
新規なスチレンモノオキシゲナーゼ、その製造方法、およびこれを利用する光学活性なスチレンオキシドの製造方法
【課題】スチレンに酸素を付加してスチレンオキシドを生成するスチレンモノオキシゲナーゼをコードする新規なDNA、該DNAによりコードされるスチレンモノオキシゲナーゼ、該スチレンモノオキシゲナーゼの製造方法などの提供を課題とする
【解決手段】ロドコッカスsp.ST-5株およびST-10株においてスチレンの分解に関与しているスチレン代謝遺伝子を単離するため、ST-5株およびST-10株のゲノムDNAに対し、縮重PCRおよびTAIL-PCRを実施した。その結果本発明者らは、これらの株より、新規なスチレンモノオキシゲナーゼ(SMO)をコードする遺伝子を単離することに成功した。
【解決手段】ロドコッカスsp.ST-5株およびST-10株においてスチレンの分解に関与しているスチレン代謝遺伝子を単離するため、ST-5株およびST-10株のゲノムDNAに対し、縮重PCRおよびTAIL-PCRを実施した。その結果本発明者らは、これらの株より、新規なスチレンモノオキシゲナーゼ(SMO)をコードする遺伝子を単離することに成功した。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、新規なスチレンモノオキシゲナーゼをコードするDNA、および該DNAによりコードされるタンパク質等に関する。また本発明は、該タンパク質を用いた光学活性なスチレンオキシドの製造方法、およびスチレンの分解方法等に関する。
【背景技術】
【0002】
スチレンは、合成高分子等の化学工業等の分野において重要な化学物質であり、ポリスチレンおよびスチレンブタジエンゴムなど多くの合成樹脂の原料として、多量に使用されている。このような合成樹脂の合成において発生するガス状スチレンやスチレン含有水の排出は、しばしば、ヒトに対する毒性および悪臭のため問題となる。
【0003】
スチレンの生物に与える影響を評価するため、哺乳動物におけるスチレン代謝の機構が研究されている(非特許文献1〜3)。哺乳動物において、スチレンは、シトクロムP-450モノオキシゲナーゼ[EC 1.14.14.1]によりスチレンオキシドへと酸化される(非特許文献4)。次いで、スチレンオキシドはいくつかの経路によって解毒される。主要な経路は、水分子の付加によるスチレングリコールの形成を介して進む。スチレングリコールの主要な代謝物は、マンデル酸、フェニルグリオキシル酸、および馬尿酸である(非特許文献5)。シュードモナスsp.、キサントバクター(Xanthobacter)sp.、およびロドコッカス(Rhodococcus)sp.のようないくつかのスチレン分解細菌が土壌から単離されており、その代謝遺伝子の特徴が報告されている(非特許文献6〜10)。
細菌によるスチレンの分解に関して、2つの異なる経路が報告されている。第一の経路は、ビニル側鎖の酸化を含む経路である。第二の経路は、芳香環の酸化を含む経路である(非特許文献11、12)。
【0004】
これまでの研究において、本発明者らは、土壌から単離されたスチレン資化細菌ロドコッカスsp.ST-5株およびST-10株を報告した。これらの細菌は、スチレン代謝に関与している酵素活性を有する(非特許文献13)。さらに、本発明者らは、ST-5株およびST-10株におけるスチレンの分解経路を、以下のように提唱した。
スチレン→スチレンオキシド→フェニルアセトアルデヒド→2-フェニルエタノールおよびフェニル酢酸。
しかしながら、これらの株からスチレン分解に関与している遺伝子は見出されておらず、代謝機構の詳細は不明なままであった。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Korn, M., R. Wodarz, W. Schoknecht, H. Weichardt, and E. Bayer. 1984. Styrene metabolism in man: gas chromatographic separation of mandelic acid enantiomers in the urine of exposed persons. Arch. Toxicol. 55:59-63.
【非特許文献2】Lof, A., E. Gullstrand, E. Lundgren, and M. B. Nordqvist. 1984. Occurrence of styrene-7,8-oxide and styrene glycol in mouse after the administration of styrene. Scand. J. Work Environ. Health 10:179-187.
【非特許文献3】Watabe, T., M. Isobe, T. Sawahata, K. Yoshikawa, S. Yamada, and E. Takabatake. 1978. Metabolism and mutagenicity of styrene. Scand. J. Work Environ. Health 4(Suppl):142-155.
【非特許文献4】Vainio, H., H. Norppa, K. Hemminki, and M. Sorsa. 1981. Metabolism and genotoxicity of styrene. Adv. Exp. Med. Biol. 136:257-274.
【非特許文献5】Sumner, S. C. J., and T. R. Fennell. 1994. Review of the metabolic fate of styrene. Crit. Rev. Toxicol. 24(Suppl): S11-33.
【非特許文献6】Beltrametti, F., A. M. Marconi, G. Bestetti, C. Colombo, E. Galli, M. Ruzzi, and E. Zennaro. 1997. Sequencing and functional analysis of styrene catabolism genes from Pseudomonas fluorescens ST. Appl. Environ. Microbiol, 63: 2232-2239.
【非特許文献7】Hartmans, S., M. J. van der Werf, and J. A. de Bont. 1990. Bacterial degradation of styrene involving a novel flavin adenine dinucleotide-dependent styrene monooxygenase. Appl. Environ. Microbiol. 56:1347-1351.
【非特許文献8】Panke, S., B. Witholt, A. Schmid, and M. G. Wubbolts. 1998. Towards a biocatalyst for (S)-styrene oxide productio: characterization of the styrene degradation pathway of Pseudomonas sp. strain VLB120. Appl. Environ. Microbiol. 64:2032-2043.
【非特許文献9】Velasco, A., S. Alonso, J. L. Garcia, J. Perera, and E. Diaz. 1998. Genetic and functional analysis of the styrene catabolic cluster of Pseudomonas sp. strain Y2. J. Bacteriol. 180:1063-1071.
【非特許文献10】Warhurst, A.M., K. F. Clarke, R. A. Hill, R. A. Holt, and C. A. Fewson. 1994. Metabolism of styrene by Rhodococcus rhodochrous NCIMB 13259. Appl. Environ. Microbiol. 60:1137-1145.
【非特許文献11】Mooney, A., P. G. Ward, and K. E. O'Connor. 2006. Microbial degradation of styrene: biochemistry, molecular genetics, and perspectives for biotechnological applications. Appl. Microbiol. Biotechnol. 72:1-10.
【非特許文献12】Warhurst, A. M., and C. A. Fewson. 1994. Microbial metabolism and biotransformations of styrene. J. Appl. Bacteriol. 77:597-606.
【非特許文献13】Itoh, N., K. Yoshida, and K. Okada. 1996. Isolation and identification of styrene-degrading Corynebacterium strains, and their styrene metabolism. Biosci. Biotech. Biochem. 60:1826-1830.
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明は、スチレンのビニル基をオキシラニル基に酸化する活性を有するスチレンモノオキシゲナーゼをコードする新規なDNAを提供することを課題とする。また本発明は、該DNAによりコードされるタンパク質、該タンパク質の製造方法等を提供する。さらに本発明はスチレンモノオキシゲナーゼをコードするDNAあるいは当該DNAを含むベクターで形質転換された形質転換細胞を利用するスチレンの分解方法、および光学活性なスチレンオキシドの製造方法を提供することを課題とする。
【課題を解決するための手段】
【0007】
本発明者らは、ロドコッカスsp.ST-5株およびST-10株においてスチレンの分解に関与しているスチレン代謝遺伝子を単離するため、ST-5株およびST-10株のゲノムDNAに対し、縮重PCRおよびTAIL-PCR(thermal asymmetric interlaced PCR)を実施した。その結果本発明者らは、ST-5株において4つのオープンリーディングフレーム(ORF)を見出すことに成功した。この4つのORFによってコードされる推定アミノ酸配列は、シュードモナス sp.におけるスチレン代謝遺伝子styABCDと類似性を示した。よって、4つのORFはそれぞれ、スチレンモノオキシゲナーゼ(SMO)、フラビンオキシドレダクターゼ、スチレンオキシドイソメラーゼ(SOI)、およびフェニルアセトアルデヒドデヒドロゲナーゼ(PAD)に対応する遺伝子であると判断された(図1および図2)。
一方ST-10株については、3つのORFを見出すことに成功した。これらのORFのうち2つは、SMOおよびフラビンオキシドレダクターゼに対応する遺伝子であると判断された(図1および図2)。なお残りの1つについては、ロドコッカス属にのみホモログが見出される機能未知タンパク質と高い類似性を示した。
本発明者らは、これらの遺伝子によってコードされるポリペプチドの作用を明らかにして本発明を完成した。すなわち本発明は、新規なスチレンモノオキシゲナーゼをコードするDNA、該DNAによりコードされるスチレンモノオキシゲナーゼ、該スチレンモノオキシゲナーゼの製造方法、および該スチレンモノオキシゲナーゼを用いるスチレンの分解方法並びに光学活性なスチレンオキシドの製造方法などに関する。
【0008】
本発明は、より詳細には以下の〔1〕から〔9〕に関する。
〔1〕以下(a)から(e)のいずれかに記載のポリヌクレオチド;
(a)配列番号:1または3に記載の塩基配列を含むポリヌクレオチド;
(b)配列番号:2または4に記載のアミノ酸配列をコードするポリヌクレオチド;
(c)配列番号:2または4に記載のアミノ酸配列において、1もしくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列を含み、かつ下記(1)および(2)に記載の理化学的性質を有するスチレンモノオキシゲナーゼをコードするポリヌクレオチド;
(d)配列番号:1または3に記載の塩基配列からなるポリヌクレオチドに対して、高度にストリンジェントな条件下でハイブリダイズし、かつ下記(1)および(2)に記載の理化学的性質を有するスチレンモノオキシゲナーゼをコードするポリヌクレオチド;および
(e)配列番号:2または4に記載のアミノ酸配列と70%以上の同一性を有し、かつ下記(1)および(2)に記載の理化学的性質を有するスチレンモノオキシゲナーゼをコードするポリヌクレオチド;
(1)作用
FADH2(還元型FAD)を補酵素として、スチレンおよび3−クロロスチレンに作用し、それぞれ、光学活性なスチレンオキシドおよび光学活性な3−クロロスチレンオキシドを生成する。
(2)分子量
ゲル濾過におけるヒスチジンを含む分子量が59,000で、SDS-PAGEによる分子量が49,000、アミノ酸組成からの推定分子量が47,000のモノマー酵素である。
〔2〕ロドコッカス(Rhodococcus)属に属する微生物由来である、〔1〕に記載のポリヌクレオチド。
〔3〕前記ロドコッカス属に属する微生物がRhodococcus opacus ST-5株またはRhodococcus erythropolis ST-10株である、〔2〕に記載のポリヌクレオチド。
〔4〕〔1〕から〔3〕のいずれかに記載のポリヌクレオチドによってコードされるタンパク質。
〔5〕〔1〕から〔3〕のいずれかに記載のポリヌクレオチドを含むベクター。
〔6〕〔1〕から〔3〕のいずれかに記載のポリヌクレオチド、または〔5〕に記載のベクターが導入された形質転換細胞。
〔7〕〔6〕に記載の形質転換細胞を培養し、その培養物から〔1〕に記載のポリヌクレオチドによってコードされるタンパク質を回収する工程を含む、〔1〕から〔3〕のいずれかに記載のポリヌクレオチドによってコードされるタンパク質の製造方法。
〔8〕〔4〕に記載のタンパク質をスチレンおよび/またはその類似体に作用させる工程、および生成される化合物を回収する工程を含む、光学活性なスチレンオキシドおよび/または光学活性なスチレン類似体の酸化物の製造方法。
〔9〕〔4〕に記載のタンパク質をスチレンおよび/またはその類似体に作用させる工程を含む、スチレンおよび/またはその類似体の分解方法。
【発明の効果】
【0009】
本発明者らにより、新規なスチレンモノオキシゲナーゼをコードするDNA、該DNAによりコードされるスチレンモノオキシゲナーゼなどが提供された。
スチレンは化学工業において重要な物質であり、ポリスチレンやスチレンブタジエンゴムなどの合成樹脂の原料として欠かせない。しかしながら、これらの合成高分子の製造において発生するスチレンガスやスチレン含有廃水は、しばしばヒトに対する毒性や環境汚染の観点から問題となっている。本発明のスチレンモノオキシゲナーゼは、このような問題の原因となるスチレンの分解や除去に有用である。
また本発明のスチレンモノオキシゲナーゼを用いることにより、医薬、農薬、電子材料等の種々の分野で必要とされる光学活性なスチレンオキシドを効率的且つ経済的に製造することができる。本発明のスチレンモノオキシゲナーゼは、このようにスチレンオキシドの製造においても有用である。
【図面の簡単な説明】
【0010】
【図1】ロドコッカスsp.ST-5株、ST-10株、およびシュードモナスsp.Y2におけるスチレン異化遺伝子の比較を示す図である。矢印は、遺伝子の転写の方向を示す。制限酵素部位を次のように示した。B、Bgl II;E、Eco RI;H、Hind III;K、Kpn I;P、Pst I;Sa、Sal I;Sp、Sph I;X、Xho I。
【図2】ロドコッカスsp.ST-5株(A)およびST-10株(B)のスチレン異化遺伝子のヌクレオチド配列および推定アミノ酸配列を示す図である。小さな矢印は、遺伝子の転写の方向を示す。推定されるリボソーム結合部位を太字で示した。下線は、推定される転写終了シグナルであるステムループ配列を示す。
【図3】スチレンの存在下(A)またはスチレンオキシドの存在下(C)における、ロドコッカスsp.ST-10株による静止菌体反応の生成物のHPLC分析結果を示す図である。(B)は(C)のコントロールであり、ST-10株の菌体の非存在下におけるHPLC分析結果を示す。STYはスチレン、SOはスチレンオキシド、ACPはアセトフェノン、PAはフェニルアセトアルデヒド、SGはスチレングリコール、PAAはフェニル酢酸を示す。
【図4】ST-5株またはST-10株におけるフェニルアセチル-CoAリガーゼの反応生成物のHPLCクロマトグラム結果を示す図である。対照は、粗細胞抽出物なしでインキュベートされた基質とした。PAAはフェニル酢酸を、PA-CoAはフェニルアセチル-CoAを示す。
【図5】ロドコッカスsp.ST-5株およびST-10株によるスチレン異化経路を示す図である。実線は酵素反応を示す。点線は自然変換を示す。
【図6】Rhodococcus sp. ST-5株 およびST-10株 両株由来のstyAおよびstyABを発現させるために構築したプラスミドを示す図である。
【発明を実施するための形態】
【0011】
スチレンモノオキシゲナーゼ
本発明のスチレンモノオキシゲナーゼは、以下の(1)および(2)の理化学的性状を有するタンパク質である。
(1)作用
FADH2(還元型FAD)を補酵素として、スチレンおよび3−クロロスチレンに作用し、それぞれ、光学活性なスチレンオキシドおよび光学活性な3−クロロスチレンオキシドを生成する。
(2)分子量
ゲル濾過におけるヒスチジンを含む分子量が59,000で、SDS-PAGEによる分子量が49,000、アミノ酸組成からの推定分子量が47,000のモノマー酵素である。
ゲル濾過カラムにおける分子量は、ヒスチジンタグを含む分子量である。アミノ酸配列から推定される、ヒスチジンタグが付加された状態における分子量は49.6 kDaである。これらから、本願のスチレンモノオキシゲナーゼはモノマーであると推測される。SDS-PAGEによるSMO分子量の分析方法、ゲル濾過におけるSMO分子量の分析方法は実施例に示した。
【0012】
スチレンは、以下の化学式で表される芳香族炭化水素である。例えば、エチルベンゼンを鉄触媒等で脱水素することにより製造することができる。
【化1】

【0013】
本発明のスチレンモノオキシゲナーゼの活性測定は、例えば以下の手順で行うことができる。
ST-5およびST-10 由来SMO の精製酵素と基質、補酵素類を50 mM MES緩衝液中に以下の濃度で添加し、反応液とする。次に、この反応液を30℃、2,500rpmで30分間インキュベーションする。反応終了後、反応液に500μlの酢酸エチルを添加し5分間ボルテックスを行う。15,000 rpm で5分間遠心し、分離した酢酸エチル相を新しいチューブに移す。再度500μl の酢酸エチルを水相に添加し、5分間ボルテックス後、再度遠心により酢酸エチル相を回収する。合計1000μl の回収した酢酸エチル相に適量の硫酸ナトリウムを添加し、2分間ボルテックスを行い、脱水処理を行う。脱水処理を行った抽出サンプルを、Hewllet Packerd HP6890 ガスクロマトグラフィーシステムを用いて、スチレンモノオキシゲナーゼ活性を分析することができる。
【0014】
反応溶液は、例えば以下の組成とすることができる。
50 mM MES Buffer (pH 7.0)
20 mM ギ酸ナトリウム
100μM NADH
200μM FAD
1 U ギ酸脱水素酵素
10 U catalase
5 mM substrate(基質変換反応時は、10 mM の基質で反応を実施)
StyA 150μl (基質変換反応時は、StyAB 共発現菌体の菌体破砕液上清を使用)
StyB 100μl
total 1000μl
【0015】
ガスクロマトグラフィーによる分析は、例えば次のように行うことができる。
酢酸エチルにより抽出された試料1μl をVarian 社製 CP-Cyclodextrin-b-2,3,6-M-19-25 キャピラリーカラム(25 m, 内径0.25 mm)に注入する。 キャリアガス(99.999% 高純度ヘリウムガス)流速は2.8 ml/min、スプリット比は50、線形流速は46 cm/s とすることができる。検出はFID(水素炎イオン化検出器)を用いて行うことができる。気化室温度は250℃、検出器温度は250℃とすることができる。各物質の同定および定量は、各標準化合物を1 mM, 5 mM, 10 mM の濃度に調製したサンプルを同条件で分析し、その検出時間およびピーク面積から検量線を作成して求めることができる。
【0016】
酵素活性1 U は、1分間に1μmol の基質を産物に変換する酵素活性と定義することができる。
本発明のスチレンモノオキシゲナーゼの酵素活性は、好ましくは1U/mg以上、さらに好ましくは5U/mg以上、特に好ましくは10U/mg以上である。
【0017】
上記のような理化学的性状を有するスチレンモノオキシゲナーゼは、例えば、ロドコッカスsp.の培養物から精製して得ることができる。ロドコッカスsp.としては、ロドコッカスsp.ST-5株およびST-10株などが挙げられるがこれらに限定されない。これらの微生物は、かつて、コリネバクテリウム(Corynebacterium)sp.ST-5株およびST-10株として同定されていた微生物である。しかし最近になり、ST-10株はロドコッカスsp.ST-10株として再同定された。ロドコッカスsp.ST-5株およびST-10株は、次のようにして取得することができる。
各地で採取した土壌サンプルに滅菌水を添加し、ボルテックスを行う。懸濁液を無機塩培地(0.3(w/v)%(NH4)2SO4、0.3% KH2PO4、0.1% NaCl、0.02% MgSO4 7H2O、および1.5%寒天、pH7.0)に塗り広げ、スチレンガス雰囲気下で培養する。例えば30℃で3〜7日間培養し、生育したコロニーを4 ml のLB 培地に接種して培養する。得られた培養液のうち例えば10μlを再度無機塩培地に塗り広げ、スチレンガス雰囲気下、30℃で培養する。生育したコロニーを再度LB 培地で培養し、スチレン資化性菌としてストックすることができる。
【0018】
ロドコッカスsp.ST-5株およびST-10株は、例えばPY培地(0.5%酵母抽出物、0.5%バクトペプトン、pH7.0)等、真菌の培養に用いられる一般的な培地で培養することができる。
これらの培地を用いて培養した菌体を回収し、無機塩培地(0.3(w/v)%(NH4)2SO4、0.3% KH2PO4、0.1% NaCl、0.02% MgSO4 7H2O、および1.5%寒天、pH7.0)に再度接種する。この菌体をスチレンガス雰囲気下において生育し回収することにより、高いSMO活性を有する菌体を得ることができる。
通常は、培養開始時のpHを2〜9、好ましくは4〜7に調節し、15〜40℃、好ましくは20〜35℃の温度条件下で培養することが望ましい。培養時間は通常は1日から7日、好ましくは1日から3日である。
【0019】
培養した菌体から無細胞抽出液を調製し、さらに、無細胞抽出液を精製することにより、本発明のスチレンモノオキシゲナーゼを得ることができる。
【0020】
具体的には、上述のようにして得られた菌体から、2−メルカプトエタノール(2−mercaptoethanol)等の還元剤や、フェニルメタンスルホニルフルオリド(phenylmethansulfonyl fluoride;PMFS)やエチレンジアミン4酢酸(以下、EDTAと略す)、ペプスタチン、ロイペプチン、ホスホラミドンのようなプロテアーゼ阻害剤を加えた緩衝液中でガラスビーズとの物理的な衝撃やミニラボ、フレンチプレスなどの高圧を利用するなどして破砕し、無細胞抽出液を得ることができる。
【0021】
無細胞抽出液から、蛋白質の溶解度による分画(アセトンやジメチルスルホキシドなどの有機溶媒による沈澱や硫安などによる塩析など)や、陽イオン交換、陰イオン交換、ゲル濾過、疎水性クロマトグラフィーや、キレート、色素、抗体などを用いたアフィニティークロマトグラフィーなどを適宜組み合わせることにより本発明の酵素を精製することができる。例えば、無細胞抽出液をブルー−セファロース、フェニル−セファロース、Resource Q(いずれもGEヘルスケア製)などのカラムクロマトグラフィーを組み合わせることにより、電気泳動的にほぼ単一バンドにまで精製することができる。
なお、アミノ酸配列から推定されるST-5株由来のStyAの分子量は46,941 Daである。また、アミノ酸配列から推定されるST-10株由来のStyAの分子量は47,290 Daである。ST-5株由来のStyA、ST-10株由来のStyAいずれも、モノマーであると推定される。
【0022】
本発明のスチレンモノオキシゲナーゼを産生する本発明者らが単離したロドコッカスsp.ST-5株、ロドコッカスsp.ST-10株は、本発明者らによって、独立行政法人製品評価技術基盤機構特許微生物寄託センターに寄託された。以下に、寄託を特定する内容を記載する。
ロドコッカスsp.ST-5株
(1)寄託機関名:独立行政法人製品評価技術基盤機構
(2)連絡先:〒292−0818千葉県木更津市かずさ鎌足2−5−8
(3)受託番号:NITE P-974
(4)識別の表示:ST-5
(5)寄託日:2010年8月27日
ロドコッカスsp.ST-10株
(1)寄託機関名:独立行政法人製品評価技術基盤機構
(2)連絡先:〒292−0818千葉県木更津市かずさ鎌足2−5−8
(3)受託番号:NITE P-975
(4)識別の表示:ST-10
(5)寄託日:2010年8月27日
【0023】
ポリヌクレオチド
本発明において、ポリヌクレオチドは、DNAおよびRNA等の天然のポリヌクレオチドに加え、人工的なヌクレオチド誘導体を含む人工的な分子であることもできる。また本発明のポリヌクレオチドは、DNA-RNAのキメラ分子であることもできる。また、本発明のポリヌクレオチドは、スチレンモノオキシゲナーゼをコードするコード鎖のみからなる一本鎖であっても、該コード鎖とその相補鎖とからなる二本鎖構造を持つものであってもよい。
【0024】
本発明のスチレンモノオキシゲナーゼをコードするポリヌクレオチドは、微生物から得ることができる。微生物としては、ロドコッカス(Rhodococcus)属に属する微生物が好ましい。より具体的には、Rhodococcus opacusやRhodococcus erythropolisは、本発明のポリヌクレオチドを得るための微生物として好ましい。例えば、配列番号:1に記載の塩基配列を含むポリヌクレオチドは、Rhodococcus opacus ST-5株からクローニングされた。配列番号:1に示す塩基配列は、配列番号:2に示すアミノ酸配列からなるタンパク質をコードしている。また、例えば、配列番号:3に記載の塩基配列を含むポリヌクレオチドは、Rhodococcus erythropolis ST-10株からクローニングされた。配列番号:3に示す塩基配列は、配列番号:4に示すアミノ酸配列からなるタンパク質をコードしている。配列番号:2または4のアミノ酸配列を含むタンパク質をコードするポリヌクレオチドは、本発明における好ましいポリヌクレオチドである。
【0025】
本発明のポリヌクレオチドには、Rhodococcus opacus ST-5株、あるいはRhodococcus erythropolis ST-10株からクローニングされた遺伝子に加え、該遺伝子のホモログも含まれる。このような遺伝子のホモログとしては、配列番号:1または3に記載の塩基配列を含むポリヌクレオチドが挙げられる。その他、スチレンモノオキシゲナーゼをコードするポリヌクレオチドに加え、その読み枠に影響しないよう適当な制御配列を含むポリヌクレオチド、適当なペプチド配列により修飾されたスチレンモノオキシゲナーゼの融合タンパク質をコードするポリヌクレオチドを、好適な例として挙げることができる。
【0026】
また、本発明のポリヌクレオチドに含まれるホモログとしては、配列番号:2または4に記載のアミノ酸配列を含むタンパク質をコードするポリヌクレオチドが挙げられる。このようなポリヌクレオチドには、配列番号:1または3に示す塩基配列を含む遺伝子に加え、遺伝暗号の縮重により配列番号:2または4に記載のアミノ酸配列をコードするが、配列番号:1または3の塩基配列とは異なる塩基配列からなるポリヌクレオチドが包含される。
【0027】
今回、配列番号:1または3の塩基配列を有するスチレンモノオキシゲナーゼをコードする遺伝子が得られた。ホモログは、例えば、配列番号:1または3の塩基配列を有するポリヌクレオチドをプローブとしてスクリーニングすることができる。あるいは、配列番号:1または3の塩基配列から選択された塩基配列をプローブとすることもできる。プローブを使ったスクリーニング方法として、次のような公知のハイブリダイゼーション法を利用することができる。
