
新規なヒト抗R7V抗体及びその使用

この発明は特異的にHIVのR7Vエピトープに結合することができる新規なヒト抗体に関する。これらの抗体は全てのヒトCDRを有し、エスケープ変異株を含むすべてのHIV株を特異的に中和することができる。これらの抗体はHIV感染治療に有益であり、特に高活性抗レトロウィルス療法(HAART)に効果のない患者の治療に有益である。
【発明の詳細な説明】
【技術分野】
【0001】
この発明は特異的にHIVのR7Vエピトープに結合することができる新規なヒト抗体に関する。これらの抗体は全てのヒトCDRを有し、エスケープ変異株を含むすべてのHIV株を特異的に中和することができる。これらの抗体はHIV感染治療に有益であり、特に高活性抗レトロウィルス療法(HAART)に効果のない患者の治療に有益である。
【背景技術】
【0002】
HIV感染は今なお世界的に流行する伝染病である。感染後のHIV複製や毒性を薬剤療法で抑制する一方、予防処置や治療処置は今なお有効とはいえない。さらに高活性抗レトロウィルス療法(HAART)は、エイズを抑制する一方で複数の副作用を伴い、薬剤耐性ウィルスの出現を促すので、HIVに対しては追加の治療法が強く望まれている。しかしノンプログレッサとして認定されているHIV患者でも感染後10年、15年以上もエイズを発症しない場合もあり、HIV疾患は弱毒化ウィルス(文献1)、欠損ウィルス(文献2)、HIVコレセプター変異株(文献3、4)あるいは中和抗体(文献5)の存在等により、様々な方法により発症を遅らせることができることを実証している。従来のエンベロップベースの抗体誘導ワクチンはいずれもウィルスの高い突然変異率及び弱い免疫原性に限界があった。先の研究(文献6)で、我々はノンプログレッサの血清から精製された広域スペクトル中和抗R7V抗体の潜在力を実証した。これらの免疫グロブリンは、β2-ミクログロブリンから抽出され、出芽中にHIVの外被に組み込まれるR7V(RTPKIQV アミノ酸配列)と呼ばれる細胞エピトープに抗するものであった(文献7、8)。我々の目的は、ノンプログレッサの患者のB-リンパ球から対応する遺伝子を、バキュロウィルスベクターを介して分離し、組み換え型抗R7V 抗体を生成することにあった。
【0003】
バキュロウィルス技術により、正しくアセンブルされグリコシル化された免疫グロブリンを生成、分泌することができる(文献9)。これら組み換え型抗体は親の免疫グロブリンの全ての機能特性を有し(文献10、11)、(i)補体成分C1q(文献12、13)やC3(文献14)と(ii)細胞傷害に対する抗体を誘導するのに必要なIgGFc レセプタ(文献15、16、13)とを結合するような効率の良いエフェクタ機能を示す。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】明細書の末尾の一覧表を参照のこと。
【発明の概要】
【0005】
この発明に関連して、ウィルスの出芽中にHIVにより獲得される細胞エピトープR7Vに抗する組み換え型抗体を作った。抗R7V抗体の可変領域をエンコードするc-DNAはノンプログレッサの患者のBリンパ球からクローニングされている。この抗体の重鎖及び軽鎖の完全なコード配列を含む2つのトランスファーベクターが作られ、組み換え型バキュロウィルスがバキュロウィルスDNAとその2つのトランスファーベクター間の2重の組み換えにより生成された。このバキュロウィルスに感染された昆虫細胞から完全なヒト抗R7V免疫グロブリンを生成した。我々は、R7Vペプチドに特異的である我々の組み換え型抗体が、耐性ウィルスを含むあらゆるHIV1のクレードを認識し中和することを解明した。これによりHIV療法に新たな展望を開くことになる。
【0006】
つまり、第1の実施例によれば、この発明の主題は単離された抗体であり、あるいはその機能を有するフラグメントの1つであり、その抗体あるいはフラグメントの1つは特にR7Vエピトープ( RTPKIQV 配列番号 11 )と結合することができ、HIV株を中和できるものであり、それは、
i)アミノ酸配列 配列番号1 (QSVLYSSNNKNY), 配列番号2 (WAS), 及び配列番号3 (QQYYSTPQT) を有する相補性決定領域CDR, あるいはその配列が、配列配列番号1, 2, 又は3 との最適アライメントの後で、少なくとも80%、好ましくは90%の同一性を持つCDRを含む軽鎖、及び
ii)アミノ酸配列配列番号6 (GGSISSYY), 配列番号7 (IYYSGST) 及び配列番号8 (ARGRSWFSY) を有するCDR, あるいはその配列が、配列配列番号6,7, 又は8 との最適アライメントの後で、少なくとも80%、好ましくは90%の同一性を持つCDRを含む重鎖を備えている。
この「発明の概要」の中で用いる用語、ポリペプチド、ポリペプチド配列、ペプチド及び抗体化合物又はその配列に結合するタンパク質は置換可能である。
【0007】
この発明は天然の抗体に関するものではなく、すなわちこの発明の抗体は自然環境に存在するものではなく、自然源から精製により単離、取得できたものであり、又は遺伝子組み換えにより取得できたものであり、あるいは化学合成により取得できたものである。それゆえにさらに説明するように、それは非天然アミノ酸を含むものであることが理解されるべきである。
【0008】
CDR領域又はCDRにより、Kabatらによって定義された 免疫グロブリンの重鎖及び軽鎖の超可変領域を示すことが意図されている。(Kabat et al. Sequences of proteins of immunological interest、 第5版、U.S. Department of Health and Human Services、NIH、1991 及び続版) 3つの重鎖CDR, 及び3つの軽鎖CDRが存在する。用語CDR 又はCDRsはここではケースに応じてこれら領域の1つまたはこれら領域の複数あるいは全体を示すために用いられ、これらの領域は、抗体の親和性によって認識した抗原又はエピトープへの結合に関与するアミノ酸残基の大多数を含んでいる。
【0009】
この発明の2つの核酸又はアミノ酸配列間の“同一性のパーセント”により、ベストアライメント(最適アライメント)後に得られる、比較される2つの配列間のヌクレオチド又は同一のアミノ酸残基のパーセントを示すことが意図されている。このパーセントは純粋に統計に基づいており、2つの配列間の差はランダムに分布しそれらの全体長を超える。従来2つの核酸又はアミノ酸配列間の比較は最適な方法によるアライメント後これらの配列を比較することにより行っている。この比較は区分毎に、あるいは“比較窓”を通して行うことができる。比較のための配列の最適アライメントはマニュアル操作に加え、Smith and Waterman (1981) [Ad. App. Math. 2:482]の局所相同性アルゴリズムにより、Neddleman and Wunsch (1970) [J. Mol. Biol. 48:443] の局所相同性アルゴリズムにより、Pearson and Lipman (1988) [Proc. Natl. Acad. Sci. USA 85:2444] の類似性サーチ法により、これらのアルゴリズムを用いたコンピュータソフトウェア(GAP, BESTFIT, FASTA and TFASTA in the Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, WI, or else by BLAST N or BLAST P comparison software ) により実行することができる。
【0010】
2つの核酸又はアミノ酸配列間の同一性のパーセントは、最適方法により整列(アライメント)された2つの配列を比較することにより決定され、比較される核酸又はアミノ酸配列はこれら2つの配列間の最適アライメントのために基準配列に対し付加あるいは削除を行うことができる。同一性のパーセントは2つの配列間で核酸又はアミノ酸残基が同一となる場合の同一位置の数を決定することにより、比較窓の中の位置の全体数でこの同一位置の数を割ることにより、及びこれら2つの配列間の同一性パーセントを得るために、得られた結果に100を掛けることにより計算される。
【0011】
例えば、サイトhttp://www.ncbi.nlm.nih.gov/gorf/b12.htmlで利用可能なBLAST プログラム “BLAST 2 sequences” (Tatusova et al, “Blast 2 sequences - タンパク質とヌクレオチド配列を比較する新規ツール”, FEMS Microbiol Lett. 174:247-250) を使うことが可能である。用いるパラメータは初期設定により得られ(具体的にはパラメータ“open gap penalty”に対し: 5及び“extension gap penalty”に対し: 2; 選択されるマトリクスはプログラムにより提案される例えばマトリクス“BLOSUM 62”)、比較される2つの配列の同一性パーセントは直接プログラムにより計算される。
【0012】
基準アミノ酸配列に対して少なくとも80パーセントの、好ましくは85%、90%、95%及び98%の同一性を持つアミノ酸配列により、これらは基準アミノ酸配列に対して特定の組み換え、具体的には少なくとも1つのアミノ酸の削除付加あるいは置換を行うが、トランケーションやエロンゲーションが好ましい。1つ又は複数の連続あるいは非連続アミノ酸の置換の場合、置換されるアミノ酸が“均等の(equivalent)”アミノ酸に置き換えられる置換が好ましい。“均等のアミノ酸”の表現は、ここでは基底構造のアミノ酸の1つにより置換され得るあらゆるアミノ酸を指すことを意図している。ただし、対応する抗体の生物活動を基本的に変更することのない置換であり、それは後述の実施例の中で具体的に記述する。
【0013】
これら均等アミノ酸は置換するアミノ酸との構造的相同性、あるいは実行可能な異なった抗体間の生物活動の比較試験の結果のいずれかに基づいて決定できる。
【0014】
例として、変更される対応の抗体の生物活動に大幅な変更が生じないように置換を実行できる可能性について記述する。例えばロイシンをバリン又はイソロイシンで、アスパラギン酸をグルタミン酸で、グルタミンをアスパラギンで、アルギニンをリシンで置換可能であり、同一条件でその逆の置換も当然予想できる。
【0015】
この発明に係る抗体は好ましくは完全ヒト単クローン抗体あるいはその機能的フラグメントである。
【0016】
具体的実施例において、この発明の抗体は、図3Bの配列番号4で表わされるアミノ酸配列との、最適アライメント後、少なくとも80%、好ましくは90%の同一性を持つアミノ酸配列を含む軽鎖、あるいは図3Aの配列番号5で表わされる配列を含むヌクレオチド配列により、又は配列番号5との、最適アライメント後、少なくとも80%、好ましくは90%の同一性を有する配列によりエンコードされる軽鎖を特徴とする。
【0017】
他の具体的実施例において、この発明の抗体は、図3Dの配列番号9で表わされるアミノ酸配列との、最適アライメント後、少なくとも80%、好ましくは90%の同一性を持つアミノ酸配列を含む重鎖、あるいは図3Cの配列番号10で表わされる配列を含むヌクレオチド配列により、又は配列番号10との、最適アライメント後、少なくとも80%、好ましくは90%の同一性を持つ配列によりエンコードされる重鎖を特徴とする。さらに他の実施例において、この発明に係る抗体は、図3Bの配列番号4で表わされるアミノ酸配列を含む軽鎖又は図3Aの配列番号5で表わされる配列を含むヌクレオチド配列によりエンコードされる軽鎖と、図3Dの配列番号 9により表わされるアミノ酸配列を含む重鎖又は図3Cの配列番号10により表わされる配列を含むヌクレオチド配列によりエンコードされる重鎖とを備える。
【0018】
この発明に係る抗体の機能的フラグメントにより、具体的にはFv, scFv (scは単一鎖), Fab, F (ab')2, Fab', scFv-Fc フラグメント又はダイアボディズ(diabodies) のような抗体フラグメント、あるいは化学修飾により又はリポソ−ムへの取り込みにより半減期が増加するあらゆるフラグメントを示すことが意図されている。化学修飾とは例えば、ポリ(エチレン)グリコール (“PEGylation”ペジレーション) (Fv-PEG, scFv-PEG, Fab-PEG, F(ab')2-PEG又はFab'-PEGと呼ばれるペグフラグメント) (“PEG”はPoly(Ethylene) Glycol ) のようなポリ(アルキレン)グリコールの添加である。このフラグメントはこの発明に係る配列番号 1, 2, 3, 6, 7 及び8 のCDR を含み、特にHIV株を中和する能力がある。
【0019】
好ましくは、前記機能フラグメントは抗体の可変重鎖又は可変軽鎖の部分配列により構成され、またはそれを含んでいる。その配列からそれらフラグメントは抽出される。前記部分配列は同一の結合特異性を保持するに十分である。好ましくは、これら機能フラグメントはFv, scFv, Fab, F (ab')2, F(ab'), scFv-Fc タイプ又はダイアボディズのフラグメントであり、これらは概してその由来する抗体と同一の結合特異性を有している。この発明によれば、抗体フラグメントはペプシンやパパインのような酵素による消化により、及び/又は化学的還元によるジスルフィド架橋の開裂により、上記のような抗体から取り込みを開始できる。他の方法としては、この発明に含まれる抗体フラグメントは当業者に良く知られている遺伝子組み換え技術により、あるいは、Applied Biosystems 社等により供給される例えば自動ペプチドシンセサイザ−によるペプチド合成によって取り込むことができる。
【0020】
より好ましい態様において、この発明は遺伝子組み換えにより、又は化学合成により得られた、この発明に係る抗体、又はその機能的フラグメントを備えている。
【0021】
好ましい態様において、この発明に係る前記機能フラグメントは、フラグメントFv, scFv, Fab, (Fab')2, Fab', scFv-Fc 又はダイアボディズ から、あるいは化学修飾、特にペジレ−ション又はリポソ−ム取り込みにより半減期が増加するあらゆる機能フラグメントから選択される。
【0022】
この発明は、配列番号5の配列との、最適アライメント後少なくとも80%、好ましくは85%、90%、95%、及び98%の同一性を持つ配列を含む単離された核酸にも関連する。
【0023】
この発明は、配列番号10の配列との、最適アライメント後少なくとも80%、好ましくは85%、90%、95%、及び98%の同一性を持つ配列を含む単離された核酸にも関連する。
【0024】
好ましい配列との、最適アライメント後少なくとも80%、好ましくは85%、90%、95%、及び98%の同一性を持つ核酸配列により、基準核酸配列に対し、具体的には欠失、トランケーション、エロンゲーション、キメラ融合、及び/又は置換、特に点置換のような特定の組み換えを行う核酸配列を示すことが意図される。それは好ましくは基準配列と同じアミノ酸配列に対する配列コードが遺伝子コードの縮退とつながりのある配列に関係し、あるいは、特に以下に説明するような好ましくは高い厳密性の条件下で、基準配列と特異的に雑種化(ハイブリッド化)を行える相補配列に関係する。
【0025】
高い厳密性条件下でのハイブリッド化は、温度条件やイオン強度条件が相補的DNAの2つのフラグメント間でのハイブリッド化が維持されるように設定されることを意図している。実例として、上述のポリヌクレオチドフラグメントを特定するためのハイブリッドステップの高い厳密性条件を次に示す。
【0026】
DNA-DNA又はDNA-RNAのハイブリッド化は2つのステップで実行される:(1)ハイブリッド化前は、温度42℃で5 × SSC (1 × SSCは0.15 M NaCl + 0.015 Mクエン酸ナトリウム液に相当)、50%のホルムアミド、7%のドデシル硫酸ナトリウム (SDS)、10 × Denhardt's、5%の硫酸デキストラン及び1%のサケ精子DNA を含有するリン酸緩衝液(20mM, pH 7.5)中に3時間;(2)実際のハイブリッド化は、プローブのサイズに応じた温度(即ち:プローブサイズ> 100 ヌクレオチドで42℃)で20時間、その後2 × SSC + 2%SDS中、温度20℃で20分間2回の洗浄、0.1 × SSC + 0.1%SDS中、温度20℃で20分間1回の洗浄。プローブサイズ> 100 ヌクレオチドには最後の洗浄が0.1 × SSC + 0.1%SDS中、温度60℃で30分間行われる。上述の限定されたサイズのポリヌクレオチドに対する高い厳密性のハイブリッド化条件は、Sambrook et al., (1989, Molecular cloning : a laboratory manual. 第2版. Cold Spring Harbor) の解説に従って、大又は小サイズのオリゴヌクレオチドの当業者が利用することができる。
【0027】
この発明は上記核酸、具体的には配列番号5及び配列番号10の核酸を有するベクターにも関係する。
【0028】
この発明は、特にクローニング及び/又はこの発明に係るヌクレオチド配列を含む発現ベクターを目的とする。例えば、上記のような、特に配列番号5及び配列番号10の核酸配列を備えるバキュロウィルストランスファーベクターを目的としている。
【0029】
