
新規なポリペプチド及びその利用
【課題】抗菌性を有するポリペプチド及びその利用を提供する。
【解決手段】特定の配列を有するイネディフェンシン様蛋白質(γ−チオニン蛋白質)のアミノ酸配列における、連続する10個以上で16個もしくは15個以下、好ましくは1番目〜10番目のアミノ酸からなる短鎖ポリペプチドであって、上記蛋白質と同様、抗菌活性を有する。
【解決手段】特定の配列を有するイネディフェンシン様蛋白質(γ−チオニン蛋白質)のアミノ酸配列における、連続する10個以上で16個もしくは15個以下、好ましくは1番目〜10番目のアミノ酸からなる短鎖ポリペプチドであって、上記蛋白質と同様、抗菌活性を有する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、抗菌性を有する新規なポリペプチド及びその利用に関するものである。
【背景技術】
【0002】
植物は外来の病原菌などの微生物に対抗するため抗菌性蛋白質を持つことが知られている。植物の持つ抗菌性蛋白質は、既に薬剤として使用されている抗菌性物質と対象微生物上の標的分子が異なるため、新たな抗菌性物質としての利用が期待されている。
【0003】
例えば、植物の持つ抗菌性蛋白質の一つとしての植物ディフェンシンは、様々な植物から単離されている。植物ディフェンシンは、一般的に、シグナルペプチドを除いた活性部位が約50前後のアミノ酸から構成され、8つのシスティン残基を有するという共通構造を持つ。そして、8つのシスティン残基により構成される4つのジスルフィド結合を分子内に形成することで保持される立体構造によって、対象微生物に対し抗菌活性を発揮する。
【0004】
植物ディフェンシンは植物体内で発現させることで、当該植物体に特定の病気に対する抵抗性を付与できることも明らかになっている。例えば、カブ(radish)のディフェンシンRs−AFP2をコードする遺伝子を導入したイネは、イネいもち病菌に抵抗性を持つことが記載されている(非特許文献1)。
【0005】
このように、新たな機能及び特性を持つ抗菌性蛋白質は、植物に特定の病原菌に抵抗性を持たせるためのツール等としての利用の可能性がある。また、精製された抗菌性蛋白質は抗菌剤としての利用の可能性がある。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Sanjay Jha and Bharat B. Chattoo, 2010. Transgenic Res 19:373-384
【非特許文献2】Samblanx et al. 1996 Pept Pes. 9(6):262-268
【発明の概要】
【発明が解決しようとする課題】
【0007】
ところで、抗菌剤等の利用を検討した場合、植物ディフェンシン等を植物体から直接に単離精製することは非常に困難で経済性も悪い。そのため、当該ディフェンシンをコードする遺伝子を取得した後、微生物による生産等によって工業的にディフェンシンを生産することが考えられる。
【0008】
しかし、ディフェンシンは立体構造の保持が抗菌活性に極めて重要であり、微生物による生産の過程もしくは蛋白質精製の過程で立体構造に損傷を受けることがあるため、安定的な活性を有する蛋白質をコンスタントに得ることは難しい。また、ディフェンシンを化学合成する方法も考えられ得るが、およそ50アミノ酸からなる分子を合成し、なおかつ抗菌活性を発揮する正しい立体構造をとらせることは、経済的、技術的に非常に困難である。
【0009】
また、上記の問題を解消するため、植物ディフェンシンを断片化してより短いペプチドとする試みもなされている。しかし、既に比較的短い配列である植物ディフェンシンを、抗菌活性を維持しつつより短くすることは困難を伴う。より具体的には、植物ディフェンシンをより短いペプチドに断片化すると、当該断片の抗菌活性は完全長と比較して大きく低下する場合が多い(非特許文献2)。特に、植物ディフェンシンを、製造が比較的容易となる20アミノ酸以下のアミノ酸からなる断片に断片化した場合には、抗菌活性の低下が顕著となる。
【0010】
本発明は、上記問題点に鑑みてなされたものであり、その目的は、比較的短いアミノ酸配列からなり、抗菌性を有する新規なポリペプチド及びその利用を提供することにある。
【課題を解決するための手段】
【0011】
本願発明者らは、イネから、抗菌活性を有するディフェンシン様の蛋白質を単離することに成功している(参考文献:提箸祥幸ら 2010. 日本農芸化学会大会講演要旨集 p239)。この蛋白質は、シグナルペプチドを除いた活性部位が約50前後のアミノ酸から構成され、4つのジスルフィド結合を分子内に形成するという、ディフェンシンに共通した構造を有している。
【0012】
本願発明者らは、上記課題を解決するためにさらに鋭意検討を行った。その結果、ディフェンシン様の上記蛋白質はユニークな抗菌スペクトルを示すこと、特異な作用機序で抗菌活性を示すと推定されること、抗菌活性を維持した状態で断片化が可能であること、及び、抗菌活性を示すドメインが複数あること等の知見を得て、本発明を完成させるに至った。
【0013】
本発明はすなわち、以下の発明を包含する。
(1) 以下の(a)又は(b)に示すポリペプチド:
(a)配列番号1に示すアミノ酸配列における連続する10個以上で16個以下のアミノ酸からなるポリペプチド
(b)配列番号2に示すアミノ酸配列における連続する10個以上で15個以下のアミノ酸からなるポリペプチド。
(2) 配列番号1に示すアミノ酸配列における1番目〜10番目のアミノ酸を含む上記(1)に記載のポリペプチド。
(3) 配列番号1に示すアミノ酸配列における1番目〜10番目のアミノ酸からなる上記(2)に記載のポリペプチド。
(4) 上記(1)から(3)の何れかに記載のポリペプチドにおいて、1から3個のアミノ酸が置換、欠失、及び/又は、付加されてなり、抗菌活性を有するポリペプチド。なお、「A及び/又はB」とは、A及びB、並びに、A又はBを意図する。
(5) 上記(1)から(3)の何れかに記載のポリペプチドにおいて、1又は2個のアミノ酸が置換されてなる、及び/又は、末端側に位置する1又は2個のアミノ酸が欠失されてなり、抗菌活性を有するポリペプチド。
(6) 上記(1)から(5)の何れかに記載のポリペプチドをコードしているポリヌクレオチド。
(7) 上記(6)に記載のポリヌクレオチドを含んでいるベクター。
(8) 上記(6)に記載のポリヌクレオチド、又は上記(7)に記載のベクターが発現可能に導入されている細胞、又は複数個の当該細胞により構成された形質転換体。
(9) 上記(7)に記載のベクターを用いて形質転換されている植物。
(10) 上記(1)から(5)の何れかに記載のポリペプチドを活性成分として含んでいる抗菌剤。
(11) 以下の(a)及び(b)に示すポリペプチドの両方を活性成分として含んでいる上記(10)に記載の抗菌剤:
(a)配列番号1に示すアミノ酸配列における連続する10個以上で16個以下のアミノ酸からなるポリペプチド
(b)配列番号2に示すアミノ酸配列における連続する10個以上で15個以下のアミノ酸からなるポリペプチド。
【発明の効果】
【0014】
本発明によれば、抗菌性を示す新規なポリペプチド等を提供することが出来るという効果を奏する。
【図面の簡単な説明】
【0015】
【図1】イネγ−チオニン蛋白質の抗菌活性部位(AK121915(Def))の抗菌活性を示す図である。
【図2】AK121915(Def)及びこれを断片化した短鎖ペプチドの抗菌活性を示す図である。
【図3】カラシナのディフェンシンBj-AFP1及びこれを断片化した短鎖ペプチドの抗菌活性を示す図である。
【発明を実施するための形態】
【0016】
〔1.本発明に係るポリペプチド〕
本発明に係るポリペプチドは、イネのディフェンシン様蛋白質であるγ−チオニン蛋白質の配列に基づき設計された、抗菌活性を有するポリペプチドである。
【0017】
これまで、植物ディフェンシンは、その抗菌活性の発揮には、分子内S−S結合により形成される立体構造が重要とされ、断片化して立体構造を壊すと活性が低下するとの報告があった(非特許文献2)。従って、製造及び取扱性をより向上させるための短鎖化は非常に困難であった。しかし、本発明に係るポリペプチドは、γ−チオニン蛋白質の活性部位(49アミノ酸から構成される)を約1/3程度以下、さらには約1/5程度以下に短鎖化したものに相当しながら、抗菌活性が維持されているという、極めて特異な性質を示す。
【0018】
また、一般に、植物ディフェンシンはその多くが糸状菌に抗菌スペクトルがあり、その他の微生物に対して抗菌活性を有するものは少ない。しかし、本発明に係るポリペプチドは、少なくとも糸状菌、及び酵母に対して抗菌スペクトルを示す。
【0019】
本発明に係るポリペプチドはさらに、一般的に植物由来のディフェンシンが標的としていると推定される細胞膜上のグルコシルセラミド、及びマンノシル・ジイノシトール・ホスホリル・セラミド以外を標的とすると推定される(実施例参照)。従って、本発明に係るポリペプチドは、新規な作用機序により従来とは異なる抗菌性を示す抗菌剤の開発等に貢献すると考えられる。
【0020】
本発明に係るポリペプチドの一例は、以下の(a)又は(b)に示すものである。
(a)配列番号1に示すアミノ酸配列における連続する10個以上で16個以下のアミノ酸からなるポリペプチド;(b)配列番号2に示すアミノ酸配列における連続する10個以上で15個以下のアミノ酸からなるポリペプチド。
【0021】
製造及び取扱性の容易さという観点では、ポリペプチドを構成するアミノ酸の個数が少ない程好ましい。すなわち、製造及び取扱性の容易さという観点で、上記(a)又は(b)に示すポリペプチドは、好ましくは14個以下又は13個以下のアミノ酸からなり、より好ましくは12個以下又は11個以下のアミノ酸からなり、さらに好ましくは10個のアミノ酸からなる。
【0022】
また、抗菌活性により優れるという観点では、本発明に係るポリペプチドは、配列番号1に示すアミノ酸配列における連続する10個以上で16個以下のアミノ酸からなり、かつ1番目〜10番目のアミノ酸を含むものが好ましい。特に好ましいポリペプチドは、配列番号1に示すアミノ酸配列における1番目〜10番目のアミノ酸からなるものである。
【0023】
なお、本発明に係るポリペプチドの製造方法は特に限定されず、固相合成法等のペプチド化学合成法により製造してもよく、当該ポリペプチドをコードするポリヌクレオチドを細胞に導入し当該細胞に大量に産生させてもよく、又は細胞抽出液を用いた無細胞ペプチド合成系により産生させてもよい。本発明に係るポリペプチドは、比較的短鎖であり、かつ特別な修飾等を行わずとも抗菌活性を示すため、ペプチド化学合成法による製造も容易である。
【0024】
また、本発明に係るポリペプチドは、抗菌活性を有する限りにおいて、アミノ酸の置換、付加、及び/又は欠失が生じていてもよい。ここで許容されるアミノ酸の置換、付加、又は欠失とは、具体的には以下に示すものである。なお、対象となる一つのポリペプチドにおいて、置換、付加、又は欠失が生じるアミノ酸の総数は好ましくは3個以下である。
【0025】
アミノ酸の置換とは、抗菌活性を失わせることのない1又は2個のアミノ酸の置換であり、好ましくは1個のアミノ酸の置換である。具体的には例えば、上記(a)又は(b)に示すポリペプチドを構成する塩基性アミノ酸を他の塩基性アミノ酸に置換すること、酸性アミノ酸を他の酸性アミノ酸に置換すること、中性アミノ酸を性質の類似する他の中性アミノ酸に置換すること、アルギニン以外のアミノ酸をアルギニンに置換すること、等を意図する。さらに具体的には例えば、上記(a)又は(b)に示すポリペプチドにおいて、以下の1)〜7)のいずれかの置換が生じていること等を意図する。