コロニーハイブリダイゼーション、
プラークハイブリダイゼーション、
サザンブロット法
スクリーニングには、酵素生産株であるRhodococcus opacus、Rhodococcus erythropolisなどロドコッカス属の微生物、あるいはその他の生物種等の染色体DNA、またはcDNAライブラリーを利用することができる。
【0028】
また、酵素生産株の染色体DNAまたはcDNAライブラリーを鋳型としたPCRによって、本発明のポリヌクレオチドを得ることもできる。PCR用のプライマーは、配列番号:1または3の塩基配列を元にデザインすることができる。PCRによって得られたDNAが断片であれば、DNA断片の塩基配列に基づいてさらにその全長配列を決定することができる。例えば、インバースPCR(inverse PCR; Genetics(1988)120: 621-3)を利用して、断片配列情報に基づいて、全長配列を決定することができる。インバースPCRは、部分的に塩基配列が不明なDNAを適当な制限酵素で消化後、自己環化反応により環化されたDNAを鋳型として利用するPCRである。断片配列内部の既知領域に特異的なプライマーを利用して、その前後に位置する塩基配列が未知の領域を増幅することができる。
あるいはRACE法(Rapid Amplification of cDNA End;「PCR実験マニュアル」HBJ出版局,p25-33)によって本発明のポリヌクレオチドを得ることもできる。
【0029】
また、本発明におけるポリヌクレオチドのホモログは、配列番号:1または3に示される塩基配列からなるポリヌクレオチドと高度にストリンジェントな条件下でハイブリダイズできるポリヌクレオチドであって、かつ、スチレンモノオキシゲナーゼ活性を有するタンパク質をコードするポリヌクレオチドも含む。
【0030】
本発明において、高度にストリンジェントな条件とは、例えばECL direct nucleic acid labeling and detection system(Amersham Pharmacia Biotech社製)を用いて、マニュアルに記載の条件(例えば、wash:42℃、0.5×SSCを含むprimary wash buffer)においてハイブリダイズすることを言う。より具体的な「高度にストリンジェントな条件」とは、例えば、通常、42℃、2×SSC、 0.1%SDSの条件であり、好ましくは50℃、2×SSC、0.1%SDSの条件であり、さらに好ましくは、65℃、0.1×SSCおよび0.1% SDSの条件である。これら温度、塩濃度に加えて、プローブ濃度、プローブの長さ、反応時間を含む複数の要素がハイブリダイゼーションのストリンジェンシーに影響する。当業者であればこれら要素を適宜選択することでストリンジェンシーを調節することができる。
【0031】
例えばハイブリダイゼーションに使用するプローブの塩基配列は、配列番号:1または3に記載の塩基配列から選択することができる。例えば、少なくとも20個、好ましくは少なくとも30個、例えば40、60または100個の連続した塩基配列を有するポリヌクレオチドをプローブとすることができる。あるいは、配列番号:1または3に記載した塩基配列の全長を有するポリヌクレオチドをプローブとすることもできる。
ハイブリダイゼーション法の詳細な手順については、「Molecular Cloning, A Laboratory Manual 2nd ed.」(Cold Spring Harbor Press(1989)、特にSection9.47-9.58)、「Current Protocols in Molecular Biology」(John Wiley&Sons(1987-1997)、特にSection6.3-6.4)、「DNA Cloning 1: Core Techniques, A Practical Approach 2nd ed.」(Oxford University(1995)、特にSection2.10)等を参照することができる。
【0032】
高度にストリンジェントな条件下でハイブリダイズできるポリヌクレオチドは、配列番号:2または4に記載のアミノ酸配列と高い同一性(相同性)を有するアミノ酸配列をコードしている可能性が高い。具体的には、配列番号:2または4に示されるアミノ酸配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは90%以上、さらに好ましくは95%あるいは98%、特に好ましくは99%以上の同一性を有するアミノ酸配列をコードするポリヌクレオチドは、本発明における好ましいポリヌクレオチドに含まれる。このような高い同一性を有するタンパク質同士は、同じまたは類似した活性を有する可能性が高い。
タンパク質の同一性は、アミノ酸配列に関するデータベースを利用して検索することができる。例えばSWISS-PROT、PIR、DAD等のタンパク質のアミノ酸に配列情報を蓄積したデータベースを利用することができる。あるいはDNAの同一性は、塩基配列情報を蓄積したデータベースを利用して検索することができる。DDBJ、EMBLまたはGenBank等のDNAに関するデータベースが公知である。これらのデータベースにおいては、DNAの塩基配列を元に予想されたアミノ酸配列情報を利用することもできる。各種の配列情報は、これらのデータベース等を対象に、BLAST、FASTA等のプログラムを利用して検索することができる。ここに例示したデータベースは、いずれもインターネット(例えば、 http://www.ncbi.nlm.nih.gov.)を通じて利用することができる。
【0033】
さらに、本発明のポリヌクレオチドのホモログとしては、配列番号:2または4に記載のアミノ酸配列において1もしくは複数のアミノ酸が欠失、置換、挿入および/または付加されたアミノ酸配列からなり、かつ、スチレンモノオキシゲナーゼ活性を有するタンパク質をコードするポリヌクレオチドを挙げることができる。当業者は、ある塩基配列を元に、適宜置換、欠失、挿入、および/または付加変異を導入することにより、このようなポリヌクレオチドのホモログを得ることができる。例えば、配列番号:1または3に記載の塩基配列からなるポリヌクレオチドに、部位特異的変異導入法(Nucleic Acids Res (1982) 10:6487;Methods in Enzymol (1983) 100:448;Molecular Cloning 2nd Edt., Cold Spring Harbor Laboratory Press(1989);PCR A Practical Approach, IRL Press(1991) pp.200)などを用いて、任意の変異を導入することができる。
【0034】
タンパク質においてアミノ酸残基を置換する場合、特に、側鎖の化学的性質が類似したアミノ酸による置換、いわゆる保存的なアミノ酸置換を行うことが好ましい。アミノ酸は、それらの側鎖の化学的性質に従い、例えば、次のように分類される:
(1)中性疎水性側鎖(アラニン、トリプトファン、バリン、フェニルアラニン、プロリン、メチオニン、ロイシン);
(2)中性極性側鎖(アスパラギン、グリシン、グルタミン、システイン、セリン、チロシン、トレオニン);
(3)塩基性側鎖(アルギニン、ヒスチジン、リシン);
(4)酸性側鎖(アスパラギン酸、グルタミン酸);
(5)脂肪族側鎖(アラニン、イソロイシン、グリシン、バリン、ロイシン);
(6)脂肪族水酸基側鎖(セリン、トレオニン);
(7)アミン含有側鎖(アスパラギン、アルギニン、グルタミン、ヒスチジン、リシン);
(8)芳香族側鎖(チロシン、トリプトファン、フェニルアラニン);および
(9)硫黄含有側鎖(システイン、メチオニン)。
【0035】
すなわち、これらの各群を構成するアミノ酸残基の相互の置換を、保存的置換と言う。本発明において、「1もしくは複数のアミノ酸が置換、欠失、挿入、および/または付加」するアミノ酸の数や場所は、上記DNAがコードするタンパク質がスチレンモノオキシゲナーゼ活性を有する限り、制限されない。変異が許容されるアミノ酸残基の数は、典型的には全アミノ酸の10%以内、好ましくは全アミノ酸の5%、さらに好ましくは全アミノ酸の1%以内である。より具体的には、配列番号:2または4のアミノ酸配列において、通常50以内、例えば20以内、より好ましくは5以内のアミノ酸残基の変異は、許容される。
【0036】
本発明のポリヌクレオチドのホモログには、上述のように、配列番号:2または4に記載のアミノ酸配列において1もしくは複数のアミノ酸が欠失されたアミノ酸配列からなり、かつ、スチレンモノオキシゲナーゼ活性を有するタンパク質をコードするポリヌクレオチドが含まれる。このような欠失を含むポリヌクレオチドには、配列番号:2または4のアミノ酸配列からなるタンパク質の「一部分」をコードするポリヌクレオチドが包含される。
【0037】
ポリヌクレオチドによってコードされるタンパク質が、元のタンパク質の生物学的活性(スチレンモノオキシゲナーゼ活性)を維持している限り、アミノ酸配列におけるアミノ酸残基の欠失・置換・付加および/または挿入は許容される。ここで、生物学的活性の維持とは、元の酵素が触媒する少なくとも一つの反応を触媒する能力が維持されることを言う。「活性の維持」には、同じ活性レベルのみならず、より高い活性も含まれる。また、活性のレベルが低下する場合であっても、実質的に同等の活性であれば、活性の維持に含まれる。実質的に同等とは、元の活性に対して、例えば50%〜100%、通常70〜100%、好ましくは80〜100%、より好ましくは90、あるいは95〜100%の活性を言う。本発明におけるタンパク質の生物学的活性、すなわちスチレンモノオキシゲナーゼ活性の評価方法は既に述べた。
【0038】
本発明のポリヌクレオチドには、天然よりクローニングされたゲノムDNA、およびcDNAの他、合成によって得られるポリヌクレオチドも含まれる。本発明のポリヌクレオチドは、例えば、配列番号:1または3に記載の配列情報を元に、周知の手法により合成することができる。
【0039】
また、本発明の酵素は遺伝子工学的な手法、あるいは化学的な合成法によっても得ることができる。例えば、細胞を含まない試験管内でのタンパク質の製造方法としてin vitroトランスレーション(Dasso and Jackson, Nucleic Acids Res(1989)17:3129-44)が知られている。また、本発明のポリヌクレオチドを適当な発現ベクターに組込み、該発現ベクターが発現される宿主に形質転換し、該宿主細胞より所望のタンパク質を得ることもできる。このような宿主-ベクター系によりタンパク質を産生する方法については、以下の「組換えベクターおよび形質転換体」の項において具体的に述べる。これら公知のタンパク質の製造方法は、本発明のタンパク質を得るための方法として利用することができる。
【0040】
遺伝子工学的な手法により製造されたタンパク質は、当該タンパク質を含む生物材料から回収される。例えば、タンパク質が宿主細胞外に分泌される場合には、当該細胞を培養した培地からタンパク質が回収される。宿主がトランスジェニック生物の場合にはその体液から目的のタンパク質を回収できる。あるいは細胞内に産生される場合には細胞を溶解した溶解物よりタンパク質を回収する。
【0041】
回収されたタンパク質は、該タンパク質を天然において産生する細胞から精製する場合と同様の手段により精製することができる。すなわち、公知の塩析、蒸留、各種クロマトグラフィー、ゲル電気泳動、ゲル濾過、限外濾過、再結晶、酸抽出、透析、免疫沈降、溶媒沈澱、溶媒抽出、硫安またはエタノール沈澱等の精製手法を組み合わせて、目的とするタンパク質を精製することができる。当業者は、例えば次のような各種のクロマトグラフィーを組み合わせて利用することができる。これらのクロマトグラフィーには、HPLCおよびFPLC等の液相クロマトグラフィーシステムを用いることができる。
アフィニティークロマトグラフィー、
アニオンまたはカチオン交換等のイオン交換クロマトグラフィー、
逆相クロマトグラフィー、
吸着クロマトグラフィー、
ゲル濾過、
疎水性クロマトグラフィー、
ヒドロキシアパタイトクロマトグラフィー
【0042】
また、本発明のタンパク質(スチレンモノオキシゲナーゼ)をタグとの融合タンパク質として発現させれば、タグに結合するカラムを利用して回収することができる。例えばGSTタグを有する融合タンパク質は、グルタチオンカラムを用いて容易に分離することができる。あるいはヒスチジンタグとの融合タンパク質は、ニッケルカラムを用いて精製することができる。タグとタンパク質(スチレンモノオキシゲナーゼ)の間にプロテアーゼ認識配列を挿入することができる。プロテアーゼには、例えばトロンビンやファクターXa等を利用することができる。融合タンパク質をカラムに結合させた後、必要に応じてカラムを洗浄する。次いでこれらのプロテアーゼを作用させると、目的とするタンパク質がタグから切断される。その後、切断されたタンパク質を回収することによって、目的とするタンパク質を容易に精製することができる。
【0043】
組換えベクターおよび形質転換体
本発明のポリヌクレオチドを公知の発現ベクターに挿入することにより、スチレンモノオキシゲナーゼ発現ベクターが提供される。即ち本発明は、本発明のポリヌクレオチドを含む組換えベクターに関する。適当なベクターとして、プラスミド、コスミド、ウイルス、バクテリオファージ等の種々のベクターを挙げることができる(Molecular Cloning, A Laboratory Manual 2nd ed., Cold Spring Harbor Press(1989); Current Protocols in Molecular Biology, John Wiley&Sons(1987)参照)。本発明の好ましいベクターとしては、これに限定されるわけではないが、例えば、大腸菌における発現ベクターpETにスチレンモノオキシゲナーゼをコードする遺伝子を発現可能に挿入したE.coli.BL21(DE3)等が挙げられる。
【0044】
本発明の組換えベクターは、分子生物学、生物工学および遺伝子工学の分野において慣用されている技術に準じて構築することができる(Molecular Cloning, A Laboratory Manual 2nd ed., Cold Spring Harbor Press(1989); Current Protocols in Molecular Biology, John Wiley&Sons(1987)参照)。微生物等を宿主として、本発明のポリヌクレオチドを発現させるためには、まず、当該微生物中において安定に存在するプラスミドベクターまたはファージベクター中に該DNAを導入し、その遺伝情報を転写・翻訳させる必要がある。そのためには、通常、プロモーターを本発明のポリヌクレオチドの5’側上流に配置する。プロモーターは、転写・翻訳を制御するユニットである。
【0045】
そして本発明のポリヌクレオチドの3’側下流には、ターミネーターを配置するのが好ましい。プロモーターおよびターミネーターとしては、宿主として利用する微生物中において機能するものを選択することができる。各種微生物において利用可能なベクター、プロモーターおよびターミネーター等の制御配列は、「微生物学基礎講座8遺伝子工学」(共立出版)などで参照することができる。また特に酵母の制御配列について、Adv Biochem Eng (1990) 43:75-102およびYeast (1992) 8:423-88等に詳細に記述されている。その他、必要に応じて、エンハンサー、オペレーター配列、開始シグナル、ポリアデニル化シグナル、リボソーム結合部位等の転写および/または翻訳に必要な制御配列を組込むことができる。
本発明のベクターは、好ましくは、挿入された本発明のポリヌクレオチドの発現に必要とされる制御配列の全ての構成成分を含むものである。さらに、本発明のベクターは、該ベクターが導入された宿主細胞を選択するための選択マーカーを含むことができる。
【0046】
本発明においては、本発明のポリヌクレオチドに、シグナルペプチドをコードする配列を付加することもできる。シグナルペプチドの付加によって、宿主細胞内で発現されたタンパク質を小胞体内腔に移行させることができる。あるいは、グラム陰性菌を宿主とする場合には、シグナルペプチドによって、宿主細胞内で発現されたタンパク質を、ペリプラズム内、または細胞外へと移行させることができる。利用する宿主細胞において機能することができる任意のシグナルペプチドを利用することができる。従って、宿主にとって異種由来のシグナルペプチドを利用することもできる。さらに必要に応じ、本発明のポリヌクレオチドのベクターへの導入に当たって、リンカー、開始コドン(ATG)、終止コドン(TAA、TAGまたはTGA)等を付加することもできる。
【0047】
本発明のポリヌクレオチドを発現させるための宿主は、該ポリヌクレオチドを含む組換えベクターによって形質転換することができ、かつスチレンモノオキシゲナーゼ活性を有するタンパク質を発現することができる生物であれば特に制限されない。本発明は、本発明のポリヌクレオチド、または本発明のベクターにより形質転換された形質転換体(形質転換細胞)を提供する。本発明の形質転換体の対象となる微生物としては、例えば以下のような微生物を示すことができる。
(1)宿主ベクター系の開発されている細菌
・エシェリヒア(Escherichia)属
・バチルス(Bacillus)属
・シュードモナス(Pseudomonas)属
・セラチア(Serratia)属
・ブレビバクテリウム(Brevibacterium)属
・コリネバクテリイウム(Corynebacterium)属
・ストレプトコッカス(Streptococcus)属
・ラクトバチルス(Lactobacillus)属など
(2)宿主ベクター系の開発されている放線菌
・ロドコッカス(Rhodococcus)属
・ストレプトマイセス(Streptomyces)属など
(3)宿主ベクター系の開発されている酵母
・サッカロマイセス(Saccharomyces)属
・クライベロマイセス(Kluyveromyces)属
・シゾサッカロマイセス(Schizosaccharomyces)属
・チゴサッカロマイセス(Zygosaccharomyces)属
・ヤロウイア(Yarrowia)属
・トリコスポロン(Trichosporon)属
・ロドスポリジウム(Rhodosporidium)属
・ピキア(Pichia)属
・キャンディダ(Candida)属など
(4)宿主ベクター系の開発されているカビ
・ノイロスポラ(Neurospora)属
・アスペルギルス(Aspergillus)属
・セファロスポリウム(Cephalosporium)属
・トリコデルマ(Trichoderma)属など
【0048】
例えばエシェリヒア属、特に大腸菌(Escherichia coli) においては、プラスミドベクターとして、pET、pBR、pUC系プラスミドを利用でき、lac(β-ガラクトシダーゼ)、trp(トリプトファンオペロン)、 tac、trc (lac、trpの融合)、λファージ PL、PR等に由来するプロモーターが利用できる。また、ターミネーターとしては、trpA由来、ファージ由来、rrnBリボソーマルRNA由来のターミネーターを用いることができる。特に、市販のpSE420(Invitrogen製)のマルチクローニングサイトを一部改変したベクターpSE420D (特開2000-189170に記載)は、エシェリヒア属細菌を宿主とした場合の好適なベクターである。
【0049】
バチルス属においては、ベクターとしてpUB110系プラスミド、pC194系プラスミド等が利用可能であり、これらのベクターを利用した場合、本発明のポリヌクレオチドを宿主染色体にインテグレートすることもできる。また、プロモーター、ターミネーターとしては、apr(アルカリプロテアーゼ)、 npr(中性プロテアーゼ)、amy(α-アミラーゼ)等が利用できる。
【0050】
シュードモナス属においては、シュードモナス・プチダ(Pseudomonas putida)、シュードモナス・セパシア(Pseudomonas cepacia) 等で宿主ベクター系が開発されている。トルエン化合物の分解に関与するプラスミドTOLプラスミドを基本にした広宿主域ベクター(RSF1010等に由来する自律的複製に必要な遺伝子を含む)pKT240等が利用可能であり、プロモーター・ターミネーターとして、リパーゼ(特開平5-284973)遺伝子由来のものが利用できる。
【0051】
ブレビバクテリウム属、特にブレビバクテリウム・ラクトファーメンタム(Brevibacterium lactofermentum)においては、pAJ43(Gene (1985) 39:281)等のプラスミドベクターが利用可能である。プロモーター・ターミネーターとしては、大腸菌で使用されているプロモーター、ターミネーターがそのまま利用可能である。
【0052】
コリネバクテリウム属、特にコリネバクテリウム・グルタミカム(Corynebacterium glutamicum)においては、pCS11(特開昭57-183799)、pCB101(Mol Gen Genet (1984)196:175)等のプラスミドベクターが利用可能である。
【0053】
ストレプトコッカス属においては、pHV1301(FEMS Microbiol Lett (1985) 26:239)、pGK1(Appl Environ Microbiol (1985) 50:94)等がプラスミドベクターとして利用可能である。
【0054】
ラクトバチルス属においては、ストレプトコッカス属用に開発されたpAMβ1(J Bacteriol (1979) 137: 614)等がベクターとして利用可能であり、プロモーターとしては、大腸菌で利用されているものが利用可能である。
【0055】
ロドコッカス属においては、Rhodococcus opacusやRhodococcus erythropolisに加え、ロドコッカス・ロドクロウス(Rhodococcus rhodochrous)から単離されたプラスミドベクターが使用可能である (J Gen Microbiol (1992) 138:1003)。
【0056】
ストレプトマイセス属においては、HopwoodらのGenetic Manipulation of Streptomyces: A Laboratory Manual Cold Spring Harbor Laboratories (1985)に記載の方法に従って、プラスミドを構築することができる。特に、ストレプトマイセス・リビダンス(Streptomyces lividans) においては、pIJ486 (Mol Gen Genet (1986) 203: 468-78)、pKC1064(Gene (1991) 103:97-9)、pUWL-KS (Gene (1995) 165:149-50)が使用できる。また、ストレプトマイセス・バージニア(Streptomyces virginiae)においても、同様のプラスミドを使用することができる(Actinomycetol (1997) 11:46-53)。
【0057】
サッカロマイセス属、特にサッカロマイセス・セレビジアエ(Saccharomyces cerevisiae) においては、YRp系、YEp系、YCp系、YIp系プラスミドが利用可能である。また、染色体内に多コピー存在するリボソームDNAとの相同組換えを利用したインテグレーションベクター(EP537456等)は、多コピーで遺伝子を導入でき、かつ安定に遺伝子を保持できるため極めて有用である。また、 ADH(アルコール脱水素酵素)、GAPDH(グリセルアルデヒド-3-リン酸脱水素酵素)、PHO(酸性フォスファターゼ)、GAL(β-ガラクトシダーゼ)、PGK(ホスホグリセレートキナーゼ)、ENO(エノラーゼ)等のプロモーター・ターミネーターが利用可能である。
【0058】
クライベロマイセス属、特にクライベロマイセス・ラクティス(Kluyveromyces lactis)においては、サッカロマイセス・セレビジアエ由来2μm系プラスミド、pKD1系プラスミド(J Bacteriol (1981) 145:382-90)、キラー活性に関与するpGKl1由来プラスミド、クライベロマイセス属における自律増殖遺伝子KARS系プラスミド、リボソーム DNA等との相同組換えにより染色体中にインテグレート可能なベクタープラスミド(EP537456等)等が利用可能である。また、ADH、PGK等に由来するプロモーター・ターミネーターが利用可能である。
【0059】
シゾサッカロマイセス属においては、シゾサッカロマイセス・ポンベ(Schizosaccharomyces pombe) 由来のARS (自律複製に関与する遺伝子)およびサッカロマイセス・セレビジアエ由来の栄養要求性を相補する選択マーカーを含むプラスミドベクターが利用可能である(Mol Cell Biol (1986) 6:80)。また、シゾサッカロマイセス・ポンベ由来のADHプロモーター等が利用できる(EMBO J (1987) 6:729)。特に、pAUR224は、宝酒造から市販されており容易に利用できる。
【0060】
チゴサッカロマイセス属においては、チゴサッカロマイセス・ロウキシ(Zygosaccharomyces rouxii) 由来のpSB3(Nucleic Acids Res. 13:4267(1985))等に由来するプラスミドベクターが利用可能であり、サッカロマイセス・セレビジアエ由来 PHO5 プロモーター、およびチゴサッカロマイセス・ロウキシ由来 GAP-Zr(グリセルアルデヒド-3-リン酸脱水素酵素)のプロモーター(Agri Biol Chem (1990) 54:2521)等が利用可能である。
【0061】
ピキア属においては、ピキア・アンガスタ(Pichia angusta;旧名:ハンゼヌラ・ポリモルファ(Hansenula polymorpha))を用いた宿主ベクター系が開発されている。ベクターとしては、ピキア・アンガスタ由来自律複製に関与する遺伝子(HARS1、HARS2)も利用可能であるが、比較的不安定であるため、染色体への多コピーインテグレーションが有効である(Yeast (1991) 7:431-43)。また、メタノール等で誘導されるAOX(アルコールオキシダーゼ)、FDH(ギ酸脱水素酵素)のプロモーター等が利用可能である。また、ピキア・パストリス(Pichia pastoris)等について、ピキア由来自律複製に関与する遺伝子 (PARS1、 PARS2)等を利用した宿主ベクター系が開発されており(Mol Cell Biol (1985) 5:3376)、高濃度培養とメタノールで誘導可能なAOX等の強いプロモーターが利用できる(Nucleic Acids Res (1987) 15:3859)。
【0062】
キャンディダ属においては、キャンディダ・マルトーサ(Candida maltosa)、キャンディダ・アルビカンス(Candida albicans)、キャンディダ・トロピカリス(Candida tropicalis)、キャンディダ・ウチルス(Candida utilis) 等について宿主ベクター系が開発されている。キャンディダ・マルトーサにおいては、キャンディダ・マルトーサ由来ARSがクローニングされ(Agri Biol Chem (1987) 51:1587)、これを利用したベクターが開発されている。また、キャンディダ・ウチルスにおいては、染色体インテグレートタイプのベクターでは、強力なプロモーターが開発されている(特開平08-173170)。
【0063】
アスペルギルス属においては、アスペルギルス・ニガー(Aspergillus niger)、アスペルギルス・オリジー(Aspergillus oryzae) 等が最もよく研究されている。これらを宿主とするプラスミドが利用可能であり、染色体への所望遺伝子のインテグレーションを行うことができる。菌体外プロテアーゼおよびアミラーゼ由来のプロモーターも利用可能である(Trends in Biotechnology (1989) 7:283-7)。
【0064】
トリコデルマ属においては、トリコデルマ・リーゼイ(Trichoderma reesei)を利用した宿主ベクター系が開発されており、菌体外セルラーゼ遺伝子由来プロモーター等が利用できる(Biotechnology (1989) 7:596-603)。
【0065】
また、微生物以外でも、植物および動物を宿主とする様々な宿主ベクター系が開発されている。例えば、大量に異種タンパク質を発現させる系として、蚕を用いた昆虫ベクター系(Nature (1985) 315:592-4)、並びに、菜種、トウモロコシおよびジャガイモ等の植物ベクター系が開発されており、好適に利用できる。
【0066】
