
改良された薬物動態を有するFc−エリスロポエチン融合タンパク質
【課題】Fc部分に加えEPO部分に改変を含み、改善された薬物動態を有する高度にシアル酸化されたFc-EPO融合タンパク質を提供する。
【解決手段】ヒンジ領域、CH2及びCH3ドメインを含むヒトIgG分子の二量体Fc部分並びにヒトエリスロポエチン(EPO)から本質的になる精製された二量体融合タンパク質であって、前記二量体Fc部分の各鎖はそのC-末端でEPO分子のN-末端へ直接又はリンカーペプチドを介して連結しており、下記の特徴を有する前記二量体融合タンパク質:
(i) 15〜28個のシアル酸残基を有することにより高度にシアル酸化されている;
(ii) CH2領域がヒトIgG2に由来し、前記CH2領域中のGln-Phe-Asn-Ser配列内のPheおよびAsnがAlaおよびAsnで置換され、その結果前記CH2領域内に配列Gln-Ala-Gln-Serが形成されることによって前記CH2領域が改変されている;および、
(iii)CH3ドメインのC-末端近傍のLeu-Ser-Leu-Serアミノ酸配列が、Ala-Thr-Ala-Thrで置換されている。
【解決手段】ヒンジ領域、CH2及びCH3ドメインを含むヒトIgG分子の二量体Fc部分並びにヒトエリスロポエチン(EPO)から本質的になる精製された二量体融合タンパク質であって、前記二量体Fc部分の各鎖はそのC-末端でEPO分子のN-末端へ直接又はリンカーペプチドを介して連結しており、下記の特徴を有する前記二量体融合タンパク質:
(i) 15〜28個のシアル酸残基を有することにより高度にシアル酸化されている;
(ii) CH2領域がヒトIgG2に由来し、前記CH2領域中のGln-Phe-Asn-Ser配列内のPheおよびAsnがAlaおよびAsnで置換され、その結果前記CH2領域内に配列Gln-Ala-Gln-Serが形成されることによって前記CH2領域が改変されている;および、
(iii)CH3ドメインのC-末端近傍のLeu-Ser-Leu-Serアミノ酸配列が、Ala-Thr-Ala-Thrで置換されている。
【発明の詳細な説明】
【技術分野】
【0001】
発明の技術分野
本発明は、改良された薬物動態を有する新規な、通例として高度にシアル酸化されたFc-EPO融合タンパク質を提供する。詳細に述べると、Fc-EPOタンパク質は延長された血清半減期及び増大したin vivo効能を有する。BHK細胞において合成されたFc-EPO融合タンパク質は、例えば、NS/0細胞などの他の細胞株において産生された対応するFc-EPO融合タンパク質と比較した場合に、劇的に延長された血清半減期及び増加したin vivo効能を有する。本発明はFc-EPOにも関し、Fc部分に加えEPO部分の種々の改変が、更に改良された特性を伴う各分子を得るために実行される。
【背景技術】
【0002】
背景
エリスロポエチンは、赤血球形成前駆細胞の赤血球への成熟に必要な糖タンパク質ホルモンである。これは、腎臓で生成され、循環内の赤血球細胞のレベルの調節に不可欠である。低レベルの組織酸素シグナルにより特徴付けられる状態は、エリスロポエチン生成を増大し、これは次に赤血球形成を刺激する。循環内のエリスロポエチンレベルは、赤血球が長期酸素欠乏にのみ反応して作られることを確実にするよう、厳密に調節されている。エリスロポエチンの70%は、受容体-媒介型エンドサイトーシスにより消失される。エリスロポエチンがその受容体へ結合する場合、この複合体はエンドサイトーシスされ、分解され、その結果シグナル伝達の範囲が制限される。エリスロポエチンの残余は、腎濾過を通じ尿中へクリアランスされる。結果的に、エリスロポエチンは、比較的短い血清半減期を有する。
【0003】
哺乳動物細胞において生成される天然のヒトエリスロポエチン又は組換えエリスロポエチンは、3個のN-結合型及び1個のO-結合型オリゴ糖鎖を含む。N-結合グリコシル化は、24、38及び83位に位置したアスパラギン残基で生じるのに対し、O-結合グリコシル化は、126位に位置したセリン残基で生じる(Laiら、J. Biol. Chem.、261:3116 (1986);Broudyら、Arch. Biochem. Biophys.、265:329 (1988))。オリゴ糖鎖は、末端シアル酸残基で改変されることが示されている。典型的にはN-結合型鎖は、1個の鎖あたり最大4個のシアル酸を有し、及びO-結合型鎖は、最大2個のシアル酸を有する。従ってエリスロポエチンポリペプチドは、合計最大14個のシアル酸を収容することができる。糖は、細胞からのエリスロポエチンの分泌、エリスロポエチン溶解度の増大、及びエリスロポエチンのin vivo生物学的活性に必要であることが示されている(Dubeら、J. Biol. Chem.、263:17516 (1988);DeLormeら、Biochemistry、31:9871-9876 (1992))。
【0004】
組換えヒトエリスロポエチンの投与は、例えば腎不全、HIV感染症、失血及び慢性疾患に関連したものを含む、様々な形の貧血のような、造血障害又は欠損の治療において有効である。エリスロポエチンは典型的には静脈内注射により投与される。エリスロポエチンは比較的短い血清半減期を有するので、循環内で治療的有効レベルのエリスロポエチン維持するためには、頻繁な静脈内注射が必要である。天然の又は組換えのヒトエリスロポエチンを含有する医薬組成物は典型的には、投与量約25〜100単位/kgで週3回投与される。この型のエリスロポエチン療法は、極めて有効であるが、非常に高価であり、かつ静脈内投与は頻繁に診療所又は病院を来院することを必要とするので不便である。現在、新規赤血球形成刺激タンパク質(NESP)である、高度にグリコシル化された組換えヒトエリスロポエチンアナログは、貧血の治療のための商標Aranesp(登録商標)(Amgen Inc.、サザンオークス、CA)として入手可能である。Aranesp(登録商標)は、同じ生物学的反応を得るために、通常のエリスロポエチンよりも少ない頻度で投与することができる。
【0005】
別の投与経路は皮下注射である。この投与形態は患者が自宅で行うことができ、及び投与部位からのより遅い吸収を提供し、その結果持続放出作用を引き起こす徐放製剤により適合している。しかし有意に低い循環レベルが皮下注射により実現され、その結果望ましい治療作用を実現するためには頻繁な注射が必要である。更にタンパク質製剤の皮下投与は一般に静脈内投与よりもより免疫原性である。その理由は、感染の主障壁としての皮膚は樹状細胞が多い免疫臓器であり、擦過傷及び異物を同定しかつ反応するための感度の高い機構を有するからである。Casadevallらは最近、エリスロポエチンを皮下投与された患者は、抗-エリスロポエチン抗体を発生したことを報告した(Casadevallら、N Engl. J. Med.、346(7): 469-75 (2002))。
従って、投与頻度の少ないより有効なエリスロポエチン療法が必要である。
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明は、様々な態様において、野生型もしくは天然のエリスロポエチン、組換えエリスロポエチン、又は高グリコシル化されたエリスロポエチンアナログNESP(PCT国際公開公報第00/24893号)と比べ、改善された薬物動態を有するエリスロポエチン融合タンパク質を提供する。従って、エリスロポエチン療法を簡略化し、並びに造血障害又は欠損又は他のエリスロポエチン投与の適応症を伴うヒト又は他の哺乳動物の治療に関する経費を削減することは、本発明の目的である。
【0007】
詳細に述べると本発明は、延長された血清半減期及び増加したin vivo効能を有する、生物学的活性のあるFc-エリスロポエチン(Fc-EPO)融合タンパク質を提供する。
本明細書において「Fc-EPO融合タンパク質」は、Fc部分及びエリスロポエチン部分を有するポリペプチドを含むタンパク質を意味する。本明細書において「Fc部分」は、免疫グロブリン、好ましくはヒト免疫グロブリンの定常領域に由来したドメインを包含し、これは定常領域の断片、アナログ、変種、変異体又は誘導体を含む。本明細書において「エリスロポエチン部分」は、ヒト及び他の種由来の野生型又は天然のエリスロポエチン、組換えエリスロポエチン、及びエリスロポエチン-様分子を包含し、これは生物学的活性のあるエリスロポエチン断片、エリスロポエチンのアナログ、変種、変異体又は誘導体を含む。
【課題を解決するための手段】
【0008】
本発明は、以下のことに関する:
ヒンジ領域、CH2及びCH3ドメインを含むヒトIgG分子の二量体Fc部分、並びにヒトエリスロポエチン(EPO)から本質的になる精製された二量体融合タンパク質であって、前記二量体Fc部分の各鎖がそのC-末端においてEPO分子のN-末端へ直接又はリンカーペプチドを介して連結しており、下記の特徴を有する、前記融合タンパク質:(i) 15〜28個のシアル酸残基を含むことにより、高度にシアル酸化されている;(ii)CH2ドメインがヒトIgG2に由来し、CH2ドメインの Gln-Phe-Asn-Ser配列内のアミノ酸残基Phe及びAsnが、Ala及びAsnで置換され、その結果CH2ドメイン内に配列Gln-Ala-Gln-Serが形成されることによってCH2ドメインが改変されている;および、(iii)CH3ドメインのC-末端近傍のLeu-Ser-Leu-Serアミノ酸配列がAla-Thr-Ala-Thrで置換されている。
【図面の簡単な説明】
【0009】
【図1A】ヒトIgG1、IgG2及びIgG4の定常領域のアミノ酸配列のアラインメントを示す。
【図1B】図1A続き。
【図2】図2は、α相において、Fc-EPO投与量とFc-EPO血清濃度の減少量の間の相関関係を示すマウスにおける薬物動態実験を示す。この実験においては、NS/0細胞において合成された過小(under)シアル酸化されたFc-EPO変種が使用された。
【図3】図3は、Fc-EPO融合タンパク質の可能性のある消失経路、及びこれらの経路を変調する可能性のある融合タンパク質への改変を示す。
【図4】図4は、Fcg2h(FN>AQ)-EPO投与後の、マウスにおけるヘマトクリット反応の例を示す。
【図5】図5は、BHK細胞で生成されたFcg2h-EPO、Fcg2h-EPO(NDS)、Fcg4h-EPO、及びFcg4h(N>Q)-EPOタンパク質投与後の、ラットにおけるヘマトクリット反応の例を示す。
【図6】図6は、BHK細胞で生成されたFcg2h-EPO(NDS)、NS/0細胞で生成されたFcg2h-EPO(NDS)、及びNESP(すなわちAranesp(登録商標))投与後の、マウスにおけるヘマトクリット反応の例を示す。
【図7】図7は、成熟型Fc-EPOタンパク質をコードしている核酸配列の例を示す。
【図8】図8は、マウスにおけるBHK細胞から生成されたFcg2h(N>Q)-EPO及びNS/0細胞から生成されたFcg2h(N>Q)-EPOの薬物動態プロファイルを示す。
【図9】図9は、マウスにおけるBHK細胞から生成されたFcg2h-EPO(NDS)及びNS/0細胞から生成されたFcg2h-EPO(NDS)の薬物動態プロファイルを示す。
【図10】図10は、マウスにおけるBHK-21細胞、PERC6細胞、及び293細胞において生成されたFcg2h-EPO(NDS)タンパク質の薬物動態プロファイルを示す。
【図11】図11は、BHK細胞において合成されたFcg2h(FN→AQ)-EPOタンパク質による処置後の、ビーグル犬におけるヘマトクリット反応を示す。
【発明を実施するための形態】
【0010】
ひとつの局面において、本発明は、BHK細胞において合成されたFc-EPOタンパク質を提供する。本発明のBHK細胞において合成されたFc-EPO融合タンパク質は、例えばNS/0、PerC6、又は293細胞などの、他の細胞株において生成された対応するFc-EPO融合タンパク質と比較した場合に、劇的に延長された血清半減期及び増加したin vivo効能が明らかにされている。本発明は、哺乳動物への投与に適した、高度にシアル酸化されたFc-EPO融合タンパク質の集団も提供する。高度にシアル酸化されたFc-EPO融合タンパク質は、様々な態様において、野生型又は天然のエリスロポエチン、組換えエリスロポエチン、高グリコシル化されたエリスロポエチンアナログNESP、又はNS/0、PerC6、もしくは293細胞で合成された同じアミノ酸配列のFc-EPO融合タンパク質と比較して、より長い血清半減期及び増加したin vivo効能を有する。本発明に従い、Fc-EPO融合タンパク質は、一般にFc融合タンパク質の血清半減期を延長するようなFc部分のアミノ酸改変を含むことができる。例えば、このようなアミノ酸改変は、Fc受容体結合活性又は補体結合活性を実質的に減少又は除去する変異を含む。加えて、Fc-EPO融合タンパク質は、EPO受容体-媒介したエンドサイトーシスを減少するか又はエリスロポエチンの生物学的活性を増加するようなエリスロポエチン部分のアミノ酸改変も含む。様々な態様において、本発明は、免疫グロブリン融合タンパク質、Fc部分及びエリスロポエチン部分のアミノ酸改変、並びにBHK細胞における生成(例えば、高レベルのシアル酸化)によりもたらされる恩恵を組合せている。これらの組合せた恩恵は、驚くほど延長された血清半減期及び増加したin vivo効能を伴うFc-EPO融合タンパク質を生じる、相加作用又は相乗作用を有する。
【0011】
従って本発明は、ひとつの局面において、Fc-EPO融合タンパク質をコードしている核酸配列を含むBHK細胞に関する。ひとつの態様において、本発明のBHK細胞は、無タンパク質培地における増殖に適合している。別の態様において、BHK細胞は、懸濁液における増殖に適合している。更に別の態様において、BHK細胞は、無タンパク質培地中で懸濁液における増殖に適合している。無タンパク質培地中で増殖したBHK細胞から生成されたFc-EPO融合タンパク質は、他の培地において増殖されたBHK細胞から生成されたFc-EPO融合タンパク質と比べ、驚くほど増加した及びより均質なシアル酸化を示すことがわかっている。好ましい態様において、核酸はBHK細胞において安定して維持される。本明細書において「安定して維持される核酸」とは、抗生物質-ベースの選択のような核酸を維持するための選択圧の非存在下で、母細胞から娘細胞への喪失率が3%未満の核酸を意味する。従って核酸を安定して維持する細胞が分裂する場合、得られる細胞の少なくとも97%(より好ましくは、98%よりも多く、99%よりも多く、又は99.5%よりも多く)がその核酸を含む。その核酸を含む得られる細胞が分裂する場合、(2回目の)分裂から得られる細胞の少なくとも97%がその核酸を含むであろう。更に1個の細胞当たりの核酸のコピー数は、繰返しの細胞分裂により実質的に低下しない。好ましい態様において、安定して維持される核酸配列はBHK細胞の染色体へ組込まれる。
【0012】
核酸配列は、様々な配置のいずれでもFc-EPO融合タンパク質をコードすることができる。好ましい態様において、核酸配列はFc-EPO融合タンパク質のN-末端側のFc部分及びFc-EPO融合タンパク質のC-末端側のエリスロポエチン部分を含むFc-EPO融合タンパク質をコードしている。Fc部分は一般に、免疫グロブリンの定常領域由来の領域を包含し、これは定常領域の断片、アナログ、変種、変異体又は誘導体を含む。好ましい態様において、Fc部分は、ヒト免疫グロブリン重鎖、例えば、IgG1、IgG2、IgG3、IgG4、又は他のクラスに由来する。一部の態様において、本Fc-EPO融合タンパク質は免疫グロブリンの可変領域を含まない。ひとつの態様において、Fc部分は、CH2ドメインを含む。別の態様において、Fc部分はCH2及びCH3ドメインを含む。
【0013】
好ましい態様において、Fc部分は、Fc受容体への親和性を低下するか、又はFcエフェクター機能を低下させる変異を含む。例えばFc部分は、IgG重鎖のFc部分内のグリコシル化部位を除去する変異を含むことができる。一部の態様において、Fc部分はIgG1のLeu234、Leu235、Gly236、Gly237、Asn297、又はPro331に対応するアミノ酸位置での変異、欠失、又は挿入を含む(アミノ酸は、EU命名法に従い番号付けられる)。好ましい態様において、Fc部分は、IgG1のAsn297に対応するアミノ酸位置に変異を含む。別の態様において、Fc部分はIgG1のLeu281、Leu282、Gly283、Gly284、Asn344、又はPro378に対応するアミノ酸位置に変異、欠失、又は挿入を含む。
【0014】
一部の態様において、Fc部分はヒトIgG2又はIgG4重鎖由来のCH2ドメインを含む。好ましくは、CH2ドメインは、CH2ドメイン内のグリコシル化部位を除去する変異を含む。ひとつの態様において、変異は、IgG2又はIgG4重鎖のCH2ドメイン内のGln-Phe-Asn-Serアミノ酸配列内のアスパラギンを変更する。好ましくは、変異は、アスパラギンをグルタミンへ変更する。あるいは、変異は、Gln-Phe-Asn-Serアミノ酸配列内のフェニルアラニン及びアスパラギンの両方を変更する。ひとつの態様において、Gln-Phe-Asn-Serアミノ酸配列はGln-Ala-Gln-Serアミノ酸配列で置換される。
【0015】
Gln-Phe-Asn-Serアミノ酸配列内のアスパラギンは、IgG1のAsn297に対応している。IgG2又はIgG4のGln-Phe-Asn-Serアミノ酸配列内のアスパラギンの変異(すなわちIgG1のAsn297に対応)も同じくFc-EPO融合タンパク質のEPO受容体への結合を驚くほど低下させることがわかっている。理論に拘束されることを欲するものではないが、IgG2又はIgG4のGln-Phe-Asn-Serアミノ酸配列内のアスパラギンの変異(すなわちIgG1のAsn297に対応)は、Fc-EPO融合タンパク質の全般的立体構造の変化を誘導し、これは劇的に改善された薬物動態特性につながるのであろう。
【0016】
別の態様において、Fc部分は、CH2ドメイン及びヒンジ領域の少なくとも一部を含む。ヒンジ領域は、免疫グロブリン重鎖、例えば、IgG1、IgG2、IgG3、IgG4、又は他のクラスに由来することができる。好ましくは、ヒンジ領域は、ヒトIgG1、IgG2、IgG3、IgG4、又は他の適当なクラスに由来する。より好ましくは、ヒンジ領域は、ヒトIgG1重鎖に由来する。ひとつの態様において、IgG1ヒンジ領域のPro-Lys-Ser-Cys-Asp-Lysアミノ酸配列内のシステインが改変される。好ましい態様において、Pro-Lys-Ser-Cys-Asp-Lysアミノ酸配列がPro-Lys-Ser-Ser-Asp-Lysアミノ酸配列で置換される。ひとつの態様において、Fc部分は、第一の抗体アイソタイプ由来のCH2ドメイン及び第二の抗体アイソタイプ由来のヒンジ領域を含む。特定の態様において、CH2ドメインは、ヒトIgG2又はIgG4重鎖に由来するが、ヒンジ領域は改変されたヒトIgG1重鎖に由来する。
【0017】
好ましい態様において、Fc部分は、定常領域のC-末端近傍のLeu-Ser-Leu-Serアミノ酸配列が、接合部の潜在的なT-細胞エピトープを除去するように変更されている、IgG配列に由来する。例えばひとつの態様において、Leu-Ser-Leu-Serアミノ酸配列はAla-Thr-Ala-Thrアミノ酸配列で置換される。別の態様において、Fc部分は、C-末端リジン残基が置換されたIgG配列に由来する。好ましくは、IgG配列のC-末端リジンは、アラニンのような非-リジンアミノ酸により交換され、Fc融合タンパク質の血清半減期を更に増加する。
【0018】
本発明に従い、Fc部分は、本明細書に記載された1個又は複数の変異を含むことができる。Fc部分の変異の組合せは一般に、Fc-EPO融合タンパク質の延長された血清半減期及び増加されたin vivo効能に対して相加作用又は相乗作用を有する。従って態様の一例において、Fc部分は、(i)Lys-Ser-Lys-Serアミノ酸配列がAla-Thr-Ala-Thrアミノ酸配列に置換されたIgG配列由来の領域;(ii)リジンの代わりにC-末端アラニン残基;(iii)異なる抗体アイソタイプ由来の、CH2ドメイン及びヒンジ領域、例えばIgG2 CH2ドメイン及び代わりのIgG1ヒンジ領域;(iv)IgG2-由来のCH2ドメイン内のグリコシル化部位を除去する変異、例えばIgG2-由来のCH2ドメイン内のGln-Phe-Asn-Serアミノ酸配列の代わりにGln-Ala-Gln-Serアミノ酸配列:を含む。
【0019】
Fc-EPO融合タンパク質のエリスロポエチン部分は、完全長の野生型又は天然のエリスロポエチン、組換えエリスロポエチン、又はエリスロポエチン-様分子、例えば生物学的活性のあるエリスロポエチン断片、エリスロポエチンのアナログ、変種、変異体又は誘導体であることができる。好ましくは、エリスロポエチン部分は、ヒトエリスロポエチンに由来する。一部の態様において、エリスロポエチン部分は、EPO受容体への結合親和性を低下するか又はエリスロポエチンの生物学的活性を増加するようなアミノ酸改変を含むことができる。一部の態様において、エリスロポエチン部分は、以下の変異を少なくとも1種含む:Arg131→Glu及びArg139→Glu(アミノ酸番号付けは、成熟型ヒトエリスロポエチン配列を基にしている)。別の態様において、エリスロポエチン部分は、以下の変異を少なくとも1種含む:His32→Gly、Ser34→Arg、及びPro90→Ala。更に別の態様において、エリスロポエチン部分は、ヒトエリスロポエチンとは異なるジスルフィド結合のパターンを有する。例えばエリスロポエチン部分は、以下のアミノ酸置換の1種又は複数を含むことができる:29位の非-システイン残基、33位の非-システイン残基、88位のシステイン残基、及び139位のシステイン残基。ひとつの態様において、エリスロポエチン部分は、7、29、88、及び161位にシステイン残基を含む。別の態様において、エリスロポエチン部分は、更に、下記の置換を1個又は複数含む:His32→Gly、Cys33→Pro、及びPro90→Ala。本発明に従い、エリスロポエチン部分は、本明細書に説明された変異のいずれかの組合せを含むことができる。
【0020】
一部の態様において、Fc-EPO融合タンパク質はFc部分とエリスロポエチン部分の間にリンカーを含む。リンカーが含まれるならば、これは一般に、1〜25個のアミノ酸を含み、プロテアーゼ切断部位を含まないことが好ましい。このリンカーは、タンパク質分解を阻害するために、N-結合型された又はO-結合型されたグリコシル化部位を含むことができる。例えばひとつの態様において、リンカーは、Asn-Ala-Thrアミノ酸配列を含む。
【0021】
本発明は更に、Fc-EPO融合タンパク質を生産する方法にも関する。この方法は、Fc-EPO融合タンパク質をコードしている核酸配列を含むBHK細胞を、コードされたFc-EPO融合タンパク質の発現に適した条件下で維持する工程、及び発現されたFc-EPO融合タンパク質を回収する工程を含む。ひとつの態様において、BHK細胞は、無タンパク質培地中で培養される。別の態様において、BHK細胞は懸濁物として培養される。更に別の態様において、BHK細胞は無タンパク質培地中で懸濁培養される。一部の態様において、この核酸は、BHK細胞において安定して維持される。一般に、BHK細胞において生成されたFc-EPO融合タンパク質は、例えば、NS/0、PerC6、もしくは293細胞などの他の細胞株において生成された対応するFc-EPO融合タンパク質よりも、より長い血清半減期を有する。
【0022】
本発明は、BHK細胞において生成されたFc-EPO融合タンパク質を含有する医薬組成物を提供する。好ましい態様において、医薬組成物において使用されるFc-EPO融合タンパク質は、シアル酸残基を除去するようには処理されない。この医薬組成物は、医薬として許容できる担体も含む。本発明は、医薬組成物を哺乳動物へ投与することにより、哺乳動物を治療する方法も提供する。一部の態様において、治療される哺乳動物は、造血障害又は欠損を有する。本発明のFc-EPO融合タンパク質は、増加したin vivo効能及び延長された血清半減期を有するので、Fc-EPO融合タンパク質を含有する医薬組成物は一般に、他の細胞において生成された天然のもしくは組換えエリスロポエチン又は対応するFc-EPO融合タンパク質を含有する医薬組成物と比べ、より少ない投与頻度しか必要としない。好ましい態様において、この医薬組成物は、週3回よりも少ない頻度(例えば、週2回、毎週、又は多くて10日に1回、例えば2週間に1回、1ヶ月に1回、又は2ヶ月に1回など)で投与される。
【0023】
別の局面において、本発明は、Fc部分及びエリスロポエチン部分を含む融合タンパク質をコードしている核酸を安定して維持するBHK細胞を選択する方法を提供する。この方法は、ハイグロマイシンBをコードしている核酸配列及びこの融合タンパク質をコードしている核酸配列をBHK細胞へ導入する工程;および、ハイグロマイシンBの存在下で前記BHK細胞を培養する工程を含む。ひとつの態様において、ハイグロマイシンBをコードしている核酸配列及び融合タンパク質をコードしている核酸配列は、単一の核酸中に存在する。別の態様において、ハイグロマイシンBをコードしている核酸配列及びこの融合タンパク質をコードしている核酸配列は、2つの個別の核酸内に存在する。
【0024】
別の局面において、本発明は、哺乳動物への投与に適した精製されたFc-EPO融合タンパク質の集団を提供する。好ましい態様において、Fc-EPO融合タンパク質は、Fc-EPO融合タンパク質のN-末端側にFc部分、及びFc-EPO融合タンパク質のC-末端側にエリスロポエチン部分を含む。より好ましい態様において、精製されたFc-EPO融合タンパク質の集団は、高度にシアル酸化され、すなわち精製されたFc-EPO融合タンパク質あたり平均11〜28個のシアル酸残基を有する。好ましいFc-EPO融合タンパク質の高度にシアル酸化された集団は、精製されたFc-EPO融合タンパク質あたり平均13〜28、15〜28、17〜28、19〜28、又は21〜28個のシアル酸残基を有する。例えばひとつの好ましいFc-EPO融合タンパク質の高度にシアル酸化された集団は、精製されたFc-EPO融合タンパク質あたり平均20〜22個のシアル酸残基を有する。好ましい態様において、精製されたFc-EPO融合タンパク質はBHK細胞において合成される。ひとつの態様において、BHK細胞は、懸濁物としての増殖に適応している。別の態様において、BHK細胞は無タンパク質培地中の増殖に適応している。更に別の態様において、BHK細胞は無タンパク質培地中で懸濁物としての増殖に適応している。本発明により提供される精製されたFc-EPO融合タンパク質の高度にシアル酸化された集団は、例えば、NS/0、PerC6、もしくは293細胞などの細胞において生成された対応するFc-EPO融合タンパク質の集団と比べ、より長い血清半減期を有する。本発明に従い、精製されたFc-EPO融合タンパク質のFc部分及びエリスロポエチン部分は、本明細書に記載されたような1個又は複数の変異又は改変を含み、増強されたシアル酸化による相加作用又は相乗作用により、延長された血清半減期及び増加したin vivo効能を提供することができる。
【0025】
本発明は、本明細書に記載された精製されたFc-EPO融合タンパク質の高度にシアル酸化された集団を含有する医薬組成物も提供する。好ましい医薬組成物は更に医薬として許容できる担体を含む。本発明は更に、哺乳動物へ、精製されたFc-EPO融合タンパク質の高度にシアル酸化された集団を含有する医薬組成物を投与することを含む、哺乳動物を治療する方法を提供する。好ましい態様において、本医薬組成物は1週間に3回よりも少なく投与される(例えば、週2回、週1回、又は多くて10日に1回、例えば2週間に1回、1ヶ月に1回又は2ヶ月に1回など)。
【0026】
まとめると、本発明は、以下のことに関する:
・ヒンジ領域、CH2及びCH3ドメインを含むヒトIgG分子の二量体Fc部分、並びにヒトエリスロポエチン(EPO)から本質的になる精製された二量体融合タンパク質であって、前記二量体Fc部分の各鎖がそのC-末端においてEPO分子のN-末端へ直接又はリンカーペプチドを介して連結しており、下記の特徴を有する、前記融合タンパク質:(i) 15〜28個のシアル酸残基を含むことにより、高度にシアル酸化されている;(ii)CH2ドメインがヒトIgG2に由来し、CH2ドメインの Gln-Phe-Asn-Ser配列内のアミノ酸残基Phe及びAsnが、Ala及びAsnで置換され、その結果CH2ドメイン内に配列Gln-Ala-Gln-Serが形成されることによってCH2ドメインが改変されている;および、(iii)CH3ドメインのC-末端近傍のLeu-Ser-Leu-Serアミノ酸配列がAla-Thr-Ala-Thrで置換されている。
【0027】
・更にCH3ドメインのC-末端Lys残基がAlaに置換されている、各二量体Fc-EPO融合タンパク質。
・ヒンジ領域がヒトIgG1に由来する、各二量体Fc-EPO融合タンパク質。
・IgG1ヒンジ領域の Pro-Lys-Ser-Cys-Asp-Lys配列内のCysアミノ酸残基がSer残基によって置換され、その結果前記ヒンジ領域内に配列Pro-Lys-Ser-Ser-Asp-Lysが形成されることによってIgG1ヒンジ領域が改変されている、各二量体Fc-EPO融合タンパク質。
【0028】
・エリスロポエチン部分が以下のアミノ酸置換の少なくとも1種を含む、各二量体Fc-EPO融合タンパク質:
(i)EPO分子の29位の非-システイン残基、
(ii)EPO分子の33位の非-システイン残基、
(iii)EPO分子の88位のシステイン残基、及び
(iv)EPO分子の139位のシステイン残基。
【0029】
・EPO分子の元のCys残基の代わりにEPO分子の33位に非-Cysアミノ酸残基があり、元のEPO分子の88位にTrp残基の代わりCys残基があり、その結果融合タンパク質内のEPO部分が、Cys29-Cys88ジスルフィド結合を形成することが可能である、各二量体Fc-EPO融合タンパク質。
・33位の非-Cysアミノ酸残基がProである、各二量体Fc-EPO融合タンパク質。
・EPO部分が以下の群から選択される1種又は複数の変異を含む、各二量体Fc-EPO融合タンパク質:
(i)Arg131→Glu131
(ii)Arg139→Glu139
(iii)His32→Gly32
(iv)Ser34→Arg34
(v)Pro90→Ala90。
【0030】
・リンカーペプチドがグリコシル化部位を含む、各二量体Fc-EPO融合タンパク質。
・グリコシル化部位がAsn-Ala-Thrアミノ酸配列を含む、各二量体Fc-EPO融合タンパク質。
・更にCH1ドメインを含む、各二量体Fc-EPO融合タンパク質。
・CH2、CH3及び場合によりCH1を含む完全なIgG分子がIgG2に由来し、ヒンジ領域がIgG1に由来する、各Fc-EPO融合タンパク質。
・CH2、CH3及びヒンジ領域および場合によりCH1を含む完全なIgG分子がIgG1に由来する、
各Fc-EPO融合タンパク質。
・18〜24個、好ましくは20〜22個のシアル酸残基を有する、各二量体Fc-EPO融合タンパク質。
【0031】
・以下の配列を含む、二量体Fc-EPO融合タンパク質:
EPKSSDKTHTCPPCPAPPVAGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVQFNWYVDGVEVHNAKTKPREEQAQSTFRVVSVLTVVHQDWLNGKEYKCKVSNKGLPAPIEKTISKTKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPMLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSATATPGAAPPRLICDSRVLERYLLEAKEAENITTGCAEHCSLNENITVPDTKVNFYAWKRMEVGQQAVEVWQGLALLSEAVLRGQALLVNSSQPWEPLQLHVDKAVSGLRSLTTLLRALGAQKEAISPPDAASAAPLRTITADTFRKLFRVYSNFLRGKLKLYTGEACRTGDR (配列番号:14)。
・以下の配列を含む、二量体Fc-EPO融合タンパク質:
EPKSSDKTHTCPPCPAPPVAGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVQFNWYVDGVEVHNAKTKPREEQAQSTFRVVSVLTVVHQDWLNGKEYKCKVSNKGLPAPIEKTISKTKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPMLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSATATPGAAPPRLICDSRVLERYLLEAKEAENITTGCAEGPSLNENITVPDTKVNFYAWKRMEVGQQAVEVWQGLALLSEAVLRGQALLVNSSQPCEALQLHVDKAVSGLRSLTTLLRALGAQKEAISPPDAASAAPLRTITADTFRKLFRVYSNFLRGKLKLYTGEACRTGDR (配列番号:15)。
【0032】
・先に特定した融合タンパク質をコードするDNA分子。
・先に特定したか又は請求項のFc-EPO融合タンパク質を有効量含み、場合により医薬として許容できる担体、希釈剤又は賦形剤を含有する、哺乳動物における造血障害または欠損の治療に適した医薬組成物。
・Fc-EPO融合タンパク質のN-末端側にFc部分、及びFc-EPO融合タンパク質のC-末端側にエリスロポエチン部分を含むFc-EPO融合タンパク質の集団であって、精製されたFc-EPO融合タンパク質あたり平均15〜28個のシアル酸残基を有し、各Fc-EPO融合タンパク質をコードするDNA分子をBHK細胞へ導入し、対応するFc-EOP融合タンパク質を発現、単離及び精製することによって得られ、NS/0、PerC6、又は293細胞において合成された対応するFc-EPO融合タンパク質の集団と比較して、より長い血清半減期を有する、哺乳動物への投与に適した精製された高度にシアル酸化されたFc-EPO融合タンパク質の集団。
【0033】
・精製されたFc-EPO融合タンパク質あたり平均20〜22個のシアル酸残基を有する、前記精製されたFc-EPO融合タンパク質の対応する集団。
・BHK細胞が、無タンパク質培地又は懸濁液中で増殖するのに適応している、精製されたFc-EPO融合タンパク質の対応する集団。
【0034】
・Fc-EPO融合タンパク質のN-末端側にFc部分、及びFc-EPO融合タンパク質のC-末端側にエリスロポエチン部分を含む、高度にシアル酸化された精製された組換えFc-EPO融合タンパク質の集団を作製する方法であって:
(i)Fc-EPO融合タンパク質をコードしているDNA分子を構築する工程;
(ii)無タンパク質培地又は懸濁液中において該DNA分子でBHK細胞を形質転換する工程;
(iii)該DNA分子によりコードされたFc-融合タンパク質の集団を発現する工程、
(iv)Fc-EPO融合タンパク質の集団を収集、単離及び精製する工程、
を含む前記方法。
【0035】
・前記合成された融合タンパク質の集団が、精製されたFc-EPO融合タンパク質あたり平均15〜28、好ましくは15〜25、より好ましくは20〜22個のシアル酸残基を有する、対応する方法。
・Fc部分及びエリスロポエチン部分を含むFc-EPO融合タンパク質をコードする核酸配列を安定して維持しているBHK細胞を選択する方法であって:
(a)ハイグロマイシンBをコードしている核酸配列及びFc-EPO融合タンパク質をコードしている核酸配列をBHK細胞へ導入する工程;及び
(b)ハイグロマイシンBの存在下で前記BHK細胞を培養する工程、
を含む前記方法。
・ハイグロマイシンBをコードしている核酸配列及びFc-EPO融合タンパク質をコードしている核酸配列が単一のDNA分子内に存在する、対応する方法。
【0036】
発明の詳細な説明
本発明は、改善された薬物動態を伴う、Fc-EPO融合タンパク質を提供する。特に、本発明により提供されるFc-EPOタンパク質は、延長された血清半減期及び増加したin vivo効能を有する。ひとつの局面において、本発明は、BHK細胞において合成されたFc-EPO融合タンパク質を提供する。BHK細胞において合成されたFc-EPO融合タンパク質は、例えばNS/0、PerC6、又は293細胞などの他の細胞株において生成された対応するFc-EPO融合タンパク質と比べた場合に、劇的に延長された血清半減期及び増加したin vivo効能を有する。別の局面において、本発明は、高度にシアル酸化されたFc-EPO融合タンパク質の集団を提供する。高度にシアル酸化されたFc-EPO融合タンパク質の集団は、より少ないレベルのシアル酸化を伴う対応するFc-EPO融合タンパク質の集団と比べ、より長い血清半減期を有する。本発明に従い、Fc-EPO融合タンパク質は、Fc受容体結合活性の実質的減少もしくは除去又は補体結合活性を低下する改変などにより、Fc融合タンパク質の血清半減期を延長するFc部分のアミノ酸改変を含むことができる。加えてFc-EPO融合タンパク質は、EPO受容体-媒介型エンドサイトーシスを減少するか又はエリスロポエチンの生物学的活性を増加するエリスロポエチン部分のアミノ酸改変も含む。
【0037】
Fc-EPO融合タンパク質
本明細書において「Fc-EPO融合タンパク質」は、そのポリペプチドに通常は存在しない少なくともふたつの部分、すなわちFc部分及びエリスロポエチン部分を有するポリペプチドを含むタンパク質を意味する。本発明の好ましい態様において、Fc部分及びエリスロポエチン部分を有するポリペプチドは、ホモ二量体を形成し;従って、Fc-EPO融合タンパク質は一般に、1個又は複数のジスルフィド結合により一緒に保持された二量体タンパク質であり、各ポリペプチド鎖は、Fc部分及びエリスロポエチン部分を含む。しかし本発明のFc-EPO融合タンパク質は、エリスロポエチン部分が、Fc部分と安定して会合することを可能にしつつ、エリスロポエチン活性を維持するようないかなる配置をも有し得る。例えばこのような配置は、ふたつのFc部分及びふたつのエリスロポエチン部分を含む単一のポリペプチド、ふたつのFc部分及びひとつのエリスロポエチン部分を含む単一のポリペプチド、Fc部分及びエリスロポエチン部分を含むひとつのポリペプチドと、Fc部分を含む別のポリペプチドを含むヘテロ二量体タンパク質、並びに他の適当な配置を含むが、これらに限定されるものではない。
【0038】
エリスロポエチン部分は、様々な配置で直接又は間接にFc部分へ連結することができる。ひとつの態様において、エリスロポエチン部分は、Fc部分へ、共有結合を介して直接連結される。例えばエリスロポエチン部分は、そのC-末端又はそのN-末端のいずれかで、Fc部分へ直接融合することができる。ひとつの態様において、Fc部分のC-末端は、エリスロポエチン部分のN-末端へ融合されており、すなわち、Nterm-Fc-Cterm-Nterm-EPO-Ctermである。この配置において、Fc部分は、Fc-EPO融合タンパク質のN-末端側にあり、エリスロポエチン部分はC-末端側にある。別の態様において、エリスロポエチンのC-末端は、Fc部分のN-末端へ融合され、すなわち、Nterm-EPO-Cterm-Nterm-Fc-Ctermである。この配置において、エリスロポエチン部分は、Fc-EPO融合タンパク質のN-末端側にあり、Fc部分はC-末端側にある。
【0039】
別の態様において、エリスロポエチン部分はFc部分へ間接的に連結される。例えば、Fc-EPO融合タンパク質は、Fc部分とエリスロポエチン部分の間に、リンカー(L)を含むことができる。直接融合と同様に、エリスロポエチン部分は好ましくは、Fc部分のC-末端へリンカーを介して融合され、すなわち、Nterm-Fc-Cterm-L-Nterm-EPO-Ctermである。従ってFc部分は、Fc-EPO融合タンパク質のN-末端へ側にあり、C-末端側のエリスロポエチン部分からリンカーにより隔てられている。あるいは、エリスロポエチン部分は、Fc部分のN-末端へリンカーを介して融合されており、すなわち、Nterm-EPO-Cterm-L-Nterm-Fc-Ctermである。
【0040】
Fc部分
本明細書において、「Fc部分」は免疫グロブリン、好ましくはヒト免疫グロブリンの定常領域由来のドメインを包含し、これは定常領域の断片、アナログ、変種、変異体又は誘導体を含む。適当な免疫グロブリンは、IgG1、IgG2、IgG3、IgG4、及び他のクラスを含む。免疫グロブリンの定常領域は、免疫グロブリンC-末端領域と相同な天然又は合成により生成されたポリペプチドと定義され、これはCH1ドメイン、ヒンジ、CH2ドメイン、CH3ドメイン、又はCH4ドメインを、個別に又はいずれかの組合せで含むことができる。ヒトIgG1、IgG2及びIgG4の定常領域の配列アラインメントを、図1A及び1Bに示している。Paulの論文(Fundamental Immunology、第4版、Lippincott-Raven、(1999))によると、CH1ドメインは、アミノ酸118-215を含み;ヒンジ領域は、アミノ酸216-230を含み;CH2ドメインは、アミノ酸231-340を含み;及び、CH3ドメインはアミノ酸341-447を含む(アミノ酸位置はIgG1配列を基にしている)。ヒンジ領域はCH1ドメインをCH2及びCH3ドメインへ連結する。
【0041】
本発明において、Fc部分は典型的には、少なくともCH2ドメインを含む。例えばFc部分はヒンジ-CH2-CH3を含む。あるいはFc部分は、ヒンジ領域、CH2ドメイン及び/又はCH3ドメインの全て又は一部を含むことができる。
免疫グロブリンの定常領域は、Fc受容体(FcR)結合及び補体結合を含む多くの重要な抗体機能に関与する。重鎖定常領域には5種の主なクラスがあり、これはIgA、IgG、IgD、IgE、及びIgMに分類され、各々、アイソタイプにより指定される特徴的エフェクター機能を伴う。例えば、IgGは4種のγサブクラス:γ1、γ2、γ3、及びγ4に分類され、これらは各々、IgG1、IgG2、IgG3、及びIgG4としても知られている。
【0042】
IgG分子は、抗体のIgGクラスに特異的なFcγ受容体(FcγR)の3種のクラス(すなわちFcγRI、FcγRII、及びFcγRIII)を含む、細胞受容体の複数のクラスと相互作用する。IgGのFcγR受容体への結合に重要な配列は、CH2及びCH3ドメインに局在することが報告されている。抗体の血清半減期は、その抗体がFc受容体(FcR)へ結合する能力により影響を受ける。同様に、免疫グロブリン融合タンパク質の血清半減期も、そのような受容体に結合する能力の影響を受ける(Gilliesら、Cancer Res.、59:2159-66 (1999))。IgG1と比べ、IgG2及びIgG4のCH2及びCH3ドメインは、生化学的に検出不可能であるか又は低下したFc受容体への結合親和性を有する。IgG2又はIgG4のCH2及びCH3ドメインを含む免疫グロブリン融合タンパク質は、IgG1のCH2及びCH3ドメインを含む対応する融合タンパク質と比べ、より長い血清半減期を有することが報告されている(米国特許第5,541,087号;Loら、Protein Engineering、11: 495-500 (1998))。従って本発明に好ましいCH2及びCH3ドメインは、例えば、IgG2又はIgG4のような、低下した受容体結合親和性及びエフェクター機能を伴う、抗体アイソタイプに由来する。より好ましいCH2及びCH3ドメインは、IgG2に由来する。
【0043】
