
改質されたインターロイキン−11とその製造方法
【課題】IL-11の生物活性と構造との相関を明らかにし、IL-11の生物活性に関与する部位を改変することで機能的に改質されたIL-11を提供する。
【解決手段】アミノ酸又はペプチドを付加することにより改質されたIL-11である。ここで、本発明にかかる改質されたIL-11は、改質対象のIL-11に対して1個のアミノ酸を付加したものであっても良いし、改質対象のIL-11に対して複数個のアミノ酸からなるペプチド(例えば、3〜8個のアミノ酸残基)を付加したものであっても良い。
【解決手段】アミノ酸又はペプチドを付加することにより改質されたIL-11である。ここで、本発明にかかる改質されたIL-11は、改質対象のIL-11に対して1個のアミノ酸を付加したものであっても良いし、改質対象のIL-11に対して複数個のアミノ酸からなるペプチド(例えば、3〜8個のアミノ酸残基)を付加したものであっても良い。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、造血幹細胞の増殖を誘導する機能等、様々な生理活性を有するインターロイキン-11(IL-11)及びその製造方法、さらにそれを含有する医薬組成物に関する。
【背景技術】
【0002】
IL-11は、アミノ酸178残基からなる糖鎖を持たない単純タンパク質で、マカクザルの骨髄間質細胞株PU34が産生する形質細胞腫増殖刺激因子として1990年に発見され(Paul, S. R., et al., Proc. Natl. Acad. Sci. USA, 82, 7512-7516, 1990〔非特許文献1〕)、また、前脂肪細胞から脂肪細胞への分化を抑制する脂肪細胞分化抑制因子(Kawasima, I., et al., FEBS Lett. 283, 199-202, 1991〔非特許文献2〕)として見出された。ヒトIL-11遺伝子は全長7kbにわたり、5つのエクソンと4つのイントロンより構成されている。mRNAサイズは、1.5kb及び2.5kbであるが、これらはpoly(A)付加シグナルの認識の違いにより生じており、同一の蛋白質をコードしている。IL-11は、元来IL-6様活性をもとにクローニングされたにも拘わらず、DNAレベルあるいは蛋白質レベルのいずれにおいてもIL-6とのホモロジーが認められない。これに対して、IL-11レセプターは、IL-6、LIF、CNTF、OSMなどの共通のシグナルトランスジューサーであるgp130(Yin,T., et al., J. Immunol., 151, 2555-2561, 1993〔非特許文献3〕)及びIL-11レセプターα鎖から構成されている(Hilton, D. J., et al., EMBO J., 13, 4765-4775, 1994〔非特許文献4〕)。細胞内シグナル伝達はJAKチロシンキナーゼ(JAK1, JAK2及びTYK2)、STAT及びRasが関与することが知られている(Taga,T., Annu. Rev. Immunol., 15, 797-819, 1997〔非特許文献5〕)。
【0003】
IL-11の生物学的作用としては、IL-3、-4、-7、-12、-13、SCF(stem cell factor)、Flt3-ligand、GM-CSFと共同で造血幹細胞の増殖を誘導すること、IL-3、TPO(thrombopoietin)あるいはSCFとともに骨髄細胞に作用し、巨核球及び血小板の産生を促進すること、単独あるいはIL-3、SCF、EPO(erythropoietin)と協調して赤血球造血を促進すること、骨髄前駆細胞の分化、成熟を促進すること、SCF, IL-4と協調してB細胞分化を促進すること、また、骨髄微小環境において、オートクラインあるいはパラクラインされる増殖因子として働くことが知られている(Du, X. X., and Williams, D. A., Blood, 89, 3897-3908, 1997〔非特許文献6〕)。また、肺胞及び気管上皮は特に炎症に伴い多量のIL-11を分泌することから、IL-11の生物学的作用として、呼吸器での炎症反応との関係も示唆されている。そのほかに、肝癌細胞株で急性期タンパク質を誘導させる作用(Baumann, H and Schendel, P., Biol. Chem., 266, 20424-20427, 1991〔非特許文献7〕)や骨芽細胞がIL-11を分泌し、骨芽細胞/破骨細胞両者に働き骨代謝を調節しているとの報告もある。In vitroの生物活性の指標には、サルのPU34細胞株培養上清からIL-11の発見をもたらしたマウスプラズマサイトーマ細胞株(T1165、Nordan, R. P and Potter, M., Science, 233, 566-569, 1986 〔非特許文献8〕)に対する増殖活性がよく用いられている。T1165細胞はIL-6及びIL-11に対し、特異的増殖活性を示す。
【0004】
このように、多くの生理活性を有するIL-11は、医薬への期待が高まり、近年、悪性腫瘍に対する化学療法で問題視されている血小板減少症に対し、血小板増殖因子としてあるいは炎症性腸疾患などを対象に米国、日本で開発が進められている。米国Genetics Institute社は、1997年に重度血小板減少症の危険が高い、非骨髄性腫瘍患者における骨髄抑制化学療法後の重度血小板減少症の予防と血小板輸血量の減量の適応で組み換えIL-11製剤の販売認可を獲得し、現在、小児を対象とした臨床試験が進行中である。また、Wyeth社では組み換えIL-11の経口製剤を炎症性腸疾患を対象に臨床試験を進めている。また、日本では、アステラス製薬がGenetics Institute社から導入した組み換え(大腸菌)IL-11の血小板減少症に対する臨床試験に着手している。
【0005】
このように医薬への期待が高まり、臨床試験もフェーズIIあるいはフェーズIIIに入り、その有効性が確認されると共にいくつかの副作用についても報告されるようになった(Smith, J.W. 2nd, Oncology, 14(9 Suppl 8),41, 7, 2000[非特許文献9])。例えば、浮腫、呼吸困難、胸膜からの漿液の滲出、結膜充血、うっ血性心房性不整脈などが報告された。深刻な血小板減少症に対し有効な医薬が望まれている折から、これらIL-11の投与に伴う副作用への対応は重要な課題になっている。また、IL-11は多くのサイトカインと異なり、システイン残基を欠き、N-グリコシレーションサイトが無いことが特徴的である。このことは、構造的不安定さを示唆しており、血中における安定性にも影響を及ぼすことが考えられている。これに対応して、血中における安定性を向上させるための研究が世界でスタートしている。米国のBolder BioTechnology社は、ポリエチレングリコール化技術を用いて数週間に一回の投与で済むIL-11誘導体の開発について米国国立衛生研究所から中小企業革新研究補助金を得て研究開発に着手している。また、エリスロポイエチンの例に見るように、糖鎖付加により安定性が増強されることも考えられる。しかしながら、血中安定性を増すなどIL-11の望ましい生物活性を十分に発揮するための多くの研究が今後の課題として残されている。そのためには、生物活性にかかわる部位の特定は必須のテーマであり、それにより安定型IL-11の創出やIL-11拮抗作用物質を見つけることが可能になる。しかし、現在までにIL-11の構造と生物活性との相関を見出したという報告は見つかっていない。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Paul, S. R., et al., Proc. Natl. Acad. Sci. USA, 82, 7512-7516, 1990
【非特許文献2】Kawasima, I., et al., FEBS Lett. 283, 199-202, 1991
【非特許文献3】Yin,T., et al., J. Immunol., 151, 2555-2561, 1993
【非特許文献4】Hilton, D. J., et al., EMBO J., 13, 4765-4775, 1994
【非特許文献5】Taga,T., Annu. Rev. Immunol., 15, 797-819, 1997
【非特許文献6】Du, X. X., and Williams, D. A., Blood, 89, 3897-3908, 1997
【非特許文献7】Baumann, H and Schendel, P., Biol. Chem., 266, 20424-20427, 1991
【非特許文献8】Nordan, P. N., and Potter, M, Science, 233, 566-569, 1986
【非特許文献9】Smith, J.W., 2nd, Oncology, 14(9 Suppl 8),41, 7, 2000
【発明の概要】
【発明が解決しようとする課題】
【0007】
そこで、本発明は、IL-11の生物活性と構造との相関を明らかにし、IL-11の生物活性に関与する部位を改変することで機能的に改質されたIL-11及びその製造方法を提供するとともに、改質されたIL-11を含む医薬組成物を提供することを目的としている。
【課題を解決するための手段】
【0008】
本発明者らは上記課題を解決すべくIL-11の生物活性にかかわる部位の同定に鋭意研究を重ね、ペプチド鎖の挿入によりIL-11の生物活性が低減或いは失われる部位を見つけることができた。本発明はこのような知見に基づいて完成されたものである。
【0009】
すなわち、本発明は、アミノ酸を付加させることにより改質されたIL-11を提供する。ここで、本発明にかかる改質されたIL-11は、改質対象のIL-11に対して1個のアミノ酸を付加したものであっても良いし、改質対象のIL-11に対して複数個のアミノ酸からなるペプチド(例えば、3〜8個のアミノ酸残基)を付加したものであっても良い。
【0010】
特に、本発明にかかる改質されたIL-11は、野生型IL-11の一次構造から立体構造予測プログラムによってコイルと推定された部位に、上記アミノ酸又は上記ペプチドを付加することが望ましい。上記コイルは、当該プログラムによって推定される6個のコイルのうちN末端側から2番目に位置するコイルである。そして、上記アミノ酸又は上記ペプチドを付加する領域としては、上記コイルを構成する領域のうち、野生型IL-11のアミノ酸配列における68番目(グリシン)〜72番目(ロイシン)の間、好ましくは、70番目(ヒスチジン)〜71番目(アスパラギン)の間であることが好ましい。
【0011】
また、本発明は、アミノ酸を付加することにより改質されたIL-11の製造方法で以下の工程を含むことを特徴とする。
(1)付加するアミノ酸あるいはペプチドをコードするDNA断片とIL-11をコードするcDNAを連結する工程
(2)当該連結cDNAを発現ベクターに組み込む工程
