
新規のIgEエピトープの同定
本発明は、IgEに特異的に結合する高親和性抗体によって認識される、IgEのCH3ドメインに由来する新規のペプチドエピトープに関する。これらの新規のペプチドは、被験体においてこれらのペプチドを投与して高親和性抗体を産生させることによる、被験体の能動免疫のため、ならびに被験体の受動免疫のために、非ヒト宿主においてIgEのこれらの領域に特異的に結合する高親和性抗IgE抗体を産生するための、両方に使用され得る。
【発明の詳細な説明】
【技術分野】
【0001】
(相互関連出願)
本出願は、PCT出願番号PCT/US04/02892およびPCT/US04/02894(2004年2月2日出願)の優先権を主張する。上記出願の全ては、本明細書において参考として援用される。
【背景技術】
【0002】
(発明の背景)
アレルギーは、外来の因子(例えば、アレルゲン)に対する過大な免疫応答によって誘導される過敏性状態である。即時(I型)過敏症は、アレルゲンとの接触直後のアレルギー性応答によって特徴付けられ、これは、B細胞を介して媒介され、そして抗原−抗体反応に基づく。遅延性過敏症は、T細胞を介して媒介され、そして細胞免疫の機構に基づく。近年、用語「アレルギー」は、よりI型過敏症と同義になってきている。
【0003】
即時過敏症は、アレルゲンに暴露されて抗体分泌性プラスマ細胞に分化するB細胞による、免疫グロブリンクラスEの抗体(IgE抗体)の産生に基づく反応である。IgEに誘導される反応は、身体へのアレルゲン侵入部位で(すなわち、粘膜表面および/または局所リンパ節において)生じる局所的事象である。局所的に産生されたIgEは、最初に局所性肥満細胞を感作する。すなわち、IgE抗体は、定常領域を用いて、肥満細胞の表面上のFcεレセプターに結合し、次いで、「溢出」IgEが循環に侵入し、全身にわたって循環性好塩基球および組織固定(tissue−fixed)肥満細胞の両方のレセプターに結合する。その結合したIgEが、その後アレルゲンに接触する場合、Fcεレセプターは、アレルゲンと結合することによって交差結合され、細胞を脱顆粒させ、多くのアナフィラキシー性媒介物質(例えば、ヒスタミン、プロスタグランジン、ロイコトリエンなど)を放出させる。即時過敏症に典型的な臨床症状(すなわち、気道または腸の平滑筋の収縮、小血管の拡張、ならびに水および血漿タンパク質に対する透過性の増大、粘液分泌(これらは、例えば、アレルギー性鼻炎、アトピー性湿疹(excema)、および喘息を引き起こす)、そして皮膚内に終わる神経の刺激(これは、かゆみおよび痛みを引き起こす))の原因となるのは、これらの物質の放出である。さらに、アレルゲンとの2回目の接触における反応は、IgEを細胞表面上に発現することによるアレルゲンとの最初の接触の後に、一部のB細胞が表面IgE陽性B細胞(sIgE+B細胞)の「記憶プール」を形成することに起因して、強化される。
【0004】
IgEに対する2つの主要なレセプターが存在する。抗親和性レセプターFcεRIおよび低親和性レセプターFcεRIIである。FcεRIは、肥満細胞および好塩基球の表面上に優先的に発現されるが、ヒトランゲルハンス細胞、樹状細胞、および単球上にもまた、低レベルのFcεRIが見出され、ここで、FcεRIは、IgE媒介性アレルゲン提示において機能する。さらに、FcεRIは、ヒト好酸球および血小板上でも報告されている(非特許文献1)。FcεRIは、B細胞、T細胞または好中球の表面上には見いだされない。ランゲルハンス細胞および皮膚樹状細胞でのFcεRIの発現は、機能的におよび生物学的に、アレルギーの個体におけるIgE結合抗原提示に関して重要である(非特許文献2)。
【0005】
低親和性レセプターFcεRII(CD23)は、細胞質膜からの長いらせん形のコイル状の柄から伸びる頭部構造を有する3つの同一なサブユニットを備えるレクチン様分子である(非特許文献3)。IgEへの結合のとき、FcεRIIは、IgEの合成の調節に関与するB細胞上のCD21と会合する(非特許文献4、非特許文献5)。FccRIIは、アレルゲン提示に関して長く認識されてきた。(非特許文献6)。上皮細胞上のFcεRIIに結合したIgEは、特異的かつ迅速なアレルゲン提示を引き起こす(非特許文献7)。FcεRIIは、数種の細胞型(B細胞、好酸球、血小板、ナチュラルキラー細胞、T細胞、濾胞樹状細胞、およびランゲルハンス細胞が挙げられる)に存在する。
【0006】
FcεRIおよびFcεRIIと相互作用するIgE分子の構造の実体もまた同定されている。突然変異誘発研究は、CH3ドメインが、FcεRI(非特許文献8;非特許文献9)およびFcεRII(非特許文献6;非特許文献10)の両方とのIgEの相互作用を媒介することを示した。高親和性レセプターおよび低親和レセプターの両方に対する結合部位は、2つのCH3ドメインを通る中心回転軸に沿って対称的に位置する。FcεRI結合部位は、CH3ドメインの、CH2ドメインの接合部近くの外側に位置し、一方FcεRII結合部位は、CH3のカルボキシル末端にある。
【0007】
アレルギー処置のための有望な考え方は、IgEアイソタイプ特異的であり、したがってIgEに結合し得る、モノクローナル抗体の適用に関する。このアプローチは、IgE免疫応答(アレルギーの誘導における最も初期の反応であり、そしてアレルギー状態を維持させる)をダウンレギュレートすることによる、アレルギー性反応の阻害に基づく。他の抗体クラスの応答が影響を与えない場合、アレルギー症状に対する即時効果および長期持続効果の両方が達成される。ヒト好塩基球密度の初期の研究は、患者の血漿中のIgEレベルと好塩基球あたりのFcεRIレセプターの数との間の相関を示した(非特許文献11)。彼らは、アレルギーの人物と非アレルギーの人物とにおけるFcεRI密度が、好塩基球あたり104〜106レセプターの範囲に及ぶことを示した。後に、抗IgEによるアレルギー性疾患の処置が、循環性IgEの量を処置前のレベルの1%まで減少させることが示された(非特許文献12)。MacGlashanは、患者の血清中を循環する遊離IgEに結合する、全抗IgE抗体で処置された患者から得られた血清を分析した。彼らは、患者中の循環性IgEのレベルを低下させることは、好塩基球上に存在するレセプターの数を減少させる結果となることを報告した。したがって、彼らは、好塩基球および肥満細胞上のFcεRI密度が循環性IgE抗体のレベルによって直接的もしくは間接的に調節される、という仮説を立てた。
【0008】
より最近では、特許文献1は、FcεRIおよびFcεRIIのIgE結合部位に結合してIgEのレセプターへの結合をブロックする、IgE分子およびフラグメントの使用を開示した。しかし、これらのアレルギー性疾患の管理に対する有害な副作用のない効果的な治療は、限られている。アレルギー性疾患の処置に対する1つの治療アプローチは、ヒト化抗IgE抗体を使用して、アレルギー性鼻炎および喘息を処置することに関する(非特許文献13;非特許文献14;非特許文献15;非特許文献16;非特許文献17)。これらの臨床的データは、IgEのそのレセプターへの結合の阻害が、アレルギー性疾患を処置するための効果的なアプローチであることを実証する。
【0009】
抗アレルギー剤として適切な抗体は、IgE産生性プラスマ細胞へと分化する表面IgE陽性B細胞と反応すべきであり、これによってこれらは、機能的にそれらのB細胞を消失させるために使用され得る。しかし、IgEに対する抗体は、基本的に、Fcεレセプターと交差結合することによって、IgE感作された肥満細胞から放出される媒介物質を誘導し得る。したがって、血清IgEレベルおよびsIgE+B細胞レベルに対して発揮される有利な効果を拮抗する。抗IgE治療を発展させ得ることに伴う潜在的に危険な問題の1つは、既に抗親和性レセプターに結合しているIgEに結合する治療用抗体によって引き起こされ、そしてヒスタミン放出を誘発し、潜在的にアナフィラキシー性の反応を引き起こす、IgE交差結合の可能性である。
【0010】
したがって、アレルギーの治療のために適用可能な抗体は、感作された肥満細胞および好塩基球に結合したIgEと反応してはいけないが、sIgE+B細胞を認識する能力を保持しているべきである。このようなIgEアイソタイプ特異的抗体は、例えば、非特許文献18)によって、特許文献2において、および数件の米国特許(例えば、特許文献3)において、記載されている。
【0011】
抗IgE抗体を産生するために使用されるペプチドはまた、アナフィラキシー性抗体を誘導する危険の可能性を被る。能動的ワクチン接種の間の抗IgE抗体の産生は、免疫の間に産生された抗体が、高親和性IgEレセプターに結合したIgEに結合する場合、または他の機構によって、受動的に投与された抗IgE抗体と同様の方法で、ヒスタミン放出を誘発し得る。
【特許文献1】国際公開第99/62550号パンフレット
【特許文献2】欧州特許第0407392号明細書
【特許文献3】米国特許第5,449,760号明細書
【非特許文献1】Hasegawa,S.ら,Hematopoiesis,1999年,93:2543−2551
【非特許文献2】Klubal R.ら,J.Invest.Dermatol.1997年,108(3):336−42
【非特許文献3】Dierks,A.E.ら,J.Immunol.1993年,150:2372−2382
【非特許文献4】Sanon,A.ら,J.Allergy Clin.Immunol.1990年,86:333−344
【非特許文献5】Bonnefoy,J.ら,Eur.Resp.J.1996年,9:63s−66s
【非特許文献6】SuttonおよびGould,Nature,1993年,366:421−428
【非特許文献7】Yang,P.P.,J.Clin.Invest.,2000年,106:879−886
【非特許文献8】Prestaら,J.Biol.Chem.1994年,269:26368−26373
【非特許文献9】Henry A.J.ら,Biochemistry,1997年,36:15568−15578
【非特許文献10】Shi,J.ら,Biochemistry,1997年,36:2112−2122
【非特許文献11】Malveauxら,J.Clin.Invest.,1978年,62:176
【非特許文献12】MacGlashanら,J.Immunol.,1997年,158:1438−1445
【非特許文献13】Corne,J.ら,J.Clin.Invest.1997年,99:879−887
【非特許文献14】Racine−Poon,A.ら,Clin.Pharmcol.Ther.1997年,62:675−690
【非特許文献15】Fahy,J.V.ら,Am.J.Resp.Crit.Care Med.1997年,155:1824−1834
【非特許文献16】Boulet,L.P.ら,Am.J.Resp.Crit.Care Med.,1997年,155:1835−1840
【非特許文献17】Milgrom,E.ら,N.Engl.J.Med.,1999年,341:1966−1973
【非特許文献18】Changら(Biotechnology 1990年,8,122−126
【発明の開示】
【発明が解決しようとする課題】
【0012】
したがって、IgEに特異的に結合するが、既にその高親和性レセプターに結合したIgEには結合しない、高親和性の非アナフィラキシー性抗体に対する必要性、ならびに、アナフィラキシー性抗体を誘導しない、能動免疫のためのペプチドに対する必要性が存在する。本発明者らは、肥満細胞または好塩基球上のIgEに結合することなく、抗体の高親和性結合を提供する、IgEの特異的エピトープを同定した。これらの特異的エピトープは、次いで、レセプターに結合するIgEの領域に結合するのみである、IgEに対する抗体を産生する能動免疫のための、特異的ペプチドを産生するために使用され得る。このことは、この抗体が、既にレセプターに結合したIgEに交差結合しないこと、したがって非アナフィラキシー性であることを保証する。
【課題を解決するための手段】
【0013】
(発明の要旨)
本発明は、IgEのCH3ドメインに由来する新規のペプチドエピトープに関する。これらのペプチドエピトープは、IgEに特異的に結合する高親和性抗体によって認識される。これらの新規のペプチドは、哺乳動物においてこれらのペプチドを投与して高親和性抗体を産生することによる、哺乳動物の能動免疫のために使用され得る。このペプチドエピトープはまた、非ヒト宿主において、IgEのこれらの領域に特異的に結合する高親和性抗IgE抗体を産生すること、および得られた抗体を哺乳動物の受動免疫のために使用することに使用され得る。
【0014】
本発明の一免疫原(エピトープA、図11)は、以下のアミノ酸配列を含む:
Asn Pro Arg Gly Val Ser Xaa Tyr Xaa Xaa Arg Xaa(配列番号72)
エピトープAの1つの例は、以下である:
Asn Pro Arg Gly Val Ser Ala Tyr Leu Ser Arg Pro(配列番号73)
別の免疫原(エピトープB、図11)は、以下のアミノ酸配列を含む:
Leu Pro Arg Ala Leu Xaa Arg Ser Xaa(配列番号74)
エピトープBの例としては、以下が挙げられる:
Leu Pro Arg Ala Leu Met Arg Ser Thr(配列番号75)
His Pro His Leu Pro Arg Ala Leu Met Arg Ser Thr(配列番号76)
Leu Pro Arg Ala Leu Met Arg Ser Thr Thr Lys Thr(配列番号77)