ヒンジ領域は通常、重鎖定常領域のCH1ドメインのC-末端に位置している。IgGアイソタイプにおいて、ジスルフィド結合は典型的にはこのヒンジ領域において生じ、最終の四量体抗体分子を形成することを可能にする。この領域は、プロリン、セリン及びトレオニンが支配的である。本発明に含まれる場合、ヒンジ領域は典型的には、ふたつのFc部分を連結するジスルフィド結合を形成するためにシステイン残基を含む天然の免疫グロブリン領域と少なくとも相同である。ヒト及びマウス免疫グロブリンのヒンジ領域の代表的配列は、Borrebaeck C. A. K.編集の「Antibody Engineering, A Practical Guide」(1992)、W. H. Freeman and Co.に認められる。本発明に適したヒンジ領域は、IgG1、IgG2、IgG3、IgG4、及び他の免疫グロブリンクラスに由来することができる。IgG1ヒンジ領域は、3個のシステインを有し、その2個は免疫グロブリンの2本の重鎖間のジスルフィド結合に関連している。これらの同じシステインは、Fc部分の間に有効かつ一貫したジスルフィド結合をもたらす。従って、本発明の好ましいヒンジ領域は、IgG1に由来し、より好ましくはヒトIgG1に由来する。一部の態様において、ヒトIgG1ヒンジ領域内の第一のシステインは、好ましくは別のアミノ酸、好ましくはセリンへ変異される。IgG2アイソタイプヒンジ領域は、組換えシステムにおける分泌時にオリゴマー化及び恐らくは不正確なジスルフィド結合を促進する傾向がある4個のジスルフィド結合を有する。適当なヒンジ領域は、IgG2ヒンジに由来することができ;最初の2個のシステインは、好ましくは各々別のアミノ酸へ変異される。IgG4のヒンジ領域は、非効率的に鎖内ジスルフィド結合を形成することがわかっている。しかし本発明に適したヒンジ領域は、IgG4ヒンジ領域に由来し、好ましくは重鎖-由来の部分の間のジスルフィド結合の正確な形成を増強する変異を含む(Angal S.らMol. Immunol.、30: 105-8 (1993))。
【0044】
本発明に従い、Fc部分は、異なる抗体アイソタイプに由来する、CH2及び/又はCH3及びヒンジ領域、すなわちハイブリッドFc部分を含むことができる。例えばひとつの態様において、Fc部分は、IgG2又はIgG4に由来したCH2及び/又はCH3ドメイン並びにIgG1に由来した変異体ヒンジ領域を含む。あるいは、別のIgGサブクラス由来の変異体ヒンジ領域が、ハイブリッドFc部分において使用される。例えば、2本の重鎖の間の効率的ジスルフィド結合を可能にするIgG4ヒンジの変異体型を使用することができる。変異体ヒンジは、最初の2個のシステインが各々別のアミノ酸に変異されているIgG2ヒンジに由来することもできる。このようなハイブリッドFc部分は高-レベル発現を促進し、及びFc-EPO融合タンパク質の正確なアッセンブルを改善する。そのようなハイブリッドFc部分の集成は、米国特許公開番号第20030044423号に開示されており、(すなわち米国特許出願第10/093,958号)、その開示は本明細書に引用により取り込まれる。
【0045】
一部の態様において、Fc部分は、一般にFc融合タンパク質の血清半減期を延長するアミノ酸改変を含む。そのようなアミノ酸改変は、Fc受容体結合又は補体結合活性を実質的に減少又は除去する変異を含む。例えば免疫グロブリン重鎖のFc部分内のグリコシル化部位を除去することができる。IgG1において、このグリコシル化部位はAsn297である。別の免疫グロブリンアイソタイプにおいて、グリコシル化部位はIgG1のAsn297に相当している。例えばIgG2及びIgG4において、グリコシル化部位はアミノ酸配列Gln-Phe-Asn-Ser内のアスパラギンである。従ってIgG1のAsn297の変異はIgG1由来のFc部分のグリコシル化部位を除去する。ひとつの態様において、Asn297はGlnに置換される。同様に、IgG2又はIgG4において、アミノ酸配列Gln-Phe-Asn-Ser内のアスパラギンの変異は、IgG2又はIgG4重鎖由来のFc部分のグリコシル化部位を除去する。ひとつの態様において、アスパラギンはグルタミンに置換される。別の態様において、アミノ酸配列Gln-Phe-Asn-Ser内のフェニルアラニンは、アスパラギン変異から生じる可能性のある非-自己T-細胞エピトープを除去するように更に変異される。例えば、IgG2又はIgG4重鎖内のアミノ酸配列Gln-Phe-Asn-SerはGln-Ala-Gln-Serアミノ酸配列に置換することができる。
【0046】
Fc部分及び非-Fc部分の接合部近傍のアミノ酸の変更は、Fc融合タンパク質の血清半減期を劇的に増加することができることも観察される。(PCT国際公開公報第01/58957号、その開示は本明細書に参照として組入れられている)。従って本発明のFc-EPO融合タンパク質の接合部領域は、免疫グロブリン重鎖及びエリスロポエチンの天然の配列に対して、好ましくは接合点のアミノ酸約10個以内に位置する変更を含むことができる。これらのアミノ酸変化は、例えば、Fc部分のC-末端リジンをアラニン又はロイシンのような疎水性アミノ酸へ変えることにより、疎水性の増加を引き起こすことができる。
【0047】
別の態様において、Fc部分は免疫グロブリン重鎖のFc部分のC-末端近傍のLeu-Ser-Leu-Serセグメントのアミノ酸変更を含む。Leu-Ser-Leu-Serセグメントのアミノ酸置換は可能性のある接合部T-細胞エピトープを除去する。ひとつの態様において、Fc部分のC-末端近傍のLeu-Ser-Leu-Serアミノ酸配列はAla-Thr-Ala-Thrアミノ酸配列に置換される。別の態様において、Leu-Ser-Leu-Serセグメント内のアミノ酸は、グリシン又はプロリンなどの別のアミノ酸に置換される。IgG1、IgG2、IgG3、IgG4、又は他の免疫グロブリンクラス分子のC-末端近傍のLeu-Ser-Leu-Serセグメントのアミノ酸置換を生じる詳細な方法は、米国特許公開番号第20030166877号(すなわち、米国特許出願第10/112,582号)に開示されており、その開示は本明細書に参照として組入れられている。
【0048】
エリスロポエチン部分
本明細書において「エリスロポエチン部分」は、ヒト及び他の種に由来する野生型又は天然のエリスロポエチン、組換えエリスロポエチン、及び生物学的活性のあるエリスロポエチン断片、エリスロポエチンのアナログ、変種、変異体もしくは誘導体を含む、エリスロポエチン-様分子を包含している。
野生型又は天然のエリスロポエチンは、エリスロポエチン前駆細胞からの赤血球の増殖及び発達を刺激する、34KD糖タンパク質ホルモンである。野生型又は天然のエリスロポエチンは、低酸素症(例えば、貧血による赤血球喪失)に応答して腎臓で産生され、その同族細胞受容体との相互作用を介して、赤血球の増殖及び分化を調節する。野生型又は天然のエリスロポエチンは、血液から(Miyake T.ら、J. Biol. Chem.、252: 5558-5564 (1977))、又は血漿から(Goldwasser, E.ら、Proc. Natl. Acad. Sci. U.S.A.、68: 697-698 (1971))、又は尿から単離及び精製することができる。
【0049】
組換えにより又は化学的に合成されたエリスロポエチンは、当業者に周知の技術を用い、生成することができる。組換えヒトエリスロポエチン(rHuEPO)のふたつの型が市販されている:AmgenからEPOGEN(登録商標)、及びJohnson & JohnsonからPROCRIT(登録商標)。
【0050】
本明細書において、エリスロポエチンの生物学的活性は、エリスロポエチン受容体との相互作用を介した細胞増殖を刺激する能力として定義される。エリスロポエチンの機能アッセイは、in vitro又はin vivoにおいて行うことができる。例えば、エリスロポエチンのin vitro活性は、細胞-ベースのアッセイにおいて試験することができる。特にエリスロポエチン活性は、TF-1細胞増殖アッセイを基に決定することができる。TF-1細胞はEPO受容体を発現する。トリチウム標識されたチミジンの組込みにより決定されるTF-1細胞の増殖はエリスロポエチン活性の関数である(Hammerlling ら、J. Pharmaceutical and Biomedical Analysis、14: 1455 (1996);Kitamura ら、J. Cellular Physiol.、140: 323 (1989))。In vitro細胞-ベースのアッセイは、実施例6においてより詳細に説明されている。In vivoアッセイは典型的には、例えばマウス及びラットなどの動物モデルにおいて行われる。In vivoアッセイの例は、ヘマトクリット(HCT)アッセイ、及び網状赤血球アッセイを含むが、これらに限定されるものではない。HCTアッセイは、エリスロポエチン-処置した動物から採取した血液試料から赤血球容積を測定し、及び毛細管中の血液を遠心し、沈降した赤血球により占拠された総容積の割合を測定することにより行う。In vivo HCTアッセイについては、実施例8により詳細に説明されている。網状赤血球アッセイは、前駆細胞から分化したばかりでまだ前駆細胞に特徴的な核酸を残している、網状赤血球としても知られている新生赤血球を測定するものである。網状赤血球は、アクリジンオレンジ又はチアゾールオレンジなどの核酸-染色色素により染色した後、フローサイトメーターにおける赤血球の選別、及び陽性に染色された網状赤血球画分の計測により測定される。
【0051】
生物学的活性又は機能活性のあるエリスロポエチン-様分子は典型的には、野生型又は天然のエリスロポエチンの対応する配列と、実質的アミノ酸配列類似性又は同一性(例えば、少なくとも約55%、約65%、約75%の同一性、典型的には少なくとも約80%、及び最も典型的には約90〜95%の同一性)を共有し、それらの野生型エリスロポエチンの1種又は複数の機能を持っている。
【0052】
従って本発明のエリスロポエチンは、野生型エリスロポエチンの配列に類似したアミノ酸配列を有するエリスロポエチンポリペプチドを特に含むと理解される。このようなタンパク質は、本明細書においてエリスロポエチンアナログと定義される。「アナログ」は、本明細書において、そのタンパク質の生物学的活性を保持するために野生型エリスロポエチンのアミノ酸配列と十分な類似性を伴うアミノ酸配列を意味すると定義される。例えば、エリスロポエチンアナログは、野生型エリスロポエチンのアミノ酸配列中に1個又は複数のアミノ酸変更を含むことができ、依然例えば赤血球形成又は成熟を刺激する能力を保持する。このようなアミノ酸変更の例は、アミノ酸残基の付加、欠失又は置換を含む。本発明のエリスロポエチンは、米国特許第5,614,184号に開示されたような、野生型エリスロポエチンよりも、より大きい又はより少ない生物学的活性を示す変異体タンパク質も包含している。
【0053】
本発明のエリスロポエチンは、エリスロポエチンの生物学的活性のある断片も包含している。このような断片は、エリスロポエチンの完全長アミノ酸配列の一部のみを含むが、依然生物学的活性を有する。本明細書において「生物学的活性のある断片」は、完全長タンパク質に類似した生物学的作用を発揮する断片を意味する。このような断片は、アミノ-及びカルボキシ-末端の欠失に加え、内部の欠失により作出され得る。これらは、エリスロポエチンの切断型及びハイブリッド型も含む。「短縮」型は、より短いエリスロポエチンの型であり、例えばアミノ末端、又はカルボキシ末端の残基が取り除かれている。
【0054】
エリスロポエチン配列の変動
EPO受容体への結合親和性を低下するため;タンパク質安定性を増強するため;正確な活性のある立体構造の選定を強化するため;薬物動態特性を増強するため;合成を増強するため;又は、他の有利な特徴を提供するために、本発明のエリスロポエチン部分にアミノ酸改変が導入される。例えば、EPO受容体-媒介したエンドサイトーシスは、エリスロポエチンとEPO受容体の間の結合親和性により決定される。ヒトエリスロポエチンとEPO受容体の複合体の三次元構造は、エリスロポエチンのその受容体への結合がエリスロポエチン表面上の正電荷及びEPO受容体上の負電荷により支配されることを明らかにしている。Syedら、Nature、395: 511 (1998)。結合の速度(on-rate)を低下させるために、変異を導入し、エリスロポエチン-EPO受容体接触表面近傍に位置する正に荷電したアミノ酸を置換することができる。例えばひとつの態様において、ヒトエリスロポエチンのArg131及びArg139の一方又は両方を置換することができる(EPO配列のアミノ酸番号付けは、成熟型ヒトEPOを基にしている)。好ましくは、Arg131及びArg139は、グルタミン酸、アスパラギン酸、又は他の非-正荷電したアミノ酸に置換される。ヒトエリスロポエチンのArg131及びArg139に対応するアミノ酸を置換するように、他の種のエリスロポエチンに変異を導入することができる。しかし、EPOアミノ酸配列において改変を行う場合、EPO生物学的活性を保存するために、EPO-EPO受容体相互作用の中心に位置する残基は避けなければならない。
【0055】
あるいは、アラニン走査突然変異導入により、アミノ酸置換を容認する領域又は位置を経験的に決定することができる(Cunninghamら、Science、244 1081-1085 (1989))。この方法において選択されるアミノ酸残基は、生物学的活性に対する効果を決定するために、中性アミノ酸(例えば、アラニン)と個々に置換される。
【0056】
ひとつの態様において、エリスロポエチン部分は以下の変異の少なくともひとつを含む:His32→Gly及び/又はSer34→Arg、及びPro90→Ala。別の態様において、システイン-システインジスルフィド結合のパターンを変更し、新規ジスルフィド結合形成を生じるために、システイン置換がエリスロポエチンへ導入される(「NDS変異」)。哺乳動物エリスロポエチンの中でも独自性を示す天然のヒトエリスロポエチンは、7、29、33及び161位に正確に4個のシステインを有し、これらは2個のジスルフィド結合を形成する。エリスロポエチン部分のこれらのシステイン残基の1個又は複数を改変することができる。改変されたジスルフィド結合を作製するために、1個のシステイン残基が、構造的に互換性のあるアミノ酸、例えばアラニン又はセリンへ変異され、及び三次元構造において隣接する第二のアミノ酸がシステインへ変異される。例えば、アミノ酸Gln86、Pro87、Trp88、Glu89、及びLeu91のひとつは、Cysにより置き換えることができる。Trp88がCysにより置き換えられ及びCys33が別のアミノ酸により置き換えられる場合、そのエリスロポエチン部分は、ヒトEPOにおいては認められないCys29-Cys88ジスルフィド結合を形成するであろう。この結合は、典型的Cys29-Cys33ジスルフィド結合を伴う融合タンパク質よりもより大きい活性を有する融合タンパク質を生じる。加えてCys29-Cys88融合タンパク質は、融合タンパク質のエリスロポエチン部分における他の変異の存在下で、Cys29-Cys33融合タンパク質と比べ、活性の際だった増加を示す。従って本発明のひとつの態様において、エリスロポエチン部分は、以下のアミノ酸置換の少なくともひとつを含む:29位の非-システイン残基、33位の非-システイン残基、88位のシステイン残基、及び139位のシステイン残基。ひとつの態様において、エリスロポエチン部分は、7、29、88、及び161位にシステインを含む。別の態様において、エリスロポエチン部分は更に、以下の置換を1個又は複数含む:His32→Gly、Cys33→Pro、及びPro90→Ala。別の態様において、全体の新たなジスルフィド結合が、2個のアミノ酸のシステインへの変異により、このタンパク質に付加される。これらのCys変異が引き起こし得る構造内の可能性のある緊張を補償するために、本発明の好ましいCys-操作された態様において、エリスロポエチン部分は更に、これらの可能性のある緊張を緩和するように設計された変異を含む。
【0057】
システイン置換に関連する更なる態様は、PCT国際公開公報第01/36489号(すなわち、米国特許出願第09/708,506号)に開示されており、その開示は本明細書に引用により取り込まれるものとする。
エリスロポエチンに変異を導入する方法は、当該技術分野においてよく知られている。例えば変異は、位置指定突然変異導入技術により導入することができる。広範な部位特異的突然変異導入技術を、利用することができ、及び同様の結果を実現するための代替として使用することができることに注目することは重要である。他の技術は、ランダム及びセミ-ランダム突然変異誘発を含むが、これらに限定されるものではない。
【0058】
リンカー
本発明のFc-EPO融合タンパク質は、Fc部分とエリスロポエチン部分の間に、リンカー分子、好ましくはペプチドリンカーを含むことができる。リンカーを伴う融合タンパク質は、増加した生物学的活性のような、改善された特性を有することができる。リンカーは一般に、1〜25個のアミノ酸(例えば、5〜25個又は10〜20個のアミノ酸)を含む。リンカーは、プロテアーゼ切断部位を含まないように設計することができる。更にリンカーは、タンパク質分解を立体的に阻害するために、N-結合型された又はO-結合型されたグリコシル化部位を含むことができる。従ってひとつの態様において、リンカーは、Asn-Ala-Thrアミノ酸配列を含む。
追加の適当なリンカーは、Robinsonら、Proc. Natl. Acad. Sci. USA、95 5929 (1998);及び、米国特許出願第09/708,506号に開示されている。
【0059】
グリコシル化
天然のヒトエリスロポエチン及び哺乳動物細胞において発現された組換えエリスロポエチンは、3個のN-結合型及び1個のO-結合型オリゴ糖鎖を含む。N-結合グリコシル化は、24、38及び83位に位置したアスパラギン残基で生じるのに対し、O-結合グリコシル化は、126位のセリン残基で生じる(Laiら、J. Biol. Chem.、261: 3116 (1986);Broudyら、Arch. Biochem. Biophys.、265: 329 (1988))。オリゴ糖鎖は、末端シアル酸残基により改変されることが示されている。N-結合型鎖は典型的には、1本の鎖あたり最大4個のシアル酸を有し、O-結合型鎖は典型的には、最大2個のシアル酸を有する。従ってエリスロポエチンポリペプチドは合計最大14個のシアル酸を収容することができる。
【0060】
シアル酸は、N-結合型又はO-結合型オリゴ糖上の末端糖である。シアル酸化の程度は、位置毎、タンパク質毎に変動し、使用される細胞培養条件、細胞型、及び具体的細胞クローンによって左右される。BHK細胞において合成された本発明のFc-EPO融合タンパク質は、高度にシアル酸化されることがわかっている。Fc-EPO融合タンパク質のシアル酸化の程度は、無タンパク質培地中、懸濁液中、又は無タンパク質培地中の懸濁物としての増殖に関するBHK細胞の適応により、更に増強され得ることもわかっている。ある種のその他の通常使用される細胞株、例えばNS/0、PerC6、又は293細胞は、標準の培養条件下で、高度にシアル酸化されたFc-EPO融合タンパク質を生成することができない。種々の細胞株から生成されたFc-EPO融合タンパク質のシアル酸化の程度は、それらの高度に負に荷電したシアル酸残基に基づく、等電点(IEF)ゲル電気泳動により決定することができ;IEFゲル電気泳動の詳細は、実施例5Bに説明されている。種々の細胞株において生成されたFc-EPO融合タンパク質のシアル酸化の程度は、当業者に良く知られた方法を用いるレクチン-結合試験により定性的に確認することができる。レクチン-結合アッセイの例は、実施例5Bに説明されている。
【0061】
典型的には、本発明の高度にシアル酸化された精製されたFc-EPO融合タンパク質の集団は、精製されたFc-EPO融合タンパク質あたり平均11〜28個のシアル酸残基を有する。好ましい高度にシアル酸化されたFc-EPO融合タンパク質の集団は、精製されたFc-EPO融合タンパク質あたり平均13〜28、15〜28、17〜28、19〜28、又は21〜28個のシアル酸残基を有する。例えば、ひとつの好ましい高度にシアル酸化されたFc-EPO融合タンパク質集団は、精製されたFc-EPO融合タンパク質あたり平均20〜22個のシアル酸残基を有する。別のFc-EPO融合タンパク質の好ましい集団は、精製されたFc-EPO融合タンパク質あたり平均23〜28個のシアル酸残基を有する。
【0062】
シアル酸化されたFc-EPO融合タンパク質の薬物動態
赤血球形成-刺激剤のin vivo生物学的活性を決定する最も重要な因子のひとつは、タンパク質の血清濃度が、赤血球形成に必要な閾値を上回り続ける時間の長さであり、これは赤血球形成-刺激剤の薬物動態により決定される。高度にシアル酸化されたFc-EPO融合タンパク質の薬物動態プロファイルは、天然の又は組換えエリスロポエチンのプロファイルとは異なる。大きな差異は、高度にシアル酸化されたFc-EPO融合タンパク質は、はるかに長い血清半減期及び遅いクリアランスを有し、これは増加したin vivo生物学的効能につながることである。理論に拘束されることを欲するものではないが、シアル酸残基はエリスロポエチン分子上の負電荷を増加し、負に荷電したEPO受容体結合の速度減少及びEPO受容体に媒介されたエンドサイトーシスの減少を生じさせ、血清半減期を長期化すると考えられている。更にシアル酸は、エリスロポエチンタンパク質が、露出したガラクトース残基に糖タンパク質を結合するアシアロ糖タンパク質受容体によりエンドサイトーシスを受けることも阻害する。
一般にエリスロポエチンのような、治療的分子のほとんどの薬物動態プロファイルは、血清濃度の初期低下(α相)と、それに続く投与後のより段階的な低下(β相)を示す。
【0063】
α相に影響を及ぼす因子
小型-分子の薬物動態理論に従い、α相は血液の外側のコンパートメントへ分子がどのように分配されるかを記述する、分布の容積を規定する。α相において認められる減衰は、種々の細胞株において合成された種々のFc-EPO融合タンパク質について広範に変動する。理論的には、この差異は、分布容積の変動又はコンパートメント間の通行の変動が原因である。しかし、シアル酸化の程度とマウスにおけるFc-EPOタンパク質の薬物動態挙動の間には相関関係が存在することが認められている。例えば、BHK細胞において合成されたFc-EPO融合タンパク質は、高度にシアル酸化され、最良の薬物動態プロファイルを示す。NS/0細胞において合成されたFc-EPO融合タンパク質は、若干シアル酸化され、中間の薬物動態プロファイルを有する。293及びPerC6細胞において合成されたFc-EPO融合タンパク質は、ほとんど又は全くシアル酸化されず、最初の30分間における約1/100の血清濃度減衰で特徴付けられる劣った薬物動態プロファイルを有する。従って特定のFc-EPO融合タンパク質のα相に影響を及ぼす重要な因子は、グリコシル化種の分布及びシアル酸化のレベルである。少なくシアル酸化されているFc-EPO融合タンパク質は、迅速に消失する。
【0064】
加えて図2に示されるように、α相の間のFc-EPO血清濃度の減衰の程度は投与量に従い変動し、この挙動は飽和させることが可能で、受容体-媒介性であろうことを示している。α相減衰を媒介する受容体は、EPO受容体でもFc受容体でもなく、アシアロ糖タンパク質受容体などの別の受容体であり得る。Aranesp(登録商標)は、正常なヒトエリスロポエチンと比べ、EPO受容体に対して低下した結合親和性を有しているが、その理由は、Aranesp(登録商標)は、追加のN-結合グリコシル化部位の結果として増加した負電荷を有するからである。しかしAranesp(登録商標)及び正常なヒトエリスロポエチンは、α相の間に同様の減衰を示す。加えて、一般に赤血球前駆細胞の細胞表面上のEPO受容体の数は、わずかに約200個であり、これらの受容体は図2において使用されたものよりもはるかに低用量のエリスロポエチンで完全に飽和されるであろう。グリコシル化部位を除去する変異、例えばIgG1のAsn297に対応するアミノ酸の変異を伴うFc-EPO融合タンパク質は依然としてα相における急勾配の減衰を示すことができるので、Fc受容体がα相における劇的減衰を媒介していることは恐らくありそうもない。加えて、凝集しない場合、IgG2 CH2領域は一般にFc受容体に結合しないが、IgG2 CH2領域を含むFc-EPOタンパク質は依然としてα相の間に有意な減衰を示す。
【0065】
理論に拘束されることを欲するものではないが、α相におけるFc-EPO融合タンパク質の血清濃度の減衰は、アシアロ糖タンパク質-受容体-媒介型エンドサイトーシスを介してアシアロ糖タンパク質-受容体により媒介され得る。少なくシアル酸化されたFc-EPO融合タンパク質は、アシアロ糖タンパク質受容体により結合されてアシアロ糖タンパク質-受容体-媒介型エンドサイトーシスを生じる、露出したガラクトース残基を含む。結果的に少なくシアル酸化されたFc-EPO融合タンパク質は迅速に消失する。
【0066】
β相に影響する因子
β相におけるFc-EPO融合タンパク質の血清濃度の減衰は、α相における減衰と比べて緩やかな勾配である。例えばマウスにおいて、投与後8〜24時間の間に、Fc-EPO融合タンパク質の血清濃度の1/2〜1/3倍の減衰が認められる。β相の間の減衰の差異も、異なる細胞株で合成された異なるFc-EPOタンパク質間では劇的さが少ない。しかしα相におけるように、シアル酸化の程度はβ相における薬物動態挙動と相関している。例えば、BHK細胞において合成されたFc-EPO融合タンパク質は、NS/0細胞で合成されたその他は同じであるFc-EPOタンパク質と比べて有意に改善されたβ相を有する。EPO受容体-媒介エンドサイトーシスは、β相の間のFc-EPO融合タンパク質の血清濃度の減衰に少なくとも部分的に寄与しているように見える。正常なヒトエリスロポエチンと比べ、EPO受容体への結合親和性が低下したAranesp(登録商標)は、同様のα相プロファイルにも関わらず、正常なヒトエリスロポエチンと比べて有意に改善されたβ相を有する。
【0067】
本発明のFc-EPO融合タンパク質は一般に、天然の又は組換えエリスロポエチンと比べ改善されたβ相を示しており、このことはFc部分の追加がβ相の間の血清濃度の低下を有意に遅延することを示唆している。Fc部分又はエリスロポエチン部分のある種のアミノ酸改変は、β相を有意に改善することも認められる。例えば、Fc部分のグリコシル化部位を除去する変異は、Fc-EPO融合タンパク質のβ相を改善する。エリスロポエチン部分の安定性を増加する変異、例えばエリスロポエチン部分においてジスルフィド結合を操作する変異(例えば、NDS変異)は、Fc-EPO融合タンパク質のβ相を有意に改善する。一般に改善されたβ相は、Fc-EPO融合タンパク質の終末血清半減期を延長する。
【0068】
Fc-EPO融合タンパク質の消失経路
エリスロポエチンタンパク質分子を体から排出するいくつかの可能性のある経路が存在する。野生型又は天然のエリスロポエチンタンパク質分子は、腎濾過及び受容体-媒介されたエンドサイトーシスにより体から排出される。エンドサイトーシスされたエリスロポエチンは、効率的に分解される。図3に示したように、Fc部分のエリスロポエチン部分への追加は、本質的にFc-EPO融合タンパク質の腎臓を通る排出を除去することが期待される。結果的に受容体-媒介エンドサイトーシスがFc-EPO融合タンパク質の排出の主要経路となる。更にFc部分のエリスロポエチン部分への追加は、インターナリゼーション後の分解を低下することも予想される。その理由は、FcRnエンドソーム受容体はこの融合タンパク質を細胞の外側へ再循環させると予想されるからである。
【0069】
原理上は、少なくとも3種の受容体、すなわちFc-受容体、EPO受容体、及びアシアロ糖タンパク質受容体がFc-EPO融合タンパク質のクリアランスを媒介することができる。Fc-EPO融合タンパク質のFc受容体を介したクリアランスは、Fc部分においてIgG1-由来のCH2の代わりにIgG2-由来のCH2ドメインを使用することにより有意に低下するはずである。IgG2-由来のCH2ドメインは、IgG1-由来のCH2ドメインと比べてFcγRI(IgG類への最高の親和性を有する)に対して約1/100低い親和性を有する。IgG2-由来のCH2とFcγRIの間の相互作用は、ほとんどの結合アッセイにおいて検出不能である。しかしIgG2-由来のCH2ドメインの残留FcγR-結合活性は、Fc-EPO融合タンパク質のクリアランスにおいて依然役割を果たす。その理由はCH2ドメイン内のグリコシル化部位を除去するアスパラギン変異は更にFc-受容体結合を低下させ、及びFc-EPO融合タンパク質の薬物動態を改善するからである。
NDS変異は、エリスロポエチン構造を安定化する作用を有し、その結果、インターナリゼーション後のFc-EPO融合タンパク質の分解を低下すると予想される。NDS変異を含むFc-EPO融合タンパク質は薬物動態特性を改善し、血清半減期を延長する。
【0070】
シアル酸化はFc-EPO融合タンパク質の負電荷を増加し、Fc-EPO融合タンパク質のEPO受容体への結合親和性を低下させる。シアル酸化は同じく、Fc-EPO融合タンパク質上の露出したガラクトース残基の数を減少し、これはFc-EPO融合タンパク質のアシアロ糖タンパク質受容体への結合親和性を低下させる。従って図3に示したように、シアル酸化はEPO受容体-媒介されたエンドサイトーシス及びアシアロ糖タンパク質受容体-媒介されたエンドサイトーシスの両方を減少させる。従って高度にシアル酸化されたFc-EPO融合タンパク質は、劇的に遅延されたクリアランス率を有し、その結果血清半減期が有意に増加する。
Fc部分の追加、Fc及びエリスロポエチン部分の置換、並びにシアル酸化は、各々、Fc-EPO融合タンパク質のクリアランスを低下させる。クリアランス及び血清半減期に対する組合せ作用は相加的又は相乗的である。
【0071】
Fc-EPO融合タンパク質のin vitro活性及びin vivo効能
Fc-EPOタンパク質のin vitro活性は、細胞-ベースのアッセイにおいて試験することができる。特に、Fc-EPOとEPO受容体の間の相互作用は、TF-1細胞増殖アッセイを基に行うことができる。TF-1細胞はEPO受容体を発現し、その結果、トリチウム標識されたチミジンの組込みにより決定されるTF-1細胞の増殖はエリスロポエチン活性の関数である(Hammerllingら、J. Pharmaceutical and Biomedical Analysis、14: 1455 (1996);Kitamuraら、J. Cellular Physiol.、140: 323 (1989))。本発明において、TF-1細胞の増殖は、エリスロポエチン部分とEPO受容体の間の相互作用の関数である。特にFc-EPO融合タンパク質のエリスロポエチン部分がEPO受容体に対して低下した結合速度を有する場合、Fc-EPOタンパク質は一般に、細胞-ベースのアッセイにおいて低下した活性を有する(増加したED50値により示される)。
【0072】
入手が比較的容易である細胞-ベースのアッセイのデータは、一般にFc-EPOタンパク質の薬物動態及びin vivo効能と相関する。In vitro活性の低下は、EPO受容体に対する低下した結合速度を示すが、これは一般に改善された薬物動態特性及び増強されたin vivo効能と相関する。対照的に、増加したin vitro活性(減少したED50値により示される)は、EPO受容体に対する増加された結合速度を示すが、これは一般に、貧弱な薬物動態特性及び低下したin vivo効能に相関する。
【0073】
Fc-EPO融合タンパク質のin vivo生物学的活性は、例えばマウス及びラットなどの動物モデルにおいて行われるアッセイにより測定することができる。In vivoアッセイの例は、ヘマトクリット(HCT)アッセイ及び網状赤血球アッセイを含むが、これらに限定されるものではない。HCTアッセイは、赤血球(RBC)により占拠された血液体積を測定し、毛細管内の血液を遠心し、沈積したRBCにより占拠された総容積の割合を測定することにより簡便に行われる。網状赤血球は、前駆細胞から新たに分化されるRBCであり、前駆細胞由来の核酸の名残を含むことを特徴としている。網状赤血球は、例えばアクリジンオレンジ又はチアゾールオレンジなどの、核酸-染色色素による染色後のフローサイトメーターにおける赤血球ソーティング、及び染色画分の計測により測定される。典型的には、ヘマトクリット及び網状赤血球は、週2回測定される。
【0074】
網状赤血球データは、ある意味で、ヘマトクリットデータの最初の派生物である。網状赤血球数は、赤血球の生成速度の測定値であるが、ヘマトクリットは、総赤血球を測定する。典型的実験において、Fc-EPO融合タンパク質を投与された動物のヘマトクリットは増加し、その後ベースラインに戻る。ヘマトクリットが高く、投与されたFc-EPOタンパク質が動物の循環系から消失した場合、赤血球形成が抑制されるので、網状赤血球数は、ベースラインを下回る。
網状赤血球は通常、前駆体がRBCとなるべく運命づけられた後、4日目に骨髄から出現する。しかし高レベルのエリスロポエチンの存在下では、網状赤血球は投与後1〜3日で骨髄から離れることが多いであろう。
【0075】
動物においてFc-EPOタンパク質の注射に応答してヘマトクリット読み値は増加し、一定で有り続け、その後ベースラインに戻る。そのようなヘマトクリット反応の例を図4-6に示した。最大減少率は、マウスにおいて週1回で血液体積の約7%であり、これはマウスにおける約45日間のRBC生存期間に相当し、及びラットにおいて週1回で血液体積の約5%であり、これはラットにおける約65日間のRBC生存期間に相当する。この最大減少率は、新たな合成の存在しない場合のRBCの破壊を恐らく表しているであろう。生物学的活性のあるFc-EPOタンパク質が赤血球形成の閾値を上回る濃度でその系内に存在し続ける場合、ヘマトクリットレベルは高く有り続け、生物学的活性のあるFc-EPOのレベルが薬物動態実験において検出不可能である場合であっても低下しない。
【0076】
Fc-EPOタンパク質の薬物動態特性は、このタンパク質のin vivo効能と相関することがわかった。先に説明されたようにFc-EPO融合タンパク質の薬物動態を増強する本発明の特徴は全て動物実験におけるin vivo効能も増強する。表1に示されたように、そのような特徴は例えばFc部分の追加、Fc部分内のグリコシル化部位の除去(例えば、IgG1のAsn297に相当する位置でのN→Q置換)、NDS変異のエリスロポエチン部分への導入、及びBHK細胞におけるFc-EPOタンパク質の合成による高レベルのシアル酸化を含む。
【0077】
表1
Fc-EPOタンパク質の薬物動態及び生物学的活性に影響を及ぼす因子

【0078】
エリスロポエチン部分について、BHK細胞で生成されたFcg2h(FN→AQ)-Epo及びFcg2h-EPO(NDS)は、最良の薬物動態及び最も強力なin vivo生物学的活性を示すことがわかっている。Fcg2h(FN→AQ)-Epo及びFcg2h-EPO(NDS)は各々、Aranesp(登録商標)よりもエリスロポエチン部分について、より長い血清半減期及びより強力なin vivo活性を有する。
【0079】
Fc-EPO融合タンパク質の合成
本発明のFc-EPO融合タンパク質は、ヒト又は他の哺乳動物細胞株のような適当な細胞又は細胞株において生成することができる。適当な細胞株は、ベビーハムスター腎(BHK)細胞、チャイニーズハムスター卵巣(CHO)細胞(ジヒドロ葉酸還元酵素(DHFR)-欠損細胞を含む)、及びCOS細胞を含むが、これらに限定されるものではない。好ましい態様において、BHK細胞が使用される。
【0080】
適当な宿主細胞(例えばBHK細胞)においてFc-EPO融合タンパク質を発現するために、まずFc-EPO融合タンパク質をコードしている核酸配列が当業者に周知の標準組換え分子技術を用いて発現ベクターに導入される。エリスロポエチン部分をコードしている配列は、高レベルの発現のためにコドン-最適化されることが好ましい。コドン-最適化されたヒトエリスロポエチンは、PCT国際公開公報第01/36489号(すなわち米国特許出願第09/708,506号)に開示されており、その開示は本明細書に参照として組入れられている。エリスロポエチン部分をコードしている核酸配列の例を配列番号:1に示す:
GCCCCACCACGCCTCATCTGTGACAGCCGAGTGCTGGAGAGGTACCTCTTGGAGGCCAAGGAGGCCGAGAATATCACGACCGGCTGTGCTGAACACTGCAGCTTGAATGAGAACATCACCGTGCCTGACACCAAAGTGAATTTCTATGCCTGGAAGAGGATGGAGGTTGGCCAGCAGGCCGTAGAAGTGTGGCAGGGCCTGGCCCTGCTGTCGGAAGCTGTCCTGCGGGGCCAGGCCCTGTTGGTCAACTCTTCCCAGCCGTGGGAGCCCCTGCAACTGCATGTGGATAAAGCCGTGAGTGGCCTTCGCAGCCTCACCACTCTGCTTCGGGCTCTGGGAGCCCAGAAGGAAGCCATCTCCCCTCCAGATGCGGCCTCAGCTGCTCCCCTCCGCACAATCACTGCTGACACTTTCCGCAAACTCTTCCGAGTCTACTCCAATTTCCTCCGGGGAAAGCTGAAGCTGTACACAGGGGAGGCCTGCCGGACAGGGGACAGATGA (配列番号:1)
【0081】
好ましいFc部分、例えばIgG2由来の CH2ドメイン及びIgG1由来のヒンジ領域を含むFc部分をコードしている核酸配列の例は、米国特許公開番号第20030044423号(すなわち米国特許出願第10/093,958号)に開示されており、その開示は本明細書に引用により取り込まれるものとする。
一般に、Fc-EPO融合タンパク質をコードしている核酸配列はシグナルペプチドをコードしている核酸配列(リーダー配列)を含む。このリーダー配列は分泌プロセッシング時に切断される。リーダー配列を伴わない成熟型Fc-EPOタンパク質をコードしている核酸配列の例(配列番号:2)を図7に示している。
【0082】
適当なベクターは、哺乳動物宿主細胞における発現に適したものを含む。これらのベクターは、例えば、プラスミド又はウイルスであることができる。ベクターは典型的には、以下のエレメントを含む:哺乳動物宿主細胞における使用に適合したプロモーター及び他の「上流」調節エレメント、複製起点、リボソーム結合部位、転写終結部位、ポリリンカー部位、及び選択マーカー。ベクターは、更に真核宿主細胞において増殖及び維持を可能にするエレメントも含む。本発明に適したベクターは、pdCs-Fc-X及びそれに由来するベクター、並びにphC10-Fc-X及びそれに由来するベクターを含むが、これらに限定されるものではない。
【0083】
Fc-EPOタンパク質をコードしているベクターは、トランスフェクション及びウイルス技術を含む、標準の細胞生物学的技術を用いて宿主細胞へ導入される。トランスフェクションは、単離されたDNA、RNA、又は合成ヌクレオチドポリマーを使用する、遺伝子情報の細胞への移動を意味する。適当なトランスフェクション法は、いくつか例を挙げると、リン酸カルシウム-媒介型共-沈殿(Sambrookら、Molecular Cloning: A Laboratory Manual、第2版、Cold Spring Harbor Laboratory Press、(1989))、リポフェクション(例えば、Life Technologies(ロックビル、ML)から得られるLipofectamine Plus)、DEAE-デキストラン-媒介型トランスフェクション技術、リゾチーム融合又は赤血球融合、スクレーピング(scraping)、直接取込み、浸透圧もしくはショ糖ショック、直接マイクロインジェクション、赤血球-媒介技術を介した間接マイクロインジェクション、プロトプラスト融合、又は宿主細胞の電流への曝露(例えばエレクトロポレーション)を含むが、これらに限定されるものではない。前述のトランスフェクション法のリストは網羅的であるとは考えられず、細胞へ遺伝子情報を導入する他の手法が開発されることは疑いもない。
【0084】
Fc-EPO融合タンパク質をコードしている核酸を含む宿主細胞の選択を促進するために、Fc-EPO融合タンパク質をコードしている核酸には典型的には選択マーカーが導入される。選択マーカーは、Fc-EPO融合タンパク質をコードしている同じ発現ベクター上に存在する核酸配列によりコードされていてもよい。あるいは、選択マーカーは異なるベクター上に存在する核酸配列によりコードされていてもよい。後者の場合、これら2種のベクターは、同時トランスフェクション又は同時-形質導入のいずれかにより、宿主細胞へ同時に導入される。適当な選択マーカーは、例えば、ハイグロマイシンB(Hyg B)及びジヒドロ葉酸還元酵素(DHFR)を含む。
一過性発現は、Fc-EPO融合タンパク質の小規模タンパク質生成及び迅速な分析に有用である。Fc-EPO融合タンパク質をコードしている核酸配列を含む宿主細胞は、コードされたFc-EPO融合タンパク質の発現に適した条件下で維持される。標準の細胞培養の方法、条件及び培地を、Fc-EPO融合タンパク質を発現している宿主細胞を維持するために使用することができる。
【0085】
安定的にトランスフェクションされた細胞は、大規模生成、高レベル発現、及び他の目的に好ましいことが多い。安定して維持される核酸は、宿主細胞内において様々な形態のいずれかで存在することができる。例えばひとつの態様において、安定して維持される核酸配列は、宿主細胞の染色体に組込まれる。別の態様において、安定して維持される核酸配列は、染色体外アレイとして、人工染色体として、又は別の適当な形態で存在することができる。
【0086】
ひとつの態様において、Fc-EPO融合タンパク質を合成するためにBHK細胞が使用される。安定的にトランスフェクションされたBHK細胞を得るために、融合タンパク質をコードしている核酸配列及び選択マーカーをコードしている核酸配列が、好ましくはエレクトロポレーション、プロトプラスト融合又はリポフェクション法を用いてBHK細胞へ導入される。融合タンパク質をコードしている核酸配列及び選択マーカーをコードしている核酸配列は、同じ発現ベクター上に存在することができる。あるいは、融合タンパク質をコードしている核酸配列及び選択マーカーをコードしている核酸配列は、個別のベクター上に存在することができる。安定したBHK細胞を確立するのに好ましい選択マーカーは、HygBである。DHFRなどの他の選択マーカーも使用することができる。安定的にトランスフェクションされたクローンは、単離され、適当な濃度(例えば、200、250、又は300μg/ml)のHygBの存在下、例えば、Life Technologiesから入手できるMEM+FBS、DMEM/F-12培地、もしくはVP-SFM、及び他の適当な培地などの標準組織培養培地中でのそれらの増殖により増やされる。Fc-EPO融合タンパク質の発現レベルは、上清及び培養培地からの試料に対する、例えばELISA試験、ウェスタンブロット、ドットブロット、又は他の適当なアッセイなどの、標準タンパク質-検出アッセイによりモニタリングすることができる。高発現クローンが、選択され、大規模に繁殖される。
【0087】
