
新規のプラスチドターゲティング核酸配列、新規のβ−アミラーゼ配列、刺激反応性プロモーター、およびその使用
【課題】新規の葉緑体に標的化される、新規のβ−アミラーゼ配列(ctβ−アミラーゼ)、新規の葉緑体標的化核酸配列、および新規のβ−アミラーゼ配列及び、光または糖の刺激により独立して刺激される誘導性プロモーターもまた提供する。
【解決手段】これらの配列を用いて植物を形質転換する方法、ならびに形質転換植物細胞、形質転換植物、およびその種子、ならびに前記配列を含むキメラ遺伝子。植物中のデンプン量の改善、ならびにデンプン生合成経路または分解経路の遺伝子の標的化、病害抵抗性または有害生物抵抗性、あるいは刺激による遺伝子発現の変化した形質転換植物細胞、形質転換植物、およびその種子。
【解決手段】これらの配列を用いて植物を形質転換する方法、ならびに形質転換植物細胞、形質転換植物、およびその種子、ならびに前記配列を含むキメラ遺伝子。植物中のデンプン量の改善、ならびにデンプン生合成経路または分解経路の遺伝子の標的化、病害抵抗性または有害生物抵抗性、あるいは刺激による遺伝子発現の変化した形質転換植物細胞、形質転換植物、およびその種子。
【発明の詳細な説明】
【技術分野】
【0001】
植物中でデンプンが合成および分解される正確な機構は、このプロセスに関与すると推定される多数の酵素が単離および特徴付けられているのにも関わらず、知られていない。
【0002】
デンプンは、日中、葉の葉緑体に蓄積され、夜間、植物がエネルギーおよび生合成を必要とするのを満たすために使用される。このいわゆる一過性デンプンが動員される機構は十分に理解されていないが、合成酵素および分解酵素の活性を協調して調節するのに違いない。葉組織における主要な分解経路は、加リン酸分解活性および加水分解活性、特に、α−グルコシダーゼ(E.C.3.2.1.3.)(NielsonおよびStitt,1997)が関与することが考えられる。
【0003】
デンプンはまた、種子、果実、および塊茎などの貯蔵器官中のアミロプラストに蓄積される。この場合、デンプンは長期間にわたって貯蔵され、デンプンの動員に付随して、貯蔵器官組織の退化ならびにデンプン分解活性および加リン酸分解活性の増加が起こる。しかしながら、デンプン代謝回転は貯蔵器官のアミロプラストでも起こっていることを示唆する証拠がある(Sweetloveら,1996)。これもまた、合成酵素活性および分解酵素活性を協調して調節することを必要とする。
【0004】
葉緑体およびアミロプラストは両方ともプロプラスチドに由来し、従って、それぞれ葉および貯蔵器官におけるデンプン合成の場であることの他に、共通の特徴が多数ある。葉緑体は、アミロプラストおよび他の型のプラスチドに変わることができる(ThomsonおよびWhatley,1980)。
【0005】
デンプンは、2種類の多糖:α−1,4−グリコシド結合により連結したグリコシル単位からなる直鎖であるアミロースと、α−1,6グリコシド結合により結合した多数のα−1,4−ポリグルカン直鎖からなるアミロペクチンの混合物である。
【0006】
デンプン合成に関与する酵素は、ADPGピロホスホリラーゼ(E.C.2.7.7.21)、デンプン合成酵素(E.C.2.4.1.21)、および分枝酵素(E.C.2.4.1.18)である。ADPGピロホスホリラーゼは基質ADPGの供給を担い、この分子は、デンプン合成酵素(α−1,4結合)および分枝酵素(α−1,6結合)の共同作用により結合されるグルコースモノマー供与体として働く。
【0007】
デンプン粒の不溶性結晶構造は、幅の広い、らせん状の、枝分かれしたアミロペクチン分子と、空間を充填する直線状アミロース分子とが密に束ねられることにより形成されると考えられる。
【0008】
α−アミラーゼ(E.C.3.2.1.1)、イソアミラーゼ(E.C.3.2.1.68)、β−アミラーゼ(E.C.3.2.1.2)、α−グルコシダーゼ(E.C.3.2.1.3)、デンプンホスホリラーゼ(E.C.2.4.1.1)、および不均化酵素(E.C.2.4.1.25)を含む、多種のデンプン分解酵素活性が報告されている。これらの酵素活性の多くは植物において複数形態で存在し、デンプン合成に関与すると考えられているものもある。恐らく、ある程度、全ての酵素がデンプン動員プロセスに関与しているが、それらの正確な役割および可能な相互作用はまだ決定されていない。様々な酵素の役割を考える難しさは、植物におけるデンプン分解を主に担うと考えられる2つの酵素活性:デンプンホスホリラーゼとアミラーゼを言及することで最もよく例示される。
【0009】
デンプンホスホリラーゼは、α−1,4−グルカンからのグルコース−1−リン酸の可逆的放出を触媒する。2つの形態のデンプンホスホリラーゼが植物組織に見出される。Pho1すなわちL型はプラスチド内にあり、マルトデキストリンに対する親和性が高い。Pho2すなわちH型はサイトゾルにあり、グリコーゲンなどの大きな、非常に枝分かれしたポリグルカンに対する親和性が高い。プラスチド型Pho1酵素は、デンプン動員に関与する有望な候補であるが、この葉酵素活性がアンチセンスにより阻害されても、トランスジェニックジャガイモ植物の葉におけるデンプン蓄積には影響しなかった(Sonnewaldら,1995)。別の研究では、細胞質Pho2がアンチセンスにより阻害されると、トランスジェニックジャガイモ塊茎の発芽行動に影響したが、デンプン蓄積および分解には影響しなかった(Duwenigら、1997)。
【0010】
アミラーゼには2つの主要なグループがあり、両方とも、アミロースおよびアミロペクチンのα−1,4−グルコシド結合を加水分解する。α−アミラーゼは非末端結合に対してランダムに作用するが、β−アミラーゼは、ポリグルカン鎖の非還元末端から始まるマルトース単位を放出するように作用する。植物細胞のアポプラスト空間におけるα−アミラーゼの亜細胞位置は、この酵素が正常に分泌されたことを反映していると考えられる。しかしながら、多数のα−アミラーゼタンパク質のアミノ末端シグナル配列がプラスチド膜ではなくER膜を通るタンパク質移動を示すという知見にもかかわらず(Chenら,1994)、イネ(Chenら,1994)およびテンサイ(Liら,1992)などの多数の植物では、この酵素は葉緑体およびアミロプラスト内にも位置する。イネα−アミラーゼ遺伝子のプロモーターおよびシグナル配列を細菌GUS遺伝子と融合し、アグロバクテリウム媒介形質転換を用いてイネ、タバコ、およびジャガイモに導入した研究(Chenら,1994)では、発現したGUS融合タンパク質は最初に小胞体に輸送され、次いで、トランスジェニック細胞からなる懸濁培養物の培地に出されることが証明された。α−アミラーゼは未変性デンプン分子を分解することが多数の研究で示されている。
【0011】
対照に、インビトロ研究から、他の酵素により未変性デンプン粒が前消化されなければ、β−アミラーゼは未変性デンプン粒を分解しないことがわかっている。活性β−アミラーゼを欠いているか、または微量の活性しかないライムギ変異体(Daussantら,1981)およびダイズ変異体(HildebrandおよびHymowitz,1981)はそれぞれ、見たところ正常な成長および発育を示す。さらに、β−アミラーゼレベルが著しく減少したトランスジェニックシロイヌナズナ植物は、ひどい成長異常を示さない(Mitaら,1997)。植物におけるβ−アミラーゼの正確な生理学的役割を明らかにする試みは、亜細胞位置に関する不確かなデータにより妨げられている。ある研究(Kakefudaら,1986)によりエンドウ葉緑体には2種類のβ−アミラーゼが存在することが報告されたが、ソラマメ、オオムギ、コムギ、ダイズ、サツマイモ、およびエンドウなどの種に関係する大多数の研究では、全てではないがほとんどのβ−アミラーゼ活性は葉緑体外にある(Nakamuraら,1991)。この考えは、現在までクローニングされた全てのβ−アミラーゼ遺伝子が、アミノ末端葉緑体トランジットペプチド配列を欠くタンパク質をコードするということにより支持される。
【0012】
穀類では3つの型のβ−アミラーゼ:穎果成熟中に蓄積する内胚乳特異的形態;発芽中にイネおよびトウモロコシのアリューロン細胞においてデノボ合成される形態(Wangら,1996;1997);ならびに栄養器官に遍在するβ−アミラーゼが記載されている。シロイヌナズナでは、この遍在形態はロゼット葉の総デンプン分解活性の約80%を占める。現在までクローニングされた他の全てのβ−アミラーゼと共通して、遍在シロイヌナズナ β−アミラーゼの遺伝子は、亜細胞ターゲティングシグナルを有するタンパク質をコードせず、従って、この酵素はサイトゾルに位置するらしい。
【0013】
植物の生存能力に悪影響を及ぼすことなく分解活性を除去することができるという多数の研究からの知見と、プラスチド外にあるデンプン分解酵素の亜細胞位置は驚くべきことである。β−アミラーゼ活性の予想される主要な最終産物、すなわちマルトースが、単離葉緑体においてデンプン分解産物として同定されたことを考慮すれば、β−アミラーゼ活性がプラスチドに明らかにないことは特に驚くべきことである(Peaveyら,1977)。さらに最近では、グルコースとマルトースが、デンプン動員プロセス中に、単離されたカリフラワー芽アミロプラストから出されることが示された(Neuhausら,1995)。
【0014】
葉または貯蔵器官のプラスチド中のデンプン量を操作できることは、植物デンプンを利用する様々な工業プロセスに非常に有利であろう。例えば、ジャガイモ塊茎のデンプン含有量を増加させようとして、E.coli ADPG PPase glgC16をトランスジェニックジャガイモ塊茎において過剰発現させると、デンプンへの炭素フラックスは増加するが、デンプンの正味蓄積は少ししか増加しないことが、以前に示された(Sweetloveら,1997)。過剰発現系統における酵素活性の分析から、ADPG PPaseが変化した以外に、アミラーゼ(特に、β−アミラーゼ)活性もまた変化したことがわかった。このデータから、glgC16タンパク質を過剰発現する塊茎におけるデンプン蓄積は、新たに合成されたデンプンの分解により妨げられる、すなわち、デンプンが代謝回転されることが示唆される。
【0015】
別の例では、麦芽製造工程中のデンプン利用率は、植物、特に、貯蔵器官中の分解酵素活性の型および量と密接に相関する。前記作物の分解能が増加すると、穀粒の麦芽形成、あるいは塊茎または他の貯蔵器官に由来するデンプンからアルコールへの変換がより効率的かつ生産的になる。
【0016】
貯蔵器官に存在するデンプンの型は、ADPGピロホスホリラーゼ、デンプン合成酵素、分枝酵素、および存在する分解酵素の形態および活性による。様々な酵素間の相互作用もまた重要である。
【0017】
植物において新規のデンプンを産生することへの関心がかなりある。なぜなら、この新規デンプン産生により、食品、製紙、製薬、接着剤、油、および繊維などの様々な産業で使用する前にデンプンを加工および改良する費用が削減されるからである。以下の例は、どのくらいデンプン加水分解活性が、インビボでのデンプン構造の改変に重要であるのかを示す。
【0018】
トウモロコシ穀粒にsugaryl変異があると、デンプンのα−1,6−グルコシル結合を加水分解する脱分枝酵素がなくなることが示されている(Jamesら,1995)。この変異は、アミロペクチン濃度と、非常に枝分かれのあるグルコポリサッカライドであるフィトグリコーゲンの蓄積の低下をもたらす。
【0019】
エンドウでは、マルトースで始まりマルトヘプトースまで連続グルコース単位を付加する短いオリゴ糖分子は、アミロース合成を担う主要酵素であると一般に認められているデンプン粒結合型デンプン合成酵素I(GBSSI)(Denyerら,1996)の活性を特異的に刺激することが示されている(例えば、van der Leijら,1991;Hyltonら,1995;Ainsworthら,1993)。マルト−オリゴ糖の供給制御によるGBSSI活性の操作は、最近の特許(WO97/16554)の主題であり、分解酵素、すなわち、α−アミラーゼ、β−アミラーゼ、不均化酵素、脱分枝酵素、およびデンプンホスホリラーゼを導入することにより、マルト−オリゴ糖濃度を増加することができ、従って、デンプン中のアミロースとアミロペクチンの比を増加することができることを示唆する。特許WO97/16554はまた、これらの酵素のプラスチド型アイソフォームがクローニングされたことについて述べている。しかしながら、上記のように、現在まで単離されたβ−アミラーゼ遺伝子は、タンパク質ターゲティング配列を有するβ−アミラーゼ酵素をコードせず、さらに、α−アミラーゼが初めからプラスチドに標的化されたことについては疑問がある(Chenら,1994;Chanら,1994)。WO97/16554のもっと後になって、プラスチドターゲティング配列を付加するための適切なβ−アミラーゼcDNA配列の操作が言及される。
【0020】
貯蔵器官中のデンプンの産業用途に加えて、葉中のデンプン量は、作物の農学上、非常に重要である。デンプンは、昼間、葉において、光合成中に固定された炭素から合成される。デンプンは葉緑体に貯蔵され、夜間に分解されて、植物のエネルギー源および代謝中間体になる。ソース−シンク関係が制御される機構は現在未知であるが、葉プラスチド中のデンプンの量および利用率が、植物生産性(バイオマスおよび収穫量)にかなり影響を及ぼすことは明らかである。
【0021】
葉中のデンプン量はまた、葉が主要な植物商品である作物(例えば、タバコ)に重要である。デンプン含有量は、喫煙した時のタバコの最終的なフレーバーに影響を及ぼすことが知られている。タバコ葉中のデンプン量を操作する手段の条項は、タバコ産業が関心を持つ可能性がある。
【0022】
本発明者らは、本明細書中で初めて、新規のターゲティング配列によりプラスチドに標的化される新規のβ−アミラーゼ酵素(今後、葉緑体標的化(ct)β−アミラーゼとして知られる)をコードするcDNAの単離について述べる。この全コード配列の単離は驚くべきことである。なぜなら、β−アミラーゼは、プラスチドから細胞質への移動または膜破壊により、さらに小さなポリグルカン断片を放出したデンプンの加水分解のみに関与すると一般に考えられていたからである。この酵素がプラスチドに存在するので、ct β−アミラーゼは、葉緑体にある一過性デンプンとアミロプラストにある貯蔵デンプンの分解に関与している意外な可能性がある。
【0023】
葉緑体とアミロプラストとの特徴の類似性(ThomsonおよびWhatley,1980)は本発明と関連している。なぜなら、葉緑体標的化ポリペプチドに由来するトランジットペプチドが異種ポリペプチドをアミロプラストに運ぶことができ、逆もまた同様であることが示されたからである。例えば、トウモロコシデンプン粒結合型デンプン合成酵素に由来するトランジットペプチドは、E.coli β−グルクロニダーゼ(GUS)タンパク質に融合された場合、GUSタンパク質をアミロプラストだけではなく葉緑体にも運ぶ(KlosgenおよびWeil,1991)。
【0024】
さらに、本発明者らは、シロイヌナズナにおけるct−Bmy遺伝子の発現、およびトランスジェニックタバコにおけるct−Bmyプロモーター:GUS融合の発現は、光およびスクロースにより独立して調節することができることを示す。このことは、シロイヌナズナ ATβ−Amyの密接に結びついた光および糖誘導反応を考えると、驚くべきことである(Mitaら,1995)。
【0025】
本発明は、本明細書では配列番号1として知られ、1〜294のヌクレオチドであり、かつ前記ヌクレオチド中に、さらなるコード配列を植物プラスチドに標的化することができる配列を有する核酸配列、または開示された配列である配列番号1と少なくとも65%以上相同であり、かつ同じ標的化能力を有する配列を提供する。
【0026】
前記核酸配列は、約94のアミノ酸残基をコードすることが好ましく、約85のアミノ酸残基をコードすることがより好ましい。
【0027】
本発明はまた、本明細書では配列番号2として知られ、1〜1642のヌクレオチドであり、かつ前記ヌクレオチド中に、β−アミラーゼをコードすることができる配列を有する核酸配列、または配列番号2内の開示された配列と少なくとも65%以上相同であり、かつ同じコード能力を有する配列を提供する。
【0028】
本発明はまた、本明細書では配列番号3として知られ、1〜1953のヌクレオチドであり、かつ前記ヌクレオチド中に、葉緑体標的化β−アミラーゼをコードすることができる配列を有する核酸配列、または配列番号3内の開示された配列と少なくとも65%以上相同であり、かつ同じコード能力を有する配列を提供する。
【0029】
相同配列として、中程度にストリンジェントな条件下で(65℃の2×SSCで洗浄)、配列番号1、配列番号2、または配列番号3にハイブリダイズする配列もまた挙げられる。
【0030】
前記核酸配列はmRNAまたはcDNAの配列が好ましいが、ゲノムDNAでもよい。
【0031】
本発明はまた、デンプン生合成経路または分解経路の酵素の活性を植物において増加または減少させる方法を提供する。前記方法は、プラスチドターゲティング配列をコードする核酸配列とデンプン生合成経路または分解経路の酵素のコード配列とを含むキメラ遺伝子を植物ゲノムに安定に組み込ませるステップと、改変したゲノムを有する植物を再生するステップを含む。
