
新規なバチルスmHKcelセルラーゼ

本発明は新規なセルラーゼ核酸配列、指定のmHKcel、及び対応するmHKcelアミノ酸配列を提供する。また、本発明はmHKcelをエンコードする核酸配列を含む発現ベクター及び宿主細胞、組換えmHKcelタンパク質及びこれらを生成する方法を提供する。
【発明の詳細な説明】
【発明の詳細な説明】
【0001】
関連出願の相互参照
本出願は、ここにその全体を引用する、2003年4月30日に出願された米国仮出願60/467,315号(代理人整理番号、GC797P)に基づく優先権を主張するものである。
【0002】
連邦政府後援の研究開発の下、行われた発明の権利に関する声明
適用なし。
【0003】
発明の分野
本発明はここでmHKcelとして言及する新規なセルラーゼに関する。また、セルラーゼをエンコードする核酸、前記セルラーゼを含む組成物、新規なセルラーゼを同定する方法、及び前記組成物を用いる方法についても記載する。好ましくは、当該セルラーゼはバチルス種から単離され、好ましくはB.agaradhaerensである。本発明はさらに、洗剤組成物、セルロース含有繊維処理、パルプ及び紙の処理、及び高フルクトース・コーンシロップまたはエタノールの生成用のデンプン処理に添加剤として、当業界で有利にセルラーゼが添加されるものとして認識される、新規なセルラーゼの組成物中での使用に関する。
【0004】
発明の背景
セルロース及びヘミセルロースは光合成により最も豊富に生成される植物材料である。これらは高分子基材を単糖類に加水分解できる細胞外酵素を生成する多数の微生物、例えば細菌、酵母及び真菌によりエネルギー源として分解及び使用される(Aro、他、2001)。非再生源の限界が近づいているため、セルロースが主な再生可能エネルギー源となる可能性は非常に大きい(Krishna、他、2001)。生物プロセスを通じたセルロースの効果的な活用は食料、飼料及び燃料不足の克服手段のひとつである(Ohmiya、他、1997)。
【0005】
セルラーゼはセルロース(β−1,4−グルカンまたはβ−D−グルコシド結合)を加水分解してグルコース、セロビオース、セロオリゴ糖等の形成を生じる酵素である。セルラーゼは従来から3つの主な分類に分けられる:エンドグルカナーゼ(EC 3.2.1.4)(“EG”)、エキソグルカナーゼまたはセロビオヒドロラーゼ(EC 3.2.1.91)(“CBH”)、及びβ−グルコシダーゼ([β]−D−グルコシドグルコヒドロラーゼ;EC 3.2.1.21)(“BG”)(Knowles、他、1987;Shulein、1988)。エンドグルカナーゼは主にセルロース繊維のアモルファス部分で作用するのに対し、セロビオヒドロラーゼは結晶セルロースを分解することもできる(Nevalainen and Penttila、1995)。従って、結晶セルロースを効果的に可溶化するために、セルラーゼ系中にセロビオヒドロラーゼの存在が必要である(Suurnakki、他、2000)。β−グルコシダーゼはセロビース、セロ−オリゴ糖、及びその他のグルコシド由来D−グルコース単位の遊離に作用する(Freer、1993)。
【0006】
結晶セルロースをグルコースに効果的に変換するためには、結晶セルロースの加水分解にあまり効果がない単離成分とともにCBH、EG及びBGの各分類由来の成分を含む完全なセルラーゼ系が必要である(Filho、他、1996)。それぞれの分類由来のセルラーゼ成分間で相乗関係が観測されている。特に、EG型セルラーゼ及びCBH型セルラーゼは相乗的に相互作用してより効果的にセルロースを分解する。例えば、Wood、1985を参照されたい。
【0007】
従来からセルラーゼ組成物について記載されているが、家庭用洗剤、ストーンウォッシュ組成物または洗濯用洗剤等に使用するための新規で改善されたセルラーゼ組成物の必要性が依然としてある。改善された性能を示すセルラーゼは特に関心が高い。
【0008】
発明の概要
本発明の目的は、洗剤、織物処理、バイオマス転換及びパルプ及び紙製造への使用に有益な性質を有する新規なセルラーゼを提供することである。
【0009】
本発明の目的は、セルロース分解活性を有するポリペプチド及び当該ポリペプチドをエンコードするポリヌクレオチドを提供することである。当該ポリペプチドは細胞壁材料、例えばセルロース及び/またはヘミセルロースの分解を改善する。また、当該ポリペプチドは、例えばバイオマスなどの植物細胞壁材料の分解に関係するその他の酵素の安定性または活性も改善する。
【0010】
本発明の目的は、新規なセルラーゼ及びその誘導体、当該セルラーゼを生成する方法、及びこのような新規なセルラーゼを含む組成物を提供することである。本発明はさらに、当業界でセルラーゼの添加が有利なものとして認識される、新規なセルラーゼ及びその誘導体の組成物中での使用に関し、例えば、洗剤組成物、有用なセルロース含有布及び繊維などの織物処理での添加剤としての使用、動物飼料添加物としての使用、バイオマス転換、パルプ及び紙の処理及び高フルクトース・コーンシロップまたはエタノールの生成用のデンプン処理への添加剤としての使用である。
【0011】
本発明のさらなる目的は、組換え宿主細胞から異種発現により新規なセルラーゼを生成する方法を提供することである。
【0012】
さらに本発明の目的は、本発明のセルラーゼをエンコードする核酸配列を提供することである。1の側面において、当該核酸及びアミノ酸配列は本発明の新規なセルラーゼ及びセルラーゼ組成物の商業生産を容易にする。
【0013】
さらに本発明の目的は、洗剤、織物処理、飼料補助剤及びパルプ及び紙製造において使用するための優れた性質を有する新規なセルラーゼを提供することである。さらなる側面において、当該セルラーゼはバイオマス転換において有用である。
【0014】
第1の側面において、本発明はmHKcelをエンコードする配列、mHKcel遺伝子コード配列に相補的な配列を有する単離ポリヌクレオチド及び当該ポリヌクレオチドを含む組成物を含む。当該ポリヌクレオチドは、mRNA、DNA、cDNA、ゲノムDNAまたはそれらのアンチセンス類似体である。
【0015】
1の実施態様において、mHKcelポリヌクレオチドは、中から高ストリンジェンシー条件下で配列番号2として表される核酸の相補体にハイブリダイズする単離核酸分子を含み、ここで核酸分子はセルロース結合活性を示すmHKcelポリペプチドをエンコードする。
【0016】
配列番号2として表される配列に少なくとも80%、85%、90%、95%、98%またはそれ以上の配列同一性を有するポリヌクレオチドはmHKcelタンパク質をエンコードできる。特定の実施態様において、ポリヌクレオチドは配列番号2に実質的に同一な配列を含む。本発明はポリヌクレオチド断片、好ましくは少なくとも約15−30のヌクレオチド長さの断片も含む。
【0017】
第2の側面において、新規なセルラーゼまたは誘導体はバチルスから得られる。好ましくは、本発明のセルラーゼは図3(配列番号3)に従うアミノ酸配列、断片またはそれらの誘導体を含み、活性部分に対して90%以上の配列同一性、好ましくは95%以上の配列同一性、及びより好ましくは97%以上の配列同一性を有する。
【0018】
第3の側面において、本発明は、本発明のポリペプチドをエンコードし、適当な宿主中ポリペプチド生成を目的とする1以上の制御配列に動作可能に連結した、ヌクレオチド配列を含む核酸構築体に関する。
【0019】
本発明はさらに、mHKcelまたは断片またはそれらのスプライス変異体をエンコードする核酸配列を含む組換え発現ベクターを提供し、当該核酸配列は選択宿主中タンパク質の発現に効果的な調節因子に動作可能に連結する。関連する側面において、本発明は当該ベクターを含む宿主細胞を含む。
【0020】
第4の側面において、本発明は本発明の核酸構築体を含む組換え発現ベクターに関する。
【0021】
第5の側面において、本発明は本発明の核酸構築体を含む組換え宿主細胞に関する。
【0022】
本発明は、さらに、組換え技術、mHKcelをエンコードする核酸配列を含む原核または真核組換え宿主細胞を当該タンパク質の発現を促進するために効果的な条件下で培養し、続いて宿主細胞または細胞培養基からタンパク質を回収することにより、mHKcelを生成する方法を含む。
【0023】
第6の側面において、本発明は本発明のポリペプチドを生成する方法に関し、当該方法は以下を含む:(a)微生物を培養し、ここで当該微生物は野生型の形態で前記ポリペプチドを生成でき、前記ポリペプチドを生成する工程;及び(b)前記ポリペプチドを回収する工程。
【0024】
第7の側面において、本発明はセルロースをエタノールに変換するために有用な酵素組成物を提供する。好ましい実施態様において、酵素組成物はmHKcelを含む。当該組成物はさらに、エンドグルカナーゼ及び/またはセロビオヒドロラーゼなどの他のセルラーゼ酵素を含んでもよい。当該組成物はmHKcelを豊富に含むことができる。
【0025】
1の実施態様において、本発明は、全微生物集団DNAを周囲から単離し、大腸菌においてゲノムDNAライブラリーを構築し、セルラーゼ活性の発現についてライブラリーをスクリーニングし、セルラーゼ陽性クローン中のセルラーゼ遺伝子を同定し、新規なセルラーゼ酵素をキャラクタライズすることによる、新規な酵素を同定する方法を提供する。
【0026】
さらに、ここに提供するmHKcel核酸及びmHKcelタンパク質を検出する分析方法も本発明の一部である。
【0027】
本発明のさらに別の実施態様において、本発明に従うセルラーゼをエンコードする核酸配列を用いて適当な微生物を形質転換する方法を提供する。前記形質転換微生物から本発明に従うセルラーゼを生成する方法を提供する。
【0028】
本発明のさらなる目的は、ストレプトマイセスに特に効果的な発現ベクターを提供することである。ストレプトマイセスは種々のタンパク質生成のための中間宿主細胞として働き、セルラーゼの発現及び生成について、特に高細胞密度で細胞が成長する場合、バチルス宿主細胞を超える多数の利点を提供する。好ましい発現ベクターは、アクチノプラーネスのグルコースイソメラーゼ遺伝子由来のプロモーター配列など調節ポリヌクレオチド配列、ストレプトマイセスセルラーゼ遺伝子由来のシグナル配列及びセルラーゼ、特に本発明のセルラーゼをエンコードするDNA配列を含む。
【0029】
本発明の好ましい実施態様において、完全長セルラーゼはバチルスから得ることができる。
【0030】
本発明のその他の目的、特徴及び利点は以下の詳細な説明から明らかである。しかしながら、詳細な説明及び具体的な実施例は、本発明の好ましい実施態様を示すが、説明目的のため与えられるものであり、本発明の範囲及び概念内で種々の変化及び修正がこの詳細な説明から当業者に明らかであることが当然に理解される。
【0031】
発明の詳細な説明
本発明を以下の定義及び実施例を用いて参照することにより詳細を説明する。ここに参照する全ての特許及び刊行物、及び当該特許及び刊行物内に開示される全ての配列は明示的に引用するものとする。
【0032】
特段に示す場合を除いて、ここで用いる全ての技術及び科学用語は本発明の属する技術の当業者にとって理解される意味と同じ意味を有する。Singleton、他、Dictionary of Microbiology and Molecular Biology(微生物学及び分子生物学の辞典)、第2版、John Wiley and Sons、New York(1994)、及びHale&Marham,The Harper Collins Dictionary of Biology,Harper Perennial,NY(1991)は当業者に本発明で用いる多くの用語の一般的な辞書を提供するものである。ここに記載の方法及び物質に類似または同等のいかなる方法及び物質も本発明を実施及び試験する際に用いることができるが、好ましい方法及び物質を記載する。数値範囲は範囲を定める数字を含める。別に示す場合を除いて、それぞれ核酸は左から右に5’〜3’方向;アミノ酸配列は左から右にアミノからカルボキシ方向に記載する。実務者は特にSambrook et al.、1989、及びAusubel FM et al.、1993を当業界の定義及び用語のために参照できる。本発明は記載された特定の方法、手順及び試薬に限定されないことは、これらが種々多様であることから当然に理解される。
【0033】
ここに提供する表題は本発明の種々の側面または実施態様を限定するものではなく、明細書を全体として参照することにより理解できるものである。従って、下記に定義する用語は明細書全体を参照することにより、さらに十分に定義される。
【0034】
ここに引用する全ての文献は、本発明に関連し得る組成物及び方法を記載及び開示する目的のため参照することによりここに明示的に引用する。
【0035】
定義
“セルラーゼ”、“セルロース分解酵素”、または“セルラーゼ酵素”は、ここに記載の本発明の細菌性エンドグルカナーゼを意味する。これらの異なる種類のセルラーゼ酵素はセルロース及びその誘導体をグルコースに変換するために相乗的に作用する。
【0036】
“セルラーゼ”の語はセルロースポリマーをより短いセロ−オリゴ糖オリゴマー、セロビオース及び/またはグルコースに加水分解できる酵素分類をいう。多数のセルラーゼの例として、例えば、エキソグルカナーゼ、エキソセロビオヒドロラーゼ、エンドグルカナーゼ及びグルコシダーゼがあり、セルロース分解性生物、特に真菌及び細菌から得られる。これらの微生物から作られる酵素は、セルロースからグルコースへの転換に有用な3種類の作用を有するタンパク質の混合物である:エンドグルカナーゼ(EG)、セロビオヒドロラーゼ(CBH)、及びβ−グルコシダーゼ。これらの3種の異なるセルラーゼ酵素はセルロース及びその誘導体をグルコースに変換するために相乗的に作用する。
【0037】
多くの微生物はセルロースを加水分解する酵素を作り、例えば、木材腐朽菌トリコデルマ、土壌細菌Thermomonospora、バチルス、及びセルロモナス;ストレプトマイセス;及び真菌フミコーラ、アルペルギルス及びフサリウムなどがある。
【0038】
“宿主細胞”の語はベクターを含み、複製及び/または転写または発現構築体の転写及び翻訳(発現)を助ける細胞を意味する。本発明で用いる宿主細胞は大腸菌などの原核細胞または酵母などの真核細胞、植物、昆虫、両生類または哺乳類細胞などの細胞である。本発明の1の実施態様において、“宿主細胞”はバチルス属の細胞を意味する。本発明の他の好ましい実施態様において、“宿主細胞”はストレプトマイセスの細胞を意味する。ストレプトマイセスは、Buchanan他、The Shorter Bergey’s Manual For Determinative Bacteriology(Williams&Wilkens 1982)において分類されるストレプトマイセス属のひとつである任意の細菌株を意味する。特に好ましいストレプトマイセス株はS.lividens、S.rubiginosus、及びS.corlicolorである。S.lividensはLomovskaya、他、J.Virology 9:258(1972)に記載されている。しかしながら、当業者であれば任意の適当な宿主細胞、例えば、細菌、真菌、真核及び植物細胞が使用できることが理解できる。
【0039】
細胞、または核酸、タンパク質、またはベクターに関して用いる“組換え体”の語は、該細胞、核酸、タンパク質またはベクターが、異種核酸またはタンパク質の挿入により、または天然核酸またはタンパク質の変異により修飾されたこと、または該細胞がそのように修飾された細胞由来であることを示す。従って、例えば、組換え細胞は当該細胞の天然(非組換え)型内では見られない遺伝子を発現し、または発現、または全く発現しない条件下で、通常は異常発現される天然遺伝子を発現する。
【0040】
“分泌シグナル配列”の語は、大ポリペプチドの構成要素として、ポリペプチドが合成される細胞の分泌経路から当該大ポリペプチドに導かれるポリペプチド(“分泌ペプチド”または“分泌シグナルペプチド”)をエンコードするDNA配列を示す。当該大ペプチドは普通は分割されて、分泌経路を通過する際に分泌ペプチドが取り除かれ、分泌シグナルペプチド及び通常、成熟ポリペプチドと呼ばれるより小さいペプチドを生じる。
【0041】
ここで用いる、“セルラーゼ調製物全体”及び“セルラーゼ組成物全体”の語句は交換可能に用いられ、天然及び非天然組成物の両方をいう。“天然”組成物は天然源により生成した組成物であり、例えば、1以上のセロビオヒドロラーゼ型、1以上のエンドグルカナーゼ型、及び1以上のβ−グルコシダーゼ成分を含み、これらの各成分は供給源より生じた比率で見られる。特定の真菌は、エキソセロビオヒドロラーゼまたはCBH型セルラーゼ、エンドグルカナーゼまたはEG型セルラーゼ及びβ−グルコシダーゼまたはBG型セルラーゼを含む完全なセルラーゼ系を生じる(Schulein、1988)。しかしながら、これらの系はCBH型セルラーゼを欠く場合があり、また、細菌性セルラーゼは通常CBH型セルラーゼを含まないか、またはほとんど含まない。天然組成物はセルロース分解酵素に関して非修飾の微生物により生成されたものであり、従って当該成分酵素の比率は天然微生物により生成されるものから変更されていない。
【0042】
“非天然”組成物は、(1)天然比率または非天然、すなわち変更された比率で成分をセルロース分解酵素と混合;または(2)1以上のセルロース分解酵素を過剰発現または過少発現するように微生物を修飾;または(3)少なくとも1のセルロース分解酵素が欠失するように微生物を修飾;または(4)異種成分セルロース分解酵素を発現するように微生物を修飾することにより生成したこれらの組成物を包含する。
【0043】
ここで用いる、“プロモーター”の語は下流遺伝子の転写を導くように作用する核酸配列をいう。プロモーターは通常、標的遺伝子が発現する宿主細胞に適したものである。プロモーターはその他の転写及び翻訳調節核酸配列(“制御配列”とも呼ぶ)と一緒に所定の遺伝子を発現するために必要である。通常、転写及び翻訳調節配列は、限定されないが、プロモーター配列、リボソーム結合部位、転写開始及び停止配列、翻訳開始及び停止配列及びエンハンサーまたは活性化配列を含む。プロモーターは通常、下流遺伝子に関連するプロモーターまたは異種のものであり、すなわち、遺伝子に対して機能する限り他の遺伝子または他の微生物由来のものであってもよい。形質転換宿主細胞がバチルスの場合の好ましいプロモーターはaprEプロモーターである。1の側面において、プロモーターは導入可能プロモーターである。1の側面において、宿主細胞は糸状菌であり、プロモーターはアクセッション番号D86235でジェンバンク(GenBank)に寄託されているT.リーゼイ(reesei)cbh1プロモーターである。他の側面において、プロモーターはT.リーゼイ由来のcbhIIまたはキシラナーゼプロモーターである。
【0044】
核酸は、他の核酸配列と機能的な関係に位置する場合、“動作可能に連結”する。例えば、分泌リーダー(leader)をエンコードするDNA、すなわちシグナルペプチドは、ポリペプチドの分泌に関与するプレタンパク質として発現する場合、ポリペプチドのDNAに動作可能に連結し;プロモーターまたはエンハンサーは、配列の転写に影響する場合、コード配列に動作可能に連結し;またはリボソーム結合部位は、翻訳を促進するために位置する場合、コード配列に動作可能に連結する。通常、“動作可能に連結する”とは、連結DNA配列が隣接しており、及び分泌リーダーの場合、隣接し、かつリーディングフレーム内にあることを意味する。しかし、エンハンサーは隣接している必要はない。連結は都合のよい制限部位で連結反応により達成される。そのような部位が存在しない場合、合成オリゴヌクレオチド・アダプタまたはリンカーを従来実務に従って用いる。
【0045】
“DNA構築体”または“DNAベクター”とは、新規なセルラーゼをエンコードする1以上のDNA断片を含むヌクレオチド配列を意味する。“DNAベクター”には“発現ベクター”が含まれる。通常の発現ベクターは調節配列を含み、例えば、転写及び翻訳ターミネーター、転写及び翻訳開始配列、シグナル配列、及び特定の核酸の発現の調製に有用なプロモーターである。“プロモーター”の語は、タンパク質をエンコードするポリヌクレオチド配列の転写開始の制御に関係するポリヌクレオチド配列を指す通常の意味において用いられる。“シグナル配列”は、所望のタンパク質の宿主からの輸送を生物活性形態で導くことが可能なタンパク質のシグナルペプチドまたは部分をいう。細胞外タンパク質の成熟形態は分泌プロセス中開裂されるシグナル配列を欠く。本発明を限定する趣旨ではないが、シグナルペプチド中のアミノ酸残基の数は約5〜約100アミノ酸残基がよい。シグナル配列は、DNAの連結またはセルラーゼをエンコードするDNAの挿入を可能にするクローニング部位を提供するように修飾できる。ベクターは任意で、少なくとも1の独立ターミネーター配列、原核、真核または両方(例えば、シャトルベクター)においてカセットの複製を可能にする配列、及び原核及び真核系両方用の選択マーカーを含むジェネリック発現カセットを含む。ベクターは原核、真核または両方において複製及び統合するために適している。Giliman and Smith,Gene 8:81−97(1979);Robert、他、Nature 328:731−734(1987);Berger and Kimmel,Guide to Molecular Cloning Techniques,Methods In Enzymology,Vol 152、Academic Press,Inc.,サンディエゴ、カリフォルニア州(“Berger”);Scheider,B.et al.,Protein Expr.Purif 6435:10(1995);Sambrook、他、Molecular Cloning−A Laboratory Manual(第2版)Vol.1−3、Cold Springs Harbor Publishing(1989)(“Sambrook”);及びCurrent Protocols In Molecular Biology,Ausubel、他、(編集)、Current Protocols,Greene Publishing Associates,Inc.とJohn Wiley&Sons,Incとの共同事業(1997、補足)(“Ausubel”)を参照されたい。ストレプトマイセスに有用なクローニングベクターは公知であり、米国特許第4,338,397号、第4,411,994号、第4,513,085号、第4,513,086号、第4,745,056号、第5,514,590号及び第5,622,866号及びWO88/07079を参照できる。
【0046】
ここで用いる“遺伝子”の語は、ポリペプチド鎖の生成に関与するDNA断片を意味し、コード領域の先行及び後の領域、例えば5’非翻訳(5’UTR)または“リーダー”配列及び3’UTRまたは“トレーラー”配列、及び個々のコード断片(エキソン)間の介在配列(イントロン)などを含んでも含まなくてもよい。
【0047】
核酸の一部に言及して用いる“異種”の語は、当該核酸が、通常は同じ関係において天然では見られない2以上のサブ配列を含むことを示す。例えば、当該核酸は通常、遺伝子組換えにより生成し、例えば新しい機能的核酸を作るために配置した関連性のない遺伝子から2以上の配列を有し、例えば、1の供給源からプロモーターを有し、他の供給源からコード領域を有する。同様に、異種タンパク質は同じ関係でお互いに天然では見られない2以上のサブ配列をいうことも多い(例えば、融合タンパク質)。
