
改良されたRNAポリメラーゼ
【課題】熱安定性が高く、しかもより高い温度域で反応することができるRNAポリメラーゼを提供する。
【解決手段】RNAポリメラーゼ活性を有するタンパク質を構成するアミノ酸配列の少なくとも1個のアミノ酸を付加、欠失、挿入もしくは置換により変異させたタンパク質であって、改変前の該タンパク質に比して熱安定性もしくは高温域での比活性が向上したことを特徴とする改変型RNAポリメラーゼ。
【解決手段】RNAポリメラーゼ活性を有するタンパク質を構成するアミノ酸配列の少なくとも1個のアミノ酸を付加、欠失、挿入もしくは置換により変異させたタンパク質であって、改変前の該タンパク質に比して熱安定性もしくは高温域での比活性が向上したことを特徴とする改変型RNAポリメラーゼ。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、RNAポリメラーゼ活性を有するタンパク質を遺伝子工学的手法により改変することにより得られる、熱安定性あるいは高温度域での比活性が改変前の野生型RNAポリメラーゼに比して向上した改変型RNAポリメラーゼ、該RNAポリメラーゼをコ−ドする遺伝子および該遺伝子を使用する該RNAポリメラーゼの製造方法及びその用途に関する。
【背景技術】
【0002】
従来から、RNAポリメラーゼは遺伝子の転写発現に関するキー酵素として多くの研究がなされ、様々な機能、性質が知られてきている。なかでもT7、T3、K11及びSP6といったバクテリオファージ由来のRNAポリメラーゼについては、大腸菌や高等生物のRNAポリメラーゼとは異なり、単一のポリペプチド鎖のみで活性を発現することから、転写メカニズムを解析するための格好の材料とされている。また、プロモーター配列を含む2本鎖DNAを鋳型に、NTPを基質として、プロモーター下流の鋳型DNAに相補的な一本鎖RNAを合成するという性質及びその際のプロモーター配列に対する認識が非常に厳密であるという性質から、これらファージ由来のRNAポリメラーゼは、イン・ヴィトロ(invitro)でRNAを合成するための酵素としても汎用されている。これにより合成されたRNAは、多くの分子生物学的手法、例えばノーザンハイブリダイゼーション、サザンハイブリダイゼーション時に用いるRNAプローブ(リボプローブ)として、あるいは細胞もしくは無細胞タンパク合成(翻訳)系に加えて、翻訳される鋳型RNAとして利用されている。
【0003】
さらに近年になって、RNAポリメラーゼの持つ、一定温度下にて鋳型DNAから多くのRNAコピーを作り出す特性すなわち転写活性を利用して、例えば、NASBA法(Nucleicacid sequene‐based amplification method)、TMA法(Transcription mediated amplification method)と呼ばれる転写に基づく核酸増幅方法や転写に基づいたシークエンス方法といった、より複雑な系の方法が開発されている。
【0004】
NASBA法、TMA法などの転写に基づく核酸増幅方法は、RNAポリメラーゼ及び逆転写酵素さらに必要によりRNA−DNA2本鎖のRNA部分のみを分解するRNaseHを共存させ、これらの反応をカップリングさせることにより、微量のRNAを増幅、あるいは特定のRNAの有無を検出する方法である。この方法は、DNAポリメラーゼを利用した核酸増幅反応、いわゆるPCR法とは異なり、温度サイクルをかけることなく一定温度にて、従って特別な機器を用いることなく、短時間に目的核酸を増幅することが可能であり、これらの特徴から臨床診断用として広く用いられることが期待されている。
【0005】
一方、転写に基づくシークエンス方法(特許文献1)は、従来のいわゆるサンガー法あるいはダイデオキシ法と呼ばれるシークエンス方法において、DNAポリメラーゼ、2’−デオキシリボヌクレオシド−5’−トリフォスフェイト(2’−dNTPs)及び2’、3’−ジデオキシリボヌクレオシド−5’−トリフォスフェイト(2’、3’−ddNTPs)の代わりにRNAポリメラーゼ、リボヌクレオシド5’−トリフォスフェイト(rNTPs)及び3’−デオキシリボヌクレオシド−5’−トリフォスフェイト(3’−dNTPs)を用いてシークエンスを行う方法である。従来のDNAポリメラーゼを用いるシークエンス方法では、PCR反応で得られた生成物(DNA断片)をシークエンス反応に供する場合は、その反応溶液中に存在する、取り込まれなかったプライマー及び2’−dNTPsを除去することが必要であった。しかしながら、このRNAポリメラーゼを用いた転写に基づくシークエンス反応では、上述のプライマー及び2’−dNTPsは反応の基質とはなり得ないため、PCR生成物を直接シークエンス反応の鋳型として用いることができる。こうした理由から、この転写に基づくシークエンス法は、PCR反応からシークエンス反応までの工程を自動化できる可能性を持つ画期的な方法として期待されている。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】国際公開 WO1996/14434号公報
【発明の概要】
【発明が解決しようとする課題】
【0007】
しかしながら、このようにRNAポリメラーゼが使用される反応系が複雑化し、かつ用途が広がるなかで、RNAポリメラーゼの熱安定性が低いこと及び/又は高温度域で反応することができないことが問題となっていた。
【0008】
NASBA反応の場合では、反応温度を上げることによりターゲットに対する特異性ひいては検出感度の向上に繋がることが期待されるが、RNAポリメラーゼが失活してしまうために不可能であった。また診断に用いるキットの一構成物としての酵素の熱安定性の向上は、輸送を容易にし、結果の再現性を向上させる意味からも重要である。
【0009】
一方、転写に基づくシークエンス反応においてもRNAポリメラーゼの熱安定性が低いために、PCR生成物をシークエンス決定する際には、PCR反応後にRNAポリメラーゼを追加することが必要となり、自動機器を用いる場合における試薬の長期保存の観点から、自動化する際の大きな障壁となりうることが考えられる。
【0010】
以上のようなことから、従来のものよりも熱安定性が高く、しかもより高い温度域で反応できるRNAポリメラーゼが待ち望まれていた。
【課題を解決するための手段】
【0011】
本発明者らは、上記目的を達成するためにT7RNAポリメラーゼにおいて熱安定性の向上した改変型RNAポリメラーゼを構築することを目的として鋭意検討した結果、野生型RNAポリメラーゼの少なくとも1つのアミノ酸を遺伝子工学的手法により改変することで熱安定性を向上させることができることを見出し、本発明を完成するに至った。
【0012】
すなわち本発明は、以下のような構成からなる。
(1)RNAポリメラーゼ活性を有するタンパク質を構成するアミノ酸配列の少なくとも1個のアミノ酸を付加、欠失、挿入もしくは置換により変異させたタンパク質であって、改変前の該タンパク質に比して熱安定性もしくは高温域での比活性が向上したことを特徴とする改変型RNAポリメラーゼ。
(2)RNAポリメラーゼ活性を有するタンパク質が、T7ファージ、T3ファージおよびK11ファージよりなる群から選択されるファージ由来のタンパク質である(1)の改変型RNAポリメラーゼ。
(3)配列表・配列番号1に記載されるアミノ酸配列の少なくとも1個のアミノ酸を付加、欠失、挿入もしくは置換により変異させてなる(1)の改変型RNAポリメラーゼ。
(4)T7ファージ由来のRNAポリメラーゼであって、430番目のアミノ酸残基セリン、849番目のアミノ酸残基フェニルアラニンおよび880番目のアミノ酸残基フェニルアラニンよりなる群から選ばれた少なくとも1箇所のアミノ酸が他のアミノ酸に置換されたアミノ酸配列を有する(1)〜(3)のいずれかの改変型RNAポリメラーゼ。
(5)T7ファージ由来のRNAポリメラーゼであって、少なくとも430番目のアミノ酸残基セリンがプロリンに置換されたアミノ酸配列を有する(4)の改変型RNAポリメラーゼ。
(6)T7ファージ由来のRNAポリメラーゼであって、少なくとも849番目のアミノ酸残基フェニルアラニンがイソロイシンに置換されたアミノ酸配列を有する(4)の改変型RNAポリメラーゼ。
(7)T7ファージ由来のRNAポリメラーゼであって、少なくとも880番目のアミノ酸残基フェニルアラニンがチロシンに置換されたアミノ酸配列を有する(4)の改変型RNAポリメラーゼ。
(8)T7ファージ由来のRNAポリメラーゼであって、少なくとも430番目のアミノ酸残基セリン、849番目のアミノ酸残基フェニルアラニン、880番目のアミノ酸残基フェニルアラニンがそれぞれプロリン、イソロイシン、チロシンに置換されたアミノ酸配列を有する(4)の改変型RNAポリメラーゼ。
(9)T7ファージ由来のRNAポリメラーゼであって、430番目のアミノ酸残基セリン、849番目のアミノ酸残基フェニルアラニン、880番目のアミノ酸残基フェニルアラニンがそれぞれプロリン、イソロイシン、チロシンに置換されたアミノ酸配列を有する(4)の改変型RNAポリメラーゼ。
(10)T3ファージ由来のRNAポリメラーゼであって、850番目のアミノ酸残基フェニルアラニンもしくは881番目のアミノ酸残基フェニルアラニンのうちの少なくとも1箇所のアミノ酸が他のアミノ酸に置換されたアミノ酸配列を有する(1)または(2)の改変型RNAポリメラーゼ。
(11)T3ファージ由来のRNAポリメラーゼであって、少なくとも850番目のアミノ酸残基フェニルアラニンがイソロイシンに置換されたアミノ酸配列を有する(10)の改変型RNAポリメラーゼ。
(12)T3ファージ由来のRNAポリメラーゼであって、少なくとも881番目のアミノ酸残基フェニルアラニンがチロシンに置換されたアミノ酸配列を有する(10)の改変型RNAポリメラーゼ。
