
新規の非タンパク分解起源の可溶性EPCRタンパク質およびその使用
本発明は、選択的スプライシング機構に由来する新規の可溶形EPCRに関し、より詳細には、配列番号1の配列の残基201〜256または前記配列のフラグメントを含むポリペプチドに関する。本発明はまた、前記ポリペプチドを含む組み換え型または単離されたEPCRタンパク質、前記ポリペプチドをコードする単離されたポリヌクレオチド、前記ポリペプチドを含むタンパク質をコードするmRNA、および疾患の検出におけるこれらの使用に関する。本発明はさらに、前記ポリヌクレオチドを含む発現ベクターまたは宿主細胞、発現ベクターと前記細胞を含む発現系、前記ポリペプチドに対して特異的な単離された抗体、および生体試料における配列番号1の配列を含むEPCRタンパク質の選択的なインビトロ検出のための方法およびキットに関する。
【発明の詳細な説明】
【技術分野】
【0001】
[発明の背景]
最近記載されているプロテインCシステム(PC)のタンパク質は、内皮プロテインC/活性化プロテインC受容体(EPCR)である(1、2)。EPCRは、内皮細胞の膜上に発現され、高親和性でPCおよび活性化プロテインC(APC)と結合する(Kd〜30nM)。その生理学的使命は、内皮表面上のPCを濃縮し、これをトロンビン−トロンボモジュリン複合体に提示し、それによって、PCの効率的な活性化を助けることである(3)。ヒト以外の霊長類に関して実施された研究では、モノクローナル抗体を用いてEPCRをブロックすることは、APCの生理学的産生を90%減少させることが実証されており、また、最近の研究では、EPCRの遺伝的欠陥が、静脈および動脈の血栓形成を患う素因になる可能性があることが示唆されている(4〜7)。これらの事実は、凝固調節において、また血栓形成において、EPCRが果たす可能性がある重要な役割を実証する。さらに、EPCRは、PCの強力な抗炎症作用を担う分子の1つであるように思われる。
【0002】
EPCRとPC/APCの相互作用は、グラム陰性細菌による感染中に現れる凝血異常性および炎症性の反応の調節の際に、重要な役割を果たす可能性がある(8)。近年では、APCは、内皮細胞上の抗炎症および抗アポトーシス作用を有すること、また、この作用が現れるためにはEPCRの存在が不可欠であることが実証されている(9、10)。
【0003】
EPCRは、約46kDaの糖タンパク質である。成熟タンパク質は、17残基からなるシグナルペプチドの排除後の、221アミノ酸からなる(11)。これは、3つのイントロンによって分断された4つのエキソンによって形成される、染色体20(20q11.2)のロングアーム中に位置する遺伝子によってコードされる(12)。エキソンIは、5’非翻訳領域およびシグナルペプチドをコードし、エキソンIVは、膜貫通ドメイン、3残基の細胞質内テール、および3’非翻訳領域をコードする。エキソンIIおよびIIIは、クラスI主要組織適合遺伝子複合体のCD1スーパーファミリーに属するタンパク質のドメインα1およびα2との相同性を有する、EPCRの細胞外領域の大部分をコードする。近年では、EPCRが、ポケット(そこには、分子を安定させるリン脂質が収容される)を形成する、折り畳まれたβシート(その上にはαヘリックス構造を2つ有する)によって形成されるプラットフォームを、どのように有するかを示す三次元構造が、首尾よく結晶化され、分析されている(13)。同様に、アラニンスキャニングによって、PC/APCとの結合に関与する主要な残基(カルシウムおよびマグネシウムイオンの存在下で、そのGLAドメインを介して結合する)が特定されている(14)。
【0004】
しかし、内皮細胞膜上に固定されるEPCRに加えて、トロンビンなどの刺激によって誘導されるメタロプロテイナーゼによるEPCRの消化に少なくとも部分的に由来する、血漿可溶形(sEPCR)が存在する(15、16)。
【0005】
論文(15)「J.Clin.Invest.Vol.100」、no.2、1997年7月、411〜418は、血漿中に見られるEPCRの可溶形に関する。この論文には、血漿中のEPCRは、独特の種であるように思われ、精製されたタンパク質のアミノ末端フラグメントの配列決定では、血漿EPCRの特有の配列が示され、これは、組換えsEPCRのアミノ末端配列と一致したことが記載されている。この論文には、前記可溶性EPCRの起源は、他の受容体の場合と同様に、タンパク分解性の破断または選択的切断−スプライシング機構である可能性があり、特に、ウシEPCRのゲノム構造は、タンパク質をコードする選択的切断部位を含有する(ここでは、膜貫通ドメインが、エキソンIIIの後に位置しているイントロンによってコードされる配列によって置き換えられる)ことが記載されている(17)。この文献は、選択的切断から得られるEPCRの形の可能性を証明するために、より多くの研究が必要であることを示している。
【0006】
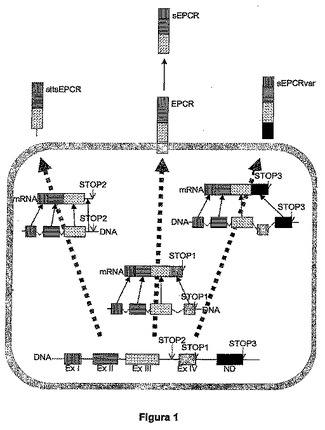
さらに、国際公開第−9900673号には、ヒト血漿におけるsEPCRの単離および特徴づけに関し、sEPCRが、タンパク質加水分解に由来する(ここでは、EPCRの細胞外ドメインは遊離され、膜に結合した残りのタンパク質から離れる)、あるいは、mRNAの選択的切断−スプライシング機構(これは、altsEPCRの配列を生じさせる)に由来する可能性があることが記載されている。本出願の図1は、タンパク分解起源の可溶性EPCR(psEPCR)と、選択的切断−スプライシングによって得られる国際公開第−9900673号に開示されたEPCR(altsEPCR)と、本発明の新規のEPCR目的物(sEPCRvar)との間の違いを見ることができる図を示す。
【0007】
したがって、本出願によって、少なくとも、ヒトEPCRは、選択的切断−スプライシング・プロセスを受け、前記選択的切断−スプライシング機構から得られる可溶性のEPCR(sEPCRvar)中にのみ存在する特有のインサートを含む、可溶性の切断型EPCRを生じる可能性があることが示される。前記sEPCRvarは、大血管における、またガンにおける疾患プロセスマーカーとして働く可能性がある。
【0008】
APCが、sEPCRに結合する場合、その抗凝血特性を失うが、その酵素的能力は維持されるため、sEPCRが、APC特異性を変化させ、おそらく、これを基質(現在は未知である)に方向付け、これが、APCに起因する抗炎症特性につながる可能性があると考えられる(18)。sEPCRは、膜上に存在するEPCRと、プロテインCに対して競合し、このようにしてAPC産生を抑制する。sEPCRは、プロテイナーゼ−3(PR3)(19)、好中球顆粒およびMac−1(CD11b/CD18)中に存在するプロテイナーゼ、好中球および単球の表面に存在する誘導性の発現のインテグリンを介して、活性化された好中球と相互作用することが可能であることも記載されている。PR3は、おそらく、膜結合型TNF−アルファおよびIL−1ベータ前駆体に作用する組織破壊メカニズムに関与する。Mac−1は、細胞シグナル伝達および細胞間接着現象に介在し、内皮ICAM−1などの接着分子と相互作用し、その理由で、炎症性プロセス中の好中球の補充において役割を果たす。PR3およびMac−1へのsEPCRの結合は、おそらく、その活性を低下させるであろう。
【発明の開示】
【発明が解決しようとする課題】
【0009】
本発明者らは、ヒトゲノムDNA(cDNA)から開始して、新規のタンパク質をクローン化し、発現させることを可能にした。前記タンパク質は、その配列の、他の既知の配列との分析および比較、ならびにその機能の特徴づけによって、選択的スプライシング・プロセスによって産生される可溶性のEPCRタンパク質の未知の形であることが示された。一方、前記タンパク質は、示差的な特徴として、現在知られているEPCR形には存在しない、当初EPCR遺伝子3’非翻訳領域と考えられていた領域によってコードされるポリペプチドの領域を有する。
【課題を解決するための手段】
【0010】
[発明の詳細な説明]
本発明は第一に、新規の可溶形のEPCR(これを、本発明者らは、可溶性のEPCR変異体(sEPCRvar)と呼ぶ)に、より詳細には、配列番号1の配列の残基201〜256を含む、内皮プロテインC受容体または前記配列の残基のフラグメントから得られるポリペプチドに関する。ある特定の実施形態では、前記フラグメントは、少なくとも9アミノ酸を有する。本発明の特定の実施形態によれば、前記フラグメントは、配列番号1の残基213〜227、229〜242、212〜226、227〜241、242〜256を含むフラグメントから選択される。
【0011】
ある特定の実施形態では、前記フラグメントは免疫原特性を有し、これは特に、特異的抗体を産生するための手順で有用である。
【0012】
本発明はさらに、配列番号1の配列の残基201〜256から形成されるポリペプチドを含む、単離されたあるいは組み換え型のタンパク質に関する。前記タンパク質は、EPCRタンパク質である。
【0013】
本発明の好ましい実施形態によれば、前記単離されたあるいは組み換え型のEPCRタンパク質は、配列番号1の配列を含む。
【0014】
本発明のさらなる目的は、配列番号1の配列の残基201〜256から形成されるポリペプチドまたは前記配列の残基のフラグメントを含む組み換え型タンパク質である。
【0015】
本発明のさらなる目的は、配列番号1の配列の残基201〜256によって形成されるポリペプチドまたは前記配列の残基のフラグメントをコードする配列を含むポリヌクレオチドである。ある特定の実施形態では、前記ポリヌクレオチドは、配列番号2に相当するcDNA配列に由来する。このcDNAは、mRNAから逆転写されたDNAに関する。
【0016】
本発明の特定の実施形態では、本発明のポリヌクレオチドは、DNA構築物中に含有される可能性がある。したがって、本発明のさらなる目的は、配列番号1の配列の残基201〜256を含むタンパク質またはポリペプチドまたは前記配列の残基のフラグメントをコードするDNA構築物である。好ましくは、前記フラグメントは、少なくとも9アミノ酸を有する。前記構築物のある特定の実施形態では、これは、配列番号2に相当するcDNA配列に由来するポリヌクレオチドを含む。前記DNA構築物は、前記タンパク質またはポリペプチドに作動可能に結合される制御配列を組み込む可能性がある。核酸またはポリヌクレオチドに関して、「作動可能に結合される」は、あるヌクレオチド領域が、別のヌクレオチド配列と、機能的関係に置かれることを意味する。「制御配列」は、機能(例えばある種のコード配列の転写および翻訳など)を調節する、ある種の宿主細胞によって特に認識される発現シグナルである。プロモーター、発現エンハンサー、転写ターミネーター、リボソーム結合部位などは制御配列である。DNA構築物を形成するための所望の配列のスプライシングは、適切な制限部位における切断およびスプライシングによって行うことができる。これらの結合部位が存在しない場合、アダプターまたはスペーサーとして合成のオリゴヌクレオチドを使用して、従来の遺伝子工学的方法によってこれらを産生することが可能である。
【0017】
本発明のポリヌクレオチドまたはDNA構築物は、しばしば実験室マニュアルに書き留められている従来の遺伝子工学的方法によって得ることができる。
【0018】
本発明のポリヌクレオチドまたは構築物は、適切な発現ベクター中に挿入させることができる。したがって、本発明のさらなる目的は、前記ポリヌクレオチドまたはDNA構築物を含む発現ベクターである。本発明の様々な実施形態における発現ベクターの選択は、前記発現ベクターが挿入されることとなる宿主細胞に依存する。例えば、本発明のポリヌクレオチドまたはDNA構築物が挿入されるベクターは、プラスミドまたはウイルス(これらは、一旦宿主細胞中に導入された場合に、細胞のゲノムに組み込まれる可能性もあるし、組み込まれない可能性もある)である可能性がある。この発現ベクターはまた、従来方法によって得ることができる。
【0019】
本発明のさらなる目的は、配列番号1の配列の残基201〜256含むポリペプチドまたは前記配列の残基の少なくとも9アミノ酸を有するフラグメントををコードするポリヌクレオチドまたはDNA構築物を含む、形質転換された宿主細胞である。本発明によれば、形質転換された宿主細胞は、原核生物細胞(例えば大腸菌)または真核生物(例えば酵母、特にピキアパストリス(Pichia Pastoris)およびサッカロミセスセレビシエ(Saccharomyces cerevisiae))、昆虫細胞または哺乳類細胞株であり得る。
【0020】
本発明のある特定の実施形態では、配列番号2を有する本発明のポリヌクレオチドは、本発明において開示される選択的切断およびスプライシングによって産生される新規の形のsEPCR(sEPCRvar)に相当する、配列番号1の組み換え型のsEPCRタンパク質を産生するために、ピキアパストリス(Pichia Pastoris)中に導入される。組み換え型タンパク質の産生に必要なDNA構築物、発現ベクター、および形質転換された宿主細胞を産生するために使用される方法は、その精製のためのプロセスを含めて、後ほど詳述する。
【0021】
本発明のある特定の実施形態では、本発明のポリヌクレオチドまたはDNA構築物を含む発現ベクターは、組成物中でのその使用、およびインビボでの遺伝子導入または治療のプロセスのために設計される。より具体的な実施形態では、この発現ベクターは、ウイルスベクターである。この目的に適したウイルスベクターとしては、アデノウイルス、アデノ随伴ウイルス、レトロウイルス、レンチウイルス、アルファウイルス、ヘルペスウイルス、コロナウイルス類縁体(derivative)ベクターなどが挙げられる。
【0022】
本発明のさらなる目的は、配列番号1の配列の残基201〜256を含むポリペプチドまたは前記配列の残基のフラグメントをコードするポリヌクレオチドを含む発現ベクターと、前記ポリヌクレオチドまたは前記構築物で形質転換される宿主細胞を含む発現系である。好ましくは、前記フラグメントは、少なくとも9アミノ酸を含む。
【0023】
本発明のさらなる目的は、配列番号1の残基201〜256を含むタンパク質またはポリペプチド、または前記配列の残基の好ましくは少なくとも9アミノ酸を有するフラグメントを産生する方法であって、前記方法が、前記タンパク質、ポリペプチド、またはフラグメントの発現を可能にする条件で、本発明のポリヌクレオチドまたはDNA構築物を含む宿主細胞を培養することを含むことを特徴とする方法である。培養を最適化する条件は、使用される宿主細胞に依存する。所望により、この方法はまた、発現されたタンパク質またはポリペプチドを単離および精製するためのいくつかのステップを含む可能性がある。
【0024】
あるいは、本発明のタンパク質またはポリペプチドは、他の従来方法によって、例えば固相技術に関する化学合成、高速液体クロマトグラフィー(HPLC)による精製によって得ることができ、所望により、これらを、従来の技術、例えば配列決定および質量分析、アミノ酸分析、核磁気共鳴などによって分析することもできる。
【0025】
本発明のさらなる目的は、配列番号1の配列を含むタンパク質または前記配列の残基のフラグメントをコードするmRNAである。
【0026】
本発明のさらなる目的は、配列番号1の残基201〜256を含むポリペプチドまたは前記配列の残基の免疫原特性を有するフラグメントに対する、特定の単離された抗体である。ある特定の実施形態では、前記フラグメントは、少なくとも9アミノ酸を有する。
【0027】
ある特定の実施形態では、前記抗体は、その配列が残基201〜256、213〜227、229〜242、212〜226、227〜241、および242〜256(すべて配列番号1に関する)から選択されるポリペプチドまたはフラグメントに対して特異的である。
【0028】
別の特定の実施形態では、前記抗体は、配列番号1に対して特異的であり、これが領域201〜256に結合しない場合には領域1〜200を認識しない。
【0029】
好ましい実施形態によれば、前記抗体は、モノクローナル抗体である。
【0030】
本発明のさらなる目的は、生体試料における、配列番号1の配列の残基201〜256を含むEPCRタンパク質(例えば配列番号1の新規のEPCRタンパク質)、または前記配列の残基のフラグメントの選択的インビトロ検出のための方法であって、
−生体試料を得ること、
−配列番号1の配列の残基201〜256を含むEPCRタンパク質または前記配列の残基のフラグメントの量を分析すること、
を含む方法である。
【0031】
前記生体試料は、生体材料を含む任意の試料である可能性がある。好ましい実施形態では、前記生体試料は、尿、血漿、血清、組織、または間質液試料である。
【0032】
前記方法は、例えば、質量分析、イムノアッセイ、化学アッセイ、液体クロマトグラフィー、および様々な間接的および直接的光度測定方法を含めた、任意の既知の方法であり得る。例えば、いくつかの分析法は、質量分析を使用するマーカーを定量化するために適用される可能性がある。一般的に言うと、マーカーは、例えば液体クロマトグラフィーまたは二次元ゲル電気泳動によって、生体試料から単離される。マーカーは、質量分析、例えば、液体クロマトグラフィー(LC−MS)を伴うタンデム質量分析、MALDI−TOF−MS(「マトリクス支援レーザー脱離/イオン化質量分析・飛行時間型MS」)などによって定量化される。標的マーカーは、既知量の精製されたマーカー標準と比較することによって、あるいは、健康な対照から得られる同じタイプの生体試料中に存在する前記マーカーの量と比較することによって定量化することができる。
