
断片化耐性IgG1Fc−コンジュゲート
本発明は、フリーラジカル媒介断片化および凝集に対して耐性の、ヒトIgG1およびIgG3 Fc−コンジュゲートに関する組成物および方法を提供する。本発明はまた、本発明のFc−コンジュゲートを作成するための組成物および方法を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
関連出願の相互参照
[0001] 本出願は、2009年4月21日に出願された35U.S.C. 119(e)の米国特許出願番号61/171,393の利益を主張し、当該出願は参照により本明細書に援用される。
【0002】
技術分野
[0002] 本発明は、活性酸素種(reactive oxygen species)による断片化に対して耐性である、療法的および診断的適用に用いるための免疫グロブリンに関する。
【背景技術】
【0003】
本発明の背景
[0003] ヒト免疫グロブリン(IgG)分子は軽鎖(LC)および重鎖(HC)の2つの同一のコピーからなる。LCおよびHCの間の鎖間ジスルフィド結合はそれらを結びつけ、半抗体を形成する;半抗体の二つの同一のコピーのHCは、いわゆるヒンジ配列においてジスルフィド結合で結びつけられて、天然の抗体を形成する。ヒトIgG1ヒンジ配列は、2つの別々のジスルフィド結合を形成できる、システイン(Cys)残基の2つの組を含む。しかしながら、単一のヒンジジスルフィドのみが、補体に仲介される溶解ならびに抗体依存性細胞介在性細胞障害およびファゴサイトーシスのために必要であることが示唆されてきた。Michaelsen, T. E. ら、 Proc. Natl. Acad. Sci. USA 91:9243-9247, 1994。IgG1 b12の結晶構造において、単一の重鎖間ジスルフィド結合のみが観察されてきた −著者らは壊れたジスルフィド結合は、動力学的であるかまたはシンクロトロン照射損傷の結果でありうると示唆した。Stanfield, R. ら、Science 248: 712-719, 1990; Saphire, E.ら、J. Mol. Biol. 319:9-18, 2002; Weik, M.ら、Proc. Natl. Acad. Sci. USA 97:623-628, 2000。事実、結晶構造における溶媒に露出した一つのシステイン対についての酸化型および還元型コンフォメーションの両方が記述されてきている。Burling, F. T.ら、Science 271:72-77, 1996。IgG1において、LCのC末端のCys残基は、最初にHCのヒンジのCys残基へと連結する;しかしながら、LCおよびHCは、それらの間の会合定数は〜1010M−1 と推定されたので、ジスルフィド結合なしでもなお強く一緒に会合することができる。Bigelow. C.ら、Biochemistry 13:4602-4609, 1978; Horne, C.ら、J. Biol. Chem., 129:660-664, 1982。総合すれば、これらの知見は、IgG1におけるジスルフィド結合は特定の攻撃に対して脆弱であり、そして関連するシステイン残基は対をつくらないままでいることができることを示唆している。
【0004】
[0004] 活性酸素種(ROS)は、酸化的ストレスの主な原因である。ROS、例えば過酸化水素類およびアルキルヒドロペルオキシド類は、タンパク質の生物学的機能を調節することができる。Poole, L. B.ら、Annu. Rev. Pharmacol. Toxicol. 44:325-347, 2004;Philip, E., Free Rad. Biol. Med. 40: 1889-1899, 2006;Salmeen A.ら、Nature 423: 769-773, 2003;Claiborne A.ら、Adv. Protein Chem. 58: 215-276, 2001;Paget, M. S. B.およびButtner, M. J., Annu. Rev. Genet. 37: 91-121, 2003。H2O2により調節されるタンパク質は、酸化に敏感な特徴的なシステインを有する、なぜならそれらの環境はチオール基(Cys−SH)のチオレートアニオン(Cys−S−)(これはCys−SHよりもスルフェン酸(Cys−SOH)へとより容易に酸化される)へのイオン化を促進するからである。Rhee, S. G.ら、(2000) Sci. STKE 200, pe1;Kim, J. R.ら、Anal. Biochem. 283:214, 2000。スルフェン酸は不安定であり、いずれかの接近可能なチオールと反応してジスルフィドを形成するか、さらなる酸化を経てスルフィン酸(Cys−SO2H)またはスルホン酸(Cys−SO3H)となる。Kice, J. L., Adv. Phys. Org. Chem. 17:65, 1980;Claiborne, A., Biochemistry 38: 15407-15412, 1999。
【0005】
[0005] システインに基づくラジカルは、短範囲水素原子引き抜き反応、または一電子移動反応によって形成されることができる。Giles, N. M.ら、Chemistry & Biology 10: 677-693, 2003;Garrison, W. M., Chem. Rev., 87:381-398, 1987;Bonifacic, M. ら、J. Chem. Soc. Perkin Trans., 2: 675-685, 1975;Elliot, A. J.ら、J. Phys. Chem. 85: 68-75, 1981;Jacob, C.ら、Biol. Chem. 387: 1385-1397, 2006。チイル(RS)、スルフィニル(RSO)およびスルホニル(RSOO)ラジカルは、酸化的ストレスの間、存在することが見出されてきている。Harman, L. S.ら、J. Biol. Chem. 259: 5606-5611, 1984;Giles, G. I.およびJacob, C, Biol. Chem. 383: 375-388, 2002;Witting, P. K.,およびMauk, A. G., J. Biol. Chem. 276: 16540-16547, 2001;Stadtman. E. R.およびLevine, R. L., Amino Acids. 25: 207-218, 2003;Berlett, B. S.およびStadtman, E. R., J. Biol. Chem. 272: 20313- 20316, 1997。Cysラジカルおよび他の残基の間の電子移動は、ミオグロビンのオリゴメリック産物形成に関与することが決定されてきている(Witting, P. K.およびMauk, A. G., J. Biol. Chem. 276: 16540-16547, 2001)、一方ProおよびHis残基は、BSAおよびコラーゲンの断片化を生じることとなるROS攻撃のターゲットであることが見出された。Garrison, W. M., Chem. Rev. 87: 381-398, 1987;Davies, M. J.およびDean, R. T., 1997, Radical mediated protein oxidation. Oxford University press, pp 50-120;Zhang, N.ら、J. Phys. Chem. 95: 4718-4722, 1991;Zhang, H.ら、J. Biol. Chem. 280: 40684-40698, 2005;Uchida, K.およびKawakishi, S., Biochem. Biophys. Res. Commun. 138: 659-665, 1986;Dean, R. T.ら、Free Radical Res. Commun. 7: 97-103, 1989。しかしながら、Cysに基づくラジカルがペプチド結合の切断に関与するかどうかについては依然として不明である。StamlerおよびHausladen(Stamler, J. S.およびHausladen A., Nat. Struct. Biol. 5: 247-251, 1998)は、一方で重要な生物学的シグナル事象を構成し、そして他方で酸化的ストレスの不可逆的な特徴である、H2O2に仲介される修飾の連続を提唱した。
【0006】
[0006] 多くの異なる生理学的および環境的プロセスが、活性酸素種(ROS)のインビトロおよびインビボでの形成につながる。細胞内のROSのレベルは、その年齢および生理学的状態に依存し、そしてプロテアーゼ、ビタミン類(A、C、およびE)、および酸化還元金属イオンのような因子の関数である。Bigelow, C.ら、Biochemistry 13: 4602-4609, 1978。ミトコンドリアは、細胞におけるROS生成の重大な源である。Salmeen, A.ら、Nature 423: 769-773, 2003。単離されたミトコンドリアにおけるH2O2産生速度は、生理学的条件下での総酸素取り込みの約2%である。Salmeen, A.ら、Nature 423: 769-773, 2003;Claiborne, A.ら、Adv. Protein Chem. 58: 215-276, 2001;Paget, M.およびButtner, M., Annu. Rev. Genet. 37: 91-121, 2003。
【0007】
[0007] ROSは、インビトロならびにインビボでのラジカルにより媒介されるタンパク質の断片化および凝集を招き得る。これらの酸化的修飾は、療法的および診断的製品の製造量を低下させ、ならびにそれらの効率を低下させ得る。抗体は、療法的および診断的タンパク質の特に有用なクラスとして証明されてきた。しかしながら、抗体のFcヒンジ領域は酸化的修飾を受けやすい。ラジカル攻撃に対するこの脆弱性は、抗体候補ならびに一般的なFc−コンジュゲート化化合物の療法的および診断的開発について、Fcヒンジ領域の安定化を優先課題とする。
【先行技術文献】
【非特許文献】
【0008】
【非特許文献1】Michaelsen, T. E. ら、 Proc. Natl. Acad. Sci. USA 91:9243-9247, 1994
【非特許文献2】Stanfield, R. ら、Science 248: 712-719, 1990
【非特許文献3】Saphire, E.ら、J. Mol. Biol. 319:9-18, 2002
【非特許文献4】Weik, M.ら、Proc. Natl. Acad. Sci. USA 97:623-628, 2000
【非特許文献5】Burling, F. T.ら、Science 271:72-77, 1996。
【非特許文献6】Bigelow. C.ら、Biochemistry 13:4602-4609, 1978
【非特許文献7】Horne, C.ら、J. Biol. Chem., 129:660-664, 1982
【非特許文献8】Poole, L. B.ら、Annu. Rev. Pharmacol. Toxicol. 44:325-347, 2004
【非特許文献9】Philip, E., Free Rad. Biol. Med. 40: 1889-1899, 2006
【非特許文献10】Salmeen A.ら、Nature 423: 769-773, 2003
【非特許文献11】Claiborne A.ら、Adv. Protein Chem. 58: 215-276, 2001
【非特許文献12】Paget, M. S. B.およびButtner, M. J., Annu. Rev. Genet. 37: 91-121, 2003
【非特許文献13】Rhee, S. G.ら、(2000) Sci. STKE 200, pe1
【非特許文献14】Kim, J. R.ら、Anal. Biochem. 283:214, 2000
【非特許文献15】Kice, J. L., Adv. Phys. Org. Chem. 17:65, 1980
【非特許文献16】Claiborne, A., Biochemistry 38: 15407-15412, 1999
【非特許文献17】Giles, N. M.ら、Chemistry & Biology 10: 677-693, 2003
【非特許文献18】Garrison, W. M., Chem. Rev., 87:381-398, 1987
【非特許文献19】Bonifacic, M. ら、J. Chem. Soc. Perkin Trans., 2: 675-685, 1975
【非特許文献20】Elliot, A. J.ら、J. Phys. Chem. 85: 68-75, 1981
【非特許文献21】Jacob, C.ら、Biol. Chem. 387: 1385-1397, 2006
【非特許文献22】Harman, L. S.ら、J. Biol. Chem. 259: 5606-5611, 1984
【非特許文献23】Giles, G. I.およびJacob, C, Biol. Chem. 383: 375-388, 2002
【非特許文献24】Witting, P. K.,およびMauk, A. G., J. Biol. Chem. 276: 16540-16547, 2001
【非特許文献25】Stadtman. E. R.およびLevine, R. L., Amino Acids. 25: 207-218, 2003
【非特許文献26】Berlett, B. S.およびStadtman, E. R., J. Biol. Chem. 272: 20313- 20316, 1997
【非特許文献27】Davies, M. J.およびDean, R. T., 1997, Radical mediated protein oxidation. Oxford University press, pp 50-120
【非特許文献28】Zhang, N.ら、J. Phys. Chem. 95: 4718-4722, 1991
【非特許文献29】Zhang, H.ら、J. Biol. Chem. 280: 40684-40698, 2005
【非特許文献30】Uchida, K.およびKawakishi, S., Biochem. Biophys. Res. Commun. 138: 659-665, 1986
【非特許文献31】Dean, R. T.ら、Free Radical Res. Commun. 7: 97-103, 1989
【非特許文献32】Stamler, J. S.およびHausladen A., Nat. Struct. Biol. 5: 247-251, 1998
【発明の開示】
【課題を解決するための手段】
【0009】
[0008] 本発明は、ラジカル媒介断片化に耐性の、IgG1またはIgG3クラスのヒンジ配列を含むイムノグロブリンFcを提供する。断片化耐性は、そうでなければ2つの半抗体を生じるジスルフィド結合切断の減少により、ならびにこれらの半抗体のそれぞれを作り上げているポリペプチド内の断片化事象の減少により、現れる。一態様において本発明は、FcがヒトIgG1またはIgG3 Fcである、Fc−コンジュゲートである。当該IgG1およびIgG3 Fcは、アミノ酸一文字表記でTHTCPXCPであり、ここでXはRまたはP残基を示す、ヒンジコア配列を含む。本発明において、天然型IgG1またはIgG3 Fcのヒンジコア配列におけるH(ヒスチジン)残基は、Ser(セリン)、Gln(グルタミン)、Asn(アスパラギン)、またはThr(スレオニン)残基で置換される。いくつかの態様において、Fc−コンジュゲートは、薬学的に許容可能な担体中にある。
【0010】
[0009] 本発明はまた、本発明のFcまたはFc−コンジュゲートをコードするポリヌクレオチドを含む単離された核酸、ならびに当該単離された核酸を含む発現ベクター、および前述の発現ベクターを含む宿主細胞に関する。よって、本発明はまた、本発明の核酸を含む発現ベクターを適切な宿主細胞中で、当該核酸の発現に適した条件下で培養し、そして当該宿主細胞から発現したFcまたはFc−コンジュゲートを単離するすることを必然的に含み得る、本発明のFcまたはFc−コンジュゲートを作成する組成物および方法をも含む。
【図面の簡単な説明】
【0011】
【図1】[0010] 図1は、H2O2と実施例において詳述した追加的な試薬の組み合わせによって生じる、IgG1抗体のラジカル媒介断片化の程度を示す。
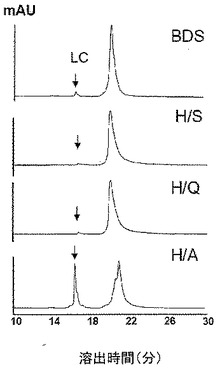
【図2】[0011] 図2は、実施例において詳述した種々のIgG1ヒンジ配列置換変異体の鎖間ジスルフィド結合切断について、ミリ吸光度単位(mAU)で測定したラジカル媒介断片化の程度を示す。
【発明を実施するための態様】
【0012】
発明の詳細な説明
[0012] 本発明は、天然型IgG1またはIgG3 Fcよりもラジカル媒介断片化に対して耐性であるように修飾された、ヒトIgG1およびIgG3 FcおよびFc−コンジュゲートに関する組成物および方法を提供する。断片化耐性IgG1およびIgG3 Fcは、例えば、インビトロまたはインビボ断片化または凝集に対してより高い耐性を有する療法的および診断的使用のための抗体の製造において、使用することができる。本発明の組成物は、以下を含む:Fc−コンジュゲート、本発明のFcまたはFc−コンジュゲートをコードする核酸を含むポリヌクレオチド、これらの核酸を含むベクター、これらのベクターを含む宿主細胞およびこれらのベクターを発現する宿主細胞、および医薬組成物。これらの組成物のそれぞれを製造するおよび使用する方法もまた、提供する。
【0013】
[0013] 単位、接頭辞、および記号はSIで認められた形式で表示されうる。特に言及しなければ、核酸は5’から3’方向へ、左から右へと記載される;アミノ酸配列は、アミノからカルボキシ方向へ、左から右へと記載される。本明細書において記載される数値範囲は、当該範囲を規定する数字を含むものであり、そして定められた範囲内のそれぞれの整数を含むおよび支持する。アミノ酸は本明細書において、IUPAC−IUBMB命名法委員会が推奨する、一般的に知られたそれらの三文字記号または一文字記号のいずれかによって言及されてもよい。同様に、核酸も一般的に認められているそれらの一文字コードによって言及されてもよい。特に言及しない限り、「a」または「an」の語は、「少なくとも1つの」を意味するものとして解釈される。本明細書で用いるセクション見出しは、整理の目的のためにのみ用いられるものであって、記載された主題を限定するものとして解釈されてはならない。
【0014】
A.定義
