
新たなクラスのウイロソーム粒子
本発明は、少なくとも一つの脂質、及び包膜ウイルスの包膜蛋白質を備えるウイロソーム膜と、ウイロソームの内側及び外側に位置し、前記包膜蛋白質に付着した前記包膜ウイルスのヌクレオカプシド粒子を備えるウイロソームを提供する。更に本発明は、本発明のウイロソームを備えるワクチンと、本発明のウイロソームを産生する方法を提供する。更に本発明は、本発明のウイロソームを、例えばHBV感染に関連した疾患の予防又は緩和のためのワクチンを調製するために使用する方法、及び被験者にワクチンを接種する方法を提供する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、少なくとも一つの脂質及び包膜ウイルスの包膜蛋白質を備えるウイロソーム膜、及びウイロソームの内側及び外側に位置し、前記包膜蛋白質に付着した前記包膜ウイルスのヌクレオカプシド粒子を備えるウイロソームに関する。更に本発明は、本発明のウイロソームを備えるワクチン、及び本発明のウイロソームの生成のための方法に関する。更に本発明は、本発明のウイロソームをワクチンの調製のために、例えばHBV感染に関連した疾患の予防又は緩和のために使用する方法、及び被験者にワクチンを接種する方法に関する。
【0002】
本願明細書のテキスト全般を通じていくつかの文献が引用されている。本願明細書中で引用されている各文献(いずれの製造業者の仕様書、指示書等も含む)の開示内容は参照することにより本願明細書に援用される。
【背景技術】
【0003】
新規であり、安全性がより高いワクチンの開発には、特性がよく知られた抗原、特に高度に精製された組み換え蛋白質又は合成ペプチドが頻繁に利用されている。いくつかの成果に関わらず、そのような抗原は単独で投与されたときには免疫原性が低いことが多いという事実により、このアプローチは妨げられる。この事実により、所定の抗原の免疫原性を促進する能力を有している適切なアジュバント及びキャリアシステムの開発が余儀なくさせる。一つの可能なアプローチは抗原を、例えばウイルス様の粒子など、より高度な構造中に組み込むことである。単一の粒子中で全てのワクチン成分が物理的に結合することは、それらが個々の免疫細胞と同時に相互作用することを保証し、そしてそれによって、相乗的な潜在力が最大に利用される。これは、免疫刺激性又は免疫調節性の成分(アジュバント)が調製物に含まれている場合には特に関連する。更に、粒子構造それ自身が免疫刺激性の効果を有すことができ、そして、個々の成分の安定性及び免疫原性の両者を増加させることができる。
【0004】
そこで問題は関連する抗原を産業上利用可能な調整物中に混ぜる適切なアプローチを見出すということであり、それは効率的な予防及び/又は治療上の適用へ至る。
【0005】
宿主細胞中でウイルスが複製する間にウイルスゲノムのコピー作られ、細胞内の設備基盤を利用しながらウイルス蛋白質が会合して成熟したビリオンとなるが、その前にウイルス蛋白質が発現してプロセスされる。共通の基礎は、ウイルスの複製と子孫の会合には、生きている宿主細胞の環境、および、ウイルスの核酸、ウイルスと宿主細胞の蛋白質と脂質膜の間の、順序が決まった一連の特異的な相互作用が必要であるということであり、それによって高分子のビリオン構造の分別と会合へと至る。多数の異なった分子が必用とされ、加えて、関連している細胞構造は、いずれのビリオンの会合も非常に複雑であることを示す。異なったウイルスクラス、特に包膜ウイルスと非包膜ウイルスの間には大きな差がある。全ての包膜ウイルスに共通しているのは、ウイルスの蛋白質が組み込まれた脂質膜からなるウイルスの外部殻、および、その結果としての、成熟した包膜ウイルスが会合するための、膜に結合し可溶性のウイルス蛋白質又は蛋白質に基づいた構造(例えばヌクレオカプシド)の間の必用な相互作用である。非包膜ウイルスは脂質に基づいた膜を欠き、蛋白質及び核酸分子のみから組みたてられている。
【0006】
インビトロとインビボでウイルス粒子を再構成しようとする数多くのアプロ−チが文献中に述べられており、異なったカテゴリーに区分けできる。
【0007】
(a)ウイルス包膜のインビトロ再構成
ウイルス由来の又は組み換えられた包膜蛋白質を精製し、追加の脂質の存在下又は非存在下でプロテオリポソームへ調製される。この純粋なインビトロのアプローチにより、包膜ウイルスの外殻、包膜の生成を達成できるが、これはウイルスの核、ヌクレオカプシドを含んでいない。異なったウイルス由来の包膜蛋白質を組み込んだ、キメラのウイロソーム構造の例がある。再構成されたウイルス包膜も、遺伝子(DNA又はRNA)の移入に成功裏に使用されているが、これらの方法は機能を有し、蛋白質に基づいたヌクレオカプシドのパッケージングには依存しておらず、むしろ、核酸と再構成された包膜との直接的な結合に依存している。
【0008】
(b)一つ又はそれ以上のウイルス蛋白質の非相同発現
単離された組み換えウイルス蛋白質は自己会合し、ウイルス様の構造(VLPs): HPV(酵母、バキュロ)、HCV(バキュロ)、HBs抗原(酵母、CHO)、HBc(大腸菌)となることができる。これらのアプロ−チの全てに共通することとして、自己会合は異種の細胞発現システム中で起こり、引き続いてウイルス様の粒子が精製される。そこで会合はインビトロでは起こらないが、細胞システムを頼りにする。VLPsはワクチン及びワクチン担体として使用されてきた(Pumpens, P.;Grens, E. (2001) Interviology 44 (2-3); 98-114; Noad R, Roy P. (2003) Trnds Microbiol. 11 (9): 438-44)。
【0009】
(c)インビトロでの非包膜(20面体の)ウイルス又はウイルス様粒子の再構成
このアプローチは、別々に精製された成分に基づいている。脂質膜に基づいた包膜が存在しないために、非包膜ウイルスは構造においてより単純であり、もし全ての必要な成分が正しい化学量論で存在していれば、インビトロである条件下において自己会合することができる。同様に包膜ウイルスの内部核、脂質がないヌクレオカプシド、又はそれらのサブユニットが、例えばインフルエンザウイルスの精製された組み換え成分からインビトロで再構成されている(Martin-Benito J. et al. (2001) EMBO Rep. 2 (4): 313-7)。
【0010】
(d)ウイルスのヌクレオカプシドの精製
多くの異なった型のウイルスからヌクレオカプシドが抽出され単離され、それらの組成の特性解析が行われてきた。ウイルスのレスキューを目的として、これらの調製物を感染しやすい細胞のトランスフェクションのために使用することもできる。成功裏のウイルスレスキューは、機能を有するヌクレオカプシドが単離され、宿主細胞の細胞質へ運ばれることを意味する。しかしこれは、機能を有する包膜ウイルスの再構成の成功を示すものではない。なぜならば、機能を有する包膜に依存する天然の感染経路は形質転換体の使用によってバイパスされ、後者は宿主細胞の細胞質へヌクレオカプシドを直接的に運搬することを仲介している。
【0011】
(e)細胞培養システム中における包膜ウイルス及びウイルスベクターの偽タイピング
このインビボのアプローチは、実験室規模のキメラウイルス又はベクター(例えば、レトロウイルス、レンチウイルス、及びAAV)の産生において、広範かつ成功裏に使用されている。偽タイピングされたウイルスの産生の鍵となる要素は、ビリオン中へ組み込まれるべき全ての蛋白質を共発現し、ビリオンの組み立てを仲介するヘルパーウイルスである。対照的に、包膜ウイルスのインビトロの会合は、規定された、別々に産生及び精製がなされた成分に基づいており、物理的な結合はインビトロで制御された条件の下で行われる(Sandrin V. et al. (2003) Curr Rop Microbiol Immunol.; 281: 137-78)。
【0012】
ウイルス様の粒子の特異的な型として、ウイロソームは臨床的に証明されたワクチン担体/アジュバントシステムであり、ヒトにおいて素晴らしい安全性と耐性プロファイルを有している。ウイロソ−ム担体が、外来性及び内在性の経路の両者を通じて抗原プロセシングを仲介する能力により、このシステムは治療用ワクチンのための良い候補となっている。
【0013】
ウイロソームの基本的な概念はインビトロでの空のウイルス包膜の、あるいはより一般的には、球状の脂質二重層中へ組み込まれたウイルスの、包膜蛋白質の再構成を備える。ウイロソームは多数のウイルスから作製されている(Y Kaneda, (2000) Adv. Drug Derivery Rev. 43, 197-205; Drummond DC et al. (2000) Prog Lipid Res. 39 (5): 409-60)。2つの異なったウイルスに由来する包膜を含んでいるキメラのウイロソームを作製する可能性が示されている(Bagai S, Saeler DP. (1994) FEBS Lett. 353 (3): 332-6)。
【0014】
全ての場合において、興味の対象であるウイルス蛋白質膜は貫通型又は膜に固定された(anchored)構造を有しており、それは自然に発生する組み込みの必要条件である。
【0015】
ウイロソーム液体膜と直接的に相互作用しない分子のウイロソームの調製を達成することは、ずっと困難である。分子をウイロソーム構造と連結させるという考えは先に提唱されており(WO 95/32706, INEX)、安定であって効率的な調製物を達成する技術的な障害は膨大であり、興味の対象である分子の生化学的な性質に依存している。陽性に荷電した脂質を使用することを介して核酸を結合させてウイロソーム構造にすることができる(WO 98/52603, Berna)。その機能のための二次構造及び三次構造を欠いている小分子(ペプチド、薬物)を生化学的に改変し、結合、組み込み、又は被包化を可能とする。ウイロソームの調製物、特に小分子(Walti et al, (2002) Canc Res.)又は合成粒子(Jana et al. (2002) FEBS Lett.; 515 (1-3: 184-188)の被包化について、多数の方法が述べられている。これらの方法は、より大きな蛋白質の真正の立体構造に影響するであろう化学的な条件下においてのみ機能するであろうし、ウイルスのヌクレオカプシドなど多量体蛋白質複合体の完全性(integrity)はもっとそうであろう(例えばHBc抗原粒子)。露出した親油性領域を欠いている一つ又はそれ以上の大きな蛋白質をウイロソーム膜中へ結合させるという、これまで述べられてきた方法は、各々の蛋白質を脂質膜中へ組み込むために、例えば脂質分子との共有結合など、蛋白質の生化学的な修飾を必要とする(Hunziker IP. et al. (2002) Int Immunol. 14 (6): 615-26)。架橋された抗体を介して特定の細胞型のウイロソ−ムを再標的化するのに、この方法は有効であると証明された(Mastrobattista E. et al. (2001) FEBS Lett. 509 (1): 71-6, Walti et al, Canc. Res. 2002)。しかし、生化学的な修飾には、非共有結合的に連結した多量体構造(例えばウイルスのヌクレオカプシド)が解離するような条件(例えば、反応性の側鎖の基を活性化するための酸化的条件)が必要である。加えてそのような条件は問題となっている蛋白質分子の立体構造も変化させる可能性があり、その結果としてその免疫原性に、そして最終的にはワクチンの効力に影響を与える。更に架橋の手段は調製に必要とされる工程の数と、抗原の損失の両者を増加させる。生化学的な修飾なしに成功裏にウイロソームと結合した多量体蛋白質について、一つの例があるのみであり、すなわちA型肝炎ワクチンのエパキサール(Epaxal:登録商標)である(Gruck R., 1995, J. of Liposome Research 1995, 5(3), 467-479)。しかしこのワクチンにおいて、ウイロソーム膜とウイルス粒子の間の静電相互作用に起因し、インフルエンザのウイロソームを調製した後にのみ、抗原は外部表層と連結している。その結果、ウイロソームの水性の内部に位置している抗原は無いが、そこは治療用ワクチンに必要とされる、効率的な細胞質送達及びCD8に基づいた細胞反応の誘起に好適な位置である(Bungener L et al. (2002) J Liposome Res. 12 (1-2): 155-63; Bungener L. et al. (2002) Vaccine. 20 (17-18): 2287-95)。
【0016】
感染疾患の予防と治療のためのアプローチの可能性と限界について、HBVを例として本願明細書において下記で議論する。特に生命を脅かす後期合併症のために、HBV感染は地球規模の大きな健康問題となっている。世界保健機関(WHO)は、現在約4億人が慢性的なHBVキャリアであると推定している。慢性的なHBV感染に罹患している患者は広範囲の症状を示し、臨床的に現れないものから重篤な肝疾患まであるが、それでも長期的な肝疾患の危険性は全ての慢性的なHBVキャリアにおいて劇的に増加する(感染後20から30年以内に25%の発生率)。全ての慢性的な患者に共通することとして、多量の抗原が慢性の感染の間に循環しているという事実に関わらず、原因物質であるHBV、特にHBVコア蛋白質に対する免疫応答が貧しい。対照的に、自然発症的な又は治療による慢性B型肝炎の治癒のみならず、急性B型肝炎の治癒は、HBV抗原に対する広範かつ活発な免疫応答と密接に連関している。インターフェロン又は抗ウイルス薬による治療により慢性肝炎をコントロ−ルするなど、従来の治療のアプローチは部分的にしか成功していないが、それでも費用がかかり、顕著な副作用を伴う。そこでHBV関連の慢性肝炎を有する患者は、この持続的なウイルス感染を制御できる治療ワクチンから大きな利益を受けるであろう。
【0017】
現在におけるHBV発病機序と免疫学の理解によると、ワクチン接種治療の成功の鍵は、慢性のキャリアにはHBV特異的な免疫学的な応答性がないことを克服することである。この目的を達成するために、存在している無効なTh2型(体液性)免疫が、強力かつ持続的なTh1型(細胞性)の反応に変わり、同時に、Th2型反応を高める(ブースト)するような方法で、関連抗原(HBcとHBs)が患者の免疫機構に提示されなければならない。
【0018】
免疫回避変異体のウイルスを防ぐために、関連抗原に対する免疫反応は広範であり、かつ、同時に多くの異なったエピトープに対して向けられていなければならない。そのような変異体は単一エピトープに対して向けられた選択的な圧力の下で発達することが示されている。更に、ワクチンの抗原として全長の蛋白質を使用することは、抗原のプロセシングとMHC遺伝子型依存的なエピトープ選択に関する患者の多様性を考慮に入れる。
【0019】
M.Hillemanの総説(Vaccine 32 (2003) 4626-4649)にあるように、過去においてHBVの治療用ワクチンの開発に多くの努力がなされてきた。多くの臨床試験において、従来のHBsに基づいた予防用ワクチンが慢性のHBV患者に使用されてきたが、持続的であって前向きである効果はこれまで観察されていない。ペプチドに基づいたワクチンは、少数の関連エピトープへの免疫応答に焦点を当てることを意図していた(Englerらによる総説、Mol Immunol. 2001 Dec; 38 (6): 457-65)。このアプローチは前臨床研究において期待をもてる結果を収めたが、ヒトにおけるものではなかった。より最近になって、二つの関連したHBV抗原を一つのワクチンに組み合わせる試みとして、表面上にHBsの単一エピトープを有する組み換えHBc粒子が作製された(Chen et al, Vaccine. 2004 Jan 2; 22 (3-4): 439-46)。
【0020】
これらのアプローチは主として、免疫システムの体液性応答を目的としている。より進んだアプローチでは、HBVに対する細胞性応答(Th1型)は治療上の免疫付与の成功における鍵となる要素である、という概念に従っている。
【0021】
HBV抗原、特にHBcに対するTh-1型の免疫応答を誘導することは、治療用のHBVワクチンの最終的なゴールである。HBsは単独でTh1応答をある程度引き起こすことができるが、HBc単独ではできない。そこでこれらの抗原は単独では、治療効果に必用とされる適切な免疫応答を誘導することができない。これは、HBV抗原をTh-1支持アジュバント又はキャリアシステムと組み合わせることのみにより達成することができる。対照的に、現在ヒトのワクチンにおけるアジュバントとして最も広く使用されているアルミニウム塩は、Th1応答を止めてTh2応答を支持することがよく知られている。アルミニウム塩のこのような性質により、それは高いタイターの保護抗体の誘導を第一の目的とする予防用ワクチンのための非常に魅力的なアジュバントとなる。治療の場面において持続的なTh1応答は決定的な役割を果すが、Th1エフェクター細胞はウイルス複製の制御とウイルスに感染した細胞の除去を仲介するからである。
【0022】
細胞応答を主として促進することが知られているDNAワクチン(HBVコア又はS遺伝子をコードするプラスミドDNA)を作る試みがなされている。DNAワクチンがマウスのモデルにおいて非常によく働くという事実に関わらず、HBVの分野のみならず、多くの臨床上の試みはヒトにおける原理の証拠を提供することができない。同様に、細胞性応答の促進を目的としたHBV抗原を発現しているウイルスベクター(例えばワクチニア)を使用しても、ヒトにおいて顕著で持続的な応答を誘導することはできなかった。
【0023】
治療用のHBVワクチンについて種々のアプローチが試みられたが、これらのいずれも有意義な治療用のワクチンへ至ることはなかった。
【0024】
2つの主要な構造的なHBV蛋白質であるHBsとHBcは、いくつかの異種のシステム:大腸菌、酵母、及び哺乳動物細胞株の中で、個々に発現させることができる。両者の抗原は典型的なウイルス様粒子構造(それぞれ、HBs粒子とHBc粒子)を形成するが、それらは感染性であって、包膜を有し、ヌクレオカプシドを含有しているHBVビリオンとは明らかに異なっている。
【0025】
組み換えB型肝炎コア抗原(HBc)は、この蛋白質は糖化されていないために、細菌又は酵母に基づいた発現システムの中で生成することができる。HBVコアモノマーは約30nmの直径を有するウイルス様の粒子へと自己会合し、プロデューサー細胞からこの型で精製できる。真性のHBVヌクレオカプシドと非常に類似して、HBc粒子は180または240モノマーコア分子のいずれかより構成され、粒子構造へ自己会合する。HBc粒子は脂質を含まない。HBVコアモノマーは183から185アミノ酸残基から構成される(長さは単離物に依存している)。C末端30アミノ酸は核酸結合領域であることを特徴とし、精製されたHBc粒子の調製物中には、好ましくない混入である、かなりの量の核酸が存在する(主としてHBVゲノムの非存在下で発現している細胞からもたらされたRNA)という結果となる。HBcはC末端において切断されて144アミノ酸でとなることができ、それは粒子構造を保ちながら核酸の含量を99%減少させる。144アミノ酸よりも短い調製物はもはや粒子を形成しない
【0026】
組み換えHBc粒子の生成は多くの変異体において述べられている。改変されたHBcの変異体は異種抗原のためのキャリアシステムとして使用されてきた(Pumpens, P.; Grens, E (2001) Intervirology 44 (2-3); 98-114)。このアプローチにおいて、外来性のアミノ酸配列が蛋白鎖のその領域(アミノ酸70−90)の中に挿入され、それは多量体粒子からみて外部表層へ露出している。しかし、遺伝子工学的に挿入される外来性抗原の配列の大きさは非常に限定されており、それは、モノマーが粒子へと自己会合する能力を保持する必要性からである。動物モデルにおけるワクチンとして使用されたとき、HBc粒子は単独で明らかなホルモン性応答を誘導するが、治療効果に必須であると考えられているHBc特異的なCD8型の細胞性応答を産生する能力を欠いている。
【0027】
WO 00/32635(ビオゲン)には、免疫原、エピトープを備え、多価のB型肝炎コア粒子を生じる可能性を有しているB型肝炎コア粒子が述べられている。
【0028】
既に臨床的な試行に入っているアプローチは、マラリアの予防のために、熱帯熱マラリア原虫の複数のエピトープを含んでいる修飾されたB型肝炎コア粒子を構築することである(Birkett A., et al, Infection and Immunity 2002, p686-6870)。