【0033】
本発明の別の実施形態では、この方法は、イムノアッセイであり得る。一般に、イムノアッセイには、1種または複数のマーカーリガンドが使用される。本発明で使用される「リガンド」は、標的マーカーに特異的に結合することが可能な任意の化合物または分子である。これらのリガンドは、別々に、あるいは組み合わせて使用することができる(例えば、抗体は、アプタマーと組み合わせて使用することができる)。
【0034】
本発明の一つの実施形態では、イムノアッセイは、均一(homogenous)アッセイ、不均一(heterogeneous)アッセイ、酵素免疫測定法(EIA、ELISA)、競合アッセイ、イムノメトリックアッセイ(サンドイッチ)、濁度アッセイ、比濁アッセイ、または他の同様のアッセイである可能性がある。同様に、イムノアッセイは、手動で、あるいは自動分析器を用いて行われる可能性がある。
【0035】
本発明の方法では、前記タンパク質の量は、血管損傷に関連する炎症性疾患、炎症、凝固異常に関連する疾患、およびガンから選択される疾患に関連する。前記疾患はまた、狼瘡などの自己免疫性疾患、敗血症、ショック、子癇前症、糖尿病、移植のモニタリングの際の不安定狭心症、再狭窄、血管形成、および肝臓または腎臓病である可能性もある。
【0036】
本発明の方法によれば、検出されるEPCRタンパク質の量はまた、較正標準と比較することができる。
【0037】
本発明のさらなる目的は、配列番号1の配列の残基201〜256を含むEPCRタンパク質、例えば、配列番号1の新規のEPCRタンパク質、または前記配列の残基の好ましくは少なくとも9アミノ酸のフラグメントの検出および定量化のための試験キットであって、
a)配列番号1の配列の残基201〜256を含むポリペプチドまたは前記配列の残基のフラグメントに特異的な抗体、即ち、領域201〜256に結合しない場合には領域1〜200を認識しない配列番号1のタンパク質に特異的な抗体、
b)抗体a)と、生体試料中に存在する配列番号1の配列の残基201〜256を含むEPCRタンパク質または前記配列の残基のフラグメントとの間の反応を検出する試薬、
を含むキットである。
【0038】
好ましくは、このキットはまた、配列番号1の配列の残基201〜256を含むEPCRタンパク質の正常および異常なレベルと、産生される反応物の量を相関させるための標準を含む。
【0039】
本発明のさらなる目的は、配列番号1の配列の残基201〜256を含むEPCRタンパク質、例えば配列番号1の新規のEPCRタンパク質、または前記配列の残基のフラグメントに相当するmRNAの検出および定量化のための、配列番号1の配列の残基201〜256を含むポリペプチドまたは前記配列の残基のフラグメント(好ましくは、前記フラグメントは、少なくとも9アミノ酸を有する)をコードするポリヌクレオチドの使用である。前記EPCRタンパク質は、血管損傷に関連する炎症性疾患、炎症、ガン、および凝固異常に関連する疾患から選択される疾患に関連する量で存在する。
【0040】
さらに、非タンパク分解起源の新規の可溶形のEPCRは、この形の可溶性のEPCR(sEPCRvar)の存在を、単独的かつ独占的に検出するのに選択的である分析方法であって、他の形の可溶性のEPCRまたは膜EPCRが検出される方法と重複する前記方法を用いずに、特定の疾患を検出できるという利点を有する分析方法の開発のために役割を果たす可能性がある。
【0041】
[図面の簡単な説明]
図1は、RNAがプロセッシングされ、成熟したメッセンジャーRNA(mRNA)が産生される方式に依存する、異なるタイプのEPCRを示す図である。3つの別の形のmRNAが産生される可能性があり、これらは、続いて、3つの異なる形のタンパク質を産生することとなる:1:膜EPCR(mEPCR)として翻訳されるタンパク質とは異なる4つのエキソンを伴うmRNA。2.エキソンEx I、Ex II、およびEx IIIのみを含み、その後にイントロン3およびこのイントロンの後の全配列が続く、国際公開第9900673号パンフレット中にエスモン(Esmon)によって開示された可溶形のEPCR(altsESPC)として翻訳されることとなるmRNA。3.本発明に開示された新規の形の選択的切断およびスプライシングによって、エキソンEx I、Ex II、およびEx IIIを含むmRNAが産生されることとなるが、プロセッシング中に、エキソンEx IVは失われ、この理由から、膜貫通ドメインも細胞質のテールも転写されず、その場所では、エキソンIIIの後、3’非翻訳領域に位置する潜在性(cryptic)エキソンであるであろう配列が出現し、この場合、mRNAは、本発明の新規の可溶性EPCR形(sEPCRvar)として翻訳される。可溶性EPCRはまた、おそらくメタロプロテアーゼ(まだ特徴づけられていない)から、タンパク分解活性によって産生することもできる。メタロプロテアーゼは、膜高さでEPCRを切断し、その結果、細胞外受容体の画分からなり、膜貫通ドメインおよび細胞質内テールを欠く、新規の可溶形のEPCR(sEPCR)が産生される。
【0042】
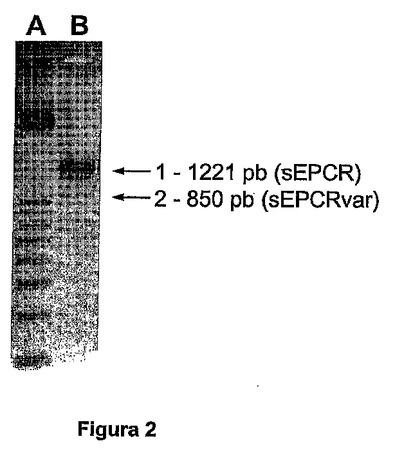
図2は、特異的なEPCR遺伝子プライマーを用いるHUVEC cDNAの増幅の後に得られるバンドパターンである:野生型EPCR cDNA(1221bp、矢印1)に相当するバンドに加えて、選択的切断−スプライシングによって産生された新規の変異体アイソフォームのcDNAに相当する新規の増幅断片が認められる(矢印2)。
【0043】
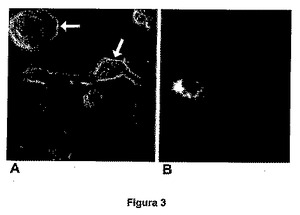
図3は、どちらも緑色蛍光タンパク質と融合させた、野生型EPCR(パネルA)を用いて、あるいはアイソフォーム(パネルB)を用いて形質移入されたCOS−7細胞である。パネルAは、野生型タンパク質が、細胞膜(矢印)上に蓄積する傾向があることを示すのに対して、アイソフォーム3を用いた場合、この局在化パターンは認められない(パネルB)。
【0044】
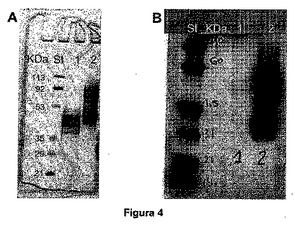
図4は、ピキアパストリス(Pichia Pastoris)におけるsEPCRvar発現を説明する図である。本明細書に示すように、sEPCRvarは、安定に形質転換されたP.パストリス(P.pastoris)細胞の上清から精製された。パネルA:SDS−PAGEによって分離され、GELCODE Blue染料を使用して検出された様々なタンパク質を示す。ストリート St:発現される各バンドの分子量をkDaで示す、分子量マーカー、ストリート 1:組み換え型のsEPCR。約42kDaのブロードなバンド(タンパク質の強いグリコシル化に起因する)が観察される。ストリート 2:sEPCRvar。47kDaのブロードなバンド(タンパク質の強いグリコシル化に起因する)が観察される。パネルB:モノクローナル抗−EPCR抗体を用いるウェスタンブロットによって検出されるタンパク質、ストリート St:発現される各バンドの分子量をkDaで示す、分子量マーカー、ストリート 1:ブランク、ストリート 2:sEPCRvar。約47kDaのブロードなバンド(様々なタンパク質のグリコシル化に起因し、35から60kDaまでの間に広がる)が観察される。
【0045】
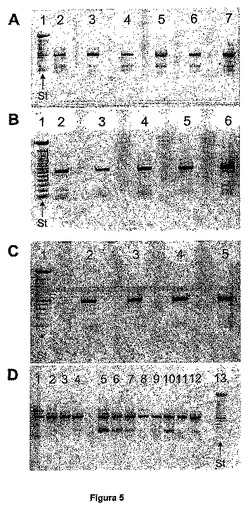
図5は、異なる組織および腫瘍由来のcDNAからのEPCR遺伝子の増幅である。パネルA)1:分子量マーカー、2:心臓、3:肝臓、4:腎臓、5:膵臓、6:肺、7:胎盤。パネルB)1:分子量マーカー、2:骨格筋、3:脳、4:胸腺、5:小腸、6:脾臓。パネルC)1:マーカー、2:前立腺、3:睾丸、4:卵巣、5:大腸。パネルD)は、肺腫瘍の腫瘍株:2:H 446、3:H 510、4:H 1264、5:H 549、6:H 441、7:HTB 51、8:H 676、9:H 727、10:H 720、11:H 385、12:H 1299からのサンプルである。分子量マーカーを含有するストリートは、図中に略語St(「standard(標準)」より)で示す。マーカーの下方のバンドは、100bpサイズに相当し、各新しいバンドは、その前のバンドに対して100bpの増分(100bp、200bp、300bp・・・)に相当する。
【発明を実施するための最良の形態】
【0046】
内皮細胞cDNAからのEPCR cDNAの増幅
HUVEC細胞を、ヒトの臍帯静脈から得て、標準の手順に従って培養し(ジャッフェ EA(Jaffe EA)、ナフマン RL(Nachman RL)、ベッカー CG(Becker CG)、ミニック CR(Minick CR)「Culture of human endothelian cells derived from umbilical veins.Identification by morphologic and immunologic criteria.」、「J Clin Inves.」、1973年11月、52(11):2745〜56)、全RNAを、(ペレス−ルイス A(Perez−Ruiz A)、モンテス R(Montes R)、ベラスコ F(Velasco F)、ロペス−ペドレラ C(Lopez−Pedrera C)、アントニオパラモ J(Antonio Paramo J)、オルベ J(Orbe J)、ハーミダ J(Hermida J)、ローシャ E(Rocha E)、「Regulation by nitric oxide of endotoxin−induced tissue factor and plasminogen activator inhibitor−1 in endothelial cells.」、「Thromb Haemost.」、2002年12月、88(6):1060〜5)に記載される通り、標準の技術に従って抽出した。得られたRNAは、転写酵素を用いて37℃で60分間インキュベートし、cDNAに逆転写させ、それに続いて酵素失活のために65℃で5分間インキュベートした。これを行うために、120単位(U)のM−MuLV逆転写酵素(ギブコBRL(Gibco BRL))を、RNA1μgに加え、10μLの最終体積にした。これは、4μLの逆転写酵素緩衝液、2mmol/Lのデオキシヌクレオチド(dNTP、インビトロジェン(Invitrogen))、0.3μg/mLのランダムヘキサマー(ギブコBRL(Gibco BRL))、0.5mmol/Lのジオチオトレイトール(diothiotreitol)、および35UのRnase阻害剤(ヒト胎盤由来のRNAsin 48,000U/mL、RNAガード、GEヘルスケアバイオサイエンス(GE Healthcare Bio−Science))を含有していた。
【0047】
EPCR遺伝子の発現を、ポリメラーゼ連鎖反応(PCR)によって分析した。使用されるプライマー(ジェンセット(Genset)、フランス)は、配列番号3の配列に相当する5’プライマー、および配列番号4の配列に相当する3’プライマーであった。
【0048】
PCRは、25μLの体積で行われた。1μgのRNAの逆転写によってあらかじめ得られた一定分量のcDNAが使用され、それに、1Uのtaq DNAポリメラーゼ(ロシュ(Roche))、2.5μLの反応緩衝液(ロシュ(Roche))、0.5μLの各プライマー(10μmol/L溶液から)、0.25μLのデオキシヌクレオチド(dNTP、GEヘルスケアバイオサイエンス(GE Healthcare Bio−Science))、および0.25μlの1mg/mL BSAが加えられた。この反応混合物を、水で25μLに調節した。増幅サイクルは、94℃で30秒の変性相と、別のアニーリングと、72℃で3.5分の伸張相からなる。このサイクルを5回繰り返し、その後、同じ変性相と、別のアニーリングと、68℃で3分の伸張相の32サイクルを実施する。最終の伸張期間として、混合物を5分間68℃に維持して、増幅を終結させた。PCR増幅されたセグメントは、1221塩基対(bp)の長さを有する。EPCR遺伝子がこれらの条件で増幅された場合、さらなるバンドが、約850塩基対の高さで出現する(図2)。
【0049】
増幅されたEPCRのさらなるバンドの配列決定
バンド2は、鋳型として図2の850bpバンドから開始すること以外は、同じ増幅条件を使用して増幅した。アガロースゲル電気泳動の後、製造業者の指示書に従って市販のキット(QIAquick Gel Extraction Kit)を使用して、増幅産物を精製した。このようにして精製された産物を、配列決定反応(ABIプリズム(ABI Prism)(登録商標)、BigDyeTM Terminator Cycle Sequencing Ready Reaction Kit)のための鋳型として使用した。配列番号5および配列番号6の配列に相当するプライマーを使用して、2つの配列決定反応(1つは5’−3’方向、もう1つは逆方向)を実施した。
【0050】
反応の結果は、自動シーケンサー、ABI Prism 377 DNA Sequencerで分析した。結論としては、新規のバンドは、配列番号7の配列によって定義されるフラグメントに相当する。
【0051】
その配列が配列番号2であるこのフラグメントの中央部分は、エキソン4および3’非翻訳領域の部分を欠く新規のEPCRのcDNAに相当する。このcDNAの起源は、おそらく、EPCR RNAの選択的切断−スプライシング現象である。
【0052】
この新種のcDNAは、アミノ酸の配列が配列番号1であるポリペプチドをコードする。前記配列の残基1〜17は、シグナルペプチドに相当し、残基18〜200は、EPCRの共通の細胞外ドメインに相当し、残基201〜256は、この新規の形のEPCRの新規の示差的なドメインに相当し、EPCRの膜貫通ドメインに代わるものである。この新規のドメインは、膜貫通ドメインであるための基準を満たさない。
【0053】
緑色蛍光タンパク質と融合させたEPCRおよびsEPCRvarの哺乳類細胞における発現
EPCRアイソフォームの細胞内の局在化を研究するために、そのcDNAを、pcDNA3.1/CT−GFP−TOPOベクター中にクローニングし、それを緑色蛍光タンパク質との融合タンパク質として発現させた。このようにして調製されるベクターを使用して、Lipofectamine 2000(インビトロジェン(Invitrogen))を使用する培養で、哺乳類COS−7細胞を一過的に形質転換させた。対照として、同じベクターを使用し、そこでは野生型EPCRのcDNAをクローニングし、緑色蛍光タンパク質と融合させた。野生型EPCRおよびアイソフォームの細胞内局在化を、蛍光顕微鏡検査によって検出した。図3に示さているように、野生型EPCRは、主に細胞膜上に局在化する(ここでは緑色蛍光が認められる)が、sEPCRvarアイソフォームは、本発明者らの仮定と一致して、細胞膜上には局在化しない。
【0054】
EPCRアイソフォームのクローニング、発現、精製、および特徴付け
クローニングおよび発現
同定されたEPCR変異体(EPCRvar)を発現させるために、sEPCR var配列を、cDNA鋳型として内皮細胞を使用して、イニシエーター
5’−agcttggcatatcgattagccaagacgcctcagatg−3’および
5’−agctatcgtagcggccgcctaccctattatatcagc−3’
