
新規な発現ベクター
【課題】翻訳後検定及び転写後検定における、改良されたベクター及びシステム、並びに遺伝子発現における変化のより一層実時間測定を可能にする検定法を提供する。
【解決手段】ヌクレオチド配列を導入するための複数クローニンブ部位、レポーター遺伝子、概転写可能なポリヌクレオチドの発現を調節するためのプロモーター及び/またはエンハンサー、ポリアデニル化配列、選択可能マーカー遺伝子、及び複製起点からなる群より選択される一つ以上の構成員を含む、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列からなる該転写可能なポリヌクレオチド発現ベクターからなる。
【解決手段】ヌクレオチド配列を導入するための複数クローニンブ部位、レポーター遺伝子、概転写可能なポリヌクレオチドの発現を調節するためのプロモーター及び/またはエンハンサー、ポリアデニル化配列、選択可能マーカー遺伝子、及び複製起点からなる群より選択される一つ以上の構成員を含む、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列からなる該転写可能なポリヌクレオチド発現ベクターからなる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は一般的に遺伝子発現又は遺伝子調節検定におけるベクター及びその使用に関する。より具体的には、本発明はタンパク質発現の動力学にプロモーター活性との改良された時的相関関係を与える発現ベクター及び/又はレポーターベクターを提供する。本発明はとりわけ遺伝子発現を調節するために有用な、ヒト疾病における治療的介入のため調節配列、新たな標的及び薬物を同定し分析するため及び薬物スクリーニングのために有用な新規なベクター及び細胞系統を提供する。
【背景技術】
【0002】
本発明者らが本明細書で言及した出版物の文献学的詳細は本明細書の末尾に集めてある。
【0003】
一以上の任意の先行技術文献を含む本明細書における先行技術についての言及は、該先行技術が当分野の技術水準であることを認め又は示唆するものと取られるべきではない。
【0004】
組換えDNA技術の精緻化の急速な進展は医学及び関連する厚生分野における研究及び発展を大いに促進している。とりわけ重要な研究領域は遺伝子発現を研究するための発現ベクターの利用である。しかしながら、今日まで、遺伝子発現の実時間分析は適切に設計されたベクターの不存在により制限されてきた。
【0005】
レポーター検定は、例えば、DNA配列、転写因子、RNA配列、RNA結合タンパク質、シグナル変換経路、及び特異的刺激などの何が目的の遺伝子の発現を制御するのかを理解させる。
【0006】
さらに、レポーター検定はヒト疾病における治療的介入のための新たな標的として役立つ遺伝子調節の局面を同定するために使用できる。レポーター検定は遺伝子発現を調節する能力について薬物をスクリーニングするために使用できる可能性がある。しかしながら、現在のレポーター検定システムに必要な費用と時間は、応答時間の長いことと併せ、その適用を妨げてきた。
【0007】
ゲノム配列は、一般にコード領域の上流にプロモーター配列を有し、それは細胞の特異性と転写の誘導可能性を規定し、それによりタンパク質産物の発現レベルに影響を与える。
【0008】
典型的にはヌクレオチド塩基A及びUが豊富であり、時には遺伝子の3’UTRに位置する特異的な配列因子は、mRNAの安定性に影響を与え、それによりタンパク質産物の発現レベルに影響を与える。RNA結合タンパク質は特定のmRNA配列に結合し、それによりmRNAの安定性及びタンパク質の発現を調節する。他の配列は翻訳効率を調節する。
【0009】
遺伝子レポーター検定の普通の適用は転写を調節するDNA配列の研究である。通常、これらの配列は転写開始部位の5’側、プロモーター領域に位置している。このようなDNA因子は、転写、従ってレポータータンパク質の発現をを駆動及び/又は調節するようにレポータープラスミド内の類似の部位の中にそれらをクローニングすることにより試験される。このレポータータンパク質は内因性タンパク質から識別可能であり、容易に定量されるべきである。種々のレポータータンパク質が使用され、最も普通なのはルシフェラーゼ、クロラムフェニコールトランスフェラーゼ(CAT)及びβガラクトシダーゼ(β-gal) である。
【0010】
レポータータンパク質は適当な検定法で定量され、しばしば、例えばプロモーター SV40 などのどこにでもあるプロモーターにより駆動される対照レポーターのレベルと比較して表される。この対照レポーターは、テストレポーターとは識別可能でなければならず、テストベクターと共トランスフェクトされる別のベクターに含まれていなければならず、トランスフェクション効率を制御するために使用されねばならない。このような検定は細胞が両方のベクターを量に比例して等しく摂取するという前提に基づいている。プラスミドベクターの一過性のトランスフェクションは最も普通に使用される。
【0011】
うえの検定はプロモーター内のプロモーター領域又は特定の因子を同定するために用いられる。または、これらの検定はプロモーター又は調節因子の種々の刺激に対する応答を研究するために用いられる。一部の適用では、レポーター構築物又はトランスフェクトされた細胞はイン・ビボにおけるプロモーター機能を研究するため生物中に配置される。
【0012】
これらのレポーター検定の別の適用は特定のプロモーターの上流のシグナル変換経路の研究又は測定である。例えば、転写のためMAPキナーゼ(MAPK) に依存するあるプロモーターを、レポーター構築物に連結して細胞中のMAPK活性化(又はMAPK依存性転写)のレベルを測定するために使用することができる。この技術は様々な情報を与えるプロモーター又はエンハンサーと共に利用することができ、そして細胞又はトランスジェニックマウスなどの生きている生物に適用できる。例えば、フォトンカメラは目的のプロモーターに連結したルシフェラーゼレポーターを含む完全マウスにおけるルシフェラーゼレポーター活性を測定するために使用できる(特許文献1を参照)。
【0013】
ルシフェラーゼはこれまでイン・ビトロ系のレポーター検定に最も普通に使用されている。二重ルシフェラーゼ検定(DLA,プロメガ, マジソン, WI, USA)は、テストレポーターと対照レポーターの両方が本質的に同一検定で測定され得るという点で他のルシフェラーゼ系システムに優る改良がなされている。現在の使用の一例として、典型的なDLA法は次のように行なわれる。
【0014】
プロモーター因子と推定されるものを、蛍のルシフェラーゼレポーター遺伝子の上流にクローニングし、それがその発現を駆動するようにする。このプラスミドを SV40 プロモーターにより駆動されるウミシイタケ(renilla)のルシフェラーゼ遺伝子を含む対照のプラスミドと共に細胞系統中に一時的にトランスフェクトする。約2〜50%の細胞がプラスミドを取込み、約3日間このレポーターを発現する。発現の動力学は、ルシフェラーゼタンパク質が蓄積する最初の約24時間の間の増加、及びそれに続き細胞内に維持されたプラスミドの数が減少する約48時間以降の低下を含む。トランスフェクションから24〜48時間の後、細胞を収穫し溶解する。細胞の溶解液を蛍ルシフェラーゼに特異的な基質と共にインキュベートし、活性(光の放射)をルミノメータ(96穴プレート又は個々の試料) を用いて測定する。次に、蛍のルシフェラーゼを失活させるがウミシイタケのルシフェラーゼに光を形成させる追加の基質を添加する。ウミシイタケのルシフェラーゼの活性を次に測定する。
【0015】
蛍のルシフェラーゼ活性のレベルはプロモーター活性のみでなくトランスフェクション効率にも依存する。これは、DNAの量、DNA調製物の品質、及び細胞の状態に応じて大きく変化する。共トランスフェクトされた対照のプラスミド( SV40 プロモーターにより駆動されるウミシイタケのルシフェラーゼ)は、ウミシイタケのルシフェラーゼの活性が細胞によって取り込まれた蛍のルシフェラーゼプラスミドの量に比例するという前提に基づいてこれらの変数を補正するために使用される。データは蛍ルシフェラーゼ活性/ウミシイタケのルシフェラーゼ活性として表される。
【0016】
二重ルシフェラーゼ検定の欠点は
(i) 試薬が高価であり、変質しやすく、調製して直ぐに使用しなければならない、
(ii) 一般的にこの検定は細胞溶解液の調製物を含む。これは時間がかかり、不正確さを加える。例えば、溶解の操作中の細胞の損失、ピペット操作の誤差、残存緩衝液/培地の容量が変化する、
(iii) 個々の試料は細胞集団の全活性である唯一つのデータのみを生ずる。レポーターを発現する細胞のパーセンテージに関する情報も、細胞当たりの発現の量に関する情報も得られない、
(iv) トランスフェクション対照(ウミシイタケ)はトランスフェクション効率における大きな変動を常に補正するとは限らない。何故なら
a. 特定のDNA調製物は僅かしかトランスフェクト/発現せず(おそらくスーパーコイルDNAの割合の減少により)、共トランスフェクトさせる対照プラスミドの量は相当する減少を惹き起こさないこと、
b. 二つのプラスミドのプロモーターの間に、対照レポーターの活性が共トランスフェクトされる構築物に依存する、例えば、ウミシイタケのルシフェラーゼの発現が強力なプロモーターを含むプラスミドと共トランスフェクトされた場合に最高となるようにみえる、というような、クロス−トークの証拠があること。プロモーターの間の干渉も、テストレポーターと対照レポーターの両方を発現する1個のプラスミドの使用を、阻害はしないとしても、制限したということ、
c. 転写及び転写後の両方の研究における普通の適用は、種々の刺激(例えば、PMA 、EGF 、ホルモン) による活性化/抑圧を測定することである。不幸にも、SV40、RSV 、TK 及びおそらく他の多数の普遍的に発現されるプロモーターが様々な刺激により活性化される。これらのプロモーターはトランスフェクション対照レポーター(ウミシイタケ)の発現を駆動するために使用されるから、これらのレポーターはこのような処置の後のトランスフェクション効率の真の反映を与えない(非特許文献1を参照)こと、
d. 蛍対ウミシイタケのルシフェラーゼタンパク質及びおそらくmRNAの半減期の相違は系全体を極めて時間感受性とすること、
e. とりわけウミシイタケのルシフェラーゼにおける急速に減少する光放射は、測定のタイミングにおける絶対的精度を必要とすること、
f. ルシフェラーゼタンパク質及びmRNAの比較的長い半減期は、転写における時間的変化を効果的に覆い隠す(例えば、種々の刺激又は処理の後)こと。
である。
【0017】
現行の転写後/mRNA安定性レポーター検定では、mRNA安定性に影響を与えると考えられた候補因子を SV40 又は RSVなどの構成プロモーターにより駆動されるレポーターベクター(例えば、蛍のルシフェラーゼ)の対応する領域にクローニングする。空のベクター(目的の因子を含まない同じベクター)と比較した場合の発現の変化は改変されたmRNA安定性の結果であると仮定される。予備的に記述されたトランスフェクション検定については、トランスフェクション対照プラスミド(例えば、 SV40 又は RSVなどの構成プロモーターにより駆動されるウミシイタケルシフェラーゼ)を共トランスフェクトさせて、トランスフェクション効率について補正させる。これらの検定は
(i) 現存するベクターは転写後研究のために設計されておらず、転写を切り換えるための手段を持たない、
(ii) これらのプロトコルの目的は候補mRNA因子の転写後の効果を研究することであるが、これらの因子はDNAのレベルでレポーターの転写に影響を与えることもでき、さらに、目的の遺伝子の内因性プロモーターは使用されないから、見られる如何なる転写効果も生理学的関連性はほとんどない、
というさらなる不利益を有する。
【0018】
mRNA安定性を研究するための系は存在するが、タンパク質レポーターではなくmRNAの直接測定を含む。mRNAを定量化するためのプロトコル類の労働集約的性質のため、このような系はさらに一層時間がかかる。
【0019】
例えば、一つの系は血清誘導に応答して転写の短時間の爆発をもたらす c-fosプロモーターを利用する。不安定因子と思われるものを、血清誘導性の(c-fos) プロモーターの制御下で極めて安定なmRNAを発現するベータグロビン(BBB) 構築物の 3-UTR中にクローニングする。トランスフェクトされた細胞(一般に NIH 3T3細胞) をまず血清から断絶させ、ついで血清を含む培地に曝露する。この転写の応答の簡単な性質のため、レポーターmRNAの分解の動力学を経時的に追跡することが可能となる。これらの検定は
(i) この検定は極めて時間がかかり、従って迅速なスクリーニングに適用できない、
(ii) c-fos プロモーターの血清誘導性を支える細胞中でしか使用できない。例えば、多数の腫瘍細胞系統は血清の非存在下でc-fos プロモーター活性を維持する、
(iii) 所望の血清応答を有するNIH 3T3 細胞などの細胞では、血清涸渇は細胞周期を阻止する原因となり、その後血清を添加すると、同時に、この阻止から細胞を解き放つ。それ故、mRNAの安定性は細胞周期の特定の期でしか測定できない、
(iv) c-fos プロモーターの活性化に加え、血清は他の経路の多くを活性化する。これらの経路は望ましくない変数を導入し、より特異的な刺激の研究を阻害する、
という不利益を有する。
【0020】
別の検定では、細胞は全ての遺伝子からの転写を阻害するアクチノマイシンDなどの薬物で処理される。mRNAの分解速度を決定するため、そのmRNAレベルが経時的に測定される。この系は内因性遺伝子を研究するために用いられるが、
(i) 転写阻害剤は、mRNA安定性がストレスを与えた又は死にかかっている細胞でしばしば測定される場合に必要な用量では極めて毒性である、
(ii) 転写阻害剤は特定のmRNAの安定化などの無数の望ましくない活性を有する、
(iii) このプロセスは全ての遺伝子からの転写を阻止し、多数のシグナル変換カスケードが阻止される一方、他のカスケードは活性化される。従って、結果は生理学的に重要であるように思われない、
(iv) この技術は極端に労働集約的である、
(v) この技術は検定内及び検定間で極めて変動し易い、
(vi) この技術は一過性のトランスフェクションレポーター検定においてはしばしば感度が十分でなく、特に低いトランスフェクション効率を持つ細胞中ではそうである。
という欠点を有する。
【先行技術文献】
【特許文献】
【0021】
【特許文献1】コンタグら,米国特許第5,650,135号公報。
【非特許文献】
【0022】
【非特許文献1】イブラヒムら,内部対照プラスミドの落とし穴:ウミシイタケ・ルシフェラーゼプラスミド(pRL-TK) のジヒドロテストステロン及びデキサメタソンへの応答, Biotechniques 2000; 29: 682-684 。
【発明の概要】
【発明が解決しようとする課題】
【0023】
従って、遺伝子発現検定を行なうための、とりわけ翻訳後検定及び転写後検定における、改良されたベクター及びシステム、並びに遺伝子発現における変化のより一層実時間測定を可能にする検定法を開発する必要がある。
【課題を解決するための手段】
【0024】
発明の概要
本明細書を通じて、文脈が別意を要求しない限り、用語「含む(comprise) 」及び「comprises 」や「comprising」などのその変形は、述べられた整数若しくは工程又は整数若しくは工程の群を含むことを意味するが、他の整数若しくは工程又は整数若しくは工程の群の排除を意味するものではないと理解される。本発明は、例示の目的のみを意図する本明細書に記載された具体的な実施態様によりその範囲が制限されるべきではない。機能的に等価な産物、組成物及び方法は、本明細書に記載されたような本発明の範囲内にあることは明らかである。
【0025】
本発明によれば、本発明者はとりわけ転写物の安定性の調節及び測定及び/又は遺伝子発現の実時間測定の改良を可能とする一連のベクター及び方法を開発した。
【0026】
ヌクレオチド配列は配列識別子数(配列番号:)により言及される。この配列番号:は配列識別子<400>1、<400>2などに数値で対応している。配列表は請求の範囲の後ろに提示する。
【0027】
本発明の一つの側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む発現ベクターに向けられている。本明細書で用いられるとき転写物の安定性とはこの転写物の半減期に相当すると考えてよい。
【0028】
本発明の別の一側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節し且つ該転写物の安定性を低減させる不安定化因子であるRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む発現ベクターを意図する。
【0029】
関連する実施態様では、本発明は転写可能なポリヌクレオチドに対応する転写物の安定性を調節し且つ該転写物の安定性を増加させる安定化因子であるRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む発現ベクターを意図する。
【0030】
本発明の別の一側面は、RNA因子をコードするヌクレオチド配列を含む転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写可能なポリヌクレオチドに対応する転写物の安定性を低減させる不安定化因子であり、該転写可能なポリヌクレオチドが目的のポリヌクレオチドを含み且つポリペプチドをコードするものである発現ベクターを意図する。
【0031】
本発明のさらに別の一側面は、RNA因子をコードするヌクレオチド配列を含む転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写可能なポリヌクレオチドに対応する転写物の安定性を低減させる不安定化因子であり、該転写可能なポリヌクレオチドが目的のポリヌクレオチドを含み且つレポーターポリペプチドをコードするものである発現ベクターを意図する。
【0032】
本発明のさらなる別の一側面は、RNA因子をコードするヌクレオチド配列を含む転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写可能なポリヌクレオチドに対応する転写物の安定性を低減させる不安定化因子であり、該転写可能なポリヌクレオチドがレポーターポリペプチドをコードするヌクレオチド配列を含むものである発現ベクターを意図する。
【0033】
本発明のなお別の一側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写可能なポリヌクレオチドに対応する転写物の安定性を低減させる不安定化因子であり、該ベクターが
(i) ヌクレオチド配列を導入するための複数クローニング部位、
(ii) レポーター遺伝子、
(iii) 該転写可能なポリヌクレオチドの発現を調節するためのプロモーター及び/又はエンハンサー、
(iv) ポリアデニル化配列、
(v) 選択可能なマーカー遺伝子、及び
(vi) 複製起点、
からなる群より選択される一つ以上の構成員を含むものである、発現ベクターに向けられている。
【0034】
本発明のさらに別の一側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む本発明のベクターを含む細胞を意図する。
【0035】
本発明のさらに別の一側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む本発明のベクターを含む細胞であって、該RNA因子が該転写物の安定性を低減する不安定化因子である細胞を意図する。
【0036】
本発明の関連する側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む遺伝的に改変された非ヒト生物であって、該RNA因子が該転写物の安定性を低減させる不安定化因子である生物を意図する。
【0037】
本発明のさらなる実施態様は、目的のポリヌクレオチドの発現を測定する方法であって、RNA合成及びタンパク質合成が起こるのに十分な時間及び条件の下で発現ベクターから該目的のポリヌクレオチドを発現させる工程、この工程では、該ベクターが目的の該ポリヌクレオチド及び転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含むものであり、且つ該発現ベクターが
(i) ヌクレオチド配列を導入するための複数クローニング部位、
(ii) レポーター遺伝子、
(iii) 該転写可能なポリヌクレオチドの発現を調節するためのプロモーター及び/又はエンハンサー、
(iv) ポリアデニル化配列、
(v) 選択可能なマーカー遺伝子、及び
(vi) 複製起点、
から成る群より選択される一つ以上の構成員を含むものである、及び転写可能なポリヌクレオチドの発現産物のレベル及び/又は機能的活性を対照と比べ経時的に測定する工程、
この工程では該因子が該転写可能なポリヌクレオチドに機能しうるように連結されたプロモーター及び/又はエンハンサーの活性と該発現産物のレベル及び/又は機能的活性の間の時的相関関係を強化するものである、を含む方法を意図する。
【0038】
さらに別の一実施態様では、本発明はRNA転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を同定する方法であって、発現ベクター中にテストヌクレオチド配列を導入することにより該ヌクレオチド配列をレポータータンパク質をコードするポリヌクレオチドに連結させプロモーター及び/又はエンハンサーに機能しうるように連結された転写可能なポリヌクレオチドを形成させる工程、RNA合成及びタンパク質合成が起こるのに十分な時間及び条件の下で該転写可能なポリヌクレオチドを発現させる工程、
この工程で該発現ベクターは
(i) 該テストヌクレオチド配列を導入するための複数クローニング部位、
(ii) ポリアデニル化配列、
(iii) 選択可能なマーカー遺伝子、及び
(iv) 複製起点、
から成る群より選択される一つ以上の構成員を含むものである、及び該ヌクレオチド配列の非存在下における対照ベクターのそれと比べ該転写可能なポリヌクレオチドの発現産物のレベル及び/又は機能的活性を経時的に測定する工程、この工程でその時間にわたって対照ベクターのそれと異なるレベル及び/又は機能的活性は該RNA因子をコードするヌクレオチド配列の存在を示す、を含む方法を意図する。
【図面の簡単な説明】
【0039】
【図1】図1は、不安定化mRNAをコードする発現ベクターの図式的表示である。
【図2】図2は、転写レポーターベクターの図式的表示である。図2aはベクター系2、図2bはベクター系3、そして図2cはベクター系4を示す。
【図3】図3は、双方向性転写レポーターベクターの図式的表示である。図3aはベクター系5を、そして図3bはベクター系6を示す。
【図4】図4は、転写後調節を研究するためのレポーターベクターの図式的表示である。図4aはベクター系7を、そして図4bはベクター系8を示す。
【図5】図5は、トランスフェクトされたDNAの量に対する尺度としてのレポーター活性を示すグラフ的表示である。蛍ルシフェラーゼをコードするプラスミドの一本鎖DNA調製物をウミシイタケルシフェラーゼをコードする別のプラスミドと30:1の比率で混合する。両方のDNA調製物は分光測光法(OD260/280)とエチジウムブロマイド染色アガロース・ゲル(データは示していない)において、正常に見えた。この混合物をDNAの総量が1、2又は3マイクログラムになるように細胞内に量を変えてトランスフェクトしたが、蛍プラスミドとウミシイタケプラスミドの比率は同じままであった。 図5Aはウミシイタケルシフェラーゼ活性がトランスフェクトされたDNAの量に依存することを示すグラフである。しかしながら、蛍ルシフェラーゼ活性(図5B)はDNAの量を増やしても増加しなかった。これは恐らく、蛍DNA調製物の品質が悪かったためであろう。従って、通常蛍プロモーター活性の尺度として用いられる蛍/ウミシイタケ比率(図5C)は、使用されたDNAの量に応じてかなり変わった。これらのデータはウミシイタケプラスミドとの共トランスフェクションが蛍プラスミドのトランスフェクション効率を適切に制御しないことを示す。
【図6】図6は二重ルシフェラーゼ検定を用いて得られた様々なプロモーター系に対するレポーター活性を示すグラフ的表示である。六つの異なるプロモーター断片(1−6と番号を付す)を、pGL3蛍ルシフェラーゼプラスミドにクローニングした。1マイクログラムの各クローンをSV40プロモーターで駆動されたウミシイタケ(トランスフェクション対照)プラスミドの30ngと共トランスフェクトし、蛍ルシフェラーゼとウミシイタケルシフェラーゼの活性を二重ルシフェラーゼ検定(プロメガ、米国、マディソン群、ウィスコンシン)で測定した。その結果がウミシイタケルシフェラーゼ活性(A)、蛍ルシフェラーゼ活性(B)、及びウミシイタケで割った蛍の活性(C)として表す。それぞれの構築物の少なくとも二つの異なる調製物を用いた複数の実験で同様の結果が見られた。 ウミシイタケルシフェラーゼ活性(図6A)はトランスフェクションの対照を意図したものであり、この結果を単独で分析すると異常に高い変動がトランスフェクション効率にあること示唆するであろう。例えば、ウミシイタケルシフェラーゼ活性は構築物3と共トランスフェクトした場合と比べて、構築物4と共トランスフェクトした場合のほうが3.5倍高い。DNA品質のばらつき又はDNAの定量の際の誤差のばらつきは、別のDNA調製物のセットでも同じパターンが見られた(データ非提示)ことから、エラーの原因であるとは思われない。 蛍ルシフェラーゼ活性(図6B)はトランスフェクション効率とプロモーター1〜6の間の差異の両方の影響を受ける。この差異のパターンは、ウミシイタケで見られる(図6A)ものと類似している。例えば、3と6は低く、一方4と5は高い。しかしながら、構築物間のこのような差異は、蛍で一層顕著であり(例えば、構築物4は構築物3より12倍高い)、プロモーター1〜6の活性がウミシイタケの発現に何らかの形で影響を与えている(またはその逆)ことを示唆する。 蛍/ウミシイタケ(図6C)は、トランスフェクション効率(ウミシイタケ)を修正した後に真の蛍プロモーター活性(1〜6)の尺度になると考えられる。ここでも実際に3と6が最も弱いプロモーターで、一方4と5が最強であることを示唆する類似のパターンが見られる。プロモーター活性(図6C)がトランスフェクション効率(6A)と偶然に相関したという可能性もあるが、多数の異なる構築物及び同じ構築物の複数の異なる調製物で類似の結果が得られたことを考えれば、この可能性はまずあり得ない。ウミシイタケルシフェラーゼの発現レベルがそれと共トランスフェクトされるプロモーター構築物の強さの影響を受けるという方が可能性が大きいであろう。従って、プロモーター1〜6の間の見掛けの差異は、真の差異を過小評価している可能性もある。
【図7】図7は転写の阻害後のBTL、BTG2、BTG1及びBTG1N4発現ベクターに対する異なるレポーターレベルを経時的に示すグラフ的表示である。Tet-オフ・ヒーラ細胞をそれぞれがレポーター遺伝子と連結したTREプロモーターを含んでいる次のレポータープラスミドでトランスフェクトした。即ち、 BTL(ルシフェラーゼ)、BTG2(d2EGFP)、BTG1(d1EGFP)、及びBTG1N4(BTG1と同じであるが、3’UTR−コード領域に存在する9量体UUAUUUAUUの4つのコピーを持つ)。トランスフェクションから10時間後、それぞれのフラスコの細胞を複数の小皿に分けた。ドキシサイクリン(1□g/ml)をトランスフェクション(タイム・ゼロ)の24時間後に添加し、レポーター遺伝子の転写を阻害した。実施例14に記載したように、レポーターレベル(蛍光又は発光)をこの時点及びその後の時点で測定し、タイム・ゼロのパーセンテージとして表した。10時間経過中にルシフェラーゼ活性(BTL)の減少は見られなかった。2時間半減期のEGFP構築物(BTG2)はドキシサイクリンで誘導される転写の阻害に対して穏やかな反応を示し、1時間半減期のEGFP(BTG1)ではより早い反応が見られた。しかしながら、9量体を含む構築物(BTG1N4)では、転写の阻害に対して群を抜いて速い反応を示した。
【図8】図8は、図7で使用したデータを線状スケールで表したもの示すグラフ的表示である。ドキシサイクリンで誘導される転写の阻害はBTG1で約6.5時間後のレポーターレベルで50%の阻害として検出できる。しかしながら、これは9量体(BTG1N4)を含めることにより3時間にまで減少する。
【図9】図9は、異なる数(1、2又は4)の9量体RNA不安定化因子の効果を示すグラフ的表示である。時間経過は、ゼロ時間をこの薬物の活性の遅延効果を除去するためにドキシサイクリンを添加して4時間後と定義したことを除き、図7に記載した通り実施した。1個の9量体の存在(BTG1N1)は「減衰の有効速度」を増加するのに十分であり、2個の9量体(BTG1N2)及び4個の9量体(BTG1N4)でその効果が徐々に強くなることが見られた。後者の構築物は約1時間20分の「有効半減期」を示したが、それはタンパク質単独の1時間半減期と大差はない。
【図10】図10は、転写の阻害がない状態で時間と共に変化するレポーターレベルを示すグラフ的表示である。時間の経過は図7に記載した通り実施した。しかしながら、示したデータはドキシサイクリンで処置されておらず、トランスフェクションの24時間後(開始)又はトランスフェクションの34時間後(終了)に測定された試料を表している。一貫した発現レベルはBTG1N4でしか見られなかった。
【図11】図11は、転写の阻害がない状態で時間と共に変化するレポーターレベルを示すグラフ的表示である。時間の経過は図7に記載したとおり実施した。BTG1 fosは c-fos ARE を含む。これらのデータは別の種類のmRNA不安定化因子が同じ効果を得るために使用できることを証明する。
【図12】図12は、ルシフェラーゼレポータータンパク質が使用された場合にも、RNA不安定化因子は発現を測定するのに役立つということを示すグラフ的表示である。ルシフェラーゼレポータータンパク質をタンパク質不安定化因子と共に使用すれば、更なる増強が期待できるであろう。時間経過は、二つのルシフェラーゼ発現構築物を用いて図7に記載したとおり実施した。BTLは標準的な蛍ルシフェラーゼのコード領域と3’UTR(pGL3−ベーシック、プロメガ)を含み、一方 BTLN6は3’UTRに9量体UUAUUUAUUの6個のコピーを含む。
【図13】図13は、RNA不安定化因子及びタンパク質不安定化因子により不安定化されたDsRedを用いたレポーターレベルの時間経過を示すグラフ的表示である。時間経過は図7及び実施例14に記載したとおり実施した。使用した構築物は、3’UTRに4個の UUAUUUAUU9量体(BRT1N4)を含むDsRed2(BTR)、DsRed-MODC(BTR1)及びDsRed-MODCであった。ドキシサイクリンで転写を阻害した後、赤色蛍光が全ての構築物で増え続けた。これはタンパク質不安定化因子により実質的に減少し、mRNA不安定化因子によってさらに減少する。
【図14】図14は、時間経過が図7に記載されたとおりに実施されたことを示すグラフ的表示である。テストしたmRNA不安定化因子は全て、対照(BTG1)と比べて減衰速度の増加に非常に効果的であった。これらのデータは、c-myc AREが効果的な不安定化因子(BTG1myc)であり、不安定化活性の緩やかな増加が myc AREと4個の9量体を結合(BTG14myc)させることにより得られるということを示す。6個の9量体(BTG1N6)もまた4個の9量体(BTG1N4)よりも幾分強く不安定化するようであった。
【発明を実施するための形態】
【0040】
好ましい実施態様の詳細な説明
定義
「3’UTR」とは、ポリヌクレオチドのタンパク質コード領域の終止コドンの下流のポリヌクレオチド領域、これはタンパク質の生産のために翻訳されることはない、ことを意味する。
「5’UTR」とは、mRNAの5’(上流)側の翻訳されない領域を意味する。mRNAの5’UTRをコードするDNA領域を指すためにも使用される。
「約」とは、参照する数量、レベル、値、寸法、大きさ、又は量に対し 30%まで、好ましくは 20%まで、より好ましくは10% まで変動する数量、レベル、値、寸法、大きさ、又は量を意味する。
【0041】
「ARE」とは、mRNAにおけるAU−豊富因子、即ちアデニン及びウラシルヌクレオチドの高い割合を含む配列を意味する。このようなmRNA因子をコードするDNA領域を指すためにも用いられる。
【0042】
「生物学的に活性な断片」とは、参照ポリヌクレオチド又はポリペプチドの活性をそれぞれ保持する全長参照ポリヌクレオチド又はポリペプチドの断片を意味する。
「c-fos 」とは、有糸分裂シグナルにより簡単に誘導される中間初期遺伝子を意味する。
「CAT 」とは、クロラムフェニコール・アセチルトランスフェラーゼを意味する。レポーターとしてしばしば用いられる細菌の酵素である。
「d1EGFP」とは、突然変異した PEST 配列に融合し従って約1時間の半減期しか持たない EGFP の変異型を意味する。同様に、d1ECFP及び d1EYFP も入手可能である。DsRed の不安定化変異型は同様な方法で作成できるであろう。今後は d1DsRedと呼ばれる。
【0043】
「d2EGFP」とは、 PEST 配列に融合し従って約2時間の半減期しか持たない EGFP 変異型の突然変異型を意味する。同様に、d2ECFP (藍色) 及び d2EYFP(黄色) も入手可能である。DsRed の不安定化変異型はおそらく同様にして作成できるであろう。今後はd2DsRed と呼ばれる。
「dEGFP 」とは、生成した EGFP(全ての色を含む) の全ての不安定化変異型(リーら)に対する一般的用語を意味する。
「DNA」とは、デオキシリボ核酸を意味する。
「誘導体」とは、例えば、他の化合物部分と共役させ若しくは複合体形成させることにより又は当分野で理解されているような転写後若しくは翻訳後修飾により参照ポリヌクレオチド又はポリペプチドからそれぞれ誘導されたポリヌクレオチド又はポリペプチドを意味する。
「DsRed 」とは、インドパシフィックシー・アネモネ近似のディスコソマ・スピーシーズから単離された赤色蛍光タンパク質を意味する。
「ECFP」とは、藍色の光の蛍光を発する、改変された励起/放射スペクトルを持つ EGFP の突然変異型を意味する。
【0044】
「EGF 」とは、表皮成長因子を意味する。
「EGFP」とは、強化された緑色蛍光タンパク質を意味する。強化された蛍光を持つ GFPの突然変異型である(コルマックら)。
「ELISA 」とは、酵素結合イムノソルベント検定法を意味する。
「ErbB2 」とは、表皮成長因子受容体ファミリーの第2の構成員を意味する。HER-2 としても知られている。
「エキソン」とは、メッセンジャーRNA分子又はこの配列をコードするDNAの一部であるRNA一次転写物の配列を意味する。一次転写物では、隣接するエキソンはイントロンにより隔てられている。
「発現ベクター」とは、DNAのクローニングされたセグメントを細胞内で発現させるベクターを意味する。
「EYFP」とは、黄色の光の蛍光を発する改変された励起/放射スペクトルを有する EGFP の突然変異型を意味する。
【0045】
「蛍ルシフェラーゼ」とは、蛍からクローニングされた luc遺伝子から誘導される酵素を意味する。酸素と Mg++の存在下で D- ルシフェリン及び ATPを用いて反応を触媒し結果として光の放射を生ずる。しばしばレポーターとして用いられる。
「フローサイトメトリー」とは、生きた又は固定した細胞の懸濁液を、懸濁液の細胞と結合した検出可能な標識の活性又は性質を個々に測定するフローサイトメーターにかける方法を意味する。細胞の標識化は、例えば蛍光性化合物を介して又は特定の蛍光性化合物に共有結合させた抗体により行なうことができる。異なる幾つかの励起/放射波長を同時にテストし異なるタイプの蛍光を測定することができる。所望の特性(蛍光、細胞の大きさ)を持つ細胞の部分集団をゲートにかけ、ゲートを通った細胞だけをさらなる統計分析にかけることができる。細胞選別機オプションを備えたフローサイトメーターは所望の蛍光を持つ細胞を物理的に分離し、最初の細胞集団の残りからこれらの(生きた)細胞を管中に回収することができる。FACS(蛍光標示式細胞分取法)とも呼ばれる。
「遺伝子」とは、RNA分子をコードするDNAのセグメントを意味する。「遺伝子」という用語は、時には、常にではないが、プロモーター領域を含むことがある。
「GFP 」とは、蛍光タンパク質(ツィエンら)を意味する。これはクラゲ、エクオリア・ヴィクトリアから単離されうるものであり、レポータータンパク質として使用することができる。GFP をコードするDNA構築物は哺乳動物細胞中で発現でき、特定の波長で励起すると細胞が緑色の蛍光を発するようにすることができる。この用語「GFP 」は色の変異型及びエクオリア・ヴィクトリア以外の生物から誘導される蛍光タンパク質(例えば、DsRed,クローンテック、hrGFP,ストラタジーン)を含む全ての相同体及び類似体を指すために本明細書で用いられる。
「半減期」とは、分子の活性、量若しくは数の半分が消失するのに必要な時間を意味する。
「イントロン」とは、遺伝子、又はその一次転写物内の非コード配列を意味する。これは該一次転写物から除去され、対応するメッセンジャーRNA中に存在しない。
【0046】
「ルシフェラーゼ」とは、光の放射をもたらす反応を触媒する普通に使用されるレポーター酵素を意味する。外因性基質を添加し、ルミノメーターを用いてこの反応を定量化する。蛍及びウミシイタケのルシフェラーゼに対する基質の要件は異なっているので、二重ルシフェラーゼ検定法(プロメガ,マディソン,WI, USA)では両者を識別することができる。
「MAPK」とは、有糸分裂活性化タンパク質キナーゼを意味する。成長又はアポトーシス(細胞死)に至る細胞内シグナル変換経路に関与する幾つかの異なるキナーゼを含む。用語「MAPK」はしばしば二つの特定の MAPKs、Erk1及びErk2(細胞外の調節キナーゼ1及び2)を指すために用いられる。
「MCMV」とは、最小 CMVプロモーターを意味する。それ自体の転写を活性化しないが、TRE に連結されるとテトラサイクリン(及びドクシサイクリン) 依存性の転写を行なわせることができる。
【0047】
「MCS 」とは、複数クローニング部位を意味する。その中にDNA断片を挿入できるユニークな制限酵素認識部位を含むDNAベクターの領域である。用語「MCS 」は本明細書で用いられるとき、このベクター中へのDNA断片の挿入を助ける任意の他の部位をも含む。例えば、Tオーバーハング(プロメガ,マディソン,WI, USA)である。これはポリメラーゼ連鎖反応(PCR)により形成された断片の直接挿入を可能とする。
「mRNA」とは、メッセンジャーRNAを意味する。DNAを鋳型として用い細胞内で生産される、それ自体はタンパク質をコードする「転写物」である。mRNAは通常5’UTR、タンパク質をコードする領域及び3’UTRから構成される。mRNAは細胞内で限定された半減期を有し、それはとりわけ3’UTR内にある、しかし5’UTR及びタンパク質をコードする領域にもある安定性因子により部分的に決定される。
「MODC」とは、マウス・オルニチン・デカルボキシラーゼ又は PEST 配列を含むその部分及び/又は誘導体を意味する。
「調節する」とは、目的の分子の安定性を、直接的若しくは間接的のいずれかで増加又は低減させることを意味する。
【0048】
