
改良免疫糖タンパク質のための物質および方法
抗体を含み、改良されたADCC及び代わりにグリコシル化パターンを伴う免疫糖タンパク質を提供する。該免疫糖タンパク質はカスタノスペルミンを含む培地でホスト細胞成長によって生産される。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、抗体を含む、特性が改良された免疫糖タンパク質に関し、それには、抗体依存性細胞傷害性およびグリコシル化パターン、細胞培養方法およびそのような免疫糖タンパク質を生成するための培地、並びに病気の処置における当該免疫糖タンパク質の使用が含まれる。
【0002】
なお、本件出願は、2006年10月24日に出願された米国仮出願第60/853,944号の利益を享受し、その内容を援用することで全体は組み入れられているものである。
【背景技術】
【0003】
免疫薬でターゲットとする細胞数を削減するのは、適応症のいくつかにおいては重要な治療処置である。そのように、ターゲットとする細胞の削減を達成するために、免疫薬が用いているメカニズムは、補体が媒体となる細胞の溶解、細胞消滅の信号を送る動作経路の活性化、生存のために必要な信号を送る経路の遮断、およびFc依存性細胞傷害性とも称される、抗体依存性細胞傷害性(ADCC)を含み得る。ADCCは、効力のあるメカニズムであり、多くの免疫薬の有効性にとって重要であると考えられている。
【0004】
ADCCの活性化のためのメカニズムには、ターゲット細胞の表面に結合される免疫薬分子へのFc受容体の結合が必要である。免疫薬へのFc受容体の結合は、CH2および/またはCH3ドメインのような、免疫グロブリンの一定領域内のドメインを媒体とすることができる。異なる種類の一定領域が、異なるFc受容体と結合する。例として含まれるのは、IgG1Fcドメインの、同種のFc受容体CD16(FcγRIII)、CD32(FcγRII‐B1および‐B2)およびCD64(FcγRI)への結合、IgAFcドメインの、同種のFc受容体CD89(FcαRI)への結合、およびIgEドメインの、同種のFc受容体FcεR1およびCD23への結合である。
【0005】
Fc受容体の結合を高めた免疫薬組成物は、ADCCにおいてより大きな有効性が呈示される。IgGFcドメインでこれを達成する方法として報告されているものには、アミノ酸の変化の導入や糖類構造の修飾 (modification) が含まれる。Fcドメインにおけるアミノ酸の変化は、免疫薬組成の免疫原性を高めるので、糖類構造の修飾が好ましい。免疫グロブリン分子については、N‐リンクされた糖類の、CH2ドメインのAsn−297への付着がADCC機能にとっては重大であるということが示されている。酵素で、またはN‐リンクされた共通サイトの突然変異を通してそれを取り除くと、結果的に、ADCC機能がほとんどまたは全く無くなってしまう。免疫グロブリン分子についてのADCC機能のレベルもまた、糖類の構造に依存すると報告している研究もあるが、実際の糖類の、ADCCに関する部位または構造は、いまだ解明されていない。まして、非免疫グロブリンFc融合タンパク質のADCCに最適な糖類の構造については、なおさら知られていない。
【0006】
糖タンパク質において、糖類は、トリペプチドのモチーフAsn‐X‐Thr/Serにおけるアスパラギンの側鎖でアミド窒素原子に付着する。N‐リンクされたグリコシル化と称されるこの種のグリコシル化は、複数の単糖類がリン酸ドリコールに加わって14‐残基側鎖の糖類複合体を形成している小胞体(ER)において開始する。そして、この糖類複合体は、オリゴサッカリルトランスフェラーゼ(OST)複合体によってタンパク質へと転移される。糖タンパク質がERの内腔を離れる前に、3つのグルコース分子が、14‐残基オリゴ糖から取り除かれる。酵素ERグルコシダーゼI、ERグルコシダーゼIIおよびERマノシダーゼがER処理に関係している。
【0007】
続いて、ポリペプチドがゴルジ複合体へと運ばれ、そこではN‐リンクされた糖鎖が異なる多くの方法で修飾される。ゴルジ複合体のシスおよび中間区画において、元の14‐糖N‐リンクされた複合体は、マンノース(Man)残基を取り除いて切り詰められ、N‐アセチルグルコサミン(GlcNac)および/またはフコース(Fuc)残基を加えることで伸長される。N‐リンクの糖類の形は様々であるが、一般には、3つのマンノース残基と2つのN‐アセチルグルコサミン残基からなる五糖(ペンタサッカライド)を共有する。最後に、トランスゴルジにおいて、他のGlcNac残基を加え、それに続いてガラクトース(Gal)および末端シアル酸(Sial)を加えることができる。ゴルジ複合体における糖処理は、「末端グリコシル化」と呼ばれ、ERにおいて起こる「コアグリコシル化」とは区別される。最終の複合糖ユニットは、多くの形態および構造を取り得るものである、その中には、2本、3本または4本の側鎖(バイアンテナリー、トリアンテナリーまたはテトラアンテナリーと称する)を有するものもある。ゴルジ処理には、複数の酵素が関係しており、それらには、ゴルジマノシダーゼIA、IBおよびIC、GlcNAc‐トランスフェラーゼI、ゴルジマノシダーゼII、GlcNAc‐トランスフェラーゼII、ガラクトシルトランスフェラーゼおよびシアリルトランスフェラーゼが含まれている。
【0008】
FcγRIIIa受容体が媒体となるADCC機能の重大な糖の決定要素は、コアN‐リンクの構造に加えられるアルファ‐1、6‐フコース部の欠如であることを、ある報告が示唆している(シンカワら、J Biol Chem. 2003年1月31日;278(5):3466−73;また、シールズら、J Biol Chem. 2002年7月26日;277(30):26733−40も参照)。別のレベルのグリコフォーム、すなわち、二等分されたN‐リンクの糖類もまたADCCを増大することができるものとして提案されている(ウマナら、Nat Biotechnol. 1999年2月;17(2):176−80)が、それに反する証拠もある(シンカワら、J Biol Chem. 2003年1月31日;278(5):3466−73)。この反対の証拠が解明される可能性もあることが示唆されており、それは、ホスト細胞においてGnTIIIが増加することで、二等分された糖が増加しているのみならず、コアフコースの修飾が欠如している免疫グロブリンが生成されるという発見による(フェラーラら、Biotechnol Bioeng. 2006年4月5日;93(5):851‐61)。これは、フコースのみがADCCの有効性を変える主要な役割を果たしており、他で見られる二等分された糖との関連は、ホスト細胞における2つの修飾の結び付きを反映しているという提言と一致している。しかしながら、GnTIIIを備えるリタクサン抗体およびハーセプチン抗体の、二等分された糖を増やすためのインビトロの処置が、結果としてADCCを増大させたという別の報告があり、二等分された糖の直接の影響が示唆されている(ホドニクツキーら、Biotechnol. Prog., 2005年11月−12月;21(6):1644−52)。しかしながら、非常に高いレベルでのGntIIIの過大な発現は、細胞に対して有毒である(ウマナら、Biotechnol. Prog., 1998年3月−4月;14(2):189−92)。
【0009】
フコース含有量の低い免疫グロブリンを生成する方法が提案されているが、治療の指標として最適なADCC機能を備えるバイオ医薬品の製造にとって大きな障害があるものもある。例えば、フコース残基(フコーシダーゼ)を取り除く酵素を備える免疫グロブリンの処置には、重大な経済上の、および薬品の安定性上の危険が潜在していて、費用のかかる製造工程を追加する必要がある。フコシル化糖タンパク質の合成に関係する主要酵素を破壊するための細胞系(line)の分子的な処理には、特別なホストの負担が必要であり、現在のプラクティスにおいては、治療上の使用のために有効性および安全性が最適化されるようADCCの有効性を変えると、薬品の「調節可能な」生成が可能ではなくなる。ADCCの高められていない比較生成物を生成するには、費用および時間がかかる。RNAiまたはアンチセンス分子で細胞系(line)を処置してこれらの主要酵素のレベルを破壊すると、予想できない的外れの効果が得られ、また、生産を行う規模で実施される程度には実用的であるとしても、費用がかかるであろう。
このように、ADCCの高められた免疫薬を調合するのに有効な方法、またそれによって生成され、治療上使用される改良された免疫薬が切望されている。
【先行技術文献】
【非特許文献】
【0010】
【非特許文献1】J Biol Chem、2003年1月31日、278(5)、3466−73
【非特許文献2】J Biol Chem、 2002年7月26日、277(30)、26733−40
【非特許文献3】J Biol Chem、 2003年1月31日、278(5)、3466−73
【非特許文献4】Nat Biotechnol、 1999年2月、17(2)、176−80
【非特許文献5】Biotechnol Bioeng、 2006年4月5日、93(5)、851−61
【非特許文献6】Biotechnol. Prog、 2005年11月−12月、21(6)、1644−52
【非特許文献7】Biotechnol. Prog、 1998年3月−4月、14(2)、189−92
【発明の概要】
【発明が解決しようとする課題】
【0011】
本発明は、培地(培養媒体)および免疫糖タンパク質の特性を改良する大規模細胞培養方法を提供するが、これには、ADCCのようなエフェクター機能および/またはフコース含有量の低減のようなグリコシル化パターンが含まれる。本発明はさらに、上記方法によって生成される、改良された免疫糖タンパク質、および病気の治療におけるそのような免疫糖タンパク質の使用を提供する。
【課題を解決するための手段】
【0012】
本発明は、ホスト細胞により生成された免疫糖タンパク質分子の抗体依存傷害性(ADCC)を増大する方法を提供するが、それは、濃度が、約25μM〜約800μMの間、または約100μM〜約500μMの間、または約100μM〜約400μMの間、または約100μM〜約300μMのカスタノスペルミンを備える培地において、ホスト細胞を成長させることによるものである。代表的な実施例において、ADCCは、少なくとも2倍、3倍、4倍または5倍に増大する。
【0013】
また本発明は、ホスト細胞により生成された免疫糖タンパク質分子のCD16結合を増大する方法を提供するが、それは、濃度が、約25μMと約800μMの間、または約100μMと約500μMの間、または約100μMと約400μMの間、または約100μMと約300μMの間にあるカスタノスペルミンを備える培地において、ホスト細胞を成長させることによるものである。代表的な実施例において、CD16結合は、少なくとも50%、75%、100%、125%、150%、175%または200%増大する。
【0014】
本発明の方法において、細胞の成長、生存度および/または密度は大きな影響を受けない(例えば、処置されていない細胞の少なくとも80%以上に留まる)。培地における免疫糖タンパク質の生成のレベルは、少なくとも100μg/mL、125μg/mLまたは150μg/mLである。
【0015】
前記本発明において、培地は実質的に漿液が含まれておらず、第二の糖修飾因子が含まれている。
【0016】
本発明では、上記方法で生成された免疫糖タンパク質分子を、必要に応じて無菌の薬学的に受け入れ可能な担体または希釈剤とともに含む組成物もまた意図されている。かかる組成物は、前記免疫糖タンパク質分子が結合した分子を表面に発現する癌細胞を死滅させる、あるいは、その成長を抑制する方法、または前記免疫糖タンパク質分子が結合する分子を表面に発現する細胞を枯渇させる方法において投与される。
【0017】
本発明の方法は、一般に、適当な濃度の糖修飾因子、例えば、カスタノスペルミンを含む培地において、免疫糖タンパク質を生成するホスト細胞を培養することを伴い、細胞の成長またはタンパク質生成レベルに大きな影響を与えることなく、エフェクター機能を向上させるという利点がある。本発明の方法を用いて製造することのできる代表的な免疫糖タンパク質には、免疫グロブリンおよび小さなモジュラー免疫薬(SMIPTM)生成物が含まれる。本発明の方法により調製されるそのような結合分子には、ターゲットへの結合特性を実質的に同じに保ち、その結果、生物的活性が直接得られ、エフェクターを媒体とする機能の向上が呈示されるという利点がある。
【0018】
本発明は、抗体依存傷害性(ADCC)および/またはホスト細胞により生成される免疫糖タンパク質のFc受容体結合を向上させる方法を提供する。当該方法は、糖修飾因子、例えば、カスタノスペルミンを、ホスト細胞が生成した免疫糖タンパク質分子の組成物のADCC活性および/またはFc受容体結合を増大させるような濃度で備え、少なくとも体積が750mL、1L、2L、3L、4L、5L、10L、15L、20Lまたはそれ以上の培地において、ホスト細胞を成長させることを伴っている。かかる糖修飾因子、例えば、カスタノスペルミンの最適な濃度は、糖修飾因子の有効性および所望のADCCの関連する調節(modulation)に依存するが、培地における糖修飾因子の代表的な最終濃度は、800μM未満または750、700、650、600、550、500、450、400、350、300、250、200、150、125、100、90、80、70、60、50、40、30、20または10μM未満である。
【0019】
それに関係した、ADCCへの影響は、細胞培養に適用される糖修飾因子、例えば、カスタノスペルミンの濃度または持続時間を変えることで調整されるが、グリコシル化を変えることでADCCを向上させる従来の方法と比較すると、顕著な利点がある。ADCC活性は、当業界で知られているアッセイを用いて測定され表されるが、代表的な実施例においては、少なくとも10%、20%、30%、40%、50%、60%、70%、80%、90%、2倍、3倍、4倍、5倍、6倍、7倍、8倍、9倍、10倍、15倍または20倍増大する。
【0020】
グリコシル化および糖含有量は、ADCC、CDCおよび循環半減期を含む、免疫グロブリンエフェクターを媒介とする様々な機能に影響を及ぼすことが知られている。ここに記述されるデータでは、本発明の方法によって、驚くことに、CDCまたは半減期に影響を及ぼすことなく、ADCCが増大することを示す免疫糖タンパク質が提供されるということが示されている。このように、代表的な実施例において、免疫糖タンパク質分子組成物のADCCは増大するが、補体依存傷害性(CDC)および/または長くなった循環半減期のような他の免疫グロブリン型エフェクター機能は、同様であるか大きくは影響を受けない(例えば、2倍未満の増大または減少、あるいは50%、40%、30%、20%または10%未満の増大または減少)。
【0021】
免疫糖タンパク質分子の組成物のFc受容体結合は、糖修飾因子で処置された免疫糖タンパク質分子対CD16に結合する処置されていない免疫糖タンパク質分子という相対比として測定される。代表的なアッセイが、以下の例で記述される。代表的な実施例におけるFc受容体結合は、少なくとも10%、20%、30%、40%、50%、60%、70%、80%、90%、2倍、3倍、4倍、5倍または6倍増大する。本発明により、糖修飾因子、例えば、カスタノスペルミンで処置されたホスト細胞が生成した免疫糖タンパク質組成は、CD16(高または低親和形態、すなわちアミノ酸158でVまたはF)および/またはCD32aまたはbおよび/またはCD64に、そのように処置されていないホスト細胞が生成する免疫糖タンパク質組成物よりもFcR結合検査において大きな親和力で結合する。Fc受容体結合親和力がこのように増大するのは、ADCC機能の向上と相関していることがここに示されている。
【0022】
本発明によって、糖含有量/グリコシル化パターンを変え、および/または免疫糖タンパク質のフコース含有量を低減する方法もまた提供されるが、該方法は、総フコース含有量を低減し、および/またはホスト細胞が生成する免疫糖タンパク質分子の組成物のグリコシル化パターンを変えるような濃度で、糖修飾因子、例えばカスタノスペルミンを備え、体積が、少なくとも750mL、1L、2L、3L、4L、5L、10L、15L、20Lまたはそれ以上である培地において、ホスト細胞を成長させることによるものである。培地における糖修飾因子の代表的な最終濃度は、800μM未満または750、700、650、600、550、500、450、400、350、300、250、200、150、125、100、90、80、70、60、50、40、30、20または10μM未満である。
【0023】
フコース含有量への影響もまた、細胞培養に適用される糖修飾因子、例えば、カスタノスペルミンの濃度または持続時間を変えることによって調整される。組成物の総フコース含有量は、フコース化されていない免疫糖タンパク質分子の、組成物における免疫糖タンパク質分子の総数に対する相対比またはパーセンテージとして表される。代表的な組成物は、少なくとも30%、40%、50%、60%、65%、70%、75%、80%、85%、90%、95%またはそれ以上のフコース化されていない分子を含む。本発明により、糖修飾因子、例えば、カスタノスペルミンで処置されたホスト細胞が生成する免疫糖タンパク質組成物のフコース含有量は、そのように処置されなかったホスト細胞が生成する組成物と比べて、少なくとも2倍、3倍、4倍、5倍、6倍、7倍、8倍、9倍または10倍以上低減される。
【0024】
前述の本発明の方法のいずれにおいても、ホスト細胞は、糖修飾因子、例えば、カスタノスペルミンにさらされている間、高いレベルでの成長を呈示している。例えば、免疫糖タンパク質を生成するCHO細胞の代表的な倍化時間は、約24時間であるが、本発明による糖修飾因子の濃度(例えば、ADCCを増大するのに有効な濃度)によって、そのような倍化時間が低減されるとの期待はない。理想的には、糖修飾因子、例えば、カスタノスペルミンの有効な濃度では、糖修飾因子を加えてから72時間後の時点で、10、20、30、40、50、60または70%を超えるほどには、細胞の成長は低減されない。
【0025】
前述の方法のいずれにおいても、ホスト細胞は、糖修飾因子、例えば、カスタノスペルミンにさらされている間、高いレベルでのタンパク質生成を呈示している。例えば、糖修飾因子、例えば、カスタノスペルミンが有効な濃度であるときのタンパク質生成レベルは、約50μg/mL以上であり、または約75、100、125または150μg/mL以上である。好ましくは、ホスト細胞は、高いレベルでの成長および高いレベルでのタンパク質生成の双方を呈示する。
【0026】
実質的に漿液を含まない培地を含む、当業界で知られている任意の培地を用いても良い。バッチ培養、流加培養、連続培養、その他の業界で知られている培養方法を本発明の方法で用いることができる。糖修飾因子が、急速な成長段階後に、シーズ列に添加され、すなわち、最初のバッチ培地に添加されるか、又は連続して培地と一緒に(例えば、連続して供給する間に)添加される。例えば、糖修飾因子は、10Xまたは100Xの濃度で初期のシーズ列または原料に添加されるが、次いで培地を加えると、糖修飾因子の濃度が希釈されるが、組み換え生成物のADCCを増大させるのになおも有効なレベルになるようにする。あるいは、糖修飾因子が、細胞に添加されるあらゆる培地に有効な濃度で含まれ、希釈する必要は無いものとする。いずれにしても、糖修飾因子は、細胞培養プロセスにおいて比較的早期に加えられ、免疫糖タンパク質の均質性を最適化するために、培養処理の間有効な濃度が維持される。糖修飾因子の効果は、長期に続くものである、糖修飾因子を一度加えると、少なくとも11乃至12日間は持続して認められる。
【0027】
代表的な糖修飾因子には、コアグリコシル化抑制剤、端末グリコシル化抑制剤、マノシダーゼ抑制剤および/または初期段階糖修飾因子が含まれており、選択的にフコシル化に特有の抑制剤が含まれていても含まれていなくてもよく、さらなる詳細は以下に記述される。本発明におていは、2つ以上あるいは3つ以上の糖修飾因子を組み合わせることでさらに利点が得られるということが意図されている。カスタノスペルミンは、特に有効な1つの糖修飾因子である。
【0028】
さらに、本発明によって、前述の方法のいずれかで生成された免疫糖タンパク質分子を含み、好ましくは、ターゲット分子について少なくとも107M−1または少なくとも108M−1または109M−1の結合親和力Kdを有する組成物が提供される。そのような組成物は、無菌の、薬学的に受け入れ可能な担体または希釈剤を1つ以上含んでいてもよい。
【0029】
さらに、本発明によって、上記組成物の被体への投与を伴っている治療方法が提供されるが、その被体は、当該投与から効果を得ることができるもので、例えば、ターゲット分子を発現する細胞が媒体となる障害を患っているか、癌細胞が表面にターゲット分子を発現するある種の癌を患っている。本発明によって、上記組成物を、ターゲット分子を表面に発現している細胞を枯渇する方法において用いることもまた意図されている。ターゲットがCD37であるところでは、本発明によって、癌細胞の成長を抑制するか、癌細胞を破壊する方法が特に意図されており、それは、本発明の方法により生成されたアンチCD37SMIP生成物を含む組成物を被体に投与する工程を備えている。同様に、ターゲットがCD20であるところでは、本発明によって、癌細胞の成長を抑制するか癌細胞を破壊する方法が特に意図されており、それは、本発明の方法により生成されたアンチCD20SMIP生成物を含む組成物を被体に投与する工程を備えている。好適な形態において、癌の進行を停止させるか反転させることを伴って癌を治療する方法が意図されている。本発明によって、本発明の方法により生成されたアンチCD37またはアンチCD20SMIP生成物を投与することで、自己免疫性または炎症性の疾患を治療する方法がさらに提供される。さらに、本発明において、ここに記述される疾患または障害のいずれかを治療する薬剤を調製するための、無菌又は滅菌された担体または希釈剤を選択的に有する、本発明の糖タンパク質組成物の使用が意図されている。
【0030】
免疫糖タンパク質
「免疫糖タンパク質」という語は、ターゲット分子と結合するグリコシル化ポリペプチドのことを称し、免疫グロブリンの一定領域から得られる十分なアミノ酸配列を含んで、好ましくは、ADCCおよび/またはCDCというエフェクター機能を提供する。代表的な分子には、免疫グロブリンのCH2ドメインから得られるシーケンス、または1つ以上の免疫グロブリンから得られるCH2およびCH3ドメインが含有される。本発明によって生成することが意図された免疫糖タンパク質の特定の部分集合には、ヒンジにおける共有結合性の、または非共有結合性の結合および/またはCH3ドメインを通して選択的に二量体になる単鎖タンパク質が含まれる。単鎖タンパク質のこの部分集合には、免疫グロブリンの典型的な四量体の形態は含まれない(軽鎖が無いことによる)が、Fc配位子またはFc可溶の受容体融合が含まれる。単鎖タンパク質の特定の例には、SMIP生成物が含まれる。
【0031】
SMIP生成物とそれらを生成する方法は、共同で所有される米国出願第10/627,556号および米国特許公報2003/133939、米国特許公報2003/0118592および米国特許公報2005/0136049において以前記述されており、それらは各々、援用することでその内容全体をここに組み入れる。エフェクター機能を備える単鎖多価結合タンパク質が、2007年6月12日に出願された国際特許出願第PCT/US07/71052号に記述されている(2006年6月12日に出願されたU.S.S.N.60/813,261および2006年10月20日に出願されたU.S.S.N.60/853,287の利益を主張する)が、それらは各々、援用することでその内容全体をここに組み入れる。SMIP生成物は、抗原、反対受容体等の同種の構造についての結合ドメインを特徴とする新規の結合ドメイン‐免疫グロブリン融合タンパク質であり、すなわち、IgG1、IgAまたはIgEヒンジ領域ポリペプチド、または0、1または2のシスティン残基を有する突然変異IgG1ヒンジ領域ポリペプチドおよび免疫グロブリンCH2およびCH3ドメインである。一例においては、結合ドメイン分子は、1つまたは2つのシスティン残基を有する。また他の例においては、結合ドメイン分子が2つのシスティン残基を備えるとき、典型的には重鎖可変領域と軽鎖可変領域との間の結合に携わる第一のシスティンが削除されたり、アミノ酸で置換されたりすることの無いように意図されている。SMIP生成物は、ADCCおよび/またはCDCが可能であるが、二硫化物に結合した多量体を形成する能力で折り合っている。代表的なSMIP生成物は、免疫グロブリンから得られる可変軽鎖および/または可変重鎖結合領域の免疫グロブリン上科要素の結合領域のような1つ以上の結合領域を有する。代表的な例において、これらの領域は、リンカーペプチドで分離されるが、そのリンカーペプチドは、ドメインまたは領域接合と適合性のあることが当業界で知られているいずれのものであっても良い。代表的なリンカーは、(Gly4Ser)n、ただしn=3〜5、のようなGly4Serリンカーモチーフに基づくリンカーである。本発明により生成できる代表的なSMIP生成物には、CD20またはCD37を結合する生成物が含まれている。CD20またはCD37を結合し、特定の結合シーケンスおよび/またはアミノ酸の修飾を備えるSMIP生成物は、共同で出願された米国出願10/627,556および米国出願11/493,132に記述されており、これらの各々の内容は、ここに援用することでその全体をここに組み入れる。
【0032】