【0029】
真性のHBV包膜蛋白質(HBs)は、ずれた3つの翻訳開始部位から発現している、L(大)、M(中)、S(小)の3つの型で存在している。多量体の型の3つの型は全てHBVビリオンの包膜中に存在している。C末端のプレS1(大)およびS2(中)領域は、感染の間にHBVが細胞へ結合することに関連し、プレS1領域に対する抗体はHBVを中和できる。酵母または哺乳動物細胞中で組み換え蛋白質として発現したとき、HBsは35から45nmの直径を有するミセル粒子構造の型で分泌され、それはかなりの量(60% w/w)の細胞内膜脂質も含んでいる(Satoh O. et al. (2000) J.Biochem; 127(4): 543-50)。発現コンストラクトに依存し、組み換え粒子はS単独を含むか、またはC末端のプレS1および/またはプレS2領域を含むかのいずれかである。
【0030】
現在のHBVに対する予防的なワクチンは全て、アルミニウム塩と共に調製された組み換えHBV包膜(HBs)蛋白質に基づいている。多くの産物は酵母の発現に基づいている。より最近の産物である所謂第三世代のHBVワクチンは、哺乳細胞からもたらされる。これらのワクチンはプレS領域を含み、それに加えて、真性の哺乳類の糖化パターンと哺乳類の脂質組成の両者は、免疫反応のために有用であると考えられている。
【0031】
HBs粒子をワクチンの担体として使用することはWO 99/39736(Yissum)中にクレームされているが、そのシステムは単量体抗原に限定されており、それによってHBc粒子又は他のヌクレオカプシド型の構造と共に調製することが除外される。加えて前記システムは、キャリア粒子のインビトロでの破壊と再会合の進行を予測するものではない。
【0032】
近年の出版物(Ponsel and Bruss, (2003) JV 77 416-422)には、HBsとHBc両者の抗原のための発現プラスミドで同時トランスフェクションを行った哺乳細胞から、HBsとHBcの両者を含んでいるHBV粒子を生成し、分泌することが記載されている。しかし、HBsとHBc両者の抗原を含んでいる包膜粒子をインビトロで再構成することに関する報告はない。
【0033】
米国特許6,020,167(Medeva)は、プレS1又はHBVコア由来の一つまたはそれ以上のT細胞活性化エピトープ、及びそのポリペプチドを提示することが可能なキャリアを備える組成物を投与することによるB型肝炎の治療方法をクレームする。その発明におけるキャリアはHBs抗原粒子でもよい。
【0034】
ウイルス様粒子の特定の型として、ウイロソームは臨床的な実績があるワクチンの担体/アジュバントのシステムであり、ヒトにおいて優れた安全性と許容性のプロファイルを有する。ウイロソームの担体が、外在性および内在性の経路の両者を介して抗原のプロセシングを仲介する能力により、このシステムは治療用ワクチンの良い候補となる。
【0035】
ウイロソームの基本的な概念はインビトロでの空のウイルス包膜の、あるいはより一般的には、球状の脂質二重層中へ組み込まれたウイルスの、包膜蛋白質の再構成を備える。ウイロソームは多数のウイルスから作製されている(Y Kaneda, (2000) Adv. Drug Derivery Rev. 43, 197-205; Drummond DC et al. (2000) Prog Lipid Res. 39 (5): 409-60)。2つの異なったウイルスに由来する包膜を含んでいるキメラのウイロソームを作製する可能性が示されている(Bagai S, Saeler DP. (1994) FEBS Lett. 353 (3): 332-6)。
【0036】
全ての場合において、興味の対象であるウイルス蛋白質膜は貫通型又は膜に固定された(anchored)構造を有しており、それは自然に発生する組み込みの必要条件である
【0037】
ウイロソームの脂質膜と直接的に相互作用しない分子のウイロソームの調製を達成することはずっと困難である。分子をウイロソーム構造と結合させるという考えは先に提唱されており(WO 95/32706, INEX)、安定であって効果的な調製を達成するための技術的なハードルは膨大であり、興味の対象である分子の生化学的な性質に依存する。陽性に荷電した脂質を使用することを介して、核酸をウイロソ−ム構造と連結させることができる(WO 98/52603, Berna)。その機能のための二次構造及び三次構造を欠いている小分子(ペプチド、薬物)を生化学的に改変し、結合、組み込み、又は被包化を可能とする。ウイロソームの調製、特に小分子(Walti et al, (2002) Canc Res.)又は合成粒子(Jana et al. (2002) FEBS Lett.; 515 (1-3: 184-188)の被包化について、多数の方法が述べられている。これらの方法は、より大きな蛋白質の真正の立体構造に影響するであろう化学的な条件下においてのみ機能し、ウイルスのヌクレオカプシドなど多量体蛋白質複合体の完全性(integrity)はもっとそうであろう(例えばHBc抗原粒子)。露出した親油性領域を欠いている一つ又はそれ以上の大きな蛋白質をウイロソーム膜中へ結合させるという、これまで述べられてきた方法は、各々の蛋白質を脂質膜中へ組み込むために、例えば脂質分子との共有結合など、蛋白質の生化学的な修飾を必要とする(Hunziker IP. et al. (2002) Int Immunol. 14 (6): 615-26)。この方法は架橋された抗体を介して、特定の細胞型のウイロソ−ムを再標的化するのに有効であると証明された(Mastrobattista E. et al. (2001) FEBS Lett. 509 (1): 71-6, Walti et al, Canc. Res. 2002)。しかし、生化学的な修飾には、非共有結合的に連結した多量体構造(例えばウイルスのヌクレオカプシド)が解離するような条件(例えば、反応性の側鎖の基を活性化するための酸化的条件)が必要である。加えてそのような条件は問題となっている蛋白質分子の立体構造も変化させる可能性があり、その結果としてその免疫原性に、そして最終的にはワクチンの効力に影響を与える。更に架橋の手段は調製に必要とされる工程の数と、抗原の損失の両者を増加させる。生化学的な修飾なしに成功裏にウイロソームと結合した多量体蛋白質について、一つの例があるのみであり、すなわちA型肝炎ワクチンのエパキサール(Epaxal:登録商標)である(Gruck R., 1995, J. of Liposome Research 1995, 5(3), 467-479)。しかしこのワクチンにおいて、ウイロソーム膜とウイルス粒子の間の静電相互作用に起因し、インフルエンザのウイロソームを調製した後にのみ、抗原は外部表層と連結している。その結果、ウイロソームの水性の内部に位置している抗原は無いが、そこは治療用ワクチンに必要とされる、効率的な細胞質送達及びCD8に基づいた細胞反応の誘起に好適な位置である(Bungener L et al. (2002) J Liposome Res. 12 (1-2): 155-63; Bungener L. et al. (2002) Vaccine. 20 (17-18): 2287-95)。
【0038】
インフルンザのウイロソーム(WO 92/19267 WO 98/52603, Berna)、及び他の包膜ウイルス由来(例えばセンダイウイルス)のウイロソ−ム様構造について、多くの特許が出願され、特許が付与されてきた。これらの方法は、ウイルスの包膜を可溶化し、ウイルスゲノムを含んでいるヌクレオカプシドを除去し、続いて「空の」ウイルスの包膜を再構築することを備える。更に、付加的な抗原は、それらをウイロソーム膜の中に固定するために、既成のウイロソーム(エパキサール(Epaxal:登録商標))に付着されているか、あるいは脂質分子と架橋されている。
【発明の開示】
【発明が解決しようとする課題】
【0039】
ウイルス感染に対するワクチン接種の上記に述べた制限を鑑みて、本発明の基礎となる技術上の課題は、ウイルス感染の予防、軽減、又は治療のために、被験者にワクチンを接種するための改良された手段と方法を提供することである。
【課題を解決するための手段】
【0040】
前記の技術的な課題の解決は、特許請求の範囲中に特性が示された態様を提供することによって達成される。
【発明を実施するための最良の形態】
【0041】
従って、本発明は
(a)少なくとも1つの脂質、及び包膜ウイルスの包膜蛋白質を備えるウイロソーム膜と、
(b)ウイロソームの内側及び外側に位置し、前記包膜蛋白質に付着した前記包膜ウイルスのヌクレオカプシド粒子
を備えるウイロソームを提供する。
【0042】
「ウイロソーム」という言葉はウイルス様粒子(VLP)の特定の形態を定義する。ウイロソームはウイルス粒子由来の半合成複合体であり、インビトロの方法で生成される。これらは本質的に再構成されたウイルス被覆であるが、ウイルスのヌクレオカプシドが任意の化合物で置換されている。ウイロソームは融合活性を保持しており、それ故組込まれた化合物(抗原、薬物、遺伝子)を標的細胞の内側に送達する。これらはワクチン、薬物送達、遺伝子導入に使用することが出来る。
【0043】
VLPとは、形態と粒度に関して親ウイルスと類似している、又は見分けがつかない粒子構造であるが、感染し宿主細胞中で複製する能力を欠いている。VLPはウイルス蛋白質(真正、又は改変されたその変異体)から構成される多量体構造である。それに加え、VLPは核酸や脂質を含んでも含まなくてもよく、脂質膜構造を含んでも含まなくてもよい。1つのウイルス(HBV)に由来する2つの典型的で、かつ大幅に異なるVLPの例は、HBsとHBc粒子である。
【0044】
本発明の文脈中において「ウイロソーム膜」という言葉は、人工的に再構成され脂質2重膜とそれに組込まれたウイルス包膜蛋白質から構成される球形の膜構造を定義する。
【0045】
本発明の文脈中において「包膜蛋白質」という言葉は、元来の形態においてウイルス脂質膜と直接相互作用する、包膜ウイルスによってコードされる蛋白質を意味することを意図している。
【0046】
本発明に従って、広範囲の脂質を前記ウイロソーム膜は備えることが出来る。脂質の集団は中性又は荷電したリン脂質、ステロイド由来の脂質、中性又は荷電した合成脂質を備える。調製物に加えられる精製した脂質に加え、例えばインフルエンザウイルスや他の任意の包含された包膜ウイルスに由来する脂質、又は調合物に含まれるVLP(例えばHBs粒子等)を囲む脂質に由来する脂質といった、ウイルス成分に含まれる脂質も同様に最終的な調製物に含めることが出来る。これらのウイルス由来の脂質は不均一であり、ウイルス産生細胞又は組換え発現細胞の脂質組成を反映している。調合物の複雑性を最小限にするため、好適な調合物はリン脂質のみを基にしている。添付した実施例に記述しているHBウイロソームに使用されているリン脂質は、通常GMPグレードであり、登録されたワクチンであるインフレキサール(Inflexal:登録商標)とエパキサール(Epaxal:登録商標)に使用されている材料と同一であることが好適である。
【0047】
本発明の文脈中において「包膜ウイルス」という言葉は成熟したビリオン構造において宿主細胞由来の脂質膜を含むウイルスを定義する。包膜ウイルスの分類は表1に列記してある。
【0048】
本発明の文脈中において「ヌクレオカプシド粒子」という言葉はウイルスカプシド蛋白質で構成される粒子構造を意味することを意図している。この粒子構造は、(1つまたは多数の組換えウイルスカプシド蛋白質で構成される)VLP、又は親ウイルスから精製されたヌクレオカプシド複合体とすることが出来る。ヌクレオカプシド粒子が核酸を含むか否かは粒子の形成と関係がない。
【0049】
本発明のウイロソーム(キメラのウイルス様粒子)は1つの粒子中で物理的に結合した上記の性質を持つ分子を備える。本発明のウイロソーム中の包膜蛋白質は、対応するヌクレオカプシド粒子と相互作用する側をウイロソームの内側に向ける本来の方向で、又は対応するヌクレオカプシド粒子との相互作用する側をウイロソームの外側に向ける人工的な方向で、ウイロソームの表面に組込むことが出来る。そのようなウイロソームの一例を図1に描写している。この新しいクラスのウイロソームの構造は、上記の個々の成分の粒状の構造、又は元来のウイルスと全く異なる。この型の粒子は天然には存在せず、最先端においても今のところ人工的に生成される構造として記述も提案もされていない。つまりこの粒子構造は、分離された成分から完全に人工的に再組立させた最初の包膜ウイルス様粒子を表している。
【0050】
当技術分野で公知であり本明細書において上記で説明しているウイロソームは、細胞に基づいたシステムで産生する。そのような細胞を基にしたシステムでは、全ての成分を同じ細胞内で同時に産生しなければならず、発現システムの選択を大幅に制限し、収量と大量生産性に関しての妥協を強いることとなる。制御の難しい生物学的発現システムと代謝過程が、例えば成分間の比率といった、結果として生じるウイルス様粒子の組成を決定する。更に、有用な標本を得るために、細胞システムで産生されたウイルス様粒子は、粒子構造や組成に影響を与えずに、次に抽出し精製しなければならない。
【0051】
対照的に、また本明細書において下記で述べるように、本発明のウイロソームの人工的な調製物の組成は、加える材料と選択する生物学的変数によって制御することが出来る。過程の単純性はその安定性を保証する。結果として生じる調合液は更なる精製を必要としない。個々の成分を事前に別々の細胞を基にしたシステム(例;大腸菌、哺乳類細胞、酵母菌)で産生し、それぞれの成分に関して収量、大量生産性、及び純度に関して最適なシステムを選択することが出来る。
【0052】
本発明におけるウイロソームの調製過程は、ウイロソーム型構造に容易に組込まれるウイルス蛋白質(膜付着蛋白質、包膜蛋白質)と、それ自身は膜に結合しない蛋白質との相互作用を利用している。宿主細胞中での天然の複製過程における大半の包膜ウイルスの組立においてこの相互作用は必要であり有効であるが、薬学的産物の人工的調製過程においてこの性質を使用することは新しい。驚くべきことに、全く違う条件下ではあるが、細胞内のウイルスの組立は本当に人工的に模倣することが出来る。
【0053】
ウイロソームは少なくとも1つの脂質が少なくとも1つのリン脂質を備えるウイロソームであることが好適である。前記のリン脂質がフォスファチジルコリン、フォスファチジルエタノールアミン、フォスファチジルセリンを備えると更に好適である。
【0054】
本発明で同様に想定するのは、更に好適な実施例において、前記の包膜蛋白質が第1と第2の包膜ウイルスの包膜蛋白質でありヌクレオカプシド粒子が前記の第2の包膜ウイルスのヌクレオカプシド粒子であるウイロソームであることである。
【0055】
前記の第1の包膜ウイルスは任意の包膜ウイルスから選択することが出来る。本発明に特に好適なのはインフルエンザウイルスである。
【0056】
本発明の更に好適な実施例においては前記の第1の包膜ウイルスの包膜蛋白質はヘマグルチニン(HA)及び/又はノイラミニダーゼ(NA)である。
【0057】
本発明のウイロソームのインフルエンザ成分である、ヘマグルチニン(HA)及びノイラミニダーゼ(NA)は、確立され特許化されたインフルエンザウイロソームの調製物(エパキサール(Epaxal:登録商標) WO 92/19267, WO 92/19268, Gluck R., 1995, Journal of Liposome Research 5(3), 467-479)と同様に、不活化されたインフルエンザウイルス(例えばシンガポールA型)から精製することが出来る。インフルエンザ由来の蛋白質又は前記の第1の包膜ウイルスの蛋白質を、構造的や機械的というよりは機能的な理由で含めることが出来るが、これは本発明のウイロソームはインフルエンザ蛋白質又は前記の第1の包膜ウイルスの蛋白質が無くても同様に調製することが出来るためである。本発明のウイロソームの、ウイロソーム様担体の免疫学的性質を強化するために、インフルエンザ成分を含めることが出来る。
【0058】
同様に本発明に従って、前記の第2の包膜ウイルスは、(好適である)B型肝炎ウイルス(HBV)、C型肝炎ウイルス(HCV)、任意の他のフラビウイルス、及びヒト免疫不全ウイルス(HIV)から成る包膜ウイルスの一群から選択すると好適である。
【0059】
本発明の更に好適な実施例においては、ヌクレオカプシド粒子はHBc蛋白質を備える。
【0060】
HBc粒子は、全長のアミノ酸配列又は短縮型を含む形で、大腸菌で産生することが出来る。全長の産物と、短縮した144個のアミノ酸の構築物をHBウイロソームに調製することに成功している。代わりに、HBウイロソームに更に短い(非特異的な)HBV核を組込むことも考えられる。対応する技術は当技術分野で公知であり、付属の例で説明している。
【0061】
前記の第2の包膜ウイルスの包膜蛋白質がHBs蛋白質であると、同様に好適である。
【0062】
HBs粒子はS蛋白質単独、又はプレSとSの結合蛋白質を含むことが出来る。前記の粒子の産生の方法は当技術分野に公知である。粒子は例えば酵母又は哺乳類細胞で産生することが出来る。本発明のウイロソームを備えるワクチンにおけるプレSドメインの存在は、より広範でより効果的な免疫反応に寄与する可能性が高いが、調合過程に何の影響も与えない。HBウイロソームはいずれの供給源からのHBsも産生することが出来る。1つのHBウイロソーム中において、異なる供給源からの異なるHBs型、又は異なる血清型を組み合わせることさえも、本明細書で説明している方法を用いて行われてきた。本発明は上記の好適な実施例においてインフルエンザウイロソームの分類に当てはまる。しかし、同じウイルスのヌクレオカプシド蛋白質(HBc)をウイロソーム構造に繋げるために替わりに使用する、全く関係のないウイルス(HBV)の包膜蛋白質(HBsAg)を組込むことは、全く新しい。
【0063】
本発明の代替的な実施例は本発明のウイロソームを備えるワクチンに関連している。
【0064】
本発明の文脈において「ワクチン」という言葉は、ウイルス病の予防のために対象者に投与される予防的組成物を定義するものと理解される。代わりに、もしくはそれに加え、この言葉はウイルス病の緩和又は治療のために対象者に投与される薬学的組成物を意味することを意図する。
【0065】
本発明に従って、「予防的組成物」及び「薬学的組成物」という言葉は患者、好ましくは人間の患者への投与のための組成物に関連している。好適な実施例においては、前記組成物は非経口、経皮、腔内、動脈内、髄腔内投与、もしくは組織への直接注入のための組成物を備える。前記組成物が輸液もしくは注射によって患者に投与されることは特に想定される。適切な組成物の投与は、例えば静脈中、腹膜中、皮下、筋中、局所、又は皮内注射等の、異なる経路によって影響される。本発明のワクチン/組成物は更に薬学的に適切な補形薬を備えることが出来る。本発明に従って使用される補形薬は、例えばカプセル、賦形剤、保護剤、着色剤、崩壊剤、結合剤、乳化剤、可溶化剤、湿潤剤、溶媒、緩衝剤、ゲル形成剤、増粘剤、膜形成剤、潤滑剤、流動促進剤、形状分離剤、流量調整剤、吸着剤、及び抗酸化剤や味と匂いの矯正剤等の添加物等の、担体、添加物、及び希釈剤を備える。適切な薬学的担体の例は当技術分野において周知であり、リン酸緩衝生理食塩水、水、例えば油水乳剤等の乳剤、さまざまな種類の湿潤剤、無菌溶液等を含む。そのような担体を備える組成物は周知の従来の方法により調合することが出来る。これらの組成物は対象者に対し適切な用量で投与することが出来る。投薬量は関与する医師と臨床的因子により決定される。医療技術分野において周知のように、どのような一人の患者に対する投薬でも、患者の大きさ、体表面積、年齢、投与される特定の化合物、性別、時間、投与経路、全般的健康状態、及び同時に投与されている他の薬等の、多くの因子によって決定される。投与の好適な用量は一回の使用あたり1ngから1mgの範囲であることが多い。
【0066】
