
新品種
【課題】従来よりも北の地域でも栽培可能なイネの新品種の提供。
【解決手段】受領番号がFERM AP−22175である、イネ品種コシヒカリかずさ6号(Oryza sativa L.cultivar Koshihikari−kazusa6 gou)、並びに、前記記載の品種の個体及び前記記載の品種の個体の後代個体からなる群より選択される2個体を交配して得られる後代個体。
【解決手段】受領番号がFERM AP−22175である、イネ品種コシヒカリかずさ6号(Oryza sativa L.cultivar Koshihikari−kazusa6 gou)、並びに、前記記載の品種の個体及び前記記載の品種の個体の後代個体からなる群より選択される2個体を交配して得られる後代個体。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、非遺伝子組み換え法により作出された新品種、当該新品種の鑑別方法、及びイネ個体を早生化する方法に関する。
【背景技術】
【0002】
同一生物種に属するが、遺伝的構成が異なるために、ある形質において他の集団と異なる集団を品種という。すなわち、同じ種類の植物であったとしても、品種により、栽培の難易性や病虫害に対する抵抗性、収量、品質等が異なる。このため、農作物、特にイネやムギ類等の主要な作物においては、より優良な品種を得るための品種改良が古くから行われており、近年では、種苗会社等のみならず、国や県等の公的機関においても積極的に行われてきている。
【0003】
近年の核酸解析技術等の進歩に伴い、シロイヌナズナ、イネ、コムギ等の様々な植物の遺伝子が解析され、得られた遺伝子情報が開示されている。これらの開示された遺伝子情報を利用して、遺伝子組み換え法による外来種の遺伝子を導入する品種改良も多く行われている。例えば、植物の感光性を増加させる機能を有する植物由来のタンパク質をコードするHd1遺伝子、及びHd1遺伝子を導入した形質転換植物の作製方法等が開示されている(例えば、特許文献1参照。)。しかしながら、遺伝子組み換え法による品種改良は、通常は交配不可能な遠縁種が有する形質を導入し得るという利点はあるものの、その安全性に対する検証は必ずしも十分ではないという問題点がある。
【0004】
このため、イネをはじめとする食用植物においては、非遺伝子組み換え法による新品種の作出が多く行われている。例えば特許文献2には、非遺伝子組み換え法により、外来の有用な染色体断片で置換する場合に、導入される外来品種由来の染色体断片による置換領域をコントロールし、元品種が有する好ましい形質を変更することなく、標的形質を有する新品種を作製するための方法が開示されている。同じく特許文献2には、この新品種を作製するための方法により、ハバタキのHd1遺伝子を含む領域のみをコシヒカリに導入した新品種イネ品種コシヒカリえいち3号が記載されている。
【0005】
特に、イネでは、より栽培可能地域が広いコシヒカリが望まれている。コシヒカリは、他の品種よりも食味に優れており、消費者に好まれている。このため米農家は、必ずしもコシヒカリの栽培に適しているとはいえない地域であっても、コシヒカリを好んで栽培している。しかしながら、コシヒカリを南の地域で栽培した場合には出穂期が早すぎ、充分な収穫量が望めない。その上、出穂期に高温が続くために食味が落ちてしまう。一方で、コシヒカリを北の地域で栽培した場合には晩生となり、たとえ出穂が出来たとしても、低温により登熟が不良となり、米を収穫することができない。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】特許第3660967号公報
【特許文献2】特許第4409610号公報
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明者による研究の結果、コシヒカリはおおよそ、日本の北緯35.5度から38.5度までの範囲内でしか栽培に適していないことがわかった。北緯38.5度よりも北の地域でも栽培可能なコシヒカリがあれば、例えば北海道等の従来では栽培不可能であった地域でもコシヒカリを収穫することができる。
【0008】
本発明は、従来よりも北の地域でも栽培可能なイネの新品種、及びイネ個体を早生化する方法を提供することを目的とする。
【課題を解決するための手段】
【0009】
本発明者は、上記課題を解決すべく鋭意研究した結果、イネ品種ハバタキの第3染色体上に存在する特定の領域の染色体断片とイネ品種ハバタキの第6染色体上に存在する特定の領域の染色体断片とを、イネ品種コシヒカリに置換することにより、従来よりも北の地域でも栽培可能なほど十分に早生化することが可能であることを見出し、本発明を完成させた。
【0010】
すなわち、本発明は、
(1) 受領番号がFERM AP−22175である、イネ品種コシヒカリかずさ6号(Oryza sativa L.cultivar Koshihikari−kazusa6 gou)、
(2) 前記(1)記載の品種の個体及び前記(1)記載の品種の個体の後代個体からなる群より選択される2個体を交配して得られる後代個体、
を、提供するものである。
【発明の効果】
【0011】
本発明の新品種であるイネ品種コシヒカリかずさ6号は、コシヒカリよりも非常に早生化されており、北緯38.5度よりも北の地域でも栽培し、米を収穫することができる。また、イネ品種コシヒカリかずさ6号は、品質や収穫量等の収穫期以外の特性はコシヒカリとほぼ同等な新品種である。
【図面の簡単な説明】
【0012】
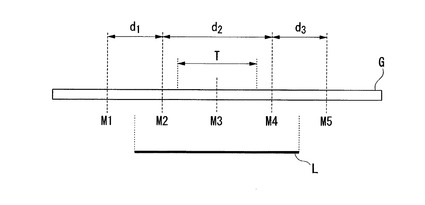
【図1】元品種の染色体G上における標的領域T、置換された外来品種由来染色体断片L、及びDNAマーカーM1〜M5を示した図である。
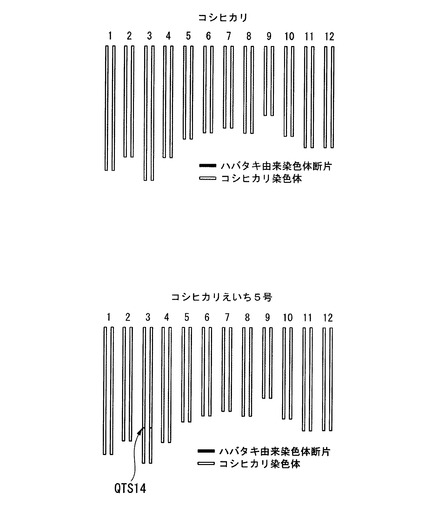
【図2】コシヒカリ、及びコシヒカリえいち5号のゲノムを模式的に表した図である。
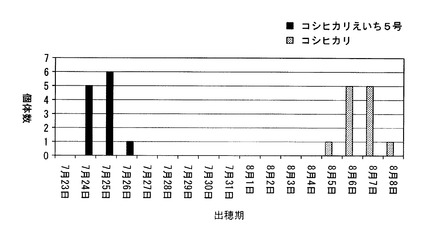
【図3】千葉県において、コシヒカリ、及びコシヒカリえいち5号の出穂期を調べた結果を示した図である。
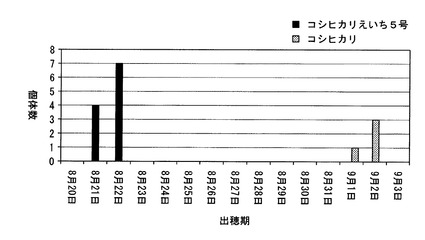
【図4】北海道において、コシヒカリ、及びコシヒカリえいち5号の出穂期を調べた結果を示した図である。
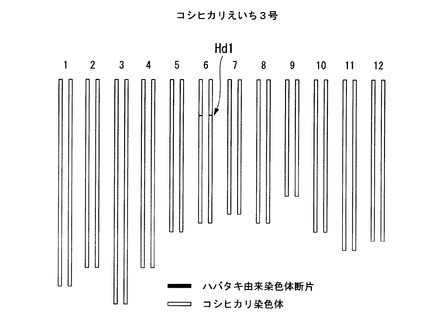
【図5】コシヒカリえいち3号のゲノムを模式的に表した図である。
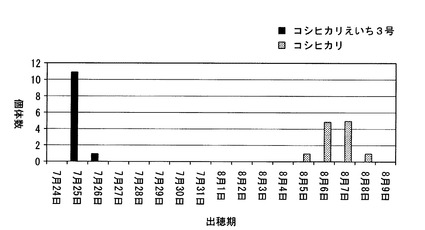
【図6】千葉県において、コシヒカリ、及びコシヒカリえいち3号の出穂期を調べた結果を示した図である。
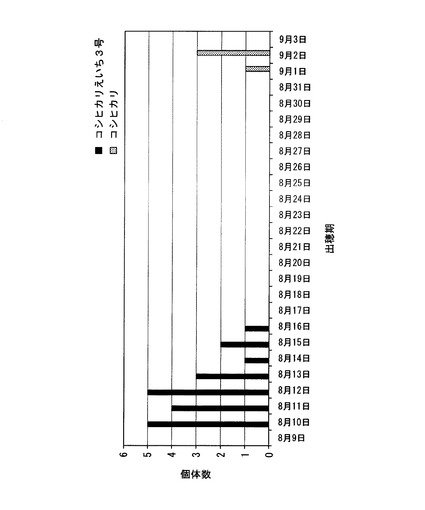
【図7】北海道において、コシヒカリ、及びコシヒカリえいち3号の出穂期を調べた結果を示した図である。
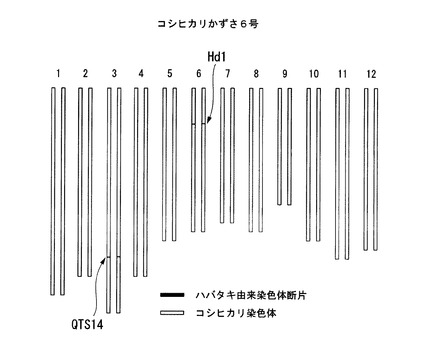
【図8】コシヒカリかずさ6号のゲノムを模式的に表した図である。
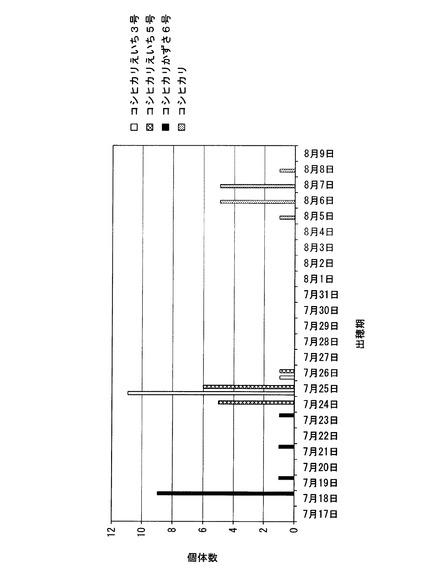
【図9】千葉県において、コシヒカリかずさ6号の出穂期を調べた結果を、コシヒカリ、コシヒカリえいち5号、及びコシヒカリえいち3号の結果とともに示した図である。
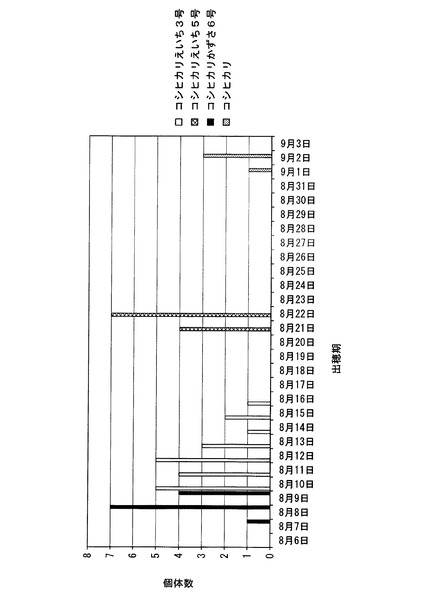
【図10】北海道において、コシヒカリかずさ6号の出穂期を調べた結果を、コシヒカリ、コシヒカリえいち5号、及びコシヒカリえいち3号の結果とともに示した図である。
【発明を実施するための形態】
【0013】
本発明において染色体断片置換系統とは、元品種の染色体の一部のみが外来品種由来の染色体断片に置換されている系統を意味する。ここで、外来品種は、元品種以外の品種であれば特に限定されるものではなく、元品種と同一種の植物の品種であってもよく、元品種と異なる種の植物の品種であってもよく、動物等の植物以外の品種であってもよい。なお、本発明において品種とは、同一種の植物であって、遺伝的構成が異なるために、ある形質において同種内の他品種から明確に識別し得る集団を意味する。
【0014】
本発明においてDNAマーカーは、元品種由来の染色体と外来品種由来の染色体を識別し得る染色体上のDNA配列の差異を検出し得るものであれば、特に限定されるものではなく、遺伝子解析分野で通常用いられているDNAマーカーを用いることができる。該DNAマーカーとして、例えば、SNP(Single Nucleotide Polymorphism、一遺伝子多型)やSSR(Simple Sequence Repeats、単純反覆配列)の繰り返し数の違い等の遺伝子多型を検出し得るマーカーであってもよく、RFLP(Restriction Fragment Length Polymorphism、制限酵素断片長多型)マーカーであってもよい。なお、これらのDNAマーカーによる、元品種由来アレルと外来品種由来アレルとの識別は、常法により行うことができる。例えば、各個体から抽出したDNAを鋳型とし、特定のSNPやSSRと特異的にハイブリダイズし得るプライマー等を用いてPCRを行い、電気泳動法等を用いてPCR産物の有無を検出し、各多型を識別することができる。また、各個体から抽出したDNAを制限酵素処理した後、電気泳動法等を用いてDNA断片のパターンを検出し、各多型を識別することができる。なお、特定のSNPやSSRと特異的にハイブリダイズし得るプライマー等は、該SNPやSSRの塩基配列に応じて、汎用されているプライマー設計ツール等を用いて常法により設計することができる。また、設計されたプライマー等は、当該技術分野においてよく知られている方法のいずれを用いても合成することができる。
【0015】
