
放出されたピロリン酸を検出することによるオリゴヌクレオチド連結アッセイ
本発明は、核酸サンプルにおける遺伝因子(例えば、ヌクレオチド反復、または微生物型決定のためのマーカーなど)の存在を明らかにするための新規な方法に関する。本発明の方法は、連結反応を行うことに基づいており、そこでは連結副生成物が検出されて、所望によりヌクレオチド反復を含む核酸サンプル中のヌクレオチド反復単位の数を決定するために使用される。本発明は、本発明の方法を行うためのキット、および本発明の方法を行うための成分を含む組成物にも関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、連結反応を行い、連結副生成物を検出することによって核酸サンプルにおける遺伝因子(例えば、ヌクレオチド反復、または微生物型決定のためのマーカーなど)の存在を明らかにするための方法に関する。
【背景技術】
【0002】
遺伝的変異は疾患に関係づけられることが多く、遺伝的変異の同定は臨床的診断における重要な手段である。遺伝的変異は、一塩基のレベルでの違い(塩基の変化、挿入または欠失を伴う突然変異およびSNP)、数塩基のレベルでの違い(典型的にはコドン欠失を伴う)、または、遺伝子全体の増加までの様々な長さの反復した配列の変化する数のレベルでの違いを伴う。
【0003】
ヌクレオチド反復の拡大は種々の遺伝病に関係づけられることが示されている。例としては、脆弱X(Webb,TP他,1986年、Am.J.Med.Genet.23:573;Gustavson,KH他,1986年、Am.J.Med.Genet.23:581)、筋緊張性ジストロフィー(Harper,PS他,1989年、Myotonic Dystrophy(第2版)、London、England、WB Saunders Co.1989年)およびハンチントン病(MacDonald,ME他,1993年、Cell.72:971)が含まれる。これらのヌクレオチド反復は、例えば、PCRに基づくアッセイおよびサザンブロッティング技術を使用して分析されている。さらに、ジヌクレオチド反復、トリヌクレオチド反復、テトラヌクレオチド反復、ペンタヌクレオチド反復およびヘキサヌクレオチド反復が、遺伝子マッピングプロジェクトにおいて、また、父子鑑定のような種々の法医学的適用において広範囲に使用されている。
【0004】
パイロシーケンス(商標)(Pyrosequencing)は、ピロリン酸の放出の検出および酵素によるヌクレオチド分解に基づく、王立技術研究所(ストックホルム)で開発された、合成による配列決定の方法である(米国特許第6210891号および米国特許第6258568号)。合成による配列決定のこの方法では、従来のサンガー配列決定とは対照的に、ヌクレオチドが配列決定反応時に1つずつ加えられる。使用される酵素混合物は4つの異なる酵素(DNAポリメラーゼ、ATP−スルフリラーゼ、ルシフェラーゼおよびアピラーゼ)からなる。ヌクレオチドが、使用者によって、テンプレートに依存する指定された順序に従って順次加えられ、決定される。加えられたヌクレオチドがテンプレートと一致するならば、DNAポリメラーゼがそのヌクレオチドを成長途中のDNA鎖に取り込み、ピロリン酸(PPi)が放出される。ATP−スルフリラーゼにより、PPiがATPに変換され、第3の酵素(ルシフェラーゼ)により、ATPが光シグナルに転換される。これらの反応の後、第4の酵素(アピラーゼ)により、過剰なヌクレオチドおよびATPが分解され、テンプレートが、その時点で次の反応サイクル(すなわち、別のヌクレオチド添加)のために使用できる状態になる。ヌクレオチドが取り込まれない限り、PPiが放出されないので、光シグナルは、正しいヌクレオチドが取り込まれたときにだけ生じることになる。パイロシーケンスは、合成による配列決定に基づくリアルタイムのDNA配列決定法である。この方法は、SNP(単一ヌクレオチド多型)スコア化、より短いDNA領域(識別タグ)の配列決定、および対立遺伝子頻度の評価のための迅速かつ正確な方法であることが判明している(Ronaghi,M.,Uhlen,M.,Nyren,P.,リアルタイムのピロリン酸に基づく配列決定法(1998年)、Science.281、363〜365;Alderborn,A.,Kristofferson,A.,Hammerling,U.リアルタイムのピロリン酸DNA配列決定による単一ヌクレオチド多型の決定(2000年)、Genome Res.10:1249〜1258)。
【0005】
パイロシーケンスの形態での合成による配列決定は遺伝的変異の分析において多くの利点を有する。例えば、SNP分析のために使用されている他の技術(例えば、ハイブリダイゼーション技術、ミニ配列分析、RFLPおよびSSCPなど)と比較した場合、合成による配列決定では、周りの配列を示すことによって、多型位置だけでなく正しいSNP位置を調査できることが確認される。さらに、パイロシーケンスは、分離工程を伴わず、かつ自動化可能な様式でのリアルタイム分析を提供する迅速な技術であり、このことは、SSCPおよびRFLPと比較して有益であり、従って、パイロシーケンスは遺伝子分析のための魅力的な方法である。
【0006】
しかしながら、パイロシーケンスは、1塩基〜100塩基の範囲での短いヌクレオチド範囲を分析するときに最も効果的である。この方法は、一致しないプライマー伸長(いわゆる「シフト」)の生成物の蓄積によって主に制限される。シフトは、例えば、最適でないレベルのヌクレオチドの存在に起因するプライマーの不完全な伸長または過度な伸長の結果である。均一なパイロシーケンス反応では、正しいヌクレオチドを取り込むDNAポリメラーゼと、取り込まれなかったヌクレオチドを分解するアピラーゼとの間での競合が伴う。正しいヌクレオチドのレベルが、取り込み工程時においてDNAポリメラーゼについての最適値よりも低くなる場合、一部のプライマーのさらなる伸長が不完全となる。その結果は、正しい完全に伸長したプライマーよりも1塩基または数塩基短い伸長されたプライマー分子の集団である(いわゆるマイナスシフト)。他方で、アピラーゼが、ヌクレオチドを分解することにおいて十分に効果的でない場合、過剰な取り込まれていないヌクレオチドが反応液に残留し得るし、また、次の正しいヌクレオチドが与えられ、かつ取り込まれ、配列により、所望されないヌクレオチドバックグラウンドを使用するさらなる伸長が可能になる特定の状況では取り込まれ得る(いわゆるプラスシフト)。さらなる現象では、DNAポリメラーゼの活性を妨げ、従って、不完全な取り込みおよびマイナスシフトを生じさせる複雑な二次構造を有するテンプレートが伴う。これらの誤りは、当然のことではあるが、プライマー伸長の数が増大するにつれて増大する。従って、ヌクレオチド反復数を推定するための一ヌクレオチド伸長による核酸の延長に依存する技術の正確性は、ヌクレオチド反復数および長さが増大するにつれて低下する。
【0007】
米国特許第6309829号には、ヌクレオチド反復数を分析するための方法が記載され、この方法では、プライマーがテンプレート核酸にアニーリングされ、一ヌクレオチドずつ伸長し、かつ、ヌクレオチドは標識され、伸長が起こったことの目安としての測定可能なシグナルを与える。最後に加えられたヌクレオチドの標識が、その後、次のヌクレオチドが加えられる前に除かれる。1つのサイクルでのシグナルが前回の反応でのシグナルよりも実質的に小さくなるまで、伸長が続けられる。
【0008】
連結反応に基づく方法は近年、遺伝的変異の分析における重要な手段になっている。米国特許第5695933号には、ヌクレオチド反復の拡大を検出するための、RED(反復拡大検出)と呼ばれる方法が記載され、この方法では、反復配列に対して相補的なオリゴヌクレオチドがリガーゼの存在下で反復配列にアニーリングされる。その後、リガーゼは、並置されたオリゴヌクレオチドをつないで、連結されたオリゴヌクレオチドの多量体を生じさせる。多量体および標的ヌクレオチドの複合体が、その後、変性し、アニーリング、連結および変性の新たな一連の操作が開始される。多量体の長さが最終的には、ゲル分離を伴う技術によって決定される。生じた多量体の長さにより、拡大したオリゴヌクレオチド反復の長さが示され、これにより、次に、サンプリングされた個体の遺伝子状態を示すことができる。この方法では、より長い核酸範囲が分析されることになる場合でさえ、反復数を推定することが可能になる。しかしながら、この分析方法では、放射能標識されたDNAのゲルに基づく分離、それに続くハイブリダイゼーションによる検出、または、放射能標識されたDNAの直接的な検出を伴う。このような検出は、時間および手間が非常にかかり、リアルタイムの分析ができない。さらに、ゲルに基づく分離は、より大きなDNAフラグメントの分解能が低く、このことは、反復数に関して不明瞭な結果をもたらすことになる。
【0009】
まとめると、核酸サンプルにおけるヌクレオチド反復数を含めて、遺伝的変異の迅速かつ正確な分析を可能にする新しい方法が依然として求められている。
【発明の開示】
【0010】
発明の概要
本発明者らは、驚くべきことに、連結反応を行い、連結副生成物を検出することによって、ヌクレオチド反復の数を推定するだけでなく、他の遺伝因子(例えば、微生物型決定のためのマーカーなど)の存在を明らかにする新しい方法を見出している。第1の実施形態において、本発明は、下記の工程を含む、核酸サンプルにおける遺伝因子(例えば、ヌクレオチド反復または微生物型決定のためのマーカーなど)の存在を明らかにするための方法に関連する:
a)遺伝因子を含む核酸サンプルを提供する工程;
b)この核酸サンプルの遺伝因子を含む領域に対して完全または部分的に相補的なオリゴヌクレオチドを提供する工程;
c)このオリゴヌクレオチドをこの核酸サンプルにアニーリングさせる工程;
d)この核酸サンプルにアニーリングしたこのオリゴヌクレオチドを、リガーゼ酵素を使用して互いに連結する工程;および
e)連結副生成物を、連結反応が起こっているかどうかを明らかにするために、遺伝因子の存在の指標として検出する工程;
ただし、工程a)〜工程e)は、同時に、または連続して、または任意の組合せの連続する工程で行われる。
【0011】
連結副生成物の量は、起こった連結反応の数に比例しているので、連結反応の数を、どのくらいの連結副生成物が生じているかを明らかにすることによって計算することができる。あるいは、本発明の1つの実施形態において、連結サイクルが続いて行われ、1つの連結反応の存在が、その次の反応が開始される前に検出される。この場合、存在する遺伝因子(例えば、反復した核酸配列における反復ユニットなど)の数を決定するために、(生じた連結副生成物の実際の量ではなく)連結反応が起こっているか否かを検出することが必要であるだけである。本発明はまた、本発明の方法を行うためのキット、および、本発明の方法を行うために必要な成分を含む組成物に関連する。本発明において、連結生成物そのものではなく、連結副生成物が分析されるので、本発明は、ヌクレオチド反復の数を決定する新しい方法を開拓している。本発明は、ヌクレオチド反復配列におけるヌクレオチド反復ユニットの数、または、微生物型決定のためのマーカーを、様々な遺伝因子(例えば、ヌクレオチド反復など)を含む非常に長い配列の場合でさえ、これまでに可能であったよりも迅速に、かつ、手間がかからない方法で、かつ、正確に明らかにすることを可能にする。加えて、分析をリアルタイムで行うことができる。本発明はまた、必要な場合には、ヘテロ接合サンプルにおけるヌクレオチド反復などの遺伝因子の数を決定することを可能にする。
【0012】
図面の簡単な記述
図1は本発明による、いわゆる「終点」分析を説明する図である。
図2は本発明による、いわゆる「段階的」分析を説明する図である。
図3は熱サイクル処理による連結反応を用いた遺伝子内の可変位置における2つのオリゴヌクレオチドの連結、それに続く生物発光検出を示す図である。
図4は生物発光検出に連係した連結反応を用いた遺伝子内の可変位置における2つのオリゴヌクレオチドの連結(プレアニーリングされたプライマー/テンプレート複合体に対するリガーゼ工程およびPPDK工程の組合せ)を示す図である。
図5はCTG反復数の違いの検出を示す図である。
図6aは、遺伝子内の可変位置における2つのオリゴヌクレオチドの連結を示す図であり、ATP依存性リガーゼ(T4DNAリガーゼ)と、補因子としてのATPとの使用を示す図である。アピラーゼが、過剰な補因子を減少させるために使用される。図6bは、2つの連結方法(ATP依存性リガーゼおよびNAD+依存性リガーゼ)を使用する遺伝子内の可変位置における2つのオリゴヌクレオチドの連結の比較を示す図である。
図7は連鎖球菌属の数多くの種から得られたrnpB遺伝子の領域に由来する配列のアライメントを示す。
図8は異なる種の領域に対するプローブのアニーリングを示す図である。
【0013】
(定義)
「遺伝因子」は、核酸分子の検出可能な特徴、例えば、ヌクレオチド反復または微生物型決定のためのマーカーなどを意味する。
【0014】
「遺伝因子を含む領域」は、核酸分子におけるヌクレオチド配列を意味する。そのような領域は遺伝因子(例えば、ヌクレオチド反復など)と同一であってもよく、あるいは、1つ以上の遺伝因子(例えば、微生物型決定のためのマーカーなど)を含んでもよい。典型的には、遺伝因子を含む領域は、本発明の方法において使用されるようなオリゴヌクレオチドに対応する領域である。
【0015】
「微生物型決定のためのマーカー」は、特定の微生物型について特徴的であり、従って、核酸サンプルの型を決定するために使用される核酸サンプルにおける1つ以上のヌクレオチド位置を意味する。
【0016】
「連結パターン」は、連結事象のパターン(すなわち、連結されるオリゴヌクレオチドの数および/またはオリゴヌクレオチドの同一性)を意味する。典型的には、サンプルの連結パターンはサンプルの型について特徴的である。従って、「参照パターン」は、既知の参照サンプルに対する連結事象のパターンを意味する。
【0017】
「ヌクレオチド反復」は、2個、3個、4個から、数千個までのヌクレオチドの特定配列の連続した繰り返しを意味する。「反復ユニット」は1つのヌクレオチド反復配列である。反復ユニットが2ヌクレオチド〜6ヌクレオチドの間であるならば、ヌクレオチド反復はミクロサテライトと呼ばれる;反復ユニットが7ヌクレオチド〜約30ヌクレオチドの間であるならば、ヌクレオチド反復はミニサテライトと呼ばれる;反復ユニットが30を超えるならば、ヌクレオチド反復はサテライトと呼ばれる。用語「ヌクレオチド反復」はまた、短いタンデム反復および一定しない数のタンデム反復(VNTR)の用語を包含する。一部の遺伝的障害では、反復ユニットが数千回繰り返され得る。線状染色体を有するヒトおよび他の生物は、数千回反復される短い反復ユニットから構成されるテロメアをその染色体末端に有する。
【0018】
「隣接配列」は、ヌクレオチド反復の外側に位置する任意のヌクレオチド配列を意味する。この配列は、多くの状況において、増幅工程におけるプライマーのために使用される。
【0019】
「相補的」は、オリゴヌクレオチドが、選ばれたアニーリング条件のもとでテンプレート核酸にアニーリングすることができるための十分な相補性(すなわち、塩基対形成するための能力)を有することを意味する。従って、「相補的」により、すべてのヌクレオチドが、すべての位置において、テンプレート核酸に対して相補的でなければならないことは意味されず、オリゴヌクレオチドが、選ばれたアニーリング条件のもとでテンプレート核酸にアニーリングするための十分な相補性を有することのみを意味する。
【0020】
「連結副生成物」は、連結された核酸の一部を構成しないが、連結反応時に放出される生成物を意味する。
【0021】
「ルシフェラーゼ」は、本発明では、ATPに依存する酵素反応において光を生成できる任意の酵素を意味する。従って、任意の種から得られるそのような酵素が、その変異体および組換え変異体を含めて、本発明による「ルシフェラーゼ」の定義に含まれる。ルシフェラーゼに基づくアッセイにより、ルシフェラーゼ酵素を用いるアッセイを意味する。
【0022】
「光度測定法アッセイ」は、酵素の活性または化合物のレベルが光放射反応においてモニターされ、それにより、光の放射が酵素反応の結果(すなわち、ルシフェラーゼ酵素の活性の結果)であるアッセイである。
【発明を実施するための最良の形態】
【0023】
一部の生物は、生物発光をもたらす発エルゴン反応で光を生成できる。生物発光反応に関与する酵素に対する一般名はルシフェラーゼである。この酵素の種々の変異体が、細菌、昆虫および渦鞭毛藻類などの様々な生物において見出されている。
【0024】
ホタルルシフェラーゼ(EC.1.13.12.7)は、ATP、マグネシウムおよび酸素の存在下でのD−ルシフェリンの酸化を触媒する。生成物のオキシルシフェリンが、光子の放射を伴って基底状態に崩壊する励起された状態で生じる。ホタルルシフェラーゼは、特に、ATPの効率的な検出および定量のために、また、遺伝子の調節および発現の研究のためのレポーター酵素として、分子生物学および細胞生物学において広範囲に使用されている(Gould,S.J.およびSubramani,S.(1988年)、分子生物学および細胞生物学における手段としてのホタルルシフェラーゼ、Anal.Biochem.175、5〜13)。ATP変換反応に関与するすべての酵素および代謝産物をホタルルシフェラーゼ系によって分析することができる(Kricka,L.J.(2000年)生物医学科学における生物発光および化学発光の適用、Methods Enzymol.305、333〜345;Kricka,L.J.(1988年)、ルシフェラーゼおよびルシフェリンの臨床的適用および生化学的適用、Anal.Biochem.175,14〜21;Kricka,L.J.(1991年)、化学発光技術および生物発光技術、Clin.Chem.37、1472〜1481)。
【0025】
その上、様々なルシフェラーゼ系が様々な生物発光アッセイにおいて使用されている。例えば、ルシフェラーゼおよび別のタンパク質(例えば、RNA同定のためのRNA結合タンパク質など)を含む様々な融合タンパク質、あるいは、ホタルルシフェラーゼと、ビオチンアクセプターペプチドまたは免疫アッセイのための単鎖抗体との融合コンジュゲートが知られている。真核生物のルシフェラーゼ酵素は、光を産生する反応においてATPを用いる。光を検出することができる簡便性のために、ルシフェラーゼに基づく分析は多くの適用のための注目される分析手段である。
【0026】
第1の実施形態において、本発明は、下記の工程を含む、核酸サンプルにおける遺伝因子(例えば、ヌクレオチド反復、または微生物型決定のためのマーカーなど)の存在を明らかにするための方法に関連する:
a)遺伝因子を含む核酸サンプルを提供する工程;
b)この核酸サンプルの遺伝因子を含む領域に対して完全または部分的に相補的なオリゴヌクレオチドを提供する工程;
c)このオリゴヌクレオチドをこの核酸サンプルにアニーリングさせる工程;
d)この核酸サンプルにアニーリングしたこのオリゴヌクレオチドを、リガーゼ酵素を使用して互いに連結する工程;および
e)連結副生成物を、連結反応が起こっているかどうかを明らかにするために、遺伝因子の存在の指標として検出する工程;
ただし、工程a)〜工程e)は、同時に、すなわち、すべての成分を一緒に混合することによって行われるか、または連続して、すなわち、それぞれの工程のために必要な成分のみを混合し、さらなる成分を続いて加えることによって行われる。あるいは、一部の工程を同時に行うことができ、一部の工程を連続して行うことができる。これは、例えば、異なる工程で、実施されるための異なる条件(例えば、異なる緩衝液、温度など)が要求されるときには好ましくなり得る。1つの成分を除いて反応のために必要なすべての成分はまた、分析に先だって混合することができ、この一連の反応を、最後の成分を加えることによって開始することができる。
【0027】
本発明は、連結生成物そのものの代わりに連結副生成物の検出に関する。これは、連結事象が起こっているかどうかを検出する新しい方法を開拓しており、これにより、先行技術と比較して、より迅速で、より高感度な分析が可能になる。
【0028】
核酸サンプルは、当業者に既知の標準的な手段によって、必ずしも必要ではないが、好ましくは単離される生物学的サンプル(例えば、組織、細胞または血液など)の任意のDNAサンプルまたはRNAサンプルであり得る。好ましくは、核酸サンプルは、ヒト対象から得られる核酸サンプルである。好ましくは、核酸サンプルはDNAである。好ましくは、核酸サンプルは、例えば、PCRによって増幅される。
【0029】
本発明による方法において、核酸サンプルは、分析される特定の反復した配列に対して相補的なオリゴヌクレオチドと混合され、アニーリングされる。アニーリング工程に先だって、核酸を変性させなければならない。当業者は、核酸を変性させるための好適な条件を選択する方法を知っている。アニーリングのための条件は、アニーリングされる特定のオリゴヌクレオチドに依存して変化させなければならない。当業者は、種々の適用ためのアニーリング時におけるストリンジェンシーを選択する方法を知っている。オリゴヌクレオチドの長さおよび同一性を、検出されることになる遺伝因子(例えば、ヌクレオチド反復など)を含むそれぞれの特定の領域について調節しなければならない。すべてのオリゴヌクレオチドは、連結反応において機能するために、リン酸を5’末端に必要とする。
【0030】
今日、その起源をトリヌクレオチド反復展開部に有する、ヒトに影響を及ぼす少なくとも14個の障害が報告されている。それらはポリグルタミン疾患(8個の障害)および非ポリグルタミン疾患(6個の疾患)に分類することができる。
【0031】
8個の障害(その1つがハンチントン病である)はすべてが、その原因としての同じ反復したコドン(CAG)をともに有する。CAGは、グルタミンと呼ばれるアミノ酸をコードするので、これら8個のトリヌクレオチド反復障害はまとめてポリグルタミン疾患として知られている。
【0032】
ポリグルタミン疾患は多くの共通点を有する:ポリグルタミン疾患のそれぞれが、身体の特定部分における神経細胞の進行性の変性によって特徴づけられ、これに対して、6個の非ポリグルタミン疾患はそれらの症状発現において非常に不均一である。下記は、疾患、トリヌクレオチド反復、および、知られている場合には、罹患者における反復数に関する表である。
【表1】

【0033】
現在、これらの反復展開部のすべてが、PCR、およびそれに続く電気泳動工程によって分析されている。この分析は時間がかかり、より長い反復した配列については、感度が問題となる場合がある。
【0034】
第2の実施形態において、本発明は、下記の工程を含む、核酸サンプルにおけるヌクレオチド反復の数を分析するための方法に関連する:
a)ヌクレオチド反復を含む可能性がある核酸サンプルを提供する工程;
b)このヌクレオチド反復に対して相補的なオリゴヌクレオチドを提供する工程;
c)このオリゴヌクレオチドをこの核酸サンプルにアニーリングさせる工程;
d)この核酸サンプルにアニーリングしたこのオリゴヌクレオチドを、リガーゼ酵素を使用して互いに連結する工程;および
e)この連結副生成物を、連結反応が起こっているかどうかを明らかにするために検出する工程;
ただし、工程a)〜工程e)は、同時に、すなわち、すべての成分を一緒に混合することによって行われるか、または連続して、すなわち、それぞれの工程のために必要な成分のみを混合し、さらなる成分を続いて加えることによって行われる。
