
新規な組換えヘルペスウイルスおよび変異体ヘルペスウイルス
【課題】動物において活性な免疫を誘導するための方法および試薬の提供。
【解決手段】ヘルペスウイルスに対する免疫を誘導し得る外来DNAを有する組換えヘルペスウイルス。その外来DNAの供給源。それらの欠失したゲノムの部分を有する変異体ヘルペスウイルス。鳥類ヘルペスウイルスのUL54.5オープンリーディングフレームまたはマレク病ウイルスのUL43オープンリーディングフレームにおいて、外来性DNAが導入されるか、またはゲノムの一部が欠失される、組換えヘルペスウイルス。
【解決手段】ヘルペスウイルスに対する免疫を誘導し得る外来DNAを有する組換えヘルペスウイルス。その外来DNAの供給源。それらの欠失したゲノムの部分を有する変異体ヘルペスウイルス。鳥類ヘルペスウイルスのUL54.5オープンリーディングフレームまたはマレク病ウイルスのUL43オープンリーディングフレームにおいて、外来性DNAが導入されるか、またはゲノムの一部が欠失される、組換えヘルペスウイルス。
【発明の詳細な説明】
【技術分野】
【0001】
(発明の分野)
本発明は、動物のワクチン接種のためのウイルスベクターに関する。詳細には、本発明は、外来DNAの導入のための遺伝子挿入部位を有するウイルスベクターに関する。
【背景技術】
【0002】
(発明の背景)
マレク病は、マレク病ウイルス(MDV)によって引き起こされるニワトリのリンパ球増殖性疾患である。MDV(天然に存在するヘルペスウイルス)は、ニワトリの嚢由来リンパ球および胸腺由来リンパ球に感染し、そして引き続き胸腺由来リンパ球のリンパ腫を誘導し得る。MDVは、鳥類ヘルペスウイルスファミリーの1つの名称である。例えば、MDV1は、ニワトリにおけるヘルペスウイルスの病原性株であり、MDV2は、ニワトリにおける自然に弱毒化されたヘルペスウイルス株であり、そしてMDV3は、シチメンチョウの非病原性ヘルペスウイルスである。
【0003】
マレク病は、伝染病なので、特に大規模な飼育環境(例えば、養鶏業)において、そのウイルスはニワトリの重要な病原体になっている。現在、マレク病は、ふ卵の17〜19日目の胚または1日齢ニワトリ(one−day−old chick)のワクチン接種によって制御される。
【0004】
一般的に、動物ウイルスに対する組換えDNA技術の適用は、最近の歴史を有する。操作されるべき第1のウイルスは、最も小さなゲノムを有するウイルスである。例えば、パポバウイルスの場合において、これらのウイルスは、たいへん小さく、そして多くの余分なDNAを収容し得ないので、遺伝子工学におけるそれらの用途は、欠損レプリコンとしてであった。従って、これらのウイルスに由来する外来DNA配列発現は、野生型のヘルパーウイルスを必要とし、そして細胞培養系に制限される。他方では、アデノウイルスについて、外来配列によって置換され得る少量の非必須DNAが存在する。この技術はまた、鳥類のヘルペスウイルスにおけるヘルペスウイルスゲノムの部分に適用されている(Cochranらに対する米国特許第5,853,733号を参照のこと)。
【0005】
ヘルペスウイルスへの遺伝子の欠失または挿入という実例が、組換えDNA技術によって、ヘルペスウイルスゲノムを遺伝子的に操作することが可能であるということを実証する。過去において、遺伝子を挿入するために使用されてきた方法は、プラスミド内にクローン化されたウイルスDNAと、同じ動物細胞内にトランスフェクトされた精製されたウイルスDNAとの間の相同組換えを含む。しかし、欠失の位置および外来DNA配列の挿入のための部位を一般化し得る程度が、これらのこれまでの研究から公知ではない。
【0006】
鳥類のヘルペスウイルスにおける適切なDNA配列挿入部位の同定が、新たなワクチンの開発のために役立つ。しかし、(i)適切なウイルスおよび(ii)外来DNA配列発現のためのベクターを作製するための挿入部位として使用するためのゲノムの特定部分の選択は、重要な課題をもたらす。詳細には、その挿入部位は、ウイルスの生存可能な複製ならびに組織培養およびインビボにおけるその作動のために非必須でなくてはならない。さらに、その挿入部位は、新たな遺伝子材料を受容し得なければならないのと同時に、ウイルスが複製し続けることを保証し得なければならない。
【0007】
必要とされることは、新規なウイルスおよび新規なウイルスベクターの作製のための遺伝子挿入部位の同定である。
【発明の概要】
【課題を解決するための手段】
【0008】
(発明の要旨)
本発明は、ヘルペスウイルスゲノム内の部位に挿入された外来DNA配列を含む変異体および組換えヘルペスウイルスを提供する。1つの実施形態において、その部位は、ウイルス複製のために非必須である。好ましい実施形態において、外来DNA配列は、組換えヘルペスウイルスおよびその発現物で感染された宿主細胞において発現され得る。特に好ましい実施形態において、外来DNA配列はまた、その外来DNA配列の上流に位置するプロモーターの制御下にある。本発明は、挿入または欠失のための特定部位に限定されない。1つの実施形態において、欠失および/または挿入は、マレク病ウイルスのUL54.5オープンリーディングフレーム内に存在する。別の実施形態において、欠失および/または挿入は、マレク病ウイルスのUL43オープンリーディングフレーム内に存在する。好ましい実施形態において、挿入物は、1型マレク病ウイルスのゲノム内に存在する。
【0009】
挿入されたDNAの特定の型に限定されないが、本発明の1つの実施形態において、ヘルペスウイルスゲノム内に挿入された外来DNA配列はポリペプチドをコードする。好ましくは、そのポリペプチドは、組換えヘルペスウイルスが導入される動物に対して免疫原性である。好ましくは、この免疫原性ポリペプチドは、抗体を産生するように動物を刺激する、10個より多くのアミノ酸がペプチド結合により連結された線状ポリマーである。好ましい実施形態において、この外来DNA配列はまた、検出マーカーをコードする。好ましくは、検出マーカーは、E.coli B−ガラクトシダーゼである。
【0010】
好ましい実施形態において、この組換えヘルペスウイルスは、ニワトリ貧血ウイルス(CAV)、伝染性ファルビキウス病ウイルス(IBDV)、マレク病ウイルス(MDV)、ニューカッスル病ウイルス(NDV)、伝染性喉頭気管炎ウイルス(ILTV)または伝染性気管支炎ウイルス(IBV)、そのフラグメントおよび/または実質的に相同な配列に由来する免疫原性ポリペプチドをコードする外来DNA配列を含む。別の好ましい実施形態において、この外来DNAは、サイトカインをコードする。本発明はまた、1つの抗原または複数の抗原をコードする1つよりも多くの外来DNA配列を有する組換えヘルペスウイルスを意図する。
【0011】
本発明の組換えヘルペスウイルスの外来DNA配列が、伝染性ファルビキウス病ウイルス(IBDV)由来の免疫原性ポリペプチドをコードする場合、免疫原性ポリペプチドが、IBVD、VP2、VP3もしくはVP4タンパク質、そのフラグメントおよび/または実質的に相同な配列であることが好ましい。外来DNA配列が、MDV由来の免疫原性ポリペプチドをコードする場合、好ましくは、その免疫原性ポリペプチドは、MDV糖タンパク質B(gB)、糖タンパク質D(gD)、もしくは糖タンパク質A(gA)、そのフラグメントおよび/または実質的に相同な配列である。
【0012】
外来DNA配列が、ニューカッスル病ウイルス(NDV)由来の免疫原性ポリペプチドをコードする場合、免疫原性ポリペプチドは、NDV融合(F)タンパク質またはNDV血球凝集素ノイラミニダーゼ(HN)、そのフラグメントおよび/または実質的に相同な配列であることが好ましい。
【0013】
外来DNA配列が、伝染性喉頭気管炎ウイルス(ILTV)由来の免疫原性ポリペプチドをコードする場合、免疫原性ポリペプチドは、ILTV糖タンパク質「B」(gB)、ILTV糖タンパク質D(gD)、もしくはILTV糖タンパク質I(gI)、そのフラグメントおよび/または実質的に相同な配列であることが好ましい。
【0014】
外来DNA配列が、伝染性気管支炎ウイルス(IBV)由来の免疫原性ポリペプチドをコードする場合、免疫原性ポリペプチドは、IBVスパイクタンパク質、IBVマトリックスタンパク質、ヌクレオカプシドタンパク質、そのフラグメントおよび/または実質的に相同な配列であることが好ましい。
【0015】
挿入された外来DNA配列の発現は、外来DNA配列の上流に位置するプロモーターに制御され得る。好ましくは、そのプロモーターは、ヘルペスウイルスプロモーターである。より好ましくは、そのプロモーターは、仮性狂犬病ウイルス(PRV)gXプロモーター、MDV gBプロモーター、MDV gAプロモーター、MDV gDプロモーター、ILTV gBプロモーター、ILTV gDプロモーター、ILTV gIプロモーター、ヒトサイトメガロウイルスウイルス(HCMV)最初期(immediate early)プロモーターおよび/または実質的に相同な配列からなる群から選択される。
【0016】
本発明は、ヘルペスウイルスゲノム内に外来DNA配列を挿入することによって組換えヘルペスウイルスを産生するための相同ベクターをさらに提供する。1つの実施形態において、相同ベクターは、本質的に二本鎖外来DNA配列からなる二本鎖DNA分子を含み、その外来DNA配列の1つの末端において、ヘルペスウイルスゲノムの非必須部位の一方の側に位置するゲノムDNAと相同である二本鎖DNAを有し、そしてその外来DNA配列のもう1つの末端において、同じ部位のもう一方の側に位置するヘルペスウイルスゲノムDNA配列と相同である二本鎖DNAを有する。そのような実施形態において、二本鎖DNAは、1型マレク病ウイルスのBamHI「B」ゲノムフラグメント内に含まれる、3212塩基対のSacI〜BglIIサブフラグメント内に存在するDNA配列と相同であり得る。好ましくは、プロモーターに対応するDNA配列は、その外来DNA配列の上流に位置し、その発現を制御する。同様に、外来DNA配列は、免疫原性ポリペプチドをコードすることが好ましい(例えば、上記のポリペプチド)。
【0017】
本発明の1つの実施形態において、二本鎖ヘルペスウイルスDNAは、MDVヘルペスウイルスゲノムのBamHI「B」フラグメント内に存在するDNA配列と相同である。好ましくは、その二本鎖ヘルペスウイルスDNAは、ヘルペスウイルスゲノムのUL43タンパク質をコードするオープンリーディングフレーム内に存在するDNA配列と相同である。本発明の別の実施形態において、二本鎖ヘルペスウイルスDNAは、ヘルペスウイルスゲノムのBamHI「M」フラグメント内に存在するDNA配列と相同である。好ましくは、二本鎖ヘルペスウイルスDNAは、ヘルペスウイルスゲノムのUL54.5遺伝子コード領域内に存在するDNA配列と相同である。
【0018】
本発明は、免疫化に効果的な量の本発明の組換えヘルペスウイルスまたは変異体ヘルペスウイルスおよび適切なキャリアを含むワクチンをさらに提供する。
【0019】
本発明は、動物を免疫化する方法をさらに提供する。免疫化されるべき好ましい動物は、ニワトリである。
【0020】
本発明はまた、インビボにおいてニワトリを免疫化する方法を提供する。本発明の目的のために、これは伝染性ファルビキウス病ウイルス、マレク病ウイルス、ニューカッスル病ウイルス、伝染性喉頭気管炎ウイルスまたは伝染性気管支炎ウイルスに対して、ニワトリを免疫化することを含む。好ましくは、その方法は、効果的に免疫化する本発明のワクチンの用量をニワトリに投与する工程を包含する。そのワクチンは、当業者に周知の方法(例えば、筋肉内注射、皮下注射、腹腔内注射または静脈内注射による)のいずれかによって投与され得る。あるいは、そのワクチンは、鼻腔内、経口または眼内的に投与され得る。
【0021】
本発明はまた、本発明の組換えヘルペスウイルスで感染された宿主細胞を提供する。好ましくは、その宿主細胞は、鳥類の細胞である。
本発明はさらに、以下の項目を提供する。
(項目1)マレク病ウイルス(MDV)のUL54.5オープンリーディングフレームに挿入された外来性DNA配列を含む、組換え鳥類ヘルペスウイルス。
(項目2)前記外来性DNAがポリペプチドをコードする、項目1に記載の組換えヘルペスウイルス。
(項目3)前記ポリペプチドが10より多いアミノ酸を含有する、項目2に記載の組換えヘルペスウイルス。
(項目4)前記ポリペプチドが抗原性である、項目2に記載の組換えヘルペスウイルス。
(項目5)前記鳥類ヘルペスウイルスが、I型マレク病ウイルス(MDV−1)を含む、項目1に記載の組換えヘルペスウイルス。
(項目6)項目5に記載の組換えヘルペスウイルスであって、ここで前記外来性DNA配列が、該外来性DNA配列の上流に位置するヘルペスウイルスプロモーターの制御下にあり、そして、該外来性DNA配列が、PRV gXプロモーター、MDV gBプロモーター、MDV gAプロモーター、MDV gDプロモーター、ILTV gBプロモーター、ILTV gIプロモーター、HCMV最初期プロモーター、および実質的に相同な配列からなる群より選択される、組換えヘルペスウイルス。
(項目7)項目5に記載の組換えヘルペスウイルスであって、ここで前記外来性DNA配列が、ニワトリ貧血ウイルス、伝染性ファルビキウス病ウイルス、マレク病ウイルス、ニューカッスル病ウイルス、伝染性喉頭気管炎ウイルス、伝染性気管支炎ウイルス、および実質的に相同な配列からなる群より選択されるウイルス由来の抗原性ポリペプチドをコードする、組換えヘルペスウイルス、
(項目8)項目7に記載の組換えヘルペスウイルスであって、前記外来性DNA配列が、VP2、VP3、VP4、および実質的に相同な配列からなる群より選択される、伝染性ファルビキウス病ウイルス由来の抗原性ポリペプチドをコードするDNA配列を含む、組換えヘルペスウイルス。
(項目9)項目7に記載の組換えヘルペスウイルスであって、前記外来性DNAが、B糖タンパク質、D糖タンパク質、A糖タンパク質、および実質的に相同な配列からなる群より選択される、マレク病ウイルス由来の抗原性ポリペプチドをコードするDNA配列を含む、組換えヘルペスウイルス。
(項目10)項目7に記載の組換えヘルペスウイルスであって、前記外来性DNA配列が、F、HN、および実質的に相同な配列からなる群より選択される、ニューカッスル病ウイルス由来の抗原性ポリペプチドをコードするDNA配列を含む、組換えヘルペスウイルス。
(項目11)項目7に記載の組換えヘルペスウイルスであって、前記外来性DNAが、スパイクタンパク質、ヌクレオカプシドタンパク質、マトリックスタンパク質、および実質的に相同な配列からなる群より選択される、伝染性気管支炎ウイルス由来の抗原性ポリペプチドをコードするDNA配列を含む、組換えヘルペスウイルス。
(項目12)マレク病ウイルス(MDV)のUL54.5オープンリーディングフレームの少なくとも一部の欠失を含む、変異体鳥類ヘルペスウイルス。
(項目13)前記UL54.5オープンリーディングフレームが完全に欠失している、項目12に記載の変異体ヘルペスウイルス。
(項目14)マレク病ウイルスのUL43オープンリーディングフレームに挿入されている外来性DNA配列を含む、組換え鳥類ヘルペスウイルス。
(項目15)前記外来性DNAがポリペプチドをコードする、項目14に記載の組換えヘルペスウイルス。
(項目16)前記ポリペプチドが10より多いアミノ酸を含む、項目15に記載の組換えヘルペスウイルス。
(項目17)前記ポリペプチドが抗原性である、項目15に記載の組換えヘルペスウイルス。
(項目18)前記鳥類ヘルペスウイルスがI型マレク病ウイルス(MDV−1)を含む、項目17に記載の組換えヘルペスウイルス。
(項目19)項目14に記載の組換えヘルペスウイルスであって、ここで前記外来性DNA配列が、該外来性DNA配列の上流に位置するヘルペスウイルスプロモーターの制御下にあり、そして、該外来性DNA配列は、PRV gXプロモーター、MDV gBプロモーター、MDV gAプロモーター、MDV gDプロモーター、ILTV gBプロモーター、ILTV gIプロモーター、HCMV最初期プロモーター、および実質的に相同な配列からなる群より選択される、組換えヘルペスウイルス。
(項目20)項目19に記載の組換えヘルペスウイルスであって、ここで前記外来性DNA配列が、ニワトリ貧血ウイルス、伝染性ファルビキウス病ウイルス、マレク病ウイルス、ニューカッスル病ウイルス、伝染性喉頭気管炎ウイルス、伝染性気管支炎ウイルス、および実質的に相同な配列からなる群より選択される、ウイルス由来の抗原性ポリペプチドをコードする、組換えヘルペスウイルス。
【0022】
(定義)
本発明の目的では、「宿主細胞」は、ベクターおよびその挿入物を増幅するために使用される細胞である。この細胞の感染は、当業者に周知の方法(例えば、以下のDNA Transfection For Generating Recombinant Herpesvirus 11)に記載されるような方法)によって達成され得る。
【0023】
用語「動物」とは、動物界における生物のことをいう。従って、この用語は、ヒトならびに他の生物を含む。好ましくは、この用語は、脊椎動物をいう。より好ましくは、この用語は鳥類である動物をいう。
【0024】
本発明の組換えヘルペスウイルスの「免疫化有効量」は、102〜109プラーク形成単位(PFU)/用量の範囲内である。
【0025】
本発明の目的では、「相同性ベクター」は、ヘルペスウイルスのゲノム上の特定の部位において外来DNA配列を挿入するために構築されたプラスミドである。
【0026】
「外来DNA配列」は、組換え技術を使用して別のDNA分子に結合されているか、または結合されるDNAのセグメントであり、ここでこの特定のDNAセグメントは、天然において他のDNA分子に関連して見出されていない。このような外来DNAの供給源は、このDNAが配置される生物とは、別の生物由来であってもよいし、そうでなくてもよい。この外来DNAはまた、ネイティブ遺伝子とは異なるコドンを有する合成配列であり得る。組換え技術の例としては、DNAを繋ぎ合わせるための制限酵素およびリガーゼの使用が挙げられるが、これらに限定されない。
【0027】
「挿入部位」は、DNA分子における、外来DNAが挿入され得る制限部位である。
【0028】
「複製可能ウイルス」は、野生型のこの生物において見出されているように、遺伝物質にウイルスの複製のために必要なDNA配列またはRNA配列の全てが含まれるウイルスである。従って、複製可能ウイルスは、複製をするためにウイルスで欠損しているものまたはウイルスから欠けているものを補充する第2のウイルスも細胞株も必要としない。「ヘルペスウイルスゲノムにおける非必須部位」は、ヘルペスウイルスゲノムにおいて、その領域のポリペプチド産物が、ウイルスの感染にも複製にも必要ではない領域を意味する。
【0029】
「ベクター」は、別のDNA配列が、結合したDNA配列の発現を達成するように結合され得る、プラスミド、ファージ、コスミドまたはウイルスのようなレプリコンである。
【0030】
「二本鎖DNA分子」とは、その正常な二本鎖ヘリックスにおけるデオキシリボヌクレオチド(アデニン、グアニン、チミン、またはシトシン)のポリマー形態をいう。この用語は、この分子の一次構造および二次構造のみをいい、そして任意の特定の3次形態には限定されない。従って、この用語は、直鎖状DNA分子において見出される二本鎖DNA(例えば、ウイルス、プラスミドおよび染色体由来のDNAの制限フラグメント)を含む。
【0031】
DNA「コード配列」は、適切な調節配列の制御下に置かれる場合、インビボで転写およびポリペプチドに翻訳されるDNA配列である。コード配列の境界は、5’(アミノ)末端の開始コドンおよび3’(カルボキシ)末端の翻訳終止コドンによって決定される。コード配列は、原核生物配列、真核生物のmRNA由来のcDNA、真核生物(例えば、哺乳動物)のDNA由来のゲノムDNA配列、ウイルスDNA、およびさらに合成DNA配列を含み得るが、これらに限定されない。ポリアデニル化シグナルおよび転写終結配列は、コード配列に対して3’に配置され得る。
【0032】
「プロモーター配列」は、細胞においてRNAポリメラーゼまたは補助タンパク質を結合し得、かつ下流(3’方向)コード配列の転写を開始し得るDNA調節領域である。本発明の定義の目的で、プロモーター配列は、コード配列の翻訳開始コドン(ATG)が3’末端に近接しており、そしてバックグラウンドよりも上で検出可能なレベルでの転写を容易にするのに必要な最少の塩基数またはエレメントを含むように上流(5’方向)に広がっている。プロモーター配列内には、転写開始部位、およびRNAポリメラーゼの結合を担うタンパク質結合ドメイン(コンセンサス配列)が見出される。真核生物プロモーターはしばしば(しかし、いつもではない)、「TATA」ボックスおよび「CAAT」ボックスを含み、これらは、多くの真核生物のプロモーター領域において見出されている保存配列である。
【0033】
RNAポリメラーゼが、プロモーター配列と直接的または間接的に相互作用し、そしてコード配列のmRNAへの転写を生じ、次いで、mRNAが、そのコード配列によってコードされるポリペプチドへと翻訳される場合、コード配列は、細胞において制御配列に「作動可能に連結」されているかまたは制御配列の「制御下」にある。
【0034】
2つのポリペプチド配列は、この分子の規定された長さにわたって少なくとも約80%(好ましくは、少なくとも約90%、そして最も好ましくは少なくとも約95%)のアミノ酸が一致する場合、「実質的に相同」である。
【0035】
2つのDNA配列は、これらが同一であるか、またはそのヌクレオチドの40%より多く、より好ましくは、そのヌクレオチドの約20%より多く、そして最も好ましくはそのヌクレオチドの約10%より多くにおいて異ならない場合、「実質的に相同」である。
【0036】
外来DNA配列がそのゲノムに挿入されているウイルスは、「組換えウイルス」であるが、一方、そのゲノムの一部が故意の欠失(例えば、遺伝子操作)によって除去されているウイルスは、「変異ウイルス」である。
【0037】
用語「ポリペプチド」は、その最も広い意味で使用される(すなわち、ペプチド結合によって結合されたアミノ酸の任意のポリマー(ジペプチド以上))。従って、用語「ポリペプチド」としては、タンパク質、オリゴペプチド、タンパク質フラグメント、アナログ、ムテイン、融合タンパク質などが挙げられる。
【0038】
「抗原性の」とは、1つ以上のエピトープを含む分子の、動物免疫系またはヒト免疫系を刺激して体液性抗原特異的応答および/または細胞性抗原特異的応答を起こす能力をいう。「抗原」は、抗原性ポリペプチドである。
【0039】
組成物またはワクチンに対する「免疫学的応答」は、目的の組成物またはワクチンに対する宿主における細胞媒介免疫応答および/または抗体媒介免疫応答の発生である。通常、このような応答は、目的の組成物またはワクチンに含まれる抗原に特異的に指向される抗体、B細胞、ヘルパーT細胞、サプレッサーT細胞、および/または細胞傷害性T細胞が被験体によって産生されることからなる。
【0040】
用語「免疫原性ポリペプチド」および「免疫原性アミノ酸配列」とは、ウイルス感染性を中和する、および/または抗体補体依存性細胞傷害性もしくは抗体依存性細胞傷害性を媒介して免疫された宿主の防御を与える抗体を惹起するそれぞれポリペプチドまたはアミノ酸配列をいう。本明細書中で使用される場合、「免疫原性ポリペプチド」は、所望のタンパク質またはその免疫原性フラグメントの全長(または全長に近い)配列を含む。
【0041】
