
新規なアミノペプチダーゼ及びその製造法
【発明の詳細な説明】
【0001】
【産業上の利用分野】本発明は、シュードモナス属に属する細菌により生産される、ペプチドのN末端からアミノ酸を順次遊離する作用を有し、基質特異性の広い特性を有する新規なアミノペプチダーゼ及びその製造法に関する。
【0002】この酵素は、上記の特性を有することから、蛋白系食品加工における呈味付与、増強にも役立ち、消化剤としての医薬品への利用や、また種々のタンパク質の一次構造決定における生化学試薬としても利用し得るものである。
【0003】
【従来の技術および発明が解決しようとする問題点】従来、アミノペプチダーゼとしては、高等動物の臓器から多くの報告がなされている。哺乳動物の腎臓や小腸のミクロソーム画分に存在する膜酵素として、Wachsmuthらによって(Biochemistry 5,169,1966)およびKimらによって(J.Biol.Chem.,251,3199,1976)また、家兎骨格筋中に存在する酵素として、Otsukaらによって(Agric.Biol.Chem.,44,1617,1980)、サル脳からShimamuraらによって(Biochim.Biophys.Acta,756,223,1983)さらに豚の肝臓からKawataらによって(J.Biochem.,92,1093,1982)アミノペプチダーゼが報告されている。また、市販されているアミノペプチダーゼとしては、シグマ社からロイシンアミノペプチダーゼ、ミクロソーマル(Leucine aminopeptidase,Microsomal)〔EC.3.4.11.2〕が豚腎臓由来のアミノペプチダーゼとしてある。しかし、これらの動物起源の酵素は、その原料の入手の制約、分離、採取の操作、収量などの面で種々の制約を受ける点からも、これに代わる微生物起源の酵素を供給することが望ましい。
【0004】また、微生物起源のアミノペプチダーゼとして、中台らが(Agric.Biol.Chem.,37,157,1973)(Agric.Biol.Chem.,37,767,1973)(Agric.Biol.Chem.,37,775,1973)アスペルギルス・オリゼ(Aspergillus oryzae)の生産するロイシンアミノペプチダーゼI,II,III(分子量26,500;61,000;56,000)を報告している。またこれらと同じ起源であるアスペルギルス・オリゼ由来のプロテアーゼ酵素剤が市販されている。これらの酵素は、N末端のアミノ酸がロイシンであるペプチドのN末端に特によく作用するアミノペプチダーゼ活性を有するところの、基質特異性の狭い(高い)とされているものが殆どであり、特に市販のプロテアーゼ製剤で、種々のアミノ酸を導入したナフチルアミド誘導体を基質として使用して、その酵素活性をみてみると、そのアミノ酸がロイシンのものに特によく作用するような基質特異性の狭いものが殆んどである。このように、種々の用途に幅広く使用し得るアミノペプチダーゼとして、広い基質特異性を有する酵素が望まれているにも拘わらず、従来のものは必ずしも満足し得ないのが、実情である。
【0005】
【問題点を解決するための手段】そこで本発明者らは、基質特異性の広いアミノペプチダーゼを生産する微生物を探索すべく研究を重ねた結果、シュードモナス(Pseudomonas)属に属する細菌が目的とする酵素を生産することを見いだし、本発明を完成した。
【0006】すなわち本発明は、ペプチドのN末端からアミノ酸を順次遊離する作用を有し、基質特異性の広い特性を有する特定のアミノペプチダーゼならびにシュードモナス属に属し、該アミノペプチダーゼを産生する能力のある特定の細菌を培養して該アミノペプチダーゼを製造する方法を提供するものである。
【0007】本発明のアミノペプチダーゼは、シュードモナス属に属し、該酵素生産能を有する特定の細菌を栄養培地に培養し、培養液もしくは、培養上澄液から該酵素を採取することによって製造することができる。
【0008】目的とするアミノペプチダーゼを生産する能力のある微生物としては、シュードモナス・クロロラフィス(Pseudomonas chlororaphis)(IAM12354t )(この菌は、日本微生物株目録(JFCC CATALOGUE OF CULTURES)第4版1987、日本微生物株保存連盟(The Japan Federation for Culture Collections)、昭和62年3月1日発行のPseudomonaschlororaphisの項にIAM(type strain)として記載されている)がある。
【0009】栄養培地は、上記微生物が十分に生育し、目的とする酵素を生産しうるものであればよく、例えば、ペプトン、肉エキス等に塩化ナトリウムなどの無機塩類等を加えたものが好適に用いられる。
【0010】培養は、本発明の場合、静置培養法によって行うことが好ましい。一般には、中性付近のpHに調整し、20〜30℃の温度で2〜16日間程度培養を行い、アミノペプチダーゼを十分に生成せしめることができる。この酵素は、主として菌体外の培養液中に生成される。