[0014] 本明細書おいて使用される用語「抗体」は、免疫化によって、組換え技術を介して、インビトロ合成手段の手法によって、またはその他の方法によって、完全にまたは部分的に、製造される抗体であるかどうかにかかわらず、ヒト(例えばCDRグラフトされた)、ヒト化、キメラ、多重特異性、モノクローナル、ポリクローナル、およびそのオリゴマーを含む、グリコシル化されたおよびグリコシル化されていない、いずれかのアイソタイプまたはサブクラスのイムノグロブリンの双方への言及を含む。よって、用語「抗体」は、例えば(a)ヒトイムノグロブリン遺伝子について遺伝子導入した動物(例えばマウス)またはそれから調製したハイブリドーマから単離した抗体、(b)当該抗体を発現するようにトランスフェクトされた宿主細胞から(例えば、トランスフェクトーマから)単離された抗体、(c)組換え、コンビナトリアル抗体ライブラリーから単離された抗体、および(d)イムノグロブリン遺伝子配列の他のDNA配列へのスプライシングを伴う他のいずれかの手段により調製され、発現され、創造され、または単離された抗体、のような、組換え手段により調製され、発現され、創造され、または単離されたものを含む。そのような抗体は、動物の2つの別個の種の生殖系列イムノグロブリン配列に由来する可変および定常領域を有する。しかしながら、特定の態様において、そのような抗体はインビトロ突然変異誘発(または、ヒトイムノグロブリン遺伝子について遺伝子導入した動物を用いる場合、インビボ体細胞突然変異誘発)を受けうるので、該抗体のVHおよびVL領域のアミノ酸配列は、特定の種(例えばヒト)の生殖系列VHおよびVL配に由来しおよび関連するが、当該種のインビボ抗体生殖系列レパートリーには天然に存在しない可能性がある配列である。
【0015】
[0015] 本明細書において、「コンジュゲート」は、Fcへとコンジュゲート化した場合に診断的または療法的機能を発揮する、いずれかの化学的または生物学的部分を意味する。コンジュゲートは、直接的にまたは間接的に(すなわち、化学的スペーサーを通して)共有結合的に付着されることができる。例示的なコンジュゲートは、以下を含む:細胞傷害性薬物または細胞分裂阻害剤(例えば、抗腫瘍剤または抗血管新生剤)、ポリエチレングリコール、脂質、および受容体または細胞表面受容体の細胞外ドメインのような受容体フラグメント。
【0016】
[0016] 「宿主細胞」は、核酸、例えば本願発明の核酸、を発現するために用いることができる細胞である。宿主細胞は、原核生物、例えば大腸菌(E. coli)であることもでき、あるいは真核生物、例えば単細胞真核生物(例えば酵母もしくは他の真菌類)、植物細胞(例えば、タバコもしくはトマト植物細胞)、動物細胞(例えばヒト細胞、サル細胞、ハムスター細胞、ラット細胞、マウス細胞、もしくは昆虫細胞)、またはハイブリドーマであることができる。宿主細胞の例は、サル腎臓細胞のCOS−7系列(ATCC CRL 1651)(Gluzmanら、Cell 23:175, 1981を参照)、L細胞、C127細胞、3T3細胞(ATCC CCL 163)、チャイニーズハムスター卵巣(CHO)細胞またはそれらの派生物であるVeggie CHOおよび無血清培地中で増殖する関連する細胞系列(Rasmussenら、Cytotechnology 28: 31, 1998)、または、DHFR欠損であるCHO DX−B11株(Urlaubら、Proc. Natl. Acad. Sci. USA 77: 4216-4220, 1980)、を含む。
【0017】
[0017] 典型的には、宿主細胞は、ポリペプチドをコードする核酸でトランスフェクトされることができる培養細胞である、そして当該核酸は当該宿主細胞中で発現されることができる。「組換え宿主細胞」の句は、発現されるヌクレオチドでトランスフェクトされている宿主細胞を示すものとして用いることがでいる。典型的には、宿主細胞は核酸を含むが、制御配列が該核酸と機能可能に連結されるように該宿主細胞に制御配列が導入されない限り、該核酸を評価できるレベルで発現しない。宿主細胞という用語は、特定の対象細胞のみに言及するものではなく、そのような細胞の後代または潜在的な後代にも言及する。突然変異または環境の影響のいずれかのために続く世代においてある一定の修飾が生じ得るので、そのような後代は、実際に、親細胞と同一ではない可能性があるが、それでもなお本明細書において用いられる用語の範囲内に含められる。
【0018】
[0018] 用語「ヒト抗体」は、定常領域およびフレームワークが完全にまたは実質的にヒト配列からなり、そのためヒト抗体はヒト宿主に投与したときにそれ自体に対する免疫原性反応、好ましくは検出可能な免疫原性反応を実質的に誘発しない、抗体を意味する。
【0019】
[0019] 用語「ヒト化抗体」は、定常領域のすべてが、ヒトイムノグロブリンに由来するか対応する一方、1またはそれより多くの可変領域のすべてまたは一部が他の種、例えばマウスに由来する、抗体を意味する。
【0020】
[0020] 本明細書において、核酸について「単離された」というときは、ヒトの直接的介入の結果物としてのDNAまたはRNAを意味する:1)天然には見出されないゲノムの座に組み込まれたもの、2)天然では機能可能に連結しない核酸と機能可能に連結したもの、3)天然の状態では混合された状態にある細胞構成物から実質的に精製されたもの(例えば、少なくとも70%、80%、または90%)。
【0021】
[0021] 本明細書において、FcまたはFc−コンジュゲートについて「単離された」というときは以下を意味する:(1)存在する優勢な種である発現された状態では混合された状態にある細胞構成物から実質的に精製されたもの(例えば、少なくとも60%、70%、80%、または90%)、(2)天然では連結されていないポリペプチドまたはその他の部分へとコンジュゲート化されたもの、(3)より大きなポリペプチド配列の一部として天然には生じないもの、(4)詳細に明らかにされた組成物において異なる特異性を有する他の化学的または生物学的な剤と組み合わされたもの、または(5)天然に見出されない限りにおいてヒトの操作された配列を含むもの。
【0022】
[0022] 本明細書において用いられる用語「モノクローナル抗体」または「モノクローナル抗体組成物」は、抗体分子の単一分子組成物の調製物を意味し、典型的には同一の核酸分子によってコードされる。モノクローナル抗体組成物は、特定のエピトープについて単一結合特異性および親和性を示す。ある態様において、モノクローナル抗体は、単一のハイブリドーマまたは他の細胞株(例えば、トランスフェクトーマ)、あるいはトランスジェニック哺乳動物によって、産生される。用語「モノクローナル」は、抗体の製造について特定の方法に限定するものではない。
【0023】
[0023] 本明細書において用いられる「核酸」および「ポリヌクレオチド」は、デオキシリボヌクレオチドもしくはボヌクレオチドポリマー、またはそのキメラ、への言及を含み、そして他に限定しない限り、言及した配列の相補鎖を包含する。
【0024】
[0024] 制御配列が核酸配列の発現(例えば、発現のレベル、タイミング、または位置)に影響を及ぼす場合、該核酸配列は制御配列に「機能可能に連結している」。「制御配列」は、第二の核酸の発現(例えば発現のレベル、タイミング、または位置)に影響を及ぼす核酸である。よって、制御配列と第二の配列の間の機能的連結が、該第二の配列に対応するDNA配列の転写を該制御配列開始または仲介するようなものである場合、制御配列および第二の配列は機能可能に連結している。制御配列の例は、プロモーター、エンハンサー及び他の発現制御因子(例えばポリアデニル化シグナル)を含む。制御配列のさらなる例は、例えば、Goeddel, 1990, Gene Expression Technology: Methods in Enzymology, 185, Academic Press, San Diego, CA およびBaronら、Nucleic Acids Res. 23: 3605-3606, 1995に記載されている。
【0025】
[0025] 用語「ペプチド」、「ポリペプチド」および「タンパク質」は、本明細書を通して互換的に用いられ、そしてペプチド結合によりお互いに結合した2以上のアミノ酸残基を含む分子に言及するものである。用語「ポリペプチド」、「ペプチド」および「タンパク質」はまた、グリコシル化、脂質付加、硫酸化、グルタミン酸残基のガンマ−カルボキシル化、ヒドロキシル化、およびADP−リボシル化、を含むがこれらに限定されない修飾を含めたものである。
【0026】
[0026] 用語「ポリヌクレオチド」、「オリゴヌクレオチド」および「核酸」は、本明細書を通して互換的に用いられ、そしてDNA分子(例えばcDNAまたはゲノムDNA)、RNA分子(例えばmRNA)およびそれらのハイブリッドを含む。核酸分子は一本鎖または二本鎖であることができる。
【0027】
[0027] 本明細書において使用される「特異的な結合をする」または「特異的な結合」または「特異的に結合する」は、異種起源のタンパク質および他の生体物質の集団の存在下での、標的(例えば、タンパク質)の存在について決定的な結合反応に言及するものである。よって、指定されたイムノアッセイ条件下で、抗体もしくはペプチボディのような特定されたFc−コンジュゲートまたは他の結合ポリペプチドは、特定のタンパク質に結合し、そして試料中に存在する他のタンパク質へは統計学的に有意な量で結合しない。典型的には、Fc−コンジュゲート(例えば、抗体、ペプチボディ)は、スクリーニング方法(例えば、ファージディスプレイ)によって、またはタンパク質またはそのエピトープを用いる免疫化によって、タンパク質へ特異的に結合する能力について選択される。特異的結合を決定するために用いることができるイムノアッセイの形式についての記載は、HarlowおよびLane (1998), Antibodies, A Laboratory Manual, Cold Spring Harbor Publications, New Yorkを参照のこと。例えば、固相ELISAイムノアッセイは特異的結合を決定するのに用いることができる。特異的結合は、少なくとも約1×107M−1、およびしばしば少なくとも1×108M−1、1×109M−1、1×1010M−1、の結合定数を生じる。
【0028】
[0028] 本明細書において、「ベクター」は、本発明のポリヌクレオチドの宿主細胞への導入に用いる核酸への言及を含む。ベクターはしばしばレプリコンである。発現ベクターは、適切な宿主細胞中で、または適切なインビトロ条件下で存在するときに、その中に挿入された核酸の転写を許容する。
【0029】
B.Fc−コンジュゲート
[0029] 本発明は、天然のIgG1またはIgG3 Fcと比較して断片化および/または凝集に対して耐性である、単離されたIgG1およびIgG3 FcおよびFc−コンジュゲート、ならびにこれらの組成物を製造し使用する方法を提供する。理論に拘束されるわけではないが、フリーラジカル仲介断片化の機構は、Fcの断片化において、IgG1イムノグロブリンのヒンジコア配列に存在するヒスチジン残基に関係する。IgG1およびIgG3Fc中のヒンジコア配列ヒスチジン残基の適切な置換または欠失は、修飾されていないFcまたはFc−コンジュゲートに対して、ラジカル仲介断片化および/または凝集の度合いを減少させることができる。
【0030】
[0030] 本発明は活性酸素種による断片化および/または凝集に対して耐性にする修飾を有する、単離されたFcおよびFc−コンジュゲートを提供する。哺乳類イムノグロブリンのFc(結晶化可能フラグメント)は、「ヒンジコア配列」を有するヒンジ領域を含む、よく特徴付けされた構造である。表1は、ヒトIgGサブタイプにおいて見出され、一文字アミノ酸コードで表示された、ヒンジコア配列のリストを示す。Edelmanら(Proc. Natl. Acad. Sci. USA 63:78-85, 1969)の番号付けシステムにおいて、IgG1のヒンジコア配列はIgG1重鎖の残基216〜230に対応する一方、IgG3のヒンジコア配列はIgG3重鎖の残基214〜230に対応する。本発明において、表1に示されるように、IgG1またはIgG3ヒンジコア配列中に(224残基において)存在するヒスチジン残基(「H」)は、水素結合を形成することができる極性アミノ酸残基に置換されている。IgG1およびIgG3のヒンジコア配列中のヒスチジン残基について置換可能なアミノ酸残基の特定の例は、Ser、Gln、Asn、またはThr残基である。あるいは、当該ヒスチジン残基はヒンジコア配列から欠失されていてもよい。
【0031】
[0031]
【0032】
【表1】

【0033】
[0032] 典型的には、ラジカル媒介断片化に耐性のFcを生じる置換または欠失に供された本発明のFcまたはFc−コンジュゲートは、ヒトのIgG1またはIgG3 Fcであろう。しかしながら、IgGサブタイプの特徴を保持しながら、ヒトIgG1またはIgG3 Fcへの限定された数の置換、付加または欠失を行うことができる。よって、例えば、IgG1またはIgG3 Fcの0、1、2、3、4、5、6、7、8、9、または10アミノ酸を修飾することができ、そしてそれらもなお本願発明の範囲内である。よって、修飾されたIgG1またはIgG3 Fcは、天然型のヒトIgG1またはIgG3 Fcに対して95%、96%、97%、98%、または99%同一であろう。いくつかの態様において、(表1に示したような)本発明のIgG1またはIgG3ヒンジコア配列への唯一の修飾は、上述のようにヒンジコア配列中のヒスチジン残基の置換である。Fc−コンジュゲートは一価または二価構造であることができる。二価Fc−コンジュゲートのそれぞれのコンジュゲートは、同一または異なるコンジュゲートであることができる。
【0034】
[0033] Fcに共有結合または非共有結合で結合してFc−コンジュゲートを形成するコンジュゲートは、化学療法剤のような薬剤、放射標識のような診断標識、ヒト細胞表面受容体の細胞外ドメインのようなタンパク質を含むまたはそれらからなることができる。いくつかの態様において、当該コンジュゲートは、Fc−コンジュゲートがIgG1またはIgG3抗体であるように、Fab抗体セグメントを含むまたはそれからなる。抗体はポリクローナルまたはモノクローナルであることができる。いくつかの態様において、Fc−コンジュゲートは、完全なヒトモノクローナル、または、そうでなければ完全なヒトIgG1またはIgG3上に非ヒト供給源(例えば、ネズミ)由来のCDR(相補性決定領域)がグラフトされた、ヒト化モノクローナルである。抗体はアゴニストまたはアンタゴニスト抗体であることができ、それにより該抗体は受容体活性化を活性化または阻害する。いくつかの態様において、受容体はヒト細胞表面受容体であり、ここでFc−コンジュゲートは当該細胞表面受容体の細胞外ドメインに特異的に結合する。他の態様において、Fc−コンジュゲートはヒト細胞表面受容体のリガンドに特異的に結合して、該リガンドが該受容体に結合するのを妨げる。Fc−コンジュゲートが結合できるヒト細胞表面受容体の例は、デスレセプター4(TRAIL受容体−1)、デスレセプター5(TRAIL受容体−2)、VEGF(血管内皮増殖因子)、TNFR(腫瘍壊死因子受容体)、RANK(核内因子カッパb活性化受容体)受容体、またはTie−1及びTie−2受容体を含む。他の態様において、Fc−コンジュゲートのコンジュゲートは、望ましい標的に特異的に結合するペプチド(「ペプチボディ」)である。ペプチボディは、公開番号WO 2000/24782(引用により本明細書に援用する)を有する国際出願において教示されている。
【0035】
C.核酸
[0034] 本発明はまた、本発明のFcまたはFc−コンジュゲートをコードする核酸を含む単離されたポリヌクレオチドに向けられる。都合の良いことには、Fc−コンジュゲートのコンジュゲートがタンパク質であるとき(Fc−タンパク質コンジュゲート)であり、そして、例えば抗体、ペプチボディ、またはFc−細胞表面受容体融合体(またはその断片)をコードする場合、本発明の核酸はFc−タンパク質コンジュゲートを全体としてコードすることができる。
【0036】
[0035] 本発明のFcおよびFc−タンパク質コンジュゲートを製造するための組換え方法は、一般的に本発明のIgG1またはIgG3 Fcをコードする単離された核酸を含むポリヌクレオチドを使用する。本発明のFc−タンパク質コンジュゲートをコードする核酸は、当該技術分野において公知のインビトロオリゴヌクレオチド合成の方法により直接的に合成することができる。あるいは、より小さいフラグメントを合成し、当該技術分野で公知の組換え法を用いてより大きなフラグメントを形成するように連結することができる。いくつかの態様において、望ましいヒンジコア配列置換または欠失を伴う核酸プライマーをPCRに基づくインビトロ突然変異誘発に用いて、本発明のFcまたはFc−コンジュゲートを創出する。本発明のポリヌクレオチドはまた、インビトロ合成手段(例えば、固相ホスホロアミダイト合成)、またはその組み合わせ、を通じて構築することもできる。そのような方法は、当業者に周知である。例えば、Current Protocols in Molecular Biology, Ausubelら編、Greene Publishing and Wiley-Interscience, New York (1995)を参照されたい。
【0037】
D.Fc−コンジュゲートの構築
[0036] 本発明の単離されたFcまたはFc−タンパク質コンジュゲートを発現するために、これらの組成をコードする単離されたDNAを、標準的な分子生物学的技術(例えば、PCR増幅、部位特異的突然変異誘発)によって得ることができ、そしてそれを、当該遺伝子が転写および翻訳制御配列に機能可能に連結されるように、発現ベクターに挿入することができる。
【0038】
[0037] よって、本発明は本発明の核酸を含む発現ベクター(ポリヌクレオチド)を含む。発現ベクターは、プラスミド、レトロウイルス、コスミド、YAC、EBV由来エピソーム等を含む。発現ベクターは、本願発明のFcまたはFc−タンパク質コンジュゲートの宿主細胞からの分泌を促進するシグナルペプチドをコードすることができる。FcまたはFc−タンパク質コンジュゲート遺伝子は、シグナルペプチドがFc/Fc−タンパク質コンジュゲート遺伝子のアミノ末端に対してインフレームで(in-frame)連結されるように、ベクター中へクローニングすることができる。シグナルペプチドは、イムノグロブリンシグナルペプチドまたは異種性のシグナルペプチド(すなわち、イムノグロブリンではないタンパク質由来のシグナルペプチド)であることができる。
【0039】