「免疫原性フラグメント」によって、1つ以上のエピトープを含み、従って、ウイルス感染性を中和する、および/または抗体補体依存性細胞傷害性もしくは抗体依存性細胞傷害性を媒介して免疫された宿主の防御を与えるポリペプチドのフラグメントを意味する。このようなフラグメントは通常、少なくとも約5アミノ酸長、そして好ましくは少なくとも約10〜15アミノ酸長である。このフラグメントの長さには、臨界的な上限はなく、このフラグメントは、タンパク質配列のほぼ全長を含み得るか、または2つ以上の抗原のフラグメントを含む融合タンパク質さえ含み得る。
【0042】
「感染性の」によって、細胞へウイルスゲノムを送達する能力を有することを意味する。
【0043】
用語「オープンリーディングフレーム」または「ORF」は、発現される場合、特定の遺伝子についての、完全タンパク質および特定のポリペプチド鎖タンパク質を産生し得る遺伝子コード領域として定義される。
【0044】
用語「鳥類ヘルペスウイルス」は、鳥類宿主において複製し得、かつ他の宿主動物において自然には複製しない、ヘルペスウイルスを意味する。
【図面の簡単な説明】
【0045】
【図1】図1は、MDVゲノムのBamHI制限酵素地図であり、特に「M」フラグメントの位置を示している。
【図2】図2は、MDVゲノムのBamHI「M」フラグメントにおけるオープンリーディングフレームを示す地図である。
【図3】図3は、MDVゲノムのBamHI制限酵素地図であり、特にSacI−BglIIIフラグメントの位置を示している。
【図4】図4は、MDVゲノムのSacI−BglIIIフラグメントにおけるオープンリーディングフレームを示す地図である。
【図5】図5は、MDVゲノムのBamHI制限酵素地図であり、特に「G」フラグメントの位置を示している。
【図6】図6は、MDVゲノムのBamHI「G」フラグメントにおけるオープンリーディングフレームの地図である。
【発明を実施するための形態】
【0046】
(発明の詳細な説明)
この開示を通じて、種々の刊行物、特許および特許出願を参考とした。これらの刊行物、特許および特許出願の開示は、本明細書中で参考として援用される。
本発明の方法および組成物は、挿入、欠失、1つまたは複数の塩基の変化によって、種々の原核生物供給源および真核生物供給源由来のクローン化DNA配列を改変する工程、および引き続くこれらの改変した配列をヘルペスウイルスのゲノムに挿入する工程を包含する。1つの例は、ヘルペスウイルスDNAの一部を細菌におけるプラスミドにクローニングする工程、ウイルスDNAが特定の配列の欠失を含むようにクローン化状態においてウイルスDNAを再構築する工程、および/またはその上、その欠失の代わりか、もしくはその欠失以外の部位かのいずれかに外来DNA配列を付加する工程を包含する。本発明の方法および組成物としてはまた、変異ウイルスを産生する、ヘルペスウイルスのゲノムの一部の欠失を含む。
【0047】
一般的に、外来遺伝子構築物は、全ヘルペスウイルスゲノムの一部だけに相当するヌクレオチド配列にクローン化される。このヌクレオチド配列は、1つ以上の適切な欠失を有し得る。このキメラDNA配列は通常、この配列の複数コピーを産生する好首尾なクローニングを可能にするプラスミド中する。次いで、このクローン化された外来遺伝子構築物は、例えば、DNA媒介同時トランスフェクション技術後のインビボ組換えによって完全ウイルスゲノムに含まれ得る。複数のコピーのコード配列または1つより多くのコード配列は、組換えベクターが、1つより多くの外来タンパク質を発現し得るように挿入され得る。外来遺伝子は、付加、欠失または置換を有して、発現したタンパク質の発現および/または免疫学的効果を増強し得る。
【0048】
遺伝子の好首尾な発現を起こすために、遺伝子は、エンハンサーエレメントおよびポリアデニル化配列を含む適切なプロモーターとともに発現ベクターに挿入され得る。哺乳動物細胞における外来遺伝子の好首尾な発現を提供する多数の真核生物プロモーターおよびポリアデニル化配列ならびに発現カセットを構築する方法は、当該分野で、例えば、米国特許第5,151,267号において公知である。このプロモーターは、免疫原性タンパク質の最適な発現を与えるように選択され、この免疫原性タンパク質は、次に公知の基準に従って体液性免疫応答、細胞媒介性免疫応答および粘膜免疫応答を満足に引き起こす。
【0049】
組換えウイルス感染細胞におけるインビボ発現によって産生される外来タンパク質は、それ自体が免疫原性であり得る。1つより多くの外来遺伝子をウイルスゲノムに挿入し、1つより多くの効果的なタンパク質の好首尾な産生が得られ得る。
【0050】
従って、変異ヘルペスウイルスの使用または外来DNA配列のヘルペスウイルスのゲノムへの付加の1つの有用性は、動物を予防接種することである。例えば、変異ウイルスを動物に導入して、その変異ウイルスに対する免疫応答を誘発し得る。
【0051】
あるいは、ポリペプチドをコードするゲノムに挿入された外来DNA配列を有する組換えヘルペスウイルスはまた、動物において外来DNA配列、外来DNA配列によってコードされるポリペプチド、および/またはヘルペスウイルスに対する免疫応答を誘発することに作用し得る。このようなウイルスをまた使用して、外来DNAおよびその産物を宿主動物に導入して、宿主動物における欠損ゲノム状態を軽減し得る。これらの組換えヘルペスウイルスは、宿主動物において外来DNAを保有し得るウイルスである場合、ウイルスベクターといわる。
【0052】
本発明は、特定のヘルペスベクターの使用に限定されない。ウイルスベクターとしての使用のために適切な1つの鳥類ヘルペスウイルスはMDVである。マレク病に対するベクターおよびワクチンとしてMDVを提供するために、MDVゲノム内に、ウイルスの複製および機能のために必須でなく、かつその中にMDV抗原をコードする1つ以上の内因性遺伝子を挿入して、コードされた抗原に対する免疫応答をさらに刺激し得る部位を配置することが望ましい。他方で、多価ワクチンとして使用するためのウイルスベクターまたは発現ベクターとしてのMDVを提供するために、MDVゲノム内に、ウイルスの複製および機能のために必須でなく、かつその中にMDV以外の家きん類病原体の抗原をコードする1つ以上の内因性遺伝子を挿入して、MDVおよびこのような他の家きん類病原体に対する免疫応答をさらに刺激し得る部位を配置することが望ましい。あるいは、内因性遺伝子のコピーおよび内因性遺伝子のコピーの組み合わせは、このようなウイルスベクターの非必須的な領域に挿入され得る。
【0053】
MDVゲノムが使用される場合、弱毒化1型MDV株を使用することが好ましい。Rispens CVI−988は、MDVの非常に有毒な株に対する防御を提供するために使用され得る、弱毒化1血清型MDVワクチン株である。
【0054】
このMDVゲノムは、2つの独特な領域(独特な短い領域(US)、および独特な長い領域(UL))からなる直鎖状180kb塩基対の二本鎖分子である。各々の独特な領域は、逆方向反復(ULについては、長い末端反復(TRL)および内部の長い逆方向反復(IRL)、そしてUSについては、短い内部の逆方向反復(IRS)および短い末端反復(TRS))に隣接している。
【0055】
本発明は、特定のDNA欠失部位および/またはDNA挿入部位に限定されないが、一方、鳥類ヘルペスウイルスのUL43領域およびUL54.5領域は、欠失および挿入のために適切な部位を含むことが発見されている。例えば、鳥類ヘルペスウイルスのUL43領域内、特にMDVゲノム内にXhoI部位が存在する。UL54領域およびUL55領域に隣接しているオープンリーディングフレーム(ORF)もまた存在する。UL54.5と称されるこのORFは、欠失および挿入のために適切なNdeI部位を含む。
【0056】
詳細には、1型マレク病ウイルスのBamHI「B」ゲノムフラグメント内に含まれる3212塩基対のSacI〜BglIIの部分フラグメントがある。BamHI「B」ゲノムフラグメント内に含まれる3212塩基対のSacI〜BglIIの部分フラグメント内の好ましい欠失部位および/または挿入部位は、ヘルペスウイルスUL43をコードするオープンリーディングフレーム内に位置し、そしてそのオープンリーディングフレーム内の好ましい挿入部分は、XhoI制限エンドヌクレアーゼ部位である。
【0057】
同様に、欠失および/または挿入は、ヘルペスウイルスゲノムのBamHI「M」ゲノムフラグメント内に位置され得る。BamHI「M」ゲノムフラグメント内の好ましい挿入部位は、ヘルペスウイルスUL54.5をコードするオープンリーディングフレーム内に位置し、そしてそのオープンリーディングフレーム内の好ましい挿入部位は、NdeI制限エンドヌクレアーゼ部位である。特に好ましい実施形態において、UL54.5オープンリーディングフレームの産物は、ウイルスの複製のために必須ではない。
【0058】
種々の外来DNA配列またはコード配列(ウイルス、原核生物、および真核生物)を、本発明に従ってヘルペスウイルスヌクレオチド配列(例えば、DNA)において挿入して、特に広範の疾患に対する防御を提供し得る。そして多くのこのような遺伝子は、当該分野ですでに公知である。任意の特定の外来DNA配列に限定されないが、代表的には、目的の外来DNA配列は、家きん産業に経済的な影響を有する疾患を鳥類において引き起こす病原体由来である。この遺伝子は、既存のワクチンが存在する生物由来であり得、そしてベクター化(vectoring)技術の新規の利点のために、このヘルペスウイルス由来ワクチンは優れている。また、目的の遺伝子は、現在ワクチンはないが、疾患の制御が必要とされている病原体由来であり得る。代表的には、目的の遺伝子は、病原体の免疫原性ポリペプチドをコードし、そして表面タンパク質、分泌タンパク質および構造タンパク質を示し得る。
【0059】
ヘルペスウイルスベクター化のための標的である関連鳥類病原体は、伝染性喉頭気管炎ウイルス(ILTV)である。ILTVは、ヘルペスウイルス科のメンバーであり、そしてこの病原体は、呼吸低下、伝染性気管支炎および観血性滲出液の喀出によって特徴付けられるニワトリの急性疾患を引き起こす。
【0060】
ヘルペスウイルスベクター化アプローチの別の標的は、ニューカッスル病である。ニューカッスル病は、高伝染性でかつ消耗性の感染性疾患であり、ニューカッスル病ウイルス(NDV)によって引き起こされる。NDVは、パラミクソウイルス科の一本鎖RNAウイルスである。NDVの種々の病原型(短潜伏期性、亜病原性、レント原性)は、この疾患の重篤度、特異性、および症状に関しては異なるが、ほとんどの型は、呼吸系および神経系に感染するようである。NDV科は、ニワトリ、シチメンチョウおよび他の鳥類種に感染する。
【0061】
本発明はまた、生物由来の特定のDNA配列の使用には限定されない。しばしば、ヘルペスウイルスゲノムへの挿入のための外来DNA配列の選択は、それがコードするタンパク質に基づく。好ましくは、外来DNA配列は、免疫原性ポリペプチドをコードする。本発明のウイルス系によって発現される好ましい免疫原性ポリペプチドは、抗原をコードする全長(または全長に近い)配列を含む。あるいは、免疫原性である(すなわち、1つ以上のエピトープをコードする)より短い配列が使用され得る。より短い配列は、中和エピトープをコードし得、このエピトープは、インビトロアッセイにおいてウイルス感染性を中和する抗体を誘発し得るエピトープとして定義される。好ましくは、このペプチドは、宿主において防御免疫応答(すなわち、感染から免疫された宿主を保護する抗体媒介性免疫応答および/または細胞媒介性免疫応答)を惹起し得る防御エピトープをコードするべきである。いくつかの場合、特定の抗原の遺伝子は、多数のイントロンを含み得るか、またはRNAウイルス由来であり得、これらの場合、相補DNAコピー(cDNA)が、使用され得る。
【0062】
野生型生物において見出されたような完全配列ではなく、遺伝子のヌクレオチド配列のフラグメントのみを、使用し得ることも可能である(これらが、防御免疫応答を生成するのに十分である場合)。入手可能である場合、合成遺伝子またはそのフラグメントもまた、使用され得る。しかし、本発明は、広範な種々の遺伝子、フラグメントなどを用いて使用され得、そして本明細書中に記載の遺伝子、フラグメントなどには、限定されない。
【0063】
従って、本発明において使用される外来DNA配列によってコードされる抗原は、ネイティブまたは組換えの免疫原性ポリペプチドまたはフラグメントのいずれかであり得る。それらは、部分配列、全長配列、または融合体(例えば、組換え宿主に適切なリーダー配列を有するか、または別の病原体についてのさらなる抗原配列を有する)でさえあり得る。
【0064】
好ましい実施形態において、本発明の変異ウイルスおよびウイルスベクターは、複製可能である。このように、ヘルペスウイルスゲノムからの欠失および/またはこのゲノムへの挿入は、ヘルペスウイルスの複製能力を破壊しない。しかし、欠失および/または挿入が、ヘルペスウイルスの複製能力を破壊または十分に阻害する場合、本発明は、本発明のヘルペスウイルスを含む発現カセットを構築し、そしてこのカセットで宿主細胞を形質転換して欠失または破壊されたDNA配列によってコードされるタンパク質を発現する細胞株または培養物を提供することによる組換え細胞株の使用を包含する。
【0065】
これらの組換え細胞株は、複製コンピテントではない組換えヘルペスウイルスが、その組換えヘルペスウイルス内にコードされる所望の外来DNA配列またはそのフラグメントを複製および発現可能にすることができる。これらの細胞株はまた、インビボ組換え、続くDNA媒介同時トランスフェクションによって組換えヘルペスウイルスを作製することに非常に有用である。
【0066】
本発明の方法および組成物がワクチン接種に使用される場合、任意の特定の投与に制限されない。1つの例は、非経口投与である。非経口投与される場合、ワクチンはワクチンキャリアの使用を含み得る。ワクチンキャリアは、当該分野で周知である:例えば、ウシ血清アルブミン(BSA)、ヒト血清アルブミン(HAS)およびキーホールリンペットヘモシアニン(HLH)。好ましいキャリアタンパク質であるロタウイスルVP6は、欧州特許公開第0259149号に開示される。
【0067】
ワクチンはまた、腸溶性投薬形態のような適切な経口キャリア中で、経口投与され得る。経口処方物としては、例えば、医薬品グレードのマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、サッカリンセルロースナトリウム(sodium saccharin cellulose)、炭酸マグネシウムなどのような通常使用される賦形剤が挙げられる。経口ワクチン組成物は、溶液、懸濁液、錠剤、ピル、カプセル、徐放性処方物または粉末の形態をとり得、約10%〜約95%の活性成分、好ましくは約25%〜約70%の活性成分を含む。経口ワクチンは、全身性免疫と組み合わせた粘膜免疫を惹起することに好ましくあり得、これは、病原体の胃腸管感染に対する防御に重要な役割を果たす。
【0068】
さらに、ワクチンは坐剤へと処方される。坐剤に関して、ワクチン組成物はポリアルカリグリコールまたはトリグリセリドのような従来の結合剤およびキャリアを含む。そのような坐剤は、約0.5%〜約10%(w/w)、好ましくは約1%〜約2%の範囲の活性成分を含む混合物から形成され得る。
【0069】
本発明のワクチン組成物を動物に投与するためのプロトコールは、本開示を考慮して当業者の範囲内である。当業者は、免疫原性フラグメントに対して抗体および/またはT細胞媒介性免疫応答を誘発するのに有効な用量でワクチン組成物の濃度を選択する。広い範囲において、この投薬量は、重要であると考えられていない。
【0070】
代表的に、ワクチン組成物は、好都合な体積のビヒクル(例えば、約1〜10cc)中にて約1から約1000μgの間のサブユニット抗原を送達する様式で投与される。好ましくは、単一免疫における投薬量は、約1から約500μgのサブユニット抗原、より好ましくは約5〜10から約100〜200μg(例えば、5〜200μg)のサブユニット抗原を送達する。
【0071】
投与の時期もまた、重要であり得る。例えば、一次接種は、好ましくは、必要な場合、後のブースター接種が続き得る。必要に応じてではあるが、二次のブースター免疫を、最初の免疫の数週間から数ヶ月間後に、動物に投与することもまた、好ましくあり得る。疾患に対する高レベルの防御の維持を確実にするためには、一定の間隔(例えば、数年毎に1回)で動物にブースター免疫を再投与することが有益であり得る。あるいは、最初の用量は経口投与され、続いて後に接種され得るか、またはその逆もあり得る。好ましいワクチン接種プロトコールは、慣用的なワクチン接種プロトコールの実験を介して確立され得る。
【0072】
本発明の組換えヘルペスウイルスはまた、本発明のワクチンによってワクチン接種された動物を、天然に存在する野生型伝染性ヘルペスウイルスまたは他の病原体によって感染された動物から区別する方法を提供し得る。これは、組換えヘルペスウイルスが、対応する病原体に対する防御免疫を与えるのに必要である上記のウイルス由来の限定数の抗原をコードする外来DNAを含むので、可能である。結果的に、これらの組換えヘルペスウイルスを用いてワクチン接種された宿主動物は、野生型伝染性ファルビキウス病ウイルス(bursal disease virus)、マレク病ウイルス、ニューカッスル病ウイルス、伝染性喉頭気管炎ウイルスまたは伝染性気管支炎ウイルスによって感染された宿主動物から、野生型ウイルスに通常存在する抗原が存在しないことによって、区別され得る。さらに、ヘルペスウイルスベクターが、免疫原性ポリペプチドをコードするそのゲノムの部分の欠失を含む場合、欠失された部分の産物に対してワクチン接種された動物からの免疫応答が欠失することによって、ワクチン接種された動物が示される。
【0073】
本発明はまた、遺伝子欠損を制御するための遺伝子治療が必要である動物に、遺伝子治療を提供するための方法を含み、この方法は、この動物に、生組換えヘルペスウイルス(これは、非欠失形態の上記遺伝子をコードする外来ヌクレオチド配列を含む)を、組換えウイルスベクターのゲノムが、上記の哺乳動物のゲノム中に組み込まれるかまたは独立して染色体外的に保持されるかして、標的器官または組織において目的の遺伝子の発現を与える条件下で、投与する工程を包含する。
【0074】
欠失遺伝子またはその部分を置換するためのこれらの種類の技術は、当業者によって使用される。例えば、Andersonら(米国特許第5,399,346号)は、遺伝子治療に関する技術を記載する。さらに、好都合な遺伝子治療に使用するために組み込まれ得る、外来DNA配列のヌクレオチド配列またはその部分の例としては、嚢胞性線維症膜貫通調節遺伝子(CFTR遺伝子(cystic fibrosis transmenbrane conductance regulator gene))、ヒトミニジストロフィン遺伝子、α−抗トリプシン遺伝子などが挙げられる。
【0075】
組換えヘルペスウイルスの構築、選択および精製に関する方法は、以下の実施例に詳述される。以下の実施例は、特定の好ましい実施形態および本発明の局面を示すために役立ち、本発明の範囲を制限するを解釈されるべきではない。
【実施例】
【0076】
(実施例1)
(マレク病ウイルス(MDV−1)ストックの調製)
マレク病ウイルスストックサンプルは、2mMグルタミン、100ユニット/ml ペニシリン、100ユニット/ml ストレプトマイシンを含むHAM’S F10および199培地の1:1混合物(これらの成分は、Sigmaまたは同等の供給者から得、そしてこれより後は、完全DME培地として呼ぶ)(1%ウシ胎仔血清を含む)中、0.01PFU/細胞の感染多重度で組織培養細胞を感染させることによって調製した。細胞変性効果が完了した後、培地および細胞を回収し、そして細胞を、臨床遠心分離機中、3000rpm、5分間でペレットにした。感染された細胞は、20%ウシ胎仔血清、10% DMSOを含む完全培地中に再懸濁し、そして−70℃で凍結保存した。
【0077】
(実施例2)
(マレク病ウイルス(MDV−1)DNAの調製)
マレク病ウイルスの全ての操作は、菌株GA5(ATCC #624)またはRispens CVI−988(Vineland Labs)を用いて行った。感染された細胞の細胞質からのMDVウイルスDNAの調製に関して、初代ニワトリ胚線維芽細胞を、細胞の過剰増殖前に、広範な細胞変性効果が起こるに十分なMOIで感染させた。全てのインキュベーションは、5% CO2空気中の加湿インキュベーターにおいて、39℃で行った。最高のDNA収量は、最大に感染されたが、不完全な細胞溶解を示す単層を収集することによって得た(代表的に5〜7日)。感染された細胞は、細胞スクレーパを用いて培地中に細胞をこすり取ることによって収集した。この細胞懸濁液を、GS−3ローター中、3000rpm、10分間、5℃で遠心分離した。
【0078】
生じたペレットを、冷PBS(20ml/ローターボトル)中に再懸濁し、そして冷やしながら3000rpmで10分間さらに遠心分離に供した。PBSをデカントした後、その細胞ペレットをRSB緩衝液(10mM Tris pH7.5、1mM EDTAおよび1.5mM MgCl2)の4ml/ローターボトル中に再懸濁した。NP40(Nonidet P−40;Sigma)を、時々混合しながら0.5%の最終濃度までサンプルに添加した。そのサンプルを、冷やしながら10分間、3000rpmで遠心分離し、核をペレットにし、そして細胞の細片を除去した。上清の流体を注意深く15mlのCorex遠心分離チューブに移した。EDTA(0.5M pH 8.0)およびSDS(ドデシル硫酸ナトリウム;20%保存)の両方を、それぞれ、最終濃度が5mMおよび1%までサンプルに添加した。100μlのプロテイナーゼ−K(10mg/ml;Boehringer Mannheim)をサンプル4ml当たりに添加し、混合し、そして45℃で1〜2時間インキュベートした。この期間の後、等量の飽和フェノールをサンプルに添加し、そして手で穏やかに混合した。このサンプルを臨床遠心分離機中、5分間、3000rpmで回転させ、相を分離させた。酢酸ナトリウム(保存溶液は3M pH5.2)を0.3Mの最終濃度で水相に添加し、そして核酸を2.5容量の冷無水エタノールを添加した後に−70℃で30分間沈澱させた。サンプル中のDNAは、HB−4ローター中5℃で、20分間、8000rpmで回転させることによってペレットにした。上清を注意深く除去し、そしてDNAペレットを25mlの80%エタノールを用いて1回洗浄した。DNAペレットを真空によって簡単に乾燥させ(2〜3分間)、そして50μl/感染された細胞のローターボトルのTE緩衝液(10mM Tris pH7.5、1mM EDTA)中に再懸濁した。代表的には、ウイルスDNAの収量は、5〜10μg/感染された細胞のローターボトルの間の範囲であった。全てのウイルスDNAは、約10℃で保存した。
【0079】
(実施例3)
(DNA配列決定)
DNA配列決定は、ABI PRISM Dye Terminator Cycle Sequencing Ready Reaction KitおよびAmplitaq DNAポリメラーゼ、FS(Perkin−Elmer;製造者らの指示書に従って)を用いた蛍光標識ジデオキシ配列決定反応をし、そして製造者らの指示書に従ってPerkin−Elmer/Applied Biosystems自動DNAシークエンサーModel 373A上で電気泳動することによって、実施した。dGTP混合物およびdITP混合物の両方を用いた反応を、圧縮の領域を明確にするために実施した。あるいは、圧縮領域は、ホルムアミドゲル上で分離させた。テンプレートは、二本鎖プラスミドサブクローンまたは一本鎖M13サブクローンであり、そしてプライマーは、配列決定される挿入物のちょうど外側のベクターか、または以前に得られた配列のいずれかに対して作製した。得られた配列を集め、そしてDNAStarソフトウェアを用いて比較した。