【0035】
本発明に従ってヌクレオチド反復を分析するためには2つのタイプのオリゴヌクレオチドが好ましい。一方の好ましいタイプのオリゴヌクレオチドは、反復ユニットそのものの単量体、二量体または多量体である(すなわち、オリゴヌクレオチドは、反復した配列の1つ以上の反復からなる)。もう一方の好ましいタイプは、ヌクレオチド反復ユニットの単量体、二量体または多量体(n−mer体)と等しい長さを有しない「一致しない」オリゴヌクレオチドである(すなわち、このオリゴヌクレオチドは、ヌクレオチド反復ユニットそのもののn−mer体よりも1塩基または数塩基だけ短いか、または長い)。「一致しない」場合におけるオリゴヌクレオチドは一つずつ加えられ、連結させられ、連結副生成物が、その次のオリゴヌクレオチドが加えられる前に検出される。ヌクレオチド反復の開始を、反復した配列のすぐ外側において、すなわち、ヌクレオチド反復に隣接する配列の1つにおいてアニーリングするように適合化されたオリゴヌクレオチドの使用によって開始することが好ましい。これらの異なるタイプのオリゴヌクレオチド、および、そのようなオリゴヌクレオチドにより行うことが可能である異なるタイプの分析が下記においてより詳しく記載される。
【0036】
下記には、ポリグルタミン障害における反復の数の計算のために使用することができる本発明のために好適なオリゴヌクレオチドの例がいくつか示される。
【表2】
【0037】
変異体1により、2つの反復したユニットを分析するためのオリゴヌクレオチドの3つの異なるタイプの二量体(それぞれ通常の書体、太字および下線)が記述される。
【0038】
変異体2により、4つの反復したユニットを分析するためのオリゴヌクレオチドの3つの異なるタイプの4mer体(それぞれ通常の書体、太字および下線)が記述される。
【0039】
変異体3により、5つの反復したユニットを分析するためのオリゴヌクレオチドの3つの異なるタイプの5mer体(それぞれ通常の書体、太字および下線)が記述される。
【0040】
変異体4により、7つの反復したユニットを分析するためのオリゴヌクレオチドの3つの異なるタイプの7mer体(それぞれ通常の書体、太字および下線)が記述される。
【0041】
下記には、脆弱X症候群における反復の数の計算のために使用することができる本発明のために好適なオリゴヌクレオチドの例がいくつか示される。
【表3】
【0042】
酵素による連結反応が、リガーゼ活性を有する数多くの異なる酵素によって行われる。そのなかでも、好ましい例が下記に詳しく記載される。リガーゼ活性を有する酵素により、核酸鎖内の末端間で、または、異なる核酸鎖との間でホスホジエステル結合を形成する能力を有する酵素を意味する。リガーゼ酵素の組換え変異体もまた本発明において意図される。
【0043】
DNAリガーゼは2つの大きな種類に分けられる(DohertyおよびSuh,2000年、Nucleic Acid Res.28、21:4051を参照のこと):ATPを補因子として要求するDNAリガーゼ、および、NAD+を要求するDNAリガーゼ。真核生物、ウイルスおよび古細菌のコードされる酵素はすべてがATPを要求する。しかしながら、ATPはまた、(下記に記載されるように)真核生物のルシフェラーゼ反応における基質でもあるので、ルシフェラーゼに基づく分析は、ATP依存性DNAリガーゼが使用されたときには、連結反応を検出することが以前では事実上不可能であった。ATP依存性リガーゼを使用するための1つの可能性は、(1)何らかの手段によって、過剰なATPを、(例えば、アピラーゼを使用することなどによって)減少させることができるか、あるいは、(2)ルシフェラーゼに対する基質として機能しないか、または、ATPよりも著しく低い程度にルシフェラーゼに対する基質として機能するATPアナログ(例えば、dATPなど)が使用されるか、あるいは、(3)(1)および(2)の組合せであると考えられる。
【0044】
1つの実施形態において、本発明では、NAD+依存性DNAリガーゼが、サンプル核酸にアニーリングさせられたDNA−オリゴヌクレオチドを連結するために使用される。この結果、エネルギー源として、ATPの代わりに、NAD+に依存するDNAリガーゼが使用される。NAD+依存性DNAリガーゼは、下記の反応(式中、Eはリガーゼであり、pAはAMPであり、pはリン酸基を示す)を触媒する、もっぱら真核生物において見出される70kD〜80kDの非常に相同的な単量体タンパク質である:
【0045】
T4RNAリガーゼもまた、DNA−オリゴヌクレオチド、または、RNA−オリゴヌクレオチド、または、DNA−オリゴヌクレオチドおよびRNA−オリゴヌクレオチドの組合せが互いに連結されるときには本発明において好ましく用いられる。例えば、T4RNAリガーゼはRNA−RNA連結のための下記の反応を触媒する:
【0046】
従って、この反応における最初の工程では、ATPを使用するRNAのアデニル化が伴う。しかしながら、いくつかの実施形態では、これは、連結反応を検出するための好ましい分析方法の1つが使用されるとき(すなわち、ルシフェラーゼに基づくアッセイが使用されるとき)には好ましくないので、別の方法が必要である。数多くの研究者がAdoPP5’RNA中間体の合成を報告している(例えば、Sninsky JJ他,等モル濃度のアクセプター分子およびドナー分子を含有するRNAリガーゼ反応におけるオリゴヌクレオチドの特異的な結合のための末端ブロック基の使用(1976年)、Nucleic Acids Res.3(11):3157〜3166;England,TE他,ジヌクレシドピロリン酸はT4誘導型RNAリガーゼに対する基質である(1977年)、Proc.Natl.Acad.Sci.USA.74(11):4839〜4842)。従って、ある方法が、そのような中間体(AdoPP5’RNAまたはAdoPP5’DNA)を本方法においてそのまま利用するために考案されており、これにより、そうでない場合には検出を妨げるATPの存在が回避されている。従って、T4RNAリガーゼもまた本発明のために好ましい。
【0047】
代わりの連結方法として、別の態様は、ATP依存性リガーゼをATPアナログ(例えば、dATPまたは2−アミノプリンリボシド三リン酸など)との組合せで使用することである。首尾よく機能するために、アナログは、まず連結反応を助けなければならない。次に、アナログは理想的には、過剰なルシフェラーゼ基質の存在に起因するバックグラウンドを最小限に抑えるために、少なくともATPによって達成されるのと同じ程度に、ルシフェラーゼ反応において基質として機能すべきではない。加えて、連結反応の生成物は好ましくはピロリン酸でなければならず、その結果、このピロリン酸は、その後、スルフリラーゼによってATPに変換することができ、次いでこのATPをルシフェラーゼ反応によって検出することができる。そのような補因子アナログは、過剰なアナログの存在から生じる低いバックグラウンドシグナルを超える連結反応の検出を容易にする。改善された方法では、検出に先だって、アナログそのもののその後の除去を伴うことができる。
【0048】
例えば、実験により、dATPが、ATP依存性リガーゼのT4DNAリガーゼに対する効果的な補因子であることが明らかにされている(Kinoshita他,J.Biochem.122:205〜211(1997年)を参照のこと)。しかしながら、dATPでさえ、ルシフェラーゼによる著しいバックグラウンドシグナルを生じさせるが、これは、ATPによって引き起こされるバックグラウンドシグナルよりも低い。代わりの方法は、dATPの使用を、アピラーゼを用いた連結後処理と組み合わせ、その結果、過剰なdATPが、ルシフェラーゼを使用する光シグナルの検出に先立って分解されるようにすることである。この組合せは、実施例の節に記載されるように、例えば、DNA反復の長さの変動を検出するための驚くほど効果的な方法をもたらしている。その結果はまた、NAD+依存性リガーゼをPPDKとの組合せで使用して得られた結果と匹敵し得る。
【0049】
さらに、アピラーゼは基質(例えば、dATP)についてATP依存性リガーゼと競合するので、アピラーゼ、リガーゼおよび基質の濃度は特定の状況について最適化されなければならない。従って、過剰量のDNAリガーゼ基質を減少させるために、アピラーゼを、連結反応の後だけでなく、連結反応の前またはその期間中に加えることもまた可能である。
【0050】
従って、別の実施形態では、連結工程(工程d)が、ATP依存性リガーゼを用いて行われ、アピラーゼが、過剰量のDNAリガーゼ基質を減少させるために、連結前に、または連結期間中に、または連結後に工程d)の連結混合物に加えられる。
【0051】
好ましい実施形態において、ATP依存性リガーゼはT4DNAリガーゼである。
【0052】
別の好ましい実施形態において、dATPが、連結工程(工程d)においてATP依存性リガーゼに対する基質として使用される。
【0053】
本発明の特許請求項に記載される方法の最後の工程として、連結副生成物が、連結反応が起こっているかどうかを明らかにするために検出される。生じる連結副生成物の量は、起こった連結反応の数に比例している。下記に記載されるように、場合により、ヌクレオチド反復ユニットの数を推定することを可能にするために、連結反応が起こっているか否かを検出することが十分である。当業者は、最終生成物のさらなる操作が望ましいことを明らかにすることができ、従って、本発明では、本発明の方法の使用に対するこのような調節が意図される。
【0054】
本発明のさらに別の実施形態は、下記の工程を含む、核酸サンプルにおけるヌクレオチド反復の数を分析するための方法に関連する:
a)ヌクレオチド反復を含む可能性がある核酸サンプルを提供する工程;
b)このヌクレオチド反復に対して相補的なオリゴヌクレオチドを提供する工程;
c)このオリゴヌクレオチドをこの核酸サンプルにアニーリングさせる工程;
d)この核酸サンプルにアニーリングしたこのオリゴヌクレオチドを、リガーゼ酵素を使用して互いに連結する工程;
e)連結副生成物をATPに変換する工程;および
f)連結反応が起こっているかどうかを明らかにするために、このATPを検出する工程;
ただし、工程a)〜工程f)は、同時に、すなわち、すべての成分を一緒に混合することによって行われるか、または連続して、すなわち、それぞれの工程のために必要な成分のみを混合し、さらなる成分を続いて加えることによって行われる。あるいは、一部の工程を同時に行うことができ、一部の工程を連続して行うことができる。これは、例えば、異なる工程で、実施されるための異なる条件(例えば、異なる緩衝液、温度など)が要求されるときには好ましくなり得る。1つの成分を除いて反応のために必要なすべての成分はまた、分析に先だって混合することができ、この一連の反応を、最後の成分を加えることによって開始することができる。
【0055】
この実施形態において、連結副生成物の1つ(すなわち、AMPもしくは修飾AMPまたはPPi(ピロリン酸))がATPに変換される。
【0056】
1つの好ましい実施形態において、連結副生成物はAMPである。
【0057】
本発明による連結反応において生じたAMPは好ましくは、ATPに変換される。1つの好ましい実施形態において、本発明による連結反応において生じたAMPは、下記のようにピルビン酸リン酸ジキナーゼ(PPDK)を使用する一段階の酵素反応によってATPに変換される:
【0058】
AMPに対するKmはATPに対するKmの1/250である。従って、この反応はAMP−>ATPの方向に強く動かされる。ピルビン酸リン酸ジキナーゼは、6.5〜7のpH至適、6〜11の範囲内でのpH安定性、55℃〜60℃の至適温度を有しており、55℃未満の温度で安定である。この酵素の特筆すべき利点は、高い温度(50℃〜60℃)でのその安定性および高活性である。多くの場合、この範囲での温度は、好適なストリンジェンシーを有するアニーリング条件を提供するために好ましいので、このことは利点である。また、PPDKが、ホタルルシフェラーゼを使用する生物発光検出(これは、下記に詳しく記載される好ましい検出アッセイである)が容易に行われる緩衝液において使用される。
【0059】
あるいは、連結反応の別の副生成物(すなわち、PPi)が好ましくはATPに変化される。上記で議論されたように、連結反応における基質(ATP)は、(例えば、上記で概略されたように、アピラーゼが使用されない場合)、ルシフェラーゼ反応のための基質と同じであるので、ATP依存性DNAリガーゼは、いくつかの実施形態では、本発明のために好適ではない。しかしながら、修飾されたATP分子(例えば、dATPまたは2−アミノプリンリボシド三リン酸など)を使用することができるDNAリガーゼ(Kinoshita,YおよびNishigaki,K、ATP要求酵素におけるATPの予想外の一般的代替可能性、J.Biochem.122:205〜211(1997年))が本発明では用いられる。好ましいそのようなDNAリガーゼは下記の反応を触媒する(式中、A*は、上記で定義されるような修飾されたアデノシン分子を示す):
【0060】
修飾されたATP分子を使用することの利点の1つは、修飾されたATP分子が、(下記に記載される)好ましい、ルシフェラーゼに基づくアッセイを、ATPが妨害するのと同じ程度に妨害しないことであり、従って、修飾されたATP分子を、下記に記載される好ましい、ルシフェラーゼに基づくアッセイの期間中、存在させることができることである。リガーゼによって要求される補因子のタイプおよび濃度は、連結事象のルシフェラーゼ媒介による検出を成功させるための重要な要素である。この補因子は効果的な連結を助けなければならないが、(例えば、上記で概略されたように、アピラーゼが使用されない場合)検出工程を妨害してはならない。
【0061】
本発明による連結反応において生じるPPiは、その後、好ましくは、ATP−スルフリラーゼを用いる下記の反応によってATPに変換される(式中、APSはアデノシン5’−ホスホ硫酸を示す):
【0062】
さらに別の実施形態において、本発明は、下記の工程を含む、核酸サンプルにおけるヌクレオチド反復の数を分析するための方法に関連する:
a)ヌクレオチド配列を含む可能性がある核酸サンプルを提供する工程;
b)このヌクレオチド配列に対して相補的なオリゴヌクレオチドを提供する工程;
c)このオリゴヌクレオチドをこの核酸サンプルにアニーリングさせる工程;
d)この核酸サンプルにアニーリングしたこのオリゴヌクレオチドを、リガーゼ酵素を使用して互いに連結する工程;
e)連結副生成物をATPに変換する工程;および
f)連結反応が起こっているかどうかの指標として、ルシフェラーゼに基づくアッセイによってこのATPを検出する工程;
ただし、工程a)〜工程f)は、同時に、すなわち、すべての成分を一緒に混合することによって行われるか、または連続して、すなわち、それぞれの工程のために必要な成分のみを混合し、さらなる成分を続いて加えることによって行われる。あるいは、一部の工程を同時に行うことができ、一部の工程を連続して行うことができる。これは、例えば、異なる工程で、実施されるための異なる条件(例えば、異なる緩衝液、温度など)が要求されるときには好ましくなり得る。1つの成分を除いて反応のために必要なすべての成分はまた、分析に先だって混合することができ、この一連の反応を、最後の成分を加えることによって開始することができる。
【0063】
生じたATP(例えば、連結副生成物をATPに変換するための上記の反応のいずれかによって生じたATP)は、好ましくは、この実施形態に従って、ルシフェラーゼに基づくアッセイにおいて基質として使用される。ATPの量は、生じた連結副生成物(AMPまたはPPi)の量に、すなわち、起こった連結反応の数に比例しているので、また、ATPの量がルシフェラーゼ反応のために限定されているという条件では、すなわち、ルシフェリンおよびO2が過剰に存在するという条件では、光の発生は連結反応の数に対応する。本発明のために好適なルシフェラーゼは下記の反応を触媒する:
【0064】
放射される光の波長は、用いられるルシフェラーゼの起源に依存する。
【0065】
さらに別の実施形態において、本発明は、下記の工程を含む、核酸サンプルの微生物型決定のための方法に関連する:
a)微生物型決定のための少なくとも1つのマーカーを含む核酸サンプルを提供する工程;
b)この核酸サンプルの微生物型決定のためのマーカーを含む領域に対して完全または部分的に相補的なオリゴヌクレオチドを提供する工程;
c)このオリゴヌクレオチドをこの核酸サンプルにアニーリングさせる工程;
d)この核酸サンプルにアニーリングしたこのオリゴヌクレオチドを、リガーゼ酵素を使用して互いに連結する工程;および
e)連結反応が起こっているかどうかを明らかにするために、連結副生成物を検出する工程;
f)微生物型を決定するために、サンプルの連結パターンを参照パターンと比較する工程;
ただし、工程a)〜工程e)は同時に行われるか、または連続して行われるか、または任意の組合せの連続する工程で行われる。
【0066】
本発明により、様々な異なる種のDNA、例えば、様々な異なる微生物種に由来するDNAが、好適な長さおよび組成のプローブを選ぶことによって解明される。例えば、細菌における比較配列分析のための最も広範囲に使用されている標的遺伝子は、30Sのリボソームスモールサブユニットの構造的一部をコードする16SrRNA遺伝子である。16SrRNA遺伝子のDNA配列決定は系統発生的研究のための重要な手段であり、また、微生物同定のためにも使用されている。
【0067】
上記反応を触媒することができる、すべての異なる微生物に由来するすべてのルシフェラーゼ酵素が、その組換え変異体を含めて、本発明において意図される。熱活性なルシフェラーゼを、種々の種に由来する野生型酵素、変異体および組換え変異体を含めて、入手することができ、また、より高い温度でルシフェラーゼ反応を行うことが好ましいときには使用することができる。ルシフェラーゼ反応を、反応のために必要な試薬を加えることによって、連結反応および変換反応に続いて、これらの反応を生じさせることが可能になった後で行うことができる。あるいは、連結のリアルタイム測定を可能にするために連結反応および変換反応を進行させることができるので、発光反応のために必要な試薬を同時に存在させることができる。別の実施形態において、より低い温度で活性である熱安定性のルシフェラーゼ、例えば、Kajiyama NおよびNakano E,1993年(217位における一アミノ酸置換によるホタルルシフェラーゼの熱安定化)、Biochemistry.32(50 13795〜9))に記載されるルシフェラーゼなどを用いることができ、また、2段階で行われる酵素反応に共役させることができる。すなわち、最初に、連結反応および変換反応が、特異的な適用のために好適なより高い温度で行われ、その後、温度が下げられ、ルシフェラーゼ活性が測定される。
【0068】
天然のホタルルシフェラーゼは、反応混合物におけるMg2+、ATP、ルシフェリンおよび分子状酸素の存在に完全に依存している(Ford,SRおよびLeach,FR、ホタルルシフェラーゼアッセイの適用における改善、Methods in Molecular Biology,第102巻:生物発光方法およびプロトコル(3頁〜20頁、R.A.LaRossa編、Humana Press Inc.1998年)を参照のこと)。ジチオスレイトール(DTT)およびエチレンジアミン四酢酸(EDTA)が、システイン残基の酸化または金属イオンによる反応の阻害を防止するために反応混合物にそれぞれ加えられる。従って、ルシフェラーゼ反応は、好ましくは、約10mMのMg2+(例えば、10mMの酢酸Mg)およびDTT(例えば、0.2mM)を含む緩衝液において行われる。至適温度は25℃である。30℃を超える温度(特に、35℃を超える温度)は野生型酵素の迅速な不活性化を引き起こす。従って、ルシフェラーゼ反応は、好ましくは、pH7.5〜8(より好ましくはpH7.8)において、2mM〜10mMのMg2+、0.5mM〜2mMのDTTおよび0.5mM〜2mMのEDTAの存在下、20℃〜30℃の温度で行われる。しかしながら、ルシフェラーゼの変異体は、例えば、上記に記載されるような温度に関して、他の要求を有することがある。重要なことは、塩化物イオンはルシフェラーゼ反応を阻害することがあり、従って、塩化物イオンをルシフェラーゼ反応の期間中に存在させることは次善となることがあるということである。
【0069】
多くの方法を、発光反応の結果としての光の検出のために利用することができ、例えば、写真フィルム、X線フィルムにおける検出、光電子増倍管(PMT)または電荷結合デバイス(CCD)カメラの使用による検出などのために利用することができる。本発明のために、PMTまたはCCDカメラによる光度測定法検出アッセイが好ましい。そのような方法は光放射の定量的な測定を可能にするからである。本発明において提供される方法による大きな利点は、この方法が、連結反応をリアルタイムで検出することを可能にすることである。
【0070】
連結のリアルタイム検出は、ルシフェリン/ルシフェラーゼ反応による光産生を、初期レベルが検出された後で「停止する」酵素を含めることによってさらに改善することができる。「初期レベル」は、光放出が検出されるために必要な光のレベルを示す。このレベルは、使用される検出装置に関連して当業者によって容易に決定される。この「停止」原理はまた、ルシフェラーゼ酵素の低い「バーンレート」に関しても好都合であり得る。そのような酵素は連結事象間の応答時間を増大させる。好適な酵素は、上記で記載されたように、ATPをADPに分解するが、AMPを分解しないアピラーゼである。この酵素は、従来のピロシーケンシング(米国特許第6258568号)において非常に都合良く使用されている。
【0071】
従って、さらに別の実施形態において、ルシフェリン/ルシフェラーゼ反応における光産生が、産生された光の初期レベルに到達した後、酵素的に停止される。好ましくは、これはアピラーゼの添加によって行われる。
【0072】
本発明の方法による連結副生成物の検出は2種類の情報をもたらす:1)連結副生成物が生じるならば、連結反応が起こっている。場合により、(下記に記載される)、この情報は十分である。2)上記で述べられたように、連結副生成物の量が連結反応の数に比例しており、従って、核酸サンプルの量が既知であるならば、核酸サンプルにおける反復したヌクレオチド配列の反復数、または微生物型決定のためのマーカーを計算するために使用することができる。
【0073】
PPDKを使用して、AMPをATPに変換し、次いで、そのATPを使用して、ルシフェリン−ルシフェラーゼに基づくシグナルを生じさせることが、Sakakibara他(Sakakibara T他,Anal Biochenm.1999年、268(1):94〜101)およびItoh他(Itoh他,Anal.Sci.2003年,19:105〜109))によって報告されている。しかしながら、これらの適用において、この方法は、RNAの分解によって生じたAMPを測定して、食品の衛生状態に対する指標を提供するために(Sakakibara他,1999年)、または、二重免疫学的反応において(Itoh他,2003年)、それぞれ使用されていた。
【0074】
