
新型インフルエンザウイルスのヘマグルチニンに結合するアプタマー
【課題】新型インフルエンザウイルス、特に、A/California/04/2009(H1N1)、A/Vietnam/1203/2004(H5N1)、及びA/HongKong/1073/1999(H9N2)のHAタンパク質をターゲットとするアプタマーを取得し、新型インフルエンザウイルスの検査薬を提供する。
【解決手段】特定の塩基配列、又は当該塩基配列において、1若しくは数個の塩基が欠失、置換若しくは付加された塩基配列を含むRNA、又はさらに短鎖化されたRNAからなる、新型インフルエンザのHAタンパク質に対する結合能を有するアプタマーRNA。
【解決手段】特定の塩基配列、又は当該塩基配列において、1若しくは数個の塩基が欠失、置換若しくは付加された塩基配列を含むRNA、又はさらに短鎖化されたRNAからなる、新型インフルエンザのHAタンパク質に対する結合能を有するアプタマーRNA。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、新型インフルエンザウイルスのヘマグルチニンに結合能を有するアプタマーに関する。
【背景技術】
【0002】
インフルエンザウイルスにはA、B、及びCの3つの型があり、季節性インフルエンザとして毎年流行を起こすのはA型とB型であることが知られている。このうちA型インフルエンザウイルスは、ウイルス表面に突出したタンパク質であるヘマグルチニン(以下HAとする)とノイラミニダーゼ(以下NAとする)の組み合わせによって多くの亜型に分類されている。HAには、H1からH16の亜型、NAにはN1からN9の亜型が存在し、この組み合わせによって、例えばH1N1、H2N2、H3N2等と表現される。
【0003】
季節性インフルエンザウイルスとして毎年流行を繰り返しているのは、A型インフルエンザウイルスのH3N2亜型(香港型)、H1N1亜型(ソ連型)とB型インフルエンザウイルスであるが、これらは高い死亡率を示すほどの病原性は持たない。
【0004】
一方、季節性のインフルエンザウイルスとは別に、近年、新型インフルエンザの世界的流行が懸念されている。新型インフルエンザは、人類がこれまで感染したことのない抗原タンパク質を持っているため、多くの人々が免疫を持たず、高い確率で感染、発症し、パンデミックを引き起こす可能性がある。
新型インフルエンザの例として、1997年に香港で発生したH5N1亜型の高病原性鳥インフルエンザウイルスがあり、家禽類だけでなくヒトへと感染して死亡者をだして以来、H5N1亜型の鳥インフルエンザウイルスのヒトへの感染は現在まで続いている。また、H9N2亜型の鳥インフルエンザウイルスについても、ヒトへの感染が確認されている。今後、これらのウイルスがヒトからヒトへ容易に感染しうるウイルスへ変異する可能性もあり、その対策が急がれている。実際に、2009年にパンデミックを引き起こした、豚由来の新型インフルエンザウイルス(H1N1)は、ヒトからヒトへ感染し、その伝播力は季節性インフルエンザウイルスよりも強い。幸い病原性は高くはなかったが、将来高病原性へ変異する危険性をもっている。
【0005】
これらの新型インフルエンザウイルスが従来の季節性インフルエンザウイルスと大きく異なる点は、本来豚インフルエンザ、鳥インフルエンザとして、豚又は鳥というそれぞれの種内のみで流行を引き起こしていたウイルスが、種の壁を乗り越えてヒトへと感染し、ヒトとヒトとの間で感染するか、あるいは感染が懸念される点である。また、新型インフルエンザに対して免疫を持たない人類にとっては、ウイルスに暴露すると高い確率で発症することが考えられる。このような点において、新型インフルエンザウイルスは季節性インフルエンザウイルスとは大きく異なる。
また、インフルエンザウイルスにおいても、種々の型・亜型が存在して、それぞれ異なる性質を持ち、同じ亜型に属するウイルスでも株の違いによって抗原性が異なる場合もあるため、インフルエンザの予防対策や治療にとって、ウイルスの型・亜型・株の識別は非常に重要な課題である。
【0006】
現在、使用されている迅速診断キットは、インフルエンザウイルスの抗原を検出するキットが主流になっているが、A型かB型かの型の識別のみである。新型かどうかを区別するには、亜型・株の識別を行う必要がある。これにはインフルエンザウイルスの遺伝子を検出するRT-PCR法を用いた診断が必要であるが、設備や手間および費用の問題から日常臨床での使用には向いていない。したがって、詳しい検査が行われないまま発見が遅れると初期の段階での封じ込めを損ない、広い範囲にわたって流行が拡大するので、常にインフルエンザウイルスの有無と共に、型、亜型、株をルーチンでチェックできるようなシステムが求められる。
【0007】
一方、アプタマー、特にRNAアプタマーは複雑な3次構造をとることによって、標的化合物を高度に識別して結合することが可能である(非特許文献1)。したがって、アプタマーの高識別機能は近縁のタンパク質どうしや、あるいは微生物やウイルスの型、亜型や株どうしを識別するために大変価値のある道具として使える可能性がある。
アプタマーとは、機能性核酸のことであり、抗体と類似の機能をもつが、標的化合物として、イオンのような小さいものから、ウイルスのような巨大複合体まで対象となるので、抗体よりも利用範囲が広い。
【0008】
本発明者は、既に季節性インフルエンザであるインフルエンザA(H3N2)およびBのHAタンパク質に対するアプタマーを取得しており、これらのアプタマーは他の型のHAタンパク質のみならず株の違いをも識別することを報告し、季節性インフルエンザの有無と共に型、亜型、株を迅速かつ正確に判定できる診断手法を提供することができた。さらに該インフルエンザウイルスの宿主細胞表面への結合を妨げる作用をもつことも報告している。(非特許文献2,3)(特許文献1)。
【0009】
したがって、新型インフルエンザに対しても、同様の迅速かつ正確な診断手法を提供するための優れた識別能を有するアプタマーの開発が強く望まれていた。
【先行技術文献】
【特許文献】
【0010】
【特許文献1】特許第4441618号公報
【非特許文献】
【0011】
【非特許文献1】Jenison R.D.,Gill,S.C.,Pardi,A., and Polisky,B.1994.High-resolution molecular discrimination by RNA.Science 263,1425-1429.
【非特許文献2】Gopinath,S.C.B.,Misono,T.,Kawasaki,K.,Mizuno,T.,Imai,M.,Odagiri,T., and Kumar,P.K.R.2006.An RNA aptamer that distinguishes between closely related human influenza viruses and inhibits hemagglutinin-mediated membrane fusion.J.Gen.Virol.87,479-487.
【非特許文献3】Gopinath,S.C.B.,Sakamaki,Y.,Kawasaki,K. and Kumar,P.K.R.2006.An efficient RNA aptamer against human influenza B virus hemagglutinin.J.Biochem.139,837-846.
【非特許文献4】Kumar,P.K.R.,Machida,K.,Urvil,P.T.,Kakiuchi,P.N.,Vishnuvardhan,D.,Shimotohno,K.,Taira,K., and Nishikawa, S. 1997.Isolation of RNA aptamers specific to the NS3 protein of hepatitis C virus from a pool of completely random RNA.Virology 237,270-282.
【非特許文献5】Kumarevel,T.S.,Gopinath,S.C.B.,Nishikawa,S.,Mizuno,H., and Kumar,P.K.R.2004.Identification of important chemical groups of the hut mRNA for HutP interactions that regulate the hut operon in Bacillus subtilis.Nucleic Acids Res.32,3904-3912.
【発明の概要】
【発明が解決しようとする課題】
【0012】
本発明は、新型インフルエンザウイルス、特に、A/California/04/2009(H1N1)、A/Vietnam/1203/2004(H5N1)、及びA/HongKong/1073/1999(H9N2)のHAタンパク質をターゲットとするアプタマーを取得し、新型インフルエンザウイルスの検査薬を新たに提供しようとするものである。
【課題を解決するための手段】
【0013】
本発明者らは、上記課題を解決するために鋭意研究を行った結果、新型インフルエンザウイルスのA/California/04/2009(H1N1)、A/Vietnam/1203/2004(H5N1)、及びA/HongKong/1073/1999(H9N2)のHAタンパク質に結合するアプタマーを取得した。
得られたアプタマーと各ターゲットとの親和性及び相互作用解析、亜型または株との識別には、フィルター結合定量法、及び表面プラズモン共鳴装置(GEヘルスケアバイオサイエンス社製BiacoreT100)を用いた。
【0014】
A/California/04/2009(H1N1)をターゲットとした選別では、アプタマーD12とD26を取得した。アプタマーD12及びD26は、A/California/04/2009に対して高い親和性を示したが、
A/California/04/2009と同じ亜型H1N1に属するA/NewCaledonia/20/1999(H1N1)とA/Brevig mission/1/1918(H1N1)に対してはいずれも結合能を示さなかった。
【0015】
A/Vietnam/1203/2004(H5N1)をターゲットとした選別では、アプタマー8-1、8-3、及び8-10を取得した。アプタマー8-1、8-3、及び8-10は、A/Vietnam/1203/2004(H5N1)に対し、高い親和性を示した。さらに、同じ亜型(H5N1)に属するA/Indonesia/05/2005(H5N1)及びA/ Anhui/1/2005(H5N1)への親和性も調べたところ、アプタマー8-1、8-3、及び8-10は、これらの株にも結合能を有することが確認された。
これに対して、H5N1型以外の亜型との親和性を確認したところ、アプタマー8-3は、A/NewYork/55/2004(H3N2)、A/Netherlands/219/2003(H7N7)、及びA/HongKong/1073/1999(H9N2)に対しては結合しないか、結合したとしてもごくわずかであり、A/California/04/2009(H1N1)に対しては親和性を示したものの、そのKd値は30nMであり、A/Vietnam/1203/2004(H5N1)に対するKd値、50pMの600倍であった。すなわち、A/California/04/2009(H1N1)への親和性は、A/Vietnam/1203/2004(H5N1)に対するよりも600分の1程度であることが示された。
アプタマー8-1及び8-10においても、H5N1型以外の亜型との親和性は、いずれもA/California/04/2009(H1N1)、A/NewYork/55/2004(H3N2)、A/Netherlands/219/2003(H7N7)及びA/HongKong/1073/1999(H9N2)には結合しないか、結合したとしてもごくわずかであった。
【0016】
A/HongKong/1073/1999(H9N2) をターゲットとした選別では、アプタマー10-1を取得した。アプタマー10-1は、A/HongKong/1073/1999(H9N2)に対して高い親和性を示したが、他の亜型との親和性を調べたところ、A/NewYork/55/2004(H3N2)、A/Vietnam/1203/2004(H5N1)、A/Indonesia/05/2005(H5N1)、及びA/Netherlands/219/2003(H7N7)のいずれにも結合能を示さなかった。
そして、本発明のアプタマーを取得した方法を上記以外の新型インフルエンザウイルスあるいは今後出現してくる新型インフルエンザウイルスに対して適用することで、他の新型インフルエンザウイルスに特異的なアプタマーを取得することができる。
【0017】
さらに、本発明者らはアプタマー8-3を短鎖化して、アプタマー8-3Sを得た。これらはターゲットとするA/Vietnam/1203/2004(H5N1)のHAタンパク質に対し高い親和性を保持していた。
【0018】
また、アプタマーD12、D26、8-3SのRNAのリボース基の2’-OHをフルオロ基(2’-F)に置き換えることによって、より安定なアプタマーD12-2’FC&U、D26-2’FC&U、及び8-3S-2’FCを得ることができた。
以上の知見を得て本発明を完成した。
【0019】
すなわち、本発明は以下の発明を含むものである。
〔1〕 配列番号4〜10のいずれかで示される塩基配列、又は当該塩基配列において、1若しくは数個の塩基が欠失、置換若しくは付加された塩基配列を含むRNAからなることを特徴とする、新型インフルエンザのHAタンパク質に対する結合能を有するアプタマーRNA。
〔2〕 配列番号4〜9のいずれかで示される塩基配列、又は当該塩基配列において、1若しくは数個の塩基が欠失、置換若しくは付加された塩基配列を含むRNAが構成する2次構造において、HAタンパク質に対する結合能を有する先端領域部の構造を保持するように短鎖化された塩基配列からなるアプタマーRNA。
〔3〕 アプタマーRNAを構成するヌクレオチドのリボース部位の少なくとも1箇所が、化学修飾されていることを特徴とする、前記〔1〕及び〔2〕のいずれかに記載のアプタマーRNA。
〔4〕 前記化学修飾が、リボース部位の2’位に対するフルオロ基(2’-F)もしくはメトキシ基(2’-OMe)による修飾、又は水素による置換(2’-deoxy)であることを特徴とする、前記〔1〕及び〔2〕のいずれかに記載のアプタマーRNA。
〔5〕 アプタマーRNAの3’および/または5’末端がインバーストデオキシチミジン(idT)あるいはポリエチレングリコール(PEG)により修飾されていることを特徴とする、前記〔1〕〜〔4〕のいずれかに記載のアプタマーRNA。
〔6〕 前記〔1〕又は〔2〕に記載のアプタマーRNAと同一の塩基配列又は当該塩基配列と相補的な塩基配列を含み、かつ前記〔1〕又は〔2〕に記載のアプタマーに変換可能な一本鎖DNA、二本鎖DNA又は相補的RNA。
〔7〕 前記〔1〕〜〔5〕のいずれかに記載のアプタマーRNAを有効成分として含有する、新型インフルエンザウイルスの検出剤。
〔8〕 前記〔1〕〜〔5〕のいずれかに記載のアプタマーRNAを有効成分として含有する、新型インフルエンザウイルス用診断剤。
〔9〕 前記〔1〕〜〔5〕のいずれかに記載のアプタマーRNAを有効成分として含有する、新型インフルエンザウイルスのHAタンパク質の亜型又は株を特定するための診断用キット。
〔10〕 前記〔1〕〜〔5〕のいずれかに記載のアプタマーRNAを有効成分として用いることを特徴とする新型インフルエンザウイルスの検出方法。
〔11〕 前記〔1〕〜〔5〕のいずれかに記載のアプタマーRNAを有効成分として用いることを特徴とする、新型インフルエンザウイルスのHAタンパク質の亜型又は株を特定するための方法。
【発明の効果】
【0020】
本発明により、新型インフルエンザウイルスの型・亜型・株の同定のための検査薬として用いることができるアプタマーを提供できた。これらアプタマーを用いた検出法は、今までにはなかった新型インフルエンザウイルスに対する正確で迅速なルーチン検査を可能とするものである。また、本発明のアプタマーは、新型インフルエンザウイルスの宿主細胞表面への結合を妨げる作用をもつと考えられるので、新型インフルエンザウイルスの予防及び治療薬となる可能性が高い。