【0080】
(実施例4)
(分子生物学的技術)
細菌およびDNAの操作に関する技術(制限エンドヌクレアーゼを用いた消化、ゲル電気泳動、ゲルからのDNAの抽出、連結、キナーゼを用いたリン酸化、ホスファターゼを用いた処理、細菌培養物の増殖、DNAを用いた細菌の形質転換、および他の分子生物学的方法のような手順を含む)は、J.Sambrookら、Molecular Cloning A Laboratory Manual 第2版、Cold Spring Harbor Press、1989およびCurrent Protocols in Molecular Biology(1992)John Wiley & Son’s,Inc.によって記載される。記載される以外、これらは、少し改変して使用した。
【0081】
(実施例5)
(ポリメラーゼフィルイン反応)
DNAを、50mM Tris pH 7.4、50mM KCl、5mM MgCl2および400μMの4つのデオキシヌクレオチド各々を含む緩衝液中に再懸濁した。10ユニットのKlenow DNAポリメラーゼ(BRL)を添加し、そして反応を15分間室温で進行させた。次いで、このDNAをフェノール抽出し、そして上記のようにエタノール沈殿した。
【0082】
(実施例6)
(ポリメラーゼ連鎖反応を用いたクローニング)
ポリメラーゼ連鎖反応(PCR)を使用して、種々のDNAの操作に簡便な制限部位を導入した。使用した手順は、M.A.Innisら(PCR Protocols A Guide to Methods and Applications、84−91、Academic Press,Inc.,San Diego,1990)によって記載される。一般的に、増幅されたフラグメントは、サイズが500塩基対未満であり、そして増幅されたフラグメントの重要な領域は、DNA配列決定によって確認した。各場合に使用したプライマーは、以下の相同性ベクターの構築の説明に詳述する。
【0083】
(実施例7)
(感染された細胞の溶解物の調製)
25cm2フラスコまたは60mmペトリ皿中の、二次(secondary)ニワトリ胚線維芽細胞のコンフルエントな単層を、100μlのウイルスサンプルを用いて感染させた。細胞変性効果が完全になった後、培地および細胞を収集し、そして細胞を、臨床遠心分離機中、3000rpm、5分間でペレットにした。この細胞ペレットを250μlの破壊緩衝液(2% ドデシル硫酸ナトリウム、2% β−メルカプトエタノール)中に、再懸濁した。このサンプルを、氷上で30秒間超音波処理し、そして−20℃で保存した。
【0084】
(実施例8)
(ウエスタンブロット手順)
溶解物のサンプルおよびタンパク質標準物質を、Laemnli,U.K.(1970)Nature 277:680の手順に従ってポリアクリルアミドゲル上で泳動した。ゲルを電気泳動した後、このタンパク質を、Sambrookら(1989)に従って、トランスファーし、そして処理した。一次抗体は、Tris−塩化ナトリウムおよびアジ化ナトリウム(TSA:1lのH2O当たり6.61g Tris−HCl、0.97g Tris塩基、9.0gNaClおよび2.0g アジ化ナトリウム)中の5%脱脂粉乳を用いて1:100希釈した。二次抗体は、アルカリホスファターゼ結合体化し、そしてTSAを用いて1:1000希釈した。
【0085】
(実施例9)
(cDNAクローニング手順)
cDNAクローニングとは、最新技術の手順に従ってRNA分子をDNA分子に変換するために使用される方法をいう。出願人の方法は、(U.GublerおよびB.J Hoffman,Gene 25,263−269)に記載される。Bethesda Research Laboratories(Gaithersburg,Md.)は、出願人によって使用された手順と非常に類似し、そして我々の結果の再現に使用され得る試薬および手順のセットを含むcDNAクローニングキットを設計した。
【0086】
ウイルスmRNA種のクローニングに関して、ウイルスによる感染に感受性な宿主細胞株を、細胞当たり5〜10プラーク形成単位で感染させた。細胞変性効果が明白な場合であるが全て破壊する前に、培地を除去し、そして細胞を10mlの溶解緩衝液(4M グアニジンチオシアネート、0.1% アンチフォームA(antifoam A)、25mM クエン酸ナトリウム pH7.0、0.5%N−ラウリルサルコシン、0.1M β−メルカプトエタノール)中に溶解した。細胞溶解物を滅菌したDounce型ホモジナイザーに注ぎ、そして溶液が均質になるまで、氷上で8〜10分間ホモジナイズした。RNA精製に関して、8mlの細胞溶解液を、Beckman SW41遠心分離チューブ中、3.5mlのCsCl溶液(5.7M CsCl、25mM クエン酸ナトリウム pH7.0)上に穏やかに重層した。サンプルを、Beckman SW41ローター中、20℃で36000rpm、18時間遠心分離した。このチューブを氷上に置き、そしてチューブからの上清を注意深く吸引によって除去し、RNAペレットを乱さないようにした。ペレットを400μlのガラス蒸留水中に再懸濁し、そして2.6mlのグアニジン溶液(7.5M グアニジン−HCL、25mM クエン酸ナトリウム pH7.0、5mM ジチオトレイトール)に添加した。0.37容量の1M酢酸を添加し、続いて0.75容量の冷エタノールを添加し、そしてサンプルを−20℃に18時間置いてRNAを沈澱させた。沈澱物は、SS34ローター中、Sorvall遠心分離機で10分間、4℃、10000rpmで遠心分離することによって収集した。そのペレットを1.0mlの滅菌水に溶解し、13000rpmで再び遠心分離し、そして上清を保存した。RNAをそのペレットから上記のように0.5mlの滅菌水を用いてさらに2回再抽出し、そして、上清をプールした。0.1容量の2M 酢酸カリウム溶液をサンプルに添加し、続いて2倍の冷エタノールを添加し、そしてサンプルを−20℃で18時間置いた。沈澱したRNAをSS34ローター中、40℃、10分間、10000rpmで遠心分離することによって収集した。そのペレットを1mlの滅菌水に溶解し、そして濃度は、A260/280の吸収によって得た。このRNAは、−70℃で保存した。
【0087】
ポリアデニル化テイル(poly−A)を含むmRNAを、オリゴdTセルロース(Pharmacia#27 5543−0)を用いて選択した。3mgの総RNAを煮沸し、そして冷却し、結合緩衝液(0.1M Tris pH 7.5、0.5M LiCl、5mM EDTA pH 8.0、0.1% ドデシル硫酸リチウム)中、100mgオリゴdTセルロースカラムに適用した。保持されたポリ−A RNAを、溶出緩衝液(5mM Tris pH 7.5、1mM EDTA pH8.0、0.1% ドデシル硫酸ナトリウム)を用いてカラムから溶出した。このmRNAを結合緩衝液中、オリゴdTカラムに再び適用し、そして溶出緩衝液を用いて再び溶出した。このサンプルを200mM酢酸ナトリウムおよび2容量の冷エタノールを用いて−20℃、18時間で沈澱させた。このRNAは、50μlの滅菌水に再懸濁した。
【0088】
10μgのポリ−A RNAを、20mMの水酸化メチル水銀中、6分間、22℃で変性させた。β−メルカプトエタノールを75mMまで添加し、そしてそのサンプルを5分間、22℃でインキュベートした。第1鎖のcDNA合成のための0.25mlの反応混合物は、1μg オリゴ−dTプライマー(P−L Biochemicals)もしくは1μgの合成プライマー、28ユニットの胎盤リボヌクレアーゼインヒビター(Bethesda Research Labs #5518SA)、100mM Tris pH8.3、140mM KCl、10mM MgCl2、0.8mM DATP、dCTP、dGTPおよびdTTP(Pharmacia)、100マイクロキューリーの32P−標識dCTP(New England Nuclear #NEG−013H)ならびに180ユニットのAMVリバーストランスクリプターゼ(Molecular Genetics Resources #MG 101)を含んだ。この反応液を42℃で90分間インキュベートし、次いで20mM EDTA pH8.0を用いて終了させた。このサンプルを、等量のフェノール/クロロホルム(1:1)を用いて抽出し、そして2M 酢酸アンモニウムおよび2容量の冷エタノールを用いて−20℃、3時間で沈澱させた。沈澱および遠心分離後、このペレットを100μlの滅菌水に溶解した。サンプルを、緩衝液(100mM Tris pH 7.5、1mM EDTA pH 8.0、100mM NaCl)中、15ml G−100 Sephadexカラム(Pharmacia)上にロードした。抽出されたDNA画分の立ち上がり(leading edge)をプールし、そしてDNAを、体積が約100μlになるまで凍結乾燥することによって濃縮し、次いでDNAを上記のように酢酸アンモニウムおよびエタノールを用いて沈澱させた。
【0089】
第1鎖のサンプル全体を、50μg/ml dNTP、5.4ユニットのDNAポリメラーゼI(Boerhinger Mannheim #642−711)、および100ユニット/ml E.coli DNAリガーゼ(New England Biolabs #205)(総体積 50μl)を使用する以外はGublerおよびHoffman(前出)の方法に従う、第2鎖反応に使用した。第2鎖合成の後、そのcDNAをフェノール/クロロホルム抽出し、そして沈澱させた。DNAを10μlの滅菌水に再懸濁し、1μg RNase Aを用いて10分間、22℃で処理し、そして40mM Tris−アセテート pH 6.85中、アガロースゲル(シグマ Type IIアガロース)を介して電気泳動させた。そのゲルをエチジウムブロマイドを用いて染色し、そして予想のサイズの範囲のDNAをゲルから切除し、そして8mM Tris−アセテート pH 6.85中で電気溶出した。電気溶出したDNAを約100μlまで凍結乾燥し、そして酢酸アンモニウムおよびエタノールを用いて上記のように沈澱させた。このDNAを20μlの水に再懸濁した。
【0090】
オリゴdCテールを、クローニングを容易にするために、そのDNAに付加した。この反応は、そのDNA、100mMカコジル酸カルシウム(pH7.2)、0.2mM ジチオスレイトール、2mM CaCl2、80μmol dCTP、および25単位の末端デオキシヌクレオチジルトランスフェラーゼ(Molecular Genetic Resources #S1001)を、50μl中に含んだ。37℃で30分後、この反応を10mM EDTAで終了させ、そしてそのサンプルを、上記のようにフェノール/クロロホルム抽出し、そして沈殿させた。
【0091】
dCテール化DNAサンプルを、200μlの0.01M Tris pH7.5、0.1M NaCl、1mM EDTA pH8.0中で、オリゴdGテール(Bethesda Research Labs #5355 SA/SB)を含む200ngのプラスミドベクターpBR322に、65℃で2分間、次いで57℃で2時間、アニーリングさせた。D.Hanahan、Molecular Biology 166、557〜580、1983により記載されるように、新鮮なコンピテントE.coli DH−1細胞を調製し、そして200μlの細胞アリコート20個においてこのアニールしたcDNAサンプルの半分を使用して、形質転換した。形質転換した細胞を、10μg/mlテトラサイクリンを含むLブロス寒天プレート上に配置した。コロニーを、Ampscreen(Bethesda Research Labs #5537 UA)を使用して、アンピシリン遺伝子へのインサートの存在についてスクリーニングし、そして陽性コロニーを分析用に拾った。
【0092】
(実施例10)
(組換えマレク病ウイルスを作製するためのDNAトランスフェクション)
この方法は、以下の改変を加えた、KawaiおよびNishizawa、Mol.and Cell.Biol.4:1172〜1174(1984)のポリブレン−DMSO手順に基づく。組換えMDVウイルスの生成は、MDVウイルスDNAと、適切なヘルペスウイルスのクローン配列が隣接する所望の外来DNAを含むプラスミド相同性ベクターとの間の、相同組換えに依存する。トランスフェクションを、50%コンフルエントな一次ニワトリ胚線維芽(CFF)細胞の6cmプレート(Corning plastic)にて実行した。この細胞は、4μg/mlポリブレン(1×HBSS中4mg/mlのストック)を含む、CEF増殖培地(1×F10/199、5%ウシ胎仔血清、2%グルタミン、1%非必須アミノ酸、および2%ペニシリン/ストレプトマイシン)中に前日に配置した。CEF細胞への同時トランスフェクションのために、5μgのインタクトMDV DNAを、30μg/mlポリブレン(1×HBSS中4mg/mlのストック)を含む1mlのCEF培地中に懸濁した。次いで、このDNA−ポリブレン懸濁物(1ml)を、培地を吸引したCEF細胞の6cmプレートに添加し、そして39℃で30分間インキュベートした。このプレートをこの時間の間周期的に揺り動かし、接種物を再分布させた。この期間の後、4mlのCEF増殖培地を、プレートを洗浄するために直接添加し、そして39℃でさらに2.5時間インキュベートした。この時点で、この培地を各プレートから除去し、そして細胞に、1×HBSS中30%のDMSO(ジメチルスルホキシド、J.T.Baker Chemical Co.、Phillipsburg、NJ)2mlを用いて、室温で4分間ショックを与えた。この30% DMSOを注意深く除去し、そして単層を、室温にて、1×HBSSで1回洗浄した。次いで、細胞に、5mlのCEF増殖培地を添加した後、39℃でこの細胞をインキュベートした。翌日、培地を交換して、DMSOの最後の残渣まで取り除き、そして細胞増殖を刺激した。ウイルスからの細胞変性効果は、6日間以内に明らかになる。高力価ストック(80%〜90% CPE)の生成は、通常は、この日から1週間以内に行い得る。MDVストックサンプルを、20%ウシ胎仔血清、10% DMSOを含むCEF増殖培地中に感染した細胞を再懸濁することによって調製し、そして−70℃にて貯蔵した。
【0093】
(実施例11)
(β−ガラクトシダーゼを発現する組換えマレク病ウイルスについてのスクリーニング(BluogalおよびCPRCアッセイ)またはβ−グルクロニダーゼを発現する組換えマレク病ウイルスについてのスクリーニング(X−Glucアッセイ))
E.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子を組換えウイルスに組み込んだ場合、その組換え体を含むプラークを、簡単な2つの方法のうちの1つにより視覚化した。第1の方法において、化学物質BluogalTM(Life Sciences Technology、Bethesda、MD)を、プラークアッセイの間にアガロース重層物に組み込んだ(200μg/ml)。そして活性なβ−ガラクトシダーゼを発現するプラークは、青色に変化した。次いで、この青色プラークを新鮮なCEF細胞へと拾い、そしてさらなる青色プラーク単離により精製した。第2の方法において、CPRG(Boehringer Mannheim)を、プラークアッセイの間にアガロース重層物に組み込んだ(400μg/ml)。活性なβ−ガラクトシダーゼを発現するプラークは、赤色に変化した。次いで、この赤いプラークを新鮮なCEF細胞へと拾い、そしてさらなる赤色プラーク単離によって精製した。両方の場合において、代表的には、ウイルスを、3回〜4回のプラーク精製により精製した。
【0094】
E.coli β−グルクロニダーゼ(uidA)マーカー遺伝子を組換えウイルスに組み込んだ場合、この組換えウイルスを含むプラークを、発色性基質X−β−D−gluUA CHX(X−GLUC;5−ブロモ−4−クロロ−3−インドキシル−β−D−グルクロン酸、シクロヘキシルアンモニウム塩;Biosynth AG;Switzerland)を、プラークアッセイの間にアガロース重層物に組み込んだ(200μg/ml)。そして活性なβ−グルクロニダーゼを発現するプラークは、青色に変化した。次いで、その青色プラークを新鮮なCEF細胞へと拾い、そしてさらなる青色プラーク単離によって精製した。
【0095】
(実施例12)
(黒色プラークアッセイを使用する、組換えマレク病ウイルスにおける外来DNA配列発現についてのスクリーニング)
組換えMDVウイルスにより発現される外来抗原の発現を分析するために、CEF細胞の単層を組換えMDVに感染させ、栄養アガロース培地を重層し、そして39℃で4〜5日間インキュベートする。一旦プラークが発色すると、アガロース重層物をディッシュから除去し、単層を1×PBSでリンスし、室温で10分間100%メタノールで固定し、そして細胞を風乾する。プレートをPBSで再水和した後、一次抗体を適切な希釈物までPBSで希釈し、そして室温にて2時間〜一晩、細胞単層とともにインキュベートする。次いで、非結合抗体を、PBSによる室温にて3回の洗浄によって細胞から除去する。アルカリホスファターゼ結合体化二次抗体をPBSで希釈し、そして室温にて2時間細胞とともにインキュベートする。次いで、非結合二次抗体を、室温にてPBSを用いて3回細胞を洗浄することによって除去する。次いで、単層を発色緩衝液(100mM Tris pH9.5/100mM NaCl/5mM MgCl2)中でリンスし、次いで、新たに調製した基質溶液(発色緩衝液中、0.3mg/ml Nitro Blueテトラゾリウム+0.15mg/ml 5−ブロモ−4−クロロ−3−インドリルホスファターゼ)とともに、10分間〜一晩、室温でインキュベートする。最後に、反応を、基質溶液をTE(10mM Tris、pH7.5/1mM EDTA)に置換することによって停止する。正しい抗原を発現するプラークは、黒色に染まる。
【0096】
(実施例13)
(1型マレク病ウイルスのオープンリーディングフレームUL54.5に外来DNAを挿入されたプラスミド)
プラスミド440−29.2を、1型マレク病ウイルス(MDV−1)に外来DNAを挿入する目的のために構築した。このプラスミドは、1型マレク病ウイルスの、約2596塩基対のBamHI「M」ゲノムフラグメント(配列番号1)を含む。このBamHI「M」フラグメント中の3つのオープンリーディングフレームは、UL54(ICP27)ORF(配列番号1の1位〜1353位)、以前に未同定であったORF(従ってUL54.5と名付けた)(配列番号1の2187位〜1483位)およびUL55(配列番号1の2459位〜2593位)のヘルペスウイルスホモログである(表1および表2を参照のこと)。MDV−1 UL54(ICP27)遺伝子に広がるDNA配列(1492塩基対)は、公開されている(Virology 1994年10月;204(1):242〜50)。MDV ICP27(HSV−1 ICP27に対する有意な類似性に基づく)は、1419ヌクレオチド長であり、かつ473アミノ酸(54.5kDa)をコードする。UL55 ORFは短縮型であり、そしてこのタンパク質の最初の49アミノ酸のみしか含まない。UL5とUL55との間に位置する潜在的ORFを同定した。このORFは、705塩基対長であり、そしておそらく、サイズが235アミノ酸のタンパク質をコードする。このUL54.5アミノ酸配列を使用してのタンパク質データベースのBLAST検索によって、MDV−2中に類似の遺伝子を同定した(Virology 1994年5月15日;201(1):142〜6)。このMDV−2遺伝子が、1型ウマヘルペスウイルスの第1のオープンリーディングフレーム(ORF−1)と低い相同性を共有することが注目された。MDV−1およびMDV−2由来のUL54.5タンパク質の類似性指標は、170アミノ酸のコンセンサス長にわたって、63%である。UL54.5は、UL54およびUL55に対して、反対の方向で転写される。
【0097】
UL54.5 ORFは、非必須であり、そしてこのORF中またはこれらのORF間の遺伝子間領域に、外来DNAが挿入される。この領域中のすべての制限部位が、外来DNAのための挿入部位として有用である。唯一ではないこの領域中の制限酵素部位は、その部位を唯一の制限酵素認識配列へと変換するDNAリンカーの挿入によって変更する。好ましくは、外来DNAの挿入のために使用する制限酵素部位は、この2596塩基対のBamHI「M」ゲノムフラグメント中約ヌクレオチド2596にある、NdeI部位である。この挿入部位は、UL54.5 ORF中で、このオープンリーディングフレームのアミノ酸4とアミノ酸5との間にある。このプラスミドベクターは、pSP64(Promega)の約3045塩基対のBamHI制限フラグメントに由来した。フラグメント1は、1型マレク病ウイルスの約2596塩基対の、BamHI「M」フラグメントである。プラスミド440−29.2を使用して、組換えマレク病ウイルス中への外来DNAの挿入のための相同性ベクターを作製した。
【0098】
(実施例14)
(1型マレク病ウイルスのオープンリーディングフレームUL54.5に伝染性喉頭気管炎ウイルスDNAが挿入されたプラスミド)
プラスミド980−85.1を、組換え1型マレク病ウイルス(MDV−1)中に外来DNAを挿入する目的のために構築した。このプラスミドは、MDV−1 DNAが隣接する、ILTウイルスgD遺伝子およびgI遺伝子、ならびにE.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子を組み込む。これらの遺伝子を、合成DNAリンカーを使用してPacI部位へと変換した唯一のNdeI部位に挿入した。この外来DAN配列の上流は、MDV DNAの約422塩基対フラグメントである。この外来DNA配列の下流は、MDV DNAの約2174塩基対フラグメントである。ILTウイルスgD遺伝子およびgI遺伝子ならびにE.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子の転写の方向は、MDVのUL54 ORFおよびUL55 ORFの転写の方向と反対である。このプラスミドを、組換えマレク病ウイルスを生成するためのDNAトランスフェクション(実施例10)、ならびにβ−ガラクトシダーゼを発現する組換えマレク病ウイルスについてのスクリーニング(BluogalおよびCPRCアッセイ)またはβ−グルクロニダーゼを発現する組換えマレク病ウイルスについてのスクリーニング(X−Glucアッセイ)(実施例11)に従って使用する場合、外来DNA配列をコードするDNAを含むウイルスが生じる。ILTV gD遺伝子およびgI遺伝子は、それらの遺伝子自身の各内因性ILTVプロモーターからの重複転写物として発現され、そしてそれらの自身の内因性ポリアデニル化シグナルを共有し、そしてE.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子は、PRV gXプロモーターから転写され、それにPRV gXポリアデニル化シグナルが続く。
【0099】