1)フェニルアラニン(F)、チロシン(Y)、及びトリプトファン(W)からなる群から選択される1アミノ酸から、当該群中の他のアミノ酸への置換。
2)リシン(K)、アルギニン(R)、及びヒスチジン(H)からなる群から選択される1アミノ酸から、当該群中の他のアミノ酸への置換。
3)アスパラギン(N)、及びグルタミン(Q)からなる群から選択される1アミノ酸から、当該群中の他のアミノ酸への置換。
4)アスパラギン酸(D)、及びグルタミン酸(E)からなる群から選択される1アミノ酸から、当該群中の他のアミノ酸への置換。
5)アラニン(A)、バリン(V)、ロイシン(L)、イソロイシン(I)、及びプロリン(P)からなる群から選択される1アミノ酸から、当該群中の他のアミノ酸への置換。
6)セリン(S)、及びトレオニン(T)からなる群から選択される1アミノ酸から、当該群中の他のアミノ酸への置換。
7)アルギニン以外のアミノ酸からアルギニンへの置換。なお、アルギニン以外のアミノ酸は、酸性又は中性アミノ酸であることが好ましい。特に限定されないが、配列番号1に示すアミノ酸配列における3番目のアミノ酸、及び/又は、10番目のアミノ酸を、アルギニンに置換することが挙げられる。
【0026】
アミノ酸の欠失とは、抗菌活性を失わせることのない1又は2個のアミノ酸の欠失であり、好ましくは1個のアミノ酸の欠失である。より具体的には例えば、上記(a)又は(b)に示すポリペプチドを構成する末端側に位置するアミノ酸を欠失させることを意図する。すなわち、アミノ末端側に位置する1又は2個のアミノ酸を欠失させる、カルボキシ末端側に位置する1又は2個のアミノ酸を欠失させる、或いは、アミノ末端側に位置する1個のアミノ酸とカルボキシ末端側に位置する1個のアミノ酸とを欠失させる、ことを意図する。或いは、末端側に限定されずに1又は2個のアミノ酸(好ましくは1個のアミノ酸)を欠失させることを意図する。特に限定されないが、配列番号1に示すアミノ酸配列における3番目のアミノ酸、及び/又は、10番目のアミノ酸を、欠失させることが挙げられる。
【0027】
アミノ酸の付加とは、原則として、抗菌活性を失わせることのない1又は2個のアミノ酸の付加であり、好ましくは1個のアミノ酸の付加である。より具体的には例えば、上記(a)又は(b)に示すポリペプチドを構成する末端側に1又は2個のアミノ酸を付加することを意図する。すなわち、アミノ末端側に1又は2個のアミノ酸を付加する、カルボキシ末端側に1又は2個のアミノ酸を付加する、或いは、アミノ末端側とカルボキシ末端側とにぞれぞれ1個のアミノ酸を付加する、ことを意図する。
【0028】
上記したアミノ酸の置換、欠失、及び/又は付加は、公知の方法で行うことができる。公知の方法とは、例えば、Kunkel法等の部位特異的な変異導入法を利用してイネγ−チオニン蛋白質の部分ペプチドに所望の変異を導入する方法が挙げられる。或いは、上記したアミノ酸の置換、欠失、及び/又は付加をした配列を、ペプチド合成法により合成することが挙げられる。
【0029】
なお、本発明に係るポリペプチドに、例えば、シグナルペプチド、マーカー蛋白質、ペプチドリンカー等、及び/又は他の機能性蛋白質を融合して融合蛋白質を構成することもできる。また、本発明に係るポリペプチドを複数個、ペプチドリンカーを介して又は介さずにタンデムに結合して、抗菌活性が増強された融合蛋白質を構成することもできる。但し、これらの融合蛋白質は、イネγ−チオニン蛋白質の活性部位の全長(49アミノ酸)を意図するものではない。
【0030】
〔2.本発明に係るポリヌクレオチド、ベクター〕
(ポリヌクレオチド)
本発明に係るポリヌクレオチドは、上記した本発明に係るポリペプチドをコードしている。当該ポリヌクレオチドは、DNA又はRNAであり、一本鎖又は二本鎖である。当該ポリヌクレオチドを構成する塩基の配列は、コードすべきポリペプチドのアミノ酸配列に応じて適宜決定すればよい。
【0031】
配列番号3は、イネγ−チオニン蛋白質の活性部位をコードするcDNAの塩基配列(コード領域のみ)を示す。本発明に係るポリヌクレオチドの塩基配列は、例えば、配列番号3に示す塩基配列を参照して適宜決定すればよい。例えば、配列番号1に示すアミノ酸配列における連続する10個以上で16個以下のアミノ酸からなるポリペプチドは、配列番号3に示す塩基配列における1番目〜48番目の塩基を有するポリヌクレオチドの全長又は部分配列によりコードされる。また、配列番号2に示すアミノ酸配列における連続する10個以上で15個以下のアミノ酸配列からなるポリペプチドは、配列番号3に示す塩基配列における103番目〜147番目の塩基を有するポリヌクレオチドの全長又は部分配列によりコードされる。
【0032】
上記ポリヌクレオチドは、細胞中又は無細胞合成系での転写及び/又は翻訳を容易とするため、プロモータ、エンハンサ等の調節領域;開始コドン及び終止コドン;その他の転写及び/又は翻訳関連配列;が機能可能に付加されていることが好ましい。さらに、必要に応じて、上記ポリヌクレオチドには、選択マーカーをコードする配列等が付加されていてもよい。これらの配列が付加された本発明に係るポリヌクレオチドの一例は、後述するベクターに組み込まれた形態のものである。
【0033】
(ベクター)
本発明に係るベクターは、本発明に係る上記ポリヌクレオチドを含んでいる。このベクターは、本発明に係るポリペプチドを産生するため、又は宿主細胞に抗菌活性を付与するために使用される。本発明に係るベクターは、細胞抽出液等を用いた無細胞ペプチド合成系に用いるベクターであっても、組換え発現に用いる発現ベクターであってもよい。
【0034】
組換え発現に用いる発現ベクターの作製方法としては、プラスミド、ファージ、コスミド、またはウイルスなどを用いる方法が挙げられるが特に限定されない。本発明に係るポリヌクレオチドは比較的短いものであるため、プラスミドを用いる方法が好ましい。
【0035】
組換え発現に用いる発現ベクターの具体的な種類は特に限定されず、宿主細胞中で発現可能なベクターを適宜選択すればよい。すなわち、宿主細胞の種類に応じて、本発明に係るポリヌクレオチドを発現させるために適当なプロモータ配列等を選択し、これと本発明に係るポリヌクレオチドとを各種プラスミド等に組み込んで発現ベクターを構築すればよい。例えば、宿主細胞が植物細胞である場合は、構成的発現用のプロモータ(カリフラワーモザイクウイルスの35Sプロモータ等)を有するベクター、または外的な刺激によって誘導性に活性化されるプロモータを有するベクターが挙げられる。或いは、抗菌性を有する植物の作出を目的とする場合には、例えば、表皮近傍に特異的な発現を誘導するプロモータ(例えば、ATML1プロモータ等)を有するベクターが挙げられる。
【0036】
また、抗菌性を有する植物の作出を目的とする場合には、例えば、細胞表層又は細胞間隙に局在する蛋白質との融合蛋白質として本発明のポリペプチドを発現させるという方法も取りうる。その他、植物病原菌の感染メカニズムに応じて、本発明のポリペプチドを、植物体内の適切な場所に局所的に発現させるようにしてもよい。
【0037】
発現ベクターは、使用される宿主細胞の種類に応じて、少なくとも1つの選択マーカーを含んでいてもよい。このようなマーカーとしては、宿主細胞が真核細胞の場合は、ジヒドロ葉酸レダクターゼの遺伝子、ネオマイシン耐性遺伝子、及びGFP(Green Fluorescent Protein)その他の蛍光/発光蛋白質をコードする遺伝子等、が挙げられる。
【0038】
〔3.本発明に係る細胞、及び形質転換体〕
本発明に係る形質転換細胞、及び形質転換体には、本発明に係る上記ポリヌクレオチド、又は、本発明に係る上記ベクターが発現可能に導入されている。本発明において形質転換体とは、複数個の形質転換細胞により構成された単位(multicellular form)を指し、細胞とは区別される。形質転換体とは、例えば、組織または器官だけでなく、多細胞生物の個体がその範疇に含まれることを意図する。
【0039】
上記の細胞、及び形質転換体の調製に用いる宿主細胞は、特に限定されるものではないが、産生されるポリペプチドが示す抗菌活性の影響を受け難いものがより好ましい。好ましいものとして、具体的には例えば、細菌類、動物細胞(例えば、昆虫細胞、両生類細胞、哺乳類細胞等)、及び植物細胞(高等植物の細胞、高等植物のプロトプラスト等)から選択される何れかが挙げられる。なお、糸状菌類、及び真菌類等の真核菌類を宿主細胞として用いても、本発明のポリペプチドの産生は可能である。これら宿主細胞のための適切な培養培地および培養条件は当分野で周知である。
【0040】
本発明に係る上記ポリヌクレオチド、又は上記発現ベクターを宿主細胞等に導入する方法、すなわち形質転換法も特に限定されるものではなく、PEG法、電気穿孔法、リン酸カルシウム法、リポソーム法、DEAEデキストラン法、パーティクルガン法、アグロバクテリウム法(植物細胞に限る)等の従来公知の方法を好適に用いることができる。また、例えば、本発明に係るポリペプチドを昆虫で転移発現させる場合には、バキュロウイルスを用いた発現系を用いればよい。
【0041】
本発明に係る形質転換体は、上記形質転換細胞から分化誘導させて得られる。或いは、複数個の細胞により構成された単位(multicellular form)に対して、本発明に係る上記ポリヌクレオチド、又は、本発明に係る上記ベクターを発現可能に導入して得てもよい。以下の説明では、複数個の植物細胞により構成された単位(以下、多細胞植物単位と称する)を形質転換して、本発明に係る形質転換体を得る方法を説明する。
【0042】
多細胞植物単位とは、植物体全体、植物器官(例えば葉、花弁、茎、根、種子など)、植物組織(例えば表皮、師部、柔組織、木部、維管束、柵状組織、海綿状組織など)、カルスなどのいずれをも意味する。植物の種類は特に限定されず、単子葉植物綱または双子葉植物綱に属する植物のいずれでもよい。また、多細胞植物単位への遺伝子の導入には、上記宿主細胞への遺伝子導入と同様、当業者に公知の形質転換方法が用いられる。
【0043】
形質転換の結果得られるカルスやシュート、毛状根などは、そのまま細胞培養、組織培養または器官培養に用いることが可能であり、また従来知られている植物組織培養法を用い、適当な濃度の植物ホルモン(オーキシン、サイトカイニン、ジベレリン、アブシジン酸、エチレン、ブラシノライドなど)の投与などによって植物体に再生させることができる。
【0044】
本発明に係るポリヌクレオチド又はベクターがゲノム内に組み込まれた形質転換植物体(個体)がいったん得られれば、当該植物体の有性生殖または無性生殖によって子孫を得ることができる。また、当該植物体またはその子孫、あるいはこれらのクローンから、例えば、種子、果実、切穂、塊茎、塊根、株、カルス、プロトプラストなどを得て、それらを基に当該植物体を量産することができる。したがって、本発明に係る形質転換体の範疇には、本発明に係るポリヌクレオチドが発現可能に導入された植物体、もしくは、当該植物体と同一の性質を有する当該植物体の子孫、またはこれら由来の組織も含まれる。
【0045】
本発明に係る形質転換体のより具体的な例示は以下の通りであるが、本発明は特にこれらの例示に限定されるものではない。
1)本発明に係るポリペプチドをコードする遺伝子を発現する遺伝子組換えイネ。当該イネは、糸状菌等に代表されるイネ病原菌に対する抵抗性を示すようになる。
2)本発明に係るポリペプチドをコードする遺伝子を発現する、イネ以外の遺伝子組換え植物体。当該植物体は、糸状菌等に代表される植物病原菌に対する抵抗性を示すようになる。