を用いる(これらは、それぞれ5’および3’末端にClaIおよび別のNotI制限部位を付加されている)ポリメラーゼ連鎖反応(PCR)によって、そのシグナルペプチドを伴わずに増幅させた(残基17〜256)。こうした修飾により、サッカロミセスセレビシエのα因子分泌シグナルの後の、pPICZαC(インビトロジェン(Invitrogen))プラスミドのClaIおよびNotI部位に、EPCR変異体配列を結合することが可能になり、酵母から細胞外培地への多くのタンパク質の効率的な分泌が可能になった。クローニングプロセスにより、セリン残基および別のイソロイシン残基が、sEPCRvarのアミノ末端に加えられた。直接的配列決定を使用して、挿入配列およびベクターが適切であることが確認されることが確認された。あらかじめ調製された発現ベクターを用いて、PmeI制限酵素を用いるそれらの直鎖化の後、ピキアパストリス(Pichia Pastoris)細胞を、化学的方法を使用して形質転換させ(Easy Comp、インビトロジェン(Invitrogen))、その結果、メタノール反応内因性プロモーターにおける、sEPCRvarコード配列の相同的組換えによる組込みが示された。ゼオシンの存在下で形質転換産物を培養し、sEPCRvarコード配列を含有する、言い換えるとゼオシン抵抗性遺伝子を含有するベクターで形質転換されたP.パストリス(P.Pastoris)コロニーを選択した。簡単に言うと、形質転換された酵母を、1%(v/v)のグリセロール(BMGY)が補充された4mlのBMY培地[1%(w/v)の酵母抽出物、2%(w/v)のペプトン、100mMリン酸カリウム(pH6,0)、硫酸アンモニウムを含む酵母窒素源1.34%(w/v)、4×10−5%(w/v)のビオチン]中で培養し、撹拌しながら約18時間28〜30℃でインキュベートした。細胞は、室温で5分間、2,000gで遠心分離することによって収集した。上清を捨て、18時間、1%メタノールを用いて、sEPCRvar発現を誘導した。これを行うために、細胞を、0.5%(w/v)メタノールが補充された3mlのMBYに再懸濁し、勢いよく攪拌しながら、約28〜30℃で18時間インキュベートした。誘導後、条件培地からのサンプルを、12% NuPAGE Bis−Trisゲル(インビトロジェン(Invitrogen)、カリフォルニア州カールズバッド(Carlsbad,CA))にローディングし、RCR−2モノクローナル抗体(フクドメケンジ(Kenji Fukodome)博士、佐賀大学(Saga University)、日本(Japan)によって快く提供された)を使用するウェスタンブロットによってsEPCRvarを検出した(図4)。大規模産生については、最も高濃度のsEPCRvarを分泌したコロニーを選択した。その高いsEPCRvar産生能力に起因して選択されたコロニーにおいて、そのメタノール代謝(新陳代謝が早いものまたは遅いもの)を研究し、これによって、最も適切なコロニーのための最適発現条件が確立された。一旦培養条件およびメタノール誘導が最適化されたら、大量のsEPCRvarを産生するために規模を増大した。
【0055】
精製
P.パストリス(P.Pastoris)培養上清を濃縮し、NaClを含まない20mM Tris−HCl(pH7.6)に対して透析した後、2つの連続する精製ステップ:イオン交換クロマトグラフィーおよびゲル濾過を実施した。イオン交換精製では、Resource Qカラム(GEヘルスケアバイオサイエンス(GE Healthcare Bio−Science))を使用し、溶出は、20カラム分に匹敵する体積の、0.0〜300mMのNaClグラジエントを用いて実施した。sEPCRvarを含有する溶出分画を合わせ、遠心限外濾過によって濃縮し、その後、Superdex 75−HR10/30カラム(GEヘルスケアバイオサイエンス(GE Healthcare Bio−Science))にかけてゲル濾過を実施した。精製されたタンパク質の濃度を、BCA総タンパク質アッセイ(ピエール(Pierre)、イリノイ州ロックフォード(Rockford,IL))およびBSA標準を使用して決定した。精製されたsEPCRvarを検出するために、サンプルを、12% NuPAGE Bis−Trisゲル(インビトロジェン(Invitrogen)、カリフォルニア州カールズバッド(Carlsbad,CA))にローディングし、還元条件下で電気泳動を実施し、続いて、クマシーブルーで染色した。電気泳動ゲルをエレクトロブロッティングにかけ、RCR−2モノクローナル抗体を用いてsEPCRvarを検出した。sEPCRvarの分子量を評価するために、各ゲル電気泳動には分子量標準を含ませた。
【0056】
新規の可溶性EPCR変異体の活性を特徴づけるための生化学的分析
PCに対するsEPCRvarの親和性
培養された内皮細胞におけるAPCの産生。使用される細胞株は、EA.hy926(トロンボモジュリンおよびEPCRを発現する能力を保持している、形質転換されたヒトの内皮細胞株)であった(スターンズ−クロサワDJ(Stearns−Kurosawa DJ)、クロサワ S(Kurosawa S)、モリカ JS(Mollica JS)、フェレル GL(Ferrell GL)、エスモン CT(Esmon CT)、「The endothelial cell protein C receptor augments protein C activation by the thrombin−thrombomodulin complex.」「Proc Natl Acad Sci U S A.」1996年、93:10212〜6)。96ウエルプレート上の5×104細胞/ウェルを、0.02U/mlのトロンビン(0.17nM)(ERL、イギリス、スウォンジー(Swansea,United Kingdom))と共にインキュベートし、150mM NaCl、5mM CaCl2、0.6mM MgCl2、1% BSA、0.001% Tween−20および0.02% NaN3を補充された、20mMトリス緩衝液(pH7.4)中で、PC(バクスター(Baxter)、米国イリノイ州ディアフィールド(Deerfield,IL.USA))の濃度を50nMと1,000nMとの間で変化させ、室温にて45分後、トロンビンを阻害するために、レピルジン(シェーリングAG(Schering AG)、ドイツ、ベルリン(Berlin,Germany))を、0.2μlの最終濃度で加え、3〜4分後に、APCによるそのタンパク質加水分解をモニタリングする目的で、発色基質S−2366(クロモジェニックス(Chromogenix)、イタリア、ミラノ(Milan,Italy)を0.4mMの最終濃度で加えた。405nmでの吸光度の上昇を、マイクロプレートリーダー(iEMS Reader、ラボシステムズ(Labsystems)、フィンランド(Finland)で動力学的に記録した。ミハエリス・メンテンの式に対する曲線データの調整を、これらの条件でのPC活性化Km値を算出するEnzfitterプログラム(バイオソフト(Biosoft)、イギリス、ケンブリッジ(Cambridge,United Kingdom))を使用して実施した。
【0057】
sEPCRvarの阻害効果を研究するために、2μmol/Lを、トロンビンおよびPCと同時に加えた。このようにして、PC活性化に対するsEPCRvarの効果を分析し、PCに対するその親和性を反映する、sEPCRが活性化に対して有する効果と比較することができた。
【0058】
表1は、sEPCRおよびsEPCRvarの存在下または非存在下での、培養された内皮細胞の表面上の、トロンビンによるプロテインC活性化の生化学的特性を示す。
【0059】
表1
【表1】

【0060】
PCに対するsEPCRおよびsEPCRvarの親和性
活性化部分卜ロンボプラスチン時間の決定。健康な対象からの5血漿の混合物を使用し、ダイアグノスティカ・スタゴ・ベーリンガーマンハイム(Diagnostica Stago Boehringer Mannheim)STA Compact装置(ロシュ(Roche))、およびパトロンチン(Pathromtin)試薬(デイドベーリング(Dade Behring)、米国(USA))を使用して、活性化部分卜ロンボプラスチン時間(APTT)を決定した。周知のように、正常な血漿の混合物中のAPCの存在は、APTTを延長させる。使用される実験条件では、APCが存在しない場合のAPTTは、33.1±0.4s(平均±SD)であったのに対して、APCが1nmol/Lの最終濃度で加えられた場合、APTTは、43.9±0.4sに延長された。sEPCRが加えられた場合、APCの効果は低下し、1mmol/LのsEPCRが存在する場合には、APTTは、37.2±0.2sであった。APCの抗凝血効果に対するsEPCRのこの阻害効果は、用量依存的である。正常な血漿の混合物に加えられたsEPCRは、APTTに対する直接的な効果を有しなかった(sEPCRが存在する場合の33.9±0.4sと比較して、sEPCRが存在しない場合には33.1±0.4s)。すべての実験を4回繰り返した。sEPCRではなく、同じ濃度のsEPCRvarが加えられた場合、結果は、sEPCRを用いて得られるものと重ねることができ、sEPCRと同様に、sEPCRvarもAPCと結合することが実証された。
【0061】
表2は、APC抗凝血作用に対するsEPCRおよびsEPCRvarの効果を示す。それに1nmol/LのAPCおよび様々な濃度のsEPCRが加えられた、正常な血漿の混合物の血液凝固時間(APTT)。4つの実験の平均±SDを示す。APCが存在しない場合のAPTTは、33.1±0.4秒であった。
【0062】
表2
【表2】
【0063】
様々な組織および腫瘍株におけるsEPCRvarの発現分析
sEPCRvarが発現される組織を同定するために、EPCRおよびsEPCRvarに相当するcDNAの同時増幅を可能にするプライマーを使用するPCRによって、様々なヒト組織のcDNAライブラリ(Multiple tissue cDNA(MTC)パネルIおよびII、バイオサイエンス(Bioscience)、米国(USA))を研究した。使用されるプライマー(ジェンセット(Genset、フランス)は、以下の通りであった:
プライマー5’:5’−GCAGTATGTGCAGAAACATATTTCCGC−3’および
プライマー3’:5’−CATCCCAAGTCTGACACACCTGGAAGT−3’
【0064】
PCRは、25μLの体積で実施した。cDNAのアリコットを使用し、これに、1 Uのtaq DNAポリメラーゼ(ロシュ(Roche))、2.5μLの反応緩衝液(ロシュ(Roche))、0.5μLの各プライマー(10μmol/L溶液から)、0.25μLのデオキシヌクレオチド(dNTP、GEヘルスケアバイオサイエンス(GE Healthcare Bio−Science))、および0.25μlの1mg/mL BSAを加えた。反応混合物を、水で25μLに調節した。増幅サイクルは、94℃で30秒の変性段階、56℃で1分の別のアニーリング段階、72℃で1.5分の伸張からなる。このサイクルを35回繰り返す。最終の伸張期間として72℃で5分間混合物を維持して、増幅を終結させた。増幅されたEPCRのcDNAセグメントは、578bp長であったのに対し、sEPCRvarのcDNAセグメントは、189bpであった(図5)。sEPCRvar形は、膵臓、心臓、肝臓、腎臓、肺、胎盤、胸腺、小腸、脾臓、および大腸に多量に発現された。sEPCRvar形は、いくつかの腫瘍株に、特にH 549、H 441、およびH 720に多量に発現された。いくつかの腫瘍株、特にH 446およびH 676は、まだ同定されていない別の形(これは、その電気泳動移動度によれば、1000bpを有する)をさらに発現した。
【0065】
抗体産生
免疫化
生後少なくとも1ヵ月のBALB/C雄を、以下のパターンに従って、sEPCRvarで免疫化させた:フロイント・アジュバントまたは別のアジュバント中の10μgと200μgの間の量の免疫原を用いる、少なくとも2週間間隔での3回の皮下、皮内、または腹腔内の免疫化、あるいはこれらの組み合わせ、3回目の免疫化の2週間後、2日間隔で、同じ量の免疫原(ただし、生理食塩水血清(saline serum)中)を使用して、2回の新規の追加免疫を投与した。他の動物も、配列番号1のペプチド213〜227、229〜242、212〜226、227〜241、および242〜256の混合物を用いて免疫化させた。これらのペプチドは、カイト J.(Kyte J.)、ドリトル R.F.(Doolittle R.F.)のアルゴリズム(「J.Mol.Biol.」、1982年、157:105〜132)に従うProtscaleプログラムによると非常に親水性であることから選択された。
【0066】
融合
最後の追加免疫投与の2日後に、ハイブリドーマを産生した。ポリエチレングリコール1500または4000の存在下で、動物の脾細胞を骨髄腫細胞(SP2/O−Ag14、P3X63−Ag8.6.5.3、P3−NS−1−Ag4−1、その他)と融合させることからなる技術を用いた。この薬剤は、膜融合を可能にし、その結果、様々な細胞型との間のハイブリッドが産生されることとなる。動物Bリンパ球と骨髄腫細胞との間のハイブリッドは、新規の細胞型、ハイブリドーマを形成し、これは、抗体を産生し、また、培養中に無限に増殖する。これらのハイブリドーマを選択し、残りのハイブリッドを排除するために、細胞培養物にHAT培地を加えた。HATは、アミノプテリン、すなわちグアノシンの新規の合成経路(新生経路)の阻害剤を含有する。骨髄腫細胞は、核酸合成サルベージ経路に必要な機能性のHPRT酵素を欠いているので、唯一の利用可能な経路(新生経路)が、アミノプテリンでブロックされる場合、これらは、核酸を合成することができない。したがって、培養の2週後、培養中で生き残る唯一の細胞は、脾細胞−骨髄腫ハイブリッドである。脾細胞は、ハイブリッドに代替の経路によるグアノシン合成能力を付与し、骨髄腫細胞に、インビトロの培養において際限なく培養される能力を付与する。
【0067】
抗sEPCRvarおよび抗sEPCR抗体を産生するハイブリドーマの検出
HAT培地中の細胞を、少なくとも10の96ウェル培養マイクロプレートに分配する。融合の9日後と14日後との間に、コロニーのサイズは、上清中の抗体の存在を分析するのに十分となる。目的の抗体、すなわちsEPCRvarに対する抗体を分泌するハイブリドーマを選択するために、クローンを含有する5枚のマイクロプレートのすべてのウェルから上清のサンプルを取り出し、これをイムノアッセイにかけた。ELISAおよびウェスタンブロットアッセイを実施した。ELISAについては、プレートを、sEPCRvar(0.3μg/ウェル)で被覆し、4℃での15時間のインキュベーション、および適切なタンパク質による対応するブロッキングの後、培養上清を加え、相応な洗浄を実施し、次いで、二次マウス抗免疫グロブリン抗体を加える。このようにして、ウェルの洗浄およびペルオキシダーゼおよびo−フェニレンジアミンを用いる発色の後、着色または吸光度(492nm)の上昇が検出されたウェルは、sEPCRvar抗体を分泌するハイブリドーマのクローンを含有する可能性がある。ウェスタンブロットについては、組み換え型タンパク質および/またはsEPCRvarタンパク質を発現する組織または細胞抽出物を、ポリアクリルアミドゲルで泳動した。これを、上清中の特異的抗体の存在を証明するために、格子型(grille−type)装置によって独立した領域に分けられたニトロセルロースまたはPVDF膜にトランスファーした。ブロッキングおよび特異的抗体とのインキュベーションを行った後、これを3回洗浄し、ペルオキシダーゼ酵素に結合した二次抗体と共にインキュベートした。発色は、酵素によって形質転換される際、特異的な抗体に結合された場所で光を産生する基質を使用して実施した。発光バンドが、予測分子量を有するタンパク質に相当する場合、その上清が由来するウェルからの抗体産生細胞を増殖させ、液体窒素中に凍結させることとなる。
【0068】
sEPCRvarは、全sEPCR部分を含有するので、sEPCRvarに対して陽性であるハイブリドーマの、組み換え型EPCRの細胞外領域との反応性が分析されることとなる。このようにして、sEPCRvarの特定の領域に対する抗体を産生するハイブリドーマを選択することができる。
【0069】
ハイブリドーマの単クローン性
抗sEPCRvar抗体を分泌する各細胞培養物がモノクローナルであることを確認するために、クローニングまたは限界希釈技術が適用される。第1のウェスタンブロットまたはELISA試験で陽性であった最初の培養物からの、あるいは幹細胞からの、単離された細胞を、新しい培養マイクロプレート上で増殖させる。1種または複数の細胞から生じている新規のコロニーが、一旦十分なサイズを獲得したら、これから再び上清を取り出し、これを新規のELISAまたはウェスタンブロットにかけることになる。再び陽性であったコロニーを限界希釈にかけ、このプロセスを、第3の限界希釈の後に分析された上清の100%が、sEPCRvarに対する抗体を含有するようになるまで繰り返した。
【0070】
抗体精製
より大きな抗体濃度を達成するために、ハイブリドーマを特別な条件で培養した後(インテグラバイオサイエンス(Integra Biosciences)からのCELlineシステムを備えた培養ビン、またはベクトンディッキンソン(Becton Dickinson)からのCELlineシステム)、IgGを含有する上清を、以下の方法のうちの1つまたは複数を用いる液体クロマトグラフィーによる精製にかけることとなる:AKTA FPLC装置、GEヘルスケアバイオサイエンス(GE Healthcare Bio−Science)を使用する、免疫親和性クロマトグラフィー、アフィニティークロマトグラフィー(プロテインA/プロテインG/プロテインL、固定化金属、バイオアフィニティ)、陽イオン交換、ヒドロキシアパタイト、疎水的相互作用、ゲル濾過、など。
【0071】
[参考文献]
1. Bangalore N, Drohan WN, Orthner CL. High affinity binding sites for activated protein C and protein C on cultured human umbilical vein endothelial cells. Independent of protein S and distinct from known ligands. Thromb Haemost 1994; 72: 465-74.