この発明に係るベクターは、好ましくは決定された宿主細胞のヌクレオチド配列の発現及び/又は分泌を可能とするエレメントを含む。そのベクターは、それ故、プロモータ遺伝子、開始のシグナル及び翻訳の終止、並びに転写の適切な制限領域を含まねばならない。それは宿主細胞の中で、安定した状態で維持されなければならないし、翻訳されるタンパク質の分泌を指定する具体的なシグナルを随意持つことができなければならない。これら異なるエレメントは使用される宿主細胞の機能として、当業者により選択され最適化される。この趣旨で、この発明に係るヌクレオチド配列は選択された宿主細胞の自律複製ベクターへ導入でき、あるいは、選択された宿主細胞の統合的ベクターへ導入できる。
【0030】
そのようなベクターは当業者が現在使っている方法で形成され、結果として生じるクローンはリポフェクション、エレクトロポレーション、熱衝撃、又は化学的手法といった標準方法で適当な宿主細胞へ導入される。
【0031】
この発明に係るベクターは、例えば、プラスミドあるいはウィルス性のベクターである。それらは、この発明に係るヌクレオチド配列のクローンを作るため、あるいは発現するために宿主細胞を形質転換するうえで有用である。
【0032】
同様にこの発明は、この発明にかかるベクターにより形質転換される、あるいはこの発明にかかるベクターを有する宿主細胞を備えている。
【0033】
宿主細胞は原核細胞系又は真核細胞系から選択でき、例えば細菌性細胞から選択できるが、同様に酵母細胞や動物細胞、具体的には哺乳類細胞から選択できる。昆虫細胞や植物細胞を使うことも同様に可能である。
【0034】
従って、別の態様によれば、この発明は上記の抗R7Vヒト抗体を分泌する細胞株に関係する。例えば、上記抗体はEVB不死化Bリンパ球により、バキュロウィルスベクターを使ってSf9細胞のような昆虫細胞により、あるいは他の抗体生成細胞株、たとえばCHO (ATCC number CCL-61)、低フコシル化抗体を生成する遺伝子組み換えされたCHO、あるいはYB2/0 (ATCC CRL-1662) 細胞株により得られる。
【0035】
さらに別の態様によれば、この発明はこの発明に係る抗体あるいは機能フラグメントの1つの生成方法を目的とし、
a) この発明に係る宿主細胞の培地と適当な培養条件の中で培養し、及び
b) 前記培養細胞の培地から前記抗体を抽出する
ステップを備えるものである。
上記方法により獲得することができる抗体は、あるいはその機能フラグメントの1つは、この発明の範囲に含まれる。
【0036】
さらに他の態様によれば、この発明は医薬品として上記抗体、あるいはその機能フラグメントの1つに関係する。この発明はさらに、有効成分としてこの発明に係る抗体あるいはそのフラグメントの1つと、賦形剤及び/又は薬学的に受け入れられる媒体とを含む医薬組成物に関係する。
【0037】
さらに他の実施例において、この発明は、同時、別々、あるいは順次使用の併用製品として現在エイズ療法に使われている少なくとも1つの薬剤と上記に係る抗体をさらに含む上述の組成物を対象にしている。“同時使用”とはこの発明に係る組成物の2つの化合物を単一かつ同一の医薬品形態で投与するものとして理解される。“別々の使用”とはこの発明に係る組成物の2つの化合物を異なった医薬品形態で同時に投与するものとして理解される。“順次使用”とはこの発明に係る組成物の2つの化合物をそれぞれが異なる医薬品形態で順次投与するものとして理解される。
例えば、抗R7V抗体の投与を:
エファビレンツ + ジドブジン + ラミブジン
エファビレンツ + テノホビル + エムトリシタビン
スタブジン + ラミブジン + ネビラピン
リトナビルによりブーストされたロピナビル+ ジドブジン + ラミブジン
リトナビルによりブーストされたロピナビル+ テノホビル + エムトリシタビン
と組み合わせることが可能である。
【0038】
この発明は、特にHIV感染、エイズ治療に対して、例えばHAART療法の患者、具体的にはHAART療法の効果がない患者の治療に対して医薬品の調合のためにここに記載する抗体の使用を含む。
【0039】
この発明を以下の図面及び事例を参照してさらに説明する。
【図面の簡単な説明】
【0040】
【図1A】抗R7V抗体の発現に使う免疫グロブリン特異トランスファーベクターの概略図。pVT-Ckの概略図。軽鎖の発現を許すトランスファーベクター。
【図1B】抗R7V抗体の発現に使う免疫グロブリン特異トランスファーベクターの概略図。pVT-Cγ1の概略図。重鎖の発現を許すトランスファーベクター。
【図2A】R7V抗原上で選択された不死化Bリンパ球から抽出された全体RNAから合成されるc-DNA上に存在するVH配列のPCR増幅。
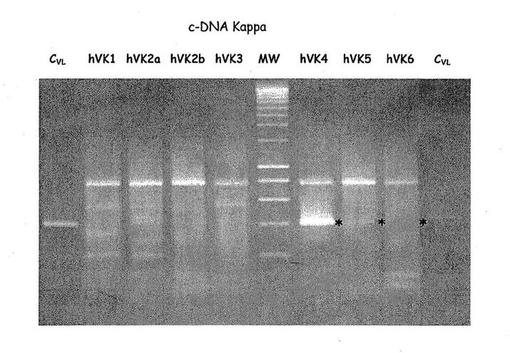
【図2B】R7V抗原上で選択された不死化Bリンパ球から抽出された全体RNAから合成されるc-DNA上に存在するVL配列のPCR増幅。
【図3A】最相同生殖細胞遺伝子と比較される不死化Bリンパ球に発現する抗体の軽鎖(K4) の可変領域のヌクレオチド配列。
【図3B】最相同生殖細胞遺伝子と比較される不死化Bリンパ球に発現する抗体の軽鎖(K4) の可変領域のアミノ酸配列。
【図3C】最相同生殖細胞遺伝子と比較される不死化Bリンパ球に発現する抗体の重鎖(M4) の可変領域のヌクレオチド配列。
【図3D】最相同生殖細胞遺伝子と比較される不死化Bリンパ球に発現する抗体の重鎖(M4) の可変領域のアミノ酸配列。
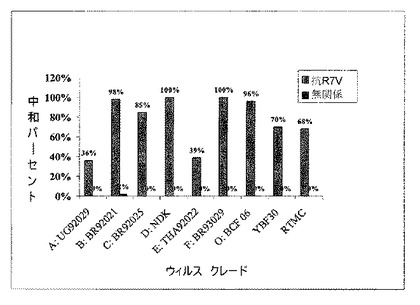
【図4】抗R7Vまたは無関係の抗体50 μg/ml によるHIV1クレードの中和。
【発明を実施するための形態】
【0041】
図1:抗R7V抗体の発現に使う免疫グロブリン特異トランスファーベクターの概略図である。
図1A:pVT-Ckの概略図−軽鎖の発現を許すトランスファーベクター。
図1B:pVT-Cγ1の概略図−重鎖の発現を許すトランスファーベクター。
図2:R7V抗原上で選択された不死化Bリンパ球から抽出された全体RNAから合成されるc-DNA上に存在するVH (図2A) 配列又はVL (図2B) 配列のPCR増幅。増幅は特定のVH 又はVL遺伝子ファミリーに特異的な適合定数3' プライマー及び5'プライマーのセットで、1.1材料と方法に報告されるように実行された。PCR反応の20μlは1.5%のアガロースゲルで分別されエチジウムブロマイドにより染色された。Lane CVH: コントロールVH配列。Lane CVL: コントロールVL配列。Lane MW: スマートラダー分子量マーカー(Eurogentec): 200, 400, 600, 800, 1000, 1500, 2000, 2500, 3000, 4000, 5000, 6000, 8000, 10000 bp.
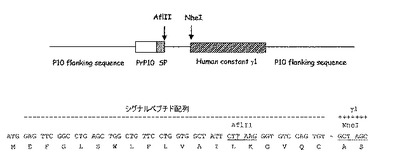
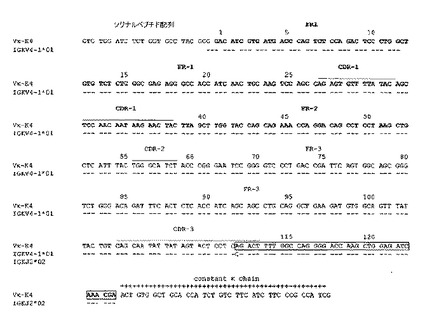
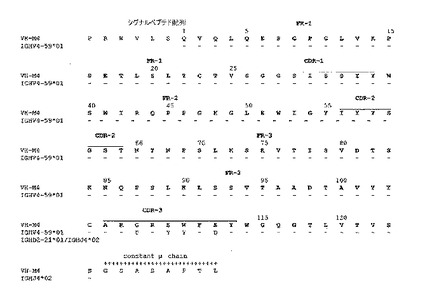
図3:最相同生殖細胞遺伝子と比較される不死化Bリンパ球に発現する抗体の軽鎖(K4) 及び重鎖(M4) の可変領域のヌクレオチド配列(図3A及び図3C)及びアミノ酸配列(図3B及び図3D)。アミノ酸配列は1文字コードで与えられている。使用される番号方式はIMGT (http://imgt.cines.fr) のルールに基づいている。VH及びVL配列の相補性決定領域 (CDR) は強調されている。配列のダッシュはトップラインに与えられる残基との一致を示している。IGHJ, IGHD, 及びIGKJ 遺伝子はボックスに収まっている。
【実施例】
【0042】
実施例1:有効ヒト組み換え型抗R7V抗体の単離生成
【0043】
1.1材料と方法
【0044】
1.1.1細胞とウィルス
ヒト末梢血単核細胞(PBMC)は血清反応陰性の健康なドナーの未使用の K2E-EDTA 血液サンプルからFicoll- Paque(Amersham)勾配遠心分離により分離された。 培養される細胞は完全RPMI培地の中で1 × 106 cells/ml の密度で次の組成で成長した:10%の加熱不活性化したウシ胎仔血清 (GIBCO) , 1%のペニシリン/グルタミン(GIBCO) , 10 UI/ml IL2 (Euromedex), 最初の3日間10 μg/ml PHA-P (Difco) , 及び2 μg/ml ポリブレン (Biowhittaker), を補充された RPMI 1640 (Biowhittaker) 。
CEM細胞株は0.5 × 106 cells/mlでRPMI-10%の培地 (10%の加熱不活性化したウシ胎仔血清、1%のペニシリン/グルタミン、2 μg/ml ポリブレンを含むRPMI 1640) の中で培養された。
NDK (クレードD) 及びAZT耐性RTMC (クレードB) ウィルスは感染CEM細胞で生成した。92UG029 (クレードA), 92BR021 (クレードB), 92BR025 (クレードC), 及び93BR029 (クレードF) ウィルスは当初AIDS Research and Reference Reagent Program, Division of AIDS, NAID, NIHにより提供され、PBMCにより生成された。ウィルスBCF06 (クレードO), 及びYBF30 (古いクレード) はF. Barre-Sinoussi (パスツール研、フランス) よりご好意で頂いた。感染細胞の上澄みで漸増したウィルスのサンプルはマイナス80℃で凍結保存された。
Sf9細胞は5%の加熱不活性化したウシ胎仔血清 (GIBCO)を補充されたTC100 培地(GIBCO)の中で28℃で維持された。野生型のAutographa californica 複合核多角体病 (AcMNPV) ウィルスクローン1.2(文献17)及び組み換え型バキュロウィルスがSf9細胞の中で繁殖した。
【0045】
1.1.2 ノンプログレッサー患者からの末梢血単核細胞 (PBMC) の単離
インフォームドコンセントを行った血清反応陽性のノンプログレッサの患者が我々の研究に登録され、Ficoll-Paque 密度勾配法による単離により未使用のK2E-EDTA静脈血からPBMCを精製した。これらのPBMCは、15%の加熱不活性化したFCSと1%のペニシリン/グルタミンを補充したRPMI 1640 培地の中で不死化前のBリンパ球の成長を助けるためにIL2及びPHA無しで2日間予備培養した。
【0046】
1.1.3 Epstein-Barr ウィルス(EBV)によるBリンパ球の不死化
その後Bリンパ球は2ml のB-95.8 培養上澄み(EBV生成細胞株)を、加熱不活性化した3ml 10 % FCSの中で予備培養した9 × 106 PBMC, 50 ml の円錐管の1%のペニシリン/グルタミンRPMI と混合することにより不死化された。37℃の水槽で2時間の培養後、10%の加熱不活性化したFCS, 1μg/μlのシクロスポリンA(Calibiochem)及び1%のペニシリン/グルタミンを 補充された 5 mlのRPMI 1640を加えた。その10-mlの細胞懸濁液は25 cm2の組織培養フラスコへ移され、37℃で加湿されCO2 濃度5%の培養器の中でそのまま4週間培養された。4週間の培養の最後には、EVB-不死化細胞が肉眼で見える塊を形成した。この細胞株をRPMI-20%、106 cells/mlで週に2回リフィーディングすることで維持した。
【0047】
1.1.4 Bリンパ球を分泌する抗R7V抗体の単離
R7Vペプチドのアミノヘキサン酸形による磁気ビーズのコーティング:10 μg のR-8-Ahx ペプチド (Neosystem) を107磁気トシル活性ビーズ(Dynal Dynabead(登録商標) M450) により低速傾斜回転、37℃で16-24時間培養した。ビーズは製造者の手順に従って洗浄し、PBS pH 7.4の中、4×108 beads/mlで再懸濁した。
Bリンパ球を分泌する抗R7V抗体の磁気選別:1 mlの無菌PBSに107 のEBV-不死化されたBリンパ球は24x106 R-8-Ahx でコートされたビーズと4℃で20分混合され、細胞がビーズに固着しなくなるまで3回繰り返される。ロゼット細胞は管を2分間磁石に入れておくことで単離された。ビーズはそのままにして上澄みを取り除き、細胞はPBS洗浄緩衝液に再懸濁された。ビーズに固着した細胞を37℃でRPMI-20% FCS 37℃, 5% CO2 で培養する前に、洗浄ステップを3回繰り返した。1日後細胞はビ−ズから分離し、106 cells/mlに成長した。
この磁気選別は2週間の培養後同じ手順で、Bリンパ球を分泌するこれらの予め選択した抗R7V抗体に対し繰り返された。
【0048】
1.1.5 ELISA 手順
抗R7V抗体は製造者が示す抗R7V ELISA試験 (Anti R7VTM IVR 96000, IVAGEN, France) により検出された。簡潔に言えば、陽性対照、陰性対照、遮断キャリブレータ及び希釈抗体( 100 μl/well)が、R7Vをコーティングする試験板に添加され、室温で30分培養された。結合された抗R7V抗体は西洋ワサビペルオキシダーゼ抱合抗ヒトIgG抗体により検出された。
【0049】
1.1.6 中和試験
ウィルスのストックは、以下の希釈に対応して1回の試験(文献18)で100 TCID50を持つように前もって滴定された:HIV-1NDK (希釈10-5), HIV-1RTMC AZT-耐性(希釈 5×10-5), 92UG029 (希釈10-2), 92BR021 (希釈10-3), 92BR025 (希釈10-2), THA92022 (希釈10-2), 93BR029 (希釈10-2), BCF06 (希釈10-4), 及びHIV-1YBF30 (希釈10-3)。ウィルス(50 μl) の希釈液は100 μg /mlの抗体(最終濃度 50 μg/ml)を含む50 μl RPMI-0%で96- wellマイクロタイタープレートで、加湿された37℃、5%CO2 の培養器の中で1時間予備培養された。PBMC (50 μl中1 ×10 6) はウィルス‐抗体混合液に1時間37℃で添加され、細胞は培地で3回洗浄され、106 cells/ml、 24- well マイクロタイタープレートで、50 μg/ml 抗体完全RPMI-10 %の存在下、最初の3日間培養された。 培養液は10日間成長し、3日毎にリフィーディングされた。同じ試験がウィルス対照 (抗体無しのHIV感染細胞)、細胞対照 (抗体無しの非感染細胞) 及び抗体対照 (非HIV 関連エピトープに対抗する無関係の抗体) に対して行われた。それぞれのサンプルのウィルス複製を測定するため、逆トランクリプタス酵素が以下のように定量化された。3日毎に採取した無細胞の1 ml の上澄みサンプルは4℃ で5分間 95,000 rpm (TL 100 Beckman) の超遠心分離機にかけた。ウィルスペレットは0.1 % Triton X-100 NTE (NaCl 100mM, Tris 10mM, EDTA 1mM) 緩衝液10ml中で再懸濁され、ウィルス酵素を解放した。酵素反応はTris 50mM, pH 7.8; MgCl2 20mM; KCl 20mM; ジチオスレイトール (DTT) 2mM ; オリゴ dT 0.25 OD/ml; ポリrA 0.25 OD/ml 及び3H dTTP 50 μCi/ml を含む50 μl の反応混合液で行われた。37℃で1時間後、抗原抗体反応は5%TCA中、1 ml のピロリン酸ナトリウムで停止し、合成されたDNA生産物が20%のトリクロロ酢酸で沈殿され、ミリポア0.45 μm でろ過により採取され、パッカードシンチレーションカウンターによりdpm/mlでβ放射能が測定された。中和のパーセントは以下のように表わされた:
[100 - (サンプルの逆転写酵素活性/ウィルスの逆転写酵素活性) × 100]
【0050】
1.1.7 抗R7V 特異性を発現する抗体の可変領域の単離とクローニング
手順はマウス可変抗体領域の増幅のための技術を応用した。(文献19)全てのRNAはRNeasy キット(Qiagen) を使って約5×106不死化Bリンパ球から抽出された。簡潔に言えば、細胞は600 μl のRLTTM/βメルカプトエタノール緩衝液で溶解され、20 ゲージ針連続継代により均質化された。600 μl の70%エタノールを添加後、混合液はRNeasy カラムに沈着され、15秒間12,000 rpm (Biofuge, Heraeus) で遠心分離機にかけた。カラムは700 μl RW1TM 緩衝液及び500 μl RPETM 緩衝液で連続洗浄を行った。 RNAは50 μl RNAse フリーの水で溶出され、使用するまでマイナス80℃で保存された。