プラスミド980−85.1を、標準的組換えDNA技術を使用して、以下の供給源由来の制限フラグメントを合成DNA配列と結合することによって、構築した。ILT gD、gI、およびE.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子を、合成DNAリンカーを使用してPacI部位へと変換した唯一のNdeI部位に、相同性ベクター440−29.2へのカセットとして挿入した。このプラスミドベクターは、pSP64(Promega)の約3045塩基対のHindIII制限フラグメントに由来した。フラグメント1は、MDV BamHI制限フラグメントMの、約418塩基対のBamHI〜NdeI制限サブフラグメントである。フラグメント2は、ILT Asp 7181ゲノムフラグメント#8(10.6kb)の、約3556塩基対のSalI〜HindIII制限サブフラグメントである。フラグメント3は、PRV BamHI制限フラグメント#10の、約413塩基対のSalI〜BamHI制限サブフラグメントである。フラグメント4は、プラスミドpJF751(11)の、約3010塩基対のBamHI〜PvuII制限サブフラグメントである。フラグメント5は、PRV BamHIゲノムフラグメント#7の、約754塩基対のNdeI〜SalI制限サブフラグメントである。フラグメント6は、MDV BamHI制限フラグメントMの、約2174塩基対のNdeI〜BamHI制限サブフラグメントである。
【0100】
(実施例15)
(1型マレク病ウイルスのオープンリーディングフレームUL54.5にニューカッスル病ウイルスDNAが挿入されたプラスミド)
プラスミド980−46.74を、組換え1型マレク病ウイルス(MDV−1)中に外来DNAを挿入する目的のために構築した。このプラスミドは、MDV DNAが隣接する、E.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子ならびにニューカッスル病ウイルス(NDV)F遺伝子を組み込む。E.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子ならびにNDV F遺伝子を、合成DNAリンカーを使用してPacI部位へと変換した唯一のNdeI部位に、相同性ベクター440−29.2(実施例13)へのカセットとして挿入した。
【0101】
この外来DAN配列の上流は、MDV DNAの約422塩基対フラグメントである。この外来DNA配列の下流は、MDV DNAの約2174塩基対フラグメントである。E.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子ならびにNDV F遺伝子の転写の方向は、MDVのUL54 ORFおよびUL55 ORFの転写の方向と反対である。このプラスミドを、組換えマレク病ウイルスを生成するためのDNAトランスフェクション(実施例10)、ならびにβ−ガラクトシダーゼを発現する組換えマレク病ウイルスについてのスクリーニング(BluogalおよびCPRCアッセイ)またはβ−グルクロニダーゼを発現する組換えマレク病ウイルスについてのスクリーニング(X−Glucアッセイ)(実施例11)に従って使用する場合、外来DNA配列をコードするDNAを含むウイルスが生じる。NDV F遺伝子は、HCMV最初期プロモーターの制御下にあり、それにHSV TKポリアデニル化シグナルが続く。E.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子は、PRV gXプロモーターから転写され、PRV gXポリアデニル化シグナルが続く。
【0102】
プラスミド980−85.1を、標準的組換えDNA技術を使用して、以下の供給源由来の制限フラグメントを合成DNA配列と結合することによって、構築した。E.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子ならびにNDV F遺伝子を、合成DNAリンカーを使用してPacI部位へと変換した唯一のNdeI部位に、相同性ベクター440−29.2(実施例13)へのカセットとして挿入した。このプラスミドベクターは、pSP64(Promega)の約3045塩基対のHindIII制限フラグメントに由来した。フラグメント1は、MDV BamHI制限フラグメントMの、約418塩基対のBamHI〜NdeI制限サブフラグメントである。フラグメント2は、PRV BamHI制限フラグメント#10の、約413塩基対のSalI〜BamHI制限サブフラグメントである。フラグメント3は、プラスミドpJF751の、約3010塩基対のBamHI〜PvuII制限フラグメントである。フラグメント4は、PRV BamHI制限フラグメント#7の、約754塩基対のNdeI〜SalI制限サブフラグメントである。フラグメント5は、HCMVゲノムのXbaI Eフラグメントの、約1191塩基対のPstI〜AvaII制限サブフラグメントである。フラグメント6は、全長NDV F cDNAクローン(B1株)の、約1812塩基対のBamHI〜PstI制限フラグメントである。フラグメント7は、HSV BamHI制限フラグメントQの、約784塩基対のSmaI〜SmaI制限サブフラグメントである。最後のフラグメントは、MDV BamHI制限フラグメント「M」の、約2174塩基対のNdeI〜BamHI制限サブフラグメントである。
【0103】
(実施例16)
(1型マレク病ウイルスのオープンリーディングフレームUL43に挿入された外来DNAを有するプラスミド)
プラスミド962−80.1を、1型マレク病ウイルス(MDV−1)中に外来DNAを挿入する目的のために構築した。これは、1型マレク病ウイルスのBamHI「B」ゲノムフラグメント内に含まれる、およそ3212塩基対のSacI〜BglIIサブフラグメント(配列番号2)を含む。3212塩基対のSacI〜BglIIサブフラグメント内の3つのオープンリーディングフレームは、UL42 ORF(配列番号2の35位〜1144位)、UL43(配列番号2の1304位〜2566位)、およびUL44(gC)(配列番号2の2786位〜3220位)のヘルペスウイルスホモログである(図3および図4を参照のこと)。MDV−1 UL44(gC)遺伝子およびプロモーター領域にわたるDNA配列(732塩基対)が、公開されている(Virus Genes 3,125〜137(1989))。MDV−2 UL42、UL43およびUL44遺伝子のDNA配列が、公開されている(J.Gen.Virol.79(Pt 8),1997〜2001(1998))。MDV−1およびMDV−2由来のUL42タンパク質の類似性指数は、369アミノ酸のコンセンサス長に対して74パーセントである。MDV−1およびMDV−2由来のUL43タンパク質の類似性指数は、401アミノ酸のコンセンサス長に対して54パーセントである。MDV−1およびMDV−2由来のUL44タンパク質の類似性指数は、145アミノ酸のコンセンサス長に対して45パーセントである。MDV−1 UL43 ORFは、非必須であり、そして外来DNAを、このORF内に、またはORF間の遺伝子間領域中に挿入する。この領域内の任意の制限部位は、外来DNAの挿入部位として有用である。唯一ではないこの領域内の制限酵素部位を、DNAリンカーの挿入によって変更する。このDNAリンカーによって、この部位が唯一の制限酵素認識配列に変換される。好ましくは、外来DNAの挿入に使用される制限酵素部位は、1型マレク病ウイルスのBamHI「B」ゲノムフラグメント内に含まれる3212塩基対のSacI〜BglIIサブフラグメントにおけるおよそヌクレオチド1386でのXhoI部位である。この挿入部位は、UL43 ORF内に、このオープンリーディングフレームのアミノ酸29と30との間にある。このプラスミドベクターは、pSP64(Promega)のおよそ3045塩基対のBamHI制限フラグメントに由来した。フラグメント1は、1型マレク病ウイルスのBamHI「B」ゲノムフラグメント内に含まれる、およそ3212塩基対のSacI〜BglIIサブフラグメントである。プラスミド962−80.1を使用して、組換えマレク病ウイルス中への外来DNAの挿入のための相同ベクターを作製した。
【0104】
(実施例17)
(1型マレク病ウイルスのオープンリーディングフレームUL43に挿入された感染性喉頭気管炎ウイルスDNAを有するプラスミド)
プラスミド980−85.22を、組換え1型マレク病ウイルス(MDV−1)に外来DNAを挿入する目的のために構築した。これは、ILTウイルスgD遺伝子およびgI遺伝子ならびにE.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子を、MDV−1 DNAに隣接して組み込んでいる。これらの遺伝子を、合成DNAリンカーを使用して、PacI部位に変換した唯一のXhoI部位に挿入した。外来DNA配列の上流は、MDV DNAのおよそ1386塩基対のフラグメントである。外来DNA配列の下流は、MDV DNAのおよそ1826塩基対のフラグメントである。ILTウイルスgD遺伝子およびgI遺伝子ならびにE.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子の転写方向は、MDV UL42およびUL43 ORFと同じ転写方向である。このプラスミドを、組換えマレク病ウイルスを作製するためのDNAトランスフェクション(実施例10)およびβ−ガラクトシダーゼを発現する組換えマレク病ウイルスのスクリーニング(BluogalおよびCprgアッセイ)、またはβ−グルクロニダーゼを発現する組換えマレク病ウイルスのスクリーニング(X−Glucアッセイ)(実施例11)に従って使用する場合、外来DNA配列をコードするDNAを含むウイルスが生じる。ILTV gD遺伝子およびgI遺伝子は、それら独自のそれぞれの内因性ILTVプロモーターから重複転写物として発現され、そしてそれら独自の内因性ポリアデニル化シグナルを共有し、そしてE.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子を、PRV gXプロモーターから転写して、そしてPRV gXポリアデニル化シグナルが続く。
【0105】
プラスミド980−85.22を、標準的な組換えDNA技術を利用して、以下の供給源からの制限フラグメントを合成DNA配列と連結することによって構築した。ILT gD、gI、およびE.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子を、唯一のXhoI部位(これは、合成DNAリンカーを使用してPacI部位に変換された)にて、相同ベクター962−80.1に、カセットとして挿入した。このプラスミドベクターは、pSP64(Promega,Madison,WI)のおよそ3045塩基対のHindIII制限フラグメントに由来した。フラグメント1は、1型マレク病ウイルスのBamHI「B」ゲノムフラグメント内に含まれる、およそ1386塩基対のSacI〜XhoI制限サブフラグメントである。フラグメント2は、ILTAsp718Iゲノムフラグメント番号8(10.6キロベース)のうちの、およそ3556塩基対のSalI〜HindIII制限サブフラグメントである。フラグメント3は、PRV BamHI制限フラグメント番号10のうちの、およそ413塩基対のSalI〜BamHI制限サブフラグメントである。フラグメント4は、プラスミドpJF751のうちの、およそ3010塩基対のBamHI〜PvuII制限フラグメントである(11)。フラグメント5は、PRV BamHI制限フラグメント番号7のうちの、およそ754塩基対のNdeI〜SalI制限サブフラグメントである。フラグメント6は、1型マレク病ウイルスのBamHI「B」ゲノムフラグメント内に含まれる、およそ1826塩基対のXhoI〜BglII制限サブフラグメントである。
【0106】
(実施例18)
(オープンリーディングフレームUL43に挿入された感染性喉頭気管炎ウイルスDNAを有する組換え1型マレク病ウイルス)
S−MDV−006は、3つの外来DNA配列を発現する1型マレク病ウイルスである。ILTウイルスgDおよびgIの遺伝子、ならびにE.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子を、唯一のPacI制限部位(1型マレク病ウイルスのBamHI「B」ゲノムフラグメント内に含まれる、およそ3212塩基対のSacI〜BglIIサブフラグメントのUL43 ORF中の独特なXhoI制限部位に挿入されたPacIリンカー)に挿入した。ILTV gD遺伝子およびgI遺伝子は、それらの独自のそれぞれの内因性ILTVプロモーターから重複転写物として発現され、そしてそれら独自の内因性ポリアデニル化シグナルを共有し、そしてE.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子を、PRV gXプロモーターから転写させ、そしてPRV gXポリアデニル化シグナルが続く。S−MDV−006は、S−MDV−002(MDV−1;CVI−988 Rispens)由来であった。組換えマレク病ウイルスを作製するためのDNAトランスフェクション手順(実施例10)において、相同ベクター980−85.22およびウイルスS−MDV−002を利用して、これを達成した。同時トランスフェクションストックを、β−グルクロニダーゼによってスクリーニングした(X−Glucアッセイ)(実施例11)。赤色プラーク精製の最終的な結果は、S−MDV−006と命名される組換えウイルスであった。このウイルスを、実施例11に記載されるように青色プラークアッセイによってモニターされるβ−ガラクトシダーゼの発現、純度、および複数の継代によるインサートの安定性についてアッセイした。最初の4回の精製後に、観察された全てのプラークは、青色であり、このことは、このウイルスが純粋であり、安定であり、そして外来DNA配列を発現していることを示す。
【0107】
S−MDV−006を、黒色プラークアッセイを使用する組換えマレク病ウイルスにおける外来DNA配列の発現についてのスクリーニング(実施例12)を使用して、ILT特異的抗原の発現についてアッセイした。ポリクローナルニワトリ抗ILT血清(SPAFAS)は、S−MDV−006プラークと特異的に反応し、そしてS−MDV−002陰性コントロールプラークと特異的に反応しなかったことが示された。全てのS−MDV−006の観察されたプラークは、ポリクローナル血清と反応し、このことは、このウイルスが、ILT外来DNA配列を安定に発現していたことを示す。本明細書に記載されるアッセイを、CEF細胞で行い、このことにより、CEF細胞は、MDV組換えワクチンの産生に適切な培養基であることが示された。
【0108】
S−MDV−006は、ILT gDタンパク質およびgIタンパク質を発現し、かつILT感染におけるワクチンとして有用である、組換え1型マレク病ウイルスである。S−MDV−006もまた、ILT gDタンパク質およびgIタンパク質の発現に有用である。
【0109】
(実施例19)
(1型マレク病ウイルスのオープンリーディングフレームUL43に挿入されたニューカッスル病ウイルスDNAを有するプラスミド)
プラスミド980−60.02を、組換え1型マレク病ウイルス(MDV−1)に外来DNAを挿入する目的のために構築した。これは、E.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子およびニューカッスル病ウイルス(NDV)F遺伝子を、MDV DNAに隣接して組み込んでいる。このE.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子およびNDV F遺伝子を、合成DNAリンカーを使用してPacI部位に変換された唯一のXhoI部位に、相同性ベクター962−80.1へのカセットとして挿入した。
【0110】
外来DNA配列の上流は、MDV DNAのおよそ1386塩基対のフラグメントである。外来DNA配列の下流は、MDV DNAのおよそ1826塩基対のフラグメントである。E.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子およびNDV F遺伝子の転写方向は、MDV UL42およびUL43 ORFと同じ転写方向である。このプラスミドを、組換えマレク病ウイルスを作製するためのDNAトランスフェクション(実施例10)およびβ−ガラクトシダーゼを発現する組換えマレク病ウイルスのスクリーニング(BluogalおよびCprgアッセイ)、またはβ−グルクロニダーゼを発現する組換えマレク病ウイルスのスクリーニング(X−Glucアッセイ)(実施例11)に従って使用する場合、外来DNA配列をコードするDNAを含むウイルスが生じる。NDV F遺伝子は、HCMV最初期プロモーターの制御下にあり、そしてHSV TKポリアデニル化シグナルが続く。E.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子を、PRV gXプロモーターから転写させ、そしてPRV gXポリアデニル化シグナルが続く。
【0111】
プラスミド980−85.1を、標準的な組換えDNA技術を利用して、以下の供給源からの制限フラグメントを合成DNA配列と連結することによって構築した。E.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子およびNDV F遺伝子を、合成DNAリンカーを使用してPacI部位に変換した唯一のXhoI部位にて、相同ベクター962−80.1中にカセットとして挿入した。このプラスミドベクターは、pSP64(Promega)のおよそ3045塩基対のHindIII制限フラグメントに由来した。
【0112】
フラグメント1は、1型マレク病ウイルスのBamHI「B」ゲノムフラグメント内に含まれる、およそ1386塩基対のSacI〜XhoI制限サブフラグメントである。
【0113】
フラグメント2は、PRV BamHI制限フラグメント番号10のうちの、およそ413塩基対のSalI〜BamHI制限サブフラグメントである。フラグメント3は、プラスミドpJF751のうちの、およそ3010塩基対のBamHI〜PvuII制限フラグメントである。フラグメント4は、PRV BamHI制限フラグメント番号7のうちの、およそ754塩基対のNdeI〜SalI制限サブフラグメントである。フラグメント5は、HCMVゲノムXbaI Eフラグメントのうちの、およそ1191塩基対のPstI〜AvaII制限サブフラグメントである。フラグメント6は、全長NDV F cDNAクローン(B1株)のうちの、およそ1812塩基対のBamHI〜PstI制限フラグメントである。フラグメント7は、HSV BamHI制限フラグメントQのうちの、およそ784塩基対のSmaI〜SmaI制限サブフラグメントである。最後のフラグメントは、1型マレク病ウイルスのBamHI「B」ゲノムフラグメント内に含まれる、およそ1826塩基対のXhoI〜BglII制限サブフラグメントである。
【0114】
(実施例20)
(オープンリーディングフレームUL43に挿入されたニューカッスル病ウイルスDNAを有する組換え1型マレク病ウイルス)
S−MDV−004は、2つの外来DNA配列を発現する1型マレク病ウイルスである。ニューカッスル病ウイルス融合物(F)の遺伝子、およびE.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子を、唯一のPacI制限部位(1型マレク病ウイルスのBamHI「B」ゲノムフラグメント内に含まれるおよそ3212塩基対のSacI〜BglIIサブフラグメントのUL43 ORF中の唯一のXhoI制限部位に挿入したPacIリンカー)中に挿入する。このNDV F遺伝子は、HCMV最初期プロモーターの制御下にあり、そしてE.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子を、PRV gXプロモーターから転写させ、そしてPRV gXポリアデニル化シグナルが続く。S−MDV−004は、S−MDV−002(MDV−1;CVI−988 Rispens)に由来する。組換えマレク病ウイルスを作製するためのDNAトランスフェクション(実施例10)において、相同ベクター980−60.02およびウイルスS−MDV−002を利用して、これを達成する。同時トランスフェクションストックを、β−ガラクトシダーゼを発現する組換えマレク病ウイルスのスクリーニング(BluogalおよびCprgアッセイ)、またはβ−グルクロニダーゼを発現する組換えマレク病ウイルスのスクリーニング(X−Glucアッセイ)(実施例11)によってスクリーニングした。赤色プラーク精製の最終的な結果は、S−MDV−004と命名される組換えウイルスであった。このウイルスを、実施例11に記載されるように青色プラークアッセイによってモニターされるβ−ガラクトシダーゼの発現、純度、および複数の継代によるインサートの安定性についてアッセイする。最初の4回の精製後に、観察された全てのプラークは、青色であり、このことは、このウイルスが純粋であり、安定であり、そして外来DNA配列を発現していることを示す。
【0115】
S−MDV−004を、黒色プラークアッセイを使用する組換えマレク病ウイルスにおける外来DNA配列の発現についてのスクリーニング(実施例12)を使用して、NDV特異的抗原の発現についてアッセイする。NDV Fに特異的なモノクローナル抗体は、S−MDV−004プラークと特異的に反応し、そしてS−MDV−002陰性コントロールプラークと特異的に反応しないことが示される。全てのS−MDV−004の観察されたプラークは、ポリクローナル血清と反応し、このことは、このウイルスが、NDV F遺伝子を安定に発現していることを示す。本明細書に記載されるアッセイを、CEF細胞で行い、このことにより、CEF細胞は、MDV組換えワクチンの産生に適切な培養基であることが示される。
【0116】
S−MDV−004は、NDV Fタンパク質を発現し、NDV感染におけるワクチンとして有用である、組換え1型マレク病ウイルスである。S−MDV−004もまた、Fタンパク質の発現に有用である。
【0117】
(実施例21)
(1型マレク病ウイルスのオープンリーディングフレームUL7、ならびに/あるいはオープンリーディングフレームUL8およびUL7の間に挿入された外来DNAを有するプラスミド)
プラスミドを、1型マレク病ウイルス(MDV−1)中に外来DNAを挿入する目的のために構築する。これは、1型マレク病ウイルスのBamHI「G」ゲノムフラグメント内に含まれる、およそ7316塩基対のサブフラグメント(配列番号3)を含む。7316塩基対のサブフラグメント内の5つのオープンリーディングフレームは、UL9 ORF(配列番号3の1位〜425位)、UL8(配列番号3の439位〜2748位)、UL7(gC)(配列番号3の3699位〜2782位)、UL6(配列番号3の5704位〜3536位)、およびUL5(配列番号3の5772位〜7316位)のヘルペスウイルスホモログである(図5および図6を参照のこと)。ORF UL8およびUL7(配列番号3の2749位〜2781位)と、ORF UL6と重複しないOFR7の部分(配列番号3の2782位〜3535位)との間の領域は、ウイルス複製に必須ではなく、そして変異ウイルスおよび/または組換えウイルスを作製するために使用され得る。
【0118】
本発明の多くの改変およびバリエーションは、当業者に明らかなように、その精神および範囲から逸脱することなくなされ得る。本明細書中に記載される特定の実施形態は、例示のみの目的で提供され、そして本発明は、添付の特許請求の範囲が権利を付す均等物の全範囲とともに、このような特許請求の範囲によってのみ制限されるべきである。
[配列表]
【数1】

【数2】
【数3】
【数4】
【数5】
【数6】
【数7】
【数8】
【技術分野】
【0001】
(発明の分野)
本発明は、動物のワクチン接種のためのウイルスベクターに関する。詳細には、本発明は、外来DNAの導入のための遺伝子挿入部位を有するウイルスベクターに関する。
【背景技術】
【0002】
(発明の背景)