【0048】
ここで用いる“単離”または“精製”の語は、天然に結合する少なくとも1の成分から除去された核酸またはアミノ酸をいう。
【0049】
本発明の内容において、“実質的に純粋なポリペプチド”とは、天然に関連するその他のポリペプチド物質を多くとも10重量%で含む、ポリペプチド調製を意味する(その他のポリペプチド物質のより低い割合は、好ましくは、例えば、多くとも8重量%、多くとも6重量%、多くとも5重量%、多くとも4重量%、多くとも3重量%、多くとも2重量%、多くとも1重量%、及び多くとも1/2重量%である)。従って、実質的に純粋なポリペプチドは少なくとも92%純粋であることが好ましく、すなわち、当該ポリペプチドは調製剤中に存在する少なくとも92重量%の合計ポリペプチド物質を構成し、及びより高い割合は好ましく、例えば、少なくとも94%純粋、少なくとも95%純粋、少なくとも96%純粋、少なくとも96%純粋、少なくとも97%純粋、少なくとも98%純粋、少なくとも99%純粋及び多くとも99.5%純粋である。ここに開示するポリペプチドは好ましくは実質的に純粋な形態である。特に、ここに開示するポリペプチドが“本質的に純粋形態”であることが好ましく、すなわち、ポリペプチド調製物が天然に関連するその他のポリペプチド物質を本質的に含まないことが好ましい。これは例えば、公知の組換え方法手段によりポリペプチドを調製することにより達成できる。ここで、“実質的に純粋なポリペプチド”の語は、“単離ポリペプチド”及び“単離形態のポリペプチド”と同義である。
【0050】
通常、mHKcelをエンコードする核酸分子を中から高ストリンジェンシー条件下で配列番号2(mHKcel)としてここに提供する配列に対してハイブリダイズする。しかしながら、場合によっては、実質的に異なるコドン使用を有するmHKcel−エンコードヌクレオチド配列を用いるが、当該mHKcel−エンコードヌクレオチド配列によりエンコードされたタンパク質は天然タンパク質と同じまたは実質的に同じアミノ酸配列を有する。例えば、コード配列は特定の原核または真核発現系においてmHKcelのより速い発現を促進するために、宿主によって利用される特定のコドン頻度に従って修飾できる。例えば、Te’o、他(2000)は糸状菌における遺伝子発現の最適化について説明している。
【0051】
核酸配列は、2つの配列が特異的に中程度から高ストリンジェンシーなハイブリダイゼーション及び洗浄条件下でお互いにハイブリダイズする場合、対照核酸配列に“選択的にハイブリダイズ”すると考えられる。ハイブリダイゼーション条件は核酸結合複合体またはプローブの融点(Tm)に基づく。例えば、“最大ストリンジェンシー”は通常、約Tm−5℃で起こり(プローブのTmより5℃低い);“高ストリンジェンシー”はTmより約5−10℃低く;“中間”または“中ストリンジェンシー”はプローブのTmより約10−20℃低く;及び“低ストリンジェンシー”はTmより約20−25℃低い。機能的に、最大ストリンジェンシー条件はハイブリダイゼーションプローブと厳密に同一またはほぼ厳密に同一な配列を同定するために用いることができ、高ストリンジェンシー条件はプローブと約80%以上の配列同一性を有する配列を同定するために用いる。
【0052】
中から高ストリンジェンシーハイブリダイゼーション条件は当業界に公知である(例えば、明示的にここに引用するSambrook、他、1989、第9章及び11章、及びAusubel、他、1993を参照)。高ストリンジェンシー条件の例としては、50%ホルムアミド、5X SSC、5X Denhardt’s溶液、0.5% SDS及び100μg/ml変性キャリアDNA中、約42℃でのハイブリダイゼーション、及び続いて2X SSC及び0.5% SDS中、室温で2回洗浄及びさらに0.1X SSC及び0.5% SDS、42℃で2回洗浄を含む。
【0053】
ここで用いる細胞に関する“形質転換”、“安定に形質転換された”または“トランスジェニック”の語は、細胞が、ゲノム内に統合された、または複数の世代を通して維持されるエピソームプラスミドとして非天然(異種)核酸配列を有することを意味する。
【0054】
ここで用いる“発現”の語は、ポリペプチドが遺伝子の核酸配列に基づいて生成されるプロセスをいう。当該プロセスは転写及び翻訳の両方を含む。
【0055】
細胞への核酸配列の挿入に関する“導入”の語は“トランスフェクション”または“形質転換”または“形質導入”を意味し、原核または真核細胞内への核酸配列の組込みに関する場合を含み、ここで核酸配列は細胞のゲノム内(例えば、染色体、プラスミド、色素体またはミトコンドリアDNA)に組み込まれることができ、自己レプリコンに変換され、または一時的に発現される(例えば、トランスフェクトされたmRNA)。
【0056】
“mHKcel発現”の語はmHKcelセルラーゼ遺伝子の転写及び翻訳、前駆体RNA、mRNA、ポリペプチド、翻訳後プロセッシングポリペプチドの生成物ということになる。例として、mHKcel発現の分析は、mHKcelタンパク質のウエスタンブロット、CBH1 mRNAのノーザンブロット分析及び逆転写ポリメラーゼ連鎖反応(RT−PCR)分析、及びShoemaker S.P.及びBrown R.D.Jr.(Biochem.Biophys.Acta,1978,523:133−146)及びSchulein(1988)に記載のエンドグルカナーゼ活性分析を含む。
【0057】
ここで用いる“界面活性剤”の語は、表面活性特性を有するものとして当業界で通常認識されている任意の化合物をいう。従って、例えば、界面活性剤は陰イオン性、陽イオン性及び非イオン性界面活性剤を含み、例えば洗剤において通常見られるものである。陰イオン性界面活性剤は、直鎖または分岐鎖アルキルベンゼンスルフォネート;直鎖または分岐鎖アルキル基またはアルケニル基を有するアルキルまたはアルケニルエーテル硫酸塩;アルキルまたはアルケニル硫酸塩;オレフィンスルフォネート;及びアルカンスルフォネートが挙げられる。両性界面活性剤は第4級アンモニウム塩スルフォネート及びベタイン型両性界面活性剤が挙げられる。当該両性界面活性剤は同じ分子内に正及び負電荷の両方の基を有する。非イオン性界面活性剤はポリオキシアルキレンエーテル及び高脂肪酸アルカノールアミドまたはその酸化アルキレン付加、脂肪酸グリセリンモノエステル等を含むことができる。
【0058】
ここで用いる“セルロース含有繊維”の語は、セルロースを含有する綿または非綿、またはセルロースブレンド、例えば天然セルロース誘導体及び人工セルロース誘導体(例えば、ジュート(黄麻)、亜麻布、ラミー、レーヨン及びリヨセル)を含有する綿または非綿から作られた任意の織布または不織布、糸または繊維をいう。
【0059】
ここで用いる、“綿含有繊維”の語は、純粋な綿または綿ブレンドから作られた織布または不織布、糸または繊維をいい、綿織物、綿ニット、綿デニム、綿糸、原綿等を含む。
【0060】
ここで用いる、“ストーンウォッシュ組成物”の語は、セルロース含有繊維をストーンウォッシュする際に用いる製剤をいう。ストーンウォッシュ組成物は販売前、すなわち製造プロセス時にセルロース含有繊維を修正するために用いる。これに対し、洗剤組成物は汚れた衣類の洗浄を目的とするものであり、製造プロセス時には用いられない。
【0061】
ここで用いる“洗剤組成物”の語は汚れたセルロース含有繊維を洗濯するための洗浄手段で用いることを意図した混合物をいう。本発明において、当該組成物はセルラーゼ及び界面活性剤に加えて、さらに加水分解酵素、ビルダー、漂白剤、漂白活性化剤、青味剤及び蛍光色素、ケーキング防止剤、マスキング剤、セルラーゼ活性化剤、酸化防止剤及び可溶化剤を含むことができる。
【0062】
ここで用いる“活性”及び“生物活性”の語は、特定タンパク質に関する生物活性をいい、ここで交換可能に用いられる。例えばプロテアーゼに関する酵素活性はタンパク質分解性であり、従って、プロテアーゼはタンパク質分解活性を有する。従って、所定タンパク質の生物活性は通常、当業者とって当該タンパク質に起因する任意の生物活性をいうことになる。
【0063】
酵素溶液中で用いる場合、mHKcel成分は通常、バイオマスからの可溶性糖の放出速度を最高にするために十分な量で加える。加えるmHKcel成分の量は、糖化されるバイオマスの種類に依存し、当業者により容易に測定できる。しかしながら、使用時の、セルラーゼ組成物中に存在するその他のセルラーゼ型成分と比較した、mHKcel成分の重量%は、好ましくは約1、好ましくは約5、好ましくは約10、好ましくは約15、または好ましくは約20重量%〜好ましくは約25、好ましくは約30、好ましくは約35、好ましくは約40、好ましくは約45、または好ましくは約50重量%である。さらに、好ましい範囲は約0.5〜約15重量%、約0.5〜約20重量%、約1〜約10重量%、約1〜約15重量%、約1〜約20重量%、約1〜約25重量%、約5〜約20重量%、約5〜約25重量%、約5〜約30重量%、約5〜約35重量%、約5〜約40重量%、約5〜約45重量%、約5〜約50重量%、約10〜約20重量%、約10〜約25重量%、約10〜約30重量%、約10〜約35重量%、約10〜約40重量%、約10〜約45重量%、約10〜約50重量%、約15〜約20重量%、約15〜約25重量%、約15〜約30重量%、約15〜約35重量%、約15〜約40重量%、約15〜約45重量%、約15〜約50重量%である。
【0064】
II.分子生物学
本発明は組換え遺伝学の分野における一般的な技術に依拠する。本発明で使用する一般的な方法を開示する基本テキストは、Sambrook、他、Molecular Cloning(分子クローニング)、A Laboratory Manual(第2版、1989);Kriegler、Gene Transfer and Expression(遺伝子導入及び発現):A Laboratory Manual(1990);及びAusubel、他、編集。Current Protocols in Molecular Biology(1994)が挙げられる。
【0065】
高レベル発現でクローン遺伝子を得るために、異種遺伝子はプロモーターからの距離が、好ましくは天然セルラーゼ遺伝子内の場合とほぼ同じ距離で位置する。当業界に公知の通り、しかしながら、ある程度の距離の変動はプロモーター機能を損なうことなく適応させることができる。
【0066】
天然プロモーターを1以上のヌクレオチドの置換、付加または欠失により、その機能を変化させることなく修飾できることは当業者であれば知っていることである。本発明の実施はプロモーターに対するそのような変異を包含するが、それに拘束されない。
【0067】
発現ベクター/構築体は通常、異種配列の発現に必要なその他全ての要素を含む転写ユニットまたは発現カセットを含む。従って、一般的な発現カセットは異種核酸配列に動作可能に連結するプロモーター及び転写、リボソーム結合部位及び翻訳停止の効率的なポリアデニル化に必要なシグナルを含む。当該カセットの他の要素は、ゲノムDNAを構造遺伝子、機能的スプライス供与体及び受容体部位を含むイントロンとして用いる場合、エンハンサーを含んでもよい。
【0068】
本発明の実施は、遺伝子構築体中のプロモーター選択により制限されない。プロモーター選択についての唯一の制限は、使用される宿主細胞において機能的であることである。形質転換宿主細胞がバチルスである場合の好ましいプロモーターは、aprEプロモーターである。
【0069】
プロモーター配列に加え、発現カセットは、効果的に停止できるように構造遺伝子の下流に転写停止領域も含むべきである。停止領域はプロモーター配列と同じ遺伝子から得られ、または異なる遺伝子から得ることもできる。
【0070】
遺伝子情報を細胞内に輸送するために用いる特定の発現ベクターは特に重要ではない。真核または原核細胞内で発現するために使用される任意の従来ベクターが使用できる。標準的な細菌性発現ベクターは、バクテリオファージλ及びM13、及びプラスミド、例えばpBR322ベースプラスミド、pSKF、pET23D、及び融合発現系、例えばMBP、GST及びLacZが挙げられる。エピトープ標識も単離方法を容易にするために組換えタンパク質に加えることができ、例えば、c−mycである。
【0071】
また、発現ベクター内に通常含まれる要素は、レプリコン、組換え体プラスミドを含む細菌選択を可能にする抗生物質耐性をエンコードする遺伝子、及び異種配列の挿入を可能にするプラスミドの非必須領域における固有制限部位も含む。特定の抗生物質耐性遺伝子の選択は重要でなく、当業者に公知の任意の多くの耐性遺伝子が適している。
【0072】
本発明の形質転換方法は、形質転換ベクターの全部または一部を糸状菌のゲノム内に安定して統合することができる。しかしながら、自己複製の細胞外形質転換ベクターの維持を生じる形質転換も考えられる。
【0073】
本発明のセルラーゼをエンコードする遺伝子はλ−ファージ(発現)ベクター及び大腸菌宿主細胞を用いてクローンできる。(もしくは、保存ドメイン上で設計されたコンセンサスプライマーを用いるPCRクローニングも使用できる。)出願人は、本発明のセルラーゼをエンコードする遺伝子の形質転換及び大腸菌内での発現により、活性タンパク質を生じることを見出した。大腸菌内での第1のクローニング工程後、本発明のセルラーゼ遺伝子はバチルスまたはストレプトマイセス種などのより好ましい工業用発現宿主、アスペルギルスまたはトリコデルマなどの糸状菌、またはサッカロマイセスなどの酵母に移動できる。これらの宿主微生物内で得られる高レベル発現及び分泌により、発酵培地中、セルラーゼの蓄積が可能となり、その後その培地から回収できる。
【0074】
プロテアーゼ欠失バチルス株の好ましい一般的な形質転換及び発現手順は、ここに引用するFerrari、他、米国特許第5,264,366号に開示されている。アスペルギルスにおける形質転換及び発現は、例えば、ここに引用する、Berka、他、米国特許第5,364,700号に記載されている。
【0075】
多くの標準的なトランスフェクション方法が、異種タンパク質を大量に発現するトリコデルマ・リーゼイ細胞株を生成するために使用できる。DNA構築体をトリコデルマのセルラーゼ生成株内に導入するための開示方法のいくつかは、Lorito,Hayes,DiPietro及びHarman,1993、Curr.Genet.24:349−356;Goldman,VanMontagu及びHerreta−Estrella,1990,Curr.Genet.17:169−174;Penttila,Nevalainen,Ratto,Salminen及びKnowles,1987,Gene 6:155−164、アスペルギルスについて、Yelton,Hamer及びTimberlake,1984,Proc.Natl.Acad.Sci.USA 81:1470−1474、フサリウム・Bajar,Podila及びKolattukudy,1991、Proc.Natl.Acad.Sci.USA 88:8202−8212、ストレプトマイセスについて、Hopwood、他、1985、The John Innes Foundation,Norwich,UK及びバチルスについて、Brigidi,DeRossi,Bertarni,Riccardi及びMatteuzzi,1990,FEMS Microbiol.Lett.55:135−138)が挙げられ、全てここに引用する。
【0076】
しかしながら、外来ヌクレオチド配列を宿主細胞に導入する任意の公知手段を用いることができる。これらは、リン酸カルシウムトランスフェクション、ポリブレン、原形質融合、電気泳動法、バイオリスティック、リポソーム、マイクロインジェクション、プラズマベクター、ウイルスベクター及びクローンゲノムDNA、cDNA、合成DNAまたはその他の外来ゲノム物質を宿主細胞内に導入するその他の公知方法の使用を含む(例えば、Sambrook.他、上記を参照)。また、米国特許第6,255,115号に記載のアグロバクテリウム媒介トランスフェクション方法も使用される。必要なのは、使用する特定の遺伝子工学手順が少なくとも1の遺伝子を異種遺伝子を発現できる宿主細胞内にうまく導入できるということだけである。
【0077】
発現ベクターが細胞内に導入された後、トランスフェクト細胞はセルラーゼ遺伝子プロモーター配列の制御下で遺伝子発現に有利な条件の下、培養される。形質転換細胞の大バッチは以下に記載の通り培養できる。最終的に、生成物は標準技術を用いて培養基から回収する。
【0078】
従って、本発明は本発明のセルラーゼの発現及び高められた分泌を提供し、発現は天然セルラーゼ遺伝子、融合DNA配列及び種々の異種構築体を含むセルラーゼ遺伝子プロモーター配列に制御される。また、本発明は本発明のセルラーゼを高レベルで発現及び分泌する方法も提供する。
【0079】
III.核酸及びエンコードされたタンパク質配列の同定
バチルス・agaradhaerans(DSM8721)由来のゲノムライブラリーを当業者に公知の標準技術を用いて調製した。この微生物はアルカリ性セルラーゼ(エンド−1,4−ベータ−グルカナーゼ)を生成し、これはグリコシル・ヒドロラーゼ、エンドグルカナーゼ5A、EC3.2.1.4、Swiss−Prot:085465、エントリー名GUN5_BACAG.EBIアクセッション番号(AF067428)のセルラーゼファミリー5に属し、その遺伝子は1203bp長である(Davies、他、1998)。セルラーゼ陽性クローンはプレート分析において発生率1/3000で検出された。本発明の側面に従う遺伝子を単離する方法において、この酵素のコード配列に基づく変性プライマーを用いた。しかしながら、意外なことに、公知のB.agaradhaeransセルラーゼを増幅することが知られるプライマーを用いてもPCR生成物は得られなかった。従って、セルラーゼをコードする挿入の完全な配列はプライマーウォーキングにより測定した。
【0080】
本発明の第2の側面に従う遺伝子を単離する方法は、配列リストに示す配列番号2のヌクレオチド配列の全部または一部を含むヌクレオチド配列に対する相同性を利用する。そのような方法の例としては以下を含む:
a)mHKcel遺伝子を推定上含む遺伝子ライブラリーをプローブとして当該ヌクレオチド配列を用いてスクリーニングする工程。
【0081】
b)ヌクレオチド配列情報に基づいてプライマーを調製し、次にテンプレートとしてmHKcel遺伝子を推定上含むサンプルを用いてPCRを行う工程。
【0082】
より具体的には、上記a)工程は、a)セルラーゼ遺伝子を推定上含む遺伝子ライブラリーを調製し、配列リストに示す配列番号2のヌクレオチド配列の全部または一部を含むヌクレオチド配列を用いて遺伝子ライブラリーをスクリーニングし、当該遺伝子ライブラリー由来の配列リストに示す配列番号2のヌクレオチド配列の全部または一部を含むヌクレオチド配列とハイブリダイズする配列を選択し、次に、選択された配列を単離し、前記遺伝子ライブラリーから選択及び単離された配列由来のmHKcel遺伝子を単離する工程、を含む。
【0083】
遺伝子ライブラリーはゲノムDNAライブラリーまたはcDNAライブラリーであってもよく、公知の手順に従って調製できる。
【0084】
IV.タンパク質発現
本発明のタンパク質は、発現がプロモーター配列に制御される本発明のセルラーゼ遺伝子を含む発現ベクターを用いて形質転換された細胞を培養することにより生成される。本発明は特にタンパク質の細胞内及び/または細胞外での生成を高めるために有用である。タンパク質は同種または異種である。
【0085】
また、本発明のタンパク質は、他の、異種ポリペプチドまたはアミノ酸配列に融合された目的タンパク質を含むキメラ分子を形成するための方法で修飾できる。1の実施態様において、このようなキメラ分子は目的タンパク質と抗標識抗体が選択的に結合できるエピトープを備える標識ポリペプチドとの融合を含む。エピトープ標識は通常、目的タンパク質のアミノ−、またはカルボキシ末端に位置する。
【0086】
種々の標識ポリペプチド及びそれぞれの抗体は当業界で公知である。例としては、ポリ−ヒスチジン(poly−his)またはポリ−ヒスチジン−グリシン(poly−his−gly)標識;HIS6及び金属キレート標識、flu HA標識ポリペプチド及びその抗体12CA5(Field、他、Mol.Cell.Biol.8:2159−2165(1988));c−myc標識及びその8F9、3C7、6E10、G4、B7及び9E10抗体(Evan、他、Molecular and Cellular Biology 5:3610−3616(1985));及び単純ヘルペスウイルス性糖タンパク質D(gD)標識及びその抗体(Paborsky、他、Ptotein Engineering 3(6):547−553(1990))が挙げられる。その他の標識ポリペプチドは、FLAG−ペプチド(Hopp、他、Bio Technology 6:1204−1210(1988));KT3エピトープペプチド(Martin、他、Science 255:192−194(1992);チューブリン・エピトープペプチド(Skinner、他、J.Biol.Chem.266:15163−15166(1991));及びT7遺伝子10タンパク質ペプチド標識(Lutz−Freyermuth、他、Proc.Natl.Acad.Sci.USA 87:6393−6397(1990))が挙げられる。
【0087】
前記mHKcel遺伝子の発現に適当な条件は、本発明のセルラーゼの成長及び/または発現に必要な成分を培養物に提供する工程を含む。タンパク質の生成に最適な条件は宿主細胞の選択に伴って、及び発現するタンパク質の選択に伴って変化する。そのような条件は、一般的な実験または最適化により当業者により容易に確かめられる。
【0088】
目的タンパク質は通常、発現後に精製または単離される。目的タンパク質は、サンプル中に存在する他の成分に応じて、当業者に公知の種々の方法で単離または精製できる。標準精製方法は電気泳動、分子、免疫学及びクロマトグラフィー技術を含み、イオン交換、疎水性、親和性及び逆相HPLCクロマトグラフィー及びクロマト分画を含む。例えば、目的タンパク質は目的抗体カラムの標準抗タンパク質を用いて精製できる。また限外ろ過及びダイアフィルトレーション技術も、タンパク濃度と併せて、有用である。適当な精製技術に関する一般的な手引きは、Scopes,Ptotein Purification(1982)を参照されたい。必要な精製の程度は目的タンパク質の使用に応じて変化する。時には精製が必要ない場合もある。
【0089】
V.セルラーゼの有用性
本発明に従う織物処理は、本発明のセルラーゼを含む組成物を用いた織物加工または織物洗浄が考えられる。そのような処理は、限定されないが、ストーンウォッシュ、セルロース含有繊維の生地、手触り及び/または外観の修正、またはセルロース含有繊維の製造または洗浄/修繕時に用いられるその他の技術を含む。さらに、本発明の文脈中の処理とは、セルロース系布または繊維から“未成熟”綿または“死”綿を取除くことも考えられる。未成熟綿は成熟綿よりも著しくアモルファスが多く、例えば不規則な染色を原因とする。本発明の組成物は、汚れた製造時のセルロース含有布を洗浄する際に使用するセルラーゼ成分をさらに含む。例えば、本発明のセルラーゼは洗濯物を洗う洗剤組成物に使用できる。本発明に有用な洗剤組成物は前洗浄用、前含浸用(pre−soak)、及び家庭用色修復組成物などの特定の剤形を含む。そのような処理組成物はここに記載するように、希釈が必要な濃縮形態または希釈溶液形態またはセルロース含有布に直接適用できる形態でもよい。織物をセルラーゼ処理するための一般的な処理技術は例えば、欧州出願第220 016及びGB出願第1,368,599号及び2,095,275号に記載されている。