[0038] 発現ベクターおよび発現制御配列は、使用する発現宿主細胞に適合性であるように選択される。適合性のベクターおよび宿主細胞系は、例えば、抗体であるFc−コンジュゲートの可変重鎖及び可変軽鎖の共発現および会合を許容することができる。発現のために適切な系は、当業者によって決定されることができる。いくつかの態様において、発現ベクターは分断(split)DHFRベクター、PDC323またはPDC324である;McGrew, J. T.およびBianchi, A. A. (2002) “Selection of cells expressing heteromeric proteins”, 米国特許出願第20030082735号;および、Bianchi, A. A.およびMcGrew, J. T., “High-level expression of full antibodies using trans-complementing expression vectors,” Bioengineering and Biotechnology 84(4): 439-444, 2003を参照のこと。Fc−コンジュゲートが抗体である場合、本願発明の可変重鎖核酸及び抗体可変軽鎖核酸は別々のベクター中へと挿入される、あるいは、頻繁に、両遺伝子は同じ発現ベクター中へと挿入されることができる。標準的な方法によって、核酸を発現ベクター中に挿入することができる(例えば、抗体核酸フラグメントおよびベクター上の相補的な制限酵素部位のライゲーション、または制限酵素部位が存在しない場合には平滑末端ライゲーション)。
【0040】
[0039] 本発明の核酸および発現ベクターは、トランスフェクションを介して宿主細胞へと導入することができる。用語「トランスフェクション」の様々な形は、原核生物または真核生物の宿主細胞への外来性DNAの導入のために一般的に用いられる多種多様の技術(例えば、エレクトロポレーション、リン酸カルシウム沈殿法、DEAE−デキストラントランスフェクション、など)を包含することを意図する。本発明のFc−コンジュゲートを原核生物宿主細胞または真核生物宿主細胞のいずれかにおいて発現することが理論的には可能であるが、真核生物細胞(そして最も好ましくは哺乳動物宿主細胞)における抗体の発現、が最も典型的である;なぜなら、そのような真核生物細胞(そして特に哺乳動物細胞において)は、原核生物細胞よりも、適切にフォールディングされ、そして免疫学的に活性な抗へと会合し、それを分泌する可能性が高いからである。
【0041】
[0040] 本願発明の発現ベクターは、宿主細胞において配列の発現を制御する調節配列を保有する。そのような調節配列は、例えばGoeddel; Gene Expression Technology. Methods in Enzymology 185, Academic Press, カリフォルニア州サンディエゴ(1990)に記載されている。調節配列の選択を含む発現ベクターの設計は、形質転換する宿主細胞の選択、望むタンパク質の発現レベル、等の要因に依存してもよいことを当業者は理解するであろう。哺乳動物宿主細胞発現のための望ましい調節配列は、哺乳動物細胞において高レベルのタンパク質発現を指示するウイルスの成分、例えば、サイトメガロウイルス(CMV)、サル・ウイルス40(SV40)、アデノウイルス、(例えばアデノウイルス主要後期プロモーター(AdMLP))およびポリオーマ由来のプロモーターおよび/またはエンハンサー、を含む。あるいは、非ウイルス性調節配列、例えばユビキチンプロモーターまたはベータ・グロブリンプロモーターを用いてもよい。
【0042】
[0041] 本発明の発現ベクターは付加的な配列、例えば宿主細胞中のベクターの複製を調節する配列(例えば、複製起点)および選択可能マーカー遺伝子、を保有してもよい。選択可能マーカー遺伝子は、ベクターが導入された宿主細胞の選択を容易にする(例えば、すべてAxelらによる、米国特許第4,399,216号、第4,364,665号及び第5,179,017号を参照されたい)。例えば、典型的には選択可能マーカー遺伝子は、ベクターが導入された宿主細胞に、G418、ハイグロマイシンまたはメトトレキセートのような薬物に対する耐性を付与する。好ましい選択可能マーカー遺伝子は、(dhfr− 宿主細胞において、メトトレキセート選択/増幅に用いるための)ジヒドロ葉酸還元酵素(DHFR)遺伝子、および(G418選択のための)neo遺伝子を含む。
【0043】
[0042] 本発明のFcまたはFc−コンジュゲートを発現するための好ましい哺乳動物宿主細胞は、チャイニーズハムスター卵巣(CHO細胞)(例えばKaufman, R. J.およびSharp, P. A., Mol. Biol. 159:601-621, 1982に記載されるDHFR選択可能マーカーとともに使用される、UrlaubおよびChasin, Proc. Natl. Acad. Sci. USA 77: 4216-4220, 1980に記載されるdhfr− CHO細胞を含む)、NS/0骨髄腫細胞、COS細胞およびSP2.0細胞を含む。特に、NS/0骨髄腫細胞を伴う使用について、別の好ましい発現系は、WO 87/04462, WO 89/01036およびEP 338841に開示されるGS遺伝子発現系である。本発明の発現ベクターが哺乳動物宿主細胞に導入される場合、FcおよびFc−コンジュゲートは、宿主細胞を適切な培養培地中で、宿主細胞におけるそれらの発現を可能にさせるのに十分な時間で培養することにより、より好ましくは当該宿主細胞が増殖した培養培地へのFcまたはFc−コンジュゲートの分泌により、産生される。
【0044】
[0043] ひとたび発現されたなら、FcまたはFc−コンジュゲートは、HPLC精製、分画カラムクロマトグラフィー、ゲル電気泳動等の当該技術分野の標準的な方法に従って単離のために精製することができる(例えば、Scopes, Protein Purification, Springer-Verlag, NY, 1982を参照のこと)。特定の態様において、ポリペプチドはクロマトグラフィーまたは電気泳動技術を用いて精製される。例示的な精製方法は、非限定的に、硫酸アンモニウムでの沈殿;PEGでの沈殿;免疫沈降;熱変性に続く遠心分離;アフィニティークロマトグラフィー(例えば、プロテインA−セファロース)、イオン交換クロマトグラフィー、排除クロマトグラフィー、および逆相クロマトグラフィーを含むがこれらに限定されないクロマトグラフィー;ゲル濾過;ヒドロキシアパタイト・クロマトグラフィー;等電点電気泳動;ポリアクリルアミドゲル電気泳動;ならびに、それらおよび他の技術の組み合わせ;を含む。特定の態様において、ポリペプチドは高速タンパク質液体クロマトグラフィーまたは高速液体クロマトグラフィー(HPLC)によって精製される。
【0045】
E.医薬組成物
[0044] 本発明は、薬学的に許容されうる担体と共に処方された本発明のFc及びFc−コンジュゲートを含む医薬組成物を提供する。いくつかの態様において、当該医薬組成物はヒト対象における投与に適している。本明細書において「薬学的に許容されうる担体」は、特定の対象に投与したときに生理学的に適合しうる、溶媒、分散媒、コーティング、抗菌薬および抗真菌薬、等張剤および吸収遅延剤、等のいずれか及びすべてを含む。典型的には、医薬組成物は、製造および保管の条件下で無菌であり、そして安定でなければならない。
【0046】
[0045] 本発明の医薬組成物は、組み合わせ療法において投与する、すなわち他の剤との組み合わせることができる。剤は、インビトロで合成的に調製した化学的組成物、抗体、抗原結合領域、放射性核種、ならびに、それらの組み合わせおよびコンジュゲート、を含むがこれらに限定されない。
【0047】
[0046] 用量投与計画は、望ましい最適な応答(例えば、療法的応答)を提供するように調整される。例えば、単一ボーラスを投与してもよく、いくつかの分割された用量が時間をかけて投与されてもよく、または用量は療法的状況の緊急性が示すところにより、比例的に減少または増加させてもよい。投与の容易さおよび用量の均一性のために、用量単位剤形(dosage unit form)で非経口組成物を処方することが特に有利である。本明細書で用いる用量単位剤形は、治療される対象のための単位用量に適した物理的に別個の単位を意味する;それぞれの単位は、薬学的担体を伴って、望ましい療法的効果を生じるよう計算された、予め決定された量の活性化合物を含有する。本発明の用量単位剤形の内訳は、(a)活性化合物の独特の特徴および達成される特定の療法的効果、および(b)患者における感受性の治療のための、そのような活性化合物を混ぜ合わせる技術分野に固有の制限、によって指図され、そしてそれらに直接的に依存する。
【0048】
E.療法的おより診断的コンジュゲート
[0047]療法的および/または診断的利点を向上する、本明細書に記載した種々の療法的部分は、本発明のFcへ直接的または間接的(例えば、「連結基」を介して)に共有結合により連結し、Fc−コンジュゲートを生じることができる。連結基は任意である。リンカーはしばしば、ペプチド結合でともに連結されたアミノ酸で作られる。当業者によく理解されているように、1またはそれより多くのこれらのアミノ酸はグリコシル化されていてもよい。非ペプチドリンカーもまた、可能である。非ペプチドリンカーの例は、PEG(ポリエチレングリコール)リンカーである。
【0049】
[0048] そのような療法的部分を抗体にコンジュゲート化する技術は周知である、例えばArnonら、 “Monoclonal Antibodies For Immunotargeting Of Drugs In Cancer Therapy”, Monoclonal Antibodies And Cancer Therapy中, Reisfeldら(編), pp.243-256 (Alan R. Liss, Inc. 1985); Hellstromら、“Antibodies For Drug Delivery”, Controlled Drug Delivery (2nd Ed.)中, Robinsonら(編), pp. 623-653 (Marcel Dekker, Inc. 1987); Thorpe, “Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review”, Monoclonal Antibodies '84: Biological And Clinical Applications中, Pincheraら(編), pp. 475-506 (1985); “Analysis, Results, And Future Prospective Of The Therapeutic Use Of Radiolabeled Antibody In Cancer Therapy”, Monoclonal Antibodies For Cancer Detection And Therapy中, Baldwinら(編), pp. 303-316 (Academic Press 1985), およびThorpeら、 “The Preparation And Cytotoxic Properties Of Antibody-Toxin Conjugates”, Immunol. Rev. 62: 119-158, 1982、を参照のこと。本発明の組成物は、放射性核種とカップリングしてもよい、例えば、Goldenberg, D. M.ら、Cancer Res. 41 : 4354-4360, 1981、及びEP 0365 997に記載されるような、131I、90Y、105Rh、インジウム−111等。
【実施例】
【0050】
[0049] 実施した実験および達成した結果を含む以下の実施例は、例証の目的のためにのみ提供されるものであり、本発明を限定する物として解釈されるものではない。
実施例1
[0050] 本実施例は、一つのFabドメインの損失および部分的分子の形成につながるH2O2媒介ラジカル切断による、ヒトIgG1抗体の特定のヒンジ断片化の結果を示すものである。IgG1に対するH2O2攻撃は、ヒンジ領域中の位置226に位置する二つのシステイン残基(Cys226)の間の鎖間ジスルフィド結合の破壊、そして続いて、スルフェン酸(Cys226SOH)およびチイルラジカル(Cys226S・)の形成を生じ、これは上部ヒンジ領域への電子移動を開始し、ラジカル媒介ポリペプチド骨格断片化につながる。
【0051】
[0051] 用いた抗体は、IgG1サブクラスの組換え完全ヒト抗体であった。当該分子はCHO細胞において発現され、そして慣習的な技術を用いてクロマトグラフィーにより精製した。抗体フラグメントはサイズ排除クロマトグラフィー(SEC)により分離した。抗体の切断は、部分的分子の割合(C1およびC2)によって測定した。
【0052】
[0052] 簡略的に、緩衝液中の2mgから10mgのIgG1抗体を含有する反応混合物(1.0mL)を、変化させたH2O2濃度を伴ってインキュベートした。H2O2を除去するために、フィルターユニットおける遠心分離によりサンプルの緩衝液交換を行った。精製された部分的分子(〜1mg/mL)を還元し、アルキル化した。アルキル化は暗所において室温で行い、そして0.5M DTTストック溶液を加えてアルキル化を停止した。逆相高速液体クロマトグラフィー(RP−HPLC)、続いてエレクトロスプレーイオン化(ESI)飛行時間(TOF)質量分析(MS)を行った。
【0053】
[0053] 精製されたバルクの抗体を、サイズ排除クロマトグラフィー(SEC)により分析し、そして〜0.9%の部分分子を示した(P1)。これは一つのロットからの単一のケースではなく、0.9〜1.1%の範囲でいくつかのランにおいて示された。P1種は、SEQにより95%より大きい純度にさらに精製され、そしてRP−HPLC−TOF/MSにより分析した。結果は、P1は一つのFabドメインを喪失した重度に酸化された部分抗体であることを示した。
【0054】
[0054] H2O2は、タンパク質に対して酸化および損傷を生じることができるものとして公知である。酸化的ストレスが切断を生じるかどうか探るために、IgG1を処理するためにH2O2を用い、そしてSECによりその影響を測定した。5〜20mM H2O2の範囲を通じて、最初の8時間のインキュベーションでは特筆すべき切断は見出されなかった。20mM H2O2でのインキュベーションの48時間後にのみ、二つの部分フラグメント、−C1およびC2−、が観察された。これらの二つのフラグメントの量は、インキュベーションの長さに対して直接比例して増加した。この断片化は、抗体濃度およびpH条件にも依存する。加えて、切断は、8週間までの間にも、顕著な安定相を伴わずに進行した。二つの産物(C1およびC2)のみが観察されたという事実は、切断は特異的であり、そしておそらく特定の機構により駆動されたものであることを示唆した。P1の重度に酸化された性質を示す続いての研究は、ヒンジ断片化はCHO細胞による抗体産生の間の酸化的ストレスによるものであり得ることを示唆する。P1およびC1の類似性、特に長期のH2O2処理において観察されたより高い酸化レベルは、酸化的ストレスがヒンジ断片化を生じさせたことを示す。
【0055】
[0055] C1およびP1のRP−HPLC−TOF/MS分析は、それらが同じ種であり、それらのそれぞれはFabドメインを失っている重度に酸化された部分分子である、特に、Fcドメインの単一の相補的HCは、上部ヒンジ領域におけるN末端残基Asp221、Lys222、Thr223およびThr225の独特の「ラダー(ladder)」を含む、ことを示した。加えて、45Daおよび71Daの二つの付加物が同じFc断片において観察された、これらは公知の修飾とは一致しないために通常の付加物ではない。
【0056】
[0056] C2断片のRP−HPLC−TOF/MS分析は、これがIgG1のFabドメインであり、そして重度に酸化されていることを明らかにした。C2のLCは、C1について対応するものに対して類似のプロフィールを示した。C2におけるHC(Fd)のFab部分は二つの構成成分を有し、それらの双方は1つまたは3つの酸素付加を伴って重度に酸化されていた。より重度に酸化された構成成分はC末端残基Asp221、Lys222、Thr223、His224およびThr225のラダーを含有した;より軽度に酸化されたFd構成成分は、Ser218からThr225のC末端残基からなる、より幅広いラダーを持っていた。これらの結果は、H2O2処理は、IgG1のLCおよびHCの双方におけるヒンジ切断および顕著なレベルの酸化という結果に帰することを示した。
【0057】
[0057] これらの付加物の性質およびその位置を組み合わせると、データはラジカル切断はヒンジ断片化の原因であることを示唆する。過酸化水素は、ラジカルにより誘導される酸化経路を通してタンパク質の生物学的機能を制御することができる。ヒドロキシルラジカルを伴う反応は、結果的にタンパク質の分解に導く種々の化学反応をもたらすことができる(Garrison, W. M., Chem. Rev. 87: 381-398, 1987; Davies, M. J. および Dean, R. T., 1997, Radical mediated protein oxidation. Oxford University press, pp 50-120; Berlett, B. S. およびStadtman, E. R., J. Biol. Chem. 272: 20313-20316, 1997)。
【0058】
[0058] OHラジカルがヒンジ断片化に関与するかを調べるため、そして当該切断に影響を及ぼしてもよいいくつかの因子を評価するため、いくつかの前処理の後にIgG1をH2O2攻撃に供した。これらは、H2O2処理の前に対を形成していないCys残基をブロキングするためのN−エチル−マレイミド(NEM)前処理、または反応系へとカタラーゼもしくはエチレン−ジアミン−テトラー酢酸(EDTA)を添加することを含む。SECを影響を測定するために実施した。カタラーゼはほぼ完全に切断をブロックしたことが見出され、OHラジカルは切断について重要であることを強く示した。フリーのチオール基の総量は、Ellmanのリガンド、5,5’−ジチオビス(2−ニトロ安息香酸)(DTNB)を用いる4M GdnHClの存在下の変性条件下で、〜0.28モル/モル抗体であると測定された。H2O2処理の前に、IgG1はNEMとともに、pH5.0、37℃で、3時間インキュベートした。NEMでブロックされた試料は、切断において〜7%の現象を示す一方、NEM処理によってフリーのチオール(−SH)基は完全にブロックされた。よって、この結果は、対を形成していないCys残基は切断について重大な意味を持たないことを示唆した。
【0059】