マレク病は、マレク病ウイルス(MDV)によって引き起こされるニワトリのリンパ球増殖性疾患である。MDV(天然に存在するヘルペスウイルス)は、ニワトリの嚢由来リンパ球および胸腺由来リンパ球に感染し、そして引き続き胸腺由来リンパ球のリンパ腫を誘導し得る。MDVは、鳥類ヘルペスウイルスファミリーの1つの名称である。例えば、MDV1は、ニワトリにおけるヘルペスウイルスの病原性株であり、MDV2は、ニワトリにおける自然に弱毒化されたヘルペスウイルス株であり、そしてMDV3は、シチメンチョウの非病原性ヘルペスウイルスである。
【0003】
マレク病は、伝染病なので、特に大規模な飼育環境(例えば、養鶏業)において、そのウイルスはニワトリの重要な病原体になっている。現在、マレク病は、ふ卵の17〜19日目の胚または1日齢ニワトリ(one−day−old chick)のワクチン接種によって制御される。
【0004】
一般的に、動物ウイルスに対する組換えDNA技術の適用は、最近の歴史を有する。操作されるべき第1のウイルスは、最も小さなゲノムを有するウイルスである。例えば、パポバウイルスの場合において、これらのウイルスは、たいへん小さく、そして多くの余分なDNAを収容し得ないので、遺伝子工学におけるそれらの用途は、欠損レプリコンとしてであった。従って、これらのウイルスに由来する外来DNA配列発現は、野生型のヘルパーウイルスを必要とし、そして細胞培養系に制限される。他方では、アデノウイルスについて、外来配列によって置換され得る少量の非必須DNAが存在する。この技術はまた、鳥類のヘルペスウイルスにおけるヘルペスウイルスゲノムの部分に適用されている(Cochranらに対する米国特許第5,853,733号を参照のこと)。
【0005】
ヘルペスウイルスへの遺伝子の欠失または挿入という実例が、組換えDNA技術によって、ヘルペスウイルスゲノムを遺伝子的に操作することが可能であるということを実証する。過去において、遺伝子を挿入するために使用されてきた方法は、プラスミド内にクローン化されたウイルスDNAと、同じ動物細胞内にトランスフェクトされた精製されたウイルスDNAとの間の相同組換えを含む。しかし、欠失の位置および外来DNA配列の挿入のための部位を一般化し得る程度が、これらのこれまでの研究から公知ではない。
【0006】
鳥類のヘルペスウイルスにおける適切なDNA配列挿入部位の同定が、新たなワクチンの開発のために役立つ。しかし、(i)適切なウイルスおよび(ii)外来DNA配列発現のためのベクターを作製するための挿入部位として使用するためのゲノムの特定部分の選択は、重要な課題をもたらす。詳細には、その挿入部位は、ウイルスの生存可能な複製ならびに組織培養およびインビボにおけるその作動のために非必須でなくてはならない。さらに、その挿入部位は、新たな遺伝子材料を受容し得なければならないのと同時に、ウイルスが複製し続けることを保証し得なければならない。
【0007】
必要とされることは、新規なウイルスおよび新規なウイルスベクターの作製のための遺伝子挿入部位の同定である。
【発明の概要】
【課題を解決するための手段】
【0008】
(発明の要旨)
本発明は、ヘルペスウイルスゲノム内の部位に挿入された外来DNA配列を含む変異体および組換えヘルペスウイルスを提供する。1つの実施形態において、その部位は、ウイルス複製のために非必須である。好ましい実施形態において、外来DNA配列は、組換えヘルペスウイルスおよびその発現物で感染された宿主細胞において発現され得る。特に好ましい実施形態において、外来DNA配列はまた、その外来DNA配列の上流に位置するプロモーターの制御下にある。本発明は、挿入または欠失のための特定部位に限定されない。1つの実施形態において、欠失および/または挿入は、マレク病ウイルスのUL54.5オープンリーディングフレーム内に存在する。別の実施形態において、欠失および/または挿入は、マレク病ウイルスのUL43オープンリーディングフレーム内に存在する。好ましい実施形態において、挿入物は、1型マレク病ウイルスのゲノム内に存在する。
【0009】
挿入されたDNAの特定の型に限定されないが、本発明の1つの実施形態において、ヘルペスウイルスゲノム内に挿入された外来DNA配列はポリペプチドをコードする。好ましくは、そのポリペプチドは、組換えヘルペスウイルスが導入される動物に対して免疫原性である。好ましくは、この免疫原性ポリペプチドは、抗体を産生するように動物を刺激する、10個より多くのアミノ酸がペプチド結合により連結された線状ポリマーである。好ましい実施形態において、この外来DNA配列はまた、検出マーカーをコードする。好ましくは、検出マーカーは、E.coli B−ガラクトシダーゼである。
【0010】
好ましい実施形態において、この組換えヘルペスウイルスは、ニワトリ貧血ウイルス(CAV)、伝染性ファルビキウス病ウイルス(IBDV)、マレク病ウイルス(MDV)、ニューカッスル病ウイルス(NDV)、伝染性喉頭気管炎ウイルス(ILTV)または伝染性気管支炎ウイルス(IBV)、そのフラグメントおよび/または実質的に相同な配列に由来する免疫原性ポリペプチドをコードする外来DNA配列を含む。別の好ましい実施形態において、この外来DNAは、サイトカインをコードする。本発明はまた、1つの抗原または複数の抗原をコードする1つよりも多くの外来DNA配列を有する組換えヘルペスウイルスを意図する。
【0011】
本発明の組換えヘルペスウイルスの外来DNA配列が、伝染性ファルビキウス病ウイルス(IBDV)由来の免疫原性ポリペプチドをコードする場合、免疫原性ポリペプチドが、IBVD、VP2、VP3もしくはVP4タンパク質、そのフラグメントおよび/または実質的に相同な配列であることが好ましい。外来DNA配列が、MDV由来の免疫原性ポリペプチドをコードする場合、好ましくは、その免疫原性ポリペプチドは、MDV糖タンパク質B(gB)、糖タンパク質D(gD)、もしくは糖タンパク質A(gA)、そのフラグメントおよび/または実質的に相同な配列である。
【0012】
外来DNA配列が、ニューカッスル病ウイルス(NDV)由来の免疫原性ポリペプチドをコードする場合、免疫原性ポリペプチドは、NDV融合(F)タンパク質またはNDV血球凝集素ノイラミニダーゼ(HN)、そのフラグメントおよび/または実質的に相同な配列であることが好ましい。
【0013】
外来DNA配列が、伝染性喉頭気管炎ウイルス(ILTV)由来の免疫原性ポリペプチドをコードする場合、免疫原性ポリペプチドは、ILTV糖タンパク質「B」(gB)、ILTV糖タンパク質D(gD)、もしくはILTV糖タンパク質I(gI)、そのフラグメントおよび/または実質的に相同な配列であることが好ましい。
【0014】
外来DNA配列が、伝染性気管支炎ウイルス(IBV)由来の免疫原性ポリペプチドをコードする場合、免疫原性ポリペプチドは、IBVスパイクタンパク質、IBVマトリックスタンパク質、ヌクレオカプシドタンパク質、そのフラグメントおよび/または実質的に相同な配列であることが好ましい。
【0015】
挿入された外来DNA配列の発現は、外来DNA配列の上流に位置するプロモーターに制御され得る。好ましくは、そのプロモーターは、ヘルペスウイルスプロモーターである。より好ましくは、そのプロモーターは、仮性狂犬病ウイルス(PRV)gXプロモーター、MDV gBプロモーター、MDV gAプロモーター、MDV gDプロモーター、ILTV gBプロモーター、ILTV gDプロモーター、ILTV gIプロモーター、ヒトサイトメガロウイルスウイルス(HCMV)最初期(immediate early)プロモーターおよび/または実質的に相同な配列からなる群から選択される。
【0016】
本発明は、ヘルペスウイルスゲノム内に外来DNA配列を挿入することによって組換えヘルペスウイルスを産生するための相同ベクターをさらに提供する。1つの実施形態において、相同ベクターは、本質的に二本鎖外来DNA配列からなる二本鎖DNA分子を含み、その外来DNA配列の1つの末端において、ヘルペスウイルスゲノムの非必須部位の一方の側に位置するゲノムDNAと相同である二本鎖DNAを有し、そしてその外来DNA配列のもう1つの末端において、同じ部位のもう一方の側に位置するヘルペスウイルスゲノムDNA配列と相同である二本鎖DNAを有する。そのような実施形態において、二本鎖DNAは、1型マレク病ウイルスのBamHI「B」ゲノムフラグメント内に含まれる、3212塩基対のSacI〜BglIIサブフラグメント内に存在するDNA配列と相同であり得る。好ましくは、プロモーターに対応するDNA配列は、その外来DNA配列の上流に位置し、その発現を制御する。同様に、外来DNA配列は、免疫原性ポリペプチドをコードすることが好ましい(例えば、上記のポリペプチド)。
【0017】
本発明の1つの実施形態において、二本鎖ヘルペスウイルスDNAは、MDVヘルペスウイルスゲノムのBamHI「B」フラグメント内に存在するDNA配列と相同である。好ましくは、その二本鎖ヘルペスウイルスDNAは、ヘルペスウイルスゲノムのUL43タンパク質をコードするオープンリーディングフレーム内に存在するDNA配列と相同である。本発明の別の実施形態において、二本鎖ヘルペスウイルスDNAは、ヘルペスウイルスゲノムのBamHI「M」フラグメント内に存在するDNA配列と相同である。好ましくは、二本鎖ヘルペスウイルスDNAは、ヘルペスウイルスゲノムのUL54.5遺伝子コード領域内に存在するDNA配列と相同である。
【0018】
本発明は、免疫化に効果的な量の本発明の組換えヘルペスウイルスまたは変異体ヘルペスウイルスおよび適切なキャリアを含むワクチンをさらに提供する。
【0019】
本発明は、動物を免疫化する方法をさらに提供する。免疫化されるべき好ましい動物は、ニワトリである。
【0020】
本発明はまた、インビボにおいてニワトリを免疫化する方法を提供する。本発明の目的のために、これは伝染性ファルビキウス病ウイルス、マレク病ウイルス、ニューカッスル病ウイルス、伝染性喉頭気管炎ウイルスまたは伝染性気管支炎ウイルスに対して、ニワトリを免疫化することを含む。好ましくは、その方法は、効果的に免疫化する本発明のワクチンの用量をニワトリに投与する工程を包含する。そのワクチンは、当業者に周知の方法(例えば、筋肉内注射、皮下注射、腹腔内注射または静脈内注射による)のいずれかによって投与され得る。あるいは、そのワクチンは、鼻腔内、経口または眼内的に投与され得る。
【0021】
本発明はまた、本発明の組換えヘルペスウイルスで感染された宿主細胞を提供する。好ましくは、その宿主細胞は、鳥類の細胞である。
本発明はさらに、以下の項目を提供する。
(項目1)マレク病ウイルス(MDV)のUL54.5オープンリーディングフレームに挿入された外来性DNA配列を含む、組換え鳥類ヘルペスウイルス。
(項目2)前記外来性DNAがポリペプチドをコードする、項目1に記載の組換えヘルペスウイルス。
(項目3)前記ポリペプチドが10より多いアミノ酸を含有する、項目2に記載の組換えヘルペスウイルス。
(項目4)前記ポリペプチドが抗原性である、項目2に記載の組換えヘルペスウイルス。
(項目5)前記鳥類ヘルペスウイルスが、I型マレク病ウイルス(MDV−1)を含む、項目1に記載の組換えヘルペスウイルス。
(項目6)項目5に記載の組換えヘルペスウイルスであって、ここで前記外来性DNA配列が、該外来性DNA配列の上流に位置するヘルペスウイルスプロモーターの制御下にあり、そして、該外来性DNA配列が、PRV gXプロモーター、MDV gBプロモーター、MDV gAプロモーター、MDV gDプロモーター、ILTV gBプロモーター、ILTV gIプロモーター、HCMV最初期プロモーター、および実質的に相同な配列からなる群より選択される、組換えヘルペスウイルス。
(項目7)項目5に記載の組換えヘルペスウイルスであって、ここで前記外来性DNA配列が、ニワトリ貧血ウイルス、伝染性ファルビキウス病ウイルス、マレク病ウイルス、ニューカッスル病ウイルス、伝染性喉頭気管炎ウイルス、伝染性気管支炎ウイルス、および実質的に相同な配列からなる群より選択されるウイルス由来の抗原性ポリペプチドをコードする、組換えヘルペスウイルス、
(項目8)項目7に記載の組換えヘルペスウイルスであって、前記外来性DNA配列が、VP2、VP3、VP4、および実質的に相同な配列からなる群より選択される、伝染性ファルビキウス病ウイルス由来の抗原性ポリペプチドをコードするDNA配列を含む、組換えヘルペスウイルス。
(項目9)項目7に記載の組換えヘルペスウイルスであって、前記外来性DNAが、B糖タンパク質、D糖タンパク質、A糖タンパク質、および実質的に相同な配列からなる群より選択される、マレク病ウイルス由来の抗原性ポリペプチドをコードするDNA配列を含む、組換えヘルペスウイルス。
(項目10)項目7に記載の組換えヘルペスウイルスであって、前記外来性DNA配列が、F、HN、および実質的に相同な配列からなる群より選択される、ニューカッスル病ウイルス由来の抗原性ポリペプチドをコードするDNA配列を含む、組換えヘルペスウイルス。
(項目11)項目7に記載の組換えヘルペスウイルスであって、前記外来性DNAが、スパイクタンパク質、ヌクレオカプシドタンパク質、マトリックスタンパク質、および実質的に相同な配列からなる群より選択される、伝染性気管支炎ウイルス由来の抗原性ポリペプチドをコードするDNA配列を含む、組換えヘルペスウイルス。
(項目12)マレク病ウイルス(MDV)のUL54.5オープンリーディングフレームの少なくとも一部の欠失を含む、変異体鳥類ヘルペスウイルス。
(項目13)前記UL54.5オープンリーディングフレームが完全に欠失している、項目12に記載の変異体ヘルペスウイルス。
(項目14)マレク病ウイルスのUL43オープンリーディングフレームに挿入されている外来性DNA配列を含む、組換え鳥類ヘルペスウイルス。
(項目15)前記外来性DNAがポリペプチドをコードする、項目14に記載の組換えヘルペスウイルス。
(項目16)前記ポリペプチドが10より多いアミノ酸を含む、項目15に記載の組換えヘルペスウイルス。
(項目17)前記ポリペプチドが抗原性である、項目15に記載の組換えヘルペスウイルス。
(項目18)前記鳥類ヘルペスウイルスがI型マレク病ウイルス(MDV−1)を含む、項目17に記載の組換えヘルペスウイルス。
(項目19)項目14に記載の組換えヘルペスウイルスであって、ここで前記外来性DNA配列が、該外来性DNA配列の上流に位置するヘルペスウイルスプロモーターの制御下にあり、そして、該外来性DNA配列は、PRV gXプロモーター、MDV gBプロモーター、MDV gAプロモーター、MDV gDプロモーター、ILTV gBプロモーター、ILTV gIプロモーター、HCMV最初期プロモーター、および実質的に相同な配列からなる群より選択される、組換えヘルペスウイルス。
(項目20)項目19に記載の組換えヘルペスウイルスであって、ここで前記外来性DNA配列が、ニワトリ貧血ウイルス、伝染性ファルビキウス病ウイルス、マレク病ウイルス、ニューカッスル病ウイルス、伝染性喉頭気管炎ウイルス、伝染性気管支炎ウイルス、および実質的に相同な配列からなる群より選択される、ウイルス由来の抗原性ポリペプチドをコードする、組換えヘルペスウイルス。
【0022】
(定義)
本発明の目的では、「宿主細胞」は、ベクターおよびその挿入物を増幅するために使用される細胞である。この細胞の感染は、当業者に周知の方法(例えば、以下のDNA Transfection For Generating Recombinant Herpesvirus 11)に記載されるような方法)によって達成され得る。
【0023】
用語「動物」とは、動物界における生物のことをいう。従って、この用語は、ヒトならびに他の生物を含む。好ましくは、この用語は、脊椎動物をいう。より好ましくは、この用語は鳥類である動物をいう。
【0024】
本発明の組換えヘルペスウイルスの「免疫化有効量」は、102〜109プラーク形成単位(PFU)/用量の範囲内である。
【0025】
本発明の目的では、「相同性ベクター」は、ヘルペスウイルスのゲノム上の特定の部位において外来DNA配列を挿入するために構築されたプラスミドである。
【0026】
「外来DNA配列」は、組換え技術を使用して別のDNA分子に結合されているか、または結合されるDNAのセグメントであり、ここでこの特定のDNAセグメントは、天然において他のDNA分子に関連して見出されていない。このような外来DNAの供給源は、このDNAが配置される生物とは、別の生物由来であってもよいし、そうでなくてもよい。この外来DNAはまた、ネイティブ遺伝子とは異なるコドンを有する合成配列であり得る。組換え技術の例としては、DNAを繋ぎ合わせるための制限酵素およびリガーゼの使用が挙げられるが、これらに限定されない。
【0027】
「挿入部位」は、DNA分子における、外来DNAが挿入され得る制限部位である。
【0028】
「複製可能ウイルス」は、野生型のこの生物において見出されているように、遺伝物質にウイルスの複製のために必要なDNA配列またはRNA配列の全てが含まれるウイルスである。従って、複製可能ウイルスは、複製をするためにウイルスで欠損しているものまたはウイルスから欠けているものを補充する第2のウイルスも細胞株も必要としない。「ヘルペスウイルスゲノムにおける非必須部位」は、ヘルペスウイルスゲノムにおいて、その領域のポリペプチド産物が、ウイルスの感染にも複製にも必要ではない領域を意味する。
【0029】
「ベクター」は、別のDNA配列が、結合したDNA配列の発現を達成するように結合され得る、プラスミド、ファージ、コスミドまたはウイルスのようなレプリコンである。
【0030】
「二本鎖DNA分子」とは、その正常な二本鎖ヘリックスにおけるデオキシリボヌクレオチド(アデニン、グアニン、チミン、またはシトシン)のポリマー形態をいう。この用語は、この分子の一次構造および二次構造のみをいい、そして任意の特定の3次形態には限定されない。従って、この用語は、直鎖状DNA分子において見出される二本鎖DNA(例えば、ウイルス、プラスミドおよび染色体由来のDNAの制限フラグメント)を含む。
【0031】
DNA「コード配列」は、適切な調節配列の制御下に置かれる場合、インビボで転写およびポリペプチドに翻訳されるDNA配列である。コード配列の境界は、5’(アミノ)末端の開始コドンおよび3’(カルボキシ)末端の翻訳終止コドンによって決定される。コード配列は、原核生物配列、真核生物のmRNA由来のcDNA、真核生物(例えば、哺乳動物)のDNA由来のゲノムDNA配列、ウイルスDNA、およびさらに合成DNA配列を含み得るが、これらに限定されない。ポリアデニル化シグナルおよび転写終結配列は、コード配列に対して3’に配置され得る。
【0032】
「プロモーター配列」は、細胞においてRNAポリメラーゼまたは補助タンパク質を結合し得、かつ下流(3’方向)コード配列の転写を開始し得るDNA調節領域である。本発明の定義の目的で、プロモーター配列は、コード配列の翻訳開始コドン(ATG)が3’末端に近接しており、そしてバックグラウンドよりも上で検出可能なレベルでの転写を容易にするのに必要な最少の塩基数またはエレメントを含むように上流(5’方向)に広がっている。プロモーター配列内には、転写開始部位、およびRNAポリメラーゼの結合を担うタンパク質結合ドメイン(コンセンサス配列)が見出される。真核生物プロモーターはしばしば(しかし、いつもではない)、「TATA」ボックスおよび「CAAT」ボックスを含み、これらは、多くの真核生物のプロモーター領域において見出されている保存配列である。
【0033】
RNAポリメラーゼが、プロモーター配列と直接的または間接的に相互作用し、そしてコード配列のmRNAへの転写を生じ、次いで、mRNAが、そのコード配列によってコードされるポリペプチドへと翻訳される場合、コード配列は、細胞において制御配列に「作動可能に連結」されているかまたは制御配列の「制御下」にある。
【0034】
2つのポリペプチド配列は、この分子の規定された長さにわたって少なくとも約80%(好ましくは、少なくとも約90%、そして最も好ましくは少なくとも約95%)のアミノ酸が一致する場合、「実質的に相同」である。
【0035】
2つのDNA配列は、これらが同一であるか、またはそのヌクレオチドの40%より多く、より好ましくは、そのヌクレオチドの約20%より多く、そして最も好ましくはそのヌクレオチドの約10%より多くにおいて異ならない場合、「実質的に相同」である。
【0036】
外来DNA配列がそのゲノムに挿入されているウイルスは、「組換えウイルス」であるが、一方、そのゲノムの一部が故意の欠失(例えば、遺伝子操作)によって除去されているウイルスは、「変異ウイルス」である。
【0037】
用語「ポリペプチド」は、その最も広い意味で使用される(すなわち、ペプチド結合によって結合されたアミノ酸の任意のポリマー(ジペプチド以上))。従って、用語「ポリペプチド」としては、タンパク質、オリゴペプチド、タンパク質フラグメント、アナログ、ムテイン、融合タンパク質などが挙げられる。
【0038】
「抗原性の」とは、1つ以上のエピトープを含む分子の、動物免疫系またはヒト免疫系を刺激して体液性抗原特異的応答および/または細胞性抗原特異的応答を起こす能力をいう。「抗原」は、抗原性ポリペプチドである。
【0039】
組成物またはワクチンに対する「免疫学的応答」は、目的の組成物またはワクチンに対する宿主における細胞媒介免疫応答および/または抗体媒介免疫応答の発生である。通常、このような応答は、目的の組成物またはワクチンに含まれる抗原に特異的に指向される抗体、B細胞、ヘルパーT細胞、サプレッサーT細胞、および/または細胞傷害性T細胞が被験体によって産生されることからなる。
【0040】
用語「免疫原性ポリペプチド」および「免疫原性アミノ酸配列」とは、ウイルス感染性を中和する、および/または抗体補体依存性細胞傷害性もしくは抗体依存性細胞傷害性を媒介して免疫された宿主の防御を与える抗体を惹起するそれぞれポリペプチドまたはアミノ酸配列をいう。本明細書中で使用される場合、「免疫原性ポリペプチド」は、所望のタンパク質またはその免疫原性フラグメントの全長(または全長に近い)配列を含む。
【0041】
「免疫原性フラグメント」によって、1つ以上のエピトープを含み、従って、ウイルス感染性を中和する、および/または抗体補体依存性細胞傷害性もしくは抗体依存性細胞傷害性を媒介して免疫された宿主の防御を与えるポリペプチドのフラグメントを意味する。このようなフラグメントは通常、少なくとも約5アミノ酸長、そして好ましくは少なくとも約10〜15アミノ酸長である。このフラグメントの長さには、臨界的な上限はなく、このフラグメントは、タンパク質配列のほぼ全長を含み得るか、または2つ以上の抗原のフラグメントを含む融合タンパク質さえ含み得る。
【0042】
「感染性の」によって、細胞へウイルスゲノムを送達する能力を有することを意味する。
【0043】
用語「オープンリーディングフレーム」または「ORF」は、発現される場合、特定の遺伝子についての、完全タンパク質および特定のポリペプチド鎖タンパク質を産生し得る遺伝子コード領域として定義される。
【0044】
用語「鳥類ヘルペスウイルス」は、鳥類宿主において複製し得、かつ他の宿主動物において自然には複製しない、ヘルペスウイルスを意味する。
【図面の簡単な説明】
【0045】
【図1】図1は、MDVゲノムのBamHI制限酵素地図であり、特に「M」フラグメントの位置を示している。
【図2】図2は、MDVゲノムのBamHI「M」フラグメントにおけるオープンリーディングフレームを示す地図である。
【図3】図3は、MDVゲノムのBamHI制限酵素地図であり、特にSacI−BglIIIフラグメントの位置を示している。
【図4】図4は、MDVゲノムのSacI−BglIIIフラグメントにおけるオープンリーディングフレームを示す地図である。