配列番号72または配列番号74のいずれにおいても、Xaaは、任意のアミノ酸であり得る。
【0015】
これらのペプチドは、上記ペプチドの少なくとも1つ、および生理学的に受容可能なキャリア、希釈剤、安定剤、もしくは賦形剤、ならびに免疫原性キャリアを含む組成物中に含まれ得る。上記免疫原性キャリアは、例えば、BSA、KLH、破傷風トキソイド、およびジフテリアトキソイドであり得る。本発明はまた、配列番号72〜77をコードするポリヌクレオチド、このポリヌクレオチドを含むベクター、およびこのベクターを有する細胞に関する。
【0016】
本発明はまた、エピトープAおよび/またはエピトープBに特異的に結合する抗体に関する。本発明はまた、エピトープAおよび/またはエピトープBに特異的に結合する抗体を作製する方法に関する。
【0017】
本発明は、配列番号72および/または配列番号74を含むペプチドの、IgE媒介性疾患もしくはIgE媒介性状態に罹患する被験体への投与に関する。
【0018】
本発明は、配列番号72および/または配列番号74を含むペプチドを用いて産生された高親和性抗体の、IgE媒介性疾患もしくはIgE媒介性状態に罹患する哺乳動物への投与に関する。この高親和性抗体は、ヒト抗体、ヒト化抗体、またはキメラ抗体であり得る。この抗体は、ポリクローナル抗体またはモノクローナル抗体であり得る。このようなIgE媒介性疾患もしくはIgE媒介性状態としては、例えば、喘息、アトピー性皮膚炎、蕁麻疹、アレルギー性鼻炎、および湿疹が挙げられる。
【0019】
(発明の詳細な説明)
(定義)
本出願を通して使用される用語は、当業者にとって通常かつ典型的な意味で解釈されるべきである。しかし、本出願人らは、以下の用語が以下に定義されるような特定の定義を与えられることを所望する。
【0020】
抗体鎖ポリペプチド配列に関する用語「実質的に同一」は、参照ポリペプチド配列に対して、少なくとも70%、もしくは少なくとも80%、もしくは少なくとも90%、もしくは少なくとも95%の配列同一性を示す抗体鎖として解釈され得る。核酸配列に関するこの用語は、参照核酸配列に対して、少なくとも約85%、もしくは少なくとも約90%、もしくは少なくとも約95%、もしくは少なくとも約97%の配列同一性を示すヌクレオチドの配列として解釈され得る。
【0021】
用語「同一性」または「相同性」は、配列全体に対する最大パーセント同一性を達成することを必要し、そして配列同一性の一部として任意の保存的置換を考慮しない場合、配列をアラインメントさせ、そしてギャップを導入した後に比較される、対応する配列の残基と同一な候補配列中のアミノ酸残基のパーセンテージを意味すると解釈されるべきである。N末端またはC末端の伸張も、それらの挿入も、同一性または相同性の低減として解釈されるべきではない。アラインメントのための方法およびコンピュータプログラムは、当該分野で周知である。配列同一性は、配列分析ソフトウェアを使用して測定され得る。
【0022】
用語「抗体」は、最も広い意味で使用され、そして具体的には、モノクローナル抗体(全長モノクローナル抗体が挙げられる)、ポリクローナル抗体、および多重特異性抗体(例えば、二重特異性抗体)を網羅する。抗体(Ab)および免疫グロブリン(Ig)は、同じ構造特性を有する、糖タンパク質である。抗体が特定の標的に対する結合特異性を示す一方で、免疫グロブリンは、抗体および標的特異性を欠く他の抗体様分子の両方を含む。ネイティブな抗体および免疫グロブリンは、通常、約150,000ダルトンのヘテロテトラマー糖タンパク質であり、2つの同一な軽(L)鎖と2つの同一な重(H)鎖から構成される。各重鎖は、一端に可変ドメイン(VH)と、それに続く多数の定常ドメインを有する。各軽鎖は、一方の端に可変ドメイン(VL)を有し、そして他方の端に定常ドメインを有する。「高親和性」抗体とは、少なくとも10−10、好ましくは10−12の結合親和性を有するこれらの抗体をいう。
【0023】
本明細書で使用される場合、「抗ヒトIgE抗体」とは、このようなIgEの高親和性レセプターFcεRIへの結合を、阻害または実質的に低下させるような様式で、ヒトIgEを結合する抗体を意味する。
【0024】
抗体の可変ドメインに関連する用語「可変」とは、可変ドメインの特定の部分が、抗体間で配列がかなり異なり、そして各特定の抗体の、その特定の標的に対する結合および特異性において使用されるという事実を意味する。しかし、可変性は、抗体の可変ドメインを通して均一に分布していない。これは、軽鎖可変ドメインと重鎖可変ドメインとの両方において相補性決定領域(CDR)と呼ばれる(または超可変領域としても知られる)3つのセグメントに集中されている。可変ドメインのより高度に保存された部分は、フレームワーク(FR)と呼ばれる。ネイティブな重鎖および軽鎖の可変ドメインは、各々、4つのFR領域を含み、その多くはβ−シート構造を採用し、3つのCDRによって結合され、これらはループを形成し、上記β−シート構造を結合し、いくつかの場合はその一部を形成する。各鎖のCDRは、FR領域によって、他の鎖からのCDRと一緒に近接して保持され、抗体の標的結合部位の形成に寄与する(Kabatらを参照のこと)。本明細書において使用される場合、免疫グロブリンアミノ酸残基の番号付けは、他に示されない限り、Kabatら(Sequences of Proteins of Immunological Interest,National Institute of Health,Bethesda,Md.1987)の免疫グロブリンアミノ酸残基の番号付けシステムに従って行われる。
【0025】
用語「抗体フラグメント」とは、全長抗体の一部(一般的には、標的結合領域または可変領域)をいう。抗体フラグメントの例としては、Fab、Fab’、F(ab’)2およびFvフラグメントが挙げられる。語句、抗体の「機能的フラグメントまたはアナログ」は、全長抗体に共通の定性的生物学的活性を有する化合物である。例えば、抗IgE抗体の機能的フラグメントまたはアナログは、IgE免疫グロブリンに対して、このような分子の能力を、高親和性レセプターFcεRIに結合する能力を有することから防ぐかまたは実質的に低減させる様式で、結合し得るものをいう。本明細書において使用される場合、抗体に関して、「機能的フラグメント」とは、Fv、F(ab)およびF(ab’)2フラグメントをいう。「Fv」フラグメントは、完全な標的認識部位および完全な標的結合部位を含む、最小抗体フラグメントである。この領域は、1つの重鎖可変領域と1つの軽鎖可変領域との強い、非共有結合性結合のダイマー(VH−VLダイマー)からなる。これは、各可変ドメインの3つのCDRがVH−VLダイマーの表面における標的結合部位を規定するために相互作用する構造にある。集合的に、6つのCDRが、抗体に対する標的結合特異性を与える。しかし、単一の可変ドメイン(または、標的に関して特異的な3つのCDRのみを含む、Fvの半分)でさえ、全結合部位よりも低い親和性ではあるが、標的に認識および結合する能力を有する。「単鎖Fv」抗体フラグメントまたは「sFv」抗体フラグメントは、抗体のVHドメインおよびVLドメインを含み、これらのドメインは、単一のポリペプチド鎖中に存在する。一般的に、Fvポリペプチドはさらに、VHドメインとVLドメインとの間に、標的結合に関する所望の構造を形成するsFvを形成し得るポリペプチドリンカーを含む。
【0026】
Fabフラグメントは、軽鎖の定常ドメインおよび重鎖の第一定常ドメイン(CH1)を含む。Fab’フラグメントは、重鎖CH1ドメインのカルボキシル末端における数個の残基(抗体ヒンジ領域からの1つ以上のシステインを含む)の付加によってFabフラグメントと異なる。F(ab’)フラグメントは、F(ab’)2ペプシン消化産物のヒンジシステインにおけるジスルフィド結合の切断によって生成される。抗体フラグメントのさらなる化学結合は、当業者に公知である。
【0027】
用語「モノクローナル抗体」は、本明細書において使用される場合、実質的に均一な抗体の集団から得られた抗体をいう(すなわち、この集団を構成する個々の抗体は、少数の量で存在し得る、可能性のある天然に存在する変異を除いて、同一である)。モノクローナル抗体は、高度に特異的であり、単一の標的部位に対して指向する。さらに、種々の決定基(エピトープ)に対して指向する種々の抗体を代表的に含む従来の(ポリクローナル)抗体調製物に対して、各モノクローナル抗体は、標的上の単一の決定基に対して指向する。それらの特異性に加えて、モノクローナル抗体は、ハイブリドーマ培養によって合成され得、他の免疫グロブリンによって汚染されない可能性があるという点で有利である。修飾語「モノクローナル」は、実質的に均一の抗体集団から得られるような抗体の特徴を示し、そして任意の特定の方法による抗体の産生を必要とすると解釈されるべきではない。例えば、本発明で使用するためのモノクローナル抗体は、周知の技術を使用して、ファージ抗体ライブラリーから単離され得る。本発明によって使用されるべき親モノクローナル抗体は、KohlerおよびMilstein,Nature 256,495(1975)によって最初に記載されたハイブリドーマ法によって作製され得るか、または組換え法によって作製され得る。
【0028】
非ヒト(例えば、マウス)抗体の「ヒト化」形態は、非ヒト免疫グロブリンに由来する最低の配列を含む、キメラ免疫グロブリン、キメラ免疫グロブリン鎖、またはそれらのフラグメント(例えば、Fv、Fab、Fab’、F(ab’)2または他の抗体の標的結合配列)である。一般に、ヒト化抗体は、少なくとも1つ、そして代表的には2つの可変ドメインの実質的に全てを含む。ここで、CDR領域の全てまたは実質的に全ては、非ヒト免疫グロブリンに対応し、そしてFR領域の全てまたは実質的に全ては、ヒト免疫グロブリンコンセンサス配列のそれらの領域である。非ヒト抗体はまた、免疫グロブリン定常領域(Fc)(代表的に、選択されたヒト免疫グロブリンテンプレートの免疫グロブリン定常領域)の少なくとも一部を含み得る。
【0029】
用語「細胞」、「細胞株」および「細胞培養物」は、子孫を含む。全ての子孫は、意図的な変異または不慮の変異に起因して、DNA内容物において正確には同一でない可能性があることが、また理解される。最初に形質転換された細胞についてスクリーニングされる場合、同じ機能特性または生物学的特性を有する改変体子孫が含まれる。本発明において使用される「宿主細胞」は、一般的に、原核生物宿主または真核生物宿主である。
【0030】
DNAによる細胞生物体の「形質転換」とは、生物体にDNAを導入し、そのDNAが、染色体外エレメントとして、または染色体組み込みによって複製されることを意味する。DNAによる細胞生物体の「トランスフェクション」とは、任意のコード配列が実際に発現されていてもいなくとも、細胞または生物体によるDNA(例えば、発現ベクター)の取り込みをいう。用語「トランスフェクトされた宿主細胞」および「形質転換された」とは、DNAが導入された細胞をいう。この細胞は、「宿主細胞」と称され、原核生物または真核生物のいずれかであり得る。代表的な原核生物宿主細胞としては、E.coliの種々の系統が挙げられる。代表的な真核生物宿主細胞としては、哺乳動物(例えば、チャイニーズハムスター卵巣またはヒト起原の細胞)が挙げられる。導入されるDNA配列は、宿主細胞と同種由来、または(of)宿主細胞と異なる種であり得るか、あるいは、一部の外来DNAおよび一部の同種DNAを含む、ハイブリッドDNA配列であり得る。
【0031】
用語「ベクター」とは、適切な宿主中におけるDNAの発現を生じ得る適切な制御配列に作動可能に連結されたDNA配列を含む、DNA構築物を意味する。このような制御配列としては、転写を引き起こすプロモーター、このような転写を制御する任意のオペレーター配列、適切なmRNAリボゾーム結合部位をコードする配列、ならびに転写および翻訳の終止を制御する配列が挙げられる。ベクターは、プラスミド、ファージ粒子、または、単に潜在的ゲノムインサートであり得る。一旦適切な宿主中へと形質転換された場合、ベクターは、複製して宿主ゲノムと独立して機能し得るか、またはいくつかの例では、ゲノムそのものへと一体化し得る。本明細書において、「プラスミド」および「ベクター」は、時として、交換可能に使用される。なぜなら、プラスミドは最も一般的に使用されるベクターの形態だからである。しかし、本発明は、当該分野で公知であるか、公知となる機能と等価な機能を果たすような他のベクターの形態を包含することを意図する。
【0032】
発現「制御配列」とは、特定の宿主生物において作動可能に連結されたコード配列の発現に必要なDNA配列をいう。原核生物に適切な制御配列としては、例えば、プロモーター、必要に応じて、オペレーター配列、およびリボゾーム結合部位が挙げられる。真核生物細胞は、プロモーター、ポリアデニル化シグナル、およびエンハンサーを利用することが知られている。前配列(presequence)または分泌リーダーのためのDNAは、それがポリペプチドの分泌に関わる前タンパク質(preprotein)として発現される場合、そのポリペプチドのためのDNAと作動可能に連結されている可能性がある;プロモーターまたはエンハンサーは、それがコード配列の転写に影響する場合、そのコード配列に作動可能に連結されている;あるいは、リボゾーム結合部位は、それが翻訳を促進するように配置されている場合、コード配列に作動可能に連結されている。一般的に、「操作可能に連結された」とは、連結されたDNA配列が、隣接しており、そして分泌リーダーの場合、隣接していてリーディングフェーズにあることを意味する。しかし、エンハンサーは、連続している必要はない。