本発明のワクチン/組成物は局所的もしくは全身的に投与することが出来る。投与は例えばバイオリスティック法による内部又は外部の場所への送達のように、非経口的であることが望ましい。非経口投与のための調製液は無菌的な水性又は非水性溶液、懸濁液、及び乳液を含む。水性の担体は生理食塩水及び緩衝媒体を含む水、乳液又は懸濁液を含む。非経口の賦形剤は食塩溶液、デキストロースリンゲル液、デキストロース及び食塩の溶液、又は乳酸リンゲル液を含む。静脈中の賦形剤は液体及び栄養補充剤、電解質補充剤(例えばデキストロースリンゲル液に基づいた物等)、及びそれに類する物を含む。例えば抗菌剤、抗酸化剤、キレート剤、不活性化ガス、及びそれに類する物等の、保存料及び他の添加物が存在してもよい。組成物の使用意図に応じて、本発明のワクチン/組成物が、本発明のウイロソームに加え、更に生物学的活性のある薬剤を備えることが想定される。そのような薬剤はアジュバントとすることが出来る。アジュバントとは、ワクチンに含まれる抗原に対する免疫反応を増強もしくは制御するためにワクチンの調製に加えられる物質のことである。様々な種類の異なるアジュバントが当技術分野において知られており、それらは脂質、蛋白質、炭水化物、界面活性剤、塩、もしくはこれらの組み合わせから構成される。
【0067】
添付された実施例において説明しているように、ウイロソームの調製物は本当に抗原に対する細胞反応を改善することが示されている。これはマウスにHBウイロソームを接種した後のHBcに対する細胞反応の検出により特に証明されている。例えばHBc等のウイルス核蛋白質に対する細胞反応、特にCD8/TH1タイプ反応の持続的な誘導は、有用であり、本発明のワクチンの患者への接種の特に好適な結果である。
【0068】
随意に、本発明のワクチンは更に薬学的に適切な担体または希釈剤及び/又はアジュバントを備えることが出来る。
【0069】
薬学的に適切な担体又は希釈剤は本明細書において下記で説明している。前記ワクチンに含まれる抗原に対する免疫反応を更に増強又は調整するために、アジュバントと呼ばれる免疫刺激物質を本発明の調合に加えることが出来る。アジュバントの性質を持つ多数の化合物が当技術分野において知られている。そのような化合物の一群は蛋白質、脂質、炭水化物、核酸、及びそれらの組み合わせを備える。化合物は合成することも、生物学的に産生することも出来る。アジュバントは事前に調製したウイロソームに加えること、又は共に調製したウイロソーム構造中に組込むことが出来る。アジュバントの生化学的性質によりウイロソーム成分のいずれかと相互作用するのであれば、後者は可能である。アジュバントがRC529(Corixa)であることが特に好適である。
【0070】
本発明の好適なHBウイロソームは、本明細書において上記で述べたように、インフルエンザとHBVの抗原が1粒子中で物理的に結合している、安定して均一なウイロソームの共調合である。ウイロソーム担体に抗原が結合していることは、ウイロソーム抗原担体/アジュバントシステムの免疫刺激作用の十分な研究のための定評のある必須条件である(Moser C et al. (2003) Expert Rev Vaccines, 2 (2): 189-96で概説している)。
・ 抗原のHAを介したMHC−1提示、及び抗原に対するTh1免疫反応
・ 反復したウイルス様構造の中の抗原の提示
・ 抗原提示細胞の標的化
・ 細胞外分解からの保護
【0071】
治療的HBVワクチンに関しては、ウイロソームのHAを介したMHC−1提示、及び樹状細胞の標的化がウイロソームワクチン担体の最重要の特徴である。それ故、抗原提示細胞の細胞質に送達されるためにHBc抗原の少なくとも一部がウイロソーム中に被包されていることが好ましく、それによって抗原特異的なCD8T細胞が誘導される(Bungener L et al. (2002) J Liposome Res. 12 (1-2) 155-63)。より最近の研究ではウイロソームがCD4T細胞活性化を通してMHCクラス1制限的CTLを増強したことが証明された(Schumacher et al. Vaccine 22 (2004): 714-723)。実験的な試作品の調製の段階で、無修正のHBsとHBc抗原の物理的な組込み及び結合はそれぞれ大きな技術的障壁となっており、産業的なcGMP規模では更に難しかった。前記の障壁は本発明によって克服されている。調製に使用するHBV及びインフルエンザ抗原は異なる組換え発現システム(哺乳類細胞、酵母菌、又は大腸菌)で別々に産生され精製され、これらの精製された抗原はそれら自身で特徴的な粒子を形成する。
【0072】
更なる実施例においては、本発明は:
(a) 界面活性剤溶液中において脂質存在下で包膜ウイルスの包膜蛋白質を可溶化し;
(b) 溶液中の界面活性剤の濃度を低下させ;
(c) 前記包膜ウイルスのヌクレオカプシド粒子を工程(b)で取得した溶液に加え;
(d) ウイロソームが産生されるように界面活性剤又はレシチンを除去する;
という工程を含むウイロソームの産生方法を提供する。
本発明の文脈中において「濃度を低下させ」という言葉は、前記の界面活性剤が入っていない溶液を加えること、又は前記の界面活性剤の濃度が工程(a)において取得した溶液に比べて低い溶液を加えることを含むと理解される。
【0073】
本発明の文脈中において、「界面活性剤を除去する」という言葉は透析、膜分離、又はクロマトグラフィー等の過程を含むと理解される。界面活性剤が基質(例えばビーズ、レシン等)への吸着によって除去される、後者のクロマトグラフィーが好適である。
【0074】
本発明のウイロソームの調整工程は、苦も無くGMP産生の方法に適合させることが出来る。HBウイロソームの例で説明しているように、HBc蛋白質(図2)を含む新しい多成分の粒子構造の均一で効率的な調製を得るためには、生化学及び量的関係の条件に関する調整が必要であり、本明細書の教唆に基づいて過度の負担なしに行うことが出来る。
【0075】
本発明の方法の根底にある概念は、ウイルスの複製過程の間の細胞内のウイルス組立で起こるような、ウイルス包膜蛋白質と対応するヌクレオカプシド又はヌクレオカプシド様粒子の間の特異的な相互作用を利用した、ウイロソームの完全に人工的な組立である。例においてはHBsとHBcはそれぞれ典型的で好適な包膜蛋白質とヌクレオカプシド複合体であるが、本発明はそれらに限定したものではなく任意の包膜ウイルスに適用出来ることを理解するべきである。一般的な原則として、人工的な調製において選択する生化学的な条件は、包膜蛋白質(env)とヌクレオカプシド(nc)成分の相互作用を可能にするものでなければならない。調製における重要な生化学的変数は、界面活性剤濃度、pH値、浸透圧、及びキレート剤、特定の塩、緩衝分子の存在を含む。
【0076】
最初の材料(ビリオン又はVLPの包膜)及び脂質成分を溶解するために、界面活性剤の存在が必要である。界面活性剤の種類及び濃度の範囲は本明細書において下記で説明している。
【0077】
相互作用の低下又は他の成分との非特異的な相互作用の危険性を下げるために、pHと浸透圧は技術的に可能な限り生理的な条件(pH 7.4、150 mEq)に近づけるのが好適である。他の包膜ウイルスに関してはpHが5〜10の範囲、また浸透圧が10〜400 mEqであることが好適である。塩、キレート剤、緩衝液は同様に蛋白質間の相互作用、つまりenv / nc の調合の効率に同様に影響する。添付した実施例で説明しているプロトコルでは、生理的な塩組成を模倣し、NaCl及びリン酸塩という生理的な緩衝システムを備えるリン酸緩衝生理食塩水(PBS)を使用している。しかし、他の包膜ウイルスの調合には修正した緩衝システム(例えばTrisに基づいた緩衝液又は炭酸塩に基づいた緩衝液)をNaCl、MgCl、KCl、及びCaCl塩と組み合わせて使用することが必要になる可能性がある。本発明の好適な実施例においては、望ましくない酵素活性を不活化するために、キレート剤(例えばEDTA、EGTA)を含むことを想定してもよい。
【0078】
成功した調合物とは、個々の最初の材料(精製されたウイルス又はVLP)のそれぞれと粒度、内容、又は生化学的性質において全く異なるウイルス様粒子の単一かつ均一な集団として最もよく説明される。図4において図4のHBウイロソームに関して図示しているように、粒度の違いは光子相関法(photon correlation spectroscopy、PCS)によって簡単に検出することが出来る。解析方法は実施例4(p33)で説明している。
【0079】
ヌクレオカプシドが(HBcでそうであるように)単一の組換え蛋白質のサブユニットで構成されているのか、又は核酸に結合している複数の異なるウイルス蛋白質を含むのかは、方法の原理に関しては重要でない。例えばHBs等のウイルス包膜蛋白質成分は、それ自身では膜構造と効率的な相互作用又は結合をしないヌクレオカプシド粒子(例えばHBc)と、再構成されたウイロソーム膜との間の架橋として働く。効率的なHBsとHBcの相互作用を可能にするために、根本的には異なるものの、人工的な調整工程はある程度HBV感染細胞の内部の条件を模倣しなければならない。今回の人工的な方法では、巨大分子的な細胞構造のない条件で、組立が界面活性剤の除去の間に起こる。対照的に、HBVの複製の間は、HBVヌクレオカプシドと相互作用しながら、HBs分子は細胞膜に固定されている。驚くべきことに、根本的に異なる条件にも関わらず、ウイルス組立、つまりウイルス包膜中へのヌクレオカプシド粒子の被包は我々の調整工程において高い効率で起こった。両方の抗原をウイロソーム構造につなげるために2つの工程が同時に起こる必要がある:
(i) 膜貫通蛋白質、インフルエンザ包膜(HAとNA)及びHBs(HBV包膜)がウイロソーム膜中に組込まれなければならず、また
(ii) HBc粒子が膜に固定されたHBsと効率的に結合しなければならない。
【0080】
従って、効率的なHBウイロソーム組立は、最適化された生化学的条件と個々の成分の正しい量的関係の下でのみ起こり得る。同じことがインフルエンザウイルスとHBV以外のウイルスからの蛋白質を備えるウイロソームの組立に関しても言える。複雑なヌクレオカプシド構造の完全性が、膜に固定された包膜蛋白質との相互作用にとっての必須条件である。
【0081】
本実施例においては、異なる供給源からの3つの異なる生物学的粒子構造が、最初の構造いずれとも明らかに異なり自然には存在しない、1つの新しい種類の合成ウイルス様粒子に人工的に変形される。
【0082】
本発明の方法の上記の実施例において述べた脂質が、少なくとも1つのリン脂質を備えることが好適である。前記のリン脂質がフォスファチジルコリン、フォスファチジルエタノールアミン、フォスファチジルセリンを備えると更に好適である。
【0083】
本発明の方法において、前記の包膜蛋白質が第1と第2の包膜ウイルスの包膜蛋白質でありヌクレオカプシド粒子が前記の第2の包膜ウイルスのヌクレオカプシド粒子であると好適である。
【0084】
本発明の更に好適な実施例では、前記の第1の包膜ウイルスはインフルエンザウイルスである。前記の包膜蛋白質がヘマグルチニン(HA)及び/又はノイラミニダーゼ(NA)であると更に好適である。
【0085】
本発明の方法において、前記の第2の包膜ウイルスがB型肝炎ウイルス(HBV)であると更に好適である。ヌクレオカプシド粒子がHBc蛋白質を備えると更に好適である。包膜蛋白質がHBs蛋白質であると同様に好適である。
【0086】
本発明の方法に従って、工程(a)における可溶化の段階の前に、包膜蛋白質の溶液を遠心し、取得した沈殿を脂質の存在下で界面活性剤又はレシチン溶液に可溶化することが好適である。本発明の方法において使用する可能性のある脂質は本明細書において上記で更に詳細に定義してある。
【0087】
工程(a)が遠心の前に溶液の超音波処理を含むと更に好適である。
【0088】
遠心を100000g、4℃で少なくとも2時間行い、及び/又は超音波処理を37℃のウォーターバス中で少なくとも2分間行うと更に好適である。
【0089】
本発明の方法の1つの変法において、前記の方法は更に工程(b)に続いて段階工程(b’)を行うことを含むことが出来る:
(b’)工程(b)で得られた希釈液の無菌濾過。
溶液の無菌濾過の手段と方法は当技術分野において公知である。無菌濾過は添付した実施例で説明するように例えば0.22μmのフィルターを通して段階工程(b)で取得した溶液を濾過することを含むことが出来る。
【0090】
上記で説明した本発明の方法の工程(d)における界面活性剤の除去は、
(i)Bio-Beads SM-2を加え回転させながらインキュベーションすること;及び
(ii)Bio-Beads SM-2を溶液から除去すること
によって達成することが出来る。
【0091】
前記の工程(i)と工程(ii)を新しいBio-Beads SM-2を使用して少なくとも1回、好ましくは少なくとも2回は繰り返すことが好適である。
【0092】
工程(i)と工程(ii)を室温で行うことが同様に好適である。更に、工程(i)でのインキュベーションが少なくとも30分間であると同様に好適である。
【0093】
本発明の方法が更に工程(e)を含むことが同様に想定される:
(e) 工程(d)で得られた希釈液の無菌濾過。
【0094】
本明細書において上記で述べた界面活性剤が非イオン性界面活性剤であることが好適である。本発明に従った非イオン性界面活性剤の例は、例えばオクタエチレングリコールモノ(N-ドデシル)エーテル(OEG)、Triton X-100、Triton X-114、NP40、Tween 20/80、及びレシチン等の界面活性剤を含む。界面活性剤は好ましくは0.1から15% (v/v)の濃度の範囲で使用することが出来る。界面活性剤の性質により、好適な濃度は一般的に(v/v)で与えられる。しかし、OEGは粉末の形状で取得し、それ故その濃度はmM単位で与えられることが好ましい。例えば100mMのOEGの濃度は大体5.5% (v/v)のOEGの濃度に相当する。
【0095】
非イオン性界面活性剤がオクタエチレングリコールモノ(N-ドデシル)エーテル(OEG)であると特に好適である。
【0096】
本発明の方法においてOEGが20mMから100mMまでの濃度の範囲で工程(b)に適用されることが同様に好適である。前記の濃度が50mM(約2.75% OEG(v/v))であると最も好適である。
【0097】
本発明の方法の好適な実施例においては、適用される脂質:蛋白質比が1:10から20:1の間の範囲である。前記の脂質:蛋白質比が約5:1であると更に好適である。
【0098】
本発明の方法においてフォスファチジルコリンのウイロソームにおける割合が22%であると好適である。
【0099】
本発明の方法においてHA:ウイルス包膜蛋白質:ウイルスカプシド蛋白質の比が1:1:1であると同様に好適である。
【0100】
更に、本発明の方法が工程(d)においてウイロソームを産生する前にアジュバントを加えることを含むと好適である。
【0101】
更に代替的な本発明の実施例は、本発明のウイロソーム又は本発明の方法によって産生されたウイロソームの、ワクチン産生のための使用に関係している。
【0102】
更に代替的な本発明の態様は、本発明のウイロソーム又は本発明の方法によって産生されたウイロソームの、HBV感染の予防、緩和、又は治療のためのワクチン産生のための使用に関係している。
【0103】
本発明は同様に、本発明のウイロソーム又は本発明の方法によって産生されたウイロソームの、それを必要としている対象者への投与を含む、対象者への接種の方法に関係している。本明細書において上記で薬学的組成物について一般的に説明したように、ウイロソームを投与することが出来る。
【0104】
本発明の更なる態様は本発明のウイロソーム又は本発明の方法によって産生されたウイロソームの、それを必要としている対象者への投与を含む、HBV感染の予防、緩和、又は治療のための対象者への接種の方法に関係している。随意に、ウイロソームは薬学的に適切な担体又は希釈剤及び/又はアジュバントと組み合わせて投与することが出来る。
上記の対象者が人間であると特に好適である。
【0105】
【表1】
図が示しているのは:
【0106】
図1:
個々の成分と結果として生じるHBウイロソームの構造を描いた概略図が図1に示してある。分析的データは提案された構造と一致している。
【0107】
図2:
調製過程のフローチャートが図2に示してある。HBウイロソームの詳細な調合プロトコルは添付した実施例に記述してある。
【0108】
図3:
図3はHBウイロソームのSDS-PAGE分析の結果を示している。SDS-PAGE分析に基づくHBウイロソーム及び最初の材料(インフルエンザ、HBs、及びHBc)の蛋白質組成が図3に示してある。
HBウイロソームの予測される物理的構造は以下の分析データによって確認されている(図4−7)。
【0109】
図4:
図4は個々の成分の粒度と比較したHBウイロソームの粒度の分析の結果を示している。HBウイロソームは個々の成分とは全く異なる単一種類の粒子から構成され、光子相関分析の粒子の粒度の分布において1つの狭いピークを形成している。
【0110】
図5:
図5はHBウイロソームの勾配分画分析の結果を示している。ショ糖勾配中でHBウイロソームを超遠心した後、全ての抗原が同一の勾配分画で見え、それらの物理的な結合を示唆している。
【0111】
図6:
図6はHBウイロソームのSDS-PAGE/ウェスタンブロット分析の結果を示している。抗HA又は抗HBs抗体のいずれかを使用したHBウイロソームの免疫沈降では、両方の場合で全ての抗原(HBs、HBc、及びHA)の共沈降が起こった。界面活性剤の添加によってHBウイロソーム構造が破壊されると、抗体によって直接認識される抗原しか沈降されない(それぞれHA又はHBs)。この所見は全ての抗原が1粒子中で結合していることを確かめている。
【0112】
図7:
図7はHBウイロソームのSDS-PAGE/ウェスタンブロット分析の結果を示している。HBウイロソームがトリプシン処理に晒された場合、両方のHBV抗原が部分的に保護された(ウェスタンブロットによれば50%)。トリプシン中でインキュベーションする前に界面活性剤の添加によってウイロソーム構造が破壊されると、HBV抗原は短時間のうちに完全に分解された。
【0113】
図8:
図8はマウス免疫化の後のHBウイロソームに対する抗体免疫反応の試験の結果を示している。この時点ではHBウイロソームの免疫原性に関して予備的なデータのみ利用可能である。付加的なアジュバント無しのHBウイロソームを用いてマウスで実験を行った。マウスは異なる量のHBウイロソームの筋内注射によって免疫化され、高い抗体力価が異なる3つの抗原に対して検出された。
【0114】
図9:
図9はマウス免疫化の後のHBウイロソームに対する細胞免疫反応の試験の結果を示している。3回目の増強の後、免疫化された動物より胸腺細胞が精製され、それぞれの抗原を用いた試験管内での再刺激の後、インターフェロンγに対するELISPOTによって細胞反応を決定した。注目すべきはこの方法はCD4型とCD8型反応を区別しないことである。
【0115】
図10:
要約表はHBウイロソームによる免疫化の後これまでに取得した免疫学的データを示している。免疫化した動物においてHBs及びHBcの両方に対する体液性と細胞性の応答が検出可能であった。
【0116】
これより本発明は以下の例を参照することによって説明され、この例は単に例証であり本発明の範囲の制限と解釈されるべきではない。
【実施例】
【0117】
実施例1:HBs抗原の酵母における発現と精製
Schaefer S. et al., in Hansenula polymorpha: Biology and Applications, WILEY-VCH Verlag, Weinheim, 2002, Recombinant hepatitis B vaccines- disease characterization and vaccine production (p187 - p193)
【0118】
この過程は発現カセットとベクターの作製、Hansenula polymorphaという酵母の形質転換、菌株の選択及び評価、発酵、細胞の回収、細胞の破壊、洗浄、吸着、イオン交換クロマトグラフィー、限外濾過、超遠心、ゲル濾過クロマトグラフィー、及び最終産物の液体の濾過滅菌という段階を含む。
【0119】
最終産物の品質は、Lowry法、SDS/PAGE法、ウェスタンブロット分析、又はAUSZYME等の種々の生化学的分析により確認している。
【0120】
実施例2:HBcの大腸菌における発現
Zheng et al, Journal of Biological Chemistry 1992, 13:9422-9429
The structure of Hepadnaviral core antigens
Preikschat el al., Journal of General Virology 1999, 80: 1777-1788
Expression, assembly competence and antigenic properties of hepatitis B virus core gene deletion variants from infected liver cells.