(13)T3ファージ由来のRNAポリメラーゼであって、少なくとも850番目のアミノ酸残基フェニルアラニンがイソロイシンに置換され、かつ881番目のアミノ酸残基フェニルアラニンがチロシンに置換されたアミノ酸配列を有する(10)の改変型RNAポリメラーゼ。
(14)T3ファージ由来のRNAポリメラーゼであって、850番目のアミノ酸残基フェニルアラニンがイソロイシンに置換され、かつ881番目のアミノ酸残基フェニルアラニンがチロシンに置換されたアミノ酸配列を有する(10)の改変型RNAポリメラーゼ。
(15)K11ファージ由来のRNAポリメラーゼであって、453番目のアミノ酸残基セリン、872番目のアミノ酸残基フェニルアラニンおよび903番目のアミノ酸残基フェニルアラニンよりなる群から選ばれた少なくとも1箇所のアミノ酸が他のアミノ酸に置換されたアミノ酸配列を有する(1)または(2)の改変型RNAポリメラーゼ。
(16)K11ファージ由来のRNAポリメラーゼであって、少なくとも453番目のアミノ酸残基セリンがプロリンに置換されたアミノ酸配列を有する(15)の改変型RNAポリメラーゼ。
(17)K11ファージ由来のRNAポリメラーゼであって、少なくとも872番目のアミノ酸残基フェニルアラニンがイソロイシンに置換されたアミノ酸配列を有する(15)の改変型RNAポリメラーゼ。
(18)K11ファージ由来のRNAポリメラーゼであって、少なくとも903番目のアミノ酸残基フェニルアラニンがチロシンに置換されたアミノ酸配列を有する(15)の改変型RNAポリメラーゼ。
(19)K11ファージ由来のRNAポリメラーゼであって、少なくとも453番目のアミノ酸残基セリン、872番目のアミノ酸残基フェニルアラニン、903番目のアミノ酸残基フェニルアラニンがそれぞれプロリン、イソロイシン、チロシンに置換されたアミノ酸配列を有する(15)の改変型RNAポリメラーゼ。
(20)K11ファージ由来のRNAポリメラーゼであって、453番目のアミノ酸残基セリン、872番目のアミノ酸残基フェニルアラニン、903番目のアミノ酸残基フェニルアラニンがそれぞれプロリン、イソロイシン、チロシンに置換されたアミノ酸配列を有する(15)の改変型RNAポリメラーゼ。
(21)(1)〜(20)のいずれかの改変型RNAポリメラーゼの少なくとも一部をコードする塩基配列を含有することを特徴とするDNAフラグメント。
(22)(21)のDNAをベクタ−に挿入したことを特徴とするDNA組換えベクタ−。
(23)(22)のDNA組換え発現ベクタ−を用いて形質転換されたことを特徴とする組換え宿主細胞。
(24)宿主細胞が大腸菌(Escherichia coli)である(23)の組換え宿主細胞。
(25)(23)または(24)の組換え宿主細胞を培養しRNAポリメラーゼを採取することを特徴とする改変型RNAポリメラーゼの製造方法。
(26)(1)〜(20)のいずれかの改変型RNAポリメラーゼを用いて恒温核酸増幅反応によりRNAを増幅せしめることを特徴とするRNAを合成する方法。
(27)(1)〜(18)のいずれかの改変型RNAポリメラーゼを含むことを特徴とする核酸増幅用試薬キット。
【0013】
尚、アミノ酸残基については、以下、慣例により使用されている一文字表記法を用いて説明する。本発明において改変されるアミノ酸のみを記述すると、セリン(S)、プロリン(P、フェニルアラニン(F)、チロシン(Y)、イソロイシン(I)である。また、例えば、S430Pの表記は、430番目のアミノ酸残基SをPに置換させたことあるいは置換させた変異体(変異型酵素)を意味し、さらにS430P+F849I+F880Yは、S430P、F849IおよびF880Yの3種類の置換を含む三重変異体(三重変異型酵素)を意味する。
【発明の効果】
【0014】
上述したように、本発明により提供される改変型RNAポリメラーゼは、野生型に比べて熱安定性及び特に高温度域での比活性が向上しており、従来の野生型RNAポリメラーゼでは使用できなかった高温度下における反応に用いることができる。
【図面の簡単な説明】
【0015】
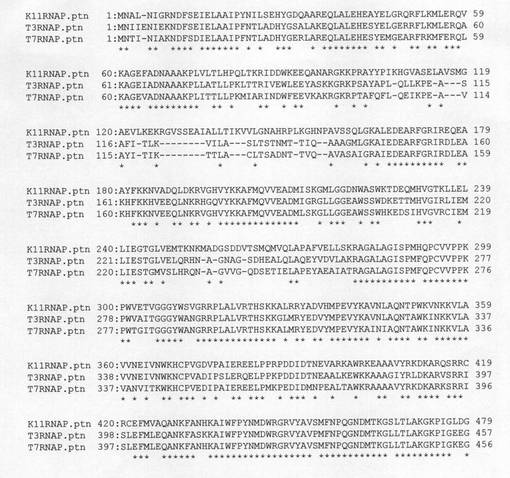
【図1】K11ファージ、T3ファージ及びT7ファージ由来のRNAポリメラーゼのアミノ酸配列を比較した図である(前半部)。各段の最下行のアスタリスク(*)は、3種類のファージ由来のRNAポリメラーゼに共通しているアミノ酸であることを示している。
【図2】K11ファージ、T3ファージ及びT7ファージ由来のRNAポリメラーゼのアミノ酸配列を比較した図である(後半部)。各段の最下行のアスタリスク(*)は、3種類のファージ由来のRNAポリメラーゼに共通しているアミノ酸であることを示している。
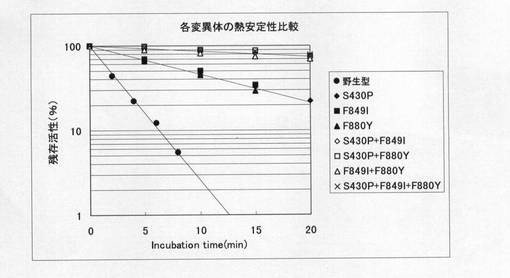
【図3】野生型T7 RNAポリメラーゼ酵素及び各変異型酵素の熱安定性を比較した図である。野生型及び各変異型酵素を48℃にてインキュベートし、経時的にサンプリングして残存活性を測定したものである。
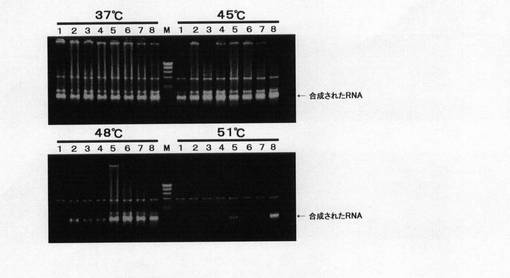
【図4】野生型T7 RNAポリメラーゼ酵素及び各変異型酵素のin vitro転写活性を反応温度がそれぞれ37、45、48及び51℃の条件で測定したものである。T7プロモーターを含むプラスミドに野生型及び各変異型酵素を加え、各温度にて1時間反応させた後、合成(転写)されたRNAを0.7%アガロースゲル電気泳動により調べたものである。
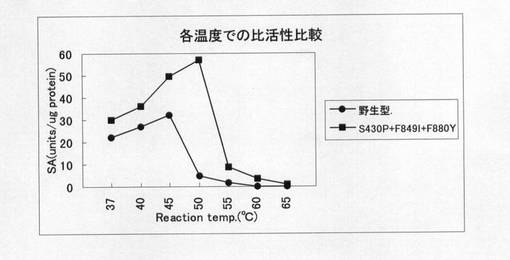
【図5】野生型T7 RNAポリメラーゼ酵素と変異型酵素(S430P+F849I+F880Y)の37〜65℃での比活性を比較した図である。
【発明を実施するための形態】
【0016】
本発明において、「野生型RNAポリメラーゼ」とは、天然に存在する全てのRNAポリメラーゼを意味する。さらに、「野生型RNAポリメラーゼ」は、対応する野生型RNAポリメラーゼの能力と比較して、熱安定性を向上させることを目的とする改変以外のアミノ酸の置換、挿入または欠失を、さらに有するものであることもできる。即ち、野生型RNAポリメラーゼを人為的に上記以外の目的で改変したRNAポリメラーゼも、上記「野生型RNAポリメラーゼ」に含まれる。但し、そのようなアミノ酸の置換、挿入または欠失は、RNAポリメラーゼとしての活性を維持する範囲で、行われたものであることが適当である。さらに「野生型RNAポリメラーゼ」としては、例えば、T7ファージ、T3ファージ、K11ファージに由来するRNAポリメラーゼを挙げることができる。但し、これらのRNAポリメラーゼに限定されるものではない。
【0017】
本発明のRNAポリメラーゼは、対応する野生型RNAポリメラーゼの少なくとも1つのアミノ酸が改変された熱安定性もしくは高温域での比活性の向上したRNAポリメラーゼである。ここで、「熱安定性」とは、例えば、0℃から100℃の温度下で、ある特定の時間インキュベーションした場合に、インキュベーション後の酵素が熱により失活せずに活性が残存していること、あるいは残存している酵素活性のインキュベーション前に対する割合をいうものである。また、「高温域での比活性」とは、例えば、37℃から100℃の温度下で酵素活性を測定した場合における単位タンパク質重量当たりの酵素活性をいうものである。
【0018】
本発明において、アミノ酸残基の変異は公知の方法により実施することができる。具体的には、例えば、付加によるタンパク質の機能を改変する方法としては、NatureBiotechnology第17巻、第58〜61頁(1999年)に記載されている。また、欠失による方法としては、Biochem.Biophys.Res.Commun.第248巻、第2号、第372〜377頁、挿入による方法としては、FEBS Lett.第442巻、第241〜245頁(1999年)、置換による方法としては、NucleicAcidsReserch 第26巻、第2号、第681〜683頁(1998年)にそれぞれ記載されている。
【0019】
本発明者らは、まず、野生型T7 RNAポリメラーゼ遺伝子を挿入した発現プラスミドpKKT7RNAPを構築し、次に、この発現プラスミドpKKT7RNAPを基にT7RNAポリメラーゼの変異体を作製した。即ち、430番目のアミノ酸残基SをPに置換させた変異体S430Pを、また、同様に849番目のアミノ酸残基FをIに置換させた変異体F849Iを、さらに同様に880番目のアミノ酸残基FをYに置換させた変異体F880Yを作製した。次にこれらの変異型酵素を精製し、その熱安定性について調べた。
【0020】
本明細書における、野生型T7RNAポリメラーゼのアミノ酸配列は、遺伝子配列データベースであるGeneBankより、accessionNo. M38308で登録されているT7RNAポリメラーゼ遺伝子配列を若干訂正した配列表・配列番号1及び配列番号2を基礎としている。配列番号1及び配列番号2において右端の数字は、アミノ酸の場合、T7RNAポリメラーゼの最初のM(メチオニン)を1として全長883アミノ酸残基からなっていることを示し、また塩基配列の場合、開始コドンATGのAを1として全長2652塩基よりなることを示す。従って、本発明における野生型T7RNAポリメラーゼのアミノ酸配列及び各アミノ酸に付された番号は、この配列番号1及び配列番号2に示される配列及び番号のことである。
【0021】
さらに、上述のように、上記野生型T7RNAポリメラーゼは、本発明で目的とする改変以外のアミノ酸の置換、挿入または欠失をさらに有するものであってもよい。従って、本発明の目的に基づいて変異を導入すべき野生型RNAポリメラーゼが、野生型T7RNAポリメラーゼに別の変異を導入したものである場合、特にそのような変異が、アミノ酸の挿入または欠失である場合、それに応じて上記アミノ酸番号は変動し、図1及び図2に示す番号とは異なったとしても、T7RNAポリメラーゼ活性を維持している限り、そのような挿入または欠失を有するT7RNAポリメラーゼも本発明の目的とする変異を導入する野生型T7RNAポリメラーゼの範疇に含まれる。