【0033】
処置目的のための「哺乳動物」とは、哺乳動物として分類される任意の動物をいい、ヒト、家畜動物(domestic animal)および家畜動物(farm animal)、非ヒト霊長類、および動物園の動物、競技用動物、またはペット用動物(例えば、イヌ、ウマ、ネコ、ウシなど)が挙げられる。
【0034】
用語「エピトープタグ化」とは、ポリペプチドに関して本明細書において使用される場合、「エピトープタグ」に融合されたポリペプチドをいう。エピトープタグポリペプチドは、そのエピトープに対して抗体が産生され得るエピトープを提供するのに十分な残基を有するが、ポリペプチドの活性を干渉しないように十分短い。エピトープタグはまた、好ましくは、かなり独特であり、したがって抗体は、他のエピトープと実質的に交差反応しない。適切なタグポリペプチドは、一般的に、少なくとも6アミノ酸残基を有し、通常、約8〜50アミノ酸残基(好ましくは、約9〜30残基)の間である。例としては、以下が挙げられる:インフルエンザHAタグポリペプチドおよびその抗体12CA5(Fieldら,Mol Cell.Biol.8:2159−2165(1988));c−mycタグならびにそれに対する8F9抗体、3C7抗体、6E10抗体、G4抗体、B7抗体および9E10抗体(Evanら,Mol Cell.Biol.5(12):3610−3616(1985));そして単純ヘルペスウイルス糖タンパク質D(gD)タグおよびその抗体(Paborskyら,Protein Engineering 3(6):547−553(1990))。特定の実施形態において、エピトープタグは、IgG分子(例えば、IgG1、IgG2、IgG3またはIgG4)のインビボ血清半減期の延長を担う、IgG分子のFc領域のエピトープであり得る。
【0035】
語「標識」とは、本明細書において使用される場合、分子またはタンパク質(例えば、抗体)と直接的または間接的に結合され得る検出可能な化合物または組成物をいう。標識は、それ自身検出可能(例えば、放射性同位体標識または蛍光標識)であり得るか、または酵素標識の場合、検出可能な基質化合物または組成物の化学的変化を触媒し得る。
【0036】
本明細書において使用される場合、「固相」とは、本発明の抗体が接着し得る非水性マトリックスを意味する。本明細書に包含される固相の例としては、部分的もしくは全体的に、ガラス(例えば、制御された多孔質ガラス)、多糖類(例えば、アガロース)、ポリアクリルアミド、ポリスチレン、ポリビニルアルコール、およびシリコーンから形成されたものが挙げられる。特定の実施形態において、状況に依存して、固相は、アッセイプレートのウェルを包含し得;他方、それは精製カラム(例えば、アフィニティークロマトグラフィーカラム)である。
【0037】
本明細書中で使用される場合、用語「IgE媒介性障害」とは、免疫グロブリンIgEの過剰生成および/または免疫グロブリンIgEに対する過剰な感受性によって特徴付けられる状態または疾患を意味する。具体的には、アナフィラキシー性過敏症ならびにアトピー性アレルギー(例えば、喘息、アレルギー性鼻炎、およびアレルギー性結膜炎(花粉症)、湿疹、蕁麻疹、アトピー性皮膚炎、および食物アレルギーが挙げられる)に関する状態を包含すると解釈される。例えば、蜂に刺されること、蛇にかまれること、食物、または医薬によって引き起こされるアナフィラキシーショックの重篤な生理学的状態もまた、本用語の範囲下に包含される。
【0038】
(抗体の生成)
出発抗体または「親」抗体は、このような抗体を生成するために当該分野で利用可能な技術を用いて調製され得る。これらの技術は、当該分野で周知である。出発抗体を生成するための例示的方法は、以下の節においてより詳細に記載される。これらの記載は、親抗体を作製または選択するための考えられる代替案であり、このような分子が生成され得る方法を決して限定しない。
【0039】
抗体の結合親和性は、本発明の高親和性抗体を生成する前に決定される。また、この抗体は、他の生物学的に活性なアッセイに(例えば、治療剤としての有効性を評価するために)供され得る。このようなアッセイは、当該分野で公知であり、そして抗体に対する標的および意図される用途に依存する。
【0040】
特定のエピトープに結合する抗体(例えば、IgEの、その高親和性レセプターへの結合をブロックするもの)についてスクリーニングするために、慣習的な交差ブロックアッセイ(例えば、Antibodies:A Laboratory Manual」(Cold Spring Harbor Laboratory,Ed HarlowおよびDavid Lane(1988))に記載されるようなアッセイ)が実施される。あるいは、エピトープマッピングは、抗体が目的のエピトープにどこで結合するかを決定するために実施され得る。必要に応じて、抗体を生成するために使用される標的のホモログに対する抗体の結合親和性(ここで、このホモログは、異なる種に由来する)は、当該分野で公知の技術を用いて評価され得る。一実施形態において、他の種は、前臨床研究において抗体が投与される非ヒト動物である。したがって、この種は、アカゲザル、カニクイザル、ヒヒ、チンパンジー、およびマカクのような非ヒト動物であり得る。他の実施形態において、この種は、例えば、げっ歯類、ネコ、またはイヌであり得る。
【0041】
親抗体は、標的に対して親抗体よりもより高いかもしくはより強い結合親和性を有する抗体を生成するように、本発明に従って変化される。抗体特異性は、抗体とその標的との間に形成される独特の界面から生じる;この表面は、独特のはめ合いを生じるように互いに補完する(Jones,S.およびThornton,J.M.(1996)Proc.Natl.Acad.Sci.USA 93:13−20)。その界面に沿った接触をさらに改善することによって、結合パートナーの会合を助けるために必要とされるエネルギーコストがより低いことの結果として、全体的な親和性が増加し得る。
【0042】
抗体の結合表面は、一般的に、6つの相補性決定領域(CDR)から構成される。これらは、コアから外に伸びるループである。CDRは、特異的標的への結合のために独特の配列を有するアミノ酸から構成される。抗体の、その抗原に対する親和性を増加させるために、これらのアミノ酸の周辺の環境は、種々の非共有結合力を導入または改善し、最終的に、相互作用のエネルギー特性を低下させ、その結果としてより高い親和性をもたらすことによって、より都合よくならねばならない。
【0043】
ファンデルワールス力は、2つの電気的に中性な分子の間で生じる非共有結合性相互作用である(Voet,D.およびVoet,J.G.(1990)Biochemistry John Wiley and Sons,NY,NY)。会合は、永久双極子または誘起双極子から生じる静電気的相互作用に由来して、2つの表面の間で起こり得る。これらの双極子は、αヘリックスの端に沿って存在し得るか、または極性アミノ酸の近くに存在し得る。結合界面に沿ったファンデルワールス力の数の増加によって、より都合よい会合が生じる。
【0044】
水素結合の導入もまた、抗体とその抗原との間の相互作用の特異性を増加させる。水素結合に関与する一般的なドナーおよびアクセプターは、主にアミノ酸を構成する窒素原子、酸素原子および硫黄原子である(上述のVoetらを参照のこと)。水素結合は、短い距離のみにわたる傾向にあり(通常2.7〜3.1Å)、そのため、その結合パートナーは、これらの相互作用が生じるためには密接しなければならない。したがって、親和性を改善し得る1つの様式は、潜在的ドナー分子と潜在的アクセプター分子とを密接して接触させて、水素結合を確立させることである。
【0045】
最後に、疎水性相互作用の改善もまた、2つの結合パートナーの間の有利なエネルギー特性を増加させる。結合表面に密接して位置する非極性残基は、他の非極性残基によって囲まれるべきであり、それによって有利な環境に存在する。非極性側鎖の囲み(the burial of nonpolar side chains)を満足することによって、相互作用のエネルギー特性は、強い結合界面のために有利となる。
【0046】
タンパク質−タンパク質界面を安定化させる相互作用は、これらの接触を維持するエネルギーコストを低下させ、したがって、全体的親和性を増加させる。結合界面近くの個々のアミノ酸の周りの環境を改善することによって、より有利な環境が生成され、結果としてより高い親和性が得られる。したがって、有利な接触を導入し、そしてさらなる相補性を介して界面を改善することによって、抗体と抗原との間の全体的な結合相互作用は大きく改善される。
【0047】
得られた抗親和性抗体は、好ましくは、親抗体の標的に対する結合親和性よりも少なくとも約10倍高い、または少なくとも約20倍高い、または少なくとも約500倍高い、または1000倍〜5000倍高くあり得る、標的に対する結合親和性を有する。必要な、もしくは所望される、結合親和性における増強の程度は、親抗体の最初の結合親和性に依存する。
【0048】
一般的に、親抗体由来の高親和性抗体を作製するための方法は、以下の工程を包含する。
【0049】
1.目的の標的に結合する、軽鎖可変ドメインおよび重鎖可変ドメインを含む親抗体を得るかまたか選択する工程。これは、伝統的なハイブリドーマ技術、ファージディスプレイ技術、または標的特異的抗体を生成する任意の他の方法によって行われ得る。
【0050】
2.配列が親フレームワーク配列(好ましくはヒトテンプレート配列)に近いフレームワーク配列を選択する工程。このテンプレートは、例えば、その比較される全体の長さ、CDRのサイズ、フレームワークとCDRとの間の接合部に位置するアミノ酸残基、全体的な相同性などに基づいて選択され得る。選択されるテンプレートは、1より多くの配列の混合物であり得るか、またはコンセンサステンプレートであり得る。
【0051】
3.考えられるCDR位置の各々および全てにおいてランダムアミノ酸置換を作製することによってクローンのライブラリーを生成する工程。これは、例えば、CDRに隣接するか、または全ての考えられるアミノ酸と結合するか、それをフォールディングすることに影響を及ぼすヒトフレームワークテンプレート中のアミノ酸をランダムに置換し得、フレームワーク置換のライブラリーを生成する。これらのフレームワーク置換は、標的結合および抗体フォールディングに対するそれらの潜在的効果について評価され得る。フレームワーク中のアミノ酸の置換は、CDR中のアミノ酸の置換と同時にか、または順次にかのいずれかで行われ得る。改変体のライブラリーを生成するための1つの方法は、オリゴヌクレオチド合成による。
【0052】
4.工程(3)において生成された重鎖および/または軽鎖改変体を含む発現ベクターを構築する工程。上記改変体は、以下の式を含み得る:FRH1−CDRH1−FRH2−CDRH2−FRH3−CDRH3−FRH4(I)およびFRL1−CDRL1−FRL2−CDRL2−FRL3−CDRL3−FRL4(II)、ここで、FRL1、FRL2、FRL3、FRL4、FRH1、FRH2、FRH3およびFRH4は、工程3において選択されたフレームワークテンプレートの軽鎖配列および重鎖配列の改変体を表し、そしてCDRは、親抗体CDRの改変体CDRを表す。このような重鎖配列および軽鎖配列を含むベクターの例は、図1に示される。
【0053】
5.特異的標的に対するクローンのライブラリーをスクリーニングする工程。標的に結合するこれらのクローンは、改善された結合親和性についてスクリーニングされる。親分子より大きな親和性で結合するこれらのクローンが選択され得る。最適な高親和性候補は、親抗体と比較して、(好ましくは、20倍より大きい、100倍より大きい、1000倍より大きい、または5000倍より大きい)最大の結合親和性の可能性を有する。選択された改変体が、望ましくない特定のアミノ酸(例えば、導入されたグリコシル化部位または潜在的に免疫原性の部位)を含む場合、それらのアミノ酸は、より有利なアミノ酸残基で置き換えられ得、そして結合親和性が再評価され得る。
【0054】
また、この方法を使用して、完全ヒト親抗体から、CDR領域のみをランダムに置換することによって、ヒトフレームワーク領域をインタクトなままにして、高親和性抗体を生成し得る。
【0055】
改善されたハイスループットスクリーニング技術および図1に記載されたようなベクターによって、当業者は、所与のCDR中および/またはフレームワーク領域中の全ての部位における置換の包括的なライブラリーを、迅速かつ効果的にスクリーニングし得る。全てのアミノ酸を全ての位置において同時に、ランダムに置換することによって、個々の置換によっては予測または同定できない(例えば、相乗作用に起因する)親和性を有意に増加させる可能性のある組合せをスクリーニングすることができる。
【0056】
(親抗体の調製)
(標的の調製)
可溶性の標的またはそれらのフラグメントは、抗体を生成するための免疫原として使用され得る。この抗体は、目的の標的に対して指向する。好ましくは、この標的は、生物学的に重要なポリペプチドであり、疾患または障害に罹患する哺乳動物へのこの抗体の投与は、哺乳動物に治療的利益をもたらし得る。しかし、抗体は、非ポリペプチド標的に対して指向し得る。標的がポリペプチドである場合、それは、膜貫通分子(例えば、レセプター)またはリガンド(例えば、成長因子)であり得る。本発明の1つの標的は、IgEである。全細胞が抗体を作製するための免疫原として使用され得る。標的は、組み換え的に使用され得るか、または合成法を用いて使用され得る。標的はまた、天然の供給源から単離され得る。