これらのDNAマーカーは、公知のDNAマーカーを適宜用いることができる。また、新規に作製したDNAマーカーであってもよい。公知のDNAマーカーとして、例えば、イネにおいては、国際公開第2003/070934号パンフレット等において開示されているSNPマーカーや、Rice Genome Research Program(RGP:http://rgp.dna.affrc.go.jp/publicdata.html)において公開されているDNAマーカーを用いることができる。
【0016】
なお、各品種の遺伝子情報等は、例えば、国際的な塩基配列データベースであるNCBI(National center for Biotechnology Information)やDDBJ(DNA Data Bank of Japan)等において入手することができる。特にイネの各品種の遺伝子情報は、KOME(Knowledge−based Oryza Molecular biological Encyclopedia、http://cdna01.dna.affrc.go.jp/cDNA/)等において入手することができる。
【0017】
本発明及び本願明細書において「イネ品種日本晴の染色体のX番目の塩基からY番目の塩基までの領域」は、RGBにおいて公開されているイネ品種日本晴のゲノムDNAの塩基配列(バージョン4;IRGSP−build4−06/04/21)に基づいて決定される領域である。
【0018】
また、本発明及び本願明細書において、「イネ品種日本晴の染色体のX番目の塩基からY番目の塩基までの領域に相当する領域」とは、イネ個体の染色体中のイネ品種日本晴の染色体中の当該領域と相同性の高い領域であり、イネ品種日本晴の公知のゲノムDNAと当該イネ個体のゲノムDNAの塩基配列を、最もホモロジーが高くなるようにアラインメントすることにより決定することができる。また、イネ品種日本晴以外のイネ個体中の「イネ品種日本晴のSNPに相当するSNP」は、当該SNPを含む領域において、イネ品種日本晴の公知のゲノムDNAと当該イネ個体のゲノムDNAの塩基配列を、最もホモロジーが高くなるようにアラインメントした場合に、当該SNPに対応する位置にある塩基を意味する。
【0019】
従来の品種よりも北の地域で栽培可能な新品種を育種するため、本発明の発明者は、まず、出穂期に関して、イネ品種ハバタキとイネ品種コシヒカリとを交配して、分離集団でQTL(Quantitative Trait Locus) 解析を行った。この結果、イネ品種ハバタキの第3染色体の長腕のQTS14領域〔イネ品種日本晴の第3染色体の31,720,064番目の塩基から31,724,043番目の塩基までの領域に相当する領域〕に、出穂期を早め、早生とするQTLが存在していることもわかった。そこで、本発明者は、コシヒカリの当該領域に含まれている遺伝子をハバタキ由来の遺伝子に置換した新品種の作出を行った。当該新品種は、元品種コシヒカリよりも早生のイネであると予想された。
【0020】
非遺伝子組み換え法により植物の品種改良を行う場合において、導入される外来品種由来の染色体断片が大きすぎる場合には、目的の形質遺伝子以外の機能不明な他の遺伝子を多数導入してしまうおそれや、元品種が有する好ましい形質を損なうおそれがある。そこで、本発明者は、導入される外来品種由来の染色体断片による置換領域をコントロールし、元品種が有する好ましい形質を変更することなく、標的形質を有する新品種を作製するため、特許文献2に記載の方法により、新品種の作出を行った。
【0021】
特許文献2に記載の新品種の作出方法は、具体的には以下の通りである。まず、公知のイネの遺伝子情報に基づき、図1に示すような位置関係にある5種類のDNAマーカーを設定した。すなわち、標的領域Tの上流側末端又はその上流にDNAマーカーM2を、DNAマーカーM2の上流にDNAマーカーM1を、標的領域Tの下流側末端又はその下流にDNAマーカーM4を、DNAマーカーM4の下流にDNAマーカーM5を、標的領域T中にDNAマーカーM3を、それぞれ設定する。次いで、コシヒカリの染色体のうち、標的領域Tを含む一部のみがハバタキ由来の染色体断片に置換されている染色体断片置換系統に対して戻し交配を行い、得られた交雑集団から前記5種類のDNAマーカーM1〜M5に基づいて好ましい個体を選抜する。その後、当該個体に対して適宜自家交配又は戻し交配を行い、同様にDNAマーカーM1〜M5に基づいて好ましい個体を選抜することを適宜繰り返すことにより、ハバタキ由来の染色体断片により置換される領域の上流側末端がDNAマーカーM1とM2の間、該領域の下流側末端がDNAマーカーM4とM5の間にあるような後代個体を得ることができる。図1に示すように、当該後代個体は、DNAマーカーM1及びM5が元品種と同じタイプであり、DNAマーカーM2、M3、及びM4が、外来品種(本発明では、ハバタキ)と同じタイプである。
【0022】
ここで、特許文献2に記載の新品種の製造方法では、DNAマーカーM1とM2の距離d1が長ければ、外来品種由来染色体断片(本願では、ハバタキ由来染色体断片)Lの上流側末端が存在し得る範囲が広く、導入されるハバタキ由来染色体断片Lの長さが確定しにくくなる。一方、距離d1が短ければ、ハバタキ由来染色体断片Lの上流側末端が存在し得る範囲が狭く、導入されるハバタキ由来染色体断片Lの長さが確定しやすくなる。同様に、DNAマーカーM4とM5の距離d3が長ければ、ハバタキ由来染色体断片Lの下流側末端が存在し得る範囲が広く、導入されるハバタキ由来染色体断片Lの長さが確定しにくくなり、距離d3が短ければ、ハバタキ由来染色体断片Lの下流側末端が存在し得る範囲が狭く、導入されるハバタキ由来染色体断片Lの長さが確定しやすくなる。
【0023】
具体的には、表1に示すDNAマーカーM1〜M5のセット、すなわち、イネ品種日本晴の第3染色体の31,521,442番目のSNP(一塩基多型)に相当するSNP(イネ品種コシヒカリではA、イネ品種ハバタキではC)をDNAマーカーM1(DNAマーカーM1−Ac(QTS14))とし、イネ品種日本晴の第3染色体の31,689,690番目のSNPに相当するSNP(イネ品種コシヒカリではC、イネ品種ハバタキではT)をDNAマーカーM2(DNAマーカーM2−Ct(QTS14))とし、イネ品種日本晴の第3染色体の32,208,924番目のSNPに相当するSNP(イネ品種コシヒカリではA、イネ品種ハバタキではG)をDNAマーカーM3(DNAマーカーM3−Ag(QTS14))とし、イネ品種日本晴の第3染色体の32,298,686番目のSNPに相当するSNP(イネ品種コシヒカリではG、イネ品種ハバタキではC)をDNAマーカーM4(DNAマーカーM4−Gc(QTS14))とし、イネ品種日本晴の第3染色体の32,363,157番目のSNPに相当するSNP(イネ品種コシヒカリではA、イネ品種ハバタキではT)をDNAマーカーM5(DNAマーカーM5−At(QTS14))としてそれぞれ用いて、特許文献2に記載の新品種の作出方法によって新品種を作出した。これらの結果から、イネ個体の第3染色体中の、DNAマーカーM2−Ct(QTS14)からDNAマーカーM4−Gc(QTS14)までの領域(すなわち、イネ品種日本晴の第3染色体中の31,689,690番目の塩基から32,298,686番目の塩基までの領域に相当する領域)を含む領域が、ハバタキ由来の染色体断片によって置換された新品種が作出された。当該新品種では、ハバタキ由来の染色体断片の上流端は、DNAマーカーM1−Ac(QTS14)よりも下流であってDNAマーカーM2−Ct(QTS14)までの領域(すなわち、イネ品種日本晴の第3染色体中の31,521,443番目の塩基から31,689,690番目の塩基までの領域に相当する領域)に存在し、当該染色体断片の下流端が、DNAマーカーM4−Gc(QTS14)からDNAマーカーM5−At(QTS14)よりも上流までの領域(すなわち、イネ品種日本晴の第3染色体中の32,298,686番目の塩基から32,363,156番目の塩基までを含む領域に相当する領域)に存在する。
【0024】
【表1】

【0025】
本発明者はこの新品種を「コシヒカリえいち5号」と命名した。図2は、コシヒカリ及びコシヒカリえいち5号のゲノムを模式的に表した図である。千葉県にある圃場において、コシヒカリえいち5号の出穂期を測定したところ(種まき日:2010年5月6日、移植日: 2010年6月1日)、図3に示すように、コシヒカリの出穂期が8月5日〜8月8日であったのに対して、コシヒカリえいち5号は7月24日〜7月26日であった。つまり、元品種のコシヒカリよりも、コシヒカリえいち5号は明らかに早生であることが判明した。
【0026】
さらに、北緯38.5度よりも北の北海道にある圃場において、コシヒカリえいち5号の出穂期を測定した(種まき日:2010年4月28日、移植日: 2010年6月7日)。この結果、図4に示すように、コシヒカリの出穂期が9月1日〜9月2日であったのに対して、コシヒカリえいち5号は8月21日〜8月22日であった。つまり、北海道で栽培した場合にも、元品種のコシヒカリよりも、コシヒカリえいち5号は明らかに早生であることが判明した。但し、コシヒカリとコシヒカリえいち5号は、いずれも出穂はしたものの、充分に成熟せず、米は収穫できなかった。
【0027】
本発明の発明者は、コシヒカリえいち5号に、さらに早生化の機能を有する外来染色体断片を積み重ねることにより、北の地域でも栽培可能なほど十分に早生であるコシヒカリが得られるのではないかと考えた。そこで、コシヒカリえいち5号とイネ品種コシヒカリえいち3号を交配した。
【0028】