【0022】
また、本明細書におけるT7RNAポリメラーゼ以外のRNAポリメラーゼについてのアミノ酸配列及び各アミノ酸に付された番号は、図1及び図2に示されている配列及び番号である。さらに、本発明で目的とする改変以外のアミノ酸の置換、挿入または欠失を、さらに有するものであることもできる。従って、これらのアミノ酸配列及びその番号に付いても、T7RNAポリメラーゼの場合と同様であり、アミノ酸の挿入または欠失による変異がある場合、それに応じて上記アミノ酸番号は変動するが、そのような一部に変異を有する野生型のRNAポリメラーゼも本発明において本発明の目的とする変異を導入する野生型RNAポリメラーゼの範疇に含まれる。
【0023】
T7RNAポリメラーゼ遺伝子を含む発現ベクターは、T7ファージゲノミックDNA(シグマ製)を鋳型にして、T7 RNAポリメラーゼ遺伝子の開始コドン領域に特異的なプライマーと終始コドン領域に特異的なプライマーを用いてPCRを実施し、この増幅産物を制限酵素にて消化した後、発現ベクターpKK223‐3(ファルマシア製)に導入することにより構築した。この発現ベクターを用いて、大腸菌JM109に形質転換することにより、T7RNAポリメラーゼ蛋白質を大量に発現させることができる。
【0024】
発現ベクターpKKT7RNAPを持つ大腸菌エシェリヒア・コリ(Escherichia coli)から精製したT7 RNAポリメラーゼは、in vitroでT7プロモーターを含んだDNA存在下で充分なRNA合成活性を有していた。この発現プラスミドpKKT7RNAPをベースにして変異型T7RNAポリメラーゼとして、上述のS430P、F849I、F880Y及びそれらの置換を複数含んだ多重変異体を構築した。これらの変異型T7RNAポリメラーゼを大腸菌組換え宿主より精製し、各変異型酵素の熱安定性を野生型T7RNAポリメラーゼと比較した。結果を表1に示す。表1に示すように、これらの変異型酵素では、熱安定性の一つの指標である活性半減期が野生型酵素に比べて大幅に向上していた。
【0025】
ここで上記3種のファージ、即ち、T7、T3及びK11ファージ由来のRNAポリメラーゼにおいては、それらのアミノ酸配列は互いに極めて高いホモロジーを有していることが知られている。図1及び図2に、これら3種類のファージ由来RNAポリメラーゼのアミノ酸配列を比較して示す。このように、T7、T3、K11由来のRNAポリメラーゼは互いに極めて類似しており、T7RNAポリメラーゼにおいて得られた結果を、アミノ酸配列の類似する他のRNAポリメラーゼに適用することは比較的容易にできると考えられる。即ち、図1及び図2から、T7ファージ由来のRNAポリメラーゼの430、849及び880番目のアミノ酸残基に対応するアミノ酸残基は、T3ファージ由来のRNAポリメラーゼではそれぞれ431、850及び881番目のアミノ酸残基であり、またK11ファージ由来のRNAポリメラーゼではそれぞれ453、872及び903番目のアミノ酸残基であり、従って、T3RNAポリメラーゼにおけるF850I及びF881Y変異体、さらにK11 RNAポリメラーゼにおけるS453P、F872I及びF903Y変異体も、T7RNAポリメラーゼにおける変異体と同様に野生型に比べてより高い熱安定性を有していることは容易に類推できる。
【0026】
本発明は、上記改変型RNAポリメラーゼを製造する方法であって、RNAポリメラーゼをコードする核酸分子を用意し、次いでその塩基配列内の1つまたはそれ以上の部位における塩基を変異させるように該核酸分子を改変し、次いで変異させた核酸分子により発現される改変されたRNAポリメラーゼを回収することを含む方法を包含する。RNAポリメラーゼをコードする核酸分子の調製、核酸分子への突然変異の導入、改変されたRNAポリメラーゼの精製はいずれも、公知の手法を用いて行うことが出来る。
【0027】
即ち、変異型T7RNAポリメラーゼは、野生型T7RNAポリメラーゼ遺伝子をベースとして適当な塩基を改変して変異型T7RNAポリメラーゼ遺伝子を作製し、続いてこの遺伝子をベクターに導入し、発現ベクターを構築する。次に、この発現ベクターを用いて、宿主細胞を形質転換することにより変異型T7RNAポリメラーゼ蛋白質を大量に発現させることができる。
【0028】
本発明における核酸分子への変異の導入は、当業者がなし得る方法であればいかなる方法でもよいが、例えばサイトディレクテッドミュータジェネシス法がある。
【0029】
本発明において使用するベクターは、RNAポリメラーゼのクローニング及び発現を可能とするものであればいかなるものでもよく、例えばファージ及びプラスミドが挙げられる。プラスミドとしては、pUC118,pUC18、pBR322、pBluescript、pLED−M1、p73、pGW7などが挙げられる。ファージとしては、例えばλgt11、λZAPIIなどが挙げられる。
【0030】
本発明において使用する宿主細胞としては、大腸菌、酵母、バチルス(Bacillus)属細菌などが挙げられる。大腸菌としては、例えばエシェリヒア・コリDH5α、JM109、HB101、XL1Blue、PR1、BL21などが挙げられる。本発明においては、上記の熱安定性の向上した改変型RNAポリメラーゼをコードする遺伝子を上記ベクターに挿入して組換え発現ベクターとし、さらにこの組換え発現ベクターにて宿主細胞を形質転換する。
【0031】
本発明の製造方法においては、上記組換え宿主細胞を培養して、熱安定性の向上した改変型RNAポリメラーゼを発現させる。組換え宿主細胞の培養に使用する培地ならびに条件は常法に従う。具体例としては、熱安定性の向上した改変型RNAポリメラーゼ遺伝子を含むプラスミドにより形質転換された大腸菌を、例えばTB培地にて培養する。
【0032】
上記培養により得られた改変型RNAポリメラーゼの精製法としては、(a)組換え宿主を集めた後、破砕して細胞抽出物を調製し、(b)宿主細胞由来の不純タンパク質を除去する工程を含む。組換え宿主細胞より産出された熱安定性の向上した改変型RNAポリメラーゼは、宿主菌体を培地で培養後、培養液から遠心分離等にて分離、回収する。該菌体を緩衝液に再懸濁した後、超音波処理、ダイノミル・フレンチプレンス等により菌体を破砕する。次いでカラムクロマトグラフィーを実施し、熱安定性の向上した改変型RNAポリメラーゼを回収する。カラムクロマトグラフィーは、陽イオン交換体、例えば、フォスフォセルロース、あるいは陰イオン交換体、例えば、DEAEセファロース、あるいはアフィニティー吸着体ヘパリンセファロースなどが好ましい。
【0033】
本発明における改変型RNAポリメラーゼは、例えば恒温核酸増幅反応により特定のRNAを増幅せしめるのに特に有用である。ここで、恒温核酸増幅反応とは、NASBA法、TMA法、3SR法(Self‐sustainedsequence replication method)、TAS法(Transcription based amplification system method)などが挙げられる。
【実施例】
【0034】
以下、本発明を実施例により具体的に説明する。なお、以下の実施例において、T7 RNAポリメラーゼ活性の測定は以下のように行なった。すなわち、活性測定用緩衝液(40mMトリス−塩酸緩衝液(pH8.0)、20mM塩化マグネシウム、5mMジチオスレイトール、0.5mM rNTP、T7ゲノムDNA(シグマD4931)、50μg/ml BSA、[3H]rUTP(370kBq/μl))中で、酵素を37℃,10分間反応させたのち、合成されたRNAを酸不溶性沈殿として回収し、シンチレーションカウンターにて取り込まれた3H量を測定した。酵素活性の1単位(U)は、この条件下で60分間に1ナノモルのrNTPを酸不溶性沈殿画分に取り込ませる酵素量とした。
【0035】
実施例1 野生型T7 RNAポリメラーゼ遺伝子のクローニングと発現プラスミドの構築
野生型RNAポリメラーゼの発現プラスミドは以下のようにして構築した。すなわち、T7ファージゲノミックDNA(シグマ製)を鋳型にして、T7RNAポリメラーゼ遺伝子の開始コドン領域に特異的なプライマー(T7EcoATG : 5’−CGC GAA TTCATG AAC ACGATT AAC ATCGCT ‐3’)、及び終始コドン領域に特異的なプライマー(T7TerPstI: 5’−TTT CTG CAG TGG CGT TAC GCG AAC GCG AAG−3’)を用いてPCRを実施し、T7 RNAポリメラーゼをコードする遺伝子を増幅した。次に、この増幅産物を制限酵素EcoRIとPstIにて消化した後、発現プラスミドpKK223‐3(ファルマシア製)の同制限酵素サイトに導入しT7RNAポリメラーゼの発現プラスミドを作製した。これをpKKT7RNAPと名付けた。なお、得られたT7 RNAポリメラーゼ遺伝子の塩基配列に関しては、発現プラスミド中の得られた遺伝子をシークエンス決定することにより、PCRにより増幅した際の意図しない塩基置換が入っていないことを確認した。
【0036】
実施例2 改変型T7 RNAポリメラーゼ遺伝子の作製
改変型T7 RNAポリメラーゼ遺伝子は、上記実施例1で得られた野生型T7 RNAポリメラーゼ遺伝子に点変異を導入することにより行った。点変異の導入はクイックチェインジミュータジェネシスキット(ストラタジーン社製)および点変異導入用オリゴヌクレオチドを用いて、説明書の指示に従い行った。S430P、F849I及びF880Yのための点変異の導入は、変異導入用のプライマーを変えること以外は基本的に同じ条件にて実施できる。以下に一例として、T7RNAポリメラーゼ遺伝子の1288番目の塩基であるTをCに置換した(その結果、430番目のアミノ酸残基SがPに置換される)変異型遺伝子S430Pを作製した際の条件を示す。