【0057】
本発明の抗体を生成するために使用される抗原は、本発明のポリペプチドおよびポリペプチドフラグメント(エピトープAおよび/またはエピトープBが挙げられる)を包含し得る。動物を免疫するために使用されるポリペプチドは、標準的な組み換え法、化学合成法、または精製法によって得ることができる。当該分野で周知であるように、免疫原性を増加させるため、抗原は、キャリアタンパク質に結合され得る。一般的に使用されるキャリアとしては、キーホールリンペットヘモシアニン(KLH)、サイログロブリン、ウシ血清アルブミン(BSA)および破傷風トキソイドが挙げられるが、これらに限定されない。次いで、結合されたペプチドは、動物(例えば、マウス、ラット、もしくはウサギ)を免疫するために使用される。このようなキャリアに加えて、周知のアジュバントが抗原とともに投与されて、強力な免疫応答の誘導を促進し得る。
【0058】
(ポリクローナル抗体)
ポリクローナル抗体は、通常、非ヒト哺乳動物において、アジュバントと組み合わせての関連する標的の複数回の皮下(sc)注射または腹腔内(ip)注射によって生成される。免疫応答を誘起し得る多くの因子は当該分野において周知である。
【0059】
動物は、(各々、ウサギまたはマウスについての)タンパク質または結合体をフロイント完全アジュバントと組み合わせて、そしてその溶液を皮内に注射することによって、標的、免疫原性結合体、または誘導体に対して免疫される。一ヵ月後、その動物は、フロイント不完全アジュバント中の最初の量の1/5〜1/10のペプチドまたは結合体を用いて、複数部位への皮下注射によって、追加免疫される。7日〜14日後、この動物は、採血されて、その血清が抗体力価についてアッセイされる。動物は、力価がプラトーに達するまで追加免疫される。
【0060】
選択された哺乳動物抗体は、通常、標的に対して十分に強力な結合親和性を有する。例えば、この抗体は、ヒト抗IgE標的と、約1×10−8Mの結合親和性(Kd)の値で結合し得る。抗体親和性は、飽和結合;酵素結合イムノソルベントアッセイ(ELISA);および競合アッセイ(例えば、ラジオイムノアッセイ)によって決定され得る。
【0061】
目的の標的に結合する抗体をスクリーニングするため、慣習的な交差結合アッセイ(例えば、Antibodies,A Laboratory Manual,Cold Spring Harbor Laboratory,Ed HarlowおよびDavid Lane(1988)に記載されるようなアッセイ)が行われ得る。あるいは、エピトープマッピング(例えば、Champeら,J.Biol.Chem.270:1388−1394(1995)に記載される)が行われて、結合を決定し得る。
【0062】
(モノクローナル抗体)
モノクローナル抗体は、単一の抗原部位を認識する抗体である。これらの均一な特異性は、モノクローナル抗体を、種々の異なる抗原部位を認識する抗体を通常含むポリクローナル抗体よりもずっと有用にしている。モノクローナル抗体は、Kohlerら,Nature,256:495(1975)によって最初に記載されたハイブリドーマ法を用いて作製され得るか、または組み換えDNA法によって作製され得る。
【0063】
ハイブリドーマ法において、マウスまたは他の適切な宿主動物(例えば、げっ歯類)は、本明細書において上記されるように免疫されて、免疫のために使用されたタンパク質に特異的に結合する抗体を産生するか、または産生し得るリンパ球を誘起される。あるいは、リンパ球は、インビトロで免疫され得る。次いで、リンパ球は、ハイブリドーマ細胞を形成するために適切な融合因子(例えば、ポリエチレングリコール)を用いて、骨髄腫細胞と融合される(Goding,Monoclonal Antibodies:Principals and Practice,pp.590−103(Academic Press,1986))。
【0064】
このようにして調製されたハイブリドーマ細胞は、好ましくは非融合の親骨髄腫細胞の成長または生存を阻害する1つ以上の物質を含む適切な培養培地中に播種され、そして増殖する。例えば、親骨髄腫細胞が酵素ヒポキサンチングアニンホスホリボシルトランスフェラーゼ(HGPRTまたはHPRT)を欠く場合、ハイブリドーマのための培養培地は、代表的に、ヒポキサンチン、アミノプテリンおよびチミジン(HGPRT欠損細胞の成長を妨害する物質)を含む(HAT培地)。好ましい骨髄腫細胞は、効果的に融合し、選択された抗体産生細胞による抗体の安定な高レベル産生を支持し、そしてHAT培地のような培地に対する感受性のある細胞である。ヒト骨髄腫細胞株およびマウス−ヒトヘテロ骨髄腫細胞株は、ヒトモノクローナル抗体の産生に関して記載されている(Kozbar,J.Immunol.133:3001(1984);Brodeurら,Monoclonal Antibody Production Techniques and Applications,pp.51−63(Marcel Dekker,Inc.,New York,1987))。
【0065】
所望の特異性、親和性、および/または活性の抗体を産生するハイブリドーマ細胞が同定された後、クローンが、限界希釈手順によってサブクローニングされ得、そして標準的方法によって増殖され得る(Goding,Monoclonal Antibodies:Principals and Practice,pp.59−103,Academic Press,1986)。この目的のための適切な培養培地は含む。サブクローンによって分泌されるモノクローナル抗体は、培養培地から従来の免疫グロブリン精製手順(例えば、プロテインAセファロース、ヒドロキシアパタイトクロマトグラフィー、ゲル電気泳動、透析、またはアフィニティークロマトグラフィー)によって適切に分離される。
【0066】
モノクローナル抗体をコードするDNAは、従来の手順を用いて(例えば、そのモノクローナル抗体の重鎖および軽鎖をコードする遺伝子に特異的に結合し得るオリゴヌクレオチドプローブを用いることによって)容易に単離され、配列決定される。ハイブリドーマ細胞は、このようなDNAの供給源として役立つ。一旦単離された場合、このDNAは、発現ベクター中に配置され得る。次いでこの発現ベクターは、宿主細胞(例えば、E.coli細胞、NS0細胞、チャイニーズハムスター卵巣(CHO)細胞、または骨髄腫細胞)に移送されて、この組み換え宿主細胞におけるモノクローナル抗体の合成を得る。このDNAはまた、例えば、相同なマウス配列の代わりに、ヒト重鎖および軽鎖の定常ドメインについてのコード配列を置換することによって(米国特許第4,816,567号;Morrisonら,Proc.Natl Acad.Sci.USA 81:6851(1984))、または免疫グロブリンポリペプチドに共有結合性に結合することによって、改変され得る。
【0067】
(ヒト化抗体)
ヒト化は、キメラ抗体を作製するための技術であり、キメラ抗体においては、インタクトなヒト可変ドメインよりもずっとわずかが、対応する非ヒト種由来の配列によって置換されている。ヒト化抗体は、非ヒトである供給源からその中に導入された1つ以上のアミノ酸残基を有する。これらの非ヒトアミノ酸残基は、多くの場合、「移入(import)」残基と称され、これは代表的に、「移入」可変ドメインから取られる。ヒト化は、本質的に、Winterおよび共同研究者(Jonesら,Nature 321:522−525(1986);Riechmanら,Nature 332:323−327(1988);Verhoeyensら,Science 239:1534−1536(1988))の方法に従って、非ヒトCDRまたはヒト抗体中の対応する配列についてのCDR配列で置換することによって行われる(例えば、米国特許第4,816,567号を参照のこと)。本発明において実施される場合、ヒト化抗体は、マウス抗体における類似部位由来の残基によって置換された、いくつかのCDR残基およびいくつかのFR残基を有し得る。
【0068】
ヒト化抗体を作製するのに使用されるべきヒト可変ドメイン(軽鎖および重鎖の両方)の選択は、抗原性を低減させるために非常に重要である。いわゆる「ベストフィット(best fit)」法に従って、非ヒト抗体の可変ドメインの配列は、公知のヒト可変ドメイン配列のライブラリーと比較される。非ヒト親抗体の配列に最も近いヒト配列が、ヒト化抗体についてのヒトフレームワークとして受容される(Simsら,J.Immunol.151:2296(1993);Chothiaら,J.Mol.Biol.196:901(1987))。別の方法は、軽鎖または重鎖の特定のサブグループの全てのヒト抗体のコンセンサス配列に由来する、特定のフレームワークを使用する。同じフレームワークが、数種の異なるヒト化抗体に対して使用され得る(Carterら,Proc.Natl.Acad.Sci.USA,89:4285(1992);Prestaら,J.Immunol.151:2623(1993))。
【0069】
(抗体フラグメント)
種々の技術が、抗体フラグメントの生成のために開発されている。伝統的には、これらのフラグメントは、インタクトな抗体のタンパク質分解性消化を介して得られた(例えば、Morimotoら,Journal of Biochemical and Biophysical Methods 24:107−117(1992)およびBrennanら,Science 229:81(1985))。しかし、これらのフラグメントは、現在、組み換え宿主細胞によって直接的に生成され得る。例えば、抗体フラグメントは、抗体ファージライブラリーから単離され得る。あるいは、F(ab’)2−SHフラグメントが、E.coliから直接的に回収され得、化学的に結合されてF(ab’)2フラグメントを形成し得る(Carterら,Bio/Technology 10:163−167(1992))。別のアプローチに従うと、F(ab’)2フラグメントは、組み換え宿主細胞培養物から直接的に単離され得る。抗体フラグメントを生成するための他の技術は、当業者に明らかである。別の実施形態において、最適な抗体は、単鎖Fvフラグメントである(scFv)(PCT特許出願 国際公開番号93/16185)。
【0070】
(高親和性抗体の調製)
一旦親抗体が同定され、単離された場合、親抗体の可変領域の1つ以上において、1つ以上のアミノ酸残基が変化され得る。あるいは、または加えて、フレームワーク残基の1つ以上の置換が、親抗体中に導入され得、これらは、抗体の(例えば、ヒトIgEに対する)結合親和性に改善をもたらす。改変するフレームワーク領域の残基の例としては、以下が挙げられる:直接的に標的に非共有結合性に結合するもの(Amitら,Science 233:747−753(1986));CDRの高次構造と相互作用するもの/CDRの高次構造に影響するもの(Chothiaら,J.Mol.Biol.196:901−917(1987));および/またはVL−VH界面に(EP 239 400 B1)関与するもの。特定の実施形態において、このようなフレームワーク領域の残基の1つ以上の改変は、目的の標的に対する抗体の結合親和性の増強をもたらす。
【0071】
抗体の生物学的特性の改変は、例えば、以下を維持することに対するそれらの影響において有意に異なる置換を選択することによって達成され得る:(a)置換の範囲におけるポリペプチド骨格の構造(例えば、シート構造もしくはヘリックス構造);(b)標的部位における分子の電荷もしくは疎水性、または(c)側鎖の大きさ(bulk)。非保存的置換は、これらの分類の1つのメンバーと別の分類との交換を伴う。
【0072】
アミノ酸配列改変体をコードする核酸分子は、当該分野で公知の種々の方法によって調製される。これらの方法としては、より早期に調製された、種依存性抗体の改変体または非改変バージョンの、オリゴヌクレオチド媒介性(または部位指向性)突然変異誘発、PCR突然変異誘発、およびカセット突然変異誘発が挙げられるが、これらに限定されない。改変体を生成するための好ましい方法は、オリゴヌクレオチド媒介性合成である。特定の実施形態において、抗体改変体は、例えば、約2〜約15の超可変領域置換を置換した、単一の超可変領域残基を有するのみである。
【0073】
改変体のライブラリーを生成するための1つの方法は、図2に示されるスキームに従うオリゴヌクレオチド媒介性合成による方法である。各々約100ヌクレオチドの3つのオリゴヌクレオチドが、全軽鎖可変領域または全重鎖可変領域にわたって合成され得る。各オリゴヌクレオチドは、以下を含む:(1)トリプレット(NNK)20によって生成された60アミノ酸伸長(ここで、Nは任意のヌクレオチドであり、KはGまたはTである)、および(2)隣のオリゴか、または各端のベクター配列とのおよそ15〜30ヌクレオチドの重複。PCR反応におけるこれら3つのオリゴヌクレオチドのアニーリングのとき、ポリメラーゼは、反対側の鎖を充填し、完全二重鎖化された重鎖可変領域配列または軽鎖可変領域配列を生成する。トリプレットの数は、任意の反復長に調整され得、そして、オリゴヌクレオチド中のそれらの位置は、所与のCDRまたはフレームワーク領域におけるアミノ酸のみを置換するように選択され得る。(NNK)を用いることによって、コードされた改変体中の各々の位置において、全20アミノ酸が考えられる。5〜10アミノ酸(15〜30ヌクレオチド)の重複配列は、置換されないが、これはフレームワークの積み重なり(stacking)領域内に含まれるように選択され得るか、または合成の別のラウンドまたはその後のラウンドで置換され得る。オリゴヌクレオチドを合成するための方法は、当該分野で周知であり、そしてまた市販されている。これらのオリゴヌクレオチドから抗体改変体を生成するための方法もまた、当該分野で周知である(例えば、PCR)。