【0090】
本発明に従うセルロース系材料の処理は、当業界に公知の目的に関して動物飼料、パルプ及び/または紙、食品及び穀物を処理することがさらに考えられる。例えば、セルラーゼは動物飼料の価値を増加させることが知られ、また、木材パルプの水切れ能を改善し、食品製造を高め、穀物湿式粉砕法または乾式製粉法時に穀物中の繊維を減少することが知られる。
【0091】
本発明に従う処理は、効果的な量でセルラーゼまたはセルラーゼと一緒にその他の任意の成分、例えば緩衝液、界面活性剤、及び/または精練剤との組み合わせを含む水溶液を調製することを含む。効果的な量のセルラーゼ酵素組成物とは、意図した目的のために十分なセルラーゼ酵素の濃度である。従って、例えば、本発明に従うストーンウオッシュ組成物における“効果的な量”のセルラーゼは、所望の効果を与える量であり、例えば、縫い目及びはぎ布上の擦り切れた及び色あせた外観を生じる。同様に、セルロース−含有布の手触り及び/または外観の改善を意図した組成物中のセルラーゼの“効果的な量”は、適度に手触りの改善を生じる量であり、例えば、布の滑らかさまたは外観の改善、例えば、布の外観の鮮明さを減少する傾向にある毛玉及び繊維を取除くことである。また、使用するセルラーゼの量は、使用する設備、使用するプロセス・パラメーター(セルラーゼ処理溶液の温度、セルラーゼ溶液への曝露時間等)及びセルラーゼ活性(例えば、特定の溶液は低濃度のセルラーゼが必要であり、より活性の低いセルラーゼ組成物と比較して、より活性の高いセルラーゼ組成物を使用する)に依存する。処理布に添加する処理水溶液中の正確なセルラーゼ濃度は上記の要件及び所望の結果に基づいて当業者により容易に測定できる。ストーンウォッシュ工程において、通常、セルラーゼの濃度は処理水溶液中、約0.5〜5,000ppmが好ましく、及び最も好ましくは約10〜200ppm合計タンパク質である。セルロース含有布の手触り及び/または外観を改善するための組成物において、処理水溶液中に存在するセルラーゼ濃度は、約0.1〜2000ppmが好ましく、及び最も好ましくは約0.5〜200ppm合計タンパク質である。
【0092】
好ましい処理実施態様において、緩衝液は処理組成物中で、使用するセルラーゼが活性を示す範囲内で緩衝液の濃度が溶液のpHを維持するのに十分な程度で用いる。セルラーゼが活性を示すpHは用いるセルラーゼの性質に依存する。使用する緩衝液の正確な濃度は当業者が用意に考慮に入れることができるいくつかの要素に依存する。例えば、好ましい実施態様において、緩衝液及び緩衝液濃度は、最適なセルラーゼ活性に必要なpHの範囲内で最終的なセルラーゼ溶液のpHを維持するために選択される。本発明のセルラーゼの最適なpH範囲の決定は公知の技術に従って確認できる。セルラーゼの活性範囲内のpHでの適当な緩衝液も当業者に公知である。
【0093】
セルラーゼ及び緩衝液に加えて、処理組成物は任意で界面活性剤を含むことができる。適当な界面活性剤としては、利用するセルラーゼ及び布と適合性のある界面活性剤を含み、例えば、陰イオン性、非イオン性、及び両イオン性界面活性剤が挙げられる。適当な陰イオン性界面活性剤は、限定されないが、直鎖または分岐鎖アルキルベンゼンスルフォネート;直鎖または分岐鎖アルキル基またはアルケニル基を有するアルキルまたはアルケニルエーテル硫酸塩;アルキルまたはアルケニル硫酸塩;オレフィンスルフォネート;アルカンスルフォネート等が挙げられる。陰イオン性界面活性剤に関する適当な対イオンは、限定されないが、アルカリ金属イオン、例えばナトリウム及びカリウム;アルカリ土類金属イオン、例えば、カルシウム及びマグネシウム;アンモニウムイオン;及び炭素数2または3の1〜3のアルカノール基を有するアルカノールアミンが挙げられる。両性界面活性剤は、例えば、第4級アンモニウム塩スルフォネート及びベタイン型両性界面活性剤が挙げられる。当該両性界面活性剤は同じ分子内に正及び負電荷の両方の基を有する。非イオン性界面活性剤は通常、ポリオキシアルキレンエーテル及び高脂肪酸アルカノールアミドまたはその酸化アルキレン付加、及び脂肪酸グリセリンモノエステルを含むことができる。界面活性剤の混合物も当業者に公知の手段で用いることができる。
【0094】
濃縮セルラーゼ組成物はここに記載の方法において使用するために調製できる。このような濃縮物は濃縮した量の上述のセルラーゼ組成物、緩衝液及び界面活性剤を、好ましくは水溶液中に含む。そのように処方された場合、セルラーゼ濃縮物は、必要な濃度で各成分を含むセルラーゼ調製物を素早く、正確に調製するために水で容易に希釈できる。水性濃縮物を処方した場合、これらの濃縮物は希釈でき、上述のセルラーゼ溶液中において必要な成分濃度に達することができる。明らかなことであるが、このようなセルラーゼ濃縮物により、セルラーゼ溶液の容易な処方が可能となり、使用場所への組成物の輸送が実現可能なものとなる。処理濃縮物は当業者が認識する任意の形態をとることができ、例えば、液体、乳液、ゲルまたはペーストである。このような形態は当業者に公知である。
【0095】
固体セルラーゼ濃縮物を用いる場合、セルラーゼ組成物は顆粒、粉末、凝集体または固体ディスクである。顆粒は、洗浄媒体内での顆粒溶解速度を減少させる物質を含むように処方できる。当該物質及び顆粒は、その全体をここに引用する米国特許第5,254,283号に開示されている。
【0096】
その他の物質も所望により本発明のセルラーゼ組成物と一緒に使用または添加でき、例えば、組成物の最終的な用途に応じて石、軽石、充填剤、溶媒、酵素活性化剤、及び抗再沈殿剤が挙げられる。
【0097】
例として、ストーンウオッシュ法を詳細に説明するが、記載のパラメーターはその他の用途のため、すなわち、布の感触及び/外観改善のために、当業者により容易に修正される。セルロース含有布は、処理組成物とストーンウオッシュ組成物を混合することにより、効果的な量のセルラーゼを含むストーンウオッシュ組成物を含有したセルラーゼと接触させ、従って、セルラーゼ酵素を布の近くまで持っていく。次に、セルラーゼ及び布を含む水溶液を攪拌する。処理組成物が水溶液の場合、布を当該水溶液中に直接含浸できる。同様に、ストーンウオッシュ組成物が濃縮物の場合、濃縮物はセルロース含有布と一緒に水槽内で希釈する。ストーンウオッシュ組成物が固体形態の場合、例えば、前洗浄ゲルまたは固体スティックの場合、ストーンウオッシュ組成物は組成物を布または洗浄溶液につけながら直接接触してもよい。
【0098】
セルロース含有布は、酵素活性がセルロース含有布にストーンウオッシュ外観を与えることができる効果的な条件下でストーンウオッシュ溶液を用いて培養される。例えば、ストーンウオッシュ中のpH、溶液比、温度及び反応時間はストーンウオッシュ組成物が作用する条件を最適化するために調節できる。“効果的な条件”とは必然的に、セルラーゼ酵素が効果的にセルロース含有布と反応することが可能なpH、溶液比、及び温度をいい、この場合、ストーンウオッシュ効果を生じる。本発明に従ってストーンウオッシュ組成物を用いる条件を最大化することは当業者の能力の範囲内である。
【0099】
ここで用いるストーンウオッシュ中の溶液比、すなわち、ストーンウオッシュ組成物溶液の重量(すなわち、洗浄溶液)と布重量との比は通常、デニム生地に所望のストーンウオッシュ効果を達成するために十分な量であり、使用する方法に依存する。好ましくは、溶液比は約4:1〜約50:1であり、より好ましくは約5:1〜約20:1、及び最も好ましくは約10:1〜約15:1である。
【0100】
本発明のストーンウオッシュ組成物を用いるストーンウオッシュ時の反応温度は2つの競合因子に支配される。まず、高温に対応して、通常、反応速度が高められ、すなわち、反応が速まり、低温の場合に必要な反応時間と比較して反応時間を減少させることができる。従って、反応温度は通常、少なくとも約10℃以上である。第2に、セルラーゼは所定の反応温度以上で活性を失うタンパク質であり、当該温度は使用するセルラーゼの性質に依存する。従って、反応温度が高くなりすぎた場合、セルロース分解活性はセルラーゼ変性の結果として失われる。当業界において標準的なセルラーゼ使用温度は通常、35℃〜65℃の範囲であり、これらの条件が本発明のセルラーゼにも適したものであると考えられるが、最適な温度条件は使用される特定のセルラーゼに関して公知技術に従い、確認するべきである。
【0101】
反応時間はストーンウオッシュが生じる特定の条件に依存する。例えば、pH、温度及びセルラーゼ濃度はすべて最適な反応時間に影響する。通常、反応時間は約5分〜約5時間であり、好ましくは約10分〜約3時間、及び最も好ましくは約20分〜約1時間である。
【0102】
本発明のさらに好ましい実施態様に従い、本発明のセルラーゼは洗剤組成物中に用いてもよい。本発明に従う洗剤組成物は前洗浄組成物、前含浸(pre−soak)組成物として、または標準洗浄またはすすぎサイクル間の洗浄に有用である。好ましくは、本発明の洗剤組成物は効果的な量のセルラーゼ、界面活性剤を含み、及び任意で下記のその他の成分を含む。
【0103】
本発明の洗剤組成物で使用される効果的な量のセルラーゼは、セルロース含有布にセルラーゼにより生じることが知られる所望の効果、例えば毛羽取り、柔軟化、毛羽防止、表面繊維除去、抗灰色化及び洗浄効果を与えるために十分な量である。好ましくは、洗剤組成物中のセルラーゼは洗剤の約10ppm〜約20,000ppm濃度で用いる。
【0104】
洗剤組成物中で使用されるセルラーゼ酵素の濃度は好ましくは、水媒体中へ希釈して、セルラーゼ酵素の濃度が約0.01〜約1000ppm、好ましくは約0.02ppm〜約500ppm、及び最も好ましくは約0.5ppm〜約250ppm合計タンパク質になるように選択される。洗剤組成物中で使用されるセルラーゼ酵素の量は、洗浄溶液を作るために水を添加することにより洗剤が希釈される程度に依存する。
【0105】
本発明の洗剤組成物は当業者が認識する任意の形態であり、例えば、液体、顆粒、乳液、ゲル、ペーストである。このような形態は当業者に公知である。固体洗剤組成物を使用する場合、セルラーゼは顆粒として処方するのが好ましい。好ましくは、顆粒はセルラーゼ保護剤をさらに含むように処方できる。顆粒は水媒体への顆粒の溶解速度を減少させる物質を含むように処方することもできる。このような物質及び顆粒は、その全体をここに引用する米国特許第5,254,283号に開示されている。
【0106】
本発明の洗剤組成物は、表面活性剤、すなわち界面活性剤、例えば洗剤組成物での使用が公知の陰イオン性、非イオン性、及び両イオン性界面活性剤を用いる。
【0107】
本発明の洗剤組成物での使用が適した陰イオン性界面活性剤は、直鎖または分岐鎖アルキルベンゼンスルフォネート;直鎖または分岐鎖アルキル基またはアルケニル基を有するアルキルまたはアルケニルエーテル硫酸塩;アルキルまたはアルケニル硫酸塩;オレフィンスルフォネート;及びアルカンスルフォネートが挙げられる。陰イオン性界面活性剤に関する適当な対イオンは、アルカリ金属イオン、例えばナトリウム及びカリウム;アルカリ土類金属イオン、例えば、カルシウム及びマグネシウム;アンモニウムイオン;及び炭素数2または3の1〜3のアルカノール基を有するアルカノールアミンが挙げられる。両性界面活性剤は第4級アンモニウム塩スルフォネート及びベタイン型両性界面活性剤が挙げられる。当該両性界面活性剤は同じ分子内に正及び負電荷の両方の基を有する。非イオン性界面活性剤は通常、ポリオキシアルキレンエーテル及び高脂肪酸アルカノールアミドまたはその酸化アルキレン付加、脂肪酸グリセリンモノエステル等を含むことができる。本発明での使用に適した界面活性剤は英国特許出願番号第2 094 826Aに開示されており、その開示の内容をここに引用する。このような界面活性剤の混合物も使用することができる。界面活性剤または界面活性剤の混合物は通常、本発明の洗剤組成物において合計洗剤組成物の約1重量%〜約95重量%の量、好ましくは合計洗剤組成物の約5重量%〜約45重量%で使用される。セルラーゼ組成物及び界面活性剤に加え、本発明の洗剤組成物は任意で1以上の以下の成分を含むことができる。
【0108】
セルラーゼ以外のヒドロラーゼ
適当なヒドロラーゼは、エステル結合上に作用するカルボキシレートエステルヒドロラーゼ、チオエステルヒドロラーゼ、ホスフェートモノエステルヒドロラーゼ、及びホスフェートジエステルヒドロラーゼ;グリコシル成分上に作用するグリコシドヒドロラーゼ;N−グリコシル成分を加水分解する酵素;エーテル結合上に作用するチオエーテルヒドロラーゼ;ペプチド結合上に作用するa−アミノ−アシル−ペプチドヒドロラーゼ、ペプチジル−アミノ酸ヒドロラーゼ、アシル−アミノ酸ヒドロラーゼ、ジペプチドヒドロラーゼ及びペプチジル−ペプチドヒドロラーゼが挙げられる。この中で好ましいものは、カルボキシレートエステルヒドロヒドロラーゼ、グリコシドヒドロラーゼ、及びペプチジル−ペプチドヒドロラーゼである。適当なヒドロラーゼは、(1)ペプチジル−ペプチドヒドロラーゼに属するプロテアーゼ、例えば、ペプシン、ペプシンB、レンニン、トリプシン、キモトリプシンA、キモトリプシンB、エラスターゼ、エンテロキナーゼ、カテプシンC、パパイン、キモパパイン、フィシン、トロンビン、フィブリノリジン、レニン、スブチリシン、アスペルギロペプチダーゼA(aspergillopeptidase A)、コラゲナーゼ、クロストリジオペプチダーゼB(clostridiopeptidase B)、カリクレイン、ガストリシン、カテプシンD、ブロメリン、ケラチナーゼ、キモトリプシンンC、ペプシンC、アスペルギロペプチダーゼB、ウロキナーゼ、カルボキシペプチダーゼA及びB、及びアミノペプチダーゼ;(2)グリコシドヒドロラーゼ(必須成分であるセルラーゼはこの群から除く)α−アミラーゼ、β−アミラーゼ、グルコアミラーゼ、インベルターゼ、リゾチーム、ペクチナーゼ、キチナーゼ及びデキストラナーゼを含む。この中で好ましくは、α−アミラーゼ及びβ−アミラーゼである。これらは酸から中性の系で機能するが、細菌から得られるものはアルカリ性の系において高い活性を示す;(3)カルボキシレートエステルヒドロラーゼ、例えばカルボキシルエステラーゼ、リパーゼ、ペクチンエステラーゼ、及びクロロフィラーゼなど。この中で特に効果的なものはリパーゼである。
【0109】
セルラーゼ以外のヒドロラーゼは本目的に従って必要とされる量だけ洗剤組成物内に組み込まれる。好ましくは、精製タンパク質に関して、0.001〜5重量%、及びより好ましくは0.02〜3重量%の量で組み込まれるべきである。この酵素は、未精製酵素だけで作られた、またはその他の成分と組合わせて、顆粒形態で洗剤組成物中で使用すべきである。未精製酵素の顆粒は、顆粒中、精製酵素が0.001〜50重量%となるような量で使用される。当該顆粒は0.002〜20及び好ましくは0.1〜10重量%の量で用いる。セルラーゼと同様、これらの顆粒は酵素保護剤及び分解遅延剤を含むように処方できる。
【0110】
陽イオン界面活性剤及び長鎖脂肪酸塩
当該陽イオン界面活性剤及び長鎖脂肪酸塩は、飽和または不飽和脂肪酸塩、アルキルまたはアルケニルエーテルカルボン酸塩、α−スルホ脂肪酸塩またはエステル、アミノ酸型界面活性剤、リン酸エステル界面活性剤、4級アンモニウム塩、例えば3〜4のアルキル置換基及び1以下のフェニル置換アルキル置換基を有するものを含む。適当な陽イオン界面活性剤及び長鎖脂肪酸塩は英国特許出願番号第2 094 826Aに開示されており、その開示の内容をここに引用する。本組成物は約1〜約20重量%の当該陽イオン界面活性剤及び長鎖脂肪酸塩を含むことができる。
【0111】
ビルダー
A.二価金属イオン封鎖剤(sequestering agents)
本組成物は約0〜約50重量%の1以上のビルダー成分を含むことができ、以下の化合物のアルカリ金属塩及びアルカノールアミン塩からなる群より選択される:リン酸塩、ホスホン酸塩、ホスホノカルボキシレート、アミノ酸塩、アミノポリアセテート高分子量電解質、非分散ポリマー、ジカルボン酸塩、及びアルミノケイ酸塩。適当な二価金属イオン封鎖剤は米国特許出願番号第2 094 826Aに開示されており、その開示の内容をここに引用する。
【0112】
B.アルカリまたは無機電解質
本組成物は当該組成物に基づいて、約1〜約50重量%、好ましくは約5〜約30重量%の1以上の以下の化合物のアルカリ金属塩をアルカリまたは無機電解質として含むことができる:ケイ酸塩、炭酸塩及び硫酸塩及び有機アルカリ、例えば、トリエタノールアミン、ジエタノールアミン、モノエタノールアミン及びトリイソプロパノールアミン。
【0113】
再付着防止剤
本組成物は約0.1〜約5重量%の1以上の以下の化合物を再付着防止剤として含むことができる:ポリエチレングリコール、ポリビニルアルコール、ポリビニルピロリドン、及びカルボキシメチルセルロース。
【0114】
この中でも、カルボキシメチル−セルロース及び/またはポリエチレングリコールと本発明のセルラーゼ組成物との組み合わせは特に有用な汚れ除去組成物を提供する。
【0115】
漂白剤
本発明のセルラーゼと漂白剤とを組み合わせた使用、例えば漂白剤はモノ過硫酸カリウム、過炭酸ナトリウム、過ホウ酸ナトリウム、硫酸ナトリウム/過酸化水素付加化合物及び塩化ナトリウム/過酸化水素付加化合物または/及び光過敏漂白剤、例えば、スルホン化フタロシアニンの亜鉛またはアルミニウム塩は、さらに洗剤効果を改善する。同様に、EP684 304に記載の漂白剤及び漂白触媒も用いることができる。
【0116】
青味剤及び蛍光染料
種々の青味剤及び蛍光染料は必要に応じて本組成物に組み込むことができる。適当な青味剤及び蛍光染料は英国特許出願第2 094 826Aに開示されており、その開示の内容をここに引用する。
【0117】
凝結防止剤
以下の凝結防止剤は粉末洗剤中に組み込むことができる:p−トルエンスルホン酸塩、キシレンスルホン酸塩、酢酸塩、スルホ琥珀酸塩、タルク、微細粉末シリカ、アモルファスシリカ、粘土、ケイ酸カルシウム(例えば、Johns Manville Co.のマイクロ−セル(Micro−Cell))、炭酸カルシウム及び酸化マグネシウム。
【0118】
酸化防止剤
酸化防止剤は、例えば、tert−ブチル−ヒドロキシトルエン、4,4’−ブチリデンビス(6−tert−ブチル−3−メチルフェノール)、2,2’−ブチリデンビス(6−tert−ブチル−4−メチルフェノール)、モノスチレン化クレゾール、ジスチレン化クレゾール、モノスチレン化フェノール、ジスチレン化フェノール及び1,1−ビス(4−ヒドロキシ−フェニル)シクロヘキサンが挙げられる。
【0119】
可溶化剤
可溶化剤は例えば、低級アルコール、例えば、エタノール、ベンゼンスルホン酸塩、低級アルキルベンゼンスルホン酸塩、例えば、p−トルエンスルホン酸塩、グリコール、例えば、プロピレングリコール、アセチルベンゼン−スルホン酸塩、アセトアミド、ピリジンジカルボン酸アミド、安息香酸塩及び尿素が挙げられる。
【0120】
本発明の洗剤組成物は酸性からアルカリ性pHまで広いpH範囲で用いることができる。好ましい実施態様において、本発明の洗剤組成物は、5以上から約12以下のpHを有する弱酸性、中性またはアルカリ性洗浄媒体中で用いることができる。
【0121】
上記成分に加えて、香料、緩衝剤、保存料、染料等を所望により本発明の洗剤組成物と一緒に用いることができる。当該成分は従来当業界で用いられてきた量で適宜用いられる。
【0122】
本発明で使用される洗剤基材が粉末形態の場合、公知の調製方法、例えば噴霧乾燥法及び造粒法により調製されるものである。特に噴霧乾燥法、凝集法、乾燥混合法または非塔(non−tower)ルート法により得られる洗剤基材は好ましい。噴霧乾燥法により得られる洗剤基材は調製条件については制限されない。噴霧乾燥法により得られる洗剤基材は、熱耐性成分、例えば表面活性剤及びビルダーの水性スラリーを温空間内に噴霧することにより得られる空洞の顆粒である。噴霧乾燥後、香料、酵素、漂白剤、無機アルカリ性ビルダーを加えることができる。噴霧乾燥造粒または凝集法などにより得られた高い密度の顆粒洗剤基材と一緒に、種々の成分も基材の調製後に加えることできる。
【0123】
洗剤基材が液体の場合、均質溶液または不均質分散液であってもよい。洗剤中のセルラーゼによるカルボキシメチルセルロースの分解を除去するために、カルボキシメチルセルラーゼは組成物に組み込む前に顆粒化またはコーティングされていることが望ましい。
【0124】
本発明の洗剤組成物はセルロース含有布、例えば、汚れた布と一緒に工業的及び家庭用途においてこれらの環境で適宜用いられる温度、反応時間及び溶液比で培養できる。
【0125】
本発明に従う洗剤は、十分な活性が存在し、所望の改善、軟化、毛羽取り、毛羽立ち防止、表面繊維除去、または洗浄が得られる、中性pHで適当な溶液中で前洗浄としてさらに処方できる。洗剤組成物が液体、スプレー、ゲルまたはペースト組成物としての前含浸(pre−soak)(例えば、前洗浄または前処理)組成物の場合、セルラーゼ酵素は通常、前含浸または前処理組成物の全重量に基づいて約0.0001〜約1重量%で用いられる。このような組成物において、界面活性剤を任意で用いることができ、用いる場合、通常、前含浸の全重量に基づいて、約0.005〜約20重量%の濃度で存在する。組成物の残りは前含浸において用いられる従来成分を含み、すなわち、従来濃度での希釈剤、緩衝剤、その他の酵素(プロテアーゼ)等である。
【0126】
ここに記載のセルラーゼを含む組成物は、色あせた布の色修復(例えば、ここにその全体を引用する米国特許第4,738,682号を参照)及びシミ取り及び毛羽取り及び抗毛羽立ち(毛羽立ち防止)のための使用に適した単独組成物として家庭用で用いることができると考えられる。
【0127】
本発明に従うセルラーゼの使用は飼料添加剤及びパルプ及び紙処理において特に効果的である。これらのさらなる工業的用途については例えば、PCT国際公開番号第95/16360号及びフィンランド発行特許第87372号にそれぞれ記載されている。
【0128】
本発明及びその利点についてさらに説明するために、以下に具体的な実施例を与えるが、これらは本発明を説明するために提供するものであり、その範囲を限定するものとしていかなる方法においても解釈すべきでないことが理解される。
【0129】
実施例
以下の実施例を与えるが、本発明を限定するものではない。
【0130】
実施例1 サンプル収集及び処理
この実施例はcDNAライブラリーを作成するために十分なDNAを得るためのサンプルの収集法及び処理法について説明する。
【0131】
水サンプル(250ml)をSonachi湖(クレーター)、ケニアの沿岸帯から、柔軟伸展性1−m棒の先端に取り付けたステンレススチール・ビーカーを用いて収集し、室温で研究所に輸送するため密封可能なプラスティック容器(Whirlpak)内に納めた。水の表面温度は28℃、pH10及び伝導率は7.23mScm−1(27℃で)であった。
【0132】