全てのRNAと5つの特異的プライマーがヒト免疫グロブリンの定常部でハイブリッド化を行い、hCLa, hCLb, hCK, hCG, 及びhCM (表3) が使われて、ラムダ、カッパ、ガンマ1及びミューmRNAそれぞれに対応する第1ストランドc-DNAs を合成した。逆転写は次のように実行された:全RNA 1μg, 4μlの10X RTTM 緩衝液(Qiagen), 4μl の5mM 各dNTP (Qiagen), 10 pMoles/μl 20 ユニットの RNAse インヒビター (Roche) での4μl の特異的プライマー及び最終容積40 μlのオムニスクリプト逆転写酵素 (Qiagen) 8ユニット。混合液は1時間37℃で培養された。逆転写活動は93℃で5分間加熱不活性化された。VH 配列、VL 配列の全長は、ヒト免疫グロブリン(表3) の重鎖及び軽鎖のシグナルペプチド配列にデザインされる特異的プライマー及び充填剤としてラムダ、カッパ、ガンマ又はミュー第1ストランドc-DNA を使って、PCRによって増幅された。PCR 反応は、2 μl の10X Vent DNA ポリメラーゼ(Biolabs), 2 μl の10mM各dNTP (Biolabs), 各プライマーの20 pMoles, 1.5 μlの25 mM MgSO4, 1ユニットのVent DNA ポリメラーゼ(Biolabs), 0.5 μl の逆転写混合液を含む20 μl の最終容積で実行された。30サイクルの増幅が95℃で30秒間、55℃で45秒間、72℃で1分間実行された。72℃で10分間延長の後、PCR生成物が1.5% のアガロースゲル (SeaKem, FMC) で分別されエチジウムブロマイドで染色された。
PCR生成物はゲル洗浄され、Advantage Taq ポリメラーゼ(Clonetech)で増幅され、プラスミド pGemT easy (Promega) でクローニングされた。PCR増幅(MWG Biotech) に使われる3'及び5' プライマを使って、両方のストランド上に導入の配列が決定された。可変領域の配列比較、生殖細胞遺伝子分析がBLAST(文献20)及びIMGT(文献21)データベースを使って実行された。
【0051】
1.1.8 抗R7V抗体発現組み換え型バキュロウィルスの生成
VH, VL 配列は, ヒト免疫グロブリンシグナルペプチド配列、2つのユニークな制限酵素認識部位及びヒトガンマ1とカッパ定常部をそれぞれエンコードする配列を含む特異的トランスファーベクター pVTCγ1 及びpVTCκ (図1)に導入された。pVTCγ1 ベクターはシグナルペプチド配列の中のユニークなAfl II部位及びガンマ1配列の2つの最初のコドン有するNhel 部位を含み、一方pVTCκはシグナルペプチド配列の中にユニークなBssH II部位及びJ領域の最後の保存アミノ酸及び定常カッパ領域の最初のアミノ酸がオーバラップするBsiW I部位を含む。
適切な制限酵素認識部位が以下のプライマーを使うPCRによりVH, VL 配列の5' 及び3'末尾に導入された:
FOR-M4: CCATCTTAAGGGTGTCCAGTGTCAGGTGCAGCTGCAGGAGTCGGGCCCAGGACTGGTGAAGC (SEQ ID N°16),
BAC-M4: GCATGCTAGCTGAGGAGACGGTGACCAGGGT (SEQ ID N°17),
FOR-K4: CGATGCGCGCTGTGACATCGTGATGACCCAGTCT (SEQ ID N°18) 及び
BAC-K4: CGATCGTACGTTTGATCTCCAGCTTGGTCCCCTGGCC (SEQ ID N°19)。
VHに対してはAfl II-Nhel, VLに対してはBssH II-BsiWIにより消化されたPCR生成物は精製されて個々のトランスファーベクターpVTCγ1 及びpVTCκに導入された。最終作成物pVT Cγ1-M4及びpVTCκ-K4はシークエンシングにより制御された。
抗体発現組み換え型バキュロウィルスは前述するように(文献22、10、11)Sf9細胞の共トランスフェクション後に生成された。生殖能力のあるクローンをELISAによりスクリーニングした(文献23)。簡潔に示せば、100 μl の1 μg/ml 抗ヒト重鎖 Fdγ1 ポリクローナル抗体 (The Binding Site) をコーティングしたマイクロタイタープレートが細胞培養上澄みの連続希釈により37℃で2時間培養された。結合した組み換え型IgGが西洋ワサビペルオキシダーゼ標識抗ヒトカッパ軽鎖抗体 (Sigma) を使って検出された。
組み換えウィルスのゲノムはサザンブロットにより制御された。7 mlの細胞培養上澄み中のウィルス粒子は35,000 rpm 40 分間で沈殿させた(TL 100.4, Beckman)。ペレットはTEK 緩衝液の1 ml (0.1 M Tris, 0.1M Na2EDTA 2 H2O, 0.2 M KCl, pH 7.5)に10 μlのプロテイナーゼ K 20 mg/ml水溶液(Roche) 及び10 μl のN ラウリルサルコシン(Sigma) 10 % (w/v) 水溶液を加えて再懸濁し、一晩50℃で培養した。ウィルスDNAがフェノールとクロロホルムイソアミルアルコール(24:1 v/v) により連続的に抽出され、エタノールで沈殿させた。水で再懸濁した後、Hind IIIでDNAは消化された。制限されたDNAは1% アガロースゲルで電気泳動により分析され、ニトラン膜 (Schleicher and Schull)へ移された。ヒト定常部γ1 及びκ領域をそれぞれエンコードするc-DNAは製造者が推奨するようにジゴキシゲニンで標識され、ハイブリダイゼーション プローブとして使われた。洗浄後、ブロットはアルカリホスタファーゼ ( Roche, dilution 1: 10,000)に接合する抗ジゴキシン抗体により培養された。標識されたDNAの検出は、化学発光基質CSPD(Roche)により行った。
【0052】
1.1.9 組み換え型抗体の生成と精製
Sf9 細胞は密度500,000 cells/ml, 400ml のローラボトルの無血清培地に蒔かれ、2 per cell の感染多重度で感染させた。28℃で4日間の培養後、上澄が採取され、製造者が示すように、分泌した組み換え型抗体がタンパク質A セファロース (Amersham) 上に精製された。精製されたIgGの量がELISAによって測定された(文献23)。
組み換え型抗R7V抗体が同様な条件でCHO 発現システムによっても生成された。
【0053】
1.2 結果
【0054】
1.2.1 B リンパ球を分泌する抗R7V抗体の選択
B リンパ球を生成する抗R7V抗体はR7Vをコーティングした磁気ビーズを使ってノンプログレッサーのHIV 感染患者から選択された。B リンパ球を分泌する27%の抗R7V抗体を最初の選択で取得し、事前に選択したB リンパを分泌する抗R7V抗体により2回目の選択で14%を取得した。B 細胞培養上澄みにはフリーな抗R7V抗体は抗R7V ELISA によって検出されず、抗体は分泌するB リンパ球膜に結合したか、ELISA試験の検出限界以下であったかいずれかを示唆している。
【0055】
1.2.2 選択した不死化B リンパ球により発現したVH 及びVL配列の単離及びクローニング
選択されたB リンパ球により発現した抗体のVH, VL 領域の増幅は、我々が以前マウス免疫グロブリンに対し記述した(文献19)RT-PCRによって行われた。
図2に示すように、プライマーの僅かな組合せのみが割り当てられたサイズ、VHに対しては約450 bp, VL に対しては 400 bp でフラグメントの増幅をもたらした。hCG/hVH5, hCM/hVH2 及び hCM/hVH3 を使って僅かのしまが観察されるに過ぎなかったが、hCM/hVH4ではより多くの物質が観察できた。主な生成物はhCK/hVK4でも合成された。しかしながら、hCLa 及び hCLb プライマー(図示せず)を使ういかなる結合も増幅は検出できなかった。PCR生成物の配列解析及びBLAST 解析はそれらのうち2つのみ、M4 フラグメント (hCG/hVH4) 及び K4 フラグメント(hCK/hVK4) のみがヒト重鎖、軽鎖の可変領域にそれぞれ対応することを示した。これらの結果は選択された不死化B リンパ球の集団がおそらくは単クローン抗体で、膜IGMカッパ抗体を発現することを示している。これら配列とIMGTデータベースとの比較によりVH-M4 重鎖可変領域の配列がIGHV-4-59*01(文献24), IGHD2-21*01(文献25)及び IGHJ4*02(文献26) 生殖細胞遺伝子(図3C)の転位から生じることを示している。それに対応するVκ-K4はIGKV4-1*01(文献27)/ IGKJ2*02(文献28), 軽鎖可変領域(図3A)の転位を示している。興味深いことに、この抗体はカッパ軽鎖レパートリーから最もJ-proximal なIGKV4-1遺伝子を使った。このような軽鎖領域は主として変異株はなく、唯一の変異株が相補性決定領域3のIGKV/IGKJ 接合に存在した(図3B)。一方、相補性決定領域3において4つのアミノ酸変異株をもたらした7つのヌクレオチド置換がVH-M4 配列に観察される中、ただ2つのサイレントヌクレオチド置換がフレームワーク領域に認められた(図3C及び3D)。
【0056】
1.2.3 バキュロウィルス発現システムにおける抗R7V抗体の発現
抗R7V抗体の可変領域をエンコードする配列は、重鎖及び軽鎖のカセットバキュロウィルストランスファーベクター (i)ポリへドリン遺伝子座の中で再結合するpVT-CK 及び(ii)P10 遺伝子座の中で再結合するpVT-Cγ1に導入された。これらの生成物で、重鎖及び軽鎖遺伝子は合成P10 プロモータ、P'10及びP10プロモ−タのそれぞれに制御される(図1)。特異的プライマーはK4及びM4フラグメントを増幅するよう設計されており、図1に示す免疫グロブリンシグナルペプチド配列及び定常領域の枠組みでそれらの直接クロ−ニングを行う。この2つの最終生成物pVT-Cκ-K4及びpVT-Cγ1-M4は配列決定により制御され、精製したウィルスDNAのもとでSF9細胞を同時遺伝子導入するために使われた。2重組み換え型ウィルスは前記(文献10、11)のように2回の組み換えの後に取得された。組み換え型ウィルスは血小板洗浄され増幅された。感染された細胞の細胞培養上静の抗体の存在が抗ヒト抗体ELISAによって分析された。4つの繁殖力のあるクローンのゲノムはヒトγ1及び κ定常部DNAをプローブとして使ってサザンブロット法により制御された。AcR7VI/K4-M4と名付けられた1つのウィルスクローンがさらなる実験のために選択された。
【0057】
1.2.4 組み換え型抗R7V抗体の特異性
組み換え型抗R7V抗体 はウェルの0.625μgの抗体濃度に相当する6.25 μg/ml においてもIVAGEN 抗R7V ELISA キットにより陽性であった。無関係な抗体はその濃度に関係なく陰性であった。
ノンプログレッサの患者から精製した抗R7V抗体について先に報告したように、組み換え型単クローン抗体はフローサイトメトリー分析(データは示していない)が実証するようにどの細胞とも結合しない。
【0058】
1.2.5 HIV-1の複数クレードに対する中和試験
患者から精製した抗R7V抗体は幅広い中和スペクトルを表わすことが説明された。そこでこの抗R7V単クローン抗体はいくつかのクレードに対し同一条件下で試験された。治療抗体としての使用を確かめるため、中和試験は薬物耐性ウィルス(RTMC)に対してもなされた。抗R7V組み換え型抗体の中和効果を測定するために、50 μg/ml で希釈された抗体が細胞に感染する前のいくつかのHIV-1のクレードと混合された。抗R7V抗体はHIV-1の8つのクレードとAZT耐性クレードB RTMCウィルスを中和した(図4)。バキュロウィルスシステムで発現し同一の条件下で対照として使われた無関係の抗体に対しては中和は観察されなかった。85 % 以上の中和が5つのクレード(B, C, D, F, O)に対して得られた。50 μg/mlの抗R7V組み換え型抗体については、異なるウィルスに応じて異なるパーセントの中和が得られた。この異種の結果は、HIV ノンプログレッサ患者から精製した抗R7V抗体の結果と同じであり、おそらくはウィルスにより様々の量のR7Vエピトープによりもたらされたからであろう。
【0059】
CHO 発現システムにおける組み換え型抗R7V抗体の作成
【0060】
1) K4M4 ロット番号13.11.06: 40 μg/mlの抗R7V ELISA 陽性結果
【表1】

【0061】
2) K4M4 ロット番号 28.02.07: 50 μg/mlの抗R7V ELISA 陽性結果
【表2】
【0062】
1.3 結論
我々はここにバキュロウィルス発現システムによる組み換え型ヒト抗R7V抗体の生成に関する結果を報告する。このシステムは機能的組み換え型抗体を大量に生成するうえで迅速かつ効率的である(文献10、11、29)。鱗翅目細胞に発現する組み換えタンパクには、哺乳類細胞で観測される全ての翻訳後修飾が見られる。しかしながら、N-結合型オリゴ糖は短く、実質的には高マンノース又は少数マンノース型である(文献30、31)。抗体の生物活動は免疫グロブリンのCH2 定常領域のAsn-292に結合するN型グリカンに大きく依存している(文献32、33)。未完成の糖鎖付加パターンではあるが、Sf9 細胞で発現された組み換え型抗体は、Clq 及びFcγR 結合による補体依存性及び抗体依存性の細胞媒介性細胞傷害のような特異的生物活動を示した(文献14、16、13)。
HIV-1ノンプログレッサー患者により生成された抗R7V抗体を単離し特性を示すために、EBV ウィルスにより不死化したR7V-反応性B 細胞が患者の一人から選択され、IgG 又はIgM 免疫グロブリンの可変領域をエンコードするc-DNAがRT-PCR を使って特異的に増幅された。この目的のため、V 遺伝子ファミリーに関わりない、ヒトVH 及びVL 領域の特異的増幅ためのコンセンサスプライマーの3つの原型がデザインされた。
そのシグナル配列でハイブリッド化するこれらプライマーはヒト定常領域γ, μ, κ, λ それぞれに向けられる3' プライマーのセットと併せて使われた。体細胞変異を受ける“FR”増幅戦略で目標となるフレームワーク 1の領域とは対照的に(文献34、35)、シグナル配列の突然変異の頻度は非常に低く、そのためこの領域のプライミングは突然変異無しで全体配列の増幅が可能となる。
【0063】
これらc-DNAs の配列の分析により、これらの不死化細胞がおそらく唯一の細胞膜IgM カッパ抗体を発現する単クローン抗体であることが分かった。VL 配列の大部分がVJ ジャンクションで唯一のサイレント変異を伴い突然変異がないのに対し、FR3 でただ2つのサイレント変異を伴うVH領域のVDJ ジャンクションで6つの突然変異アミノ酸がCDR3 の中に見つかっている。その可変領域で低い突然変異率が観察されるにもかかわらず、この組み換え型抗体はフローサイトメトリー分析に従ってどんな細胞にも反応しないのでポリリアクティブではない。
【0064】
HIV-1 亜型 A, B, C, D, E, F, N, O ウィルス及び抗レトロウィルス治療耐性ウィルスに対するこの完全ヒト組み換え型抗体の中和能力は、ノンプログレッサ患者から得る多クローン性抗体と明らかに同じである。これらの結果は、全てのHIV-1 変異株が、細胞により生成されたR7V エピトープを捕捉することを裏付けている。50 μg/mlの抗R7V抗体により異なった中和のパーセントが得られたのは、おそらくウィルスに存在するR7V の様々の量に関連している。増え続ける抗体はそれぞれのクレードに対し100 % の中和を達成するように試験されねばならない。
【0065】
単クローン抗体にとってHIV感染患者の治療薬として作用するために最も重要な特徴の1つは、中和のその幅広いスペクトルである。HIVは感染時間とともに及び抗レトロウィルス治療(エスケープ突然変異体の出現)とともに1つの個体から他へと絶え間なく変化し、免疫システムがウィルスの複製を制御することの難しさを物語っている。今日、抗R7V抗体とは別に、幅広い中和を行う他の4つの単クローン抗体は、全てHIV-1 亜型B に抗するものであるが、そのような潜在力を示している。面gp 120 上に結合するCD4 に抗するIgG1b12は無症状のHIV 陽性個人からファージ提示技術によって生成されている(文献36、37、38)。2F5 及び4E10 抗体はgp41(文献39、40)の定常部を認識し、2G12 はgp120上エピトープに抗する(文献41、42)。これら4つの抗体は幅広く中和する抗体として報告されているが、最も効果を発揮するのはそれらが組み合わされた時であった(文献43、44)。
【0066】
我々が得た結果は、組み換え型抗R7V抗体がHIV-1 亜型C単離株を中和することを示した。この亜型では、単クローン抗体2F5 及び2F12 は効果が無く、IgG1b12 は部分的に効果があり、そして4E10 のみが顕著な活動を示す(文献45)。
【0067】
従って、抗R7V抗体は今日まで報告されているHIV-1に対し最も幅広く効果のあるMab (単クローン抗体) の1つであると思われる。その細胞起源にもかかわらず、抗R7V抗体を生成するどの患者も自己免疫疾患のいかなる臨床的症状も示さないので、R7V エピトープは自己免疫反応には関与しない(文献5)。このことは、この抗R7V抗体はHIV 感染患者の治療にとって強力な候補となることを裏付けている。
【0068】
【表3】
【0069】
参考文献一覧
1. Oelrichs R, Tsykin A, Rhodes D, Solomon A, Ellett A, McPhee D, et al: Genomic sequences of HIV type 1 from four members of the Sydney Blood Ban Cohort of long-term nonprogressors. AIDS Res Hum Retrovir 1998 ;14:811-814.