「機能しうるように連結される」とは、機能的な関係の中でポリヌクレオチド因子を連結することを意味する。核酸は、それが別の核酸配列と機能的な関係に置かれる場合に「機能しうるように連結される」。例えば、プロモーター又はエンハンサーは、それがコード配列の転写に影響を与える場合、コード配列に機能しうるように連結されている。機能しうるように連結されるとは、連結されている核酸配列が通常隣接している、そして二つのタンパク質コード領域を結合する必要がある場合は、隣接し且つ読み取り枠内にあることを意味する。RNAポリメラーゼが二つのコード配列を一つのmRNAに転写する場合コード配列は別のコード配列に「機能しうるように連結されている」。mRNAは次いで両方のコード配列から誘導されるアミノ酸を持つ一本のポリペプチドに翻訳される。このコード配列は発現された配列が最終的にプロセスされ所望のタンパク質を生産する限り相互に隣接している必要はない。転写可能なポリヌクレオチドにプロモーターを「機能しうるように連結する」とは、転写可能なポリヌクレオチド(例えば、タンパク質をコードするポリヌクレオチド又は他の転写物)をプロモーターの調節的制御の下に置くことを意味する。このプロモーターは次いで該ポリヌクレオチドの転写を制御し、任意選択的に翻訳を制御する。プロモーター/構造遺伝子の異質の組み合わせの構築では、転写可能なポリヌクレオチドの転写開始部位から離れた位置にプロモーター又はその変異型を配置することが好ましい。この距離はその天然の設定、即ち、プロモーターがそれから由来した遺伝子における、プロモーターとそれが制御する遺伝子の間の距離とほぼ同じである。当分野で知られているように、この距離のある程度の変化は機能の損失無しに順応できる。同様に、その制御の下に置かれる転写可能なポリヌクレオチドに関する調節配列因子(例えば、オペレーター、エンハンサーなど)の好ましい配置は、天然の設定、即ちそれが由来した遺伝子における該因子の位置により規定される。
【0049】
「pA」とは、ポリアデニル化部位を示すために本明細書のダイアグラム中で用いられる、転写を停止させるための部位として及び未成熟mRNAにポリA尾を添加するために働くDNA配列を意味する。SV40ウイルス遺伝子又はβガラクトシダーゼ遺伝子又は合成ポリアデニル化部位を含む他の供給源からの様々な pA 配列を、この目的で発現ベクター中で使用できる。
「PEST」は、アミノ酸プロリン(P)、グルタミン酸(E)、セリン(S)、及びスレオニン(T)の豊富なアミノ酸配列を指す。PEST配列を含むタンパク質は半減期が短い。
「プラスミド」とは、環状DNAベクターを意味する。プラスミドは宿主細胞DNA中にプラスミドを組み込むことなしに細菌細胞(しばしば真核細胞)中でプラスミドの多数のコピーを生産させる複製起点を含む。
「ポリヌクレオチド」又は「核酸」とは、DNA又はRNAを含むヌクレオチドの線状配列を意味する。これは二本鎖でも一本鎖でもよい。
【0050】
「ポリペプチド」、「ぺプチド」又は「タンパク質」とは、特定の配列でぺプチド結合により結合されたアミノ酸のポリマーを意味する。
「プロモーター」とは、一般にmRNAをコードする領域の上流(5’)のDNA領域を意味する。これは転写の開始及び転写レベルを制御する。この用語はその範囲内で誘導プロモーター、抑圧プロモーター、及び構成プロモーターをも含む。
「PMA 」とは、ホルボール・ミリストロイックアシッドを意味する。
「ウミシイタケのルシフェラーゼ」とは、シーパンジー(レニッラ・レニホルミス)から誘導され、酸素とコエレンテレートルシフェリン(コエレンテラジン)を利用して光放射を形成する酵素を意味する。
「レポーターベクター」とは、容易に検定できるタンパク質又はポリペプチド(又はmRNA)をコードする「レポーター遺伝子」を含む発現ベクターを意味する。このレポーター遺伝子はその機能又は活性がテストされる調節配列に連結されるのが通常である。
【0051】
「レポーター」とは、レポーター遺伝子によりコードされそしてレポーター検定法で測定される、通常タンパク質又はポリペプチドである分子を意味する。現在のシステムは一般に酵素レポーターを利用しそしてレポーター活性を測定する。
「RNA」とは、リボ核酸を意味する。
「rtTA」とは、TRE に結合しテトラサイクリン又はドキシサイクリンの存在下でのみ転写を活性化する逆 tTA (下記を参照) を意味する。
「SEAP」とは、分泌されるアルカリホスファターゼレポーター遺伝子を意味する。
「SKBR3 」とは、ErbB2 を過剰発現するヒト乳癌細胞系統を意味する。
【0052】
「ストリンジェント条件」とは、高頻度の相補的塩基を有するヌクレオチド配列のみがハイブリッドを形成する温度及びイオン条件を意味する。要求されるストリンジェンシーはヌクレオチド配列依存性でありハイブリダイゼーションの間に存在する様々な成分及びその後の洗浄、及びこれらの処理にかける時間に依存する。一般に、ハイブリダイゼーション速度を最大にするため、熱的融点(Tm)よりも約20〜25℃低い、非ストリンジェント・ハイブリダイゼーション条件が選択さる。このTmは規定されたイオン強度及びpHで溶液中で完全に相補的なプローブに特定の標的配列の50%がハイブリッド形成をする温度である。一般に、ハイブリダイズされる配列の少なくとも約85%のヌクレオチド相補性を要求するためには、Tmよりも約5〜15℃低い高ストリンジェント洗浄条件が選択される。ハイブリダイズされる配列の少なくとも約70%のヌクレオチド相補性を要求するためには、Tmよりも約15〜30℃低い中程度のストリンジェント洗浄条件が選択される。高度に寛容な(低ストリンジェンシー)洗浄条件はTmより50℃も低いものであり、ハイブリダイズされる配列の間の高レベルの不適合を許容する。当業者はハイブリダイゼーション及び洗浄の段階で、標的配列とプローブ配列の間の相同性の特定のレベルからの検出可能なハイブリダイゼーションシグナルの結果に影響を与えるように他の物理的及び化学的パラメータを変更することもできることを認識するはずである。
【0053】
「SV40/CMV/RSV」は、サルウイルス、サイトメガロウイルス及びラウス肉腫ウイルスからそれぞれ誘導されたプロモーター因子を指す。一般に、これらのプロモーターは哺乳動物細胞で構成的に活性であると考えられている。
「TetO」とは、大腸菌のテトラサイクリン耐性オペロンから誘導されたTet オペレーターDNA配列を意味する。
「Tet-Off 細胞系統」とは、テトラサイクリン又はドキシサイクリンが TREプロモーターからの転写を遮断するように tTAを安定に発現する細胞系統を意味する。
「Tet-On細胞系統」とは、テトラサイクリン又はドキシサイクリンが TREプロモーターからの転写を点火するように rtTA を安定に発現する細胞系統を意味する。
「転写」とは、DNA鋳型に相補的なRNA分子を合成するプロセスを意味する。
【0054】
「トランスフェクション」とは、プラスミド又はDNA断片が真核細胞の中に挿入されるプロセスを意味する。通常、2〜50%の細胞がプラスミドを取込み、細胞の染色体にプラスミドDNAを組み込むことなしに(一過性のトランスフェクション)約3日間タンパク質産物を発現する。これらの細胞の小部分はついにはそれらのゲノム中にプラスミドDNAを組み込み、該タンパク質産物を永久に発現する(安定なトランスフェクション)。
「翻訳」とは、mRNA分子がタンパク質合成のための鋳型として使用されるプロセスを意味する。
「TRE 」とは、本明細書では、転写が外因性の因子(例えば、tTA 又は rtTA)の TREへの結合を介してのみ起こるように、一般に最小のプロモーターと結合された任意のテトラサイクリン応答因子(ゴッセンら)を規定する。本発明の好ましい実施態様では、最小CMVプロモーター(mCMV)(クローンテック・ラボラトリーズ社,Palo Alto, CA, USA) に連結した tetO 配列の7個の反復から構成されるTRE を利用する。
「tTA 」とは、テトラサイクリンにより制御されたトランスアクチベーターを意味する。
これは、TRE に結合し、テトラサイクリン又はドキシサイクリンの非存在下でのみ転写を活性化するように、Tet 抑制タンパク質(TetR)及び VP16 活性化ドメインから構成される。
【0055】
「TS」とは、トロンボキサンシンターゼのプロモーターを意味する。
「変異型」とは、参照ポリヌクレオチド又はポリペプチドとそれぞれ実質的な配列同一性を示すポリヌクレオチド又はポリペプチドを意味する。変異型ポリヌクレオチドはストリンジェント条件下で参照配列とハイブリッドを形成するポリヌクレオチドをも含む。これらの用語は参照配列と少なくとも一つのヌクレオチドの付加、欠失又は置換だけ異なるポリヌクレオチドをも包含する。この点で、突然変異を含む特定の改変、付加、欠失及び置換が参照ポリヌクレオチドに成され得ることそしてそれにより改変されたポリヌクレオチドが参照ポリヌクレオチドの生物学的機能又は活性を保持することが当分野でよく知られている。「ポリヌクレオチド変異型」及び「変異型」という用語は天然に生ずる対立遺伝子変異型をも含む。変異型ポリペプチドに関しては、例えば一部のアミノ酸はポリペプチドの活性の本質を変更することなく幅広く類似の性質を持つ他のアミノ酸に変更しうること(同類置換)が当分野で良く理解されている。
【0056】
「ベクター」とは、宿主細胞に外来性DNA配列を挿入し、該ベクターの複製を支える細胞中でこのDNA配列を増幅させるビークルを意味する。最も普通にはプラスミドであるが、ファージミド、バクテリオファージ、アデノウイルス又はレトロウイルスであってもよい。
「vEGFP 」、「EGFP変異型」又は「EGFPの変異型」とは、EGFPの異なる色の変異型及び/又は異なる半減期の変異型を意味する。
「vGFP」とは、GFP の全ての変異型を意味する。これには DsRedなどの相同体や類似体、EGFP変異型又は不安定化された GFP変異型をも含まれる。
【0057】
本発明はとりわけ転写物の安定性、従って該ベクターにより生産されるタンパク質の量、を調節する発現ベクターを提供する。転写物の安定性を増加させる発現ベクターが本発明により包含されることは明らかであるが、特に好ましい実施態様は不安定化されている転写物に的を絞っている。この場合、転写物の安定性は、転写可能なポリヌクレオチドに一つ以上の不安定化因子を添加することにより、又は該ポリヌクレオチドから一つ以上の安定性因子(例えば、ポリA尾)を除去することにより低減させることができる。現存する発現ベクターと比べ、本発明のベクターは、例えば減少したプロモーター活性と対応する発現産物のレベルの減少との間の時間差を減少させることにより、該プロモーター活性に対する改良された時的相関関係を持つタンパク質発現の動力学を提供する。
【0058】
従って、本発明の一つの側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む発現ベクターに向けられる。
【0059】
転写物の安定性の文脈における用語「調節する」とは、転写物の安定性を増加又は減少させることを意味し、調節の最適量は具体的な適用に応じて変化する。本発明を如何なる特定の理論又は作用様式に限定するものではないが、RNA因子が転写物を不安定化するヌクレオチド配列である場合、該因子は転写物を直接的又は間接的に標的化してこれを分解するものと考える。
【0060】
本明細書で用いるとき用語「不安定化因子」とは、細胞内で、タンパク質又は転写物の半減期をそれぞれ低減させるアミノ酸配列又はヌクレオチド配列を意味する。従って、「RNAを不安定化する因子」はRNA転写物の細胞内半減期を低減させるヌクレオチド配列を含み、そして「タンパク質を不安定化する因子」はタンパク質の細胞内半減期を低減させるアミノ酸配列を含む。
【0061】
求められる低減の程度は具体的適用に応じて変化する。好ましい実施態様では、RNA不安定化の程度は、不安定化因子を含まないベクターと比べ、プロモーター活性と発現ベクター中のレポーターのレベル又は活性との間の時的相関関係を有意に改良する。転写物の安定性を増加させることと関連して、安定性の最適レベルはここでも具体的適用に応じて変化する。
【0062】
「RN59定化因子」は細胞内半減期を増加させるヌクレオチド配列である。
【0063】
「RNA」分子には、mRNA、ヘテロ核RNA(hnRNA) 、核内低分子RNA(snRNA) 、核小体RNA(snoRNA)、細胞質低分子RNA(scRNA) 、リボソームRNA(rRNA)、翻訳調節RNA(tcRNA) 、トランスファーRNA(tRNA)、eRNA、メッセンジャーRNA−干渉性相補RNA(micRNA)又は干渉性RNA(iRNA)及びミトコンドリアRNA(mtRNA) などの全てのRNA分子が含まれる。
メッセンジャーRNA(mRNA)はRNAの好ましい形態である。
【0064】
RNA転写物又は目的のコードされたタンパク質から選択された分子の細胞内半減期を低減させることと関連して、(a)一つ以上の不安定化因子及び/又は(b)一つ以上の安定化因子は通常該分子の分解レベルを増加させるように選択され、それにより該分子の細胞内半減期を、約24時間未満が適当であり、約10時間未満がより好ましく、約5時間未満がさらに好ましく、約3時間未満がより一層好ましく、約1時間未満がさらに一層好ましく、約30分未満がより好ましく、約15分未満がより好ましく、約10分未満がより好ましく、約5分未満がさらに好ましく、約3分未満がさらに一層好ましい半減期まで低減させる。RNA転写物又は目的のコードされたタンパク質の半減期はは該転写物又はコードされたタンパク質の最低検出可能レベルの少なくとも10倍の定常状態発現レベルを与える最低半減期に相当することが好ましい。
【0065】
細胞内又は細胞内類似の条件はその細胞型の生理的条件であることが好ましい。細胞内又は細胞内類似の条件の温度はその細胞型にとっての生理的な温度であることが好ましい。哺乳動物細胞の典型的温度は約30℃から約42℃までであることが適当であり、約35℃から約37℃までであることが好ましい。
【0066】
最低でも、RNA転写物又はポリペプチドのリボ核酸分解又はタンパク質分解のそれぞれの強化は、不安定化因子(単数又は複数)の非存在下で又は安定化因子(単数又は複数)の存在下でのRNA転写物又はポリペプチドの分解より、少なくとも約5%、好ましくは少なくとも約10%、より好ましくは少なくとも約20%、さらにより好ましくは少なくとも約40%、さらに一層好ましくは少なくとも約50%、さらに好ましくは少なくとも約60%、さらに好ましくは少なくとも約70%、さらに好ましくは少なくとも約80%、さらに好ましくは少なくとも約90%、さらに好ましくは少なくとも約100%、さらに好ましくは少なくとも約150%、さらに好ましくは少なくとも約200%、さらに好ましくは少なくとも約400%、さらに好ましくは少なくとも約600%、さらに好ましくは少なくとも約1000%、さらに好ましくは少なくとも約2000%、さらに好ましくは少なくとも約4000%、さらに好ましくは少なくとも約6000%、さらに好ましくは少なくとも約8000%、さらに好ましくは少なくとも約10000%、さらに好ましくは少なくとも約12000%大きいリボ核酸分解又はタンパク質分解のレベルを指す。RNA分解を測定する検定法は当業者に知られている。例えば、RNA分解は、例えば転写阻害剤(アクチノマイシンD、DRB、コルディセピン、アルファ−アマニチン)、パルス標識化(放射活性ヌクレオシド)、無細胞減衰法(ポリソーム、細胞質ゾル、又は網状赤血球)、又は短期間プロモーター活性化(fos プロモーター、以下を参照) の使用に基づくロス,ジェイ(1995) 又はリュー,ジェイら(JBC 2000) により開示された一連の検定法を用いて測定できる。タンパク質の分解を測定する検定法も当業者に知られている。例えば、タンパク質分解はバックメアーら(米国特許第5,646,017 号) の網状赤血球溶解液検定法を含む、これに限定されないが、哺乳動物細胞溶解液検定法を用いてイン・ビトロで測定しうる。別法として、タンパク質分解は、例えばヴァズハピリ,アールとズッヒェル,エヌ(2002) 又はサイトウ, ティーら(1998)により開示されたシクロヘキシイミド又はパルス−チェイス法を用いてイン・ビボで測定しうる。
【0067】
このRNA不安定化因子は、しばしば不安定化配列を含む如何なる供給源からも、とりわけ短寿命mRNAの3’UTR又は5’UTR領域から誘導することができる。本明細書で用いるとき、用語「から誘導される」は、特定のもの (integer)又はものの群が特定された種に起源を有するが、必ずしもその特定の供給源から直接得られるとは限らないことを示すものと解すべきである。
【0068】
RNA不安定化配列は、例えば c-fos、c-jun 、c-myc 、GM-CSF、 IL-3 、TNF-アルファ、IL-2、IL-6、IL-8、IL-10 、ウロキナーゼ、bcl-2 、SGLT1 (Na(+)- 共役グルコース・トランスポーター) 、Cox-2 ( シクロオキシゲナーゼ 2) 、IL-8、PAI-2 (2型プラスミノーゲン・アクチベーター・インヒビター) 、アドレナリン性ベータ1受容体、GAP43 (5’UTR及び3’UTR)などの短寿命RNA(単数又は複数)からクローニングしうる。
【0069】
AU豊富因子(AREs) 及び/又はU豊富因子(UREs)は9量体UUAUUUA(U/A)(U/A)(U/AはA又はUのいずれかである)(ラグナドら 1994)及び/又は5量体AUUUA(スーら 1997)及び/又は4量体AUUU(ズビアーガら 1995)の単一コピー、縦列コピー又は複数コピー又は重なり合うコピーを含むが、これらに限定されない。
【0070】
RNA不安定化因子は、例えば、ホスホエノールピルビン酸カルボキシキナーゼmRNA(PEPCK)、ショウジョウバエのビコイド遺伝子、ヒトチオレドキシン遺伝子、熱安定抗原及び大豆10A5遺伝子からも記載されてきた。
【0071】
鉄応答因子及び鉄調節タンパク質結合部位も本ベクター中に有利に組み込まれてRNA安定性及びとりわけ翻訳効率を調節しうる。ヒストンのRNA、とりわけ3’UTRは細胞周期に依存した様式でRNA安定性を調節するのに特に有用である。
【0072】
上に列挙した因子の修飾又は並べ替えも意図される。用語「縦列コピー」は外側のヌクレオチドの一つ以上を二重にする場合及び/又は二重にしない場合の両方を含む。例えば、5量体AUUUAの縦列コピーはAUUUAAUUUAAUUUAだけでなくAUUUAUUUAUUUAなどの配列も含む。
【0073】
RNAを不安定化する因子が同定され、そしてそれの改変がコンピュータ手法及びデータベース分析を用いて(ダンデカール,ティーら)行なわれた。
【0074】
従って、参照を不安定化する因子の生物学的に活性な断片並びにその変異型及び誘導体は本発明に包含される。
【0075】
真核生物の発現ベクターが意図される。
【0076】
関連する実施態様において、本発明は転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写物の安定性を増加させる安定化因子である発現ベクターを意図する。
【0077】
別の関連する実施態様では、本発明は転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写物の安定性を増加させる安定化因子であり、該安定化因子が長寿命mRNAの例であるアルファ2グロビン、アルファ1グロビン、ベータグロビン、又は成長ホルモンであるか又はこれらから誘導されるものである発現ベクターを意図する。本明細書で用いられるとき、遺伝子の名前の下線又はイタリック体は遺伝子を示し、下線又はイタリックにより強調されていない遺伝子の名前で示されるのがそのタンパク質産物であることと対照的である。例えば、「アルファ2グロビン」はアルファ2グロビン遺伝子を意味し、一方、「アルファ2グロビン」は「アルファ2グロビン」遺伝子のタンパク質産物を示す。
【0078】
転写物を不安定化し細胞により生産されるタンパク質の量を低減させる能力は広範囲の適用に対して有用であることが明らかである。
【0079】
本発明の別の一側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写物の安定性を低減させる不安定化因子である発現ベクターを意図する。
【0080】
別の一側面では、本発明は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写物の安定性を低減させる不安定化因子であり、該不安定化因子をコードするヌクレオチド配列が c-fos、c-jun 、c-myc 、GM-CSF、 IL-3 、TNF-アルファ、IL-2、IL-6、IL-8、IL-10 、ウロキナーゼ、bcl-2 、SGLT1 (Na(+)- 共役グルコース・トランスポーター) 、Cox-2 ( シクロオキシゲナーゼ 2) 、IL-8、PAI-2 (2型プラスミノーゲン・アクチベーター・インヒビター) 、アドレナリン性ベータ1受容体又はGAP43 から選択される遺伝子であり、又は選択される遺伝子から誘導されるものである発現ベクターを意図する。
【0081】
一つの特定の実施態様では、RNAを不安定化する因子をコードするポリヌクレオチド配列を、目的のタンパク質をコードする配列に連結し、これを次いで好ましくは調節可能な(即ち、誘導性又は抑圧性の)目的のプロモーターに連結して、発現の調節のオン−オフを行なうようにする。この適用では、RNA不安定化因子は通常機能的なmRNA又はタンパク質の発現の期間を短縮するのに役立つ。これはイン・ビトロ又はイン・ビボで適用しうる。例えば、細胞周期に特異的なプロモーターをRNA不安定化因子と組み合わせて、細胞周期の特定の期でのみ目的のタンパク質を発現させることができよう。目的のタンパク質は機能的なタンパク質であってもよく、又はレポータータンパク質であってもよい。後者の例では、レポーターのレベルは細胞周期の期又は細胞の増殖の指標として使用することができる。
【0082】
本発明のさらに別の一側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写物の安定性を低減させる不安定化因子であり、該不安定化因子をコードするヌクレオチド配列が配列番号:1〜配列番号:23のいずれか一つ、又はそれらの生物学的に活性な断片、又はこれらの変異体若しくは誘導体から選択されるものである、発現ベクターを意図する。
【0083】
本発明のさらに別の一側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写物の安定性を低減させる不安定化因子であり、該不安定化因子をコードするヌクレオチド配列が配列番号:1、配列番号:2又は配列番号:22、又はその生物学的に活性な断片、変異体又は誘導体に記載されているものである、発現ベクターを意図する。
【0084】
本発明の別の一側面はRNA因子をコードするヌクレオチド配列を含む転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写物の安定性を低減させる不安定化因子であり、該転写可能なポリヌクレオチドが目的のポリヌクレオチドを含みそしてポリペプチドをコードするものである発現ベクターを意図する。
【0085】
一つの具体的な適用は遺伝子発現を測定する分野においてである。具体的には、細胞内で生産される転写物の量を低減させることにより、プロモーター活性又はエンハンサー活性をより正確に測定することが可能である。この適用では、レポーター遺伝子を用いてプロモーター活性を、直接的にか、又はその発現がベクター内の調節因子により調節される別のポリペプチドとの融合タンパク質として間接的に測定する。
【0086】
一つの実施態様では、このRNA不安定化配列をレポーターmRNAの3’UTRをコードする領域に組み込む。代わりに又は加えて、不安定化因子を5’UTR及び/又はコードされたタンパク質の選択された活性に好ましくは必要でない又はそれを妨害しないタンパク質コード領域に組み込む。
【0087】
関連する実施態様では、このRNA不安定化配列を用いて、例えばその発現を正確にモニターし又は低減させる必要がある場合、目的の遺伝子を不安定化する。通常、この適用では、RNA不安定化因子はレポータータンパク質不安定化因子と一緒に使用される。
【0088】
本発現ベクターは、mRNA又はタンパク質の発現の期間が短いことが好ましい場合、又はプロモーター活性の変化とそれによって生ずるmRNA/タンパク質のレベルの変化の間の時間差を最小にすることが望ましい場合に、様々な遺伝子発現系において適用がある。
【0089】
従って、本発明のさらに別の一側面は、RNA因子をコードするヌクレオチド配列を含む転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写物の安定性を低減させる不安定化因子であり、該転写可能なポリヌクレオチドが目的のポリヌクレオチドを含み且つレポーターポリペプチドをコードするものである発現ベクターを意図する。
【0090】
本発明のさらに別の一側面はRNA因子をコードするヌクレオチド配列を含む転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写物の安定性を低減させる不安定化因子であり、且つ該転写可能なポリヌクレオチドがレポーターポリペプチドをコードするヌクレオチド配列を含むものである発現ベクターを意図する。
【0091】
この発現ベクターは真核細胞系で使用するように設計される。しかしながら、RNAを不安定化する因子は酵母、昆虫、線虫、魚、鳥又は哺乳動物として定義された細胞、組織又は完全生物を含む広範囲の真核系及び/又は植物系で使用しうることに留意すべきである。植物で使用する場合には、異なるプロモーター及びおそらく異なるレポーター及びRNAを不安定化する因子(例えば、DST 配列) が使用されうる。
【0092】
本発明の発現ベクターは標準的なタンパク質レポーター分子又は不安定化されたレポータータンパク質分子を組み込むことが意図される。標準的レポーター分子は当分野で良く知られている。
【0093】
本発明の別の一側面は、タンパク質を不安定化する因子(例えば、目的のポリペプチドのアミノ末端にある不安定化アミノ酸から選択されうる細胞内タンパク質分解シグナル又はデグロンをコードするDNA/RNA配列、PEST領域又はユビキチン) とmRNAを不安定化する因子(例えば、9量体のUUAUUUAUUの複数のコピー)を、mRNAとタンパク質の両方が不安定化されるように組み合わせることを意図する。例えば、一つのこのような実施態様は、 PEST 配列を翻訳停止コドンの直ぐ上流に、そして4個の9量体を該停止コドンの下流(好ましくは停止コドンから20ヌクレオチド又はそれ以上)に配置して発現ベクターに組み込む。
【0094】
このようにして、レポータータンパク質はタンパク質レベル及びmRNAレベルの両方で不安定化されうる。
【0095】
不安定化されたレポータータンパク質は任意の適切なタンパク質であってよい。例えば、不安定化された GFPタンパク質は、例えば、MODCのd1突然変異体を含むd1EGFP、d1EYFP及び d1ECFP などには適当である。不安定化ルシフェラーゼタンパク質はレクラーク,ジー.らにより記載された。 MODC PEST配列が使用された。d1EGFP由来の MODC も意図される。
【0096】
目的のポリペプチドを不安定化する方法はすべて本発明で意図される。例えば、目的のポリペプチドは、そのように修飾されたタンパク質が、例えばバックメアら(米国特許第 5,093,242号) 及びヴァルシャヴスキーら (米国特許第 5,122,463号) により開示されたようなN末端ルール経路に従うように、不安定化するアミノ酸をそのアミノ末端に含むように修飾することができる。この種の好ましい実施態様では、不安定化するアミノ酸はイソロイシン及びグルタミン酸から、より好ましくはヒスチジン、チロシン及びグルタミンから、さらに一層好ましくはアスパラギン酸、アスパラギン、フェニルアラニン、ロイシン、トリプトファン及びリジンから選択される。とりわけ好ましい実施態様では、不安定化アミノ酸はアルギニンである。一部のタンパク質では、アミノ末端はタンパク質のコンホーメーション(即ち、その三次元又は四次元構造)の結果として曖昧になる。これらの場合には、アミノ末端のより広範な改変がN末端ルール経路に該タンパク質を従わせるためには必要であるかも知れない。例えば、単一のアミノ末端残基の単純な付加又は置換ではアミノ末端への接近不能のために不十分である場合、(ユビキチンの基質タンパク質に結合する部位であるリジンを含め)幾つかのアミノ酸を元のアミノ末端に添加して工学操作されたアミノ末端の接近可能性及び/又はセグメントの移動性を相加させてもよい。
【0097】
タンパク質のアミノ末端の修飾又は設計は遺伝子レベルで行なうこともできる。単離され又は合成された抗原をコードするポリヌクレオチドの5’末端に適当なコドンを付加又は置換するための部位特異的突然変異誘発という通常の技術はコードされたタンパク質の所望のアミノ末端構造を作るために使用できる。例えば、発現されたタンパク質がそのアミノ末端に所望のアミノ酸を有するように、不安定化アミノ酸の適当なコドンをタンパク質コード配列のアミノ末端中に挿入又は構築できる。必要な場合、タンパク質のアミノ末端領域をコードする核酸配列を修飾して、適当な前後関係でリジン残基を導入することができる。これは「汎用不安定化セグメント」をコードするDNA構築物を使用することにより最も便利に達成できる。汎用不安定化セグメントは、好ましくはセグメントとして移動可能な、一つ以上のリジン残基を含むポリペプチド構造をコードする核酸構築物を含み、該構築物が抗原をコードするポリヌクレオチドのコード配列中に挿入される場合、該リジン残基が完全なアミノ末端分解シグナルの第2の決定子として役立つようにコードされたタンパク質のアミノ末端に空間的に十分に近くに存在するように、リジン残基のコドンが該構築物内に配置されている。このような構築物を抗原をコードするポリヌクレオチドの5’部位に挿入すると、不安定化のための適当な前後関係でリジン残基(単数又は複数)を有するコードされたタンパク質がえられるであろう。
【0098】
別の一実施態様では、目的のポリペプチドは PEST 領域を含むように修飾される。この領域はプロリン、グルタミン酸、セリン及びスレオニンから選択されるアミノ酸を豊富に含み、任意選択的に正の電荷を有する側鎖を含むアミノ酸が両側を挟んでいる。この点で、例えば、ロジャーズら(1986, Science234(4774): 364-368)により示されたように、2時間未満の細胞内半減期を持つタンパク質のアミノ酸配列はプロリン(P)、グルタミン酸(E)、セリン(S)、及びスレオニン(T)の豊富な領域を一つ以上含むことが知られている。さらに別の実施態様では、目的のポリペプチドをユビキチン又はその生物学的に活性な断片と結合させて、細胞内タンパク質分解の速度が無修飾のポリペプチドと比べ増加、向上又は他の方法で上昇する修飾されたポリペプチドを作成する。
【0099】
本発明のさらに別の側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写物の安定性を低減させる不安定化因子であり、該レポータータンパク質がルシフェラーゼ、緑色蛍光タンパク質、赤色蛍光タンパク質、SEAP、CAT 、又はそれらの生物学的に活性な断片、又はこれらの変異体若しくは誘導体から選択されるものである発現ベクターを意図する。
【0100】
このようなベクターは該プロモーターの活性を変化させる薬物又は処理をスクリーニングするために使用できる。現存するレポーターベクターと比べ、薬物の作用の近「実時間」測定を行なうことができる。
【0101】
本発明のさらに別の側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写物の安定性を低減させる不安定化因子であり、
(i) ヌクレオチド配列を導入するための複数クローニング部位であって、その部位が好ましくは酵素により又は他の生化学的方法により切断されて PCR増幅産物をそれに直接クローニングできる線状化されたベクターを生じうる部位(例えば、Ec1HK1部位) 、
(ii) レポーター遺伝子、
(iii) 該転写可能なポリヌクレオチドの発現を調節するためのプロモーター及び/又はエンハンサー、
(iv) ポリアデニル化配列、
(v) 選択可能なマーカー遺伝子、及び
(vi) 複製起点、
から成る群より選択される一つ以上の構成員を任意の順序で含む発現ベクターに向けられる。
【0102】
本発明の別の一側面は、遺伝子発現(例えば、プロモーター活性)の研究又は測定又は監視における適用でのベクター又はベクター群、とりわけプラスミド、しかしこれに限らないが、を意図する。例えば、ウイルス、人工染色体及び他の非プラスミドベクターなどの多くの他のベクターも使用できる。
【0103】
一つの実施態様はプラスミドの対又はセットを含む。そのそれぞれは、例えば d1EGFP 、d1EYFP、d1ECFP又は d1DsRedなどの不安定化されたレポータータンパク質をコードする構築物中に組み込まれた上述のmRNAを不安定化する配列を一つ以上含む。各対又はセットからの一つのプラスミド(対照)はレポーターをコードする領域のプロモーター 5’ を含む。このプロモーターは外因性処理(例えば、mCMVなどの最小プロモーターと結合したTRE、図2cを参照)により調節される(即ち、誘導又は抑圧されうる)因子から構成される。または、TS、SV40、CMV、TK又はRSVなどの構成的に活性なプロモーターが使用される(図2b参照)。植物系では、トップ−テン・プロモーターがTREに取って代わり、カリフラワー・モザイク・ウイルスの35SプロモーターはSV40等に取って代わることができる。アグロバクテリウム・ツメファシエンスは遺伝子移送を容易にするために植物で使用できる。対又はセットでの他のプラスミド(単数又は複数)は、クローニング部位(MCS)がプロモーターに取って代わり、レポーターをコードする領域が対照レポーターに類似するが識別可能なレポーターをコードする(図2a参照)点を除き、対照のプラスミドと同一である。好ましい実施態様では、対照プラスミドはEGFPの不安定化変異体(例えばd1EGFP、d1EYFP又はd1ECFP)をコードし、他のベクター(テストベクター)はそれぞれ同じリスト(同じタンパク質半減期)からの異なる色変異体をコードする。
【0104】
他の実施態様では、対照及びテストレポーターの一つが、例えば双方向性プラスミドなどの1個のベクターの中に組み込まれる(図3参照)。
【0105】
上の実施態様では、対照及びテストプラスミドの両方が不安定化されたmRNAをコードしており、これらが今度は不安定化されたタンパク質をコードする。こうして、低減したプロモーター活性と低減したレポータータンパク質レベルの間の時間差は現存する構築物についての時間差と比べ有意に減少する。同様に、プロモーター活性の増加は先に存在するmRNA及びタンパク質のレベルの減少により、より容易かつ迅速に検出可能である。誤りを招き得る、対照とテスト構築物の間の他の相違は、相互に二、三の小さな突然変異しか異ならない蛍光タンパク質を用いることにより最小にされる。ルシフェラーゼ又は他の酵素に基づく検定法と比べ、ここに記載した蛍光レポーターは、下記の
a)数個の異なるレポーターを同一の細胞/試料の中で測定できる、
b)生きている細胞を測定でき、同一試料の複数の時点又は測定後のさらなる操作、例えば薬物処理の前後の同一細胞の測定を可能とする、
c)巧くトランスフェクトされた細胞は蛍光顕微鏡で可視化できる。従って、不十分なトランスフェクションは顕微鏡下で細胞を見ることにより簡単に確認でき、さらなる資源の投入を必要としない、
d)基質は必要でない、それ故この方法は技術的な要求が少なく、迅速で、安価であり、且つ精度も高い、
e)対照の発現及びテストレポーターの発現の両方をフローサイトメトリーにより同時に測定できる(下記のフローサイトメトリーの利点を参照)、
f)対照プロモーターとしてTREを用いる実施態様はTet−オン又はTet−オフの細胞系統で使用できるだけであるが、他の対照プロモーターと比較すると、テストプロモーターによる又はそれへの妨害を示し難く、そしてテストプロモーターの誘導性を検査するために用いられる種々の刺激により影響され難い。従って、これらにより、トランスフェクション効率及び関連するテストプロモーター活性はより正確に測定される。必要な場合、対照レポーターの発現はスイッチをオン・オフでき、プロモーターのクロストークの欠如を確認するため又は存在するときはそれを補償するために用いることができる、
g)別の実施態様では、上記の対照及びテストレポーターの一つは、両方とも単一のベクター、好ましくは双方向プラスミド、の中に組み込まれる。この二つのプロモーターの間の干渉は、これは先の二重プロモーターベクターの主要な欠点であるが、該対照プロモーター中のTREを用いることにより最小にする、このような単一ベクター系は共トランスフェクション研究の不正確さを防止する、
という幾つかの他の利点を有する。
【0106】
本発明は、情報を与えるプロモーター又はプロモーター断片がレポーターをコードする領域の上流に置かれるベクターをも提供する。本発明は、レポーターとしてルシフェラーゼ又は他の酵素とは全く異なってEGFP変異型を利用する場合、このような検定のためのより単純で、より迅速で、より費用効果的なレポーター系を提供する。さらに、mRNA不安定化因子の組み込みはほぼ実時間分析を可能とする。
【0107】
情報を与えるプロモーターには、細胞周期依存性のプロモーター(例えば、サイクリンA,B,D1、ヒストン又はトポイソメラーゼIのプロモーター類)、アポトーシス(細胞死)経路により活性化されるプロモーター類、及び有糸分裂シグナルに連結したプロモーター/断片類が含まれる(表1)が、これらに限定されない。情報を与えるエンハンサーの使用可能な例にはクローンテックのマーキュリ・パスウエー・プロファイリング・システムズで使用されるもののいずれかが含まれる。クローンテックのマーキュリ・イン・ビボ・キナーゼ・アッセイ・キッツは本発明の使用方法の別の一例を示す。この例では、プロモーター因子は、特定のキナーゼが活性であり融合タンパク質のトランスアクチベータードメインをリン酸化できる場合のみTREからの転写を可能とするキメラのTerR−トランスアクチベータータンパク質と細胞内で結合するTREである。従って、本発明は特定のキナーゼ活性のより実時間に近い測定を行なうために使用することができる。
【0108】
本発明のさらに別の側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む転写可能なポリヌクレオチドを含む本発明のベクターでトランスフェクト又は形質導入された細胞を意図する。
【0109】
一部の適用では、発現ベクター又はレポーター構築物を発現する細胞が生物に挿入され、イン・ビボでレポーター活性を測定することを可能とする。これらの適用の一部では、不安定化されたEGFP変異型ではなく不安定化されたルシフェラーゼが好ましいレポーターでありうる。例えば、情報を与えるプロモーターの制御下で不安定化されたルシフェラーゼを発現するトランスジェニックマウスは、ホトンカメラを用いて(ホトンカメラ分析はコンタグらにより記述されている、1997) 生きているマウスの組織中でプロモーターの活性を測定するために使用できる。mRNA不安定化配列はプロモーター活性とレポーターレベルの間の時的相関関係を改良するのに役立ち、こうして、プロモーター活性のほぼ実時間測定から利益を得る薬物スクリーニングなどの適用に顕著な改良を与える。
【0110】