なお、イネ以外の植物の種類は特に限定されないが、具体的には例えば、イネ科(コムギ、オオムギ、ライ麦、トウモロコシ等でイネを除く)、ウリ科(例えば、キュウリ、メロン、スイカ、カボチャ、ニガウリ)、ナス科(例えば、トマト、ナス、ピーマン、パプリカ、トウガラシ、ジャガイモ、ペチュニア)、マメ科(例えば、インゲン、ソラマメ、アズキ、ダイズ、ササゲ、エンドウ)、キク科(例えば、ゴボウ、シュンギク、レタス、キク、ガーベラ、シネラリア、マリーゴールド)、アブラナ科(例えば、ハクサイ、キャベツ、ダイコン、カブ、コマツナ、ブロッコリー、チンゲンサイ、ストック)、アオイ科(例えば、オクラ、ワタ)、セリ科(例えば、セロリ、ニンジン、パセリ)、ユリ科(例えば、タマネギ、ニラ、アスパラガス、ネギ)、ナデシコ科(例えば、カスミソウ、カーネーション)、ミカン科(例えば、ミカン)、バラ科(例えば、バラ、イチゴ、リンゴ、ナシ、洋ナシ、モモ、ネクタリン)、アカザ科(例えば、ホウレンソウ)、等に属する植物が挙げられる。
【0046】
〔4.本発明に係るポリペプチドの用途の例示〕
(抗菌剤、及びその具体的用途)
本発明に係る抗菌剤は、本発明に係る少なくとも一種のポリペプチドを抗菌活性成分として含んでいる。本発明に係る抗菌剤は、必要に応じて、当該ポリペプチドを安定化させる安定化剤、酸化防止剤、等を含んだ組成物であってもよい。本発明に係る抗菌剤は、必要に応じて、製剤化のための、担体、等を含んだ組成物であってもよい。本発明に係る抗菌剤は、抗菌活性を増強する目的、或いは抗菌スペクトルをより拡張する目的で、本発明に係るポリペプチド以外の他の抗菌成分をさらに含んだ組成物であってもよい。
【0047】
本発明に係る抗菌剤は、原核菌類(亜界Procaryomycota)及び/又は真核菌類(亜界Eucaryomycota)に抗菌活性を示すことが好ましい。原核菌類の中では特に細菌門に属する菌類に対して抗菌活性を示すことが好ましい。また、真核菌類の中では特に糸状菌類、酵母類(カンジダ等)に対して抗菌活性を示すことが好ましい。
【0048】
本発明に係る抗菌剤の、より具体的な用途は特に限定されないが、例えば、糸状菌、及びカンジダに対する抗菌性を示しうるため、当該抗菌剤を、例えば、農薬又は医薬の分野で利用することもできる。また、本発明に係る抗菌剤を含む洗剤等を提供することもできる。
【0049】
本発明に係る抗菌剤のより具体的な一例は、以下の(a)及び(b)に示す本発明に係るポリペプチドの一方、又は両方を活性成分として含むものである。
(a)配列番号1に示すアミノ酸配列における連続する10個以上で16個以下のアミノ酸からなるポリペプチド;
(b)配列番号2に示すアミノ酸配列における連続する10個以上で15個以下のアミノ酸からなるポリペプチド。
【0050】
イネγ−チオニン蛋白質は、他の一般的な植物ディフェンシンとは異なり、複数箇所(上記(a)及び(b)に示すポリペプチドに相当)に抗菌活性ドメインを有するという特性を持つ。従って、上記(a)及び(b)に示すポリペプチドの両方を活性成分として含む抗菌剤は、イネγ−チオニン蛋白質により近い特性を示すと推定される。
【0051】
(殺菌又は制菌の方法)
本発明はまた殺菌又は制菌の方法を提供する。すなわち、当該方法は、殺菌又は制菌の対象となる原核菌類及び真核菌類に、本発明に係るポリペプチドを接触させることで、当該原核菌類及び真核菌類を殺菌又は制菌する。なお、「制菌」とは、本発明に係るポリペプチドを接触させない状態と比較して、原核菌類及び/又は真核菌類の活動が抑制されることを指す。なお、原核菌類及び/又は真核菌類の活動が抑制されたか否かは、例えば、増殖速度の低下、又は増殖の停止(静菌)等を指標にして判断することが出来る。
【0052】
(その他のより具体的な応用)
本発明に係るポリペプチドは何れも小分子で、その抗菌活性に立体構造の影響を受けない。そのため、他の蛋白質分子に融合させることで、融合蛋白質分子に新たに抗菌性を付与することが出来る。
【0053】
植物繊維又は工業的に素材となる蛋白質に、本発明に係るポリペプチドを融合させることで、抗菌性の素材を開発することが出来る。一例では、本発明に係るポリペプチドが絹糸を構成する蛋白質に融合された融合蛋白質を産生するようにカイコの遺伝子組換えを行い、抗菌性を有する絹糸を得ること等が考えられる。
【0054】
本発明に係るポリペプチドと比較して細菌に抗菌スペクトルを示す抗菌性蛋白質と、本発明に係るポリペプチドとを融合することで、細菌と糸状菌とに強い抗菌活性を有する蛋白質を作製出来る。
【0055】
本発明は上述した各実施形態に限定されるものではなく、請求項に示した範囲で種々の変更が可能であり、異なる実施形態にそれぞれ開示された技術的手段を適宜組み合わせて得られる実施形態についても本発明の技術的範囲に含まれる。
【実施例】
【0056】
以下、本発明を実施例、比較例及び参考例によりさらに詳細に説明するが、本発明はこの例に限定されるものではない。
【0057】
〔参考例1:イネディフェンシン様蛋白質(γ−チオニン蛋白質)の製造、及び抗菌活性〕
(1)γ−チオニン蛋白質の活性部位(「AK121915(Def)」)の製造
イネ由来のγ−チオニン蛋白質は、抗菌活性に寄与する49個のアミノ酸から構成される活性部位(成熟領域)と、31個のアミノ酸から構成されるシグナルペプチド部位とからなる。本参考例では、以下の方法に従い、上記活性部位のみを製造した。
【0058】
初めに、イネ(品種:日本晴)のゲノムDNAを鋳型として、プライマー1(CGCGGATCCAGGCACTGCCTGTCGCAGAGC)とプライマー2(CCGGAATTCCTAGCAGACCTTCTTGCAGAAG)とを用いた定法のPCR(Polymerase Chain Reaction)により、上記活性部位をコードする遺伝子断片を増幅した。
【0059】
PCRの後、増幅された遺伝子断片に対してBamHI及びEcoRIによる制限酵素処理を行った。次いで、制限酵素処理後の遺伝子断片を、BamHI及びEcoRIで同様に制限酵素処理したpGEX6P−1ベクター(GEヘルスケア社)に挿入し、当該ベクターを用いて大腸菌Rosetta-gamiB(DE3)pLysS株(タカラバイオ社)に形質転換を行った。得られた形質転換体を培養し、IPTG(Isopropyl-thio-β-D-galactoside)添加により誘導的な蛋白質発現を行った。発現蛋白質の精製はGSTrapTMFF column (GEヘルスケア社)及びHiTrapTM CM FF column (GEヘルスケア社)を用いて行い、精製蛋白質からのGST(Glutathion S transferase)タグの除去はPreScission Protease (GEヘルスケア社)を用いて行った。この一連のプロセスにより、γ−チオニン蛋白質の活性部位(AK121915(Def))を得た。
【0060】
(2)AK121915(Def)が有する抗菌活性の測定
図1に示す複数種の糸状菌、酵母(真菌)、及び複数種の細菌に関して、AK121915(Def)が抗菌活性を示すか否かを確認した。結果を図1に示す。図1に示す通り、糸状菌及び酵母に対する抗菌活性が顕著であることが確認された。なお、糸状菌及び酵母に対する抗菌活性の試験方法は、後述する実施例2及び3に示す通りである。
【0061】
また、細菌に対する抗菌活性の測定も、Broekaertらの方法を参考にして行った(参考文献:Broekaert et al. 1990 FEMS. Microbiol. Lett., 69, 55-60.)。測定開始時の菌体濃度(濁度)はOD600が0.1となるように調整した。抗菌活性の測定は、96穴のマイクロプレートを用い、ポリペプトン培地(1L作製につき、ポリペプトン:10g、酵母エキス:2g、MgSO4/7H2O:1g、pH7.0)を1/4濃度に希釈したものを細菌培養用培地として使用した。
【0062】
次いで、AK121915(Def)を含む蛋白質溶液を適当な濃度に希釈して、マイクロプレートの各ウェルに添加し、経時的に濁度測定を行うことで、蛋白質溶液に依存的な細菌に対する生育阻害を測定した。生育阻害の測定温度は25℃とした。なお、図1に示すように、細菌では、25μl/ml以下の濃度では抗菌活性が見られなかったため、IC50値は算出していない。
【0063】
〔実施例1:短鎖ペプチドの合成〕
AK121915(Def)のアミノ酸配列に基づき、短鎖ペプチドを合成した。短鎖ペプチドは北海道システムサイエンス社の受託ペプチド合成により合成した。純度は98%以上のものとした。
【0064】
合成した短鎖ペプチドのアミノ酸配列は以下のとおりである。なお、各配列の後に括弧書きで、AK121915(Def)のアミノ末端側から数えて何番目のアミノ酸に相当するかを示している。
No.1: RHCLSQSHRF(1−10番目のアミノ酸)
No.2: VSSNNCANV(15−23番目のアミノ酸)
No.3: RTESFPDGE(25−33番目のアミノ酸)
No.4: KSHGLERKCF(35−44番目のアミノ酸)
No.5: SHRFKGMCVS(7−16番目のアミノ酸)
No.6: SNNCANVCRTE(17−27番目のアミノ酸)
No.7: FPDGECKSHG(29−38番目のアミノ酸)
No.8: ERKCFCKKVC(40−49番目のアミノ酸)
なお、上記No.1〜No.8の短鎖ペプチドを、順に、AK121915(1-10)、AK121915(15-23)、AK121915(25-33)、AK121915(35-44)、AK121915(7-16)、AK121915(17-27)、AK121915(29-38)、AK121915(40-49)、と称する場合もある。
【0065】
〔実施例2:短鎖ペプチドの抗菌活性試験(糸状菌)〕
参考例1で得たAK121915(Def)、及び実施例1で得た短鎖ペプチドを用いて、イネいもち病菌(Magnaporthe grisea)に対する抗菌活性の測定を行った。
【0066】
抗菌活性の測定は、Broekaertらの方法を参考にして行った(参考文献:Broekaert et al. 1990 FEMS. Microbiol. Lett., 69, 55-60.)。また、イネいもち病菌としてMagnaporthe grisea Guy11株(独立行政法人農業生物資源研究所の西村麻里江博士から新潟薬科大学高久洋暁博士へ分譲されたものを使用)を使用した。抗菌活性の測定は、96穴のマイクロプレートを用い、培地は1/2濃度のpotato dextrose broth (Difco)を使用し、イネいもち病菌の胞子は20,000個/mLとなるように調整した。
【0067】
次いで、AK121915(Def)又は何れかの上記短鎖ペプチドを含む蛋白質溶液を適当な濃度に希釈して、マイクロプレートの各ウェルに添加し、経時的に濁度測定を行うことで、蛋白質溶液に依存的なイネいもち病菌に対する生育阻害を測定した。生育阻害の測定温度は25℃とした。そして、蛋白質濃度と生育阻害率とから各蛋白質のIC50値を算出した。また、対照試験としてWakoのカスガマイシン塩酸塩水和物を用いた。カスガマイシンは、イネいもち病菌等に対する抗菌剤である。
【0068】
抗菌活性試験の結果は、図2及び表1に纏めて示す。なお、カスガマイシンのIC50値は2.5(μg/ml)であった。
【0069】
【表1】

【0070】
〔実施例3:短鎖ペプチドの抗菌活性試験(酵母)〕
参考例1で得たAK121915(Def)、及び実施例1で得た短鎖ペプチドを用いて、カンジダ(Candida albicans)に対する抗菌活性の測定を行った。
【0071】