2. Sanchez G, Xu X, Chermann JC and Hirsch I: Accumulation of defective viral genomes in peripherical blood mononuclear cells of Human Immunodeficiency Virus type 1-infected individual. J Virol 1997;71 :2233-2240.
3. Connor RI, Sheridan KE, Ceradini D, Choe S, Landau NR: Change in coreceptor use correlates with disease progression in HIV 1-infected individuals. J Exp Med 1997;185 : 621-628.
4. De Roda Husman AM, Schuitemaker H: Chemokine receptors and the clinical course of HIV-I infection. Trends Microbiol 1998;6:244-249.
5. Haslin C, Chermann JC: Anti-R7V antibodies as therapeutics for HIV-infected patients in failure of HAART. Curr Opin Biotechnol 2002;13(6):621-624.
6. Galea P, Le Contel C, Coutton C, Chermann JC: Rationale for a vaccine using cellular-derived epitope presented by HIV isolates. Vaccine 1999 ;17:1700-1705.
7. Arthur LO, Bess JW Jr, Sowder RC 2nd, Benveniste RE, Mann DL, Chermann JC, et al.: Cellular proteins bound to immunodeficiency viruses: implications for pathogenesis and vaccines. Science 1992; 258:1935-1938.
8. Le Contel C, Galea P, Silvy F, Hirsch I, Chermann JC: Identification of the beta- 2-microglobulin-derivated epitope responsible for neutralisation of HIV isolates. Cell Pharmacol 1996;3:68-73.
9. Hasemann CA, Capra D: High-level production of a functional immunoglobulin heterodimer in a baculovirus expression system. Proc Natl Acad Sci USA 1990;87: 3942-3946.
10. Poul MA, Cerutti M, Chaabihi H, Ticchioni M, Deramoult FX, Bernard A, et al. : Cassette baculovirus vectors for the production of chimeric, humanized, or human antibodies in insect cells. Eur J Immunol 1995 ; 25:2005-2009.
11. Poul MA, Cerutti M, Chaabihi H, Devauchelle G, Kaczoreck M, Lefranc MP : Design of cassette baculovirus vectors for the production of therapeutic antibodies in insect cells. Immunotechnology 1995 ;1 : 189-196.
12. zu Putlitz J, Kubasek WL, Duchene M, Marget M, von Specht BU, Domdey H: Antibody production in Baculovirus-infected insect cells. Biotechnology (New York) 1990;8:651-654.
13. Troadec S, Chentouf M, Cerutti M, Nguyen B, Olive D, Bes C, Chardes T: In vitro anti-tumoral activity of baculo virus-expressed chimeric recombinant anti-CD4 antibody 13B8.2 on T-cell lymphomas. J Immunother (in press) 2006.
14. Carayannopoulos L, Max EE, Capra JD : Recombinant human IgA expressed in insect cells. Proc Natl Acad Sci USA 1994; 91 :8348-8352.
15. Nesbit M, Fu ZF, McDonald-Smith J, Steplewski Z, Curtis PJ: Production of a functional monoclonal antibody recognizing human colorectal carcinoma cells from a baculovirus expression system. J Immunol Methods 1992;151 : 201-208.
16. Edelman L, Margaritte C, Chaabihi H, Monchatre E, Blanchard D, Cardona A, et al.: Obtaining a functional recombinant anti-rhesus (D) antibody using the baculo virus-insect cell expression system. Immunology 1997;91 :13-19.
17. Croizier G, Croizier L, Quiot J-M, Lereclus D: Recombination of Autographa californica and Rachiplusia ou nuclear polyhedrosis viruses in Galleria mellonella L. J Gen Virol 1988;69: 177-185.
18. Rey F, Donker G, Hirsch I, and Chermann JC: Productive infection of CD4+ cells by selected HIV strains is not inhibited by anti-CD4 monoclonal antibodies. Virology 1991;181 : 165-171.
19. Chardes T, Villard S, Ferrieres G, Piechaczyk M, Cerutti M, Devauchelle G, et al. : Efficient amplification and direct sequencing of mouse variable regions from any immunoglobulin gene family. FEBS Lett 1999 ;452:386-394.
20. Altschul SF, Madden TL, Schaeffer AA, Zhang J, Zhang Z, Miller W, et al.: Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res 1997;25:3389-3402.
21. Lefranc MP, Giudicelli V, Kaasl Q, Duprat E, Jabado-Michaloud J, Scaviner D: IMGT, the international ImMunoGeneTics information system. Nucleic Acids Research 2005;33:D593-D597.
22. Cerutti M, Chaabihi H, Devauchelle G, Gautier L, Kaczorek M, Lefranc MP, et al.: Recombinant Baculovirus and use thereof in the production of monoclonal antibodies. INRA-CNRS Patent, France, FR 94/01015 1994.
23. Bes C, Briant-Longuet L, Cerutti M, Heitz F, Troadec S, Pugniere M, et al. :
Mapping the paratope of the anti-CD4 recombinant Fab 13B8.2 by combining parallel peptide synthesis and site-directed mutagenesis. J Biol Chem 2003;278: 14265-14273.
24. Van der Maarel S, van Dijk KW, Alexander CM, Sasso EH, Bull A, Milner EC: Chromosomal organization of the human VH4 gene family. Location of individual gene segments. J Immunol 1993;150:2858-2868.
25. Siebenlist U, Ravetch JV, Korsmeyer S, Waldmann T, Leder P: Human immunoglobulin D segments encoded in tandem multigenic families. Nature 1981;294:631-635.
26. Mattila PS, Schugk J, Wu H and Makela O: Extensive allelic sequence variation in the J region of the human immunoglobulin heavy chain gene locus. Eur J Immunol 1995 ;25(9):2578-2582.
27. Klobeck HG, Bornkamm GW, Combriato G, Mocikat R, Pohlenz HD, Zachau HG : Subgroup IV of human immunoglobulin K light chains is encoded by a single germline gene. Nucleic Acids Res 1985;13:6515-6529.
28. Sahota SS, Leo R, Hamblin TJ, Stevenson FK: Myeloma VL gene sequences reveal somatic hypermutation with intraclonal homogeneity, and a role for VL in antigen selection (Unpublished, access number : Z70260) 1996.
29. Lieby P, Soley A, Levallois H, Hugel B, Freyssinet JM, Cerutti M: The clonal analysis of anticardiolipin antibodies in a single patient with primary antiphospho lipid syndrome reveals an extreme antibody heterogeneity. Blood 2001;97:3820-3828.
30. Marchal I, Mir AM, Kmiecik D, Verbert A, Cacan R: Use of inhibitors to characterize intermediates in the processing of N-glycans synthesized by insect cells: a metabolic study with Sf9 cell line. Glycobiology 1999;9: 645-654.
31. Marchal I, Jarvis DL, Cacan R, Verbert A: Glycoproteins from insect cells: sialylated or not ? Biol Chem 2001;382:151-159.
32. Boyd PN, Lines AC, Patel AK: The effect of the removal of sialic acid, galactose and total carbohydrate on the functional activity of Campath-1H. MoI Immunol 1995;32:1311-1318.
33. Shinkawa T, Nakamura K, Yamane N, Shoji-Hosaka E, Kanda Y, Sakurada M, et al: The absence of fucose but not the presence of galactose or bisecting N- acetylglucosamine of human IgGl complex-type oligosaccharides shows the critical role of enhancing antibody-dependent cellular cytotoxicity. J Biol Chem 2003;278:3466-3473.
34. Kettleborough CA, Saldanha J, Ansell KH, Bendig MM: Optimization of primers for cloning libraries of mouse immunoglobulin genes using the polymerase chain reaction. Eur J Immunol 1993 ;23:206-211.
35. Orlandi R, Gussow, DH, Jones PT, Winter G: Cloning immunoglobulin variable domains for expression by the polymerase chain reaction. Proc Natl Acad Sci USA 1989;86:3833-3837.
36. Burton DR, Barbas CF 3rd, Persson MA, Koenig S, Chanock RM, Lerner RA: A large array of human monoclonal antibodies to type 1 human immunodeficiency virus from combinatorial libraries of asymptomatic seropositive individuals. Proc Natl Acad Sci U S A 1991;88(22):10134-10137.
37. Burton DR, Pyati J, Koduri R, Sharp SJ, Thornton GB, Parren PW, et al.: Efficient neutralization of primary isolates of HIV-I by a recombinant human monoclonal antibody. Science 1994;266 (5187): 1024- 1027.
38. Roben P, Moore JP, Thali M, Sodroski J, Barbas CF 3rd, Burton DR: Recognition properties of a panel of human recombinant Fab fragments to the CD4 binding site of gpl20 that show differing abilities to neutralize human immunodeficiency virus type 1. J Virol 1994;68(8):4821-4828.
39. Muster T, Steindl F, Purtscher M, Trkola A, Klima A, Himmler G, et al.: A conserved neutralizing epitope on gp41 of human immunodeficiency virus type 1. J Virol 1993;67(l l):6642-6647.
40. Zwick MB, Labrijn AF, Wang M, Spenlehauer C, Saphire EO, Binley JM, et al: Broadly neutralizing antibodies targeted to the membrane-proximal external region of human immunodeficiency virus type 1 glycoprotein gp41. J Virol 2001;75(22): 10892- 10905.
41. Trkola A, Purtscher M, Muster T, Ballaun C, Buchacher A, Sullivan N, et al. : Human monoclonal antibody 2Gl 2 defines a distinctive neutralization epitope on the gpl20 glycoprotein of human immunodeficiency virus type 1. J Virol 1996;70(2): 1100- 1108.
42. Scanlan CN, Pantophlet R, Wormald MR, Ollmann Saphire E, Stanfield R, Wilson IA, et al.: The broadly neutralizing anti-human immunodeficiency virus type 1 antibody 2G12 recognizes a cluster of alphal~>2 mannose residues on the outer face of gpl20. J Virol 2002;76(14):7306-7321. 43. Binley JM, Wrin T, Korber B, Zwick MB, Wang M, Chappey C, et al.: Comprehensive cross-clade neutralization analysis of a panel of anti-human immunodeficiency virus type 1 monoclonal antibodies. J Virol 2004;78(23): 13232- 13252.
44. Mehandru S, Wrin T, Galovich J, Stiegler G, Vcelar B, Hurley A, et al.: Neutralization of newly transmitted human immunodeficiency virus type 1 by monoclonal antibodies 2G12, 2F5 and 4E10. J Virol 2004 ;78:14039-14042.
45. Gray ES, Meyers T, Gray G, Montefiori DC, Morris L: Insensitivity of Paediatric HIV-I Subtype C Viruses to Broadly Neutralising Monoclonal Antibodies Raised against Subtype. B PLoS Med 2006;18;3(7):e255.