免疫糖タンパク質の他の例には、結合ドメイン‐Ig融合体が含まれており、ここで、結合ドメインは、非自然発生のペプチドまたは自然発生のリガンドまたはレセプターのフラグメントである。レセプターの場合、細胞外ドメインのフラグメント(断片)が好ましい。免疫グロブリンまたはFc領域を備える代表的な融合体には、Fc領域を備えるsTNFRIIの融合タンパク質であるエタナーセプト(米国特許第5,605,690号)、Fc領域を備える抗原提示細胞で示されるLFA‐3の融合タンパク質であるアレファセプト(米国特許第5,914,111号)、Fc領域を備える細胞毒性Tリンパ球関連の抗原‐4(CTLA‐4)の融合タンパク質[J. Exp. Med. 181、1869(1995)]、Fc領域を備えるインターロイキン15の融合タンパク質[J. Immunol. 160、5742(1998)]、Fc領域を備える因子VIIの融合タンパク質[Proc. Natl. Acad. Sci. USA, 98,12180(2001)]、Fc領域を備えるインターロイキン10の融合タンパク質[J. Immunol. 154、5590(1995)]、Fc領域を備えるインターロイキン2の融合タンパク質[J. Immunol. 146、915(1991)]、Fc領域を備えるCD40の融合タンパク質[サージェリー、132、149(2002)]、抗体Fc領域を備えるFlt‐3(fms様チロシンキナーゼ)の融合タンパク質[Acta. Haemato., 95、218(1996)]、抗体Fc領域を備えるOX40の融合タンパク質[J. Leu. Biol., 72、522(2002)]、他のCD分子[例えば、CD2、CD30(TNFRSF8)、CD95(Fas)、CD106(VCAM‐1)、CD137]、接着分子[例えば、ALCAM(活性化白血球細胞接着分子)、カドヘリン、ICAM(細胞間接着分子)‐1、ICAM‐2、ICAM‐3]、サイトカイン受容体[例えば、インターロイキン‐4R、インターロイキン‐5R、インターロイキン‐6R、インターロイキン‐9R、インターロイキン‐10R、インターロイキン‐12R、インターロイキン‐13Rα1、インターロイキン‐13Rα2、インターロイキン‐15R、インターロイキン‐21R]、ケモカイン、細胞死誘発信号分子[例えば、B7‐H1、DR6(死受容体6)、PD‐1(プログラム化死‐1)、TRAIL R1]、同時刺激分子[例えば、B7‐1、B7‐2、B7‐H2、ICOS(誘発性共同刺激因子)]、成長要因[例えば、ErbB2、ErbB3、ErbB4、HGFR]、分化誘発因子(例えば、B7‐H3)、活性化要因(例えば、NKG2D)、信号転送分子(例えば、gp130)が含まれてる。
【0033】
免疫糖タンパク質のさらに別の例としては、抗体が含まれる。ここで「抗体」という語は、完全に組み立てられた抗体、単クローン抗体、多クローン抗体、多重特異性抗体(例えば、二重特異性抗体)、抗原を結合できる抗体断片(例えば、Fab’、F’(ab)2、Fv、単鎖抗体、ダイアボディ)、および所望の抗原結合活性を呈示する限りにおいて前述のいずれかを備える組み換えペプチドを含むように定義される。化学的に誘導される抗体を含んで、損傷のない完全な分子および/または断片の多量体または集合体が意図されている。IgG、IgM、IgD、IgA、IgE、IgG1、IgG2,IgG3、IgG4、IgA1およびIgA2を含む、あらゆるアイソタイプの分類または下位分類の抗体が意図されている。異なるアイソタイプは異なるエフェクター機能を有し、例えばIgG1およびIgG3アイソタイプは、抗体依存細胞傷害性(ADCC)活性を有する。
【0034】
「免疫グロブリン」または「自然のままの抗体」は、同一の二組のポリペプチド鎖(二つの「軽」鎖および二つの「重」鎖)からなる四量体糖タンパク質である。各鎖のアミノ端子部には、抗原を認識する主要な役割を果たす約100乃至110またはそれ以上のアミノ酸の「可変」(「V」)領域が含まれている。この可変領域内では、「超可変性の」領域または「相補性判別領域」(CDR)は、[カバトらが免疫学に関係する一連のタンパク質(Sequences of Proteins of Immunological Interest)第5版、保健サービス、国立衛生研究所、ベセズダ、メリーランド州(Public Health Service, National Institutes of Health, Bethesda, Md. )(1991) で説明したような]軽鎖可変ドメインにおける残基24−34(L1)、50−56(L2)および89−97(L3)および重鎖可変ドメインにおける31−35(H1)、50−65(H2)および95−102(H3)、および/または[チョシアらがJ. Mol. Biol. 196: 901-917 (1987) で説明したような]超可変性のループからのこれらの残基(すなわち、軽鎖可変ドメインにおける残基26−32(L1)、50−52(L2)および91−96(L3)および重鎖可変ドメインにおける26−32(H1)、53−55(H2)および96−101(H3))からなる。
【0035】
各鎖のカルボキシ端部には、一定領域が含まれる。軽鎖は、その一定領域内に単一ドメインを有する。そうして軽鎖は、可変領域を1つと一定領域ドメインを1つ有する。重鎖は、一定領域内にいくつかのドメインを有する。IgG、IgAおよびIgD抗体中の重鎖は、一定領域ドメインを3つ有しており、それらはCH1、CH2およびCH3と称され、IgMとIgE抗体における重鎖は、一定領域ドメイン4つと、CH1、CH2、CH3およびCH4を有する。従って、重鎖は、可変領域を1つと一定領域を3乃至4つ有する。
【0036】
免疫グロブリンの重鎖はまた、3つの機能領域、Fd領域(VHおよびCH1、すなわち、重鎖のN−端末領域を2つ有する断片)、ヒンジ領域およびFc領域(一定領域から得られ、ペプシン消化の後に形成される「断片結晶化」領域)に分けることができる。軽鎖と組み合わせられるFd領域が、Fabを形成する(「断片抗原結合」)。抗原は、各Fabのアミノ端末で抗原結合領域と立体化学的に反応するので、IgG分子は二価であり、すなわち、それは、抗原分子2つと結合する。Fc領域は、細胞で免疫グロブリン受容体、および補体カスケードの最初の要素と相互作用するドメインを含有する。従って、Fc断片は一般に、補体固定やFc受容体への結合のような免疫グロブリンのエフェクター機能の原因であると考えられている。
【0037】
ここで用いられている「単一クローン性抗体」という語は、実質的に均一な抗体の母集団から得られる抗体のことを称し、すなわち、その母集団に備わる個々の抗体は同一であるが、雑種細胞から生成されても、組み換えDNA技術から生成されても、可能性として自然に起こる突然変異、あるいは少量において現れる翻訳後の修飾単一クローン性抗体は除外される。単一クローン性抗体の例には、制限が無く、マウス、キメラ、ヒト化またはヒトの抗体またはそれらの変種または誘導体が含まれる。
【0038】
抗体シーケンスをよりヒト様であるようにヒト化または変更することは、例えば、ジョーンズらによる ネイチャー321: 522 525 (1986); モリソンらによるProc. Natl. Acad. Sci., U.S.A., 81: 6851 6855 (1984); モリソンおよびオオイによるAdv. Immunol., 44: 65 92 (1988); バーホエヤらによるサイエンス239: 1534 1536 (1988); パドランによる Molec. Immun. 28: 489 498 (1991); パドランによる Molec. Immunol. 31(3): 169 217 (1994); ケトルボロウ, C. A. らによるProtein Eng. 4(7): 773 83 (1991);コウ, M. S. らによる(1994), J. Immunol. 152, 2968-2976); スタドニッカらによるプロテインエンジニアリング7: 805-814 (1994) に記載されており、それら各々は、ここで援用することでその内容をここに組み入れる。
【0039】
ヒトの単一クローン性抗体を分離する1つの方法は、ファージディスプレイ技術を用いることである。ファージディスプレイは、例えば、ダワーらによるWO 91/17271, マキャファーティらによるWO 92/01047 およびケイトンとコプロウスキーによるProc. Natl. Acad. Sci. USA 87: 6450-6454 (1990) に記載されており、それら各々の内容は、援用することでここに組み入れる。ヒトの単一クローン性抗体を分離する別の方法は、内生的に免疫グロブリンを生成せず、ヒト免疫グロブリン遺伝子座を含有するよう操作されるトランスジェニックの動物を用いることである。例えば、ヤコボビッツらによるProc. Natl. Acad. Sci. USA, 90: 2551 (1993); ヤコボビッツらによるネイチャー, 362: 255-258 (1993); ブラガーマンらによるYear in Immuno., 7: 33 (1993); WO 91/10741、WO 96/34096、WO 98/24893または米国特許公報第20030194404号、米国特許公報第20030031667号、米国特許公報第20020199213号が参照され、それら各々の内容は、援用することでここに組み入れる。
【0040】
抗体フラグメント(断片)は、組み換えDNA技法、または完全な抗体を酵素でまたは化学的に分割することで生成される。「抗体断片」は、全長を保った完全な抗体の一部、好ましくは、抗原結合または完全な抗体の可変領域を備え、抗体断片から形成される多重特異性(二重特異性、三重特異性等)抗体を含んでいる。抗体断片の例には制限が無く、Fab、Fab’、F(ab’)2、Fv[可変領域]ドメイン抗体(dAb)[ワードらによるネイチャー341: 544-546, 1989]、相補性判別領域(CDR)断片、単鎖抗体(scFv)[バードらによるサイエンス242: 423-426, 1988、およびハスチョンらによるProc. Natl. Acad. Sci. USA 85: 5879-5883, 1988、選択的にポリペプチドリンカーを含む;また選択的に多重特異性である、グルバーらによるJ. Immunol. 152: 5368 (1994)]単鎖抗体断片、ダイアボディ[EP 404, 097; WO 93/11161; ホリンガーらによるProc. Natl. Acad. Sci. USA, 90: 6444-6448 (1993)]、トライアボディ、テトラボディ、ミニボディ[オラフセンらによるProtein Eng Des Sel. 2004 Apr; 17(4): 315-23]、線形抗体[ザパタらによるProtein Eng. 8(10): 1057-1062 (1995)];キレート化組み換え抗体[ネリらによるJ Mol Biol. 246: 367-73, 1995]、トライボディまたはバイボディ[スクーンジャンズらによる J Immunol. 165: 7050-57, 2000; ウィレムズらによるJ Chromatogr B Analyt Technol Biomed Life Sci. 786: 161-76, 2003]、イントラボディ[ビオッカらによるEMBO J. 9: 101-108, 1990; コルビーらによるProc Natl Acad Sci USA. 101: 17616-21, 2004]、ナノボディ[コルテツレタモゾらによるキャンサーリサーチ64: 2853-57, 2004]、抗原結合ドメイン免疫グロブリン融合タンパク質、キャメル化抗体[デスミターらによるJ. Biol. Chem. 276: 26285-90, 2001; エワートらによるバイオケミストリー41: 3628-36, 2002、米国特許公報第20050136049号および米国特許公報第20050037421号]、VHH含有抗体、ミメチボディ[米国特許公報第20050095700号および米国特許公報第20060127404号; WO 04/002424 A2; WO 05/081687 A2]またはそれらの変種または誘導体および、抗体が所望の抗原結合活性を保持する限りにおいて、CDRシーケンスのような、ポリペプチドへの特定の抗原結合を起こすのに十分な免疫グロブリンの少なくとも一部を含むポリペプチドが含まれてる。
【0041】
「変種」という語は、抗体との関係で用いられるとき、その変種が、所望のターゲット結合親和力または生物学的活性を保持するとすれば、可変領域または可変領域と等価な部分に少なくとも1つのアミノ酸が置き換えられるか削除されるか挿入されるかしている抗体のペプチドシーケンスのことを称する。加えて、本発明の抗体は、一定領域においてアミノ酸を変更して、半減期またはクリアランス、ADCCおよび/またはCDC活性を含めて抗体のエフェクター機能を変更する。そのような変更は、薬物動態を高めるか、例えば癌の治療をする上で抗体の有効性を高める。IgG1の場合、一定領域、とりわけヒンジまたはCH2領域の変更は、ADCCおよび/またはCDC活性を含めてエフェクター機能を増大または減少させる。他の例においては、IgG2一定領域が変更されて、抗体−抗原集合体の形成が低減される。IgG4の場合、一定領域、とりわけヒンジ領域の変更によって、半抗体の形成が低減される。
【0042】
「誘導体」という語は、抗体との関係で用いられるとき、治療または診断試薬への共役、(例えば、放射性核種または種々の酵素での)ラベリング、ペジレーション(ポリエチレングリコールでの誘導体化)のような共有結合高分子付着、および人工アミノ酸の化学合成による挿入または置き換えによって共有結合で変更される抗体のことを称する。本発明の誘導体は、本発明の、誘導されていない分子の結合特性を保持する。癌をターゲットとする抗体の細胞毒性試薬、例えば、放射性同位体(例えば、I131、I125、Y90およびRe186)、化学療法剤、または毒素への共役は、癌細胞の破壊を高める。
【0043】
ターゲット分子にとって「特定の」免疫糖タンパク質は、他のどのターゲットよりも大きな親和力で、そのターゲットに結合する。本発明の免疫糖タンパク質は、Kaが少なくとも約104M−1またはその代わりに少なくとも約105M−1、106M−1、107M−1、108M―1、109M−1または1010M−1というターゲットについて親和力を有する。そのような親和力は、通常の技法、例えば、BIAコアインスツルメントを用いることまたは放射能標識のついたターゲット抗原を用いる放射免疫測定によって容易に測定される。親和力データは、例えば、スキャッチャードらによるAnn N.Y. Acad. Sci., 51: 660 (1949) の方法によって分析される。
【0044】
糖修飾因子
「糖修飾因子」は、好ましくは、分子量が<1000ドルトンの、ポリペプチドに付着する炭水化物(糖)の一部である糖の付加、除去または修飾に関与する酵素の活性を抑制する小さな有機化合物である。グリコシル化は、小胞体(「コアグリコシル化」)およびゴルジ体(「端末グリコシル化」)で起こる極めて複雑なプロセスである。
【0045】
本発明においては、初期段階の糖修飾因子の活性を抑制するRNAiまたはアンチセンスを含む、グリコシル化酵素の、ポリペプチドベースまたはポリヌクレオチドベースの抑制体が有用であるが、「糖修飾因子」の定義からは除外する。
【0046】
ここで用いられるように、「初期段階の糖修飾因子」は、N‐アセチルグルコサミンをマンノースに加える前の1つ以上のグリコシル化工程のインヒビターのことを称し、ERグルコシダーゼI、ERグルコシダーゼII、ERマンノシダーゼ、ゴルジマンノシダーゼIA、ゴルジマンノシダーゼIB、ゴルジマンノシダーゼICおよびGlcNAc‐トランスフェラーゼIが含まれる。
【0047】
それに続くグリコシル化工程には、ゴルジマンノシダーゼII、GlcNAc‐トランスフェラーゼII、ガラクトシルトランスフェラーゼおよびシアリルトランスフェラーゼ、フコシルトランスフェラーゼおよびフコキナーゼが含まれる。
【0048】
代表的な糖修飾因子には、以下のいずれかが含まれる。カスタノスペルミンは、グルコシダーゼIおよびIIのインヒビターであると考えられている。デオキシフコノジリミシンは、フコシダーゼのインヒビターである。6‐メチル‐テトラヒドロ‐ピラン‐2H‐2、3、4‐トリオールは、インビトロで、L‐フコースのリン酸化、すなわち、GDP‐L‐フコースの生合成の第一工程を阻害することが報告されている。6、8a‐ジエピカスタノスペルミンは、フコシルトランスフェラーゼのインヒビターであると報告されている。1‐N‐イミュノシュガーAおよびB(それぞれ、1‐ブチル‐5‐メチル‐ピペリジン‐3、4‐ジオール塩酸塩および5‐メチル‐ピペリジン‐3、4‐ジオール塩酸塩としても知られている)は、フコシルトランスフェラーゼのインヒビターであることが報告されている。デオキシマンノジリミシン(DMJ)は、ERマンノシダーゼIのインヒビターである。キフネンシン(Kf)は、ERマンノシダーゼIのインヒビターである。スワインゾニン(Sw)は、ERマンノシダーゼIIのインヒビターである。モネンシン(Mn)は、コアオリゴ糖の伸長に干渉するERとゴルジとの間の細胞内タンパク質輸送のインヒビターである。
【0049】
本明細書に記述されているデータが示すように、さまざまなグリコシダーゼおよび/またはマンノシダーゼのインヒビターによって、ADCC活性を増大する1つ以上の所望の効果が得られ、Fc受容体の結合が増大され、グリコシル化パターンが変えられている。
【0050】
代表的な例においては、カスタノスペルミン(MW189.21)が、培地に添加され、最終の濃度を約200μM(約37.8μg/mLに相当する)とするか、濃度範囲を約10、20、30、40、50、60、70、80、90、100、110、120、130、140または150μMより大きく、約300、275、250、225、200、175、150、125、100、75、60または50μg/mLまでとする。例えば、10−50か、50−200か、50−300か、100−300か、150−250μMの範囲が意図されている。
【0051】
他の代表的な例において、DMJ、例えば、DMJ−HCl(MW199.6)が培地に加えられて、最終の濃度を約200μM(約32.6μgDMJ/mLに相当する)とするか、濃度範囲を約10、20、30、40、50、60、70、80、90、100、110、120、130、140または150μMより大きく、約300、275、250、225、200、175、150、125、100、75、60または50μg/mLまでとする。例えば、10−50か、50−200か、50−300か、100−300か、150−250μMの範囲が意図されている。
【0052】
他の代表的な例において、キフネンシン(MW232.2)が培地に加えられて、最終の濃度を約10μM(約2.3μg/mLに相当する)とするか、濃度範囲を約0.5、1、2、3、4、5、6、7、8、9または10μMより大きく、約50、45、40、35、30、25、20、19、18、17、16、15、14、13、12または11μMまでとする。例えば、1−10か、1−25か、1−50か、5−10か、5−25か、5−15μMの範囲が意図されている。
【0053】
組み換え構造物、細胞および培養方法
ここで用いられるように、「ホスト細胞」は、蚕食性ハイブリドーマを特に除外するが、グリコシル化(すなわち、ポリペプチドのアミノ酸への糖の付加)が可能であり、かつ組み換え手段を通して修飾され、レベルの増大したタンパク質プロダクトを発現させる他の細胞を含む。組み換え修飾とタンパク質プロダクトを発現する能力を保持するホスト細胞の子孫も「ホスト細胞」という語の中に含まれる。
【0054】
発現ベクトルまたは規定シーケンスの代表的な要素には、複製の元になるもの、プロモーター、オペレーターまたは転写および翻訳の媒体となるその他の要素が含まれる。プロモーターは、構成性であるか活性であり、かつさらに細胞種特異であるか、組織特有であるか、個々の細胞に特異有であるか、イベント特異であるか、一時的特異であるか、誘導性である。イベントに特異なプロモーターは、イベントが起こるときのみに活性であるか上方制御される。プロモーターに加えて、抑制体シーケンス、負の調節遺伝子または組織に特有のサイレンサーが挿入されて特異でない発現を低減する。他の要素には、内部リボゾーム結合サイト、ポリアデニル化シーケンスを含む転写ターミネータシーケンス、スプライスドナーおよび受容体サイト、エンハンサー、選択可能なマーカー等が含まれる。
【0055】
培地には、当業界で知られている必要なまたは望ましい成分をいずれも含み得るが、例えば、グルコースを含む糖類、必須および/または非必須アミノ酸、脂質および脂質前駆体、核酸前駆体、ビタミン、無機塩、希少金属を含む微量要素および/または細胞成長因子が含まれる。培地は、化学的に定義され、漿液、植物加水分解物またはその他の誘導された物質が含まれる。培地は、実質的にまたは完全に漿液や動物要素が含まれない。「実質的に漿液が含まれない」は、媒体には漿液が欠如しているか、わずかな量の漿液しか含んでいないことを意味する。細胞培養の間に枯渇される代表的な補遺のアミノ酸には、アスパラギン、アスパラギン酸、システイン、シスチン、イソロイシン、ロイシン、トロプトファンおよびバリンが含まれる。
【0056】
商業的に入手できる脂質および/または脂質前駆体には、コリン、エタノールアミンまたはホスホエタノールアミン、コレステロール、オレイン酸のような脂肪酸、リノール酸、リノレン酸、メチルエステル、例えば、酢酸塩の形でのD‐アルファ‐トコフェノール、ステアリン酸、ミリスチン酸、パルミチン酸、パルミトオレイン酸、またはアラキノン酸が含まれる。必須アミノ酸には、アルギニン、ヒスチジン、イソロイシン、ロイシン、リジン、メチオニン、フェニルアラニン、スレオニン、トリプトファンおよびバリンが含まれる。非必須アミノ酸には、アラニン、アスパラギン、アスパラギン酸塩、システイン、グルタミン酸塩、グルタミン、グリシン、プロリン、セリンおよびチロシンが含まれる。適当な塩として供給され、商業的に入手できる無機または微量要素には、ナトリウム、カルシウム、カリウム、マグネシウム、銅、鉄、亜鉛、セレニウム、モリブデン、バナジウム、マンガン、ニッケル、ケイ素、錫、アルミニウム、バリウム、カドミウム、クロム、コバルト、ゲルマニウム、カリウム、銀、ルビジウム、ジルコニウム、フッ化物、臭化物、ヨウ化物および塩化物が含まれる。媒体にはまた、混合や通気から細胞が保護されるように、非イオン性界面活性剤または表面活性剤が選択的に含まれる。培地はまた、重炭酸ソーダ、一塩基性および二塩基性リン酸塩、HEPSおよび/またはトリスのような緩衝剤を含む。培地はまた、酪酸ナトリウムまたはカフェインのようなタンパク質生成の誘発剤を含む。
【0057】
本発明によって、免疫糖タンパク質を生成する方法もまた提供されるが、任意の培地において、またはここに記述されるいずれかの条件のもとで、ホスト細胞を培養することを含むものである。当該方法はさらに、ホスト細胞または培地から免疫糖タンパク質を調製する工程を含む。糖修飾因子は、最初の培地に含まれているか、または最初の成長段階またはその後の段階で添加される。組み換えタンパク質が媒体中に分泌されるとき、培地を周期的に取り出すことができ、また何回かの取り出しサイクルを経て新しい培地と置き換えることができる。
【0058】
治療用のタンパク質生成に広く用いられているCHO細胞が好ましいけれども、グリコシル化タンパク質を生成することが当業界で知られているいずれのホスト細胞を用いてもよく、これには、酵母細胞、植物細胞、植物、昆虫細胞およびほ乳類の細胞が含まれる。代表的な酵母細胞には、ピキア属(Pichia)、例えば、P. パストリス(Pastoris)およびサッカロミセス属(Saccharomyces)、例えば、S. セレビシエ(cerevisiae) 並びにシゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)、 クリベロミセス属(Kluyveromyces)、 K. ザクティス(Zactis)、 K. フラギリス(fragilis)、 K.ブルガリカス( bulgaricus)、 K. ウィッケラミイ(wickeramii)、 K. ワルティイ(waltii)、 K. ドロソフィララム(drosophilarum)、 K.(サルノトレランス) thernotolerans、 K.(マルキアヌス) marxianus、 K. (ヤルロウィア)yarrowia、 トリコデルマ・リージア(Trichoderma reesia)、 アカバンカビ(Neurospora crassa)、シュワンニオミセス( Schwanniomyces)、 シュワンニオミセス オッシデンタリス(Schwanniomyces occidentalis)、ニューロスポラ( Neurospora)、 ペニシリウム属(Penicillium)、 トチポクラディウム(Totypocladium)、 アスペルギウス属(Aspergillus)、 アスペルギウスニジュランス(A. nidulans)、 クロコウジカビ(A. niger)、 ハンゼヌラ(Hansenula)、 カンジダ属(Candida)、 クロエケラ属(Kloeckera)、トルロプシス( Torulopsis)およびロドトルラ属(Rhodotorula)が含まれる。代表的な昆虫細胞には、オートグラファカリフォルニカ(Autographa californica)およびスポドプトーラフルギペルダ( Spodoptola frugiperda)並びにショウジョウバエ属(Drosophila)が含まれる。代表的なほ乳類の細胞には、CHO、BHK、HEK-293、NS0、YB2/3、SP2/0の変種およびPER-C6またはHT1080のようなヒト細胞、並びにVERO、HeLa、COS、MDCK、NIH3T3、Jurkat、Saos、PC-12、HCT 116、 L929 、Ltk-、WI38、CV1、TM4、W138、Hep G2、MMT白血病細胞系、胚幹細胞または受精卵細胞が含まれる。
【0059】
細胞は、当業界で知られているいかなる培養システムにおいても、かついかなる方法で培養されてもよく、それには、T‐フラスコ、スピナーおよび振とうフラスコ、ローラーボトル、撹拌タンクバイオリアクターが含まれる。付着依存性細胞もまた、撹拌タンクバイオリアクターに懸濁して保持されるマイクロキャリア、例えば、高分子球体で培養することができる。あるいは、細胞は、単細胞懸濁液において成長させることができる。培地が添加されるのは、バッチ処理であって、例えば、培地が単一のバッチで細胞に一度に添加されても良いし、培地の小さなバッチが周期的に添加される流加バッチ処理であっても良い。培地は、培養の終わりに回収されるか、培養中に何度かにわたって取り出される。継続的に潅流させる生成処理もまた当業界で知られており、新しい培地を、同じ量でリアクターから継続的に引き出しながら、培養に継続的に供給することが伴われる。潅流による培養は、一般にバッチによる培養よりも細胞密度が高く、繰り返して回収しながら何週間もまたは何ヶ月も維持できる。