【図5】図5は、MDVゲノムのBamHI制限酵素地図であり、特に「G」フラグメントの位置を示している。
【図6】図6は、MDVゲノムのBamHI「G」フラグメントにおけるオープンリーディングフレームの地図である。
【発明を実施するための形態】
【0046】
(発明の詳細な説明)
この開示を通じて、種々の刊行物、特許および特許出願を参考とした。これらの刊行物、特許および特許出願の開示は、本明細書中で参考として援用される。
本発明の方法および組成物は、挿入、欠失、1つまたは複数の塩基の変化によって、種々の原核生物供給源および真核生物供給源由来のクローン化DNA配列を改変する工程、および引き続くこれらの改変した配列をヘルペスウイルスのゲノムに挿入する工程を包含する。1つの例は、ヘルペスウイルスDNAの一部を細菌におけるプラスミドにクローニングする工程、ウイルスDNAが特定の配列の欠失を含むようにクローン化状態においてウイルスDNAを再構築する工程、および/またはその上、その欠失の代わりか、もしくはその欠失以外の部位かのいずれかに外来DNA配列を付加する工程を包含する。本発明の方法および組成物としてはまた、変異ウイルスを産生する、ヘルペスウイルスのゲノムの一部の欠失を含む。
【0047】
一般的に、外来遺伝子構築物は、全ヘルペスウイルスゲノムの一部だけに相当するヌクレオチド配列にクローン化される。このヌクレオチド配列は、1つ以上の適切な欠失を有し得る。このキメラDNA配列は通常、この配列の複数コピーを産生する好首尾なクローニングを可能にするプラスミド中する。次いで、このクローン化された外来遺伝子構築物は、例えば、DNA媒介同時トランスフェクション技術後のインビボ組換えによって完全ウイルスゲノムに含まれ得る。複数のコピーのコード配列または1つより多くのコード配列は、組換えベクターが、1つより多くの外来タンパク質を発現し得るように挿入され得る。外来遺伝子は、付加、欠失または置換を有して、発現したタンパク質の発現および/または免疫学的効果を増強し得る。
【0048】
遺伝子の好首尾な発現を起こすために、遺伝子は、エンハンサーエレメントおよびポリアデニル化配列を含む適切なプロモーターとともに発現ベクターに挿入され得る。哺乳動物細胞における外来遺伝子の好首尾な発現を提供する多数の真核生物プロモーターおよびポリアデニル化配列ならびに発現カセットを構築する方法は、当該分野で、例えば、米国特許第5,151,267号において公知である。このプロモーターは、免疫原性タンパク質の最適な発現を与えるように選択され、この免疫原性タンパク質は、次に公知の基準に従って体液性免疫応答、細胞媒介性免疫応答および粘膜免疫応答を満足に引き起こす。
【0049】
組換えウイルス感染細胞におけるインビボ発現によって産生される外来タンパク質は、それ自体が免疫原性であり得る。1つより多くの外来遺伝子をウイルスゲノムに挿入し、1つより多くの効果的なタンパク質の好首尾な産生が得られ得る。
【0050】
従って、変異ヘルペスウイルスの使用または外来DNA配列のヘルペスウイルスのゲノムへの付加の1つの有用性は、動物を予防接種することである。例えば、変異ウイルスを動物に導入して、その変異ウイルスに対する免疫応答を誘発し得る。
【0051】
あるいは、ポリペプチドをコードするゲノムに挿入された外来DNA配列を有する組換えヘルペスウイルスはまた、動物において外来DNA配列、外来DNA配列によってコードされるポリペプチド、および/またはヘルペスウイルスに対する免疫応答を誘発することに作用し得る。このようなウイルスをまた使用して、外来DNAおよびその産物を宿主動物に導入して、宿主動物における欠損ゲノム状態を軽減し得る。これらの組換えヘルペスウイルスは、宿主動物において外来DNAを保有し得るウイルスである場合、ウイルスベクターといわる。
【0052】
本発明は、特定のヘルペスベクターの使用に限定されない。ウイルスベクターとしての使用のために適切な1つの鳥類ヘルペスウイルスはMDVである。マレク病に対するベクターおよびワクチンとしてMDVを提供するために、MDVゲノム内に、ウイルスの複製および機能のために必須でなく、かつその中にMDV抗原をコードする1つ以上の内因性遺伝子を挿入して、コードされた抗原に対する免疫応答をさらに刺激し得る部位を配置することが望ましい。他方で、多価ワクチンとして使用するためのウイルスベクターまたは発現ベクターとしてのMDVを提供するために、MDVゲノム内に、ウイルスの複製および機能のために必須でなく、かつその中にMDV以外の家きん類病原体の抗原をコードする1つ以上の内因性遺伝子を挿入して、MDVおよびこのような他の家きん類病原体に対する免疫応答をさらに刺激し得る部位を配置することが望ましい。あるいは、内因性遺伝子のコピーおよび内因性遺伝子のコピーの組み合わせは、このようなウイルスベクターの非必須的な領域に挿入され得る。
【0053】
MDVゲノムが使用される場合、弱毒化1型MDV株を使用することが好ましい。Rispens CVI−988は、MDVの非常に有毒な株に対する防御を提供するために使用され得る、弱毒化1血清型MDVワクチン株である。
【0054】
このMDVゲノムは、2つの独特な領域(独特な短い領域(US)、および独特な長い領域(UL))からなる直鎖状180kb塩基対の二本鎖分子である。各々の独特な領域は、逆方向反復(ULについては、長い末端反復(TRL)および内部の長い逆方向反復(IRL)、そしてUSについては、短い内部の逆方向反復(IRS)および短い末端反復(TRS))に隣接している。
【0055】
本発明は、特定のDNA欠失部位および/またはDNA挿入部位に限定されないが、一方、鳥類ヘルペスウイルスのUL43領域およびUL54.5領域は、欠失および挿入のために適切な部位を含むことが発見されている。例えば、鳥類ヘルペスウイルスのUL43領域内、特にMDVゲノム内にXhoI部位が存在する。UL54領域およびUL55領域に隣接しているオープンリーディングフレーム(ORF)もまた存在する。UL54.5と称されるこのORFは、欠失および挿入のために適切なNdeI部位を含む。
【0056】
詳細には、1型マレク病ウイルスのBamHI「B」ゲノムフラグメント内に含まれる3212塩基対のSacI〜BglIIの部分フラグメントがある。BamHI「B」ゲノムフラグメント内に含まれる3212塩基対のSacI〜BglIIの部分フラグメント内の好ましい欠失部位および/または挿入部位は、ヘルペスウイルスUL43をコードするオープンリーディングフレーム内に位置し、そしてそのオープンリーディングフレーム内の好ましい挿入部分は、XhoI制限エンドヌクレアーゼ部位である。
【0057】
同様に、欠失および/または挿入は、ヘルペスウイルスゲノムのBamHI「M」ゲノムフラグメント内に位置され得る。BamHI「M」ゲノムフラグメント内の好ましい挿入部位は、ヘルペスウイルスUL54.5をコードするオープンリーディングフレーム内に位置し、そしてそのオープンリーディングフレーム内の好ましい挿入部位は、NdeI制限エンドヌクレアーゼ部位である。特に好ましい実施形態において、UL54.5オープンリーディングフレームの産物は、ウイルスの複製のために必須ではない。
【0058】
種々の外来DNA配列またはコード配列(ウイルス、原核生物、および真核生物)を、本発明に従ってヘルペスウイルスヌクレオチド配列(例えば、DNA)において挿入して、特に広範の疾患に対する防御を提供し得る。そして多くのこのような遺伝子は、当該分野ですでに公知である。任意の特定の外来DNA配列に限定されないが、代表的には、目的の外来DNA配列は、家きん産業に経済的な影響を有する疾患を鳥類において引き起こす病原体由来である。この遺伝子は、既存のワクチンが存在する生物由来であり得、そしてベクター化(vectoring)技術の新規の利点のために、このヘルペスウイルス由来ワクチンは優れている。また、目的の遺伝子は、現在ワクチンはないが、疾患の制御が必要とされている病原体由来であり得る。代表的には、目的の遺伝子は、病原体の免疫原性ポリペプチドをコードし、そして表面タンパク質、分泌タンパク質および構造タンパク質を示し得る。
【0059】
ヘルペスウイルスベクター化のための標的である関連鳥類病原体は、伝染性喉頭気管炎ウイルス(ILTV)である。ILTVは、ヘルペスウイルス科のメンバーであり、そしてこの病原体は、呼吸低下、伝染性気管支炎および観血性滲出液の喀出によって特徴付けられるニワトリの急性疾患を引き起こす。
【0060】
ヘルペスウイルスベクター化アプローチの別の標的は、ニューカッスル病である。ニューカッスル病は、高伝染性でかつ消耗性の感染性疾患であり、ニューカッスル病ウイルス(NDV)によって引き起こされる。NDVは、パラミクソウイルス科の一本鎖RNAウイルスである。NDVの種々の病原型(短潜伏期性、亜病原性、レント原性)は、この疾患の重篤度、特異性、および症状に関しては異なるが、ほとんどの型は、呼吸系および神経系に感染するようである。NDV科は、ニワトリ、シチメンチョウおよび他の鳥類種に感染する。
【0061】
本発明はまた、生物由来の特定のDNA配列の使用には限定されない。しばしば、ヘルペスウイルスゲノムへの挿入のための外来DNA配列の選択は、それがコードするタンパク質に基づく。好ましくは、外来DNA配列は、免疫原性ポリペプチドをコードする。本発明のウイルス系によって発現される好ましい免疫原性ポリペプチドは、抗原をコードする全長(または全長に近い)配列を含む。あるいは、免疫原性である(すなわち、1つ以上のエピトープをコードする)より短い配列が使用され得る。より短い配列は、中和エピトープをコードし得、このエピトープは、インビトロアッセイにおいてウイルス感染性を中和する抗体を誘発し得るエピトープとして定義される。好ましくは、このペプチドは、宿主において防御免疫応答(すなわち、感染から免疫された宿主を保護する抗体媒介性免疫応答および/または細胞媒介性免疫応答)を惹起し得る防御エピトープをコードするべきである。いくつかの場合、特定の抗原の遺伝子は、多数のイントロンを含み得るか、またはRNAウイルス由来であり得、これらの場合、相補DNAコピー(cDNA)が、使用され得る。
【0062】
野生型生物において見出されたような完全配列ではなく、遺伝子のヌクレオチド配列のフラグメントのみを、使用し得ることも可能である(これらが、防御免疫応答を生成するのに十分である場合)。入手可能である場合、合成遺伝子またはそのフラグメントもまた、使用され得る。しかし、本発明は、広範な種々の遺伝子、フラグメントなどを用いて使用され得、そして本明細書中に記載の遺伝子、フラグメントなどには、限定されない。
【0063】
従って、本発明において使用される外来DNA配列によってコードされる抗原は、ネイティブまたは組換えの免疫原性ポリペプチドまたはフラグメントのいずれかであり得る。それらは、部分配列、全長配列、または融合体(例えば、組換え宿主に適切なリーダー配列を有するか、または別の病原体についてのさらなる抗原配列を有する)でさえあり得る。
【0064】
好ましい実施形態において、本発明の変異ウイルスおよびウイルスベクターは、複製可能である。このように、ヘルペスウイルスゲノムからの欠失および/またはこのゲノムへの挿入は、ヘルペスウイルスの複製能力を破壊しない。しかし、欠失および/または挿入が、ヘルペスウイルスの複製能力を破壊または十分に阻害する場合、本発明は、本発明のヘルペスウイルスを含む発現カセットを構築し、そしてこのカセットで宿主細胞を形質転換して欠失または破壊されたDNA配列によってコードされるタンパク質を発現する細胞株または培養物を提供することによる組換え細胞株の使用を包含する。
【0065】
これらの組換え細胞株は、複製コンピテントではない組換えヘルペスウイルスが、その組換えヘルペスウイルス内にコードされる所望の外来DNA配列またはそのフラグメントを複製および発現可能にすることができる。これらの細胞株はまた、インビボ組換え、続くDNA媒介同時トランスフェクションによって組換えヘルペスウイルスを作製することに非常に有用である。
【0066】
本発明の方法および組成物がワクチン接種に使用される場合、任意の特定の投与に制限されない。1つの例は、非経口投与である。非経口投与される場合、ワクチンはワクチンキャリアの使用を含み得る。ワクチンキャリアは、当該分野で周知である:例えば、ウシ血清アルブミン(BSA)、ヒト血清アルブミン(HAS)およびキーホールリンペットヘモシアニン(HLH)。好ましいキャリアタンパク質であるロタウイスルVP6は、欧州特許公開第0259149号に開示される。
【0067】
ワクチンはまた、腸溶性投薬形態のような適切な経口キャリア中で、経口投与され得る。経口処方物としては、例えば、医薬品グレードのマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、サッカリンセルロースナトリウム(sodium saccharin cellulose)、炭酸マグネシウムなどのような通常使用される賦形剤が挙げられる。経口ワクチン組成物は、溶液、懸濁液、錠剤、ピル、カプセル、徐放性処方物または粉末の形態をとり得、約10%〜約95%の活性成分、好ましくは約25%〜約70%の活性成分を含む。経口ワクチンは、全身性免疫と組み合わせた粘膜免疫を惹起することに好ましくあり得、これは、病原体の胃腸管感染に対する防御に重要な役割を果たす。
【0068】
さらに、ワクチンは坐剤へと処方される。坐剤に関して、ワクチン組成物はポリアルカリグリコールまたはトリグリセリドのような従来の結合剤およびキャリアを含む。そのような坐剤は、約0.5%〜約10%(w/w)、好ましくは約1%〜約2%の範囲の活性成分を含む混合物から形成され得る。
【0069】
本発明のワクチン組成物を動物に投与するためのプロトコールは、本開示を考慮して当業者の範囲内である。当業者は、免疫原性フラグメントに対して抗体および/またはT細胞媒介性免疫応答を誘発するのに有効な用量でワクチン組成物の濃度を選択する。広い範囲において、この投薬量は、重要であると考えられていない。
【0070】
代表的に、ワクチン組成物は、好都合な体積のビヒクル(例えば、約1〜10cc)中にて約1から約1000μgの間のサブユニット抗原を送達する様式で投与される。好ましくは、単一免疫における投薬量は、約1から約500μgのサブユニット抗原、より好ましくは約5〜10から約100〜200μg(例えば、5〜200μg)のサブユニット抗原を送達する。
【0071】
投与の時期もまた、重要であり得る。例えば、一次接種は、好ましくは、必要な場合、後のブースター接種が続き得る。必要に応じてではあるが、二次のブースター免疫を、最初の免疫の数週間から数ヶ月間後に、動物に投与することもまた、好ましくあり得る。疾患に対する高レベルの防御の維持を確実にするためには、一定の間隔(例えば、数年毎に1回)で動物にブースター免疫を再投与することが有益であり得る。あるいは、最初の用量は経口投与され、続いて後に接種され得るか、またはその逆もあり得る。好ましいワクチン接種プロトコールは、慣用的なワクチン接種プロトコールの実験を介して確立され得る。
【0072】
本発明の組換えヘルペスウイルスはまた、本発明のワクチンによってワクチン接種された動物を、天然に存在する野生型伝染性ヘルペスウイルスまたは他の病原体によって感染された動物から区別する方法を提供し得る。これは、組換えヘルペスウイルスが、対応する病原体に対する防御免疫を与えるのに必要である上記のウイルス由来の限定数の抗原をコードする外来DNAを含むので、可能である。結果的に、これらの組換えヘルペスウイルスを用いてワクチン接種された宿主動物は、野生型伝染性ファルビキウス病ウイルス(bursal disease virus)、マレク病ウイルス、ニューカッスル病ウイルス、伝染性喉頭気管炎ウイルスまたは伝染性気管支炎ウイルスによって感染された宿主動物から、野生型ウイルスに通常存在する抗原が存在しないことによって、区別され得る。さらに、ヘルペスウイルスベクターが、免疫原性ポリペプチドをコードするそのゲノムの部分の欠失を含む場合、欠失された部分の産物に対してワクチン接種された動物からの免疫応答が欠失することによって、ワクチン接種された動物が示される。
【0073】
本発明はまた、遺伝子欠損を制御するための遺伝子治療が必要である動物に、遺伝子治療を提供するための方法を含み、この方法は、この動物に、生組換えヘルペスウイルス(これは、非欠失形態の上記遺伝子をコードする外来ヌクレオチド配列を含む)を、組換えウイルスベクターのゲノムが、上記の哺乳動物のゲノム中に組み込まれるかまたは独立して染色体外的に保持されるかして、標的器官または組織において目的の遺伝子の発現を与える条件下で、投与する工程を包含する。
【0074】
欠失遺伝子またはその部分を置換するためのこれらの種類の技術は、当業者によって使用される。例えば、Andersonら(米国特許第5,399,346号)は、遺伝子治療に関する技術を記載する。さらに、好都合な遺伝子治療に使用するために組み込まれ得る、外来DNA配列のヌクレオチド配列またはその部分の例としては、嚢胞性線維症膜貫通調節遺伝子(CFTR遺伝子(cystic fibrosis transmenbrane conductance regulator gene))、ヒトミニジストロフィン遺伝子、α−抗トリプシン遺伝子などが挙げられる。
【0075】
組換えヘルペスウイルスの構築、選択および精製に関する方法は、以下の実施例に詳述される。以下の実施例は、特定の好ましい実施形態および本発明の局面を示すために役立ち、本発明の範囲を制限するを解釈されるべきではない。
【実施例】
【0076】
(実施例1)
(マレク病ウイルス(MDV−1)ストックの調製)
マレク病ウイルスストックサンプルは、2mMグルタミン、100ユニット/ml ペニシリン、100ユニット/ml ストレプトマイシンを含むHAM’S F10および199培地の1:1混合物(これらの成分は、Sigmaまたは同等の供給者から得、そしてこれより後は、完全DME培地として呼ぶ)(1%ウシ胎仔血清を含む)中、0.01PFU/細胞の感染多重度で組織培養細胞を感染させることによって調製した。細胞変性効果が完了した後、培地および細胞を回収し、そして細胞を、臨床遠心分離機中、3000rpm、5分間でペレットにした。感染された細胞は、20%ウシ胎仔血清、10% DMSOを含む完全培地中に再懸濁し、そして−70℃で凍結保存した。
【0077】
(実施例2)
(マレク病ウイルス(MDV−1)DNAの調製)
マレク病ウイルスの全ての操作は、菌株GA5(ATCC #624)またはRispens CVI−988(Vineland Labs)を用いて行った。感染された細胞の細胞質からのMDVウイルスDNAの調製に関して、初代ニワトリ胚線維芽細胞を、細胞の過剰増殖前に、広範な細胞変性効果が起こるに十分なMOIで感染させた。全てのインキュベーションは、5% CO2空気中の加湿インキュベーターにおいて、39℃で行った。最高のDNA収量は、最大に感染されたが、不完全な細胞溶解を示す単層を収集することによって得た(代表的に5〜7日)。感染された細胞は、細胞スクレーパを用いて培地中に細胞をこすり取ることによって収集した。この細胞懸濁液を、GS−3ローター中、3000rpm、10分間、5℃で遠心分離した。
【0078】
生じたペレットを、冷PBS(20ml/ローターボトル)中に再懸濁し、そして冷やしながら3000rpmで10分間さらに遠心分離に供した。PBSをデカントした後、その細胞ペレットをRSB緩衝液(10mM Tris pH7.5、1mM EDTAおよび1.5mM MgCl2)の4ml/ローターボトル中に再懸濁した。NP40(Nonidet P−40;Sigma)を、時々混合しながら0.5%の最終濃度までサンプルに添加した。そのサンプルを、冷やしながら10分間、3000rpmで遠心分離し、核をペレットにし、そして細胞の細片を除去した。上清の流体を注意深く15mlのCorex遠心分離チューブに移した。EDTA(0.5M pH 8.0)およびSDS(ドデシル硫酸ナトリウム;20%保存)の両方を、それぞれ、最終濃度が5mMおよび1%までサンプルに添加した。100μlのプロテイナーゼ−K(10mg/ml;Boehringer Mannheim)をサンプル4ml当たりに添加し、混合し、そして45℃で1〜2時間インキュベートした。この期間の後、等量の飽和フェノールをサンプルに添加し、そして手で穏やかに混合した。このサンプルを臨床遠心分離機中、5分間、3000rpmで回転させ、相を分離させた。酢酸ナトリウム(保存溶液は3M pH5.2)を0.3Mの最終濃度で水相に添加し、そして核酸を2.5容量の冷無水エタノールを添加した後に−70℃で30分間沈澱させた。サンプル中のDNAは、HB−4ローター中5℃で、20分間、8000rpmで回転させることによってペレットにした。上清を注意深く除去し、そしてDNAペレットを25mlの80%エタノールを用いて1回洗浄した。DNAペレットを真空によって簡単に乾燥させ(2〜3分間)、そして50μl/感染された細胞のローターボトルのTE緩衝液(10mM Tris pH7.5、1mM EDTA)中に再懸濁した。代表的には、ウイルスDNAの収量は、5〜10μg/感染された細胞のローターボトルの間の範囲であった。全てのウイルスDNAは、約10℃で保存した。
【0079】
(実施例3)
(DNA配列決定)
DNA配列決定は、ABI PRISM Dye Terminator Cycle Sequencing Ready Reaction KitおよびAmplitaq DNAポリメラーゼ、FS(Perkin−Elmer;製造者らの指示書に従って)を用いた蛍光標識ジデオキシ配列決定反応をし、そして製造者らの指示書に従ってPerkin−Elmer/Applied Biosystems自動DNAシークエンサーModel 373A上で電気泳動することによって、実施した。dGTP混合物およびdITP混合物の両方を用いた反応を、圧縮の領域を明確にするために実施した。あるいは、圧縮領域は、ホルムアミドゲル上で分離させた。テンプレートは、二本鎖プラスミドサブクローンまたは一本鎖M13サブクローンであり、そしてプライマーは、配列決定される挿入物のちょうど外側のベクターか、または以前に得られた配列のいずれかに対して作製した。得られた配列を集め、そしてDNAStarソフトウェアを用いて比較した。
【0080】
(実施例4)
(分子生物学的技術)
細菌およびDNAの操作に関する技術(制限エンドヌクレアーゼを用いた消化、ゲル電気泳動、ゲルからのDNAの抽出、連結、キナーゼを用いたリン酸化、ホスファターゼを用いた処理、細菌培養物の増殖、DNAを用いた細菌の形質転換、および他の分子生物学的方法のような手順を含む)は、J.Sambrookら、Molecular Cloning A Laboratory Manual 第2版、Cold Spring Harbor Press、1989およびCurrent Protocols in Molecular Biology(1992)John Wiley & Son’s,Inc.によって記載される。記載される以外、これらは、少し改変して使用した。
【0081】
(実施例5)
(ポリメラーゼフィルイン反応)
DNAを、50mM Tris pH 7.4、50mM KCl、5mM MgCl2および400μMの4つのデオキシヌクレオチド各々を含む緩衝液中に再懸濁した。10ユニットのKlenow DNAポリメラーゼ(BRL)を添加し、そして反応を15分間室温で進行させた。次いで、このDNAをフェノール抽出し、そして上記のようにエタノール沈殿した。
【0082】
(実施例6)
(ポリメラーゼ連鎖反応を用いたクローニング)