典型的にはBHK細胞は接着細胞株であり、通常はMEM+10%熱-失活したウシ胎仔血清(FBS)のような血清-含有培地において増殖させる。しかしBHK細胞は懸濁物として無血清培地、例えば、VP-SFM(Invitrogen Corp.、カタログ番号11681-020)又はOpti-Pro SFM(Invitrogen Corp.、カタログ番号12309)などの無血清培地中で増殖するために適合ささることができる。適合プロセスの例は実施例3に説明されている。無血清培地における増殖に適応したBHK細胞は、更に例えばDMEM/F-12(Invitrogen Corp.、カタログ番号11039-021)のような無タンパク質培地における増殖に適合させる。適合法の一例は実施例3に説明されている。DMEM/F-12には、適当なアミノ酸及び他の成分、例えばグルタミン、HyPep 4601(Quest International、カタログ番号5Z10419)及びHyPep 1510(Quest International、カタログ番号5X59053)などのタンパク質加水分解産物、Ethanolamine(Sigma、カタログ番号E0135)、及びTropolone(Sigma、カタログ番号T7387)が補充されることが好ましい。当業者は、各補充物の適当な濃度を慣習的な実験により経験的に決定することができる。
【0088】
無タンパク質培地において増殖されたBHK細胞において合成されたFc-EPO融合タンパク質は、血清-含有培地(例えばMEM+FBS)、又は無血清だが無タンパク質でない培地(例えばVP-SFM)において増殖させた細胞において合成された対応するタンパク質よりも、より多くシアル酸化され、より均質なシアル酸化を示す。加えてこうして得られたF c-EPOタンパク質は実質的に非-凝集性、すなわち総収量の約98%が非-凝集性である。無タンパク質培地において増殖されたBHK細胞から得たタンパク質収量は、血清-含有培地中で増殖されたBHK細胞からのものに類似しており、すなわち10μg/mLを上回る。従って、懸濁液及び/又は無タンパク質培地における増殖は多くの利点をもたらす。これには1)増加したシアル酸化の結果としてFc-EPO融合タンパク質の薬物動態が改善されること;および、2)タンパク質は懸濁態様及びタンパク質を欠いた培地において増殖された細胞から精製することができるので、下流の精製工程が容易になること、が含まれる。
【0089】
精製
Fc-EPOの精製は、当業者に公知の標準GMP手法に従い行われる。このタンパク質は一般に、均質又はほぼ均質に精製される。カラムクロマトグラフィーに関連するもののような、クロマトグラフィー精製が一般に好ましい。一般にFc-EPO融合タンパク質の精製スキームは、最初のタンパク質捕獲工程;ウイルス失活工程;一又は複数のポリッシュ(polishing)工程;ウイルス除去工程;及び、タンパク質濃縮及び/又は製剤工程を含むが、これらに限定されるものではない。例えば融合タンパク質のFc部分に結合するクロマトグラフィー用の樹脂材料を使用し、Fc-EPOタンパク質を捕獲することができる。適当な樹脂材料は、プロテインAに結合された樹脂を含むが、これらに限定されるものではない。夾雑成分を除去するためにポリッシュ工程を含んでもよい。例えば、ヒドロキシアパタイトクロマトグラフィー、Sepharose Qクロマトグラフィー、サイズ排除クロマトグラフィー、又は疎水相互作用クロマトグラフィーを使用し、夾雑物を除去することができる。Fc部分に結合し及びFc-EPO融合タンパク質を精製するためのプロテインA-ベースのカラムクロマトグラフィーを使用するひとつの精製法は、ウイルス失活及び除去の任意的方法として実施例12に説明されている。精製されたタンパク質は一般に、限外濾過;適当な配合の緩衝液への透析;滅菌濾過;及び、バイアルへの分注を用いて所望の濃度へ濃縮される。
【0090】
投与
医薬組成物及び投与経路
本発明は、本発明に従い作製されたFc-EPOタンパク質を含有する医薬組成物も提供する。これらの医薬組成物は、赤血球形成を刺激し、並びに貧血を予防及び治療するために使用することができる。本発明により治療可能な状態は中でも、腎機能の低下又は喪失(慢性腎不全)に随伴した貧血、化学療法もしくは抗-ウイルス薬(AZTなど)のような骨髄抑制療法に随伴した貧血、非-骨髄性癌の進行に随伴した貧血、ウイルス感染症(例えばHIV)に随伴した貧血、及び慢性疾患の貧血を含む。手術時の予想外の失血のような、それ以外は健康な個人において貧血につながり得る状態も同じく治療可能である。一般にrHuEpoにより治療可能ないずれの状態も、本発明のFc-EPO融合タンパク質により治療することができる。
【0091】
Fc-EPOタンパク質含有製剤
一般に製剤は、液体又は固体の剤形で、Fc-EPOタンパク質、緩衝液及び界面活性剤を含有する。固形製剤は、凍結乾燥された、噴霧凍結乾燥された、又は噴霧乾燥された製剤も含むが、これらに限定されるものではない。液体製剤は、好ましくは水を基剤にしているが、いくつか例を挙げると、エタノール、プロパノール、プロパンジオール、又はグリセロールなどの他の成分を含むことができる。
Fc-EPOタンパク質は、当業者に公知の標準GMP手法に従い、水溶液中に製剤される。一般に製剤は、適当な成分を適当な濃度で含む指定された容量の水溶液の混合により、作製される。例えば製剤は典型的には、Fc-EPOタンパク質を濃度0.1〜200mg/ml、好ましくは0.2〜10mg/ml、より好ましくは0.5〜6mg/mlで含有する。
【0092】
緩衝液成分は、pHを調節することが可能である任意の生理的に許容できる物質、例えば、クエン酸塩、酢酸塩、ヒスチジン塩、コハク酸塩、マレイン酸塩、リン酸塩、乳酸塩、それらの各酸もしくは塩基、又はそれらの混合物を含む。一般に使用される緩衝液成分は、クエン酸塩及び/又はそれらの遊離酸である。製剤は典型的には、緩衝液成分を濃度10〜100mmol/l、好ましくは2〜20mmol/l、より好ましくは10mmol/l含む。
Fc-EPO製剤のための界面活性剤は、医薬組成物において界面活性剤として使用される任意の賦形剤であることができ、好ましくはポリエチレン-ソルビタン-エステル(Tweens(登録商標))、例えばポリオキシエチレン(20)-ソルビタンモノラウレート、ポリオキシエチレン(20)-ソルビタンモノパルミテート、及びポリオキシエチレン(20)-ソルビタンモノステアレート、並びにポリオキシエチレン-ポリオキシプロピレン-コポリマーである。製剤は典型的には、界面活性剤を濃度0.001〜1.0%w/v、好ましくは0.005〜0.1%w/v、より好ましくは0.01〜0.5%w/vで含有する。
【0093】
製剤は、1種又は複数のアミノ酸も含有することができる。適当なアミノ酸は、アルギニン、ヒスチジン、オルニチン、リジン、グリシン、メチオニン、イソロイシン、ロイシン、アラニン、フェニルアラニン、チロシン及びトリプトファンを含むが、これらに限定されるものではない。ひとつの態様において、Fc-EPO製剤は、グリシンを含有する。好ましいアミノ酸は、塩の形、例えば塩酸塩で使用される。使用可能なアミノ酸濃度は、2〜200mmol/L、又は50〜150mmol/Lの範囲である。
加えて製剤は、ショ糖、トレハロース、ソルビトールなどの、糖;アスコルビン酸又はグルタチオンなどの、抗酸化剤;フェノール、m-クレゾール、メチル-もしくはプロピルパラベンなどの、保存剤;クロロブタノール;チメロサール;塩化ベンザルコニウム;ポリエチレングリコール;シクロデキストリン及び他の適当な成分を含むことができる。
【0094】
Fc-EPO製剤は等張であることが望ましい。例えば製剤の浸透圧は、150〜450mOsmol/kgの範囲であることができる。医薬製剤は、望ましい貯蔵温度、例えば2〜8℃、又は室温で、望ましい貯蔵期間安定していなければならない。Fc-EPOタンパク質を含有する有用な製剤は、生理的に忍容性がよく、製造が容易であり、正確に投与することができ、及び2℃〜8℃又は25℃の貯蔵期間、複数の凍結解凍サイクルの間、更には40℃で少なくとも3ヶ月の貯蔵のような他のストレス時に安定したものである。Fc-EPO製剤の安定性はストレス試験において試験することができる。ストレス試験の例は実施例13に記載されている。
【0095】
投与
本発明に従い製造されたFc-EPO融合タンパク質を含有する治療的組成物は、哺乳動物宿主へ任意の経路により投与することができる。従って、適当ならば、投与は、経口又は非経口(例えば、i.v.、i.a.、s.c.、i.m.)であってよく、これは静脈内及び腹腔内投与経路を含む。加えて投与は、ボーラスの治療薬の定期的注射によるか、又は外部にある貯蔵庫(例えば静注バッグ)からの静脈内又は腹腔内投与によりより連続して行うことができる。ある態様において、本発明の治療薬は、医薬-等級であることができる。すなわちある態様は、ヒトへの投与に必要な純度及び品質管理基準に従う。獣医学用途も、本明細書に使用される意図された意味に含まれる。
【0096】
本発明の治療薬の獣医学及びヒト医療用途の両方のための製剤は、典型的には医薬として許容できる担体及び任意に他の成分(複数)と会合している治療薬を含む。担体(複数)は、製剤の他の成分と共存でき、及びそれらのレシピエントにとって有害ではないという意味で「許容できる」。これに関して、医薬として許容できる担体は、医薬投与に適合可能ないずれか及び全ての溶媒、分散媒、コーティング、抗菌剤及び抗真菌剤、等張剤及び吸収遅延剤などを含むことが意図される。医薬活性物質のためのこのような媒体及び物質の使用は、当該技術分野においてよく知られている。従来の媒体又は物質が活性化合物と共存できないような場合を除いて、それらの本組成物における使用が企図されている。補助的活性化合物も、この組成物へ混入することができる。これらの製剤は、都合の良いことに単位剤形で存在することができ、及び薬学/微生物学の技術分野において周知の方法により調製することができる。一般に、一部の製剤は治療薬を液体担体又は細かく粉砕された固形担体又はそれら両方と会合し、その後生成物を必要ならば所望の製剤へ成形することにより調製される。
【0097】
本発明の医薬組成物は、その意図された投与経路と適合可能であるように製剤される。投与経路の例は、経口又は非経口、例えば、静脈内、皮下、吸入(例えばネブライザーによる)、経皮(局所的)、経粘膜、鼻腔内、口腔内、及び直腸投与を含む。非経口、皮内、又は皮下適用のために使用される液剤又は懸濁剤は以下の成分を含むことができる:注射用水、生理食塩水、不揮発油、ポリエチレングリコール、グリセリン、プロピレングリコール又は他の合成溶媒のような、無菌の希釈剤;ベンジルアルコール又はメチルパラベンなどの、抗菌剤;アスコルビン酸又は亜硫酸水素ナトリウムなどの、抗酸化剤;エチレンジアミン四酢酸などの、キレート剤;酢酸塩、クエン酸塩又はリン酸塩などの緩衝剤、及び塩化ナトリウム又はデキストロースなどの張性調節剤。pHは、塩酸又は水酸化ナトリウムなどの、酸又は塩基により調節することができる。
【0098】
本発明のFc-EPOタンパク質製品の好ましい投与法は、非経口(例えば、IV、IM、SC、又はIP)経路によるものであり、並びに投与される組成物は通常、治療有効量の生成物を、許容できる希釈剤、担体及び/又は補助剤と組合せて含有する。有効量は実質的に治療される状態にかなり左右されると予想されるが、治療量は活性物質0.2〜2μg/kg体重の範囲であると現在予想されている。ヒト血清アルブミンのような標準希釈剤は生理食塩水のような標準担体として本発明の医薬組成物について企図されている。本発明の組成物における使用に適した補助剤物質は、テストステロン、前駆細胞刺激剤、インスリン-様増殖因子、プロスタグランジン、セロトニン、cAMP、プロラクチン及びトリヨードチロシンなどの赤血球形成刺激作用について独立して注目される化合物、更には一般に再生不良性貧血の治療に使用される物質、例えばメテノレン(methenolene)、スタノゾロール及びナンドロンなどを含む。例えば、Resegottiら、Panminerva Medics、23: 243-248 (1981);Mgonigleら、Kidney Int., 25(2): 437-444 (1984);Pavlovic-Kanteraら、Expt. Hematol.、8(Supp. 8): 283-291 (1980);及び、Kurtz、FEBS Letters、14a(1): 105-108 (1982)を参照のこと。
【0099】
補助剤として、Fc-EPOの作用を増強又はこれと相乗作用することが報告された物質、例えばアドレナリン作動薬、甲状腺ホルモン、アンドロゲン及びBPA、更には「肝赤血球形成性因子」(Naughtonら、Acta. Haemat.、69: 171-179 (1983)参照)及び「エリスロトロピン(erythrotropins)」(Congoteら、Abstract 364、Proceedings 7th International Congress of Endocrinology、ケベック市、ケベック州、1984年7月1-7日;Congote、Biochem. Biophys. Res. Comm.、115(2): 447-483 (1983);及び、Congote、Anal. Biochem.、140: 428-433 (1984)により説明)、及び「エリスロゲニン(erythrogenins)」(Rothmanら、J. Surg. Oncol.、20: 105-108 (1982)により説明)と称される化合物のクラスも企図される。
【0100】
経口又は非経口投与に有用な溶液は、例えば「レミントン薬科学」(Gennaro, A.編集)、Mack Pub.(1990)に記載されたような、医薬技術分野において周知の方法のいずれかにより調製され得る。非経口投与用の製剤は、口腔内投与のためのグリココール酸、直腸投与のためのメトキシサリチル酸、又は膣内投与のためのクエン酸も含むことができる。非経口用調製物は、ガラス又はプラスチック製のアンプル、使い捨て注射筒、又は反復投与用バイアル中に封入することができる。直腸投与用の坐薬も同じく、この薬剤をココアバター、他のグリセリド又は室温で固形であり体温で液体である他の組成物などの非-刺激性賦形剤と共に混合することにより調製することができる。製剤は、例えばポリエチレングリコールなどのポリアルキレングリコール、植物起源の油、水素化されたナフタレンなども含むことができる。直接投与するための製剤はグリセロール及び高粘度の他の組成物を含むことができる。これらの治療薬のためのその他の可能性のある有用な非経口担体は、エチレン-酢酸ビニルコポリマー粒子、浸透圧ポンプ、埋込み可能な注入システム、及びリポソームを含む。吸入投与するための製剤は、賦形剤として、例えば乳糖を含むか、又は例えば、ポリオキシエチレン-9-ラウリルエーテル、グリココール酸及びデオキシコール酸を含有する水溶液、又は点鼻薬の形の投与用の油性溶液、又は鼻腔内塗布用のゲル剤である。持続浣腸(retention enemas)も、直腸送達に使用することができる。
【0101】
注射用途に適した医薬組成物は、無菌の液剤(ここで水溶性)又は分散剤、及び無菌の注射用液剤もしくは分散剤を即時調製するための無菌の散剤を含む。静脈内投与に関して、適当な担体は、生理食塩水、静菌水、Cremophor ELTM(BASF、ピスカタウェイ、NJ)又はリン酸緩衝生理食塩水(PBS)を含む。全ての場合において、この組成物は無菌であり、及び容易に注射筒内に入れることができる程度に流体であることができる。これは、製造及び貯蔵の条件下では安定することができ、並びに細菌及び真菌などの微生物の混入作用に対し保存することができる。この担体は、例えば、水、エタノール、ポリオール(例えば、グリセロール、プロピレングリコール、及び液体ポリエチレングリコールなど)、並びにそれらの適当な混合物を含有する溶媒又は分散媒であることができる。例えば、レシチンのようなコーティングの使用によるか、分散剤の場合必要な粒子サイズの維持によるか、及び界面活性剤の使用により適当な流動性が維持される。微生物の作用の防止は、例えばパラベン、クロロブタノール、フェノール、アスコルビン酸、チメロサールなどのような、様々な抗菌剤及び抗真菌剤により実現される。多くの場合、組成物中に例えば、糖、ポリアルコール、例えばマンニトール、ソルビトール、及び塩化ナトリウムなどの等張化剤を含むことが好ましい。注射用組成物の持続吸収は、組成物中に吸収を遅延させる物質、例えばモノステアリン酸アルミニウム及びゼラチンを含有することによりもたらされる。
【0102】
無菌の注射用液剤は、必要量の活性化合物を適当な溶媒中に、必要ならば先に列記した成分の1種又は組合せと共に混入し、その後濾過滅菌することにより調製することができる。一般に分散剤は、活性化合物を、基本の分散媒及び先に列記されたものから必要な他の成分を含む無菌のビヒクルに混入することにより調製される。無菌の注射用液剤の調製のための無菌の散剤の場合、調製法は、活性成分に加え追加の所望の成分の散剤をそれらの先に濾過滅菌された溶液から得る、真空乾燥及び凍結乾燥を含む。
【0103】
ひとつの態様において、治療薬は、インプラント及び微小カプセル化された送達システムを含む、制御された放出製剤のような、迅速な体からの消失に対し保護する担体により調製される。エチレン-酢酸ビニル、ポリ無水物、ポリグリコール酸、コラーゲン、ポリオルトエステル、及びポリ乳酸などの、生分解性、生体適合性ポリマーを使用することができる。このような製剤の調製法は、当業者に明らかであろう。これらの材料は、Alza Corporation及びNova Pharmaceuticals, Inc.から市販されている。リポソーム懸濁液も、医薬として許容できる担体として使用することができる。これらは、当業者に公知の方法、例えば米国特許第4,522,811号に開示された方法に従い調製することができる。マイクロソーム及びマイクロ粒子も使用することができる。
【0104】
経口又は非経口組成物を、投与に容易でかつ均質な用量の単位剤形に製剤することができる。単位剤形は、治療される対象にとって単位用量として適した物理的に個別の単位を意味し;各単位は、所望の治療作用を生じるように計算された、予め決められた量の活性化合物を、必要な医薬担体と会合して含有する。本発明の単位剤形の特定は、活性化合物の独自の特性及び達成されるべき具体的な治療作用により指示されかつ直接左右され、並びにこれらの制限は、個体の治療に関するそのような活性化合物の混和の技術分野において固有のものである。
【0105】
Fc-EPOの治療有効量及び投与頻度の決定
一般に、本発明に従い作製されたFc-EPO融合タンパク質を含有する治療薬は、例えば治療有効量、すなわち望ましい作用を誘導するのに十分な時間、標的組織へ薬剤の適当な濃度を提供する量でヒト又は他の哺乳動物へ非経口投与又は経口投与用に製剤することができる。より詳細に述べると、本明細書において「治療有効量」は、目標ヘマトクリット、又は患者へ恩恵をもたらす目標ヘマトクリット範囲までのヘマトクリットの増大をもたらすか、あるいは目標ヘマトクリットに、もしくは目標ヘマトクリット範囲内に患者を維持するような、Fc-EPO融合タンパク質の量を意味する。この量は、個人毎に変動し、並びに患者の全身の生理的条件、貧血の重症度及び基礎となる原因、並びに個々の患者に関する最終的な目標ヘマトクリットを含む、多くの要因に左右されるであろう。目標ヘマトクリットは、典型的には少なくとも約30%、又は30%〜38%の範囲であり、好ましくは38%を上回り、より好ましくは40%〜45%である。rHuEpoに関する目標ヘマトクリット範囲に関する全般的指針は、1996年12月23日付けのEPOGEN(登録商標)の添付文書においても認められ、これはそこに記されているように、30%〜36%、あるいは32%〜38%である。そのような目標は個人毎に変動し、その結果、所定の患者について実際の目標ヘマトクリットの決定においては医師の判断が適当であろうことが理解される。しかしながら、目標ヘマトクリットの決定は十分に当業者の能力範囲内である。
【0106】
Fc-EPOタンパク質の治療有効量は、当業者により容易に確定され得る。実施例15は、週1回、2週に1回、及び1ヶ月に1回の投与の、Fc-EPOの治療有効量を決定するためのひとつの目標としての臨床プロトコールを示している。例えば、週1回又は2週に1回の投与量範囲は、約0.075〜約4.5μg Fc-EPO/kg/投与である。1ヶ月に1回の投与量範囲は、0.45〜4.5μg Fc-EPO/kg/投与である。
【0107】
治療的組成物中で送達される本発明のFc-EPO融合タンパク質の有効濃度は、投与される薬剤の最終の望ましい用量及び投与経路を含む、多くの要因に応じて変動するであろう。投与される好ましい用量は、恐らく治療される疾患又は適応症の種類及び程度、特定の患者の全身の健康状態、送達される治療薬の相対的生物学的効能(例えば、シアル酸化のレベル)、治療薬の製剤、製剤中の賦形剤の存在及び種類、並びに投与経路のような変数により決まるであろう。一部の態様において、本発明の治療薬は、非-ヒト霊長類及び齧歯類を使用する哺乳動物試験から推定された典型的単位投与量を用い、個体へ提供される。先に説明されたように、用量単位は、患者へ投与することが可能であり、容易に取扱い及び包装することができ、そのまま又はそれの固形もしくは液体の医薬希釈剤もしくは担体との混合物として治療薬を含有する物理的及び生物学的に安定した単位投与量として維持される単位投与量を意味する。
【0108】
Fc-EPO融合タンパク質を含有する治療薬の投与頻度は、治療される状態及び目標のヘマトクリットに応じて変動するが、一般には週3回未満であろう。投与頻度は、ほぼ週1回又は2回であってよい。投与頻度は、週に約1回未満、例えば2週間に約1回(14日間に約1回)、1ヶ月に1回又は2ヶ月に1回であることもできる。エリスロポエチン及びそのアナログに対する異なる個体による反応の変動のために、実際に使用される投与頻度は、本明細書に明らかにされた頻度から若干変動することができることが理解され;用語「約」は、そのような変動を反映することが意図されている。
【0109】
本発明は、治療時に増加した赤血球形成を維持するための、治療有効量の鉄分の投与も提供する。投与される量は、rHuEpoによる治療を基に当業者により容易に決定され得る。加えて本発明の治療薬は、単独で、又は特定の疾患又は関心のある適応症に有益な作用を有することがわかっている他の分子と組合せて投与することができる。単なる例として、有用な補因子は、防腐薬、抗生物質、抗ウイルス薬、及び抗真菌薬、並びに鎮痛薬及び麻酔薬などの、症状を緩和する補因子を含む。
【0110】
プロドラッグ
本発明の治療薬は、「プロドラッグ」誘導体も含む。用語プロドラッグは、活性成分を放出もしくは活性化するために、生体内で自発的又は酵素的のいずれかの生体転換を必要とする、親分子の薬理学的に不活性(又は部分的に不活性)の誘導体を意味する。プロドラッグは、代謝条件下で切断可能な基を有する、本発明の治療薬の変種又は誘導体である。プロドラッグは、生理的条件下で加溶媒分解を受けるか又は酵素分解を受ける場合に、in vivoにおいて医薬活性のある本発明の治療薬となり始める。本発明のプロドラッグは、生体内での活性のある薬剤成分の放出又は活性化に必要な生体転換工程の数に応じてシングル、ダブル、トリプルなどと称され、これは前駆体型に存在する官能基の数を示す。プロドラッグ型は、哺乳動物生体において、溶解度、組織適合性、又は遅延した放出の利点を提供することが多い(Bundgard、Design of Prodrugs、7-9頁、21-24頁(1985)、Elsevier, Amsterdam;Silverman、The Organic Chemistry of Drug Design and Drug Action、352-401頁(1992)、Academic Press、サンディエゴ、CA)。更に本発明のプロドラッグ誘導体は、バイオアベイラビリティを増強するために、他の特徴と組合せることができる。
【0111】
In vivo発現
本発明のFc-EPO融合タンパク質はin vivo発現法により提供することができる。例えば、Fc-EPO融合タンパク質をコードしている核酸は、有利なことに造血障害又は欠損を罹患した患者へ直接投与することができるか、又は細胞へex vivoで提供し、その後生存細胞を患者へ投与することができる。当該技術分野において公知であるin vivo遺伝子療法は、精製されたDNA(例えば、プラスミド内)を提供すること、ウイルスベクター内のDNAを提供すること、又はリポソームもしくは他のベシクル内のDNAを提供することを含む(例えば、遺伝子療法において使用する脂質担体を開示している、米国特許第5,827,703号、及び遺伝子療法において有用なアデノウイルスベクターを提供する、米国特許第6,281,010号を参照のこと)。
【0112】
組換えタンパク質を発現するように改変されている細胞を移植することにより疾患を治療する方法も、公知である。例えばヒトへ導入するために、核酸を初代ヒト細胞へ導入する方法を開示している、米国特許第5,399,346号を参照のこと。
In vivo発現法は、精製することなく、タンパク質の標的組織又は細胞コンパートメントへの直接の送達に特に有用である。本発明において、Fc-EPOをコードしている配列を使用する遺伝子療法は、貧血を含む、血液系の不調の様々な病態、障害及び状態における、特に慢性腎不全などに随伴した型の貧血の調整における用途が認められる。Fc-EPO融合タンパク質をコードしている核酸配列は、適当な転写又は発現カセットに挿入し、宿主哺乳動物へ裸のDNAとして又は適当な担体と複合して導入することができる。活性Fc-EPOタンパク質の生成のモニタリングは、核酸ハイブリダイゼーション、ELISA、ウェスタンハイブリダイゼーション、及び当業者に公知の他の適当な方法により行うことができる。
【0113】
導入遺伝子の全身投与によって複数の組織を形質転換することができることがわかっている。哺乳動物宿主へのカチオン性脂質担体/外来性DNA複合体の静脈内注射後の外来性DNAの発現は、Tリンパ球、細網内皮系、心内皮細胞、肺細胞、及び骨髄細胞、例えば骨髄由来の造血細胞を含む、複数の組織において示されている。
米国特許第6,627,615号に説明されたin vivo遺伝子療法の送達技術は、動物において無毒であり、並びに導入遺伝子発現は、単回投与後少なくとも60日間最後まで示される。導入遺伝子は、サザン分析により測定して、in vivoにおいて検出可能なレベルで、宿主細胞DNAへ組込まれるようには見えず、このことは、遺伝子治療に関するこの技術は、癌を惹起する癌遺伝子を活性化するか、又は癌を予防する腫瘍抑制遺伝子を停止するように、正常な細胞遺伝子の発現を変更することにより、宿主哺乳動物に問題を引き起こさないことを示唆している。
【0114】
本発明の具体的態様を以下に列挙する。
(1) ヒンジ領域、CH2及びCH3ドメインを含むヒトIgG分子の二量体Fc部分、並びにヒトエリスロポエチン(EPO)から本質的になる、精製された二量体融合タンパク質であって、前記二量体Fc部分の各鎖は、そのC-末端でEPO分子のN-末端へ直接又はリンカーペプチドを介して連結しており、下記の特徴を有する前記二量体融合タンパク質:
(i) 15〜28個のシアル酸残基を有することにより高度にシアル酸化されている;;
(ii) 前記CH2領域がヒトIgG2に由来し、前記CH2領域中のGln-Phe-Asn-Ser配列内のPheおよびAsnがAlaおよびAsnで置換され、その結果前記CH2領域内に配列Gln-Ala-Gln-Serが形成されることによって前記CH2領域が改変されている;および、
(iii)CH3ドメインのC-末端近傍のLeu-Ser-Leu-Serアミノ酸配列が、Ala-Thr-Ala-Thrで置換されている。
(2) 更に、CH3ドメインのC-末端Lys残基がAlaに置換されている、(1)記載の二量体Fc-EPO融合タンパク質。
(3) ヒンジ領域がヒトIgG1に由来する、(1)又は(2)記載の二量体Fc-EPO融合タンパク質。
(4) IgG1ヒンジ領域中のPro-Lys-Ser-Cys-Asp-Lys配列内のCysアミノ酸残基がSer残基に置換され、前記ヒンジ領域中に配列Pro-Lys-Ser-Ser-Asp-Lysが形成されることによって、I前記ヒンジ領域が改変されている(3)記載の二量体Fc-EPO融合タンパク質。
(5) エリスロポエチン部分が少なくとも以下のアミノ酸置換の一つを含む、(1)〜(4)のいずれかに記載のFc-EPO融合タンパク質:
(i)EPO分子の29位における非システイン残基;
(ii)EPO分子の33位における非システイン残基;
(iii)EPO分子の88位におけるシステイン残基;
(iv)EPO分子の139位におけるシステイン残基。
(6) EPO分子の33位のCys残基の代わりに非-Cysアミノ酸残基が存在し、EPO分子の88位のTrpの代わりにCys残基が存在し、それによって融合タンパク質内のEPO 部分がCys29-Cys88ジスルフィド結合を形成することが可能である、(5)記載の二量体Fc-EPO融合タンパク質。
(7) 33位の非-Cysアミノ酸残基がProである、(6)記載の二量体Fc-EPO融合タンパク質。
(8) EPO部分が以下から選ばれる1以上の変異を含む(1)〜(7)のいずれかに記載の二量体Fc-EPO融合タンパク質:
(i)Arg131→Glu131;
(ii)Arg139→Glu139;
(iii)His32→Gly32
(iv)Ser34→Arg34;
(v)Pro90→Ala90。
(9) リンカーペプチドがグリコシル化部位を含む、(1)〜(8)のいずれかに記載の二量体Fc-EPO融合タンパク質。
(10) グリコシル化部位がAsn-Ala-Thrアミノ酸配列を含む、(9)記載の二量体Fc-EPO融合タンパク質。
(11) 完全なIgG分子がIgG2に由来し、ヒンジ領域がIgG1に由来する、(1)〜(10)のいずれかに記載の二量体Fc-EPO融合タンパク質。
(12) 更にCH1ドメインを含む、(1)〜(11)のいずれかに記載の二量体Fc-EPO融合タンパク質。
(13) 融合タンパク質が、20〜22個のシアル酸残基を有する、(1)〜(12)のいずれかに記載の二量体Fc-EPO融合タンパク質。
(14) 配列:
EPKSSDKTHTCPPCPAPPVAGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVQFNWYVDGVEVHNAKTKPREEQAQSTFRVVSVLTVVHQDWLNGKEYKCKVSNKGLPAPIEKTISKTKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPMLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSATATPGAAPPRLICDSRVLERYLLEAKEAENITTGCAEHCSLNENITVPDTKVNFYAWKRMEVGQQAVEVWQGLALLSEAVLRGQALLVNSSQPWEPLQLHVDKAVSGLRSLTTLLRALGAQKEAISPPDAASAAPLRTITADTFRKLFRVYSNFLRGKLKLYTGEACRTGDR (配列番号:14)
を含む、二量体Fc-EPO融合タンパク質。
(15) 配列:
EPKSSDKTHTCPPCPAPPVAGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVQFNWYVDGVEVHNAKTKPREEQAQSTFRVVSVLTVVHQDWLNGKEYKCKVSNKGLPAPIEKTISKTKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPMLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSATATPGAAPPRLICDSRVLERYLLEAKEAENITTGCAEGPSLNENITVPDTKVNFYAWKRMEVGQQAVEVWQGLALLSEAVLRGQALLVNSSQPCEALQLHVDKAVSGLRSLTTLLRALGAQKEAISPPDAASAAPLRTITADTFRKLFRVYSNFLRGKLKLYTGEACRTGDR (配列番号:15)
を含む、二量体Fc-EPO融合タンパク質。
(16) (1)〜(15)のいずれかに記載の融合タンパク質をコードするDNA分子。
(17) (1)〜(15)のいずれかに記載の融合タンパク質を効果的量で含み、場合により製薬的に許容される単体、希釈剤または賦形剤とを含む、哺乳動物の造血疾患または不全の治療に適した医薬組成物。
(18) N-末端側にFc部分を含みC-末端側にエリスロポエチンを含む、哺乳動物への投与に適した高度にシアル酸化された精製されたFc-EPO融合タンパク質の集団であって、前記精製されたFc-EPO融合タンパク質1分子あたり平均15〜28個のシアル酸残基を有し、それぞれのFc-EPO融合タンパク質をコードするDNA分子をBHK細胞に導入し、対応する前記Fc-EPO融合タンパク質を発現、単離及び精製することによって得られ、NS/0、PerC6または293細胞中で合成された対応するFcーEPO融合タンパク質の集団に比較して長い血清半減期を有する前記Fc-EPO融合タンパク質の集団。
(19) 精製されたFc-EPO融合タンパク質1分子あたり平均20〜22個のシアル酸残基を有する、(18)に記載のFc-EPO融合タンパク質の集団。
(20) BHK細胞が無タンパク質培地又は懸濁液中での増殖に適応している、(18)又は(19)に記載の精製されたFc-EPO融合タンパク質の集団。
(21) 以下の工程を含む、N-末端側にFc部分を含みC-末端側にエリスロポエチンを含む、哺乳動物への投与に適した高度にシアル酸化された精製されたFc-EPO融合タンパク質の集団を作製する方法:
(i)Fc-EPO融合タンパク質をコードするDNA分子を構築する工程;
(ii)無タンパク質培地中または懸濁状態で、前記DNA分子でBHK細胞を形質転換する工程;
(iii)前記DNA分子よってコードされるFc-EPO融合タンパク質の集団を発現させる工程;
(iv)前記Fc-EPO融合タンパク質の集団を回収、単離および精製する工程。
(22) 作製された融合タンパク質の集団が、精製されたFc-EPO融合タンパク質あたり平均15〜28個のシアル酸残基を有する、(21)記載の方法。
(23) 作製された融合タンパク質の集団が、精製されたFc-EPO融合タンパク質あたり平均20〜22個のシアル酸残基を有する、(22)記載の方法。
(24) 以下の工程を含む、Fc部分およびエリスロポエチン部分を含むFc-EPO融合タンパク質をコードする核酸配列を安定に維持するBHK細胞を選抜する方法:
(a)ハイグロマイシンBをコードする核酸配列および前記Fc-EPO融合タンパク質をコードする核酸配列をBHK細胞に導入する工程;
(b)前記BHK細胞をハイグロマイシンBの存在下で培養する工程。
(25) ハイグロマイシンBをコードしている核酸配列及びFc-EPO融合タンパク質をコードする核酸配列が単一のDNA分子中に存在する、(24)記載の方法。
【実施例1】
【0115】
実施例1. Fc-EPO融合タンパク質をコードしている構築物
正常なエリスロポエチン部分を含むFc-EPO融合タンパク質をコードしているプラスミドphC10-Fcg2h(FN→AQ)-M1-EPO及びNDS変異を伴うFc-EPO融合タンパク質をコードしているプラスミド phC10-Fcg2h(FN→AQ)-M1-EPO(NDS)を以下のように構築した。
ヒトエリスロポエチンをコードしている核酸配列を、哺乳動物細胞における高発現のためにコドン-最適化した。例えば配列番号:3は翻訳を最適化するように改変されたコドンを伴う、成熟型ヒトエリスロポエチンのコード配列の例を示している。5'末端の配列は、サブクローニングを容易にするためにSmaI部位を含むように改変した。
【0116】
配列番号:3
CCCGGGtGCCCCACCACGCCTCATCTGTGACAGCCGAGTgCTGGAGAGGTACCTCTTGGAGGCCAAGGAGGCCGAGAATATCACGACcGGCTGTGCTGAACACTGCAGCTTGAATGAGAAcATCACcGTgCCtGACACCAAAGTgAATTTCTATGCCTGGAAGAGGATGGAGGTtGGcCAGCAGGCCGTAGAAGTgTGGCAGGGCCTGGCCCTGCTGTCGGAAGCTGTCCTGCGGGGCCAGGCCCTGTTGGTCAACTCTTCCCAGCCGTGGGAGCCCCTGCAaCTGCATGTGGATAAAGCCGTgAGTGGCCTTCGCAGCCTCACCACTCTGCTTCGGGCTCTGgGAGCCCAGAAGGAAGCCATCTCCCCTCCAGATGCGGCCTCAGCTGCTCCcCTCCGcACAATCACTGCTGACACTTTCCGCAAACTCTTCCGAGTCTACTCCAATTTCCTCCGGGGAAAGCTGAAGCTGTACACAGGGGAGGCCTgcCGGACAGGGGACAGATGActcgag
(小文字は、野生型ヒトエリスロポエチンコード配列と異なる塩基を示す。これらの変更は、哺乳動物細胞における発現レベルを増加させると期待されるが、発現されたタンパク質配列を変化させない。)
【0117】
NDS変異は、こその開示は本明細書に参照として組入れられている、PCT国際公開公報第01/36489号に開示された位置指定突然変異誘発により、エリスロポエチン部分へ導入した。例えば、国際公開公報第01/36489号に開示された、アミノ酸置換His32Gly、Cys33Pro、Trp88Cys、及びPro90Alaを生じる変異を伴うヒトエリスロポエチンコード配列の形を含むXmaI-XhoI DNA断片を使用した。対応するタンパク質配列を配列番号:4に示した。
【0118】
APPRLICDSRVLERYLLEAKEAENITTGCAEGPSLNENITVPDTKVNFYAWKRMEVGQQAVEVWQGLALLSEAVLRGQALLVNSSQPCEGLQLHVDKAVSGLRSLTTLLRALGAQKEAISPPDAASAAPLRTITADTFRKLFRVYSNFLRGKLKLYTGEACRTGDR (配列番号:4)
【0119】
IgG2-由来のCH2ドメイン及びIgG1-由来のヒンジ領域を含むハイブリッドFc部分を、米国特許公報第20020147311号、及び例えば国際公開公報第01/058957号に開示されたように構築した。
ある形のエリスロポエチンをコードしているXmaI-XhoI DNA断片を、プラスミドベクター、例えばpdCs-Fc-Xへ挿入したが、このベクターは、CH3 C-末端領域におけるアミノ酸置換を生じる2セットの変異(ここではM1セット変異と称される)が存在することを除き、IgG1由来の変更されたヒンジ領域並びにIgG2由来のCH2及びCH3領域をコードしており、その結果CH3 C-末端及びEPO N-末端の接合部の配列は、以下である:
【0120】
. . . . TQKSATATPGA-APPRLI . . . .(配列番号:5)
【0121】
IgG2 CH3領域の配列KSLSLSPG (配列番号:6)をKSATATPG (配列番号:7)へ変更する変異の第一のセットは、国際公開公報第02/079232号に開示されている。Leu-Ser-Leu-Ser (配列番号:6の3位から6位)のAla-Thr-Ala-Thr (配列番号:7の3位から6位)による置換の効果は、ヒトFcとヒトエリスロポエチンの間の接合部が非-自己ペプチド配列を含むことによって生じる可能性のあるヒト非-自己T-細胞エピトープを除去することである。CH3領域のC-末端アミノ酸の単一アミノ酸置換KからAを含む第二のセットは、国際公開公報第01/58957号に開示されている。
【0122】
Fc融合タンパク質の発現のための発現ベクターpdCs-Fc-Xは、Loらの論文(Protein Engineering、11: 495 (1998))に記載されている。プラスミドphC10-Fc-Xを、pdCs-Fc-Xから、メトトレキセート耐性を付与するジヒドロ葉酸還元酵素(DHFR)遺伝子のコード領域を、ハイグロマイシンBに対する耐性を付与する遺伝子と置換することにより、構築した。NheI/NsiIハイグロマイシンB DNA断片は、プライマー5'-GCTAGCTTGGTGCCCTCATGAAAAAGCCTGAACTC-3' (配列番号:8)及び5'-ATGCATTCAGTTAGCCTCCCCCATC-3' (配列番号:9)を使用する、鋳型プラスミドpCEP4 (Invitrogen)からのハイグロマイシンB遺伝子のPCR増幅により得た。PCR断片をTAクローニングベクターpCR2.1 (Invitrogen)にクローニングし、その配列を確認した。
【0123】
プラスミドphC10-Fcg2h-M1-EPO(NDS)を、pdCs-Fcg2h-M1-EPO(NDS)由来のNheI/AflI及びAflII/NsiI DNA断片並びにNheI/NseIハイグロマイシンB断片のトリプルライゲーションにより作製した。
更に、PCR突然変異誘発により、IgG2重鎖のCH2ドメイン内のGln-Phe-Asn-Serアミノ酸配列内に可能性のあるT-細胞エピトープ及びFc部分内のN-結合グリコシル化を除去する二重アミノ酸置換「FN>AQ」を生じさせる変異を導入した。変異導入プライマー5'-AGCAGGCCCAGAGCACGTTCCGTGTGGT-3' (配列番号:10)及び5'-GAACGTGCTCTGGGCCTGCTCCTCCCGT-3' (配列番号:11)を、各々、SacII部位を含む下流プライマー5'-CCCCGCGGGTCCCACCTTTGG-3' (配列番号:12)及びPvuII部位を含む上流プライマー5'-CCCAGCTGGGTGCTGACACGT-3' (配列番号:13)と組み合わせ、ふたつの重複するDNA断片を鋳型DNA pdC10-Fcg2h-M1-EPO(NDS)から増幅した。第二の増幅ラウンドにおいて、変異(FN→AQ)を含むPvuII/SacII断片を、第一の増幅ラウンドのPCR産物から、上流プライマー(配列番号:13)及び下流プライマー(配列番号:12)を用いて増幅した。PvuII/SacII断片を、TAベクターpCR2.1 (Invitrogen)へクローニングし、その配列を検証した。構築物pdC10-Fcg2h(FN>AQ)-M1-EPO(NDS)を、pdC10-Fcg2h-M1-EPO由来のPvuII/SacII断片、XhoI/SacII断片、並びにpdC10-Fcg2h-M1-EPO(NDS)由来のXhoI/PvuII断片のトリプルライゲーションから作製した。
【0124】
FN>AQ変異をプラスミドphC10-Fcg2h-M1-EPOへ導入するために、phC10-Fcg2h-M1-EPO及びpdC10-Fcg2h(FN→AQ)-M1-EPO由来の適当なDNA断片を組合せた。phC10-Fcg2h-M1-EPO及びpdC10-Fcg2h(FN→AQ)-M1-EPOの両構築物をXhoI及びXbaIで消化し、5.7kbのXhoI/XbaI phC10-Fcg2h-M1-EPO(NDS)断片を、1.9kbのpdC10-Fcg2h(FN→AQ)-M1-EPO断片にライゲーションし、phC10-Fcg2h(FN→AQ)-M1-EPOを作出した。
【0125】
FN→AQ変異をプラスミドphC10-Fcg2h-M1-EPO(NDS)へ導入するために、phC10-Fcg2h-M1-EPO(NDS)及びphC10-Fcg2h(FN→AQ)-M1-EPO由来の2種の適当なXhoI/SmaI消化した断片を互いにライゲーションし、phC10-Fcg2h(FN→AQ)-M1-EPO(NDS)を作出した。
【0126】
pdC10-huFcg2h(FN→AQ)-M1-EPOによりコードされたFc-EPOのアミノ酸配列を、配列番号:14に示した。
[EPKSSDKTHTCPPCP]APPVAGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVQFNWYVDGVEVHNAKTKPREEQAQSTFRVVSVLTVVHQDWLNGKEYKCKVSNKGLPAPIEKTISKTKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPMLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSATATPGAAPPRLICDSRVLERYLLEAKEAENITTGCAEHCSLNENITVPDTKVNFYAWKRMEVGQQAVEVWQGLALLSEAVLRGQALLVNSSQPWEPLQLHVDKAVSGLRSLTTLLRALGAQKEAISPPDAASAAPLRTITADTFRKLFRVYSNFLRGKLKLYTGEACRTGDR (配列番号:14)