【0060】
免疫糖タンパク質の使用
本発明の免疫糖タンパク質は、ターゲット分子が媒体となる疾患を処置する治療術として有用であり、例えば、発現したまたは細胞表面と結びついたターゲット分子を有する癌細胞を死滅させるための細胞崩壊剤として有用である。
【0061】
「処置」または「処置する」は、治療または予防または防止処置のことを言う。治療は処置を受ける被体の疾患の少なくとも1つの症状が良好となるか、被体において進行する疾患の悪化を遅延させ、あるいは関連する疾患が新たに発症することを防止する。改善された効果は、疾患状態について、当業界で周知の臨床基準を評価することで、査定される。
【0062】
免疫糖タンパク質の「治療で有効なドース」または「有効なドース」は、処置される疾患の1つ以上の症状が結果的に良くなるのに十分な化合物の量のことを称する。個人に活性成分を単独で投与するとき、治療に有効なドースは、その成分のみについて称する。組み合わせて投与されるとき、治療で有効なドースは、連続して投与されようとも同時に投与されようとも、結果的に治療の効果が得られる活性成分を合わせた量のことを称する。ドースは、患者の体重に基づいて投与され、例えば0.01乃至50mg/kgのドースであって、日単位または週単位で、あるいは二週間毎、三週間毎または月に一度投与される。
【0063】
本発明の免疫糖タンパク質をヒトまたは実験動物に投与するのに、組成物が非経口的な投与のためのものであるならば、1つ以上の薬学的に許容され得る担体または希釈剤、好ましくは、殺菌されたまたは無菌の担体また希釈剤を有する組成物において、分子を調製するのが好ましい。「薬学的にまたは薬理学的に受け入れ可能な若しくは許容され得る」という文言は、以下に記述されるような当業界で周知の経路を用いて投与されるとき、アレルギーまたはその他の有害な反応を引き起こさない分子状態および組成物のことを称する。「薬学的に受け入れ可能な担体」には、臨床的に有用な溶剤、分散媒体、被覆、抗菌性剤および抗真菌性剤、等張剤および吸収遅滞剤等が全て含まれる。一般に、組成物にはまた、発熱物質並びに被体に有害であり得るその他の不純物は実質的に含まれない。
【0064】
免疫糖タンパク質が投与されるのは、経口であっても、局部的であっても、経皮であっても、非経口であっても、吸入スプレーによっても、経膣であっても、頭蓋内注射によってでも良い。ここで用いられる非経口という語には、皮下注射、静脈内、筋肉内、嚢内への注射または注入技法が含まれる。静脈内、皮膚内、筋肉内、乳房内、腹腔内、髄膜内、眼球後方、肺内への注射および/または特定の場所への手術での注入による投与も同様に意図されている。
【0065】
1つの例において、処置を必要とする癌または病気にかかった組織の箇所に、その箇所への直接の注射によるか、製剤を内部に放出できる、持続的な放出法または持続的な吐出メカニズムを介して投与が行われる。例えば、組成を持続して放出可能な生分解性の微小球またはカプセルその他の生分解性高分子構成物(例えば、可溶ポリペプチド、抗体または小分子)は、癌の近辺に注入される本発明の製剤に含ませることができる。
【0066】
また治療組成物を、患者の複数の箇所に放出しても良い。複数の投与が同時に行われても、また継続した期間内に投与されても良い。
【0067】
特に、水溶液の注入が好ましい。水性の組成物は、真空状態で凍結乾燥して貯蔵され、使用に先立って適切な担体で復元できる。この技法は、従来の免疫グロブリンで有効であることが示されている。真空状態で凍結乾燥し、復元する技法は、適切であれば任意のものを用いることができる。当業者には、真空状態で凍結乾燥して復元すると、活性の損失の度合いがさまざまであり、それを補償するために使用のレベルを調節しなければならないということが理解される。
【0068】
あらゆる場合において、製剤は、殺菌され、かつ注射器での扱いが容易である程度に液化されていなければならない。例えば、レシチンのような被覆を用いること、分散している場合には必要な粒子のサイズを維持すること、および界面活性剤を用いることによって、適切な流動性を維持することができる。また、製造および貯蔵する条件のもとで安定していなければならず、バクテリアや菌類のような微生物の汚染作用を防いで保存される。微生物の作用は、さまざまな抗菌性剤および抗真菌性剤、例えば、パラベン、クロロブタノール、フェノール、ソルビン酸、チメロサール等によって防ぐことができる。多くの場合、等張剤、例えば、砂糖または塩化ナトリウムを含むことが望ましい。
【0069】
さらに、本発明での使用が意図されている組成物は、親水性と疎水性のバランスが良好であり、これによりインビトロおよびインビボの使用、特にインビボでの使用について実用性が高いが、他の組成物は、このようなバランスを欠いており実用性が相当に低い。とりわけ、本発明での使用が意図されている組成物は、身体での吸収および生物学的利用を可能とする水性媒体に可溶である度合いが適切であり、また一方、化合物が、細胞膜を横切って作用すると推定される箇所に至ることを可能とする程度に脂質にも可溶である。
【0070】
また、本発明で意図されているのは、免疫糖タンパク質組成物を第二の薬剤と一緒に投与することである。
【0071】
さらに、本発明には、本発明の方法を実施するために、1つ以上の化合物または組成物が容易に使用できるようにパッケージとなった状態で、それらを備えるキットまたは製品が含まれる。1つの例において、そのようなキットには、選択的に第二の治療薬と一緒に、密封された瓶または器のような容器にパッケージされた状態で、本発明における免疫糖タンパク質が含まれており、その容器にはラベルが貼られているか、該方法を実施する上での、化合物または組成物の使用について記述するパッケージに納められている。好ましくは、その化合物または組成物は、ドース単位の形でパッケージされている。そのキットにはさらに、特定の投与経路によりその組成物を投与するのに適切な、またはスクリーニングアッセイを行うのに適切な装置が含まれる。好ましくは、当該キットには、該組成物の使用について記述されたラベルが入っている。
【0072】
さらに本発明では、免疫糖タンパク質が結びつくターゲットにより特徴付けられるか媒介される被体における、疾患、病状または障害を抑制、防止または処置するための医薬の製造で、本発明の免疫糖タンパク質を用いることが意図されている。
【図面の簡単な説明】
【0073】
【図1】図1は、様々な濃度のカスタノスペルミンを備える細胞媒体において成長するTRU‐016を発現するCHO細胞の成長を、cells/mlという細胞数で示す。
【図2】図2は、様々な濃度のカスタノスペルミンを備える細胞媒体において成長するTRU‐016を発現するCHO細胞の生存度を、生存細胞の%で示す。
【図3】図3は、様々な濃度のカスタノスペルミンの存在で培養される細胞が生成するTRU‐015のCD16結合を示し、かつ幾何学的に示された平均蛍光強度対カスタノスペルミン濃度を示す。
【図4】図4は、様々な濃度の6、8a‐ジエピカスタノスペルミン、スワインソニンまたはデオキシマンノジリミシン(DMJ)の存在で培養される細胞が生成するTRU‐016のCD16結合を、幾何学的に示された平均蛍光強度で示す。
【図5】図5は、様々な濃度のキフネンシンの存在で培養される細胞が生成するTRU‐016のCD16結合を、蛍光強度の平均で示す。
【図6】図6は、様々な濃度のカスタノスペルミンの存在で培養される細胞が生成するタンパク質Aが取り除かれたTRU‐016のCD16結合を、蛍光強度の平均で示す。
【図7】図7は、高親和性および低親和性ドナーのPBMCを用いて測定したTRU‐15のADCCを示し、添加TRU‐015の濃度対%特有の致死量をプロットした図である。
【図8】図8は、高親和性および低親和性ドナーのPBMCを用いて測定したTRU‐15のADCCを示し、添加TRU‐015の濃度対%特有の致死量をプロットした図である。
【図9】図9は、様々な濃度のカスタノスペルミンの存在で培養される細胞が生成するTRU‐016のADCCを示し、%特有の致死量対添加TRU‐016の濃度をプロットした図である。
【図10】図10は、様々な糖修飾因子の存在で培養される細胞が生成するTRU‐016のADCCを示し、%特有の致死量対添加TRU‐016の濃度をプロットした図である。
【図11】図11は、様々な糖修飾因子の存在で培養される細胞が生成するTRU‐016を投与されたマウスにおける薬物動態データを示す。
【図12】図12は、様々な糖修飾因子で処置した細胞が生成するTRU‐016を投与されたマウスの漿液におけるTRU‐016のCD16結合を示す。
【図13】図13は、腫瘍細胞をインプランとされ、様々な糖修飾因子で処置した細胞または処置していない細胞から生成されるTRU‐016を投与されたマウスにおいて8日後の腫瘍の相対容量を示す。
【図14】図14は、腫瘍細胞をインプラントされ、様々な糖修飾更因子で処置した細胞または処置していない細胞から生成されるTRU‐016を投与されたマウスの生存率%を示す。
【図15】図15は、カスタノスペルミンの存在で培養された細胞が生成するTRU‐015のCDCを示し、ヨウ化プロピジウムでポジティブ(正)の%(死亡細胞)対TRU‐015実験タンパク質の濃度をプロットした図である。
【図16】図16は、様々な糖修飾因子の存在で培養された細胞が生成するTRU‐016のCDCを示し、ヨウ化プロピジウムでポジティブ(正)の%(死亡細胞)対TRU‐016実験タンパク質の濃度をプロットした図である。
【図17】図17は、カスタノスペルミン濃度範囲に対する、TRU‐016に特異なタンパク質生成を示す。
【図18】図18は、カスタノスペルミン濃度範囲に対する、TRU‐016のCD37およびFcγRIIIa(CD16)への同時結合についてのアッセイ結果を示す。
【図19】図19は、カスタノスペルミン濃度範囲に対する、TRU‐016からCD37を発現する細胞の用量反応結合曲線を示す。
【図20】図20は、カスタノスペルミン濃度範囲に対する、TRU‐016のADCC活性曲線を示す。
【発明を実施するための形態】
【0074】
例
例1
SMIPプロダクトの製造
TRU‐016
CD37‐特有のSMIPが、共同で所有されている米国特許出願第10/627,556号および米国特許公開第2003/133939号、米国特許公開第2003/0118592号および米国特許公開第2005/0136049号に記述されており、それら各々は、援用することでその全体をここに組み入れられる。代表的なSMIP、すなわちTRU‐016は、以下に記載されるようにして生成される。
【0075】
TRU‐016[G28-1 scFv VH11S(SSC-P)H WCH2 WCH3]は、CD37抗原に結合する組み換え単鎖タンパク質である。TRU‐016のヌクレオチドおよびアミノ酸配列は、それぞれSEQ ID NOS: 1および2で並べられている。結合ドメインは、前段落に列挙されている特許公報において以前開示されたG28‐1抗体シーケンスに基づいた。結合ドメインは、モディファイされたヒンジ領域を通して、エフェクタードメイン、すなわち、ヒトIgG1のCH2およびCH3ドメインに接続されている。TRU‐016は、溶液において二量体として存在する。
【0076】
TRU‐016は、チャイニーズハムスター卵巣(CHO)ほ乳類細胞出現システムにおいて、組み換えDNA技術によって生成される。TRU‐016SMIPは、タンパク質A親和クロマトグラフィーでCHO培養上清から精製される。dPBSを用いると、50mLのrタンパク質A FFセファロースカラム(GEヘルスケアrタンパク質AセファロースFF、カタログ#17‐0974‐04)は、1.5カラム体積(CV)について5.0mls/min(150cm/hr)で平衡となる。AKTAエクスプローラ100Air(GEヘルスケアAKTAエクスプローラ100Air、カタログ#18‐1403‐00)を用いて、培養上清が1.7mls/minという流速でrタンパク質AセファロースFFカラムに加えられ、組み換えTRU‐016を捕捉する。そのカラムは、5カラム容量(CV)分のdPBSで洗浄され、そして1.0M NaCl、20mMリン酸ナトリウム、pH6.0で、さらに25mM NaCl、20mM NaOAc、pH5.0で洗浄される。これらの洗浄工程により、不特定に結合されたCHOホスト細胞タンパク質が、溶出後の生成物沈殿に貢献するrタンパク質Aカラムから取り除かれる。
【0077】
組み換えTRU‐016は、100mMグリシン、pH3.5を備えるカラムから溶出される。その溶出された生成物の一部である10mLが回収され、そしてその溶出された生成物が、0.5M 2‐(N‐モルフォリノ)エタンスルフォン酸(MES)pH6.0の溶出した容量の20%で、pH5.0とした。この溶出された生成物は、約25mg/mLのTRU‐016に濃縮されて濾過殺菌される。
【0078】
そして、精製されたタンパク質がGPCサイズ排除クロマトグラフィー(SEC)にかけられ、より高い分子量の集合体からTRU‐016(二量体)分子をさらに精製する。dPBSを用いて、1Lのスーパーデックス200FFセファロースを含むXK50/100カラム(GEヘルスケアXK50/100からのクロマトグラフィーカラム、カタログ#18‐8753‐01)が、1.5カラム容量(CV)について12.6mls/min(38cm/hr)で平衡となる。サンプルの54mls(3%CV)という最大容量が、カラムに適用される。カラムは、12.6ml/minで操作され、容出されたタンパク質は、40mlの画分に分けられる。各画分は、分析HPLCを用いて生成物品質について分析され、そして溶出された部分は、>95%POI(集合体になっていない)TRU‐016についてプールされる。この得られたプール物が、0.22μmで濾過殺菌される。そして、物質が、20mMリン酸ナトリウムと240mMショ糖で、pH6.0で濃縮されて形成される。
【0079】
グリコバリアントを精製する代わりの方法は、以下の通りである。TRU‐016が、タンパク質A親和クロマトグラフィーによってCHO培養上清から精製される。dPBSを用いて、1mL MabSelect親和クロマトグラフィーカラム(GEヘルスケアHitrap MabSelect、カタログ#28‐4082‐53)が、7カラム容量(CV)について1.0mL/minで平衡とされる。培養上清が、Aktaエクスプローラ100Air(GEヘルスケア、Aktaエクスプローラ100Air、カタログ#18‐1403‐00)を用いて流速1.0mL/minでMabSelectカラムに加えられ、組み換えTRU‐016を捕捉する。そのカラムは、20CV分のdPBSで洗浄され、そして20mMリン酸ナトリウム、5CV分の1.0M NaCl、pH7.0でそして3CV分のdPBSで洗浄する。
【0080】
組み換えTRU‐016が、10mMクエン酸塩(シトレート)、pH3.5でカラムから溶出され、そのカラムは、8CV分の10mMシトレート3.0でストリップされる。ストリップに続いてそのカラムはdPBSで5CV分再平衡とされる。タンパク質は、溶出の間、各画分に集められて、それらは吸着度に基づいてプールされ、このプールされた物質は、溶出物5mLにつき約400μLの0.55M 2‐(N‐モルフォリン)エタンスルフォン酸(MES)、pH6.0を加えてpH5.0とされる。この中和された溶出物は、濾過殺菌されて、活性アッセイとプロセス分析アッセイの双方に課される。
【0081】
親抗体のCD37細胞表面受容体への結合特異性がTRU‐016において保存されるということを確認するために実験が行われた。ヒトPBMCが、LSM密度勾配を持って単離され、非抱合TRU‐016およびPE抱合抗ヒトCD19でインキュベートされる。細胞は洗浄され、45分間氷上にて1:100FITC GAH IgG(Fc特異)でインキュベートされる。細胞は、洗浄され、セルクエストソフトウェアを用いるFACsCaliburインスツルメントで二色フロー血球計算によって分析される。細胞は、CD19染色によって、Bリンパ球または非Bリンパ球について振り分けられる。
【0082】
TRU‐016の濃度が増大すると、Bリンパ球でのFITC信号(CD19正ゲート)が、0.01‐1.0μg/mlから、約1μg/mLでの飽和または平均蛍光強度(MFI)1000に至るまで急速に増大する。反対に、非Bリンパ球数は、染色が検出可能であるが非常に低いので、scFvIgの濃度が増大すると、ゆっくりと増大する。
【0083】
TRU‐015
CD20‐特異のSMIPが同様に調製される。CD20‐特異のSMIPは、共同で所有される米国特許公報2003/133939、2003/0118592および2005/0136049において記述され、各々は、援用することでその全体をここに組み入れられる。代表的なSMIP、すなわち、TRU‐015を以下に記載する。
【0084】
TRU‐015は、CD20抗原に結合する組み換え単鎖タンパク質である。TRU‐15のヌクレオチドとアミノ酸シーケンスは、それぞれSEQ ID NOS:3および4で並べられる。結合ドメインは、公的に入手可能なヒトCD20抗体シーケンスに基づいた。結合ドメインは、修飾されたCSSヒンジ領域を通して、エフェクタードメイン、すなわち、ヒトIgG1のCH2およびCH3ドメインに接続される。TRU‐015は、溶液において二量体として存在する。
【0085】
TRU‐015は、SEQ ID NO:4のアミノ酸1−23からの2e12リーダーペプチドクーロン化シーケンスと、SEQ ID NO:4におけるポジション34で反映される、可変領域における残基11でリジンからセリン(VHL11S)アミノ酸が置き換えられた2H7ネズミ抗ヒトCD20軽鎖可変領域と、SEQ ID NO:4の残基129で始まるasp-gly3-ser-(gly4ser)2リンカーと、重鎖領域の端にセリン残基が無く、すなわち、VTVSSからVTVSに変化した2H7ネズミ抗ヒトCD20重鎖可変領域と、(CSS)シーケンスとワイルド型CH2およびCH3ドメインを備えるモディファイされたヒンジ領域を含むヒトIgG1 Fcドメインとを備える。
【0086】
例2
糖修飾因子(モディファイア)を有するホスト細胞の培養
TRU‐016またはTRU‐015 cDNAがトランスフェクトされたCHO細胞は、一般に以下に記述される手順によって、種々の糖修飾因子(modifier)について濃度を変えて、振とうフラスコまたは揺動バッグで培養された。
【0087】
振とうフラスコでの作業については、ログ位相ホスト細胞がテストすべき濃度の糖修飾因子とともに100,000 cells/mlで接種され、また選択的に50nMのメトトレキセートとともに接種された。
【0088】
細胞は、t=0で1350mLのEx-Cell 302培地(SATCバイオサイエンス;全てがインビトロゲンからの非必須アミノ酸、ピルケート、L‐グルタミン、ペン/ストレップ、HTサプリメントおよびインシュリンを加えて)に3x10E6/mLで接種され、T>=72時間で合計容量5Lとされた。細胞は、37℃でおよび5%の二酸化炭素で培養され、6−7日目から毎日成長および生存度についてモニターした。上清は、典型的には10−12日目に、細胞の生存度が60%未満に落ちたとき回収された。
【0089】
Naアジドを加えて0.02%とし、細胞を遠心で取り除き、かつ上清を0.22μMフィルターを通して濾過殺菌した。他の例に記述されるアッセイは、示された通りに、上清において行われるものもあり、また他のアッセイは、さらなるタンパク質A精製を経た材料に対して行われるものもあった。揺動バッグの作業について、ログ位相ホスト細胞が、テストされるべき濃度の糖修飾因子(モディファイア)ともに10−20%という条件のEx-Cell 302媒体(SATCバイオサイエンス;全てがインビトロゲンからの非必須アミノ酸、ピルケート、L‐グルタミン、ペン/ストレップ、HTサプリメントおよびインシュリンを加えて)において、100,000−200,000 cells/mLで5L揺動バッグ中に接種された。細胞は、37℃でかつ5%の二酸化炭素で培養され、成長と生存度について毎日モニターされた。上清は、典型的には、11−12日目に、または細胞生存率が50%未満に低下したとき回収された。
【0090】
ソルバールレジェンド(Sorvall Legend)における3000rpm(1932rcf)での20分間の遠心によって細胞が取り除かれ、上清は濾過殺菌された。他の例に記述されるアッセイは、示されたように、上清において行われたものもあるが、他のアッセイは、さらなるタンパク質A精製を受けた材料において行われた。
【0091】
種々の糖修飾因子の濃度を変えて培養される細胞により生成されるTRU‐016は、以下に記述されるように、CD16結合、ADCC、CDC、薬物動態パラメータ、およびインビボの活性についてアッセイされる。
【0092】
図1および図2は、典型例を示しており、1000μMまでの濃度の糖修飾因子カスタノスペルミンでの処置は、サンプルを採取した期間全体を通して(144時間まで)、細胞数やパーセント細胞生存度に影響を与えなかったということが示されている。
【0093】
例3
FcRsへの結合
例2により生成される免疫糖タンパク質を、受容体の細胞外ドメインがネズミIgG2a Fcに融合する、Fcγ受容体の可溶性Ig‐融合バージョンへの結合について、インビトロでアッセイした。
【0094】
可溶性Fcγ受容体材料は、Fcγ受容体I(Genbank Acc. No. BC032634)、IIa(Genbank Acc. No. NM_021642)、IIb(Genbank Acc. No. BC031992)およびIII‐V158(高親和対立遺伝子)(Genbank Acc. No. X07934)およびIII‐F158(低親和対立遺伝子)それぞれの細胞外ドメインを、残基238(MIgG2aP238S)にPro乃至Serの突然変異があるネズミIgG2a Fcに融合することによって生成された。FcγRIII(CD16)の双方の形について、HE4リーダーは、CD16アミノ酸1‐178でクローン化され、そしてMIgG2aP238Sに融合される。
【0095】
アッセイは以下のように行われた。1%ウシ胎仔漿液(FBS)を含むリン酸塩で緩衝された生理食塩水(PBS)において、45分間、5μg/mlのTRU‐015かTRU‐016かを入れたコスター96ウェルプレートにおいて氷上で500,000のWIL2‐S細胞(表面上においてCD37並びにCD20を発現するBリンパ腫細胞系(line))が培養された。結合されていないTRU‐015またはTRU‐016は、細胞をスピンし、希釈剤(PBS+1%FBS)で洗浄し、ソルバールレジェンド RTにおいて1200rpmで再度スピンすることで取り除かれた。そして細胞は、45分間、氷上で1μg/mlの濃度の同じ希釈剤において、所望のFcγR‐MIgと融合して培養された。
【0096】
そして、錯体(WIL2-S細胞/SMIP/FcγR-MIg)が、1:100の希釈の、PE抱合AffiniPure F(Ab’)2 ヤギ抗マウスIgG[ジャクソンイミュノリサーチ(Jackson Immunoresearch)](ヒトFcと最小の交差反応性を有するマウスFc‐特異の抗体)で培養された。細胞は、セルクエストソフトウェア(ベクトンディキンソン)を用いるFACsCaliburでの一色フロー血球計算によって分析された。
【0097】
このアッセイで、精製されたTRU‐016タンパク質の代わりに例2のTRU‐016上清が用いられるときは、上清のSMIP濃度は、TRU‐016基準に沿って、希釈上清でWIL2‐S細胞を直接染色することによって定量化された。TRU‐016は、1:50希釈のFITC抱合F(Ab’)2ヤギ抗ヒト(ガンマ)[Caltag H10101]で染色することによって検出された。
【0098】
低親和対立遺伝子か高親和対立遺伝子への結合が、ADCC活性に同じように相関するように測定された。CD16(低または高親和対立遺伝子)結合の増大がADCC活性の増大と相関していた。
【0099】
典型的な結果を、図3乃至図6に示す。
【0100】
0、2、5、10、30または100μg/mLのカスタノスペルミンを含む媒体において培養されたCHO細胞により生成されるTRU‐015精製のタンパク質がCD16結合についてテストされた(低親和対立遺伝子)。幾何平均蛍光強度の典型的な結果が図3に示されており、培地におけるカスタノスペルミンの濃度が増大すると、CD16結合が、用量依存的に増大することが示されている。
【0101】
50または250μMという濃度で6、8a‐ジエピカスタノスペルミンを、50または250μMという濃度でスワインソニンをまたは50または250μMという濃度でデオキシマンノジリミシン(DMJ)を含む培地で培養されたCHO細胞が生成したTRU‐016上清が、CD16結合のためにテストされた。平均蛍光強度の典型的な結果が図4に示されており、DMJのいずれの濃度もCD16結合を増大させたことが示されている。これらの濃度では、6,8a‐ジエピカスタノスペルミンまたはスワインソニンについては効果が見られないが、精製されたタンパク質でさらなるテストが行われて効果を測定する。
【0102】
0、0.5、1、3、5または10μMという濃度のキフネンシンを含む培地で培養されたCHO細胞が生成するTRU‐016上清が、CD16結合についてテストされた。平均蛍光強度の典型的な結果が図5に示されており、キフネンシンは、CD16結合を増大させるのにDMJよりずっと大きな効力があり、濃度が最低の0.5μMであったときでさえ、CD16結合を大きく増大させたことが示されている。
【0103】
0、10、25、50、100または200μMのカスタノスペルミンを含む培地で培養された、CHO細胞により生成されるタンパク質A精製TRU‐016が、CD16結合についてテストされた。平均蛍光強度の典型的な結果が図6に示されており、培地におけるカスタノスペルミンの濃度が増大するとCD16結合が用量依存的に増大することが示されている。
【0104】
例4
ADCC活性