【0121】
この過程は発現カセットとベクターの作製、大腸菌の形質転換、菌株の選択及び評価、発酵、細胞の回収、細胞の破壊、洗浄、吸着、クロマトグラフィー、ゲル濾過、及び最終産物の液体の濾過滅菌という段階を含む。
【0122】
最終産物の品質は、Lowry法、SDS/PAGE法、又はウェスタンブロット分析等の種々の生化学的分析により確認している。
【0123】
実施例3:インフルエンザウイロソームの再構成
Gluck R., 1995, Journal of Liposome Research 1995 5(3), 467-479:
Liposomal Hepatitis A Vaccine and Liposomal multiantigen combination vaccines
【0124】
インフルエンザウイロソームは、リン脂質と、孵化鶏卵と細胞培地のいずれかで培養されたインフルエンザウイルスから産生される。ウイルスを回収し、精製し、一回又はそれ以上の遠心段階で濃縮し、次にベータプロピオラクトン(BPL)で処理することにより不活化した。
【0125】
不活化されたウイルスを超遠心により沈殿にし、界面活性剤(100mMのOEGを含むPBS)に再懸濁し、それによってウイルスの外殻、つまりウイルス包膜を溶解するが、ウイルスの内部、つまりヌクレオカプシドは蛋白質と残存核酸の複合体として残る。同時に、脂質(レシチン他)を同じ界面活性剤(100mMのOEGを含むPBS)に溶解する。その後脂質と溶解したインフルエンザウイルスを混合し、随意に完全に分離するために超音波パルスで処理する。次に、混合物中の全ての不溶性物質を沈殿させ除去するために、超遠心を行う。この不溶性物質の大部分はウイルスヌクレオカプシド複合体を含む。超遠心の後の上清は、別々に加えたウイルス包膜蛋白質及び脂質とリン脂質といった、将来のウイロソームの全ての成分を含む。最終段階においては、SM-2 Bio-Beadsを用いたバッチクロマトグラフィーによって上清から界面活性剤を取り除く。この連続的な界面活性剤の除去が、厳密な組成と脂質:蛋白質比に依存して平均直径が100から200nmとなる、ウイロソームベシクルの均一な集団の自発的な形成をもたらす。
【0126】
実施例4:HBウイロソームの調製及び分析論
HBウイロソームへのHBs及びHBc成分の定量的な組込みを達成するために、インフルエンザウイロソームに関して確立された調製過程は大幅に修正され、多数の変数に関して最適化された。標準的なインフルエンザウイロソームのプロトコルを用いると、HBV抗原の組込み率及び過程の再現性が不満足なものとなるとことが分かった。
【0127】
4.1 図3-11に示しているようなHBウイロソームの詳細かつ基本的な調合プロトコル
共にリン酸緩衝生理食塩水(PBS)に入った、2mgのHAを含む不活化されたインフルエンザウイルス(シンガポールA型)及び2mgの精製された組換えHBs抗原を混合し、100000g、4℃で2時間遠心した。結果として生じる沈殿を1mlの100mM PBS-OEGを含むPBS中に可溶化した。
【0128】
卵由来の粉末状のリン脂質(16.5mgのフォスファチジルコリン及び4.5mgのフォスファチジルエタノールアミン)を1mlの100mM PBS-OEG中に溶解した。
【0129】
その後リン脂質とHA-HBs抗原溶液を混合し、完全に溶解するために37℃のウォーターバス中で2分間超音波処理した。不溶性の残存物質は100000g、4℃で2時間遠心することにより除去した。結果として生じる上清(2ml)を回収し、PBSで最終容量が3.5mlとなるように希釈した。
【0130】
HBc抗原を4mg/mlとなるようにPBSで希釈し、希釈液の0.5mlをPBS-OEG中にHA、HBs抗原、及びリン脂質を含む溶液に加え、結果的に最終OEG濃度が50mMとなった。
【0131】
調製液を0.22マイクロメートルフィルター(Millipore)を通して濾過し、界面活性剤除去手順を行った。混合液を1.2g(乾燥質量)のBio-Beads SM-2(BioRad)に加え、回転させながら室温で30分インキュベーションした。次に、懸濁液を0.8gの新しいBio-Beadsに移し同じ条件下で30分インキュベーションし、その後0.8gの新しいBio-Beadsと同じ条件下で3回目のインキュベーションを行った。結果として生じるHBウイロソームをその後濾過滅菌(0.22マイクロメートル)し、使用までガラスアンプル中に4℃で保存した。
【0132】
4.2 HBウイロソームの組成の可能な修正
異なる供給源(CHO由来のHBs、酵母由来のHBs)、サブタイプ(ayw 及びadv HBV核粒子)、又は変異体(HBV核の全長又は短縮型)からのHBV抗原を含むHBウイロソームを同様の方法で調製した。上記で説明した再構成HBVウイロソームの製剤は、粒度と抗原組込み率に関しての重要な変数を同定するために行った一連の調製の結果であり、最終濾過滅菌に適した均一な粒度の粒子への抗原の良好な組込みを可能にした。異なるリン脂質組成(異なる比率のフォスファチジルエタノールアミンとフォスファチジルコリン)による調合は、粒度とフォスファチジルエタノールアミン量の間の逆相関の関係を示した。蛋白質に対する脂質の比(2.5、5、6、7.5)の粒度と抗原組込みに対する影響は一連の調製の中で調べている。異なる抗原の相対量が組込みに影響することが示されており、調合において(HBc抗原無しで)HAの濃度を増やすことがHBs抗原の組込みの増加につながり、1対1の比率の80%まで届いた。以下の変数を系統的に試験し最適化した:
【0133】
脂質:蛋白質比
我々の手では最適な脂質:蛋白質比は5:1であったが、リン脂質を使用するなら抗原組込みが最大となるのは20:1から1:10の範囲であることは考えられる。他の脂質(合成脂質、ステロイド型脂質)又は異なる脂質の組み合わせを使用するなら、脂質:蛋白質比は更に異なる可能性がある。
【0134】
リン脂質組成(PC、PE、他の脂質)
PCのみのHBウイロソームを産生することはでき、我々の手では粒度と均一性に関してPEが22%であることが最適であった。もう一度言うが、他の脂質を使用するならば、これらの比は大幅に異なる可能性がある。
【0135】
抗原間の比率(HA:HBs:HBc)
我々の手では1:1:1という比が最適であると分かった。しかし、修正した脂質組成と脂質:蛋白質比では、抗原組込みが最大となる最適量は変わる可能性が高い。それに加え、HAの一切無い調製物も同様に所望の粒子構造を作り出した。
【0136】
界面活性剤
選択した界面活性剤は50mMの濃度のOEGである。しかし、組成や比率を修正する時には20から100mMの範囲が適切である可能性がある。他の非イオン性、イオン性、又は双極性の性質の界面活性剤をOEGの代わりに調製過程で使用してもよい。
【0137】
【表2】
【0138】
4.3 ウイロソーム調製物の分析論
HBウイロソームの徹底した物理化学的な分析は、調製過程の最適化と将来の製品の品質管理に必須の要素である。それ故、HBウイロソームの内容と構造を調べる測定法の開発に莫大な努力が費やされた。アジュバント効果(MHC-1提示)がHBウイロソームの物理的構造に直接依存するため、ワクチンのHBV成分をウイロソーム担体に物理的に結合させる単一の粒子型の証明に特に重点を置いた。
【0139】
成分の定量:
蛋白質(SDS-PAGE)
HBs(ELISA、ウェスタンブロット)
HBc(ELISA、ウェスタンブロット)
HA(SRD)
リン脂質(酵素測定法)
ウイロソーム構造:
光子相関法
免疫共沈降法
密度勾配超遠心法
トリプシン処理
電子顕微鏡(計画中)
【0140】
全蛋白質濃度の決定
蛋白質濃度は260、280、320nmの波長のUV吸光度によって測定し、以下の公式に従って計算した:1.55×(A280 - A320) - 0.76×(A260 - A320)。結果はmg / mlで表示した。
【0141】
HBs及びHBc抗原の定量
HBウイロソームに組込まれたHBs及びHBc抗原の量は定量的Elisa測定法により決定した。抗原に対する最大限の接触をもたらすために、基準抗原とHBウイロソーム試料を最初の希釈時にPBS-OEGに溶解し、次に連続する希釈はPBSにて行った。HBs測定は市販のHBs Elisa detection kit (DADE Behring)を使用して行い、同じ測定法にて測定する精製したHBs抗原の希釈系列が、HBsウイロソーム試料中の抗原の量の決定を可能にした。定量的HBc ELISAでは、マイクロタイタープレートを、50mM Na2CO3 (PH 9.6) に入ったHBcに対するモノクローナル抗体(mAb)(クローン7E6、Biogenesis、希釈率1:1000)によってコートした。その後プレートを1% BSA、5%ショ糖、0.05% NaN3入りのPBSで室温で少なくとも1時間ブロックし、0.05% Tween20 (v/v)入りのPBS(洗浄バッファー)で洗浄した。試料(0.1ml)を載せ、室温で1時間インキュベートした。第2の、HBcに対するビオチン化されたmAb(クローン4H5、Biogenesis)を0.05% Tween20、0.1% BSA入りのPBS(希釈バッファー)中で1:1000に希釈し、プレートに加え(1ウェルあたり0.1ml)、室温で1時間インキュベートした。4回の洗浄の後、プレートをストレプトアビジン(希釈バッファー中で1:5000)の存在下で室温で1時間インキュベートした。更なる4回の洗浄の後、TMB(1ウェルあたり0.075ml)を30分インキュベートし、発色反応を1M H2SO4(1ウェルあたり75μl)で停止し、450nmにおける光学密度を測定した。
【0142】
HAの定量
HBウイロソームのHAの量は標準的な一元免疫拡散(SRD)法を使用して決定した。この試験はウイロソームワクチンの分析に関して有効な測定法であり、エパキサール(Epaxal:登録商標)に関するそれぞれの標準操作手順に従ってBerna Biotch Ltd. QC departmentによって行われた。
【0143】
ゲル電気泳動、ウェスタンブロット、銀染色
HA、HBs又はHBc抗原のHBウイロソームにおける存在を証明するために、調製物、勾配分画、プロテアーゼ処理、又は免疫共沈降法からの試料をMESバッファーを用いNuPage Bis-Tris SDS-PAGE pre-cast gel(Invitrogen)上で分離し、製造者の使用説明書に従いニトロセルロースメンブレンに転写した。メンブレンを5%ミルク入りのPBST(1% Tween 20入りのPBS)でブロックし、抗原特異的な抗体の1:1000の希釈液中で室温で1時間インキュベートした。メンブレンを洗浄し、その後ペルオキシダーゼ標識の抗ウサギ又は抗ヒツジ免疫グロブリンの1:10000の希釈液中で1時間インキュベートした。蛋白質をECL Plus 基質溶液 (Amersham Pharmacia, Piscataway, N.J.)を用いて可視化した。
ゲルの銀染色は製造者の使用説明書(Invitrogen)に従って行った。
【0144】
粒度の測定:光子相関法
最初の材料及び調製したHBウイロソームの流体力学的径、多分散性係数、及び統計的粒度分布を光子相関法又は動的光散乱によって決定した。この方法は、光散乱の時間変動として測定される、ブラウン運動の粒度依存的な速度に基づいている。生データ、つまり光強度の変動からの変数の計算のためのソフトウェアを含め、Malvern Zetasizer 1000HS(Malvern Ltd, Malvern, UK)をこの目的に使用した。試料を測定のためにPBS中に十分に希釈し、1mlの希釈液を25℃の標準条件下で分析した。
【0145】
ショ糖勾配
個々の成分の異なる密度に基づいて、HBウイロソーム構造への抗原の組込みを推定するための分析的方法として、不連続ショ糖勾配を通した超遠心を行った。PBS中のHBウイロソーム調製物の一部を、PBS中の20-60% (w/v)不連続ショ糖勾配の上に載せ、4℃で24時間100000gで遠心した。次に回収した分画を密度と蛋白質量に関して分析し、また異なる抗原の存在に関してウェスタンブロットを用いて分析した。
【0146】
トリプシン処理
HBウイロソーム粒子の抗原の組込み及び最後の皮膜化を部分トリプシン処理によって調査した。次に処理後の試料を、蛋白質分解抵抗性の断片又は保護された全長蛋白質に関して、ウェスタンブロットによって分析した。HBウイロソームをPBSバッファー(無傷の粒子=自然条件)、又は0.5% デオキシコール酸ナトリウム及び1% Triton X-100入りのPBS(ウイロソーム構造の破壊=変性条件)のいずれかで希釈した。プロテアーゼ処理は5%トリプシン(w/w 蛋白質)を用いて、室温で0、2、5、及び10時間行った。トリプシン処理に対する個々の成分の効率的な接触可能性のコントロールとして、同濃度の異なる抗原の混合物を同じ条件下で処理した。反応は4×SDS-PAGEサンプルバッファーを加えることにより停止した。95℃での10分間の変性の後、試料をSDS-PAGE電気泳動、及びHBsとHBcに対する免疫ブロット分析にかけた。
【0147】
免疫共沈降法
HBウイロソーム構造中でのHA、HBs、HBc抗原の物理的結合は、(トリプシン処理に関して説明したような)天然及び変性条件下におけるそれぞれの抗原に対する別々の(個別の)免疫共沈降法、及びそれに続くウェスタンブロット分析による免疫共沈降された抗原の同定によって証明した。HBウイロソーム調合の免疫共沈降法は、HA、HBc、HBsに対する特異的抗体の存在下に、同時平行の実験で行われた。次に免疫複合体をプロテインGセファロースビーズ(Promega)と共に4時間インキュベーションし、遠心した。結果として生じる沈殿をPBSで5回洗浄した。免疫沈降された蛋白質をサンプルバッファー中で10分間煮沸することにより再懸濁し、SDS-PAGEとウェスタンブロットによって分析した。それぞれの抗原の存在はそれぞれの特異的抗体と共にインキュベーションすることにより調査した。
【0148】
ウイロソームの調製物:マウスモデルにおける免疫原性
HBウイロソームの免疫原性に関してはこの時点で予備的なデータのみが利用可能である。HBウイロソームに加え、付加的なアジュバントRC529(Corixa)がある条件と無い条件で、マウスにおける実験を行った。マウスは異なる量のHBウイロソームの筋内注射によって免疫化され、全ての3つの抗原に対する高い抗体力価が検出された(図8)。3回目の増強の後、免疫化動物から脾臓細胞を精製し、それぞれの抗体を用いた試験管内での再刺激の後、インターフェロンγに対するELISPOTによって細胞反応を決定した(図9)。注目すべきはこの方法はCD4型とCD8型反応を区別しないことである。それに加え、免疫化動物からの脾細胞をFACSによって分析した(図10)。新しい脾細胞はHBc特異的5量体によって抗CD8抗体と共に直接染色された。別個に、脾細胞はCD8特異的ペプチド又は全蛋白質によって刺激され、次に細胞内インターフェロンγに対して抗CD4と抗CD8抗体のいずれかと共に染色された。
【0149】
実施例5:他のウイルスシステムに対する調製原理の適用
異なるウイルス蛋白質間の相互作用はビリオンの組立に重要であり、全てのウイルスに見られる一般原則である。包膜ウイルス粒子は組立に関して細胞膜構造に依存している。核酸と蛋白質の複合体であるヌクレオカプシドは、成熟したウイルス構造を形成するために、膜に結合したウイルス蛋白質と結合する。我々はB型肝炎ウイルスに関して、インビトロで、細胞膜構造のない条件で、異なる組換え供給源からのウイルス蛋白質を利用して、この過程が模倣できることを示した。ウイルス抗原の組換え供給源が量と質に関して好適であるが、インフルエンザHAに関して本明細書で適用したように、ウイルス蛋白質は同様に元のウイルスに由来してもよい。インビトロの調製物の柔軟性は異なる供給源からの多数の抗原の包含を可能にする。(HBVとインフルエンザに関して示したように)複数のウイルスからの蛋白質を含む多面的調製物は、この原理に基づいて物理的安定性、免疫学的性質、又はワクチンの防御範囲を改善する可能性がある。
【0150】
条件をそれぞれの病原体に合わせ、必要な成分が十分な量で利用可能ならば、任意の他の包膜ウイルスに対して同じ原理が適用出来ることが想定される。インビトロの調製物の原則が適用でき、また魅力的で切迫したワクチンの標的である、多数の候補を下記に列挙してある。しかし、そのような憶測上のウイルス様粒子によって誘導された免疫反応が防御的、又は更に治療的な効果までを持つかどうかは、予測することが出来ない。
【0151】
5.1 C型肝炎ウイルス
C型肝炎を引き起こす原因としてのHCVは世界的な健康問題となっている。世界人口の内 %がこのウイルスに感染していると推定されている。予防的又は治療的用途のための、HCVに対するワクチンは現在存在しない。細胞システムにおけるウイルス様粒子の組立は実現されている(Baumert et al, Journal of Virology 1998, 75: 3827-3838; Hepatitis C virus structural proteins assemble into virus-like particles in isect cells)。HBVのように、核蛋白質の1量体が結合して膜に結合した包膜蛋白質(E1、E2)と相互作用する20面体のヌクレオカプシドとなる。
【0152】
5.2 他のフラビウイルス
HCVとは別に、多数の重要なヒト病原体がフラビウイルス科ファミリーに含まれる(ウェストナイルウイルス、クンジンウイルス、日本脳炎ウイルス、デングウイルス、黄熱ウイルス、及びダニ媒介脳炎ウイルス)。HCV(及びHBV)との構造レベルでの類似性は高く、それ故ウイルス様粒子のインビトロの再組み立ては可能だと思われる。
【0153】
5.3 HIV
HIVはレトロウイルス科ファミリーの中で最も著名なメンバーである。繰り返すが、切迫した医療需要があるにも関わらず、効果的なワクチンは存在しない。これらのウイルスは更に複雑なヌクレオカプシドを形成するが、同じ原理に基づいた多抗原ワクチンのインビトロの調製は可能だと思われる。
【図面の簡単な説明】
【0154】
【図1】個々の成分と結果として生じるHBウイロソームの構造を描いた概略図が図1に示してある。
【図2】調製過程のフローチャートが図2に示してある。
【図3】図3はHBウイロソームのSDS-PAGE分析の結果を示している。SDS-PAGE分析に基づくHBウイロソーム及び最初の材料(インフルエンザ、HBs、及びHBc)の蛋白質組成が図3に示してある。
【図4】図4は個々の成分の粒度と比較したHBウイロソームの粒度の分析の結果を示している。
【図5】図5はHBウイロソームの勾配分画分析の結果を示している。
【図6】図6はHBウイロソームのSDS-PAGE/ウェスタンブロット分析の結果を示している。
【図7】図7はHBウイロソームのSDS-PAGE/ウェスタンブロット分析の結果を示している。
【図8】図8はマウス免疫化の後のHBウイロソームに対する抗体免疫反応の試験の結果を示している。
【図9】図9はマウス免疫化の後のHBウイロソームに対する細胞免疫反応の試験の結果を示している。
【図10】要約表はHBウイロソームによる免疫化の後これまでに取得した免疫学的データを示している。
【技術分野】
【0001】
本発明は、少なくとも一つの脂質及び包膜ウイルスの包膜蛋白質を備えるウイロソーム膜、及びウイロソームの内側及び外側に位置し、前記包膜蛋白質に付着した前記包膜ウイルスのヌクレオカプシド粒子を備えるウイロソームに関する。更に本発明は、本発明のウイロソームを備えるワクチン、及び本発明のウイロソームの生成のための方法に関する。更に本発明は、本発明のウイロソームをワクチンの調製のために、例えばHBV感染に関連した疾患の予防又は緩和のために使用する方法、及び被験者にワクチンを接種する方法に関する。
【0002】
本願明細書のテキスト全般を通じていくつかの文献が引用されている。本願明細書中で引用されている各文献(いずれの製造業者の仕様書、指示書等も含む)の開示内容は参照することにより本願明細書に援用される。
【背景技術】
【0003】
新規であり、安全性がより高いワクチンの開発には、特性がよく知られた抗原、特に高度に精製された組み換え蛋白質又は合成ペプチドが頻繁に利用されている。いくつかの成果に関わらず、そのような抗原は単独で投与されたときには免疫原性が低いことが多いという事実により、このアプローチは妨げられる。この事実により、所定の抗原の免疫原性を促進する能力を有している適切なアジュバント及びキャリアシステムの開発が余儀なくさせる。一つの可能なアプローチは抗原を、例えばウイルス様の粒子など、より高度な構造中に組み込むことである。単一の粒子中で全てのワクチン成分が物理的に結合することは、それらが個々の免疫細胞と同時に相互作用することを保証し、そしてそれによって、相乗的な潜在力が最大に利用される。これは、免疫刺激性又は免疫調節性の成分(アジュバント)が調製物に含まれている場合には特に関連する。更に、粒子構造それ自身が免疫刺激性の効果を有すことができ、そして、個々の成分の安定性及び免疫原性の両者を増加させることができる。
【0004】
そこで問題は関連する抗原を産業上利用可能な調整物中に混ぜる適切なアプローチを見出すということであり、それは効率的な予防及び/又は治療上の適用へ至る。
【0005】
宿主細胞中でウイルスが複製する間にウイルスゲノムのコピー作られ、細胞内の設備基盤を利用しながらウイルス蛋白質が会合して成熟したビリオンとなるが、その前にウイルス蛋白質が発現してプロセスされる。共通の基礎は、ウイルスの複製と子孫の会合には、生きている宿主細胞の環境、および、ウイルスの核酸、ウイルスと宿主細胞の蛋白質と脂質膜の間の、順序が決まった一連の特異的な相互作用が必要であるということであり、それによって高分子のビリオン構造の分別と会合へと至る。多数の異なった分子が必用とされ、加えて、関連している細胞構造は、いずれのビリオンの会合も非常に複雑であることを示す。異なったウイルスクラス、特に包膜ウイルスと非包膜ウイルスの間には大きな差がある。全ての包膜ウイルスに共通しているのは、ウイルスの蛋白質が組み込まれた脂質膜からなるウイルスの外部殻、および、その結果としての、成熟した包膜ウイルスが会合するための、膜に結合し可溶性のウイルス蛋白質又は蛋白質に基づいた構造(例えばヌクレオカプシド)の間の必用な相互作用である。非包膜ウイルスは脂質に基づいた膜を欠き、蛋白質及び核酸分子のみから組みたてられている。