数多くのリボザイムが、リガーゼ活性を有することもまた報告されており、これらもまた本発明において意図される(Glasner,ME他(2002年)、RNAリガーゼリボザイムの構造および触媒作用のための金属イオン要求、Biochemistry,41,8103〜8112;McGinness,K.E.およびJoyce,G.F.(2002年)、外部RNAテンプレートにおけるRNA触媒によるRNA連結、Chemistry and Biology9,297〜307)。連結は、基質に直接に連結するリボザイム(すなわち、クラスIリガーゼ、Glasner他,2002年)を伴い、また、相補的なテンプレートにおいて隣接位置で結合される2つのオリゴヌクレオチドの連結さえ伴う(hcリガーゼリボザイム、McGinnessおよびJoyce,2002年)。それぞれの場合において、連結は、一方の分子の3’−OH基を別の分子の5’−三リン酸に結合すること、およびピロリン酸の放出を伴う。ATPなどの補因子はこの反応に関与しておらず、従って、リボザイムは、連結反応の好ましい検出が、ルシフェラーゼに基づくアッセイの使用を伴うとき、本発明のために好適である。
【0075】
本開示の他所で言及されたリガーゼ酵素に加えて、本発明のための好適なリガーゼ酵素が下記に例示されるが、下記のリガーゼ酵素に限定されない:ATP依存性リガーゼ酵素(例えば、T4DNAリガーゼ、T7DNAリガーゼ、BstDNAリガーゼ、TfiDNAリガーゼなど)、NAD+依存性リガーゼ酵素(例えば、TthDNAリガーゼ、TflDNAリガーゼ、TaqDNAリガーゼ、大腸菌DNAリガーゼなど)、およびRNAリガーゼ酵素(例えば、tRNAリガーゼなど)。
【0076】
本発明の方法が行われるとき、2つのタイプの分析が可能である。特定の核酸サンプルにおける反復の数を、既知の数の反復した配列を有する既知量のテンプレートDNAを使用することにより連結副生成物の検出の標準曲線を構築することによって決定することができる。それぞれの連結反応が1つの連結副生成物をもたらすので、連結副生成物の量の決定により、ヌクレオチド反復の数を決定することが可能になる。分析される核酸サンプルにおけるテンプレートDNAの量が既知であるならば、反復の数を、標準曲線を使用して連結副生成物の量を反復の数に関連づけることによって決定することができる。
【0077】
別の好ましい実施形態において、「一致しない」オリゴヌクレオチドが、上記で記載されたように使用される。この場合、例えば、上記で記載されたように、テンプレートDNAの全範囲が連結オリゴヌクレオチドで満たされるまで、さらなるオリゴヌクレオチドのそれぞれの添加において検出可能である連結副生成物が生じる。従って、この場合、テンプレートの量が既知である必要はない。反復ユニットが2である場合、それぞれの一致しないオリゴヌクレオチドは、奇数の数を有する3つ以上のヌクレオチドを有しなければならない。反復数が3である場合、オリゴヌクレオチドは、3の倍数を除いて、2ヌクレオチド以上から、反復したユニットの配列の全長までであり得る。例えば、三重反復の検出の場合、4mer体、5mer体、7mer体、8mer体、10mer体、11mer体などの組合せを使用することができる。一致しないオリゴヌクレオチドが使用されるとき、ヌクレオチド反復配列のすぐ外側でアニーリングするように適合化されているオリゴヌクレオチドが好ましくは最初に加えられる。重要なことは、オリゴヌクレオチドは、それぞれのオリゴヌクレオチドが、1つの場所で、(すなわち、現在の連結サイクルの直前の連結サイクルにおいて加えられたオリゴヌクレオチドの隣りで)連結することができるだけであるように構築されなければならないということである。
【0078】
一致しないオリゴヌクレオチドが使用されるとき、1つの連結サイクルに由来するアニーリングしていないオリゴヌクレオチド(すなわち、アニーリングまたは連結されないオリゴヌクレオチド)が好ましくは、その次の連結サイクルが開始される前に除かれる。これは種々の方法で達成される。1つの方法は、連結反応時に存在するアニーリングしていないオリゴヌクレオチドを消化する一本鎖DNA依存性エキソヌクレアーゼを使用することである。例えば、エキソヌクレアーゼI(ExoI)(大腸菌のsbcB遺伝子の産物)は、一本鎖(ss)DNAを3’から5’の方向で消化するエキソデオキシリボヌクレアーゼである。酵素活性は自由な3’−ヒドロキシル末端の存在を絶対的に要求する。この場合、テンプレートを、3’末端の修飾によって、例えば、ターミナルデオキシヌクレオチジルトランスフェラーゼを使用するジデオキシヌクレオチドの取り込みなどによって保護することが必要である。あるいは、本発明の別の好ましい実施形態において、アニーリングしていないオリゴヌクレオチドを、5’リン酸をオリゴヌクレオチドから除くことによって不活性化する酵素が連結反応工程の期間中または反応工程の後に加えられる。この酵素はホスファターゼであり得るか、または、5’末端リン酸に依存する5’末端特異的エキソヌクレアーゼであり得る。ホスファターゼは、エビのアルカリホスファターゼSAP、子ウシ腸アルカリホスファターゼCIP、または任意の細菌アルカリホスファターゼからなる群から選択することができる。上記で記載されたようなアニーリングしていないオリゴヌクレオチドの除去はまた、反復ユニットのn−mer体がオリゴヌクレオチドとして使用されるときに利用され得ることが当業者によって容易に理解される。
【0079】
本発明の別の好ましい実施形態において、核酸サンプルが担体に固定化される。固定化は共有結合性または非共有結合性のいずれかであり得る。この分野で使用されている一般的な非共有結合性の方法はビオチン−ストレプトアビジン結合対の利用であり、この場合、ビオチンがプライマーに結合され、ストレプトアビジンが担体に結合される。頻繁に使用されている共有結合性の結合方法は、プライマーに結合されたアミノリンカーの使用であり、これは、エポキシシラン処理された表面と容易に反応する。担体は数タイプのものが可能である:例えば、ビーズ(非中空または多孔性)、またはチップもしくはファイバーの表面である。別の方法は、相補的であるプライマーを、遺伝因子(例えば、ヌクレオチド反復、または微生物型決定のための1つのマーカーなど)を含む領域の隣接配列に、例えば、ビオチン−ストレプトアビジン結合対を介して結合することである。固定化された核酸サンプルまたはプライマーの使用による1つの利点は、過剰な酵素、オリゴヌクレオチドおよび/または分解生成物が、1つの好ましい実施形態では、1つの連結−検出サイクルの後、その次が開始される前に、洗浄工程によって容易に除くことができることである。
【0080】
上記で記載されたこれら2つのタイプの分析が、連結副生成物がATPに変換され、このATPが、連結反応が起こったことを検出するための、ルシフェラーゼに基づくアッセイにおいて使用されるときの本発明の変形とともに図1および図2に例示される。図1は、反復ユニットそのものの単量体または多量体であるオリゴヌクレオチドがオリゴヌクレオチドとして使用される「終点」分析を示す。それぞれの連結反応が特定量の光を放射させ、その総量が定量される。放射された光の量は、サンプル核酸の量が既知であるならば、上記で記載されたような標準曲線を使用して、反復数に関連づけることができる。図2において、続いて加えられ、連結することを可能にする一致しないオリゴヌクレオチドが使用され、1つの連結反応において放射された光が、その次のオリゴヌクレオチドが加えられ、サイクル(すなわち、添加、連結、および光放射の検出)がもう一度開始される前に「段階的」様式で検出される。これは、それ以上の光が放射されなくなるまで繰り返される。すなわち、これは、サンプル核酸における反復した配列の全範囲が連結オリゴヌクレオチドで満たされるときには当てはまる。従って、反復した配列における反復ユニットの数を、サンプル核酸の量を知ることなく決定することができる。
【0081】
オリゴヌクレオチドの長さおよび配列の選択に依存して、この方法は、2塩基〜数千塩基の範囲における反復配列について使用することができる。
【0082】
連結反応は、リガーゼ、および、連結されることになる核酸のタイプの選択に依存して、広範囲の温度にわたって行われる。例えば、ハイブリダイゼーションさせられるテンプレート核酸に対する大きな程度の相補性を有するオリゴヌクレオチドプローブが使用されるとき、相補性が低い場合よりも高い温度を、熱に安定なリガーゼを使用することによって連結反応時に使用することができる。緩衝液の選択、および、連結反応時におけるリガーゼの濃度は、それぞれの特異的なリガーゼについて最適化されなければならず、また、当業者の能力の範囲内である。
【0083】
好ましい実施形態において、連結反応は、NAD+依存性DNAリガーゼを使用して行われ、生じたAMP連結副生成物が、PPDK酵素を使用してATPに転換される。生じたATPが、その後、好ましくは、ルシフェラーゼに基づくアッセイを使用して検出される。
【0084】
本発明の好ましい実施形態において、熱安定性リガーゼ(例えば、TaqDNAリガーゼ)が、(熱に不安定な試薬を伴う)検出工程に関係なく、連結反応を循環させるために使用され、反応の生成物が検出試薬に移される。あるいは、反応液を冷却し、その後、熱に不安定な成分が反応チャンバーに導入される。
【0085】
本発明の1つの実施形態において、PCR反応が、標的核酸の量を連結反応の前に増幅するために核酸サンプルに対して行われる。これは、ヌクレオチド反復について分析されるテンプレート核酸の量が低いならば好まれる。また、これは、反復した配列に隣接する対立遺伝子特異的な配列に対して特異的なPCRプライマーが使用されるならば、対立遺伝子特異性が得られることを可能にする。
【0086】
ヒトなどの生物では、ヌクレオチド反復の2つの対立遺伝子の様々な異なる対立遺伝子形態を見つけることは非常に一般的であり、このことは、犯罪を解決するために法医学においてしばしば使用される。本発明は、連結副生成物の量が連結反応の数に比例しているので、異なる数の反復が2つの対立遺伝子に存在する状況を解決する。短い方の対立遺伝子がオリゴヌクレオチドプローブで満たされるとき、連結副生成物の数が50%減少する。例えば、連結反応が、上記で記載されるような、ルシフェラーゼに基づくアッセイを使用して、放射された光によって検出されるとき、光の産出は約50%低下することになり、これは、光度測定法によって容易にモニターすることができる差である。従って、それぞれの対立遺伝子における反復ユニットの数を本発明によって決定することができる。
【0087】
本発明はまた、本発明による方法を行うためのキットに関連し、そのようなキットは、別個のバイアルにおいて、リガーゼ酵素と、連結副生成物をATPに変換するための酵素とを含む。好ましくは、リガーゼ酵素はNAD+依存性DNAリガーゼである。好ましくは、連結副生成物を変換するための酵素はPPDK酵素である。本発明はまた、別個のバイアルにおいて、ルシフェラーゼ酵素をさらに含むキットに関連する。本発明によるキットは、好ましくは、下記の疾患群から選択される疾患に関連した遺伝因子(例えば、ヌクレオチド反復など)を含む領域に対して相補的なオリゴヌクレオチド(必要な場合にはAdoPP5’修飾を有する)を含むことができる:歯状核赤核淡蒼球ルイ体萎縮症(DRPLA)、脆弱X症候群、脆弱部位FRAXE、ハンチントン病、ケネディ病、マシャド・ジョセフ病、筋緊張性ジストロフィー、フリードライヒ運動失調症、1型脊髄小脳運動失調、2型脊髄小脳運動失調、3型脊髄小脳運動失調、6型脊髄小脳運動失調、8型脊髄小脳運動失調および12型脊髄小脳運動失調。さらに、1つの実施形態において、キットはさらに、別個のバイアルにおいて、アピラーゼを含む。
【0088】
本発明による別のキットは、好ましくは、下記の群に由来する、微生物種の同定のための情報を提供する遺伝子領域に対して相補的なオリゴヌクレオチド(必要な場合にはAdoPP5’修飾を有する)を含むことができる:16SrRNA遺伝子、23SrRNA遺伝子、groEL、gyrB、rpoB、rnpBおよびgroEL、ミクロサテライト配列およびミニサテライト配列、VNTR、核リボソームDNA(rDNA)アレイ、すなわち、スモールサブユニット(SSU)(18S様)、ラージサブユニット(LSU)(23S、26Sまたは28S様)、5.8SrRNA遺伝子、ならびに内部転写リボソームDNA(rDNA)スペーサー(ITS1およびITS2)。
【0089】
本発明によるキットにおけるバイアルはまた、キットが少なくとも2つのバイアルを含むならば、キットの成分の2つ以上を含むことができる。
【0090】
本発明はまた、本発明による方法を行うための組成物に関連し、そのような組成物は、リガーゼ酵素と、連結副生成物をATPに変換するための酵素とを含む。好ましくは、リガーゼ酵素はNAD+依存性DNAリガーゼである。好ましくは、連結副生成物を変換するための酵素はPPDK酵素である。本発明はまた、ルシフェラーゼ酵素をさらに含む組成物に関連する。本発明による組成物は、好ましくは、下記の疾患群から選択される疾患に関連した遺伝因子(例えば、ヌクレオチド反復など)を含む領域に対して相補的なオリゴヌクレオチド(必要な場合にはAdoPP5’修飾を有する)を含むことができる:歯状核赤核淡蒼球ルイ体萎縮症(DRPLA)、脆弱X症候群、脆弱部位FRAXE、ハンチントン病、ケネディ病、マシャド・ジョセフ病、筋緊張性ジストロフィー、フリードライヒ運動失調症、1型脊髄小脳運動失調、2型脊髄小脳運動失調、3型脊髄小脳運動失調、6型脊髄小脳運動失調、8型脊髄小脳運動失調および12型脊髄小脳運動失調。
【0091】
本発明による別の組成物は、好ましくは、下記の群に由来する、微生物種の同定のための情報を提供する遺伝子領域に対して相補的なオリゴヌクレオチド(必要な場合にはAdoPP5’修飾を有する)を含むことができる:16SrRNA遺伝子、23SrRNA遺伝子、groEL、gyrB、rpoB、rnpBおよびgroEL、ミクロサテライト配列およびミニサテライト配列、VNTR、核リボソームDNA(rDNA)アレイ、すなわち、スモールサブユニット(SSU)(18S様)、ラージサブユニット(LSU)(23S、26Sまたは28S様)、5.8SrRNA遺伝子、ならびに内部転写リボソームDNA(rDNA)スペーサー(ITS1およびITS2)。
【0092】
米国特許第4988617号には、「上流側」オリゴヌクレオチドの3’末端が多型位置を覆って置かれるように設計された特異的なオリゴヌクレオチドを使用して、連結反応の検出を伴うSNPを分析するための方法が記載される。「下流側」オリゴヌクレオチドが、その5’末端が上流側プライマーに並置されて置かれる。下流側プライマーの5’末端が、下流側オリゴヌクレオチドの3’−OH末端に対する連結を可能にするためにリン酸化される。上流側プライマーの1つの型は、その3’末端において特定のテンプレート分子に関して相補的であり、一方で、別の遺伝子型のテンプレート分子とのミスマッチを形成する。このミスマッチはDNAリガーゼの特異性のために連結反応を妨げ、従って、連結または連結の欠如を使用して、多型を検出することができる。その後、プローブが標的ヌクレオチド配列から分離され、連結された生成物の存在または非存在が検出される。この検出では、固定化されたプローブを使用することによってゲルでのヌクレオチド範囲の分離を伴うことができる。
【0093】
本発明の別の好ましい実施形態において、本発明の方法は、SNPを分析するために使用される。ヒトゲノムにおける大多数の変異は単一ヌクレオチド多型(SNP)であり、それらは多くの場合、遺伝病に関係づけられ得る。従って、そのようなSNPを検出するための確実な方法を有することは重要である。SNPを検出するための現在利用することができる方法には、ミスマッチしたプローブの異なる変性に依存する方法、例えば、対立遺伝子特異的なオリゴヌクレオチドのハイブリダイゼーション(Wallece,RB他,1979年、Nucl.Acid Res.6:3553)および変性勾配ゲル電気泳動(Myers,RM他,1985年、Nature 313:495)などにおける方法がある。あるいは、制限断片長多型(RFLP)(Geever他,1987年、Proc.Natl.Acad.Sci.78:508)を使用することができ、この場合、ヌクレオチド配列の違いの結果である別の消化パターンが検出される。しかしながら、ヒトゲノムにおけるヌクレオチドの約1/3のみがこの技術によって分析され得るにすぎない。RFLPでは、SNPの約1/2を分析することが可能になる。本発明は、任意の植物、動物または微生物から得られる任意のDNA含有サンプル(例えば、血液、細胞または組織など)におけるSNPを分析するために使用することができる。テンプレートDNA上において互いにすぐ隣接してアニーリングする様々なオリゴヌクレオチドが構築される。これらのオリゴヌクレオチドは、オリゴヌクレオチドのうちの1つの末端ヌクレオチドが、もう一方のプローブに隣接する末端で、その位置においてテンプレート核酸のヌクレオチドの正常なヌクレオチドまたは異常なヌクレオチドのいずれかに対して相補的であるヌクレオチドを有するように構築される。これらのオリゴヌクレオチドは、テンプレート核酸にアニーリングすることができ、その後、本発明の方法の各工程を上記のように行うことができる。テンプレート核酸の多型部位におけるヌクレオチドがオリゴヌクレオチドのこの位置におけるヌクレオチドに対して相補的であるならば、連結反応が起こり、連結副生成物が放出され、この連結副生成物を、光シグナルを生じさせるためにATPに変換することができ、最終的には光シグナルを検出することができる。オリゴヌクレオチドとテンプレート核酸との間に相補性が存在しないならば、連結は起こらず、従って、光放射が起こらない。従って、多型部位におけるヌクレオチドの同一性を明らかにすることができる。
【0094】
本発明はまた、約8個〜10個のヌクレオチドからなるヌクレオチド反復であるテロメアの長さを決定するために好適である。ヒトの老化がテロメアの短縮化に関連づけられ、また、ガンの種々の形態もまたテロメアの短縮化に依存しているかもしれないことが示唆されている。
【0095】
本発明はまた、種々の法医学的検査および父子鑑定においてしばしば使用されるミクロサテライトの長さを決定するために好適である。最も頻繁に使用されているミクロサテライトはCA反復である。
【0096】
連続したプローブをアニーリングし、その後、どのプローブがアニーリングしているかを、ルシフェラーゼ反応に連係した連結事象によって明らかにする方法を、多くの他のタイプの遺伝子分析に適用することができる。一例が、そのための分子的方法が標的配列フラグメントの増幅にしばしば基づいている細菌の同定である。その後、アンプリコンが、特定の菌株、種または群に関連し得る、有り/無しの答え(DNAハイブリダイゼーション、種特異的なPCR)またはパターン(断片多型、DNA配列決定)を得るために分析される。細菌における比較配列分析のための最も広範に使用されている標的遺伝子は、30Sリボソームスモールサブユニットの構造的一部をコードする16SrRNA遺伝子である。16SrRNA遺伝子のDNA配列決定は系統発生的研究のための重要な手段であり、また、微生物同定のためにも使用されている。16S遺伝子はかなり大きく(約1500塩基)、多くの場合、近縁菌株の分類のためにはあまりにも少ない変動を含有する。さらに、多くの属において、16S遺伝子は、配列不均一性をもたらし得る多コピー遺伝子である。これらの欠点のために、さらなる標的遺伝子が調べられており、これらには、groEL、gyrB、rpoB、rnpBおよびgrpELが含まれる。23S遺伝子もまた、好適な代わりになりつつある。ミクロサテライト配列およびミニサテライト配列は1塩基対〜10塩基対および10塩基対〜100塩基対の範囲での反復をそれぞれ伴うが、これらもまた、細菌同定において使用される。多くがVNTR(変動数のタンデム反復)として示される。さらに、菌類などの真核生物のなかには、核リボソームDNA(rDNA)アレイ、すなわち、スモールサブユニット(SSU)(18様)、ラージサブユニット(LSU)(23S、26Sまたは28S様)、5.8SrRNA遺伝子、ならびに内部転写リボソームDNA(rDNA)スペーサー(ITS1およびITS2)が、分子的同定のための理想的な標的を提供する。菌類において、23S様遺伝子の埋め込まれた拡大セグメント(ES)領域は非常に変わりやすく、従って、診断的標的配列として役立つ。DNAウイルスおよびRNAウイルスを、数多くの特異的な遺伝子領域を使用して検出し、型決定することができる(例えば、ヒト乳頭腫ウイルス(HPV)を型決定するためのL1、E6、E6/E7、E7/E1およびE1の各領域)。上述の可変領域は、連結媒介型方法による分析のための好適な標的であり得る。
【0097】
従って、本発明はまた、微生物種におけるヌクレオチド反復の長さを決定するために、または、そうでない場合には、微生物種における遺伝子の変動性を特徴づけるために好適であり、このことは、菌株および単離体を同定するための有用な手段である。
【0098】
さらに、本発明の1つの実施形態において、連結反応から得られたシグナルが、テンプレート分子または代表的な代わりのDNA鎖における別の位置において行われた別個の連結事象またはプライマー伸長事象により生じたシグナルを使用することによって、本発明の方法を行う前に正規化/校正される。
【0099】
また、別の実施形態では、多重反応で同時または逐次的に鎖を分析するために、分析される種々のDNA鎖が同じ反応に含められ、好適な連結可能なプローブが加えられる。
下記において、本発明が実施例として記載されるが、そのような実施例は、本発明を例示することが意図され、本発明の範囲をいかなる点においても限定することは意図されない。
【実施例】
【0100】
実施例1:熱サイクルによる連結反応を用いた遺伝子内の可変位置における2つのオリゴヌクレオチドの連結、それに続く生物発光検出
これらのオリゴヌクレオチドは、鎌状赤血球貧血を引き起こす点突然変異を含有するβ−グロビン遺伝子の領域の合成体を表すために設計された(Barany,F.(1991年)、クローン化された熱安定性リガーゼを使用する遺伝病の検出およびDNA増幅、Proc.Natl.Acad.Sci.USA 88,189〜193を参照のこと)。
【0101】
オリゴヌクレオチドは、アニーリング後、下記の複合体を形成する:
【0102】
下記を200μLのPCRチューブにおいて50μLの最終体積で混合した:20mMのTris−酢酸塩緩衝液(pH7.6);10mMの酢酸マグネシウム;10mMのジチオスレイトール;1mMのNAD+;20UのTaqDNAリガーゼ(New England Biolabs);0pmol、2pmol、4pmolまたは10pmolのテンプレートBGL−T、40pmolの各オリゴヌクレオチド(BGL−1およびBGL−2)。リガーゼ酵素を省いたコントロールが含められた。チューブを、下記の方法を使用してサーモサイクラー(MJ Research Tetrad)でインキュベーションした:
【0103】
連結の成功が、ストレプトアビジン−セファロースビーズでの連結生成物(ビオチン−BGL1−BGL2)の捕獲、それに続く、変性、特異的な配列決定用プライマーBG−S1のアニーリング、および、パイロシーケンスによる完全な配列の確認を伴う並行実験において確認された。
【0104】