【0011】培養液からのアミノペプチダーゼの採取は、適宜既知の手法を組み合わせて行えばよく、その一例を示すと、培養終了後、培養液から遠心分離等によって菌体を除去し、得られた上澄液を適当な緩衝液で透析して粗酵素液を得る。
【0012】次いで必要に応じ、粗酵素液をイオン交換クロマト、ゲル濾過、ハイドロキシルアパタイトクロマト、疎水クロマトなどの酵素精製に用いられる通常の操作を適宜組み合わせることによって、精製または、部分精製されたアミノペプチダーゼを得ることができる。本発明の場合、透析した粗酵素液を直接カラムクロマトに供することが、好ましい。
【0013】次に、本発明の酵素の性質を以下に示す。本発明の酵素は、その分子量が、98,000で、その至適pHは、pH8.0付近にあり、至適温度は、35℃付近にあり、その熱安定性は、37℃以下では、安定であり、図3及び図4で例証されるように50℃より高い温度では相対活性が実質上0である。より具体的には、40℃、10分間の加熱で約60%の活性が残存し、1mMのCo2+イオンでその活性が、約1.3倍に上昇し、ベスタチンに著しく阻害され、また、ロイペプチン、EDTAにも若干阻害されるが、その基質特異性は、アミノ酸−2−ナフチルアミドの基質においてN末端が、アラニン、リジン、ロイシン、メチオニンといったアミノ酸であるものによく作用するなど広い基質特異性が認められ、さらに、トリ、テトラペプチドにも良好な活性を有し、それらのN末端から良好にアミノ酸を遊離するアミノペプチダーゼである。
【0014】なお、酵素活性の測定は、実施例に示した精製酵素を用いて以下の方法で行った。
1)アミノ酸−2−ナフチルアミドに対する活性の測定アミノ酸−2−ナフチルアミドに対する活性の測定は、Matsutaniら(J.Med.Technol.,11,300,1967)の方法に準じて行った。酵素を0.4mlの1mM DTT−0.1M リン酸緩衝液(pH8.0)の存在下、0.5mM のアミノ酸−2−ナフチルアミドに37℃で一定時間作用させた後、0.4mlの0.23N HCl−エタノール溶液を加えて反応を停止させた。さらに0.4mlの0.06%p−ジメチルアミノシンナムアルデヒド・エタノール溶液を加えて30分間放置後、生成したシッフ塩基色素の量を540nmにおける吸光値を分光々度計で測定し、遊離した2−ナフチルアミンを定量した。また、アミノ酸の種類によっては、その基質濃度、基質溶液を変えて行った。酵素活性の単位は、pH8.0、37℃において酵素が、アミノ酸−2−ナフチルアミドから1分間に1μmolの2−ナフチルアミンを遊離させる酵素量を1単位(U)とした。
【0015】2)一般の合成ペプチドに対する活性の測定一般の合成ペプチドに対する活性の測定は、Yemm & Cocking(Analyst,80,209,1975)の方法に準じて行った。酵素を終濃度0.5mM の基質と、0.4mlの1mM DTT−0.1M リン酸緩衝液(pH8.0)の存在下、一定時間作用させた後、氷冷した0.2M クエン酸緩衝液(pH5.0)0.2mlを加えて反応を停止した。さらに1M のモノヨード酢酸(中和済)を0.1ml加えて後、0.4mlのニンヒドリン試薬を加えて、100℃で15分間煮沸した後、流水で5分間冷却した。さらに60%エタノール2mlで希釈した後、570nmにおける吸光値を分光々度計で測定した。
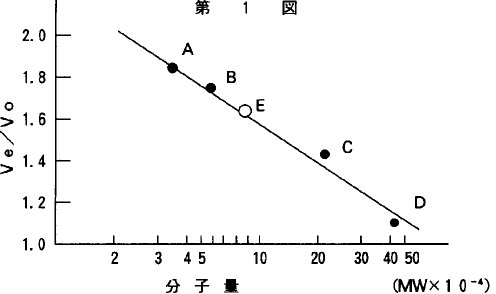
【0016】1)分子量本酵素の分子量は、Andrewsの方法(Biochem.J.,96,595,1965)に準じ、セファデックスG−200によるゲル濾過カラム(φ2.4×105cm)に精製酵素および標準タンパク質を供して1mM NaN3−0.1%MCE−10mM Tris−HCl緩衝液(pH7.2)で、流速0.25ml/min.にて溶出して求めた。精製酵素および標準タンパク質の溶出位置と分子量の関係を第1図に示した。その結果、本酵素の分子量は、98,000と推定した。
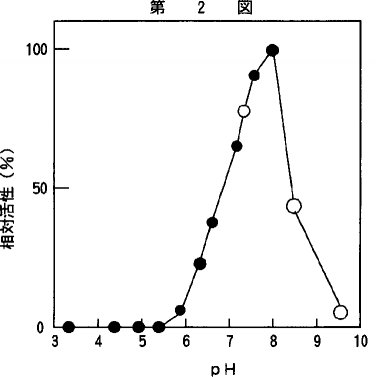
【0017】2)至適作用pH本酵素の至適作用pHは、pH3.0〜10.0の範囲で、アラニン−2−ナフチルアミドを基質として37℃、10分間の反応条件で測定した。緩衝液は、各pHに調製したMcIlvaine緩衝液(pH3〜8)、0.1M リン酸緩衝液(pH8〜10)を使用した。その結果を第2図に示した。至適作用pHは、8.0付近であった。pH8.5よりアルカリ性側、pH7.0より酸性側では、相対的活性は、低下した。
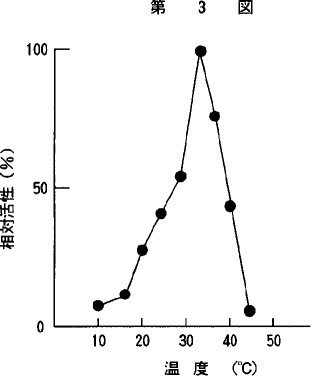
【0018】3)至適作用温度精製酵素の至適作用温度は、10℃〜45℃の温度で、アラニン−2−ナフチルアミドを基質としてpH8.0、0.1mM DTT−0.1M リン酸緩衝液を用いて10分間の反応で測定した。その結果を第3図に示した。本酵素は、35℃付近に至適作用温度を示し、40℃では、相対的活性で約45%、30℃では、約55%であった。
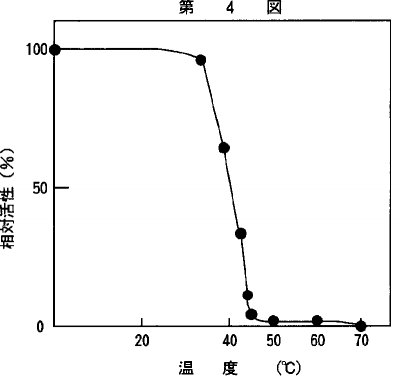