抗菌活性の測定は、Broekaertらの方法を参考にして行った(参考文献:Broekaert et al. 1990 FEMS. Microbiol. Lett., 69, 55-60.)。また、カンジダとしてCandida albicans CAI4株(千葉大学真菌医学研究センターから新潟薬科大学高久洋暁博士に分譲されたものを使用)を使用した。抗菌活性の測定は、96穴のマイクロプレートを用い、培地は1/2濃度のpotato dextrose broth (Difco)を使用し、測定開始時のカンジダ菌体の濁度はOD600が0.01となるように調整した。
【0072】
次いで、AK121915(Def)又は何れかの上記短鎖ペプチドを含む蛋白質溶液を適当な濃度に希釈して、マイクロプレートの各ウェルに添加し、経時的に濁度測定を行うことで、蛋白質溶液に依存的なカンジダに対する生育阻害を測定した。生育阻害の測定温度は30℃とした。そして、蛋白質濃度と生育阻害率とから各蛋白質のIC50値を算出した。
【0073】
抗菌活性試験の結果は、図2に纏めて示す。
【0074】
〔実施例4:AK121915(Def)、及び短鎖ペプチドの標的分子の探索〕
これまで、植物ディフェンシンが抗菌活性を発揮するための微生物上の標的分子として、グルコシルセラミド及びマンノシル・ジイノシトール・ホスホリル・セラミド(mannosyl diinositolphosphoryl ceramide)が候補とされてきた(参考文献:Thevissen et al. 2005 FEBS Let. 579 : 1973-1977)。
【0075】
そして、標的分子の候補がグルコシルセラミドとされている植物ディフェンシンとしては、カブのディフェンシンRs-AFP2が、標的分子の候補がマンノシル・ジイノシトール・ホスホリル・セラミドとされている植物ディフェンシンとしては、ダリアのディフェンシンDm−AMP1が、これまで比較的詳細に解析されてきた。
【0076】
本実施例では、AK121915(Def)、及びその短鎖ペプチドAK121915(1-10)の標的分子が、グルコシルセラミド又はマンノシル・ジイノシトール・ホスホリル・セラミドである可能性について以下の検証をした。
【0077】
(1) 標的分子がグルコシルセラミドである可能性の検証
カラシナディフェンシンBj-AFP1、イネディフェンシンAK121915(Def)、及びAK121915(1-10)の三種の蛋白質及びペプチドを用い、Candida albicans CAI4株、C. albicans CAI4 △gcs株(新潟薬科大学高久洋暁博士がCAI4株より作製)、Saccharomyces cerevisiae W303-1A株(ATCCより入手可)に対する抗菌活性を測定した。抗菌活性の測定は、何れも実施例3の記載に従い行った。測定の結果は、表2に纏めて示す。表2中の○(抗菌活性有)はIC50値が15μg/mL以下であることを示し、×(抗菌活性無)はIC50値が25μg/mL以上であることを示す。
【0078】
なお、Bj-AFP1はカブ由来のディフェンシンRs-AFP2と高い相同性を有する。そのため、Rs-AFP2と同様にBj-AFP1もグルコシルセラミドが抗菌活性を発揮するための標的分子と推定されている。また、カンジダ(C. albicans CAI4株)は、細胞膜成分としてグルコシルセラミドを有する。カンジダのグルコシルセラミド合成系の遺伝子gcs (Glucosylceramide Synthase)を破壊した△gcs株はグルコシルセラミドを合成できない。また、出芽酵母(S. cerevisiae W303-1A株)は細胞膜成分としてのグルコシルセラミドを持たない。
【0079】
【表2】
【0080】
表2に示す通り、Bj-AFP1はグルコシルセラミドが存在する株にのみ抗菌活性を示したことから、予想通りグルコシルセラミドが標的分子である(もしくは関与している)可能性が高い。一方、AK121915(Def)及びその短鎖ペプチドAK121915(1-10)はグルコシルセラミドの有無に関係なく全ての株に対し抗菌活性を示したことから、グルコシルセラミドがターゲットの候補にはならない。すなわち、AK121915(Def)及びその短鎖ペプチドはBj-AFP1をはじめとする植物ディフェンシンと作用機序が異なる可能性が示唆される。
【0081】
(2) 標的分子がマンノシル・ジイノシトール・ホスホリル・セラミドである可能性の検証
カラシナディフェンシンBj-AFP1、イネディフェンシンAK121915(Def)、及びAK121915(1-10)の三種の蛋白質及びペプチドを用い、S. cerevisiae W303-1A株、S. cerevisiae W303-1A△skn1△ipt1株(新潟薬科大学高久洋暁博士がW303-1A株より作製)、C. albicans CAI4株に対する抗菌活性を測定した。抗菌活性の測定は、何れも実施例3の記載に従い行った。測定の結果は、表3に纏めて示す。表3中の○(抗菌活性有)はIC50値が15μg/mL以下であることを示し、×(抗菌活性無)はIC50値が25μg/mL以上であることを示す。
【0082】
なお、出芽酵母(S. cerevisiae W303-1A株)は細胞膜成分としてマンノシル・ジイノシトール・ホスホリル・セラミドを有する。一方、カンジダ(C. albicans CAI4株)は細胞膜成分としてマンノシル・ジイノシトール・ホスホリル・セラミドを持たない。また、S. cerevisiae W303-1A株においてskn1遺伝子及びipt1遺伝子を破壊したS. cerevisiae W303-1A△skn1△ipt1株は、マンノシル・ジイノシトール・ホスホリル・セラミドを合成できない。
【0083】
【表3】
【0084】
表3に示す通り、AK121915(Def)及びその短鎖ペプチドAK121915(1-10)は、マンノシル・ジイノシトール・ホスホリル・セラミド分子の有無に関係なくS. cerevisiae及びC. albicansに対し抗菌活性を示した。このことからAK121915(Def)及びその短鎖ペプチドの標的分子は、ダリアのディフェンシンDm−AMP1ともタイプの異なる抗菌性蛋白質であることが示唆された。
【0085】
〔参考例2:カラシナディフェンシンBj-AFP1由来の短鎖ペプチドの抗菌活性〕
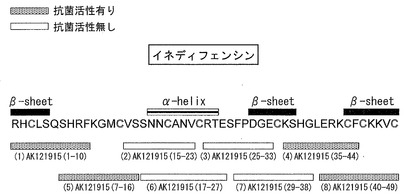
図3に示すように、Bj-AFP1はAK121915(Def)とほぼ同様の位置に8つのシスティン残基を有するという、ディフェンシンに特有の共通構造を持つ。そこで、Bj-AFP1の配列に基づいて、実施例1に示す短鎖ペプチドAK121915(1-10)、AK121915(15-23)、AK121915(25-33)、及びAK121915(35-44)に対応する短鎖ペプチドを合成した。合成した短鎖ペプチドは順に、Bj-AFP1(1-10)、Bj-AFP1(16-24)、Bj-AFP1(26-35)、及びBj-AFP1(37-46)として図3中に示す。なお、これら短鎖ペプチドは、北海道システムサイエンス社の受託ペプチド合成により合成した。純度は98%以上のものとした。
【0086】
その上で、実施例2及び3に記載した方法に従い、Bj-AFP1に基づく上記短鎖ペプチド、及びBj-AFP1の活性部位(全長)の、イネいもち病菌、及びカンジダに対する抗菌活性を測定した。その結果を図3中に纏めて示す。結果から明らかなように、カラシナディフェンシンBj-AFP1では、AK121915(1-10)に対応する短鎖ペプチドBj-AFP1(1-10)でさえ、抗菌活性を示さないことが明らかとなった。
【0087】
〔実施例5:AK121915(Def)の短鎖ペプチドAK121915(1-10)の改変体〕
AK121915(Def)のアミノ酸配列情報から、1―10番目のアミノ酸からなる短鎖ペプチドAK121915(1-10)(表4のNo.1。実施例1も参照)をベースに、アミノ酸の置換、欠失、及び付加を組み合わせた5種類の改変短鎖ペプチドを合成した。改変短鎖ペプチドは北海道システムサイエンス社の受託ペプチド合成により合成した。純度は98%以上のものとした。
【0088】
合成した改変短鎖ペプチドNo.2〜No.6のアミノ酸配列は、表4に示す。
【0089】
【表4】
【0090】
表4に示す改変短鎖ペプチドNo.2は、短鎖ペプチドAK121915(1-10)の10番目のアミノ酸Fを欠失させたものである。改変短鎖ペプチドNo.3は、短鎖ペプチドAK121915(1-10)の3番目のアミノ酸Cと、10番目のアミノ酸Fとを欠失させたものである。改変短鎖ペプチドNo.4は、短鎖ペプチドAK121915(1-10)の3番目のアミノ酸Cを欠失させ、10番目のアミノ酸Fをアミノ酸R(アルギニン)に置換し、かつC末端にアミノ酸Rを付加したものである。改変短鎖ペプチドNo.5は、短鎖ペプチドAK121915(1-10)の10番目のアミノ酸Fをアミノ酸Rに置換したものである。改変短鎖ペプチドNo.6は、短鎖ペプチドAK121915(1-10)の3番目のアミノ酸C及び10番目のアミノ酸Fを何れもアミノ酸Rに置換したものである。
【0091】
次いで、実施例3と同様の方法で、表4のNo.1〜6で示した短鎖ペプチドを用いてカンジダ(Candida albicans)に対する生育阻害を測定し、IC50値(単位はμg/ml)を算出した。結果を表4に示した。特に、短鎖ペプチドAK121915(1-10)の10番目のアミノ酸Fの欠失やアミノ酸Rへの置換、或いは、3番目のアミノ酸Cの欠失やアミノ酸Rへの置換を行っても、その抗菌活性は維持又は向上することが示された。
【産業上の利用可能性】
【0092】
本発明に係るポリペプチド等は抗菌剤、抗菌性付与植物の作出等に利用できる。
【技術分野】
【0001】
本発明は、抗菌性を有する新規なポリペプチド及びその利用に関するものである。
【背景技術】
【0002】
植物は外来の病原菌などの微生物に対抗するため抗菌性蛋白質を持つことが知られている。植物の持つ抗菌性蛋白質は、既に薬剤として使用されている抗菌性物質と対象微生物上の標的分子が異なるため、新たな抗菌性物質としての利用が期待されている。
【0003】
例えば、植物の持つ抗菌性蛋白質の一つとしての植物ディフェンシンは、様々な植物から単離されている。植物ディフェンシンは、一般的に、シグナルペプチドを除いた活性部位が約50前後のアミノ酸から構成され、8つのシスティン残基を有するという共通構造を持つ。そして、8つのシスティン残基により構成される4つのジスルフィド結合を分子内に形成することで保持される立体構造によって、対象微生物に対し抗菌活性を発揮する。
【0004】
植物ディフェンシンは植物体内で発現させることで、当該植物体に特定の病気に対する抵抗性を付与できることも明らかになっている。例えば、カブ(radish)のディフェンシンRs−AFP2をコードする遺伝子を導入したイネは、イネいもち病菌に抵抗性を持つことが記載されている(非特許文献1)。
【0005】
このように、新たな機能及び特性を持つ抗菌性蛋白質は、植物に特定の病原菌に抵抗性を持たせるためのツール等としての利用の可能性がある。また、精製された抗菌性蛋白質は抗菌剤としての利用の可能性がある。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Sanjay Jha and Bharat B. Chattoo, 2010. Transgenic Res 19:373-384
【非特許文献2】Samblanx et al. 1996 Pept Pes. 9(6):262-268
【発明の概要】
【発明が解決しようとする課題】
【0007】
ところで、抗菌剤等の利用を検討した場合、植物ディフェンシン等を植物体から直接に単離精製することは非常に困難で経済性も悪い。そのため、当該ディフェンシンをコードする遺伝子を取得した後、微生物による生産等によって工業的にディフェンシンを生産することが考えられる。