【0127】
pdC10-huFcg2h(FN→AQ)-M1-EPO(NDS)によりコードされたFc-EPO(NDS)のアミノ酸配列を、配列番号:15に示した。
[EPKSSDKTHTCPPCP]APPVAGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVQFNWYVDGVEVHNAKTKPREEQAQSTFRVVSVLTVVHQDWLNGKEYKCKVSNKGLPAPIEKTISKTKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPMLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSATATPGAAPPRLICDSRVLERYLLEAKEAENITTGCAEGPSLNENITVPDTKVNFYAWKRMEVGQQAVEVWQGLALLSEAVLRGQALLVNSSQPCEALQLHVDKAVSGLRSLTTLLRALGAQKEAISPPDAASAAPLRTITADTFRKLFRVYSNFLRGKLKLYTGEACRTGDR (配列番号:15)
下線を付けた配列トラックはEPO部分を表し、下線を付けた[]内配列は、IgG1ヒンジを表し、下線を付けていない配列は改変されたIgG鎖のCH2及びCH3ドメインを表し、ここで点線を付した配列はCH3ドメインを表す。
【実施例2】
【0128】
実施例2. 様々な細胞株におけるFc-EPOの発現
融合タンパク質の即時分析のために、標準の一過性トランスフェクション法、例えば、リン酸カルシウム-媒介型DNA共沈法(Sambrookら、Molecular Cloning: A Laboratory Manual、第2版、Cold Spring Harbor Laboratory Press、 (1989))、又はLipofectamine Plus (Life Technologies)を製造業者のプロトコールに従い使用するリポフェクション法により、プラスミドphC10-Fcg2h(FN→AQ)-M1-EPO(NDS)又はphC10-Fcg2h(FN→AQ)-M1-EPOを、適当な組織培養細胞へ導入した。
【0129】
安定的にトランスフェクションされたBHK-21細胞を得るために、プラスミドphC10-Fcg2h(FN→AQ)-M1-EPO(NDS)又はphC10-Fcg2h(FN→AQ)-M1-EPOをエレクトロポレーションによりBHK-21細胞へ導入した。高効率のエレクトロポレーションのために、BHK-21細胞はMEM培地(アメリカン・タイプ・カルチャー・コレクション(ATCC)の推奨により、非-必須アミノ酸及びピルビン酸ナトリウムを補充)中で増殖させ、PBSで1回洗浄し;並びに、約5x106個細胞を、0.5ml PBS中に再懸濁し、0.4cm電極間隙を備えたGene Pulser(商標)キュベット(BioRad, ハーキュレス, CA)中で、直鎖状プラスミドDNA 10μgと共に、氷上で、10分間インキュベーションした。エレクトロポレーションは、0.25V及び500μFに設定したGene Pulser(商標)(BioRad, ハーキュレス, CA)を用いて行った。細胞を氷上で10分間回復させ、増殖培地中に再懸濁し、2個の96ウェルプレート上に播種した。ハイグロマイシンB(Hyg B)を、トランスフェクション後2日間、濃度300μg/mlで増殖培地へ添加した。これらの細胞は、3日毎に2〜3回培地供給し(fed)、Hyg B耐性安定クローンが、2〜3週間で出現した。
【0130】
高レベルFc-EPO融合タンパク質を産生している安定クローンを同定するために、クローン上清を抗-Fc抗体によるELISAによりアッセイした。高-産生クローンを単離し、300μg/ml Hyg Bを含有する増殖培地において増殖させた。タンパク質産生の目的では、BHK-21細胞を、補充したDMEM/F-12培地又はVP-SFM (Life Technologies)などの別の適当な培地においてルーチン的に生育させた。Fc-EPO融合タンパク質を、標準的ノーマル-フロー濾過により、コンディション培地から収集し、清澄化した材料を更なる精製まで4℃で貯蔵した。典型的には、ローラーボトル産生モードでFc-EPOタンパク質6〜12μg/mlがBHK-21細胞から得られた。
【0131】
Fc-EPO融合タンパク質は、NS/0細胞においても発現され、これを回収した。プラスミドpdC10-Fcg2h(FN→AQ)-M1-EPO又はpdC10-Fcg2h(FN→AQ)-M1-EPO(NDS)を安定して維持しているNS/0クローンを、先にPCT国際公開公報第01/36489号に開示された方法により確立した。この文献の開示は本明細書に引用により取り込まれるものとする。典型的には、NS/0細胞からFc-EPOタンパク質50〜100μg/mlが得られた。
【実施例3】
【0132】
実施例3. 懸濁物及び/又は無タンパク質培地中の増殖へのBHK細胞の適応
BHKは、一般に例えばMEM+10%熱-失活したウシ胎仔血清(FBS)のような血清-含有培地において増殖される接着細胞株である。BHK細胞を維持および拡張(増殖)するために、これらは定期的に(例えば4日間隔で)、典型的にはトリプシン-EDTA溶液の作用によりそれらの基質から剥離し、新鮮培地に希釈し、適当な容器に再播種される。しかしBHK細胞は、以下の手順により、懸濁液並びに無血清及び/又は無タンパク質培地における増殖に適応させることができる。
【0133】
典型的適応プロセスにおいて、BHK細胞をまずMEM+FBS:標的培地の75:25(v/v)混合培地中で指数増殖期まで培養し、続いて当初の培地:標的培地の50:50(v/v)、25:75(v/v)、及び最後に0:100(v/v)の中で、適当な細胞密度でサブクローニングした。適応プロセスの間、BHK細胞の増殖を、目視検査によりモニタリングした。以下の無血清培地を、それらに対する適応性について試験した:293 SFM II (Invitrogen Corp., カタログ番号11686-929)、CHO-S-SFM II (Invitrogen Corp., カタログ番号12052-098)、VP-SFM (Invitrogen Corp., カタログ番号11681-020)、Opti-Pro SFM (Invitrogen Corp., カタログ番号12309)、CDハイブリドーマ(Invitrogen Corp., カタログ番号11279-023)、及びH-SFM (Invitrogen Corp., カタログ番号12045-076)。
【0134】
この適応プロセスの間にBHK細胞を接着細胞株から懸濁細胞株へ転換するために、この培養混合液を各継代前に静置し、細胞懸濁液の上層25%を取り出して新鮮培地へ希釈した。凝集した細胞は、単独細胞及び対細胞(doublet cell)よりもより迅速に培養容器の底に沈降するので、上層25%の細胞懸濁液は一般に凝集の最低量を示す細胞を含む。従って各継代は凝集しにくいBHK細胞を拡張および濃縮し、Fc-EPOタンパク質を発現しているBHKクローンの浮遊細胞株がこの方法で樹立された。
【0135】
Fc-EPOタンパク質を発現しているBHK細胞は、VP-SFM又はOpti-PRO SFM無血清培地における増殖に適応し、浮遊培養物が得られたことがわかった。Fc-EPO融合タンパク質を発現しているBHK細胞は、以下の無血清培地において増殖することはできなかった:293 SFM II、CHO-S-SFM II、CDハイブリドーマ、及びH-SFM。
無血清培地VP-SFMに適応されたBHK細胞は、75:25(v/v)、50:50(v/v)、25:75(v/v)、最後に0:100(v/v)のVP-SFM:DMEM/F-12混合液中で、BHK細胞を適当な細胞密度で逐次培養することにより、無タンパク質培地、例えばDMEM/F-12 (Invitrogen Corp., カタログ番号11039-021)における増殖に更に適応した。無タンパク質培地DMEM/F-12には、グルタミン(最終6mM)、2g/l HyPep 4601 (Quest International, シカゴ, IL, カタログ番号5Z10419)、2g/l HyPep 1510 (Quest International, シカゴ, IL, カタログ番号5X59053)、10μl/l(v/v)エタノールアミン(Sigma, カタログ番号E0135)、及び5μM Tropolone (Sigma, カタログ番号T7387)を補充した。補充したDMEM/F-12における増殖に対しコンピテントなFc-EPO融合タンパク質を安定して発現しているBHK細胞株をこの方法により得、高い細胞生存率で維持した。
【実施例4】
【0136】
実施例4. 凝集状態のタンパク質の精製及び特徴決定
分析のために、Fc-EPO融合タンパク質を、細胞-培養上清から、Fc部分のプロテインAへの親和性を基に、プロテインAクロマトグラフィーにより精製した。Fc-EPOタンパク質を発現している細胞の馴化した上清を、予め平衡化したファストフロー・プロテインA Sepharoseカラムに装加した。カラムをリン酸ナトリウム緩衝液(150mMリン酸ナトリウム、100mM NaCl、中性pH)で十分洗浄した。結合したタンパク質を低pH(pH2.5〜3)のリン酸ナトリウム緩衝液(前記組成)で溶離し、溶離された画分を即座に中和した。
【0137】
異なる細胞株により産生されたFc-EPO融合タンパク質の凝集状態を評価するために、プロテインA精製された試料を分析用サイズ排除クロマトグラフィー(SEC)により分析した。試料を、HPLC-SEC(例えば、Super 3000 SW, TosoHaas, Montgomeryville, PA)により、流量0.35ml/分で15分間流し、分画した。BHK細胞から産生したFc-EPOタンパク質のかなりの部分(例えば、総収量の最大90%〜100%)は凝集しなかった。更にFc-EPO融合タンパク質の試料を、還元SDS-PAGE(予め成形されたNuPAGE 4%〜12%ゲル、NuPAGE, Novex)により分析し、実質的に1本バンドであることを明らかにした。このことはこれらの生成物が、標準操作手法を行った場合、分解に抵抗性があることを示している。
【0138】
懸濁液、無血清培地、及び/又は無タンパク質培地において増殖したBHK細胞から精製されたFc-EPO融合タンパク質も前述のSDS-PAGE及び分析用SECにより特徴決定した。これらのタンパク質は、血清-含有培地において増殖したBHK細胞において合成されたタンパク質同様、実質的に凝集せず、分解されないことがわかった。
【実施例5】
【0139】
実施例5A. グリコシル化パターンの特徴決定
ヒトエリスロポエチンのセリン126は、O-グリコシル化と適合性のある配列内であり、全ての哺乳動物エリスロポエチンタンパク質において保存されている。しかしセリン126は、そのタンパク質の残余に対し密にパックされない「フロッピー-ループ(floppy loop)」内にある。O-グリコシル化が存在しない場合、エリスロポエチンのこの領域はタンパク質分解に特に感受性がある。
【0140】
異なる細胞株で産生されたFc-EPOタンパク質のSer126でのO-グリコシル化の状態を、逆相HPLCにより試験した。試料は変性及び還元し、0.1%トリフルオロ酢酸(TFA)に希釈し、逆相HPLCカラム(例えば、Vydac C4カラム, Grace Vydac)へ注入した。アセトニトリル中0.085%TFAまでの勾配を適用し、タンパク質試料の保持時間を記録した。BHK-21細胞において合成されたFc-EPO及びFc-g2h(FN→AQ)-EPOは、ふたつの部分的に重複する大きいピークを生じたことがわかった(ピーク#1及びピーク#2)。これらのピーク画分は更に、ペプチドマッピングにより分析した。ピーク#1は、特徴的ペプチド(ペプチド#36)の非存在により示されるように、Ser126でグリコシル化されたFc-EPOの形態に対応するのに対し、ピーク#2は、特徴的ペプチド(ペプチド#36)の存在により示されるように、Ser126でグリコシル化されないFc-EPOの形態に対応することがわかった。Ser126は、BHK細胞から産生されたFc-EPO分子の約60%においてO-グリコシル化により修飾されることが明らかになる、これは天然のEPOについて報告されているものと一致する。更に、補充した無タンパク質DMEM/F-12培地中でBHK細胞を増殖させることは、O-グリコシル化の頻度に正の効果を有した。
【0141】
実施例5B. シアル酸化パターンの特徴決定
NS/0、BHK、293、及びPerC6細胞において合成されたFc-EPO融合タンパク質のシアル酸化の程度を等電点(IEF)ゲル電気泳動により比較した。簡単に述べると、2mg/mlに濃縮し及び必要ならば脱塩した試料を、等量のIEF試料緩衝液(pH3-7)に添加し、垂直の予め成形されたNovex pH 3-7 IEFゲル(Novex, カタログ番号EC6655B/B2)上で2.5時間、最初の1時間は100V、次の1時間は200Vで及び最後の30分間は500Vで流した。その後ゲルを固定、染色及び脱染色した。
【0142】
ひとつの特定の実験において、以下の試料を比較した(試料は、血清-含有培地において増殖した細胞に由来した):
1. NS/0由来のFcg2h-EPO(NDS)
2. BHK-21由来のFcg2h-EPO(NDS)
3. BHK-21由来のFcg2h-EPO
4. BHK-21由来のFcg2h(“Delta Lys”)-EPO
5. BHK-21由来のFcg4h(FN→AQ “Delta Lys”)-EPO
6. BHK-21由来のFcg4h(“Delta Lys”)-EPO
【0143】
この群において、「Delta Lys」は、FcドメインのC-末端におけるリジンの欠失を意味する(試料4-6)。試料1-3は、このC-末端リジンのアラニンへの変異を有する。従ってこのC-末端リジンは全ての試料において存在せず、試料間に生じる電荷の差は存在しない。全ての細胞は、血清-含有培地において接着細胞として増殖させた。
【0144】
試料を、pH3-7 IEFゲルに載せ、pH3.5、4.2、4.5、5.2、5.3、6.0、及び6.9に焦点のある標準 (Serva Electrophoresis, 独国)と比較した。第一の試料のNS/0由来のFcg2h-EPO(NDS)は、等電点約pH5.3〜6.5のバンド分布として移動し;最も強いバンドは、pH6.0-6.1に存在した。第二の試料のBHK-21由来のFcg2h-EPO(NDS)は、等電点約pH4.6〜pH5.0に強いバンド、pH5.0〜約pH6.0に弱いバンドの分布として泳動され;最も強いバンドは、pH4.8-4.9に存在した。第三及び第四の試料のBHK-21由来のFcg2h-EPO及びBHK-21由来のFcg2h(“Delta Lys”)-EPOは、各々、両方とも約pH4.7〜6.0にバンド分布を有し、最も強いバンドは約pH5.3に焦点があった。第五及び第六の試料のBHK-21由来のFcg4h(FN→AQ “Delta Lys”)-EPO及びBHK-21由来のFcg4h(“Delta Lys”)-EPOは、各々、第二の試料に類似した収束パターンを有していた。すなわち、これらは等電点約pH4.6〜pH5.0の強いバンド、pH5.0〜約pH6.0の弱いバンドの分布として泳動された。これらの結果は、BHK細胞におけるFc-EPO融合タンパク質の合成は、一般にNS/0細胞において合成された同じ又は類似の生成物よりも、有意により酸性の生成物を生じることを示している。
【0145】
別の実験において、BHK細胞由来のFcg2h-M1-EPO(NDS)試料をノイラミニダーゼにより処理し、オリゴ糖からシアル酸を除去した。得られたノイラミニダーゼ-処理した試料は、IEFゲル上で泳動し、pH6.9又はそれより大きいところに数個のバンドがフォーカシングすることが認められた。ノイラミニダーゼ-処理を伴う又は伴わないBHK細胞由来の試料並びにNS/0細胞由来の試料のバンド形成パターンを比較し、約27種の個別のシアル酸化された種を同定した。これらの27種は、ホモ二量体配置のFc-EPO融合タンパク質のシアル酸化の程度の変動により生じ得る予想される28の異なる種と良く対応している。この分析に従い、4〜5個のシアル酸残基を伴うFcg2h-EPOはpH6.9マーカーと共に収束し、及び11〜12個のシアル酸残基を伴うFcg2h-EPOはpH6.0マーカーと共に収束した。BHK細胞において合成されたFcg2h-EPOタンパク質の集団は、タンパク質1分子あたり平均21個のシアル酸残基を有することがわかった。対照的に、NS/0細胞において合成されたFc(g2h)-EPOタンパク質の集団は、タンパク質1分子あたり平均約10個のシアル酸残基を有するように見えた。
【0146】
引き続きの実験において、Fc-EPOタンパク質を発現しているBHK細胞を、無血清増殖条件及び大規模生成に適した条件、例えば懸濁条件に適応させた。無血清かつ懸濁液中で増殖させたBHK細胞において産生されたFc-EPOタンパク質を、前述のようにIEFゲル電気泳動により分析した。増殖条件のこれらの変更は、最強のバンドの等電点について最大でも0.1〜0.3 pH単位のシフトしか生じさせなかった。
補充したDMEM/F-12無タンパク質培地において合成されたFc-EPO融合タンパク質の試料も、IEFゲル電気泳動により同様に特徴決定した。タンパク質生成物は、VP-SFMのような無血清培地において増殖された細胞から得られた対応する生成物よりも、より程度が大きくシアル酸化され、かつ、より均質なシアル酸化を示したことがわかった。
【0147】
異なる細胞株で生成されたFc-EPOタンパク質のシアル酸化の程度は更に、レクチン-結合試験により定量的に確認した。例えばFc-EPO融合タンパク質を最初に、標準SDSゲル電気泳動により分離し、ブロッティングし、その後異なる糖質部分を認識する改変されたレクチン(例えばRoche Applied Science, インディアナポリス, INから市販されている)でプロービングし、結合したレクチンを視認した。適当なレクチンは、特定の結合を伴うシアル酸を認識するSambucus nigraアグルチニン(SNA)又はMaackia amurensisアグルチニン(MAA)、並びにO-グリカンコアのような糖鎖部分の他の領域を認識するDatura stramoniumアグルチニン(DAA)、Peanutアグルチニン(PNA)及びジャカリンを含むが、これらに限定されるものではない。レクチン結合アッセイを基に、異なる細胞株で生成されたFc-EPO融合タンパク質のシアル酸化レベルを決定した。
【実施例6】
【0148】
実施例6. Fc-EPO変種のin vitro生物学的活性
種々のFc-EPOタンパク質のin vitro活性を細胞-ベースのアッセイにおいて試験した。TF-1細胞株はEPO受容体を発現しており、従って適当な培養条件下におけるそのトリチウム標識されたチミジンの取込みは、EPO又はEPO-様タンパク質活性の関数である(Hammerllingら、J. Pharmaceutical and Biomedical Analysis、14: 1455 (1996);Kitamuraら、J. Cellular Physiol.、140: 323 (1989))。具体的には、活発な対数相にあるTF-1細胞をEPOを含まない培地において2回洗浄し、マイクロタイタープレートへ約104個細胞/ウェルで播種した。その後これらの細胞を、Fc-EPO変種の滴定した連続希釈を伴う培地において、48時間インキュベーションした。3H-チミジン0.3μCiをウェルに添加し、その10時間後に細胞増殖をアッセイした。対照として、TF-1細胞を、組換えヒトEPO、及び高グリコシル化されたEPOアナログAranesp(登録商標)の存在下でもインキュベーションした。放射性チミジンの取込みは、総TCA-沈降可能な数として測定した。表2に示したように、Fcg2h-M1-EPO分子の活性は組換えヒトEPOの活性と同等であった。
【0149】
いくつかの全般的結論をこのデータから引き出すことができる。先に報告された結果と一致して、CHO細胞から生成されたEPOはED50 約0.7ng/mlを有し;これは、NIBSC EPO標準、R&D SystemsからのEPO、及び市販のProcrit(登録商標)を含む。Aranesp(登録商標)はin vitroにおいて有意に活性が低く、これは恐らくその増加した負電荷に起因する低下した結合速度を反映しているであろう。同様にBHK細胞から生成されたFc-EPOはNS/0細胞から産生されたFc-EPOよりも少ない活性を有し、これはBHK細胞により生成されたFc-EPOタンパク質は、より高度にシアル酸化され、タンパク質上に増加した負電荷を生じるという知見と一致している。
【0150】
表2
【実施例7】
【0151】
実施例7. Fc-EPO変種の薬物動態分析
様々な細胞株において合成された様々なFc-EPOタンパク質の薬物動態プロファイルを、以下のin vivo実験を基に特徴付けた。ひとつの実験において、図8に示したように、NS/0細胞及びBHK細胞において合成されたFcg2h(N>Q)-EPOタンパク質約14μgを、Swiss-Websterマウスへ静脈内投与した。投与後様々な時点(例えば、T=投与後0、1/2、1、2、4、8、及び24時間)で、血液試料を採取し、血清を遠心により調製した。Fc-EPOの血清濃度を、抗-Fc抗体を用いるELISAにより決定した。図8に示したように、投与後24時間で、BHK-由来のFc-EPOの初期血清濃度の10%より多くが血清中に残存したのに対し、NS/0-由来のFc-EPOの初期血清濃度の0.1%未満しか血清中に残存しなかった。
【0152】
同様の実験を、NS/0細胞及びBHK細胞で合成されたFcg2h-EPO(NDS)タンパク質で行った。NS/0細胞及びBHK細胞において合成されたFcg2h-EPO(NDS)タンパク質約14μgを、Swiss-Websterマウスへ静脈内投与した。血液試料を投与後、T=0、1/2、1、2、4、8、24及び36時間で採取し、血清中のFcg2h-EPO(NDS)濃度を、抗-Fc抗体を用いるELISAにより決定した。図9に示したように、投与後24時間で、BHK-由来のFcg2h-EPO(NDS)の初期血清濃度の10%より多くが血清中に残存したのに対し、NS/0-由来のFcg2h-EPO(NDS)の初期血清濃度の0.1%未満が血清中に残存した。
【0153】
BHK-21細胞、PERC6細胞、及び293細胞において生成されたFcg2h-EPO(NDS)の薬物動態プロファイルも比較した。特にFcg2h-Epo(NDS)を発現するプラスミドを、BHK、293、及びPERC6細胞へ一過性にトランスフェクションした。発現されたFcg2h-Epo(NDS)融合タンパク質を、異なる細胞株で精製し、Swiss-Websterマウスへ、濃度1.7μg/マウスで、静脈内注射した。血液試料を、T=0、1/2、1、2、4、8、24、48、及び72時間に採取し、血清中のFcg2h-Epo(NDS)濃度を抗-Fc ELISAにより測定した。図10に示したように、投与後24時間で、BHK-由来のFcg2h-EPO(NDS)の初期血清濃度の10%より多くが血清中に残存したのに対し、293細胞由来のFcg2h-EPO(NDS)の初期血清濃度の1%未満が血清中に残存し、並びにPerC6細胞-由来のFcg2h-EPO(NDS)は血清中でほとんど検出不能であった。BHK、PerC6、及び293細胞において産生されたFcg2h(N→Q)-EPOタンパク質において同様の結果が得られた。
【0154】
Fcg2h(N→Q)-EPO、Fcg2h-EPO(NDS)、Fcg2h-EPO、及びAranesp(登録商標)(すなわち、NESP)の薬物動態プロファイルを比較するために、同様の実験をマウスにおいて行った。ここで使用したFc-EPO変種は、BHK細胞から合成した。投与後48時間で、Aranesp(登録商標)の初期血清濃度の10%未満が血清中に残存したのに対し、Fcg2h(N→Q)-EPO及びFcg2h-EPO(NDS)の両方の初期血清濃度の10%よりも多くが血清中に残存したことが観察された。これらの結果は、BHK-21細胞から生成されたFcg2h(N→Q)-EPO及びFcg2h-EPO(NDS)タンパク質は、Aranesp(登録商標)よりもはるかに長い血清半減期を有したことを示している。
【実施例8】
【0155】
実施例8. Fc-EPO変種のin vivo効能
様々なFc-EPO変種のin vivo生物学的活性を、マウス及びラットにおけるヘマトクリット(HCT)アッセイ及び網状赤血球アッセイにより測定した。
ひとつのHCT実験において、CD1マウスに、BHK細胞において合成されたFcg2h(FN→AQ)-EPOタンパク質を投与量20μg/kg及び10μg/kgで腹腔内注射した。血液試料をマウスから第4、7、11及び14日目に採取し、毛細管で遠心した。沈降したRBCの量を総容積の割合として測定した。図4に示したように、Fcg2h(FN>AQ)-EPOタンパク質注射に応答してヘマトクリットは初めに劇的に増加し、その後定常状態を維持し、最後に漸減した。
【0156】
別の実験において、BHK細胞において合成された以下のタンパク質を、Sprague-Dawleyラットに腹腔内注射した。全ての動物に42.5μg/kgで投与した。
1. Fcg2h-EPO
2. Fcg2h-EPO(NDS)
3. Fcg4h-EPO
4. Fcg4h(N>Q)-EPO
HCTアッセイを、前述のように注射したマウスから採取した血液試料について行った。図5に示したように、Fcg2h-EPO(NDS)及びFcg2h-EPOに応答して、注射したラットにおけるヘマトクリットの量は延長された期間定常状態を維持した。このことは、Fcg2h-EPO(NDS)及びFcg2h-EPOの両タンパク質は延長された血清半減期及び強力なin vivo生物学的活性を有することを示している。図5に示されたように、Fcg4h-EPO及びFcg4h(N→Q)-EPOは、Fcg2h-EPO(NDS)及びFcg2h-EPOタンパク質と比べてより短い定常状態の期間及びより迅速な血清濃度の低下を示すことも認められた。
【0157】
別の実験において、CD1マウスに、以下の試料を腹腔内投与した:
1. BHK細胞由来のFcg2h-EPO(NDS)、投与量85μg/kg、42.5μg/kg、及び21.25μg/kg
2. NS/0細胞由来のFcg2h-EPO(NDS)、投与量85μg/kg、42.5μg/kg、及び21.25μg/kg
3. Aranesp(登録商標)(すなわち、NESP)、投与量50μg/kg、25μg/kg、及び12.5μg/kg
これらのタンパク質の量は、糖鎖を伴わないタンパク質分子量を基に算出した。この実験において、Fcg2h-EPO(NDS)タンパク質の分子量は単量体ポリペプチドを基にしている。従ってFcg2h-EPO(NDS)のNESPに対する分子量の比は、約1.71〜1であった。従って、この実験において各タンパク質の投与量範囲はほぼ等しかった。
【0158】
図6に示されたように、BHK細胞において合成されたFcg2h-EPO(NDS)タンパク質は、効能及び作用期間に関して最良のヘマトクリットプロファイルを示し、このことは、BHK細胞由来のFcg2h-EPO(NDS)タンパク質は、NS/0細胞由来のFcg2h-EPO(NDS)及びNESPの両方と比べ、より長い血清半減期及びより強力なin vivo活性を有することを示している。NS/0細胞由来のFcg2h-EPO(NDS)及びNESPのヘマトクリットプロファイルは、同等である。
【実施例9】
【0159】
実施例9. IgG2及びIgG4由来のCH2-CH3ドメインを伴うFc-EPOタンパク質の比較
様々なFc-EPOタンパク質の細胞-ベースのエリスロポエチン活性の比較は、IgG4由来のCH2及びCH3ドメインを伴う融合タンパク質は一般に、IgG2由来のCH2及びCH3ドメインを伴う対応するタンパク質よりも活性が低いことを明らかにした。この結論は、少なくとも3種のFc-EPOタンパク質、すなわち、エリスロポエチン部分にNDS変異を伴い、及びNS/0細胞において合成されたタンパク質(表3)、BHK細胞において合成されたNDS変異を伴うタンパク質(表4)、及びBHK細胞において合成された正常なエリスロポエチンを伴うタンパク質(表5)について、当てはまった。
【0160】
下記表3から5で比較したタンパク質は全て、IgG1由来の改変されたヒンジ、及びFc部分のC-末端のM1セット変異を有する。これらのタンパク質の活性を、実施例6に説明された標準手法による、タンパク質により刺激されたTF-1細胞へのトリチウム標識されたチミジンの取込みの測定により決定した。活性は、エリスロポエチン部分のED50(ng/ml)として示した。
【0161】
表3:NDS変異を伴う、NS/0細胞において合成されたFc-EPO融合タンパク質の細胞-ベースの活性
【0162】
表4:NDS変異を伴う、BHK細胞において合成されたFc-EPO融合タンパク質の細胞-ベースの活性
【0163】
表5:野生型EPOを有する、BHK細胞において合成されたFc-EPO融合タンパク質の細胞-ベースの活性
【0164】
In vitro細胞-ベースのアッセイからの活性データは通常、エリスロポエチン-含有タンパク質の薬物動態プロファイル及びin vivo効能を示すことができる。一般に細胞-ベースのアッセイにおける減少したin vitro活性は、EPO受容体の低下したon-rateを示し、これは改善された薬物動態特性(例えば延長された半減期)及び増強されたin vivo活性に相関している。しかしIgG4-由来のCH2及びCH3ドメインを伴うFc-EPO融合タンパク質の減少したin vitro活性は、改善した薬物動態及び増強されたin vivo生物学的活性とは相関しない。IgG4-由来のCH2及びCH3ドメインを伴うFc-EPO融合タンパク質の薬物動態プロファイルは一般に、IgG2-由来のCH2及びCH3ドメインを伴う対応するタンパク質とは識別不能であることがわかった。IgG4-由来のCH2及びCH3ドメインを伴うFc-EPO融合タンパク質は一般に、IgG2-由来のCH2及びCH3ドメインを伴う対応するタンパク質と比べ、in vivoにおいてより少ない活性を有することもわかった(図5参照)。
【実施例10】
【0165】
実施例10. Fc部分のグリコシル化部位の除去の効果
Fc部分のグリコシル化部位の除去の、in vitro活性、薬物動態、及びin vivo効能に対する作用を試験するために実験を行った。特にIgG2-由来のCH2及びCH3ドメイン又はIgG4-由来のCH2及びCH3ドメインのいずれかを含むFc-EPO融合タンパク質を試験した。IgG1のAsn297に相当する、IgG2又はIgG4のGln-Phe-Asn-Serアミノ酸配列内のアスパラギンをグルタミンに置換した。ほとんどの実験において、Gln-Phe-Asn-Serアミノ酸配列のフェニルアラニンをアラニンに置換し、アスパラギンの変異の結果として生ずる可能性のある非-自己T-細胞エピトープを除去した。表6に示されるように、細胞-ベースのin vitroアッセイにおいて、Fc部分のN-結合グリコシル化部位を除去するFN>AQ変異を伴うFc-EPOタンパク質のED50値は、一般にこの変異を伴わないFc-EPOタンパク質の約1/5倍であり、このことはN-結合グリコシル化部位の除去が細胞-ベースのアッセイにおいてin vitro活性を減少させることを示した。
【0166】
N-結合グリコシル化の除去の薬物動態及びin vivo効能に対する作用を試験するために実験を行った。CD1マウスを、BHK細胞で合成したFcg2h-M1-EPO、Fcg2h-M1-EPO(NDS)、及びFcg2h(N>Q)-M1-EPOタンパク質で、それぞれ投与量42μg/kgで処置した。Fcg2h(N→Q)-M1-EPOタンパク質は、NRQ変異を伴わない対応するタンパク質よりも、より良い薬物動態プロファイルを示したことが認められた。従って、IgG2-由来のFc部分においてN-結合グリコシル化を除去するN>Q変異は、改善された薬物動態(例えば、延長された血清半減期)を生じた。延長された血清半減期は、Fc受容体への結合に対する効果により説明することはできない。その理由は、IgG2-由来のCH2及びCH3ドメインはそもそも本質的に検出不能なFc-受容体結合を有しているからである。
【0167】
表6:Fc部分のグリコシル化部位の除去はFc-EPO融合タンパク質のin vitro細胞-ベースの活性を低下させる
【0168】
IgG2及びIgG4由来のFc部分のN-結合グリコシル化の除去により引き起こされる効果は、エリスロポエチン受容体についての低下した結合速度とほとんど一致するので、これらの効果は予想外で驚くべきことである。理論に拘束されることを欲するものではないが、IgG2及びIgG4由来のFc部分のN-結合グリコシル化の除去は、Fc-EPO融合タンパク質の全体的な立体構造の変化を引き起こすかもしれない。
【実施例11】
【0169】
実施例11. BHK細胞において合成されたFc-EPO融合タンパク質によるビーグル犬の処置
Fc-EPO融合タンパク質のヘマトクリット、網状赤血球数、及び他の血液パラメーターに対する作用を試験するために、Fc-EPO融合タンパク質をビーグル犬に投与した。具体的には、Fcg2h(FN→AQ)-EPOタンパク質を、2種の個別に安定的にトランスフェクションされたBHK細胞株であるクローン65及びクローン187から精製し、ビーグル犬に静脈内投与した。1匹のオス及び1匹のメスのビーグル犬に、以下のスケジュールで各調製物を注射した:
0日目:3μg/kg
16日目:10μg/kg
23日目:100μg/kg
【0170】
各投与後異なる時点で、血液約2mlを収集し、ヘマトクリット、網状赤血球数及び他の血液パラメーターなどの血液パラメーターを測定した。
処置後のヘマトクリット反応を、図11に示した。Fc-EPO融合タンパク質3μg/kgの投与後、血液パラメーターは正常範囲から増加しなかった。10μg/kgの投与後1週間以内に、網状赤血球数は、4匹の動物中3匹において、総血液体積の3%を超えて増加し、ヘマトクリットは1匹において51に増加した。他の血液パラメーターは正常な範囲から増加しなかった。100μg/kgの投与後、ヘマトクリット数は急激に上昇し、57〜62のピークレベルに到達し、5〜6週間にわたり正常な範囲を上回り続けた。網状赤血球数は、2〜3週間上昇し続けた。
各動物について、1dl当たりのg数で測定した血液1μl当たりの赤血球数及びヘモグロビン数はヘマトクリット量に比例した。これらの結果は、Fc-EPOタンパク質は正常なヘモグロビン含量の正常なサイズの赤血球形成を刺激することを示した。
【実施例12】
【0171】
実施例12. 臨床使用のためのFc-EPOタンパク質の精製
Fc-EPOタンパク質は、当業者に公知の標準GMP手法に従い精製した。バンクで作出したクローンからのBHK-21細胞を、追加の2.5mM L-グルタミン(Invitrogen)、2g/lの各HyPep1501及びHyPep 4601(Quest International, シカゴ, IL)、10μl/lエタノールアミン(Sigma)、及び5μM Tropolone(Sigma)を補充したDMEM/F-12培地(Invitrogen)において、7〜10日間、バッチ培養し、その間高細胞生存率(例えば、80%を上回る)を維持した。馴化培地を収集し、ノーマル-フロー-濾過により清澄化し、予め-平衡としたプロテインA Sepharoseファスト-フローカラム(Pharmacia)に載せた(Fc部分に関するプロテインAの親和性を基に融合タンパク質を捕獲する)。このカラムを中性pHの150mMリン酸ナトリウム及び100mM NaClを含有するリン酸ナトリウム緩衝液15カラム容量で十分に洗浄した。結合したタンパク質を、更に150mMリン酸ナトリウム及び100mM NaClを含有する酸性リン酸ナトリウム緩衝液(pH2.5〜3)15カラム容量により、低pHで溶離した。
【0172】
ウイルス失活のため、プールされたピーク画分のpHをpH3.8に調節し、更に30分間室温でインキュベーションした。30分間インキュベーションした後、プールした画分を中和し、滅菌濾過し、その後Q-Sepharoseファスト-フローアニオン交換カラム(Pharmacia)に装加した。これにより、その高度なシアル酸化の結果としてのFc-EPOタンパク質の酸性pIを利用してFc-EPOタンパク質と同時-溶離した可能性のある夾雑物が効果的に除去される。具体的には、中和した画分を、pH5.0でQ-Sepharoseファスト-フローアニオン交換カラム(Pharmacia)上に載せ、NaCl溶液の勾配で溶離した。その後Fc-EPOの画分を収集し、引き続きの分析及び更なる精製プロセスのためにプールした。例えば、Q-Sepharoseカラムからの高塩ストリップを逆相クロマトグラフィーカラムに載せ、過剰なNaClを除去した。希釈した逆相カラムからの溶離液を更に第二のQ-Sepharoseファストフロー(Pharmacia, 3cm X 9cm)カラムに載せた。
【0173】
その後、ナノ-濾過(例えば、MilliporeのViresolve)により、このプールから存在し得るウイルス粒子を除去した。場合により、ヒドロキシアパタイトカラム又はフェニル-ホウ酸カラム(cis-ジオールに結合)などの更なる精製工程を使用することができる。最後に、精製されたタンパク質を、限外濾過を用いて望ましい濃度に濃縮し、その後適当な配合の緩衝液へ透析した。この材料を最終的に滅菌濾過し、所定の体積でバイアルに分注した。
【実施例13】
【0174】
実施例13. Fc-EPOタンパク質製剤の安定性を決定するためのストレス試験
例証のための試料Fc-EPO製剤又は参照Fc-EPO製剤を含有するバイアルを、40℃で相対湿度75%で、指定された貯蔵期間(例えば、0週、4週、8週など)貯蔵した。一定の貯蔵期間の後、各バイアルからアリコート試料を採取し、分析した。これらの試料は、濁度について、冷光源による直接照射下で目視により評価した。更に、濁度を吸光度350nm及び550nmで測定することにより決定した。加えて試料中のFc-EPOタンパク質の条件及びタンパク質分解産物の存在を、分析用サイズ排除クロマトグラフィー(HPLC-SEC)により分析した。0.5mg/ml Fc-EPO、10mMクエン酸塩(pH6.2)、100mMグリシン、100mM NaCl、0.01%w/vポリソルベート20を含有する製剤は、参照溶液と比べて安定性を有意に増加したことがわかった。
【実施例14】
【0175】
実施例14. ヒトにおけるFcg2h(FN>AQ)-M1-EPO融合タンパク質の第I相試験
ヒトにおけるFcg2h(FN>AQ)-M1-EPO融合タンパク質の第I相臨床試験は、下記のように実施されている。薬物動態パラメーターは、本質的にAranesp(登録商標)について、MacDougallら(J. Am. Soc. Nephrol.、10: 2392-2395 (1999))により説明されたように決定したが、この論文の教示は本明細書に引用により取り込まれるものとする。ヒトに静脈内注射されたFcg2h(FN→AQ)-M1-EPO融合タンパク質(用量1μg/kg)の終末血清半減期は、約20〜30時間であることがわかった。従って、用量1μg/kg、又は成人貧血患者における約70μgは、初期血清濃度約10ng/mlをもたらした。正常なヒトエリスロポエチン濃度は、約0.04〜0.25ng/mlであるので(Cazzolaら、Blood、91: 2139-2145 (1998))、Fc-EPOタンパク質の薬理学的に活性なレベルは患者の全身において少なくとも5〜10日間維持される。
【実施例15】
【0176】
実施例15. Fcg2h(FN→AQ)-M1-EPO融合タンパク質の第II相用量設定及び投与スケジュール決定試験
Fcg2h(FN>AQ)-M1-EPO融合タンパク質について、透析を受けている慢性腎不全(CRF)患者へ皮下又は静脈内注射により投与した場合の最適用量及び投与スケジュールを調べるために、多施設無作為化逐次用量漸増試験を開始する。
臨床の実践において、以下の指針に従い、Fcg2h(FN→AQ)-M1-EPO融合タンパク質の投与を個々の貧血患者に合わせて調節することは一般に都合がよい。初回量を投与し、ヘマトクリット、ヘモグロビン、網状赤血球数、及び血小板数などの血液パラメーターをモニタリングする。初回量は、典型的には約0.3〜3μg/kgである。都合の良い初回量は1μg/kgである。治療の8週間後にヘマトクリットの増加が血液体積の5〜6%未満である場合には、用量が増加されなければならない。2週間のヘマトクリットの増加が血液体積の4%を上回るか、又はヘマトクリットが36%に達する場合、用量を減少させなければならない。
【0177】
投与スケジュールの例は、以下である:
週1回投与:0.075、0.225、0.45、0.75、1.5及び4.5μg/kg/投与。
2週に1回投与:0.075、0.225、0.45、0.75、1.5及び4.5μg/kg/投与。
1ヶ月に1回投与:0.45、0.75、1.5及び4.5μg/kg/投与。
【0178】
試験は、2部において行われる。第一の部は、週1回、2週に1回、1ヶ月に1回のいずれかで投与される、Fcg2h(FN→AQ)-M1-EPO融合タンパク質の用量を評価するためにデザインされた、用量-漸増試験であり、これは4週間にわたり、最適速度でヘモグロビンを増加する(1g/dLよりも大きい又はこれと等しいが、3g/dL未満である)。各試験の第二の部は、治療目標にヘマトクリットを維持するために必要な用量を決定するためにデザインされている(静脈内又は皮下のいずれかの投与経路で、週1回、2週に1回、1ヶ月に1回投与される場合)。
【技術分野】
【0001】
発明の技術分野
本発明は、改良された薬物動態を有する新規な、通例として高度にシアル酸化されたFc-EPO融合タンパク質を提供する。詳細に述べると、Fc-EPOタンパク質は延長された血清半減期及び増大したin vivo効能を有する。BHK細胞において合成されたFc-EPO融合タンパク質は、例えば、NS/0細胞などの他の細胞株において産生された対応するFc-EPO融合タンパク質と比較した場合に、劇的に延長された血清半減期及び増加したin vivo効能を有する。本発明はFc-EPOにも関し、Fc部分に加えEPO部分の種々の改変が、更に改良された特性を伴う各分子を得るために実行される。
【背景技術】
【0002】
背景
エリスロポエチンは、赤血球形成前駆細胞の赤血球への成熟に必要な糖タンパク質ホルモンである。これは、腎臓で生成され、循環内の赤血球細胞のレベルの調節に不可欠である。低レベルの組織酸素シグナルにより特徴付けられる状態は、エリスロポエチン生成を増大し、これは次に赤血球形成を刺激する。循環内のエリスロポエチンレベルは、赤血球が長期酸素欠乏にのみ反応して作られることを確実にするよう、厳密に調節されている。エリスロポエチンの70%は、受容体-媒介型エンドサイトーシスにより消失される。エリスロポエチンがその受容体へ結合する場合、この複合体はエンドサイトーシスされ、分解され、その結果シグナル伝達の範囲が制限される。エリスロポエチンの残余は、腎濾過を通じ尿中へクリアランスされる。結果的に、エリスロポエチンは、比較的短い血清半減期を有する。
【0003】
