
新規なヘモポエチン受容体およびそれをコードする遺伝子配列
【課題】新規ヘモポヘチン受容体およびそれをコードする核酸の提供。また、上記受容体のリガンドの検出及び受容体とリガンドの相互作用の解明により機能するアゴニスト、アンタゴニスト、治療剤等の提供。
【解決手段】マウス由来の新規ヘモポエチン受容体NR4。上記受容体のリガンドとしてのインターロイキン13(IL−13)及びインターロイキン13と構造的類似性の高いインターロイキン4(IL−4)とその受容体との複合体と上記ヘモポエチン受容体との間の相互作用に係わる核酸分子、該核酸分子を含む遺伝子構築物、組換えポリペプチド。
【解決手段】マウス由来の新規ヘモポエチン受容体NR4。上記受容体のリガンドとしてのインターロイキン13(IL−13)及びインターロイキン13と構造的類似性の高いインターロイキン4(IL−4)とその受容体との複合体と上記ヘモポエチン受容体との間の相互作用に係わる核酸分子、該核酸分子を含む遺伝子構築物、組換えポリペプチド。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は新規なヘモポエチン受容体またはその成分またはその部分そしてそれをコードする遺伝子配列に広く関係する。本発明のこのような受容体分子およびそれらの成分および/または部分そしてそれをコードする遺伝子配列はその受容体とリガンド相互作用により機能するアゴニスト、アンタゴニスト、治療剤および診断試薬などの広範囲のものの開発に有用である。
【背景技術】
【0002】
本明細書で数値により言及する出版物の文献的詳細は明細書の末尾に集めてある。本明細書で言及したヌクレオチド配列およびアミノ酸配列の配列番号は文献の次に記載してある。
【0003】
本明細書およびそれに続く請求の範囲を通じて、文脈上他の意味を必要とする場合を除き、「コンプライズ(含む)」という用語、あるいは「コンプライジーズ」または「コンプライジング」などのそれらの変形は、述べられた実体または一群の実体の包含を意味し、他の実体または他の一群の実体の排除を意味するものではないと理解すべきである。
【0004】
本発明の好ましいヘモポエチン受容体は本明細書では「NR4」と言及される。このNR4受容体はインターロイキン−13(IL−13)と相互作用する、従って本明細書ではIL−13受容体、より具体的にはIL−13受容体α−鎖(IL−13Rα)と呼ばれる。これらの用語は特定の主題について互換的に使用される。特定のNR4の起源となる種は1個の縮小文字で与えられる。例えば、ねずみ(murine)は「M」そしてヒトは「H」である。組換え型は接頭辞「r」が付されることがある。
【0005】
組換えDNA技術の精密化の迅速な発展により、医学分野や類似の保健衛生分野への研究が大いに促進された。サイトカインの研究は特に重要である。とりわけ、多様な細胞の増殖、分化および機能を調節するこれらの分子は重要である。組換えサイトカインの投与やサイトカインの機能および/または合成の調節がある範囲の疾病状態の治療のための医学的研究の焦点となる場合が増加しつつある。
【0006】
ある範囲のサイトカインや他の分泌される細胞機能調節物質の発見にもかかわらず、比較的僅かなサイトカインしか治療分野で直接使用されたり標的とされたりしていない。この理由の一つは多数のサイトカインが多面的な性質を持っていることである。例えば、インターロイキン(IL)−11は機能的に多面的な分子であり(1,2)、IL−6依存性の形質細胞腫の細胞株、T1165の増殖を刺激する能力によって特徴付けられたのが最初である(3)。IL−11の他の生物学的作用には、多機能性ヘモポエチン原始体の細胞増殖の誘導(4,5,6)、巨核球や血小板の形成の増強(7、8、9、10)、急性タンパク合成の刺激(11)および脂肪細胞リポタンパクリパーゼ活性の阻害(12、13)が含まれる。
【0007】
インターロイキン−13(IL−13)はインターロイキン−4(IL−4)と幾つかの構造的特徴を共有するもう一つの重要なサイトカインである(14および15を参照)。IL−4の遺伝子とIL−13の遺伝子とは関連のあるイントロン/エキソン構造を持っておりそしてヒト染色体5の上に共に近くに局在しており、またネズミでも染色体11のシンテニー領域に存在する(14、15)。タンパクレベルでは、IL−4とIL−13は4個のシステイン残基を含め約30%のアミノ酸同一性を共有している。生物学的にも、IL−13とIL−4は類似しており、活性化されたT−細胞により産生されそして、例えば、マクロファージに作用して分化を誘発しそして炎症性サイトカインの産生を抑制する。さらに、ヒトIL−13はB−細胞増殖に対する共−刺激シグナルとして作用しそして免疫グロブリン・イソタイプ・スイッチングに影響を与えることができる(14、15)。IL−13および他のヘモポエチンサイトカインの多様かつ多面的な機能はこのグループを重要な研究対象とし、特にサイトカインとその受容体との相互作用のレベルでの研究が重要である。サイトカイン受容体およびサイトカイン−受容体相互作用の操作および制御は多くの治療分野で、とりわけ標的サイトカインが機能的に多面的でありそして標的サイトカインのある機能を阻止したいがそのすべての機能を阻止したくはない場合には、潜在的に極めて重要である。
【非特許文献1】デュー,エックス.エックス.およびウィリアムス,ディー.エイ.(1994)Blood 83 : 2023-2030。
【非特許文献2】ヤング,ワイ.シー.およびイン,ティー.(1992)Biofactors 4 : 15-21。
【非特許文献3】ポール,エス.アール.,ベネット,エフ.,カルベッティ,ジェイ.エイ.,ケレハー,ケイ., ウッド,シー.アール.,オオハラ,アール.ジェイ.ジェイ.,レアリー,エイ.シー.,シブリー,ビー.,クラーク,エス.シー.,ウィリアムス,ディー.エイ.およびヤング,ワイ.シー.(1990)Proc.Natl.Acad.Sci.USA 87: 7512。
【非特許文献4】ムサシ,エム.,クラーク,エス.シー.,スドー,ティー.,ウーダル,ディー.エル.およびオガワ,エム.(1991)Blood 78: 1448-1451。
【非特許文献5】シブラー,ケイ.アール.,ヤング,ワイ.シー.およびクリステンセン,アール.ディー.(1992)Blood 80: 900-3。
【非特許文献6】ツジ,ケイ.,リーマン,エス.ディー.,スドー ,ティー.,クラーク,エス.シー.およびオガワ,エム.(1992)Blood 79: 2855-60。
【非特許文献7】バーンスタイン,エス.エイ.,メイ,アール.エル.,ヘンソーン,ジェイ.,フリース,ピー.およびターナー,ケイ.(1992)J.Cell.Physiol.153 : 305-12。
【非特許文献8】ハンゴク,ジー.,イン,ティー.,クーパー,エス.,シェンデル,ピー.,ヤング,ワイ.シー.およびブロックスメイヤー,エイチ.イー.(1993)Blood 81: 965-72。
【非特許文献9】テラムラ,エム.コバヤシ,エス.,ホシノ,エス.,オシミ,ケイ.およびミゾクチ,エイチ.(1992)Blood 79: 327-31。
【非特許文献10】ヨネムラ,ワイ.,カワキタ,エム.,マスダ,ティー.,フジモト,ケイ.,カトウ,ケイ.およびタカツキ,ケイ.(1992)Exp.Hematol.20: 1011-6。
【非特許文献11】バウマン,エイチ.およびシェンデル,ピー.(1991)J.Biol.Chem.266 : 20424-7。
【非特許文献12】カワシマ,アイ.,オオスミ,ジェイ.,ミタ−ホンジョウ,ケイ.,シモダ−タカノ,ケイ.,イシカワ,エイチ.,サカキバラ,エス.,ミヤダイ,ケイ.およびタキグチ,ワイ.(1991)Febs.Lett. 283: 199-202。
【非特許文献13】ケラー,ディー.シー., デュー,エックス.エックス.,スローアー,イー.エフ.,ホフマン,アール.およびウィリアムス,ディー.エイ.(1993)Blood 82 : 1428-35。
【非特許文献14】マッケンジー,エイ.エヌ.ジェイ.およびズラウスキー,ジー.(1994)サイトカインおよびその受容体へのガイドブック,オクスフォード大学出版部,オクスフォード。
【非特許文献15】ズラウスキー,ジー.およびド フリース,ジェイ.イー.(1994)Immunol.Today 15: 19-26。
【発明の開示】
【0008】
IL−13およびその受容体に関する研究はIL−13受容体のすべてまたは部分をコードする遺伝子配列をクローニングすることができなかったため妨げられてきた。本発明によれば、IL−13受容体α−鎖、すなわちIL−4受容体とも共有されている受容体サブユニットをコードする遺伝子配列が今やクローニングされたのである。これらの遺伝子配列の利用可能性により、構造または機能のレベルにおいてIL−13の活性並びにIL−13に関連するサイトカインの活性を調節または監視することができる一群の治療薬および診断薬の開発が可能となる。本発明によれば、構造および機能においてIL−13に関連するサイトカインの1例はIL−4である。
【0009】
従って、本発明の一つの側面は動物由来のヘモポエチン受容体または該受容体の誘導体をコードするヌクレオチド配列またはコードする配列に相補的なヌクレオチド配列を含む核酸分子に関する。
【0010】
より具体的には、本発明は動物ヘモポエチン受容体−該受容体はIL−13またはIL−13の誘導体と相互作用をすることができるものであるか−またはそれらの誘導体をコードするヌクレオチド配列またはコードする配列に相補的なヌクレオチド配列を含む単離された核酸分子を提供する。
【0011】
関連する一つの態様で、本発明は、該受容体が、
(i)IL−13またはその誘導体と相互作用をすることができ、そして
(ii)IL−4とIL−4受容体α−鎖との間でできる複合体と相互作用をすることができる、
ものである、動物ヘモポエチン受容体またはそれらの誘導体をコードするヌクレオチド配列またはコードする配列に相補的なヌクレオチド配列を含む単離された核酸分子を提供する。
【0012】
これらの態様により、IL−13の誘導体には、アゴニスト、アンタゴニスト、抗体および擬症物質(mimetics)が含まれることになる。
【0013】
本発明は動物IL−13受容体α−鎖またはその誘導体をコードするヌクレオチド配列またはコードする配列に相補的なヌクレオチド配列を含む核酸分子にも関する。
【0014】
関連する一つの態様では、本発明は動物IL−4受容体またはその誘導体の1成分をコードするヌクレオチド配列またはコードする配列に相補的なヌクレオチド配列を含む核酸分子を包含する。
【0015】
この動物は哺乳類または鳥の1種であることが好ましい。とりわけ好ましい動物としては、ヒト、実験室テスト動物(例えば、霊長類、マウス、ウサギ、ハムスター、モルモット)、家畜(例えば、羊、山羊、馬、豚、牛、ロバ)、愛玩動物(例えば、犬、猫)、捕獲した野生の動物(例えば、キツネ、カンガルー、ディンゴー)および家禽(例えば、鶏、鵞鳥、鴨)およびゲーム鳥(例えば、エミュー、ダチョウ)などが挙げられる。本発明はマウスやヒトを例としてあげるが、主題である本発明の範囲はすべての動物および鳥類に及ぶものである。
【0016】
本発明は配列類似性に基づきヘモポエチン受容体ファミリーのメンバーを同定する能力について部分的に予言する。このアプローチにより、ヘモポエチン受容体をコードする一つの遺伝子配列が本発明により同定された。発現された遺伝子配列は本明細書で「NR4」と呼ばれる。本発明によれば、NR4はトランスフェクトされたCOS細胞により合成されたとき、約50,000〜約70,000ダルトン、より好ましくは約55,000〜約65,000ダルトンの見掛けの分子量を持っている。NR4はIL−13と特異的にかつ低親和性をもって結合する、それ故、IL−13受容体α−鎖であると考えられる。従って、「NR4」と「IL−13受容体α−鎖」(または「IL−13Rα」)とは本明細書を通して互換可能なものとして使用される。さらに、IL−13のその受容体への結合はIL−4受容体α−鎖を発現する細胞内でIL−4またはその成分により競争的に阻害されることが見出された、そしてこれはIL−13と受容体との相互作用を制御する方法を提供できるものであり、そして擬症物質の調製および構築のための基礎をも提供するものである。
【0017】
本発明の別の側面では、配列番号:2に記載されたアミノ酸配列を持つIL−13受容体α−鎖またはその全部または一部に少なくとも約50%の類似性を有するアミノ酸配列をコードするヌクレオチド配列を含む核酸分子が提供される。この類似性パーセンテージは少なくとも約60%が好ましく、少なくとも約70%がより好ましく、少なくとも約80〜85%がさらに好ましく、そして少なくとも約90〜95%またはそれ以上がさらに一層好ましい。配列の全部または一部に言及しているのは、二つの受容体の各部分を含むハイブリッド分子を定義して含めることを意図している。単一アミノ酸を包含することは意図していない。
【0018】
本発明のさらなる態様では、IL−13受容体α−鎖をコードしそして配列番号:1に実質的に記載されたヌクレオチド配列を有するヌクレオチド配列またはその全部または一部と少なくとも約50%の類似性を有するヌクレオチド配列を含む核酸分子が提供される。この類似性パーセンテージは少なくとも約60%が好ましく、少なくとも約70%がより好ましく、少なくとも約80〜85%がさらに好ましく、そして少なくとも約90〜95%またはそれ以上がさらに一層好ましい。
【0019】
本発明のさらに別の側面では、配列番号:4に記載されたアミノ酸配列を有するIL−13受容体α−鎖またはその全部または一部と少なくとも約50%の類似性を有するアミノ酸配列をコードするヌクレオチド配列を含む核酸分子が提供される。この類似性パーセンテージは少なくとも約60%が好ましく、少なくとも約70%がより好ましく、少なくとも約80〜85%がさらに好ましく、そして少なくとも約90〜95%またはそれ以上がさらに一層好ましい。
【0020】
本発明のさらなる態様では、IL−13受容体α−鎖をコードしそして配列番号:3に記載されたようなヌクレオチド配列を実質的に有するヌクレオチド配列またはその全部または一部に少なくとも約50%の類似性を有するヌクレオチド配列を含む核酸分子が提供される。この類似性パーセンテージは少なくとも約60%が好ましく、少なくとも約70%がより好ましく、少なくとも約80〜85%がさらに好ましく、そして少なくとも約90〜95%またはそれ以上がさらに一層好ましい。
【0021】
従って、本発明は配列番号:1または配列番号:3に記載されたヌクレオチド配列または配列番号:2または配列番号:4に記載されたアミノ酸配列またはこれらに対し1個または複数のヌクレオチドまたはアミノ酸の置換、欠失および/または付加がなされたものに及ぶことになる。
【0022】
本発明はさらに、低厳格条件下で配列番号:1または配列番号:3に記載されたヌクレオチド配列またはそれらの相補型とハイブリッドを形成することができる核酸分子に及ぶことになる。
【0023】
本発明は組換えヘモポエチン受容体および特に組換えNR4およびNR4を含む組換えハイブリッドに及ぶ。組換えポリペプチドはIL−13と低親和性で相互作用することが好ましく、高い親和性で相互作用すればより一層好ましい。
【0024】
とりわけ好ましい態様では、ポリペプチドは下記の特性のうちの少なくとも二つを有するものである。
(i)配列番号:2または配列番号:4に実質的に記載されたアミノ酸配列またはそれらの全部または一部と少なくとも約50%の類似性を有するアミノ酸配列を含むこと、
(ii)配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列またはそれらの全部または一部と少なくとも約50%の類似性を有するヌクレオチド配列によりコードされるものであること、
(iii)IL−13またはその誘導体と少なくとも低親和性で相互作用すること、および
(iv)COS細胞で発現させたとき、ウエスタン・ブロット分析により測定すると約50,000から約70,000ダルトンの分子量を有すること。
【0025】
関連する態様では、このポリペプチドは下記の特性のうちの少なくとも三つを有する。
(i)配列番号:2または配列番号:4に実質的に記載されたアミノ酸配列またはそれらの全部または一部と少なくとも約50%の類似性を有するアミノ酸配列を含むこと、
(ii)配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列またはそれらの全部または一部と少なくとも約50%の類似性を有するヌクレオチド配列によりコードされるものであること、
(iii)IL−13またはその誘導体と少なくとも低親和性で相互作用すること、および
(iv)COS細胞で発現させたとき、ウエスタン・ブロット分析により測定すると約50,000から約70,000ダルトンの分子量を有すること、
(v)IL−4受容体α−鎖由来のアミノ酸配列を含むこと、そして
(vi)IL−13と相互作用をすることができ、かつその相互作用がIL−4受容体α−鎖を発現する細胞中でIL−4により競争的に阻害されること。
【0026】
本明細書で「組換えヘモポエチン受容体」、「NR4」、「IL−13受容体」または「IL−13受容体α−鎖」と言及するときは、それらの一部、断片、部分、相同体、ハイブリッドまたはアナログなどのそれらの誘導体への言及も含まれる。誘導体は機能的であってもなくてもよく、または非機能的ではあるが受容体の全部または一部に対する抗体と免疫学的に相互作用し得るものであってもよい。受容体の誘導体は受容体−リガンド相互作用のアゴニストまたはアンタゴニストをも包含する。機能とは、IL−13またはその誘導体と相互作用するNR4の能力あるいは可溶性NR4の場合にはIL−13で誘発されたある細胞の活性と競合する能力により便宜的に定義される。
【0027】
本発明により提供される特に好ましい誘導体には、高親和性でIL−13とまたは高親和性でIL−13およびIL−4と結合することができるIL−13受容体α−鎖の誘導体が含まれ、また誘導体にはIL−13受容体α−鎖と、例えば、IL−13とも高親和性で結合するIL−4受容体α−鎖の間などのキメラ分子が包含される。
【0028】
本発明により提供される他の融合分子またはキメラ分子には、NR4とヘモポエチン受容体ファミリーのメンバー、受容体チロシンキナーゼ、TNF/NGF受容体およびGタンパク質−共役受容体の間のものが含まれる。例えば、キメラにはNR4とIL−13結合タンパク、IL−4受容体α−鎖、IL−2受容体γ−鎖またはIL−5などの気管支喘息やアレルギーに関与する他のサイトカインに対する受容体との間に生じ得る。他の重要なキメラにはNR4と免疫グロブリンまたは特定の細胞または組織にNR4を標的化させる他の分子、NR4と毒素、およびNR4と成長因子が含まれる。
【0029】
本明細書において42℃の低厳格性というときは、ハイブリッド形成条件においては少なくとも約1%v/vから少なくとも約15%v/vまでのホルムアミドおよび少なくとも約1Mから少なくとも約2Mまでの塩を、そして洗浄条件においては少なくとも約1Mから少なくとも約2Mまでの塩を範囲として包含する。必要なときは、中度の厳格性などの別の厳格条件を適用することもできる。中度の厳格性は、ハイブリッド形成条件においては少なくとも約16%v/vから少なくとも約30%v/vまでのホルムアミドおよび少なくとも約0.5Mから少なくとも約0.9Mの塩を、そして洗浄条件においては少なくとも約0.5Mから少なくとも約0.9Mの塩を範囲として包含し、あるいは高厳格性は、ハイブリッド形成条件においては少なくとも約31%v/vから少なくとも約50%v/vまでのホルムアミドおよび少なくとも約0.01Mから少なくとも約0.15Mの塩を、そして洗浄条件においては少なくとも約0.01Mから少なくとも約0.15Mの塩を範囲として包含する。
【0030】
本発明のもっと別の側面では、IL−13受容体α−鎖をコードするヌクレオチド配列またはコードする配列に相補的なヌクレオチド配列を含む核酸分子であって、配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列を有するものあるいは構造的に類似するIL−13受容体α−鎖またはその誘導体をコードしそして配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列またはその相補型に低厳格性条件下でハイブリッドを形成することができる核酸分子が提供される。
【0031】
本発明のさらに別の側面では、配列番号:2または配列番号:4に実質的に記載されたアミノ酸配列を持つIL−13受容体α−鎖をコードするヌクレオチド配列またはコードする配列に相補的なヌクレオチド配列を含む核酸分子が、または配列番号:2または配列番号:4に記載された配列に少なくとも約50%の類似性を有するアミノ酸配列をコードするヌクレオチド配列であって、かつ配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列に低厳格性条件下でハイブリッドを形成することができるものを含む核酸分子が提供される。
【0032】
本発明により提供される核酸分子は一般に単離された形のものであり、一本鎖でも二本鎖でも、直鎖状でも閉じた環状のDNA(例えば、ゲノムDNA)でも、cDNAまたはmRNAでも、あるいはDNA:RNAハイブリッドの形などのそれらの結合物でもよい。特に好ましい態様では、核酸分子はベクターの形であり、特定の宿主細胞内で発現させることができる発現ベクターが最も好ましい。特に有用な宿主細胞としては原核細胞、哺乳類細胞、酵母細胞および昆虫細胞が挙げられる。この細胞は細胞株の形であってもよい。
【0033】
本発明のこの側面により、本明細書で先に述べたIL−13受容体α−鎖をコードする核酸分子を含む発現ベクターが提供される。該発現ベクターは特定の宿主細胞内で発現することができる。
【0034】
本発明の別の側面では、配列番号:2または配列番号:4に実質的に記載されたアミノ酸配列またはその全部または一部と少なくとも約50%の類似性を有するアミノ酸配列を含む組換えポリペプチドが提供される。この類似性パーセンテージは少なくとも約60%が好ましく、少なくとも約70%がより好ましく、少なくとも約80〜85%がさらに好ましく、そして少なくとも約90〜95%またはそれ以上がさらに一層好ましい。
【0035】
本発明により提供される組換えポリペプチドは、従って、IL−13受容体α−鎖の成分、部分、断片、誘導体、相同体、またはアナログを含み、そして配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列、またはその全部または一部と少なくとも約50%の類似性を有する分子、または配列番号:1または配列番号:3に記載されたヌクレオチド配列またはそれらの相補型とハイブリッドを形成することができる分子によりコードされることが好ましい。この組換え分子はグリコシル化されていてもいなくてもよい。グリコシル化型であるときは、グリコシル化は天然に生ずるIL−13受容体α−鎖と実質的に同じであってもよく、またはグリコシル化の修飾型であってもよい。グリコシル化の変更または相違はIL−13受容体α−鎖の結合活性に影響を与えることも与えないこともある。
【0036】
組換えIL−13受容体α−鎖は可溶性の形であることも、細胞表面上に発現されることもまたは固体支持体または別の分子に結合したりまたは融合したりしていることもある。
【0037】
本発明はさらに下記の特性のうちの少なくとも二つを有する組換えポリペプチドを製造する方法を提供する。
(i)配列番号:2または配列番号:4に実質的に記載されたアミノ酸配列またはそれに少なくとも約50%の類似性を有するアミノ酸配列を含むこと、
(ii)配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列またはそれに少なくとも約50%の類似性を有するヌクレオチド配列によりコードされるものであること、
(iii)IL−13またはその誘導体と低親和性で相互作用すること、そして
(iv)COS細胞中で発現させるとき、ウエスタン・ブロット分析により測定すると約50,000から約70,000ダルトンの分子量を持つこと。
【0038】
該方法は、本発明の遺伝子構築物を含む細胞を、該遺伝子構築物中の核酸分子を発現して組換えポリペプチドを産生するのに十分な時間および条件の下で培養する工程、そして該組換えポリペプチドを単離する工程を含んで成る方法である。
【0039】
別の態様は下記の特性のうちの少なくとも三つを有する組換えポリペプチドを製造する方法を提供する。
(i)配列番号:2または配列番号:4に実質的に記載されたアミノ酸配列またはその全部または一部に少なくとも約50%の類似性を有するアミノ酸配列を含むこと、
(ii)配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列またはその全部または一部に少なくとも約50%の類似性を有するヌクレオチド配列によりコードされるものであること、
(iii)IL−13またはその誘導体と少なくとも低親和性で相互作用すること、
(iv)COS細胞中で発現させたとき、ウエスタン・ブロット分析により測定すると約50,000から約70,000ダルトンの分子量を持つこと、
(v)IL−4受容体α−鎖由来のアミノ酸配列を含むこと、そして
(vi)IL−13と相互作用をすることができること、かつ、その相互作用がIL−4受容体α−鎖を発現する細胞内でIL−4により競争的に阻害されること。
【0040】
該方法は、本発明の融合遺伝子構築物を含む細胞を、該融合遺伝子構築物内の核酸分子が発現し組み換えポリペプチドを産生するのに十分な時間および条件の下で培養する工程および該組換えポリペプチドを単離する工程を含んで成る方法である。
【0041】
本発明はさらに上記の組換えポリペプチドを発現する動物細胞などの細胞にまで及ぶ。
【0042】
本発明の別の態様では、組換えIL−13受容体α−鎖の化学的アナログが提供される。
【0043】