精製されたTRU‐016のADCC活性を測定するために、標識化したBJAB B細胞をターゲットとして用い、ヒト末梢血単核細胞(PBMC)をエフェクター細胞として用いた。IMDM/10%FBSにおいて37℃で二時間、BJAB B細胞(107個の細胞)が、500μCi/mL51Crクロム酸ナトリウムで標識付けされた。PBMCは、リンパ球分離メディア(LSM、ICNバイオメディカル)勾配について分画することで、ヘパリン処理したヒト全血から単離された。10%FBSをともなってRPMI培地に試薬サンプルが加えられ、各試薬の一連の希釈物が調製された。51Crで標識付けされたBJABを、2x104cells/wellで加えられた。そして、PBMCが、25:1のエフェクター(PBMC):ターゲット(BJAB)の最終比について5x105cells/wellだけ加えられた。反応は96ウェルプレートの四つ組ウェルで提供された。TRU‐016の一連の希釈物が、図に示されるように、10ng/mLから20μg/mLの範囲の最終濃度でウェルに加えられた。回収したり計数したりするのに先立って、5%CO2において37℃で6時間反応を進行させた。放出されたCMPが50μlの乾燥した培養上清からPackard TopCountNXTで測定された。パーセント特有の死亡率が減算(サンプルのcpm[4倍のサンプルの平均]‐cpm自発放出)/(cpm最大放出‐cpm自発放出)x100によって計算され、データが%特有の死亡率対TRU‐016濃度としてプロットされた。
【0105】
典型的な結果が、図7乃至図10に示されている。
【0106】
0、2、5、10、30または100μg/mLのカスタノスペルミンを含む培地で培養されたCHO細胞が生成するTRU‐015精製タンパク質が、高親和(V/V158)および低親和(F/F158)CD16ドナーからのPBMCを用いて測定されたADCCについてテストされた。%特有の死亡率の典型的な結果が、図7および図8(それぞれ、高親和ドナーおよび低親和ドナー)に示されており、培養媒体におけるカスタノスペルミンの濃度が増大すると、ADCC活性が用量依存的に増大することが示されている。
【0107】
0、10、25、50、100または200μMのカスタノスペルミンを含む媒体で培養されたCHO細胞が生成するTRU‐016精製のタンパク質が、ADCCについてテストされた。%特有の死亡率の典型的な結果が図9に示されており、培地におけるカスタノスペルミンの濃度が増大するとき、ADCC活性が用量依存的に増大することが示されている。
【0108】
200μMのDMJ、10μMのキフネンシンまたは200μMのカスタノスペルミンを含む培地で培養されたCHO細胞により生成されるTRU‐016精製のタンパク質が、ADCCについてテストされた。%特有の死亡率の典型的な結果が図10に示されており、糖修飾因子の、これらの濃度の全てについて、CHO細胞が生成する免疫糖タンパク質のADCCが改良されたということが示されている。
【0109】
例5
CDC活性
例2により生成されるTRU‐016精製のタンパク質のCDC活性を測定するために、ラモスB細胞が、75μlに5x105cells/wellでイスコベス(ギブコ/インビトロゲン、グランドアイランド、NY)に懸濁された。指示された二倍の濃度でTRU‐016(75μl)が細胞に加えられた。漿液を含まないイスコベスにおいて遠心分離および洗浄を行うのに先立ち、45分間結合反応を進行させた。種々の濃度のヒト漿液(補体を含む)を備えるイスコベスにおいて細胞が再懸濁された。細胞は、37℃で60分間培養された。細胞は、遠心分離によって洗浄され、0.5μg/mlのヨウ化プロピジウムを備える染色媒体において再懸濁された。FACsCaliburおよびCellQuestソフトウェアを用いるフロー血球計算による分析に先立ち、暗所において室温で15分間サンプルが培養された(ベクトンディキンソン)。
【0110】
未処置のCHO細胞が生成したTR‐015精製タンパク質または30μg/mlのカスタノスペルミンで処置されたCHO細胞が、CDC活性についてテストされた。結果を図15に示す。
【0111】
未処置のCHO細胞が生成するTRU‐016精製のタンパク質、または200μMのDMJ、10μMのキフネンシンまたは200μMのカスタノスペルミンを含む媒体で培養されたCHO細胞が、CDC活性についてテストされた。結果を図16に示す。
【0112】
これらの結果により、糖修飾TRU‐015またはTRU‐016についてのCDCは、未処置のCHO細胞が生成する、対応するタンパク質のCDCに類似していたことが示され、ホスト細胞の培地に糖修飾因子が存在することで、ホスト細胞が生成する免疫糖タンパク質のCDCに大きな影響が及ぼされなかったことを示している。
【0113】
例6
薬物動態プロフィール
雌のBALB/cマウスに200μgのTRU‐016テストタンパク質(未処置のCHO細胞が生成したTRU‐016または200μMのDMJ、10μMのキフネンシンまたは200μMのカスタノスペルミンで処置したCHO細胞が生成したTRU‐016)を時刻0で静脈注射した。注射後15分、2、6、24、48、72、96および192時間で漿液のサンプルが集められた(各時点でマウス3匹)。
【0114】
TRU‐016テストサンプルの各々の漿液濃度を、CD37+ラモスヒト細胞ラインを用いるFACSに基づく結合アッセイで測定した。CD37+ラモス細胞(5x105cells/well)がテストすべき漿液サンプルと一緒に底の平らな96ウェルプレートで培養された。スパイク漿液サンプルを用いて標準曲線とした。細胞は、4℃で1時間培養され、検出抗体を添加する前に洗浄した。TRU‐016テストタンパク質のCD37+ラモス細胞への結合は、フルオレッセイン結合ヤギ抗ヒトIgG Fcγフラグメン特異抗体を用いて検出された。標準曲線を用いて、抗原濃度の関数として結合曲線を描いた。要するに、標準曲線は、FACS緩衝剤で1:20に希釈された通常のマウス漿液に加えられたTRU‐016テストタンパク質の周知の様々な濃度からなる。標準曲線は、各プレートにおいて二重に引かれていた。平均蛍光強度(MFI)が、FACS分析からソフトマックスプロソフトウェアに移入され、TRU‐016テストタンパク質の漿液濃度を計算するのに用いられた。
【0115】
薬物動態研究の結果、200μMのDMJ、10μMのキフネンシンまたは200μMのカスタノスペルミンを含む媒体で培養されたCHO細胞が生成したTRU‐016(図11に示されている)が、マウスに投与されたとき、未処置のCHO細胞が生成したTRU‐016に類似する薬物動態プロフィールを呈示するということが示され、ホスト細胞の培地における糖修飾因子は、半減期またはその他の薬物動態パラメータに大きな影響を及ぼさないということが示された。
【0116】
TRU‐016の投与後48、72、96および192時間でマウスから得られる、TRU‐016を含む漿液についてCD16アッセイを繰り返すことで、テストのあらゆる時点で、増大したCD16結合活性が漿液によって保持されるということが示された。結果を図12に示す。
【0117】
例7
インビボの糖修飾免疫糖タンパク質活性
ヌードマウスが、0日目に、5x106ラモス細胞を皮下に投与され、0、2、4、6および8日目に、200μgの対照ヒトIgGまたは200μMのDMJ、10μMのキフネンシンまたは200μMのカスタノスペルミンで処置されたCHO細胞が生成したTRU‐016テストタンパク質を静脈に注射された。典型的には、マウスは6日以内に腫瘍が発達し、その後すぐに死亡する。腫瘍は、デジタルカリパスおよびラブキャットソフトウェアで週に3度測定され、腫瘍の体積は、1/2[長さx(幅)]2で計算される。また、週に1度体重を測定する。
【0118】
マウスは、腫瘍が1500mm3(金曜日に1200mm3)の大きさに到達するとき、死亡する。マウスはまた、腫瘍から潰瘍が形成されると死亡し、体重が20%以上失われてしまうと、腫瘍によって動物としての動きが抑制されてしまう。
【0119】
研究が開始されてから8日目の相対的な腫瘍の体積について平均の結果を図13に示す。研究開始後の生存率%についてのデータが、図14および以下の表1に示される。
【0120】
【表1】
【0121】
このインビボ研究の結果、200μMのDMJ、10μMのキフネンシンまたは200μMのカスタノスペルミンで処置されたCHO細胞が生成したTRU‐016は、癌の動物モデルにおいて腫瘍の体積を低減し、平均生存時間を増大させることができた。
【0122】
例8
様々な濃度のカスタノスペルミンのタンパク質生成への影響
さらなる実験を行って、カスタノスペルミンの濃度の、TRU‐016の細胞生存度、密度および特定のタンパク質生成への影響を測定した。
【0123】
実験の開始に先立って、TRU‐016をトランスフェクトしたCHO細胞が、加湿された培養器において37℃かつ5%の二酸化炭素で、1x非必須アミノ酸(メディアテック)、1xピルビン酸ナトリウム(メディアテック)、4mMのL−グルタミン(メディアテック)、500nMのメトトレキセート(MPバイオメディカルズ)および1mg/Lの組み換えインシュリン(リコンビュリン‐ジブコ/インビトロゲン社)を補ったEx-CellTM 302というCHO漿液の含まれない媒体(SAFCバイオサイエンス)の入った振とうフラスコで成長された。殺菌されたカスタノスペルミンを、蒸留した/イオンを取り除いた水(メディアテック)で希釈し、0.2μmHTタフリン膜を備える13mmのアクロディスク(R)(ポール社)を通して濾過することで、カスタノスペルミンの200mMの蓄積濃度(アレクシスバイオケミカルズ)が調節された。蓄積溶液を、殺菌され、Oリングのついた0.5mLのマイクロ遠心分離チューブ(フィッシャーブランド、フィッシャーサイエンティフィック)に取り分け、−20℃で凍らせた。実験を開始する約1時間前に、取り分けたサンプルの必要なものを室温に解凍し、ボルテックスして各小瓶の中身を良く混ぜた。
【0124】
各実験では、対数期成長の細胞が前述の培地に接種され、250mLの振とうフラスコにおける合計容量を200,000 cells/mLという密度で60mLとし、テストすべき濃度でCSを付加した。800μM、400μM、200μM、100μM、50μM、25μMおよび0μMという最終のCS濃度が各々、対のフラスコでテストされた。加湿された培養器において37℃および5%二酸化炭素で、全ての培養がなされ、生存細胞密度および全体の細胞の生存度について少なくとも1日おきにモニターした。
【0125】
8日目に全体の細胞の生存度が50%乃至70%(実験1)および30%乃至50%(実験2)であったときに、培養物が回収された。細胞および細胞のデブリが、3000rpmのソルバールスーパー(Sorvall Super)T21で20分間の遠心分離によって取り除かれ、その後、上清が、0.22μmミリポアエクスプレスプラス膜を備えるミリポアステリフリップ装置を通して殺菌濾過され、精製されるまで2℃乃至8℃で蓄積された。
【0126】
各サンプルの細胞領域全体(ICA)、表2、が指示するように、細胞の生存度と成長は、大きく影響を受けなかったように見えるが、カスタノスペルミンの濃度が増大すると免疫糖タンパク質の生成が低減した。結果を、図17および以下の表2に示す。400μmおよび800μmのCSの濃度が、TRU‐016タンパク質の生成をそれぞれ約40%乃至55%低減することが示されている。
【0127】
【表2】
【0128】
例9
TRU‐016のCD37およびFcγRIIIa(CD16)への同時結合についてのアッセイ
【0129】
カスタノスペルミン濃度の、TRU‐016の機能活性への影響を測定するために実験が行われ、それがFcγRIIIaに結合することおよびそれがターゲットの抗原CD37に結合することにより測定された。
【0130】
例8において記述されるように生成されるTRU‐016は、以下のアッセイにおいてテストされたが、それによって同時に、TRU‐016結合ドメインの、CD37発現ターゲット細胞に結合する能力および、TRU‐016SMIPのFc部の、ヒトCD16およびネズミIgGFcの融合タンパク質に結合する能力が評価される。
【0131】
使用されるターゲット細胞は、ダウディ(ATCC CRL‐213)細胞系(line)である。ダウディ細胞は、バーキットリンパ腫から得られるヒトB‐リンパ芽球状細胞系であり、高レベルのCD37を発現する。カスタム可溶性CD16:MuIgGFc融合タンパク質は、ネズミIgGFcに結び付いたヒトCD16(低親和多形性)である。
【0132】
適当な数のダウディ細胞(350,000/ウェルのウェル数倍)がアリコートされて、250xgで5分間、15℃で遠心分離される。上清は取り除かれる。USB(USB US19943)1:4からの4%原料をFACS緩衝剤で希釈することによって、1%の冷たいパラホルムアルデヒドが調製される。FACD緩衝剤は、2%FBS(ギブコ)をダルベッコPBS(インビトロジェン)(v/v)に加え、0.22μmフィルターで殺菌濾過することによって調製される。FACS緩衝剤は貯蔵され4℃で使用される。細胞は、1%パラホルムアルデヒド(50μL/ウェルに等しい体積のウェル数倍)に再懸濁され、底の丸い96ウェルプレートで培養される。細胞は、4℃で30分間培養される。この培養に続いて、細胞は、150μLのFACS緩衝剤を各ウェルに加え、250xgで3分間の遠心分離を15℃で行い、上清を取り除くことによって洗浄される。細胞は、50μLのFACS緩衝剤に再懸濁される。TRU‐016がFACS緩衝剤において、飽和からバックグラウンドのレベルの範囲にある濃度(24μg/mL乃至0.011μg/mL)に希釈され、適当なウェル、50μL/well、に加えられ、そして細胞は4℃で25分間培養される。CD16:MuIgGFc融合タンパク質は、FACS緩衝剤において飽和レベル(20μg/ml)に希釈され、アッセイ(50μL/ウェル)に加えられ、4℃でさらに30分間培養されて、細胞表面に結合しているTRU‐016と複合体を形成する。結合していない試薬は、250xgで3分間の遠心分離を15℃で行い、上清を取り除いて、そして200μL/ウェルのFACS緩衝剤で三回洗浄することによって取り除かれる。そして、細胞は、ネズミFc特異の(かつヒトFcには最小限しか反応しないように選択された)、蛍光物質(R‐フィコエリトリン、ジャクソン115‐116‐071)でタグを付けたF(ab’)2抗体で培養される。この抗体は、CD16:MuIgGFc融合タンパク質のMuIgGFc部に結合する。抗体は、FACS緩衝剤において1:200に希釈され、各ウェルに100μLが加えられる。プレートは、暗所において4℃で45分間培養される。結合していないR‐PEは、150μLのFACS緩衝剤を各ウェルに加え、250xgで3分間の遠心分離を15℃で行い、それに続いて上清を取り除くことで取り除かれる。これに続いて、200μL/wellのFACS緩衝剤で二回目の洗浄を行い、250xgで3分間の遠心分離を15℃で行い、そして上清を取り除く。細胞は、200μL/wellの1%パラホルムアルデヒドで再懸濁され、4℃で一晩静置される。
【0133】
各サンプルに結合した蛍光は、BD FACSキャリバーフロー血球計算システムで測定され、セルクエストプロソフトウェア(ベクトンディキンソンver5.2)で分析される。各サンプルのGeoMean蛍光強度がTRU‐016の濃度に対してプロットされる。用量反応が生成されて、SoftMax Proソフトウェア(モルキュラーデバイスver5.0.1)を用いて4パラメータ対数(4‐PL)曲線に当てはめられる。TRU‐016の滴定を用いて、テスト物質および比較のための参照物質の用量反応曲線が求められる。処置したおよび未処置のサンプルを比較するために、「D」‐パラメータ(最大曲線漸近線)を参照として用いる。「D」値の増大は、対応するサンプルについての結合活性の増大を表す。
【0134】
実験の結果を図18に示し、CSの濃度に対する、用量依存的結合反応が400μMまで示され、この点で、結合が横ばいになっている。
【0135】
CSで処置されたTRU‐016のサンプルのCD16への結合が高められても、それは、部分的には分子のCD37への結合が高められることによらないということを示すために、前記のアッセイを繰り返したが、処置されたまたは未処置のTRU‐016サンプルをアッセイプレートに加えて培養した後、250xgで3分間遠心分離を15℃で行い、上清を取り除き、そして200μL/ウェルのFACS緩衝剤で三度洗浄することによって、結合していないTRU‐016をウェルから取り除くことは除外した。そして、細胞は、FITC抱合ヤギ抗ヒトIgGFc特定抗体(カルタグH10501)で培養される。この抗体は、細胞に結合したTRU‐016のヒトIgG鎖のFc領域に結合する。抗体は、FACS緩衝剤において1:50に希釈され、各ウェルに100μL加えられる。プレートは暗所において4℃で45分間培養される。100μLのFACS緩衝剤を各ウェルに加え、250xgで3分間の遠心分離を15℃で行い、続いて上清を取り除くことによって、未結合のFITCラベル化された抗体を取り除く。これに続いて、200μL/ウェルのFACS緩衝剤で二度目の洗浄を行う。細胞は、200μL/ウェルの2%パラホルムアルデヒドで再懸濁され、4℃で一晩静置される。各サンプルに結合した蛍光は、BD FACSキャリバーフロー血球計算システムで測定され、セルクエストプロソフトウェア(ベクトンディキンソンver5.2)を用いて分析される。各サンプルのGeoMean蛍光強度が、TRU‐016の濃度に対してプロットされる。用量反応曲線が生成され、SoftMax Proソフトウェア(モルキュラーデバイスver5.0.1)を用いて4パラメータ対数(4‐PL)曲線に当てはめられる。TRU‐016の滴定を用いて、未処置の対照サンプルおよび比較のためにCSで処置されたサンプルの用量反応曲線が求められる。
【0136】
図19に示されるように、CSで処置されたサンプル全てについて、CD37発現細胞に対する用量反応結合曲線は、相互に、および未処置のTRU‐016サンプルと実質的に同一であって、CSでの処置によっても、TRU‐016の、その特定のターゲット抗原への結合は変わらなかったということが示されている。
【0137】
例10
抗体依存性細胞障害性(ADCC)のアッセイ
カスタノスペルミン濃度の、TRU‐016の機能活性に対する影響を測定するために実験が行われ、それはADCC活性によって測定された。
【0138】
例8に記述されるように生成されたTRU‐016は、プライマリーヒト末梢血リンパ球(PBL)エフェクター細胞とともにCD37発現ダウディ癌B細胞系で培養されて、ADCC活性が評価される。
【0139】
ダウディターゲット細胞(5x106)が、15mlの円錐チューブに加えられ、そして250xgで5分間の遠心分離が20℃で行われ、上清が取り除かれた。細胞ペレットは、0.3mCiクロム‐51(51Cr、GEヘルスケア、CJ51)を加えることで再懸濁される。細胞は、5%のCO2において、37℃で75分間培養され、細胞が放射性同位元素を取り込むことが可能となる。そして細胞は、三度洗浄され、取り込まれなかった51Crが取り除かれる。これが行われるのは、10mLの完全培地‐10%FBS(ギブコ)を備えるIMDM(ギブコ)‐をチューブに加え、250xgで5分間の遠心分離を20℃で行い、それに続いて上清を取り除くことによってである。最終の再懸濁は、11.5mLの完全培地においてである。TRU‐016は、完全培地において、最大からバックグラウンドレベルまでの細胞溶解(500ng/mL乃至0.005ng/mL)を生成することができる濃度に希釈される。これらの滴定は、底の丸い96ウェルプレートにおいて、50μL/ウェルで行われる。51Crのラベルを付けられたターゲット細胞が、50μL/ウェルおよび対照ウェル(TRU‐016の無い対照媒体)でTRU‐016の用量滴定に加えられる。プロトコル(LSM、MPバイオメディカル、50494/36427)に従う、リンパ球セパレーションメディアを用いての密度勾配遠心分離によって、ヘパリン処理した新鮮な全血からPBLが分離される。PBLエフェクター細胞は、25:1乃至30:1の割合でウェルに、100mL/ウェル加えられる(エフェクター:ターゲット)。5%のCO2にてアッセイは37℃で4.5乃至5時間培養される。エフェクター細胞は、TRU‐016の濃度に対してターゲット細胞を溶解させ、比例した量の51Crをアッセイの上清に放出する。培養に続いて、250xgで3分間、プレートの遠心分離が20℃で行われる。25μL容量の細胞を含まない上清が、全てのウェルから取り除かれてシンチレーションプレート(パーキンエルマー6005185)に入れられ、一晩乾燥される。シンチレーションプレートの各ウェルにおける51Cr同位元素の量は、トップカウントプレートリーダー(パーキンエルマー、C9904VO)を用いて測定される。データは、特定の放出のパーセントとして表される。特定の放出は、以下のように計算される。
【0140】
(サンプル値‐自発値)/(最大値‐自発値)*100%
自発=ターゲット細胞のみから放出される51Crの量
最大放出=洗浄溶解剤で処置されたターゲットから放出される51Crの量
バックグラウンド対照=ターゲット細胞+エフェクター細胞(No TRU‐016)から放出される51Crの量
【0141】
用量反応が形成され、SoftMax Proソフトウェア(モルキュラーデバイスver5.0.1)を用いて4パラメータ対数曲線に当てはめられる。TRU‐016の滴定を用いて、テスト物質および比較のための参照物質の用量反応曲線が求められる。処置したものについてのEC50値が、処置されていない対照(CSでない)と比較され、ADCC活性のパーセント増大が測定される。以下の表に、図20に示されるデータがまとめられている。データでは、100μM乃至800μMの最終濃度の範囲についてCSで処置されたTRU‐016のADCC活性が、未処置のTRU‐016に対して大きく増大していることが示されている。
【0142】
【表3】
【0143】
本発明の構成および方法は、前述の代表的な実施例について記述されているが、発明の概念、思想および範囲から逸脱することなく、ここに記述されている構成および/または方法および工程または方法における一連の工程には、変形を施してもよいことが当業者には明らかである。さらに詳しくは、化学的にも生理学的にも関係のある試薬をここに記述されている試薬と置き換えても、同じまたは同様の結果が達成されるということも明らかである。当業者に明らかなそのような類似の置き換えおよび変形は全て、添付の請求の範囲に定義される本発明の思想、範囲および概念内のものと看做される。
【0144】
本明細書の至る所に引用され、当該説明を例示によって詳細に増補する参考文献は全て、援用することでここに組み入れることを明示する。
【技術分野】
【0001】
本発明は、抗体を含む、特性が改良された免疫糖タンパク質に関し、それには、抗体依存性細胞傷害性およびグリコシル化パターン、細胞培養方法およびそのような免疫糖タンパク質を生成するための培地、並びに病気の処置における当該免疫糖タンパク質の使用が含まれる。
【0002】
なお、本件出願は、2006年10月24日に出願された米国仮出願第60/853,944号の利益を享受し、その内容を援用することで全体は組み入れられているものである。
【背景技術】
【0003】
免疫薬でターゲットとする細胞数を削減するのは、適応症のいくつかにおいては重要な治療処置である。そのように、ターゲットとする細胞の削減を達成するために、免疫薬が用いているメカニズムは、補体が媒体となる細胞の溶解、細胞消滅の信号を送る動作経路の活性化、生存のために必要な信号を送る経路の遮断、およびFc依存性細胞傷害性とも称される、抗体依存性細胞傷害性(ADCC)を含み得る。ADCCは、効力のあるメカニズムであり、多くの免疫薬の有効性にとって重要であると考えられている。
【0004】
ADCCの活性化のためのメカニズムには、ターゲット細胞の表面に結合される免疫薬分子へのFc受容体の結合が必要である。免疫薬へのFc受容体の結合は、CH2および/またはCH3ドメインのような、免疫グロブリンの一定領域内のドメインを媒体とすることができる。異なる種類の一定領域が、異なるFc受容体と結合する。例として含まれるのは、IgG1Fcドメインの、同種のFc受容体CD16(FcγRIII)、CD32(FcγRII‐B1および‐B2)およびCD64(FcγRI)への結合、IgAFcドメインの、同種のFc受容体CD89(FcαRI)への結合、およびIgEドメインの、同種のFc受容体FcεR1およびCD23への結合である。
【0005】
Fc受容体の結合を高めた免疫薬組成物は、ADCCにおいてより大きな有効性が呈示される。IgGFcドメインでこれを達成する方法として報告されているものには、アミノ酸の変化の導入や糖類構造の修飾 (modification) が含まれる。Fcドメインにおけるアミノ酸の変化は、免疫薬組成の免疫原性を高めるので、糖類構造の修飾が好ましい。免疫グロブリン分子については、N‐リンクされた糖類の、CH2ドメインのAsn−297への付着がADCC機能にとっては重大であるということが示されている。酵素で、またはN‐リンクされた共通サイトの突然変異を通してそれを取り除くと、結果的に、ADCC機能がほとんどまたは全く無くなってしまう。免疫グロブリン分子についてのADCC機能のレベルもまた、糖類の構造に依存すると報告している研究もあるが、実際の糖類の、ADCCに関する部位または構造は、いまだ解明されていない。まして、非免疫グロブリンFc融合タンパク質のADCCに最適な糖類の構造については、なおさら知られていない。
【0006】
糖タンパク質において、糖類は、トリペプチドのモチーフAsn‐X‐Thr/Serにおけるアスパラギンの側鎖でアミド窒素原子に付着する。N‐リンクされたグリコシル化と称されるこの種のグリコシル化は、複数の単糖類がリン酸ドリコールに加わって14‐残基側鎖の糖類複合体を形成している小胞体(ER)において開始する。そして、この糖類複合体は、オリゴサッカリルトランスフェラーゼ(OST)複合体によってタンパク質へと転移される。糖タンパク質がERの内腔を離れる前に、3つのグルコース分子が、14‐残基オリゴ糖から取り除かれる。酵素ERグルコシダーゼI、ERグルコシダーゼIIおよびERマノシダーゼがER処理に関係している。
【0007】