【図面の簡単な説明】
【0021】
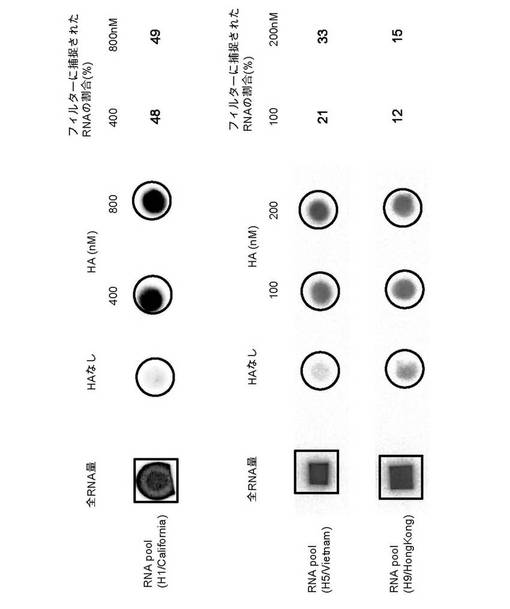
【図1】フィルター結合定量法で調べた第8回目の選別サイクル目におけるRNAプールと、各ターゲットHAタンパク質との結合能力を示す。四角及び丸で囲まれた内側の黒化度は標識RNAの濃度を示す。括弧内はターゲットタンパク質を示す。
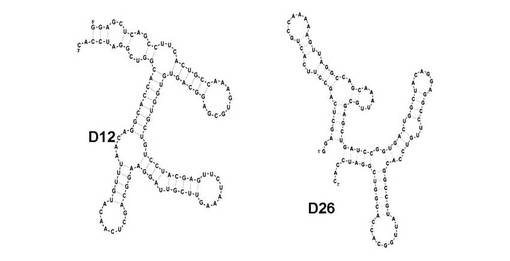
【図2】第10回目の選別サイクルで得られたA/California/04/2009(H1N1)に対するアプタマーの二次構造モデルを示す図である。
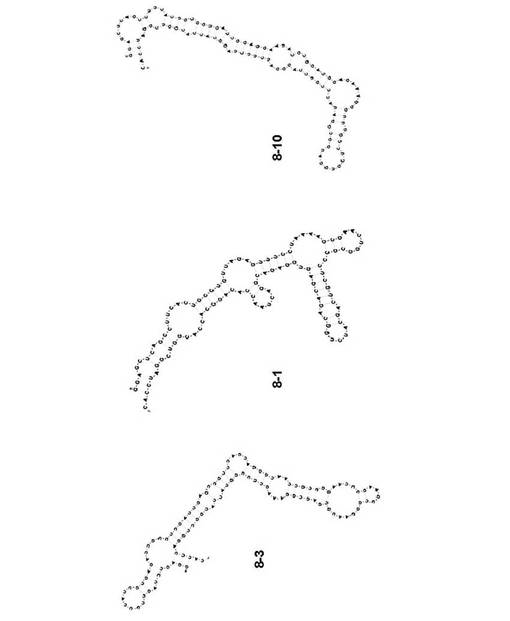
【図3】第10回目の選別サイクルで得られたA/Vietnam/1203/2004(H5N1)に対するアプタマーの二次構造モデルを示す図である。
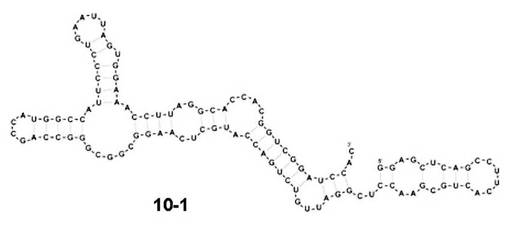
【図4】第10回目の選別サイクルで得られたA/HongKong/1073/1999(H9N2)に対するアプタマーの二次構造モデルを示す図である。
【図5】フィルター結合定量法で調べた、アプタマーD12及びD26とA/California/04/2009(H1N1)のHAタンパク質との親和性の解析図である。四角及び丸で囲まれた内側の黒化度は標識RNAの濃度を示す。
【図6】フィルター結合定量法で調べた、アプタマー8-1、8-3及び8-10とA/Vietnam/1203/2004(H5N1)のHAタンパク質との親和性の解析図である。四角及び丸で囲まれた内側の黒化度は標識RNAの濃度を示す。
【図7】フィルター結合定量法で調べた、アプタマー10-1とA/HongKong/1073/1999(H9N2)のHAタンパク質との親和性の解析図である。四角及び丸で囲まれた内側の黒化度は標識RNAの濃度を示す。
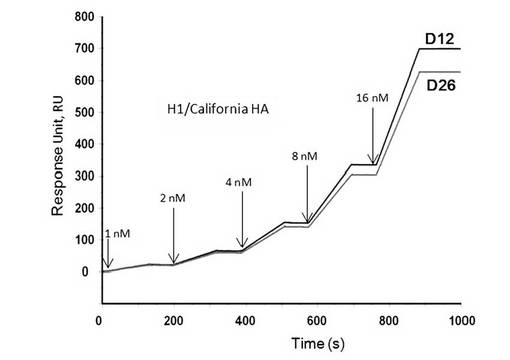
【図8】表面プラズモン共鳴法(シングルサイクルカイネティクス法)によって測定したアプタマーD12及びD26とA/California/04/2009(H1N1)のHAタンパク質との相互作用の解析図である。
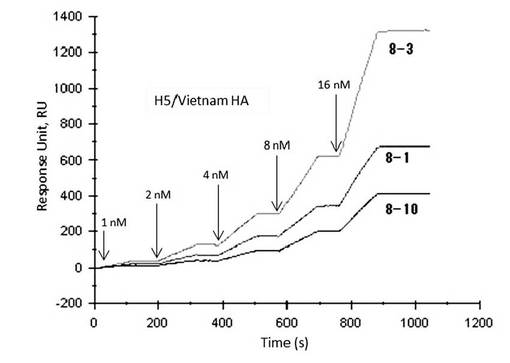
【図9】表面プラズモン共鳴法(シングルサイクルカイネティクス法)によって測定したアプタマー8-1、8-3、及び8-10とA/Vietnam/1203/2004(H5N1)のHAタンパク質との相互作用の解析図である。
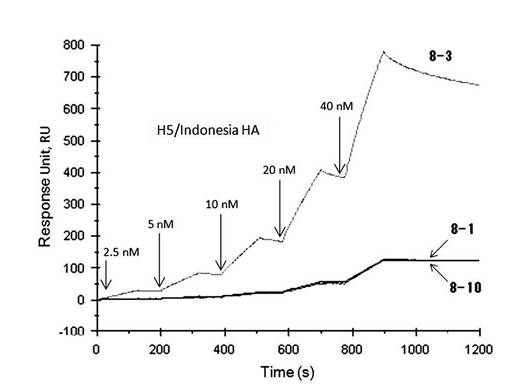
【図10】表面プラズモン共鳴法(シングルサイクルカイネティクス法)によって測定したアプタマー8-1、8-3、及び8-10とA/Indonesia/05/2005(H5N1)のHAタンパク質との相互作用の解析図である。
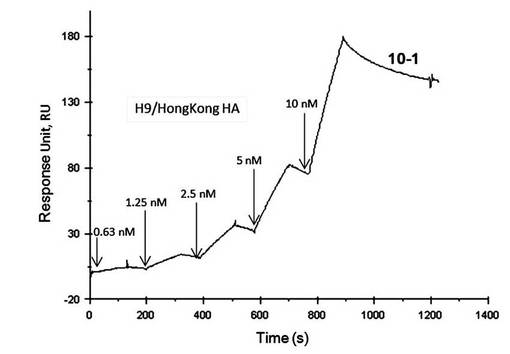
【図11】表面プラズモン共鳴法(シングルサイクルカイネティクス法)によって測定したアプタマー10-1とA/HongKong/1073/1999(H9N2)のHAタンパク質との相互作用の解析図である。
【図12】短鎖化アプタマー8-3Sの二次構造モデルを示す図である。
【図13】表面プラズモン共鳴法(シングルサイクルカイネティクス法)によって測定したアプタマー8-3S及び8-3とA/Vietnam/1203/2004(H5N1)のHAタンパク質との相互作用の解析図である。
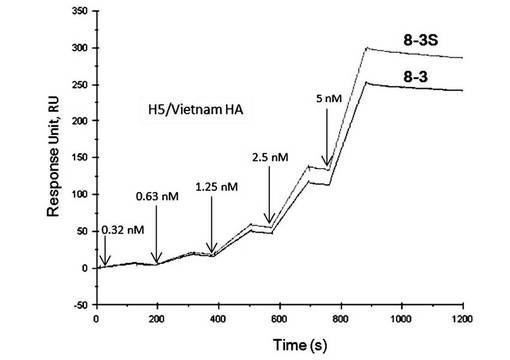
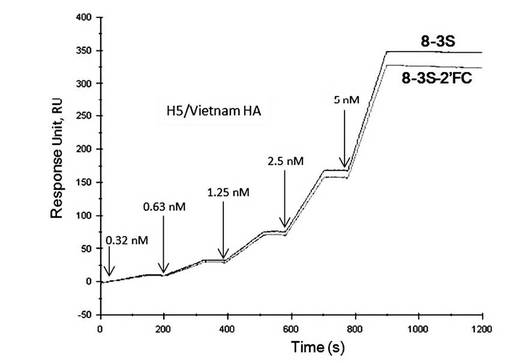
【図14】表面プラズモン共鳴法(シングルサイクルカイネティクス法)によって測定した、2’フルオロ化したアプタマー8-3S-2’FCとA/Vietnam/1203/2004(H5N1)のHAタンパク質との相互作用の解析図である。
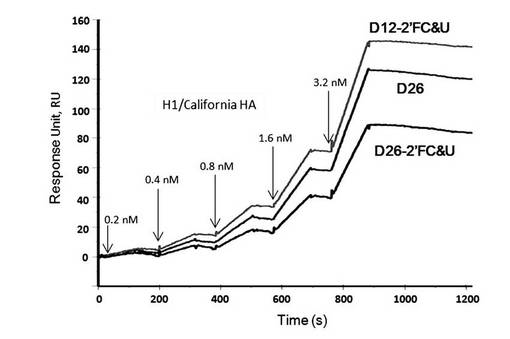
【図15】表面プラズモン共鳴法(シングルサイクルカイネティクス法)によって測定した、2’フルオロ化したアプタマーD12-2’FC&U及びD26-2’FC&UとA/California/04/2009(H1N1)のHAタンパク質との相互作用の解析図である。
【発明を実施するための形態】
【0022】
1.本発明のアプタマーについて
(1)本発明の「アプタマー」とは
「アプタマー」とは、一般に、ある標的、例えばウイルス、タンパク質、ペプチド、糖類、金属イオン、小分子等に特異的に結合するように人工的に創製された核酸リガンドをいうが、本発明において用いる「アプタマー」は、新型インフルエンザウイルスのHAタンパク質に特異的な結合能を有する。核酸としてはRNAが好ましく、RNAを構成するリボース基が化学的に修飾されている場合も含む。
本発明のアプタマーは、周知のSELEX法(systematic evolution of ligands by exponential enrichment)を適用し、中央の74塩基部分をランダム化したRNAライブラリーから得られたクローン由来のものであって、典型的には配列番号4〜10に示される塩基配列からなるRNAが挙げられる。本発明においては、ランダム領域を比較的長い74塩基配列としたことによって、より複雑な3次構造ができ、高度な識別が可能なアプタマーを提供することができたものと考えられる。
【0023】
(2)「アプタマー」の構造
本発明のアプタマーの2次構造モデルは、例えば図2〜4などに示されるように、先端のループ構造とステム構造とループ構造の繰り返しが続いたステム・ループ構造となっている。実際にインフルエンザウイルスのHAタンパク質抗原と結合する部位は、ランダム領域からなる領域で、主にこのうちの先端ループを含むステム領域であると考えられるため、当該先端部領域の構造を壊さない範囲でRNA鎖を短縮化することもできる。
【0024】
(3)本発明のアプタマーを構成する核酸
本発明においては、上記配列番号で示されるRNAに限らず、これらRNAと対応する配列を有する、DNA及び、これらDNAと相補のDNA、あるいはこれら相補のDNA鎖同士が2本鎖を構成したdsDNAを包含する。さらに、上記RNAと相補の塩基配列を有するRNAも包含する。これらはそれ自体慣用の遺伝子操作手段により、たやすく本発明のアプタマーRNAに変換可能であり、本発明のアプタマー製造用中間体である。また、本発明においては、上述のように、典型的な配列番号4〜10に示された塩基配列と共に、当該塩基配列中のヌクレオチド残基を1若しくは数個欠失、置換、あるいは付加させた改変アプタマーであっても、上記HAタンパク質と結合するものであればこれを包含する。また、後述のように、RNAを構成するリボース基を化学修飾させた修飾アプタマーも包含される。
【0025】
(4)本発明のアプタマーの認識する部位とその親和性
新型インフルエンザウイルス粒子表面も季節性のA型インフルエンザウイルス粒子表面と同様、それぞれ約900コピーのヘマグルチニン(HA)と約300コピーのノイラミニダーゼ(NA)が存在すると言われている。本発明のアプタマーRNAには新型インフルエンザウイルスのHAタンパク質に結合する能力がある。
本発明のアプタマーが認識する部位は、新型インフルエンザのHAタンパク質の表面に突出した領域であり、且つ、アミノ酸の変異が認められる領域であると考えられる。
また、親和性に関する実験は、後述で示すフィルター結合定量法、及び表面プラズモン共鳴装置(GEヘルスケアバイオサイエンス社製BiacoreT100)を用いて行い、本発明のアプタマーの、新型インフルエンザHAタンパク質に対する親和性は高く、且つ特異的である。(図5−11、図13−15)。
【0026】
2.本発明のアプタマーRNAの製造方法
(1)SELEX法
本発明のアプタマーは、それ自体周知のSELEX法により得られたものである。SELEX法は、ランダム配列の核酸ライブラリーの作成から出発し、標的とするタンパク質との結合性を指標に選別して、PCR増幅するサイクルを複数回繰り返すもので、これにより、標的とするタンパク質に対し高い結合能を有する核酸のみを選別することができる。上記選別及びPCR増幅はそれぞれ、自然界における「淘汰」及び「増殖」に相当し、自然界における進化を試験管内で短時間に再現させるものである。
【0027】
本発明においては、試験管内選択法によって合計10サイクルの選別を行い、その結果得られた特異的なウイルスに結合するRNAプールについてクローニングを行った。選別を経たRNAプールには各ターゲットに高い親和性を有するアプタマーが含まれていた。
具体的には、配列中央の74塩基をランダム領域とするDNAライブラリー(配列番号1)を合成し、PCR増幅した後、転写してRNAプールとし、このRNAプールに対し、上記HAタンパク質と結合するRNAを選別、増幅し、このサイクルを複数回繰り返すことにより、該HAタンパク質に対して特異性及び結合性の高い、配列番号4から9に示されるアプタマーを得たものであり、本発明においては配列番号6に示すアプタマー(アプタマー8-3)をさらに短鎖化して、配列番号10に示される短鎖化アプタマー(アプタマー8-3S)を得ており、これはHAタンパク質に対する結合能を保持している。
【0028】
(2)その他の方法
本発明のアプタマーは、上記したとおり、基本的にはSELEX法により得たものではあるが、本発明においては、アプタマーの塩基配列を明らかにしており、SELEX法によらずとも、この塩基配列から本発明のアプタマーを合成することができる。これには、例えば、それ自体周知の以下の方法を用いることができる。
インビトロ転写法:合成目的のアプタマーRNAに対応する上記DNAを化学合成し、これをPCR増幅し、増幅されたDNAからRNAポリメラーゼによる転写反応によりアプタマーRNAを合成する。例えば、T7プロモーターの下流側に上記DNAを連結してPCR増幅し2本鎖DNAを得て、該2本鎖DNAを鋳型として、T7RNAポリメラーゼ、およびATP、GTP、CTP及びUTPからなる該RNAポリメラーゼ基質を含む反応溶液中で、RNA伸長反応を行うことにより、アプタマーRNAを得ることができる。
【0029】
この方法においては、T7プロモーター配列を含むプラスミドに上記dsDNAを連結したもの、あるいは化学合成したDNAも鋳型として用いることができる。また、上記DNAの合成においては従来公知の種々の方法で合成することが可能である。例えば、DNA合成機を用いて、dNTPを材料として、末端塩基から化学合成することができる。
化学合成法:RNAの合成には、リボース部位の2’水酸基の保護が必要であり、保護基として種々のアミダイトが開発されてきており、2-cyanoethoxymethyl(CEM)基を用いるとRNA合成時の鎖長伸長反応の効率が高くなり、100merを超える長鎖RNAの合成も可能となる。本発明のアプタマーRNAはこのような化学合成法を用いて合成することができる。
また、このほか、上記アプタマーRNAと相補のRNAからは、RNA依存RNAポリメラーゼを使用することにより本発明のアプタマーRNAを得ることも可能である。
【0030】
3.アプタマーRNAの短縮化
アプタマーの短縮化の主な目的は、アプタマーRNAの製造コストを削減することである。本発明のアプタマーは、HAタンパク質との結合に関与するコア領域となるステム・ループ構造を残して、HAタンパク質との結合活性を保持したまま短鎖化することができた(アプタマー8-3S;図12)。
本発明のアプタマーの短鎖化は、配列番号4〜9に示されるアプタマーに対して、一般的なアプタマーの短鎖化方法であるマッピング法(非特許文献2、特許文献1)に従って行うことができる。また、本発明の実施例5において短鎖化の対象とした配列番号6のアプタマー8-3(図3)のように、比較的単純な2次構造をとっていて、先端部のHA結合領域の予測がつきやすい場合は、マッピング法実験を省略することができる。
【0031】
4.アプタマーRNAの化学修飾
本発明のアプタマーRNAにおいては、リボヌクレアーゼ耐性を付加するために、その配列中に化学修飾(modified)したヌクレオチドを有していても良い。このようなアプタマーRNAは、例えば、アプタマーRNA中のヌクレオチドのリボース部位の2’-OH基を、常法によりフロオロ基(2’-F)にまたはメトキシ基(2’-OMe)に、あるいは同リボース部位の2’位を水素に置換(2’-deoxy)することにより得られる。この化学修飾は、ピリミジンヌクレオチド部位がリボヌクレアーゼにより分解されやすいので、ピリミジンヌクレオチドに対してより有効である。
このような化学修飾は、ヘマグルチニンとの結合に関与する部位のヌクレオチドに対しては行わない。それ以外のヌクレオチドであれば全て修飾することも可能であるが、一般的にはループ領域のピリミジンヌクレオチドのリボース基について行う。
【0032】
本発明においては、さらにアプタマーRNAの3’及び/又は5’末端をインバーストデオキシチミジン(idT)あるいはポリエチレングリコール(PEG)により修飾することもでき、これらにより、アプタマーRNAのリボヌクレアーゼ耐性はさらに向上し、生体中での分解による活性の低下を抑制することができる。され、生体内で有効な抗ウイルス作用を発揮させることが可能となる。
【0033】
5.アプタマーRNAを用いた新型インフルエンザウイルスの検出法
(1)標識化されたアプタマーRNAを用いる方法
本発明のアプタマーRNAは新型インフルエンザウイルスの検査薬の検出剤として用いられる。例えば、本発明のアプタマーRNAをフルオロセイン、ローダミン、テキサスレッド等の蛍光色素で標識し、被験試料と接触、結合反応を行い、結合しなかったものを除去した後、蛍光あるいはその強度を検出、測定することにより、新型インフルエンザウイルスの検出、あるいはその量を測定できる。この際、例えば、標識アプタマーRNAあるいは被験試料のいずれかを固定化した基板を用いて行うことができる。
また、結合時に蛍光発光する物質を用いて、本発明のアプタマー及び被験試料のいずれかを標識して、結合時の蛍光あるいはその強度を検出、測定することにより行うことも可能である。このような蛍光色素としては、例えば、フルオロセイン、ローダミン、テキサスレッド等が挙げられる。
【0034】
(2)表面プラズモン共鳴法(SPR)
また、本発明においては、蛍光色素を標識せずに、新型インフルエンザウイルスを検出することも可能である。これには、例えば、表面プラズモン共鳴法(SPR)がある。すなわち、分子間の結合反応を、センサー表面でおこる分子間の結合時の微細な質量変化を表面プラズモン共鳴とよばれる光学現象を採用することで測定することができる。SPR法を利用した装置として、GEヘルスケアバイオサイエンス社製BiacoreT100がある。測定はセンサーチップの金薄膜表面に本発明のアプタマーRNAあるいは被験試料のいずれかを周知の方法で固定化し、これに被験試料あるいはアプタマーRNAを供給し、結合反応をリアルタイムでモニタリングしながら行う。
【0035】
6.本発明のアプタマーを用いた診断方法及び診断用キット
本発明は、新型インフルエンザウイルスで発現するHAタンパク質に結合するアプタマーを、新型インフルエンザウイルスを含有するか、または含有する可能性のあるサンプルと接触させることによって、サンプル中に目的の新型インフルエンザウイルスが存在すること、あるいは存在しないことが確認できる。
また、本発明においては、新型インフルエンザウイルスの特定の亜型及び/又は株で発現するHAタンパク質と特異的に結合するアプタマーを提供することができたので、これらアプタマーを用いて、新型インフルエンザウイルスの特定の亜型及び/又は株を識別し、判定することができる。
そして、本発明は、新型インフルエンザウイルスの特定の亜型及び/又は株で発現するHAタンパク質と特異的に結合するアプタマーを備えた新型インフルエンザウイルス診断用キットを提供する。
診断剤、診断用キットを製造するに際しては、適宜、周知の薬学的に許容可能な希釈剤、安定化剤、その他の担体などと組み合わせて用いる。
本発明の診断用キットの対象となるサンプルとしては、例えば、被験者の咽頭拭い液、鼻汁等の採取検体等や、それら検体を培養細胞やニワトリ受精卵に与えて感染・増殖の工程によって得た溶液等が挙げられる。