【0019】4)熱安定性酵素活性の熱安定性は、精製酵素をpH8.0、1mM DTT−0.1Mリン酸緩衝液を用いて0℃〜70℃の範囲で10分間加熱処理を行い、加熱後に残存する活性は、pH8.0でアラニン−2−ナフチルアミドを基質として、37℃、5分間の反応で測定し、未加熱の酵素活性を基準として相対活性値を示した。得られた結果を第4図に示した。40℃、10分間の加熱で、約60%、44℃、10分間の加熱で約10%の活性が残存した。
【0020】5)金属イオンの影響本酵素の活性におよぼす各種金属イオンを測定した結果を第1表に示した。各種金属イオンをそれぞれ酵素溶液に0.5mM となるように加え、アラニン−2−ナフチルアミドを基質としてpH8.0、37℃10分間の反応を行った。本酵素は、Co2+に対して若干活性化される傾向であった。またZn2+に対してある程度影響を受けた。
【0021】
【表1】

【0022】6)試薬の影響本酵素の活性におよぼす各種試薬の影響を測定した結果を第2表に示した。酵素溶液にそれぞれの濃度の各試薬の存在下で、アラニン−2−ナフチルアミドを基質としてpH8.0、37℃、10分間の反応を行った。本酵素の活性は0.1mM ベスタチンで著しく阻害された。ロイペプチン、EDTAに対して若干阻害された。
【0023】
【表2】
【0024】7)Km値およびVmax.
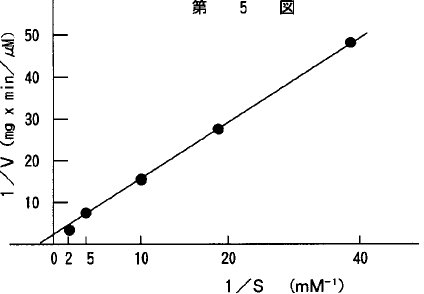
本酵素のKm値およびVmax.は、アラニン−2−ナフチルアミドを基質として求めた。各種濃度の基質をpH8.0の0.1M リン酸緩衝液で調製して、37℃、10分間で酵素反応を行った。得られた結果からLineweaver−Burkの逆数プロットを第5図に示した。Km値は、0.63mMであり、Vmax.は0.63μM/min./mg proteinであった。
【0025】8)基質特異性(1).各種アミノ酸−2−ナフチルアミドに対する基質特異性本酵素の各種アミノ酸−2−ナフチルアミドに対する基質特異性を測定し、アラニン−2−ナフチルアミドを基準とした相対活性値を第3表に示した。酵素反応は、pH8.0で37℃、10分間行った。本酵素は、アミノ酸が、アラニン、リジン、ロイシン、メチオニンであるアミノ酸−2−ナフチルアミドによく作用し、広い基質特異性を示した。
【0026】
【表3】
【0027】(2).各種ペプチドに対する基質特異性本酵素の各種ペプチドに対する基質特異性を測定し、Met−Glyを基準とした相対活性値を第4表に示した。酵素反応は、pH8.0で37℃、30分間行った。C末端をGlyに揃えた13種のジペプチドに作用させた場合、N末端がLys,Leu,Phe,Trp,Metのジペプチドによく作用した。その他、Ala,Ser,Gly,のペプチドにも作用した。ジおよびトリペプチドよりテトラペプチドへの作用が大きかった。テトラペプチドでは、Gly,Ala,Met,LysがN末端にあるものによく作用し、広い基質特異性を示した。
【0028】
【表4】
【0029】10)ペプチドに対する加水分解作用本酵素のLeu−Gly−Gly,Met−Gly−Phe−Alaに対する加水分解作用を測定した。酵素反応は、pH8.0、1mM DTT−0.1Mリン酸緩衝液を用いて、37℃で、作用時間を変えて作用させ、終濃度80%になるようにエタノールを加えて反応を停止した。反応液を濾過後、濾液を蒸発乾固して、その後蒸留水で一定量にしてアミノ酸分析計で、遊離したアミノ酸を定量した。その結果、作用時間の経過とともに、それぞれ遊離されるLeu及びMetがLeuで0.1μM/ml,30min.;0.18μM/ml,60min.、Metで0.2μM/ml,15min.;0.34μM/ml,30min.というように増加することが示され、本酵素が、アミノペプチダーゼ活性を有する事を確認した。
【0030】本発明は、上記したような特性を有するものであれば、すべて本発明のアミノペプチダーゼとして含むとしてよいものである。
【0031】次に、本発明を実施例により詳しく説明する。
【実施例】ペプトン30g、肉エキス15g、NaCl 15gを脱イオン蒸留水3lに溶解し、121℃、20分間滅菌を行ったものを培地とした。この培地にシュードモナス・クロロラフィス(Pseudomonas chlororaphis)(IAM12354t)を接種し、25℃で8日間静置培養した。培養後、培養液から遠心分離して菌体を除去した。得られた上澄液を1mM NaN3−0.1%MCE−10mM Tris−HCl緩衝液(pH7.2)(以下緩衝液Aと略す)に透析して菌体外粗酵素液とした。次いで、以下の方法により酵素を精製した。
【0032】1)DEAE−celluloseカラムクロマトグラフィー菌体外粗酵素液2.5lをあらかじめ緩衝液Aで平衡化しておいたDEAE−celluloseカラム(φ3.0×25cm)にかけた。緩衝液Aで非吸着タンパク質を除去した後、0M〜0.4M NaClまでの食塩濃度傾斜法で溶出した。溶出液は、1画分20mlずつ分画した。分画した各画分のタンパク質量(280nmの吸光値)およびアラニン−2−ナフチルアミドに対する加水分解活性を測定し、アミノペプチダーゼ活性を含む溶出画分No.49〜52を集め、限外濾過膜で濃縮を行った後に、DEAE−celluloseカラムクロマトグラフィー酵素画分とした。