ベクターへの本発明のポリヌクレオチドの導入は、制限酵素サイトを利用したリガーゼ反応により行うことができる(Current Protocols in Molecular Biology, John Wiley & Sons(1987) Section 11.4-11.11; Molecular Cloning, A Laboratory Manual 2nd ed., Cold Spring Harbor Press(1989) Section 5.61-5.63)。また、使用する宿主のコドン使用頻度を考慮し、必要に応じ本発明のポリヌクレオチド配列の改変を行い、発現効率の高いベクターを設計するようにしてもよい(Grantham et al., Nucleic Acids Res (1981) 9:r43-74)。
【0067】
上述のように、様々な細胞が宿主細胞株として確立されている。そして、各細胞株に適した発現ベクターの導入法も公知であり、当業者であれば、各選択した宿主細胞に好適な導入法を選択することができる。例えば、原核細胞については、カルシウム処理、エレクトポレーションによる形質転換等が知られている。また、植物細胞については、アグロバクテリウムを用いた方法が公知であり、哺乳動物細胞についてはリン酸カルシウム沈降法を例示することができる。本発明は特にこれらの方法に限定されるわけではなく、選択した宿主に応じ、その他公知の核マイクロインジェクション、プロトプラスト融合、DEAE-デキストラン法、細胞融合、電気パルス穿孔法、リポフェクタミン法(GIBCO BRL)、FuGENE6試薬(Boehringer-Mannheim)を用いた方法をはじめとする種々の公知の方法により発現ベクターの導入を行うことができる。
【0068】
以上のようにして本発明のポリヌクレオチドを含む組換えベクターにより形質転換された形質転換体を培養することにより、本発明のスチレンモノオキシゲナーゼ活性を有するタンパク質を製造することができる。よって、本発明の好ましい一態様として、本発明の上記形質転換体を培養し、その培養物から前記ポリヌクレオチドによってコードされる蛋白質を回収する工程を含む、光学活性なスチレンモノオキシゲナーゼの製造方法が提供される。形質転換体の培養方法は特に限定されず、選択した各宿主細胞の生育に適し、かつ、本発明の酵素の生産に最も適した培地、温度、時間等の条件を選択することが望ましい。例えば実施例に記載の条件が挙げられるがこれに限定されない。
【0069】
スチレンオキシドおよび/またはスチレン類似体の酸化物の製造方法
本発明は光学活性なスチレンオキシドおよび/または光学活性なスチレン類似体の酸化物の製造方法に関する。当該方法は、スチレンモノオキシゲナーゼにより、スチレンおよび/またはその類似体のビニル基がオキシラニル基に酸化されることを特徴とする。本発明のスチレンオキシドおよび/またはスチレン類似体の酸化物の製造方法は、スチレンモノオキシゲナーゼをスチレンおよび/またはその類似体に作用させる工程、および生成される酸化物を回収する工程を含む。
【0070】
スチレンモノオキシゲナーゼと基質は、基質の酸化が可能な条件下で作用させられる。具体的には、本発明の上記酸化反応では、FADH2(または還元型FAD)を補酵素として加えることが必要である。以下に、本発明の酸化反応を示す。本発明の酸化反応においては、補酵素FADH2が酸化されFADが生成すると同時に、水が生じる。
【化2】
【0071】
スチレンモノオキシゲナーゼの使用形態は、酵素分子、その処理物、酵素分子を含む培養物、酵素を生成する形質転換体等の微生物またはその処理物の形態で使用できる。
スチレンモノオキシゲナーゼを生成する形質転換体の微生物の処理物には、具体的には界面活性剤やトルエンなどの有機溶媒処理によって細胞膜の透過性を変化させた微生物、あるいはガラスビーズや酵素処理によって菌体を破砕した無細胞抽出液やそれを部分精製したもの、高度に精製したもの、各種固定化担体に固定化したものなどが含まれる。
【0072】
このようなスチレンモノオキシゲナーゼを反応溶液中、スチレンおよび/またはその類似体と作用させることにより、目的とする酵素反応を行わせ、スチレンオキシドおよび/またはスチレン類似体の酸化物を得ることができる。なお、酵素と反応溶液の作用形態はこれらの具体例に限定されるものではない。また本発明において「作用」は「接触」と表現することもできる。
【0073】
反応溶液は、好ましくは、基質や酵素反応に必要な補酵素であるFADH2を酵素活性の発現に望ましい環境を与える適当な溶媒に溶解したものである。このような溶媒としては、例えば、水中もしくは水に溶解しにくい有機溶媒、例えば、酢酸エチル、酢酸ブチル、トルエン、クロロホルム、n−ヘキサン、メチルイソブチルケトン、メチルターシャリーブチルエステルなどの有機溶媒、もしくは、水性媒体との2相系、もしくは水に溶解する有機溶媒、例えば、メタノール、エタノール、イソプロピルアルコール、アセトニトリル、アセトン、ジメチルスルホキシド、ジメチルホルムアミドなどとの混合系が挙げられる。
【0074】
本発明におけるスチレン類似体としては、例えば、スチレンの2位、3位または4位の少なくともいずれかがハロゲンに置換された化合物が挙げられる。ハロゲンとしてはフッ素、塩素、臭素などが挙げられるがこれらに限定されない。このようなスチレン類似体として以下の化合物を例示することができる。
・2-フルオロスチレン、3-フルオロスチレン、4−フルオロスチレン
・2-クロロスチレン、3-クロロスチレン、4−クロロスチレン
・2-ブロモスチレン、3-ブロモスチレン、4−ブロモスチレン
あるいは、本発明におけるスチレン類似体のその他の例として、以下の化合物を挙げることもできる。
・インデン
・p-アミノスチレン
・p-メトキシスチレン
・trans-β-メチルスチレン
・4-ビニル-1-クロロヘキセン
・N,N-ジメチル-1-ナフチルアミン
・桂皮アルデヒド
このようなスチレン類似体を本発明のスチレンモノオキシゲナーゼと接触させることによって酸化することにより、スチレン類似体の酸化物を得ることができる。具体的には、例えば2−フルオロスチレンを本発明のスチレンモノオキシゲナーゼによって酸化することにより、2−フルオロスチレンオキシドを得ることができる。あるいは、例えばインデンを本発明のスチレンモノオキシゲナーゼによって酸化することにより、インデンオキシドを得ることができる。
【0075】
酸化反応に付随してFADH2から生成するFADの、FADH2への再生は、微生物の持つFAD還元能(呼吸活性など)を用いて行うことができる。また、FADからFADH2を生成する能力を有する微生物やその処理物、酵素を反応系に添加することによっても行うことができる。例えば、フラビンオキシドレダクターゼを含む微生物、その処理物、ならびに部分精製もしくは精製酵素を用いてFADH2の再生を行うことができる。これらのFADH2再生に必要な反応を構成する成分は、本発明によるスチレンオキシドまたはスチレン類似体の酸化物の製造のための反応系に添加する、固定化したものを添加する、あるいはFAD+の交換が可能な膜を介して接触させることができる。
あるいは、フラビンオキシドレダクターゼをコードするDNAでトランスフェクトされた微生物やその処理物を、本発明の酸化反応のための反応系に添加することも可能である。
【0076】
本発明の反応は、固定化酵素、膜リアクターなどを利用して行うことも可能である。
【0077】
スチレンオキシドまたはスチレン類似体の酸化物の製造方法においては、反応温度が好ましくは4〜60℃、さらに好ましくは15〜37℃、pHが好ましくは4〜11、さらに好ましくはpH5〜9、基質濃度が好ましくは0.01〜90%、さらに好ましくは0.1〜30%で行うことができる。
【0078】
反応系には必要に応じて補酵素FADH2を、好ましくは0.001mM〜100mM、さらに好ましくは、0.01〜10mM添加できる。
また、基質は反応開始時に一括して添加することも可能であるが、反応液中の基質濃度が高くなりすぎないように連続的、もしくは非連続的に添加することが望ましい。基質濃度は、例えば0.01g/Lから500g/L、好ましくは0.1g/Lから100g/L、より好ましくは0.5g/Lから10g/Lとすることができるがこれらに限定されない。
【0079】
FADH2再生用の酵素、例えば、フラビンオキシドレダクターゼなどは、本発明の酸化酵素に比較して酵素活性で好ましくは0.1〜100倍、さらに好ましくは0.5〜20倍程度添加することができる。
【0080】
本発明の酸化反応により生成するスチレンオキシドまたはスチレン類似体の酸化物の回収は遠心分離や膜濾過による菌体や蛋白質の除去、膜処理等による蛋白質の分離、酢酸エチル、酢酸ブチル、クロロホルム、メチル−t−ブチルエーテル、n−ブタノール、sec−ブタノール、t−ブタノール、イソブタノール、n−ペンタノール、sec−ペンタノール、t−ペンタノール、イソペンタノールなどによる溶媒抽出、蒸留、酢酸エチル、酢酸ブチル、キシレン、トルエン、メシチレン、クロルベンゼン、クロロホルム、n−ヘキサン、イソオクタン、ジイソプロピルエーテル、メチル−t−ブチルエーテルなどによる結晶化、等を適当に組み合わせることにより行うことができる。あるいは、抽出溶媒液に塩酸を滴下することで生成する塩酸塩として回収することもできる。目的物は、必要によりシリカゲルカラムクロマトグラフィー、蒸留によりさらに精製することができる。
【0081】
スチレンまたはその類似体の分解方法
さらに本発明は、本発明のスチレンモノオキシゲナーゼをスチレンまたはその類似体と接触させる工程を含む、スチレンまたはその類似体の分解方法に関する。上述のように本発明のスチレンモノオキシゲナーゼは、スチレンまたはその誘導体のビニル基をオキシラニル基に酸化する。従って本発明のスチレンモノオキシゲナーゼは、スチレンまたはその類似体を分解することができる。
本発明において分解とは、元の化合物がこれとは異なる化合物に変換されることを意味する。変換としては、例えば酸化などが挙げられる。従って本発明において分解方法は、酸化する方法と表現することもできる。あるいは、スチレンまたはその類似体の量あるいは濃度を減少させる方法と表現することもできる。
スチレン類似体ついては上述のものが挙げられる。またスチレンモノオキシゲナーゼと基質との接触の形態についても既に述べた。
なお本明細書において引用された全ての文献は、参照として本明細書に組み入れられる。
【実施例】
【0082】
以下、実施例により本発明をさらに具体的に説明する。但し、本発明は以下の実施例に限定されるものではない。
材料および方法
(1)細菌株およびベクター
スチレン同化細菌ST-5株およびST-10株を、スチレン代謝遺伝子の単離のための染色体DNAの起源として使用した。これらの微生物は、以前はコリネバクテリウム(Corynebacterium)sp.ST-5株およびST-10株として同定されていた(Itoh, N., K. Yoshida, and K. Okada. 1996. Isolation and identification of styrene-degrading Corynebacterium strains, and their styrene metabolism. Biosci. Biotech. Biochem. 60:1826-1830.)。最近、Makinoらによって、ST-10株はロドコッカスsp.ST-10株として再同定された(Makino, Y., K. Inoue, T. Dairi, and N. Itoh. 2005. Engineering of phenylacetaldehyde reductase for efficient substrate conversion in concentrated 2-propanol. Appl. Environ. Microbiol. 71:4713-4720)。本発明者らも、Makinoらによって記載された方法により、ST-5株の再同定を実施した。結果は、ST-5株がロドコッカスの典型的な特性を有することを示した。従って、本発明者らは、この微生物がロドコッカスsp.ST-5株として同定されるべきであるとの結論を下した。大腸菌JM109株およびプラスミドベクターpGEM-T Easy(Plomega)をDNA操作において使用した。
【0083】
(2)培養条件
ロドコッカスsp.ST-5株およびST-10株を、30℃で2日間、振とうしながら、PY培地(0.5%酵母抽出物、0.5%バクトペプトン、pH7.0)中で培養した。10μlの培養液を、寒天プレート(0.3(w/v)%(NH4)2SO4、0.3% KH2PO4、0.1% NaCl、0.02% MgSO4 7H2O、および1.5%寒天、pH7.0)へ播き、スチレン雰囲気下において30℃で4日間培養した(Itoh, N., K. Yoshida, and K. Okada. 1996. Isolation and identification of styrene-degrading Corynebacterium strains, and their styrene metabolism. Biosci. Biotech. Biochem. 60:1826-1830)。
【0084】
(3)スチレン代謝遺伝子のクローニング
DNA操作には標準的な技術を使用した(Sambrook, J., and W. D. Russell. 2001. "Molecular cloning, a laboratory manual" 3rd ed. Cold Spring Harbor Laboratory Press.)。採集されたST-5株 およびST-10株 の細胞をリゾチームにより処理した後、Eulbergら(Eulberg, D., L. A. Golovleva, and M. Schlomann. 1997. Characterization of catechol catabolic genes from Rhodococcus erythropolis 1CP. J. Bacteriol. 179:370-381)によって記載された方法によってゲノムDNAを調製した。調製されたゲノムDNAを、PCRの鋳型およびサザンブロット分析に使用した。
styA遺伝子およびstyC遺伝子をコードする部分断片を、縮重PCRにより増幅した。styAおよびstyCの増幅のために使用されたプライマーを表1に示した。これらのオリゴヌクレオチドプライマーは、それぞれ、スチレンモノオキシゲナーゼおよびスチレンオキシドイソメラーゼにおいて保存されている部分アミノ酸配列に基づき設計された。スチレン代謝に関与している遺伝子の全長を入手するために、明らかになったstyA遺伝子またはstyC遺伝子の部分塩基配列を用いて、TAIL(thermal asymmetric interlaced)PCRを実施した。TAIL-PCRは、Liu らによって報告された方法(Liu, Y. G., and R. F. Whittier. 1995. Thermal asymmetric interlaced PCR: automatable amplification and sequencing of insert end fragments from P1 and YAC clones for chromosome walking. Genomics 25:674-681)に従い実施された。TAIL-PCRのために使用されたアダプタープライマーおよび既知配列特異的プライマーを表1に示した。一次および二次のPCR産物を希釈し、それぞれ二次および三次のPCRのための鋳型として使用した。増幅された断片は、アガロースゲル電気泳動による分離および抽出により精製された。精製された断片を、pGEM-T Easyへとクローニングし、キャピラリーDNAシーケンサー310(Applied Biosystems,Tokyo,Japan)を使用して塩基配列を決定した。
【0085】
【表1】
【0086】
(4)サザンブロット分析
ロドコッカスsp.ST-10株のゲノムDNAを、いくつかの制限酵素により消化し、アガロースゲル電気泳動によって分離した。分離されたDNAを、Hybond N+ナイロンメンブラン(GE Healthcare UK Ltd,Buckinghamshire,UK)へ転写し、80℃で2時間ベーキングした。ST-10株由来のスチレンモノオキシゲナーゼ遺伝子断片(1.2 kbp)、およびST-5株由来のスチレンオキシドイソメラーゼ遺伝子断片(0.6 kbp)をPCRにより増幅し、プローブとしてハイブリダイゼーションのために使用した。各断片を、Random Primer DNA Labeling Kit Ver.2(TaKaRa,Shiga,Japan)を使用してα-32P-dCTPにより標識した。標識操作は製品プロトコルに従って行った。プレハイブリダイゼーションおよびハイブリダイゼーションを、5×SSC、0.1%SDS、5×デンハルツ液、40%ホルムアミドからなるハイブリダイゼーション緩衝液において実施した。標識されたプローブDNA 1ngをメンブランに添加し、45℃で一夜インキュベートした。メンブランを、45℃で10分間2×SSC緩衝液により2回洗浄し、次いで、45℃で10分間、0.1%SDSを含有している2×SSCにより2回洗浄した。メンブランにハイブリダイズした標識DNAの検出は、BAS-3000画像分析装置(Fujifilm,Tokyo,Japan)を使用して行った。
【0087】
(5)静止菌体反応による代謝物の抽出
スチレン雰囲気下でプレート培養した菌体を、50mMリン酸カリウム緩衝液(pH7.0)2mlに懸濁し、遠心分離(8000×g、20分)によって収集した。収集した菌体を同緩衝液に再懸濁した。次いで、菌体懸濁液(OD600=1)4mlおよび各基質50mgからなる反応混合物を使用して、静止菌体反応を実施した。反応混合物を22ml のバイアルに添加後シリコンセプタムを用いて密栓し、30℃、20時間、170rpmでインキュベーションした。インキュベーション後、反応混合物に酢酸エチル2mlを添加・混合し、反応生成物を抽出した。抽出産物をガスクロマトグラフィー質量分析(GC-MS)および高速液体クロマトグラフィ(HPLC)によって分析した。
【0088】
(6)GC-MSおよびHPLCの分析条件
静止菌体反応による生成物のGC-MS分析は、GC-17AガスクロマトグラフィーシステムおよびShimadzu QP-5000質量分析計を使用して実施された。酢酸エチルにより抽出された試料1μlを、HP-1MSキャピラリーカラム(J & W Scientific Inc.;30m×内径0.25mm、フィルム幅0.25μm)に注入した。キャリアガス(He)流速は1.7ml/min(100kPa)、スプリット比は26、線形速度は47.4cm/sであった。カラムオーブンの温度プログラムは:50℃、5分間;10℃/minで220℃まで上昇; 220℃、5分間で行った。質量スペクトルは、電子衝撃イオン源(EI、200℃)を使用して、1.5kVで取得した。各反応中間体の同定は、標準化合物の保持時間および質量電荷比(m/z)との比較によって行った。
静止菌体反応による生成物のHPLC分析は、移動相Aとして10mM酢酸アンモニウム、移動相Bとしてアセトニトリルを用い、流速0.5ml/分、カラム温度は40℃で分析を行った。試料(10μl)を注入した後、移動相Bを8分間10%に維持し、次いで20分かけて90%に増加させ、10分間維持した。254nmで生成物をモニタリングした。
フェニルアセチル-CoAリガーゼの反応生成物の同定は、Shimadzu Prominence GE HPLCシステムを使用して実施された。分析にはCadenza CD-C18カラム(Imtakt,Kyoto,Japan)を用いた。移動相には10% (v/v)イソプロピルアルコールを含有する0.2M KPB(pH4.5)を用いた。移動相の流速は0.5ml/分、カラムは40℃に維持された。254nmで生成物をモニタリングした。
【0089】
(7)無細胞粗抽出物の調製および酵素アッセイ
ガス状スチレンを単一炭素源として培養された菌体から、無細胞粗抽出物を入手した。採集された菌体を、50mMリン酸カリウム緩衝液(pH7.0)に懸濁し、2回洗浄した。菌体を、10%グリセロールを含有している50mMリン酸カリウム緩衝液(pH7.0)に再懸濁した。菌体懸濁液は、各1分間の超音波処理を5回行い、遠心分離(16,000×g、10分、4℃)した。上清を同緩衝液に対して透析し、酵素アッセイのために使用した。フェニルアセチル-CoAリガーゼ(PCL)活性を、以前に報告された方法(Martinez-Blanco, H., A. Reglero, L. B. Rodriguez-Aparicio, and J. M. Luengo. 1990. Purification and biochemical characterization of phenylacetyl-CoA ligase from Pseudomonas putida. J. Biol. Chem. 265:7084-7090)に従って測定した。
【0090】
(8)化学物質
スチレンおよびスチレンオキシドはNacalai Tesque Inc.(Kyoto,Japan)より購入した。フェニルアセトアルデヒドはToyotama International Inc.(Tokyo,Japan)より供給された。その他の化学物質は、全て、Wako Pure Chemical Industries Ltd.(Osaka,Japan)より購入した。
【0091】
〔実施例1〕ロドコッカスsp.ST-5株およびST-10株のスチレン代謝遺伝子の遺伝子構造
代謝遺伝子を単離し解析するため、本発明者らは、データベース上に登録されたスチレンモノオキシゲナーゼおよびスチレンオキシドイソメラーゼのアミノ酸配列から高度に保存されている領域を選択し、縮重PCR用のオリゴヌクレオチドプライマーを設計した。ST-5株のゲノムDNAを鋳型として縮重PCRを行い、styCの一部をコードする260bpのサイズの断片を入手することができた。しかし、styAに対応する断片は増幅されなかった。他方、ST-10株のゲノムDNAを鋳型として縮重PCRを行った場合には、styAをコードする部分断片が増幅されたが、styCをコードする遺伝子は増幅されなかった。これらの断片から推定されたアミノ酸配列は、シュードモナスのスチレン分解酵素と有意な類似性を示した。
スチレン代謝遺伝子の全長を入手するため、本発明者らは、ST-5株およびST-10株のゲノムDNAに対してTAIL-PCRを実施した。TAIL-PCRのため使用されたプライマーを表1に示した。その結果、本発明者らは、それぞれST-5株の染色体の約4.7kbpの領域およびST-10株の4.2kbpの領域のヌクレオチド配列を決定することができた。ST-5株およびST-10株の決定された染色体領域には、それぞれ、4つおよび3つのオープンリーディングフレーム(ORF)が存在した(図1および図2)。推定アミノ酸配列がシュードモナスのstyABCDと類似性を示したため、これらの遺伝子がスチレン代謝遺伝子であると推定した。ST-5株は、SMO、フラビンオキシドレダクターゼ、SOI、およびフェニルアセトアルデヒドデヒドロゲナーゼ(PAD)に対応する4つの連続する遺伝子を有していた。他方、ST-10株はSMOおよびフラビンオキシドレダクターゼに対応する2つの遺伝子を有していた。styBの下流に存在したorf3は、ロドコッカス・ジョスティ(jostii)RHA1のようなロドコッカス属にのみホモログが見出される機能未知タンパク質に対して高い類似性を示した。さらに、転写ターミネーターとして作用することが予測されるステムループ構造が、ST-5株のstyCおよびST-10株のstyBの下流にそれぞれ存在した(図1)。
【0092】
〔実施例2〕ST-10株ゲノムDNAのサザンブロット分析
SOI遺伝子の存在を確認するため、本発明者らは、ST-10株の染色体に対してサザンブロット解析を実施した。ST-10株のstyA遺伝子およびST-5株のstyC遺伝子に対応する各々1.2kbpおよび0.8kbpのプローブを、α-32P-dCTPにより標識し、様々な制限酵素により消化されたST-10株ゲノムDNAにハイブリダイズさせた。その結果、styA遺伝子に対応するプローブにハイブリダイズする有意な断片を見出すことができたが、styC遺伝子をプローブとして使用した場合には、検出可能なシグナルは見出されなかった。
【0093】
〔実施例3〕ST-5株およびST-10株のスチレン代謝遺伝子の構造的特性
ST-5株およびST-10株に由来するstyA遺伝子は427および428アミノ酸残基をコードし、推定アミノ酸配列は、シュードモナスsp.Y2由来のSMOの配列との有意な類似性を示した(60.6%および60.4%)(表2)。
【0094】
【表2】
【0095】
ST-5株およびST-10株に由来するStyAの推定アミノ酸配列は、フラビンおよびニコチンアミドアデニンジヌクレオチド(NAD+/NADH)との結合に関与していることが公知であるSMOにおいて保存されているいくつかの配列を含有していた。しかしながら、ST-5株およびST-10株のStyAは、SMOおよびフラビンオキシドレダクターゼの融合タンパク質として報告されたロドコッカス・オパカス(opacus)1CPのstyA2B(Tischler, D., D. Eulberg, S. Lankner, S. R. Kaschabek, W. J. H. van Berkel, and M. Schlomann. 2009. Identification of a novel self-sufficient styrene monooxygenase from Rhodococcus opacus 1CP. J. Bacteriol. 191:4996-5009)のStyA部分に対しては比較的低い類似性を示した(29.2%および28.0%)。ロドコッカス・オパカス1CPは、styCまたはstyDに対応するスチレン代謝遺伝子をstyA2Bの近傍に有していないことが報告された。これらの事実は、既報のロドコッカスのスチレン代謝遺伝子が本発明の遺伝子とは異なることを示している。
【0096】
ST-5株およびST-10株のstyB遺伝子は、それぞれ179および183アミノ酸残基をコードしており、そのアミノ酸配列から計算される推定分子量は19,284Daおよび19,543Daであった。ST-5株およびST-10株に由来するStyBの推定アミノ酸配列は、フラビンオキシドレダクターゼおよび様々なモノオキシゲナーゼにおいて高度に保存されているいくつかのモチーフを含有している。相同性検索の結果から、ST-5株およびST-10株に由来するstyBが、シュードモナスsp.Y2 StyB(53.8%および50.6%)、バチルス(Bacillus)sp.JF8 MocB(36.5%および36.8%)、ゲオバチルス・サーモグルコシダシウス(Geobacillus thermoglucosidasius)A7 PheA2(33.6%および35.1%)、ならびにロドコッカス・オパカス1CP StyA2B(27.6%および33.3%、B部分)を含む、NAD(P)H:フラビンオキシドレダクターゼのファミリーに属しているいくつかのタンパク質との相同性をそれぞれ示すことが明らかとなった(表2)。この結果より、ST-5株およびST-10株のstyBが、NAD(P)H:フラビンオキシドレダクターゼのメンバーであり、StyAと共に2成分からなるモノオキシゲナーゼ系を構成していることが示された。
【0097】
styC遺伝子およびstyD遺伝子はST-5株にのみ観察された。styC遺伝子は168アミノ酸残基をコードし、推定アミノ酸配列は、シュードモナスsp.Y2のSOIとの有意な類似性(69.5%)、およびシュードモナス・オーレオファシエンス(aureofaciens)のp-シメンメチルヒドロキシラーゼとの中程度の類似性(35.0%)を示した(表2)。styDをコードするオープンリーディングフレームには、2ヶ所の推定開始コドンが3068塩基目および3086塩基目に存在した(図2A)。このことから、styD がコードするポリペプチドは502アミノ酸ないし496アミノ酸であることが示唆された。このタンパク質は、ロドコッカス・ジョスティRHA1 ALDH(78.6%)、バークホルデリア(Burkholderia)sp.383 BADH(54.4%)、アゾアルカス(Azoarcus)sp.BH72 ThmS2(54.5%)、シュードモナスsp.Y2 StyD(49.9%)、および大腸菌W3110 PadA(40.1%)のような様々なアルデヒドデヒドロゲナーゼとの高い類似性を示した(表2)。StyDは、様々なアルデヒドデヒドロゲナーゼにおいて保存されているいくつかの補因子結合モチーフを有する(Hempel, J., H. Nicholas, and R. Lindahl. 1993. Aldehyde dehydrogenases: Widespread structural and functional diversity within a shared framework. Protein Sci. 2:1890-1900、Liu, Z. J., Y. J. Sun, J. Rose, Y. J. Chung, C. D. Hsial, W. R. Chang, I. Kuo, J. Perozich, R. Lindahl, J. Hempel, and B. C. Wang. 1997. The first structure of an aldehyde dehydrogenase reveals novel interactions between NAD and the Rossmann fold. Nat. Struct. Biol. 4:317-326)。