【0032】
本発明はまた、タンパク質または酵素を植物プラスチドに標的化する方法を提供する。前記方法は、プラスチドターゲティング配列をコードする核酸配列とタンパク質または酵素のコード配列とを含むキメラ遺伝子を植物ゲノムに安定に組み込ませるステップと、改変したゲノムを有する植物を再生するステップを含む。前記タンパク質または酵素は、1以上の下記の群:脂質合成、光合成、アミノ酸代謝、窒素固定、炭素固定、または炭水化物ポリマー合成の経路であるか、あるいは前記植物にある特徴を付与することができる。前記特徴は、1以上の下記の群:除草剤抵抗性および有害生物抵抗性(例えば、真菌抵抗性、細菌抵抗性、またはウイルス抵抗性を含む)から選択される。
【0033】
本発明はまた、プロモーターと、プラスチドターゲティング配列をコードする核酸コード配列と、ターミネーターとを含むキメラ遺伝子を有する植物を提供する。前記配列は、デンプン生合成経路または分解経路の酵素のコード配列を植物プラスチドに標的化することができる。
【0034】
本発明は、産物の発現を指向することができる核酸配列をさらに提供する。前記産物は、前記核酸配列に作動可能に連結されたコード配列によりコードされる。前記核酸配列は、本明細書では配列番号8として知られるか、あるいは配列番号8と少なくとも65%相同であり、かつ配列番号8と実質的に同じ機能を有する。前記核酸配列は刺激反応性であり、前記産物の発現レベルは、前記核酸配列に適用された刺激に応じて変化する。
【0035】
本発明は、産物の発現レベルを変化させる方法をさらに提供する。前記産物は、植物において産物の発現を指向することができる核酸配列に作動可能に連結されたコード配列によりコードされる。前記方法は、産物の発現を指向することができる核酸配列を含むキメラ遺伝子を植物ゲノムに安定に組み込ませるステップを含む。前記産物は、前記核酸配列に作動可能に連結されたコード配列によりコードされる。前記核酸配列は、配列番号8の配列を実質的に有するか、または配列番号8と少なくとも65%相同であり、かつ配列番号8と実質的に同じ機能を有し、刺激反応性である。
【0036】
前記刺激は、光および/または様々なレベルの糖があることか、またはないことが好ましい。あるいは、前記刺激は、発育段階により制御される刺激である。
【0037】
前記糖は、1以上のスクロースまたはグルコースであることが有利である。
【0038】
前記糖はスクロースであることが好ましい。
【0039】
前記誘導性プロモーターまたは植物において産物の発現を指向することができる核酸配列は、光をうけていないが糖が存在する条件下で作動可能であるか、または糖は存在しないが光をうけている条件下で作動可能であることが有利である。光をうけていないが糖が存在する植物組織は、地下器官またはシンク器官が適切であり得る。地下器官は、例えば、塊茎、根茎、または根でもよいが、他のシンク器官は若い葉または種子でもよい。
【0040】
糖は存在しないが光をうけている植物組織は、古い葉(糖は輸送されない)、花の部分、または発芽中の種子である。
【0041】
上記のDNA構造特徴を有する構築物およびキメラ遺伝子もまた本発明の態様である。
【0042】
上記のプラスチドターゲティング配列をコードする核酸配列とデンプン生合成経路または分解経路の酵素の核酸コード配列とを含むキメラ遺伝子を含む植物細胞、さらなるコード配列の発現を指向することができる核酸配列を含むキメラ遺伝子を含む植物細胞、または刺激反応性の上記の核酸配列とコード配列とを含むキメラ遺伝子を含み、前記コード配列の発現レベルが前記核酸配列に適用された刺激に応じて変化する植物細胞、本発明による1以上のキメラ遺伝子を含む形質転換植物の種子もまた、本発明の態様である。
【0043】
前記プラスチドターゲティング配列は配列番号1の配列であることが有利である。
【0044】
本発明の第1の態様では、上記の方法を用いて、デンプンが葉に蓄積されるか、または葉から動員されるように葉の代謝を改変することができる。このプロセスは、全体的に植物内のソース−シンク関係を改変する。これは、適切なプロモーターの制御下にある、ターゲティング配列とデンプン生合成経路または分解経路の酵素の核酸コード配列を作成することにより達成することができる。適切なプロモーターを選択すると、葉中のデンプン量が増加または減少した植物(例えば、これはタバコ産業で有用であり得る)が得られるか、あるいは、植物のソース−シンク関係を改善した後に、様々な他の植物組織(例えば、塊茎、果実、および根)のデンプン収量が変化する。
【0045】
本発明の本実施態様では、適切なプロモーターは、プラスチドターゲティング配列とデンプン生合成経路酵素または分解経路の酵素のコード配列との発現を植物全体で指向するか(いわゆる構成的発現)、あるいは特異的に葉に指向する。これらの変化は著しい影響を及ぼし、植物器官のデンプン含有量および/または収量がかなり変化する。
【0046】
全ての植物組織全体にわたって発現を指向することができる好ましいプロモーターは、カリフラワーモザイクウイルス35S遺伝子から得られるプロモーターである。葉での発現に好ましいプロモーターは、リブロースビスリン酸カルボキシラーゼ小サブユニット遺伝子またはエンドウプラストシアニン遺伝子から得ることができる。当業者であれば、構成的発現および葉特異的発現に適切な他のプロモーター(例えば、それぞれ、ノパリン合成酵素プロモーターおよびクロロフィルa/b結合タンパク質プロモーター)を認識するであろう。
【0047】
デンプン生合成経路または分解経路の酵素のコード配列またはその一部を、正常なリーディングフレーム方向(すなわち、センス)で配置してもよいし、逆のリーディングフレーム方向(すなわち、アンチセンス)で配置してもよい。センス、またはアンチセンス、またはコサプレッション技術(最後の技術は、DNAPにより欧州特許第0465572号および同第0647715号において説明される)を用いた、植物における酵素活性のアップレギュレーションまたはダウンレギュレーションを用いて、植物デンプンを変えることができる。
【0048】
本発明の第2の態様では、本発明の方法を用いて、デンプン含有量が増加し、かつ/またはデンプンが特定の工業プロセスに必要とされる適切な形態で得られるように、貯蔵器官におけるデンプン代謝を変えることができる。このようなプロセスとして、製紙;医薬品、繊維、染料、および建築用品の製造;天火焼き製品、乳製品、およびスナック食品の供給;缶詰製品、乾燥食品、またはインスタント食品の製造;穀粒の麦芽形成ならびにシロップおよびアルコールの製造が挙げられる。
【0049】
本方法の第1または第2の態様では、本方法のキメラ遺伝子で使用するために選択される酵素は、デンプン分解経路に由来する酵素(すなわち、デンプン分解酵素)でもよい。キメラ遺伝子は、葉緑体標的化β−アミラーゼ(以下、ct β−アミラーゼとして知られる)を含むことが有利であり、シロイヌナズナに由来するct β−アミラーゼ(以下、At ct β−アミラーゼとして知られる)(配列番号3を参照のこと)を含むことがより好ましい。ジャガイモ、タバコ、コムギ、トウモロコシ、およびオオムギなどの他の植物供給源から得ることができる、At ct β−アミラーゼと相同な配列もまた使用することができる。ハイブリダイゼーションまたはポリメラーゼ連鎖反応(PCR)技術によるクローニングの標準的な方法、例えば、Sambrookら(1989)に記載される技術およびInnesら(1990)に記載されるPCR技術などの分子クローニング技術を用いて、このような生物から配列を単離することができる。他のデンプン分解酵素として、α−アミラーゼ、不均化酵素、脱分枝酵素、デンプンホスホリラーゼ、α−グルコシダーゼ、および非プラスチド型β−アミラーゼが挙げられる。前記酵素の1以上のコード配列がプラスチドターゲティング配列との併用に適する。
【0050】
本方法の第2の態様では、植物の貯蔵器官に発現を指向する好ましいプロモーターを、例えば、以下のリストからの遺伝子から選択することができる:コムギ内胚乳高分子量グルテニン遺伝子、コムギ内胚乳α,β−グリアジン遺伝子、オオムギ内胚乳ホルデイン遺伝子、またはジャガイモ塊茎パタチン遺伝子。他の適切なプロモーターは当業者に周知である。
【0051】
本発明のどちらの態様でも、組織代謝の変化あるいはデンプンの型または特徴の変化を、本明細書に記載の誘導性プロモーター(配列番号8)を用いることによって、刺激反応性、すなわち、誘導性にすることができる。例えば、光誘導性の誘導性プロモーターは、Barnase(特許WO98/10081に例示される)などの遺伝子の誘導により花粉発育に影響を及ぼすように種子を操作するために、または果実追熟および種子発芽などの光依存性プロセスにおいて非光反応性遺伝子に影響を及ぼすために使用することができる。光誘導性プロモーターはまた、葉での二次代謝産生(例えば、アルカロイド産生)に影響を及ぼす遺伝子を活性化するために使用することができる。光誘導性プロモーターはまた、葉または他の光合成組織においてデンプン生合成酵素遺伝子を操作するために、あるいは塊茎を例えば暗所貯蔵から取り出した後に遺伝子を活性化するために使用することができる。糖誘導性の誘導性プロモーターは、例えば、発育中の塊茎または他の非光合成組織において、有害生物耐性遺伝子および/または収穫後の作物の品質に影響を及ぼすことが可能な遺伝子などの遺伝子を調節するために使用することができる。ジャガイモでは、胴枯れ病、黒脚病、および乾腐病に対する耐性遺伝子は特に有益であり、糖誘導性プロモーターを有する組換え遺伝子にクローニングできることが最も有利である。あるいは、糖誘導性の誘導性プロモーターは、組織培養プロセスにおいて選択マーカー遺伝子を発現させるために使用することができる。
【0052】
当業者は、欠失試験などの周知の技術を用いることにより、配列番号8から糖誘導性反応性エレメントおよび/または光誘導性反応性エレメントを容易に示すことができる。PweeおよびGray(1993)は、エンドウプラストシアニン遺伝子の作動可能な領域を決定するために、この遺伝子内での、マーカー遺伝子を用いたこのような欠失試験について記載している。
【0053】
遺伝子配列をクローニングし、それらを適切なキャリア(ベクターまたはプラスミドなど)に挿入するための、本明細書に記載の方法、あるいは、例えば、Sambrookら(1989)ならびにGelvinおよびStanton(1995)による実験マニュアルに記載の方法は、このような概念を実行に移す当業者に周知の技術である。上記の1以上のキメラ遺伝子を単独で導入してもよいし、1以上の他のキメラ遺伝子(例えば、1以上の上記の他の遺伝子)と共に導入してもよい。デンプン分解経路の酵素をコードする第1のキメラ遺伝子を用いる上記の実施態様の場合、第2のキメラ遺伝子は、例えば、デンプン生合成経路の酵素をコードする核酸配列(この配列もまた、適切なプロモーターおよび適切なターミネーターの制御下にある)を含んでもよい。第2のキメラ遺伝子のプロモーターおよび/またはターミネーターは、第1のキメラ遺伝子のプロモーターおよび/またはターミネーターと同じでもよいし、異なっていてもよい。デンプン生合成経路の酵素をコードする適切な配列は、スクロース合成酵素、ADPGピロホスホリラーゼ、デンプン合成酵素の核酸配列であり、分枝酵素、α−アミラーゼ、イソアミラーゼ、非プラスチド型β−アミラーゼ、α−グルコシダーゼ、デンプンホスホリラーゼ、および不均化酵素も含み得る。
【0054】
2以上のキメラ遺伝子を植物に導入する方法は説明されており、1つの分子内に結合したキメラ遺伝子を有するバイナリーベクターを構築すること、例えば、異なるキメラ遺伝子を含む異なるバイナリベクターを有する2以上の異なるアグロバクレリウム細胞を用いて同時形質転換するか、または、キメラ遺伝子を既に有する植物を第2の異なるキメラ遺伝子で形質転換(すなわち、再形質転換)することを含む。後者の場合、第2のキメラ遺伝子を導入した後の、トランスジェニック植物の選択方法は、第1のキメラ遺伝子の導入に用いた選択方法とは異ならなければならない。適切な選択マーカーとして、ハイグロマイシン、カナマイシン、スルホンアミド、およびBasta耐性マーカーが挙げられる。各植物が単一のキメラ遺伝子を含む2つの植物を交配するなど、生物学的方法もまた使用することができる。
【0055】
第1の酵素活性が有意に増加し、結果としてデンプン合成が変化した既に形質転換されている植物のデンプン含有量を変えるために、2つのキメラ遺伝子構築物を使用することができる。
【0056】
従って、本発明は、さらなる酵素をアップレギュレートまたはダウンレギュレートし、それにより再形質転換植物により産生されるデンプン量を増加または減少させるために、トランスジェニック植物(この植物では、遺伝的形質転換の結果としてある酵素活性が既に増加または減少している)において、さらなる酵素を変える方法をさらに提供する。
【0057】
第1の形質転換植物は、デンプン生合成経路の酵素活性が増加した植物が有利である。植物のデンプン含有量を増加させる試みの例は、ADPG−PPase遺伝子、例えば、glgC16(例えば、WO91/19806を参照のこと)で形質転換されたトランスジェニックジャガイモである。このような植物のデンプン増加量は比較的少ない。この第1の形質転換植物は、デンプン分解酵素のキメラ遺伝子(例えば、At ct β−アミラーゼを含むことが適する)で再形質転換されることが有利である。glgC16タンパク質は第1の形質転換塊茎で発現し、ADPG−PPase活性増加およびデンプンへの炭素フラックスの増加をもたらす。再形質転換塊茎においてキメラAt ct β−アミラーゼ遺伝子またはその一部を発現させると、ct β−アミラーゼ活性がダウンレギュレートされ(すなわち、コサプレッションまたはアンチセンス技術)、従って、デンプン蓄積が増加する。
【0058】
第2の酵素の発現は塊茎に指向されることが好ましい。At ct β−アミラーゼキメラ遺伝子の発現を塊茎に指向するのに適したプロモーターは、パタチン遺伝子に由来するプロモーターである。
【0059】
glgC16を発現する第1の形質転換ジャガイモ植物はカナマイシン耐性であり、従って、At ct β−アミラーゼキメラ遺伝子用のバイナリーベクター構築物は異なる耐性遺伝子を有し、例えば、スルホンアミド耐性遺伝子が適切である。ジャガイモ乾物が1%増加すると製品が4%増加するので、ジャガイモ塊茎のデンプン産生増加は、例えば、ポテトチップス製造業者に有益であろう。
【0060】
ポテトチップス製造はまた、本発明の別の利益を例示するのに役立つ。ジャガイモ塊茎を8℃以下の温度で貯蔵すると、デンプン分解により還元糖であるグルコースおよびフルクトースが蓄積する。このジャガイモをポテトチップスにするために油で揚げると、メイラード反応により還元糖はアミノ酸と反応して、製品は茶色になり、味がまずくなる。デンプン破壊を妨げ、従って、還元糖の蓄積を妨げるキメラ遺伝子をジャガイモ植物に導入することは、スナック食品産業に有益であろう。このキメラ遺伝子は、コサプレッションまたはアンチセンス構築物中に、適切なプロモーターおよびターミネーターで駆動される、ct β−アミラーゼのコード配列またはその配列の一部を含むことが好ましい。適切なプロモーターは、ジャガイモ塊茎パタチン遺伝子から得られる。有利にも、ct β−アミラーゼの代わりに、上記の他のどのデンプン分解酵素も使用できる。
【0061】
発育中の葉および発育中の塊茎での協調した発現が必要であれば、パタチンプロモーターがスクロース誘導性でもあるので(Rocha−Sosaら(1989))、配列番号8の誘導性プロモーターもまた前記構築物に使用することができる。同様に、配列番号1の葉緑体ターゲティングポリペプチド配列もまた、自身のターゲティング配列を欠き、かつプラスチドに指向されることが必要な他の任意の遺伝子と併用することができる。
【0062】
上記の例は、本発明を使用する可能な利益を例示するのに役立つ。当業者であれば、遺伝子の組み合わせと本発明を適用することができる植物とが多数あることを認識するであろう。
【0063】
遺伝子の組み合わせとして、好ましくは、ct β−アミラーゼと、1以上のスクロース合成酵素、ADPGピロホスホリラーゼ、デンプン合成酵素、分枝酵素、α−アミラーゼ、イソアミラーゼ、非プラスチド型β−アミラーゼ、α−グルコシダーゼ、デンプンホスホリラーゼ、および不均化酵素の遺伝子とが挙げられる。上記遺伝子の配列は当業者に周知である。あるいは、ct β−アミラーゼに由来するターゲティング配列を、1以上の上記遺伝子と併用してもよい。
【0064】
形質転換することができる植物のリストには、好ましくは、ジャガイモ、コムギ、トウモロコシ、オオムギ、トマト、イネ、エンドウ、ダイズ、ラッカセイ、キャッサバ、ヤムイモ、バナナ、およびタバコが含まれる。
【0065】
本発明を、一例として、シロイヌナズナに由来するct β−アミラーゼcDNAを単離し、このcDNAをタバコおよびジャガイモ植物に組み込ませる実施態様に関して説明する。刺激反応性プロモーターおよびトランスジェニック植物におけるその活性に関する実施例もまた示す。
【0066】
本発明を容易に実行に移すことができるために、一例として、以下の図面が参照されよう。
【0067】