また、本発明の方法により、新型インフルエンザウイルスの特定の亜型及び/又は株を識別し、判定することができるから、例えばヒトへの感染やパンデミックが発生する前に、豚や鳥類において実際に流行しているインフルエンザウイルスの亜型及び/又は株をモニタリングし、将来のヒトへの感染やパンデミック予測に用いることができる。
【0036】
本発明におけるアプタマーは、抗体に匹敵する特異性を有し、現在新型インフルエンザウイルスの検出のために使用されている抗体を用いた迅速診断キットと同様の用途に使用することができる。臨床上、新型インフルエンザであるか、又は単なる季節性インフルエンザ、感冒等のその他のかぜ症候群の疾患であるかを迅速に診断することは、治療上、また予防上においても非常に重要である。また、インフルエンザであっても、ウイルスの型により、また亜型により、症状が異なり、種類が異なることでワクチンの効果が得られない場合もあり、治療に用いる医薬の決定、ワクチンの選択等においても、その識別は重要な意味を有する。更に、新型インフルエンザの流行状況をモニタリングすることで、将来の流行予測、ワクチンの製造の上でも有用な情報を得ることができ、更に研究用の試薬としても利用することができる。
【0037】
使用の際には、上記のようにして得られた1種のアプタマーを単独で用いても良いが、場合によっては、同じウイルスまたはタンパク質に結合し得るアプタマー、または異なるウイルスまたはタンパク質に結合し得るアプタマーを複数混合して用いても良い。
【0038】
7.抗ウイルス剤、又は新型インフルエンザウイルスの予防もしくは治療用組成物
本発明のアプタマーは、新型インフルエンザウイルスのHAタンパク質の表面にある突出領域に特異的に結合し、ウイルスの細胞への結合を阻害すると考えられる。特に化学修飾されたアプタマーはヌクレアーゼ抵抗性で、血中半減期も長いばかりか、生体内への抗原性も無視できるので、各種の抗ウイルス製剤として、また新型インフルエンザの予防用または治療用の医薬組成物として利用する事も可能である。投与対象はヒトのみならず、鳥や豚などに適用することができる。
抗ウイルス製剤、または医薬組成物とする場合には、経口、非経口のいずれでもよく、適宜、周知の薬学的に許容可能な無毒性の担体、希釈剤と組み合わせて用いる。非経口投与としては、典型的には注射剤であるが、噴霧剤などと共に吸入による投与も可能である。
また、直接ヒトなどに投与せずに、抗ウイルス剤として、マスクや看護用の衣服などに含浸させて用いることもできる。
【実施例】
【0039】
以下に本発明の一実施形態として、新型インフルエンザウイルスのうちで、A/California/04/2009(H1N1)、A/Vietnam/1203/2004(H5N1)、及びA/HongKong/1073/1999(H9N2)に結合するアプタマーの選別例を挙げ、下記実施例において具体的に説明するが、本発明は以下の実施例に限定されるものではない。
なお、本発明で使用されている技術的用語は、別途定義されていない限り、当業者により普通に理解されている意味を持つ。本発明の実施例で用いた遺伝子組換え技術、PCR法、その他の手法などの具体的な手順や条件は、特に断らない限り、Sambrook and Russell,Molecular Cloning:A Laboratory Manual,3rd Edition.Cold Spring Harbor Laboratory Press,Plainview,NY(2001)、Cold Spring Harbor Laboratory Press,Cold Spring Harbor,New York(1989);D.M.Glover et al.ed.,"DNA Cloning",2nd ed.,Vol.1 to 4,(The Practical Approach Series),IRL Press,Oxford University Press(1995);Ausubel,F.M.et al.,Current Protocols in Molecular Biology,John Wiley & Sons,New York,N.Y,1995;日本生化学会編、「続生化学実験講座1、遺伝子研究法II」、東京化学同人 (1986);日本生化学会編、「新生化学実験講座2、核酸III(組換えDNA技術)」、東京化学同人(1992);R.Wu ed.,"Methods in Enzymology",Vol.68(Recombinant DNA),Academic Press,New York(1980);R.Wu et al.ed.,"Methods in Enzymology",Vol.100(Recombinant DNA,PartB) & 101(Recombinant DNA,Part C),Academic Press,New York(1983);R.Wu et al.ed.,"Methods in Enzymology",Vol.153(Recombinant DNA,Part D),154(Recombinant DNA,Part E) & 155(Recombinant DNA,Part F),Academic Press,New York(1987)などに記載の方法あるいはそこで引用された文献記載の方法またはそれらと実質的に同様な方法により行うことができる。
また、本発明で引用した先行文献又は特許出願明細書の記載内容は、参照して本明細書の記載として組み入れるものとする。
【0040】
(実施例1) A/California/04/2009(H1N1)、A/Vietnam/1203/2004(H5N1)、及びA/HongKong/1073/1999(H9N2)に特異的なアプタマーのインビトロでの選別
(1−1)RNAランダムプールの作成
以下に示す、中央の74塩基をランダム領域とする一本鎖DNA(ssDNA)のライブラリーを合成してテンプレートとし(配列番号1)、5’末端プライマー(配列番号2)、及び3’末端プライマー(配列番号3)を用いてPCRを行った。
〔テンプレート〕
5'-GGAGCTCAGCCTTCACTGC-(N)74-GGCACCACGGTCGGATCCAC-3'(配列番号1)
〔5’末端プライマー〕
5'-TCTAATACGACTCACTATAGGAGCTCAGCCTTCACTGC-3'(配列番号2)
〔3’末端プライマー〕
5'-GTGGATCCGACCGTGGTGCC-3'(配列番号3)
【0041】
次いでT7 Ampliscribe kit(Epicentre Technologies社製)を用いて、インビトロでの転写を行い、増幅されたDNAライブラリーをRNAライブラリーに変換した。
【0042】
(1−2)インビトロにおける選別
上記(1)で得られたRNAライブラリー(16μg)を結合用緩衝液(0.01M HEPES,0.15M NaCl,pH7.4)に溶解させ、474≒3.57×1044通りの異なるRNA配列のコンフォメーションの平衡を促進するために、95℃で2分間処理して変性させた後に、室温で10分間冷却させた(アニーリング)。非特異的に結合するRNAを除去するためコンペティターとしてtRNA(E.coliのトータルtRNA(Boehringer-Mannheim社製))を加えた後、ターゲットとなるHAタンパク質を加えた。ターゲットとして選別に用いたHAタンパク質は、A/California/04/2009(H1N1)、A/Vietnam/1203/2004(H5N1)、及びA/HongKong/1073/1999(H9N2)である。各選別サイクルで用いたRNA、タンパク質、tRNAの分子比は表1に示すとおり。
【0043】
これらの混合液、50μLを室温で10分間インキュベートした後、”Pop-top”フィルターホルダー(Nucleopore社製)に装着した湿潤済みニトロセルロース・アセテート・フィルター(HA WPフィルター,0.45μm,径13.0mm,Millipore)に通し、タンパク質に結合したRNAをフィルター上に捕捉させて、フィルターを1mlの結合用緩衝液で洗浄した。フィルター上に残留したタンパク質結合RNAは溶出用緩衝液(0.01M HEPES,0.15M NaCl,7M Urea,pH7.4)で回収し、エタノールで沈殿精製後、50mM Tris-HCl(pH8.3),50mM KCl,10mM MgCl2,0.5mM Spermidine,10mM DTT,0.4μMプライマー(配列番号3),0.4mM dNTPs,25U AMV逆転写酵素(Wako社製)を含む20μlの反応液中で逆転写した。dNTPsと逆転写酵素は、変性及びアニーリングステップ(95℃で2分間処理後、室温で5分間インキュベート)の後、加えた。逆転写は37℃で45分間行った。
【0044】
<表1>

【0045】
得られたcDNAはPCRにより増幅され、次の選別ラウンドにおけるRNAを得るための鋳型として用いられた。PCRによる増幅のために、逆転写後の混合液(cDNA反応液)20μlを80μlのPCR用混合液(PrimeSTAR Max Premix(TaKaRa社製),1μMプライマー,1μMプライマー)で希釈した。反応液は95℃で30秒間加熱後、95℃、15秒;-54℃、10秒;-72℃、10秒のサイクルを、適正サイズで産物のバンドが得られるまでの回数(12-18サイクル)繰り返した。得られたPCR産物はエタノールで沈殿させて転写に用いた。試験管内転写反応はT7 Ampliscribeキットを用いて、37℃、3時間またはオーバーナイトで行った。RNA合成とDNase I処理の後、反応液を8%変性ポリアクリルアミドゲルで分画した。RNAをゲルから抽出し、エタノール沈殿により精製した後に定量し、次回の選別及び増幅サイクルに用いた。
【0046】
(1−3)選別及び増幅サイクル
以下においては、HAタンパク質に対する特異性と高い親和性を有するRNAを得るために、各選別サイクルを行った。表1に示したようにRNAとタンパク質比、コンペティター濃度は各選別サイクルごとに修正した。
非特異的に結合するRNAの濃縮を避けるために、第2、第4、第6及び第8の各選別サイクルでは、フィルターではなく96穴タイタープレート(Xenobind plate;xenopore社製)を使用した。Xenobind選別のために、最初に結合用緩衝液1mlあたり上記HAタンパク質10μgで各ウェルをコートし、残りの部分をBSA(0.1% stock solution)でブロックした。次いで、各ウェルを洗浄し、選別に使用した。
【0047】
前のサイクルで得たRNAプールを、結合用緩衝液中で95℃、2分間変性させた。続いて、室温で10分間冷却した後、tRNAを加え、HAタンパク質でコートされたウェルに充填した。10分間インキュベートした後、結合用緩衝液400μl(第6及び第8回目の選別サイクルでは1mL)で洗浄し、未吸着のRNAを除いた。その後、タンパク質結合RNAを、加熱した溶出用緩衝液(0.01M HEPES,0.15M NaCl,7M Urea,pH7.4)で回収し、エタノールで沈殿精製後、逆転写、PCR及びインビトロにおける転写により再生した。
【0048】
高親和性のアプタマーが濃縮されていく進行状況とその特異性を評価するため、第4回目の選別サイクルと第8回目の選別サイクルにおけるRNAプールの結合活性をフィルター結合定量法で解析した。[α-32P]ATPを使用して放射性標識(ラベル)したRNAを調製し、以前報告した方法(非特許文献3,4,5)に従ってフィルター結合分析を行った。結合反応では、モル濃度で10倍過剰の大腸菌tRNAを非特異的競争阻害剤として加え、RNAとHAタンパク質を1対4のモル比で混合し結合させた後、フィルターを通した。フィルターは1mlの上記結合用緩衝液で洗浄し、空気中で乾燥し、フィルターに残留した標識RNAの放射活性を画像解析装置(BAS2500,富士フイルム社製)で定量した。結合活性は、反応液に入れたRNA全量の内、フィルター結合したRNA量のパーセンテージで表した(図1)。第8回目の選別サイクル後のフィルターに捕捉されたRNAの割合は、A/California/04/2009(H1N1)のHAタンパク質をターゲットとした場合、HAタンパク質濃度400nM及び800nMにおいて、48%及び49%、A/Vietnam/1203/2004(H5N1)のHAタンパク質をターゲットとした場合、HAタンパク質濃度100nM及び200nMにおいて21%及び33%、さらに、A/HongKong/1073/1999(H9N2)の場合では、HAタンパク質濃度100nM及び200nMにおいて12%及び15%であった。
【0049】
(1−4)アプタマーの分析
個別のアプタマーを得るために、第10回目の選別サイクルで得たPCR産物をTAクローニング・ベクター(Invitrogen社製)に導入し、サブクローニングした後、大腸菌にトランスフォームした。プラスミドDNAの個々のクローンをプラスミド精製キット(Promega社製)で単離し、DNA塩基配列をDye Terminator Sequencing kit(Applied Biosystems Inc.社製)とDNAシークエンサー(Model 373A,Applied Biosystems Inc.社製)を用いて配列を解読し、そのDNA塩基配列に相当するRNA塩基配列を求めた。配列が解読されたRNAのクローンは配列によって幾つかのクラスに分類された。
【0050】
A/California/04/2009(H1N1)をターゲットとして選別されたRNAの塩基配列は、3つの主要なクラスに分類された。このうち、アプタマーD12(配列番号4)は、全体の19%を占めるクラス2に分類され、また、アプタマーD26(配列番号5)は、全体の9%を占めるクラス3に分類された。二次構造解析ソフト「BayesFold」(version 1.01、http://bayes.colorado.edu/Bayes/上で解析可能)を用いて得られたアプタマーの二次構造予測を行ったところ、それぞれ異なる二次構造の形成が予測された(図2)。
【0051】
A/Vietnam/1203/2004(H5N1)をターゲットとして選別されたRNAの塩基配列は、大きく3つのクラスに分類された。クラス1は、全体の42.9%を占め、配列番号6で示す塩基配列を得た(アプタマー8-3)。また、クラス2は全体の25.7%、クラス3は11.4%を占め、各塩基配列を、アプタマー8-1(配列番号7)及び8-10(配列番号8)とした。BayesFoldを用いてアプタマー8-1、8-3、及び8-10の二次構造予測を行ったところ、それぞれ異なる二次構造の形成が予測された(図3)。
【0052】
A/HongKong/1073/1999(H9N2)をターゲットとして選別されたRNAの塩基配列は、3つの主要なクラスを含み、このうち最も大きい割合を占めたクラス1は、全体の26.5%であり、この塩基配列をアプタマー10-1(配列番号9)とした。BayesFoldを用いてアプタマー10-1の二次構造予測を行ったところ、図4に示す二次構造が得られた。
【0053】
(実施例2) フィルター結合定量法によるアプタマーと各ターゲットHAタンパク質との親和性解析
実施例1で選別されたアプタマーとHAタンパク質との結合効率をフィルター結合定量法(非特許文献2に記載の方法による。)によって求めた。
【0054】
A/California/04/2009(H1N1)をターゲットとして選別されたアプタマーD12及びD26は、A/California/04/2009(H1N1)のHAタンパク質濃度に依存した結合がみられた。HAタンパク質濃度100nM及び200nMにおける、フィルターに捕捉されたRNAの割合は、D12が40%及び58%、D26が39%及び66%であった(図5)。
【0055】
A/Vietnam/1203/2004(H5N1)をターゲットとして選別されたアプタマー8-1、8-3及び8-10はA/Vietnam/1203/2004(H5N1)のHAタンパク質濃度に依存した結合がみられた。HAタンパク質濃度100nM及び200nMにおける、フィルターに捕捉されたRNAの割合は、8-1が12%及び25%、8-3が13%及び25%、8-10が23%及び41%であった(図6)。
【0056】