微生物叢を収集するために、Sonachi(クレーター)湖、ケニアからの水(750ml)を、減少した細孔サイズの硝酸セルロースまたは酢酸セルロースからなる一連の殺菌薄膜フィルター(直径47mm)を通して、水流が全て止まるまで(手動吸引ポンプを用いて)現地でろ過した。一連のフィルターは8μm、3μm及び0.22μmであった。個々の薄膜フィルターを10mM Tris HCl、pH8.0;1mM EDTA及び5%w/v NaClを含む30ml殺菌プラスティック・ユニバーサル菅内の10mlの冷、殺菌細胞安定化緩衝剤(TES)中に速やかに加え、冷蔵された冷却ボックス内の氷上に、さらに処理するときまで、通常はサンプル採取4時間以内で保管した。フィルター上の細菌物質は、殺菌ガラスビーズ(5ml)及び微量遠心菅内で5分間、13,000gで遠心することにより沈殿した細胞を混合しながら激しくかき混ぜることにより分散させた。細菌物質を、推定で108〜109の等量の細菌細胞を含む体積で微量遠心菅に等分し、合計12個の菅を得た。DNAをGenomicPrep(登録商標)Cells及びTissue DNA単離キット(アマシャム・ファルマシア・バイオテック(Amersham Pharmacia Biotech)、Piscataway,ニュージャージー州、米国)を用いて取扱説明書に従って抽出した。各菅中の細胞は所定の600μlの細胞溶解液中に再懸濁し、80℃、5分間で培養し、細胞を溶解させた。この方法により調製したサンプルは室温で少なくとも18ヶ月間安定であり、この形態で研究室に戻した。DNA抽出を製造業者手順に従って、RNAアーゼ処理、タンパク質沈殿及びDNAイソプロパノール沈殿により完結させた。各DNAペレットを100μl殺菌Tris緩衝液10mM pH8.5中に溶解させた。
【0133】
DNA収量は5μlのサンプルを0.5% w/vアガロースゲル上で処理し、細菌性ゲノムDNAの公知量と比較することにより推測した。サンプルをプールし、合計約20μgのDNAを得た。収量が低かったので、on−site物質として同時に収集した水サンプルから抽出した約30%の余分物質で物質を補充し、必要なときまで4℃で研究室で保存した。約30μgの量のDNAは、予備実験が示した開始物質の量であり、試験及びバルク制限消化及びサイズ分画を行うために必要であり、ライブラリー構築のために十分な物質を与えた。
【0134】
実施例2
ライブラリー構築
以下の実施例は、大腸菌内の新規な配列のスクリーニング及び検出に使用するためのDNAライブラリーをどのように調製するかについて詳述する。
【0135】
DNA調製
プールしたDNAをゲノムDNAライブラリーの構築のために用いた。精製DNAをSau3A1を用いて部分的に消化し、約5kbの平均断片サイズを得た。制限DNAを電気泳動法によりTAEにおいて0.5%アガロース上でサイズ分画した(0.04M Tris−アセテート、0.001M EDTA pH8.0)。1.5〜10kb範囲の物質を切り取り、アガロースゲルの未使用部分をカットした同じサイズのウェル中に置き換え、逆電流により狭いバンドに濃縮させた。DNAバンドを切断し、キアゲン(QIAGEN)(Crawley,UK)QIAEXIIゲル抽出キットを用いて、取扱説明書に従ってDNA抽出した。溶出DNAをエタノールを用いて沈殿させ、10mM Tris HCl緩衝液、pH8.5中に再懸濁した。
【0136】
ラムダライブラリーの調製
制限DNAをラムダベクター内に、ZAP−エクスプレス(登録商標)ベクターキット(BamH1で前消化、及びアルカリホスファターゼ処理)及びGigapak(登録商標)IIIゴールド・パッケージング抽出(ストラタジーン(Stratagene)、アムステルダム、オランダ)を用いて製造業者手順に従いクローンした。最初のライブラリーを手順通りに、〜5×104pfuを含むアリコートを宿主大腸菌株XL1−ブルーMRF’で150mmペトリ皿にプレートし、緩衝液中のファージを溶出させることにより増幅させた。液体窒素中で凍結させた後、増幅ライブラリーを7% v/vジメチルスルホキシド中、−80℃で保存した。最初の滴定量の合計は1.8×106pfuであり、増幅後は6.8×109pfu ml−1であった。
【0137】
ライブラリー特質の評価
ファージミドベクターpBK−CMVをExAssistヘルパーファージ(ストラタジーン)を用いて製造業者の説明書通りにラムダZAPライブラリーから切り取り、大腸菌株XLOLRを感染させるために用いた。プラスミド含有クローンを50μg ml−1カナマイシンを含むルリア栄養(LB)寒天上にプレートすることにより単離した。Xgal[5−ブロモ−4−クロロ−3−インドイル−β−D−ガラクトシド]及びIPTG[イソプロピルチオ−β−D−ガラクトシド]の存在下、青白スクリーニングを用いてクローニング効率を測定した。DNAがラムダベクター内にクローンされなかった場合、β−ガラクトシド遺伝子が誘導因子IPTGの存在下で発現され、基質類似体Xgalの分裂を生じ、コロニー内に青色着色を生じる。しかしながら、ゲノムDNAの断片がうまくラムダベクター内にクローンされた場合、遺伝子が阻害され、従って、酵素は生成されず、コロニーは白色のままである。従って、青色と白色コロニーの比は挿入を含むクローンの割合を計算するために用いることができる。このライブラリーに関して、青白スクリーニングは青7、白286のコロニー比を生じ、97%のコロニーがゲノムDNA挿入を含んでいたことを示した。無作為に24のコロニーを選択し、BamH1クローニング部位に隣接するPst1及びHindIIIを用いるWizard(登録商標)Plus SV Miniprep DNA精製システム(Promega UK、サウサンプトン(Sauthampton))制限分析を用い、続いてアガロースゲル電気泳動法により調製したプラスミドDNAを使用して挿入サイズを測定した。24個の中、1個のクローンは検出可能な挿入を含まなかった。残りは1.5kb〜8.0kbの範囲の挿入を有した。
【0138】
実施例3
セルラーゼのためのライブラリースクリーニング
pBK−CMVファージミド中のDNAライブラリーを大腸菌クローンのプレート分析におけるセルラーゼ活性に関して、スクリーニングした。セルラーゼ活性を検出するために、ゲノムライブラリーをカナマイシン、0.5% w/vカルボキシメチルセルロース(低粘度ナトリウム塩;シグマ(Sigma)、Poole,UK)及びIPTG(直径7cmのペトリ皿内で寒天の表面上に広げた15μlの0.5M溶液)を含むLB寒天上にプレートした。37℃で一晩中成長させた後、当該コロニーを水に溶かして50℃まで冷やした3ml溶融0.7% w/vアガロースでかぶせた。これが終わると、当該プレートを0.1% w/v コンゴレッド(Congo Red)溶液で30分間水浸しにし、続いて1M NaClで2回洗浄した。細胞外セルラーゼ活性を示す陽性クローンは赤背景に対してイエロー・ハロ(yellow halo)で囲まれた(R.Teather and P.J.Wood,Applied&Environmental Microbiology,43:770−780,1982)。
【0139】
110,000の大腸菌pBK−CMVクローンのスクリーニングにより4領域のクリアリング指示潜在性(clearing indicating potential)のセルラーゼ生成コロニーを生じた。これらの3つは、除去領域(cleared zone)から取除いた寒天プラグを均質化し、単一のコロニーの筋をつけて、コンゴレッド試験により表現型を確認した後、セルラーゼ生成クローンとしてうまく回収された。
【0140】
実施例4
セルラーゼ陽性クローンのキャラクタリゼーション
プラスミドDNAを3つのセルラーゼ陽性クローンから単離し、挿入サイズを上述の通り制限消化により測定した。ゲル電気泳動により測定して消化後、3つ全てが同じサイズ(約3.5kb)であり、及び同じサイズの断片を有した。このことは、3つ全ての単離体が同一であり、単一のクローンの増幅により生じたことを示す。これは、プラスミドDNAのシーケンシングの第1ラウンドにより確認された(pBKCMVプラスミド中のプライマー部位を用いて)。これはレスター(Leicester)大学でタンパク質及び核酸化学研究室により、パーキンエルマー(Perkin Elmer)‘BigDye’ターミネーター・ケミストリー及び377モデルABI自動DNAシーケンサーを用いて行われた。配列の完全な範囲が挿入の5’及び3’の両末端から‘プライマーウォーキング’により得られた。当該配列はアプライド・バイオシステムス(Applied Biosystems)マルチ配列編集Seqed(登録商標)バージョン1.0.3を用いて編集した。配列はレスター大学から入手可能なGCGウィスコンシン(Wisconsin)パッケージ、バージョン10.2−UNIXにおけるプログラムを用いて組み立てた。これにより3796ヌクレオチド塩基の環境DNAの挿入を同定した(図1)。
【0141】
実施例5
セルラーゼ遺伝子の同定
クローンmHKcelの挿入環境DNAのヌクレオチド配列における可能なオープンリーディングフレーム(ORF)をMapDrawプログラム(DNASTAR、Brighton、マサチューセッツ州、米国)のORF Find設備またはベクターNTI式(Suite)のプログラムからのORFサーチ(InforMax(登録商標、North Bethesda,メリーランド州、米国)を用いて同定した。
【0142】
これにより、571アミノ酸のタンパク質に対応した1716ヌクレオチドからなるORFが同定され、挿入配列の位置923から開始して位置2638で終わった。このORFの配列はEditSeq(DNASTAR)を用いて切り取り、BLASTプログラムにより測定した。
【0143】
このORFのヌクレオチド配列を図2に示す。
【0144】
BLASTnプログラムを用いたヌクレオチド配列の調査は、重複のないヌクレオチド配列データベースに対してヌクレオチド質問配列を比較するものであり、公知配列に対してバチルス・ハロデュランス(halodurans)ゲノムの一部と同一性を示し、特にエンド−ベータ−1,4−グルカナーゼ(“セルラーゼB”)(GenBankアクセッション AP001509)に対応する1585ヌクレオチド領域に68%同一性を示した。
【0145】
BLASTxプログラムを用いるヌクレオチド配列の調査は、ヌクレオチド質問配列(両ストランド)の6フレーム概念的翻訳産物をタンパク質配列データベースに対して比較するものであり、多数の細菌性エンドセルラーゼに著しい類似性を示した。最も高い配列スコアは、574アミノ酸を含む酵素であり、通性好アルカリ性(facultative alkaliphilic)細菌バチルス・ハロデュランス株C125(タンパク質ID BAB04322、アクセッション AP001509)のエンド−β−グルカナーゼ(セルラーゼB)の553アミノ酸領域に対して67%同一性(372アミノ酸)を示した。この新しい同定遺伝子はバチルス・ハロデュランスセルラーゼに67%同一性を有する571アミノ酸のタンパク質をコードした。
【0146】
571個のアミノ酸からなる翻訳タンパク質を図3に示す。
【0147】
実施例6
酵素キャラクタリゼーション
塩の影響
mHKcel遺伝子を含む大腸菌pBK−CMV細胞を5ml緩衝液(20mM TRIS−HCl、pH8.0;500mM NaCl;0.1mM EDTA;0.1%Triton X−100)中に懸濁し、氷上で超音波処理することにより分解させる。超音波分解した抽出物をカルボキシメチルセルロース(CMC)上で寒天拡散分析により、種々のNaCl濃度で調べる。超音波分解抽出物(100μL)及び10個中1個の希釈物を種々の量のNaClを含むCMC−寒天プレート中に穴を開けたウェル内に配置する。当該プレートを37℃、16時間で培養し、セルロース加水分解を示す生じた除去(clearing)領域をミリメーターで測定する。結果は、セルラーゼmHKcelは0〜25% w/v NaClの範囲で活性だが、25% w/v NaClでの活性は0% NaClでの活性の約50%しかないということを示した(図4)。
【0148】
pHの影響
セルラーゼ活性におけるpHの影響をGrant&Tindall(Isolation of alkaliphilic bacteria(アルカリ性細菌の単離),In:Microbial Growth and Survival in Extreme Environments,Academic Press,ロンドン、1980、第27〜36頁)に記載のpH勾配プレート法を用いて調査した。CMCを含む寒天培地を四角形ペトリ皿の深さ1cmまで注ぎ、設置する。幅1cmの均一な鉢をプレートの端から切り取り、20% w/v Na2CO3・10H2O及び0.2M NaOHを含む寒天(同体積の殺菌0.4M NaOH/40% w/v Na2CO3・10H2O及び4% w/v寒天を60℃で混合することにより調製したもの)を鉢の中に注いだ。当該プレートを37℃で一晩中成長させ、均一にpH12〜pH7勾配を形成させる。mHKcelセルラーゼのpH耐性を調べるために、鉢を狭く(寒天)勾配により元の鉢に対して直角に切り取り、1mlの超音波分解した細胞抽出物で満たした。当該プレートを37℃で一晩中成長させた。当該プレートをコンゴレッドで30分間処理し、セルロース加水分解の領域を視覚化する。図5に示す結果は、mHKcelセルラーゼが約pH11.5まで活性であることを示す。
【0149】
ここに記載される実施例及び実施態様は説明目的のためのみであり、これに照らして種々の修正または変形が当業者に示唆され、本発明の概念及び精神及び請求の範囲の範囲内であることが理解される。ここに記載の全ての文献、特許及び特許出願はその全体を全ての目的においてここに引用するものとする。
【図面の簡単な説明】
【0150】
【図1A】環境(environmental)ヌクレオチド配列(配列番号1)を示す。
【図1B】環境(environmental)ヌクレオチド配列(配列番号1)を示す。
【図2】図2は新規なセルラーゼをエンコードする核酸配列を示す(配列番号2)。
【図3】図3は本発明のセルラーゼの推定アミノ酸配列を示す(配列番号3)。
【図4】図4は増加した塩化ナトリウム濃度を有する酵素活性を描いたグラフである。
【図5】図5はpH7〜12の範囲でのmHKcel活性へのpHの影響を描いたグラフである。

【発明の詳細な説明】
【0001】
関連出願の相互参照
本出願は、ここにその全体を引用する、2003年4月30日に出願された米国仮出願60/467,315号(代理人整理番号、GC797P)に基づく優先権を主張するものである。
【0002】
連邦政府後援の研究開発の下、行われた発明の権利に関する声明
適用なし。
【0003】
発明の分野
本発明はここでmHKcelとして言及する新規なセルラーゼに関する。また、セルラーゼをエンコードする核酸、前記セルラーゼを含む組成物、新規なセルラーゼを同定する方法、及び前記組成物を用いる方法についても記載する。好ましくは、当該セルラーゼはバチルス種から単離され、好ましくはB.agaradhaerensである。本発明はさらに、洗剤組成物、セルロース含有繊維処理、パルプ及び紙の処理、及び高フルクトース・コーンシロップまたはエタノールの生成用のデンプン処理に添加剤として、当業界で有利にセルラーゼが添加されるものとして認識される、新規なセルラーゼの組成物中での使用に関する。
【0004】
発明の背景
セルロース及びヘミセルロースは光合成により最も豊富に生成される植物材料である。これらは高分子基材を単糖類に加水分解できる細胞外酵素を生成する多数の微生物、例えば細菌、酵母及び真菌によりエネルギー源として分解及び使用される(Aro、他、2001)。非再生源の限界が近づいているため、セルロースが主な再生可能エネルギー源となる可能性は非常に大きい(Krishna、他、2001)。生物プロセスを通じたセルロースの効果的な活用は食料、飼料及び燃料不足の克服手段のひとつである(Ohmiya、他、1997)。
【0005】
セルラーゼはセルロース(β−1,4−グルカンまたはβ−D−グルコシド結合)を加水分解してグルコース、セロビオース、セロオリゴ糖等の形成を生じる酵素である。セルラーゼは従来から3つの主な分類に分けられる:エンドグルカナーゼ(EC 3.2.1.4)(“EG”)、エキソグルカナーゼまたはセロビオヒドロラーゼ(EC 3.2.1.91)(“CBH”)、及びβ−グルコシダーゼ([β]−D−グルコシドグルコヒドロラーゼ;EC 3.2.1.21)(“BG”)(Knowles、他、1987;Shulein、1988)。エンドグルカナーゼは主にセルロース繊維のアモルファス部分で作用するのに対し、セロビオヒドロラーゼは結晶セルロースを分解することもできる(Nevalainen and Penttila、1995)。従って、結晶セルロースを効果的に可溶化するために、セルラーゼ系中にセロビオヒドロラーゼの存在が必要である(Suurnakki、他、2000)。β−グルコシダーゼはセロビース、セロ−オリゴ糖、及びその他のグルコシド由来D−グルコース単位の遊離に作用する(Freer、1993)。
【0006】
結晶セルロースをグルコースに効果的に変換するためには、結晶セルロースの加水分解にあまり効果がない単離成分とともにCBH、EG及びBGの各分類由来の成分を含む完全なセルラーゼ系が必要である(Filho、他、1996)。それぞれの分類由来のセルラーゼ成分間で相乗関係が観測されている。特に、EG型セルラーゼ及びCBH型セルラーゼは相乗的に相互作用してより効果的にセルロースを分解する。例えば、Wood、1985を参照されたい。
【0007】
従来からセルラーゼ組成物について記載されているが、家庭用洗剤、ストーンウォッシュ組成物または洗濯用洗剤等に使用するための新規で改善されたセルラーゼ組成物の必要性が依然としてある。改善された性能を示すセルラーゼは特に関心が高い。
【0008】
発明の概要
本発明の目的は、洗剤、織物処理、バイオマス転換及びパルプ及び紙製造への使用に有益な性質を有する新規なセルラーゼを提供することである。
【0009】
本発明の目的は、セルロース分解活性を有するポリペプチド及び当該ポリペプチドをエンコードするポリヌクレオチドを提供することである。当該ポリペプチドは細胞壁材料、例えばセルロース及び/またはヘミセルロースの分解を改善する。また、当該ポリペプチドは、例えばバイオマスなどの植物細胞壁材料の分解に関係するその他の酵素の安定性または活性も改善する。
【0010】
本発明の目的は、新規なセルラーゼ及びその誘導体、当該セルラーゼを生成する方法、及びこのような新規なセルラーゼを含む組成物を提供することである。本発明はさらに、当業界でセルラーゼの添加が有利なものとして認識される、新規なセルラーゼ及びその誘導体の組成物中での使用に関し、例えば、洗剤組成物、有用なセルロース含有布及び繊維などの織物処理での添加剤としての使用、動物飼料添加物としての使用、バイオマス転換、パルプ及び紙の処理及び高フルクトース・コーンシロップまたはエタノールの生成用のデンプン処理への添加剤としての使用である。
【0011】
本発明のさらなる目的は、組換え宿主細胞から異種発現により新規なセルラーゼを生成する方法を提供することである。
【0012】
さらに本発明の目的は、本発明のセルラーゼをエンコードする核酸配列を提供することである。1の側面において、当該核酸及びアミノ酸配列は本発明の新規なセルラーゼ及びセルラーゼ組成物の商業生産を容易にする。
【0013】
さらに本発明の目的は、洗剤、織物処理、飼料補助剤及びパルプ及び紙製造において使用するための優れた性質を有する新規なセルラーゼを提供することである。さらなる側面において、当該セルラーゼはバイオマス転換において有用である。
【0014】
第1の側面において、本発明はmHKcelをエンコードする配列、mHKcel遺伝子コード配列に相補的な配列を有する単離ポリヌクレオチド及び当該ポリヌクレオチドを含む組成物を含む。当該ポリヌクレオチドは、mRNA、DNA、cDNA、ゲノムDNAまたはそれらのアンチセンス類似体である。
【0015】
1の実施態様において、mHKcelポリヌクレオチドは、中から高ストリンジェンシー条件下で配列番号2として表される核酸の相補体にハイブリダイズする単離核酸分子を含み、ここで核酸分子はセルロース結合活性を示すmHKcelポリペプチドをエンコードする。
【0016】
配列番号2として表される配列に少なくとも80%、85%、90%、95%、98%またはそれ以上の配列同一性を有するポリヌクレオチドはmHKcelタンパク質をエンコードできる。特定の実施態様において、ポリヌクレオチドは配列番号2に実質的に同一な配列を含む。本発明はポリヌクレオチド断片、好ましくは少なくとも約15−30のヌクレオチド長さの断片も含む。
【0017】
第2の側面において、新規なセルラーゼまたは誘導体はバチルスから得られる。好ましくは、本発明のセルラーゼは図3(配列番号3)に従うアミノ酸配列、断片またはそれらの誘導体を含み、活性部分に対して90%以上の配列同一性、好ましくは95%以上の配列同一性、及びより好ましくは97%以上の配列同一性を有する。
【0018】
第3の側面において、本発明は、本発明のポリペプチドをエンコードし、適当な宿主中ポリペプチド生成を目的とする1以上の制御配列に動作可能に連結した、ヌクレオチド配列を含む核酸構築体に関する。
【0019】
本発明はさらに、mHKcelまたは断片またはそれらのスプライス変異体をエンコードする核酸配列を含む組換え発現ベクターを提供し、当該核酸配列は選択宿主中タンパク質の発現に効果的な調節因子に動作可能に連結する。関連する側面において、本発明は当該ベクターを含む宿主細胞を含む。
【0020】
第4の側面において、本発明は本発明の核酸構築体を含む組換え発現ベクターに関する。
【0021】
第5の側面において、本発明は本発明の核酸構築体を含む組換え宿主細胞に関する。
【0022】
本発明は、さらに、組換え技術、mHKcelをエンコードする核酸配列を含む原核または真核組換え宿主細胞を当該タンパク質の発現を促進するために効果的な条件下で培養し、続いて宿主細胞または細胞培養基からタンパク質を回収することにより、mHKcelを生成する方法を含む。
【0023】
第6の側面において、本発明は本発明のポリペプチドを生成する方法に関し、当該方法は以下を含む:(a)微生物を培養し、ここで当該微生物は野生型の形態で前記ポリペプチドを生成でき、前記ポリペプチドを生成する工程;及び(b)前記ポリペプチドを回収する工程。
【0024】
第7の側面において、本発明はセルロースをエタノールに変換するために有用な酵素組成物を提供する。好ましい実施態様において、酵素組成物はmHKcelを含む。当該組成物はさらに、エンドグルカナーゼ及び/またはセロビオヒドロラーゼなどの他のセルラーゼ酵素を含んでもよい。当該組成物はmHKcelを豊富に含むことができる。
【0025】
1の実施態様において、本発明は、全微生物集団DNAを周囲から単離し、大腸菌においてゲノムDNAライブラリーを構築し、セルラーゼ活性の発現についてライブラリーをスクリーニングし、セルラーゼ陽性クローン中のセルラーゼ遺伝子を同定し、新規なセルラーゼ酵素をキャラクタライズすることによる、新規な酵素を同定する方法を提供する。
【0026】
さらに、ここに提供するmHKcel核酸及びmHKcelタンパク質を検出する分析方法も本発明の一部である。
【0027】
本発明のさらに別の実施態様において、本発明に従うセルラーゼをエンコードする核酸配列を用いて適当な微生物を形質転換する方法を提供する。前記形質転換微生物から本発明に従うセルラーゼを生成する方法を提供する。
【0028】