[0059] 5’5−ジメチルー1−ピロリン N−オキシド(DMPO)スピントラップは、多くの生物学的反応におけるフリーのラジカルの関与、特にOHラジカル、についての証拠を提供するために広く用いられている。DMPOはミオグロビンおよび他の分子におけるラジカル損傷へと曝露されたラジカル部位を同定するのに用いられてきた。よって、IgG1をDMPOの存在下で1週間H2O2で処理し、そしてSECによって断片化を監視した。50:1から5:1のDMPO:H2O2のモル比範囲で、DMPOは2週間のインキュベーションの時間経過を通して断片化を完全にブロックした。
【0060】
[0060] ラジカル形成部位を同定するために、Lys−Cペプチドマッピングを行った。Cys231−SO3Hを含有する無傷のヒンジペプチドが観察される一方、HC Cys231のみがDMPO付加物を含有するものとして見出された。上部ヒンジ残基においてラジカル形成が見出されなかったことは、質量分析の感度の問題またはOHラジカルと上部ヒンジ残基の間の反応率によるものではなさそうである。OHラジカルはCysとの速度定数、3.4×1010M−1s−1を有し、これはHis(1.3×1010M−1s−1)、Thr(5.1×108M−1s−1)、Asp(7.5×107M−1s−1)、およびLys(3.5×107M−1s−1)よりもずっと速い(Davies, M. J. およびDean, R. T., 1997, Radical mediated protein oxidation. Oxford University press, pp 50-120)。よって、これらの結果は、分子あたりのラジカル切断に導く、HC Cys231から上部ヒンジの残基への電子伝達の必要性を立証した。電子がHisについて6.4×107M−1s−1、Thrについて2.0×107M−1s−1、Lysについて2.0×107M−1s−1、Aspについて1.8×107M−1s−1の反応速度定数を有することが決定され(Davies, M. J.およびDean, R. T., 1997, Radical mediated protein oxidation, Oxford University press, pp.50-120)、このことはこれらの残基はラジカル誘導骨格切断を進めるために電子を局在化させることができることを示している。この機構は、FabフラグメントのC末端残基(C2)および部分抗体のFcのN末端残基(C1)の相補性を形成する特異的ヒンジ断片化を説明する。
【0061】
実施例2
[0061] この実施例はIgG1 Fcのラジカル媒介断片化の結果をまとめたものである。
1.IgG1バルク抗体は〜1%の切断型抗体(P1)を含有し、これは、Fabドメインの一つを欠損した、重度に酸化された型と決定された。
2.IgG1バルク薬剤物質(BDS)とH2O2の反応は、切断型分子、そして重鎖のFabドメイン(Fd)におけるC末端残基のラダー(Cys220−Asp221−Lys222−Thr223−His224−Thr225)およびFcドメインにおける相補的N末端残基のラダーの形成に帰する、ヒンジ領域における特異的切断による一つのフリーのFabドメイン断片を生じた。
3.H2O2で処理した試料において、無傷のまたは切断された分子の大部分において、Cys226残基間の鎖間ジスルフィド結合は無傷であることが見出された。
4.BDS試料において、潜在的な対形成していないCysのN−エチルマレイミド(NEM)によるプレブロッキング後に行った天然型Lys−Cペプチドマップによって観察される、ヒンジ領域中の対形成していないジスルフィド結合は存在しなかった。
5.LC−MS/MS分析は、Cys226における少量のCys−SO3Hを、無傷のヒンジペプチド(THTCys226PPCAPELLGGPSVFLFPPKPK)(配列番号5)および切断型ヒンジペプチド(Cys226PPCAPELLGGPSVFLFPPKPK)(配列番号6)の両方において同定した。
6.切断された抗体において、FcドメインのN−末端ヒンジ領域において、それぞれ45または71Daの付加質量を導入するイソシアネートまたはN−α−ケトアシル誘導体のいずれかとして、付加物が同定された。
7.IgG1は〜0.28モル/モル抗体の対形成していないCys残基を含有し、これは、すべての対形成していないCys残基のブロッキングは断片化においてまったく効果を示さないか、ごくわずかな効果しか生じないという事実によって示されるように、切断反応には重大なものではない。
8.広く用いられるラジカルスピントラップ 5,5’−ジメチル−1−ピロリン N−オキシド(DMPO)は、それのCys226への結合のために、ヒンジ断片化をブロックすることができることが見出された。しかしながら、DMPO結合はCys226−SO3Hの形成をブロックしなかった。
【0062】
実施例3
[0062] 本実施例は、Cu2+ではなくヒドロキシルラジカルがヒンジ断片化を誘発することを示す。過酸化水素はタンパク質の生物学的機能を、ラジカルに誘導される酸化経路を通して制御することができる。加えて、ヒドロキシルラジカルを伴う反応は、タンパク質の分解に帰する、種々の化学反応を導くことができる。OHラジカルがヒンジ断片化に関与するか試験し、そして当該切断に影響しうるいくつかの因子を評価するために、IgG1をH2O2攻撃にさらした。図1に示すように、H2O2が誘導する断片化はカタラーゼによって完全にブロックされ、これはOHラジカルは当該切断の原因であることを示している。フリーのチオール基の全体は、Ellman試薬、5,5’−ジチオビス(2−ニトロ安息香酸)(DTNB)を用いる4M GdnHClの存在下の変性条件下で、〜0.28モル/モル抗体であると決定された。H2O2処理の前に、IgG1をNEMとともにpH5.0で3時間、37℃でインキュベートした。NEMでブロックした試料は、〜7%の切断の減少しか示さなかった一方、NEM処理によってフリーのチオール(−SH)基は完全にブロックされたことが見出され、対形成していないCys残基は切断に重要ではないことが示唆された。
【0063】
[0063] 加えて、EDTAとのプレインキュベーションは、IgG1のH2O2に誘導される切断の〜90%を阻害することが見出され、反応における遷移金属の関与が示唆される。しかしながら、そのような前処理はH2O2による切断を完全にブロックすることはなく、より遅い反応速度を有するものではあったが、依然としてIgG1を切断することが可能であった。これらの結果はOHラジカルがヒンジ断片化に重要であり、そしてこの反応は、OHラジカルを生成する金属により触媒される反応によって、促進されうることを示唆した。この仮説は、10μM 酢酸銅(Cu(OAc)2)の存在下でのH2O2を伴う処理が、H2O2処理単独よりもおよそ4倍多くの切断となる一方で、10μM Cu(OAc)2単独では5日間のインキュベーションの間にごくわずかの切断しか生じなかったという結果により支持された。
【0064】
[0064] Smithらは、いくつかの合成ペプチドを試験することにより、中性または塩基性pHでの1mMのCuSO4による、IgG1の上部ヒンジDKTHT(配列番号7)領域中のK−T結合の切断を報告した(Smith, M. A.ら、Int. J. Pept. Protein Res., 48:48-55, 1996)。本明細書に記載される実験条件下(pH5.2、および25℃でのインキュベーション)では、上部ヒンジ領域(例えば、His、Lys)へのCu2+の結合は中性または塩基性pHの場合よりも有望ではなく、そしてヒンジ断片化の〜30%増加という結果となった。溶媒またはタンパク質中に微量の遷移金属イオンが存在するので、それらの濃度はラジカル誘発ヒンジ断片化のための触媒として機能するのに十分であり得た。この結論は、いくつかの遷移金属(例えばCu2+およびFe3+)が、タンパク質に結合するかまたは溶媒中に留まることのいずれかにより、部位特異的ラジカル攻撃における重要な役割を果たすという理論に一致する。両方の場合において、当該金属は、Fenton様反応を通してヒドロキシルラジカルの生成を触媒することにより、反応を促進する。まとめると、これらの事実はラジカル誘発ヒンジ断片化機構を独立して確認した。
【0065】
実施例4
[0065] 本実施例は、ラジカル媒介Fc断片化の機構を提案する。我々のヒトIgG1の研究の実験結果は、このヒトIgG1抗体におけるラジカル媒介ヒンジ断片化を明らかにした。
【0066】
[0066] 微量の遷移金属は反応系におけるOHラジカルの生成を触媒する。IgG1抗体とOHラジカルとの反応は、抗体のヒンジ領域(Cys226−Pro−Pro−Cys−Pro)における226位に位置する二つのシステイン残基(Cys226)の間の鎖間ジスルフィド結合の切断という結果となった。そのジスルフィド結合切断の後、スルフェン酸(Cys226−SOH)およびチイルラジカル(Cys226−S・)の形成が続いた。酸素の存在下でのこれらの種の続く反応は、主要な産物としてスルフィン酸(Cys226−SO2H)およびスルホン酸(Cys226−SO3H)の形成という結果となった。一方チイルラジカルは、ヒンジポリペプチド骨格に沿って、電子移動の上流側(electron transfer upstream)を開始する。この電子移動は、ラジカル媒介ポリペプチド骨格断片化を導き、これは、いくつかの隣接するヒンジ残基(Asp221、Lys222、Thr223、His224、およびThr225)における切断のために生じる、重鎖のFabドメイン(Fd)におけるC末端残基のラダーによって特徴づけされる。我々は、広く用いられているラジカルスピントラップである5,5’−ジメチル−1−ピロリン N−オキシド(DMPO)の結合を、Cys226でのみ観察し、これはヒンジ断片化をブロックした。DMPOのCys226のみへの特異的結合は、IgG分子を通じて高度に保存された配列モチーフCys−Pro−X−Cys−Pro(X=Pro、Arg、およびSer)(表1)である、CPPCP配列においてCys226のみにラジカルが存在することが確認された。
【0067】
[0067] +45Da付加物の決定は、ジアミド経路を通じてFcのN末端にイソシアネート構造(MW=28Da)を生じるラジカル切断機構を示唆した。イソシアネート基は、その不安定な性質により、カルボン酸(+45Da付加物)へと加水分解する。一方、特定のアミノ酸の側鎖のγ−炭素位におけるOHラジカル攻撃は、側鎖としてβ−CH2のみを保持する、デヒドロペプチドの不飽和産物の形成へとつながる酸化的分解という結果になりうる。この化合物は容易に加水分解されて、+71Da付加物(N−ピルビル基)である、アミドおよびケト官能基を生じることができる。そのために、Thr225のN末端において観察された+71Da付加物は、His224の酸化的分解によって生じたものであり得た。一方、これらの不安定な中間体の加水分解は、それらをリサイクルする別の方法である可能性があり、そしてこの過程は通常のN末端残基を含有するいくつかの切断されたヒンジペプチドをもたらした。まとめると、上部ヒンジ領域のN末端残基における+45Daおよび+71Da付加物は、それぞれタンパク質骨格のα−炭素およびアミノ酸側鎖のγ−炭素位におけるラジカル切断の産物であり、このことはタンパク質骨格切断についてのラジカル媒介機構を確認するものである。
【0068】
実施例5
[0068] 本実施例は、ヒンジコア配列中のHisおよびLys残基の変異によるラジカル媒介断片化への耐性を実証する。調査は、ヒト野生型IgG1との比較において、His224およびLys222を変異させている効果について決定するために行った。野生型IgG1および7つの変異体をH2O2とともにインキュベートし、部分的分子の形成および特にFabドメイン断片の放出をSECにより監視した。7つの変異体は以下である:Lys222Ser(K/S)、Lys222Gln(K/Q)、Lys222Ala(K/A)、His224Ser(H/S)、His224Gln(H/Q)、His224Ala(H/A)、およびLys222Ser/His224Ser(K/S+H/S)。これらの変異体の中で、HisをGlnまたはSerで置換したものは、Fabドメイン(C2)および部分的分子C1の放出につながるOHラジカルに誘発される断片化をほぼ完全にブロックした(>97%)。His/Ala変異は8日間のインキュベーション期間で〜6%の断片化を示し、これに対して天然型IgG1については〜15%であった。対照的に、すべてのLys単独変異体は、31〜33%で切断を促進した。より重要なことには、二重変異体であるK/S+H/Sは>97%の断片化阻害を示し、これはHis/SerまたはHis/Gln単独変異体について測定されたのと同じ割合であり、このことは断片化におけるHis残基の重要性を示している。
【0069】
[0069] His/Ala変異体は切断を示したが、当該変異体が同じ構造的分解を含むかどうかは知られていなかった。LCおよびHCは、それらを連結している鎖間ジスルフィド結合なしに、強く会合したままであることが記載されていた(Bigelow. C.ら、Biochemistry, 13: 4602-4609, 1978)。よって、LCおよびHCが、鎖間ジスルフィド結合なしに共に保持されること、およびFabドメイン断片と同様のSECプロフィールを示すことは可能である。よって、変異体を、H2O2処理の1日後に、非還元条件下でRP−HPLC−TOF/MSによってさらに試験した。これらの条件下では、共有結合で結合していない成分のみが主要な種から分離されることが期待される。図2に示すように、〜21分で溶出される主要ピークのほかに、16.5分の保持時間で移動してくる一つの成分がすべての変異体について観察された。特に、H/A変異体はこの種をH/SおよびH/Q変異体よりもおよそ15倍多く放出した。TOF/MS分析は、この種について23,437.5Daの分子量を決定し、これはLCについての理論質量である23,389.0Daよりも+48Da重い。このLys−CペプチドマップのRP−HPLC−MS/MS分析は、この種はLC Cys215のスルホン酸(+48Da)への完全な転換を示すものと確認し、このことはH2O2攻撃による鎖間ジスルフィド結合の切断はLCの酸化を導いたことを示唆するものである。これらの結果はOH基の除去は、側鎖におけるH−結合形成の可能性を消滅させ、この残基がラジカル攻撃に耐える能力を不利に損なったことを示唆した。
【0070】
[0070] IgG1の上部ヒンジと同じ配列(DKTHT)(配列番号7)を含む合成ペプチド(FDKTHTY)(配列番号8)を用いて、Allenら(Allen, G.およびCampbell, R., Int. J. Peptide Protein Res. 48: 265-273, 1996)はHis/Ala置換は、Cu2+(1mM)が誘導するペプチドの切断を阻止することを見出した。しかしながら、我々の結果はHis/Ala変異体は、LCおよびHCの間の鎖間ジスルフィド結合のH2O2誘導切断によるLCの放出を阻止しなかったことを明らかに示した。LCの喪失は、IgGの機能を破壊するであろう。特に、二つのヒンジ鎖間ジスルフィド結合がFabドメインへと連結している上部ヒンジ(DKTHT)(配列番号7)で二つのHCを連結しているヒンジ領域は、溶液中の合成ペプチドのコンフォメーションとは非常に異なる可能性が最も高いコンフォメーションをヒンジが採用するよう制限する、二重鎖構造である。したがって、ペプチドから得られた結果は、同じまたは類似の配列を含有するタンパク質に適用する場合には、注意深く取り扱う必要がある。まとめると、我々の結果は、His/Ala変異体ではなく、His/SerおよびHis/Gln変異体が、OHラジカル媒介切断を阻害したことを明らかに示した。
【0071】
[0071] His、Gln、Ser、AlaおよびLysの側鎖の性質を前提とすると、これらの変異体の解析結果は、His残基の側鎖のγ−炭素よりはむしろイミダゾール環が、ヒンジ切断の原因であると我々は結論付けさせる。この仮定は、Glnはその側鎖にγ−炭素を有するが、His/Gln変異体はラジカル誘導切断を阻害したという観察結果によって支持された。電子付加の主要な部位は、プロトン化されるpH値におけるHis224のイミダゾール環であるようである。OHラジカルはイミダゾール環のC−2、C−4およびC−5位に付着することが知られている。IgG1の公知の三次元構造におけるこれらのヒンジ領域の配置、およびヒンジ周囲の水素結合ネットワークに基づいて、続くこれらの種への酸素の付加が、最初のラジカルに塩基に触媒される水の喪失を被らせ、高度に安定化したビスアリルラジカルを与えることを我々は提案する。His残基は電子を局在化させる中心的な標的として機能し、そして続いて隣接する残基からプロトンを抽出し、ジアミドおよびα−アミド化経路によりラジカル誘発切断に導いた。まとめると、この結果は、合理的設計を用いてヒンジ断片化を防止する可能性を立証した。
【技術分野】
【0001】
関連出願の相互参照
[0001] 本出願は、2009年4月21日に出願された35U.S.C. 119(e)の米国特許出願番号61/171,393の利益を主張し、当該出願は参照により本明細書に援用される。
【0002】
技術分野
[0002] 本発明は、活性酸素種(reactive oxygen species)による断片化に対して耐性である、療法的および診断的適用に用いるための免疫グロブリンに関する。
【背景技術】
【0003】
本発明の背景
[0003] ヒト免疫グロブリン(IgG)分子は軽鎖(LC)および重鎖(HC)の2つの同一のコピーからなる。LCおよびHCの間の鎖間ジスルフィド結合はそれらを結びつけ、半抗体を形成する;半抗体の二つの同一のコピーのHCは、いわゆるヒンジ配列においてジスルフィド結合で結びつけられて、天然の抗体を形成する。ヒトIgG1ヒンジ配列は、2つの別々のジスルフィド結合を形成できる、システイン(Cys)残基の2つの組を含む。しかしながら、単一のヒンジジスルフィドのみが、補体に仲介される溶解ならびに抗体依存性細胞介在性細胞障害およびファゴサイトーシスのために必要であることが示唆されてきた。Michaelsen, T. E. ら、 Proc. Natl. Acad. Sci. USA 91:9243-9247, 1994。IgG1 b12の結晶構造において、単一の重鎖間ジスルフィド結合のみが観察されてきた −著者らは壊れたジスルフィド結合は、動力学的であるかまたはシンクロトロン照射損傷の結果でありうると示唆した。Stanfield, R. ら、Science 248: 712-719, 1990; Saphire, E.ら、J. Mol. Biol. 319:9-18, 2002; Weik, M.ら、Proc. Natl. Acad. Sci. USA 97:623-628, 2000。事実、結晶構造における溶媒に露出した一つのシステイン対についての酸化型および還元型コンフォメーションの両方が記述されてきている。Burling, F. T.ら、Science 271:72-77, 1996。IgG1において、LCのC末端のCys残基は、最初にHCのヒンジのCys残基へと連結する;しかしながら、LCおよびHCは、それらの間の会合定数は〜1010M−1 と推定されたので、ジスルフィド結合なしでもなお強く一緒に会合することができる。Bigelow. C.ら、Biochemistry 13:4602-4609, 1978; Horne, C.ら、J. Biol. Chem., 129:660-664, 1982。総合すれば、これらの知見は、IgG1におけるジスルフィド結合は特定の攻撃に対して脆弱であり、そして関連するシステイン残基は対をつくらないままでいることができることを示唆している。