2. Fukodome K, Esmon CT. Identification, cloning and regulation of a novel endothelial cell protein C/activated protein C receptor. J Biol Chem 1994; 269: 26486-91.
3. Stearns-Kurosawa DJ, Kurosawa S, Mollica JS. Ferrell GL, Esmon CT. The endothelial cell protein C receptor augments protein C activation by the thrombin-thrombomodulin complex. Proc. Natl Acad Sci USA 1996; 93: 10212-6.
4. Taylor FB Jr, Peer GT, Lockhart MS, Ferrell G, Esmon CT. Endothelial cell protein C receptor plays an important role in protein C activation in vivo. Blood 2001; 97; 1685-8.
5. Biguzzi E, Merati G, Liaw PC, Bucciarelli P, Oganesyan N, Qu D, Gu JM, Fetiveau R, Esmon CT, msnnucci PM, Faioni EM. A 23bp insertion in the endothelial protein C receptor (EPCR) gene impairs EPCR function. Thromb Haemost 2001; 86: 945-8.
6. Saposnik B, Reny JL, Gaussem P, Emmerich J, Aiach M, Gandrille S: A haplotype of
the EPCR gene is associated with increased plasma levels of sEPCR and is a candidate risk factor for thrombosis. Blood 2004: 103; 1311-1318.
7. Uitte de Willige S, Van Marion V, Rosendaal FR, Vos HL, de Visser MCH, Bertina RM.
Haplotypes of the EPCR gene, plasma sEPCR levels and the risk of deep venous thrombosis. J. Thromb Haemost. 2004; 2: 1305-10.
8. Taylor FB Jr, Stearns-Kurosawa DJ, Kurosawa S, Ferrell G, Chang AC, Laszik Z, Kosanke S, Peer. G, Esmon CT: The endothelial cell protein C receptor aids in host defense against Escherichia coli sepsis. Blood 2000; 95: 1680-6.
9. Joyce DE, Gelbert L, Ciaccia A, DeHoff B, Grinnell BW. Gene expression profile
of antithrombotic protein C defines new mechanisms modulating inflammation and apoptosis. J Biol Chem. 2001; 276: i1199-203.
10. Riewald M, Petrovan RJ, Donner A, Mueller BM, Ruf W. Activation of endothelial
cell protease activated receptor 1 by the protein C pathway. Science 2002; 296: 1880-2.
11. Fukudome K, Kurosawa S, Stearns-Kurosawa DJ, He X, Rezaie AR, Esmon CT.
The endothelial cell protein C receptor. Cell surface expression and direct ligand binding by the soluble receptor. J Biol Chem 1996; 271: 17491-8.
12. Simmonds, R E, Lane, DA. Structural and functional implications of the intron/exon organisation of the human endothelial cell protein C/activated protein C receptor (EPCR) gene: comparison with the structure of CD1/major histocompatibility complex alpha-1 and alpha-2 domains. Blood 1999; 94: 632-41.
13. Oganesyan V, Oganesyan N, Terzyan S, Qu D, Dauter Z, Esmon NL, Esmon CT. The crystal structure of the endothelial protein C receptor and a bound phospholipid. J Biol Chem 2002; 277: 24851-4.
14. Liaw PC, Mather T, Oganesyan N, Ferrell GL, and Esmon CT. Identification of the protein C/activated protein C binding sites on the endothelial cell protein C receptor. Implications for a novel mode of ligand recognition by a major histocompatibility complex class 1-type receptor. J Biol Chem 2001; 276: 8364-70.
15. Kurosawa S, Stearns-Kurosawa DJ, Hidari N, Esmon CT. Identification of functional endothelial protein C receptor in human plasma. J Clin Invest 1997; 100: 411-8.
16. 8, Xu J, Qu D, Esmon NL, Esmon CT. Metalloproteolytic release of endothelial cell protein C receptor. J Biol Chem 1999; 275: 603844.
17. Fukudome K, Esmon CT. Molecular cloning and expression of murine and bovine endothelial cell protein C/activated protein C receptor (EPCR). The structural and functional conservation in human, bovine, and murine EPCR. J Biol Chem. 1995;270:5571-7.
18. Liaw PCY, Neuenschwander PF, Smirnov M, Esmon CT. (2000) Mechanisms by which soluble endothelial cell protein C modulates protein C and activated protein C function. J Biol Chem. 2000; 275:5447-52.
19. Kurosawa S, Esmon CT, Stearns-Kurosawa DJ. The soluble endothelial protein C receptor binds to activated neutrophils: involvement of proteinase-3 and CD11b/CD18. J Immunol 2000; 165: 4697-4703.
【図面の簡単な説明】
【0072】
【図1】図1は、RNAがプロセッシングされ、成熟したメッセンジャーRNA(mRNA)が産生される方式に依存する、異なるタイプのEPCRを示す図である。3つの別の形のmRNAが産生される可能性があり、これらは、続いて、3つの異なる形のタンパク質を産生することとなる:1:膜EPCR(mEPCR)として翻訳されるタンパク質とは異なる4つのエキソンを伴うmRNA。2.エキソンEx I、Ex II、およびEx IIIのみを含み、その後にイントロン3およびこのイントロンの後の全配列が続く、国際公開第9900673号パンフレット中にエスモン(Esmon)によって開示された可溶形のEPCR(altsESPC)として翻訳されることとなるmRNA。3.本発明に開示された新規の形の選択的切断およびスプライシングによって、エキソンEx I、Ex II、およびEx IIIを含むmRNAが産生されることとなるが、プロセッシング中に、エキソンEx IVは失われ、この理由から、膜貫通ドメインも細胞質のテールも転写されず、その場所では、エキソンIIIの後、3’非翻訳領域に位置する潜在性(cryptic)エキソンであるであろう配列が出現し、この場合、mRNAは、本発明の新規の可溶性EPCR形(sEPCRvar)として翻訳される。可溶性EPCRはまた、おそらくメタロプロテアーゼ(まだ特徴づけられていない)から、タンパク分解活性によって産生することもできる。メタロプロテアーゼは、膜高さでEPCRを切断し、その結果、細胞外受容体の画分からなり、膜貫通ドメインおよび細胞質内テールを欠く、新規の可溶形のEPCR(sEPCR)が産生される。
【図2】図2は、特異的なEPCR遺伝子プライマーを用いるHUVEC cDNAの増幅の後に得られるバンドパターンである:野生型EPCR cDNA(1221bp、矢印1)に相当するバンドに加えて、選択的切断−スプライシングによって産生された新規の変異体アイソフォームのcDNAに相当する新規の増幅断片が認められる(矢印2)。
【図3】図3は、どちらも緑色蛍光タンパク質と融合させた、野生型EPCR(パネルA)を用いて、あるいはアイソフォーム(パネルB)を用いて形質移入されたCOS−7細胞である。パネルAは、野生型タンパク質が、細胞膜(矢印)上に蓄積する傾向があることを示すのに対して、アイソフォーム3を用いた場合、この局在化パターンは認められない(パネルB)。
【図4】図4は、ピキアパストリス(Pichia Pastoris)におけるsEPCRvar発現を説明する図である。本明細書に示すように、sEPCRvarは、安定に形質転換されたP.パストリス(P.pastoris)細胞の上清から精製された。パネルA:SDS−PAGEによって分離され、GELCODE Blue染料を使用して検出された様々なタンパク質を示す。ストリート St:発現される各バンドの分子量をkDaで示す、分子量マーカー、ストリート 1:組み換え型のsEPCR。約42kDaのブロードなバンド(タンパク質の強いグリコシル化に起因する)が観察される。Street 2:sEPCRvar。47kDaのブロードなバンド(タンパク質の強いグリコシル化に起因する)が観察される。パネルB:モノクローナル抗−EPCR抗体を用いるウェスタンブロットによって検出されるタンパク質、ストリート St:発現される各バンドの分子量をkDaで示す、分子量マーカー、ストリート 1:ブランク、ストリート 2:sEPCRvar。約47kDaのブロードなバンド(様々なタンパク質のグリコシル化に起因し、35から60kDaまでの間に広がる)が観察される。
【図5】図5は、異なる組織および腫瘍由来のcDNAからのEPCR遺伝子の増幅である。パネルA)1:分子量マーカー、2:心臓、3:肝臓、4:腎臓、5:膵臓、6:肺、7:胎盤。パネルB)1:分子量マーカー、2:骨格筋、3:脳、4:胸腺、5:小腸、6:脾臓。パネルC)1:マーカー、2:前立腺、3:睾丸、4:卵巣、5:大腸。パネルD)は、肺腫瘍の腫瘍株:2:H 446、3:H 510、4:H 1264、5:H 549、6:H 441、7:HTB 51、8:H 676、9:H 727、10:H 720、11:H 385、12:H 1299からのサンプルである。分子量マーカーを含有するストリートは、図中に略語St(「standard(標準)」より)で示す。マーカーの下方のバンドは、100bpサイズに相当し、各新しいバンドは、その前のバンドに対して100bpの増分(100bp、200bp、300bp・・・)に相当する。
【技術分野】
【0001】
[発明の背景]
最近記載されているプロテインCシステム(PC)のタンパク質は、内皮プロテインC/活性化プロテインC受容体(EPCR)である(1、2)。EPCRは、内皮細胞の膜上に発現され、高親和性でPCおよび活性化プロテインC(APC)と結合する(Kd〜30nM)。その生理学的使命は、内皮表面上のPCを濃縮し、これをトロンビン−トロンボモジュリン複合体に提示し、それによって、PCの効率的な活性化を助けることである(3)。ヒト以外の霊長類に関して実施された研究では、モノクローナル抗体を用いてEPCRをブロックすることは、APCの生理学的産生を90%減少させることが実証されており、また、最近の研究では、EPCRの遺伝的欠陥が、静脈および動脈の血栓形成を患う素因になる可能性があることが示唆されている(4〜7)。これらの事実は、凝固調節において、また血栓形成において、EPCRが果たす可能性がある重要な役割を実証する。さらに、EPCRは、PCの強力な抗炎症作用を担う分子の1つであるように思われる。
【0002】
EPCRとPC/APCの相互作用は、グラム陰性細菌による感染中に現れる凝血異常性および炎症性の反応の調節の際に、重要な役割を果たす可能性がある(8)。近年では、APCは、内皮細胞上の抗炎症および抗アポトーシス作用を有すること、また、この作用が現れるためにはEPCRの存在が不可欠であることが実証されている(9、10)。
【0003】
EPCRは、約46kDaの糖タンパク質である。成熟タンパク質は、17残基からなるシグナルペプチドの排除後の、221アミノ酸からなる(11)。これは、3つのイントロンによって分断された4つのエキソンによって形成される、染色体20(20q11.2)のロングアーム中に位置する遺伝子によってコードされる(12)。エキソンIは、5’非翻訳領域およびシグナルペプチドをコードし、エキソンIVは、膜貫通ドメイン、3残基の細胞質内テール、および3’非翻訳領域をコードする。エキソンIIおよびIIIは、クラスI主要組織適合遺伝子複合体のCD1スーパーファミリーに属するタンパク質のドメインα1およびα2との相同性を有する、EPCRの細胞外領域の大部分をコードする。近年では、EPCRが、ポケット(そこには、分子を安定させるリン脂質が収容される)を形成する、折り畳まれたβシート(その上にはαヘリックス構造を2つ有する)によって形成されるプラットフォームを、どのように有するかを示す三次元構造が、首尾よく結晶化され、分析されている(13)。同様に、アラニンスキャニングによって、PC/APCとの結合に関与する主要な残基(カルシウムおよびマグネシウムイオンの存在下で、そのGLAドメインを介して結合する)が特定されている(14)。
【0004】
しかし、内皮細胞膜上に固定されるEPCRに加えて、トロンビンなどの刺激によって誘導されるメタロプロテイナーゼによるEPCRの消化に少なくとも部分的に由来する、血漿可溶形(sEPCR)が存在する(15、16)。
【0005】
論文(15)「J.Clin.Invest.Vol.100」、no.2、1997年7月、411〜418は、血漿中に見られるEPCRの可溶形に関する。この論文には、血漿中のEPCRは、独特の種であるように思われ、精製されたタンパク質のアミノ末端フラグメントの配列決定では、血漿EPCRの特有の配列が示され、これは、組換えsEPCRのアミノ末端配列と一致したことが記載されている。この論文には、前記可溶性EPCRの起源は、他の受容体の場合と同様に、タンパク分解性の破断または選択的切断−スプライシング機構である可能性があり、特に、ウシEPCRのゲノム構造は、タンパク質をコードする選択的切断部位を含有する(ここでは、膜貫通ドメインが、エキソンIIIの後に位置しているイントロンによってコードされる配列によって置き換えられる)ことが記載されている(17)。この文献は、選択的切断から得られるEPCRの形の可能性を証明するために、より多くの研究が必要であることを示している。
【0006】
さらに、国際公開第−9900673号には、ヒト血漿におけるsEPCRの単離および特徴づけに関し、sEPCRが、タンパク質加水分解に由来する(ここでは、EPCRの細胞外ドメインは遊離され、膜に結合した残りのタンパク質から離れる)、あるいは、mRNAの選択的切断−スプライシング機構(これは、altsEPCRの配列を生じさせる)に由来する可能性があることが記載されている。本出願の図1は、タンパク分解起源の可溶性EPCR(psEPCR)と、選択的切断−スプライシングによって得られる国際公開第−9900673号に開示されたEPCR(altsEPCR)と、本発明の新規のEPCR目的物(sEPCRvar)との間の違いを見ることができる図を示す。
【0007】