配列表中では、以下の通りである。
【0068】
配列番号1は、コード配列を植物プラスチド、特に葉緑体に標的化することができる核酸である。
【0069】
配列番号2は、β−アミラーゼをコードする核酸である。
【0070】
配列番号3は、葉緑体標的化(ct)β−アミラーゼの完全配列である。
【0071】
配列番号4および5は、実施例3の増幅プロセスに用いられたプライマーである。
【0072】
配列番号6および7は、実施例4の増幅プロセスに用いられたプライマーである。
【0073】
配列番号8は、刺激反応性、特に光および/または糖に反応性の核酸である。
実施例1
シロイヌナズナ葉緑体標的化β−アミラーゼの単離および特徴
pBmy81内にあるcDNA挿入物の配列決定
【0074】
コスミドG16599(Bevanら,1988)内にある37kb シロイヌナズナ 4番染色体DNAフラグメントのヌクレオチド配列のBLASTNデータベース検索から、シロイヌナズナ、オオムギ、トウモロコシ、イネ、ダイズ、およびイネの葉緑体外β−アミラーゼと有意な相同性を有する遺伝子が存在することがわかった。この検索から、いくつかの3’末端EST配列もまた同定された。これらの1つであるEST 81E10T7(Newmanら,1995)(以下、pBmy81と呼ぶ)は約300ヌクレオチドにわたって同一であった。クローンEST 81E10T7は、Arabidopsis Biological Resource Center(ABRC)DNA Stock Center(Ohio University,USA)により供給された。pBmy81内のcDNA挿入物にまたがるBal31欠失サブクローンのネステッドセットを、蛍光色素標識ユニバーサルプライマーを用いた二本鎖PCRサイクルシークエンシング反応でのDNA鋳型として使用した。シークエンシング反応を、Apllied Biosystems Model373A自動化シーケンサーにより分析した。pBmy81の前記cDNA挿入物のヌクレオチド配列を配列番号3に示す。特許手続上の微生物の寄託の国際的承認に関するブタペスト条約により、Advanced Technologies(Cambridge)Limited,210 Cambridge Science Park,Cambridge CB4 4WAは、構築物pBmy81を、1998年8月4日にアクセッション番号NCIMB40964で、National Collection of Indurtrial and Marine Bacteria(NCIMB),23 St.Machar Street,Aberdeen Scotlandに寄託した。
推定葉緑体ターゲティングシグナルの同定
【0075】
pBmy81 cDNA挿入物は、5’末端に36の非翻訳ヌクレオチド、548アミノ酸からなるタンパク質をコードするオープンリーディングフレーム(ORF)、および232bpの3’非翻訳領域(UTR)を含む。pBmy81 cDNA挿入物によりコードされるタンパク質は、61kDaの推定分子量を有し、トウモロコシ、イネ、オオムギ、ダイズ、およびサツマイモに由来する植物葉緑体外β−アミラーゼと高いアミノ酸類似性を有する。しかしながら、pBmy81によりコードされるタンパク質は、葉緑体ターゲティングシグナルの特徴(すなわち、高含有量のセリン(16%)、スレオニン(10%)、および正に荷電したアミノ酸残基(15%))(BaierおよびDietz,1997)を有する独特のN末端伸長を含む点で、現在まで報告された他の全てのβ−アミラーゼとは異なる。葉緑体ターゲティングシグナルの特徴を識別する3つのドメインである、非荷電アミノ末端ドメイン、ヒドロキシル化アミノ酸リッチな中心ドメイン、および脂肪族β鎖を形成し得るカルボキシ末端ドメイン(SchatzおよびDobberstein,1996)を、このシグナル配列内で同定した。
pBmy81内のcDNA挿入物は、葉緑体標的化β−アミラーゼをコードする
【0076】
無傷の葉緑体を、パーコール段階勾配を用いて、50〜60gのエンドウシュート(Pisum sativum L.var Feltham First)から単離した。植物材料を成長させ、MouldおよびGray(1997a)の方法に従って葉緑体を単離した。
【0077】
pBmy81プラスミドを、NotIを用いた制限消化により線状化し、T7RNAポリメラーゼを用いてインビトロ転写した。放射性標識前駆体タンパク質を、本質的にMouldおよびGray(1997b)に記載のように、pBmy81 cDNAの転写物から35Sメチオニンおよび35Sシステインを含むコムギ胚翻訳系において合成した。
【0078】
放射性標識インビトロ翻訳産物の移入を、MouldおよびGray(1997b)に記載のように行った。移入インキュベーションの後、無傷の葉緑体を、氷上で30分間、サーモリシン(移入緩衝液中で最終濃度0.2mg/ml)を用いて処理し、次いで、プロテアーゼ反応を、50mM EDTAを移入緩衝液に添加することで止めた。葉緑体を、移入緩衝液に溶かした40%パーコールの緩衝物により再単離し、次いで、移入緩衝液で洗浄した(MouldおよびGray,1997b)。サーモリシン処理した葉緑体試料のアリコート(1/10)を分析のために採取し、残りを、本質的にSchnellおよびBlobel(1993)に記載のように分画した。サーモリシン処理葉緑体、ストロマ画分、チラコイド、およびサーモリシン処理チラコイドの試料を、SDS−PAGE、続いてクマシーブルー染色、染色タンパク質バンドのデンシトメータースキャニングにより定量した(リブロースビスリン酸カルボキシラーゼおよび集光性複合体タンパク質のサブユニットを標準として使用した)。等量のこれらの画分(パーコール勾配から回収した葉緑体の2%にほぼ等しい)と回収した内包膜および外包膜画分の505とを、SDS存在下での10%ポリアクリルアミドゲルによる電気泳動、続いてフルオログラフィーにより分析した。結果(図1)から、主要な翻訳産物(レーンTr)が約58kDaであることがわかる。単離した無傷のエンドウ葉緑体を、ATPの存在下で放射性標識タンパク質とインキュベートすると、約50kDaおよび48kDaのポリペプチドが産生された(レーンC)。外から添加したサーモリシンによる分解に対する耐性から、これらのポリペプチドは、放射性標識タンパク質移入の産物であることがわかる。無傷のサーモリシン処理葉緑体のストロマ、洗浄チラコイド、サーモリシン処理チラコイド、内包膜および外包膜への分画により、2つの放射性標識ポリペプチドがストロマ画分にあることが証明された。
実施例2
シロイヌナズナ ct β−アミラーゼ遺伝子のスクロースおよび光誘導
【0079】
明所でのct b−アミラーゼ誘導を証明するために、シロイヌナズナ生態型Landsberg植物を、18時間明期、6時間暗期、18℃で、温室において成長させた。5週間後、2トレイの芽生えを真っ暗闇に移し、2トレイの芽生えを連続光で成長させた。2日後、1トレイの暗所適応した芽生えと1トレイの明所で成長させた芽生えを総RNA単離のために使用し、それぞれの2つ目のトレイを、さらに3日間、連続光に暴露した。
【0080】
スクロース−明−暗を組み合わせた処理のために、Landsberg生態型の種子を表面滅菌し、1%スクロースを含むMS寒天培地上に置き、18時間明期、6時間暗期で培養室において成長させた。5週齢の芽生えを、滅菌蒸留水上に、あるいは5%スクロース水溶液またはグルコース水溶液上に移した。この芽生えを、3日間、連続して明所または暗所で維持した。総RNAを各試験の芽生えから調製し、Eggermontら(1996)により記載されるようにノザンブロット分析により分析した。ノザンを、FeinbergおよびVogelstein(1983)により記載されるように、32P−dCTPでランダム標識した後に、pBmy81内のゲル精製したcDNA挿入物を用いてプローブした。
【0081】
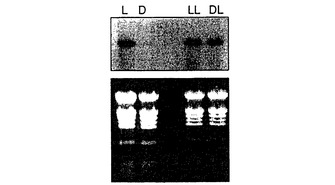
図2Aに示す結果から、ct β−アミラーゼ遺伝子転写物は光誘導性であることがわかる。
【0082】
図2Bに示す結果から、ct β−アミラーゼ転写物は、暗所において5%スクロースにより誘導され、それより少ない程度で5%グルコースにより誘導されることがわかる。この誘導は、明所で、糖の存在下でさらに高まった。これらの結果から、光および糖の効果は互いに独立していることがわかる。
実施例3
ct β−アミラーゼプロモーター−GUS融合の構築
【0083】
プロモーターフラグメントを、制限酵素消化により、コスミドG16599(Bevenら,1998)内にあるct β−アミラーゼ遺伝子から単離した。このプロモーター内にある便利な制限部位は、ヌクレオチド−1662bp位のHindIII(配列番号8のマイナス鎖の19179bpで開始する)、−1127bpのSalI、および−371bpのPstIであり、ct β−アミラーゼ開始メチオニンの下流にある+21bp位に位置するXhoI部位を用いて、3つの異なる長さのプロモーターとトランジットペプチド配列を単離した(翻訳開始メチオニンATGのAに+1の番号をつけた)。
【0084】
コスミドG16599(Bevenら,1998)内にあるct β−アミラーゼ遺伝子の294bp(配列番号1)フラグメントを、以下のオリゴヌクレオチドプライマー:
配列番号4
P1:(5’−ATT TCC TCG AGT TCT CTT ATC−3’)および
配列番号5
P2:(5’−cgg gAT CCC TGA CAT TGT TAC−3’)
を用いて増幅した。
【0085】
プライマーP1の下線を付けた塩基は、+21bp位に位置するXhoI部位を示す。プライマー2の小文字の塩基は、BamHI部位を作成するために付加したヌクレオチドを示す。
【0086】
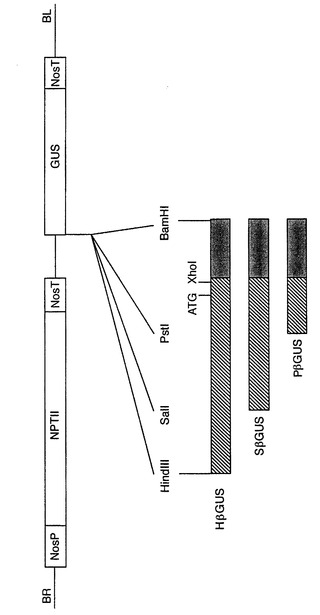
プロモーターフラグメントと、XhoIおよびBamHIで消化したPCR結合フラグメントと、HindIII−BamHI、SalI−BamHI、またはPstI−BamHIで消化したGUSベクターpBI101(Jeffersonら,1987)のトリプルライゲーションにより、キメラct β−アミラーゼプロモーター−GUS遺伝子を作成した(図3)。構築物を、それぞれ、HβGUS、SβGUS、およびPβGUSと名づけた。
【0087】
キメラ遺伝子構築物を、トリペアレンタルメイティング(Bevan,1984)によりアグロバクテリウム−ツメファシエンス LBA4404に移し、リーフディスク形質転換法(Horschら,1985)によりNicotiana tabacum var Samsumに導入した。
実施例3A
タバコ芽生えにおけるキメラシロイヌナズナ ct β−アミラーゼプロモーター−GUS遺伝子のスクロースおよび光誘導
【0088】
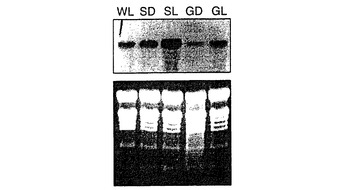
HβGUSおよびPβGUS構築物を含む植物は高レベルのGUS活性を発現し、この系統のF1芽生え子孫を用いて、キメラ遺伝子の光およびスクロース誘導性発現を調べた。F1タバコ種子を表面滅菌し、1%スクロースを含むMS寒天培地上に置き、18時間明期、6時間暗期で培養室において成長させた。2〜3週齢の芽生えを5%スクロース溶液または蒸留水上に移し、3日間、連続して明所または暗所で維持した。Jeffersonら(1987)に記載のように、10〜14個の芽生えプールからの総タンパク質抽出物を、蛍光基質4−メチルウンベリフェリル−グルクロニド(4−MUG)を用いてGUS活性について分析した。両構築物を用いた、スクロースの非存在下で連続光に暴露された芽生えのGUS活性レベルは、光の非存在下でスクロースに暴露された芽生えのGUS活性レベルと類似した(図4)。しかしながら、芽生えを連続光とスクロースとの両方に暴露すると、GUS活性レベルは約2〜3倍増加した。これらの結果は、光誘導性とスクロース誘導性が独立したプロセスであることを示した、ct b−アミラーゼ遺伝子自体を用いた実験からの結果と大まかに一致する。
【0089】
組織化学的GUS染色から、活性は2週齢芽生えの子葉で検出され、第1本葉または茎および根の活性は全くといってよいくらいないことがわかった。4週齢の芽生えでは、さらなるGUS活性が第1本葉全体で示され、茎でも示された。GUS染色は、木部放射組織間にある、および木部と茎内部師部を構成する師部維管束との間にある、葉緑体が豊富な柔組織(葉緑組織)細胞と特に関連していた。
実施例4
タバコおよびジャガイモの葉の形質転換に使用するためのct β−アミラーゼプラスミドの構築
【0090】
位置指定突然変異誘発を用いて、pBmy81プラスミドの2302bp位に位置するKpnI部位をBamHI部位に変換した。オリゴヌクレオチドプライマー:
配列番号6
P3:(5’−GCT GGT ACG CCT GCA GGA TCC GGT CCG GAA TTC CC−3’)および
配列番号7
P4:(5’−GGG AAT TCC GGA CCG GAT CCT GCA GGC GTA CCA GC−3’)
を設計し、Quick Change位置指定突然変異誘発キット(Promega)と共に使用した。プロトコールは、製造業者により概説された通りであった。
【0091】
完全長ct β−アミラーゼコード配列を、BamHI切断により突然変異pBmy81プラスミドから切り出し、次いで、GeneClean(BIO101)を用いて精製した。BamHIフラグメントを、供与体ベクターpDV35S(SK)V(図5を参照のこと)およびpDV02000(図6を参照のこと)のBamHI部位に連結した。pDV35S(SK)Vは、35S CaMVプロモーター−35Sターミネーターを有するpBluescript(Stratagene)からなり、類似する構築物が当該分野で周知である(例えば、Odellら,1985)。pDV02000は、1.4kbパタチンプロモーター−ノパリン合成酵素ターミネーターを有するpBluescriptからなる。当業者は、既知の配列から類似する構築物を作成することができる(例えば、Liuら,1990)。プロモーターに対してセンス方向とアンチセンス方向のコード配列を有するプラスミドを単離し、ct β−アミラーゼキメラ遺伝子を、供与体ベクターからバイナリーベクターpBinPlus(van Engelenら,1995)にサブクローニングした。プラスミドマップを図7〜10に示す。
実施例5
植物の形質転換または再形質転換
【0092】
ジャガイモ植物を、本質的にHorsch(1985)に記載のように、リーフディスク共培養法を用いて形質転換した。上記のバイナリーベクターを、エレクトロポレーション法を用いてアグロバクテリウム−ツメファシエンス LBA4404に移し、このアグテバクテリア培養物を、再生した植物が実施例4に記載のキメラ遺伝子を有するように形質転換に使用した。
【0093】
パタチンプロモーター−ct β−アミラーゼ−ノパリン合成酵素ターミネーターキメラ遺伝子バイナリープラスミドは、リーフディスク共培養法により、E.coli ADPG−PPase glgC16キメラ遺伝子を既に有するジャガイモ植物を形質転換するために使用することができる。
実施例6
AT ct β−アミラーゼのターゲティングペプチドを有するプラスミドの構築
【0094】
AT ct β−アミラーゼのプラスチドターゲティング配列は、配列番号1に等しい294bpフラグメント内に含まれる。PCR増幅または制限酵素消化を用いて、DNAフラグメントを実施例3に記載のプラスミドから単離することができる。すなわち、フラグメントは、35S CaMVプロモーター+プラスチドターゲティング配列またはパタチンプロモーター+プラスチドターゲティング配列からなる。キメラ遺伝子は、翻訳融合として、タンパク質または酵素のコード配列とトランジットペプチド配列とを連結することにより構築することができる。翻訳されたタンパク質はプラスチドに運ばれて、新規の活性をもたらすか、または代謝経路に影響を及ぼす。
参考文献
Ainsworth,C.,Clark,J.およびBalsdon,J.(1993).Plant.Mol.Biol.,12,67−82.
Baier,M.およびDietz,X.J.,(1997)Plant J.12,179−190
Bevan,M.W.(1984)Nucl.Acids.Res.,12,8711−8721
Bevan,M.W.ら(1998).Nature,391,485−488.
Chan,MT.,Chao,YC.およびYu,SM.(1994).J.Biol.Chem.,269,17635−17641.
Chen,NH.,Liu,LF.,Chen,YR.,Wu,HK.およびYu,SM.(1994).Plant J.,6,625−636.
Daussant,J.,Zbaszyniak,B.,Sadowaki,J.およびWiatroszak,I.(1981).Planta,151,176−179.
Denyer,K.,Clarke,B.,Hylton,C.,Tatge,H;およびSmith,A.M.(1996).Plant J.,10,1135−1143.
Duwenig,E.,Steup,M.,Willmitzer,L.およびKossmann,J.(1997).Plant J.,12,323−333.