(3)当該発現ベクターを宿主細胞に導入する工程。
(4)当該宿主細胞において、アミノ酸又はペプチドが付加されたIL-11を発現させる工程
(5)当該宿主細胞において発現された、アミノ酸又はペプチドが付加されたIL-11を単離・精製する工程。
【0012】
また、本発明は、アミノ酸又はペプチドが付加されることにより改質されたIL-11の製造方法であって、アミノ酸又はペプチドを化学的結合法によりIL-11に結合させる工程を含むことを特徴とする。さらに、本発明は、アミノ酸又はペプチドを付加させることにより改質された前記のIL-11を有効成分として含有する医薬組成物を提供する。また、本発明の技術的思想は、IL-11と類似した立体構造が予測されるIL-6ファミリーサイトカインであるIL-6、CNTF(ciliary neurotrophic factor)、LIF(leukemia inhibitory factor)などにも適用される幅広い応用が可能である。
【発明の効果】
【0013】
本発明によれば、IL-11が本来的に有している様々な生理活性が改質されたインターロイキン-11を提供することができる。また、本発明によれば、改質されたIL-11を有効成分として含み、従来のIL-11を有効成分として含有する医薬組成物より優れた薬効を示したり、あるいはIL-11を有効成分として含有する医薬組成物の薬効をより有効なものにする効果を期待できる医薬組成物を提供することができる。
【0014】
さらに、本発明によれば、IL-11が本来的に有している様々な生理活性が改質されたインターロイキン-11の製造方法を提供することができる。
【図面の簡単な説明】
【0015】
【図1】哺乳類細胞発現用pSRαhIL-11プラスミドのマップ示す。
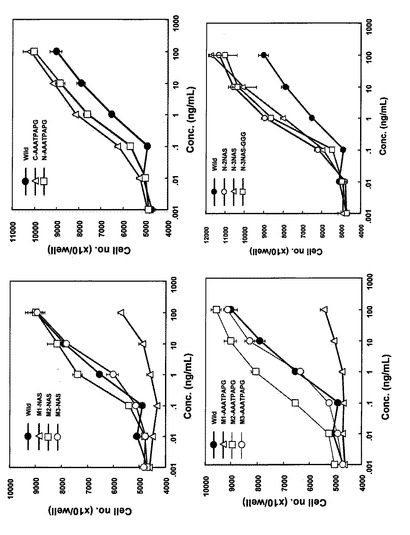
【図2】COS7細胞由来各種改変IL-11のT1165細胞に対する細胞増殖促進活性を示す特性図である。
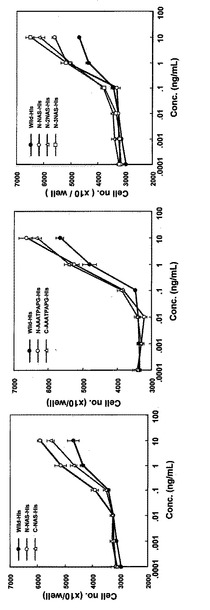
【図3】COS7細胞由来Hisタグ付き改変IL-11のT1165細胞に対する細胞増殖促進活性を示す特性図である。
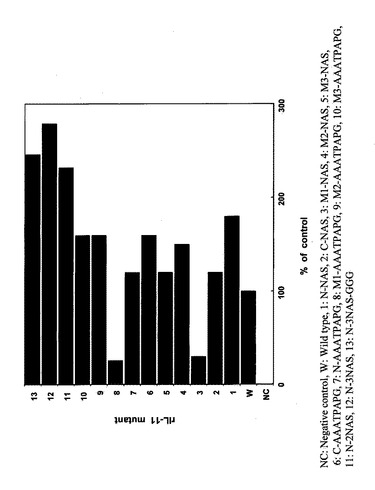
【図4】COS7細胞由来各種改変IL-11のT1165細胞に対する細胞増殖促進活性を野性型IL-11と比較検討した結果を示す特性図である。
【図5】野生型IL-11のアミノ酸配列における70番目と71番目のアミノ酸の間にペプチドを付加することにより細胞増殖促進活性が低下することを示す特性図である。
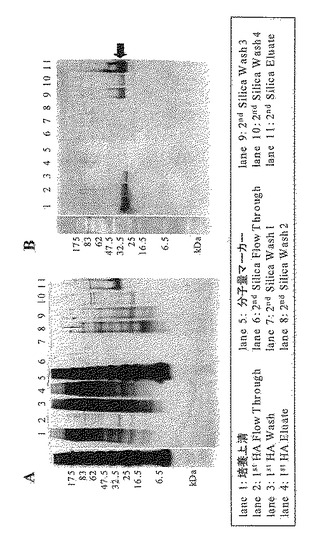
【図6】改質されたIL-11(M1-AAATPAPG)の精製画分の銀染色(図6のA)とウエスタンブロッテイング解析(図6のB)の結果を示す写真である。
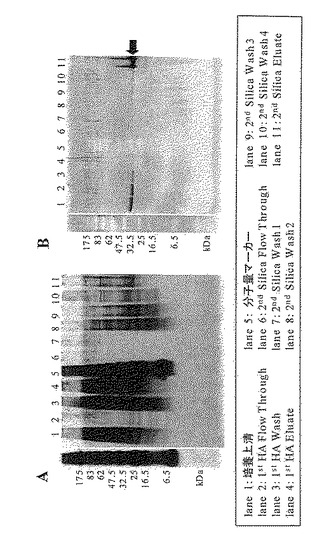
【図7】野性型IL-11の精製画分の銀染色(図7のA)とウエスタンブロッテイング解析(図7のB)の結果を示す写真である。
【発明を実施するための形態】
【0016】
以下、本発明を詳細に説明する。
本発明に係る改質されたインターロイキン-11(以下、改質されたIL-11)は、IL-11の所定の領域にアミノ酸又はペプチドが付加され、IL-11が本来有する生理活性が低下或いは欠損したものである。すなわち、本発明において、改質されたとは、IL-11が本来的に有する生物活性が低下或いは欠損したことを意味する。
【0017】
本発明において、改質対象のIL-11としては、IL-11として知られている蛋白質であれば何ら限定することなく適用することができる。IL-11の一例として、ヒト由来IL-11のアミノ酸配列を配列番号2、ヒト由来IL-11をコードするcDNAの塩基配列を配列番号1に挙げる。なお、本発明に係る高機能化IL-11は、配列番号2に記載されたアミノ酸配列からなる蛋白質に限定されず、糖鎖の付加を受けることができるペプチド及びシグナルペプチドを有する蛋白質であっても良い。なお、シグナルペプチドとは、所定の蛋白質のアミノ末端に存在する分泌のためのペプチド配列であって、当該蛋白質が細胞から分泌される際に機能する。本発明に係る改質されたIL-11は、初めからシグナルペプチドを欠損する形で製造してもその有用性には変化がない。なお、野生型IL-11のcDNAは、日本DNAデータバンク(DDBJ)など遺伝子バンクに登録された配列から適当なプライマーを設計し、当該動物の当該組織のmRNAよりRT-PCRを行うことにより取得できる。
【0018】
また、IL-11としては、配列番号2のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換、付加若しくは修飾されたアミノ酸配列からなり、IL-11活性を有する蛋白質も含まれる。ここで、数個のアミノ酸とは、2〜50個のアミノ酸、好ましくは2〜25個のアミノ酸、より好ましくは2〜10個のアミノ酸を意味する。
【0019】
さらに、改質対象のIL-11としては、野生型のIL-11に対して、N型糖鎖、O型糖鎖及びN型O型並びにこれらの組み合わせの糖鎖のいずれかの糖鎖を付加する機能を有するペプチドが導入された変異型IL-11であってもよい。
【0020】
ここで、N型糖鎖とは、蛋白質の一次構造に対するアスパラギン残基に結合したN-アセチルガラクトサミンを起点として伸長する多様な糖鎖構造を総称するものであり、例えば、Asn-X-Ser/ThrのAsnNH2基にC-N結合を介して結合したGlcNAcを還元末端とする糖鎖を挙げることができる。すなわち、N型糖鎖の付加を受けることが知られているペプチドの例として、Asn-X-Thr、Asn-X-Ser(配列中、Xはプロリン以外の任意のアミノ酸である。)が挙げられる。また、O型糖鎖とは、蛋白質の一次構造に存在するセリン残基又はスレオニン残基に結合したN-アセチルガラクトサミンを起点として伸長する多様な糖鎖構造を総称するものであり、例えば、Ser又はThrの水酸基にGalNAcがC-O結合を介して結合した糖鎖を挙げることができる。すなわち、O型糖鎖の付加を受けることが知られているペプチドの例として、Ala-Thr-Pro-Ala-Proが挙げられる。
【0021】
本発明にかかる改質されたIL-11は、改質対象のIL-11(例えば、野生型IL-11)の一次構造から立体構造予測プログラムによりコイルと推定された領域に、アミノ酸又はペプチドを付加させた一次構造を有している。ここで立体構造予測プログラムとしては、特に限定されず、例えば、GENETYX ver. 7(ゼネテイックス)を例示することができる。当該プログラムを使用し、Chou-Fasman法(Chou, P. Y. and Fasman, G. D., Enzymology, 47, 45-147, 1978)に従ってアミノ酸配列から立体構造におけるコイルを予測することができる。例えば、野生型IL-11を当該プログラムにより解析すると、シグナルペプチドを含むアミノ酸配列のN末端から順に、30番目(アルギニン)と31番目(バリン)の間、68番目(グリシン)と72番目(アスパラギン)の間、99番目(アラニン)と100番目(アスパラギン)の間、114番目(アラニン)、156番目(プロリン)と162番目(アラニン)の間、167番目(アラニン)と170番目(グリシン)の間の6箇所にコイル構造が予測される。
【0022】
本発明にかかる改質されたIL-11は、N末端側から2番目に位置するコイルにアミノ酸又はペプチドを付加することが好ましい。具体的には、本発明にかかる改質されたIL-11は、N末端側から2番目に位置するコイルを立体的に破壊するようにアミノ酸又はペプチドを付加することが好ましい。例えば、野生型IL-11のシグナルペプチドを含むアミノ酸配列における68番目(グリシン)〜72番目(ロイシン)の間にアミノ酸又はペプチドを付加することによって、ここに位置するコイルを立体的に破壊することができる。より好ましくは、野生型IL-11のアミノ酸配列における70番目(ヒスチジン)〜71番目(アスパラギン)の間にアミノ酸又はペプチドを付加することによって、最も効果的にN末端側から2番目に位置するコイルを立体的に破壊することができる。
【0023】
なお、上述したアミノ酸残基の位置を示す数値は、野生型IL-11のシグナルペプチドを含むアミノ酸配列(配列番号2)を基準とした値である。したがって、改変対象のIL-11が野生型IL-11と異なるアミノ酸配列である場合には、上述したアミノ酸残基の位置を示す数値は同じ値にならないことに留意すべきである。
【0024】
改質されたIL-11において、付加するアミノ酸の種類及びペプチドの配列には限定されず、いかなる種類のアミノ酸を付加しても良いし、いかなる配列及びいかなる長さのペプチドを付加しても良い。具体的にペプチドとしては、3〜8個の長さのペプチドを挙げることができる。好ましくは、改変対象IL-11全体の立体構造に大きな影響を与えないアミノ酸あるいはペプチドを付加することが好ましい。例えば、グリシン単独あるいはグリシンのみからなるペプチドを挙げることができる。
【0025】
改質されたIL-11を製造するには、まず、付加するアミノ酸あるいはペプチドをコードするDNA断片をIL-11をコードするcDNAに組み込んだ発現ベクターを構築する。次に、当該発現ベクターを適当な宿主細胞に導入して、IL-11を発現させれば良い。発現ベクターは、プラスミド・ベクターを基本として構築しても良いし、ウィルス・ベクターを基本として構築しても良い。
【0026】
より具体的には、まず、分泌シグナル及び付加するアミノ酸又はペプチドをコードするオリゴヌクレオチドを合成し、あるいはPCR反応によって増幅し、これをIL-11をコードするcDNAの所望の位置(例えば、N末端側から2番目に位置するコイルを構成するアミノ酸配列をコードするコドン)に組み込む。分泌シグナルとしては、典型的な分泌型糖蛋白質のアミノ末端を利用することができる。このIL-11をコードするcDNAを組み込むプラスミドとしては、宿主内で複製保持されるものであれば、いずれも使用することができるが、例えば大腸菌由来のpBR322、pUC18及びこれらを基に構築されたpET-3cなどを挙げることができる。
【0027】
IL-11をコードするcDNAに上記オリゴヌクレオチドを組み込む方法としては、例えばT.Maniatisらにより紹介された方法(Molecular Cloning, Cold Spring Harbor Laboratory, p239, 1982)がある。上記のようにして作製したベクターから、分泌シグナル、付加するアミノ酸又はペプチド及びIL-11をコードする塩基配列を含む領域を切り出し、これを発現に適したベクター中のプロモーターの下流に連結することにより、発現型ベクターを得ることができる。
【0028】