したがって、本出願によって、少なくとも、ヒトEPCRは、選択的切断−スプライシング・プロセスを受け、前記選択的切断−スプライシング機構から得られる可溶性のEPCR(sEPCRvar)中にのみ存在する特有のインサートを含む、可溶性の切断型EPCRを生じる可能性があることが示される。前記sEPCRvarは、大血管における、またガンにおける疾患プロセスマーカーとして働く可能性がある。
【0008】
APCが、sEPCRに結合する場合、その抗凝血特性を失うが、その酵素的能力は維持されるため、sEPCRが、APC特異性を変化させ、おそらく、これを基質(現在は未知である)に方向付け、これが、APCに起因する抗炎症特性につながる可能性があると考えられる(18)。sEPCRは、膜上に存在するEPCRと、プロテインCに対して競合し、このようにしてAPC産生を抑制する。sEPCRは、プロテイナーゼ−3(PR3)(19)、好中球顆粒およびMac−1(CD11b/CD18)中に存在するプロテイナーゼ、好中球および単球の表面に存在する誘導性の発現のインテグリンを介して、活性化された好中球と相互作用することが可能であることも記載されている。PR3は、おそらく、膜結合型TNF−アルファおよびIL−1ベータ前駆体に作用する組織破壊メカニズムに関与する。Mac−1は、細胞シグナル伝達および細胞間接着現象に介在し、内皮ICAM−1などの接着分子と相互作用し、その理由で、炎症性プロセス中の好中球の補充において役割を果たす。PR3およびMac−1へのsEPCRの結合は、おそらく、その活性を低下させるであろう。
【発明の開示】
【発明が解決しようとする課題】
【0009】
本発明者らは、ヒトゲノムDNA(cDNA)から開始して、新規のタンパク質をクローン化し、発現させることを可能にした。前記タンパク質は、その配列の、他の既知の配列との分析および比較、ならびにその機能の特徴づけによって、選択的スプライシング・プロセスによって産生される可溶性のEPCRタンパク質の未知の形であることが示された。一方、前記タンパク質は、示差的な特徴として、現在知られているEPCR形には存在しない、当初EPCR遺伝子3’非翻訳領域と考えられていた領域によってコードされるポリペプチドの領域を有する。
【課題を解決するための手段】
【0010】
[発明の詳細な説明]
本発明は第一に、新規の可溶形のEPCR(これを、本発明者らは、可溶性のEPCR変異体(sEPCRvar)と呼ぶ)に、より詳細には、配列番号1の配列の残基201〜256を含む、内皮プロテインC受容体または前記配列の残基のフラグメントから得られるポリペプチドに関する。ある特定の実施形態では、前記フラグメントは、少なくとも9アミノ酸を有する。本発明の特定の実施形態によれば、前記フラグメントは、配列番号1の残基213〜227、229〜242、212〜226、227〜241、242〜256を含むフラグメントから選択される。
【0011】
ある特定の実施形態では、前記フラグメントは免疫原特性を有し、これは特に、特異的抗体を産生するための手順で有用である。
【0012】
本発明はさらに、配列番号1の配列の残基201〜256から形成されるポリペプチドを含む、単離されたあるいは組み換え型のタンパク質に関する。前記タンパク質は、EPCRタンパク質である。
【0013】
本発明の好ましい実施形態によれば、前記単離されたあるいは組み換え型のEPCRタンパク質は、配列番号1の配列を含む。
【0014】
本発明のさらなる目的は、配列番号1の配列の残基201〜256から形成されるポリペプチドまたは前記配列の残基のフラグメントを含む組み換え型タンパク質である。
【0015】
本発明のさらなる目的は、配列番号1の配列の残基201〜256によって形成されるポリペプチドまたは前記配列の残基のフラグメントをコードする配列を含むポリヌクレオチドである。ある特定の実施形態では、前記ポリヌクレオチドは、配列番号2に相当するcDNA配列に由来する。このcDNAは、mRNAから逆転写されたDNAに関する。
【0016】
本発明の特定の実施形態では、本発明のポリヌクレオチドは、DNA構築物中に含有される可能性がある。したがって、本発明のさらなる目的は、配列番号1の配列の残基201〜256を含むタンパク質またはポリペプチドまたは前記配列の残基のフラグメントをコードするDNA構築物である。好ましくは、前記フラグメントは、少なくとも9アミノ酸を有する。前記構築物のある特定の実施形態では、これは、配列番号2に相当するcDNA配列に由来するポリヌクレオチドを含む。前記DNA構築物は、前記タンパク質またはポリペプチドに作動可能に結合される制御配列を組み込む可能性がある。核酸またはポリヌクレオチドに関して、「作動可能に結合される」は、あるヌクレオチド領域が、別のヌクレオチド配列と、機能的関係に置かれることを意味する。「制御配列」は、機能(例えばある種のコード配列の転写および翻訳など)を調節する、ある種の宿主細胞によって特に認識される発現シグナルである。プロモーター、発現エンハンサー、転写ターミネーター、リボソーム結合部位などは制御配列である。DNA構築物を形成するための所望の配列のスプライシングは、適切な制限部位における切断およびスプライシングによって行うことができる。これらの結合部位が存在しない場合、アダプターまたはスペーサーとして合成のオリゴヌクレオチドを使用して、従来の遺伝子工学的方法によってこれらを産生することが可能である。
【0017】
本発明のポリヌクレオチドまたはDNA構築物は、しばしば実験室マニュアルに書き留められている従来の遺伝子工学的方法によって得ることができる。
【0018】
本発明のポリヌクレオチドまたは構築物は、適切な発現ベクター中に挿入させることができる。したがって、本発明のさらなる目的は、前記ポリヌクレオチドまたはDNA構築物を含む発現ベクターである。本発明の様々な実施形態における発現ベクターの選択は、前記発現ベクターが挿入されることとなる宿主細胞に依存する。例えば、本発明のポリヌクレオチドまたはDNA構築物が挿入されるベクターは、プラスミドまたはウイルス(これらは、一旦宿主細胞中に導入された場合に、細胞のゲノムに組み込まれる可能性もあるし、組み込まれない可能性もある)である可能性がある。この発現ベクターはまた、従来方法によって得ることができる。
【0019】
本発明のさらなる目的は、配列番号1の配列の残基201〜256含むポリペプチドまたは前記配列の残基の少なくとも9アミノ酸を有するフラグメントををコードするポリヌクレオチドまたはDNA構築物を含む、形質転換された宿主細胞である。本発明によれば、形質転換された宿主細胞は、原核生物細胞(例えば大腸菌)または真核生物(例えば酵母、特にピキアパストリス(Pichia Pastoris)およびサッカロミセスセレビシエ(Saccharomyces cerevisiae))、昆虫細胞または哺乳類細胞株であり得る。
【0020】
本発明のある特定の実施形態では、配列番号2を有する本発明のポリヌクレオチドは、本発明において開示される選択的切断およびスプライシングによって産生される新規の形のsEPCR(sEPCRvar)に相当する、配列番号1の組み換え型のsEPCRタンパク質を産生するために、ピキアパストリス(Pichia Pastoris)中に導入される。組み換え型タンパク質の産生に必要なDNA構築物、発現ベクター、および形質転換された宿主細胞を産生するために使用される方法は、その精製のためのプロセスを含めて、後ほど詳述する。
【0021】
本発明のある特定の実施形態では、本発明のポリヌクレオチドまたはDNA構築物を含む発現ベクターは、組成物中でのその使用、およびインビボでの遺伝子導入または治療のプロセスのために設計される。より具体的な実施形態では、この発現ベクターは、ウイルスベクターである。この目的に適したウイルスベクターとしては、アデノウイルス、アデノ随伴ウイルス、レトロウイルス、レンチウイルス、アルファウイルス、ヘルペスウイルス、コロナウイルス類縁体(derivative)ベクターなどが挙げられる。
【0022】
本発明のさらなる目的は、配列番号1の配列の残基201〜256を含むポリペプチドまたは前記配列の残基のフラグメントをコードするポリヌクレオチドを含む発現ベクターと、前記ポリヌクレオチドまたは前記構築物で形質転換される宿主細胞を含む発現系である。好ましくは、前記フラグメントは、少なくとも9アミノ酸を含む。
【0023】
本発明のさらなる目的は、配列番号1の残基201〜256を含むタンパク質またはポリペプチド、または前記配列の残基の好ましくは少なくとも9アミノ酸を有するフラグメントを産生する方法であって、前記方法が、前記タンパク質、ポリペプチド、またはフラグメントの発現を可能にする条件で、本発明のポリヌクレオチドまたはDNA構築物を含む宿主細胞を培養することを含むことを特徴とする方法である。培養を最適化する条件は、使用される宿主細胞に依存する。所望により、この方法はまた、発現されたタンパク質またはポリペプチドを単離および精製するためのいくつかのステップを含む可能性がある。
【0024】
あるいは、本発明のタンパク質またはポリペプチドは、他の従来方法によって、例えば固相技術に関する化学合成、高速液体クロマトグラフィー(HPLC)による精製によって得ることができ、所望により、これらを、従来の技術、例えば配列決定および質量分析、アミノ酸分析、核磁気共鳴などによって分析することもできる。
【0025】
本発明のさらなる目的は、配列番号1の配列を含むタンパク質または前記配列の残基のフラグメントをコードするmRNAである。
【0026】
本発明のさらなる目的は、配列番号1の残基201〜256を含むポリペプチドまたは前記配列の残基の免疫原特性を有するフラグメントに対する、特定の単離された抗体である。ある特定の実施形態では、前記フラグメントは、少なくとも9アミノ酸を有する。
【0027】
ある特定の実施形態では、前記抗体は、その配列が残基201〜256、213〜227、229〜242、212〜226、227〜241、および242〜256(すべて配列番号1に関する)から選択されるポリペプチドまたはフラグメントに対して特異的である。
【0028】
別の特定の実施形態では、前記抗体は、配列番号1に対して特異的であり、これが領域201〜256に結合しない場合には領域1〜200を認識しない。
【0029】
好ましい実施形態によれば、前記抗体は、モノクローナル抗体である。
【0030】
本発明のさらなる目的は、生体試料における、配列番号1の配列の残基201〜256を含むEPCRタンパク質(例えば配列番号1の新規のEPCRタンパク質)、または前記配列の残基のフラグメントの選択的インビトロ検出のための方法であって、
−生体試料を得ること、
−配列番号1の配列の残基201〜256を含むEPCRタンパク質または前記配列の残基のフラグメントの量を分析すること、
を含む方法である。
【0031】
前記生体試料は、生体材料を含む任意の試料である可能性がある。好ましい実施形態では、前記生体試料は、尿、血漿、血清、組織、または間質液試料である。
【0032】
前記方法は、例えば、質量分析、イムノアッセイ、化学アッセイ、液体クロマトグラフィー、および様々な間接的および直接的光度測定方法を含めた、任意の既知の方法であり得る。例えば、いくつかの分析法は、質量分析を使用するマーカーを定量化するために適用される可能性がある。一般的に言うと、マーカーは、例えば液体クロマトグラフィーまたは二次元ゲル電気泳動によって、生体試料から単離される。マーカーは、質量分析、例えば、液体クロマトグラフィー(LC−MS)を伴うタンデム質量分析、MALDI−TOF−MS(「マトリクス支援レーザー脱離/イオン化質量分析・飛行時間型MS」)などによって定量化される。標的マーカーは、既知量の精製されたマーカー標準と比較することによって、あるいは、健康な対照から得られる同じタイプの生体試料中に存在する前記マーカーの量と比較することによって定量化することができる。
【0033】
本発明の別の実施形態では、この方法は、イムノアッセイであり得る。一般に、イムノアッセイには、1種または複数のマーカーリガンドが使用される。本発明で使用される「リガンド」は、標的マーカーに特異的に結合することが可能な任意の化合物または分子である。これらのリガンドは、別々に、あるいは組み合わせて使用することができる(例えば、抗体は、アプタマーと組み合わせて使用することができる)。
【0034】
本発明の一つの実施形態では、イムノアッセイは、均一(homogenous)アッセイ、不均一(heterogeneous)アッセイ、酵素免疫測定法(EIA、ELISA)、競合アッセイ、イムノメトリックアッセイ(サンドイッチ)、濁度アッセイ、比濁アッセイ、または他の同様のアッセイである可能性がある。同様に、イムノアッセイは、手動で、あるいは自動分析器を用いて行われる可能性がある。
【0035】
本発明の方法では、前記タンパク質の量は、血管損傷に関連する炎症性疾患、炎症、凝固異常に関連する疾患、およびガンから選択される疾患に関連する。前記疾患はまた、狼瘡などの自己免疫性疾患、敗血症、ショック、子癇前症、糖尿病、移植のモニタリングの際の不安定狭心症、再狭窄、血管形成、および肝臓または腎臓病である可能性もある。
【0036】
本発明の方法によれば、検出されるEPCRタンパク質の量はまた、較正標準と比較することができる。
【0037】
本発明のさらなる目的は、配列番号1の配列の残基201〜256を含むEPCRタンパク質、例えば、配列番号1の新規のEPCRタンパク質、または前記配列の残基の好ましくは少なくとも9アミノ酸のフラグメントの検出および定量化のための試験キットであって、
a)配列番号1の配列の残基201〜256を含むポリペプチドまたは前記配列の残基のフラグメントに特異的な抗体、即ち、領域201〜256に結合しない場合には領域1〜200を認識しない配列番号1のタンパク質に特異的な抗体、
b)抗体a)と、生体試料中に存在する配列番号1の配列の残基201〜256を含むEPCRタンパク質または前記配列の残基のフラグメントとの間の反応を検出する試薬、
を含むキットである。
【0038】
好ましくは、このキットはまた、配列番号1の配列の残基201〜256を含むEPCRタンパク質の正常および異常なレベルと、産生される反応物の量を相関させるための標準を含む。
【0039】
本発明のさらなる目的は、配列番号1の配列の残基201〜256を含むEPCRタンパク質、例えば配列番号1の新規のEPCRタンパク質、または前記配列の残基のフラグメントに相当するmRNAの検出および定量化のための、配列番号1の配列の残基201〜256を含むポリペプチドまたは前記配列の残基のフラグメント(好ましくは、前記フラグメントは、少なくとも9アミノ酸を有する)をコードするポリヌクレオチドの使用である。前記EPCRタンパク質は、血管損傷に関連する炎症性疾患、炎症、ガン、および凝固異常に関連する疾患から選択される疾患に関連する量で存在する。
【0040】
さらに、非タンパク分解起源の新規の可溶形のEPCRは、この形の可溶性のEPCR(sEPCRvar)の存在を、単独的かつ独占的に検出するのに選択的である分析方法であって、他の形の可溶性のEPCRまたは膜EPCRが検出される方法と重複する前記方法を用いずに、特定の疾患を検出できるという利点を有する分析方法の開発のために役割を果たす可能性がある。
【0041】
[図面の簡単な説明]
図1は、RNAがプロセッシングされ、成熟したメッセンジャーRNA(mRNA)が産生される方式に依存する、異なるタイプのEPCRを示す図である。3つの別の形のmRNAが産生される可能性があり、これらは、続いて、3つの異なる形のタンパク質を産生することとなる:1:膜EPCR(mEPCR)として翻訳されるタンパク質とは異なる4つのエキソンを伴うmRNA。2.エキソンEx I、Ex II、およびEx IIIのみを含み、その後にイントロン3およびこのイントロンの後の全配列が続く、国際公開第9900673号パンフレット中にエスモン(Esmon)によって開示された可溶形のEPCR(altsESPC)として翻訳されることとなるmRNA。3.本発明に開示された新規の形の選択的切断およびスプライシングによって、エキソンEx I、Ex II、およびEx IIIを含むmRNAが産生されることとなるが、プロセッシング中に、エキソンEx IVは失われ、この理由から、膜貫通ドメインも細胞質のテールも転写されず、その場所では、エキソンIIIの後、3’非翻訳領域に位置する潜在性(cryptic)エキソンであるであろう配列が出現し、この場合、mRNAは、本発明の新規の可溶性EPCR形(sEPCRvar)として翻訳される。可溶性EPCRはまた、おそらくメタロプロテアーゼ(まだ特徴づけられていない)から、タンパク分解活性によって産生することもできる。メタロプロテアーゼは、膜高さでEPCRを切断し、その結果、細胞外受容体の画分からなり、膜貫通ドメインおよび細胞質内テールを欠く、新規の可溶形のEPCR(sEPCR)が産生される。
【0042】
図2は、特異的なEPCR遺伝子プライマーを用いるHUVEC cDNAの増幅の後に得られるバンドパターンである:野生型EPCR cDNA(1221bp、矢印1)に相当するバンドに加えて、選択的切断−スプライシングによって産生された新規の変異体アイソフォームのcDNAに相当する新規の増幅断片が認められる(矢印2)。
【0043】
図3は、どちらも緑色蛍光タンパク質と融合させた、野生型EPCR(パネルA)を用いて、あるいはアイソフォーム(パネルB)を用いて形質移入されたCOS−7細胞である。パネルAは、野生型タンパク質が、細胞膜(矢印)上に蓄積する傾向があることを示すのに対して、アイソフォーム3を用いた場合、この局在化パターンは認められない(パネルB)。
【0044】