【0037】
pKKT7RNAP5ng、変異導入用プライマー(5’‐GTT TAC GCT GTG CCAATG TTCAAC CCGCAA‐3’及び 5’‐TTG CGG GTT GAACAT TGG CACAGC GTA AAC‐3’)各125ng、上記キット添付の反応用緩衝液5μl、10mM dNTPmix 1μl、Pfu DNAポリメラーゼ2.5Uを含む50μlの溶液を、95℃で30秒間インキュベートした後、95℃、30秒/55℃、1分/68℃、12分の温度サイクルを12サイクル実施した。温度サイクル終了後、反応液に制限酵素DpnI10Uを添加し、37℃で1時間インキュベートした。次にこの反応液1μlをエシェリヒア・コリJM109株コンピテントセル100μlに加え、30分間氷冷した後、42℃で30秒間インキュベートし、900μlのSOC培地を加え37℃で1時間インキュベートした。これに50μg/mlのアンピシリンを含むLB寒天培地上にスプレッドし、37℃で一晩インキュベートした。得られたコロニーをLB培地2.5mlにて一晩培養した後、定法に従いプラスミドを抽出した。最後に、これらのプラスミドの当該領域のシークエンスを行い、1288番目のTがCに置換されているクローンを取得し、pKKT7S430Pと名付けた。
【0038】
また、同様の方法で、変異導入用プライマーとして5’‐TTC TAC GAC CAG ATCGCT GAC CAGTTG CAC‐3’及び 5’‐GTG CAA CTG GTCAGC GAT CTGGTC GTA GAA‐3’を用いることにより2545番目のTをAに置換した(結果として、849番目のFがIに置換される)遺伝子を含む発現プラスミド(pKKT7F849I)、5’‐TCT TAG AGT CGGACT ACG CGTTCG CGT AAC‐3’及び 5’‐GTT ACG CGAACG CGT AGT CCG ACT CTA AGA‐3’を用いることにより2639番目のTをAに置換した(結果として、880番目のFがYに置換される)遺伝子を含む発現プラスミド(pKKT7F880Y)を作製した。さらに、これらの3種類の変異を複数含む変異型遺伝子に関しては、変異を含む領域を制限酵素にて切り出し、他の変異を含む発現プラスミドの同領域と入れ替えることにより作製した。
【0039】
実施例3 形質転換体の作製
実施例1及び実施例2で得られたプラスミド1ngをJM109コンピテントセル100μlに加え、30分間氷冷した後、42℃で30秒間インキュベートし、900μlのSOC培地を加え、37℃で1時間インキュベートした。これを50μg/mlのアンピシリンを含むLB寒天培地上にて37℃で一晩インキュベートし、形質転換体を得た。
【0040】
実施例4 野生型及び各種改変型T7RNAポリメラーゼの精製
野生型及び各種改変型T7RNAポリメラーゼは基本的に同じ方法にて精製、調製することができる。実施例3で得られた各形質転換体を100μg/mlのアンピシリンを含むTB培地100mlに植菌し、37℃で一晩インキュベートした。得られた菌体を12,000回転/分で5分間遠心することにより回収した。菌体約10gをバッファー1(20mMトリス−塩酸(pH8.0),1mM EDTA、1mMジチオスレイトール, 5%グリセロール)20mlに懸濁し、これを超音波破砕機で破砕した後、12000回転/分で20分間遠心することにより沈殿を分離した。得られた上清に0.6%ポリエチレンイミン溶液を0.4ml添加し、30分間攪拌した。これを12000回転/分で10分間遠心することにより沈殿を分離し、上清を回収した。この液に硫酸アンモニウムを4.56g加え、30分間攪拌した後、これを12000回転/分で10分間遠心することにより沈殿を分離し回収した。得られた沈殿をバッファー2(20mMトリス−塩酸(pH8.0),1mM EDTA、1mMジチオスレイトール、20mM NaCl、5%グリセロール)5mlに溶解し、100mlのバッファー2に対して透析した。次に、これをDEAEセファロースカラム(5ml)にチャージし、10mlのバッファー2で洗浄後、NaClの0〜500mMのグラジエントにより溶出した。得られたフラクションのうち、RNAポリメラーゼ活性を含む画分をプールし、100mlのバッファー2に対して透析した。次に、これをフォスフォセルロースカラム(5ml)にチャージし、15mlのバッファー2で洗浄後、バッファー2をベースとしてNaCl濃度の100〜400mMのグラジェントにより溶出した。得られたフラクションのうち、RNAポリメラーゼ活性を含む画分を回収した。以上の操作で、SDS−PAGEによりほぼ単一なバンドを示す10mgのT7RNAポリメラーゼタンパク質を得た。
【0041】
実施例5 野生型及び各変異体の活性半減期の測定
熱安定性の指標の一つである活性半減期(ある温度でインキュベートした際に活性が半減するまでの時間)の測定は以下のようにして実施した。すなわち40mMトリス−塩酸緩衝液(pH8.0)、20mM塩化マグネシウム、5mMジチオスレイトール、70mM KCl、100μg/ml BSAからなる緩衝液中、酵素を48℃にてインキュベートし、一定の時間間隔ごとにサンプリングを行い、各サンプリング時点での残存活性を測定した。野生型および各種改変型T7RNAポリメラーゼを48℃でインキュベートした際の残存活性を経時的に測定した結果を図3に示す。また、図3から導き出される野生型及び各変異型酵素の活性半減期を表1に示す。
【0042】
表1の数値は、48℃で酵素をインキュベートした場合に、酵素活性が半減するまでの時間(min)を示しており、数値が大きいほどその酵素の熱安定性が高いことを示している。図3及び表1から、各変異型酵素はいずれも野生型酵素に比してより高い熱安定性を有していることがわかる。また今回作製した変異型酵素の中では、S430P、F849I及びF880Yの3箇所の変異を全て含む三重変異型酵素(S430P+F849I+F880Y)が最も熱安定性に優れている。
【0043】
【表1】

【0044】
実施例6 野生型及び各変異型酵素のin vitro転写反応
野生型及び各変異型酵素のin vitro転写反応は以下のように実施した。即ち、野生型及び各変異型酵素各50Uを、40mMトリス−塩酸緩衝液(pH8.0)、20mM塩化マグネシウム、5mMジチオスレイトール、0.4mMrNTP、0.1%BSA、0.5μg pUCT7−1からなる転写反応バッファー50μlに加え、37、45、48、51℃の各温度にて1時間反応させた。ここでpUCT7−1は、T7プロモーターの下流に約1.5kbpのDNAフラグメントが連結されたプラスミドDNAであり、T7RNAポリメラーゼにより転写されると約1.5KbpのRNAが合成されるものである。1時間の反応の後、反応液のうちの3μlを0.7%アガロースゲル電気泳動に供し、合成されたRNAの有無、量を測定した。その0.7%アガロースゲル電気泳動の結果を図4に示す。なお各レーンに付されているナンバーは上記表1における各変異体のナンバーに相当する。図4より、変異型酵素、特にNo.8の三重変異体(S430P+F849I+F880Y)は、より高い反応温度でも合成された転写産物(RNA)のバンドを確認でき、野生型に比してより高い温度下でも活性を有していることが確認できる。
【0045】
実施例7 野生型及び三重変異型酵素の各温度での比活性測定
各温度での比活性の測定は以下のようにして実施した。すなわち、野生型及び改変型酵素を、予め以下の温度に保った活性測定緩衝液(40mMトリス−塩酸緩衝液(pH8.0)、20mM塩化マグネシウム、5mMジチオスレイトール、0.5mM rNTP、T7ファージゲノムDNA(シグマD4931)、50ug/mlBSA、[3H]rUTP(370kBq/ul)に加え、それぞれ37℃、40℃、45℃、50℃、55℃、60℃、65℃にて10分間反応させ、各温度における活性を測定した。また、各酵素のタンパク質濃度は、プロテインアッセイキット(バイオラッド社製)を用いて、キット付属のイムノグロブリンガンマ(IgG)を標準タンパク質として用いて測定した。比活性(SA)は、単位タンパク質重量当たりの酵素活性のことであり、活性量を酵素タンパク量で割ることにより算出できる。野生型および三重変異型酵素(S430P+F849I+F880Y)の各温度での比活性を測定した結果を図5に示す。図5より、改変型酵素は、野生型酵素に比べて各温度全般にわたって高い比活性を有しており、特に高温域での比活性に優れており、とりわけ50℃での比活性は野生型の約12倍もの値を有していることが判る。
【技術分野】
【0001】
本発明は、RNAポリメラーゼ活性を有するタンパク質を遺伝子工学的手法により改変することにより得られる、熱安定性あるいは高温度域での比活性が改変前の野生型RNAポリメラーゼに比して向上した改変型RNAポリメラーゼ、該RNAポリメラーゼをコ−ドする遺伝子および該遺伝子を使用する該RNAポリメラーゼの製造方法及びその用途に関する。
【背景技術】
【0002】
従来から、RNAポリメラーゼは遺伝子の転写発現に関するキー酵素として多くの研究がなされ、様々な機能、性質が知られてきている。なかでもT7、T3、K11及びSP6といったバクテリオファージ由来のRNAポリメラーゼについては、大腸菌や高等生物のRNAポリメラーゼとは異なり、単一のポリペプチド鎖のみで活性を発現することから、転写メカニズムを解析するための格好の材料とされている。また、プロモーター配列を含む2本鎖DNAを鋳型に、NTPを基質として、プロモーター下流の鋳型DNAに相補的な一本鎖RNAを合成するという性質及びその際のプロモーター配列に対する認識が非常に厳密であるという性質から、これらファージ由来のRNAポリメラーゼは、イン・ヴィトロ(invitro)でRNAを合成するための酵素としても汎用されている。これにより合成されたRNAは、多くの分子生物学的手法、例えばノーザンハイブリダイゼーション、サザンハイブリダイゼーション時に用いるRNAプローブ(リボプローブ)として、あるいは細胞もしくは無細胞タンパク合成(翻訳)系に加えて、翻訳される鋳型RNAとして利用されている。