上記IL-11をコードするcDNAを含む領域は、5’末端に翻訳開始コドンとしてのATGを有し、また、3’末端には翻訳終止コドンとしてのTAA、TGA又はTAGを有しても良い。さらに、該コーデイング領域にコードされている蛋白質を発現させるには、その上流にプロモーターを接続する必要がある。本発明で用いられるプロモーターとしては、遺伝子の発現に用いる宿主に対応した適切なプロモーターであればいかなるものでも用いることが出来る。形質転換する宿主が枯草菌である場合には、SP01、SP02、penPプロモーターなどがあり、酵母ではPH05、PGK、GAP、ADHプロモーターなどが挙げられる。また、動物細胞ではSV40由来のプロモーター、レトロウイルスのプロモーターが挙げられる。
【0029】
宿主細胞としては、いかなるものでも良く、枯草菌(例えば、Bacillus subtilis DB105)、酵母(例えば、Pichia pasto ris、Saccharomyces cerevisiae)、動物細胞(例えば、COS、CHO、KB、293、BHK、NIH3T3、BALB/c3T3、臍帯静脈内皮細胞)、昆虫細胞(例えば、Sf9、Tn細胞)などが挙げられるが、本発明に係る改質されたIL-11を医薬組成物としてヒトに投与する目的で大量培養をおこなう場合、動物細胞を用いて製造することが好ましい。
【0030】
上記の形質転換は、それぞれの宿主について一般的に行われている方法あるいは適応可能な方法であれば良い。例えば、宿主が酵母であれば、リチウム法などで作製したコンピテント細胞に組み換えDNAを含むベクターを温度ショック法あるいはエレクトロポレーション法により導入する。宿主が動物細胞であれば、増殖期等の細胞に組み換えDNAを含むベクターをリン酸カルシウム法、リポフェクション法あるいはエレクトロポレーション法により導入する。
【0031】
このようにして得られた形質転換体は、それぞれの宿主に一般的に用いられている培地、あるいは適用可能な培養液を用いて培養することにより、改質されたIL-11を産生することができる。宿主が酵母であればYPD培地などを用い、動物細胞であればDulbecco's MEMにウシ胎児血清を加えた培養液を用いる。培養は、それぞれの宿主について一般的に用いられている条件あるいは適用可能な条件であれば良い。例えば、宿主が酵母であれば、約25〜37℃で、約12時間〜2週間行い、必要により、通気や撹拌を加えることができる。動物細胞の場合は、37℃で、5%炭酸ガス、100%湿度の条件で約24時間〜2週間行い、必要により、気相の条件を変えたり撹拌を加えることが出来る。
【0032】
細胞から得られた、本発明にかかる改質されたIL-11の生物活性は、IL-11及びIL-6により特異的な増殖を示すマウスプラズマサイトーマ細胞株T1165細胞あるいはマウスハイブリドーマ細胞株7TD細胞を用いてその増殖活性を調べることにより測定することが出来る。具体的には、24穴プレートや96穴プレートに接種したT1165や7TD1細胞に、改質されたIL-11標品などを処理し、3〜5日後の細胞数をコールターカウンターでそのまま測定するかMTT法をなどにより吸光度測定から細胞増殖活性を測定することができる。
【0033】
本発明にかかる改質されたIL-11を医薬に活用するには、ヒトに投与して抗原性を持たないことが重要であり、ヒト正常細胞に近い糖鎖付加機能を有した動物細胞を宿主に用いるのが好ましい。動物細胞は発現効率が良く、糖鎖を付加した有用蛋白質の生産にはCHO細胞が好ましく用いられているが、これに限定されない。CHO細胞を用いる場合には、染色体に遺伝子を組み込み、構成的に生産させる方法が好ましく活用されている。
【0034】
動物細胞を用いた、改質されたIL-11の医薬のための商品化には、動物細胞の大量高密度培養が必須となる。動物細胞の高密度細胞培養方法には、マイクロキャリアー培養法、細胞固定用デイスクを用いたバイオリアクター、多段式培養装置やローラーボトルを用いる方法、中空糸モジュールを用いた培養システム、浮遊細胞のサスペンジョンカルチャー、マイクロカプセルに細胞を固定化培養する方法などがあるが、本発明にかかる改質されたIL-11遺伝子を導入した細胞の大量培養には、接着依存性細胞の大量培養に最も適したマイクロキャリアー法を用いるのが望ましい。
【0035】
マイクロキャリアー培養では、200mLスケールの培養瓶1本で内径100mmのシャーレの100枚分に相当する細胞数が得られ、しかも単位液量当たりの細胞数は約4倍の高密度培養を達成できる接着依存性細胞の大量培養化に理想的な方法である。マイクロキャリアーとしては、マトリックス素材はコラーゲン、ゼラチン、セルロース、架橋デキストラン、ポリスチレンのような合成樹脂からなり、荷電基としてジメチルアミノプロピル、ジメチルアミノエチル、トリメチルハイドロキシアミノプロピル、負電荷が付加されているものが好ましく用いられる。また、マトリックス素材をコラーゲンやゼラチンでコートしたものも使用される。市販品としては、架橋デキストランにジメチルアミノエチルを付加した“Cytodex-1、ファルマシア社”、“Cytodex-3、ファルマシア社”がある。
【0036】
中空糸としては、修飾セルロースを使用したものがある(”Vitafiber”、アミコン社)。マイクロカプセルは水透過性のあるゲルを形成するコラーゲンやアルギン酸ソーダを用いて、内部に細胞を包埋して作製する方法が知られている(A.Klausner,Bio/Technol.,1,736, 1983)。
【0037】
マイクロキャリアーの小スケール培養は、500mL用スピナーフラスコに0.3w/v%のマイクロキャリアーを含む200mLのPBS(-)を入れ、高圧蒸気滅菌(121℃、20分)したあと、200mLの培養液(10%ウシ胎児血清添加)に培地交換し、細胞を約2×105個/mL接種して培養を開始する。二日毎に培地交換し、細胞をマイクロキャリアー上にコンフルエントに増殖させる。改質されたIL-11を構成的に産生するように染色体に遺伝子を組み込んだ組み換えCHO細胞では、細胞がコンフルエントに増殖後、無血清培地あるいは低血清培地に培地交換し、改質されたIL-11を含む培養上清を2〜3日毎に回収し、細胞がマイクロキャリアーから脱落するまで培養を継続する。一過性の発現を行うには、COS7、KB、CHO、293細胞などをマイクロキャリアー上に増殖させ、無血清条件下でリポフェクション試薬と混合したDNAを4時間程度処理し、無血清培地に培地交換して産生のための培養を開始する。培養液は2〜3日毎に回収し、2〜3回のハーベストが可能である。
【0038】
改質されたIL-11蛋白質は、培養液中に放出されたものを、遠心分離後、上清を精製することにより、上記のような遺伝子導入細胞の培養液から単離することができる。精製には公知の分離・精製法を適切に組み合わせて行うことができる。これらの方法としては、塩析、溶媒沈殿、透析、限外濾過、ゲル濾過、SDS-ポリアクリルアミドゲル電気泳動、イオン交換クロマトグラフィー、アフィニテイークロマトグラフィー、逆相高速液体クロマトグラフィー、等電点電気泳動などがある。本発明に係る改質されたIL-11の精製初段には、操作が簡便で大量原液にも対応でき、回収率及び精製純度も上げられる担体を用いるのが好ましく、ハイドロキシアパタイト担体、シリカ担体、ブルーセファロース担体などを用いることができるがこれらに限定されない。さらにこれら担体や陽イオン交換担体、ゲル濾過などを組み合わせることにより、改質されたIL-11の精製標品を高純度に、かつ高収率で得ることができる。
【0039】
このようにして得られた改質されたIL-11標品は、透析、凍結乾燥を行い、乾燥粉末にすることもできる。さらに、容器への吸着を防ぐ担体として、血清アルブミンなどを添加したり、標品の酸化を防止するために精製過程、保存過程に微量の還元剤(β-メルカプトエタノール、ジチオスレイトール、グルタチオンなど)を添加することも可能である。
【0040】
本発明にかかる改質されたIL-11は医薬として利用可能である。IL-11には、他のサイトカイン(IL-3、-4、-7、-12、-13、SCF、Flt-3 ligand、GM-CSFなど)と協調して造血幹細胞の増殖を誘導する作用や、IL-3、TPO、SCFなどと共に骨髄細胞に作用し、巨核球及び血小板の産生を促進する作用、赤芽球系前駆細胞に作用して赤血球造血を促進する作用、骨髄前駆細胞の分化、成熟を促進する作用、SCFやIL-4と協調してB細胞分化を促進する作用など造血系に多くの作用を有している。また、肺胞や気管の上皮系細胞の炎症反応に深い関わりがあることや骨代謝や神経系にも細胞増殖や代謝を調節していることが報告されている。これら作用はいずれも医薬への展開の可能性を示しているが、悪性腫瘍への化学療法で引き起こされる血小板減少症に対する臨床試験では、浮腫やうっ血性心房性不整脈など深刻な副作用も報告されてきた。
【0041】
本発明にかかる改質されたIL-11は、野生型のIL-11が有する上記生物活性が野生型IL-11と比較して低減しているか、又は欠損しているといった特徴を有している。この特徴によれば、本発明にかかる改質されたIL-11は、高機能化されるだけでなく上述した副作用を軽減することにも寄与することが考えられる。すなわち、本発明にかかる改質されたIL-11は、野生型IL-11に対するアンタゴニストとして幅広く利用できる。
【0042】
上記のようにして得られた本発明にかかる改質されたIL-11は、医薬的に許容できる溶剤、賦形剤、担体、補助剤などを使用し、製剤化の常法に従って液剤、ローション剤、エアゾール剤、注射剤、散剤、顆粒剤、錠剤、座剤、腸溶剤及びカプセル剤などの医薬組成物としても良い。該医薬組成物は、ヒト、マウス、ラット、ウサギ、イヌ、ネコなどの哺乳動物に対して非経口的あるいは経口的に安全に投与することができる。本医薬組成物の投与量は、剤形、投与ルート、症状などにより適宜変更しうる。
【0043】
より具体的に、本発明にかかる改質されたIL-11を有効成分として含有する医薬組成物は、癌疾患における化学療法剤投与後の骨髄抑制に伴う重度血小板減少症、放射線療法後や骨髄移植後の血小板回復、再生不良性貧血、あるいは骨形成促進夜薬などといった疾患に対するIL-11製剤の治療をより効果的にする。また、本発明にかかる改質されたIL-11を有効成分として含有する医薬組成物は、野生型IL-11に対するアンタゴニストとして使用できることから、血小板増多症の血栓予防剤として使用することができる。
【実施例】
【0044】
以下、本発明を実施例を用いてより詳細に説明するが、本発明の技術的範囲は以下の実施例に限定されるものではない。
【0045】
〔実施例1〕
ヒトIL-11をコードする遺伝子断片をPCRにより増幅し、BstX1リンカーを付加した。これをBstX1制限酵素処理により切断したプラスミド・ベクターSRα BX3とライゲーションさせた。得られたプラスミドを用いてコンピテントセルJM109を形質転換し、アンピシリンによるスクリーニングを経てpSRα hIL-11を得た(図1)。なお、本例で使用したヒトIL-11をコードする遺伝子の塩基配列を配列番号1に、当該遺伝子によりコードされるヒトIL-11のアミノ酸配列を配列番号2に示す。