図4は、ピキアパストリス(Pichia Pastoris)におけるsEPCRvar発現を説明する図である。本明細書に示すように、sEPCRvarは、安定に形質転換されたP.パストリス(P.pastoris)細胞の上清から精製された。パネルA:SDS−PAGEによって分離され、GELCODE Blue染料を使用して検出された様々なタンパク質を示す。ストリート St:発現される各バンドの分子量をkDaで示す、分子量マーカー、ストリート 1:組み換え型のsEPCR。約42kDaのブロードなバンド(タンパク質の強いグリコシル化に起因する)が観察される。ストリート 2:sEPCRvar。47kDaのブロードなバンド(タンパク質の強いグリコシル化に起因する)が観察される。パネルB:モノクローナル抗−EPCR抗体を用いるウェスタンブロットによって検出されるタンパク質、ストリート St:発現される各バンドの分子量をkDaで示す、分子量マーカー、ストリート 1:ブランク、ストリート 2:sEPCRvar。約47kDaのブロードなバンド(様々なタンパク質のグリコシル化に起因し、35から60kDaまでの間に広がる)が観察される。
【0045】
図5は、異なる組織および腫瘍由来のcDNAからのEPCR遺伝子の増幅である。パネルA)1:分子量マーカー、2:心臓、3:肝臓、4:腎臓、5:膵臓、6:肺、7:胎盤。パネルB)1:分子量マーカー、2:骨格筋、3:脳、4:胸腺、5:小腸、6:脾臓。パネルC)1:マーカー、2:前立腺、3:睾丸、4:卵巣、5:大腸。パネルD)は、肺腫瘍の腫瘍株:2:H 446、3:H 510、4:H 1264、5:H 549、6:H 441、7:HTB 51、8:H 676、9:H 727、10:H 720、11:H 385、12:H 1299からのサンプルである。分子量マーカーを含有するストリートは、図中に略語St(「standard(標準)」より)で示す。マーカーの下方のバンドは、100bpサイズに相当し、各新しいバンドは、その前のバンドに対して100bpの増分(100bp、200bp、300bp・・・)に相当する。
【発明を実施するための最良の形態】
【0046】
内皮細胞cDNAからのEPCR cDNAの増幅
HUVEC細胞を、ヒトの臍帯静脈から得て、標準の手順に従って培養し(ジャッフェ EA(Jaffe EA)、ナフマン RL(Nachman RL)、ベッカー CG(Becker CG)、ミニック CR(Minick CR)「Culture of human endothelian cells derived from umbilical veins.Identification by morphologic and immunologic criteria.」、「J Clin Inves.」、1973年11月、52(11):2745〜56)、全RNAを、(ペレス−ルイス A(Perez−Ruiz A)、モンテス R(Montes R)、ベラスコ F(Velasco F)、ロペス−ペドレラ C(Lopez−Pedrera C)、アントニオパラモ J(Antonio Paramo J)、オルベ J(Orbe J)、ハーミダ J(Hermida J)、ローシャ E(Rocha E)、「Regulation by nitric oxide of endotoxin−induced tissue factor and plasminogen activator inhibitor−1 in endothelial cells.」、「Thromb Haemost.」、2002年12月、88(6):1060〜5)に記載される通り、標準の技術に従って抽出した。得られたRNAは、転写酵素を用いて37℃で60分間インキュベートし、cDNAに逆転写させ、それに続いて酵素失活のために65℃で5分間インキュベートした。これを行うために、120単位(U)のM−MuLV逆転写酵素(ギブコBRL(Gibco BRL))を、RNA1μgに加え、10μLの最終体積にした。これは、4μLの逆転写酵素緩衝液、2mmol/Lのデオキシヌクレオチド(dNTP、インビトロジェン(Invitrogen))、0.3μg/mLのランダムヘキサマー(ギブコBRL(Gibco BRL))、0.5mmol/Lのジオチオトレイトール(diothiotreitol)、および35UのRnase阻害剤(ヒト胎盤由来のRNAsin 48,000U/mL、RNAガード、GEヘルスケアバイオサイエンス(GE Healthcare Bio−Science))を含有していた。
【0047】
EPCR遺伝子の発現を、ポリメラーゼ連鎖反応(PCR)によって分析した。使用されるプライマー(ジェンセット(Genset)、フランス)は、配列番号3の配列に相当する5’プライマー、および配列番号4の配列に相当する3’プライマーであった。
【0048】
PCRは、25μLの体積で行われた。1μgのRNAの逆転写によってあらかじめ得られた一定分量のcDNAが使用され、それに、1Uのtaq DNAポリメラーゼ(ロシュ(Roche))、2.5μLの反応緩衝液(ロシュ(Roche))、0.5μLの各プライマー(10μmol/L溶液から)、0.25μLのデオキシヌクレオチド(dNTP、GEヘルスケアバイオサイエンス(GE Healthcare Bio−Science))、および0.25μlの1mg/mL BSAが加えられた。この反応混合物を、水で25μLに調節した。増幅サイクルは、94℃で30秒の変性相と、別のアニーリングと、72℃で3.5分の伸張相からなる。このサイクルを5回繰り返し、その後、同じ変性相と、別のアニーリングと、68℃で3分の伸張相の32サイクルを実施する。最終の伸張期間として、混合物を5分間68℃に維持して、増幅を終結させた。PCR増幅されたセグメントは、1221塩基対(bp)の長さを有する。EPCR遺伝子がこれらの条件で増幅された場合、さらなるバンドが、約850塩基対の高さで出現する(図2)。
【0049】
増幅されたEPCRのさらなるバンドの配列決定
バンド2は、鋳型として図2の850bpバンドから開始すること以外は、同じ増幅条件を使用して増幅した。アガロースゲル電気泳動の後、製造業者の指示書に従って市販のキット(QIAquick Gel Extraction Kit)を使用して、増幅産物を精製した。このようにして精製された産物を、配列決定反応(ABIプリズム(ABI Prism)(登録商標)、BigDyeTM Terminator Cycle Sequencing Ready Reaction Kit)のための鋳型として使用した。配列番号5および配列番号6の配列に相当するプライマーを使用して、2つの配列決定反応(1つは5’−3’方向、もう1つは逆方向)を実施した。
【0050】
反応の結果は、自動シーケンサー、ABI Prism 377 DNA Sequencerで分析した。結論としては、新規のバンドは、配列番号7の配列によって定義されるフラグメントに相当する。
【0051】
その配列が配列番号2であるこのフラグメントの中央部分は、エキソン4および3’非翻訳領域の部分を欠く新規のEPCRのcDNAに相当する。このcDNAの起源は、おそらく、EPCR RNAの選択的切断−スプライシング現象である。
【0052】
この新種のcDNAは、アミノ酸の配列が配列番号1であるポリペプチドをコードする。前記配列の残基1〜17は、シグナルペプチドに相当し、残基18〜200は、EPCRの共通の細胞外ドメインに相当し、残基201〜256は、この新規の形のEPCRの新規の示差的なドメインに相当し、EPCRの膜貫通ドメインに代わるものである。この新規のドメインは、膜貫通ドメインであるための基準を満たさない。
【0053】
緑色蛍光タンパク質と融合させたEPCRおよびsEPCRvarの哺乳類細胞における発現
EPCRアイソフォームの細胞内の局在化を研究するために、そのcDNAを、pcDNA3.1/CT−GFP−TOPOベクター中にクローニングし、それを緑色蛍光タンパク質との融合タンパク質として発現させた。このようにして調製されるベクターを使用して、Lipofectamine 2000(インビトロジェン(Invitrogen))を使用する培養で、哺乳類COS−7細胞を一過的に形質転換させた。対照として、同じベクターを使用し、そこでは野生型EPCRのcDNAをクローニングし、緑色蛍光タンパク質と融合させた。野生型EPCRおよびアイソフォームの細胞内局在化を、蛍光顕微鏡検査によって検出した。図3に示さているように、野生型EPCRは、主に細胞膜上に局在化する(ここでは緑色蛍光が認められる)が、sEPCRvarアイソフォームは、本発明者らの仮定と一致して、細胞膜上には局在化しない。
【0054】
EPCRアイソフォームのクローニング、発現、精製、および特徴付け
クローニングおよび発現
同定されたEPCR変異体(EPCRvar)を発現させるために、sEPCR var配列を、cDNA鋳型として内皮細胞を使用して、イニシエーター
5’−agcttggcatatcgattagccaagacgcctcagatg−3’および
5’−agctatcgtagcggccgcctaccctattatatcagc−3’
を用いる(これらは、それぞれ5’および3’末端にClaIおよび別のNotI制限部位を付加されている)ポリメラーゼ連鎖反応(PCR)によって、そのシグナルペプチドを伴わずに増幅させた(残基17〜256)。こうした修飾により、サッカロミセスセレビシエのα因子分泌シグナルの後の、pPICZαC(インビトロジェン(Invitrogen))プラスミドのClaIおよびNotI部位に、EPCR変異体配列を結合することが可能になり、酵母から細胞外培地への多くのタンパク質の効率的な分泌が可能になった。クローニングプロセスにより、セリン残基および別のイソロイシン残基が、sEPCRvarのアミノ末端に加えられた。直接的配列決定を使用して、挿入配列およびベクターが適切であることが確認されることが確認された。あらかじめ調製された発現ベクターを用いて、PmeI制限酵素を用いるそれらの直鎖化の後、ピキアパストリス(Pichia Pastoris)細胞を、化学的方法を使用して形質転換させ(Easy Comp、インビトロジェン(Invitrogen))、その結果、メタノール反応内因性プロモーターにおける、sEPCRvarコード配列の相同的組換えによる組込みが示された。ゼオシンの存在下で形質転換産物を培養し、sEPCRvarコード配列を含有する、言い換えるとゼオシン抵抗性遺伝子を含有するベクターで形質転換されたP.パストリス(P.Pastoris)コロニーを選択した。簡単に言うと、形質転換された酵母を、1%(v/v)のグリセロール(BMGY)が補充された4mlのBMY培地[1%(w/v)の酵母抽出物、2%(w/v)のペプトン、100mMリン酸カリウム(pH6,0)、硫酸アンモニウムを含む酵母窒素源1.34%(w/v)、4×10−5%(w/v)のビオチン]中で培養し、撹拌しながら約18時間28〜30℃でインキュベートした。細胞は、室温で5分間、2,000gで遠心分離することによって収集した。上清を捨て、18時間、1%メタノールを用いて、sEPCRvar発現を誘導した。これを行うために、細胞を、0.5%(w/v)メタノールが補充された3mlのMBYに再懸濁し、勢いよく攪拌しながら、約28〜30℃で18時間インキュベートした。誘導後、条件培地からのサンプルを、12% NuPAGE Bis−Trisゲル(インビトロジェン(Invitrogen)、カリフォルニア州カールズバッド(Carlsbad,CA))にローディングし、RCR−2モノクローナル抗体(フクドメケンジ(Kenji Fukodome)博士、佐賀大学(Saga University)、日本(Japan)によって快く提供された)を使用するウェスタンブロットによってsEPCRvarを検出した(図4)。大規模産生については、最も高濃度のsEPCRvarを分泌したコロニーを選択した。その高いsEPCRvar産生能力に起因して選択されたコロニーにおいて、そのメタノール代謝(新陳代謝が早いものまたは遅いもの)を研究し、これによって、最も適切なコロニーのための最適発現条件が確立された。一旦培養条件およびメタノール誘導が最適化されたら、大量のsEPCRvarを産生するために規模を増大した。
【0055】
精製
P.パストリス(P.Pastoris)培養上清を濃縮し、NaClを含まない20mM Tris−HCl(pH7.6)に対して透析した後、2つの連続する精製ステップ:イオン交換クロマトグラフィーおよびゲル濾過を実施した。イオン交換精製では、Resource Qカラム(GEヘルスケアバイオサイエンス(GE Healthcare Bio−Science))を使用し、溶出は、20カラム分に匹敵する体積の、0.0〜300mMのNaClグラジエントを用いて実施した。sEPCRvarを含有する溶出分画を合わせ、遠心限外濾過によって濃縮し、その後、Superdex 75−HR10/30カラム(GEヘルスケアバイオサイエンス(GE Healthcare Bio−Science))にかけてゲル濾過を実施した。精製されたタンパク質の濃度を、BCA総タンパク質アッセイ(ピエール(Pierre)、イリノイ州ロックフォード(Rockford,IL))およびBSA標準を使用して決定した。精製されたsEPCRvarを検出するために、サンプルを、12% NuPAGE Bis−Trisゲル(インビトロジェン(Invitrogen)、カリフォルニア州カールズバッド(Carlsbad,CA))にローディングし、還元条件下で電気泳動を実施し、続いて、クマシーブルーで染色した。電気泳動ゲルをエレクトロブロッティングにかけ、RCR−2モノクローナル抗体を用いてsEPCRvarを検出した。sEPCRvarの分子量を評価するために、各ゲル電気泳動には分子量標準を含ませた。
【0056】
新規の可溶性EPCR変異体の活性を特徴づけるための生化学的分析
PCに対するsEPCRvarの親和性
培養された内皮細胞におけるAPCの産生。使用される細胞株は、EA.hy926(トロンボモジュリンおよびEPCRを発現する能力を保持している、形質転換されたヒトの内皮細胞株)であった(スターンズ−クロサワDJ(Stearns−Kurosawa DJ)、クロサワ S(Kurosawa S)、モリカ JS(Mollica JS)、フェレル GL(Ferrell GL)、エスモン CT(Esmon CT)、「The endothelial cell protein C receptor augments protein C activation by the thrombin−thrombomodulin complex.」「Proc Natl Acad Sci U S A.」1996年、93:10212〜6)。96ウエルプレート上の5×104細胞/ウェルを、0.02U/mlのトロンビン(0.17nM)(ERL、イギリス、スウォンジー(Swansea,United Kingdom))と共にインキュベートし、150mM NaCl、5mM CaCl2、0.6mM MgCl2、1% BSA、0.001% Tween−20および0.02% NaN3を補充された、20mMトリス緩衝液(pH7.4)中で、PC(バクスター(Baxter)、米国イリノイ州ディアフィールド(Deerfield,IL.USA))の濃度を50nMと1,000nMとの間で変化させ、室温にて45分後、トロンビンを阻害するために、レピルジン(シェーリングAG(Schering AG)、ドイツ、ベルリン(Berlin,Germany))を、0.2μlの最終濃度で加え、3〜4分後に、APCによるそのタンパク質加水分解をモニタリングする目的で、発色基質S−2366(クロモジェニックス(Chromogenix)、イタリア、ミラノ(Milan,Italy)を0.4mMの最終濃度で加えた。405nmでの吸光度の上昇を、マイクロプレートリーダー(iEMS Reader、ラボシステムズ(Labsystems)、フィンランド(Finland)で動力学的に記録した。ミハエリス・メンテンの式に対する曲線データの調整を、これらの条件でのPC活性化Km値を算出するEnzfitterプログラム(バイオソフト(Biosoft)、イギリス、ケンブリッジ(Cambridge,United Kingdom))を使用して実施した。
【0057】
sEPCRvarの阻害効果を研究するために、2μmol/Lを、トロンビンおよびPCと同時に加えた。このようにして、PC活性化に対するsEPCRvarの効果を分析し、PCに対するその親和性を反映する、sEPCRが活性化に対して有する効果と比較することができた。
【0058】
表1は、sEPCRおよびsEPCRvarの存在下または非存在下での、培養された内皮細胞の表面上の、トロンビンによるプロテインC活性化の生化学的特性を示す。
【0059】
表1
【表1】
【0060】
PCに対するsEPCRおよびsEPCRvarの親和性
活性化部分卜ロンボプラスチン時間の決定。健康な対象からの5血漿の混合物を使用し、ダイアグノスティカ・スタゴ・ベーリンガーマンハイム(Diagnostica Stago Boehringer Mannheim)STA Compact装置(ロシュ(Roche))、およびパトロンチン(Pathromtin)試薬(デイドベーリング(Dade Behring)、米国(USA))を使用して、活性化部分卜ロンボプラスチン時間(APTT)を決定した。周知のように、正常な血漿の混合物中のAPCの存在は、APTTを延長させる。使用される実験条件では、APCが存在しない場合のAPTTは、33.1±0.4s(平均±SD)であったのに対して、APCが1nmol/Lの最終濃度で加えられた場合、APTTは、43.9±0.4sに延長された。sEPCRが加えられた場合、APCの効果は低下し、1mmol/LのsEPCRが存在する場合には、APTTは、37.2±0.2sであった。APCの抗凝血効果に対するsEPCRのこの阻害効果は、用量依存的である。正常な血漿の混合物に加えられたsEPCRは、APTTに対する直接的な効果を有しなかった(sEPCRが存在する場合の33.9±0.4sと比較して、sEPCRが存在しない場合には33.1±0.4s)。すべての実験を4回繰り返した。sEPCRではなく、同じ濃度のsEPCRvarが加えられた場合、結果は、sEPCRを用いて得られるものと重ねることができ、sEPCRと同様に、sEPCRvarもAPCと結合することが実証された。