上に述べたように、本発明はさらにNR4のある範囲の誘導体を提供する。誘導体には、NR4ポリペプチドおよび対応する遺伝子配列の断片、一部、部分、変異体、ハイブリッド(融合分子やキメラ分子を含む)、相同体およびアナログが含まれる。一つの好ましい態様では、この誘導体は高親和性でもってIL−13と結合する。他の好ましい誘導体はアゴニスト、アンタゴニストまたは擬症物質として作用する。これらの誘導体には、NR4に1個または複数のアミノ酸が置換、欠失、および/または付加されたもの、およびNR4をコードする遺伝子配列に1個または複数のヌクレオチドが置換、欠失、および/または付加されたものも含まれる。アミノ酸配列またはヌクレオチド配列への「付加」には、他のペプチド、ポリペプチド、またはタンパク質との融合またはヌクレオチド配列への融合も含まれる。本明細書で「NR4」と言及するときは、機能的誘導体や「NR4」と免疫学的に相互作用し得る誘導体も含めそのすべての誘導体を指している。本発明はネズミNR4、ヒトNR4またはそれらの誘導体の間などのハイブリッド分子にも及ぶ。特に好ましいハイブリッドはNR4とIL−4受容体α−鎖を含むものである。
【0044】
本明細書において提供されるNR4の類似体には、側鎖の修飾、ペプチド合成、ポリペプチド合成またはタンパク合成の間における非天然型アミノ酸および/またはそれらの誘導体の取り込み、および架橋剤や、タンパク性分子またはそれらのアナログにコンホーメーション上の制約を加える他の方法の使用などが含まれるが、これらに限定されるものではない。
【0045】
本発明によって提供される側鎖の修飾の例としては、アルデヒドとの反応による還元的アルキル化に続くNaBH4による還元、メチルアセチミデートを用いるアミジノ化(amidination)、無水酢酸を用いるアシル化、シアネートを用いるアミノ基のカルバモイル化、2,4,6−トリニトロベンゼンスルホン酸(TNBS)を用いるアミノ基のトリニトロベンジル化、無水コハク酸および無水テトラハイドロフタル酸を用いるアミノ基のアシル化、およびピリドキサル−5−リン酸に続いてNaBH4で還元することによるリシンのピリドキシレーションなどのアミノ基の修飾が挙げられる。
【0046】
アルギニン残基のグアニジン基は2,3−ブタンジオン、フェニルグリオキサルおよびグリオキサルなどの試薬を用いて複素環縮合産物の形成により修飾することができる。
【0047】
カルボキシル基はO−アシルイソウレア形成を経るカルボジイミド活性化それに続く、例えば対応するアミドへの誘導により修飾することができる。
【0048】
スルフヒドリル基はヨード酢酸またはヨードアセトアミドを用いるカルボキシメチレーション、システイン酸への過ギ酸酸化、他のチオール化合物との混合ジスルフィドの形成、マレイミド、無水マレイン酸または他の置換マレイミドとの反応、4−クロロマーキュリベンゾエート、4−クロロマーキュリフェニルスルホン酸、フェニルマーキュリクロライド、2−クロロマーキュリ−4−ニトロフェノールおよび他の水銀化合物を用いる水銀誘導体の形成、アルカリ性pHでシアネートを用いるカルバモイル化などの諸方法により修飾することができる。
【0049】
トリプトファン残基は、例えば、N−ブロモスクシンイミドを用いる酸化または2−ヒドロキシ−5−ニトロベンジルブロマイドまたはスルフェニルハライドを用いるインドール環のアルキル化により修飾することができる。チロジン残基は、他方、テトラニトロメタンを用いる硝酸化により改変し3−ニトロチロジン誘導体を形成することができる。
【0050】
ヒスチジン残基のイミダゾール環の修飾はヨード酢酸誘導体を用いるアルキル化またはジエチルピロカーボネートを用いるN−カルボエトキシレーションにより達成することができる。
【0051】
ペプチド合成の間に非天然型アミノ酸および誘導体を取り込む例としては、ノルロイシン、4−アミノ酪酸、4−アミノ−3−ヒドロキシ−5−フェニルペンタン酸、6−アミノヘキサン酸、t−ブチルグリシン、ノルバリン、フェニルグリシン、オルニチン、ザルコシン、4−アミノ−3−ヒドロキシ−6−メチルヘプタン酸、2−チエニルアラニンおよび/またはアミノ酸のD−異性体が挙げられるが、これらに限定されるものではない。本明細書で提供される非天然型アミノ酸のリストを表1に示す。
【0052】
架橋剤は、例えば、3Dコンホーメーションを安定化するために使用することができ、n=1からn=6までのスペーサー基(CH2)nを持つ二官能性イミドエステル、グルタルアルデヒド、N−ヒドロキシスクシンイミドエステルなどのホモ−二官能性架橋剤および通常N−ヒドロキシスクシンイミドなどのアミノ−反応性部分とマレイミドまたはジチオ部分(SH)またはカルボジイミド(COOH)などの別の基の特異的反応性部分とを持つヘテロ−二官能性試薬を使用する。また、ペプチドのコンホーメーションは、例えば、CαおよびNα−メチルアミノ酸の取り込み、アミノ酸のCαとCβ原子の間に二重結合の導入、およびN末端とC末端の間、二つの側鎖の間または側鎖とN末端またはC末端との間にアマイド結合を形成させるなどして共有結合を導入することにより環状ペプチドまたは環状アナログの形成により制約することができる。
【0053】
これらのタイプの修飾は個体に投与するとき、あるいは診断試薬として使用するために、NR4を安定化するために重要である。
【0054】
本発明はさらにNR4のアンタゴニストまたはアゴニストとして作用することができる、すなわちNR4の機能的アナログとして作用することができるNR4の化学的アナログを提供する。化学的アナログは必ずしもNR4から誘導されるとは限らず、ある種のコンホーメーションの類似性を共有するものであってもよい。また、化学的アナログはNR4のある種の物理化学的性質を真似するように特別に設計することもできる。化学的アナログは化学的に合成することができまたは例えば天然物のスクリーニングにより検出することができる。
【0055】
NR4の同定により、NR4の発現を調節したりNR4の活性を調節したりすることができる1群の治療に有用な分子の誕生を可能とする。本発明により提供される調節物質にはNR4遺伝子発現またはNR4タンパク活性のアゴニストおよびアンタゴニストが含まれる。NR4遺伝子発現のアンタゴニストにはアンチセンス分子、リボザイムおよび共−抑制分子(co-suppression molecules)が含まれる。アゴニストにはプロモーター能を増強したりまたは負の調節機構を妨害したりする分子が含まれる。NR4タンパクのアゴニストには抗体、リガンドおよび擬症物質が含まれる。NR4のアンタゴニストには抗体および阻害ペプチド断片が含まれる。細胞がNR4とIL−4受容体α−鎖を共−発現するときは、アゴニストおよびアンタゴニストはこのIL−4受容体α−鎖を標的とすることができる。
【0056】
【表1】

【0057】
本発明により提供される他の誘導体には、完全に脱グリコシル化された分子から修飾されたグリコシル化分子までのある範囲のグリコシル化変異型が含まれる。改変されたグリコシレーションパターンは異なる宿主細胞中での組換え分子の発現から生ずることがある。
【0058】
本発明の別の態様では、ヒトにおけるNR4遺伝子の発現を調節する方法が提供される。この方法はNR4をコードするNR4遺伝子をNR4発現の調節物質の有効量と、NR2の発現を高く調節しまたは低く調節しまたは別の仕方で調節するのに十分な時間および条件の下で、接触させることを含んでなる方法である。NR4をコードする核酸分子またはその誘導体を細胞内に導入して、例えば、欠陥を持つまたは一つ以上の望ましくない変異を持つ内因性NR4遺伝子配列の置換を含め、その細胞のNR4関連活性を増強したり改変したりすることができる。逆に、オリゴヌクレオチドなどのNR4アンチセンス配列(または共−抑制のためのセンス配列)を導入して、内因性のNR4遺伝子を発現しているどの細胞のNR4関連活性をも減少させることができる。リボザイムも使用することができる。
【0059】
本発明の別の側面は、ヒトにおけるNR4の活性を調節する方法を提供する。この方法はNR4活性を増加または減少させるのに十分な時間および条件の下である分子の調節有効量を該哺乳類に投与することを含んでなる方法である。この分子はタンパク性の分子であっても化学的実体であってもよくそしてNR4の誘導体またはそのリガンドまたは化学的アナログまたはNR4の切断変異体またはそのリガンドであってもよい。
【0060】
例えば、IL−13とIL−4は免疫応答の調節に関与し、そして喘息などのアレルギー性またはアトピー性疾患と関連する免疫グロブリン・イソタイプであるIgEの産生に関与している。IL−13/IL−4とそれらの受容体との相互作用を調節することはアレルギー状態などの炎症状態を治療するのに重要であり得る。IL−4/IL−13およびIgEのレベルの上昇はネフローゼ症候群、春季カタルおよび角結膜炎などの疾病にも重要である。本明細書でその治療法が提供される他の疾病としては、気管支喘息、鼻炎(perennial rhinitis)およびアトピー性皮膚炎が挙げられる。IL−13−受容体相互作用の調節が重要となり得る他の病態としては、IL−13がサイトカイン形成を誘発し、今度はこのサイトカインが疾病の開始、進行および/または重篤度に関与するような病態が含まれる。同様に、IL−4−受容体相互作用の調節も疾病状態を制御するのに重要である。例えば、一部の癌はサイトカインIL−13またはIL−4により悪化させられることがある。これらは免疫抑制効果または免疫抑制効果分子を誘発し、今度はこれが癌の成長に応答する身体の能力を減退させるのである。
【0061】
従って、本発明はNR4またはその誘導体またはNR4発現またはNR4活性の調節物質および1以上の薬学的に許容され得る担体および/または希釈剤を含んでなる薬学的組成物を提供する。これらの成分は「活性成分」と呼ばれる。
【0062】
これについては、前に記載した組換えヘモポエチン受容体またはそのリガンド(例えば、IL−13)結合性部分および1以上の薬学的に許容され得る担体および/または希釈剤を含んでなる薬学的組成物が提供される。
【0063】
別の態様では、前に述べた組換えヘモポエチン受容体に対するリガンド(例えば、IL−13)および1以上の薬学的に許容され得る担体および/または希釈剤を含んでなる薬学的組成物が提供される。
【0064】
本発明のさらなる側面は、動物の治療方法を提供する。この方法は前に記載した組換えヘモポエチン受容体またはそのリガンド結合性部分または該ヘモポエチン受容体に対するリガンド(例えば、IL−13)の治療有効量を、該治療が実質的に達成されるのに十分な時間および条件の下であるいは実質的に改善される条件の下で、該動物に投与することを含むものである。
【0065】
注射可能な用途に適する薬学的剤形としては、滅菌水溶液(水溶性の場合)または分散剤および滅菌した注射可能な溶液または分散剤の用時調製のための滅菌粉末が挙げられ、あるいはクリームの形状であってもまたは局所適用に適した他の剤形であってもよい。それは製造および貯蔵の条件下で安定でなければならず、細菌や糸状菌などの微生物の汚染作用に対して保護されなければならない。担体は溶媒または分散剤であることができ、例えば、水、エタノール、ポリオール(例えば、グリセロール、プロピレングリコールおよび液体ポリエチレングリコール、およびその他のもの)、それらの適当な混合物、および植物性油などを含む。例えば、リシチン(licithin)などの皮膜の使用により、分散剤の場合には必要な粒子サイズの維持によりそしてスーパーファクタントの使用により、適当な流動性を維持することができる。微生物の作用の防止は種々の抗細菌物質および抗かび物質、例えば、パラベン、クロロブタノール、フェノール、ソルビン酸、チルメロサル(thirmerosal)およびその他により可能となる。多くの場合、等張物質、例えば、砂糖または塩化ナトリウムを含めることが好ましい。注射可能な組成物の長期間吸収は吸収を遅延させる物質、例えばアルミニウムモノステアレートおよびゼラチンを組成物の中で使用することにより可能とすることができる。
【0066】
注射可能な滅菌溶液は必要な量の活性化合物を必要な場合は上に挙げた他の様々な成分と共に適当な溶媒の中に溶解し次いで無菌ろ過を行うことにより調製される。一般に、分散剤は様々な滅菌された活性成分を、基本的な分散媒体および上に挙げたものの中の必要な他の成分を含む滅菌担体の中に取り込むことにより調製される。
【0067】
注射可能な滅菌溶液を調製するための滅菌粉末の場合には、好ましい調製方法は真空乾燥法および凍結−乾燥法である。これらは予めそれらの無菌ろ過された溶液から、活性成分プラス付加的な必要成分の粉末を生ずる。
【0068】
活性成分を適当に保護するときは、例えば、不活性な希釈剤または資化し得る食用の担体と共に経口投与することができ、あるいはそれを固いまたは軟らかい殻のゼラチンカプセルの中に閉じ込めることができ、あるいはそれを錠剤中に圧縮することができ、あるいは食事の食品と共に直接摂取することができる。治療のための経口投与を行うためには、活性化合物は賦形剤と混ぜ、消化可能な錠剤、バッカル錠剤、トローチ、カプセル、エリクサル、懸濁剤、シロップ、ウェハース、その他の形で使用される。このような組成物および調製物は少なくとも1重量%の活性化合物を含むべきである。この組成物および調製物のパーセンテージは、もちろん、変動するものであり、単位重量の約5%から約80%までの間にあるのが便宜である。治療上有用な組成物の中の活性化合物の量は適当な投与量が得られるようなものである。本発明による組成物または調製物は経口投与の単位形態が活性化合物を約0.1μgと2000mgの間で含むように調製されることが好ましい。
【0069】
錠剤、トローチ、ピル、カプセルなども後にリストされる成分を含むことができる。ガム、アカシア、コーンスターチまたはゼラチンなどのバインダー、ジカルシウムホスフェートなどの賦形剤、コーンスターチ、ポテトスターチ、アルギン酸、その他の崩壊剤、ステアリン酸マグネシウムなどの滑剤、およびスクロース、ラクトースまたはサッカリンなどの甘味剤またはペパーミント、サリチル酸メチル(oil of wintergreen)、または桜の芳香などの芳香剤を添加することができる。投与単位の形態がカプセルのときは、上のタイプの物質に加えて液体の担体を含めることができる。投与単位の物理的形態を被覆あるいは別の仕方で修飾するための様々な物質がほかにもある。例えば、錠剤、ピル、またはカプセルはシェラック、砂糖または両方で被覆することができる。シロップまたはエリクサルは活性化合物、甘味剤として砂糖、保存剤としてメチルおよびプロピルパラベン、色素および桜またはオレンジなどの芳香剤を含むことができる。もちろん、いかなる投与単位形態を作る場合に使用されるどんな物質も薬学的に純粋でありそしてその使用量では実質的に無毒であるべきである。さらに、活性化合物(単数または複数)は制御放出(sustained-release)用の調製物や処方に加えることもできる。
【0070】
本発明はクリーム、ローションおよびゲルなどの局所適用に適した形態をも包含する。
【0071】
薬学的に許容され得る担体および/または希釈剤としては、溶媒、分散媒体、被覆剤、抗細菌および抗かび剤、等張物質および吸収遅延物質などのいずれかおよびすべてが挙げられる。このような媒体や物質を薬学的に活性な物質に使用することは当技術分野では良く知られている。通常の媒体または物質が活性成分と配合禁忌である場合を除き、治療用組成物にそれらを使用することは本発明に包含される。補充的な活性成分もこの組成物の中に取り込むことができる。
【0072】
投与の容易および投与量の均一性のため投与単位の形態で非経口的組成物を処方することはとりわけ有利である。本明細書で使用されるとき投与単位形態とは治療すべき対象哺乳類に対し1回投与に適した物理的に区別された単位を指し、各単位は必要な薬学的担体との組み合わせで望まれる治療効果を生じさせるように計算された活性物質の予め計測された量を含んでいる。本発明の新規な投与単位形態の仕様は、(a)活性物質のユニークな特性および達成すべき具体的治療効果、および(b)身体の健康が損なわれている疾病状態を持つ生物対象における疾病の治療のためにこのような活性物質を混ぜ合わせる技術上の固有の制約、により支配されそして直接それらに依存している。
【0073】
主要な活性成分は便利で効果的な投与のために、前に開示したような投与単位形態で、その有効量が薬学的に許容され得る適当な担体と混ぜ合わされる。単位の投与形態は、例えば、0.5μgから約2000mgにわたる量で主要な活性化合物を含むことができる。比率で表すと、活性化合物は一般に担体の1ml当たり約0.5μgから約2000mgまで存在する。補充的活性成分を含む組成物の場合は、投与量は該成分の通常の投与量および投与方法を参照して決定する。
【0074】
薬学的組成物は標的細胞をトランスフェクトすることができるベクターのような遺伝子分子を含むこともでき、該ベクターはNR4発現またはNR4活性を調節することができる核酸分子を担っている。このベクターは例えばウイルスベクターであることができる。
【0075】
本発明のさらに別の側面はNR4に対する抗体およびその誘導体またはそのリガンド(例えば、IL−13)に関する。このような抗体はモノクローン抗体またはポリクローン抗体であることができ、そして天然に生ずるNR4への抗体から選択することができ、あるいはNR4またはその誘導体に対し特異的に高めることもできる。後者の場合は、NR4またはその誘導体はまず担体分子と結合することが必要になることがある。本発明の抗体および/または組換えNR4またはその誘導体は治療剤または診断試薬として特に有用である。
【0076】
例えば、NR4およびその誘導体はNR4への天然に生ずる抗体のスクリーニングに使用することができる。これらは、例えば、自己免疫疾患の一部で生ずる。また、NR4をスクリーニングするために特異的抗体を使用することができる。このような試験の技法は当技術分野で良く知られており、例えば、サンドイッチ試験やELISAが含まれる。NR4レベルおよび/またはIL−13レベルの知識はある種の癌または癌の素因の診断のためまたはある種の治療プロトコルの監視のために重要であり得る。特に、IgE応答またはIL−13またはIL−4またはその両方を監視することが重要である。これらは、今度は免疫系に影響を及ぼすからである。
【0077】
本発明のNR4に対する抗体はモノクローンでもポリクローンでもよい。また、Fab断片などの抗体の断片も使用することができる。さらに、本発明は組換え抗体および合成抗体に及びそして抗体ハイブリッドにまで及ぶ。「合成抗体」は本明細書では抗体の断片および抗体のハイブリッドを含むと考える。本発明のこの側面の抗体は免疫治療法にとりわけ有用であり、受容体または受容体−リガンド相互作用の評価または治療計画のプログラムの監視のための診断手段として使用することもできる。
【0078】
例えば、特異的抗体をNR4タンパクのスクリーニングに使用することができる。後者は、例えば、細胞抽出液または他の生物学的液体中のNR4レベルをスクリーニングするための手段としてまたは培養物上清から組換え手段で調製されたNR4を精製するための手段として重要であろう。本明細書で提供される試験のための技法は当技術分野で知られており、例えば、サンドイッチ試験やELISAが含まれる。
【0079】
上に論じた最初に述べた抗体に対するいかなる第2の抗体(モノクローン抗体、ポリクローン抗体または抗体断片または合成抗体)も本発明の範囲に含まれるべきものである。第1および第2の抗体は両方とも検出試験に使用することができ、あるいは第1の抗体は市場で入手可能な抗−免疫グロブリン抗体と共に使用することができる。本明細書で提供される抗体にはNR4のどの領域に特異的な抗体であってもすべて含まれる。
【0080】
ポリクローン抗体およびモノクローン抗体は両方とも受容体で免疫化することにより取得することができ、そしていずれのタイプも免疫試験に利用することができる。両方のタイプの血清を得る方法は当技術分野で良く知られている。ポリクローン血清はあまり好ましくないが、適当な実験室動物にNR4またはその抗原部分の有効量を注射し、その動物から血清を集め、そして既知の免疫吸着法のいずれかにより特異的な血清を単離することにより、比較的容易に調製される。この方法により製造された抗体は事実上どのタイプの免疫試験にも利用可能であるが、この生産物の潜在的不均質性のため一般的にはあまり好ましくない。
【0081】
免疫試験でのモノクローン抗体の使用は、大量にそれを作る能力があることおよび生産物の均質性のためとりわけ好ましい。不死の細胞株と免疫原調製物に対して感作させたリンパ球を融合させて作成したモノクローン抗体生産用のハイブリドーマ細胞株の調製は当技術分野で熟練した者に良く知られた技法により行うことができる。
【0082】
本発明の別の側面は主体からの生物試料内のNR4を検出する方法を提供する。該方法は該生物試料をNR4またはその誘導体または同族体に特異的な抗体と、抗体−NR4複合体が形成するのに十分な時間および条件の下で接触させ、そして該複合体を検出することを含んで成る。
【0083】
NR4が存在することはウエスタン・ブロッティングやELISAによるなどの幾つかの方法で確認できる。広い範囲の免疫試験法が利用できることは米国特許第4,016,043号、第4,424,279号および第4,018,653号を参照することにより見ることができる。これらは、もちろん、従来の競争的結合試験におけると同様に非−競争的タイプの1部位試験および2部位試験の両方または「サンドイッチ」試験を含んでいる。これらの試験は標的への標識化抗体の直接結合をも含んでいる。
【0084】
サンドイッチ試験は中でも最も有用でありそして普通に使用されている試験であって、本発明における使用にとっても好ましい。サンドイッチ試験法は幾つかの変法があり、それらのすべてが本発明に包含されるものである。簡単に言えば、典型的なホワード試験(forward assay)では、非標識化抗体が固体基質上に固定化されておりそして試験対象の試料が結合した分子と接触させられる。適当な時間、すなわち抗体−抗原複合体の形成を行わせるのに十分な時間、インキュベートした後、抗原に特異的な第2の抗体であって検出可能なシグナルを形成することができるレポーター分子で標識された抗体を次に添加し、抗体−抗原−標識化抗体の別の複合体が形成するのに十分な時間インキュベートする。未反応物質をすべて洗い流し、そしてレポーター分子が形成したシグナルの観察により抗原の存在が測定される。この結果は可視的なシグナルの簡単な観察による定性的なものであることもあり、また既知の量のハプテンを含む対照試料と比較することにより定量化することもできる。ホワード試験の変法には試料と標識化抗体の両方が結合抗体に同時に添加される同時試験が含まれる。これらの技法は当技術分野の熟練した者に良く知られており、容易に分かるような僅かな修飾を包含するものである。本発明によれば、試料は細胞抽出物、組織生検またはおそらく血清、唾液、粘液分泌物、リンパ液、組織液および呼吸液などのNR4を含むかも知れないものである。試料は、従って、一般に生物学的液体、細胞抽出液、骨髄またはリンパ液、組織抽出物(例えば、腎臓、肝臓、脾臓、など)、醗酵液および細胞培養物や細胞条件化培地からの上清を含む生物試料である。
【0085】
典型的なホワードサンドイッチ試験では、NR4またはその抗原部分に特異性を有する第1の抗体は固体表面に共有結合または受動的に結合している。固体表面はガラスまたはポリマーが代表的であり、最も普通に使用されるポリマーはセルローズ、ポリアクリルアミド、ナイロン、ポリスチレン、ポリ塩化ビニルまたはポリプロピレンである。固体支持体は管、ビーズ、ミクロプレートの平円盤の形、または免疫試験を行うのに適した表面であればなんでもよい。結合工程は当技術分野で良く知られており、一般に架橋共有結合または物理的吸着からなり、ポリマー−抗体複合体は試験試料の調製の際に洗浄される。試験対象の試料の一部をとり、ついで固相複合体に加え、そして抗体中に存在するサブユニットのいずれかと結合するのに十分な時間(例えば、2〜40分)および適当な条件(例えば、25℃)の下でインキュベートする。インキュベーション期間の後に、抗体サブユニット固相を洗浄し乾燥し、そしてこのハプテンの一部に特異的な第2の抗体とインキュベートする。第2の抗体はレポーター分子に結合しており、この分子が第2の抗体のハプテンへの結合を示すために用いられる。
【0086】
別の方法は生物試料中の標的分子を固定化し、ついでレポーター分子で標識されまたはされていない特異的抗体にこの固定化された標的を曝す。標的の量およびレポーター分子シグナルの強度に依存して、結合した標的は抗体による直接の標識化により検出可能となる。また、第1の抗体に特異的な第2の標識化抗体を標的−第1の抗体複合体と接触させ、標的−第1抗体−第2抗体の三次複合体を形成させる。この複合体はレポーター分子が発するシグナルにより検出される。
【0087】
本明細書で使用されるとき「レポーター分子」は、その化学的性質により抗原−結合抗体の検出を可能とする、分析的に同定可能なシグナルを供給する分子を意味する。検出は定性的にも定量的にも行うことができる。このタイプの試験で最も普通に用いられるレポーター分子は、酵素、発蛍光団または放射性核種を含む分子(すなわち、放射性同位元素)および化学発光性分子である。酵素免疫試験の場合には、酵素を通常グルタルアルデヒドまたは過ヨウ素酸により第2の抗体に結合させる。しかし、容易に分かるように、熟練した技術者に容易に利用可能な多様な異なる結合手段が存在する。通常使用される酵素としては、中でも、ホースラディッシュペルオキシダーゼ、グルコーズオキシダーゼ、ベーターガラクトシダーゼおよびアルカリホスファターゼか挙げられる。特異的酵素と共に使用されるべき基質は一般に対応する酵素による加水分解の際の検出可能な色変化の生成に対して選択される。適当な酵素の例としては、アルカリホスファターゼおよびペルオキシダーゼが挙げられる。発蛍光性基質を採用することも可能である。これは上に述べた発色原性基質よりもむしろ蛍光性の生産物を生ずる。全ての場合に、酵素−標識化抗体を第1抗体ハプテン複合体に添加して、結合させ、ついで過剰の試薬を洗い流す。ついで、適当な基質を含む溶液を抗体−抗原−抗体の複合体に添加する。この基質は第2の抗体に結合した酵素と反応し、定性的な視覚的シグナルを生じ、これは通常分光学的に定量化することができ、試料中に存在したハプテンの量の指標を与える。「レポーター分子」はラテックスビーズなどの上での赤血球などの細胞凝集または凝集の阻害の利用にも広がっている。
【0088】