A/HongKong/1073/1999(H9N2)をターゲットとして選別されたアプタマー10-1はA/HongKong/1073/1999(H9N2)のHAタンパク質濃度に依存した結合がみられ、HAタンパク質濃度100nM及び200nMにおける、フィルターに捕捉されたRNAの割合は、それぞれ4%及び9%であった(図7)。
【0057】
(実施例3) BiacoreT100による各アプタマーとターゲットHAタンパク質との相互作用解析
実施例1で選別されたアプタマーとHAタンパク質との親和性や特異性をBiacoreT100(GEヘルスケアバイオサイエンス社製)によって測定した。
【0058】
HBS-P+緩衝液(0.01M HEPES,0.15M NaCl,0.05% v/v Surfactant P20,pH7.4)に溶解した5μMのビオチン化オリゴdT(24ヌクレオチド長)[5'-(dT)24-3']を、ストレプトアビジンでコートされたセンサーチップ(Series S Sensor Chip SA Certified,GEヘルスケアバイオサイエンス社製)に流速10μl/minで、2分間添加し、固定化を行った。次いで、3'-端に24ヌクレオチド長のオリゴ・アデニン(A)を延長させたアプタマー(30-60μM)を流速2μl/minで10分間添加し、オリゴdTに結合させた。
【0059】
上記のSAチップへ0.2-40nMの濃度範囲のHAタンパク質溶液を流速30μl/minで2分間添加し、固定化されたアプタマーとの相互作用解析を行った。
【0060】
A/California/04/2009(H1N1)をターゲットとして選別されたアプタマーD12及びD26はA/California/04/2009(H1N1)のHAタンパク質濃度に依存した結合が見られ、いずれも高い親和性を示した(図8)。
【0061】
A/Vietnam/1203/2004(H5N1)をターゲットとして選別されたアプタマー8-1、8-3、及び8-10は、A/Vietnam/1203/2004(H5N1)のHAタンパク質濃度に依存した結合が見られ、いずれも高い親和性を示した(図9)。
【0062】
さらに、アプタマー8-1、8-3、及び8-10は、A/Vietnam/1203/2004(H5N1)と同じ亜型に属するA/Indonesia/05/2005(H5N1)(図10)及びA/Anhui/1/2005(H5N1)にもまた結合することが確認された。
【0063】
A/HongKong/1073/1999(H9N2)をターゲットとして選別されたアプタマー10-1は、A/HongKong/1073/1999(H9N2)のHAタンパク質濃度に依存した結合が見られ、高い親和性を示した(図11)。
【0064】
また、各アプタマーの結合は、糖鎖を除去していないHAタンパク質に対しても糖鎖を除去したHAタンパク質に対してもほぼ同等の親和性を示したので、HAタンパク質の糖鎖はこのRNAアプタマーとの結合には関与していないことが示された。
本研究において選別されたアプタマーの種類と各ターゲットとの解離定数(Kd)値は表2に示す通りである。
【0065】
<表2>
【0066】
(実施例4) BiacoreT100による各アプタマーの特異性解析
(4−1)異なる亜型との相互作用解析
選別されたアプタマーと、そのターゲットとは異なる亜型のHAタンパク質との相互作用を調べるために、BiacoreT100を利用して解析を行った(表3)。
【0067】
A/Vietnam/1203/2004(H5N1)をターゲットとして選別されたアプタマー8-3と、ターゲットとは異なる亜型のHAタンパク質との相互作用を調べたところ、A/NewYork/55/2004(H3N2)、A/Netherlands/219/2003(H7N7)及びA/HongKong/1073/1999(H9N2)に対しては、結合しないか、結合したとしてもごくわずかであった。一方、A/California/04/2009(H1N1)に対しては親和性を示したものの、そのKd値は30nMであり、A/Vietnam/1203/2004(H5N1)に対するKd値、50pMの600倍であった。すなわち、A/California/04/2009 (H1N1)への親和性は、A/Vietnam/1203/2004(H5N1)に対するよりも600分の1程度であることが示された。
【0068】
同様に、A/Vietnam/1203/2004(H5N1)をターゲットとして選別されたアプタマー8-1及び8-10は、A/California/04/2009(H1N1)、A/NewYork/55/2004(H3N2)A/Netherlands/219/2003(H7N7)及びA/HongKong/1073/1999(H9N2)には結合しないか、結合したとしてもごくわずかであった。
【0069】
A/HongKong/1073/1999(H9N2)をターゲットとして選別されたアプタマー10-1は、ターゲットとは異なる亜型、A/California/04/2009(H1N1)、A/NewYork/55/2004(H3N2)、A/Vietnam/1203/2004(H5N1)、A/ Indonesia/05/2005(H5N1)及びA/Netherlands/219/2003(H7N7)のいずれにも結合能を示さなかった。
<表3>
【0070】
(4−2)異なる株との相互作用解析
選別されたアプタマーと、そのターゲットとは異なる株のHAタンパク質との相互作用を調べるために、BiacoreT100を利用して解析を行った。
A/California/04/2009(H1N1)をターゲットとして選別されたアプタマーD12及びD26と、それらのターゲットと同じ亜型(H1N1)に属し、異なる株であるA/New Caledonia/20/1999(H1N1)とA/Brevig mission/1/1918(H1N1)のHAタンパク質との相互作用解析を行った。その結果、アプタマーD12及びD26は、同じ亜型であってもA/New Caledonia/20/1999(H1N1)とA/Brevig mission/1/1918(H1N1)には結合せず、株間のわずかな変異も識別できることが示された(表4)。
<表4>
【0071】
(実施例5) 短鎖化アプタマーの作製
実施例1で選別を行った各アプタマーは74ヌクレオチドの長さを持つが、合成コストの面や様々な解析のためにはより短い方が有用であるので、HAタンパク質への結合を保持したまま、より短いアプタマーの誘導体を作製することを目標とした。本発明においてはアプタマー8-3を短鎖化して、アプタマー8-3Sを作製した。
アプタマー8-3(配列番号6)の48-91塩基の3’末端に“C”の1塩基を加えた45merの短鎖化アプタマー8-3S:5’-GGGCAACCGCUGGAACUUGAAGUCGGUAAUGCGAGCGGAAAGCCC-3’(配列番号10、図12)を、以下のテンプレートとプライマーを用いてPCR、インビトロ転写反応により調製した。
【0072】
・テンプレート:5’-GGAGCTCAGCCTTCACTGCGAGATTGTTCTGACCGAGTTGCCTAGCAGGGCAACCGCTGGAACTTGAAGTCGGTAATGCGAGCGGAAAGCCTTGGCACCACGGTCGGATCCAC-3’(配列番号11)
・プライマー8-3S-F:
5’-AGTAATACGACTCACTATAGGGCAACCGCTGGAACTTGAA-3’(配列番号12)
・プライマー8-3S-R:
5’-GGGCTTTCCGCTCGCATTAC-3’(配列番号13)
【0073】
得られたPCR産物はエタノール沈殿で精製させた後、T7 Ampliscribeキットを用いて、37℃、3時間またはオーバーナイトでインビトロ転写反応を行った。RNA合成とDNaseI処理の後、反応液を8%変性ポリアクリルアミドゲルで分画した。RNAをゲルから抽出し、エタノール沈殿により精製した。
【0074】
(実施例6) BiacoreT100による短鎖化アプタマー8-3SとA/Vietnam/1203/2004(H5N1)との相互作用解析
実施例5で得られたアプタマー8-3Sについて、実施例3と同様にして、BiacoreT100により、A/Vietnam/1203/2004(H5N1)のHAタンパク質との相互作用を測定した。
図13に示すように、アプタマー8-3Sは、A/Vietnam/1203/2004(H5N1)のHAタンパク質に対して、そのタンパク質量に依存した結合が見られた。また、フィルター結合定量法によっても同様の結果を得た。このことから、短縮化アプタマー8-3Sが、A/Vietnam/1203/2004 (H5N1)のHAタンパク質に対し、高い親和性を維持していることが示された。
【0075】
(実施例7) アプタマー8-3Sの安定化
より安定なアプタマーを得るため、アプタマー8-3Sのシチジンの2’-OH 基を、転写工程において2’-フルオロ基(2’-F)で修飾されるよう調製した(アプタマー8-3S-2’FCとする)。
実施例5と同様の方法でPCRを実施し、得られたdsDNAを、T7 Durascribe kit (Epicentre Technologies社製)を使用して、37℃、オーバーナイトでインキュベーションを行い、インビトロ転写により安定なRNAに変換した。次いで、精製されたアプタマー8-3S-2’FCとA/Vietnam/1203/2004(H5N1)のHAタンパク質との相互作用解析をBiacoreT100を用いて行った。
A/Vietnam/1203/2004(H5N1)に対するアプタマー8-3S-2’FCの親和性は8-3Sとほぼ同程度の値を示した(図14)。すなわち、アプタマー8-3S-2’FCはA/Vietnam/1203/2004(H5N1)に対する結合能を維持していることが示された。
【0076】
(実施例8) アプタマーD12、D26の安定化
実施例7と同様にアプタマーD12及びD26の安定化を行った。D12及びD26に対しては、シチジン及びウリジンの2’-OH基を2’-フルオロ基で修飾した(アプタマーD12-2’FC&U及びD26-2’FC&Uとする)。
PCRには、選別に用いた配列番号2のプライマーとオリゴdTを持つ配列番号3のプライマーを使用した。得られたdsDNAを、T7 Durascribe kitを使用して、37℃、オーバーナイトでインキュベーションを行い、インビトロ転写により安定なRNAに変換した。次いで、精製されたアプタマーD12-2’FC&U及びD26-2’FC&Uと、A/California/04/2009(H1N1)のHAタンパク質との相互作用解析をBiacoreT100を用いて行った。
アプタマーD12-2’FC&U及びD26-2’FC&UはいずれもA/California/04/2009(H1N1)に対する結合能を維持していた(図15)。
【配列表フリーテキスト】
【0077】
1.(配列番号1)RNAランダムプール用テンプレート:
5'-GGAGCTCAGCCTTCACTGC-(N)74-GGCACCACGGTCGGATCCAC-3'
2.(配列番号2)RNAランダムプール用5’末端プライマー:
5'-TCTAATACGACTCACTATAGGAGCTCAGCCTTCACTGC-3'
3.(配列番号3)RNAランダムプール用3’末端プライマー:
5'-GTGGATCCGACCGTGGTGCC-3'
4.(配列番号4)アプタマーD12:
5'-GGAGCUCAGCCUUCACUGCCAAAGUGCGAGGCAGUGUGGUGCUGUCCUACGAGUUCUAAAGUUCGUUAGGAAGGCAGCUCAACAUGUUUAACAGGCACCACGGUCGGAUCCAC-3’
5.(配列番号5)アプタマーD26:
5'-GGAGCUCAGCCUUCACUGCCAAAAAGUUAGGCCAGCAAAUUGCGAGCUGAUCCGGUGACUGGCUACAGGAGGCCUUGUCCACGGCCGUAUUGGCACCACGGUCGGAUCCAC-3’
6.(配列番号6)アプタマー8-3:
5'-GGAGCUCAGCCUUCACUGCGAGAUUGUUCUGACCGAGUUGCCUAGCAGGGCAACCGCUGGAACUUGAAGUCGGUAAUGCGAGCGGAAAGCCUUGGCACCACGGUCGGAUCCAC-3’
7.(配列番号7)アプタマー8-1:
5'-GGAGCUCAGCCUUCACUGCCUGUUAGAGUUUCCUAAAAGCGAACUGGCGCCCUCGUCAGCAUCUGGCAGACGAGUGGAGACGGACUAACCACAGGCACCACGGUCGGAUCCAC-3’
8.(配列番号8)アプタマー8-10:
5'-GGAGCUCAGCCUUCACUGCGGUGACCGGAGGAAUACGCGGACGGAGAAAGGGUUGGCCUCGUGGAUGGCGGAUACCCGUCCAGGGAUCUUCUAGGCACCACGGUCGGAUCCAC-3’
9.(配列番号9)アプタマー10-1:
5'-GGAGCUCAGCCUUCACUGCGAACCUCGGAUUGUCUGACCAUGCUCAAGGCGGCGGGCCAGCCAUGGCCAUUCCCUGAAUUAGUGGAAACCUUAGGCACCACGGUCGGAUCCAC-3’
10.(配列番号10)短鎖化アプタマー8-3S:
5’-GGGCAACCGCUGGAACUUGAAGUCGGUAAUGCGAGCGGAAAGCCC-3’
11.(配列番号11)短鎖化アプタマー8-3S用テンプレート:5’-GGAGCTCAGCCTTCACTGCGAGATTGTTCTGACCGAGTTGCCTAGCAGGGCAACCGCTGGAACTTGAAGTCGGTAATGCGAGCGGAAAGCCTTGGCACCACGGTCGGATCCAC-3’
12.(配列番号12)プライマー8-3S-F:
5’-AGTAATACGACTCACTATAGGGCAACCGCTGGAACTTGAA-3’
13.(配列番号13)プライマー8-3S-R:
5’-GGGCTTTCCGCTCGCATTAC-3’
【技術分野】
【0001】
本発明は、新型インフルエンザウイルスのヘマグルチニンに結合能を有するアプタマーに関する。
【背景技術】
【0002】
インフルエンザウイルスにはA、B、及びCの3つの型があり、季節性インフルエンザとして毎年流行を起こすのはA型とB型であることが知られている。このうちA型インフルエンザウイルスは、ウイルス表面に突出したタンパク質であるヘマグルチニン(以下HAとする)とノイラミニダーゼ(以下NAとする)の組み合わせによって多くの亜型に分類されている。HAには、H1からH16の亜型、NAにはN1からN9の亜型が存在し、この組み合わせによって、例えばH1N1、H2N2、H3N2等と表現される。
【0003】
季節性インフルエンザウイルスとして毎年流行を繰り返しているのは、A型インフルエンザウイルスのH3N2亜型(香港型)、H1N1亜型(ソ連型)とB型インフルエンザウイルスであるが、これらは高い死亡率を示すほどの病原性は持たない。
【0004】
一方、季節性のインフルエンザウイルスとは別に、近年、新型インフルエンザの世界的流行が懸念されている。新型インフルエンザは、人類がこれまで感染したことのない抗原タンパク質を持っているため、多くの人々が免疫を持たず、高い確率で感染、発症し、パンデミックを引き起こす可能性がある。
新型インフルエンザの例として、1997年に香港で発生したH5N1亜型の高病原性鳥インフルエンザウイルスがあり、家禽類だけでなくヒトへと感染して死亡者をだして以来、H5N1亜型の鳥インフルエンザウイルスのヒトへの感染は現在まで続いている。また、H9N2亜型の鳥インフルエンザウイルスについても、ヒトへの感染が確認されている。今後、これらのウイルスがヒトからヒトへ容易に感染しうるウイルスへ変異する可能性もあり、その対策が急がれている。実際に、2009年にパンデミックを引き起こした、豚由来の新型インフルエンザウイルス(H1N1)は、ヒトからヒトへ感染し、その伝播力は季節性インフルエンザウイルスよりも強い。幸い病原性は高くはなかったが、将来高病原性へ変異する危険性をもっている。
【0005】