一部の適用では、レポーター分子及び機能性の遺伝子産物の両方を、イン・ビトロの細胞系システム又はイン・ビボの哺乳動物システムのいずれかで発現させることが望ましい。これは、それぞれがmRNAを不安定化する因子を含む二つの別々のmRNAを含む。または、mRNAを不安定化する因子を、(例えば、内部リポソームエントリ部位,IRESを用いて)二つの別々のタンパク質を生ずる単一の不安定化された転写物又はレポーター及び機能性遺伝子産物から構成される融合タンパク質の中に組み込む。
【0111】
本発明はこれらのベクター(対照と共に又は対照なしで)を安定に発現する細胞系統をも提供する。このような細胞は薬物スクリーニングなどの分野で適用がある。例えば、MAPK−依存性レポーターベクターを含む細胞は、これらの細胞においてMAPK、又はMAPK−依存性転写の上流の任意の経路を阻害するように設計された薬物の効力をテストするための迅速で且つ安価な方法を提供する。SKBR3ヒト乳癌細胞では、例えば、MAPK活性は過剰発現したErbB2タンパク質からのシグナル発信に依存する。従って、ErbB2を阻害する薬物はこのような構築物を含むSKBR3細胞の蛍光の減少を惹起するが、ErbB2を欠く細胞ではそうはならないであろう。または、細胞は、該経路の阻害をモニターするために、異なる経路を経てMAPK活性化をもたらす、薬物の有無及び特異的リガンド若しくは処理の有無をテストできるであろう。細胞周期により調節されるプロモーターに連結したベクターを安定に発現する細胞系統(又は生物)は細胞周期の進行又は細胞の増殖を測定するための極めて迅速な、単純なそして安価な手段として使用できる。このような細胞系統は薬物のスクリーニングにおいて明らかに有用であり、本発明で意図される。細胞周期により調節されるプロモーターの例は容易に入手可能である。例えば(リー,エイチら,1995) 、(シュタイン,ジェイら,1996) 及び(ユエ,エックスら,1996) がある。
【0112】
本発明の別の実施態様には、転写後調節、とりわけmRNAの安定性、の研究のためのベクターが含まれる。このレポーターは、例えば、EGFPの不安定化変異型(例えば、d1EGFP、d1EYFP、d1ECFP)であり、それぞれ別のベクターに異なる色の変異型がある。(mCMVなどの最小プロモーターに連結した)このTREはレポーターコード領域の5’にあり、テトラサイクリン(又はドキシサイクリン)依存性の様式で転写を駆動する。他の誘導性プロモーター系も使用することができる。
【0113】
一つの実施態様では、上述したmRNA不安定化因子は含まれておらず、その代わりにMCSが主として3’UTRに(図4aを参照)しかし5’UTRにも、及び/又は一部の具体的実施態様ではコード領域に配置される。mRNAの安定性に影響を与えると考えられる配列は、それを一つの色の変異型を含むベクターの適当なクローニング部位にクローニングし、テトラサイクリン又はドキシサイクリンで転写を阻止した後レポーターレベルの減少速度を測定する(図7を参照)ことによりテストすることができる。所望ならば、減衰の速度を同一細胞中で「テストベクター」と「対照ベクター」(異なる色のEGFP変異型をコードし且つテストされる配列を含まない)との間で比較することができる。
【0114】
MCSはオーバーハングを有するPCR産物の直接クローニングを可能とする制限エンドヌクレアーゼ部位を含め又は該部位と一緒に作用させると有用である(以下を参照)。
【0115】
本発明の別の関連する実施態様では、mRNAを安定化する因子を特に探索している科学者を援助するため一つ以上のmRNA不安定化因子(単数又は複数)が含められる。同様に、他の実施態様では、mRNAを不安定化する因子を特に探索している科学者を援助するためmRNAを安定化する因子が含められる。
【0116】
他の実施態様では、対照及びテストレポーターの一つが両方とも単一のベクター、好ましくは双方向プラスミド中に組み込まれる(図4bを参照)。
【0117】
安定化因子は、例えば、高レベルのタンパク質が要求され又はプロモーターが弱い場合、タンパク質精製の過程で発現するタンパク質のレベルを増加させるのに有用である。
【0118】
二つのプロモーターの間の干渉、さらに因子又はテストされた種々の刺激の転写効果の間の干渉は、両方のレポーターを駆動するためTRE又は類似の因子を用いることにより、そしてベクターからの転写を阻止するドキシサイクリン(又はテトラサイクリン)を添加した後レポーターレベルを測定することにより回避される。
【0119】
本発明のさらに別の側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む本発明のベクターでトランスフェクトされ又は形質導入された細胞であって、該RNA因子が該転写物の安定性を低減させる不安定化因子である細胞を意図する。
【0120】
本発明の関連する側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む遺伝的に改変された非ヒト生物であって、該RNA因子が該転写物の安定性を低減させる不安定化因子である生物を意図する。
【0121】
本発明のさらなる実施態様は、目的のポリヌクレオチドの発現を測定する方法であって、RNA及びタンパク質の合成が起こるのに十分な時間及び条件の下でレポーター発現ベクターから目的の該ポリヌクレオチドを発現させる工程であり、該ベクターが転写される因子をコードするヌクレオチド配列及び該目的のポリヌクレオチドを含む転写可能なポリヌクレオチドを含むものであり、該転写される因子が該転写可能なポリヌクレオチドに対応する転写物の安定性を調節するものであり、且つ該発現ベクターが
(i) 目的の該ポリヌクレオチドを導入するための複数クローニング部位、
(ii) レポーター遺伝子、
(iii) 該目的のポリヌクレオチドの発現を調節するためのプロモーター及び/又はレポーター遺伝子であって、該プロモーターが好ましくは調節可能であるもの(例えば,テトラサイクリン応答因子(TRE)を使用)、
(iv) ポリアデニル化配列、
(v) 選択可能マーカー遺伝子、及び
(vi) 複製起点、
から成る群より選択される一つ以上の構成員を任意の順序で含むものである工程、及び対照と比べレポーターのレベル又は活性を経時的に測定する工程であり、該不安定化因子がプロモーター又はエンハンサーの活性とレポーターのレベル若しくは活性の間の時的相関関係を増強するものである工程を含む方法を意図する。
【0122】
組み合わせで作用する、異なるRNA不安定化因子の組み合わせが本明細書で意図される。
【0123】
本発明は非限定的なさらなる実施例によりさらに説明される。
【実施例1】
【0124】
DNA因子のベクターへのクローニング
クローニングは、既存の手法に従い、ベクターがMCS内で「Tオーバーハング」を持つ場合は、MCSの制限酵素部位又はPCR産物の直接連結を用いて実施した。しかしながら転写後レポーターベクターについては、3’−UTR又は他の領域にMCSを含むことは、転写又はその他の研究用に設計されこれらの位置に便利なクローニング部位を含まない現行のベクターからの顕著な改良である。
【実施例2】
【0125】
トランスフェクション
対照ベクターとテストベクターの共トランスフェクションは、一つのベクターのみを必要とし、それ故に共トランスフェクションに伴う不正確さを排除する上述の単一(例えば、双方向性)ベクター系の場合を除き、既存の手法(例えば、Fugene [ベーリンゲル・マンハイム、マンハイム、ドイツ]又は電気穿孔法)により実施した。
【実施例3】
【0126】
レポーター発現の測定
vGFPシステムの直接の利点は、単に組織培養プレート又はフラスコを蛍光顕微鏡下で見るだけで、生きた細胞内でレポーター発現を直接目に見えるようにすることができることである。従って、無駄な時間を費やす前に、劣悪なトランスフェクションを確認して破棄することができる。定量的測定は、蛍光光度計(例えば、96穴プレートフォーマット)を用いて実施することができ、生きた細胞を測定できるため、同一の資料を繰り返し、例えば経時的に、測定することができる。
【0127】
ルシフェラーゼ分析、及び他の酵素に基づく分析と比べてさらなる利点は、レポーターレベルを測定するのにフローサイトメトリーも使用できることである。
【実施例4】
【0128】
レポートレベルの測定にフローサイトメトリーを使用する利点
i. 二つ以上のレポーター(対照及びテスト)並びに追加のパラメータを、全ての細胞において秒速2,000細胞より大の速度で個々に測定することができる。従ってこれを適用すると、既存のルシフェラーゼ検定法の一つのデータポイントに対し、この手法は試料当たり数千から数十万のデータポイントを生ずる。
【0129】
ii.トランスフェクション効率の正確な測定:これはトランスフェクションの手順を最適化するのに役立つ。異なる手法の比較を可能にするのに加え、細胞当たりの発現と細胞発現の比率の双方を測定することもできる。これは研究者が何らかの問題の原因を割り出すのに役立つ。
【0130】
iii.共トランスフェクションのエラーの同定:共トランスフェクション研究は、細胞がそれを取込みそして同じ細胞に取り込まれるテストプラスミドの量に比例するある量の対照レポーターを発現するという前提に基づいている。これは必ずしもそうなるとは限らない。本明細書に記載したフローサイトメトリー法を使用することにより、同一試料中の異なる細胞でのテスト発現レベルと対照発現レベルを相関させることができる。テストレポーターレベルと対照レポーターレベルの間に良好な直線関係が欠如していれば、無効な試料であると同定できる。このようなエラーは現行の手法では見過ごされてしまう。
【0131】
iv.追加パラメータの同時測定:蛍光標識された抗体を用いて細胞ごとの特定のタンパク質を定量することができ、そしてこれをレポーターレベルと相関させてそのタンパク質がレポーター構築物にクローニングされた因子を介して遺伝子発現に影響を与えるかどうかを決定することができる。または、目的のタンパク質を適切な(誘導可能な又は誘導不可能な)発現ベクターのトランスフェクションを介して、vGFP融合タンパク質(GFP変異体と融合した目的タンパク質)として発現させることができる。次いで、特定のタンパク質のレベルを目的の(共トランスフェクトした又は異なる時点でトランスフェクトした)調節因子に連結した異なるGFP変異体の発現と相関させることができる。第三の適用では、vGFPレポーターを細胞周期特異的であると考えられている調節因子(例えば、プロモーター)と連結させる。トランスフェクトした細胞をDNA内容物を測定するためにヨウ化プロピジウムなどの蛍光DNA色素で着色し、次いでそれをレポーター発現と相関させる。原則として、本明細書で記載したDNA構築物の幾つかは、それぞれが異なるvGFPを含んでいるが、共に発現し個々に測定できるであろう。さらに、他の蛍光標識をこれらのベクター(単一または複数)と併せて使用することができるであろう。
【0132】
v. セルソーティング:セルソーターを使用すると、生存能力のある vGFP 発現細胞を非発現細胞から単離することができる。この技術は、検定の開始に先立って、安定して発現する細胞を選別したり、又は発現しない細胞を除去するのに使用できる。同様に、極めて低レベル及び/又は極めて高レベルの vGFP を発現する細胞を除去することができる。この技術は、もっと均質な集団を形成するために使用することができ、及び/又は生理学的に適切でないであろう、又は通常の細胞機能を乱すであろう、及び/又は本明細書に記載したDNAベクターから得たデータにその他の方法で悪影響を及ぼすであろう程に高いレベルで発現する細胞を除去するために使用することができる。
【0133】
発現ベクターの一過性のトランスフェクション及び安定なトランスフェクションは極めて不均質な発現レベルを有する細胞集団を結果として生ずることに注目することが大切である。一般的に、最高の発現細胞と最低の発現細胞の間で千倍の差があるというのは珍しいことではない。本発明は必要なときに均質な集団を選択する方法を提供する(上述のvを参照)だけでなく、科学者の利益のために不均質性を利用することもできる。例えば、共トランスフェクションのエラーの同定である。これの別の例は上述の(iv)に関する。タンパク質XがプロモーターYからの転写に影響を与えるかどうかを決定するために、細胞を、プロモーターYの制御の下、d1EGFPを発現するレポーター構築物でトランスフェクトする。必要に応じて、セルソーティングを用いて適正レベルの d1EGFPを一時的に又は安定して発現する細胞を単離することができる。これらの細胞を今度はタンパク質X-EYFP融合タンパク質を発現するベクターで一時的にトランスフェクトする。フローサイトメトリーの間、EGFPを一つの軸上にプロットし、EYFPを他方の軸上にプロットする。正相関はタンパク質XがプロモーターYからの転写を増加させることを示し、負の相関はタンパク質XがプロモーターYからの転写を阻害することを示すであろう。
【0134】
現在では、このような相関関係を確立しようとしている科学者らは、タンパク質Xを高度に発現するもの対低度に発現するものの幾つかの異なるクローンを選択するであろう。それぞれのクローンを、次いでプロモーターY−ルシフェラーゼ構築物で別々にトランスフェクトし、ルシフェラーゼ活性を測定するであろう。細胞クローンの使用には、調製に何ヶ月も必要であり、宿主細胞とベクター組み込みの種々の部位の間に事前に存在する不均質性を含め様々な変数を導入する(ベクターDNAは組み込み部位の特定の遺伝子を妨害し、この部位は全てのクローンで異なる)。さらに、このような手法ではそれぞれが異なるクローンの異なるトランスフェクションから得られる極めてわずかなデータポイントしか得られない。従って、この新しいシステムは既存の手法よりも用途が広いだけでなく、より迅速で且つより正確である。
【実施例5】
【0135】
レーザー走査サイトメトリー(LSC)
フローサイトメトリーとは異なり、LSCはスライド上で細胞の複数色の蛍光と光散乱を測定し、分析した各細胞についてその計測位置と時刻を記録する。この技法はフローサイトメトリーと同等のデータを提供するが、顕微鏡用スライドに基づいているという利点がある(ダルジンキーヴィッツら,1999、カメンツキーら,1997)。GFPとその変異体の蛍光のため、フローサイトメトリー用に記述された技術もLSCに適用できる。
【実施例6】
【0136】
転写後分析のための特異的方法
転写後分析のための特異的方法は、特定の3’−UTR断片がmRNAの安定性に影響を与えるかどうかの測定を目標とした研究例
(i) 3’−UTR断片をテストベクターの3’−UTRクローニング部位中に連結し、対照ベクターと共にTet−オフの細胞系統に共トランスフェクトされる。双方向性ベクターの場合は、対照ベクターは必要でない。実際、典型的な適用では、同一トランスフェクションの範囲内からの試料で減衰速度を測定できるため、対照レポーターまたはベクターを必要としない。5’−UTR断片は5’−UTRクローニング部位を持つベクター中に挿入することによりテストできる、
(ii) 細胞をドキシサイクリン(又はテトラサイクリン)の非存在下で6〜48時間増殖させ、両方のベクターを発現させる。または、細胞を低用量のドキシサイクリン(又はテトラサイクリン)と共に6〜48時間増殖させて転写を阻止し、次いでドキシサイクリン(又はテトラサイクリン)なしの培地に切替えて2〜12時間培養し、短時間で転写を一気に爆発させる、
(iii)次いで、両方のベクターからの転写を遮断するため、高用量のドキシサイクリン(又はテトラサイクリン)を適用する、
(iv) 両レポーターの蛍光を経時的に測定し(フローサイトメトリー、蛍光高度法、又はLSCにより)、続いてドキシサイクリン(又はテトラサイクリン)を添加する、
という研究例を用いると最適にまとめられる。この例は一過性の発現の一つであるが、安定なトランスフェクションにも使用できるであろう。
【0137】
クローニングされた因子が mRNA を不安定にするのであれば、同一細胞又は同一試料の「対照」蛍光と比べて「テスト」蛍光でより急激な減少が見られるはずである。同様の研究が、異なる細胞内若しくはRNA結合タンパク質のような特定のタンパク質を異なる量で発現する細胞内での特定の刺激又はその効果に対する mRNA 因子の応答をテストするのに使用することができる。ドキシサイクリンの後に該刺激を適用すると、事前に存在した転写物が刺激により影響されたかどうかが決定される。該因子を異なる位置(例えば、5’−UTR、3’−UTR)に挿入すると、その機能が位置に依存するものかどうかが決定される。タンパク質/ポリペプチドのコード配列を(フレームを揃えて)該ベクターのレポーターコード領域内に挿入すると、mRNAに対するその配列の効果とタンパク質の安定性を測定するために使用することができる。
【0138】
RNAはトランスフェクトされた細胞から抽出することができ、レポーターmRNAを直接測定するために使用することができる。
【実施例7】
【0139】
転写レポーターベクター
該ベクターは大腸菌内での増幅、及び真核細胞における蛍光レポーターの発現に適したプラスミドである。該プラスミドはセットで使用しうる。それぞれのセットは一つ以上の「対照」ベクターと一つ以上の「テスト」ベクターから成る。セット内のどのベクターも同様に不安定化された mRNA 及び同様に不安定化された蛍光レポータータンパク質を発現する。このようなプラスミドの標準特性(アンピシリン耐性、複製起点など)に加えて、各プラスミドは次の構築物を含む(図2及び図3をも参照)。
5’---MCS/プロモーター---転写開始部位---5’UTR---ATG---vEGFPコード領域--停止コドン---mRNA不安定化因子を伴う3’UTR---ポリアデニル化シグナル
【0140】
上式中、
MCS/プロモーターは、複数クローニング部位(テストベクター、図2a参照)又はSV40などの構成的に活性なプロモーター(対照ベクター、図2b参照)、又は TRE-mCMV のような誘導可能なプロモーター(対照ベクター、図2c参照)のいずれかを意味する。
ATGは、翻訳開始コドンを意味する。
停止コドンは、翻訳停止コドンを意味する。
5’UTRは、5’非翻訳領域を意味する。
mRNA不安定化因子を伴う3’UTRは、概説したmRNA不安定化因子を一つ以上含む3’非翻訳領域を意味する。
vEGFPは、EGFPの不安定化変異体を意味する。各種の不安定化修飾(例えば、1時間半減期、2時間半減期)に対してプラスミドの一つのセットが提供される。プラスミドの各セット内で、それぞれ色の異なる変異体に対して一つのベクターが提供される。例えば、一つのセットはd1EGFP、d1EYFP、d1ECFPを発現するベクターを含み、もう一つのセットは d2 変異体を発現する。
【0141】
他の実施例では、上述の対照とテストレポーターの一つは共に単一ベクター、好ましくは双方向性プラスミドに組み込まれる(図3参照)。
【実施例8】
【0142】
転写後レポーターベクター
3’−UTR内の mRNA 不安定化因子がMCSに置き換わっている(図4a参照)点を除けば、TRE-mCMVプロモーターを含む転写レポーター「対照」ベクターと類似する。幾つかの実施態様では、MCSは5’UTR及び/又はコドン領域にも配置される。
【0143】
このような構築物は、本明細書で概説した転写後検定のための「テスト」ベクター又は「対照」ベクターとして使用することができる。
【0144】
他の実施例では、上述の対照レポーターとテストレポーターの一つは共に単一ベクター、好ましくは双方向性プラスミドに組み込まれる(図4b参照)。
【実施例9】
【0145】
特定の経路を検定するレポーターベクター
調節因子を研究又は測定する目的で、該調節因子が MCS 中に挿入された本明細書記載のものに類似するベクター。例えば、MCS内に表1で言及するものなどの経路に敏感なプロモーター又はプロモーター因子(単数または複数)又はエンハンサー、及び/又はクローンテックズ・マーキュリー・パスウェイ・プロファイリング・システムに記載されたようなシスで作用するエンハンサー(AP1、CRE、E2F、GRE、HSE、ISRE、Myc、NFAT、NFkB、p53、Rb、SRE)のいずれかを含むエンハンサー(単数または複数)を含むという点を除けば、本明細書で概説した転写レポータープラスミドに類似するプラスミド。該レポーターは、GFP、ルシフェラーゼ又は SEAP の不安定化変異体であることが好ましい。
【0146】
特定の経路を検定する細胞系統及びマウス
本明細書記載のベクターの一つ以上を安定して発現する細胞系統又は遺伝的に改変されたマウス。
【実施例10】
【0147】
使用方法
本発明で記載するベクターは、本明細書で記載する新しい方法以外は、本発明で記載するベクターに取って代わられる既存のベクターと本質的には同じ方法で実験に使用する。
【0148】
構築方法
本明細書で概説するベクター及び DNA構築物は標準的なクローニング技術を用いて組み立てる。本明細書記載の SV40 及び TRE-mCMV プロモーター並びにプラスミドベクターのより標準的な構成要素(例えば、複製起点、抗生物質耐性又はその他の選択用遺伝子)は、様々な普通のベクターですぐに入手可能である。EGFPの不安定化変異型(例えば、d1EGFP、d1EYFP、d1ECFP及びd2EGFP、d2EYFP、d2ECFP)をコードするDNA配列がクローンテック(クローンテック・ラボラトリーズ・インク、パロアルト、カリフォルニア、米国)から入手可能である。不安定化 DsRed 変異型をコードするDNA配列は、DsRedコード領域の3’末端に、短寿命のタンパク質由来の分解ドメイン(又はその突然変異体)をコードする配列を融合することにより構築される。例えば、PEST配列を含むマウス・オルニチンデカルボキシラーゼ由来のアミノ酸422-461である。このような配列はおそらく既存のdEGFP変異体から誘導できるであろう。
【実施例11】
【0149】
概要
概要では本発明のベクターと方法が入手可能である。
【0150】
1)比較的短寿命な mRNA を供するために一つ以上の mRNA 不安定化因子を組み込む発現ベクター又はその部分。既存の発現ベクターと比べて、本発明で請求されたベクターはプロモーター活性とより密接に相関するタンパク質発現の動力学を供する。例えば、減少したプロモーター活性、及び減少したmRNAとタンパク質レベルの間の時間差は、実質的に減少した。
【0151】
2)不安定化mRNAをコードする発現ベクター又はその部分。このmRNAは今度は不安定化したタンパク質をコードする。既存のベクターと比べて、本発明で請求されたベクターはプロモーター活性とより密接に相関するタンパク質発現の動力学を提供する。
【0152】
3)その中のmRNA不安定化因子がc-fosのような短寿命mRNAからクローニングされた配列を含むものである発現ベクター又はその部分。該短寿命mRNAの例には、c-fos、c-myc、GM-CSF、IL-3、TNF-alpha、IL-2、IL-6、IL-8、ウロキナーゼ、bcl-2、SGLT1 (Na(+)結合ブドウ糖輸送担体)、Cox-2(シクロオキシゲナーゼ2)、IL8、PAI-2(2型プラスミノーゲン・アクチベータ・インヒビター)、ベータ1−アドレナリン受容体、GAP43(5’UTR及び3’UTR)、AU豊富因子(AREs)及び/又はU豊富因子が含まれ、9量体UUAUUUA(U/A)(U/A)(U/AはAまたはUのいずれか)(ラグナードら 1994)及び/又は5量体AUUUA(スーら 997)及び/又は4量体AUUU(ズビアーガら 1995)の単一コピー、縦列コピー又は複数コピー又は重なりコピーが含まれるがこれらに限定されない。上述のリストの因子の小さな改変又は並べ替えも含まれる。「縦列コピー」という用語は、外側のヌクレオチド(複数)の一つ以上の重複及び/又は非重複の両方を許す。例えば、5量体AUUAの縦列コピーは、AUUUAAUUUAAUUUAだけでなく、AUUUAUUUAUUUAのような配列を含む。短寿命 mRNA の3’UTR又は5’UTR領域は、しばしば不安定化配列を含む。
【0153】
4)その中のmRNA不安定化因子が本明細書記載のベクターを用いて同定又は検証された発現ベクター又はその部分。これらはこのような因子を同定するための実質的に改良された手法を提供する。
【0154】
5)その中の不安定化mRNAがEGFP又はルシフェラーゼの不安定化変異体などの短寿命レポータータンパク質をコードするものである、発現ベクター又はその部分。既存のレポーターベクターと比べて、本発明で請求されるベクターは、プロモーター活性とより密接に相関するレポーター発現の動力学を提供する。例えば、減少したプロモーター活性、及び減少したmRNAとタンパク質レベルの間の時間差は、実質的に減少する。
【0155】
6)同様に不安定化されたmRNA(同じ集合の他のベクターと同様)をコードするレポーターベクター又はその部分の集合。これらのmRNAは、今度は同様に(同じ集合の他のベクターと同様)不安定化されたEGFP若しくはDsRedの変異体、又は他の蛍光標識をコードする。各セット内の一つ以上のベクター(対照ベクター)は一つの構成的プロモーター(例えば、SV40、CMV、RSV、TK、TS。図2b参照)又は一つの誘導可能なプロモーター(例えば、TRE-mCMV。図2c参照)を含み、一方各セット内の他のベクター(テストベクター)はプロモーターの代わりにクローニング部位(例えば、MCS)を含む(例えば、図2a参照)。これらのベクターの適用は、プロモーター活性の研究又は測定を含むが、これらに限定されない。例えば、目的のプロモーター因子は d1EGFP をコードするテストベクターの MCS中にクローングでき、そしてレポーターの発現は d1EYFP を発現する対照ベクターのレポーター発現と比較して測定できる。同一のベクター内に一つのテストレポーター構築物及び一つの対照レポーター構築物を組み込む双方向性ベクター又は他の単一ベクター系(例えば、図3a及び3b)として詳しく記載した個々のベクターのそれぞれも請求される。既存のレポーターベクターの集合と比べ、本発明で請求されるベクターは
a) プロモーター活性の実時間により近い測定、
b) 対照構築物とテスト構築物の類似性が高くなったことによるエラーの減少、
c) テストプロモーターと対照プロモーターの間のクロストークに起因するエラーの減少。対照ベクターの誘導可能プロモーターを使用することにより、このようなクロストークは誘導あり及び誘導なしの測定により最小限に抑えられ、及び/又は同定され、そして修正される、
d) 記載されたフローサイトメトリー/LSC法と併せて使用できる、
という利点を有する。
【0156】
7)誘導可能なプロモーター、テトラサイクリン応答因子(TRE)が好ましいがこれに限定しない、を利用して、不安定化した蛍光性レポータータンパク質(不安定化EGFP変異体が好ましいがこれに限定しない)の発現を駆動するレポーターベクター又はレポーターベクターの集合、又はその部分。このようなベクターは、調節因子又は調節因子であると推定されるものが1色の蛍光レポーターを発現するベクター中にクローニングできるように、そして必要であれば、異なる色のレポーターを発現し且つ目的の因子を含まない対照ベクターと比較できるように、3’−UTR(例えば、図4a)及び/又は5’−UTR及び/又はレポーターコード領域にクローニング部位を含む。このようなベクターは、転写が誘導可能なプロモーターにより要望どおりに転写を阻止できるため、転写後調節の研究又は測定に適用がある。これらのベクターがもたらす利点はb〜dにリストした利点、即ち、転写後効率を転写効率と切り離すという利点、並びに
a)他のベクターには存在しない便利なクローニング部位の組み込み、及び
b)この技術は、既存の方法よりも速い、
という利点をも含む。
【0157】
8)本質的に一つのテスト構築物と対照構築物を連結する、記載された単一ベクター系(例えば、図4b)。テストレポーターと対照レポーターは共に誘導可能なプロモーターで駆動され、クローニング部位は調節因子をテスト構築物にだけ連結させる。概説したベクターの利点に加え、単一ベクター系は別々のテストベクターと対照ベクターの共トランスフェクションに伴う諸問題と不正確さを解決する。
【0158】
9)概説したベクターを介して発現される2以上の蛍光レポーターのレベルを測定するためのフローサイトメトリーまたはLSCの使用。この適用においては、既存の酵素に基づく検定では1サンプルにつき1データポイントであるのに対し、該手法では1サンプルにつき数千から数十万のデータポイントが得られる。二つ以上のレポーター(対照及びテスト)だけでなく、さらなるパラメータ(例えば、DNA含量、他のタンパク質のレベル)も、全てのセルで個々に測定できる。同一試料内の複数の細胞の2以上のレベルを相関させるフローサイトメトリーの使用、及びトランスフェクション手順を最適化するため及び/または共トランスフェクションに伴う諸問題を特定するためにこのようなデータを利用することをも包含される。例えば、無効なサンプルはテストレポーターレベルと対照レポーターレベルの間に良好な線状関係が欠けていることで特定できる。このような誤差は、現行手法では認識されないままになってしまう。
【0159】
10)請求される転写後レポーターベクターを利用する方法。これらの方法は特定の3’−UTR断片がmRNAの安定性に影響を与えるかどうかの測定を目的とした研究例を用いて最も良く要約できる。この例、即ち
(i) 3’−UTR断片を、テストベクターの3’−UTRクローニング部位に連結し、対照ベクターと共にTet−オフ細胞系統中に共トランスフェクトする。単一ベクター系の場合、対照ベクターは必要ない。5’−UTR断片は5’−UTRクローニング部位を持つベクター中にそれを挿入することによりテストできる、
(ii) 細胞はドキシサイクリン(又はテトラサイクリン)の非存在下で6〜48時間増殖させて両方のベクターを発現させる。または、細胞を低用量のドキシサイクリン(又はテトラサイクリン)と共に6〜48時間増殖させて転写を阻止し、次にドキシサイクリン(又はテトラサイクリン)を含まない培地に切替えて2〜12時間培養し、転写を短時間で爆発させる、
(iii)次に高用量のドキシサイクリン(又はテトラサイクリン)を適用して両方のベクターからの転写を遮断する、
(iv) ドキシサイクリン(又はテトラサイクリン)の添加に続き、両方のレポーターの蛍光を(フローサイトメトリー、フルオロメトリー、又はLSCにより)経時的に測定する、
は一過性発現の例であるが、安定なトランスフェクションにも使用できるであろう。
クローニングされた因子がmRNAを不安定化すれば、同一の細胞又は試料の「対照」の蛍光と比べ、「テスト」蛍光ではより急速な減少が見られる。同様の研究は、異なる細胞又はRNA-結合タンパク質などの特定のタンパク質を異なる量で発現する細胞における特定の刺激又はその効果に対するmRNA因子の反応をテストするために使用することができる。ドキシサイクリンの後に該刺激を適用することで事前に存在した転写がその刺激の影響を受けたかどうかを測定することができる。該因子を別の位置(例えば、5’−UTR、3’−UTR)に挿入すると、その機能が位置に依存するものかどうかを決定できる。タンパク質/ポリペプチドコード配列を(フレームを揃えて)該ベクターのレポータータンパク質コード領域内に挿入する手法は、その配列がmRNA及びタンパク質の安定性に及ぼす効果を決定するために使用することができる。
RNAはトランスフェクトした細胞から抽出でき、レポーターmRNAを直接測定するのに使用できる。
【0160】
11)請求される、発現構築物又はその部分の一つ以上を一時的に又は安定して発現する細胞系統。
【0161】
12)請求される、発現構築物又はその部分の一つ以上を一時的に又は安定して発現する細胞系統であって、該発現構築物がヒトの病気及び/又は薬物治療に対する反応に関わるシグナル変換経路の活性化のマーカーとしての機能を果たす調節因子を含むものである細胞系統。このような経路は表1のリストと本明細書のどこかに記載されているもの(例えば、CRE、SRE、AP1、サイクリンA、B、及びD1のプロモーター)を含むが、それらに限定されない。
【0162】
13)請求される発現構築物又はその部分の一つ以上を発現するトランスジェニックマウス、遺伝子削除マウス、又は他の遺伝的に改変されたマウス。
【0163】
14)請求される、発現構築物又はその部分の一つ以上を発現する、トランスジェニックマウス、遺伝子導入(knock-in) マウス、又は他の遺伝子的に改変されたマウスであって、該発現構築物がヒトの病気及び/又は薬物治療に対する反応に関わるシグナル変換経路の活性化のマーカーとして機能する調節因子を含むものであるマウス。このような経路は、表1のリストを含むが、これらに限定されない。
【0164】
15)DsRedの不安定化変異体又はDsRed1-E5の突然変異体。これらはDsRedのC-末端に、様々な不安定なタンパク質からの分解ドメイン(又はその突然変異体)を融合して構築することができる。例えば、PEST配列を含むマウス・オルニチンデカルボキシラーゼのアミノ酸422〜461である(リーら 1998)。さらなる分解因子もまた追加できる。DsRedの不安定化変異体をコードするDNA構築物も意図される。
【0165】
16)概説したmRNA不安定化因子をも含むようなベクターを含む、概説したDsRedの不安定化変異体をコードするベクター。
【0166】
17)Tet−オフ又はTet−オン細胞系統を創出する以下の方法。
tTA又はrtTA発現ベクター、好ましくはレトロウイルス、アデノウイルス、又はプラスミドは、標準技術を使用すると目的の細胞系統で安定して発現し、発現する細胞は薬物耐性マーカーを介して単離される。次に、これらの細胞をTRE-vGFP構築物で一時的にトランスフェクトし、フローサイトメトリーによる何回りかのセルソーティング(細胞分取)にかける。例えば、良好なTet−オフ細胞はドキシサイクリンの存在下で蛍光を発しないであろうし、そのように分取される。ドキシサイクリンなしでさらに5〜48時間経過した後、緑色細胞が分取される。最終的に、該細胞をドキシサイクリンなしで1週間以上増殖させ、最後に分取して、安定にトランスフェクトした(緑色)細胞を除去する。
【実施例12】
【0167】
mRNA及びタンパク質の不安定化因子を組み込んだベクター
目的のコーディング領域(例えば、EGFP又はルシフェラーゼのようなレポーター)はタンパク質不安定化因子(例えば、MODCのd1突然変異体(クローンテック)、但し他のPEST配列又はユビキチン化部位などの他のタンパク質不安定化因子をも含む)とmRNA不安定化因子(例えば、AU豊富因子)の結合した配列を含むことができるであろう。
【0168】
例えば、ルシフェラーゼ及びDsRedの停止コドンはHind3部位(AAGCTT)で置換され、次の配列、
AAGCTTAGCCATGGCTTCCCGCCGGCGGTGGCGGCGCAGGATGATGGCACGCTGCCCATG
TCTTGTGCCCAGGAGAGCGGGATGGACCGTCACCCTGCAGCCTGTGCTTCTGCTAGGATC
AATGTGTAG
の付加を可能にする。
これは、EGFPに1時間の半減期を与えるクローンテックのMODCのd1突然変異体である。これにはリンカー(3’UTRの一部となる)が後続する。次に、4個の9量体を含み、親ベクター(pGL3、プロメガ)のXba1部位に連結する
UUAUUUAUU GGCGG UUAUUUAUU CGGCG UUAUUUAUU GCGCG UUAUUUAUU ACTAG
も3’UTR中に、ただしさらに下流において後続する。
【実施例13】
【0169】
PCR産物の直接連結
二つに分かれているが近くの RE 認識部位をベクターのMCSに包含させること。このRE(単数又は複数)で切断すると、残りのベクターの両端に単一のTヌクレオチドの3’オーバーハングが残る。例えば、Ec1HK1の認識配列はGACNNN、NNGTC(3番目のNと4番目のNの間を5’から切り離して両端に単一のNという3’オーバーハングを残す)である。これらの部位のうち二つは、その二つの間の短い領域がEclHK1での消化により放出されて、両端に単一のNの3’オーバーハングを持つ線状ベクターが残るようにMCS中に組み込まれる。この例では、上流認識配列は 5’GACNNTNNGTC3’で、下流認識配列は5’GACNNANNGTC3’でなければならない。Ec1HK1で切断した後、大きなベクター断片は両端に単一の3’Tオーバーハングを含むことになる(プロメガのpGEM-Tイージーベクターと同様)。これは5’Aオーバーハングを生ずるTaqのようなポリメラーゼで製造されるPCR産物の直接連結を容易にする。これは、RE部位のPCRプライマーへの包含とその後のPCR産物の消化なしでは PCR産物の直接連結をサポートしない標準的なMCSに対する顕著な改良点となる。これはまた、増幅(線状として供給される)できず、サブクローニングにしか役立たない(即ち、PCR産物は通常は pGEM-T イージーに連結され、増幅され、次いでRE消化によって切り出され、続いて目的の発現ベクターにクローニングされる)pGEM-イージーベクターに対する顕著な改良でもある。従って、この MCS は、PCR産物をRE(しばしば問題となる)で消化したり、又は中間ベクターにそれらをサブクローニングする必要なく、PCR産物の直接連結を可能にする。
【実施例14】
【0170】
不安定化レポーターモデルは改良された実時間分析を示す
プラスミドレポーターベクターは、標準的なクローニング技術を使用してpGL3−ベーシック(プロメガ)バックボーン(アンピシリン耐性遺伝子など)に組み込んだ。クローンテックの pTRE-d2EGFPベクターから誘導したテトラサイクリン応答因子を MCSに挿入した。一部の構築物では、クローンテックが定めたようにルシフェラーゼ−コード領域を d1EGFP −コード配列又は d2EGFP −コード配列(コザック配列を含む)で置き換えた。この工程は便利な5’隣接 RE 部位を持つ適切なプライマーを用いたPCRにより実行した。一部の構築物では、mRNA不安定化因子の特定の例が3’UTR−コード領域にクローニングされた。通常、これらの配列はセンス配列及びアンチセンス配列を合成し、ハイブリッド形成させることにより調製した。両側を挟む配列は、3’UTR−コード領域が特定の制限酵素で切断される場合に生成するもの適合する「付着末端」のオーバーハングを提供した。これらの酵素による該ベクターの消化とその後の精製の後、ハイブリッド形成するオリゴマーを標準的な技術を用いて該ベクターに連結した。適切な供給源から得たゲノムDNA又はcDNAのPCRをc-myc-AREのようなより大きい不安定化因子を得るための代替手法として使用した。極めて小さな因子(例えば、1又は2個の9量体)を、一つの5’隣接RE部位とベクター鋳型中の既存の3’UTRに相補的な一つの3’隣接領域を含む逆向きPCRプライマーに組み込んだ。適切な前向きプライマー(タンパク質コード領域に相補的であり且つ内因性RE部位と重なる)を用いる PCRの後、該PCR産物を適切なRE部位で消化し、元のベクターに連結した。
【0171】
用語:
B=プロメガのpGL3- ベイシックから誘導されたベクターバックボーン。
T=テトラサイクリン応答因子(TRE)。クローンテックの pTRE-d2EGFPベクターから誘導され、レポーターの転写を駆動するプロモーターとして用いられる。
G1=レポーターとして用いられる1時間の半減期を持つGFP、即ち、クローンテックにより定義されたような d1EGFP タンパク質をコードする配列。
G2=レポーターとして用いられる2時間の半減期を持つGFP、即ち、クローンテックにより定義されたような d2EGFP タンパク質をコードする配列。
L=レポーターとして用いられるルシフェラーゼ、即ち、pGL3-ベイシック(プロメガ)由来の蛍ルシフェラーゼをコードする配列。