また、フルオレッセインやローダミンなどの蛍光性化合物は抗体の結合活性を変えることなく抗体に化学的に結合させることができる。特定波長の光を照射することにより活性化すると、フルオロクロム−標識化抗体が光エネルギーを吸収し、分子内で励起状態を誘発し、続いて光学顕微鏡で視覚的に検出可能な特徴的な色の光を発射する。EIAの場合のように、蛍光性標識化抗体は第1抗体−ハプテン複合体に結合させられる。未結合試薬を洗い流した後、残りの三次複合体を次に適当な波長の光に曝すと、観察される蛍光は問題のハプテンの存在の証拠となる。免疫蛍光法およびEIA法は両方とも当技術分野で十分に確立されており、本発明の方法にとってとりわけ好ましい。しかしながら、放射性同位元素、化学発光性分子または生物発光性分子などの他のレポーター分子も採用することができる。
【0089】
NR4およびIL−4受容体α−鎖を発現することができる細胞の場合は別の形の試験ができる。例えば、IL−4受容体α−鎖とNR4がCOS細胞などの細胞上に共−発現されるときは、IL4の存在下でIL−13は高親和性でNR4に結合する。
【0090】
一つの理論または一つの作用様式に本発明を制限する積もりはないが、NR4とIL−4受容体とが同一細胞中で発現されるとき、これらはIL−4受容体およびIL−13受容体両方の形成に寄与する。IL−4の場合には、結合はまずIL−4受容体α−鎖を介して起こり、ついでこの複合体にNR4が相互作用する。IL−13の場合には、結合はまずNR4に対して起こり、ついでこの複合体とIL−4受容体α−鎖が相互作用してシグナル伝達を可能とする高親和性受容体を形成する。NR4とIL−4受容体α−鎖との共−発現の結果は、IL−4とIL−13がIL−4受容体α−鎖およびNR4と結合するために相互に競争することになることである。
【0091】
この挙動に基づけば、IL−4またはIL−13が両受容体成分を含む細胞表面複合体を形成するのを阻止するタンパク質または低分子はアンタゴニストとなり得るということであろう。このような分子はこのサイトカインとその低親和性受容体との相互作用を阻止することができる。例えば、可溶性IL−13BPはIL−13とNR4との相互作用を阻止することができる。同様に、可溶性IL−4受容体α−鎖はIL−4の細胞表面IL−4受容体α−鎖への結合を阻止することができる。これらの作用物質はIL−4またはIL−13に特異的なアンタゴニストとなるであろう。
【0092】
敷衍すると、可溶性NR4はその低親和性のためIL−13の極めて無能なアンタゴニストである。もしIL−4に結合しそしてIL−13とも高親和性で結合する可溶性NR4突然変異体が選択されるならば、これはIL−4とIL−13の両方の有用なアンタゴニストとなるであろう。
【0093】
可溶性受容体の利用のもう一つは、NR4に対するモノクローン抗体のパネルを作ることである。機能的IL−4受容体と機能的IL−13受容体の両方の形成における決定的な事象である、NR4とIL−4受容体α−鎖との相互作用を阻止する抗体が得られるならば、そのときは、両方のサイトカインの作用がさらに阻止される。
【0094】
一つの特殊な態様においては、本発明は生物学的試料中のIL−4のレベルを監視する方法を提供する。該方法はNR4とIL−4受容体α−鎖を発現する細胞と該生物学的試料を、IL−4がその受容体と結合するのを競争的に阻害するのに有効な量のIL−13と共にインキュベートする工程および競争的阻害の程度を測定する工程とを含んで成る方法である。
【0095】
本発明の関連する態様においては、生物学的試料中のIL−13のレベルを監視する方法を提供する。該方法はNR4とIL−4受容体α−鎖を発現する細胞と該生物学的試料を、IL−13がその受容体と結合するのを競争的に阻害するのに有効な量のIL−4と共にインキュベートする工程および競争的阻害の程度を測定する工程とを含んで成る方法である。
【0096】
このサイトカインは上述のようにレポーター分子で標識することが好ましい。
【0097】
この生物学的試料には、血液、血清、血漿、組織液、組織抽出物、リンパ液、T細胞またはその抽出物、培養上清および培養条件化培地などが含まれるが、これらに限定されるものではない。
【0098】
本発明はNR4遺伝子またはその誘導体を検出するためPCR分析を含むような遺伝子試験をも提供する。別の方法または関連して用いられる方法には、一本鎖コンホーメーション多形性分析(SSCP)、特異的オリゴヌクレオチドハイブリッド形成、直接タンパク切断テストのような方法などの直接ヌクレオチド配列決定法または突然変異スキャニングが含まれる。このような遺伝子テストは、例えば、疾病状態をもたらすNR4の非−発現または実質的な発現の欠如または突然変異型の発現について動物(例えば、ヒト)の遺伝子をスクリーニングする場合に重要である。
【0099】
本発明の核酸分子はDNAでもRNAでもよい。核酸分子がDNA型であるときは、それはゲノムDNAまたはcDNAであり得る。本発明の核酸分子のRNA型は一般にmRNAである。
【0100】
本発明の核酸分子は一般に単離型であるが、それはベクター分子そして特に発現ベクター分子などの他の遺伝子分子中に組み込まれたりまたはそれに連結されたりまたはそれと別な仕方で融合したりまたはそれと集合状態にあったりすることができる。ベクターおよび発現ベクターは一般に複製することができ、そして適用可能なときは、原核細胞または真核細胞の一方または両方の中で発現する。原核細胞は大腸菌、バチルス・スピーシーズおよびシュードモナス・スピーシーズを含むことが好ましい。真核細胞は酵母、カビ、哺乳類および昆虫の細胞を含むことが好ましい。
【0101】
従って、本発明の別の側面はベクター部分と哺乳類の、より具体的にはヒトのNR4遺伝子部分を含む遺伝子構築物を提供する。このNR4遺伝子部分はNR4ポリペプチドまたは機能的または免疫学的に相互作用するその誘導体をコードすることができるものである。
【0102】
遺伝子構築物のNR4遺伝子部分は、適当な細胞中でプロモーターが該NR4遺伝子部分の発現を指令することができるようにベクター上の該プロモーターに機能的に連結させることが好ましい。
【0103】
また、遺伝子構築物のNR4遺伝子部分はグルタチオン−S−トランスフェラーゼまたはその部分またはサイトカインまたは別のヘモポエチン受容体をコードするヌクレオチド配列などの別の遺伝子配列と融合した遺伝子の全部または一部を含むことができる。ハイブリッド受容体分子は多機能性治療薬および診断薬の開発において特に有用である。
【0104】
本発明はこのような遺伝子構築物を包含しそして該構築物を含む原核細胞または真核細胞をも包含する。
【0105】
本発明はNR4の変異体、一部、断片、部分、同族体およびアナログを含むNR4のすべての誘導体またはそれらをコードする遺伝子配列に及ぶ。この遺伝子配列には、天然に生ずるヌクレオチド配列またはアミノ酸配列において1個または多重のヌクレオチドまたはアミノ酸が置換、付加および/または欠失することにより生ずる遺伝子配列も包含される。
【0106】
本発明のNR4およびその遺伝子配列はある範囲の治療薬および診断薬の創薬に有用でありそして対応するリガンドの検出にとりわけ有用である。例えば、組換えNR4は同定可能なシグナルを形成することができるレポーター分子に結合または融合し、リガンドを含むと考えられる生物試料と接触させ、そして標識化NR4のリガンドへの結合についてスクリーニングをすることができる。また、標識化NR4は、リガンド遺伝子と考えられるものまたはその機能的部分の発現ライブラリーをスクリーニングするために使用することができる。
【0107】
別の態様では、NR4がまず固定化される。この態様によれば、リガンドと考えられるものを含む生物試料を、固体支持体に固定化されたヘモポエチン受容体またはそのリガンド結合部分と、該生物試料中にリガンドが存在するときは該受容体と該リガンドとの間で複合体が形成されるのに十分な時間および条件の下で接触させる工程、および結合したリガンドを溶出しそしてそれを単離する工程を含んで成る方法が提供される。
【0108】
可溶性NR4ポリペプチドも種々のハイブリッドと同様に神経系における疾病、障害または異常の治療において有用であると考えられる。例えば、中枢神経系または末梢神経系に関して脳性麻痺、外傷により誘発される麻痺、発作と関連する血管虚血症、ニューロン腫瘍、運動ニューロン病、パーキンソン病、ハンチントン病、アルツハイマー病、多発性硬化症、糖尿病と関連する末梢神経症、重金属またはアルコール毒性、腎不全、およびヘルペス、風疹、麻疹、チキンポックス、HIVまたはHTLV−1などの感染症の治療である。NR4ポリペプチドおよびハイブリッドはサイトカイン活性を調節したり、造血を調節したりするのにも重要である。これらは慢性関節リューマチなどの他の炎症性症状と同様にアレルギー性またはアトピー性症状の治療にも重要である。
【0109】
上に述べたように、本発明のNR4またはそのリガンドまたはその機能的誘導体は1以上の薬学的に許容され得る担体および/または希釈剤と共に薬学的組成物中に提供され得る。さらに、本発明は本発明のNR4の有効量の投与を含む治療方法を提供する。本発明はNR4のアンタゴニストおよびアゴニストおよび/またはそのリガンドをも包含しそして治療用の組成物および治療の方法論におけるそれらの使用をも包含する。
【0110】
本発明のさらなる側面はNR4に起因する欠陥状態または欠損状態の治療用の医薬品の製造におけるNR4またはその機能的誘導体の使用を提供する。
アミノ酸残基に対する下記の1文字略号および3文字略号が本明細書で使用される。
本発明については下記の非−制限的な図および実施例を参照してさらに記載する。
【0111】
実施例1
NR4をコードするゲノムDNAおよびcDNAの単離
胚幹細胞株から抽出されたApoI消化ゲノムDNAをλZAPIIバクテリオファージ(ストラタジーン、LaJolla,CA)の中にクローニングした。このライブラリーから得た約106プラークを配列Trp−Ser−Asp−Trp−Ser(16)に対応する32P標識オリゴヌクレオチドでスクリーニングした。ハイブリッド形成陽性のクローンを自動化DNAシークェンサー(アプライド・バイオシステムズ、Foster City、CA)を用い製造者の指示に従って配列決定を行った。一つのクローンがヘモポエチン受容体ファミリーの新規なメンバーの一部をコードするように見えた。このゲノムDNA配列に基づいてオリゴヌクレオチドを設計しそしてマウス腹膜マクロファージ(クロンテク・ラボラトリーズ、PaloAlto、CA)の、マウス皮膚の、マウス肺の、マウス腎のそしてWEHI−3B(ストラタジーン、LaJolla、CA)のλ−バクテリオファージcDNAライブラリーからクローンを単離するために通常の方法で使用した。
【0112】
実施例2
発現ベクターの構築および細胞のトランスフェクション
PCRを使用して、NR4の成熟コード領域(Thr27からPro424まで、図1)の前にあるIL−3シグナル配列(配列番号:5)およびN−末端FLAGエピトープ−標識(配列番号:6)をコードするNR4cDNAの誘導体を作成した。このPCR産物を哺乳類発現ベクターpEF−BOS(17)の中にクローニングした。使用に先立ち、構築物の全体の配列決定を行った。細胞をトランスフェクトし、そして既に述べられたように選択した(16、18)。
【0113】
実施例3
ノーザン・ブロット
ノーザン・ブロットは既に記載されたように行った(16)。ハイブリッド形成用プローブの起源は次のものである。すなわち、NR4については、ヌクレオチド32〜984(図1)からのPCR産物であり、そしてGAPDHに対しては、cDNA断片スパンニングヌクレオチド(19)である。
【0114】
実施例4
サイトカイン類と放射性ヨード化サイトカインの使用
IL−2、IL−4、IL−7、IL−9、IL−13、およびIL−15は市販品を使用した(R & Dシステムズ、Minneapolis MN)。放射性ヨード化のために、10mMリン酸ナトリウム、150mM塩化ナトリウム(PBS)、0.02%v/vトゥイーン20および0.02%w/vアジ化ナトリウム、pH7.4に、サイトカインを100μg/mlの濃度で溶解した。IL−13の2μgをヨードモノクロリド法(20、21)を用いて放射性ヨード化し、一方IL−4の2μgをジ−ヨード−ボルトン−ハンター試薬(16)を用いて放射標識した。結合の研究および比放射活性の測定および標識化サイトカインの結合可能性については既に記載されたように行った(2)。
【0115】
架橋実験のために、組換えネズミIL−13をFLAG標識化タンパクとしてピチア・パストリス中で製造した。
【0116】
架橋試験のために、精製した可溶性IL−13Rα(NR4)の一定量を競争剤の存在下または非存在下に最終容量20μlで少なくとも30分間40℃で125I−IL−13とインキュベートした。次いで、0.02%v/vトゥイーン20を含むPBS中のBS3(スベリン酸ビス(スルホスクシミジル)の12mM溶液の5μlを添加し、そして混合物を4℃で30分間インキュベートした。試料を4×SDS試料緩衝液の8μlと混合しそして13%w/vSDS−PAGEにより非還元条件下で分析した。ゲルを乾燥しそしてオートラジオグラフィーまたはホスフォイメージャー(PhosphoImager)のいずれかで可視化した。
【0117】
実施例5
増殖試験
サイトカインに応じて得られるBa/F3細胞およびCTLL細胞の増殖をラックス60ミクロ穴HL−Aプレート(ヌンク・インク、IL、USA)中で測定した。細胞を、20%v/v新生児ウシ血清を含むDMEM中で3回洗浄し、同培地に2×104細胞/mlの濃度で再懸濁した。この細胞懸濁液の10μl部分を5μlの種々の濃度の精製組換えサイトカインと共に培養穴の中に入れた。空気中10%v/vCO2を含む完全加湿インキュベーター中で37℃で2日間インキュベートした後、生存細胞を倒立顕微鏡を用いて計測した。
【0118】
実施例6
ネズミNR4のクローニングと特性決定
胚幹細胞由来のゲノムDNAのApoI消化物を用いてλZAPII中に一つのライブラリーを構築しそしてヘモポエチン受容体ファミリーの多くのメンバーに見出されるアミノ酸配列Trp−Ser−Asp−Trp−Serをコードする32P標識化オリゴヌクレオチドのプールでスクリーニングした。ハイブリッドを形成する一つのバクテリオファージクローンがヘモポエチン受容体ファミリーの新規なメンバーの一部をコードするように見える配列を含むことが見出された。この受容体はNR4という機能的名称が与えられた。このゲノムクローンの配列を使用して、WEHI−3B細胞の、腹膜マクロファージの、骨髄の、皮膚の、そして腎のライブラリーからNR4をコードするcDNAを単離した。これらのcDNAの合成物であるヌクレオチド配列(配列番号:1)および予言されたアミノ酸配列(配列番号:2)を図1に示す。このNR4cDNAは424のアミノ酸残基からなるタンパクをコードすることを予言する。このタンパクの中には、シグナル配列および膜貫通ドメインと思われるものが含まれる。このタンパクの細胞外領域は、保存された4個のシステイン残基と特徴的なTrp−Ser−Asp−Trp−Serモチーフ(図1、WSXWSとして太字で記載)を含む典型的なヘモポエチン受容体ドメイン(アミノ酸118〜340)に加えて、免疫グロブリン−様ドメイン(アミノ酸27〜117)を含んでいた。この新規な受容体の細胞質テール(cytoplasmic tail)は長さが60アミノ酸であった。
【0119】
実施例7
NR4cDNAの発現パターン
NR4mRNAの発現パターンをノーザン分析で検討した。5.2kbおよび2.2kbの2個のハイブリッド形成物を多くの組織由来のmRNA中に検出した(図2)。NR4mRNAは骨格筋には検出できなかった(図2)。図8はマウス組織内のNR4の発現を示す。
【0120】
実施例8
NR4はIL−13受容体の特異的結合サブユニットであるIL−13受容体α−鎖(IL−13Rα)をコードする
COS細胞で発現したNR4を抗−FLAG抗体を用いるウエスタン・ブロットから評価すると、見掛けの分子量は約50,000から約70,000ダルトンであり、より具体的には約55,000から約65,000である。これはNR4がIL−13受容体の結合性サブユニットをコードするかも知れないということを示唆した。この可能性をテストするため、NR4をCOS細胞で発現させた。トランスフェクトされていないCOS細胞は比較的低レベルのIL−4受容体およびIL−13受容体を発現した。NR4cDNAを含むプラスミドをトランスフェクトすると、COS細胞によって発現されるIL−13の数は劇的に増加した(図3A、細胞当たり100,000から500,000まで)が、IL−4受容体の数は増加しなかった。COS細胞により発現されたNR4に対するIL−13の親和性は低かった(KDは約2〜10nM)そしてこのNR4は非標識IL−13と競争できた(図4A)がIL−2、IL−4、IL−7、IL−9またはIL−15を含む他のサイトカインとは競争できなかったから、その結合は特異的であった。これらの結果から、NR4がIL−13受容体α−鎖(IL−13Rα)であることが示唆された。
【0121】
実施例9
IL−13Rα(NR4)およびIL−4RαはIL−4受容体およびIL−13受容体の成分を共有する。
IL−4受容体とIL−13受容体の間の関係を研究するため、IL−4に応答する細胞株CTLLを検討した。親のCTLL細胞は1個のクラスのIL−4受容体(KD約660pM、約3600受容体/細胞)を発現したがIL−13受容体は検出されなかった(図3B)。CTLL細胞によって発現されたIL−4受容体は、125I−IL−4の結合が非標識IL−4と競争できたがIL−13とは競争できなかったから(図4B)、特異的であるように見えた。CTLL細胞中でIL−13Rα(NR4)が発現する際、IL−4受容体の数および親和性には変化がみられなかったが、一方、高親和性IL−13受容体の1個のクラスが検出された(図3C、KD約75pM、1350受容体/細胞)。CTLL細胞内で発現したIL−13Rα(NR4)に対するIL−13の親和性はCOS細胞中よりも高かった。これは前者がIL−13Rα(NR4)と相互作用しIL−13に対する親和性を増強することができるあるタンパクを発現したことを示唆する。以前の研究に基づく同様な候補はIL−4Rαである。この可能性を解明するため、IL−13Rα(NR4)を発現するCTLL細胞への結合に対し125I−IL−13と競争するIL−4の能力を評価した。図4Bは125I−IL−13の結合に対する競争ではIL−4とIL−13は同等に効果的であった(IC50は約300pM、図4C)のであり、そしてさらにこれらは125I−IL−4とも結合に対して競争できたのであった(IC50は約300pM、図4D)。
【0122】
実施例10
IL−13Rα(NR4)はIL−13による増殖性シグナルの形質導入に必要である。
CTLL細胞は生存および増殖に外因性サイトカイン類の添加を必要とする。
IL−2はCTLL細胞に対し強力な増殖性刺激を有する(EC50は約100〜200pM)一方、IL−4は比較的弱く(EC50は2〜7nM)そしてIL−13は不活性であった(図5A)。CTLL細胞中にIL−13Rα(NR4)が発現することにより、IL−13に応答して(EC50は約700pM)弱く生き残りそして増殖する能力が、そしてIL−4に応答して(EC50は約700pM、図5B)親細胞よりもややより強く増殖する能力が生じた。
【0123】
実施例11
ヒトIL−13Rα(NR4)のクローニング
ネズミIL−13Rα(NR4)に相同な遺伝子が他の脊椎動物種に存在するか否かを決定するため、ネズミIL−13Rα(NR4)のヌクレオチド840〜1270を包含するプローブを、EcoRIで消化した種々の種由来のゲノムDNAとハイブリッド形成させた。ハイブリッド形成は500mMリン酸水素二ナトリウム(約5×SSC)中50℃で一晩行った。フィルターを40mMリン酸水素二ナトリウム(約0.2×SSC)中50℃で2時間洗浄し、オートラジオグラフィー様フィルムに48時間曝した。図6はヒトを含む種々の種由来のゲノムDNAの中に比較的少数(1〜5)のハイブリッド形成バンドが観察されることを示す。これはネズミcDNAプローブを使用してヒトIL−13Rα(NR4)をクローニングすることが容易であることを示唆する。従って、λZAPIIバクテリオファージ中のヒト骨髄cDNAライブラリーのクローンをネズミIL−13Rα(NR4)cDNA由来の二つのプローブ(ヌクレオチド82〜840およびヌクレオチド840〜1270)でスクリーニングした。ハイブリッド形成は6×SSC、0.1%w/vSDS中42℃で一晩行った。フィルターを2×SSC:0.1%w/vSDS中50℃で2時間洗浄しそしオートラジオグラフィー様フィルムに48時間曝した。両方のネズミIL−13Rα(NR4)プローブとハイブリッドを形成したプラークを釣り上げ、通常の方法で精製した。ハイブリッドを形成するバクテリオファージ由来のこのcDNA挿入物をpBluescript プラスミドの中に切り取り、そしてABI自動化シークェンサーを用いてその全体の配列決定を行った。図7は単離されたクローンの配列の合成体を示し、そしてこれらのクローンがネズミIL−13Rα(NR4)と高度の配列類似性を共有する一つのタンパクをコードすることを明らかにする。これらのクローンはこのタンパクの全コード領域をコードする。高度の配列類似性(320/425アミノ酸、約75%)はこのcDNAがネズミIL−13Rα(NR4)のヒト同族体であることを予言する。ヌクレオチド配列は配列番号:3として表されそしてそのアミノ酸配列は配列番号:4である。
【0124】
実施例12
可溶性ネズミIL−13Rα(NR4)はIL−13と結合する
N−末端「FLAG」エピトープ(インターナショナル・バイオテクノロジーズ/イーストマン・コダック、New Haven CT)を持つNR4の可溶性バージョンを発現するような構築物を構築した。まず、哺乳類発現ベクターpEF−BOSの誘導体を、それがネズミIL−3のシグナル配列(MVLASSTTSIHTMLLLLLMLFHLGLQASIS(配列番号:5))をコードするDNAおよびFLAGエピトープ(DYKDDDDK(配列番号:6))を含みそれに続いてユニークなXbaIクローニング部位を含むように作成した。このベクターをpEF/IL3SIG/FLAGと名付けた。成熟NR4コード領域(Thr27〜Thr344)の細胞外部分はプライマー1478および1480を用いるPCRによって作成した。生じた産物をXbaIで消化しそしてpEF/IL3SIG/FLAGのXbaI部位にクローニングしてpEF/IL3SIG/FLAG/solNR4を得た。この構築物の同定はジデオキシ配列決定法により確認された。
オリゴ 1478 5'AGCTTCTAGAACAGAAGTTCAGCCACCTGTG3'〔配列番号:7〕、
オリゴ 1480 5'AACTCCACCTTCTACACCACCTGATCTAGA3'〔配列番号:8〕
【0125】
CHO細胞中にトランスフェクトし、発現させた後、抗−FLAG抗体(M2)アフィニティカラム上でCHO細胞コンディション化培地から可溶性NR4を吸着させ遊離のFLAGペプチド(サイエンス・イメージング・システムズ)で溶出することにより精製した。
【0126】
COS細胞により発現されたNR4に対しIL−13の親和性が低いことと一致して、精製した可溶性NR4はゲルろ過クロマトグラフィーで評価するとIL−13と結合することができないようであった。しかしながら、感度の高い架橋試験を用いると、可溶性IL−13Rα(NR4)のIL−13に結合する能力が証明された(図8、レーン1)。この相互作用は非標識IL−13により競争されたが非標識IL−4によっては競争されることはなかった(図8、レーン2およびレーン3)。
【0127】
実施例13
可溶性IL−13Rα(NR4)に対するポリクローン抗血清
精製した可溶性NR4をウサギに注射し3ヵ月後に採血してNR4に対するポリクローン抗血清を調製した。この抗血清は125I−IL−13の可溶性NR4との架橋産物を免疫沈降させた(図9、レーン11)が、免疫化前(pre-immune)の血清では免疫沈降は観察されなかった(図9、レーン9)。複合体の免疫沈降はFLAGペプチドによっては阻害されなかった(図9、レーン10)。
【0128】
免疫沈降反応試験は次のように行われた。
【0129】
架橋反応はトリス−塩酸、pH7.5を最終濃度40mMになるまで添加することにより終了させた。次いで、試料を1:50希釈の対照ウサギ血清またはFLAGペプチドの存在下にまたは非存在下に予めインキュベートしておいた抗−NR4血清と混合した。4℃で30分間インキュベートした後、混合物に50%v/vタンパク質G−セファロースゲルスラリー(ファルマシア)の40μlを添加しそして4℃で30分間インキュベートした。試料を遠心分離しタンパク質Gビーズを3×0.5mlのPBSで洗浄し、2×濃縮SDS−PAGE試料緩衝液の40μlと混ぜ、95℃で2分間加熱した。この上清を非還元条件下で13%w/vSDS−PAGEにより分析した。
【0130】
実施例14
NR4のN−末端アミノ酸配列
NR4のN−末端アミノ酸配列を決定した。これを図10に示す。
【0131】
実施例15
IL−13の試験
IL−13は喘息などのアレルギー性疾患において重要な免疫グロブリンイソタイプであるIgEの産生に関与するサイトカインである。IL−13のレベルを監視することは、従って、重要な診断学的意味がある。IL−4とIL−13は多くの生物学的効果を共有しているから、これらのサイトカインを識別する試験法を作成することも重要である。
【0132】
COS細胞で発現したNR4は125I−IL−13と結合する。この結合はウシ血清やヒト血清などの無関係のタンパクの大量の存在下で非標識IL−13により投与量依存的に阻害される。IL−4はこの状況下では125I−IL−13結合に対して競争する能力を示さない、従って、この試験はIL−13に対して特異的であるように見える。
【0133】
この試験法は可溶性NR4をELISAプレート上に被覆し、そして、例えば、蛍光標識IL−13をプローブとして使用することにより組立てられる。次いで、試験用試料中の非標識IL−13の存在が蛍光シグナルの減少として記録される。
【0134】
NR4およびIL−4受容体α−鎖を発現する細胞を用いることにより、IL−4およびIL−13の両方を測定する同様な試験法が組み立てられる。これらの細胞には正常にIL−4受容体α−鎖を発現しそしてNR4を発現するように工学的に処理されるCTLL細胞が含まれる。125I−IL−13または125I−IL−4の結合はIL−4およびIL−13の両方の非標識型により阻害することができる。