続いて、ポリペプチドがゴルジ複合体へと運ばれ、そこではN‐リンクされた糖鎖が異なる多くの方法で修飾される。ゴルジ複合体のシスおよび中間区画において、元の14‐糖N‐リンクされた複合体は、マンノース(Man)残基を取り除いて切り詰められ、N‐アセチルグルコサミン(GlcNac)および/またはフコース(Fuc)残基を加えることで伸長される。N‐リンクの糖類の形は様々であるが、一般には、3つのマンノース残基と2つのN‐アセチルグルコサミン残基からなる五糖(ペンタサッカライド)を共有する。最後に、トランスゴルジにおいて、他のGlcNac残基を加え、それに続いてガラクトース(Gal)および末端シアル酸(Sial)を加えることができる。ゴルジ複合体における糖処理は、「末端グリコシル化」と呼ばれ、ERにおいて起こる「コアグリコシル化」とは区別される。最終の複合糖ユニットは、多くの形態および構造を取り得るものである、その中には、2本、3本または4本の側鎖(バイアンテナリー、トリアンテナリーまたはテトラアンテナリーと称する)を有するものもある。ゴルジ処理には、複数の酵素が関係しており、それらには、ゴルジマノシダーゼIA、IBおよびIC、GlcNAc‐トランスフェラーゼI、ゴルジマノシダーゼII、GlcNAc‐トランスフェラーゼII、ガラクトシルトランスフェラーゼおよびシアリルトランスフェラーゼが含まれている。
【0008】
FcγRIIIa受容体が媒体となるADCC機能の重大な糖の決定要素は、コアN‐リンクの構造に加えられるアルファ‐1、6‐フコース部の欠如であることを、ある報告が示唆している(シンカワら、J Biol Chem. 2003年1月31日;278(5):3466−73;また、シールズら、J Biol Chem. 2002年7月26日;277(30):26733−40も参照)。別のレベルのグリコフォーム、すなわち、二等分されたN‐リンクの糖類もまたADCCを増大することができるものとして提案されている(ウマナら、Nat Biotechnol. 1999年2月;17(2):176−80)が、それに反する証拠もある(シンカワら、J Biol Chem. 2003年1月31日;278(5):3466−73)。この反対の証拠が解明される可能性もあることが示唆されており、それは、ホスト細胞においてGnTIIIが増加することで、二等分された糖が増加しているのみならず、コアフコースの修飾が欠如している免疫グロブリンが生成されるという発見による(フェラーラら、Biotechnol Bioeng. 2006年4月5日;93(5):851‐61)。これは、フコースのみがADCCの有効性を変える主要な役割を果たしており、他で見られる二等分された糖との関連は、ホスト細胞における2つの修飾の結び付きを反映しているという提言と一致している。しかしながら、GnTIIIを備えるリタクサン抗体およびハーセプチン抗体の、二等分された糖を増やすためのインビトロの処置が、結果としてADCCを増大させたという別の報告があり、二等分された糖の直接の影響が示唆されている(ホドニクツキーら、Biotechnol. Prog., 2005年11月−12月;21(6):1644−52)。しかしながら、非常に高いレベルでのGntIIIの過大な発現は、細胞に対して有毒である(ウマナら、Biotechnol. Prog., 1998年3月−4月;14(2):189−92)。
【0009】
フコース含有量の低い免疫グロブリンを生成する方法が提案されているが、治療の指標として最適なADCC機能を備えるバイオ医薬品の製造にとって大きな障害があるものもある。例えば、フコース残基(フコーシダーゼ)を取り除く酵素を備える免疫グロブリンの処置には、重大な経済上の、および薬品の安定性上の危険が潜在していて、費用のかかる製造工程を追加する必要がある。フコシル化糖タンパク質の合成に関係する主要酵素を破壊するための細胞系(line)の分子的な処理には、特別なホストの負担が必要であり、現在のプラクティスにおいては、治療上の使用のために有効性および安全性が最適化されるようADCCの有効性を変えると、薬品の「調節可能な」生成が可能ではなくなる。ADCCの高められていない比較生成物を生成するには、費用および時間がかかる。RNAiまたはアンチセンス分子で細胞系(line)を処置してこれらの主要酵素のレベルを破壊すると、予想できない的外れの効果が得られ、また、生産を行う規模で実施される程度には実用的であるとしても、費用がかかるであろう。
このように、ADCCの高められた免疫薬を調合するのに有効な方法、またそれによって生成され、治療上使用される改良された免疫薬が切望されている。
【先行技術文献】
【非特許文献】
【0010】
【非特許文献1】J Biol Chem、2003年1月31日、278(5)、3466−73
【非特許文献2】J Biol Chem、 2002年7月26日、277(30)、26733−40
【非特許文献3】J Biol Chem、 2003年1月31日、278(5)、3466−73
【非特許文献4】Nat Biotechnol、 1999年2月、17(2)、176−80
【非特許文献5】Biotechnol Bioeng、 2006年4月5日、93(5)、851−61
【非特許文献6】Biotechnol. Prog、 2005年11月−12月、21(6)、1644−52
【非特許文献7】Biotechnol. Prog、 1998年3月−4月、14(2)、189−92
【発明の概要】
【発明が解決しようとする課題】
【0011】
本発明は、培地(培養媒体)および免疫糖タンパク質の特性を改良する大規模細胞培養方法を提供するが、これには、ADCCのようなエフェクター機能および/またはフコース含有量の低減のようなグリコシル化パターンが含まれる。本発明はさらに、上記方法によって生成される、改良された免疫糖タンパク質、および病気の治療におけるそのような免疫糖タンパク質の使用を提供する。
【課題を解決するための手段】
【0012】
本発明は、ホスト細胞により生成された免疫糖タンパク質分子の抗体依存傷害性(ADCC)を増大する方法を提供するが、それは、濃度が、約25μM〜約800μMの間、または約100μM〜約500μMの間、または約100μM〜約400μMの間、または約100μM〜約300μMのカスタノスペルミンを備える培地において、ホスト細胞を成長させることによるものである。代表的な実施例において、ADCCは、少なくとも2倍、3倍、4倍または5倍に増大する。
【0013】
また本発明は、ホスト細胞により生成された免疫糖タンパク質分子のCD16結合を増大する方法を提供するが、それは、濃度が、約25μMと約800μMの間、または約100μMと約500μMの間、または約100μMと約400μMの間、または約100μMと約300μMの間にあるカスタノスペルミンを備える培地において、ホスト細胞を成長させることによるものである。代表的な実施例において、CD16結合は、少なくとも50%、75%、100%、125%、150%、175%または200%増大する。
【0014】
本発明の方法において、細胞の成長、生存度および/または密度は大きな影響を受けない(例えば、処置されていない細胞の少なくとも80%以上に留まる)。培地における免疫糖タンパク質の生成のレベルは、少なくとも100μg/mL、125μg/mLまたは150μg/mLである。
【0015】
前記本発明において、培地は実質的に漿液が含まれておらず、第二の糖修飾因子が含まれている。
【0016】
本発明では、上記方法で生成された免疫糖タンパク質分子を、必要に応じて無菌の薬学的に受け入れ可能な担体または希釈剤とともに含む組成物もまた意図されている。かかる組成物は、前記免疫糖タンパク質分子が結合した分子を表面に発現する癌細胞を死滅させる、あるいは、その成長を抑制する方法、または前記免疫糖タンパク質分子が結合する分子を表面に発現する細胞を枯渇させる方法において投与される。
【0017】
本発明の方法は、一般に、適当な濃度の糖修飾因子、例えば、カスタノスペルミンを含む培地において、免疫糖タンパク質を生成するホスト細胞を培養することを伴い、細胞の成長またはタンパク質生成レベルに大きな影響を与えることなく、エフェクター機能を向上させるという利点がある。本発明の方法を用いて製造することのできる代表的な免疫糖タンパク質には、免疫グロブリンおよび小さなモジュラー免疫薬(SMIPTM)生成物が含まれる。本発明の方法により調製されるそのような結合分子には、ターゲットへの結合特性を実質的に同じに保ち、その結果、生物的活性が直接得られ、エフェクターを媒体とする機能の向上が呈示されるという利点がある。
【0018】
本発明は、抗体依存傷害性(ADCC)および/またはホスト細胞により生成される免疫糖タンパク質のFc受容体結合を向上させる方法を提供する。当該方法は、糖修飾因子、例えば、カスタノスペルミンを、ホスト細胞が生成した免疫糖タンパク質分子の組成物のADCC活性および/またはFc受容体結合を増大させるような濃度で備え、少なくとも体積が750mL、1L、2L、3L、4L、5L、10L、15L、20Lまたはそれ以上の培地において、ホスト細胞を成長させることを伴っている。かかる糖修飾因子、例えば、カスタノスペルミンの最適な濃度は、糖修飾因子の有効性および所望のADCCの関連する調節(modulation)に依存するが、培地における糖修飾因子の代表的な最終濃度は、800μM未満または750、700、650、600、550、500、450、400、350、300、250、200、150、125、100、90、80、70、60、50、40、30、20または10μM未満である。
【0019】
それに関係した、ADCCへの影響は、細胞培養に適用される糖修飾因子、例えば、カスタノスペルミンの濃度または持続時間を変えることで調整されるが、グリコシル化を変えることでADCCを向上させる従来の方法と比較すると、顕著な利点がある。ADCC活性は、当業界で知られているアッセイを用いて測定され表されるが、代表的な実施例においては、少なくとも10%、20%、30%、40%、50%、60%、70%、80%、90%、2倍、3倍、4倍、5倍、6倍、7倍、8倍、9倍、10倍、15倍または20倍増大する。
【0020】
グリコシル化および糖含有量は、ADCC、CDCおよび循環半減期を含む、免疫グロブリンエフェクターを媒介とする様々な機能に影響を及ぼすことが知られている。ここに記述されるデータでは、本発明の方法によって、驚くことに、CDCまたは半減期に影響を及ぼすことなく、ADCCが増大することを示す免疫糖タンパク質が提供されるということが示されている。このように、代表的な実施例において、免疫糖タンパク質分子組成物のADCCは増大するが、補体依存傷害性(CDC)および/または長くなった循環半減期のような他の免疫グロブリン型エフェクター機能は、同様であるか大きくは影響を受けない(例えば、2倍未満の増大または減少、あるいは50%、40%、30%、20%または10%未満の増大または減少)。
【0021】
免疫糖タンパク質分子の組成物のFc受容体結合は、糖修飾因子で処置された免疫糖タンパク質分子対CD16に結合する処置されていない免疫糖タンパク質分子という相対比として測定される。代表的なアッセイが、以下の例で記述される。代表的な実施例におけるFc受容体結合は、少なくとも10%、20%、30%、40%、50%、60%、70%、80%、90%、2倍、3倍、4倍、5倍または6倍増大する。本発明により、糖修飾因子、例えば、カスタノスペルミンで処置されたホスト細胞が生成した免疫糖タンパク質組成は、CD16(高または低親和形態、すなわちアミノ酸158でVまたはF)および/またはCD32aまたはbおよび/またはCD64に、そのように処置されていないホスト細胞が生成する免疫糖タンパク質組成物よりもFcR結合検査において大きな親和力で結合する。Fc受容体結合親和力がこのように増大するのは、ADCC機能の向上と相関していることがここに示されている。
【0022】
本発明によって、糖含有量/グリコシル化パターンを変え、および/または免疫糖タンパク質のフコース含有量を低減する方法もまた提供されるが、該方法は、総フコース含有量を低減し、および/またはホスト細胞が生成する免疫糖タンパク質分子の組成物のグリコシル化パターンを変えるような濃度で、糖修飾因子、例えばカスタノスペルミンを備え、体積が、少なくとも750mL、1L、2L、3L、4L、5L、10L、15L、20Lまたはそれ以上である培地において、ホスト細胞を成長させることによるものである。培地における糖修飾因子の代表的な最終濃度は、800μM未満または750、700、650、600、550、500、450、400、350、300、250、200、150、125、100、90、80、70、60、50、40、30、20または10μM未満である。
【0023】
フコース含有量への影響もまた、細胞培養に適用される糖修飾因子、例えば、カスタノスペルミンの濃度または持続時間を変えることによって調整される。組成物の総フコース含有量は、フコース化されていない免疫糖タンパク質分子の、組成物における免疫糖タンパク質分子の総数に対する相対比またはパーセンテージとして表される。代表的な組成物は、少なくとも30%、40%、50%、60%、65%、70%、75%、80%、85%、90%、95%またはそれ以上のフコース化されていない分子を含む。本発明により、糖修飾因子、例えば、カスタノスペルミンで処置されたホスト細胞が生成する免疫糖タンパク質組成物のフコース含有量は、そのように処置されなかったホスト細胞が生成する組成物と比べて、少なくとも2倍、3倍、4倍、5倍、6倍、7倍、8倍、9倍または10倍以上低減される。
【0024】
前述の本発明の方法のいずれにおいても、ホスト細胞は、糖修飾因子、例えば、カスタノスペルミンにさらされている間、高いレベルでの成長を呈示している。例えば、免疫糖タンパク質を生成するCHO細胞の代表的な倍化時間は、約24時間であるが、本発明による糖修飾因子の濃度(例えば、ADCCを増大するのに有効な濃度)によって、そのような倍化時間が低減されるとの期待はない。理想的には、糖修飾因子、例えば、カスタノスペルミンの有効な濃度では、糖修飾因子を加えてから72時間後の時点で、10、20、30、40、50、60または70%を超えるほどには、細胞の成長は低減されない。
【0025】
前述の方法のいずれにおいても、ホスト細胞は、糖修飾因子、例えば、カスタノスペルミンにさらされている間、高いレベルでのタンパク質生成を呈示している。例えば、糖修飾因子、例えば、カスタノスペルミンが有効な濃度であるときのタンパク質生成レベルは、約50μg/mL以上であり、または約75、100、125または150μg/mL以上である。好ましくは、ホスト細胞は、高いレベルでの成長および高いレベルでのタンパク質生成の双方を呈示する。
【0026】
実質的に漿液を含まない培地を含む、当業界で知られている任意の培地を用いても良い。バッチ培養、流加培養、連続培養、その他の業界で知られている培養方法を本発明の方法で用いることができる。糖修飾因子が、急速な成長段階後に、シーズ列に添加され、すなわち、最初のバッチ培地に添加されるか、又は連続して培地と一緒に(例えば、連続して供給する間に)添加される。例えば、糖修飾因子は、10Xまたは100Xの濃度で初期のシーズ列または原料に添加されるが、次いで培地を加えると、糖修飾因子の濃度が希釈されるが、組み換え生成物のADCCを増大させるのになおも有効なレベルになるようにする。あるいは、糖修飾因子が、細胞に添加されるあらゆる培地に有効な濃度で含まれ、希釈する必要は無いものとする。いずれにしても、糖修飾因子は、細胞培養プロセスにおいて比較的早期に加えられ、免疫糖タンパク質の均質性を最適化するために、培養処理の間有効な濃度が維持される。糖修飾因子の効果は、長期に続くものである、糖修飾因子を一度加えると、少なくとも11乃至12日間は持続して認められる。
【0027】
代表的な糖修飾因子には、コアグリコシル化抑制剤、端末グリコシル化抑制剤、マノシダーゼ抑制剤および/または初期段階糖修飾因子が含まれており、選択的にフコシル化に特有の抑制剤が含まれていても含まれていなくてもよく、さらなる詳細は以下に記述される。本発明におていは、2つ以上あるいは3つ以上の糖修飾因子を組み合わせることでさらに利点が得られるということが意図されている。カスタノスペルミンは、特に有効な1つの糖修飾因子である。
【0028】
さらに、本発明によって、前述の方法のいずれかで生成された免疫糖タンパク質分子を含み、好ましくは、ターゲット分子について少なくとも107M−1または少なくとも108M−1または109M−1の結合親和力Kdを有する組成物が提供される。そのような組成物は、無菌の、薬学的に受け入れ可能な担体または希釈剤を1つ以上含んでいてもよい。
【0029】
さらに、本発明によって、上記組成物の被体への投与を伴っている治療方法が提供されるが、その被体は、当該投与から効果を得ることができるもので、例えば、ターゲット分子を発現する細胞が媒体となる障害を患っているか、癌細胞が表面にターゲット分子を発現するある種の癌を患っている。本発明によって、上記組成物を、ターゲット分子を表面に発現している細胞を枯渇する方法において用いることもまた意図されている。ターゲットがCD37であるところでは、本発明によって、癌細胞の成長を抑制するか、癌細胞を破壊する方法が特に意図されており、それは、本発明の方法により生成されたアンチCD37SMIP生成物を含む組成物を被体に投与する工程を備えている。同様に、ターゲットがCD20であるところでは、本発明によって、癌細胞の成長を抑制するか癌細胞を破壊する方法が特に意図されており、それは、本発明の方法により生成されたアンチCD20SMIP生成物を含む組成物を被体に投与する工程を備えている。好適な形態において、癌の進行を停止させるか反転させることを伴って癌を治療する方法が意図されている。本発明によって、本発明の方法により生成されたアンチCD37またはアンチCD20SMIP生成物を投与することで、自己免疫性または炎症性の疾患を治療する方法がさらに提供される。さらに、本発明において、ここに記述される疾患または障害のいずれかを治療する薬剤を調製するための、無菌又は滅菌された担体または希釈剤を選択的に有する、本発明の糖タンパク質組成物の使用が意図されている。
【0030】
免疫糖タンパク質
「免疫糖タンパク質」という語は、ターゲット分子と結合するグリコシル化ポリペプチドのことを称し、免疫グロブリンの一定領域から得られる十分なアミノ酸配列を含んで、好ましくは、ADCCおよび/またはCDCというエフェクター機能を提供する。代表的な分子には、免疫グロブリンのCH2ドメインから得られるシーケンス、または1つ以上の免疫グロブリンから得られるCH2およびCH3ドメインが含有される。本発明によって生成することが意図された免疫糖タンパク質の特定の部分集合には、ヒンジにおける共有結合性の、または非共有結合性の結合および/またはCH3ドメインを通して選択的に二量体になる単鎖タンパク質が含まれる。単鎖タンパク質のこの部分集合には、免疫グロブリンの典型的な四量体の形態は含まれない(軽鎖が無いことによる)が、Fc配位子またはFc可溶の受容体融合が含まれる。単鎖タンパク質の特定の例には、SMIP生成物が含まれる。
【0031】
SMIP生成物とそれらを生成する方法は、共同で所有される米国出願第10/627,556号および米国特許公報2003/133939、米国特許公報2003/0118592および米国特許公報2005/0136049において以前記述されており、それらは各々、援用することでその内容全体をここに組み入れる。エフェクター機能を備える単鎖多価結合タンパク質が、2007年6月12日に出願された国際特許出願第PCT/US07/71052号に記述されている(2006年6月12日に出願されたU.S.S.N.60/813,261および2006年10月20日に出願されたU.S.S.N.60/853,287の利益を主張する)が、それらは各々、援用することでその内容全体をここに組み入れる。SMIP生成物は、抗原、反対受容体等の同種の構造についての結合ドメインを特徴とする新規の結合ドメイン‐免疫グロブリン融合タンパク質であり、すなわち、IgG1、IgAまたはIgEヒンジ領域ポリペプチド、または0、1または2のシスティン残基を有する突然変異IgG1ヒンジ領域ポリペプチドおよび免疫グロブリンCH2およびCH3ドメインである。一例においては、結合ドメイン分子は、1つまたは2つのシスティン残基を有する。また他の例においては、結合ドメイン分子が2つのシスティン残基を備えるとき、典型的には重鎖可変領域と軽鎖可変領域との間の結合に携わる第一のシスティンが削除されたり、アミノ酸で置換されたりすることの無いように意図されている。SMIP生成物は、ADCCおよび/またはCDCが可能であるが、二硫化物に結合した多量体を形成する能力で折り合っている。代表的なSMIP生成物は、免疫グロブリンから得られる可変軽鎖および/または可変重鎖結合領域の免疫グロブリン上科要素の結合領域のような1つ以上の結合領域を有する。代表的な例において、これらの領域は、リンカーペプチドで分離されるが、そのリンカーペプチドは、ドメインまたは領域接合と適合性のあることが当業界で知られているいずれのものであっても良い。代表的なリンカーは、(Gly4Ser)n、ただしn=3〜5、のようなGly4Serリンカーモチーフに基づくリンカーである。本発明により生成できる代表的なSMIP生成物には、CD20またはCD37を結合する生成物が含まれている。CD20またはCD37を結合し、特定の結合シーケンスおよび/またはアミノ酸の修飾を備えるSMIP生成物は、共同で出願された米国出願10/627,556および米国出願11/493,132に記述されており、これらの各々の内容は、ここに援用することでその全体をここに組み入れる。
【0032】
免疫糖タンパク質の他の例には、結合ドメイン‐Ig融合体が含まれており、ここで、結合ドメインは、非自然発生のペプチドまたは自然発生のリガンドまたはレセプターのフラグメントである。レセプターの場合、細胞外ドメインのフラグメント(断片)が好ましい。免疫グロブリンまたはFc領域を備える代表的な融合体には、Fc領域を備えるsTNFRIIの融合タンパク質であるエタナーセプト(米国特許第5,605,690号)、Fc領域を備える抗原提示細胞で示されるLFA‐3の融合タンパク質であるアレファセプト(米国特許第5,914,111号)、Fc領域を備える細胞毒性Tリンパ球関連の抗原‐4(CTLA‐4)の融合タンパク質[J. Exp. Med. 181、1869(1995)]、Fc領域を備えるインターロイキン15の融合タンパク質[J. Immunol. 160、5742(1998)]、Fc領域を備える因子VIIの融合タンパク質[Proc. Natl. Acad. Sci. USA, 98,12180(2001)]、Fc領域を備えるインターロイキン10の融合タンパク質[J. Immunol. 154、5590(1995)]、Fc領域を備えるインターロイキン2の融合タンパク質[J. Immunol. 146、915(1991)]、Fc領域を備えるCD40の融合タンパク質[サージェリー、132、149(2002)]、抗体Fc領域を備えるFlt‐3(fms様チロシンキナーゼ)の融合タンパク質[Acta. Haemato., 95、218(1996)]、抗体Fc領域を備えるOX40の融合タンパク質[J. Leu. Biol., 72、522(2002)]、他のCD分子[例えば、CD2、CD30(TNFRSF8)、CD95(Fas)、CD106(VCAM‐1)、CD137]、接着分子[例えば、ALCAM(活性化白血球細胞接着分子)、カドヘリン、ICAM(細胞間接着分子)‐1、ICAM‐2、ICAM‐3]、サイトカイン受容体[例えば、インターロイキン‐4R、インターロイキン‐5R、インターロイキン‐6R、インターロイキン‐9R、インターロイキン‐10R、インターロイキン‐12R、インターロイキン‐13Rα1、インターロイキン‐13Rα2、インターロイキン‐15R、インターロイキン‐21R]、ケモカイン、細胞死誘発信号分子[例えば、B7‐H1、DR6(死受容体6)、PD‐1(プログラム化死‐1)、TRAIL R1]、同時刺激分子[例えば、B7‐1、B7‐2、B7‐H2、ICOS(誘発性共同刺激因子)]、成長要因[例えば、ErbB2、ErbB3、ErbB4、HGFR]、分化誘発因子(例えば、B7‐H3)、活性化要因(例えば、NKG2D)、信号転送分子(例えば、gp130)が含まれてる。
【0033】
免疫糖タンパク質のさらに別の例としては、抗体が含まれる。ここで「抗体」という語は、完全に組み立てられた抗体、単クローン抗体、多クローン抗体、多重特異性抗体(例えば、二重特異性抗体)、抗原を結合できる抗体断片(例えば、Fab’、F’(ab)2、Fv、単鎖抗体、ダイアボディ)、および所望の抗原結合活性を呈示する限りにおいて前述のいずれかを備える組み換えペプチドを含むように定義される。化学的に誘導される抗体を含んで、損傷のない完全な分子および/または断片の多量体または集合体が意図されている。IgG、IgM、IgD、IgA、IgE、IgG1、IgG2,IgG3、IgG4、IgA1およびIgA2を含む、あらゆるアイソタイプの分類または下位分類の抗体が意図されている。異なるアイソタイプは異なるエフェクター機能を有し、例えばIgG1およびIgG3アイソタイプは、抗体依存細胞傷害性(ADCC)活性を有する。
【0034】
「免疫グロブリン」または「自然のままの抗体」は、同一の二組のポリペプチド鎖(二つの「軽」鎖および二つの「重」鎖)からなる四量体糖タンパク質である。各鎖のアミノ端子部には、抗原を認識する主要な役割を果たす約100乃至110またはそれ以上のアミノ酸の「可変」(「V」)領域が含まれている。この可変領域内では、「超可変性の」領域または「相補性判別領域」(CDR)は、[カバトらが免疫学に関係する一連のタンパク質(Sequences of Proteins of Immunological Interest)第5版、保健サービス、国立衛生研究所、ベセズダ、メリーランド州(Public Health Service, National Institutes of Health, Bethesda, Md. )(1991) で説明したような]軽鎖可変ドメインにおける残基24−34(L1)、50−56(L2)および89−97(L3)および重鎖可変ドメインにおける31−35(H1)、50−65(H2)および95−102(H3)、および/または[チョシアらがJ. Mol. Biol. 196: 901-917 (1987) で説明したような]超可変性のループからのこれらの残基(すなわち、軽鎖可変ドメインにおける残基26−32(L1)、50−52(L2)および91−96(L3)および重鎖可変ドメインにおける26−32(H1)、53−55(H2)および96−101(H3))からなる。