25マイクロリットルの反応液をPSQ96プレートに移した。20マイクロリットルの検出混合物#1を加えた(これは、20mMのTris−酢酸塩緩衝液(pH7.6);10mMの酢酸マグネシウム;0.625mMのホスホエノールピルビン酸;0.375mMのピロリン酸ナトリウム;50μgのPPDKを含有した)。プレートをPSQ96パイロシーケンス装置に移した。装置内で、5μLの検出混合物#2(20mMのTris−酢酸塩緩衝液(pH7.6);10mMの酢酸マグネシウム;13μgのルシフェラーゼ;7.5μgのルシフェリン)が、37℃のインキュベーション温度とともに、PSQ96装置の分注カセットを使用して加えられた。光の発生が、PSQ96装置においてCCDカメラを使用して追跡された。最初の反応における0pmol、2pmol、4pmolおよび10pmolのテンプレートBGL−Tについての結果が図3に示される。
【0105】
シグナルが、反応混合物におけるテンプレートの量とともに明らかに増大している。このことは、連結、放出されたAMPのATPへの変換、次いで光への変換の連係が機能していることを示している。
【0106】
実施例2:生物発光検出に連係した連結反応を用いた遺伝子内の可変位置における2つのオリゴヌクレオチドの連結。プレアニーリングされたプライマー/テンプレート複合体に対するリガーゼ工程およびPPDK工程の組合せ。
下記を、三連ウエルで、200μLのPCRチューブにおいて20μLの最終体積で混合した:20mMのTris−酢酸塩緩衝液、pH7.6;10mMの酢酸マグネシウム;5pmolのテンプレートBGL−T;20pmolの各オリゴヌクレオチド(BGL−1およびBGL−2)。オリゴヌクレオチドを、80℃で5分間インキュベーションし、その後、室温に冷却することによってテンプレートにアニーリングさせた。アニーリングした反応液を、PSQ96において使用されるマイクロタイタープレートに移した。コントロールは緩衝液のみを含んだ。その後、連結、および、放出されたAMPのATPへの変換のための試薬を含有する混合物の20マイクロリットルを加えた。この混合物は下記を含有した:20mMのTris−酢酸塩緩衝液(pH7.6);10mMの酢酸マグネシウム;2.5mMのNAD+;25mMのDTT;0.625mMのホスホエノールピルビン酸;0.375mMのピロリン酸ナトリウム;50μgのPPDK;20UのTaqDNAリガーゼ(コントロールでは省かれた)。反応液を45℃で10分間インキュベーションした。プレートをPSQ96に移し、5μLの検出混合物#2(20mMのTris−酢酸塩緩衝液(pH7.6);10mMの酢酸マグネシウム;13μgのルシフェラーゼ;7.5μgのルシフェリン)が、28℃のインキュベーション温度とともに、PSQ96装置の分注カセットを使用して加えられた。光の発生が、PSQ96装置においてCCDカメラを使用して追跡された(図4参照)。
【0107】
実施例3:CTG反復の数の差の検出
本実験は、数多くのポリグルタミン疾患(上記の表を参照のこと)に関与するトリヌクレオチド反復(CAG/CTG)に基づいた。(CTG)20または(CTG)10の配列を有するオリゴヌクレオチドテンプレートの1ピコモルを、96ウエルのPSQ96プレートにおいて、20μLのアニーリング緩衝液(20mMのTris−酢酸塩緩衝液(pH7.6);10mMの酢酸マグネシウム;20mMの酢酸カリウム)中で、40ピコモルの相補的な5’−リン酸化オリゴヌクレオチド(CAG)3と混合した。短いリン酸化オリゴヌクレオチドを、80℃で5分間インキュベーションし、その後、室温に冷却することによって、長い方のオリゴヌクレオチドテンプレートにアニ−リングさせた。連結を、5μLの連結混合物(アニーリング緩衝液において20UのTaqDNAリガーゼ、6.25mMのNAD+、および62.5mMのジチオスレイトール)を加え、45℃で30分間インキュベーションすることによって行った。リガーゼを含まない連結混合物を用いたコントロールもまた処理された。連結反応によって放出されたAMPを、15μLのPPDK混合物(アニーリング緩衝液において50μgのPPDK、0.8mMのPEP、および0.5mMのピロリン酸ナトリウム)を加え、45℃で10分間インキュベーションすることによってATPに変換した。生じたATPの量を明らかにするために、PSQ96プレートをPSQ96パイロシーケンス装置に移した。装置内で、5μLの検出混合物(アニーリング緩衝液において13μgのルシフェラーゼおよび7.5μgのルシフェリン)が装置によって分注され、生じた光放出が検出された。四連についての結果が、コントロール(リガーゼを含まない)からのシグナルが引かれて、図5に示される。これらの結果は、CTG反復の数が増大するとき、シグナルの増大を明瞭に示している(図5)。
【0108】
実施例4:ヌクレオチド反復の数を決定するためのずれたオリゴヌクレオチドの使用
本実施例では、サンプルDNAにおける反復した配列が、本発明によって分析されることになる反復したユニットよりも長いオリゴヌクレオチドまたは短いオリゴヌクレオチド(すなわち、ずれたオリゴヌクレオチド)の段階的な連結工程を用いてどのように分析されるかが記載される。
【0109】
最初に、プライマー−テンプレート複合体を、ヌクレオチド反復配列に隣接する配列に対して相補的なオリゴヌクレオチドを、アニーリングが起こることを可能にする条件のもとで加えることによって形成させる:
【0110】
次に、反復したユニットよりも長い(または短い)リン酸化オリゴヌクレオチド(この場合には3)を好適な連結緩衝液においてリガーゼおよび連結補因子と一緒に加える:
【0111】
その後、連結反応において放出された連結副生成物を、連結反応が起こっていることの目安として検出する。例えば、放出されたAMPまたはPPiを、上記で記載されたような、ルシフェラーゼに基づくアッセイで検出することができるATPに変換することによって。
【0112】
その後、過剰なリン酸化オリゴヌクレオチドを、その次のリン酸化オリゴヌクレオチド(pCCGC)が、上記で記載されたように、リガーゼおよび連結補因子と一緒に加えられ、アニーリング、連結および検出の別の一連操作が開始される前に除く。
【0113】
その後、アニーリング、連結および検出のこのサイクルを、ヌクレオチド反復の全範囲が連結オリゴヌクレオチドで満たされるまで繰り返す。起こった連結反応の数は既知であるので、テンプレート核酸における反復ユニットの数を計算することができる。
【0114】
実施例5:過剰な補因子のアピラーゼ媒介による除去との組合せでのATP依存性リガーゼの使用
本実験は、数多くのポリグルタミン疾患(上記の表を参照のこと)に関与するトリヌクレオチド反復(CAG/CTG)に基づいた。(CTG)10または(CTG)20の配列を有するオリゴヌクレオチドテンプレートの1ピコモルを、96ウエルのPSQ96プレートにおいて、15μLのアニーリング緩衝液(20mMのTris−酢酸塩緩衝液(pH7.6);2mMの酢酸マグネシウム)中で、40ピコモルの相補的な5’−リン酸化オリゴヌクレオチド(CAG)3と混合した。短いリン酸化オリゴヌクレオチドを、80℃で5分間インキュベーションし、その後、室温に冷却することによって、長い方のオリゴヌクレオチドテンプレートにアニ−リングさせた。連結を、15μLの連結混合物(アニーリング緩衝液において200UのT4DNAリガーゼ、2mMのdATP、および2mMのジチオスレイトール)を加え、37℃で30分間インキュベーションすることによって行った。リガーゼおよび(CAG)3を含まない連結混合物を用いたコントロールもまた処理された。連結反応液の25マイクロリットルを、50mUのアピラーゼを15μlのアニーリング緩衝液において加え、室温(約25℃)で25分間インキュベーションすることによってアピラーゼで処理して、過剰なdATPを消化した。連結反応によって生じたピロリン酸の量を明らかにするために、PSQ96プレートをPSQ96パイロシーケンス装置に移した。装置内で、5μlの酵素混合物(アニーリング緩衝液において25mUのスルフリラーゼおよび0.5μgのルシフェラーゼ)および5μLの基質混合物(アニーリング緩衝液において280pmolのAPSおよび7.5μgのルシフェリン)が装置によって分注され、生じた光放射が検出された。その後、既知量のピロリン酸およびATPから得られるシグナルを、5ピコモルのピロリン酸をそれぞれのウエルに分注し、続いて5ピコモルのATPを分注することによって測定した。三連についての結果が、コントロール(リガーゼおよびリン酸化オリゴヌクレオチドを含まない)からのシグナルとともに、図6aに示される。
【0115】
図6aにおける結果は、dATPとの組合せでT4DNAリガーゼから得られたシグナルが、連結されたオリゴヌクレオチドの非存在(コントロール)下で測定されるようなバックグラウンドを上回る連結シグナル(試験)の良好な分離を伴って、DNA反復領域の長さに依存していることを明瞭に示している。
【0116】
図6bにおいて、NAD+依存性TaqDNAリガーゼ+PPDK(実施例3)およびATP依存性T4DNAリガーゼ(dATPを基質として使用する;本実施例)から得られた結果(「観測シグナル」)が、テンプレート分子あたりの連結の最大数から得ることができるシグナル(これは連結の理論的最大数から計算される)、および、5pmolのピロリン酸から得られるシグナル(「最大予想シグナル」)と比較される。これらの結果は、(1)予想された結果と、観測された結果との間における明瞭な相関、および、(2)2つの独立した連結方法の間における明瞭な相関さえも示している。
【0117】
実施例6:微生物の型決定
本実施例では、本発明の連結法が、細菌の型を決定するためにどのように使用され得るかが明らかにされる。図7は、連鎖球菌属の数多くの種に由来するrnpB遺伝子の領域から得られた配列のアラインメントを示す。認められ得るように、共通する領域(陰影部)と、種間で変化する領域との両方が存在する。異なる種または種群の連結媒介による検出が下記のように行われる。長さおよび組成が異なるプローブが、種に依存して大きい効率または低い効率でそのような領域の連続した部分に対して同時または連続的のいずれかでアニーリングすることができるように設計される(図8参照)。アニーリングのレベルは、例えば、温度、塩濃度、または当業者に既知の他の薬剤(例えば、ジメチルスルホキシドなど)とともにストリンジェントな条件を適用することによって制御される。アニーリングの成功が、得られるシグナルが、どのくらい多くのプローブがアニーリングしているかの目安であるように連結反応によってモニターされ、従って、アニーリングの成功はテンプレートの同一性の目安を与える。アニーリング工程および検出工程が、適用に依存して、その後の検出を伴って、すべてのプローブを同時に使用するか、または、プローブを連続して加えることによるかのいずれかで行われる。これは、種の異なる群などを同定するための簡便なスクリーニング方法として使用することができ、その後、必要ならば、配列決定によって詳しく分析することができる。この原理は、当然ながら、数塩基を伴う任意の遺伝的変異に対して適用することができ、また、微生物の型決定に限定されない。本発明の方法ではさらに、アニーリングし、かつ、従って、(1)反応が機能していることを確認し、また(2)他の連結事象からのその後のシグナルを正規化するための標準シグナルをもたらすシグナルを与えることが常に予想され得る保存された領域におけるプローブを含むことができる。
【図面の簡単な説明】
【0118】
【図1】本発明による、いわゆる「終点」分析を説明する図である。
【図2】本発明による、いわゆる「段階的」分析を説明する図である。
【図3】熱サイクル処理による連結反応を用いた遺伝子内の可変位置における2つのオリゴヌクレオチドの連結、それに続く生物発光検出を示す図である。
【図4】生物発光検出に連係した連結反応を用いた遺伝子内の可変位置における2つのオリゴヌクレオチドの連結(プレアニーリングされたプライマー/テンプレート複合体に対するリガーゼ工程およびPPDK工程の組合せ)を示す図である。
【図5】CTG反復数の違いの検出を示す図である。
【図6】図6aは、遺伝子内の可変位置における2つのオリゴヌクレオチドの連結を示し、ATP依存性リガーゼ(T4DNAリガーゼ)と、補因子としてのATPとの使用を示す図である。アピラーゼが、過剰な補因子を減少させるために使用される。図6bは、2つの連結方法(ATP依存性リガーゼおよびNAD+依存性リガーゼ)を使用する遺伝子内の可変位置における2つのオリゴヌクレオチドの連結の比較を示す図である。
【図7】連鎖球菌属の数多くの種から得られたrnpB遺伝子の領域に由来する配列のアライメントを示す。
【図8】異なる種の領域に対するプローブのアニーリングを示す図である。
【配列表フリーテキスト】
【0119】
配列番号1〜19は人工的な配列である。
【技術分野】
【0001】
本発明は、連結反応を行い、連結副生成物を検出することによって核酸サンプルにおける遺伝因子(例えば、ヌクレオチド反復、または微生物型決定のためのマーカーなど)の存在を明らかにするための方法に関する。
【背景技術】
【0002】
遺伝的変異は疾患に関係づけられることが多く、遺伝的変異の同定は臨床的診断における重要な手段である。遺伝的変異は、一塩基のレベルでの違い(塩基の変化、挿入または欠失を伴う突然変異およびSNP)、数塩基のレベルでの違い(典型的にはコドン欠失を伴う)、または、遺伝子全体の増加までの様々な長さの反復した配列の変化する数のレベルでの違いを伴う。
【0003】
ヌクレオチド反復の拡大は種々の遺伝病に関係づけられることが示されている。例としては、脆弱X(Webb,TP他,1986年、Am.J.Med.Genet.23:573;Gustavson,KH他,1986年、Am.J.Med.Genet.23:581)、筋緊張性ジストロフィー(Harper,PS他,1989年、Myotonic Dystrophy(第2版)、London、England、WB Saunders Co.1989年)およびハンチントン病(MacDonald,ME他,1993年、Cell.72:971)が含まれる。これらのヌクレオチド反復は、例えば、PCRに基づくアッセイおよびサザンブロッティング技術を使用して分析されている。さらに、ジヌクレオチド反復、トリヌクレオチド反復、テトラヌクレオチド反復、ペンタヌクレオチド反復およびヘキサヌクレオチド反復が、遺伝子マッピングプロジェクトにおいて、また、父子鑑定のような種々の法医学的適用において広範囲に使用されている。
【0004】
パイロシーケンス(商標)(Pyrosequencing)は、ピロリン酸の放出の検出および酵素によるヌクレオチド分解に基づく、王立技術研究所(ストックホルム)で開発された、合成による配列決定の方法である(米国特許第6210891号および米国特許第6258568号)。合成による配列決定のこの方法では、従来のサンガー配列決定とは対照的に、ヌクレオチドが配列決定反応時に1つずつ加えられる。使用される酵素混合物は4つの異なる酵素(DNAポリメラーゼ、ATP−スルフリラーゼ、ルシフェラーゼおよびアピラーゼ)からなる。ヌクレオチドが、使用者によって、テンプレートに依存する指定された順序に従って順次加えられ、決定される。加えられたヌクレオチドがテンプレートと一致するならば、DNAポリメラーゼがそのヌクレオチドを成長途中のDNA鎖に取り込み、ピロリン酸(PPi)が放出される。ATP−スルフリラーゼにより、PPiがATPに変換され、第3の酵素(ルシフェラーゼ)により、ATPが光シグナルに転換される。これらの反応の後、第4の酵素(アピラーゼ)により、過剰なヌクレオチドおよびATPが分解され、テンプレートが、その時点で次の反応サイクル(すなわち、別のヌクレオチド添加)のために使用できる状態になる。ヌクレオチドが取り込まれない限り、PPiが放出されないので、光シグナルは、正しいヌクレオチドが取り込まれたときにだけ生じることになる。パイロシーケンスは、合成による配列決定に基づくリアルタイムのDNA配列決定法である。この方法は、SNP(単一ヌクレオチド多型)スコア化、より短いDNA領域(識別タグ)の配列決定、および対立遺伝子頻度の評価のための迅速かつ正確な方法であることが判明している(Ronaghi,M.,Uhlen,M.,Nyren,P.,リアルタイムのピロリン酸に基づく配列決定法(1998年)、Science.281、363〜365;Alderborn,A.,Kristofferson,A.,Hammerling,U.リアルタイムのピロリン酸DNA配列決定による単一ヌクレオチド多型の決定(2000年)、Genome Res.10:1249〜1258)。
【0005】
パイロシーケンスの形態での合成による配列決定は遺伝的変異の分析において多くの利点を有する。例えば、SNP分析のために使用されている他の技術(例えば、ハイブリダイゼーション技術、ミニ配列分析、RFLPおよびSSCPなど)と比較した場合、合成による配列決定では、周りの配列を示すことによって、多型位置だけでなく正しいSNP位置を調査できることが確認される。さらに、パイロシーケンスは、分離工程を伴わず、かつ自動化可能な様式でのリアルタイム分析を提供する迅速な技術であり、このことは、SSCPおよびRFLPと比較して有益であり、従って、パイロシーケンスは遺伝子分析のための魅力的な方法である。
【0006】
しかしながら、パイロシーケンスは、1塩基〜100塩基の範囲での短いヌクレオチド範囲を分析するときに最も効果的である。この方法は、一致しないプライマー伸長(いわゆる「シフト」)の生成物の蓄積によって主に制限される。シフトは、例えば、最適でないレベルのヌクレオチドの存在に起因するプライマーの不完全な伸長または過度な伸長の結果である。均一なパイロシーケンス反応では、正しいヌクレオチドを取り込むDNAポリメラーゼと、取り込まれなかったヌクレオチドを分解するアピラーゼとの間での競合が伴う。正しいヌクレオチドのレベルが、取り込み工程時においてDNAポリメラーゼについての最適値よりも低くなる場合、一部のプライマーのさらなる伸長が不完全となる。その結果は、正しい完全に伸長したプライマーよりも1塩基または数塩基短い伸長されたプライマー分子の集団である(いわゆるマイナスシフト)。他方で、アピラーゼが、ヌクレオチドを分解することにおいて十分に効果的でない場合、過剰な取り込まれていないヌクレオチドが反応液に残留し得るし、また、次の正しいヌクレオチドが与えられ、かつ取り込まれ、配列により、所望されないヌクレオチドバックグラウンドを使用するさらなる伸長が可能になる特定の状況では取り込まれ得る(いわゆるプラスシフト)。さらなる現象では、DNAポリメラーゼの活性を妨げ、従って、不完全な取り込みおよびマイナスシフトを生じさせる複雑な二次構造を有するテンプレートが伴う。これらの誤りは、当然のことではあるが、プライマー伸長の数が増大するにつれて増大する。従って、ヌクレオチド反復数を推定するための一ヌクレオチド伸長による核酸の延長に依存する技術の正確性は、ヌクレオチド反復数および長さが増大するにつれて低下する。
【0007】
米国特許第6309829号には、ヌクレオチド反復数を分析するための方法が記載され、この方法では、プライマーがテンプレート核酸にアニーリングされ、一ヌクレオチドずつ伸長し、かつ、ヌクレオチドは標識され、伸長が起こったことの目安としての測定可能なシグナルを与える。最後に加えられたヌクレオチドの標識が、その後、次のヌクレオチドが加えられる前に除かれる。1つのサイクルでのシグナルが前回の反応でのシグナルよりも実質的に小さくなるまで、伸長が続けられる。
【0008】
連結反応に基づく方法は近年、遺伝的変異の分析における重要な手段になっている。米国特許第5695933号には、ヌクレオチド反復の拡大を検出するための、RED(反復拡大検出)と呼ばれる方法が記載され、この方法では、反復配列に対して相補的なオリゴヌクレオチドがリガーゼの存在下で反復配列にアニーリングされる。その後、リガーゼは、並置されたオリゴヌクレオチドをつないで、連結されたオリゴヌクレオチドの多量体を生じさせる。多量体および標的ヌクレオチドの複合体が、その後、変性し、アニーリング、連結および変性の新たな一連の操作が開始される。多量体の長さが最終的には、ゲル分離を伴う技術によって決定される。生じた多量体の長さにより、拡大したオリゴヌクレオチド反復の長さが示され、これにより、次に、サンプリングされた個体の遺伝子状態を示すことができる。この方法では、より長い核酸範囲が分析されることになる場合でさえ、反復数を推定することが可能になる。しかしながら、この分析方法では、放射能標識されたDNAのゲルに基づく分離、それに続くハイブリダイゼーションによる検出、または、放射能標識されたDNAの直接的な検出を伴う。このような検出は、時間および手間が非常にかかり、リアルタイムの分析ができない。さらに、ゲルに基づく分離は、より大きなDNAフラグメントの分解能が低く、このことは、反復数に関して不明瞭な結果をもたらすことになる。
【0009】
まとめると、核酸サンプルにおけるヌクレオチド反復数を含めて、遺伝的変異の迅速かつ正確な分析を可能にする新しい方法が依然として求められている。
【発明の開示】
【0010】
発明の概要
本発明者らは、驚くべきことに、連結反応を行い、連結副生成物を検出することによって、ヌクレオチド反復の数を推定するだけでなく、他の遺伝因子(例えば、微生物型決定のためのマーカーなど)の存在を明らかにする新しい方法を見出している。第1の実施形態において、本発明は、下記の工程を含む、核酸サンプルにおける遺伝因子(例えば、ヌクレオチド反復または微生物型決定のためのマーカーなど)の存在を明らかにするための方法に関連する:
a)遺伝因子を含む核酸サンプルを提供する工程;
b)この核酸サンプルの遺伝因子を含む領域に対して完全または部分的に相補的なオリゴヌクレオチドを提供する工程;
c)このオリゴヌクレオチドをこの核酸サンプルにアニーリングさせる工程;
d)この核酸サンプルにアニーリングしたこのオリゴヌクレオチドを、リガーゼ酵素を使用して互いに連結する工程;および
e)連結副生成物を、連結反応が起こっているかどうかを明らかにするために、遺伝因子の存在の指標として検出する工程;
ただし、工程a)〜工程e)は、同時に、または連続して、または任意の組合せの連続する工程で行われる。
【0011】
連結副生成物の量は、起こった連結反応の数に比例しているので、連結反応の数を、どのくらいの連結副生成物が生じているかを明らかにすることによって計算することができる。あるいは、本発明の1つの実施形態において、連結サイクルが続いて行われ、1つの連結反応の存在が、その次の反応が開始される前に検出される。この場合、存在する遺伝因子(例えば、反復した核酸配列における反復ユニットなど)の数を決定するために、(生じた連結副生成物の実際の量ではなく)連結反応が起こっているか否かを検出することが必要であるだけである。本発明はまた、本発明の方法を行うためのキット、および、本発明の方法を行うために必要な成分を含む組成物に関連する。本発明において、連結生成物そのものではなく、連結副生成物が分析されるので、本発明は、ヌクレオチド反復の数を決定する新しい方法を開拓している。本発明は、ヌクレオチド反復配列におけるヌクレオチド反復ユニットの数、または、微生物型決定のためのマーカーを、様々な遺伝因子(例えば、ヌクレオチド反復など)を含む非常に長い配列の場合でさえ、これまでに可能であったよりも迅速に、かつ、手間がかからない方法で、かつ、正確に明らかにすることを可能にする。加えて、分析をリアルタイムで行うことができる。本発明はまた、必要な場合には、ヘテロ接合サンプルにおけるヌクレオチド反復などの遺伝因子の数を決定することを可能にする。