【0135】
実施例16
IL−4およびIL−13の修飾
機能的に重要であると予測される分子の領域に突然変異を導入する。NR4の場合には、この領域は、このサイトカインがIL−4受容体α−鎖に結合するときは、IL−13と相互作用する領域、IL−4受容体α−鎖と相互作用する領域またはIL−4と相互作用する領域を含む。これらの領域は直接の実験、例えば、NR4の構造またはIL−4、IL−13およびIL−4受容体α−鎖のような他のタンパクとNR4との複合体の構造を解明することにより、またはその構造に関する情報が存在する類似のタンパク、例えば、成長ホルモン/成長ホルモン受容体の複合体に基づきこれらのタンパクをモデル化することにより決定される。次いで、得られるNR4突然変異体を機能の改善の有無につき個々にテストする。
【0136】
別の方法では、分子内に無差別突然変異が作られる。適当な技術としては、正しくないdNTPの取り込みを促進するようなポリメラーゼや反応条件を使用するNR4cDNAの合成や、DNAシャフリングと呼ばれる技術が挙げられる(23、24、25、26)。
【0137】
目的のcDNAの無差別突然変異体を作成した後、潜在的に有用な突然変異体を選別する。NR4の場合には、一つの試験法は、もしNR4がIL−4受容体α−鎖を欠如している細胞(例えば、COS細胞)の中で発現されるときは、IL−4とは如何なる検出感度でも結合できないがIL−13とは低親和性で結合する細胞が得られるという知識に基づくものである。このようにして、もしCOS細胞をNR4でトランスフェクトし、そしてFITC−結合IL−4およびフィコエリトリン−結合IL−13と結合させ、未結合リガンドを洗い流すものとすれば、IL−4は全く結合せずそして結合したIL−13は洗浄の間に解離するであろう。
【0138】
これらの細胞をFACS−選別するときは、FITC−チャンネルにもPE−チャンネルにもシグナルはほとんどあるいは全く得られないであろう。COS細胞をNR4の無差別突然変異体106〜107でトランスフェクトしそして結合試験のために処理する。野生のNR4でトランスフェクトされたものよりも強くサイトカインに結合するとして選別される細胞はすべてFACSで選別することができる。これらの「改良された」NR4cDNAを含むプラスミドを回収し、大腸菌で増やし、そして再度COS細胞を用いてその改良度を確認する。一貫して改良された突然変異体は次にさらにもう一度無差別変化の導入のために使用してさらに改良された分子を得ることができる。この反復プロセスを数回繰り返してもよい。
【0139】
当技術分野で熟練した者は本明細書に記載された発明が具体的に記載されたこと以外の変更や修正を受け得るものであることを認めるであろう。本発明はそのような変更や修正のすべてを包含することが理解されるべきである。本発明はまた本明細書で個々にあるいはまとめて言及しあるいは指摘した工程、特徴、組成物および化合物のすべてを、そして該工程あるいは特徴のいかなる2以上の組み合わせをも包含する。
【0140】
引用文献
1.デュー,エックス.エックス.およびウィリアムス,ディー.エイ.(1994)Blood 83 : 2023-2030。
2.ヤング,ワイ.シー.およびイン,ティー.(1992)Biofactors 4 : 15-21。
3.ポール,エス.アール.,ベネット,エフ.,カルベッティ,ジェイ.エイ.,ケレハー,ケイ., ウッド,シー.アール.,オオハラ,アール.ジェイ.ジェイ.,レアリー,エイ.シー.,シブリー,ビー.,クラーク,エス.シー.,ウィリアムス,ディー.エイ.およびヤング,ワイ.シー.(1990)Proc.Natl.Acad.Sci.USA 87: 7512。
4.ムサシ,エム.,クラーク,エス.シー.,スドー,ティー.,ウーダル,ディー.エル.およびオガワ,エム.(1991)Blood 78: 1448-1451。
5.シブラー,ケイ.アール.,ヤング,ワイ.シー.およびクリステンセン,アール.ディー.(1992)Blood 80: 900-3。
6.ツジ,ケイ.,リーマン,エス.ディー.,スドー ,ティー.,クラーク,エス.シー.およびオガワ,エム.(1992)Blood 79: 2855-60。
7.バーンスタイン,エス.エイ.,メイ,アール.エル.,ヘンソーン,ジェイ.,フリース,ピー.およびターナー,ケイ.(1992)J.Cell.Physiol.153 : 305-12。
8.ハンゴク,ジー.,イン,ティー.,クーパー,エス.,シェンデル,ピー.,ヤング,ワイ.シー.およびブロックスメイヤー,エイチ.イー.(1993)Blood 81: 965-72。
9.テラムラ,エム.コバヤシ,エス.,ホシノ,エス.,オシミ,ケイ.およびミゾクチ,エイチ.(1992)Blood 79: 327-31。
10.ヨネムラ,ワイ.,カワキタ,エム.,マスダ,ティー.,フジモト,ケイ.,カトウ,ケイ.およびタカツキ,ケイ.(1992)Exp.Hematol.20: 1011-6。
11.バウマン,エイチ.およびシェンデル,ピー.(1991)J.Biol.Chem.266 : 20424-7。
12.カワシマ,アイ.,オオスミ,ジェイ.,ミタ−ホンジョウ,ケイ.,シモダ−タカノ,ケイ.,イシカワ,エイチ.,サカキバラ,エス.,ミヤダイ,ケイ.およびタキグチ,ワイ.(1991)Febs.Lett. 283: 199-202。
13.ケラー,ディー.シー., デュー,エックス.エックス.,スローアー,イー.エフ.,ホフマン,アール.およびウィリアムス,ディー.エイ.(1993)Blood 82 : 1428-35。
14.マッケンジー,エイ.エヌ.ジェイ.およびズラウスキー,ジー.(1994)サイトカインおよびその受容体へのガイドブック,オクスフォード大学出版部,オクスフォード。
15.ズラウスキー,ジー.およびド フリース,ジェイ.イー.(1994)Immunol.Today 15: 19-26。
16.ヒルトン,ディー.ジェイ.,ヒルトン,エイ.エイ.,ライセビック,エイ.,ラカー,エス.,ハリソン−スミス,エム.,ゴウ,エヌ.エム.,ベグリー,シー.ジー.,メトカーフ,ディー.,ニコラ,エヌ.エイ.およびウイルソン,ティー.エイ.(1994)EMBO J.13: 4765-4777。
17.ミズシマ,エス.およびナガタ,エス.(1990)Nucleic Acids Research18; 5322。
18.ロック,ピー.,メトカーフ,ディー.およびニコラ,エヌ.エイ.(1994)Proc.Natl.Acad.Sci.USA 91: 252-256。
19.デゥガイシク,エイ.ら(1983) Biochemistry 22 : 1605-1613。
20.コントレラス,エム.エイ.,ベイル,ダブリュー.エフ.およびスパー,アイ.エル.(1983)Methods in Enzymol.92 : 277-292。
21.ヒルトン,ディ.ジェイ.およびニコラ,エヌ.エイ.(1992)J.Biol Chem.267 : 10238-10247
22.サンブルック,ジェイ.,フリッチュ,イー.エフ.およびマニアティス,ティー.(1989)モレキュラー クローニング:ア ラボラトリー マニュアル,コールド・スプリング・ハーバー・ラボラトリー,コールド・スプリング・ハーバー,ニューヨーク。
23.ステンマー,ダブリュー.ピー.シー.,(1994)Nature 370: 389-391。
24.ステンマー,ダブリュー.ピー.シー.,(1995)Biotechnology 13 : 549-553。
25.ガスマンら(1995)Proc.Natl.Acad.Sci.USA 92 : 1292-1296。
26.グラメリら(1996)Nature Biotechnology 14 : 315-319。
【図面の簡単な説明】
【0141】
【図1−1】図1はネズミNR4の配列番号:1に記載のヌクレオチド配列および配列番号:2に記載の予言されるアミノ酸配列を示す。非翻訳部分は下方の箱の中にそして翻訳部分は上方の箱の中に示してある。アミノ酸については、通常の1文字符号が略号が用いられ、グリコシレーション部位にリンクすると考えられるアスパラギンには下線を付し、保存されているシステイン残基とヘモポエチン受容体ファミリー・メンバーのWSXWSモチーフは太字で示してある。予想されるシグナル配列は太字で下線を付し、一方膜貫通ドメインはダッシュで下線を付している。示した配列は3種のライブラリー由来の8個のcDNAクローンの分析から得られたものの合成物である。この配列の5’−末端(ヌクレオチド−60から351まで)は1個のcDNAクローンから由来するが、単離されたゲノムDNAクローン中にも存在する。箱領域は:典型的なヘモポエチン受容体ドメインであり、アミノ酸118〜340である。
【図1−2】図1−2は図1−1の続きを示す図である。
【図1−3】図1−3は図1−2の続きを示す図である。
【図1−4】図1−4は図1−3の続きを示す図である。
【図1−5】図1−5は図1−4の続きを示す図である。
【図1−6】図1−6は図1−5の続きを示す図である。
【図1−7】図1−7は図1−6の続きを示す図である。
【図2】図2は選択された組織および器官でのネズミNR4mRNA発現のノーザン分析を示す写真である。
【図3】図3は125I−IL−13および125I−IL−4結合の飽和等温線、すなわち、IL−4(○)およびIL−13(●)の(A)IL−13Rα(NR4)を発現するCOS細胞への結合、(B)CTLL細胞への結合、そして(C)IL−13Rα(NR4)を発現するCTLL細胞への結合のスカチャードプロットとして図示された飽和等温線を示すグラフである。データは1×104COS細胞および1×106CTLL細胞に規格化し、そして結合は2〜4時間氷上で行った。
【図4】図4はIL−4とIL−13の結合、すなわち、IL−4(○)およびIL−13(●)が125I−IL−13の(A)IL−13Rα(NR4)を発現するCOS細胞への結合および(C)IL−13Rα(NR4)を発現するCTLL細胞への結合と競争する能力、または125I−IL−4の(B)CTLL細胞への結合および(D)IL−13Rα(NR4)を発現するCTLL細胞への結合と競争する能力、の特異性を示すグラフである。結合は4℃で2〜4時間行われそしてデータは競争物質(△)の横座標で観察された特異的結合のパーセンテージとして表された。
【図5】図5はNR4を発現する細胞の因子依存性増殖を示すグラフである。(A)CTLL細胞または(B)IL−13Rα(NR4)を発現するCTLL細胞の200個をサイトカインの非存在下にまたは種々の濃度のIL−2(□)、IL−4(○)またはIL−13(●)と共にインキュベートした。48時間後に生存細胞を計測し、データをIL−2の最高濃度で観察された生存数のパーセンテージとして表した。
【図6】図6はNR4(IL−13Rα)遺伝子の種間保存を示す写真である。
【図7−1】図7はネズミおよびヒトのNR4(IL−13Rα)遺伝子のヌクレオチド配列と対応するアミノ酸配列の表示である。ヒト(H)およびネズミ(M)のIL−13Rα(NR4)のヌクレオチド配列および予言されるアミノ酸配列を眼で配列しそして配列が最適となるようにギャップ(−)を挿入して配列した。ネズミクローンについての数字付けは、コード領域の部分を形成するヌクレオチドは上方の箱に、一方非翻訳領域のヌクレオチドは下方の箱に示してある。ネズミの予言されたタンパクとヒトの予言されたタンパクの間で同一なアミノ酸は(*)で指摘してある。ネズミのシグナル配列をコードするDNAは下線を付してあり、成熟タンパクの予言された第1アミノ酸はA26またはT27である。
【図7−2】図7−2は図7−1の続きを示す図である。
【図7−3】図7−3は図7−2の続きを示す図である。
【図7−4】図7−4は図7−3の続きを示す図である。
【図7−5】図7−5は図7−4の続きを示す図である。
【図7−6】図7−6は図7−5の続きを示す図である。
【図7−7】図7−7は図7−6の続きを示す図である。
【図7−8】図7−8は図7−7の続きを示す図である。
【図7−9】図7−9は図7−8の続きを示す図である。
【図7−10】図7−10は図7−9の続きを示す図である。
【図7−11】図7−11は図7−10の続きを示す図である。
【図8】図8は可溶性NR4に架橋している125I−IL−13を示す写真である。レーン:125I−IL−13(100,000cpm)+2μg/ml可溶性NR4、レーン2:過剰の非標識IL−13の存在下における125I−IL−13(100,000cpm)+2μg/ml可溶性NR4、レーン3:過剰の非標識IL−4の存在下における125I−IL−13(100,000cpm)+2μg/ml可溶性NR4。
【図9】図9はIL−13Rα(NR4)と架橋している125I−IL−13の抗−NR4ポリクローン抗血清による免疫沈降反応の写真である。レーン9〜11:125I−IL−13(750,000cpm)に架橋している可溶性IL−13Rα(3μg/mlの30μl)、およびそれぞれ、対照のウサギ血清で沈降させたもの、または100μg/m1FLAGペプチドの存在下または非存在下において抗−NR4ポリクローン抗血清で沈降させたものである。レーン12〜14:0.5μg/mlの非標識IL−13の存在下における125I−IL−13(750,000cpm)に架橋している可溶性IL−13Rα(NR4)(3μg/mlの30μl)、およびそれぞれ100μg/mlのFLAGペプチドの存在下または非存在下に抗−IL−13Rα(NR4)ポリクローン抗血清で沈降させたものである。
【図10】図10はネズミNR4のN−末端アミノ酸配列の表示である。
【技術分野】
【0001】
本発明は新規なヘモポエチン受容体またはその成分またはその部分そしてそれをコードする遺伝子配列に広く関係する。本発明のこのような受容体分子およびそれらの成分および/または部分そしてそれをコードする遺伝子配列はその受容体とリガンド相互作用により機能するアゴニスト、アンタゴニスト、治療剤および診断試薬などの広範囲のものの開発に有用である。
【背景技術】
【0002】
本明細書で数値により言及する出版物の文献的詳細は明細書の末尾に集めてある。本明細書で言及したヌクレオチド配列およびアミノ酸配列の配列番号は文献の次に記載してある。
【0003】
本明細書およびそれに続く請求の範囲を通じて、文脈上他の意味を必要とする場合を除き、「コンプライズ(含む)」という用語、あるいは「コンプライジーズ」または「コンプライジング」などのそれらの変形は、述べられた実体または一群の実体の包含を意味し、他の実体または他の一群の実体の排除を意味するものではないと理解すべきである。
【0004】
本発明の好ましいヘモポエチン受容体は本明細書では「NR4」と言及される。このNR4受容体はインターロイキン−13(IL−13)と相互作用する、従って本明細書ではIL−13受容体、より具体的にはIL−13受容体α−鎖(IL−13Rα)と呼ばれる。これらの用語は特定の主題について互換的に使用される。特定のNR4の起源となる種は1個の縮小文字で与えられる。例えば、ねずみ(murine)は「M」そしてヒトは「H」である。組換え型は接頭辞「r」が付されることがある。
【0005】
組換えDNA技術の精密化の迅速な発展により、医学分野や類似の保健衛生分野への研究が大いに促進された。サイトカインの研究は特に重要である。とりわけ、多様な細胞の増殖、分化および機能を調節するこれらの分子は重要である。組換えサイトカインの投与やサイトカインの機能および/または合成の調節がある範囲の疾病状態の治療のための医学的研究の焦点となる場合が増加しつつある。
【0006】
ある範囲のサイトカインや他の分泌される細胞機能調節物質の発見にもかかわらず、比較的僅かなサイトカインしか治療分野で直接使用されたり標的とされたりしていない。この理由の一つは多数のサイトカインが多面的な性質を持っていることである。例えば、インターロイキン(IL)−11は機能的に多面的な分子であり(1,2)、IL−6依存性の形質細胞腫の細胞株、T1165の増殖を刺激する能力によって特徴付けられたのが最初である(3)。IL−11の他の生物学的作用には、多機能性ヘモポエチン原始体の細胞増殖の誘導(4,5,6)、巨核球や血小板の形成の増強(7、8、9、10)、急性タンパク合成の刺激(11)および脂肪細胞リポタンパクリパーゼ活性の阻害(12、13)が含まれる。
【0007】
インターロイキン−13(IL−13)はインターロイキン−4(IL−4)と幾つかの構造的特徴を共有するもう一つの重要なサイトカインである(14および15を参照)。IL−4の遺伝子とIL−13の遺伝子とは関連のあるイントロン/エキソン構造を持っておりそしてヒト染色体5の上に共に近くに局在しており、またネズミでも染色体11のシンテニー領域に存在する(14、15)。タンパクレベルでは、IL−4とIL−13は4個のシステイン残基を含め約30%のアミノ酸同一性を共有している。生物学的にも、IL−13とIL−4は類似しており、活性化されたT−細胞により産生されそして、例えば、マクロファージに作用して分化を誘発しそして炎症性サイトカインの産生を抑制する。さらに、ヒトIL−13はB−細胞増殖に対する共−刺激シグナルとして作用しそして免疫グロブリン・イソタイプ・スイッチングに影響を与えることができる(14、15)。IL−13および他のヘモポエチンサイトカインの多様かつ多面的な機能はこのグループを重要な研究対象とし、特にサイトカインとその受容体との相互作用のレベルでの研究が重要である。サイトカイン受容体およびサイトカイン−受容体相互作用の操作および制御は多くの治療分野で、とりわけ標的サイトカインが機能的に多面的でありそして標的サイトカインのある機能を阻止したいがそのすべての機能を阻止したくはない場合には、潜在的に極めて重要である。
【非特許文献1】デュー,エックス.エックス.およびウィリアムス,ディー.エイ.(1994)Blood 83 : 2023-2030。
【非特許文献2】ヤング,ワイ.シー.およびイン,ティー.(1992)Biofactors 4 : 15-21。
【非特許文献3】ポール,エス.アール.,ベネット,エフ.,カルベッティ,ジェイ.エイ.,ケレハー,ケイ., ウッド,シー.アール.,オオハラ,アール.ジェイ.ジェイ.,レアリー,エイ.シー.,シブリー,ビー.,クラーク,エス.シー.,ウィリアムス,ディー.エイ.およびヤング,ワイ.シー.(1990)Proc.Natl.Acad.Sci.USA 87: 7512。
【非特許文献4】ムサシ,エム.,クラーク,エス.シー.,スドー,ティー.,ウーダル,ディー.エル.およびオガワ,エム.(1991)Blood 78: 1448-1451。
【非特許文献5】シブラー,ケイ.アール.,ヤング,ワイ.シー.およびクリステンセン,アール.ディー.(1992)Blood 80: 900-3。
【非特許文献6】ツジ,ケイ.,リーマン,エス.ディー.,スドー ,ティー.,クラーク,エス.シー.およびオガワ,エム.(1992)Blood 79: 2855-60。
【非特許文献7】バーンスタイン,エス.エイ.,メイ,アール.エル.,ヘンソーン,ジェイ.,フリース,ピー.およびターナー,ケイ.(1992)J.Cell.Physiol.153 : 305-12。
【非特許文献8】ハンゴク,ジー.,イン,ティー.,クーパー,エス.,シェンデル,ピー.,ヤング,ワイ.シー.およびブロックスメイヤー,エイチ.イー.(1993)Blood 81: 965-72。
【非特許文献9】テラムラ,エム.コバヤシ,エス.,ホシノ,エス.,オシミ,ケイ.およびミゾクチ,エイチ.(1992)Blood 79: 327-31。
【非特許文献10】ヨネムラ,ワイ.,カワキタ,エム.,マスダ,ティー.,フジモト,ケイ.,カトウ,ケイ.およびタカツキ,ケイ.(1992)Exp.Hematol.20: 1011-6。
【非特許文献11】バウマン,エイチ.およびシェンデル,ピー.(1991)J.Biol.Chem.266 : 20424-7。
【非特許文献12】カワシマ,アイ.,オオスミ,ジェイ.,ミタ−ホンジョウ,ケイ.,シモダ−タカノ,ケイ.,イシカワ,エイチ.,サカキバラ,エス.,ミヤダイ,ケイ.およびタキグチ,ワイ.(1991)Febs.Lett. 283: 199-202。
【非特許文献13】ケラー,ディー.シー., デュー,エックス.エックス.,スローアー,イー.エフ.,ホフマン,アール.およびウィリアムス,ディー.エイ.(1993)Blood 82 : 1428-35。
【非特許文献14】マッケンジー,エイ.エヌ.ジェイ.およびズラウスキー,ジー.(1994)サイトカインおよびその受容体へのガイドブック,オクスフォード大学出版部,オクスフォード。
【非特許文献15】ズラウスキー,ジー.およびド フリース,ジェイ.イー.(1994)Immunol.Today 15: 19-26。
【発明の開示】
【0008】
IL−13およびその受容体に関する研究はIL−13受容体のすべてまたは部分をコードする遺伝子配列をクローニングすることができなかったため妨げられてきた。本発明によれば、IL−13受容体α−鎖、すなわちIL−4受容体とも共有されている受容体サブユニットをコードする遺伝子配列が今やクローニングされたのである。これらの遺伝子配列の利用可能性により、構造または機能のレベルにおいてIL−13の活性並びにIL−13に関連するサイトカインの活性を調節または監視することができる一群の治療薬および診断薬の開発が可能となる。本発明によれば、構造および機能においてIL−13に関連するサイトカインの1例はIL−4である。
【0009】
従って、本発明の一つの側面は動物由来のヘモポエチン受容体または該受容体の誘導体をコードするヌクレオチド配列またはコードする配列に相補的なヌクレオチド配列を含む核酸分子に関する。
【0010】
より具体的には、本発明は動物ヘモポエチン受容体−該受容体はIL−13またはIL−13の誘導体と相互作用をすることができるものであるか−またはそれらの誘導体をコードするヌクレオチド配列またはコードする配列に相補的なヌクレオチド配列を含む単離された核酸分子を提供する。
【0011】
関連する一つの態様で、本発明は、該受容体が、
(i)IL−13またはその誘導体と相互作用をすることができ、そして
(ii)IL−4とIL−4受容体α−鎖との間でできる複合体と相互作用をすることができる、
ものである、動物ヘモポエチン受容体またはそれらの誘導体をコードするヌクレオチド配列またはコードする配列に相補的なヌクレオチド配列を含む単離された核酸分子を提供する。
【0012】
これらの態様により、IL−13の誘導体には、アゴニスト、アンタゴニスト、抗体および擬症物質(mimetics)が含まれることになる。
【0013】
本発明は動物IL−13受容体α−鎖またはその誘導体をコードするヌクレオチド配列またはコードする配列に相補的なヌクレオチド配列を含む核酸分子にも関する。
【0014】
関連する一つの態様では、本発明は動物IL−4受容体またはその誘導体の1成分をコードするヌクレオチド配列またはコードする配列に相補的なヌクレオチド配列を含む核酸分子を包含する。
【0015】
この動物は哺乳類または鳥の1種であることが好ましい。とりわけ好ましい動物としては、ヒト、実験室テスト動物(例えば、霊長類、マウス、ウサギ、ハムスター、モルモット)、家畜(例えば、羊、山羊、馬、豚、牛、ロバ)、愛玩動物(例えば、犬、猫)、捕獲した野生の動物(例えば、キツネ、カンガルー、ディンゴー)および家禽(例えば、鶏、鵞鳥、鴨)およびゲーム鳥(例えば、エミュー、ダチョウ)などが挙げられる。本発明はマウスやヒトを例としてあげるが、主題である本発明の範囲はすべての動物および鳥類に及ぶものである。
【0016】
本発明は配列類似性に基づきヘモポエチン受容体ファミリーのメンバーを同定する能力について部分的に予言する。このアプローチにより、ヘモポエチン受容体をコードする一つの遺伝子配列が本発明により同定された。発現された遺伝子配列は本明細書で「NR4」と呼ばれる。本発明によれば、NR4はトランスフェクトされたCOS細胞により合成されたとき、約50,000〜約70,000ダルトン、より好ましくは約55,000〜約65,000ダルトンの見掛けの分子量を持っている。NR4はIL−13と特異的にかつ低親和性をもって結合する、それ故、IL−13受容体α−鎖であると考えられる。従って、「NR4」と「IL−13受容体α−鎖」(または「IL−13Rα」)とは本明細書を通して互換可能なものとして使用される。さらに、IL−13のその受容体への結合はIL−4受容体α−鎖を発現する細胞内でIL−4またはその成分により競争的に阻害されることが見出された、そしてこれはIL−13と受容体との相互作用を制御する方法を提供できるものであり、そして擬症物質の調製および構築のための基礎をも提供するものである。
【0017】
本発明の別の側面では、配列番号:2に記載されたアミノ酸配列を持つIL−13受容体α−鎖またはその全部または一部に少なくとも約50%の類似性を有するアミノ酸配列をコードするヌクレオチド配列を含む核酸分子が提供される。この類似性パーセンテージは少なくとも約60%が好ましく、少なくとも約70%がより好ましく、少なくとも約80〜85%がさらに好ましく、そして少なくとも約90〜95%またはそれ以上がさらに一層好ましい。配列の全部または一部に言及しているのは、二つの受容体の各部分を含むハイブリッド分子を定義して含めることを意図している。単一アミノ酸を包含することは意図していない。
【0018】
本発明のさらなる態様では、IL−13受容体α−鎖をコードしそして配列番号:1に実質的に記載されたヌクレオチド配列を有するヌクレオチド配列またはその全部または一部と少なくとも約50%の類似性を有するヌクレオチド配列を含む核酸分子が提供される。この類似性パーセンテージは少なくとも約60%が好ましく、少なくとも約70%がより好ましく、少なくとも約80〜85%がさらに好ましく、そして少なくとも約90〜95%またはそれ以上がさらに一層好ましい。
【0019】
本発明のさらに別の側面では、配列番号:4に記載されたアミノ酸配列を有するIL−13受容体α−鎖またはその全部または一部と少なくとも約50%の類似性を有するアミノ酸配列をコードするヌクレオチド配列を含む核酸分子が提供される。この類似性パーセンテージは少なくとも約60%が好ましく、少なくとも約70%がより好ましく、少なくとも約80〜85%がさらに好ましく、そして少なくとも約90〜95%またはそれ以上がさらに一層好ましい。