本発明のさらなる目的は、ストレプトマイセスに特に効果的な発現ベクターを提供することである。ストレプトマイセスは種々のタンパク質生成のための中間宿主細胞として働き、セルラーゼの発現及び生成について、特に高細胞密度で細胞が成長する場合、バチルス宿主細胞を超える多数の利点を提供する。好ましい発現ベクターは、アクチノプラーネスのグルコースイソメラーゼ遺伝子由来のプロモーター配列など調節ポリヌクレオチド配列、ストレプトマイセスセルラーゼ遺伝子由来のシグナル配列及びセルラーゼ、特に本発明のセルラーゼをエンコードするDNA配列を含む。
【0029】
本発明の好ましい実施態様において、完全長セルラーゼはバチルスから得ることができる。
【0030】
本発明のその他の目的、特徴及び利点は以下の詳細な説明から明らかである。しかしながら、詳細な説明及び具体的な実施例は、本発明の好ましい実施態様を示すが、説明目的のため与えられるものであり、本発明の範囲及び概念内で種々の変化及び修正がこの詳細な説明から当業者に明らかであることが当然に理解される。
【0031】
発明の詳細な説明
本発明を以下の定義及び実施例を用いて参照することにより詳細を説明する。ここに参照する全ての特許及び刊行物、及び当該特許及び刊行物内に開示される全ての配列は明示的に引用するものとする。
【0032】
特段に示す場合を除いて、ここで用いる全ての技術及び科学用語は本発明の属する技術の当業者にとって理解される意味と同じ意味を有する。Singleton、他、Dictionary of Microbiology and Molecular Biology(微生物学及び分子生物学の辞典)、第2版、John Wiley and Sons、New York(1994)、及びHale&Marham,The Harper Collins Dictionary of Biology,Harper Perennial,NY(1991)は当業者に本発明で用いる多くの用語の一般的な辞書を提供するものである。ここに記載の方法及び物質に類似または同等のいかなる方法及び物質も本発明を実施及び試験する際に用いることができるが、好ましい方法及び物質を記載する。数値範囲は範囲を定める数字を含める。別に示す場合を除いて、それぞれ核酸は左から右に5’〜3’方向;アミノ酸配列は左から右にアミノからカルボキシ方向に記載する。実務者は特にSambrook et al.、1989、及びAusubel FM et al.、1993を当業界の定義及び用語のために参照できる。本発明は記載された特定の方法、手順及び試薬に限定されないことは、これらが種々多様であることから当然に理解される。
【0033】
ここに提供する表題は本発明の種々の側面または実施態様を限定するものではなく、明細書を全体として参照することにより理解できるものである。従って、下記に定義する用語は明細書全体を参照することにより、さらに十分に定義される。
【0034】
ここに引用する全ての文献は、本発明に関連し得る組成物及び方法を記載及び開示する目的のため参照することによりここに明示的に引用する。
【0035】
定義
“セルラーゼ”、“セルロース分解酵素”、または“セルラーゼ酵素”は、ここに記載の本発明の細菌性エンドグルカナーゼを意味する。これらの異なる種類のセルラーゼ酵素はセルロース及びその誘導体をグルコースに変換するために相乗的に作用する。
【0036】
“セルラーゼ”の語はセルロースポリマーをより短いセロ−オリゴ糖オリゴマー、セロビオース及び/またはグルコースに加水分解できる酵素分類をいう。多数のセルラーゼの例として、例えば、エキソグルカナーゼ、エキソセロビオヒドロラーゼ、エンドグルカナーゼ及びグルコシダーゼがあり、セルロース分解性生物、特に真菌及び細菌から得られる。これらの微生物から作られる酵素は、セルロースからグルコースへの転換に有用な3種類の作用を有するタンパク質の混合物である:エンドグルカナーゼ(EG)、セロビオヒドロラーゼ(CBH)、及びβ−グルコシダーゼ。これらの3種の異なるセルラーゼ酵素はセルロース及びその誘導体をグルコースに変換するために相乗的に作用する。
【0037】
多くの微生物はセルロースを加水分解する酵素を作り、例えば、木材腐朽菌トリコデルマ、土壌細菌Thermomonospora、バチルス、及びセルロモナス;ストレプトマイセス;及び真菌フミコーラ、アルペルギルス及びフサリウムなどがある。
【0038】
“宿主細胞”の語はベクターを含み、複製及び/または転写または発現構築体の転写及び翻訳(発現)を助ける細胞を意味する。本発明で用いる宿主細胞は大腸菌などの原核細胞または酵母などの真核細胞、植物、昆虫、両生類または哺乳類細胞などの細胞である。本発明の1の実施態様において、“宿主細胞”はバチルス属の細胞を意味する。本発明の他の好ましい実施態様において、“宿主細胞”はストレプトマイセスの細胞を意味する。ストレプトマイセスは、Buchanan他、The Shorter Bergey’s Manual For Determinative Bacteriology(Williams&Wilkens 1982)において分類されるストレプトマイセス属のひとつである任意の細菌株を意味する。特に好ましいストレプトマイセス株はS.lividens、S.rubiginosus、及びS.corlicolorである。S.lividensはLomovskaya、他、J.Virology 9:258(1972)に記載されている。しかしながら、当業者であれば任意の適当な宿主細胞、例えば、細菌、真菌、真核及び植物細胞が使用できることが理解できる。
【0039】
細胞、または核酸、タンパク質、またはベクターに関して用いる“組換え体”の語は、該細胞、核酸、タンパク質またはベクターが、異種核酸またはタンパク質の挿入により、または天然核酸またはタンパク質の変異により修飾されたこと、または該細胞がそのように修飾された細胞由来であることを示す。従って、例えば、組換え細胞は当該細胞の天然(非組換え)型内では見られない遺伝子を発現し、または発現、または全く発現しない条件下で、通常は異常発現される天然遺伝子を発現する。
【0040】
“分泌シグナル配列”の語は、大ポリペプチドの構成要素として、ポリペプチドが合成される細胞の分泌経路から当該大ポリペプチドに導かれるポリペプチド(“分泌ペプチド”または“分泌シグナルペプチド”)をエンコードするDNA配列を示す。当該大ペプチドは普通は分割されて、分泌経路を通過する際に分泌ペプチドが取り除かれ、分泌シグナルペプチド及び通常、成熟ポリペプチドと呼ばれるより小さいペプチドを生じる。
【0041】
ここで用いる、“セルラーゼ調製物全体”及び“セルラーゼ組成物全体”の語句は交換可能に用いられ、天然及び非天然組成物の両方をいう。“天然”組成物は天然源により生成した組成物であり、例えば、1以上のセロビオヒドロラーゼ型、1以上のエンドグルカナーゼ型、及び1以上のβ−グルコシダーゼ成分を含み、これらの各成分は供給源より生じた比率で見られる。特定の真菌は、エキソセロビオヒドロラーゼまたはCBH型セルラーゼ、エンドグルカナーゼまたはEG型セルラーゼ及びβ−グルコシダーゼまたはBG型セルラーゼを含む完全なセルラーゼ系を生じる(Schulein、1988)。しかしながら、これらの系はCBH型セルラーゼを欠く場合があり、また、細菌性セルラーゼは通常CBH型セルラーゼを含まないか、またはほとんど含まない。天然組成物はセルロース分解酵素に関して非修飾の微生物により生成されたものであり、従って当該成分酵素の比率は天然微生物により生成されるものから変更されていない。
【0042】
“非天然”組成物は、(1)天然比率または非天然、すなわち変更された比率で成分をセルロース分解酵素と混合;または(2)1以上のセルロース分解酵素を過剰発現または過少発現するように微生物を修飾;または(3)少なくとも1のセルロース分解酵素が欠失するように微生物を修飾;または(4)異種成分セルロース分解酵素を発現するように微生物を修飾することにより生成したこれらの組成物を包含する。
【0043】
ここで用いる、“プロモーター”の語は下流遺伝子の転写を導くように作用する核酸配列をいう。プロモーターは通常、標的遺伝子が発現する宿主細胞に適したものである。プロモーターはその他の転写及び翻訳調節核酸配列(“制御配列”とも呼ぶ)と一緒に所定の遺伝子を発現するために必要である。通常、転写及び翻訳調節配列は、限定されないが、プロモーター配列、リボソーム結合部位、転写開始及び停止配列、翻訳開始及び停止配列及びエンハンサーまたは活性化配列を含む。プロモーターは通常、下流遺伝子に関連するプロモーターまたは異種のものであり、すなわち、遺伝子に対して機能する限り他の遺伝子または他の微生物由来のものであってもよい。形質転換宿主細胞がバチルスの場合の好ましいプロモーターはaprEプロモーターである。1の側面において、プロモーターは導入可能プロモーターである。1の側面において、宿主細胞は糸状菌であり、プロモーターはアクセッション番号D86235でジェンバンク(GenBank)に寄託されているT.リーゼイ(reesei)cbh1プロモーターである。他の側面において、プロモーターはT.リーゼイ由来のcbhIIまたはキシラナーゼプロモーターである。
【0044】
核酸は、他の核酸配列と機能的な関係に位置する場合、“動作可能に連結”する。例えば、分泌リーダー(leader)をエンコードするDNA、すなわちシグナルペプチドは、ポリペプチドの分泌に関与するプレタンパク質として発現する場合、ポリペプチドのDNAに動作可能に連結し;プロモーターまたはエンハンサーは、配列の転写に影響する場合、コード配列に動作可能に連結し;またはリボソーム結合部位は、翻訳を促進するために位置する場合、コード配列に動作可能に連結する。通常、“動作可能に連結する”とは、連結DNA配列が隣接しており、及び分泌リーダーの場合、隣接し、かつリーディングフレーム内にあることを意味する。しかし、エンハンサーは隣接している必要はない。連結は都合のよい制限部位で連結反応により達成される。そのような部位が存在しない場合、合成オリゴヌクレオチド・アダプタまたはリンカーを従来実務に従って用いる。
【0045】
“DNA構築体”または“DNAベクター”とは、新規なセルラーゼをエンコードする1以上のDNA断片を含むヌクレオチド配列を意味する。“DNAベクター”には“発現ベクター”が含まれる。通常の発現ベクターは調節配列を含み、例えば、転写及び翻訳ターミネーター、転写及び翻訳開始配列、シグナル配列、及び特定の核酸の発現の調製に有用なプロモーターである。“プロモーター”の語は、タンパク質をエンコードするポリヌクレオチド配列の転写開始の制御に関係するポリヌクレオチド配列を指す通常の意味において用いられる。“シグナル配列”は、所望のタンパク質の宿主からの輸送を生物活性形態で導くことが可能なタンパク質のシグナルペプチドまたは部分をいう。細胞外タンパク質の成熟形態は分泌プロセス中開裂されるシグナル配列を欠く。本発明を限定する趣旨ではないが、シグナルペプチド中のアミノ酸残基の数は約5〜約100アミノ酸残基がよい。シグナル配列は、DNAの連結またはセルラーゼをエンコードするDNAの挿入を可能にするクローニング部位を提供するように修飾できる。ベクターは任意で、少なくとも1の独立ターミネーター配列、原核、真核または両方(例えば、シャトルベクター)においてカセットの複製を可能にする配列、及び原核及び真核系両方用の選択マーカーを含むジェネリック発現カセットを含む。ベクターは原核、真核または両方において複製及び統合するために適している。Giliman and Smith,Gene 8:81−97(1979);Robert、他、Nature 328:731−734(1987);Berger and Kimmel,Guide to Molecular Cloning Techniques,Methods In Enzymology,Vol 152、Academic Press,Inc.,サンディエゴ、カリフォルニア州(“Berger”);Scheider,B.et al.,Protein Expr.Purif 6435:10(1995);Sambrook、他、Molecular Cloning−A Laboratory Manual(第2版)Vol.1−3、Cold Springs Harbor Publishing(1989)(“Sambrook”);及びCurrent Protocols In Molecular Biology,Ausubel、他、(編集)、Current Protocols,Greene Publishing Associates,Inc.とJohn Wiley&Sons,Incとの共同事業(1997、補足)(“Ausubel”)を参照されたい。ストレプトマイセスに有用なクローニングベクターは公知であり、米国特許第4,338,397号、第4,411,994号、第4,513,085号、第4,513,086号、第4,745,056号、第5,514,590号及び第5,622,866号及びWO88/07079を参照できる。
【0046】
ここで用いる“遺伝子”の語は、ポリペプチド鎖の生成に関与するDNA断片を意味し、コード領域の先行及び後の領域、例えば5’非翻訳(5’UTR)または“リーダー”配列及び3’UTRまたは“トレーラー”配列、及び個々のコード断片(エキソン)間の介在配列(イントロン)などを含んでも含まなくてもよい。
【0047】
核酸の一部に言及して用いる“異種”の語は、当該核酸が、通常は同じ関係において天然では見られない2以上のサブ配列を含むことを示す。例えば、当該核酸は通常、遺伝子組換えにより生成し、例えば新しい機能的核酸を作るために配置した関連性のない遺伝子から2以上の配列を有し、例えば、1の供給源からプロモーターを有し、他の供給源からコード領域を有する。同様に、異種タンパク質は同じ関係でお互いに天然では見られない2以上のサブ配列をいうことも多い(例えば、融合タンパク質)。
【0048】
ここで用いる“単離”または“精製”の語は、天然に結合する少なくとも1の成分から除去された核酸またはアミノ酸をいう。
【0049】
本発明の内容において、“実質的に純粋なポリペプチド”とは、天然に関連するその他のポリペプチド物質を多くとも10重量%で含む、ポリペプチド調製を意味する(その他のポリペプチド物質のより低い割合は、好ましくは、例えば、多くとも8重量%、多くとも6重量%、多くとも5重量%、多くとも4重量%、多くとも3重量%、多くとも2重量%、多くとも1重量%、及び多くとも1/2重量%である)。従って、実質的に純粋なポリペプチドは少なくとも92%純粋であることが好ましく、すなわち、当該ポリペプチドは調製剤中に存在する少なくとも92重量%の合計ポリペプチド物質を構成し、及びより高い割合は好ましく、例えば、少なくとも94%純粋、少なくとも95%純粋、少なくとも96%純粋、少なくとも96%純粋、少なくとも97%純粋、少なくとも98%純粋、少なくとも99%純粋及び多くとも99.5%純粋である。ここに開示するポリペプチドは好ましくは実質的に純粋な形態である。特に、ここに開示するポリペプチドが“本質的に純粋形態”であることが好ましく、すなわち、ポリペプチド調製物が天然に関連するその他のポリペプチド物質を本質的に含まないことが好ましい。これは例えば、公知の組換え方法手段によりポリペプチドを調製することにより達成できる。ここで、“実質的に純粋なポリペプチド”の語は、“単離ポリペプチド”及び“単離形態のポリペプチド”と同義である。
【0050】
通常、mHKcelをエンコードする核酸分子を中から高ストリンジェンシー条件下で配列番号2(mHKcel)としてここに提供する配列に対してハイブリダイズする。しかしながら、場合によっては、実質的に異なるコドン使用を有するmHKcel−エンコードヌクレオチド配列を用いるが、当該mHKcel−エンコードヌクレオチド配列によりエンコードされたタンパク質は天然タンパク質と同じまたは実質的に同じアミノ酸配列を有する。例えば、コード配列は特定の原核または真核発現系においてmHKcelのより速い発現を促進するために、宿主によって利用される特定のコドン頻度に従って修飾できる。例えば、Te’o、他(2000)は糸状菌における遺伝子発現の最適化について説明している。
【0051】
核酸配列は、2つの配列が特異的に中程度から高ストリンジェンシーなハイブリダイゼーション及び洗浄条件下でお互いにハイブリダイズする場合、対照核酸配列に“選択的にハイブリダイズ”すると考えられる。ハイブリダイゼーション条件は核酸結合複合体またはプローブの融点(Tm)に基づく。例えば、“最大ストリンジェンシー”は通常、約Tm−5℃で起こり(プローブのTmより5℃低い);“高ストリンジェンシー”はTmより約5−10℃低く;“中間”または“中ストリンジェンシー”はプローブのTmより約10−20℃低く;及び“低ストリンジェンシー”はTmより約20−25℃低い。機能的に、最大ストリンジェンシー条件はハイブリダイゼーションプローブと厳密に同一またはほぼ厳密に同一な配列を同定するために用いることができ、高ストリンジェンシー条件はプローブと約80%以上の配列同一性を有する配列を同定するために用いる。
【0052】
中から高ストリンジェンシーハイブリダイゼーション条件は当業界に公知である(例えば、明示的にここに引用するSambrook、他、1989、第9章及び11章、及びAusubel、他、1993を参照)。高ストリンジェンシー条件の例としては、50%ホルムアミド、5X SSC、5X Denhardt’s溶液、0.5% SDS及び100μg/ml変性キャリアDNA中、約42℃でのハイブリダイゼーション、及び続いて2X SSC及び0.5% SDS中、室温で2回洗浄及びさらに0.1X SSC及び0.5% SDS、42℃で2回洗浄を含む。
【0053】
ここで用いる細胞に関する“形質転換”、“安定に形質転換された”または“トランスジェニック”の語は、細胞が、ゲノム内に統合された、または複数の世代を通して維持されるエピソームプラスミドとして非天然(異種)核酸配列を有することを意味する。
【0054】
ここで用いる“発現”の語は、ポリペプチドが遺伝子の核酸配列に基づいて生成されるプロセスをいう。当該プロセスは転写及び翻訳の両方を含む。
【0055】
細胞への核酸配列の挿入に関する“導入”の語は“トランスフェクション”または“形質転換”または“形質導入”を意味し、原核または真核細胞内への核酸配列の組込みに関する場合を含み、ここで核酸配列は細胞のゲノム内(例えば、染色体、プラスミド、色素体またはミトコンドリアDNA)に組み込まれることができ、自己レプリコンに変換され、または一時的に発現される(例えば、トランスフェクトされたmRNA)。
【0056】
“mHKcel発現”の語はmHKcelセルラーゼ遺伝子の転写及び翻訳、前駆体RNA、mRNA、ポリペプチド、翻訳後プロセッシングポリペプチドの生成物ということになる。例として、mHKcel発現の分析は、mHKcelタンパク質のウエスタンブロット、CBH1 mRNAのノーザンブロット分析及び逆転写ポリメラーゼ連鎖反応(RT−PCR)分析、及びShoemaker S.P.及びBrown R.D.Jr.(Biochem.Biophys.Acta,1978,523:133−146)及びSchulein(1988)に記載のエンドグルカナーゼ活性分析を含む。
【0057】
ここで用いる“界面活性剤”の語は、表面活性特性を有するものとして当業界で通常認識されている任意の化合物をいう。従って、例えば、界面活性剤は陰イオン性、陽イオン性及び非イオン性界面活性剤を含み、例えば洗剤において通常見られるものである。陰イオン性界面活性剤は、直鎖または分岐鎖アルキルベンゼンスルフォネート;直鎖または分岐鎖アルキル基またはアルケニル基を有するアルキルまたはアルケニルエーテル硫酸塩;アルキルまたはアルケニル硫酸塩;オレフィンスルフォネート;及びアルカンスルフォネートが挙げられる。両性界面活性剤は第4級アンモニウム塩スルフォネート及びベタイン型両性界面活性剤が挙げられる。当該両性界面活性剤は同じ分子内に正及び負電荷の両方の基を有する。非イオン性界面活性剤はポリオキシアルキレンエーテル及び高脂肪酸アルカノールアミドまたはその酸化アルキレン付加、脂肪酸グリセリンモノエステル等を含むことができる。
【0058】
ここで用いる“セルロース含有繊維”の語は、セルロースを含有する綿または非綿、またはセルロースブレンド、例えば天然セルロース誘導体及び人工セルロース誘導体(例えば、ジュート(黄麻)、亜麻布、ラミー、レーヨン及びリヨセル)を含有する綿または非綿から作られた任意の織布または不織布、糸または繊維をいう。
【0059】
ここで用いる、“綿含有繊維”の語は、純粋な綿または綿ブレンドから作られた織布または不織布、糸または繊維をいい、綿織物、綿ニット、綿デニム、綿糸、原綿等を含む。
【0060】
ここで用いる、“ストーンウォッシュ組成物”の語は、セルロース含有繊維をストーンウォッシュする際に用いる製剤をいう。ストーンウォッシュ組成物は販売前、すなわち製造プロセス時にセルロース含有繊維を修正するために用いる。これに対し、洗剤組成物は汚れた衣類の洗浄を目的とするものであり、製造プロセス時には用いられない。
【0061】
ここで用いる“洗剤組成物”の語は汚れたセルロース含有繊維を洗濯するための洗浄手段で用いることを意図した混合物をいう。本発明において、当該組成物はセルラーゼ及び界面活性剤に加えて、さらに加水分解酵素、ビルダー、漂白剤、漂白活性化剤、青味剤及び蛍光色素、ケーキング防止剤、マスキング剤、セルラーゼ活性化剤、酸化防止剤及び可溶化剤を含むことができる。
【0062】
ここで用いる“活性”及び“生物活性”の語は、特定タンパク質に関する生物活性をいい、ここで交換可能に用いられる。例えばプロテアーゼに関する酵素活性はタンパク質分解性であり、従って、プロテアーゼはタンパク質分解活性を有する。従って、所定タンパク質の生物活性は通常、当業者とって当該タンパク質に起因する任意の生物活性をいうことになる。
【0063】
酵素溶液中で用いる場合、mHKcel成分は通常、バイオマスからの可溶性糖の放出速度を最高にするために十分な量で加える。加えるmHKcel成分の量は、糖化されるバイオマスの種類に依存し、当業者により容易に測定できる。しかしながら、使用時の、セルラーゼ組成物中に存在するその他のセルラーゼ型成分と比較した、mHKcel成分の重量%は、好ましくは約1、好ましくは約5、好ましくは約10、好ましくは約15、または好ましくは約20重量%〜好ましくは約25、好ましくは約30、好ましくは約35、好ましくは約40、好ましくは約45、または好ましくは約50重量%である。さらに、好ましい範囲は約0.5〜約15重量%、約0.5〜約20重量%、約1〜約10重量%、約1〜約15重量%、約1〜約20重量%、約1〜約25重量%、約5〜約20重量%、約5〜約25重量%、約5〜約30重量%、約5〜約35重量%、約5〜約40重量%、約5〜約45重量%、約5〜約50重量%、約10〜約20重量%、約10〜約25重量%、約10〜約30重量%、約10〜約35重量%、約10〜約40重量%、約10〜約45重量%、約10〜約50重量%、約15〜約20重量%、約15〜約25重量%、約15〜約30重量%、約15〜約35重量%、約15〜約40重量%、約15〜約45重量%、約15〜約50重量%である。
【0064】
II.分子生物学
本発明は組換え遺伝学の分野における一般的な技術に依拠する。本発明で使用する一般的な方法を開示する基本テキストは、Sambrook、他、Molecular Cloning(分子クローニング)、A Laboratory Manual(第2版、1989);Kriegler、Gene Transfer and Expression(遺伝子導入及び発現):A Laboratory Manual(1990);及びAusubel、他、編集。Current Protocols in Molecular Biology(1994)が挙げられる。