【0098】
〔実施例4〕ロドコッカスsp.ST-10株によるスチレン代謝の中間体の分析
ST-5株およびST-10株を用いて静止菌体反応を行い、生成した中間体をGC-MSによって同定した。スチレンを基質としたとき、ST-10株の反応生成物としてスチレンオキシドが検出されたが、フェニルアセトアルデヒドは検出されなかった。対照的に、ST-5株は反応生成物としてフェニルアセトアルデヒドのみを蓄積した。これらの結果は、ST-5株およびST-10株のそれぞれの遺伝子発現の差異によることが示唆された。ST-5株においては、スチレンが、StyA、StyB、およびStyCの作用によってフェニルアセトアルデヒドに変換されるが、ST-10株ではStyCを欠くために、スチレンオキシドをフェニルアセトアルデヒドに変換することができず、反応生成物中に蓄積されたと推測される。
【0099】
ST-10株のスチレン資化メカニズムを解明するため、本発明者らは、HPLCによってST-10株の静止菌体反応による生成物を詳細に調査した。スチレンを基質とした反応産物からは、スチレンオキシド、アセトフェノン、スチレングリコール、およびフェニル酢酸を含む様々な芳香族化合物が見出された(図3A)。また、コントロールとしてスチレンオキシドをST-10株菌体を含まない緩衝液中で反応させたとき、スチレングリコール、アセトフェノン、およびフェニルアセトアルデヒドが検出された(図3B)。スチレンオキシドは不安定な化合物であり、水、過酸化物、または空気の存在下で、他の化合物へと容易に変換されることが報告されている(Liu, Z., J. Michel, Z. Wang, B. Witholt, and L. Zhi. 2006. Enantioselective hydrolysis of styrene oxide with the epoxide hydrolase of Sphingomonas sp. HXN-200. Tetrahedron Asymmetry 17:47-52、Pfaffli, P., H. Vainio, and A. Hesso. 1979. Styrene and styrene oxide concentration in the air during the lamination process int the reinforced plastics industry. Scand. J. Work Environ. Health 5:158-161)。従って、アセトフェノンおよびフェニルアセトアルデヒドは、自発的な反応によってスチレンオキシドから生成したと推測される。さらに、ST-10株の菌体と共にインキュベートしたとき、フェニルアセトアルデヒドは消失し、代わりにフェニル酢酸が増加した(図3C)。
【0100】
〔実施例5〕フェニル酢酸の代謝に関与している酵素活性の同定
フェニル酢酸の分解経路を同定するため、ST-5株およびST-10株のフェニルアセチル-CoAリガーゼ(PCL)活性を測定した。基質としてフェニル酢酸、CoA、ATPを含む緩衝液にST-5株およびST-10株の無細胞粗抽出物を添加し、反応生成物をHPLCにより分析した。いずれの株についても、無細胞粗抽出物の存在下で、フェニルアセチル-CoAに対応するピークが8.1分の位置に検出された(図4)。これらの結果は、いずれの株もPCL活性を有し、シュードモナス属細菌で報告されているフェニル酢酸(PAA)分解経路と同様の経路を介してフェニル酢酸を分解し得ることを示す。
【0101】
以下に、実施例1〜5で得られた結果について考察する。
ロドコッカスsp.ST-5株およびST-10株(以前はコリネバクテリアsp.)が、ガス状スチレンおよびスチレンオキシドを単一炭素源として資化できること、またこれらの菌株がシュードモナスと同様のスチレン代謝経路を有し、スチレンおよびスチレンオキシドを資化していることが示唆されていた(Itoh, N., K. Yoshida, and K. Okada. 1996. Isolation and identification of styrene-degrading Corynebacterium strains, and their styrene metabolism. Biosci. Biotech. Biochem. 60:1826-1830)。
【0102】
ST-5株およびST-10株におけるスチレンの分解経路を解明するため、本発明者らはスチレン分解に関与している遺伝子群をPCR技術を利用して単離した。
【0103】
図1および図2に示されるように、本発明者らは、ST-5株の約4.7kbpの染色体領域およびST-10株の約4.2kbpの領域の塩基配列を決定した。ST-5株において決定された配列中には4つのORFが含まれ、その推定アミノ酸配列はシュードモナス属細菌において報告されているスチレン代謝遺伝子styABCDと相同性を示した。他方、ST-10株において決定された配列は、styABおよび機能未知タンパク質の3つのORFを有する。シュードモナス属細菌を含む各種微生物から単離されたスチレン代謝遺伝子は、SMO、フラビンオキシドレダクターゼ、SOI、およびフェニルアセトアルデヒドデヒドロゲナーゼを含む、4つの遺伝子から構成されている。さらに、いくつかの微生物はこれらの遺伝子の上流または下流に転写制御因子(styRS)および基質トランスポーター(styE)をコードする遺伝子を有する(Mooney, A., N. D. O'Leary, and A. D. W. Dobson. 2006. Cloning and functional characterization of the styE gene, involved in styrene transport in Pseudomonas putida CA-3. Appl. Environ. Microbiol. 72:1302-1309、Panke, S., B. Witholt, A. Schmid, and M. G. Wubbolts. 1998. Towards a biocatalyst for (S)-styrene oxide productio: characterization of the styrene degradation pathway of Pseudomonas sp. strain VLB120. Appl. Environ. Microbiol. 64:2032-2043、Velasco, A., S. Alonso, J. L. Garcia, J. Perera, and E. Diaz. 1998. Genetic and functional analysis of the styrene catabolic cluster of Pseudomonas sp. strain Y2. J. Bacteriol. 180:1063-1071)。ST-5株およびST-10株の両株はstyAの上流に制御遺伝子を有しておらず、ST-10株はstyCDを欠くことが見出された(図1)。しかしながら、これらの株をガス状スチレンまたはスチレンオキシドで培養したとき、スチレンモノオキシゲナーゼの活性が上昇したことから、シュードモナス属細菌と同様これらの遺伝子の発現を制御している制御遺伝子が存在することが示唆された。ただし、そのような遺伝子はスチレン代謝遺伝子の近隣の領域には存在しない。
【0104】
ST-10株の染色体中にスチレンオキシドイソメラーゼ遺伝子が存在しないことを確認するため、本発明者らは、ST-5株由来のstyC遺伝子をプローブとして、ST-10株ゲノムDNAに対してサザンブロット解析を実施した。その結果、styC遺伝子に相当する特異的なシグナルを見出すことができなかった。一方、コントロールとしてstyA遺伝子をプローブに用いた場合、特異的なバンドが検出された。Itohらは、ST-10株がSOI活性は有していないが、SMO活性およびPAD活性は有することを報告している(Itoh, N., K. Yoshida, and K. Okada. 1996. Isolation and identification of styrene-degrading Corynebacterium strains, and their styrene metabolism. Biosci. Biotech. Biochem. 60:1826-1830)。これらの事実は、ST-10株がSOI以外のスチレン分解酵素を有し、未知経路を介してスチレンオキシドを資化することを示唆している。
【0105】
ST-10株のスチレン資化経路を解明するため、ST-5株およびST-10株による静止菌体反応を行い、生成した中間体を同定した。GC-MS分析によって、ST-10株の反応生成物としてスチレンオキシドが蓄積され、ST-5株ではフェニルアセトアルデヒドが蓄積されることが明らかとなった。これらの現象は、ST-10株にSOIをコードする遺伝子が存在しないことと一致する。以前の報告において、ST-5株はPAD活性より高いSOI活性を示すことが見出されている(それぞれ、菌体湿重量で33U/g cellおよび1.5U/g cell)。ST-5株においては、転写ターミネーターと推定される配列がstyCとstyDとの間の領域に見出されている(図2)。従って、styDの発現レベルは他のスチレン代謝遺伝子より低いことが推測される。スチレン代謝フラックスにおいて、PAD活性が低いことが要因となりフェニルアセトアルデヒドがST-5株の代謝中間体として蓄積されていることが示唆される。対照的に、ST-10株においては、StyCが欠落しているため、スチレンオキシドが代謝中間体として蓄積している。HPLC分析により、スチレンオキシドがフェニルアセトアルデヒドおよびアセトフェノンへと自発的に異性化されること、またST-10株が自発的に異性化したフェニルアセトアルデヒドを資化していることが証明された(図3)。Itohらは、スチレン、スチレンオキシド、および2-フェニルエタノールがST-10株によって代謝されるが、アセトフェノンおよびスチレングリコールは炭素源として利用されないことを報告している。HPLC分析の結果は、これらの事実とよく一致している。スチレンまたはスチレンオキシドを基質としたとき、両株の反応生成物中に2-フェニルエタノールが観察された。図5に示されるように、フェニルアセトアルデヒドレダクターゼ(PAR)の活性によってフェニルアセトアルデヒドは2-フェニルエタノールに変換される。スチレンの代謝経路において、2-フェニルエタノールは、貯蔵化合物の役割を担っていることが示唆される。2-フェニルエタノール/フェニルアセトアルデヒドの比率は、各化合物の濃度およびNAD+/NADH比によって制御されると推測される。
【0106】
フェニル酢酸代謝経路は、シュードモナス.プチダ(putida)および大腸菌のようなグラム陰性菌においてよく解析されている(Luengo, J. M., J. L. Garcia, and E. R. Olivera. 2001. The phenylacetyl-CoA catabolon: a complex catabolic unit with broad biotechnological applications. Mol. Microbiol. 39:1434-1442)。しかしながら、グラム陽性菌の同経路に関してはほとんど情報がない。近年Llorensにより、ロドコッカスsp.RHA1がグラム陰性菌のフェニル酢酸代謝系路に対応するpaa遺伝子群を有することを報告している(Navarro-Llorens, J. M., M. A. Patrauchan, G. R. Stewart, J. E. Davies, L. D. Eltis, and W. W. Mohn. 2005. Phenylacetic acid catabolism in Rhodococcus sp. strain RHA1: a central pathway for degradation of aromatic compounds. J. Bacteriol. 187:4497-4504)。さらに、数種のロドコッカス属細菌の全ゲノム配列が解析されたことにより、そのゲノム中にpaa遺伝子群が存在することが明らかになった。ST-5株およびST-10株も、他のロドコッカス属細菌と同様にPCL活性を有する(図4)。この事実は、ST-5株およびST-10株がpaa遺伝子群を有し、他のグラム陰性菌と同様の経路を利用してフェニル酢酸を資化していることを示唆している。
ST-5株およびST-10株における推定スチレン代謝経路を図5に示した。スチレンはまずSMOによってスチレンオキシドへと酸化され、続いてSOIによってフェニルアセトアルデヒドへと変換される。ST-10株においてはSOIを欠落しているため、この反応は自発的な化学変換によって起きていると推測される。次いで、フェニルアセトアルデヒドはPAD活性によりフェニル酢酸へと酸化され、PAA分解経路により分解される。本発明者らは、ST-5株およびST-10株のゲノム中にスチレン分解遺伝子群が存在することを確認した。ST-10株のスチレン代謝経路はSOIを欠落した不完全なものであるが、ST-10株はスチレンおよびスチレンオキシドを資化することができる。
【0107】
〔実施例6〕Rhodococcus sp. ST-5株 およびST-10株 由来styAの基質特異性
(1)Rhodococcus sp. ST-5株 およびST-10株 由来styAB の大腸菌による異種宿主発現系構築および組換え酵素の発現・精製
Rhodococcus sp. ST-5株 およびST-10株 両株由来のstyAB 遺伝子を大腸菌Escherichia coli において異種宿主発現させるため、以下の手順により発現用プラスミドを構築した。発現には大腸菌株E. coli BL21 (DE3)を用いた。
1. ST-5株 およびST-10株 のstyAB の末端配列に特異的なオリゴヌクレオチドプライマー(表3)を作成し、ST-5株 およびST-10株 ゲノムDNA に対してPCRを実施した。この際N-末端およびC-末端に制限酵素サイト(Bam HI, Hind III) を付加した。
2. 増幅した約1.8 kbp の増幅断片をpGEM-T Easy vector (plomega) にクローニングし、内部配列を確認した。
3. Bam HI、Hind III 処理によりstyAB 断片を切り出し、アガロースゲル電気泳動により精製した。精製したstyAB 断片を同制限酵素によりあらかじめ処理したpET-21 (b) にライゲーションし、E. coli DH5αに導入した。
4. 抗生物質プレート上で形質転換体を選抜した。プラスミドを抽出精製し、E. coli BL21 (DE3) にエレクトロポレーション法により導入した。さらに、組換えタンパク発現を効率化させるためにシャペロン発現ベクターpG-KJE8 を重ねて導入した。
5. 形質転換体を抗生物質プレート上で選抜し、以後発現実験へ用いた。
なお、上記と同様の手法により、styAを単体で発現させるコンストラクトも作成した。
発現プラスミド構築に使用したプライマー配列を表3に、構築したプラスミドを図6に示した。
【0108】
SDS-PAGEによるSMO分子量の分析は以下の条件で行った。
1. 精製したヒスチジンタグを含むStyA サンプルを2x SDS-PAGE サンプルバッファー(100 mM Tris-HCl, 20% グリセロール, 4% SDS, 0.05% ブロモフェノールブルー)に溶解した。
2. 10% ポリアクリルアミドゲルに、タンパク量にして5μg相当の上記サンプルをアプライした。
3. 150 V, 40 mAで 100 分間 泳動した。
4. 泳動したゲルをSDS-PAGE染色液(50% メタノール、10% 酢酸、0.25% クマジーブルー)に浸し、40分間振とうしながら染色を行った。
5. 染色したゲルを脱色液(30% メタノール、10% 酢酸)に浸し、適宜脱色液を交換しながら脱色を行った。
6. 得られたバンドの泳動度から、精製したStyA の分子量は約49.0 kDaであると推定した。分子量の算出には、サンプルと同時に流した分子量マーカー(Prestained APRO-Marker) を指標として用いた。
【0109】
また、ゲル濾過によるSMO分子量の分析は以下の条件で行った。
1. 分析システムとしてShimadzu LC-10 HPLC システムを、分析カラムとしてGE ヘルスケア社製Superdex 200 10/300 GL を用いた。
2. 移動相として100 mM KPB, 200 mM NaCl pH 7.0 を用い、流速0.4 ml/min で分離を行った。
3. ヒスチジンタグを含む精製StyA サンプル 100μl (500μg タンパク相当) を分析サンプルとしてアプライし、35.3 min に単一ピークとして溶出した。
4. 同様に、標準試料 (オリエンタル酵母社MW-Marker (HPLC)) を分析した。
5. 標準試料の分子量および溶出時間を以下に示す。
酵母グルタミン酸脱水素酵素 290 kDa 26.9 min
豚心筋乳酸脱水素酵素 142 kDa 31.3 min
酵母エノラーゼ 67 kDa 34.3 min
酵母ミオキナーゼ 32 kDa 39.1 min
馬心筋チトクロームc 12.4 kDa 43.0 min
6. 上記の標準試料の各タンパクの溶出時間および分子量を用いて検量線を作成した。得られた検量線に対して精製StyAの溶出時間35.3 min をプロットした結果、StyA の分子量は59.2 kDa に相当すると推測された。
【0110】
【表3】
【0111】
また、組換えStyAB の発現および精製は以下の通りに行った。
1. 発現プラスミドを含む形質転換体を、4 mlのLB medium (50μg/ml アンピシリン、20μg/ml クロラムフェニコール)で37℃、170 rpm で培養した。
2. 前培養液1 ml を100 ml LB medium (50μg/ml アンピシリン、20μg/ml クロラムフェニコール、0.5 mg/ml アラビノース、5 ng/ml テトラサイクリン)に植菌し、37℃、250 rpm で培養した。
3. OD600 が0.5 になったらIPTG を終濃度0.1 mM になるように添加し、18℃、250 rpm、24時間誘導した。
4. 菌体を遠心(18000 rpm, 4℃,10 min)により回収し、50 mM KPB (pH 7.0), 10% glycerol に懸濁、洗浄した。
5. 再度遠心により菌体を回収し、Ni-Sepharose カラムBinding Buffer (50 mM KPB, 10% glycerol, 500 mM NaCl, 10 mM imidazole, pH 7.5 )に再懸濁した。
6. 超音波破砕により菌体を破砕し、遠心(18,000 rpm, 4℃, 20 min x2回)により上清を回収した。
7. Ni-Sepharose カラムBinding Buffer で平衡化したNi-Sepharose High performance カラムに、上記の菌体破砕液上清をアプライし、目的の組換えタンパクを吸着させた。
8. カラム体積の10倍量のBinding Buffer でカラムを洗浄後、Elution Buffer (50 mM KPB, 10% glycerol, 500 mM NaCl, 500 mM imidazole, pH 7.5) により吸着させた目的タンパクを溶出させた。
9. 得られた精製タンパク溶液を透析し、50 mM KPB, 10% glycerol, pH 7.0 に置換した。
10. 透析したタンパク溶液を各種イオン交換クロマトグラフィー(DEAE-Sepharose, Resource Q, MonoQ) を用いて最終精製し、目的タンパク質の精製サンプルを得た。
【0112】
(2)SMO の活性測定
ST-5およびST-10 由来SMO の活性測定は、カラム精製した組換え酵素を用いて行った。各精製酵素、基質、補酵素を50 mM MES緩衝液中に以下の濃度で添加し、反応混合液とした。
反応混合液組成
50 mM MES Buffer (pH 7.0)
20 mM ギ酸ナトリウム
100μM NADH
200μM FAD
1 U ギ酸脱水素酵素
10 U catalase
5 mM substrate(基質変換反応時は、10 mM の基質で反応を実施)
StyA 150μl (基質変換反応時は、StyAB 共発現菌体の菌体破砕液上清を使用)
StyB 100μl
total 1000μl
この反応混合液を30℃、2,500rpmで30分間インキュベーションした。反応終了後、反応混合液に500μlの酢酸エチルを添加し5分間ボルテックスを行った。15,000 rpm で5分間遠心し、分離した酢酸エチル相を新しいチューブに移した。再度500μl の酢酸エチルを水相に添加し、5分間ボルテックス後、再度遠心により酢酸エチル相を回収した。合計1000μl の回収した酢酸エチル相に適量の硫酸ナトリウムを添加し、2分間ボルテックスを行い、脱水処理を行った。脱水処理を行った抽出サンプルを、Hewllet Packerd HP6890 ガスクロマトグラフィーシステムを用いて分析した。
1 U は、1分間に1μmol の基質を産物に変換する酵素活性と定義した。
【0113】
(3)Gas Chromatography 分析条件
ST-5 およびST-10 由来SMOの反応産物の測定は、ガスクロマトグラフィーを用いて行った。上記反応条件にて反応し、酢酸エチルで抽出した反応産物をHewllet Packard HP6890 ガスクロマトグラフィーシステムを用いて分析した。酢酸エチルにより抽出された試料1μl をVarian 社製CP-Cyclodextrin-b-2,3,6-M-19-25 キャピラリーカラム(25 m, 内径0.25 mm)に注入した。キャリアガス(99.999% 高純度ヘリウムガス)流速は2.8 ml/min、スプリット比は50、線形流速は46 cm/s であった。検出はFID(水素炎イオン化検出器)を用いた。気化室温度は250℃、検出器温度は250℃で行った。
カラムオーブンの温度プログラムは、定温プログラムを用いて行った。各分析サンプルに対するカラムオーブン温度および各物質の検出時間は表4に示した。
各物質の同定および定量は、各標準化合物を1 mM, 5 mM, 10 mM の濃度に調製したサンプルを同条件で分析し、その検出時間およびピーク面積から検量線を作成して求めた。
【0114】
【表4】
【0115】
(4)Rhodococcus sp. ST-5株 およびST-10株 由来styAの基質特異性
ST-5およびST-10由来SMOは、スチレンをはじめ様々な基質に対してエポキシ化活性を示した(表5)。その基質特異性は、スチレンを100としたときに、4−フルオロスチレン に対しておよそ2倍の活性を示した。また、生成物の鏡像体過剰率(e.e.)はスチレンや4−クロロスチレン、4−ブロモスチレン、4−フルオロスチレンに対してはやや低いものの、2−クロロスチレンや3−クロロスチレンに対しては99.9%以上の値を示し、その絶対配置は(S)-体であった。
【0116】
【表5】
反応産物の立体選択性については、styrene oxide および3-chloro styrene oxide 以外は産物の標品が市販されていないか、ラセミ体しかないため、S-体であると推測されるが確認はできていない。
【産業上の利用可能性】
【0117】
本発明は、スチレンモノオキシゲナーゼ、該酵素をコードするポリヌクレオチドを提供する。本発明のスチレンモノオキシゲナーゼを利用することにより、スチレンモノオキシゲナーゼを効率よく生産することができる。
【技術分野】
【0001】
本発明は、新規なスチレンモノオキシゲナーゼをコードするDNA、および該DNAによりコードされるタンパク質等に関する。また本発明は、該タンパク質を用いた光学活性なスチレンオキシドの製造方法、およびスチレンの分解方法等に関する。
【背景技術】
【0002】
スチレンは、合成高分子等の化学工業等の分野において重要な化学物質であり、ポリスチレンおよびスチレンブタジエンゴムなど多くの合成樹脂の原料として、多量に使用されている。このような合成樹脂の合成において発生するガス状スチレンやスチレン含有水の排出は、しばしば、ヒトに対する毒性および悪臭のため問題となる。
【0003】
スチレンの生物に与える影響を評価するため、哺乳動物におけるスチレン代謝の機構が研究されている(非特許文献1〜3)。哺乳動物において、スチレンは、シトクロムP-450モノオキシゲナーゼ[EC 1.14.14.1]によりスチレンオキシドへと酸化される(非特許文献4)。次いで、スチレンオキシドはいくつかの経路によって解毒される。主要な経路は、水分子の付加によるスチレングリコールの形成を介して進む。スチレングリコールの主要な代謝物は、マンデル酸、フェニルグリオキシル酸、および馬尿酸である(非特許文献5)。シュードモナスsp.、キサントバクター(Xanthobacter)sp.、およびロドコッカス(Rhodococcus)sp.のようないくつかのスチレン分解細菌が土壌から単離されており、その代謝遺伝子の特徴が報告されている(非特許文献6〜10)。
細菌によるスチレンの分解に関して、2つの異なる経路が報告されている。第一の経路は、ビニル側鎖の酸化を含む経路である。第二の経路は、芳香環の酸化を含む経路である(非特許文献11、12)。
【0004】
これまでの研究において、本発明者らは、土壌から単離されたスチレン資化細菌ロドコッカスsp.ST-5株およびST-10株を報告した。これらの細菌は、スチレン代謝に関与している酵素活性を有する(非特許文献13)。さらに、本発明者らは、ST-5株およびST-10株におけるスチレンの分解経路を、以下のように提唱した。
スチレン→スチレンオキシド→フェニルアセトアルデヒド→2-フェニルエタノールおよびフェニル酢酸。
しかしながら、これらの株からスチレン分解に関与している遺伝子は見出されておらず、代謝機構の詳細は不明なままであった。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Korn, M., R. Wodarz, W. Schoknecht, H. Weichardt, and E. Bayer. 1984. Styrene metabolism in man: gas chromatographic separation of mandelic acid enantiomers in the urine of exposed persons. Arch. Toxicol. 55:59-63.
【非特許文献2】Lof, A., E. Gullstrand, E. Lundgren, and M. B. Nordqvist. 1984. Occurrence of styrene-7,8-oxide and styrene glycol in mouse after the administration of styrene. Scand. J. Work Environ. Health 10:179-187.
【非特許文献3】Watabe, T., M. Isobe, T. Sawahata, K. Yoshikawa, S. Yamada, and E. Takabatake. 1978. Metabolism and mutagenicity of styrene. Scand. J. Work Environ. Health 4(Suppl):142-155.
【非特許文献4】Vainio, H., H. Norppa, K. Hemminki, and M. Sorsa. 1981. Metabolism and genotoxicity of styrene. Adv. Exp. Med. Biol. 136:257-274.
【非特許文献5】Sumner, S. C. J., and T. R. Fennell. 1994. Review of the metabolic fate of styrene. Crit. Rev. Toxicol. 24(Suppl): S11-33.
【非特許文献6】Beltrametti, F., A. M. Marconi, G. Bestetti, C. Colombo, E. Galli, M. Ruzzi, and E. Zennaro. 1997. Sequencing and functional analysis of styrene catabolism genes from Pseudomonas fluorescens ST. Appl. Environ. Microbiol, 63: 2232-2239.