【技術分野】
【0001】
この発明は特異的にHIVのR7Vエピトープに結合することができる新規なヒト抗体に関する。これらの抗体は全てのヒトCDRを有し、エスケープ変異株を含むすべてのHIV株を特異的に中和することができる。これらの抗体はHIV感染治療に有益であり、特に高活性抗レトロウィルス療法(HAART)に効果のない患者の治療に有益である。
【背景技術】
【0002】
HIV感染は今なお世界的に流行する伝染病である。感染後のHIV複製や毒性を薬剤療法で抑制する一方、予防処置や治療処置は今なお有効とはいえない。さらに高活性抗レトロウィルス療法(HAART)は、エイズを抑制する一方で複数の副作用を伴い、薬剤耐性ウィルスの出現を促すので、HIVに対しては追加の治療法が強く望まれている。しかしノンプログレッサとして認定されているHIV患者でも感染後10年、15年以上もエイズを発症しない場合もあり、HIV疾患は弱毒化ウィルス(文献1)、欠損ウィルス(文献2)、HIVコレセプター変異株(文献3、4)あるいは中和抗体(文献5)の存在等により、様々な方法により発症を遅らせることができることを実証している。従来のエンベロップベースの抗体誘導ワクチンはいずれもウィルスの高い突然変異率及び弱い免疫原性に限界があった。先の研究(文献6)で、我々はノンプログレッサの血清から精製された広域スペクトル中和抗R7V抗体の潜在力を実証した。これらの免疫グロブリンは、β2-ミクログロブリンから抽出され、出芽中にHIVの外被に組み込まれるR7V(RTPKIQV アミノ酸配列)と呼ばれる細胞エピトープに抗するものであった(文献7、8)。我々の目的は、ノンプログレッサの患者のB-リンパ球から対応する遺伝子を、バキュロウィルスベクターを介して分離し、組み換え型抗R7V 抗体を生成することにあった。
【0003】
バキュロウィルス技術により、正しくアセンブルされグリコシル化された免疫グロブリンを生成、分泌することができる(文献9)。これら組み換え型抗体は親の免疫グロブリンの全ての機能特性を有し(文献10、11)、(i)補体成分C1q(文献12、13)やC3(文献14)と(ii)細胞傷害に対する抗体を誘導するのに必要なIgGFc レセプタ(文献15、16、13)とを結合するような効率の良いエフェクタ機能を示す。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】明細書の末尾の一覧表を参照のこと。
【発明の概要】
【0005】
この発明に関連して、ウィルスの出芽中にHIVにより獲得される細胞エピトープR7Vに抗する組み換え型抗体を作った。抗R7V抗体の可変領域をエンコードするc-DNAはノンプログレッサの患者のBリンパ球からクローニングされている。この抗体の重鎖及び軽鎖の完全なコード配列を含む2つのトランスファーベクターが作られ、組み換え型バキュロウィルスがバキュロウィルスDNAとその2つのトランスファーベクター間の2重の組み換えにより生成された。このバキュロウィルスに感染された昆虫細胞から完全なヒト抗R7V免疫グロブリンを生成した。我々は、R7Vペプチドに特異的である我々の組み換え型抗体が、耐性ウィルスを含むあらゆるHIV1のクレードを認識し中和することを解明した。これによりHIV療法に新たな展望を開くことになる。
【0006】
つまり、第1の実施例によれば、この発明の主題は単離された抗体であり、あるいはその機能を有するフラグメントの1つであり、その抗体あるいはフラグメントの1つは特にR7Vエピトープ( RTPKIQV 配列番号 11 )と結合することができ、HIV株を中和できるものであり、それは、
i)アミノ酸配列 配列番号1 (QSVLYSSNNKNY), 配列番号2 (WAS), 及び配列番号3 (QQYYSTPQT) を有する相補性決定領域CDR, あるいはその配列が、配列配列番号1, 2, 又は3 との最適アライメントの後で、少なくとも80%、好ましくは90%の同一性を持つCDRを含む軽鎖、及び
ii)アミノ酸配列配列番号6 (GGSISSYY), 配列番号7 (IYYSGST) 及び配列番号8 (ARGRSWFSY) を有するCDR, あるいはその配列が、配列配列番号6,7, 又は8 との最適アライメントの後で、少なくとも80%、好ましくは90%の同一性を持つCDRを含む重鎖を備えている。
この「発明の概要」の中で用いる用語、ポリペプチド、ポリペプチド配列、ペプチド及び抗体化合物又はその配列に結合するタンパク質は置換可能である。
【0007】
この発明は天然の抗体に関するものではなく、すなわちこの発明の抗体は自然環境に存在するものではなく、自然源から精製により単離、取得できたものであり、又は遺伝子組み換えにより取得できたものであり、あるいは化学合成により取得できたものである。それゆえにさらに説明するように、それは非天然アミノ酸を含むものであることが理解されるべきである。
【0008】
CDR領域又はCDRにより、Kabatらによって定義された 免疫グロブリンの重鎖及び軽鎖の超可変領域を示すことが意図されている。(Kabat et al. Sequences of proteins of immunological interest、 第5版、U.S. Department of Health and Human Services、NIH、1991 及び続版) 3つの重鎖CDR, 及び3つの軽鎖CDRが存在する。用語CDR 又はCDRsはここではケースに応じてこれら領域の1つまたはこれら領域の複数あるいは全体を示すために用いられ、これらの領域は、抗体の親和性によって認識した抗原又はエピトープへの結合に関与するアミノ酸残基の大多数を含んでいる。
【0009】
この発明の2つの核酸又はアミノ酸配列間の“同一性のパーセント”により、ベストアライメント(最適アライメント)後に得られる、比較される2つの配列間のヌクレオチド又は同一のアミノ酸残基のパーセントを示すことが意図されている。このパーセントは純粋に統計に基づいており、2つの配列間の差はランダムに分布しそれらの全体長を超える。従来2つの核酸又はアミノ酸配列間の比較は最適な方法によるアライメント後これらの配列を比較することにより行っている。この比較は区分毎に、あるいは“比較窓”を通して行うことができる。比較のための配列の最適アライメントはマニュアル操作に加え、Smith and Waterman (1981) [Ad. App. Math. 2:482]の局所相同性アルゴリズムにより、Neddleman and Wunsch (1970) [J. Mol. Biol. 48:443] の局所相同性アルゴリズムにより、Pearson and Lipman (1988) [Proc. Natl. Acad. Sci. USA 85:2444] の類似性サーチ法により、これらのアルゴリズムを用いたコンピュータソフトウェア(GAP, BESTFIT, FASTA and TFASTA in the Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, WI, or else by BLAST N or BLAST P comparison software ) により実行することができる。
【0010】
2つの核酸又はアミノ酸配列間の同一性のパーセントは、最適方法により整列(アライメント)された2つの配列を比較することにより決定され、比較される核酸又はアミノ酸配列はこれら2つの配列間の最適アライメントのために基準配列に対し付加あるいは削除を行うことができる。同一性のパーセントは2つの配列間で核酸又はアミノ酸残基が同一となる場合の同一位置の数を決定することにより、比較窓の中の位置の全体数でこの同一位置の数を割ることにより、及びこれら2つの配列間の同一性パーセントを得るために、得られた結果に100を掛けることにより計算される。
【0011】
例えば、サイトhttp://www.ncbi.nlm.nih.gov/gorf/b12.htmlで利用可能なBLAST プログラム “BLAST 2 sequences” (Tatusova et al, “Blast 2 sequences - タンパク質とヌクレオチド配列を比較する新規ツール”, FEMS Microbiol Lett. 174:247-250) を使うことが可能である。用いるパラメータは初期設定により得られ(具体的にはパラメータ“open gap penalty”に対し: 5及び“extension gap penalty”に対し: 2; 選択されるマトリクスはプログラムにより提案される例えばマトリクス“BLOSUM 62”)、比較される2つの配列の同一性パーセントは直接プログラムにより計算される。
【0012】
基準アミノ酸配列に対して少なくとも80パーセントの、好ましくは85%、90%、95%及び98%の同一性を持つアミノ酸配列により、これらは基準アミノ酸配列に対して特定の組み換え、具体的には少なくとも1つのアミノ酸の削除付加あるいは置換を行うが、トランケーションやエロンゲーションが好ましい。1つ又は複数の連続あるいは非連続アミノ酸の置換の場合、置換されるアミノ酸が“均等の(equivalent)”アミノ酸に置き換えられる置換が好ましい。“均等のアミノ酸”の表現は、ここでは基底構造のアミノ酸の1つにより置換され得るあらゆるアミノ酸を指すことを意図している。ただし、対応する抗体の生物活動を基本的に変更することのない置換であり、それは後述の実施例の中で具体的に記述する。
【0013】
これら均等アミノ酸は置換するアミノ酸との構造的相同性、あるいは実行可能な異なった抗体間の生物活動の比較試験の結果のいずれかに基づいて決定できる。
【0014】
例として、変更される対応の抗体の生物活動に大幅な変更が生じないように置換を実行できる可能性について記述する。例えばロイシンをバリン又はイソロイシンで、アスパラギン酸をグルタミン酸で、グルタミンをアスパラギンで、アルギニンをリシンで置換可能であり、同一条件でその逆の置換も当然予想できる。
【0015】
この発明に係る抗体は好ましくは完全ヒト単クローン抗体あるいはその機能的フラグメントである。
【0016】
具体的実施例において、この発明の抗体は、図3Bの配列番号4で表わされるアミノ酸配列との、最適アライメント後、少なくとも80%、好ましくは90%の同一性を持つアミノ酸配列を含む軽鎖、あるいは図3Aの配列番号5で表わされる配列を含むヌクレオチド配列により、又は配列番号5との、最適アライメント後、少なくとも80%、好ましくは90%の同一性を有する配列によりエンコードされる軽鎖を特徴とする。
【0017】
他の具体的実施例において、この発明の抗体は、図3Dの配列番号9で表わされるアミノ酸配列との、最適アライメント後、少なくとも80%、好ましくは90%の同一性を持つアミノ酸配列を含む重鎖、あるいは図3Cの配列番号10で表わされる配列を含むヌクレオチド配列により、又は配列番号10との、最適アライメント後、少なくとも80%、好ましくは90%の同一性を持つ配列によりエンコードされる重鎖を特徴とする。さらに他の実施例において、この発明に係る抗体は、図3Bの配列番号4で表わされるアミノ酸配列を含む軽鎖又は図3Aの配列番号5で表わされる配列を含むヌクレオチド配列によりエンコードされる軽鎖と、図3Dの配列番号 9により表わされるアミノ酸配列を含む重鎖又は図3Cの配列番号10により表わされる配列を含むヌクレオチド配列によりエンコードされる重鎖とを備える。
【0018】
この発明に係る抗体の機能的フラグメントにより、具体的にはFv, scFv (scは単一鎖), Fab, F (ab')2, Fab', scFv-Fc フラグメント又はダイアボディズ(diabodies) のような抗体フラグメント、あるいは化学修飾により又はリポソ−ムへの取り込みにより半減期が増加するあらゆるフラグメントを示すことが意図されている。化学修飾とは例えば、ポリ(エチレン)グリコール (“PEGylation”ペジレーション) (Fv-PEG, scFv-PEG, Fab-PEG, F(ab')2-PEG又はFab'-PEGと呼ばれるペグフラグメント) (“PEG”はPoly(Ethylene) Glycol ) のようなポリ(アルキレン)グリコールの添加である。このフラグメントはこの発明に係る配列番号 1, 2, 3, 6, 7 及び8 のCDR を含み、特にHIV株を中和する能力がある。
【0019】
好ましくは、前記機能フラグメントは抗体の可変重鎖又は可変軽鎖の部分配列により構成され、またはそれを含んでいる。その配列からそれらフラグメントは抽出される。前記部分配列は同一の結合特異性を保持するに十分である。好ましくは、これら機能フラグメントはFv, scFv, Fab, F (ab')2, F(ab'), scFv-Fc タイプ又はダイアボディズのフラグメントであり、これらは概してその由来する抗体と同一の結合特異性を有している。この発明によれば、抗体フラグメントはペプシンやパパインのような酵素による消化により、及び/又は化学的還元によるジスルフィド架橋の開裂により、上記のような抗体から取り込みを開始できる。他の方法としては、この発明に含まれる抗体フラグメントは当業者に良く知られている遺伝子組み換え技術により、あるいは、Applied Biosystems 社等により供給される例えば自動ペプチドシンセサイザ−によるペプチド合成によって取り込むことができる。
【0020】
より好ましい態様において、この発明は遺伝子組み換えにより、又は化学合成により得られた、この発明に係る抗体、又はその機能的フラグメントを備えている。
【0021】
好ましい態様において、この発明に係る前記機能フラグメントは、フラグメントFv, scFv, Fab, (Fab')2, Fab', scFv-Fc 又はダイアボディズ から、あるいは化学修飾、特にペジレ−ション又はリポソ−ム取り込みにより半減期が増加するあらゆる機能フラグメントから選択される。
【0022】
この発明は、配列番号5の配列との、最適アライメント後少なくとも80%、好ましくは85%、90%、95%、及び98%の同一性を持つ配列を含む単離された核酸にも関連する。
【0023】
この発明は、配列番号10の配列との、最適アライメント後少なくとも80%、好ましくは85%、90%、95%、及び98%の同一性を持つ配列を含む単離された核酸にも関連する。
【0024】
好ましい配列との、最適アライメント後少なくとも80%、好ましくは85%、90%、95%、及び98%の同一性を持つ核酸配列により、基準核酸配列に対し、具体的には欠失、トランケーション、エロンゲーション、キメラ融合、及び/又は置換、特に点置換のような特定の組み換えを行う核酸配列を示すことが意図される。それは好ましくは基準配列と同じアミノ酸配列に対する配列コードが遺伝子コードの縮退とつながりのある配列に関係し、あるいは、特に以下に説明するような好ましくは高い厳密性の条件下で、基準配列と特異的に雑種化(ハイブリッド化)を行える相補配列に関係する。
【0025】
高い厳密性条件下でのハイブリッド化は、温度条件やイオン強度条件が相補的DNAの2つのフラグメント間でのハイブリッド化が維持されるように設定されることを意図している。実例として、上述のポリヌクレオチドフラグメントを特定するためのハイブリッドステップの高い厳密性条件を次に示す。
【0026】
DNA-DNA又はDNA-RNAのハイブリッド化は2つのステップで実行される:(1)ハイブリッド化前は、温度42℃で5 × SSC (1 × SSCは0.15 M NaCl + 0.015 Mクエン酸ナトリウム液に相当)、50%のホルムアミド、7%のドデシル硫酸ナトリウム (SDS)、10 × Denhardt's、5%の硫酸デキストラン及び1%のサケ精子DNA を含有するリン酸緩衝液(20mM, pH 7.5)中に3時間;(2)実際のハイブリッド化は、プローブのサイズに応じた温度(即ち:プローブサイズ> 100 ヌクレオチドで42℃)で20時間、その後2 × SSC + 2%SDS中、温度20℃で20分間2回の洗浄、0.1 × SSC + 0.1%SDS中、温度20℃で20分間1回の洗浄。プローブサイズ> 100 ヌクレオチドには最後の洗浄が0.1 × SSC + 0.1%SDS中、温度60℃で30分間行われる。上述の限定されたサイズのポリヌクレオチドに対する高い厳密性のハイブリッド化条件は、Sambrook et al., (1989, Molecular cloning : a laboratory manual. 第2版. Cold Spring Harbor) の解説に従って、大又は小サイズのオリゴヌクレオチドの当業者が利用することができる。
【0027】
この発明は上記核酸、具体的には配列番号5及び配列番号10の核酸を有するベクターにも関係する。
【0028】
この発明は、特にクローニング及び/又はこの発明に係るヌクレオチド配列を含む発現ベクターを目的とする。例えば、上記のような、特に配列番号5及び配列番号10の核酸配列を備えるバキュロウィルストランスファーベクターを目的としている。
【0029】
この発明に係るベクターは、好ましくは決定された宿主細胞のヌクレオチド配列の発現及び/又は分泌を可能とするエレメントを含む。そのベクターは、それ故、プロモータ遺伝子、開始のシグナル及び翻訳の終止、並びに転写の適切な制限領域を含まねばならない。それは宿主細胞の中で、安定した状態で維持されなければならないし、翻訳されるタンパク質の分泌を指定する具体的なシグナルを随意持つことができなければならない。これら異なるエレメントは使用される宿主細胞の機能として、当業者により選択され最適化される。この趣旨で、この発明に係るヌクレオチド配列は選択された宿主細胞の自律複製ベクターへ導入でき、あるいは、選択された宿主細胞の統合的ベクターへ導入できる。
【0030】
そのようなベクターは当業者が現在使っている方法で形成され、結果として生じるクローンはリポフェクション、エレクトロポレーション、熱衝撃、又は化学的手法といった標準方法で適当な宿主細胞へ導入される。
【0031】
この発明に係るベクターは、例えば、プラスミドあるいはウィルス性のベクターである。それらは、この発明に係るヌクレオチド配列のクローンを作るため、あるいは発現するために宿主細胞を形質転換するうえで有用である。
【0032】
同様にこの発明は、この発明にかかるベクターにより形質転換される、あるいはこの発明にかかるベクターを有する宿主細胞を備えている。
【0033】
宿主細胞は原核細胞系又は真核細胞系から選択でき、例えば細菌性細胞から選択できるが、同様に酵母細胞や動物細胞、具体的には哺乳類細胞から選択できる。昆虫細胞や植物細胞を使うことも同様に可能である。
【0034】
従って、別の態様によれば、この発明は上記の抗R7Vヒト抗体を分泌する細胞株に関係する。例えば、上記抗体はEVB不死化Bリンパ球により、バキュロウィルスベクターを使ってSf9細胞のような昆虫細胞により、あるいは他の抗体生成細胞株、たとえばCHO (ATCC number CCL-61)、低フコシル化抗体を生成する遺伝子組み換えされたCHO、あるいはYB2/0 (ATCC CRL-1662) 細胞株により得られる。
【0035】
さらに別の態様によれば、この発明はこの発明に係る抗体あるいは機能フラグメントの1つの生成方法を目的とし、
a) この発明に係る宿主細胞の培地と適当な培養条件の中で培養し、及び
b) 前記培養細胞の培地から前記抗体を抽出する
ステップを備えるものである。
上記方法により獲得することができる抗体は、あるいはその機能フラグメントの1つは、この発明の範囲に含まれる。
【0036】
さらに他の態様によれば、この発明は医薬品として上記抗体、あるいはその機能フラグメントの1つに関係する。この発明はさらに、有効成分としてこの発明に係る抗体あるいはそのフラグメントの1つと、賦形剤及び/又は薬学的に受け入れられる媒体とを含む医薬組成物に関係する。
【0037】
さらに他の実施例において、この発明は、同時、別々、あるいは順次使用の併用製品として現在エイズ療法に使われている少なくとも1つの薬剤と上記に係る抗体をさらに含む上述の組成物を対象にしている。“同時使用”とはこの発明に係る組成物の2つの化合物を単一かつ同一の医薬品形態で投与するものとして理解される。“別々の使用”とはこの発明に係る組成物の2つの化合物を異なった医薬品形態で同時に投与するものとして理解される。“順次使用”とはこの発明に係る組成物の2つの化合物をそれぞれが異なる医薬品形態で順次投与するものとして理解される。
例えば、抗R7V抗体の投与を:
エファビレンツ + ジドブジン + ラミブジン
エファビレンツ + テノホビル + エムトリシタビン
スタブジン + ラミブジン + ネビラピン
リトナビルによりブーストされたロピナビル+ ジドブジン + ラミブジン
リトナビルによりブーストされたロピナビル+ テノホビル + エムトリシタビン
と組み合わせることが可能である。
【0038】
この発明は、特にHIV感染、エイズ治療に対して、例えばHAART療法の患者、具体的にはHAART療法の効果がない患者の治療に対して医薬品の調合のためにここに記載する抗体の使用を含む。
【0039】
この発明を以下の図面及び事例を参照してさらに説明する。
【図面の簡単な説明】
【0040】
【図1A】抗R7V抗体の発現に使う免疫グロブリン特異トランスファーベクターの概略図。pVT-Ckの概略図。軽鎖の発現を許すトランスファーベクター。
【図1B】抗R7V抗体の発現に使う免疫グロブリン特異トランスファーベクターの概略図。pVT-Cγ1の概略図。重鎖の発現を許すトランスファーベクター。
【図2A】R7V抗原上で選択された不死化Bリンパ球から抽出された全体RNAから合成されるc-DNA上に存在するVH配列のPCR増幅。
【図2B】R7V抗原上で選択された不死化Bリンパ球から抽出された全体RNAから合成されるc-DNA上に存在するVL配列のPCR増幅。
【図3A】最相同生殖細胞遺伝子と比較される不死化Bリンパ球に発現する抗体の軽鎖(K4) の可変領域のヌクレオチド配列。
【図3B】最相同生殖細胞遺伝子と比較される不死化Bリンパ球に発現する抗体の軽鎖(K4) の可変領域のアミノ酸配列。
【図3C】最相同生殖細胞遺伝子と比較される不死化Bリンパ球に発現する抗体の重鎖(M4) の可変領域のヌクレオチド配列。
【図3D】最相同生殖細胞遺伝子と比較される不死化Bリンパ球に発現する抗体の重鎖(M4) の可変領域のアミノ酸配列。
【図4】抗R7Vまたは無関係の抗体50 μg/ml によるHIV1クレードの中和。
【発明を実施するための形態】
【0041】
図1:抗R7V抗体の発現に使う免疫グロブリン特異トランスファーベクターの概略図である。
図1A:pVT-Ckの概略図−軽鎖の発現を許すトランスファーベクター。
図1B:pVT-Cγ1の概略図−重鎖の発現を許すトランスファーベクター。
図2:R7V抗原上で選択された不死化Bリンパ球から抽出された全体RNAから合成されるc-DNA上に存在するVH (図2A) 配列又はVL (図2B) 配列のPCR増幅。増幅は特定のVH 又はVL遺伝子ファミリーに特異的な適合定数3' プライマー及び5'プライマーのセットで、1.1材料と方法に報告されるように実行された。PCR反応の20μlは1.5%のアガロースゲルで分別されエチジウムブロマイドにより染色された。Lane CVH: コントロールVH配列。Lane CVL: コントロールVL配列。Lane MW: スマートラダー分子量マーカー(Eurogentec): 200, 400, 600, 800, 1000, 1500, 2000, 2500, 3000, 4000, 5000, 6000, 8000, 10000 bp.