哺乳動物細胞において生成される天然のヒトエリスロポエチン又は組換えエリスロポエチンは、3個のN-結合型及び1個のO-結合型オリゴ糖鎖を含む。N-結合グリコシル化は、24、38及び83位に位置したアスパラギン残基で生じるのに対し、O-結合グリコシル化は、126位に位置したセリン残基で生じる(Laiら、J. Biol. Chem.、261:3116 (1986);Broudyら、Arch. Biochem. Biophys.、265:329 (1988))。オリゴ糖鎖は、末端シアル酸残基で改変されることが示されている。典型的にはN-結合型鎖は、1個の鎖あたり最大4個のシアル酸を有し、及びO-結合型鎖は、最大2個のシアル酸を有する。従ってエリスロポエチンポリペプチドは、合計最大14個のシアル酸を収容することができる。糖は、細胞からのエリスロポエチンの分泌、エリスロポエチン溶解度の増大、及びエリスロポエチンのin vivo生物学的活性に必要であることが示されている(Dubeら、J. Biol. Chem.、263:17516 (1988);DeLormeら、Biochemistry、31:9871-9876 (1992))。
【0004】
組換えヒトエリスロポエチンの投与は、例えば腎不全、HIV感染症、失血及び慢性疾患に関連したものを含む、様々な形の貧血のような、造血障害又は欠損の治療において有効である。エリスロポエチンは典型的には静脈内注射により投与される。エリスロポエチンは比較的短い血清半減期を有するので、循環内で治療的有効レベルのエリスロポエチン維持するためには、頻繁な静脈内注射が必要である。天然の又は組換えのヒトエリスロポエチンを含有する医薬組成物は典型的には、投与量約25〜100単位/kgで週3回投与される。この型のエリスロポエチン療法は、極めて有効であるが、非常に高価であり、かつ静脈内投与は頻繁に診療所又は病院を来院することを必要とするので不便である。現在、新規赤血球形成刺激タンパク質(NESP)である、高度にグリコシル化された組換えヒトエリスロポエチンアナログは、貧血の治療のための商標Aranesp(登録商標)(Amgen Inc.、サザンオークス、CA)として入手可能である。Aranesp(登録商標)は、同じ生物学的反応を得るために、通常のエリスロポエチンよりも少ない頻度で投与することができる。
【0005】
別の投与経路は皮下注射である。この投与形態は患者が自宅で行うことができ、及び投与部位からのより遅い吸収を提供し、その結果持続放出作用を引き起こす徐放製剤により適合している。しかし有意に低い循環レベルが皮下注射により実現され、その結果望ましい治療作用を実現するためには頻繁な注射が必要である。更にタンパク質製剤の皮下投与は一般に静脈内投与よりもより免疫原性である。その理由は、感染の主障壁としての皮膚は樹状細胞が多い免疫臓器であり、擦過傷及び異物を同定しかつ反応するための感度の高い機構を有するからである。Casadevallらは最近、エリスロポエチンを皮下投与された患者は、抗-エリスロポエチン抗体を発生したことを報告した(Casadevallら、N Engl. J. Med.、346(7): 469-75 (2002))。
従って、投与頻度の少ないより有効なエリスロポエチン療法が必要である。
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明は、様々な態様において、野生型もしくは天然のエリスロポエチン、組換えエリスロポエチン、又は高グリコシル化されたエリスロポエチンアナログNESP(PCT国際公開公報第00/24893号)と比べ、改善された薬物動態を有するエリスロポエチン融合タンパク質を提供する。従って、エリスロポエチン療法を簡略化し、並びに造血障害又は欠損又は他のエリスロポエチン投与の適応症を伴うヒト又は他の哺乳動物の治療に関する経費を削減することは、本発明の目的である。
【0007】
詳細に述べると本発明は、延長された血清半減期及び増加したin vivo効能を有する、生物学的活性のあるFc-エリスロポエチン(Fc-EPO)融合タンパク質を提供する。
本明細書において「Fc-EPO融合タンパク質」は、Fc部分及びエリスロポエチン部分を有するポリペプチドを含むタンパク質を意味する。本明細書において「Fc部分」は、免疫グロブリン、好ましくはヒト免疫グロブリンの定常領域に由来したドメインを包含し、これは定常領域の断片、アナログ、変種、変異体又は誘導体を含む。本明細書において「エリスロポエチン部分」は、ヒト及び他の種由来の野生型又は天然のエリスロポエチン、組換えエリスロポエチン、及びエリスロポエチン-様分子を包含し、これは生物学的活性のあるエリスロポエチン断片、エリスロポエチンのアナログ、変種、変異体又は誘導体を含む。
【課題を解決するための手段】
【0008】
本発明は、以下のことに関する:
ヒンジ領域、CH2及びCH3ドメインを含むヒトIgG分子の二量体Fc部分、並びにヒトエリスロポエチン(EPO)から本質的になる精製された二量体融合タンパク質であって、前記二量体Fc部分の各鎖がそのC-末端においてEPO分子のN-末端へ直接又はリンカーペプチドを介して連結しており、下記の特徴を有する、前記融合タンパク質:(i) 15〜28個のシアル酸残基を含むことにより、高度にシアル酸化されている;(ii)CH2ドメインがヒトIgG2に由来し、CH2ドメインの Gln-Phe-Asn-Ser配列内のアミノ酸残基Phe及びAsnが、Ala及びAsnで置換され、その結果CH2ドメイン内に配列Gln-Ala-Gln-Serが形成されることによってCH2ドメインが改変されている;および、(iii)CH3ドメインのC-末端近傍のLeu-Ser-Leu-Serアミノ酸配列がAla-Thr-Ala-Thrで置換されている。
【図面の簡単な説明】
【0009】
【図1A】ヒトIgG1、IgG2及びIgG4の定常領域のアミノ酸配列のアラインメントを示す。
【図1B】図1A続き。
【図2】図2は、α相において、Fc-EPO投与量とFc-EPO血清濃度の減少量の間の相関関係を示すマウスにおける薬物動態実験を示す。この実験においては、NS/0細胞において合成された過小(under)シアル酸化されたFc-EPO変種が使用された。
【図3】図3は、Fc-EPO融合タンパク質の可能性のある消失経路、及びこれらの経路を変調する可能性のある融合タンパク質への改変を示す。
【図4】図4は、Fcg2h(FN>AQ)-EPO投与後の、マウスにおけるヘマトクリット反応の例を示す。
【図5】図5は、BHK細胞で生成されたFcg2h-EPO、Fcg2h-EPO(NDS)、Fcg4h-EPO、及びFcg4h(N>Q)-EPOタンパク質投与後の、ラットにおけるヘマトクリット反応の例を示す。
【図6】図6は、BHK細胞で生成されたFcg2h-EPO(NDS)、NS/0細胞で生成されたFcg2h-EPO(NDS)、及びNESP(すなわちAranesp(登録商標))投与後の、マウスにおけるヘマトクリット反応の例を示す。
【図7】図7は、成熟型Fc-EPOタンパク質をコードしている核酸配列の例を示す。
【図8】図8は、マウスにおけるBHK細胞から生成されたFcg2h(N>Q)-EPO及びNS/0細胞から生成されたFcg2h(N>Q)-EPOの薬物動態プロファイルを示す。
【図9】図9は、マウスにおけるBHK細胞から生成されたFcg2h-EPO(NDS)及びNS/0細胞から生成されたFcg2h-EPO(NDS)の薬物動態プロファイルを示す。
【図10】図10は、マウスにおけるBHK-21細胞、PERC6細胞、及び293細胞において生成されたFcg2h-EPO(NDS)タンパク質の薬物動態プロファイルを示す。
【図11】図11は、BHK細胞において合成されたFcg2h(FN→AQ)-EPOタンパク質による処置後の、ビーグル犬におけるヘマトクリット反応を示す。
【発明を実施するための形態】
【0010】
ひとつの局面において、本発明は、BHK細胞において合成されたFc-EPOタンパク質を提供する。本発明のBHK細胞において合成されたFc-EPO融合タンパク質は、例えばNS/0、PerC6、又は293細胞などの、他の細胞株において生成された対応するFc-EPO融合タンパク質と比較した場合に、劇的に延長された血清半減期及び増加したin vivo効能が明らかにされている。本発明は、哺乳動物への投与に適した、高度にシアル酸化されたFc-EPO融合タンパク質の集団も提供する。高度にシアル酸化されたFc-EPO融合タンパク質は、様々な態様において、野生型又は天然のエリスロポエチン、組換えエリスロポエチン、高グリコシル化されたエリスロポエチンアナログNESP、又はNS/0、PerC6、もしくは293細胞で合成された同じアミノ酸配列のFc-EPO融合タンパク質と比較して、より長い血清半減期及び増加したin vivo効能を有する。本発明に従い、Fc-EPO融合タンパク質は、一般にFc融合タンパク質の血清半減期を延長するようなFc部分のアミノ酸改変を含むことができる。例えば、このようなアミノ酸改変は、Fc受容体結合活性又は補体結合活性を実質的に減少又は除去する変異を含む。加えて、Fc-EPO融合タンパク質は、EPO受容体-媒介したエンドサイトーシスを減少するか又はエリスロポエチンの生物学的活性を増加するようなエリスロポエチン部分のアミノ酸改変も含む。様々な態様において、本発明は、免疫グロブリン融合タンパク質、Fc部分及びエリスロポエチン部分のアミノ酸改変、並びにBHK細胞における生成(例えば、高レベルのシアル酸化)によりもたらされる恩恵を組合せている。これらの組合せた恩恵は、驚くほど延長された血清半減期及び増加したin vivo効能を伴うFc-EPO融合タンパク質を生じる、相加作用又は相乗作用を有する。
【0011】
従って本発明は、ひとつの局面において、Fc-EPO融合タンパク質をコードしている核酸配列を含むBHK細胞に関する。ひとつの態様において、本発明のBHK細胞は、無タンパク質培地における増殖に適合している。別の態様において、BHK細胞は、懸濁液における増殖に適合している。更に別の態様において、BHK細胞は、無タンパク質培地中で懸濁液における増殖に適合している。無タンパク質培地中で増殖したBHK細胞から生成されたFc-EPO融合タンパク質は、他の培地において増殖されたBHK細胞から生成されたFc-EPO融合タンパク質と比べ、驚くほど増加した及びより均質なシアル酸化を示すことがわかっている。好ましい態様において、核酸はBHK細胞において安定して維持される。本明細書において「安定して維持される核酸」とは、抗生物質-ベースの選択のような核酸を維持するための選択圧の非存在下で、母細胞から娘細胞への喪失率が3%未満の核酸を意味する。従って核酸を安定して維持する細胞が分裂する場合、得られる細胞の少なくとも97%(より好ましくは、98%よりも多く、99%よりも多く、又は99.5%よりも多く)がその核酸を含む。その核酸を含む得られる細胞が分裂する場合、(2回目の)分裂から得られる細胞の少なくとも97%がその核酸を含むであろう。更に1個の細胞当たりの核酸のコピー数は、繰返しの細胞分裂により実質的に低下しない。好ましい態様において、安定して維持される核酸配列はBHK細胞の染色体へ組込まれる。
【0012】
核酸配列は、様々な配置のいずれでもFc-EPO融合タンパク質をコードすることができる。好ましい態様において、核酸配列はFc-EPO融合タンパク質のN-末端側のFc部分及びFc-EPO融合タンパク質のC-末端側のエリスロポエチン部分を含むFc-EPO融合タンパク質をコードしている。Fc部分は一般に、免疫グロブリンの定常領域由来の領域を包含し、これは定常領域の断片、アナログ、変種、変異体又は誘導体を含む。好ましい態様において、Fc部分は、ヒト免疫グロブリン重鎖、例えば、IgG1、IgG2、IgG3、IgG4、又は他のクラスに由来する。一部の態様において、本Fc-EPO融合タンパク質は免疫グロブリンの可変領域を含まない。ひとつの態様において、Fc部分は、CH2ドメインを含む。別の態様において、Fc部分はCH2及びCH3ドメインを含む。
【0013】
好ましい態様において、Fc部分は、Fc受容体への親和性を低下するか、又はFcエフェクター機能を低下させる変異を含む。例えばFc部分は、IgG重鎖のFc部分内のグリコシル化部位を除去する変異を含むことができる。一部の態様において、Fc部分はIgG1のLeu234、Leu235、Gly236、Gly237、Asn297、又はPro331に対応するアミノ酸位置での変異、欠失、又は挿入を含む(アミノ酸は、EU命名法に従い番号付けられる)。好ましい態様において、Fc部分は、IgG1のAsn297に対応するアミノ酸位置に変異を含む。別の態様において、Fc部分はIgG1のLeu281、Leu282、Gly283、Gly284、Asn344、又はPro378に対応するアミノ酸位置に変異、欠失、又は挿入を含む。
【0014】
一部の態様において、Fc部分はヒトIgG2又はIgG4重鎖由来のCH2ドメインを含む。好ましくは、CH2ドメインは、CH2ドメイン内のグリコシル化部位を除去する変異を含む。ひとつの態様において、変異は、IgG2又はIgG4重鎖のCH2ドメイン内のGln-Phe-Asn-Serアミノ酸配列内のアスパラギンを変更する。好ましくは、変異は、アスパラギンをグルタミンへ変更する。あるいは、変異は、Gln-Phe-Asn-Serアミノ酸配列内のフェニルアラニン及びアスパラギンの両方を変更する。ひとつの態様において、Gln-Phe-Asn-Serアミノ酸配列はGln-Ala-Gln-Serアミノ酸配列で置換される。
【0015】
Gln-Phe-Asn-Serアミノ酸配列内のアスパラギンは、IgG1のAsn297に対応している。IgG2又はIgG4のGln-Phe-Asn-Serアミノ酸配列内のアスパラギンの変異(すなわちIgG1のAsn297に対応)も同じくFc-EPO融合タンパク質のEPO受容体への結合を驚くほど低下させることがわかっている。理論に拘束されることを欲するものではないが、IgG2又はIgG4のGln-Phe-Asn-Serアミノ酸配列内のアスパラギンの変異(すなわちIgG1のAsn297に対応)は、Fc-EPO融合タンパク質の全般的立体構造の変化を誘導し、これは劇的に改善された薬物動態特性につながるのであろう。
【0016】
別の態様において、Fc部分は、CH2ドメイン及びヒンジ領域の少なくとも一部を含む。ヒンジ領域は、免疫グロブリン重鎖、例えば、IgG1、IgG2、IgG3、IgG4、又は他のクラスに由来することができる。好ましくは、ヒンジ領域は、ヒトIgG1、IgG2、IgG3、IgG4、又は他の適当なクラスに由来する。より好ましくは、ヒンジ領域は、ヒトIgG1重鎖に由来する。ひとつの態様において、IgG1ヒンジ領域のPro-Lys-Ser-Cys-Asp-Lysアミノ酸配列内のシステインが改変される。好ましい態様において、Pro-Lys-Ser-Cys-Asp-Lysアミノ酸配列がPro-Lys-Ser-Ser-Asp-Lysアミノ酸配列で置換される。ひとつの態様において、Fc部分は、第一の抗体アイソタイプ由来のCH2ドメイン及び第二の抗体アイソタイプ由来のヒンジ領域を含む。特定の態様において、CH2ドメインは、ヒトIgG2又はIgG4重鎖に由来するが、ヒンジ領域は改変されたヒトIgG1重鎖に由来する。
【0017】
好ましい態様において、Fc部分は、定常領域のC-末端近傍のLeu-Ser-Leu-Serアミノ酸配列が、接合部の潜在的なT-細胞エピトープを除去するように変更されている、IgG配列に由来する。例えばひとつの態様において、Leu-Ser-Leu-Serアミノ酸配列はAla-Thr-Ala-Thrアミノ酸配列で置換される。別の態様において、Fc部分は、C-末端リジン残基が置換されたIgG配列に由来する。好ましくは、IgG配列のC-末端リジンは、アラニンのような非-リジンアミノ酸により交換され、Fc融合タンパク質の血清半減期を更に増加する。
【0018】
本発明に従い、Fc部分は、本明細書に記載された1個又は複数の変異を含むことができる。Fc部分の変異の組合せは一般に、Fc-EPO融合タンパク質の延長された血清半減期及び増加されたin vivo効能に対して相加作用又は相乗作用を有する。従って態様の一例において、Fc部分は、(i)Lys-Ser-Lys-Serアミノ酸配列がAla-Thr-Ala-Thrアミノ酸配列に置換されたIgG配列由来の領域;(ii)リジンの代わりにC-末端アラニン残基;(iii)異なる抗体アイソタイプ由来の、CH2ドメイン及びヒンジ領域、例えばIgG2 CH2ドメイン及び代わりのIgG1ヒンジ領域;(iv)IgG2-由来のCH2ドメイン内のグリコシル化部位を除去する変異、例えばIgG2-由来のCH2ドメイン内のGln-Phe-Asn-Serアミノ酸配列の代わりにGln-Ala-Gln-Serアミノ酸配列:を含む。
【0019】
Fc-EPO融合タンパク質のエリスロポエチン部分は、完全長の野生型又は天然のエリスロポエチン、組換えエリスロポエチン、又はエリスロポエチン-様分子、例えば生物学的活性のあるエリスロポエチン断片、エリスロポエチンのアナログ、変種、変異体又は誘導体であることができる。好ましくは、エリスロポエチン部分は、ヒトエリスロポエチンに由来する。一部の態様において、エリスロポエチン部分は、EPO受容体への結合親和性を低下するか又はエリスロポエチンの生物学的活性を増加するようなアミノ酸改変を含むことができる。一部の態様において、エリスロポエチン部分は、以下の変異を少なくとも1種含む:Arg131→Glu及びArg139→Glu(アミノ酸番号付けは、成熟型ヒトエリスロポエチン配列を基にしている)。別の態様において、エリスロポエチン部分は、以下の変異を少なくとも1種含む:His32→Gly、Ser34→Arg、及びPro90→Ala。更に別の態様において、エリスロポエチン部分は、ヒトエリスロポエチンとは異なるジスルフィド結合のパターンを有する。例えばエリスロポエチン部分は、以下のアミノ酸置換の1種又は複数を含むことができる:29位の非-システイン残基、33位の非-システイン残基、88位のシステイン残基、及び139位のシステイン残基。ひとつの態様において、エリスロポエチン部分は、7、29、88、及び161位にシステイン残基を含む。別の態様において、エリスロポエチン部分は、更に、下記の置換を1個又は複数含む:His32→Gly、Cys33→Pro、及びPro90→Ala。本発明に従い、エリスロポエチン部分は、本明細書に説明された変異のいずれかの組合せを含むことができる。
【0020】
一部の態様において、Fc-EPO融合タンパク質はFc部分とエリスロポエチン部分の間にリンカーを含む。リンカーが含まれるならば、これは一般に、1〜25個のアミノ酸を含み、プロテアーゼ切断部位を含まないことが好ましい。このリンカーは、タンパク質分解を阻害するために、N-結合型された又はO-結合型されたグリコシル化部位を含むことができる。例えばひとつの態様において、リンカーは、Asn-Ala-Thrアミノ酸配列を含む。
【0021】
本発明は更に、Fc-EPO融合タンパク質を生産する方法にも関する。この方法は、Fc-EPO融合タンパク質をコードしている核酸配列を含むBHK細胞を、コードされたFc-EPO融合タンパク質の発現に適した条件下で維持する工程、及び発現されたFc-EPO融合タンパク質を回収する工程を含む。ひとつの態様において、BHK細胞は、無タンパク質培地中で培養される。別の態様において、BHK細胞は懸濁物として培養される。更に別の態様において、BHK細胞は無タンパク質培地中で懸濁培養される。一部の態様において、この核酸は、BHK細胞において安定して維持される。一般に、BHK細胞において生成されたFc-EPO融合タンパク質は、例えば、NS/0、PerC6、もしくは293細胞などの他の細胞株において生成された対応するFc-EPO融合タンパク質よりも、より長い血清半減期を有する。
【0022】
本発明は、BHK細胞において生成されたFc-EPO融合タンパク質を含有する医薬組成物を提供する。好ましい態様において、医薬組成物において使用されるFc-EPO融合タンパク質は、シアル酸残基を除去するようには処理されない。この医薬組成物は、医薬として許容できる担体も含む。本発明は、医薬組成物を哺乳動物へ投与することにより、哺乳動物を治療する方法も提供する。一部の態様において、治療される哺乳動物は、造血障害又は欠損を有する。本発明のFc-EPO融合タンパク質は、増加したin vivo効能及び延長された血清半減期を有するので、Fc-EPO融合タンパク質を含有する医薬組成物は一般に、他の細胞において生成された天然のもしくは組換えエリスロポエチン又は対応するFc-EPO融合タンパク質を含有する医薬組成物と比べ、より少ない投与頻度しか必要としない。好ましい態様において、この医薬組成物は、週3回よりも少ない頻度(例えば、週2回、毎週、又は多くて10日に1回、例えば2週間に1回、1ヶ月に1回、又は2ヶ月に1回など)で投与される。
【0023】
別の局面において、本発明は、Fc部分及びエリスロポエチン部分を含む融合タンパク質をコードしている核酸を安定して維持するBHK細胞を選択する方法を提供する。この方法は、ハイグロマイシンBをコードしている核酸配列及びこの融合タンパク質をコードしている核酸配列をBHK細胞へ導入する工程;および、ハイグロマイシンBの存在下で前記BHK細胞を培養する工程を含む。ひとつの態様において、ハイグロマイシンBをコードしている核酸配列及び融合タンパク質をコードしている核酸配列は、単一の核酸中に存在する。別の態様において、ハイグロマイシンBをコードしている核酸配列及びこの融合タンパク質をコードしている核酸配列は、2つの個別の核酸内に存在する。
【0024】
別の局面において、本発明は、哺乳動物への投与に適した精製されたFc-EPO融合タンパク質の集団を提供する。好ましい態様において、Fc-EPO融合タンパク質は、Fc-EPO融合タンパク質のN-末端側にFc部分、及びFc-EPO融合タンパク質のC-末端側にエリスロポエチン部分を含む。より好ましい態様において、精製されたFc-EPO融合タンパク質の集団は、高度にシアル酸化され、すなわち精製されたFc-EPO融合タンパク質あたり平均11〜28個のシアル酸残基を有する。好ましいFc-EPO融合タンパク質の高度にシアル酸化された集団は、精製されたFc-EPO融合タンパク質あたり平均13〜28、15〜28、17〜28、19〜28、又は21〜28個のシアル酸残基を有する。例えばひとつの好ましいFc-EPO融合タンパク質の高度にシアル酸化された集団は、精製されたFc-EPO融合タンパク質あたり平均20〜22個のシアル酸残基を有する。好ましい態様において、精製されたFc-EPO融合タンパク質はBHK細胞において合成される。ひとつの態様において、BHK細胞は、懸濁物としての増殖に適応している。別の態様において、BHK細胞は無タンパク質培地中の増殖に適応している。更に別の態様において、BHK細胞は無タンパク質培地中で懸濁物としての増殖に適応している。本発明により提供される精製されたFc-EPO融合タンパク質の高度にシアル酸化された集団は、例えば、NS/0、PerC6、もしくは293細胞などの細胞において生成された対応するFc-EPO融合タンパク質の集団と比べ、より長い血清半減期を有する。本発明に従い、精製されたFc-EPO融合タンパク質のFc部分及びエリスロポエチン部分は、本明細書に記載されたような1個又は複数の変異又は改変を含み、増強されたシアル酸化による相加作用又は相乗作用により、延長された血清半減期及び増加したin vivo効能を提供することができる。
【0025】
本発明は、本明細書に記載された精製されたFc-EPO融合タンパク質の高度にシアル酸化された集団を含有する医薬組成物も提供する。好ましい医薬組成物は更に医薬として許容できる担体を含む。本発明は更に、哺乳動物へ、精製されたFc-EPO融合タンパク質の高度にシアル酸化された集団を含有する医薬組成物を投与することを含む、哺乳動物を治療する方法を提供する。好ましい態様において、本医薬組成物は1週間に3回よりも少なく投与される(例えば、週2回、週1回、又は多くて10日に1回、例えば2週間に1回、1ヶ月に1回又は2ヶ月に1回など)。
【0026】
まとめると、本発明は、以下のことに関する:
・ヒンジ領域、CH2及びCH3ドメインを含むヒトIgG分子の二量体Fc部分、並びにヒトエリスロポエチン(EPO)から本質的になる精製された二量体融合タンパク質であって、前記二量体Fc部分の各鎖がそのC-末端においてEPO分子のN-末端へ直接又はリンカーペプチドを介して連結しており、下記の特徴を有する、前記融合タンパク質:(i) 15〜28個のシアル酸残基を含むことにより、高度にシアル酸化されている;(ii)CH2ドメインがヒトIgG2に由来し、CH2ドメインの Gln-Phe-Asn-Ser配列内のアミノ酸残基Phe及びAsnが、Ala及びAsnで置換され、その結果CH2ドメイン内に配列Gln-Ala-Gln-Serが形成されることによってCH2ドメインが改変されている;および、(iii)CH3ドメインのC-末端近傍のLeu-Ser-Leu-Serアミノ酸配列がAla-Thr-Ala-Thrで置換されている。
【0027】
・更にCH3ドメインのC-末端Lys残基がAlaに置換されている、各二量体Fc-EPO融合タンパク質。
・ヒンジ領域がヒトIgG1に由来する、各二量体Fc-EPO融合タンパク質。
・IgG1ヒンジ領域の Pro-Lys-Ser-Cys-Asp-Lys配列内のCysアミノ酸残基がSer残基によって置換され、その結果前記ヒンジ領域内に配列Pro-Lys-Ser-Ser-Asp-Lysが形成されることによってIgG1ヒンジ領域が改変されている、各二量体Fc-EPO融合タンパク質。
【0028】
・エリスロポエチン部分が以下のアミノ酸置換の少なくとも1種を含む、各二量体Fc-EPO融合タンパク質:
(i)EPO分子の29位の非-システイン残基、
(ii)EPO分子の33位の非-システイン残基、
(iii)EPO分子の88位のシステイン残基、及び
(iv)EPO分子の139位のシステイン残基。
【0029】
・EPO分子の元のCys残基の代わりにEPO分子の33位に非-Cysアミノ酸残基があり、元のEPO分子の88位にTrp残基の代わりCys残基があり、その結果融合タンパク質内のEPO部分が、Cys29-Cys88ジスルフィド結合を形成することが可能である、各二量体Fc-EPO融合タンパク質。
・33位の非-Cysアミノ酸残基がProである、各二量体Fc-EPO融合タンパク質。
・EPO部分が以下の群から選択される1種又は複数の変異を含む、各二量体Fc-EPO融合タンパク質:
(i)Arg131→Glu131
(ii)Arg139→Glu139
(iii)His32→Gly32
(iv)Ser34→Arg34
(v)Pro90→Ala90。
【0030】
・リンカーペプチドがグリコシル化部位を含む、各二量体Fc-EPO融合タンパク質。
・グリコシル化部位がAsn-Ala-Thrアミノ酸配列を含む、各二量体Fc-EPO融合タンパク質。
・更にCH1ドメインを含む、各二量体Fc-EPO融合タンパク質。
・CH2、CH3及び場合によりCH1を含む完全なIgG分子がIgG2に由来し、ヒンジ領域がIgG1に由来する、各Fc-EPO融合タンパク質。
・CH2、CH3及びヒンジ領域および場合によりCH1を含む完全なIgG分子がIgG1に由来する、
各Fc-EPO融合タンパク質。
・18〜24個、好ましくは20〜22個のシアル酸残基を有する、各二量体Fc-EPO融合タンパク質。
【0031】
・以下の配列を含む、二量体Fc-EPO融合タンパク質:
EPKSSDKTHTCPPCPAPPVAGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVQFNWYVDGVEVHNAKTKPREEQAQSTFRVVSVLTVVHQDWLNGKEYKCKVSNKGLPAPIEKTISKTKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPMLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSATATPGAAPPRLICDSRVLERYLLEAKEAENITTGCAEHCSLNENITVPDTKVNFYAWKRMEVGQQAVEVWQGLALLSEAVLRGQALLVNSSQPWEPLQLHVDKAVSGLRSLTTLLRALGAQKEAISPPDAASAAPLRTITADTFRKLFRVYSNFLRGKLKLYTGEACRTGDR (配列番号:14)。
・以下の配列を含む、二量体Fc-EPO融合タンパク質:
EPKSSDKTHTCPPCPAPPVAGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVQFNWYVDGVEVHNAKTKPREEQAQSTFRVVSVLTVVHQDWLNGKEYKCKVSNKGLPAPIEKTISKTKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPMLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSATATPGAAPPRLICDSRVLERYLLEAKEAENITTGCAEGPSLNENITVPDTKVNFYAWKRMEVGQQAVEVWQGLALLSEAVLRGQALLVNSSQPCEALQLHVDKAVSGLRSLTTLLRALGAQKEAISPPDAASAAPLRTITADTFRKLFRVYSNFLRGKLKLYTGEACRTGDR (配列番号:15)。
【0032】
・先に特定した融合タンパク質をコードするDNA分子。
・先に特定したか又は請求項のFc-EPO融合タンパク質を有効量含み、場合により医薬として許容できる担体、希釈剤又は賦形剤を含有する、哺乳動物における造血障害または欠損の治療に適した医薬組成物。
・Fc-EPO融合タンパク質のN-末端側にFc部分、及びFc-EPO融合タンパク質のC-末端側にエリスロポエチン部分を含むFc-EPO融合タンパク質の集団であって、精製されたFc-EPO融合タンパク質あたり平均15〜28個のシアル酸残基を有し、各Fc-EPO融合タンパク質をコードするDNA分子をBHK細胞へ導入し、対応するFc-EOP融合タンパク質を発現、単離及び精製することによって得られ、NS/0、PerC6、又は293細胞において合成された対応するFc-EPO融合タンパク質の集団と比較して、より長い血清半減期を有する、哺乳動物への投与に適した精製された高度にシアル酸化されたFc-EPO融合タンパク質の集団。
【0033】
・精製されたFc-EPO融合タンパク質あたり平均20〜22個のシアル酸残基を有する、前記精製されたFc-EPO融合タンパク質の対応する集団。
・BHK細胞が、無タンパク質培地又は懸濁液中で増殖するのに適応している、精製されたFc-EPO融合タンパク質の対応する集団。
【0034】
・Fc-EPO融合タンパク質のN-末端側にFc部分、及びFc-EPO融合タンパク質のC-末端側にエリスロポエチン部分を含む、高度にシアル酸化された精製された組換えFc-EPO融合タンパク質の集団を作製する方法であって:
(i)Fc-EPO融合タンパク質をコードしているDNA分子を構築する工程;
(ii)無タンパク質培地又は懸濁液中において該DNA分子でBHK細胞を形質転換する工程;
(iii)該DNA分子によりコードされたFc-融合タンパク質の集団を発現する工程、
(iv)Fc-EPO融合タンパク質の集団を収集、単離及び精製する工程、
を含む前記方法。
【0035】
・前記合成された融合タンパク質の集団が、精製されたFc-EPO融合タンパク質あたり平均15〜28、好ましくは15〜25、より好ましくは20〜22個のシアル酸残基を有する、対応する方法。
・Fc部分及びエリスロポエチン部分を含むFc-EPO融合タンパク質をコードする核酸配列を安定して維持しているBHK細胞を選択する方法であって:
(a)ハイグロマイシンBをコードしている核酸配列及びFc-EPO融合タンパク質をコードしている核酸配列をBHK細胞へ導入する工程;及び
(b)ハイグロマイシンBの存在下で前記BHK細胞を培養する工程、
を含む前記方法。
・ハイグロマイシンBをコードしている核酸配列及びFc-EPO融合タンパク質をコードしている核酸配列が単一のDNA分子内に存在する、対応する方法。
【0036】
発明の詳細な説明
本発明は、改善された薬物動態を伴う、Fc-EPO融合タンパク質を提供する。特に、本発明により提供されるFc-EPOタンパク質は、延長された血清半減期及び増加したin vivo効能を有する。ひとつの局面において、本発明は、BHK細胞において合成されたFc-EPO融合タンパク質を提供する。BHK細胞において合成されたFc-EPO融合タンパク質は、例えばNS/0、PerC6、又は293細胞などの他の細胞株において生成された対応するFc-EPO融合タンパク質と比べた場合に、劇的に延長された血清半減期及び増加したin vivo効能を有する。別の局面において、本発明は、高度にシアル酸化されたFc-EPO融合タンパク質の集団を提供する。高度にシアル酸化されたFc-EPO融合タンパク質の集団は、より少ないレベルのシアル酸化を伴う対応するFc-EPO融合タンパク質の集団と比べ、より長い血清半減期を有する。本発明に従い、Fc-EPO融合タンパク質は、Fc受容体結合活性の実質的減少もしくは除去又は補体結合活性を低下する改変などにより、Fc融合タンパク質の血清半減期を延長するFc部分のアミノ酸改変を含むことができる。加えてFc-EPO融合タンパク質は、EPO受容体-媒介型エンドサイトーシスを減少するか又はエリスロポエチンの生物学的活性を増加するエリスロポエチン部分のアミノ酸改変も含む。
【0037】
Fc-EPO融合タンパク質
本明細書において「Fc-EPO融合タンパク質」は、そのポリペプチドに通常は存在しない少なくともふたつの部分、すなわちFc部分及びエリスロポエチン部分を有するポリペプチドを含むタンパク質を意味する。本発明の好ましい態様において、Fc部分及びエリスロポエチン部分を有するポリペプチドは、ホモ二量体を形成し;従って、Fc-EPO融合タンパク質は一般に、1個又は複数のジスルフィド結合により一緒に保持された二量体タンパク質であり、各ポリペプチド鎖は、Fc部分及びエリスロポエチン部分を含む。しかし本発明のFc-EPO融合タンパク質は、エリスロポエチン部分が、Fc部分と安定して会合することを可能にしつつ、エリスロポエチン活性を維持するようないかなる配置をも有し得る。例えばこのような配置は、ふたつのFc部分及びふたつのエリスロポエチン部分を含む単一のポリペプチド、ふたつのFc部分及びひとつのエリスロポエチン部分を含む単一のポリペプチド、Fc部分及びエリスロポエチン部分を含むひとつのポリペプチドと、Fc部分を含む別のポリペプチドを含むヘテロ二量体タンパク質、並びに他の適当な配置を含むが、これらに限定されるものではない。
【0038】
エリスロポエチン部分は、様々な配置で直接又は間接にFc部分へ連結することができる。ひとつの態様において、エリスロポエチン部分は、Fc部分へ、共有結合を介して直接連結される。例えばエリスロポエチン部分は、そのC-末端又はそのN-末端のいずれかで、Fc部分へ直接融合することができる。ひとつの態様において、Fc部分のC-末端は、エリスロポエチン部分のN-末端へ融合されており、すなわち、Nterm-Fc-Cterm-Nterm-EPO-Ctermである。この配置において、Fc部分は、Fc-EPO融合タンパク質のN-末端側にあり、エリスロポエチン部分はC-末端側にある。別の態様において、エリスロポエチンのC-末端は、Fc部分のN-末端へ融合され、すなわち、Nterm-EPO-Cterm-Nterm-Fc-Ctermである。この配置において、エリスロポエチン部分は、Fc-EPO融合タンパク質のN-末端側にあり、Fc部分はC-末端側にある。
【0039】
別の態様において、エリスロポエチン部分はFc部分へ間接的に連結される。例えば、Fc-EPO融合タンパク質は、Fc部分とエリスロポエチン部分の間に、リンカー(L)を含むことができる。直接融合と同様に、エリスロポエチン部分は好ましくは、Fc部分のC-末端へリンカーを介して融合され、すなわち、Nterm-Fc-Cterm-L-Nterm-EPO-Ctermである。従ってFc部分は、Fc-EPO融合タンパク質のN-末端へ側にあり、C-末端側のエリスロポエチン部分からリンカーにより隔てられている。あるいは、エリスロポエチン部分は、Fc部分のN-末端へリンカーを介して融合されており、すなわち、Nterm-EPO-Cterm-L-Nterm-Fc-Ctermである。
【0040】
Fc部分
本明細書において、「Fc部分」は免疫グロブリン、好ましくはヒト免疫グロブリンの定常領域由来のドメインを包含し、これは定常領域の断片、アナログ、変種、変異体又は誘導体を含む。適当な免疫グロブリンは、IgG1、IgG2、IgG3、IgG4、及び他のクラスを含む。免疫グロブリンの定常領域は、免疫グロブリンC-末端領域と相同な天然又は合成により生成されたポリペプチドと定義され、これはCH1ドメイン、ヒンジ、CH2ドメイン、CH3ドメイン、又はCH4ドメインを、個別に又はいずれかの組合せで含むことができる。ヒトIgG1、IgG2及びIgG4の定常領域の配列アラインメントを、図1A及び1Bに示している。Paulの論文(Fundamental Immunology、第4版、Lippincott-Raven、(1999))によると、CH1ドメインは、アミノ酸118-215を含み;ヒンジ領域は、アミノ酸216-230を含み;CH2ドメインは、アミノ酸231-340を含み;及び、CH3ドメインはアミノ酸341-447を含む(アミノ酸位置はIgG1配列を基にしている)。ヒンジ領域はCH1ドメインをCH2及びCH3ドメインへ連結する。
【0041】
本発明において、Fc部分は典型的には、少なくともCH2ドメインを含む。例えばFc部分はヒンジ-CH2-CH3を含む。あるいはFc部分は、ヒンジ領域、CH2ドメイン及び/又はCH3ドメインの全て又は一部を含むことができる。
免疫グロブリンの定常領域は、Fc受容体(FcR)結合及び補体結合を含む多くの重要な抗体機能に関与する。重鎖定常領域には5種の主なクラスがあり、これはIgA、IgG、IgD、IgE、及びIgMに分類され、各々、アイソタイプにより指定される特徴的エフェクター機能を伴う。例えば、IgGは4種のγサブクラス:γ1、γ2、γ3、及びγ4に分類され、これらは各々、IgG1、IgG2、IgG3、及びIgG4としても知られている。
【0042】
IgG分子は、抗体のIgGクラスに特異的なFcγ受容体(FcγR)の3種のクラス(すなわちFcγRI、FcγRII、及びFcγRIII)を含む、細胞受容体の複数のクラスと相互作用する。IgGのFcγR受容体への結合に重要な配列は、CH2及びCH3ドメインに局在することが報告されている。抗体の血清半減期は、その抗体がFc受容体(FcR)へ結合する能力により影響を受ける。同様に、免疫グロブリン融合タンパク質の血清半減期も、そのような受容体に結合する能力の影響を受ける(Gilliesら、Cancer Res.、59:2159-66 (1999))。IgG1と比べ、IgG2及びIgG4のCH2及びCH3ドメインは、生化学的に検出不可能であるか又は低下したFc受容体への結合親和性を有する。IgG2又はIgG4のCH2及びCH3ドメインを含む免疫グロブリン融合タンパク質は、IgG1のCH2及びCH3ドメインを含む対応する融合タンパク質と比べ、より長い血清半減期を有することが報告されている(米国特許第5,541,087号;Loら、Protein Engineering、11: 495-500 (1998))。従って本発明に好ましいCH2及びCH3ドメインは、例えば、IgG2又はIgG4のような、低下した受容体結合親和性及びエフェクター機能を伴う、抗体アイソタイプに由来する。より好ましいCH2及びCH3ドメインは、IgG2に由来する。
【0043】
ヒンジ領域は通常、重鎖定常領域のCH1ドメインのC-末端に位置している。IgGアイソタイプにおいて、ジスルフィド結合は典型的にはこのヒンジ領域において生じ、最終の四量体抗体分子を形成することを可能にする。この領域は、プロリン、セリン及びトレオニンが支配的である。本発明に含まれる場合、ヒンジ領域は典型的には、ふたつのFc部分を連結するジスルフィド結合を形成するためにシステイン残基を含む天然の免疫グロブリン領域と少なくとも相同である。ヒト及びマウス免疫グロブリンのヒンジ領域の代表的配列は、Borrebaeck C. A. K.編集の「Antibody Engineering, A Practical Guide」(1992)、W. H. Freeman and Co.に認められる。本発明に適したヒンジ領域は、IgG1、IgG2、IgG3、IgG4、及び他の免疫グロブリンクラスに由来することができる。IgG1ヒンジ領域は、3個のシステインを有し、その2個は免疫グロブリンの2本の重鎖間のジスルフィド結合に関連している。これらの同じシステインは、Fc部分の間に有効かつ一貫したジスルフィド結合をもたらす。従って、本発明の好ましいヒンジ領域は、IgG1に由来し、より好ましくはヒトIgG1に由来する。一部の態様において、ヒトIgG1ヒンジ領域内の第一のシステインは、好ましくは別のアミノ酸、好ましくはセリンへ変異される。IgG2アイソタイプヒンジ領域は、組換えシステムにおける分泌時にオリゴマー化及び恐らくは不正確なジスルフィド結合を促進する傾向がある4個のジスルフィド結合を有する。適当なヒンジ領域は、IgG2ヒンジに由来することができ;最初の2個のシステインは、好ましくは各々別のアミノ酸へ変異される。IgG4のヒンジ領域は、非効率的に鎖内ジスルフィド結合を形成することがわかっている。しかし本発明に適したヒンジ領域は、IgG4ヒンジ領域に由来し、好ましくは重鎖-由来の部分の間のジスルフィド結合の正確な形成を増強する変異を含む(Angal S.らMol. Immunol.、30: 105-8 (1993))。
【0044】
本発明に従い、Fc部分は、異なる抗体アイソタイプに由来する、CH2及び/又はCH3及びヒンジ領域、すなわちハイブリッドFc部分を含むことができる。例えばひとつの態様において、Fc部分は、IgG2又はIgG4に由来したCH2及び/又はCH3ドメイン並びにIgG1に由来した変異体ヒンジ領域を含む。あるいは、別のIgGサブクラス由来の変異体ヒンジ領域が、ハイブリッドFc部分において使用される。例えば、2本の重鎖の間の効率的ジスルフィド結合を可能にするIgG4ヒンジの変異体型を使用することができる。変異体ヒンジは、最初の2個のシステインが各々別のアミノ酸に変異されているIgG2ヒンジに由来することもできる。このようなハイブリッドFc部分は高-レベル発現を促進し、及びFc-EPO融合タンパク質の正確なアッセンブルを改善する。そのようなハイブリッドFc部分の集成は、米国特許公開番号第20030044423号に開示されており、(すなわち米国特許出願第10/093,958号)、その開示は本明細書に引用により取り込まれる。
【0045】
一部の態様において、Fc部分は、一般にFc融合タンパク質の血清半減期を延長するアミノ酸改変を含む。そのようなアミノ酸改変は、Fc受容体結合又は補体結合活性を実質的に減少又は除去する変異を含む。例えば免疫グロブリン重鎖のFc部分内のグリコシル化部位を除去することができる。IgG1において、このグリコシル化部位はAsn297である。別の免疫グロブリンアイソタイプにおいて、グリコシル化部位はIgG1のAsn297に相当している。例えばIgG2及びIgG4において、グリコシル化部位はアミノ酸配列Gln-Phe-Asn-Ser内のアスパラギンである。従ってIgG1のAsn297の変異はIgG1由来のFc部分のグリコシル化部位を除去する。ひとつの態様において、Asn297はGlnに置換される。同様に、IgG2又はIgG4において、アミノ酸配列Gln-Phe-Asn-Ser内のアスパラギンの変異は、IgG2又はIgG4重鎖由来のFc部分のグリコシル化部位を除去する。ひとつの態様において、アスパラギンはグルタミンに置換される。別の態様において、アミノ酸配列Gln-Phe-Asn-Ser内のフェニルアラニンは、アスパラギン変異から生じる可能性のある非-自己T-細胞エピトープを除去するように更に変異される。例えば、IgG2又はIgG4重鎖内のアミノ酸配列Gln-Phe-Asn-SerはGln-Ala-Gln-Serアミノ酸配列に置換することができる。