【非特許文献7】Hartmans, S., M. J. van der Werf, and J. A. de Bont. 1990. Bacterial degradation of styrene involving a novel flavin adenine dinucleotide-dependent styrene monooxygenase. Appl. Environ. Microbiol. 56:1347-1351.
【非特許文献8】Panke, S., B. Witholt, A. Schmid, and M. G. Wubbolts. 1998. Towards a biocatalyst for (S)-styrene oxide productio: characterization of the styrene degradation pathway of Pseudomonas sp. strain VLB120. Appl. Environ. Microbiol. 64:2032-2043.
【非特許文献9】Velasco, A., S. Alonso, J. L. Garcia, J. Perera, and E. Diaz. 1998. Genetic and functional analysis of the styrene catabolic cluster of Pseudomonas sp. strain Y2. J. Bacteriol. 180:1063-1071.
【非特許文献10】Warhurst, A.M., K. F. Clarke, R. A. Hill, R. A. Holt, and C. A. Fewson. 1994. Metabolism of styrene by Rhodococcus rhodochrous NCIMB 13259. Appl. Environ. Microbiol. 60:1137-1145.
【非特許文献11】Mooney, A., P. G. Ward, and K. E. O'Connor. 2006. Microbial degradation of styrene: biochemistry, molecular genetics, and perspectives for biotechnological applications. Appl. Microbiol. Biotechnol. 72:1-10.
【非特許文献12】Warhurst, A. M., and C. A. Fewson. 1994. Microbial metabolism and biotransformations of styrene. J. Appl. Bacteriol. 77:597-606.
【非特許文献13】Itoh, N., K. Yoshida, and K. Okada. 1996. Isolation and identification of styrene-degrading Corynebacterium strains, and their styrene metabolism. Biosci. Biotech. Biochem. 60:1826-1830.
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明は、スチレンのビニル基をオキシラニル基に酸化する活性を有するスチレンモノオキシゲナーゼをコードする新規なDNAを提供することを課題とする。また本発明は、該DNAによりコードされるタンパク質、該タンパク質の製造方法等を提供する。さらに本発明はスチレンモノオキシゲナーゼをコードするDNAあるいは当該DNAを含むベクターで形質転換された形質転換細胞を利用するスチレンの分解方法、および光学活性なスチレンオキシドの製造方法を提供することを課題とする。
【課題を解決するための手段】
【0007】
本発明者らは、ロドコッカスsp.ST-5株およびST-10株においてスチレンの分解に関与しているスチレン代謝遺伝子を単離するため、ST-5株およびST-10株のゲノムDNAに対し、縮重PCRおよびTAIL-PCR(thermal asymmetric interlaced PCR)を実施した。その結果本発明者らは、ST-5株において4つのオープンリーディングフレーム(ORF)を見出すことに成功した。この4つのORFによってコードされる推定アミノ酸配列は、シュードモナス sp.におけるスチレン代謝遺伝子styABCDと類似性を示した。よって、4つのORFはそれぞれ、スチレンモノオキシゲナーゼ(SMO)、フラビンオキシドレダクターゼ、スチレンオキシドイソメラーゼ(SOI)、およびフェニルアセトアルデヒドデヒドロゲナーゼ(PAD)に対応する遺伝子であると判断された(図1および図2)。
一方ST-10株については、3つのORFを見出すことに成功した。これらのORFのうち2つは、SMOおよびフラビンオキシドレダクターゼに対応する遺伝子であると判断された(図1および図2)。なお残りの1つについては、ロドコッカス属にのみホモログが見出される機能未知タンパク質と高い類似性を示した。
本発明者らは、これらの遺伝子によってコードされるポリペプチドの作用を明らかにして本発明を完成した。すなわち本発明は、新規なスチレンモノオキシゲナーゼをコードするDNA、該DNAによりコードされるスチレンモノオキシゲナーゼ、該スチレンモノオキシゲナーゼの製造方法、および該スチレンモノオキシゲナーゼを用いるスチレンの分解方法並びに光学活性なスチレンオキシドの製造方法などに関する。
【0008】
本発明は、より詳細には以下の〔1〕から〔9〕に関する。
〔1〕以下(a)から(e)のいずれかに記載のポリヌクレオチド;
(a)配列番号:1または3に記載の塩基配列を含むポリヌクレオチド;
(b)配列番号:2または4に記載のアミノ酸配列をコードするポリヌクレオチド;
(c)配列番号:2または4に記載のアミノ酸配列において、1もしくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列を含み、かつ下記(1)および(2)に記載の理化学的性質を有するスチレンモノオキシゲナーゼをコードするポリヌクレオチド;
(d)配列番号:1または3に記載の塩基配列からなるポリヌクレオチドに対して、高度にストリンジェントな条件下でハイブリダイズし、かつ下記(1)および(2)に記載の理化学的性質を有するスチレンモノオキシゲナーゼをコードするポリヌクレオチド;および
(e)配列番号:2または4に記載のアミノ酸配列と70%以上の同一性を有し、かつ下記(1)および(2)に記載の理化学的性質を有するスチレンモノオキシゲナーゼをコードするポリヌクレオチド;
(1)作用
FADH2(還元型FAD)を補酵素として、スチレンおよび3−クロロスチレンに作用し、それぞれ、光学活性なスチレンオキシドおよび光学活性な3−クロロスチレンオキシドを生成する。
(2)分子量
ゲル濾過におけるヒスチジンを含む分子量が59,000で、SDS-PAGEによる分子量が49,000、アミノ酸組成からの推定分子量が47,000のモノマー酵素である。
〔2〕ロドコッカス(Rhodococcus)属に属する微生物由来である、〔1〕に記載のポリヌクレオチド。
〔3〕前記ロドコッカス属に属する微生物がRhodococcus opacus ST-5株またはRhodococcus erythropolis ST-10株である、〔2〕に記載のポリヌクレオチド。
〔4〕〔1〕から〔3〕のいずれかに記載のポリヌクレオチドによってコードされるタンパク質。
〔5〕〔1〕から〔3〕のいずれかに記載のポリヌクレオチドを含むベクター。
〔6〕〔1〕から〔3〕のいずれかに記載のポリヌクレオチド、または〔5〕に記載のベクターが導入された形質転換細胞。
〔7〕〔6〕に記載の形質転換細胞を培養し、その培養物から〔1〕に記載のポリヌクレオチドによってコードされるタンパク質を回収する工程を含む、〔1〕から〔3〕のいずれかに記載のポリヌクレオチドによってコードされるタンパク質の製造方法。
〔8〕〔4〕に記載のタンパク質をスチレンおよび/またはその類似体に作用させる工程、および生成される化合物を回収する工程を含む、光学活性なスチレンオキシドおよび/または光学活性なスチレン類似体の酸化物の製造方法。
〔9〕〔4〕に記載のタンパク質をスチレンおよび/またはその類似体に作用させる工程を含む、スチレンおよび/またはその類似体の分解方法。
【発明の効果】
【0009】
本発明者らにより、新規なスチレンモノオキシゲナーゼをコードするDNA、該DNAによりコードされるスチレンモノオキシゲナーゼなどが提供された。
スチレンは化学工業において重要な物質であり、ポリスチレンやスチレンブタジエンゴムなどの合成樹脂の原料として欠かせない。しかしながら、これらの合成高分子の製造において発生するスチレンガスやスチレン含有廃水は、しばしばヒトに対する毒性や環境汚染の観点から問題となっている。本発明のスチレンモノオキシゲナーゼは、このような問題の原因となるスチレンの分解や除去に有用である。
また本発明のスチレンモノオキシゲナーゼを用いることにより、医薬、農薬、電子材料等の種々の分野で必要とされる光学活性なスチレンオキシドを効率的且つ経済的に製造することができる。本発明のスチレンモノオキシゲナーゼは、このようにスチレンオキシドの製造においても有用である。
【図面の簡単な説明】
【0010】
【図1】ロドコッカスsp.ST-5株、ST-10株、およびシュードモナスsp.Y2におけるスチレン異化遺伝子の比較を示す図である。矢印は、遺伝子の転写の方向を示す。制限酵素部位を次のように示した。B、Bgl II;E、Eco RI;H、Hind III;K、Kpn I;P、Pst I;Sa、Sal I;Sp、Sph I;X、Xho I。
【図2】ロドコッカスsp.ST-5株(A)およびST-10株(B)のスチレン異化遺伝子のヌクレオチド配列および推定アミノ酸配列を示す図である。小さな矢印は、遺伝子の転写の方向を示す。推定されるリボソーム結合部位を太字で示した。下線は、推定される転写終了シグナルであるステムループ配列を示す。
【図3】スチレンの存在下(A)またはスチレンオキシドの存在下(C)における、ロドコッカスsp.ST-10株による静止菌体反応の生成物のHPLC分析結果を示す図である。(B)は(C)のコントロールであり、ST-10株の菌体の非存在下におけるHPLC分析結果を示す。STYはスチレン、SOはスチレンオキシド、ACPはアセトフェノン、PAはフェニルアセトアルデヒド、SGはスチレングリコール、PAAはフェニル酢酸を示す。
【図4】ST-5株またはST-10株におけるフェニルアセチル-CoAリガーゼの反応生成物のHPLCクロマトグラム結果を示す図である。対照は、粗細胞抽出物なしでインキュベートされた基質とした。PAAはフェニル酢酸を、PA-CoAはフェニルアセチル-CoAを示す。
【図5】ロドコッカスsp.ST-5株およびST-10株によるスチレン異化経路を示す図である。実線は酵素反応を示す。点線は自然変換を示す。
【図6】Rhodococcus sp. ST-5株 およびST-10株 両株由来のstyAおよびstyABを発現させるために構築したプラスミドを示す図である。
【発明を実施するための形態】
【0011】
スチレンモノオキシゲナーゼ
本発明のスチレンモノオキシゲナーゼは、以下の(1)および(2)の理化学的性状を有するタンパク質である。
(1)作用
FADH2(還元型FAD)を補酵素として、スチレンおよび3−クロロスチレンに作用し、それぞれ、光学活性なスチレンオキシドおよび光学活性な3−クロロスチレンオキシドを生成する。
(2)分子量
ゲル濾過におけるヒスチジンを含む分子量が59,000で、SDS-PAGEによる分子量が49,000、アミノ酸組成からの推定分子量が47,000のモノマー酵素である。
ゲル濾過カラムにおける分子量は、ヒスチジンタグを含む分子量である。アミノ酸配列から推定される、ヒスチジンタグが付加された状態における分子量は49.6 kDaである。これらから、本願のスチレンモノオキシゲナーゼはモノマーであると推測される。SDS-PAGEによるSMO分子量の分析方法、ゲル濾過におけるSMO分子量の分析方法は実施例に示した。
【0012】
スチレンは、以下の化学式で表される芳香族炭化水素である。例えば、エチルベンゼンを鉄触媒等で脱水素することにより製造することができる。
【化1】
【0013】
本発明のスチレンモノオキシゲナーゼの活性測定は、例えば以下の手順で行うことができる。
ST-5およびST-10 由来SMO の精製酵素と基質、補酵素類を50 mM MES緩衝液中に以下の濃度で添加し、反応液とする。次に、この反応液を30℃、2,500rpmで30分間インキュベーションする。反応終了後、反応液に500μlの酢酸エチルを添加し5分間ボルテックスを行う。15,000 rpm で5分間遠心し、分離した酢酸エチル相を新しいチューブに移す。再度500μl の酢酸エチルを水相に添加し、5分間ボルテックス後、再度遠心により酢酸エチル相を回収する。合計1000μl の回収した酢酸エチル相に適量の硫酸ナトリウムを添加し、2分間ボルテックスを行い、脱水処理を行う。脱水処理を行った抽出サンプルを、Hewllet Packerd HP6890 ガスクロマトグラフィーシステムを用いて、スチレンモノオキシゲナーゼ活性を分析することができる。
【0014】
反応溶液は、例えば以下の組成とすることができる。
50 mM MES Buffer (pH 7.0)
20 mM ギ酸ナトリウム
100μM NADH
200μM FAD
1 U ギ酸脱水素酵素
10 U catalase
5 mM substrate(基質変換反応時は、10 mM の基質で反応を実施)
StyA 150μl (基質変換反応時は、StyAB 共発現菌体の菌体破砕液上清を使用)
StyB 100μl
total 1000μl
【0015】
ガスクロマトグラフィーによる分析は、例えば次のように行うことができる。
酢酸エチルにより抽出された試料1μl をVarian 社製 CP-Cyclodextrin-b-2,3,6-M-19-25 キャピラリーカラム(25 m, 内径0.25 mm)に注入する。 キャリアガス(99.999% 高純度ヘリウムガス)流速は2.8 ml/min、スプリット比は50、線形流速は46 cm/s とすることができる。検出はFID(水素炎イオン化検出器)を用いて行うことができる。気化室温度は250℃、検出器温度は250℃とすることができる。各物質の同定および定量は、各標準化合物を1 mM, 5 mM, 10 mM の濃度に調製したサンプルを同条件で分析し、その検出時間およびピーク面積から検量線を作成して求めることができる。
【0016】
酵素活性1 U は、1分間に1μmol の基質を産物に変換する酵素活性と定義することができる。
本発明のスチレンモノオキシゲナーゼの酵素活性は、好ましくは1U/mg以上、さらに好ましくは5U/mg以上、特に好ましくは10U/mg以上である。
【0017】
上記のような理化学的性状を有するスチレンモノオキシゲナーゼは、例えば、ロドコッカスsp.の培養物から精製して得ることができる。ロドコッカスsp.としては、ロドコッカスsp.ST-5株およびST-10株などが挙げられるがこれらに限定されない。これらの微生物は、かつて、コリネバクテリウム(Corynebacterium)sp.ST-5株およびST-10株として同定されていた微生物である。しかし最近になり、ST-10株はロドコッカスsp.ST-10株として再同定された。ロドコッカスsp.ST-5株およびST-10株は、次のようにして取得することができる。
各地で採取した土壌サンプルに滅菌水を添加し、ボルテックスを行う。懸濁液を無機塩培地(0.3(w/v)%(NH4)2SO4、0.3% KH2PO4、0.1% NaCl、0.02% MgSO4 7H2O、および1.5%寒天、pH7.0)に塗り広げ、スチレンガス雰囲気下で培養する。例えば30℃で3〜7日間培養し、生育したコロニーを4 ml のLB 培地に接種して培養する。得られた培養液のうち例えば10μlを再度無機塩培地に塗り広げ、スチレンガス雰囲気下、30℃で培養する。生育したコロニーを再度LB 培地で培養し、スチレン資化性菌としてストックすることができる。
【0018】
ロドコッカスsp.ST-5株およびST-10株は、例えばPY培地(0.5%酵母抽出物、0.5%バクトペプトン、pH7.0)等、真菌の培養に用いられる一般的な培地で培養することができる。
これらの培地を用いて培養した菌体を回収し、無機塩培地(0.3(w/v)%(NH4)2SO4、0.3% KH2PO4、0.1% NaCl、0.02% MgSO4 7H2O、および1.5%寒天、pH7.0)に再度接種する。この菌体をスチレンガス雰囲気下において生育し回収することにより、高いSMO活性を有する菌体を得ることができる。
通常は、培養開始時のpHを2〜9、好ましくは4〜7に調節し、15〜40℃、好ましくは20〜35℃の温度条件下で培養することが望ましい。培養時間は通常は1日から7日、好ましくは1日から3日である。
【0019】
培養した菌体から無細胞抽出液を調製し、さらに、無細胞抽出液を精製することにより、本発明のスチレンモノオキシゲナーゼを得ることができる。
【0020】
具体的には、上述のようにして得られた菌体から、2−メルカプトエタノール(2−mercaptoethanol)等の還元剤や、フェニルメタンスルホニルフルオリド(phenylmethansulfonyl fluoride;PMFS)やエチレンジアミン4酢酸(以下、EDTAと略す)、ペプスタチン、ロイペプチン、ホスホラミドンのようなプロテアーゼ阻害剤を加えた緩衝液中でガラスビーズとの物理的な衝撃やミニラボ、フレンチプレスなどの高圧を利用するなどして破砕し、無細胞抽出液を得ることができる。
【0021】
無細胞抽出液から、蛋白質の溶解度による分画(アセトンやジメチルスルホキシドなどの有機溶媒による沈澱や硫安などによる塩析など)や、陽イオン交換、陰イオン交換、ゲル濾過、疎水性クロマトグラフィーや、キレート、色素、抗体などを用いたアフィニティークロマトグラフィーなどを適宜組み合わせることにより本発明の酵素を精製することができる。例えば、無細胞抽出液をブルー−セファロース、フェニル−セファロース、Resource Q(いずれもGEヘルスケア製)などのカラムクロマトグラフィーを組み合わせることにより、電気泳動的にほぼ単一バンドにまで精製することができる。
なお、アミノ酸配列から推定されるST-5株由来のStyAの分子量は46,941 Daである。また、アミノ酸配列から推定されるST-10株由来のStyAの分子量は47,290 Daである。ST-5株由来のStyA、ST-10株由来のStyAいずれも、モノマーであると推定される。
【0022】
本発明のスチレンモノオキシゲナーゼを産生する本発明者らが単離したロドコッカスsp.ST-5株、ロドコッカスsp.ST-10株は、本発明者らによって、独立行政法人製品評価技術基盤機構特許微生物寄託センターに寄託された。以下に、寄託を特定する内容を記載する。
ロドコッカスsp.ST-5株
(1)寄託機関名:独立行政法人製品評価技術基盤機構
(2)連絡先:〒292−0818千葉県木更津市かずさ鎌足2−5−8
(3)受託番号:NITE P-974
(4)識別の表示:ST-5
(5)寄託日:2010年8月27日
ロドコッカスsp.ST-10株
(1)寄託機関名:独立行政法人製品評価技術基盤機構
(2)連絡先:〒292−0818千葉県木更津市かずさ鎌足2−5−8
(3)受託番号:NITE P-975
(4)識別の表示:ST-10
(5)寄託日:2010年8月27日
【0023】
ポリヌクレオチド
本発明において、ポリヌクレオチドは、DNAおよびRNA等の天然のポリヌクレオチドに加え、人工的なヌクレオチド誘導体を含む人工的な分子であることもできる。また本発明のポリヌクレオチドは、DNA-RNAのキメラ分子であることもできる。また、本発明のポリヌクレオチドは、スチレンモノオキシゲナーゼをコードするコード鎖のみからなる一本鎖であっても、該コード鎖とその相補鎖とからなる二本鎖構造を持つものであってもよい。
【0024】
本発明のスチレンモノオキシゲナーゼをコードするポリヌクレオチドは、微生物から得ることができる。微生物としては、ロドコッカス(Rhodococcus)属に属する微生物が好ましい。より具体的には、Rhodococcus opacusやRhodococcus erythropolisは、本発明のポリヌクレオチドを得るための微生物として好ましい。例えば、配列番号:1に記載の塩基配列を含むポリヌクレオチドは、Rhodococcus opacus ST-5株からクローニングされた。配列番号:1に示す塩基配列は、配列番号:2に示すアミノ酸配列からなるタンパク質をコードしている。また、例えば、配列番号:3に記載の塩基配列を含むポリヌクレオチドは、Rhodococcus erythropolis ST-10株からクローニングされた。配列番号:3に示す塩基配列は、配列番号:4に示すアミノ酸配列からなるタンパク質をコードしている。配列番号:2または4のアミノ酸配列を含むタンパク質をコードするポリヌクレオチドは、本発明における好ましいポリヌクレオチドである。
【0025】
本発明のポリヌクレオチドには、Rhodococcus opacus ST-5株、あるいはRhodococcus erythropolis ST-10株からクローニングされた遺伝子に加え、該遺伝子のホモログも含まれる。このような遺伝子のホモログとしては、配列番号:1または3に記載の塩基配列を含むポリヌクレオチドが挙げられる。その他、スチレンモノオキシゲナーゼをコードするポリヌクレオチドに加え、その読み枠に影響しないよう適当な制御配列を含むポリヌクレオチド、適当なペプチド配列により修飾されたスチレンモノオキシゲナーゼの融合タンパク質をコードするポリヌクレオチドを、好適な例として挙げることができる。
【0026】
また、本発明のポリヌクレオチドに含まれるホモログとしては、配列番号:2または4に記載のアミノ酸配列を含むタンパク質をコードするポリヌクレオチドが挙げられる。このようなポリヌクレオチドには、配列番号:1または3に示す塩基配列を含む遺伝子に加え、遺伝暗号の縮重により配列番号:2または4に記載のアミノ酸配列をコードするが、配列番号:1または3の塩基配列とは異なる塩基配列からなるポリヌクレオチドが包含される。
【0027】
今回、配列番号:1または3の塩基配列を有するスチレンモノオキシゲナーゼをコードする遺伝子が得られた。ホモログは、例えば、配列番号:1または3の塩基配列を有するポリヌクレオチドをプローブとしてスクリーニングすることができる。あるいは、配列番号:1または3の塩基配列から選択された塩基配列をプローブとすることもできる。プローブを使ったスクリーニング方法として、次のような公知のハイブリダイゼーション法を利用することができる。
コロニーハイブリダイゼーション、
プラークハイブリダイゼーション、
サザンブロット法
スクリーニングには、酵素生産株であるRhodococcus opacus、Rhodococcus erythropolisなどロドコッカス属の微生物、あるいはその他の生物種等の染色体DNA、またはcDNAライブラリーを利用することができる。
【0028】
また、酵素生産株の染色体DNAまたはcDNAライブラリーを鋳型としたPCRによって、本発明のポリヌクレオチドを得ることもできる。PCR用のプライマーは、配列番号:1または3の塩基配列を元にデザインすることができる。PCRによって得られたDNAが断片であれば、DNA断片の塩基配列に基づいてさらにその全長配列を決定することができる。例えば、インバースPCR(inverse PCR; Genetics(1988)120: 621-3)を利用して、断片配列情報に基づいて、全長配列を決定することができる。インバースPCRは、部分的に塩基配列が不明なDNAを適当な制限酵素で消化後、自己環化反応により環化されたDNAを鋳型として利用するPCRである。断片配列内部の既知領域に特異的なプライマーを利用して、その前後に位置する塩基配列が未知の領域を増幅することができる。
あるいはRACE法(Rapid Amplification of cDNA End;「PCR実験マニュアル」HBJ出版局,p25-33)によって本発明のポリヌクレオチドを得ることもできる。
【0029】
また、本発明におけるポリヌクレオチドのホモログは、配列番号:1または3に示される塩基配列からなるポリヌクレオチドと高度にストリンジェントな条件下でハイブリダイズできるポリヌクレオチドであって、かつ、スチレンモノオキシゲナーゼ活性を有するタンパク質をコードするポリヌクレオチドも含む。
【0030】
本発明において、高度にストリンジェントな条件とは、例えばECL direct nucleic acid labeling and detection system(Amersham Pharmacia Biotech社製)を用いて、マニュアルに記載の条件(例えば、wash:42℃、0.5×SSCを含むprimary wash buffer)においてハイブリダイズすることを言う。より具体的な「高度にストリンジェントな条件」とは、例えば、通常、42℃、2×SSC、 0.1%SDSの条件であり、好ましくは50℃、2×SSC、0.1%SDSの条件であり、さらに好ましくは、65℃、0.1×SSCおよび0.1% SDSの条件である。これら温度、塩濃度に加えて、プローブ濃度、プローブの長さ、反応時間を含む複数の要素がハイブリダイゼーションのストリンジェンシーに影響する。当業者であればこれら要素を適宜選択することでストリンジェンシーを調節することができる。
【0031】
例えばハイブリダイゼーションに使用するプローブの塩基配列は、配列番号:1または3に記載の塩基配列から選択することができる。例えば、少なくとも20個、好ましくは少なくとも30個、例えば40、60または100個の連続した塩基配列を有するポリヌクレオチドをプローブとすることができる。あるいは、配列番号:1または3に記載した塩基配列の全長を有するポリヌクレオチドをプローブとすることもできる。
ハイブリダイゼーション法の詳細な手順については、「Molecular Cloning, A Laboratory Manual 2nd ed.」(Cold Spring Harbor Press(1989)、特にSection9.47-9.58)、「Current Protocols in Molecular Biology」(John Wiley&Sons(1987-1997)、特にSection6.3-6.4)、「DNA Cloning 1: Core Techniques, A Practical Approach 2nd ed.」(Oxford University(1995)、特にSection2.10)等を参照することができる。
【0032】
高度にストリンジェントな条件下でハイブリダイズできるポリヌクレオチドは、配列番号:2または4に記載のアミノ酸配列と高い同一性(相同性)を有するアミノ酸配列をコードしている可能性が高い。具体的には、配列番号:2または4に示されるアミノ酸配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは90%以上、さらに好ましくは95%あるいは98%、特に好ましくは99%以上の同一性を有するアミノ酸配列をコードするポリヌクレオチドは、本発明における好ましいポリヌクレオチドに含まれる。このような高い同一性を有するタンパク質同士は、同じまたは類似した活性を有する可能性が高い。
タンパク質の同一性は、アミノ酸配列に関するデータベースを利用して検索することができる。例えばSWISS-PROT、PIR、DAD等のタンパク質のアミノ酸に配列情報を蓄積したデータベースを利用することができる。あるいはDNAの同一性は、塩基配列情報を蓄積したデータベースを利用して検索することができる。DDBJ、EMBLまたはGenBank等のDNAに関するデータベースが公知である。これらのデータベースにおいては、DNAの塩基配列を元に予想されたアミノ酸配列情報を利用することもできる。各種の配列情報は、これらのデータベース等を対象に、BLAST、FASTA等のプログラムを利用して検索することができる。ここに例示したデータベースは、いずれもインターネット(例えば、 http://www.ncbi.nlm.nih.gov.)を通じて利用することができる。