図3:最相同生殖細胞遺伝子と比較される不死化Bリンパ球に発現する抗体の軽鎖(K4) 及び重鎖(M4) の可変領域のヌクレオチド配列(図3A及び図3C)及びアミノ酸配列(図3B及び図3D)。アミノ酸配列は1文字コードで与えられている。使用される番号方式はIMGT (http://imgt.cines.fr) のルールに基づいている。VH及びVL配列の相補性決定領域 (CDR) は強調されている。配列のダッシュはトップラインに与えられる残基との一致を示している。IGHJ, IGHD, 及びIGKJ 遺伝子はボックスに収まっている。
【実施例】
【0042】
実施例1:有効ヒト組み換え型抗R7V抗体の単離生成
【0043】
1.1材料と方法
【0044】
1.1.1細胞とウィルス
ヒト末梢血単核細胞(PBMC)は血清反応陰性の健康なドナーの未使用の K2E-EDTA 血液サンプルからFicoll- Paque(Amersham)勾配遠心分離により分離された。 培養される細胞は完全RPMI培地の中で1 × 106 cells/ml の密度で次の組成で成長した:10%の加熱不活性化したウシ胎仔血清 (GIBCO) , 1%のペニシリン/グルタミン(GIBCO) , 10 UI/ml IL2 (Euromedex), 最初の3日間10 μg/ml PHA-P (Difco) , 及び2 μg/ml ポリブレン (Biowhittaker), を補充された RPMI 1640 (Biowhittaker) 。
CEM細胞株は0.5 × 106 cells/mlでRPMI-10%の培地 (10%の加熱不活性化したウシ胎仔血清、1%のペニシリン/グルタミン、2 μg/ml ポリブレンを含むRPMI 1640) の中で培養された。
NDK (クレードD) 及びAZT耐性RTMC (クレードB) ウィルスは感染CEM細胞で生成した。92UG029 (クレードA), 92BR021 (クレードB), 92BR025 (クレードC), 及び93BR029 (クレードF) ウィルスは当初AIDS Research and Reference Reagent Program, Division of AIDS, NAID, NIHにより提供され、PBMCにより生成された。ウィルスBCF06 (クレードO), 及びYBF30 (古いクレード) はF. Barre-Sinoussi (パスツール研、フランス) よりご好意で頂いた。感染細胞の上澄みで漸増したウィルスのサンプルはマイナス80℃で凍結保存された。
Sf9細胞は5%の加熱不活性化したウシ胎仔血清 (GIBCO)を補充されたTC100 培地(GIBCO)の中で28℃で維持された。野生型のAutographa californica 複合核多角体病 (AcMNPV) ウィルスクローン1.2(文献17)及び組み換え型バキュロウィルスがSf9細胞の中で繁殖した。
【0045】
1.1.2 ノンプログレッサー患者からの末梢血単核細胞 (PBMC) の単離
インフォームドコンセントを行った血清反応陽性のノンプログレッサの患者が我々の研究に登録され、Ficoll-Paque 密度勾配法による単離により未使用のK2E-EDTA静脈血からPBMCを精製した。これらのPBMCは、15%の加熱不活性化したFCSと1%のペニシリン/グルタミンを補充したRPMI 1640 培地の中で不死化前のBリンパ球の成長を助けるためにIL2及びPHA無しで2日間予備培養した。
【0046】
1.1.3 Epstein-Barr ウィルス(EBV)によるBリンパ球の不死化
その後Bリンパ球は2ml のB-95.8 培養上澄み(EBV生成細胞株)を、加熱不活性化した3ml 10 % FCSの中で予備培養した9 × 106 PBMC, 50 ml の円錐管の1%のペニシリン/グルタミンRPMI と混合することにより不死化された。37℃の水槽で2時間の培養後、10%の加熱不活性化したFCS, 1μg/μlのシクロスポリンA(Calibiochem)及び1%のペニシリン/グルタミンを 補充された 5 mlのRPMI 1640を加えた。その10-mlの細胞懸濁液は25 cm2の組織培養フラスコへ移され、37℃で加湿されCO2 濃度5%の培養器の中でそのまま4週間培養された。4週間の培養の最後には、EVB-不死化細胞が肉眼で見える塊を形成した。この細胞株をRPMI-20%、106 cells/mlで週に2回リフィーディングすることで維持した。
【0047】
1.1.4 Bリンパ球を分泌する抗R7V抗体の単離
R7Vペプチドのアミノヘキサン酸形による磁気ビーズのコーティング:10 μg のR-8-Ahx ペプチド (Neosystem) を107磁気トシル活性ビーズ(Dynal Dynabead(登録商標) M450) により低速傾斜回転、37℃で16-24時間培養した。ビーズは製造者の手順に従って洗浄し、PBS pH 7.4の中、4×108 beads/mlで再懸濁した。
Bリンパ球を分泌する抗R7V抗体の磁気選別:1 mlの無菌PBSに107 のEBV-不死化されたBリンパ球は24x106 R-8-Ahx でコートされたビーズと4℃で20分混合され、細胞がビーズに固着しなくなるまで3回繰り返される。ロゼット細胞は管を2分間磁石に入れておくことで単離された。ビーズはそのままにして上澄みを取り除き、細胞はPBS洗浄緩衝液に再懸濁された。ビーズに固着した細胞を37℃でRPMI-20% FCS 37℃, 5% CO2 で培養する前に、洗浄ステップを3回繰り返した。1日後細胞はビ−ズから分離し、106 cells/mlに成長した。
この磁気選別は2週間の培養後同じ手順で、Bリンパ球を分泌するこれらの予め選択した抗R7V抗体に対し繰り返された。
【0048】
1.1.5 ELISA 手順
抗R7V抗体は製造者が示す抗R7V ELISA試験 (Anti R7VTM IVR 96000, IVAGEN, France) により検出された。簡潔に言えば、陽性対照、陰性対照、遮断キャリブレータ及び希釈抗体( 100 μl/well)が、R7Vをコーティングする試験板に添加され、室温で30分培養された。結合された抗R7V抗体は西洋ワサビペルオキシダーゼ抱合抗ヒトIgG抗体により検出された。
【0049】
1.1.6 中和試験
ウィルスのストックは、以下の希釈に対応して1回の試験(文献18)で100 TCID50を持つように前もって滴定された:HIV-1NDK (希釈10-5), HIV-1RTMC AZT-耐性(希釈 5×10-5), 92UG029 (希釈10-2), 92BR021 (希釈10-3), 92BR025 (希釈10-2), THA92022 (希釈10-2), 93BR029 (希釈10-2), BCF06 (希釈10-4), 及びHIV-1YBF30 (希釈10-3)。ウィルス(50 μl) の希釈液は100 μg /mlの抗体(最終濃度 50 μg/ml)を含む50 μl RPMI-0%で96- wellマイクロタイタープレートで、加湿された37℃、5%CO2 の培養器の中で1時間予備培養された。PBMC (50 μl中1 ×10 6) はウィルス‐抗体混合液に1時間37℃で添加され、細胞は培地で3回洗浄され、106 cells/ml、 24- well マイクロタイタープレートで、50 μg/ml 抗体完全RPMI-10 %の存在下、最初の3日間培養された。 培養液は10日間成長し、3日毎にリフィーディングされた。同じ試験がウィルス対照 (抗体無しのHIV感染細胞)、細胞対照 (抗体無しの非感染細胞) 及び抗体対照 (非HIV 関連エピトープに対抗する無関係の抗体) に対して行われた。それぞれのサンプルのウィルス複製を測定するため、逆トランクリプタス酵素が以下のように定量化された。3日毎に採取した無細胞の1 ml の上澄みサンプルは4℃ で5分間 95,000 rpm (TL 100 Beckman) の超遠心分離機にかけた。ウィルスペレットは0.1 % Triton X-100 NTE (NaCl 100mM, Tris 10mM, EDTA 1mM) 緩衝液10ml中で再懸濁され、ウィルス酵素を解放した。酵素反応はTris 50mM, pH 7.8; MgCl2 20mM; KCl 20mM; ジチオスレイトール (DTT) 2mM ; オリゴ dT 0.25 OD/ml; ポリrA 0.25 OD/ml 及び3H dTTP 50 μCi/ml を含む50 μl の反応混合液で行われた。37℃で1時間後、抗原抗体反応は5%TCA中、1 ml のピロリン酸ナトリウムで停止し、合成されたDNA生産物が20%のトリクロロ酢酸で沈殿され、ミリポア0.45 μm でろ過により採取され、パッカードシンチレーションカウンターによりdpm/mlでβ放射能が測定された。中和のパーセントは以下のように表わされた:
[100 - (サンプルの逆転写酵素活性/ウィルスの逆転写酵素活性) × 100]
【0050】
1.1.7 抗R7V 特異性を発現する抗体の可変領域の単離とクローニング
手順はマウス可変抗体領域の増幅のための技術を応用した。(文献19)全てのRNAはRNeasy キット(Qiagen) を使って約5×106不死化Bリンパ球から抽出された。簡潔に言えば、細胞は600 μl のRLTTM/βメルカプトエタノール緩衝液で溶解され、20 ゲージ針連続継代により均質化された。600 μl の70%エタノールを添加後、混合液はRNeasy カラムに沈着され、15秒間12,000 rpm (Biofuge, Heraeus) で遠心分離機にかけた。カラムは700 μl RW1TM 緩衝液及び500 μl RPETM 緩衝液で連続洗浄を行った。 RNAは50 μl RNAse フリーの水で溶出され、使用するまでマイナス80℃で保存された。
全てのRNAと5つの特異的プライマーがヒト免疫グロブリンの定常部でハイブリッド化を行い、hCLa, hCLb, hCK, hCG, 及びhCM (表3) が使われて、ラムダ、カッパ、ガンマ1及びミューmRNAそれぞれに対応する第1ストランドc-DNAs を合成した。逆転写は次のように実行された:全RNA 1μg, 4μlの10X RTTM 緩衝液(Qiagen), 4μl の5mM 各dNTP (Qiagen), 10 pMoles/μl 20 ユニットの RNAse インヒビター (Roche) での4μl の特異的プライマー及び最終容積40 μlのオムニスクリプト逆転写酵素 (Qiagen) 8ユニット。混合液は1時間37℃で培養された。逆転写活動は93℃で5分間加熱不活性化された。VH 配列、VL 配列の全長は、ヒト免疫グロブリン(表3) の重鎖及び軽鎖のシグナルペプチド配列にデザインされる特異的プライマー及び充填剤としてラムダ、カッパ、ガンマ又はミュー第1ストランドc-DNA を使って、PCRによって増幅された。PCR 反応は、2 μl の10X Vent DNA ポリメラーゼ(Biolabs), 2 μl の10mM各dNTP (Biolabs), 各プライマーの20 pMoles, 1.5 μlの25 mM MgSO4, 1ユニットのVent DNA ポリメラーゼ(Biolabs), 0.5 μl の逆転写混合液を含む20 μl の最終容積で実行された。30サイクルの増幅が95℃で30秒間、55℃で45秒間、72℃で1分間実行された。72℃で10分間延長の後、PCR生成物が1.5% のアガロースゲル (SeaKem, FMC) で分別されエチジウムブロマイドで染色された。
PCR生成物はゲル洗浄され、Advantage Taq ポリメラーゼ(Clonetech)で増幅され、プラスミド pGemT easy (Promega) でクローニングされた。PCR増幅(MWG Biotech) に使われる3'及び5' プライマを使って、両方のストランド上に導入の配列が決定された。可変領域の配列比較、生殖細胞遺伝子分析がBLAST(文献20)及びIMGT(文献21)データベースを使って実行された。
【0051】
1.1.8 抗R7V抗体発現組み換え型バキュロウィルスの生成
VH, VL 配列は, ヒト免疫グロブリンシグナルペプチド配列、2つのユニークな制限酵素認識部位及びヒトガンマ1とカッパ定常部をそれぞれエンコードする配列を含む特異的トランスファーベクター pVTCγ1 及びpVTCκ (図1)に導入された。pVTCγ1 ベクターはシグナルペプチド配列の中のユニークなAfl II部位及びガンマ1配列の2つの最初のコドン有するNhel 部位を含み、一方pVTCκはシグナルペプチド配列の中にユニークなBssH II部位及びJ領域の最後の保存アミノ酸及び定常カッパ領域の最初のアミノ酸がオーバラップするBsiW I部位を含む。
適切な制限酵素認識部位が以下のプライマーを使うPCRによりVH, VL 配列の5' 及び3'末尾に導入された:
FOR-M4: CCATCTTAAGGGTGTCCAGTGTCAGGTGCAGCTGCAGGAGTCGGGCCCAGGACTGGTGAAGC (SEQ ID N°16),
BAC-M4: GCATGCTAGCTGAGGAGACGGTGACCAGGGT (SEQ ID N°17),
FOR-K4: CGATGCGCGCTGTGACATCGTGATGACCCAGTCT (SEQ ID N°18) 及び
BAC-K4: CGATCGTACGTTTGATCTCCAGCTTGGTCCCCTGGCC (SEQ ID N°19)。
VHに対してはAfl II-Nhel, VLに対してはBssH II-BsiWIにより消化されたPCR生成物は精製されて個々のトランスファーベクターpVTCγ1 及びpVTCκに導入された。最終作成物pVT Cγ1-M4及びpVTCκ-K4はシークエンシングにより制御された。
抗体発現組み換え型バキュロウィルスは前述するように(文献22、10、11)Sf9細胞の共トランスフェクション後に生成された。生殖能力のあるクローンをELISAによりスクリーニングした(文献23)。簡潔に示せば、100 μl の1 μg/ml 抗ヒト重鎖 Fdγ1 ポリクローナル抗体 (The Binding Site) をコーティングしたマイクロタイタープレートが細胞培養上澄みの連続希釈により37℃で2時間培養された。結合した組み換え型IgGが西洋ワサビペルオキシダーゼ標識抗ヒトカッパ軽鎖抗体 (Sigma) を使って検出された。
組み換えウィルスのゲノムはサザンブロットにより制御された。7 mlの細胞培養上澄み中のウィルス粒子は35,000 rpm 40 分間で沈殿させた(TL 100.4, Beckman)。ペレットはTEK 緩衝液の1 ml (0.1 M Tris, 0.1M Na2EDTA 2 H2O, 0.2 M KCl, pH 7.5)に10 μlのプロテイナーゼ K 20 mg/ml水溶液(Roche) 及び10 μl のN ラウリルサルコシン(Sigma) 10 % (w/v) 水溶液を加えて再懸濁し、一晩50℃で培養した。ウィルスDNAがフェノールとクロロホルムイソアミルアルコール(24:1 v/v) により連続的に抽出され、エタノールで沈殿させた。水で再懸濁した後、Hind IIIでDNAは消化された。制限されたDNAは1% アガロースゲルで電気泳動により分析され、ニトラン膜 (Schleicher and Schull)へ移された。ヒト定常部γ1 及びκ領域をそれぞれエンコードするc-DNAは製造者が推奨するようにジゴキシゲニンで標識され、ハイブリダイゼーション プローブとして使われた。洗浄後、ブロットはアルカリホスタファーゼ ( Roche, dilution 1: 10,000)に接合する抗ジゴキシン抗体により培養された。標識されたDNAの検出は、化学発光基質CSPD(Roche)により行った。
【0052】
1.1.9 組み換え型抗体の生成と精製
Sf9 細胞は密度500,000 cells/ml, 400ml のローラボトルの無血清培地に蒔かれ、2 per cell の感染多重度で感染させた。28℃で4日間の培養後、上澄が採取され、製造者が示すように、分泌した組み換え型抗体がタンパク質A セファロース (Amersham) 上に精製された。精製されたIgGの量がELISAによって測定された(文献23)。
組み換え型抗R7V抗体が同様な条件でCHO 発現システムによっても生成された。
【0053】
1.2 結果
【0054】
1.2.1 B リンパ球を分泌する抗R7V抗体の選択
B リンパ球を生成する抗R7V抗体はR7Vをコーティングした磁気ビーズを使ってノンプログレッサーのHIV 感染患者から選択された。B リンパ球を分泌する27%の抗R7V抗体を最初の選択で取得し、事前に選択したB リンパを分泌する抗R7V抗体により2回目の選択で14%を取得した。B 細胞培養上澄みにはフリーな抗R7V抗体は抗R7V ELISA によって検出されず、抗体は分泌するB リンパ球膜に結合したか、ELISA試験の検出限界以下であったかいずれかを示唆している。
【0055】
1.2.2 選択した不死化B リンパ球により発現したVH 及びVL配列の単離及びクローニング
選択されたB リンパ球により発現した抗体のVH, VL 領域の増幅は、我々が以前マウス免疫グロブリンに対し記述した(文献19)RT-PCRによって行われた。