【0020】
本発明のさらなる態様では、IL−13受容体α−鎖をコードしそして配列番号:3に記載されたようなヌクレオチド配列を実質的に有するヌクレオチド配列またはその全部または一部に少なくとも約50%の類似性を有するヌクレオチド配列を含む核酸分子が提供される。この類似性パーセンテージは少なくとも約60%が好ましく、少なくとも約70%がより好ましく、少なくとも約80〜85%がさらに好ましく、そして少なくとも約90〜95%またはそれ以上がさらに一層好ましい。
【0021】
従って、本発明は配列番号:1または配列番号:3に記載されたヌクレオチド配列または配列番号:2または配列番号:4に記載されたアミノ酸配列またはこれらに対し1個または複数のヌクレオチドまたはアミノ酸の置換、欠失および/または付加がなされたものに及ぶことになる。
【0022】
本発明はさらに、低厳格条件下で配列番号:1または配列番号:3に記載されたヌクレオチド配列またはそれらの相補型とハイブリッドを形成することができる核酸分子に及ぶことになる。
【0023】
本発明は組換えヘモポエチン受容体および特に組換えNR4およびNR4を含む組換えハイブリッドに及ぶ。組換えポリペプチドはIL−13と低親和性で相互作用することが好ましく、高い親和性で相互作用すればより一層好ましい。
【0024】
とりわけ好ましい態様では、ポリペプチドは下記の特性のうちの少なくとも二つを有するものである。
(i)配列番号:2または配列番号:4に実質的に記載されたアミノ酸配列またはそれらの全部または一部と少なくとも約50%の類似性を有するアミノ酸配列を含むこと、
(ii)配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列またはそれらの全部または一部と少なくとも約50%の類似性を有するヌクレオチド配列によりコードされるものであること、
(iii)IL−13またはその誘導体と少なくとも低親和性で相互作用すること、および
(iv)COS細胞で発現させたとき、ウエスタン・ブロット分析により測定すると約50,000から約70,000ダルトンの分子量を有すること。
【0025】
関連する態様では、このポリペプチドは下記の特性のうちの少なくとも三つを有する。
(i)配列番号:2または配列番号:4に実質的に記載されたアミノ酸配列またはそれらの全部または一部と少なくとも約50%の類似性を有するアミノ酸配列を含むこと、
(ii)配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列またはそれらの全部または一部と少なくとも約50%の類似性を有するヌクレオチド配列によりコードされるものであること、
(iii)IL−13またはその誘導体と少なくとも低親和性で相互作用すること、および
(iv)COS細胞で発現させたとき、ウエスタン・ブロット分析により測定すると約50,000から約70,000ダルトンの分子量を有すること、
(v)IL−4受容体α−鎖由来のアミノ酸配列を含むこと、そして
(vi)IL−13と相互作用をすることができ、かつその相互作用がIL−4受容体α−鎖を発現する細胞中でIL−4により競争的に阻害されること。
【0026】
本明細書で「組換えヘモポエチン受容体」、「NR4」、「IL−13受容体」または「IL−13受容体α−鎖」と言及するときは、それらの一部、断片、部分、相同体、ハイブリッドまたはアナログなどのそれらの誘導体への言及も含まれる。誘導体は機能的であってもなくてもよく、または非機能的ではあるが受容体の全部または一部に対する抗体と免疫学的に相互作用し得るものであってもよい。受容体の誘導体は受容体−リガンド相互作用のアゴニストまたはアンタゴニストをも包含する。機能とは、IL−13またはその誘導体と相互作用するNR4の能力あるいは可溶性NR4の場合にはIL−13で誘発されたある細胞の活性と競合する能力により便宜的に定義される。
【0027】
本発明により提供される特に好ましい誘導体には、高親和性でIL−13とまたは高親和性でIL−13およびIL−4と結合することができるIL−13受容体α−鎖の誘導体が含まれ、また誘導体にはIL−13受容体α−鎖と、例えば、IL−13とも高親和性で結合するIL−4受容体α−鎖の間などのキメラ分子が包含される。
【0028】
本発明により提供される他の融合分子またはキメラ分子には、NR4とヘモポエチン受容体ファミリーのメンバー、受容体チロシンキナーゼ、TNF/NGF受容体およびGタンパク質−共役受容体の間のものが含まれる。例えば、キメラにはNR4とIL−13結合タンパク、IL−4受容体α−鎖、IL−2受容体γ−鎖またはIL−5などの気管支喘息やアレルギーに関与する他のサイトカインに対する受容体との間に生じ得る。他の重要なキメラにはNR4と免疫グロブリンまたは特定の細胞または組織にNR4を標的化させる他の分子、NR4と毒素、およびNR4と成長因子が含まれる。
【0029】
本明細書において42℃の低厳格性というときは、ハイブリッド形成条件においては少なくとも約1%v/vから少なくとも約15%v/vまでのホルムアミドおよび少なくとも約1Mから少なくとも約2Mまでの塩を、そして洗浄条件においては少なくとも約1Mから少なくとも約2Mまでの塩を範囲として包含する。必要なときは、中度の厳格性などの別の厳格条件を適用することもできる。中度の厳格性は、ハイブリッド形成条件においては少なくとも約16%v/vから少なくとも約30%v/vまでのホルムアミドおよび少なくとも約0.5Mから少なくとも約0.9Mの塩を、そして洗浄条件においては少なくとも約0.5Mから少なくとも約0.9Mの塩を範囲として包含し、あるいは高厳格性は、ハイブリッド形成条件においては少なくとも約31%v/vから少なくとも約50%v/vまでのホルムアミドおよび少なくとも約0.01Mから少なくとも約0.15Mの塩を、そして洗浄条件においては少なくとも約0.01Mから少なくとも約0.15Mの塩を範囲として包含する。
【0030】
本発明のもっと別の側面では、IL−13受容体α−鎖をコードするヌクレオチド配列またはコードする配列に相補的なヌクレオチド配列を含む核酸分子であって、配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列を有するものあるいは構造的に類似するIL−13受容体α−鎖またはその誘導体をコードしそして配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列またはその相補型に低厳格性条件下でハイブリッドを形成することができる核酸分子が提供される。
【0031】
本発明のさらに別の側面では、配列番号:2または配列番号:4に実質的に記載されたアミノ酸配列を持つIL−13受容体α−鎖をコードするヌクレオチド配列またはコードする配列に相補的なヌクレオチド配列を含む核酸分子が、または配列番号:2または配列番号:4に記載された配列に少なくとも約50%の類似性を有するアミノ酸配列をコードするヌクレオチド配列であって、かつ配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列に低厳格性条件下でハイブリッドを形成することができるものを含む核酸分子が提供される。
【0032】
本発明により提供される核酸分子は一般に単離された形のものであり、一本鎖でも二本鎖でも、直鎖状でも閉じた環状のDNA(例えば、ゲノムDNA)でも、cDNAまたはmRNAでも、あるいはDNA:RNAハイブリッドの形などのそれらの結合物でもよい。特に好ましい態様では、核酸分子はベクターの形であり、特定の宿主細胞内で発現させることができる発現ベクターが最も好ましい。特に有用な宿主細胞としては原核細胞、哺乳類細胞、酵母細胞および昆虫細胞が挙げられる。この細胞は細胞株の形であってもよい。
【0033】
本発明のこの側面により、本明細書で先に述べたIL−13受容体α−鎖をコードする核酸分子を含む発現ベクターが提供される。該発現ベクターは特定の宿主細胞内で発現することができる。
【0034】
本発明の別の側面では、配列番号:2または配列番号:4に実質的に記載されたアミノ酸配列またはその全部または一部と少なくとも約50%の類似性を有するアミノ酸配列を含む組換えポリペプチドが提供される。この類似性パーセンテージは少なくとも約60%が好ましく、少なくとも約70%がより好ましく、少なくとも約80〜85%がさらに好ましく、そして少なくとも約90〜95%またはそれ以上がさらに一層好ましい。
【0035】
本発明により提供される組換えポリペプチドは、従って、IL−13受容体α−鎖の成分、部分、断片、誘導体、相同体、またはアナログを含み、そして配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列、またはその全部または一部と少なくとも約50%の類似性を有する分子、または配列番号:1または配列番号:3に記載されたヌクレオチド配列またはそれらの相補型とハイブリッドを形成することができる分子によりコードされることが好ましい。この組換え分子はグリコシル化されていてもいなくてもよい。グリコシル化型であるときは、グリコシル化は天然に生ずるIL−13受容体α−鎖と実質的に同じであってもよく、またはグリコシル化の修飾型であってもよい。グリコシル化の変更または相違はIL−13受容体α−鎖の結合活性に影響を与えることも与えないこともある。
【0036】
組換えIL−13受容体α−鎖は可溶性の形であることも、細胞表面上に発現されることもまたは固体支持体または別の分子に結合したりまたは融合したりしていることもある。
【0037】
本発明はさらに下記の特性のうちの少なくとも二つを有する組換えポリペプチドを製造する方法を提供する。
(i)配列番号:2または配列番号:4に実質的に記載されたアミノ酸配列またはそれに少なくとも約50%の類似性を有するアミノ酸配列を含むこと、
(ii)配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列またはそれに少なくとも約50%の類似性を有するヌクレオチド配列によりコードされるものであること、
(iii)IL−13またはその誘導体と低親和性で相互作用すること、そして
(iv)COS細胞中で発現させるとき、ウエスタン・ブロット分析により測定すると約50,000から約70,000ダルトンの分子量を持つこと。
【0038】
該方法は、本発明の遺伝子構築物を含む細胞を、該遺伝子構築物中の核酸分子を発現して組換えポリペプチドを産生するのに十分な時間および条件の下で培養する工程、そして該組換えポリペプチドを単離する工程を含んで成る方法である。
【0039】
別の態様は下記の特性のうちの少なくとも三つを有する組換えポリペプチドを製造する方法を提供する。
(i)配列番号:2または配列番号:4に実質的に記載されたアミノ酸配列またはその全部または一部に少なくとも約50%の類似性を有するアミノ酸配列を含むこと、
(ii)配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列またはその全部または一部に少なくとも約50%の類似性を有するヌクレオチド配列によりコードされるものであること、
(iii)IL−13またはその誘導体と少なくとも低親和性で相互作用すること、
(iv)COS細胞中で発現させたとき、ウエスタン・ブロット分析により測定すると約50,000から約70,000ダルトンの分子量を持つこと、
(v)IL−4受容体α−鎖由来のアミノ酸配列を含むこと、そして
(vi)IL−13と相互作用をすることができること、かつ、その相互作用がIL−4受容体α−鎖を発現する細胞内でIL−4により競争的に阻害されること。
【0040】
該方法は、本発明の融合遺伝子構築物を含む細胞を、該融合遺伝子構築物内の核酸分子が発現し組み換えポリペプチドを産生するのに十分な時間および条件の下で培養する工程および該組換えポリペプチドを単離する工程を含んで成る方法である。
【0041】
本発明はさらに上記の組換えポリペプチドを発現する動物細胞などの細胞にまで及ぶ。
【0042】
本発明の別の態様では、組換えIL−13受容体α−鎖の化学的アナログが提供される。
【0043】
上に述べたように、本発明はさらにNR4のある範囲の誘導体を提供する。誘導体には、NR4ポリペプチドおよび対応する遺伝子配列の断片、一部、部分、変異体、ハイブリッド(融合分子やキメラ分子を含む)、相同体およびアナログが含まれる。一つの好ましい態様では、この誘導体は高親和性でもってIL−13と結合する。他の好ましい誘導体はアゴニスト、アンタゴニストまたは擬症物質として作用する。これらの誘導体には、NR4に1個または複数のアミノ酸が置換、欠失、および/または付加されたもの、およびNR4をコードする遺伝子配列に1個または複数のヌクレオチドが置換、欠失、および/または付加されたものも含まれる。アミノ酸配列またはヌクレオチド配列への「付加」には、他のペプチド、ポリペプチド、またはタンパク質との融合またはヌクレオチド配列への融合も含まれる。本明細書で「NR4」と言及するときは、機能的誘導体や「NR4」と免疫学的に相互作用し得る誘導体も含めそのすべての誘導体を指している。本発明はネズミNR4、ヒトNR4またはそれらの誘導体の間などのハイブリッド分子にも及ぶ。特に好ましいハイブリッドはNR4とIL−4受容体α−鎖を含むものである。
【0044】
本明細書において提供されるNR4の類似体には、側鎖の修飾、ペプチド合成、ポリペプチド合成またはタンパク合成の間における非天然型アミノ酸および/またはそれらの誘導体の取り込み、および架橋剤や、タンパク性分子またはそれらのアナログにコンホーメーション上の制約を加える他の方法の使用などが含まれるが、これらに限定されるものではない。
【0045】
本発明によって提供される側鎖の修飾の例としては、アルデヒドとの反応による還元的アルキル化に続くNaBH4による還元、メチルアセチミデートを用いるアミジノ化(amidination)、無水酢酸を用いるアシル化、シアネートを用いるアミノ基のカルバモイル化、2,4,6−トリニトロベンゼンスルホン酸(TNBS)を用いるアミノ基のトリニトロベンジル化、無水コハク酸および無水テトラハイドロフタル酸を用いるアミノ基のアシル化、およびピリドキサル−5−リン酸に続いてNaBH4で還元することによるリシンのピリドキシレーションなどのアミノ基の修飾が挙げられる。
【0046】
アルギニン残基のグアニジン基は2,3−ブタンジオン、フェニルグリオキサルおよびグリオキサルなどの試薬を用いて複素環縮合産物の形成により修飾することができる。
【0047】
カルボキシル基はO−アシルイソウレア形成を経るカルボジイミド活性化それに続く、例えば対応するアミドへの誘導により修飾することができる。
【0048】
スルフヒドリル基はヨード酢酸またはヨードアセトアミドを用いるカルボキシメチレーション、システイン酸への過ギ酸酸化、他のチオール化合物との混合ジスルフィドの形成、マレイミド、無水マレイン酸または他の置換マレイミドとの反応、4−クロロマーキュリベンゾエート、4−クロロマーキュリフェニルスルホン酸、フェニルマーキュリクロライド、2−クロロマーキュリ−4−ニトロフェノールおよび他の水銀化合物を用いる水銀誘導体の形成、アルカリ性pHでシアネートを用いるカルバモイル化などの諸方法により修飾することができる。
【0049】
トリプトファン残基は、例えば、N−ブロモスクシンイミドを用いる酸化または2−ヒドロキシ−5−ニトロベンジルブロマイドまたはスルフェニルハライドを用いるインドール環のアルキル化により修飾することができる。チロジン残基は、他方、テトラニトロメタンを用いる硝酸化により改変し3−ニトロチロジン誘導体を形成することができる。
【0050】
ヒスチジン残基のイミダゾール環の修飾はヨード酢酸誘導体を用いるアルキル化またはジエチルピロカーボネートを用いるN−カルボエトキシレーションにより達成することができる。
【0051】
ペプチド合成の間に非天然型アミノ酸および誘導体を取り込む例としては、ノルロイシン、4−アミノ酪酸、4−アミノ−3−ヒドロキシ−5−フェニルペンタン酸、6−アミノヘキサン酸、t−ブチルグリシン、ノルバリン、フェニルグリシン、オルニチン、ザルコシン、4−アミノ−3−ヒドロキシ−6−メチルヘプタン酸、2−チエニルアラニンおよび/またはアミノ酸のD−異性体が挙げられるが、これらに限定されるものではない。本明細書で提供される非天然型アミノ酸のリストを表1に示す。
【0052】
架橋剤は、例えば、3Dコンホーメーションを安定化するために使用することができ、n=1からn=6までのスペーサー基(CH2)nを持つ二官能性イミドエステル、グルタルアルデヒド、N−ヒドロキシスクシンイミドエステルなどのホモ−二官能性架橋剤および通常N−ヒドロキシスクシンイミドなどのアミノ−反応性部分とマレイミドまたはジチオ部分(SH)またはカルボジイミド(COOH)などの別の基の特異的反応性部分とを持つヘテロ−二官能性試薬を使用する。また、ペプチドのコンホーメーションは、例えば、CαおよびNα−メチルアミノ酸の取り込み、アミノ酸のCαとCβ原子の間に二重結合の導入、およびN末端とC末端の間、二つの側鎖の間または側鎖とN末端またはC末端との間にアマイド結合を形成させるなどして共有結合を導入することにより環状ペプチドまたは環状アナログの形成により制約することができる。
【0053】
これらのタイプの修飾は個体に投与するとき、あるいは診断試薬として使用するために、NR4を安定化するために重要である。
【0054】
本発明はさらにNR4のアンタゴニストまたはアゴニストとして作用することができる、すなわちNR4の機能的アナログとして作用することができるNR4の化学的アナログを提供する。化学的アナログは必ずしもNR4から誘導されるとは限らず、ある種のコンホーメーションの類似性を共有するものであってもよい。また、化学的アナログはNR4のある種の物理化学的性質を真似するように特別に設計することもできる。化学的アナログは化学的に合成することができまたは例えば天然物のスクリーニングにより検出することができる。
【0055】
NR4の同定により、NR4の発現を調節したりNR4の活性を調節したりすることができる1群の治療に有用な分子の誕生を可能とする。本発明により提供される調節物質にはNR4遺伝子発現またはNR4タンパク活性のアゴニストおよびアンタゴニストが含まれる。NR4遺伝子発現のアンタゴニストにはアンチセンス分子、リボザイムおよび共−抑制分子(co-suppression molecules)が含まれる。アゴニストにはプロモーター能を増強したりまたは負の調節機構を妨害したりする分子が含まれる。NR4タンパクのアゴニストには抗体、リガンドおよび擬症物質が含まれる。NR4のアンタゴニストには抗体および阻害ペプチド断片が含まれる。細胞がNR4とIL−4受容体α−鎖を共−発現するときは、アゴニストおよびアンタゴニストはこのIL−4受容体α−鎖を標的とすることができる。
【0056】
【表1】
【0057】
本発明により提供される他の誘導体には、完全に脱グリコシル化された分子から修飾されたグリコシル化分子までのある範囲のグリコシル化変異型が含まれる。改変されたグリコシレーションパターンは異なる宿主細胞中での組換え分子の発現から生ずることがある。
【0058】
本発明の別の態様では、ヒトにおけるNR4遺伝子の発現を調節する方法が提供される。この方法はNR4をコードするNR4遺伝子をNR4発現の調節物質の有効量と、NR2の発現を高く調節しまたは低く調節しまたは別の仕方で調節するのに十分な時間および条件の下で、接触させることを含んでなる方法である。NR4をコードする核酸分子またはその誘導体を細胞内に導入して、例えば、欠陥を持つまたは一つ以上の望ましくない変異を持つ内因性NR4遺伝子配列の置換を含め、その細胞のNR4関連活性を増強したり改変したりすることができる。逆に、オリゴヌクレオチドなどのNR4アンチセンス配列(または共−抑制のためのセンス配列)を導入して、内因性のNR4遺伝子を発現しているどの細胞のNR4関連活性をも減少させることができる。リボザイムも使用することができる。
【0059】
本発明の別の側面は、ヒトにおけるNR4の活性を調節する方法を提供する。この方法はNR4活性を増加または減少させるのに十分な時間および条件の下である分子の調節有効量を該哺乳類に投与することを含んでなる方法である。この分子はタンパク性の分子であっても化学的実体であってもよくそしてNR4の誘導体またはそのリガンドまたは化学的アナログまたはNR4の切断変異体またはそのリガンドであってもよい。
【0060】
例えば、IL−13とIL−4は免疫応答の調節に関与し、そして喘息などのアレルギー性またはアトピー性疾患と関連する免疫グロブリン・イソタイプであるIgEの産生に関与している。IL−13/IL−4とそれらの受容体との相互作用を調節することはアレルギー状態などの炎症状態を治療するのに重要であり得る。IL−4/IL−13およびIgEのレベルの上昇はネフローゼ症候群、春季カタルおよび角結膜炎などの疾病にも重要である。本明細書でその治療法が提供される他の疾病としては、気管支喘息、鼻炎(perennial rhinitis)およびアトピー性皮膚炎が挙げられる。IL−13−受容体相互作用の調節が重要となり得る他の病態としては、IL−13がサイトカイン形成を誘発し、今度はこのサイトカインが疾病の開始、進行および/または重篤度に関与するような病態が含まれる。同様に、IL−4−受容体相互作用の調節も疾病状態を制御するのに重要である。例えば、一部の癌はサイトカインIL−13またはIL−4により悪化させられることがある。これらは免疫抑制効果または免疫抑制効果分子を誘発し、今度はこれが癌の成長に応答する身体の能力を減退させるのである。
【0061】
従って、本発明はNR4またはその誘導体またはNR4発現またはNR4活性の調節物質および1以上の薬学的に許容され得る担体および/または希釈剤を含んでなる薬学的組成物を提供する。これらの成分は「活性成分」と呼ばれる。
【0062】
これについては、前に記載した組換えヘモポエチン受容体またはそのリガンド(例えば、IL−13)結合性部分および1以上の薬学的に許容され得る担体および/または希釈剤を含んでなる薬学的組成物が提供される。
【0063】
別の態様では、前に述べた組換えヘモポエチン受容体に対するリガンド(例えば、IL−13)および1以上の薬学的に許容され得る担体および/または希釈剤を含んでなる薬学的組成物が提供される。
【0064】
本発明のさらなる側面は、動物の治療方法を提供する。この方法は前に記載した組換えヘモポエチン受容体またはそのリガンド結合性部分または該ヘモポエチン受容体に対するリガンド(例えば、IL−13)の治療有効量を、該治療が実質的に達成されるのに十分な時間および条件の下であるいは実質的に改善される条件の下で、該動物に投与することを含むものである。
【0065】
注射可能な用途に適する薬学的剤形としては、滅菌水溶液(水溶性の場合)または分散剤および滅菌した注射可能な溶液または分散剤の用時調製のための滅菌粉末が挙げられ、あるいはクリームの形状であってもまたは局所適用に適した他の剤形であってもよい。それは製造および貯蔵の条件下で安定でなければならず、細菌や糸状菌などの微生物の汚染作用に対して保護されなければならない。担体は溶媒または分散剤であることができ、例えば、水、エタノール、ポリオール(例えば、グリセロール、プロピレングリコールおよび液体ポリエチレングリコール、およびその他のもの)、それらの適当な混合物、および植物性油などを含む。例えば、リシチン(licithin)などの皮膜の使用により、分散剤の場合には必要な粒子サイズの維持によりそしてスーパーファクタントの使用により、適当な流動性を維持することができる。微生物の作用の防止は種々の抗細菌物質および抗かび物質、例えば、パラベン、クロロブタノール、フェノール、ソルビン酸、チルメロサル(thirmerosal)およびその他により可能となる。多くの場合、等張物質、例えば、砂糖または塩化ナトリウムを含めることが好ましい。注射可能な組成物の長期間吸収は吸収を遅延させる物質、例えばアルミニウムモノステアレートおよびゼラチンを組成物の中で使用することにより可能とすることができる。
【0066】
注射可能な滅菌溶液は必要な量の活性化合物を必要な場合は上に挙げた他の様々な成分と共に適当な溶媒の中に溶解し次いで無菌ろ過を行うことにより調製される。一般に、分散剤は様々な滅菌された活性成分を、基本的な分散媒体および上に挙げたものの中の必要な他の成分を含む滅菌担体の中に取り込むことにより調製される。
【0067】
注射可能な滅菌溶液を調製するための滅菌粉末の場合には、好ましい調製方法は真空乾燥法および凍結−乾燥法である。これらは予めそれらの無菌ろ過された溶液から、活性成分プラス付加的な必要成分の粉末を生ずる。
【0068】
活性成分を適当に保護するときは、例えば、不活性な希釈剤または資化し得る食用の担体と共に経口投与することができ、あるいはそれを固いまたは軟らかい殻のゼラチンカプセルの中に閉じ込めることができ、あるいはそれを錠剤中に圧縮することができ、あるいは食事の食品と共に直接摂取することができる。治療のための経口投与を行うためには、活性化合物は賦形剤と混ぜ、消化可能な錠剤、バッカル錠剤、トローチ、カプセル、エリクサル、懸濁剤、シロップ、ウェハース、その他の形で使用される。このような組成物および調製物は少なくとも1重量%の活性化合物を含むべきである。この組成物および調製物のパーセンテージは、もちろん、変動するものであり、単位重量の約5%から約80%までの間にあるのが便宜である。治療上有用な組成物の中の活性化合物の量は適当な投与量が得られるようなものである。本発明による組成物または調製物は経口投与の単位形態が活性化合物を約0.1μgと2000mgの間で含むように調製されることが好ましい。
【0069】