【0033】
さらに、本発明のポリヌクレオチドのホモログとしては、配列番号:2または4に記載のアミノ酸配列において1もしくは複数のアミノ酸が欠失、置換、挿入および/または付加されたアミノ酸配列からなり、かつ、スチレンモノオキシゲナーゼ活性を有するタンパク質をコードするポリヌクレオチドを挙げることができる。当業者は、ある塩基配列を元に、適宜置換、欠失、挿入、および/または付加変異を導入することにより、このようなポリヌクレオチドのホモログを得ることができる。例えば、配列番号:1または3に記載の塩基配列からなるポリヌクレオチドに、部位特異的変異導入法(Nucleic Acids Res (1982) 10:6487;Methods in Enzymol (1983) 100:448;Molecular Cloning 2nd Edt., Cold Spring Harbor Laboratory Press(1989);PCR A Practical Approach, IRL Press(1991) pp.200)などを用いて、任意の変異を導入することができる。
【0034】
タンパク質においてアミノ酸残基を置換する場合、特に、側鎖の化学的性質が類似したアミノ酸による置換、いわゆる保存的なアミノ酸置換を行うことが好ましい。アミノ酸は、それらの側鎖の化学的性質に従い、例えば、次のように分類される:
(1)中性疎水性側鎖(アラニン、トリプトファン、バリン、フェニルアラニン、プロリン、メチオニン、ロイシン);
(2)中性極性側鎖(アスパラギン、グリシン、グルタミン、システイン、セリン、チロシン、トレオニン);
(3)塩基性側鎖(アルギニン、ヒスチジン、リシン);
(4)酸性側鎖(アスパラギン酸、グルタミン酸);
(5)脂肪族側鎖(アラニン、イソロイシン、グリシン、バリン、ロイシン);
(6)脂肪族水酸基側鎖(セリン、トレオニン);
(7)アミン含有側鎖(アスパラギン、アルギニン、グルタミン、ヒスチジン、リシン);
(8)芳香族側鎖(チロシン、トリプトファン、フェニルアラニン);および
(9)硫黄含有側鎖(システイン、メチオニン)。
【0035】
すなわち、これらの各群を構成するアミノ酸残基の相互の置換を、保存的置換と言う。本発明において、「1もしくは複数のアミノ酸が置換、欠失、挿入、および/または付加」するアミノ酸の数や場所は、上記DNAがコードするタンパク質がスチレンモノオキシゲナーゼ活性を有する限り、制限されない。変異が許容されるアミノ酸残基の数は、典型的には全アミノ酸の10%以内、好ましくは全アミノ酸の5%、さらに好ましくは全アミノ酸の1%以内である。より具体的には、配列番号:2または4のアミノ酸配列において、通常50以内、例えば20以内、より好ましくは5以内のアミノ酸残基の変異は、許容される。
【0036】
本発明のポリヌクレオチドのホモログには、上述のように、配列番号:2または4に記載のアミノ酸配列において1もしくは複数のアミノ酸が欠失されたアミノ酸配列からなり、かつ、スチレンモノオキシゲナーゼ活性を有するタンパク質をコードするポリヌクレオチドが含まれる。このような欠失を含むポリヌクレオチドには、配列番号:2または4のアミノ酸配列からなるタンパク質の「一部分」をコードするポリヌクレオチドが包含される。
【0037】
ポリヌクレオチドによってコードされるタンパク質が、元のタンパク質の生物学的活性(スチレンモノオキシゲナーゼ活性)を維持している限り、アミノ酸配列におけるアミノ酸残基の欠失・置換・付加および/または挿入は許容される。ここで、生物学的活性の維持とは、元の酵素が触媒する少なくとも一つの反応を触媒する能力が維持されることを言う。「活性の維持」には、同じ活性レベルのみならず、より高い活性も含まれる。また、活性のレベルが低下する場合であっても、実質的に同等の活性であれば、活性の維持に含まれる。実質的に同等とは、元の活性に対して、例えば50%〜100%、通常70〜100%、好ましくは80〜100%、より好ましくは90、あるいは95〜100%の活性を言う。本発明におけるタンパク質の生物学的活性、すなわちスチレンモノオキシゲナーゼ活性の評価方法は既に述べた。
【0038】
本発明のポリヌクレオチドには、天然よりクローニングされたゲノムDNA、およびcDNAの他、合成によって得られるポリヌクレオチドも含まれる。本発明のポリヌクレオチドは、例えば、配列番号:1または3に記載の配列情報を元に、周知の手法により合成することができる。
【0039】
また、本発明の酵素は遺伝子工学的な手法、あるいは化学的な合成法によっても得ることができる。例えば、細胞を含まない試験管内でのタンパク質の製造方法としてin vitroトランスレーション(Dasso and Jackson, Nucleic Acids Res(1989)17:3129-44)が知られている。また、本発明のポリヌクレオチドを適当な発現ベクターに組込み、該発現ベクターが発現される宿主に形質転換し、該宿主細胞より所望のタンパク質を得ることもできる。このような宿主-ベクター系によりタンパク質を産生する方法については、以下の「組換えベクターおよび形質転換体」の項において具体的に述べる。これら公知のタンパク質の製造方法は、本発明のタンパク質を得るための方法として利用することができる。
【0040】
遺伝子工学的な手法により製造されたタンパク質は、当該タンパク質を含む生物材料から回収される。例えば、タンパク質が宿主細胞外に分泌される場合には、当該細胞を培養した培地からタンパク質が回収される。宿主がトランスジェニック生物の場合にはその体液から目的のタンパク質を回収できる。あるいは細胞内に産生される場合には細胞を溶解した溶解物よりタンパク質を回収する。
【0041】
回収されたタンパク質は、該タンパク質を天然において産生する細胞から精製する場合と同様の手段により精製することができる。すなわち、公知の塩析、蒸留、各種クロマトグラフィー、ゲル電気泳動、ゲル濾過、限外濾過、再結晶、酸抽出、透析、免疫沈降、溶媒沈澱、溶媒抽出、硫安またはエタノール沈澱等の精製手法を組み合わせて、目的とするタンパク質を精製することができる。当業者は、例えば次のような各種のクロマトグラフィーを組み合わせて利用することができる。これらのクロマトグラフィーには、HPLCおよびFPLC等の液相クロマトグラフィーシステムを用いることができる。
アフィニティークロマトグラフィー、
アニオンまたはカチオン交換等のイオン交換クロマトグラフィー、
逆相クロマトグラフィー、
吸着クロマトグラフィー、
ゲル濾過、
疎水性クロマトグラフィー、
ヒドロキシアパタイトクロマトグラフィー
【0042】
また、本発明のタンパク質(スチレンモノオキシゲナーゼ)をタグとの融合タンパク質として発現させれば、タグに結合するカラムを利用して回収することができる。例えばGSTタグを有する融合タンパク質は、グルタチオンカラムを用いて容易に分離することができる。あるいはヒスチジンタグとの融合タンパク質は、ニッケルカラムを用いて精製することができる。タグとタンパク質(スチレンモノオキシゲナーゼ)の間にプロテアーゼ認識配列を挿入することができる。プロテアーゼには、例えばトロンビンやファクターXa等を利用することができる。融合タンパク質をカラムに結合させた後、必要に応じてカラムを洗浄する。次いでこれらのプロテアーゼを作用させると、目的とするタンパク質がタグから切断される。その後、切断されたタンパク質を回収することによって、目的とするタンパク質を容易に精製することができる。
【0043】
組換えベクターおよび形質転換体
本発明のポリヌクレオチドを公知の発現ベクターに挿入することにより、スチレンモノオキシゲナーゼ発現ベクターが提供される。即ち本発明は、本発明のポリヌクレオチドを含む組換えベクターに関する。適当なベクターとして、プラスミド、コスミド、ウイルス、バクテリオファージ等の種々のベクターを挙げることができる(Molecular Cloning, A Laboratory Manual 2nd ed., Cold Spring Harbor Press(1989); Current Protocols in Molecular Biology, John Wiley&Sons(1987)参照)。本発明の好ましいベクターとしては、これに限定されるわけではないが、例えば、大腸菌における発現ベクターpETにスチレンモノオキシゲナーゼをコードする遺伝子を発現可能に挿入したE.coli.BL21(DE3)等が挙げられる。
【0044】
本発明の組換えベクターは、分子生物学、生物工学および遺伝子工学の分野において慣用されている技術に準じて構築することができる(Molecular Cloning, A Laboratory Manual 2nd ed., Cold Spring Harbor Press(1989); Current Protocols in Molecular Biology, John Wiley&Sons(1987)参照)。微生物等を宿主として、本発明のポリヌクレオチドを発現させるためには、まず、当該微生物中において安定に存在するプラスミドベクターまたはファージベクター中に該DNAを導入し、その遺伝情報を転写・翻訳させる必要がある。そのためには、通常、プロモーターを本発明のポリヌクレオチドの5’側上流に配置する。プロモーターは、転写・翻訳を制御するユニットである。
【0045】
そして本発明のポリヌクレオチドの3’側下流には、ターミネーターを配置するのが好ましい。プロモーターおよびターミネーターとしては、宿主として利用する微生物中において機能するものを選択することができる。各種微生物において利用可能なベクター、プロモーターおよびターミネーター等の制御配列は、「微生物学基礎講座8遺伝子工学」(共立出版)などで参照することができる。また特に酵母の制御配列について、Adv Biochem Eng (1990) 43:75-102およびYeast (1992) 8:423-88等に詳細に記述されている。その他、必要に応じて、エンハンサー、オペレーター配列、開始シグナル、ポリアデニル化シグナル、リボソーム結合部位等の転写および/または翻訳に必要な制御配列を組込むことができる。
本発明のベクターは、好ましくは、挿入された本発明のポリヌクレオチドの発現に必要とされる制御配列の全ての構成成分を含むものである。さらに、本発明のベクターは、該ベクターが導入された宿主細胞を選択するための選択マーカーを含むことができる。
【0046】
本発明においては、本発明のポリヌクレオチドに、シグナルペプチドをコードする配列を付加することもできる。シグナルペプチドの付加によって、宿主細胞内で発現されたタンパク質を小胞体内腔に移行させることができる。あるいは、グラム陰性菌を宿主とする場合には、シグナルペプチドによって、宿主細胞内で発現されたタンパク質を、ペリプラズム内、または細胞外へと移行させることができる。利用する宿主細胞において機能することができる任意のシグナルペプチドを利用することができる。従って、宿主にとって異種由来のシグナルペプチドを利用することもできる。さらに必要に応じ、本発明のポリヌクレオチドのベクターへの導入に当たって、リンカー、開始コドン(ATG)、終止コドン(TAA、TAGまたはTGA)等を付加することもできる。
【0047】
本発明のポリヌクレオチドを発現させるための宿主は、該ポリヌクレオチドを含む組換えベクターによって形質転換することができ、かつスチレンモノオキシゲナーゼ活性を有するタンパク質を発現することができる生物であれば特に制限されない。本発明は、本発明のポリヌクレオチド、または本発明のベクターにより形質転換された形質転換体(形質転換細胞)を提供する。本発明の形質転換体の対象となる微生物としては、例えば以下のような微生物を示すことができる。
(1)宿主ベクター系の開発されている細菌
・エシェリヒア(Escherichia)属
・バチルス(Bacillus)属
・シュードモナス(Pseudomonas)属
・セラチア(Serratia)属
・ブレビバクテリウム(Brevibacterium)属
・コリネバクテリイウム(Corynebacterium)属
・ストレプトコッカス(Streptococcus)属
・ラクトバチルス(Lactobacillus)属など
(2)宿主ベクター系の開発されている放線菌
・ロドコッカス(Rhodococcus)属
・ストレプトマイセス(Streptomyces)属など
(3)宿主ベクター系の開発されている酵母
・サッカロマイセス(Saccharomyces)属
・クライベロマイセス(Kluyveromyces)属
・シゾサッカロマイセス(Schizosaccharomyces)属
・チゴサッカロマイセス(Zygosaccharomyces)属
・ヤロウイア(Yarrowia)属
・トリコスポロン(Trichosporon)属
・ロドスポリジウム(Rhodosporidium)属
・ピキア(Pichia)属
・キャンディダ(Candida)属など
(4)宿主ベクター系の開発されているカビ
・ノイロスポラ(Neurospora)属
・アスペルギルス(Aspergillus)属
・セファロスポリウム(Cephalosporium)属
・トリコデルマ(Trichoderma)属など
【0048】
例えばエシェリヒア属、特に大腸菌(Escherichia coli) においては、プラスミドベクターとして、pET、pBR、pUC系プラスミドを利用でき、lac(β-ガラクトシダーゼ)、trp(トリプトファンオペロン)、 tac、trc (lac、trpの融合)、λファージ PL、PR等に由来するプロモーターが利用できる。また、ターミネーターとしては、trpA由来、ファージ由来、rrnBリボソーマルRNA由来のターミネーターを用いることができる。特に、市販のpSE420(Invitrogen製)のマルチクローニングサイトを一部改変したベクターpSE420D (特開2000-189170に記載)は、エシェリヒア属細菌を宿主とした場合の好適なベクターである。
【0049】
バチルス属においては、ベクターとしてpUB110系プラスミド、pC194系プラスミド等が利用可能であり、これらのベクターを利用した場合、本発明のポリヌクレオチドを宿主染色体にインテグレートすることもできる。また、プロモーター、ターミネーターとしては、apr(アルカリプロテアーゼ)、 npr(中性プロテアーゼ)、amy(α-アミラーゼ)等が利用できる。
【0050】
シュードモナス属においては、シュードモナス・プチダ(Pseudomonas putida)、シュードモナス・セパシア(Pseudomonas cepacia) 等で宿主ベクター系が開発されている。トルエン化合物の分解に関与するプラスミドTOLプラスミドを基本にした広宿主域ベクター(RSF1010等に由来する自律的複製に必要な遺伝子を含む)pKT240等が利用可能であり、プロモーター・ターミネーターとして、リパーゼ(特開平5-284973)遺伝子由来のものが利用できる。
【0051】
ブレビバクテリウム属、特にブレビバクテリウム・ラクトファーメンタム(Brevibacterium lactofermentum)においては、pAJ43(Gene (1985) 39:281)等のプラスミドベクターが利用可能である。プロモーター・ターミネーターとしては、大腸菌で使用されているプロモーター、ターミネーターがそのまま利用可能である。
【0052】
コリネバクテリウム属、特にコリネバクテリウム・グルタミカム(Corynebacterium glutamicum)においては、pCS11(特開昭57-183799)、pCB101(Mol Gen Genet (1984)196:175)等のプラスミドベクターが利用可能である。
【0053】
ストレプトコッカス属においては、pHV1301(FEMS Microbiol Lett (1985) 26:239)、pGK1(Appl Environ Microbiol (1985) 50:94)等がプラスミドベクターとして利用可能である。
【0054】
ラクトバチルス属においては、ストレプトコッカス属用に開発されたpAMβ1(J Bacteriol (1979) 137: 614)等がベクターとして利用可能であり、プロモーターとしては、大腸菌で利用されているものが利用可能である。
【0055】
ロドコッカス属においては、Rhodococcus opacusやRhodococcus erythropolisに加え、ロドコッカス・ロドクロウス(Rhodococcus rhodochrous)から単離されたプラスミドベクターが使用可能である (J Gen Microbiol (1992) 138:1003)。
【0056】
ストレプトマイセス属においては、HopwoodらのGenetic Manipulation of Streptomyces: A Laboratory Manual Cold Spring Harbor Laboratories (1985)に記載の方法に従って、プラスミドを構築することができる。特に、ストレプトマイセス・リビダンス(Streptomyces lividans) においては、pIJ486 (Mol Gen Genet (1986) 203: 468-78)、pKC1064(Gene (1991) 103:97-9)、pUWL-KS (Gene (1995) 165:149-50)が使用できる。また、ストレプトマイセス・バージニア(Streptomyces virginiae)においても、同様のプラスミドを使用することができる(Actinomycetol (1997) 11:46-53)。
【0057】
サッカロマイセス属、特にサッカロマイセス・セレビジアエ(Saccharomyces cerevisiae) においては、YRp系、YEp系、YCp系、YIp系プラスミドが利用可能である。また、染色体内に多コピー存在するリボソームDNAとの相同組換えを利用したインテグレーションベクター(EP537456等)は、多コピーで遺伝子を導入でき、かつ安定に遺伝子を保持できるため極めて有用である。また、 ADH(アルコール脱水素酵素)、GAPDH(グリセルアルデヒド-3-リン酸脱水素酵素)、PHO(酸性フォスファターゼ)、GAL(β-ガラクトシダーゼ)、PGK(ホスホグリセレートキナーゼ)、ENO(エノラーゼ)等のプロモーター・ターミネーターが利用可能である。
【0058】
クライベロマイセス属、特にクライベロマイセス・ラクティス(Kluyveromyces lactis)においては、サッカロマイセス・セレビジアエ由来2μm系プラスミド、pKD1系プラスミド(J Bacteriol (1981) 145:382-90)、キラー活性に関与するpGKl1由来プラスミド、クライベロマイセス属における自律増殖遺伝子KARS系プラスミド、リボソーム DNA等との相同組換えにより染色体中にインテグレート可能なベクタープラスミド(EP537456等)等が利用可能である。また、ADH、PGK等に由来するプロモーター・ターミネーターが利用可能である。
【0059】
シゾサッカロマイセス属においては、シゾサッカロマイセス・ポンベ(Schizosaccharomyces pombe) 由来のARS (自律複製に関与する遺伝子)およびサッカロマイセス・セレビジアエ由来の栄養要求性を相補する選択マーカーを含むプラスミドベクターが利用可能である(Mol Cell Biol (1986) 6:80)。また、シゾサッカロマイセス・ポンベ由来のADHプロモーター等が利用できる(EMBO J (1987) 6:729)。特に、pAUR224は、宝酒造から市販されており容易に利用できる。
【0060】
チゴサッカロマイセス属においては、チゴサッカロマイセス・ロウキシ(Zygosaccharomyces rouxii) 由来のpSB3(Nucleic Acids Res. 13:4267(1985))等に由来するプラスミドベクターが利用可能であり、サッカロマイセス・セレビジアエ由来 PHO5 プロモーター、およびチゴサッカロマイセス・ロウキシ由来 GAP-Zr(グリセルアルデヒド-3-リン酸脱水素酵素)のプロモーター(Agri Biol Chem (1990) 54:2521)等が利用可能である。
【0061】
ピキア属においては、ピキア・アンガスタ(Pichia angusta;旧名:ハンゼヌラ・ポリモルファ(Hansenula polymorpha))を用いた宿主ベクター系が開発されている。ベクターとしては、ピキア・アンガスタ由来自律複製に関与する遺伝子(HARS1、HARS2)も利用可能であるが、比較的不安定であるため、染色体への多コピーインテグレーションが有効である(Yeast (1991) 7:431-43)。また、メタノール等で誘導されるAOX(アルコールオキシダーゼ)、FDH(ギ酸脱水素酵素)のプロモーター等が利用可能である。また、ピキア・パストリス(Pichia pastoris)等について、ピキア由来自律複製に関与する遺伝子 (PARS1、 PARS2)等を利用した宿主ベクター系が開発されており(Mol Cell Biol (1985) 5:3376)、高濃度培養とメタノールで誘導可能なAOX等の強いプロモーターが利用できる(Nucleic Acids Res (1987) 15:3859)。
【0062】
キャンディダ属においては、キャンディダ・マルトーサ(Candida maltosa)、キャンディダ・アルビカンス(Candida albicans)、キャンディダ・トロピカリス(Candida tropicalis)、キャンディダ・ウチルス(Candida utilis) 等について宿主ベクター系が開発されている。キャンディダ・マルトーサにおいては、キャンディダ・マルトーサ由来ARSがクローニングされ(Agri Biol Chem (1987) 51:1587)、これを利用したベクターが開発されている。また、キャンディダ・ウチルスにおいては、染色体インテグレートタイプのベクターでは、強力なプロモーターが開発されている(特開平08-173170)。
【0063】
アスペルギルス属においては、アスペルギルス・ニガー(Aspergillus niger)、アスペルギルス・オリジー(Aspergillus oryzae) 等が最もよく研究されている。これらを宿主とするプラスミドが利用可能であり、染色体への所望遺伝子のインテグレーションを行うことができる。菌体外プロテアーゼおよびアミラーゼ由来のプロモーターも利用可能である(Trends in Biotechnology (1989) 7:283-7)。
【0064】
トリコデルマ属においては、トリコデルマ・リーゼイ(Trichoderma reesei)を利用した宿主ベクター系が開発されており、菌体外セルラーゼ遺伝子由来プロモーター等が利用できる(Biotechnology (1989) 7:596-603)。
【0065】
また、微生物以外でも、植物および動物を宿主とする様々な宿主ベクター系が開発されている。例えば、大量に異種タンパク質を発現させる系として、蚕を用いた昆虫ベクター系(Nature (1985) 315:592-4)、並びに、菜種、トウモロコシおよびジャガイモ等の植物ベクター系が開発されており、好適に利用できる。
【0066】
ベクターへの本発明のポリヌクレオチドの導入は、制限酵素サイトを利用したリガーゼ反応により行うことができる(Current Protocols in Molecular Biology, John Wiley & Sons(1987) Section 11.4-11.11; Molecular Cloning, A Laboratory Manual 2nd ed., Cold Spring Harbor Press(1989) Section 5.61-5.63)。また、使用する宿主のコドン使用頻度を考慮し、必要に応じ本発明のポリヌクレオチド配列の改変を行い、発現効率の高いベクターを設計するようにしてもよい(Grantham et al., Nucleic Acids Res (1981) 9:r43-74)。
【0067】
上述のように、様々な細胞が宿主細胞株として確立されている。そして、各細胞株に適した発現ベクターの導入法も公知であり、当業者であれば、各選択した宿主細胞に好適な導入法を選択することができる。例えば、原核細胞については、カルシウム処理、エレクトポレーションによる形質転換等が知られている。また、植物細胞については、アグロバクテリウムを用いた方法が公知であり、哺乳動物細胞についてはリン酸カルシウム沈降法を例示することができる。本発明は特にこれらの方法に限定されるわけではなく、選択した宿主に応じ、その他公知の核マイクロインジェクション、プロトプラスト融合、DEAE-デキストラン法、細胞融合、電気パルス穿孔法、リポフェクタミン法(GIBCO BRL)、FuGENE6試薬(Boehringer-Mannheim)を用いた方法をはじめとする種々の公知の方法により発現ベクターの導入を行うことができる。
【0068】
以上のようにして本発明のポリヌクレオチドを含む組換えベクターにより形質転換された形質転換体を培養することにより、本発明のスチレンモノオキシゲナーゼ活性を有するタンパク質を製造することができる。よって、本発明の好ましい一態様として、本発明の上記形質転換体を培養し、その培養物から前記ポリヌクレオチドによってコードされる蛋白質を回収する工程を含む、光学活性なスチレンモノオキシゲナーゼの製造方法が提供される。形質転換体の培養方法は特に限定されず、選択した各宿主細胞の生育に適し、かつ、本発明の酵素の生産に最も適した培地、温度、時間等の条件を選択することが望ましい。例えば実施例に記載の条件が挙げられるがこれに限定されない。
【0069】
スチレンオキシドおよび/またはスチレン類似体の酸化物の製造方法
本発明は光学活性なスチレンオキシドおよび/または光学活性なスチレン類似体の酸化物の製造方法に関する。当該方法は、スチレンモノオキシゲナーゼにより、スチレンおよび/またはその類似体のビニル基がオキシラニル基に酸化されることを特徴とする。本発明のスチレンオキシドおよび/またはスチレン類似体の酸化物の製造方法は、スチレンモノオキシゲナーゼをスチレンおよび/またはその類似体に作用させる工程、および生成される酸化物を回収する工程を含む。
【0070】
スチレンモノオキシゲナーゼと基質は、基質の酸化が可能な条件下で作用させられる。具体的には、本発明の上記酸化反応では、FADH2(または還元型FAD)を補酵素として加えることが必要である。以下に、本発明の酸化反応を示す。本発明の酸化反応においては、補酵素FADH2が酸化されFADが生成すると同時に、水が生じる。
【化2】
【0071】
スチレンモノオキシゲナーゼの使用形態は、酵素分子、その処理物、酵素分子を含む培養物、酵素を生成する形質転換体等の微生物またはその処理物の形態で使用できる。
スチレンモノオキシゲナーゼを生成する形質転換体の微生物の処理物には、具体的には界面活性剤やトルエンなどの有機溶媒処理によって細胞膜の透過性を変化させた微生物、あるいはガラスビーズや酵素処理によって菌体を破砕した無細胞抽出液やそれを部分精製したもの、高度に精製したもの、各種固定化担体に固定化したものなどが含まれる。
【0072】
このようなスチレンモノオキシゲナーゼを反応溶液中、スチレンおよび/またはその類似体と作用させることにより、目的とする酵素反応を行わせ、スチレンオキシドおよび/またはスチレン類似体の酸化物を得ることができる。なお、酵素と反応溶液の作用形態はこれらの具体例に限定されるものではない。また本発明において「作用」は「接触」と表現することもできる。
【0073】