図2に示すように、プライマーの僅かな組合せのみが割り当てられたサイズ、VHに対しては約450 bp, VL に対しては 400 bp でフラグメントの増幅をもたらした。hCG/hVH5, hCM/hVH2 及び hCM/hVH3 を使って僅かのしまが観察されるに過ぎなかったが、hCM/hVH4ではより多くの物質が観察できた。主な生成物はhCK/hVK4でも合成された。しかしながら、hCLa 及び hCLb プライマー(図示せず)を使ういかなる結合も増幅は検出できなかった。PCR生成物の配列解析及びBLAST 解析はそれらのうち2つのみ、M4 フラグメント (hCG/hVH4) 及び K4 フラグメント(hCK/hVK4) のみがヒト重鎖、軽鎖の可変領域にそれぞれ対応することを示した。これらの結果は選択された不死化B リンパ球の集団がおそらくは単クローン抗体で、膜IGMカッパ抗体を発現することを示している。これら配列とIMGTデータベースとの比較によりVH-M4 重鎖可変領域の配列がIGHV-4-59*01(文献24), IGHD2-21*01(文献25)及び IGHJ4*02(文献26) 生殖細胞遺伝子(図3C)の転位から生じることを示している。それに対応するVκ-K4はIGKV4-1*01(文献27)/ IGKJ2*02(文献28), 軽鎖可変領域(図3A)の転位を示している。興味深いことに、この抗体はカッパ軽鎖レパートリーから最もJ-proximal なIGKV4-1遺伝子を使った。このような軽鎖領域は主として変異株はなく、唯一の変異株が相補性決定領域3のIGKV/IGKJ 接合に存在した(図3B)。一方、相補性決定領域3において4つのアミノ酸変異株をもたらした7つのヌクレオチド置換がVH-M4 配列に観察される中、ただ2つのサイレントヌクレオチド置換がフレームワーク領域に認められた(図3C及び3D)。
【0056】
1.2.3 バキュロウィルス発現システムにおける抗R7V抗体の発現
抗R7V抗体の可変領域をエンコードする配列は、重鎖及び軽鎖のカセットバキュロウィルストランスファーベクター (i)ポリへドリン遺伝子座の中で再結合するpVT-CK 及び(ii)P10 遺伝子座の中で再結合するpVT-Cγ1に導入された。これらの生成物で、重鎖及び軽鎖遺伝子は合成P10 プロモータ、P'10及びP10プロモ−タのそれぞれに制御される(図1)。特異的プライマーはK4及びM4フラグメントを増幅するよう設計されており、図1に示す免疫グロブリンシグナルペプチド配列及び定常領域の枠組みでそれらの直接クロ−ニングを行う。この2つの最終生成物pVT-Cκ-K4及びpVT-Cγ1-M4は配列決定により制御され、精製したウィルスDNAのもとでSF9細胞を同時遺伝子導入するために使われた。2重組み換え型ウィルスは前記(文献10、11)のように2回の組み換えの後に取得された。組み換え型ウィルスは血小板洗浄され増幅された。感染された細胞の細胞培養上静の抗体の存在が抗ヒト抗体ELISAによって分析された。4つの繁殖力のあるクローンのゲノムはヒトγ1及び κ定常部DNAをプローブとして使ってサザンブロット法により制御された。AcR7VI/K4-M4と名付けられた1つのウィルスクローンがさらなる実験のために選択された。
【0057】
1.2.4 組み換え型抗R7V抗体の特異性
組み換え型抗R7V抗体 はウェルの0.625μgの抗体濃度に相当する6.25 μg/ml においてもIVAGEN 抗R7V ELISA キットにより陽性であった。無関係な抗体はその濃度に関係なく陰性であった。
ノンプログレッサの患者から精製した抗R7V抗体について先に報告したように、組み換え型単クローン抗体はフローサイトメトリー分析(データは示していない)が実証するようにどの細胞とも結合しない。
【0058】
1.2.5 HIV-1の複数クレードに対する中和試験
患者から精製した抗R7V抗体は幅広い中和スペクトルを表わすことが説明された。そこでこの抗R7V単クローン抗体はいくつかのクレードに対し同一条件下で試験された。治療抗体としての使用を確かめるため、中和試験は薬物耐性ウィルス(RTMC)に対してもなされた。抗R7V組み換え型抗体の中和効果を測定するために、50 μg/ml で希釈された抗体が細胞に感染する前のいくつかのHIV-1のクレードと混合された。抗R7V抗体はHIV-1の8つのクレードとAZT耐性クレードB RTMCウィルスを中和した(図4)。バキュロウィルスシステムで発現し同一の条件下で対照として使われた無関係の抗体に対しては中和は観察されなかった。85 % 以上の中和が5つのクレード(B, C, D, F, O)に対して得られた。50 μg/mlの抗R7V組み換え型抗体については、異なるウィルスに応じて異なるパーセントの中和が得られた。この異種の結果は、HIV ノンプログレッサ患者から精製した抗R7V抗体の結果と同じであり、おそらくはウィルスにより様々の量のR7Vエピトープによりもたらされたからであろう。
【0059】
CHO 発現システムにおける組み換え型抗R7V抗体の作成
【0060】
1) K4M4 ロット番号13.11.06: 40 μg/mlの抗R7V ELISA 陽性結果
【表1】
【0061】
2) K4M4 ロット番号 28.02.07: 50 μg/mlの抗R7V ELISA 陽性結果
【表2】
【0062】
1.3 結論
我々はここにバキュロウィルス発現システムによる組み換え型ヒト抗R7V抗体の生成に関する結果を報告する。このシステムは機能的組み換え型抗体を大量に生成するうえで迅速かつ効率的である(文献10、11、29)。鱗翅目細胞に発現する組み換えタンパクには、哺乳類細胞で観測される全ての翻訳後修飾が見られる。しかしながら、N-結合型オリゴ糖は短く、実質的には高マンノース又は少数マンノース型である(文献30、31)。抗体の生物活動は免疫グロブリンのCH2 定常領域のAsn-292に結合するN型グリカンに大きく依存している(文献32、33)。未完成の糖鎖付加パターンではあるが、Sf9 細胞で発現された組み換え型抗体は、Clq 及びFcγR 結合による補体依存性及び抗体依存性の細胞媒介性細胞傷害のような特異的生物活動を示した(文献14、16、13)。
HIV-1ノンプログレッサー患者により生成された抗R7V抗体を単離し特性を示すために、EBV ウィルスにより不死化したR7V-反応性B 細胞が患者の一人から選択され、IgG 又はIgM 免疫グロブリンの可変領域をエンコードするc-DNAがRT-PCR を使って特異的に増幅された。この目的のため、V 遺伝子ファミリーに関わりない、ヒトVH 及びVL 領域の特異的増幅ためのコンセンサスプライマーの3つの原型がデザインされた。
そのシグナル配列でハイブリッド化するこれらプライマーはヒト定常領域γ, μ, κ, λ それぞれに向けられる3' プライマーのセットと併せて使われた。体細胞変異を受ける“FR”増幅戦略で目標となるフレームワーク 1の領域とは対照的に(文献34、35)、シグナル配列の突然変異の頻度は非常に低く、そのためこの領域のプライミングは突然変異無しで全体配列の増幅が可能となる。
【0063】
これらc-DNAs の配列の分析により、これらの不死化細胞がおそらく唯一の細胞膜IgM カッパ抗体を発現する単クローン抗体であることが分かった。VL 配列の大部分がVJ ジャンクションで唯一のサイレント変異を伴い突然変異がないのに対し、FR3 でただ2つのサイレント変異を伴うVH領域のVDJ ジャンクションで6つの突然変異アミノ酸がCDR3 の中に見つかっている。その可変領域で低い突然変異率が観察されるにもかかわらず、この組み換え型抗体はフローサイトメトリー分析に従ってどんな細胞にも反応しないのでポリリアクティブではない。
【0064】
HIV-1 亜型 A, B, C, D, E, F, N, O ウィルス及び抗レトロウィルス治療耐性ウィルスに対するこの完全ヒト組み換え型抗体の中和能力は、ノンプログレッサ患者から得る多クローン性抗体と明らかに同じである。これらの結果は、全てのHIV-1 変異株が、細胞により生成されたR7V エピトープを捕捉することを裏付けている。50 μg/mlの抗R7V抗体により異なった中和のパーセントが得られたのは、おそらくウィルスに存在するR7V の様々の量に関連している。増え続ける抗体はそれぞれのクレードに対し100 % の中和を達成するように試験されねばならない。
【0065】
単クローン抗体にとってHIV感染患者の治療薬として作用するために最も重要な特徴の1つは、中和のその幅広いスペクトルである。HIVは感染時間とともに及び抗レトロウィルス治療(エスケープ突然変異体の出現)とともに1つの個体から他へと絶え間なく変化し、免疫システムがウィルスの複製を制御することの難しさを物語っている。今日、抗R7V抗体とは別に、幅広い中和を行う他の4つの単クローン抗体は、全てHIV-1 亜型B に抗するものであるが、そのような潜在力を示している。面gp 120 上に結合するCD4 に抗するIgG1b12は無症状のHIV 陽性個人からファージ提示技術によって生成されている(文献36、37、38)。2F5 及び4E10 抗体はgp41(文献39、40)の定常部を認識し、2G12 はgp120上エピトープに抗する(文献41、42)。これら4つの抗体は幅広く中和する抗体として報告されているが、最も効果を発揮するのはそれらが組み合わされた時であった(文献43、44)。
【0066】
我々が得た結果は、組み換え型抗R7V抗体がHIV-1 亜型C単離株を中和することを示した。この亜型では、単クローン抗体2F5 及び2F12 は効果が無く、IgG1b12 は部分的に効果があり、そして4E10 のみが顕著な活動を示す(文献45)。
【0067】
従って、抗R7V抗体は今日まで報告されているHIV-1に対し最も幅広く効果のあるMab (単クローン抗体) の1つであると思われる。その細胞起源にもかかわらず、抗R7V抗体を生成するどの患者も自己免疫疾患のいかなる臨床的症状も示さないので、R7V エピトープは自己免疫反応には関与しない(文献5)。このことは、この抗R7V抗体はHIV 感染患者の治療にとって強力な候補となることを裏付けている。
【0068】
【表3】
【0069】
参考文献一覧
1. Oelrichs R, Tsykin A, Rhodes D, Solomon A, Ellett A, McPhee D, et al: Genomic sequences of HIV type 1 from four members of the Sydney Blood Ban Cohort of long-term nonprogressors. AIDS Res Hum Retrovir 1998 ;14:811-814.
2. Sanchez G, Xu X, Chermann JC and Hirsch I: Accumulation of defective viral genomes in peripherical blood mononuclear cells of Human Immunodeficiency Virus type 1-infected individual. J Virol 1997;71 :2233-2240.
3. Connor RI, Sheridan KE, Ceradini D, Choe S, Landau NR: Change in coreceptor use correlates with disease progression in HIV 1-infected individuals. J Exp Med 1997;185 : 621-628.
4. De Roda Husman AM, Schuitemaker H: Chemokine receptors and the clinical course of HIV-I infection. Trends Microbiol 1998;6:244-249.
5. Haslin C, Chermann JC: Anti-R7V antibodies as therapeutics for HIV-infected patients in failure of HAART. Curr Opin Biotechnol 2002;13(6):621-624.
6. Galea P, Le Contel C, Coutton C, Chermann JC: Rationale for a vaccine using cellular-derived epitope presented by HIV isolates. Vaccine 1999 ;17:1700-1705.
7. Arthur LO, Bess JW Jr, Sowder RC 2nd, Benveniste RE, Mann DL, Chermann JC, et al.: Cellular proteins bound to immunodeficiency viruses: implications for pathogenesis and vaccines. Science 1992; 258:1935-1938.
8. Le Contel C, Galea P, Silvy F, Hirsch I, Chermann JC: Identification of the beta- 2-microglobulin-derivated epitope responsible for neutralisation of HIV isolates. Cell Pharmacol 1996;3:68-73.
9. Hasemann CA, Capra D: High-level production of a functional immunoglobulin heterodimer in a baculovirus expression system. Proc Natl Acad Sci USA 1990;87: 3942-3946.
10. Poul MA, Cerutti M, Chaabihi H, Ticchioni M, Deramoult FX, Bernard A, et al. : Cassette baculovirus vectors for the production of chimeric, humanized, or human antibodies in insect cells. Eur J Immunol 1995 ; 25:2005-2009.
11. Poul MA, Cerutti M, Chaabihi H, Devauchelle G, Kaczoreck M, Lefranc MP : Design of cassette baculovirus vectors for the production of therapeutic antibodies in insect cells. Immunotechnology 1995 ;1 : 189-196.
12. zu Putlitz J, Kubasek WL, Duchene M, Marget M, von Specht BU, Domdey H: Antibody production in Baculovirus-infected insect cells. Biotechnology (New York) 1990;8:651-654.
13. Troadec S, Chentouf M, Cerutti M, Nguyen B, Olive D, Bes C, Chardes T: In vitro anti-tumoral activity of baculo virus-expressed chimeric recombinant anti-CD4 antibody 13B8.2 on T-cell lymphomas. J Immunother (in press) 2006.
14. Carayannopoulos L, Max EE, Capra JD : Recombinant human IgA expressed in insect cells. Proc Natl Acad Sci USA 1994; 91 :8348-8352.
15. Nesbit M, Fu ZF, McDonald-Smith J, Steplewski Z, Curtis PJ: Production of a functional monoclonal antibody recognizing human colorectal carcinoma cells from a baculovirus expression system. J Immunol Methods 1992;151 : 201-208.
16. Edelman L, Margaritte C, Chaabihi H, Monchatre E, Blanchard D, Cardona A, et al.: Obtaining a functional recombinant anti-rhesus (D) antibody using the baculo virus-insect cell expression system. Immunology 1997;91 :13-19.
17. Croizier G, Croizier L, Quiot J-M, Lereclus D: Recombination of Autographa californica and Rachiplusia ou nuclear polyhedrosis viruses in Galleria mellonella L. J Gen Virol 1988;69: 177-185.
18. Rey F, Donker G, Hirsch I, and Chermann JC: Productive infection of CD4+ cells by selected HIV strains is not inhibited by anti-CD4 monoclonal antibodies. Virology 1991;181 : 165-171.