R=レポーターとして用いられる DsRed2 。
R1= d1EGFP に存在するのと同じMODC突然変異体にカルボキシ末端で融合した DsRed 。
N6=3’UTR−コード領域に挿入された6個の9量体TTATTTATT(配列番号:1)。
N4=3’UTR−コード領域に挿入された4個の9量体TTATTTATT。
N2=3’UTR−コード領域に挿入された2個の9量体TTATTTATT。
N1=3’UTR−コード領域に挿入された1個の9量体TTATTTATT。
fos=シューら(1989)により定義され、3’UTR−コード領域に挿入された c-fos ARE。即ち、
5’AAAACGTTTTATTGTGTTTTTAATTTATTTATTAAGATGG
ATTCTCAGATATTTATATTTTTATTTTATTTTTTT3’ (配列番号:2)
myc=次のように定義される myc ARE である、
5’ATGCATGATCAAATGCAACCTCACAACCTTGGCTGAGTCTTGAGACTGAA
AGATTTAGCCATAATGTAAACTGCCTCAAATTGGACTTTGGGCATAAAAG
AACTTTTTTATGCTTACCATCTTTTTTTTTTCTTTAACAGATTTGTATTT
AAGAATTGTTTTTAAAAAATTTTAAGATTTACACAATGTTTCTCTGTAAA
TATTGCCATTAAATGTAAATAACTTT3’ (配列番号:21)。
【0172】
方法:
5マイクログラムのマキシプレップ品質のDNAを、フジーン(Fugene) 試薬(ロシュ)を用いて10cmフラスコの約50%コンフルーエントのヒーラのTet−オフ細胞(クローンテック)中にトランスフェクトした。約10時間後に、該フラスコの細胞をそれぞれ約12の小皿(6cm)に分け、ついで一晩(約12〜14時間)インキュベートした。この時点(通常タイムゼロ又は T0と呼ぶ)で、殆どのプレートに1マイクログラム/mlの最終濃度でドキシサイクリンを添加した。細胞をトリプシン処理し、この時点及び後続の時点で回収した。構築物を発現するGFPのために、これらの試料を標準的なFITCフィルターを用いたフローサイトメトリーで分析した。(背景蛍光のみの)非トランスフェクト細胞を遮断し、次いで細胞当たりの平均蛍光に陽性細胞数を掛けることにより、GFPの総蛍光を測定した。RFPの蛍光(DsRed)を同様に適切なフィルターを用いて測定した。ルシフェラーゼ−をコードするベクターでトランスフェクトされた細胞を溶解し、プロメガの二重ルシフェラーゼ検定法及び試薬を用いた照度計中で測定した。
【0173】
データは通常、タイムゼロと比較した場合の、残ったレポーター(蛍光又は発光)のパーセンテージとして表される。
【0174】
タイムゼロで添加されたドキシサイクリンは該レポーターの転写を阻害する原因となるため、レポーターレベルにおける減少速度は改変された転写レベルと改変されたレポーター/タンパク質レベルの間の時間差を示す。本発明の主な目的はこの時間差を減少させることであり、図7、8、9、及び11〜14はその目的が達成されたことを示す。
【0175】
本発明の使用例として、製薬会社は疾病に関与する遺伝子の転写を減少させる薬物をスクリーニングしたいと思うかもしれない。TREプロモーター由来のテトラサイクリン/ドキシサイクリンが誘導する転写の阻害はこのようなシステムのモデルである。図7と図8は、標準的なルシフェラーゼレポーターベクターでは、(ドキシサイクリンによる)転写の完全な阻害さえも、10時間以内のルシフェラーゼ活性の減少として検出されることはないということを示している。不安定化EGFP突然変異体は、転写の完全阻害が11時間以内(d2EGFP; BTG2)又は7時間以内(d1EGFP; BTG1)にEGFP蛍光の50%減少として検出できるという点で改良を表す。しかしながら、後者のレポーターが4個の9量体UUAUUUAUU(BTG1N4)のようなmRNA不安定化因子と結合した場合、レポーターレベルの50%の減少が3時間以内に検出できる。このことから、転写の増加も不安定化因子を含む構築物を用いてより早く検出されうるであろうと結論される(ロス、1995)。
【0176】
もちろん、ドキシサイクリンの作用はすぐに起こるわけではなく、それ故に時間差の一部はこの薬物が100%の転写阻害を誘導するのに必要な時間に起因する。従って、「減衰の有効速度」はドキシサイクリンを添加した4時間後の時点に続くデータポイントをその時点と比較してプロットすることにより測定した(図9)。減衰の有効速度は、従って薬物作用の遅れを排除し、且つタンパク質の半減期とmRNAの半減期の総合的な効果である。図9は、1個、2個、又は4個の9量体を含む構築物による減衰の有効速度を示す。これらのデータから、4個の9量体が2個の9量体よりも効果的であり、2個の9量体は1個の9量体よりも効果的であることが分かる。さらに、これらのデータは1時間の半減期のタンパク質(d1EGFP)を4個の9量体と結合させることにより、およそ1時間20分という減衰の有効速度が達成できることを示す。これは該タンパク質の1時間半減期に極めて近く、mRNAの半減期が極めて短いことを例証する。二つ以上の異なるmRNA不安定化因子を結合することで、更なる減衰を達成することができるであろう(図13)。しかしながら、これは殆どの適用で必要とされないと思われる。より適度な不安定化因子を必要とする適用では4個ではなく、1個又は2個の9量体を利用できるであろう。
【0177】
標準的なルシフェラーゼレポーターにより、発光はドキシサイクリンの添加後に実際に増加した。これはデータを均等目盛り上に表した場合に最も明らかであり(図8)、部分的にはドキシサイクリンの作用の遅れによって説明できる。しかしながら、4時間先ですら減衰が見られず、時間を掛けて転写の変化を測定するにはこのレポーターは不適当であることを示している。このベクターのさらなる問題を図10に示す。これらのデータは、何らの処置又は薬物も加えない状態での、長時間(トランスフェクション後24〜34時間)にわたるレポーターレベルの変化に関する。レポーターレベルは一般的には該プラスミドが細胞に入り発現を始める、トランスフェクションの24時間後に増加する。減少は一般的には該プラスミドが細胞から排出される約48時間から見られる。従って、測定は通常は24時間と48時間の間に行われる。薬物又は処置の非存在下で、不安定化因子を含んでいる新しいベクター(BTG1N4)はレポーターレベルの優れた安定性を示す。対照的に、ルシフェラーゼベクターは、明らかに発現レベルを依然として増加させている。緩やかな安定性を持つ構築物(例えば、BTG1)は中間的な結果を示した。mRNAとタンパク質のより長い半減期をもつレポーターは、図10に示すようにより長い増加相を示すことが明らかである。24〜34時間の重要な期間に新しい構築物で見られたより安定な発現レベルは正確な測定を容易にし、本発明の別の利点を表す。
【0178】
レポーターレベルのこの減少速度は、それらのレポーターmRNAの配列(例えば、3’UTRの配列)では異なるが、同一のタンパク質又は同じ半減期を有する異なるタンパク質(例えば、d2EGFP、d2EYFP)をコードする二つ以上の構築物の間で比較できる。この状況では、減衰の速度の差異は、mRNA安定性に対する改変されたmRNA配列の効果を示す。例えば、3’UTR内における DNA TTATTTATT (配列番号:1)(図7〜9)又は c-fos ARE(図11)(配列番号:2)としての4個の UUAUUUAUU9量体の存在は、mRNA減衰の速度を有意に増大させた。これらの因子の有効性の実証に加えて、使用した手法とベクターは、他のシスに作用する mRNA の安定化/不安定化因子を検出するための実質的に改良されたシステムをも表し、このプロセスも本発明に包含される。
【0179】
図12から14に示すように、mRNA不安定化因子はルシフェラーゼ、GFP及びDsRedと共に働く。これらのレポーター間の相同性のレベルが低いにもかかわらずである。DsREdは EGFP と23%の相同性しかない。図14に示すように、myc ARE(配列番号:21)は効果的であり、異なる不安定化因子との組み合わせでも効果的である。
【実施例15】
【0180】
mRNA不安定化因子
本発明の RNA不安定化因子は、とりわけ次の遺伝子の3’UTRから誘導できる。殆どの場合、全長の3’UTRが用いられる。しかしながら、U豊富因子及び/又はAU豊富因子はしばしば単独で用いられる。
【0181】
a)ラテルツァ, オウエフらにより記載されたホスホエノールピルビン酸カルボキシキナーゼ(PEPCK)mRNA不安定化因子。JW6及びJW7と称される3’UTRの3’側の半分以内の領域、即ち、
GTATGTTTAAATTATTTTTATACACTGCCCTTTCTTACCT
TTCTTTACATAATTGAAATAGGTATCCTGACCA (配列番号:4)。
【0182】
b)ショウジョウバエメラノガスター(melanogaster)由来のビコイド遺伝子は、3’UTRの最初の43ヌクレオチド内にmRNA不安定化因子(サーデイ, ピーら)を含む。このような因子は特に昆虫細胞のmRNAを不安定化するために使用できる。
【0183】
c)ヒト・チオレドキシン還元酵素遺伝子(ガスダスカ、ジェイアールら)。3’UTR全体。ヌクレオチド1933〜3690(6個のAU豊富因子を含む)。3個の上流AU反復(ヌクレオチド1975〜3360)を含むセグメント。ヌクレオチド 1933〜2014には非AU豊富不安定化因子もある。
【0184】
d)ジョウ, キューらにより記載された熱安定性抗原(HSA)遺伝子。例えば、3’UTRにあるヌクレオチド1465〜1625。
【0185】
e)チー−イイング,エイらにより記載された顆粒球マクロファージコロニー刺激因子(GM-CSF)ARE。AGUAAUAUUUAUAUAUUUAUAUUUUUAA AAUAUUUAUUUAUUUAUUUAUUUAA 即ち、
DNAとして:AGTAAUATTTATATATTTATATTTTTAAAATATTTATTTATTTATTTATTTAA(配列番号:5)。
【0186】
f)c-fosの3’UTR全長又はその部分、又はシュウらにより記載されたARE、
5’AAAACGTTTTATTGTGTTTTTAATTTATTTATTAAGATGG
ATTCTCAGATATTTATATTTTTATTTTATTTTTTT3’ (配列番号:2)。
又は、ペング, エスらによる
5’TTTTATTGTGTTTTTAATTTATTTATTAAGATGGATTCTC
AGATATTTATATTTTTATTTTATTTTTTTT3’(配列番号:6)。
【0187】
g)ペング, エスらにより記載されたようなc-jun ARE。
5’UUUCGUUAACUGUGUAUGUACAUAUAUAUAUUUUUUAAUUUGAUUAAAGC
UGAUUACUGUGAAUAAACAGCUUCAUGCCUUUGUAAGUU3’ 。
DNAとしての配列:
5’TTTCGTTAACTGTGTATGTACATATATATATTTTTTAATTTGATTAAAGC
TGATTACTGTGAATAAACAGCTTCATGCCTTTGTAAGTT3’(配列番号:7)。
又は、ポリアデニル化(AAUAAA)シグナルを含まないその突然変異型、即ち、
5’UUUCGUUAACUGUGUAUGUACAUAUAUAUAUUUUUUAAUUUGAUUAAAGC
UGAUUACUGUGgAUccACAGCUUCAUGCCUUUGUAAGUU3’ 又は、DNAとして
5’TTTCGTTAACTGTGTATGTACATATATATATTTTTTAATTTGATTAAAGC
TGATTACTGTGgATccACAGCTTCATGCCTTTGTAAGTT3’ (配列番号:8)。
【0188】
h)次の遺伝子由来の配列。これらはへニックス, ティーらにより記載されたように、それぞれの ARE構成成分を含む。
IFN-γ ARE
5’UCUAUUUAUUAAUAUUUAACAUUAUUUAUAUAUGGG3’又は、DNAとして
5’TCTATTTATTAATATTTAACATTATTTATATATGGG3’(配列番号:9)。
【0189】
i)IL-2 ARE
5’CUCUAUUUAUUUAAAUAUUUAACUUUAAUUUAUUUUUGGAUGUAUUGU
UUACUAACUUUUAGUGCUUCCCACUUAAAACAUAUCAGGCUUCUAUUU
AUUUAAAUAUUUAAAUUUUAUAUUUAUU3’又は、DNAとして
5’CTCTATTTATTTAAATATTTAACTTTAATTTATTTTTGGATGTATTGT
TTACTAACTTTTAGTGCTTCCCACTTAAAACATATCAGGCTTCTATTT
ATTTAAATATTTAAATTTTATATTTATT3’(配列番号:10)。
【0190】
j)c-myc ARE(配列番号:21も参照)
5’AUAAACCCUAAUUUUUUUUAUUUAAGUACAUUUUGCUUUUAAAGUU3’又は、DNAとして
5’ATAAACCCTAATTTTTTTTATTTAAGTACATTTTGCTTTTAAAGTT3’(配列番号:11)。
【0191】
k)IL-10
5’UAGAAUAUUUAUUACCUCUGAUACCUCAACCCCCAUUUCUAUUUAUUUAC
UGAGCUUCUCUGUGAACGAUUUAGAAAGAAGCCCAAUAUUAUAAUUUUUU
UCAAUAUUUAUUAUUUUCA3’ 又は、DNAとして
5’TAGAATATTTATTACCTCTGATACCTCAACCCCCATTTCTATTTATTTAC
TGAGCTTCTCTGTGAACGATTTAGAAAGAAGCCCAATATTATAATTTTTT
TCAATATTTATTATTTTCA3’(配列番号:12)。
【0192】
l)bcl-2
スキアヴォーン, エヌらにより記載されたようなbcl-2 AREの全部又は一部を含むbcl-2 の3’UTR由来の配列。
5’UCAGCUAUUUACUGCCAAAGGGAAAUAUCAUUUAUUUUUUACAUUAUUAAGAAAAAAGAU
UUAUUUAUUUAAGACAGUCCCAUCAAAACUCCGUCUUUGGAAAUC3’
(M13994のヌクレオチド 2371〜2475)又は、DNAとして
5’TCAGCTATTTACTGCCAAAGGGAAATATCATTTATTTTTTACATTATTAAGAAAAAAGAT
TTATTTATTTAAGACAGTCCCATCAAAACTCCGTCTTTGGAAATC3’
(配列番号:13)。
【0193】
m)スー、エヌらにより記載されたTNF ARE。
5’AUUAUUUAUUAUUUAUUUAUUAUUUAUUUAUUUA3’又は、DNAとして
5’ATTATTTATTATTTATTTATTATTTATTTATTTA3’(配列番号:14)。
【0194】
n)スー、エヌらにより記載されたIL3ARE。
5’UAUUUUAUUCCAUUAAGGCUAUUUAUUUAUGUAUUUAUGUAUUUAUUUAUUUAUU3’ 又は、
DNAとして
5’TATTTT ATTCCATTAAGGCTATTTATTTATGTATTTATGTATTTATTTATTTATT3’
(配列番号:15)
【0195】
o)ズビアーガ, エイらにより記載されたような、9量体 UUAUUUAUU。DNAとして
TTATTTATT(配列番号:1)。
【0196】
p)ラグナード、シーらにより記載されたような9量体UUAUUUAUU(U/A)(U/A) 。DNAとして TTATTTA(T/A)(T/A)( 配列番号:3)。
【0197】
q)スー、エヌらにより記載されたような5量体 AUUUA。又はDNAとして
ATTTA(配列番号:16)。
【0198】
r)4量体 AUUU 、又はDNAとして ATTT (配列番号:17)。
AU豊富因子(ARE)は、チェン、シー及びシュー、エイにより記載されたような、一般的にはクラスIとクラスIIの両方のものである。
植物は、不安定化因子として作用するDST(下流配列)を有する。DST配列は、ニューマン、ティーらに規定されている。提案されたコンセンサスDST配列は
GGAgN2-9cATAGATTaN3-8(A/C)(T/A)(A/T)TttGTA(T/C)
である。
【0199】
s)上記のDST配列は9つの異なったDST配列の比較に基づくものである。
下線部=9/9の遺伝子で保存されている。
大文字=少なくとも7/9の遺伝子で保存されている。
N2-9=2〜9個のヌクレオチドの範囲で長さが可変の領域。平均=5。
N3-8=3〜8個のヌクレオチドの範囲で長さが可変の領域。平均=6。
停止コドンからの距離=19〜83ヌクレオチド。
DST配列の更なる例は、次のものを含む。
大豆の10A5遺伝子:
5’GGAGN5CATAGATTAN8AAATTTGTAC3’(配列番号:18)。
アラビドプシスSURAC1遺伝子:
5’GGAAN9CATAGATCGN8CAATGCGTAT3’(配列番号:19)。
DST配列は植物に使用するためのAU豊富因子の代替物である。AU豊富因子とDST配列は共に植物における転写を不安定化する。
【0200】
t)鉄応答因子(IRE)
トムソン、エイら、1999。
IREはヘアピン・ループ内にコンセンサス CAGUGを含む。
例:
フェリチンIRE:
GUUCUUGCUUCAACAGUGUUUGAACGGAAC 又は、
DNAとして、GTTCTTGCTTCAACAGTGTTTGAACGGAAC(配列番号:20)。
トランスフェリン受容体のIRE:
GAUUAUCGGGAGCAGUGUCUUCCAUAAUC 又は、
DNAとして、GATTATCGGGAGCAGTGTCTTCCATAATC(配列番号:21)。
【0201】
鉄調節タンパク質(IRP、例えば、IRP1及びIRP2)は鉄に依存した形でIREと結合する。結合は又、様々な他の刺激及び処置(例えば、酸化的ストレス、一酸化窒素、エリスロポエチン、甲状腺ホルモン、及びPKCによるリン酸化反応)によって調節される。
【0202】
IREは、翻訳効率及びmRNA安定性の双方を調節することができる。例えば、フェリチンmRNA内の5’UTR IREは、IRPと結合した場合のみ翻訳を阻止する。トランスフェリン受容体mRNAの3’UTR内のIREは、IRPと結合した場合にmRNAの減衰を阻害する。従って、IREを発現ベクターの5’UTR又は3’UTRに挿入して、鉄レベルまたは他の刺激によって制御されうる発現を提供することができる。
【0203】
不安定化因子はクローンテックのマーキュリー経路プロファイリングベクター及びインビボ・キナーゼ検定キットと共に使用できる。クローンテックは3つの異なるタンパク質不安定化因子を製造しており、全てがPEST配列を含み、全てがMODC遺伝子から誘導される。EGFPのカルボキシ末端に配置された異なる変異体MODCは、1時間、2時間、及び4時間のタンパク質半減期を提供する。本発明のmRNA不安定化因子はこれらの不安定化因子と他の如何なるタンパク質不安定化因子(例えば、ユビキチン・シグナル)とも一緒に使用できる。
【0204】
u)c-myc AREは、次のように定義されうる:
5’ATGCATGATCAAATGCAACCTCACAACCTTGGCTGAGTCTTGAGACTGAA
AGATTTAGCCATAATGTAAACTGCCTCAAATTGGACTTTGGGCATAAAAG
AACTTTTTTATGCTTACCATCTTTTTTTTTTCTTTAACAGATTTGTATTT
AAGAATTGTTTTTAAAAAATTTTAAGATTTACACAATGTTTCTCTGTAAA
TATTGCCATTAAATGTAAATAACTTT3’ (配列番号:22)。
【0205】
別の役に立つmRNA因子は、ヒストンmRNAから得ることができる。とりわけ、コンセンサス・ステム・ループ(consensus stem loop)構造を含む3’UTR配列が、ガリー、ディーらに記載されている:
TGA-N20-40-CCAAAGGYYYUUYUNARRRCCACCCA、この式で、Y=ピリミジン、R=プリン、N=何らかのヌクレオチドである。
又は、DNAとして、TGA-N20-40-CCAAAGGYYYTTYTNARRRCCACCCA(配列番号:23)。
【0206】
このような配列は、翻訳効率を増大できる。さらに、このような配列は、特にS期(DNA合成期)の外側でmRNAの減衰を指示することができる。細胞周期特異的プロモーターを含むレポーター構築物は、mRNA不安定化因子と共に、細胞周期特異的発現(例えば、レポーターの発現)を指示するための道具として本発明で意図される。ヒストンの3’UTR因子は、S期と比べてG2期でmRNAの減衰の増大を指示し、それ故にタンパク質発現をS期にさらに制限するので、S期又は後期G1特異的プロモーターと共に使用するための代替物を提供する。
【0207】
発現ベクターにおける3’UTR因子の更に別の用途は、キメラmRNAを特異的に局在化するためのものである。例えば、ユトロフィン(utrophin)の3’UTRはレポーターmRNAを細胞骨格結合ポリソームに導くことができる。mRNA不安定化因子も、この3’UTRに含まれる(グラモリーニ、エイら)。
【実施例16】
【0208】
mRNA安定化因子と安定化されたmRNAをコードする発現ベクター
安定化配列はCT豊富因子及び/又は長寿命mRNA(特に3’UTR領域)から誘導された配列を含む。
【0209】
CT豊富因子は、ホルシックとリーブハーバー、1997により記載されたような
(C/U)CCANxCCC(U/A)PyxUC(C/U)CCを含む。
【0210】
CT豊富因子は次の因子を含む。
CCTCCTGCCCGCTGGGCCTCCCAACGGGCCCTCCTCCCCTCCTTGC
又は、CCTCC、CCTCCTGCC、又はCCCTCCTCCCCTGGを含むその部分。
【0211】
ユーとラッセルにより記載されたヒトのベータ−グロビンの3’UTRに由来する14ヌクレオチドのピリミジン豊富領域も安定化因子としての使用が意図される。
【0212】
そこから安定化因子が誘導されるであろう長寿命mRNAの例には、ヒト、マウス、ウサギ、又は他の種からのアルファ2グロビン、アルファ1グロビン、ベータグロビン、牛成長ホルモン3’UTRが含まれる。
【0213】
本明細書記載のmRNA不安定化因子は、一般的にはキメラ遺伝子を不安定化するために優性な形で作用する。従って、mRNA安定化因子はしばしば劣性に作用しているということになる。例えば、c-fos AREをウサギのベータグロビン遺伝子に挿入すると、mRNA安定性因子が持続的に存在するにも関わらず不安定化された転写という結果になる(シュー、エイら、1989)。アルファ−及びベータ−グロビンmRNAは両方ともそれぞれの3’UTRに配置された安定化因子を含み、一方ゼータグロビンmRNAはこれらの因子を欠いており、安定性が低い。ゼータグロビン3’UTRをアルファグロビンmRNAの3’UTRに置き換えるとmRNA安定性が殆ど2倍になる(ラッセル、ジェイら、1998)。しかしながら、このような因子は全ての転写を安定化するわけではない。従って、安定性mRNAを発現する発現ベクターを生成するための必要条件は、安定化の対象となる元のmRNAによって異なる。このようなベクターを創り出すためには、一般的にはアルファ−又はベータ−グロビンなどの安定な遺伝子に由来する大きなセグメントを含めることが好ましい。これらの例では、このようなセグメントは、好ましくは内因性3’UTRに替えてグロビン3’UTR全体を含むべきである。ゼータ−グロビンで例証したように、これで十分な場合も多い。しかしながら、タンパク質コード配列及び/又は5’UTR配列を更に組み込むことがしばしば必要となる。一般的に、優性の不安定化因子(これらは本明細書記載の技術を用いて同定できる)として作用するであろう内因性のAU又はU豊富領域を置き換えることが好ましい。5’UTR又は3’UTR内のこのような領域は、同じ関連位置からのアルファ−又はベータ−グロビン配列で単に置き換えられる。コード領域からの不安定化因子は同義のコドンへ突然変異させることにより非機能的とすることができる。グロビンタンパク質をコードする領域を目的の遺伝子のコード領域に組み込んで、N末端又はC末端融合タンパク質を創り出すことができる。しかしながら、これは望ましくないことが多く、一般的にはグロビンタンパク質をコードする領域(及び3’UTR)をキメラ遺伝子の3’UTRに配置すれば十分である。これはより安定な転写物から所望のタンパク質を発現させることを可能にし、従って該タンパク質のレベルは顕著に増加する。所望のタンパク質がレポーターであり、又はレポーターに融合しており、又は内因性タンパク質から容易に識別できる場合、本明細書記載(図7参照)のTREベクター系は、キメラ構築物のmRNA安定性のテストを大幅に容易にする。
【0214】
当業者は本明細書に記載した発明が特に記載されたもの以外の変化及び修飾を受けることを承知している。本明細書記載の発明は、そのような変化及び修飾の全てを含むものであると理解されるべきである。本発明は又、この明細書で言及又は示唆したそのような工程、特性、組成物及び化合物の全てを個々に又は集合的に含み、該工程又は特性の二つ以上のありとあらゆる組み合わせを含む。
【0215】
【表1】

【0216】
引用文献
チェン,シー.とシュー,エイ.、Trends Biochem. Sci. vol 20: 465-470, 1995。
チー−ユィング,エイ.ら、MCB vol 15(10): 5777-5788, 1995。
コンタグら、哺乳類における光放射性コンジュゲートの非侵襲性局在化,米国特許第 5,650,135号(1997年 7月22日) 。
コーマックら、緑色蛍光タンパク質 (GFP)の FACS 最適化変異型、米国特許第5,804,387 号( 1998年 9月 8日) 。
ダンデカー、ティー.ら、Bioinformation14(3): 271-278, 1998。
【0217】
ダーツィンキーウイック、レーザー走査サイトメトリー:様々な適用を持つ新たな装置,Exp Cell Res 249: 1-12,1999。
ガリー,ディー.ら、Nucleic Acids Res. vol 24(10): 1954-1962,1996。
ガスダスカ,ジェイアール.ら、JBC vol 274(36): 25379-25385,1999。
ゴッセンら、テトラサイクリン応答プロモーターによる真核細胞中での遺伝子発現の厳格な制御,米国特許第 5,464,758号(1995年11月 7日)。
グラモリーニ,エイ.ら、JBC vol 154(6): 1173-1183, 2001。
【0218】
へニックス,ティー.ら、JBC vol 274(24): 17318-17324,1999。
ホルシックとリブハーバー、PNAS vol 94: 2410-2414, 1997。
ユエ,エックス.ら、Mol Cell Biol. vol 16(7): 3789-3798, 1996。
イブラヒムら、内部制御プラスミドの落とし穴:ウミシイタケルシフェラーゼ(pRL-TK)プラスミドのジヒドロテストステロンとデキサメタゾンに対する反応,Biotechniques 29: 682-684, 2000。
カメンツキーら、スライドベースのレーザー走査サイトメトリー,Acta Cytol 41: 123-43,1997。
【0219】
リーら、転写レポーターとしての不安定化緑色蛍光タンパク質の形成,J Biol Chem 273: 34970-34975,1998。
ラグナード,シー.ら、MCB14: 7984-7995, 1994。
ラテルツァ,オーエフ.ら、Am J Physiol Renal Physiol. vol 279(5): F866-F873, 2000。
ルクレール,ジー.ら、Biotechniques29(3): 590-595, 2000。
リー,エイチ.ら、Gene Expr vol 4(3): 95-109, 1995。
【0220】
リーら、GFP−融合タンパク質の急速分解,米国特許第 6,130,313号(2000年10月10日)。
リュー,ジェイ.ら、JBC vol 275(16): 11846-11851, 2000。
ニューナン,ティー.ら、The Plant Cell vol 5: 701-714, 1993。
ペング,エス.ら、MCB vol 6(4): 1490-1499, 1996。
ロス,ジェイ.、Microbiological Reviews. vol 59(3): 423-450, 1995。
【0221】
サイトウ,ティー.ら、Biochem Biophys Res Commun. vol 252(3): 775-778, 1998。
スキアヴォーン,エヌ.ら、FASEB J. vol 14: 174-184, 2000。
シューら、Genes & Dev. vol 3: 60-72, 1989。
シュタイン,ジェイ.ら、Int J Obes Relat Metab Disord. Suupl 3:S84-90, (1996年3月20日)。
サーデュ,ピー.とジェーコブス−ロレーナ、M. MCB vol 18(5): 2892-2900, 1998。
【0222】
トムソン,エイ.ら、Int. J Biochem Cell Biol. vol 31: 1139-1152, 1999。
ツィエンら、改変された緑色蛍光タンパク質,米国特許第 5,625,048号(1997年 4月29日)。
ツィエンら、改変された緑色蛍光タンパク質,米国特許第 5,777,079号(1998年 6月 7日)。
ヴァズハピリー,アール.とサッチャー,エヌ.、Neurosci Lett. vol 318(3): 153-157, 2002。
スー,ダブリュー.ら、MCB17(8): 4611-4621, 1997。
ユーとラッセル、MCB : 5879-88, Sept 2001 。
ジョウ,キュー.ら、MCB vol 18(2): 815-826, 1998。
ツビアガら、MCB 15: 2219-30, 1995。
【技術分野】
【0001】
本発明は一般的に遺伝子発現又は遺伝子調節検定におけるベクター及びその使用に関する。より具体的には、本発明はタンパク質発現の動力学にプロモーター活性との改良された時的相関関係を与える発現ベクター及び/又はレポーターベクターを提供する。本発明はとりわけ遺伝子発現を調節するために有用な、ヒト疾病における治療的介入のため調節配列、新たな標的及び薬物を同定し分析するため及び薬物スクリーニングのために有用な新規なベクター及び細胞系統を提供する。
【背景技術】
【0002】
本発明者らが本明細書で言及した出版物の文献学的詳細は本明細書の末尾に集めてある。
【0003】
一以上の任意の先行技術文献を含む本明細書における先行技術についての言及は、該先行技術が当分野の技術水準であることを認め又は示唆するものと取られるべきではない。
【0004】
組換えDNA技術の精緻化の急速な進展は医学及び関連する厚生分野における研究及び発展を大いに促進している。とりわけ重要な研究領域は遺伝子発現を研究するための発現ベクターの利用である。しかしながら、今日まで、遺伝子発現の実時間分析は適切に設計されたベクターの不存在により制限されてきた。
【0005】
レポーター検定は、例えば、DNA配列、転写因子、RNA配列、RNA結合タンパク質、シグナル変換経路、及び特異的刺激などの何が目的の遺伝子の発現を制御するのかを理解させる。
【0006】
さらに、レポーター検定はヒト疾病における治療的介入のための新たな標的として役立つ遺伝子調節の局面を同定するために使用できる。レポーター検定は遺伝子発現を調節する能力について薬物をスクリーニングするために使用できる可能性がある。しかしながら、現在のレポーター検定システムに必要な費用と時間は、応答時間の長いことと併せ、その適用を妨げてきた。
【0007】
ゲノム配列は、一般にコード領域の上流にプロモーター配列を有し、それは細胞の特異性と転写の誘導可能性を規定し、それによりタンパク質産物の発現レベルに影響を与える。
【0008】
典型的にはヌクレオチド塩基A及びUが豊富であり、時には遺伝子の3’UTRに位置する特異的な配列因子は、mRNAの安定性に影響を与え、それによりタンパク質産物の発現レベルに影響を与える。RNA結合タンパク質は特定のmRNA配列に結合し、それによりmRNAの安定性及びタンパク質の発現を調節する。他の配列は翻訳効率を調節する。
【0009】
遺伝子レポーター検定の普通の適用は転写を調節するDNA配列の研究である。通常、これらの配列は転写開始部位の5’側、プロモーター領域に位置している。このようなDNA因子は、転写、従ってレポータータンパク質の発現をを駆動及び/又は調節するようにレポータープラスミド内の類似の部位の中にそれらをクローニングすることにより試験される。このレポータータンパク質は内因性タンパク質から識別可能であり、容易に定量されるべきである。種々のレポータータンパク質が使用され、最も普通なのはルシフェラーゼ、クロラムフェニコールトランスフェラーゼ(CAT)及びβガラクトシダーゼ(β-gal) である。
【0010】
レポータータンパク質は適当な検定法で定量され、しばしば、例えばプロモーター SV40 などのどこにでもあるプロモーターにより駆動される対照レポーターのレベルと比較して表される。この対照レポーターは、テストレポーターとは識別可能でなければならず、テストベクターと共トランスフェクトされる別のベクターに含まれていなければならず、トランスフェクション効率を制御するために使用されねばならない。このような検定は細胞が両方のベクターを量に比例して等しく摂取するという前提に基づいている。プラスミドベクターの一過性のトランスフェクションは最も普通に使用される。
【0011】
うえの検定はプロモーター内のプロモーター領域又は特定の因子を同定するために用いられる。または、これらの検定はプロモーター又は調節因子の種々の刺激に対する応答を研究するために用いられる。一部の適用では、レポーター構築物又はトランスフェクトされた細胞はイン・ビボにおけるプロモーター機能を研究するため生物中に配置される。
【0012】
これらのレポーター検定の別の適用は特定のプロモーターの上流のシグナル変換経路の研究又は測定である。例えば、転写のためMAPキナーゼ(MAPK) に依存するあるプロモーターを、レポーター構築物に連結して細胞中のMAPK活性化(又はMAPK依存性転写)のレベルを測定するために使用することができる。この技術は様々な情報を与えるプロモーター又はエンハンサーと共に利用することができ、そして細胞又はトランスジェニックマウスなどの生きている生物に適用できる。例えば、フォトンカメラは目的のプロモーターに連結したルシフェラーゼレポーターを含む完全マウスにおけるルシフェラーゼレポーター活性を測定するために使用できる(特許文献1を参照)。
【0013】
ルシフェラーゼはこれまでイン・ビトロ系のレポーター検定に最も普通に使用されている。二重ルシフェラーゼ検定(DLA,プロメガ, マジソン, WI, USA)は、テストレポーターと対照レポーターの両方が本質的に同一検定で測定され得るという点で他のルシフェラーゼ系システムに優る改良がなされている。現在の使用の一例として、典型的なDLA法は次のように行なわれる。
【0014】
プロモーター因子と推定されるものを、蛍のルシフェラーゼレポーター遺伝子の上流にクローニングし、それがその発現を駆動するようにする。このプラスミドを SV40 プロモーターにより駆動されるウミシイタケ(renilla)のルシフェラーゼ遺伝子を含む対照のプラスミドと共に細胞系統中に一時的にトランスフェクトする。約2〜50%の細胞がプラスミドを取込み、約3日間このレポーターを発現する。発現の動力学は、ルシフェラーゼタンパク質が蓄積する最初の約24時間の間の増加、及びそれに続き細胞内に維持されたプラスミドの数が減少する約48時間以降の低下を含む。トランスフェクションから24〜48時間の後、細胞を収穫し溶解する。細胞の溶解液を蛍ルシフェラーゼに特異的な基質と共にインキュベートし、活性(光の放射)をルミノメータ(96穴プレート又は個々の試料) を用いて測定する。次に、蛍のルシフェラーゼを失活させるがウミシイタケのルシフェラーゼに光を形成させる追加の基質を添加する。ウミシイタケのルシフェラーゼの活性を次に測定する。
【0015】
蛍のルシフェラーゼ活性のレベルはプロモーター活性のみでなくトランスフェクション効率にも依存する。これは、DNAの量、DNA調製物の品質、及び細胞の状態に応じて大きく変化する。共トランスフェクトされた対照のプラスミド( SV40 プロモーターにより駆動されるウミシイタケのルシフェラーゼ)は、ウミシイタケのルシフェラーゼの活性が細胞によって取り込まれた蛍のルシフェラーゼプラスミドの量に比例するという前提に基づいてこれらの変数を補正するために使用される。データは蛍ルシフェラーゼ活性/ウミシイタケのルシフェラーゼ活性として表される。
【0016】
二重ルシフェラーゼ検定の欠点は
(i) 試薬が高価であり、変質しやすく、調製して直ぐに使用しなければならない、
(ii) 一般的にこの検定は細胞溶解液の調製物を含む。これは時間がかかり、不正確さを加える。例えば、溶解の操作中の細胞の損失、ピペット操作の誤差、残存緩衝液/培地の容量が変化する、
(iii) 個々の試料は細胞集団の全活性である唯一つのデータのみを生ずる。レポーターを発現する細胞のパーセンテージに関する情報も、細胞当たりの発現の量に関する情報も得られない、
(iv) トランスフェクション対照(ウミシイタケ)はトランスフェクション効率における大きな変動を常に補正するとは限らない。何故なら
a. 特定のDNA調製物は僅かしかトランスフェクト/発現せず(おそらくスーパーコイルDNAの割合の減少により)、共トランスフェクトさせる対照プラスミドの量は相当する減少を惹き起こさないこと、
b. 二つのプラスミドのプロモーターの間に、対照レポーターの活性が共トランスフェクトされる構築物に依存する、例えば、ウミシイタケのルシフェラーゼの発現が強力なプロモーターを含むプラスミドと共トランスフェクトされた場合に最高となるようにみえる、というような、クロス−トークの証拠があること。プロモーターの間の干渉も、テストレポーターと対照レポーターの両方を発現する1個のプラスミドの使用を、阻害はしないとしても、制限したということ、
c. 転写及び転写後の両方の研究における普通の適用は、種々の刺激(例えば、PMA 、EGF 、ホルモン) による活性化/抑圧を測定することである。不幸にも、SV40、RSV 、TK 及びおそらく他の多数の普遍的に発現されるプロモーターが様々な刺激により活性化される。これらのプロモーターはトランスフェクション対照レポーター(ウミシイタケ)の発現を駆動するために使用されるから、これらのレポーターはこのような処置の後のトランスフェクション効率の真の反映を与えない(非特許文献1を参照)こと、
d. 蛍対ウミシイタケのルシフェラーゼタンパク質及びおそらくmRNAの半減期の相違は系全体を極めて時間感受性とすること、
e. とりわけウミシイタケのルシフェラーゼにおける急速に減少する光放射は、測定のタイミングにおける絶対的精度を必要とすること、
f. ルシフェラーゼタンパク質及びmRNAの比較的長い半減期は、転写における時間的変化を効果的に覆い隠す(例えば、種々の刺激又は処理の後)こと。
である。
【0017】