【0046】
Fc部分及び非-Fc部分の接合部近傍のアミノ酸の変更は、Fc融合タンパク質の血清半減期を劇的に増加することができることも観察される。(PCT国際公開公報第01/58957号、その開示は本明細書に参照として組入れられている)。従って本発明のFc-EPO融合タンパク質の接合部領域は、免疫グロブリン重鎖及びエリスロポエチンの天然の配列に対して、好ましくは接合点のアミノ酸約10個以内に位置する変更を含むことができる。これらのアミノ酸変化は、例えば、Fc部分のC-末端リジンをアラニン又はロイシンのような疎水性アミノ酸へ変えることにより、疎水性の増加を引き起こすことができる。
【0047】
別の態様において、Fc部分は免疫グロブリン重鎖のFc部分のC-末端近傍のLeu-Ser-Leu-Serセグメントのアミノ酸変更を含む。Leu-Ser-Leu-Serセグメントのアミノ酸置換は可能性のある接合部T-細胞エピトープを除去する。ひとつの態様において、Fc部分のC-末端近傍のLeu-Ser-Leu-Serアミノ酸配列はAla-Thr-Ala-Thrアミノ酸配列に置換される。別の態様において、Leu-Ser-Leu-Serセグメント内のアミノ酸は、グリシン又はプロリンなどの別のアミノ酸に置換される。IgG1、IgG2、IgG3、IgG4、又は他の免疫グロブリンクラス分子のC-末端近傍のLeu-Ser-Leu-Serセグメントのアミノ酸置換を生じる詳細な方法は、米国特許公開番号第20030166877号(すなわち、米国特許出願第10/112,582号)に開示されており、その開示は本明細書に参照として組入れられている。
【0048】
エリスロポエチン部分
本明細書において「エリスロポエチン部分」は、ヒト及び他の種に由来する野生型又は天然のエリスロポエチン、組換えエリスロポエチン、及び生物学的活性のあるエリスロポエチン断片、エリスロポエチンのアナログ、変種、変異体もしくは誘導体を含む、エリスロポエチン-様分子を包含している。
野生型又は天然のエリスロポエチンは、エリスロポエチン前駆細胞からの赤血球の増殖及び発達を刺激する、34KD糖タンパク質ホルモンである。野生型又は天然のエリスロポエチンは、低酸素症(例えば、貧血による赤血球喪失)に応答して腎臓で産生され、その同族細胞受容体との相互作用を介して、赤血球の増殖及び分化を調節する。野生型又は天然のエリスロポエチンは、血液から(Miyake T.ら、J. Biol. Chem.、252: 5558-5564 (1977))、又は血漿から(Goldwasser, E.ら、Proc. Natl. Acad. Sci. U.S.A.、68: 697-698 (1971))、又は尿から単離及び精製することができる。
【0049】
組換えにより又は化学的に合成されたエリスロポエチンは、当業者に周知の技術を用い、生成することができる。組換えヒトエリスロポエチン(rHuEPO)のふたつの型が市販されている:AmgenからEPOGEN(登録商標)、及びJohnson & JohnsonからPROCRIT(登録商標)。
【0050】
本明細書において、エリスロポエチンの生物学的活性は、エリスロポエチン受容体との相互作用を介した細胞増殖を刺激する能力として定義される。エリスロポエチンの機能アッセイは、in vitro又はin vivoにおいて行うことができる。例えば、エリスロポエチンのin vitro活性は、細胞-ベースのアッセイにおいて試験することができる。特にエリスロポエチン活性は、TF-1細胞増殖アッセイを基に決定することができる。TF-1細胞はEPO受容体を発現する。トリチウム標識されたチミジンの組込みにより決定されるTF-1細胞の増殖はエリスロポエチン活性の関数である(Hammerlling ら、J. Pharmaceutical and Biomedical Analysis、14: 1455 (1996);Kitamura ら、J. Cellular Physiol.、140: 323 (1989))。In vitro細胞-ベースのアッセイは、実施例6においてより詳細に説明されている。In vivoアッセイは典型的には、例えばマウス及びラットなどの動物モデルにおいて行われる。In vivoアッセイの例は、ヘマトクリット(HCT)アッセイ、及び網状赤血球アッセイを含むが、これらに限定されるものではない。HCTアッセイは、エリスロポエチン-処置した動物から採取した血液試料から赤血球容積を測定し、及び毛細管中の血液を遠心し、沈降した赤血球により占拠された総容積の割合を測定することにより行う。In vivo HCTアッセイについては、実施例8により詳細に説明されている。網状赤血球アッセイは、前駆細胞から分化したばかりでまだ前駆細胞に特徴的な核酸を残している、網状赤血球としても知られている新生赤血球を測定するものである。網状赤血球は、アクリジンオレンジ又はチアゾールオレンジなどの核酸-染色色素により染色した後、フローサイトメーターにおける赤血球の選別、及び陽性に染色された網状赤血球画分の計測により測定される。
【0051】
生物学的活性又は機能活性のあるエリスロポエチン-様分子は典型的には、野生型又は天然のエリスロポエチンの対応する配列と、実質的アミノ酸配列類似性又は同一性(例えば、少なくとも約55%、約65%、約75%の同一性、典型的には少なくとも約80%、及び最も典型的には約90〜95%の同一性)を共有し、それらの野生型エリスロポエチンの1種又は複数の機能を持っている。
【0052】
従って本発明のエリスロポエチンは、野生型エリスロポエチンの配列に類似したアミノ酸配列を有するエリスロポエチンポリペプチドを特に含むと理解される。このようなタンパク質は、本明細書においてエリスロポエチンアナログと定義される。「アナログ」は、本明細書において、そのタンパク質の生物学的活性を保持するために野生型エリスロポエチンのアミノ酸配列と十分な類似性を伴うアミノ酸配列を意味すると定義される。例えば、エリスロポエチンアナログは、野生型エリスロポエチンのアミノ酸配列中に1個又は複数のアミノ酸変更を含むことができ、依然例えば赤血球形成又は成熟を刺激する能力を保持する。このようなアミノ酸変更の例は、アミノ酸残基の付加、欠失又は置換を含む。本発明のエリスロポエチンは、米国特許第5,614,184号に開示されたような、野生型エリスロポエチンよりも、より大きい又はより少ない生物学的活性を示す変異体タンパク質も包含している。
【0053】
本発明のエリスロポエチンは、エリスロポエチンの生物学的活性のある断片も包含している。このような断片は、エリスロポエチンの完全長アミノ酸配列の一部のみを含むが、依然生物学的活性を有する。本明細書において「生物学的活性のある断片」は、完全長タンパク質に類似した生物学的作用を発揮する断片を意味する。このような断片は、アミノ-及びカルボキシ-末端の欠失に加え、内部の欠失により作出され得る。これらは、エリスロポエチンの切断型及びハイブリッド型も含む。「短縮」型は、より短いエリスロポエチンの型であり、例えばアミノ末端、又はカルボキシ末端の残基が取り除かれている。
【0054】
エリスロポエチン配列の変動
EPO受容体への結合親和性を低下するため;タンパク質安定性を増強するため;正確な活性のある立体構造の選定を強化するため;薬物動態特性を増強するため;合成を増強するため;又は、他の有利な特徴を提供するために、本発明のエリスロポエチン部分にアミノ酸改変が導入される。例えば、EPO受容体-媒介したエンドサイトーシスは、エリスロポエチンとEPO受容体の間の結合親和性により決定される。ヒトエリスロポエチンとEPO受容体の複合体の三次元構造は、エリスロポエチンのその受容体への結合がエリスロポエチン表面上の正電荷及びEPO受容体上の負電荷により支配されることを明らかにしている。Syedら、Nature、395: 511 (1998)。結合の速度(on-rate)を低下させるために、変異を導入し、エリスロポエチン-EPO受容体接触表面近傍に位置する正に荷電したアミノ酸を置換することができる。例えばひとつの態様において、ヒトエリスロポエチンのArg131及びArg139の一方又は両方を置換することができる(EPO配列のアミノ酸番号付けは、成熟型ヒトEPOを基にしている)。好ましくは、Arg131及びArg139は、グルタミン酸、アスパラギン酸、又は他の非-正荷電したアミノ酸に置換される。ヒトエリスロポエチンのArg131及びArg139に対応するアミノ酸を置換するように、他の種のエリスロポエチンに変異を導入することができる。しかし、EPOアミノ酸配列において改変を行う場合、EPO生物学的活性を保存するために、EPO-EPO受容体相互作用の中心に位置する残基は避けなければならない。
【0055】
あるいは、アラニン走査突然変異導入により、アミノ酸置換を容認する領域又は位置を経験的に決定することができる(Cunninghamら、Science、244 1081-1085 (1989))。この方法において選択されるアミノ酸残基は、生物学的活性に対する効果を決定するために、中性アミノ酸(例えば、アラニン)と個々に置換される。
【0056】
ひとつの態様において、エリスロポエチン部分は以下の変異の少なくともひとつを含む:His32→Gly及び/又はSer34→Arg、及びPro90→Ala。別の態様において、システイン-システインジスルフィド結合のパターンを変更し、新規ジスルフィド結合形成を生じるために、システイン置換がエリスロポエチンへ導入される(「NDS変異」)。哺乳動物エリスロポエチンの中でも独自性を示す天然のヒトエリスロポエチンは、7、29、33及び161位に正確に4個のシステインを有し、これらは2個のジスルフィド結合を形成する。エリスロポエチン部分のこれらのシステイン残基の1個又は複数を改変することができる。改変されたジスルフィド結合を作製するために、1個のシステイン残基が、構造的に互換性のあるアミノ酸、例えばアラニン又はセリンへ変異され、及び三次元構造において隣接する第二のアミノ酸がシステインへ変異される。例えば、アミノ酸Gln86、Pro87、Trp88、Glu89、及びLeu91のひとつは、Cysにより置き換えることができる。Trp88がCysにより置き換えられ及びCys33が別のアミノ酸により置き換えられる場合、そのエリスロポエチン部分は、ヒトEPOにおいては認められないCys29-Cys88ジスルフィド結合を形成するであろう。この結合は、典型的Cys29-Cys33ジスルフィド結合を伴う融合タンパク質よりもより大きい活性を有する融合タンパク質を生じる。加えてCys29-Cys88融合タンパク質は、融合タンパク質のエリスロポエチン部分における他の変異の存在下で、Cys29-Cys33融合タンパク質と比べ、活性の際だった増加を示す。従って本発明のひとつの態様において、エリスロポエチン部分は、以下のアミノ酸置換の少なくともひとつを含む:29位の非-システイン残基、33位の非-システイン残基、88位のシステイン残基、及び139位のシステイン残基。ひとつの態様において、エリスロポエチン部分は、7、29、88、及び161位にシステインを含む。別の態様において、エリスロポエチン部分は更に、以下の置換を1個又は複数含む:His32→Gly、Cys33→Pro、及びPro90→Ala。別の態様において、全体の新たなジスルフィド結合が、2個のアミノ酸のシステインへの変異により、このタンパク質に付加される。これらのCys変異が引き起こし得る構造内の可能性のある緊張を補償するために、本発明の好ましいCys-操作された態様において、エリスロポエチン部分は更に、これらの可能性のある緊張を緩和するように設計された変異を含む。
【0057】
システイン置換に関連する更なる態様は、PCT国際公開公報第01/36489号(すなわち、米国特許出願第09/708,506号)に開示されており、その開示は本明細書に引用により取り込まれるものとする。
エリスロポエチンに変異を導入する方法は、当該技術分野においてよく知られている。例えば変異は、位置指定突然変異導入技術により導入することができる。広範な部位特異的突然変異導入技術を、利用することができ、及び同様の結果を実現するための代替として使用することができることに注目することは重要である。他の技術は、ランダム及びセミ-ランダム突然変異誘発を含むが、これらに限定されるものではない。
【0058】
リンカー
本発明のFc-EPO融合タンパク質は、Fc部分とエリスロポエチン部分の間に、リンカー分子、好ましくはペプチドリンカーを含むことができる。リンカーを伴う融合タンパク質は、増加した生物学的活性のような、改善された特性を有することができる。リンカーは一般に、1〜25個のアミノ酸(例えば、5〜25個又は10〜20個のアミノ酸)を含む。リンカーは、プロテアーゼ切断部位を含まないように設計することができる。更にリンカーは、タンパク質分解を立体的に阻害するために、N-結合型された又はO-結合型されたグリコシル化部位を含むことができる。従ってひとつの態様において、リンカーは、Asn-Ala-Thrアミノ酸配列を含む。
追加の適当なリンカーは、Robinsonら、Proc. Natl. Acad. Sci. USA、95 5929 (1998);及び、米国特許出願第09/708,506号に開示されている。
【0059】
グリコシル化
天然のヒトエリスロポエチン及び哺乳動物細胞において発現された組換えエリスロポエチンは、3個のN-結合型及び1個のO-結合型オリゴ糖鎖を含む。N-結合グリコシル化は、24、38及び83位に位置したアスパラギン残基で生じるのに対し、O-結合グリコシル化は、126位のセリン残基で生じる(Laiら、J. Biol. Chem.、261: 3116 (1986);Broudyら、Arch. Biochem. Biophys.、265: 329 (1988))。オリゴ糖鎖は、末端シアル酸残基により改変されることが示されている。N-結合型鎖は典型的には、1本の鎖あたり最大4個のシアル酸を有し、O-結合型鎖は典型的には、最大2個のシアル酸を有する。従ってエリスロポエチンポリペプチドは合計最大14個のシアル酸を収容することができる。
【0060】
シアル酸は、N-結合型又はO-結合型オリゴ糖上の末端糖である。シアル酸化の程度は、位置毎、タンパク質毎に変動し、使用される細胞培養条件、細胞型、及び具体的細胞クローンによって左右される。BHK細胞において合成された本発明のFc-EPO融合タンパク質は、高度にシアル酸化されることがわかっている。Fc-EPO融合タンパク質のシアル酸化の程度は、無タンパク質培地中、懸濁液中、又は無タンパク質培地中の懸濁物としての増殖に関するBHK細胞の適応により、更に増強され得ることもわかっている。ある種のその他の通常使用される細胞株、例えばNS/0、PerC6、又は293細胞は、標準の培養条件下で、高度にシアル酸化されたFc-EPO融合タンパク質を生成することができない。種々の細胞株から生成されたFc-EPO融合タンパク質のシアル酸化の程度は、それらの高度に負に荷電したシアル酸残基に基づく、等電点(IEF)ゲル電気泳動により決定することができ;IEFゲル電気泳動の詳細は、実施例5Bに説明されている。種々の細胞株において生成されたFc-EPO融合タンパク質のシアル酸化の程度は、当業者に良く知られた方法を用いるレクチン-結合試験により定性的に確認することができる。レクチン-結合アッセイの例は、実施例5Bに説明されている。
【0061】
典型的には、本発明の高度にシアル酸化された精製されたFc-EPO融合タンパク質の集団は、精製されたFc-EPO融合タンパク質あたり平均11〜28個のシアル酸残基を有する。好ましい高度にシアル酸化されたFc-EPO融合タンパク質の集団は、精製されたFc-EPO融合タンパク質あたり平均13〜28、15〜28、17〜28、19〜28、又は21〜28個のシアル酸残基を有する。例えば、ひとつの好ましい高度にシアル酸化されたFc-EPO融合タンパク質集団は、精製されたFc-EPO融合タンパク質あたり平均20〜22個のシアル酸残基を有する。別のFc-EPO融合タンパク質の好ましい集団は、精製されたFc-EPO融合タンパク質あたり平均23〜28個のシアル酸残基を有する。
【0062】
シアル酸化されたFc-EPO融合タンパク質の薬物動態
赤血球形成-刺激剤のin vivo生物学的活性を決定する最も重要な因子のひとつは、タンパク質の血清濃度が、赤血球形成に必要な閾値を上回り続ける時間の長さであり、これは赤血球形成-刺激剤の薬物動態により決定される。高度にシアル酸化されたFc-EPO融合タンパク質の薬物動態プロファイルは、天然の又は組換えエリスロポエチンのプロファイルとは異なる。大きな差異は、高度にシアル酸化されたFc-EPO融合タンパク質は、はるかに長い血清半減期及び遅いクリアランスを有し、これは増加したin vivo生物学的効能につながることである。理論に拘束されることを欲するものではないが、シアル酸残基はエリスロポエチン分子上の負電荷を増加し、負に荷電したEPO受容体結合の速度減少及びEPO受容体に媒介されたエンドサイトーシスの減少を生じさせ、血清半減期を長期化すると考えられている。更にシアル酸は、エリスロポエチンタンパク質が、露出したガラクトース残基に糖タンパク質を結合するアシアロ糖タンパク質受容体によりエンドサイトーシスを受けることも阻害する。
一般にエリスロポエチンのような、治療的分子のほとんどの薬物動態プロファイルは、血清濃度の初期低下(α相)と、それに続く投与後のより段階的な低下(β相)を示す。
【0063】
α相に影響を及ぼす因子
小型-分子の薬物動態理論に従い、α相は血液の外側のコンパートメントへ分子がどのように分配されるかを記述する、分布の容積を規定する。α相において認められる減衰は、種々の細胞株において合成された種々のFc-EPO融合タンパク質について広範に変動する。理論的には、この差異は、分布容積の変動又はコンパートメント間の通行の変動が原因である。しかし、シアル酸化の程度とマウスにおけるFc-EPOタンパク質の薬物動態挙動の間には相関関係が存在することが認められている。例えば、BHK細胞において合成されたFc-EPO融合タンパク質は、高度にシアル酸化され、最良の薬物動態プロファイルを示す。NS/0細胞において合成されたFc-EPO融合タンパク質は、若干シアル酸化され、中間の薬物動態プロファイルを有する。293及びPerC6細胞において合成されたFc-EPO融合タンパク質は、ほとんど又は全くシアル酸化されず、最初の30分間における約1/100の血清濃度減衰で特徴付けられる劣った薬物動態プロファイルを有する。従って特定のFc-EPO融合タンパク質のα相に影響を及ぼす重要な因子は、グリコシル化種の分布及びシアル酸化のレベルである。少なくシアル酸化されているFc-EPO融合タンパク質は、迅速に消失する。
【0064】
加えて図2に示されるように、α相の間のFc-EPO血清濃度の減衰の程度は投与量に従い変動し、この挙動は飽和させることが可能で、受容体-媒介性であろうことを示している。α相減衰を媒介する受容体は、EPO受容体でもFc受容体でもなく、アシアロ糖タンパク質受容体などの別の受容体であり得る。Aranesp(登録商標)は、正常なヒトエリスロポエチンと比べ、EPO受容体に対して低下した結合親和性を有しているが、その理由は、Aranesp(登録商標)は、追加のN-結合グリコシル化部位の結果として増加した負電荷を有するからである。しかしAranesp(登録商標)及び正常なヒトエリスロポエチンは、α相の間に同様の減衰を示す。加えて、一般に赤血球前駆細胞の細胞表面上のEPO受容体の数は、わずかに約200個であり、これらの受容体は図2において使用されたものよりもはるかに低用量のエリスロポエチンで完全に飽和されるであろう。グリコシル化部位を除去する変異、例えばIgG1のAsn297に対応するアミノ酸の変異を伴うFc-EPO融合タンパク質は依然としてα相における急勾配の減衰を示すことができるので、Fc受容体がα相における劇的減衰を媒介していることは恐らくありそうもない。加えて、凝集しない場合、IgG2 CH2領域は一般にFc受容体に結合しないが、IgG2 CH2領域を含むFc-EPOタンパク質は依然としてα相の間に有意な減衰を示す。
【0065】
理論に拘束されることを欲するものではないが、α相におけるFc-EPO融合タンパク質の血清濃度の減衰は、アシアロ糖タンパク質-受容体-媒介型エンドサイトーシスを介してアシアロ糖タンパク質-受容体により媒介され得る。少なくシアル酸化されたFc-EPO融合タンパク質は、アシアロ糖タンパク質受容体により結合されてアシアロ糖タンパク質-受容体-媒介型エンドサイトーシスを生じる、露出したガラクトース残基を含む。結果的に少なくシアル酸化されたFc-EPO融合タンパク質は迅速に消失する。
【0066】
β相に影響する因子
β相におけるFc-EPO融合タンパク質の血清濃度の減衰は、α相における減衰と比べて緩やかな勾配である。例えばマウスにおいて、投与後8〜24時間の間に、Fc-EPO融合タンパク質の血清濃度の1/2〜1/3倍の減衰が認められる。β相の間の減衰の差異も、異なる細胞株で合成された異なるFc-EPOタンパク質間では劇的さが少ない。しかしα相におけるように、シアル酸化の程度はβ相における薬物動態挙動と相関している。例えば、BHK細胞において合成されたFc-EPO融合タンパク質は、NS/0細胞で合成されたその他は同じであるFc-EPOタンパク質と比べて有意に改善されたβ相を有する。EPO受容体-媒介エンドサイトーシスは、β相の間のFc-EPO融合タンパク質の血清濃度の減衰に少なくとも部分的に寄与しているように見える。正常なヒトエリスロポエチンと比べ、EPO受容体への結合親和性が低下したAranesp(登録商標)は、同様のα相プロファイルにも関わらず、正常なヒトエリスロポエチンと比べて有意に改善されたβ相を有する。
【0067】
本発明のFc-EPO融合タンパク質は一般に、天然の又は組換えエリスロポエチンと比べ改善されたβ相を示しており、このことはFc部分の追加がβ相の間の血清濃度の低下を有意に遅延することを示唆している。Fc部分又はエリスロポエチン部分のある種のアミノ酸改変は、β相を有意に改善することも認められる。例えば、Fc部分のグリコシル化部位を除去する変異は、Fc-EPO融合タンパク質のβ相を改善する。エリスロポエチン部分の安定性を増加する変異、例えばエリスロポエチン部分においてジスルフィド結合を操作する変異(例えば、NDS変異)は、Fc-EPO融合タンパク質のβ相を有意に改善する。一般に改善されたβ相は、Fc-EPO融合タンパク質の終末血清半減期を延長する。
【0068】
Fc-EPO融合タンパク質の消失経路
エリスロポエチンタンパク質分子を体から排出するいくつかの可能性のある経路が存在する。野生型又は天然のエリスロポエチンタンパク質分子は、腎濾過及び受容体-媒介されたエンドサイトーシスにより体から排出される。エンドサイトーシスされたエリスロポエチンは、効率的に分解される。図3に示したように、Fc部分のエリスロポエチン部分への追加は、本質的にFc-EPO融合タンパク質の腎臓を通る排出を除去することが期待される。結果的に受容体-媒介エンドサイトーシスがFc-EPO融合タンパク質の排出の主要経路となる。更にFc部分のエリスロポエチン部分への追加は、インターナリゼーション後の分解を低下することも予想される。その理由は、FcRnエンドソーム受容体はこの融合タンパク質を細胞の外側へ再循環させると予想されるからである。
【0069】
原理上は、少なくとも3種の受容体、すなわちFc-受容体、EPO受容体、及びアシアロ糖タンパク質受容体がFc-EPO融合タンパク質のクリアランスを媒介することができる。Fc-EPO融合タンパク質のFc受容体を介したクリアランスは、Fc部分においてIgG1-由来のCH2の代わりにIgG2-由来のCH2ドメインを使用することにより有意に低下するはずである。IgG2-由来のCH2ドメインは、IgG1-由来のCH2ドメインと比べてFcγRI(IgG類への最高の親和性を有する)に対して約1/100低い親和性を有する。IgG2-由来のCH2とFcγRIの間の相互作用は、ほとんどの結合アッセイにおいて検出不能である。しかしIgG2-由来のCH2ドメインの残留FcγR-結合活性は、Fc-EPO融合タンパク質のクリアランスにおいて依然役割を果たす。その理由はCH2ドメイン内のグリコシル化部位を除去するアスパラギン変異は更にFc-受容体結合を低下させ、及びFc-EPO融合タンパク質の薬物動態を改善するからである。
NDS変異は、エリスロポエチン構造を安定化する作用を有し、その結果、インターナリゼーション後のFc-EPO融合タンパク質の分解を低下すると予想される。NDS変異を含むFc-EPO融合タンパク質は薬物動態特性を改善し、血清半減期を延長する。
【0070】
シアル酸化はFc-EPO融合タンパク質の負電荷を増加し、Fc-EPO融合タンパク質のEPO受容体への結合親和性を低下させる。シアル酸化は同じく、Fc-EPO融合タンパク質上の露出したガラクトース残基の数を減少し、これはFc-EPO融合タンパク質のアシアロ糖タンパク質受容体への結合親和性を低下させる。従って図3に示したように、シアル酸化はEPO受容体-媒介されたエンドサイトーシス及びアシアロ糖タンパク質受容体-媒介されたエンドサイトーシスの両方を減少させる。従って高度にシアル酸化されたFc-EPO融合タンパク質は、劇的に遅延されたクリアランス率を有し、その結果血清半減期が有意に増加する。
Fc部分の追加、Fc及びエリスロポエチン部分の置換、並びにシアル酸化は、各々、Fc-EPO融合タンパク質のクリアランスを低下させる。クリアランス及び血清半減期に対する組合せ作用は相加的又は相乗的である。
【0071】
Fc-EPO融合タンパク質のin vitro活性及びin vivo効能
Fc-EPOタンパク質のin vitro活性は、細胞-ベースのアッセイにおいて試験することができる。特に、Fc-EPOとEPO受容体の間の相互作用は、TF-1細胞増殖アッセイを基に行うことができる。TF-1細胞はEPO受容体を発現し、その結果、トリチウム標識されたチミジンの組込みにより決定されるTF-1細胞の増殖はエリスロポエチン活性の関数である(Hammerllingら、J. Pharmaceutical and Biomedical Analysis、14: 1455 (1996);Kitamuraら、J. Cellular Physiol.、140: 323 (1989))。本発明において、TF-1細胞の増殖は、エリスロポエチン部分とEPO受容体の間の相互作用の関数である。特にFc-EPO融合タンパク質のエリスロポエチン部分がEPO受容体に対して低下した結合速度を有する場合、Fc-EPOタンパク質は一般に、細胞-ベースのアッセイにおいて低下した活性を有する(増加したED50値により示される)。
【0072】
入手が比較的容易である細胞-ベースのアッセイのデータは、一般にFc-EPOタンパク質の薬物動態及びin vivo効能と相関する。In vitro活性の低下は、EPO受容体に対する低下した結合速度を示すが、これは一般に改善された薬物動態特性及び増強されたin vivo効能と相関する。対照的に、増加したin vitro活性(減少したED50値により示される)は、EPO受容体に対する増加された結合速度を示すが、これは一般に、貧弱な薬物動態特性及び低下したin vivo効能に相関する。
【0073】
Fc-EPO融合タンパク質のin vivo生物学的活性は、例えばマウス及びラットなどの動物モデルにおいて行われるアッセイにより測定することができる。In vivoアッセイの例は、ヘマトクリット(HCT)アッセイ及び網状赤血球アッセイを含むが、これらに限定されるものではない。HCTアッセイは、赤血球(RBC)により占拠された血液体積を測定し、毛細管内の血液を遠心し、沈積したRBCにより占拠された総容積の割合を測定することにより簡便に行われる。網状赤血球は、前駆細胞から新たに分化されるRBCであり、前駆細胞由来の核酸の名残を含むことを特徴としている。網状赤血球は、例えばアクリジンオレンジ又はチアゾールオレンジなどの、核酸-染色色素による染色後のフローサイトメーターにおける赤血球ソーティング、及び染色画分の計測により測定される。典型的には、ヘマトクリット及び網状赤血球は、週2回測定される。
【0074】
網状赤血球データは、ある意味で、ヘマトクリットデータの最初の派生物である。網状赤血球数は、赤血球の生成速度の測定値であるが、ヘマトクリットは、総赤血球を測定する。典型的実験において、Fc-EPO融合タンパク質を投与された動物のヘマトクリットは増加し、その後ベースラインに戻る。ヘマトクリットが高く、投与されたFc-EPOタンパク質が動物の循環系から消失した場合、赤血球形成が抑制されるので、網状赤血球数は、ベースラインを下回る。
網状赤血球は通常、前駆体がRBCとなるべく運命づけられた後、4日目に骨髄から出現する。しかし高レベルのエリスロポエチンの存在下では、網状赤血球は投与後1〜3日で骨髄から離れることが多いであろう。
【0075】
動物においてFc-EPOタンパク質の注射に応答してヘマトクリット読み値は増加し、一定で有り続け、その後ベースラインに戻る。そのようなヘマトクリット反応の例を図4-6に示した。最大減少率は、マウスにおいて週1回で血液体積の約7%であり、これはマウスにおける約45日間のRBC生存期間に相当し、及びラットにおいて週1回で血液体積の約5%であり、これはラットにおける約65日間のRBC生存期間に相当する。この最大減少率は、新たな合成の存在しない場合のRBCの破壊を恐らく表しているであろう。生物学的活性のあるFc-EPOタンパク質が赤血球形成の閾値を上回る濃度でその系内に存在し続ける場合、ヘマトクリットレベルは高く有り続け、生物学的活性のあるFc-EPOのレベルが薬物動態実験において検出不可能である場合であっても低下しない。
【0076】
Fc-EPOタンパク質の薬物動態特性は、このタンパク質のin vivo効能と相関することがわかった。先に説明されたようにFc-EPO融合タンパク質の薬物動態を増強する本発明の特徴は全て動物実験におけるin vivo効能も増強する。表1に示されたように、そのような特徴は例えばFc部分の追加、Fc部分内のグリコシル化部位の除去(例えば、IgG1のAsn297に相当する位置でのN→Q置換)、NDS変異のエリスロポエチン部分への導入、及びBHK細胞におけるFc-EPOタンパク質の合成による高レベルのシアル酸化を含む。
【0077】
表1
Fc-EPOタンパク質の薬物動態及び生物学的活性に影響を及ぼす因子
【0078】
エリスロポエチン部分について、BHK細胞で生成されたFcg2h(FN→AQ)-Epo及びFcg2h-EPO(NDS)は、最良の薬物動態及び最も強力なin vivo生物学的活性を示すことがわかっている。Fcg2h(FN→AQ)-Epo及びFcg2h-EPO(NDS)は各々、Aranesp(登録商標)よりもエリスロポエチン部分について、より長い血清半減期及びより強力なin vivo活性を有する。
【0079】
Fc-EPO融合タンパク質の合成
本発明のFc-EPO融合タンパク質は、ヒト又は他の哺乳動物細胞株のような適当な細胞又は細胞株において生成することができる。適当な細胞株は、ベビーハムスター腎(BHK)細胞、チャイニーズハムスター卵巣(CHO)細胞(ジヒドロ葉酸還元酵素(DHFR)-欠損細胞を含む)、及びCOS細胞を含むが、これらに限定されるものではない。好ましい態様において、BHK細胞が使用される。
【0080】
適当な宿主細胞(例えばBHK細胞)においてFc-EPO融合タンパク質を発現するために、まずFc-EPO融合タンパク質をコードしている核酸配列が当業者に周知の標準組換え分子技術を用いて発現ベクターに導入される。エリスロポエチン部分をコードしている配列は、高レベルの発現のためにコドン-最適化されることが好ましい。コドン-最適化されたヒトエリスロポエチンは、PCT国際公開公報第01/36489号(すなわち米国特許出願第09/708,506号)に開示されており、その開示は本明細書に参照として組入れられている。エリスロポエチン部分をコードしている核酸配列の例を配列番号:1に示す:
GCCCCACCACGCCTCATCTGTGACAGCCGAGTGCTGGAGAGGTACCTCTTGGAGGCCAAGGAGGCCGAGAATATCACGACCGGCTGTGCTGAACACTGCAGCTTGAATGAGAACATCACCGTGCCTGACACCAAAGTGAATTTCTATGCCTGGAAGAGGATGGAGGTTGGCCAGCAGGCCGTAGAAGTGTGGCAGGGCCTGGCCCTGCTGTCGGAAGCTGTCCTGCGGGGCCAGGCCCTGTTGGTCAACTCTTCCCAGCCGTGGGAGCCCCTGCAACTGCATGTGGATAAAGCCGTGAGTGGCCTTCGCAGCCTCACCACTCTGCTTCGGGCTCTGGGAGCCCAGAAGGAAGCCATCTCCCCTCCAGATGCGGCCTCAGCTGCTCCCCTCCGCACAATCACTGCTGACACTTTCCGCAAACTCTTCCGAGTCTACTCCAATTTCCTCCGGGGAAAGCTGAAGCTGTACACAGGGGAGGCCTGCCGGACAGGGGACAGATGA (配列番号:1)
【0081】
好ましいFc部分、例えばIgG2由来の CH2ドメイン及びIgG1由来のヒンジ領域を含むFc部分をコードしている核酸配列の例は、米国特許公開番号第20030044423号(すなわち米国特許出願第10/093,958号)に開示されており、その開示は本明細書に引用により取り込まれるものとする。
一般に、Fc-EPO融合タンパク質をコードしている核酸配列はシグナルペプチドをコードしている核酸配列(リーダー配列)を含む。このリーダー配列は分泌プロセッシング時に切断される。リーダー配列を伴わない成熟型Fc-EPOタンパク質をコードしている核酸配列の例(配列番号:2)を図7に示している。
【0082】
適当なベクターは、哺乳動物宿主細胞における発現に適したものを含む。これらのベクターは、例えば、プラスミド又はウイルスであることができる。ベクターは典型的には、以下のエレメントを含む:哺乳動物宿主細胞における使用に適合したプロモーター及び他の「上流」調節エレメント、複製起点、リボソーム結合部位、転写終結部位、ポリリンカー部位、及び選択マーカー。ベクターは、更に真核宿主細胞において増殖及び維持を可能にするエレメントも含む。本発明に適したベクターは、pdCs-Fc-X及びそれに由来するベクター、並びにphC10-Fc-X及びそれに由来するベクターを含むが、これらに限定されるものではない。
【0083】
Fc-EPOタンパク質をコードしているベクターは、トランスフェクション及びウイルス技術を含む、標準の細胞生物学的技術を用いて宿主細胞へ導入される。トランスフェクションは、単離されたDNA、RNA、又は合成ヌクレオチドポリマーを使用する、遺伝子情報の細胞への移動を意味する。適当なトランスフェクション法は、いくつか例を挙げると、リン酸カルシウム-媒介型共-沈殿(Sambrookら、Molecular Cloning: A Laboratory Manual、第2版、Cold Spring Harbor Laboratory Press、(1989))、リポフェクション(例えば、Life Technologies(ロックビル、ML)から得られるLipofectamine Plus)、DEAE-デキストラン-媒介型トランスフェクション技術、リゾチーム融合又は赤血球融合、スクレーピング(scraping)、直接取込み、浸透圧もしくはショ糖ショック、直接マイクロインジェクション、赤血球-媒介技術を介した間接マイクロインジェクション、プロトプラスト融合、又は宿主細胞の電流への曝露(例えばエレクトロポレーション)を含むが、これらに限定されるものではない。前述のトランスフェクション法のリストは網羅的であるとは考えられず、細胞へ遺伝子情報を導入する他の手法が開発されることは疑いもない。
【0084】
Fc-EPO融合タンパク質をコードしている核酸を含む宿主細胞の選択を促進するために、Fc-EPO融合タンパク質をコードしている核酸には典型的には選択マーカーが導入される。選択マーカーは、Fc-EPO融合タンパク質をコードしている同じ発現ベクター上に存在する核酸配列によりコードされていてもよい。あるいは、選択マーカーは異なるベクター上に存在する核酸配列によりコードされていてもよい。後者の場合、これら2種のベクターは、同時トランスフェクション又は同時-形質導入のいずれかにより、宿主細胞へ同時に導入される。適当な選択マーカーは、例えば、ハイグロマイシンB(Hyg B)及びジヒドロ葉酸還元酵素(DHFR)を含む。
一過性発現は、Fc-EPO融合タンパク質の小規模タンパク質生成及び迅速な分析に有用である。Fc-EPO融合タンパク質をコードしている核酸配列を含む宿主細胞は、コードされたFc-EPO融合タンパク質の発現に適した条件下で維持される。標準の細胞培養の方法、条件及び培地を、Fc-EPO融合タンパク質を発現している宿主細胞を維持するために使用することができる。
【0085】
安定的にトランスフェクションされた細胞は、大規模生成、高レベル発現、及び他の目的に好ましいことが多い。安定して維持される核酸は、宿主細胞内において様々な形態のいずれかで存在することができる。例えばひとつの態様において、安定して維持される核酸配列は、宿主細胞の染色体に組込まれる。別の態様において、安定して維持される核酸配列は、染色体外アレイとして、人工染色体として、又は別の適当な形態で存在することができる。
【0086】
ひとつの態様において、Fc-EPO融合タンパク質を合成するためにBHK細胞が使用される。安定的にトランスフェクションされたBHK細胞を得るために、融合タンパク質をコードしている核酸配列及び選択マーカーをコードしている核酸配列が、好ましくはエレクトロポレーション、プロトプラスト融合又はリポフェクション法を用いてBHK細胞へ導入される。融合タンパク質をコードしている核酸配列及び選択マーカーをコードしている核酸配列は、同じ発現ベクター上に存在することができる。あるいは、融合タンパク質をコードしている核酸配列及び選択マーカーをコードしている核酸配列は、個別のベクター上に存在することができる。安定したBHK細胞を確立するのに好ましい選択マーカーは、HygBである。DHFRなどの他の選択マーカーも使用することができる。安定的にトランスフェクションされたクローンは、単離され、適当な濃度(例えば、200、250、又は300μg/ml)のHygBの存在下、例えば、Life Technologiesから入手できるMEM+FBS、DMEM/F-12培地、もしくはVP-SFM、及び他の適当な培地などの標準組織培養培地中でのそれらの増殖により増やされる。Fc-EPO融合タンパク質の発現レベルは、上清及び培養培地からの試料に対する、例えばELISA試験、ウェスタンブロット、ドットブロット、又は他の適当なアッセイなどの、標準タンパク質-検出アッセイによりモニタリングすることができる。高発現クローンが、選択され、大規模に繁殖される。
【0087】
典型的にはBHK細胞は接着細胞株であり、通常はMEM+10%熱-失活したウシ胎仔血清(FBS)のような血清-含有培地において増殖させる。しかしBHK細胞は懸濁物として無血清培地、例えば、VP-SFM(Invitrogen Corp.、カタログ番号11681-020)又はOpti-Pro SFM(Invitrogen Corp.、カタログ番号12309)などの無血清培地中で増殖するために適合ささることができる。適合プロセスの例は実施例3に説明されている。無血清培地における増殖に適応したBHK細胞は、更に例えばDMEM/F-12(Invitrogen Corp.、カタログ番号11039-021)のような無タンパク質培地における増殖に適合させる。適合法の一例は実施例3に説明されている。DMEM/F-12には、適当なアミノ酸及び他の成分、例えばグルタミン、HyPep 4601(Quest International、カタログ番号5Z10419)及びHyPep 1510(Quest International、カタログ番号5X59053)などのタンパク質加水分解産物、Ethanolamine(Sigma、カタログ番号E0135)、及びTropolone(Sigma、カタログ番号T7387)が補充されることが好ましい。当業者は、各補充物の適当な濃度を慣習的な実験により経験的に決定することができる。
【0088】
無タンパク質培地において増殖されたBHK細胞において合成されたFc-EPO融合タンパク質は、血清-含有培地(例えばMEM+FBS)、又は無血清だが無タンパク質でない培地(例えばVP-SFM)において増殖させた細胞において合成された対応するタンパク質よりも、より多くシアル酸化され、より均質なシアル酸化を示す。加えてこうして得られたF c-EPOタンパク質は実質的に非-凝集性、すなわち総収量の約98%が非-凝集性である。無タンパク質培地において増殖されたBHK細胞から得たタンパク質収量は、血清-含有培地中で増殖されたBHK細胞からのものに類似しており、すなわち10μg/mLを上回る。従って、懸濁液及び/又は無タンパク質培地における増殖は多くの利点をもたらす。これには1)増加したシアル酸化の結果としてFc-EPO融合タンパク質の薬物動態が改善されること;および、2)タンパク質は懸濁態様及びタンパク質を欠いた培地において増殖された細胞から精製することができるので、下流の精製工程が容易になること、が含まれる。
【0089】
精製
Fc-EPOの精製は、当業者に公知の標準GMP手法に従い行われる。このタンパク質は一般に、均質又はほぼ均質に精製される。カラムクロマトグラフィーに関連するもののような、クロマトグラフィー精製が一般に好ましい。一般にFc-EPO融合タンパク質の精製スキームは、最初のタンパク質捕獲工程;ウイルス失活工程;一又は複数のポリッシュ(polishing)工程;ウイルス除去工程;及び、タンパク質濃縮及び/又は製剤工程を含むが、これらに限定されるものではない。例えば融合タンパク質のFc部分に結合するクロマトグラフィー用の樹脂材料を使用し、Fc-EPOタンパク質を捕獲することができる。適当な樹脂材料は、プロテインAに結合された樹脂を含むが、これらに限定されるものではない。夾雑成分を除去するためにポリッシュ工程を含んでもよい。例えば、ヒドロキシアパタイトクロマトグラフィー、Sepharose Qクロマトグラフィー、サイズ排除クロマトグラフィー、又は疎水相互作用クロマトグラフィーを使用し、夾雑物を除去することができる。Fc部分に結合し及びFc-EPO融合タンパク質を精製するためのプロテインA-ベースのカラムクロマトグラフィーを使用するひとつの精製法は、ウイルス失活及び除去の任意的方法として実施例12に説明されている。精製されたタンパク質は一般に、限外濾過;適当な配合の緩衝液への透析;滅菌濾過;及び、バイアルへの分注を用いて所望の濃度へ濃縮される。
【0090】
投与
医薬組成物及び投与経路
本発明は、本発明に従い作製されたFc-EPOタンパク質を含有する医薬組成物も提供する。これらの医薬組成物は、赤血球形成を刺激し、並びに貧血を予防及び治療するために使用することができる。本発明により治療可能な状態は中でも、腎機能の低下又は喪失(慢性腎不全)に随伴した貧血、化学療法もしくは抗-ウイルス薬(AZTなど)のような骨髄抑制療法に随伴した貧血、非-骨髄性癌の進行に随伴した貧血、ウイルス感染症(例えばHIV)に随伴した貧血、及び慢性疾患の貧血を含む。手術時の予想外の失血のような、それ以外は健康な個人において貧血につながり得る状態も同じく治療可能である。一般にrHuEpoにより治療可能ないずれの状態も、本発明のFc-EPO融合タンパク質により治療することができる。