【0074】
重鎖改変体および軽鎖改変体のライブラリーは、それらの配列のランダムな位置において異なり、任意の発現ベクター(例えば、バクテリオファージ、具体的には、図1のベクター)中で構築され得る。これらのベクターの各々は、特定の重鎖改変体および軽鎖改変体をコードするDNAを含む。
【0075】
抗体改変体の生成に続いて、親抗体に比較した改変体の生物学的活性が決定される。上記のように、これは、改変体の標的に対する結合親和性を決定する工程を包含する。多くのハイスループット法が、抗体改変体を、目的の標的と結合するそれらの能力に関して迅速にスクリーニングするために存在する。
【0076】
次いで、この最初のスクリーニングから選択された1つ以上の抗体改変体が、親抗体に対して増強された結合親和性についてスクリーニングされ得る。結合親和性を決定するための1つの一般的な方法は、BIAcoreTM表面プラズモン共鳴システム(BIAcore,Inc.)を用いて会合速度定数および解離速度定数を評価することによる方法である。バイオセンサーチップは、製造元(BIAcore)の指示に従って、標的の共有結合のために活性化される。次いで、標的は、希釈され、そしてチップ上に注入されて、固定材料の反応ユニット(RU)におけるシグナルを得る。RUのシグナルは、固定材料の質量に比例するので、このシグナルは、マトリックス上の固定された標的密度の範囲を示す。解離データは、一部位モデル(one−site model)に適合されて、koff +/− s.d.(測定の標準偏差)を得る。偽第一次速度定数(ks)は、各会合曲線について計算されて、タンパク質濃度の関数としてプロットされてkon +/− s.e.(適合の標準誤差)を得る。結合の平衡解離定数KDは、koff/konとしてSPR測定から計算される。平衡解離定数KDは、koffに反比例するので、親和性改善の推定は、会合速度(kon)が全改変体について一定であると仮定して行われ得る。
【0077】
得られた高親和性を有する候補は、必要に応じて、1つ以上のさらなる生物学的活性アッセイに供されて、増強された結合親和性を有する抗体改変体が、さらに所望される治療特性を保有することを確認し得る。例えば、抗IgE抗体の場合、IgEのレセプターへの結合をブロックし、ヒスタミンの放出を阻害するものについてスクリーニングし得る。最適な抗体改変体は、親抗体よりも有意に高い結合親和性で標的に結合する能力を保有する。
【0078】
そのようにして選択された抗体改変体は、多くの場合抗体の意図された用途に依存して、さらなる改変に供され得る。このような改変は、アミノ酸配列のさらなる変化、異種ポリペプチドとの融合、および/または以下に詳述されるような共有結合性の改変を含む。例えば、抗体の適切な高次構造の維持に関与しない任意のシステイン残基が、一般的にセリンによって置換されて、分子の酸化的安定性を改善し得、そして異常な交差結合を防止し得る。逆に、システイン結合が抗体に付加されて、(特に、この抗体が、Fvフラグメントのような抗体フラグメントである場合)その安定性を改善し得る。
【0079】
(ベクター)
本発明はまた、本明細書において開示されるような抗体改変体をコードする単離された核酸、この核酸を含むベクターおよび宿主細胞、ならびに上記抗体改変体の生成のための組み換え技術を提供する。抗体改変体の組み換え生成のため、それをコードする核酸が単離され、そしてさらなるクローニング(DNAの増幅)または発現のため、複製可能なベクターに挿入される。抗体改変体をコードするDNAは、容易に単離され、従来の手順を用いて(例えば、この抗体改変体の重鎖および軽鎖をコードする遺伝子に対して特異的に結合し得るオリゴヌクレオチドプローブを用いることによって)配列決定される。
【0080】
多くのベクターが利用可能である。ベクター構成要素としては、一般的に以下の1つ以上が挙げられるが、これらに限定されない:シグナル配列、複製起点、1つ以上のマーカー遺伝子、エンハンサーエレメント、プロモーター、および転写終止配列。
【0081】
図1に示されるファージ発現ベクターは、一般的に使用されるM13ベクターと、迅速な分泌のためおよび適切な結合特異性および最低の親和性基準に関して改変体Fabをスクリーニングするため、M13の自身の遺伝子IIIウイルス分泌シグナルとから構成される。このベクターは、全遺伝子III配列を使用せず、したがって細菌細胞の表面上に表示が存在しないが、Fabはペリプラズム空間に分泌される。あるいは、Fabは、細胞質中で発現され得、そして単離され得る。重鎖および軽鎖の各々は、それら自身の分泌シグナルを有するが、単一の強力な誘導プロモーターに依存して発現される。
【0082】
図1におけるベクターはまた、容易な精製および検出のために、Hisタグおよびmycタグを提供する。当業者は、Fabが、別個のプロモーターから独立して発現され得るか、または分泌シグナルが選択されたウイルス配列である必要はないが、選択された宿主細胞からの抗体フラグメントの分泌に適切な原核生物もしくは真核生物のシグナル配列であり得ることを認識する、重鎖および軽鎖が異なるベクター上に位置していてもよいことが、また認識されるべきである。
【0083】
(A.シグナル配列構成要素)
本発明の抗体改変体は、組み換え的に生成され得る。この改変体はまた、異種ポリペプチド(好ましくは、シグナル配列、または成熟タンパク質または成熟ポリペプチドのN末端に特異的切断部位を有する他のポリペプチド)と融合された融合ポリペプチドとして発現され得る。好んで選択される異種シグナル配列は、宿主細胞によって認識およびプロセシングされる(すなわち、シグナルペプチダーゼによって切断される)配列である。ネイティブ抗体シグナル配列を認識およびプロセシングしない原核生物宿主細胞に対しては、このシグナル配列は、例えば、アルカリホスファターゼ、ペニシリナーゼ、Ippまたは熱安定性エンテロトキシンIIリーダーの群から選択された原核生物シグナル配列によって置換され得る。あるいは、図1のベクターの場合、選択されたシグナル配列は、遺伝子III由来のウイルスシグナル配列であった。酵母分泌については、ネイティブシグナル配列は、例えば、酵母インベルターゼリーダー、α因子リーダー(SaccharomycesおよびKluyveromyces α因子リーダーが挙げられる)もしくは酸性ホスファターゼリーダー、C.albicansグルコアミラーゼリーダー、または、例えば国際公開番号90/13646において記載されるシグナルによって置換され得る。哺乳動物細胞発現においては、哺乳動物シグナル配列およびウイルス分泌リーダー(例えば、単純ヘルペスgDシグナル)が利用可能である。このような前駆領域のためのDNAは、リーディングフレームにおいて、抗体改変体をコードするDNAに連結される。
【0084】
(B.複製起点の構成要素)
ベクターは、通常、ベクターが1種以上の選択された宿主細胞において複製することを可能にする核酸配列を含む。一般的に、この配列は、ベクターが宿主の染色体DNAと独立に複製することを可能にする配列であり、そして複製起点または自立的に複製する配列を含む。このような配列は、種々の細菌、酵母、およびウイルスに関して周知である。プラスミドpBR322由来の複製起点は、ほとんどのグラム陰性細菌に対して適切であり、2μプラスミド起点は、酵母に対して適切であり、そして種々のウイルス起点(SV40、ポリオーマ、アデノウイルス、VSVもしくはBPV)は、哺乳動物細胞中のベクターに対して有用である。一般的に、複製起点の構成要素は、哺乳動物発現ベクターにとって必要ではない(SV40起点は、初期プロモーターを含むので、代表的に唯一使用され得る)。
【0085】
(C.選択遺伝子構成要素)
ベクターは、選択遺伝子(選択マーカーとも称される)を含み得る。代表的な選択遺伝子は、(a)抗生物質もしくは他の毒素(例えば、アンピシリン、ネオマイシン、メトトレキサート、もしくはテトラサイクリン)に対する耐性を与えるか、(b)栄養素要求性欠損を補完するか、または(c)複合培地から得られない重要な栄養素を供給する、タンパク質をコードする(例えば、Bacillus属にとってのD−アラニンラセマーゼをコードする遺伝子)。
【0086】
選択スキームの1つの例は、宿主細胞の増殖を阻止する薬物を利用する。異種遺伝子によって首尾よく形質転換されたこれらの細胞は、薬物耐性を与えるタンパク質を産生し、したがって選択レジメンを生き延びる。このような優性選択の例は、薬物(ネオマイシン、ミコフェノール酸およびハイグロマイシン)を使用する。
【0087】
哺乳動物細胞のための適切な選択マーカーの別の例は、抗体核酸(例えば、DHFR、チミジンキナーゼ、メタロチオネイン−Iおよび−II、好ましくは、霊長類メタロチオネイン遺伝子、アデノシンデアミナーゼ、オルニチンデカルボキシラーゼなど)を取り込む能力のある細胞の同定を可能にするものである。
【0088】
例えば、DHFR選択遺伝子によって形質転換された細胞は、全ての形質転換体をメトトレキサート(Mtx)(DHFRの競合的アンタゴニスト)を含む培養培地中で培養することによって、最初に同定される。野生型DHFRが利用される場合、適切な宿主細胞は、DHFR活性が欠損したチャイニーズハムスター卵巣(CHO)細胞株である。
【0089】
あるいは、抗体、野生型DHFRタンパク質およびアミノグリコシド3’−ホスホトランスフェラーゼ(APH)のような別の選択マーカーをコードするDNA配列によって形質転換または同時形質転換された宿主細胞(特に、内因性DHFRを含む野生型宿主)は、上記選択マーカーに対する選択因子(例えば、アミノグリコシド系抗生物質(例えば、カナマイシン、ネオマイシンまたはG418(米国特許第4,965,199号)))を含む培地中での細胞増殖によって選択され得る。
【0090】
酵母中での使用のための適切な選択遺伝子は、酵母プラスミドYrp7に存在するtrp1遺伝子である(Stinchcombら,Nature 282:39(1979))。trp1遺伝子は、トリプトファン中で増殖する能力を欠く酵母の改変体系統(例えば、ATCC No.44076もしくはPEP4−1)に対する選択マーカーを提供する(Jones,Genetics 85:12(1977))。次いで、酵母宿主細胞ゲノムにおけるtrp1障害の存在は、トリプトファンの非存在下での増殖によって形質転換を検出するための有効な環境を提供する。同様に、Leu2−欠損性酵母系統(ATCC 20,622もしくは38,626)は、Leu2遺伝子を保有する公知のプラスミドによって補完される。
【0091】
(D.プロモーター構成要素)
発現ベクターおよびクローニングベクターは、通常、宿主生物体によって認識され、かつ抗体核酸に作動可能に連結されたプロモーターを含む。原核生物宿主とともに使用するために適切なプロモーターとしては、phoAプロモーター系、β−ラクタマーゼプロモーター系およびラクトースプロモーター系、アルカリホスファターゼプロモーター系、トリプトファン(trp)プロモーター系、ならびにハイブリッドプロモーター(例えば、tacプロモーター)が挙げられる。しかし、他の公知の細菌プロモーターが適切である。細菌系において使用するためのプロモーターはまた、抗体をコードするDNAに作動可能に連結された、シャイン−ダルガーノ(S.D.)配列を含む。
【0092】
プロモーター配列は、真核生物に関して知られている。実質的に全ての真核生物遺伝子は、転写が開始される部位から約25〜30塩基上流に位置するATリッチな領域を有する。多くの遺伝子の転写の開始部から70〜80塩基上流に見出される別の配列は、CNCAAT領域である(ここで、Nは、任意のヌクレオチドであり得る)。ほとんどの真核生物遺伝子の3’末端には、コード配列の3’末端へのポリAテイルの付加のためのシグナルであり得るAATAAA配列がある。これらの配列の全ては、真核生物発現ベクター中に適切に挿入される。
【0093】
酵母宿主とともに使用するための適切なプロモーター配列の例としては、3−ホスホグリセリン酸キナーゼまたは他の糖分解酵素(例えば、エノラーゼ、グリセルアルデヒド3−リン酸デヒドロゲナーゼ、ヘキソキナーゼ、ピルビン酸デカルボキシラーゼ、ホスホフルクトキナーゼ、グルコース6−リン酸イソメラーゼ、3−ホスホグリセリン酸ムターゼ、ピルビン酸キナーゼ、トリオースリン酸イソメラーゼ、グルコースリン酸イソメラーゼおよびグルコキナーゼ)に対するプロモーターが挙げられる。
【0094】
他の酵母プロモーター(増殖条件によって制御される転写というさらなる利点を有する誘導プロモーター)は、アルコールデヒドロゲナーゼ2、イソシトクロムC、酸性ホスファターゼ、窒素代謝に関連する分解酵素、メタロチオネイン、グルセルアルデヒド3−リン酸デヒドロゲナーゼ、ならびにマルトース利用およびガラクトース利用を担う酵素に対するプロモーター領域である。酵母発現において使用するための適切なベクターおよびプロモーターは、EP 73,657にさらに記載されている。酵母エンハンサーもまた、酵母プロモーターとともに有利に使用される。
【0095】