これらの新型インフルエンザウイルスが従来の季節性インフルエンザウイルスと大きく異なる点は、本来豚インフルエンザ、鳥インフルエンザとして、豚又は鳥というそれぞれの種内のみで流行を引き起こしていたウイルスが、種の壁を乗り越えてヒトへと感染し、ヒトとヒトとの間で感染するか、あるいは感染が懸念される点である。また、新型インフルエンザに対して免疫を持たない人類にとっては、ウイルスに暴露すると高い確率で発症することが考えられる。このような点において、新型インフルエンザウイルスは季節性インフルエンザウイルスとは大きく異なる。
また、インフルエンザウイルスにおいても、種々の型・亜型が存在して、それぞれ異なる性質を持ち、同じ亜型に属するウイルスでも株の違いによって抗原性が異なる場合もあるため、インフルエンザの予防対策や治療にとって、ウイルスの型・亜型・株の識別は非常に重要な課題である。
【0006】
現在、使用されている迅速診断キットは、インフルエンザウイルスの抗原を検出するキットが主流になっているが、A型かB型かの型の識別のみである。新型かどうかを区別するには、亜型・株の識別を行う必要がある。これにはインフルエンザウイルスの遺伝子を検出するRT-PCR法を用いた診断が必要であるが、設備や手間および費用の問題から日常臨床での使用には向いていない。したがって、詳しい検査が行われないまま発見が遅れると初期の段階での封じ込めを損ない、広い範囲にわたって流行が拡大するので、常にインフルエンザウイルスの有無と共に、型、亜型、株をルーチンでチェックできるようなシステムが求められる。
【0007】
一方、アプタマー、特にRNAアプタマーは複雑な3次構造をとることによって、標的化合物を高度に識別して結合することが可能である(非特許文献1)。したがって、アプタマーの高識別機能は近縁のタンパク質どうしや、あるいは微生物やウイルスの型、亜型や株どうしを識別するために大変価値のある道具として使える可能性がある。
アプタマーとは、機能性核酸のことであり、抗体と類似の機能をもつが、標的化合物として、イオンのような小さいものから、ウイルスのような巨大複合体まで対象となるので、抗体よりも利用範囲が広い。
【0008】
本発明者は、既に季節性インフルエンザであるインフルエンザA(H3N2)およびBのHAタンパク質に対するアプタマーを取得しており、これらのアプタマーは他の型のHAタンパク質のみならず株の違いをも識別することを報告し、季節性インフルエンザの有無と共に型、亜型、株を迅速かつ正確に判定できる診断手法を提供することができた。さらに該インフルエンザウイルスの宿主細胞表面への結合を妨げる作用をもつことも報告している。(非特許文献2,3)(特許文献1)。
【0009】
したがって、新型インフルエンザに対しても、同様の迅速かつ正確な診断手法を提供するための優れた識別能を有するアプタマーの開発が強く望まれていた。
【先行技術文献】
【特許文献】
【0010】
【特許文献1】特許第4441618号公報
【非特許文献】
【0011】
【非特許文献1】Jenison R.D.,Gill,S.C.,Pardi,A., and Polisky,B.1994.High-resolution molecular discrimination by RNA.Science 263,1425-1429.
【非特許文献2】Gopinath,S.C.B.,Misono,T.,Kawasaki,K.,Mizuno,T.,Imai,M.,Odagiri,T., and Kumar,P.K.R.2006.An RNA aptamer that distinguishes between closely related human influenza viruses and inhibits hemagglutinin-mediated membrane fusion.J.Gen.Virol.87,479-487.
【非特許文献3】Gopinath,S.C.B.,Sakamaki,Y.,Kawasaki,K. and Kumar,P.K.R.2006.An efficient RNA aptamer against human influenza B virus hemagglutinin.J.Biochem.139,837-846.
【非特許文献4】Kumar,P.K.R.,Machida,K.,Urvil,P.T.,Kakiuchi,P.N.,Vishnuvardhan,D.,Shimotohno,K.,Taira,K., and Nishikawa, S. 1997.Isolation of RNA aptamers specific to the NS3 protein of hepatitis C virus from a pool of completely random RNA.Virology 237,270-282.
【非特許文献5】Kumarevel,T.S.,Gopinath,S.C.B.,Nishikawa,S.,Mizuno,H., and Kumar,P.K.R.2004.Identification of important chemical groups of the hut mRNA for HutP interactions that regulate the hut operon in Bacillus subtilis.Nucleic Acids Res.32,3904-3912.
【発明の概要】
【発明が解決しようとする課題】
【0012】
本発明は、新型インフルエンザウイルス、特に、A/California/04/2009(H1N1)、A/Vietnam/1203/2004(H5N1)、及びA/HongKong/1073/1999(H9N2)のHAタンパク質をターゲットとするアプタマーを取得し、新型インフルエンザウイルスの検査薬を新たに提供しようとするものである。
【課題を解決するための手段】
【0013】
本発明者らは、上記課題を解決するために鋭意研究を行った結果、新型インフルエンザウイルスのA/California/04/2009(H1N1)、A/Vietnam/1203/2004(H5N1)、及びA/HongKong/1073/1999(H9N2)のHAタンパク質に結合するアプタマーを取得した。
得られたアプタマーと各ターゲットとの親和性及び相互作用解析、亜型または株との識別には、フィルター結合定量法、及び表面プラズモン共鳴装置(GEヘルスケアバイオサイエンス社製BiacoreT100)を用いた。
【0014】
A/California/04/2009(H1N1)をターゲットとした選別では、アプタマーD12とD26を取得した。アプタマーD12及びD26は、A/California/04/2009に対して高い親和性を示したが、
A/California/04/2009と同じ亜型H1N1に属するA/NewCaledonia/20/1999(H1N1)とA/Brevig mission/1/1918(H1N1)に対してはいずれも結合能を示さなかった。
【0015】
A/Vietnam/1203/2004(H5N1)をターゲットとした選別では、アプタマー8-1、8-3、及び8-10を取得した。アプタマー8-1、8-3、及び8-10は、A/Vietnam/1203/2004(H5N1)に対し、高い親和性を示した。さらに、同じ亜型(H5N1)に属するA/Indonesia/05/2005(H5N1)及びA/ Anhui/1/2005(H5N1)への親和性も調べたところ、アプタマー8-1、8-3、及び8-10は、これらの株にも結合能を有することが確認された。
これに対して、H5N1型以外の亜型との親和性を確認したところ、アプタマー8-3は、A/NewYork/55/2004(H3N2)、A/Netherlands/219/2003(H7N7)、及びA/HongKong/1073/1999(H9N2)に対しては結合しないか、結合したとしてもごくわずかであり、A/California/04/2009(H1N1)に対しては親和性を示したものの、そのKd値は30nMであり、A/Vietnam/1203/2004(H5N1)に対するKd値、50pMの600倍であった。すなわち、A/California/04/2009(H1N1)への親和性は、A/Vietnam/1203/2004(H5N1)に対するよりも600分の1程度であることが示された。
アプタマー8-1及び8-10においても、H5N1型以外の亜型との親和性は、いずれもA/California/04/2009(H1N1)、A/NewYork/55/2004(H3N2)、A/Netherlands/219/2003(H7N7)及びA/HongKong/1073/1999(H9N2)には結合しないか、結合したとしてもごくわずかであった。
【0016】
A/HongKong/1073/1999(H9N2) をターゲットとした選別では、アプタマー10-1を取得した。アプタマー10-1は、A/HongKong/1073/1999(H9N2)に対して高い親和性を示したが、他の亜型との親和性を調べたところ、A/NewYork/55/2004(H3N2)、A/Vietnam/1203/2004(H5N1)、A/Indonesia/05/2005(H5N1)、及びA/Netherlands/219/2003(H7N7)のいずれにも結合能を示さなかった。
そして、本発明のアプタマーを取得した方法を上記以外の新型インフルエンザウイルスあるいは今後出現してくる新型インフルエンザウイルスに対して適用することで、他の新型インフルエンザウイルスに特異的なアプタマーを取得することができる。
【0017】
さらに、本発明者らはアプタマー8-3を短鎖化して、アプタマー8-3Sを得た。これらはターゲットとするA/Vietnam/1203/2004(H5N1)のHAタンパク質に対し高い親和性を保持していた。
【0018】
また、アプタマーD12、D26、8-3SのRNAのリボース基の2’-OHをフルオロ基(2’-F)に置き換えることによって、より安定なアプタマーD12-2’FC&U、D26-2’FC&U、及び8-3S-2’FCを得ることができた。
以上の知見を得て本発明を完成した。
【0019】
すなわち、本発明は以下の発明を含むものである。
〔1〕 配列番号4〜10のいずれかで示される塩基配列、又は当該塩基配列において、1若しくは数個の塩基が欠失、置換若しくは付加された塩基配列を含むRNAからなることを特徴とする、新型インフルエンザのHAタンパク質に対する結合能を有するアプタマーRNA。
〔2〕 配列番号4〜9のいずれかで示される塩基配列、又は当該塩基配列において、1若しくは数個の塩基が欠失、置換若しくは付加された塩基配列を含むRNAが構成する2次構造において、HAタンパク質に対する結合能を有する先端領域部の構造を保持するように短鎖化された塩基配列からなるアプタマーRNA。
〔3〕 アプタマーRNAを構成するヌクレオチドのリボース部位の少なくとも1箇所が、化学修飾されていることを特徴とする、前記〔1〕及び〔2〕のいずれかに記載のアプタマーRNA。
〔4〕 前記化学修飾が、リボース部位の2’位に対するフルオロ基(2’-F)もしくはメトキシ基(2’-OMe)による修飾、又は水素による置換(2’-deoxy)であることを特徴とする、前記〔1〕及び〔2〕のいずれかに記載のアプタマーRNA。
〔5〕 アプタマーRNAの3’および/または5’末端がインバーストデオキシチミジン(idT)あるいはポリエチレングリコール(PEG)により修飾されていることを特徴とする、前記〔1〕〜〔4〕のいずれかに記載のアプタマーRNA。
〔6〕 前記〔1〕又は〔2〕に記載のアプタマーRNAと同一の塩基配列又は当該塩基配列と相補的な塩基配列を含み、かつ前記〔1〕又は〔2〕に記載のアプタマーに変換可能な一本鎖DNA、二本鎖DNA又は相補的RNA。
〔7〕 前記〔1〕〜〔5〕のいずれかに記載のアプタマーRNAを有効成分として含有する、新型インフルエンザウイルスの検出剤。
〔8〕 前記〔1〕〜〔5〕のいずれかに記載のアプタマーRNAを有効成分として含有する、新型インフルエンザウイルス用診断剤。
〔9〕 前記〔1〕〜〔5〕のいずれかに記載のアプタマーRNAを有効成分として含有する、新型インフルエンザウイルスのHAタンパク質の亜型又は株を特定するための診断用キット。
〔10〕 前記〔1〕〜〔5〕のいずれかに記載のアプタマーRNAを有効成分として用いることを特徴とする新型インフルエンザウイルスの検出方法。
〔11〕 前記〔1〕〜〔5〕のいずれかに記載のアプタマーRNAを有効成分として用いることを特徴とする、新型インフルエンザウイルスのHAタンパク質の亜型又は株を特定するための方法。
【発明の効果】
【0020】
本発明により、新型インフルエンザウイルスの型・亜型・株の同定のための検査薬として用いることができるアプタマーを提供できた。これらアプタマーを用いた検出法は、今までにはなかった新型インフルエンザウイルスに対する正確で迅速なルーチン検査を可能とするものである。また、本発明のアプタマーは、新型インフルエンザウイルスの宿主細胞表面への結合を妨げる作用をもつと考えられるので、新型インフルエンザウイルスの予防及び治療薬となる可能性が高い。
【図面の簡単な説明】
【0021】
【図1】フィルター結合定量法で調べた第8回目の選別サイクル目におけるRNAプールと、各ターゲットHAタンパク質との結合能力を示す。四角及び丸で囲まれた内側の黒化度は標識RNAの濃度を示す。括弧内はターゲットタンパク質を示す。
【図2】第10回目の選別サイクルで得られたA/California/04/2009(H1N1)に対するアプタマーの二次構造モデルを示す図である。
【図3】第10回目の選別サイクルで得られたA/Vietnam/1203/2004(H5N1)に対するアプタマーの二次構造モデルを示す図である。
【図4】第10回目の選別サイクルで得られたA/HongKong/1073/1999(H9N2)に対するアプタマーの二次構造モデルを示す図である。
【図5】フィルター結合定量法で調べた、アプタマーD12及びD26とA/California/04/2009(H1N1)のHAタンパク質との親和性の解析図である。四角及び丸で囲まれた内側の黒化度は標識RNAの濃度を示す。
【図6】フィルター結合定量法で調べた、アプタマー8-1、8-3及び8-10とA/Vietnam/1203/2004(H5N1)のHAタンパク質との親和性の解析図である。四角及び丸で囲まれた内側の黒化度は標識RNAの濃度を示す。
【図7】フィルター結合定量法で調べた、アプタマー10-1とA/HongKong/1073/1999(H9N2)のHAタンパク質との親和性の解析図である。四角及び丸で囲まれた内側の黒化度は標識RNAの濃度を示す。
【図8】表面プラズモン共鳴法(シングルサイクルカイネティクス法)によって測定したアプタマーD12及びD26とA/California/04/2009(H1N1)のHAタンパク質との相互作用の解析図である。
【図9】表面プラズモン共鳴法(シングルサイクルカイネティクス法)によって測定したアプタマー8-1、8-3、及び8-10とA/Vietnam/1203/2004(H5N1)のHAタンパク質との相互作用の解析図である。
【図10】表面プラズモン共鳴法(シングルサイクルカイネティクス法)によって測定したアプタマー8-1、8-3、及び8-10とA/Indonesia/05/2005(H5N1)のHAタンパク質との相互作用の解析図である。
【図11】表面プラズモン共鳴法(シングルサイクルカイネティクス法)によって測定したアプタマー10-1とA/HongKong/1073/1999(H9N2)のHAタンパク質との相互作用の解析図である。
【図12】短鎖化アプタマー8-3Sの二次構造モデルを示す図である。
【図13】表面プラズモン共鳴法(シングルサイクルカイネティクス法)によって測定したアプタマー8-3S及び8-3とA/Vietnam/1203/2004(H5N1)のHAタンパク質との相互作用の解析図である。
【図14】表面プラズモン共鳴法(シングルサイクルカイネティクス法)によって測定した、2’フルオロ化したアプタマー8-3S-2’FCとA/Vietnam/1203/2004(H5N1)のHAタンパク質との相互作用の解析図である。
【図15】表面プラズモン共鳴法(シングルサイクルカイネティクス法)によって測定した、2’フルオロ化したアプタマーD12-2’FC&U及びD26-2’FC&UとA/California/04/2009(H1N1)のHAタンパク質との相互作用の解析図である。