【0065】
高レベル発現でクローン遺伝子を得るために、異種遺伝子はプロモーターからの距離が、好ましくは天然セルラーゼ遺伝子内の場合とほぼ同じ距離で位置する。当業界に公知の通り、しかしながら、ある程度の距離の変動はプロモーター機能を損なうことなく適応させることができる。
【0066】
天然プロモーターを1以上のヌクレオチドの置換、付加または欠失により、その機能を変化させることなく修飾できることは当業者であれば知っていることである。本発明の実施はプロモーターに対するそのような変異を包含するが、それに拘束されない。
【0067】
発現ベクター/構築体は通常、異種配列の発現に必要なその他全ての要素を含む転写ユニットまたは発現カセットを含む。従って、一般的な発現カセットは異種核酸配列に動作可能に連結するプロモーター及び転写、リボソーム結合部位及び翻訳停止の効率的なポリアデニル化に必要なシグナルを含む。当該カセットの他の要素は、ゲノムDNAを構造遺伝子、機能的スプライス供与体及び受容体部位を含むイントロンとして用いる場合、エンハンサーを含んでもよい。
【0068】
本発明の実施は、遺伝子構築体中のプロモーター選択により制限されない。プロモーター選択についての唯一の制限は、使用される宿主細胞において機能的であることである。形質転換宿主細胞がバチルスである場合の好ましいプロモーターは、aprEプロモーターである。
【0069】
プロモーター配列に加え、発現カセットは、効果的に停止できるように構造遺伝子の下流に転写停止領域も含むべきである。停止領域はプロモーター配列と同じ遺伝子から得られ、または異なる遺伝子から得ることもできる。
【0070】
遺伝子情報を細胞内に輸送するために用いる特定の発現ベクターは特に重要ではない。真核または原核細胞内で発現するために使用される任意の従来ベクターが使用できる。標準的な細菌性発現ベクターは、バクテリオファージλ及びM13、及びプラスミド、例えばpBR322ベースプラスミド、pSKF、pET23D、及び融合発現系、例えばMBP、GST及びLacZが挙げられる。エピトープ標識も単離方法を容易にするために組換えタンパク質に加えることができ、例えば、c−mycである。
【0071】
また、発現ベクター内に通常含まれる要素は、レプリコン、組換え体プラスミドを含む細菌選択を可能にする抗生物質耐性をエンコードする遺伝子、及び異種配列の挿入を可能にするプラスミドの非必須領域における固有制限部位も含む。特定の抗生物質耐性遺伝子の選択は重要でなく、当業者に公知の任意の多くの耐性遺伝子が適している。
【0072】
本発明の形質転換方法は、形質転換ベクターの全部または一部を糸状菌のゲノム内に安定して統合することができる。しかしながら、自己複製の細胞外形質転換ベクターの維持を生じる形質転換も考えられる。
【0073】
本発明のセルラーゼをエンコードする遺伝子はλ−ファージ(発現)ベクター及び大腸菌宿主細胞を用いてクローンできる。(もしくは、保存ドメイン上で設計されたコンセンサスプライマーを用いるPCRクローニングも使用できる。)出願人は、本発明のセルラーゼをエンコードする遺伝子の形質転換及び大腸菌内での発現により、活性タンパク質を生じることを見出した。大腸菌内での第1のクローニング工程後、本発明のセルラーゼ遺伝子はバチルスまたはストレプトマイセス種などのより好ましい工業用発現宿主、アスペルギルスまたはトリコデルマなどの糸状菌、またはサッカロマイセスなどの酵母に移動できる。これらの宿主微生物内で得られる高レベル発現及び分泌により、発酵培地中、セルラーゼの蓄積が可能となり、その後その培地から回収できる。
【0074】
プロテアーゼ欠失バチルス株の好ましい一般的な形質転換及び発現手順は、ここに引用するFerrari、他、米国特許第5,264,366号に開示されている。アスペルギルスにおける形質転換及び発現は、例えば、ここに引用する、Berka、他、米国特許第5,364,700号に記載されている。
【0075】
多くの標準的なトランスフェクション方法が、異種タンパク質を大量に発現するトリコデルマ・リーゼイ細胞株を生成するために使用できる。DNA構築体をトリコデルマのセルラーゼ生成株内に導入するための開示方法のいくつかは、Lorito,Hayes,DiPietro及びHarman,1993、Curr.Genet.24:349−356;Goldman,VanMontagu及びHerreta−Estrella,1990,Curr.Genet.17:169−174;Penttila,Nevalainen,Ratto,Salminen及びKnowles,1987,Gene 6:155−164、アスペルギルスについて、Yelton,Hamer及びTimberlake,1984,Proc.Natl.Acad.Sci.USA 81:1470−1474、フサリウム・Bajar,Podila及びKolattukudy,1991、Proc.Natl.Acad.Sci.USA 88:8202−8212、ストレプトマイセスについて、Hopwood、他、1985、The John Innes Foundation,Norwich,UK及びバチルスについて、Brigidi,DeRossi,Bertarni,Riccardi及びMatteuzzi,1990,FEMS Microbiol.Lett.55:135−138)が挙げられ、全てここに引用する。
【0076】
しかしながら、外来ヌクレオチド配列を宿主細胞に導入する任意の公知手段を用いることができる。これらは、リン酸カルシウムトランスフェクション、ポリブレン、原形質融合、電気泳動法、バイオリスティック、リポソーム、マイクロインジェクション、プラズマベクター、ウイルスベクター及びクローンゲノムDNA、cDNA、合成DNAまたはその他の外来ゲノム物質を宿主細胞内に導入するその他の公知方法の使用を含む(例えば、Sambrook.他、上記を参照)。また、米国特許第6,255,115号に記載のアグロバクテリウム媒介トランスフェクション方法も使用される。必要なのは、使用する特定の遺伝子工学手順が少なくとも1の遺伝子を異種遺伝子を発現できる宿主細胞内にうまく導入できるということだけである。
【0077】
発現ベクターが細胞内に導入された後、トランスフェクト細胞はセルラーゼ遺伝子プロモーター配列の制御下で遺伝子発現に有利な条件の下、培養される。形質転換細胞の大バッチは以下に記載の通り培養できる。最終的に、生成物は標準技術を用いて培養基から回収する。
【0078】
従って、本発明は本発明のセルラーゼの発現及び高められた分泌を提供し、発現は天然セルラーゼ遺伝子、融合DNA配列及び種々の異種構築体を含むセルラーゼ遺伝子プロモーター配列に制御される。また、本発明は本発明のセルラーゼを高レベルで発現及び分泌する方法も提供する。
【0079】
III.核酸及びエンコードされたタンパク質配列の同定
バチルス・agaradhaerans(DSM8721)由来のゲノムライブラリーを当業者に公知の標準技術を用いて調製した。この微生物はアルカリ性セルラーゼ(エンド−1,4−ベータ−グルカナーゼ)を生成し、これはグリコシル・ヒドロラーゼ、エンドグルカナーゼ5A、EC3.2.1.4、Swiss−Prot:085465、エントリー名GUN5_BACAG.EBIアクセッション番号(AF067428)のセルラーゼファミリー5に属し、その遺伝子は1203bp長である(Davies、他、1998)。セルラーゼ陽性クローンはプレート分析において発生率1/3000で検出された。本発明の側面に従う遺伝子を単離する方法において、この酵素のコード配列に基づく変性プライマーを用いた。しかしながら、意外なことに、公知のB.agaradhaeransセルラーゼを増幅することが知られるプライマーを用いてもPCR生成物は得られなかった。従って、セルラーゼをコードする挿入の完全な配列はプライマーウォーキングにより測定した。
【0080】
本発明の第2の側面に従う遺伝子を単離する方法は、配列リストに示す配列番号2のヌクレオチド配列の全部または一部を含むヌクレオチド配列に対する相同性を利用する。そのような方法の例としては以下を含む:
a)mHKcel遺伝子を推定上含む遺伝子ライブラリーをプローブとして当該ヌクレオチド配列を用いてスクリーニングする工程。
【0081】
b)ヌクレオチド配列情報に基づいてプライマーを調製し、次にテンプレートとしてmHKcel遺伝子を推定上含むサンプルを用いてPCRを行う工程。
【0082】
より具体的には、上記a)工程は、a)セルラーゼ遺伝子を推定上含む遺伝子ライブラリーを調製し、配列リストに示す配列番号2のヌクレオチド配列の全部または一部を含むヌクレオチド配列を用いて遺伝子ライブラリーをスクリーニングし、当該遺伝子ライブラリー由来の配列リストに示す配列番号2のヌクレオチド配列の全部または一部を含むヌクレオチド配列とハイブリダイズする配列を選択し、次に、選択された配列を単離し、前記遺伝子ライブラリーから選択及び単離された配列由来のmHKcel遺伝子を単離する工程、を含む。
【0083】
遺伝子ライブラリーはゲノムDNAライブラリーまたはcDNAライブラリーであってもよく、公知の手順に従って調製できる。
【0084】
IV.タンパク質発現
本発明のタンパク質は、発現がプロモーター配列に制御される本発明のセルラーゼ遺伝子を含む発現ベクターを用いて形質転換された細胞を培養することにより生成される。本発明は特にタンパク質の細胞内及び/または細胞外での生成を高めるために有用である。タンパク質は同種または異種である。
【0085】
また、本発明のタンパク質は、他の、異種ポリペプチドまたはアミノ酸配列に融合された目的タンパク質を含むキメラ分子を形成するための方法で修飾できる。1の実施態様において、このようなキメラ分子は目的タンパク質と抗標識抗体が選択的に結合できるエピトープを備える標識ポリペプチドとの融合を含む。エピトープ標識は通常、目的タンパク質のアミノ−、またはカルボキシ末端に位置する。
【0086】
種々の標識ポリペプチド及びそれぞれの抗体は当業界で公知である。例としては、ポリ−ヒスチジン(poly−his)またはポリ−ヒスチジン−グリシン(poly−his−gly)標識;HIS6及び金属キレート標識、flu HA標識ポリペプチド及びその抗体12CA5(Field、他、Mol.Cell.Biol.8:2159−2165(1988));c−myc標識及びその8F9、3C7、6E10、G4、B7及び9E10抗体(Evan、他、Molecular and Cellular Biology 5:3610−3616(1985));及び単純ヘルペスウイルス性糖タンパク質D(gD)標識及びその抗体(Paborsky、他、Ptotein Engineering 3(6):547−553(1990))が挙げられる。その他の標識ポリペプチドは、FLAG−ペプチド(Hopp、他、Bio Technology 6:1204−1210(1988));KT3エピトープペプチド(Martin、他、Science 255:192−194(1992);チューブリン・エピトープペプチド(Skinner、他、J.Biol.Chem.266:15163−15166(1991));及びT7遺伝子10タンパク質ペプチド標識(Lutz−Freyermuth、他、Proc.Natl.Acad.Sci.USA 87:6393−6397(1990))が挙げられる。
【0087】
前記mHKcel遺伝子の発現に適当な条件は、本発明のセルラーゼの成長及び/または発現に必要な成分を培養物に提供する工程を含む。タンパク質の生成に最適な条件は宿主細胞の選択に伴って、及び発現するタンパク質の選択に伴って変化する。そのような条件は、一般的な実験または最適化により当業者により容易に確かめられる。
【0088】
目的タンパク質は通常、発現後に精製または単離される。目的タンパク質は、サンプル中に存在する他の成分に応じて、当業者に公知の種々の方法で単離または精製できる。標準精製方法は電気泳動、分子、免疫学及びクロマトグラフィー技術を含み、イオン交換、疎水性、親和性及び逆相HPLCクロマトグラフィー及びクロマト分画を含む。例えば、目的タンパク質は目的抗体カラムの標準抗タンパク質を用いて精製できる。また限外ろ過及びダイアフィルトレーション技術も、タンパク濃度と併せて、有用である。適当な精製技術に関する一般的な手引きは、Scopes,Ptotein Purification(1982)を参照されたい。必要な精製の程度は目的タンパク質の使用に応じて変化する。時には精製が必要ない場合もある。
【0089】
V.セルラーゼの有用性
本発明に従う織物処理は、本発明のセルラーゼを含む組成物を用いた織物加工または織物洗浄が考えられる。そのような処理は、限定されないが、ストーンウォッシュ、セルロース含有繊維の生地、手触り及び/または外観の修正、またはセルロース含有繊維の製造または洗浄/修繕時に用いられるその他の技術を含む。さらに、本発明の文脈中の処理とは、セルロース系布または繊維から“未成熟”綿または“死”綿を取除くことも考えられる。未成熟綿は成熟綿よりも著しくアモルファスが多く、例えば不規則な染色を原因とする。本発明の組成物は、汚れた製造時のセルロース含有布を洗浄する際に使用するセルラーゼ成分をさらに含む。例えば、本発明のセルラーゼは洗濯物を洗う洗剤組成物に使用できる。本発明に有用な洗剤組成物は前洗浄用、前含浸用(pre−soak)、及び家庭用色修復組成物などの特定の剤形を含む。そのような処理組成物はここに記載するように、希釈が必要な濃縮形態または希釈溶液形態またはセルロース含有布に直接適用できる形態でもよい。織物をセルラーゼ処理するための一般的な処理技術は例えば、欧州出願第220 016及びGB出願第1,368,599号及び2,095,275号に記載されている。
【0090】
本発明に従うセルロース系材料の処理は、当業界に公知の目的に関して動物飼料、パルプ及び/または紙、食品及び穀物を処理することがさらに考えられる。例えば、セルラーゼは動物飼料の価値を増加させることが知られ、また、木材パルプの水切れ能を改善し、食品製造を高め、穀物湿式粉砕法または乾式製粉法時に穀物中の繊維を減少することが知られる。
【0091】
本発明に従う処理は、効果的な量でセルラーゼまたはセルラーゼと一緒にその他の任意の成分、例えば緩衝液、界面活性剤、及び/または精練剤との組み合わせを含む水溶液を調製することを含む。効果的な量のセルラーゼ酵素組成物とは、意図した目的のために十分なセルラーゼ酵素の濃度である。従って、例えば、本発明に従うストーンウオッシュ組成物における“効果的な量”のセルラーゼは、所望の効果を与える量であり、例えば、縫い目及びはぎ布上の擦り切れた及び色あせた外観を生じる。同様に、セルロース−含有布の手触り及び/または外観の改善を意図した組成物中のセルラーゼの“効果的な量”は、適度に手触りの改善を生じる量であり、例えば、布の滑らかさまたは外観の改善、例えば、布の外観の鮮明さを減少する傾向にある毛玉及び繊維を取除くことである。また、使用するセルラーゼの量は、使用する設備、使用するプロセス・パラメーター(セルラーゼ処理溶液の温度、セルラーゼ溶液への曝露時間等)及びセルラーゼ活性(例えば、特定の溶液は低濃度のセルラーゼが必要であり、より活性の低いセルラーゼ組成物と比較して、より活性の高いセルラーゼ組成物を使用する)に依存する。処理布に添加する処理水溶液中の正確なセルラーゼ濃度は上記の要件及び所望の結果に基づいて当業者により容易に測定できる。ストーンウォッシュ工程において、通常、セルラーゼの濃度は処理水溶液中、約0.5〜5,000ppmが好ましく、及び最も好ましくは約10〜200ppm合計タンパク質である。セルロース含有布の手触り及び/または外観を改善するための組成物において、処理水溶液中に存在するセルラーゼ濃度は、約0.1〜2000ppmが好ましく、及び最も好ましくは約0.5〜200ppm合計タンパク質である。
【0092】
好ましい処理実施態様において、緩衝液は処理組成物中で、使用するセルラーゼが活性を示す範囲内で緩衝液の濃度が溶液のpHを維持するのに十分な程度で用いる。セルラーゼが活性を示すpHは用いるセルラーゼの性質に依存する。使用する緩衝液の正確な濃度は当業者が用意に考慮に入れることができるいくつかの要素に依存する。例えば、好ましい実施態様において、緩衝液及び緩衝液濃度は、最適なセルラーゼ活性に必要なpHの範囲内で最終的なセルラーゼ溶液のpHを維持するために選択される。本発明のセルラーゼの最適なpH範囲の決定は公知の技術に従って確認できる。セルラーゼの活性範囲内のpHでの適当な緩衝液も当業者に公知である。
【0093】
セルラーゼ及び緩衝液に加えて、処理組成物は任意で界面活性剤を含むことができる。適当な界面活性剤としては、利用するセルラーゼ及び布と適合性のある界面活性剤を含み、例えば、陰イオン性、非イオン性、及び両イオン性界面活性剤が挙げられる。適当な陰イオン性界面活性剤は、限定されないが、直鎖または分岐鎖アルキルベンゼンスルフォネート;直鎖または分岐鎖アルキル基またはアルケニル基を有するアルキルまたはアルケニルエーテル硫酸塩;アルキルまたはアルケニル硫酸塩;オレフィンスルフォネート;アルカンスルフォネート等が挙げられる。陰イオン性界面活性剤に関する適当な対イオンは、限定されないが、アルカリ金属イオン、例えばナトリウム及びカリウム;アルカリ土類金属イオン、例えば、カルシウム及びマグネシウム;アンモニウムイオン;及び炭素数2または3の1〜3のアルカノール基を有するアルカノールアミンが挙げられる。両性界面活性剤は、例えば、第4級アンモニウム塩スルフォネート及びベタイン型両性界面活性剤が挙げられる。当該両性界面活性剤は同じ分子内に正及び負電荷の両方の基を有する。非イオン性界面活性剤は通常、ポリオキシアルキレンエーテル及び高脂肪酸アルカノールアミドまたはその酸化アルキレン付加、及び脂肪酸グリセリンモノエステルを含むことができる。界面活性剤の混合物も当業者に公知の手段で用いることができる。
【0094】
濃縮セルラーゼ組成物はここに記載の方法において使用するために調製できる。このような濃縮物は濃縮した量の上述のセルラーゼ組成物、緩衝液及び界面活性剤を、好ましくは水溶液中に含む。そのように処方された場合、セルラーゼ濃縮物は、必要な濃度で各成分を含むセルラーゼ調製物を素早く、正確に調製するために水で容易に希釈できる。水性濃縮物を処方した場合、これらの濃縮物は希釈でき、上述のセルラーゼ溶液中において必要な成分濃度に達することができる。明らかなことであるが、このようなセルラーゼ濃縮物により、セルラーゼ溶液の容易な処方が可能となり、使用場所への組成物の輸送が実現可能なものとなる。処理濃縮物は当業者が認識する任意の形態をとることができ、例えば、液体、乳液、ゲルまたはペーストである。このような形態は当業者に公知である。
【0095】
固体セルラーゼ濃縮物を用いる場合、セルラーゼ組成物は顆粒、粉末、凝集体または固体ディスクである。顆粒は、洗浄媒体内での顆粒溶解速度を減少させる物質を含むように処方できる。当該物質及び顆粒は、その全体をここに引用する米国特許第5,254,283号に開示されている。
【0096】
その他の物質も所望により本発明のセルラーゼ組成物と一緒に使用または添加でき、例えば、組成物の最終的な用途に応じて石、軽石、充填剤、溶媒、酵素活性化剤、及び抗再沈殿剤が挙げられる。
【0097】
例として、ストーンウオッシュ法を詳細に説明するが、記載のパラメーターはその他の用途のため、すなわち、布の感触及び/外観改善のために、当業者により容易に修正される。セルロース含有布は、処理組成物とストーンウオッシュ組成物を混合することにより、効果的な量のセルラーゼを含むストーンウオッシュ組成物を含有したセルラーゼと接触させ、従って、セルラーゼ酵素を布の近くまで持っていく。次に、セルラーゼ及び布を含む水溶液を攪拌する。処理組成物が水溶液の場合、布を当該水溶液中に直接含浸できる。同様に、ストーンウオッシュ組成物が濃縮物の場合、濃縮物はセルロース含有布と一緒に水槽内で希釈する。ストーンウオッシュ組成物が固体形態の場合、例えば、前洗浄ゲルまたは固体スティックの場合、ストーンウオッシュ組成物は組成物を布または洗浄溶液につけながら直接接触してもよい。
【0098】
セルロース含有布は、酵素活性がセルロース含有布にストーンウオッシュ外観を与えることができる効果的な条件下でストーンウオッシュ溶液を用いて培養される。例えば、ストーンウオッシュ中のpH、溶液比、温度及び反応時間はストーンウオッシュ組成物が作用する条件を最適化するために調節できる。“効果的な条件”とは必然的に、セルラーゼ酵素が効果的にセルロース含有布と反応することが可能なpH、溶液比、及び温度をいい、この場合、ストーンウオッシュ効果を生じる。本発明に従ってストーンウオッシュ組成物を用いる条件を最大化することは当業者の能力の範囲内である。
【0099】
ここで用いるストーンウオッシュ中の溶液比、すなわち、ストーンウオッシュ組成物溶液の重量(すなわち、洗浄溶液)と布重量との比は通常、デニム生地に所望のストーンウオッシュ効果を達成するために十分な量であり、使用する方法に依存する。好ましくは、溶液比は約4:1〜約50:1であり、より好ましくは約5:1〜約20:1、及び最も好ましくは約10:1〜約15:1である。
【0100】
本発明のストーンウオッシュ組成物を用いるストーンウオッシュ時の反応温度は2つの競合因子に支配される。まず、高温に対応して、通常、反応速度が高められ、すなわち、反応が速まり、低温の場合に必要な反応時間と比較して反応時間を減少させることができる。従って、反応温度は通常、少なくとも約10℃以上である。第2に、セルラーゼは所定の反応温度以上で活性を失うタンパク質であり、当該温度は使用するセルラーゼの性質に依存する。従って、反応温度が高くなりすぎた場合、セルロース分解活性はセルラーゼ変性の結果として失われる。当業界において標準的なセルラーゼ使用温度は通常、35℃〜65℃の範囲であり、これらの条件が本発明のセルラーゼにも適したものであると考えられるが、最適な温度条件は使用される特定のセルラーゼに関して公知技術に従い、確認するべきである。
【0101】
反応時間はストーンウオッシュが生じる特定の条件に依存する。例えば、pH、温度及びセルラーゼ濃度はすべて最適な反応時間に影響する。通常、反応時間は約5分〜約5時間であり、好ましくは約10分〜約3時間、及び最も好ましくは約20分〜約1時間である。
【0102】
本発明のさらに好ましい実施態様に従い、本発明のセルラーゼは洗剤組成物中に用いてもよい。本発明に従う洗剤組成物は前洗浄組成物、前含浸(pre−soak)組成物として、または標準洗浄またはすすぎサイクル間の洗浄に有用である。好ましくは、本発明の洗剤組成物は効果的な量のセルラーゼ、界面活性剤を含み、及び任意で下記のその他の成分を含む。
【0103】
本発明の洗剤組成物で使用される効果的な量のセルラーゼは、セルロース含有布にセルラーゼにより生じることが知られる所望の効果、例えば毛羽取り、柔軟化、毛羽防止、表面繊維除去、抗灰色化及び洗浄効果を与えるために十分な量である。好ましくは、洗剤組成物中のセルラーゼは洗剤の約10ppm〜約20,000ppm濃度で用いる。
【0104】