哺乳動物宿主細胞におけるベクターからの抗体転写は、例えば、ウイルス(例えば、ポリオーマウイルス、トリポックスウイルス、アデノウイルス2のようなアデノウイルス、ウシパピローマウイルス、トリ肉腫ウイルス、サイトメガロウイルス、レトロウイルス、B型肝炎ウイルス、および最も好ましくは、シミアンウイルス40(SV40))のゲノムから得られたプロモーター、異種哺乳動物プロモーター(例えば、アクチンプロモーターまたは免疫グロブリンプロモーター)、熱ショックプロモーターによって制御される(ただし、このようなプロモーターが宿主細胞系と適合することを条件とする)。
【0096】
SV40ウイルスの初期プロモーターおよび後期プロモーターは、SV40ウイルス複製起点もまた含むSV40制限フラグメントとして、都合よく得られる。ヒトサイトメガロウイルスの極初期プロモーターは、HindIII E制限フラグメントとして都合よく得られる。ウシパピローマウイルスをベクターとして使用して、哺乳動物宿主においてDNAを発現するための系は、米国特許第4,419,446号に開示されている。この系の改変形態は、米国特許第4,601,978号に記載されている。あるいは、ヒトβ−インターフェロンcDNAは、マウス細胞において、単純ヘルペスウイルス由来のチミジンキナーゼプロモーターの制御下で発現された。あるいは、ラウス肉腫ウイルスの長い末端反復(long terminal repeat)が、プロモーターとして使用され得る。
【0097】
(E.エンハンサーエレメント構成要素)
高等真核生物による本発明の抗体をコードするDNAの転写は、多くの場合、ベクター中にエンハンサー配列を挿入することによって増大する。多くのエンハンサー配列が、現在、哺乳動物遺伝子から知られている(グロブリン、エラスターゼ、アルブミン、α−フェトプロテイン、およびインシュリン)。しかし代表的には、真核生物細胞ウイルス由来のエンハンサーを使用する。例としては、複製起点の後期側におけるSV40エンハンサー(bp100〜270)、サイトメガロウイルス初期プロモーターエンハンサー、複製起点の後期側におけるポリオーマエンハンサー、およびアデノウイルスエンハンサーが挙げられる。真核生物プロモーターの活性化のためのエンハンサーエレメントに関する、Yaniv,Nature 297:17−18(1982)もまた参照のこと。エンハンサーは、抗体をコードする配列に対して5’位置または3’位置において、ベクター中にスプライシングされ得るが、好ましくはプロモーターの5’部位に位置する。
【0098】
(F.転写終止構成要素)
真核生物宿主細胞(酵母、真菌、昆虫、植物、動物、ヒト、もしくは他の多細胞生物由来の有核細胞)において使用される発現ベクターはまた、転写の終止のためおよびmRNAを安定化するために必要な配列を含み得る。このような配列は、一般的に、真核生物またはウイルスのDNAもしくはcDNAの、5’非翻訳領域から、そしてときには3’非翻訳領域から得られる。これらの領域は、抗体をコードするmRNAの非翻訳部分においてポリアデニル化フラグメントとして転写される、ヌクレオチドセグメントを含む。1つの有用な転写終止構成要素は、ウシ成長ホルモンポリアデニル化領域である。例えば、国際公開番号94/11026を参照のこと。
【0099】
(宿主細胞の選択および形質転換)
本明細書中に記載のベクターにDNAをクローニングまたは発現させるのに適した宿主細胞は、原核生物、酵母または高等真核生物の細胞である。この目的に適した原核生物としては、グラム陰性生物およびグラム陽性生物(例えば、E.coli、Enterobacter属、エルウィニア属、Klebsiella属、Proteus属、サルモネラ属、Serratia属およびShigella属などの腸内細菌、ならびにバチルス属、シュードモナス属およびストレプトミセス属)の両方が挙げられる。一つの好ましいE.coliクローニング宿主は、E.coli294(ATCC 31,446)であるが、E.coli B、E.coli X1776(ATCC 31,537)およびE.coli W3110(ATCC 27,325)のような他の菌株も適している。これらの例は、限定的ではなく、むしろ例示的である。
【0100】
原核生物に加えて真核生物微生物(例えば、糸状菌または酵母)は、抗体をコードするベクターのための適したクローニング宿主または発現宿主である。Saccharomyces cerevisiaeは、下等真核宿主微生物の内で最も一般的に使用される。しかし、例えば、Schizosaccharomyces pombe;Kluyveromyces属;カンジダ属;Trichoderma属;Neurospora crassaおよび糸状菌(例えば、アカパンカビ属、ペニシリウム属、イトクズグサ属およびコウジカビ属(例えば、A.nidulansおよびA.niger)の宿主などの他の多数の属、種および菌株が、本明細書中で一般的に利用可能であり、そして有用である。
【0101】
グリコシル化抗体の発現のために適した宿主細胞は、多細胞生物に由来する。第一に、脊椎動物培養または無脊椎動物培養に由来しようと、任意の高等真核細胞培養が機能し得る。無脊椎動物細胞の例としては、植物細胞および昆虫細胞が挙げられる(Luckowら、Bio/Technology 6、47〜55(1988);Millerら、Genetic Engineering、Setlowら編、第8巻、p.277−279(Plenam publishing 1986);Msedaら、Nature 第315巻、592〜594(1985))。多数のバキュロウイルス株および改変体ならびに宿主由来の対応する許容される昆虫宿主細胞(例えば、Spodoptera frugiperda(毛虫)、ヤブカ属(蚊)、Drosophila melanogaster(ショウジョウバエ)およびBombyx mori)が同定されている。トランスフェクションのための種々のウイルス株(例えば、Autographa californica NPVのL−1改変体、Bombyx mori NPVのBm−5株)が、公けに利用可能であり、このようなウイルスが、本発明による本明細書中のウイルスとして、特にSpodoptera frugiperda細胞のトランスフェクションに使用され得る。さらに、綿、トウモロコシ、ジャガイモ、大豆、ペチュニア、トマトおよびタバコの植物細胞培養物もまた宿主として利用される。
【0102】
脊椎動物細胞および培養(組織培養)における脊椎動物細胞の増殖は、慣例的になっている。Tissue Culture、Academic Press、KruseおよびPatterson編(1973)を参照のこと。有用な哺乳動物宿主細胞系統の例は、サル腎臓系統、ヒト胚腎臓系統、生まれたばかりのハムスターの腎臓細胞;チャイニーズハムスターの卵巣細胞/−DHFR(CHO、Urlaubら、Proc.Natl.Acad.Sci.USA 77:4216(1980);マウスセルトリ細胞;ヒト子宮頸部癌細胞(HELA);イヌ腎臓細胞;ヒト肺細胞;ヒト肝細胞;マウス乳腺癌;およびNS0細胞である。
【0103】
宿主細胞は、抗体産生のために上記ベクターで形質転換され、プロモーターを含むため、形質転換体を選択するため、または所望の配列をコードする遺伝子を増幅するために適切なものとして改変された通常の栄養培地で培養される。
【0104】
本発明の抗体改変体を産生するために使用される宿主細胞は、種々の培地で培養され得る。例えば、Ham’s F10(Sigma)、最小必須培地(MEM、Sigma)、RPMI−1640(Sigma)およびダルベッコ改変イーグル培地(DMEM、Sigma)などの市販の培地が、宿主細胞の培養に適している。さらに、Hamら、Meth.Enzymol.58:44(1979)、Barnesら、Anal.Biochem.102:255(1980)、米国特許第4,767,704号、同第4,657,866号;同第4,560,655号;同第5,122,469号;同第5,712,163号または同第6,048,728号に記載される任意の培地が、宿主細胞のための培養培地として使用され得る。これらの培地のいずれもが、必要な場合、ホルモンおよび/または他の増殖因子(例えば、インスリン、トランスフェリン、または表皮増殖因子)、塩(例えば、X塩化物、ここで、Xは、ナトリウム、カルシウム、マグネシウムである;およびリン酸塩)、緩衝液(例えば、HEPES)、ヌクレオチド(例えば、アデノシンおよびチミジン)、抗生物質(例えば、GENTAMYCIN.TM.薬物)、痕跡元素(通常マイクロモル範囲の最終濃度で存在する無機化合物として定義される)、およびグルコースまたは同等のエネルギー源を補充され得る。任意の他の必要な補充物はまた、当業者に公知である適切な濃度で含まれ得る。培養条件(例えば、温度、pHなど)は、発現のために選択された宿主細胞にこれまで用いられてきたものであり、当業者に明らかである。
【0105】
(抗体精製)
組換え技術を使用する場合、抗体改変体は、細胞内で産生され得るか、ペリプラズム空間で産生され得るか、または培地に直接分泌され得る。抗体改変体が、細胞内で産生される場合、第一の工程として、粒状の破片である、宿主細胞の断片または溶解した断片のいずれかは、例えば、遠心分離または限外濾過により取り除かれ得る。Carterら、Bio/Technology 10:163〜167(1992)は、E.coliのペリプラズム空間に分泌される抗体を単離するための手順を記述する。手短かには、細胞ペーストを、酢酸ナトリウム(pH3.5)、EDTAおよびフェニルメチルスルホニルフルオリド(PMSF)の存在下で約30分間にわたって溶解する。細胞の破片は、遠心分離によって取り除かれ得る。抗体改変体が、培地中に分泌される場合、そのような発現系由来の上清は、一般的には最初に市販のタンパク質濃縮フィルター(例えば、Amicon限外濾過ユニットまたはMillipore Pellicon限外濾過ユニット)を使用して濃縮される。タンパク質分解を阻害するために、前の工程のいずれかに例えば、PMSFのようなプロテアーゼインヒビターが含まれてもよく、外来性の夾雑物の増殖を防止するために抗生物質が含まれてもよい。
【0106】
細胞から調製された抗体組成物は、例えば、ヒドロキシアパタイトクロマトグラフィー、ゲル電気泳動、透析およびアフィニティークロマトグラフィーを使用して精製され得、アフィニティークロマトグラフィーが好ましい精製技術である。親和性リガンドとしてのプロテインAの適合性は、抗体改変体に存在するいずれかの免疫グロブリンFcドメインの種およびアイソタイプに依存する。プロテインAは、ヒトIgG1重鎖、ヒトIgG2重鎖またはヒトIgG4重鎖に基づく抗体を精製するために使用され得る(Lindmarkら、J.Immunol Meth.62:1〜13(1983))。プロテインGは、全てのマウスアイソタイプおよびヒトIgG3に対して推奨される(Gussら、EMBO J.5:1567〜1575(1986))。親和性リガンドが取り付けられるマトリクスは、多くの場合アガロースであるが、他のマトリクスも利用可能である。機械的に安定なマトリクス(例えば、調節された多孔ガラスまたはポリ(スチレンジビニル)ベンゼン)は、アガロースを用いて達成され得るより早い流速および短いプロセス時間を可能にする。抗体改変体が、CH3ドメインを含む場合、Bakerbond ABXTM樹脂(J.T.Baker、Phillipsburg,N.J.)が、精製に有用である。イオン交換カラムによる分画、エタノール沈殿、逆相HPLC、シリカによるクロマトグラフィー、アニオン交換樹脂もしくはカチオン交換樹脂(例えば、ポリアスパラギン酸カラム)上のヘパリンSEPHAROSETMクロマトグラフィーによるクロマトグラフィー、等電点電気泳動、SDS−PAGEおよび硫酸アンモニウム沈殿などのタンパク質精製のための技術がまた回収されるべき抗体改変体によって利用可能である。
【0107】
任意の予備精製工程の後、目的の抗体改変体および夾雑物を含む混合物は、約2.5〜4.5の間のpHで、好ましくは低塩濃度(例えば、約0〜0.25Mの塩)で、溶出緩衝液を使用して、低pHの疎水性相互作用クロマトグラフィーに供され得る。
【0108】
(薬学的処方物)
ポリペプチドまたは抗体の治療処方物は、凍結乾燥処方物または水溶液として保存するために、所望の程度の精製度を有するポリペプチドを、当該分野で代表的に利用される任意の「薬学的に受容可能な」キャリア、賦形剤または安定化剤(これらは全て「賦形剤」と称される)と混合することにより調製され得る。例えば、緩衝剤、安定化剤、保存剤、等張化剤、非イオン性洗浄剤、抗酸化剤および他の種々の添加剤である(Remington’s Pharmaceutical Sciences、第16版、A.Osol編(1980)を参照のこと)。このような添加剤は、用いられる投薬量および濃度では、レシピエントに対して無毒性でなければならない。
【0109】
緩衝剤は、生理学的条件に近い範囲にpHを維持するのを補助する。それらは、好ましくは約2mM〜約50mMの範囲の濃度で存在する。本発明に使用するための適した緩衝剤としては、有機酸とその塩および無機酸とその塩の両方が挙げられ、例えば、クエン酸緩衝液(例えば、クエン酸一ナトリウム−クエン酸二ナトリウムの混合物、クエン酸三ナトリウムの混合物、クエン酸−クエン酸一ナトリウムの混合物など)、コハク酸緩衝液(例えば、コハク酸−コハク酸一ナトリウムの混合物、コハク酸−水酸化ナトリウムの混合物、コハク酸−コハク酸二ナトリウムの混合物など)、酒石酸緩衝液(例えば、酒石酸−酒石酸ナトリウムの混合物、酒石酸−酒石酸カリウムの混合物、酒石酸−水酸化ナトリウムの混合物など)、フマル酸緩衝液(例えば、フマル酸−フマル酸一ナトリウムの混合物など)、フマル酸緩衝液(例えば、フマル酸−フマル酸一ナトリウムの混合物、フマル酸−フマル酸二ナトリウムの混合物、フマル酸一ナトリウム−フマル酸二ナトリウムの混合物など)、グルコン酸緩衝液(例えば、グルコン酸−グルコン酸ナトリウム混合物、グルコン酸−水酸化ナトリウム混合物、グルコン酸−グルコン酸カリウム混合物など)、シュウ酸緩衝液(例えば、シュウ酸−シュウ酸ナトリウム混合物、シュウ酸−水酸化ナトリウム混合物、シュウ酸−シュウ酸カリウム混合物など)、乳酸緩衝液(例えば、乳酸−乳酸ナトリウム混合物、乳酸−水酸化ナトリウム混合物、乳酸−乳酸カリウム混合物など)および酢酸緩衝液(例えば、酢酸−酢酸ナトリウム混合物、酢酸−水酸化ナトリウム混合物など)が挙げられる。さらに、リン酸緩衝液、ヒスチジン緩衝液およびトリメチルアミン塩(例えば、Tris)が言及され得る。