【0006】
インビトロとインビボでウイルス粒子を再構成しようとする数多くのアプロ−チが文献中に述べられており、異なったカテゴリーに区分けできる。
【0007】
(a)ウイルス包膜のインビトロ再構成
ウイルス由来の又は組み換えられた包膜蛋白質を精製し、追加の脂質の存在下又は非存在下でプロテオリポソームへ調製される。この純粋なインビトロのアプローチにより、包膜ウイルスの外殻、包膜の生成を達成できるが、これはウイルスの核、ヌクレオカプシドを含んでいない。異なったウイルス由来の包膜蛋白質を組み込んだ、キメラのウイロソーム構造の例がある。再構成されたウイルス包膜も、遺伝子(DNA又はRNA)の移入に成功裏に使用されているが、これらの方法は機能を有し、蛋白質に基づいたヌクレオカプシドのパッケージングには依存しておらず、むしろ、核酸と再構成された包膜との直接的な結合に依存している。
【0008】
(b)一つ又はそれ以上のウイルス蛋白質の非相同発現
単離された組み換えウイルス蛋白質は自己会合し、ウイルス様の構造(VLPs): HPV(酵母、バキュロ)、HCV(バキュロ)、HBs抗原(酵母、CHO)、HBc(大腸菌)となることができる。これらのアプロ−チの全てに共通することとして、自己会合は異種の細胞発現システム中で起こり、引き続いてウイルス様の粒子が精製される。そこで会合はインビトロでは起こらないが、細胞システムを頼りにする。VLPsはワクチン及びワクチン担体として使用されてきた(Pumpens, P.;Grens, E. (2001) Interviology 44 (2-3); 98-114; Noad R, Roy P. (2003) Trnds Microbiol. 11 (9): 438-44)。
【0009】
(c)インビトロでの非包膜(20面体の)ウイルス又はウイルス様粒子の再構成
このアプローチは、別々に精製された成分に基づいている。脂質膜に基づいた包膜が存在しないために、非包膜ウイルスは構造においてより単純であり、もし全ての必要な成分が正しい化学量論で存在していれば、インビトロである条件下において自己会合することができる。同様に包膜ウイルスの内部核、脂質がないヌクレオカプシド、又はそれらのサブユニットが、例えばインフルエンザウイルスの精製された組み換え成分からインビトロで再構成されている(Martin-Benito J. et al. (2001) EMBO Rep. 2 (4): 313-7)。
【0010】
(d)ウイルスのヌクレオカプシドの精製
多くの異なった型のウイルスからヌクレオカプシドが抽出され単離され、それらの組成の特性解析が行われてきた。ウイルスのレスキューを目的として、これらの調製物を感染しやすい細胞のトランスフェクションのために使用することもできる。成功裏のウイルスレスキューは、機能を有するヌクレオカプシドが単離され、宿主細胞の細胞質へ運ばれることを意味する。しかしこれは、機能を有する包膜ウイルスの再構成の成功を示すものではない。なぜならば、機能を有する包膜に依存する天然の感染経路は形質転換体の使用によってバイパスされ、後者は宿主細胞の細胞質へヌクレオカプシドを直接的に運搬することを仲介している。
【0011】
(e)細胞培養システム中における包膜ウイルス及びウイルスベクターの偽タイピング
このインビボのアプローチは、実験室規模のキメラウイルス又はベクター(例えば、レトロウイルス、レンチウイルス、及びAAV)の産生において、広範かつ成功裏に使用されている。偽タイピングされたウイルスの産生の鍵となる要素は、ビリオン中へ組み込まれるべき全ての蛋白質を共発現し、ビリオンの組み立てを仲介するヘルパーウイルスである。対照的に、包膜ウイルスのインビトロの会合は、規定された、別々に産生及び精製がなされた成分に基づいており、物理的な結合はインビトロで制御された条件の下で行われる(Sandrin V. et al. (2003) Curr Rop Microbiol Immunol.; 281: 137-78)。
【0012】
ウイルス様の粒子の特異的な型として、ウイロソームは臨床的に証明されたワクチン担体/アジュバントシステムであり、ヒトにおいて素晴らしい安全性と耐性プロファイルを有している。ウイロソ−ム担体が、外来性及び内在性の経路の両者を通じて抗原プロセシングを仲介する能力により、このシステムは治療用ワクチンのための良い候補となっている。
【0013】
ウイロソームの基本的な概念はインビトロでの空のウイルス包膜の、あるいはより一般的には、球状の脂質二重層中へ組み込まれたウイルスの、包膜蛋白質の再構成を備える。ウイロソームは多数のウイルスから作製されている(Y Kaneda, (2000) Adv. Drug Derivery Rev. 43, 197-205; Drummond DC et al. (2000) Prog Lipid Res. 39 (5): 409-60)。2つの異なったウイルスに由来する包膜を含んでいるキメラのウイロソームを作製する可能性が示されている(Bagai S, Saeler DP. (1994) FEBS Lett. 353 (3): 332-6)。
【0014】
全ての場合において、興味の対象であるウイルス蛋白質膜は貫通型又は膜に固定された(anchored)構造を有しており、それは自然に発生する組み込みの必要条件である。
【0015】
ウイロソーム液体膜と直接的に相互作用しない分子のウイロソームの調製を達成することは、ずっと困難である。分子をウイロソーム構造と連結させるという考えは先に提唱されており(WO 95/32706, INEX)、安定であって効率的な調製物を達成する技術的な障害は膨大であり、興味の対象である分子の生化学的な性質に依存している。陽性に荷電した脂質を使用することを介して核酸を結合させてウイロソーム構造にすることができる(WO 98/52603, Berna)。その機能のための二次構造及び三次構造を欠いている小分子(ペプチド、薬物)を生化学的に改変し、結合、組み込み、又は被包化を可能とする。ウイロソームの調製物、特に小分子(Walti et al, (2002) Canc Res.)又は合成粒子(Jana et al. (2002) FEBS Lett.; 515 (1-3: 184-188)の被包化について、多数の方法が述べられている。これらの方法は、より大きな蛋白質の真正の立体構造に影響するであろう化学的な条件下においてのみ機能するであろうし、ウイルスのヌクレオカプシドなど多量体蛋白質複合体の完全性(integrity)はもっとそうであろう(例えばHBc抗原粒子)。露出した親油性領域を欠いている一つ又はそれ以上の大きな蛋白質をウイロソーム膜中へ結合させるという、これまで述べられてきた方法は、各々の蛋白質を脂質膜中へ組み込むために、例えば脂質分子との共有結合など、蛋白質の生化学的な修飾を必要とする(Hunziker IP. et al. (2002) Int Immunol. 14 (6): 615-26)。架橋された抗体を介して特定の細胞型のウイロソ−ムを再標的化するのに、この方法は有効であると証明された(Mastrobattista E. et al. (2001) FEBS Lett. 509 (1): 71-6, Walti et al, Canc. Res. 2002)。しかし、生化学的な修飾には、非共有結合的に連結した多量体構造(例えばウイルスのヌクレオカプシド)が解離するような条件(例えば、反応性の側鎖の基を活性化するための酸化的条件)が必要である。加えてそのような条件は問題となっている蛋白質分子の立体構造も変化させる可能性があり、その結果としてその免疫原性に、そして最終的にはワクチンの効力に影響を与える。更に架橋の手段は調製に必要とされる工程の数と、抗原の損失の両者を増加させる。生化学的な修飾なしに成功裏にウイロソームと結合した多量体蛋白質について、一つの例があるのみであり、すなわちA型肝炎ワクチンのエパキサール(Epaxal:登録商標)である(Gruck R., 1995, J. of Liposome Research 1995, 5(3), 467-479)。しかしこのワクチンにおいて、ウイロソーム膜とウイルス粒子の間の静電相互作用に起因し、インフルエンザのウイロソームを調製した後にのみ、抗原は外部表層と連結している。その結果、ウイロソームの水性の内部に位置している抗原は無いが、そこは治療用ワクチンに必要とされる、効率的な細胞質送達及びCD8に基づいた細胞反応の誘起に好適な位置である(Bungener L et al. (2002) J Liposome Res. 12 (1-2): 155-63; Bungener L. et al. (2002) Vaccine. 20 (17-18): 2287-95)。
【0016】
感染疾患の予防と治療のためのアプローチの可能性と限界について、HBVを例として本願明細書において下記で議論する。特に生命を脅かす後期合併症のために、HBV感染は地球規模の大きな健康問題となっている。世界保健機関(WHO)は、現在約4億人が慢性的なHBVキャリアであると推定している。慢性的なHBV感染に罹患している患者は広範囲の症状を示し、臨床的に現れないものから重篤な肝疾患まであるが、それでも長期的な肝疾患の危険性は全ての慢性的なHBVキャリアにおいて劇的に増加する(感染後20から30年以内に25%の発生率)。全ての慢性的な患者に共通することとして、多量の抗原が慢性の感染の間に循環しているという事実に関わらず、原因物質であるHBV、特にHBVコア蛋白質に対する免疫応答が貧しい。対照的に、自然発症的な又は治療による慢性B型肝炎の治癒のみならず、急性B型肝炎の治癒は、HBV抗原に対する広範かつ活発な免疫応答と密接に連関している。インターフェロン又は抗ウイルス薬による治療により慢性肝炎をコントロ−ルするなど、従来の治療のアプローチは部分的にしか成功していないが、それでも費用がかかり、顕著な副作用を伴う。そこでHBV関連の慢性肝炎を有する患者は、この持続的なウイルス感染を制御できる治療ワクチンから大きな利益を受けるであろう。
【0017】
現在におけるHBV発病機序と免疫学の理解によると、ワクチン接種治療の成功の鍵は、慢性のキャリアにはHBV特異的な免疫学的な応答性がないことを克服することである。この目的を達成するために、存在している無効なTh2型(体液性)免疫が、強力かつ持続的なTh1型(細胞性)の反応に変わり、同時に、Th2型反応を高める(ブースト)するような方法で、関連抗原(HBcとHBs)が患者の免疫機構に提示されなければならない。
【0018】
免疫回避変異体のウイルスを防ぐために、関連抗原に対する免疫反応は広範であり、かつ、同時に多くの異なったエピトープに対して向けられていなければならない。そのような変異体は単一エピトープに対して向けられた選択的な圧力の下で発達することが示されている。更に、ワクチンの抗原として全長の蛋白質を使用することは、抗原のプロセシングとMHC遺伝子型依存的なエピトープ選択に関する患者の多様性を考慮に入れる。
【0019】
M.Hillemanの総説(Vaccine 32 (2003) 4626-4649)にあるように、過去においてHBVの治療用ワクチンの開発に多くの努力がなされてきた。多くの臨床試験において、従来のHBsに基づいた予防用ワクチンが慢性のHBV患者に使用されてきたが、持続的であって前向きである効果はこれまで観察されていない。ペプチドに基づいたワクチンは、少数の関連エピトープへの免疫応答に焦点を当てることを意図していた(Englerらによる総説、Mol Immunol. 2001 Dec; 38 (6): 457-65)。このアプローチは前臨床研究において期待をもてる結果を収めたが、ヒトにおけるものではなかった。より最近になって、二つの関連したHBV抗原を一つのワクチンに組み合わせる試みとして、表面上にHBsの単一エピトープを有する組み換えHBc粒子が作製された(Chen et al, Vaccine. 2004 Jan 2; 22 (3-4): 439-46)。
【0020】
これらのアプローチは主として、免疫システムの体液性応答を目的としている。より進んだアプローチでは、HBVに対する細胞性応答(Th1型)は治療上の免疫付与の成功における鍵となる要素である、という概念に従っている。
【0021】
HBV抗原、特にHBcに対するTh-1型の免疫応答を誘導することは、治療用のHBVワクチンの最終的なゴールである。HBsは単独でTh1応答をある程度引き起こすことができるが、HBc単独ではできない。そこでこれらの抗原は単独では、治療効果に必用とされる適切な免疫応答を誘導することができない。これは、HBV抗原をTh-1支持アジュバント又はキャリアシステムと組み合わせることのみにより達成することができる。対照的に、現在ヒトのワクチンにおけるアジュバントとして最も広く使用されているアルミニウム塩は、Th1応答を止めてTh2応答を支持することがよく知られている。アルミニウム塩のこのような性質により、それは高いタイターの保護抗体の誘導を第一の目的とする予防用ワクチンのための非常に魅力的なアジュバントとなる。治療の場面において持続的なTh1応答は決定的な役割を果すが、Th1エフェクター細胞はウイルス複製の制御とウイルスに感染した細胞の除去を仲介するからである。
【0022】
細胞応答を主として促進することが知られているDNAワクチン(HBVコア又はS遺伝子をコードするプラスミドDNA)を作る試みがなされている。DNAワクチンがマウスのモデルにおいて非常によく働くという事実に関わらず、HBVの分野のみならず、多くの臨床上の試みはヒトにおける原理の証拠を提供することができない。同様に、細胞性応答の促進を目的としたHBV抗原を発現しているウイルスベクター(例えばワクチニア)を使用しても、ヒトにおいて顕著で持続的な応答を誘導することはできなかった。
【0023】
治療用のHBVワクチンについて種々のアプローチが試みられたが、これらのいずれも有意義な治療用のワクチンへ至ることはなかった。
【0024】
2つの主要な構造的なHBV蛋白質であるHBsとHBcは、いくつかの異種のシステム:大腸菌、酵母、及び哺乳動物細胞株の中で、個々に発現させることができる。両者の抗原は典型的なウイルス様粒子構造(それぞれ、HBs粒子とHBc粒子)を形成するが、それらは感染性であって、包膜を有し、ヌクレオカプシドを含有しているHBVビリオンとは明らかに異なっている。
【0025】
組み換えB型肝炎コア抗原(HBc)は、この蛋白質は糖化されていないために、細菌又は酵母に基づいた発現システムの中で生成することができる。HBVコアモノマーは約30nmの直径を有するウイルス様の粒子へと自己会合し、プロデューサー細胞からこの型で精製できる。真性のHBVヌクレオカプシドと非常に類似して、HBc粒子は180または240モノマーコア分子のいずれかより構成され、粒子構造へ自己会合する。HBc粒子は脂質を含まない。HBVコアモノマーは183から185アミノ酸残基から構成される(長さは単離物に依存している)。C末端30アミノ酸は核酸結合領域であることを特徴とし、精製されたHBc粒子の調製物中には、好ましくない混入である、かなりの量の核酸が存在する(主としてHBVゲノムの非存在下で発現している細胞からもたらされたRNA)という結果となる。HBcはC末端において切断されて144アミノ酸でとなることができ、それは粒子構造を保ちながら核酸の含量を99%減少させる。144アミノ酸よりも短い調製物はもはや粒子を形成しない
【0026】
組み換えHBc粒子の生成は多くの変異体において述べられている。改変されたHBcの変異体は異種抗原のためのキャリアシステムとして使用されてきた(Pumpens, P.; Grens, E (2001) Intervirology 44 (2-3); 98-114)。このアプローチにおいて、外来性のアミノ酸配列が蛋白鎖のその領域(アミノ酸70−90)の中に挿入され、それは多量体粒子からみて外部表層へ露出している。しかし、遺伝子工学的に挿入される外来性抗原の配列の大きさは非常に限定されており、それは、モノマーが粒子へと自己会合する能力を保持する必要性からである。動物モデルにおけるワクチンとして使用されたとき、HBc粒子は単独で明らかなホルモン性応答を誘導するが、治療効果に必須であると考えられているHBc特異的なCD8型の細胞性応答を産生する能力を欠いている。
【0027】
WO 00/32635(ビオゲン)には、免疫原、エピトープを備え、多価のB型肝炎コア粒子を生じる可能性を有しているB型肝炎コア粒子が述べられている。
【0028】
既に臨床的な試行に入っているアプローチは、マラリアの予防のために、熱帯熱マラリア原虫の複数のエピトープを含んでいる修飾されたB型肝炎コア粒子を構築することである(Birkett A., et al, Infection and Immunity 2002, p686-6870)。
【0029】
真性のHBV包膜蛋白質(HBs)は、ずれた3つの翻訳開始部位から発現している、L(大)、M(中)、S(小)の3つの型で存在している。多量体の型の3つの型は全てHBVビリオンの包膜中に存在している。C末端のプレS1(大)およびS2(中)領域は、感染の間にHBVが細胞へ結合することに関連し、プレS1領域に対する抗体はHBVを中和できる。酵母または哺乳動物細胞中で組み換え蛋白質として発現したとき、HBsは35から45nmの直径を有するミセル粒子構造の型で分泌され、それはかなりの量(60% w/w)の細胞内膜脂質も含んでいる(Satoh O. et al. (2000) J.Biochem; 127(4): 543-50)。発現コンストラクトに依存し、組み換え粒子はS単独を含むか、またはC末端のプレS1および/またはプレS2領域を含むかのいずれかである。
【0030】
現在のHBVに対する予防的なワクチンは全て、アルミニウム塩と共に調製された組み換えHBV包膜(HBs)蛋白質に基づいている。多くの産物は酵母の発現に基づいている。より最近の産物である所謂第三世代のHBVワクチンは、哺乳細胞からもたらされる。これらのワクチンはプレS領域を含み、それに加えて、真性の哺乳類の糖化パターンと哺乳類の脂質組成の両者は、免疫反応のために有用であると考えられている。
【0031】
HBs粒子をワクチンの担体として使用することはWO 99/39736(Yissum)中にクレームされているが、そのシステムは単量体抗原に限定されており、それによってHBc粒子又は他のヌクレオカプシド型の構造と共に調製することが除外される。加えて前記システムは、キャリア粒子のインビトロでの破壊と再会合の進行を予測するものではない。
【0032】
近年の出版物(Ponsel and Bruss, (2003) JV 77 416-422)には、HBsとHBc両者の抗原のための発現プラスミドで同時トランスフェクションを行った哺乳細胞から、HBsとHBcの両者を含んでいるHBV粒子を生成し、分泌することが記載されている。しかし、HBsとHBc両者の抗原を含んでいる包膜粒子をインビトロで再構成することに関する報告はない。
【0033】
米国特許6,020,167(Medeva)は、プレS1又はHBVコア由来の一つまたはそれ以上のT細胞活性化エピトープ、及びそのポリペプチドを提示することが可能なキャリアを備える組成物を投与することによるB型肝炎の治療方法をクレームする。その発明におけるキャリアはHBs抗原粒子でもよい。
【0034】
ウイルス様粒子の特定の型として、ウイロソームは臨床的な実績があるワクチンの担体/アジュバントのシステムであり、ヒトにおいて優れた安全性と許容性のプロファイルを有する。ウイロソームの担体が、外在性および内在性の経路の両者を介して抗原のプロセシングを仲介する能力により、このシステムは治療用ワクチンの良い候補となる。
【0035】
ウイロソームの基本的な概念はインビトロでの空のウイルス包膜の、あるいはより一般的には、球状の脂質二重層中へ組み込まれたウイルスの、包膜蛋白質の再構成を備える。ウイロソームは多数のウイルスから作製されている(Y Kaneda, (2000) Adv. Drug Derivery Rev. 43, 197-205; Drummond DC et al. (2000) Prog Lipid Res. 39 (5): 409-60)。2つの異なったウイルスに由来する包膜を含んでいるキメラのウイロソームを作製する可能性が示されている(Bagai S, Saeler DP. (1994) FEBS Lett. 353 (3): 332-6)。
【0036】
全ての場合において、興味の対象であるウイルス蛋白質膜は貫通型又は膜に固定された(anchored)構造を有しており、それは自然に発生する組み込みの必要条件である
【0037】