【0035】
各鎖のカルボキシ端部には、一定領域が含まれる。軽鎖は、その一定領域内に単一ドメインを有する。そうして軽鎖は、可変領域を1つと一定領域ドメインを1つ有する。重鎖は、一定領域内にいくつかのドメインを有する。IgG、IgAおよびIgD抗体中の重鎖は、一定領域ドメインを3つ有しており、それらはCH1、CH2およびCH3と称され、IgMとIgE抗体における重鎖は、一定領域ドメイン4つと、CH1、CH2、CH3およびCH4を有する。従って、重鎖は、可変領域を1つと一定領域を3乃至4つ有する。
【0036】
免疫グロブリンの重鎖はまた、3つの機能領域、Fd領域(VHおよびCH1、すなわち、重鎖のN−端末領域を2つ有する断片)、ヒンジ領域およびFc領域(一定領域から得られ、ペプシン消化の後に形成される「断片結晶化」領域)に分けることができる。軽鎖と組み合わせられるFd領域が、Fabを形成する(「断片抗原結合」)。抗原は、各Fabのアミノ端末で抗原結合領域と立体化学的に反応するので、IgG分子は二価であり、すなわち、それは、抗原分子2つと結合する。Fc領域は、細胞で免疫グロブリン受容体、および補体カスケードの最初の要素と相互作用するドメインを含有する。従って、Fc断片は一般に、補体固定やFc受容体への結合のような免疫グロブリンのエフェクター機能の原因であると考えられている。
【0037】
ここで用いられている「単一クローン性抗体」という語は、実質的に均一な抗体の母集団から得られる抗体のことを称し、すなわち、その母集団に備わる個々の抗体は同一であるが、雑種細胞から生成されても、組み換えDNA技術から生成されても、可能性として自然に起こる突然変異、あるいは少量において現れる翻訳後の修飾単一クローン性抗体は除外される。単一クローン性抗体の例には、制限が無く、マウス、キメラ、ヒト化またはヒトの抗体またはそれらの変種または誘導体が含まれる。
【0038】
抗体シーケンスをよりヒト様であるようにヒト化または変更することは、例えば、ジョーンズらによる ネイチャー321: 522 525 (1986); モリソンらによるProc. Natl. Acad. Sci., U.S.A., 81: 6851 6855 (1984); モリソンおよびオオイによるAdv. Immunol., 44: 65 92 (1988); バーホエヤらによるサイエンス239: 1534 1536 (1988); パドランによる Molec. Immun. 28: 489 498 (1991); パドランによる Molec. Immunol. 31(3): 169 217 (1994); ケトルボロウ, C. A. らによるProtein Eng. 4(7): 773 83 (1991);コウ, M. S. らによる(1994), J. Immunol. 152, 2968-2976); スタドニッカらによるプロテインエンジニアリング7: 805-814 (1994) に記載されており、それら各々は、ここで援用することでその内容をここに組み入れる。
【0039】
ヒトの単一クローン性抗体を分離する1つの方法は、ファージディスプレイ技術を用いることである。ファージディスプレイは、例えば、ダワーらによるWO 91/17271, マキャファーティらによるWO 92/01047 およびケイトンとコプロウスキーによるProc. Natl. Acad. Sci. USA 87: 6450-6454 (1990) に記載されており、それら各々の内容は、援用することでここに組み入れる。ヒトの単一クローン性抗体を分離する別の方法は、内生的に免疫グロブリンを生成せず、ヒト免疫グロブリン遺伝子座を含有するよう操作されるトランスジェニックの動物を用いることである。例えば、ヤコボビッツらによるProc. Natl. Acad. Sci. USA, 90: 2551 (1993); ヤコボビッツらによるネイチャー, 362: 255-258 (1993); ブラガーマンらによるYear in Immuno., 7: 33 (1993); WO 91/10741、WO 96/34096、WO 98/24893または米国特許公報第20030194404号、米国特許公報第20030031667号、米国特許公報第20020199213号が参照され、それら各々の内容は、援用することでここに組み入れる。
【0040】
抗体フラグメント(断片)は、組み換えDNA技法、または完全な抗体を酵素でまたは化学的に分割することで生成される。「抗体断片」は、全長を保った完全な抗体の一部、好ましくは、抗原結合または完全な抗体の可変領域を備え、抗体断片から形成される多重特異性(二重特異性、三重特異性等)抗体を含んでいる。抗体断片の例には制限が無く、Fab、Fab’、F(ab’)2、Fv[可変領域]ドメイン抗体(dAb)[ワードらによるネイチャー341: 544-546, 1989]、相補性判別領域(CDR)断片、単鎖抗体(scFv)[バードらによるサイエンス242: 423-426, 1988、およびハスチョンらによるProc. Natl. Acad. Sci. USA 85: 5879-5883, 1988、選択的にポリペプチドリンカーを含む;また選択的に多重特異性である、グルバーらによるJ. Immunol. 152: 5368 (1994)]単鎖抗体断片、ダイアボディ[EP 404, 097; WO 93/11161; ホリンガーらによるProc. Natl. Acad. Sci. USA, 90: 6444-6448 (1993)]、トライアボディ、テトラボディ、ミニボディ[オラフセンらによるProtein Eng Des Sel. 2004 Apr; 17(4): 315-23]、線形抗体[ザパタらによるProtein Eng. 8(10): 1057-1062 (1995)];キレート化組み換え抗体[ネリらによるJ Mol Biol. 246: 367-73, 1995]、トライボディまたはバイボディ[スクーンジャンズらによる J Immunol. 165: 7050-57, 2000; ウィレムズらによるJ Chromatogr B Analyt Technol Biomed Life Sci. 786: 161-76, 2003]、イントラボディ[ビオッカらによるEMBO J. 9: 101-108, 1990; コルビーらによるProc Natl Acad Sci USA. 101: 17616-21, 2004]、ナノボディ[コルテツレタモゾらによるキャンサーリサーチ64: 2853-57, 2004]、抗原結合ドメイン免疫グロブリン融合タンパク質、キャメル化抗体[デスミターらによるJ. Biol. Chem. 276: 26285-90, 2001; エワートらによるバイオケミストリー41: 3628-36, 2002、米国特許公報第20050136049号および米国特許公報第20050037421号]、VHH含有抗体、ミメチボディ[米国特許公報第20050095700号および米国特許公報第20060127404号; WO 04/002424 A2; WO 05/081687 A2]またはそれらの変種または誘導体および、抗体が所望の抗原結合活性を保持する限りにおいて、CDRシーケンスのような、ポリペプチドへの特定の抗原結合を起こすのに十分な免疫グロブリンの少なくとも一部を含むポリペプチドが含まれてる。
【0041】
「変種」という語は、抗体との関係で用いられるとき、その変種が、所望のターゲット結合親和力または生物学的活性を保持するとすれば、可変領域または可変領域と等価な部分に少なくとも1つのアミノ酸が置き換えられるか削除されるか挿入されるかしている抗体のペプチドシーケンスのことを称する。加えて、本発明の抗体は、一定領域においてアミノ酸を変更して、半減期またはクリアランス、ADCCおよび/またはCDC活性を含めて抗体のエフェクター機能を変更する。そのような変更は、薬物動態を高めるか、例えば癌の治療をする上で抗体の有効性を高める。IgG1の場合、一定領域、とりわけヒンジまたはCH2領域の変更は、ADCCおよび/またはCDC活性を含めてエフェクター機能を増大または減少させる。他の例においては、IgG2一定領域が変更されて、抗体−抗原集合体の形成が低減される。IgG4の場合、一定領域、とりわけヒンジ領域の変更によって、半抗体の形成が低減される。
【0042】
「誘導体」という語は、抗体との関係で用いられるとき、治療または診断試薬への共役、(例えば、放射性核種または種々の酵素での)ラベリング、ペジレーション(ポリエチレングリコールでの誘導体化)のような共有結合高分子付着、および人工アミノ酸の化学合成による挿入または置き換えによって共有結合で変更される抗体のことを称する。本発明の誘導体は、本発明の、誘導されていない分子の結合特性を保持する。癌をターゲットとする抗体の細胞毒性試薬、例えば、放射性同位体(例えば、I131、I125、Y90およびRe186)、化学療法剤、または毒素への共役は、癌細胞の破壊を高める。
【0043】
ターゲット分子にとって「特定の」免疫糖タンパク質は、他のどのターゲットよりも大きな親和力で、そのターゲットに結合する。本発明の免疫糖タンパク質は、Kaが少なくとも約104M−1またはその代わりに少なくとも約105M−1、106M−1、107M−1、108M―1、109M−1または1010M−1というターゲットについて親和力を有する。そのような親和力は、通常の技法、例えば、BIAコアインスツルメントを用いることまたは放射能標識のついたターゲット抗原を用いる放射免疫測定によって容易に測定される。親和力データは、例えば、スキャッチャードらによるAnn N.Y. Acad. Sci., 51: 660 (1949) の方法によって分析される。
【0044】
糖修飾因子
「糖修飾因子」は、好ましくは、分子量が<1000ドルトンの、ポリペプチドに付着する炭水化物(糖)の一部である糖の付加、除去または修飾に関与する酵素の活性を抑制する小さな有機化合物である。グリコシル化は、小胞体(「コアグリコシル化」)およびゴルジ体(「端末グリコシル化」)で起こる極めて複雑なプロセスである。
【0045】
本発明においては、初期段階の糖修飾因子の活性を抑制するRNAiまたはアンチセンスを含む、グリコシル化酵素の、ポリペプチドベースまたはポリヌクレオチドベースの抑制体が有用であるが、「糖修飾因子」の定義からは除外する。
【0046】
ここで用いられるように、「初期段階の糖修飾因子」は、N‐アセチルグルコサミンをマンノースに加える前の1つ以上のグリコシル化工程のインヒビターのことを称し、ERグルコシダーゼI、ERグルコシダーゼII、ERマンノシダーゼ、ゴルジマンノシダーゼIA、ゴルジマンノシダーゼIB、ゴルジマンノシダーゼICおよびGlcNAc‐トランスフェラーゼIが含まれる。
【0047】
それに続くグリコシル化工程には、ゴルジマンノシダーゼII、GlcNAc‐トランスフェラーゼII、ガラクトシルトランスフェラーゼおよびシアリルトランスフェラーゼ、フコシルトランスフェラーゼおよびフコキナーゼが含まれる。
【0048】
代表的な糖修飾因子には、以下のいずれかが含まれる。カスタノスペルミンは、グルコシダーゼIおよびIIのインヒビターであると考えられている。デオキシフコノジリミシンは、フコシダーゼのインヒビターである。6‐メチル‐テトラヒドロ‐ピラン‐2H‐2、3、4‐トリオールは、インビトロで、L‐フコースのリン酸化、すなわち、GDP‐L‐フコースの生合成の第一工程を阻害することが報告されている。6、8a‐ジエピカスタノスペルミンは、フコシルトランスフェラーゼのインヒビターであると報告されている。1‐N‐イミュノシュガーAおよびB(それぞれ、1‐ブチル‐5‐メチル‐ピペリジン‐3、4‐ジオール塩酸塩および5‐メチル‐ピペリジン‐3、4‐ジオール塩酸塩としても知られている)は、フコシルトランスフェラーゼのインヒビターであることが報告されている。デオキシマンノジリミシン(DMJ)は、ERマンノシダーゼIのインヒビターである。キフネンシン(Kf)は、ERマンノシダーゼIのインヒビターである。スワインゾニン(Sw)は、ERマンノシダーゼIIのインヒビターである。モネンシン(Mn)は、コアオリゴ糖の伸長に干渉するERとゴルジとの間の細胞内タンパク質輸送のインヒビターである。
【0049】
本明細書に記述されているデータが示すように、さまざまなグリコシダーゼおよび/またはマンノシダーゼのインヒビターによって、ADCC活性を増大する1つ以上の所望の効果が得られ、Fc受容体の結合が増大され、グリコシル化パターンが変えられている。
【0050】
代表的な例においては、カスタノスペルミン(MW189.21)が、培地に添加され、最終の濃度を約200μM(約37.8μg/mLに相当する)とするか、濃度範囲を約10、20、30、40、50、60、70、80、90、100、110、120、130、140または150μMより大きく、約300、275、250、225、200、175、150、125、100、75、60または50μg/mLまでとする。例えば、10−50か、50−200か、50−300か、100−300か、150−250μMの範囲が意図されている。
【0051】
他の代表的な例において、DMJ、例えば、DMJ−HCl(MW199.6)が培地に加えられて、最終の濃度を約200μM(約32.6μgDMJ/mLに相当する)とするか、濃度範囲を約10、20、30、40、50、60、70、80、90、100、110、120、130、140または150μMより大きく、約300、275、250、225、200、175、150、125、100、75、60または50μg/mLまでとする。例えば、10−50か、50−200か、50−300か、100−300か、150−250μMの範囲が意図されている。
【0052】
他の代表的な例において、キフネンシン(MW232.2)が培地に加えられて、最終の濃度を約10μM(約2.3μg/mLに相当する)とするか、濃度範囲を約0.5、1、2、3、4、5、6、7、8、9または10μMより大きく、約50、45、40、35、30、25、20、19、18、17、16、15、14、13、12または11μMまでとする。例えば、1−10か、1−25か、1−50か、5−10か、5−25か、5−15μMの範囲が意図されている。
【0053】
組み換え構造物、細胞および培養方法
ここで用いられるように、「ホスト細胞」は、蚕食性ハイブリドーマを特に除外するが、グリコシル化(すなわち、ポリペプチドのアミノ酸への糖の付加)が可能であり、かつ組み換え手段を通して修飾され、レベルの増大したタンパク質プロダクトを発現させる他の細胞を含む。組み換え修飾とタンパク質プロダクトを発現する能力を保持するホスト細胞の子孫も「ホスト細胞」という語の中に含まれる。
【0054】
発現ベクトルまたは規定シーケンスの代表的な要素には、複製の元になるもの、プロモーター、オペレーターまたは転写および翻訳の媒体となるその他の要素が含まれる。プロモーターは、構成性であるか活性であり、かつさらに細胞種特異であるか、組織特有であるか、個々の細胞に特異有であるか、イベント特異であるか、一時的特異であるか、誘導性である。イベントに特異なプロモーターは、イベントが起こるときのみに活性であるか上方制御される。プロモーターに加えて、抑制体シーケンス、負の調節遺伝子または組織に特有のサイレンサーが挿入されて特異でない発現を低減する。他の要素には、内部リボゾーム結合サイト、ポリアデニル化シーケンスを含む転写ターミネータシーケンス、スプライスドナーおよび受容体サイト、エンハンサー、選択可能なマーカー等が含まれる。
【0055】
培地には、当業界で知られている必要なまたは望ましい成分をいずれも含み得るが、例えば、グルコースを含む糖類、必須および/または非必須アミノ酸、脂質および脂質前駆体、核酸前駆体、ビタミン、無機塩、希少金属を含む微量要素および/または細胞成長因子が含まれる。培地は、化学的に定義され、漿液、植物加水分解物またはその他の誘導された物質が含まれる。培地は、実質的にまたは完全に漿液や動物要素が含まれない。「実質的に漿液が含まれない」は、媒体には漿液が欠如しているか、わずかな量の漿液しか含んでいないことを意味する。細胞培養の間に枯渇される代表的な補遺のアミノ酸には、アスパラギン、アスパラギン酸、システイン、シスチン、イソロイシン、ロイシン、トロプトファンおよびバリンが含まれる。
【0056】
商業的に入手できる脂質および/または脂質前駆体には、コリン、エタノールアミンまたはホスホエタノールアミン、コレステロール、オレイン酸のような脂肪酸、リノール酸、リノレン酸、メチルエステル、例えば、酢酸塩の形でのD‐アルファ‐トコフェノール、ステアリン酸、ミリスチン酸、パルミチン酸、パルミトオレイン酸、またはアラキノン酸が含まれる。必須アミノ酸には、アルギニン、ヒスチジン、イソロイシン、ロイシン、リジン、メチオニン、フェニルアラニン、スレオニン、トリプトファンおよびバリンが含まれる。非必須アミノ酸には、アラニン、アスパラギン、アスパラギン酸塩、システイン、グルタミン酸塩、グルタミン、グリシン、プロリン、セリンおよびチロシンが含まれる。適当な塩として供給され、商業的に入手できる無機または微量要素には、ナトリウム、カルシウム、カリウム、マグネシウム、銅、鉄、亜鉛、セレニウム、モリブデン、バナジウム、マンガン、ニッケル、ケイ素、錫、アルミニウム、バリウム、カドミウム、クロム、コバルト、ゲルマニウム、カリウム、銀、ルビジウム、ジルコニウム、フッ化物、臭化物、ヨウ化物および塩化物が含まれる。媒体にはまた、混合や通気から細胞が保護されるように、非イオン性界面活性剤または表面活性剤が選択的に含まれる。培地はまた、重炭酸ソーダ、一塩基性および二塩基性リン酸塩、HEPSおよび/またはトリスのような緩衝剤を含む。培地はまた、酪酸ナトリウムまたはカフェインのようなタンパク質生成の誘発剤を含む。
【0057】
本発明によって、免疫糖タンパク質を生成する方法もまた提供されるが、任意の培地において、またはここに記述されるいずれかの条件のもとで、ホスト細胞を培養することを含むものである。当該方法はさらに、ホスト細胞または培地から免疫糖タンパク質を調製する工程を含む。糖修飾因子は、最初の培地に含まれているか、または最初の成長段階またはその後の段階で添加される。組み換えタンパク質が媒体中に分泌されるとき、培地を周期的に取り出すことができ、また何回かの取り出しサイクルを経て新しい培地と置き換えることができる。
【0058】
治療用のタンパク質生成に広く用いられているCHO細胞が好ましいけれども、グリコシル化タンパク質を生成することが当業界で知られているいずれのホスト細胞を用いてもよく、これには、酵母細胞、植物細胞、植物、昆虫細胞およびほ乳類の細胞が含まれる。代表的な酵母細胞には、ピキア属(Pichia)、例えば、P. パストリス(Pastoris)およびサッカロミセス属(Saccharomyces)、例えば、S. セレビシエ(cerevisiae) 並びにシゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)、 クリベロミセス属(Kluyveromyces)、 K. ザクティス(Zactis)、 K. フラギリス(fragilis)、 K.ブルガリカス( bulgaricus)、 K. ウィッケラミイ(wickeramii)、 K. ワルティイ(waltii)、 K. ドロソフィララム(drosophilarum)、 K.(サルノトレランス) thernotolerans、 K.(マルキアヌス) marxianus、 K. (ヤルロウィア)yarrowia、 トリコデルマ・リージア(Trichoderma reesia)、 アカバンカビ(Neurospora crassa)、シュワンニオミセス( Schwanniomyces)、 シュワンニオミセス オッシデンタリス(Schwanniomyces occidentalis)、ニューロスポラ( Neurospora)、 ペニシリウム属(Penicillium)、 トチポクラディウム(Totypocladium)、 アスペルギウス属(Aspergillus)、 アスペルギウスニジュランス(A. nidulans)、 クロコウジカビ(A. niger)、 ハンゼヌラ(Hansenula)、 カンジダ属(Candida)、 クロエケラ属(Kloeckera)、トルロプシス( Torulopsis)およびロドトルラ属(Rhodotorula)が含まれる。代表的な昆虫細胞には、オートグラファカリフォルニカ(Autographa californica)およびスポドプトーラフルギペルダ( Spodoptola frugiperda)並びにショウジョウバエ属(Drosophila)が含まれる。代表的なほ乳類の細胞には、CHO、BHK、HEK-293、NS0、YB2/3、SP2/0の変種およびPER-C6またはHT1080のようなヒト細胞、並びにVERO、HeLa、COS、MDCK、NIH3T3、Jurkat、Saos、PC-12、HCT 116、 L929 、Ltk-、WI38、CV1、TM4、W138、Hep G2、MMT白血病細胞系、胚幹細胞または受精卵細胞が含まれる。
【0059】
細胞は、当業界で知られているいかなる培養システムにおいても、かついかなる方法で培養されてもよく、それには、T‐フラスコ、スピナーおよび振とうフラスコ、ローラーボトル、撹拌タンクバイオリアクターが含まれる。付着依存性細胞もまた、撹拌タンクバイオリアクターに懸濁して保持されるマイクロキャリア、例えば、高分子球体で培養することができる。あるいは、細胞は、単細胞懸濁液において成長させることができる。培地が添加されるのは、バッチ処理であって、例えば、培地が単一のバッチで細胞に一度に添加されても良いし、培地の小さなバッチが周期的に添加される流加バッチ処理であっても良い。培地は、培養の終わりに回収されるか、培養中に何度かにわたって取り出される。継続的に潅流させる生成処理もまた当業界で知られており、新しい培地を、同じ量でリアクターから継続的に引き出しながら、培養に継続的に供給することが伴われる。潅流による培養は、一般にバッチによる培養よりも細胞密度が高く、繰り返して回収しながら何週間もまたは何ヶ月も維持できる。
【0060】
免疫糖タンパク質の使用
本発明の免疫糖タンパク質は、ターゲット分子が媒体となる疾患を処置する治療術として有用であり、例えば、発現したまたは細胞表面と結びついたターゲット分子を有する癌細胞を死滅させるための細胞崩壊剤として有用である。
【0061】
「処置」または「処置する」は、治療または予防または防止処置のことを言う。治療は処置を受ける被体の疾患の少なくとも1つの症状が良好となるか、被体において進行する疾患の悪化を遅延させ、あるいは関連する疾患が新たに発症することを防止する。改善された効果は、疾患状態について、当業界で周知の臨床基準を評価することで、査定される。
【0062】
免疫糖タンパク質の「治療で有効なドース」または「有効なドース」は、処置される疾患の1つ以上の症状が結果的に良くなるのに十分な化合物の量のことを称する。個人に活性成分を単独で投与するとき、治療に有効なドースは、その成分のみについて称する。組み合わせて投与されるとき、治療で有効なドースは、連続して投与されようとも同時に投与されようとも、結果的に治療の効果が得られる活性成分を合わせた量のことを称する。ドースは、患者の体重に基づいて投与され、例えば0.01乃至50mg/kgのドースであって、日単位または週単位で、あるいは二週間毎、三週間毎または月に一度投与される。
【0063】
本発明の免疫糖タンパク質をヒトまたは実験動物に投与するのに、組成物が非経口的な投与のためのものであるならば、1つ以上の薬学的に許容され得る担体または希釈剤、好ましくは、殺菌されたまたは無菌の担体また希釈剤を有する組成物において、分子を調製するのが好ましい。「薬学的にまたは薬理学的に受け入れ可能な若しくは許容され得る」という文言は、以下に記述されるような当業界で周知の経路を用いて投与されるとき、アレルギーまたはその他の有害な反応を引き起こさない分子状態および組成物のことを称する。「薬学的に受け入れ可能な担体」には、臨床的に有用な溶剤、分散媒体、被覆、抗菌性剤および抗真菌性剤、等張剤および吸収遅滞剤等が全て含まれる。一般に、組成物にはまた、発熱物質並びに被体に有害であり得るその他の不純物は実質的に含まれない。
【0064】
免疫糖タンパク質が投与されるのは、経口であっても、局部的であっても、経皮であっても、非経口であっても、吸入スプレーによっても、経膣であっても、頭蓋内注射によってでも良い。ここで用いられる非経口という語には、皮下注射、静脈内、筋肉内、嚢内への注射または注入技法が含まれる。静脈内、皮膚内、筋肉内、乳房内、腹腔内、髄膜内、眼球後方、肺内への注射および/または特定の場所への手術での注入による投与も同様に意図されている。
【0065】
1つの例において、処置を必要とする癌または病気にかかった組織の箇所に、その箇所への直接の注射によるか、製剤を内部に放出できる、持続的な放出法または持続的な吐出メカニズムを介して投与が行われる。例えば、組成を持続して放出可能な生分解性の微小球またはカプセルその他の生分解性高分子構成物(例えば、可溶ポリペプチド、抗体または小分子)は、癌の近辺に注入される本発明の製剤に含ませることができる。
【0066】
また治療組成物を、患者の複数の箇所に放出しても良い。複数の投与が同時に行われても、また継続した期間内に投与されても良い。
【0067】
特に、水溶液の注入が好ましい。水性の組成物は、真空状態で凍結乾燥して貯蔵され、使用に先立って適切な担体で復元できる。この技法は、従来の免疫グロブリンで有効であることが示されている。真空状態で凍結乾燥し、復元する技法は、適切であれば任意のものを用いることができる。当業者には、真空状態で凍結乾燥して復元すると、活性の損失の度合いがさまざまであり、それを補償するために使用のレベルを調節しなければならないということが理解される。
【0068】
あらゆる場合において、製剤は、殺菌され、かつ注射器での扱いが容易である程度に液化されていなければならない。例えば、レシチンのような被覆を用いること、分散している場合には必要な粒子のサイズを維持すること、および界面活性剤を用いることによって、適切な流動性を維持することができる。また、製造および貯蔵する条件のもとで安定していなければならず、バクテリアや菌類のような微生物の汚染作用を防いで保存される。微生物の作用は、さまざまな抗菌性剤および抗真菌性剤、例えば、パラベン、クロロブタノール、フェノール、ソルビン酸、チメロサール等によって防ぐことができる。多くの場合、等張剤、例えば、砂糖または塩化ナトリウムを含むことが望ましい。