【0091】
Fc-EPOタンパク質含有製剤
一般に製剤は、液体又は固体の剤形で、Fc-EPOタンパク質、緩衝液及び界面活性剤を含有する。固形製剤は、凍結乾燥された、噴霧凍結乾燥された、又は噴霧乾燥された製剤も含むが、これらに限定されるものではない。液体製剤は、好ましくは水を基剤にしているが、いくつか例を挙げると、エタノール、プロパノール、プロパンジオール、又はグリセロールなどの他の成分を含むことができる。
Fc-EPOタンパク質は、当業者に公知の標準GMP手法に従い、水溶液中に製剤される。一般に製剤は、適当な成分を適当な濃度で含む指定された容量の水溶液の混合により、作製される。例えば製剤は典型的には、Fc-EPOタンパク質を濃度0.1〜200mg/ml、好ましくは0.2〜10mg/ml、より好ましくは0.5〜6mg/mlで含有する。
【0092】
緩衝液成分は、pHを調節することが可能である任意の生理的に許容できる物質、例えば、クエン酸塩、酢酸塩、ヒスチジン塩、コハク酸塩、マレイン酸塩、リン酸塩、乳酸塩、それらの各酸もしくは塩基、又はそれらの混合物を含む。一般に使用される緩衝液成分は、クエン酸塩及び/又はそれらの遊離酸である。製剤は典型的には、緩衝液成分を濃度10〜100mmol/l、好ましくは2〜20mmol/l、より好ましくは10mmol/l含む。
Fc-EPO製剤のための界面活性剤は、医薬組成物において界面活性剤として使用される任意の賦形剤であることができ、好ましくはポリエチレン-ソルビタン-エステル(Tweens(登録商標))、例えばポリオキシエチレン(20)-ソルビタンモノラウレート、ポリオキシエチレン(20)-ソルビタンモノパルミテート、及びポリオキシエチレン(20)-ソルビタンモノステアレート、並びにポリオキシエチレン-ポリオキシプロピレン-コポリマーである。製剤は典型的には、界面活性剤を濃度0.001〜1.0%w/v、好ましくは0.005〜0.1%w/v、より好ましくは0.01〜0.5%w/vで含有する。
【0093】
製剤は、1種又は複数のアミノ酸も含有することができる。適当なアミノ酸は、アルギニン、ヒスチジン、オルニチン、リジン、グリシン、メチオニン、イソロイシン、ロイシン、アラニン、フェニルアラニン、チロシン及びトリプトファンを含むが、これらに限定されるものではない。ひとつの態様において、Fc-EPO製剤は、グリシンを含有する。好ましいアミノ酸は、塩の形、例えば塩酸塩で使用される。使用可能なアミノ酸濃度は、2〜200mmol/L、又は50〜150mmol/Lの範囲である。
加えて製剤は、ショ糖、トレハロース、ソルビトールなどの、糖;アスコルビン酸又はグルタチオンなどの、抗酸化剤;フェノール、m-クレゾール、メチル-もしくはプロピルパラベンなどの、保存剤;クロロブタノール;チメロサール;塩化ベンザルコニウム;ポリエチレングリコール;シクロデキストリン及び他の適当な成分を含むことができる。
【0094】
Fc-EPO製剤は等張であることが望ましい。例えば製剤の浸透圧は、150〜450mOsmol/kgの範囲であることができる。医薬製剤は、望ましい貯蔵温度、例えば2〜8℃、又は室温で、望ましい貯蔵期間安定していなければならない。Fc-EPOタンパク質を含有する有用な製剤は、生理的に忍容性がよく、製造が容易であり、正確に投与することができ、及び2℃〜8℃又は25℃の貯蔵期間、複数の凍結解凍サイクルの間、更には40℃で少なくとも3ヶ月の貯蔵のような他のストレス時に安定したものである。Fc-EPO製剤の安定性はストレス試験において試験することができる。ストレス試験の例は実施例13に記載されている。
【0095】
投与
本発明に従い製造されたFc-EPO融合タンパク質を含有する治療的組成物は、哺乳動物宿主へ任意の経路により投与することができる。従って、適当ならば、投与は、経口又は非経口(例えば、i.v.、i.a.、s.c.、i.m.)であってよく、これは静脈内及び腹腔内投与経路を含む。加えて投与は、ボーラスの治療薬の定期的注射によるか、又は外部にある貯蔵庫(例えば静注バッグ)からの静脈内又は腹腔内投与によりより連続して行うことができる。ある態様において、本発明の治療薬は、医薬-等級であることができる。すなわちある態様は、ヒトへの投与に必要な純度及び品質管理基準に従う。獣医学用途も、本明細書に使用される意図された意味に含まれる。
【0096】
本発明の治療薬の獣医学及びヒト医療用途の両方のための製剤は、典型的には医薬として許容できる担体及び任意に他の成分(複数)と会合している治療薬を含む。担体(複数)は、製剤の他の成分と共存でき、及びそれらのレシピエントにとって有害ではないという意味で「許容できる」。これに関して、医薬として許容できる担体は、医薬投与に適合可能ないずれか及び全ての溶媒、分散媒、コーティング、抗菌剤及び抗真菌剤、等張剤及び吸収遅延剤などを含むことが意図される。医薬活性物質のためのこのような媒体及び物質の使用は、当該技術分野においてよく知られている。従来の媒体又は物質が活性化合物と共存できないような場合を除いて、それらの本組成物における使用が企図されている。補助的活性化合物も、この組成物へ混入することができる。これらの製剤は、都合の良いことに単位剤形で存在することができ、及び薬学/微生物学の技術分野において周知の方法により調製することができる。一般に、一部の製剤は治療薬を液体担体又は細かく粉砕された固形担体又はそれら両方と会合し、その後生成物を必要ならば所望の製剤へ成形することにより調製される。
【0097】
本発明の医薬組成物は、その意図された投与経路と適合可能であるように製剤される。投与経路の例は、経口又は非経口、例えば、静脈内、皮下、吸入(例えばネブライザーによる)、経皮(局所的)、経粘膜、鼻腔内、口腔内、及び直腸投与を含む。非経口、皮内、又は皮下適用のために使用される液剤又は懸濁剤は以下の成分を含むことができる:注射用水、生理食塩水、不揮発油、ポリエチレングリコール、グリセリン、プロピレングリコール又は他の合成溶媒のような、無菌の希釈剤;ベンジルアルコール又はメチルパラベンなどの、抗菌剤;アスコルビン酸又は亜硫酸水素ナトリウムなどの、抗酸化剤;エチレンジアミン四酢酸などの、キレート剤;酢酸塩、クエン酸塩又はリン酸塩などの緩衝剤、及び塩化ナトリウム又はデキストロースなどの張性調節剤。pHは、塩酸又は水酸化ナトリウムなどの、酸又は塩基により調節することができる。
【0098】
本発明のFc-EPOタンパク質製品の好ましい投与法は、非経口(例えば、IV、IM、SC、又はIP)経路によるものであり、並びに投与される組成物は通常、治療有効量の生成物を、許容できる希釈剤、担体及び/又は補助剤と組合せて含有する。有効量は実質的に治療される状態にかなり左右されると予想されるが、治療量は活性物質0.2〜2μg/kg体重の範囲であると現在予想されている。ヒト血清アルブミンのような標準希釈剤は生理食塩水のような標準担体として本発明の医薬組成物について企図されている。本発明の組成物における使用に適した補助剤物質は、テストステロン、前駆細胞刺激剤、インスリン-様増殖因子、プロスタグランジン、セロトニン、cAMP、プロラクチン及びトリヨードチロシンなどの赤血球形成刺激作用について独立して注目される化合物、更には一般に再生不良性貧血の治療に使用される物質、例えばメテノレン(methenolene)、スタノゾロール及びナンドロンなどを含む。例えば、Resegottiら、Panminerva Medics、23: 243-248 (1981);Mgonigleら、Kidney Int., 25(2): 437-444 (1984);Pavlovic-Kanteraら、Expt. Hematol.、8(Supp. 8): 283-291 (1980);及び、Kurtz、FEBS Letters、14a(1): 105-108 (1982)を参照のこと。
【0099】
補助剤として、Fc-EPOの作用を増強又はこれと相乗作用することが報告された物質、例えばアドレナリン作動薬、甲状腺ホルモン、アンドロゲン及びBPA、更には「肝赤血球形成性因子」(Naughtonら、Acta. Haemat.、69: 171-179 (1983)参照)及び「エリスロトロピン(erythrotropins)」(Congoteら、Abstract 364、Proceedings 7th International Congress of Endocrinology、ケベック市、ケベック州、1984年7月1-7日;Congote、Biochem. Biophys. Res. Comm.、115(2): 447-483 (1983);及び、Congote、Anal. Biochem.、140: 428-433 (1984)により説明)、及び「エリスロゲニン(erythrogenins)」(Rothmanら、J. Surg. Oncol.、20: 105-108 (1982)により説明)と称される化合物のクラスも企図される。
【0100】
経口又は非経口投与に有用な溶液は、例えば「レミントン薬科学」(Gennaro, A.編集)、Mack Pub.(1990)に記載されたような、医薬技術分野において周知の方法のいずれかにより調製され得る。非経口投与用の製剤は、口腔内投与のためのグリココール酸、直腸投与のためのメトキシサリチル酸、又は膣内投与のためのクエン酸も含むことができる。非経口用調製物は、ガラス又はプラスチック製のアンプル、使い捨て注射筒、又は反復投与用バイアル中に封入することができる。直腸投与用の坐薬も同じく、この薬剤をココアバター、他のグリセリド又は室温で固形であり体温で液体である他の組成物などの非-刺激性賦形剤と共に混合することにより調製することができる。製剤は、例えばポリエチレングリコールなどのポリアルキレングリコール、植物起源の油、水素化されたナフタレンなども含むことができる。直接投与するための製剤はグリセロール及び高粘度の他の組成物を含むことができる。これらの治療薬のためのその他の可能性のある有用な非経口担体は、エチレン-酢酸ビニルコポリマー粒子、浸透圧ポンプ、埋込み可能な注入システム、及びリポソームを含む。吸入投与するための製剤は、賦形剤として、例えば乳糖を含むか、又は例えば、ポリオキシエチレン-9-ラウリルエーテル、グリココール酸及びデオキシコール酸を含有する水溶液、又は点鼻薬の形の投与用の油性溶液、又は鼻腔内塗布用のゲル剤である。持続浣腸(retention enemas)も、直腸送達に使用することができる。
【0101】
注射用途に適した医薬組成物は、無菌の液剤(ここで水溶性)又は分散剤、及び無菌の注射用液剤もしくは分散剤を即時調製するための無菌の散剤を含む。静脈内投与に関して、適当な担体は、生理食塩水、静菌水、Cremophor ELTM(BASF、ピスカタウェイ、NJ)又はリン酸緩衝生理食塩水(PBS)を含む。全ての場合において、この組成物は無菌であり、及び容易に注射筒内に入れることができる程度に流体であることができる。これは、製造及び貯蔵の条件下では安定することができ、並びに細菌及び真菌などの微生物の混入作用に対し保存することができる。この担体は、例えば、水、エタノール、ポリオール(例えば、グリセロール、プロピレングリコール、及び液体ポリエチレングリコールなど)、並びにそれらの適当な混合物を含有する溶媒又は分散媒であることができる。例えば、レシチンのようなコーティングの使用によるか、分散剤の場合必要な粒子サイズの維持によるか、及び界面活性剤の使用により適当な流動性が維持される。微生物の作用の防止は、例えばパラベン、クロロブタノール、フェノール、アスコルビン酸、チメロサールなどのような、様々な抗菌剤及び抗真菌剤により実現される。多くの場合、組成物中に例えば、糖、ポリアルコール、例えばマンニトール、ソルビトール、及び塩化ナトリウムなどの等張化剤を含むことが好ましい。注射用組成物の持続吸収は、組成物中に吸収を遅延させる物質、例えばモノステアリン酸アルミニウム及びゼラチンを含有することによりもたらされる。
【0102】
無菌の注射用液剤は、必要量の活性化合物を適当な溶媒中に、必要ならば先に列記した成分の1種又は組合せと共に混入し、その後濾過滅菌することにより調製することができる。一般に分散剤は、活性化合物を、基本の分散媒及び先に列記されたものから必要な他の成分を含む無菌のビヒクルに混入することにより調製される。無菌の注射用液剤の調製のための無菌の散剤の場合、調製法は、活性成分に加え追加の所望の成分の散剤をそれらの先に濾過滅菌された溶液から得る、真空乾燥及び凍結乾燥を含む。
【0103】
ひとつの態様において、治療薬は、インプラント及び微小カプセル化された送達システムを含む、制御された放出製剤のような、迅速な体からの消失に対し保護する担体により調製される。エチレン-酢酸ビニル、ポリ無水物、ポリグリコール酸、コラーゲン、ポリオルトエステル、及びポリ乳酸などの、生分解性、生体適合性ポリマーを使用することができる。このような製剤の調製法は、当業者に明らかであろう。これらの材料は、Alza Corporation及びNova Pharmaceuticals, Inc.から市販されている。リポソーム懸濁液も、医薬として許容できる担体として使用することができる。これらは、当業者に公知の方法、例えば米国特許第4,522,811号に開示された方法に従い調製することができる。マイクロソーム及びマイクロ粒子も使用することができる。
【0104】
経口又は非経口組成物を、投与に容易でかつ均質な用量の単位剤形に製剤することができる。単位剤形は、治療される対象にとって単位用量として適した物理的に個別の単位を意味し;各単位は、所望の治療作用を生じるように計算された、予め決められた量の活性化合物を、必要な医薬担体と会合して含有する。本発明の単位剤形の特定は、活性化合物の独自の特性及び達成されるべき具体的な治療作用により指示されかつ直接左右され、並びにこれらの制限は、個体の治療に関するそのような活性化合物の混和の技術分野において固有のものである。
【0105】
Fc-EPOの治療有効量及び投与頻度の決定
一般に、本発明に従い作製されたFc-EPO融合タンパク質を含有する治療薬は、例えば治療有効量、すなわち望ましい作用を誘導するのに十分な時間、標的組織へ薬剤の適当な濃度を提供する量でヒト又は他の哺乳動物へ非経口投与又は経口投与用に製剤することができる。より詳細に述べると、本明細書において「治療有効量」は、目標ヘマトクリット、又は患者へ恩恵をもたらす目標ヘマトクリット範囲までのヘマトクリットの増大をもたらすか、あるいは目標ヘマトクリットに、もしくは目標ヘマトクリット範囲内に患者を維持するような、Fc-EPO融合タンパク質の量を意味する。この量は、個人毎に変動し、並びに患者の全身の生理的条件、貧血の重症度及び基礎となる原因、並びに個々の患者に関する最終的な目標ヘマトクリットを含む、多くの要因に左右されるであろう。目標ヘマトクリットは、典型的には少なくとも約30%、又は30%〜38%の範囲であり、好ましくは38%を上回り、より好ましくは40%〜45%である。rHuEpoに関する目標ヘマトクリット範囲に関する全般的指針は、1996年12月23日付けのEPOGEN(登録商標)の添付文書においても認められ、これはそこに記されているように、30%〜36%、あるいは32%〜38%である。そのような目標は個人毎に変動し、その結果、所定の患者について実際の目標ヘマトクリットの決定においては医師の判断が適当であろうことが理解される。しかしながら、目標ヘマトクリットの決定は十分に当業者の能力範囲内である。
【0106】
Fc-EPOタンパク質の治療有効量は、当業者により容易に確定され得る。実施例15は、週1回、2週に1回、及び1ヶ月に1回の投与の、Fc-EPOの治療有効量を決定するためのひとつの目標としての臨床プロトコールを示している。例えば、週1回又は2週に1回の投与量範囲は、約0.075〜約4.5μg Fc-EPO/kg/投与である。1ヶ月に1回の投与量範囲は、0.45〜4.5μg Fc-EPO/kg/投与である。
【0107】
治療的組成物中で送達される本発明のFc-EPO融合タンパク質の有効濃度は、投与される薬剤の最終の望ましい用量及び投与経路を含む、多くの要因に応じて変動するであろう。投与される好ましい用量は、恐らく治療される疾患又は適応症の種類及び程度、特定の患者の全身の健康状態、送達される治療薬の相対的生物学的効能(例えば、シアル酸化のレベル)、治療薬の製剤、製剤中の賦形剤の存在及び種類、並びに投与経路のような変数により決まるであろう。一部の態様において、本発明の治療薬は、非-ヒト霊長類及び齧歯類を使用する哺乳動物試験から推定された典型的単位投与量を用い、個体へ提供される。先に説明されたように、用量単位は、患者へ投与することが可能であり、容易に取扱い及び包装することができ、そのまま又はそれの固形もしくは液体の医薬希釈剤もしくは担体との混合物として治療薬を含有する物理的及び生物学的に安定した単位投与量として維持される単位投与量を意味する。
【0108】
Fc-EPO融合タンパク質を含有する治療薬の投与頻度は、治療される状態及び目標のヘマトクリットに応じて変動するが、一般には週3回未満であろう。投与頻度は、ほぼ週1回又は2回であってよい。投与頻度は、週に約1回未満、例えば2週間に約1回(14日間に約1回)、1ヶ月に1回又は2ヶ月に1回であることもできる。エリスロポエチン及びそのアナログに対する異なる個体による反応の変動のために、実際に使用される投与頻度は、本明細書に明らかにされた頻度から若干変動することができることが理解され;用語「約」は、そのような変動を反映することが意図されている。
【0109】
本発明は、治療時に増加した赤血球形成を維持するための、治療有効量の鉄分の投与も提供する。投与される量は、rHuEpoによる治療を基に当業者により容易に決定され得る。加えて本発明の治療薬は、単独で、又は特定の疾患又は関心のある適応症に有益な作用を有することがわかっている他の分子と組合せて投与することができる。単なる例として、有用な補因子は、防腐薬、抗生物質、抗ウイルス薬、及び抗真菌薬、並びに鎮痛薬及び麻酔薬などの、症状を緩和する補因子を含む。
【0110】
プロドラッグ
本発明の治療薬は、「プロドラッグ」誘導体も含む。用語プロドラッグは、活性成分を放出もしくは活性化するために、生体内で自発的又は酵素的のいずれかの生体転換を必要とする、親分子の薬理学的に不活性(又は部分的に不活性)の誘導体を意味する。プロドラッグは、代謝条件下で切断可能な基を有する、本発明の治療薬の変種又は誘導体である。プロドラッグは、生理的条件下で加溶媒分解を受けるか又は酵素分解を受ける場合に、in vivoにおいて医薬活性のある本発明の治療薬となり始める。本発明のプロドラッグは、生体内での活性のある薬剤成分の放出又は活性化に必要な生体転換工程の数に応じてシングル、ダブル、トリプルなどと称され、これは前駆体型に存在する官能基の数を示す。プロドラッグ型は、哺乳動物生体において、溶解度、組織適合性、又は遅延した放出の利点を提供することが多い(Bundgard、Design of Prodrugs、7-9頁、21-24頁(1985)、Elsevier, Amsterdam;Silverman、The Organic Chemistry of Drug Design and Drug Action、352-401頁(1992)、Academic Press、サンディエゴ、CA)。更に本発明のプロドラッグ誘導体は、バイオアベイラビリティを増強するために、他の特徴と組合せることができる。
【0111】
In vivo発現
本発明のFc-EPO融合タンパク質はin vivo発現法により提供することができる。例えば、Fc-EPO融合タンパク質をコードしている核酸は、有利なことに造血障害又は欠損を罹患した患者へ直接投与することができるか、又は細胞へex vivoで提供し、その後生存細胞を患者へ投与することができる。当該技術分野において公知であるin vivo遺伝子療法は、精製されたDNA(例えば、プラスミド内)を提供すること、ウイルスベクター内のDNAを提供すること、又はリポソームもしくは他のベシクル内のDNAを提供することを含む(例えば、遺伝子療法において使用する脂質担体を開示している、米国特許第5,827,703号、及び遺伝子療法において有用なアデノウイルスベクターを提供する、米国特許第6,281,010号を参照のこと)。
【0112】
組換えタンパク質を発現するように改変されている細胞を移植することにより疾患を治療する方法も、公知である。例えばヒトへ導入するために、核酸を初代ヒト細胞へ導入する方法を開示している、米国特許第5,399,346号を参照のこと。
In vivo発現法は、精製することなく、タンパク質の標的組織又は細胞コンパートメントへの直接の送達に特に有用である。本発明において、Fc-EPOをコードしている配列を使用する遺伝子療法は、貧血を含む、血液系の不調の様々な病態、障害及び状態における、特に慢性腎不全などに随伴した型の貧血の調整における用途が認められる。Fc-EPO融合タンパク質をコードしている核酸配列は、適当な転写又は発現カセットに挿入し、宿主哺乳動物へ裸のDNAとして又は適当な担体と複合して導入することができる。活性Fc-EPOタンパク質の生成のモニタリングは、核酸ハイブリダイゼーション、ELISA、ウェスタンハイブリダイゼーション、及び当業者に公知の他の適当な方法により行うことができる。
【0113】
導入遺伝子の全身投与によって複数の組織を形質転換することができることがわかっている。哺乳動物宿主へのカチオン性脂質担体/外来性DNA複合体の静脈内注射後の外来性DNAの発現は、Tリンパ球、細網内皮系、心内皮細胞、肺細胞、及び骨髄細胞、例えば骨髄由来の造血細胞を含む、複数の組織において示されている。
米国特許第6,627,615号に説明されたin vivo遺伝子療法の送達技術は、動物において無毒であり、並びに導入遺伝子発現は、単回投与後少なくとも60日間最後まで示される。導入遺伝子は、サザン分析により測定して、in vivoにおいて検出可能なレベルで、宿主細胞DNAへ組込まれるようには見えず、このことは、遺伝子治療に関するこの技術は、癌を惹起する癌遺伝子を活性化するか、又は癌を予防する腫瘍抑制遺伝子を停止するように、正常な細胞遺伝子の発現を変更することにより、宿主哺乳動物に問題を引き起こさないことを示唆している。
【0114】
本発明の具体的態様を以下に列挙する。
(1) ヒンジ領域、CH2及びCH3ドメインを含むヒトIgG分子の二量体Fc部分、並びにヒトエリスロポエチン(EPO)から本質的になる、精製された二量体融合タンパク質であって、前記二量体Fc部分の各鎖は、そのC-末端でEPO分子のN-末端へ直接又はリンカーペプチドを介して連結しており、下記の特徴を有する前記二量体融合タンパク質:
(i) 15〜28個のシアル酸残基を有することにより高度にシアル酸化されている;;
(ii) 前記CH2領域がヒトIgG2に由来し、前記CH2領域中のGln-Phe-Asn-Ser配列内のPheおよびAsnがAlaおよびAsnで置換され、その結果前記CH2領域内に配列Gln-Ala-Gln-Serが形成されることによって前記CH2領域が改変されている;および、
(iii)CH3ドメインのC-末端近傍のLeu-Ser-Leu-Serアミノ酸配列が、Ala-Thr-Ala-Thrで置換されている。
(2) 更に、CH3ドメインのC-末端Lys残基がAlaに置換されている、(1)記載の二量体Fc-EPO融合タンパク質。
(3) ヒンジ領域がヒトIgG1に由来する、(1)又は(2)記載の二量体Fc-EPO融合タンパク質。
(4) IgG1ヒンジ領域中のPro-Lys-Ser-Cys-Asp-Lys配列内のCysアミノ酸残基がSer残基に置換され、前記ヒンジ領域中に配列Pro-Lys-Ser-Ser-Asp-Lysが形成されることによって、I前記ヒンジ領域が改変されている(3)記載の二量体Fc-EPO融合タンパク質。
(5) エリスロポエチン部分が少なくとも以下のアミノ酸置換の一つを含む、(1)〜(4)のいずれかに記載のFc-EPO融合タンパク質:
(i)EPO分子の29位における非システイン残基;
(ii)EPO分子の33位における非システイン残基;
(iii)EPO分子の88位におけるシステイン残基;
(iv)EPO分子の139位におけるシステイン残基。
(6) EPO分子の33位のCys残基の代わりに非-Cysアミノ酸残基が存在し、EPO分子の88位のTrpの代わりにCys残基が存在し、それによって融合タンパク質内のEPO 部分がCys29-Cys88ジスルフィド結合を形成することが可能である、(5)記載の二量体Fc-EPO融合タンパク質。
(7) 33位の非-Cysアミノ酸残基がProである、(6)記載の二量体Fc-EPO融合タンパク質。
(8) EPO部分が以下から選ばれる1以上の変異を含む(1)〜(7)のいずれかに記載の二量体Fc-EPO融合タンパク質:
(i)Arg131→Glu131;
(ii)Arg139→Glu139;
(iii)His32→Gly32
(iv)Ser34→Arg34;
(v)Pro90→Ala90。
(9) リンカーペプチドがグリコシル化部位を含む、(1)〜(8)のいずれかに記載の二量体Fc-EPO融合タンパク質。
(10) グリコシル化部位がAsn-Ala-Thrアミノ酸配列を含む、(9)記載の二量体Fc-EPO融合タンパク質。
(11) 完全なIgG分子がIgG2に由来し、ヒンジ領域がIgG1に由来する、(1)〜(10)のいずれかに記載の二量体Fc-EPO融合タンパク質。
(12) 更にCH1ドメインを含む、(1)〜(11)のいずれかに記載の二量体Fc-EPO融合タンパク質。
(13) 融合タンパク質が、20〜22個のシアル酸残基を有する、(1)〜(12)のいずれかに記載の二量体Fc-EPO融合タンパク質。
(14) 配列:
EPKSSDKTHTCPPCPAPPVAGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVQFNWYVDGVEVHNAKTKPREEQAQSTFRVVSVLTVVHQDWLNGKEYKCKVSNKGLPAPIEKTISKTKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPMLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSATATPGAAPPRLICDSRVLERYLLEAKEAENITTGCAEHCSLNENITVPDTKVNFYAWKRMEVGQQAVEVWQGLALLSEAVLRGQALLVNSSQPWEPLQLHVDKAVSGLRSLTTLLRALGAQKEAISPPDAASAAPLRTITADTFRKLFRVYSNFLRGKLKLYTGEACRTGDR (配列番号:14)
を含む、二量体Fc-EPO融合タンパク質。
(15) 配列:
EPKSSDKTHTCPPCPAPPVAGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVQFNWYVDGVEVHNAKTKPREEQAQSTFRVVSVLTVVHQDWLNGKEYKCKVSNKGLPAPIEKTISKTKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPMLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSATATPGAAPPRLICDSRVLERYLLEAKEAENITTGCAEGPSLNENITVPDTKVNFYAWKRMEVGQQAVEVWQGLALLSEAVLRGQALLVNSSQPCEALQLHVDKAVSGLRSLTTLLRALGAQKEAISPPDAASAAPLRTITADTFRKLFRVYSNFLRGKLKLYTGEACRTGDR (配列番号:15)
を含む、二量体Fc-EPO融合タンパク質。
(16) (1)〜(15)のいずれかに記載の融合タンパク質をコードするDNA分子。
(17) (1)〜(15)のいずれかに記載の融合タンパク質を効果的量で含み、場合により製薬的に許容される単体、希釈剤または賦形剤とを含む、哺乳動物の造血疾患または不全の治療に適した医薬組成物。
(18) N-末端側にFc部分を含みC-末端側にエリスロポエチンを含む、哺乳動物への投与に適した高度にシアル酸化された精製されたFc-EPO融合タンパク質の集団であって、前記精製されたFc-EPO融合タンパク質1分子あたり平均15〜28個のシアル酸残基を有し、それぞれのFc-EPO融合タンパク質をコードするDNA分子をBHK細胞に導入し、対応する前記Fc-EPO融合タンパク質を発現、単離及び精製することによって得られ、NS/0、PerC6または293細胞中で合成された対応するFcーEPO融合タンパク質の集団に比較して長い血清半減期を有する前記Fc-EPO融合タンパク質の集団。
(19) 精製されたFc-EPO融合タンパク質1分子あたり平均20〜22個のシアル酸残基を有する、(18)に記載のFc-EPO融合タンパク質の集団。
(20) BHK細胞が無タンパク質培地又は懸濁液中での増殖に適応している、(18)又は(19)に記載の精製されたFc-EPO融合タンパク質の集団。
(21) 以下の工程を含む、N-末端側にFc部分を含みC-末端側にエリスロポエチンを含む、哺乳動物への投与に適した高度にシアル酸化された精製されたFc-EPO融合タンパク質の集団を作製する方法:
(i)Fc-EPO融合タンパク質をコードするDNA分子を構築する工程;
(ii)無タンパク質培地中または懸濁状態で、前記DNA分子でBHK細胞を形質転換する工程;
(iii)前記DNA分子よってコードされるFc-EPO融合タンパク質の集団を発現させる工程;
(iv)前記Fc-EPO融合タンパク質の集団を回収、単離および精製する工程。
(22) 作製された融合タンパク質の集団が、精製されたFc-EPO融合タンパク質あたり平均15〜28個のシアル酸残基を有する、(21)記載の方法。
(23) 作製された融合タンパク質の集団が、精製されたFc-EPO融合タンパク質あたり平均20〜22個のシアル酸残基を有する、(22)記載の方法。
(24) 以下の工程を含む、Fc部分およびエリスロポエチン部分を含むFc-EPO融合タンパク質をコードする核酸配列を安定に維持するBHK細胞を選抜する方法:
(a)ハイグロマイシンBをコードする核酸配列および前記Fc-EPO融合タンパク質をコードする核酸配列をBHK細胞に導入する工程;
(b)前記BHK細胞をハイグロマイシンBの存在下で培養する工程。
(25) ハイグロマイシンBをコードしている核酸配列及びFc-EPO融合タンパク質をコードする核酸配列が単一のDNA分子中に存在する、(24)記載の方法。
【実施例1】
【0115】
実施例1. Fc-EPO融合タンパク質をコードしている構築物
正常なエリスロポエチン部分を含むFc-EPO融合タンパク質をコードしているプラスミドphC10-Fcg2h(FN→AQ)-M1-EPO及びNDS変異を伴うFc-EPO融合タンパク質をコードしているプラスミド phC10-Fcg2h(FN→AQ)-M1-EPO(NDS)を以下のように構築した。
ヒトエリスロポエチンをコードしている核酸配列を、哺乳動物細胞における高発現のためにコドン-最適化した。例えば配列番号:3は翻訳を最適化するように改変されたコドンを伴う、成熟型ヒトエリスロポエチンのコード配列の例を示している。5'末端の配列は、サブクローニングを容易にするためにSmaI部位を含むように改変した。
【0116】
配列番号:3
CCCGGGtGCCCCACCACGCCTCATCTGTGACAGCCGAGTgCTGGAGAGGTACCTCTTGGAGGCCAAGGAGGCCGAGAATATCACGACcGGCTGTGCTGAACACTGCAGCTTGAATGAGAAcATCACcGTgCCtGACACCAAAGTgAATTTCTATGCCTGGAAGAGGATGGAGGTtGGcCAGCAGGCCGTAGAAGTgTGGCAGGGCCTGGCCCTGCTGTCGGAAGCTGTCCTGCGGGGCCAGGCCCTGTTGGTCAACTCTTCCCAGCCGTGGGAGCCCCTGCAaCTGCATGTGGATAAAGCCGTgAGTGGCCTTCGCAGCCTCACCACTCTGCTTCGGGCTCTGgGAGCCCAGAAGGAAGCCATCTCCCCTCCAGATGCGGCCTCAGCTGCTCCcCTCCGcACAATCACTGCTGACACTTTCCGCAAACTCTTCCGAGTCTACTCCAATTTCCTCCGGGGAAAGCTGAAGCTGTACACAGGGGAGGCCTgcCGGACAGGGGACAGATGActcgag
(小文字は、野生型ヒトエリスロポエチンコード配列と異なる塩基を示す。これらの変更は、哺乳動物細胞における発現レベルを増加させると期待されるが、発現されたタンパク質配列を変化させない。)
【0117】
NDS変異は、こその開示は本明細書に参照として組入れられている、PCT国際公開公報第01/36489号に開示された位置指定突然変異誘発により、エリスロポエチン部分へ導入した。例えば、国際公開公報第01/36489号に開示された、アミノ酸置換His32Gly、Cys33Pro、Trp88Cys、及びPro90Alaを生じる変異を伴うヒトエリスロポエチンコード配列の形を含むXmaI-XhoI DNA断片を使用した。対応するタンパク質配列を配列番号:4に示した。
【0118】
APPRLICDSRVLERYLLEAKEAENITTGCAEGPSLNENITVPDTKVNFYAWKRMEVGQQAVEVWQGLALLSEAVLRGQALLVNSSQPCEGLQLHVDKAVSGLRSLTTLLRALGAQKEAISPPDAASAAPLRTITADTFRKLFRVYSNFLRGKLKLYTGEACRTGDR (配列番号:4)
【0119】
IgG2-由来のCH2ドメイン及びIgG1-由来のヒンジ領域を含むハイブリッドFc部分を、米国特許公報第20020147311号、及び例えば国際公開公報第01/058957号に開示されたように構築した。
ある形のエリスロポエチンをコードしているXmaI-XhoI DNA断片を、プラスミドベクター、例えばpdCs-Fc-Xへ挿入したが、このベクターは、CH3 C-末端領域におけるアミノ酸置換を生じる2セットの変異(ここではM1セット変異と称される)が存在することを除き、IgG1由来の変更されたヒンジ領域並びにIgG2由来のCH2及びCH3領域をコードしており、その結果CH3 C-末端及びEPO N-末端の接合部の配列は、以下である:
【0120】
. . . . TQKSATATPGA-APPRLI . . . .(配列番号:5)
【0121】
IgG2 CH3領域の配列KSLSLSPG (配列番号:6)をKSATATPG (配列番号:7)へ変更する変異の第一のセットは、国際公開公報第02/079232号に開示されている。Leu-Ser-Leu-Ser (配列番号:6の3位から6位)のAla-Thr-Ala-Thr (配列番号:7の3位から6位)による置換の効果は、ヒトFcとヒトエリスロポエチンの間の接合部が非-自己ペプチド配列を含むことによって生じる可能性のあるヒト非-自己T-細胞エピトープを除去することである。CH3領域のC-末端アミノ酸の単一アミノ酸置換KからAを含む第二のセットは、国際公開公報第01/58957号に開示されている。
【0122】
Fc融合タンパク質の発現のための発現ベクターpdCs-Fc-Xは、Loらの論文(Protein Engineering、11: 495 (1998))に記載されている。プラスミドphC10-Fc-Xを、pdCs-Fc-Xから、メトトレキセート耐性を付与するジヒドロ葉酸還元酵素(DHFR)遺伝子のコード領域を、ハイグロマイシンBに対する耐性を付与する遺伝子と置換することにより、構築した。NheI/NsiIハイグロマイシンB DNA断片は、プライマー5'-GCTAGCTTGGTGCCCTCATGAAAAAGCCTGAACTC-3' (配列番号:8)及び5'-ATGCATTCAGTTAGCCTCCCCCATC-3' (配列番号:9)を使用する、鋳型プラスミドpCEP4 (Invitrogen)からのハイグロマイシンB遺伝子のPCR増幅により得た。PCR断片をTAクローニングベクターpCR2.1 (Invitrogen)にクローニングし、その配列を確認した。
【0123】
プラスミドphC10-Fcg2h-M1-EPO(NDS)を、pdCs-Fcg2h-M1-EPO(NDS)由来のNheI/AflI及びAflII/NsiI DNA断片並びにNheI/NseIハイグロマイシンB断片のトリプルライゲーションにより作製した。
更に、PCR突然変異誘発により、IgG2重鎖のCH2ドメイン内のGln-Phe-Asn-Serアミノ酸配列内に可能性のあるT-細胞エピトープ及びFc部分内のN-結合グリコシル化を除去する二重アミノ酸置換「FN>AQ」を生じさせる変異を導入した。変異導入プライマー5'-AGCAGGCCCAGAGCACGTTCCGTGTGGT-3' (配列番号:10)及び5'-GAACGTGCTCTGGGCCTGCTCCTCCCGT-3' (配列番号:11)を、各々、SacII部位を含む下流プライマー5'-CCCCGCGGGTCCCACCTTTGG-3' (配列番号:12)及びPvuII部位を含む上流プライマー5'-CCCAGCTGGGTGCTGACACGT-3' (配列番号:13)と組み合わせ、ふたつの重複するDNA断片を鋳型DNA pdC10-Fcg2h-M1-EPO(NDS)から増幅した。第二の増幅ラウンドにおいて、変異(FN→AQ)を含むPvuII/SacII断片を、第一の増幅ラウンドのPCR産物から、上流プライマー(配列番号:13)及び下流プライマー(配列番号:12)を用いて増幅した。PvuII/SacII断片を、TAベクターpCR2.1 (Invitrogen)へクローニングし、その配列を検証した。構築物pdC10-Fcg2h(FN>AQ)-M1-EPO(NDS)を、pdC10-Fcg2h-M1-EPO由来のPvuII/SacII断片、XhoI/SacII断片、並びにpdC10-Fcg2h-M1-EPO(NDS)由来のXhoI/PvuII断片のトリプルライゲーションから作製した。
【0124】
FN>AQ変異をプラスミドphC10-Fcg2h-M1-EPOへ導入するために、phC10-Fcg2h-M1-EPO及びpdC10-Fcg2h(FN→AQ)-M1-EPO由来の適当なDNA断片を組合せた。phC10-Fcg2h-M1-EPO及びpdC10-Fcg2h(FN→AQ)-M1-EPOの両構築物をXhoI及びXbaIで消化し、5.7kbのXhoI/XbaI phC10-Fcg2h-M1-EPO(NDS)断片を、1.9kbのpdC10-Fcg2h(FN→AQ)-M1-EPO断片にライゲーションし、phC10-Fcg2h(FN→AQ)-M1-EPOを作出した。
【0125】
FN→AQ変異をプラスミドphC10-Fcg2h-M1-EPO(NDS)へ導入するために、phC10-Fcg2h-M1-EPO(NDS)及びphC10-Fcg2h(FN→AQ)-M1-EPO由来の2種の適当なXhoI/SmaI消化した断片を互いにライゲーションし、phC10-Fcg2h(FN→AQ)-M1-EPO(NDS)を作出した。
【0126】
pdC10-huFcg2h(FN→AQ)-M1-EPOによりコードされたFc-EPOのアミノ酸配列を、配列番号:14に示した。
[EPKSSDKTHTCPPCP]APPVAGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVQFNWYVDGVEVHNAKTKPREEQAQSTFRVVSVLTVVHQDWLNGKEYKCKVSNKGLPAPIEKTISKTKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPMLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSATATPGAAPPRLICDSRVLERYLLEAKEAENITTGCAEHCSLNENITVPDTKVNFYAWKRMEVGQQAVEVWQGLALLSEAVLRGQALLVNSSQPWEPLQLHVDKAVSGLRSLTTLLRALGAQKEAISPPDAASAAPLRTITADTFRKLFRVYSNFLRGKLKLYTGEACRTGDR (配列番号:14)
【0127】
pdC10-huFcg2h(FN→AQ)-M1-EPO(NDS)によりコードされたFc-EPO(NDS)のアミノ酸配列を、配列番号:15に示した。
[EPKSSDKTHTCPPCP]APPVAGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVQFNWYVDGVEVHNAKTKPREEQAQSTFRVVSVLTVVHQDWLNGKEYKCKVSNKGLPAPIEKTISKTKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPMLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSATATPGAAPPRLICDSRVLERYLLEAKEAENITTGCAEGPSLNENITVPDTKVNFYAWKRMEVGQQAVEVWQGLALLSEAVLRGQALLVNSSQPCEALQLHVDKAVSGLRSLTTLLRALGAQKEAISPPDAASAAPLRTITADTFRKLFRVYSNFLRGKLKLYTGEACRTGDR (配列番号:15)
下線を付けた配列トラックはEPO部分を表し、下線を付けた[]内配列は、IgG1ヒンジを表し、下線を付けていない配列は改変されたIgG鎖のCH2及びCH3ドメインを表し、ここで点線を付した配列はCH3ドメインを表す。
【実施例2】
【0128】
実施例2. 様々な細胞株におけるFc-EPOの発現
融合タンパク質の即時分析のために、標準の一過性トランスフェクション法、例えば、リン酸カルシウム-媒介型DNA共沈法(Sambrookら、Molecular Cloning: A Laboratory Manual、第2版、Cold Spring Harbor Laboratory Press、 (1989))、又はLipofectamine Plus (Life Technologies)を製造業者のプロトコールに従い使用するリポフェクション法により、プラスミドphC10-Fcg2h(FN→AQ)-M1-EPO(NDS)又はphC10-Fcg2h(FN→AQ)-M1-EPOを、適当な組織培養細胞へ導入した。
【0129】
安定的にトランスフェクションされたBHK-21細胞を得るために、プラスミドphC10-Fcg2h(FN→AQ)-M1-EPO(NDS)又はphC10-Fcg2h(FN→AQ)-M1-EPOをエレクトロポレーションによりBHK-21細胞へ導入した。高効率のエレクトロポレーションのために、BHK-21細胞はMEM培地(アメリカン・タイプ・カルチャー・コレクション(ATCC)の推奨により、非-必須アミノ酸及びピルビン酸ナトリウムを補充)中で増殖させ、PBSで1回洗浄し;並びに、約5x106個細胞を、0.5ml PBS中に再懸濁し、0.4cm電極間隙を備えたGene Pulser(商標)キュベット(BioRad, ハーキュレス, CA)中で、直鎖状プラスミドDNA 10μgと共に、氷上で、10分間インキュベーションした。エレクトロポレーションは、0.25V及び500μFに設定したGene Pulser(商標)(BioRad, ハーキュレス, CA)を用いて行った。細胞を氷上で10分間回復させ、増殖培地中に再懸濁し、2個の96ウェルプレート上に播種した。ハイグロマイシンB(Hyg B)を、トランスフェクション後2日間、濃度300μg/mlで増殖培地へ添加した。これらの細胞は、3日毎に2〜3回培地供給し(fed)、Hyg B耐性安定クローンが、2〜3週間で出現した。