【0012】
図面の簡単な記述
図1は本発明による、いわゆる「終点」分析を説明する図である。
図2は本発明による、いわゆる「段階的」分析を説明する図である。
図3は熱サイクル処理による連結反応を用いた遺伝子内の可変位置における2つのオリゴヌクレオチドの連結、それに続く生物発光検出を示す図である。
図4は生物発光検出に連係した連結反応を用いた遺伝子内の可変位置における2つのオリゴヌクレオチドの連結(プレアニーリングされたプライマー/テンプレート複合体に対するリガーゼ工程およびPPDK工程の組合せ)を示す図である。
図5はCTG反復数の違いの検出を示す図である。
図6aは、遺伝子内の可変位置における2つのオリゴヌクレオチドの連結を示す図であり、ATP依存性リガーゼ(T4DNAリガーゼ)と、補因子としてのATPとの使用を示す図である。アピラーゼが、過剰な補因子を減少させるために使用される。図6bは、2つの連結方法(ATP依存性リガーゼおよびNAD+依存性リガーゼ)を使用する遺伝子内の可変位置における2つのオリゴヌクレオチドの連結の比較を示す図である。
図7は連鎖球菌属の数多くの種から得られたrnpB遺伝子の領域に由来する配列のアライメントを示す。
図8は異なる種の領域に対するプローブのアニーリングを示す図である。
【0013】
(定義)
「遺伝因子」は、核酸分子の検出可能な特徴、例えば、ヌクレオチド反復または微生物型決定のためのマーカーなどを意味する。
【0014】
「遺伝因子を含む領域」は、核酸分子におけるヌクレオチド配列を意味する。そのような領域は遺伝因子(例えば、ヌクレオチド反復など)と同一であってもよく、あるいは、1つ以上の遺伝因子(例えば、微生物型決定のためのマーカーなど)を含んでもよい。典型的には、遺伝因子を含む領域は、本発明の方法において使用されるようなオリゴヌクレオチドに対応する領域である。
【0015】
「微生物型決定のためのマーカー」は、特定の微生物型について特徴的であり、従って、核酸サンプルの型を決定するために使用される核酸サンプルにおける1つ以上のヌクレオチド位置を意味する。
【0016】
「連結パターン」は、連結事象のパターン(すなわち、連結されるオリゴヌクレオチドの数および/またはオリゴヌクレオチドの同一性)を意味する。典型的には、サンプルの連結パターンはサンプルの型について特徴的である。従って、「参照パターン」は、既知の参照サンプルに対する連結事象のパターンを意味する。
【0017】
「ヌクレオチド反復」は、2個、3個、4個から、数千個までのヌクレオチドの特定配列の連続した繰り返しを意味する。「反復ユニット」は1つのヌクレオチド反復配列である。反復ユニットが2ヌクレオチド〜6ヌクレオチドの間であるならば、ヌクレオチド反復はミクロサテライトと呼ばれる;反復ユニットが7ヌクレオチド〜約30ヌクレオチドの間であるならば、ヌクレオチド反復はミニサテライトと呼ばれる;反復ユニットが30を超えるならば、ヌクレオチド反復はサテライトと呼ばれる。用語「ヌクレオチド反復」はまた、短いタンデム反復および一定しない数のタンデム反復(VNTR)の用語を包含する。一部の遺伝的障害では、反復ユニットが数千回繰り返され得る。線状染色体を有するヒトおよび他の生物は、数千回反復される短い反復ユニットから構成されるテロメアをその染色体末端に有する。
【0018】
「隣接配列」は、ヌクレオチド反復の外側に位置する任意のヌクレオチド配列を意味する。この配列は、多くの状況において、増幅工程におけるプライマーのために使用される。
【0019】
「相補的」は、オリゴヌクレオチドが、選ばれたアニーリング条件のもとでテンプレート核酸にアニーリングすることができるための十分な相補性(すなわち、塩基対形成するための能力)を有することを意味する。従って、「相補的」により、すべてのヌクレオチドが、すべての位置において、テンプレート核酸に対して相補的でなければならないことは意味されず、オリゴヌクレオチドが、選ばれたアニーリング条件のもとでテンプレート核酸にアニーリングするための十分な相補性を有することのみを意味する。
【0020】
「連結副生成物」は、連結された核酸の一部を構成しないが、連結反応時に放出される生成物を意味する。
【0021】
「ルシフェラーゼ」は、本発明では、ATPに依存する酵素反応において光を生成できる任意の酵素を意味する。従って、任意の種から得られるそのような酵素が、その変異体および組換え変異体を含めて、本発明による「ルシフェラーゼ」の定義に含まれる。ルシフェラーゼに基づくアッセイにより、ルシフェラーゼ酵素を用いるアッセイを意味する。
【0022】
「光度測定法アッセイ」は、酵素の活性または化合物のレベルが光放射反応においてモニターされ、それにより、光の放射が酵素反応の結果(すなわち、ルシフェラーゼ酵素の活性の結果)であるアッセイである。
【発明を実施するための最良の形態】
【0023】
一部の生物は、生物発光をもたらす発エルゴン反応で光を生成できる。生物発光反応に関与する酵素に対する一般名はルシフェラーゼである。この酵素の種々の変異体が、細菌、昆虫および渦鞭毛藻類などの様々な生物において見出されている。
【0024】
ホタルルシフェラーゼ(EC.1.13.12.7)は、ATP、マグネシウムおよび酸素の存在下でのD−ルシフェリンの酸化を触媒する。生成物のオキシルシフェリンが、光子の放射を伴って基底状態に崩壊する励起された状態で生じる。ホタルルシフェラーゼは、特に、ATPの効率的な検出および定量のために、また、遺伝子の調節および発現の研究のためのレポーター酵素として、分子生物学および細胞生物学において広範囲に使用されている(Gould,S.J.およびSubramani,S.(1988年)、分子生物学および細胞生物学における手段としてのホタルルシフェラーゼ、Anal.Biochem.175、5〜13)。ATP変換反応に関与するすべての酵素および代謝産物をホタルルシフェラーゼ系によって分析することができる(Kricka,L.J.(2000年)生物医学科学における生物発光および化学発光の適用、Methods Enzymol.305、333〜345;Kricka,L.J.(1988年)、ルシフェラーゼおよびルシフェリンの臨床的適用および生化学的適用、Anal.Biochem.175,14〜21;Kricka,L.J.(1991年)、化学発光技術および生物発光技術、Clin.Chem.37、1472〜1481)。
【0025】
その上、様々なルシフェラーゼ系が様々な生物発光アッセイにおいて使用されている。例えば、ルシフェラーゼおよび別のタンパク質(例えば、RNA同定のためのRNA結合タンパク質など)を含む様々な融合タンパク質、あるいは、ホタルルシフェラーゼと、ビオチンアクセプターペプチドまたは免疫アッセイのための単鎖抗体との融合コンジュゲートが知られている。真核生物のルシフェラーゼ酵素は、光を産生する反応においてATPを用いる。光を検出することができる簡便性のために、ルシフェラーゼに基づく分析は多くの適用のための注目される分析手段である。
【0026】
第1の実施形態において、本発明は、下記の工程を含む、核酸サンプルにおける遺伝因子(例えば、ヌクレオチド反復、または微生物型決定のためのマーカーなど)の存在を明らかにするための方法に関連する:
a)遺伝因子を含む核酸サンプルを提供する工程;
b)この核酸サンプルの遺伝因子を含む領域に対して完全または部分的に相補的なオリゴヌクレオチドを提供する工程;
c)このオリゴヌクレオチドをこの核酸サンプルにアニーリングさせる工程;
d)この核酸サンプルにアニーリングしたこのオリゴヌクレオチドを、リガーゼ酵素を使用して互いに連結する工程;および
e)連結副生成物を、連結反応が起こっているかどうかを明らかにするために、遺伝因子の存在の指標として検出する工程;
ただし、工程a)〜工程e)は、同時に、すなわち、すべての成分を一緒に混合することによって行われるか、または連続して、すなわち、それぞれの工程のために必要な成分のみを混合し、さらなる成分を続いて加えることによって行われる。あるいは、一部の工程を同時に行うことができ、一部の工程を連続して行うことができる。これは、例えば、異なる工程で、実施されるための異なる条件(例えば、異なる緩衝液、温度など)が要求されるときには好ましくなり得る。1つの成分を除いて反応のために必要なすべての成分はまた、分析に先だって混合することができ、この一連の反応を、最後の成分を加えることによって開始することができる。
【0027】
本発明は、連結生成物そのものの代わりに連結副生成物の検出に関する。これは、連結事象が起こっているかどうかを検出する新しい方法を開拓しており、これにより、先行技術と比較して、より迅速で、より高感度な分析が可能になる。
【0028】
核酸サンプルは、当業者に既知の標準的な手段によって、必ずしも必要ではないが、好ましくは単離される生物学的サンプル(例えば、組織、細胞または血液など)の任意のDNAサンプルまたはRNAサンプルであり得る。好ましくは、核酸サンプルは、ヒト対象から得られる核酸サンプルである。好ましくは、核酸サンプルはDNAである。好ましくは、核酸サンプルは、例えば、PCRによって増幅される。
【0029】
本発明による方法において、核酸サンプルは、分析される特定の反復した配列に対して相補的なオリゴヌクレオチドと混合され、アニーリングされる。アニーリング工程に先だって、核酸を変性させなければならない。当業者は、核酸を変性させるための好適な条件を選択する方法を知っている。アニーリングのための条件は、アニーリングされる特定のオリゴヌクレオチドに依存して変化させなければならない。当業者は、種々の適用ためのアニーリング時におけるストリンジェンシーを選択する方法を知っている。オリゴヌクレオチドの長さおよび同一性を、検出されることになる遺伝因子(例えば、ヌクレオチド反復など)を含むそれぞれの特定の領域について調節しなければならない。すべてのオリゴヌクレオチドは、連結反応において機能するために、リン酸を5’末端に必要とする。
【0030】
今日、その起源をトリヌクレオチド反復展開部に有する、ヒトに影響を及ぼす少なくとも14個の障害が報告されている。それらはポリグルタミン疾患(8個の障害)および非ポリグルタミン疾患(6個の疾患)に分類することができる。
【0031】
8個の障害(その1つがハンチントン病である)はすべてが、その原因としての同じ反復したコドン(CAG)をともに有する。CAGは、グルタミンと呼ばれるアミノ酸をコードするので、これら8個のトリヌクレオチド反復障害はまとめてポリグルタミン疾患として知られている。
【0032】
ポリグルタミン疾患は多くの共通点を有する:ポリグルタミン疾患のそれぞれが、身体の特定部分における神経細胞の進行性の変性によって特徴づけられ、これに対して、6個の非ポリグルタミン疾患はそれらの症状発現において非常に不均一である。下記は、疾患、トリヌクレオチド反復、および、知られている場合には、罹患者における反復数に関する表である。
【表1】
【0033】
現在、これらの反復展開部のすべてが、PCR、およびそれに続く電気泳動工程によって分析されている。この分析は時間がかかり、より長い反復した配列については、感度が問題となる場合がある。
【0034】
第2の実施形態において、本発明は、下記の工程を含む、核酸サンプルにおけるヌクレオチド反復の数を分析するための方法に関連する:
a)ヌクレオチド反復を含む可能性がある核酸サンプルを提供する工程;
b)このヌクレオチド反復に対して相補的なオリゴヌクレオチドを提供する工程;
c)このオリゴヌクレオチドをこの核酸サンプルにアニーリングさせる工程;
d)この核酸サンプルにアニーリングしたこのオリゴヌクレオチドを、リガーゼ酵素を使用して互いに連結する工程;および
e)この連結副生成物を、連結反応が起こっているかどうかを明らかにするために検出する工程;
ただし、工程a)〜工程e)は、同時に、すなわち、すべての成分を一緒に混合することによって行われるか、または連続して、すなわち、それぞれの工程のために必要な成分のみを混合し、さらなる成分を続いて加えることによって行われる。
【0035】
本発明に従ってヌクレオチド反復を分析するためには2つのタイプのオリゴヌクレオチドが好ましい。一方の好ましいタイプのオリゴヌクレオチドは、反復ユニットそのものの単量体、二量体または多量体である(すなわち、オリゴヌクレオチドは、反復した配列の1つ以上の反復からなる)。もう一方の好ましいタイプは、ヌクレオチド反復ユニットの単量体、二量体または多量体(n−mer体)と等しい長さを有しない「一致しない」オリゴヌクレオチドである(すなわち、このオリゴヌクレオチドは、ヌクレオチド反復ユニットそのもののn−mer体よりも1塩基または数塩基だけ短いか、または長い)。「一致しない」場合におけるオリゴヌクレオチドは一つずつ加えられ、連結させられ、連結副生成物が、その次のオリゴヌクレオチドが加えられる前に検出される。ヌクレオチド反復の開始を、反復した配列のすぐ外側において、すなわち、ヌクレオチド反復に隣接する配列の1つにおいてアニーリングするように適合化されたオリゴヌクレオチドの使用によって開始することが好ましい。これらの異なるタイプのオリゴヌクレオチド、および、そのようなオリゴヌクレオチドにより行うことが可能である異なるタイプの分析が下記においてより詳しく記載される。
【0036】
下記には、ポリグルタミン障害における反復の数の計算のために使用することができる本発明のために好適なオリゴヌクレオチドの例がいくつか示される。
【表2】
【0037】
変異体1により、2つの反復したユニットを分析するためのオリゴヌクレオチドの3つの異なるタイプの二量体(それぞれ通常の書体、太字および下線)が記述される。
【0038】
変異体2により、4つの反復したユニットを分析するためのオリゴヌクレオチドの3つの異なるタイプの4mer体(それぞれ通常の書体、太字および下線)が記述される。
【0039】
変異体3により、5つの反復したユニットを分析するためのオリゴヌクレオチドの3つの異なるタイプの5mer体(それぞれ通常の書体、太字および下線)が記述される。
【0040】
変異体4により、7つの反復したユニットを分析するためのオリゴヌクレオチドの3つの異なるタイプの7mer体(それぞれ通常の書体、太字および下線)が記述される。
【0041】
下記には、脆弱X症候群における反復の数の計算のために使用することができる本発明のために好適なオリゴヌクレオチドの例がいくつか示される。
【表3】
【0042】
酵素による連結反応が、リガーゼ活性を有する数多くの異なる酵素によって行われる。そのなかでも、好ましい例が下記に詳しく記載される。リガーゼ活性を有する酵素により、核酸鎖内の末端間で、または、異なる核酸鎖との間でホスホジエステル結合を形成する能力を有する酵素を意味する。リガーゼ酵素の組換え変異体もまた本発明において意図される。
【0043】
DNAリガーゼは2つの大きな種類に分けられる(DohertyおよびSuh,2000年、Nucleic Acid Res.28、21:4051を参照のこと):ATPを補因子として要求するDNAリガーゼ、および、NAD+を要求するDNAリガーゼ。真核生物、ウイルスおよび古細菌のコードされる酵素はすべてがATPを要求する。しかしながら、ATPはまた、(下記に記載されるように)真核生物のルシフェラーゼ反応における基質でもあるので、ルシフェラーゼに基づく分析は、ATP依存性DNAリガーゼが使用されたときには、連結反応を検出することが以前では事実上不可能であった。ATP依存性リガーゼを使用するための1つの可能性は、(1)何らかの手段によって、過剰なATPを、(例えば、アピラーゼを使用することなどによって)減少させることができるか、あるいは、(2)ルシフェラーゼに対する基質として機能しないか、または、ATPよりも著しく低い程度にルシフェラーゼに対する基質として機能するATPアナログ(例えば、dATPなど)が使用されるか、あるいは、(3)(1)および(2)の組合せであると考えられる。
【0044】
1つの実施形態において、本発明では、NAD+依存性DNAリガーゼが、サンプル核酸にアニーリングさせられたDNA−オリゴヌクレオチドを連結するために使用される。この結果、エネルギー源として、ATPの代わりに、NAD+に依存するDNAリガーゼが使用される。NAD+依存性DNAリガーゼは、下記の反応(式中、Eはリガーゼであり、pAはAMPであり、pはリン酸基を示す)を触媒する、もっぱら真核生物において見出される70kD〜80kDの非常に相同的な単量体タンパク質である:
【0045】
T4RNAリガーゼもまた、DNA−オリゴヌクレオチド、または、RNA−オリゴヌクレオチド、または、DNA−オリゴヌクレオチドおよびRNA−オリゴヌクレオチドの組合せが互いに連結されるときには本発明において好ましく用いられる。例えば、T4RNAリガーゼはRNA−RNA連結のための下記の反応を触媒する:
【0046】
従って、この反応における最初の工程では、ATPを使用するRNAのアデニル化が伴う。しかしながら、いくつかの実施形態では、これは、連結反応を検出するための好ましい分析方法の1つが使用されるとき(すなわち、ルシフェラーゼに基づくアッセイが使用されるとき)には好ましくないので、別の方法が必要である。数多くの研究者がAdoPP5’RNA中間体の合成を報告している(例えば、Sninsky JJ他,等モル濃度のアクセプター分子およびドナー分子を含有するRNAリガーゼ反応におけるオリゴヌクレオチドの特異的な結合のための末端ブロック基の使用(1976年)、Nucleic Acids Res.3(11):3157〜3166;England,TE他,ジヌクレシドピロリン酸はT4誘導型RNAリガーゼに対する基質である(1977年)、Proc.Natl.Acad.Sci.USA.74(11):4839〜4842)。従って、ある方法が、そのような中間体(AdoPP5’RNAまたはAdoPP5’DNA)を本方法においてそのまま利用するために考案されており、これにより、そうでない場合には検出を妨げるATPの存在が回避されている。従って、T4RNAリガーゼもまた本発明のために好ましい。
【0047】
代わりの連結方法として、別の態様は、ATP依存性リガーゼをATPアナログ(例えば、dATPまたは2−アミノプリンリボシド三リン酸など)との組合せで使用することである。首尾よく機能するために、アナログは、まず連結反応を助けなければならない。次に、アナログは理想的には、過剰なルシフェラーゼ基質の存在に起因するバックグラウンドを最小限に抑えるために、少なくともATPによって達成されるのと同じ程度に、ルシフェラーゼ反応において基質として機能すべきではない。加えて、連結反応の生成物は好ましくはピロリン酸でなければならず、その結果、このピロリン酸は、その後、スルフリラーゼによってATPに変換することができ、次いでこのATPをルシフェラーゼ反応によって検出することができる。そのような補因子アナログは、過剰なアナログの存在から生じる低いバックグラウンドシグナルを超える連結反応の検出を容易にする。改善された方法では、検出に先だって、アナログそのもののその後の除去を伴うことができる。
【0048】
例えば、実験により、dATPが、ATP依存性リガーゼのT4DNAリガーゼに対する効果的な補因子であることが明らかにされている(Kinoshita他,J.Biochem.122:205〜211(1997年)を参照のこと)。しかしながら、dATPでさえ、ルシフェラーゼによる著しいバックグラウンドシグナルを生じさせるが、これは、ATPによって引き起こされるバックグラウンドシグナルよりも低い。代わりの方法は、dATPの使用を、アピラーゼを用いた連結後処理と組み合わせ、その結果、過剰なdATPが、ルシフェラーゼを使用する光シグナルの検出に先立って分解されるようにすることである。この組合せは、実施例の節に記載されるように、例えば、DNA反復の長さの変動を検出するための驚くほど効果的な方法をもたらしている。その結果はまた、NAD+依存性リガーゼをPPDKとの組合せで使用して得られた結果と匹敵し得る。
【0049】
さらに、アピラーゼは基質(例えば、dATP)についてATP依存性リガーゼと競合するので、アピラーゼ、リガーゼおよび基質の濃度は特定の状況について最適化されなければならない。従って、過剰量のDNAリガーゼ基質を減少させるために、アピラーゼを、連結反応の後だけでなく、連結反応の前またはその期間中に加えることもまた可能である。
【0050】
従って、別の実施形態では、連結工程(工程d)が、ATP依存性リガーゼを用いて行われ、アピラーゼが、過剰量のDNAリガーゼ基質を減少させるために、連結前に、または連結期間中に、または連結後に工程d)の連結混合物に加えられる。
【0051】
好ましい実施形態において、ATP依存性リガーゼはT4DNAリガーゼである。
【0052】
別の好ましい実施形態において、dATPが、連結工程(工程d)においてATP依存性リガーゼに対する基質として使用される。
【0053】
本発明の特許請求項に記載される方法の最後の工程として、連結副生成物が、連結反応が起こっているかどうかを明らかにするために検出される。生じる連結副生成物の量は、起こった連結反応の数に比例している。下記に記載されるように、場合により、ヌクレオチド反復ユニットの数を推定することを可能にするために、連結反応が起こっているか否かを検出することが十分である。当業者は、最終生成物のさらなる操作が望ましいことを明らかにすることができ、従って、本発明では、本発明の方法の使用に対するこのような調節が意図される。
【0054】
本発明のさらに別の実施形態は、下記の工程を含む、核酸サンプルにおけるヌクレオチド反復の数を分析するための方法に関連する:
a)ヌクレオチド反復を含む可能性がある核酸サンプルを提供する工程;
b)このヌクレオチド反復に対して相補的なオリゴヌクレオチドを提供する工程;
c)このオリゴヌクレオチドをこの核酸サンプルにアニーリングさせる工程;
d)この核酸サンプルにアニーリングしたこのオリゴヌクレオチドを、リガーゼ酵素を使用して互いに連結する工程;
e)連結副生成物をATPに変換する工程;および
f)連結反応が起こっているかどうかを明らかにするために、このATPを検出する工程;
ただし、工程a)〜工程f)は、同時に、すなわち、すべての成分を一緒に混合することによって行われるか、または連続して、すなわち、それぞれの工程のために必要な成分のみを混合し、さらなる成分を続いて加えることによって行われる。あるいは、一部の工程を同時に行うことができ、一部の工程を連続して行うことができる。これは、例えば、異なる工程で、実施されるための異なる条件(例えば、異なる緩衝液、温度など)が要求されるときには好ましくなり得る。1つの成分を除いて反応のために必要なすべての成分はまた、分析に先だって混合することができ、この一連の反応を、最後の成分を加えることによって開始することができる。
【0055】
この実施形態において、連結副生成物の1つ(すなわち、AMPもしくは修飾AMPまたはPPi(ピロリン酸))がATPに変換される。
【0056】
1つの好ましい実施形態において、連結副生成物はAMPである。
【0057】
本発明による連結反応において生じたAMPは好ましくは、ATPに変換される。1つの好ましい実施形態において、本発明による連結反応において生じたAMPは、下記のようにピルビン酸リン酸ジキナーゼ(PPDK)を使用する一段階の酵素反応によってATPに変換される:
【0058】
AMPに対するKmはATPに対するKmの1/250である。従って、この反応はAMP−>ATPの方向に強く動かされる。ピルビン酸リン酸ジキナーゼは、6.5〜7のpH至適、6〜11の範囲内でのpH安定性、55℃〜60℃の至適温度を有しており、55℃未満の温度で安定である。この酵素の特筆すべき利点は、高い温度(50℃〜60℃)でのその安定性および高活性である。多くの場合、この範囲での温度は、好適なストリンジェンシーを有するアニーリング条件を提供するために好ましいので、このことは利点である。また、PPDKが、ホタルルシフェラーゼを使用する生物発光検出(これは、下記に詳しく記載される好ましい検出アッセイである)が容易に行われる緩衝液において使用される。
【0059】
あるいは、連結反応の別の副生成物(すなわち、PPi)が好ましくはATPに変化される。上記で議論されたように、連結反応における基質(ATP)は、(例えば、上記で概略されたように、アピラーゼが使用されない場合)、ルシフェラーゼ反応のための基質と同じであるので、ATP依存性DNAリガーゼは、いくつかの実施形態では、本発明のために好適ではない。しかしながら、修飾されたATP分子(例えば、dATPまたは2−アミノプリンリボシド三リン酸など)を使用することができるDNAリガーゼ(Kinoshita,YおよびNishigaki,K、ATP要求酵素におけるATPの予想外の一般的代替可能性、J.Biochem.122:205〜211(1997年))が本発明では用いられる。好ましいそのようなDNAリガーゼは下記の反応を触媒する(式中、A*は、上記で定義されるような修飾されたアデノシン分子を示す):
【0060】
修飾されたATP分子を使用することの利点の1つは、修飾されたATP分子が、(下記に記載される)好ましい、ルシフェラーゼに基づくアッセイを、ATPが妨害するのと同じ程度に妨害しないことであり、従って、修飾されたATP分子を、下記に記載される好ましい、ルシフェラーゼに基づくアッセイの期間中、存在させることができることである。リガーゼによって要求される補因子のタイプおよび濃度は、連結事象のルシフェラーゼ媒介による検出を成功させるための重要な要素である。この補因子は効果的な連結を助けなければならないが、(例えば、上記で概略されたように、アピラーゼが使用されない場合)検出工程を妨害してはならない。
【0061】
本発明による連結反応において生じるPPiは、その後、好ましくは、ATP−スルフリラーゼを用いる下記の反応によってATPに変換される(式中、APSはアデノシン5’−ホスホ硫酸を示す):
【0062】
さらに別の実施形態において、本発明は、下記の工程を含む、核酸サンプルにおけるヌクレオチド反復の数を分析するための方法に関連する:
a)ヌクレオチド配列を含む可能性がある核酸サンプルを提供する工程;
b)このヌクレオチド配列に対して相補的なオリゴヌクレオチドを提供する工程;
c)このオリゴヌクレオチドをこの核酸サンプルにアニーリングさせる工程;
d)この核酸サンプルにアニーリングしたこのオリゴヌクレオチドを、リガーゼ酵素を使用して互いに連結する工程;
e)連結副生成物をATPに変換する工程;および
f)連結反応が起こっているかどうかの指標として、ルシフェラーゼに基づくアッセイによってこのATPを検出する工程;
ただし、工程a)〜工程f)は、同時に、すなわち、すべての成分を一緒に混合することによって行われるか、または連続して、すなわち、それぞれの工程のために必要な成分のみを混合し、さらなる成分を続いて加えることによって行われる。あるいは、一部の工程を同時に行うことができ、一部の工程を連続して行うことができる。これは、例えば、異なる工程で、実施されるための異なる条件(例えば、異なる緩衝液、温度など)が要求されるときには好ましくなり得る。1つの成分を除いて反応のために必要なすべての成分はまた、分析に先だって混合することができ、この一連の反応を、最後の成分を加えることによって開始することができる。
【0063】
生じたATP(例えば、連結副生成物をATPに変換するための上記の反応のいずれかによって生じたATP)は、好ましくは、この実施形態に従って、ルシフェラーゼに基づくアッセイにおいて基質として使用される。ATPの量は、生じた連結副生成物(AMPまたはPPi)の量に、すなわち、起こった連結反応の数に比例しているので、また、ATPの量がルシフェラーゼ反応のために限定されているという条件では、すなわち、ルシフェリンおよびO2が過剰に存在するという条件では、光の発生は連結反応の数に対応する。本発明のために好適なルシフェラーゼは下記の反応を触媒する:
【0064】
放射される光の波長は、用いられるルシフェラーゼの起源に依存する。
【0065】
さらに別の実施形態において、本発明は、下記の工程を含む、核酸サンプルの微生物型決定のための方法に関連する:
a)微生物型決定のための少なくとも1つのマーカーを含む核酸サンプルを提供する工程;
b)この核酸サンプルの微生物型決定のためのマーカーを含む領域に対して完全または部分的に相補的なオリゴヌクレオチドを提供する工程;
c)このオリゴヌクレオチドをこの核酸サンプルにアニーリングさせる工程;
d)この核酸サンプルにアニーリングしたこのオリゴヌクレオチドを、リガーゼ酵素を使用して互いに連結する工程;および
e)連結反応が起こっているかどうかを明らかにするために、連結副生成物を検出する工程;
f)微生物型を決定するために、サンプルの連結パターンを参照パターンと比較する工程;
ただし、工程a)〜工程e)は同時に行われるか、または連続して行われるか、または任意の組合せの連続する工程で行われる。
【0066】
本発明により、様々な異なる種のDNA、例えば、様々な異なる微生物種に由来するDNAが、好適な長さおよび組成のプローブを選ぶことによって解明される。例えば、細菌における比較配列分析のための最も広範囲に使用されている標的遺伝子は、30Sのリボソームスモールサブユニットの構造的一部をコードする16SrRNA遺伝子である。16SrRNA遺伝子のDNA配列決定は系統発生的研究のための重要な手段であり、また、微生物同定のためにも使用されている。
【0067】
上記反応を触媒することができる、すべての異なる微生物に由来するすべてのルシフェラーゼ酵素が、その組換え変異体を含めて、本発明において意図される。熱活性なルシフェラーゼを、種々の種に由来する野生型酵素、変異体および組換え変異体を含めて、入手することができ、また、より高い温度でルシフェラーゼ反応を行うことが好ましいときには使用することができる。ルシフェラーゼ反応を、反応のために必要な試薬を加えることによって、連結反応および変換反応に続いて、これらの反応を生じさせることが可能になった後で行うことができる。あるいは、連結のリアルタイム測定を可能にするために連結反応および変換反応を進行させることができるので、発光反応のために必要な試薬を同時に存在させることができる。別の実施形態において、より低い温度で活性である熱安定性のルシフェラーゼ、例えば、Kajiyama NおよびNakano E,1993年(217位における一アミノ酸置換によるホタルルシフェラーゼの熱安定化)、Biochemistry.32(50 13795〜9))に記載されるルシフェラーゼなどを用いることができ、また、2段階で行われる酵素反応に共役させることができる。すなわち、最初に、連結反応および変換反応が、特異的な適用のために好適なより高い温度で行われ、その後、温度が下げられ、ルシフェラーゼ活性が測定される。
【0068】
天然のホタルルシフェラーゼは、反応混合物におけるMg2+、ATP、ルシフェリンおよび分子状酸素の存在に完全に依存している(Ford,SRおよびLeach,FR、ホタルルシフェラーゼアッセイの適用における改善、Methods in Molecular Biology,第102巻:生物発光方法およびプロトコル(3頁〜20頁、R.A.LaRossa編、Humana Press Inc.1998年)を参照のこと)。ジチオスレイトール(DTT)およびエチレンジアミン四酢酸(EDTA)が、システイン残基の酸化または金属イオンによる反応の阻害を防止するために反応混合物にそれぞれ加えられる。従って、ルシフェラーゼ反応は、好ましくは、約10mMのMg2+(例えば、10mMの酢酸Mg)およびDTT(例えば、0.2mM)を含む緩衝液において行われる。至適温度は25℃である。30℃を超える温度(特に、35℃を超える温度)は野生型酵素の迅速な不活性化を引き起こす。従って、ルシフェラーゼ反応は、好ましくは、pH7.5〜8(より好ましくはpH7.8)において、2mM〜10mMのMg2+、0.5mM〜2mMのDTTおよび0.5mM〜2mMのEDTAの存在下、20℃〜30℃の温度で行われる。しかしながら、ルシフェラーゼの変異体は、例えば、上記に記載されるような温度に関して、他の要求を有することがある。重要なことは、塩化物イオンはルシフェラーゼ反応を阻害することがあり、従って、塩化物イオンをルシフェラーゼ反応の期間中に存在させることは次善となることがあるということである。
【0069】
多くの方法を、発光反応の結果としての光の検出のために利用することができ、例えば、写真フィルム、X線フィルムにおける検出、光電子増倍管(PMT)または電荷結合デバイス(CCD)カメラの使用による検出などのために利用することができる。本発明のために、PMTまたはCCDカメラによる光度測定法検出アッセイが好ましい。そのような方法は光放射の定量的な測定を可能にするからである。本発明において提供される方法による大きな利点は、この方法が、連結反応をリアルタイムで検出することを可能にすることである。
【0070】
連結のリアルタイム検出は、ルシフェリン/ルシフェラーゼ反応による光産生を、初期レベルが検出された後で「停止する」酵素を含めることによってさらに改善することができる。「初期レベル」は、光放出が検出されるために必要な光のレベルを示す。このレベルは、使用される検出装置に関連して当業者によって容易に決定される。この「停止」原理はまた、ルシフェラーゼ酵素の低い「バーンレート」に関しても好都合であり得る。そのような酵素は連結事象間の応答時間を増大させる。好適な酵素は、上記で記載されたように、ATPをADPに分解するが、AMPを分解しないアピラーゼである。この酵素は、従来のピロシーケンシング(米国特許第6258568号)において非常に都合良く使用されている。
【0071】
従って、さらに別の実施形態において、ルシフェリン/ルシフェラーゼ反応における光産生が、産生された光の初期レベルに到達した後、酵素的に停止される。好ましくは、これはアピラーゼの添加によって行われる。
【0072】
本発明の方法による連結副生成物の検出は2種類の情報をもたらす:1)連結副生成物が生じるならば、連結反応が起こっている。場合により、(下記に記載される)、この情報は十分である。2)上記で述べられたように、連結副生成物の量が連結反応の数に比例しており、従って、核酸サンプルの量が既知であるならば、核酸サンプルにおける反復したヌクレオチド配列の反復数、または微生物型決定のためのマーカーを計算するために使用することができる。
【0073】
PPDKを使用して、AMPをATPに変換し、次いで、そのATPを使用して、ルシフェリン−ルシフェラーゼに基づくシグナルを生じさせることが、Sakakibara他(Sakakibara T他,Anal Biochenm.1999年、268(1):94〜101)およびItoh他(Itoh他,Anal.Sci.2003年,19:105〜109))によって報告されている。しかしながら、これらの適用において、この方法は、RNAの分解によって生じたAMPを測定して、食品の衛生状態に対する指標を提供するために(Sakakibara他,1999年)、または、二重免疫学的反応において(Itoh他,2003年)、それぞれ使用されていた。
【0074】
数多くのリボザイムが、リガーゼ活性を有することもまた報告されており、これらもまた本発明において意図される(Glasner,ME他(2002年)、RNAリガーゼリボザイムの構造および触媒作用のための金属イオン要求、Biochemistry,41,8103〜8112;McGinness,K.E.およびJoyce,G.F.(2002年)、外部RNAテンプレートにおけるRNA触媒によるRNA連結、Chemistry and Biology9,297〜307)。連結は、基質に直接に連結するリボザイム(すなわち、クラスIリガーゼ、Glasner他,2002年)を伴い、また、相補的なテンプレートにおいて隣接位置で結合される2つのオリゴヌクレオチドの連結さえ伴う(hcリガーゼリボザイム、McGinnessおよびJoyce,2002年)。それぞれの場合において、連結は、一方の分子の3’−OH基を別の分子の5’−三リン酸に結合すること、およびピロリン酸の放出を伴う。ATPなどの補因子はこの反応に関与しておらず、従って、リボザイムは、連結反応の好ましい検出が、ルシフェラーゼに基づくアッセイの使用を伴うとき、本発明のために好適である。
【0075】
本開示の他所で言及されたリガーゼ酵素に加えて、本発明のための好適なリガーゼ酵素が下記に例示されるが、下記のリガーゼ酵素に限定されない:ATP依存性リガーゼ酵素(例えば、T4DNAリガーゼ、T7DNAリガーゼ、BstDNAリガーゼ、TfiDNAリガーゼなど)、NAD+依存性リガーゼ酵素(例えば、TthDNAリガーゼ、TflDNAリガーゼ、TaqDNAリガーゼ、大腸菌DNAリガーゼなど)、およびRNAリガーゼ酵素(例えば、tRNAリガーゼなど)。
【0076】
本発明の方法が行われるとき、2つのタイプの分析が可能である。特定の核酸サンプルにおける反復の数を、既知の数の反復した配列を有する既知量のテンプレートDNAを使用することにより連結副生成物の検出の標準曲線を構築することによって決定することができる。それぞれの連結反応が1つの連結副生成物をもたらすので、連結副生成物の量の決定により、ヌクレオチド反復の数を決定することが可能になる。分析される核酸サンプルにおけるテンプレートDNAの量が既知であるならば、反復の数を、標準曲線を使用して連結副生成物の量を反復の数に関連づけることによって決定することができる。
【0077】
別の好ましい実施形態において、「一致しない」オリゴヌクレオチドが、上記で記載されたように使用される。この場合、例えば、上記で記載されたように、テンプレートDNAの全範囲が連結オリゴヌクレオチドで満たされるまで、さらなるオリゴヌクレオチドのそれぞれの添加において検出可能である連結副生成物が生じる。従って、この場合、テンプレートの量が既知である必要はない。反復ユニットが2である場合、それぞれの一致しないオリゴヌクレオチドは、奇数の数を有する3つ以上のヌクレオチドを有しなければならない。反復数が3である場合、オリゴヌクレオチドは、3の倍数を除いて、2ヌクレオチド以上から、反復したユニットの配列の全長までであり得る。例えば、三重反復の検出の場合、4mer体、5mer体、7mer体、8mer体、10mer体、11mer体などの組合せを使用することができる。一致しないオリゴヌクレオチドが使用されるとき、ヌクレオチド反復配列のすぐ外側でアニーリングするように適合化されているオリゴヌクレオチドが好ましくは最初に加えられる。重要なことは、オリゴヌクレオチドは、それぞれのオリゴヌクレオチドが、1つの場所で、(すなわち、現在の連結サイクルの直前の連結サイクルにおいて加えられたオリゴヌクレオチドの隣りで)連結することができるだけであるように構築されなければならないということである。
【0078】
一致しないオリゴヌクレオチドが使用されるとき、1つの連結サイクルに由来するアニーリングしていないオリゴヌクレオチド(すなわち、アニーリングまたは連結されないオリゴヌクレオチド)が好ましくは、その次の連結サイクルが開始される前に除かれる。これは種々の方法で達成される。1つの方法は、連結反応時に存在するアニーリングしていないオリゴヌクレオチドを消化する一本鎖DNA依存性エキソヌクレアーゼを使用することである。例えば、エキソヌクレアーゼI(ExoI)(大腸菌のsbcB遺伝子の産物)は、一本鎖(ss)DNAを3’から5’の方向で消化するエキソデオキシリボヌクレアーゼである。酵素活性は自由な3’−ヒドロキシル末端の存在を絶対的に要求する。この場合、テンプレートを、3’末端の修飾によって、例えば、ターミナルデオキシヌクレオチジルトランスフェラーゼを使用するジデオキシヌクレオチドの取り込みなどによって保護することが必要である。あるいは、本発明の別の好ましい実施形態において、アニーリングしていないオリゴヌクレオチドを、5’リン酸をオリゴヌクレオチドから除くことによって不活性化する酵素が連結反応工程の期間中または反応工程の後に加えられる。この酵素はホスファターゼであり得るか、または、5’末端リン酸に依存する5’末端特異的エキソヌクレアーゼであり得る。ホスファターゼは、エビのアルカリホスファターゼSAP、子ウシ腸アルカリホスファターゼCIP、または任意の細菌アルカリホスファターゼからなる群から選択することができる。上記で記載されたようなアニーリングしていないオリゴヌクレオチドの除去はまた、反復ユニットのn−mer体がオリゴヌクレオチドとして使用されるときに利用され得ることが当業者によって容易に理解される。
【0079】
本発明の別の好ましい実施形態において、核酸サンプルが担体に固定化される。固定化は共有結合性または非共有結合性のいずれかであり得る。この分野で使用されている一般的な非共有結合性の方法はビオチン−ストレプトアビジン結合対の利用であり、この場合、ビオチンがプライマーに結合され、ストレプトアビジンが担体に結合される。頻繁に使用されている共有結合性の結合方法は、プライマーに結合されたアミノリンカーの使用であり、これは、エポキシシラン処理された表面と容易に反応する。担体は数タイプのものが可能である:例えば、ビーズ(非中空または多孔性)、またはチップもしくはファイバーの表面である。別の方法は、相補的であるプライマーを、遺伝因子(例えば、ヌクレオチド反復、または微生物型決定のための1つのマーカーなど)を含む領域の隣接配列に、例えば、ビオチン−ストレプトアビジン結合対を介して結合することである。固定化された核酸サンプルまたはプライマーの使用による1つの利点は、過剰な酵素、オリゴヌクレオチドおよび/または分解生成物が、1つの好ましい実施形態では、1つの連結−検出サイクルの後、その次が開始される前に、洗浄工程によって容易に除くことができることである。
【0080】
上記で記載されたこれら2つのタイプの分析が、連結副生成物がATPに変換され、このATPが、連結反応が起こったことを検出するための、ルシフェラーゼに基づくアッセイにおいて使用されるときの本発明の変形とともに図1および図2に例示される。図1は、反復ユニットそのものの単量体または多量体であるオリゴヌクレオチドがオリゴヌクレオチドとして使用される「終点」分析を示す。それぞれの連結反応が特定量の光を放射させ、その総量が定量される。放射された光の量は、サンプル核酸の量が既知であるならば、上記で記載されたような標準曲線を使用して、反復数に関連づけることができる。図2において、続いて加えられ、連結することを可能にする一致しないオリゴヌクレオチドが使用され、1つの連結反応において放射された光が、その次のオリゴヌクレオチドが加えられ、サイクル(すなわち、添加、連結、および光放射の検出)がもう一度開始される前に「段階的」様式で検出される。これは、それ以上の光が放射されなくなるまで繰り返される。すなわち、これは、サンプル核酸における反復した配列の全範囲が連結オリゴヌクレオチドで満たされるときには当てはまる。従って、反復した配列における反復ユニットの数を、サンプル核酸の量を知ることなく決定することができる。
【0081】
オリゴヌクレオチドの長さおよび配列の選択に依存して、この方法は、2塩基〜数千塩基の範囲における反復配列について使用することができる。
【0082】
連結反応は、リガーゼ、および、連結されることになる核酸のタイプの選択に依存して、広範囲の温度にわたって行われる。例えば、ハイブリダイゼーションさせられるテンプレート核酸に対する大きな程度の相補性を有するオリゴヌクレオチドプローブが使用されるとき、相補性が低い場合よりも高い温度を、熱に安定なリガーゼを使用することによって連結反応時に使用することができる。緩衝液の選択、および、連結反応時におけるリガーゼの濃度は、それぞれの特異的なリガーゼについて最適化されなければならず、また、当業者の能力の範囲内である。
【0083】
好ましい実施形態において、連結反応は、NAD+依存性DNAリガーゼを使用して行われ、生じたAMP連結副生成物が、PPDK酵素を使用してATPに転換される。生じたATPが、その後、好ましくは、ルシフェラーゼに基づくアッセイを使用して検出される。
【0084】
本発明の好ましい実施形態において、熱安定性リガーゼ(例えば、TaqDNAリガーゼ)が、(熱に不安定な試薬を伴う)検出工程に関係なく、連結反応を循環させるために使用され、反応の生成物が検出試薬に移される。あるいは、反応液を冷却し、その後、熱に不安定な成分が反応チャンバーに導入される。
【0085】
本発明の1つの実施形態において、PCR反応が、標的核酸の量を連結反応の前に増幅するために核酸サンプルに対して行われる。これは、ヌクレオチド反復について分析されるテンプレート核酸の量が低いならば好まれる。また、これは、反復した配列に隣接する対立遺伝子特異的な配列に対して特異的なPCRプライマーが使用されるならば、対立遺伝子特異性が得られることを可能にする。
【0086】
ヒトなどの生物では、ヌクレオチド反復の2つの対立遺伝子の様々な異なる対立遺伝子形態を見つけることは非常に一般的であり、このことは、犯罪を解決するために法医学においてしばしば使用される。本発明は、連結副生成物の量が連結反応の数に比例しているので、異なる数の反復が2つの対立遺伝子に存在する状況を解決する。短い方の対立遺伝子がオリゴヌクレオチドプローブで満たされるとき、連結副生成物の数が50%減少する。例えば、連結反応が、上記で記載されるような、ルシフェラーゼに基づくアッセイを使用して、放射された光によって検出されるとき、光の産出は約50%低下することになり、これは、光度測定法によって容易にモニターすることができる差である。従って、それぞれの対立遺伝子における反復ユニットの数を本発明によって決定することができる。
【0087】
本発明はまた、本発明による方法を行うためのキットに関連し、そのようなキットは、別個のバイアルにおいて、リガーゼ酵素と、連結副生成物をATPに変換するための酵素とを含む。好ましくは、リガーゼ酵素はNAD+依存性DNAリガーゼである。好ましくは、連結副生成物を変換するための酵素はPPDK酵素である。本発明はまた、別個のバイアルにおいて、ルシフェラーゼ酵素をさらに含むキットに関連する。本発明によるキットは、好ましくは、下記の疾患群から選択される疾患に関連した遺伝因子(例えば、ヌクレオチド反復など)を含む領域に対して相補的なオリゴヌクレオチド(必要な場合にはAdoPP5’修飾を有する)を含むことができる:歯状核赤核淡蒼球ルイ体萎縮症(DRPLA)、脆弱X症候群、脆弱部位FRAXE、ハンチントン病、ケネディ病、マシャド・ジョセフ病、筋緊張性ジストロフィー、フリードライヒ運動失調症、1型脊髄小脳運動失調、2型脊髄小脳運動失調、3型脊髄小脳運動失調、6型脊髄小脳運動失調、8型脊髄小脳運動失調および12型脊髄小脳運動失調。さらに、1つの実施形態において、キットはさらに、別個のバイアルにおいて、アピラーゼを含む。