【0046】
また、得られたpSRα hIL-11プラスミドのIL-11の5’末端から22番目(プロリン)、70番目(ヒスチジン)、114番目(アラニン)、160番目(プロリン)、199番目(ロイシン)のアミノ酸残基をコードするコドンの次に、ペプチドNASあるいはAAATPAPG配列をコードした塩基配列をPCRにより挿入し、各種発現ベクターを得た。構築された野生型および改変蛋白質の発現ベクターとそれら由来の改変蛋白質名を表1にまとめた。
【0047】
【表1】

【0048】
なお、表1「ペプチド挿入位置」の欄には、野生型IL-11に対してペプチドを付加した位置と、ペプチドのアミノ酸配列が記載されている。例えば、野生型IL-11のアミノ酸配列における70番目と71番目の間に3個のグリシンを付加した場合、この欄には「70-71 (GGG)」と記載されている。
【0049】
〔実施例2〕
本実施例では、実施例1で構築された各種プラスミドを用いてCOS7細胞を形質転換し、一過性発現で得られた各種改変蛋白質を用いて細胞増殖促進活性を比較検討した。形質転換は以下のようにして行った。先ず、6穴プレートを用いてCOS7細胞をダルベッコーMEMにウシ胎児血清10%を含む培養液を用いて80%コンフルエントになるように細胞を接種し、翌日、無血清培地Opti-MEM-1(GIBCO)に培地交換した。次に、5μgの各種プラスミドを15μLのリポフェクション試薬(Lipofectoamine 2000、Invitrogen)を用いて細胞に遺伝子導入し、4時間後に1%ウシ胎児血清を含むOpti MEM-1に培地交換した。このとき、2日毎に各種改変IL-11を含む培養液をハーベストし、産生量をELISAキット(R&D Systems)で測定した。これらを用いてT1165細胞の増殖に対する作用を調べた。
【0050】
細胞増殖活性は24穴プレートにRPMI 1640培地に10%FCSおよび10 ng/mLの各種改変IL-11を添加した培養液で培養してきた細胞をIL-11不含の培地で2回洗浄後の細胞を用いて測定した。ウェルあたり1万個の細胞を接種し、Hisタグを付加しない改変体については100 ng/mLから、Hisタグの付加した改変体は10 ng/mLから10倍希釈した各IL-11改変体希釈液を細胞に処理した。4日間培養し、増殖した細胞数をコールターカウンター(コールター)で測定した。結果を図2、図3に示した。1希釈に付き4ウェルを使用し、その平均値と標準誤差をグラフに示した。これらの結果から、アミノ酸配列の70番目(ヒスチジン)と71番目(アスパラギン)の間に3残基(NAS)又は8残基(AAATPAPG)付加した改変体を除き、全ての改変体は野生型と同等あるいはそれ以上の活性を示すことが明らかになった。Hisタグの付加は活性に影響を与えないことも確認された。
【0051】
本実施例の結果より、IL-11におけるN末端側から2番目に位置するコイルを立体的に破壊することによって、IL-11の有する生物活性が低下するか又は欠損することが明かになった。
【0052】
〔実施例3〕
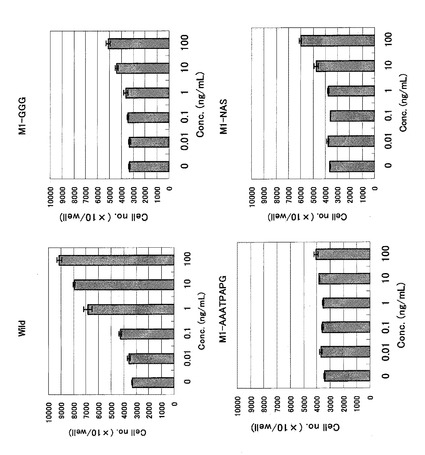
本実施例では、実施例2で得られたCOS7由来の各種改変体(Hisタグなし)を10ng/mLの濃度で細胞に処理し、増殖した4日後の細胞数を野生型と比較検討した。具体的には、まず、24穴プレートにウェルあたり1万個の細胞を接種し、10ng/mLの各種改変IL-11を添加し、4日後の細胞数をコールターカウンターで測定した。各改変IL-11につき4ウェルを使用し、野生型(Wild type)を100%にした時の各各改変IL-11の平均値の相対的な増殖の割合をグラフに示した。結果を図4に示す。
【0053】
なお、図4において、横軸には改変IL-11に付した番号、野生型(WT)及び無添加区(NC)を示した。図4の横軸において、1はN-NASであり、2はC-NASであり、3はM1-NASであり、4はM2-NASであり、5はM3-NASであり、6はC-AAATPAPGであり、7はN-AAATPAPGであり、8はM1-AAATPAPGであり、9はM2-AAATPAPGであり、10はM3-AAATPAPGであり、11はN-2NASであり、12はN-3NASであり、13はN-3NAS-GGGである。
【0054】
この結果は実施例2で得られた結果と同様、野生型IL-11のアミノ酸配列における70番目(ヒスチジン)と71番目(アスパラギン)の間にペプチド(具体的には3残基(NAS)又は8残基(AAATPAPG))を挿入した改変IL-11のみが著しい生物活性低下を示した。これらの結果は、アミノ酸配列の70番目と71番目近傍がIL-11の生物活性に大きく関与していることを示唆している。この知見は、IL-11に関する新規な知見である。実施例2及び3に示した結果から、野生型IL-11のアミノ酸配列における70番目(ヒスチジン)と71番目(アスパラギン)の間にアミノ酸又はペプチドを付加することによって、IL-11のアンタゴニストが創出できることが示された。
【0055】
〔実施例4〕
実施例2及び実施例3で得られた結果は、野生型IL-11のアミノ酸配列の70番目と71番目の間にNAS又はAAATPAPG配列を挿入して得られた結果である。これらの配列はN型又はO型糖鎖の付加を受けるペプチド配列であり、図2、図3、図4で観察された細胞増殖促進活性の低下は、これら配列に付加した糖鎖の影響である可能性が考えられた。
【0056】
そこで、本実施例ではこれら配列に代わり、GGGの3残基をコードする遺伝子を組み込んだ発現ベクターを新たに構築し、COS7細胞に導入して得た改変胃IL-11の細胞増殖促進活性を調べた。なお、GGGの3残基からなるペプチドは糖鎖の付加を受けないものである。
24穴プレートに接種した細胞にアミノ酸配列の70番目と71番目の間にNAS、GGG又はAAATPAPGのペプチドを付加した改変IL-11と野生型IL-11を100ng/mLから希釈した各希釈液を処理し、培養後4日目の細胞数をコールターカウンターで測定した。結果として、それぞれ4ウェルの平均値と標準誤差をグラフ(図5)にした。図5の結果から、糖鎖付加配列に代わりGGGを挿入した改変IL-11も細胞増殖促進活性の低下が同様に起きることが明らかになった。このことは、野生型IL-11のアミノ酸配列における70番目と71番目の近傍にIL-11の生物活性に深く関わる立体構造があることを示しており、この部位へのアミノ酸又はペプチドの挿入により生物活性を持たない又は生物活性が低減された、改質されたIL-11を作製できることが証明された。
【0057】
〔実施例5〕
本実施例では改質されたIL-11の生化学的、物理学的、構造科学的解析や動物を用いた評価用サンプルの調製に向けて、改変IL-11を構成的に産生する組換えCHO細胞を樹立し、得られた培養上清を用いて確立した精製法について示した。実施例1で得られたプラスミドpSRα-M1-AAATPAPG-hIL-11およびpSRα-hIL-11をマウスdhfr-遺伝子が挿入されたプラスミドpAdD26SV(A)-3と共に、Scahillらの方法(Proc. Natl. Acad. Sci. USA, 80, 4654-4658, 1983)に従ってCHOdhfr-細胞に遺伝子導入した。10%FCSを添加した核酸不含のα−MEMで細胞のスクリーニングを行い、0.01-1μMのメソトレキセートで遺伝子の増幅を行い、野性型IL-11および改変体M1-AAATPAPGを構成的に産生するCHO細胞株を得た。これらの細胞を培養フラスコおよびマイクロキャリアー法を用いて大量培養し、それぞれ精製原液10リットルを得た。
【0058】
初段の精製として、Bio-Gel HT (Hydrated) Hydroxyapatite(BIO-RAD)担体(HA)を詰めたXK26/20 column(amersham)をAKTA explorer(amersham)に接続し、CHO細胞由来培養上清1000 mLを13.4 mL/minの流速で送液した。Flow Throughを回収した後、0.1 M PB(pH 6.8)を洗浄し、0.2 M PB(pH 6.8)で溶出を行った。
【0059】
次に二段目の精製として、カラムをMicro Bead Silica Gel (FUJI-DAVISON CHEMICAL)担体(Silica)を詰めたXK26/20 columnに交換し、1000mLの1st HA Flow Throughを6.7 mL/minの流速で送液した。Flow Throughを回収した後、20 mM PB(pH 6.8)、20 mM PB-40%EG(pH 6.8)さらに50 mM AB-40%EG(pH 4.0)、15 mM HCl-20%EG(pH 1.2)でカラムを洗浄し、15 mM HCl-50%EGを用いて溶出した。また、溶出液回収時には、酸性の溶出バッファーにて溶出された精製サンプルを出来るだけ素早く1 M Tris-HCl(pH8.0)で中和した。表2および表3にHydroxyapatiteおよびSilica gelを用いたM1-AAATPAPGおよび野性型IL-11の精製の結果を示した。
【0060】
【表2】
【0061】
【表3】
【0062】
また、これらの精製で得られた各画分をSDS-PAGE後銀染色、または抗IL-11抗体を用いたウエスタンブロット解析した結果を図6および図7に示した。以上のように、本発明にかかる改変されたIL-11を培養上清から精製する方法を確立することができた。本方法を使用することによって、生化学的、物理学的、構造科学的解析に使用するためのサンプルや、動物を用いた実験系の評価用サンプルとして使用するための改質されたIL-11を大量に取得することが可能となった。
【技術分野】
【0001】
本発明は、造血幹細胞の増殖を誘導する機能等、様々な生理活性を有するインターロイキン-11(IL-11)及びその製造方法、さらにそれを含有する医薬組成物に関する。
【背景技術】
【0002】