【発明を実施するための形態】
【0022】
1.本発明のアプタマーについて
(1)本発明の「アプタマー」とは
「アプタマー」とは、一般に、ある標的、例えばウイルス、タンパク質、ペプチド、糖類、金属イオン、小分子等に特異的に結合するように人工的に創製された核酸リガンドをいうが、本発明において用いる「アプタマー」は、新型インフルエンザウイルスのHAタンパク質に特異的な結合能を有する。核酸としてはRNAが好ましく、RNAを構成するリボース基が化学的に修飾されている場合も含む。
本発明のアプタマーは、周知のSELEX法(systematic evolution of ligands by exponential enrichment)を適用し、中央の74塩基部分をランダム化したRNAライブラリーから得られたクローン由来のものであって、典型的には配列番号4〜10に示される塩基配列からなるRNAが挙げられる。本発明においては、ランダム領域を比較的長い74塩基配列としたことによって、より複雑な3次構造ができ、高度な識別が可能なアプタマーを提供することができたものと考えられる。
【0023】
(2)「アプタマー」の構造
本発明のアプタマーの2次構造モデルは、例えば図2〜4などに示されるように、先端のループ構造とステム構造とループ構造の繰り返しが続いたステム・ループ構造となっている。実際にインフルエンザウイルスのHAタンパク質抗原と結合する部位は、ランダム領域からなる領域で、主にこのうちの先端ループを含むステム領域であると考えられるため、当該先端部領域の構造を壊さない範囲でRNA鎖を短縮化することもできる。
【0024】
(3)本発明のアプタマーを構成する核酸
本発明においては、上記配列番号で示されるRNAに限らず、これらRNAと対応する配列を有する、DNA及び、これらDNAと相補のDNA、あるいはこれら相補のDNA鎖同士が2本鎖を構成したdsDNAを包含する。さらに、上記RNAと相補の塩基配列を有するRNAも包含する。これらはそれ自体慣用の遺伝子操作手段により、たやすく本発明のアプタマーRNAに変換可能であり、本発明のアプタマー製造用中間体である。また、本発明においては、上述のように、典型的な配列番号4〜10に示された塩基配列と共に、当該塩基配列中のヌクレオチド残基を1若しくは数個欠失、置換、あるいは付加させた改変アプタマーであっても、上記HAタンパク質と結合するものであればこれを包含する。また、後述のように、RNAを構成するリボース基を化学修飾させた修飾アプタマーも包含される。
【0025】
(4)本発明のアプタマーの認識する部位とその親和性
新型インフルエンザウイルス粒子表面も季節性のA型インフルエンザウイルス粒子表面と同様、それぞれ約900コピーのヘマグルチニン(HA)と約300コピーのノイラミニダーゼ(NA)が存在すると言われている。本発明のアプタマーRNAには新型インフルエンザウイルスのHAタンパク質に結合する能力がある。
本発明のアプタマーが認識する部位は、新型インフルエンザのHAタンパク質の表面に突出した領域であり、且つ、アミノ酸の変異が認められる領域であると考えられる。
また、親和性に関する実験は、後述で示すフィルター結合定量法、及び表面プラズモン共鳴装置(GEヘルスケアバイオサイエンス社製BiacoreT100)を用いて行い、本発明のアプタマーの、新型インフルエンザHAタンパク質に対する親和性は高く、且つ特異的である。(図5−11、図13−15)。
【0026】
2.本発明のアプタマーRNAの製造方法
(1)SELEX法
本発明のアプタマーは、それ自体周知のSELEX法により得られたものである。SELEX法は、ランダム配列の核酸ライブラリーの作成から出発し、標的とするタンパク質との結合性を指標に選別して、PCR増幅するサイクルを複数回繰り返すもので、これにより、標的とするタンパク質に対し高い結合能を有する核酸のみを選別することができる。上記選別及びPCR増幅はそれぞれ、自然界における「淘汰」及び「増殖」に相当し、自然界における進化を試験管内で短時間に再現させるものである。
【0027】
本発明においては、試験管内選択法によって合計10サイクルの選別を行い、その結果得られた特異的なウイルスに結合するRNAプールについてクローニングを行った。選別を経たRNAプールには各ターゲットに高い親和性を有するアプタマーが含まれていた。
具体的には、配列中央の74塩基をランダム領域とするDNAライブラリー(配列番号1)を合成し、PCR増幅した後、転写してRNAプールとし、このRNAプールに対し、上記HAタンパク質と結合するRNAを選別、増幅し、このサイクルを複数回繰り返すことにより、該HAタンパク質に対して特異性及び結合性の高い、配列番号4から9に示されるアプタマーを得たものであり、本発明においては配列番号6に示すアプタマー(アプタマー8-3)をさらに短鎖化して、配列番号10に示される短鎖化アプタマー(アプタマー8-3S)を得ており、これはHAタンパク質に対する結合能を保持している。
【0028】
(2)その他の方法
本発明のアプタマーは、上記したとおり、基本的にはSELEX法により得たものではあるが、本発明においては、アプタマーの塩基配列を明らかにしており、SELEX法によらずとも、この塩基配列から本発明のアプタマーを合成することができる。これには、例えば、それ自体周知の以下の方法を用いることができる。
インビトロ転写法:合成目的のアプタマーRNAに対応する上記DNAを化学合成し、これをPCR増幅し、増幅されたDNAからRNAポリメラーゼによる転写反応によりアプタマーRNAを合成する。例えば、T7プロモーターの下流側に上記DNAを連結してPCR増幅し2本鎖DNAを得て、該2本鎖DNAを鋳型として、T7RNAポリメラーゼ、およびATP、GTP、CTP及びUTPからなる該RNAポリメラーゼ基質を含む反応溶液中で、RNA伸長反応を行うことにより、アプタマーRNAを得ることができる。
【0029】
この方法においては、T7プロモーター配列を含むプラスミドに上記dsDNAを連結したもの、あるいは化学合成したDNAも鋳型として用いることができる。また、上記DNAの合成においては従来公知の種々の方法で合成することが可能である。例えば、DNA合成機を用いて、dNTPを材料として、末端塩基から化学合成することができる。
化学合成法:RNAの合成には、リボース部位の2’水酸基の保護が必要であり、保護基として種々のアミダイトが開発されてきており、2-cyanoethoxymethyl(CEM)基を用いるとRNA合成時の鎖長伸長反応の効率が高くなり、100merを超える長鎖RNAの合成も可能となる。本発明のアプタマーRNAはこのような化学合成法を用いて合成することができる。
また、このほか、上記アプタマーRNAと相補のRNAからは、RNA依存RNAポリメラーゼを使用することにより本発明のアプタマーRNAを得ることも可能である。
【0030】
3.アプタマーRNAの短縮化
アプタマーの短縮化の主な目的は、アプタマーRNAの製造コストを削減することである。本発明のアプタマーは、HAタンパク質との結合に関与するコア領域となるステム・ループ構造を残して、HAタンパク質との結合活性を保持したまま短鎖化することができた(アプタマー8-3S;図12)。
本発明のアプタマーの短鎖化は、配列番号4〜9に示されるアプタマーに対して、一般的なアプタマーの短鎖化方法であるマッピング法(非特許文献2、特許文献1)に従って行うことができる。また、本発明の実施例5において短鎖化の対象とした配列番号6のアプタマー8-3(図3)のように、比較的単純な2次構造をとっていて、先端部のHA結合領域の予測がつきやすい場合は、マッピング法実験を省略することができる。
【0031】
4.アプタマーRNAの化学修飾
本発明のアプタマーRNAにおいては、リボヌクレアーゼ耐性を付加するために、その配列中に化学修飾(modified)したヌクレオチドを有していても良い。このようなアプタマーRNAは、例えば、アプタマーRNA中のヌクレオチドのリボース部位の2’-OH基を、常法によりフロオロ基(2’-F)にまたはメトキシ基(2’-OMe)に、あるいは同リボース部位の2’位を水素に置換(2’-deoxy)することにより得られる。この化学修飾は、ピリミジンヌクレオチド部位がリボヌクレアーゼにより分解されやすいので、ピリミジンヌクレオチドに対してより有効である。
このような化学修飾は、ヘマグルチニンとの結合に関与する部位のヌクレオチドに対しては行わない。それ以外のヌクレオチドであれば全て修飾することも可能であるが、一般的にはループ領域のピリミジンヌクレオチドのリボース基について行う。
【0032】
本発明においては、さらにアプタマーRNAの3’及び/又は5’末端をインバーストデオキシチミジン(idT)あるいはポリエチレングリコール(PEG)により修飾することもでき、これらにより、アプタマーRNAのリボヌクレアーゼ耐性はさらに向上し、生体中での分解による活性の低下を抑制することができる。され、生体内で有効な抗ウイルス作用を発揮させることが可能となる。
【0033】
5.アプタマーRNAを用いた新型インフルエンザウイルスの検出法
(1)標識化されたアプタマーRNAを用いる方法
本発明のアプタマーRNAは新型インフルエンザウイルスの検査薬の検出剤として用いられる。例えば、本発明のアプタマーRNAをフルオロセイン、ローダミン、テキサスレッド等の蛍光色素で標識し、被験試料と接触、結合反応を行い、結合しなかったものを除去した後、蛍光あるいはその強度を検出、測定することにより、新型インフルエンザウイルスの検出、あるいはその量を測定できる。この際、例えば、標識アプタマーRNAあるいは被験試料のいずれかを固定化した基板を用いて行うことができる。
また、結合時に蛍光発光する物質を用いて、本発明のアプタマー及び被験試料のいずれかを標識して、結合時の蛍光あるいはその強度を検出、測定することにより行うことも可能である。このような蛍光色素としては、例えば、フルオロセイン、ローダミン、テキサスレッド等が挙げられる。
【0034】
(2)表面プラズモン共鳴法(SPR)
また、本発明においては、蛍光色素を標識せずに、新型インフルエンザウイルスを検出することも可能である。これには、例えば、表面プラズモン共鳴法(SPR)がある。すなわち、分子間の結合反応を、センサー表面でおこる分子間の結合時の微細な質量変化を表面プラズモン共鳴とよばれる光学現象を採用することで測定することができる。SPR法を利用した装置として、GEヘルスケアバイオサイエンス社製BiacoreT100がある。測定はセンサーチップの金薄膜表面に本発明のアプタマーRNAあるいは被験試料のいずれかを周知の方法で固定化し、これに被験試料あるいはアプタマーRNAを供給し、結合反応をリアルタイムでモニタリングしながら行う。
【0035】
6.本発明のアプタマーを用いた診断方法及び診断用キット
本発明は、新型インフルエンザウイルスで発現するHAタンパク質に結合するアプタマーを、新型インフルエンザウイルスを含有するか、または含有する可能性のあるサンプルと接触させることによって、サンプル中に目的の新型インフルエンザウイルスが存在すること、あるいは存在しないことが確認できる。
また、本発明においては、新型インフルエンザウイルスの特定の亜型及び/又は株で発現するHAタンパク質と特異的に結合するアプタマーを提供することができたので、これらアプタマーを用いて、新型インフルエンザウイルスの特定の亜型及び/又は株を識別し、判定することができる。
そして、本発明は、新型インフルエンザウイルスの特定の亜型及び/又は株で発現するHAタンパク質と特異的に結合するアプタマーを備えた新型インフルエンザウイルス診断用キットを提供する。
診断剤、診断用キットを製造するに際しては、適宜、周知の薬学的に許容可能な希釈剤、安定化剤、その他の担体などと組み合わせて用いる。
本発明の診断用キットの対象となるサンプルとしては、例えば、被験者の咽頭拭い液、鼻汁等の採取検体等や、それら検体を培養細胞やニワトリ受精卵に与えて感染・増殖の工程によって得た溶液等が挙げられる。
また、本発明の方法により、新型インフルエンザウイルスの特定の亜型及び/又は株を識別し、判定することができるから、例えばヒトへの感染やパンデミックが発生する前に、豚や鳥類において実際に流行しているインフルエンザウイルスの亜型及び/又は株をモニタリングし、将来のヒトへの感染やパンデミック予測に用いることができる。
【0036】
本発明におけるアプタマーは、抗体に匹敵する特異性を有し、現在新型インフルエンザウイルスの検出のために使用されている抗体を用いた迅速診断キットと同様の用途に使用することができる。臨床上、新型インフルエンザであるか、又は単なる季節性インフルエンザ、感冒等のその他のかぜ症候群の疾患であるかを迅速に診断することは、治療上、また予防上においても非常に重要である。また、インフルエンザであっても、ウイルスの型により、また亜型により、症状が異なり、種類が異なることでワクチンの効果が得られない場合もあり、治療に用いる医薬の決定、ワクチンの選択等においても、その識別は重要な意味を有する。更に、新型インフルエンザの流行状況をモニタリングすることで、将来の流行予測、ワクチンの製造の上でも有用な情報を得ることができ、更に研究用の試薬としても利用することができる。
【0037】
使用の際には、上記のようにして得られた1種のアプタマーを単独で用いても良いが、場合によっては、同じウイルスまたはタンパク質に結合し得るアプタマー、または異なるウイルスまたはタンパク質に結合し得るアプタマーを複数混合して用いても良い。
【0038】
7.抗ウイルス剤、又は新型インフルエンザウイルスの予防もしくは治療用組成物
本発明のアプタマーは、新型インフルエンザウイルスのHAタンパク質の表面にある突出領域に特異的に結合し、ウイルスの細胞への結合を阻害すると考えられる。特に化学修飾されたアプタマーはヌクレアーゼ抵抗性で、血中半減期も長いばかりか、生体内への抗原性も無視できるので、各種の抗ウイルス製剤として、また新型インフルエンザの予防用または治療用の医薬組成物として利用する事も可能である。投与対象はヒトのみならず、鳥や豚などに適用することができる。
抗ウイルス製剤、または医薬組成物とする場合には、経口、非経口のいずれでもよく、適宜、周知の薬学的に許容可能な無毒性の担体、希釈剤と組み合わせて用いる。非経口投与としては、典型的には注射剤であるが、噴霧剤などと共に吸入による投与も可能である。
また、直接ヒトなどに投与せずに、抗ウイルス剤として、マスクや看護用の衣服などに含浸させて用いることもできる。
【実施例】
【0039】
以下に本発明の一実施形態として、新型インフルエンザウイルスのうちで、A/California/04/2009(H1N1)、A/Vietnam/1203/2004(H5N1)、及びA/HongKong/1073/1999(H9N2)に結合するアプタマーの選別例を挙げ、下記実施例において具体的に説明するが、本発明は以下の実施例に限定されるものではない。
なお、本発明で使用されている技術的用語は、別途定義されていない限り、当業者により普通に理解されている意味を持つ。本発明の実施例で用いた遺伝子組換え技術、PCR法、その他の手法などの具体的な手順や条件は、特に断らない限り、Sambrook and Russell,Molecular Cloning:A Laboratory Manual,3rd Edition.Cold Spring Harbor Laboratory Press,Plainview,NY(2001)、Cold Spring Harbor Laboratory Press,Cold Spring Harbor,New York(1989);D.M.Glover et al.ed.,"DNA Cloning",2nd ed.,Vol.1 to 4,(The Practical Approach Series),IRL Press,Oxford University Press(1995);Ausubel,F.M.et al.,Current Protocols in Molecular Biology,John Wiley & Sons,New York,N.Y,1995;日本生化学会編、「続生化学実験講座1、遺伝子研究法II」、東京化学同人 (1986);日本生化学会編、「新生化学実験講座2、核酸III(組換えDNA技術)」、東京化学同人(1992);R.Wu ed.,"Methods in Enzymology",Vol.68(Recombinant DNA),Academic Press,New York(1980);R.Wu et al.ed.,"Methods in Enzymology",Vol.100(Recombinant DNA,PartB) & 101(Recombinant DNA,Part C),Academic Press,New York(1983);R.Wu et al.ed.,"Methods in Enzymology",Vol.153(Recombinant DNA,Part D),154(Recombinant DNA,Part E) & 155(Recombinant DNA,Part F),Academic Press,New York(1987)などに記載の方法あるいはそこで引用された文献記載の方法またはそれらと実質的に同様な方法により行うことができる。