【0088】
本発明による別のキットは、好ましくは、下記の群に由来する、微生物種の同定のための情報を提供する遺伝子領域に対して相補的なオリゴヌクレオチド(必要な場合にはAdoPP5’修飾を有する)を含むことができる:16SrRNA遺伝子、23SrRNA遺伝子、groEL、gyrB、rpoB、rnpBおよびgroEL、ミクロサテライト配列およびミニサテライト配列、VNTR、核リボソームDNA(rDNA)アレイ、すなわち、スモールサブユニット(SSU)(18S様)、ラージサブユニット(LSU)(23S、26Sまたは28S様)、5.8SrRNA遺伝子、ならびに内部転写リボソームDNA(rDNA)スペーサー(ITS1およびITS2)。
【0089】
本発明によるキットにおけるバイアルはまた、キットが少なくとも2つのバイアルを含むならば、キットの成分の2つ以上を含むことができる。
【0090】
本発明はまた、本発明による方法を行うための組成物に関連し、そのような組成物は、リガーゼ酵素と、連結副生成物をATPに変換するための酵素とを含む。好ましくは、リガーゼ酵素はNAD+依存性DNAリガーゼである。好ましくは、連結副生成物を変換するための酵素はPPDK酵素である。本発明はまた、ルシフェラーゼ酵素をさらに含む組成物に関連する。本発明による組成物は、好ましくは、下記の疾患群から選択される疾患に関連した遺伝因子(例えば、ヌクレオチド反復など)を含む領域に対して相補的なオリゴヌクレオチド(必要な場合にはAdoPP5’修飾を有する)を含むことができる:歯状核赤核淡蒼球ルイ体萎縮症(DRPLA)、脆弱X症候群、脆弱部位FRAXE、ハンチントン病、ケネディ病、マシャド・ジョセフ病、筋緊張性ジストロフィー、フリードライヒ運動失調症、1型脊髄小脳運動失調、2型脊髄小脳運動失調、3型脊髄小脳運動失調、6型脊髄小脳運動失調、8型脊髄小脳運動失調および12型脊髄小脳運動失調。
【0091】
本発明による別の組成物は、好ましくは、下記の群に由来する、微生物種の同定のための情報を提供する遺伝子領域に対して相補的なオリゴヌクレオチド(必要な場合にはAdoPP5’修飾を有する)を含むことができる:16SrRNA遺伝子、23SrRNA遺伝子、groEL、gyrB、rpoB、rnpBおよびgroEL、ミクロサテライト配列およびミニサテライト配列、VNTR、核リボソームDNA(rDNA)アレイ、すなわち、スモールサブユニット(SSU)(18S様)、ラージサブユニット(LSU)(23S、26Sまたは28S様)、5.8SrRNA遺伝子、ならびに内部転写リボソームDNA(rDNA)スペーサー(ITS1およびITS2)。
【0092】
米国特許第4988617号には、「上流側」オリゴヌクレオチドの3’末端が多型位置を覆って置かれるように設計された特異的なオリゴヌクレオチドを使用して、連結反応の検出を伴うSNPを分析するための方法が記載される。「下流側」オリゴヌクレオチドが、その5’末端が上流側プライマーに並置されて置かれる。下流側プライマーの5’末端が、下流側オリゴヌクレオチドの3’−OH末端に対する連結を可能にするためにリン酸化される。上流側プライマーの1つの型は、その3’末端において特定のテンプレート分子に関して相補的であり、一方で、別の遺伝子型のテンプレート分子とのミスマッチを形成する。このミスマッチはDNAリガーゼの特異性のために連結反応を妨げ、従って、連結または連結の欠如を使用して、多型を検出することができる。その後、プローブが標的ヌクレオチド配列から分離され、連結された生成物の存在または非存在が検出される。この検出では、固定化されたプローブを使用することによってゲルでのヌクレオチド範囲の分離を伴うことができる。
【0093】
本発明の別の好ましい実施形態において、本発明の方法は、SNPを分析するために使用される。ヒトゲノムにおける大多数の変異は単一ヌクレオチド多型(SNP)であり、それらは多くの場合、遺伝病に関係づけられ得る。従って、そのようなSNPを検出するための確実な方法を有することは重要である。SNPを検出するための現在利用することができる方法には、ミスマッチしたプローブの異なる変性に依存する方法、例えば、対立遺伝子特異的なオリゴヌクレオチドのハイブリダイゼーション(Wallece,RB他,1979年、Nucl.Acid Res.6:3553)および変性勾配ゲル電気泳動(Myers,RM他,1985年、Nature 313:495)などにおける方法がある。あるいは、制限断片長多型(RFLP)(Geever他,1987年、Proc.Natl.Acad.Sci.78:508)を使用することができ、この場合、ヌクレオチド配列の違いの結果である別の消化パターンが検出される。しかしながら、ヒトゲノムにおけるヌクレオチドの約1/3のみがこの技術によって分析され得るにすぎない。RFLPでは、SNPの約1/2を分析することが可能になる。本発明は、任意の植物、動物または微生物から得られる任意のDNA含有サンプル(例えば、血液、細胞または組織など)におけるSNPを分析するために使用することができる。テンプレートDNA上において互いにすぐ隣接してアニーリングする様々なオリゴヌクレオチドが構築される。これらのオリゴヌクレオチドは、オリゴヌクレオチドのうちの1つの末端ヌクレオチドが、もう一方のプローブに隣接する末端で、その位置においてテンプレート核酸のヌクレオチドの正常なヌクレオチドまたは異常なヌクレオチドのいずれかに対して相補的であるヌクレオチドを有するように構築される。これらのオリゴヌクレオチドは、テンプレート核酸にアニーリングすることができ、その後、本発明の方法の各工程を上記のように行うことができる。テンプレート核酸の多型部位におけるヌクレオチドがオリゴヌクレオチドのこの位置におけるヌクレオチドに対して相補的であるならば、連結反応が起こり、連結副生成物が放出され、この連結副生成物を、光シグナルを生じさせるためにATPに変換することができ、最終的には光シグナルを検出することができる。オリゴヌクレオチドとテンプレート核酸との間に相補性が存在しないならば、連結は起こらず、従って、光放射が起こらない。従って、多型部位におけるヌクレオチドの同一性を明らかにすることができる。
【0094】
本発明はまた、約8個〜10個のヌクレオチドからなるヌクレオチド反復であるテロメアの長さを決定するために好適である。ヒトの老化がテロメアの短縮化に関連づけられ、また、ガンの種々の形態もまたテロメアの短縮化に依存しているかもしれないことが示唆されている。
【0095】
本発明はまた、種々の法医学的検査および父子鑑定においてしばしば使用されるミクロサテライトの長さを決定するために好適である。最も頻繁に使用されているミクロサテライトはCA反復である。
【0096】
連続したプローブをアニーリングし、その後、どのプローブがアニーリングしているかを、ルシフェラーゼ反応に連係した連結事象によって明らかにする方法を、多くの他のタイプの遺伝子分析に適用することができる。一例が、そのための分子的方法が標的配列フラグメントの増幅にしばしば基づいている細菌の同定である。その後、アンプリコンが、特定の菌株、種または群に関連し得る、有り/無しの答え(DNAハイブリダイゼーション、種特異的なPCR)またはパターン(断片多型、DNA配列決定)を得るために分析される。細菌における比較配列分析のための最も広範に使用されている標的遺伝子は、30Sリボソームスモールサブユニットの構造的一部をコードする16SrRNA遺伝子である。16SrRNA遺伝子のDNA配列決定は系統発生的研究のための重要な手段であり、また、微生物同定のためにも使用されている。16S遺伝子はかなり大きく(約1500塩基)、多くの場合、近縁菌株の分類のためにはあまりにも少ない変動を含有する。さらに、多くの属において、16S遺伝子は、配列不均一性をもたらし得る多コピー遺伝子である。これらの欠点のために、さらなる標的遺伝子が調べられており、これらには、groEL、gyrB、rpoB、rnpBおよびgrpELが含まれる。23S遺伝子もまた、好適な代わりになりつつある。ミクロサテライト配列およびミニサテライト配列は1塩基対〜10塩基対および10塩基対〜100塩基対の範囲での反復をそれぞれ伴うが、これらもまた、細菌同定において使用される。多くがVNTR(変動数のタンデム反復)として示される。さらに、菌類などの真核生物のなかには、核リボソームDNA(rDNA)アレイ、すなわち、スモールサブユニット(SSU)(18様)、ラージサブユニット(LSU)(23S、26Sまたは28S様)、5.8SrRNA遺伝子、ならびに内部転写リボソームDNA(rDNA)スペーサー(ITS1およびITS2)が、分子的同定のための理想的な標的を提供する。菌類において、23S様遺伝子の埋め込まれた拡大セグメント(ES)領域は非常に変わりやすく、従って、診断的標的配列として役立つ。DNAウイルスおよびRNAウイルスを、数多くの特異的な遺伝子領域を使用して検出し、型決定することができる(例えば、ヒト乳頭腫ウイルス(HPV)を型決定するためのL1、E6、E6/E7、E7/E1およびE1の各領域)。上述の可変領域は、連結媒介型方法による分析のための好適な標的であり得る。
【0097】
従って、本発明はまた、微生物種におけるヌクレオチド反復の長さを決定するために、または、そうでない場合には、微生物種における遺伝子の変動性を特徴づけるために好適であり、このことは、菌株および単離体を同定するための有用な手段である。
【0098】
さらに、本発明の1つの実施形態において、連結反応から得られたシグナルが、テンプレート分子または代表的な代わりのDNA鎖における別の位置において行われた別個の連結事象またはプライマー伸長事象により生じたシグナルを使用することによって、本発明の方法を行う前に正規化/校正される。
【0099】
また、別の実施形態では、多重反応で同時または逐次的に鎖を分析するために、分析される種々のDNA鎖が同じ反応に含められ、好適な連結可能なプローブが加えられる。
下記において、本発明が実施例として記載されるが、そのような実施例は、本発明を例示することが意図され、本発明の範囲をいかなる点においても限定することは意図されない。
【実施例】
【0100】
実施例1:熱サイクルによる連結反応を用いた遺伝子内の可変位置における2つのオリゴヌクレオチドの連結、それに続く生物発光検出
これらのオリゴヌクレオチドは、鎌状赤血球貧血を引き起こす点突然変異を含有するβ−グロビン遺伝子の領域の合成体を表すために設計された(Barany,F.(1991年)、クローン化された熱安定性リガーゼを使用する遺伝病の検出およびDNA増幅、Proc.Natl.Acad.Sci.USA 88,189〜193を参照のこと)。
【0101】
オリゴヌクレオチドは、アニーリング後、下記の複合体を形成する:
【0102】
下記を200μLのPCRチューブにおいて50μLの最終体積で混合した:20mMのTris−酢酸塩緩衝液(pH7.6);10mMの酢酸マグネシウム;10mMのジチオスレイトール;1mMのNAD+;20UのTaqDNAリガーゼ(New England Biolabs);0pmol、2pmol、4pmolまたは10pmolのテンプレートBGL−T、40pmolの各オリゴヌクレオチド(BGL−1およびBGL−2)。リガーゼ酵素を省いたコントロールが含められた。チューブを、下記の方法を使用してサーモサイクラー(MJ Research Tetrad)でインキュベーションした:
【0103】
連結の成功が、ストレプトアビジン−セファロースビーズでの連結生成物(ビオチン−BGL1−BGL2)の捕獲、それに続く、変性、特異的な配列決定用プライマーBG−S1のアニーリング、および、パイロシーケンスによる完全な配列の確認を伴う並行実験において確認された。
【0104】
25マイクロリットルの反応液をPSQ96プレートに移した。20マイクロリットルの検出混合物#1を加えた(これは、20mMのTris−酢酸塩緩衝液(pH7.6);10mMの酢酸マグネシウム;0.625mMのホスホエノールピルビン酸;0.375mMのピロリン酸ナトリウム;50μgのPPDKを含有した)。プレートをPSQ96パイロシーケンス装置に移した。装置内で、5μLの検出混合物#2(20mMのTris−酢酸塩緩衝液(pH7.6);10mMの酢酸マグネシウム;13μgのルシフェラーゼ;7.5μgのルシフェリン)が、37℃のインキュベーション温度とともに、PSQ96装置の分注カセットを使用して加えられた。光の発生が、PSQ96装置においてCCDカメラを使用して追跡された。最初の反応における0pmol、2pmol、4pmolおよび10pmolのテンプレートBGL−Tについての結果が図3に示される。
【0105】
シグナルが、反応混合物におけるテンプレートの量とともに明らかに増大している。このことは、連結、放出されたAMPのATPへの変換、次いで光への変換の連係が機能していることを示している。
【0106】
実施例2:生物発光検出に連係した連結反応を用いた遺伝子内の可変位置における2つのオリゴヌクレオチドの連結。プレアニーリングされたプライマー/テンプレート複合体に対するリガーゼ工程およびPPDK工程の組合せ。
下記を、三連ウエルで、200μLのPCRチューブにおいて20μLの最終体積で混合した:20mMのTris−酢酸塩緩衝液、pH7.6;10mMの酢酸マグネシウム;5pmolのテンプレートBGL−T;20pmolの各オリゴヌクレオチド(BGL−1およびBGL−2)。オリゴヌクレオチドを、80℃で5分間インキュベーションし、その後、室温に冷却することによってテンプレートにアニーリングさせた。アニーリングした反応液を、PSQ96において使用されるマイクロタイタープレートに移した。コントロールは緩衝液のみを含んだ。その後、連結、および、放出されたAMPのATPへの変換のための試薬を含有する混合物の20マイクロリットルを加えた。この混合物は下記を含有した:20mMのTris−酢酸塩緩衝液(pH7.6);10mMの酢酸マグネシウム;2.5mMのNAD+;25mMのDTT;0.625mMのホスホエノールピルビン酸;0.375mMのピロリン酸ナトリウム;50μgのPPDK;20UのTaqDNAリガーゼ(コントロールでは省かれた)。反応液を45℃で10分間インキュベーションした。プレートをPSQ96に移し、5μLの検出混合物#2(20mMのTris−酢酸塩緩衝液(pH7.6);10mMの酢酸マグネシウム;13μgのルシフェラーゼ;7.5μgのルシフェリン)が、28℃のインキュベーション温度とともに、PSQ96装置の分注カセットを使用して加えられた。光の発生が、PSQ96装置においてCCDカメラを使用して追跡された(図4参照)。
【0107】
実施例3:CTG反復の数の差の検出
本実験は、数多くのポリグルタミン疾患(上記の表を参照のこと)に関与するトリヌクレオチド反復(CAG/CTG)に基づいた。(CTG)20または(CTG)10の配列を有するオリゴヌクレオチドテンプレートの1ピコモルを、96ウエルのPSQ96プレートにおいて、20μLのアニーリング緩衝液(20mMのTris−酢酸塩緩衝液(pH7.6);10mMの酢酸マグネシウム;20mMの酢酸カリウム)中で、40ピコモルの相補的な5’−リン酸化オリゴヌクレオチド(CAG)3と混合した。短いリン酸化オリゴヌクレオチドを、80℃で5分間インキュベーションし、その後、室温に冷却することによって、長い方のオリゴヌクレオチドテンプレートにアニ−リングさせた。連結を、5μLの連結混合物(アニーリング緩衝液において20UのTaqDNAリガーゼ、6.25mMのNAD+、および62.5mMのジチオスレイトール)を加え、45℃で30分間インキュベーションすることによって行った。リガーゼを含まない連結混合物を用いたコントロールもまた処理された。連結反応によって放出されたAMPを、15μLのPPDK混合物(アニーリング緩衝液において50μgのPPDK、0.8mMのPEP、および0.5mMのピロリン酸ナトリウム)を加え、45℃で10分間インキュベーションすることによってATPに変換した。生じたATPの量を明らかにするために、PSQ96プレートをPSQ96パイロシーケンス装置に移した。装置内で、5μLの検出混合物(アニーリング緩衝液において13μgのルシフェラーゼおよび7.5μgのルシフェリン)が装置によって分注され、生じた光放出が検出された。四連についての結果が、コントロール(リガーゼを含まない)からのシグナルが引かれて、図5に示される。これらの結果は、CTG反復の数が増大するとき、シグナルの増大を明瞭に示している(図5)。
【0108】
実施例4:ヌクレオチド反復の数を決定するためのずれたオリゴヌクレオチドの使用
本実施例では、サンプルDNAにおける反復した配列が、本発明によって分析されることになる反復したユニットよりも長いオリゴヌクレオチドまたは短いオリゴヌクレオチド(すなわち、ずれたオリゴヌクレオチド)の段階的な連結工程を用いてどのように分析されるかが記載される。
【0109】
最初に、プライマー−テンプレート複合体を、ヌクレオチド反復配列に隣接する配列に対して相補的なオリゴヌクレオチドを、アニーリングが起こることを可能にする条件のもとで加えることによって形成させる:
【0110】
次に、反復したユニットよりも長い(または短い)リン酸化オリゴヌクレオチド(この場合には3)を好適な連結緩衝液においてリガーゼおよび連結補因子と一緒に加える:
【0111】
その後、連結反応において放出された連結副生成物を、連結反応が起こっていることの目安として検出する。例えば、放出されたAMPまたはPPiを、上記で記載されたような、ルシフェラーゼに基づくアッセイで検出することができるATPに変換することによって。
【0112】
その後、過剰なリン酸化オリゴヌクレオチドを、その次のリン酸化オリゴヌクレオチド(pCCGC)が、上記で記載されたように、リガーゼおよび連結補因子と一緒に加えられ、アニーリング、連結および検出の別の一連操作が開始される前に除く。
【0113】
その後、アニーリング、連結および検出のこのサイクルを、ヌクレオチド反復の全範囲が連結オリゴヌクレオチドで満たされるまで繰り返す。起こった連結反応の数は既知であるので、テンプレート核酸における反復ユニットの数を計算することができる。
【0114】
実施例5:過剰な補因子のアピラーゼ媒介による除去との組合せでのATP依存性リガーゼの使用
本実験は、数多くのポリグルタミン疾患(上記の表を参照のこと)に関与するトリヌクレオチド反復(CAG/CTG)に基づいた。(CTG)10または(CTG)20の配列を有するオリゴヌクレオチドテンプレートの1ピコモルを、96ウエルのPSQ96プレートにおいて、15μLのアニーリング緩衝液(20mMのTris−酢酸塩緩衝液(pH7.6);2mMの酢酸マグネシウム)中で、40ピコモルの相補的な5’−リン酸化オリゴヌクレオチド(CAG)3と混合した。短いリン酸化オリゴヌクレオチドを、80℃で5分間インキュベーションし、その後、室温に冷却することによって、長い方のオリゴヌクレオチドテンプレートにアニ−リングさせた。連結を、15μLの連結混合物(アニーリング緩衝液において200UのT4DNAリガーゼ、2mMのdATP、および2mMのジチオスレイトール)を加え、37℃で30分間インキュベーションすることによって行った。リガーゼおよび(CAG)3を含まない連結混合物を用いたコントロールもまた処理された。連結反応液の25マイクロリットルを、50mUのアピラーゼを15μlのアニーリング緩衝液において加え、室温(約25℃)で25分間インキュベーションすることによってアピラーゼで処理して、過剰なdATPを消化した。連結反応によって生じたピロリン酸の量を明らかにするために、PSQ96プレートをPSQ96パイロシーケンス装置に移した。装置内で、5μlの酵素混合物(アニーリング緩衝液において25mUのスルフリラーゼおよび0.5μgのルシフェラーゼ)および5μLの基質混合物(アニーリング緩衝液において280pmolのAPSおよび7.5μgのルシフェリン)が装置によって分注され、生じた光放射が検出された。その後、既知量のピロリン酸およびATPから得られるシグナルを、5ピコモルのピロリン酸をそれぞれのウエルに分注し、続いて5ピコモルのATPを分注することによって測定した。三連についての結果が、コントロール(リガーゼおよびリン酸化オリゴヌクレオチドを含まない)からのシグナルとともに、図6aに示される。
【0115】
図6aにおける結果は、dATPとの組合せでT4DNAリガーゼから得られたシグナルが、連結されたオリゴヌクレオチドの非存在(コントロール)下で測定されるようなバックグラウンドを上回る連結シグナル(試験)の良好な分離を伴って、DNA反復領域の長さに依存していることを明瞭に示している。
【0116】
図6bにおいて、NAD+依存性TaqDNAリガーゼ+PPDK(実施例3)およびATP依存性T4DNAリガーゼ(dATPを基質として使用する;本実施例)から得られた結果(「観測シグナル」)が、テンプレート分子あたりの連結の最大数から得ることができるシグナル(これは連結の理論的最大数から計算される)、および、5pmolのピロリン酸から得られるシグナル(「最大予想シグナル」)と比較される。これらの結果は、(1)予想された結果と、観測された結果との間における明瞭な相関、および、(2)2つの独立した連結方法の間における明瞭な相関さえも示している。
【0117】
実施例6:微生物の型決定
本実施例では、本発明の連結法が、細菌の型を決定するためにどのように使用され得るかが明らかにされる。図7は、連鎖球菌属の数多くの種に由来するrnpB遺伝子の領域から得られた配列のアラインメントを示す。認められ得るように、共通する領域(陰影部)と、種間で変化する領域との両方が存在する。異なる種または種群の連結媒介による検出が下記のように行われる。長さおよび組成が異なるプローブが、種に依存して大きい効率または低い効率でそのような領域の連続した部分に対して同時または連続的のいずれかでアニーリングすることができるように設計される(図8参照)。アニーリングのレベルは、例えば、温度、塩濃度、または当業者に既知の他の薬剤(例えば、ジメチルスルホキシドなど)とともにストリンジェントな条件を適用することによって制御される。アニーリングの成功が、得られるシグナルが、どのくらい多くのプローブがアニーリングしているかの目安であるように連結反応によってモニターされ、従って、アニーリングの成功はテンプレートの同一性の目安を与える。アニーリング工程および検出工程が、適用に依存して、その後の検出を伴って、すべてのプローブを同時に使用するか、または、プローブを連続して加えることによるかのいずれかで行われる。これは、種の異なる群などを同定するための簡便なスクリーニング方法として使用することができ、その後、必要ならば、配列決定によって詳しく分析することができる。この原理は、当然ながら、数塩基を伴う任意の遺伝的変異に対して適用することができ、また、微生物の型決定に限定されない。本発明の方法ではさらに、アニーリングし、かつ、従って、(1)反応が機能していることを確認し、また(2)他の連結事象からのその後のシグナルを正規化するための標準シグナルをもたらすシグナルを与えることが常に予想され得る保存された領域におけるプローブを含むことができる。
【図面の簡単な説明】
【0118】