【0003】
さらに近年になって、RNAポリメラーゼの持つ、一定温度下にて鋳型DNAから多くのRNAコピーを作り出す特性すなわち転写活性を利用して、例えば、NASBA法(Nucleicacid sequene‐based amplification method)、TMA法(Transcription mediated amplification method)と呼ばれる転写に基づく核酸増幅方法や転写に基づいたシークエンス方法といった、より複雑な系の方法が開発されている。
【0004】
NASBA法、TMA法などの転写に基づく核酸増幅方法は、RNAポリメラーゼ及び逆転写酵素さらに必要によりRNA−DNA2本鎖のRNA部分のみを分解するRNaseHを共存させ、これらの反応をカップリングさせることにより、微量のRNAを増幅、あるいは特定のRNAの有無を検出する方法である。この方法は、DNAポリメラーゼを利用した核酸増幅反応、いわゆるPCR法とは異なり、温度サイクルをかけることなく一定温度にて、従って特別な機器を用いることなく、短時間に目的核酸を増幅することが可能であり、これらの特徴から臨床診断用として広く用いられることが期待されている。
【0005】
一方、転写に基づくシークエンス方法(特許文献1)は、従来のいわゆるサンガー法あるいはダイデオキシ法と呼ばれるシークエンス方法において、DNAポリメラーゼ、2’−デオキシリボヌクレオシド−5’−トリフォスフェイト(2’−dNTPs)及び2’、3’−ジデオキシリボヌクレオシド−5’−トリフォスフェイト(2’、3’−ddNTPs)の代わりにRNAポリメラーゼ、リボヌクレオシド5’−トリフォスフェイト(rNTPs)及び3’−デオキシリボヌクレオシド−5’−トリフォスフェイト(3’−dNTPs)を用いてシークエンスを行う方法である。従来のDNAポリメラーゼを用いるシークエンス方法では、PCR反応で得られた生成物(DNA断片)をシークエンス反応に供する場合は、その反応溶液中に存在する、取り込まれなかったプライマー及び2’−dNTPsを除去することが必要であった。しかしながら、このRNAポリメラーゼを用いた転写に基づくシークエンス反応では、上述のプライマー及び2’−dNTPsは反応の基質とはなり得ないため、PCR生成物を直接シークエンス反応の鋳型として用いることができる。こうした理由から、この転写に基づくシークエンス法は、PCR反応からシークエンス反応までの工程を自動化できる可能性を持つ画期的な方法として期待されている。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】国際公開 WO1996/14434号公報
【発明の概要】
【発明が解決しようとする課題】
【0007】
しかしながら、このようにRNAポリメラーゼが使用される反応系が複雑化し、かつ用途が広がるなかで、RNAポリメラーゼの熱安定性が低いこと及び/又は高温度域で反応することができないことが問題となっていた。
【0008】
NASBA反応の場合では、反応温度を上げることによりターゲットに対する特異性ひいては検出感度の向上に繋がることが期待されるが、RNAポリメラーゼが失活してしまうために不可能であった。また診断に用いるキットの一構成物としての酵素の熱安定性の向上は、輸送を容易にし、結果の再現性を向上させる意味からも重要である。
【0009】
一方、転写に基づくシークエンス反応においてもRNAポリメラーゼの熱安定性が低いために、PCR生成物をシークエンス決定する際には、PCR反応後にRNAポリメラーゼを追加することが必要となり、自動機器を用いる場合における試薬の長期保存の観点から、自動化する際の大きな障壁となりうることが考えられる。
【0010】
以上のようなことから、従来のものよりも熱安定性が高く、しかもより高い温度域で反応できるRNAポリメラーゼが待ち望まれていた。
【課題を解決するための手段】
【0011】
本発明者らは、上記目的を達成するためにT7RNAポリメラーゼにおいて熱安定性の向上した改変型RNAポリメラーゼを構築することを目的として鋭意検討した結果、野生型RNAポリメラーゼの少なくとも1つのアミノ酸を遺伝子工学的手法により改変することで熱安定性を向上させることができることを見出し、本発明を完成するに至った。
【0012】
すなわち本発明は、以下のような構成からなる。
(1)RNAポリメラーゼ活性を有するタンパク質を構成するアミノ酸配列の少なくとも1個のアミノ酸を付加、欠失、挿入もしくは置換により変異させたタンパク質であって、改変前の該タンパク質に比して熱安定性もしくは高温域での比活性が向上したことを特徴とする改変型RNAポリメラーゼ。
(2)RNAポリメラーゼ活性を有するタンパク質が、T7ファージ、T3ファージおよびK11ファージよりなる群から選択されるファージ由来のタンパク質である(1)の改変型RNAポリメラーゼ。
(3)配列表・配列番号1に記載されるアミノ酸配列の少なくとも1個のアミノ酸を付加、欠失、挿入もしくは置換により変異させてなる(1)の改変型RNAポリメラーゼ。
(4)T7ファージ由来のRNAポリメラーゼであって、430番目のアミノ酸残基セリン、849番目のアミノ酸残基フェニルアラニンおよび880番目のアミノ酸残基フェニルアラニンよりなる群から選ばれた少なくとも1箇所のアミノ酸が他のアミノ酸に置換されたアミノ酸配列を有する(1)〜(3)のいずれかの改変型RNAポリメラーゼ。
(5)T7ファージ由来のRNAポリメラーゼであって、少なくとも430番目のアミノ酸残基セリンがプロリンに置換されたアミノ酸配列を有する(4)の改変型RNAポリメラーゼ。
(6)T7ファージ由来のRNAポリメラーゼであって、少なくとも849番目のアミノ酸残基フェニルアラニンがイソロイシンに置換されたアミノ酸配列を有する(4)の改変型RNAポリメラーゼ。
(7)T7ファージ由来のRNAポリメラーゼであって、少なくとも880番目のアミノ酸残基フェニルアラニンがチロシンに置換されたアミノ酸配列を有する(4)の改変型RNAポリメラーゼ。
(8)T7ファージ由来のRNAポリメラーゼであって、少なくとも430番目のアミノ酸残基セリン、849番目のアミノ酸残基フェニルアラニン、880番目のアミノ酸残基フェニルアラニンがそれぞれプロリン、イソロイシン、チロシンに置換されたアミノ酸配列を有する(4)の改変型RNAポリメラーゼ。
(9)T7ファージ由来のRNAポリメラーゼであって、430番目のアミノ酸残基セリン、849番目のアミノ酸残基フェニルアラニン、880番目のアミノ酸残基フェニルアラニンがそれぞれプロリン、イソロイシン、チロシンに置換されたアミノ酸配列を有する(4)の改変型RNAポリメラーゼ。
(10)T3ファージ由来のRNAポリメラーゼであって、850番目のアミノ酸残基フェニルアラニンもしくは881番目のアミノ酸残基フェニルアラニンのうちの少なくとも1箇所のアミノ酸が他のアミノ酸に置換されたアミノ酸配列を有する(1)または(2)の改変型RNAポリメラーゼ。
(11)T3ファージ由来のRNAポリメラーゼであって、少なくとも850番目のアミノ酸残基フェニルアラニンがイソロイシンに置換されたアミノ酸配列を有する(10)の改変型RNAポリメラーゼ。
(12)T3ファージ由来のRNAポリメラーゼであって、少なくとも881番目のアミノ酸残基フェニルアラニンがチロシンに置換されたアミノ酸配列を有する(10)の改変型RNAポリメラーゼ。
(13)T3ファージ由来のRNAポリメラーゼであって、少なくとも850番目のアミノ酸残基フェニルアラニンがイソロイシンに置換され、かつ881番目のアミノ酸残基フェニルアラニンがチロシンに置換されたアミノ酸配列を有する(10)の改変型RNAポリメラーゼ。
(14)T3ファージ由来のRNAポリメラーゼであって、850番目のアミノ酸残基フェニルアラニンがイソロイシンに置換され、かつ881番目のアミノ酸残基フェニルアラニンがチロシンに置換されたアミノ酸配列を有する(10)の改変型RNAポリメラーゼ。
(15)K11ファージ由来のRNAポリメラーゼであって、453番目のアミノ酸残基セリン、872番目のアミノ酸残基フェニルアラニンおよび903番目のアミノ酸残基フェニルアラニンよりなる群から選ばれた少なくとも1箇所のアミノ酸が他のアミノ酸に置換されたアミノ酸配列を有する(1)または(2)の改変型RNAポリメラーゼ。
(16)K11ファージ由来のRNAポリメラーゼであって、少なくとも453番目のアミノ酸残基セリンがプロリンに置換されたアミノ酸配列を有する(15)の改変型RNAポリメラーゼ。
(17)K11ファージ由来のRNAポリメラーゼであって、少なくとも872番目のアミノ酸残基フェニルアラニンがイソロイシンに置換されたアミノ酸配列を有する(15)の改変型RNAポリメラーゼ。
(18)K11ファージ由来のRNAポリメラーゼであって、少なくとも903番目のアミノ酸残基フェニルアラニンがチロシンに置換されたアミノ酸配列を有する(15)の改変型RNAポリメラーゼ。
(19)K11ファージ由来のRNAポリメラーゼであって、少なくとも453番目のアミノ酸残基セリン、872番目のアミノ酸残基フェニルアラニン、903番目のアミノ酸残基フェニルアラニンがそれぞれプロリン、イソロイシン、チロシンに置換されたアミノ酸配列を有する(15)の改変型RNAポリメラーゼ。
(20)K11ファージ由来のRNAポリメラーゼであって、453番目のアミノ酸残基セリン、872番目のアミノ酸残基フェニルアラニン、903番目のアミノ酸残基フェニルアラニンがそれぞれプロリン、イソロイシン、チロシンに置換されたアミノ酸配列を有する(15)の改変型RNAポリメラーゼ。