【0008】
しかし、ディフェンシンは立体構造の保持が抗菌活性に極めて重要であり、微生物による生産の過程もしくは蛋白質精製の過程で立体構造に損傷を受けることがあるため、安定的な活性を有する蛋白質をコンスタントに得ることは難しい。また、ディフェンシンを化学合成する方法も考えられ得るが、およそ50アミノ酸からなる分子を合成し、なおかつ抗菌活性を発揮する正しい立体構造をとらせることは、経済的、技術的に非常に困難である。
【0009】
また、上記の問題を解消するため、植物ディフェンシンを断片化してより短いペプチドとする試みもなされている。しかし、既に比較的短い配列である植物ディフェンシンを、抗菌活性を維持しつつより短くすることは困難を伴う。より具体的には、植物ディフェンシンをより短いペプチドに断片化すると、当該断片の抗菌活性は完全長と比較して大きく低下する場合が多い(非特許文献2)。特に、植物ディフェンシンを、製造が比較的容易となる20アミノ酸以下のアミノ酸からなる断片に断片化した場合には、抗菌活性の低下が顕著となる。
【0010】
本発明は、上記問題点に鑑みてなされたものであり、その目的は、比較的短いアミノ酸配列からなり、抗菌性を有する新規なポリペプチド及びその利用を提供することにある。
【課題を解決するための手段】
【0011】
本願発明者らは、イネから、抗菌活性を有するディフェンシン様の蛋白質を単離することに成功している(参考文献:提箸祥幸ら 2010. 日本農芸化学会大会講演要旨集 p239)。この蛋白質は、シグナルペプチドを除いた活性部位が約50前後のアミノ酸から構成され、4つのジスルフィド結合を分子内に形成するという、ディフェンシンに共通した構造を有している。
【0012】
本願発明者らは、上記課題を解決するためにさらに鋭意検討を行った。その結果、ディフェンシン様の上記蛋白質はユニークな抗菌スペクトルを示すこと、特異な作用機序で抗菌活性を示すと推定されること、抗菌活性を維持した状態で断片化が可能であること、及び、抗菌活性を示すドメインが複数あること等の知見を得て、本発明を完成させるに至った。
【0013】
本発明はすなわち、以下の発明を包含する。
(1) 以下の(a)又は(b)に示すポリペプチド:
(a)配列番号1に示すアミノ酸配列における連続する10個以上で16個以下のアミノ酸からなるポリペプチド
(b)配列番号2に示すアミノ酸配列における連続する10個以上で15個以下のアミノ酸からなるポリペプチド。
(2) 配列番号1に示すアミノ酸配列における1番目〜10番目のアミノ酸を含む上記(1)に記載のポリペプチド。
(3) 配列番号1に示すアミノ酸配列における1番目〜10番目のアミノ酸からなる上記(2)に記載のポリペプチド。
(4) 上記(1)から(3)の何れかに記載のポリペプチドにおいて、1から3個のアミノ酸が置換、欠失、及び/又は、付加されてなり、抗菌活性を有するポリペプチド。なお、「A及び/又はB」とは、A及びB、並びに、A又はBを意図する。
(5) 上記(1)から(3)の何れかに記載のポリペプチドにおいて、1又は2個のアミノ酸が置換されてなる、及び/又は、末端側に位置する1又は2個のアミノ酸が欠失されてなり、抗菌活性を有するポリペプチド。
(6) 上記(1)から(5)の何れかに記載のポリペプチドをコードしているポリヌクレオチド。
(7) 上記(6)に記載のポリヌクレオチドを含んでいるベクター。
(8) 上記(6)に記載のポリヌクレオチド、又は上記(7)に記載のベクターが発現可能に導入されている細胞、又は複数個の当該細胞により構成された形質転換体。
(9) 上記(7)に記載のベクターを用いて形質転換されている植物。
(10) 上記(1)から(5)の何れかに記載のポリペプチドを活性成分として含んでいる抗菌剤。
(11) 以下の(a)及び(b)に示すポリペプチドの両方を活性成分として含んでいる上記(10)に記載の抗菌剤:
(a)配列番号1に示すアミノ酸配列における連続する10個以上で16個以下のアミノ酸からなるポリペプチド
(b)配列番号2に示すアミノ酸配列における連続する10個以上で15個以下のアミノ酸からなるポリペプチド。
【発明の効果】
【0014】
本発明によれば、抗菌性を示す新規なポリペプチド等を提供することが出来るという効果を奏する。
【図面の簡単な説明】
【0015】
【図1】イネγ−チオニン蛋白質の抗菌活性部位(AK121915(Def))の抗菌活性を示す図である。
【図2】AK121915(Def)及びこれを断片化した短鎖ペプチドの抗菌活性を示す図である。
【図3】カラシナのディフェンシンBj-AFP1及びこれを断片化した短鎖ペプチドの抗菌活性を示す図である。
【発明を実施するための形態】
【0016】
〔1.本発明に係るポリペプチド〕
本発明に係るポリペプチドは、イネのディフェンシン様蛋白質であるγ−チオニン蛋白質の配列に基づき設計された、抗菌活性を有するポリペプチドである。
【0017】
これまで、植物ディフェンシンは、その抗菌活性の発揮には、分子内S−S結合により形成される立体構造が重要とされ、断片化して立体構造を壊すと活性が低下するとの報告があった(非特許文献2)。従って、製造及び取扱性をより向上させるための短鎖化は非常に困難であった。しかし、本発明に係るポリペプチドは、γ−チオニン蛋白質の活性部位(49アミノ酸から構成される)を約1/3程度以下、さらには約1/5程度以下に短鎖化したものに相当しながら、抗菌活性が維持されているという、極めて特異な性質を示す。
【0018】
また、一般に、植物ディフェンシンはその多くが糸状菌に抗菌スペクトルがあり、その他の微生物に対して抗菌活性を有するものは少ない。しかし、本発明に係るポリペプチドは、少なくとも糸状菌、及び酵母に対して抗菌スペクトルを示す。
【0019】
本発明に係るポリペプチドはさらに、一般的に植物由来のディフェンシンが標的としていると推定される細胞膜上のグルコシルセラミド、及びマンノシル・ジイノシトール・ホスホリル・セラミド以外を標的とすると推定される(実施例参照)。従って、本発明に係るポリペプチドは、新規な作用機序により従来とは異なる抗菌性を示す抗菌剤の開発等に貢献すると考えられる。
【0020】
本発明に係るポリペプチドの一例は、以下の(a)又は(b)に示すものである。
(a)配列番号1に示すアミノ酸配列における連続する10個以上で16個以下のアミノ酸からなるポリペプチド;(b)配列番号2に示すアミノ酸配列における連続する10個以上で15個以下のアミノ酸からなるポリペプチド。
【0021】
製造及び取扱性の容易さという観点では、ポリペプチドを構成するアミノ酸の個数が少ない程好ましい。すなわち、製造及び取扱性の容易さという観点で、上記(a)又は(b)に示すポリペプチドは、好ましくは14個以下又は13個以下のアミノ酸からなり、より好ましくは12個以下又は11個以下のアミノ酸からなり、さらに好ましくは10個のアミノ酸からなる。
【0022】
また、抗菌活性により優れるという観点では、本発明に係るポリペプチドは、配列番号1に示すアミノ酸配列における連続する10個以上で16個以下のアミノ酸からなり、かつ1番目〜10番目のアミノ酸を含むものが好ましい。特に好ましいポリペプチドは、配列番号1に示すアミノ酸配列における1番目〜10番目のアミノ酸からなるものである。
【0023】
なお、本発明に係るポリペプチドの製造方法は特に限定されず、固相合成法等のペプチド化学合成法により製造してもよく、当該ポリペプチドをコードするポリヌクレオチドを細胞に導入し当該細胞に大量に産生させてもよく、又は細胞抽出液を用いた無細胞ペプチド合成系により産生させてもよい。本発明に係るポリペプチドは、比較的短鎖であり、かつ特別な修飾等を行わずとも抗菌活性を示すため、ペプチド化学合成法による製造も容易である。
【0024】
また、本発明に係るポリペプチドは、抗菌活性を有する限りにおいて、アミノ酸の置換、付加、及び/又は欠失が生じていてもよい。ここで許容されるアミノ酸の置換、付加、又は欠失とは、具体的には以下に示すものである。なお、対象となる一つのポリペプチドにおいて、置換、付加、又は欠失が生じるアミノ酸の総数は好ましくは3個以下である。
【0025】
アミノ酸の置換とは、抗菌活性を失わせることのない1又は2個のアミノ酸の置換であり、好ましくは1個のアミノ酸の置換である。具体的には例えば、上記(a)又は(b)に示すポリペプチドを構成する塩基性アミノ酸を他の塩基性アミノ酸に置換すること、酸性アミノ酸を他の酸性アミノ酸に置換すること、中性アミノ酸を性質の類似する他の中性アミノ酸に置換すること、アルギニン以外のアミノ酸をアルギニンに置換すること、等を意図する。さらに具体的には例えば、上記(a)又は(b)に示すポリペプチドにおいて、以下の1)〜7)のいずれかの置換が生じていること等を意図する。
1)フェニルアラニン(F)、チロシン(Y)、及びトリプトファン(W)からなる群から選択される1アミノ酸から、当該群中の他のアミノ酸への置換。
2)リシン(K)、アルギニン(R)、及びヒスチジン(H)からなる群から選択される1アミノ酸から、当該群中の他のアミノ酸への置換。
3)アスパラギン(N)、及びグルタミン(Q)からなる群から選択される1アミノ酸から、当該群中の他のアミノ酸への置換。
4)アスパラギン酸(D)、及びグルタミン酸(E)からなる群から選択される1アミノ酸から、当該群中の他のアミノ酸への置換。
5)アラニン(A)、バリン(V)、ロイシン(L)、イソロイシン(I)、及びプロリン(P)からなる群から選択される1アミノ酸から、当該群中の他のアミノ酸への置換。
6)セリン(S)、及びトレオニン(T)からなる群から選択される1アミノ酸から、当該群中の他のアミノ酸への置換。
7)アルギニン以外のアミノ酸からアルギニンへの置換。なお、アルギニン以外のアミノ酸は、酸性又は中性アミノ酸であることが好ましい。特に限定されないが、配列番号1に示すアミノ酸配列における3番目のアミノ酸、及び/又は、10番目のアミノ酸を、アルギニンに置換することが挙げられる。
【0026】
アミノ酸の欠失とは、抗菌活性を失わせることのない1又は2個のアミノ酸の欠失であり、好ましくは1個のアミノ酸の欠失である。より具体的には例えば、上記(a)又は(b)に示すポリペプチドを構成する末端側に位置するアミノ酸を欠失させることを意図する。すなわち、アミノ末端側に位置する1又は2個のアミノ酸を欠失させる、カルボキシ末端側に位置する1又は2個のアミノ酸を欠失させる、或いは、アミノ末端側に位置する1個のアミノ酸とカルボキシ末端側に位置する1個のアミノ酸とを欠失させる、ことを意図する。或いは、末端側に限定されずに1又は2個のアミノ酸(好ましくは1個のアミノ酸)を欠失させることを意図する。特に限定されないが、配列番号1に示すアミノ酸配列における3番目のアミノ酸、及び/又は、10番目のアミノ酸を、欠失させることが挙げられる。
【0027】
アミノ酸の付加とは、原則として、抗菌活性を失わせることのない1又は2個のアミノ酸の付加であり、好ましくは1個のアミノ酸の付加である。より具体的には例えば、上記(a)又は(b)に示すポリペプチドを構成する末端側に1又は2個のアミノ酸を付加することを意図する。すなわち、アミノ末端側に1又は2個のアミノ酸を付加する、カルボキシ末端側に1又は2個のアミノ酸を付加する、或いは、アミノ末端側とカルボキシ末端側とにぞれぞれ1個のアミノ酸を付加する、ことを意図する。
【0028】
上記したアミノ酸の置換、欠失、及び/又は付加は、公知の方法で行うことができる。公知の方法とは、例えば、Kunkel法等の部位特異的な変異導入法を利用してイネγ−チオニン蛋白質の部分ペプチドに所望の変異を導入する方法が挙げられる。或いは、上記したアミノ酸の置換、欠失、及び/又は付加をした配列を、ペプチド合成法により合成することが挙げられる。