【図1】本発明による、いわゆる「終点」分析を説明する図である。
【図2】本発明による、いわゆる「段階的」分析を説明する図である。
【図3】熱サイクル処理による連結反応を用いた遺伝子内の可変位置における2つのオリゴヌクレオチドの連結、それに続く生物発光検出を示す図である。
【図4】生物発光検出に連係した連結反応を用いた遺伝子内の可変位置における2つのオリゴヌクレオチドの連結(プレアニーリングされたプライマー/テンプレート複合体に対するリガーゼ工程およびPPDK工程の組合せ)を示す図である。
【図5】CTG反復数の違いの検出を示す図である。
【図6】図6aは、遺伝子内の可変位置における2つのオリゴヌクレオチドの連結を示し、ATP依存性リガーゼ(T4DNAリガーゼ)と、補因子としてのATPとの使用を示す図である。アピラーゼが、過剰な補因子を減少させるために使用される。図6bは、2つの連結方法(ATP依存性リガーゼおよびNAD+依存性リガーゼ)を使用する遺伝子内の可変位置における2つのオリゴヌクレオチドの連結の比較を示す図である。
【図7】連鎖球菌属の数多くの種から得られたrnpB遺伝子の領域に由来する配列のアライメントを示す。
【図8】異なる種の領域に対するプローブのアニーリングを示す図である。
【配列表フリーテキスト】
【0119】
配列番号1〜19は人工的な配列である。
【特許請求の範囲】
【請求項1】
以下の工程を含む、核酸サンプルにおける、ヌクレオチド反復または微生物型決定のためのマーカーの如き遺伝因子の存在を明らかにするための方法:
a)遺伝因子を含む核酸サンプルを提供する工程;
b)前記核酸サンプルの遺伝因子を含む領域に対して完全または部分的に相補的なオリゴヌクレオチドを提供する工程;
c)前記オリゴヌクレオチドを前記核酸サンプルにアニーリングさせる工程;
d)前記核酸サンプルにアニーリングした前記オリゴヌクレオチドを、リガーゼ酵素を使用して互いに連結する工程;および
e)連結副生成物を、連結反応が起こっているかどうかを明らかにするために、遺伝因子の存在の指標として検出する工程;
ただし、工程a)〜工程e)は、同時に、または連続して、または任意の組合せの連続する工程で行われる。
【請求項2】
以下の工程を含む、核酸サンプルにおけるヌクレオチド反復の数を分析するための方法:
a)ヌクレオチド反復を含む可能性がある核酸サンプルを提供する工程;
b)前記ヌクレオチド反復に対して相補的なオリゴヌクレオチドを提供する工程;
c)前記オリゴヌクレオチドを前記核酸サンプルにアニーリングさせる工程;
d)前記核酸サンプルにアニーリングした前記オリゴヌクレオチドを、リガーゼ酵素を使用して互いに連結する工程;および
e)連結副生成物を、連結反応が起こっているかどうかを明らかにするために検出する工程;
ただし、工程a)〜工程e)は、同時に、または連続して、または任意の組合せの連続する工程で行われる。
【請求項3】
以下の工程を含む、核酸サンプルにおけるヌクレオチド反復の数を分析するための方法:
a)ヌクレオチド反復を含む可能性がある核酸サンプルを提供する工程;
b)前記ヌクレオチド反復に対して相補的なオリゴヌクレオチドを提供する工程;
c)前記オリゴヌクレオチドを前記核酸サンプルにアニーリングさせる工程;
d)前記核酸サンプルにアニーリングした前記オリゴヌクレオチドを、リガーゼ酵素を使用して互いに連結する工程;
e)連結副生成物をATPに変換する工程;および
f)連結反応が起こっているかどうかを明らかにするために、前記ATPを検出する工程;
ただし、工程a)〜工程f)は、同時に、または連続して、または任意の組合せの連続する工程で行われる。
【請求項4】
以下の工程を含む、核酸サンプルにおけるヌクレオチド反復の数を分析するための方法:
a)ヌクレオチド反復を含む可能性がある核酸サンプルを提供する工程;
b)前記ヌクレオチド反復に対して相補的なオリゴヌクレオチドを提供する工程;
c)前記オリゴヌクレオチドを前記核酸サンプルにアニーリングさせる工程;
d)前記核酸サンプルにアニーリングした前記オリゴヌクレオチドを、リガーゼ酵素を使用して互いに連結する工程;
e)連結副生成物をATPに変換する工程;および
f)連結反応が起こっているかどうかの指標として、ルシフェラーゼに基づくアッセイによって前記ATPを検出する工程;
ただし、工程a)〜工程f)は、同時に、または連続して、または任意の組合せの連続する工程で行われる。
【請求項5】
以下の工程を含む、核酸サンプルの微生物型決定のための方法:
a)微生物型決定のための少なくとも1つのマーカーを含む核酸サンプルを提供する工程;
b)前記核酸サンプルの微生物型決定のためのマーカーを含む領域に対して完全または部分的に相補的なオリゴヌクレオチドを提供する工程;
c)前記オリゴヌクレオチドを前記核酸サンプルにアニーリングさせる工程;
d)前記核酸サンプルにアニーリングした前記オリゴヌクレオチドを、リガーゼ酵素を使用して互いに連結する工程;および
e)連結反応が起こっているかどうかを明らかにするために、連結副生成物を検出する工程;
f)微生物型を決定するために、サンプルの連結パターンを参照パターンと比較する工程;
ただし、工程a)〜工程e)は同時に、または連続して、または任意の組合せの連続する工程で行われる。
【請求項6】
工程b)におけるオリゴヌクレオチドの一つが、反復配列のすぐ外側でアニーリングするように適合されている請求項1〜5のいずれか一項に記載の方法。
【請求項7】
連結副生成物がAMPである請求項1〜6のいずれか一項に記載の方法。
【請求項8】
工程d)がNAD+依存性DNAリガーゼを使用して行われる請求項1〜7のいずれか一項に記載の方法。
【請求項9】
工程e)がピルビン酸リン酸ジキナーゼを使用して行われる請求項1〜8のいずれか一項に記載の方法。
【請求項10】
工程d)が、ATP依存性リガーゼを用いて行われ、アピラーゼが、過剰量のDNAリガーゼ基質を減少させるために、連結前に、または連結期間中に、または連結後に工程d)の連結混合物に加えられる請求項1〜6のいずれか一項に記載の方法。
【請求項11】
ATP依存性リガーゼはT4DNAリガーゼである請求項10に記載の方法。
【請求項12】
dATPが、工程d)においてATP依存性リガーゼに対する基質として使用される請求項10または11に記載の方法。
【請求項13】
連結副生成物がPPiである請求項1〜6または10〜12のいずれか一項に記載の方法。
【請求項14】
工程e)がATP−スルフリラーゼを使用して行われる請求項1〜6または10〜13のいずれか一項に記載の方法。
【請求項15】
使用されたオリゴヌクレオチドが反復そのものの単量体、二量体または多量体である請求項1〜14のいずれか一項に記載の方法。
【請求項16】
オリゴヌクレオチドが前記ヌクレオチド反復に相補的であるが、前記ヌクレオチド反復とはずれている請求項1〜14のいずれか一項に記載の方法。
【請求項17】
アニーリングしていないオリゴヌクレオチドが検出の後でエキソヌクレアーゼの使用によって除かれる工程をさらに含む請求項16に記載の方法。
【請求項18】
アニーリングしていないオリゴヌクレオチドが検出の後でホスファターゼの使用によって不活性化される工程をさらに含む請求項16に記載の方法。
【請求項19】
核酸サンプルが担体に固定化される請求項1〜18のいずれか一項に記載の方法。
【請求項20】
アニーリングしていないオリゴヌクレオチドが検出の後で洗浄によって除かれる工程をさらに含む請求項19に記載の方法。
【請求項21】
核酸サンプルが増幅される工程によって先行される請求項1〜20のいずれか一項に記載の方法。
【請求項22】
ルシフェラーゼに基づくアッセイが光度測定法アッセイである請求項1〜21のいずれか一項に記載の方法。
【請求項23】
ルシフェラーゼ反応で産生された光が、産生された光の初期レベルが達成された後、酵素的に停止される請求項1〜22のいずれか一項に記載の方法。
【請求項24】
光産生が、アピラーゼの添加によって停止される請求項23に記載の方法。
【請求項25】
分析されるべき領域の外側に相補的なオリゴヌクレオチドが、分析されるべき領域から得られるシグナルを正規化するために使用されることができるシグナルを、連結またはプライマー伸長によって生成するために使用される請求項1〜24のいずれか一項に記載の方法。
【請求項26】
別個のバイアルにおいて、リガーゼ酵素と、連結副生成物をATPに変換するための酵素とを含む、請求項1〜25のいずれか一項に記載の方法を行うためのキット。
【請求項27】
別個のバイアルにおいて、ルシフェラーゼ酵素をさらに含む請求項26に記載のキット。
【請求項28】
別個のバイアルにおいて、アピラーゼをさらに含む請求項26または27に記載のキット。
【請求項29】
以下の疾患群から選択される疾患に関連したヌクレオチド反復に対して相補的なオリゴヌクレオチド(必要な場合にはAdoPP5’修飾を有する)をさらに含む請求項26〜28のいずれか一項に記載のキット:歯状核赤核淡蒼球ルイ体萎縮症(DRPLA)、脆弱X症候群、脆弱部位FRAXE、ハンチントン病、ケネディ病、マシャド・ジョセフ病、筋緊張性ジストロフィー、フリードライヒ運動失調症、1型脊髄小脳運動失調、2型脊髄小脳運動失調、3型脊髄小脳運動失調、6型脊髄小脳運動失調、8型脊髄小脳運動失調および12型脊髄小脳運動失調。
【請求項30】
以下の群に由来する、微生物種の同定のための情報を提供する遺伝子領域に対して相補的なオリゴヌクレオチド(必要な場合にはAdoPP5’修飾を有する)をさらに含む請求項26〜28のいずれか一項に記載のキット:16SrRNA遺伝子、23SrRNA遺伝子、groEL、gyrB、rpoB、rnpBおよびgroEL、ミクロサテライト配列およびミニサテライト配列、VNTR、核リボソームDNA(rDNA)アレイ、すなわち、スモールサブユニット(SSU)(18S様)、ラージサブユニット(LSU)(23S、26Sまたは28S様)、5.8SrRNA遺伝子、ならびに内部転写リボソームDNA(rDNA)スペーサー(ITS1およびITS2)。
【請求項31】
リガーゼ酵素と、連結副生成物をATPに変換するための酵素とを含む組成物。
【請求項32】
ルシフェラーゼ酵素をさらに含む請求項31に記載の組成物。
【請求項33】
以下の疾患群から選択される疾患に関連したヌクレオチド反復に対して相補的なオリゴヌクレオチド(必要な場合にはAdoPP5’修飾を有する)をさらに含む請求項31または32に記載の組成物:歯状核赤核淡蒼球ルイ体萎縮症(DRPLA)、脆弱X症候群、脆弱部位FRAXE、ハンチントン病、ケネディ病、マシャド・ジョセフ病、筋緊張性ジストロフィー、フリードライヒ運動失調症、1型脊髄小脳運動失調、2型脊髄小脳運動失調、3型脊髄小脳運動失調、6型脊髄小脳運動失調、8型脊髄小脳運動失調および12型脊髄小脳運動失調。
【請求項34】
以下の群に由来する、微生物種の同定のための情報を提供する遺伝子領域に対して相補的なオリゴヌクレオチド(必要な場合にはAdoPP5’修飾を有する)をさらに含む請求項31または32に記載の組成物:16SrRNA遺伝子、23SrRNA遺伝子、groEL、gyrB、rpoB、rnpBおよびgroEL、ミクロサテライト配列およびミニサテライト配列、VNTR、核リボソームDNA(rDNA)アレイ、すなわち、スモールサブユニット(SSU)(18S様)、ラージサブユニット(LSU)(23S、26Sまたは28S様)、5.8SrRNA遺伝子、ならびに内部転写リボソームDNA(rDNA)スペーサー(ITS1およびITS2)。
【請求項1】
以下の工程を含む、核酸サンプルにおける、ヌクレオチド反復または微生物型決定のためのマーカーの如き遺伝因子の存在を明らかにするための方法:
a)遺伝因子を含む核酸サンプルを提供する工程;
b)前記核酸サンプルの遺伝因子を含む領域に対して完全または部分的に相補的なオリゴヌクレオチドを提供する工程;
c)前記オリゴヌクレオチドを前記核酸サンプルにアニーリングさせる工程;
d)前記核酸サンプルにアニーリングした前記オリゴヌクレオチドを、リガーゼ酵素を使用して互いに連結する工程;および
e)連結副生成物を、連結反応が起こっているかどうかを明らかにするために、遺伝因子の存在の指標として検出する工程;
ただし、工程a)〜工程e)は、同時に、または連続して、または任意の組合せの連続する工程で行われる。
【請求項2】
以下の工程を含む、核酸サンプルにおけるヌクレオチド反復の数を分析するための方法:
a)ヌクレオチド反復を含む可能性がある核酸サンプルを提供する工程;
b)前記ヌクレオチド反復に対して相補的なオリゴヌクレオチドを提供する工程;
c)前記オリゴヌクレオチドを前記核酸サンプルにアニーリングさせる工程;
d)前記核酸サンプルにアニーリングした前記オリゴヌクレオチドを、リガーゼ酵素を使用して互いに連結する工程;および
e)連結副生成物を、連結反応が起こっているかどうかを明らかにするために検出する工程;
ただし、工程a)〜工程e)は、同時に、または連続して、または任意の組合せの連続する工程で行われる。
【請求項3】
以下の工程を含む、核酸サンプルにおけるヌクレオチド反復の数を分析するための方法:
a)ヌクレオチド反復を含む可能性がある核酸サンプルを提供する工程;
b)前記ヌクレオチド反復に対して相補的なオリゴヌクレオチドを提供する工程;
c)前記オリゴヌクレオチドを前記核酸サンプルにアニーリングさせる工程;
d)前記核酸サンプルにアニーリングした前記オリゴヌクレオチドを、リガーゼ酵素を使用して互いに連結する工程;
e)連結副生成物をATPに変換する工程;および
f)連結反応が起こっているかどうかを明らかにするために、前記ATPを検出する工程;
ただし、工程a)〜工程f)は、同時に、または連続して、または任意の組合せの連続する工程で行われる。
【請求項4】
以下の工程を含む、核酸サンプルにおけるヌクレオチド反復の数を分析するための方法:
a)ヌクレオチド反復を含む可能性がある核酸サンプルを提供する工程;
b)前記ヌクレオチド反復に対して相補的なオリゴヌクレオチドを提供する工程;
c)前記オリゴヌクレオチドを前記核酸サンプルにアニーリングさせる工程;
d)前記核酸サンプルにアニーリングした前記オリゴヌクレオチドを、リガーゼ酵素を使用して互いに連結する工程;
e)連結副生成物をATPに変換する工程;および
f)連結反応が起こっているかどうかの指標として、ルシフェラーゼに基づくアッセイによって前記ATPを検出する工程;
ただし、工程a)〜工程f)は、同時に、または連続して、または任意の組合せの連続する工程で行われる。
【請求項5】
以下の工程を含む、核酸サンプルの微生物型決定のための方法:
a)微生物型決定のための少なくとも1つのマーカーを含む核酸サンプルを提供する工程;
b)前記核酸サンプルの微生物型決定のためのマーカーを含む領域に対して完全または部分的に相補的なオリゴヌクレオチドを提供する工程;
c)前記オリゴヌクレオチドを前記核酸サンプルにアニーリングさせる工程;
d)前記核酸サンプルにアニーリングした前記オリゴヌクレオチドを、リガーゼ酵素を使用して互いに連結する工程;および
e)連結反応が起こっているかどうかを明らかにするために、連結副生成物を検出する工程;
f)微生物型を決定するために、サンプルの連結パターンを参照パターンと比較する工程;
ただし、工程a)〜工程e)は同時に、または連続して、または任意の組合せの連続する工程で行われる。
【請求項6】
工程b)におけるオリゴヌクレオチドの一つが、反復配列のすぐ外側でアニーリングするように適合されている請求項1〜5のいずれか一項に記載の方法。
【請求項7】
連結副生成物がAMPである請求項1〜6のいずれか一項に記載の方法。
【請求項8】
工程d)がNAD+依存性DNAリガーゼを使用して行われる請求項1〜7のいずれか一項に記載の方法。
【請求項9】
工程e)がピルビン酸リン酸ジキナーゼを使用して行われる請求項1〜8のいずれか一項に記載の方法。
【請求項10】
工程d)が、ATP依存性リガーゼを用いて行われ、アピラーゼが、過剰量のDNAリガーゼ基質を減少させるために、連結前に、または連結期間中に、または連結後に工程d)の連結混合物に加えられる請求項1〜6のいずれか一項に記載の方法。
【請求項11】
ATP依存性リガーゼはT4DNAリガーゼである請求項10に記載の方法。
【請求項12】
dATPが、工程d)においてATP依存性リガーゼに対する基質として使用される請求項10または11に記載の方法。
【請求項13】
連結副生成物がPPiである請求項1〜6または10〜12のいずれか一項に記載の方法。
【請求項14】
工程e)がATP−スルフリラーゼを使用して行われる請求項1〜6または10〜13のいずれか一項に記載の方法。
【請求項15】
使用されたオリゴヌクレオチドが反復そのものの単量体、二量体または多量体である請求項1〜14のいずれか一項に記載の方法。
【請求項16】
オリゴヌクレオチドが前記ヌクレオチド反復に相補的であるが、前記ヌクレオチド反復とはずれている請求項1〜14のいずれか一項に記載の方法。
【請求項17】
アニーリングしていないオリゴヌクレオチドが検出の後でエキソヌクレアーゼの使用によって除かれる工程をさらに含む請求項16に記載の方法。
【請求項18】
アニーリングしていないオリゴヌクレオチドが検出の後でホスファターゼの使用によって不活性化される工程をさらに含む請求項16に記載の方法。
【請求項19】
核酸サンプルが担体に固定化される請求項1〜18のいずれか一項に記載の方法。
【請求項20】
アニーリングしていないオリゴヌクレオチドが検出の後で洗浄によって除かれる工程をさらに含む請求項19に記載の方法。
【請求項21】
核酸サンプルが増幅される工程によって先行される請求項1〜20のいずれか一項に記載の方法。
【請求項22】
ルシフェラーゼに基づくアッセイが光度測定法アッセイである請求項1〜21のいずれか一項に記載の方法。
【請求項23】
ルシフェラーゼ反応で産生された光が、産生された光の初期レベルが達成された後、酵素的に停止される請求項1〜22のいずれか一項に記載の方法。
【請求項24】
光産生が、アピラーゼの添加によって停止される請求項23に記載の方法。
【請求項25】
分析されるべき領域の外側に相補的なオリゴヌクレオチドが、分析されるべき領域から得られるシグナルを正規化するために使用されることができるシグナルを、連結またはプライマー伸長によって生成するために使用される請求項1〜24のいずれか一項に記載の方法。
【請求項26】
別個のバイアルにおいて、リガーゼ酵素と、連結副生成物をATPに変換するための酵素とを含む、請求項1〜25のいずれか一項に記載の方法を行うためのキット。
【請求項27】
別個のバイアルにおいて、ルシフェラーゼ酵素をさらに含む請求項26に記載のキット。
【請求項28】
別個のバイアルにおいて、アピラーゼをさらに含む請求項26または27に記載のキット。
【請求項29】
以下の疾患群から選択される疾患に関連したヌクレオチド反復に対して相補的なオリゴヌクレオチド(必要な場合にはAdoPP5’修飾を有する)をさらに含む請求項26〜28のいずれか一項に記載のキット:歯状核赤核淡蒼球ルイ体萎縮症(DRPLA)、脆弱X症候群、脆弱部位FRAXE、ハンチントン病、ケネディ病、マシャド・ジョセフ病、筋緊張性ジストロフィー、フリードライヒ運動失調症、1型脊髄小脳運動失調、2型脊髄小脳運動失調、3型脊髄小脳運動失調、6型脊髄小脳運動失調、8型脊髄小脳運動失調および12型脊髄小脳運動失調。
【請求項30】
以下の群に由来する、微生物種の同定のための情報を提供する遺伝子領域に対して相補的なオリゴヌクレオチド(必要な場合にはAdoPP5’修飾を有する)をさらに含む請求項26〜28のいずれか一項に記載のキット:16SrRNA遺伝子、23SrRNA遺伝子、groEL、gyrB、rpoB、rnpBおよびgroEL、ミクロサテライト配列およびミニサテライト配列、VNTR、核リボソームDNA(rDNA)アレイ、すなわち、スモールサブユニット(SSU)(18S様)、ラージサブユニット(LSU)(23S、26Sまたは28S様)、5.8SrRNA遺伝子、ならびに内部転写リボソームDNA(rDNA)スペーサー(ITS1およびITS2)。
【請求項31】
リガーゼ酵素と、連結副生成物をATPに変換するための酵素とを含む組成物。
【請求項32】
ルシフェラーゼ酵素をさらに含む請求項31に記載の組成物。
【請求項33】
以下の疾患群から選択される疾患に関連したヌクレオチド反復に対して相補的なオリゴヌクレオチド(必要な場合にはAdoPP5’修飾を有する)をさらに含む請求項31または32に記載の組成物:歯状核赤核淡蒼球ルイ体萎縮症(DRPLA)、脆弱X症候群、脆弱部位FRAXE、ハンチントン病、ケネディ病、マシャド・ジョセフ病、筋緊張性ジストロフィー、フリードライヒ運動失調症、1型脊髄小脳運動失調、2型脊髄小脳運動失調、3型脊髄小脳運動失調、6型脊髄小脳運動失調、8型脊髄小脳運動失調および12型脊髄小脳運動失調。
【請求項34】
以下の群に由来する、微生物種の同定のための情報を提供する遺伝子領域に対して相補的なオリゴヌクレオチド(必要な場合にはAdoPP5’修飾を有する)をさらに含む請求項31または32に記載の組成物:16SrRNA遺伝子、23SrRNA遺伝子、groEL、gyrB、rpoB、rnpBおよびgroEL、ミクロサテライト配列およびミニサテライト配列、VNTR、核リボソームDNA(rDNA)アレイ、すなわち、スモールサブユニット(SSU)(18S様)、ラージサブユニット(LSU)(23S、26Sまたは28S様)、5.8SrRNA遺伝子、ならびに内部転写リボソームDNA(rDNA)スペーサー(ITS1およびITS2)。
【図1】

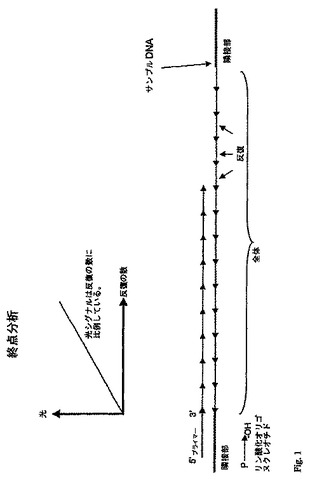
【図2】

【図3】
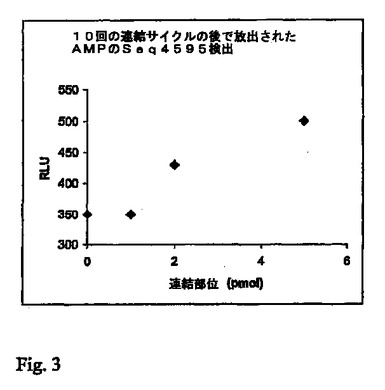
【図4】
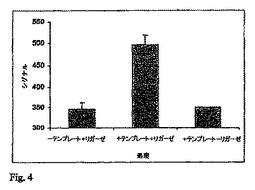
【図5】
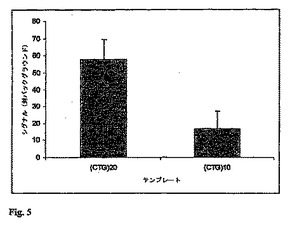
【図6】
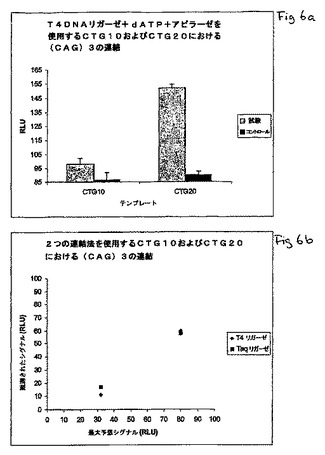
【図7】

【図8】
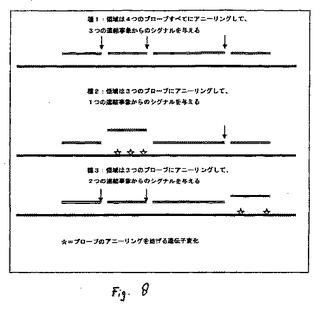
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【公表番号】特表2009−513105(P2009−513105A)
【公表日】平成21年4月2日(2009.4.2)
【国際特許分類】
【出願番号】特願2006−518071(P2006−518071)
【出願日】平成16年6月30日(2004.6.30)
【国際出願番号】PCT/EP2004/007090
【国際公開番号】WO2005/003386
【国際公開日】平成17年1月13日(2005.1.13)
【出願人】(504371561)バイオタージ アクチボラゲット (1)
【Fターム(参考)】
【公表日】平成21年4月2日(2009.4.2)
【国際特許分類】
【出願日】平成16年6月30日(2004.6.30)
【国際出願番号】PCT/EP2004/007090
【国際公開番号】WO2005/003386
【国際公開日】平成17年1月13日(2005.1.13)
【出願人】(504371561)バイオタージ アクチボラゲット (1)
【Fターム(参考)】
[ Back to top ]