現行の転写後/mRNA安定性レポーター検定では、mRNA安定性に影響を与えると考えられた候補因子を SV40 又は RSVなどの構成プロモーターにより駆動されるレポーターベクター(例えば、蛍のルシフェラーゼ)の対応する領域にクローニングする。空のベクター(目的の因子を含まない同じベクター)と比較した場合の発現の変化は改変されたmRNA安定性の結果であると仮定される。予備的に記述されたトランスフェクション検定については、トランスフェクション対照プラスミド(例えば、 SV40 又は RSVなどの構成プロモーターにより駆動されるウミシイタケルシフェラーゼ)を共トランスフェクトさせて、トランスフェクション効率について補正させる。これらの検定は
(i) 現存するベクターは転写後研究のために設計されておらず、転写を切り換えるための手段を持たない、
(ii) これらのプロトコルの目的は候補mRNA因子の転写後の効果を研究することであるが、これらの因子はDNAのレベルでレポーターの転写に影響を与えることもでき、さらに、目的の遺伝子の内因性プロモーターは使用されないから、見られる如何なる転写効果も生理学的関連性はほとんどない、
というさらなる不利益を有する。
【0018】
mRNA安定性を研究するための系は存在するが、タンパク質レポーターではなくmRNAの直接測定を含む。mRNAを定量化するためのプロトコル類の労働集約的性質のため、このような系はさらに一層時間がかかる。
【0019】
例えば、一つの系は血清誘導に応答して転写の短時間の爆発をもたらす c-fosプロモーターを利用する。不安定因子と思われるものを、血清誘導性の(c-fos) プロモーターの制御下で極めて安定なmRNAを発現するベータグロビン(BBB) 構築物の 3-UTR中にクローニングする。トランスフェクトされた細胞(一般に NIH 3T3細胞) をまず血清から断絶させ、ついで血清を含む培地に曝露する。この転写の応答の簡単な性質のため、レポーターmRNAの分解の動力学を経時的に追跡することが可能となる。これらの検定は
(i) この検定は極めて時間がかかり、従って迅速なスクリーニングに適用できない、
(ii) c-fos プロモーターの血清誘導性を支える細胞中でしか使用できない。例えば、多数の腫瘍細胞系統は血清の非存在下でc-fos プロモーター活性を維持する、
(iii) 所望の血清応答を有するNIH 3T3 細胞などの細胞では、血清涸渇は細胞周期を阻止する原因となり、その後血清を添加すると、同時に、この阻止から細胞を解き放つ。それ故、mRNAの安定性は細胞周期の特定の期でしか測定できない、
(iv) c-fos プロモーターの活性化に加え、血清は他の経路の多くを活性化する。これらの経路は望ましくない変数を導入し、より特異的な刺激の研究を阻害する、
という不利益を有する。
【0020】
別の検定では、細胞は全ての遺伝子からの転写を阻害するアクチノマイシンDなどの薬物で処理される。mRNAの分解速度を決定するため、そのmRNAレベルが経時的に測定される。この系は内因性遺伝子を研究するために用いられるが、
(i) 転写阻害剤は、mRNA安定性がストレスを与えた又は死にかかっている細胞でしばしば測定される場合に必要な用量では極めて毒性である、
(ii) 転写阻害剤は特定のmRNAの安定化などの無数の望ましくない活性を有する、
(iii) このプロセスは全ての遺伝子からの転写を阻止し、多数のシグナル変換カスケードが阻止される一方、他のカスケードは活性化される。従って、結果は生理学的に重要であるように思われない、
(iv) この技術は極端に労働集約的である、
(v) この技術は検定内及び検定間で極めて変動し易い、
(vi) この技術は一過性のトランスフェクションレポーター検定においてはしばしば感度が十分でなく、特に低いトランスフェクション効率を持つ細胞中ではそうである。
という欠点を有する。
【先行技術文献】
【特許文献】
【0021】
【特許文献1】コンタグら,米国特許第5,650,135号公報。
【非特許文献】
【0022】
【非特許文献1】イブラヒムら,内部対照プラスミドの落とし穴:ウミシイタケ・ルシフェラーゼプラスミド(pRL-TK) のジヒドロテストステロン及びデキサメタソンへの応答, Biotechniques 2000; 29: 682-684 。
【発明の概要】
【発明が解決しようとする課題】
【0023】
従って、遺伝子発現検定を行なうための、とりわけ翻訳後検定及び転写後検定における、改良されたベクター及びシステム、並びに遺伝子発現における変化のより一層実時間測定を可能にする検定法を開発する必要がある。
【課題を解決するための手段】
【0024】
発明の概要
本明細書を通じて、文脈が別意を要求しない限り、用語「含む(comprise) 」及び「comprises 」や「comprising」などのその変形は、述べられた整数若しくは工程又は整数若しくは工程の群を含むことを意味するが、他の整数若しくは工程又は整数若しくは工程の群の排除を意味するものではないと理解される。本発明は、例示の目的のみを意図する本明細書に記載された具体的な実施態様によりその範囲が制限されるべきではない。機能的に等価な産物、組成物及び方法は、本明細書に記載されたような本発明の範囲内にあることは明らかである。
【0025】
本発明によれば、本発明者はとりわけ転写物の安定性の調節及び測定及び/又は遺伝子発現の実時間測定の改良を可能とする一連のベクター及び方法を開発した。
【0026】
ヌクレオチド配列は配列識別子数(配列番号:)により言及される。この配列番号:は配列識別子<400>1、<400>2などに数値で対応している。配列表は請求の範囲の後ろに提示する。
【0027】
本発明の一つの側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む発現ベクターに向けられている。本明細書で用いられるとき転写物の安定性とはこの転写物の半減期に相当すると考えてよい。
【0028】
本発明の別の一側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節し且つ該転写物の安定性を低減させる不安定化因子であるRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む発現ベクターを意図する。
【0029】
関連する実施態様では、本発明は転写可能なポリヌクレオチドに対応する転写物の安定性を調節し且つ該転写物の安定性を増加させる安定化因子であるRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む発現ベクターを意図する。
【0030】
本発明の別の一側面は、RNA因子をコードするヌクレオチド配列を含む転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写可能なポリヌクレオチドに対応する転写物の安定性を低減させる不安定化因子であり、該転写可能なポリヌクレオチドが目的のポリヌクレオチドを含み且つポリペプチドをコードするものである発現ベクターを意図する。
【0031】
本発明のさらに別の一側面は、RNA因子をコードするヌクレオチド配列を含む転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写可能なポリヌクレオチドに対応する転写物の安定性を低減させる不安定化因子であり、該転写可能なポリヌクレオチドが目的のポリヌクレオチドを含み且つレポーターポリペプチドをコードするものである発現ベクターを意図する。
【0032】
本発明のさらなる別の一側面は、RNA因子をコードするヌクレオチド配列を含む転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写可能なポリヌクレオチドに対応する転写物の安定性を低減させる不安定化因子であり、該転写可能なポリヌクレオチドがレポーターポリペプチドをコードするヌクレオチド配列を含むものである発現ベクターを意図する。
【0033】
本発明のなお別の一側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写可能なポリヌクレオチドに対応する転写物の安定性を低減させる不安定化因子であり、該ベクターが
(i) ヌクレオチド配列を導入するための複数クローニング部位、
(ii) レポーター遺伝子、
(iii) 該転写可能なポリヌクレオチドの発現を調節するためのプロモーター及び/又はエンハンサー、
(iv) ポリアデニル化配列、
(v) 選択可能なマーカー遺伝子、及び
(vi) 複製起点、
からなる群より選択される一つ以上の構成員を含むものである、発現ベクターに向けられている。
【0034】
本発明のさらに別の一側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む本発明のベクターを含む細胞を意図する。
【0035】
本発明のさらに別の一側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む本発明のベクターを含む細胞であって、該RNA因子が該転写物の安定性を低減する不安定化因子である細胞を意図する。
【0036】
本発明の関連する側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む遺伝的に改変された非ヒト生物であって、該RNA因子が該転写物の安定性を低減させる不安定化因子である生物を意図する。
【0037】
本発明のさらなる実施態様は、目的のポリヌクレオチドの発現を測定する方法であって、RNA合成及びタンパク質合成が起こるのに十分な時間及び条件の下で発現ベクターから該目的のポリヌクレオチドを発現させる工程、この工程では、該ベクターが目的の該ポリヌクレオチド及び転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含むものであり、且つ該発現ベクターが
(i) ヌクレオチド配列を導入するための複数クローニング部位、
(ii) レポーター遺伝子、
(iii) 該転写可能なポリヌクレオチドの発現を調節するためのプロモーター及び/又はエンハンサー、
(iv) ポリアデニル化配列、
(v) 選択可能なマーカー遺伝子、及び
(vi) 複製起点、
から成る群より選択される一つ以上の構成員を含むものである、及び転写可能なポリヌクレオチドの発現産物のレベル及び/又は機能的活性を対照と比べ経時的に測定する工程、
この工程では該因子が該転写可能なポリヌクレオチドに機能しうるように連結されたプロモーター及び/又はエンハンサーの活性と該発現産物のレベル及び/又は機能的活性の間の時的相関関係を強化するものである、を含む方法を意図する。
【0038】
さらに別の一実施態様では、本発明はRNA転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を同定する方法であって、発現ベクター中にテストヌクレオチド配列を導入することにより該ヌクレオチド配列をレポータータンパク質をコードするポリヌクレオチドに連結させプロモーター及び/又はエンハンサーに機能しうるように連結された転写可能なポリヌクレオチドを形成させる工程、RNA合成及びタンパク質合成が起こるのに十分な時間及び条件の下で該転写可能なポリヌクレオチドを発現させる工程、
この工程で該発現ベクターは
(i) 該テストヌクレオチド配列を導入するための複数クローニング部位、
(ii) ポリアデニル化配列、
(iii) 選択可能なマーカー遺伝子、及び
(iv) 複製起点、
から成る群より選択される一つ以上の構成員を含むものである、及び該ヌクレオチド配列の非存在下における対照ベクターのそれと比べ該転写可能なポリヌクレオチドの発現産物のレベル及び/又は機能的活性を経時的に測定する工程、この工程でその時間にわたって対照ベクターのそれと異なるレベル及び/又は機能的活性は該RNA因子をコードするヌクレオチド配列の存在を示す、を含む方法を意図する。
【図面の簡単な説明】
【0039】
【図1】図1は、不安定化mRNAをコードする発現ベクターの図式的表示である。
【図2】図2は、転写レポーターベクターの図式的表示である。図2aはベクター系2、図2bはベクター系3、そして図2cはベクター系4を示す。
【図3】図3は、双方向性転写レポーターベクターの図式的表示である。図3aはベクター系5を、そして図3bはベクター系6を示す。
【図4】図4は、転写後調節を研究するためのレポーターベクターの図式的表示である。図4aはベクター系7を、そして図4bはベクター系8を示す。
【図5】図5は、トランスフェクトされたDNAの量に対する尺度としてのレポーター活性を示すグラフ的表示である。蛍ルシフェラーゼをコードするプラスミドの一本鎖DNA調製物をウミシイタケルシフェラーゼをコードする別のプラスミドと30:1の比率で混合する。両方のDNA調製物は分光測光法(OD260/280)とエチジウムブロマイド染色アガロース・ゲル(データは示していない)において、正常に見えた。この混合物をDNAの総量が1、2又は3マイクログラムになるように細胞内に量を変えてトランスフェクトしたが、蛍プラスミドとウミシイタケプラスミドの比率は同じままであった。 図5Aはウミシイタケルシフェラーゼ活性がトランスフェクトされたDNAの量に依存することを示すグラフである。しかしながら、蛍ルシフェラーゼ活性(図5B)はDNAの量を増やしても増加しなかった。これは恐らく、蛍DNA調製物の品質が悪かったためであろう。従って、通常蛍プロモーター活性の尺度として用いられる蛍/ウミシイタケ比率(図5C)は、使用されたDNAの量に応じてかなり変わった。これらのデータはウミシイタケプラスミドとの共トランスフェクションが蛍プラスミドのトランスフェクション効率を適切に制御しないことを示す。
【図6】図6は二重ルシフェラーゼ検定を用いて得られた様々なプロモーター系に対するレポーター活性を示すグラフ的表示である。六つの異なるプロモーター断片(1−6と番号を付す)を、pGL3蛍ルシフェラーゼプラスミドにクローニングした。1マイクログラムの各クローンをSV40プロモーターで駆動されたウミシイタケ(トランスフェクション対照)プラスミドの30ngと共トランスフェクトし、蛍ルシフェラーゼとウミシイタケルシフェラーゼの活性を二重ルシフェラーゼ検定(プロメガ、米国、マディソン群、ウィスコンシン)で測定した。その結果がウミシイタケルシフェラーゼ活性(A)、蛍ルシフェラーゼ活性(B)、及びウミシイタケで割った蛍の活性(C)として表す。それぞれの構築物の少なくとも二つの異なる調製物を用いた複数の実験で同様の結果が見られた。 ウミシイタケルシフェラーゼ活性(図6A)はトランスフェクションの対照を意図したものであり、この結果を単独で分析すると異常に高い変動がトランスフェクション効率にあること示唆するであろう。例えば、ウミシイタケルシフェラーゼ活性は構築物3と共トランスフェクトした場合と比べて、構築物4と共トランスフェクトした場合のほうが3.5倍高い。DNA品質のばらつき又はDNAの定量の際の誤差のばらつきは、別のDNA調製物のセットでも同じパターンが見られた(データ非提示)ことから、エラーの原因であるとは思われない。 蛍ルシフェラーゼ活性(図6B)はトランスフェクション効率とプロモーター1〜6の間の差異の両方の影響を受ける。この差異のパターンは、ウミシイタケで見られる(図6A)ものと類似している。例えば、3と6は低く、一方4と5は高い。しかしながら、構築物間のこのような差異は、蛍で一層顕著であり(例えば、構築物4は構築物3より12倍高い)、プロモーター1〜6の活性がウミシイタケの発現に何らかの形で影響を与えている(またはその逆)ことを示唆する。 蛍/ウミシイタケ(図6C)は、トランスフェクション効率(ウミシイタケ)を修正した後に真の蛍プロモーター活性(1〜6)の尺度になると考えられる。ここでも実際に3と6が最も弱いプロモーターで、一方4と5が最強であることを示唆する類似のパターンが見られる。プロモーター活性(図6C)がトランスフェクション効率(6A)と偶然に相関したという可能性もあるが、多数の異なる構築物及び同じ構築物の複数の異なる調製物で類似の結果が得られたことを考えれば、この可能性はまずあり得ない。ウミシイタケルシフェラーゼの発現レベルがそれと共トランスフェクトされるプロモーター構築物の強さの影響を受けるという方が可能性が大きいであろう。従って、プロモーター1〜6の間の見掛けの差異は、真の差異を過小評価している可能性もある。
【図7】図7は転写の阻害後のBTL、BTG2、BTG1及びBTG1N4発現ベクターに対する異なるレポーターレベルを経時的に示すグラフ的表示である。Tet-オフ・ヒーラ細胞をそれぞれがレポーター遺伝子と連結したTREプロモーターを含んでいる次のレポータープラスミドでトランスフェクトした。即ち、 BTL(ルシフェラーゼ)、BTG2(d2EGFP)、BTG1(d1EGFP)、及びBTG1N4(BTG1と同じであるが、3’UTR−コード領域に存在する9量体UUAUUUAUUの4つのコピーを持つ)。トランスフェクションから10時間後、それぞれのフラスコの細胞を複数の小皿に分けた。ドキシサイクリン(1□g/ml)をトランスフェクション(タイム・ゼロ)の24時間後に添加し、レポーター遺伝子の転写を阻害した。実施例14に記載したように、レポーターレベル(蛍光又は発光)をこの時点及びその後の時点で測定し、タイム・ゼロのパーセンテージとして表した。10時間経過中にルシフェラーゼ活性(BTL)の減少は見られなかった。2時間半減期のEGFP構築物(BTG2)はドキシサイクリンで誘導される転写の阻害に対して穏やかな反応を示し、1時間半減期のEGFP(BTG1)ではより早い反応が見られた。しかしながら、9量体を含む構築物(BTG1N4)では、転写の阻害に対して群を抜いて速い反応を示した。
【図8】図8は、図7で使用したデータを線状スケールで表したもの示すグラフ的表示である。ドキシサイクリンで誘導される転写の阻害はBTG1で約6.5時間後のレポーターレベルで50%の阻害として検出できる。しかしながら、これは9量体(BTG1N4)を含めることにより3時間にまで減少する。
【図9】図9は、異なる数(1、2又は4)の9量体RNA不安定化因子の効果を示すグラフ的表示である。時間経過は、ゼロ時間をこの薬物の活性の遅延効果を除去するためにドキシサイクリンを添加して4時間後と定義したことを除き、図7に記載した通り実施した。1個の9量体の存在(BTG1N1)は「減衰の有効速度」を増加するのに十分であり、2個の9量体(BTG1N2)及び4個の9量体(BTG1N4)でその効果が徐々に強くなることが見られた。後者の構築物は約1時間20分の「有効半減期」を示したが、それはタンパク質単独の1時間半減期と大差はない。
【図10】図10は、転写の阻害がない状態で時間と共に変化するレポーターレベルを示すグラフ的表示である。時間の経過は図7に記載した通り実施した。しかしながら、示したデータはドキシサイクリンで処置されておらず、トランスフェクションの24時間後(開始)又はトランスフェクションの34時間後(終了)に測定された試料を表している。一貫した発現レベルはBTG1N4でしか見られなかった。
【図11】図11は、転写の阻害がない状態で時間と共に変化するレポーターレベルを示すグラフ的表示である。時間の経過は図7に記載したとおり実施した。BTG1 fosは c-fos ARE を含む。これらのデータは別の種類のmRNA不安定化因子が同じ効果を得るために使用できることを証明する。
【図12】図12は、ルシフェラーゼレポータータンパク質が使用された場合にも、RNA不安定化因子は発現を測定するのに役立つということを示すグラフ的表示である。ルシフェラーゼレポータータンパク質をタンパク質不安定化因子と共に使用すれば、更なる増強が期待できるであろう。時間経過は、二つのルシフェラーゼ発現構築物を用いて図7に記載したとおり実施した。BTLは標準的な蛍ルシフェラーゼのコード領域と3’UTR(pGL3−ベーシック、プロメガ)を含み、一方 BTLN6は3’UTRに9量体UUAUUUAUUの6個のコピーを含む。
【図13】図13は、RNA不安定化因子及びタンパク質不安定化因子により不安定化されたDsRedを用いたレポーターレベルの時間経過を示すグラフ的表示である。時間経過は図7及び実施例14に記載したとおり実施した。使用した構築物は、3’UTRに4個の UUAUUUAUU9量体(BRT1N4)を含むDsRed2(BTR)、DsRed-MODC(BTR1)及びDsRed-MODCであった。ドキシサイクリンで転写を阻害した後、赤色蛍光が全ての構築物で増え続けた。これはタンパク質不安定化因子により実質的に減少し、mRNA不安定化因子によってさらに減少する。
【図14】図14は、時間経過が図7に記載されたとおりに実施されたことを示すグラフ的表示である。テストしたmRNA不安定化因子は全て、対照(BTG1)と比べて減衰速度の増加に非常に効果的であった。これらのデータは、c-myc AREが効果的な不安定化因子(BTG1myc)であり、不安定化活性の緩やかな増加が myc AREと4個の9量体を結合(BTG14myc)させることにより得られるということを示す。6個の9量体(BTG1N6)もまた4個の9量体(BTG1N4)よりも幾分強く不安定化するようであった。
【発明を実施するための形態】
【0040】
好ましい実施態様の詳細な説明
定義
「3’UTR」とは、ポリヌクレオチドのタンパク質コード領域の終止コドンの下流のポリヌクレオチド領域、これはタンパク質の生産のために翻訳されることはない、ことを意味する。
「5’UTR」とは、mRNAの5’(上流)側の翻訳されない領域を意味する。mRNAの5’UTRをコードするDNA領域を指すためにも使用される。
「約」とは、参照する数量、レベル、値、寸法、大きさ、又は量に対し 30%まで、好ましくは 20%まで、より好ましくは10% まで変動する数量、レベル、値、寸法、大きさ、又は量を意味する。
【0041】
「ARE」とは、mRNAにおけるAU−豊富因子、即ちアデニン及びウラシルヌクレオチドの高い割合を含む配列を意味する。このようなmRNA因子をコードするDNA領域を指すためにも用いられる。
【0042】
「生物学的に活性な断片」とは、参照ポリヌクレオチド又はポリペプチドの活性をそれぞれ保持する全長参照ポリヌクレオチド又はポリペプチドの断片を意味する。
「c-fos 」とは、有糸分裂シグナルにより簡単に誘導される中間初期遺伝子を意味する。
「CAT 」とは、クロラムフェニコール・アセチルトランスフェラーゼを意味する。レポーターとしてしばしば用いられる細菌の酵素である。
「d1EGFP」とは、突然変異した PEST 配列に融合し従って約1時間の半減期しか持たない EGFP の変異型を意味する。同様に、d1ECFP及び d1EYFP も入手可能である。DsRed の不安定化変異型は同様な方法で作成できるであろう。今後は d1DsRedと呼ばれる。
【0043】
「d2EGFP」とは、 PEST 配列に融合し従って約2時間の半減期しか持たない EGFP 変異型の突然変異型を意味する。同様に、d2ECFP (藍色) 及び d2EYFP(黄色) も入手可能である。DsRed の不安定化変異型はおそらく同様にして作成できるであろう。今後はd2DsRed と呼ばれる。
「dEGFP 」とは、生成した EGFP(全ての色を含む) の全ての不安定化変異型(リーら)に対する一般的用語を意味する。
「DNA」とは、デオキシリボ核酸を意味する。
「誘導体」とは、例えば、他の化合物部分と共役させ若しくは複合体形成させることにより又は当分野で理解されているような転写後若しくは翻訳後修飾により参照ポリヌクレオチド又はポリペプチドからそれぞれ誘導されたポリヌクレオチド又はポリペプチドを意味する。
「DsRed 」とは、インドパシフィックシー・アネモネ近似のディスコソマ・スピーシーズから単離された赤色蛍光タンパク質を意味する。
「ECFP」とは、藍色の光の蛍光を発する、改変された励起/放射スペクトルを持つ EGFP の突然変異型を意味する。
【0044】
「EGF 」とは、表皮成長因子を意味する。
「EGFP」とは、強化された緑色蛍光タンパク質を意味する。強化された蛍光を持つ GFPの突然変異型である(コルマックら)。
「ELISA 」とは、酵素結合イムノソルベント検定法を意味する。
「ErbB2 」とは、表皮成長因子受容体ファミリーの第2の構成員を意味する。HER-2 としても知られている。
「エキソン」とは、メッセンジャーRNA分子又はこの配列をコードするDNAの一部であるRNA一次転写物の配列を意味する。一次転写物では、隣接するエキソンはイントロンにより隔てられている。
「発現ベクター」とは、DNAのクローニングされたセグメントを細胞内で発現させるベクターを意味する。
「EYFP」とは、黄色の光の蛍光を発する改変された励起/放射スペクトルを有する EGFP の突然変異型を意味する。
【0045】
「蛍ルシフェラーゼ」とは、蛍からクローニングされた luc遺伝子から誘導される酵素を意味する。酸素と Mg++の存在下で D- ルシフェリン及び ATPを用いて反応を触媒し結果として光の放射を生ずる。しばしばレポーターとして用いられる。
「フローサイトメトリー」とは、生きた又は固定した細胞の懸濁液を、懸濁液の細胞と結合した検出可能な標識の活性又は性質を個々に測定するフローサイトメーターにかける方法を意味する。細胞の標識化は、例えば蛍光性化合物を介して又は特定の蛍光性化合物に共有結合させた抗体により行なうことができる。異なる幾つかの励起/放射波長を同時にテストし異なるタイプの蛍光を測定することができる。所望の特性(蛍光、細胞の大きさ)を持つ細胞の部分集団をゲートにかけ、ゲートを通った細胞だけをさらなる統計分析にかけることができる。細胞選別機オプションを備えたフローサイトメーターは所望の蛍光を持つ細胞を物理的に分離し、最初の細胞集団の残りからこれらの(生きた)細胞を管中に回収することができる。FACS(蛍光標示式細胞分取法)とも呼ばれる。
「遺伝子」とは、RNA分子をコードするDNAのセグメントを意味する。「遺伝子」という用語は、時には、常にではないが、プロモーター領域を含むことがある。
「GFP 」とは、蛍光タンパク質(ツィエンら)を意味する。これはクラゲ、エクオリア・ヴィクトリアから単離されうるものであり、レポータータンパク質として使用することができる。GFP をコードするDNA構築物は哺乳動物細胞中で発現でき、特定の波長で励起すると細胞が緑色の蛍光を発するようにすることができる。この用語「GFP 」は色の変異型及びエクオリア・ヴィクトリア以外の生物から誘導される蛍光タンパク質(例えば、DsRed,クローンテック、hrGFP,ストラタジーン)を含む全ての相同体及び類似体を指すために本明細書で用いられる。
「半減期」とは、分子の活性、量若しくは数の半分が消失するのに必要な時間を意味する。
「イントロン」とは、遺伝子、又はその一次転写物内の非コード配列を意味する。これは該一次転写物から除去され、対応するメッセンジャーRNA中に存在しない。
【0046】
「ルシフェラーゼ」とは、光の放射をもたらす反応を触媒する普通に使用されるレポーター酵素を意味する。外因性基質を添加し、ルミノメーターを用いてこの反応を定量化する。蛍及びウミシイタケのルシフェラーゼに対する基質の要件は異なっているので、二重ルシフェラーゼ検定法(プロメガ,マディソン,WI, USA)では両者を識別することができる。
「MAPK」とは、有糸分裂活性化タンパク質キナーゼを意味する。成長又はアポトーシス(細胞死)に至る細胞内シグナル変換経路に関与する幾つかの異なるキナーゼを含む。用語「MAPK」はしばしば二つの特定の MAPKs、Erk1及びErk2(細胞外の調節キナーゼ1及び2)を指すために用いられる。
「MCMV」とは、最小 CMVプロモーターを意味する。それ自体の転写を活性化しないが、TRE に連結されるとテトラサイクリン(及びドクシサイクリン) 依存性の転写を行なわせることができる。
【0047】
「MCS 」とは、複数クローニング部位を意味する。その中にDNA断片を挿入できるユニークな制限酵素認識部位を含むDNAベクターの領域である。用語「MCS 」は本明細書で用いられるとき、このベクター中へのDNA断片の挿入を助ける任意の他の部位をも含む。例えば、Tオーバーハング(プロメガ,マディソン,WI, USA)である。これはポリメラーゼ連鎖反応(PCR)により形成された断片の直接挿入を可能とする。
「mRNA」とは、メッセンジャーRNAを意味する。DNAを鋳型として用い細胞内で生産される、それ自体はタンパク質をコードする「転写物」である。mRNAは通常5’UTR、タンパク質をコードする領域及び3’UTRから構成される。mRNAは細胞内で限定された半減期を有し、それはとりわけ3’UTR内にある、しかし5’UTR及びタンパク質をコードする領域にもある安定性因子により部分的に決定される。
「MODC」とは、マウス・オルニチン・デカルボキシラーゼ又は PEST 配列を含むその部分及び/又は誘導体を意味する。
「調節する」とは、目的の分子の安定性を、直接的若しくは間接的のいずれかで増加又は低減させることを意味する。
【0048】
「機能しうるように連結される」とは、機能的な関係の中でポリヌクレオチド因子を連結することを意味する。核酸は、それが別の核酸配列と機能的な関係に置かれる場合に「機能しうるように連結される」。例えば、プロモーター又はエンハンサーは、それがコード配列の転写に影響を与える場合、コード配列に機能しうるように連結されている。機能しうるように連結されるとは、連結されている核酸配列が通常隣接している、そして二つのタンパク質コード領域を結合する必要がある場合は、隣接し且つ読み取り枠内にあることを意味する。RNAポリメラーゼが二つのコード配列を一つのmRNAに転写する場合コード配列は別のコード配列に「機能しうるように連結されている」。mRNAは次いで両方のコード配列から誘導されるアミノ酸を持つ一本のポリペプチドに翻訳される。このコード配列は発現された配列が最終的にプロセスされ所望のタンパク質を生産する限り相互に隣接している必要はない。転写可能なポリヌクレオチドにプロモーターを「機能しうるように連結する」とは、転写可能なポリヌクレオチド(例えば、タンパク質をコードするポリヌクレオチド又は他の転写物)をプロモーターの調節的制御の下に置くことを意味する。このプロモーターは次いで該ポリヌクレオチドの転写を制御し、任意選択的に翻訳を制御する。プロモーター/構造遺伝子の異質の組み合わせの構築では、転写可能なポリヌクレオチドの転写開始部位から離れた位置にプロモーター又はその変異型を配置することが好ましい。この距離はその天然の設定、即ち、プロモーターがそれから由来した遺伝子における、プロモーターとそれが制御する遺伝子の間の距離とほぼ同じである。当分野で知られているように、この距離のある程度の変化は機能の損失無しに順応できる。同様に、その制御の下に置かれる転写可能なポリヌクレオチドに関する調節配列因子(例えば、オペレーター、エンハンサーなど)の好ましい配置は、天然の設定、即ちそれが由来した遺伝子における該因子の位置により規定される。
【0049】
「pA」とは、ポリアデニル化部位を示すために本明細書のダイアグラム中で用いられる、転写を停止させるための部位として及び未成熟mRNAにポリA尾を添加するために働くDNA配列を意味する。SV40ウイルス遺伝子又はβガラクトシダーゼ遺伝子又は合成ポリアデニル化部位を含む他の供給源からの様々な pA 配列を、この目的で発現ベクター中で使用できる。
「PEST」は、アミノ酸プロリン(P)、グルタミン酸(E)、セリン(S)、及びスレオニン(T)の豊富なアミノ酸配列を指す。PEST配列を含むタンパク質は半減期が短い。
「プラスミド」とは、環状DNAベクターを意味する。プラスミドは宿主細胞DNA中にプラスミドを組み込むことなしに細菌細胞(しばしば真核細胞)中でプラスミドの多数のコピーを生産させる複製起点を含む。
「ポリヌクレオチド」又は「核酸」とは、DNA又はRNAを含むヌクレオチドの線状配列を意味する。これは二本鎖でも一本鎖でもよい。
【0050】
「ポリペプチド」、「ぺプチド」又は「タンパク質」とは、特定の配列でぺプチド結合により結合されたアミノ酸のポリマーを意味する。
「プロモーター」とは、一般にmRNAをコードする領域の上流(5’)のDNA領域を意味する。これは転写の開始及び転写レベルを制御する。この用語はその範囲内で誘導プロモーター、抑圧プロモーター、及び構成プロモーターをも含む。
「PMA 」とは、ホルボール・ミリストロイックアシッドを意味する。
「ウミシイタケのルシフェラーゼ」とは、シーパンジー(レニッラ・レニホルミス)から誘導され、酸素とコエレンテレートルシフェリン(コエレンテラジン)を利用して光放射を形成する酵素を意味する。
「レポーターベクター」とは、容易に検定できるタンパク質又はポリペプチド(又はmRNA)をコードする「レポーター遺伝子」を含む発現ベクターを意味する。このレポーター遺伝子はその機能又は活性がテストされる調節配列に連結されるのが通常である。
【0051】
「レポーター」とは、レポーター遺伝子によりコードされそしてレポーター検定法で測定される、通常タンパク質又はポリペプチドである分子を意味する。現在のシステムは一般に酵素レポーターを利用しそしてレポーター活性を測定する。
「RNA」とは、リボ核酸を意味する。
「rtTA」とは、TRE に結合しテトラサイクリン又はドキシサイクリンの存在下でのみ転写を活性化する逆 tTA (下記を参照) を意味する。
「SEAP」とは、分泌されるアルカリホスファターゼレポーター遺伝子を意味する。
「SKBR3 」とは、ErbB2 を過剰発現するヒト乳癌細胞系統を意味する。
【0052】
「ストリンジェント条件」とは、高頻度の相補的塩基を有するヌクレオチド配列のみがハイブリッドを形成する温度及びイオン条件を意味する。要求されるストリンジェンシーはヌクレオチド配列依存性でありハイブリダイゼーションの間に存在する様々な成分及びその後の洗浄、及びこれらの処理にかける時間に依存する。一般に、ハイブリダイゼーション速度を最大にするため、熱的融点(Tm)よりも約20〜25℃低い、非ストリンジェント・ハイブリダイゼーション条件が選択さる。このTmは規定されたイオン強度及びpHで溶液中で完全に相補的なプローブに特定の標的配列の50%がハイブリッド形成をする温度である。一般に、ハイブリダイズされる配列の少なくとも約85%のヌクレオチド相補性を要求するためには、Tmよりも約5〜15℃低い高ストリンジェント洗浄条件が選択される。ハイブリダイズされる配列の少なくとも約70%のヌクレオチド相補性を要求するためには、Tmよりも約15〜30℃低い中程度のストリンジェント洗浄条件が選択される。高度に寛容な(低ストリンジェンシー)洗浄条件はTmより50℃も低いものであり、ハイブリダイズされる配列の間の高レベルの不適合を許容する。当業者はハイブリダイゼーション及び洗浄の段階で、標的配列とプローブ配列の間の相同性の特定のレベルからの検出可能なハイブリダイゼーションシグナルの結果に影響を与えるように他の物理的及び化学的パラメータを変更することもできることを認識するはずである。
【0053】
「SV40/CMV/RSV」は、サルウイルス、サイトメガロウイルス及びラウス肉腫ウイルスからそれぞれ誘導されたプロモーター因子を指す。一般に、これらのプロモーターは哺乳動物細胞で構成的に活性であると考えられている。
「TetO」とは、大腸菌のテトラサイクリン耐性オペロンから誘導されたTet オペレーターDNA配列を意味する。
「Tet-Off 細胞系統」とは、テトラサイクリン又はドキシサイクリンが TREプロモーターからの転写を遮断するように tTAを安定に発現する細胞系統を意味する。
「Tet-On細胞系統」とは、テトラサイクリン又はドキシサイクリンが TREプロモーターからの転写を点火するように rtTA を安定に発現する細胞系統を意味する。
「転写」とは、DNA鋳型に相補的なRNA分子を合成するプロセスを意味する。
【0054】
「トランスフェクション」とは、プラスミド又はDNA断片が真核細胞の中に挿入されるプロセスを意味する。通常、2〜50%の細胞がプラスミドを取込み、細胞の染色体にプラスミドDNAを組み込むことなしに(一過性のトランスフェクション)約3日間タンパク質産物を発現する。これらの細胞の小部分はついにはそれらのゲノム中にプラスミドDNAを組み込み、該タンパク質産物を永久に発現する(安定なトランスフェクション)。
「翻訳」とは、mRNA分子がタンパク質合成のための鋳型として使用されるプロセスを意味する。
「TRE 」とは、本明細書では、転写が外因性の因子(例えば、tTA 又は rtTA)の TREへの結合を介してのみ起こるように、一般に最小のプロモーターと結合された任意のテトラサイクリン応答因子(ゴッセンら)を規定する。本発明の好ましい実施態様では、最小CMVプロモーター(mCMV)(クローンテック・ラボラトリーズ社,Palo Alto, CA, USA) に連結した tetO 配列の7個の反復から構成されるTRE を利用する。
「tTA 」とは、テトラサイクリンにより制御されたトランスアクチベーターを意味する。
これは、TRE に結合し、テトラサイクリン又はドキシサイクリンの非存在下でのみ転写を活性化するように、Tet 抑制タンパク質(TetR)及び VP16 活性化ドメインから構成される。
【0055】
「TS」とは、トロンボキサンシンターゼのプロモーターを意味する。
「変異型」とは、参照ポリヌクレオチド又はポリペプチドとそれぞれ実質的な配列同一性を示すポリヌクレオチド又はポリペプチドを意味する。変異型ポリヌクレオチドはストリンジェント条件下で参照配列とハイブリッドを形成するポリヌクレオチドをも含む。これらの用語は参照配列と少なくとも一つのヌクレオチドの付加、欠失又は置換だけ異なるポリヌクレオチドをも包含する。この点で、突然変異を含む特定の改変、付加、欠失及び置換が参照ポリヌクレオチドに成され得ることそしてそれにより改変されたポリヌクレオチドが参照ポリヌクレオチドの生物学的機能又は活性を保持することが当分野でよく知られている。「ポリヌクレオチド変異型」及び「変異型」という用語は天然に生ずる対立遺伝子変異型をも含む。変異型ポリペプチドに関しては、例えば一部のアミノ酸はポリペプチドの活性の本質を変更することなく幅広く類似の性質を持つ他のアミノ酸に変更しうること(同類置換)が当分野で良く理解されている。
【0056】
「ベクター」とは、宿主細胞に外来性DNA配列を挿入し、該ベクターの複製を支える細胞中でこのDNA配列を増幅させるビークルを意味する。最も普通にはプラスミドであるが、ファージミド、バクテリオファージ、アデノウイルス又はレトロウイルスであってもよい。
「vEGFP 」、「EGFP変異型」又は「EGFPの変異型」とは、EGFPの異なる色の変異型及び/又は異なる半減期の変異型を意味する。
「vGFP」とは、GFP の全ての変異型を意味する。これには DsRedなどの相同体や類似体、EGFP変異型又は不安定化された GFP変異型をも含まれる。
【0057】
本発明はとりわけ転写物の安定性、従って該ベクターにより生産されるタンパク質の量、を調節する発現ベクターを提供する。転写物の安定性を増加させる発現ベクターが本発明により包含されることは明らかであるが、特に好ましい実施態様は不安定化されている転写物に的を絞っている。この場合、転写物の安定性は、転写可能なポリヌクレオチドに一つ以上の不安定化因子を添加することにより、又は該ポリヌクレオチドから一つ以上の安定性因子(例えば、ポリA尾)を除去することにより低減させることができる。現存する発現ベクターと比べ、本発明のベクターは、例えば減少したプロモーター活性と対応する発現産物のレベルの減少との間の時間差を減少させることにより、該プロモーター活性に対する改良された時的相関関係を持つタンパク質発現の動力学を提供する。
【0058】
従って、本発明の一つの側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む発現ベクターに向けられる。
【0059】