【0061】
表2は、APC抗凝血作用に対するsEPCRおよびsEPCRvarの効果を示す。それに1nmol/LのAPCおよび様々な濃度のsEPCRが加えられた、正常な血漿の混合物の血液凝固時間(APTT)。4つの実験の平均±SDを示す。APCが存在しない場合のAPTTは、33.1±0.4秒であった。
【0062】
表2
【表2】
【0063】
様々な組織および腫瘍株におけるsEPCRvarの発現分析
sEPCRvarが発現される組織を同定するために、EPCRおよびsEPCRvarに相当するcDNAの同時増幅を可能にするプライマーを使用するPCRによって、様々なヒト組織のcDNAライブラリ(Multiple tissue cDNA(MTC)パネルIおよびII、バイオサイエンス(Bioscience)、米国(USA))を研究した。使用されるプライマー(ジェンセット(Genset、フランス)は、以下の通りであった:
プライマー5’:5’−GCAGTATGTGCAGAAACATATTTCCGC−3’および
プライマー3’:5’−CATCCCAAGTCTGACACACCTGGAAGT−3’
【0064】
PCRは、25μLの体積で実施した。cDNAのアリコットを使用し、これに、1 Uのtaq DNAポリメラーゼ(ロシュ(Roche))、2.5μLの反応緩衝液(ロシュ(Roche))、0.5μLの各プライマー(10μmol/L溶液から)、0.25μLのデオキシヌクレオチド(dNTP、GEヘルスケアバイオサイエンス(GE Healthcare Bio−Science))、および0.25μlの1mg/mL BSAを加えた。反応混合物を、水で25μLに調節した。増幅サイクルは、94℃で30秒の変性段階、56℃で1分の別のアニーリング段階、72℃で1.5分の伸張からなる。このサイクルを35回繰り返す。最終の伸張期間として72℃で5分間混合物を維持して、増幅を終結させた。増幅されたEPCRのcDNAセグメントは、578bp長であったのに対し、sEPCRvarのcDNAセグメントは、189bpであった(図5)。sEPCRvar形は、膵臓、心臓、肝臓、腎臓、肺、胎盤、胸腺、小腸、脾臓、および大腸に多量に発現された。sEPCRvar形は、いくつかの腫瘍株に、特にH 549、H 441、およびH 720に多量に発現された。いくつかの腫瘍株、特にH 446およびH 676は、まだ同定されていない別の形(これは、その電気泳動移動度によれば、1000bpを有する)をさらに発現した。
【0065】
抗体産生
免疫化
生後少なくとも1ヵ月のBALB/C雄を、以下のパターンに従って、sEPCRvarで免疫化させた:フロイント・アジュバントまたは別のアジュバント中の10μgと200μgの間の量の免疫原を用いる、少なくとも2週間間隔での3回の皮下、皮内、または腹腔内の免疫化、あるいはこれらの組み合わせ、3回目の免疫化の2週間後、2日間隔で、同じ量の免疫原(ただし、生理食塩水血清(saline serum)中)を使用して、2回の新規の追加免疫を投与した。他の動物も、配列番号1のペプチド213〜227、229〜242、212〜226、227〜241、および242〜256の混合物を用いて免疫化させた。これらのペプチドは、カイト J.(Kyte J.)、ドリトル R.F.(Doolittle R.F.)のアルゴリズム(「J.Mol.Biol.」、1982年、157:105〜132)に従うProtscaleプログラムによると非常に親水性であることから選択された。
【0066】
融合
最後の追加免疫投与の2日後に、ハイブリドーマを産生した。ポリエチレングリコール1500または4000の存在下で、動物の脾細胞を骨髄腫細胞(SP2/O−Ag14、P3X63−Ag8.6.5.3、P3−NS−1−Ag4−1、その他)と融合させることからなる技術を用いた。この薬剤は、膜融合を可能にし、その結果、様々な細胞型との間のハイブリッドが産生されることとなる。動物Bリンパ球と骨髄腫細胞との間のハイブリッドは、新規の細胞型、ハイブリドーマを形成し、これは、抗体を産生し、また、培養中に無限に増殖する。これらのハイブリドーマを選択し、残りのハイブリッドを排除するために、細胞培養物にHAT培地を加えた。HATは、アミノプテリン、すなわちグアノシンの新規の合成経路(新生経路)の阻害剤を含有する。骨髄腫細胞は、核酸合成サルベージ経路に必要な機能性のHPRT酵素を欠いているので、唯一の利用可能な経路(新生経路)が、アミノプテリンでブロックされる場合、これらは、核酸を合成することができない。したがって、培養の2週後、培養中で生き残る唯一の細胞は、脾細胞−骨髄腫ハイブリッドである。脾細胞は、ハイブリッドに代替の経路によるグアノシン合成能力を付与し、骨髄腫細胞に、インビトロの培養において際限なく培養される能力を付与する。
【0067】
抗sEPCRvarおよび抗sEPCR抗体を産生するハイブリドーマの検出
HAT培地中の細胞を、少なくとも10の96ウェル培養マイクロプレートに分配する。融合の9日後と14日後との間に、コロニーのサイズは、上清中の抗体の存在を分析するのに十分となる。目的の抗体、すなわちsEPCRvarに対する抗体を分泌するハイブリドーマを選択するために、クローンを含有する5枚のマイクロプレートのすべてのウェルから上清のサンプルを取り出し、これをイムノアッセイにかけた。ELISAおよびウェスタンブロットアッセイを実施した。ELISAについては、プレートを、sEPCRvar(0.3μg/ウェル)で被覆し、4℃での15時間のインキュベーション、および適切なタンパク質による対応するブロッキングの後、培養上清を加え、相応な洗浄を実施し、次いで、二次マウス抗免疫グロブリン抗体を加える。このようにして、ウェルの洗浄およびペルオキシダーゼおよびo−フェニレンジアミンを用いる発色の後、着色または吸光度(492nm)の上昇が検出されたウェルは、sEPCRvar抗体を分泌するハイブリドーマのクローンを含有する可能性がある。ウェスタンブロットについては、組み換え型タンパク質および/またはsEPCRvarタンパク質を発現する組織または細胞抽出物を、ポリアクリルアミドゲルで泳動した。これを、上清中の特異的抗体の存在を証明するために、格子型(grille−type)装置によって独立した領域に分けられたニトロセルロースまたはPVDF膜にトランスファーした。ブロッキングおよび特異的抗体とのインキュベーションを行った後、これを3回洗浄し、ペルオキシダーゼ酵素に結合した二次抗体と共にインキュベートした。発色は、酵素によって形質転換される際、特異的な抗体に結合された場所で光を産生する基質を使用して実施した。発光バンドが、予測分子量を有するタンパク質に相当する場合、その上清が由来するウェルからの抗体産生細胞を増殖させ、液体窒素中に凍結させることとなる。
【0068】
sEPCRvarは、全sEPCR部分を含有するので、sEPCRvarに対して陽性であるハイブリドーマの、組み換え型EPCRの細胞外領域との反応性が分析されることとなる。このようにして、sEPCRvarの特定の領域に対する抗体を産生するハイブリドーマを選択することができる。
【0069】
ハイブリドーマの単クローン性
抗sEPCRvar抗体を分泌する各細胞培養物がモノクローナルであることを確認するために、クローニングまたは限界希釈技術が適用される。第1のウェスタンブロットまたはELISA試験で陽性であった最初の培養物からの、あるいは幹細胞からの、単離された細胞を、新しい培養マイクロプレート上で増殖させる。1種または複数の細胞から生じている新規のコロニーが、一旦十分なサイズを獲得したら、これから再び上清を取り出し、これを新規のELISAまたはウェスタンブロットにかけることになる。再び陽性であったコロニーを限界希釈にかけ、このプロセスを、第3の限界希釈の後に分析された上清の100%が、sEPCRvarに対する抗体を含有するようになるまで繰り返した。
【0070】
抗体精製
より大きな抗体濃度を達成するために、ハイブリドーマを特別な条件で培養した後(インテグラバイオサイエンス(Integra Biosciences)からのCELlineシステムを備えた培養ビン、またはベクトンディッキンソン(Becton Dickinson)からのCELlineシステム)、IgGを含有する上清を、以下の方法のうちの1つまたは複数を用いる液体クロマトグラフィーによる精製にかけることとなる:AKTA FPLC装置、GEヘルスケアバイオサイエンス(GE Healthcare Bio−Science)を使用する、免疫親和性クロマトグラフィー、アフィニティークロマトグラフィー(プロテインA/プロテインG/プロテインL、固定化金属、バイオアフィニティ)、陽イオン交換、ヒドロキシアパタイト、疎水的相互作用、ゲル濾過、など。
【0071】
[参考文献]
1. Bangalore N, Drohan WN, Orthner CL. High affinity binding sites for activated protein C and protein C on cultured human umbilical vein endothelial cells. Independent of protein S and distinct from known ligands. Thromb Haemost 1994; 72: 465-74.
2. Fukodome K, Esmon CT. Identification, cloning and regulation of a novel endothelial cell protein C/activated protein C receptor. J Biol Chem 1994; 269: 26486-91.
3. Stearns-Kurosawa DJ, Kurosawa S, Mollica JS. Ferrell GL, Esmon CT. The endothelial cell protein C receptor augments protein C activation by the thrombin-thrombomodulin complex. Proc. Natl Acad Sci USA 1996; 93: 10212-6.
4. Taylor FB Jr, Peer GT, Lockhart MS, Ferrell G, Esmon CT. Endothelial cell protein C receptor plays an important role in protein C activation in vivo. Blood 2001; 97; 1685-8.
5. Biguzzi E, Merati G, Liaw PC, Bucciarelli P, Oganesyan N, Qu D, Gu JM, Fetiveau R, Esmon CT, msnnucci PM, Faioni EM. A 23bp insertion in the endothelial protein C receptor (EPCR) gene impairs EPCR function. Thromb Haemost 2001; 86: 945-8.
6. Saposnik B, Reny JL, Gaussem P, Emmerich J, Aiach M, Gandrille S: A haplotype of
the EPCR gene is associated with increased plasma levels of sEPCR and is a candidate risk factor for thrombosis. Blood 2004: 103; 1311-1318.
7. Uitte de Willige S, Van Marion V, Rosendaal FR, Vos HL, de Visser MCH, Bertina RM.
Haplotypes of the EPCR gene, plasma sEPCR levels and the risk of deep venous thrombosis. J. Thromb Haemost. 2004; 2: 1305-10.
8. Taylor FB Jr, Stearns-Kurosawa DJ, Kurosawa S, Ferrell G, Chang AC, Laszik Z, Kosanke S, Peer. G, Esmon CT: The endothelial cell protein C receptor aids in host defense against Escherichia coli sepsis. Blood 2000; 95: 1680-6.
9. Joyce DE, Gelbert L, Ciaccia A, DeHoff B, Grinnell BW. Gene expression profile
of antithrombotic protein C defines new mechanisms modulating inflammation and apoptosis. J Biol Chem. 2001; 276: i1199-203.
10. Riewald M, Petrovan RJ, Donner A, Mueller BM, Ruf W. Activation of endothelial
cell protease activated receptor 1 by the protein C pathway. Science 2002; 296: 1880-2.
11. Fukudome K, Kurosawa S, Stearns-Kurosawa DJ, He X, Rezaie AR, Esmon CT.
The endothelial cell protein C receptor. Cell surface expression and direct ligand binding by the soluble receptor. J Biol Chem 1996; 271: 17491-8.
12. Simmonds, R E, Lane, DA. Structural and functional implications of the intron/exon organisation of the human endothelial cell protein C/activated protein C receptor (EPCR) gene: comparison with the structure of CD1/major histocompatibility complex alpha-1 and alpha-2 domains. Blood 1999; 94: 632-41.
13. Oganesyan V, Oganesyan N, Terzyan S, Qu D, Dauter Z, Esmon NL, Esmon CT. The crystal structure of the endothelial protein C receptor and a bound phospholipid. J Biol Chem 2002; 277: 24851-4.
14. Liaw PC, Mather T, Oganesyan N, Ferrell GL, and Esmon CT. Identification of the protein C/activated protein C binding sites on the endothelial cell protein C receptor. Implications for a novel mode of ligand recognition by a major histocompatibility complex class 1-type receptor. J Biol Chem 2001; 276: 8364-70.
15. Kurosawa S, Stearns-Kurosawa DJ, Hidari N, Esmon CT. Identification of functional endothelial protein C receptor in human plasma. J Clin Invest 1997; 100: 411-8.
16. 8, Xu J, Qu D, Esmon NL, Esmon CT. Metalloproteolytic release of endothelial cell protein C receptor. J Biol Chem 1999; 275: 603844.
17. Fukudome K, Esmon CT. Molecular cloning and expression of murine and bovine endothelial cell protein C/activated protein C receptor (EPCR). The structural and functional conservation in human, bovine, and murine EPCR. J Biol Chem. 1995;270:5571-7.
18. Liaw PCY, Neuenschwander PF, Smirnov M, Esmon CT. (2000) Mechanisms by which soluble endothelial cell protein C modulates protein C and activated protein C function. J Biol Chem. 2000; 275:5447-52.
19. Kurosawa S, Esmon CT, Stearns-Kurosawa DJ. The soluble endothelial protein C receptor binds to activated neutrophils: involvement of proteinase-3 and CD11b/CD18. J Immunol 2000; 165: 4697-4703.