【0033】2)ウルトロゲルAcA34カラムクロマトグラフィー先の酵素液をあらかじめ緩衝液Aで平衡化しておいたウルトロゲルAcA34カラム(φ2.5×103cm)にかけ、同緩衝液で溶出した。溶出液は、1画分10mlずつ分画した。分画した各画分のタンパク質量(280nmの吸光値)およびアラニン−2−ナフチルアミドに対する加水分解活性を測定し、アミノペプチダーゼ活性を含む溶出画分No.24〜27を集め、0.1%MCE−1mM NaN3−1mM リン酸緩衝液(pH7.2)で透析を行った後に、ウルトロゲルAcA34カラムクロマトグラフィー酵素画分とした。
【0034】3)ハイドロキシルアパタイトカラムクロマトグラフィー先の酵素液をあらかじめ0.1%MCE−1mM NaN3−1mM リン酸緩衝液(pH7.2)で平衡化しておいたハイドロキシルアパタイトカラム(φ1.8×10.5cm)にかけた。同緩衝液で非吸着タンパク質を除去した後、リン酸の濃度を10mM に上げて酵素を溶出させた。溶出液は、1画分3mlずつ分画した。分画した各画分のタンパク質量(280nmの吸光値)およびアラニン−2−ナフチルアミドに対する加水分解活性を測定し、アミノペプチダーゼ活性を含む溶出画分No.30〜33を集め、0.1%MCE−1.5M (NH4)2SO4−5mM NaN3−50mM Tris−HCl緩衝液(pH7.2)で透析を行った後に、ハイドロキシルアパタイトカラムクロマトグラフィー酵素画分とした。
【0035】4)フェニルセファロースCL−4Bカラムクロマトグラフィー先の酵素液をあらかじめ0.1%MCE−1.5M (NH4)2SO4−5mM NaN3−50mM Tris−HCl緩衝液(pH7.2)で平衡化しておいたフェニルセファロースCL−4Bカラム(φ1.8×5.5cm)にかけた。非吸着タンパク質を除去した後、硫安濃度を1.5Mから0Mまで濃度傾斜法で下げることにより、吸着タンパク質を溶出した。溶出液は、1画分3mlずつ分画した。分画した各画分のタンパク質量(280nmの吸光値)およびアラニン−2−ナフチルアミドに対する加水分解活性を測定し、アミノペプチダーゼ活性を含む溶出画分No.72〜75を集め、緩衝液Aで透析を行った後に、精製酵素溶液とした。
【0036】以上の精製工程における結果を第5表に要約した。精製酵素は、アラニン−2−ナフチルアミドを基質として、菌体外粗酵素液と比較すると比活性で2161倍に精製され、活性の収率は、14%であった。また、得られた精製酵素の蛋白質濃度(280nmの吸光値)は、0.047mg/mlであった。また、得られた精製酵素の総活性は、5.0557Uであった。活性測定法は、前記した方法に準じて行った。上記フェニルセファロースCL−4B処理して得られた酵素を用いると、前記した種々の酵素の物性及び活性の特性結果が得られた。
【0037】
【表5】
【0038】
【発明の効果】本発明のアミノペプチダーゼは、広い基質特異性を有している。このように広い基質特異性とペプチドのN末端からアミノ酸を順次遊離するアミノペプチダーゼ作用を有するので、本酵素は、タンパク質の構造や機能の研究、分析用試薬、食品工業への応用などに広く利用されるベく、期待される。また、本酵素は、微生物を用いて効率よく製造することができる。
【図面の簡単な説明】
【図1】本発明の酵素および標準タンパク質のゲル濾過カラムにおける溶出位置と分子量の関係を示すグラフであり、図中のAは、オボアルブミン(分子量45,000)、Bは、牛血清アルブミン(分子量67,000)、Cは、カタラーゼ(分子量240,000)、Dは、フェリチン(分子量450,000)、Eは、本発明の酵素を示す。
【図2】pHと本発明の酵素の活性発現の影響についての関係を示すグラフであり、pH3〜8は、マックルバインの緩衝液(黒丸)、pH8〜10は、0.1Mリン酸塩緩衝液(白丸)を用いた。
【図3】温度と本発明の酵素の活性発現の影響についての関係を示すグラフである。
【図4】温度と本発明の酵素の一定時間後の残存活性を示すグラフである。
【図5】本酵素のKm値とVmax.を示すグラフである。
【0001】
【産業上の利用分野】本発明は、シュードモナス属に属する細菌により生産される、ペプチドのN末端からアミノ酸を順次遊離する作用を有し、基質特異性の広い特性を有する新規なアミノペプチダーゼ及びその製造法に関する。
【0002】この酵素は、上記の特性を有することから、蛋白系食品加工における呈味付与、増強にも役立ち、消化剤としての医薬品への利用や、また種々のタンパク質の一次構造決定における生化学試薬としても利用し得るものである。
【0003】