Eggermont,K.,Goderis,I.J.およびBroekaert,W.F.(1996).Plant Mol.Biol.Rep.14,273−279.
Feinberg,A.P.およびVogelstein,B.(1983).Anal.Biochem.132,6−13.
Gelvin,S.B.およびSchilperoort,R.A.(1995).Plant Molecular Biology Manual.第2版.Kluwer Academic Publishers,The Netherlands.
Hildebrand,D.F.およびHymowitz,T.(1981).Physiol.Plant.,53,429−434.
Horsch,R.B.,Fry,J.E.,Hoffman,N.L.,Eichholtz,D.,Rogers,S.G.およびSwaley,R.T.(1985)Science,227.1229−1231.
Hylton,C.M.,Denyer,K.,Keeling,P.L.,Chang,MT.およびSmith,A.M.(1995).Planta,198,230−237.
Innes,M.A.,Gelfand,D.H.,Sninsky,J.J.およびWhite,T.J.(1990).PCR Protocols.Publisher:Academic Press.
James,M.G.,Robertson,D.S.およびMyers,A.M.(1995).Plant Cell,7,417−429.
Jefferson,R.A.,Kavanagh,T.A.およびBevan,M.W.(1987)EMBO,J.6.3901−3907.
Kakefuda,G.,Duke,S.H.およびHostak,M.H.(1986).Planta,168,175−182.
Klosgen,R.B.およびWeil,J.H.(1991)Mol.Gen.Genet.,225,297−304
Li,B.,Servaites,J.C.およびGeiger,D.R.(1992).Plant Physiol.,98,1277−1284.
Liu,X.J.,Prat,S.,Willmitzer,L.およびFrommer,W.B.(1990)Mol.Gen.Genet.,223,401−406.
Mita,S.,Suzuki−Fujii,K.およびNakarmura,K.(1995)Plant Physiol.107,895−904.
Mita,S.,Murano,N.,Akaike,M.およびNakamura,K.(1997).Plant J.,11,841−851.
Mould,R.M.およびGray,J.C.(1997a).In Cell Biology:A Laboratory Handbook,第2版,2巻(Cells,J.E.編).New York:Academic Press,81−86頁.
Mould,R.M.およびGray,J.C.(1997b).In Cell Biology:A Laboratory Handbook,第2版,2巻(Cells,J.E.編).New York:Academic Press,286−292頁.
Nakamura,K.,Ohto,M.,Yoshida,N.およびNakamura,K.(1991).Plant Physiol.,96,902−909.
Neuhaus,H.E.,Henrichs,G.およびSchiebe,R.(1995).Planta,194,454−460.
Newman,T.ら(1994).Plant Physiol.,106,1241−1255.
Nielson,T.H.,Deiting,U.およびStitt,M.(1997).Plant Physiol.,113,503−510.
Odell,J.T.Nagy,F.およびChua,N.H.(1985)Nature,313,810−812.
Peavey,D.G.,Steup,M.およびGibbs,M.(1977).Plant Physiol.,60,305−308.
Pwee,K−H.およびGray,J.C.(1993)Plant J.3,437−449.
Rocha−Sosa,M.,Sonnewald,U.,Frommer,W.,Stratmann,M.,Schell,J.およびWillmitzer,L.(1989)EMBO,8,23−29.
Sambrook,J.,Fritsch,E.F.およびManiatis,T.(1989).Molecular Cloning.Publisher:Cold Spring Harbour.
Schatz,G.およびDobberstein,B.(1996).Science,271,1519−1526.
Schnell,D.J.およびBlobel,G.(1993).J.Cell.Biol.,120,103−115.
Sonnewald,U.,Basner,A.,Greve,B.およびSteup,M.(1995).Plant.Mol.Biol.,27,567−576
Sweetlove,L.J.,Burrell,M.M.およびap Rees,T.(1996).Biochem.J.,320,493−498.
Thomson,W.W.およびWhatley,J.M.(1980)Ann.Rev.Plant Physiol.31,375−394.
van Engelen,F.A.,Molthoff,J.W.,Conner,A.J.,Nap,J−P.,Pereira,A.およびStiekema,W.J.(1995).Transgenic Res.4,288−290.
van der Leij,F.R.,Visser,R.G.F.,Ponstein,A.S.,Jacobsen,E.およびFeenstra,W.J.(1991).Mol.Gen.Genet.,228,240−248.
Wang,SM.,Lue,WL.およびChen,J.(1996).Plant Mol.Biol.,31,975−982.
Wang,SM.,Lue,WL.,Huang,HW.およびChen,J.(1997).Plant Physiol.,113,403−409.
【図面の簡単な説明】
【0095】
【図1】SDS−PAGEゲル、続いてフルオログラフィーにより示された放射性標識インビトロ移入翻訳産物の結果を示す図である。説明:分子量マーカー(レーンM);翻訳産物(レーンTr);移入インキュベーション後に再単離およびサーモリシン処理された葉緑体(レーンC);ストロマ画分(レーンS);洗浄したチラコイド(レーンT);サーモリシン処理チラコイド(レーンtT);内包膜画分(レーンI);外包膜画分(レーンO)。それぞれ、β−アミラーゼの推定前駆体(P)、中間体(I)、および成熟(M)形態。キロダルトン(K)。
【図2】シロイヌナズナ芽生えにおける、ct β−アミラーゼ転写物発現に対する光の効果ならびに光および糖の効果を示す図である。図2Aは、土壌で成長させ、2日連続光(L)、2日連続暗所(D)、2日連続光の後に3日連続光(LL)、または2日暗所の後に3日連続光(DL)に暴露された5週齢シロイヌナズナ植物の総RNAノザンブロット分析を示す。図2Bは、インビトロで成長させ、水上に移して3日連続光に暴露された(WL)、あるいは5%スクロース上に移して3日暗所(SD)または3日連続光(SL)に暴露された、あるいは5%グルコース上に移して3日暗所(GD)または3日連続光(GL)に暴露された、5週齢シロイヌナズナ植物の総RNAノザンブロット分析を示す。ノザンブロットを、放射性標識ct−Bmy cDNA挿入物とハイブリダイズさせ、オートラジオグラフィーにかけた(上部パネル)。対応する臭化エチジウム染色ホルムアルデヒド−アガロースゲルを下部パネルに示す。
【図3】実施例3で構築されるキメラct β−アミラーゼプロモーター−GUS遺伝子からなるT−DNAを示す図である。ここで、NosPはノパリン合成酵素プロモーターを示す。NosTはノパリン合成酵素ターミネーターを示す。BRはライトボーダー逆方向反復であり、BLは、pBI101 T−DNAのレフトボーダ逆方向反復である。NPTIIは、ネオマイシンホスホトランスフェラーゼIIコード配列を示す。GUSは、β−グルクロニダーゼコード配列を示す。ct β−アミラーゼプロモーターフラグメントを、斜線を引いた長方形で示す。PCR増幅したXhoI−BamHI架橋フラグメントを黒色の長方形で示す。
【図4】タバコ芽生えにおけるct Bmyプロモーター−GUSキメラ遺伝子から発現したGUS活性に対する光およびスクロースの効果を示す図である。
【図5】供与体ベクターpDV35S(SK)Vのプラスミドマップを示す図である。
【図6】供与体ベクターpDV02000のプラスミドマップを示す図である。
【図7】バイナリープラスミドpBNP10431のプラスミドマップを示す図である。ここで35Spは、CaMV 35Sプロモーターを示す。ct bamyは完全長ct β−アミラーゼcDNAを示す。35StはCaMV 35Sターミネーターを示す。RBはバイナリーベクターpBinPlusのライトボーダーを示す。colE1oriはcolE1細菌複製起点を示す。RKoriは、RK2プラスミドのoriV複製起点を示す。nptIIIは、細菌カナマイシン耐性のためのネオマイシンホスホトランスフェラーゼ遺伝子を示す。LBは、バイナリーベクターのレフトボーダー配列を示す。kanは、植物カナマイシン耐性に必要とされる植物ネオマイシンホスホトランスフェラーゼ組換え遺伝子を示す。
【図8】バイナリープラスミドpBNP10432のプラスミドマップを示す図である。略語は図7の通りである。
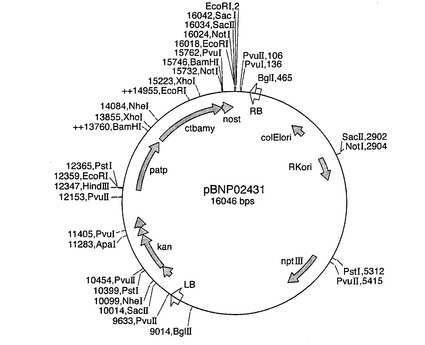
【図9】バイナリープラスミドpBNP02431のプラスミドマップを示す図である。patpが図6のベクターpDV02000に由来するパタチンクラスIプロモーターを示し、nostがノパリン合成酵素ターミネーターを示す以外は、略語は実施例7の通りである。
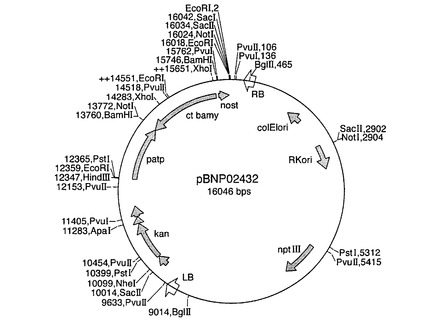
【図10】バイナリープラスミドpBNP02432のプラスミドマップを示す図である。略語は図9の通りである。
【技術分野】
【0001】
植物中でデンプンが合成および分解される正確な機構は、このプロセスに関与すると推定される多数の酵素が単離および特徴付けられているのにも関わらず、知られていない。
【0002】
デンプンは、日中、葉の葉緑体に蓄積され、夜間、植物がエネルギーおよび生合成を必要とするのを満たすために使用される。このいわゆる一過性デンプンが動員される機構は十分に理解されていないが、合成酵素および分解酵素の活性を協調して調節するのに違いない。葉組織における主要な分解経路は、加リン酸分解活性および加水分解活性、特に、α−グルコシダーゼ(E.C.3.2.1.3.)(NielsonおよびStitt,1997)が関与することが考えられる。
【0003】
デンプンはまた、種子、果実、および塊茎などの貯蔵器官中のアミロプラストに蓄積される。この場合、デンプンは長期間にわたって貯蔵され、デンプンの動員に付随して、貯蔵器官組織の退化ならびにデンプン分解活性および加リン酸分解活性の増加が起こる。しかしながら、デンプン代謝回転は貯蔵器官のアミロプラストでも起こっていることを示唆する証拠がある(Sweetloveら,1996)。これもまた、合成酵素活性および分解酵素活性を協調して調節することを必要とする。
【0004】
葉緑体およびアミロプラストは両方ともプロプラスチドに由来し、従って、それぞれ葉および貯蔵器官におけるデンプン合成の場であることの他に、共通の特徴が多数ある。葉緑体は、アミロプラストおよび他の型のプラスチドに変わることができる(ThomsonおよびWhatley,1980)。
【0005】
デンプンは、2種類の多糖:α−1,4−グリコシド結合により連結したグリコシル単位からなる直鎖であるアミロースと、α−1,6グリコシド結合により結合した多数のα−1,4−ポリグルカン直鎖からなるアミロペクチンの混合物である。
【0006】
デンプン合成に関与する酵素は、ADPGピロホスホリラーゼ(E.C.2.7.7.21)、デンプン合成酵素(E.C.2.4.1.21)、および分枝酵素(E.C.2.4.1.18)である。ADPGピロホスホリラーゼは基質ADPGの供給を担い、この分子は、デンプン合成酵素(α−1,4結合)および分枝酵素(α−1,6結合)の共同作用により結合されるグルコースモノマー供与体として働く。
【0007】
デンプン粒の不溶性結晶構造は、幅の広い、らせん状の、枝分かれしたアミロペクチン分子と、空間を充填する直線状アミロース分子とが密に束ねられることにより形成されると考えられる。
【0008】
α−アミラーゼ(E.C.3.2.1.1)、イソアミラーゼ(E.C.3.2.1.68)、β−アミラーゼ(E.C.3.2.1.2)、α−グルコシダーゼ(E.C.3.2.1.3)、デンプンホスホリラーゼ(E.C.2.4.1.1)、および不均化酵素(E.C.2.4.1.25)を含む、多種のデンプン分解酵素活性が報告されている。これらの酵素活性の多くは植物において複数形態で存在し、デンプン合成に関与すると考えられているものもある。恐らく、ある程度、全ての酵素がデンプン動員プロセスに関与しているが、それらの正確な役割および可能な相互作用はまだ決定されていない。様々な酵素の役割を考える難しさは、植物におけるデンプン分解を主に担うと考えられる2つの酵素活性:デンプンホスホリラーゼとアミラーゼを言及することで最もよく例示される。
【0009】
デンプンホスホリラーゼは、α−1,4−グルカンからのグルコース−1−リン酸の可逆的放出を触媒する。2つの形態のデンプンホスホリラーゼが植物組織に見出される。Pho1すなわちL型はプラスチド内にあり、マルトデキストリンに対する親和性が高い。Pho2すなわちH型はサイトゾルにあり、グリコーゲンなどの大きな、非常に枝分かれしたポリグルカンに対する親和性が高い。プラスチド型Pho1酵素は、デンプン動員に関与する有望な候補であるが、この葉酵素活性がアンチセンスにより阻害されても、トランスジェニックジャガイモ植物の葉におけるデンプン蓄積には影響しなかった(Sonnewaldら,1995)。別の研究では、細胞質Pho2がアンチセンスにより阻害されると、トランスジェニックジャガイモ塊茎の発芽行動に影響したが、デンプン蓄積および分解には影響しなかった(Duwenigら、1997)。
【0010】
アミラーゼには2つの主要なグループがあり、両方とも、アミロースおよびアミロペクチンのα−1,4−グルコシド結合を加水分解する。α−アミラーゼは非末端結合に対してランダムに作用するが、β−アミラーゼは、ポリグルカン鎖の非還元末端から始まるマルトース単位を放出するように作用する。植物細胞のアポプラスト空間におけるα−アミラーゼの亜細胞位置は、この酵素が正常に分泌されたことを反映していると考えられる。しかしながら、多数のα−アミラーゼタンパク質のアミノ末端シグナル配列がプラスチド膜ではなくER膜を通るタンパク質移動を示すという知見にもかかわらず(Chenら,1994)、イネ(Chenら,1994)およびテンサイ(Liら,1992)などの多数の植物では、この酵素は葉緑体およびアミロプラスト内にも位置する。イネα−アミラーゼ遺伝子のプロモーターおよびシグナル配列を細菌GUS遺伝子と融合し、アグロバクテリウム媒介形質転換を用いてイネ、タバコ、およびジャガイモに導入した研究(Chenら,1994)では、発現したGUS融合タンパク質は最初に小胞体に輸送され、次いで、トランスジェニック細胞からなる懸濁培養物の培地に出されることが証明された。α−アミラーゼは未変性デンプン分子を分解することが多数の研究で示されている。
【0011】
対照に、インビトロ研究から、他の酵素により未変性デンプン粒が前消化されなければ、β−アミラーゼは未変性デンプン粒を分解しないことがわかっている。活性β−アミラーゼを欠いているか、または微量の活性しかないライムギ変異体(Daussantら,1981)およびダイズ変異体(HildebrandおよびHymowitz,1981)はそれぞれ、見たところ正常な成長および発育を示す。さらに、β−アミラーゼレベルが著しく減少したトランスジェニックシロイヌナズナ植物は、ひどい成長異常を示さない(Mitaら,1997)。植物におけるβ−アミラーゼの正確な生理学的役割を明らかにする試みは、亜細胞位置に関する不確かなデータにより妨げられている。ある研究(Kakefudaら,1986)によりエンドウ葉緑体には2種類のβ−アミラーゼが存在することが報告されたが、ソラマメ、オオムギ、コムギ、ダイズ、サツマイモ、およびエンドウなどの種に関係する大多数の研究では、全てではないがほとんどのβ−アミラーゼ活性は葉緑体外にある(Nakamuraら,1991)。この考えは、現在までクローニングされた全てのβ−アミラーゼ遺伝子が、アミノ末端葉緑体トランジットペプチド配列を欠くタンパク質をコードするということにより支持される。