また、本発明で引用した先行文献又は特許出願明細書の記載内容は、参照して本明細書の記載として組み入れるものとする。
【0040】
(実施例1) A/California/04/2009(H1N1)、A/Vietnam/1203/2004(H5N1)、及びA/HongKong/1073/1999(H9N2)に特異的なアプタマーのインビトロでの選別
(1−1)RNAランダムプールの作成
以下に示す、中央の74塩基をランダム領域とする一本鎖DNA(ssDNA)のライブラリーを合成してテンプレートとし(配列番号1)、5’末端プライマー(配列番号2)、及び3’末端プライマー(配列番号3)を用いてPCRを行った。
〔テンプレート〕
5'-GGAGCTCAGCCTTCACTGC-(N)74-GGCACCACGGTCGGATCCAC-3'(配列番号1)
〔5’末端プライマー〕
5'-TCTAATACGACTCACTATAGGAGCTCAGCCTTCACTGC-3'(配列番号2)
〔3’末端プライマー〕
5'-GTGGATCCGACCGTGGTGCC-3'(配列番号3)
【0041】
次いでT7 Ampliscribe kit(Epicentre Technologies社製)を用いて、インビトロでの転写を行い、増幅されたDNAライブラリーをRNAライブラリーに変換した。
【0042】
(1−2)インビトロにおける選別
上記(1)で得られたRNAライブラリー(16μg)を結合用緩衝液(0.01M HEPES,0.15M NaCl,pH7.4)に溶解させ、474≒3.57×1044通りの異なるRNA配列のコンフォメーションの平衡を促進するために、95℃で2分間処理して変性させた後に、室温で10分間冷却させた(アニーリング)。非特異的に結合するRNAを除去するためコンペティターとしてtRNA(E.coliのトータルtRNA(Boehringer-Mannheim社製))を加えた後、ターゲットとなるHAタンパク質を加えた。ターゲットとして選別に用いたHAタンパク質は、A/California/04/2009(H1N1)、A/Vietnam/1203/2004(H5N1)、及びA/HongKong/1073/1999(H9N2)である。各選別サイクルで用いたRNA、タンパク質、tRNAの分子比は表1に示すとおり。
【0043】
これらの混合液、50μLを室温で10分間インキュベートした後、”Pop-top”フィルターホルダー(Nucleopore社製)に装着した湿潤済みニトロセルロース・アセテート・フィルター(HA WPフィルター,0.45μm,径13.0mm,Millipore)に通し、タンパク質に結合したRNAをフィルター上に捕捉させて、フィルターを1mlの結合用緩衝液で洗浄した。フィルター上に残留したタンパク質結合RNAは溶出用緩衝液(0.01M HEPES,0.15M NaCl,7M Urea,pH7.4)で回収し、エタノールで沈殿精製後、50mM Tris-HCl(pH8.3),50mM KCl,10mM MgCl2,0.5mM Spermidine,10mM DTT,0.4μMプライマー(配列番号3),0.4mM dNTPs,25U AMV逆転写酵素(Wako社製)を含む20μlの反応液中で逆転写した。dNTPsと逆転写酵素は、変性及びアニーリングステップ(95℃で2分間処理後、室温で5分間インキュベート)の後、加えた。逆転写は37℃で45分間行った。
【0044】
<表1>
【0045】
得られたcDNAはPCRにより増幅され、次の選別ラウンドにおけるRNAを得るための鋳型として用いられた。PCRによる増幅のために、逆転写後の混合液(cDNA反応液)20μlを80μlのPCR用混合液(PrimeSTAR Max Premix(TaKaRa社製),1μMプライマー,1μMプライマー)で希釈した。反応液は95℃で30秒間加熱後、95℃、15秒;-54℃、10秒;-72℃、10秒のサイクルを、適正サイズで産物のバンドが得られるまでの回数(12-18サイクル)繰り返した。得られたPCR産物はエタノールで沈殿させて転写に用いた。試験管内転写反応はT7 Ampliscribeキットを用いて、37℃、3時間またはオーバーナイトで行った。RNA合成とDNase I処理の後、反応液を8%変性ポリアクリルアミドゲルで分画した。RNAをゲルから抽出し、エタノール沈殿により精製した後に定量し、次回の選別及び増幅サイクルに用いた。
【0046】
(1−3)選別及び増幅サイクル
以下においては、HAタンパク質に対する特異性と高い親和性を有するRNAを得るために、各選別サイクルを行った。表1に示したようにRNAとタンパク質比、コンペティター濃度は各選別サイクルごとに修正した。
非特異的に結合するRNAの濃縮を避けるために、第2、第4、第6及び第8の各選別サイクルでは、フィルターではなく96穴タイタープレート(Xenobind plate;xenopore社製)を使用した。Xenobind選別のために、最初に結合用緩衝液1mlあたり上記HAタンパク質10μgで各ウェルをコートし、残りの部分をBSA(0.1% stock solution)でブロックした。次いで、各ウェルを洗浄し、選別に使用した。
【0047】
前のサイクルで得たRNAプールを、結合用緩衝液中で95℃、2分間変性させた。続いて、室温で10分間冷却した後、tRNAを加え、HAタンパク質でコートされたウェルに充填した。10分間インキュベートした後、結合用緩衝液400μl(第6及び第8回目の選別サイクルでは1mL)で洗浄し、未吸着のRNAを除いた。その後、タンパク質結合RNAを、加熱した溶出用緩衝液(0.01M HEPES,0.15M NaCl,7M Urea,pH7.4)で回収し、エタノールで沈殿精製後、逆転写、PCR及びインビトロにおける転写により再生した。
【0048】
高親和性のアプタマーが濃縮されていく進行状況とその特異性を評価するため、第4回目の選別サイクルと第8回目の選別サイクルにおけるRNAプールの結合活性をフィルター結合定量法で解析した。[α-32P]ATPを使用して放射性標識(ラベル)したRNAを調製し、以前報告した方法(非特許文献3,4,5)に従ってフィルター結合分析を行った。結合反応では、モル濃度で10倍過剰の大腸菌tRNAを非特異的競争阻害剤として加え、RNAとHAタンパク質を1対4のモル比で混合し結合させた後、フィルターを通した。フィルターは1mlの上記結合用緩衝液で洗浄し、空気中で乾燥し、フィルターに残留した標識RNAの放射活性を画像解析装置(BAS2500,富士フイルム社製)で定量した。結合活性は、反応液に入れたRNA全量の内、フィルター結合したRNA量のパーセンテージで表した(図1)。第8回目の選別サイクル後のフィルターに捕捉されたRNAの割合は、A/California/04/2009(H1N1)のHAタンパク質をターゲットとした場合、HAタンパク質濃度400nM及び800nMにおいて、48%及び49%、A/Vietnam/1203/2004(H5N1)のHAタンパク質をターゲットとした場合、HAタンパク質濃度100nM及び200nMにおいて21%及び33%、さらに、A/HongKong/1073/1999(H9N2)の場合では、HAタンパク質濃度100nM及び200nMにおいて12%及び15%であった。
【0049】
(1−4)アプタマーの分析
個別のアプタマーを得るために、第10回目の選別サイクルで得たPCR産物をTAクローニング・ベクター(Invitrogen社製)に導入し、サブクローニングした後、大腸菌にトランスフォームした。プラスミドDNAの個々のクローンをプラスミド精製キット(Promega社製)で単離し、DNA塩基配列をDye Terminator Sequencing kit(Applied Biosystems Inc.社製)とDNAシークエンサー(Model 373A,Applied Biosystems Inc.社製)を用いて配列を解読し、そのDNA塩基配列に相当するRNA塩基配列を求めた。配列が解読されたRNAのクローンは配列によって幾つかのクラスに分類された。
【0050】
A/California/04/2009(H1N1)をターゲットとして選別されたRNAの塩基配列は、3つの主要なクラスに分類された。このうち、アプタマーD12(配列番号4)は、全体の19%を占めるクラス2に分類され、また、アプタマーD26(配列番号5)は、全体の9%を占めるクラス3に分類された。二次構造解析ソフト「BayesFold」(version 1.01、http://bayes.colorado.edu/Bayes/上で解析可能)を用いて得られたアプタマーの二次構造予測を行ったところ、それぞれ異なる二次構造の形成が予測された(図2)。
【0051】
A/Vietnam/1203/2004(H5N1)をターゲットとして選別されたRNAの塩基配列は、大きく3つのクラスに分類された。クラス1は、全体の42.9%を占め、配列番号6で示す塩基配列を得た(アプタマー8-3)。また、クラス2は全体の25.7%、クラス3は11.4%を占め、各塩基配列を、アプタマー8-1(配列番号7)及び8-10(配列番号8)とした。BayesFoldを用いてアプタマー8-1、8-3、及び8-10の二次構造予測を行ったところ、それぞれ異なる二次構造の形成が予測された(図3)。
【0052】
A/HongKong/1073/1999(H9N2)をターゲットとして選別されたRNAの塩基配列は、3つの主要なクラスを含み、このうち最も大きい割合を占めたクラス1は、全体の26.5%であり、この塩基配列をアプタマー10-1(配列番号9)とした。BayesFoldを用いてアプタマー10-1の二次構造予測を行ったところ、図4に示す二次構造が得られた。
【0053】
(実施例2) フィルター結合定量法によるアプタマーと各ターゲットHAタンパク質との親和性解析
実施例1で選別されたアプタマーとHAタンパク質との結合効率をフィルター結合定量法(非特許文献2に記載の方法による。)によって求めた。
【0054】
A/California/04/2009(H1N1)をターゲットとして選別されたアプタマーD12及びD26は、A/California/04/2009(H1N1)のHAタンパク質濃度に依存した結合がみられた。HAタンパク質濃度100nM及び200nMにおける、フィルターに捕捉されたRNAの割合は、D12が40%及び58%、D26が39%及び66%であった(図5)。
【0055】
A/Vietnam/1203/2004(H5N1)をターゲットとして選別されたアプタマー8-1、8-3及び8-10はA/Vietnam/1203/2004(H5N1)のHAタンパク質濃度に依存した結合がみられた。HAタンパク質濃度100nM及び200nMにおける、フィルターに捕捉されたRNAの割合は、8-1が12%及び25%、8-3が13%及び25%、8-10が23%及び41%であった(図6)。
【0056】
A/HongKong/1073/1999(H9N2)をターゲットとして選別されたアプタマー10-1はA/HongKong/1073/1999(H9N2)のHAタンパク質濃度に依存した結合がみられ、HAタンパク質濃度100nM及び200nMにおける、フィルターに捕捉されたRNAの割合は、それぞれ4%及び9%であった(図7)。
【0057】
(実施例3) BiacoreT100による各アプタマーとターゲットHAタンパク質との相互作用解析
実施例1で選別されたアプタマーとHAタンパク質との親和性や特異性をBiacoreT100(GEヘルスケアバイオサイエンス社製)によって測定した。
【0058】
HBS-P+緩衝液(0.01M HEPES,0.15M NaCl,0.05% v/v Surfactant P20,pH7.4)に溶解した5μMのビオチン化オリゴdT(24ヌクレオチド長)[5'-(dT)24-3']を、ストレプトアビジンでコートされたセンサーチップ(Series S Sensor Chip SA Certified,GEヘルスケアバイオサイエンス社製)に流速10μl/minで、2分間添加し、固定化を行った。次いで、3'-端に24ヌクレオチド長のオリゴ・アデニン(A)を延長させたアプタマー(30-60μM)を流速2μl/minで10分間添加し、オリゴdTに結合させた。
【0059】
上記のSAチップへ0.2-40nMの濃度範囲のHAタンパク質溶液を流速30μl/minで2分間添加し、固定化されたアプタマーとの相互作用解析を行った。
【0060】
A/California/04/2009(H1N1)をターゲットとして選別されたアプタマーD12及びD26はA/California/04/2009(H1N1)のHAタンパク質濃度に依存した結合が見られ、いずれも高い親和性を示した(図8)。
【0061】
A/Vietnam/1203/2004(H5N1)をターゲットとして選別されたアプタマー8-1、8-3、及び8-10は、A/Vietnam/1203/2004(H5N1)のHAタンパク質濃度に依存した結合が見られ、いずれも高い親和性を示した(図9)。
【0062】
さらに、アプタマー8-1、8-3、及び8-10は、A/Vietnam/1203/2004(H5N1)と同じ亜型に属するA/Indonesia/05/2005(H5N1)(図10)及びA/Anhui/1/2005(H5N1)にもまた結合することが確認された。
【0063】
A/HongKong/1073/1999(H9N2)をターゲットとして選別されたアプタマー10-1は、A/HongKong/1073/1999(H9N2)のHAタンパク質濃度に依存した結合が見られ、高い親和性を示した(図11)。
【0064】
また、各アプタマーの結合は、糖鎖を除去していないHAタンパク質に対しても糖鎖を除去したHAタンパク質に対してもほぼ同等の親和性を示したので、HAタンパク質の糖鎖はこのRNAアプタマーとの結合には関与していないことが示された。
本研究において選別されたアプタマーの種類と各ターゲットとの解離定数(Kd)値は表2に示す通りである。
【0065】
<表2>
【0066】
(実施例4) BiacoreT100による各アプタマーの特異性解析
(4−1)異なる亜型との相互作用解析
選別されたアプタマーと、そのターゲットとは異なる亜型のHAタンパク質との相互作用を調べるために、BiacoreT100を利用して解析を行った(表3)。
【0067】
A/Vietnam/1203/2004(H5N1)をターゲットとして選別されたアプタマー8-3と、ターゲットとは異なる亜型のHAタンパク質との相互作用を調べたところ、A/NewYork/55/2004(H3N2)、A/Netherlands/219/2003(H7N7)及びA/HongKong/1073/1999(H9N2)に対しては、結合しないか、結合したとしてもごくわずかであった。一方、A/California/04/2009(H1N1)に対しては親和性を示したものの、そのKd値は30nMであり、A/Vietnam/1203/2004(H5N1)に対するKd値、50pMの600倍であった。すなわち、A/California/04/2009 (H1N1)への親和性は、A/Vietnam/1203/2004(H5N1)に対するよりも600分の1程度であることが示された。