コシヒカリえいち3号は、特許文献2に記載の新品種の作出方法により、コシヒカリの染色体中、第6染色体のHd1遺伝子を含む領域のみがハバタキ由来の遺伝子断片に置換された品種である。表2に、コシヒカリえいち3号の作製に使用した5種類のDNAマーカーを示す。DNAマーカーM1−Ctは、イネ品種日本晴の第6染色体の8,757,818番目のSNPに相当するSNP(イネ品種コシヒカリではC、イネ品種ハバタキではT)であり、DNAマーカーM2−Agは、イネ品種日本晴の第6染色体の8,940,503番目のSNPに相当するSNP(イネ品種コシヒカリではA、イネ品種ハバタキではG)であり、DNAマーカーM3−Cgは、イネ品種日本晴の第6染色体の9,325,062番目のSNPに相当するSNP(イネ品種コシヒカリではC、イネ品種ハバタキではG)であり、DNAマーカーM4−Gcは、イネ品種日本晴の第6染色体の9,533,057番目のSNPに相当するSNP(イネ品種コシヒカリではG、イネ品種ハバタキではC)であり、DNAマーカーM5−Atは、イネ品種日本晴の第6染色体の9,777,196番目のSNPに相当するSNP(イネ品種コシヒカリではA、イネ品種ハバタキではT)である。すなわち、コシヒカリえいち3号は、イネ品種コシヒカリの第6染色体中のDNAマーカーM2−Ag(Hd1)からDNAマーカーM4−Gc(Hd1)までの領域(すなわち、イネ品種日本晴の第6染色体中の8,940,503番目の塩基から9,533,057番目の塩基までの領域に相当する領域)を含む領域がハバタキ由来の染色体断片によって置換された新品種である。当該新品種では、ハバタキ由来の染色体断片の上流端は、DNAマーカーM1−Ct(Hd1)よりも下流であってDNAマーカーM2−Ag(Hd1)までの領域(すなわち、イネ品種コシヒカリの第6染色体中の8,757,819番目の塩基から8,940,503番目の塩基までの領域に相当する領域)に存在し、当該染色体断片の下流端が、DNAマーカーM4−Gc(Hd1)からDNAマーカーM5−At(Hd1)よりも上流までの領域(すなわち、9,533,057番目の塩基から9,777,195番目の塩基までを含む領域に相当する領域)に存在する。
【0029】
【表2】
【0030】
図5は、コシヒカリ及びコシヒカリえいち3号のゲノムを模式的に表した図である。千葉県にある圃場において、コシヒカリえいち3号の出穂期を測定したところ(種まき日:2010年5月6日、移植日: 2010年6月1日)、図6に示すように、コシヒカリの出穂期が8月5日〜8月8日であったのに対して、コシヒカリえいち3号は7月25日〜7月26日であった。つまり、千葉県で栽培した場合には、コシヒカリえいち3号は、コシヒカリえいち5号と同程度に早生であった。
【0031】
さらに、北緯38.5度よりも北の北海道にある圃場において、コシヒカリえいち3号の出穂期を測定した(種まき日:2010年4月28日、移植日: 2010年6月7日)。この結果、図7に示すように、コシヒカリの出穂期が9月1日〜9月2日であり、米は収穫できなかった。これに対して、コシヒカリえいち3号の出穂期は8月10日〜8月16日であり、その後、米を収穫することができた。つまり、コシヒカリえいち3号は、北緯38.5度よりも北の地域で栽培した場合には、コシヒカリえいち5号よりも顕著に早生化するため、北海道でも栽培可能なイネであることが明らかとなった。Hd1領域をハバタキ由来の染色体断片に置換することによる早生化効果の程度が栽培地によって異なること、より北の地域、例えば北緯38.5度よりも北の地域で栽培した場合のほうが、従来のコシヒカリの栽培地域(北緯35.5度から38.5度)で栽培した場合よりも早生化効果が高いことは、本発明者によって初めて見出された知見である。
【0032】
本発明者は、後記実施例1に示すように、コシヒカリえいち5号とコシヒカリえいち3号とを交配し、コシヒカリの染色体中、QTS14領域とHd1領域のみがハバタキ由来の染色体断片に置換された新品種を作製した。本発明者はこの新品種を「コシヒカリかずさ6号」と命名した。図8は、コシヒカリかずさ6号のゲノムを模式的に表した図である。コシヒカリかずさ6号の出穂期を測定したところ、コシヒカリえいち3号よりも早生であり、北緯38.5度よりも北の北海道で栽培した場合でも、米を収穫することができた。また、コシヒカリかずさ6号の表現形質をコシヒカリと比較したところ、実際の圃場試験においても、出穂期以外のその他の形質はコシヒカリとほぼ同等であった。
【0033】
コシヒカリかずさ6号は、特許文献2に記載の方法により作出された新品種であり、ゲノム構成は99%以上がコシヒカリと同じであるように設計され、育成された。コシヒカリかずさ6号は、従来コシヒカリは栽培不可能であった北海道でも栽培可能なほど早生であるにもかかわらず、コシヒカリが有する味等の優良形質を維持しているという非常に優れた品種である。そこで、出願人は、コシヒカリかずさ6号を、独立行政法人産業技術総合研究所特許生物寄託センター(茨城県つくば市東1−1−1 つくばセンター中央第6)に新規植物として寄託した(寄託日:平成23年9月20日)。受領番号はFERM AP−22175である。
【0034】
イネ品種コシヒカリかずさ6号は、元品種コシヒカリと同様の手法により、栽培し、自家交配や人工交配により米を収穫することができる。また、イネ品種コシヒカリかずさ6号及びその後代個体は、元品種コシヒカリと同様に、新品種育成の親個体とすることができる。例えば、イネ品種コシヒカリかずさ6号の個体と別の品種の個体とを交配し、得られた後代個体を、イネ品種コシヒカリかずさ6号の個体と戻し交配することにより、新品種の育種を試みることもできる。
【0035】
また、表1に記載の5種類のDNAマーカー(DNAマーカーM1−Ac(QTS14)、DNAマーカーM2−Ct(QTS14)、DNAマーカーM3−Ag(QTS14)、DNAマーカーM4−Gc(QTS14)は、イネ品種コシヒカリかずさ6号及びイネ品種コシヒカリえいち5号に特有のゲノム情報である。したがって、イネ品種コシヒカリかずさ6号及びイネ品種コシヒカリえいち5号は、これらの5種類のDNAマーカーを適宜用いて鑑別することができる。
【0036】
具体的には、品種を鑑別する対象のイネ個体に対してゲノム解析を行い、DNAマーカーM1−Ac(QTS14)、DNAマーカーM2−Ct(QTS14)、DNAマーカーM3−Ag(QTS14)、DNAマーカーM4−Gc(QTS14)、及びDNAマーカーM5−At(QTS14)からなる群より選択される1以上のDNAマーカーをタイピングし、得られたタイピング結果が、イネ品種コシヒカリかずさ6号の結果と一致する場合には、当該イネ個体がイネ品種コシヒカリかずさ6号又はイネ品種コシヒカリえいち5号であると鑑別することができる。
【0037】
また、表2に記載の5種類のDNAマーカー(DNAマーカーM1−Ct(Hd1)、DNAマーカーM2−Ag(Hd1)、DNAマーカーM3−Cg(Hd1)、DNAマーカーM4−Gc(Hd1)、及びDNAマーカーM5−At(Hd1))は、イネ品種コシヒカリかずさ6号及びイネ品種コシヒカリえいち3号に特有のゲノム情報である。したがって、イネ品種コシヒカリかずさ6号及びイネ品種コシヒカリえいち3号は、これらの5種類のDNAマーカーを適宜用いて鑑別することができる。
【0038】
具体的には、品種を鑑別する対象のイネ個体に対してゲノム解析を行い、DNAマーカーM1−Ct(Hd1)、DNAマーカーM2−Ag(Hd1)、DNAマーカーM3−Cg(Hd1)、DNAマーカーM4−Gc(Hd1)、及びDNAマーカーM5−At(Hd1)からなる群より選択される1以上のDNAマーカーをタイピングし、得られたタイピング結果が、イネ品種コシヒカリかずさ6号の結果と一致する場合には、当該イネ個体がイネ品種コシヒカリかずさ6号又はイネ品種コシヒカリえいち3号であると鑑別することができる。
【0039】
ここで、品種の鑑別には、DNAマーカーM1〜M5の全てを用いてもよく、5個のDNAマーカーのうちの幾つかを用いてもよい。例えば、上流側の組み換えポイントであるDNAマーカーM1とM2のみを用いてもよく、下流側の組み換えポイントであるDNAマーカーM4とM5のみを用いてもよく、DNAマーカーM2とM4のみを用いてもよい。複数のDNAマーカーを適宜組み合わせることにより、より厳密な品種鑑別が可能となる。
【0040】
これらの結果から、イネ個体の第3染色体中のQTS14領域、具体的には、少なくともDNAマーカーM2−Ct(QTS14)からDNAマーカーM4−Gc(QTS14)までの領域(すなわち、イネ品種日本晴の第3染色体中の31,689,690番目の塩基から32,298,686番目の塩基までを含む領域に相当する領域)を、イネ品種ハバタキの当該領域からなる染色体断片に置換することにより、当該イネ個体を元品種よりも早生化することができることが明らかである。なお、イネ品種コシヒカリかずさ6号及びイネ品種コシヒカリえいち5号の当該領域は、イネ品種ハバタキの当該領域からなる染色体断片により構成されているため、イネ品種コシヒカリかずさ6号若しくはイネ品種コシヒカリえいち5号の当該領域からなる染色体断片によって置換してもよい。イネ品種ハバタキの当該領域からなる染色体断片を導入することにより早生化するイネ個体は、当該領域がイネ品種コシヒカリと同一若しくは近似した塩基配列を有している品種であればよく、イネ品種コシヒカリに限定されるものではないが、消費者の嗜好性等から、イネ品種コシヒカリ又はそれを親品種として作出された新品種であることが好ましい。
【0041】
また、DNAマーカーM2−Ct(QTS14)からDNAマーカーM4−Gc(QTS14)までの領域を含むイネ品種ハバタキ由来(若しくはイネ品種コシヒカリかずさ6号等由来)の染色体断片の上流端が、DNAマーカーM1−Ac(QTS14)よりも下流であってDNAマーカーM2−Ct(QTS14)までの領域(すなわち、イネ品種日本晴の第3染色体中の31,521,443番目の塩基から31,689,690番目の塩基までを含む領域に相当する領域)に存在し、当該染色体断片の下流端が、DNAマーカーM4−Gc(QTS14)からDNAマーカーM5−At(QTS14)よりも上流までの領域(すなわち、イネ品種日本晴の第3染色体中の32,298,686番目の塩基から32,363,156番目の塩基までを含む領域に相当する領域)に存在するように、当該染色体断片をイネ個体の第3染色体中に導入することにより、出穂期以外の形質に明らかな影響を及ぼすことなく、当該イネ個体を、元品種よりも早生化することができる。