【0110】
保存剤は、微生物の増殖を妨げるために添加され得、0.2%〜1%(w/v)の範囲の量で添加され得る。本発明に使用するための適切な保存剤としては、フェノール、ベンジルアルコール、メタ−クレゾール、メチルパラベン、プロピルパラベン、オクタデシルジメチルベンジルアンモニウムクロライド、ベンザルコニウムハロゲン化物(例えば、塩化物、臭化物、ヨウ化物)、塩化ヘキサメトニウム、アルキルパラベン(例えば、メチルパラベンまたはプロピルパラベン)、カテコール、レゾルシノール、シクロヘキサノールおよび3−ペンタノールが挙げられる。
【0111】
「安定化剤」として公知であることもある等張化剤は、本発明の液体組成物の等張性を確実にするために添加され得、多価糖アルコール、好ましくは三価アルコールまたはそれより多価の糖アルコール(例えば、グリセリン、エリスリトール、アラビトール、キシリトール、ソルビトールおよびマンニトール)が挙げられる。
【0112】
安定化剤は、治療剤を溶解させるか、または変性もしくは容器の壁への付着を防止するのを補助する充填剤から添加剤の機能の範囲であり得る広い範疇の賦形剤をいう。代表的な安定化剤は、多価糖アルコール(上記で列挙した);アミノ酸(例えば、アルギニン、リジン、グリシン、グルタミン、アスパラギン、ヒスチジン、アラニン、オルニチン、L−ロイシン、2−フェニルアラニン、グルタミン酸、スレオニンなど)、有機糖または糖アルコール(例えば、ラクトース、トレハロース、スタキオース、マンニトール、ソルビトール、キシリトール、リビトール、ミオイノシトール、ガラクティトール、グリセロールなど、イノシトールなどのシクリトールを含む);ポリエチレングリコール;アミノ酸ポリマー;硫黄含有還元剤(例えば、尿素、グルタチオン、チオクト酸、チオグリコール酸ナトリウム、チオグリセロール、α−モノチオグリセロールおよびチオ硫酸ナトリウム);低分子量ポリペプチド(すなわち、10残基未満);タンパク質(例えば、ヒト血清アルブミン、ウシ血清アルブミン、ゼラチンまたは免疫グロブリン)、親水性ポリマー(例えば、ポリビニルピロリドン)、単糖(例えば、キシロース、マンノース、フルクトース、グルコース);二糖(例えば、ラクトース、マルトース、スクロース)および三糖(例えば、ラフィノース);多糖(例えば、デキストラン)であり得る。安定化剤は、活性タンパク質の重量部あたり0.1〜10,000の重量の範囲で存在し得る。
【0113】
非イオン性界面活性剤または洗浄剤(「湿潤剤」としても公知である)は、治療剤の可溶化を補助するため、および治療タンパク質を攪拌に誘導される凝集に対して保護するために添加され得、また処方物が、タンパク質の変性を引き起こすことなく、圧力を加えた剪断表面に露出されることを可能にする。適切な非イオン性界面活性剤としては、ポリソルベート(20、80など)、ポロキサマー(184、188など)、Pluronic(登録商標)ポリオール、ポリオキシエチレンソルビタンモノエーテル(TWEEN(登録商標)−20、TWEEN(登録商標)−80など)が挙げられる。非イオン性界面活性剤は、約0.05mg/ml〜約1.0mg/mlの範囲、好ましくは約0.07mg/ml〜約0.2mg/mlの範囲で存在し得る。
【0114】
さらなる種々の賦形剤としては、充填剤(例えば、デンプン)、キレート剤(例えば、EDTA)、抗酸化剤(例えば、アスコルビン酸、メチオニン、ビタミンE)および共溶媒が挙げられる。本明細書中の処方物はまた、必要な場合、処置される特定の適応症に必要な一種より多い活性化合物、好ましくは互いに不利に影響しない相補的活性を有する化合物を含有し得る。例えば、さらに免疫抑制剤を提供することが望ましくあり得る。このような分子は、意図される目的に対して有効である量で、組合せて適切に存在する。活性成分はまた、例えば、コアセルベーション技術により、または例えばヒドロキシメチルセルロースまたはゼラチンマイクロカプセルおよびポリ(メチルメタクリレート)マイクロカプセルそれぞれのコロイド状薬物送達系(例えば、リポソーム、アルブミンマイクロフェア、マイクロエマルジョン、ナノ粒子およびナノカプセル)またはマクロエマルジョン)中での界面のポリマー化により調製されたマイクロカプセルに閉じ込められ得る。このような技術は、Remington’s Pharmaeutical Sciences、第16版、A.Osal編(1980)に開示される。
【0115】
インビボ投与のために使用される処方物は、無菌性でなければならない。このことは、例えば、滅菌濾過膜を通す濾過によって容易に達成される。徐放調製物が調製され得る。徐放調製物の適切な例としては、抗体改変体を含む固体疎水性ポリマーの半透性マトリクスが挙げられ、このマトリクスは、成形した物品(例えば、フィルムまたはマイクロカプセル)の形態である。徐放マトリクスの例としては、ポリエステル、ヒドロゲル(例えば、ポリ(2−ヒドロキシエチルメタクリレート)またはポリ(ビニルアルコール)、ポリラクチド(米国特許第3,773,919号)、L−グルタミン酸とエチル−L−グルタメートのコポリマー、非分解酢酸ビニルエチレン、分解可能乳酸−グリコール酸コポリマー(例えば、LUPRON DEPOTTM(乳酸−グリコール酸のコポリマーと酢酸ロイプロリドからなる注射可能マイクロスフェア)およびポリ−D−(−)−3−ヒドロキシ酪酸が挙げられる。例えば、酢酸ビニルエチレンと乳酸−グリコール酸ポリマーは、100日間に亘って分子の放出を可能にし、特定のヒドロゲルは、より短い期間タンパク質を放出する。封じ込められた抗体が、体内に長い期間保持される場合、それらは、37℃にて湿気に曝露された結果として変性し得るか、または凝集し得、生物学的活性の喪失および免疫原性における可能な変化を生じる。合理的な方法は、関係する機構に依存して安定化のために、工夫され得る。例えば、凝集機構が、チオ−ジスルフィド交換を介した細胞間S−S結合形成であることが発見される場合、安定化は、スルフヒドリル残基を改変することによって、酸性溶液から凍結乾燥することによって、含水量を制御することによって、適切な添加剤を使用することによって、そして、特定のポリマーマトリクス組成物を開発することによって達成され得る。
【0116】
特定の障害または状態の処置に有効である治療ポリペプチド、抗体またはそのフラグメントの量は、障害もしくは状態の性質に依存し、そして、標準的な臨床技術により決定され得る。可能な場合、用量−応答曲線を決定し、ヒトで試験する前に、最初にインビトロで、次いで有用な動物モデル系で本発明の薬学的組成物を決定することが望ましい。
【0117】
好ましい実施形態において、治療ポリペプチド、抗体またはそのフラグメントの水溶液は、皮下注射によって投与される。各用量は、体重1kgあたり約0.5μg〜約50μgの範囲であり得るか、またはより好ましくは、体重1kgあたり約3μg〜約30μgの範囲であり得る。
【0118】
皮下投与のための投薬計画は、多数の臨床的因子(疾患の型、疾患の重篤度、および治療剤に対する患者の感受性)に依存して一ヶ月に一回から一日一回に変動し得る。
【0119】
(抗体改変体の使用)
本発明の抗体改変体は、親和性精製因子として使用され得る。このプロセスにおいて、抗体は、当該分野で周知の方法を使用して、固相(例えば、SEPHADEXTM樹脂、またはろ紙)上に固定される。固定された抗体改変体は、精製されるべき標的を含むサンプルと接触され、その後、その支持体は、精製されるべき標的を除いてサンプル中の実質的に全ての物質を取り除く適した溶媒で洗浄される。その支持体は、固定された抗体改変体結合される。最後に、その支持体は、標的を抗体改変体から放出する別の適切な溶媒(例えば、グリシン緩衝液)で洗浄される。
【0120】
改変抗体はまた、診断アッセイ(例えば、特定の細胞、組織または血清における目的の標的の発現の検出)に有用であり得る。診断適用について、抗体改変体は、代表的に検出可能部分で標識される。多数の標識が、蛍光の変化を定量するために利用可能な技術であり、上に記した。化学発光基質は、化学反応により電気的に励起され、次いで、放射光は、(例えば、化学発光計を使用して)測定され得るか、または蛍光受容体にエネルギーを供与し得る。酵素標識の例としては、ルシフェラーゼ(例えば、ホタルルシフェラーゼおよび細菌ルシフェラーゼ;米国特許第4,737,456号)、ルシフェリン、2,3−ジヒドロフタラジンジオン、リンゴ酸デヒドロゲナーゼ、ウレアーゼ、ペルオキシダーゼ(例えば、ホースラディッシュペルオキシダーゼ(HPRO)、アルカリホスファターゼ、β−ガラクトシダーゼ、グルコアミラーゼ、リゾチーム、糖類オキシダーゼ(例えば、グルコースオキシダーゼ、ガラクトースオキシダーゼおよびグルコース6リン酸デヒドロゲナーゼ)、複素環式オキシダーゼ(例えば、ウリカーゼおよびキサンチンオキシダーゼ)、ラクトペルオキシダーゼ、ミクロペルオキシダーゼなどが挙げられる。酵素を抗体に結合するための技術は、Methods in Enzym.(J.LangoneおよびH.Van Vunakis編)、Academic press、New York、73:147〜166(1981)中のO’Sullivanら、Methods for the Preparation of Enzyme−Antibody Conjugates for Use in Enzyme Immunoassayに記載されている。
【0121】
標識は、抗体改変体と間接的に結合されることもある。当業者は、このことを達成するための種々の技術を知っている。例えば、抗体改変体は、ビオチンと結合され得、上記の3種の広範な範疇の標識のいずれかが、アビジンと結合され得、逆もまた同様である。ビオチンは、アビジンに選択的に結合し、従って標識は、この間接的な様式で抗体改変体と結合され得る。あるいは、標識の抗体改変体との間接的な結合を達成するために、抗体改変体は、小型のハプテン(例えば、ジグロキシン(digloxin))と結合され、上記の異なる型の標識の一つが、抗ハプテン抗体改変体(例えば、抗ジグロキシン抗体)と結合される。従って、標識の抗体改変体との間接的な結合は、達成され得る。
【0122】
本発明の別の実施形態において、抗体改変体は、標識される必要はなく、その存在は、抗体改変体に結合する標識された抗体を使用して検出され得る。
【0123】
本発明の抗体は、任意の公知のアッセイ方法(例えば、競合結合アッセイ、直接サンドイッチアッセイおよび間接サンドイッチアッセイならびに免疫沈降アッセイ)で使用され得る。Zola、Monoclonal Antibodies:A Manual of Techniques、pp.147〜158(CRC Press Inc.1987)。
【0124】
競合結合アッセイは、標識された標準が、限られた量の抗体改変体との結合について試験サンプルと競合する能力に依存する。試験サンプル中の標的の量は、抗体に結する標準の量に反比例する。結合する標準の量の決定を容易にするために、抗体は、一般的に、競合の前か後に、不溶化される。結果として、抗体に結合する標準サンプルおよび試験サンプルは、結合しないままである標準サンプルおよび試験サンプルから好都合に分離され得る。
【0125】
サンドイッチアッセイは、二つの抗体の使用を伴い、それぞれは、検出されるべき、異なる免疫原性部分またはエピトープまたはタンパク質に結合し得る。サンドイッチアッセイにおいて、分析されるべき試験サンプルは、固体支持体上に固定された一次抗体によって結合され、その後、二次抗体は、試験サンプルに結合し、それによって不溶性の3つの部分の複合体を形成する。例えば、米国特許第4,376,110号を参照のこと。二次抗体はそれ自体、検出可能な部分で標識され得る(直接サンドイッチアッセイ)か、または検出可能な部分で標識される抗免疫グロブリン抗体を使用して測定され得る(間接サンドイッチアッセイ)。例えば、一つの型のサンドイッチアッセイは、ELISAアッセイであり、その場合、検出可能な部分は、酵素である。
【0126】
免疫組織化学に関して、腫瘍サンプルは、新鮮であっても凍結されていてもよく、パラフィンに包埋されていても保存剤(例えばホルマリン)で固定されていてもよい。
【0127】
その抗体はまたインビボ診断アッセイに使用され得る。一般的に、抗体改変体は、放射性ヌクレオチド(例えば、111In、99Tc、14C、131I、3H、32Pまたは35S)で標識されて、その結果腫瘍は、免疫シンチグラフィを使用して局在化され得る。例えば、本発明の高親和性抗IgE抗体は、例えば、喘息患者の肺に存在するIgEの量を検出するために使用され得る。
【0128】