洗剤組成物中で使用されるセルラーゼ酵素の濃度は好ましくは、水媒体中へ希釈して、セルラーゼ酵素の濃度が約0.01〜約1000ppm、好ましくは約0.02ppm〜約500ppm、及び最も好ましくは約0.5ppm〜約250ppm合計タンパク質になるように選択される。洗剤組成物中で使用されるセルラーゼ酵素の量は、洗浄溶液を作るために水を添加することにより洗剤が希釈される程度に依存する。
【0105】
本発明の洗剤組成物は当業者が認識する任意の形態であり、例えば、液体、顆粒、乳液、ゲル、ペーストである。このような形態は当業者に公知である。固体洗剤組成物を使用する場合、セルラーゼは顆粒として処方するのが好ましい。好ましくは、顆粒はセルラーゼ保護剤をさらに含むように処方できる。顆粒は水媒体への顆粒の溶解速度を減少させる物質を含むように処方することもできる。このような物質及び顆粒は、その全体をここに引用する米国特許第5,254,283号に開示されている。
【0106】
本発明の洗剤組成物は、表面活性剤、すなわち界面活性剤、例えば洗剤組成物での使用が公知の陰イオン性、非イオン性、及び両イオン性界面活性剤を用いる。
【0107】
本発明の洗剤組成物での使用が適した陰イオン性界面活性剤は、直鎖または分岐鎖アルキルベンゼンスルフォネート;直鎖または分岐鎖アルキル基またはアルケニル基を有するアルキルまたはアルケニルエーテル硫酸塩;アルキルまたはアルケニル硫酸塩;オレフィンスルフォネート;及びアルカンスルフォネートが挙げられる。陰イオン性界面活性剤に関する適当な対イオンは、アルカリ金属イオン、例えばナトリウム及びカリウム;アルカリ土類金属イオン、例えば、カルシウム及びマグネシウム;アンモニウムイオン;及び炭素数2または3の1〜3のアルカノール基を有するアルカノールアミンが挙げられる。両性界面活性剤は第4級アンモニウム塩スルフォネート及びベタイン型両性界面活性剤が挙げられる。当該両性界面活性剤は同じ分子内に正及び負電荷の両方の基を有する。非イオン性界面活性剤は通常、ポリオキシアルキレンエーテル及び高脂肪酸アルカノールアミドまたはその酸化アルキレン付加、脂肪酸グリセリンモノエステル等を含むことができる。本発明での使用に適した界面活性剤は英国特許出願番号第2 094 826Aに開示されており、その開示の内容をここに引用する。このような界面活性剤の混合物も使用することができる。界面活性剤または界面活性剤の混合物は通常、本発明の洗剤組成物において合計洗剤組成物の約1重量%〜約95重量%の量、好ましくは合計洗剤組成物の約5重量%〜約45重量%で使用される。セルラーゼ組成物及び界面活性剤に加え、本発明の洗剤組成物は任意で1以上の以下の成分を含むことができる。
【0108】
セルラーゼ以外のヒドロラーゼ
適当なヒドロラーゼは、エステル結合上に作用するカルボキシレートエステルヒドロラーゼ、チオエステルヒドロラーゼ、ホスフェートモノエステルヒドロラーゼ、及びホスフェートジエステルヒドロラーゼ;グリコシル成分上に作用するグリコシドヒドロラーゼ;N−グリコシル成分を加水分解する酵素;エーテル結合上に作用するチオエーテルヒドロラーゼ;ペプチド結合上に作用するa−アミノ−アシル−ペプチドヒドロラーゼ、ペプチジル−アミノ酸ヒドロラーゼ、アシル−アミノ酸ヒドロラーゼ、ジペプチドヒドロラーゼ及びペプチジル−ペプチドヒドロラーゼが挙げられる。この中で好ましいものは、カルボキシレートエステルヒドロヒドロラーゼ、グリコシドヒドロラーゼ、及びペプチジル−ペプチドヒドロラーゼである。適当なヒドロラーゼは、(1)ペプチジル−ペプチドヒドロラーゼに属するプロテアーゼ、例えば、ペプシン、ペプシンB、レンニン、トリプシン、キモトリプシンA、キモトリプシンB、エラスターゼ、エンテロキナーゼ、カテプシンC、パパイン、キモパパイン、フィシン、トロンビン、フィブリノリジン、レニン、スブチリシン、アスペルギロペプチダーゼA(aspergillopeptidase A)、コラゲナーゼ、クロストリジオペプチダーゼB(clostridiopeptidase B)、カリクレイン、ガストリシン、カテプシンD、ブロメリン、ケラチナーゼ、キモトリプシンンC、ペプシンC、アスペルギロペプチダーゼB、ウロキナーゼ、カルボキシペプチダーゼA及びB、及びアミノペプチダーゼ;(2)グリコシドヒドロラーゼ(必須成分であるセルラーゼはこの群から除く)α−アミラーゼ、β−アミラーゼ、グルコアミラーゼ、インベルターゼ、リゾチーム、ペクチナーゼ、キチナーゼ及びデキストラナーゼを含む。この中で好ましくは、α−アミラーゼ及びβ−アミラーゼである。これらは酸から中性の系で機能するが、細菌から得られるものはアルカリ性の系において高い活性を示す;(3)カルボキシレートエステルヒドロラーゼ、例えばカルボキシルエステラーゼ、リパーゼ、ペクチンエステラーゼ、及びクロロフィラーゼなど。この中で特に効果的なものはリパーゼである。
【0109】
セルラーゼ以外のヒドロラーゼは本目的に従って必要とされる量だけ洗剤組成物内に組み込まれる。好ましくは、精製タンパク質に関して、0.001〜5重量%、及びより好ましくは0.02〜3重量%の量で組み込まれるべきである。この酵素は、未精製酵素だけで作られた、またはその他の成分と組合わせて、顆粒形態で洗剤組成物中で使用すべきである。未精製酵素の顆粒は、顆粒中、精製酵素が0.001〜50重量%となるような量で使用される。当該顆粒は0.002〜20及び好ましくは0.1〜10重量%の量で用いる。セルラーゼと同様、これらの顆粒は酵素保護剤及び分解遅延剤を含むように処方できる。
【0110】
陽イオン界面活性剤及び長鎖脂肪酸塩
当該陽イオン界面活性剤及び長鎖脂肪酸塩は、飽和または不飽和脂肪酸塩、アルキルまたはアルケニルエーテルカルボン酸塩、α−スルホ脂肪酸塩またはエステル、アミノ酸型界面活性剤、リン酸エステル界面活性剤、4級アンモニウム塩、例えば3〜4のアルキル置換基及び1以下のフェニル置換アルキル置換基を有するものを含む。適当な陽イオン界面活性剤及び長鎖脂肪酸塩は英国特許出願番号第2 094 826Aに開示されており、その開示の内容をここに引用する。本組成物は約1〜約20重量%の当該陽イオン界面活性剤及び長鎖脂肪酸塩を含むことができる。
【0111】
ビルダー
A.二価金属イオン封鎖剤(sequestering agents)
本組成物は約0〜約50重量%の1以上のビルダー成分を含むことができ、以下の化合物のアルカリ金属塩及びアルカノールアミン塩からなる群より選択される:リン酸塩、ホスホン酸塩、ホスホノカルボキシレート、アミノ酸塩、アミノポリアセテート高分子量電解質、非分散ポリマー、ジカルボン酸塩、及びアルミノケイ酸塩。適当な二価金属イオン封鎖剤は米国特許出願番号第2 094 826Aに開示されており、その開示の内容をここに引用する。
【0112】
B.アルカリまたは無機電解質
本組成物は当該組成物に基づいて、約1〜約50重量%、好ましくは約5〜約30重量%の1以上の以下の化合物のアルカリ金属塩をアルカリまたは無機電解質として含むことができる:ケイ酸塩、炭酸塩及び硫酸塩及び有機アルカリ、例えば、トリエタノールアミン、ジエタノールアミン、モノエタノールアミン及びトリイソプロパノールアミン。
【0113】
再付着防止剤
本組成物は約0.1〜約5重量%の1以上の以下の化合物を再付着防止剤として含むことができる:ポリエチレングリコール、ポリビニルアルコール、ポリビニルピロリドン、及びカルボキシメチルセルロース。
【0114】
この中でも、カルボキシメチル−セルロース及び/またはポリエチレングリコールと本発明のセルラーゼ組成物との組み合わせは特に有用な汚れ除去組成物を提供する。
【0115】
漂白剤
本発明のセルラーゼと漂白剤とを組み合わせた使用、例えば漂白剤はモノ過硫酸カリウム、過炭酸ナトリウム、過ホウ酸ナトリウム、硫酸ナトリウム/過酸化水素付加化合物及び塩化ナトリウム/過酸化水素付加化合物または/及び光過敏漂白剤、例えば、スルホン化フタロシアニンの亜鉛またはアルミニウム塩は、さらに洗剤効果を改善する。同様に、EP684 304に記載の漂白剤及び漂白触媒も用いることができる。
【0116】
青味剤及び蛍光染料
種々の青味剤及び蛍光染料は必要に応じて本組成物に組み込むことができる。適当な青味剤及び蛍光染料は英国特許出願第2 094 826Aに開示されており、その開示の内容をここに引用する。
【0117】
凝結防止剤
以下の凝結防止剤は粉末洗剤中に組み込むことができる:p−トルエンスルホン酸塩、キシレンスルホン酸塩、酢酸塩、スルホ琥珀酸塩、タルク、微細粉末シリカ、アモルファスシリカ、粘土、ケイ酸カルシウム(例えば、Johns Manville Co.のマイクロ−セル(Micro−Cell))、炭酸カルシウム及び酸化マグネシウム。
【0118】
酸化防止剤
酸化防止剤は、例えば、tert−ブチル−ヒドロキシトルエン、4,4’−ブチリデンビス(6−tert−ブチル−3−メチルフェノール)、2,2’−ブチリデンビス(6−tert−ブチル−4−メチルフェノール)、モノスチレン化クレゾール、ジスチレン化クレゾール、モノスチレン化フェノール、ジスチレン化フェノール及び1,1−ビス(4−ヒドロキシ−フェニル)シクロヘキサンが挙げられる。
【0119】
可溶化剤
可溶化剤は例えば、低級アルコール、例えば、エタノール、ベンゼンスルホン酸塩、低級アルキルベンゼンスルホン酸塩、例えば、p−トルエンスルホン酸塩、グリコール、例えば、プロピレングリコール、アセチルベンゼン−スルホン酸塩、アセトアミド、ピリジンジカルボン酸アミド、安息香酸塩及び尿素が挙げられる。
【0120】
本発明の洗剤組成物は酸性からアルカリ性pHまで広いpH範囲で用いることができる。好ましい実施態様において、本発明の洗剤組成物は、5以上から約12以下のpHを有する弱酸性、中性またはアルカリ性洗浄媒体中で用いることができる。
【0121】
上記成分に加えて、香料、緩衝剤、保存料、染料等を所望により本発明の洗剤組成物と一緒に用いることができる。当該成分は従来当業界で用いられてきた量で適宜用いられる。
【0122】
本発明で使用される洗剤基材が粉末形態の場合、公知の調製方法、例えば噴霧乾燥法及び造粒法により調製されるものである。特に噴霧乾燥法、凝集法、乾燥混合法または非塔(non−tower)ルート法により得られる洗剤基材は好ましい。噴霧乾燥法により得られる洗剤基材は調製条件については制限されない。噴霧乾燥法により得られる洗剤基材は、熱耐性成分、例えば表面活性剤及びビルダーの水性スラリーを温空間内に噴霧することにより得られる空洞の顆粒である。噴霧乾燥後、香料、酵素、漂白剤、無機アルカリ性ビルダーを加えることができる。噴霧乾燥造粒または凝集法などにより得られた高い密度の顆粒洗剤基材と一緒に、種々の成分も基材の調製後に加えることできる。
【0123】
洗剤基材が液体の場合、均質溶液または不均質分散液であってもよい。洗剤中のセルラーゼによるカルボキシメチルセルロースの分解を除去するために、カルボキシメチルセルラーゼは組成物に組み込む前に顆粒化またはコーティングされていることが望ましい。
【0124】
本発明の洗剤組成物はセルロース含有布、例えば、汚れた布と一緒に工業的及び家庭用途においてこれらの環境で適宜用いられる温度、反応時間及び溶液比で培養できる。
【0125】
本発明に従う洗剤は、十分な活性が存在し、所望の改善、軟化、毛羽取り、毛羽立ち防止、表面繊維除去、または洗浄が得られる、中性pHで適当な溶液中で前洗浄としてさらに処方できる。洗剤組成物が液体、スプレー、ゲルまたはペースト組成物としての前含浸(pre−soak)(例えば、前洗浄または前処理)組成物の場合、セルラーゼ酵素は通常、前含浸または前処理組成物の全重量に基づいて約0.0001〜約1重量%で用いられる。このような組成物において、界面活性剤を任意で用いることができ、用いる場合、通常、前含浸の全重量に基づいて、約0.005〜約20重量%の濃度で存在する。組成物の残りは前含浸において用いられる従来成分を含み、すなわち、従来濃度での希釈剤、緩衝剤、その他の酵素(プロテアーゼ)等である。
【0126】
ここに記載のセルラーゼを含む組成物は、色あせた布の色修復(例えば、ここにその全体を引用する米国特許第4,738,682号を参照)及びシミ取り及び毛羽取り及び抗毛羽立ち(毛羽立ち防止)のための使用に適した単独組成物として家庭用で用いることができると考えられる。
【0127】
本発明に従うセルラーゼの使用は飼料添加剤及びパルプ及び紙処理において特に効果的である。これらのさらなる工業的用途については例えば、PCT国際公開番号第95/16360号及びフィンランド発行特許第87372号にそれぞれ記載されている。
【0128】
本発明及びその利点についてさらに説明するために、以下に具体的な実施例を与えるが、これらは本発明を説明するために提供するものであり、その範囲を限定するものとしていかなる方法においても解釈すべきでないことが理解される。
【0129】
実施例
以下の実施例を与えるが、本発明を限定するものではない。
【0130】
実施例1 サンプル収集及び処理
この実施例はcDNAライブラリーを作成するために十分なDNAを得るためのサンプルの収集法及び処理法について説明する。
【0131】
水サンプル(250ml)をSonachi湖(クレーター)、ケニアの沿岸帯から、柔軟伸展性1−m棒の先端に取り付けたステンレススチール・ビーカーを用いて収集し、室温で研究所に輸送するため密封可能なプラスティック容器(Whirlpak)内に納めた。水の表面温度は28℃、pH10及び伝導率は7.23mScm−1(27℃で)であった。
【0132】
微生物叢を収集するために、Sonachi(クレーター)湖、ケニアからの水(750ml)を、減少した細孔サイズの硝酸セルロースまたは酢酸セルロースからなる一連の殺菌薄膜フィルター(直径47mm)を通して、水流が全て止まるまで(手動吸引ポンプを用いて)現地でろ過した。一連のフィルターは8μm、3μm及び0.22μmであった。個々の薄膜フィルターを10mM Tris HCl、pH8.0;1mM EDTA及び5%w/v NaClを含む30ml殺菌プラスティック・ユニバーサル菅内の10mlの冷、殺菌細胞安定化緩衝剤(TES)中に速やかに加え、冷蔵された冷却ボックス内の氷上に、さらに処理するときまで、通常はサンプル採取4時間以内で保管した。フィルター上の細菌物質は、殺菌ガラスビーズ(5ml)及び微量遠心菅内で5分間、13,000gで遠心することにより沈殿した細胞を混合しながら激しくかき混ぜることにより分散させた。細菌物質を、推定で108〜109の等量の細菌細胞を含む体積で微量遠心菅に等分し、合計12個の菅を得た。DNAをGenomicPrep(登録商標)Cells及びTissue DNA単離キット(アマシャム・ファルマシア・バイオテック(Amersham Pharmacia Biotech)、Piscataway,ニュージャージー州、米国)を用いて取扱説明書に従って抽出した。各菅中の細胞は所定の600μlの細胞溶解液中に再懸濁し、80℃、5分間で培養し、細胞を溶解させた。この方法により調製したサンプルは室温で少なくとも18ヶ月間安定であり、この形態で研究室に戻した。DNA抽出を製造業者手順に従って、RNAアーゼ処理、タンパク質沈殿及びDNAイソプロパノール沈殿により完結させた。各DNAペレットを100μl殺菌Tris緩衝液10mM pH8.5中に溶解させた。
【0133】
DNA収量は5μlのサンプルを0.5% w/vアガロースゲル上で処理し、細菌性ゲノムDNAの公知量と比較することにより推測した。サンプルをプールし、合計約20μgのDNAを得た。収量が低かったので、on−site物質として同時に収集した水サンプルから抽出した約30%の余分物質で物質を補充し、必要なときまで4℃で研究室で保存した。約30μgの量のDNAは、予備実験が示した開始物質の量であり、試験及びバルク制限消化及びサイズ分画を行うために必要であり、ライブラリー構築のために十分な物質を与えた。
【0134】
実施例2
ライブラリー構築
以下の実施例は、大腸菌内の新規な配列のスクリーニング及び検出に使用するためのDNAライブラリーをどのように調製するかについて詳述する。
【0135】
DNA調製
プールしたDNAをゲノムDNAライブラリーの構築のために用いた。精製DNAをSau3A1を用いて部分的に消化し、約5kbの平均断片サイズを得た。制限DNAを電気泳動法によりTAEにおいて0.5%アガロース上でサイズ分画した(0.04M Tris−アセテート、0.001M EDTA pH8.0)。1.5〜10kb範囲の物質を切り取り、アガロースゲルの未使用部分をカットした同じサイズのウェル中に置き換え、逆電流により狭いバンドに濃縮させた。DNAバンドを切断し、キアゲン(QIAGEN)(Crawley,UK)QIAEXIIゲル抽出キットを用いて、取扱説明書に従ってDNA抽出した。溶出DNAをエタノールを用いて沈殿させ、10mM Tris HCl緩衝液、pH8.5中に再懸濁した。
【0136】
ラムダライブラリーの調製
制限DNAをラムダベクター内に、ZAP−エクスプレス(登録商標)ベクターキット(BamH1で前消化、及びアルカリホスファターゼ処理)及びGigapak(登録商標)IIIゴールド・パッケージング抽出(ストラタジーン(Stratagene)、アムステルダム、オランダ)を用いて製造業者手順に従いクローンした。最初のライブラリーを手順通りに、〜5×104pfuを含むアリコートを宿主大腸菌株XL1−ブルーMRF’で150mmペトリ皿にプレートし、緩衝液中のファージを溶出させることにより増幅させた。液体窒素中で凍結させた後、増幅ライブラリーを7% v/vジメチルスルホキシド中、−80℃で保存した。最初の滴定量の合計は1.8×106pfuであり、増幅後は6.8×109pfu ml−1であった。
【0137】
ライブラリー特質の評価
ファージミドベクターpBK−CMVをExAssistヘルパーファージ(ストラタジーン)を用いて製造業者の説明書通りにラムダZAPライブラリーから切り取り、大腸菌株XLOLRを感染させるために用いた。プラスミド含有クローンを50μg ml−1カナマイシンを含むルリア栄養(LB)寒天上にプレートすることにより単離した。Xgal[5−ブロモ−4−クロロ−3−インドイル−β−D−ガラクトシド]及びIPTG[イソプロピルチオ−β−D−ガラクトシド]の存在下、青白スクリーニングを用いてクローニング効率を測定した。DNAがラムダベクター内にクローンされなかった場合、β−ガラクトシド遺伝子が誘導因子IPTGの存在下で発現され、基質類似体Xgalの分裂を生じ、コロニー内に青色着色を生じる。しかしながら、ゲノムDNAの断片がうまくラムダベクター内にクローンされた場合、遺伝子が阻害され、従って、酵素は生成されず、コロニーは白色のままである。従って、青色と白色コロニーの比は挿入を含むクローンの割合を計算するために用いることができる。このライブラリーに関して、青白スクリーニングは青7、白286のコロニー比を生じ、97%のコロニーがゲノムDNA挿入を含んでいたことを示した。無作為に24のコロニーを選択し、BamH1クローニング部位に隣接するPst1及びHindIIIを用いるWizard(登録商標)Plus SV Miniprep DNA精製システム(Promega UK、サウサンプトン(Sauthampton))制限分析を用い、続いてアガロースゲル電気泳動法により調製したプラスミドDNAを使用して挿入サイズを測定した。24個の中、1個のクローンは検出可能な挿入を含まなかった。残りは1.5kb〜8.0kbの範囲の挿入を有した。
【0138】
実施例3
セルラーゼのためのライブラリースクリーニング
pBK−CMVファージミド中のDNAライブラリーを大腸菌クローンのプレート分析におけるセルラーゼ活性に関して、スクリーニングした。セルラーゼ活性を検出するために、ゲノムライブラリーをカナマイシン、0.5% w/vカルボキシメチルセルロース(低粘度ナトリウム塩;シグマ(Sigma)、Poole,UK)及びIPTG(直径7cmのペトリ皿内で寒天の表面上に広げた15μlの0.5M溶液)を含むLB寒天上にプレートした。37℃で一晩中成長させた後、当該コロニーを水に溶かして50℃まで冷やした3ml溶融0.7% w/vアガロースでかぶせた。これが終わると、当該プレートを0.1% w/v コンゴレッド(Congo Red)溶液で30分間水浸しにし、続いて1M NaClで2回洗浄した。細胞外セルラーゼ活性を示す陽性クローンは赤背景に対してイエロー・ハロ(yellow halo)で囲まれた(R.Teather and P.J.Wood,Applied&Environmental Microbiology,43:770−780,1982)。
【0139】
110,000の大腸菌pBK−CMVクローンのスクリーニングにより4領域のクリアリング指示潜在性(clearing indicating potential)のセルラーゼ生成コロニーを生じた。これらの3つは、除去領域(cleared zone)から取除いた寒天プラグを均質化し、単一のコロニーの筋をつけて、コンゴレッド試験により表現型を確認した後、セルラーゼ生成クローンとしてうまく回収された。
【0140】
実施例4
セルラーゼ陽性クローンのキャラクタリゼーション
プラスミドDNAを3つのセルラーゼ陽性クローンから単離し、挿入サイズを上述の通り制限消化により測定した。ゲル電気泳動により測定して消化後、3つ全てが同じサイズ(約3.5kb)であり、及び同じサイズの断片を有した。このことは、3つ全ての単離体が同一であり、単一のクローンの増幅により生じたことを示す。これは、プラスミドDNAのシーケンシングの第1ラウンドにより確認された(pBKCMVプラスミド中のプライマー部位を用いて)。これはレスター(Leicester)大学でタンパク質及び核酸化学研究室により、パーキンエルマー(Perkin Elmer)‘BigDye’ターミネーター・ケミストリー及び377モデルABI自動DNAシーケンサーを用いて行われた。配列の完全な範囲が挿入の5’及び3’の両末端から‘プライマーウォーキング’により得られた。当該配列はアプライド・バイオシステムス(Applied Biosystems)マルチ配列編集Seqed(登録商標)バージョン1.0.3を用いて編集した。配列はレスター大学から入手可能なGCGウィスコンシン(Wisconsin)パッケージ、バージョン10.2−UNIXにおけるプログラムを用いて組み立てた。これにより3796ヌクレオチド塩基の環境DNAの挿入を同定した(図1)。
【0141】
実施例5
セルラーゼ遺伝子の同定
クローンmHKcelの挿入環境DNAのヌクレオチド配列における可能なオープンリーディングフレーム(ORF)をMapDrawプログラム(DNASTAR、Brighton、マサチューセッツ州、米国)のORF Find設備またはベクターNTI式(Suite)のプログラムからのORFサーチ(InforMax(登録商標、North Bethesda,メリーランド州、米国)を用いて同定した。
【0142】
これにより、571アミノ酸のタンパク質に対応した1716ヌクレオチドからなるORFが同定され、挿入配列の位置923から開始して位置2638で終わった。このORFの配列はEditSeq(DNASTAR)を用いて切り取り、BLASTプログラムにより測定した。
【0143】
このORFのヌクレオチド配列を図2に示す。
【0144】
BLASTnプログラムを用いたヌクレオチド配列の調査は、重複のないヌクレオチド配列データベースに対してヌクレオチド質問配列を比較するものであり、公知配列に対してバチルス・ハロデュランス(halodurans)ゲノムの一部と同一性を示し、特にエンド−ベータ−1,4−グルカナーゼ(“セルラーゼB”)(GenBankアクセッション AP001509)に対応する1585ヌクレオチド領域に68%同一性を示した。