【0004】
[0004] 活性酸素種(ROS)は、酸化的ストレスの主な原因である。ROS、例えば過酸化水素類およびアルキルヒドロペルオキシド類は、タンパク質の生物学的機能を調節することができる。Poole, L. B.ら、Annu. Rev. Pharmacol. Toxicol. 44:325-347, 2004;Philip, E., Free Rad. Biol. Med. 40: 1889-1899, 2006;Salmeen A.ら、Nature 423: 769-773, 2003;Claiborne A.ら、Adv. Protein Chem. 58: 215-276, 2001;Paget, M. S. B.およびButtner, M. J., Annu. Rev. Genet. 37: 91-121, 2003。H2O2により調節されるタンパク質は、酸化に敏感な特徴的なシステインを有する、なぜならそれらの環境はチオール基(Cys−SH)のチオレートアニオン(Cys−S−)(これはCys−SHよりもスルフェン酸(Cys−SOH)へとより容易に酸化される)へのイオン化を促進するからである。Rhee, S. G.ら、(2000) Sci. STKE 200, pe1;Kim, J. R.ら、Anal. Biochem. 283:214, 2000。スルフェン酸は不安定であり、いずれかの接近可能なチオールと反応してジスルフィドを形成するか、さらなる酸化を経てスルフィン酸(Cys−SO2H)またはスルホン酸(Cys−SO3H)となる。Kice, J. L., Adv. Phys. Org. Chem. 17:65, 1980;Claiborne, A., Biochemistry 38: 15407-15412, 1999。
【0005】
[0005] システインに基づくラジカルは、短範囲水素原子引き抜き反応、または一電子移動反応によって形成されることができる。Giles, N. M.ら、Chemistry & Biology 10: 677-693, 2003;Garrison, W. M., Chem. Rev., 87:381-398, 1987;Bonifacic, M. ら、J. Chem. Soc. Perkin Trans., 2: 675-685, 1975;Elliot, A. J.ら、J. Phys. Chem. 85: 68-75, 1981;Jacob, C.ら、Biol. Chem. 387: 1385-1397, 2006。チイル(RS)、スルフィニル(RSO)およびスルホニル(RSOO)ラジカルは、酸化的ストレスの間、存在することが見出されてきている。Harman, L. S.ら、J. Biol. Chem. 259: 5606-5611, 1984;Giles, G. I.およびJacob, C, Biol. Chem. 383: 375-388, 2002;Witting, P. K.,およびMauk, A. G., J. Biol. Chem. 276: 16540-16547, 2001;Stadtman. E. R.およびLevine, R. L., Amino Acids. 25: 207-218, 2003;Berlett, B. S.およびStadtman, E. R., J. Biol. Chem. 272: 20313- 20316, 1997。Cysラジカルおよび他の残基の間の電子移動は、ミオグロビンのオリゴメリック産物形成に関与することが決定されてきている(Witting, P. K.およびMauk, A. G., J. Biol. Chem. 276: 16540-16547, 2001)、一方ProおよびHis残基は、BSAおよびコラーゲンの断片化を生じることとなるROS攻撃のターゲットであることが見出された。Garrison, W. M., Chem. Rev. 87: 381-398, 1987;Davies, M. J.およびDean, R. T., 1997, Radical mediated protein oxidation. Oxford University press, pp 50-120;Zhang, N.ら、J. Phys. Chem. 95: 4718-4722, 1991;Zhang, H.ら、J. Biol. Chem. 280: 40684-40698, 2005;Uchida, K.およびKawakishi, S., Biochem. Biophys. Res. Commun. 138: 659-665, 1986;Dean, R. T.ら、Free Radical Res. Commun. 7: 97-103, 1989。しかしながら、Cysに基づくラジカルがペプチド結合の切断に関与するかどうかについては依然として不明である。StamlerおよびHausladen(Stamler, J. S.およびHausladen A., Nat. Struct. Biol. 5: 247-251, 1998)は、一方で重要な生物学的シグナル事象を構成し、そして他方で酸化的ストレスの不可逆的な特徴である、H2O2に仲介される修飾の連続を提唱した。
【0006】
[0006] 多くの異なる生理学的および環境的プロセスが、活性酸素種(ROS)のインビトロおよびインビボでの形成につながる。細胞内のROSのレベルは、その年齢および生理学的状態に依存し、そしてプロテアーゼ、ビタミン類(A、C、およびE)、および酸化還元金属イオンのような因子の関数である。Bigelow, C.ら、Biochemistry 13: 4602-4609, 1978。ミトコンドリアは、細胞におけるROS生成の重大な源である。Salmeen, A.ら、Nature 423: 769-773, 2003。単離されたミトコンドリアにおけるH2O2産生速度は、生理学的条件下での総酸素取り込みの約2%である。Salmeen, A.ら、Nature 423: 769-773, 2003;Claiborne, A.ら、Adv. Protein Chem. 58: 215-276, 2001;Paget, M.およびButtner, M., Annu. Rev. Genet. 37: 91-121, 2003。
【0007】
[0007] ROSは、インビトロならびにインビボでのラジカルにより媒介されるタンパク質の断片化および凝集を招き得る。これらの酸化的修飾は、療法的および診断的製品の製造量を低下させ、ならびにそれらの効率を低下させ得る。抗体は、療法的および診断的タンパク質の特に有用なクラスとして証明されてきた。しかしながら、抗体のFcヒンジ領域は酸化的修飾を受けやすい。ラジカル攻撃に対するこの脆弱性は、抗体候補ならびに一般的なFc−コンジュゲート化化合物の療法的および診断的開発について、Fcヒンジ領域の安定化を優先課題とする。
【先行技術文献】
【非特許文献】
【0008】
【非特許文献1】Michaelsen, T. E. ら、 Proc. Natl. Acad. Sci. USA 91:9243-9247, 1994
【非特許文献2】Stanfield, R. ら、Science 248: 712-719, 1990
【非特許文献3】Saphire, E.ら、J. Mol. Biol. 319:9-18, 2002
【非特許文献4】Weik, M.ら、Proc. Natl. Acad. Sci. USA 97:623-628, 2000
【非特許文献5】Burling, F. T.ら、Science 271:72-77, 1996。
【非特許文献6】Bigelow. C.ら、Biochemistry 13:4602-4609, 1978
【非特許文献7】Horne, C.ら、J. Biol. Chem., 129:660-664, 1982
【非特許文献8】Poole, L. B.ら、Annu. Rev. Pharmacol. Toxicol. 44:325-347, 2004
【非特許文献9】Philip, E., Free Rad. Biol. Med. 40: 1889-1899, 2006
【非特許文献10】Salmeen A.ら、Nature 423: 769-773, 2003
【非特許文献11】Claiborne A.ら、Adv. Protein Chem. 58: 215-276, 2001
【非特許文献12】Paget, M. S. B.およびButtner, M. J., Annu. Rev. Genet. 37: 91-121, 2003
【非特許文献13】Rhee, S. G.ら、(2000) Sci. STKE 200, pe1
【非特許文献14】Kim, J. R.ら、Anal. Biochem. 283:214, 2000
【非特許文献15】Kice, J. L., Adv. Phys. Org. Chem. 17:65, 1980
【非特許文献16】Claiborne, A., Biochemistry 38: 15407-15412, 1999
【非特許文献17】Giles, N. M.ら、Chemistry & Biology 10: 677-693, 2003
【非特許文献18】Garrison, W. M., Chem. Rev., 87:381-398, 1987
【非特許文献19】Bonifacic, M. ら、J. Chem. Soc. Perkin Trans., 2: 675-685, 1975
【非特許文献20】Elliot, A. J.ら、J. Phys. Chem. 85: 68-75, 1981
【非特許文献21】Jacob, C.ら、Biol. Chem. 387: 1385-1397, 2006
【非特許文献22】Harman, L. S.ら、J. Biol. Chem. 259: 5606-5611, 1984
【非特許文献23】Giles, G. I.およびJacob, C, Biol. Chem. 383: 375-388, 2002
【非特許文献24】Witting, P. K.,およびMauk, A. G., J. Biol. Chem. 276: 16540-16547, 2001
【非特許文献25】Stadtman. E. R.およびLevine, R. L., Amino Acids. 25: 207-218, 2003
【非特許文献26】Berlett, B. S.およびStadtman, E. R., J. Biol. Chem. 272: 20313- 20316, 1997
【非特許文献27】Davies, M. J.およびDean, R. T., 1997, Radical mediated protein oxidation. Oxford University press, pp 50-120
【非特許文献28】Zhang, N.ら、J. Phys. Chem. 95: 4718-4722, 1991
【非特許文献29】Zhang, H.ら、J. Biol. Chem. 280: 40684-40698, 2005
【非特許文献30】Uchida, K.およびKawakishi, S., Biochem. Biophys. Res. Commun. 138: 659-665, 1986
【非特許文献31】Dean, R. T.ら、Free Radical Res. Commun. 7: 97-103, 1989
【非特許文献32】Stamler, J. S.およびHausladen A., Nat. Struct. Biol. 5: 247-251, 1998
【発明の開示】
【課題を解決するための手段】
【0009】
[0008] 本発明は、ラジカル媒介断片化に耐性の、IgG1またはIgG3クラスのヒンジ配列を含むイムノグロブリンFcを提供する。断片化耐性は、そうでなければ2つの半抗体を生じるジスルフィド結合切断の減少により、ならびにこれらの半抗体のそれぞれを作り上げているポリペプチド内の断片化事象の減少により、現れる。一態様において本発明は、FcがヒトIgG1またはIgG3 Fcである、Fc−コンジュゲートである。当該IgG1およびIgG3 Fcは、アミノ酸一文字表記でTHTCPXCPであり、ここでXはRまたはP残基を示す、ヒンジコア配列を含む。本発明において、天然型IgG1またはIgG3 Fcのヒンジコア配列におけるH(ヒスチジン)残基は、Ser(セリン)、Gln(グルタミン)、Asn(アスパラギン)、またはThr(スレオニン)残基で置換される。いくつかの態様において、Fc−コンジュゲートは、薬学的に許容可能な担体中にある。
【0010】
[0009] 本発明はまた、本発明のFcまたはFc−コンジュゲートをコードするポリヌクレオチドを含む単離された核酸、ならびに当該単離された核酸を含む発現ベクター、および前述の発現ベクターを含む宿主細胞に関する。よって、本発明はまた、本発明の核酸を含む発現ベクターを適切な宿主細胞中で、当該核酸の発現に適した条件下で培養し、そして当該宿主細胞から発現したFcまたはFc−コンジュゲートを単離するすることを必然的に含み得る、本発明のFcまたはFc−コンジュゲートを作成する組成物および方法をも含む。
【図面の簡単な説明】
【0011】
【図1】[0010] 図1は、H2O2と実施例において詳述した追加的な試薬の組み合わせによって生じる、IgG1抗体のラジカル媒介断片化の程度を示す。
【図2】[0011] 図2は、実施例において詳述した種々のIgG1ヒンジ配列置換変異体の鎖間ジスルフィド結合切断について、ミリ吸光度単位(mAU)で測定したラジカル媒介断片化の程度を示す。
【発明を実施するための態様】
【0012】
発明の詳細な説明
[0012] 本発明は、天然型IgG1またはIgG3 Fcよりもラジカル媒介断片化に対して耐性であるように修飾された、ヒトIgG1およびIgG3 FcおよびFc−コンジュゲートに関する組成物および方法を提供する。断片化耐性IgG1およびIgG3 Fcは、例えば、インビトロまたはインビボ断片化または凝集に対してより高い耐性を有する療法的および診断的使用のための抗体の製造において、使用することができる。本発明の組成物は、以下を含む:Fc−コンジュゲート、本発明のFcまたはFc−コンジュゲートをコードする核酸を含むポリヌクレオチド、これらの核酸を含むベクター、これらのベクターを含む宿主細胞およびこれらのベクターを発現する宿主細胞、および医薬組成物。これらの組成物のそれぞれを製造するおよび使用する方法もまた、提供する。
【0013】
[0013] 単位、接頭辞、および記号はSIで認められた形式で表示されうる。特に言及しなければ、核酸は5’から3’方向へ、左から右へと記載される;アミノ酸配列は、アミノからカルボキシ方向へ、左から右へと記載される。本明細書において記載される数値範囲は、当該範囲を規定する数字を含むものであり、そして定められた範囲内のそれぞれの整数を含むおよび支持する。アミノ酸は本明細書において、IUPAC−IUBMB命名法委員会が推奨する、一般的に知られたそれらの三文字記号または一文字記号のいずれかによって言及されてもよい。同様に、核酸も一般的に認められているそれらの一文字コードによって言及されてもよい。特に言及しない限り、「a」または「an」の語は、「少なくとも1つの」を意味するものとして解釈される。本明細書で用いるセクション見出しは、整理の目的のためにのみ用いられるものであって、記載された主題を限定するものとして解釈されてはならない。
【0014】
A.定義
[0014] 本明細書おいて使用される用語「抗体」は、免疫化によって、組換え技術を介して、インビトロ合成手段の手法によって、またはその他の方法によって、完全にまたは部分的に、製造される抗体であるかどうかにかかわらず、ヒト(例えばCDRグラフトされた)、ヒト化、キメラ、多重特異性、モノクローナル、ポリクローナル、およびそのオリゴマーを含む、グリコシル化されたおよびグリコシル化されていない、いずれかのアイソタイプまたはサブクラスのイムノグロブリンの双方への言及を含む。よって、用語「抗体」は、例えば(a)ヒトイムノグロブリン遺伝子について遺伝子導入した動物(例えばマウス)またはそれから調製したハイブリドーマから単離した抗体、(b)当該抗体を発現するようにトランスフェクトされた宿主細胞から(例えば、トランスフェクトーマから)単離された抗体、(c)組換え、コンビナトリアル抗体ライブラリーから単離された抗体、および(d)イムノグロブリン遺伝子配列の他のDNA配列へのスプライシングを伴う他のいずれかの手段により調製され、発現され、創造され、または単離された抗体、のような、組換え手段により調製され、発現され、創造され、または単離されたものを含む。そのような抗体は、動物の2つの別個の種の生殖系列イムノグロブリン配列に由来する可変および定常領域を有する。しかしながら、特定の態様において、そのような抗体はインビトロ突然変異誘発(または、ヒトイムノグロブリン遺伝子について遺伝子導入した動物を用いる場合、インビボ体細胞突然変異誘発)を受けうるので、該抗体のVHおよびVL領域のアミノ酸配列は、特定の種(例えばヒト)の生殖系列VHおよびVL配に由来しおよび関連するが、当該種のインビボ抗体生殖系列レパートリーには天然に存在しない可能性がある配列である。
【0015】
[0015] 本明細書において、「コンジュゲート」は、Fcへとコンジュゲート化した場合に診断的または療法的機能を発揮する、いずれかの化学的または生物学的部分を意味する。コンジュゲートは、直接的にまたは間接的に(すなわち、化学的スペーサーを通して)共有結合的に付着されることができる。例示的なコンジュゲートは、以下を含む:細胞傷害性薬物または細胞分裂阻害剤(例えば、抗腫瘍剤または抗血管新生剤)、ポリエチレングリコール、脂質、および受容体または細胞表面受容体の細胞外ドメインのような受容体フラグメント。
【0016】