【0042】
また、第3染色体中のQTS14領域に加えて、さらに第6染色体中のHd1領域、具体的には、少なくともDNAマーカーM2−Ag(Hd1)からDNAマーカーM4−Gc(Hd1)までの領域(すなわち、イネ品種日本晴の第6染色体中の8,940,503番目の塩基から9,533,057番目の塩基までを含む領域に相当する領域)を、イネ品種ハバタキの当該領域からなる染色体断片に置換することにより、当該イネ個体を北緯38.5度よりも北の地域でも栽培可能なほど顕著に早生化することができる。なお、イネ品種コシヒカリかずさ6号及びイネ品種コシヒカリえいち3号の当該領域は、イネ品種ハバタキの当該領域からなる染色体断片により構成されているため、イネ品種コシヒカリかずさ6号若しくはイネ品種コシヒカリえいち3号の当該領域からなる染色体断片によって置換してもよい。イネ品種ハバタキの当該領域からなる染色体断片を導入することにより早生化するイネ個体は、当該領域がイネ品種コシヒカリと同一若しくは近似した塩基配列を有している品種であればよく、イネ品種コシヒカリに限定されるものではないが、消費者の嗜好性等から、イネ品種コシヒカリ又はそれを親品種として作出された新品種であることが好ましい。
【0043】
また、DNAマーカーM2−Ag(Hd1)からDNAマーカーM4−Gc(Hd1)までの領域を含むイネ品種ハバタキ由来(若しくはイネ品種コシヒカリかずさ6号等由来)の染色体断片の上流端が、DNAマーカーM1−Ct(Hd1)よりも下流であってDNAマーカーM2−Ag(Hd1)までの領域(すなわち、イネ品種日本晴の第6染色体中の8,757,819番目の塩基から8,940,503番目の塩基までを含む領域に相当する領域)に存在し、当該染色体断片の下流端が、DNAマーカーM4−Gc(Hd1)からDNAマーカーM5−At(Hd1)よりも上流までの領域(すなわち、イネ品種日本晴の第6染色体中の9,533,057番目の塩基から9,777,195番目の塩基までを含む領域に相当する領域)に存在するように、当該染色体断片をイネ個体の第6染色体中に導入することにより、出穂期以外の形質に明らかな影響を及ぼすことなく、当該イネ個体を、元品種よりも早生化することができる。
【0044】
なお、コシヒカリえいち3号やコシヒカリかずさ6号をはじめとする、第6染色体中のHd1領域(具体的には、少なくともDNAマーカーM2−Ag(Hd1)からDNAマーカーM4−Gc(Hd1)までの領域)をイネ品種ハバタキの当該領域からなる染色体断片に置換されたイネ個体は、コシヒカリが栽培可能な地域で栽培可能であるのみならず、北緯38.5度よりも北の地域でも栽培し、米を収穫することができる。これらのイネ個体は、気温や降雨量等にも影響を受けるが、例えば、北緯38.5度から43.3度までの間の地域で栽培することができる。
【0045】
これまでの研究から、Hd1領域のうち、Hd1遺伝子が早生化を引き起こす原因遺伝子と考えられている。一方で、QTS14領域に含まれる遺伝子を調べたところ、当該領域中には、phytochrome C遺伝子をコードする領域が含まれていた。当該遺伝子は、主に植物の開花時間の制御に関与していることが報告されている(米国特許第7566815号明細書)。よって、QTS14領域において早生化を引き起こす原因遺伝子はphytochrome C遺伝子であると推察される。なお、イネ品種日本晴の対立断片では、Hd1遺伝子は、第6染色体の9,335,337番目の塩基から9,337,606番目の塩基までの領域にマップされており、phytochrome C遺伝子は、第3染色体の31,720,064番目の塩基から31,724,043番目の塩基までの領域にマップされている。
【0046】
Hd1領域中の早生化の原因遺伝子やQTS14領域中の早生化の原因遺伝子を含む領域がハバタキ由来の染色体断片によって置換されていれば、コシヒカリかずさ6号に導入されたハバタキ由来染色体断片よりも短い染色体断片によって置換されているイネ個体であっても、イネ品種コシヒカリかずさ6号と同様に早生化が引き起こされると考えられる。したがって、例えば、イネ個体の第3染色体中の、イネ品種日本晴の第3染色体中の31,689,691番目の塩基から31,724,043番目の塩基までを含む領域に相当する領域を、イネ品種コシヒカリかずさ6号、イネ品種コシヒカリえいち5号、又はイネ品種ハバタキの当該領域からなる染色体断片に置換することにより、当該イネ個体を元品種よりも早生化することができると考えられる。また、この際、当該染色体断片の上流端が、イネ品種日本晴の第3染色体の31,689,690番目の塩基から31,720,064番目の塩基までを含む領域に相当する領域に存在し、かつ当該染色体断片の下流端が、イネ品種日本晴の第3染色体の31,724,043番目の塩基から32,298,685番目の塩基までを含む領域に相当する領域に存在するように、当該染色体断片をイネ個体の第3染色体中に導入することにより、出穂期以外の形質に明らかな影響を及ぼすことなく、当該イネ個体を、元品種よりも早生化することができると考えられる。
【0047】
同様に、イネ個体の第6染色体中の、イネ品種日本晴の第6染色体中の9,335,337番目の塩基から9,337,606番目の塩基までを含む領域に相当する領域を、イネ品種コシヒカリかずさ6号、イネ品種コシヒカリえいち3号、又はイネ品種ハバタキの当該領域からなる染色体断片に置換することにより、当該イネ個体を元品種よりも早生化することができると考えられる。また、この際、当該染色体断片の上流端が、イネ品種日本晴の第6染色体の8,940,504番目の塩基から9,335,337番目の塩基までを含む領域に相当する領域に存在し、かつ当該染色体断片の下流端が、イネ品種日本晴の第3染色体の9,337,606番目の塩基から9,533,056番目の塩基までを含む領域に相当する領域に存在するように、当該染色体断片をイネ個体の第3染色体中に導入することにより、出穂期以外の形質に明らかな影響を及ぼすことなく、当該イネ個体を、元品種よりも早生化することができると考えられる。
【実施例】
【0048】
次に実施例を示して本発明をさらに詳細に説明するが、本発明は以下の実施例に限定されるものではない。
【0049】
[実施例1]
コシヒカリえいち5号とコシヒカリえいち3号とを掛け合わせ、コシヒカリの染色体中、QTS14領域とHd1領域のみがハバタキ由来の染色体断片に置換された新品種を作製した。
具体的には、コシヒカリえいち3号とコシヒカリえいち5号を交配し、得られた後代個体(種子)のうち2個を栽培し、自殖(自家交配)させ、さらに後代個体である種子を100個得た。この100個の種子を全て栽培し、各後代個体のDNAマーカーを調べ、DNAマーカーM3−Cg(Hd1)とDNAマーカーM3−Ag(QTS14)の両方がハバタキ由来アレルのホモ染色体領域である栽培個体を1個体選抜した。本発明者はこの新品種を「コシヒカリかずさ6号」と命名した。
【0050】
千葉県にある圃場において、コシヒカリかずさ6号の出穂期を測定した(種まき日:2010年5月6日、移植日: 2010年6月1日)。測定結果を、コシヒカリ、コシヒカリえいち5号、及びコシヒカリえいち3号の結果とともに図9に示す。コシヒカリの出穂期が8月5日〜8月8日であり、コシヒカリえいち5号とコシヒカリえいち3号が7月24日〜7月26日であったのに対して、コシヒカリかずさ6号は7月18日〜7月23日であった。これらの結果から、千葉県で栽培した場合に、コシヒカリかずさ6号は、コシヒカリえいち3号やコシヒカリえいち5号よりも明らかに早生であった。
【0051】
さらに、北海道にある圃場(北緯43.3度)において、コシヒカリかずさ6号の出穂期を測定した(種まき日:2010年4月28日、移植日: 2010年6月7日)。測定結果を、コシヒカリ、コシヒカリえいち5号、及びコシヒカリえいち3号の結果とともに図10に示す。コシヒカリの出穂期が9月1日〜9月2日であり、コシヒカリえいち5号が8月21日から8月22日であり、コシヒカリえいち3号が8月10日〜8月16日であったのに対して、コシヒカリかずさ6号は8月7日〜8月9日であった。また、コシヒカリ及びコシヒカリえいち5号は成熟しなかったのに対して、コシヒカリかずさ6号はコシヒカリえいち3号と同様に米を収穫することができた。これらの結果から、コシヒカリかずさ6号は、北緯38.5度よりも北の北海道においても栽培可能であることが明らかである。
【0052】
コシヒカリかずさ6号とコシヒカリの形質を比較検討した(千葉県にて、2009年に実施)。形質の検討は、種苗法(平成10年法律第83号)第5条第1項に基づく品種登録出願のための特性審査に準拠して行った。検討結果を表3〜6に示す。この結果、出穂期及び成熟期のいずれも、コシヒカリかずさ6号はコシヒカリよりも2週間程度早くなった。また、コシヒカリかずさ6号はコシヒカリよりも、稈長や穂の主軸の長さ、主茎長が若干短く、穂数及び主茎粒数も少な目であったが、それ以外の形質は基本的にコシヒカリと同じであった。
【0053】
【表3】
【0054】
【表4】
【0055】
【表5】
【0056】
【表6】
【産業上の利用可能性】
【0057】
本発明の新品種であるイネ品種コシヒカリかずさ6号は、コシヒカリとほぼ同様の特性を有し、かつ従来よりも北の地域でも栽培可能であるため、特に農業の分野において利用が可能である。また、本発明のイネ個体を早生化する方法により、イネ個体を元品種よりも早生化することができるため、当該方法は、特に植物の育種の分野において利用が可能である。
【受託番号】
【0058】
FERM AP−22175
【技術分野】