【従来の技術および発明が解決しようとする問題点】従来、アミノペプチダーゼとしては、高等動物の臓器から多くの報告がなされている。哺乳動物の腎臓や小腸のミクロソーム画分に存在する膜酵素として、Wachsmuthらによって(Biochemistry 5,169,1966)およびKimらによって(J.Biol.Chem.,251,3199,1976)また、家兎骨格筋中に存在する酵素として、Otsukaらによって(Agric.Biol.Chem.,44,1617,1980)、サル脳からShimamuraらによって(Biochim.Biophys.Acta,756,223,1983)さらに豚の肝臓からKawataらによって(J.Biochem.,92,1093,1982)アミノペプチダーゼが報告されている。また、市販されているアミノペプチダーゼとしては、シグマ社からロイシンアミノペプチダーゼ、ミクロソーマル(Leucine aminopeptidase,Microsomal)〔EC.3.4.11.2〕が豚腎臓由来のアミノペプチダーゼとしてある。しかし、これらの動物起源の酵素は、その原料の入手の制約、分離、採取の操作、収量などの面で種々の制約を受ける点からも、これに代わる微生物起源の酵素を供給することが望ましい。
【0004】また、微生物起源のアミノペプチダーゼとして、中台らが(Agric.Biol.Chem.,37,157,1973)(Agric.Biol.Chem.,37,767,1973)(Agric.Biol.Chem.,37,775,1973)アスペルギルス・オリゼ(Aspergillus oryzae)の生産するロイシンアミノペプチダーゼI,II,III(分子量26,500;61,000;56,000)を報告している。またこれらと同じ起源であるアスペルギルス・オリゼ由来のプロテアーゼ酵素剤が市販されている。これらの酵素は、N末端のアミノ酸がロイシンであるペプチドのN末端に特によく作用するアミノペプチダーゼ活性を有するところの、基質特異性の狭い(高い)とされているものが殆どであり、特に市販のプロテアーゼ製剤で、種々のアミノ酸を導入したナフチルアミド誘導体を基質として使用して、その酵素活性をみてみると、そのアミノ酸がロイシンのものに特によく作用するような基質特異性の狭いものが殆んどである。このように、種々の用途に幅広く使用し得るアミノペプチダーゼとして、広い基質特異性を有する酵素が望まれているにも拘わらず、従来のものは必ずしも満足し得ないのが、実情である。
【0005】
【問題点を解決するための手段】そこで本発明者らは、基質特異性の広いアミノペプチダーゼを生産する微生物を探索すべく研究を重ねた結果、シュードモナス(Pseudomonas)属に属する細菌が目的とする酵素を生産することを見いだし、本発明を完成した。
【0006】すなわち本発明は、ペプチドのN末端からアミノ酸を順次遊離する作用を有し、基質特異性の広い特性を有する特定のアミノペプチダーゼならびにシュードモナス属に属し、該アミノペプチダーゼを産生する能力のある特定の細菌を培養して該アミノペプチダーゼを製造する方法を提供するものである。
【0007】本発明のアミノペプチダーゼは、シュードモナス属に属し、該酵素生産能を有する特定の細菌を栄養培地に培養し、培養液もしくは、培養上澄液から該酵素を採取することによって製造することができる。
【0008】目的とするアミノペプチダーゼを生産する能力のある微生物としては、シュードモナス・クロロラフィス(Pseudomonas chlororaphis)(IAM12354t )(この菌は、日本微生物株目録(JFCC CATALOGUE OF CULTURES)第4版1987、日本微生物株保存連盟(The Japan Federation for Culture Collections)、昭和62年3月1日発行のPseudomonaschlororaphisの項にIAM(type strain)として記載されている)がある。
【0009】栄養培地は、上記微生物が十分に生育し、目的とする酵素を生産しうるものであればよく、例えば、ペプトン、肉エキス等に塩化ナトリウムなどの無機塩類等を加えたものが好適に用いられる。
【0010】培養は、本発明の場合、静置培養法によって行うことが好ましい。一般には、中性付近のpHに調整し、20〜30℃の温度で2〜16日間程度培養を行い、アミノペプチダーゼを十分に生成せしめることができる。この酵素は、主として菌体外の培養液中に生成される。
【0011】培養液からのアミノペプチダーゼの採取は、適宜既知の手法を組み合わせて行えばよく、その一例を示すと、培養終了後、培養液から遠心分離等によって菌体を除去し、得られた上澄液を適当な緩衝液で透析して粗酵素液を得る。
【0012】次いで必要に応じ、粗酵素液をイオン交換クロマト、ゲル濾過、ハイドロキシルアパタイトクロマト、疎水クロマトなどの酵素精製に用いられる通常の操作を適宜組み合わせることによって、精製または、部分精製されたアミノペプチダーゼを得ることができる。本発明の場合、透析した粗酵素液を直接カラムクロマトに供することが、好ましい。