【0012】
穀類では3つの型のβ−アミラーゼ:穎果成熟中に蓄積する内胚乳特異的形態;発芽中にイネおよびトウモロコシのアリューロン細胞においてデノボ合成される形態(Wangら,1996;1997);ならびに栄養器官に遍在するβ−アミラーゼが記載されている。シロイヌナズナでは、この遍在形態はロゼット葉の総デンプン分解活性の約80%を占める。現在までクローニングされた他の全てのβ−アミラーゼと共通して、遍在シロイヌナズナ β−アミラーゼの遺伝子は、亜細胞ターゲティングシグナルを有するタンパク質をコードせず、従って、この酵素はサイトゾルに位置するらしい。
【0013】
植物の生存能力に悪影響を及ぼすことなく分解活性を除去することができるという多数の研究からの知見と、プラスチド外にあるデンプン分解酵素の亜細胞位置は驚くべきことである。β−アミラーゼ活性の予想される主要な最終産物、すなわちマルトースが、単離葉緑体においてデンプン分解産物として同定されたことを考慮すれば、β−アミラーゼ活性がプラスチドに明らかにないことは特に驚くべきことである(Peaveyら,1977)。さらに最近では、グルコースとマルトースが、デンプン動員プロセス中に、単離されたカリフラワー芽アミロプラストから出されることが示された(Neuhausら,1995)。
【0014】
葉または貯蔵器官のプラスチド中のデンプン量を操作できることは、植物デンプンを利用する様々な工業プロセスに非常に有利であろう。例えば、ジャガイモ塊茎のデンプン含有量を増加させようとして、E.coli ADPG PPase glgC16をトランスジェニックジャガイモ塊茎において過剰発現させると、デンプンへの炭素フラックスは増加するが、デンプンの正味蓄積は少ししか増加しないことが、以前に示された(Sweetloveら,1997)。過剰発現系統における酵素活性の分析から、ADPG PPaseが変化した以外に、アミラーゼ(特に、β−アミラーゼ)活性もまた変化したことがわかった。このデータから、glgC16タンパク質を過剰発現する塊茎におけるデンプン蓄積は、新たに合成されたデンプンの分解により妨げられる、すなわち、デンプンが代謝回転されることが示唆される。
【0015】
別の例では、麦芽製造工程中のデンプン利用率は、植物、特に、貯蔵器官中の分解酵素活性の型および量と密接に相関する。前記作物の分解能が増加すると、穀粒の麦芽形成、あるいは塊茎または他の貯蔵器官に由来するデンプンからアルコールへの変換がより効率的かつ生産的になる。
【0016】
貯蔵器官に存在するデンプンの型は、ADPGピロホスホリラーゼ、デンプン合成酵素、分枝酵素、および存在する分解酵素の形態および活性による。様々な酵素間の相互作用もまた重要である。
【0017】
植物において新規のデンプンを産生することへの関心がかなりある。なぜなら、この新規デンプン産生により、食品、製紙、製薬、接着剤、油、および繊維などの様々な産業で使用する前にデンプンを加工および改良する費用が削減されるからである。以下の例は、どのくらいデンプン加水分解活性が、インビボでのデンプン構造の改変に重要であるのかを示す。
【0018】
トウモロコシ穀粒にsugaryl変異があると、デンプンのα−1,6−グルコシル結合を加水分解する脱分枝酵素がなくなることが示されている(Jamesら,1995)。この変異は、アミロペクチン濃度と、非常に枝分かれのあるグルコポリサッカライドであるフィトグリコーゲンの蓄積の低下をもたらす。
【0019】
エンドウでは、マルトースで始まりマルトヘプトースまで連続グルコース単位を付加する短いオリゴ糖分子は、アミロース合成を担う主要酵素であると一般に認められているデンプン粒結合型デンプン合成酵素I(GBSSI)(Denyerら,1996)の活性を特異的に刺激することが示されている(例えば、van der Leijら,1991;Hyltonら,1995;Ainsworthら,1993)。マルト−オリゴ糖の供給制御によるGBSSI活性の操作は、最近の特許(WO97/16554)の主題であり、分解酵素、すなわち、α−アミラーゼ、β−アミラーゼ、不均化酵素、脱分枝酵素、およびデンプンホスホリラーゼを導入することにより、マルト−オリゴ糖濃度を増加することができ、従って、デンプン中のアミロースとアミロペクチンの比を増加することができることを示唆する。特許WO97/16554はまた、これらの酵素のプラスチド型アイソフォームがクローニングされたことについて述べている。しかしながら、上記のように、現在まで単離されたβ−アミラーゼ遺伝子は、タンパク質ターゲティング配列を有するβ−アミラーゼ酵素をコードせず、さらに、α−アミラーゼが初めからプラスチドに標的化されたことについては疑問がある(Chenら,1994;Chanら,1994)。WO97/16554のもっと後になって、プラスチドターゲティング配列を付加するための適切なβ−アミラーゼcDNA配列の操作が言及される。
【0020】
貯蔵器官中のデンプンの産業用途に加えて、葉中のデンプン量は、作物の農学上、非常に重要である。デンプンは、昼間、葉において、光合成中に固定された炭素から合成される。デンプンは葉緑体に貯蔵され、夜間に分解されて、植物のエネルギー源および代謝中間体になる。ソース−シンク関係が制御される機構は現在未知であるが、葉プラスチド中のデンプンの量および利用率が、植物生産性(バイオマスおよび収穫量)にかなり影響を及ぼすことは明らかである。
【0021】
葉中のデンプン量はまた、葉が主要な植物商品である作物(例えば、タバコ)に重要である。デンプン含有量は、喫煙した時のタバコの最終的なフレーバーに影響を及ぼすことが知られている。タバコ葉中のデンプン量を操作する手段の条項は、タバコ産業が関心を持つ可能性がある。
【0022】
本発明者らは、本明細書中で初めて、新規のターゲティング配列によりプラスチドに標的化される新規のβ−アミラーゼ酵素(今後、葉緑体標的化(ct)β−アミラーゼとして知られる)をコードするcDNAの単離について述べる。この全コード配列の単離は驚くべきことである。なぜなら、β−アミラーゼは、プラスチドから細胞質への移動または膜破壊により、さらに小さなポリグルカン断片を放出したデンプンの加水分解のみに関与すると一般に考えられていたからである。この酵素がプラスチドに存在するので、ct β−アミラーゼは、葉緑体にある一過性デンプンとアミロプラストにある貯蔵デンプンの分解に関与している意外な可能性がある。
【0023】
葉緑体とアミロプラストとの特徴の類似性(ThomsonおよびWhatley,1980)は本発明と関連している。なぜなら、葉緑体標的化ポリペプチドに由来するトランジットペプチドが異種ポリペプチドをアミロプラストに運ぶことができ、逆もまた同様であることが示されたからである。例えば、トウモロコシデンプン粒結合型デンプン合成酵素に由来するトランジットペプチドは、E.coli β−グルクロニダーゼ(GUS)タンパク質に融合された場合、GUSタンパク質をアミロプラストだけではなく葉緑体にも運ぶ(KlosgenおよびWeil,1991)。
【0024】
さらに、本発明者らは、シロイヌナズナにおけるct−Bmy遺伝子の発現、およびトランスジェニックタバコにおけるct−Bmyプロモーター:GUS融合の発現は、光およびスクロースにより独立して調節することができることを示す。このことは、シロイヌナズナ ATβ−Amyの密接に結びついた光および糖誘導反応を考えると、驚くべきことである(Mitaら,1995)。
【0025】
本発明は、本明細書では配列番号1として知られ、1〜294のヌクレオチドであり、かつ前記ヌクレオチド中に、さらなるコード配列を植物プラスチドに標的化することができる配列を有する核酸配列、または開示された配列である配列番号1と少なくとも65%以上相同であり、かつ同じ標的化能力を有する配列を提供する。
【0026】
前記核酸配列は、約94のアミノ酸残基をコードすることが好ましく、約85のアミノ酸残基をコードすることがより好ましい。
【0027】
本発明はまた、本明細書では配列番号2として知られ、1〜1642のヌクレオチドであり、かつ前記ヌクレオチド中に、β−アミラーゼをコードすることができる配列を有する核酸配列、または配列番号2内の開示された配列と少なくとも65%以上相同であり、かつ同じコード能力を有する配列を提供する。
【0028】
本発明はまた、本明細書では配列番号3として知られ、1〜1953のヌクレオチドであり、かつ前記ヌクレオチド中に、葉緑体標的化β−アミラーゼをコードすることができる配列を有する核酸配列、または配列番号3内の開示された配列と少なくとも65%以上相同であり、かつ同じコード能力を有する配列を提供する。
【0029】
相同配列として、中程度にストリンジェントな条件下で(65℃の2×SSCで洗浄)、配列番号1、配列番号2、または配列番号3にハイブリダイズする配列もまた挙げられる。
【0030】
前記核酸配列はmRNAまたはcDNAの配列が好ましいが、ゲノムDNAでもよい。
【0031】
本発明はまた、デンプン生合成経路または分解経路の酵素の活性を植物において増加または減少させる方法を提供する。前記方法は、プラスチドターゲティング配列をコードする核酸配列とデンプン生合成経路または分解経路の酵素のコード配列とを含むキメラ遺伝子を植物ゲノムに安定に組み込ませるステップと、改変したゲノムを有する植物を再生するステップを含む。
【0032】
本発明はまた、タンパク質または酵素を植物プラスチドに標的化する方法を提供する。前記方法は、プラスチドターゲティング配列をコードする核酸配列とタンパク質または酵素のコード配列とを含むキメラ遺伝子を植物ゲノムに安定に組み込ませるステップと、改変したゲノムを有する植物を再生するステップを含む。前記タンパク質または酵素は、1以上の下記の群:脂質合成、光合成、アミノ酸代謝、窒素固定、炭素固定、または炭水化物ポリマー合成の経路であるか、あるいは前記植物にある特徴を付与することができる。前記特徴は、1以上の下記の群:除草剤抵抗性および有害生物抵抗性(例えば、真菌抵抗性、細菌抵抗性、またはウイルス抵抗性を含む)から選択される。
【0033】
本発明はまた、プロモーターと、プラスチドターゲティング配列をコードする核酸コード配列と、ターミネーターとを含むキメラ遺伝子を有する植物を提供する。前記配列は、デンプン生合成経路または分解経路の酵素のコード配列を植物プラスチドに標的化することができる。
【0034】
本発明は、産物の発現を指向することができる核酸配列をさらに提供する。前記産物は、前記核酸配列に作動可能に連結されたコード配列によりコードされる。前記核酸配列は、本明細書では配列番号8として知られるか、あるいは配列番号8と少なくとも65%相同であり、かつ配列番号8と実質的に同じ機能を有する。前記核酸配列は刺激反応性であり、前記産物の発現レベルは、前記核酸配列に適用された刺激に応じて変化する。
【0035】
本発明は、産物の発現レベルを変化させる方法をさらに提供する。前記産物は、植物において産物の発現を指向することができる核酸配列に作動可能に連結されたコード配列によりコードされる。前記方法は、産物の発現を指向することができる核酸配列を含むキメラ遺伝子を植物ゲノムに安定に組み込ませるステップを含む。前記産物は、前記核酸配列に作動可能に連結されたコード配列によりコードされる。前記核酸配列は、配列番号8の配列を実質的に有するか、または配列番号8と少なくとも65%相同であり、かつ配列番号8と実質的に同じ機能を有し、刺激反応性である。
【0036】
前記刺激は、光および/または様々なレベルの糖があることか、またはないことが好ましい。あるいは、前記刺激は、発育段階により制御される刺激である。
【0037】
前記糖は、1以上のスクロースまたはグルコースであることが有利である。
【0038】
前記糖はスクロースであることが好ましい。
【0039】
前記誘導性プロモーターまたは植物において産物の発現を指向することができる核酸配列は、光をうけていないが糖が存在する条件下で作動可能であるか、または糖は存在しないが光をうけている条件下で作動可能であることが有利である。光をうけていないが糖が存在する植物組織は、地下器官またはシンク器官が適切であり得る。地下器官は、例えば、塊茎、根茎、または根でもよいが、他のシンク器官は若い葉または種子でもよい。
【0040】
糖は存在しないが光をうけている植物組織は、古い葉(糖は輸送されない)、花の部分、または発芽中の種子である。
【0041】
上記のDNA構造特徴を有する構築物およびキメラ遺伝子もまた本発明の態様である。
【0042】
上記のプラスチドターゲティング配列をコードする核酸配列とデンプン生合成経路または分解経路の酵素の核酸コード配列とを含むキメラ遺伝子を含む植物細胞、さらなるコード配列の発現を指向することができる核酸配列を含むキメラ遺伝子を含む植物細胞、または刺激反応性の上記の核酸配列とコード配列とを含むキメラ遺伝子を含み、前記コード配列の発現レベルが前記核酸配列に適用された刺激に応じて変化する植物細胞、本発明による1以上のキメラ遺伝子を含む形質転換植物の種子もまた、本発明の態様である。
【0043】
前記プラスチドターゲティング配列は配列番号1の配列であることが有利である。
【0044】
本発明の第1の態様では、上記の方法を用いて、デンプンが葉に蓄積されるか、または葉から動員されるように葉の代謝を改変することができる。このプロセスは、全体的に植物内のソース−シンク関係を改変する。これは、適切なプロモーターの制御下にある、ターゲティング配列とデンプン生合成経路または分解経路の酵素の核酸コード配列を作成することにより達成することができる。適切なプロモーターを選択すると、葉中のデンプン量が増加または減少した植物(例えば、これはタバコ産業で有用であり得る)が得られるか、あるいは、植物のソース−シンク関係を改善した後に、様々な他の植物組織(例えば、塊茎、果実、および根)のデンプン収量が変化する。
【0045】
本発明の本実施態様では、適切なプロモーターは、プラスチドターゲティング配列とデンプン生合成経路酵素または分解経路の酵素のコード配列との発現を植物全体で指向するか(いわゆる構成的発現)、あるいは特異的に葉に指向する。これらの変化は著しい影響を及ぼし、植物器官のデンプン含有量および/または収量がかなり変化する。
【0046】
全ての植物組織全体にわたって発現を指向することができる好ましいプロモーターは、カリフラワーモザイクウイルス35S遺伝子から得られるプロモーターである。葉での発現に好ましいプロモーターは、リブロースビスリン酸カルボキシラーゼ小サブユニット遺伝子またはエンドウプラストシアニン遺伝子から得ることができる。当業者であれば、構成的発現および葉特異的発現に適切な他のプロモーター(例えば、それぞれ、ノパリン合成酵素プロモーターおよびクロロフィルa/b結合タンパク質プロモーター)を認識するであろう。
【0047】
デンプン生合成経路または分解経路の酵素のコード配列またはその一部を、正常なリーディングフレーム方向(すなわち、センス)で配置してもよいし、逆のリーディングフレーム方向(すなわち、アンチセンス)で配置してもよい。センス、またはアンチセンス、またはコサプレッション技術(最後の技術は、DNAPにより欧州特許第0465572号および同第0647715号において説明される)を用いた、植物における酵素活性のアップレギュレーションまたはダウンレギュレーションを用いて、植物デンプンを変えることができる。
【0048】
本発明の第2の態様では、本発明の方法を用いて、デンプン含有量が増加し、かつ/またはデンプンが特定の工業プロセスに必要とされる適切な形態で得られるように、貯蔵器官におけるデンプン代謝を変えることができる。このようなプロセスとして、製紙;医薬品、繊維、染料、および建築用品の製造;天火焼き製品、乳製品、およびスナック食品の供給;缶詰製品、乾燥食品、またはインスタント食品の製造;穀粒の麦芽形成ならびにシロップおよびアルコールの製造が挙げられる。
【0049】
本方法の第1または第2の態様では、本方法のキメラ遺伝子で使用するために選択される酵素は、デンプン分解経路に由来する酵素(すなわち、デンプン分解酵素)でもよい。キメラ遺伝子は、葉緑体標的化β−アミラーゼ(以下、ct β−アミラーゼとして知られる)を含むことが有利であり、シロイヌナズナに由来するct β−アミラーゼ(以下、At ct β−アミラーゼとして知られる)(配列番号3を参照のこと)を含むことがより好ましい。ジャガイモ、タバコ、コムギ、トウモロコシ、およびオオムギなどの他の植物供給源から得ることができる、At ct β−アミラーゼと相同な配列もまた使用することができる。ハイブリダイゼーションまたはポリメラーゼ連鎖反応(PCR)技術によるクローニングの標準的な方法、例えば、Sambrookら(1989)に記載される技術およびInnesら(1990)に記載されるPCR技術などの分子クローニング技術を用いて、このような生物から配列を単離することができる。他のデンプン分解酵素として、α−アミラーゼ、不均化酵素、脱分枝酵素、デンプンホスホリラーゼ、α−グルコシダーゼ、および非プラスチド型β−アミラーゼが挙げられる。前記酵素の1以上のコード配列がプラスチドターゲティング配列との併用に適する。