【0001】
本発明は、非遺伝子組み換え法により作出された新品種、当該新品種の鑑別方法、及びイネ個体を早生化する方法に関する。
【背景技術】
【0002】
同一生物種に属するが、遺伝的構成が異なるために、ある形質において他の集団と異なる集団を品種という。すなわち、同じ種類の植物であったとしても、品種により、栽培の難易性や病虫害に対する抵抗性、収量、品質等が異なる。このため、農作物、特にイネやムギ類等の主要な作物においては、より優良な品種を得るための品種改良が古くから行われており、近年では、種苗会社等のみならず、国や県等の公的機関においても積極的に行われてきている。
【0003】
近年の核酸解析技術等の進歩に伴い、シロイヌナズナ、イネ、コムギ等の様々な植物の遺伝子が解析され、得られた遺伝子情報が開示されている。これらの開示された遺伝子情報を利用して、遺伝子組み換え法による外来種の遺伝子を導入する品種改良も多く行われている。例えば、植物の感光性を増加させる機能を有する植物由来のタンパク質をコードするHd1遺伝子、及びHd1遺伝子を導入した形質転換植物の作製方法等が開示されている(例えば、特許文献1参照。)。しかしながら、遺伝子組み換え法による品種改良は、通常は交配不可能な遠縁種が有する形質を導入し得るという利点はあるものの、その安全性に対する検証は必ずしも十分ではないという問題点がある。
【0004】
このため、イネをはじめとする食用植物においては、非遺伝子組み換え法による新品種の作出が多く行われている。例えば特許文献2には、非遺伝子組み換え法により、外来の有用な染色体断片で置換する場合に、導入される外来品種由来の染色体断片による置換領域をコントロールし、元品種が有する好ましい形質を変更することなく、標的形質を有する新品種を作製するための方法が開示されている。同じく特許文献2には、この新品種を作製するための方法により、ハバタキのHd1遺伝子を含む領域のみをコシヒカリに導入した新品種イネ品種コシヒカリえいち3号が記載されている。
【0005】
特に、イネでは、より栽培可能地域が広いコシヒカリが望まれている。コシヒカリは、他の品種よりも食味に優れており、消費者に好まれている。このため米農家は、必ずしもコシヒカリの栽培に適しているとはいえない地域であっても、コシヒカリを好んで栽培している。しかしながら、コシヒカリを南の地域で栽培した場合には出穂期が早すぎ、充分な収穫量が望めない。その上、出穂期に高温が続くために食味が落ちてしまう。一方で、コシヒカリを北の地域で栽培した場合には晩生となり、たとえ出穂が出来たとしても、低温により登熟が不良となり、米を収穫することができない。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】特許第3660967号公報
【特許文献2】特許第4409610号公報
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明者による研究の結果、コシヒカリはおおよそ、日本の北緯35.5度から38.5度までの範囲内でしか栽培に適していないことがわかった。北緯38.5度よりも北の地域でも栽培可能なコシヒカリがあれば、例えば北海道等の従来では栽培不可能であった地域でもコシヒカリを収穫することができる。
【0008】
本発明は、従来よりも北の地域でも栽培可能なイネの新品種、及びイネ個体を早生化する方法を提供することを目的とする。
【課題を解決するための手段】
【0009】
本発明者は、上記課題を解決すべく鋭意研究した結果、イネ品種ハバタキの第3染色体上に存在する特定の領域の染色体断片とイネ品種ハバタキの第6染色体上に存在する特定の領域の染色体断片とを、イネ品種コシヒカリに置換することにより、従来よりも北の地域でも栽培可能なほど十分に早生化することが可能であることを見出し、本発明を完成させた。
【0010】
すなわち、本発明は、
(1) 受領番号がFERM AP−22175である、イネ品種コシヒカリかずさ6号(Oryza sativa L.cultivar Koshihikari−kazusa6 gou)、
(2) 前記(1)記載の品種の個体及び前記(1)記載の品種の個体の後代個体からなる群より選択される2個体を交配して得られる後代個体、
を、提供するものである。
【発明の効果】
【0011】
本発明の新品種であるイネ品種コシヒカリかずさ6号は、コシヒカリよりも非常に早生化されており、北緯38.5度よりも北の地域でも栽培し、米を収穫することができる。また、イネ品種コシヒカリかずさ6号は、品質や収穫量等の収穫期以外の特性はコシヒカリとほぼ同等な新品種である。
【図面の簡単な説明】
【0012】
【図1】元品種の染色体G上における標的領域T、置換された外来品種由来染色体断片L、及びDNAマーカーM1〜M5を示した図である。
【図2】コシヒカリ、及びコシヒカリえいち5号のゲノムを模式的に表した図である。
【図3】千葉県において、コシヒカリ、及びコシヒカリえいち5号の出穂期を調べた結果を示した図である。
【図4】北海道において、コシヒカリ、及びコシヒカリえいち5号の出穂期を調べた結果を示した図である。
【図5】コシヒカリえいち3号のゲノムを模式的に表した図である。
【図6】千葉県において、コシヒカリ、及びコシヒカリえいち3号の出穂期を調べた結果を示した図である。
【図7】北海道において、コシヒカリ、及びコシヒカリえいち3号の出穂期を調べた結果を示した図である。
【図8】コシヒカリかずさ6号のゲノムを模式的に表した図である。
【図9】千葉県において、コシヒカリかずさ6号の出穂期を調べた結果を、コシヒカリ、コシヒカリえいち5号、及びコシヒカリえいち3号の結果とともに示した図である。
【図10】北海道において、コシヒカリかずさ6号の出穂期を調べた結果を、コシヒカリ、コシヒカリえいち5号、及びコシヒカリえいち3号の結果とともに示した図である。
【発明を実施するための形態】
【0013】
本発明において染色体断片置換系統とは、元品種の染色体の一部のみが外来品種由来の染色体断片に置換されている系統を意味する。ここで、外来品種は、元品種以外の品種であれば特に限定されるものではなく、元品種と同一種の植物の品種であってもよく、元品種と異なる種の植物の品種であってもよく、動物等の植物以外の品種であってもよい。なお、本発明において品種とは、同一種の植物であって、遺伝的構成が異なるために、ある形質において同種内の他品種から明確に識別し得る集団を意味する。
【0014】
本発明においてDNAマーカーは、元品種由来の染色体と外来品種由来の染色体を識別し得る染色体上のDNA配列の差異を検出し得るものであれば、特に限定されるものではなく、遺伝子解析分野で通常用いられているDNAマーカーを用いることができる。該DNAマーカーとして、例えば、SNP(Single Nucleotide Polymorphism、一遺伝子多型)やSSR(Simple Sequence Repeats、単純反覆配列)の繰り返し数の違い等の遺伝子多型を検出し得るマーカーであってもよく、RFLP(Restriction Fragment Length Polymorphism、制限酵素断片長多型)マーカーであってもよい。なお、これらのDNAマーカーによる、元品種由来アレルと外来品種由来アレルとの識別は、常法により行うことができる。例えば、各個体から抽出したDNAを鋳型とし、特定のSNPやSSRと特異的にハイブリダイズし得るプライマー等を用いてPCRを行い、電気泳動法等を用いてPCR産物の有無を検出し、各多型を識別することができる。また、各個体から抽出したDNAを制限酵素処理した後、電気泳動法等を用いてDNA断片のパターンを検出し、各多型を識別することができる。なお、特定のSNPやSSRと特異的にハイブリダイズし得るプライマー等は、該SNPやSSRの塩基配列に応じて、汎用されているプライマー設計ツール等を用いて常法により設計することができる。また、設計されたプライマー等は、当該技術分野においてよく知られている方法のいずれを用いても合成することができる。
【0015】
これらのDNAマーカーは、公知のDNAマーカーを適宜用いることができる。また、新規に作製したDNAマーカーであってもよい。公知のDNAマーカーとして、例えば、イネにおいては、国際公開第2003/070934号パンフレット等において開示されているSNPマーカーや、Rice Genome Research Program(RGP:http://rgp.dna.affrc.go.jp/publicdata.html)において公開されているDNAマーカーを用いることができる。
【0016】
なお、各品種の遺伝子情報等は、例えば、国際的な塩基配列データベースであるNCBI(National center for Biotechnology Information)やDDBJ(DNA Data Bank of Japan)等において入手することができる。特にイネの各品種の遺伝子情報は、KOME(Knowledge−based Oryza Molecular biological Encyclopedia、http://cdna01.dna.affrc.go.jp/cDNA/)等において入手することができる。
【0017】
本発明及び本願明細書において「イネ品種日本晴の染色体のX番目の塩基からY番目の塩基までの領域」は、RGBにおいて公開されているイネ品種日本晴のゲノムDNAの塩基配列(バージョン4;IRGSP−build4−06/04/21)に基づいて決定される領域である。
【0018】
また、本発明及び本願明細書において、「イネ品種日本晴の染色体のX番目の塩基からY番目の塩基までの領域に相当する領域」とは、イネ個体の染色体中のイネ品種日本晴の染色体中の当該領域と相同性の高い領域であり、イネ品種日本晴の公知のゲノムDNAと当該イネ個体のゲノムDNAの塩基配列を、最もホモロジーが高くなるようにアラインメントすることにより決定することができる。また、イネ品種日本晴以外のイネ個体中の「イネ品種日本晴のSNPに相当するSNP」は、当該SNPを含む領域において、イネ品種日本晴の公知のゲノムDNAと当該イネ個体のゲノムDNAの塩基配列を、最もホモロジーが高くなるようにアラインメントした場合に、当該SNPに対応する位置にある塩基を意味する。