ウイロソームの脂質膜と直接的に相互作用しない分子のウイロソームの調製を達成することはずっと困難である。分子をウイロソーム構造と結合させるという考えは先に提唱されており(WO 95/32706, INEX)、安定であって効果的な調製を達成するための技術的なハードルは膨大であり、興味の対象である分子の生化学的な性質に依存する。陽性に荷電した脂質を使用することを介して、核酸をウイロソ−ム構造と連結させることができる(WO 98/52603, Berna)。その機能のための二次構造及び三次構造を欠いている小分子(ペプチド、薬物)を生化学的に改変し、結合、組み込み、又は被包化を可能とする。ウイロソームの調製、特に小分子(Walti et al, (2002) Canc Res.)又は合成粒子(Jana et al. (2002) FEBS Lett.; 515 (1-3: 184-188)の被包化について、多数の方法が述べられている。これらの方法は、より大きな蛋白質の真正の立体構造に影響するであろう化学的な条件下においてのみ機能し、ウイルスのヌクレオカプシドなど多量体蛋白質複合体の完全性(integrity)はもっとそうであろう(例えばHBc抗原粒子)。露出した親油性領域を欠いている一つ又はそれ以上の大きな蛋白質をウイロソーム膜中へ結合させるという、これまで述べられてきた方法は、各々の蛋白質を脂質膜中へ組み込むために、例えば脂質分子との共有結合など、蛋白質の生化学的な修飾を必要とする(Hunziker IP. et al. (2002) Int Immunol. 14 (6): 615-26)。この方法は架橋された抗体を介して、特定の細胞型のウイロソ−ムを再標的化するのに有効であると証明された(Mastrobattista E. et al. (2001) FEBS Lett. 509 (1): 71-6, Walti et al, Canc. Res. 2002)。しかし、生化学的な修飾には、非共有結合的に連結した多量体構造(例えばウイルスのヌクレオカプシド)が解離するような条件(例えば、反応性の側鎖の基を活性化するための酸化的条件)が必要である。加えてそのような条件は問題となっている蛋白質分子の立体構造も変化させる可能性があり、その結果としてその免疫原性に、そして最終的にはワクチンの効力に影響を与える。更に架橋の手段は調製に必要とされる工程の数と、抗原の損失の両者を増加させる。生化学的な修飾なしに成功裏にウイロソームと結合した多量体蛋白質について、一つの例があるのみであり、すなわちA型肝炎ワクチンのエパキサール(Epaxal:登録商標)である(Gruck R., 1995, J. of Liposome Research 1995, 5(3), 467-479)。しかしこのワクチンにおいて、ウイロソーム膜とウイルス粒子の間の静電相互作用に起因し、インフルエンザのウイロソームを調製した後にのみ、抗原は外部表層と連結している。その結果、ウイロソームの水性の内部に位置している抗原は無いが、そこは治療用ワクチンに必要とされる、効率的な細胞質送達及びCD8に基づいた細胞反応の誘起に好適な位置である(Bungener L et al. (2002) J Liposome Res. 12 (1-2): 155-63; Bungener L. et al. (2002) Vaccine. 20 (17-18): 2287-95)。
【0038】
インフルンザのウイロソーム(WO 92/19267 WO 98/52603, Berna)、及び他の包膜ウイルス由来(例えばセンダイウイルス)のウイロソ−ム様構造について、多くの特許が出願され、特許が付与されてきた。これらの方法は、ウイルスの包膜を可溶化し、ウイルスゲノムを含んでいるヌクレオカプシドを除去し、続いて「空の」ウイルスの包膜を再構築することを備える。更に、付加的な抗原は、それらをウイロソーム膜の中に固定するために、既成のウイロソーム(エパキサール(Epaxal:登録商標))に付着されているか、あるいは脂質分子と架橋されている。
【発明の開示】
【発明が解決しようとする課題】
【0039】
ウイルス感染に対するワクチン接種の上記に述べた制限を鑑みて、本発明の基礎となる技術上の課題は、ウイルス感染の予防、軽減、又は治療のために、被験者にワクチンを接種するための改良された手段と方法を提供することである。
【課題を解決するための手段】
【0040】
前記の技術的な課題の解決は、特許請求の範囲中に特性が示された態様を提供することによって達成される。
【発明を実施するための最良の形態】
【0041】
従って、本発明は
(a)少なくとも1つの脂質、及び包膜ウイルスの包膜蛋白質を備えるウイロソーム膜と、
(b)ウイロソームの内側及び外側に位置し、前記包膜蛋白質に付着した前記包膜ウイルスのヌクレオカプシド粒子
を備えるウイロソームを提供する。
【0042】
「ウイロソーム」という言葉はウイルス様粒子(VLP)の特定の形態を定義する。ウイロソームはウイルス粒子由来の半合成複合体であり、インビトロの方法で生成される。これらは本質的に再構成されたウイルス被覆であるが、ウイルスのヌクレオカプシドが任意の化合物で置換されている。ウイロソームは融合活性を保持しており、それ故組込まれた化合物(抗原、薬物、遺伝子)を標的細胞の内側に送達する。これらはワクチン、薬物送達、遺伝子導入に使用することが出来る。
【0043】
VLPとは、形態と粒度に関して親ウイルスと類似している、又は見分けがつかない粒子構造であるが、感染し宿主細胞中で複製する能力を欠いている。VLPはウイルス蛋白質(真正、又は改変されたその変異体)から構成される多量体構造である。それに加え、VLPは核酸や脂質を含んでも含まなくてもよく、脂質膜構造を含んでも含まなくてもよい。1つのウイルス(HBV)に由来する2つの典型的で、かつ大幅に異なるVLPの例は、HBsとHBc粒子である。
【0044】
本発明の文脈中において「ウイロソーム膜」という言葉は、人工的に再構成され脂質2重膜とそれに組込まれたウイルス包膜蛋白質から構成される球形の膜構造を定義する。
【0045】
本発明の文脈中において「包膜蛋白質」という言葉は、元来の形態においてウイルス脂質膜と直接相互作用する、包膜ウイルスによってコードされる蛋白質を意味することを意図している。
【0046】
本発明に従って、広範囲の脂質を前記ウイロソーム膜は備えることが出来る。脂質の集団は中性又は荷電したリン脂質、ステロイド由来の脂質、中性又は荷電した合成脂質を備える。調製物に加えられる精製した脂質に加え、例えばインフルエンザウイルスや他の任意の包含された包膜ウイルスに由来する脂質、又は調合物に含まれるVLP(例えばHBs粒子等)を囲む脂質に由来する脂質といった、ウイルス成分に含まれる脂質も同様に最終的な調製物に含めることが出来る。これらのウイルス由来の脂質は不均一であり、ウイルス産生細胞又は組換え発現細胞の脂質組成を反映している。調合物の複雑性を最小限にするため、好適な調合物はリン脂質のみを基にしている。添付した実施例に記述しているHBウイロソームに使用されているリン脂質は、通常GMPグレードであり、登録されたワクチンであるインフレキサール(Inflexal:登録商標)とエパキサール(Epaxal:登録商標)に使用されている材料と同一であることが好適である。
【0047】
本発明の文脈中において「包膜ウイルス」という言葉は成熟したビリオン構造において宿主細胞由来の脂質膜を含むウイルスを定義する。包膜ウイルスの分類は表1に列記してある。
【0048】
本発明の文脈中において「ヌクレオカプシド粒子」という言葉はウイルスカプシド蛋白質で構成される粒子構造を意味することを意図している。この粒子構造は、(1つまたは多数の組換えウイルスカプシド蛋白質で構成される)VLP、又は親ウイルスから精製されたヌクレオカプシド複合体とすることが出来る。ヌクレオカプシド粒子が核酸を含むか否かは粒子の形成と関係がない。
【0049】
本発明のウイロソーム(キメラのウイルス様粒子)は1つの粒子中で物理的に結合した上記の性質を持つ分子を備える。本発明のウイロソーム中の包膜蛋白質は、対応するヌクレオカプシド粒子と相互作用する側をウイロソームの内側に向ける本来の方向で、又は対応するヌクレオカプシド粒子との相互作用する側をウイロソームの外側に向ける人工的な方向で、ウイロソームの表面に組込むことが出来る。そのようなウイロソームの一例を図1に描写している。この新しいクラスのウイロソームの構造は、上記の個々の成分の粒状の構造、又は元来のウイルスと全く異なる。この型の粒子は天然には存在せず、最先端においても今のところ人工的に生成される構造として記述も提案もされていない。つまりこの粒子構造は、分離された成分から完全に人工的に再組立させた最初の包膜ウイルス様粒子を表している。
【0050】
当技術分野で公知であり本明細書において上記で説明しているウイロソームは、細胞に基づいたシステムで産生する。そのような細胞を基にしたシステムでは、全ての成分を同じ細胞内で同時に産生しなければならず、発現システムの選択を大幅に制限し、収量と大量生産性に関しての妥協を強いることとなる。制御の難しい生物学的発現システムと代謝過程が、例えば成分間の比率といった、結果として生じるウイルス様粒子の組成を決定する。更に、有用な標本を得るために、細胞システムで産生されたウイルス様粒子は、粒子構造や組成に影響を与えずに、次に抽出し精製しなければならない。
【0051】
対照的に、また本明細書において下記で述べるように、本発明のウイロソームの人工的な調製物の組成は、加える材料と選択する生物学的変数によって制御することが出来る。過程の単純性はその安定性を保証する。結果として生じる調合液は更なる精製を必要としない。個々の成分を事前に別々の細胞を基にしたシステム(例;大腸菌、哺乳類細胞、酵母菌)で産生し、それぞれの成分に関して収量、大量生産性、及び純度に関して最適なシステムを選択することが出来る。
【0052】
本発明におけるウイロソームの調製過程は、ウイロソーム型構造に容易に組込まれるウイルス蛋白質(膜付着蛋白質、包膜蛋白質)と、それ自身は膜に結合しない蛋白質との相互作用を利用している。宿主細胞中での天然の複製過程における大半の包膜ウイルスの組立においてこの相互作用は必要であり有効であるが、薬学的産物の人工的調製過程においてこの性質を使用することは新しい。驚くべきことに、全く違う条件下ではあるが、細胞内のウイルスの組立は本当に人工的に模倣することが出来る。
【0053】
ウイロソームは少なくとも1つの脂質が少なくとも1つのリン脂質を備えるウイロソームであることが好適である。前記のリン脂質がフォスファチジルコリン、フォスファチジルエタノールアミン、フォスファチジルセリンを備えると更に好適である。
【0054】
本発明で同様に想定するのは、更に好適な実施例において、前記の包膜蛋白質が第1と第2の包膜ウイルスの包膜蛋白質でありヌクレオカプシド粒子が前記の第2の包膜ウイルスのヌクレオカプシド粒子であるウイロソームであることである。
【0055】
前記の第1の包膜ウイルスは任意の包膜ウイルスから選択することが出来る。本発明に特に好適なのはインフルエンザウイルスである。
【0056】
本発明の更に好適な実施例においては前記の第1の包膜ウイルスの包膜蛋白質はヘマグルチニン(HA)及び/又はノイラミニダーゼ(NA)である。
【0057】
本発明のウイロソームのインフルエンザ成分である、ヘマグルチニン(HA)及びノイラミニダーゼ(NA)は、確立され特許化されたインフルエンザウイロソームの調製物(エパキサール(Epaxal:登録商標) WO 92/19267, WO 92/19268, Gluck R., 1995, Journal of Liposome Research 5(3), 467-479)と同様に、不活化されたインフルエンザウイルス(例えばシンガポールA型)から精製することが出来る。インフルエンザ由来の蛋白質又は前記の第1の包膜ウイルスの蛋白質を、構造的や機械的というよりは機能的な理由で含めることが出来るが、これは本発明のウイロソームはインフルエンザ蛋白質又は前記の第1の包膜ウイルスの蛋白質が無くても同様に調製することが出来るためである。本発明のウイロソームの、ウイロソーム様担体の免疫学的性質を強化するために、インフルエンザ成分を含めることが出来る。
【0058】
同様に本発明に従って、前記の第2の包膜ウイルスは、(好適である)B型肝炎ウイルス(HBV)、C型肝炎ウイルス(HCV)、任意の他のフラビウイルス、及びヒト免疫不全ウイルス(HIV)から成る包膜ウイルスの一群から選択すると好適である。
【0059】
本発明の更に好適な実施例においては、ヌクレオカプシド粒子はHBc蛋白質を備える。
【0060】
HBc粒子は、全長のアミノ酸配列又は短縮型を含む形で、大腸菌で産生することが出来る。全長の産物と、短縮した144個のアミノ酸の構築物をHBウイロソームに調製することに成功している。代わりに、HBウイロソームに更に短い(非特異的な)HBV核を組込むことも考えられる。対応する技術は当技術分野で公知であり、付属の例で説明している。
【0061】
前記の第2の包膜ウイルスの包膜蛋白質がHBs蛋白質であると、同様に好適である。
【0062】
HBs粒子はS蛋白質単独、又はプレSとSの結合蛋白質を含むことが出来る。前記の粒子の産生の方法は当技術分野に公知である。粒子は例えば酵母又は哺乳類細胞で産生することが出来る。本発明のウイロソームを備えるワクチンにおけるプレSドメインの存在は、より広範でより効果的な免疫反応に寄与する可能性が高いが、調合過程に何の影響も与えない。HBウイロソームはいずれの供給源からのHBsも産生することが出来る。1つのHBウイロソーム中において、異なる供給源からの異なるHBs型、又は異なる血清型を組み合わせることさえも、本明細書で説明している方法を用いて行われてきた。本発明は上記の好適な実施例においてインフルエンザウイロソームの分類に当てはまる。しかし、同じウイルスのヌクレオカプシド蛋白質(HBc)をウイロソーム構造に繋げるために替わりに使用する、全く関係のないウイルス(HBV)の包膜蛋白質(HBsAg)を組込むことは、全く新しい。
【0063】
本発明の代替的な実施例は本発明のウイロソームを備えるワクチンに関連している。
【0064】
本発明の文脈において「ワクチン」という言葉は、ウイルス病の予防のために対象者に投与される予防的組成物を定義するものと理解される。代わりに、もしくはそれに加え、この言葉はウイルス病の緩和又は治療のために対象者に投与される薬学的組成物を意味することを意図する。
【0065】
本発明に従って、「予防的組成物」及び「薬学的組成物」という言葉は患者、好ましくは人間の患者への投与のための組成物に関連している。好適な実施例においては、前記組成物は非経口、経皮、腔内、動脈内、髄腔内投与、もしくは組織への直接注入のための組成物を備える。前記組成物が輸液もしくは注射によって患者に投与されることは特に想定される。適切な組成物の投与は、例えば静脈中、腹膜中、皮下、筋中、局所、又は皮内注射等の、異なる経路によって影響される。本発明のワクチン/組成物は更に薬学的に適切な補形薬を備えることが出来る。本発明に従って使用される補形薬は、例えばカプセル、賦形剤、保護剤、着色剤、崩壊剤、結合剤、乳化剤、可溶化剤、湿潤剤、溶媒、緩衝剤、ゲル形成剤、増粘剤、膜形成剤、潤滑剤、流動促進剤、形状分離剤、流量調整剤、吸着剤、及び抗酸化剤や味と匂いの矯正剤等の添加物等の、担体、添加物、及び希釈剤を備える。適切な薬学的担体の例は当技術分野において周知であり、リン酸緩衝生理食塩水、水、例えば油水乳剤等の乳剤、さまざまな種類の湿潤剤、無菌溶液等を含む。そのような担体を備える組成物は周知の従来の方法により調合することが出来る。これらの組成物は対象者に対し適切な用量で投与することが出来る。投薬量は関与する医師と臨床的因子により決定される。医療技術分野において周知のように、どのような一人の患者に対する投薬でも、患者の大きさ、体表面積、年齢、投与される特定の化合物、性別、時間、投与経路、全般的健康状態、及び同時に投与されている他の薬等の、多くの因子によって決定される。投与の好適な用量は一回の使用あたり1ngから1mgの範囲であることが多い。
【0066】
本発明のワクチン/組成物は局所的もしくは全身的に投与することが出来る。投与は例えばバイオリスティック法による内部又は外部の場所への送達のように、非経口的であることが望ましい。非経口投与のための調製液は無菌的な水性又は非水性溶液、懸濁液、及び乳液を含む。水性の担体は生理食塩水及び緩衝媒体を含む水、乳液又は懸濁液を含む。非経口の賦形剤は食塩溶液、デキストロースリンゲル液、デキストロース及び食塩の溶液、又は乳酸リンゲル液を含む。静脈中の賦形剤は液体及び栄養補充剤、電解質補充剤(例えばデキストロースリンゲル液に基づいた物等)、及びそれに類する物を含む。例えば抗菌剤、抗酸化剤、キレート剤、不活性化ガス、及びそれに類する物等の、保存料及び他の添加物が存在してもよい。組成物の使用意図に応じて、本発明のワクチン/組成物が、本発明のウイロソームに加え、更に生物学的活性のある薬剤を備えることが想定される。そのような薬剤はアジュバントとすることが出来る。アジュバントとは、ワクチンに含まれる抗原に対する免疫反応を増強もしくは制御するためにワクチンの調製に加えられる物質のことである。様々な種類の異なるアジュバントが当技術分野において知られており、それらは脂質、蛋白質、炭水化物、界面活性剤、塩、もしくはこれらの組み合わせから構成される。
【0067】
添付された実施例において説明しているように、ウイロソームの調製物は本当に抗原に対する細胞反応を改善することが示されている。これはマウスにHBウイロソームを接種した後のHBcに対する細胞反応の検出により特に証明されている。例えばHBc等のウイルス核蛋白質に対する細胞反応、特にCD8/TH1タイプ反応の持続的な誘導は、有用であり、本発明のワクチンの患者への接種の特に好適な結果である。
【0068】
随意に、本発明のワクチンは更に薬学的に適切な担体または希釈剤及び/又はアジュバントを備えることが出来る。
【0069】
薬学的に適切な担体又は希釈剤は本明細書において下記で説明している。前記ワクチンに含まれる抗原に対する免疫反応を更に増強又は調整するために、アジュバントと呼ばれる免疫刺激物質を本発明の調合に加えることが出来る。アジュバントの性質を持つ多数の化合物が当技術分野において知られている。そのような化合物の一群は蛋白質、脂質、炭水化物、核酸、及びそれらの組み合わせを備える。化合物は合成することも、生物学的に産生することも出来る。アジュバントは事前に調製したウイロソームに加えること、又は共に調製したウイロソーム構造中に組込むことが出来る。アジュバントの生化学的性質によりウイロソーム成分のいずれかと相互作用するのであれば、後者は可能である。アジュバントがRC529(Corixa)であることが特に好適である。
【0070】
本発明の好適なHBウイロソームは、本明細書において上記で述べたように、インフルエンザとHBVの抗原が1粒子中で物理的に結合している、安定して均一なウイロソームの共調合である。ウイロソーム担体に抗原が結合していることは、ウイロソーム抗原担体/アジュバントシステムの免疫刺激作用の十分な研究のための定評のある必須条件である(Moser C et al. (2003) Expert Rev Vaccines, 2 (2): 189-96で概説している)。
・ 抗原のHAを介したMHC−1提示、及び抗原に対するTh1免疫反応
・ 反復したウイルス様構造の中の抗原の提示
・ 抗原提示細胞の標的化
・ 細胞外分解からの保護
【0071】
治療的HBVワクチンに関しては、ウイロソームのHAを介したMHC−1提示、及び樹状細胞の標的化がウイロソームワクチン担体の最重要の特徴である。それ故、抗原提示細胞の細胞質に送達されるためにHBc抗原の少なくとも一部がウイロソーム中に被包されていることが好ましく、それによって抗原特異的なCD8T細胞が誘導される(Bungener L et al. (2002) J Liposome Res. 12 (1-2) 155-63)。より最近の研究ではウイロソームがCD4T細胞活性化を通してMHCクラス1制限的CTLを増強したことが証明された(Schumacher et al. Vaccine 22 (2004): 714-723)。実験的な試作品の調製の段階で、無修正のHBsとHBc抗原の物理的な組込み及び結合はそれぞれ大きな技術的障壁となっており、産業的なcGMP規模では更に難しかった。前記の障壁は本発明によって克服されている。調製に使用するHBV及びインフルエンザ抗原は異なる組換え発現システム(哺乳類細胞、酵母菌、又は大腸菌)で別々に産生され精製され、これらの精製された抗原はそれら自身で特徴的な粒子を形成する。
【0072】
更なる実施例においては、本発明は:
(a) 界面活性剤溶液中において脂質存在下で包膜ウイルスの包膜蛋白質を可溶化し;
(b) 溶液中の界面活性剤の濃度を低下させ;
(c) 前記包膜ウイルスのヌクレオカプシド粒子を工程(b)で取得した溶液に加え;
(d) ウイロソームが産生されるように界面活性剤又はレシチンを除去する;
という工程を含むウイロソームの産生方法を提供する。
本発明の文脈中において「濃度を低下させ」という言葉は、前記の界面活性剤が入っていない溶液を加えること、又は前記の界面活性剤の濃度が工程(a)において取得した溶液に比べて低い溶液を加えることを含むと理解される。
【0073】
本発明の文脈中において、「界面活性剤を除去する」という言葉は透析、膜分離、又はクロマトグラフィー等の過程を含むと理解される。界面活性剤が基質(例えばビーズ、レシン等)への吸着によって除去される、後者のクロマトグラフィーが好適である。
【0074】
本発明のウイロソームの調整工程は、苦も無くGMP産生の方法に適合させることが出来る。HBウイロソームの例で説明しているように、HBc蛋白質(図2)を含む新しい多成分の粒子構造の均一で効率的な調製を得るためには、生化学及び量的関係の条件に関する調整が必要であり、本明細書の教唆に基づいて過度の負担なしに行うことが出来る。
【0075】
本発明の方法の根底にある概念は、ウイルスの複製過程の間の細胞内のウイルス組立で起こるような、ウイルス包膜蛋白質と対応するヌクレオカプシド又はヌクレオカプシド様粒子の間の特異的な相互作用を利用した、ウイロソームの完全に人工的な組立である。例においてはHBsとHBcはそれぞれ典型的で好適な包膜蛋白質とヌクレオカプシド複合体であるが、本発明はそれらに限定したものではなく任意の包膜ウイルスに適用出来ることを理解するべきである。一般的な原則として、人工的な調製において選択する生化学的な条件は、包膜蛋白質(env)とヌクレオカプシド(nc)成分の相互作用を可能にするものでなければならない。調製における重要な生化学的変数は、界面活性剤濃度、pH値、浸透圧、及びキレート剤、特定の塩、緩衝分子の存在を含む。
【0076】
最初の材料(ビリオン又はVLPの包膜)及び脂質成分を溶解するために、界面活性剤の存在が必要である。界面活性剤の種類及び濃度の範囲は本明細書において下記で説明している。
【0077】