【0013】次に、本発明の酵素の性質を以下に示す。本発明の酵素は、その分子量が、98,000で、その至適pHは、pH8.0付近にあり、至適温度は、35℃付近にあり、その熱安定性は、37℃以下では、安定であり、図3及び図4で例証されるように50℃より高い温度では相対活性が実質上0である。より具体的には、40℃、10分間の加熱で約60%の活性が残存し、1mMのCo2+イオンでその活性が、約1.3倍に上昇し、ベスタチンに著しく阻害され、また、ロイペプチン、EDTAにも若干阻害されるが、その基質特異性は、アミノ酸−2−ナフチルアミドの基質においてN末端が、アラニン、リジン、ロイシン、メチオニンといったアミノ酸であるものによく作用するなど広い基質特異性が認められ、さらに、トリ、テトラペプチドにも良好な活性を有し、それらのN末端から良好にアミノ酸を遊離するアミノペプチダーゼである。
【0014】なお、酵素活性の測定は、実施例に示した精製酵素を用いて以下の方法で行った。
1)アミノ酸−2−ナフチルアミドに対する活性の測定アミノ酸−2−ナフチルアミドに対する活性の測定は、Matsutaniら(J.Med.Technol.,11,300,1967)の方法に準じて行った。酵素を0.4mlの1mM DTT−0.1M リン酸緩衝液(pH8.0)の存在下、0.5mM のアミノ酸−2−ナフチルアミドに37℃で一定時間作用させた後、0.4mlの0.23N HCl−エタノール溶液を加えて反応を停止させた。さらに0.4mlの0.06%p−ジメチルアミノシンナムアルデヒド・エタノール溶液を加えて30分間放置後、生成したシッフ塩基色素の量を540nmにおける吸光値を分光々度計で測定し、遊離した2−ナフチルアミンを定量した。また、アミノ酸の種類によっては、その基質濃度、基質溶液を変えて行った。酵素活性の単位は、pH8.0、37℃において酵素が、アミノ酸−2−ナフチルアミドから1分間に1μmolの2−ナフチルアミンを遊離させる酵素量を1単位(U)とした。
【0015】2)一般の合成ペプチドに対する活性の測定一般の合成ペプチドに対する活性の測定は、Yemm & Cocking(Analyst,80,209,1975)の方法に準じて行った。酵素を終濃度0.5mM の基質と、0.4mlの1mM DTT−0.1M リン酸緩衝液(pH8.0)の存在下、一定時間作用させた後、氷冷した0.2M クエン酸緩衝液(pH5.0)0.2mlを加えて反応を停止した。さらに1M のモノヨード酢酸(中和済)を0.1ml加えて後、0.4mlのニンヒドリン試薬を加えて、100℃で15分間煮沸した後、流水で5分間冷却した。さらに60%エタノール2mlで希釈した後、570nmにおける吸光値を分光々度計で測定した。
【0016】1)分子量本酵素の分子量は、Andrewsの方法(Biochem.J.,96,595,1965)に準じ、セファデックスG−200によるゲル濾過カラム(φ2.4×105cm)に精製酵素および標準タンパク質を供して1mM NaN3−0.1%MCE−10mM Tris−HCl緩衝液(pH7.2)で、流速0.25ml/min.にて溶出して求めた。精製酵素および標準タンパク質の溶出位置と分子量の関係を第1図に示した。その結果、本酵素の分子量は、98,000と推定した。
【0017】2)至適作用pH本酵素の至適作用pHは、pH3.0〜10.0の範囲で、アラニン−2−ナフチルアミドを基質として37℃、10分間の反応条件で測定した。緩衝液は、各pHに調製したMcIlvaine緩衝液(pH3〜8)、0.1M リン酸緩衝液(pH8〜10)を使用した。その結果を第2図に示した。至適作用pHは、8.0付近であった。pH8.5よりアルカリ性側、pH7.0より酸性側では、相対的活性は、低下した。
【0018】3)至適作用温度精製酵素の至適作用温度は、10℃〜45℃の温度で、アラニン−2−ナフチルアミドを基質としてpH8.0、0.1mM DTT−0.1M リン酸緩衝液を用いて10分間の反応で測定した。その結果を第3図に示した。本酵素は、35℃付近に至適作用温度を示し、40℃では、相対的活性で約45%、30℃では、約55%であった。
【0019】4)熱安定性酵素活性の熱安定性は、精製酵素をpH8.0、1mM DTT−0.1Mリン酸緩衝液を用いて0℃〜70℃の範囲で10分間加熱処理を行い、加熱後に残存する活性は、pH8.0でアラニン−2−ナフチルアミドを基質として、37℃、5分間の反応で測定し、未加熱の酵素活性を基準として相対活性値を示した。得られた結果を第4図に示した。40℃、10分間の加熱で、約60%、44℃、10分間の加熱で約10%の活性が残存した。
【0020】5)金属イオンの影響本酵素の活性におよぼす各種金属イオンを測定した結果を第1表に示した。各種金属イオンをそれぞれ酵素溶液に0.5mM となるように加え、アラニン−2−ナフチルアミドを基質としてpH8.0、37℃10分間の反応を行った。本酵素は、Co2+に対して若干活性化される傾向であった。またZn2+に対してある程度影響を受けた。
【0021】
【表1】
【0022】6)試薬の影響本酵素の活性におよぼす各種試薬の影響を測定した結果を第2表に示した。酵素溶液にそれぞれの濃度の各試薬の存在下で、アラニン−2−ナフチルアミドを基質としてpH8.0、37℃、10分間の反応を行った。本酵素の活性は0.1mM ベスタチンで著しく阻害された。ロイペプチン、EDTAに対して若干阻害された。
【0023】
【表2】
【0024】7)Km値およびVmax.