【0019】
従来の品種よりも北の地域で栽培可能な新品種を育種するため、本発明の発明者は、まず、出穂期に関して、イネ品種ハバタキとイネ品種コシヒカリとを交配して、分離集団でQTL(Quantitative Trait Locus) 解析を行った。この結果、イネ品種ハバタキの第3染色体の長腕のQTS14領域〔イネ品種日本晴の第3染色体の31,720,064番目の塩基から31,724,043番目の塩基までの領域に相当する領域〕に、出穂期を早め、早生とするQTLが存在していることもわかった。そこで、本発明者は、コシヒカリの当該領域に含まれている遺伝子をハバタキ由来の遺伝子に置換した新品種の作出を行った。当該新品種は、元品種コシヒカリよりも早生のイネであると予想された。
【0020】
非遺伝子組み換え法により植物の品種改良を行う場合において、導入される外来品種由来の染色体断片が大きすぎる場合には、目的の形質遺伝子以外の機能不明な他の遺伝子を多数導入してしまうおそれや、元品種が有する好ましい形質を損なうおそれがある。そこで、本発明者は、導入される外来品種由来の染色体断片による置換領域をコントロールし、元品種が有する好ましい形質を変更することなく、標的形質を有する新品種を作製するため、特許文献2に記載の方法により、新品種の作出を行った。
【0021】
特許文献2に記載の新品種の作出方法は、具体的には以下の通りである。まず、公知のイネの遺伝子情報に基づき、図1に示すような位置関係にある5種類のDNAマーカーを設定した。すなわち、標的領域Tの上流側末端又はその上流にDNAマーカーM2を、DNAマーカーM2の上流にDNAマーカーM1を、標的領域Tの下流側末端又はその下流にDNAマーカーM4を、DNAマーカーM4の下流にDNAマーカーM5を、標的領域T中にDNAマーカーM3を、それぞれ設定する。次いで、コシヒカリの染色体のうち、標的領域Tを含む一部のみがハバタキ由来の染色体断片に置換されている染色体断片置換系統に対して戻し交配を行い、得られた交雑集団から前記5種類のDNAマーカーM1〜M5に基づいて好ましい個体を選抜する。その後、当該個体に対して適宜自家交配又は戻し交配を行い、同様にDNAマーカーM1〜M5に基づいて好ましい個体を選抜することを適宜繰り返すことにより、ハバタキ由来の染色体断片により置換される領域の上流側末端がDNAマーカーM1とM2の間、該領域の下流側末端がDNAマーカーM4とM5の間にあるような後代個体を得ることができる。図1に示すように、当該後代個体は、DNAマーカーM1及びM5が元品種と同じタイプであり、DNAマーカーM2、M3、及びM4が、外来品種(本発明では、ハバタキ)と同じタイプである。
【0022】
ここで、特許文献2に記載の新品種の製造方法では、DNAマーカーM1とM2の距離d1が長ければ、外来品種由来染色体断片(本願では、ハバタキ由来染色体断片)Lの上流側末端が存在し得る範囲が広く、導入されるハバタキ由来染色体断片Lの長さが確定しにくくなる。一方、距離d1が短ければ、ハバタキ由来染色体断片Lの上流側末端が存在し得る範囲が狭く、導入されるハバタキ由来染色体断片Lの長さが確定しやすくなる。同様に、DNAマーカーM4とM5の距離d3が長ければ、ハバタキ由来染色体断片Lの下流側末端が存在し得る範囲が広く、導入されるハバタキ由来染色体断片Lの長さが確定しにくくなり、距離d3が短ければ、ハバタキ由来染色体断片Lの下流側末端が存在し得る範囲が狭く、導入されるハバタキ由来染色体断片Lの長さが確定しやすくなる。
【0023】
具体的には、表1に示すDNAマーカーM1〜M5のセット、すなわち、イネ品種日本晴の第3染色体の31,521,442番目のSNP(一塩基多型)に相当するSNP(イネ品種コシヒカリではA、イネ品種ハバタキではC)をDNAマーカーM1(DNAマーカーM1−Ac(QTS14))とし、イネ品種日本晴の第3染色体の31,689,690番目のSNPに相当するSNP(イネ品種コシヒカリではC、イネ品種ハバタキではT)をDNAマーカーM2(DNAマーカーM2−Ct(QTS14))とし、イネ品種日本晴の第3染色体の32,208,924番目のSNPに相当するSNP(イネ品種コシヒカリではA、イネ品種ハバタキではG)をDNAマーカーM3(DNAマーカーM3−Ag(QTS14))とし、イネ品種日本晴の第3染色体の32,298,686番目のSNPに相当するSNP(イネ品種コシヒカリではG、イネ品種ハバタキではC)をDNAマーカーM4(DNAマーカーM4−Gc(QTS14))とし、イネ品種日本晴の第3染色体の32,363,157番目のSNPに相当するSNP(イネ品種コシヒカリではA、イネ品種ハバタキではT)をDNAマーカーM5(DNAマーカーM5−At(QTS14))としてそれぞれ用いて、特許文献2に記載の新品種の作出方法によって新品種を作出した。これらの結果から、イネ個体の第3染色体中の、DNAマーカーM2−Ct(QTS14)からDNAマーカーM4−Gc(QTS14)までの領域(すなわち、イネ品種日本晴の第3染色体中の31,689,690番目の塩基から32,298,686番目の塩基までの領域に相当する領域)を含む領域が、ハバタキ由来の染色体断片によって置換された新品種が作出された。当該新品種では、ハバタキ由来の染色体断片の上流端は、DNAマーカーM1−Ac(QTS14)よりも下流であってDNAマーカーM2−Ct(QTS14)までの領域(すなわち、イネ品種日本晴の第3染色体中の31,521,443番目の塩基から31,689,690番目の塩基までの領域に相当する領域)に存在し、当該染色体断片の下流端が、DNAマーカーM4−Gc(QTS14)からDNAマーカーM5−At(QTS14)よりも上流までの領域(すなわち、イネ品種日本晴の第3染色体中の32,298,686番目の塩基から32,363,156番目の塩基までを含む領域に相当する領域)に存在する。
【0024】
【表1】
【0025】
本発明者はこの新品種を「コシヒカリえいち5号」と命名した。図2は、コシヒカリ及びコシヒカリえいち5号のゲノムを模式的に表した図である。千葉県にある圃場において、コシヒカリえいち5号の出穂期を測定したところ(種まき日:2010年5月6日、移植日: 2010年6月1日)、図3に示すように、コシヒカリの出穂期が8月5日〜8月8日であったのに対して、コシヒカリえいち5号は7月24日〜7月26日であった。つまり、元品種のコシヒカリよりも、コシヒカリえいち5号は明らかに早生であることが判明した。
【0026】
さらに、北緯38.5度よりも北の北海道にある圃場において、コシヒカリえいち5号の出穂期を測定した(種まき日:2010年4月28日、移植日: 2010年6月7日)。この結果、図4に示すように、コシヒカリの出穂期が9月1日〜9月2日であったのに対して、コシヒカリえいち5号は8月21日〜8月22日であった。つまり、北海道で栽培した場合にも、元品種のコシヒカリよりも、コシヒカリえいち5号は明らかに早生であることが判明した。但し、コシヒカリとコシヒカリえいち5号は、いずれも出穂はしたものの、充分に成熟せず、米は収穫できなかった。
【0027】
本発明の発明者は、コシヒカリえいち5号に、さらに早生化の機能を有する外来染色体断片を積み重ねることにより、北の地域でも栽培可能なほど十分に早生であるコシヒカリが得られるのではないかと考えた。そこで、コシヒカリえいち5号とイネ品種コシヒカリえいち3号を交配した。
【0028】
コシヒカリえいち3号は、特許文献2に記載の新品種の作出方法により、コシヒカリの染色体中、第6染色体のHd1遺伝子を含む領域のみがハバタキ由来の遺伝子断片に置換された品種である。表2に、コシヒカリえいち3号の作製に使用した5種類のDNAマーカーを示す。DNAマーカーM1−Ctは、イネ品種日本晴の第6染色体の8,757,818番目のSNPに相当するSNP(イネ品種コシヒカリではC、イネ品種ハバタキではT)であり、DNAマーカーM2−Agは、イネ品種日本晴の第6染色体の8,940,503番目のSNPに相当するSNP(イネ品種コシヒカリではA、イネ品種ハバタキではG)であり、DNAマーカーM3−Cgは、イネ品種日本晴の第6染色体の9,325,062番目のSNPに相当するSNP(イネ品種コシヒカリではC、イネ品種ハバタキではG)であり、DNAマーカーM4−Gcは、イネ品種日本晴の第6染色体の9,533,057番目のSNPに相当するSNP(イネ品種コシヒカリではG、イネ品種ハバタキではC)であり、DNAマーカーM5−Atは、イネ品種日本晴の第6染色体の9,777,196番目のSNPに相当するSNP(イネ品種コシヒカリではA、イネ品種ハバタキではT)である。すなわち、コシヒカリえいち3号は、イネ品種コシヒカリの第6染色体中のDNAマーカーM2−Ag(Hd1)からDNAマーカーM4−Gc(Hd1)までの領域(すなわち、イネ品種日本晴の第6染色体中の8,940,503番目の塩基から9,533,057番目の塩基までの領域に相当する領域)を含む領域がハバタキ由来の染色体断片によって置換された新品種である。当該新品種では、ハバタキ由来の染色体断片の上流端は、DNAマーカーM1−Ct(Hd1)よりも下流であってDNAマーカーM2−Ag(Hd1)までの領域(すなわち、イネ品種コシヒカリの第6染色体中の8,757,819番目の塩基から8,940,503番目の塩基までの領域に相当する領域)に存在し、当該染色体断片の下流端が、DNAマーカーM4−Gc(Hd1)からDNAマーカーM5−At(Hd1)よりも上流までの領域(すなわち、9,533,057番目の塩基から9,777,195番目の塩基までを含む領域に相当する領域)に存在する。