【0069】
さらに、本発明での使用が意図されている組成物は、親水性と疎水性のバランスが良好であり、これによりインビトロおよびインビボの使用、特にインビボでの使用について実用性が高いが、他の組成物は、このようなバランスを欠いており実用性が相当に低い。とりわけ、本発明での使用が意図されている組成物は、身体での吸収および生物学的利用を可能とする水性媒体に可溶である度合いが適切であり、また一方、化合物が、細胞膜を横切って作用すると推定される箇所に至ることを可能とする程度に脂質にも可溶である。
【0070】
また、本発明で意図されているのは、免疫糖タンパク質組成物を第二の薬剤と一緒に投与することである。
【0071】
さらに、本発明には、本発明の方法を実施するために、1つ以上の化合物または組成物が容易に使用できるようにパッケージとなった状態で、それらを備えるキットまたは製品が含まれる。1つの例において、そのようなキットには、選択的に第二の治療薬と一緒に、密封された瓶または器のような容器にパッケージされた状態で、本発明における免疫糖タンパク質が含まれており、その容器にはラベルが貼られているか、該方法を実施する上での、化合物または組成物の使用について記述するパッケージに納められている。好ましくは、その化合物または組成物は、ドース単位の形でパッケージされている。そのキットにはさらに、特定の投与経路によりその組成物を投与するのに適切な、またはスクリーニングアッセイを行うのに適切な装置が含まれる。好ましくは、当該キットには、該組成物の使用について記述されたラベルが入っている。
【0072】
さらに本発明では、免疫糖タンパク質が結びつくターゲットにより特徴付けられるか媒介される被体における、疾患、病状または障害を抑制、防止または処置するための医薬の製造で、本発明の免疫糖タンパク質を用いることが意図されている。
【図面の簡単な説明】
【0073】
【図1】図1は、様々な濃度のカスタノスペルミンを備える細胞媒体において成長するTRU‐016を発現するCHO細胞の成長を、cells/mlという細胞数で示す。
【図2】図2は、様々な濃度のカスタノスペルミンを備える細胞媒体において成長するTRU‐016を発現するCHO細胞の生存度を、生存細胞の%で示す。
【図3】図3は、様々な濃度のカスタノスペルミンの存在で培養される細胞が生成するTRU‐015のCD16結合を示し、かつ幾何学的に示された平均蛍光強度対カスタノスペルミン濃度を示す。
【図4】図4は、様々な濃度の6、8a‐ジエピカスタノスペルミン、スワインソニンまたはデオキシマンノジリミシン(DMJ)の存在で培養される細胞が生成するTRU‐016のCD16結合を、幾何学的に示された平均蛍光強度で示す。
【図5】図5は、様々な濃度のキフネンシンの存在で培養される細胞が生成するTRU‐016のCD16結合を、蛍光強度の平均で示す。
【図6】図6は、様々な濃度のカスタノスペルミンの存在で培養される細胞が生成するタンパク質Aが取り除かれたTRU‐016のCD16結合を、蛍光強度の平均で示す。
【図7】図7は、高親和性および低親和性ドナーのPBMCを用いて測定したTRU‐15のADCCを示し、添加TRU‐015の濃度対%特有の致死量をプロットした図である。
【図8】図8は、高親和性および低親和性ドナーのPBMCを用いて測定したTRU‐15のADCCを示し、添加TRU‐015の濃度対%特有の致死量をプロットした図である。
【図9】図9は、様々な濃度のカスタノスペルミンの存在で培養される細胞が生成するTRU‐016のADCCを示し、%特有の致死量対添加TRU‐016の濃度をプロットした図である。
【図10】図10は、様々な糖修飾因子の存在で培養される細胞が生成するTRU‐016のADCCを示し、%特有の致死量対添加TRU‐016の濃度をプロットした図である。
【図11】図11は、様々な糖修飾因子の存在で培養される細胞が生成するTRU‐016を投与されたマウスにおける薬物動態データを示す。
【図12】図12は、様々な糖修飾因子で処置した細胞が生成するTRU‐016を投与されたマウスの漿液におけるTRU‐016のCD16結合を示す。
【図13】図13は、腫瘍細胞をインプランとされ、様々な糖修飾因子で処置した細胞または処置していない細胞から生成されるTRU‐016を投与されたマウスにおいて8日後の腫瘍の相対容量を示す。
【図14】図14は、腫瘍細胞をインプラントされ、様々な糖修飾更因子で処置した細胞または処置していない細胞から生成されるTRU‐016を投与されたマウスの生存率%を示す。
【図15】図15は、カスタノスペルミンの存在で培養された細胞が生成するTRU‐015のCDCを示し、ヨウ化プロピジウムでポジティブ(正)の%(死亡細胞)対TRU‐015実験タンパク質の濃度をプロットした図である。
【図16】図16は、様々な糖修飾因子の存在で培養された細胞が生成するTRU‐016のCDCを示し、ヨウ化プロピジウムでポジティブ(正)の%(死亡細胞)対TRU‐016実験タンパク質の濃度をプロットした図である。
【図17】図17は、カスタノスペルミン濃度範囲に対する、TRU‐016に特異なタンパク質生成を示す。
【図18】図18は、カスタノスペルミン濃度範囲に対する、TRU‐016のCD37およびFcγRIIIa(CD16)への同時結合についてのアッセイ結果を示す。
【図19】図19は、カスタノスペルミン濃度範囲に対する、TRU‐016からCD37を発現する細胞の用量反応結合曲線を示す。
【図20】図20は、カスタノスペルミン濃度範囲に対する、TRU‐016のADCC活性曲線を示す。
【発明を実施するための形態】
【0074】
例
例1
SMIPプロダクトの製造
TRU‐016
CD37‐特有のSMIPが、共同で所有されている米国特許出願第10/627,556号および米国特許公開第2003/133939号、米国特許公開第2003/0118592号および米国特許公開第2005/0136049号に記述されており、それら各々は、援用することでその全体をここに組み入れられる。代表的なSMIP、すなわちTRU‐016は、以下に記載されるようにして生成される。
【0075】
TRU‐016[G28-1 scFv VH11S(SSC-P)H WCH2 WCH3]は、CD37抗原に結合する組み換え単鎖タンパク質である。TRU‐016のヌクレオチドおよびアミノ酸配列は、それぞれSEQ ID NOS: 1および2で並べられている。結合ドメインは、前段落に列挙されている特許公報において以前開示されたG28‐1抗体シーケンスに基づいた。結合ドメインは、モディファイされたヒンジ領域を通して、エフェクタードメイン、すなわち、ヒトIgG1のCH2およびCH3ドメインに接続されている。TRU‐016は、溶液において二量体として存在する。
【0076】
TRU‐016は、チャイニーズハムスター卵巣(CHO)ほ乳類細胞出現システムにおいて、組み換えDNA技術によって生成される。TRU‐016SMIPは、タンパク質A親和クロマトグラフィーでCHO培養上清から精製される。dPBSを用いると、50mLのrタンパク質A FFセファロースカラム(GEヘルスケアrタンパク質AセファロースFF、カタログ#17‐0974‐04)は、1.5カラム体積(CV)について5.0mls/min(150cm/hr)で平衡となる。AKTAエクスプローラ100Air(GEヘルスケアAKTAエクスプローラ100Air、カタログ#18‐1403‐00)を用いて、培養上清が1.7mls/minという流速でrタンパク質AセファロースFFカラムに加えられ、組み換えTRU‐016を捕捉する。そのカラムは、5カラム容量(CV)分のdPBSで洗浄され、そして1.0M NaCl、20mMリン酸ナトリウム、pH6.0で、さらに25mM NaCl、20mM NaOAc、pH5.0で洗浄される。これらの洗浄工程により、不特定に結合されたCHOホスト細胞タンパク質が、溶出後の生成物沈殿に貢献するrタンパク質Aカラムから取り除かれる。
【0077】
組み換えTRU‐016は、100mMグリシン、pH3.5を備えるカラムから溶出される。その溶出された生成物の一部である10mLが回収され、そしてその溶出された生成物が、0.5M 2‐(N‐モルフォリノ)エタンスルフォン酸(MES)pH6.0の溶出した容量の20%で、pH5.0とした。この溶出された生成物は、約25mg/mLのTRU‐016に濃縮されて濾過殺菌される。
【0078】
そして、精製されたタンパク質がGPCサイズ排除クロマトグラフィー(SEC)にかけられ、より高い分子量の集合体からTRU‐016(二量体)分子をさらに精製する。dPBSを用いて、1Lのスーパーデックス200FFセファロースを含むXK50/100カラム(GEヘルスケアXK50/100からのクロマトグラフィーカラム、カタログ#18‐8753‐01)が、1.5カラム容量(CV)について12.6mls/min(38cm/hr)で平衡となる。サンプルの54mls(3%CV)という最大容量が、カラムに適用される。カラムは、12.6ml/minで操作され、容出されたタンパク質は、40mlの画分に分けられる。各画分は、分析HPLCを用いて生成物品質について分析され、そして溶出された部分は、>95%POI(集合体になっていない)TRU‐016についてプールされる。この得られたプール物が、0.22μmで濾過殺菌される。そして、物質が、20mMリン酸ナトリウムと240mMショ糖で、pH6.0で濃縮されて形成される。
【0079】
グリコバリアントを精製する代わりの方法は、以下の通りである。TRU‐016が、タンパク質A親和クロマトグラフィーによってCHO培養上清から精製される。dPBSを用いて、1mL MabSelect親和クロマトグラフィーカラム(GEヘルスケアHitrap MabSelect、カタログ#28‐4082‐53)が、7カラム容量(CV)について1.0mL/minで平衡とされる。培養上清が、Aktaエクスプローラ100Air(GEヘルスケア、Aktaエクスプローラ100Air、カタログ#18‐1403‐00)を用いて流速1.0mL/minでMabSelectカラムに加えられ、組み換えTRU‐016を捕捉する。そのカラムは、20CV分のdPBSで洗浄され、そして20mMリン酸ナトリウム、5CV分の1.0M NaCl、pH7.0でそして3CV分のdPBSで洗浄する。
【0080】
組み換えTRU‐016が、10mMクエン酸塩(シトレート)、pH3.5でカラムから溶出され、そのカラムは、8CV分の10mMシトレート3.0でストリップされる。ストリップに続いてそのカラムはdPBSで5CV分再平衡とされる。タンパク質は、溶出の間、各画分に集められて、それらは吸着度に基づいてプールされ、このプールされた物質は、溶出物5mLにつき約400μLの0.55M 2‐(N‐モルフォリン)エタンスルフォン酸(MES)、pH6.0を加えてpH5.0とされる。この中和された溶出物は、濾過殺菌されて、活性アッセイとプロセス分析アッセイの双方に課される。
【0081】
親抗体のCD37細胞表面受容体への結合特異性がTRU‐016において保存されるということを確認するために実験が行われた。ヒトPBMCが、LSM密度勾配を持って単離され、非抱合TRU‐016およびPE抱合抗ヒトCD19でインキュベートされる。細胞は洗浄され、45分間氷上にて1:100FITC GAH IgG(Fc特異)でインキュベートされる。細胞は、洗浄され、セルクエストソフトウェアを用いるFACsCaliburインスツルメントで二色フロー血球計算によって分析される。細胞は、CD19染色によって、Bリンパ球または非Bリンパ球について振り分けられる。
【0082】
TRU‐016の濃度が増大すると、Bリンパ球でのFITC信号(CD19正ゲート)が、0.01‐1.0μg/mlから、約1μg/mLでの飽和または平均蛍光強度(MFI)1000に至るまで急速に増大する。反対に、非Bリンパ球数は、染色が検出可能であるが非常に低いので、scFvIgの濃度が増大すると、ゆっくりと増大する。
【0083】
TRU‐015
CD20‐特異のSMIPが同様に調製される。CD20‐特異のSMIPは、共同で所有される米国特許公報2003/133939、2003/0118592および2005/0136049において記述され、各々は、援用することでその全体をここに組み入れられる。代表的なSMIP、すなわち、TRU‐015を以下に記載する。
【0084】
TRU‐015は、CD20抗原に結合する組み換え単鎖タンパク質である。TRU‐15のヌクレオチドとアミノ酸シーケンスは、それぞれSEQ ID NOS:3および4で並べられる。結合ドメインは、公的に入手可能なヒトCD20抗体シーケンスに基づいた。結合ドメインは、修飾されたCSSヒンジ領域を通して、エフェクタードメイン、すなわち、ヒトIgG1のCH2およびCH3ドメインに接続される。TRU‐015は、溶液において二量体として存在する。
【0085】
TRU‐015は、SEQ ID NO:4のアミノ酸1−23からの2e12リーダーペプチドクーロン化シーケンスと、SEQ ID NO:4におけるポジション34で反映される、可変領域における残基11でリジンからセリン(VHL11S)アミノ酸が置き換えられた2H7ネズミ抗ヒトCD20軽鎖可変領域と、SEQ ID NO:4の残基129で始まるasp-gly3-ser-(gly4ser)2リンカーと、重鎖領域の端にセリン残基が無く、すなわち、VTVSSからVTVSに変化した2H7ネズミ抗ヒトCD20重鎖可変領域と、(CSS)シーケンスとワイルド型CH2およびCH3ドメインを備えるモディファイされたヒンジ領域を含むヒトIgG1 Fcドメインとを備える。
【0086】
例2
糖修飾因子(モディファイア)を有するホスト細胞の培養
TRU‐016またはTRU‐015 cDNAがトランスフェクトされたCHO細胞は、一般に以下に記述される手順によって、種々の糖修飾因子(modifier)について濃度を変えて、振とうフラスコまたは揺動バッグで培養された。
【0087】
振とうフラスコでの作業については、ログ位相ホスト細胞がテストすべき濃度の糖修飾因子とともに100,000 cells/mlで接種され、また選択的に50nMのメトトレキセートとともに接種された。
【0088】
細胞は、t=0で1350mLのEx-Cell 302培地(SATCバイオサイエンス;全てがインビトロゲンからの非必須アミノ酸、ピルケート、L‐グルタミン、ペン/ストレップ、HTサプリメントおよびインシュリンを加えて)に3x10E6/mLで接種され、T>=72時間で合計容量5Lとされた。細胞は、37℃でおよび5%の二酸化炭素で培養され、6−7日目から毎日成長および生存度についてモニターした。上清は、典型的には10−12日目に、細胞の生存度が60%未満に落ちたとき回収された。
【0089】
Naアジドを加えて0.02%とし、細胞を遠心で取り除き、かつ上清を0.22μMフィルターを通して濾過殺菌した。他の例に記述されるアッセイは、示された通りに、上清において行われるものもあり、また他のアッセイは、さらなるタンパク質A精製を経た材料に対して行われるものもあった。揺動バッグの作業について、ログ位相ホスト細胞が、テストされるべき濃度の糖修飾因子(モディファイア)ともに10−20%という条件のEx-Cell 302媒体(SATCバイオサイエンス;全てがインビトロゲンからの非必須アミノ酸、ピルケート、L‐グルタミン、ペン/ストレップ、HTサプリメントおよびインシュリンを加えて)において、100,000−200,000 cells/mLで5L揺動バッグ中に接種された。細胞は、37℃でかつ5%の二酸化炭素で培養され、成長と生存度について毎日モニターされた。上清は、典型的には、11−12日目に、または細胞生存率が50%未満に低下したとき回収された。
【0090】
ソルバールレジェンド(Sorvall Legend)における3000rpm(1932rcf)での20分間の遠心によって細胞が取り除かれ、上清は濾過殺菌された。他の例に記述されるアッセイは、示されたように、上清において行われたものもあるが、他のアッセイは、さらなるタンパク質A精製を受けた材料において行われた。
【0091】
種々の糖修飾因子の濃度を変えて培養される細胞により生成されるTRU‐016は、以下に記述されるように、CD16結合、ADCC、CDC、薬物動態パラメータ、およびインビボの活性についてアッセイされる。
【0092】
図1および図2は、典型例を示しており、1000μMまでの濃度の糖修飾因子カスタノスペルミンでの処置は、サンプルを採取した期間全体を通して(144時間まで)、細胞数やパーセント細胞生存度に影響を与えなかったということが示されている。
【0093】
例3
FcRsへの結合
例2により生成される免疫糖タンパク質を、受容体の細胞外ドメインがネズミIgG2a Fcに融合する、Fcγ受容体の可溶性Ig‐融合バージョンへの結合について、インビトロでアッセイした。
【0094】
可溶性Fcγ受容体材料は、Fcγ受容体I(Genbank Acc. No. BC032634)、IIa(Genbank Acc. No. NM_021642)、IIb(Genbank Acc. No. BC031992)およびIII‐V158(高親和対立遺伝子)(Genbank Acc. No. X07934)およびIII‐F158(低親和対立遺伝子)それぞれの細胞外ドメインを、残基238(MIgG2aP238S)にPro乃至Serの突然変異があるネズミIgG2a Fcに融合することによって生成された。FcγRIII(CD16)の双方の形について、HE4リーダーは、CD16アミノ酸1‐178でクローン化され、そしてMIgG2aP238Sに融合される。
【0095】
アッセイは以下のように行われた。1%ウシ胎仔漿液(FBS)を含むリン酸塩で緩衝された生理食塩水(PBS)において、45分間、5μg/mlのTRU‐015かTRU‐016かを入れたコスター96ウェルプレートにおいて氷上で500,000のWIL2‐S細胞(表面上においてCD37並びにCD20を発現するBリンパ腫細胞系(line))が培養された。結合されていないTRU‐015またはTRU‐016は、細胞をスピンし、希釈剤(PBS+1%FBS)で洗浄し、ソルバールレジェンド RTにおいて1200rpmで再度スピンすることで取り除かれた。そして細胞は、45分間、氷上で1μg/mlの濃度の同じ希釈剤において、所望のFcγR‐MIgと融合して培養された。
【0096】
そして、錯体(WIL2-S細胞/SMIP/FcγR-MIg)が、1:100の希釈の、PE抱合AffiniPure F(Ab’)2 ヤギ抗マウスIgG[ジャクソンイミュノリサーチ(Jackson Immunoresearch)](ヒトFcと最小の交差反応性を有するマウスFc‐特異の抗体)で培養された。細胞は、セルクエストソフトウェア(ベクトンディキンソン)を用いるFACsCaliburでの一色フロー血球計算によって分析された。
【0097】
このアッセイで、精製されたTRU‐016タンパク質の代わりに例2のTRU‐016上清が用いられるときは、上清のSMIP濃度は、TRU‐016基準に沿って、希釈上清でWIL2‐S細胞を直接染色することによって定量化された。TRU‐016は、1:50希釈のFITC抱合F(Ab’)2ヤギ抗ヒト(ガンマ)[Caltag H10101]で染色することによって検出された。
【0098】
低親和対立遺伝子か高親和対立遺伝子への結合が、ADCC活性に同じように相関するように測定された。CD16(低または高親和対立遺伝子)結合の増大がADCC活性の増大と相関していた。
【0099】
典型的な結果を、図3乃至図6に示す。
【0100】
0、2、5、10、30または100μg/mLのカスタノスペルミンを含む媒体において培養されたCHO細胞により生成されるTRU‐015精製のタンパク質がCD16結合についてテストされた(低親和対立遺伝子)。幾何平均蛍光強度の典型的な結果が図3に示されており、培地におけるカスタノスペルミンの濃度が増大すると、CD16結合が、用量依存的に増大することが示されている。
【0101】
50または250μMという濃度で6、8a‐ジエピカスタノスペルミンを、50または250μMという濃度でスワインソニンをまたは50または250μMという濃度でデオキシマンノジリミシン(DMJ)を含む培地で培養されたCHO細胞が生成したTRU‐016上清が、CD16結合のためにテストされた。平均蛍光強度の典型的な結果が図4に示されており、DMJのいずれの濃度もCD16結合を増大させたことが示されている。これらの濃度では、6,8a‐ジエピカスタノスペルミンまたはスワインソニンについては効果が見られないが、精製されたタンパク質でさらなるテストが行われて効果を測定する。
【0102】
0、0.5、1、3、5または10μMという濃度のキフネンシンを含む培地で培養されたCHO細胞が生成するTRU‐016上清が、CD16結合についてテストされた。平均蛍光強度の典型的な結果が図5に示されており、キフネンシンは、CD16結合を増大させるのにDMJよりずっと大きな効力があり、濃度が最低の0.5μMであったときでさえ、CD16結合を大きく増大させたことが示されている。
【0103】
0、10、25、50、100または200μMのカスタノスペルミンを含む培地で培養された、CHO細胞により生成されるタンパク質A精製TRU‐016が、CD16結合についてテストされた。平均蛍光強度の典型的な結果が図6に示されており、培地におけるカスタノスペルミンの濃度が増大するとCD16結合が用量依存的に増大することが示されている。
【0104】
例4
ADCC活性
精製されたTRU‐016のADCC活性を測定するために、標識化したBJAB B細胞をターゲットとして用い、ヒト末梢血単核細胞(PBMC)をエフェクター細胞として用いた。IMDM/10%FBSにおいて37℃で二時間、BJAB B細胞(107個の細胞)が、500μCi/mL51Crクロム酸ナトリウムで標識付けされた。PBMCは、リンパ球分離メディア(LSM、ICNバイオメディカル)勾配について分画することで、ヘパリン処理したヒト全血から単離された。10%FBSをともなってRPMI培地に試薬サンプルが加えられ、各試薬の一連の希釈物が調製された。51Crで標識付けされたBJABを、2x104cells/wellで加えられた。そして、PBMCが、25:1のエフェクター(PBMC):ターゲット(BJAB)の最終比について5x105cells/wellだけ加えられた。反応は96ウェルプレートの四つ組ウェルで提供された。TRU‐016の一連の希釈物が、図に示されるように、10ng/mLから20μg/mLの範囲の最終濃度でウェルに加えられた。回収したり計数したりするのに先立って、5%CO2において37℃で6時間反応を進行させた。放出されたCMPが50μlの乾燥した培養上清からPackard TopCountNXTで測定された。パーセント特有の死亡率が減算(サンプルのcpm[4倍のサンプルの平均]‐cpm自発放出)/(cpm最大放出‐cpm自発放出)x100によって計算され、データが%特有の死亡率対TRU‐016濃度としてプロットされた。
【0105】
典型的な結果が、図7乃至図10に示されている。
【0106】
0、2、5、10、30または100μg/mLのカスタノスペルミンを含む培地で培養されたCHO細胞が生成するTRU‐015精製タンパク質が、高親和(V/V158)および低親和(F/F158)CD16ドナーからのPBMCを用いて測定されたADCCについてテストされた。%特有の死亡率の典型的な結果が、図7および図8(それぞれ、高親和ドナーおよび低親和ドナー)に示されており、培養媒体におけるカスタノスペルミンの濃度が増大すると、ADCC活性が用量依存的に増大することが示されている。
【0107】
0、10、25、50、100または200μMのカスタノスペルミンを含む媒体で培養されたCHO細胞が生成するTRU‐016精製のタンパク質が、ADCCについてテストされた。%特有の死亡率の典型的な結果が図9に示されており、培地におけるカスタノスペルミンの濃度が増大するとき、ADCC活性が用量依存的に増大することが示されている。
【0108】
200μMのDMJ、10μMのキフネンシンまたは200μMのカスタノスペルミンを含む培地で培養されたCHO細胞により生成されるTRU‐016精製のタンパク質が、ADCCについてテストされた。%特有の死亡率の典型的な結果が図10に示されており、糖修飾因子の、これらの濃度の全てについて、CHO細胞が生成する免疫糖タンパク質のADCCが改良されたということが示されている。
【0109】
例5
CDC活性
例2により生成されるTRU‐016精製のタンパク質のCDC活性を測定するために、ラモスB細胞が、75μlに5x105cells/wellでイスコベス(ギブコ/インビトロゲン、グランドアイランド、NY)に懸濁された。指示された二倍の濃度でTRU‐016(75μl)が細胞に加えられた。漿液を含まないイスコベスにおいて遠心分離および洗浄を行うのに先立ち、45分間結合反応を進行させた。種々の濃度のヒト漿液(補体を含む)を備えるイスコベスにおいて細胞が再懸濁された。細胞は、37℃で60分間培養された。細胞は、遠心分離によって洗浄され、0.5μg/mlのヨウ化プロピジウムを備える染色媒体において再懸濁された。FACsCaliburおよびCellQuestソフトウェアを用いるフロー血球計算による分析に先立ち、暗所において室温で15分間サンプルが培養された(ベクトンディキンソン)。
【0110】
未処置のCHO細胞が生成したTR‐015精製タンパク質または30μg/mlのカスタノスペルミンで処置されたCHO細胞が、CDC活性についてテストされた。結果を図15に示す。
【0111】
未処置のCHO細胞が生成するTRU‐016精製のタンパク質、または200μMのDMJ、10μMのキフネンシンまたは200μMのカスタノスペルミンを含む媒体で培養されたCHO細胞が、CDC活性についてテストされた。結果を図16に示す。
【0112】
これらの結果により、糖修飾TRU‐015またはTRU‐016についてのCDCは、未処置のCHO細胞が生成する、対応するタンパク質のCDCに類似していたことが示され、ホスト細胞の培地に糖修飾因子が存在することで、ホスト細胞が生成する免疫糖タンパク質のCDCに大きな影響が及ぼされなかったことを示している。
【0113】
例6
薬物動態プロフィール