【0130】
高レベルFc-EPO融合タンパク質を産生している安定クローンを同定するために、クローン上清を抗-Fc抗体によるELISAによりアッセイした。高-産生クローンを単離し、300μg/ml Hyg Bを含有する増殖培地において増殖させた。タンパク質産生の目的では、BHK-21細胞を、補充したDMEM/F-12培地又はVP-SFM (Life Technologies)などの別の適当な培地においてルーチン的に生育させた。Fc-EPO融合タンパク質を、標準的ノーマル-フロー濾過により、コンディション培地から収集し、清澄化した材料を更なる精製まで4℃で貯蔵した。典型的には、ローラーボトル産生モードでFc-EPOタンパク質6〜12μg/mlがBHK-21細胞から得られた。
【0131】
Fc-EPO融合タンパク質は、NS/0細胞においても発現され、これを回収した。プラスミドpdC10-Fcg2h(FN→AQ)-M1-EPO又はpdC10-Fcg2h(FN→AQ)-M1-EPO(NDS)を安定して維持しているNS/0クローンを、先にPCT国際公開公報第01/36489号に開示された方法により確立した。この文献の開示は本明細書に引用により取り込まれるものとする。典型的には、NS/0細胞からFc-EPOタンパク質50〜100μg/mlが得られた。
【実施例3】
【0132】
実施例3. 懸濁物及び/又は無タンパク質培地中の増殖へのBHK細胞の適応
BHKは、一般に例えばMEM+10%熱-失活したウシ胎仔血清(FBS)のような血清-含有培地において増殖される接着細胞株である。BHK細胞を維持および拡張(増殖)するために、これらは定期的に(例えば4日間隔で)、典型的にはトリプシン-EDTA溶液の作用によりそれらの基質から剥離し、新鮮培地に希釈し、適当な容器に再播種される。しかしBHK細胞は、以下の手順により、懸濁液並びに無血清及び/又は無タンパク質培地における増殖に適応させることができる。
【0133】
典型的適応プロセスにおいて、BHK細胞をまずMEM+FBS:標的培地の75:25(v/v)混合培地中で指数増殖期まで培養し、続いて当初の培地:標的培地の50:50(v/v)、25:75(v/v)、及び最後に0:100(v/v)の中で、適当な細胞密度でサブクローニングした。適応プロセスの間、BHK細胞の増殖を、目視検査によりモニタリングした。以下の無血清培地を、それらに対する適応性について試験した:293 SFM II (Invitrogen Corp., カタログ番号11686-929)、CHO-S-SFM II (Invitrogen Corp., カタログ番号12052-098)、VP-SFM (Invitrogen Corp., カタログ番号11681-020)、Opti-Pro SFM (Invitrogen Corp., カタログ番号12309)、CDハイブリドーマ(Invitrogen Corp., カタログ番号11279-023)、及びH-SFM (Invitrogen Corp., カタログ番号12045-076)。
【0134】
この適応プロセスの間にBHK細胞を接着細胞株から懸濁細胞株へ転換するために、この培養混合液を各継代前に静置し、細胞懸濁液の上層25%を取り出して新鮮培地へ希釈した。凝集した細胞は、単独細胞及び対細胞(doublet cell)よりもより迅速に培養容器の底に沈降するので、上層25%の細胞懸濁液は一般に凝集の最低量を示す細胞を含む。従って各継代は凝集しにくいBHK細胞を拡張および濃縮し、Fc-EPOタンパク質を発現しているBHKクローンの浮遊細胞株がこの方法で樹立された。
【0135】
Fc-EPOタンパク質を発現しているBHK細胞は、VP-SFM又はOpti-PRO SFM無血清培地における増殖に適応し、浮遊培養物が得られたことがわかった。Fc-EPO融合タンパク質を発現しているBHK細胞は、以下の無血清培地において増殖することはできなかった:293 SFM II、CHO-S-SFM II、CDハイブリドーマ、及びH-SFM。
無血清培地VP-SFMに適応されたBHK細胞は、75:25(v/v)、50:50(v/v)、25:75(v/v)、最後に0:100(v/v)のVP-SFM:DMEM/F-12混合液中で、BHK細胞を適当な細胞密度で逐次培養することにより、無タンパク質培地、例えばDMEM/F-12 (Invitrogen Corp., カタログ番号11039-021)における増殖に更に適応した。無タンパク質培地DMEM/F-12には、グルタミン(最終6mM)、2g/l HyPep 4601 (Quest International, シカゴ, IL, カタログ番号5Z10419)、2g/l HyPep 1510 (Quest International, シカゴ, IL, カタログ番号5X59053)、10μl/l(v/v)エタノールアミン(Sigma, カタログ番号E0135)、及び5μM Tropolone (Sigma, カタログ番号T7387)を補充した。補充したDMEM/F-12における増殖に対しコンピテントなFc-EPO融合タンパク質を安定して発現しているBHK細胞株をこの方法により得、高い細胞生存率で維持した。
【実施例4】
【0136】
実施例4. 凝集状態のタンパク質の精製及び特徴決定
分析のために、Fc-EPO融合タンパク質を、細胞-培養上清から、Fc部分のプロテインAへの親和性を基に、プロテインAクロマトグラフィーにより精製した。Fc-EPOタンパク質を発現している細胞の馴化した上清を、予め平衡化したファストフロー・プロテインA Sepharoseカラムに装加した。カラムをリン酸ナトリウム緩衝液(150mMリン酸ナトリウム、100mM NaCl、中性pH)で十分洗浄した。結合したタンパク質を低pH(pH2.5〜3)のリン酸ナトリウム緩衝液(前記組成)で溶離し、溶離された画分を即座に中和した。
【0137】
異なる細胞株により産生されたFc-EPO融合タンパク質の凝集状態を評価するために、プロテインA精製された試料を分析用サイズ排除クロマトグラフィー(SEC)により分析した。試料を、HPLC-SEC(例えば、Super 3000 SW, TosoHaas, Montgomeryville, PA)により、流量0.35ml/分で15分間流し、分画した。BHK細胞から産生したFc-EPOタンパク質のかなりの部分(例えば、総収量の最大90%〜100%)は凝集しなかった。更にFc-EPO融合タンパク質の試料を、還元SDS-PAGE(予め成形されたNuPAGE 4%〜12%ゲル、NuPAGE, Novex)により分析し、実質的に1本バンドであることを明らかにした。このことはこれらの生成物が、標準操作手法を行った場合、分解に抵抗性があることを示している。
【0138】
懸濁液、無血清培地、及び/又は無タンパク質培地において増殖したBHK細胞から精製されたFc-EPO融合タンパク質も前述のSDS-PAGE及び分析用SECにより特徴決定した。これらのタンパク質は、血清-含有培地において増殖したBHK細胞において合成されたタンパク質同様、実質的に凝集せず、分解されないことがわかった。
【実施例5】
【0139】
実施例5A. グリコシル化パターンの特徴決定
ヒトエリスロポエチンのセリン126は、O-グリコシル化と適合性のある配列内であり、全ての哺乳動物エリスロポエチンタンパク質において保存されている。しかしセリン126は、そのタンパク質の残余に対し密にパックされない「フロッピー-ループ(floppy loop)」内にある。O-グリコシル化が存在しない場合、エリスロポエチンのこの領域はタンパク質分解に特に感受性がある。
【0140】
異なる細胞株で産生されたFc-EPOタンパク質のSer126でのO-グリコシル化の状態を、逆相HPLCにより試験した。試料は変性及び還元し、0.1%トリフルオロ酢酸(TFA)に希釈し、逆相HPLCカラム(例えば、Vydac C4カラム, Grace Vydac)へ注入した。アセトニトリル中0.085%TFAまでの勾配を適用し、タンパク質試料の保持時間を記録した。BHK-21細胞において合成されたFc-EPO及びFc-g2h(FN→AQ)-EPOは、ふたつの部分的に重複する大きいピークを生じたことがわかった(ピーク#1及びピーク#2)。これらのピーク画分は更に、ペプチドマッピングにより分析した。ピーク#1は、特徴的ペプチド(ペプチド#36)の非存在により示されるように、Ser126でグリコシル化されたFc-EPOの形態に対応するのに対し、ピーク#2は、特徴的ペプチド(ペプチド#36)の存在により示されるように、Ser126でグリコシル化されないFc-EPOの形態に対応することがわかった。Ser126は、BHK細胞から産生されたFc-EPO分子の約60%においてO-グリコシル化により修飾されることが明らかになる、これは天然のEPOについて報告されているものと一致する。更に、補充した無タンパク質DMEM/F-12培地中でBHK細胞を増殖させることは、O-グリコシル化の頻度に正の効果を有した。
【0141】
実施例5B. シアル酸化パターンの特徴決定
NS/0、BHK、293、及びPerC6細胞において合成されたFc-EPO融合タンパク質のシアル酸化の程度を等電点(IEF)ゲル電気泳動により比較した。簡単に述べると、2mg/mlに濃縮し及び必要ならば脱塩した試料を、等量のIEF試料緩衝液(pH3-7)に添加し、垂直の予め成形されたNovex pH 3-7 IEFゲル(Novex, カタログ番号EC6655B/B2)上で2.5時間、最初の1時間は100V、次の1時間は200Vで及び最後の30分間は500Vで流した。その後ゲルを固定、染色及び脱染色した。
【0142】
ひとつの特定の実験において、以下の試料を比較した(試料は、血清-含有培地において増殖した細胞に由来した):
1. NS/0由来のFcg2h-EPO(NDS)
2. BHK-21由来のFcg2h-EPO(NDS)
3. BHK-21由来のFcg2h-EPO
4. BHK-21由来のFcg2h(“Delta Lys”)-EPO
5. BHK-21由来のFcg4h(FN→AQ “Delta Lys”)-EPO
6. BHK-21由来のFcg4h(“Delta Lys”)-EPO
【0143】
この群において、「Delta Lys」は、FcドメインのC-末端におけるリジンの欠失を意味する(試料4-6)。試料1-3は、このC-末端リジンのアラニンへの変異を有する。従ってこのC-末端リジンは全ての試料において存在せず、試料間に生じる電荷の差は存在しない。全ての細胞は、血清-含有培地において接着細胞として増殖させた。
【0144】
試料を、pH3-7 IEFゲルに載せ、pH3.5、4.2、4.5、5.2、5.3、6.0、及び6.9に焦点のある標準 (Serva Electrophoresis, 独国)と比較した。第一の試料のNS/0由来のFcg2h-EPO(NDS)は、等電点約pH5.3〜6.5のバンド分布として移動し;最も強いバンドは、pH6.0-6.1に存在した。第二の試料のBHK-21由来のFcg2h-EPO(NDS)は、等電点約pH4.6〜pH5.0に強いバンド、pH5.0〜約pH6.0に弱いバンドの分布として泳動され;最も強いバンドは、pH4.8-4.9に存在した。第三及び第四の試料のBHK-21由来のFcg2h-EPO及びBHK-21由来のFcg2h(“Delta Lys”)-EPOは、各々、両方とも約pH4.7〜6.0にバンド分布を有し、最も強いバンドは約pH5.3に焦点があった。第五及び第六の試料のBHK-21由来のFcg4h(FN→AQ “Delta Lys”)-EPO及びBHK-21由来のFcg4h(“Delta Lys”)-EPOは、各々、第二の試料に類似した収束パターンを有していた。すなわち、これらは等電点約pH4.6〜pH5.0の強いバンド、pH5.0〜約pH6.0の弱いバンドの分布として泳動された。これらの結果は、BHK細胞におけるFc-EPO融合タンパク質の合成は、一般にNS/0細胞において合成された同じ又は類似の生成物よりも、有意により酸性の生成物を生じることを示している。
【0145】
別の実験において、BHK細胞由来のFcg2h-M1-EPO(NDS)試料をノイラミニダーゼにより処理し、オリゴ糖からシアル酸を除去した。得られたノイラミニダーゼ-処理した試料は、IEFゲル上で泳動し、pH6.9又はそれより大きいところに数個のバンドがフォーカシングすることが認められた。ノイラミニダーゼ-処理を伴う又は伴わないBHK細胞由来の試料並びにNS/0細胞由来の試料のバンド形成パターンを比較し、約27種の個別のシアル酸化された種を同定した。これらの27種は、ホモ二量体配置のFc-EPO融合タンパク質のシアル酸化の程度の変動により生じ得る予想される28の異なる種と良く対応している。この分析に従い、4〜5個のシアル酸残基を伴うFcg2h-EPOはpH6.9マーカーと共に収束し、及び11〜12個のシアル酸残基を伴うFcg2h-EPOはpH6.0マーカーと共に収束した。BHK細胞において合成されたFcg2h-EPOタンパク質の集団は、タンパク質1分子あたり平均21個のシアル酸残基を有することがわかった。対照的に、NS/0細胞において合成されたFc(g2h)-EPOタンパク質の集団は、タンパク質1分子あたり平均約10個のシアル酸残基を有するように見えた。
【0146】
引き続きの実験において、Fc-EPOタンパク質を発現しているBHK細胞を、無血清増殖条件及び大規模生成に適した条件、例えば懸濁条件に適応させた。無血清かつ懸濁液中で増殖させたBHK細胞において産生されたFc-EPOタンパク質を、前述のようにIEFゲル電気泳動により分析した。増殖条件のこれらの変更は、最強のバンドの等電点について最大でも0.1〜0.3 pH単位のシフトしか生じさせなかった。
補充したDMEM/F-12無タンパク質培地において合成されたFc-EPO融合タンパク質の試料も、IEFゲル電気泳動により同様に特徴決定した。タンパク質生成物は、VP-SFMのような無血清培地において増殖された細胞から得られた対応する生成物よりも、より程度が大きくシアル酸化され、かつ、より均質なシアル酸化を示したことがわかった。
【0147】
異なる細胞株で生成されたFc-EPOタンパク質のシアル酸化の程度は更に、レクチン-結合試験により定量的に確認した。例えばFc-EPO融合タンパク質を最初に、標準SDSゲル電気泳動により分離し、ブロッティングし、その後異なる糖質部分を認識する改変されたレクチン(例えばRoche Applied Science, インディアナポリス, INから市販されている)でプロービングし、結合したレクチンを視認した。適当なレクチンは、特定の結合を伴うシアル酸を認識するSambucus nigraアグルチニン(SNA)又はMaackia amurensisアグルチニン(MAA)、並びにO-グリカンコアのような糖鎖部分の他の領域を認識するDatura stramoniumアグルチニン(DAA)、Peanutアグルチニン(PNA)及びジャカリンを含むが、これらに限定されるものではない。レクチン結合アッセイを基に、異なる細胞株で生成されたFc-EPO融合タンパク質のシアル酸化レベルを決定した。
【実施例6】
【0148】
実施例6. Fc-EPO変種のin vitro生物学的活性
種々のFc-EPOタンパク質のin vitro活性を細胞-ベースのアッセイにおいて試験した。TF-1細胞株はEPO受容体を発現しており、従って適当な培養条件下におけるそのトリチウム標識されたチミジンの取込みは、EPO又はEPO-様タンパク質活性の関数である(Hammerllingら、J. Pharmaceutical and Biomedical Analysis、14: 1455 (1996);Kitamuraら、J. Cellular Physiol.、140: 323 (1989))。具体的には、活発な対数相にあるTF-1細胞をEPOを含まない培地において2回洗浄し、マイクロタイタープレートへ約104個細胞/ウェルで播種した。その後これらの細胞を、Fc-EPO変種の滴定した連続希釈を伴う培地において、48時間インキュベーションした。3H-チミジン0.3μCiをウェルに添加し、その10時間後に細胞増殖をアッセイした。対照として、TF-1細胞を、組換えヒトEPO、及び高グリコシル化されたEPOアナログAranesp(登録商標)の存在下でもインキュベーションした。放射性チミジンの取込みは、総TCA-沈降可能な数として測定した。表2に示したように、Fcg2h-M1-EPO分子の活性は組換えヒトEPOの活性と同等であった。
【0149】
いくつかの全般的結論をこのデータから引き出すことができる。先に報告された結果と一致して、CHO細胞から生成されたEPOはED50 約0.7ng/mlを有し;これは、NIBSC EPO標準、R&D SystemsからのEPO、及び市販のProcrit(登録商標)を含む。Aranesp(登録商標)はin vitroにおいて有意に活性が低く、これは恐らくその増加した負電荷に起因する低下した結合速度を反映しているであろう。同様にBHK細胞から生成されたFc-EPOはNS/0細胞から産生されたFc-EPOよりも少ない活性を有し、これはBHK細胞により生成されたFc-EPOタンパク質は、より高度にシアル酸化され、タンパク質上に増加した負電荷を生じるという知見と一致している。
【0150】
表2
【実施例7】
【0151】
実施例7. Fc-EPO変種の薬物動態分析
様々な細胞株において合成された様々なFc-EPOタンパク質の薬物動態プロファイルを、以下のin vivo実験を基に特徴付けた。ひとつの実験において、図8に示したように、NS/0細胞及びBHK細胞において合成されたFcg2h(N>Q)-EPOタンパク質約14μgを、Swiss-Websterマウスへ静脈内投与した。投与後様々な時点(例えば、T=投与後0、1/2、1、2、4、8、及び24時間)で、血液試料を採取し、血清を遠心により調製した。Fc-EPOの血清濃度を、抗-Fc抗体を用いるELISAにより決定した。図8に示したように、投与後24時間で、BHK-由来のFc-EPOの初期血清濃度の10%より多くが血清中に残存したのに対し、NS/0-由来のFc-EPOの初期血清濃度の0.1%未満しか血清中に残存しなかった。
【0152】
同様の実験を、NS/0細胞及びBHK細胞で合成されたFcg2h-EPO(NDS)タンパク質で行った。NS/0細胞及びBHK細胞において合成されたFcg2h-EPO(NDS)タンパク質約14μgを、Swiss-Websterマウスへ静脈内投与した。血液試料を投与後、T=0、1/2、1、2、4、8、24及び36時間で採取し、血清中のFcg2h-EPO(NDS)濃度を、抗-Fc抗体を用いるELISAにより決定した。図9に示したように、投与後24時間で、BHK-由来のFcg2h-EPO(NDS)の初期血清濃度の10%より多くが血清中に残存したのに対し、NS/0-由来のFcg2h-EPO(NDS)の初期血清濃度の0.1%未満が血清中に残存した。
【0153】
BHK-21細胞、PERC6細胞、及び293細胞において生成されたFcg2h-EPO(NDS)の薬物動態プロファイルも比較した。特にFcg2h-Epo(NDS)を発現するプラスミドを、BHK、293、及びPERC6細胞へ一過性にトランスフェクションした。発現されたFcg2h-Epo(NDS)融合タンパク質を、異なる細胞株で精製し、Swiss-Websterマウスへ、濃度1.7μg/マウスで、静脈内注射した。血液試料を、T=0、1/2、1、2、4、8、24、48、及び72時間に採取し、血清中のFcg2h-Epo(NDS)濃度を抗-Fc ELISAにより測定した。図10に示したように、投与後24時間で、BHK-由来のFcg2h-EPO(NDS)の初期血清濃度の10%より多くが血清中に残存したのに対し、293細胞由来のFcg2h-EPO(NDS)の初期血清濃度の1%未満が血清中に残存し、並びにPerC6細胞-由来のFcg2h-EPO(NDS)は血清中でほとんど検出不能であった。BHK、PerC6、及び293細胞において産生されたFcg2h(N→Q)-EPOタンパク質において同様の結果が得られた。
【0154】
Fcg2h(N→Q)-EPO、Fcg2h-EPO(NDS)、Fcg2h-EPO、及びAranesp(登録商標)(すなわち、NESP)の薬物動態プロファイルを比較するために、同様の実験をマウスにおいて行った。ここで使用したFc-EPO変種は、BHK細胞から合成した。投与後48時間で、Aranesp(登録商標)の初期血清濃度の10%未満が血清中に残存したのに対し、Fcg2h(N→Q)-EPO及びFcg2h-EPO(NDS)の両方の初期血清濃度の10%よりも多くが血清中に残存したことが観察された。これらの結果は、BHK-21細胞から生成されたFcg2h(N→Q)-EPO及びFcg2h-EPO(NDS)タンパク質は、Aranesp(登録商標)よりもはるかに長い血清半減期を有したことを示している。
【実施例8】
【0155】
実施例8. Fc-EPO変種のin vivo効能
様々なFc-EPO変種のin vivo生物学的活性を、マウス及びラットにおけるヘマトクリット(HCT)アッセイ及び網状赤血球アッセイにより測定した。
ひとつのHCT実験において、CD1マウスに、BHK細胞において合成されたFcg2h(FN→AQ)-EPOタンパク質を投与量20μg/kg及び10μg/kgで腹腔内注射した。血液試料をマウスから第4、7、11及び14日目に採取し、毛細管で遠心した。沈降したRBCの量を総容積の割合として測定した。図4に示したように、Fcg2h(FN>AQ)-EPOタンパク質注射に応答してヘマトクリットは初めに劇的に増加し、その後定常状態を維持し、最後に漸減した。
【0156】
別の実験において、BHK細胞において合成された以下のタンパク質を、Sprague-Dawleyラットに腹腔内注射した。全ての動物に42.5μg/kgで投与した。
1. Fcg2h-EPO
2. Fcg2h-EPO(NDS)
3. Fcg4h-EPO
4. Fcg4h(N>Q)-EPO
HCTアッセイを、前述のように注射したマウスから採取した血液試料について行った。図5に示したように、Fcg2h-EPO(NDS)及びFcg2h-EPOに応答して、注射したラットにおけるヘマトクリットの量は延長された期間定常状態を維持した。このことは、Fcg2h-EPO(NDS)及びFcg2h-EPOの両タンパク質は延長された血清半減期及び強力なin vivo生物学的活性を有することを示している。図5に示されたように、Fcg4h-EPO及びFcg4h(N→Q)-EPOは、Fcg2h-EPO(NDS)及びFcg2h-EPOタンパク質と比べてより短い定常状態の期間及びより迅速な血清濃度の低下を示すことも認められた。
【0157】
別の実験において、CD1マウスに、以下の試料を腹腔内投与した:
1. BHK細胞由来のFcg2h-EPO(NDS)、投与量85μg/kg、42.5μg/kg、及び21.25μg/kg
2. NS/0細胞由来のFcg2h-EPO(NDS)、投与量85μg/kg、42.5μg/kg、及び21.25μg/kg
3. Aranesp(登録商標)(すなわち、NESP)、投与量50μg/kg、25μg/kg、及び12.5μg/kg
これらのタンパク質の量は、糖鎖を伴わないタンパク質分子量を基に算出した。この実験において、Fcg2h-EPO(NDS)タンパク質の分子量は単量体ポリペプチドを基にしている。従ってFcg2h-EPO(NDS)のNESPに対する分子量の比は、約1.71〜1であった。従って、この実験において各タンパク質の投与量範囲はほぼ等しかった。
【0158】
図6に示されたように、BHK細胞において合成されたFcg2h-EPO(NDS)タンパク質は、効能及び作用期間に関して最良のヘマトクリットプロファイルを示し、このことは、BHK細胞由来のFcg2h-EPO(NDS)タンパク質は、NS/0細胞由来のFcg2h-EPO(NDS)及びNESPの両方と比べ、より長い血清半減期及びより強力なin vivo活性を有することを示している。NS/0細胞由来のFcg2h-EPO(NDS)及びNESPのヘマトクリットプロファイルは、同等である。
【実施例9】
【0159】
実施例9. IgG2及びIgG4由来のCH2-CH3ドメインを伴うFc-EPOタンパク質の比較
様々なFc-EPOタンパク質の細胞-ベースのエリスロポエチン活性の比較は、IgG4由来のCH2及びCH3ドメインを伴う融合タンパク質は一般に、IgG2由来のCH2及びCH3ドメインを伴う対応するタンパク質よりも活性が低いことを明らかにした。この結論は、少なくとも3種のFc-EPOタンパク質、すなわち、エリスロポエチン部分にNDS変異を伴い、及びNS/0細胞において合成されたタンパク質(表3)、BHK細胞において合成されたNDS変異を伴うタンパク質(表4)、及びBHK細胞において合成された正常なエリスロポエチンを伴うタンパク質(表5)について、当てはまった。
【0160】
下記表3から5で比較したタンパク質は全て、IgG1由来の改変されたヒンジ、及びFc部分のC-末端のM1セット変異を有する。これらのタンパク質の活性を、実施例6に説明された標準手法による、タンパク質により刺激されたTF-1細胞へのトリチウム標識されたチミジンの取込みの測定により決定した。活性は、エリスロポエチン部分のED50(ng/ml)として示した。
【0161】
表3:NDS変異を伴う、NS/0細胞において合成されたFc-EPO融合タンパク質の細胞-ベースの活性
【0162】
表4:NDS変異を伴う、BHK細胞において合成されたFc-EPO融合タンパク質の細胞-ベースの活性
【0163】
表5:野生型EPOを有する、BHK細胞において合成されたFc-EPO融合タンパク質の細胞-ベースの活性
【0164】
In vitro細胞-ベースのアッセイからの活性データは通常、エリスロポエチン-含有タンパク質の薬物動態プロファイル及びin vivo効能を示すことができる。一般に細胞-ベースのアッセイにおける減少したin vitro活性は、EPO受容体の低下したon-rateを示し、これは改善された薬物動態特性(例えば延長された半減期)及び増強されたin vivo活性に相関している。しかしIgG4-由来のCH2及びCH3ドメインを伴うFc-EPO融合タンパク質の減少したin vitro活性は、改善した薬物動態及び増強されたin vivo生物学的活性とは相関しない。IgG4-由来のCH2及びCH3ドメインを伴うFc-EPO融合タンパク質の薬物動態プロファイルは一般に、IgG2-由来のCH2及びCH3ドメインを伴う対応するタンパク質とは識別不能であることがわかった。IgG4-由来のCH2及びCH3ドメインを伴うFc-EPO融合タンパク質は一般に、IgG2-由来のCH2及びCH3ドメインを伴う対応するタンパク質と比べ、in vivoにおいてより少ない活性を有することもわかった(図5参照)。
【実施例10】
【0165】
実施例10. Fc部分のグリコシル化部位の除去の効果
Fc部分のグリコシル化部位の除去の、in vitro活性、薬物動態、及びin vivo効能に対する作用を試験するために実験を行った。特にIgG2-由来のCH2及びCH3ドメイン又はIgG4-由来のCH2及びCH3ドメインのいずれかを含むFc-EPO融合タンパク質を試験した。IgG1のAsn297に相当する、IgG2又はIgG4のGln-Phe-Asn-Serアミノ酸配列内のアスパラギンをグルタミンに置換した。ほとんどの実験において、Gln-Phe-Asn-Serアミノ酸配列のフェニルアラニンをアラニンに置換し、アスパラギンの変異の結果として生ずる可能性のある非-自己T-細胞エピトープを除去した。表6に示されるように、細胞-ベースのin vitroアッセイにおいて、Fc部分のN-結合グリコシル化部位を除去するFN>AQ変異を伴うFc-EPOタンパク質のED50値は、一般にこの変異を伴わないFc-EPOタンパク質の約1/5倍であり、このことはN-結合グリコシル化部位の除去が細胞-ベースのアッセイにおいてin vitro活性を減少させることを示した。
【0166】
N-結合グリコシル化の除去の薬物動態及びin vivo効能に対する作用を試験するために実験を行った。CD1マウスを、BHK細胞で合成したFcg2h-M1-EPO、Fcg2h-M1-EPO(NDS)、及びFcg2h(N>Q)-M1-EPOタンパク質で、それぞれ投与量42μg/kgで処置した。Fcg2h(N→Q)-M1-EPOタンパク質は、NRQ変異を伴わない対応するタンパク質よりも、より良い薬物動態プロファイルを示したことが認められた。従って、IgG2-由来のFc部分においてN-結合グリコシル化を除去するN>Q変異は、改善された薬物動態(例えば、延長された血清半減期)を生じた。延長された血清半減期は、Fc受容体への結合に対する効果により説明することはできない。その理由は、IgG2-由来のCH2及びCH3ドメインはそもそも本質的に検出不能なFc-受容体結合を有しているからである。
【0167】
表6:Fc部分のグリコシル化部位の除去はFc-EPO融合タンパク質のin vitro細胞-ベースの活性を低下させる
【0168】
IgG2及びIgG4由来のFc部分のN-結合グリコシル化の除去により引き起こされる効果は、エリスロポエチン受容体についての低下した結合速度とほとんど一致するので、これらの効果は予想外で驚くべきことである。理論に拘束されることを欲するものではないが、IgG2及びIgG4由来のFc部分のN-結合グリコシル化の除去は、Fc-EPO融合タンパク質の全体的な立体構造の変化を引き起こすかもしれない。
【実施例11】
【0169】
実施例11. BHK細胞において合成されたFc-EPO融合タンパク質によるビーグル犬の処置
Fc-EPO融合タンパク質のヘマトクリット、網状赤血球数、及び他の血液パラメーターに対する作用を試験するために、Fc-EPO融合タンパク質をビーグル犬に投与した。具体的には、Fcg2h(FN→AQ)-EPOタンパク質を、2種の個別に安定的にトランスフェクションされたBHK細胞株であるクローン65及びクローン187から精製し、ビーグル犬に静脈内投与した。1匹のオス及び1匹のメスのビーグル犬に、以下のスケジュールで各調製物を注射した:
0日目:3μg/kg
16日目:10μg/kg
23日目:100μg/kg
【0170】
各投与後異なる時点で、血液約2mlを収集し、ヘマトクリット、網状赤血球数及び他の血液パラメーターなどの血液パラメーターを測定した。
処置後のヘマトクリット反応を、図11に示した。Fc-EPO融合タンパク質3μg/kgの投与後、血液パラメーターは正常範囲から増加しなかった。10μg/kgの投与後1週間以内に、網状赤血球数は、4匹の動物中3匹において、総血液体積の3%を超えて増加し、ヘマトクリットは1匹において51に増加した。他の血液パラメーターは正常な範囲から増加しなかった。100μg/kgの投与後、ヘマトクリット数は急激に上昇し、57〜62のピークレベルに到達し、5〜6週間にわたり正常な範囲を上回り続けた。網状赤血球数は、2〜3週間上昇し続けた。
各動物について、1dl当たりのg数で測定した血液1μl当たりの赤血球数及びヘモグロビン数はヘマトクリット量に比例した。これらの結果は、Fc-EPOタンパク質は正常なヘモグロビン含量の正常なサイズの赤血球形成を刺激することを示した。
【実施例12】
【0171】
実施例12. 臨床使用のためのFc-EPOタンパク質の精製
Fc-EPOタンパク質は、当業者に公知の標準GMP手法に従い精製した。バンクで作出したクローンからのBHK-21細胞を、追加の2.5mM L-グルタミン(Invitrogen)、2g/lの各HyPep1501及びHyPep 4601(Quest International, シカゴ, IL)、10μl/lエタノールアミン(Sigma)、及び5μM Tropolone(Sigma)を補充したDMEM/F-12培地(Invitrogen)において、7〜10日間、バッチ培養し、その間高細胞生存率(例えば、80%を上回る)を維持した。馴化培地を収集し、ノーマル-フロー-濾過により清澄化し、予め-平衡としたプロテインA Sepharoseファスト-フローカラム(Pharmacia)に載せた(Fc部分に関するプロテインAの親和性を基に融合タンパク質を捕獲する)。このカラムを中性pHの150mMリン酸ナトリウム及び100mM NaClを含有するリン酸ナトリウム緩衝液15カラム容量で十分に洗浄した。結合したタンパク質を、更に150mMリン酸ナトリウム及び100mM NaClを含有する酸性リン酸ナトリウム緩衝液(pH2.5〜3)15カラム容量により、低pHで溶離した。
【0172】
ウイルス失活のため、プールされたピーク画分のpHをpH3.8に調節し、更に30分間室温でインキュベーションした。30分間インキュベーションした後、プールした画分を中和し、滅菌濾過し、その後Q-Sepharoseファスト-フローアニオン交換カラム(Pharmacia)に装加した。これにより、その高度なシアル酸化の結果としてのFc-EPOタンパク質の酸性pIを利用してFc-EPOタンパク質と同時-溶離した可能性のある夾雑物が効果的に除去される。具体的には、中和した画分を、pH5.0でQ-Sepharoseファスト-フローアニオン交換カラム(Pharmacia)上に載せ、NaCl溶液の勾配で溶離した。その後Fc-EPOの画分を収集し、引き続きの分析及び更なる精製プロセスのためにプールした。例えば、Q-Sepharoseカラムからの高塩ストリップを逆相クロマトグラフィーカラムに載せ、過剰なNaClを除去した。希釈した逆相カラムからの溶離液を更に第二のQ-Sepharoseファストフロー(Pharmacia, 3cm X 9cm)カラムに載せた。
【0173】
その後、ナノ-濾過(例えば、MilliporeのViresolve)により、このプールから存在し得るウイルス粒子を除去した。場合により、ヒドロキシアパタイトカラム又はフェニル-ホウ酸カラム(cis-ジオールに結合)などの更なる精製工程を使用することができる。最後に、精製されたタンパク質を、限外濾過を用いて望ましい濃度に濃縮し、その後適当な配合の緩衝液へ透析した。この材料を最終的に滅菌濾過し、所定の体積でバイアルに分注した。
【実施例13】
【0174】
実施例13. Fc-EPOタンパク質製剤の安定性を決定するためのストレス試験
例証のための試料Fc-EPO製剤又は参照Fc-EPO製剤を含有するバイアルを、40℃で相対湿度75%で、指定された貯蔵期間(例えば、0週、4週、8週など)貯蔵した。一定の貯蔵期間の後、各バイアルからアリコート試料を採取し、分析した。これらの試料は、濁度について、冷光源による直接照射下で目視により評価した。更に、濁度を吸光度350nm及び550nmで測定することにより決定した。加えて試料中のFc-EPOタンパク質の条件及びタンパク質分解産物の存在を、分析用サイズ排除クロマトグラフィー(HPLC-SEC)により分析した。0.5mg/ml Fc-EPO、10mMクエン酸塩(pH6.2)、100mMグリシン、100mM NaCl、0.01%w/vポリソルベート20を含有する製剤は、参照溶液と比べて安定性を有意に増加したことがわかった。
【実施例14】
【0175】
実施例14. ヒトにおけるFcg2h(FN>AQ)-M1-EPO融合タンパク質の第I相試験
ヒトにおけるFcg2h(FN>AQ)-M1-EPO融合タンパク質の第I相臨床試験は、下記のように実施されている。薬物動態パラメーターは、本質的にAranesp(登録商標)について、MacDougallら(J. Am. Soc. Nephrol.、10: 2392-2395 (1999))により説明されたように決定したが、この論文の教示は本明細書に引用により取り込まれるものとする。ヒトに静脈内注射されたFcg2h(FN→AQ)-M1-EPO融合タンパク質(用量1μg/kg)の終末血清半減期は、約20〜30時間であることがわかった。従って、用量1μg/kg、又は成人貧血患者における約70μgは、初期血清濃度約10ng/mlをもたらした。正常なヒトエリスロポエチン濃度は、約0.04〜0.25ng/mlであるので(Cazzolaら、Blood、91: 2139-2145 (1998))、Fc-EPOタンパク質の薬理学的に活性なレベルは患者の全身において少なくとも5〜10日間維持される。
【実施例15】
【0176】
実施例15. Fcg2h(FN→AQ)-M1-EPO融合タンパク質の第II相用量設定及び投与スケジュール決定試験
Fcg2h(FN>AQ)-M1-EPO融合タンパク質について、透析を受けている慢性腎不全(CRF)患者へ皮下又は静脈内注射により投与した場合の最適用量及び投与スケジュールを調べるために、多施設無作為化逐次用量漸増試験を開始する。
臨床の実践において、以下の指針に従い、Fcg2h(FN→AQ)-M1-EPO融合タンパク質の投与を個々の貧血患者に合わせて調節することは一般に都合がよい。初回量を投与し、ヘマトクリット、ヘモグロビン、網状赤血球数、及び血小板数などの血液パラメーターをモニタリングする。初回量は、典型的には約0.3〜3μg/kgである。都合の良い初回量は1μg/kgである。治療の8週間後にヘマトクリットの増加が血液体積の5〜6%未満である場合には、用量が増加されなければならない。2週間のヘマトクリットの増加が血液体積の4%を上回るか、又はヘマトクリットが36%に達する場合、用量を減少させなければならない。
【0177】
投与スケジュールの例は、以下である:
週1回投与:0.075、0.225、0.45、0.75、1.5及び4.5μg/kg/投与。
2週に1回投与:0.075、0.225、0.45、0.75、1.5及び4.5μg/kg/投与。
1ヶ月に1回投与:0.45、0.75、1.5及び4.5μg/kg/投与。
【0178】
試験は、2部において行われる。第一の部は、週1回、2週に1回、1ヶ月に1回のいずれかで投与される、Fcg2h(FN→AQ)-M1-EPO融合タンパク質の用量を評価するためにデザインされた、用量-漸増試験であり、これは4週間にわたり、最適速度でヘモグロビンを増加する(1g/dLよりも大きい又はこれと等しいが、3g/dL未満である)。各試験の第二の部は、治療目標にヘマトクリットを維持するために必要な用量を決定するためにデザインされている(静脈内又は皮下のいずれかの投与経路で、週1回、2週に1回、1ヶ月に1回投与される場合)。
【特許請求の範囲】
【請求項1】
ヒンジ領域、CH2およびCH3ドメインおよび、ヒトエリスロポエチン(EPO)を含む精製された二量体組換えFc-EPO融合タンパク質であって、前記二量体Fc部分の各鎖がそのC末端でEPO分子のN末端に接続しており、無タンパク質培地中でBHK-12細胞にて産生され、かつ、以下から選択される、前記融合タンパク:
(i)配列番号14に記載されたFcg2h(FN→AQ)-M1-EPO;および、
(ii)配列番号15に記載されたFcg2h(FN→AQ)-M1-EPO(NDS)。
【請求項2】
DMEM/F-12培地で産生される、請求項1記載の精製された二量体組換えFc-EPO融合タンパク質。
【請求項3】
アミノ酸、タンパク質加水分解産物、エタノールアミンおよびトロポロンが補充されたDMEM/F-12培地で産生された、請求項1記載の精製された二量体組換えFc-EPO融合タンパク質。
【請求項4】
請求項1〜3のいずれか1項記載のFc-EPO融合タンパク質の有効量を含み、場合により製薬的に許容できる単体、希釈剤または賦形剤を含む、医薬組成物。
【請求項5】
哺乳動物の造血疾患または不全の治療用の、請求項1〜3のいずれか1項記載の精製された二量体Fc-EPO融合タンパク質または4記載の医薬組成物。
【請求項1】
ヒンジ領域、CH2およびCH3ドメインおよび、ヒトエリスロポエチン(EPO)を含む精製された二量体組換えFc-EPO融合タンパク質であって、前記二量体Fc部分の各鎖がそのC末端でEPO分子のN末端に接続しており、無タンパク質培地中でBHK-12細胞にて産生され、かつ、以下から選択される、前記融合タンパク:
(i)配列番号14に記載されたFcg2h(FN→AQ)-M1-EPO;および、
(ii)配列番号15に記載されたFcg2h(FN→AQ)-M1-EPO(NDS)。
【請求項2】
DMEM/F-12培地で産生される、請求項1記載の精製された二量体組換えFc-EPO融合タンパク質。
【請求項3】
アミノ酸、タンパク質加水分解産物、エタノールアミンおよびトロポロンが補充されたDMEM/F-12培地で産生された、請求項1記載の精製された二量体組換えFc-EPO融合タンパク質。
【請求項4】
請求項1〜3のいずれか1項記載のFc-EPO融合タンパク質の有効量を含み、場合により製薬的に許容できる単体、希釈剤または賦形剤を含む、医薬組成物。
【請求項5】
哺乳動物の造血疾患または不全の治療用の、請求項1〜3のいずれか1項記載の精製された二量体Fc-EPO融合タンパク質または4記載の医薬組成物。
【図1A】

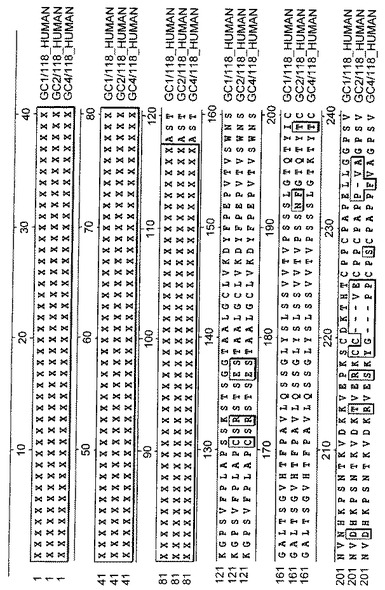
【図1B】
【図2】
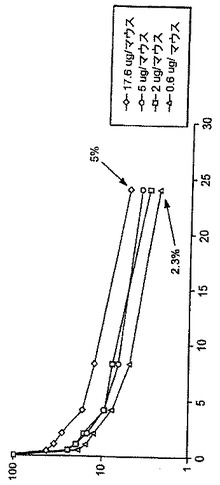
【図3】
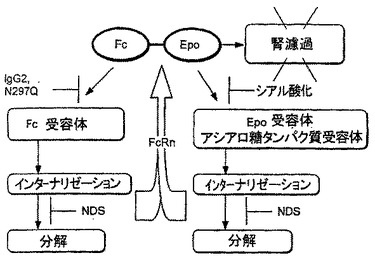
【図4】
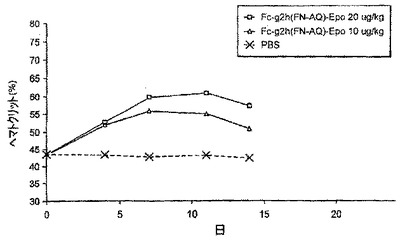
【図5】
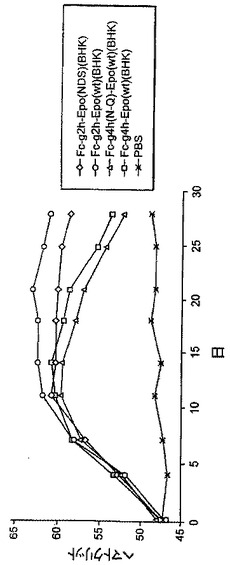
【図6】
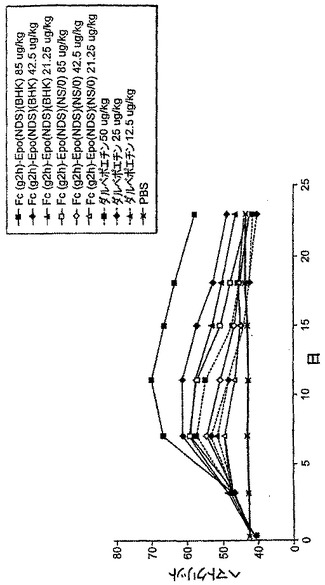
【図7】

【図8】
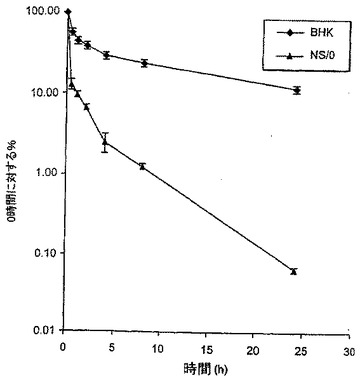
【図9】
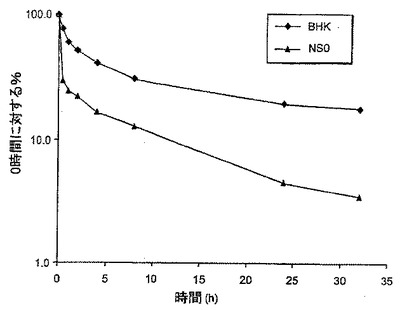
【図10】
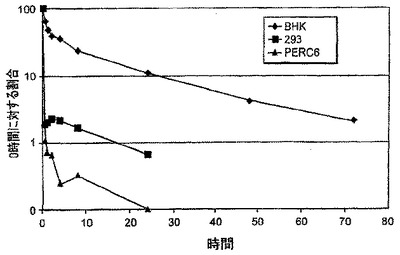
【図11】
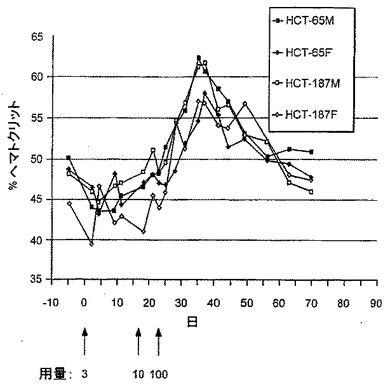
【図1B】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【公開番号】特開2012−107042(P2012−107042A)
【公開日】平成24年6月7日(2012.6.7)
【国際特許分類】
【出願番号】特願2012−32557(P2012−32557)
【出願日】平成24年2月17日(2012.2.17)
【分割の表示】特願2006−546058(P2006−546058)の分割
【原出願日】平成16年12月22日(2004.12.22)
【出願人】(591032596)メルク パテント ゲゼルシャフト ミット ベシュレンクテル ハフツング (1,043)
【氏名又は名称原語表記】Merck Patent Gesellschaft mit beschraenkter Haftung
【住所又は居所原語表記】Frankfurter Str. 250,D−64293 Darmstadt,Federal Republic of Germany
【Fターム(参考)】
【公開日】平成24年6月7日(2012.6.7)
【国際特許分類】
【出願日】平成24年2月17日(2012.2.17)
【分割の表示】特願2006−546058(P2006−546058)の分割
【原出願日】平成16年12月22日(2004.12.22)
【出願人】(591032596)メルク パテント ゲゼルシャフト ミット ベシュレンクテル ハフツング (1,043)
【氏名又は名称原語表記】Merck Patent Gesellschaft mit beschraenkter Haftung
【住所又は居所原語表記】Frankfurter Str. 250,D−64293 Darmstadt,Federal Republic of Germany
【Fターム(参考)】
[ Back to top ]