転写物の安定性の文脈における用語「調節する」とは、転写物の安定性を増加又は減少させることを意味し、調節の最適量は具体的な適用に応じて変化する。本発明を如何なる特定の理論又は作用様式に限定するものではないが、RNA因子が転写物を不安定化するヌクレオチド配列である場合、該因子は転写物を直接的又は間接的に標的化してこれを分解するものと考える。
【0060】
本明細書で用いるとき用語「不安定化因子」とは、細胞内で、タンパク質又は転写物の半減期をそれぞれ低減させるアミノ酸配列又はヌクレオチド配列を意味する。従って、「RNAを不安定化する因子」はRNA転写物の細胞内半減期を低減させるヌクレオチド配列を含み、そして「タンパク質を不安定化する因子」はタンパク質の細胞内半減期を低減させるアミノ酸配列を含む。
【0061】
求められる低減の程度は具体的適用に応じて変化する。好ましい実施態様では、RNA不安定化の程度は、不安定化因子を含まないベクターと比べ、プロモーター活性と発現ベクター中のレポーターのレベル又は活性との間の時的相関関係を有意に改良する。転写物の安定性を増加させることと関連して、安定性の最適レベルはここでも具体的適用に応じて変化する。
【0062】
「RN59定化因子」は細胞内半減期を増加させるヌクレオチド配列である。
【0063】
「RNA」分子には、mRNA、ヘテロ核RNA(hnRNA) 、核内低分子RNA(snRNA) 、核小体RNA(snoRNA)、細胞質低分子RNA(scRNA) 、リボソームRNA(rRNA)、翻訳調節RNA(tcRNA) 、トランスファーRNA(tRNA)、eRNA、メッセンジャーRNA−干渉性相補RNA(micRNA)又は干渉性RNA(iRNA)及びミトコンドリアRNA(mtRNA) などの全てのRNA分子が含まれる。
メッセンジャーRNA(mRNA)はRNAの好ましい形態である。
【0064】
RNA転写物又は目的のコードされたタンパク質から選択された分子の細胞内半減期を低減させることと関連して、(a)一つ以上の不安定化因子及び/又は(b)一つ以上の安定化因子は通常該分子の分解レベルを増加させるように選択され、それにより該分子の細胞内半減期を、約24時間未満が適当であり、約10時間未満がより好ましく、約5時間未満がさらに好ましく、約3時間未満がより一層好ましく、約1時間未満がさらに一層好ましく、約30分未満がより好ましく、約15分未満がより好ましく、約10分未満がより好ましく、約5分未満がさらに好ましく、約3分未満がさらに一層好ましい半減期まで低減させる。RNA転写物又は目的のコードされたタンパク質の半減期はは該転写物又はコードされたタンパク質の最低検出可能レベルの少なくとも10倍の定常状態発現レベルを与える最低半減期に相当することが好ましい。
【0065】
細胞内又は細胞内類似の条件はその細胞型の生理的条件であることが好ましい。細胞内又は細胞内類似の条件の温度はその細胞型にとっての生理的な温度であることが好ましい。哺乳動物細胞の典型的温度は約30℃から約42℃までであることが適当であり、約35℃から約37℃までであることが好ましい。
【0066】
最低でも、RNA転写物又はポリペプチドのリボ核酸分解又はタンパク質分解のそれぞれの強化は、不安定化因子(単数又は複数)の非存在下で又は安定化因子(単数又は複数)の存在下でのRNA転写物又はポリペプチドの分解より、少なくとも約5%、好ましくは少なくとも約10%、より好ましくは少なくとも約20%、さらにより好ましくは少なくとも約40%、さらに一層好ましくは少なくとも約50%、さらに好ましくは少なくとも約60%、さらに好ましくは少なくとも約70%、さらに好ましくは少なくとも約80%、さらに好ましくは少なくとも約90%、さらに好ましくは少なくとも約100%、さらに好ましくは少なくとも約150%、さらに好ましくは少なくとも約200%、さらに好ましくは少なくとも約400%、さらに好ましくは少なくとも約600%、さらに好ましくは少なくとも約1000%、さらに好ましくは少なくとも約2000%、さらに好ましくは少なくとも約4000%、さらに好ましくは少なくとも約6000%、さらに好ましくは少なくとも約8000%、さらに好ましくは少なくとも約10000%、さらに好ましくは少なくとも約12000%大きいリボ核酸分解又はタンパク質分解のレベルを指す。RNA分解を測定する検定法は当業者に知られている。例えば、RNA分解は、例えば転写阻害剤(アクチノマイシンD、DRB、コルディセピン、アルファ−アマニチン)、パルス標識化(放射活性ヌクレオシド)、無細胞減衰法(ポリソーム、細胞質ゾル、又は網状赤血球)、又は短期間プロモーター活性化(fos プロモーター、以下を参照) の使用に基づくロス,ジェイ(1995) 又はリュー,ジェイら(JBC 2000) により開示された一連の検定法を用いて測定できる。タンパク質の分解を測定する検定法も当業者に知られている。例えば、タンパク質分解はバックメアーら(米国特許第5,646,017 号) の網状赤血球溶解液検定法を含む、これに限定されないが、哺乳動物細胞溶解液検定法を用いてイン・ビトロで測定しうる。別法として、タンパク質分解は、例えばヴァズハピリ,アールとズッヒェル,エヌ(2002) 又はサイトウ, ティーら(1998)により開示されたシクロヘキシイミド又はパルス−チェイス法を用いてイン・ビボで測定しうる。
【0067】
このRNA不安定化因子は、しばしば不安定化配列を含む如何なる供給源からも、とりわけ短寿命mRNAの3’UTR又は5’UTR領域から誘導することができる。本明細書で用いるとき、用語「から誘導される」は、特定のもの (integer)又はものの群が特定された種に起源を有するが、必ずしもその特定の供給源から直接得られるとは限らないことを示すものと解すべきである。
【0068】
RNA不安定化配列は、例えば c-fos、c-jun 、c-myc 、GM-CSF、 IL-3 、TNF-アルファ、IL-2、IL-6、IL-8、IL-10 、ウロキナーゼ、bcl-2 、SGLT1 (Na(+)- 共役グルコース・トランスポーター) 、Cox-2 ( シクロオキシゲナーゼ 2) 、IL-8、PAI-2 (2型プラスミノーゲン・アクチベーター・インヒビター) 、アドレナリン性ベータ1受容体、GAP43 (5’UTR及び3’UTR)などの短寿命RNA(単数又は複数)からクローニングしうる。
【0069】
AU豊富因子(AREs) 及び/又はU豊富因子(UREs)は9量体UUAUUUA(U/A)(U/A)(U/AはA又はUのいずれかである)(ラグナドら 1994)及び/又は5量体AUUUA(スーら 1997)及び/又は4量体AUUU(ズビアーガら 1995)の単一コピー、縦列コピー又は複数コピー又は重なり合うコピーを含むが、これらに限定されない。
【0070】
RNA不安定化因子は、例えば、ホスホエノールピルビン酸カルボキシキナーゼmRNA(PEPCK)、ショウジョウバエのビコイド遺伝子、ヒトチオレドキシン遺伝子、熱安定抗原及び大豆10A5遺伝子からも記載されてきた。
【0071】
鉄応答因子及び鉄調節タンパク質結合部位も本ベクター中に有利に組み込まれてRNA安定性及びとりわけ翻訳効率を調節しうる。ヒストンのRNA、とりわけ3’UTRは細胞周期に依存した様式でRNA安定性を調節するのに特に有用である。
【0072】
上に列挙した因子の修飾又は並べ替えも意図される。用語「縦列コピー」は外側のヌクレオチドの一つ以上を二重にする場合及び/又は二重にしない場合の両方を含む。例えば、5量体AUUUAの縦列コピーはAUUUAAUUUAAUUUAだけでなくAUUUAUUUAUUUAなどの配列も含む。
【0073】
RNAを不安定化する因子が同定され、そしてそれの改変がコンピュータ手法及びデータベース分析を用いて(ダンデカール,ティーら)行なわれた。
【0074】
従って、参照を不安定化する因子の生物学的に活性な断片並びにその変異型及び誘導体は本発明に包含される。
【0075】
真核生物の発現ベクターが意図される。
【0076】
関連する実施態様において、本発明は転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写物の安定性を増加させる安定化因子である発現ベクターを意図する。
【0077】
別の関連する実施態様では、本発明は転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写物の安定性を増加させる安定化因子であり、該安定化因子が長寿命mRNAの例であるアルファ2グロビン、アルファ1グロビン、ベータグロビン、又は成長ホルモンであるか又はこれらから誘導されるものである発現ベクターを意図する。本明細書で用いられるとき、遺伝子の名前の下線又はイタリック体は遺伝子を示し、下線又はイタリックにより強調されていない遺伝子の名前で示されるのがそのタンパク質産物であることと対照的である。例えば、「アルファ2グロビン」はアルファ2グロビン遺伝子を意味し、一方、「アルファ2グロビン」は「アルファ2グロビン」遺伝子のタンパク質産物を示す。
【0078】
転写物を不安定化し細胞により生産されるタンパク質の量を低減させる能力は広範囲の適用に対して有用であることが明らかである。
【0079】
本発明の別の一側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写物の安定性を低減させる不安定化因子である発現ベクターを意図する。
【0080】
別の一側面では、本発明は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写物の安定性を低減させる不安定化因子であり、該不安定化因子をコードするヌクレオチド配列が c-fos、c-jun 、c-myc 、GM-CSF、 IL-3 、TNF-アルファ、IL-2、IL-6、IL-8、IL-10 、ウロキナーゼ、bcl-2 、SGLT1 (Na(+)- 共役グルコース・トランスポーター) 、Cox-2 ( シクロオキシゲナーゼ 2) 、IL-8、PAI-2 (2型プラスミノーゲン・アクチベーター・インヒビター) 、アドレナリン性ベータ1受容体又はGAP43 から選択される遺伝子であり、又は選択される遺伝子から誘導されるものである発現ベクターを意図する。
【0081】
一つの特定の実施態様では、RNAを不安定化する因子をコードするポリヌクレオチド配列を、目的のタンパク質をコードする配列に連結し、これを次いで好ましくは調節可能な(即ち、誘導性又は抑圧性の)目的のプロモーターに連結して、発現の調節のオン−オフを行なうようにする。この適用では、RNA不安定化因子は通常機能的なmRNA又はタンパク質の発現の期間を短縮するのに役立つ。これはイン・ビトロ又はイン・ビボで適用しうる。例えば、細胞周期に特異的なプロモーターをRNA不安定化因子と組み合わせて、細胞周期の特定の期でのみ目的のタンパク質を発現させることができよう。目的のタンパク質は機能的なタンパク質であってもよく、又はレポータータンパク質であってもよい。後者の例では、レポーターのレベルは細胞周期の期又は細胞の増殖の指標として使用することができる。
【0082】
本発明のさらに別の一側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写物の安定性を低減させる不安定化因子であり、該不安定化因子をコードするヌクレオチド配列が配列番号:1〜配列番号:23のいずれか一つ、又はそれらの生物学的に活性な断片、又はこれらの変異体若しくは誘導体から選択されるものである、発現ベクターを意図する。
【0083】
本発明のさらに別の一側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写物の安定性を低減させる不安定化因子であり、該不安定化因子をコードするヌクレオチド配列が配列番号:1、配列番号:2又は配列番号:22、又はその生物学的に活性な断片、変異体又は誘導体に記載されているものである、発現ベクターを意図する。
【0084】
本発明の別の一側面はRNA因子をコードするヌクレオチド配列を含む転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写物の安定性を低減させる不安定化因子であり、該転写可能なポリヌクレオチドが目的のポリヌクレオチドを含みそしてポリペプチドをコードするものである発現ベクターを意図する。
【0085】
一つの具体的な適用は遺伝子発現を測定する分野においてである。具体的には、細胞内で生産される転写物の量を低減させることにより、プロモーター活性又はエンハンサー活性をより正確に測定することが可能である。この適用では、レポーター遺伝子を用いてプロモーター活性を、直接的にか、又はその発現がベクター内の調節因子により調節される別のポリペプチドとの融合タンパク質として間接的に測定する。
【0086】
一つの実施態様では、このRNA不安定化配列をレポーターmRNAの3’UTRをコードする領域に組み込む。代わりに又は加えて、不安定化因子を5’UTR及び/又はコードされたタンパク質の選択された活性に好ましくは必要でない又はそれを妨害しないタンパク質コード領域に組み込む。
【0087】
関連する実施態様では、このRNA不安定化配列を用いて、例えばその発現を正確にモニターし又は低減させる必要がある場合、目的の遺伝子を不安定化する。通常、この適用では、RNA不安定化因子はレポータータンパク質不安定化因子と一緒に使用される。
【0088】
本発現ベクターは、mRNA又はタンパク質の発現の期間が短いことが好ましい場合、又はプロモーター活性の変化とそれによって生ずるmRNA/タンパク質のレベルの変化の間の時間差を最小にすることが望ましい場合に、様々な遺伝子発現系において適用がある。
【0089】
従って、本発明のさらに別の一側面は、RNA因子をコードするヌクレオチド配列を含む転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写物の安定性を低減させる不安定化因子であり、該転写可能なポリヌクレオチドが目的のポリヌクレオチドを含み且つレポーターポリペプチドをコードするものである発現ベクターを意図する。
【0090】
本発明のさらに別の一側面はRNA因子をコードするヌクレオチド配列を含む転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写物の安定性を低減させる不安定化因子であり、且つ該転写可能なポリヌクレオチドがレポーターポリペプチドをコードするヌクレオチド配列を含むものである発現ベクターを意図する。
【0091】
この発現ベクターは真核細胞系で使用するように設計される。しかしながら、RNAを不安定化する因子は酵母、昆虫、線虫、魚、鳥又は哺乳動物として定義された細胞、組織又は完全生物を含む広範囲の真核系及び/又は植物系で使用しうることに留意すべきである。植物で使用する場合には、異なるプロモーター及びおそらく異なるレポーター及びRNAを不安定化する因子(例えば、DST 配列) が使用されうる。
【0092】
本発明の発現ベクターは標準的なタンパク質レポーター分子又は不安定化されたレポータータンパク質分子を組み込むことが意図される。標準的レポーター分子は当分野で良く知られている。
【0093】
本発明の別の一側面は、タンパク質を不安定化する因子(例えば、目的のポリペプチドのアミノ末端にある不安定化アミノ酸から選択されうる細胞内タンパク質分解シグナル又はデグロンをコードするDNA/RNA配列、PEST領域又はユビキチン) とmRNAを不安定化する因子(例えば、9量体のUUAUUUAUUの複数のコピー)を、mRNAとタンパク質の両方が不安定化されるように組み合わせることを意図する。例えば、一つのこのような実施態様は、 PEST 配列を翻訳停止コドンの直ぐ上流に、そして4個の9量体を該停止コドンの下流(好ましくは停止コドンから20ヌクレオチド又はそれ以上)に配置して発現ベクターに組み込む。
【0094】
このようにして、レポータータンパク質はタンパク質レベル及びmRNAレベルの両方で不安定化されうる。
【0095】
不安定化されたレポータータンパク質は任意の適切なタンパク質であってよい。例えば、不安定化された GFPタンパク質は、例えば、MODCのd1突然変異体を含むd1EGFP、d1EYFP及び d1ECFP などには適当である。不安定化ルシフェラーゼタンパク質はレクラーク,ジー.らにより記載された。 MODC PEST配列が使用された。d1EGFP由来の MODC も意図される。
【0096】
目的のポリペプチドを不安定化する方法はすべて本発明で意図される。例えば、目的のポリペプチドは、そのように修飾されたタンパク質が、例えばバックメアら(米国特許第 5,093,242号) 及びヴァルシャヴスキーら (米国特許第 5,122,463号) により開示されたようなN末端ルール経路に従うように、不安定化するアミノ酸をそのアミノ末端に含むように修飾することができる。この種の好ましい実施態様では、不安定化するアミノ酸はイソロイシン及びグルタミン酸から、より好ましくはヒスチジン、チロシン及びグルタミンから、さらに一層好ましくはアスパラギン酸、アスパラギン、フェニルアラニン、ロイシン、トリプトファン及びリジンから選択される。とりわけ好ましい実施態様では、不安定化アミノ酸はアルギニンである。一部のタンパク質では、アミノ末端はタンパク質のコンホーメーション(即ち、その三次元又は四次元構造)の結果として曖昧になる。これらの場合には、アミノ末端のより広範な改変がN末端ルール経路に該タンパク質を従わせるためには必要であるかも知れない。例えば、単一のアミノ末端残基の単純な付加又は置換ではアミノ末端への接近不能のために不十分である場合、(ユビキチンの基質タンパク質に結合する部位であるリジンを含め)幾つかのアミノ酸を元のアミノ末端に添加して工学操作されたアミノ末端の接近可能性及び/又はセグメントの移動性を相加させてもよい。
【0097】
タンパク質のアミノ末端の修飾又は設計は遺伝子レベルで行なうこともできる。単離され又は合成された抗原をコードするポリヌクレオチドの5’末端に適当なコドンを付加又は置換するための部位特異的突然変異誘発という通常の技術はコードされたタンパク質の所望のアミノ末端構造を作るために使用できる。例えば、発現されたタンパク質がそのアミノ末端に所望のアミノ酸を有するように、不安定化アミノ酸の適当なコドンをタンパク質コード配列のアミノ末端中に挿入又は構築できる。必要な場合、タンパク質のアミノ末端領域をコードする核酸配列を修飾して、適当な前後関係でリジン残基を導入することができる。これは「汎用不安定化セグメント」をコードするDNA構築物を使用することにより最も便利に達成できる。汎用不安定化セグメントは、好ましくはセグメントとして移動可能な、一つ以上のリジン残基を含むポリペプチド構造をコードする核酸構築物を含み、該構築物が抗原をコードするポリヌクレオチドのコード配列中に挿入される場合、該リジン残基が完全なアミノ末端分解シグナルの第2の決定子として役立つようにコードされたタンパク質のアミノ末端に空間的に十分に近くに存在するように、リジン残基のコドンが該構築物内に配置されている。このような構築物を抗原をコードするポリヌクレオチドの5’部位に挿入すると、不安定化のための適当な前後関係でリジン残基(単数又は複数)を有するコードされたタンパク質がえられるであろう。
【0098】
別の一実施態様では、目的のポリペプチドは PEST 領域を含むように修飾される。この領域はプロリン、グルタミン酸、セリン及びスレオニンから選択されるアミノ酸を豊富に含み、任意選択的に正の電荷を有する側鎖を含むアミノ酸が両側を挟んでいる。この点で、例えば、ロジャーズら(1986, Science234(4774): 364-368)により示されたように、2時間未満の細胞内半減期を持つタンパク質のアミノ酸配列はプロリン(P)、グルタミン酸(E)、セリン(S)、及びスレオニン(T)の豊富な領域を一つ以上含むことが知られている。さらに別の実施態様では、目的のポリペプチドをユビキチン又はその生物学的に活性な断片と結合させて、細胞内タンパク質分解の速度が無修飾のポリペプチドと比べ増加、向上又は他の方法で上昇する修飾されたポリペプチドを作成する。
【0099】
本発明のさらに別の側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写物の安定性を低減させる不安定化因子であり、該レポータータンパク質がルシフェラーゼ、緑色蛍光タンパク質、赤色蛍光タンパク質、SEAP、CAT 、又はそれらの生物学的に活性な断片、又はこれらの変異体若しくは誘導体から選択されるものである発現ベクターを意図する。
【0100】
このようなベクターは該プロモーターの活性を変化させる薬物又は処理をスクリーニングするために使用できる。現存するレポーターベクターと比べ、薬物の作用の近「実時間」測定を行なうことができる。
【0101】
本発明のさらに別の側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む発現ベクターであって、該RNA因子が該転写物の安定性を低減させる不安定化因子であり、
(i) ヌクレオチド配列を導入するための複数クローニング部位であって、その部位が好ましくは酵素により又は他の生化学的方法により切断されて PCR増幅産物をそれに直接クローニングできる線状化されたベクターを生じうる部位(例えば、Ec1HK1部位) 、
(ii) レポーター遺伝子、
(iii) 該転写可能なポリヌクレオチドの発現を調節するためのプロモーター及び/又はエンハンサー、
(iv) ポリアデニル化配列、
(v) 選択可能なマーカー遺伝子、及び
(vi) 複製起点、
から成る群より選択される一つ以上の構成員を任意の順序で含む発現ベクターに向けられる。
【0102】
本発明の別の一側面は、遺伝子発現(例えば、プロモーター活性)の研究又は測定又は監視における適用でのベクター又はベクター群、とりわけプラスミド、しかしこれに限らないが、を意図する。例えば、ウイルス、人工染色体及び他の非プラスミドベクターなどの多くの他のベクターも使用できる。
【0103】
一つの実施態様はプラスミドの対又はセットを含む。そのそれぞれは、例えば d1EGFP 、d1EYFP、d1ECFP又は d1DsRedなどの不安定化されたレポータータンパク質をコードする構築物中に組み込まれた上述のmRNAを不安定化する配列を一つ以上含む。各対又はセットからの一つのプラスミド(対照)はレポーターをコードする領域のプロモーター 5’ を含む。このプロモーターは外因性処理(例えば、mCMVなどの最小プロモーターと結合したTRE、図2cを参照)により調節される(即ち、誘導又は抑圧されうる)因子から構成される。または、TS、SV40、CMV、TK又はRSVなどの構成的に活性なプロモーターが使用される(図2b参照)。植物系では、トップ−テン・プロモーターがTREに取って代わり、カリフラワー・モザイク・ウイルスの35SプロモーターはSV40等に取って代わることができる。アグロバクテリウム・ツメファシエンスは遺伝子移送を容易にするために植物で使用できる。対又はセットでの他のプラスミド(単数又は複数)は、クローニング部位(MCS)がプロモーターに取って代わり、レポーターをコードする領域が対照レポーターに類似するが識別可能なレポーターをコードする(図2a参照)点を除き、対照のプラスミドと同一である。好ましい実施態様では、対照プラスミドはEGFPの不安定化変異体(例えばd1EGFP、d1EYFP又はd1ECFP)をコードし、他のベクター(テストベクター)はそれぞれ同じリスト(同じタンパク質半減期)からの異なる色変異体をコードする。
【0104】
他の実施態様では、対照及びテストレポーターの一つが、例えば双方向性プラスミドなどの1個のベクターの中に組み込まれる(図3参照)。
【0105】
上の実施態様では、対照及びテストプラスミドの両方が不安定化されたmRNAをコードしており、これらが今度は不安定化されたタンパク質をコードする。こうして、低減したプロモーター活性と低減したレポータータンパク質レベルの間の時間差は現存する構築物についての時間差と比べ有意に減少する。同様に、プロモーター活性の増加は先に存在するmRNA及びタンパク質のレベルの減少により、より容易かつ迅速に検出可能である。誤りを招き得る、対照とテスト構築物の間の他の相違は、相互に二、三の小さな突然変異しか異ならない蛍光タンパク質を用いることにより最小にされる。ルシフェラーゼ又は他の酵素に基づく検定法と比べ、ここに記載した蛍光レポーターは、下記の
a)数個の異なるレポーターを同一の細胞/試料の中で測定できる、
b)生きている細胞を測定でき、同一試料の複数の時点又は測定後のさらなる操作、例えば薬物処理の前後の同一細胞の測定を可能とする、
c)巧くトランスフェクトされた細胞は蛍光顕微鏡で可視化できる。従って、不十分なトランスフェクションは顕微鏡下で細胞を見ることにより簡単に確認でき、さらなる資源の投入を必要としない、
d)基質は必要でない、それ故この方法は技術的な要求が少なく、迅速で、安価であり、且つ精度も高い、
e)対照の発現及びテストレポーターの発現の両方をフローサイトメトリーにより同時に測定できる(下記のフローサイトメトリーの利点を参照)、
f)対照プロモーターとしてTREを用いる実施態様はTet−オン又はTet−オフの細胞系統で使用できるだけであるが、他の対照プロモーターと比較すると、テストプロモーターによる又はそれへの妨害を示し難く、そしてテストプロモーターの誘導性を検査するために用いられる種々の刺激により影響され難い。従って、これらにより、トランスフェクション効率及び関連するテストプロモーター活性はより正確に測定される。必要な場合、対照レポーターの発現はスイッチをオン・オフでき、プロモーターのクロストークの欠如を確認するため又は存在するときはそれを補償するために用いることができる、
g)別の実施態様では、上記の対照及びテストレポーターの一つは、両方とも単一のベクター、好ましくは双方向プラスミド、の中に組み込まれる。この二つのプロモーターの間の干渉は、これは先の二重プロモーターベクターの主要な欠点であるが、該対照プロモーター中のTREを用いることにより最小にする、このような単一ベクター系は共トランスフェクション研究の不正確さを防止する、
という幾つかの他の利点を有する。
【0106】
本発明は、情報を与えるプロモーター又はプロモーター断片がレポーターをコードする領域の上流に置かれるベクターをも提供する。本発明は、レポーターとしてルシフェラーゼ又は他の酵素とは全く異なってEGFP変異型を利用する場合、このような検定のためのより単純で、より迅速で、より費用効果的なレポーター系を提供する。さらに、mRNA不安定化因子の組み込みはほぼ実時間分析を可能とする。
【0107】
情報を与えるプロモーターには、細胞周期依存性のプロモーター(例えば、サイクリンA,B,D1、ヒストン又はトポイソメラーゼIのプロモーター類)、アポトーシス(細胞死)経路により活性化されるプロモーター類、及び有糸分裂シグナルに連結したプロモーター/断片類が含まれる(表1)が、これらに限定されない。情報を与えるエンハンサーの使用可能な例にはクローンテックのマーキュリ・パスウエー・プロファイリング・システムズで使用されるもののいずれかが含まれる。クローンテックのマーキュリ・イン・ビボ・キナーゼ・アッセイ・キッツは本発明の使用方法の別の一例を示す。この例では、プロモーター因子は、特定のキナーゼが活性であり融合タンパク質のトランスアクチベータードメインをリン酸化できる場合のみTREからの転写を可能とするキメラのTerR−トランスアクチベータータンパク質と細胞内で結合するTREである。従って、本発明は特定のキナーゼ活性のより実時間に近い測定を行なうために使用することができる。
【0108】
本発明のさらに別の側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む転写可能なポリヌクレオチドを含む本発明のベクターでトランスフェクト又は形質導入された細胞を意図する。
【0109】
一部の適用では、発現ベクター又はレポーター構築物を発現する細胞が生物に挿入され、イン・ビボでレポーター活性を測定することを可能とする。これらの適用の一部では、不安定化されたEGFP変異型ではなく不安定化されたルシフェラーゼが好ましいレポーターでありうる。例えば、情報を与えるプロモーターの制御下で不安定化されたルシフェラーゼを発現するトランスジェニックマウスは、ホトンカメラを用いて(ホトンカメラ分析はコンタグらにより記述されている、1997) 生きているマウスの組織中でプロモーターの活性を測定するために使用できる。mRNA不安定化配列はプロモーター活性とレポーターレベルの間の時的相関関係を改良するのに役立ち、こうして、プロモーター活性のほぼ実時間測定から利益を得る薬物スクリーニングなどの適用に顕著な改良を与える。
【0110】
一部の適用では、レポーター分子及び機能性の遺伝子産物の両方を、イン・ビトロの細胞系システム又はイン・ビボの哺乳動物システムのいずれかで発現させることが望ましい。これは、それぞれがmRNAを不安定化する因子を含む二つの別々のmRNAを含む。または、mRNAを不安定化する因子を、(例えば、内部リポソームエントリ部位,IRESを用いて)二つの別々のタンパク質を生ずる単一の不安定化された転写物又はレポーター及び機能性遺伝子産物から構成される融合タンパク質の中に組み込む。
【0111】
本発明はこれらのベクター(対照と共に又は対照なしで)を安定に発現する細胞系統をも提供する。このような細胞は薬物スクリーニングなどの分野で適用がある。例えば、MAPK−依存性レポーターベクターを含む細胞は、これらの細胞においてMAPK、又はMAPK−依存性転写の上流の任意の経路を阻害するように設計された薬物の効力をテストするための迅速で且つ安価な方法を提供する。SKBR3ヒト乳癌細胞では、例えば、MAPK活性は過剰発現したErbB2タンパク質からのシグナル発信に依存する。従って、ErbB2を阻害する薬物はこのような構築物を含むSKBR3細胞の蛍光の減少を惹起するが、ErbB2を欠く細胞ではそうはならないであろう。または、細胞は、該経路の阻害をモニターするために、異なる経路を経てMAPK活性化をもたらす、薬物の有無及び特異的リガンド若しくは処理の有無をテストできるであろう。細胞周期により調節されるプロモーターに連結したベクターを安定に発現する細胞系統(又は生物)は細胞周期の進行又は細胞の増殖を測定するための極めて迅速な、単純なそして安価な手段として使用できる。このような細胞系統は薬物のスクリーニングにおいて明らかに有用であり、本発明で意図される。細胞周期により調節されるプロモーターの例は容易に入手可能である。例えば(リー,エイチら,1995) 、(シュタイン,ジェイら,1996) 及び(ユエ,エックスら,1996) がある。
【0112】
本発明の別の実施態様には、転写後調節、とりわけmRNAの安定性、の研究のためのベクターが含まれる。このレポーターは、例えば、EGFPの不安定化変異型(例えば、d1EGFP、d1EYFP、d1ECFP)であり、それぞれ別のベクターに異なる色の変異型がある。(mCMVなどの最小プロモーターに連結した)このTREはレポーターコード領域の5’にあり、テトラサイクリン(又はドキシサイクリン)依存性の様式で転写を駆動する。他の誘導性プロモーター系も使用することができる。
【0113】
一つの実施態様では、上述したmRNA不安定化因子は含まれておらず、その代わりにMCSが主として3’UTRに(図4aを参照)しかし5’UTRにも、及び/又は一部の具体的実施態様ではコード領域に配置される。mRNAの安定性に影響を与えると考えられる配列は、それを一つの色の変異型を含むベクターの適当なクローニング部位にクローニングし、テトラサイクリン又はドキシサイクリンで転写を阻止した後レポーターレベルの減少速度を測定する(図7を参照)ことによりテストすることができる。所望ならば、減衰の速度を同一細胞中で「テストベクター」と「対照ベクター」(異なる色のEGFP変異型をコードし且つテストされる配列を含まない)との間で比較することができる。
【0114】
MCSはオーバーハングを有するPCR産物の直接クローニングを可能とする制限エンドヌクレアーゼ部位を含め又は該部位と一緒に作用させると有用である(以下を参照)。
【0115】
本発明の別の関連する実施態様では、mRNAを安定化する因子を特に探索している科学者を援助するため一つ以上のmRNA不安定化因子(単数又は複数)が含められる。同様に、他の実施態様では、mRNAを不安定化する因子を特に探索している科学者を援助するためmRNAを安定化する因子が含められる。
【0116】
他の実施態様では、対照及びテストレポーターの一つが両方とも単一のベクター、好ましくは双方向プラスミド中に組み込まれる(図4bを参照)。
【0117】
安定化因子は、例えば、高レベルのタンパク質が要求され又はプロモーターが弱い場合、タンパク質精製の過程で発現するタンパク質のレベルを増加させるのに有用である。
【0118】
二つのプロモーターの間の干渉、さらに因子又はテストされた種々の刺激の転写効果の間の干渉は、両方のレポーターを駆動するためTRE又は類似の因子を用いることにより、そしてベクターからの転写を阻止するドキシサイクリン(又はテトラサイクリン)を添加した後レポーターレベルを測定することにより回避される。
【0119】
本発明のさらに別の側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む本発明のベクターでトランスフェクトされ又は形質導入された細胞であって、該RNA因子が該転写物の安定性を低減させる不安定化因子である細胞を意図する。
【0120】
本発明の関連する側面は、転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む遺伝的に改変された非ヒト生物であって、該RNA因子が該転写物の安定性を低減させる不安定化因子である生物を意図する。
【0121】
本発明のさらなる実施態様は、目的のポリヌクレオチドの発現を測定する方法であって、RNA及びタンパク質の合成が起こるのに十分な時間及び条件の下でレポーター発現ベクターから目的の該ポリヌクレオチドを発現させる工程であり、該ベクターが転写される因子をコードするヌクレオチド配列及び該目的のポリヌクレオチドを含む転写可能なポリヌクレオチドを含むものであり、該転写される因子が該転写可能なポリヌクレオチドに対応する転写物の安定性を調節するものであり、且つ該発現ベクターが
(i) 目的の該ポリヌクレオチドを導入するための複数クローニング部位、
(ii) レポーター遺伝子、
(iii) 該目的のポリヌクレオチドの発現を調節するためのプロモーター及び/又はレポーター遺伝子であって、該プロモーターが好ましくは調節可能であるもの(例えば,テトラサイクリン応答因子(TRE)を使用)、
(iv) ポリアデニル化配列、
(v) 選択可能マーカー遺伝子、及び
(vi) 複製起点、
から成る群より選択される一つ以上の構成員を任意の順序で含むものである工程、及び対照と比べレポーターのレベル又は活性を経時的に測定する工程であり、該不安定化因子がプロモーター又はエンハンサーの活性とレポーターのレベル若しくは活性の間の時的相関関係を増強するものである工程を含む方法を意図する。
【0122】
組み合わせで作用する、異なるRNA不安定化因子の組み合わせが本明細書で意図される。
【0123】
本発明は非限定的なさらなる実施例によりさらに説明される。
【実施例1】
【0124】
DNA因子のベクターへのクローニング
クローニングは、既存の手法に従い、ベクターがMCS内で「Tオーバーハング」を持つ場合は、MCSの制限酵素部位又はPCR産物の直接連結を用いて実施した。しかしながら転写後レポーターベクターについては、3’−UTR又は他の領域にMCSを含むことは、転写又はその他の研究用に設計されこれらの位置に便利なクローニング部位を含まない現行のベクターからの顕著な改良である。
【実施例2】
【0125】
トランスフェクション
対照ベクターとテストベクターの共トランスフェクションは、一つのベクターのみを必要とし、それ故に共トランスフェクションに伴う不正確さを排除する上述の単一(例えば、双方向性)ベクター系の場合を除き、既存の手法(例えば、Fugene [ベーリンゲル・マンハイム、マンハイム、ドイツ]又は電気穿孔法)により実施した。
【実施例3】
【0126】
レポーター発現の測定
vGFPシステムの直接の利点は、単に組織培養プレート又はフラスコを蛍光顕微鏡下で見るだけで、生きた細胞内でレポーター発現を直接目に見えるようにすることができることである。従って、無駄な時間を費やす前に、劣悪なトランスフェクションを確認して破棄することができる。定量的測定は、蛍光光度計(例えば、96穴プレートフォーマット)を用いて実施することができ、生きた細胞を測定できるため、同一の資料を繰り返し、例えば経時的に、測定することができる。
【0127】
ルシフェラーゼ分析、及び他の酵素に基づく分析と比べてさらなる利点は、レポーターレベルを測定するのにフローサイトメトリーも使用できることである。
【実施例4】
【0128】
レポートレベルの測定にフローサイトメトリーを使用する利点
i. 二つ以上のレポーター(対照及びテスト)並びに追加のパラメータを、全ての細胞において秒速2,000細胞より大の速度で個々に測定することができる。従ってこれを適用すると、既存のルシフェラーゼ検定法の一つのデータポイントに対し、この手法は試料当たり数千から数十万のデータポイントを生ずる。
【0129】
ii.トランスフェクション効率の正確な測定:これはトランスフェクションの手順を最適化するのに役立つ。異なる手法の比較を可能にするのに加え、細胞当たりの発現と細胞発現の比率の双方を測定することもできる。これは研究者が何らかの問題の原因を割り出すのに役立つ。
【0130】
iii.共トランスフェクションのエラーの同定:共トランスフェクション研究は、細胞がそれを取込みそして同じ細胞に取り込まれるテストプラスミドの量に比例するある量の対照レポーターを発現するという前提に基づいている。これは必ずしもそうなるとは限らない。本明細書に記載したフローサイトメトリー法を使用することにより、同一試料中の異なる細胞でのテスト発現レベルと対照発現レベルを相関させることができる。テストレポーターレベルと対照レポーターレベルの間に良好な直線関係が欠如していれば、無効な試料であると同定できる。このようなエラーは現行の手法では見過ごされてしまう。
【0131】
iv.追加パラメータの同時測定:蛍光標識された抗体を用いて細胞ごとの特定のタンパク質を定量することができ、そしてこれをレポーターレベルと相関させてそのタンパク質がレポーター構築物にクローニングされた因子を介して遺伝子発現に影響を与えるかどうかを決定することができる。または、目的のタンパク質を適切な(誘導可能な又は誘導不可能な)発現ベクターのトランスフェクションを介して、vGFP融合タンパク質(GFP変異体と融合した目的タンパク質)として発現させることができる。次いで、特定のタンパク質のレベルを目的の(共トランスフェクトした又は異なる時点でトランスフェクトした)調節因子に連結した異なるGFP変異体の発現と相関させることができる。第三の適用では、vGFPレポーターを細胞周期特異的であると考えられている調節因子(例えば、プロモーター)と連結させる。トランスフェクトした細胞をDNA内容物を測定するためにヨウ化プロピジウムなどの蛍光DNA色素で着色し、次いでそれをレポーター発現と相関させる。原則として、本明細書で記載したDNA構築物の幾つかは、それぞれが異なるvGFPを含んでいるが、共に発現し個々に測定できるであろう。さらに、他の蛍光標識をこれらのベクター(単一または複数)と併せて使用することができるであろう。