反応溶液は、好ましくは、基質や酵素反応に必要な補酵素であるFADH2を酵素活性の発現に望ましい環境を与える適当な溶媒に溶解したものである。このような溶媒としては、例えば、水中もしくは水に溶解しにくい有機溶媒、例えば、酢酸エチル、酢酸ブチル、トルエン、クロロホルム、n−ヘキサン、メチルイソブチルケトン、メチルターシャリーブチルエステルなどの有機溶媒、もしくは、水性媒体との2相系、もしくは水に溶解する有機溶媒、例えば、メタノール、エタノール、イソプロピルアルコール、アセトニトリル、アセトン、ジメチルスルホキシド、ジメチルホルムアミドなどとの混合系が挙げられる。
【0074】
本発明におけるスチレン類似体としては、例えば、スチレンの2位、3位または4位の少なくともいずれかがハロゲンに置換された化合物が挙げられる。ハロゲンとしてはフッ素、塩素、臭素などが挙げられるがこれらに限定されない。このようなスチレン類似体として以下の化合物を例示することができる。
・2-フルオロスチレン、3-フルオロスチレン、4−フルオロスチレン
・2-クロロスチレン、3-クロロスチレン、4−クロロスチレン
・2-ブロモスチレン、3-ブロモスチレン、4−ブロモスチレン
あるいは、本発明におけるスチレン類似体のその他の例として、以下の化合物を挙げることもできる。
・インデン
・p-アミノスチレン
・p-メトキシスチレン
・trans-β-メチルスチレン
・4-ビニル-1-クロロヘキセン
・N,N-ジメチル-1-ナフチルアミン
・桂皮アルデヒド
このようなスチレン類似体を本発明のスチレンモノオキシゲナーゼと接触させることによって酸化することにより、スチレン類似体の酸化物を得ることができる。具体的には、例えば2−フルオロスチレンを本発明のスチレンモノオキシゲナーゼによって酸化することにより、2−フルオロスチレンオキシドを得ることができる。あるいは、例えばインデンを本発明のスチレンモノオキシゲナーゼによって酸化することにより、インデンオキシドを得ることができる。
【0075】
酸化反応に付随してFADH2から生成するFADの、FADH2への再生は、微生物の持つFAD還元能(呼吸活性など)を用いて行うことができる。また、FADからFADH2を生成する能力を有する微生物やその処理物、酵素を反応系に添加することによっても行うことができる。例えば、フラビンオキシドレダクターゼを含む微生物、その処理物、ならびに部分精製もしくは精製酵素を用いてFADH2の再生を行うことができる。これらのFADH2再生に必要な反応を構成する成分は、本発明によるスチレンオキシドまたはスチレン類似体の酸化物の製造のための反応系に添加する、固定化したものを添加する、あるいはFAD+の交換が可能な膜を介して接触させることができる。
あるいは、フラビンオキシドレダクターゼをコードするDNAでトランスフェクトされた微生物やその処理物を、本発明の酸化反応のための反応系に添加することも可能である。
【0076】
本発明の反応は、固定化酵素、膜リアクターなどを利用して行うことも可能である。
【0077】
スチレンオキシドまたはスチレン類似体の酸化物の製造方法においては、反応温度が好ましくは4〜60℃、さらに好ましくは15〜37℃、pHが好ましくは4〜11、さらに好ましくはpH5〜9、基質濃度が好ましくは0.01〜90%、さらに好ましくは0.1〜30%で行うことができる。
【0078】
反応系には必要に応じて補酵素FADH2を、好ましくは0.001mM〜100mM、さらに好ましくは、0.01〜10mM添加できる。
また、基質は反応開始時に一括して添加することも可能であるが、反応液中の基質濃度が高くなりすぎないように連続的、もしくは非連続的に添加することが望ましい。基質濃度は、例えば0.01g/Lから500g/L、好ましくは0.1g/Lから100g/L、より好ましくは0.5g/Lから10g/Lとすることができるがこれらに限定されない。
【0079】
FADH2再生用の酵素、例えば、フラビンオキシドレダクターゼなどは、本発明の酸化酵素に比較して酵素活性で好ましくは0.1〜100倍、さらに好ましくは0.5〜20倍程度添加することができる。
【0080】
本発明の酸化反応により生成するスチレンオキシドまたはスチレン類似体の酸化物の回収は遠心分離や膜濾過による菌体や蛋白質の除去、膜処理等による蛋白質の分離、酢酸エチル、酢酸ブチル、クロロホルム、メチル−t−ブチルエーテル、n−ブタノール、sec−ブタノール、t−ブタノール、イソブタノール、n−ペンタノール、sec−ペンタノール、t−ペンタノール、イソペンタノールなどによる溶媒抽出、蒸留、酢酸エチル、酢酸ブチル、キシレン、トルエン、メシチレン、クロルベンゼン、クロロホルム、n−ヘキサン、イソオクタン、ジイソプロピルエーテル、メチル−t−ブチルエーテルなどによる結晶化、等を適当に組み合わせることにより行うことができる。あるいは、抽出溶媒液に塩酸を滴下することで生成する塩酸塩として回収することもできる。目的物は、必要によりシリカゲルカラムクロマトグラフィー、蒸留によりさらに精製することができる。
【0081】
スチレンまたはその類似体の分解方法
さらに本発明は、本発明のスチレンモノオキシゲナーゼをスチレンまたはその類似体と接触させる工程を含む、スチレンまたはその類似体の分解方法に関する。上述のように本発明のスチレンモノオキシゲナーゼは、スチレンまたはその誘導体のビニル基をオキシラニル基に酸化する。従って本発明のスチレンモノオキシゲナーゼは、スチレンまたはその類似体を分解することができる。
本発明において分解とは、元の化合物がこれとは異なる化合物に変換されることを意味する。変換としては、例えば酸化などが挙げられる。従って本発明において分解方法は、酸化する方法と表現することもできる。あるいは、スチレンまたはその類似体の量あるいは濃度を減少させる方法と表現することもできる。
スチレン類似体ついては上述のものが挙げられる。またスチレンモノオキシゲナーゼと基質との接触の形態についても既に述べた。
なお本明細書において引用された全ての文献は、参照として本明細書に組み入れられる。
【実施例】
【0082】
以下、実施例により本発明をさらに具体的に説明する。但し、本発明は以下の実施例に限定されるものではない。
材料および方法
(1)細菌株およびベクター
スチレン同化細菌ST-5株およびST-10株を、スチレン代謝遺伝子の単離のための染色体DNAの起源として使用した。これらの微生物は、以前はコリネバクテリウム(Corynebacterium)sp.ST-5株およびST-10株として同定されていた(Itoh, N., K. Yoshida, and K. Okada. 1996. Isolation and identification of styrene-degrading Corynebacterium strains, and their styrene metabolism. Biosci. Biotech. Biochem. 60:1826-1830.)。最近、Makinoらによって、ST-10株はロドコッカスsp.ST-10株として再同定された(Makino, Y., K. Inoue, T. Dairi, and N. Itoh. 2005. Engineering of phenylacetaldehyde reductase for efficient substrate conversion in concentrated 2-propanol. Appl. Environ. Microbiol. 71:4713-4720)。本発明者らも、Makinoらによって記載された方法により、ST-5株の再同定を実施した。結果は、ST-5株がロドコッカスの典型的な特性を有することを示した。従って、本発明者らは、この微生物がロドコッカスsp.ST-5株として同定されるべきであるとの結論を下した。大腸菌JM109株およびプラスミドベクターpGEM-T Easy(Plomega)をDNA操作において使用した。
【0083】
(2)培養条件
ロドコッカスsp.ST-5株およびST-10株を、30℃で2日間、振とうしながら、PY培地(0.5%酵母抽出物、0.5%バクトペプトン、pH7.0)中で培養した。10μlの培養液を、寒天プレート(0.3(w/v)%(NH4)2SO4、0.3% KH2PO4、0.1% NaCl、0.02% MgSO4 7H2O、および1.5%寒天、pH7.0)へ播き、スチレン雰囲気下において30℃で4日間培養した(Itoh, N., K. Yoshida, and K. Okada. 1996. Isolation and identification of styrene-degrading Corynebacterium strains, and their styrene metabolism. Biosci. Biotech. Biochem. 60:1826-1830)。
【0084】
(3)スチレン代謝遺伝子のクローニング
DNA操作には標準的な技術を使用した(Sambrook, J., and W. D. Russell. 2001. "Molecular cloning, a laboratory manual" 3rd ed. Cold Spring Harbor Laboratory Press.)。採集されたST-5株 およびST-10株 の細胞をリゾチームにより処理した後、Eulbergら(Eulberg, D., L. A. Golovleva, and M. Schlomann. 1997. Characterization of catechol catabolic genes from Rhodococcus erythropolis 1CP. J. Bacteriol. 179:370-381)によって記載された方法によってゲノムDNAを調製した。調製されたゲノムDNAを、PCRの鋳型およびサザンブロット分析に使用した。
styA遺伝子およびstyC遺伝子をコードする部分断片を、縮重PCRにより増幅した。styAおよびstyCの増幅のために使用されたプライマーを表1に示した。これらのオリゴヌクレオチドプライマーは、それぞれ、スチレンモノオキシゲナーゼおよびスチレンオキシドイソメラーゼにおいて保存されている部分アミノ酸配列に基づき設計された。スチレン代謝に関与している遺伝子の全長を入手するために、明らかになったstyA遺伝子またはstyC遺伝子の部分塩基配列を用いて、TAIL(thermal asymmetric interlaced)PCRを実施した。TAIL-PCRは、Liu らによって報告された方法(Liu, Y. G., and R. F. Whittier. 1995. Thermal asymmetric interlaced PCR: automatable amplification and sequencing of insert end fragments from P1 and YAC clones for chromosome walking. Genomics 25:674-681)に従い実施された。TAIL-PCRのために使用されたアダプタープライマーおよび既知配列特異的プライマーを表1に示した。一次および二次のPCR産物を希釈し、それぞれ二次および三次のPCRのための鋳型として使用した。増幅された断片は、アガロースゲル電気泳動による分離および抽出により精製された。精製された断片を、pGEM-T Easyへとクローニングし、キャピラリーDNAシーケンサー310(Applied Biosystems,Tokyo,Japan)を使用して塩基配列を決定した。
【0085】
【表1】
【0086】
(4)サザンブロット分析
ロドコッカスsp.ST-10株のゲノムDNAを、いくつかの制限酵素により消化し、アガロースゲル電気泳動によって分離した。分離されたDNAを、Hybond N+ナイロンメンブラン(GE Healthcare UK Ltd,Buckinghamshire,UK)へ転写し、80℃で2時間ベーキングした。ST-10株由来のスチレンモノオキシゲナーゼ遺伝子断片(1.2 kbp)、およびST-5株由来のスチレンオキシドイソメラーゼ遺伝子断片(0.6 kbp)をPCRにより増幅し、プローブとしてハイブリダイゼーションのために使用した。各断片を、Random Primer DNA Labeling Kit Ver.2(TaKaRa,Shiga,Japan)を使用してα-32P-dCTPにより標識した。標識操作は製品プロトコルに従って行った。プレハイブリダイゼーションおよびハイブリダイゼーションを、5×SSC、0.1%SDS、5×デンハルツ液、40%ホルムアミドからなるハイブリダイゼーション緩衝液において実施した。標識されたプローブDNA 1ngをメンブランに添加し、45℃で一夜インキュベートした。メンブランを、45℃で10分間2×SSC緩衝液により2回洗浄し、次いで、45℃で10分間、0.1%SDSを含有している2×SSCにより2回洗浄した。メンブランにハイブリダイズした標識DNAの検出は、BAS-3000画像分析装置(Fujifilm,Tokyo,Japan)を使用して行った。
【0087】
(5)静止菌体反応による代謝物の抽出
スチレン雰囲気下でプレート培養した菌体を、50mMリン酸カリウム緩衝液(pH7.0)2mlに懸濁し、遠心分離(8000×g、20分)によって収集した。収集した菌体を同緩衝液に再懸濁した。次いで、菌体懸濁液(OD600=1)4mlおよび各基質50mgからなる反応混合物を使用して、静止菌体反応を実施した。反応混合物を22ml のバイアルに添加後シリコンセプタムを用いて密栓し、30℃、20時間、170rpmでインキュベーションした。インキュベーション後、反応混合物に酢酸エチル2mlを添加・混合し、反応生成物を抽出した。抽出産物をガスクロマトグラフィー質量分析(GC-MS)および高速液体クロマトグラフィ(HPLC)によって分析した。
【0088】
(6)GC-MSおよびHPLCの分析条件
静止菌体反応による生成物のGC-MS分析は、GC-17AガスクロマトグラフィーシステムおよびShimadzu QP-5000質量分析計を使用して実施された。酢酸エチルにより抽出された試料1μlを、HP-1MSキャピラリーカラム(J & W Scientific Inc.;30m×内径0.25mm、フィルム幅0.25μm)に注入した。キャリアガス(He)流速は1.7ml/min(100kPa)、スプリット比は26、線形速度は47.4cm/sであった。カラムオーブンの温度プログラムは:50℃、5分間;10℃/minで220℃まで上昇; 220℃、5分間で行った。質量スペクトルは、電子衝撃イオン源(EI、200℃)を使用して、1.5kVで取得した。各反応中間体の同定は、標準化合物の保持時間および質量電荷比(m/z)との比較によって行った。
静止菌体反応による生成物のHPLC分析は、移動相Aとして10mM酢酸アンモニウム、移動相Bとしてアセトニトリルを用い、流速0.5ml/分、カラム温度は40℃で分析を行った。試料(10μl)を注入した後、移動相Bを8分間10%に維持し、次いで20分かけて90%に増加させ、10分間維持した。254nmで生成物をモニタリングした。
フェニルアセチル-CoAリガーゼの反応生成物の同定は、Shimadzu Prominence GE HPLCシステムを使用して実施された。分析にはCadenza CD-C18カラム(Imtakt,Kyoto,Japan)を用いた。移動相には10% (v/v)イソプロピルアルコールを含有する0.2M KPB(pH4.5)を用いた。移動相の流速は0.5ml/分、カラムは40℃に維持された。254nmで生成物をモニタリングした。
【0089】
(7)無細胞粗抽出物の調製および酵素アッセイ
ガス状スチレンを単一炭素源として培養された菌体から、無細胞粗抽出物を入手した。採集された菌体を、50mMリン酸カリウム緩衝液(pH7.0)に懸濁し、2回洗浄した。菌体を、10%グリセロールを含有している50mMリン酸カリウム緩衝液(pH7.0)に再懸濁した。菌体懸濁液は、各1分間の超音波処理を5回行い、遠心分離(16,000×g、10分、4℃)した。上清を同緩衝液に対して透析し、酵素アッセイのために使用した。フェニルアセチル-CoAリガーゼ(PCL)活性を、以前に報告された方法(Martinez-Blanco, H., A. Reglero, L. B. Rodriguez-Aparicio, and J. M. Luengo. 1990. Purification and biochemical characterization of phenylacetyl-CoA ligase from Pseudomonas putida. J. Biol. Chem. 265:7084-7090)に従って測定した。
【0090】
(8)化学物質
スチレンおよびスチレンオキシドはNacalai Tesque Inc.(Kyoto,Japan)より購入した。フェニルアセトアルデヒドはToyotama International Inc.(Tokyo,Japan)より供給された。その他の化学物質は、全て、Wako Pure Chemical Industries Ltd.(Osaka,Japan)より購入した。
【0091】
〔実施例1〕ロドコッカスsp.ST-5株およびST-10株のスチレン代謝遺伝子の遺伝子構造
代謝遺伝子を単離し解析するため、本発明者らは、データベース上に登録されたスチレンモノオキシゲナーゼおよびスチレンオキシドイソメラーゼのアミノ酸配列から高度に保存されている領域を選択し、縮重PCR用のオリゴヌクレオチドプライマーを設計した。ST-5株のゲノムDNAを鋳型として縮重PCRを行い、styCの一部をコードする260bpのサイズの断片を入手することができた。しかし、styAに対応する断片は増幅されなかった。他方、ST-10株のゲノムDNAを鋳型として縮重PCRを行った場合には、styAをコードする部分断片が増幅されたが、styCをコードする遺伝子は増幅されなかった。これらの断片から推定されたアミノ酸配列は、シュードモナスのスチレン分解酵素と有意な類似性を示した。
スチレン代謝遺伝子の全長を入手するため、本発明者らは、ST-5株およびST-10株のゲノムDNAに対してTAIL-PCRを実施した。TAIL-PCRのため使用されたプライマーを表1に示した。その結果、本発明者らは、それぞれST-5株の染色体の約4.7kbpの領域およびST-10株の4.2kbpの領域のヌクレオチド配列を決定することができた。ST-5株およびST-10株の決定された染色体領域には、それぞれ、4つおよび3つのオープンリーディングフレーム(ORF)が存在した(図1および図2)。推定アミノ酸配列がシュードモナスのstyABCDと類似性を示したため、これらの遺伝子がスチレン代謝遺伝子であると推定した。ST-5株は、SMO、フラビンオキシドレダクターゼ、SOI、およびフェニルアセトアルデヒドデヒドロゲナーゼ(PAD)に対応する4つの連続する遺伝子を有していた。他方、ST-10株はSMOおよびフラビンオキシドレダクターゼに対応する2つの遺伝子を有していた。styBの下流に存在したorf3は、ロドコッカス・ジョスティ(jostii)RHA1のようなロドコッカス属にのみホモログが見出される機能未知タンパク質に対して高い類似性を示した。さらに、転写ターミネーターとして作用することが予測されるステムループ構造が、ST-5株のstyCおよびST-10株のstyBの下流にそれぞれ存在した(図1)。
【0092】
〔実施例2〕ST-10株ゲノムDNAのサザンブロット分析
SOI遺伝子の存在を確認するため、本発明者らは、ST-10株の染色体に対してサザンブロット解析を実施した。ST-10株のstyA遺伝子およびST-5株のstyC遺伝子に対応する各々1.2kbpおよび0.8kbpのプローブを、α-32P-dCTPにより標識し、様々な制限酵素により消化されたST-10株ゲノムDNAにハイブリダイズさせた。その結果、styA遺伝子に対応するプローブにハイブリダイズする有意な断片を見出すことができたが、styC遺伝子をプローブとして使用した場合には、検出可能なシグナルは見出されなかった。
【0093】
〔実施例3〕ST-5株およびST-10株のスチレン代謝遺伝子の構造的特性
ST-5株およびST-10株に由来するstyA遺伝子は427および428アミノ酸残基をコードし、推定アミノ酸配列は、シュードモナスsp.Y2由来のSMOの配列との有意な類似性を示した(60.6%および60.4%)(表2)。
【0094】
【表2】
【0095】
ST-5株およびST-10株に由来するStyAの推定アミノ酸配列は、フラビンおよびニコチンアミドアデニンジヌクレオチド(NAD+/NADH)との結合に関与していることが公知であるSMOにおいて保存されているいくつかの配列を含有していた。しかしながら、ST-5株およびST-10株のStyAは、SMOおよびフラビンオキシドレダクターゼの融合タンパク質として報告されたロドコッカス・オパカス(opacus)1CPのstyA2B(Tischler, D., D. Eulberg, S. Lankner, S. R. Kaschabek, W. J. H. van Berkel, and M. Schlomann. 2009. Identification of a novel self-sufficient styrene monooxygenase from Rhodococcus opacus 1CP. J. Bacteriol. 191:4996-5009)のStyA部分に対しては比較的低い類似性を示した(29.2%および28.0%)。ロドコッカス・オパカス1CPは、styCまたはstyDに対応するスチレン代謝遺伝子をstyA2Bの近傍に有していないことが報告された。これらの事実は、既報のロドコッカスのスチレン代謝遺伝子が本発明の遺伝子とは異なることを示している。
【0096】
ST-5株およびST-10株のstyB遺伝子は、それぞれ179および183アミノ酸残基をコードしており、そのアミノ酸配列から計算される推定分子量は19,284Daおよび19,543Daであった。ST-5株およびST-10株に由来するStyBの推定アミノ酸配列は、フラビンオキシドレダクターゼおよび様々なモノオキシゲナーゼにおいて高度に保存されているいくつかのモチーフを含有している。相同性検索の結果から、ST-5株およびST-10株に由来するstyBが、シュードモナスsp.Y2 StyB(53.8%および50.6%)、バチルス(Bacillus)sp.JF8 MocB(36.5%および36.8%)、ゲオバチルス・サーモグルコシダシウス(Geobacillus thermoglucosidasius)A7 PheA2(33.6%および35.1%)、ならびにロドコッカス・オパカス1CP StyA2B(27.6%および33.3%、B部分)を含む、NAD(P)H:フラビンオキシドレダクターゼのファミリーに属しているいくつかのタンパク質との相同性をそれぞれ示すことが明らかとなった(表2)。この結果より、ST-5株およびST-10株のstyBが、NAD(P)H:フラビンオキシドレダクターゼのメンバーであり、StyAと共に2成分からなるモノオキシゲナーゼ系を構成していることが示された。
【0097】
styC遺伝子およびstyD遺伝子はST-5株にのみ観察された。styC遺伝子は168アミノ酸残基をコードし、推定アミノ酸配列は、シュードモナスsp.Y2のSOIとの有意な類似性(69.5%)、およびシュードモナス・オーレオファシエンス(aureofaciens)のp-シメンメチルヒドロキシラーゼとの中程度の類似性(35.0%)を示した(表2)。styDをコードするオープンリーディングフレームには、2ヶ所の推定開始コドンが3068塩基目および3086塩基目に存在した(図2A)。このことから、styD がコードするポリペプチドは502アミノ酸ないし496アミノ酸であることが示唆された。このタンパク質は、ロドコッカス・ジョスティRHA1 ALDH(78.6%)、バークホルデリア(Burkholderia)sp.383 BADH(54.4%)、アゾアルカス(Azoarcus)sp.BH72 ThmS2(54.5%)、シュードモナスsp.Y2 StyD(49.9%)、および大腸菌W3110 PadA(40.1%)のような様々なアルデヒドデヒドロゲナーゼとの高い類似性を示した(表2)。StyDは、様々なアルデヒドデヒドロゲナーゼにおいて保存されているいくつかの補因子結合モチーフを有する(Hempel, J., H. Nicholas, and R. Lindahl. 1993. Aldehyde dehydrogenases: Widespread structural and functional diversity within a shared framework. Protein Sci. 2:1890-1900、Liu, Z. J., Y. J. Sun, J. Rose, Y. J. Chung, C. D. Hsial, W. R. Chang, I. Kuo, J. Perozich, R. Lindahl, J. Hempel, and B. C. Wang. 1997. The first structure of an aldehyde dehydrogenase reveals novel interactions between NAD and the Rossmann fold. Nat. Struct. Biol. 4:317-326)。
【0098】
〔実施例4〕ロドコッカスsp.ST-10株によるスチレン代謝の中間体の分析
ST-5株およびST-10株を用いて静止菌体反応を行い、生成した中間体をGC-MSによって同定した。スチレンを基質としたとき、ST-10株の反応生成物としてスチレンオキシドが検出されたが、フェニルアセトアルデヒドは検出されなかった。対照的に、ST-5株は反応生成物としてフェニルアセトアルデヒドのみを蓄積した。これらの結果は、ST-5株およびST-10株のそれぞれの遺伝子発現の差異によることが示唆された。ST-5株においては、スチレンが、StyA、StyB、およびStyCの作用によってフェニルアセトアルデヒドに変換されるが、ST-10株ではStyCを欠くために、スチレンオキシドをフェニルアセトアルデヒドに変換することができず、反応生成物中に蓄積されたと推測される。
【0099】
ST-10株のスチレン資化メカニズムを解明するため、本発明者らは、HPLCによってST-10株の静止菌体反応による生成物を詳細に調査した。スチレンを基質とした反応産物からは、スチレンオキシド、アセトフェノン、スチレングリコール、およびフェニル酢酸を含む様々な芳香族化合物が見出された(図3A)。また、コントロールとしてスチレンオキシドをST-10株菌体を含まない緩衝液中で反応させたとき、スチレングリコール、アセトフェノン、およびフェニルアセトアルデヒドが検出された(図3B)。スチレンオキシドは不安定な化合物であり、水、過酸化物、または空気の存在下で、他の化合物へと容易に変換されることが報告されている(Liu, Z., J. Michel, Z. Wang, B. Witholt, and L. Zhi. 2006. Enantioselective hydrolysis of styrene oxide with the epoxide hydrolase of Sphingomonas sp. HXN-200. Tetrahedron Asymmetry 17:47-52、Pfaffli, P., H. Vainio, and A. Hesso. 1979. Styrene and styrene oxide concentration in the air during the lamination process int the reinforced plastics industry. Scand. J. Work Environ. Health 5:158-161)。従って、アセトフェノンおよびフェニルアセトアルデヒドは、自発的な反応によってスチレンオキシドから生成したと推測される。さらに、ST-10株の菌体と共にインキュベートしたとき、フェニルアセトアルデヒドは消失し、代わりにフェニル酢酸が増加した(図3C)。
【0100】