相互作用の低下又は他の成分との非特異的な相互作用の危険性を下げるために、pHと浸透圧は技術的に可能な限り生理的な条件(pH 7.4、150 mEq)に近づけるのが好適である。他の包膜ウイルスに関してはpHが5〜10の範囲、また浸透圧が10〜400 mEqであることが好適である。塩、キレート剤、緩衝液は同様に蛋白質間の相互作用、つまりenv / nc の調合の効率に同様に影響する。添付した実施例で説明しているプロトコルでは、生理的な塩組成を模倣し、NaCl及びリン酸塩という生理的な緩衝システムを備えるリン酸緩衝生理食塩水(PBS)を使用している。しかし、他の包膜ウイルスの調合には修正した緩衝システム(例えばTrisに基づいた緩衝液又は炭酸塩に基づいた緩衝液)をNaCl、MgCl、KCl、及びCaCl塩と組み合わせて使用することが必要になる可能性がある。本発明の好適な実施例においては、望ましくない酵素活性を不活化するために、キレート剤(例えばEDTA、EGTA)を含むことを想定してもよい。
【0078】
成功した調合物とは、個々の最初の材料(精製されたウイルス又はVLP)のそれぞれと粒度、内容、又は生化学的性質において全く異なるウイルス様粒子の単一かつ均一な集団として最もよく説明される。図4において図4のHBウイロソームに関して図示しているように、粒度の違いは光子相関法(photon correlation spectroscopy、PCS)によって簡単に検出することが出来る。解析方法は実施例4(p33)で説明している。
【0079】
ヌクレオカプシドが(HBcでそうであるように)単一の組換え蛋白質のサブユニットで構成されているのか、又は核酸に結合している複数の異なるウイルス蛋白質を含むのかは、方法の原理に関しては重要でない。例えばHBs等のウイルス包膜蛋白質成分は、それ自身では膜構造と効率的な相互作用又は結合をしないヌクレオカプシド粒子(例えばHBc)と、再構成されたウイロソーム膜との間の架橋として働く。効率的なHBsとHBcの相互作用を可能にするために、根本的には異なるものの、人工的な調整工程はある程度HBV感染細胞の内部の条件を模倣しなければならない。今回の人工的な方法では、巨大分子的な細胞構造のない条件で、組立が界面活性剤の除去の間に起こる。対照的に、HBVの複製の間は、HBVヌクレオカプシドと相互作用しながら、HBs分子は細胞膜に固定されている。驚くべきことに、根本的に異なる条件にも関わらず、ウイルス組立、つまりウイルス包膜中へのヌクレオカプシド粒子の被包は我々の調整工程において高い効率で起こった。両方の抗原をウイロソーム構造につなげるために2つの工程が同時に起こる必要がある:
(i) 膜貫通蛋白質、インフルエンザ包膜(HAとNA)及びHBs(HBV包膜)がウイロソーム膜中に組込まれなければならず、また
(ii) HBc粒子が膜に固定されたHBsと効率的に結合しなければならない。
【0080】
従って、効率的なHBウイロソーム組立は、最適化された生化学的条件と個々の成分の正しい量的関係の下でのみ起こり得る。同じことがインフルエンザウイルスとHBV以外のウイルスからの蛋白質を備えるウイロソームの組立に関しても言える。複雑なヌクレオカプシド構造の完全性が、膜に固定された包膜蛋白質との相互作用にとっての必須条件である。
【0081】
本実施例においては、異なる供給源からの3つの異なる生物学的粒子構造が、最初の構造いずれとも明らかに異なり自然には存在しない、1つの新しい種類の合成ウイルス様粒子に人工的に変形される。
【0082】
本発明の方法の上記の実施例において述べた脂質が、少なくとも1つのリン脂質を備えることが好適である。前記のリン脂質がフォスファチジルコリン、フォスファチジルエタノールアミン、フォスファチジルセリンを備えると更に好適である。
【0083】
本発明の方法において、前記の包膜蛋白質が第1と第2の包膜ウイルスの包膜蛋白質でありヌクレオカプシド粒子が前記の第2の包膜ウイルスのヌクレオカプシド粒子であると好適である。
【0084】
本発明の更に好適な実施例では、前記の第1の包膜ウイルスはインフルエンザウイルスである。前記の包膜蛋白質がヘマグルチニン(HA)及び/又はノイラミニダーゼ(NA)であると更に好適である。
【0085】
本発明の方法において、前記の第2の包膜ウイルスがB型肝炎ウイルス(HBV)であると更に好適である。ヌクレオカプシド粒子がHBc蛋白質を備えると更に好適である。包膜蛋白質がHBs蛋白質であると同様に好適である。
【0086】
本発明の方法に従って、工程(a)における可溶化の段階の前に、包膜蛋白質の溶液を遠心し、取得した沈殿を脂質の存在下で界面活性剤又はレシチン溶液に可溶化することが好適である。本発明の方法において使用する可能性のある脂質は本明細書において上記で更に詳細に定義してある。
【0087】
工程(a)が遠心の前に溶液の超音波処理を含むと更に好適である。
【0088】
遠心を100000g、4℃で少なくとも2時間行い、及び/又は超音波処理を37℃のウォーターバス中で少なくとも2分間行うと更に好適である。
【0089】
本発明の方法の1つの変法において、前記の方法は更に工程(b)に続いて段階工程(b’)を行うことを含むことが出来る:
(b’)工程(b)で得られた希釈液の無菌濾過。
溶液の無菌濾過の手段と方法は当技術分野において公知である。無菌濾過は添付した実施例で説明するように例えば0.22μmのフィルターを通して段階工程(b)で取得した溶液を濾過することを含むことが出来る。
【0090】
上記で説明した本発明の方法の工程(d)における界面活性剤の除去は、
(i)Bio-Beads SM-2を加え回転させながらインキュベーションすること;及び
(ii)Bio-Beads SM-2を溶液から除去すること
によって達成することが出来る。
【0091】
前記の工程(i)と工程(ii)を新しいBio-Beads SM-2を使用して少なくとも1回、好ましくは少なくとも2回は繰り返すことが好適である。
【0092】
工程(i)と工程(ii)を室温で行うことが同様に好適である。更に、工程(i)でのインキュベーションが少なくとも30分間であると同様に好適である。
【0093】
本発明の方法が更に工程(e)を含むことが同様に想定される:
(e) 工程(d)で得られた希釈液の無菌濾過。
【0094】
本明細書において上記で述べた界面活性剤が非イオン性界面活性剤であることが好適である。本発明に従った非イオン性界面活性剤の例は、例えばオクタエチレングリコールモノ(N-ドデシル)エーテル(OEG)、Triton X-100、Triton X-114、NP40、Tween 20/80、及びレシチン等の界面活性剤を含む。界面活性剤は好ましくは0.1から15% (v/v)の濃度の範囲で使用することが出来る。界面活性剤の性質により、好適な濃度は一般的に(v/v)で与えられる。しかし、OEGは粉末の形状で取得し、それ故その濃度はmM単位で与えられることが好ましい。例えば100mMのOEGの濃度は大体5.5% (v/v)のOEGの濃度に相当する。
【0095】
非イオン性界面活性剤がオクタエチレングリコールモノ(N-ドデシル)エーテル(OEG)であると特に好適である。
【0096】
本発明の方法においてOEGが20mMから100mMまでの濃度の範囲で工程(b)に適用されることが同様に好適である。前記の濃度が50mM(約2.75% OEG(v/v))であると最も好適である。
【0097】
本発明の方法の好適な実施例においては、適用される脂質:蛋白質比が1:10から20:1の間の範囲である。前記の脂質:蛋白質比が約5:1であると更に好適である。
【0098】
本発明の方法においてフォスファチジルコリンのウイロソームにおける割合が22%であると好適である。
【0099】
本発明の方法においてHA:ウイルス包膜蛋白質:ウイルスカプシド蛋白質の比が1:1:1であると同様に好適である。
【0100】
更に、本発明の方法が工程(d)においてウイロソームを産生する前にアジュバントを加えることを含むと好適である。
【0101】
更に代替的な本発明の実施例は、本発明のウイロソーム又は本発明の方法によって産生されたウイロソームの、ワクチン産生のための使用に関係している。
【0102】
更に代替的な本発明の態様は、本発明のウイロソーム又は本発明の方法によって産生されたウイロソームの、HBV感染の予防、緩和、又は治療のためのワクチン産生のための使用に関係している。
【0103】
本発明は同様に、本発明のウイロソーム又は本発明の方法によって産生されたウイロソームの、それを必要としている対象者への投与を含む、対象者への接種の方法に関係している。本明細書において上記で薬学的組成物について一般的に説明したように、ウイロソームを投与することが出来る。
【0104】
本発明の更なる態様は本発明のウイロソーム又は本発明の方法によって産生されたウイロソームの、それを必要としている対象者への投与を含む、HBV感染の予防、緩和、又は治療のための対象者への接種の方法に関係している。随意に、ウイロソームは薬学的に適切な担体又は希釈剤及び/又はアジュバントと組み合わせて投与することが出来る。
上記の対象者が人間であると特に好適である。
【0105】
【表1】
図が示しているのは:
【0106】
図1:
個々の成分と結果として生じるHBウイロソームの構造を描いた概略図が図1に示してある。分析的データは提案された構造と一致している。
【0107】
図2:
調製過程のフローチャートが図2に示してある。HBウイロソームの詳細な調合プロトコルは添付した実施例に記述してある。
【0108】
図3:
図3はHBウイロソームのSDS-PAGE分析の結果を示している。SDS-PAGE分析に基づくHBウイロソーム及び最初の材料(インフルエンザ、HBs、及びHBc)の蛋白質組成が図3に示してある。
HBウイロソームの予測される物理的構造は以下の分析データによって確認されている(図4−7)。
【0109】
図4:
図4は個々の成分の粒度と比較したHBウイロソームの粒度の分析の結果を示している。HBウイロソームは個々の成分とは全く異なる単一種類の粒子から構成され、光子相関分析の粒子の粒度の分布において1つの狭いピークを形成している。
【0110】
図5:
図5はHBウイロソームの勾配分画分析の結果を示している。ショ糖勾配中でHBウイロソームを超遠心した後、全ての抗原が同一の勾配分画で見え、それらの物理的な結合を示唆している。
【0111】
図6:
図6はHBウイロソームのSDS-PAGE/ウェスタンブロット分析の結果を示している。抗HA又は抗HBs抗体のいずれかを使用したHBウイロソームの免疫沈降では、両方の場合で全ての抗原(HBs、HBc、及びHA)の共沈降が起こった。界面活性剤の添加によってHBウイロソーム構造が破壊されると、抗体によって直接認識される抗原しか沈降されない(それぞれHA又はHBs)。この所見は全ての抗原が1粒子中で結合していることを確かめている。
【0112】
図7:
図7はHBウイロソームのSDS-PAGE/ウェスタンブロット分析の結果を示している。HBウイロソームがトリプシン処理に晒された場合、両方のHBV抗原が部分的に保護された(ウェスタンブロットによれば50%)。トリプシン中でインキュベーションする前に界面活性剤の添加によってウイロソーム構造が破壊されると、HBV抗原は短時間のうちに完全に分解された。
【0113】
図8:
図8はマウス免疫化の後のHBウイロソームに対する抗体免疫反応の試験の結果を示している。この時点ではHBウイロソームの免疫原性に関して予備的なデータのみ利用可能である。付加的なアジュバント無しのHBウイロソームを用いてマウスで実験を行った。マウスは異なる量のHBウイロソームの筋内注射によって免疫化され、高い抗体力価が異なる3つの抗原に対して検出された。
【0114】
図9:
図9はマウス免疫化の後のHBウイロソームに対する細胞免疫反応の試験の結果を示している。3回目の増強の後、免疫化された動物より胸腺細胞が精製され、それぞれの抗原を用いた試験管内での再刺激の後、インターフェロンγに対するELISPOTによって細胞反応を決定した。注目すべきはこの方法はCD4型とCD8型反応を区別しないことである。
【0115】
図10:
要約表はHBウイロソームによる免疫化の後これまでに取得した免疫学的データを示している。免疫化した動物においてHBs及びHBcの両方に対する体液性と細胞性の応答が検出可能であった。
【0116】
これより本発明は以下の例を参照することによって説明され、この例は単に例証であり本発明の範囲の制限と解釈されるべきではない。
【実施例】
【0117】
実施例1:HBs抗原の酵母における発現と精製
Schaefer S. et al., in Hansenula polymorpha: Biology and Applications, WILEY-VCH Verlag, Weinheim, 2002, Recombinant hepatitis B vaccines- disease characterization and vaccine production (p187 - p193)
【0118】
この過程は発現カセットとベクターの作製、Hansenula polymorphaという酵母の形質転換、菌株の選択及び評価、発酵、細胞の回収、細胞の破壊、洗浄、吸着、イオン交換クロマトグラフィー、限外濾過、超遠心、ゲル濾過クロマトグラフィー、及び最終産物の液体の濾過滅菌という段階を含む。
【0119】
最終産物の品質は、Lowry法、SDS/PAGE法、ウェスタンブロット分析、又はAUSZYME等の種々の生化学的分析により確認している。
【0120】
実施例2:HBcの大腸菌における発現
Zheng et al, Journal of Biological Chemistry 1992, 13:9422-9429
The structure of Hepadnaviral core antigens
Preikschat el al., Journal of General Virology 1999, 80: 1777-1788
Expression, assembly competence and antigenic properties of hepatitis B virus core gene deletion variants from infected liver cells.
【0121】
この過程は発現カセットとベクターの作製、大腸菌の形質転換、菌株の選択及び評価、発酵、細胞の回収、細胞の破壊、洗浄、吸着、クロマトグラフィー、ゲル濾過、及び最終産物の液体の濾過滅菌という段階を含む。
【0122】
最終産物の品質は、Lowry法、SDS/PAGE法、又はウェスタンブロット分析等の種々の生化学的分析により確認している。
【0123】
実施例3:インフルエンザウイロソームの再構成
Gluck R., 1995, Journal of Liposome Research 1995 5(3), 467-479:
Liposomal Hepatitis A Vaccine and Liposomal multiantigen combination vaccines
【0124】
インフルエンザウイロソームは、リン脂質と、孵化鶏卵と細胞培地のいずれかで培養されたインフルエンザウイルスから産生される。ウイルスを回収し、精製し、一回又はそれ以上の遠心段階で濃縮し、次にベータプロピオラクトン(BPL)で処理することにより不活化した。
【0125】
不活化されたウイルスを超遠心により沈殿にし、界面活性剤(100mMのOEGを含むPBS)に再懸濁し、それによってウイルスの外殻、つまりウイルス包膜を溶解するが、ウイルスの内部、つまりヌクレオカプシドは蛋白質と残存核酸の複合体として残る。同時に、脂質(レシチン他)を同じ界面活性剤(100mMのOEGを含むPBS)に溶解する。その後脂質と溶解したインフルエンザウイルスを混合し、随意に完全に分離するために超音波パルスで処理する。次に、混合物中の全ての不溶性物質を沈殿させ除去するために、超遠心を行う。この不溶性物質の大部分はウイルスヌクレオカプシド複合体を含む。超遠心の後の上清は、別々に加えたウイルス包膜蛋白質及び脂質とリン脂質といった、将来のウイロソームの全ての成分を含む。最終段階においては、SM-2 Bio-Beadsを用いたバッチクロマトグラフィーによって上清から界面活性剤を取り除く。この連続的な界面活性剤の除去が、厳密な組成と脂質:蛋白質比に依存して平均直径が100から200nmとなる、ウイロソームベシクルの均一な集団の自発的な形成をもたらす。
【0126】
実施例4:HBウイロソームの調製及び分析論
HBウイロソームへのHBs及びHBc成分の定量的な組込みを達成するために、インフルエンザウイロソームに関して確立された調製過程は大幅に修正され、多数の変数に関して最適化された。標準的なインフルエンザウイロソームのプロトコルを用いると、HBV抗原の組込み率及び過程の再現性が不満足なものとなるとことが分かった。
【0127】
4.1 図3-11に示しているようなHBウイロソームの詳細かつ基本的な調合プロトコル
共にリン酸緩衝生理食塩水(PBS)に入った、2mgのHAを含む不活化されたインフルエンザウイルス(シンガポールA型)及び2mgの精製された組換えHBs抗原を混合し、100000g、4℃で2時間遠心した。結果として生じる沈殿を1mlの100mM PBS-OEGを含むPBS中に可溶化した。
【0128】
卵由来の粉末状のリン脂質(16.5mgのフォスファチジルコリン及び4.5mgのフォスファチジルエタノールアミン)を1mlの100mM PBS-OEG中に溶解した。
【0129】
その後リン脂質とHA-HBs抗原溶液を混合し、完全に溶解するために37℃のウォーターバス中で2分間超音波処理した。不溶性の残存物質は100000g、4℃で2時間遠心することにより除去した。結果として生じる上清(2ml)を回収し、PBSで最終容量が3.5mlとなるように希釈した。
【0130】
HBc抗原を4mg/mlとなるようにPBSで希釈し、希釈液の0.5mlをPBS-OEG中にHA、HBs抗原、及びリン脂質を含む溶液に加え、結果的に最終OEG濃度が50mMとなった。
【0131】
調製液を0.22マイクロメートルフィルター(Millipore)を通して濾過し、界面活性剤除去手順を行った。混合液を1.2g(乾燥質量)のBio-Beads SM-2(BioRad)に加え、回転させながら室温で30分インキュベーションした。次に、懸濁液を0.8gの新しいBio-Beadsに移し同じ条件下で30分インキュベーションし、その後0.8gの新しいBio-Beadsと同じ条件下で3回目のインキュベーションを行った。結果として生じるHBウイロソームをその後濾過滅菌(0.22マイクロメートル)し、使用までガラスアンプル中に4℃で保存した。
【0132】
4.2 HBウイロソームの組成の可能な修正
異なる供給源(CHO由来のHBs、酵母由来のHBs)、サブタイプ(ayw 及びadv HBV核粒子)、又は変異体(HBV核の全長又は短縮型)からのHBV抗原を含むHBウイロソームを同様の方法で調製した。上記で説明した再構成HBVウイロソームの製剤は、粒度と抗原組込み率に関しての重要な変数を同定するために行った一連の調製の結果であり、最終濾過滅菌に適した均一な粒度の粒子への抗原の良好な組込みを可能にした。異なるリン脂質組成(異なる比率のフォスファチジルエタノールアミンとフォスファチジルコリン)による調合は、粒度とフォスファチジルエタノールアミン量の間の逆相関の関係を示した。蛋白質に対する脂質の比(2.5、5、6、7.5)の粒度と抗原組込みに対する影響は一連の調製の中で調べている。異なる抗原の相対量が組込みに影響することが示されており、調合において(HBc抗原無しで)HAの濃度を増やすことがHBs抗原の組込みの増加につながり、1対1の比率の80%まで届いた。以下の変数を系統的に試験し最適化した:
【0133】
脂質:蛋白質比
我々の手では最適な脂質:蛋白質比は5:1であったが、リン脂質を使用するなら抗原組込みが最大となるのは20:1から1:10の範囲であることは考えられる。他の脂質(合成脂質、ステロイド型脂質)又は異なる脂質の組み合わせを使用するなら、脂質:蛋白質比は更に異なる可能性がある。
【0134】
リン脂質組成(PC、PE、他の脂質)
PCのみのHBウイロソームを産生することはでき、我々の手では粒度と均一性に関してPEが22%であることが最適であった。もう一度言うが、他の脂質を使用するならば、これらの比は大幅に異なる可能性がある。
【0135】
抗原間の比率(HA:HBs:HBc)
我々の手では1:1:1という比が最適であると分かった。しかし、修正した脂質組成と脂質:蛋白質比では、抗原組込みが最大となる最適量は変わる可能性が高い。それに加え、HAの一切無い調製物も同様に所望の粒子構造を作り出した。
【0136】
界面活性剤
選択した界面活性剤は50mMの濃度のOEGである。しかし、組成や比率を修正する時には20から100mMの範囲が適切である可能性がある。他の非イオン性、イオン性、又は双極性の性質の界面活性剤をOEGの代わりに調製過程で使用してもよい。
【0137】
【表2】
【0138】
4.3 ウイロソーム調製物の分析論
HBウイロソームの徹底した物理化学的な分析は、調製過程の最適化と将来の製品の品質管理に必須の要素である。それ故、HBウイロソームの内容と構造を調べる測定法の開発に莫大な努力が費やされた。アジュバント効果(MHC-1提示)がHBウイロソームの物理的構造に直接依存するため、ワクチンのHBV成分をウイロソーム担体に物理的に結合させる単一の粒子型の証明に特に重点を置いた。
【0139】
成分の定量:
蛋白質(SDS-PAGE)
HBs(ELISA、ウェスタンブロット)
HBc(ELISA、ウェスタンブロット)
HA(SRD)
リン脂質(酵素測定法)
ウイロソーム構造:
光子相関法
免疫共沈降法
密度勾配超遠心法
トリプシン処理
電子顕微鏡(計画中)
【0140】
全蛋白質濃度の決定
蛋白質濃度は260、280、320nmの波長のUV吸光度によって測定し、以下の公式に従って計算した:1.55×(A280 - A320) - 0.76×(A260 - A320)。結果はmg / mlで表示した。
【0141】
HBs及びHBc抗原の定量