【0132】
v. セルソーティング:セルソーターを使用すると、生存能力のある vGFP 発現細胞を非発現細胞から単離することができる。この技術は、検定の開始に先立って、安定して発現する細胞を選別したり、又は発現しない細胞を除去するのに使用できる。同様に、極めて低レベル及び/又は極めて高レベルの vGFP を発現する細胞を除去することができる。この技術は、もっと均質な集団を形成するために使用することができ、及び/又は生理学的に適切でないであろう、又は通常の細胞機能を乱すであろう、及び/又は本明細書に記載したDNAベクターから得たデータにその他の方法で悪影響を及ぼすであろう程に高いレベルで発現する細胞を除去するために使用することができる。
【0133】
発現ベクターの一過性のトランスフェクション及び安定なトランスフェクションは極めて不均質な発現レベルを有する細胞集団を結果として生ずることに注目することが大切である。一般的に、最高の発現細胞と最低の発現細胞の間で千倍の差があるというのは珍しいことではない。本発明は必要なときに均質な集団を選択する方法を提供する(上述のvを参照)だけでなく、科学者の利益のために不均質性を利用することもできる。例えば、共トランスフェクションのエラーの同定である。これの別の例は上述の(iv)に関する。タンパク質XがプロモーターYからの転写に影響を与えるかどうかを決定するために、細胞を、プロモーターYの制御の下、d1EGFPを発現するレポーター構築物でトランスフェクトする。必要に応じて、セルソーティングを用いて適正レベルの d1EGFPを一時的に又は安定して発現する細胞を単離することができる。これらの細胞を今度はタンパク質X-EYFP融合タンパク質を発現するベクターで一時的にトランスフェクトする。フローサイトメトリーの間、EGFPを一つの軸上にプロットし、EYFPを他方の軸上にプロットする。正相関はタンパク質XがプロモーターYからの転写を増加させることを示し、負の相関はタンパク質XがプロモーターYからの転写を阻害することを示すであろう。
【0134】
現在では、このような相関関係を確立しようとしている科学者らは、タンパク質Xを高度に発現するもの対低度に発現するものの幾つかの異なるクローンを選択するであろう。それぞれのクローンを、次いでプロモーターY−ルシフェラーゼ構築物で別々にトランスフェクトし、ルシフェラーゼ活性を測定するであろう。細胞クローンの使用には、調製に何ヶ月も必要であり、宿主細胞とベクター組み込みの種々の部位の間に事前に存在する不均質性を含め様々な変数を導入する(ベクターDNAは組み込み部位の特定の遺伝子を妨害し、この部位は全てのクローンで異なる)。さらに、このような手法ではそれぞれが異なるクローンの異なるトランスフェクションから得られる極めてわずかなデータポイントしか得られない。従って、この新しいシステムは既存の手法よりも用途が広いだけでなく、より迅速で且つより正確である。
【実施例5】
【0135】
レーザー走査サイトメトリー(LSC)
フローサイトメトリーとは異なり、LSCはスライド上で細胞の複数色の蛍光と光散乱を測定し、分析した各細胞についてその計測位置と時刻を記録する。この技法はフローサイトメトリーと同等のデータを提供するが、顕微鏡用スライドに基づいているという利点がある(ダルジンキーヴィッツら,1999、カメンツキーら,1997)。GFPとその変異体の蛍光のため、フローサイトメトリー用に記述された技術もLSCに適用できる。
【実施例6】
【0136】
転写後分析のための特異的方法
転写後分析のための特異的方法は、特定の3’−UTR断片がmRNAの安定性に影響を与えるかどうかの測定を目標とした研究例
(i) 3’−UTR断片をテストベクターの3’−UTRクローニング部位中に連結し、対照ベクターと共にTet−オフの細胞系統に共トランスフェクトされる。双方向性ベクターの場合は、対照ベクターは必要でない。実際、典型的な適用では、同一トランスフェクションの範囲内からの試料で減衰速度を測定できるため、対照レポーターまたはベクターを必要としない。5’−UTR断片は5’−UTRクローニング部位を持つベクター中に挿入することによりテストできる、
(ii) 細胞をドキシサイクリン(又はテトラサイクリン)の非存在下で6〜48時間増殖させ、両方のベクターを発現させる。または、細胞を低用量のドキシサイクリン(又はテトラサイクリン)と共に6〜48時間増殖させて転写を阻止し、次いでドキシサイクリン(又はテトラサイクリン)なしの培地に切替えて2〜12時間培養し、短時間で転写を一気に爆発させる、
(iii)次いで、両方のベクターからの転写を遮断するため、高用量のドキシサイクリン(又はテトラサイクリン)を適用する、
(iv) 両レポーターの蛍光を経時的に測定し(フローサイトメトリー、蛍光高度法、又はLSCにより)、続いてドキシサイクリン(又はテトラサイクリン)を添加する、
という研究例を用いると最適にまとめられる。この例は一過性の発現の一つであるが、安定なトランスフェクションにも使用できるであろう。
【0137】
クローニングされた因子が mRNA を不安定にするのであれば、同一細胞又は同一試料の「対照」蛍光と比べて「テスト」蛍光でより急激な減少が見られるはずである。同様の研究が、異なる細胞内若しくはRNA結合タンパク質のような特定のタンパク質を異なる量で発現する細胞内での特定の刺激又はその効果に対する mRNA 因子の応答をテストするのに使用することができる。ドキシサイクリンの後に該刺激を適用すると、事前に存在した転写物が刺激により影響されたかどうかが決定される。該因子を異なる位置(例えば、5’−UTR、3’−UTR)に挿入すると、その機能が位置に依存するものかどうかが決定される。タンパク質/ポリペプチドのコード配列を(フレームを揃えて)該ベクターのレポーターコード領域内に挿入すると、mRNAに対するその配列の効果とタンパク質の安定性を測定するために使用することができる。
【0138】
RNAはトランスフェクトされた細胞から抽出することができ、レポーターmRNAを直接測定するために使用することができる。
【実施例7】
【0139】
転写レポーターベクター
該ベクターは大腸菌内での増幅、及び真核細胞における蛍光レポーターの発現に適したプラスミドである。該プラスミドはセットで使用しうる。それぞれのセットは一つ以上の「対照」ベクターと一つ以上の「テスト」ベクターから成る。セット内のどのベクターも同様に不安定化された mRNA 及び同様に不安定化された蛍光レポータータンパク質を発現する。このようなプラスミドの標準特性(アンピシリン耐性、複製起点など)に加えて、各プラスミドは次の構築物を含む(図2及び図3をも参照)。
5’---MCS/プロモーター---転写開始部位---5’UTR---ATG---vEGFPコード領域--停止コドン---mRNA不安定化因子を伴う3’UTR---ポリアデニル化シグナル
【0140】
上式中、
MCS/プロモーターは、複数クローニング部位(テストベクター、図2a参照)又はSV40などの構成的に活性なプロモーター(対照ベクター、図2b参照)、又は TRE-mCMV のような誘導可能なプロモーター(対照ベクター、図2c参照)のいずれかを意味する。
ATGは、翻訳開始コドンを意味する。
停止コドンは、翻訳停止コドンを意味する。
5’UTRは、5’非翻訳領域を意味する。
mRNA不安定化因子を伴う3’UTRは、概説したmRNA不安定化因子を一つ以上含む3’非翻訳領域を意味する。
vEGFPは、EGFPの不安定化変異体を意味する。各種の不安定化修飾(例えば、1時間半減期、2時間半減期)に対してプラスミドの一つのセットが提供される。プラスミドの各セット内で、それぞれ色の異なる変異体に対して一つのベクターが提供される。例えば、一つのセットはd1EGFP、d1EYFP、d1ECFPを発現するベクターを含み、もう一つのセットは d2 変異体を発現する。
【0141】
他の実施例では、上述の対照とテストレポーターの一つは共に単一ベクター、好ましくは双方向性プラスミドに組み込まれる(図3参照)。
【実施例8】
【0142】
転写後レポーターベクター
3’−UTR内の mRNA 不安定化因子がMCSに置き換わっている(図4a参照)点を除けば、TRE-mCMVプロモーターを含む転写レポーター「対照」ベクターと類似する。幾つかの実施態様では、MCSは5’UTR及び/又はコドン領域にも配置される。
【0143】
このような構築物は、本明細書で概説した転写後検定のための「テスト」ベクター又は「対照」ベクターとして使用することができる。
【0144】
他の実施例では、上述の対照レポーターとテストレポーターの一つは共に単一ベクター、好ましくは双方向性プラスミドに組み込まれる(図4b参照)。
【実施例9】
【0145】
特定の経路を検定するレポーターベクター
調節因子を研究又は測定する目的で、該調節因子が MCS 中に挿入された本明細書記載のものに類似するベクター。例えば、MCS内に表1で言及するものなどの経路に敏感なプロモーター又はプロモーター因子(単数または複数)又はエンハンサー、及び/又はクローンテックズ・マーキュリー・パスウェイ・プロファイリング・システムに記載されたようなシスで作用するエンハンサー(AP1、CRE、E2F、GRE、HSE、ISRE、Myc、NFAT、NFkB、p53、Rb、SRE)のいずれかを含むエンハンサー(単数または複数)を含むという点を除けば、本明細書で概説した転写レポータープラスミドに類似するプラスミド。該レポーターは、GFP、ルシフェラーゼ又は SEAP の不安定化変異体であることが好ましい。
【0146】
特定の経路を検定する細胞系統及びマウス
本明細書記載のベクターの一つ以上を安定して発現する細胞系統又は遺伝的に改変されたマウス。
【実施例10】
【0147】
使用方法
本発明で記載するベクターは、本明細書で記載する新しい方法以外は、本発明で記載するベクターに取って代わられる既存のベクターと本質的には同じ方法で実験に使用する。
【0148】
構築方法
本明細書で概説するベクター及び DNA構築物は標準的なクローニング技術を用いて組み立てる。本明細書記載の SV40 及び TRE-mCMV プロモーター並びにプラスミドベクターのより標準的な構成要素(例えば、複製起点、抗生物質耐性又はその他の選択用遺伝子)は、様々な普通のベクターですぐに入手可能である。EGFPの不安定化変異型(例えば、d1EGFP、d1EYFP、d1ECFP及びd2EGFP、d2EYFP、d2ECFP)をコードするDNA配列がクローンテック(クローンテック・ラボラトリーズ・インク、パロアルト、カリフォルニア、米国)から入手可能である。不安定化 DsRed 変異型をコードするDNA配列は、DsRedコード領域の3’末端に、短寿命のタンパク質由来の分解ドメイン(又はその突然変異体)をコードする配列を融合することにより構築される。例えば、PEST配列を含むマウス・オルニチンデカルボキシラーゼ由来のアミノ酸422-461である。このような配列はおそらく既存のdEGFP変異体から誘導できるであろう。
【実施例11】
【0149】
概要
概要では本発明のベクターと方法が入手可能である。
【0150】
1)比較的短寿命な mRNA を供するために一つ以上の mRNA 不安定化因子を組み込む発現ベクター又はその部分。既存の発現ベクターと比べて、本発明で請求されたベクターはプロモーター活性とより密接に相関するタンパク質発現の動力学を供する。例えば、減少したプロモーター活性、及び減少したmRNAとタンパク質レベルの間の時間差は、実質的に減少した。
【0151】
2)不安定化mRNAをコードする発現ベクター又はその部分。このmRNAは今度は不安定化したタンパク質をコードする。既存のベクターと比べて、本発明で請求されたベクターはプロモーター活性とより密接に相関するタンパク質発現の動力学を提供する。
【0152】
3)その中のmRNA不安定化因子がc-fosのような短寿命mRNAからクローニングされた配列を含むものである発現ベクター又はその部分。該短寿命mRNAの例には、c-fos、c-myc、GM-CSF、IL-3、TNF-alpha、IL-2、IL-6、IL-8、ウロキナーゼ、bcl-2、SGLT1 (Na(+)結合ブドウ糖輸送担体)、Cox-2(シクロオキシゲナーゼ2)、IL8、PAI-2(2型プラスミノーゲン・アクチベータ・インヒビター)、ベータ1−アドレナリン受容体、GAP43(5’UTR及び3’UTR)、AU豊富因子(AREs)及び/又はU豊富因子が含まれ、9量体UUAUUUA(U/A)(U/A)(U/AはAまたはUのいずれか)(ラグナードら 1994)及び/又は5量体AUUUA(スーら 997)及び/又は4量体AUUU(ズビアーガら 1995)の単一コピー、縦列コピー又は複数コピー又は重なりコピーが含まれるがこれらに限定されない。上述のリストの因子の小さな改変又は並べ替えも含まれる。「縦列コピー」という用語は、外側のヌクレオチド(複数)の一つ以上の重複及び/又は非重複の両方を許す。例えば、5量体AUUAの縦列コピーは、AUUUAAUUUAAUUUAだけでなく、AUUUAUUUAUUUAのような配列を含む。短寿命 mRNA の3’UTR又は5’UTR領域は、しばしば不安定化配列を含む。
【0153】
4)その中のmRNA不安定化因子が本明細書記載のベクターを用いて同定又は検証された発現ベクター又はその部分。これらはこのような因子を同定するための実質的に改良された手法を提供する。
【0154】
5)その中の不安定化mRNAがEGFP又はルシフェラーゼの不安定化変異体などの短寿命レポータータンパク質をコードするものである、発現ベクター又はその部分。既存のレポーターベクターと比べて、本発明で請求されるベクターは、プロモーター活性とより密接に相関するレポーター発現の動力学を提供する。例えば、減少したプロモーター活性、及び減少したmRNAとタンパク質レベルの間の時間差は、実質的に減少する。
【0155】
6)同様に不安定化されたmRNA(同じ集合の他のベクターと同様)をコードするレポーターベクター又はその部分の集合。これらのmRNAは、今度は同様に(同じ集合の他のベクターと同様)不安定化されたEGFP若しくはDsRedの変異体、又は他の蛍光標識をコードする。各セット内の一つ以上のベクター(対照ベクター)は一つの構成的プロモーター(例えば、SV40、CMV、RSV、TK、TS。図2b参照)又は一つの誘導可能なプロモーター(例えば、TRE-mCMV。図2c参照)を含み、一方各セット内の他のベクター(テストベクター)はプロモーターの代わりにクローニング部位(例えば、MCS)を含む(例えば、図2a参照)。これらのベクターの適用は、プロモーター活性の研究又は測定を含むが、これらに限定されない。例えば、目的のプロモーター因子は d1EGFP をコードするテストベクターの MCS中にクローングでき、そしてレポーターの発現は d1EYFP を発現する対照ベクターのレポーター発現と比較して測定できる。同一のベクター内に一つのテストレポーター構築物及び一つの対照レポーター構築物を組み込む双方向性ベクター又は他の単一ベクター系(例えば、図3a及び3b)として詳しく記載した個々のベクターのそれぞれも請求される。既存のレポーターベクターの集合と比べ、本発明で請求されるベクターは
a) プロモーター活性の実時間により近い測定、
b) 対照構築物とテスト構築物の類似性が高くなったことによるエラーの減少、
c) テストプロモーターと対照プロモーターの間のクロストークに起因するエラーの減少。対照ベクターの誘導可能プロモーターを使用することにより、このようなクロストークは誘導あり及び誘導なしの測定により最小限に抑えられ、及び/又は同定され、そして修正される、
d) 記載されたフローサイトメトリー/LSC法と併せて使用できる、
という利点を有する。
【0156】
7)誘導可能なプロモーター、テトラサイクリン応答因子(TRE)が好ましいがこれに限定しない、を利用して、不安定化した蛍光性レポータータンパク質(不安定化EGFP変異体が好ましいがこれに限定しない)の発現を駆動するレポーターベクター又はレポーターベクターの集合、又はその部分。このようなベクターは、調節因子又は調節因子であると推定されるものが1色の蛍光レポーターを発現するベクター中にクローニングできるように、そして必要であれば、異なる色のレポーターを発現し且つ目的の因子を含まない対照ベクターと比較できるように、3’−UTR(例えば、図4a)及び/又は5’−UTR及び/又はレポーターコード領域にクローニング部位を含む。このようなベクターは、転写が誘導可能なプロモーターにより要望どおりに転写を阻止できるため、転写後調節の研究又は測定に適用がある。これらのベクターがもたらす利点はb〜dにリストした利点、即ち、転写後効率を転写効率と切り離すという利点、並びに
a)他のベクターには存在しない便利なクローニング部位の組み込み、及び
b)この技術は、既存の方法よりも速い、
という利点をも含む。
【0157】
8)本質的に一つのテスト構築物と対照構築物を連結する、記載された単一ベクター系(例えば、図4b)。テストレポーターと対照レポーターは共に誘導可能なプロモーターで駆動され、クローニング部位は調節因子をテスト構築物にだけ連結させる。概説したベクターの利点に加え、単一ベクター系は別々のテストベクターと対照ベクターの共トランスフェクションに伴う諸問題と不正確さを解決する。
【0158】
9)概説したベクターを介して発現される2以上の蛍光レポーターのレベルを測定するためのフローサイトメトリーまたはLSCの使用。この適用においては、既存の酵素に基づく検定では1サンプルにつき1データポイントであるのに対し、該手法では1サンプルにつき数千から数十万のデータポイントが得られる。二つ以上のレポーター(対照及びテスト)だけでなく、さらなるパラメータ(例えば、DNA含量、他のタンパク質のレベル)も、全てのセルで個々に測定できる。同一試料内の複数の細胞の2以上のレベルを相関させるフローサイトメトリーの使用、及びトランスフェクション手順を最適化するため及び/または共トランスフェクションに伴う諸問題を特定するためにこのようなデータを利用することをも包含される。例えば、無効なサンプルはテストレポーターレベルと対照レポーターレベルの間に良好な線状関係が欠けていることで特定できる。このような誤差は、現行手法では認識されないままになってしまう。
【0159】
10)請求される転写後レポーターベクターを利用する方法。これらの方法は特定の3’−UTR断片がmRNAの安定性に影響を与えるかどうかの測定を目的とした研究例を用いて最も良く要約できる。この例、即ち
(i) 3’−UTR断片を、テストベクターの3’−UTRクローニング部位に連結し、対照ベクターと共にTet−オフ細胞系統中に共トランスフェクトする。単一ベクター系の場合、対照ベクターは必要ない。5’−UTR断片は5’−UTRクローニング部位を持つベクター中にそれを挿入することによりテストできる、
(ii) 細胞はドキシサイクリン(又はテトラサイクリン)の非存在下で6〜48時間増殖させて両方のベクターを発現させる。または、細胞を低用量のドキシサイクリン(又はテトラサイクリン)と共に6〜48時間増殖させて転写を阻止し、次にドキシサイクリン(又はテトラサイクリン)を含まない培地に切替えて2〜12時間培養し、転写を短時間で爆発させる、
(iii)次に高用量のドキシサイクリン(又はテトラサイクリン)を適用して両方のベクターからの転写を遮断する、
(iv) ドキシサイクリン(又はテトラサイクリン)の添加に続き、両方のレポーターの蛍光を(フローサイトメトリー、フルオロメトリー、又はLSCにより)経時的に測定する、
は一過性発現の例であるが、安定なトランスフェクションにも使用できるであろう。
クローニングされた因子がmRNAを不安定化すれば、同一の細胞又は試料の「対照」の蛍光と比べ、「テスト」蛍光ではより急速な減少が見られる。同様の研究は、異なる細胞又はRNA-結合タンパク質などの特定のタンパク質を異なる量で発現する細胞における特定の刺激又はその効果に対するmRNA因子の反応をテストするために使用することができる。ドキシサイクリンの後に該刺激を適用することで事前に存在した転写がその刺激の影響を受けたかどうかを測定することができる。該因子を別の位置(例えば、5’−UTR、3’−UTR)に挿入すると、その機能が位置に依存するものかどうかを決定できる。タンパク質/ポリペプチドコード配列を(フレームを揃えて)該ベクターのレポータータンパク質コード領域内に挿入する手法は、その配列がmRNA及びタンパク質の安定性に及ぼす効果を決定するために使用することができる。
RNAはトランスフェクトした細胞から抽出でき、レポーターmRNAを直接測定するのに使用できる。
【0160】
11)請求される、発現構築物又はその部分の一つ以上を一時的に又は安定して発現する細胞系統。
【0161】
12)請求される、発現構築物又はその部分の一つ以上を一時的に又は安定して発現する細胞系統であって、該発現構築物がヒトの病気及び/又は薬物治療に対する反応に関わるシグナル変換経路の活性化のマーカーとしての機能を果たす調節因子を含むものである細胞系統。このような経路は表1のリストと本明細書のどこかに記載されているもの(例えば、CRE、SRE、AP1、サイクリンA、B、及びD1のプロモーター)を含むが、それらに限定されない。
【0162】
13)請求される発現構築物又はその部分の一つ以上を発現するトランスジェニックマウス、遺伝子削除マウス、又は他の遺伝的に改変されたマウス。
【0163】
14)請求される、発現構築物又はその部分の一つ以上を発現する、トランスジェニックマウス、遺伝子導入(knock-in) マウス、又は他の遺伝子的に改変されたマウスであって、該発現構築物がヒトの病気及び/又は薬物治療に対する反応に関わるシグナル変換経路の活性化のマーカーとして機能する調節因子を含むものであるマウス。このような経路は、表1のリストを含むが、これらに限定されない。
【0164】
15)DsRedの不安定化変異体又はDsRed1-E5の突然変異体。これらはDsRedのC-末端に、様々な不安定なタンパク質からの分解ドメイン(又はその突然変異体)を融合して構築することができる。例えば、PEST配列を含むマウス・オルニチンデカルボキシラーゼのアミノ酸422〜461である(リーら 1998)。さらなる分解因子もまた追加できる。DsRedの不安定化変異体をコードするDNA構築物も意図される。
【0165】
16)概説したmRNA不安定化因子をも含むようなベクターを含む、概説したDsRedの不安定化変異体をコードするベクター。
【0166】
17)Tet−オフ又はTet−オン細胞系統を創出する以下の方法。
tTA又はrtTA発現ベクター、好ましくはレトロウイルス、アデノウイルス、又はプラスミドは、標準技術を使用すると目的の細胞系統で安定して発現し、発現する細胞は薬物耐性マーカーを介して単離される。次に、これらの細胞をTRE-vGFP構築物で一時的にトランスフェクトし、フローサイトメトリーによる何回りかのセルソーティング(細胞分取)にかける。例えば、良好なTet−オフ細胞はドキシサイクリンの存在下で蛍光を発しないであろうし、そのように分取される。ドキシサイクリンなしでさらに5〜48時間経過した後、緑色細胞が分取される。最終的に、該細胞をドキシサイクリンなしで1週間以上増殖させ、最後に分取して、安定にトランスフェクトした(緑色)細胞を除去する。
【実施例12】
【0167】
mRNA及びタンパク質の不安定化因子を組み込んだベクター
目的のコーディング領域(例えば、EGFP又はルシフェラーゼのようなレポーター)はタンパク質不安定化因子(例えば、MODCのd1突然変異体(クローンテック)、但し他のPEST配列又はユビキチン化部位などの他のタンパク質不安定化因子をも含む)とmRNA不安定化因子(例えば、AU豊富因子)の結合した配列を含むことができるであろう。
【0168】
例えば、ルシフェラーゼ及びDsRedの停止コドンはHind3部位(AAGCTT)で置換され、次の配列、
AAGCTTAGCCATGGCTTCCCGCCGGCGGTGGCGGCGCAGGATGATGGCACGCTGCCCATG
TCTTGTGCCCAGGAGAGCGGGATGGACCGTCACCCTGCAGCCTGTGCTTCTGCTAGGATC
AATGTGTAG
の付加を可能にする。
これは、EGFPに1時間の半減期を与えるクローンテックのMODCのd1突然変異体である。これにはリンカー(3’UTRの一部となる)が後続する。次に、4個の9量体を含み、親ベクター(pGL3、プロメガ)のXba1部位に連結する
UUAUUUAUU GGCGG UUAUUUAUU CGGCG UUAUUUAUU GCGCG UUAUUUAUU ACTAG
も3’UTR中に、ただしさらに下流において後続する。
【実施例13】
【0169】
PCR産物の直接連結
二つに分かれているが近くの RE 認識部位をベクターのMCSに包含させること。このRE(単数又は複数)で切断すると、残りのベクターの両端に単一のTヌクレオチドの3’オーバーハングが残る。例えば、Ec1HK1の認識配列はGACNNN、NNGTC(3番目のNと4番目のNの間を5’から切り離して両端に単一のNという3’オーバーハングを残す)である。これらの部位のうち二つは、その二つの間の短い領域がEclHK1での消化により放出されて、両端に単一のNの3’オーバーハングを持つ線状ベクターが残るようにMCS中に組み込まれる。この例では、上流認識配列は 5’GACNNTNNGTC3’で、下流認識配列は5’GACNNANNGTC3’でなければならない。Ec1HK1で切断した後、大きなベクター断片は両端に単一の3’Tオーバーハングを含むことになる(プロメガのpGEM-Tイージーベクターと同様)。これは5’Aオーバーハングを生ずるTaqのようなポリメラーゼで製造されるPCR産物の直接連結を容易にする。これは、RE部位のPCRプライマーへの包含とその後のPCR産物の消化なしでは PCR産物の直接連結をサポートしない標準的なMCSに対する顕著な改良点となる。これはまた、増幅(線状として供給される)できず、サブクローニングにしか役立たない(即ち、PCR産物は通常は pGEM-T イージーに連結され、増幅され、次いでRE消化によって切り出され、続いて目的の発現ベクターにクローニングされる)pGEM-イージーベクターに対する顕著な改良でもある。従って、この MCS は、PCR産物をRE(しばしば問題となる)で消化したり、又は中間ベクターにそれらをサブクローニングする必要なく、PCR産物の直接連結を可能にする。
【実施例14】
【0170】
不安定化レポーターモデルは改良された実時間分析を示す
プラスミドレポーターベクターは、標準的なクローニング技術を使用してpGL3−ベーシック(プロメガ)バックボーン(アンピシリン耐性遺伝子など)に組み込んだ。クローンテックの pTRE-d2EGFPベクターから誘導したテトラサイクリン応答因子を MCSに挿入した。一部の構築物では、クローンテックが定めたようにルシフェラーゼ−コード領域を d1EGFP −コード配列又は d2EGFP −コード配列(コザック配列を含む)で置き換えた。この工程は便利な5’隣接 RE 部位を持つ適切なプライマーを用いたPCRにより実行した。一部の構築物では、mRNA不安定化因子の特定の例が3’UTR−コード領域にクローニングされた。通常、これらの配列はセンス配列及びアンチセンス配列を合成し、ハイブリッド形成させることにより調製した。両側を挟む配列は、3’UTR−コード領域が特定の制限酵素で切断される場合に生成するもの適合する「付着末端」のオーバーハングを提供した。これらの酵素による該ベクターの消化とその後の精製の後、ハイブリッド形成するオリゴマーを標準的な技術を用いて該ベクターに連結した。適切な供給源から得たゲノムDNA又はcDNAのPCRをc-myc-AREのようなより大きい不安定化因子を得るための代替手法として使用した。極めて小さな因子(例えば、1又は2個の9量体)を、一つの5’隣接RE部位とベクター鋳型中の既存の3’UTRに相補的な一つの3’隣接領域を含む逆向きPCRプライマーに組み込んだ。適切な前向きプライマー(タンパク質コード領域に相補的であり且つ内因性RE部位と重なる)を用いる PCRの後、該PCR産物を適切なRE部位で消化し、元のベクターに連結した。
【0171】
用語:
B=プロメガのpGL3- ベイシックから誘導されたベクターバックボーン。
T=テトラサイクリン応答因子(TRE)。クローンテックの pTRE-d2EGFPベクターから誘導され、レポーターの転写を駆動するプロモーターとして用いられる。
G1=レポーターとして用いられる1時間の半減期を持つGFP、即ち、クローンテックにより定義されたような d1EGFP タンパク質をコードする配列。
G2=レポーターとして用いられる2時間の半減期を持つGFP、即ち、クローンテックにより定義されたような d2EGFP タンパク質をコードする配列。
L=レポーターとして用いられるルシフェラーゼ、即ち、pGL3-ベイシック(プロメガ)由来の蛍ルシフェラーゼをコードする配列。
R=レポーターとして用いられる DsRed2 。
R1= d1EGFP に存在するのと同じMODC突然変異体にカルボキシ末端で融合した DsRed 。
N6=3’UTR−コード領域に挿入された6個の9量体TTATTTATT(配列番号:1)。
N4=3’UTR−コード領域に挿入された4個の9量体TTATTTATT。
N2=3’UTR−コード領域に挿入された2個の9量体TTATTTATT。
N1=3’UTR−コード領域に挿入された1個の9量体TTATTTATT。
fos=シューら(1989)により定義され、3’UTR−コード領域に挿入された c-fos ARE。即ち、
5’AAAACGTTTTATTGTGTTTTTAATTTATTTATTAAGATGG
ATTCTCAGATATTTATATTTTTATTTTATTTTTTT3’ (配列番号:2)
myc=次のように定義される myc ARE である、
5’ATGCATGATCAAATGCAACCTCACAACCTTGGCTGAGTCTTGAGACTGAA
AGATTTAGCCATAATGTAAACTGCCTCAAATTGGACTTTGGGCATAAAAG
AACTTTTTTATGCTTACCATCTTTTTTTTTTCTTTAACAGATTTGTATTT
AAGAATTGTTTTTAAAAAATTTTAAGATTTACACAATGTTTCTCTGTAAA
TATTGCCATTAAATGTAAATAACTTT3’ (配列番号:21)。
【0172】
方法:
5マイクログラムのマキシプレップ品質のDNAを、フジーン(Fugene) 試薬(ロシュ)を用いて10cmフラスコの約50%コンフルーエントのヒーラのTet−オフ細胞(クローンテック)中にトランスフェクトした。約10時間後に、該フラスコの細胞をそれぞれ約12の小皿(6cm)に分け、ついで一晩(約12〜14時間)インキュベートした。この時点(通常タイムゼロ又は T0と呼ぶ)で、殆どのプレートに1マイクログラム/mlの最終濃度でドキシサイクリンを添加した。細胞をトリプシン処理し、この時点及び後続の時点で回収した。構築物を発現するGFPのために、これらの試料を標準的なFITCフィルターを用いたフローサイトメトリーで分析した。(背景蛍光のみの)非トランスフェクト細胞を遮断し、次いで細胞当たりの平均蛍光に陽性細胞数を掛けることにより、GFPの総蛍光を測定した。RFPの蛍光(DsRed)を同様に適切なフィルターを用いて測定した。ルシフェラーゼ−をコードするベクターでトランスフェクトされた細胞を溶解し、プロメガの二重ルシフェラーゼ検定法及び試薬を用いた照度計中で測定した。
【0173】
データは通常、タイムゼロと比較した場合の、残ったレポーター(蛍光又は発光)のパーセンテージとして表される。
【0174】
タイムゼロで添加されたドキシサイクリンは該レポーターの転写を阻害する原因となるため、レポーターレベルにおける減少速度は改変された転写レベルと改変されたレポーター/タンパク質レベルの間の時間差を示す。本発明の主な目的はこの時間差を減少させることであり、図7、8、9、及び11〜14はその目的が達成されたことを示す。
【0175】
本発明の使用例として、製薬会社は疾病に関与する遺伝子の転写を減少させる薬物をスクリーニングしたいと思うかもしれない。TREプロモーター由来のテトラサイクリン/ドキシサイクリンが誘導する転写の阻害はこのようなシステムのモデルである。図7と図8は、標準的なルシフェラーゼレポーターベクターでは、(ドキシサイクリンによる)転写の完全な阻害さえも、10時間以内のルシフェラーゼ活性の減少として検出されることはないということを示している。不安定化EGFP突然変異体は、転写の完全阻害が11時間以内(d2EGFP; BTG2)又は7時間以内(d1EGFP; BTG1)にEGFP蛍光の50%減少として検出できるという点で改良を表す。しかしながら、後者のレポーターが4個の9量体UUAUUUAUU(BTG1N4)のようなmRNA不安定化因子と結合した場合、レポーターレベルの50%の減少が3時間以内に検出できる。このことから、転写の増加も不安定化因子を含む構築物を用いてより早く検出されうるであろうと結論される(ロス、1995)。
【0176】
もちろん、ドキシサイクリンの作用はすぐに起こるわけではなく、それ故に時間差の一部はこの薬物が100%の転写阻害を誘導するのに必要な時間に起因する。従って、「減衰の有効速度」はドキシサイクリンを添加した4時間後の時点に続くデータポイントをその時点と比較してプロットすることにより測定した(図9)。減衰の有効速度は、従って薬物作用の遅れを排除し、且つタンパク質の半減期とmRNAの半減期の総合的な効果である。図9は、1個、2個、又は4個の9量体を含む構築物による減衰の有効速度を示す。これらのデータから、4個の9量体が2個の9量体よりも効果的であり、2個の9量体は1個の9量体よりも効果的であることが分かる。さらに、これらのデータは1時間の半減期のタンパク質(d1EGFP)を4個の9量体と結合させることにより、およそ1時間20分という減衰の有効速度が達成できることを示す。これは該タンパク質の1時間半減期に極めて近く、mRNAの半減期が極めて短いことを例証する。二つ以上の異なるmRNA不安定化因子を結合することで、更なる減衰を達成することができるであろう(図13)。しかしながら、これは殆どの適用で必要とされないと思われる。より適度な不安定化因子を必要とする適用では4個ではなく、1個又は2個の9量体を利用できるであろう。
【0177】
標準的なルシフェラーゼレポーターにより、発光はドキシサイクリンの添加後に実際に増加した。これはデータを均等目盛り上に表した場合に最も明らかであり(図8)、部分的にはドキシサイクリンの作用の遅れによって説明できる。しかしながら、4時間先ですら減衰が見られず、時間を掛けて転写の変化を測定するにはこのレポーターは不適当であることを示している。このベクターのさらなる問題を図10に示す。これらのデータは、何らの処置又は薬物も加えない状態での、長時間(トランスフェクション後24〜34時間)にわたるレポーターレベルの変化に関する。レポーターレベルは一般的には該プラスミドが細胞に入り発現を始める、トランスフェクションの24時間後に増加する。減少は一般的には該プラスミドが細胞から排出される約48時間から見られる。従って、測定は通常は24時間と48時間の間に行われる。薬物又は処置の非存在下で、不安定化因子を含んでいる新しいベクター(BTG1N4)はレポーターレベルの優れた安定性を示す。対照的に、ルシフェラーゼベクターは、明らかに発現レベルを依然として増加させている。緩やかな安定性を持つ構築物(例えば、BTG1)は中間的な結果を示した。mRNAとタンパク質のより長い半減期をもつレポーターは、図10に示すようにより長い増加相を示すことが明らかである。24〜34時間の重要な期間に新しい構築物で見られたより安定な発現レベルは正確な測定を容易にし、本発明の別の利点を表す。
【0178】
レポーターレベルのこの減少速度は、それらのレポーターmRNAの配列(例えば、3’UTRの配列)では異なるが、同一のタンパク質又は同じ半減期を有する異なるタンパク質(例えば、d2EGFP、d2EYFP)をコードする二つ以上の構築物の間で比較できる。この状況では、減衰の速度の差異は、mRNA安定性に対する改変されたmRNA配列の効果を示す。例えば、3’UTR内における DNA TTATTTATT (配列番号:1)(図7〜9)又は c-fos ARE(図11)(配列番号:2)としての4個の UUAUUUAUU9量体の存在は、mRNA減衰の速度を有意に増大させた。これらの因子の有効性の実証に加えて、使用した手法とベクターは、他のシスに作用する mRNA の安定化/不安定化因子を検出するための実質的に改良されたシステムをも表し、このプロセスも本発明に包含される。
【0179】
図12から14に示すように、mRNA不安定化因子はルシフェラーゼ、GFP及びDsRedと共に働く。これらのレポーター間の相同性のレベルが低いにもかかわらずである。DsREdは EGFP と23%の相同性しかない。図14に示すように、myc ARE(配列番号:21)は効果的であり、異なる不安定化因子との組み合わせでも効果的である。
【実施例15】
【0180】
mRNA不安定化因子
本発明の RNA不安定化因子は、とりわけ次の遺伝子の3’UTRから誘導できる。殆どの場合、全長の3’UTRが用いられる。しかしながら、U豊富因子及び/又はAU豊富因子はしばしば単独で用いられる。
【0181】
a)ラテルツァ, オウエフらにより記載されたホスホエノールピルビン酸カルボキシキナーゼ(PEPCK)mRNA不安定化因子。JW6及びJW7と称される3’UTRの3’側の半分以内の領域、即ち、
GTATGTTTAAATTATTTTTATACACTGCCCTTTCTTACCT
TTCTTTACATAATTGAAATAGGTATCCTGACCA (配列番号:4)。
【0182】
b)ショウジョウバエメラノガスター(melanogaster)由来のビコイド遺伝子は、3’UTRの最初の43ヌクレオチド内にmRNA不安定化因子(サーデイ, ピーら)を含む。このような因子は特に昆虫細胞のmRNAを不安定化するために使用できる。
【0183】
c)ヒト・チオレドキシン還元酵素遺伝子(ガスダスカ、ジェイアールら)。3’UTR全体。ヌクレオチド1933〜3690(6個のAU豊富因子を含む)。3個の上流AU反復(ヌクレオチド1975〜3360)を含むセグメント。ヌクレオチド 1933〜2014には非AU豊富不安定化因子もある。
【0184】
d)ジョウ, キューらにより記載された熱安定性抗原(HSA)遺伝子。例えば、3’UTRにあるヌクレオチド1465〜1625。
【0185】
e)チー−イイング,エイらにより記載された顆粒球マクロファージコロニー刺激因子(GM-CSF)ARE。AGUAAUAUUUAUAUAUUUAUAUUUUUAA AAUAUUUAUUUAUUUAUUUAUUUAA 即ち、
DNAとして:AGTAAUATTTATATATTTATATTTTTAAAATATTTATTTATTTATTTATTTAA(配列番号:5)。
【0186】
f)c-fosの3’UTR全長又はその部分、又はシュウらにより記載されたARE、
5’AAAACGTTTTATTGTGTTTTTAATTTATTTATTAAGATGG
ATTCTCAGATATTTATATTTTTATTTTATTTTTTT3’ (配列番号:2)。
又は、ペング, エスらによる
5’TTTTATTGTGTTTTTAATTTATTTATTAAGATGGATTCTC
AGATATTTATATTTTTATTTTATTTTTTTT3’(配列番号:6)。
【0187】
g)ペング, エスらにより記載されたようなc-jun ARE。