【0050】
本方法の第2の態様では、植物の貯蔵器官に発現を指向する好ましいプロモーターを、例えば、以下のリストからの遺伝子から選択することができる:コムギ内胚乳高分子量グルテニン遺伝子、コムギ内胚乳α,β−グリアジン遺伝子、オオムギ内胚乳ホルデイン遺伝子、またはジャガイモ塊茎パタチン遺伝子。他の適切なプロモーターは当業者に周知である。
【0051】
本発明のどちらの態様でも、組織代謝の変化あるいはデンプンの型または特徴の変化を、本明細書に記載の誘導性プロモーター(配列番号8)を用いることによって、刺激反応性、すなわち、誘導性にすることができる。例えば、光誘導性の誘導性プロモーターは、Barnase(特許WO98/10081に例示される)などの遺伝子の誘導により花粉発育に影響を及ぼすように種子を操作するために、または果実追熟および種子発芽などの光依存性プロセスにおいて非光反応性遺伝子に影響を及ぼすために使用することができる。光誘導性プロモーターはまた、葉での二次代謝産生(例えば、アルカロイド産生)に影響を及ぼす遺伝子を活性化するために使用することができる。光誘導性プロモーターはまた、葉または他の光合成組織においてデンプン生合成酵素遺伝子を操作するために、あるいは塊茎を例えば暗所貯蔵から取り出した後に遺伝子を活性化するために使用することができる。糖誘導性の誘導性プロモーターは、例えば、発育中の塊茎または他の非光合成組織において、有害生物耐性遺伝子および/または収穫後の作物の品質に影響を及ぼすことが可能な遺伝子などの遺伝子を調節するために使用することができる。ジャガイモでは、胴枯れ病、黒脚病、および乾腐病に対する耐性遺伝子は特に有益であり、糖誘導性プロモーターを有する組換え遺伝子にクローニングできることが最も有利である。あるいは、糖誘導性の誘導性プロモーターは、組織培養プロセスにおいて選択マーカー遺伝子を発現させるために使用することができる。
【0052】
当業者は、欠失試験などの周知の技術を用いることにより、配列番号8から糖誘導性反応性エレメントおよび/または光誘導性反応性エレメントを容易に示すことができる。PweeおよびGray(1993)は、エンドウプラストシアニン遺伝子の作動可能な領域を決定するために、この遺伝子内での、マーカー遺伝子を用いたこのような欠失試験について記載している。
【0053】
遺伝子配列をクローニングし、それらを適切なキャリア(ベクターまたはプラスミドなど)に挿入するための、本明細書に記載の方法、あるいは、例えば、Sambrookら(1989)ならびにGelvinおよびStanton(1995)による実験マニュアルに記載の方法は、このような概念を実行に移す当業者に周知の技術である。上記の1以上のキメラ遺伝子を単独で導入してもよいし、1以上の他のキメラ遺伝子(例えば、1以上の上記の他の遺伝子)と共に導入してもよい。デンプン分解経路の酵素をコードする第1のキメラ遺伝子を用いる上記の実施態様の場合、第2のキメラ遺伝子は、例えば、デンプン生合成経路の酵素をコードする核酸配列(この配列もまた、適切なプロモーターおよび適切なターミネーターの制御下にある)を含んでもよい。第2のキメラ遺伝子のプロモーターおよび/またはターミネーターは、第1のキメラ遺伝子のプロモーターおよび/またはターミネーターと同じでもよいし、異なっていてもよい。デンプン生合成経路の酵素をコードする適切な配列は、スクロース合成酵素、ADPGピロホスホリラーゼ、デンプン合成酵素の核酸配列であり、分枝酵素、α−アミラーゼ、イソアミラーゼ、非プラスチド型β−アミラーゼ、α−グルコシダーゼ、デンプンホスホリラーゼ、および不均化酵素も含み得る。
【0054】
2以上のキメラ遺伝子を植物に導入する方法は説明されており、1つの分子内に結合したキメラ遺伝子を有するバイナリーベクターを構築すること、例えば、異なるキメラ遺伝子を含む異なるバイナリベクターを有する2以上の異なるアグロバクレリウム細胞を用いて同時形質転換するか、または、キメラ遺伝子を既に有する植物を第2の異なるキメラ遺伝子で形質転換(すなわち、再形質転換)することを含む。後者の場合、第2のキメラ遺伝子を導入した後の、トランスジェニック植物の選択方法は、第1のキメラ遺伝子の導入に用いた選択方法とは異ならなければならない。適切な選択マーカーとして、ハイグロマイシン、カナマイシン、スルホンアミド、およびBasta耐性マーカーが挙げられる。各植物が単一のキメラ遺伝子を含む2つの植物を交配するなど、生物学的方法もまた使用することができる。
【0055】
第1の酵素活性が有意に増加し、結果としてデンプン合成が変化した既に形質転換されている植物のデンプン含有量を変えるために、2つのキメラ遺伝子構築物を使用することができる。
【0056】
従って、本発明は、さらなる酵素をアップレギュレートまたはダウンレギュレートし、それにより再形質転換植物により産生されるデンプン量を増加または減少させるために、トランスジェニック植物(この植物では、遺伝的形質転換の結果としてある酵素活性が既に増加または減少している)において、さらなる酵素を変える方法をさらに提供する。
【0057】
第1の形質転換植物は、デンプン生合成経路の酵素活性が増加した植物が有利である。植物のデンプン含有量を増加させる試みの例は、ADPG−PPase遺伝子、例えば、glgC16(例えば、WO91/19806を参照のこと)で形質転換されたトランスジェニックジャガイモである。このような植物のデンプン増加量は比較的少ない。この第1の形質転換植物は、デンプン分解酵素のキメラ遺伝子(例えば、At ct β−アミラーゼを含むことが適する)で再形質転換されることが有利である。glgC16タンパク質は第1の形質転換塊茎で発現し、ADPG−PPase活性増加およびデンプンへの炭素フラックスの増加をもたらす。再形質転換塊茎においてキメラAt ct β−アミラーゼ遺伝子またはその一部を発現させると、ct β−アミラーゼ活性がダウンレギュレートされ(すなわち、コサプレッションまたはアンチセンス技術)、従って、デンプン蓄積が増加する。
【0058】
第2の酵素の発現は塊茎に指向されることが好ましい。At ct β−アミラーゼキメラ遺伝子の発現を塊茎に指向するのに適したプロモーターは、パタチン遺伝子に由来するプロモーターである。
【0059】
glgC16を発現する第1の形質転換ジャガイモ植物はカナマイシン耐性であり、従って、At ct β−アミラーゼキメラ遺伝子用のバイナリーベクター構築物は異なる耐性遺伝子を有し、例えば、スルホンアミド耐性遺伝子が適切である。ジャガイモ乾物が1%増加すると製品が4%増加するので、ジャガイモ塊茎のデンプン産生増加は、例えば、ポテトチップス製造業者に有益であろう。
【0060】
ポテトチップス製造はまた、本発明の別の利益を例示するのに役立つ。ジャガイモ塊茎を8℃以下の温度で貯蔵すると、デンプン分解により還元糖であるグルコースおよびフルクトースが蓄積する。このジャガイモをポテトチップスにするために油で揚げると、メイラード反応により還元糖はアミノ酸と反応して、製品は茶色になり、味がまずくなる。デンプン破壊を妨げ、従って、還元糖の蓄積を妨げるキメラ遺伝子をジャガイモ植物に導入することは、スナック食品産業に有益であろう。このキメラ遺伝子は、コサプレッションまたはアンチセンス構築物中に、適切なプロモーターおよびターミネーターで駆動される、ct β−アミラーゼのコード配列またはその配列の一部を含むことが好ましい。適切なプロモーターは、ジャガイモ塊茎パタチン遺伝子から得られる。有利にも、ct β−アミラーゼの代わりに、上記の他のどのデンプン分解酵素も使用できる。
【0061】
発育中の葉および発育中の塊茎での協調した発現が必要であれば、パタチンプロモーターがスクロース誘導性でもあるので(Rocha−Sosaら(1989))、配列番号8の誘導性プロモーターもまた前記構築物に使用することができる。同様に、配列番号1の葉緑体ターゲティングポリペプチド配列もまた、自身のターゲティング配列を欠き、かつプラスチドに指向されることが必要な他の任意の遺伝子と併用することができる。
【0062】
上記の例は、本発明を使用する可能な利益を例示するのに役立つ。当業者であれば、遺伝子の組み合わせと本発明を適用することができる植物とが多数あることを認識するであろう。
【0063】
遺伝子の組み合わせとして、好ましくは、ct β−アミラーゼと、1以上のスクロース合成酵素、ADPGピロホスホリラーゼ、デンプン合成酵素、分枝酵素、α−アミラーゼ、イソアミラーゼ、非プラスチド型β−アミラーゼ、α−グルコシダーゼ、デンプンホスホリラーゼ、および不均化酵素の遺伝子とが挙げられる。上記遺伝子の配列は当業者に周知である。あるいは、ct β−アミラーゼに由来するターゲティング配列を、1以上の上記遺伝子と併用してもよい。
【0064】
形質転換することができる植物のリストには、好ましくは、ジャガイモ、コムギ、トウモロコシ、オオムギ、トマト、イネ、エンドウ、ダイズ、ラッカセイ、キャッサバ、ヤムイモ、バナナ、およびタバコが含まれる。
【0065】
本発明を、一例として、シロイヌナズナに由来するct β−アミラーゼcDNAを単離し、このcDNAをタバコおよびジャガイモ植物に組み込ませる実施態様に関して説明する。刺激反応性プロモーターおよびトランスジェニック植物におけるその活性に関する実施例もまた示す。
【0066】
本発明を容易に実行に移すことができるために、一例として、以下の図面が参照されよう。
【0067】
配列表中では、以下の通りである。
【0068】
配列番号1は、コード配列を植物プラスチド、特に葉緑体に標的化することができる核酸である。
【0069】
配列番号2は、β−アミラーゼをコードする核酸である。
【0070】
配列番号3は、葉緑体標的化(ct)β−アミラーゼの完全配列である。
【0071】
配列番号4および5は、実施例3の増幅プロセスに用いられたプライマーである。
【0072】
配列番号6および7は、実施例4の増幅プロセスに用いられたプライマーである。
【0073】
配列番号8は、刺激反応性、特に光および/または糖に反応性の核酸である。
実施例1
シロイヌナズナ葉緑体標的化β−アミラーゼの単離および特徴
pBmy81内にあるcDNA挿入物の配列決定
【0074】
コスミドG16599(Bevanら,1988)内にある37kb シロイヌナズナ 4番染色体DNAフラグメントのヌクレオチド配列のBLASTNデータベース検索から、シロイヌナズナ、オオムギ、トウモロコシ、イネ、ダイズ、およびイネの葉緑体外β−アミラーゼと有意な相同性を有する遺伝子が存在することがわかった。この検索から、いくつかの3’末端EST配列もまた同定された。これらの1つであるEST 81E10T7(Newmanら,1995)(以下、pBmy81と呼ぶ)は約300ヌクレオチドにわたって同一であった。クローンEST 81E10T7は、Arabidopsis Biological Resource Center(ABRC)DNA Stock Center(Ohio University,USA)により供給された。pBmy81内のcDNA挿入物にまたがるBal31欠失サブクローンのネステッドセットを、蛍光色素標識ユニバーサルプライマーを用いた二本鎖PCRサイクルシークエンシング反応でのDNA鋳型として使用した。シークエンシング反応を、Apllied Biosystems Model373A自動化シーケンサーにより分析した。pBmy81の前記cDNA挿入物のヌクレオチド配列を配列番号3に示す。特許手続上の微生物の寄託の国際的承認に関するブタペスト条約により、Advanced Technologies(Cambridge)Limited,210 Cambridge Science Park,Cambridge CB4 4WAは、構築物pBmy81を、1998年8月4日にアクセッション番号NCIMB40964で、National Collection of Indurtrial and Marine Bacteria(NCIMB),23 St.Machar Street,Aberdeen Scotlandに寄託した。
推定葉緑体ターゲティングシグナルの同定
【0075】
pBmy81 cDNA挿入物は、5’末端に36の非翻訳ヌクレオチド、548アミノ酸からなるタンパク質をコードするオープンリーディングフレーム(ORF)、および232bpの3’非翻訳領域(UTR)を含む。pBmy81 cDNA挿入物によりコードされるタンパク質は、61kDaの推定分子量を有し、トウモロコシ、イネ、オオムギ、ダイズ、およびサツマイモに由来する植物葉緑体外β−アミラーゼと高いアミノ酸類似性を有する。しかしながら、pBmy81によりコードされるタンパク質は、葉緑体ターゲティングシグナルの特徴(すなわち、高含有量のセリン(16%)、スレオニン(10%)、および正に荷電したアミノ酸残基(15%))(BaierおよびDietz,1997)を有する独特のN末端伸長を含む点で、現在まで報告された他の全てのβ−アミラーゼとは異なる。葉緑体ターゲティングシグナルの特徴を識別する3つのドメインである、非荷電アミノ末端ドメイン、ヒドロキシル化アミノ酸リッチな中心ドメイン、および脂肪族β鎖を形成し得るカルボキシ末端ドメイン(SchatzおよびDobberstein,1996)を、このシグナル配列内で同定した。
pBmy81内のcDNA挿入物は、葉緑体標的化β−アミラーゼをコードする
【0076】
無傷の葉緑体を、パーコール段階勾配を用いて、50〜60gのエンドウシュート(Pisum sativum L.var Feltham First)から単離した。植物材料を成長させ、MouldおよびGray(1997a)の方法に従って葉緑体を単離した。
【0077】
pBmy81プラスミドを、NotIを用いた制限消化により線状化し、T7RNAポリメラーゼを用いてインビトロ転写した。放射性標識前駆体タンパク質を、本質的にMouldおよびGray(1997b)に記載のように、pBmy81 cDNAの転写物から35Sメチオニンおよび35Sシステインを含むコムギ胚翻訳系において合成した。
【0078】
放射性標識インビトロ翻訳産物の移入を、MouldおよびGray(1997b)に記載のように行った。移入インキュベーションの後、無傷の葉緑体を、氷上で30分間、サーモリシン(移入緩衝液中で最終濃度0.2mg/ml)を用いて処理し、次いで、プロテアーゼ反応を、50mM EDTAを移入緩衝液に添加することで止めた。葉緑体を、移入緩衝液に溶かした40%パーコールの緩衝物により再単離し、次いで、移入緩衝液で洗浄した(MouldおよびGray,1997b)。サーモリシン処理した葉緑体試料のアリコート(1/10)を分析のために採取し、残りを、本質的にSchnellおよびBlobel(1993)に記載のように分画した。サーモリシン処理葉緑体、ストロマ画分、チラコイド、およびサーモリシン処理チラコイドの試料を、SDS−PAGE、続いてクマシーブルー染色、染色タンパク質バンドのデンシトメータースキャニングにより定量した(リブロースビスリン酸カルボキシラーゼおよび集光性複合体タンパク質のサブユニットを標準として使用した)。等量のこれらの画分(パーコール勾配から回収した葉緑体の2%にほぼ等しい)と回収した内包膜および外包膜画分の505とを、SDS存在下での10%ポリアクリルアミドゲルによる電気泳動、続いてフルオログラフィーにより分析した。結果(図1)から、主要な翻訳産物(レーンTr)が約58kDaであることがわかる。単離した無傷のエンドウ葉緑体を、ATPの存在下で放射性標識タンパク質とインキュベートすると、約50kDaおよび48kDaのポリペプチドが産生された(レーンC)。外から添加したサーモリシンによる分解に対する耐性から、これらのポリペプチドは、放射性標識タンパク質移入の産物であることがわかる。無傷のサーモリシン処理葉緑体のストロマ、洗浄チラコイド、サーモリシン処理チラコイド、内包膜および外包膜への分画により、2つの放射性標識ポリペプチドがストロマ画分にあることが証明された。
実施例2
シロイヌナズナ ct β−アミラーゼ遺伝子のスクロースおよび光誘導
【0079】