〔実施例5〕フェニル酢酸の代謝に関与している酵素活性の同定
フェニル酢酸の分解経路を同定するため、ST-5株およびST-10株のフェニルアセチル-CoAリガーゼ(PCL)活性を測定した。基質としてフェニル酢酸、CoA、ATPを含む緩衝液にST-5株およびST-10株の無細胞粗抽出物を添加し、反応生成物をHPLCにより分析した。いずれの株についても、無細胞粗抽出物の存在下で、フェニルアセチル-CoAに対応するピークが8.1分の位置に検出された(図4)。これらの結果は、いずれの株もPCL活性を有し、シュードモナス属細菌で報告されているフェニル酢酸(PAA)分解経路と同様の経路を介してフェニル酢酸を分解し得ることを示す。
【0101】
以下に、実施例1〜5で得られた結果について考察する。
ロドコッカスsp.ST-5株およびST-10株(以前はコリネバクテリアsp.)が、ガス状スチレンおよびスチレンオキシドを単一炭素源として資化できること、またこれらの菌株がシュードモナスと同様のスチレン代謝経路を有し、スチレンおよびスチレンオキシドを資化していることが示唆されていた(Itoh, N., K. Yoshida, and K. Okada. 1996. Isolation and identification of styrene-degrading Corynebacterium strains, and their styrene metabolism. Biosci. Biotech. Biochem. 60:1826-1830)。
【0102】
ST-5株およびST-10株におけるスチレンの分解経路を解明するため、本発明者らはスチレン分解に関与している遺伝子群をPCR技術を利用して単離した。
【0103】
図1および図2に示されるように、本発明者らは、ST-5株の約4.7kbpの染色体領域およびST-10株の約4.2kbpの領域の塩基配列を決定した。ST-5株において決定された配列中には4つのORFが含まれ、その推定アミノ酸配列はシュードモナス属細菌において報告されているスチレン代謝遺伝子styABCDと相同性を示した。他方、ST-10株において決定された配列は、styABおよび機能未知タンパク質の3つのORFを有する。シュードモナス属細菌を含む各種微生物から単離されたスチレン代謝遺伝子は、SMO、フラビンオキシドレダクターゼ、SOI、およびフェニルアセトアルデヒドデヒドロゲナーゼを含む、4つの遺伝子から構成されている。さらに、いくつかの微生物はこれらの遺伝子の上流または下流に転写制御因子(styRS)および基質トランスポーター(styE)をコードする遺伝子を有する(Mooney, A., N. D. O'Leary, and A. D. W. Dobson. 2006. Cloning and functional characterization of the styE gene, involved in styrene transport in Pseudomonas putida CA-3. Appl. Environ. Microbiol. 72:1302-1309、Panke, S., B. Witholt, A. Schmid, and M. G. Wubbolts. 1998. Towards a biocatalyst for (S)-styrene oxide productio: characterization of the styrene degradation pathway of Pseudomonas sp. strain VLB120. Appl. Environ. Microbiol. 64:2032-2043、Velasco, A., S. Alonso, J. L. Garcia, J. Perera, and E. Diaz. 1998. Genetic and functional analysis of the styrene catabolic cluster of Pseudomonas sp. strain Y2. J. Bacteriol. 180:1063-1071)。ST-5株およびST-10株の両株はstyAの上流に制御遺伝子を有しておらず、ST-10株はstyCDを欠くことが見出された(図1)。しかしながら、これらの株をガス状スチレンまたはスチレンオキシドで培養したとき、スチレンモノオキシゲナーゼの活性が上昇したことから、シュードモナス属細菌と同様これらの遺伝子の発現を制御している制御遺伝子が存在することが示唆された。ただし、そのような遺伝子はスチレン代謝遺伝子の近隣の領域には存在しない。
【0104】
ST-10株の染色体中にスチレンオキシドイソメラーゼ遺伝子が存在しないことを確認するため、本発明者らは、ST-5株由来のstyC遺伝子をプローブとして、ST-10株ゲノムDNAに対してサザンブロット解析を実施した。その結果、styC遺伝子に相当する特異的なシグナルを見出すことができなかった。一方、コントロールとしてstyA遺伝子をプローブに用いた場合、特異的なバンドが検出された。Itohらは、ST-10株がSOI活性は有していないが、SMO活性およびPAD活性は有することを報告している(Itoh, N., K. Yoshida, and K. Okada. 1996. Isolation and identification of styrene-degrading Corynebacterium strains, and their styrene metabolism. Biosci. Biotech. Biochem. 60:1826-1830)。これらの事実は、ST-10株がSOI以外のスチレン分解酵素を有し、未知経路を介してスチレンオキシドを資化することを示唆している。
【0105】
ST-10株のスチレン資化経路を解明するため、ST-5株およびST-10株による静止菌体反応を行い、生成した中間体を同定した。GC-MS分析によって、ST-10株の反応生成物としてスチレンオキシドが蓄積され、ST-5株ではフェニルアセトアルデヒドが蓄積されることが明らかとなった。これらの現象は、ST-10株にSOIをコードする遺伝子が存在しないことと一致する。以前の報告において、ST-5株はPAD活性より高いSOI活性を示すことが見出されている(それぞれ、菌体湿重量で33U/g cellおよび1.5U/g cell)。ST-5株においては、転写ターミネーターと推定される配列がstyCとstyDとの間の領域に見出されている(図2)。従って、styDの発現レベルは他のスチレン代謝遺伝子より低いことが推測される。スチレン代謝フラックスにおいて、PAD活性が低いことが要因となりフェニルアセトアルデヒドがST-5株の代謝中間体として蓄積されていることが示唆される。対照的に、ST-10株においては、StyCが欠落しているため、スチレンオキシドが代謝中間体として蓄積している。HPLC分析により、スチレンオキシドがフェニルアセトアルデヒドおよびアセトフェノンへと自発的に異性化されること、またST-10株が自発的に異性化したフェニルアセトアルデヒドを資化していることが証明された(図3)。Itohらは、スチレン、スチレンオキシド、および2-フェニルエタノールがST-10株によって代謝されるが、アセトフェノンおよびスチレングリコールは炭素源として利用されないことを報告している。HPLC分析の結果は、これらの事実とよく一致している。スチレンまたはスチレンオキシドを基質としたとき、両株の反応生成物中に2-フェニルエタノールが観察された。図5に示されるように、フェニルアセトアルデヒドレダクターゼ(PAR)の活性によってフェニルアセトアルデヒドは2-フェニルエタノールに変換される。スチレンの代謝経路において、2-フェニルエタノールは、貯蔵化合物の役割を担っていることが示唆される。2-フェニルエタノール/フェニルアセトアルデヒドの比率は、各化合物の濃度およびNAD+/NADH比によって制御されると推測される。
【0106】
フェニル酢酸代謝経路は、シュードモナス.プチダ(putida)および大腸菌のようなグラム陰性菌においてよく解析されている(Luengo, J. M., J. L. Garcia, and E. R. Olivera. 2001. The phenylacetyl-CoA catabolon: a complex catabolic unit with broad biotechnological applications. Mol. Microbiol. 39:1434-1442)。しかしながら、グラム陽性菌の同経路に関してはほとんど情報がない。近年Llorensにより、ロドコッカスsp.RHA1がグラム陰性菌のフェニル酢酸代謝系路に対応するpaa遺伝子群を有することを報告している(Navarro-Llorens, J. M., M. A. Patrauchan, G. R. Stewart, J. E. Davies, L. D. Eltis, and W. W. Mohn. 2005. Phenylacetic acid catabolism in Rhodococcus sp. strain RHA1: a central pathway for degradation of aromatic compounds. J. Bacteriol. 187:4497-4504)。さらに、数種のロドコッカス属細菌の全ゲノム配列が解析されたことにより、そのゲノム中にpaa遺伝子群が存在することが明らかになった。ST-5株およびST-10株も、他のロドコッカス属細菌と同様にPCL活性を有する(図4)。この事実は、ST-5株およびST-10株がpaa遺伝子群を有し、他のグラム陰性菌と同様の経路を利用してフェニル酢酸を資化していることを示唆している。
ST-5株およびST-10株における推定スチレン代謝経路を図5に示した。スチレンはまずSMOによってスチレンオキシドへと酸化され、続いてSOIによってフェニルアセトアルデヒドへと変換される。ST-10株においてはSOIを欠落しているため、この反応は自発的な化学変換によって起きていると推測される。次いで、フェニルアセトアルデヒドはPAD活性によりフェニル酢酸へと酸化され、PAA分解経路により分解される。本発明者らは、ST-5株およびST-10株のゲノム中にスチレン分解遺伝子群が存在することを確認した。ST-10株のスチレン代謝経路はSOIを欠落した不完全なものであるが、ST-10株はスチレンおよびスチレンオキシドを資化することができる。
【0107】
〔実施例6〕Rhodococcus sp. ST-5株 およびST-10株 由来styAの基質特異性
(1)Rhodococcus sp. ST-5株 およびST-10株 由来styAB の大腸菌による異種宿主発現系構築および組換え酵素の発現・精製
Rhodococcus sp. ST-5株 およびST-10株 両株由来のstyAB 遺伝子を大腸菌Escherichia coli において異種宿主発現させるため、以下の手順により発現用プラスミドを構築した。発現には大腸菌株E. coli BL21 (DE3)を用いた。
1. ST-5株 およびST-10株 のstyAB の末端配列に特異的なオリゴヌクレオチドプライマー(表3)を作成し、ST-5株 およびST-10株 ゲノムDNA に対してPCRを実施した。この際N-末端およびC-末端に制限酵素サイト(Bam HI, Hind III) を付加した。
2. 増幅した約1.8 kbp の増幅断片をpGEM-T Easy vector (plomega) にクローニングし、内部配列を確認した。
3. Bam HI、Hind III 処理によりstyAB 断片を切り出し、アガロースゲル電気泳動により精製した。精製したstyAB 断片を同制限酵素によりあらかじめ処理したpET-21 (b) にライゲーションし、E. coli DH5αに導入した。
4. 抗生物質プレート上で形質転換体を選抜した。プラスミドを抽出精製し、E. coli BL21 (DE3) にエレクトロポレーション法により導入した。さらに、組換えタンパク発現を効率化させるためにシャペロン発現ベクターpG-KJE8 を重ねて導入した。
5. 形質転換体を抗生物質プレート上で選抜し、以後発現実験へ用いた。
なお、上記と同様の手法により、styAを単体で発現させるコンストラクトも作成した。
発現プラスミド構築に使用したプライマー配列を表3に、構築したプラスミドを図6に示した。
【0108】
SDS-PAGEによるSMO分子量の分析は以下の条件で行った。
1. 精製したヒスチジンタグを含むStyA サンプルを2x SDS-PAGE サンプルバッファー(100 mM Tris-HCl, 20% グリセロール, 4% SDS, 0.05% ブロモフェノールブルー)に溶解した。
2. 10% ポリアクリルアミドゲルに、タンパク量にして5μg相当の上記サンプルをアプライした。
3. 150 V, 40 mAで 100 分間 泳動した。
4. 泳動したゲルをSDS-PAGE染色液(50% メタノール、10% 酢酸、0.25% クマジーブルー)に浸し、40分間振とうしながら染色を行った。
5. 染色したゲルを脱色液(30% メタノール、10% 酢酸)に浸し、適宜脱色液を交換しながら脱色を行った。
6. 得られたバンドの泳動度から、精製したStyA の分子量は約49.0 kDaであると推定した。分子量の算出には、サンプルと同時に流した分子量マーカー(Prestained APRO-Marker) を指標として用いた。
【0109】
また、ゲル濾過によるSMO分子量の分析は以下の条件で行った。
1. 分析システムとしてShimadzu LC-10 HPLC システムを、分析カラムとしてGE ヘルスケア社製Superdex 200 10/300 GL を用いた。
2. 移動相として100 mM KPB, 200 mM NaCl pH 7.0 を用い、流速0.4 ml/min で分離を行った。
3. ヒスチジンタグを含む精製StyA サンプル 100μl (500μg タンパク相当) を分析サンプルとしてアプライし、35.3 min に単一ピークとして溶出した。
4. 同様に、標準試料 (オリエンタル酵母社MW-Marker (HPLC)) を分析した。
5. 標準試料の分子量および溶出時間を以下に示す。
酵母グルタミン酸脱水素酵素 290 kDa 26.9 min
豚心筋乳酸脱水素酵素 142 kDa 31.3 min
酵母エノラーゼ 67 kDa 34.3 min
酵母ミオキナーゼ 32 kDa 39.1 min
馬心筋チトクロームc 12.4 kDa 43.0 min
6. 上記の標準試料の各タンパクの溶出時間および分子量を用いて検量線を作成した。得られた検量線に対して精製StyAの溶出時間35.3 min をプロットした結果、StyA の分子量は59.2 kDa に相当すると推測された。
【0110】
【表3】
【0111】
また、組換えStyAB の発現および精製は以下の通りに行った。
1. 発現プラスミドを含む形質転換体を、4 mlのLB medium (50μg/ml アンピシリン、20μg/ml クロラムフェニコール)で37℃、170 rpm で培養した。
2. 前培養液1 ml を100 ml LB medium (50μg/ml アンピシリン、20μg/ml クロラムフェニコール、0.5 mg/ml アラビノース、5 ng/ml テトラサイクリン)に植菌し、37℃、250 rpm で培養した。
3. OD600 が0.5 になったらIPTG を終濃度0.1 mM になるように添加し、18℃、250 rpm、24時間誘導した。
4. 菌体を遠心(18000 rpm, 4℃,10 min)により回収し、50 mM KPB (pH 7.0), 10% glycerol に懸濁、洗浄した。
5. 再度遠心により菌体を回収し、Ni-Sepharose カラムBinding Buffer (50 mM KPB, 10% glycerol, 500 mM NaCl, 10 mM imidazole, pH 7.5 )に再懸濁した。
6. 超音波破砕により菌体を破砕し、遠心(18,000 rpm, 4℃, 20 min x2回)により上清を回収した。
7. Ni-Sepharose カラムBinding Buffer で平衡化したNi-Sepharose High performance カラムに、上記の菌体破砕液上清をアプライし、目的の組換えタンパクを吸着させた。
8. カラム体積の10倍量のBinding Buffer でカラムを洗浄後、Elution Buffer (50 mM KPB, 10% glycerol, 500 mM NaCl, 500 mM imidazole, pH 7.5) により吸着させた目的タンパクを溶出させた。
9. 得られた精製タンパク溶液を透析し、50 mM KPB, 10% glycerol, pH 7.0 に置換した。
10. 透析したタンパク溶液を各種イオン交換クロマトグラフィー(DEAE-Sepharose, Resource Q, MonoQ) を用いて最終精製し、目的タンパク質の精製サンプルを得た。
【0112】
(2)SMO の活性測定
ST-5およびST-10 由来SMO の活性測定は、カラム精製した組換え酵素を用いて行った。各精製酵素、基質、補酵素を50 mM MES緩衝液中に以下の濃度で添加し、反応混合液とした。
反応混合液組成
50 mM MES Buffer (pH 7.0)
20 mM ギ酸ナトリウム
100μM NADH
200μM FAD
1 U ギ酸脱水素酵素
10 U catalase
5 mM substrate(基質変換反応時は、10 mM の基質で反応を実施)
StyA 150μl (基質変換反応時は、StyAB 共発現菌体の菌体破砕液上清を使用)
StyB 100μl
total 1000μl
この反応混合液を30℃、2,500rpmで30分間インキュベーションした。反応終了後、反応混合液に500μlの酢酸エチルを添加し5分間ボルテックスを行った。15,000 rpm で5分間遠心し、分離した酢酸エチル相を新しいチューブに移した。再度500μl の酢酸エチルを水相に添加し、5分間ボルテックス後、再度遠心により酢酸エチル相を回収した。合計1000μl の回収した酢酸エチル相に適量の硫酸ナトリウムを添加し、2分間ボルテックスを行い、脱水処理を行った。脱水処理を行った抽出サンプルを、Hewllet Packerd HP6890 ガスクロマトグラフィーシステムを用いて分析した。
1 U は、1分間に1μmol の基質を産物に変換する酵素活性と定義した。
【0113】
(3)Gas Chromatography 分析条件
ST-5 およびST-10 由来SMOの反応産物の測定は、ガスクロマトグラフィーを用いて行った。上記反応条件にて反応し、酢酸エチルで抽出した反応産物をHewllet Packard HP6890 ガスクロマトグラフィーシステムを用いて分析した。酢酸エチルにより抽出された試料1μl をVarian 社製CP-Cyclodextrin-b-2,3,6-M-19-25 キャピラリーカラム(25 m, 内径0.25 mm)に注入した。キャリアガス(99.999% 高純度ヘリウムガス)流速は2.8 ml/min、スプリット比は50、線形流速は46 cm/s であった。検出はFID(水素炎イオン化検出器)を用いた。気化室温度は250℃、検出器温度は250℃で行った。
カラムオーブンの温度プログラムは、定温プログラムを用いて行った。各分析サンプルに対するカラムオーブン温度および各物質の検出時間は表4に示した。
各物質の同定および定量は、各標準化合物を1 mM, 5 mM, 10 mM の濃度に調製したサンプルを同条件で分析し、その検出時間およびピーク面積から検量線を作成して求めた。
【0114】
【表4】
【0115】
(4)Rhodococcus sp. ST-5株 およびST-10株 由来styAの基質特異性
ST-5およびST-10由来SMOは、スチレンをはじめ様々な基質に対してエポキシ化活性を示した(表5)。その基質特異性は、スチレンを100としたときに、4−フルオロスチレン に対しておよそ2倍の活性を示した。また、生成物の鏡像体過剰率(e.e.)はスチレンや4−クロロスチレン、4−ブロモスチレン、4−フルオロスチレンに対してはやや低いものの、2−クロロスチレンや3−クロロスチレンに対しては99.9%以上の値を示し、その絶対配置は(S)-体であった。
【0116】
【表5】
反応産物の立体選択性については、styrene oxide および3-chloro styrene oxide 以外は産物の標品が市販されていないか、ラセミ体しかないため、S-体であると推測されるが確認はできていない。
【産業上の利用可能性】
【0117】
本発明は、スチレンモノオキシゲナーゼ、該酵素をコードするポリヌクレオチドを提供する。本発明のスチレンモノオキシゲナーゼを利用することにより、スチレンモノオキシゲナーゼを効率よく生産することができる。
【特許請求の範囲】
【請求項1】
以下(a)から(e)のいずれかに記載のポリヌクレオチド;
(a)配列番号:1または3に記載の塩基配列を含むポリヌクレオチド;
(b)配列番号:2または4に記載のアミノ酸配列をコードするポリヌクレオチド;
(c)配列番号:2または4に記載のアミノ酸配列において、1もしくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列を含み、かつ下記(1)および(2)に記載の理化学的性質を有するスチレンモノオキシゲナーゼをコードするポリヌクレオチド;
(d)配列番号:1または3に記載の塩基配列からなるポリヌクレオチドに対して、高度にストリンジェントな条件下でハイブリダイズし、かつ下記(1)および(2)に記載の理化学的性質を有するスチレンモノオキシゲナーゼをコードするポリヌクレオチド;および
(e)配列番号:2または4に記載のアミノ酸配列と70%以上の同一性を有し、かつ下記(1)および(2)に記載の理化学的性質を有するスチレンモノオキシゲナーゼをコードするポリヌクレオチド;
(1)作用
FADH2(還元型FAD)を補酵素として、スチレンおよび3−クロロスチレンに作用し、それぞれ、光学活性なスチレンオキシドおよび光学活性な3−クロロスチレンオキシドを生成する。
(2)分子量
ゲル濾過におけるヒスチジンを含む分子量が59,000で、SDS-PAGEによる分子量が49,000、アミノ酸組成からの推定分子量が47,000のモノマー酵素である。
【請求項2】
ロドコッカス(Rhodococcus)属に属する微生物由来である、請求項1に記載のポリヌクレオチド。
【請求項3】
前記ロドコッカス属に属する微生物がRhodococcus opacus ST-5株またはRhodococcus erythropolis ST-10株である、請求項2に記載のポリヌクレオチド。
【請求項4】
請求項1から3のいずれかに記載のポリヌクレオチドによってコードされるタンパク質。
【請求項5】
請求項1から3のいずれかに記載のポリヌクレオチドを含むベクター。
【請求項6】
請求項1から3のいずれかに記載のポリヌクレオチド、または請求項5に記載のベクターが導入された形質転換細胞。
【請求項7】
請求項6に記載の形質転換細胞を培養し、その培養物から請求項1に記載のポリヌクレオチドによってコードされるタンパク質を回収する工程を含む、請求項1から3のいずれかに記載のポリヌクレオチドによってコードされるタンパク質の製造方法。
【請求項8】
請求項4に記載のタンパク質をスチレンおよび/またはその類似体に作用させる工程、および生成される化合物を回収する工程を含む、光学活性なスチレンオキシドおよび/または光学活性なスチレン類似体の酸化物の製造方法。
【請求項9】
請求項4に記載のタンパク質をスチレンおよび/またはその類似体に作用させる工程を含む、スチレンおよび/またはその類似体の分解方法。
【請求項1】
以下(a)から(e)のいずれかに記載のポリヌクレオチド;
(a)配列番号:1または3に記載の塩基配列を含むポリヌクレオチド;
(b)配列番号:2または4に記載のアミノ酸配列をコードするポリヌクレオチド;
(c)配列番号:2または4に記載のアミノ酸配列において、1もしくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列を含み、かつ下記(1)および(2)に記載の理化学的性質を有するスチレンモノオキシゲナーゼをコードするポリヌクレオチド;
(d)配列番号:1または3に記載の塩基配列からなるポリヌクレオチドに対して、高度にストリンジェントな条件下でハイブリダイズし、かつ下記(1)および(2)に記載の理化学的性質を有するスチレンモノオキシゲナーゼをコードするポリヌクレオチド;および
(e)配列番号:2または4に記載のアミノ酸配列と70%以上の同一性を有し、かつ下記(1)および(2)に記載の理化学的性質を有するスチレンモノオキシゲナーゼをコードするポリヌクレオチド;
(1)作用
FADH2(還元型FAD)を補酵素として、スチレンおよび3−クロロスチレンに作用し、それぞれ、光学活性なスチレンオキシドおよび光学活性な3−クロロスチレンオキシドを生成する。
(2)分子量
ゲル濾過におけるヒスチジンを含む分子量が59,000で、SDS-PAGEによる分子量が49,000、アミノ酸組成からの推定分子量が47,000のモノマー酵素である。
【請求項2】
ロドコッカス(Rhodococcus)属に属する微生物由来である、請求項1に記載のポリヌクレオチド。
【請求項3】
前記ロドコッカス属に属する微生物がRhodococcus opacus ST-5株またはRhodococcus erythropolis ST-10株である、請求項2に記載のポリヌクレオチド。
【請求項4】
請求項1から3のいずれかに記載のポリヌクレオチドによってコードされるタンパク質。
【請求項5】
請求項1から3のいずれかに記載のポリヌクレオチドを含むベクター。
【請求項6】
請求項1から3のいずれかに記載のポリヌクレオチド、または請求項5に記載のベクターが導入された形質転換細胞。
【請求項7】
請求項6に記載の形質転換細胞を培養し、その培養物から請求項1に記載のポリヌクレオチドによってコードされるタンパク質を回収する工程を含む、請求項1から3のいずれかに記載のポリヌクレオチドによってコードされるタンパク質の製造方法。
【請求項8】
請求項4に記載のタンパク質をスチレンおよび/またはその類似体に作用させる工程、および生成される化合物を回収する工程を含む、光学活性なスチレンオキシドおよび/または光学活性なスチレン類似体の酸化物の製造方法。
【請求項9】
請求項4に記載のタンパク質をスチレンおよび/またはその類似体に作用させる工程を含む、スチレンおよび/またはその類似体の分解方法。
【図1】

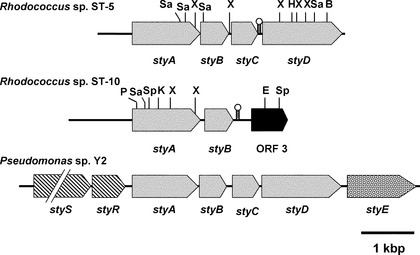
【図2】

【図3】
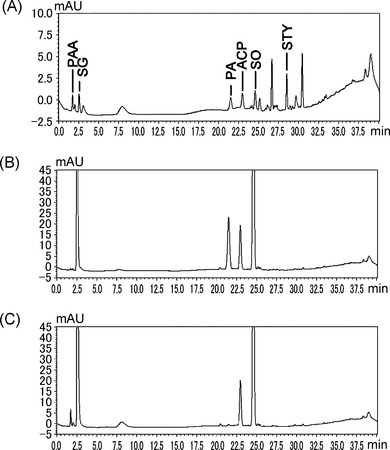
【図4】
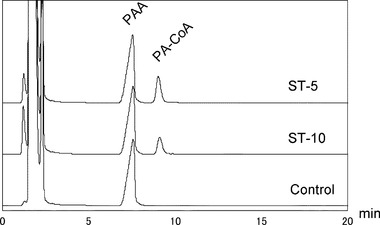
【図5】
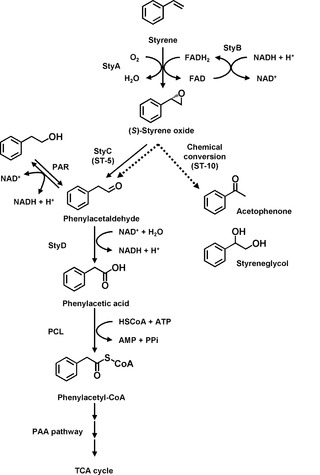
【図6】
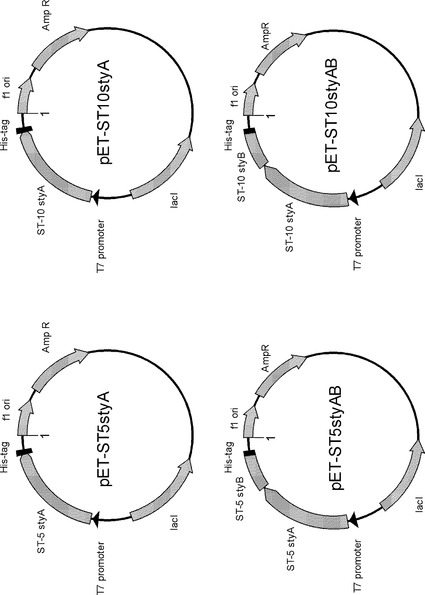
【図2】
【図3】
【図4】
【図5】
【図6】
【公開番号】特開2012−60913(P2012−60913A)
【公開日】平成24年3月29日(2012.3.29)
【国際特許分類】
【出願番号】特願2010−206789(P2010−206789)
【出願日】平成22年9月15日(2010.9.15)
【出願人】(000002901)株式会社ダイセル (1,236)
【出願人】(000236920)富山県 (197)
【Fターム(参考)】
【公開日】平成24年3月29日(2012.3.29)
【国際特許分類】
【出願日】平成22年9月15日(2010.9.15)
【出願人】(000002901)株式会社ダイセル (1,236)
【出願人】(000236920)富山県 (197)
【Fターム(参考)】
[ Back to top ]