ポリメラーゼ連鎖反応(PCR)を使用して、種々のDNAの操作に簡便な制限部位を導入した。使用した手順は、M.A.Innisら(PCR Protocols A Guide to Methods and Applications、84−91、Academic Press,Inc.,San Diego,1990)によって記載される。一般的に、増幅されたフラグメントは、サイズが500塩基対未満であり、そして増幅されたフラグメントの重要な領域は、DNA配列決定によって確認した。各場合に使用したプライマーは、以下の相同性ベクターの構築の説明に詳述する。
【0083】
(実施例7)
(感染された細胞の溶解物の調製)
25cm2フラスコまたは60mmペトリ皿中の、二次(secondary)ニワトリ胚線維芽細胞のコンフルエントな単層を、100μlのウイルスサンプルを用いて感染させた。細胞変性効果が完全になった後、培地および細胞を収集し、そして細胞を、臨床遠心分離機中、3000rpm、5分間でペレットにした。この細胞ペレットを250μlの破壊緩衝液(2% ドデシル硫酸ナトリウム、2% β−メルカプトエタノール)中に、再懸濁した。このサンプルを、氷上で30秒間超音波処理し、そして−20℃で保存した。
【0084】
(実施例8)
(ウエスタンブロット手順)
溶解物のサンプルおよびタンパク質標準物質を、Laemnli,U.K.(1970)Nature 277:680の手順に従ってポリアクリルアミドゲル上で泳動した。ゲルを電気泳動した後、このタンパク質を、Sambrookら(1989)に従って、トランスファーし、そして処理した。一次抗体は、Tris−塩化ナトリウムおよびアジ化ナトリウム(TSA:1lのH2O当たり6.61g Tris−HCl、0.97g Tris塩基、9.0gNaClおよび2.0g アジ化ナトリウム)中の5%脱脂粉乳を用いて1:100希釈した。二次抗体は、アルカリホスファターゼ結合体化し、そしてTSAを用いて1:1000希釈した。
【0085】
(実施例9)
(cDNAクローニング手順)
cDNAクローニングとは、最新技術の手順に従ってRNA分子をDNA分子に変換するために使用される方法をいう。出願人の方法は、(U.GublerおよびB.J Hoffman,Gene 25,263−269)に記載される。Bethesda Research Laboratories(Gaithersburg,Md.)は、出願人によって使用された手順と非常に類似し、そして我々の結果の再現に使用され得る試薬および手順のセットを含むcDNAクローニングキットを設計した。
【0086】
ウイルスmRNA種のクローニングに関して、ウイルスによる感染に感受性な宿主細胞株を、細胞当たり5〜10プラーク形成単位で感染させた。細胞変性効果が明白な場合であるが全て破壊する前に、培地を除去し、そして細胞を10mlの溶解緩衝液(4M グアニジンチオシアネート、0.1% アンチフォームA(antifoam A)、25mM クエン酸ナトリウム pH7.0、0.5%N−ラウリルサルコシン、0.1M β−メルカプトエタノール)中に溶解した。細胞溶解物を滅菌したDounce型ホモジナイザーに注ぎ、そして溶液が均質になるまで、氷上で8〜10分間ホモジナイズした。RNA精製に関して、8mlの細胞溶解液を、Beckman SW41遠心分離チューブ中、3.5mlのCsCl溶液(5.7M CsCl、25mM クエン酸ナトリウム pH7.0)上に穏やかに重層した。サンプルを、Beckman SW41ローター中、20℃で36000rpm、18時間遠心分離した。このチューブを氷上に置き、そしてチューブからの上清を注意深く吸引によって除去し、RNAペレットを乱さないようにした。ペレットを400μlのガラス蒸留水中に再懸濁し、そして2.6mlのグアニジン溶液(7.5M グアニジン−HCL、25mM クエン酸ナトリウム pH7.0、5mM ジチオトレイトール)に添加した。0.37容量の1M酢酸を添加し、続いて0.75容量の冷エタノールを添加し、そしてサンプルを−20℃に18時間置いてRNAを沈澱させた。沈澱物は、SS34ローター中、Sorvall遠心分離機で10分間、4℃、10000rpmで遠心分離することによって収集した。そのペレットを1.0mlの滅菌水に溶解し、13000rpmで再び遠心分離し、そして上清を保存した。RNAをそのペレットから上記のように0.5mlの滅菌水を用いてさらに2回再抽出し、そして、上清をプールした。0.1容量の2M 酢酸カリウム溶液をサンプルに添加し、続いて2倍の冷エタノールを添加し、そしてサンプルを−20℃で18時間置いた。沈澱したRNAをSS34ローター中、40℃、10分間、10000rpmで遠心分離することによって収集した。そのペレットを1mlの滅菌水に溶解し、そして濃度は、A260/280の吸収によって得た。このRNAは、−70℃で保存した。
【0087】
ポリアデニル化テイル(poly−A)を含むmRNAを、オリゴdTセルロース(Pharmacia#27 5543−0)を用いて選択した。3mgの総RNAを煮沸し、そして冷却し、結合緩衝液(0.1M Tris pH 7.5、0.5M LiCl、5mM EDTA pH 8.0、0.1% ドデシル硫酸リチウム)中、100mgオリゴdTセルロースカラムに適用した。保持されたポリ−A RNAを、溶出緩衝液(5mM Tris pH 7.5、1mM EDTA pH8.0、0.1% ドデシル硫酸ナトリウム)を用いてカラムから溶出した。このmRNAを結合緩衝液中、オリゴdTカラムに再び適用し、そして溶出緩衝液を用いて再び溶出した。このサンプルを200mM酢酸ナトリウムおよび2容量の冷エタノールを用いて−20℃、18時間で沈澱させた。このRNAは、50μlの滅菌水に再懸濁した。
【0088】
10μgのポリ−A RNAを、20mMの水酸化メチル水銀中、6分間、22℃で変性させた。β−メルカプトエタノールを75mMまで添加し、そしてそのサンプルを5分間、22℃でインキュベートした。第1鎖のcDNA合成のための0.25mlの反応混合物は、1μg オリゴ−dTプライマー(P−L Biochemicals)もしくは1μgの合成プライマー、28ユニットの胎盤リボヌクレアーゼインヒビター(Bethesda Research Labs #5518SA)、100mM Tris pH8.3、140mM KCl、10mM MgCl2、0.8mM DATP、dCTP、dGTPおよびdTTP(Pharmacia)、100マイクロキューリーの32P−標識dCTP(New England Nuclear #NEG−013H)ならびに180ユニットのAMVリバーストランスクリプターゼ(Molecular Genetics Resources #MG 101)を含んだ。この反応液を42℃で90分間インキュベートし、次いで20mM EDTA pH8.0を用いて終了させた。このサンプルを、等量のフェノール/クロロホルム(1:1)を用いて抽出し、そして2M 酢酸アンモニウムおよび2容量の冷エタノールを用いて−20℃、3時間で沈澱させた。沈澱および遠心分離後、このペレットを100μlの滅菌水に溶解した。サンプルを、緩衝液(100mM Tris pH 7.5、1mM EDTA pH 8.0、100mM NaCl)中、15ml G−100 Sephadexカラム(Pharmacia)上にロードした。抽出されたDNA画分の立ち上がり(leading edge)をプールし、そしてDNAを、体積が約100μlになるまで凍結乾燥することによって濃縮し、次いでDNAを上記のように酢酸アンモニウムおよびエタノールを用いて沈澱させた。
【0089】
第1鎖のサンプル全体を、50μg/ml dNTP、5.4ユニットのDNAポリメラーゼI(Boerhinger Mannheim #642−711)、および100ユニット/ml E.coli DNAリガーゼ(New England Biolabs #205)(総体積 50μl)を使用する以外はGublerおよびHoffman(前出)の方法に従う、第2鎖反応に使用した。第2鎖合成の後、そのcDNAをフェノール/クロロホルム抽出し、そして沈澱させた。DNAを10μlの滅菌水に再懸濁し、1μg RNase Aを用いて10分間、22℃で処理し、そして40mM Tris−アセテート pH 6.85中、アガロースゲル(シグマ Type IIアガロース)を介して電気泳動させた。そのゲルをエチジウムブロマイドを用いて染色し、そして予想のサイズの範囲のDNAをゲルから切除し、そして8mM Tris−アセテート pH 6.85中で電気溶出した。電気溶出したDNAを約100μlまで凍結乾燥し、そして酢酸アンモニウムおよびエタノールを用いて上記のように沈澱させた。このDNAを20μlの水に再懸濁した。
【0090】
オリゴdCテールを、クローニングを容易にするために、そのDNAに付加した。この反応は、そのDNA、100mMカコジル酸カルシウム(pH7.2)、0.2mM ジチオスレイトール、2mM CaCl2、80μmol dCTP、および25単位の末端デオキシヌクレオチジルトランスフェラーゼ(Molecular Genetic Resources #S1001)を、50μl中に含んだ。37℃で30分後、この反応を10mM EDTAで終了させ、そしてそのサンプルを、上記のようにフェノール/クロロホルム抽出し、そして沈殿させた。
【0091】
dCテール化DNAサンプルを、200μlの0.01M Tris pH7.5、0.1M NaCl、1mM EDTA pH8.0中で、オリゴdGテール(Bethesda Research Labs #5355 SA/SB)を含む200ngのプラスミドベクターpBR322に、65℃で2分間、次いで57℃で2時間、アニーリングさせた。D.Hanahan、Molecular Biology 166、557〜580、1983により記載されるように、新鮮なコンピテントE.coli DH−1細胞を調製し、そして200μlの細胞アリコート20個においてこのアニールしたcDNAサンプルの半分を使用して、形質転換した。形質転換した細胞を、10μg/mlテトラサイクリンを含むLブロス寒天プレート上に配置した。コロニーを、Ampscreen(Bethesda Research Labs #5537 UA)を使用して、アンピシリン遺伝子へのインサートの存在についてスクリーニングし、そして陽性コロニーを分析用に拾った。
【0092】
(実施例10)
(組換えマレク病ウイルスを作製するためのDNAトランスフェクション)
この方法は、以下の改変を加えた、KawaiおよびNishizawa、Mol.and Cell.Biol.4:1172〜1174(1984)のポリブレン−DMSO手順に基づく。組換えMDVウイルスの生成は、MDVウイルスDNAと、適切なヘルペスウイルスのクローン配列が隣接する所望の外来DNAを含むプラスミド相同性ベクターとの間の、相同組換えに依存する。トランスフェクションを、50%コンフルエントな一次ニワトリ胚線維芽(CFF)細胞の6cmプレート(Corning plastic)にて実行した。この細胞は、4μg/mlポリブレン(1×HBSS中4mg/mlのストック)を含む、CEF増殖培地(1×F10/199、5%ウシ胎仔血清、2%グルタミン、1%非必須アミノ酸、および2%ペニシリン/ストレプトマイシン)中に前日に配置した。CEF細胞への同時トランスフェクションのために、5μgのインタクトMDV DNAを、30μg/mlポリブレン(1×HBSS中4mg/mlのストック)を含む1mlのCEF培地中に懸濁した。次いで、このDNA−ポリブレン懸濁物(1ml)を、培地を吸引したCEF細胞の6cmプレートに添加し、そして39℃で30分間インキュベートした。このプレートをこの時間の間周期的に揺り動かし、接種物を再分布させた。この期間の後、4mlのCEF増殖培地を、プレートを洗浄するために直接添加し、そして39℃でさらに2.5時間インキュベートした。この時点で、この培地を各プレートから除去し、そして細胞に、1×HBSS中30%のDMSO(ジメチルスルホキシド、J.T.Baker Chemical Co.、Phillipsburg、NJ)2mlを用いて、室温で4分間ショックを与えた。この30% DMSOを注意深く除去し、そして単層を、室温にて、1×HBSSで1回洗浄した。次いで、細胞に、5mlのCEF増殖培地を添加した後、39℃でこの細胞をインキュベートした。翌日、培地を交換して、DMSOの最後の残渣まで取り除き、そして細胞増殖を刺激した。ウイルスからの細胞変性効果は、6日間以内に明らかになる。高力価ストック(80%〜90% CPE)の生成は、通常は、この日から1週間以内に行い得る。MDVストックサンプルを、20%ウシ胎仔血清、10% DMSOを含むCEF増殖培地中に感染した細胞を再懸濁することによって調製し、そして−70℃にて貯蔵した。
【0093】
(実施例11)
(β−ガラクトシダーゼを発現する組換えマレク病ウイルスについてのスクリーニング(BluogalおよびCPRCアッセイ)またはβ−グルクロニダーゼを発現する組換えマレク病ウイルスについてのスクリーニング(X−Glucアッセイ))
E.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子を組換えウイルスに組み込んだ場合、その組換え体を含むプラークを、簡単な2つの方法のうちの1つにより視覚化した。第1の方法において、化学物質BluogalTM(Life Sciences Technology、Bethesda、MD)を、プラークアッセイの間にアガロース重層物に組み込んだ(200μg/ml)。そして活性なβ−ガラクトシダーゼを発現するプラークは、青色に変化した。次いで、この青色プラークを新鮮なCEF細胞へと拾い、そしてさらなる青色プラーク単離により精製した。第2の方法において、CPRG(Boehringer Mannheim)を、プラークアッセイの間にアガロース重層物に組み込んだ(400μg/ml)。活性なβ−ガラクトシダーゼを発現するプラークは、赤色に変化した。次いで、この赤いプラークを新鮮なCEF細胞へと拾い、そしてさらなる赤色プラーク単離によって精製した。両方の場合において、代表的には、ウイルスを、3回〜4回のプラーク精製により精製した。
【0094】
E.coli β−グルクロニダーゼ(uidA)マーカー遺伝子を組換えウイルスに組み込んだ場合、この組換えウイルスを含むプラークを、発色性基質X−β−D−gluUA CHX(X−GLUC;5−ブロモ−4−クロロ−3−インドキシル−β−D−グルクロン酸、シクロヘキシルアンモニウム塩;Biosynth AG;Switzerland)を、プラークアッセイの間にアガロース重層物に組み込んだ(200μg/ml)。そして活性なβ−グルクロニダーゼを発現するプラークは、青色に変化した。次いで、その青色プラークを新鮮なCEF細胞へと拾い、そしてさらなる青色プラーク単離によって精製した。
【0095】
(実施例12)
(黒色プラークアッセイを使用する、組換えマレク病ウイルスにおける外来DNA配列発現についてのスクリーニング)
組換えMDVウイルスにより発現される外来抗原の発現を分析するために、CEF細胞の単層を組換えMDVに感染させ、栄養アガロース培地を重層し、そして39℃で4〜5日間インキュベートする。一旦プラークが発色すると、アガロース重層物をディッシュから除去し、単層を1×PBSでリンスし、室温で10分間100%メタノールで固定し、そして細胞を風乾する。プレートをPBSで再水和した後、一次抗体を適切な希釈物までPBSで希釈し、そして室温にて2時間〜一晩、細胞単層とともにインキュベートする。次いで、非結合抗体を、PBSによる室温にて3回の洗浄によって細胞から除去する。アルカリホスファターゼ結合体化二次抗体をPBSで希釈し、そして室温にて2時間細胞とともにインキュベートする。次いで、非結合二次抗体を、室温にてPBSを用いて3回細胞を洗浄することによって除去する。次いで、単層を発色緩衝液(100mM Tris pH9.5/100mM NaCl/5mM MgCl2)中でリンスし、次いで、新たに調製した基質溶液(発色緩衝液中、0.3mg/ml Nitro Blueテトラゾリウム+0.15mg/ml 5−ブロモ−4−クロロ−3−インドリルホスファターゼ)とともに、10分間〜一晩、室温でインキュベートする。最後に、反応を、基質溶液をTE(10mM Tris、pH7.5/1mM EDTA)に置換することによって停止する。正しい抗原を発現するプラークは、黒色に染まる。
【0096】
(実施例13)
(1型マレク病ウイルスのオープンリーディングフレームUL54.5に外来DNAを挿入されたプラスミド)
プラスミド440−29.2を、1型マレク病ウイルス(MDV−1)に外来DNAを挿入する目的のために構築した。このプラスミドは、1型マレク病ウイルスの、約2596塩基対のBamHI「M」ゲノムフラグメント(配列番号1)を含む。このBamHI「M」フラグメント中の3つのオープンリーディングフレームは、UL54(ICP27)ORF(配列番号1の1位〜1353位)、以前に未同定であったORF(従ってUL54.5と名付けた)(配列番号1の2187位〜1483位)およびUL55(配列番号1の2459位〜2593位)のヘルペスウイルスホモログである(表1および表2を参照のこと)。MDV−1 UL54(ICP27)遺伝子に広がるDNA配列(1492塩基対)は、公開されている(Virology 1994年10月;204(1):242〜50)。MDV ICP27(HSV−1 ICP27に対する有意な類似性に基づく)は、1419ヌクレオチド長であり、かつ473アミノ酸(54.5kDa)をコードする。UL55 ORFは短縮型であり、そしてこのタンパク質の最初の49アミノ酸のみしか含まない。UL5とUL55との間に位置する潜在的ORFを同定した。このORFは、705塩基対長であり、そしておそらく、サイズが235アミノ酸のタンパク質をコードする。このUL54.5アミノ酸配列を使用してのタンパク質データベースのBLAST検索によって、MDV−2中に類似の遺伝子を同定した(Virology 1994年5月15日;201(1):142〜6)。このMDV−2遺伝子が、1型ウマヘルペスウイルスの第1のオープンリーディングフレーム(ORF−1)と低い相同性を共有することが注目された。MDV−1およびMDV−2由来のUL54.5タンパク質の類似性指標は、170アミノ酸のコンセンサス長にわたって、63%である。UL54.5は、UL54およびUL55に対して、反対の方向で転写される。
【0097】
UL54.5 ORFは、非必須であり、そしてこのORF中またはこれらのORF間の遺伝子間領域に、外来DNAが挿入される。この領域中のすべての制限部位が、外来DNAのための挿入部位として有用である。唯一ではないこの領域中の制限酵素部位は、その部位を唯一の制限酵素認識配列へと変換するDNAリンカーの挿入によって変更する。好ましくは、外来DNAの挿入のために使用する制限酵素部位は、この2596塩基対のBamHI「M」ゲノムフラグメント中約ヌクレオチド2596にある、NdeI部位である。この挿入部位は、UL54.5 ORF中で、このオープンリーディングフレームのアミノ酸4とアミノ酸5との間にある。このプラスミドベクターは、pSP64(Promega)の約3045塩基対のBamHI制限フラグメントに由来した。フラグメント1は、1型マレク病ウイルスの約2596塩基対の、BamHI「M」フラグメントである。プラスミド440−29.2を使用して、組換えマレク病ウイルス中への外来DNAの挿入のための相同性ベクターを作製した。
【0098】
(実施例14)
(1型マレク病ウイルスのオープンリーディングフレームUL54.5に伝染性喉頭気管炎ウイルスDNAが挿入されたプラスミド)
プラスミド980−85.1を、組換え1型マレク病ウイルス(MDV−1)中に外来DNAを挿入する目的のために構築した。このプラスミドは、MDV−1 DNAが隣接する、ILTウイルスgD遺伝子およびgI遺伝子、ならびにE.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子を組み込む。これらの遺伝子を、合成DNAリンカーを使用してPacI部位へと変換した唯一のNdeI部位に挿入した。この外来DAN配列の上流は、MDV DNAの約422塩基対フラグメントである。この外来DNA配列の下流は、MDV DNAの約2174塩基対フラグメントである。ILTウイルスgD遺伝子およびgI遺伝子ならびにE.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子の転写の方向は、MDVのUL54 ORFおよびUL55 ORFの転写の方向と反対である。このプラスミドを、組換えマレク病ウイルスを生成するためのDNAトランスフェクション(実施例10)、ならびにβ−ガラクトシダーゼを発現する組換えマレク病ウイルスについてのスクリーニング(BluogalおよびCPRCアッセイ)またはβ−グルクロニダーゼを発現する組換えマレク病ウイルスについてのスクリーニング(X−Glucアッセイ)(実施例11)に従って使用する場合、外来DNA配列をコードするDNAを含むウイルスが生じる。ILTV gD遺伝子およびgI遺伝子は、それらの遺伝子自身の各内因性ILTVプロモーターからの重複転写物として発現され、そしてそれらの自身の内因性ポリアデニル化シグナルを共有し、そしてE.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子は、PRV gXプロモーターから転写され、それにPRV gXポリアデニル化シグナルが続く。
【0099】
プラスミド980−85.1を、標準的組換えDNA技術を使用して、以下の供給源由来の制限フラグメントを合成DNA配列と結合することによって、構築した。ILT gD、gI、およびE.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子を、合成DNAリンカーを使用してPacI部位へと変換した唯一のNdeI部位に、相同性ベクター440−29.2へのカセットとして挿入した。このプラスミドベクターは、pSP64(Promega)の約3045塩基対のHindIII制限フラグメントに由来した。フラグメント1は、MDV BamHI制限フラグメントMの、約418塩基対のBamHI〜NdeI制限サブフラグメントである。フラグメント2は、ILT Asp 7181ゲノムフラグメント#8(10.6kb)の、約3556塩基対のSalI〜HindIII制限サブフラグメントである。フラグメント3は、PRV BamHI制限フラグメント#10の、約413塩基対のSalI〜BamHI制限サブフラグメントである。フラグメント4は、プラスミドpJF751(11)の、約3010塩基対のBamHI〜PvuII制限サブフラグメントである。フラグメント5は、PRV BamHIゲノムフラグメント#7の、約754塩基対のNdeI〜SalI制限サブフラグメントである。フラグメント6は、MDV BamHI制限フラグメントMの、約2174塩基対のNdeI〜BamHI制限サブフラグメントである。
【0100】
(実施例15)
(1型マレク病ウイルスのオープンリーディングフレームUL54.5にニューカッスル病ウイルスDNAが挿入されたプラスミド)
プラスミド980−46.74を、組換え1型マレク病ウイルス(MDV−1)中に外来DNAを挿入する目的のために構築した。このプラスミドは、MDV DNAが隣接する、E.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子ならびにニューカッスル病ウイルス(NDV)F遺伝子を組み込む。E.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子ならびにNDV F遺伝子を、合成DNAリンカーを使用してPacI部位へと変換した唯一のNdeI部位に、相同性ベクター440−29.2(実施例13)へのカセットとして挿入した。