【0068】
同様に、A/Vietnam/1203/2004(H5N1)をターゲットとして選別されたアプタマー8-1及び8-10は、A/California/04/2009(H1N1)、A/NewYork/55/2004(H3N2)A/Netherlands/219/2003(H7N7)及びA/HongKong/1073/1999(H9N2)には結合しないか、結合したとしてもごくわずかであった。
【0069】
A/HongKong/1073/1999(H9N2)をターゲットとして選別されたアプタマー10-1は、ターゲットとは異なる亜型、A/California/04/2009(H1N1)、A/NewYork/55/2004(H3N2)、A/Vietnam/1203/2004(H5N1)、A/ Indonesia/05/2005(H5N1)及びA/Netherlands/219/2003(H7N7)のいずれにも結合能を示さなかった。
<表3>
【0070】
(4−2)異なる株との相互作用解析
選別されたアプタマーと、そのターゲットとは異なる株のHAタンパク質との相互作用を調べるために、BiacoreT100を利用して解析を行った。
A/California/04/2009(H1N1)をターゲットとして選別されたアプタマーD12及びD26と、それらのターゲットと同じ亜型(H1N1)に属し、異なる株であるA/New Caledonia/20/1999(H1N1)とA/Brevig mission/1/1918(H1N1)のHAタンパク質との相互作用解析を行った。その結果、アプタマーD12及びD26は、同じ亜型であってもA/New Caledonia/20/1999(H1N1)とA/Brevig mission/1/1918(H1N1)には結合せず、株間のわずかな変異も識別できることが示された(表4)。
<表4>
【0071】
(実施例5) 短鎖化アプタマーの作製
実施例1で選別を行った各アプタマーは74ヌクレオチドの長さを持つが、合成コストの面や様々な解析のためにはより短い方が有用であるので、HAタンパク質への結合を保持したまま、より短いアプタマーの誘導体を作製することを目標とした。本発明においてはアプタマー8-3を短鎖化して、アプタマー8-3Sを作製した。
アプタマー8-3(配列番号6)の48-91塩基の3’末端に“C”の1塩基を加えた45merの短鎖化アプタマー8-3S:5’-GGGCAACCGCUGGAACUUGAAGUCGGUAAUGCGAGCGGAAAGCCC-3’(配列番号10、図12)を、以下のテンプレートとプライマーを用いてPCR、インビトロ転写反応により調製した。
【0072】
・テンプレート:5’-GGAGCTCAGCCTTCACTGCGAGATTGTTCTGACCGAGTTGCCTAGCAGGGCAACCGCTGGAACTTGAAGTCGGTAATGCGAGCGGAAAGCCTTGGCACCACGGTCGGATCCAC-3’(配列番号11)
・プライマー8-3S-F:
5’-AGTAATACGACTCACTATAGGGCAACCGCTGGAACTTGAA-3’(配列番号12)
・プライマー8-3S-R:
5’-GGGCTTTCCGCTCGCATTAC-3’(配列番号13)
【0073】
得られたPCR産物はエタノール沈殿で精製させた後、T7 Ampliscribeキットを用いて、37℃、3時間またはオーバーナイトでインビトロ転写反応を行った。RNA合成とDNaseI処理の後、反応液を8%変性ポリアクリルアミドゲルで分画した。RNAをゲルから抽出し、エタノール沈殿により精製した。
【0074】
(実施例6) BiacoreT100による短鎖化アプタマー8-3SとA/Vietnam/1203/2004(H5N1)との相互作用解析
実施例5で得られたアプタマー8-3Sについて、実施例3と同様にして、BiacoreT100により、A/Vietnam/1203/2004(H5N1)のHAタンパク質との相互作用を測定した。
図13に示すように、アプタマー8-3Sは、A/Vietnam/1203/2004(H5N1)のHAタンパク質に対して、そのタンパク質量に依存した結合が見られた。また、フィルター結合定量法によっても同様の結果を得た。このことから、短縮化アプタマー8-3Sが、A/Vietnam/1203/2004 (H5N1)のHAタンパク質に対し、高い親和性を維持していることが示された。
【0075】
(実施例7) アプタマー8-3Sの安定化
より安定なアプタマーを得るため、アプタマー8-3Sのシチジンの2’-OH 基を、転写工程において2’-フルオロ基(2’-F)で修飾されるよう調製した(アプタマー8-3S-2’FCとする)。
実施例5と同様の方法でPCRを実施し、得られたdsDNAを、T7 Durascribe kit (Epicentre Technologies社製)を使用して、37℃、オーバーナイトでインキュベーションを行い、インビトロ転写により安定なRNAに変換した。次いで、精製されたアプタマー8-3S-2’FCとA/Vietnam/1203/2004(H5N1)のHAタンパク質との相互作用解析をBiacoreT100を用いて行った。
A/Vietnam/1203/2004(H5N1)に対するアプタマー8-3S-2’FCの親和性は8-3Sとほぼ同程度の値を示した(図14)。すなわち、アプタマー8-3S-2’FCはA/Vietnam/1203/2004(H5N1)に対する結合能を維持していることが示された。
【0076】
(実施例8) アプタマーD12、D26の安定化
実施例7と同様にアプタマーD12及びD26の安定化を行った。D12及びD26に対しては、シチジン及びウリジンの2’-OH基を2’-フルオロ基で修飾した(アプタマーD12-2’FC&U及びD26-2’FC&Uとする)。
PCRには、選別に用いた配列番号2のプライマーとオリゴdTを持つ配列番号3のプライマーを使用した。得られたdsDNAを、T7 Durascribe kitを使用して、37℃、オーバーナイトでインキュベーションを行い、インビトロ転写により安定なRNAに変換した。次いで、精製されたアプタマーD12-2’FC&U及びD26-2’FC&Uと、A/California/04/2009(H1N1)のHAタンパク質との相互作用解析をBiacoreT100を用いて行った。
アプタマーD12-2’FC&U及びD26-2’FC&UはいずれもA/California/04/2009(H1N1)に対する結合能を維持していた(図15)。
【配列表フリーテキスト】
【0077】
1.(配列番号1)RNAランダムプール用テンプレート:
5'-GGAGCTCAGCCTTCACTGC-(N)74-GGCACCACGGTCGGATCCAC-3'
2.(配列番号2)RNAランダムプール用5’末端プライマー:
5'-TCTAATACGACTCACTATAGGAGCTCAGCCTTCACTGC-3'
3.(配列番号3)RNAランダムプール用3’末端プライマー:
5'-GTGGATCCGACCGTGGTGCC-3'
4.(配列番号4)アプタマーD12:
5'-GGAGCUCAGCCUUCACUGCCAAAGUGCGAGGCAGUGUGGUGCUGUCCUACGAGUUCUAAAGUUCGUUAGGAAGGCAGCUCAACAUGUUUAACAGGCACCACGGUCGGAUCCAC-3’
5.(配列番号5)アプタマーD26:
5'-GGAGCUCAGCCUUCACUGCCAAAAAGUUAGGCCAGCAAAUUGCGAGCUGAUCCGGUGACUGGCUACAGGAGGCCUUGUCCACGGCCGUAUUGGCACCACGGUCGGAUCCAC-3’
6.(配列番号6)アプタマー8-3:
5'-GGAGCUCAGCCUUCACUGCGAGAUUGUUCUGACCGAGUUGCCUAGCAGGGCAACCGCUGGAACUUGAAGUCGGUAAUGCGAGCGGAAAGCCUUGGCACCACGGUCGGAUCCAC-3’
7.(配列番号7)アプタマー8-1:
5'-GGAGCUCAGCCUUCACUGCCUGUUAGAGUUUCCUAAAAGCGAACUGGCGCCCUCGUCAGCAUCUGGCAGACGAGUGGAGACGGACUAACCACAGGCACCACGGUCGGAUCCAC-3’
8.(配列番号8)アプタマー8-10:
5'-GGAGCUCAGCCUUCACUGCGGUGACCGGAGGAAUACGCGGACGGAGAAAGGGUUGGCCUCGUGGAUGGCGGAUACCCGUCCAGGGAUCUUCUAGGCACCACGGUCGGAUCCAC-3’
9.(配列番号9)アプタマー10-1:
5'-GGAGCUCAGCCUUCACUGCGAACCUCGGAUUGUCUGACCAUGCUCAAGGCGGCGGGCCAGCCAUGGCCAUUCCCUGAAUUAGUGGAAACCUUAGGCACCACGGUCGGAUCCAC-3’
10.(配列番号10)短鎖化アプタマー8-3S:
5’-GGGCAACCGCUGGAACUUGAAGUCGGUAAUGCGAGCGGAAAGCCC-3’
11.(配列番号11)短鎖化アプタマー8-3S用テンプレート:5’-GGAGCTCAGCCTTCACTGCGAGATTGTTCTGACCGAGTTGCCTAGCAGGGCAACCGCTGGAACTTGAAGTCGGTAATGCGAGCGGAAAGCCTTGGCACCACGGTCGGATCCAC-3’
12.(配列番号12)プライマー8-3S-F:
5’-AGTAATACGACTCACTATAGGGCAACCGCTGGAACTTGAA-3’
13.(配列番号13)プライマー8-3S-R:
5’-GGGCTTTCCGCTCGCATTAC-3’
【特許請求の範囲】
【請求項1】
配列番号4〜10のいずれかで示される塩基配列、又は当該塩基配列において、1若しくは数個の塩基が欠失、置換若しくは付加された塩基配列を含むRNAからなることを特徴とする、新型インフルエンザのHAタンパク質に対する結合能を有するアプタマーRNA。
【請求項2】
配列番号4〜9のいずれかで示される塩基配列、又は当該塩基配列において、1若しくは数個の塩基が欠失、置換若しくは付加された塩基配列を含むRNAが構成する2次構造において、HAタンパク質に対する結合能を有する先端領域部の構造を保持するように短鎖化された塩基配列からなるアプタマーRNA。
【請求項3】
アプタマーRNAを構成するヌクレオチドのリボース部位の少なくとも1箇所が、化学修飾されていることを特徴とする、請求項1及び2のいずれかに記載のアプタマーRNA。
【請求項4】
前記化学修飾が、リボース部位の2’位に対するフルオロ基(2’-F)もしくはメトキシ基(2’-OMe)による修飾、又は水素による置換(2’-deoxy)であることを特徴とする、請求項1及び2のいずれかに記載のアプタマーRNA。
【請求項5】
アプタマーRNAの3’および/または5’末端がインバーストデオキシチミジン(idT)あるいはポリエチレングリコール(PEG)により修飾されていることを特徴とする、請求項1〜4のいずれかに記載のアプタマーRNA。
【請求項6】
請求項1又は2に記載のアプタマーRNAと同一の塩基配列又は当該塩基配列と相補的な塩基配列を含み、かつ請求項1又は2に記載のアプタマーに変換可能な一本鎖DNA、二本鎖DNA又は相補的RNA。
【請求項7】
請求項1〜5のいずれかに記載のアプタマーRNAを有効成分として含有する、新型インフルエンザウイルスの検出剤。
【請求項8】
請求項1〜5のいずれかに記載のアプタマーRNAを有効成分として含有する、新型インフルエンザウイルス用診断剤。
【請求項9】
請求項1〜5のいずれかに記載のアプタマーRNAを有効成分として含有する、新型インフルエンザウイルスのHAタンパク質の亜型又は株を特定するための診断用キット。
【請求項10】
請求項1〜5のいずれかに記載のアプタマーRNAを有効成分として用いることを特徴とする新型インフルエンザウイルスの検出方法。
【請求項11】
請求項1〜5のいずれかに記載のアプタマーRNAを有効成分として用いることを特徴とする、新型インフルエンザウイルスのHAタンパク質の亜型又は株を特定するための方法。
【請求項1】
配列番号4〜10のいずれかで示される塩基配列、又は当該塩基配列において、1若しくは数個の塩基が欠失、置換若しくは付加された塩基配列を含むRNAからなることを特徴とする、新型インフルエンザのHAタンパク質に対する結合能を有するアプタマーRNA。
【請求項2】
配列番号4〜9のいずれかで示される塩基配列、又は当該塩基配列において、1若しくは数個の塩基が欠失、置換若しくは付加された塩基配列を含むRNAが構成する2次構造において、HAタンパク質に対する結合能を有する先端領域部の構造を保持するように短鎖化された塩基配列からなるアプタマーRNA。
【請求項3】
アプタマーRNAを構成するヌクレオチドのリボース部位の少なくとも1箇所が、化学修飾されていることを特徴とする、請求項1及び2のいずれかに記載のアプタマーRNA。
【請求項4】
前記化学修飾が、リボース部位の2’位に対するフルオロ基(2’-F)もしくはメトキシ基(2’-OMe)による修飾、又は水素による置換(2’-deoxy)であることを特徴とする、請求項1及び2のいずれかに記載のアプタマーRNA。
【請求項5】
アプタマーRNAの3’および/または5’末端がインバーストデオキシチミジン(idT)あるいはポリエチレングリコール(PEG)により修飾されていることを特徴とする、請求項1〜4のいずれかに記載のアプタマーRNA。
【請求項6】
請求項1又は2に記載のアプタマーRNAと同一の塩基配列又は当該塩基配列と相補的な塩基配列を含み、かつ請求項1又は2に記載のアプタマーに変換可能な一本鎖DNA、二本鎖DNA又は相補的RNA。
【請求項7】
請求項1〜5のいずれかに記載のアプタマーRNAを有効成分として含有する、新型インフルエンザウイルスの検出剤。
【請求項8】
請求項1〜5のいずれかに記載のアプタマーRNAを有効成分として含有する、新型インフルエンザウイルス用診断剤。
【請求項9】
請求項1〜5のいずれかに記載のアプタマーRNAを有効成分として含有する、新型インフルエンザウイルスのHAタンパク質の亜型又は株を特定するための診断用キット。
【請求項10】
請求項1〜5のいずれかに記載のアプタマーRNAを有効成分として用いることを特徴とする新型インフルエンザウイルスの検出方法。
【請求項11】
請求項1〜5のいずれかに記載のアプタマーRNAを有効成分として用いることを特徴とする、新型インフルエンザウイルスのHAタンパク質の亜型又は株を特定するための方法。
【図1】

【図2】
【図3】
【図4】
【図5】

【図6】

【図7】

【図8】
【図9】
【図10】
【図11】
【図12】
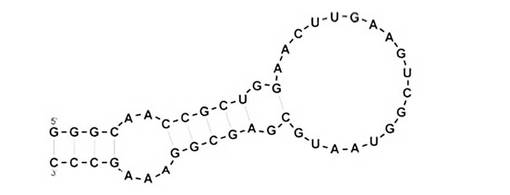
【図13】
【図14】
【図15】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【公開番号】特開2012−100636(P2012−100636A)
【公開日】平成24年5月31日(2012.5.31)
【国際特許分類】
【出願番号】特願2010−254565(P2010−254565)
【出願日】平成22年11月15日(2010.11.15)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成22年度、独立行政法人科学技術振興機構「ラベル不要の高機能性バイオセンサシステムの開発」委託研究、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
【公開日】平成24年5月31日(2012.5.31)
【国際特許分類】
【出願日】平成22年11月15日(2010.11.15)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成22年度、独立行政法人科学技術振興機構「ラベル不要の高機能性バイオセンサシステムの開発」委託研究、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
[ Back to top ]