【0029】
【表2】
【0030】
図5は、コシヒカリ及びコシヒカリえいち3号のゲノムを模式的に表した図である。千葉県にある圃場において、コシヒカリえいち3号の出穂期を測定したところ(種まき日:2010年5月6日、移植日: 2010年6月1日)、図6に示すように、コシヒカリの出穂期が8月5日〜8月8日であったのに対して、コシヒカリえいち3号は7月25日〜7月26日であった。つまり、千葉県で栽培した場合には、コシヒカリえいち3号は、コシヒカリえいち5号と同程度に早生であった。
【0031】
さらに、北緯38.5度よりも北の北海道にある圃場において、コシヒカリえいち3号の出穂期を測定した(種まき日:2010年4月28日、移植日: 2010年6月7日)。この結果、図7に示すように、コシヒカリの出穂期が9月1日〜9月2日であり、米は収穫できなかった。これに対して、コシヒカリえいち3号の出穂期は8月10日〜8月16日であり、その後、米を収穫することができた。つまり、コシヒカリえいち3号は、北緯38.5度よりも北の地域で栽培した場合には、コシヒカリえいち5号よりも顕著に早生化するため、北海道でも栽培可能なイネであることが明らかとなった。Hd1領域をハバタキ由来の染色体断片に置換することによる早生化効果の程度が栽培地によって異なること、より北の地域、例えば北緯38.5度よりも北の地域で栽培した場合のほうが、従来のコシヒカリの栽培地域(北緯35.5度から38.5度)で栽培した場合よりも早生化効果が高いことは、本発明者によって初めて見出された知見である。
【0032】
本発明者は、後記実施例1に示すように、コシヒカリえいち5号とコシヒカリえいち3号とを交配し、コシヒカリの染色体中、QTS14領域とHd1領域のみがハバタキ由来の染色体断片に置換された新品種を作製した。本発明者はこの新品種を「コシヒカリかずさ6号」と命名した。図8は、コシヒカリかずさ6号のゲノムを模式的に表した図である。コシヒカリかずさ6号の出穂期を測定したところ、コシヒカリえいち3号よりも早生であり、北緯38.5度よりも北の北海道で栽培した場合でも、米を収穫することができた。また、コシヒカリかずさ6号の表現形質をコシヒカリと比較したところ、実際の圃場試験においても、出穂期以外のその他の形質はコシヒカリとほぼ同等であった。
【0033】
コシヒカリかずさ6号は、特許文献2に記載の方法により作出された新品種であり、ゲノム構成は99%以上がコシヒカリと同じであるように設計され、育成された。コシヒカリかずさ6号は、従来コシヒカリは栽培不可能であった北海道でも栽培可能なほど早生であるにもかかわらず、コシヒカリが有する味等の優良形質を維持しているという非常に優れた品種である。そこで、出願人は、コシヒカリかずさ6号を、独立行政法人産業技術総合研究所特許生物寄託センター(茨城県つくば市東1−1−1 つくばセンター中央第6)に新規植物として寄託した(寄託日:平成23年9月20日)。受領番号はFERM AP−22175である。
【0034】
イネ品種コシヒカリかずさ6号は、元品種コシヒカリと同様の手法により、栽培し、自家交配や人工交配により米を収穫することができる。また、イネ品種コシヒカリかずさ6号及びその後代個体は、元品種コシヒカリと同様に、新品種育成の親個体とすることができる。例えば、イネ品種コシヒカリかずさ6号の個体と別の品種の個体とを交配し、得られた後代個体を、イネ品種コシヒカリかずさ6号の個体と戻し交配することにより、新品種の育種を試みることもできる。
【0035】
また、表1に記載の5種類のDNAマーカー(DNAマーカーM1−Ac(QTS14)、DNAマーカーM2−Ct(QTS14)、DNAマーカーM3−Ag(QTS14)、DNAマーカーM4−Gc(QTS14)は、イネ品種コシヒカリかずさ6号及びイネ品種コシヒカリえいち5号に特有のゲノム情報である。したがって、イネ品種コシヒカリかずさ6号及びイネ品種コシヒカリえいち5号は、これらの5種類のDNAマーカーを適宜用いて鑑別することができる。
【0036】
具体的には、品種を鑑別する対象のイネ個体に対してゲノム解析を行い、DNAマーカーM1−Ac(QTS14)、DNAマーカーM2−Ct(QTS14)、DNAマーカーM3−Ag(QTS14)、DNAマーカーM4−Gc(QTS14)、及びDNAマーカーM5−At(QTS14)からなる群より選択される1以上のDNAマーカーをタイピングし、得られたタイピング結果が、イネ品種コシヒカリかずさ6号の結果と一致する場合には、当該イネ個体がイネ品種コシヒカリかずさ6号又はイネ品種コシヒカリえいち5号であると鑑別することができる。
【0037】
また、表2に記載の5種類のDNAマーカー(DNAマーカーM1−Ct(Hd1)、DNAマーカーM2−Ag(Hd1)、DNAマーカーM3−Cg(Hd1)、DNAマーカーM4−Gc(Hd1)、及びDNAマーカーM5−At(Hd1))は、イネ品種コシヒカリかずさ6号及びイネ品種コシヒカリえいち3号に特有のゲノム情報である。したがって、イネ品種コシヒカリかずさ6号及びイネ品種コシヒカリえいち3号は、これらの5種類のDNAマーカーを適宜用いて鑑別することができる。
【0038】
具体的には、品種を鑑別する対象のイネ個体に対してゲノム解析を行い、DNAマーカーM1−Ct(Hd1)、DNAマーカーM2−Ag(Hd1)、DNAマーカーM3−Cg(Hd1)、DNAマーカーM4−Gc(Hd1)、及びDNAマーカーM5−At(Hd1)からなる群より選択される1以上のDNAマーカーをタイピングし、得られたタイピング結果が、イネ品種コシヒカリかずさ6号の結果と一致する場合には、当該イネ個体がイネ品種コシヒカリかずさ6号又はイネ品種コシヒカリえいち3号であると鑑別することができる。
【0039】
ここで、品種の鑑別には、DNAマーカーM1〜M5の全てを用いてもよく、5個のDNAマーカーのうちの幾つかを用いてもよい。例えば、上流側の組み換えポイントであるDNAマーカーM1とM2のみを用いてもよく、下流側の組み換えポイントであるDNAマーカーM4とM5のみを用いてもよく、DNAマーカーM2とM4のみを用いてもよい。複数のDNAマーカーを適宜組み合わせることにより、より厳密な品種鑑別が可能となる。
【0040】
これらの結果から、イネ個体の第3染色体中のQTS14領域、具体的には、少なくともDNAマーカーM2−Ct(QTS14)からDNAマーカーM4−Gc(QTS14)までの領域(すなわち、イネ品種日本晴の第3染色体中の31,689,690番目の塩基から32,298,686番目の塩基までを含む領域に相当する領域)を、イネ品種ハバタキの当該領域からなる染色体断片に置換することにより、当該イネ個体を元品種よりも早生化することができることが明らかである。なお、イネ品種コシヒカリかずさ6号及びイネ品種コシヒカリえいち5号の当該領域は、イネ品種ハバタキの当該領域からなる染色体断片により構成されているため、イネ品種コシヒカリかずさ6号若しくはイネ品種コシヒカリえいち5号の当該領域からなる染色体断片によって置換してもよい。イネ品種ハバタキの当該領域からなる染色体断片を導入することにより早生化するイネ個体は、当該領域がイネ品種コシヒカリと同一若しくは近似した塩基配列を有している品種であればよく、イネ品種コシヒカリに限定されるものではないが、消費者の嗜好性等から、イネ品種コシヒカリ又はそれを親品種として作出された新品種であることが好ましい。
【0041】
また、DNAマーカーM2−Ct(QTS14)からDNAマーカーM4−Gc(QTS14)までの領域を含むイネ品種ハバタキ由来(若しくはイネ品種コシヒカリかずさ6号等由来)の染色体断片の上流端が、DNAマーカーM1−Ac(QTS14)よりも下流であってDNAマーカーM2−Ct(QTS14)までの領域(すなわち、イネ品種日本晴の第3染色体中の31,521,443番目の塩基から31,689,690番目の塩基までを含む領域に相当する領域)に存在し、当該染色体断片の下流端が、DNAマーカーM4−Gc(QTS14)からDNAマーカーM5−At(QTS14)よりも上流までの領域(すなわち、イネ品種日本晴の第3染色体中の32,298,686番目の塩基から32,363,156番目の塩基までを含む領域に相当する領域)に存在するように、当該染色体断片をイネ個体の第3染色体中に導入することにより、出穂期以外の形質に明らかな影響を及ぼすことなく、当該イネ個体を、元品種よりも早生化することができる。
【0042】
また、第3染色体中のQTS14領域に加えて、さらに第6染色体中のHd1領域、具体的には、少なくともDNAマーカーM2−Ag(Hd1)からDNAマーカーM4−Gc(Hd1)までの領域(すなわち、イネ品種日本晴の第6染色体中の8,940,503番目の塩基から9,533,057番目の塩基までを含む領域に相当する領域)を、イネ品種ハバタキの当該領域からなる染色体断片に置換することにより、当該イネ個体を北緯38.5度よりも北の地域でも栽培可能なほど顕著に早生化することができる。なお、イネ品種コシヒカリかずさ6号及びイネ品種コシヒカリえいち3号の当該領域は、イネ品種ハバタキの当該領域からなる染色体断片により構成されているため、イネ品種コシヒカリかずさ6号若しくはイネ品種コシヒカリえいち3号の当該領域からなる染色体断片によって置換してもよい。イネ品種ハバタキの当該領域からなる染色体断片を導入することにより早生化するイネ個体は、当該領域がイネ品種コシヒカリと同一若しくは近似した塩基配列を有している品種であればよく、イネ品種コシヒカリに限定されるものではないが、消費者の嗜好性等から、イネ品種コシヒカリ又はそれを親品種として作出された新品種であることが好ましい。
【0043】