明所でのct b−アミラーゼ誘導を証明するために、シロイヌナズナ生態型Landsberg植物を、18時間明期、6時間暗期、18℃で、温室において成長させた。5週間後、2トレイの芽生えを真っ暗闇に移し、2トレイの芽生えを連続光で成長させた。2日後、1トレイの暗所適応した芽生えと1トレイの明所で成長させた芽生えを総RNA単離のために使用し、それぞれの2つ目のトレイを、さらに3日間、連続光に暴露した。
【0080】
スクロース−明−暗を組み合わせた処理のために、Landsberg生態型の種子を表面滅菌し、1%スクロースを含むMS寒天培地上に置き、18時間明期、6時間暗期で培養室において成長させた。5週齢の芽生えを、滅菌蒸留水上に、あるいは5%スクロース水溶液またはグルコース水溶液上に移した。この芽生えを、3日間、連続して明所または暗所で維持した。総RNAを各試験の芽生えから調製し、Eggermontら(1996)により記載されるようにノザンブロット分析により分析した。ノザンを、FeinbergおよびVogelstein(1983)により記載されるように、32P−dCTPでランダム標識した後に、pBmy81内のゲル精製したcDNA挿入物を用いてプローブした。
【0081】
図2Aに示す結果から、ct β−アミラーゼ遺伝子転写物は光誘導性であることがわかる。
【0082】
図2Bに示す結果から、ct β−アミラーゼ転写物は、暗所において5%スクロースにより誘導され、それより少ない程度で5%グルコースにより誘導されることがわかる。この誘導は、明所で、糖の存在下でさらに高まった。これらの結果から、光および糖の効果は互いに独立していることがわかる。
実施例3
ct β−アミラーゼプロモーター−GUS融合の構築
【0083】
プロモーターフラグメントを、制限酵素消化により、コスミドG16599(Bevenら,1998)内にあるct β−アミラーゼ遺伝子から単離した。このプロモーター内にある便利な制限部位は、ヌクレオチド−1662bp位のHindIII(配列番号8のマイナス鎖の19179bpで開始する)、−1127bpのSalI、および−371bpのPstIであり、ct β−アミラーゼ開始メチオニンの下流にある+21bp位に位置するXhoI部位を用いて、3つの異なる長さのプロモーターとトランジットペプチド配列を単離した(翻訳開始メチオニンATGのAに+1の番号をつけた)。
【0084】
コスミドG16599(Bevenら,1998)内にあるct β−アミラーゼ遺伝子の294bp(配列番号1)フラグメントを、以下のオリゴヌクレオチドプライマー:
配列番号4
P1:(5’−ATT TCC TCG AGT TCT CTT ATC−3’)および
配列番号5
P2:(5’−cgg gAT CCC TGA CAT TGT TAC−3’)
を用いて増幅した。
【0085】
プライマーP1の下線を付けた塩基は、+21bp位に位置するXhoI部位を示す。プライマー2の小文字の塩基は、BamHI部位を作成するために付加したヌクレオチドを示す。
【0086】
プロモーターフラグメントと、XhoIおよびBamHIで消化したPCR結合フラグメントと、HindIII−BamHI、SalI−BamHI、またはPstI−BamHIで消化したGUSベクターpBI101(Jeffersonら,1987)のトリプルライゲーションにより、キメラct β−アミラーゼプロモーター−GUS遺伝子を作成した(図3)。構築物を、それぞれ、HβGUS、SβGUS、およびPβGUSと名づけた。
【0087】
キメラ遺伝子構築物を、トリペアレンタルメイティング(Bevan,1984)によりアグロバクテリウム−ツメファシエンス LBA4404に移し、リーフディスク形質転換法(Horschら,1985)によりNicotiana tabacum var Samsumに導入した。
実施例3A
タバコ芽生えにおけるキメラシロイヌナズナ ct β−アミラーゼプロモーター−GUS遺伝子のスクロースおよび光誘導
【0088】
HβGUSおよびPβGUS構築物を含む植物は高レベルのGUS活性を発現し、この系統のF1芽生え子孫を用いて、キメラ遺伝子の光およびスクロース誘導性発現を調べた。F1タバコ種子を表面滅菌し、1%スクロースを含むMS寒天培地上に置き、18時間明期、6時間暗期で培養室において成長させた。2〜3週齢の芽生えを5%スクロース溶液または蒸留水上に移し、3日間、連続して明所または暗所で維持した。Jeffersonら(1987)に記載のように、10〜14個の芽生えプールからの総タンパク質抽出物を、蛍光基質4−メチルウンベリフェリル−グルクロニド(4−MUG)を用いてGUS活性について分析した。両構築物を用いた、スクロースの非存在下で連続光に暴露された芽生えのGUS活性レベルは、光の非存在下でスクロースに暴露された芽生えのGUS活性レベルと類似した(図4)。しかしながら、芽生えを連続光とスクロースとの両方に暴露すると、GUS活性レベルは約2〜3倍増加した。これらの結果は、光誘導性とスクロース誘導性が独立したプロセスであることを示した、ct b−アミラーゼ遺伝子自体を用いた実験からの結果と大まかに一致する。
【0089】
組織化学的GUS染色から、活性は2週齢芽生えの子葉で検出され、第1本葉または茎および根の活性は全くといってよいくらいないことがわかった。4週齢の芽生えでは、さらなるGUS活性が第1本葉全体で示され、茎でも示された。GUS染色は、木部放射組織間にある、および木部と茎内部師部を構成する師部維管束との間にある、葉緑体が豊富な柔組織(葉緑組織)細胞と特に関連していた。
実施例4
タバコおよびジャガイモの葉の形質転換に使用するためのct β−アミラーゼプラスミドの構築
【0090】
位置指定突然変異誘発を用いて、pBmy81プラスミドの2302bp位に位置するKpnI部位をBamHI部位に変換した。オリゴヌクレオチドプライマー:
配列番号6
P3:(5’−GCT GGT ACG CCT GCA GGA TCC GGT CCG GAA TTC CC−3’)および
配列番号7
P4:(5’−GGG AAT TCC GGA CCG GAT CCT GCA GGC GTA CCA GC−3’)
を設計し、Quick Change位置指定突然変異誘発キット(Promega)と共に使用した。プロトコールは、製造業者により概説された通りであった。
【0091】
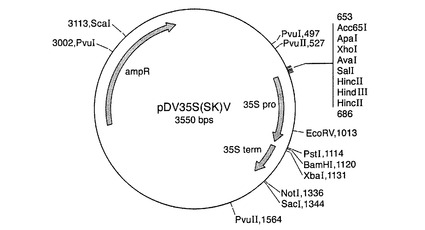
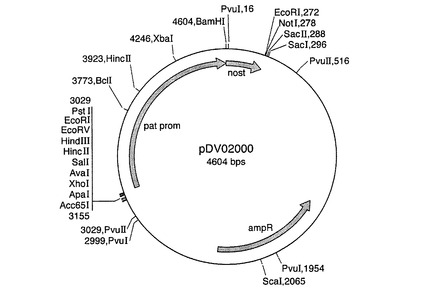
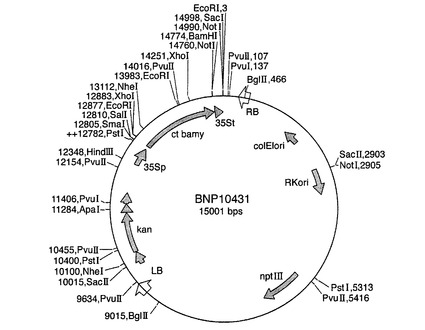
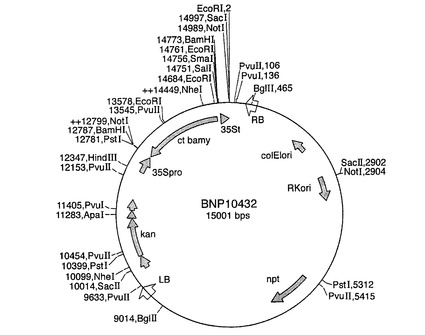
完全長ct β−アミラーゼコード配列を、BamHI切断により突然変異pBmy81プラスミドから切り出し、次いで、GeneClean(BIO101)を用いて精製した。BamHIフラグメントを、供与体ベクターpDV35S(SK)V(図5を参照のこと)およびpDV02000(図6を参照のこと)のBamHI部位に連結した。pDV35S(SK)Vは、35S CaMVプロモーター−35Sターミネーターを有するpBluescript(Stratagene)からなり、類似する構築物が当該分野で周知である(例えば、Odellら,1985)。pDV02000は、1.4kbパタチンプロモーター−ノパリン合成酵素ターミネーターを有するpBluescriptからなる。当業者は、既知の配列から類似する構築物を作成することができる(例えば、Liuら,1990)。プロモーターに対してセンス方向とアンチセンス方向のコード配列を有するプラスミドを単離し、ct β−アミラーゼキメラ遺伝子を、供与体ベクターからバイナリーベクターpBinPlus(van Engelenら,1995)にサブクローニングした。プラスミドマップを図7〜10に示す。
実施例5
植物の形質転換または再形質転換
【0092】
ジャガイモ植物を、本質的にHorsch(1985)に記載のように、リーフディスク共培養法を用いて形質転換した。上記のバイナリーベクターを、エレクトロポレーション法を用いてアグロバクテリウム−ツメファシエンス LBA4404に移し、このアグテバクテリア培養物を、再生した植物が実施例4に記載のキメラ遺伝子を有するように形質転換に使用した。
【0093】
パタチンプロモーター−ct β−アミラーゼ−ノパリン合成酵素ターミネーターキメラ遺伝子バイナリープラスミドは、リーフディスク共培養法により、E.coli ADPG−PPase glgC16キメラ遺伝子を既に有するジャガイモ植物を形質転換するために使用することができる。
実施例6
AT ct β−アミラーゼのターゲティングペプチドを有するプラスミドの構築
【0094】
AT ct β−アミラーゼのプラスチドターゲティング配列は、配列番号1に等しい294bpフラグメント内に含まれる。PCR増幅または制限酵素消化を用いて、DNAフラグメントを実施例3に記載のプラスミドから単離することができる。すなわち、フラグメントは、35S CaMVプロモーター+プラスチドターゲティング配列またはパタチンプロモーター+プラスチドターゲティング配列からなる。キメラ遺伝子は、翻訳融合として、タンパク質または酵素のコード配列とトランジットペプチド配列とを連結することにより構築することができる。翻訳されたタンパク質はプラスチドに運ばれて、新規の活性をもたらすか、または代謝経路に影響を及ぼす。
参考文献
Ainsworth,C.,Clark,J.およびBalsdon,J.(1993).Plant.Mol.Biol.,12,67−82.
Baier,M.およびDietz,X.J.,(1997)Plant J.12,179−190
Bevan,M.W.(1984)Nucl.Acids.Res.,12,8711−8721
Bevan,M.W.ら(1998).Nature,391,485−488.
Chan,MT.,Chao,YC.およびYu,SM.(1994).J.Biol.Chem.,269,17635−17641.
Chen,NH.,Liu,LF.,Chen,YR.,Wu,HK.およびYu,SM.(1994).Plant J.,6,625−636.
Daussant,J.,Zbaszyniak,B.,Sadowaki,J.およびWiatroszak,I.(1981).Planta,151,176−179.
Denyer,K.,Clarke,B.,Hylton,C.,Tatge,H;およびSmith,A.M.(1996).Plant J.,10,1135−1143.
Duwenig,E.,Steup,M.,Willmitzer,L.およびKossmann,J.(1997).Plant J.,12,323−333.
Eggermont,K.,Goderis,I.J.およびBroekaert,W.F.(1996).Plant Mol.Biol.Rep.14,273−279.
Feinberg,A.P.およびVogelstein,B.(1983).Anal.Biochem.132,6−13.
Gelvin,S.B.およびSchilperoort,R.A.(1995).Plant Molecular Biology Manual.第2版.Kluwer Academic Publishers,The Netherlands.
Hildebrand,D.F.およびHymowitz,T.(1981).Physiol.Plant.,53,429−434.
Horsch,R.B.,Fry,J.E.,Hoffman,N.L.,Eichholtz,D.,Rogers,S.G.およびSwaley,R.T.(1985)Science,227.1229−1231.
Hylton,C.M.,Denyer,K.,Keeling,P.L.,Chang,MT.およびSmith,A.M.(1995).Planta,198,230−237.
Innes,M.A.,Gelfand,D.H.,Sninsky,J.J.およびWhite,T.J.(1990).PCR Protocols.Publisher:Academic Press.
James,M.G.,Robertson,D.S.およびMyers,A.M.(1995).Plant Cell,7,417−429.
Jefferson,R.A.,Kavanagh,T.A.およびBevan,M.W.(1987)EMBO,J.6.3901−3907.
Kakefuda,G.,Duke,S.H.およびHostak,M.H.(1986).Planta,168,175−182.
Klosgen,R.B.およびWeil,J.H.(1991)Mol.Gen.Genet.,225,297−304
Li,B.,Servaites,J.C.およびGeiger,D.R.(1992).Plant Physiol.,98,1277−1284.
Liu,X.J.,Prat,S.,Willmitzer,L.およびFrommer,W.B.(1990)Mol.Gen.Genet.,223,401−406.
Mita,S.,Suzuki−Fujii,K.およびNakarmura,K.(1995)Plant Physiol.107,895−904.
Mita,S.,Murano,N.,Akaike,M.およびNakamura,K.(1997).Plant J.,11,841−851.
Mould,R.M.およびGray,J.C.(1997a).In Cell Biology:A Laboratory Handbook,第2版,2巻(Cells,J.E.編).New York:Academic Press,81−86頁.
Mould,R.M.およびGray,J.C.(1997b).In Cell Biology:A Laboratory Handbook,第2版,2巻(Cells,J.E.編).New York:Academic Press,286−292頁.
Nakamura,K.,Ohto,M.,Yoshida,N.およびNakamura,K.(1991).Plant Physiol.,96,902−909.
Neuhaus,H.E.,Henrichs,G.およびSchiebe,R.(1995).Planta,194,454−460.
Newman,T.ら(1994).Plant Physiol.,106,1241−1255.
Nielson,T.H.,Deiting,U.およびStitt,M.(1997).Plant Physiol.,113,503−510.
Odell,J.T.Nagy,F.およびChua,N.H.(1985)Nature,313,810−812.
Peavey,D.G.,Steup,M.およびGibbs,M.(1977).Plant Physiol.,60,305−308.
Pwee,K−H.およびGray,J.C.(1993)Plant J.3,437−449.
Rocha−Sosa,M.,Sonnewald,U.,Frommer,W.,Stratmann,M.,Schell,J.およびWillmitzer,L.(1989)EMBO,8,23−29.
Sambrook,J.,Fritsch,E.F.およびManiatis,T.(1989).Molecular Cloning.Publisher:Cold Spring Harbour.
Schatz,G.およびDobberstein,B.(1996).Science,271,1519−1526.
Schnell,D.J.およびBlobel,G.(1993).J.Cell.Biol.,120,103−115.
Sonnewald,U.,Basner,A.,Greve,B.およびSteup,M.(1995).Plant.Mol.Biol.,27,567−576
Sweetlove,L.J.,Burrell,M.M.およびap Rees,T.(1996).Biochem.J.,320,493−498.
Thomson,W.W.およびWhatley,J.M.(1980)Ann.Rev.Plant Physiol.31,375−394.
van Engelen,F.A.,Molthoff,J.W.,Conner,A.J.,Nap,J−P.,Pereira,A.およびStiekema,W.J.(1995).Transgenic Res.4,288−290.
van der Leij,F.R.,Visser,R.G.F.,Ponstein,A.S.,Jacobsen,E.およびFeenstra,W.J.(1991).Mol.Gen.Genet.,228,240−248.
Wang,SM.,Lue,WL.およびChen,J.(1996).Plant Mol.Biol.,31,975−982.
Wang,SM.,Lue,WL.,Huang,HW.およびChen,J.(1997).Plant Physiol.,113,403−409.