[0016] 「宿主細胞」は、核酸、例えば本願発明の核酸、を発現するために用いることができる細胞である。宿主細胞は、原核生物、例えば大腸菌(E. coli)であることもでき、あるいは真核生物、例えば単細胞真核生物(例えば酵母もしくは他の真菌類)、植物細胞(例えば、タバコもしくはトマト植物細胞)、動物細胞(例えばヒト細胞、サル細胞、ハムスター細胞、ラット細胞、マウス細胞、もしくは昆虫細胞)、またはハイブリドーマであることができる。宿主細胞の例は、サル腎臓細胞のCOS−7系列(ATCC CRL 1651)(Gluzmanら、Cell 23:175, 1981を参照)、L細胞、C127細胞、3T3細胞(ATCC CCL 163)、チャイニーズハムスター卵巣(CHO)細胞またはそれらの派生物であるVeggie CHOおよび無血清培地中で増殖する関連する細胞系列(Rasmussenら、Cytotechnology 28: 31, 1998)、または、DHFR欠損であるCHO DX−B11株(Urlaubら、Proc. Natl. Acad. Sci. USA 77: 4216-4220, 1980)、を含む。
【0017】
[0017] 典型的には、宿主細胞は、ポリペプチドをコードする核酸でトランスフェクトされることができる培養細胞である、そして当該核酸は当該宿主細胞中で発現されることができる。「組換え宿主細胞」の句は、発現されるヌクレオチドでトランスフェクトされている宿主細胞を示すものとして用いることがでいる。典型的には、宿主細胞は核酸を含むが、制御配列が該核酸と機能可能に連結されるように該宿主細胞に制御配列が導入されない限り、該核酸を評価できるレベルで発現しない。宿主細胞という用語は、特定の対象細胞のみに言及するものではなく、そのような細胞の後代または潜在的な後代にも言及する。突然変異または環境の影響のいずれかのために続く世代においてある一定の修飾が生じ得るので、そのような後代は、実際に、親細胞と同一ではない可能性があるが、それでもなお本明細書において用いられる用語の範囲内に含められる。
【0018】
[0018] 用語「ヒト抗体」は、定常領域およびフレームワークが完全にまたは実質的にヒト配列からなり、そのためヒト抗体はヒト宿主に投与したときにそれ自体に対する免疫原性反応、好ましくは検出可能な免疫原性反応を実質的に誘発しない、抗体を意味する。
【0019】
[0019] 用語「ヒト化抗体」は、定常領域のすべてが、ヒトイムノグロブリンに由来するか対応する一方、1またはそれより多くの可変領域のすべてまたは一部が他の種、例えばマウスに由来する、抗体を意味する。
【0020】
[0020] 本明細書において、核酸について「単離された」というときは、ヒトの直接的介入の結果物としてのDNAまたはRNAを意味する:1)天然には見出されないゲノムの座に組み込まれたもの、2)天然では機能可能に連結しない核酸と機能可能に連結したもの、3)天然の状態では混合された状態にある細胞構成物から実質的に精製されたもの(例えば、少なくとも70%、80%、または90%)。
【0021】
[0021] 本明細書において、FcまたはFc−コンジュゲートについて「単離された」というときは以下を意味する:(1)存在する優勢な種である発現された状態では混合された状態にある細胞構成物から実質的に精製されたもの(例えば、少なくとも60%、70%、80%、または90%)、(2)天然では連結されていないポリペプチドまたはその他の部分へとコンジュゲート化されたもの、(3)より大きなポリペプチド配列の一部として天然には生じないもの、(4)詳細に明らかにされた組成物において異なる特異性を有する他の化学的または生物学的な剤と組み合わされたもの、または(5)天然に見出されない限りにおいてヒトの操作された配列を含むもの。
【0022】
[0022] 本明細書において用いられる用語「モノクローナル抗体」または「モノクローナル抗体組成物」は、抗体分子の単一分子組成物の調製物を意味し、典型的には同一の核酸分子によってコードされる。モノクローナル抗体組成物は、特定のエピトープについて単一結合特異性および親和性を示す。ある態様において、モノクローナル抗体は、単一のハイブリドーマまたは他の細胞株(例えば、トランスフェクトーマ)、あるいはトランスジェニック哺乳動物によって、産生される。用語「モノクローナル」は、抗体の製造について特定の方法に限定するものではない。
【0023】
[0023] 本明細書において用いられる「核酸」および「ポリヌクレオチド」は、デオキシリボヌクレオチドもしくはボヌクレオチドポリマー、またはそのキメラ、への言及を含み、そして他に限定しない限り、言及した配列の相補鎖を包含する。
【0024】
[0024] 制御配列が核酸配列の発現(例えば、発現のレベル、タイミング、または位置)に影響を及ぼす場合、該核酸配列は制御配列に「機能可能に連結している」。「制御配列」は、第二の核酸の発現(例えば発現のレベル、タイミング、または位置)に影響を及ぼす核酸である。よって、制御配列と第二の配列の間の機能的連結が、該第二の配列に対応するDNA配列の転写を該制御配列開始または仲介するようなものである場合、制御配列および第二の配列は機能可能に連結している。制御配列の例は、プロモーター、エンハンサー及び他の発現制御因子(例えばポリアデニル化シグナル)を含む。制御配列のさらなる例は、例えば、Goeddel, 1990, Gene Expression Technology: Methods in Enzymology, 185, Academic Press, San Diego, CA およびBaronら、Nucleic Acids Res. 23: 3605-3606, 1995に記載されている。
【0025】
[0025] 用語「ペプチド」、「ポリペプチド」および「タンパク質」は、本明細書を通して互換的に用いられ、そしてペプチド結合によりお互いに結合した2以上のアミノ酸残基を含む分子に言及するものである。用語「ポリペプチド」、「ペプチド」および「タンパク質」はまた、グリコシル化、脂質付加、硫酸化、グルタミン酸残基のガンマ−カルボキシル化、ヒドロキシル化、およびADP−リボシル化、を含むがこれらに限定されない修飾を含めたものである。
【0026】
[0026] 用語「ポリヌクレオチド」、「オリゴヌクレオチド」および「核酸」は、本明細書を通して互換的に用いられ、そしてDNA分子(例えばcDNAまたはゲノムDNA)、RNA分子(例えばmRNA)およびそれらのハイブリッドを含む。核酸分子は一本鎖または二本鎖であることができる。
【0027】
[0027] 本明細書において使用される「特異的な結合をする」または「特異的な結合」または「特異的に結合する」は、異種起源のタンパク質および他の生体物質の集団の存在下での、標的(例えば、タンパク質)の存在について決定的な結合反応に言及するものである。よって、指定されたイムノアッセイ条件下で、抗体もしくはペプチボディのような特定されたFc−コンジュゲートまたは他の結合ポリペプチドは、特定のタンパク質に結合し、そして試料中に存在する他のタンパク質へは統計学的に有意な量で結合しない。典型的には、Fc−コンジュゲート(例えば、抗体、ペプチボディ)は、スクリーニング方法(例えば、ファージディスプレイ)によって、またはタンパク質またはそのエピトープを用いる免疫化によって、タンパク質へ特異的に結合する能力について選択される。特異的結合を決定するために用いることができるイムノアッセイの形式についての記載は、HarlowおよびLane (1998), Antibodies, A Laboratory Manual, Cold Spring Harbor Publications, New Yorkを参照のこと。例えば、固相ELISAイムノアッセイは特異的結合を決定するのに用いることができる。特異的結合は、少なくとも約1×107M−1、およびしばしば少なくとも1×108M−1、1×109M−1、1×1010M−1、の結合定数を生じる。
【0028】
[0028] 本明細書において、「ベクター」は、本発明のポリヌクレオチドの宿主細胞への導入に用いる核酸への言及を含む。ベクターはしばしばレプリコンである。発現ベクターは、適切な宿主細胞中で、または適切なインビトロ条件下で存在するときに、その中に挿入された核酸の転写を許容する。
【0029】
B.Fc−コンジュゲート
[0029] 本発明は、天然のIgG1またはIgG3 Fcと比較して断片化および/または凝集に対して耐性である、単離されたIgG1およびIgG3 FcおよびFc−コンジュゲート、ならびにこれらの組成物を製造し使用する方法を提供する。理論に拘束されるわけではないが、フリーラジカル仲介断片化の機構は、Fcの断片化において、IgG1イムノグロブリンのヒンジコア配列に存在するヒスチジン残基に関係する。IgG1およびIgG3Fc中のヒンジコア配列ヒスチジン残基の適切な置換または欠失は、修飾されていないFcまたはFc−コンジュゲートに対して、ラジカル仲介断片化および/または凝集の度合いを減少させることができる。
【0030】
[0030] 本発明は活性酸素種による断片化および/または凝集に対して耐性にする修飾を有する、単離されたFcおよびFc−コンジュゲートを提供する。哺乳類イムノグロブリンのFc(結晶化可能フラグメント)は、「ヒンジコア配列」を有するヒンジ領域を含む、よく特徴付けされた構造である。表1は、ヒトIgGサブタイプにおいて見出され、一文字アミノ酸コードで表示された、ヒンジコア配列のリストを示す。Edelmanら(Proc. Natl. Acad. Sci. USA 63:78-85, 1969)の番号付けシステムにおいて、IgG1のヒンジコア配列はIgG1重鎖の残基216〜230に対応する一方、IgG3のヒンジコア配列はIgG3重鎖の残基214〜230に対応する。本発明において、表1に示されるように、IgG1またはIgG3ヒンジコア配列中に(224残基において)存在するヒスチジン残基(「H」)は、水素結合を形成することができる極性アミノ酸残基に置換されている。IgG1およびIgG3のヒンジコア配列中のヒスチジン残基について置換可能なアミノ酸残基の特定の例は、Ser、Gln、Asn、またはThr残基である。あるいは、当該ヒスチジン残基はヒンジコア配列から欠失されていてもよい。
【0031】
[0031]
【0032】
【表1】
【0033】
[0032] 典型的には、ラジカル媒介断片化に耐性のFcを生じる置換または欠失に供された本発明のFcまたはFc−コンジュゲートは、ヒトのIgG1またはIgG3 Fcであろう。しかしながら、IgGサブタイプの特徴を保持しながら、ヒトIgG1またはIgG3 Fcへの限定された数の置換、付加または欠失を行うことができる。よって、例えば、IgG1またはIgG3 Fcの0、1、2、3、4、5、6、7、8、9、または10アミノ酸を修飾することができ、そしてそれらもなお本願発明の範囲内である。よって、修飾されたIgG1またはIgG3 Fcは、天然型のヒトIgG1またはIgG3 Fcに対して95%、96%、97%、98%、または99%同一であろう。いくつかの態様において、(表1に示したような)本発明のIgG1またはIgG3ヒンジコア配列への唯一の修飾は、上述のようにヒンジコア配列中のヒスチジン残基の置換である。Fc−コンジュゲートは一価または二価構造であることができる。二価Fc−コンジュゲートのそれぞれのコンジュゲートは、同一または異なるコンジュゲートであることができる。
【0034】
[0033] Fcに共有結合または非共有結合で結合してFc−コンジュゲートを形成するコンジュゲートは、化学療法剤のような薬剤、放射標識のような診断標識、ヒト細胞表面受容体の細胞外ドメインのようなタンパク質を含むまたはそれらからなることができる。いくつかの態様において、当該コンジュゲートは、Fc−コンジュゲートがIgG1またはIgG3抗体であるように、Fab抗体セグメントを含むまたはそれからなる。抗体はポリクローナルまたはモノクローナルであることができる。いくつかの態様において、Fc−コンジュゲートは、完全なヒトモノクローナル、または、そうでなければ完全なヒトIgG1またはIgG3上に非ヒト供給源(例えば、ネズミ)由来のCDR(相補性決定領域)がグラフトされた、ヒト化モノクローナルである。抗体はアゴニストまたはアンタゴニスト抗体であることができ、それにより該抗体は受容体活性化を活性化または阻害する。いくつかの態様において、受容体はヒト細胞表面受容体であり、ここでFc−コンジュゲートは当該細胞表面受容体の細胞外ドメインに特異的に結合する。他の態様において、Fc−コンジュゲートはヒト細胞表面受容体のリガンドに特異的に結合して、該リガンドが該受容体に結合するのを妨げる。Fc−コンジュゲートが結合できるヒト細胞表面受容体の例は、デスレセプター4(TRAIL受容体−1)、デスレセプター5(TRAIL受容体−2)、VEGF(血管内皮増殖因子)、TNFR(腫瘍壊死因子受容体)、RANK(核内因子カッパb活性化受容体)受容体、またはTie−1及びTie−2受容体を含む。他の態様において、Fc−コンジュゲートのコンジュゲートは、望ましい標的に特異的に結合するペプチド(「ペプチボディ」)である。ペプチボディは、公開番号WO 2000/24782(引用により本明細書に援用する)を有する国際出願において教示されている。
【0035】
C.核酸
[0034] 本発明はまた、本発明のFcまたはFc−コンジュゲートをコードする核酸を含む単離されたポリヌクレオチドに向けられる。都合の良いことには、Fc−コンジュゲートのコンジュゲートがタンパク質であるとき(Fc−タンパク質コンジュゲート)であり、そして、例えば抗体、ペプチボディ、またはFc−細胞表面受容体融合体(またはその断片)をコードする場合、本発明の核酸はFc−タンパク質コンジュゲートを全体としてコードすることができる。
【0036】
[0035] 本発明のFcおよびFc−タンパク質コンジュゲートを製造するための組換え方法は、一般的に本発明のIgG1またはIgG3 Fcをコードする単離された核酸を含むポリヌクレオチドを使用する。本発明のFc−タンパク質コンジュゲートをコードする核酸は、当該技術分野において公知のインビトロオリゴヌクレオチド合成の方法により直接的に合成することができる。あるいは、より小さいフラグメントを合成し、当該技術分野で公知の組換え法を用いてより大きなフラグメントを形成するように連結することができる。いくつかの態様において、望ましいヒンジコア配列置換または欠失を伴う核酸プライマーをPCRに基づくインビトロ突然変異誘発に用いて、本発明のFcまたはFc−コンジュゲートを創出する。本発明のポリヌクレオチドはまた、インビトロ合成手段(例えば、固相ホスホロアミダイト合成)、またはその組み合わせ、を通じて構築することもできる。そのような方法は、当業者に周知である。例えば、Current Protocols in Molecular Biology, Ausubelら編、Greene Publishing and Wiley-Interscience, New York (1995)を参照されたい。
【0037】
D.Fc−コンジュゲートの構築
[0036] 本発明の単離されたFcまたはFc−タンパク質コンジュゲートを発現するために、これらの組成をコードする単離されたDNAを、標準的な分子生物学的技術(例えば、PCR増幅、部位特異的突然変異誘発)によって得ることができ、そしてそれを、当該遺伝子が転写および翻訳制御配列に機能可能に連結されるように、発現ベクターに挿入することができる。
【0038】
[0037] よって、本発明は本発明の核酸を含む発現ベクター(ポリヌクレオチド)を含む。発現ベクターは、プラスミド、レトロウイルス、コスミド、YAC、EBV由来エピソーム等を含む。発現ベクターは、本願発明のFcまたはFc−タンパク質コンジュゲートの宿主細胞からの分泌を促進するシグナルペプチドをコードすることができる。FcまたはFc−タンパク質コンジュゲート遺伝子は、シグナルペプチドがFc/Fc−タンパク質コンジュゲート遺伝子のアミノ末端に対してインフレームで(in-frame)連結されるように、ベクター中へクローニングすることができる。シグナルペプチドは、イムノグロブリンシグナルペプチドまたは異種性のシグナルペプチド(すなわち、イムノグロブリンではないタンパク質由来のシグナルペプチド)であることができる。
【0039】
[0038] 発現ベクターおよび発現制御配列は、使用する発現宿主細胞に適合性であるように選択される。適合性のベクターおよび宿主細胞系は、例えば、抗体であるFc−コンジュゲートの可変重鎖及び可変軽鎖の共発現および会合を許容することができる。発現のために適切な系は、当業者によって決定されることができる。いくつかの態様において、発現ベクターは分断(split)DHFRベクター、PDC323またはPDC324である;McGrew, J. T.およびBianchi, A. A. (2002) “Selection of cells expressing heteromeric proteins”, 米国特許出願第20030082735号;および、Bianchi, A. A.およびMcGrew, J. T., “High-level expression of full antibodies using trans-complementing expression vectors,” Bioengineering and Biotechnology 84(4): 439-444, 2003を参照のこと。Fc−コンジュゲートが抗体である場合、本願発明の可変重鎖核酸及び抗体可変軽鎖核酸は別々のベクター中へと挿入される、あるいは、頻繁に、両遺伝子は同じ発現ベクター中へと挿入されることができる。標準的な方法によって、核酸を発現ベクター中に挿入することができる(例えば、抗体核酸フラグメントおよびベクター上の相補的な制限酵素部位のライゲーション、または制限酵素部位が存在しない場合には平滑末端ライゲーション)。
【0040】
[0039] 本発明の核酸および発現ベクターは、トランスフェクションを介して宿主細胞へと導入することができる。用語「トランスフェクション」の様々な形は、原核生物または真核生物の宿主細胞への外来性DNAの導入のために一般的に用いられる多種多様の技術(例えば、エレクトロポレーション、リン酸カルシウム沈殿法、DEAE−デキストラントランスフェクション、など)を包含することを意図する。本発明のFc−コンジュゲートを原核生物宿主細胞または真核生物宿主細胞のいずれかにおいて発現することが理論的には可能であるが、真核生物細胞(そして最も好ましくは哺乳動物宿主細胞)における抗体の発現、が最も典型的である;なぜなら、そのような真核生物細胞(そして特に哺乳動物細胞において)は、原核生物細胞よりも、適切にフォールディングされ、そして免疫学的に活性な抗へと会合し、それを分泌する可能性が高いからである。
【0041】