錠剤、トローチ、ピル、カプセルなども後にリストされる成分を含むことができる。ガム、アカシア、コーンスターチまたはゼラチンなどのバインダー、ジカルシウムホスフェートなどの賦形剤、コーンスターチ、ポテトスターチ、アルギン酸、その他の崩壊剤、ステアリン酸マグネシウムなどの滑剤、およびスクロース、ラクトースまたはサッカリンなどの甘味剤またはペパーミント、サリチル酸メチル(oil of wintergreen)、または桜の芳香などの芳香剤を添加することができる。投与単位の形態がカプセルのときは、上のタイプの物質に加えて液体の担体を含めることができる。投与単位の物理的形態を被覆あるいは別の仕方で修飾するための様々な物質がほかにもある。例えば、錠剤、ピル、またはカプセルはシェラック、砂糖または両方で被覆することができる。シロップまたはエリクサルは活性化合物、甘味剤として砂糖、保存剤としてメチルおよびプロピルパラベン、色素および桜またはオレンジなどの芳香剤を含むことができる。もちろん、いかなる投与単位形態を作る場合に使用されるどんな物質も薬学的に純粋でありそしてその使用量では実質的に無毒であるべきである。さらに、活性化合物(単数または複数)は制御放出(sustained-release)用の調製物や処方に加えることもできる。
【0070】
本発明はクリーム、ローションおよびゲルなどの局所適用に適した形態をも包含する。
【0071】
薬学的に許容され得る担体および/または希釈剤としては、溶媒、分散媒体、被覆剤、抗細菌および抗かび剤、等張物質および吸収遅延物質などのいずれかおよびすべてが挙げられる。このような媒体や物質を薬学的に活性な物質に使用することは当技術分野では良く知られている。通常の媒体または物質が活性成分と配合禁忌である場合を除き、治療用組成物にそれらを使用することは本発明に包含される。補充的な活性成分もこの組成物の中に取り込むことができる。
【0072】
投与の容易および投与量の均一性のため投与単位の形態で非経口的組成物を処方することはとりわけ有利である。本明細書で使用されるとき投与単位形態とは治療すべき対象哺乳類に対し1回投与に適した物理的に区別された単位を指し、各単位は必要な薬学的担体との組み合わせで望まれる治療効果を生じさせるように計算された活性物質の予め計測された量を含んでいる。本発明の新規な投与単位形態の仕様は、(a)活性物質のユニークな特性および達成すべき具体的治療効果、および(b)身体の健康が損なわれている疾病状態を持つ生物対象における疾病の治療のためにこのような活性物質を混ぜ合わせる技術上の固有の制約、により支配されそして直接それらに依存している。
【0073】
主要な活性成分は便利で効果的な投与のために、前に開示したような投与単位形態で、その有効量が薬学的に許容され得る適当な担体と混ぜ合わされる。単位の投与形態は、例えば、0.5μgから約2000mgにわたる量で主要な活性化合物を含むことができる。比率で表すと、活性化合物は一般に担体の1ml当たり約0.5μgから約2000mgまで存在する。補充的活性成分を含む組成物の場合は、投与量は該成分の通常の投与量および投与方法を参照して決定する。
【0074】
薬学的組成物は標的細胞をトランスフェクトすることができるベクターのような遺伝子分子を含むこともでき、該ベクターはNR4発現またはNR4活性を調節することができる核酸分子を担っている。このベクターは例えばウイルスベクターであることができる。
【0075】
本発明のさらに別の側面はNR4に対する抗体およびその誘導体またはそのリガンド(例えば、IL−13)に関する。このような抗体はモノクローン抗体またはポリクローン抗体であることができ、そして天然に生ずるNR4への抗体から選択することができ、あるいはNR4またはその誘導体に対し特異的に高めることもできる。後者の場合は、NR4またはその誘導体はまず担体分子と結合することが必要になることがある。本発明の抗体および/または組換えNR4またはその誘導体は治療剤または診断試薬として特に有用である。
【0076】
例えば、NR4およびその誘導体はNR4への天然に生ずる抗体のスクリーニングに使用することができる。これらは、例えば、自己免疫疾患の一部で生ずる。また、NR4をスクリーニングするために特異的抗体を使用することができる。このような試験の技法は当技術分野で良く知られており、例えば、サンドイッチ試験やELISAが含まれる。NR4レベルおよび/またはIL−13レベルの知識はある種の癌または癌の素因の診断のためまたはある種の治療プロトコルの監視のために重要であり得る。特に、IgE応答またはIL−13またはIL−4またはその両方を監視することが重要である。これらは、今度は免疫系に影響を及ぼすからである。
【0077】
本発明のNR4に対する抗体はモノクローンでもポリクローンでもよい。また、Fab断片などの抗体の断片も使用することができる。さらに、本発明は組換え抗体および合成抗体に及びそして抗体ハイブリッドにまで及ぶ。「合成抗体」は本明細書では抗体の断片および抗体のハイブリッドを含むと考える。本発明のこの側面の抗体は免疫治療法にとりわけ有用であり、受容体または受容体−リガンド相互作用の評価または治療計画のプログラムの監視のための診断手段として使用することもできる。
【0078】
例えば、特異的抗体をNR4タンパクのスクリーニングに使用することができる。後者は、例えば、細胞抽出液または他の生物学的液体中のNR4レベルをスクリーニングするための手段としてまたは培養物上清から組換え手段で調製されたNR4を精製するための手段として重要であろう。本明細書で提供される試験のための技法は当技術分野で知られており、例えば、サンドイッチ試験やELISAが含まれる。
【0079】
上に論じた最初に述べた抗体に対するいかなる第2の抗体(モノクローン抗体、ポリクローン抗体または抗体断片または合成抗体)も本発明の範囲に含まれるべきものである。第1および第2の抗体は両方とも検出試験に使用することができ、あるいは第1の抗体は市場で入手可能な抗−免疫グロブリン抗体と共に使用することができる。本明細書で提供される抗体にはNR4のどの領域に特異的な抗体であってもすべて含まれる。
【0080】
ポリクローン抗体およびモノクローン抗体は両方とも受容体で免疫化することにより取得することができ、そしていずれのタイプも免疫試験に利用することができる。両方のタイプの血清を得る方法は当技術分野で良く知られている。ポリクローン血清はあまり好ましくないが、適当な実験室動物にNR4またはその抗原部分の有効量を注射し、その動物から血清を集め、そして既知の免疫吸着法のいずれかにより特異的な血清を単離することにより、比較的容易に調製される。この方法により製造された抗体は事実上どのタイプの免疫試験にも利用可能であるが、この生産物の潜在的不均質性のため一般的にはあまり好ましくない。
【0081】
免疫試験でのモノクローン抗体の使用は、大量にそれを作る能力があることおよび生産物の均質性のためとりわけ好ましい。不死の細胞株と免疫原調製物に対して感作させたリンパ球を融合させて作成したモノクローン抗体生産用のハイブリドーマ細胞株の調製は当技術分野で熟練した者に良く知られた技法により行うことができる。
【0082】
本発明の別の側面は主体からの生物試料内のNR4を検出する方法を提供する。該方法は該生物試料をNR4またはその誘導体または同族体に特異的な抗体と、抗体−NR4複合体が形成するのに十分な時間および条件の下で接触させ、そして該複合体を検出することを含んで成る。
【0083】
NR4が存在することはウエスタン・ブロッティングやELISAによるなどの幾つかの方法で確認できる。広い範囲の免疫試験法が利用できることは米国特許第4,016,043号、第4,424,279号および第4,018,653号を参照することにより見ることができる。これらは、もちろん、従来の競争的結合試験におけると同様に非−競争的タイプの1部位試験および2部位試験の両方または「サンドイッチ」試験を含んでいる。これらの試験は標的への標識化抗体の直接結合をも含んでいる。
【0084】
サンドイッチ試験は中でも最も有用でありそして普通に使用されている試験であって、本発明における使用にとっても好ましい。サンドイッチ試験法は幾つかの変法があり、それらのすべてが本発明に包含されるものである。簡単に言えば、典型的なホワード試験(forward assay)では、非標識化抗体が固体基質上に固定化されておりそして試験対象の試料が結合した分子と接触させられる。適当な時間、すなわち抗体−抗原複合体の形成を行わせるのに十分な時間、インキュベートした後、抗原に特異的な第2の抗体であって検出可能なシグナルを形成することができるレポーター分子で標識された抗体を次に添加し、抗体−抗原−標識化抗体の別の複合体が形成するのに十分な時間インキュベートする。未反応物質をすべて洗い流し、そしてレポーター分子が形成したシグナルの観察により抗原の存在が測定される。この結果は可視的なシグナルの簡単な観察による定性的なものであることもあり、また既知の量のハプテンを含む対照試料と比較することにより定量化することもできる。ホワード試験の変法には試料と標識化抗体の両方が結合抗体に同時に添加される同時試験が含まれる。これらの技法は当技術分野の熟練した者に良く知られており、容易に分かるような僅かな修飾を包含するものである。本発明によれば、試料は細胞抽出物、組織生検またはおそらく血清、唾液、粘液分泌物、リンパ液、組織液および呼吸液などのNR4を含むかも知れないものである。試料は、従って、一般に生物学的液体、細胞抽出液、骨髄またはリンパ液、組織抽出物(例えば、腎臓、肝臓、脾臓、など)、醗酵液および細胞培養物や細胞条件化培地からの上清を含む生物試料である。
【0085】
典型的なホワードサンドイッチ試験では、NR4またはその抗原部分に特異性を有する第1の抗体は固体表面に共有結合または受動的に結合している。固体表面はガラスまたはポリマーが代表的であり、最も普通に使用されるポリマーはセルローズ、ポリアクリルアミド、ナイロン、ポリスチレン、ポリ塩化ビニルまたはポリプロピレンである。固体支持体は管、ビーズ、ミクロプレートの平円盤の形、または免疫試験を行うのに適した表面であればなんでもよい。結合工程は当技術分野で良く知られており、一般に架橋共有結合または物理的吸着からなり、ポリマー−抗体複合体は試験試料の調製の際に洗浄される。試験対象の試料の一部をとり、ついで固相複合体に加え、そして抗体中に存在するサブユニットのいずれかと結合するのに十分な時間(例えば、2〜40分)および適当な条件(例えば、25℃)の下でインキュベートする。インキュベーション期間の後に、抗体サブユニット固相を洗浄し乾燥し、そしてこのハプテンの一部に特異的な第2の抗体とインキュベートする。第2の抗体はレポーター分子に結合しており、この分子が第2の抗体のハプテンへの結合を示すために用いられる。
【0086】
別の方法は生物試料中の標的分子を固定化し、ついでレポーター分子で標識されまたはされていない特異的抗体にこの固定化された標的を曝す。標的の量およびレポーター分子シグナルの強度に依存して、結合した標的は抗体による直接の標識化により検出可能となる。また、第1の抗体に特異的な第2の標識化抗体を標的−第1の抗体複合体と接触させ、標的−第1抗体−第2抗体の三次複合体を形成させる。この複合体はレポーター分子が発するシグナルにより検出される。
【0087】
本明細書で使用されるとき「レポーター分子」は、その化学的性質により抗原−結合抗体の検出を可能とする、分析的に同定可能なシグナルを供給する分子を意味する。検出は定性的にも定量的にも行うことができる。このタイプの試験で最も普通に用いられるレポーター分子は、酵素、発蛍光団または放射性核種を含む分子(すなわち、放射性同位元素)および化学発光性分子である。酵素免疫試験の場合には、酵素を通常グルタルアルデヒドまたは過ヨウ素酸により第2の抗体に結合させる。しかし、容易に分かるように、熟練した技術者に容易に利用可能な多様な異なる結合手段が存在する。通常使用される酵素としては、中でも、ホースラディッシュペルオキシダーゼ、グルコーズオキシダーゼ、ベーターガラクトシダーゼおよびアルカリホスファターゼか挙げられる。特異的酵素と共に使用されるべき基質は一般に対応する酵素による加水分解の際の検出可能な色変化の生成に対して選択される。適当な酵素の例としては、アルカリホスファターゼおよびペルオキシダーゼが挙げられる。発蛍光性基質を採用することも可能である。これは上に述べた発色原性基質よりもむしろ蛍光性の生産物を生ずる。全ての場合に、酵素−標識化抗体を第1抗体ハプテン複合体に添加して、結合させ、ついで過剰の試薬を洗い流す。ついで、適当な基質を含む溶液を抗体−抗原−抗体の複合体に添加する。この基質は第2の抗体に結合した酵素と反応し、定性的な視覚的シグナルを生じ、これは通常分光学的に定量化することができ、試料中に存在したハプテンの量の指標を与える。「レポーター分子」はラテックスビーズなどの上での赤血球などの細胞凝集または凝集の阻害の利用にも広がっている。
【0088】
また、フルオレッセインやローダミンなどの蛍光性化合物は抗体の結合活性を変えることなく抗体に化学的に結合させることができる。特定波長の光を照射することにより活性化すると、フルオロクロム−標識化抗体が光エネルギーを吸収し、分子内で励起状態を誘発し、続いて光学顕微鏡で視覚的に検出可能な特徴的な色の光を発射する。EIAの場合のように、蛍光性標識化抗体は第1抗体−ハプテン複合体に結合させられる。未結合試薬を洗い流した後、残りの三次複合体を次に適当な波長の光に曝すと、観察される蛍光は問題のハプテンの存在の証拠となる。免疫蛍光法およびEIA法は両方とも当技術分野で十分に確立されており、本発明の方法にとってとりわけ好ましい。しかしながら、放射性同位元素、化学発光性分子または生物発光性分子などの他のレポーター分子も採用することができる。
【0089】
NR4およびIL−4受容体α−鎖を発現することができる細胞の場合は別の形の試験ができる。例えば、IL−4受容体α−鎖とNR4がCOS細胞などの細胞上に共−発現されるときは、IL4の存在下でIL−13は高親和性でNR4に結合する。
【0090】
一つの理論または一つの作用様式に本発明を制限する積もりはないが、NR4とIL−4受容体とが同一細胞中で発現されるとき、これらはIL−4受容体およびIL−13受容体両方の形成に寄与する。IL−4の場合には、結合はまずIL−4受容体α−鎖を介して起こり、ついでこの複合体にNR4が相互作用する。IL−13の場合には、結合はまずNR4に対して起こり、ついでこの複合体とIL−4受容体α−鎖が相互作用してシグナル伝達を可能とする高親和性受容体を形成する。NR4とIL−4受容体α−鎖との共−発現の結果は、IL−4とIL−13がIL−4受容体α−鎖およびNR4と結合するために相互に競争することになることである。
【0091】
この挙動に基づけば、IL−4またはIL−13が両受容体成分を含む細胞表面複合体を形成するのを阻止するタンパク質または低分子はアンタゴニストとなり得るということであろう。このような分子はこのサイトカインとその低親和性受容体との相互作用を阻止することができる。例えば、可溶性IL−13BPはIL−13とNR4との相互作用を阻止することができる。同様に、可溶性IL−4受容体α−鎖はIL−4の細胞表面IL−4受容体α−鎖への結合を阻止することができる。これらの作用物質はIL−4またはIL−13に特異的なアンタゴニストとなるであろう。
【0092】
敷衍すると、可溶性NR4はその低親和性のためIL−13の極めて無能なアンタゴニストである。もしIL−4に結合しそしてIL−13とも高親和性で結合する可溶性NR4突然変異体が選択されるならば、これはIL−4とIL−13の両方の有用なアンタゴニストとなるであろう。
【0093】
可溶性受容体の利用のもう一つは、NR4に対するモノクローン抗体のパネルを作ることである。機能的IL−4受容体と機能的IL−13受容体の両方の形成における決定的な事象である、NR4とIL−4受容体α−鎖との相互作用を阻止する抗体が得られるならば、そのときは、両方のサイトカインの作用がさらに阻止される。
【0094】
一つの特殊な態様においては、本発明は生物学的試料中のIL−4のレベルを監視する方法を提供する。該方法はNR4とIL−4受容体α−鎖を発現する細胞と該生物学的試料を、IL−4がその受容体と結合するのを競争的に阻害するのに有効な量のIL−13と共にインキュベートする工程および競争的阻害の程度を測定する工程とを含んで成る方法である。
【0095】
本発明の関連する態様においては、生物学的試料中のIL−13のレベルを監視する方法を提供する。該方法はNR4とIL−4受容体α−鎖を発現する細胞と該生物学的試料を、IL−13がその受容体と結合するのを競争的に阻害するのに有効な量のIL−4と共にインキュベートする工程および競争的阻害の程度を測定する工程とを含んで成る方法である。
【0096】
このサイトカインは上述のようにレポーター分子で標識することが好ましい。
【0097】
この生物学的試料には、血液、血清、血漿、組織液、組織抽出物、リンパ液、T細胞またはその抽出物、培養上清および培養条件化培地などが含まれるが、これらに限定されるものではない。
【0098】
本発明はNR4遺伝子またはその誘導体を検出するためPCR分析を含むような遺伝子試験をも提供する。別の方法または関連して用いられる方法には、一本鎖コンホーメーション多形性分析(SSCP)、特異的オリゴヌクレオチドハイブリッド形成、直接タンパク切断テストのような方法などの直接ヌクレオチド配列決定法または突然変異スキャニングが含まれる。このような遺伝子テストは、例えば、疾病状態をもたらすNR4の非−発現または実質的な発現の欠如または突然変異型の発現について動物(例えば、ヒト)の遺伝子をスクリーニングする場合に重要である。
【0099】
本発明の核酸分子はDNAでもRNAでもよい。核酸分子がDNA型であるときは、それはゲノムDNAまたはcDNAであり得る。本発明の核酸分子のRNA型は一般にmRNAである。
【0100】
本発明の核酸分子は一般に単離型であるが、それはベクター分子そして特に発現ベクター分子などの他の遺伝子分子中に組み込まれたりまたはそれに連結されたりまたはそれと別な仕方で融合したりまたはそれと集合状態にあったりすることができる。ベクターおよび発現ベクターは一般に複製することができ、そして適用可能なときは、原核細胞または真核細胞の一方または両方の中で発現する。原核細胞は大腸菌、バチルス・スピーシーズおよびシュードモナス・スピーシーズを含むことが好ましい。真核細胞は酵母、カビ、哺乳類および昆虫の細胞を含むことが好ましい。
【0101】
従って、本発明の別の側面はベクター部分と哺乳類の、より具体的にはヒトのNR4遺伝子部分を含む遺伝子構築物を提供する。このNR4遺伝子部分はNR4ポリペプチドまたは機能的または免疫学的に相互作用するその誘導体をコードすることができるものである。
【0102】
遺伝子構築物のNR4遺伝子部分は、適当な細胞中でプロモーターが該NR4遺伝子部分の発現を指令することができるようにベクター上の該プロモーターに機能的に連結させることが好ましい。
【0103】
また、遺伝子構築物のNR4遺伝子部分はグルタチオン−S−トランスフェラーゼまたはその部分またはサイトカインまたは別のヘモポエチン受容体をコードするヌクレオチド配列などの別の遺伝子配列と融合した遺伝子の全部または一部を含むことができる。ハイブリッド受容体分子は多機能性治療薬および診断薬の開発において特に有用である。
【0104】
本発明はこのような遺伝子構築物を包含しそして該構築物を含む原核細胞または真核細胞をも包含する。
【0105】
本発明はNR4の変異体、一部、断片、部分、同族体およびアナログを含むNR4のすべての誘導体またはそれらをコードする遺伝子配列に及ぶ。この遺伝子配列には、天然に生ずるヌクレオチド配列またはアミノ酸配列において1個または多重のヌクレオチドまたはアミノ酸が置換、付加および/または欠失することにより生ずる遺伝子配列も包含される。
【0106】
本発明のNR4およびその遺伝子配列はある範囲の治療薬および診断薬の創薬に有用でありそして対応するリガンドの検出にとりわけ有用である。例えば、組換えNR4は同定可能なシグナルを形成することができるレポーター分子に結合または融合し、リガンドを含むと考えられる生物試料と接触させ、そして標識化NR4のリガンドへの結合についてスクリーニングをすることができる。また、標識化NR4は、リガンド遺伝子と考えられるものまたはその機能的部分の発現ライブラリーをスクリーニングするために使用することができる。
【0107】
別の態様では、NR4がまず固定化される。この態様によれば、リガンドと考えられるものを含む生物試料を、固体支持体に固定化されたヘモポエチン受容体またはそのリガンド結合部分と、該生物試料中にリガンドが存在するときは該受容体と該リガンドとの間で複合体が形成されるのに十分な時間および条件の下で接触させる工程、および結合したリガンドを溶出しそしてそれを単離する工程を含んで成る方法が提供される。
【0108】
可溶性NR4ポリペプチドも種々のハイブリッドと同様に神経系における疾病、障害または異常の治療において有用であると考えられる。例えば、中枢神経系または末梢神経系に関して脳性麻痺、外傷により誘発される麻痺、発作と関連する血管虚血症、ニューロン腫瘍、運動ニューロン病、パーキンソン病、ハンチントン病、アルツハイマー病、多発性硬化症、糖尿病と関連する末梢神経症、重金属またはアルコール毒性、腎不全、およびヘルペス、風疹、麻疹、チキンポックス、HIVまたはHTLV−1などの感染症の治療である。NR4ポリペプチドおよびハイブリッドはサイトカイン活性を調節したり、造血を調節したりするのにも重要である。これらは慢性関節リューマチなどの他の炎症性症状と同様にアレルギー性またはアトピー性症状の治療にも重要である。
【0109】
上に述べたように、本発明のNR4またはそのリガンドまたはその機能的誘導体は1以上の薬学的に許容され得る担体および/または希釈剤と共に薬学的組成物中に提供され得る。さらに、本発明は本発明のNR4の有効量の投与を含む治療方法を提供する。本発明はNR4のアンタゴニストおよびアゴニストおよび/またはそのリガンドをも包含しそして治療用の組成物および治療の方法論におけるそれらの使用をも包含する。
【0110】
本発明のさらなる側面はNR4に起因する欠陥状態または欠損状態の治療用の医薬品の製造におけるNR4またはその機能的誘導体の使用を提供する。
アミノ酸残基に対する下記の1文字略号および3文字略号が本明細書で使用される。
本発明については下記の非−制限的な図および実施例を参照してさらに記載する。
【0111】
実施例1
NR4をコードするゲノムDNAおよびcDNAの単離
胚幹細胞株から抽出されたApoI消化ゲノムDNAをλZAPIIバクテリオファージ(ストラタジーン、LaJolla,CA)の中にクローニングした。このライブラリーから得た約106プラークを配列Trp−Ser−Asp−Trp−Ser(16)に対応する32P標識オリゴヌクレオチドでスクリーニングした。ハイブリッド形成陽性のクローンを自動化DNAシークェンサー(アプライド・バイオシステムズ、Foster City、CA)を用い製造者の指示に従って配列決定を行った。一つのクローンがヘモポエチン受容体ファミリーの新規なメンバーの一部をコードするように見えた。このゲノムDNA配列に基づいてオリゴヌクレオチドを設計しそしてマウス腹膜マクロファージ(クロンテク・ラボラトリーズ、PaloAlto、CA)の、マウス皮膚の、マウス肺の、マウス腎のそしてWEHI−3B(ストラタジーン、LaJolla、CA)のλ−バクテリオファージcDNAライブラリーからクローンを単離するために通常の方法で使用した。
【0112】
実施例2
発現ベクターの構築および細胞のトランスフェクション
PCRを使用して、NR4の成熟コード領域(Thr27からPro424まで、図1)の前にあるIL−3シグナル配列(配列番号:5)およびN−末端FLAGエピトープ−標識(配列番号:6)をコードするNR4cDNAの誘導体を作成した。このPCR産物を哺乳類発現ベクターpEF−BOS(17)の中にクローニングした。使用に先立ち、構築物の全体の配列決定を行った。細胞をトランスフェクトし、そして既に述べられたように選択した(16、18)。
【0113】
実施例3
ノーザン・ブロット
ノーザン・ブロットは既に記載されたように行った(16)。ハイブリッド形成用プローブの起源は次のものである。すなわち、NR4については、ヌクレオチド32〜984(図1)からのPCR産物であり、そしてGAPDHに対しては、cDNA断片スパンニングヌクレオチド(19)である。
【0114】
実施例4
サイトカイン類と放射性ヨード化サイトカインの使用
IL−2、IL−4、IL−7、IL−9、IL−13、およびIL−15は市販品を使用した(R & Dシステムズ、Minneapolis MN)。放射性ヨード化のために、10mMリン酸ナトリウム、150mM塩化ナトリウム(PBS)、0.02%v/vトゥイーン20および0.02%w/vアジ化ナトリウム、pH7.4に、サイトカインを100μg/mlの濃度で溶解した。IL−13の2μgをヨードモノクロリド法(20、21)を用いて放射性ヨード化し、一方IL−4の2μgをジ−ヨード−ボルトン−ハンター試薬(16)を用いて放射標識した。結合の研究および比放射活性の測定および標識化サイトカインの結合可能性については既に記載されたように行った(2)。
【0115】
架橋実験のために、組換えネズミIL−13をFLAG標識化タンパクとしてピチア・パストリス中で製造した。
【0116】
架橋試験のために、精製した可溶性IL−13Rα(NR4)の一定量を競争剤の存在下または非存在下に最終容量20μlで少なくとも30分間40℃で125I−IL−13とインキュベートした。次いで、0.02%v/vトゥイーン20を含むPBS中のBS3(スベリン酸ビス(スルホスクシミジル)の12mM溶液の5μlを添加し、そして混合物を4℃で30分間インキュベートした。試料を4×SDS試料緩衝液の8μlと混合しそして13%w/vSDS−PAGEにより非還元条件下で分析した。ゲルを乾燥しそしてオートラジオグラフィーまたはホスフォイメージャー(PhosphoImager)のいずれかで可視化した。
【0117】
実施例5
増殖試験
サイトカインに応じて得られるBa/F3細胞およびCTLL細胞の増殖をラックス60ミクロ穴HL−Aプレート(ヌンク・インク、IL、USA)中で測定した。細胞を、20%v/v新生児ウシ血清を含むDMEM中で3回洗浄し、同培地に2×104細胞/mlの濃度で再懸濁した。この細胞懸濁液の10μl部分を5μlの種々の濃度の精製組換えサイトカインと共に培養穴の中に入れた。空気中10%v/vCO2を含む完全加湿インキュベーター中で37℃で2日間インキュベートした後、生存細胞を倒立顕微鏡を用いて計測した。
【0118】
実施例6
ネズミNR4のクローニングと特性決定