(21)(1)〜(20)のいずれかの改変型RNAポリメラーゼの少なくとも一部をコードする塩基配列を含有することを特徴とするDNAフラグメント。
(22)(21)のDNAをベクタ−に挿入したことを特徴とするDNA組換えベクタ−。
(23)(22)のDNA組換え発現ベクタ−を用いて形質転換されたことを特徴とする組換え宿主細胞。
(24)宿主細胞が大腸菌(Escherichia coli)である(23)の組換え宿主細胞。
(25)(23)または(24)の組換え宿主細胞を培養しRNAポリメラーゼを採取することを特徴とする改変型RNAポリメラーゼの製造方法。
(26)(1)〜(20)のいずれかの改変型RNAポリメラーゼを用いて恒温核酸増幅反応によりRNAを増幅せしめることを特徴とするRNAを合成する方法。
(27)(1)〜(18)のいずれかの改変型RNAポリメラーゼを含むことを特徴とする核酸増幅用試薬キット。
【0013】
尚、アミノ酸残基については、以下、慣例により使用されている一文字表記法を用いて説明する。本発明において改変されるアミノ酸のみを記述すると、セリン(S)、プロリン(P、フェニルアラニン(F)、チロシン(Y)、イソロイシン(I)である。また、例えば、S430Pの表記は、430番目のアミノ酸残基SをPに置換させたことあるいは置換させた変異体(変異型酵素)を意味し、さらにS430P+F849I+F880Yは、S430P、F849IおよびF880Yの3種類の置換を含む三重変異体(三重変異型酵素)を意味する。
【発明の効果】
【0014】
上述したように、本発明により提供される改変型RNAポリメラーゼは、野生型に比べて熱安定性及び特に高温度域での比活性が向上しており、従来の野生型RNAポリメラーゼでは使用できなかった高温度下における反応に用いることができる。
【図面の簡単な説明】
【0015】
【図1】K11ファージ、T3ファージ及びT7ファージ由来のRNAポリメラーゼのアミノ酸配列を比較した図である(前半部)。各段の最下行のアスタリスク(*)は、3種類のファージ由来のRNAポリメラーゼに共通しているアミノ酸であることを示している。
【図2】K11ファージ、T3ファージ及びT7ファージ由来のRNAポリメラーゼのアミノ酸配列を比較した図である(後半部)。各段の最下行のアスタリスク(*)は、3種類のファージ由来のRNAポリメラーゼに共通しているアミノ酸であることを示している。
【図3】野生型T7 RNAポリメラーゼ酵素及び各変異型酵素の熱安定性を比較した図である。野生型及び各変異型酵素を48℃にてインキュベートし、経時的にサンプリングして残存活性を測定したものである。
【図4】野生型T7 RNAポリメラーゼ酵素及び各変異型酵素のin vitro転写活性を反応温度がそれぞれ37、45、48及び51℃の条件で測定したものである。T7プロモーターを含むプラスミドに野生型及び各変異型酵素を加え、各温度にて1時間反応させた後、合成(転写)されたRNAを0.7%アガロースゲル電気泳動により調べたものである。
【図5】野生型T7 RNAポリメラーゼ酵素と変異型酵素(S430P+F849I+F880Y)の37〜65℃での比活性を比較した図である。
【発明を実施するための形態】
【0016】
本発明において、「野生型RNAポリメラーゼ」とは、天然に存在する全てのRNAポリメラーゼを意味する。さらに、「野生型RNAポリメラーゼ」は、対応する野生型RNAポリメラーゼの能力と比較して、熱安定性を向上させることを目的とする改変以外のアミノ酸の置換、挿入または欠失を、さらに有するものであることもできる。即ち、野生型RNAポリメラーゼを人為的に上記以外の目的で改変したRNAポリメラーゼも、上記「野生型RNAポリメラーゼ」に含まれる。但し、そのようなアミノ酸の置換、挿入または欠失は、RNAポリメラーゼとしての活性を維持する範囲で、行われたものであることが適当である。さらに「野生型RNAポリメラーゼ」としては、例えば、T7ファージ、T3ファージ、K11ファージに由来するRNAポリメラーゼを挙げることができる。但し、これらのRNAポリメラーゼに限定されるものではない。
【0017】
本発明のRNAポリメラーゼは、対応する野生型RNAポリメラーゼの少なくとも1つのアミノ酸が改変された熱安定性もしくは高温域での比活性の向上したRNAポリメラーゼである。ここで、「熱安定性」とは、例えば、0℃から100℃の温度下で、ある特定の時間インキュベーションした場合に、インキュベーション後の酵素が熱により失活せずに活性が残存していること、あるいは残存している酵素活性のインキュベーション前に対する割合をいうものである。また、「高温域での比活性」とは、例えば、37℃から100℃の温度下で酵素活性を測定した場合における単位タンパク質重量当たりの酵素活性をいうものである。
【0018】
本発明において、アミノ酸残基の変異は公知の方法により実施することができる。具体的には、例えば、付加によるタンパク質の機能を改変する方法としては、NatureBiotechnology第17巻、第58〜61頁(1999年)に記載されている。また、欠失による方法としては、Biochem.Biophys.Res.Commun.第248巻、第2号、第372〜377頁、挿入による方法としては、FEBS Lett.第442巻、第241〜245頁(1999年)、置換による方法としては、NucleicAcidsReserch 第26巻、第2号、第681〜683頁(1998年)にそれぞれ記載されている。
【0019】
本発明者らは、まず、野生型T7 RNAポリメラーゼ遺伝子を挿入した発現プラスミドpKKT7RNAPを構築し、次に、この発現プラスミドpKKT7RNAPを基にT7RNAポリメラーゼの変異体を作製した。即ち、430番目のアミノ酸残基SをPに置換させた変異体S430Pを、また、同様に849番目のアミノ酸残基FをIに置換させた変異体F849Iを、さらに同様に880番目のアミノ酸残基FをYに置換させた変異体F880Yを作製した。次にこれらの変異型酵素を精製し、その熱安定性について調べた。
【0020】
本明細書における、野生型T7RNAポリメラーゼのアミノ酸配列は、遺伝子配列データベースであるGeneBankより、accessionNo. M38308で登録されているT7RNAポリメラーゼ遺伝子配列を若干訂正した配列表・配列番号1及び配列番号2を基礎としている。配列番号1及び配列番号2において右端の数字は、アミノ酸の場合、T7RNAポリメラーゼの最初のM(メチオニン)を1として全長883アミノ酸残基からなっていることを示し、また塩基配列の場合、開始コドンATGのAを1として全長2652塩基よりなることを示す。従って、本発明における野生型T7RNAポリメラーゼのアミノ酸配列及び各アミノ酸に付された番号は、この配列番号1及び配列番号2に示される配列及び番号のことである。
【0021】
さらに、上述のように、上記野生型T7RNAポリメラーゼは、本発明で目的とする改変以外のアミノ酸の置換、挿入または欠失をさらに有するものであってもよい。従って、本発明の目的に基づいて変異を導入すべき野生型RNAポリメラーゼが、野生型T7RNAポリメラーゼに別の変異を導入したものである場合、特にそのような変異が、アミノ酸の挿入または欠失である場合、それに応じて上記アミノ酸番号は変動し、図1及び図2に示す番号とは異なったとしても、T7RNAポリメラーゼ活性を維持している限り、そのような挿入または欠失を有するT7RNAポリメラーゼも本発明の目的とする変異を導入する野生型T7RNAポリメラーゼの範疇に含まれる。
【0022】
また、本明細書におけるT7RNAポリメラーゼ以外のRNAポリメラーゼについてのアミノ酸配列及び各アミノ酸に付された番号は、図1及び図2に示されている配列及び番号である。さらに、本発明で目的とする改変以外のアミノ酸の置換、挿入または欠失を、さらに有するものであることもできる。従って、これらのアミノ酸配列及びその番号に付いても、T7RNAポリメラーゼの場合と同様であり、アミノ酸の挿入または欠失による変異がある場合、それに応じて上記アミノ酸番号は変動するが、そのような一部に変異を有する野生型のRNAポリメラーゼも本発明において本発明の目的とする変異を導入する野生型RNAポリメラーゼの範疇に含まれる。
【0023】
T7RNAポリメラーゼ遺伝子を含む発現ベクターは、T7ファージゲノミックDNA(シグマ製)を鋳型にして、T7 RNAポリメラーゼ遺伝子の開始コドン領域に特異的なプライマーと終始コドン領域に特異的なプライマーを用いてPCRを実施し、この増幅産物を制限酵素にて消化した後、発現ベクターpKK223‐3(ファルマシア製)に導入することにより構築した。この発現ベクターを用いて、大腸菌JM109に形質転換することにより、T7RNAポリメラーゼ蛋白質を大量に発現させることができる。
【0024】
発現ベクターpKKT7RNAPを持つ大腸菌エシェリヒア・コリ(Escherichia coli)から精製したT7 RNAポリメラーゼは、in vitroでT7プロモーターを含んだDNA存在下で充分なRNA合成活性を有していた。この発現プラスミドpKKT7RNAPをベースにして変異型T7RNAポリメラーゼとして、上述のS430P、F849I、F880Y及びそれらの置換を複数含んだ多重変異体を構築した。これらの変異型T7RNAポリメラーゼを大腸菌組換え宿主より精製し、各変異型酵素の熱安定性を野生型T7RNAポリメラーゼと比較した。結果を表1に示す。表1に示すように、これらの変異型酵素では、熱安定性の一つの指標である活性半減期が野生型酵素に比べて大幅に向上していた。
【0025】