また、DNAマーカーM2−Ag(Hd1)からDNAマーカーM4−Gc(Hd1)までの領域を含むイネ品種ハバタキ由来(若しくはイネ品種コシヒカリかずさ6号等由来)の染色体断片の上流端が、DNAマーカーM1−Ct(Hd1)よりも下流であってDNAマーカーM2−Ag(Hd1)までの領域(すなわち、イネ品種日本晴の第6染色体中の8,757,819番目の塩基から8,940,503番目の塩基までを含む領域に相当する領域)に存在し、当該染色体断片の下流端が、DNAマーカーM4−Gc(Hd1)からDNAマーカーM5−At(Hd1)よりも上流までの領域(すなわち、イネ品種日本晴の第6染色体中の9,533,057番目の塩基から9,777,195番目の塩基までを含む領域に相当する領域)に存在するように、当該染色体断片をイネ個体の第6染色体中に導入することにより、出穂期以外の形質に明らかな影響を及ぼすことなく、当該イネ個体を、元品種よりも早生化することができる。
【0044】
なお、コシヒカリえいち3号やコシヒカリかずさ6号をはじめとする、第6染色体中のHd1領域(具体的には、少なくともDNAマーカーM2−Ag(Hd1)からDNAマーカーM4−Gc(Hd1)までの領域)をイネ品種ハバタキの当該領域からなる染色体断片に置換されたイネ個体は、コシヒカリが栽培可能な地域で栽培可能であるのみならず、北緯38.5度よりも北の地域でも栽培し、米を収穫することができる。これらのイネ個体は、気温や降雨量等にも影響を受けるが、例えば、北緯38.5度から43.3度までの間の地域で栽培することができる。
【0045】
これまでの研究から、Hd1領域のうち、Hd1遺伝子が早生化を引き起こす原因遺伝子と考えられている。一方で、QTS14領域に含まれる遺伝子を調べたところ、当該領域中には、phytochrome C遺伝子をコードする領域が含まれていた。当該遺伝子は、主に植物の開花時間の制御に関与していることが報告されている(米国特許第7566815号明細書)。よって、QTS14領域において早生化を引き起こす原因遺伝子はphytochrome C遺伝子であると推察される。なお、イネ品種日本晴の対立断片では、Hd1遺伝子は、第6染色体の9,335,337番目の塩基から9,337,606番目の塩基までの領域にマップされており、phytochrome C遺伝子は、第3染色体の31,720,064番目の塩基から31,724,043番目の塩基までの領域にマップされている。
【0046】
Hd1領域中の早生化の原因遺伝子やQTS14領域中の早生化の原因遺伝子を含む領域がハバタキ由来の染色体断片によって置換されていれば、コシヒカリかずさ6号に導入されたハバタキ由来染色体断片よりも短い染色体断片によって置換されているイネ個体であっても、イネ品種コシヒカリかずさ6号と同様に早生化が引き起こされると考えられる。したがって、例えば、イネ個体の第3染色体中の、イネ品種日本晴の第3染色体中の31,689,691番目の塩基から31,724,043番目の塩基までを含む領域に相当する領域を、イネ品種コシヒカリかずさ6号、イネ品種コシヒカリえいち5号、又はイネ品種ハバタキの当該領域からなる染色体断片に置換することにより、当該イネ個体を元品種よりも早生化することができると考えられる。また、この際、当該染色体断片の上流端が、イネ品種日本晴の第3染色体の31,689,690番目の塩基から31,720,064番目の塩基までを含む領域に相当する領域に存在し、かつ当該染色体断片の下流端が、イネ品種日本晴の第3染色体の31,724,043番目の塩基から32,298,685番目の塩基までを含む領域に相当する領域に存在するように、当該染色体断片をイネ個体の第3染色体中に導入することにより、出穂期以外の形質に明らかな影響を及ぼすことなく、当該イネ個体を、元品種よりも早生化することができると考えられる。
【0047】
同様に、イネ個体の第6染色体中の、イネ品種日本晴の第6染色体中の9,335,337番目の塩基から9,337,606番目の塩基までを含む領域に相当する領域を、イネ品種コシヒカリかずさ6号、イネ品種コシヒカリえいち3号、又はイネ品種ハバタキの当該領域からなる染色体断片に置換することにより、当該イネ個体を元品種よりも早生化することができると考えられる。また、この際、当該染色体断片の上流端が、イネ品種日本晴の第6染色体の8,940,504番目の塩基から9,335,337番目の塩基までを含む領域に相当する領域に存在し、かつ当該染色体断片の下流端が、イネ品種日本晴の第3染色体の9,337,606番目の塩基から9,533,056番目の塩基までを含む領域に相当する領域に存在するように、当該染色体断片をイネ個体の第3染色体中に導入することにより、出穂期以外の形質に明らかな影響を及ぼすことなく、当該イネ個体を、元品種よりも早生化することができると考えられる。
【実施例】
【0048】
次に実施例を示して本発明をさらに詳細に説明するが、本発明は以下の実施例に限定されるものではない。
【0049】
[実施例1]
コシヒカリえいち5号とコシヒカリえいち3号とを掛け合わせ、コシヒカリの染色体中、QTS14領域とHd1領域のみがハバタキ由来の染色体断片に置換された新品種を作製した。
具体的には、コシヒカリえいち3号とコシヒカリえいち5号を交配し、得られた後代個体(種子)のうち2個を栽培し、自殖(自家交配)させ、さらに後代個体である種子を100個得た。この100個の種子を全て栽培し、各後代個体のDNAマーカーを調べ、DNAマーカーM3−Cg(Hd1)とDNAマーカーM3−Ag(QTS14)の両方がハバタキ由来アレルのホモ染色体領域である栽培個体を1個体選抜した。本発明者はこの新品種を「コシヒカリかずさ6号」と命名した。
【0050】
千葉県にある圃場において、コシヒカリかずさ6号の出穂期を測定した(種まき日:2010年5月6日、移植日: 2010年6月1日)。測定結果を、コシヒカリ、コシヒカリえいち5号、及びコシヒカリえいち3号の結果とともに図9に示す。コシヒカリの出穂期が8月5日〜8月8日であり、コシヒカリえいち5号とコシヒカリえいち3号が7月24日〜7月26日であったのに対して、コシヒカリかずさ6号は7月18日〜7月23日であった。これらの結果から、千葉県で栽培した場合に、コシヒカリかずさ6号は、コシヒカリえいち3号やコシヒカリえいち5号よりも明らかに早生であった。
【0051】
さらに、北海道にある圃場(北緯43.3度)において、コシヒカリかずさ6号の出穂期を測定した(種まき日:2010年4月28日、移植日: 2010年6月7日)。測定結果を、コシヒカリ、コシヒカリえいち5号、及びコシヒカリえいち3号の結果とともに図10に示す。コシヒカリの出穂期が9月1日〜9月2日であり、コシヒカリえいち5号が8月21日から8月22日であり、コシヒカリえいち3号が8月10日〜8月16日であったのに対して、コシヒカリかずさ6号は8月7日〜8月9日であった。また、コシヒカリ及びコシヒカリえいち5号は成熟しなかったのに対して、コシヒカリかずさ6号はコシヒカリえいち3号と同様に米を収穫することができた。これらの結果から、コシヒカリかずさ6号は、北緯38.5度よりも北の北海道においても栽培可能であることが明らかである。
【0052】
コシヒカリかずさ6号とコシヒカリの形質を比較検討した(千葉県にて、2009年に実施)。形質の検討は、種苗法(平成10年法律第83号)第5条第1項に基づく品種登録出願のための特性審査に準拠して行った。検討結果を表3〜6に示す。この結果、出穂期及び成熟期のいずれも、コシヒカリかずさ6号はコシヒカリよりも2週間程度早くなった。また、コシヒカリかずさ6号はコシヒカリよりも、稈長や穂の主軸の長さ、主茎長が若干短く、穂数及び主茎粒数も少な目であったが、それ以外の形質は基本的にコシヒカリと同じであった。
【0053】
【表3】
【0054】
【表4】
【0055】
【表5】
【0056】
【表6】
【産業上の利用可能性】
【0057】
本発明の新品種であるイネ品種コシヒカリかずさ6号は、コシヒカリとほぼ同様の特性を有し、かつ従来よりも北の地域でも栽培可能であるため、特に農業の分野において利用が可能である。また、本発明のイネ個体を早生化する方法により、イネ個体を元品種よりも早生化することができるため、当該方法は、特に植物の育種の分野において利用が可能である。
【受託番号】
【0058】
FERM AP−22175
【特許請求の範囲】
【請求項1】
受領番号がFERM AP−22175である、イネ品種コシヒカリかずさ6号(Oryza sativa L.cultivar Koshihikari−kazusa6 gou)。
【請求項2】
請求項1記載の品種の個体及び請求項1記載の品種の個体の後代個体からなる群より選択される2個体を交配して得られる後代個体。
【請求項1】
受領番号がFERM AP−22175である、イネ品種コシヒカリかずさ6号(Oryza sativa L.cultivar Koshihikari−kazusa6 gou)。
【請求項2】
請求項1記載の品種の個体及び請求項1記載の品種の個体の後代個体からなる群より選択される2個体を交配して得られる後代個体。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【公開番号】特開2012−210203(P2012−210203A)
【公開日】平成24年11月1日(2012.11.1)
【国際特許分類】
【出願番号】特願2011−206766(P2011−206766)
【出願日】平成23年9月22日(2011.9.22)
【特許番号】特許第4961504号(P4961504)
【特許公報発行日】平成24年6月27日(2012.6.27)
【出願人】(000005326)本田技研工業株式会社 (23,863)
【Fターム(参考)】
【公開日】平成24年11月1日(2012.11.1)
【国際特許分類】
【出願日】平成23年9月22日(2011.9.22)
【特許番号】特許第4961504号(P4961504)
【特許公報発行日】平成24年6月27日(2012.6.27)
【出願人】(000005326)本田技研工業株式会社 (23,863)
【Fターム(参考)】
[ Back to top ]