IL-11は、アミノ酸178残基からなる糖鎖を持たない単純タンパク質で、マカクザルの骨髄間質細胞株PU34が産生する形質細胞腫増殖刺激因子として1990年に発見され(Paul, S. R., et al., Proc. Natl. Acad. Sci. USA, 82, 7512-7516, 1990〔非特許文献1〕)、また、前脂肪細胞から脂肪細胞への分化を抑制する脂肪細胞分化抑制因子(Kawasima, I., et al., FEBS Lett. 283, 199-202, 1991〔非特許文献2〕)として見出された。ヒトIL-11遺伝子は全長7kbにわたり、5つのエクソンと4つのイントロンより構成されている。mRNAサイズは、1.5kb及び2.5kbであるが、これらはpoly(A)付加シグナルの認識の違いにより生じており、同一の蛋白質をコードしている。IL-11は、元来IL-6様活性をもとにクローニングされたにも拘わらず、DNAレベルあるいは蛋白質レベルのいずれにおいてもIL-6とのホモロジーが認められない。これに対して、IL-11レセプターは、IL-6、LIF、CNTF、OSMなどの共通のシグナルトランスジューサーであるgp130(Yin,T., et al., J. Immunol., 151, 2555-2561, 1993〔非特許文献3〕)及びIL-11レセプターα鎖から構成されている(Hilton, D. J., et al., EMBO J., 13, 4765-4775, 1994〔非特許文献4〕)。細胞内シグナル伝達はJAKチロシンキナーゼ(JAK1, JAK2及びTYK2)、STAT及びRasが関与することが知られている(Taga,T., Annu. Rev. Immunol., 15, 797-819, 1997〔非特許文献5〕)。
【0003】
IL-11の生物学的作用としては、IL-3、-4、-7、-12、-13、SCF(stem cell factor)、Flt3-ligand、GM-CSFと共同で造血幹細胞の増殖を誘導すること、IL-3、TPO(thrombopoietin)あるいはSCFとともに骨髄細胞に作用し、巨核球及び血小板の産生を促進すること、単独あるいはIL-3、SCF、EPO(erythropoietin)と協調して赤血球造血を促進すること、骨髄前駆細胞の分化、成熟を促進すること、SCF, IL-4と協調してB細胞分化を促進すること、また、骨髄微小環境において、オートクラインあるいはパラクラインされる増殖因子として働くことが知られている(Du, X. X., and Williams, D. A., Blood, 89, 3897-3908, 1997〔非特許文献6〕)。また、肺胞及び気管上皮は特に炎症に伴い多量のIL-11を分泌することから、IL-11の生物学的作用として、呼吸器での炎症反応との関係も示唆されている。そのほかに、肝癌細胞株で急性期タンパク質を誘導させる作用(Baumann, H and Schendel, P., Biol. Chem., 266, 20424-20427, 1991〔非特許文献7〕)や骨芽細胞がIL-11を分泌し、骨芽細胞/破骨細胞両者に働き骨代謝を調節しているとの報告もある。In vitroの生物活性の指標には、サルのPU34細胞株培養上清からIL-11の発見をもたらしたマウスプラズマサイトーマ細胞株(T1165、Nordan, R. P and Potter, M., Science, 233, 566-569, 1986 〔非特許文献8〕)に対する増殖活性がよく用いられている。T1165細胞はIL-6及びIL-11に対し、特異的増殖活性を示す。
【0004】
このように、多くの生理活性を有するIL-11は、医薬への期待が高まり、近年、悪性腫瘍に対する化学療法で問題視されている血小板減少症に対し、血小板増殖因子としてあるいは炎症性腸疾患などを対象に米国、日本で開発が進められている。米国Genetics Institute社は、1997年に重度血小板減少症の危険が高い、非骨髄性腫瘍患者における骨髄抑制化学療法後の重度血小板減少症の予防と血小板輸血量の減量の適応で組み換えIL-11製剤の販売認可を獲得し、現在、小児を対象とした臨床試験が進行中である。また、Wyeth社では組み換えIL-11の経口製剤を炎症性腸疾患を対象に臨床試験を進めている。また、日本では、アステラス製薬がGenetics Institute社から導入した組み換え(大腸菌)IL-11の血小板減少症に対する臨床試験に着手している。
【0005】
このように医薬への期待が高まり、臨床試験もフェーズIIあるいはフェーズIIIに入り、その有効性が確認されると共にいくつかの副作用についても報告されるようになった(Smith, J.W. 2nd, Oncology, 14(9 Suppl 8),41, 7, 2000[非特許文献9])。例えば、浮腫、呼吸困難、胸膜からの漿液の滲出、結膜充血、うっ血性心房性不整脈などが報告された。深刻な血小板減少症に対し有効な医薬が望まれている折から、これらIL-11の投与に伴う副作用への対応は重要な課題になっている。また、IL-11は多くのサイトカインと異なり、システイン残基を欠き、N-グリコシレーションサイトが無いことが特徴的である。このことは、構造的不安定さを示唆しており、血中における安定性にも影響を及ぼすことが考えられている。これに対応して、血中における安定性を向上させるための研究が世界でスタートしている。米国のBolder BioTechnology社は、ポリエチレングリコール化技術を用いて数週間に一回の投与で済むIL-11誘導体の開発について米国国立衛生研究所から中小企業革新研究補助金を得て研究開発に着手している。また、エリスロポイエチンの例に見るように、糖鎖付加により安定性が増強されることも考えられる。しかしながら、血中安定性を増すなどIL-11の望ましい生物活性を十分に発揮するための多くの研究が今後の課題として残されている。そのためには、生物活性にかかわる部位の特定は必須のテーマであり、それにより安定型IL-11の創出やIL-11拮抗作用物質を見つけることが可能になる。しかし、現在までにIL-11の構造と生物活性との相関を見出したという報告は見つかっていない。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Paul, S. R., et al., Proc. Natl. Acad. Sci. USA, 82, 7512-7516, 1990
【非特許文献2】Kawasima, I., et al., FEBS Lett. 283, 199-202, 1991
【非特許文献3】Yin,T., et al., J. Immunol., 151, 2555-2561, 1993
【非特許文献4】Hilton, D. J., et al., EMBO J., 13, 4765-4775, 1994
【非特許文献5】Taga,T., Annu. Rev. Immunol., 15, 797-819, 1997
【非特許文献6】Du, X. X., and Williams, D. A., Blood, 89, 3897-3908, 1997
【非特許文献7】Baumann, H and Schendel, P., Biol. Chem., 266, 20424-20427, 1991
【非特許文献8】Nordan, P. N., and Potter, M, Science, 233, 566-569, 1986
【非特許文献9】Smith, J.W., 2nd, Oncology, 14(9 Suppl 8),41, 7, 2000
【発明の概要】
【発明が解決しようとする課題】
【0007】
そこで、本発明は、IL-11の生物活性と構造との相関を明らかにし、IL-11の生物活性に関与する部位を改変することで機能的に改質されたIL-11及びその製造方法を提供するとともに、改質されたIL-11を含む医薬組成物を提供することを目的としている。
【課題を解決するための手段】
【0008】
本発明者らは上記課題を解決すべくIL-11の生物活性にかかわる部位の同定に鋭意研究を重ね、ペプチド鎖の挿入によりIL-11の生物活性が低減或いは失われる部位を見つけることができた。本発明はこのような知見に基づいて完成されたものである。
【0009】
すなわち、本発明は、アミノ酸を付加させることにより改質されたIL-11を提供する。ここで、本発明にかかる改質されたIL-11は、改質対象のIL-11に対して1個のアミノ酸を付加したものであっても良いし、改質対象のIL-11に対して複数個のアミノ酸からなるペプチド(例えば、3〜8個のアミノ酸残基)を付加したものであっても良い。
【0010】
特に、本発明にかかる改質されたIL-11は、野生型IL-11の一次構造から立体構造予測プログラムによってコイルと推定された部位に、上記アミノ酸又は上記ペプチドを付加することが望ましい。上記コイルは、当該プログラムによって推定される6個のコイルのうちN末端側から2番目に位置するコイルである。そして、上記アミノ酸又は上記ペプチドを付加する領域としては、上記コイルを構成する領域のうち、野生型IL-11のアミノ酸配列における68番目(グリシン)〜72番目(ロイシン)の間、好ましくは、70番目(ヒスチジン)〜71番目(アスパラギン)の間であることが好ましい。
【0011】
また、本発明は、アミノ酸を付加することにより改質されたIL-11の製造方法で以下の工程を含むことを特徴とする。
(1)付加するアミノ酸あるいはペプチドをコードするDNA断片とIL-11をコードするcDNAを連結する工程
(2)当該連結cDNAを発現ベクターに組み込む工程
(3)当該発現ベクターを宿主細胞に導入する工程。
(4)当該宿主細胞において、アミノ酸又はペプチドが付加されたIL-11を発現させる工程
(5)当該宿主細胞において発現された、アミノ酸又はペプチドが付加されたIL-11を単離・精製する工程。
【0012】
また、本発明は、アミノ酸又はペプチドが付加されることにより改質されたIL-11の製造方法であって、アミノ酸又はペプチドを化学的結合法によりIL-11に結合させる工程を含むことを特徴とする。さらに、本発明は、アミノ酸又はペプチドを付加させることにより改質された前記のIL-11を有効成分として含有する医薬組成物を提供する。また、本発明の技術的思想は、IL-11と類似した立体構造が予測されるIL-6ファミリーサイトカインであるIL-6、CNTF(ciliary neurotrophic factor)、LIF(leukemia inhibitory factor)などにも適用される幅広い応用が可能である。
【発明の効果】
【0013】
本発明によれば、IL-11が本来的に有している様々な生理活性が改質されたインターロイキン-11を提供することができる。また、本発明によれば、改質されたIL-11を有効成分として含み、従来のIL-11を有効成分として含有する医薬組成物より優れた薬効を示したり、あるいはIL-11を有効成分として含有する医薬組成物の薬効をより有効なものにする効果を期待できる医薬組成物を提供することができる。
【0014】
さらに、本発明によれば、IL-11が本来的に有している様々な生理活性が改質されたインターロイキン-11の製造方法を提供することができる。
【図面の簡単な説明】
【0015】
【図1】哺乳類細胞発現用pSRαhIL-11プラスミドのマップ示す。
【図2】COS7細胞由来各種改変IL-11のT1165細胞に対する細胞増殖促進活性を示す特性図である。
【図3】COS7細胞由来Hisタグ付き改変IL-11のT1165細胞に対する細胞増殖促進活性を示す特性図である。
【図4】COS7細胞由来各種改変IL-11のT1165細胞に対する細胞増殖促進活性を野性型IL-11と比較検討した結果を示す特性図である。