本酵素のKm値およびVmax.は、アラニン−2−ナフチルアミドを基質として求めた。各種濃度の基質をpH8.0の0.1M リン酸緩衝液で調製して、37℃、10分間で酵素反応を行った。得られた結果からLineweaver−Burkの逆数プロットを第5図に示した。Km値は、0.63mMであり、Vmax.は0.63μM/min./mg proteinであった。
【0025】8)基質特異性(1).各種アミノ酸−2−ナフチルアミドに対する基質特異性本酵素の各種アミノ酸−2−ナフチルアミドに対する基質特異性を測定し、アラニン−2−ナフチルアミドを基準とした相対活性値を第3表に示した。酵素反応は、pH8.0で37℃、10分間行った。本酵素は、アミノ酸が、アラニン、リジン、ロイシン、メチオニンであるアミノ酸−2−ナフチルアミドによく作用し、広い基質特異性を示した。
【0026】
【表3】
【0027】(2).各種ペプチドに対する基質特異性本酵素の各種ペプチドに対する基質特異性を測定し、Met−Glyを基準とした相対活性値を第4表に示した。酵素反応は、pH8.0で37℃、30分間行った。C末端をGlyに揃えた13種のジペプチドに作用させた場合、N末端がLys,Leu,Phe,Trp,Metのジペプチドによく作用した。その他、Ala,Ser,Gly,のペプチドにも作用した。ジおよびトリペプチドよりテトラペプチドへの作用が大きかった。テトラペプチドでは、Gly,Ala,Met,LysがN末端にあるものによく作用し、広い基質特異性を示した。
【0028】
【表4】
【0029】10)ペプチドに対する加水分解作用本酵素のLeu−Gly−Gly,Met−Gly−Phe−Alaに対する加水分解作用を測定した。酵素反応は、pH8.0、1mM DTT−0.1Mリン酸緩衝液を用いて、37℃で、作用時間を変えて作用させ、終濃度80%になるようにエタノールを加えて反応を停止した。反応液を濾過後、濾液を蒸発乾固して、その後蒸留水で一定量にしてアミノ酸分析計で、遊離したアミノ酸を定量した。その結果、作用時間の経過とともに、それぞれ遊離されるLeu及びMetがLeuで0.1μM/ml,30min.;0.18μM/ml,60min.、Metで0.2μM/ml,15min.;0.34μM/ml,30min.というように増加することが示され、本酵素が、アミノペプチダーゼ活性を有する事を確認した。
【0030】本発明は、上記したような特性を有するものであれば、すべて本発明のアミノペプチダーゼとして含むとしてよいものである。
【0031】次に、本発明を実施例により詳しく説明する。
【実施例】ペプトン30g、肉エキス15g、NaCl 15gを脱イオン蒸留水3lに溶解し、121℃、20分間滅菌を行ったものを培地とした。この培地にシュードモナス・クロロラフィス(Pseudomonas chlororaphis)(IAM12354t)を接種し、25℃で8日間静置培養した。培養後、培養液から遠心分離して菌体を除去した。得られた上澄液を1mM NaN3−0.1%MCE−10mM Tris−HCl緩衝液(pH7.2)(以下緩衝液Aと略す)に透析して菌体外粗酵素液とした。次いで、以下の方法により酵素を精製した。
【0032】1)DEAE−celluloseカラムクロマトグラフィー菌体外粗酵素液2.5lをあらかじめ緩衝液Aで平衡化しておいたDEAE−celluloseカラム(φ3.0×25cm)にかけた。緩衝液Aで非吸着タンパク質を除去した後、0M〜0.4M NaClまでの食塩濃度傾斜法で溶出した。溶出液は、1画分20mlずつ分画した。分画した各画分のタンパク質量(280nmの吸光値)およびアラニン−2−ナフチルアミドに対する加水分解活性を測定し、アミノペプチダーゼ活性を含む溶出画分No.49〜52を集め、限外濾過膜で濃縮を行った後に、DEAE−celluloseカラムクロマトグラフィー酵素画分とした。
【0033】2)ウルトロゲルAcA34カラムクロマトグラフィー先の酵素液をあらかじめ緩衝液Aで平衡化しておいたウルトロゲルAcA34カラム(φ2.5×103cm)にかけ、同緩衝液で溶出した。溶出液は、1画分10mlずつ分画した。分画した各画分のタンパク質量(280nmの吸光値)およびアラニン−2−ナフチルアミドに対する加水分解活性を測定し、アミノペプチダーゼ活性を含む溶出画分No.24〜27を集め、0.1%MCE−1mM NaN3−1mM リン酸緩衝液(pH7.2)で透析を行った後に、ウルトロゲルAcA34カラムクロマトグラフィー酵素画分とした。
【0034】3)ハイドロキシルアパタイトカラムクロマトグラフィー先の酵素液をあらかじめ0.1%MCE−1mM NaN3−1mM リン酸緩衝液(pH7.2)で平衡化しておいたハイドロキシルアパタイトカラム(φ1.8×10.5cm)にかけた。同緩衝液で非吸着タンパク質を除去した後、リン酸の濃度を10mM に上げて酵素を溶出させた。溶出液は、1画分3mlずつ分画した。分画した各画分のタンパク質量(280nmの吸光値)およびアラニン−2−ナフチルアミドに対する加水分解活性を測定し、アミノペプチダーゼ活性を含む溶出画分No.30〜33を集め、0.1%MCE−1.5M (NH4)2SO4−5mM NaN3−50mM Tris−HCl緩衝液(pH7.2)で透析を行った後に、ハイドロキシルアパタイトカラムクロマトグラフィー酵素画分とした。
【0035】4)フェニルセファロースCL−4Bカラムクロマトグラフィー先の酵素液をあらかじめ0.1%MCE−1.5M (NH4)2SO4−5mM NaN3−50mM Tris−HCl緩衝液(pH7.2)で平衡化しておいたフェニルセファロースCL−4Bカラム(φ1.8×5.5cm)にかけた。非吸着タンパク質を除去した後、硫安濃度を1.5Mから0Mまで濃度傾斜法で下げることにより、吸着タンパク質を溶出した。溶出液は、1画分3mlずつ分画した。分画した各画分のタンパク質量(280nmの吸光値)およびアラニン−2−ナフチルアミドに対する加水分解活性を測定し、アミノペプチダーゼ活性を含む溶出画分No.72〜75を集め、緩衝液Aで透析を行った後に、精製酵素溶液とした。