【図面の簡単な説明】
【0095】
【図1】SDS−PAGEゲル、続いてフルオログラフィーにより示された放射性標識インビトロ移入翻訳産物の結果を示す図である。説明:分子量マーカー(レーンM);翻訳産物(レーンTr);移入インキュベーション後に再単離およびサーモリシン処理された葉緑体(レーンC);ストロマ画分(レーンS);洗浄したチラコイド(レーンT);サーモリシン処理チラコイド(レーンtT);内包膜画分(レーンI);外包膜画分(レーンO)。それぞれ、β−アミラーゼの推定前駆体(P)、中間体(I)、および成熟(M)形態。キロダルトン(K)。
【図2】シロイヌナズナ芽生えにおける、ct β−アミラーゼ転写物発現に対する光の効果ならびに光および糖の効果を示す図である。図2Aは、土壌で成長させ、2日連続光(L)、2日連続暗所(D)、2日連続光の後に3日連続光(LL)、または2日暗所の後に3日連続光(DL)に暴露された5週齢シロイヌナズナ植物の総RNAノザンブロット分析を示す。図2Bは、インビトロで成長させ、水上に移して3日連続光に暴露された(WL)、あるいは5%スクロース上に移して3日暗所(SD)または3日連続光(SL)に暴露された、あるいは5%グルコース上に移して3日暗所(GD)または3日連続光(GL)に暴露された、5週齢シロイヌナズナ植物の総RNAノザンブロット分析を示す。ノザンブロットを、放射性標識ct−Bmy cDNA挿入物とハイブリダイズさせ、オートラジオグラフィーにかけた(上部パネル)。対応する臭化エチジウム染色ホルムアルデヒド−アガロースゲルを下部パネルに示す。
【図3】実施例3で構築されるキメラct β−アミラーゼプロモーター−GUS遺伝子からなるT−DNAを示す図である。ここで、NosPはノパリン合成酵素プロモーターを示す。NosTはノパリン合成酵素ターミネーターを示す。BRはライトボーダー逆方向反復であり、BLは、pBI101 T−DNAのレフトボーダ逆方向反復である。NPTIIは、ネオマイシンホスホトランスフェラーゼIIコード配列を示す。GUSは、β−グルクロニダーゼコード配列を示す。ct β−アミラーゼプロモーターフラグメントを、斜線を引いた長方形で示す。PCR増幅したXhoI−BamHI架橋フラグメントを黒色の長方形で示す。
【図4】タバコ芽生えにおけるct Bmyプロモーター−GUSキメラ遺伝子から発現したGUS活性に対する光およびスクロースの効果を示す図である。
【図5】供与体ベクターpDV35S(SK)Vのプラスミドマップを示す図である。
【図6】供与体ベクターpDV02000のプラスミドマップを示す図である。
【図7】バイナリープラスミドpBNP10431のプラスミドマップを示す図である。ここで35Spは、CaMV 35Sプロモーターを示す。ct bamyは完全長ct β−アミラーゼcDNAを示す。35StはCaMV 35Sターミネーターを示す。RBはバイナリーベクターpBinPlusのライトボーダーを示す。colE1oriはcolE1細菌複製起点を示す。RKoriは、RK2プラスミドのoriV複製起点を示す。nptIIIは、細菌カナマイシン耐性のためのネオマイシンホスホトランスフェラーゼ遺伝子を示す。LBは、バイナリーベクターのレフトボーダー配列を示す。kanは、植物カナマイシン耐性に必要とされる植物ネオマイシンホスホトランスフェラーゼ組換え遺伝子を示す。
【図8】バイナリープラスミドpBNP10432のプラスミドマップを示す図である。略語は図7の通りである。
【図9】バイナリープラスミドpBNP02431のプラスミドマップを示す図である。patpが図6のベクターpDV02000に由来するパタチンクラスIプロモーターを示し、nostがノパリン合成酵素ターミネーターを示す以外は、略語は実施例7の通りである。
【図10】バイナリープラスミドpBNP02432のプラスミドマップを示す図である。略語は図9の通りである。
【特許請求の範囲】
【請求項1】
本明細書では配列番号1として知られ、1〜294のヌクレオチドであり、かつ前記ヌクレオチド中に、さらなるコード配列を植物プラスチドに標的化することができる配列を有する核酸配列、または開示された配列である配列番号1と少なくとも65%以上相同であり、かつ同じ標的化能力を有する配列。
【請求項2】
前記核酸配列は約94のアミノ酸残基をコードする、請求項1に記載の核酸配列。
【請求項3】
前記配列は約85のアミノ酸残基をコードする、請求項2に記載の核酸配列。
【請求項4】
本明細書では配列番号2として知られ、1〜1642のヌクレオチドであり、かつ前記ヌクレオチド中にβ−アミラーゼをコードすることができる配列を有する核酸配列、または配列番号2内の開示された配列と少なくとも65%以上相同であり、かつ同じコード能力を有する配列。
【請求項5】
本明細書では配列番号3として知られ、1〜1953のヌクレオチドであり、かつ前記ヌクレオチド中に、葉緑体標的化β−アミラーゼをコードすることができる配列を有する核酸配列、または配列番号3内の開示された配列と少なくとも65%以上相同であり、かつ同じコード能力を有する配列。
【請求項6】
産物の発現を指向することができる核酸配列であって、前記産物は、前記核酸配列に作動可能に連結されたコード配列によりコードされ、前記核酸配列は、本明細書では配列番号8として知られるか、あるいは配列番号8と少なくとも65%相同であり、かつ配列番号8と実質的に同じ機能を有し、前記核酸配列は刺激反応性であり、前記産物の発現レベルは前記核酸配列に適用された刺激に応じて変化する、核酸配列。
【請求項7】
前記核酸配列は、mRNA、cDNA、またはゲノムDNAの配列である、前記請求項のいずれか1つに記載の核酸配列。
【請求項8】
デンプン生合成経路または分解経路の酵素の活性を植物において増加または減少させる方法であって、前記方法は、プラスチドターゲティング配列をコードする核酸配列とデンプン生合成経路または分解経路の酵素のコード配列とを含むキメラ遺伝子を植物ゲノムに安定に組み込ませるステップと、改変したゲノムを有する植物を再生するステップを含む、方法。
【請求項9】
前記核酸配列は請求項1に記載の配列を含む、請求項8に記載の方法。
【請求項10】
前記デンプン分解経路の酵素はβ−アミラーゼである、請求項8に記載の方法。
【請求項11】
前記コード配列は請求項4に記載の核酸配列であり、デンプン分解経路の酵素のコード配列である、請求項8、9、または10に記載の方法。
【請求項12】
前記核酸配列は請求項5に記載の核酸配列である、請求項8に記載の方法。
【請求項13】
請求項8〜12のいずれか1つに記載の方法であって、前記デンプン分解経路の酵素は、スクロース合成酵素、ADPGピロホスホリラーゼ、デンプン合成酵素、分枝酵素、αアミラーゼ、イソアミラーゼ、非プラスチド型β−アミラーゼ、α−グルコシダーゼ、デンプンホスホリラーゼ、および不均化酵素からなる群の1つである、方法。
【請求項14】
タンパク質または酵素を植物プラスチドに標的化する方法であって、前記方法は、プラスチドターゲティング配列をコードする核酸配列とタンパク質または酵素のコード配列とを含むキメラ遺伝子を植物ゲノムに安定に組み込ませるステップと、改変したゲノムを有する植物を再生するステップを含み、前記タンパク質または酵素は、1以上の下記の群:脂質合成、光合成、アミノ酸代謝、窒素固定、炭素固定、または炭水化物ポリマー合成の経路であるか、あるいは前記植物にある特徴を付与することができ、前記核酸配列は請求項1に記載の核酸配列である、方法。
【請求項15】
前記特徴は、1以上の下記の群:除草剤抵抗性および有害生物抵抗性から選択される、請求項14に記載の方法。
【請求項16】
産物の発現レベルを変化させる方法であって、前記産物は、植物において前記産物の発現を指向することができる核酸配列に作動可能に連結されたコード配列によりコードされ、前記方法は、前記核酸配列に作動可能に連結されたコード配列によりコードされる産物の発現を指向することができる核酸配列を含むキメラ遺伝子を植物ゲノムに安定に組み込ませるステップを含み、前記核酸配列は、配列番号8の配列を実質的に有するか、または配列番号8と少なくとも65%相同であり、かつ配列番号8と同じ機能を実質的に有し、刺激反応性である、方法。
【請求項17】
前記コード配列は、前記植物において開花中に不稔性を引き起こす遺伝子をコードし、かつ光に暴露されると前記核酸配列により発現させられる、請求項16に記載の方法。
【請求項18】
前記コード配列は、葉または他の光合成組織におけるデンプン生合成酵素遺伝子のレベルを変化させる遺伝子をコードする、請求項16に記載の方法。
【請求項19】
前記コード配列は、発育中の貯蔵器官において糖が産生され、かつ糖が前記核酸配列に作用すると、発現レベルが変化する遺伝子をコードする、請求項16に記載の方法。
【請求項20】
前記核酸配列は、光はあるが糖がない場合に作動可能である、請求項16に記載の方法。
【請求項21】
前記核酸配列は、糖はあるが光がない場合に作動可能である、請求項16に記載の方法。
【請求項22】
さらなる酵素をアップレギュレートまたはダウンレギュレートし、それにより再形質転換植物により産生されるデンプン量を増加または減少させるために、遺伝的形質転換の結果として酵素活性の増加または減少を既に示しているトランスジェニック植物において前記さらなる酵素を変える方法。
【請求項23】
前記第1の形質転換植物は、デンプン生合成経路の酵素活性が増加または減少した植物である、請求項22に記載の方法。
【請求項24】
前記第1の形質転換植物は、ADPG−PPase遺伝子で形質転換されたトランスジェニックジャガイモである、請求項23に記載の方法。
【請求項25】
前記キメラ遺伝子は、カリフラワーモザイクウイルス35Sプロモーター(完全または切断)、ルビスコプロモーター、エンドウプラストシアニンプロモーター、ノパリン合成酵素プロモーター、クロロフィルr/b結合プロモーター、高分子量グルテニンプロモーター、α,β−グリアジンプロモーター、ホルデインプロモーター、およびパタチンプロモーターからなる群から選択されるプロモーターをさらに含む、請求項8、14、または16のいずれか1つに記載の方法。
【請求項26】
前記キメラ遺伝子中の前記酵素の前記コード配列は、前記酵素の活性をアップレギュレートまたはダウンレギュレートする、請求項8、14、または16のいずれか1つに記載の方法。
【請求項27】
前記刺激は、光および/または様々なレベルの糖があることか、またはないことである、請求項16に記載の方法。
【請求項28】
前記刺激は、発育段階により制御される刺激である、請求項16に記載の方法。
【請求項29】
前記糖は、1以上のスクロースまたはグルコースである、請求項27に記載の方法。
【請求項30】
前記糖はスクロースである、請求項29に記載の方法。
【請求項31】
請求項1に記載の核酸配列を含むキメラ遺伝子。
【請求項32】
請求項1に記載のプラスチドターゲティング配列をコードする核酸配列とデンプン分解経路の酵素の核酸コード配列とを含むキメラ遺伝子であって、前記酵素は、請求項4に記載の配列によりコードされる、キメラ遺伝子。
【請求項33】
さらなるコード配列の発現を指向することができる核酸配列を含むキメラ遺伝子であって、前記コード配列は請求項4に記載の配列である、キメラ遺伝子。
【請求項34】
刺激に反応する請求項6に記載の核酸配列とコード配列とを含むキメラ遺伝子であって、前記コード配列の発現レベルは、前記核酸配列に適用された刺激に応じて変化する、キメラ遺伝子。
【請求項35】
請求項1、4、5、または6のいずれか1つに記載の核酸配列を含む植物細胞。
【請求項36】
1以上の請求項1、4、5、または6に記載の核酸配列を含む形質転換植物の種子。
【請求項37】
請求項8、14、または16のいずれか1つに記載の方法によって形質転換された植物であって、前記植物は、1以上のジャガイモ、コムギ、トウモロコシ、オオムギ、トマト、イネ、エンドウ、ラッカセイ、キャッサバ、ヤムイモ、バナナ、およびタバコからなる群である、植物。
【請求項1】
本明細書では配列番号1として知られ、1〜294のヌクレオチドであり、かつ前記ヌクレオチド中に、さらなるコード配列を植物プラスチドに標的化することができる配列を有する核酸配列、または開示された配列である配列番号1と少なくとも65%以上相同であり、かつ同じ標的化能力を有する配列。
【請求項2】
前記核酸配列は約94のアミノ酸残基をコードする、請求項1に記載の核酸配列。
【請求項3】
前記配列は約85のアミノ酸残基をコードする、請求項2に記載の核酸配列。
【請求項4】
本明細書では配列番号2として知られ、1〜1642のヌクレオチドであり、かつ前記ヌクレオチド中にβ−アミラーゼをコードすることができる配列を有する核酸配列、または配列番号2内の開示された配列と少なくとも65%以上相同であり、かつ同じコード能力を有する配列。
【請求項5】
本明細書では配列番号3として知られ、1〜1953のヌクレオチドであり、かつ前記ヌクレオチド中に、葉緑体標的化β−アミラーゼをコードすることができる配列を有する核酸配列、または配列番号3内の開示された配列と少なくとも65%以上相同であり、かつ同じコード能力を有する配列。
【請求項6】
産物の発現を指向することができる核酸配列であって、前記産物は、前記核酸配列に作動可能に連結されたコード配列によりコードされ、前記核酸配列は、本明細書では配列番号8として知られるか、あるいは配列番号8と少なくとも65%相同であり、かつ配列番号8と実質的に同じ機能を有し、前記核酸配列は刺激反応性であり、前記産物の発現レベルは前記核酸配列に適用された刺激に応じて変化する、核酸配列。
【請求項7】
前記核酸配列は、mRNA、cDNA、またはゲノムDNAの配列である、前記請求項のいずれか1つに記載の核酸配列。
【請求項8】
デンプン生合成経路または分解経路の酵素の活性を植物において増加または減少させる方法であって、前記方法は、プラスチドターゲティング配列をコードする核酸配列とデンプン生合成経路または分解経路の酵素のコード配列とを含むキメラ遺伝子を植物ゲノムに安定に組み込ませるステップと、改変したゲノムを有する植物を再生するステップを含む、方法。
【請求項9】
前記核酸配列は請求項1に記載の配列を含む、請求項8に記載の方法。
【請求項10】
前記デンプン分解経路の酵素はβ−アミラーゼである、請求項8に記載の方法。
【請求項11】
前記コード配列は請求項4に記載の核酸配列であり、デンプン分解経路の酵素のコード配列である、請求項8、9、または10に記載の方法。
【請求項12】
前記核酸配列は請求項5に記載の核酸配列である、請求項8に記載の方法。
【請求項13】
請求項8〜12のいずれか1つに記載の方法であって、前記デンプン分解経路の酵素は、スクロース合成酵素、ADPGピロホスホリラーゼ、デンプン合成酵素、分枝酵素、αアミラーゼ、イソアミラーゼ、非プラスチド型β−アミラーゼ、α−グルコシダーゼ、デンプンホスホリラーゼ、および不均化酵素からなる群の1つである、方法。
【請求項14】
タンパク質または酵素を植物プラスチドに標的化する方法であって、前記方法は、プラスチドターゲティング配列をコードする核酸配列とタンパク質または酵素のコード配列とを含むキメラ遺伝子を植物ゲノムに安定に組み込ませるステップと、改変したゲノムを有する植物を再生するステップを含み、前記タンパク質または酵素は、1以上の下記の群:脂質合成、光合成、アミノ酸代謝、窒素固定、炭素固定、または炭水化物ポリマー合成の経路であるか、あるいは前記植物にある特徴を付与することができ、前記核酸配列は請求項1に記載の核酸配列である、方法。
【請求項15】
前記特徴は、1以上の下記の群:除草剤抵抗性および有害生物抵抗性から選択される、請求項14に記載の方法。
【請求項16】
産物の発現レベルを変化させる方法であって、前記産物は、植物において前記産物の発現を指向することができる核酸配列に作動可能に連結されたコード配列によりコードされ、前記方法は、前記核酸配列に作動可能に連結されたコード配列によりコードされる産物の発現を指向することができる核酸配列を含むキメラ遺伝子を植物ゲノムに安定に組み込ませるステップを含み、前記核酸配列は、配列番号8の配列を実質的に有するか、または配列番号8と少なくとも65%相同であり、かつ配列番号8と同じ機能を実質的に有し、刺激反応性である、方法。
【請求項17】
前記コード配列は、前記植物において開花中に不稔性を引き起こす遺伝子をコードし、かつ光に暴露されると前記核酸配列により発現させられる、請求項16に記載の方法。
【請求項18】
前記コード配列は、葉または他の光合成組織におけるデンプン生合成酵素遺伝子のレベルを変化させる遺伝子をコードする、請求項16に記載の方法。
【請求項19】
前記コード配列は、発育中の貯蔵器官において糖が産生され、かつ糖が前記核酸配列に作用すると、発現レベルが変化する遺伝子をコードする、請求項16に記載の方法。
【請求項20】
前記核酸配列は、光はあるが糖がない場合に作動可能である、請求項16に記載の方法。
【請求項21】
前記核酸配列は、糖はあるが光がない場合に作動可能である、請求項16に記載の方法。
【請求項22】
さらなる酵素をアップレギュレートまたはダウンレギュレートし、それにより再形質転換植物により産生されるデンプン量を増加または減少させるために、遺伝的形質転換の結果として酵素活性の増加または減少を既に示しているトランスジェニック植物において前記さらなる酵素を変える方法。
【請求項23】
前記第1の形質転換植物は、デンプン生合成経路の酵素活性が増加または減少した植物である、請求項22に記載の方法。
【請求項24】
前記第1の形質転換植物は、ADPG−PPase遺伝子で形質転換されたトランスジェニックジャガイモである、請求項23に記載の方法。
【請求項25】
前記キメラ遺伝子は、カリフラワーモザイクウイルス35Sプロモーター(完全または切断)、ルビスコプロモーター、エンドウプラストシアニンプロモーター、ノパリン合成酵素プロモーター、クロロフィルr/b結合プロモーター、高分子量グルテニンプロモーター、α,β−グリアジンプロモーター、ホルデインプロモーター、およびパタチンプロモーターからなる群から選択されるプロモーターをさらに含む、請求項8、14、または16のいずれか1つに記載の方法。
【請求項26】
前記キメラ遺伝子中の前記酵素の前記コード配列は、前記酵素の活性をアップレギュレートまたはダウンレギュレートする、請求項8、14、または16のいずれか1つに記載の方法。
【請求項27】
前記刺激は、光および/または様々なレベルの糖があることか、またはないことである、請求項16に記載の方法。
【請求項28】
前記刺激は、発育段階により制御される刺激である、請求項16に記載の方法。
【請求項29】
前記糖は、1以上のスクロースまたはグルコースである、請求項27に記載の方法。
【請求項30】
前記糖はスクロースである、請求項29に記載の方法。
【請求項31】
請求項1に記載の核酸配列を含むキメラ遺伝子。
【請求項32】
請求項1に記載のプラスチドターゲティング配列をコードする核酸配列とデンプン分解経路の酵素の核酸コード配列とを含むキメラ遺伝子であって、前記酵素は、請求項4に記載の配列によりコードされる、キメラ遺伝子。
【請求項33】
さらなるコード配列の発現を指向することができる核酸配列を含むキメラ遺伝子であって、前記コード配列は請求項4に記載の配列である、キメラ遺伝子。
【請求項34】
刺激に反応する請求項6に記載の核酸配列とコード配列とを含むキメラ遺伝子であって、前記コード配列の発現レベルは、前記核酸配列に適用された刺激に応じて変化する、キメラ遺伝子。
【請求項35】
請求項1、4、5、または6のいずれか1つに記載の核酸配列を含む植物細胞。
【請求項36】
1以上の請求項1、4、5、または6に記載の核酸配列を含む形質転換植物の種子。
【請求項37】
請求項8、14、または16のいずれか1つに記載の方法によって形質転換された植物であって、前記植物は、1以上のジャガイモ、コムギ、トウモロコシ、オオムギ、トマト、イネ、エンドウ、ラッカセイ、キャッサバ、ヤムイモ、バナナ、およびタバコからなる群である、植物。
【図1】



【図2A】
【図2B】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図2A】
【図2B】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【公開番号】特開2010−131029(P2010−131029A)
【公開日】平成22年6月17日(2010.6.17)
【国際特許分類】
【出願番号】特願2010−28677(P2010−28677)
【出願日】平成22年2月12日(2010.2.12)
【分割の表示】特願2000−566401(P2000−566401)の分割
【原出願日】平成11年8月13日(1999.8.13)
【出願人】(301004857)アドヴァンスト・テクノロジーズ(ケンブリッジ)リミテッド (8)
【Fターム(参考)】
【公開日】平成22年6月17日(2010.6.17)
【国際特許分類】
【出願日】平成22年2月12日(2010.2.12)
【分割の表示】特願2000−566401(P2000−566401)の分割
【原出願日】平成11年8月13日(1999.8.13)
【出願人】(301004857)アドヴァンスト・テクノロジーズ(ケンブリッジ)リミテッド (8)
【Fターム(参考)】
[ Back to top ]