【0029】
なお、本発明に係るポリペプチドに、例えば、シグナルペプチド、マーカー蛋白質、ペプチドリンカー等、及び/又は他の機能性蛋白質を融合して融合蛋白質を構成することもできる。また、本発明に係るポリペプチドを複数個、ペプチドリンカーを介して又は介さずにタンデムに結合して、抗菌活性が増強された融合蛋白質を構成することもできる。但し、これらの融合蛋白質は、イネγ−チオニン蛋白質の活性部位の全長(49アミノ酸)を意図するものではない。
【0030】
〔2.本発明に係るポリヌクレオチド、ベクター〕
(ポリヌクレオチド)
本発明に係るポリヌクレオチドは、上記した本発明に係るポリペプチドをコードしている。当該ポリヌクレオチドは、DNA又はRNAであり、一本鎖又は二本鎖である。当該ポリヌクレオチドを構成する塩基の配列は、コードすべきポリペプチドのアミノ酸配列に応じて適宜決定すればよい。
【0031】
配列番号3は、イネγ−チオニン蛋白質の活性部位をコードするcDNAの塩基配列(コード領域のみ)を示す。本発明に係るポリヌクレオチドの塩基配列は、例えば、配列番号3に示す塩基配列を参照して適宜決定すればよい。例えば、配列番号1に示すアミノ酸配列における連続する10個以上で16個以下のアミノ酸からなるポリペプチドは、配列番号3に示す塩基配列における1番目〜48番目の塩基を有するポリヌクレオチドの全長又は部分配列によりコードされる。また、配列番号2に示すアミノ酸配列における連続する10個以上で15個以下のアミノ酸配列からなるポリペプチドは、配列番号3に示す塩基配列における103番目〜147番目の塩基を有するポリヌクレオチドの全長又は部分配列によりコードされる。
【0032】
上記ポリヌクレオチドは、細胞中又は無細胞合成系での転写及び/又は翻訳を容易とするため、プロモータ、エンハンサ等の調節領域;開始コドン及び終止コドン;その他の転写及び/又は翻訳関連配列;が機能可能に付加されていることが好ましい。さらに、必要に応じて、上記ポリヌクレオチドには、選択マーカーをコードする配列等が付加されていてもよい。これらの配列が付加された本発明に係るポリヌクレオチドの一例は、後述するベクターに組み込まれた形態のものである。
【0033】
(ベクター)
本発明に係るベクターは、本発明に係る上記ポリヌクレオチドを含んでいる。このベクターは、本発明に係るポリペプチドを産生するため、又は宿主細胞に抗菌活性を付与するために使用される。本発明に係るベクターは、細胞抽出液等を用いた無細胞ペプチド合成系に用いるベクターであっても、組換え発現に用いる発現ベクターであってもよい。
【0034】
組換え発現に用いる発現ベクターの作製方法としては、プラスミド、ファージ、コスミド、またはウイルスなどを用いる方法が挙げられるが特に限定されない。本発明に係るポリヌクレオチドは比較的短いものであるため、プラスミドを用いる方法が好ましい。
【0035】
組換え発現に用いる発現ベクターの具体的な種類は特に限定されず、宿主細胞中で発現可能なベクターを適宜選択すればよい。すなわち、宿主細胞の種類に応じて、本発明に係るポリヌクレオチドを発現させるために適当なプロモータ配列等を選択し、これと本発明に係るポリヌクレオチドとを各種プラスミド等に組み込んで発現ベクターを構築すればよい。例えば、宿主細胞が植物細胞である場合は、構成的発現用のプロモータ(カリフラワーモザイクウイルスの35Sプロモータ等)を有するベクター、または外的な刺激によって誘導性に活性化されるプロモータを有するベクターが挙げられる。或いは、抗菌性を有する植物の作出を目的とする場合には、例えば、表皮近傍に特異的な発現を誘導するプロモータ(例えば、ATML1プロモータ等)を有するベクターが挙げられる。
【0036】
また、抗菌性を有する植物の作出を目的とする場合には、例えば、細胞表層又は細胞間隙に局在する蛋白質との融合蛋白質として本発明のポリペプチドを発現させるという方法も取りうる。その他、植物病原菌の感染メカニズムに応じて、本発明のポリペプチドを、植物体内の適切な場所に局所的に発現させるようにしてもよい。
【0037】
発現ベクターは、使用される宿主細胞の種類に応じて、少なくとも1つの選択マーカーを含んでいてもよい。このようなマーカーとしては、宿主細胞が真核細胞の場合は、ジヒドロ葉酸レダクターゼの遺伝子、ネオマイシン耐性遺伝子、及びGFP(Green Fluorescent Protein)その他の蛍光/発光蛋白質をコードする遺伝子等、が挙げられる。
【0038】
〔3.本発明に係る細胞、及び形質転換体〕
本発明に係る形質転換細胞、及び形質転換体には、本発明に係る上記ポリヌクレオチド、又は、本発明に係る上記ベクターが発現可能に導入されている。本発明において形質転換体とは、複数個の形質転換細胞により構成された単位(multicellular form)を指し、細胞とは区別される。形質転換体とは、例えば、組織または器官だけでなく、多細胞生物の個体がその範疇に含まれることを意図する。
【0039】
上記の細胞、及び形質転換体の調製に用いる宿主細胞は、特に限定されるものではないが、産生されるポリペプチドが示す抗菌活性の影響を受け難いものがより好ましい。好ましいものとして、具体的には例えば、細菌類、動物細胞(例えば、昆虫細胞、両生類細胞、哺乳類細胞等)、及び植物細胞(高等植物の細胞、高等植物のプロトプラスト等)から選択される何れかが挙げられる。なお、糸状菌類、及び真菌類等の真核菌類を宿主細胞として用いても、本発明のポリペプチドの産生は可能である。これら宿主細胞のための適切な培養培地および培養条件は当分野で周知である。
【0040】
本発明に係る上記ポリヌクレオチド、又は上記発現ベクターを宿主細胞等に導入する方法、すなわち形質転換法も特に限定されるものではなく、PEG法、電気穿孔法、リン酸カルシウム法、リポソーム法、DEAEデキストラン法、パーティクルガン法、アグロバクテリウム法(植物細胞に限る)等の従来公知の方法を好適に用いることができる。また、例えば、本発明に係るポリペプチドを昆虫で転移発現させる場合には、バキュロウイルスを用いた発現系を用いればよい。
【0041】
本発明に係る形質転換体は、上記形質転換細胞から分化誘導させて得られる。或いは、複数個の細胞により構成された単位(multicellular form)に対して、本発明に係る上記ポリヌクレオチド、又は、本発明に係る上記ベクターを発現可能に導入して得てもよい。以下の説明では、複数個の植物細胞により構成された単位(以下、多細胞植物単位と称する)を形質転換して、本発明に係る形質転換体を得る方法を説明する。
【0042】
多細胞植物単位とは、植物体全体、植物器官(例えば葉、花弁、茎、根、種子など)、植物組織(例えば表皮、師部、柔組織、木部、維管束、柵状組織、海綿状組織など)、カルスなどのいずれをも意味する。植物の種類は特に限定されず、単子葉植物綱または双子葉植物綱に属する植物のいずれでもよい。また、多細胞植物単位への遺伝子の導入には、上記宿主細胞への遺伝子導入と同様、当業者に公知の形質転換方法が用いられる。
【0043】
形質転換の結果得られるカルスやシュート、毛状根などは、そのまま細胞培養、組織培養または器官培養に用いることが可能であり、また従来知られている植物組織培養法を用い、適当な濃度の植物ホルモン(オーキシン、サイトカイニン、ジベレリン、アブシジン酸、エチレン、ブラシノライドなど)の投与などによって植物体に再生させることができる。
【0044】
本発明に係るポリヌクレオチド又はベクターがゲノム内に組み込まれた形質転換植物体(個体)がいったん得られれば、当該植物体の有性生殖または無性生殖によって子孫を得ることができる。また、当該植物体またはその子孫、あるいはこれらのクローンから、例えば、種子、果実、切穂、塊茎、塊根、株、カルス、プロトプラストなどを得て、それらを基に当該植物体を量産することができる。したがって、本発明に係る形質転換体の範疇には、本発明に係るポリヌクレオチドが発現可能に導入された植物体、もしくは、当該植物体と同一の性質を有する当該植物体の子孫、またはこれら由来の組織も含まれる。
【0045】
本発明に係る形質転換体のより具体的な例示は以下の通りであるが、本発明は特にこれらの例示に限定されるものではない。
1)本発明に係るポリペプチドをコードする遺伝子を発現する遺伝子組換えイネ。当該イネは、糸状菌等に代表されるイネ病原菌に対する抵抗性を示すようになる。
2)本発明に係るポリペプチドをコードする遺伝子を発現する、イネ以外の遺伝子組換え植物体。当該植物体は、糸状菌等に代表される植物病原菌に対する抵抗性を示すようになる。
なお、イネ以外の植物の種類は特に限定されないが、具体的には例えば、イネ科(コムギ、オオムギ、ライ麦、トウモロコシ等でイネを除く)、ウリ科(例えば、キュウリ、メロン、スイカ、カボチャ、ニガウリ)、ナス科(例えば、トマト、ナス、ピーマン、パプリカ、トウガラシ、ジャガイモ、ペチュニア)、マメ科(例えば、インゲン、ソラマメ、アズキ、ダイズ、ササゲ、エンドウ)、キク科(例えば、ゴボウ、シュンギク、レタス、キク、ガーベラ、シネラリア、マリーゴールド)、アブラナ科(例えば、ハクサイ、キャベツ、ダイコン、カブ、コマツナ、ブロッコリー、チンゲンサイ、ストック)、アオイ科(例えば、オクラ、ワタ)、セリ科(例えば、セロリ、ニンジン、パセリ)、ユリ科(例えば、タマネギ、ニラ、アスパラガス、ネギ)、ナデシコ科(例えば、カスミソウ、カーネーション)、ミカン科(例えば、ミカン)、バラ科(例えば、バラ、イチゴ、リンゴ、ナシ、洋ナシ、モモ、ネクタリン)、アカザ科(例えば、ホウレンソウ)、等に属する植物が挙げられる。
【0046】
〔4.本発明に係るポリペプチドの用途の例示〕
(抗菌剤、及びその具体的用途)
本発明に係る抗菌剤は、本発明に係る少なくとも一種のポリペプチドを抗菌活性成分として含んでいる。本発明に係る抗菌剤は、必要に応じて、当該ポリペプチドを安定化させる安定化剤、酸化防止剤、等を含んだ組成物であってもよい。本発明に係る抗菌剤は、必要に応じて、製剤化のための、担体、等を含んだ組成物であってもよい。本発明に係る抗菌剤は、抗菌活性を増強する目的、或いは抗菌スペクトルをより拡張する目的で、本発明に係るポリペプチド以外の他の抗菌成分をさらに含んだ組成物であってもよい。
【0047】
本発明に係る抗菌剤は、原核菌類(亜界Procaryomycota)及び/又は真核菌類(亜界Eucaryomycota)に抗菌活性を示すことが好ましい。原核菌類の中では特に細菌門に属する菌類に対して抗菌活性を示すことが好ましい。また、真核菌類の中では特に糸状菌類、酵母類(カンジダ等)に対して抗菌活性を示すことが好ましい。
【0048】
本発明に係る抗菌剤の、より具体的な用途は特に限定されないが、例えば、糸状菌、及びカンジダに対する抗菌性を示しうるため、当該抗菌剤を、例えば、農薬又は医薬の分野で利用することもできる。また、本発明に係る抗菌剤を含む洗剤等を提供することもできる。
【0049】
本発明に係る抗菌剤のより具体的な一例は、以下の(a)及び(b)に示す本発明に係るポリペプチドの一方、又は両方を活性成分として含むものである。
(a)配列番号1に示すアミノ酸配列における連続する10個以上で16個以下のアミノ酸からなるポリペプチド;
(b)配列番号2に示すアミノ酸配列における連続する10個以上で15個以下のアミノ酸からなるポリペプチド。
【0050】
イネγ−チオニン蛋白質は、他の一般的な植物ディフェンシンとは異なり、複数箇所(上記(a)及び(b)に示すポリペプチドに相当)に抗菌活性ドメインを有するという特性を持つ。従って、上記(a)及び(b)に示すポリペプチドの両方を活性成分として含む抗菌剤は、イネγ−チオニン蛋白質により近い特性を示すと推定される。