ここで上記3種のファージ、即ち、T7、T3及びK11ファージ由来のRNAポリメラーゼにおいては、それらのアミノ酸配列は互いに極めて高いホモロジーを有していることが知られている。図1及び図2に、これら3種類のファージ由来RNAポリメラーゼのアミノ酸配列を比較して示す。このように、T7、T3、K11由来のRNAポリメラーゼは互いに極めて類似しており、T7RNAポリメラーゼにおいて得られた結果を、アミノ酸配列の類似する他のRNAポリメラーゼに適用することは比較的容易にできると考えられる。即ち、図1及び図2から、T7ファージ由来のRNAポリメラーゼの430、849及び880番目のアミノ酸残基に対応するアミノ酸残基は、T3ファージ由来のRNAポリメラーゼではそれぞれ431、850及び881番目のアミノ酸残基であり、またK11ファージ由来のRNAポリメラーゼではそれぞれ453、872及び903番目のアミノ酸残基であり、従って、T3RNAポリメラーゼにおけるF850I及びF881Y変異体、さらにK11 RNAポリメラーゼにおけるS453P、F872I及びF903Y変異体も、T7RNAポリメラーゼにおける変異体と同様に野生型に比べてより高い熱安定性を有していることは容易に類推できる。
【0026】
本発明は、上記改変型RNAポリメラーゼを製造する方法であって、RNAポリメラーゼをコードする核酸分子を用意し、次いでその塩基配列内の1つまたはそれ以上の部位における塩基を変異させるように該核酸分子を改変し、次いで変異させた核酸分子により発現される改変されたRNAポリメラーゼを回収することを含む方法を包含する。RNAポリメラーゼをコードする核酸分子の調製、核酸分子への突然変異の導入、改変されたRNAポリメラーゼの精製はいずれも、公知の手法を用いて行うことが出来る。
【0027】
即ち、変異型T7RNAポリメラーゼは、野生型T7RNAポリメラーゼ遺伝子をベースとして適当な塩基を改変して変異型T7RNAポリメラーゼ遺伝子を作製し、続いてこの遺伝子をベクターに導入し、発現ベクターを構築する。次に、この発現ベクターを用いて、宿主細胞を形質転換することにより変異型T7RNAポリメラーゼ蛋白質を大量に発現させることができる。
【0028】
本発明における核酸分子への変異の導入は、当業者がなし得る方法であればいかなる方法でもよいが、例えばサイトディレクテッドミュータジェネシス法がある。
【0029】
本発明において使用するベクターは、RNAポリメラーゼのクローニング及び発現を可能とするものであればいかなるものでもよく、例えばファージ及びプラスミドが挙げられる。プラスミドとしては、pUC118,pUC18、pBR322、pBluescript、pLED−M1、p73、pGW7などが挙げられる。ファージとしては、例えばλgt11、λZAPIIなどが挙げられる。
【0030】
本発明において使用する宿主細胞としては、大腸菌、酵母、バチルス(Bacillus)属細菌などが挙げられる。大腸菌としては、例えばエシェリヒア・コリDH5α、JM109、HB101、XL1Blue、PR1、BL21などが挙げられる。本発明においては、上記の熱安定性の向上した改変型RNAポリメラーゼをコードする遺伝子を上記ベクターに挿入して組換え発現ベクターとし、さらにこの組換え発現ベクターにて宿主細胞を形質転換する。
【0031】
本発明の製造方法においては、上記組換え宿主細胞を培養して、熱安定性の向上した改変型RNAポリメラーゼを発現させる。組換え宿主細胞の培養に使用する培地ならびに条件は常法に従う。具体例としては、熱安定性の向上した改変型RNAポリメラーゼ遺伝子を含むプラスミドにより形質転換された大腸菌を、例えばTB培地にて培養する。
【0032】
上記培養により得られた改変型RNAポリメラーゼの精製法としては、(a)組換え宿主を集めた後、破砕して細胞抽出物を調製し、(b)宿主細胞由来の不純タンパク質を除去する工程を含む。組換え宿主細胞より産出された熱安定性の向上した改変型RNAポリメラーゼは、宿主菌体を培地で培養後、培養液から遠心分離等にて分離、回収する。該菌体を緩衝液に再懸濁した後、超音波処理、ダイノミル・フレンチプレンス等により菌体を破砕する。次いでカラムクロマトグラフィーを実施し、熱安定性の向上した改変型RNAポリメラーゼを回収する。カラムクロマトグラフィーは、陽イオン交換体、例えば、フォスフォセルロース、あるいは陰イオン交換体、例えば、DEAEセファロース、あるいはアフィニティー吸着体ヘパリンセファロースなどが好ましい。
【0033】
本発明における改変型RNAポリメラーゼは、例えば恒温核酸増幅反応により特定のRNAを増幅せしめるのに特に有用である。ここで、恒温核酸増幅反応とは、NASBA法、TMA法、3SR法(Self‐sustainedsequence replication method)、TAS法(Transcription based amplification system method)などが挙げられる。
【実施例】
【0034】
以下、本発明を実施例により具体的に説明する。なお、以下の実施例において、T7 RNAポリメラーゼ活性の測定は以下のように行なった。すなわち、活性測定用緩衝液(40mMトリス−塩酸緩衝液(pH8.0)、20mM塩化マグネシウム、5mMジチオスレイトール、0.5mM rNTP、T7ゲノムDNA(シグマD4931)、50μg/ml BSA、[3H]rUTP(370kBq/μl))中で、酵素を37℃,10分間反応させたのち、合成されたRNAを酸不溶性沈殿として回収し、シンチレーションカウンターにて取り込まれた3H量を測定した。酵素活性の1単位(U)は、この条件下で60分間に1ナノモルのrNTPを酸不溶性沈殿画分に取り込ませる酵素量とした。
【0035】
実施例1 野生型T7 RNAポリメラーゼ遺伝子のクローニングと発現プラスミドの構築
野生型RNAポリメラーゼの発現プラスミドは以下のようにして構築した。すなわち、T7ファージゲノミックDNA(シグマ製)を鋳型にして、T7RNAポリメラーゼ遺伝子の開始コドン領域に特異的なプライマー(T7EcoATG : 5’−CGC GAA TTCATG AAC ACGATT AAC ATCGCT ‐3’)、及び終始コドン領域に特異的なプライマー(T7TerPstI: 5’−TTT CTG CAG TGG CGT TAC GCG AAC GCG AAG−3’)を用いてPCRを実施し、T7 RNAポリメラーゼをコードする遺伝子を増幅した。次に、この増幅産物を制限酵素EcoRIとPstIにて消化した後、発現プラスミドpKK223‐3(ファルマシア製)の同制限酵素サイトに導入しT7RNAポリメラーゼの発現プラスミドを作製した。これをpKKT7RNAPと名付けた。なお、得られたT7 RNAポリメラーゼ遺伝子の塩基配列に関しては、発現プラスミド中の得られた遺伝子をシークエンス決定することにより、PCRにより増幅した際の意図しない塩基置換が入っていないことを確認した。
【0036】
実施例2 改変型T7 RNAポリメラーゼ遺伝子の作製
改変型T7 RNAポリメラーゼ遺伝子は、上記実施例1で得られた野生型T7 RNAポリメラーゼ遺伝子に点変異を導入することにより行った。点変異の導入はクイックチェインジミュータジェネシスキット(ストラタジーン社製)および点変異導入用オリゴヌクレオチドを用いて、説明書の指示に従い行った。S430P、F849I及びF880Yのための点変異の導入は、変異導入用のプライマーを変えること以外は基本的に同じ条件にて実施できる。以下に一例として、T7RNAポリメラーゼ遺伝子の1288番目の塩基であるTをCに置換した(その結果、430番目のアミノ酸残基SがPに置換される)変異型遺伝子S430Pを作製した際の条件を示す。
【0037】
pKKT7RNAP5ng、変異導入用プライマー(5’‐GTT TAC GCT GTG CCAATG TTCAAC CCGCAA‐3’及び 5’‐TTG CGG GTT GAACAT TGG CACAGC GTA AAC‐3’)各125ng、上記キット添付の反応用緩衝液5μl、10mM dNTPmix 1μl、Pfu DNAポリメラーゼ2.5Uを含む50μlの溶液を、95℃で30秒間インキュベートした後、95℃、30秒/55℃、1分/68℃、12分の温度サイクルを12サイクル実施した。温度サイクル終了後、反応液に制限酵素DpnI10Uを添加し、37℃で1時間インキュベートした。次にこの反応液1μlをエシェリヒア・コリJM109株コンピテントセル100μlに加え、30分間氷冷した後、42℃で30秒間インキュベートし、900μlのSOC培地を加え37℃で1時間インキュベートした。これに50μg/mlのアンピシリンを含むLB寒天培地上にスプレッドし、37℃で一晩インキュベートした。得られたコロニーをLB培地2.5mlにて一晩培養した後、定法に従いプラスミドを抽出した。最後に、これらのプラスミドの当該領域のシークエンスを行い、1288番目のTがCに置換されているクローンを取得し、pKKT7S430Pと名付けた。
【0038】
また、同様の方法で、変異導入用プライマーとして5’‐TTC TAC GAC CAG ATCGCT GAC CAGTTG CAC‐3’及び 5’‐GTG CAA CTG GTCAGC GAT CTGGTC GTA GAA‐3’を用いることにより2545番目のTをAに置換した(結果として、849番目のFがIに置換される)遺伝子を含む発現プラスミド(pKKT7F849I)、5’‐TCT TAG AGT CGGACT ACG CGTTCG CGT AAC‐3’及び 5’‐GTT ACG CGAACG CGT AGT CCG ACT CTA AGA‐3’を用いることにより2639番目のTをAに置換した(結果として、880番目のFがYに置換される)遺伝子を含む発現プラスミド(pKKT7F880Y)を作製した。さらに、これらの3種類の変異を複数含む変異型遺伝子に関しては、変異を含む領域を制限酵素にて切り出し、他の変異を含む発現プラスミドの同領域と入れ替えることにより作製した。
【0039】
実施例3 形質転換体の作製