19. Chardes T, Villard S, Ferrieres G, Piechaczyk M, Cerutti M, Devauchelle G, et al. : Efficient amplification and direct sequencing of mouse variable regions from any immunoglobulin gene family. FEBS Lett 1999 ;452:386-394.
20. Altschul SF, Madden TL, Schaeffer AA, Zhang J, Zhang Z, Miller W, et al.: Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res 1997;25:3389-3402.
21. Lefranc MP, Giudicelli V, Kaasl Q, Duprat E, Jabado-Michaloud J, Scaviner D: IMGT, the international ImMunoGeneTics information system. Nucleic Acids Research 2005;33:D593-D597.
22. Cerutti M, Chaabihi H, Devauchelle G, Gautier L, Kaczorek M, Lefranc MP, et al.: Recombinant Baculovirus and use thereof in the production of monoclonal antibodies. INRA-CNRS Patent, France, FR 94/01015 1994.
23. Bes C, Briant-Longuet L, Cerutti M, Heitz F, Troadec S, Pugniere M, et al. :
Mapping the paratope of the anti-CD4 recombinant Fab 13B8.2 by combining parallel peptide synthesis and site-directed mutagenesis. J Biol Chem 2003;278: 14265-14273.
24. Van der Maarel S, van Dijk KW, Alexander CM, Sasso EH, Bull A, Milner EC: Chromosomal organization of the human VH4 gene family. Location of individual gene segments. J Immunol 1993;150:2858-2868.
25. Siebenlist U, Ravetch JV, Korsmeyer S, Waldmann T, Leder P: Human immunoglobulin D segments encoded in tandem multigenic families. Nature 1981;294:631-635.
26. Mattila PS, Schugk J, Wu H and Makela O: Extensive allelic sequence variation in the J region of the human immunoglobulin heavy chain gene locus. Eur J Immunol 1995 ;25(9):2578-2582.
27. Klobeck HG, Bornkamm GW, Combriato G, Mocikat R, Pohlenz HD, Zachau HG : Subgroup IV of human immunoglobulin K light chains is encoded by a single germline gene. Nucleic Acids Res 1985;13:6515-6529.
28. Sahota SS, Leo R, Hamblin TJ, Stevenson FK: Myeloma VL gene sequences reveal somatic hypermutation with intraclonal homogeneity, and a role for VL in antigen selection (Unpublished, access number : Z70260) 1996.
29. Lieby P, Soley A, Levallois H, Hugel B, Freyssinet JM, Cerutti M: The clonal analysis of anticardiolipin antibodies in a single patient with primary antiphospho lipid syndrome reveals an extreme antibody heterogeneity. Blood 2001;97:3820-3828.
30. Marchal I, Mir AM, Kmiecik D, Verbert A, Cacan R: Use of inhibitors to characterize intermediates in the processing of N-glycans synthesized by insect cells: a metabolic study with Sf9 cell line. Glycobiology 1999;9: 645-654.
31. Marchal I, Jarvis DL, Cacan R, Verbert A: Glycoproteins from insect cells: sialylated or not ? Biol Chem 2001;382:151-159.
32. Boyd PN, Lines AC, Patel AK: The effect of the removal of sialic acid, galactose and total carbohydrate on the functional activity of Campath-1H. MoI Immunol 1995;32:1311-1318.
33. Shinkawa T, Nakamura K, Yamane N, Shoji-Hosaka E, Kanda Y, Sakurada M, et al: The absence of fucose but not the presence of galactose or bisecting N- acetylglucosamine of human IgGl complex-type oligosaccharides shows the critical role of enhancing antibody-dependent cellular cytotoxicity. J Biol Chem 2003;278:3466-3473.
34. Kettleborough CA, Saldanha J, Ansell KH, Bendig MM: Optimization of primers for cloning libraries of mouse immunoglobulin genes using the polymerase chain reaction. Eur J Immunol 1993 ;23:206-211.
35. Orlandi R, Gussow, DH, Jones PT, Winter G: Cloning immunoglobulin variable domains for expression by the polymerase chain reaction. Proc Natl Acad Sci USA 1989;86:3833-3837.
36. Burton DR, Barbas CF 3rd, Persson MA, Koenig S, Chanock RM, Lerner RA: A large array of human monoclonal antibodies to type 1 human immunodeficiency virus from combinatorial libraries of asymptomatic seropositive individuals. Proc Natl Acad Sci U S A 1991;88(22):10134-10137.
37. Burton DR, Pyati J, Koduri R, Sharp SJ, Thornton GB, Parren PW, et al.: Efficient neutralization of primary isolates of HIV-I by a recombinant human monoclonal antibody. Science 1994;266 (5187): 1024- 1027.
38. Roben P, Moore JP, Thali M, Sodroski J, Barbas CF 3rd, Burton DR: Recognition properties of a panel of human recombinant Fab fragments to the CD4 binding site of gpl20 that show differing abilities to neutralize human immunodeficiency virus type 1. J Virol 1994;68(8):4821-4828.
39. Muster T, Steindl F, Purtscher M, Trkola A, Klima A, Himmler G, et al.: A conserved neutralizing epitope on gp41 of human immunodeficiency virus type 1. J Virol 1993;67(l l):6642-6647.
40. Zwick MB, Labrijn AF, Wang M, Spenlehauer C, Saphire EO, Binley JM, et al: Broadly neutralizing antibodies targeted to the membrane-proximal external region of human immunodeficiency virus type 1 glycoprotein gp41. J Virol 2001;75(22): 10892- 10905.
41. Trkola A, Purtscher M, Muster T, Ballaun C, Buchacher A, Sullivan N, et al. : Human monoclonal antibody 2Gl 2 defines a distinctive neutralization epitope on the gpl20 glycoprotein of human immunodeficiency virus type 1. J Virol 1996;70(2): 1100- 1108.
42. Scanlan CN, Pantophlet R, Wormald MR, Ollmann Saphire E, Stanfield R, Wilson IA, et al.: The broadly neutralizing anti-human immunodeficiency virus type 1 antibody 2G12 recognizes a cluster of alphal~>2 mannose residues on the outer face of gpl20. J Virol 2002;76(14):7306-7321. 43. Binley JM, Wrin T, Korber B, Zwick MB, Wang M, Chappey C, et al.: Comprehensive cross-clade neutralization analysis of a panel of anti-human immunodeficiency virus type 1 monoclonal antibodies. J Virol 2004;78(23): 13232- 13252.
44. Mehandru S, Wrin T, Galovich J, Stiegler G, Vcelar B, Hurley A, et al.: Neutralization of newly transmitted human immunodeficiency virus type 1 by monoclonal antibodies 2G12, 2F5 and 4E10. J Virol 2004 ;78:14039-14042.
45. Gray ES, Meyers T, Gray G, Montefiori DC, Morris L: Insensitivity of Paediatric HIV-I Subtype C Viruses to Broadly Neutralising Monoclonal Antibodies Raised against Subtype. B PLoS Med 2006;18;3(7):e255.
【特許請求の範囲】
【請求項1】
R7V エピトープ ( RTPKIQV 配列番号 11 ) に特異的に結合できHIV 株を中和できる単離された抗体又はその機能フラグメントの一つであって:
1) アミノ酸配列 配列番号1(QSVLYSSNNKNY), 配列番号2 (WAS) 及び配列番号3 (QQYYSTPQT) を有する相補性決定領域CDRs、又はその配列が配列番号 1, 2 又は3と、最適アライメント後に少なくとも80 %, 好ましくは90 % の同一性を持つCDRs を含む軽鎖、及び
2) アミノ酸配列配列番号6(GGSISSYY), 配列番号7 (IYYSGST) 及び配列番号8 (ARGRSWFSY) を有するCDRs、又はその配列が配列番号No 6, 7 及び8と、最適アライメント後に少なくとも80 %, 好ましくは90 % の同一性を持つCDRs を含む重鎖
を備える単離された抗体、又はその機能フラグメント。
【請求項2】
前記抗体が完全ヒト単クローン抗体又はその機能フラグメントである請求項1に記載の抗体。
【請求項3】
図3B-配列番号4で表わされるアミノ酸配列と、最適アライメント後に少なくとも80 % 好ましくは90 % の同一性を持つアミノ酸配列を含む軽鎖、又は図3 A -配列番号5 で表わされる配列又は配列番号5 と、最適アライメント後に少なくとも80 % , 好ましくは90 %の同一性を持つ配列を含むヌクレオチド配列によりエンコードされる軽鎖を備える請求項1又は2に記載の抗体。
【請求項4】
図3D -配列番号9で表わされるアミノ酸配列と、最適アライメント後に少なくとも80 % 好ましくは90 % の同一性を持つアミノ酸配列を含む重鎖、又は図3C -配列番号10 で表わされる配列又は配列番号10 と、最適アライメント後に少なくとも80 % , 好ましくは90 %の同一性を持つ配列を含むヌクレオチド配列によりエンコードされる重鎖を備える請求項1又は2に記載の抗体。
【請求項5】
請求項1に記載の抗体又はその機能フラグメントであって、図3B - 配列番号4で表わされるアミノ酸配列を含む軽鎖及び図3D - 配列番号9 で表わされるアミノ酸配列を含む重鎖を備える抗体、又はFv, scFv, Fab, F(ab')2, F(ab'), scFv-Fcタイプ又はダイアボディズから選択されたその機能フラグメント。
【請求項6】
配列 配列番号5 と、最適アライメント後に少なくとも80 %好ましくは90 %の同一性を持つ配列を含む単離された核酸。
【請求項7】
配列 配列番号10 と、最適アライメント後に少なくとも80 %好ましくは90 %の同一性を持つ配列を含む単離された核酸。
【請求項8】
請求項6または7により限定される核酸を含むベクター。
【請求項9】
請求項6で限定される核酸配列及び請求項7で限定される核酸配列を含むバキュロウィルストランスファーベクター。
【請求項10】
請求項8又は9に記載のベクターにより転換されたまたは該ベクターを含む宿主細胞。
【請求項11】
Sf9 細胞のような昆虫細胞、細菌性細胞、酵母細胞、動物細胞、具体的にはEBV 不死化B リンパ球、CHO、低フコシル化抗体を生成する遺伝子組み換えされたCHOのような哺乳類細胞又はYB2/0である請求項10に記載の宿主細胞。
【請求項12】
請求項1ないし5のいずれかによって限定される抗体、又はその機能フラグメントの1つを生成する方法であり、
a) 培地及び適切な培養条件の中で請求項10又は11に記載の宿主細胞を培養し、
b) 前記培養された細胞の前記培地から前記抗体を抽出する
ステップを備える方法。
【請求項13】
請求項1ないし5のいずれかによって限定される薬剤としての抗体、又はその機能フラグメントの1つ。
【請求項14】
請求項1ないし5のいずれかによって限定される抗体、又はその機能フラグメントの1つ、及び賦形剤及び/又は薬学的に受け入れられる媒体を含む医薬組成物。
【請求項15】
エイズ治療に現在使われている少なくとも1つの薬剤と請求項1ないし5のいずれかに記載の抗体とを含む同時、別々又は順次に使用のための併用製品。
【請求項16】
例としてHAART治療を受けている患者、具体的にはHAART治療の効果がない患者のHIV感染、エイズ治療のための請求項1ないし5のいずれかに記載の抗体の使用。
【請求項1】
R7V エピトープ ( RTPKIQV 配列番号 11 ) に特異的に結合できHIV 株を中和できる単離された抗体又はその機能フラグメントの一つであって:
1) アミノ酸配列 配列番号1(QSVLYSSNNKNY), 配列番号2 (WAS) 及び配列番号3 (QQYYSTPQT) を有する相補性決定領域CDRs、又はその配列が配列番号 1, 2 又は3と、最適アライメント後に少なくとも80 %, 好ましくは90 % の同一性を持つCDRs を含む軽鎖、及び
2) アミノ酸配列配列番号6(GGSISSYY), 配列番号7 (IYYSGST) 及び配列番号8 (ARGRSWFSY) を有するCDRs、又はその配列が配列番号No 6, 7 及び8と、最適アライメント後に少なくとも80 %, 好ましくは90 % の同一性を持つCDRs を含む重鎖
を備える単離された抗体、又はその機能フラグメント。
【請求項2】
前記抗体が完全ヒト単クローン抗体又はその機能フラグメントである請求項1に記載の抗体。
【請求項3】
図3B-配列番号4で表わされるアミノ酸配列と、最適アライメント後に少なくとも80 % 好ましくは90 % の同一性を持つアミノ酸配列を含む軽鎖、又は図3 A -配列番号5 で表わされる配列又は配列番号5 と、最適アライメント後に少なくとも80 % , 好ましくは90 %の同一性を持つ配列を含むヌクレオチド配列によりエンコードされる軽鎖を備える請求項1又は2に記載の抗体。
【請求項4】
図3D -配列番号9で表わされるアミノ酸配列と、最適アライメント後に少なくとも80 % 好ましくは90 % の同一性を持つアミノ酸配列を含む重鎖、又は図3C -配列番号10 で表わされる配列又は配列番号10 と、最適アライメント後に少なくとも80 % , 好ましくは90 %の同一性を持つ配列を含むヌクレオチド配列によりエンコードされる重鎖を備える請求項1又は2に記載の抗体。
【請求項5】
請求項1に記載の抗体又はその機能フラグメントであって、図3B - 配列番号4で表わされるアミノ酸配列を含む軽鎖及び図3D - 配列番号9 で表わされるアミノ酸配列を含む重鎖を備える抗体、又はFv, scFv, Fab, F(ab')2, F(ab'), scFv-Fcタイプ又はダイアボディズから選択されたその機能フラグメント。
【請求項6】
配列 配列番号5 と、最適アライメント後に少なくとも80 %好ましくは90 %の同一性を持つ配列を含む単離された核酸。
【請求項7】
配列 配列番号10 と、最適アライメント後に少なくとも80 %好ましくは90 %の同一性を持つ配列を含む単離された核酸。
【請求項8】
請求項6または7により限定される核酸を含むベクター。
【請求項9】
請求項6で限定される核酸配列及び請求項7で限定される核酸配列を含むバキュロウィルストランスファーベクター。
【請求項10】
請求項8又は9に記載のベクターにより転換されたまたは該ベクターを含む宿主細胞。
【請求項11】
Sf9 細胞のような昆虫細胞、細菌性細胞、酵母細胞、動物細胞、具体的にはEBV 不死化B リンパ球、CHO、低フコシル化抗体を生成する遺伝子組み換えされたCHOのような哺乳類細胞又はYB2/0である請求項10に記載の宿主細胞。
【請求項12】
請求項1ないし5のいずれかによって限定される抗体、又はその機能フラグメントの1つを生成する方法であり、
a) 培地及び適切な培養条件の中で請求項10又は11に記載の宿主細胞を培養し、
b) 前記培養された細胞の前記培地から前記抗体を抽出する
ステップを備える方法。
【請求項13】
請求項1ないし5のいずれかによって限定される薬剤としての抗体、又はその機能フラグメントの1つ。
【請求項14】
請求項1ないし5のいずれかによって限定される抗体、又はその機能フラグメントの1つ、及び賦形剤及び/又は薬学的に受け入れられる媒体を含む医薬組成物。
【請求項15】
エイズ治療に現在使われている少なくとも1つの薬剤と請求項1ないし5のいずれかに記載の抗体とを含む同時、別々又は順次に使用のための併用製品。
【請求項16】
例としてHAART治療を受けている患者、具体的にはHAART治療の効果がない患者のHIV感染、エイズ治療のための請求項1ないし5のいずれかに記載の抗体の使用。
【図1A】


【図1B】
【図3A】
【図3B】

【図3C】

【図3D】
【図4】
【図2A】

【図2B】
【図1B】
【図3A】
【図3B】
【図3C】
【図3D】
【図4】
【図2A】
【図2B】
【公表番号】特表2010−521189(P2010−521189A)
【公表日】平成22年6月24日(2010.6.24)
【国際特許分類】
【出願番号】特願2009−554021(P2009−554021)
【出願日】平成20年3月19日(2008.3.19)
【国際出願番号】PCT/EP2008/053317
【国際公開番号】WO2008/113833
【国際公開日】平成20年9月25日(2008.9.25)
【出願人】(509261946)
【Fターム(参考)】
【公表日】平成22年6月24日(2010.6.24)
【国際特許分類】
【出願日】平成20年3月19日(2008.3.19)
【国際出願番号】PCT/EP2008/053317
【国際公開番号】WO2008/113833
【国際公開日】平成20年9月25日(2008.9.25)
【出願人】(509261946)
【Fターム(参考)】
[ Back to top ]