HBウイロソームに組込まれたHBs及びHBc抗原の量は定量的Elisa測定法により決定した。抗原に対する最大限の接触をもたらすために、基準抗原とHBウイロソーム試料を最初の希釈時にPBS-OEGに溶解し、次に連続する希釈はPBSにて行った。HBs測定は市販のHBs Elisa detection kit (DADE Behring)を使用して行い、同じ測定法にて測定する精製したHBs抗原の希釈系列が、HBsウイロソーム試料中の抗原の量の決定を可能にした。定量的HBc ELISAでは、マイクロタイタープレートを、50mM Na2CO3 (PH 9.6) に入ったHBcに対するモノクローナル抗体(mAb)(クローン7E6、Biogenesis、希釈率1:1000)によってコートした。その後プレートを1% BSA、5%ショ糖、0.05% NaN3入りのPBSで室温で少なくとも1時間ブロックし、0.05% Tween20 (v/v)入りのPBS(洗浄バッファー)で洗浄した。試料(0.1ml)を載せ、室温で1時間インキュベートした。第2の、HBcに対するビオチン化されたmAb(クローン4H5、Biogenesis)を0.05% Tween20、0.1% BSA入りのPBS(希釈バッファー)中で1:1000に希釈し、プレートに加え(1ウェルあたり0.1ml)、室温で1時間インキュベートした。4回の洗浄の後、プレートをストレプトアビジン(希釈バッファー中で1:5000)の存在下で室温で1時間インキュベートした。更なる4回の洗浄の後、TMB(1ウェルあたり0.075ml)を30分インキュベートし、発色反応を1M H2SO4(1ウェルあたり75μl)で停止し、450nmにおける光学密度を測定した。
【0142】
HAの定量
HBウイロソームのHAの量は標準的な一元免疫拡散(SRD)法を使用して決定した。この試験はウイロソームワクチンの分析に関して有効な測定法であり、エパキサール(Epaxal:登録商標)に関するそれぞれの標準操作手順に従ってBerna Biotch Ltd. QC departmentによって行われた。
【0143】
ゲル電気泳動、ウェスタンブロット、銀染色
HA、HBs又はHBc抗原のHBウイロソームにおける存在を証明するために、調製物、勾配分画、プロテアーゼ処理、又は免疫共沈降法からの試料をMESバッファーを用いNuPage Bis-Tris SDS-PAGE pre-cast gel(Invitrogen)上で分離し、製造者の使用説明書に従いニトロセルロースメンブレンに転写した。メンブレンを5%ミルク入りのPBST(1% Tween 20入りのPBS)でブロックし、抗原特異的な抗体の1:1000の希釈液中で室温で1時間インキュベートした。メンブレンを洗浄し、その後ペルオキシダーゼ標識の抗ウサギ又は抗ヒツジ免疫グロブリンの1:10000の希釈液中で1時間インキュベートした。蛋白質をECL Plus 基質溶液 (Amersham Pharmacia, Piscataway, N.J.)を用いて可視化した。
ゲルの銀染色は製造者の使用説明書(Invitrogen)に従って行った。
【0144】
粒度の測定:光子相関法
最初の材料及び調製したHBウイロソームの流体力学的径、多分散性係数、及び統計的粒度分布を光子相関法又は動的光散乱によって決定した。この方法は、光散乱の時間変動として測定される、ブラウン運動の粒度依存的な速度に基づいている。生データ、つまり光強度の変動からの変数の計算のためのソフトウェアを含め、Malvern Zetasizer 1000HS(Malvern Ltd, Malvern, UK)をこの目的に使用した。試料を測定のためにPBS中に十分に希釈し、1mlの希釈液を25℃の標準条件下で分析した。
【0145】
ショ糖勾配
個々の成分の異なる密度に基づいて、HBウイロソーム構造への抗原の組込みを推定するための分析的方法として、不連続ショ糖勾配を通した超遠心を行った。PBS中のHBウイロソーム調製物の一部を、PBS中の20-60% (w/v)不連続ショ糖勾配の上に載せ、4℃で24時間100000gで遠心した。次に回収した分画を密度と蛋白質量に関して分析し、また異なる抗原の存在に関してウェスタンブロットを用いて分析した。
【0146】
トリプシン処理
HBウイロソーム粒子の抗原の組込み及び最後の皮膜化を部分トリプシン処理によって調査した。次に処理後の試料を、蛋白質分解抵抗性の断片又は保護された全長蛋白質に関して、ウェスタンブロットによって分析した。HBウイロソームをPBSバッファー(無傷の粒子=自然条件)、又は0.5% デオキシコール酸ナトリウム及び1% Triton X-100入りのPBS(ウイロソーム構造の破壊=変性条件)のいずれかで希釈した。プロテアーゼ処理は5%トリプシン(w/w 蛋白質)を用いて、室温で0、2、5、及び10時間行った。トリプシン処理に対する個々の成分の効率的な接触可能性のコントロールとして、同濃度の異なる抗原の混合物を同じ条件下で処理した。反応は4×SDS-PAGEサンプルバッファーを加えることにより停止した。95℃での10分間の変性の後、試料をSDS-PAGE電気泳動、及びHBsとHBcに対する免疫ブロット分析にかけた。
【0147】
免疫共沈降法
HBウイロソーム構造中でのHA、HBs、HBc抗原の物理的結合は、(トリプシン処理に関して説明したような)天然及び変性条件下におけるそれぞれの抗原に対する別々の(個別の)免疫共沈降法、及びそれに続くウェスタンブロット分析による免疫共沈降された抗原の同定によって証明した。HBウイロソーム調合の免疫共沈降法は、HA、HBc、HBsに対する特異的抗体の存在下に、同時平行の実験で行われた。次に免疫複合体をプロテインGセファロースビーズ(Promega)と共に4時間インキュベーションし、遠心した。結果として生じる沈殿をPBSで5回洗浄した。免疫沈降された蛋白質をサンプルバッファー中で10分間煮沸することにより再懸濁し、SDS-PAGEとウェスタンブロットによって分析した。それぞれの抗原の存在はそれぞれの特異的抗体と共にインキュベーションすることにより調査した。
【0148】
ウイロソームの調製物:マウスモデルにおける免疫原性
HBウイロソームの免疫原性に関してはこの時点で予備的なデータのみが利用可能である。HBウイロソームに加え、付加的なアジュバントRC529(Corixa)がある条件と無い条件で、マウスにおける実験を行った。マウスは異なる量のHBウイロソームの筋内注射によって免疫化され、全ての3つの抗原に対する高い抗体力価が検出された(図8)。3回目の増強の後、免疫化動物から脾臓細胞を精製し、それぞれの抗体を用いた試験管内での再刺激の後、インターフェロンγに対するELISPOTによって細胞反応を決定した(図9)。注目すべきはこの方法はCD4型とCD8型反応を区別しないことである。それに加え、免疫化動物からの脾細胞をFACSによって分析した(図10)。新しい脾細胞はHBc特異的5量体によって抗CD8抗体と共に直接染色された。別個に、脾細胞はCD8特異的ペプチド又は全蛋白質によって刺激され、次に細胞内インターフェロンγに対して抗CD4と抗CD8抗体のいずれかと共に染色された。
【0149】
実施例5:他のウイルスシステムに対する調製原理の適用
異なるウイルス蛋白質間の相互作用はビリオンの組立に重要であり、全てのウイルスに見られる一般原則である。包膜ウイルス粒子は組立に関して細胞膜構造に依存している。核酸と蛋白質の複合体であるヌクレオカプシドは、成熟したウイルス構造を形成するために、膜に結合したウイルス蛋白質と結合する。我々はB型肝炎ウイルスに関して、インビトロで、細胞膜構造のない条件で、異なる組換え供給源からのウイルス蛋白質を利用して、この過程が模倣できることを示した。ウイルス抗原の組換え供給源が量と質に関して好適であるが、インフルエンザHAに関して本明細書で適用したように、ウイルス蛋白質は同様に元のウイルスに由来してもよい。インビトロの調製物の柔軟性は異なる供給源からの多数の抗原の包含を可能にする。(HBVとインフルエンザに関して示したように)複数のウイルスからの蛋白質を含む多面的調製物は、この原理に基づいて物理的安定性、免疫学的性質、又はワクチンの防御範囲を改善する可能性がある。
【0150】
条件をそれぞれの病原体に合わせ、必要な成分が十分な量で利用可能ならば、任意の他の包膜ウイルスに対して同じ原理が適用出来ることが想定される。インビトロの調製物の原則が適用でき、また魅力的で切迫したワクチンの標的である、多数の候補を下記に列挙してある。しかし、そのような憶測上のウイルス様粒子によって誘導された免疫反応が防御的、又は更に治療的な効果までを持つかどうかは、予測することが出来ない。
【0151】
5.1 C型肝炎ウイルス
C型肝炎を引き起こす原因としてのHCVは世界的な健康問題となっている。世界人口の内 %がこのウイルスに感染していると推定されている。予防的又は治療的用途のための、HCVに対するワクチンは現在存在しない。細胞システムにおけるウイルス様粒子の組立は実現されている(Baumert et al, Journal of Virology 1998, 75: 3827-3838; Hepatitis C virus structural proteins assemble into virus-like particles in isect cells)。HBVのように、核蛋白質の1量体が結合して膜に結合した包膜蛋白質(E1、E2)と相互作用する20面体のヌクレオカプシドとなる。
【0152】
5.2 他のフラビウイルス
HCVとは別に、多数の重要なヒト病原体がフラビウイルス科ファミリーに含まれる(ウェストナイルウイルス、クンジンウイルス、日本脳炎ウイルス、デングウイルス、黄熱ウイルス、及びダニ媒介脳炎ウイルス)。HCV(及びHBV)との構造レベルでの類似性は高く、それ故ウイルス様粒子のインビトロの再組み立ては可能だと思われる。
【0153】
5.3 HIV
HIVはレトロウイルス科ファミリーの中で最も著名なメンバーである。繰り返すが、切迫した医療需要があるにも関わらず、効果的なワクチンは存在しない。これらのウイルスは更に複雑なヌクレオカプシドを形成するが、同じ原理に基づいた多抗原ワクチンのインビトロの調製は可能だと思われる。
【図面の簡単な説明】
【0154】
【図1】個々の成分と結果として生じるHBウイロソームの構造を描いた概略図が図1に示してある。
【図2】調製過程のフローチャートが図2に示してある。
【図3】図3はHBウイロソームのSDS-PAGE分析の結果を示している。SDS-PAGE分析に基づくHBウイロソーム及び最初の材料(インフルエンザ、HBs、及びHBc)の蛋白質組成が図3に示してある。
【図4】図4は個々の成分の粒度と比較したHBウイロソームの粒度の分析の結果を示している。
【図5】図5はHBウイロソームの勾配分画分析の結果を示している。
【図6】図6はHBウイロソームのSDS-PAGE/ウェスタンブロット分析の結果を示している。
【図7】図7はHBウイロソームのSDS-PAGE/ウェスタンブロット分析の結果を示している。
【図8】図8はマウス免疫化の後のHBウイロソームに対する抗体免疫反応の試験の結果を示している。
【図9】図9はマウス免疫化の後のHBウイロソームに対する細胞免疫反応の試験の結果を示している。
【図10】要約表はHBウイロソームによる免疫化の後これまでに取得した免疫学的データを示している。
【特許請求の範囲】
【請求項1】
(a)少なくとも一つの脂質、及びインフルエンザウイルスとB型肝炎ウイルス(HBV)の包膜蛋白質を備えるウイロソーム膜;及び
(b)ウイロソームの内側及び外側に位置し、前記包膜蛋白質に付着したHBVのHBc蛋白質を備えるヌクレオカプシド粒子、
を備えるウイロソーム。
【請求項2】
前記少なくとも一つの脂質が少なくとも一つのリン脂質を備える請求項1記載のウイロソーム。
【請求項3】
前記リン脂質がホスファチジルコリン及びホスファチジルエタノールアミンである請求項2記載のウイロソーム。
【請求項4】
インフルエンザウイルスの前記包膜蛋白質が、ヘマグルチニン(HA)及び/又はニューロミニダーゼ(NA)である請求項1から請求項3のいずれか1項記載のウイロソーム。
【請求項5】
前記HBVの包膜蛋白質がHBs蛋白質である請求項1から請求項4のいずれか1項記載のウイロソーム。
【請求項6】
請求項1から請求項5のいずれか1項記載のウイロソーム、及び必要に応じて、薬学的に許容される担体又は希釈剤及び/又はアジュバントを備えるワクチン。
【請求項7】
前記アジュバントがRC529(Corixa)である請求項6記載のワクチン。
【請求項8】
(a)界面活性剤溶液中において脂質存在下で包膜ウイルスであるインフルエンザウイルスとHBVの包膜蛋白質を可溶化し;
(b)溶液中の界面活性剤の濃度を低下させ;
(c)HBc蛋白質を備えるHBVのヌクレオカプシド粒子を工程(b)で取得した溶液に加え;
(d)ウイロソームが産生されるように界面活性剤を除去する;
という工程を備えるウイロソームの産生方法。
【請求項9】
前記脂質は少なくとも一つのリン脂質を備える請求項8記載の方法。
【請求項10】
前記リン脂質がホスファチジルコリン及びホスファチジルエタノールアミンである請求項9記載の方法。
【請求項11】
インフルエンザウイルスの前記包膜蛋白質が、ヘマグルチニン(HA)及び/又はニューロミニダーゼ(NA)である請求項8から請求項10のいずれか1項記載の方法。
【請求項12】
前記HBVの包膜蛋白質がHBs蛋白質である請求項8から請求項11のいずれか1項記載の方法。
【請求項13】
工程(b)に続いて行われる以下の工程(b’):
(b’)工程(b)で得られた希釈液の無菌濾過、
を更に備える請求項8から請求項12のいずれか1項記載の方法。
【請求項14】
以下の工程(e):
(e)工程(d)で得られた希釈液の無菌濾過、
を更に備える請求項8から請求項13のいずれか1項記載の方法。
【請求項15】
前記界面活性剤が非イオン性界面活性剤である請求項8から請求項14のいずれか1項記載の方法。
【請求項16】
前記非イオン性界面活性剤がオクタエチレングリコールモノ(N-ドデシル)エーテル(OEG)である請求項15記載の方法。
【請求項17】
工程(b)において、前記非イオン性界面活性剤を20mMから100mMの範囲内の濃度とする請求項15又は請求項16記載の方法。
【請求項18】
脂質:蛋白質の比が1:1と10:1の範囲内にある請求項8から請求項17のいずれか1項記載の方法。
【請求項19】
脂質:蛋白質の比が約5:1である請求項18記載の方法。
【請求項20】
ウイロソーム中のホスファチジルコリンの割合が22%である請求項10から請求項19のいずれか1項記載の方法。
【請求項21】
HA:ウイルス包膜蛋白質:ウイルスカプシド蛋白質の比が1:1:1である請求項11から請求項20のいずれか1項記載の方法。
【請求項22】
工程(d)におけるウイロソームの生成の前にアジュバントを添加することを含む請求項8から請求項21のいずれか1項記載の方法。
【請求項23】
請求項1から請求項5のいずれか1項記載のウイロソーム、又は請求項8から請求項22のいずれか1項記載の方法により得られたウイロソームを、ワクチンの調製のために使用する方法。
【請求項24】
請求項1から請求項5のいずれか1項記載のウイロソーム、又は請求項8から請求項22のいずれか1項記載の方法により得られたウイロソームを、HBV感染の予防、緩和又は治療用のワクチンの調製のために使用する方法。
【請求項25】
請求項1から請求項5のいずれか1項記載のウイロソーム、又は請求項8から請求項22のいずれか1項記載の方法により得られたウイロソームを投与する工程を備える、被験者にワクチンを接種する方法。
【請求項26】
必要に応じて薬学的に許容される担体又は希釈剤及び/又はアジュバントと組合わせて、請求項1から請求項5のいずれか1項記載のウイロソーム、又は請求項8から請求項22のいずれか1項記載の方法により得られたウイロソームを投与する工程を備える、HBV感染の予防、緩和又は治療のために被験者にワクチンを接種する方法。
【請求項1】
(a)少なくとも一つの脂質、及びインフルエンザウイルスとB型肝炎ウイルス(HBV)の包膜蛋白質を備えるウイロソーム膜;及び
(b)ウイロソームの内側及び外側に位置し、前記包膜蛋白質に付着したHBVのHBc蛋白質を備えるヌクレオカプシド粒子、
を備えるウイロソーム。
【請求項2】
前記少なくとも一つの脂質が少なくとも一つのリン脂質を備える請求項1記載のウイロソーム。
【請求項3】
前記リン脂質がホスファチジルコリン及びホスファチジルエタノールアミンである請求項2記載のウイロソーム。
【請求項4】
インフルエンザウイルスの前記包膜蛋白質が、ヘマグルチニン(HA)及び/又はニューロミニダーゼ(NA)である請求項1から請求項3のいずれか1項記載のウイロソーム。
【請求項5】
前記HBVの包膜蛋白質がHBs蛋白質である請求項1から請求項4のいずれか1項記載のウイロソーム。
【請求項6】
請求項1から請求項5のいずれか1項記載のウイロソーム、及び必要に応じて、薬学的に許容される担体又は希釈剤及び/又はアジュバントを備えるワクチン。
【請求項7】
前記アジュバントがRC529(Corixa)である請求項6記載のワクチン。
【請求項8】
(a)界面活性剤溶液中において脂質存在下で包膜ウイルスであるインフルエンザウイルスとHBVの包膜蛋白質を可溶化し;
(b)溶液中の界面活性剤の濃度を低下させ;
(c)HBc蛋白質を備えるHBVのヌクレオカプシド粒子を工程(b)で取得した溶液に加え;
(d)ウイロソームが産生されるように界面活性剤を除去する;
という工程を備えるウイロソームの産生方法。
【請求項9】
前記脂質は少なくとも一つのリン脂質を備える請求項8記載の方法。
【請求項10】
前記リン脂質がホスファチジルコリン及びホスファチジルエタノールアミンである請求項9記載の方法。
【請求項11】
インフルエンザウイルスの前記包膜蛋白質が、ヘマグルチニン(HA)及び/又はニューロミニダーゼ(NA)である請求項8から請求項10のいずれか1項記載の方法。
【請求項12】
前記HBVの包膜蛋白質がHBs蛋白質である請求項8から請求項11のいずれか1項記載の方法。
【請求項13】
工程(b)に続いて行われる以下の工程(b’):
(b’)工程(b)で得られた希釈液の無菌濾過、
を更に備える請求項8から請求項12のいずれか1項記載の方法。
【請求項14】
以下の工程(e):
(e)工程(d)で得られた希釈液の無菌濾過、
を更に備える請求項8から請求項13のいずれか1項記載の方法。
【請求項15】
前記界面活性剤が非イオン性界面活性剤である請求項8から請求項14のいずれか1項記載の方法。
【請求項16】
前記非イオン性界面活性剤がオクタエチレングリコールモノ(N-ドデシル)エーテル(OEG)である請求項15記載の方法。
【請求項17】
工程(b)において、前記非イオン性界面活性剤を20mMから100mMの範囲内の濃度とする請求項15又は請求項16記載の方法。
【請求項18】
脂質:蛋白質の比が1:1と10:1の範囲内にある請求項8から請求項17のいずれか1項記載の方法。
【請求項19】
脂質:蛋白質の比が約5:1である請求項18記載の方法。
【請求項20】
ウイロソーム中のホスファチジルコリンの割合が22%である請求項10から請求項19のいずれか1項記載の方法。
【請求項21】
HA:ウイルス包膜蛋白質:ウイルスカプシド蛋白質の比が1:1:1である請求項11から請求項20のいずれか1項記載の方法。
【請求項22】
工程(d)におけるウイロソームの生成の前にアジュバントを添加することを含む請求項8から請求項21のいずれか1項記載の方法。
【請求項23】
請求項1から請求項5のいずれか1項記載のウイロソーム、又は請求項8から請求項22のいずれか1項記載の方法により得られたウイロソームを、ワクチンの調製のために使用する方法。
【請求項24】
請求項1から請求項5のいずれか1項記載のウイロソーム、又は請求項8から請求項22のいずれか1項記載の方法により得られたウイロソームを、HBV感染の予防、緩和又は治療用のワクチンの調製のために使用する方法。
【請求項25】
請求項1から請求項5のいずれか1項記載のウイロソーム、又は請求項8から請求項22のいずれか1項記載の方法により得られたウイロソームを投与する工程を備える、被験者にワクチンを接種する方法。
【請求項26】
必要に応じて薬学的に許容される担体又は希釈剤及び/又はアジュバントと組合わせて、請求項1から請求項5のいずれか1項記載のウイロソーム、又は請求項8から請求項22のいずれか1項記載の方法により得られたウイロソームを投与する工程を備える、HBV感染の予防、緩和又は治療のために被験者にワクチンを接種する方法。
【図1】

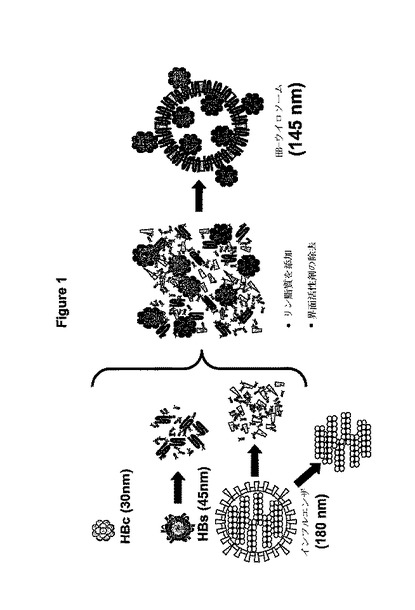
【図2】
【図3】
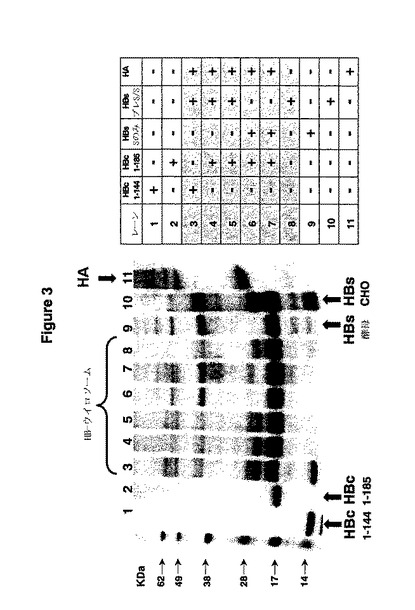
【図4】
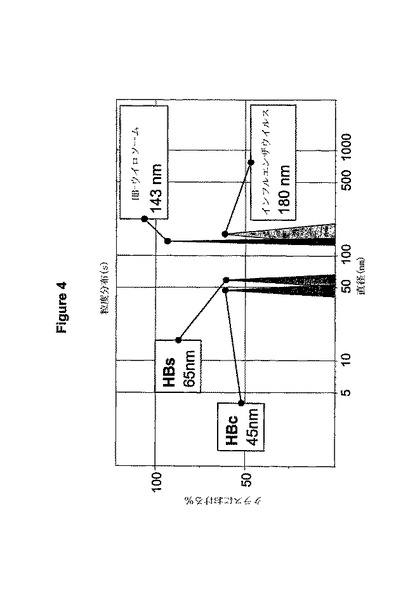
【図5】
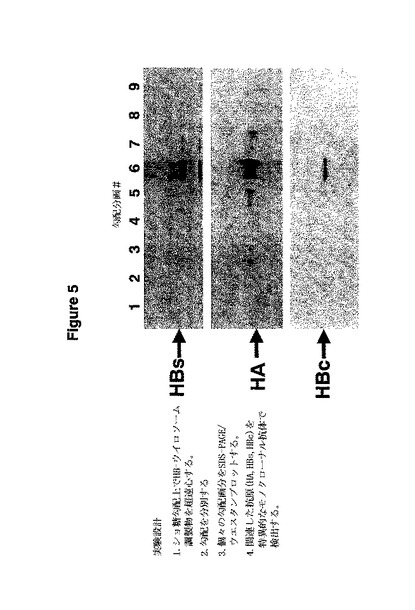
【図6】

【図7】
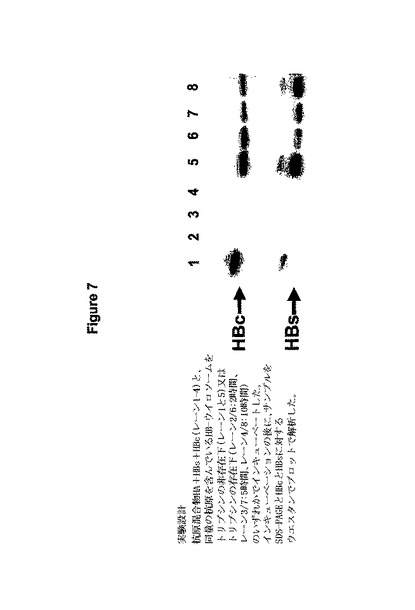
【図8】

【図9】
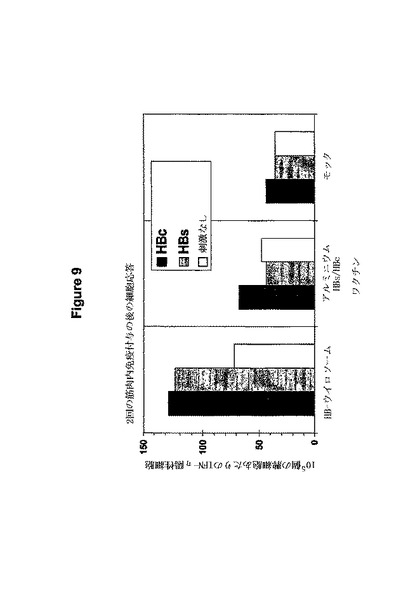
【図10】
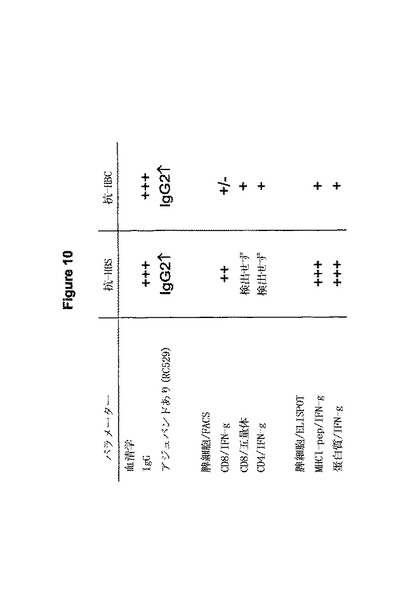
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【公表番号】特表2008−517961(P2008−517961A)
【公表日】平成20年5月29日(2008.5.29)
【国際特許分類】
【出願番号】特願2007−538311(P2007−538311)
【出願日】平成17年10月20日(2005.10.20)
【国際出願番号】PCT/EP2005/011297
【国際公開番号】WO2006/045532
【国際公開日】平成18年5月4日(2006.5.4)
【出願人】(507139579)ベルナ バイオテック アーゲー (1)
【Fターム(参考)】
【公表日】平成20年5月29日(2008.5.29)
【国際特許分類】
【出願日】平成17年10月20日(2005.10.20)
【国際出願番号】PCT/EP2005/011297
【国際公開番号】WO2006/045532
【国際公開日】平成18年5月4日(2006.5.4)
【出願人】(507139579)ベルナ バイオテック アーゲー (1)
【Fターム(参考)】
[ Back to top ]