【0051】
(殺菌又は制菌の方法)
本発明はまた殺菌又は制菌の方法を提供する。すなわち、当該方法は、殺菌又は制菌の対象となる原核菌類及び真核菌類に、本発明に係るポリペプチドを接触させることで、当該原核菌類及び真核菌類を殺菌又は制菌する。なお、「制菌」とは、本発明に係るポリペプチドを接触させない状態と比較して、原核菌類及び/又は真核菌類の活動が抑制されることを指す。なお、原核菌類及び/又は真核菌類の活動が抑制されたか否かは、例えば、増殖速度の低下、又は増殖の停止(静菌)等を指標にして判断することが出来る。
【0052】
(その他のより具体的な応用)
本発明に係るポリペプチドは何れも小分子で、その抗菌活性に立体構造の影響を受けない。そのため、他の蛋白質分子に融合させることで、融合蛋白質分子に新たに抗菌性を付与することが出来る。
【0053】
植物繊維又は工業的に素材となる蛋白質に、本発明に係るポリペプチドを融合させることで、抗菌性の素材を開発することが出来る。一例では、本発明に係るポリペプチドが絹糸を構成する蛋白質に融合された融合蛋白質を産生するようにカイコの遺伝子組換えを行い、抗菌性を有する絹糸を得ること等が考えられる。
【0054】
本発明に係るポリペプチドと比較して細菌に抗菌スペクトルを示す抗菌性蛋白質と、本発明に係るポリペプチドとを融合することで、細菌と糸状菌とに強い抗菌活性を有する蛋白質を作製出来る。
【0055】
本発明は上述した各実施形態に限定されるものではなく、請求項に示した範囲で種々の変更が可能であり、異なる実施形態にそれぞれ開示された技術的手段を適宜組み合わせて得られる実施形態についても本発明の技術的範囲に含まれる。
【実施例】
【0056】
以下、本発明を実施例、比較例及び参考例によりさらに詳細に説明するが、本発明はこの例に限定されるものではない。
【0057】
〔参考例1:イネディフェンシン様蛋白質(γ−チオニン蛋白質)の製造、及び抗菌活性〕
(1)γ−チオニン蛋白質の活性部位(「AK121915(Def)」)の製造
イネ由来のγ−チオニン蛋白質は、抗菌活性に寄与する49個のアミノ酸から構成される活性部位(成熟領域)と、31個のアミノ酸から構成されるシグナルペプチド部位とからなる。本参考例では、以下の方法に従い、上記活性部位のみを製造した。
【0058】
初めに、イネ(品種:日本晴)のゲノムDNAを鋳型として、プライマー1(CGCGGATCCAGGCACTGCCTGTCGCAGAGC)とプライマー2(CCGGAATTCCTAGCAGACCTTCTTGCAGAAG)とを用いた定法のPCR(Polymerase Chain Reaction)により、上記活性部位をコードする遺伝子断片を増幅した。
【0059】
PCRの後、増幅された遺伝子断片に対してBamHI及びEcoRIによる制限酵素処理を行った。次いで、制限酵素処理後の遺伝子断片を、BamHI及びEcoRIで同様に制限酵素処理したpGEX6P−1ベクター(GEヘルスケア社)に挿入し、当該ベクターを用いて大腸菌Rosetta-gamiB(DE3)pLysS株(タカラバイオ社)に形質転換を行った。得られた形質転換体を培養し、IPTG(Isopropyl-thio-β-D-galactoside)添加により誘導的な蛋白質発現を行った。発現蛋白質の精製はGSTrapTMFF column (GEヘルスケア社)及びHiTrapTM CM FF column (GEヘルスケア社)を用いて行い、精製蛋白質からのGST(Glutathion S transferase)タグの除去はPreScission Protease (GEヘルスケア社)を用いて行った。この一連のプロセスにより、γ−チオニン蛋白質の活性部位(AK121915(Def))を得た。
【0060】
(2)AK121915(Def)が有する抗菌活性の測定
図1に示す複数種の糸状菌、酵母(真菌)、及び複数種の細菌に関して、AK121915(Def)が抗菌活性を示すか否かを確認した。結果を図1に示す。図1に示す通り、糸状菌及び酵母に対する抗菌活性が顕著であることが確認された。なお、糸状菌及び酵母に対する抗菌活性の試験方法は、後述する実施例2及び3に示す通りである。
【0061】
また、細菌に対する抗菌活性の測定も、Broekaertらの方法を参考にして行った(参考文献:Broekaert et al. 1990 FEMS. Microbiol. Lett., 69, 55-60.)。測定開始時の菌体濃度(濁度)はOD600が0.1となるように調整した。抗菌活性の測定は、96穴のマイクロプレートを用い、ポリペプトン培地(1L作製につき、ポリペプトン:10g、酵母エキス:2g、MgSO4/7H2O:1g、pH7.0)を1/4濃度に希釈したものを細菌培養用培地として使用した。
【0062】
次いで、AK121915(Def)を含む蛋白質溶液を適当な濃度に希釈して、マイクロプレートの各ウェルに添加し、経時的に濁度測定を行うことで、蛋白質溶液に依存的な細菌に対する生育阻害を測定した。生育阻害の測定温度は25℃とした。なお、図1に示すように、細菌では、25μl/ml以下の濃度では抗菌活性が見られなかったため、IC50値は算出していない。
【0063】
〔実施例1:短鎖ペプチドの合成〕
AK121915(Def)のアミノ酸配列に基づき、短鎖ペプチドを合成した。短鎖ペプチドは北海道システムサイエンス社の受託ペプチド合成により合成した。純度は98%以上のものとした。
【0064】
合成した短鎖ペプチドのアミノ酸配列は以下のとおりである。なお、各配列の後に括弧書きで、AK121915(Def)のアミノ末端側から数えて何番目のアミノ酸に相当するかを示している。
No.1: RHCLSQSHRF(1−10番目のアミノ酸)
No.2: VSSNNCANV(15−23番目のアミノ酸)
No.3: RTESFPDGE(25−33番目のアミノ酸)
No.4: KSHGLERKCF(35−44番目のアミノ酸)
No.5: SHRFKGMCVS(7−16番目のアミノ酸)
No.6: SNNCANVCRTE(17−27番目のアミノ酸)
No.7: FPDGECKSHG(29−38番目のアミノ酸)
No.8: ERKCFCKKVC(40−49番目のアミノ酸)
なお、上記No.1〜No.8の短鎖ペプチドを、順に、AK121915(1-10)、AK121915(15-23)、AK121915(25-33)、AK121915(35-44)、AK121915(7-16)、AK121915(17-27)、AK121915(29-38)、AK121915(40-49)、と称する場合もある。
【0065】
〔実施例2:短鎖ペプチドの抗菌活性試験(糸状菌)〕
参考例1で得たAK121915(Def)、及び実施例1で得た短鎖ペプチドを用いて、イネいもち病菌(Magnaporthe grisea)に対する抗菌活性の測定を行った。
【0066】
抗菌活性の測定は、Broekaertらの方法を参考にして行った(参考文献:Broekaert et al. 1990 FEMS. Microbiol. Lett., 69, 55-60.)。また、イネいもち病菌としてMagnaporthe grisea Guy11株(独立行政法人農業生物資源研究所の西村麻里江博士から新潟薬科大学高久洋暁博士へ分譲されたものを使用)を使用した。抗菌活性の測定は、96穴のマイクロプレートを用い、培地は1/2濃度のpotato dextrose broth (Difco)を使用し、イネいもち病菌の胞子は20,000個/mLとなるように調整した。
【0067】
次いで、AK121915(Def)又は何れかの上記短鎖ペプチドを含む蛋白質溶液を適当な濃度に希釈して、マイクロプレートの各ウェルに添加し、経時的に濁度測定を行うことで、蛋白質溶液に依存的なイネいもち病菌に対する生育阻害を測定した。生育阻害の測定温度は25℃とした。そして、蛋白質濃度と生育阻害率とから各蛋白質のIC50値を算出した。また、対照試験としてWakoのカスガマイシン塩酸塩水和物を用いた。カスガマイシンは、イネいもち病菌等に対する抗菌剤である。
【0068】
抗菌活性試験の結果は、図2及び表1に纏めて示す。なお、カスガマイシンのIC50値は2.5(μg/ml)であった。
【0069】
【表1】
【0070】
〔実施例3:短鎖ペプチドの抗菌活性試験(酵母)〕
参考例1で得たAK121915(Def)、及び実施例1で得た短鎖ペプチドを用いて、カンジダ(Candida albicans)に対する抗菌活性の測定を行った。
【0071】
抗菌活性の測定は、Broekaertらの方法を参考にして行った(参考文献:Broekaert et al. 1990 FEMS. Microbiol. Lett., 69, 55-60.)。また、カンジダとしてCandida albicans CAI4株(千葉大学真菌医学研究センターから新潟薬科大学高久洋暁博士に分譲されたものを使用)を使用した。抗菌活性の測定は、96穴のマイクロプレートを用い、培地は1/2濃度のpotato dextrose broth (Difco)を使用し、測定開始時のカンジダ菌体の濁度はOD600が0.01となるように調整した。
【0072】
次いで、AK121915(Def)又は何れかの上記短鎖ペプチドを含む蛋白質溶液を適当な濃度に希釈して、マイクロプレートの各ウェルに添加し、経時的に濁度測定を行うことで、蛋白質溶液に依存的なカンジダに対する生育阻害を測定した。生育阻害の測定温度は30℃とした。そして、蛋白質濃度と生育阻害率とから各蛋白質のIC50値を算出した。
【0073】
抗菌活性試験の結果は、図2に纏めて示す。
【0074】
〔実施例4:AK121915(Def)、及び短鎖ペプチドの標的分子の探索〕
これまで、植物ディフェンシンが抗菌活性を発揮するための微生物上の標的分子として、グルコシルセラミド及びマンノシル・ジイノシトール・ホスホリル・セラミド(mannosyl diinositolphosphoryl ceramide)が候補とされてきた(参考文献:Thevissen et al. 2005 FEBS Let. 579 : 1973-1977)。
【0075】
そして、標的分子の候補がグルコシルセラミドとされている植物ディフェンシンとしては、カブのディフェンシンRs-AFP2が、標的分子の候補がマンノシル・ジイノシトール・ホスホリル・セラミドとされている植物ディフェンシンとしては、ダリアのディフェンシンDm−AMP1が、これまで比較的詳細に解析されてきた。
【0076】
本実施例では、AK121915(Def)、及びその短鎖ペプチドAK121915(1-10)の標的分子が、グルコシルセラミド又はマンノシル・ジイノシトール・ホスホリル・セラミドである可能性について以下の検証をした。
【0077】
(1) 標的分子がグルコシルセラミドである可能性の検証
カラシナディフェンシンBj-AFP1、イネディフェンシンAK121915(Def)、及びAK121915(1-10)の三種の蛋白質及びペプチドを用い、Candida albicans CAI4株、C. albicans CAI4 △gcs株(新潟薬科大学高久洋暁博士がCAI4株より作製)、Saccharomyces cerevisiae W303-1A株(ATCCより入手可)に対する抗菌活性を測定した。抗菌活性の測定は、何れも実施例3の記載に従い行った。測定の結果は、表2に纏めて示す。表2中の○(抗菌活性有)はIC50値が15μg/mL以下であることを示し、×(抗菌活性無)はIC50値が25μg/mL以上であることを示す。
【0078】