[0040] 本願発明の発現ベクターは、宿主細胞において配列の発現を制御する調節配列を保有する。そのような調節配列は、例えばGoeddel; Gene Expression Technology. Methods in Enzymology 185, Academic Press, カリフォルニア州サンディエゴ(1990)に記載されている。調節配列の選択を含む発現ベクターの設計は、形質転換する宿主細胞の選択、望むタンパク質の発現レベル、等の要因に依存してもよいことを当業者は理解するであろう。哺乳動物宿主細胞発現のための望ましい調節配列は、哺乳動物細胞において高レベルのタンパク質発現を指示するウイルスの成分、例えば、サイトメガロウイルス(CMV)、サル・ウイルス40(SV40)、アデノウイルス、(例えばアデノウイルス主要後期プロモーター(AdMLP))およびポリオーマ由来のプロモーターおよび/またはエンハンサー、を含む。あるいは、非ウイルス性調節配列、例えばユビキチンプロモーターまたはベータ・グロブリンプロモーターを用いてもよい。
【0042】
[0041] 本発明の発現ベクターは付加的な配列、例えば宿主細胞中のベクターの複製を調節する配列(例えば、複製起点)および選択可能マーカー遺伝子、を保有してもよい。選択可能マーカー遺伝子は、ベクターが導入された宿主細胞の選択を容易にする(例えば、すべてAxelらによる、米国特許第4,399,216号、第4,364,665号及び第5,179,017号を参照されたい)。例えば、典型的には選択可能マーカー遺伝子は、ベクターが導入された宿主細胞に、G418、ハイグロマイシンまたはメトトレキセートのような薬物に対する耐性を付与する。好ましい選択可能マーカー遺伝子は、(dhfr− 宿主細胞において、メトトレキセート選択/増幅に用いるための)ジヒドロ葉酸還元酵素(DHFR)遺伝子、および(G418選択のための)neo遺伝子を含む。
【0043】
[0042] 本発明のFcまたはFc−コンジュゲートを発現するための好ましい哺乳動物宿主細胞は、チャイニーズハムスター卵巣(CHO細胞)(例えばKaufman, R. J.およびSharp, P. A., Mol. Biol. 159:601-621, 1982に記載されるDHFR選択可能マーカーとともに使用される、UrlaubおよびChasin, Proc. Natl. Acad. Sci. USA 77: 4216-4220, 1980に記載されるdhfr− CHO細胞を含む)、NS/0骨髄腫細胞、COS細胞およびSP2.0細胞を含む。特に、NS/0骨髄腫細胞を伴う使用について、別の好ましい発現系は、WO 87/04462, WO 89/01036およびEP 338841に開示されるGS遺伝子発現系である。本発明の発現ベクターが哺乳動物宿主細胞に導入される場合、FcおよびFc−コンジュゲートは、宿主細胞を適切な培養培地中で、宿主細胞におけるそれらの発現を可能にさせるのに十分な時間で培養することにより、より好ましくは当該宿主細胞が増殖した培養培地へのFcまたはFc−コンジュゲートの分泌により、産生される。
【0044】
[0043] ひとたび発現されたなら、FcまたはFc−コンジュゲートは、HPLC精製、分画カラムクロマトグラフィー、ゲル電気泳動等の当該技術分野の標準的な方法に従って単離のために精製することができる(例えば、Scopes, Protein Purification, Springer-Verlag, NY, 1982を参照のこと)。特定の態様において、ポリペプチドはクロマトグラフィーまたは電気泳動技術を用いて精製される。例示的な精製方法は、非限定的に、硫酸アンモニウムでの沈殿;PEGでの沈殿;免疫沈降;熱変性に続く遠心分離;アフィニティークロマトグラフィー(例えば、プロテインA−セファロース)、イオン交換クロマトグラフィー、排除クロマトグラフィー、および逆相クロマトグラフィーを含むがこれらに限定されないクロマトグラフィー;ゲル濾過;ヒドロキシアパタイト・クロマトグラフィー;等電点電気泳動;ポリアクリルアミドゲル電気泳動;ならびに、それらおよび他の技術の組み合わせ;を含む。特定の態様において、ポリペプチドは高速タンパク質液体クロマトグラフィーまたは高速液体クロマトグラフィー(HPLC)によって精製される。
【0045】
E.医薬組成物
[0044] 本発明は、薬学的に許容されうる担体と共に処方された本発明のFc及びFc−コンジュゲートを含む医薬組成物を提供する。いくつかの態様において、当該医薬組成物はヒト対象における投与に適している。本明細書において「薬学的に許容されうる担体」は、特定の対象に投与したときに生理学的に適合しうる、溶媒、分散媒、コーティング、抗菌薬および抗真菌薬、等張剤および吸収遅延剤、等のいずれか及びすべてを含む。典型的には、医薬組成物は、製造および保管の条件下で無菌であり、そして安定でなければならない。
【0046】
[0045] 本発明の医薬組成物は、組み合わせ療法において投与する、すなわち他の剤との組み合わせることができる。剤は、インビトロで合成的に調製した化学的組成物、抗体、抗原結合領域、放射性核種、ならびに、それらの組み合わせおよびコンジュゲート、を含むがこれらに限定されない。
【0047】
[0046] 用量投与計画は、望ましい最適な応答(例えば、療法的応答)を提供するように調整される。例えば、単一ボーラスを投与してもよく、いくつかの分割された用量が時間をかけて投与されてもよく、または用量は療法的状況の緊急性が示すところにより、比例的に減少または増加させてもよい。投与の容易さおよび用量の均一性のために、用量単位剤形(dosage unit form)で非経口組成物を処方することが特に有利である。本明細書で用いる用量単位剤形は、治療される対象のための単位用量に適した物理的に別個の単位を意味する;それぞれの単位は、薬学的担体を伴って、望ましい療法的効果を生じるよう計算された、予め決定された量の活性化合物を含有する。本発明の用量単位剤形の内訳は、(a)活性化合物の独特の特徴および達成される特定の療法的効果、および(b)患者における感受性の治療のための、そのような活性化合物を混ぜ合わせる技術分野に固有の制限、によって指図され、そしてそれらに直接的に依存する。
【0048】
E.療法的おより診断的コンジュゲート
[0047]療法的および/または診断的利点を向上する、本明細書に記載した種々の療法的部分は、本発明のFcへ直接的または間接的(例えば、「連結基」を介して)に共有結合により連結し、Fc−コンジュゲートを生じることができる。連結基は任意である。リンカーはしばしば、ペプチド結合でともに連結されたアミノ酸で作られる。当業者によく理解されているように、1またはそれより多くのこれらのアミノ酸はグリコシル化されていてもよい。非ペプチドリンカーもまた、可能である。非ペプチドリンカーの例は、PEG(ポリエチレングリコール)リンカーである。
【0049】
[0048] そのような療法的部分を抗体にコンジュゲート化する技術は周知である、例えばArnonら、 “Monoclonal Antibodies For Immunotargeting Of Drugs In Cancer Therapy”, Monoclonal Antibodies And Cancer Therapy中, Reisfeldら(編), pp.243-256 (Alan R. Liss, Inc. 1985); Hellstromら、“Antibodies For Drug Delivery”, Controlled Drug Delivery (2nd Ed.)中, Robinsonら(編), pp. 623-653 (Marcel Dekker, Inc. 1987); Thorpe, “Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review”, Monoclonal Antibodies '84: Biological And Clinical Applications中, Pincheraら(編), pp. 475-506 (1985); “Analysis, Results, And Future Prospective Of The Therapeutic Use Of Radiolabeled Antibody In Cancer Therapy”, Monoclonal Antibodies For Cancer Detection And Therapy中, Baldwinら(編), pp. 303-316 (Academic Press 1985), およびThorpeら、 “The Preparation And Cytotoxic Properties Of Antibody-Toxin Conjugates”, Immunol. Rev. 62: 119-158, 1982、を参照のこと。本発明の組成物は、放射性核種とカップリングしてもよい、例えば、Goldenberg, D. M.ら、Cancer Res. 41 : 4354-4360, 1981、及びEP 0365 997に記載されるような、131I、90Y、105Rh、インジウム−111等。
【実施例】
【0050】
[0049] 実施した実験および達成した結果を含む以下の実施例は、例証の目的のためにのみ提供されるものであり、本発明を限定する物として解釈されるものではない。
実施例1
[0050] 本実施例は、一つのFabドメインの損失および部分的分子の形成につながるH2O2媒介ラジカル切断による、ヒトIgG1抗体の特定のヒンジ断片化の結果を示すものである。IgG1に対するH2O2攻撃は、ヒンジ領域中の位置226に位置する二つのシステイン残基(Cys226)の間の鎖間ジスルフィド結合の破壊、そして続いて、スルフェン酸(Cys226SOH)およびチイルラジカル(Cys226S・)の形成を生じ、これは上部ヒンジ領域への電子移動を開始し、ラジカル媒介ポリペプチド骨格断片化につながる。
【0051】
[0051] 用いた抗体は、IgG1サブクラスの組換え完全ヒト抗体であった。当該分子はCHO細胞において発現され、そして慣習的な技術を用いてクロマトグラフィーにより精製した。抗体フラグメントはサイズ排除クロマトグラフィー(SEC)により分離した。抗体の切断は、部分的分子の割合(C1およびC2)によって測定した。
【0052】
[0052] 簡略的に、緩衝液中の2mgから10mgのIgG1抗体を含有する反応混合物(1.0mL)を、変化させたH2O2濃度を伴ってインキュベートした。H2O2を除去するために、フィルターユニットおける遠心分離によりサンプルの緩衝液交換を行った。精製された部分的分子(〜1mg/mL)を還元し、アルキル化した。アルキル化は暗所において室温で行い、そして0.5M DTTストック溶液を加えてアルキル化を停止した。逆相高速液体クロマトグラフィー(RP−HPLC)、続いてエレクトロスプレーイオン化(ESI)飛行時間(TOF)質量分析(MS)を行った。
【0053】
[0053] 精製されたバルクの抗体を、サイズ排除クロマトグラフィー(SEC)により分析し、そして〜0.9%の部分分子を示した(P1)。これは一つのロットからの単一のケースではなく、0.9〜1.1%の範囲でいくつかのランにおいて示された。P1種は、SEQにより95%より大きい純度にさらに精製され、そしてRP−HPLC−TOF/MSにより分析した。結果は、P1は一つのFabドメインを喪失した重度に酸化された部分抗体であることを示した。
【0054】
[0054] H2O2は、タンパク質に対して酸化および損傷を生じることができるものとして公知である。酸化的ストレスが切断を生じるかどうか探るために、IgG1を処理するためにH2O2を用い、そしてSECによりその影響を測定した。5〜20mM H2O2の範囲を通じて、最初の8時間のインキュベーションでは特筆すべき切断は見出されなかった。20mM H2O2でのインキュベーションの48時間後にのみ、二つの部分フラグメント、−C1およびC2−、が観察された。これらの二つのフラグメントの量は、インキュベーションの長さに対して直接比例して増加した。この断片化は、抗体濃度およびpH条件にも依存する。加えて、切断は、8週間までの間にも、顕著な安定相を伴わずに進行した。二つの産物(C1およびC2)のみが観察されたという事実は、切断は特異的であり、そしておそらく特定の機構により駆動されたものであることを示唆した。P1の重度に酸化された性質を示す続いての研究は、ヒンジ断片化はCHO細胞による抗体産生の間の酸化的ストレスによるものであり得ることを示唆する。P1およびC1の類似性、特に長期のH2O2処理において観察されたより高い酸化レベルは、酸化的ストレスがヒンジ断片化を生じさせたことを示す。
【0055】
[0055] C1およびP1のRP−HPLC−TOF/MS分析は、それらが同じ種であり、それらのそれぞれはFabドメインを失っている重度に酸化された部分分子である、特に、Fcドメインの単一の相補的HCは、上部ヒンジ領域におけるN末端残基Asp221、Lys222、Thr223およびThr225の独特の「ラダー(ladder)」を含む、ことを示した。加えて、45Daおよび71Daの二つの付加物が同じFc断片において観察された、これらは公知の修飾とは一致しないために通常の付加物ではない。
【0056】
[0056] C2断片のRP−HPLC−TOF/MS分析は、これがIgG1のFabドメインであり、そして重度に酸化されていることを明らかにした。C2のLCは、C1について対応するものに対して類似のプロフィールを示した。C2におけるHC(Fd)のFab部分は二つの構成成分を有し、それらの双方は1つまたは3つの酸素付加を伴って重度に酸化されていた。より重度に酸化された構成成分はC末端残基Asp221、Lys222、Thr223、His224およびThr225のラダーを含有した;より軽度に酸化されたFd構成成分は、Ser218からThr225のC末端残基からなる、より幅広いラダーを持っていた。これらの結果は、H2O2処理は、IgG1のLCおよびHCの双方におけるヒンジ切断および顕著なレベルの酸化という結果に帰することを示した。
【0057】
[0057] これらの付加物の性質およびその位置を組み合わせると、データはラジカル切断はヒンジ断片化の原因であることを示唆する。過酸化水素は、ラジカルにより誘導される酸化経路を通してタンパク質の生物学的機能を制御することができる。ヒドロキシルラジカルを伴う反応は、結果的にタンパク質の分解に導く種々の化学反応をもたらすことができる(Garrison, W. M., Chem. Rev. 87: 381-398, 1987; Davies, M. J. および Dean, R. T., 1997, Radical mediated protein oxidation. Oxford University press, pp 50-120; Berlett, B. S. およびStadtman, E. R., J. Biol. Chem. 272: 20313-20316, 1997)。
【0058】
[0058] OHラジカルがヒンジ断片化に関与するかを調べるため、そして当該切断に影響を及ぼしてもよいいくつかの因子を評価するため、いくつかの前処理の後にIgG1をH2O2攻撃に供した。これらは、H2O2処理の前に対を形成していないCys残基をブロキングするためのN−エチル−マレイミド(NEM)前処理、または反応系へとカタラーゼもしくはエチレン−ジアミン−テトラー酢酸(EDTA)を添加することを含む。SECを影響を測定するために実施した。カタラーゼはほぼ完全に切断をブロックしたことが見出され、OHラジカルは切断について重要であることを強く示した。フリーのチオール基の総量は、Ellmanのリガンド、5,5’−ジチオビス(2−ニトロ安息香酸)(DTNB)を用いる4M GdnHClの存在下の変性条件下で、〜0.28モル/モル抗体であると測定された。H2O2処理の前に、IgG1はNEMとともに、pH5.0、37℃で、3時間インキュベートした。NEMでブロックされた試料は、切断において〜7%の現象を示す一方、NEM処理によってフリーのチオール(−SH)基は完全にブロックされた。よって、この結果は、対を形成していないCys残基は切断について重大な意味を持たないことを示唆した。
【0059】
[0059] 5’5−ジメチルー1−ピロリン N−オキシド(DMPO)スピントラップは、多くの生物学的反応におけるフリーのラジカルの関与、特にOHラジカル、についての証拠を提供するために広く用いられている。DMPOはミオグロビンおよび他の分子におけるラジカル損傷へと曝露されたラジカル部位を同定するのに用いられてきた。よって、IgG1をDMPOの存在下で1週間H2O2で処理し、そしてSECによって断片化を監視した。50:1から5:1のDMPO:H2O2のモル比範囲で、DMPOは2週間のインキュベーションの時間経過を通して断片化を完全にブロックした。
【0060】
[0060] ラジカル形成部位を同定するために、Lys−Cペプチドマッピングを行った。Cys231−SO3Hを含有する無傷のヒンジペプチドが観察される一方、HC Cys231のみがDMPO付加物を含有するものとして見出された。上部ヒンジ残基においてラジカル形成が見出されなかったことは、質量分析の感度の問題またはOHラジカルと上部ヒンジ残基の間の反応率によるものではなさそうである。OHラジカルはCysとの速度定数、3.4×1010M−1s−1を有し、これはHis(1.3×1010M−1s−1)、Thr(5.1×108M−1s−1)、Asp(7.5×107M−1s−1)、およびLys(3.5×107M−1s−1)よりもずっと速い(Davies, M. J. およびDean, R. T., 1997, Radical mediated protein oxidation. Oxford University press, pp 50-120)。よって、これらの結果は、分子あたりのラジカル切断に導く、HC Cys231から上部ヒンジの残基への電子伝達の必要性を立証した。電子がHisについて6.4×107M−1s−1、Thrについて2.0×107M−1s−1、Lysについて2.0×107M−1s−1、Aspについて1.8×107M−1s−1の反応速度定数を有することが決定され(Davies, M. J.およびDean, R. T., 1997, Radical mediated protein oxidation, Oxford University press, pp.50-120)、このことはこれらの残基はラジカル誘導骨格切断を進めるために電子を局在化させることができることを示している。この機構は、FabフラグメントのC末端残基(C2)および部分抗体のFcのN末端残基(C1)の相補性を形成する特異的ヒンジ断片化を説明する。