【図面の簡単な説明】
【0072】
【図1】図1は、RNAがプロセッシングされ、成熟したメッセンジャーRNA(mRNA)が産生される方式に依存する、異なるタイプのEPCRを示す図である。3つの別の形のmRNAが産生される可能性があり、これらは、続いて、3つの異なる形のタンパク質を産生することとなる:1:膜EPCR(mEPCR)として翻訳されるタンパク質とは異なる4つのエキソンを伴うmRNA。2.エキソンEx I、Ex II、およびEx IIIのみを含み、その後にイントロン3およびこのイントロンの後の全配列が続く、国際公開第9900673号パンフレット中にエスモン(Esmon)によって開示された可溶形のEPCR(altsESPC)として翻訳されることとなるmRNA。3.本発明に開示された新規の形の選択的切断およびスプライシングによって、エキソンEx I、Ex II、およびEx IIIを含むmRNAが産生されることとなるが、プロセッシング中に、エキソンEx IVは失われ、この理由から、膜貫通ドメインも細胞質のテールも転写されず、その場所では、エキソンIIIの後、3’非翻訳領域に位置する潜在性(cryptic)エキソンであるであろう配列が出現し、この場合、mRNAは、本発明の新規の可溶性EPCR形(sEPCRvar)として翻訳される。可溶性EPCRはまた、おそらくメタロプロテアーゼ(まだ特徴づけられていない)から、タンパク分解活性によって産生することもできる。メタロプロテアーゼは、膜高さでEPCRを切断し、その結果、細胞外受容体の画分からなり、膜貫通ドメインおよび細胞質内テールを欠く、新規の可溶形のEPCR(sEPCR)が産生される。
【図2】図2は、特異的なEPCR遺伝子プライマーを用いるHUVEC cDNAの増幅の後に得られるバンドパターンである:野生型EPCR cDNA(1221bp、矢印1)に相当するバンドに加えて、選択的切断−スプライシングによって産生された新規の変異体アイソフォームのcDNAに相当する新規の増幅断片が認められる(矢印2)。
【図3】図3は、どちらも緑色蛍光タンパク質と融合させた、野生型EPCR(パネルA)を用いて、あるいはアイソフォーム(パネルB)を用いて形質移入されたCOS−7細胞である。パネルAは、野生型タンパク質が、細胞膜(矢印)上に蓄積する傾向があることを示すのに対して、アイソフォーム3を用いた場合、この局在化パターンは認められない(パネルB)。
【図4】図4は、ピキアパストリス(Pichia Pastoris)におけるsEPCRvar発現を説明する図である。本明細書に示すように、sEPCRvarは、安定に形質転換されたP.パストリス(P.pastoris)細胞の上清から精製された。パネルA:SDS−PAGEによって分離され、GELCODE Blue染料を使用して検出された様々なタンパク質を示す。ストリート St:発現される各バンドの分子量をkDaで示す、分子量マーカー、ストリート 1:組み換え型のsEPCR。約42kDaのブロードなバンド(タンパク質の強いグリコシル化に起因する)が観察される。Street 2:sEPCRvar。47kDaのブロードなバンド(タンパク質の強いグリコシル化に起因する)が観察される。パネルB:モノクローナル抗−EPCR抗体を用いるウェスタンブロットによって検出されるタンパク質、ストリート St:発現される各バンドの分子量をkDaで示す、分子量マーカー、ストリート 1:ブランク、ストリート 2:sEPCRvar。約47kDaのブロードなバンド(様々なタンパク質のグリコシル化に起因し、35から60kDaまでの間に広がる)が観察される。
【図5】図5は、異なる組織および腫瘍由来のcDNAからのEPCR遺伝子の増幅である。パネルA)1:分子量マーカー、2:心臓、3:肝臓、4:腎臓、5:膵臓、6:肺、7:胎盤。パネルB)1:分子量マーカー、2:骨格筋、3:脳、4:胸腺、5:小腸、6:脾臓。パネルC)1:マーカー、2:前立腺、3:睾丸、4:卵巣、5:大腸。パネルD)は、肺腫瘍の腫瘍株:2:H 446、3:H 510、4:H 1264、5:H 549、6:H 441、7:HTB 51、8:H 676、9:H 727、10:H 720、11:H 385、12:H 1299からのサンプルである。分子量マーカーを含有するストリートは、図中に略語St(「standard(標準)」より)で示す。マーカーの下方のバンドは、100bpサイズに相当し、各新しいバンドは、その前のバンドに対して100bpの増分(100bp、200bp、300bp・・・)に相当する。
【特許請求の範囲】
【請求項1】
配列番号1の配列の残基201〜256、または201と256との間の前記配列の残基のフラグメントを含むことを特徴とする、大血管およびガンにおける疾患プロセスのためのマーカーとして有用な、内皮プロテインC受容体に由来するポリペプチド。
【請求項2】
配列番号1の残基213〜227、229〜242、212〜226、227〜241、242〜256を含むフラグメントであることを特徴とする、請求項1に記載のポリペプチド。
【請求項3】
配列番号1の配列の残基201〜256から形成されるポリペプチドを含むことを特徴とする、単離されたEPCRタンパク質。
【請求項4】
配列番号1の配列を含むことを特徴とする、請求項3に記載の単離されたEPCRタンパク質。
【請求項5】
請求項1〜4のいずれか一項に記載のポリペプチドを含むことを特徴とする組み換え型のタンパク質。
【請求項6】
配列番号1の配列の残基201〜256から形成されるポリペプチドまたは前記ポリペプチドのフラグメントをコードする配列を含むことを特徴とする、単離されたあるいは組み換え型のポリヌクレオチド。
【請求項7】
配列番号2に相当するcDNA配列に由来することを特徴とする、請求項6に記載のポリヌクレオチド。
【請求項8】
前記ポリヌクレオチドによってコードされるタンパク質またはポリペプチド発現を調節する作動可能に結合された制御配列をさらに含むことを特徴とする、請求項6または7のいずれか一項に記載のポリヌクレオチド。
【請求項9】
配列番号1の配列の残基201〜256によって形成されるポリペプチドまたは前記ポリペプチドのフラグメントをコードする配列を含むことを特徴とするDNA構築物。
【請求項10】
配列番号2に相当するcDNA配列から得られるポリヌクレオチドを含むことを特徴とする請求項9に記載のDNA構築物。
【請求項11】
前記ポリヌクレオチドによってコードされるタンパク質またはポリペプチドの発現を調節する作動可能に結合された制御配列をさらに含むことを特徴とする、請求項9または10のいずれか一項に記載のDNA構築物。
【請求項12】
請求項1に記載の配列番号1の配列の残基201〜256を含むポリペプチドまたは前記配列のフラグメントをコードする、請求項6〜8のいずれか一項に記載のポリヌクレオチド、または請求項9〜11のいずれか一項に記載のDNA構築物を含むことを特徴とする発現ベクター。
【請求項13】
請求項6〜8のいずれか一項に記載のポリヌクレオチド、または請求項9〜11のいずれか一項に記載のDNA構築物、または請求項12に記載の発現ベクターを含むことを特徴とする宿主細胞。
【請求項14】
前記ポリヌクレオチドまたはDNA構築物が、適切な制御配列に作動可能に結合されていることを特徴とする請求項13に記載の宿主細胞。
【請求項15】
前記細胞が、原核細胞または真核生物細胞であることを特徴とする請求項13または14のいずれか一項に記載の宿主細胞。
【請求項16】
前記細胞が、細菌大腸菌または酵母ピキアパストリス(Pichia Pastoris)であることを特徴とする請求項15に記載の宿主細胞。
【請求項17】
請求項12に記載の発現ベクターと、請求項13〜16のいずれか一項に記載の宿主細胞を含むことを特徴とする発現系。
【請求項18】
前記タンパク質、ポリペプチド、またはフラグメントの発現を可能にする条件下で、請求項13〜16のいずれか一項に記載の宿主細胞を培養することを含むことを特徴とする、請求項1〜5のいずれか一項に記載のタンパク質、ポリペプチド、または前記ポリペプチドのフラグメントを産生する方法。
【請求項19】
配列番号1の配列の残基201〜256を含むタンパク質、または前記配列の残基の少なくとも9アミノ酸のフラグメントをコードすることを特徴とするmRNA。
【請求項20】
配列番号1の配列の残基201〜256を含むポリペプチドまたは免疫原特性を有する前記配列の残基のフラグメントに対して特異的な抗体。
【請求項21】
配列番号1の残基201〜256の配列のフラグメントが、少なくとも9アミノ酸を有することを特徴とする、請求項20に記載の抗体。
【請求項22】
配列が、残基201〜256、残基213〜227、229〜242、212〜226、227〜241、242〜256(すべて配列番号1に関する)から選択される前記ポリペプチドのポリペプチドまたはフラグメントに対して特異的であることを特徴とする、請求項20に記載の抗体。
【請求項23】
配列番号1に対して特異的であり、領域201〜256に結合しない場合には領域1〜200を認識しないことを特徴とする請求項20に記載の抗体。
【請求項24】
モノクローナル抗体であることを特徴とする請求項20〜23のいずれか一項に記載の抗体。
【請求項25】
請求項1に記載の配列番号1の配列の残基201〜256を含むEPCRタンパク質、または前記配列の残基のフラグメントの、生体試料のインビトロでの選択的検出のための方法であって、
−生体試料を得ること、
−配列番号1の配列の残基201〜256を含むEPCRタンパク質または前記配列の残基のフラグメントの量を分析すること、
を含むことを特徴とする方法。
【請求項26】
前記生体試料が、尿、血漿、血清、組織、および間質液から選択されることを特徴とする請求項25に記載の方法。
【請求項27】
前記方法が、免疫学的方法、クロマトグラフィー法、および分光光度法から選択されることを特徴とする請求項25〜26のいずれか一項に記載の方法。
【請求項28】
前記タンパク質の量が、血管損傷に関連する炎症性のプロセス、炎症、ガン、および凝固異常に関連する疾患から選択される疾患に関連することを特徴とする請求項25〜27のいずれか一項に記載の方法。
【請求項29】
検出されるEPCRタンパク質の量を、較正標準と比較することによって比較することを含むことを特徴とする請求項25〜27のいずれか一項に記載の方法。
【請求項30】
配列番号1の配列の残基201〜256を含むEPCRタンパク質または前記配列の残基のフラグメントの検出および定量化のための試験キットであって、a)請求項20〜24のいずれか一項に記載の特異的な抗体、
b)生体試料中に存在する配列番号1の配列の残基201〜256を含むEPCRタンパク質と、抗体a)との間の反応を検出する試薬、
を含むことを特徴とするキット。
【請求項31】
産生される反応物の量を、配列番号1の配列の残基201〜256を含むEPCRタンパク質の正常および異常なレベルと相関させるための標準も含むことを特徴とする請求項30に記載のキット。
【請求項32】
配列番号1の配列の残基201〜256を含むEPCRタンパク質または前記配列の残基のフラグメントに相当するmRNAを検出および定量化するための、配列番号1の配列の残基201〜256を含むポリペプチドをコードするポリヌクレオチドの使用。
【請求項33】
前記EPCRタンパク質が、血管損傷に関連する炎症性の疾患、炎症、ガンおよび凝固異常に関連する疾患から選択される疾患に関連する量で存在することを特徴とする請求項32に記載のポリヌクレオチドの使用。
【請求項1】
配列番号1の配列の残基201〜256、または201と256との間の前記配列の残基のフラグメントを含むことを特徴とする、大血管およびガンにおける疾患プロセスのためのマーカーとして有用な、内皮プロテインC受容体に由来するポリペプチド。
【請求項2】
配列番号1の残基213〜227、229〜242、212〜226、227〜241、242〜256を含むフラグメントであることを特徴とする、請求項1に記載のポリペプチド。
【請求項3】
配列番号1の配列の残基201〜256から形成されるポリペプチドを含むことを特徴とする、単離されたEPCRタンパク質。
【請求項4】
配列番号1の配列を含むことを特徴とする、請求項3に記載の単離されたEPCRタンパク質。
【請求項5】
請求項1〜4のいずれか一項に記載のポリペプチドを含むことを特徴とする組み換え型のタンパク質。
【請求項6】
配列番号1の配列の残基201〜256から形成されるポリペプチドまたは前記ポリペプチドのフラグメントをコードする配列を含むことを特徴とする、単離されたあるいは組み換え型のポリヌクレオチド。
【請求項7】
配列番号2に相当するcDNA配列に由来することを特徴とする、請求項6に記載のポリヌクレオチド。
【請求項8】
前記ポリヌクレオチドによってコードされるタンパク質またはポリペプチド発現を調節する作動可能に結合された制御配列をさらに含むことを特徴とする、請求項6または7のいずれか一項に記載のポリヌクレオチド。
【請求項9】
配列番号1の配列の残基201〜256によって形成されるポリペプチドまたは前記ポリペプチドのフラグメントをコードする配列を含むことを特徴とするDNA構築物。
【請求項10】
配列番号2に相当するcDNA配列から得られるポリヌクレオチドを含むことを特徴とする請求項9に記載のDNA構築物。
【請求項11】
前記ポリヌクレオチドによってコードされるタンパク質またはポリペプチドの発現を調節する作動可能に結合された制御配列をさらに含むことを特徴とする、請求項9または10のいずれか一項に記載のDNA構築物。
【請求項12】
請求項1に記載の配列番号1の配列の残基201〜256を含むポリペプチドまたは前記配列のフラグメントをコードする、請求項6〜8のいずれか一項に記載のポリヌクレオチド、または請求項9〜11のいずれか一項に記載のDNA構築物を含むことを特徴とする発現ベクター。
【請求項13】
請求項6〜8のいずれか一項に記載のポリヌクレオチド、または請求項9〜11のいずれか一項に記載のDNA構築物、または請求項12に記載の発現ベクターを含むことを特徴とする宿主細胞。
【請求項14】
前記ポリヌクレオチドまたはDNA構築物が、適切な制御配列に作動可能に結合されていることを特徴とする請求項13に記載の宿主細胞。
【請求項15】
前記細胞が、原核細胞または真核生物細胞であることを特徴とする請求項13または14のいずれか一項に記載の宿主細胞。
【請求項16】
前記細胞が、細菌大腸菌または酵母ピキアパストリス(Pichia Pastoris)であることを特徴とする請求項15に記載の宿主細胞。
【請求項17】
請求項12に記載の発現ベクターと、請求項13〜16のいずれか一項に記載の宿主細胞を含むことを特徴とする発現系。
【請求項18】
前記タンパク質、ポリペプチド、またはフラグメントの発現を可能にする条件下で、請求項13〜16のいずれか一項に記載の宿主細胞を培養することを含むことを特徴とする、請求項1〜5のいずれか一項に記載のタンパク質、ポリペプチド、または前記ポリペプチドのフラグメントを産生する方法。
【請求項19】
配列番号1の配列の残基201〜256を含むタンパク質、または前記配列の残基の少なくとも9アミノ酸のフラグメントをコードすることを特徴とするmRNA。
【請求項20】
配列番号1の配列の残基201〜256を含むポリペプチドまたは免疫原特性を有する前記配列の残基のフラグメントに対して特異的な抗体。
【請求項21】
配列番号1の残基201〜256の配列のフラグメントが、少なくとも9アミノ酸を有することを特徴とする、請求項20に記載の抗体。
【請求項22】
配列が、残基201〜256、残基213〜227、229〜242、212〜226、227〜241、242〜256(すべて配列番号1に関する)から選択される前記ポリペプチドのポリペプチドまたはフラグメントに対して特異的であることを特徴とする、請求項20に記載の抗体。
【請求項23】
配列番号1に対して特異的であり、領域201〜256に結合しない場合には領域1〜200を認識しないことを特徴とする請求項20に記載の抗体。
【請求項24】
モノクローナル抗体であることを特徴とする請求項20〜23のいずれか一項に記載の抗体。
【請求項25】
請求項1に記載の配列番号1の配列の残基201〜256を含むEPCRタンパク質、または前記配列の残基のフラグメントの、生体試料のインビトロでの選択的検出のための方法であって、
−生体試料を得ること、
−配列番号1の配列の残基201〜256を含むEPCRタンパク質または前記配列の残基のフラグメントの量を分析すること、
を含むことを特徴とする方法。
【請求項26】
前記生体試料が、尿、血漿、血清、組織、および間質液から選択されることを特徴とする請求項25に記載の方法。
【請求項27】
前記方法が、免疫学的方法、クロマトグラフィー法、および分光光度法から選択されることを特徴とする請求項25〜26のいずれか一項に記載の方法。
【請求項28】
前記タンパク質の量が、血管損傷に関連する炎症性のプロセス、炎症、ガン、および凝固異常に関連する疾患から選択される疾患に関連することを特徴とする請求項25〜27のいずれか一項に記載の方法。
【請求項29】
検出されるEPCRタンパク質の量を、較正標準と比較することによって比較することを含むことを特徴とする請求項25〜27のいずれか一項に記載の方法。
【請求項30】
配列番号1の配列の残基201〜256を含むEPCRタンパク質または前記配列の残基のフラグメントの検出および定量化のための試験キットであって、a)請求項20〜24のいずれか一項に記載の特異的な抗体、
b)生体試料中に存在する配列番号1の配列の残基201〜256を含むEPCRタンパク質と、抗体a)との間の反応を検出する試薬、
を含むことを特徴とするキット。
【請求項31】
産生される反応物の量を、配列番号1の配列の残基201〜256を含むEPCRタンパク質の正常および異常なレベルと相関させるための標準も含むことを特徴とする請求項30に記載のキット。
【請求項32】
配列番号1の配列の残基201〜256を含むEPCRタンパク質または前記配列の残基のフラグメントに相当するmRNAを検出および定量化するための、配列番号1の配列の残基201〜256を含むポリペプチドをコードするポリヌクレオチドの使用。
【請求項33】
前記EPCRタンパク質が、血管損傷に関連する炎症性の疾患、炎症、ガンおよび凝固異常に関連する疾患から選択される疾患に関連する量で存在することを特徴とする請求項32に記載のポリヌクレオチドの使用。
【図1】

【図2】
【図3】
【図4】
【図5】
【図2】
【図3】
【図4】
【図5】
【公表番号】特表2008−532536(P2008−532536A)
【公表日】平成20年8月21日(2008.8.21)
【国際特許分類】
【出願番号】特願2008−501340(P2008−501340)
【出願日】平成18年3月17日(2006.3.17)
【国際出願番号】PCT/ES2006/000136
【国際公開番号】WO2006/097564
【国際公開日】平成18年9月21日(2006.9.21)
【出願人】(506061716)プロイェクト、デ、ビオメディシナ、シーマ、ソシエダッド、リミターダ (34)
【氏名又は名称原語表記】PROYECTO DE BIOMEDICINA CIMA, S.L.
【Fターム(参考)】
【公表日】平成20年8月21日(2008.8.21)
【国際特許分類】
【出願日】平成18年3月17日(2006.3.17)
【国際出願番号】PCT/ES2006/000136
【国際公開番号】WO2006/097564
【国際公開日】平成18年9月21日(2006.9.21)
【出願人】(506061716)プロイェクト、デ、ビオメディシナ、シーマ、ソシエダッド、リミターダ (34)
【氏名又は名称原語表記】PROYECTO DE BIOMEDICINA CIMA, S.L.
【Fターム(参考)】
[ Back to top ]