なお、Bj-AFP1はカブ由来のディフェンシンRs-AFP2と高い相同性を有する。そのため、Rs-AFP2と同様にBj-AFP1もグルコシルセラミドが抗菌活性を発揮するための標的分子と推定されている。また、カンジダ(C. albicans CAI4株)は、細胞膜成分としてグルコシルセラミドを有する。カンジダのグルコシルセラミド合成系の遺伝子gcs (Glucosylceramide Synthase)を破壊した△gcs株はグルコシルセラミドを合成できない。また、出芽酵母(S. cerevisiae W303-1A株)は細胞膜成分としてのグルコシルセラミドを持たない。
【0079】
【表2】
【0080】
表2に示す通り、Bj-AFP1はグルコシルセラミドが存在する株にのみ抗菌活性を示したことから、予想通りグルコシルセラミドが標的分子である(もしくは関与している)可能性が高い。一方、AK121915(Def)及びその短鎖ペプチドAK121915(1-10)はグルコシルセラミドの有無に関係なく全ての株に対し抗菌活性を示したことから、グルコシルセラミドがターゲットの候補にはならない。すなわち、AK121915(Def)及びその短鎖ペプチドはBj-AFP1をはじめとする植物ディフェンシンと作用機序が異なる可能性が示唆される。
【0081】
(2) 標的分子がマンノシル・ジイノシトール・ホスホリル・セラミドである可能性の検証
カラシナディフェンシンBj-AFP1、イネディフェンシンAK121915(Def)、及びAK121915(1-10)の三種の蛋白質及びペプチドを用い、S. cerevisiae W303-1A株、S. cerevisiae W303-1A△skn1△ipt1株(新潟薬科大学高久洋暁博士がW303-1A株より作製)、C. albicans CAI4株に対する抗菌活性を測定した。抗菌活性の測定は、何れも実施例3の記載に従い行った。測定の結果は、表3に纏めて示す。表3中の○(抗菌活性有)はIC50値が15μg/mL以下であることを示し、×(抗菌活性無)はIC50値が25μg/mL以上であることを示す。
【0082】
なお、出芽酵母(S. cerevisiae W303-1A株)は細胞膜成分としてマンノシル・ジイノシトール・ホスホリル・セラミドを有する。一方、カンジダ(C. albicans CAI4株)は細胞膜成分としてマンノシル・ジイノシトール・ホスホリル・セラミドを持たない。また、S. cerevisiae W303-1A株においてskn1遺伝子及びipt1遺伝子を破壊したS. cerevisiae W303-1A△skn1△ipt1株は、マンノシル・ジイノシトール・ホスホリル・セラミドを合成できない。
【0083】
【表3】
【0084】
表3に示す通り、AK121915(Def)及びその短鎖ペプチドAK121915(1-10)は、マンノシル・ジイノシトール・ホスホリル・セラミド分子の有無に関係なくS. cerevisiae及びC. albicansに対し抗菌活性を示した。このことからAK121915(Def)及びその短鎖ペプチドの標的分子は、ダリアのディフェンシンDm−AMP1ともタイプの異なる抗菌性蛋白質であることが示唆された。
【0085】
〔参考例2:カラシナディフェンシンBj-AFP1由来の短鎖ペプチドの抗菌活性〕
図3に示すように、Bj-AFP1はAK121915(Def)とほぼ同様の位置に8つのシスティン残基を有するという、ディフェンシンに特有の共通構造を持つ。そこで、Bj-AFP1の配列に基づいて、実施例1に示す短鎖ペプチドAK121915(1-10)、AK121915(15-23)、AK121915(25-33)、及びAK121915(35-44)に対応する短鎖ペプチドを合成した。合成した短鎖ペプチドは順に、Bj-AFP1(1-10)、Bj-AFP1(16-24)、Bj-AFP1(26-35)、及びBj-AFP1(37-46)として図3中に示す。なお、これら短鎖ペプチドは、北海道システムサイエンス社の受託ペプチド合成により合成した。純度は98%以上のものとした。
【0086】
その上で、実施例2及び3に記載した方法に従い、Bj-AFP1に基づく上記短鎖ペプチド、及びBj-AFP1の活性部位(全長)の、イネいもち病菌、及びカンジダに対する抗菌活性を測定した。その結果を図3中に纏めて示す。結果から明らかなように、カラシナディフェンシンBj-AFP1では、AK121915(1-10)に対応する短鎖ペプチドBj-AFP1(1-10)でさえ、抗菌活性を示さないことが明らかとなった。
【0087】
〔実施例5:AK121915(Def)の短鎖ペプチドAK121915(1-10)の改変体〕
AK121915(Def)のアミノ酸配列情報から、1―10番目のアミノ酸からなる短鎖ペプチドAK121915(1-10)(表4のNo.1。実施例1も参照)をベースに、アミノ酸の置換、欠失、及び付加を組み合わせた5種類の改変短鎖ペプチドを合成した。改変短鎖ペプチドは北海道システムサイエンス社の受託ペプチド合成により合成した。純度は98%以上のものとした。
【0088】
合成した改変短鎖ペプチドNo.2〜No.6のアミノ酸配列は、表4に示す。
【0089】
【表4】
【0090】
表4に示す改変短鎖ペプチドNo.2は、短鎖ペプチドAK121915(1-10)の10番目のアミノ酸Fを欠失させたものである。改変短鎖ペプチドNo.3は、短鎖ペプチドAK121915(1-10)の3番目のアミノ酸Cと、10番目のアミノ酸Fとを欠失させたものである。改変短鎖ペプチドNo.4は、短鎖ペプチドAK121915(1-10)の3番目のアミノ酸Cを欠失させ、10番目のアミノ酸Fをアミノ酸R(アルギニン)に置換し、かつC末端にアミノ酸Rを付加したものである。改変短鎖ペプチドNo.5は、短鎖ペプチドAK121915(1-10)の10番目のアミノ酸Fをアミノ酸Rに置換したものである。改変短鎖ペプチドNo.6は、短鎖ペプチドAK121915(1-10)の3番目のアミノ酸C及び10番目のアミノ酸Fを何れもアミノ酸Rに置換したものである。
【0091】
次いで、実施例3と同様の方法で、表4のNo.1〜6で示した短鎖ペプチドを用いてカンジダ(Candida albicans)に対する生育阻害を測定し、IC50値(単位はμg/ml)を算出した。結果を表4に示した。特に、短鎖ペプチドAK121915(1-10)の10番目のアミノ酸Fの欠失やアミノ酸Rへの置換、或いは、3番目のアミノ酸Cの欠失やアミノ酸Rへの置換を行っても、その抗菌活性は維持又は向上することが示された。
【産業上の利用可能性】
【0092】
本発明に係るポリペプチド等は抗菌剤、抗菌性付与植物の作出等に利用できる。
【特許請求の範囲】
【請求項1】
以下の(a)又は(b)に示すポリペプチド:
(a)配列番号1に示すアミノ酸配列における連続する10個以上で16個以下のアミノ酸からなるポリペプチド
(b)配列番号2に示すアミノ酸配列における連続する10個以上で15個以下のアミノ酸からなるポリペプチド。
【請求項2】
配列番号1に示すアミノ酸配列における1番目〜10番目のアミノ酸を含む請求項1に記載のポリペプチド。
【請求項3】
配列番号1に示すアミノ酸配列における1番目〜10番目のアミノ酸からなる請求項2に記載のポリペプチド。
【請求項4】
請求項1から3の何れか1項に記載のポリペプチドにおいて、1から3個のアミノ酸が置換、欠失、及び/又は、付加されてなり、抗菌活性を有するポリペプチド。
【請求項5】
請求項1から3の何れか1項に記載のポリペプチドにおいて、1又は2個のアミノ酸が置換されてなる、及び/又は、末端側に位置する1又は2個のアミノ酸が欠失されてなり、抗菌活性を有するポリペプチド。
【請求項6】
請求項1から5の何れか1項に記載のポリペプチドをコードしているポリヌクレオチド。
【請求項7】
請求項6に記載のポリヌクレオチドを含んでいるベクター。
【請求項8】
請求項6に記載のポリヌクレオチド、又は請求項7に記載のベクターが発現可能に導入されている細胞、又は複数個の当該細胞により構成された形質転換体。
【請求項9】
請求項7に記載のベクターを用いて形質転換されている植物。
【請求項10】
請求項1から5の何れか1項に記載のポリペプチドを活性成分として含んでいる抗菌剤。
【請求項11】
以下の(a)及び(b)に示すポリペプチドの両方を活性成分として含んでいる請求項10に記載の抗菌剤:
(a)配列番号1に示すアミノ酸配列における連続する10個以上で16個以下のアミノ酸からなるポリペプチド
(b)配列番号2に示すアミノ酸配列における連続する10個以上で15個以下のアミノ酸からなるポリペプチド。
【請求項1】
以下の(a)又は(b)に示すポリペプチド:
(a)配列番号1に示すアミノ酸配列における連続する10個以上で16個以下のアミノ酸からなるポリペプチド
(b)配列番号2に示すアミノ酸配列における連続する10個以上で15個以下のアミノ酸からなるポリペプチド。
【請求項2】
配列番号1に示すアミノ酸配列における1番目〜10番目のアミノ酸を含む請求項1に記載のポリペプチド。
【請求項3】
配列番号1に示すアミノ酸配列における1番目〜10番目のアミノ酸からなる請求項2に記載のポリペプチド。
【請求項4】
請求項1から3の何れか1項に記載のポリペプチドにおいて、1から3個のアミノ酸が置換、欠失、及び/又は、付加されてなり、抗菌活性を有するポリペプチド。
【請求項5】
請求項1から3の何れか1項に記載のポリペプチドにおいて、1又は2個のアミノ酸が置換されてなる、及び/又は、末端側に位置する1又は2個のアミノ酸が欠失されてなり、抗菌活性を有するポリペプチド。
【請求項6】
請求項1から5の何れか1項に記載のポリペプチドをコードしているポリヌクレオチド。
【請求項7】
請求項6に記載のポリヌクレオチドを含んでいるベクター。
【請求項8】
請求項6に記載のポリヌクレオチド、又は請求項7に記載のベクターが発現可能に導入されている細胞、又は複数個の当該細胞により構成された形質転換体。
【請求項9】
請求項7に記載のベクターを用いて形質転換されている植物。
【請求項10】
請求項1から5の何れか1項に記載のポリペプチドを活性成分として含んでいる抗菌剤。
【請求項11】
以下の(a)及び(b)に示すポリペプチドの両方を活性成分として含んでいる請求項10に記載の抗菌剤:
(a)配列番号1に示すアミノ酸配列における連続する10個以上で16個以下のアミノ酸からなるポリペプチド
(b)配列番号2に示すアミノ酸配列における連続する10個以上で15個以下のアミノ酸からなるポリペプチド。
【図1】


【図2】
【図3】

【図2】
【図3】
【公開番号】特開2013−31431(P2013−31431A)
【公開日】平成25年2月14日(2013.2.14)
【国際特許分類】
【出願番号】特願2012−138180(P2012−138180)
【出願日】平成24年6月19日(2012.6.19)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り ・刊行物名 日本農芸化学会2011年度大会講演要旨集 発行日 2011年3月5日 発行所 社団法人日本農芸化学会
【出願人】(501203344)独立行政法人農業・食品産業技術総合研究機構 (827)
【Fターム(参考)】
【公開日】平成25年2月14日(2013.2.14)
【国際特許分類】
【出願日】平成24年6月19日(2012.6.19)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り ・刊行物名 日本農芸化学会2011年度大会講演要旨集 発行日 2011年3月5日 発行所 社団法人日本農芸化学会
【出願人】(501203344)独立行政法人農業・食品産業技術総合研究機構 (827)
【Fターム(参考)】
[ Back to top ]