実施例1及び実施例2で得られたプラスミド1ngをJM109コンピテントセル100μlに加え、30分間氷冷した後、42℃で30秒間インキュベートし、900μlのSOC培地を加え、37℃で1時間インキュベートした。これを50μg/mlのアンピシリンを含むLB寒天培地上にて37℃で一晩インキュベートし、形質転換体を得た。
【0040】
実施例4 野生型及び各種改変型T7RNAポリメラーゼの精製
野生型及び各種改変型T7RNAポリメラーゼは基本的に同じ方法にて精製、調製することができる。実施例3で得られた各形質転換体を100μg/mlのアンピシリンを含むTB培地100mlに植菌し、37℃で一晩インキュベートした。得られた菌体を12,000回転/分で5分間遠心することにより回収した。菌体約10gをバッファー1(20mMトリス−塩酸(pH8.0),1mM EDTA、1mMジチオスレイトール, 5%グリセロール)20mlに懸濁し、これを超音波破砕機で破砕した後、12000回転/分で20分間遠心することにより沈殿を分離した。得られた上清に0.6%ポリエチレンイミン溶液を0.4ml添加し、30分間攪拌した。これを12000回転/分で10分間遠心することにより沈殿を分離し、上清を回収した。この液に硫酸アンモニウムを4.56g加え、30分間攪拌した後、これを12000回転/分で10分間遠心することにより沈殿を分離し回収した。得られた沈殿をバッファー2(20mMトリス−塩酸(pH8.0),1mM EDTA、1mMジチオスレイトール、20mM NaCl、5%グリセロール)5mlに溶解し、100mlのバッファー2に対して透析した。次に、これをDEAEセファロースカラム(5ml)にチャージし、10mlのバッファー2で洗浄後、NaClの0〜500mMのグラジエントにより溶出した。得られたフラクションのうち、RNAポリメラーゼ活性を含む画分をプールし、100mlのバッファー2に対して透析した。次に、これをフォスフォセルロースカラム(5ml)にチャージし、15mlのバッファー2で洗浄後、バッファー2をベースとしてNaCl濃度の100〜400mMのグラジェントにより溶出した。得られたフラクションのうち、RNAポリメラーゼ活性を含む画分を回収した。以上の操作で、SDS−PAGEによりほぼ単一なバンドを示す10mgのT7RNAポリメラーゼタンパク質を得た。
【0041】
実施例5 野生型及び各変異体の活性半減期の測定
熱安定性の指標の一つである活性半減期(ある温度でインキュベートした際に活性が半減するまでの時間)の測定は以下のようにして実施した。すなわち40mMトリス−塩酸緩衝液(pH8.0)、20mM塩化マグネシウム、5mMジチオスレイトール、70mM KCl、100μg/ml BSAからなる緩衝液中、酵素を48℃にてインキュベートし、一定の時間間隔ごとにサンプリングを行い、各サンプリング時点での残存活性を測定した。野生型および各種改変型T7RNAポリメラーゼを48℃でインキュベートした際の残存活性を経時的に測定した結果を図3に示す。また、図3から導き出される野生型及び各変異型酵素の活性半減期を表1に示す。
【0042】
表1の数値は、48℃で酵素をインキュベートした場合に、酵素活性が半減するまでの時間(min)を示しており、数値が大きいほどその酵素の熱安定性が高いことを示している。図3及び表1から、各変異型酵素はいずれも野生型酵素に比してより高い熱安定性を有していることがわかる。また今回作製した変異型酵素の中では、S430P、F849I及びF880Yの3箇所の変異を全て含む三重変異型酵素(S430P+F849I+F880Y)が最も熱安定性に優れている。
【0043】
【表1】
【0044】
実施例6 野生型及び各変異型酵素のin vitro転写反応
野生型及び各変異型酵素のin vitro転写反応は以下のように実施した。即ち、野生型及び各変異型酵素各50Uを、40mMトリス−塩酸緩衝液(pH8.0)、20mM塩化マグネシウム、5mMジチオスレイトール、0.4mMrNTP、0.1%BSA、0.5μg pUCT7−1からなる転写反応バッファー50μlに加え、37、45、48、51℃の各温度にて1時間反応させた。ここでpUCT7−1は、T7プロモーターの下流に約1.5kbpのDNAフラグメントが連結されたプラスミドDNAであり、T7RNAポリメラーゼにより転写されると約1.5KbpのRNAが合成されるものである。1時間の反応の後、反応液のうちの3μlを0.7%アガロースゲル電気泳動に供し、合成されたRNAの有無、量を測定した。その0.7%アガロースゲル電気泳動の結果を図4に示す。なお各レーンに付されているナンバーは上記表1における各変異体のナンバーに相当する。図4より、変異型酵素、特にNo.8の三重変異体(S430P+F849I+F880Y)は、より高い反応温度でも合成された転写産物(RNA)のバンドを確認でき、野生型に比してより高い温度下でも活性を有していることが確認できる。
【0045】
実施例7 野生型及び三重変異型酵素の各温度での比活性測定
各温度での比活性の測定は以下のようにして実施した。すなわち、野生型及び改変型酵素を、予め以下の温度に保った活性測定緩衝液(40mMトリス−塩酸緩衝液(pH8.0)、20mM塩化マグネシウム、5mMジチオスレイトール、0.5mM rNTP、T7ファージゲノムDNA(シグマD4931)、50ug/mlBSA、[3H]rUTP(370kBq/ul)に加え、それぞれ37℃、40℃、45℃、50℃、55℃、60℃、65℃にて10分間反応させ、各温度における活性を測定した。また、各酵素のタンパク質濃度は、プロテインアッセイキット(バイオラッド社製)を用いて、キット付属のイムノグロブリンガンマ(IgG)を標準タンパク質として用いて測定した。比活性(SA)は、単位タンパク質重量当たりの酵素活性のことであり、活性量を酵素タンパク量で割ることにより算出できる。野生型および三重変異型酵素(S430P+F849I+F880Y)の各温度での比活性を測定した結果を図5に示す。図5より、改変型酵素は、野生型酵素に比べて各温度全般にわたって高い比活性を有しており、特に高温域での比活性に優れており、とりわけ50℃での比活性は野生型の約12倍もの値を有していることが判る。
【特許請求の範囲】
【請求項1】
以下の(a)又は(b)のタンパク質からなる、T7ファージ由来のRNAポリメラーゼ。
(a)849番目のアミノ酸残基フェニルアラニン及び880番目のアミノ酸残基フェニルアラニンが、それぞれイソロイシン及びチロシンに置換された、配列番号1に記載されるアミノ酸配列からなるタンパク質
(b)アミノ酸配列(a)において1若しくは数個のアミノ酸が付加、欠失、挿入若しくは置換されたアミノ酸配列からなり、かつ改変前の該タンパク質に比して熱安定性若しくは高温域での比活性が向上したタンパク質
【請求項2】
請求項1に記載のタンパク質をコードするDNAからなる遺伝子。
【請求項3】
請求項2に記載の遺伝子を含有する組換えベクター。
【請求項4】
請求項3に記載の組換えベクターを用いて形質転換された組換え宿主細胞。
【請求項5】
宿主細胞が大腸菌(Escherichia coli)である請求項4に記載の組換え宿主細胞。
【請求項6】
請求項4又は5に記載の組換え宿主細胞を培養しRNAポリメラーゼを採取することを特徴とする改変型RNAポリメラーゼの製造方法。
【請求項7】
請求項1に記載のRNAポリメラーゼを用いて恒温核酸増幅反応によりRNAを増幅せしめることを特徴とするRNAを合成する方法。
【請求項8】
請求項1に記載のRNAポリメラーゼを含む核酸増幅用試薬キット。
【請求項1】
以下の(a)又は(b)のタンパク質からなる、T7ファージ由来のRNAポリメラーゼ。
(a)849番目のアミノ酸残基フェニルアラニン及び880番目のアミノ酸残基フェニルアラニンが、それぞれイソロイシン及びチロシンに置換された、配列番号1に記載されるアミノ酸配列からなるタンパク質
(b)アミノ酸配列(a)において1若しくは数個のアミノ酸が付加、欠失、挿入若しくは置換されたアミノ酸配列からなり、かつ改変前の該タンパク質に比して熱安定性若しくは高温域での比活性が向上したタンパク質
【請求項2】
請求項1に記載のタンパク質をコードするDNAからなる遺伝子。
【請求項3】
請求項2に記載の遺伝子を含有する組換えベクター。
【請求項4】
請求項3に記載の組換えベクターを用いて形質転換された組換え宿主細胞。
【請求項5】
宿主細胞が大腸菌(Escherichia coli)である請求項4に記載の組換え宿主細胞。
【請求項6】
請求項4又は5に記載の組換え宿主細胞を培養しRNAポリメラーゼを採取することを特徴とする改変型RNAポリメラーゼの製造方法。
【請求項7】
請求項1に記載のRNAポリメラーゼを用いて恒温核酸増幅反応によりRNAを増幅せしめることを特徴とするRNAを合成する方法。
【請求項8】
請求項1に記載のRNAポリメラーゼを含む核酸増幅用試薬キット。
【図1】

【図2】

【図3】
【図4】
【図5】
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2009−213499(P2009−213499A)
【公開日】平成21年9月24日(2009.9.24)
【国際特許分類】
【出願番号】特願2009−157706(P2009−157706)
【出願日】平成21年7月2日(2009.7.2)
【分割の表示】特願平11−230629の分割
【原出願日】平成11年8月17日(1999.8.17)
【出願人】(000003160)東洋紡績株式会社 (3,622)
【Fターム(参考)】
【公開日】平成21年9月24日(2009.9.24)
【国際特許分類】
【出願日】平成21年7月2日(2009.7.2)
【分割の表示】特願平11−230629の分割
【原出願日】平成11年8月17日(1999.8.17)
【出願人】(000003160)東洋紡績株式会社 (3,622)
【Fターム(参考)】
[ Back to top ]