5’UUUCGUUAACUGUGUAUGUACAUAUAUAUAUUUUUUAAUUUGAUUAAAGC
UGAUUACUGUGAAUAAACAGCUUCAUGCCUUUGUAAGUU3’ 。
DNAとしての配列:
5’TTTCGTTAACTGTGTATGTACATATATATATTTTTTAATTTGATTAAAGC
TGATTACTGTGAATAAACAGCTTCATGCCTTTGTAAGTT3’(配列番号:7)。
又は、ポリアデニル化(AAUAAA)シグナルを含まないその突然変異型、即ち、
5’UUUCGUUAACUGUGUAUGUACAUAUAUAUAUUUUUUAAUUUGAUUAAAGC
UGAUUACUGUGgAUccACAGCUUCAUGCCUUUGUAAGUU3’ 又は、DNAとして
5’TTTCGTTAACTGTGTATGTACATATATATATTTTTTAATTTGATTAAAGC
TGATTACTGTGgATccACAGCTTCATGCCTTTGTAAGTT3’ (配列番号:8)。
【0188】
h)次の遺伝子由来の配列。これらはへニックス, ティーらにより記載されたように、それぞれの ARE構成成分を含む。
IFN-γ ARE
5’UCUAUUUAUUAAUAUUUAACAUUAUUUAUAUAUGGG3’又は、DNAとして
5’TCTATTTATTAATATTTAACATTATTTATATATGGG3’(配列番号:9)。
【0189】
i)IL-2 ARE
5’CUCUAUUUAUUUAAAUAUUUAACUUUAAUUUAUUUUUGGAUGUAUUGU
UUACUAACUUUUAGUGCUUCCCACUUAAAACAUAUCAGGCUUCUAUUU
AUUUAAAUAUUUAAAUUUUAUAUUUAUU3’又は、DNAとして
5’CTCTATTTATTTAAATATTTAACTTTAATTTATTTTTGGATGTATTGT
TTACTAACTTTTAGTGCTTCCCACTTAAAACATATCAGGCTTCTATTT
ATTTAAATATTTAAATTTTATATTTATT3’(配列番号:10)。
【0190】
j)c-myc ARE(配列番号:21も参照)
5’AUAAACCCUAAUUUUUUUUAUUUAAGUACAUUUUGCUUUUAAAGUU3’又は、DNAとして
5’ATAAACCCTAATTTTTTTTATTTAAGTACATTTTGCTTTTAAAGTT3’(配列番号:11)。
【0191】
k)IL-10
5’UAGAAUAUUUAUUACCUCUGAUACCUCAACCCCCAUUUCUAUUUAUUUAC
UGAGCUUCUCUGUGAACGAUUUAGAAAGAAGCCCAAUAUUAUAAUUUUUU
UCAAUAUUUAUUAUUUUCA3’ 又は、DNAとして
5’TAGAATATTTATTACCTCTGATACCTCAACCCCCATTTCTATTTATTTAC
TGAGCTTCTCTGTGAACGATTTAGAAAGAAGCCCAATATTATAATTTTTT
TCAATATTTATTATTTTCA3’(配列番号:12)。
【0192】
l)bcl-2
スキアヴォーン, エヌらにより記載されたようなbcl-2 AREの全部又は一部を含むbcl-2 の3’UTR由来の配列。
5’UCAGCUAUUUACUGCCAAAGGGAAAUAUCAUUUAUUUUUUACAUUAUUAAGAAAAAAGAU
UUAUUUAUUUAAGACAGUCCCAUCAAAACUCCGUCUUUGGAAAUC3’
(M13994のヌクレオチド 2371〜2475)又は、DNAとして
5’TCAGCTATTTACTGCCAAAGGGAAATATCATTTATTTTTTACATTATTAAGAAAAAAGAT
TTATTTATTTAAGACAGTCCCATCAAAACTCCGTCTTTGGAAATC3’
(配列番号:13)。
【0193】
m)スー、エヌらにより記載されたTNF ARE。
5’AUUAUUUAUUAUUUAUUUAUUAUUUAUUUAUUUA3’又は、DNAとして
5’ATTATTTATTATTTATTTATTATTTATTTATTTA3’(配列番号:14)。
【0194】
n)スー、エヌらにより記載されたIL3ARE。
5’UAUUUUAUUCCAUUAAGGCUAUUUAUUUAUGUAUUUAUGUAUUUAUUUAUUUAUU3’ 又は、
DNAとして
5’TATTTT ATTCCATTAAGGCTATTTATTTATGTATTTATGTATTTATTTATTTATT3’
(配列番号:15)
【0195】
o)ズビアーガ, エイらにより記載されたような、9量体 UUAUUUAUU。DNAとして
TTATTTATT(配列番号:1)。
【0196】
p)ラグナード、シーらにより記載されたような9量体UUAUUUAUU(U/A)(U/A) 。DNAとして TTATTTA(T/A)(T/A)( 配列番号:3)。
【0197】
q)スー、エヌらにより記載されたような5量体 AUUUA。又はDNAとして
ATTTA(配列番号:16)。
【0198】
r)4量体 AUUU 、又はDNAとして ATTT (配列番号:17)。
AU豊富因子(ARE)は、チェン、シー及びシュー、エイにより記載されたような、一般的にはクラスIとクラスIIの両方のものである。
植物は、不安定化因子として作用するDST(下流配列)を有する。DST配列は、ニューマン、ティーらに規定されている。提案されたコンセンサスDST配列は
GGAgN2-9cATAGATTaN3-8(A/C)(T/A)(A/T)TttGTA(T/C)
である。
【0199】
s)上記のDST配列は9つの異なったDST配列の比較に基づくものである。
下線部=9/9の遺伝子で保存されている。
大文字=少なくとも7/9の遺伝子で保存されている。
N2-9=2〜9個のヌクレオチドの範囲で長さが可変の領域。平均=5。
N3-8=3〜8個のヌクレオチドの範囲で長さが可変の領域。平均=6。
停止コドンからの距離=19〜83ヌクレオチド。
DST配列の更なる例は、次のものを含む。
大豆の10A5遺伝子:
5’GGAGN5CATAGATTAN8AAATTTGTAC3’(配列番号:18)。
アラビドプシスSURAC1遺伝子:
5’GGAAN9CATAGATCGN8CAATGCGTAT3’(配列番号:19)。
DST配列は植物に使用するためのAU豊富因子の代替物である。AU豊富因子とDST配列は共に植物における転写を不安定化する。
【0200】
t)鉄応答因子(IRE)
トムソン、エイら、1999。
IREはヘアピン・ループ内にコンセンサス CAGUGを含む。
例:
フェリチンIRE:
GUUCUUGCUUCAACAGUGUUUGAACGGAAC 又は、
DNAとして、GTTCTTGCTTCAACAGTGTTTGAACGGAAC(配列番号:20)。
トランスフェリン受容体のIRE:
GAUUAUCGGGAGCAGUGUCUUCCAUAAUC 又は、
DNAとして、GATTATCGGGAGCAGTGTCTTCCATAATC(配列番号:21)。
【0201】
鉄調節タンパク質(IRP、例えば、IRP1及びIRP2)は鉄に依存した形でIREと結合する。結合は又、様々な他の刺激及び処置(例えば、酸化的ストレス、一酸化窒素、エリスロポエチン、甲状腺ホルモン、及びPKCによるリン酸化反応)によって調節される。
【0202】
IREは、翻訳効率及びmRNA安定性の双方を調節することができる。例えば、フェリチンmRNA内の5’UTR IREは、IRPと結合した場合のみ翻訳を阻止する。トランスフェリン受容体mRNAの3’UTR内のIREは、IRPと結合した場合にmRNAの減衰を阻害する。従って、IREを発現ベクターの5’UTR又は3’UTRに挿入して、鉄レベルまたは他の刺激によって制御されうる発現を提供することができる。
【0203】
不安定化因子はクローンテックのマーキュリー経路プロファイリングベクター及びインビボ・キナーゼ検定キットと共に使用できる。クローンテックは3つの異なるタンパク質不安定化因子を製造しており、全てがPEST配列を含み、全てがMODC遺伝子から誘導される。EGFPのカルボキシ末端に配置された異なる変異体MODCは、1時間、2時間、及び4時間のタンパク質半減期を提供する。本発明のmRNA不安定化因子はこれらの不安定化因子と他の如何なるタンパク質不安定化因子(例えば、ユビキチン・シグナル)とも一緒に使用できる。
【0204】
u)c-myc AREは、次のように定義されうる:
5’ATGCATGATCAAATGCAACCTCACAACCTTGGCTGAGTCTTGAGACTGAA
AGATTTAGCCATAATGTAAACTGCCTCAAATTGGACTTTGGGCATAAAAG
AACTTTTTTATGCTTACCATCTTTTTTTTTTCTTTAACAGATTTGTATTT
AAGAATTGTTTTTAAAAAATTTTAAGATTTACACAATGTTTCTCTGTAAA
TATTGCCATTAAATGTAAATAACTTT3’ (配列番号:22)。
【0205】
別の役に立つmRNA因子は、ヒストンmRNAから得ることができる。とりわけ、コンセンサス・ステム・ループ(consensus stem loop)構造を含む3’UTR配列が、ガリー、ディーらに記載されている:
TGA-N20-40-CCAAAGGYYYUUYUNARRRCCACCCA、この式で、Y=ピリミジン、R=プリン、N=何らかのヌクレオチドである。
又は、DNAとして、TGA-N20-40-CCAAAGGYYYTTYTNARRRCCACCCA(配列番号:23)。
【0206】
このような配列は、翻訳効率を増大できる。さらに、このような配列は、特にS期(DNA合成期)の外側でmRNAの減衰を指示することができる。細胞周期特異的プロモーターを含むレポーター構築物は、mRNA不安定化因子と共に、細胞周期特異的発現(例えば、レポーターの発現)を指示するための道具として本発明で意図される。ヒストンの3’UTR因子は、S期と比べてG2期でmRNAの減衰の増大を指示し、それ故にタンパク質発現をS期にさらに制限するので、S期又は後期G1特異的プロモーターと共に使用するための代替物を提供する。
【0207】
発現ベクターにおける3’UTR因子の更に別の用途は、キメラmRNAを特異的に局在化するためのものである。例えば、ユトロフィン(utrophin)の3’UTRはレポーターmRNAを細胞骨格結合ポリソームに導くことができる。mRNA不安定化因子も、この3’UTRに含まれる(グラモリーニ、エイら)。
【実施例16】
【0208】
mRNA安定化因子と安定化されたmRNAをコードする発現ベクター
安定化配列はCT豊富因子及び/又は長寿命mRNA(特に3’UTR領域)から誘導された配列を含む。
【0209】
CT豊富因子は、ホルシックとリーブハーバー、1997により記載されたような
(C/U)CCANxCCC(U/A)PyxUC(C/U)CCを含む。
【0210】
CT豊富因子は次の因子を含む。
CCTCCTGCCCGCTGGGCCTCCCAACGGGCCCTCCTCCCCTCCTTGC
又は、CCTCC、CCTCCTGCC、又はCCCTCCTCCCCTGGを含むその部分。
【0211】
ユーとラッセルにより記載されたヒトのベータ−グロビンの3’UTRに由来する14ヌクレオチドのピリミジン豊富領域も安定化因子としての使用が意図される。
【0212】
そこから安定化因子が誘導されるであろう長寿命mRNAの例には、ヒト、マウス、ウサギ、又は他の種からのアルファ2グロビン、アルファ1グロビン、ベータグロビン、牛成長ホルモン3’UTRが含まれる。
【0213】
本明細書記載のmRNA不安定化因子は、一般的にはキメラ遺伝子を不安定化するために優性な形で作用する。従って、mRNA安定化因子はしばしば劣性に作用しているということになる。例えば、c-fos AREをウサギのベータグロビン遺伝子に挿入すると、mRNA安定性因子が持続的に存在するにも関わらず不安定化された転写という結果になる(シュー、エイら、1989)。アルファ−及びベータ−グロビンmRNAは両方ともそれぞれの3’UTRに配置された安定化因子を含み、一方ゼータグロビンmRNAはこれらの因子を欠いており、安定性が低い。ゼータグロビン3’UTRをアルファグロビンmRNAの3’UTRに置き換えるとmRNA安定性が殆ど2倍になる(ラッセル、ジェイら、1998)。しかしながら、このような因子は全ての転写を安定化するわけではない。従って、安定性mRNAを発現する発現ベクターを生成するための必要条件は、安定化の対象となる元のmRNAによって異なる。このようなベクターを創り出すためには、一般的にはアルファ−又はベータ−グロビンなどの安定な遺伝子に由来する大きなセグメントを含めることが好ましい。これらの例では、このようなセグメントは、好ましくは内因性3’UTRに替えてグロビン3’UTR全体を含むべきである。ゼータ−グロビンで例証したように、これで十分な場合も多い。しかしながら、タンパク質コード配列及び/又は5’UTR配列を更に組み込むことがしばしば必要となる。一般的に、優性の不安定化因子(これらは本明細書記載の技術を用いて同定できる)として作用するであろう内因性のAU又はU豊富領域を置き換えることが好ましい。5’UTR又は3’UTR内のこのような領域は、同じ関連位置からのアルファ−又はベータ−グロビン配列で単に置き換えられる。コード領域からの不安定化因子は同義のコドンへ突然変異させることにより非機能的とすることができる。グロビンタンパク質をコードする領域を目的の遺伝子のコード領域に組み込んで、N末端又はC末端融合タンパク質を創り出すことができる。しかしながら、これは望ましくないことが多く、一般的にはグロビンタンパク質をコードする領域(及び3’UTR)をキメラ遺伝子の3’UTRに配置すれば十分である。これはより安定な転写物から所望のタンパク質を発現させることを可能にし、従って該タンパク質のレベルは顕著に増加する。所望のタンパク質がレポーターであり、又はレポーターに融合しており、又は内因性タンパク質から容易に識別できる場合、本明細書記載(図7参照)のTREベクター系は、キメラ構築物のmRNA安定性のテストを大幅に容易にする。
【0214】
当業者は本明細書に記載した発明が特に記載されたもの以外の変化及び修飾を受けることを承知している。本明細書記載の発明は、そのような変化及び修飾の全てを含むものであると理解されるべきである。本発明は又、この明細書で言及又は示唆したそのような工程、特性、組成物及び化合物の全てを個々に又は集合的に含み、該工程又は特性の二つ以上のありとあらゆる組み合わせを含む。
【0215】
【表1】
【0216】
引用文献
チェン,シー.とシュー,エイ.、Trends Biochem. Sci. vol 20: 465-470, 1995。
チー−ユィング,エイ.ら、MCB vol 15(10): 5777-5788, 1995。
コンタグら、哺乳類における光放射性コンジュゲートの非侵襲性局在化,米国特許第 5,650,135号(1997年 7月22日) 。
コーマックら、緑色蛍光タンパク質 (GFP)の FACS 最適化変異型、米国特許第5,804,387 号( 1998年 9月 8日) 。
ダンデカー、ティー.ら、Bioinformation14(3): 271-278, 1998。
【0217】
ダーツィンキーウイック、レーザー走査サイトメトリー:様々な適用を持つ新たな装置,Exp Cell Res 249: 1-12,1999。
ガリー,ディー.ら、Nucleic Acids Res. vol 24(10): 1954-1962,1996。
ガスダスカ,ジェイアール.ら、JBC vol 274(36): 25379-25385,1999。
ゴッセンら、テトラサイクリン応答プロモーターによる真核細胞中での遺伝子発現の厳格な制御,米国特許第 5,464,758号(1995年11月 7日)。
グラモリーニ,エイ.ら、JBC vol 154(6): 1173-1183, 2001。
【0218】
へニックス,ティー.ら、JBC vol 274(24): 17318-17324,1999。
ホルシックとリブハーバー、PNAS vol 94: 2410-2414, 1997。
ユエ,エックス.ら、Mol Cell Biol. vol 16(7): 3789-3798, 1996。
イブラヒムら、内部制御プラスミドの落とし穴:ウミシイタケルシフェラーゼ(pRL-TK)プラスミドのジヒドロテストステロンとデキサメタゾンに対する反応,Biotechniques 29: 682-684, 2000。
カメンツキーら、スライドベースのレーザー走査サイトメトリー,Acta Cytol 41: 123-43,1997。
【0219】
リーら、転写レポーターとしての不安定化緑色蛍光タンパク質の形成,J Biol Chem 273: 34970-34975,1998。
ラグナード,シー.ら、MCB14: 7984-7995, 1994。
ラテルツァ,オーエフ.ら、Am J Physiol Renal Physiol. vol 279(5): F866-F873, 2000。
ルクレール,ジー.ら、Biotechniques29(3): 590-595, 2000。
リー,エイチ.ら、Gene Expr vol 4(3): 95-109, 1995。
【0220】
リーら、GFP−融合タンパク質の急速分解,米国特許第 6,130,313号(2000年10月10日)。
リュー,ジェイ.ら、JBC vol 275(16): 11846-11851, 2000。
ニューナン,ティー.ら、The Plant Cell vol 5: 701-714, 1993。
ペング,エス.ら、MCB vol 6(4): 1490-1499, 1996。
ロス,ジェイ.、Microbiological Reviews. vol 59(3): 423-450, 1995。
【0221】
サイトウ,ティー.ら、Biochem Biophys Res Commun. vol 252(3): 775-778, 1998。
スキアヴォーン,エヌ.ら、FASEB J. vol 14: 174-184, 2000。
シューら、Genes & Dev. vol 3: 60-72, 1989。
シュタイン,ジェイ.ら、Int J Obes Relat Metab Disord. Suupl 3:S84-90, (1996年3月20日)。
サーデュ,ピー.とジェーコブス−ロレーナ、M. MCB vol 18(5): 2892-2900, 1998。
【0222】
トムソン,エイ.ら、Int. J Biochem Cell Biol. vol 31: 1139-1152, 1999。
ツィエンら、改変された緑色蛍光タンパク質,米国特許第 5,625,048号(1997年 4月29日)。
ツィエンら、改変された緑色蛍光タンパク質,米国特許第 5,777,079号(1998年 6月 7日)。
ヴァズハピリー,アール.とサッチャー,エヌ.、Neurosci Lett. vol 318(3): 153-157, 2002。
スー,ダブリュー.ら、MCB17(8): 4611-4621, 1997。
ユーとラッセル、MCB : 5879-88, Sept 2001 。
ジョウ,キュー.ら、MCB vol 18(2): 815-826, 1998。
ツビアガら、MCB 15: 2219-30, 1995。
【特許請求の範囲】
【請求項1】
転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む発現ベクター。
【請求項2】
該RNA因子が該転写物の安定性を低下させる不安定化因子である、請求項1記載の発現ベクター。
【請求項3】
該RNA因子が該転写物の安定性を増加させる安定化因子である、請求項1記載の発現ベクター。
【請求項4】
該転写可能なポリヌクレオチドが目的のポリヌクレオチドを含み且つあるポリペプチドをコードするものである、請求項2記載の発現ベクター。
【請求項5】
該ポリペプチドがレポータータンパク質を含むものである、請求項4記載の発現ベクター。
【請求項6】
該転写可能なポリヌクレオチドがレポータータンパク質をコードするヌクレオチド配列を含むものである、請求項2記載の発現ベクター。
【請求項7】
(i) ヌクレオチド配列を導入するための複数クローニング部位、
(ii) レポーター遺伝子、
(iii) 該転写可能なポリヌクレオチドの発現を調節するめたのプロモーター及び/又はエンハンサー、
(iv) ポリアデニル化配列、
(v) 選択可能マーカー遺伝子、及び
(iv) 複製起点、
からなる群より選択される一つ以上の構成員を含む、請求項1又は請求項2記載のベクター。
【請求項8】
酵素により又はそれ以外の生化学的方法により切断してその中にPCR増幅産物を直接クローニングできる線状化したベクターを生じうる少なくとも一つの部位をさらに含む、請求項7記載のベクター。
【請求項9】
該不安定化因子をコードするヌクレオチド配列が、c-fos 、c-jun 、c-myc 、GM-CSF、IL-3、TNF-アルファ、IL-2、IL-6、IL-8、IL-10 、ウロキナーゼ、bcl-2 、SGLT1 (Na(+)- 共役グルコーストランスポーター) 、Cox-2 ( シクロオキシゲナーゼ2)、IL-8、PAI-2 ( プラズミノーゲン・アクチベーター・インヒビター・タイプ2)、アドレナリンベータ1受容体又は GAP43から選択される遺伝子であり、又は該遺伝子から誘導されるものである、請求項2記載のベクター。
【請求項10】
該安定化因子をコードするヌクレオチド配列が、アルファ2グロビン、アルファ1グロビン、ベータグロビン、又は成長ホルモン、エリスロポエチン、リボヌクレオチドリダクターゼR1又は m1 ムスカリン性アセチルコリンから選択される遺伝子であり、又は該遺伝子から誘導されるものである、請求項3記載のベクター。
【請求項11】
該不安定化因子をコードするヌクレオチド配列が、配列番号:1〜配列番号:23のいずれか一つ、又はそれらの生物学的に活性な断片、又はこれらの変異体若しくは誘導体から選択されるものである、請求項5記載のベクター。
【請求項12】
該不安定化因子をコードするヌクレオチド配列が、配列番号:1、配列番号:2又は配列番号:22、又はそれらの生物学的に活性な断片、又はこれらの変異体若しくは誘導体から選択されるものである、請求項11記載のベクター。
【請求項13】
該レポータータンパク質がルシフェラーゼ、緑色蛍光タンパク質、赤色蛍光タンパク質、SEAP、CAT 、又はそれらの生物学的に活性な断片、又はこれらの変異体若しくは誘導体から選択されるものである、請求項5又は請求項6記載のベクター。
【請求項14】
該ポリペプチドがタンパク質を不安定化する因子を含むものである、請求項4記載のベクター。
【請求項15】
該レポータータンパク質がタンパク質を不安定化する因子を含むものである、請求項13記載のベクター。
【請求項16】
該タンパク質を不安定化する因子が PEST 配列又はユビキチン又はこれらの生物学的に活性な断片又はこれらの変異体若しくは誘導体をコードするヌクレオチド配列によりコードされるものである、請求項14又は請求項15記載のベクター。
【請求項17】
先行する請求項のいずれか一つに記載の一つ以上のベクターを含む細胞。
【請求項18】
該細胞が真核細胞である、請求項17記載の細胞。
【請求項19】
該細胞が哺乳動物の細胞である、請求項18記載の細胞。
【請求項20】
該細胞がヒトの細胞である、請求項19記載の細胞。
【請求項21】
請求項1〜請求項16いずれか1項に記載のベクターの一つ以上を含む遺伝子的に改変された非ヒト生物。
【請求項22】
目的のポリヌクレオチドの発現を測定する方法であって、RNA合成及びタンパク質合成が起こるのに十分な時間及び条件の下で発現ベクターから目的の該ポリヌクレオチドを発現させる工程であり、該ベクターが目的の該ポリヌクレオチド及び転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含むものであり、該発現ベクターが、
(i) ヌクレオチド配列を導入するための複数クローニング部位、
(ii) レポーター遺伝子、
(iii) 該転写可能なポリヌクレオチドの発現を調節するためのプロモーター及び/又はエンハンサー、
(iv) ポリアデニル化配列、
(v) 選択可能マーカー遺伝子、及び
(vi) 複製起点、
から成る群より選択される一つ以上の構成員を含むものである工程、
及び、経時的に転写可能なポリヌクレオチドの発現産物のレベル及び/又は機能的活性を対照と比べて測定する工程であり、該因子が該転写可能なポリヌクレオチドに機能しうるように連結されたプロモーター及び/又はエンハンサーの活性と該発現産物のレベル及び/又は機能的活性の間の時的相関関係を高めるものである工程、を含む方法。
【請求項23】
該RNAを不安定化する因子をコードするヌクレオチド配列を含む転写可能なポリヌクレオチドが、c-fos 、c-jun 、c-myc 、GM-CSF、IL-3、TNF-アルファ、IL-2、IL-6、IL-8、IL-10 、ウロキナーゼ、bcl-2 、SGLT1 (Na(+)- 共役グルコーストランスポーター) 、Cox-2 ( シクロオキシゲナーゼ2)、IL-8、PAI-2 ( プラズミノーゲン・アクチベーター・インヒビター・タイプ2)、アドレナリンベータ1受容体又は GAP43から選択される遺伝子であり、又は該遺伝子から誘導されるものである、請求項22記載の方法。
【請求項24】
該RNA不安定化因子をコードする転写可能なポリヌクレオチドが配列番号:1〜配列番号:23のいずれか一つに記載されたようなヌクレオチド配列、又はそれらの生物学的に活性な断片、又はこれらの変異体若しくは誘導体を含むものである、請求項22記載の方法。
【請求項25】
該RNAを不安定化する因子をコードする転写可能なポリヌクレオチドが配列番号:1、配列番号:2又は配列番号:22に記載された配列又はそれらの生物学的に活性な断片又はこれらの変異体若しくは誘導体を含むものである、請求項22記載の方法。
【請求項26】
RNA転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を同定する方法であって、テストヌクレオチド配列を発現ベクター中に導入することにより該ヌクレオチド配列をレポータータンパク質をコードするポリヌクレオチドに連結させてプロモーター及び/又はエンハンサーに機能しうるように連結されている転写可能なポリヌクレオチドを形成させる工程、RNA合成及びタンパク質合成が起こるのに十分な時間及び条件の下で該転写可能なポリヌクレオチドを発現させる工程であり、該発現ベクターが
(i) ヌクレオチド配列を導入するための複数クローニング部位、
(ii) ポリアデニル化配列、
(iii) 選択可能マーカー遺伝子、及び
(iv) 複製起点、
からなる群より選択される一つ以上の構成員を含むものである工程、及び該転写可能なポリヌクレオチドの発現産物のレベル及び/又は機能的活性を該ヌクレオチド配列の非存在下で対照ベクターのそれと比較して経時的に測定する工程であり、その時間にわたって対照ベクターのそれと異なるレベル及び/又は機能的活性が該RNA因子をコードするヌクレオチド配列を示すものである工程を含む方法。
【請求項27】
該プロモーターが調節可能なものである、請求項26記載の方法。
【請求項28】
該プロモーターが誘導可能なものである、請求項27記載の方法。
【請求項29】
該プロモーターが抑制可能なものである、請求項27記載の方法。
【請求項30】
そのレベル又は機能的活性が測定される発現産物が該レポータータンパク質である、請求項26記載の方法。
【請求項31】
該レポータータンパク質がルシフェラーゼ、緑色蛍光タンパク質、赤色蛍光タンパク質、SEAP、CAT 、又はそれらの生物学的に活性な断片、又はこれらの変異体若しくは誘導体から選択されるものである、請求項26記載の方法。
【請求項32】
該レポータータンパク質がタンパク質を不安定化する因子を含むものである、請求項31記載の方法。
【請求項33】
該タンパク質を不安定化する因子が PEST 配列又はユビキチンをコードするヌクレオチド配列又はそれらの生物学的に活性な断片又はこれらの変異体若しくは誘導体によりコードされるものである、請求項32記載の方法。
【請求項1】
転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含む発現ベクター。
【請求項2】
該RNA因子が該転写物の安定性を低下させる不安定化因子である、請求項1記載の発現ベクター。
【請求項3】
該RNA因子が該転写物の安定性を増加させる安定化因子である、請求項1記載の発現ベクター。
【請求項4】
該転写可能なポリヌクレオチドが目的のポリヌクレオチドを含み且つあるポリペプチドをコードするものである、請求項2記載の発現ベクター。
【請求項5】
該ポリペプチドがレポータータンパク質を含むものである、請求項4記載の発現ベクター。
【請求項6】
該転写可能なポリヌクレオチドがレポータータンパク質をコードするヌクレオチド配列を含むものである、請求項2記載の発現ベクター。
【請求項7】
(i) ヌクレオチド配列を導入するための複数クローニング部位、
(ii) レポーター遺伝子、
(iii) 該転写可能なポリヌクレオチドの発現を調節するめたのプロモーター及び/又はエンハンサー、
(iv) ポリアデニル化配列、
(v) 選択可能マーカー遺伝子、及び
(iv) 複製起点、
からなる群より選択される一つ以上の構成員を含む、請求項1又は請求項2記載のベクター。
【請求項8】
酵素により又はそれ以外の生化学的方法により切断してその中にPCR増幅産物を直接クローニングできる線状化したベクターを生じうる少なくとも一つの部位をさらに含む、請求項7記載のベクター。
【請求項9】
該不安定化因子をコードするヌクレオチド配列が、c-fos 、c-jun 、c-myc 、GM-CSF、IL-3、TNF-アルファ、IL-2、IL-6、IL-8、IL-10 、ウロキナーゼ、bcl-2 、SGLT1 (Na(+)- 共役グルコーストランスポーター) 、Cox-2 ( シクロオキシゲナーゼ2)、IL-8、PAI-2 ( プラズミノーゲン・アクチベーター・インヒビター・タイプ2)、アドレナリンベータ1受容体又は GAP43から選択される遺伝子であり、又は該遺伝子から誘導されるものである、請求項2記載のベクター。
【請求項10】
該安定化因子をコードするヌクレオチド配列が、アルファ2グロビン、アルファ1グロビン、ベータグロビン、又は成長ホルモン、エリスロポエチン、リボヌクレオチドリダクターゼR1又は m1 ムスカリン性アセチルコリンから選択される遺伝子であり、又は該遺伝子から誘導されるものである、請求項3記載のベクター。
【請求項11】
該不安定化因子をコードするヌクレオチド配列が、配列番号:1〜配列番号:23のいずれか一つ、又はそれらの生物学的に活性な断片、又はこれらの変異体若しくは誘導体から選択されるものである、請求項5記載のベクター。
【請求項12】
該不安定化因子をコードするヌクレオチド配列が、配列番号:1、配列番号:2又は配列番号:22、又はそれらの生物学的に活性な断片、又はこれらの変異体若しくは誘導体から選択されるものである、請求項11記載のベクター。
【請求項13】
該レポータータンパク質がルシフェラーゼ、緑色蛍光タンパク質、赤色蛍光タンパク質、SEAP、CAT 、又はそれらの生物学的に活性な断片、又はこれらの変異体若しくは誘導体から選択されるものである、請求項5又は請求項6記載のベクター。
【請求項14】
該ポリペプチドがタンパク質を不安定化する因子を含むものである、請求項4記載のベクター。
【請求項15】
該レポータータンパク質がタンパク質を不安定化する因子を含むものである、請求項13記載のベクター。
【請求項16】
該タンパク質を不安定化する因子が PEST 配列又はユビキチン又はこれらの生物学的に活性な断片又はこれらの変異体若しくは誘導体をコードするヌクレオチド配列によりコードされるものである、請求項14又は請求項15記載のベクター。
【請求項17】
先行する請求項のいずれか一つに記載の一つ以上のベクターを含む細胞。
【請求項18】
該細胞が真核細胞である、請求項17記載の細胞。
【請求項19】
該細胞が哺乳動物の細胞である、請求項18記載の細胞。
【請求項20】
該細胞がヒトの細胞である、請求項19記載の細胞。
【請求項21】
請求項1〜請求項16いずれか1項に記載のベクターの一つ以上を含む遺伝子的に改変された非ヒト生物。
【請求項22】
目的のポリヌクレオチドの発現を測定する方法であって、RNA合成及びタンパク質合成が起こるのに十分な時間及び条件の下で発現ベクターから目的の該ポリヌクレオチドを発現させる工程であり、該ベクターが目的の該ポリヌクレオチド及び転写可能なポリヌクレオチドに対応する転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を含む該転写可能なポリヌクレオチドを含むものであり、該発現ベクターが、
(i) ヌクレオチド配列を導入するための複数クローニング部位、
(ii) レポーター遺伝子、
(iii) 該転写可能なポリヌクレオチドの発現を調節するためのプロモーター及び/又はエンハンサー、
(iv) ポリアデニル化配列、
(v) 選択可能マーカー遺伝子、及び
(vi) 複製起点、
から成る群より選択される一つ以上の構成員を含むものである工程、
及び、経時的に転写可能なポリヌクレオチドの発現産物のレベル及び/又は機能的活性を対照と比べて測定する工程であり、該因子が該転写可能なポリヌクレオチドに機能しうるように連結されたプロモーター及び/又はエンハンサーの活性と該発現産物のレベル及び/又は機能的活性の間の時的相関関係を高めるものである工程、を含む方法。
【請求項23】
該RNAを不安定化する因子をコードするヌクレオチド配列を含む転写可能なポリヌクレオチドが、c-fos 、c-jun 、c-myc 、GM-CSF、IL-3、TNF-アルファ、IL-2、IL-6、IL-8、IL-10 、ウロキナーゼ、bcl-2 、SGLT1 (Na(+)- 共役グルコーストランスポーター) 、Cox-2 ( シクロオキシゲナーゼ2)、IL-8、PAI-2 ( プラズミノーゲン・アクチベーター・インヒビター・タイプ2)、アドレナリンベータ1受容体又は GAP43から選択される遺伝子であり、又は該遺伝子から誘導されるものである、請求項22記載の方法。
【請求項24】
該RNA不安定化因子をコードする転写可能なポリヌクレオチドが配列番号:1〜配列番号:23のいずれか一つに記載されたようなヌクレオチド配列、又はそれらの生物学的に活性な断片、又はこれらの変異体若しくは誘導体を含むものである、請求項22記載の方法。
【請求項25】
該RNAを不安定化する因子をコードする転写可能なポリヌクレオチドが配列番号:1、配列番号:2又は配列番号:22に記載された配列又はそれらの生物学的に活性な断片又はこれらの変異体若しくは誘導体を含むものである、請求項22記載の方法。
【請求項26】
RNA転写物の安定性を調節するRNA因子をコードするヌクレオチド配列を同定する方法であって、テストヌクレオチド配列を発現ベクター中に導入することにより該ヌクレオチド配列をレポータータンパク質をコードするポリヌクレオチドに連結させてプロモーター及び/又はエンハンサーに機能しうるように連結されている転写可能なポリヌクレオチドを形成させる工程、RNA合成及びタンパク質合成が起こるのに十分な時間及び条件の下で該転写可能なポリヌクレオチドを発現させる工程であり、該発現ベクターが
(i) ヌクレオチド配列を導入するための複数クローニング部位、
(ii) ポリアデニル化配列、
(iii) 選択可能マーカー遺伝子、及び
(iv) 複製起点、
からなる群より選択される一つ以上の構成員を含むものである工程、及び該転写可能なポリヌクレオチドの発現産物のレベル及び/又は機能的活性を該ヌクレオチド配列の非存在下で対照ベクターのそれと比較して経時的に測定する工程であり、その時間にわたって対照ベクターのそれと異なるレベル及び/又は機能的活性が該RNA因子をコードするヌクレオチド配列を示すものである工程を含む方法。
【請求項27】
該プロモーターが調節可能なものである、請求項26記載の方法。
【請求項28】
該プロモーターが誘導可能なものである、請求項27記載の方法。
【請求項29】
該プロモーターが抑制可能なものである、請求項27記載の方法。
【請求項30】
そのレベル又は機能的活性が測定される発現産物が該レポータータンパク質である、請求項26記載の方法。
【請求項31】
該レポータータンパク質がルシフェラーゼ、緑色蛍光タンパク質、赤色蛍光タンパク質、SEAP、CAT 、又はそれらの生物学的に活性な断片、又はこれらの変異体若しくは誘導体から選択されるものである、請求項26記載の方法。
【請求項32】
該レポータータンパク質がタンパク質を不安定化する因子を含むものである、請求項31記載の方法。
【請求項33】
該タンパク質を不安定化する因子が PEST 配列又はユビキチンをコードするヌクレオチド配列又はそれらの生物学的に活性な断片又はこれらの変異体若しくは誘導体によりコードされるものである、請求項32記載の方法。
【図1】

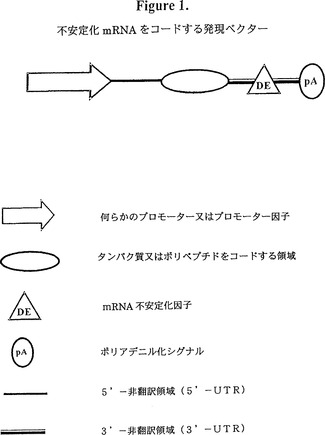
【図2】
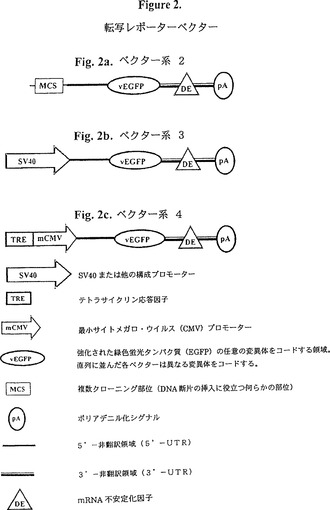
【図3】
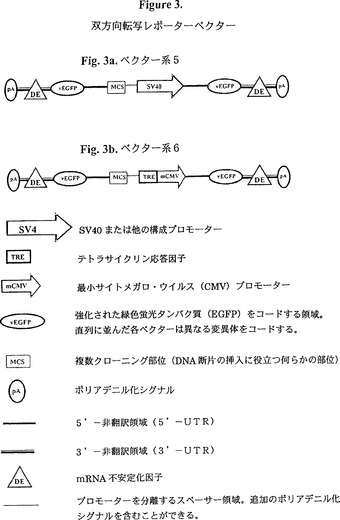
【図4】
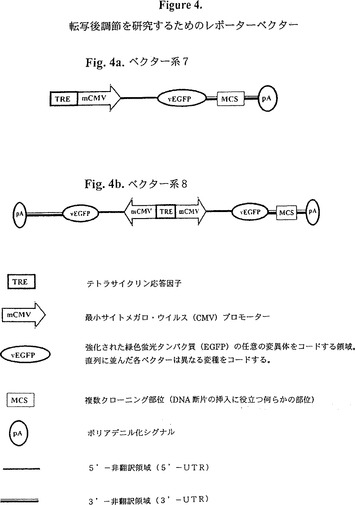
【図5】
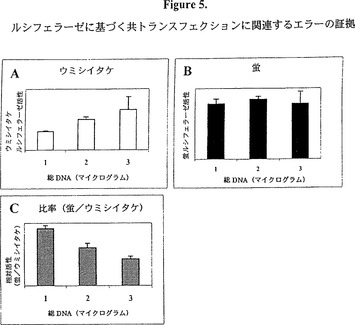
【図6】
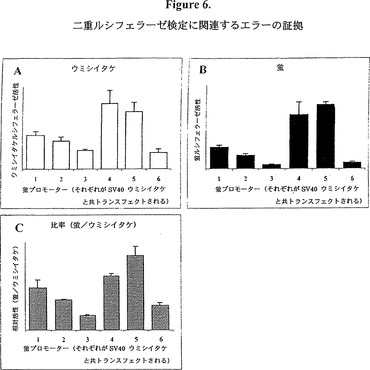
【図7】
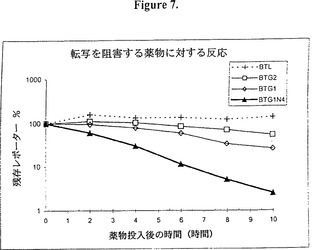
【図8】
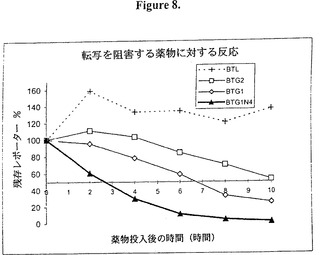
【図9】
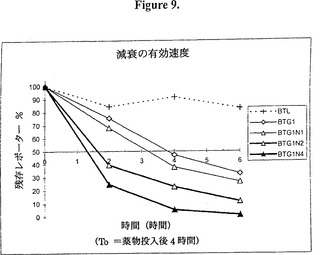
【図10】
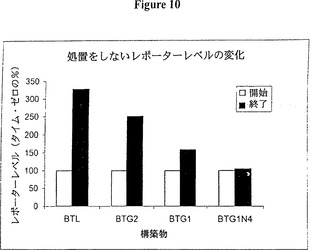
【図11】
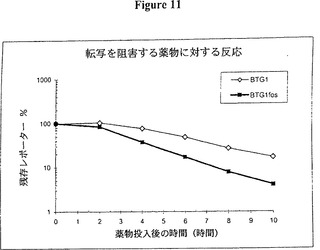
【図12】
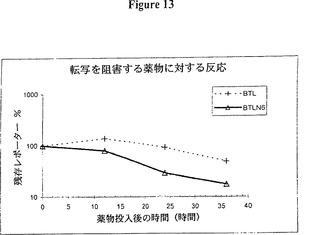
【図13】
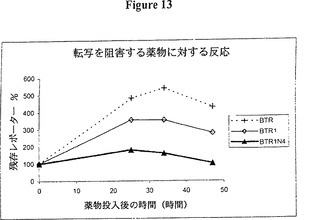
【図14】
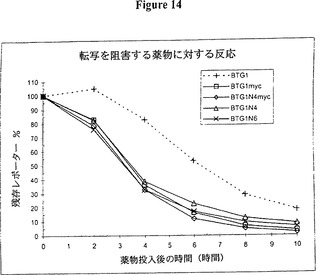
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【公開番号】特開2011−15685(P2011−15685A)
【公開日】平成23年1月27日(2011.1.27)
【国際特許分類】
【外国語出願】
【出願番号】特願2010−173310(P2010−173310)
【出願日】平成22年8月2日(2010.8.2)
【分割の表示】特願2002−571895(P2002−571895)の分割
【原出願日】平成14年3月8日(2002.3.8)
【出願人】(503323958)
【Fターム(参考)】
【公開日】平成23年1月27日(2011.1.27)
【国際特許分類】
【出願番号】特願2010−173310(P2010−173310)
【出願日】平成22年8月2日(2010.8.2)
【分割の表示】特願2002−571895(P2002−571895)の分割
【原出願日】平成14年3月8日(2002.3.8)
【出願人】(503323958)
【Fターム(参考)】
[ Back to top ]