【0036】以上の精製工程における結果を第5表に要約した。精製酵素は、アラニン−2−ナフチルアミドを基質として、菌体外粗酵素液と比較すると比活性で2161倍に精製され、活性の収率は、14%であった。また、得られた精製酵素の蛋白質濃度(280nmの吸光値)は、0.047mg/mlであった。また、得られた精製酵素の総活性は、5.0557Uであった。活性測定法は、前記した方法に準じて行った。上記フェニルセファロースCL−4B処理して得られた酵素を用いると、前記した種々の酵素の物性及び活性の特性結果が得られた。
【0037】
【表5】
【0038】
【発明の効果】本発明のアミノペプチダーゼは、広い基質特異性を有している。このように広い基質特異性とペプチドのN末端からアミノ酸を順次遊離するアミノペプチダーゼ作用を有するので、本酵素は、タンパク質の構造や機能の研究、分析用試薬、食品工業への応用などに広く利用されるベく、期待される。また、本酵素は、微生物を用いて効率よく製造することができる。
【図面の簡単な説明】
【図1】本発明の酵素および標準タンパク質のゲル濾過カラムにおける溶出位置と分子量の関係を示すグラフであり、図中のAは、オボアルブミン(分子量45,000)、Bは、牛血清アルブミン(分子量67,000)、Cは、カタラーゼ(分子量240,000)、Dは、フェリチン(分子量450,000)、Eは、本発明の酵素を示す。
【図2】pHと本発明の酵素の活性発現の影響についての関係を示すグラフであり、pH3〜8は、マックルバインの緩衝液(黒丸)、pH8〜10は、0.1Mリン酸塩緩衝液(白丸)を用いた。
【図3】温度と本発明の酵素の活性発現の影響についての関係を示すグラフである。
【図4】温度と本発明の酵素の一定時間後の残存活性を示すグラフである。
【図5】本酵素のKm値とVmax.を示すグラフである。
【特許請求の範囲】
【請求項1】 微生物により産生され、分子量が98,000で、至適pHが8.0付近にあり、至適温度35℃付近にあり、37℃以下では安定であるが50℃より高い温度では相対活性が実質上0である熱安定特性を有し、ペプチドのN末端からアミノ酸を順次遊離する作用を有し、基質特異性の広い特性を有するアミノペプチダーゼ。
【請求項2】 シュードモナス・クロロラフィス(IAM12354t )により産生され、ペプチドのN末端からアミノ酸を順次遊離する作用を有し、基質特異性の広い特性を有するアミノペプチダーゼ。
【請求項3】 ペプチドのN末端からアミノ酸を順次遊離する作用を有し、基質特異性の広い特性を有するアミノペプチダーゼを産生する能力のあるシュードモナス・クロロラフィス(IAM12354t )を栄養培地に培養して得られる培養液、もしくは培養上澄液をイオン交換クロマトグラフィー、ゲル濾過法、ハイドロキシルアパタイトクロマトグラフィー、疎水クロマトグラフィーあるいはそれらの任意の組み合わせに付することを特徴とするアミノペプチダーゼの製造法。
【請求項1】 微生物により産生され、分子量が98,000で、至適pHが8.0付近にあり、至適温度35℃付近にあり、37℃以下では安定であるが50℃より高い温度では相対活性が実質上0である熱安定特性を有し、ペプチドのN末端からアミノ酸を順次遊離する作用を有し、基質特異性の広い特性を有するアミノペプチダーゼ。
【請求項2】 シュードモナス・クロロラフィス(IAM12354t )により産生され、ペプチドのN末端からアミノ酸を順次遊離する作用を有し、基質特異性の広い特性を有するアミノペプチダーゼ。
【請求項3】 ペプチドのN末端からアミノ酸を順次遊離する作用を有し、基質特異性の広い特性を有するアミノペプチダーゼを産生する能力のあるシュードモナス・クロロラフィス(IAM12354t )を栄養培地に培養して得られる培養液、もしくは培養上澄液をイオン交換クロマトグラフィー、ゲル濾過法、ハイドロキシルアパタイトクロマトグラフィー、疎水クロマトグラフィーあるいはそれらの任意の組み合わせに付することを特徴とするアミノペプチダーゼの製造法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図2】
【図3】
【図4】
【図5】
【特許番号】特許第3076936号(P3076936)
【登録日】平成12年6月16日(2000.6.16)
【発行日】平成12年8月14日(2000.8.14)
【国際特許分類】
【出願番号】特願平3−308551
【出願日】平成3年9月12日(1991.9.12)
【公開番号】特開平5−68550
【公開日】平成5年3月23日(1993.3.23)
【審査請求日】平成10年8月5日(1998.8.5)
【出願人】(000201733)曽田香料株式会社 (56)
【参考文献】
【文献】Research Communications in Chemical Pathology and Pharmacology,Vol.39,No.3,(1983),p.463−476
【登録日】平成12年6月16日(2000.6.16)
【発行日】平成12年8月14日(2000.8.14)
【国際特許分類】
【出願日】平成3年9月12日(1991.9.12)
【公開番号】特開平5−68550
【公開日】平成5年3月23日(1993.3.23)
【審査請求日】平成10年8月5日(1998.8.5)
【出願人】(000201733)曽田香料株式会社 (56)
【参考文献】
【文献】Research Communications in Chemical Pathology and Pharmacology,Vol.39,No.3,(1983),p.463−476
[ Back to top ]