雌のBALB/cマウスに200μgのTRU‐016テストタンパク質(未処置のCHO細胞が生成したTRU‐016または200μMのDMJ、10μMのキフネンシンまたは200μMのカスタノスペルミンで処置したCHO細胞が生成したTRU‐016)を時刻0で静脈注射した。注射後15分、2、6、24、48、72、96および192時間で漿液のサンプルが集められた(各時点でマウス3匹)。
【0114】
TRU‐016テストサンプルの各々の漿液濃度を、CD37+ラモスヒト細胞ラインを用いるFACSに基づく結合アッセイで測定した。CD37+ラモス細胞(5x105cells/well)がテストすべき漿液サンプルと一緒に底の平らな96ウェルプレートで培養された。スパイク漿液サンプルを用いて標準曲線とした。細胞は、4℃で1時間培養され、検出抗体を添加する前に洗浄した。TRU‐016テストタンパク質のCD37+ラモス細胞への結合は、フルオレッセイン結合ヤギ抗ヒトIgG Fcγフラグメン特異抗体を用いて検出された。標準曲線を用いて、抗原濃度の関数として結合曲線を描いた。要するに、標準曲線は、FACS緩衝剤で1:20に希釈された通常のマウス漿液に加えられたTRU‐016テストタンパク質の周知の様々な濃度からなる。標準曲線は、各プレートにおいて二重に引かれていた。平均蛍光強度(MFI)が、FACS分析からソフトマックスプロソフトウェアに移入され、TRU‐016テストタンパク質の漿液濃度を計算するのに用いられた。
【0115】
薬物動態研究の結果、200μMのDMJ、10μMのキフネンシンまたは200μMのカスタノスペルミンを含む媒体で培養されたCHO細胞が生成したTRU‐016(図11に示されている)が、マウスに投与されたとき、未処置のCHO細胞が生成したTRU‐016に類似する薬物動態プロフィールを呈示するということが示され、ホスト細胞の培地における糖修飾因子は、半減期またはその他の薬物動態パラメータに大きな影響を及ぼさないということが示された。
【0116】
TRU‐016の投与後48、72、96および192時間でマウスから得られる、TRU‐016を含む漿液についてCD16アッセイを繰り返すことで、テストのあらゆる時点で、増大したCD16結合活性が漿液によって保持されるということが示された。結果を図12に示す。
【0117】
例7
インビボの糖修飾免疫糖タンパク質活性
ヌードマウスが、0日目に、5x106ラモス細胞を皮下に投与され、0、2、4、6および8日目に、200μgの対照ヒトIgGまたは200μMのDMJ、10μMのキフネンシンまたは200μMのカスタノスペルミンで処置されたCHO細胞が生成したTRU‐016テストタンパク質を静脈に注射された。典型的には、マウスは6日以内に腫瘍が発達し、その後すぐに死亡する。腫瘍は、デジタルカリパスおよびラブキャットソフトウェアで週に3度測定され、腫瘍の体積は、1/2[長さx(幅)]2で計算される。また、週に1度体重を測定する。
【0118】
マウスは、腫瘍が1500mm3(金曜日に1200mm3)の大きさに到達するとき、死亡する。マウスはまた、腫瘍から潰瘍が形成されると死亡し、体重が20%以上失われてしまうと、腫瘍によって動物としての動きが抑制されてしまう。
【0119】
研究が開始されてから8日目の相対的な腫瘍の体積について平均の結果を図13に示す。研究開始後の生存率%についてのデータが、図14および以下の表1に示される。
【0120】
【表1】
【0121】
このインビボ研究の結果、200μMのDMJ、10μMのキフネンシンまたは200μMのカスタノスペルミンで処置されたCHO細胞が生成したTRU‐016は、癌の動物モデルにおいて腫瘍の体積を低減し、平均生存時間を増大させることができた。
【0122】
例8
様々な濃度のカスタノスペルミンのタンパク質生成への影響
さらなる実験を行って、カスタノスペルミンの濃度の、TRU‐016の細胞生存度、密度および特定のタンパク質生成への影響を測定した。
【0123】
実験の開始に先立って、TRU‐016をトランスフェクトしたCHO細胞が、加湿された培養器において37℃かつ5%の二酸化炭素で、1x非必須アミノ酸(メディアテック)、1xピルビン酸ナトリウム(メディアテック)、4mMのL−グルタミン(メディアテック)、500nMのメトトレキセート(MPバイオメディカルズ)および1mg/Lの組み換えインシュリン(リコンビュリン‐ジブコ/インビトロゲン社)を補ったEx-CellTM 302というCHO漿液の含まれない媒体(SAFCバイオサイエンス)の入った振とうフラスコで成長された。殺菌されたカスタノスペルミンを、蒸留した/イオンを取り除いた水(メディアテック)で希釈し、0.2μmHTタフリン膜を備える13mmのアクロディスク(R)(ポール社)を通して濾過することで、カスタノスペルミンの200mMの蓄積濃度(アレクシスバイオケミカルズ)が調節された。蓄積溶液を、殺菌され、Oリングのついた0.5mLのマイクロ遠心分離チューブ(フィッシャーブランド、フィッシャーサイエンティフィック)に取り分け、−20℃で凍らせた。実験を開始する約1時間前に、取り分けたサンプルの必要なものを室温に解凍し、ボルテックスして各小瓶の中身を良く混ぜた。
【0124】
各実験では、対数期成長の細胞が前述の培地に接種され、250mLの振とうフラスコにおける合計容量を200,000 cells/mLという密度で60mLとし、テストすべき濃度でCSを付加した。800μM、400μM、200μM、100μM、50μM、25μMおよび0μMという最終のCS濃度が各々、対のフラスコでテストされた。加湿された培養器において37℃および5%二酸化炭素で、全ての培養がなされ、生存細胞密度および全体の細胞の生存度について少なくとも1日おきにモニターした。
【0125】
8日目に全体の細胞の生存度が50%乃至70%(実験1)および30%乃至50%(実験2)であったときに、培養物が回収された。細胞および細胞のデブリが、3000rpmのソルバールスーパー(Sorvall Super)T21で20分間の遠心分離によって取り除かれ、その後、上清が、0.22μmミリポアエクスプレスプラス膜を備えるミリポアステリフリップ装置を通して殺菌濾過され、精製されるまで2℃乃至8℃で蓄積された。
【0126】
各サンプルの細胞領域全体(ICA)、表2、が指示するように、細胞の生存度と成長は、大きく影響を受けなかったように見えるが、カスタノスペルミンの濃度が増大すると免疫糖タンパク質の生成が低減した。結果を、図17および以下の表2に示す。400μmおよび800μmのCSの濃度が、TRU‐016タンパク質の生成をそれぞれ約40%乃至55%低減することが示されている。
【0127】
【表2】
【0128】
例9
TRU‐016のCD37およびFcγRIIIa(CD16)への同時結合についてのアッセイ
【0129】
カスタノスペルミン濃度の、TRU‐016の機能活性への影響を測定するために実験が行われ、それがFcγRIIIaに結合することおよびそれがターゲットの抗原CD37に結合することにより測定された。
【0130】
例8において記述されるように生成されるTRU‐016は、以下のアッセイにおいてテストされたが、それによって同時に、TRU‐016結合ドメインの、CD37発現ターゲット細胞に結合する能力および、TRU‐016SMIPのFc部の、ヒトCD16およびネズミIgGFcの融合タンパク質に結合する能力が評価される。
【0131】
使用されるターゲット細胞は、ダウディ(ATCC CRL‐213)細胞系(line)である。ダウディ細胞は、バーキットリンパ腫から得られるヒトB‐リンパ芽球状細胞系であり、高レベルのCD37を発現する。カスタム可溶性CD16:MuIgGFc融合タンパク質は、ネズミIgGFcに結び付いたヒトCD16(低親和多形性)である。
【0132】
適当な数のダウディ細胞(350,000/ウェルのウェル数倍)がアリコートされて、250xgで5分間、15℃で遠心分離される。上清は取り除かれる。USB(USB US19943)1:4からの4%原料をFACS緩衝剤で希釈することによって、1%の冷たいパラホルムアルデヒドが調製される。FACD緩衝剤は、2%FBS(ギブコ)をダルベッコPBS(インビトロジェン)(v/v)に加え、0.22μmフィルターで殺菌濾過することによって調製される。FACS緩衝剤は貯蔵され4℃で使用される。細胞は、1%パラホルムアルデヒド(50μL/ウェルに等しい体積のウェル数倍)に再懸濁され、底の丸い96ウェルプレートで培養される。細胞は、4℃で30分間培養される。この培養に続いて、細胞は、150μLのFACS緩衝剤を各ウェルに加え、250xgで3分間の遠心分離を15℃で行い、上清を取り除くことによって洗浄される。細胞は、50μLのFACS緩衝剤に再懸濁される。TRU‐016がFACS緩衝剤において、飽和からバックグラウンドのレベルの範囲にある濃度(24μg/mL乃至0.011μg/mL)に希釈され、適当なウェル、50μL/well、に加えられ、そして細胞は4℃で25分間培養される。CD16:MuIgGFc融合タンパク質は、FACS緩衝剤において飽和レベル(20μg/ml)に希釈され、アッセイ(50μL/ウェル)に加えられ、4℃でさらに30分間培養されて、細胞表面に結合しているTRU‐016と複合体を形成する。結合していない試薬は、250xgで3分間の遠心分離を15℃で行い、上清を取り除いて、そして200μL/ウェルのFACS緩衝剤で三回洗浄することによって取り除かれる。そして、細胞は、ネズミFc特異の(かつヒトFcには最小限しか反応しないように選択された)、蛍光物質(R‐フィコエリトリン、ジャクソン115‐116‐071)でタグを付けたF(ab’)2抗体で培養される。この抗体は、CD16:MuIgGFc融合タンパク質のMuIgGFc部に結合する。抗体は、FACS緩衝剤において1:200に希釈され、各ウェルに100μLが加えられる。プレートは、暗所において4℃で45分間培養される。結合していないR‐PEは、150μLのFACS緩衝剤を各ウェルに加え、250xgで3分間の遠心分離を15℃で行い、それに続いて上清を取り除くことで取り除かれる。これに続いて、200μL/wellのFACS緩衝剤で二回目の洗浄を行い、250xgで3分間の遠心分離を15℃で行い、そして上清を取り除く。細胞は、200μL/wellの1%パラホルムアルデヒドで再懸濁され、4℃で一晩静置される。
【0133】
各サンプルに結合した蛍光は、BD FACSキャリバーフロー血球計算システムで測定され、セルクエストプロソフトウェア(ベクトンディキンソンver5.2)で分析される。各サンプルのGeoMean蛍光強度がTRU‐016の濃度に対してプロットされる。用量反応が生成されて、SoftMax Proソフトウェア(モルキュラーデバイスver5.0.1)を用いて4パラメータ対数(4‐PL)曲線に当てはめられる。TRU‐016の滴定を用いて、テスト物質および比較のための参照物質の用量反応曲線が求められる。処置したおよび未処置のサンプルを比較するために、「D」‐パラメータ(最大曲線漸近線)を参照として用いる。「D」値の増大は、対応するサンプルについての結合活性の増大を表す。
【0134】
実験の結果を図18に示し、CSの濃度に対する、用量依存的結合反応が400μMまで示され、この点で、結合が横ばいになっている。
【0135】
CSで処置されたTRU‐016のサンプルのCD16への結合が高められても、それは、部分的には分子のCD37への結合が高められることによらないということを示すために、前記のアッセイを繰り返したが、処置されたまたは未処置のTRU‐016サンプルをアッセイプレートに加えて培養した後、250xgで3分間遠心分離を15℃で行い、上清を取り除き、そして200μL/ウェルのFACS緩衝剤で三度洗浄することによって、結合していないTRU‐016をウェルから取り除くことは除外した。そして、細胞は、FITC抱合ヤギ抗ヒトIgGFc特定抗体(カルタグH10501)で培養される。この抗体は、細胞に結合したTRU‐016のヒトIgG鎖のFc領域に結合する。抗体は、FACS緩衝剤において1:50に希釈され、各ウェルに100μL加えられる。プレートは暗所において4℃で45分間培養される。100μLのFACS緩衝剤を各ウェルに加え、250xgで3分間の遠心分離を15℃で行い、続いて上清を取り除くことによって、未結合のFITCラベル化された抗体を取り除く。これに続いて、200μL/ウェルのFACS緩衝剤で二度目の洗浄を行う。細胞は、200μL/ウェルの2%パラホルムアルデヒドで再懸濁され、4℃で一晩静置される。各サンプルに結合した蛍光は、BD FACSキャリバーフロー血球計算システムで測定され、セルクエストプロソフトウェア(ベクトンディキンソンver5.2)を用いて分析される。各サンプルのGeoMean蛍光強度が、TRU‐016の濃度に対してプロットされる。用量反応曲線が生成され、SoftMax Proソフトウェア(モルキュラーデバイスver5.0.1)を用いて4パラメータ対数(4‐PL)曲線に当てはめられる。TRU‐016の滴定を用いて、未処置の対照サンプルおよび比較のためにCSで処置されたサンプルの用量反応曲線が求められる。
【0136】
図19に示されるように、CSで処置されたサンプル全てについて、CD37発現細胞に対する用量反応結合曲線は、相互に、および未処置のTRU‐016サンプルと実質的に同一であって、CSでの処置によっても、TRU‐016の、その特定のターゲット抗原への結合は変わらなかったということが示されている。
【0137】
例10
抗体依存性細胞障害性(ADCC)のアッセイ
カスタノスペルミン濃度の、TRU‐016の機能活性に対する影響を測定するために実験が行われ、それはADCC活性によって測定された。
【0138】
例8に記述されるように生成されたTRU‐016は、プライマリーヒト末梢血リンパ球(PBL)エフェクター細胞とともにCD37発現ダウディ癌B細胞系で培養されて、ADCC活性が評価される。
【0139】
ダウディターゲット細胞(5x106)が、15mlの円錐チューブに加えられ、そして250xgで5分間の遠心分離が20℃で行われ、上清が取り除かれた。細胞ペレットは、0.3mCiクロム‐51(51Cr、GEヘルスケア、CJ51)を加えることで再懸濁される。細胞は、5%のCO2において、37℃で75分間培養され、細胞が放射性同位元素を取り込むことが可能となる。そして細胞は、三度洗浄され、取り込まれなかった51Crが取り除かれる。これが行われるのは、10mLの完全培地‐10%FBS(ギブコ)を備えるIMDM(ギブコ)‐をチューブに加え、250xgで5分間の遠心分離を20℃で行い、それに続いて上清を取り除くことによってである。最終の再懸濁は、11.5mLの完全培地においてである。TRU‐016は、完全培地において、最大からバックグラウンドレベルまでの細胞溶解(500ng/mL乃至0.005ng/mL)を生成することができる濃度に希釈される。これらの滴定は、底の丸い96ウェルプレートにおいて、50μL/ウェルで行われる。51Crのラベルを付けられたターゲット細胞が、50μL/ウェルおよび対照ウェル(TRU‐016の無い対照媒体)でTRU‐016の用量滴定に加えられる。プロトコル(LSM、MPバイオメディカル、50494/36427)に従う、リンパ球セパレーションメディアを用いての密度勾配遠心分離によって、ヘパリン処理した新鮮な全血からPBLが分離される。PBLエフェクター細胞は、25:1乃至30:1の割合でウェルに、100mL/ウェル加えられる(エフェクター:ターゲット)。5%のCO2にてアッセイは37℃で4.5乃至5時間培養される。エフェクター細胞は、TRU‐016の濃度に対してターゲット細胞を溶解させ、比例した量の51Crをアッセイの上清に放出する。培養に続いて、250xgで3分間、プレートの遠心分離が20℃で行われる。25μL容量の細胞を含まない上清が、全てのウェルから取り除かれてシンチレーションプレート(パーキンエルマー6005185)に入れられ、一晩乾燥される。シンチレーションプレートの各ウェルにおける51Cr同位元素の量は、トップカウントプレートリーダー(パーキンエルマー、C9904VO)を用いて測定される。データは、特定の放出のパーセントとして表される。特定の放出は、以下のように計算される。
【0140】
(サンプル値‐自発値)/(最大値‐自発値)*100%
自発=ターゲット細胞のみから放出される51Crの量
最大放出=洗浄溶解剤で処置されたターゲットから放出される51Crの量
バックグラウンド対照=ターゲット細胞+エフェクター細胞(No TRU‐016)から放出される51Crの量
【0141】
用量反応が形成され、SoftMax Proソフトウェア(モルキュラーデバイスver5.0.1)を用いて4パラメータ対数曲線に当てはめられる。TRU‐016の滴定を用いて、テスト物質および比較のための参照物質の用量反応曲線が求められる。処置したものについてのEC50値が、処置されていない対照(CSでない)と比較され、ADCC活性のパーセント増大が測定される。以下の表に、図20に示されるデータがまとめられている。データでは、100μM乃至800μMの最終濃度の範囲についてCSで処置されたTRU‐016のADCC活性が、未処置のTRU‐016に対して大きく増大していることが示されている。
【0142】
【表3】
【0143】
本発明の構成および方法は、前述の代表的な実施例について記述されているが、発明の概念、思想および範囲から逸脱することなく、ここに記述されている構成および/または方法および工程または方法における一連の工程には、変形を施してもよいことが当業者には明らかである。さらに詳しくは、化学的にも生理学的にも関係のある試薬をここに記述されている試薬と置き換えても、同じまたは同様の結果が達成されるということも明らかである。当業者に明らかなそのような類似の置き換えおよび変形は全て、添付の請求の範囲に定義される本発明の思想、範囲および概念内のものと看做される。
【0144】
本明細書の至る所に引用され、当該説明を例示によって詳細に増補する参考文献は全て、援用することでここに組み入れることを明示する。
【特許請求の範囲】
【請求項1】
ホスト細胞により生成される免疫糖タンパク質分子の抗体依存細胞傷害性(ADCC)を増大させる方法であって、
前記ホスト細胞により生成される免疫糖タンパク質分子のADCCを増大させるカスタノスペルミンを、25μM〜約800μM濃度で備え、容量が少なくとも1リットルである培地で前記ホスト細胞を成長させる工程を備える、免疫糖タンパク質分子の抗体依存細胞傷害性(ADCC)を増大させる方法。
【請求項2】
ADCCが少なくとも2倍に増大される、請求項1に記載の方法。
【請求項3】
ADCCが少なくとも5倍に増大される、請求項1に記載の方法。
【請求項4】
ホスト細胞により生成される免疫糖タンパク質分子のCD16結合を増大させる方法であって、
前記ホスト細胞により生成される免疫糖タンパク質分子のCD16結合を増大させるカスタノスペルミンを、25μM〜約800μMの濃度で備え、容量が少なくとも1リットルである培地で前記ホスト細胞を成長させる工程を備える、免疫糖タンパク質分子のCD16結合を増大させる方法。
【請求項5】
CD16結合を少なくとも50%増大させる、請求項4に記載の方法。
【請求項6】
CD16結合を少なくとも2倍(200%)に増大させる、請求項4に記載の方法。
【請求項7】
培地における免疫糖タンパク質の生成レベルが少なくとも100μg/mLである、請求項1乃至7のいずれかに記載の方法。
【請求項8】
カスタノスペルミンが100μM〜400μMの濃度で存在する、請求項1乃至7のいずれかに記載の方法。
【請求項9】
培地が実質的に漿液を含んでいない、請求項1乃至8のいずれかに記載の方法。
【請求項10】
ホスト細胞がバッチ培養法で育てられる、請求項1乃至9のいずれかに記載の方法。
【請求項11】
ホスト細胞が連続培養法で育てられる、請求項1乃至9のいずれかに記載の方法。
【請求項12】
培地は第二の糖修飾因子を含む、請求項1乃至9のいずれかに記載の方法。
【請求項13】
請求項1乃至12のいずれかの方法により生成される免疫糖タンパク質分子および無菌の医薬的に受け入れ可能な担体または希釈剤を含む、組成物。
【請求項14】
請求項13の組成物を投与する工程を備える、癌細胞を死滅させるか成長を抑制する方法であって、癌細胞が前記免疫糖タンパク質分子により結合される分子を表面に発現する、癌細胞を死滅させるか成長を抑制する方法。
【請求項15】
請求項13の組成物を投与する工程を備える、細胞を枯渇させる方法であって、該枯渇される細胞が、前記免疫糖タンパク質分子により結合される分子を表面に発現する、細胞を枯渇させる方法。
【請求項1】
ホスト細胞により生成される免疫糖タンパク質分子の抗体依存細胞傷害性(ADCC)を増大させる方法であって、
前記ホスト細胞により生成される免疫糖タンパク質分子のADCCを増大させるカスタノスペルミンを、25μM〜約800μM濃度で備え、容量が少なくとも1リットルである培地で前記ホスト細胞を成長させる工程を備える、免疫糖タンパク質分子の抗体依存細胞傷害性(ADCC)を増大させる方法。
【請求項2】
ADCCが少なくとも2倍に増大される、請求項1に記載の方法。
【請求項3】
ADCCが少なくとも5倍に増大される、請求項1に記載の方法。
【請求項4】
ホスト細胞により生成される免疫糖タンパク質分子のCD16結合を増大させる方法であって、
前記ホスト細胞により生成される免疫糖タンパク質分子のCD16結合を増大させるカスタノスペルミンを、25μM〜約800μMの濃度で備え、容量が少なくとも1リットルである培地で前記ホスト細胞を成長させる工程を備える、免疫糖タンパク質分子のCD16結合を増大させる方法。
【請求項5】
CD16結合を少なくとも50%増大させる、請求項4に記載の方法。
【請求項6】
CD16結合を少なくとも2倍(200%)に増大させる、請求項4に記載の方法。
【請求項7】
培地における免疫糖タンパク質の生成レベルが少なくとも100μg/mLである、請求項1乃至7のいずれかに記載の方法。
【請求項8】
カスタノスペルミンが100μM〜400μMの濃度で存在する、請求項1乃至7のいずれかに記載の方法。
【請求項9】
培地が実質的に漿液を含んでいない、請求項1乃至8のいずれかに記載の方法。
【請求項10】
ホスト細胞がバッチ培養法で育てられる、請求項1乃至9のいずれかに記載の方法。
【請求項11】
ホスト細胞が連続培養法で育てられる、請求項1乃至9のいずれかに記載の方法。
【請求項12】
培地は第二の糖修飾因子を含む、請求項1乃至9のいずれかに記載の方法。
【請求項13】
請求項1乃至12のいずれかの方法により生成される免疫糖タンパク質分子および無菌の医薬的に受け入れ可能な担体または希釈剤を含む、組成物。
【請求項14】
請求項13の組成物を投与する工程を備える、癌細胞を死滅させるか成長を抑制する方法であって、癌細胞が前記免疫糖タンパク質分子により結合される分子を表面に発現する、癌細胞を死滅させるか成長を抑制する方法。
【請求項15】
請求項13の組成物を投与する工程を備える、細胞を枯渇させる方法であって、該枯渇される細胞が、前記免疫糖タンパク質分子により結合される分子を表面に発現する、細胞を枯渇させる方法。
【図1】


【図2】
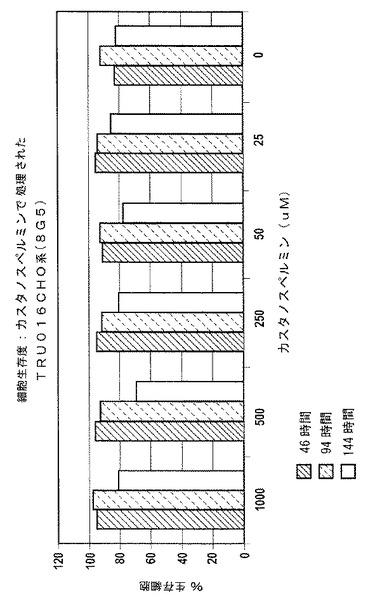
【図3】
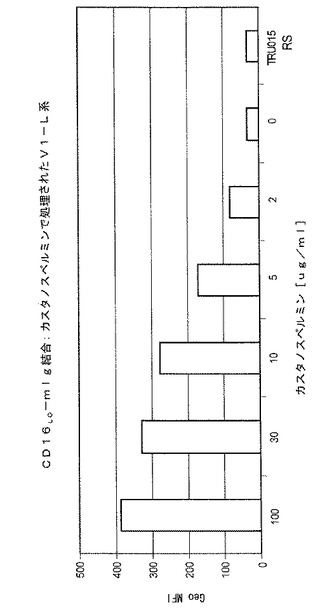
【図4】
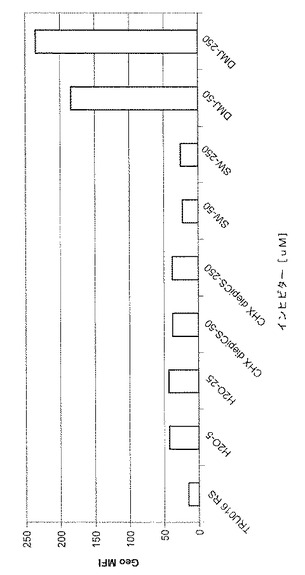
【図5】
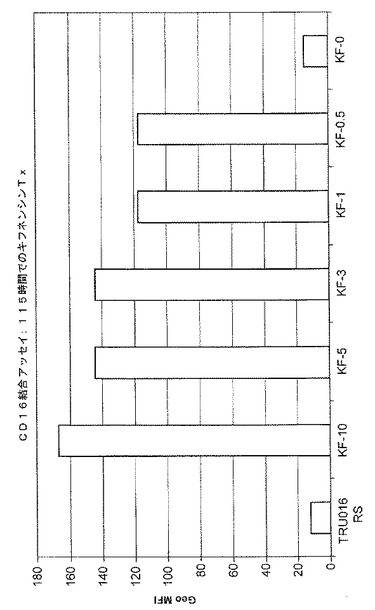
【図6】
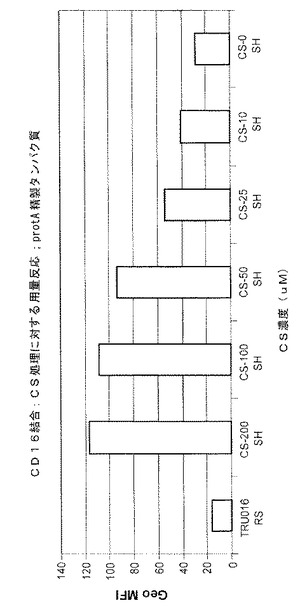
【図7】
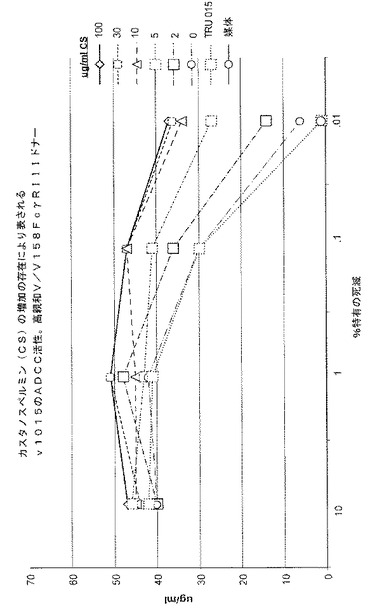
【図8】
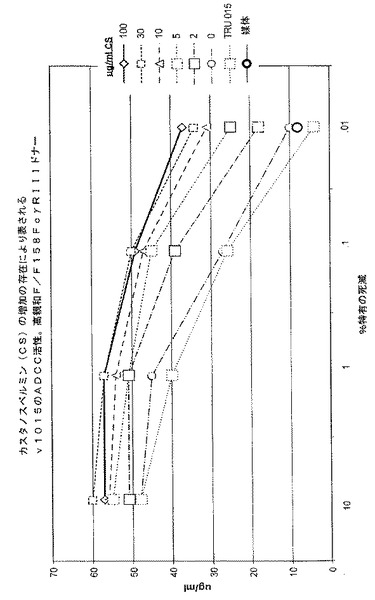
【図9】
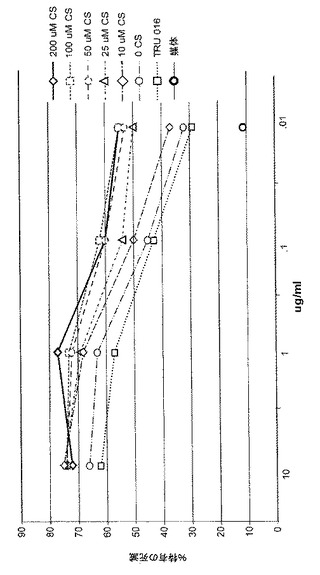
【図10】
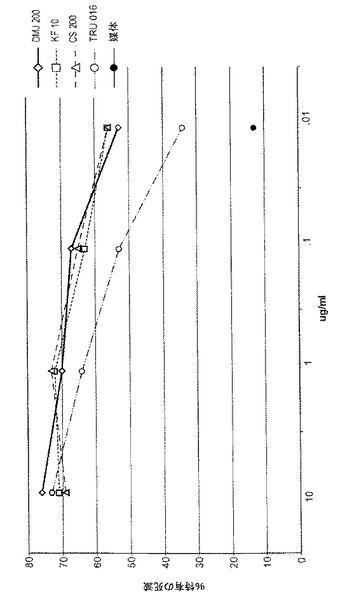
【図11】
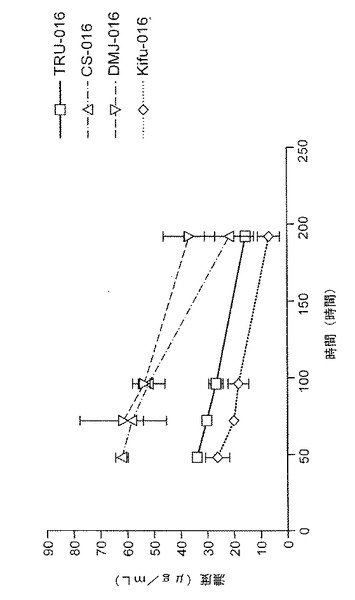
【図12】
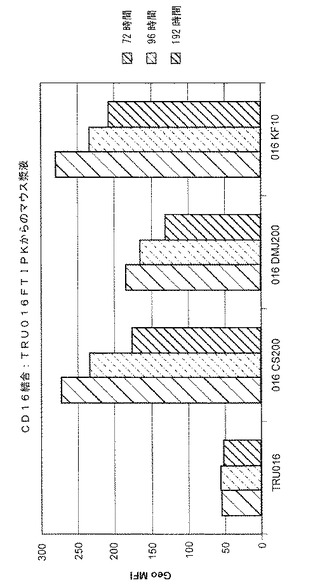
【図13】
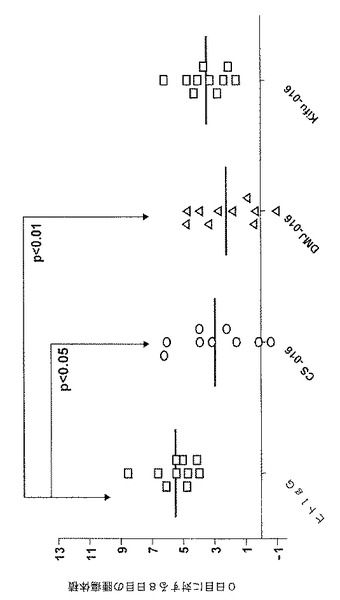
【図14】
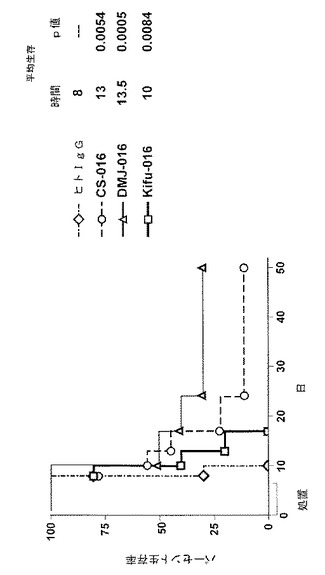
【図15】
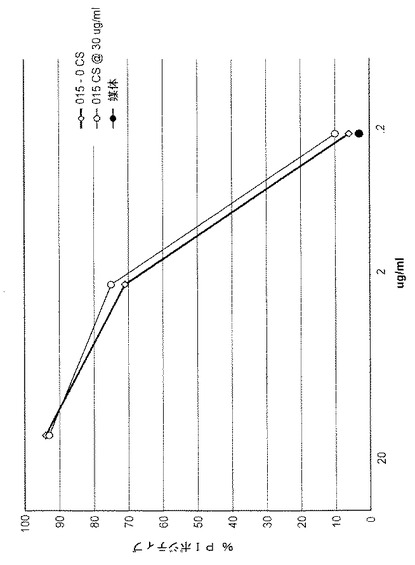
【図16】
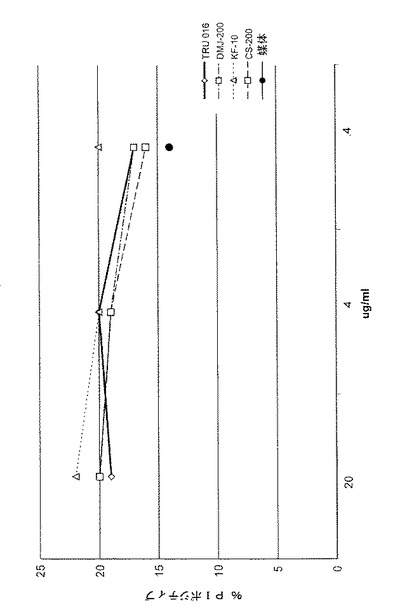
【図17】
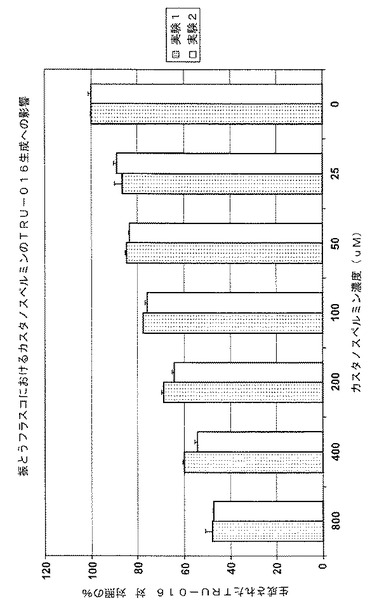
【図18】
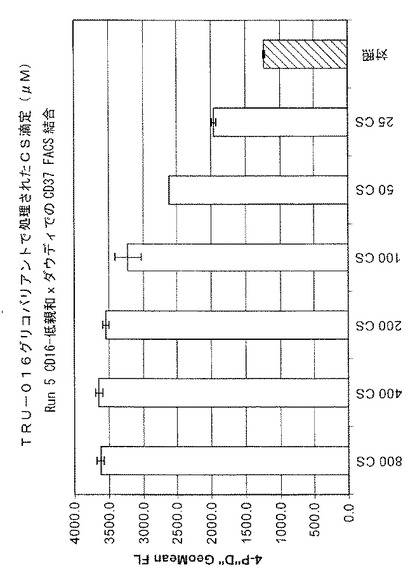
【図19】
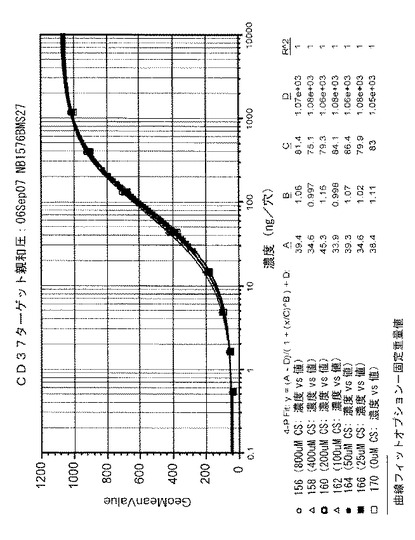
【図20】
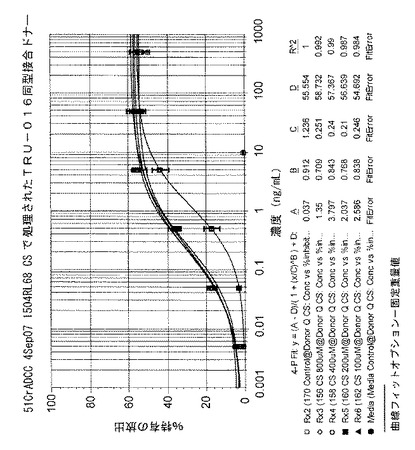
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【公表番号】特表2010−507394(P2010−507394A)
【公表日】平成22年3月11日(2010.3.11)
【国際特許分類】
【出願番号】特願2009−534837(P2009−534837)
【出願日】平成19年10月24日(2007.10.24)
【国際出願番号】PCT/US2007/082343
【国際公開番号】WO2008/052030
【国際公開日】平成20年5月2日(2008.5.2)
【出願人】(508025530)トルービオン ファーマスーティカルズ インコーポレイテッド (2)
【Fターム(参考)】
【公表日】平成22年3月11日(2010.3.11)
【国際特許分類】
【出願日】平成19年10月24日(2007.10.24)
【国際出願番号】PCT/US2007/082343
【国際公開番号】WO2008/052030
【国際公開日】平成20年5月2日(2008.5.2)
【出願人】(508025530)トルービオン ファーマスーティカルズ インコーポレイテッド (2)
【Fターム(参考)】
[ Back to top ]