【0145】
BLASTxプログラムを用いるヌクレオチド配列の調査は、ヌクレオチド質問配列(両ストランド)の6フレーム概念的翻訳産物をタンパク質配列データベースに対して比較するものであり、多数の細菌性エンドセルラーゼに著しい類似性を示した。最も高い配列スコアは、574アミノ酸を含む酵素であり、通性好アルカリ性(facultative alkaliphilic)細菌バチルス・ハロデュランス株C125(タンパク質ID BAB04322、アクセッション AP001509)のエンド−β−グルカナーゼ(セルラーゼB)の553アミノ酸領域に対して67%同一性(372アミノ酸)を示した。この新しい同定遺伝子はバチルス・ハロデュランスセルラーゼに67%同一性を有する571アミノ酸のタンパク質をコードした。
【0146】
571個のアミノ酸からなる翻訳タンパク質を図3に示す。
【0147】
実施例6
酵素キャラクタリゼーション
塩の影響
mHKcel遺伝子を含む大腸菌pBK−CMV細胞を5ml緩衝液(20mM TRIS−HCl、pH8.0;500mM NaCl;0.1mM EDTA;0.1%Triton X−100)中に懸濁し、氷上で超音波処理することにより分解させる。超音波分解した抽出物をカルボキシメチルセルロース(CMC)上で寒天拡散分析により、種々のNaCl濃度で調べる。超音波分解抽出物(100μL)及び10個中1個の希釈物を種々の量のNaClを含むCMC−寒天プレート中に穴を開けたウェル内に配置する。当該プレートを37℃、16時間で培養し、セルロース加水分解を示す生じた除去(clearing)領域をミリメーターで測定する。結果は、セルラーゼmHKcelは0〜25% w/v NaClの範囲で活性だが、25% w/v NaClでの活性は0% NaClでの活性の約50%しかないということを示した(図4)。
【0148】
pHの影響
セルラーゼ活性におけるpHの影響をGrant&Tindall(Isolation of alkaliphilic bacteria(アルカリ性細菌の単離),In:Microbial Growth and Survival in Extreme Environments,Academic Press,ロンドン、1980、第27〜36頁)に記載のpH勾配プレート法を用いて調査した。CMCを含む寒天培地を四角形ペトリ皿の深さ1cmまで注ぎ、設置する。幅1cmの均一な鉢をプレートの端から切り取り、20% w/v Na2CO3・10H2O及び0.2M NaOHを含む寒天(同体積の殺菌0.4M NaOH/40% w/v Na2CO3・10H2O及び4% w/v寒天を60℃で混合することにより調製したもの)を鉢の中に注いだ。当該プレートを37℃で一晩中成長させ、均一にpH12〜pH7勾配を形成させる。mHKcelセルラーゼのpH耐性を調べるために、鉢を狭く(寒天)勾配により元の鉢に対して直角に切り取り、1mlの超音波分解した細胞抽出物で満たした。当該プレートを37℃で一晩中成長させた。当該プレートをコンゴレッドで30分間処理し、セルロース加水分解の領域を視覚化する。図5に示す結果は、mHKcelセルラーゼが約pH11.5まで活性であることを示す。
【0149】
ここに記載される実施例及び実施態様は説明目的のためのみであり、これに照らして種々の修正または変形が当業者に示唆され、本発明の概念及び精神及び請求の範囲の範囲内であることが理解される。ここに記載の全ての文献、特許及び特許出願はその全体を全ての目的においてここに引用するものとする。
【図面の簡単な説明】
【0150】
【図1A】環境(environmental)ヌクレオチド配列(配列番号1)を示す。
【図1B】環境(environmental)ヌクレオチド配列(配列番号1)を示す。
【図2】図2は新規なセルラーゼをエンコードする核酸配列を示す(配列番号2)。
【図3】図3は本発明のセルラーゼの推定アミノ酸配列を示す(配列番号3)。
【図4】図4は増加した塩化ナトリウム濃度を有する酵素活性を描いたグラフである。
【図5】図5はpH7〜12の範囲でのmHKcel活性へのpHの影響を描いたグラフである。
【特許請求の範囲】
【請求項1】
以下からなる群より選択される単離ポリヌクレオチド:
(a)配列番号1に示す配列に少なくとも85%の配列同一性を有する核酸配列、またはその相補体;
(b)図3(配列番号3)に示すアミノ酸配列に少なくとも85%配列同一性を有するmHKcelポリペプチドをエンコードする核酸配列、または当該配列に相補的な核酸配列;
(c)図3(配列番号3)に示すアミノ酸配列に少なくとも90%配列同一性を有するmHKcelポリペプチドをエンコードする核酸配列、または当該配列に相補的な核酸配列;
(d)図3(配列番号3)に示すアミノ酸配列に少なくとも95%配列同一性を有するmHKcelポリペプチドをエンコードする核酸配列、または当該配列に相補的な核酸配列;
(e)図3(配列番号3)に示すアミノ酸配列を有するmHKcelポリペプチドをエンコードする核酸配列、または当該配列に相補的な核酸配列;
ここで、前記単離ポリヌクレオチドはセルラーゼ生物学的活性を有するポリペプチドをエンコードし、その同一性はMacVectorバージョン6.5のCLUSTAL−Wプログラムにより測定され、オープンギャップ・ペナルティ10.0、伸長ギャップ・ペナルティ0.1を含む初期設定パラメーター、及びBLOSUM 30類似マトリックスを用いて操作される。
【請求項2】
以下からなる群より選択される単離ポリヌクレオチド:
(a)配列番号1に示す核酸配列またはその相補体;
(b)高ストリンジェンシーな条件下で配列番号1に示す配列にハイブリダイズする核酸配列、またはその相補体、またはその断片;
(c)配列番号2に示す核酸配列またはその相補体;及び
(d)高ストリンジェンシーな条件下で配列番号2に示す配列にハイブリダイズする核酸配列、またはその相補体、またはその断片;
ここで、前記単離ポリヌクレオチドはセルラーゼ生物学的活性を有するポリペプチドをエンコードし、ハイブリダイゼーションは50%ホルムアミド、6X SSC、5X Denhardt’s溶液、0.5% SDS及び100μg/ml変性キャリアDNA中、42℃で行われ、続いて2X SSPE及び0.5% SDS中、室温で2回洗浄し、及びさらに0.1X SSPE及び0.5% SDS中、42℃で2回洗浄する。
【請求項3】
ヌクレオチドがmRNA、DNA、cDNA、ゲノムDNA及びそれらのアンチセンス類似体からなる群より選択される、請求項1の単離ヌクレオチド。
【請求項4】
前記ポリヌクレオチドがRNA分子である、請求項3の単離ポリヌクレオチド。
【請求項5】
セルラーゼ活性を有する酵素をエンコードし、当該酵素がトリコデルマ源から単離される、請求項1の単離ポリヌクレオチド。
【請求項6】
酵素がトリコデルマ・リーゼイから単離される、請求項5の単離ポリヌクレオチド。
【請求項7】
セルラーゼ活性を有するアミノ酸配列をエンコードするポリヌクレオチド配列を含む発現構築体であって、(i)配列番号3に示すアミノ酸配列に少なくとも85%同一性を有し、または(ii)中から高ストリンジェンシーな条件下、図2に示すヌクレオチド配列とハイブリダイズするように設計されたプローブにハイブリダイズでき、または(iii)配列番号3に示すアミノ酸配列をエンコードするヌクレオチド配列に少なくとも85%配列同一性を有するヌクレオチド配列に相補的である、ポリヌクレオチド配列を含む発現構築体であり、ここで当該同一性はMacVectorのバージョン6.5におけるCLUSTAL−Wプログラムにより測定され、オープンギャップ・ペナルティ10.0、伸長ギャップ・ペナルティ0.1を含む初期設定パラメーター、及びBLOSUM 30類似マトリックスを用いて操作される。
【請求項8】
請求項1のポリヌクレオチドを含む発現ベクター。
【請求項9】
ベクターにより形質転換された宿主細胞により認識される制御配列に動作可能に連結した、請求項1の単離ポリヌクレオチドを含む発現ベクター。
【請求項10】
アクチノプラネス(Actinoplanes)のグルコースイソメラーゼ遺伝子由来プロモーター配列、ストレプトマイセスセルラーゼ遺伝子由来のシグナル配列、及びmHKcelセルラーゼをエンコードするポリヌクレオチド配列を含む調節ポリヌクレオチド配列を含む、請求項9に従う発現ベクター。
【請求項11】
請求項8の発現構築体を含むベクター。
【請求項12】
請求項8のベクターにより形質転換された宿主細胞。
【請求項13】
原核細胞である、請求項12の宿主細胞。
【請求項14】
真核細胞である、請求項12の宿主細胞。
【請求項15】
セルラーゼ生物学的活性を有する実質的に精製されたmHKcelポリペプチドであって、以下からなる群より選択される配列を含む:
(a)図3(配列番号3)に示すアミノ酸配列に少なくとも85%の配列同一性を有するアミノ酸配列;
(b)図3(配列番号3)に示すアミノ酸配列に少なくとも90%の配列同一性を有するアミノ酸配列;
(c)図3(配列番号3)に示すアミノ酸配列に少なくとも95%の配列同一性を有するアミノ酸配列;
(d)図3(配列番号3)に示すアミノ酸配列;
(e)配列番号3に示すアミノ酸配列の実質的に精製された生物学的に活性な断片;
ここで、当該同一性はMacVectorバージョン6.5のCLUSTAL−Wプログラムにより測定され、オープンギャップ・ペナルティ10.0、伸長ギャップ・ペナルティ0.1を含む初期設定パラメーター、及びBLOSUM 30類似マトリックスを用いて操作される。
【請求項16】
バチルスから得られる、実質的に精製されたmHKcelセルラーゼポリペプチドまたは誘導体。
【請求項17】
以下の工程を含むセルラーゼの生成方法:
(a)セルラーゼを生成する適当な条件下、適した培養基中で請求項12に従う宿主細胞を培養する工程;
(b)前記の生成したセルラーゼを得る工程。
【請求項18】
宿主細胞が糸状菌または酵母細胞である、請求項17の方法。
【請求項19】
宿主細胞が細菌である、請求項17の方法。
【請求項20】
細菌がストレプトマイセスである、請求項19の方法。
【請求項21】
請求項17の方法により調製したセルラーゼ活性を有する精製酵素。
【請求項22】
mHKcel遺伝子内に欠失または挿入またはその他の変異を含み、当該遺伝子が不活性化され、mHKcelポリペプチド生成が阻害された組換え宿主細胞。
【請求項23】
配列番号3に示す配列を有するmHKcelポリペプチドをエンコードするメッセンジャーRNAに相補的なアンチセンスオリゴヌクレオチドであって、セルラーゼ生成宿主細胞に曝露すると、前記オリゴヌクレオチドは前記宿主細胞によるセルラーゼ生成を減少または阻害する、アンチセンスオリゴヌクレオチド。
【請求項24】
宿主細胞が糸状菌である、請求項23のアンチセンスオリゴヌクレオチド。
【請求項25】
以下からなる群より選択されるポリペプチドを含む洗剤組成物:
(a)図3(配列番号3)に示すアミノ酸配列に少なくとも85%配列同一性を有するアミノ酸配列;
(b)図3(配列番号3)に示すアミノ酸配列に少なくとも90%配列同一性を有するアミノ酸配列;
(c)図3(配列番号3)に示すアミノ酸配列に少なくとも95%配列同一性を有するアミノ酸配列;
(d)図3(配列番号3)に示すアミノ酸;
(e)配列番号3に示すアミノ酸配列の実質的に精製された生物学的に活性な断片;
ここで、当該同一性はMacVectorバージョン6.5のCLUSTAL−Wプログラムにより測定され、オープンギャップ・ペナルティ10.0、伸長ギャップ・ペナルティ0.1を含む初期設定パラメーター、及びBLOSUM 30類似マトリックスを用いて操作される。
【請求項26】
界面活性剤及び請求項15のセルラーゼを含む洗剤。
【請求項27】
前記洗剤が洗濯用洗剤である、請求項25の洗剤。
【請求項28】
前記洗剤が食器用洗剤である、請求項25の洗剤。
【請求項29】
請求項15のセルラーゼを含む飼料添加物。
【請求項30】
木材パルプと請求項15のセルラーゼとを接触させることを含む木材パルプの処理方法。
【請求項31】
バイオマスと請求項15のセルラーゼとを接触させることを含むバイオマスを糖に変換する方法。
【請求項32】
さらに高フルクトース・コーンシロップの生成を含む、請求項31の方法。
【請求項33】
以下の工程を含むエタノールの生成方法:
(a)バイオマス組成物とmHKcelを含む酵素組成物とを接触させて糖溶液を得る工程;
(b)前記糖溶液に発酵性微生物を加える工程;及び
(c)エタノールを生成するために十分な条件下で発酵性微生物を培養する工程。
【請求項34】
以下を含む新規な酵素を同定する方法:
(a)周囲環境から全微生物集団DNAを単離する工程;
(b)大腸菌内にゲノムDNAライブラリーを構築する工程;
(c)セルラーゼ活性の発現に関してライブラリーをスクリーニングする工程;
(d)セルラーゼ陽性クローン中のセルラーゼ遺伝子を同定する工程;及び
(e)新規なセルラーゼ酵素をキャラクタライズする工程。
【請求項1】
以下からなる群より選択される単離ポリヌクレオチド:
(a)配列番号1に示す配列に少なくとも85%の配列同一性を有する核酸配列、またはその相補体;
(b)図3(配列番号3)に示すアミノ酸配列に少なくとも85%配列同一性を有するmHKcelポリペプチドをエンコードする核酸配列、または当該配列に相補的な核酸配列;
(c)図3(配列番号3)に示すアミノ酸配列に少なくとも90%配列同一性を有するmHKcelポリペプチドをエンコードする核酸配列、または当該配列に相補的な核酸配列;
(d)図3(配列番号3)に示すアミノ酸配列に少なくとも95%配列同一性を有するmHKcelポリペプチドをエンコードする核酸配列、または当該配列に相補的な核酸配列;
(e)図3(配列番号3)に示すアミノ酸配列を有するmHKcelポリペプチドをエンコードする核酸配列、または当該配列に相補的な核酸配列;
ここで、前記単離ポリヌクレオチドはセルラーゼ生物学的活性を有するポリペプチドをエンコードし、その同一性はMacVectorバージョン6.5のCLUSTAL−Wプログラムにより測定され、オープンギャップ・ペナルティ10.0、伸長ギャップ・ペナルティ0.1を含む初期設定パラメーター、及びBLOSUM 30類似マトリックスを用いて操作される。
【請求項2】
以下からなる群より選択される単離ポリヌクレオチド:
(a)配列番号1に示す核酸配列またはその相補体;
(b)高ストリンジェンシーな条件下で配列番号1に示す配列にハイブリダイズする核酸配列、またはその相補体、またはその断片;
(c)配列番号2に示す核酸配列またはその相補体;及び
(d)高ストリンジェンシーな条件下で配列番号2に示す配列にハイブリダイズする核酸配列、またはその相補体、またはその断片;
ここで、前記単離ポリヌクレオチドはセルラーゼ生物学的活性を有するポリペプチドをエンコードし、ハイブリダイゼーションは50%ホルムアミド、6X SSC、5X Denhardt’s溶液、0.5% SDS及び100μg/ml変性キャリアDNA中、42℃で行われ、続いて2X SSPE及び0.5% SDS中、室温で2回洗浄し、及びさらに0.1X SSPE及び0.5% SDS中、42℃で2回洗浄する。
【請求項3】
ヌクレオチドがmRNA、DNA、cDNA、ゲノムDNA及びそれらのアンチセンス類似体からなる群より選択される、請求項1の単離ヌクレオチド。
【請求項4】
前記ポリヌクレオチドがRNA分子である、請求項3の単離ポリヌクレオチド。
【請求項5】
セルラーゼ活性を有する酵素をエンコードし、当該酵素がトリコデルマ源から単離される、請求項1の単離ポリヌクレオチド。
【請求項6】
酵素がトリコデルマ・リーゼイから単離される、請求項5の単離ポリヌクレオチド。
【請求項7】
セルラーゼ活性を有するアミノ酸配列をエンコードするポリヌクレオチド配列を含む発現構築体であって、(i)配列番号3に示すアミノ酸配列に少なくとも85%同一性を有し、または(ii)中から高ストリンジェンシーな条件下、図2に示すヌクレオチド配列とハイブリダイズするように設計されたプローブにハイブリダイズでき、または(iii)配列番号3に示すアミノ酸配列をエンコードするヌクレオチド配列に少なくとも85%配列同一性を有するヌクレオチド配列に相補的である、ポリヌクレオチド配列を含む発現構築体であり、ここで当該同一性はMacVectorのバージョン6.5におけるCLUSTAL−Wプログラムにより測定され、オープンギャップ・ペナルティ10.0、伸長ギャップ・ペナルティ0.1を含む初期設定パラメーター、及びBLOSUM 30類似マトリックスを用いて操作される。
【請求項8】
請求項1のポリヌクレオチドを含む発現ベクター。
【請求項9】
ベクターにより形質転換された宿主細胞により認識される制御配列に動作可能に連結した、請求項1の単離ポリヌクレオチドを含む発現ベクター。
【請求項10】
アクチノプラネス(Actinoplanes)のグルコースイソメラーゼ遺伝子由来プロモーター配列、ストレプトマイセスセルラーゼ遺伝子由来のシグナル配列、及びmHKcelセルラーゼをエンコードするポリヌクレオチド配列を含む調節ポリヌクレオチド配列を含む、請求項9に従う発現ベクター。
【請求項11】
請求項8の発現構築体を含むベクター。
【請求項12】
請求項8のベクターにより形質転換された宿主細胞。
【請求項13】
原核細胞である、請求項12の宿主細胞。
【請求項14】
真核細胞である、請求項12の宿主細胞。
【請求項15】
セルラーゼ生物学的活性を有する実質的に精製されたmHKcelポリペプチドであって、以下からなる群より選択される配列を含む:
(a)図3(配列番号3)に示すアミノ酸配列に少なくとも85%の配列同一性を有するアミノ酸配列;
(b)図3(配列番号3)に示すアミノ酸配列に少なくとも90%の配列同一性を有するアミノ酸配列;
(c)図3(配列番号3)に示すアミノ酸配列に少なくとも95%の配列同一性を有するアミノ酸配列;
(d)図3(配列番号3)に示すアミノ酸配列;
(e)配列番号3に示すアミノ酸配列の実質的に精製された生物学的に活性な断片;
ここで、当該同一性はMacVectorバージョン6.5のCLUSTAL−Wプログラムにより測定され、オープンギャップ・ペナルティ10.0、伸長ギャップ・ペナルティ0.1を含む初期設定パラメーター、及びBLOSUM 30類似マトリックスを用いて操作される。
【請求項16】
バチルスから得られる、実質的に精製されたmHKcelセルラーゼポリペプチドまたは誘導体。
【請求項17】
以下の工程を含むセルラーゼの生成方法:
(a)セルラーゼを生成する適当な条件下、適した培養基中で請求項12に従う宿主細胞を培養する工程;
(b)前記の生成したセルラーゼを得る工程。
【請求項18】
宿主細胞が糸状菌または酵母細胞である、請求項17の方法。
【請求項19】
宿主細胞が細菌である、請求項17の方法。
【請求項20】
細菌がストレプトマイセスである、請求項19の方法。
【請求項21】
請求項17の方法により調製したセルラーゼ活性を有する精製酵素。
【請求項22】
mHKcel遺伝子内に欠失または挿入またはその他の変異を含み、当該遺伝子が不活性化され、mHKcelポリペプチド生成が阻害された組換え宿主細胞。
【請求項23】
配列番号3に示す配列を有するmHKcelポリペプチドをエンコードするメッセンジャーRNAに相補的なアンチセンスオリゴヌクレオチドであって、セルラーゼ生成宿主細胞に曝露すると、前記オリゴヌクレオチドは前記宿主細胞によるセルラーゼ生成を減少または阻害する、アンチセンスオリゴヌクレオチド。
【請求項24】
宿主細胞が糸状菌である、請求項23のアンチセンスオリゴヌクレオチド。
【請求項25】
以下からなる群より選択されるポリペプチドを含む洗剤組成物:
(a)図3(配列番号3)に示すアミノ酸配列に少なくとも85%配列同一性を有するアミノ酸配列;
(b)図3(配列番号3)に示すアミノ酸配列に少なくとも90%配列同一性を有するアミノ酸配列;
(c)図3(配列番号3)に示すアミノ酸配列に少なくとも95%配列同一性を有するアミノ酸配列;
(d)図3(配列番号3)に示すアミノ酸;
(e)配列番号3に示すアミノ酸配列の実質的に精製された生物学的に活性な断片;
ここで、当該同一性はMacVectorバージョン6.5のCLUSTAL−Wプログラムにより測定され、オープンギャップ・ペナルティ10.0、伸長ギャップ・ペナルティ0.1を含む初期設定パラメーター、及びBLOSUM 30類似マトリックスを用いて操作される。
【請求項26】
界面活性剤及び請求項15のセルラーゼを含む洗剤。
【請求項27】
前記洗剤が洗濯用洗剤である、請求項25の洗剤。
【請求項28】
前記洗剤が食器用洗剤である、請求項25の洗剤。
【請求項29】
請求項15のセルラーゼを含む飼料添加物。
【請求項30】
木材パルプと請求項15のセルラーゼとを接触させることを含む木材パルプの処理方法。
【請求項31】
バイオマスと請求項15のセルラーゼとを接触させることを含むバイオマスを糖に変換する方法。
【請求項32】
さらに高フルクトース・コーンシロップの生成を含む、請求項31の方法。
【請求項33】
以下の工程を含むエタノールの生成方法:
(a)バイオマス組成物とmHKcelを含む酵素組成物とを接触させて糖溶液を得る工程;
(b)前記糖溶液に発酵性微生物を加える工程;及び
(c)エタノールを生成するために十分な条件下で発酵性微生物を培養する工程。
【請求項34】
以下を含む新規な酵素を同定する方法:
(a)周囲環境から全微生物集団DNAを単離する工程;
(b)大腸菌内にゲノムDNAライブラリーを構築する工程;
(c)セルラーゼ活性の発現に関してライブラリーをスクリーニングする工程;
(d)セルラーゼ陽性クローン中のセルラーゼ遺伝子を同定する工程;及び
(e)新規なセルラーゼ酵素をキャラクタライズする工程。
【図2】


【図3】

【図4】
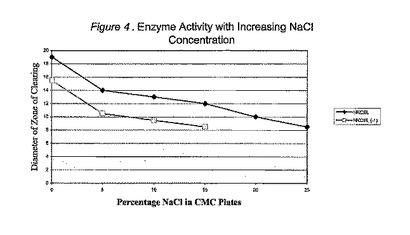
【図5】
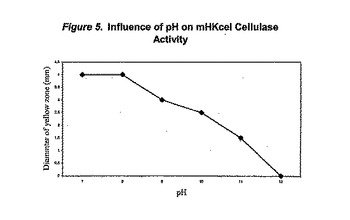
【図3】
【図4】
【図5】
【公表番号】特表2007−525180(P2007−525180A)
【公表日】平成19年9月6日(2007.9.6)
【国際特許分類】
【出願番号】特願2006−513443(P2006−513443)
【出願日】平成16年4月28日(2004.4.28)
【国際出願番号】PCT/US2004/013257
【国際公開番号】WO2004/099370
【国際公開日】平成16年11月18日(2004.11.18)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.UNIX
【出願人】(500284580)ジェネンコー・インターナショナル・インク (67)
【Fターム(参考)】
【公表日】平成19年9月6日(2007.9.6)
【国際特許分類】
【出願日】平成16年4月28日(2004.4.28)
【国際出願番号】PCT/US2004/013257
【国際公開番号】WO2004/099370
【国際公開日】平成16年11月18日(2004.11.18)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.UNIX
【出願人】(500284580)ジェネンコー・インターナショナル・インク (67)
【Fターム(参考)】
[ Back to top ]