【図5】野生型IL-11のアミノ酸配列における70番目と71番目のアミノ酸の間にペプチドを付加することにより細胞増殖促進活性が低下することを示す特性図である。
【図6】改質されたIL-11(M1-AAATPAPG)の精製画分の銀染色(図6のA)とウエスタンブロッテイング解析(図6のB)の結果を示す写真である。
【図7】野性型IL-11の精製画分の銀染色(図7のA)とウエスタンブロッテイング解析(図7のB)の結果を示す写真である。
【発明を実施するための形態】
【0016】
以下、本発明を詳細に説明する。
本発明に係る改質されたインターロイキン-11(以下、改質されたIL-11)は、IL-11の所定の領域にアミノ酸又はペプチドが付加され、IL-11が本来有する生理活性が低下或いは欠損したものである。すなわち、本発明において、改質されたとは、IL-11が本来的に有する生物活性が低下或いは欠損したことを意味する。
【0017】
本発明において、改質対象のIL-11としては、IL-11として知られている蛋白質であれば何ら限定することなく適用することができる。IL-11の一例として、ヒト由来IL-11のアミノ酸配列を配列番号2、ヒト由来IL-11をコードするcDNAの塩基配列を配列番号1に挙げる。なお、本発明に係る高機能化IL-11は、配列番号2に記載されたアミノ酸配列からなる蛋白質に限定されず、糖鎖の付加を受けることができるペプチド及びシグナルペプチドを有する蛋白質であっても良い。なお、シグナルペプチドとは、所定の蛋白質のアミノ末端に存在する分泌のためのペプチド配列であって、当該蛋白質が細胞から分泌される際に機能する。本発明に係る改質されたIL-11は、初めからシグナルペプチドを欠損する形で製造してもその有用性には変化がない。なお、野生型IL-11のcDNAは、日本DNAデータバンク(DDBJ)など遺伝子バンクに登録された配列から適当なプライマーを設計し、当該動物の当該組織のmRNAよりRT-PCRを行うことにより取得できる。
【0018】
また、IL-11としては、配列番号2のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換、付加若しくは修飾されたアミノ酸配列からなり、IL-11活性を有する蛋白質も含まれる。ここで、数個のアミノ酸とは、2〜50個のアミノ酸、好ましくは2〜25個のアミノ酸、より好ましくは2〜10個のアミノ酸を意味する。
【0019】
さらに、改質対象のIL-11としては、野生型のIL-11に対して、N型糖鎖、O型糖鎖及びN型O型並びにこれらの組み合わせの糖鎖のいずれかの糖鎖を付加する機能を有するペプチドが導入された変異型IL-11であってもよい。
【0020】
ここで、N型糖鎖とは、蛋白質の一次構造に対するアスパラギン残基に結合したN-アセチルガラクトサミンを起点として伸長する多様な糖鎖構造を総称するものであり、例えば、Asn-X-Ser/ThrのAsnNH2基にC-N結合を介して結合したGlcNAcを還元末端とする糖鎖を挙げることができる。すなわち、N型糖鎖の付加を受けることが知られているペプチドの例として、Asn-X-Thr、Asn-X-Ser(配列中、Xはプロリン以外の任意のアミノ酸である。)が挙げられる。また、O型糖鎖とは、蛋白質の一次構造に存在するセリン残基又はスレオニン残基に結合したN-アセチルガラクトサミンを起点として伸長する多様な糖鎖構造を総称するものであり、例えば、Ser又はThrの水酸基にGalNAcがC-O結合を介して結合した糖鎖を挙げることができる。すなわち、O型糖鎖の付加を受けることが知られているペプチドの例として、Ala-Thr-Pro-Ala-Proが挙げられる。
【0021】
本発明にかかる改質されたIL-11は、改質対象のIL-11(例えば、野生型IL-11)の一次構造から立体構造予測プログラムによりコイルと推定された領域に、アミノ酸又はペプチドを付加させた一次構造を有している。ここで立体構造予測プログラムとしては、特に限定されず、例えば、GENETYX ver. 7(ゼネテイックス)を例示することができる。当該プログラムを使用し、Chou-Fasman法(Chou, P. Y. and Fasman, G. D., Enzymology, 47, 45-147, 1978)に従ってアミノ酸配列から立体構造におけるコイルを予測することができる。例えば、野生型IL-11を当該プログラムにより解析すると、シグナルペプチドを含むアミノ酸配列のN末端から順に、30番目(アルギニン)と31番目(バリン)の間、68番目(グリシン)と72番目(アスパラギン)の間、99番目(アラニン)と100番目(アスパラギン)の間、114番目(アラニン)、156番目(プロリン)と162番目(アラニン)の間、167番目(アラニン)と170番目(グリシン)の間の6箇所にコイル構造が予測される。
【0022】
本発明にかかる改質されたIL-11は、N末端側から2番目に位置するコイルにアミノ酸又はペプチドを付加することが好ましい。具体的には、本発明にかかる改質されたIL-11は、N末端側から2番目に位置するコイルを立体的に破壊するようにアミノ酸又はペプチドを付加することが好ましい。例えば、野生型IL-11のシグナルペプチドを含むアミノ酸配列における68番目(グリシン)〜72番目(ロイシン)の間にアミノ酸又はペプチドを付加することによって、ここに位置するコイルを立体的に破壊することができる。より好ましくは、野生型IL-11のアミノ酸配列における70番目(ヒスチジン)〜71番目(アスパラギン)の間にアミノ酸又はペプチドを付加することによって、最も効果的にN末端側から2番目に位置するコイルを立体的に破壊することができる。
【0023】
なお、上述したアミノ酸残基の位置を示す数値は、野生型IL-11のシグナルペプチドを含むアミノ酸配列(配列番号2)を基準とした値である。したがって、改変対象のIL-11が野生型IL-11と異なるアミノ酸配列である場合には、上述したアミノ酸残基の位置を示す数値は同じ値にならないことに留意すべきである。
【0024】
改質されたIL-11において、付加するアミノ酸の種類及びペプチドの配列には限定されず、いかなる種類のアミノ酸を付加しても良いし、いかなる配列及びいかなる長さのペプチドを付加しても良い。具体的にペプチドとしては、3〜8個の長さのペプチドを挙げることができる。好ましくは、改変対象IL-11全体の立体構造に大きな影響を与えないアミノ酸あるいはペプチドを付加することが好ましい。例えば、グリシン単独あるいはグリシンのみからなるペプチドを挙げることができる。
【0025】
改質されたIL-11を製造するには、まず、付加するアミノ酸あるいはペプチドをコードするDNA断片をIL-11をコードするcDNAに組み込んだ発現ベクターを構築する。次に、当該発現ベクターを適当な宿主細胞に導入して、IL-11を発現させれば良い。発現ベクターは、プラスミド・ベクターを基本として構築しても良いし、ウィルス・ベクターを基本として構築しても良い。
【0026】
より具体的には、まず、分泌シグナル及び付加するアミノ酸又はペプチドをコードするオリゴヌクレオチドを合成し、あるいはPCR反応によって増幅し、これをIL-11をコードするcDNAの所望の位置(例えば、N末端側から2番目に位置するコイルを構成するアミノ酸配列をコードするコドン)に組み込む。分泌シグナルとしては、典型的な分泌型糖蛋白質のアミノ末端を利用することができる。このIL-11をコードするcDNAを組み込むプラスミドとしては、宿主内で複製保持されるものであれば、いずれも使用することができるが、例えば大腸菌由来のpBR322、pUC18及びこれらを基に構築されたpET-3cなどを挙げることができる。
【0027】
IL-11をコードするcDNAに上記オリゴヌクレオチドを組み込む方法としては、例えばT.Maniatisらにより紹介された方法(Molecular Cloning, Cold Spring Harbor Laboratory, p239, 1982)がある。上記のようにして作製したベクターから、分泌シグナル、付加するアミノ酸又はペプチド及びIL-11をコードする塩基配列を含む領域を切り出し、これを発現に適したベクター中のプロモーターの下流に連結することにより、発現型ベクターを得ることができる。
【0028】
上記IL-11をコードするcDNAを含む領域は、5’末端に翻訳開始コドンとしてのATGを有し、また、3’末端には翻訳終止コドンとしてのTAA、TGA又はTAGを有しても良い。さらに、該コーデイング領域にコードされている蛋白質を発現させるには、その上流にプロモーターを接続する必要がある。本発明で用いられるプロモーターとしては、遺伝子の発現に用いる宿主に対応した適切なプロモーターであればいかなるものでも用いることが出来る。形質転換する宿主が枯草菌である場合には、SP01、SP02、penPプロモーターなどがあり、酵母ではPH05、PGK、GAP、ADHプロモーターなどが挙げられる。また、動物細胞ではSV40由来のプロモーター、レトロウイルスのプロモーターが挙げられる。
【0029】
宿主細胞としては、いかなるものでも良く、枯草菌(例えば、Bacillus subtilis DB105)、酵母(例えば、Pichia pasto ris、Saccharomyces cerevisiae)、動物細胞(例えば、COS、CHO、KB、293、BHK、NIH3T3、BALB/c3T3、臍帯静脈内皮細胞)、昆虫細胞(例えば、Sf9、Tn細胞)などが挙げられるが、本発明に係る改質されたIL-11を医薬組成物としてヒトに投与する目的で大量培養をおこなう場合、動物細胞を用いて製造することが好ましい。
【0030】
上記の形質転換は、それぞれの宿主について一般的に行われている方法あるいは適応可能な方法であれば良い。例えば、宿主が酵母であれば、リチウム法などで作製したコンピテント細胞に組み換えDNAを含むベクターを温度ショック法あるいはエレクトロポレーション法により導入する。宿主が動物細胞であれば、増殖期等の細胞に組み換えDNAを含むベクターをリン酸カルシウム法、リポフェクション法あるいはエレクトロポレーション法により導入する。
【0031】
このようにして得られた形質転換体は、それぞれの宿主に一般的に用いられている培地、あるいは適用可能な培養液を用いて培養することにより、改質されたIL-11を産生することができる。宿主が酵母であればYPD培地などを用い、動物細胞であればDulbecco's MEMにウシ胎児血清を加えた培養液を用いる。培養は、それぞれの宿主について一般的に用いられている条件あるいは適用可能な条件であれば良い。例えば、宿主が酵母であれば、約25〜37℃で、約12時間〜2週間行い、必要により、通気や撹拌を加えることができる。動物細胞の場合は、37℃で、5%炭酸ガス、100%湿度の条件で約24時間〜2週間行い、必要により、気相の条件を変えたり撹拌を加えることが出来る。
【0032】