【0061】
実施例2
[0061] この実施例はIgG1 Fcのラジカル媒介断片化の結果をまとめたものである。
1.IgG1バルク抗体は〜1%の切断型抗体(P1)を含有し、これは、Fabドメインの一つを欠損した、重度に酸化された型と決定された。
2.IgG1バルク薬剤物質(BDS)とH2O2の反応は、切断型分子、そして重鎖のFabドメイン(Fd)におけるC末端残基のラダー(Cys220−Asp221−Lys222−Thr223−His224−Thr225)およびFcドメインにおける相補的N末端残基のラダーの形成に帰する、ヒンジ領域における特異的切断による一つのフリーのFabドメイン断片を生じた。
3.H2O2で処理した試料において、無傷のまたは切断された分子の大部分において、Cys226残基間の鎖間ジスルフィド結合は無傷であることが見出された。
4.BDS試料において、潜在的な対形成していないCysのN−エチルマレイミド(NEM)によるプレブロッキング後に行った天然型Lys−Cペプチドマップによって観察される、ヒンジ領域中の対形成していないジスルフィド結合は存在しなかった。
5.LC−MS/MS分析は、Cys226における少量のCys−SO3Hを、無傷のヒンジペプチド(THTCys226PPCAPELLGGPSVFLFPPKPK)(配列番号5)および切断型ヒンジペプチド(Cys226PPCAPELLGGPSVFLFPPKPK)(配列番号6)の両方において同定した。
6.切断された抗体において、FcドメインのN−末端ヒンジ領域において、それぞれ45または71Daの付加質量を導入するイソシアネートまたはN−α−ケトアシル誘導体のいずれかとして、付加物が同定された。
7.IgG1は〜0.28モル/モル抗体の対形成していないCys残基を含有し、これは、すべての対形成していないCys残基のブロッキングは断片化においてまったく効果を示さないか、ごくわずかな効果しか生じないという事実によって示されるように、切断反応には重大なものではない。
8.広く用いられるラジカルスピントラップ 5,5’−ジメチル−1−ピロリン N−オキシド(DMPO)は、それのCys226への結合のために、ヒンジ断片化をブロックすることができることが見出された。しかしながら、DMPO結合はCys226−SO3Hの形成をブロックしなかった。
【0062】
実施例3
[0062] 本実施例は、Cu2+ではなくヒドロキシルラジカルがヒンジ断片化を誘発することを示す。過酸化水素はタンパク質の生物学的機能を、ラジカルに誘導される酸化経路を通して制御することができる。加えて、ヒドロキシルラジカルを伴う反応は、タンパク質の分解に帰する、種々の化学反応を導くことができる。OHラジカルがヒンジ断片化に関与するか試験し、そして当該切断に影響しうるいくつかの因子を評価するために、IgG1をH2O2攻撃にさらした。図1に示すように、H2O2が誘導する断片化はカタラーゼによって完全にブロックされ、これはOHラジカルは当該切断の原因であることを示している。フリーのチオール基の全体は、Ellman試薬、5,5’−ジチオビス(2−ニトロ安息香酸)(DTNB)を用いる4M GdnHClの存在下の変性条件下で、〜0.28モル/モル抗体であると決定された。H2O2処理の前に、IgG1をNEMとともにpH5.0で3時間、37℃でインキュベートした。NEMでブロックした試料は、〜7%の切断の減少しか示さなかった一方、NEM処理によってフリーのチオール(−SH)基は完全にブロックされたことが見出され、対形成していないCys残基は切断に重要ではないことが示唆された。
【0063】
[0063] 加えて、EDTAとのプレインキュベーションは、IgG1のH2O2に誘導される切断の〜90%を阻害することが見出され、反応における遷移金属の関与が示唆される。しかしながら、そのような前処理はH2O2による切断を完全にブロックすることはなく、より遅い反応速度を有するものではあったが、依然としてIgG1を切断することが可能であった。これらの結果はOHラジカルがヒンジ断片化に重要であり、そしてこの反応は、OHラジカルを生成する金属により触媒される反応によって、促進されうることを示唆した。この仮説は、10μM 酢酸銅(Cu(OAc)2)の存在下でのH2O2を伴う処理が、H2O2処理単独よりもおよそ4倍多くの切断となる一方で、10μM Cu(OAc)2単独では5日間のインキュベーションの間にごくわずかの切断しか生じなかったという結果により支持された。
【0064】
[0064] Smithらは、いくつかの合成ペプチドを試験することにより、中性または塩基性pHでの1mMのCuSO4による、IgG1の上部ヒンジDKTHT(配列番号7)領域中のK−T結合の切断を報告した(Smith, M. A.ら、Int. J. Pept. Protein Res., 48:48-55, 1996)。本明細書に記載される実験条件下(pH5.2、および25℃でのインキュベーション)では、上部ヒンジ領域(例えば、His、Lys)へのCu2+の結合は中性または塩基性pHの場合よりも有望ではなく、そしてヒンジ断片化の〜30%増加という結果となった。溶媒またはタンパク質中に微量の遷移金属イオンが存在するので、それらの濃度はラジカル誘発ヒンジ断片化のための触媒として機能するのに十分であり得た。この結論は、いくつかの遷移金属(例えばCu2+およびFe3+)が、タンパク質に結合するかまたは溶媒中に留まることのいずれかにより、部位特異的ラジカル攻撃における重要な役割を果たすという理論に一致する。両方の場合において、当該金属は、Fenton様反応を通してヒドロキシルラジカルの生成を触媒することにより、反応を促進する。まとめると、これらの事実はラジカル誘発ヒンジ断片化機構を独立して確認した。
【0065】
実施例4
[0065] 本実施例は、ラジカル媒介Fc断片化の機構を提案する。我々のヒトIgG1の研究の実験結果は、このヒトIgG1抗体におけるラジカル媒介ヒンジ断片化を明らかにした。
【0066】
[0066] 微量の遷移金属は反応系におけるOHラジカルの生成を触媒する。IgG1抗体とOHラジカルとの反応は、抗体のヒンジ領域(Cys226−Pro−Pro−Cys−Pro)における226位に位置する二つのシステイン残基(Cys226)の間の鎖間ジスルフィド結合の切断という結果となった。そのジスルフィド結合切断の後、スルフェン酸(Cys226−SOH)およびチイルラジカル(Cys226−S・)の形成が続いた。酸素の存在下でのこれらの種の続く反応は、主要な産物としてスルフィン酸(Cys226−SO2H)およびスルホン酸(Cys226−SO3H)の形成という結果となった。一方チイルラジカルは、ヒンジポリペプチド骨格に沿って、電子移動の上流側(electron transfer upstream)を開始する。この電子移動は、ラジカル媒介ポリペプチド骨格断片化を導き、これは、いくつかの隣接するヒンジ残基(Asp221、Lys222、Thr223、His224、およびThr225)における切断のために生じる、重鎖のFabドメイン(Fd)におけるC末端残基のラダーによって特徴づけされる。我々は、広く用いられているラジカルスピントラップである5,5’−ジメチル−1−ピロリン N−オキシド(DMPO)の結合を、Cys226でのみ観察し、これはヒンジ断片化をブロックした。DMPOのCys226のみへの特異的結合は、IgG分子を通じて高度に保存された配列モチーフCys−Pro−X−Cys−Pro(X=Pro、Arg、およびSer)(表1)である、CPPCP配列においてCys226のみにラジカルが存在することが確認された。
【0067】
[0067] +45Da付加物の決定は、ジアミド経路を通じてFcのN末端にイソシアネート構造(MW=28Da)を生じるラジカル切断機構を示唆した。イソシアネート基は、その不安定な性質により、カルボン酸(+45Da付加物)へと加水分解する。一方、特定のアミノ酸の側鎖のγ−炭素位におけるOHラジカル攻撃は、側鎖としてβ−CH2のみを保持する、デヒドロペプチドの不飽和産物の形成へとつながる酸化的分解という結果になりうる。この化合物は容易に加水分解されて、+71Da付加物(N−ピルビル基)である、アミドおよびケト官能基を生じることができる。そのために、Thr225のN末端において観察された+71Da付加物は、His224の酸化的分解によって生じたものであり得た。一方、これらの不安定な中間体の加水分解は、それらをリサイクルする別の方法である可能性があり、そしてこの過程は通常のN末端残基を含有するいくつかの切断されたヒンジペプチドをもたらした。まとめると、上部ヒンジ領域のN末端残基における+45Daおよび+71Da付加物は、それぞれタンパク質骨格のα−炭素およびアミノ酸側鎖のγ−炭素位におけるラジカル切断の産物であり、このことはタンパク質骨格切断についてのラジカル媒介機構を確認するものである。
【0068】
実施例5
[0068] 本実施例は、ヒンジコア配列中のHisおよびLys残基の変異によるラジカル媒介断片化への耐性を実証する。調査は、ヒト野生型IgG1との比較において、His224およびLys222を変異させている効果について決定するために行った。野生型IgG1および7つの変異体をH2O2とともにインキュベートし、部分的分子の形成および特にFabドメイン断片の放出をSECにより監視した。7つの変異体は以下である:Lys222Ser(K/S)、Lys222Gln(K/Q)、Lys222Ala(K/A)、His224Ser(H/S)、His224Gln(H/Q)、His224Ala(H/A)、およびLys222Ser/His224Ser(K/S+H/S)。これらの変異体の中で、HisをGlnまたはSerで置換したものは、Fabドメイン(C2)および部分的分子C1の放出につながるOHラジカルに誘発される断片化をほぼ完全にブロックした(>97%)。His/Ala変異は8日間のインキュベーション期間で〜6%の断片化を示し、これに対して天然型IgG1については〜15%であった。対照的に、すべてのLys単独変異体は、31〜33%で切断を促進した。より重要なことには、二重変異体であるK/S+H/Sは>97%の断片化阻害を示し、これはHis/SerまたはHis/Gln単独変異体について測定されたのと同じ割合であり、このことは断片化におけるHis残基の重要性を示している。
【0069】
[0069] His/Ala変異体は切断を示したが、当該変異体が同じ構造的分解を含むかどうかは知られていなかった。LCおよびHCは、それらを連結している鎖間ジスルフィド結合なしに、強く会合したままであることが記載されていた(Bigelow. C.ら、Biochemistry, 13: 4602-4609, 1978)。よって、LCおよびHCが、鎖間ジスルフィド結合なしに共に保持されること、およびFabドメイン断片と同様のSECプロフィールを示すことは可能である。よって、変異体を、H2O2処理の1日後に、非還元条件下でRP−HPLC−TOF/MSによってさらに試験した。これらの条件下では、共有結合で結合していない成分のみが主要な種から分離されることが期待される。図2に示すように、〜21分で溶出される主要ピークのほかに、16.5分の保持時間で移動してくる一つの成分がすべての変異体について観察された。特に、H/A変異体はこの種をH/SおよびH/Q変異体よりもおよそ15倍多く放出した。TOF/MS分析は、この種について23,437.5Daの分子量を決定し、これはLCについての理論質量である23,389.0Daよりも+48Da重い。このLys−CペプチドマップのRP−HPLC−MS/MS分析は、この種はLC Cys215のスルホン酸(+48Da)への完全な転換を示すものと確認し、このことはH2O2攻撃による鎖間ジスルフィド結合の切断はLCの酸化を導いたことを示唆するものである。これらの結果はOH基の除去は、側鎖におけるH−結合形成の可能性を消滅させ、この残基がラジカル攻撃に耐える能力を不利に損なったことを示唆した。
【0070】
[0070] IgG1の上部ヒンジと同じ配列(DKTHT)(配列番号7)を含む合成ペプチド(FDKTHTY)(配列番号8)を用いて、Allenら(Allen, G.およびCampbell, R., Int. J. Peptide Protein Res. 48: 265-273, 1996)はHis/Ala置換は、Cu2+(1mM)が誘導するペプチドの切断を阻止することを見出した。しかしながら、我々の結果はHis/Ala変異体は、LCおよびHCの間の鎖間ジスルフィド結合のH2O2誘導切断によるLCの放出を阻止しなかったことを明らかに示した。LCの喪失は、IgGの機能を破壊するであろう。特に、二つのヒンジ鎖間ジスルフィド結合がFabドメインへと連結している上部ヒンジ(DKTHT)(配列番号7)で二つのHCを連結しているヒンジ領域は、溶液中の合成ペプチドのコンフォメーションとは非常に異なる可能性が最も高いコンフォメーションをヒンジが採用するよう制限する、二重鎖構造である。したがって、ペプチドから得られた結果は、同じまたは類似の配列を含有するタンパク質に適用する場合には、注意深く取り扱う必要がある。まとめると、我々の結果は、His/Ala変異体ではなく、His/SerおよびHis/Gln変異体が、OHラジカル媒介切断を阻害したことを明らかに示した。
【0071】
[0071] His、Gln、Ser、AlaおよびLysの側鎖の性質を前提とすると、これらの変異体の解析結果は、His残基の側鎖のγ−炭素よりはむしろイミダゾール環が、ヒンジ切断の原因であると我々は結論付けさせる。この仮定は、Glnはその側鎖にγ−炭素を有するが、His/Gln変異体はラジカル誘導切断を阻害したという観察結果によって支持された。電子付加の主要な部位は、プロトン化されるpH値におけるHis224のイミダゾール環であるようである。OHラジカルはイミダゾール環のC−2、C−4およびC−5位に付着することが知られている。IgG1の公知の三次元構造におけるこれらのヒンジ領域の配置、およびヒンジ周囲の水素結合ネットワークに基づいて、続くこれらの種への酸素の付加が、最初のラジカルに塩基に触媒される水の喪失を被らせ、高度に安定化したビスアリルラジカルを与えることを我々は提案する。His残基は電子を局在化させる中心的な標的として機能し、そして続いて隣接する残基からプロトンを抽出し、ジアミドおよびα−アミド化経路によりラジカル誘発切断に導いた。まとめると、この結果は、合理的設計を用いてヒンジ断片化を防止する可能性を立証した。
【特許請求の範囲】
【請求項1】
単離されたラジカル媒介断片化に耐性のFc−コンジュゲートであって、当該FcはヒトIgG1またはIgG3 Fcであり、そして当該Fcはヒンジコア配列THTCPXCP(配列番号9)を含み、ここでXはRまたはP、そして当該ヒンジコア配列中のH残基は、Ser、Gln、AsnまたはThr残基で置換されている、前記Fc−コンジュゲート。
【請求項2】
Fc−コンジュゲートが、モノクローナル抗体、ペプチボディ、またはFc−受容体融合体である、請求項1に記載のFc−コンジュゲート。
【請求項3】
モノクローナル抗体が、完全ヒト モノクローナル抗体である、請求項2に記載のFc−コンジュゲート。
【請求項4】
当該モチーフ中のヒスチジン残基が、セリンまたはグルタミン残基である、請求項3に記載のFc−コンジュゲート。
【請求項5】
薬学的に許容可能な担体中の、請求項4に記載のFc−コンジュゲート。
【請求項6】
請求項2に記載のFc−コンジュゲートをコードするポリヌクレオチドを含む、単離された核酸。
【請求項7】
請求項6に記載の単離された核酸を含む、単離された発現ベクター。
【請求項8】
請求項7に記載の発現ベクターを含む、宿主細胞。
【請求項9】
宿主細胞がCHO細胞である、請求項8に記載の宿主細胞。
【請求項10】
請求項2に記載のFc−コンジュゲートを作成する方法であって、適切な宿主細胞中で、請求項5に記載の発現ベクターを、当該ベクターを発現するのに適した条件下で培養し、そして当該宿主細胞から発現されたFc−コンジュゲートを単離することを含む、前記方法。
【請求項1】
単離されたラジカル媒介断片化に耐性のFc−コンジュゲートであって、当該FcはヒトIgG1またはIgG3 Fcであり、そして当該Fcはヒンジコア配列THTCPXCP(配列番号9)を含み、ここでXはRまたはP、そして当該ヒンジコア配列中のH残基は、Ser、Gln、AsnまたはThr残基で置換されている、前記Fc−コンジュゲート。
【請求項2】
Fc−コンジュゲートが、モノクローナル抗体、ペプチボディ、またはFc−受容体融合体である、請求項1に記載のFc−コンジュゲート。
【請求項3】
モノクローナル抗体が、完全ヒト モノクローナル抗体である、請求項2に記載のFc−コンジュゲート。
【請求項4】
当該モチーフ中のヒスチジン残基が、セリンまたはグルタミン残基である、請求項3に記載のFc−コンジュゲート。
【請求項5】
薬学的に許容可能な担体中の、請求項4に記載のFc−コンジュゲート。
【請求項6】
請求項2に記載のFc−コンジュゲートをコードするポリヌクレオチドを含む、単離された核酸。
【請求項7】
請求項6に記載の単離された核酸を含む、単離された発現ベクター。
【請求項8】
請求項7に記載の発現ベクターを含む、宿主細胞。
【請求項9】
宿主細胞がCHO細胞である、請求項8に記載の宿主細胞。
【請求項10】
請求項2に記載のFc−コンジュゲートを作成する方法であって、適切な宿主細胞中で、請求項5に記載の発現ベクターを、当該ベクターを発現するのに適した条件下で培養し、そして当該宿主細胞から発現されたFc−コンジュゲートを単離することを含む、前記方法。
【図1】


【図2】
【図2】
【公表番号】特表2012−524545(P2012−524545A)
【公表日】平成24年10月18日(2012.10.18)
【国際特許分類】
【出願番号】特願2012−507359(P2012−507359)
【出願日】平成22年4月21日(2010.4.21)
【国際出願番号】PCT/US2010/031933
【国際公開番号】WO2010/124018
【国際公開日】平成22年10月28日(2010.10.28)
【出願人】(500203709)アムジェン インコーポレイテッド (76)
【Fターム(参考)】
【公表日】平成24年10月18日(2012.10.18)
【国際特許分類】
【出願日】平成22年4月21日(2010.4.21)
【国際出願番号】PCT/US2010/031933
【国際公開番号】WO2010/124018
【国際公開日】平成22年10月28日(2010.10.28)
【出願人】(500203709)アムジェン インコーポレイテッド (76)
【Fターム(参考)】
[ Back to top ]