胚幹細胞由来のゲノムDNAのApoI消化物を用いてλZAPII中に一つのライブラリーを構築しそしてヘモポエチン受容体ファミリーの多くのメンバーに見出されるアミノ酸配列Trp−Ser−Asp−Trp−Serをコードする32P標識化オリゴヌクレオチドのプールでスクリーニングした。ハイブリッドを形成する一つのバクテリオファージクローンがヘモポエチン受容体ファミリーの新規なメンバーの一部をコードするように見える配列を含むことが見出された。この受容体はNR4という機能的名称が与えられた。このゲノムクローンの配列を使用して、WEHI−3B細胞の、腹膜マクロファージの、骨髄の、皮膚の、そして腎のライブラリーからNR4をコードするcDNAを単離した。これらのcDNAの合成物であるヌクレオチド配列(配列番号:1)および予言されたアミノ酸配列(配列番号:2)を図1に示す。このNR4cDNAは424のアミノ酸残基からなるタンパクをコードすることを予言する。このタンパクの中には、シグナル配列および膜貫通ドメインと思われるものが含まれる。このタンパクの細胞外領域は、保存された4個のシステイン残基と特徴的なTrp−Ser−Asp−Trp−Serモチーフ(図1、WSXWSとして太字で記載)を含む典型的なヘモポエチン受容体ドメイン(アミノ酸118〜340)に加えて、免疫グロブリン−様ドメイン(アミノ酸27〜117)を含んでいた。この新規な受容体の細胞質テール(cytoplasmic tail)は長さが60アミノ酸であった。
【0119】
実施例7
NR4cDNAの発現パターン
NR4mRNAの発現パターンをノーザン分析で検討した。5.2kbおよび2.2kbの2個のハイブリッド形成物を多くの組織由来のmRNA中に検出した(図2)。NR4mRNAは骨格筋には検出できなかった(図2)。図8はマウス組織内のNR4の発現を示す。
【0120】
実施例8
NR4はIL−13受容体の特異的結合サブユニットであるIL−13受容体α−鎖(IL−13Rα)をコードする
COS細胞で発現したNR4を抗−FLAG抗体を用いるウエスタン・ブロットから評価すると、見掛けの分子量は約50,000から約70,000ダルトンであり、より具体的には約55,000から約65,000である。これはNR4がIL−13受容体の結合性サブユニットをコードするかも知れないということを示唆した。この可能性をテストするため、NR4をCOS細胞で発現させた。トランスフェクトされていないCOS細胞は比較的低レベルのIL−4受容体およびIL−13受容体を発現した。NR4cDNAを含むプラスミドをトランスフェクトすると、COS細胞によって発現されるIL−13の数は劇的に増加した(図3A、細胞当たり100,000から500,000まで)が、IL−4受容体の数は増加しなかった。COS細胞により発現されたNR4に対するIL−13の親和性は低かった(KDは約2〜10nM)そしてこのNR4は非標識IL−13と競争できた(図4A)がIL−2、IL−4、IL−7、IL−9またはIL−15を含む他のサイトカインとは競争できなかったから、その結合は特異的であった。これらの結果から、NR4がIL−13受容体α−鎖(IL−13Rα)であることが示唆された。
【0121】
実施例9
IL−13Rα(NR4)およびIL−4RαはIL−4受容体およびIL−13受容体の成分を共有する。
IL−4受容体とIL−13受容体の間の関係を研究するため、IL−4に応答する細胞株CTLLを検討した。親のCTLL細胞は1個のクラスのIL−4受容体(KD約660pM、約3600受容体/細胞)を発現したがIL−13受容体は検出されなかった(図3B)。CTLL細胞によって発現されたIL−4受容体は、125I−IL−4の結合が非標識IL−4と競争できたがIL−13とは競争できなかったから(図4B)、特異的であるように見えた。CTLL細胞中でIL−13Rα(NR4)が発現する際、IL−4受容体の数および親和性には変化がみられなかったが、一方、高親和性IL−13受容体の1個のクラスが検出された(図3C、KD約75pM、1350受容体/細胞)。CTLL細胞内で発現したIL−13Rα(NR4)に対するIL−13の親和性はCOS細胞中よりも高かった。これは前者がIL−13Rα(NR4)と相互作用しIL−13に対する親和性を増強することができるあるタンパクを発現したことを示唆する。以前の研究に基づく同様な候補はIL−4Rαである。この可能性を解明するため、IL−13Rα(NR4)を発現するCTLL細胞への結合に対し125I−IL−13と競争するIL−4の能力を評価した。図4Bは125I−IL−13の結合に対する競争ではIL−4とIL−13は同等に効果的であった(IC50は約300pM、図4C)のであり、そしてさらにこれらは125I−IL−4とも結合に対して競争できたのであった(IC50は約300pM、図4D)。
【0122】
実施例10
IL−13Rα(NR4)はIL−13による増殖性シグナルの形質導入に必要である。
CTLL細胞は生存および増殖に外因性サイトカイン類の添加を必要とする。
IL−2はCTLL細胞に対し強力な増殖性刺激を有する(EC50は約100〜200pM)一方、IL−4は比較的弱く(EC50は2〜7nM)そしてIL−13は不活性であった(図5A)。CTLL細胞中にIL−13Rα(NR4)が発現することにより、IL−13に応答して(EC50は約700pM)弱く生き残りそして増殖する能力が、そしてIL−4に応答して(EC50は約700pM、図5B)親細胞よりもややより強く増殖する能力が生じた。
【0123】
実施例11
ヒトIL−13Rα(NR4)のクローニング
ネズミIL−13Rα(NR4)に相同な遺伝子が他の脊椎動物種に存在するか否かを決定するため、ネズミIL−13Rα(NR4)のヌクレオチド840〜1270を包含するプローブを、EcoRIで消化した種々の種由来のゲノムDNAとハイブリッド形成させた。ハイブリッド形成は500mMリン酸水素二ナトリウム(約5×SSC)中50℃で一晩行った。フィルターを40mMリン酸水素二ナトリウム(約0.2×SSC)中50℃で2時間洗浄し、オートラジオグラフィー様フィルムに48時間曝した。図6はヒトを含む種々の種由来のゲノムDNAの中に比較的少数(1〜5)のハイブリッド形成バンドが観察されることを示す。これはネズミcDNAプローブを使用してヒトIL−13Rα(NR4)をクローニングすることが容易であることを示唆する。従って、λZAPIIバクテリオファージ中のヒト骨髄cDNAライブラリーのクローンをネズミIL−13Rα(NR4)cDNA由来の二つのプローブ(ヌクレオチド82〜840およびヌクレオチド840〜1270)でスクリーニングした。ハイブリッド形成は6×SSC、0.1%w/vSDS中42℃で一晩行った。フィルターを2×SSC:0.1%w/vSDS中50℃で2時間洗浄しそしオートラジオグラフィー様フィルムに48時間曝した。両方のネズミIL−13Rα(NR4)プローブとハイブリッドを形成したプラークを釣り上げ、通常の方法で精製した。ハイブリッドを形成するバクテリオファージ由来のこのcDNA挿入物をpBluescript プラスミドの中に切り取り、そしてABI自動化シークェンサーを用いてその全体の配列決定を行った。図7は単離されたクローンの配列の合成体を示し、そしてこれらのクローンがネズミIL−13Rα(NR4)と高度の配列類似性を共有する一つのタンパクをコードすることを明らかにする。これらのクローンはこのタンパクの全コード領域をコードする。高度の配列類似性(320/425アミノ酸、約75%)はこのcDNAがネズミIL−13Rα(NR4)のヒト同族体であることを予言する。ヌクレオチド配列は配列番号:3として表されそしてそのアミノ酸配列は配列番号:4である。
【0124】
実施例12
可溶性ネズミIL−13Rα(NR4)はIL−13と結合する
N−末端「FLAG」エピトープ(インターナショナル・バイオテクノロジーズ/イーストマン・コダック、New Haven CT)を持つNR4の可溶性バージョンを発現するような構築物を構築した。まず、哺乳類発現ベクターpEF−BOSの誘導体を、それがネズミIL−3のシグナル配列(MVLASSTTSIHTMLLLLLMLFHLGLQASIS(配列番号:5))をコードするDNAおよびFLAGエピトープ(DYKDDDDK(配列番号:6))を含みそれに続いてユニークなXbaIクローニング部位を含むように作成した。このベクターをpEF/IL3SIG/FLAGと名付けた。成熟NR4コード領域(Thr27〜Thr344)の細胞外部分はプライマー1478および1480を用いるPCRによって作成した。生じた産物をXbaIで消化しそしてpEF/IL3SIG/FLAGのXbaI部位にクローニングしてpEF/IL3SIG/FLAG/solNR4を得た。この構築物の同定はジデオキシ配列決定法により確認された。
オリゴ 1478 5'AGCTTCTAGAACAGAAGTTCAGCCACCTGTG3'〔配列番号:7〕、
オリゴ 1480 5'AACTCCACCTTCTACACCACCTGATCTAGA3'〔配列番号:8〕
【0125】
CHO細胞中にトランスフェクトし、発現させた後、抗−FLAG抗体(M2)アフィニティカラム上でCHO細胞コンディション化培地から可溶性NR4を吸着させ遊離のFLAGペプチド(サイエンス・イメージング・システムズ)で溶出することにより精製した。
【0126】
COS細胞により発現されたNR4に対しIL−13の親和性が低いことと一致して、精製した可溶性NR4はゲルろ過クロマトグラフィーで評価するとIL−13と結合することができないようであった。しかしながら、感度の高い架橋試験を用いると、可溶性IL−13Rα(NR4)のIL−13に結合する能力が証明された(図8、レーン1)。この相互作用は非標識IL−13により競争されたが非標識IL−4によっては競争されることはなかった(図8、レーン2およびレーン3)。
【0127】
実施例13
可溶性IL−13Rα(NR4)に対するポリクローン抗血清
精製した可溶性NR4をウサギに注射し3ヵ月後に採血してNR4に対するポリクローン抗血清を調製した。この抗血清は125I−IL−13の可溶性NR4との架橋産物を免疫沈降させた(図9、レーン11)が、免疫化前(pre-immune)の血清では免疫沈降は観察されなかった(図9、レーン9)。複合体の免疫沈降はFLAGペプチドによっては阻害されなかった(図9、レーン10)。
【0128】
免疫沈降反応試験は次のように行われた。
【0129】
架橋反応はトリス−塩酸、pH7.5を最終濃度40mMになるまで添加することにより終了させた。次いで、試料を1:50希釈の対照ウサギ血清またはFLAGペプチドの存在下にまたは非存在下に予めインキュベートしておいた抗−NR4血清と混合した。4℃で30分間インキュベートした後、混合物に50%v/vタンパク質G−セファロースゲルスラリー(ファルマシア)の40μlを添加しそして4℃で30分間インキュベートした。試料を遠心分離しタンパク質Gビーズを3×0.5mlのPBSで洗浄し、2×濃縮SDS−PAGE試料緩衝液の40μlと混ぜ、95℃で2分間加熱した。この上清を非還元条件下で13%w/vSDS−PAGEにより分析した。
【0130】
実施例14
NR4のN−末端アミノ酸配列
NR4のN−末端アミノ酸配列を決定した。これを図10に示す。
【0131】
実施例15
IL−13の試験
IL−13は喘息などのアレルギー性疾患において重要な免疫グロブリンイソタイプであるIgEの産生に関与するサイトカインである。IL−13のレベルを監視することは、従って、重要な診断学的意味がある。IL−4とIL−13は多くの生物学的効果を共有しているから、これらのサイトカインを識別する試験法を作成することも重要である。
【0132】
COS細胞で発現したNR4は125I−IL−13と結合する。この結合はウシ血清やヒト血清などの無関係のタンパクの大量の存在下で非標識IL−13により投与量依存的に阻害される。IL−4はこの状況下では125I−IL−13結合に対して競争する能力を示さない、従って、この試験はIL−13に対して特異的であるように見える。
【0133】
この試験法は可溶性NR4をELISAプレート上に被覆し、そして、例えば、蛍光標識IL−13をプローブとして使用することにより組立てられる。次いで、試験用試料中の非標識IL−13の存在が蛍光シグナルの減少として記録される。
【0134】
NR4およびIL−4受容体α−鎖を発現する細胞を用いることにより、IL−4およびIL−13の両方を測定する同様な試験法が組み立てられる。これらの細胞には正常にIL−4受容体α−鎖を発現しそしてNR4を発現するように工学的に処理されるCTLL細胞が含まれる。125I−IL−13または125I−IL−4の結合はIL−4およびIL−13の両方の非標識型により阻害することができる。
【0135】
実施例16
IL−4およびIL−13の修飾
機能的に重要であると予測される分子の領域に突然変異を導入する。NR4の場合には、この領域は、このサイトカインがIL−4受容体α−鎖に結合するときは、IL−13と相互作用する領域、IL−4受容体α−鎖と相互作用する領域またはIL−4と相互作用する領域を含む。これらの領域は直接の実験、例えば、NR4の構造またはIL−4、IL−13およびIL−4受容体α−鎖のような他のタンパクとNR4との複合体の構造を解明することにより、またはその構造に関する情報が存在する類似のタンパク、例えば、成長ホルモン/成長ホルモン受容体の複合体に基づきこれらのタンパクをモデル化することにより決定される。次いで、得られるNR4突然変異体を機能の改善の有無につき個々にテストする。
【0136】
別の方法では、分子内に無差別突然変異が作られる。適当な技術としては、正しくないdNTPの取り込みを促進するようなポリメラーゼや反応条件を使用するNR4cDNAの合成や、DNAシャフリングと呼ばれる技術が挙げられる(23、24、25、26)。
【0137】
目的のcDNAの無差別突然変異体を作成した後、潜在的に有用な突然変異体を選別する。NR4の場合には、一つの試験法は、もしNR4がIL−4受容体α−鎖を欠如している細胞(例えば、COS細胞)の中で発現されるときは、IL−4とは如何なる検出感度でも結合できないがIL−13とは低親和性で結合する細胞が得られるという知識に基づくものである。このようにして、もしCOS細胞をNR4でトランスフェクトし、そしてFITC−結合IL−4およびフィコエリトリン−結合IL−13と結合させ、未結合リガンドを洗い流すものとすれば、IL−4は全く結合せずそして結合したIL−13は洗浄の間に解離するであろう。
【0138】
これらの細胞をFACS−選別するときは、FITC−チャンネルにもPE−チャンネルにもシグナルはほとんどあるいは全く得られないであろう。COS細胞をNR4の無差別突然変異体106〜107でトランスフェクトしそして結合試験のために処理する。野生のNR4でトランスフェクトされたものよりも強くサイトカインに結合するとして選別される細胞はすべてFACSで選別することができる。これらの「改良された」NR4cDNAを含むプラスミドを回収し、大腸菌で増やし、そして再度COS細胞を用いてその改良度を確認する。一貫して改良された突然変異体は次にさらにもう一度無差別変化の導入のために使用してさらに改良された分子を得ることができる。この反復プロセスを数回繰り返してもよい。
【0139】
当技術分野で熟練した者は本明細書に記載された発明が具体的に記載されたこと以外の変更や修正を受け得るものであることを認めるであろう。本発明はそのような変更や修正のすべてを包含することが理解されるべきである。本発明はまた本明細書で個々にあるいはまとめて言及しあるいは指摘した工程、特徴、組成物および化合物のすべてを、そして該工程あるいは特徴のいかなる2以上の組み合わせをも包含する。
【0140】
引用文献
1.デュー,エックス.エックス.およびウィリアムス,ディー.エイ.(1994)Blood 83 : 2023-2030。
2.ヤング,ワイ.シー.およびイン,ティー.(1992)Biofactors 4 : 15-21。
3.ポール,エス.アール.,ベネット,エフ.,カルベッティ,ジェイ.エイ.,ケレハー,ケイ., ウッド,シー.アール.,オオハラ,アール.ジェイ.ジェイ.,レアリー,エイ.シー.,シブリー,ビー.,クラーク,エス.シー.,ウィリアムス,ディー.エイ.およびヤング,ワイ.シー.(1990)Proc.Natl.Acad.Sci.USA 87: 7512。
4.ムサシ,エム.,クラーク,エス.シー.,スドー,ティー.,ウーダル,ディー.エル.およびオガワ,エム.(1991)Blood 78: 1448-1451。
5.シブラー,ケイ.アール.,ヤング,ワイ.シー.およびクリステンセン,アール.ディー.(1992)Blood 80: 900-3。
6.ツジ,ケイ.,リーマン,エス.ディー.,スドー ,ティー.,クラーク,エス.シー.およびオガワ,エム.(1992)Blood 79: 2855-60。
7.バーンスタイン,エス.エイ.,メイ,アール.エル.,ヘンソーン,ジェイ.,フリース,ピー.およびターナー,ケイ.(1992)J.Cell.Physiol.153 : 305-12。
8.ハンゴク,ジー.,イン,ティー.,クーパー,エス.,シェンデル,ピー.,ヤング,ワイ.シー.およびブロックスメイヤー,エイチ.イー.(1993)Blood 81: 965-72。
9.テラムラ,エム.コバヤシ,エス.,ホシノ,エス.,オシミ,ケイ.およびミゾクチ,エイチ.(1992)Blood 79: 327-31。
10.ヨネムラ,ワイ.,カワキタ,エム.,マスダ,ティー.,フジモト,ケイ.,カトウ,ケイ.およびタカツキ,ケイ.(1992)Exp.Hematol.20: 1011-6。
11.バウマン,エイチ.およびシェンデル,ピー.(1991)J.Biol.Chem.266 : 20424-7。
12.カワシマ,アイ.,オオスミ,ジェイ.,ミタ−ホンジョウ,ケイ.,シモダ−タカノ,ケイ.,イシカワ,エイチ.,サカキバラ,エス.,ミヤダイ,ケイ.およびタキグチ,ワイ.(1991)Febs.Lett. 283: 199-202。
13.ケラー,ディー.シー., デュー,エックス.エックス.,スローアー,イー.エフ.,ホフマン,アール.およびウィリアムス,ディー.エイ.(1993)Blood 82 : 1428-35。
14.マッケンジー,エイ.エヌ.ジェイ.およびズラウスキー,ジー.(1994)サイトカインおよびその受容体へのガイドブック,オクスフォード大学出版部,オクスフォード。
15.ズラウスキー,ジー.およびド フリース,ジェイ.イー.(1994)Immunol.Today 15: 19-26。
16.ヒルトン,ディー.ジェイ.,ヒルトン,エイ.エイ.,ライセビック,エイ.,ラカー,エス.,ハリソン−スミス,エム.,ゴウ,エヌ.エム.,ベグリー,シー.ジー.,メトカーフ,ディー.,ニコラ,エヌ.エイ.およびウイルソン,ティー.エイ.(1994)EMBO J.13: 4765-4777。
17.ミズシマ,エス.およびナガタ,エス.(1990)Nucleic Acids Research18; 5322。
18.ロック,ピー.,メトカーフ,ディー.およびニコラ,エヌ.エイ.(1994)Proc.Natl.Acad.Sci.USA 91: 252-256。
19.デゥガイシク,エイ.ら(1983) Biochemistry 22 : 1605-1613。
20.コントレラス,エム.エイ.,ベイル,ダブリュー.エフ.およびスパー,アイ.エル.(1983)Methods in Enzymol.92 : 277-292。
21.ヒルトン,ディ.ジェイ.およびニコラ,エヌ.エイ.(1992)J.Biol Chem.267 : 10238-10247
22.サンブルック,ジェイ.,フリッチュ,イー.エフ.およびマニアティス,ティー.(1989)モレキュラー クローニング:ア ラボラトリー マニュアル,コールド・スプリング・ハーバー・ラボラトリー,コールド・スプリング・ハーバー,ニューヨーク。
23.ステンマー,ダブリュー.ピー.シー.,(1994)Nature 370: 389-391。
24.ステンマー,ダブリュー.ピー.シー.,(1995)Biotechnology 13 : 549-553。
25.ガスマンら(1995)Proc.Natl.Acad.Sci.USA 92 : 1292-1296。
26.グラメリら(1996)Nature Biotechnology 14 : 315-319。
【図面の簡単な説明】
【0141】
【図1−1】図1はネズミNR4の配列番号:1に記載のヌクレオチド配列および配列番号:2に記載の予言されるアミノ酸配列を示す。非翻訳部分は下方の箱の中にそして翻訳部分は上方の箱の中に示してある。アミノ酸については、通常の1文字符号が略号が用いられ、グリコシレーション部位にリンクすると考えられるアスパラギンには下線を付し、保存されているシステイン残基とヘモポエチン受容体ファミリー・メンバーのWSXWSモチーフは太字で示してある。予想されるシグナル配列は太字で下線を付し、一方膜貫通ドメインはダッシュで下線を付している。示した配列は3種のライブラリー由来の8個のcDNAクローンの分析から得られたものの合成物である。この配列の5’−末端(ヌクレオチド−60から351まで)は1個のcDNAクローンから由来するが、単離されたゲノムDNAクローン中にも存在する。箱領域は:典型的なヘモポエチン受容体ドメインであり、アミノ酸118〜340である。
【図1−2】図1−2は図1−1の続きを示す図である。
【図1−3】図1−3は図1−2の続きを示す図である。
【図1−4】図1−4は図1−3の続きを示す図である。
【図1−5】図1−5は図1−4の続きを示す図である。
【図1−6】図1−6は図1−5の続きを示す図である。
【図1−7】図1−7は図1−6の続きを示す図である。
【図2】図2は選択された組織および器官でのネズミNR4mRNA発現のノーザン分析を示す写真である。
【図3】図3は125I−IL−13および125I−IL−4結合の飽和等温線、すなわち、IL−4(○)およびIL−13(●)の(A)IL−13Rα(NR4)を発現するCOS細胞への結合、(B)CTLL細胞への結合、そして(C)IL−13Rα(NR4)を発現するCTLL細胞への結合のスカチャードプロットとして図示された飽和等温線を示すグラフである。データは1×104COS細胞および1×106CTLL細胞に規格化し、そして結合は2〜4時間氷上で行った。
【図4】図4はIL−4とIL−13の結合、すなわち、IL−4(○)およびIL−13(●)が125I−IL−13の(A)IL−13Rα(NR4)を発現するCOS細胞への結合および(C)IL−13Rα(NR4)を発現するCTLL細胞への結合と競争する能力、または125I−IL−4の(B)CTLL細胞への結合および(D)IL−13Rα(NR4)を発現するCTLL細胞への結合と競争する能力、の特異性を示すグラフである。結合は4℃で2〜4時間行われそしてデータは競争物質(△)の横座標で観察された特異的結合のパーセンテージとして表された。
【図5】図5はNR4を発現する細胞の因子依存性増殖を示すグラフである。(A)CTLL細胞または(B)IL−13Rα(NR4)を発現するCTLL細胞の200個をサイトカインの非存在下にまたは種々の濃度のIL−2(□)、IL−4(○)またはIL−13(●)と共にインキュベートした。48時間後に生存細胞を計測し、データをIL−2の最高濃度で観察された生存数のパーセンテージとして表した。
【図6】図6はNR4(IL−13Rα)遺伝子の種間保存を示す写真である。
【図7−1】図7はネズミおよびヒトのNR4(IL−13Rα)遺伝子のヌクレオチド配列と対応するアミノ酸配列の表示である。ヒト(H)およびネズミ(M)のIL−13Rα(NR4)のヌクレオチド配列および予言されるアミノ酸配列を眼で配列しそして配列が最適となるようにギャップ(−)を挿入して配列した。ネズミクローンについての数字付けは、コード領域の部分を形成するヌクレオチドは上方の箱に、一方非翻訳領域のヌクレオチドは下方の箱に示してある。ネズミの予言されたタンパクとヒトの予言されたタンパクの間で同一なアミノ酸は(*)で指摘してある。ネズミのシグナル配列をコードするDNAは下線を付してあり、成熟タンパクの予言された第1アミノ酸はA26またはT27である。
【図7−2】図7−2は図7−1の続きを示す図である。
【図7−3】図7−3は図7−2の続きを示す図である。
【図7−4】図7−4は図7−3の続きを示す図である。
【図7−5】図7−5は図7−4の続きを示す図である。
【図7−6】図7−6は図7−5の続きを示す図である。
【図7−7】図7−7は図7−6の続きを示す図である。
【図7−8】図7−8は図7−7の続きを示す図である。
【図7−9】図7−9は図7−8の続きを示す図である。
【図7−10】図7−10は図7−9の続きを示す図である。
【図7−11】図7−11は図7−10の続きを示す図である。
【図8】図8は可溶性NR4に架橋している125I−IL−13を示す写真である。レーン:125I−IL−13(100,000cpm)+2μg/ml可溶性NR4、レーン2:過剰の非標識IL−13の存在下における125I−IL−13(100,000cpm)+2μg/ml可溶性NR4、レーン3:過剰の非標識IL−4の存在下における125I−IL−13(100,000cpm)+2μg/ml可溶性NR4。
【図9】図9はIL−13Rα(NR4)と架橋している125I−IL−13の抗−NR4ポリクローン抗血清による免疫沈降反応の写真である。レーン9〜11:125I−IL−13(750,000cpm)に架橋している可溶性IL−13Rα(3μg/mlの30μl)、およびそれぞれ、対照のウサギ血清で沈降させたもの、または100μg/m1FLAGペプチドの存在下または非存在下において抗−NR4ポリクローン抗血清で沈降させたものである。レーン12〜14:0.5μg/mlの非標識IL−13の存在下における125I−IL−13(750,000cpm)に架橋している可溶性IL−13Rα(NR4)(3μg/mlの30μl)、およびそれぞれ100μg/mlのFLAGペプチドの存在下または非存在下に抗−IL−13Rα(NR4)ポリクローン抗血清で沈降させたものである。
【図10】図10はネズミNR4のN−末端アミノ酸配列の表示である。
【特許請求の範囲】
【請求項1】
動物由来のヘモポエチン受容体または該受容体の誘導体をコードするヌクレオチド配列またはコードする配列に相補的なヌクレオチド配列を含む単離された核酸分子。