細胞から得られた、本発明にかかる改質されたIL-11の生物活性は、IL-11及びIL-6により特異的な増殖を示すマウスプラズマサイトーマ細胞株T1165細胞あるいはマウスハイブリドーマ細胞株7TD細胞を用いてその増殖活性を調べることにより測定することが出来る。具体的には、24穴プレートや96穴プレートに接種したT1165や7TD1細胞に、改質されたIL-11標品などを処理し、3〜5日後の細胞数をコールターカウンターでそのまま測定するかMTT法をなどにより吸光度測定から細胞増殖活性を測定することができる。
【0033】
本発明にかかる改質されたIL-11を医薬に活用するには、ヒトに投与して抗原性を持たないことが重要であり、ヒト正常細胞に近い糖鎖付加機能を有した動物細胞を宿主に用いるのが好ましい。動物細胞は発現効率が良く、糖鎖を付加した有用蛋白質の生産にはCHO細胞が好ましく用いられているが、これに限定されない。CHO細胞を用いる場合には、染色体に遺伝子を組み込み、構成的に生産させる方法が好ましく活用されている。
【0034】
動物細胞を用いた、改質されたIL-11の医薬のための商品化には、動物細胞の大量高密度培養が必須となる。動物細胞の高密度細胞培養方法には、マイクロキャリアー培養法、細胞固定用デイスクを用いたバイオリアクター、多段式培養装置やローラーボトルを用いる方法、中空糸モジュールを用いた培養システム、浮遊細胞のサスペンジョンカルチャー、マイクロカプセルに細胞を固定化培養する方法などがあるが、本発明にかかる改質されたIL-11遺伝子を導入した細胞の大量培養には、接着依存性細胞の大量培養に最も適したマイクロキャリアー法を用いるのが望ましい。
【0035】
マイクロキャリアー培養では、200mLスケールの培養瓶1本で内径100mmのシャーレの100枚分に相当する細胞数が得られ、しかも単位液量当たりの細胞数は約4倍の高密度培養を達成できる接着依存性細胞の大量培養化に理想的な方法である。マイクロキャリアーとしては、マトリックス素材はコラーゲン、ゼラチン、セルロース、架橋デキストラン、ポリスチレンのような合成樹脂からなり、荷電基としてジメチルアミノプロピル、ジメチルアミノエチル、トリメチルハイドロキシアミノプロピル、負電荷が付加されているものが好ましく用いられる。また、マトリックス素材をコラーゲンやゼラチンでコートしたものも使用される。市販品としては、架橋デキストランにジメチルアミノエチルを付加した“Cytodex-1、ファルマシア社”、“Cytodex-3、ファルマシア社”がある。
【0036】
中空糸としては、修飾セルロースを使用したものがある(”Vitafiber”、アミコン社)。マイクロカプセルは水透過性のあるゲルを形成するコラーゲンやアルギン酸ソーダを用いて、内部に細胞を包埋して作製する方法が知られている(A.Klausner,Bio/Technol.,1,736, 1983)。
【0037】
マイクロキャリアーの小スケール培養は、500mL用スピナーフラスコに0.3w/v%のマイクロキャリアーを含む200mLのPBS(-)を入れ、高圧蒸気滅菌(121℃、20分)したあと、200mLの培養液(10%ウシ胎児血清添加)に培地交換し、細胞を約2×105個/mL接種して培養を開始する。二日毎に培地交換し、細胞をマイクロキャリアー上にコンフルエントに増殖させる。改質されたIL-11を構成的に産生するように染色体に遺伝子を組み込んだ組み換えCHO細胞では、細胞がコンフルエントに増殖後、無血清培地あるいは低血清培地に培地交換し、改質されたIL-11を含む培養上清を2〜3日毎に回収し、細胞がマイクロキャリアーから脱落するまで培養を継続する。一過性の発現を行うには、COS7、KB、CHO、293細胞などをマイクロキャリアー上に増殖させ、無血清条件下でリポフェクション試薬と混合したDNAを4時間程度処理し、無血清培地に培地交換して産生のための培養を開始する。培養液は2〜3日毎に回収し、2〜3回のハーベストが可能である。
【0038】
改質されたIL-11蛋白質は、培養液中に放出されたものを、遠心分離後、上清を精製することにより、上記のような遺伝子導入細胞の培養液から単離することができる。精製には公知の分離・精製法を適切に組み合わせて行うことができる。これらの方法としては、塩析、溶媒沈殿、透析、限外濾過、ゲル濾過、SDS-ポリアクリルアミドゲル電気泳動、イオン交換クロマトグラフィー、アフィニテイークロマトグラフィー、逆相高速液体クロマトグラフィー、等電点電気泳動などがある。本発明に係る改質されたIL-11の精製初段には、操作が簡便で大量原液にも対応でき、回収率及び精製純度も上げられる担体を用いるのが好ましく、ハイドロキシアパタイト担体、シリカ担体、ブルーセファロース担体などを用いることができるがこれらに限定されない。さらにこれら担体や陽イオン交換担体、ゲル濾過などを組み合わせることにより、改質されたIL-11の精製標品を高純度に、かつ高収率で得ることができる。
【0039】
このようにして得られた改質されたIL-11標品は、透析、凍結乾燥を行い、乾燥粉末にすることもできる。さらに、容器への吸着を防ぐ担体として、血清アルブミンなどを添加したり、標品の酸化を防止するために精製過程、保存過程に微量の還元剤(β-メルカプトエタノール、ジチオスレイトール、グルタチオンなど)を添加することも可能である。
【0040】
本発明にかかる改質されたIL-11は医薬として利用可能である。IL-11には、他のサイトカイン(IL-3、-4、-7、-12、-13、SCF、Flt-3 ligand、GM-CSFなど)と協調して造血幹細胞の増殖を誘導する作用や、IL-3、TPO、SCFなどと共に骨髄細胞に作用し、巨核球及び血小板の産生を促進する作用、赤芽球系前駆細胞に作用して赤血球造血を促進する作用、骨髄前駆細胞の分化、成熟を促進する作用、SCFやIL-4と協調してB細胞分化を促進する作用など造血系に多くの作用を有している。また、肺胞や気管の上皮系細胞の炎症反応に深い関わりがあることや骨代謝や神経系にも細胞増殖や代謝を調節していることが報告されている。これら作用はいずれも医薬への展開の可能性を示しているが、悪性腫瘍への化学療法で引き起こされる血小板減少症に対する臨床試験では、浮腫やうっ血性心房性不整脈など深刻な副作用も報告されてきた。
【0041】
本発明にかかる改質されたIL-11は、野生型のIL-11が有する上記生物活性が野生型IL-11と比較して低減しているか、又は欠損しているといった特徴を有している。この特徴によれば、本発明にかかる改質されたIL-11は、高機能化されるだけでなく上述した副作用を軽減することにも寄与することが考えられる。すなわち、本発明にかかる改質されたIL-11は、野生型IL-11に対するアンタゴニストとして幅広く利用できる。
【0042】
上記のようにして得られた本発明にかかる改質されたIL-11は、医薬的に許容できる溶剤、賦形剤、担体、補助剤などを使用し、製剤化の常法に従って液剤、ローション剤、エアゾール剤、注射剤、散剤、顆粒剤、錠剤、座剤、腸溶剤及びカプセル剤などの医薬組成物としても良い。該医薬組成物は、ヒト、マウス、ラット、ウサギ、イヌ、ネコなどの哺乳動物に対して非経口的あるいは経口的に安全に投与することができる。本医薬組成物の投与量は、剤形、投与ルート、症状などにより適宜変更しうる。
【0043】
より具体的に、本発明にかかる改質されたIL-11を有効成分として含有する医薬組成物は、癌疾患における化学療法剤投与後の骨髄抑制に伴う重度血小板減少症、放射線療法後や骨髄移植後の血小板回復、再生不良性貧血、あるいは骨形成促進夜薬などといった疾患に対するIL-11製剤の治療をより効果的にする。また、本発明にかかる改質されたIL-11を有効成分として含有する医薬組成物は、野生型IL-11に対するアンタゴニストとして使用できることから、血小板増多症の血栓予防剤として使用することができる。
【実施例】
【0044】
以下、本発明を実施例を用いてより詳細に説明するが、本発明の技術的範囲は以下の実施例に限定されるものではない。
【0045】
〔実施例1〕
ヒトIL-11をコードする遺伝子断片をPCRにより増幅し、BstX1リンカーを付加した。これをBstX1制限酵素処理により切断したプラスミド・ベクターSRα BX3とライゲーションさせた。得られたプラスミドを用いてコンピテントセルJM109を形質転換し、アンピシリンによるスクリーニングを経てpSRα hIL-11を得た(図1)。なお、本例で使用したヒトIL-11をコードする遺伝子の塩基配列を配列番号1に、当該遺伝子によりコードされるヒトIL-11のアミノ酸配列を配列番号2に示す。
【0046】
また、得られたpSRα hIL-11プラスミドのIL-11の5’末端から22番目(プロリン)、70番目(ヒスチジン)、114番目(アラニン)、160番目(プロリン)、199番目(ロイシン)のアミノ酸残基をコードするコドンの次に、ペプチドNASあるいはAAATPAPG配列をコードした塩基配列をPCRにより挿入し、各種発現ベクターを得た。構築された野生型および改変蛋白質の発現ベクターとそれら由来の改変蛋白質名を表1にまとめた。
【0047】
【表1】
【0048】
なお、表1「ペプチド挿入位置」の欄には、野生型IL-11に対してペプチドを付加した位置と、ペプチドのアミノ酸配列が記載されている。例えば、野生型IL-11のアミノ酸配列における70番目と71番目の間に3個のグリシンを付加した場合、この欄には「70-71 (GGG)」と記載されている。
【0049】
〔実施例2〕
本実施例では、実施例1で構築された各種プラスミドを用いてCOS7細胞を形質転換し、一過性発現で得られた各種改変蛋白質を用いて細胞増殖促進活性を比較検討した。形質転換は以下のようにして行った。先ず、6穴プレートを用いてCOS7細胞をダルベッコーMEMにウシ胎児血清10%を含む培養液を用いて80%コンフルエントになるように細胞を接種し、翌日、無血清培地Opti-MEM-1(GIBCO)に培地交換した。次に、5μgの各種プラスミドを15μLのリポフェクション試薬(Lipofectoamine 2000、Invitrogen)を用いて細胞に遺伝子導入し、4時間後に1%ウシ胎児血清を含むOpti MEM-1に培地交換した。このとき、2日毎に各種改変IL-11を含む培養液をハーベストし、産生量をELISAキット(R&D Systems)で測定した。これらを用いてT1165細胞の増殖に対する作用を調べた。
【0050】
細胞増殖活性は24穴プレートにRPMI 1640培地に10%FCSおよび10 ng/mLの各種改変IL-11を添加した培養液で培養してきた細胞をIL-11不含の培地で2回洗浄後の細胞を用いて測定した。ウェルあたり1万個の細胞を接種し、Hisタグを付加しない改変体については100 ng/mLから、Hisタグの付加した改変体は10 ng/mLから10倍希釈した各IL-11改変体希釈液を細胞に処理した。4日間培養し、増殖した細胞数をコールターカウンター(コールター)で測定した。結果を図2、図3に示した。1希釈に付き4ウェルを使用し、その平均値と標準誤差をグラフに示した。これらの結果から、アミノ酸配列の70番目(ヒスチジン)と71番目(アスパラギン)の間に3残基(NAS)又は8残基(AAATPAPG)付加した改変体を除き、全ての改変体は野生型と同等あるいはそれ以上の活性を示すことが明らかになった。Hisタグの付加は活性に影響を与えないことも確認された。