【0101】
この外来DAN配列の上流は、MDV DNAの約422塩基対フラグメントである。この外来DNA配列の下流は、MDV DNAの約2174塩基対フラグメントである。E.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子ならびにNDV F遺伝子の転写の方向は、MDVのUL54 ORFおよびUL55 ORFの転写の方向と反対である。このプラスミドを、組換えマレク病ウイルスを生成するためのDNAトランスフェクション(実施例10)、ならびにβ−ガラクトシダーゼを発現する組換えマレク病ウイルスについてのスクリーニング(BluogalおよびCPRCアッセイ)またはβ−グルクロニダーゼを発現する組換えマレク病ウイルスについてのスクリーニング(X−Glucアッセイ)(実施例11)に従って使用する場合、外来DNA配列をコードするDNAを含むウイルスが生じる。NDV F遺伝子は、HCMV最初期プロモーターの制御下にあり、それにHSV TKポリアデニル化シグナルが続く。E.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子は、PRV gXプロモーターから転写され、PRV gXポリアデニル化シグナルが続く。
【0102】
プラスミド980−85.1を、標準的組換えDNA技術を使用して、以下の供給源由来の制限フラグメントを合成DNA配列と結合することによって、構築した。E.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子ならびにNDV F遺伝子を、合成DNAリンカーを使用してPacI部位へと変換した唯一のNdeI部位に、相同性ベクター440−29.2(実施例13)へのカセットとして挿入した。このプラスミドベクターは、pSP64(Promega)の約3045塩基対のHindIII制限フラグメントに由来した。フラグメント1は、MDV BamHI制限フラグメントMの、約418塩基対のBamHI〜NdeI制限サブフラグメントである。フラグメント2は、PRV BamHI制限フラグメント#10の、約413塩基対のSalI〜BamHI制限サブフラグメントである。フラグメント3は、プラスミドpJF751の、約3010塩基対のBamHI〜PvuII制限フラグメントである。フラグメント4は、PRV BamHI制限フラグメント#7の、約754塩基対のNdeI〜SalI制限サブフラグメントである。フラグメント5は、HCMVゲノムのXbaI Eフラグメントの、約1191塩基対のPstI〜AvaII制限サブフラグメントである。フラグメント6は、全長NDV F cDNAクローン(B1株)の、約1812塩基対のBamHI〜PstI制限フラグメントである。フラグメント7は、HSV BamHI制限フラグメントQの、約784塩基対のSmaI〜SmaI制限サブフラグメントである。最後のフラグメントは、MDV BamHI制限フラグメント「M」の、約2174塩基対のNdeI〜BamHI制限サブフラグメントである。
【0103】
(実施例16)
(1型マレク病ウイルスのオープンリーディングフレームUL43に挿入された外来DNAを有するプラスミド)
プラスミド962−80.1を、1型マレク病ウイルス(MDV−1)中に外来DNAを挿入する目的のために構築した。これは、1型マレク病ウイルスのBamHI「B」ゲノムフラグメント内に含まれる、およそ3212塩基対のSacI〜BglIIサブフラグメント(配列番号2)を含む。3212塩基対のSacI〜BglIIサブフラグメント内の3つのオープンリーディングフレームは、UL42 ORF(配列番号2の35位〜1144位)、UL43(配列番号2の1304位〜2566位)、およびUL44(gC)(配列番号2の2786位〜3220位)のヘルペスウイルスホモログである(図3および図4を参照のこと)。MDV−1 UL44(gC)遺伝子およびプロモーター領域にわたるDNA配列(732塩基対)が、公開されている(Virus Genes 3,125〜137(1989))。MDV−2 UL42、UL43およびUL44遺伝子のDNA配列が、公開されている(J.Gen.Virol.79(Pt 8),1997〜2001(1998))。MDV−1およびMDV−2由来のUL42タンパク質の類似性指数は、369アミノ酸のコンセンサス長に対して74パーセントである。MDV−1およびMDV−2由来のUL43タンパク質の類似性指数は、401アミノ酸のコンセンサス長に対して54パーセントである。MDV−1およびMDV−2由来のUL44タンパク質の類似性指数は、145アミノ酸のコンセンサス長に対して45パーセントである。MDV−1 UL43 ORFは、非必須であり、そして外来DNAを、このORF内に、またはORF間の遺伝子間領域中に挿入する。この領域内の任意の制限部位は、外来DNAの挿入部位として有用である。唯一ではないこの領域内の制限酵素部位を、DNAリンカーの挿入によって変更する。このDNAリンカーによって、この部位が唯一の制限酵素認識配列に変換される。好ましくは、外来DNAの挿入に使用される制限酵素部位は、1型マレク病ウイルスのBamHI「B」ゲノムフラグメント内に含まれる3212塩基対のSacI〜BglIIサブフラグメントにおけるおよそヌクレオチド1386でのXhoI部位である。この挿入部位は、UL43 ORF内に、このオープンリーディングフレームのアミノ酸29と30との間にある。このプラスミドベクターは、pSP64(Promega)のおよそ3045塩基対のBamHI制限フラグメントに由来した。フラグメント1は、1型マレク病ウイルスのBamHI「B」ゲノムフラグメント内に含まれる、およそ3212塩基対のSacI〜BglIIサブフラグメントである。プラスミド962−80.1を使用して、組換えマレク病ウイルス中への外来DNAの挿入のための相同ベクターを作製した。
【0104】
(実施例17)
(1型マレク病ウイルスのオープンリーディングフレームUL43に挿入された感染性喉頭気管炎ウイルスDNAを有するプラスミド)
プラスミド980−85.22を、組換え1型マレク病ウイルス(MDV−1)に外来DNAを挿入する目的のために構築した。これは、ILTウイルスgD遺伝子およびgI遺伝子ならびにE.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子を、MDV−1 DNAに隣接して組み込んでいる。これらの遺伝子を、合成DNAリンカーを使用して、PacI部位に変換した唯一のXhoI部位に挿入した。外来DNA配列の上流は、MDV DNAのおよそ1386塩基対のフラグメントである。外来DNA配列の下流は、MDV DNAのおよそ1826塩基対のフラグメントである。ILTウイルスgD遺伝子およびgI遺伝子ならびにE.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子の転写方向は、MDV UL42およびUL43 ORFと同じ転写方向である。このプラスミドを、組換えマレク病ウイルスを作製するためのDNAトランスフェクション(実施例10)およびβ−ガラクトシダーゼを発現する組換えマレク病ウイルスのスクリーニング(BluogalおよびCprgアッセイ)、またはβ−グルクロニダーゼを発現する組換えマレク病ウイルスのスクリーニング(X−Glucアッセイ)(実施例11)に従って使用する場合、外来DNA配列をコードするDNAを含むウイルスが生じる。ILTV gD遺伝子およびgI遺伝子は、それら独自のそれぞれの内因性ILTVプロモーターから重複転写物として発現され、そしてそれら独自の内因性ポリアデニル化シグナルを共有し、そしてE.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子を、PRV gXプロモーターから転写して、そしてPRV gXポリアデニル化シグナルが続く。
【0105】
プラスミド980−85.22を、標準的な組換えDNA技術を利用して、以下の供給源からの制限フラグメントを合成DNA配列と連結することによって構築した。ILT gD、gI、およびE.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子を、唯一のXhoI部位(これは、合成DNAリンカーを使用してPacI部位に変換された)にて、相同ベクター962−80.1に、カセットとして挿入した。このプラスミドベクターは、pSP64(Promega,Madison,WI)のおよそ3045塩基対のHindIII制限フラグメントに由来した。フラグメント1は、1型マレク病ウイルスのBamHI「B」ゲノムフラグメント内に含まれる、およそ1386塩基対のSacI〜XhoI制限サブフラグメントである。フラグメント2は、ILTAsp718Iゲノムフラグメント番号8(10.6キロベース)のうちの、およそ3556塩基対のSalI〜HindIII制限サブフラグメントである。フラグメント3は、PRV BamHI制限フラグメント番号10のうちの、およそ413塩基対のSalI〜BamHI制限サブフラグメントである。フラグメント4は、プラスミドpJF751のうちの、およそ3010塩基対のBamHI〜PvuII制限フラグメントである(11)。フラグメント5は、PRV BamHI制限フラグメント番号7のうちの、およそ754塩基対のNdeI〜SalI制限サブフラグメントである。フラグメント6は、1型マレク病ウイルスのBamHI「B」ゲノムフラグメント内に含まれる、およそ1826塩基対のXhoI〜BglII制限サブフラグメントである。
【0106】
(実施例18)
(オープンリーディングフレームUL43に挿入された感染性喉頭気管炎ウイルスDNAを有する組換え1型マレク病ウイルス)
S−MDV−006は、3つの外来DNA配列を発現する1型マレク病ウイルスである。ILTウイルスgDおよびgIの遺伝子、ならびにE.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子を、唯一のPacI制限部位(1型マレク病ウイルスのBamHI「B」ゲノムフラグメント内に含まれる、およそ3212塩基対のSacI〜BglIIサブフラグメントのUL43 ORF中の独特なXhoI制限部位に挿入されたPacIリンカー)に挿入した。ILTV gD遺伝子およびgI遺伝子は、それらの独自のそれぞれの内因性ILTVプロモーターから重複転写物として発現され、そしてそれら独自の内因性ポリアデニル化シグナルを共有し、そしてE.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子を、PRV gXプロモーターから転写させ、そしてPRV gXポリアデニル化シグナルが続く。S−MDV−006は、S−MDV−002(MDV−1;CVI−988 Rispens)由来であった。組換えマレク病ウイルスを作製するためのDNAトランスフェクション手順(実施例10)において、相同ベクター980−85.22およびウイルスS−MDV−002を利用して、これを達成した。同時トランスフェクションストックを、β−グルクロニダーゼによってスクリーニングした(X−Glucアッセイ)(実施例11)。赤色プラーク精製の最終的な結果は、S−MDV−006と命名される組換えウイルスであった。このウイルスを、実施例11に記載されるように青色プラークアッセイによってモニターされるβ−ガラクトシダーゼの発現、純度、および複数の継代によるインサートの安定性についてアッセイした。最初の4回の精製後に、観察された全てのプラークは、青色であり、このことは、このウイルスが純粋であり、安定であり、そして外来DNA配列を発現していることを示す。
【0107】
S−MDV−006を、黒色プラークアッセイを使用する組換えマレク病ウイルスにおける外来DNA配列の発現についてのスクリーニング(実施例12)を使用して、ILT特異的抗原の発現についてアッセイした。ポリクローナルニワトリ抗ILT血清(SPAFAS)は、S−MDV−006プラークと特異的に反応し、そしてS−MDV−002陰性コントロールプラークと特異的に反応しなかったことが示された。全てのS−MDV−006の観察されたプラークは、ポリクローナル血清と反応し、このことは、このウイルスが、ILT外来DNA配列を安定に発現していたことを示す。本明細書に記載されるアッセイを、CEF細胞で行い、このことにより、CEF細胞は、MDV組換えワクチンの産生に適切な培養基であることが示された。
【0108】
S−MDV−006は、ILT gDタンパク質およびgIタンパク質を発現し、かつILT感染におけるワクチンとして有用である、組換え1型マレク病ウイルスである。S−MDV−006もまた、ILT gDタンパク質およびgIタンパク質の発現に有用である。
【0109】
(実施例19)
(1型マレク病ウイルスのオープンリーディングフレームUL43に挿入されたニューカッスル病ウイルスDNAを有するプラスミド)
プラスミド980−60.02を、組換え1型マレク病ウイルス(MDV−1)に外来DNAを挿入する目的のために構築した。これは、E.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子およびニューカッスル病ウイルス(NDV)F遺伝子を、MDV DNAに隣接して組み込んでいる。このE.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子およびNDV F遺伝子を、合成DNAリンカーを使用してPacI部位に変換された唯一のXhoI部位に、相同性ベクター962−80.1へのカセットとして挿入した。
【0110】
外来DNA配列の上流は、MDV DNAのおよそ1386塩基対のフラグメントである。外来DNA配列の下流は、MDV DNAのおよそ1826塩基対のフラグメントである。E.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子およびNDV F遺伝子の転写方向は、MDV UL42およびUL43 ORFと同じ転写方向である。このプラスミドを、組換えマレク病ウイルスを作製するためのDNAトランスフェクション(実施例10)およびβ−ガラクトシダーゼを発現する組換えマレク病ウイルスのスクリーニング(BluogalおよびCprgアッセイ)、またはβ−グルクロニダーゼを発現する組換えマレク病ウイルスのスクリーニング(X−Glucアッセイ)(実施例11)に従って使用する場合、外来DNA配列をコードするDNAを含むウイルスが生じる。NDV F遺伝子は、HCMV最初期プロモーターの制御下にあり、そしてHSV TKポリアデニル化シグナルが続く。E.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子を、PRV gXプロモーターから転写させ、そしてPRV gXポリアデニル化シグナルが続く。
【0111】
プラスミド980−85.1を、標準的な組換えDNA技術を利用して、以下の供給源からの制限フラグメントを合成DNA配列と連結することによって構築した。E.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子およびNDV F遺伝子を、合成DNAリンカーを使用してPacI部位に変換した唯一のXhoI部位にて、相同ベクター962−80.1中にカセットとして挿入した。このプラスミドベクターは、pSP64(Promega)のおよそ3045塩基対のHindIII制限フラグメントに由来した。
【0112】
フラグメント1は、1型マレク病ウイルスのBamHI「B」ゲノムフラグメント内に含まれる、およそ1386塩基対のSacI〜XhoI制限サブフラグメントである。
【0113】
フラグメント2は、PRV BamHI制限フラグメント番号10のうちの、およそ413塩基対のSalI〜BamHI制限サブフラグメントである。フラグメント3は、プラスミドpJF751のうちの、およそ3010塩基対のBamHI〜PvuII制限フラグメントである。フラグメント4は、PRV BamHI制限フラグメント番号7のうちの、およそ754塩基対のNdeI〜SalI制限サブフラグメントである。フラグメント5は、HCMVゲノムXbaI Eフラグメントのうちの、およそ1191塩基対のPstI〜AvaII制限サブフラグメントである。フラグメント6は、全長NDV F cDNAクローン(B1株)のうちの、およそ1812塩基対のBamHI〜PstI制限フラグメントである。フラグメント7は、HSV BamHI制限フラグメントQのうちの、およそ784塩基対のSmaI〜SmaI制限サブフラグメントである。最後のフラグメントは、1型マレク病ウイルスのBamHI「B」ゲノムフラグメント内に含まれる、およそ1826塩基対のXhoI〜BglII制限サブフラグメントである。
【0114】
(実施例20)
(オープンリーディングフレームUL43に挿入されたニューカッスル病ウイルスDNAを有する組換え1型マレク病ウイルス)
S−MDV−004は、2つの外来DNA配列を発現する1型マレク病ウイルスである。ニューカッスル病ウイルス融合物(F)の遺伝子、およびE.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子を、唯一のPacI制限部位(1型マレク病ウイルスのBamHI「B」ゲノムフラグメント内に含まれるおよそ3212塩基対のSacI〜BglIIサブフラグメントのUL43 ORF中の唯一のXhoI制限部位に挿入したPacIリンカー)中に挿入する。このNDV F遺伝子は、HCMV最初期プロモーターの制御下にあり、そしてE.coli β−ガラクトシダーゼ(lacZ)マーカー遺伝子を、PRV gXプロモーターから転写させ、そしてPRV gXポリアデニル化シグナルが続く。S−MDV−004は、S−MDV−002(MDV−1;CVI−988 Rispens)に由来する。組換えマレク病ウイルスを作製するためのDNAトランスフェクション(実施例10)において、相同ベクター980−60.02およびウイルスS−MDV−002を利用して、これを達成する。同時トランスフェクションストックを、β−ガラクトシダーゼを発現する組換えマレク病ウイルスのスクリーニング(BluogalおよびCprgアッセイ)、またはβ−グルクロニダーゼを発現する組換えマレク病ウイルスのスクリーニング(X−Glucアッセイ)(実施例11)によってスクリーニングした。赤色プラーク精製の最終的な結果は、S−MDV−004と命名される組換えウイルスであった。このウイルスを、実施例11に記載されるように青色プラークアッセイによってモニターされるβ−ガラクトシダーゼの発現、純度、および複数の継代によるインサートの安定性についてアッセイする。最初の4回の精製後に、観察された全てのプラークは、青色であり、このことは、このウイルスが純粋であり、安定であり、そして外来DNA配列を発現していることを示す。
【0115】
S−MDV−004を、黒色プラークアッセイを使用する組換えマレク病ウイルスにおける外来DNA配列の発現についてのスクリーニング(実施例12)を使用して、NDV特異的抗原の発現についてアッセイする。NDV Fに特異的なモノクローナル抗体は、S−MDV−004プラークと特異的に反応し、そしてS−MDV−002陰性コントロールプラークと特異的に反応しないことが示される。全てのS−MDV−004の観察されたプラークは、ポリクローナル血清と反応し、このことは、このウイルスが、NDV F遺伝子を安定に発現していることを示す。本明細書に記載されるアッセイを、CEF細胞で行い、このことにより、CEF細胞は、MDV組換えワクチンの産生に適切な培養基であることが示される。
【0116】
S−MDV−004は、NDV Fタンパク質を発現し、NDV感染におけるワクチンとして有用である、組換え1型マレク病ウイルスである。S−MDV−004もまた、Fタンパク質の発現に有用である。
【0117】
(実施例21)
(1型マレク病ウイルスのオープンリーディングフレームUL7、ならびに/あるいはオープンリーディングフレームUL8およびUL7の間に挿入された外来DNAを有するプラスミド)
プラスミドを、1型マレク病ウイルス(MDV−1)中に外来DNAを挿入する目的のために構築する。これは、1型マレク病ウイルスのBamHI「G」ゲノムフラグメント内に含まれる、およそ7316塩基対のサブフラグメント(配列番号3)を含む。7316塩基対のサブフラグメント内の5つのオープンリーディングフレームは、UL9 ORF(配列番号3の1位〜425位)、UL8(配列番号3の439位〜2748位)、UL7(gC)(配列番号3の3699位〜2782位)、UL6(配列番号3の5704位〜3536位)、およびUL5(配列番号3の5772位〜7316位)のヘルペスウイルスホモログである(図5および図6を参照のこと)。ORF UL8およびUL7(配列番号3の2749位〜2781位)と、ORF UL6と重複しないOFR7の部分(配列番号3の2782位〜3535位)との間の領域は、ウイルス複製に必須ではなく、そして変異ウイルスおよび/または組換えウイルスを作製するために使用され得る。
【0118】
本発明の多くの改変およびバリエーションは、当業者に明らかなように、その精神および範囲から逸脱することなくなされ得る。本明細書中に記載される特定の実施形態は、例示のみの目的で提供され、そして本発明は、添付の特許請求の範囲が権利を付す均等物の全範囲とともに、このような特許請求の範囲によってのみ制限されるべきである。
[配列表]
【数1】
【数2】
【数3】
【数4】
【数5】
【数6】
【数7】
【数8】
【特許請求の範囲】
【請求項1】
本明細書中に記載の組換えヘルペスウイルス。
【請求項1】
本明細書中に記載の組換えヘルペスウイルス。
【図1】

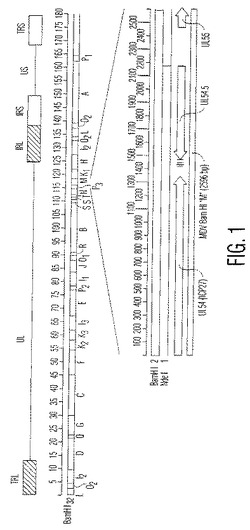
【図2】

【図3】
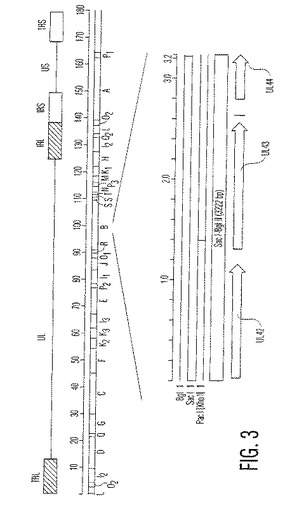
【図4】
【図5】
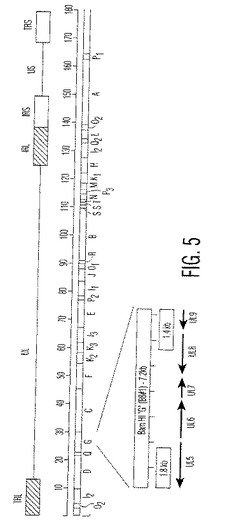
【図6】

【図2】
【図3】
【図4】
【図5】
【図6】
【公開番号】特開2010−187679(P2010−187679A)
【公開日】平成22年9月2日(2010.9.2)
【国際特許分類】
【出願番号】特願2010−63199(P2010−63199)
【出願日】平成22年3月18日(2010.3.18)
【分割の表示】特願2000−611660(P2000−611660)の分割
【原出願日】平成12年4月7日(2000.4.7)
【出願人】(503442097)シェーリング−プラウ・リミテッド (47)
【住所又は居所原語表記】Weystrasse20,Lucerne6,CH−6000,Switzerland
【Fターム(参考)】
【公開日】平成22年9月2日(2010.9.2)
【国際特許分類】
【出願日】平成22年3月18日(2010.3.18)
【分割の表示】特願2000−611660(P2000−611660)の分割
【原出願日】平成12年4月7日(2000.4.7)
【出願人】(503442097)シェーリング−プラウ・リミテッド (47)
【住所又は居所原語表記】Weystrasse20,Lucerne6,CH−6000,Switzerland
【Fターム(参考)】
[ Back to top ]