【請求項2】
動物ヘモポエチン受容体またはその誘導体をコードするヌクレオチド配列またはコードする配列に相補的なヌクレオチド配列を含む単離された核酸分子であって、該受容体が
(i)IL−13またはその誘導体と相互作用することができるものであり、かつ
(ii)IL−4とIL−4受容体α−鎖の間の複合体と相互作用することができるものである、
核酸分子。
【請求項3】
該受容体がIL−13と低親和性で相互作用することができるヘモポエチン受容体のα−鎖の誘導体を含むものである、請求項1または請求項2記載の単離された核酸分子。
【請求項4】
該受容体がIL−13と中から高親和性で相互作用するこことができるヘモポエチン受容体のα−鎖の誘導体である、請求項1または請求項2記載の単離された核酸分子。
【請求項5】
配列番号:2または配列番号:4に実質的に記載されたアミノ酸配列またはその全部または一部と少なくとも50%の類似性を持つアミノ酸配列を持つ受容体をコードする、請求項1または請求項2記載の単離された核酸分子。
【請求項6】
配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列またはその全部または一部と少なくとも約50%の類似性を持つヌクレオチド配列を含む請求項1または請求項2記載の単離された核酸分子。
【請求項7】
IL−13受容体α−鎖またはその誘導体をコードするヌクレオチド配列またはコードする配列に相補的なヌクレオチド配列を含む単離された核酸分子であって、配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列を持つものである、または機能的に類似のIL−13受容体α−鎖またはその誘導体をコードしかつ配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列またはその相補型と低厳格条件下でハイブリッドを形成することができる核酸分子を持つものである単離された核酸分子。
【請求項8】
配列番号:2または配列番号:4に実質的に記載されたアミノ酸配列を持つIL−13受容体α−鎖またはその誘導体をコードするヌクレオチド配列またはコードする配列に相補的なヌクレオチド配列を含む、または配列番号:2または配列番号:4に記載された配列に少なくとも約50%の類似性を持つアミノ酸配列をコードするヌクレオチド配列を含みかつ配列番号:1または配列番号:3に記載の配列と低厳格条件下でハイブリッドを形成することができるヌクレオチド配列を含む単離された核酸分子。
【請求項9】
IL−13またはその誘導体と相互作用することができるヘモポエチン受容体をコードする請求項1または請求項2または請求項7または請求項8記載の単離された核酸分子であって、その相互作用をIL−4受容体α−鎖を発現する細胞内でIL−4またはその誘導体により競争的に阻害することができるものである単離された核酸分子。
【請求項10】
宿主細胞中で核酸分子の発現を指令することができるプロモーターに機能的に連結された請求項1または請求項6または請求項7記載の核酸分子を含む遺伝子構築物。
【請求項11】
配列番号:2または配列番号:4に実質的に記載されたアミノ酸配列またはその全部または一部と少なくとも約50%の類似性を持つアミノ酸配列を含む組換えポリペプチドであって、IL−13またはその誘導体と相互作用することができるものである組換えポリペプチド。
【請求項12】
IL−13との相互作用がIL−4受容体α−鎖を発現する細胞内でIL−4により競争的に阻害されるものである請求項11記載の組換えポリペプチド。
【請求項13】
IL−13との相互作用が低親和性のものである請求項11記載の組換えポリペプチド。
【請求項14】
IL−13との相互作用が中から高親和性のものである請求項11記載の組換えポリペプチド。
【請求項15】
該ポリペプチドがCOS細胞で発現されたときウエスタン・ブロット分析で測定すると約50,000から約70,000ダルトンの分子量を持つものである請求項11記載の組換えポリペプチド。
【請求項16】
次の特性のうちの少なくとも二つを持つ組換えポリペプチド、
(i)配列番号:2または配列番号:4に実質的に記載されたアミノ酸配列またはその全部または一部と少なくとも約50%類似性を持つアミノ酸配列を含むこと、
(ii)配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列またはその全部または一部と少なくとも約50%の類似性を持つヌクレオチド配列によりコードされるものであること、
(iii)少なくとも低親和性でIL−13またはその誘導体と相互作用するものであること、そして
(iv)COS細胞内で発現させるときウエスタン・ブロット分析で測定すると約50,000から約70,000ダルトンの分子量を持つものであること。
【請求項17】
次の特性のうちの少なくとも三つを持つ組換えポリペプチド、
(i)配列番号:2または配列番号:4に実質的に記載されたアミノ酸配列またはその全部または一部と少なくとも約50%類似性を持つアミノ酸配列を含むこと、
(ii)配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列またはその全部または一部と少なくとも約50%の類似性を持つヌクレオチド配列によりコードされるものであること、
(iii)少なくとも低親和性でIL−13またはその誘導体と相互作用するものであること、
(iv)COS細胞内で発現させるときウエスタン・ブロット分析で測定すると約50,000から約70,000ダルトンの分子量を持つものであること、
(v)IL−4受容体α−鎖由来のアミノ酸配列を含むものであること、そして
(vi)IL−13と相互作用をすることができるものであること、かつ、その相互作用がIL−4受容体α−鎖を発現する細胞内でIL−4により競争的に阻害されるものであること。
【請求項18】
請求項16または請求項17に記載の組換えポリペプチドに対する抗体。
【請求項19】
該抗体がモノクローン抗体である、請求項16記載の抗体。
【請求項20】
少なくとも二つのサイトカインと相互作用することができるハイブリッドヘモポエチン受容体であって、該サイトカインの少なくとも一つがIL−13またはその誘導体でありそして該ハイブリッド受容体が配列番号:2または配列番号:4に記載されたアミノ酸配列の全部または一部を含むアミノ酸配列を含むものであるハイブリッドヘモポエチン受容体。
【請求項21】
少なくとも一つのサイトカインと高親和性相互作用をすることができるハイブリッドヘモポエチン受容体であって、該サイトカインの少なくとも一つがIL−13またはその誘導体でありそして該ハイブリッド受容体が配列番号:2または配列番号:4に記載されたアミノ酸配列の全部または一部を含むアミノ酸配列を含むものであるハイブリッドヘモポエチン受容体。
【請求項22】
IL−4と相互作用することができる請求項18記載のハイブリッドヘモポエチン受容体。
【請求項23】
IL−2、IL−5、IL−7、IL−9およびIL−15から選択されるサイトカインと相互作用することができる請求項21記載のハイブリッドヘモポエチン受容体。
【請求項24】
請求項16または請求項17記載の組換えポリペプチドおよび一つ以上の薬学的に許容され得る担体および/または希釈剤を含んで成る薬学的組成物。
【請求項25】
請求項1または請求項2または請求項7または請求項8に記載の核酸分子および1以上の遺伝的に許容され得る担体および/または希釈剤を含んで成る遺伝子的薬学的組成物。
【請求項26】
請求項16または請求項17記載の組換えポリペプチドの治療上有効量を動物に投与することを含む該動物の治療方法。
【請求項27】
請求項16または請求項17記載の組換えポリペプチドの治療上有効量を動物に投与することを含む該動物でのIgE産生により悪化した喘息、アレルギーまたは状態を治療する方法。
【請求項28】
次の特性のうちの少なくとも二つを持つ組換えポリペプチドを製造する方法であって、請求項10記載の遺伝子構築物を含む細胞を、該遺伝子構築物中の核酸分子を発現して組換えポリペプチドを生産するのに十分な時間および条件の下で培養する工程、および該組換えポリペプチドを単離する工程を含んで成る方法、
(i)配列番号:2または配列番号:4に実質的に記載されたアミノ酸配列またはそれに少なくとも約50%の類似性を有するアミノ酸配列を含むものであること、
(ii)配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列またはそれに少なくとも約50%の類似性を有するヌクレオチド配列によりコードされるものであること、
(iii)IL−13またはその誘導体と少なくとも低親和性で相互作用するものであること、
(iv)COS細胞で発現させたとき、ウエスタン・ブロット分析で測定すると約50,000から約70,000ダルトンの分子量を持つものであること。
【請求項29】
次の特性のうち少なくとも三つを持つ組換えポリペプチドを製造する方法であって、請求項10記載の遺伝子構築物を含む細胞を、該遺伝子構築物中の核酸分子を発現して組換えポリペプチドを生産するのに十分な時間および条件の下で培養する工程、および該組換えポリペプチドを単離する工程を含んで成る方法、
(i)配列番号:2または配列番号:4に実質的に記載されたアミノ酸配列またはそのすべてまたは一部に少なくとも約50%の類似性を有するアミノ酸配列であること、
(ii)配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列またはそのすべてまたは一部に少なくとも約50%の類似性を有するヌクレオチド配列によりコードされるものであること、
(iii)IL−13またはその誘導体と少なくとも低親和性で相互作用するものであること、
(iv)COS細胞で発現させたとき、ウエスタン・ブロット分析で測定すると約50,000から約70,000ダルトンの分子量を持つものであること、
(v)IL−4受容体α−鎖由来のアミノ酸配列を含むものであること、そして
(vi)IL−13と相互作用をすることができるものであること、かつ、その相互作用がIL−4受容体α−鎖を発現する細胞内でIL−4により競争的に阻害されるものであること。
【請求項30】
請求項28または請求項29記載の方法により製造される組換えポリペプチドを発現する動物細胞。
【請求項31】
IL−13またはその誘導体と相互作用することができる第1部分とヘモポエチン受容体、受容体チロシンキナーゼ、TNF/NGF受容体またはGタンパク質結合受容体由来の第2部分とを含んで成るキメラタンパク質。
【請求項32】
該第2部分がIL−13結合タンパク質、IL−4受容体α−鎖、IL−2受容体γ−鎖または喘息またはアレルギーに関与するサイトカインの受容体の全部または機能的部分を含むものである請求項31記載のキメラタンパク質。
【請求項33】
生物学的試料中のIL−4のレベルを監視する方法であって、NR4とIL−4受容体α−鎖とを発現する細胞およびその受容体へのIL−4の結合を競争的に阻害するのに十分な量のIL−13と該生物学的試料とをインキュベートする工程、および競争的阻害の程度を測定する工程を含んで成る方法。
【請求項34】
生物学的試料中のIL−13のレベルを監視する方法であって、NR4とIL−4受容体α−鎖とを発現する細胞およびその受容体へのIL−13の結合を競争的に阻害するのに十分な量のIL−4と該生物学的試料とをインキュベートする工程、および競争的阻害の程度を測定する工程を含んで成る方法。
【請求項35】
サイトカインがレポーター分子で標識化されているものである請求項33または請求項34記載の方法。
【請求項1】
動物由来のヘモポエチン受容体または該受容体の誘導体をコードするヌクレオチド配列またはコードする配列に相補的なヌクレオチド配列を含む単離された核酸分子。
【請求項2】
動物ヘモポエチン受容体またはその誘導体をコードするヌクレオチド配列またはコードする配列に相補的なヌクレオチド配列を含む単離された核酸分子であって、該受容体が
(i)IL−13またはその誘導体と相互作用することができるものであり、かつ
(ii)IL−4とIL−4受容体α−鎖の間の複合体と相互作用することができるものである、
核酸分子。
【請求項3】
該受容体がIL−13と低親和性で相互作用することができるヘモポエチン受容体のα−鎖の誘導体を含むものである、請求項1または請求項2記載の単離された核酸分子。
【請求項4】
該受容体がIL−13と中から高親和性で相互作用するこことができるヘモポエチン受容体のα−鎖の誘導体である、請求項1または請求項2記載の単離された核酸分子。
【請求項5】
配列番号:2または配列番号:4に実質的に記載されたアミノ酸配列またはその全部または一部と少なくとも50%の類似性を持つアミノ酸配列を持つ受容体をコードする、請求項1または請求項2記載の単離された核酸分子。
【請求項6】
配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列またはその全部または一部と少なくとも約50%の類似性を持つヌクレオチド配列を含む請求項1または請求項2記載の単離された核酸分子。
【請求項7】
IL−13受容体α−鎖またはその誘導体をコードするヌクレオチド配列またはコードする配列に相補的なヌクレオチド配列を含む単離された核酸分子であって、配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列を持つものである、または機能的に類似のIL−13受容体α−鎖またはその誘導体をコードしかつ配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列またはその相補型と低厳格条件下でハイブリッドを形成することができる核酸分子を持つものである単離された核酸分子。
【請求項8】
配列番号:2または配列番号:4に実質的に記載されたアミノ酸配列を持つIL−13受容体α−鎖またはその誘導体をコードするヌクレオチド配列またはコードする配列に相補的なヌクレオチド配列を含む、または配列番号:2または配列番号:4に記載された配列に少なくとも約50%の類似性を持つアミノ酸配列をコードするヌクレオチド配列を含みかつ配列番号:1または配列番号:3に記載の配列と低厳格条件下でハイブリッドを形成することができるヌクレオチド配列を含む単離された核酸分子。
【請求項9】
IL−13またはその誘導体と相互作用することができるヘモポエチン受容体をコードする請求項1または請求項2または請求項7または請求項8記載の単離された核酸分子であって、その相互作用をIL−4受容体α−鎖を発現する細胞内でIL−4またはその誘導体により競争的に阻害することができるものである単離された核酸分子。
【請求項10】
宿主細胞中で核酸分子の発現を指令することができるプロモーターに機能的に連結された請求項1または請求項6または請求項7記載の核酸分子を含む遺伝子構築物。
【請求項11】
配列番号:2または配列番号:4に実質的に記載されたアミノ酸配列またはその全部または一部と少なくとも約50%の類似性を持つアミノ酸配列を含む組換えポリペプチドであって、IL−13またはその誘導体と相互作用することができるものである組換えポリペプチド。
【請求項12】
IL−13との相互作用がIL−4受容体α−鎖を発現する細胞内でIL−4により競争的に阻害されるものである請求項11記載の組換えポリペプチド。
【請求項13】
IL−13との相互作用が低親和性のものである請求項11記載の組換えポリペプチド。
【請求項14】
IL−13との相互作用が中から高親和性のものである請求項11記載の組換えポリペプチド。
【請求項15】
該ポリペプチドがCOS細胞で発現されたときウエスタン・ブロット分析で測定すると約50,000から約70,000ダルトンの分子量を持つものである請求項11記載の組換えポリペプチド。
【請求項16】
次の特性のうちの少なくとも二つを持つ組換えポリペプチド、
(i)配列番号:2または配列番号:4に実質的に記載されたアミノ酸配列またはその全部または一部と少なくとも約50%類似性を持つアミノ酸配列を含むこと、
(ii)配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列またはその全部または一部と少なくとも約50%の類似性を持つヌクレオチド配列によりコードされるものであること、
(iii)少なくとも低親和性でIL−13またはその誘導体と相互作用するものであること、そして
(iv)COS細胞内で発現させるときウエスタン・ブロット分析で測定すると約50,000から約70,000ダルトンの分子量を持つものであること。
【請求項17】
次の特性のうちの少なくとも三つを持つ組換えポリペプチド、
(i)配列番号:2または配列番号:4に実質的に記載されたアミノ酸配列またはその全部または一部と少なくとも約50%類似性を持つアミノ酸配列を含むこと、
(ii)配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列またはその全部または一部と少なくとも約50%の類似性を持つヌクレオチド配列によりコードされるものであること、
(iii)少なくとも低親和性でIL−13またはその誘導体と相互作用するものであること、
(iv)COS細胞内で発現させるときウエスタン・ブロット分析で測定すると約50,000から約70,000ダルトンの分子量を持つものであること、
(v)IL−4受容体α−鎖由来のアミノ酸配列を含むものであること、そして
(vi)IL−13と相互作用をすることができるものであること、かつ、その相互作用がIL−4受容体α−鎖を発現する細胞内でIL−4により競争的に阻害されるものであること。
【請求項18】
請求項16または請求項17に記載の組換えポリペプチドに対する抗体。
【請求項19】
該抗体がモノクローン抗体である、請求項16記載の抗体。
【請求項20】
少なくとも二つのサイトカインと相互作用することができるハイブリッドヘモポエチン受容体であって、該サイトカインの少なくとも一つがIL−13またはその誘導体でありそして該ハイブリッド受容体が配列番号:2または配列番号:4に記載されたアミノ酸配列の全部または一部を含むアミノ酸配列を含むものであるハイブリッドヘモポエチン受容体。
【請求項21】
少なくとも一つのサイトカインと高親和性相互作用をすることができるハイブリッドヘモポエチン受容体であって、該サイトカインの少なくとも一つがIL−13またはその誘導体でありそして該ハイブリッド受容体が配列番号:2または配列番号:4に記載されたアミノ酸配列の全部または一部を含むアミノ酸配列を含むものであるハイブリッドヘモポエチン受容体。
【請求項22】
IL−4と相互作用することができる請求項18記載のハイブリッドヘモポエチン受容体。
【請求項23】
IL−2、IL−5、IL−7、IL−9およびIL−15から選択されるサイトカインと相互作用することができる請求項21記載のハイブリッドヘモポエチン受容体。
【請求項24】
請求項16または請求項17記載の組換えポリペプチドおよび一つ以上の薬学的に許容され得る担体および/または希釈剤を含んで成る薬学的組成物。
【請求項25】
請求項1または請求項2または請求項7または請求項8に記載の核酸分子および1以上の遺伝的に許容され得る担体および/または希釈剤を含んで成る遺伝子的薬学的組成物。
【請求項26】
請求項16または請求項17記載の組換えポリペプチドの治療上有効量を動物に投与することを含む該動物の治療方法。
【請求項27】
請求項16または請求項17記載の組換えポリペプチドの治療上有効量を動物に投与することを含む該動物でのIgE産生により悪化した喘息、アレルギーまたは状態を治療する方法。
【請求項28】
次の特性のうちの少なくとも二つを持つ組換えポリペプチドを製造する方法であって、請求項10記載の遺伝子構築物を含む細胞を、該遺伝子構築物中の核酸分子を発現して組換えポリペプチドを生産するのに十分な時間および条件の下で培養する工程、および該組換えポリペプチドを単離する工程を含んで成る方法、
(i)配列番号:2または配列番号:4に実質的に記載されたアミノ酸配列またはそれに少なくとも約50%の類似性を有するアミノ酸配列を含むものであること、
(ii)配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列またはそれに少なくとも約50%の類似性を有するヌクレオチド配列によりコードされるものであること、
(iii)IL−13またはその誘導体と少なくとも低親和性で相互作用するものであること、
(iv)COS細胞で発現させたとき、ウエスタン・ブロット分析で測定すると約50,000から約70,000ダルトンの分子量を持つものであること。
【請求項29】
次の特性のうち少なくとも三つを持つ組換えポリペプチドを製造する方法であって、請求項10記載の遺伝子構築物を含む細胞を、該遺伝子構築物中の核酸分子を発現して組換えポリペプチドを生産するのに十分な時間および条件の下で培養する工程、および該組換えポリペプチドを単離する工程を含んで成る方法、
(i)配列番号:2または配列番号:4に実質的に記載されたアミノ酸配列またはそのすべてまたは一部に少なくとも約50%の類似性を有するアミノ酸配列であること、
(ii)配列番号:1または配列番号:3に実質的に記載されたヌクレオチド配列またはそのすべてまたは一部に少なくとも約50%の類似性を有するヌクレオチド配列によりコードされるものであること、
(iii)IL−13またはその誘導体と少なくとも低親和性で相互作用するものであること、
(iv)COS細胞で発現させたとき、ウエスタン・ブロット分析で測定すると約50,000から約70,000ダルトンの分子量を持つものであること、
(v)IL−4受容体α−鎖由来のアミノ酸配列を含むものであること、そして
(vi)IL−13と相互作用をすることができるものであること、かつ、その相互作用がIL−4受容体α−鎖を発現する細胞内でIL−4により競争的に阻害されるものであること。
【請求項30】
請求項28または請求項29記載の方法により製造される組換えポリペプチドを発現する動物細胞。
【請求項31】
IL−13またはその誘導体と相互作用することができる第1部分とヘモポエチン受容体、受容体チロシンキナーゼ、TNF/NGF受容体またはGタンパク質結合受容体由来の第2部分とを含んで成るキメラタンパク質。
【請求項32】
該第2部分がIL−13結合タンパク質、IL−4受容体α−鎖、IL−2受容体γ−鎖または喘息またはアレルギーに関与するサイトカインの受容体の全部または機能的部分を含むものである請求項31記載のキメラタンパク質。
【請求項33】
生物学的試料中のIL−4のレベルを監視する方法であって、NR4とIL−4受容体α−鎖とを発現する細胞およびその受容体へのIL−4の結合を競争的に阻害するのに十分な量のIL−13と該生物学的試料とをインキュベートする工程、および競争的阻害の程度を測定する工程を含んで成る方法。
【請求項34】
生物学的試料中のIL−13のレベルを監視する方法であって、NR4とIL−4受容体α−鎖とを発現する細胞およびその受容体へのIL−13の結合を競争的に阻害するのに十分な量のIL−4と該生物学的試料とをインキュベートする工程、および競争的阻害の程度を測定する工程を含んで成る方法。
【請求項35】
サイトカインがレポーター分子で標識化されているものである請求項33または請求項34記載の方法。
【図1−1】

【図1−2】

【図1−3】

【図1−4】
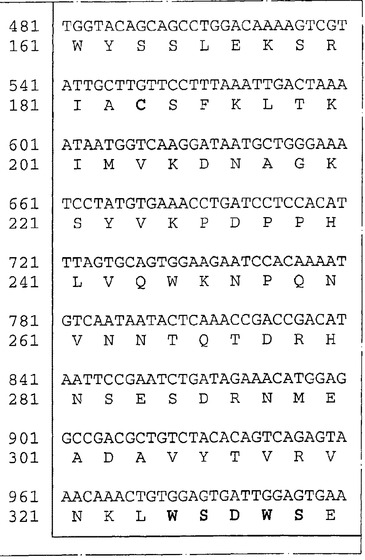
【図1−5】

【図1−6】

【図1−7】

【図2】
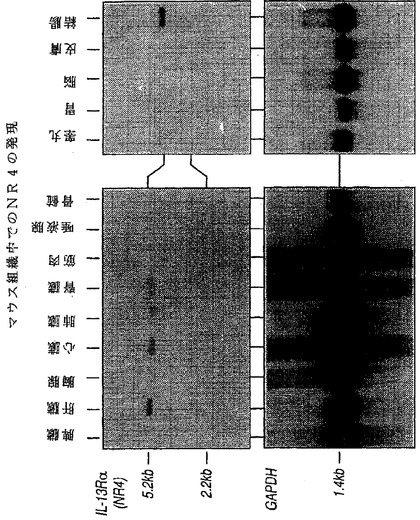
【図3】
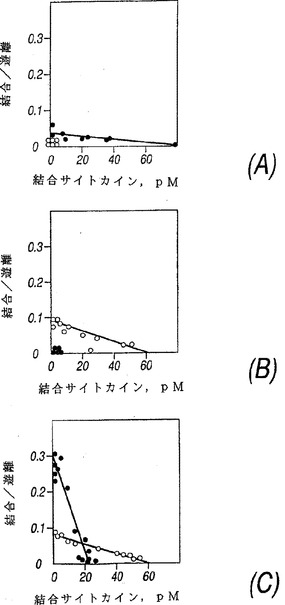
【図4】
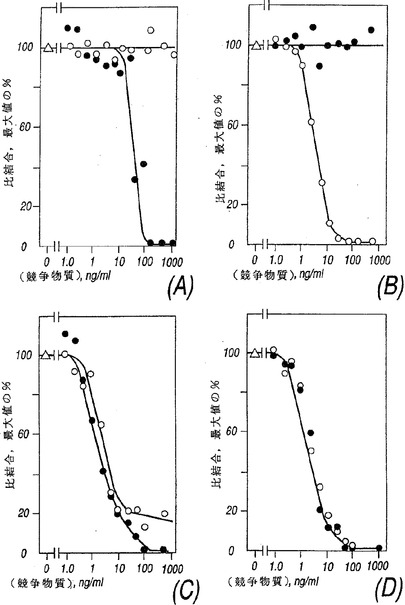
【図5】
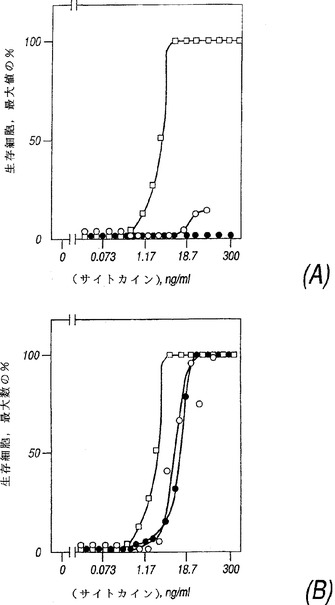
【図6】
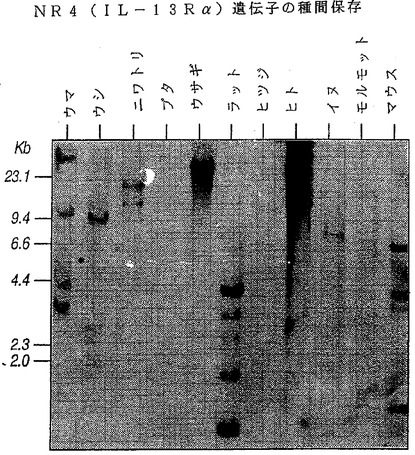
【図7−1】
【図7−2】
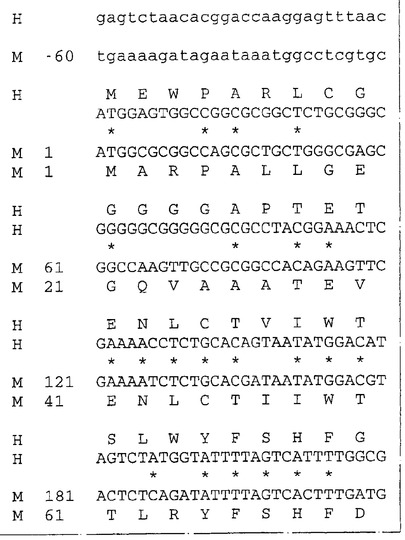
【図7−3】
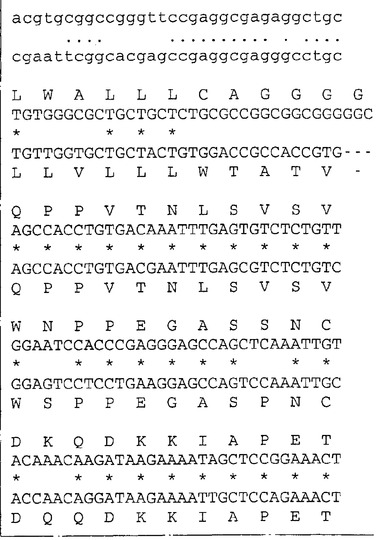
【図7−4】

【図7−5】
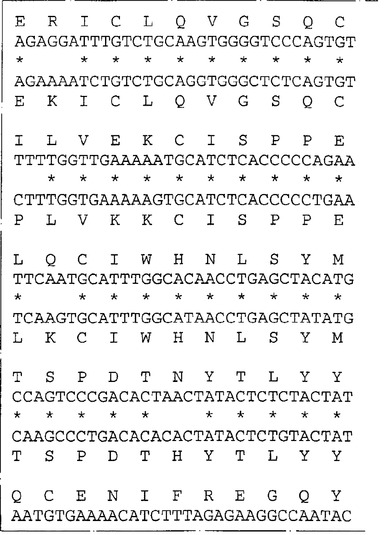
【図7−6】
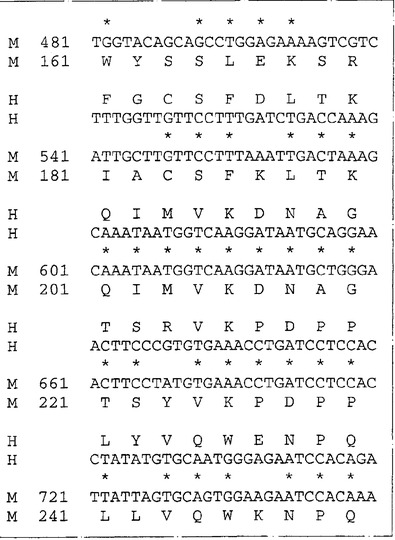
【図7−7】
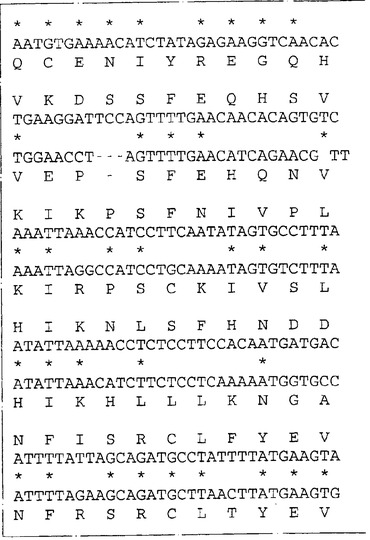
【図7−8】

【図7−9】
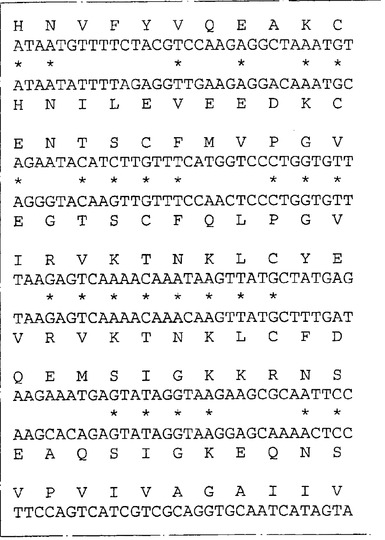
【図7−10】

【図7−11】

【図8】
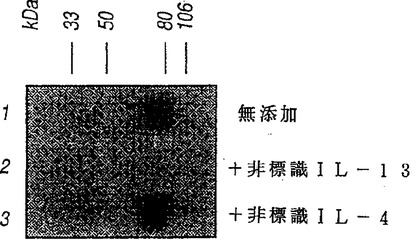
【図9】
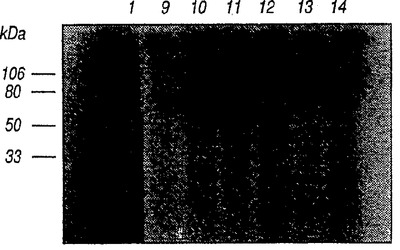
【図10】

【図1−2】
【図1−3】
【図1−4】
【図1−5】
【図1−6】
【図1−7】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7−1】
【図7−2】
【図7−3】
【図7−4】
【図7−5】
【図7−6】
【図7−7】
【図7−8】
【図7−9】
【図7−10】
【図7−11】
【図8】
【図9】
【図10】
【公開番号】特開2008−29340(P2008−29340A)
【公開日】平成20年2月14日(2008.2.14)
【国際特許分類】
【出願番号】特願2007−179629(P2007−179629)
【出願日】平成19年7月9日(2007.7.9)
【分割の表示】特願平9−516141の分割
【原出願日】平成8年10月23日(1996.10.23)
【出願人】(507231747)アムラド オペレーションズ ピーティーワイ.エルティーディー. (3)
【Fターム(参考)】
【公開日】平成20年2月14日(2008.2.14)
【国際特許分類】
【出願日】平成19年7月9日(2007.7.9)
【分割の表示】特願平9−516141の分割
【原出願日】平成8年10月23日(1996.10.23)
【出願人】(507231747)アムラド オペレーションズ ピーティーワイ.エルティーディー. (3)
【Fターム(参考)】
[ Back to top ]