【0051】
本実施例の結果より、IL-11におけるN末端側から2番目に位置するコイルを立体的に破壊することによって、IL-11の有する生物活性が低下するか又は欠損することが明かになった。
【0052】
〔実施例3〕
本実施例では、実施例2で得られたCOS7由来の各種改変体(Hisタグなし)を10ng/mLの濃度で細胞に処理し、増殖した4日後の細胞数を野生型と比較検討した。具体的には、まず、24穴プレートにウェルあたり1万個の細胞を接種し、10ng/mLの各種改変IL-11を添加し、4日後の細胞数をコールターカウンターで測定した。各改変IL-11につき4ウェルを使用し、野生型(Wild type)を100%にした時の各各改変IL-11の平均値の相対的な増殖の割合をグラフに示した。結果を図4に示す。
【0053】
なお、図4において、横軸には改変IL-11に付した番号、野生型(WT)及び無添加区(NC)を示した。図4の横軸において、1はN-NASであり、2はC-NASであり、3はM1-NASであり、4はM2-NASであり、5はM3-NASであり、6はC-AAATPAPGであり、7はN-AAATPAPGであり、8はM1-AAATPAPGであり、9はM2-AAATPAPGであり、10はM3-AAATPAPGであり、11はN-2NASであり、12はN-3NASであり、13はN-3NAS-GGGである。
【0054】
この結果は実施例2で得られた結果と同様、野生型IL-11のアミノ酸配列における70番目(ヒスチジン)と71番目(アスパラギン)の間にペプチド(具体的には3残基(NAS)又は8残基(AAATPAPG))を挿入した改変IL-11のみが著しい生物活性低下を示した。これらの結果は、アミノ酸配列の70番目と71番目近傍がIL-11の生物活性に大きく関与していることを示唆している。この知見は、IL-11に関する新規な知見である。実施例2及び3に示した結果から、野生型IL-11のアミノ酸配列における70番目(ヒスチジン)と71番目(アスパラギン)の間にアミノ酸又はペプチドを付加することによって、IL-11のアンタゴニストが創出できることが示された。
【0055】
〔実施例4〕
実施例2及び実施例3で得られた結果は、野生型IL-11のアミノ酸配列の70番目と71番目の間にNAS又はAAATPAPG配列を挿入して得られた結果である。これらの配列はN型又はO型糖鎖の付加を受けるペプチド配列であり、図2、図3、図4で観察された細胞増殖促進活性の低下は、これら配列に付加した糖鎖の影響である可能性が考えられた。
【0056】
そこで、本実施例ではこれら配列に代わり、GGGの3残基をコードする遺伝子を組み込んだ発現ベクターを新たに構築し、COS7細胞に導入して得た改変胃IL-11の細胞増殖促進活性を調べた。なお、GGGの3残基からなるペプチドは糖鎖の付加を受けないものである。
24穴プレートに接種した細胞にアミノ酸配列の70番目と71番目の間にNAS、GGG又はAAATPAPGのペプチドを付加した改変IL-11と野生型IL-11を100ng/mLから希釈した各希釈液を処理し、培養後4日目の細胞数をコールターカウンターで測定した。結果として、それぞれ4ウェルの平均値と標準誤差をグラフ(図5)にした。図5の結果から、糖鎖付加配列に代わりGGGを挿入した改変IL-11も細胞増殖促進活性の低下が同様に起きることが明らかになった。このことは、野生型IL-11のアミノ酸配列における70番目と71番目の近傍にIL-11の生物活性に深く関わる立体構造があることを示しており、この部位へのアミノ酸又はペプチドの挿入により生物活性を持たない又は生物活性が低減された、改質されたIL-11を作製できることが証明された。
【0057】
〔実施例5〕
本実施例では改質されたIL-11の生化学的、物理学的、構造科学的解析や動物を用いた評価用サンプルの調製に向けて、改変IL-11を構成的に産生する組換えCHO細胞を樹立し、得られた培養上清を用いて確立した精製法について示した。実施例1で得られたプラスミドpSRα-M1-AAATPAPG-hIL-11およびpSRα-hIL-11をマウスdhfr-遺伝子が挿入されたプラスミドpAdD26SV(A)-3と共に、Scahillらの方法(Proc. Natl. Acad. Sci. USA, 80, 4654-4658, 1983)に従ってCHOdhfr-細胞に遺伝子導入した。10%FCSを添加した核酸不含のα−MEMで細胞のスクリーニングを行い、0.01-1μMのメソトレキセートで遺伝子の増幅を行い、野性型IL-11および改変体M1-AAATPAPGを構成的に産生するCHO細胞株を得た。これらの細胞を培養フラスコおよびマイクロキャリアー法を用いて大量培養し、それぞれ精製原液10リットルを得た。
【0058】
初段の精製として、Bio-Gel HT (Hydrated) Hydroxyapatite(BIO-RAD)担体(HA)を詰めたXK26/20 column(amersham)をAKTA explorer(amersham)に接続し、CHO細胞由来培養上清1000 mLを13.4 mL/minの流速で送液した。Flow Throughを回収した後、0.1 M PB(pH 6.8)を洗浄し、0.2 M PB(pH 6.8)で溶出を行った。
【0059】
次に二段目の精製として、カラムをMicro Bead Silica Gel (FUJI-DAVISON CHEMICAL)担体(Silica)を詰めたXK26/20 columnに交換し、1000mLの1st HA Flow Throughを6.7 mL/minの流速で送液した。Flow Throughを回収した後、20 mM PB(pH 6.8)、20 mM PB-40%EG(pH 6.8)さらに50 mM AB-40%EG(pH 4.0)、15 mM HCl-20%EG(pH 1.2)でカラムを洗浄し、15 mM HCl-50%EGを用いて溶出した。また、溶出液回収時には、酸性の溶出バッファーにて溶出された精製サンプルを出来るだけ素早く1 M Tris-HCl(pH8.0)で中和した。表2および表3にHydroxyapatiteおよびSilica gelを用いたM1-AAATPAPGおよび野性型IL-11の精製の結果を示した。
【0060】
【表2】
【0061】
【表3】
【0062】
また、これらの精製で得られた各画分をSDS-PAGE後銀染色、または抗IL-11抗体を用いたウエスタンブロット解析した結果を図6および図7に示した。以上のように、本発明にかかる改変されたIL-11を培養上清から精製する方法を確立することができた。本方法を使用することによって、生化学的、物理学的、構造科学的解析に使用するためのサンプルや、動物を用いた実験系の評価用サンプルとして使用するための改質されたIL-11を大量に取得することが可能となった。
【特許請求の範囲】
【請求項1】
配列番号2に記載のアミノ酸配列からなるヒトインターロイキン−11における70番目(ヒスチジン)と71番目(アスパラギン)の間にアミノ酸配列:NASからなるペプチド又はアミノ酸配列:AAATPAPGからなるペプチドを挿入させた、インターロイキン−11。
【請求項2】
挿入するアミノ酸配列:NASからなるペプチド又はアミノ酸配列:AAATPAPGからなるペプチドをコードするDNA断片を、配列番号2に記載のアミノ酸配列からなるヒトインターロイキン−11における70番目(ヒスチジン)をコードするコドンと71番目(アスパラギン)をコードするコドンの間に組み込んだベクターを細胞に導入し、当該細胞が産生する、ペプチドが挿入されたインターロイキン−11を単離することを特徴とするインターロイキン−11の製造方法。
【請求項3】
請求項1記載のインターロイキン−11を有効成分として含有する医薬組成物。
【請求項1】
配列番号2に記載のアミノ酸配列からなるヒトインターロイキン−11における70番目(ヒスチジン)と71番目(アスパラギン)の間にアミノ酸配列:NASからなるペプチド又はアミノ酸配列:AAATPAPGからなるペプチドを挿入させた、インターロイキン−11。
【請求項2】
挿入するアミノ酸配列:NASからなるペプチド又はアミノ酸配列:AAATPAPGからなるペプチドをコードするDNA断片を、配列番号2に記載のアミノ酸配列からなるヒトインターロイキン−11における70番目(ヒスチジン)をコードするコドンと71番目(アスパラギン)をコードするコドンの間に組み込んだベクターを細胞に導入し、当該細胞が産生する、ペプチドが挿入されたインターロイキン−11を単離することを特徴とするインターロイキン−11の製造方法。
【請求項3】
請求項1記載のインターロイキン−11を有効成分として含有する医薬組成物。
【図1】

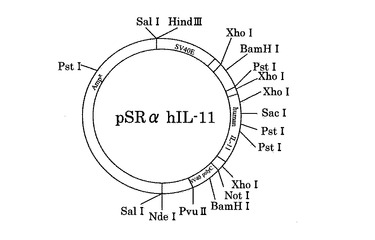
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【公開番号】特開2013−14614(P2013−14614A)
【公開日】平成25年1月24日(2013.1.24)
【国際特許分類】
【出願番号】特願2012−212096(P2012−212096)
【出願日】平成24年9月26日(2012.9.26)
【分割の表示】特願2006−144448(P2006−144448)の分割
【原出願日】平成18年5月24日(2006.5.24)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成14年度、独立行政法人新エネルギー・産業技術総合開発機構基盤技術研究促進事業委託研究、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(000003159)東レ株式会社 (7,677)
【Fターム(参考)】
【公開日】平成25年1月24日(2013.1.24)
【国際特許分類】
【出願日】平成24年9月26日(2012.9.26)
【分割の表示】特願2006−144448(P2006−144448)の分割
【原出願日】平成18年5月24日(2006.5.24)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成14年度、独立行政法人新エネルギー・産業技術総合開発機構基盤技術研究促進事業委託研究、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(000003159)東レ株式会社 (7,677)
【Fターム(参考)】
[ Back to top ]