本発明の抗体は、キット、すなわち、診断アッセイを行なうための指示書を備える、所定量の試薬のパッケージングした組合せで提供され得る。抗体改変体が、酵素で標識される場合、そのキットは、酵素に必要とされる基質および補因子(例えば、検出可能な発色団またはフルオロフォアを提供する基質前駆体)を備え得る。さらに、他の添加剤(例えば、安定化剤、緩衝液(例えば、ブロック緩衝液または溶解緩衝液)など)が含まれ得る。種々の試薬の相対量は、アッセイの感度を実質的に最適化する試薬の溶液中の濃度を提供するために、大きく変動し得る。特に、試薬は、溶解の際に、適切な濃度を有する試薬溶液を提供する賦形剤を含む、乾燥粉末、通常は、凍結乾燥した粉末として提供され得る。
【0129】
(抗体のインビボ使用)
本発明の抗体は、哺乳動物を処置するために使用され得ることが企図される。一実施形態において、この抗体は、例えば、前臨床データを得る目的で、非ヒト哺乳動物に投与される。処置されるべき例示的な非ヒト哺乳動物としては、非ヒト霊長類、イヌ、ネコ、げっ歯類、および前臨床研究が行われる他の哺乳動物が挙げられる。このような哺乳動物は、この抗体を用いて処置されるべき疾患に対する確立された動物モデルであり得るか、または、目的の抗体の毒性を研究するために使用され得る。これらの実施形態のそれぞれにおいて、用量増大研究が、哺乳動物に対して行なわれ得る。この抗体またはポリペプチドは、任意の適切な手段で投与され、その手段としては、非経口投与、皮下投与、腹腔内投与、肺内投与、鼻腔内投与が挙げられ、局所的な免疫抑制処置について所望の場合、病巣内投与が挙げられる。非経口注入としては、筋肉内投与、静脈内投与、動脈内投与、腹腔内投与または皮下投与が挙げられる。さらに、抗体改変体は、パルス注入、特に低下用量の抗体改変体を用いたパルス注入により適切に投与される。好ましくは、この用量は、注射によって与えられ、最も好ましくは、投与が、短期であるか、長期であるかに一部依存して、静脈内注射または皮下注射によって与えられる。
【0130】
疾患の予防または処置のために、適切な投薬量の抗体またはポリペプチドは、処置されるべき疾患の型、疾患の重篤度および過程、抗体改変体が、予防目的または治療目的で投与されたか否か、これまでの治療、患者の臨床履歴および抗体改変体に対する応答、および主治医の裁量に依存する。本発明の非常に高い親和性の抗ヒトIgE抗体は、患者に、一時に、または一連の処置にわたって適切に投与され得る。疾患の型および重篤度に依存して、約0.1mg/kg〜150mg/kg(例えば、0.1〜20mg/kg)の抗体が、患者に投与するための初期候補投薬量であり、この量は例えば、一回以上の別個の投与であっても、連続注入であってもよい。代表的な毎日の投薬量は、上記の因子に依存して、約1mg/kg〜100mg/kgまたはそれ以上の範囲であり得る。数日以上に亘る繰り返しの投与については、その条件に依存して、その処置は、疾患症状の所望の抑制が生じるまで維持される。しかし、他の投薬レジメンも有用であり得る。この治療の進行は、従来の技術およびアッセイにより容易にモニタリングされる。抗LFA−1抗体または抗ICAM−1抗体についての例示的な用量レジメンは、国際公開番号94/04188に開示される。抗体改変体組成物は、良好な医療行為と一致した様式で処方され、投薬され、投与される。この文脈において、考慮のための因子としては、処置される特定の障害、処置される特定の哺乳動物、個々の患者の臨床状態、障害の原因、薬剤の送達の部位、投与の方法、投与のスケジュール、および医師に公知の他の因子が挙げられる。投与されるべき抗体改変体の「治療有効量」は、このような考慮すべき事項によって決定され、疾患または障害を予防するか、改善するか、または処置するために必要である最少量である。この抗体改変体は、必ずしもではないが、必要に応じて、問題の障害を予防または処置するために現在使用されている一種以上の薬剤と一緒に処方される。このような他の薬剤の有効量は、処方物中に存在する抗体の量、障害または処置の型、および上で考察した他の因子に依存する。これらは、一般的に同じ投薬量で、上で使用した投与経路で、またはこれまでに用いた投薬量の約1〜99%で使用される。
【0131】
IgEをその標的として認識する本発明の抗体は、「IgE媒介性障害」を処置するために、使用され得る。これらとしては、例えば、ぜん息、アレルギー性鼻炎およびアレルギー性結膜炎(枯草熱)、湿疹、じんま疹、アトピー性皮膚炎および食物アレルギーのような疾患が挙げられる。例えば、蜂に刺されること、蛇にかまれること、食物または薬物によって引き起こされるアナフィラキシーショックの深刻な生理学的状態もまた本発明の範囲に包含される。
【0132】
(抗体エピトープマッピング)
用語「エピトープ」とは、B細胞および/またはT細胞が抗原に対して応答する部位をいう。B細胞エピトープは、連続するアミノ酸またはタンパク質の三次元のフォールディングによって並列する連続しないアミノ酸の両方から形成され得る。連続するアミノ酸から形成されたエピトープは、代表的に、変性溶媒に曝露されて保持されるのに対して、三次元フォールディングによって形成されるエピトープは、代表的に変性溶媒を用いた処理に対して失われる。エピトープは、代表的に、少なくとも3個、より通常は、少なくとも5個または8〜10個のアミノ酸を、固有の空間コンフォメーションに含む。同一のエピトープを認識する抗体は、ある抗体の、別の抗体の標的抗原への結合をブロックする能力を示す単純な免疫学的検定において同定され得る。
【0133】
本発明の高親和性抗体IgEに対する結合部位のエピトープマッピングは、結合についてのウエスタンブロット分析、IgEのCH3ドメインのペプチドスキャン、結合を示した領域のアラニン走査、IgG1の対応する領域からのアミノ酸置換および特定部位の変異誘発を含む。
【0134】
IgEの全CH3ドメインのペプチドスキャンは、73個の重複するペプチドを必要とした。各ペプチドを、本発明の標識した抗IgE抗体による結合に供し、IgEの、その高親和性レセプターへの結合をブロックするIgEの特異的なエピトープを決定した。ペプチドスキャンは、二つのペプチドを、IgE上の潜在的な抗IgE MAb接触部位と同定し、エピトープ「A」およびエピトープ「B」と名付けた(図11を参照のこと)。エピトープ「A」配列およびエピトープ「B」配列は、直鎖状配列で約80アミノ酸離れているが、IgEの三次元構造では互いに近くに位置している。両方とも表面に露出されていて、それらは、IgEのFcεRI結合部位に重複し、両方ともペプチドであり、Argの正に荷電した残基およびProの疎水性残基が存在する。図12は、ペプチドスキャンを使用してELISAにより決定したエピトープBの結合領域を図示する。
【0135】
どのアミノ酸残基が、これらのエピトープ内での高親和性抗体の結合に重要であるかの決定を、アラニンスキャニング変異誘発によって行なった(Cunninghamら、「High−Resolution Epitope Mapping of hGH−Receptor Interactions by Alanine−Scanning Mutagenesis」、Science、第244巻、p.1081〜1085)。エピトープAおよびエピトープBの各残基の代わりに、アラニンを用い、高親和性モノクローナル抗体の結合を決定した(実施例12の下ならびに図13および図14を参照のこと)。
【0136】
(能動免疫および受動免疫)
本発明はまた、薬学的組成物(例えば、配列番号72および/または配列番号74を含む本発明のペプチド免疫源分子、ならびに希釈剤、賦形剤、アジュバントまたはキャリアを含有するワクチン)に関する。本発明は、さらに、本発明の少なくとも一つのペプチドを、そのペプチドに対する免疫応答を誘発し得る部分と共有結合させる工程を包含する、本発明の免疫源の調製のためのプロセスに関する。
【0137】
本発明はまた、例えば、IgE媒介性疾患またはIgE媒介性状態(例えば、アレルギーおよびアトピー性皮膚炎)の処置における医薬として使用するための上に定義された免疫原性ペプチドに関する。
【0138】
本発明はさらに、上に定義する免疫原性ペプチドの治療有効量を、そのような処置を必要とする患者に投与する工程を包含する、IgE媒介性疾患またはIgE媒介性状態(例えば、アレルギーおよびアトピー性皮膚炎)に対して哺乳動物を免疫状態にする方法に関する。
【0139】
本発明の免疫原性ペプチドは、非細胞溶解性のヒスタミン放出を実質的に媒介し得ないが、エピトープAおよび/またはエピトープBの標的アミノ酸配列との血清学的な強い交差反応性を抗体に誘発し得る。
【0140】
ペプチドの初期用量(例えば、約0.2mg〜約5mg)が、例えば、筋肉内に投与され得、その後14日後〜28日後に、同じ用量で繰り返し投与される(追加抗原)。用量は、当然のことながらある程度は、患者の年齢、体重、全体的な健康に依存する。免疫処置は、「能動的」または「受動的」であり得る。「能動」免疫において、被験体は、本発明の免疫原性ペプチドを受容し、抗IgE応答が、被験体の免疫系によって能動的に誘導される。
【0141】
「能動」免疫、これは、ヒト使用について好ましいが、他の哺乳動物種(例えば、イヌ)も同様に処置され得る。用語「免疫原性キャリア」は、本明細書中では、宿主動物内で免疫原性応答を独立に誘発する特性を有し、ポリペプチドおよび免疫原性キャリア物質の対応する基の中の遊離のカルボキシル基、アミノ基またはヒドロキシル基の間のペプチド結合またはエステル結合の形成を介して、または、従来の二官能性連結基を介した結合、または融合タンパク質として直接ポリペプチドに共有結合し得る物質を包含する。
【0142】
このような免疫原性キャリアの例としては、以下:アルブミン(例えば、BSA);グロブリン;サイログロブリン;ヘモグロブリン;ヘモシアニン(特に、キーホールリンペットヘモシアニン(KLH));回虫属から抽出したタンパク質(例えば、J.Immunol.、111巻、(1973)260〜268、J.Immunol.、122巻、(1979)302〜308、J.Immun.、98巻、(1967)893〜900、Am.J.Physiol.、199巻、(1960)575〜578に記載される回虫属抽出物またはその精製産物);ポリリシン;ポリグルタミン酸;リシン−グルタミン酸コポリマー;リシンまたはオルニチンなどを含むコポリマーなどが挙げられる。ワクチンは、ジフテリアトキソイドまたは破傷風トキソイドを免疫原性キャリア物質として使用して産生され(Lepow M.L.ら、J.of Infectious Diseases、第150巻、(1984)402〜406;Coen Beuvery,E.ら、Infection and Immunity、第40巻、(1983)39〜45)、これらのトキソイド物質はまた、本発明に使用され得る。ツベルクリンの精製されたタンパク質誘導体(PPD)は、(1)それ自体ではT細胞応答を誘導せず(すなわち、それは、事実上「T細胞ハプテン」である)、なお十分にプロセスされた抗原として挙動し、従ってT細胞により認識される;(2)連結した認識様式では、最も強力なハプテン「キャリア」の一つであることが公知である;そして(3)さらなる試験を行なわずに、ヒトにおいて使用され得るので、「能動」免疫スキームにおける利用に特に好ましい。
【0143】
本発明はまた、本発明のペプチドをコードするポリヌクレオチド、このポリヌクレオチドを含むベクターおよびこのベクターを有する細胞に関する。さらに、能動免疫は、本発明のペプチドをコードするポリヌクレオチドを投与することにより達成され得る。このような治療に適したベクターは、当該分野で公知であり、例えば、アデノウイルスベクターが挙げられる。
【0144】
「受動」免疫は、本発明の抗IgE抗体を、IgE媒介性疾患またはIgE媒介性状態を罹患する患者に投与することによって達成される。これらの抗体は、本発明の免疫原性ペプチドを非ヒト哺乳動物に投与し、生じた抗血清を収集することにより調製され得る。改善された力価は、長期間にわたる繰り返しの注射によって獲得され得る。抗体を誘発するために使用され得る哺乳動物の種に特に制限はなく、一般的には、ウサギまたはモルモットを使用することが好ましいが、ウマ、ネコ、イヌ、ヤギ、ブタ、ラット、ウシ、ヒツジなども使用され得る。抗体は、最後の投与の後、1〜2週間の経過後、免疫状態にした動物から血液を収集し、血液を遠心分離し、血清を血液から単離することにより回収される。モノクローナル抗体は、例えば、ヒトまたはマウスのものであり得る。
【0145】
被験体を免疫状態にする場合、本発明の抗体は、哺乳動物に、例えば、筋肉内注射により導入され得る。しかし、抗体投与の任意の形態が使用され得る。被験体に許容され、有害な副作用を有さない任意の従来の液体または固体のビヒクルが、使用され得る。生理学的pH(例えば、pH約6.8〜7.2、好ましくはpH約7.0)のリン酸緩衝生理食塩水(PBS)が、それのみでビヒクルとして、または例えば、水酸化アルミニウムベースのアジュバントなどの適切なアジュバントと一緒にビヒクルとして使用され得る。
【0146】
以下の実施例は、限定の目的で提供されるのではなく、例示の目的で提供される。
【実施例】
【0147】
(実施例1)
(抗IgEマウスMAb TES−C21のヒト化)
マウスmAb TES−C21の重鎖可変領域(VH)および軽鎖可変領域(VL)の配列を、公開されたデータベースで利用可能なヒト抗体生殖細胞系配列と比較した。上記工程1に記載されるテンプレートを決定する場合、いくつかの基準を用い、これらの基準としては、全長、フレームワーク内の類似したCDR位置、全体の相同性、CDRの大きさなどが挙げられる。これらの基準は全てひとまとめにして、TES−C21MAb重鎖配列とTES−C21MAb軽鎖配列との間の配列アラインメント、ならびに図3Aおよび図3Bに示されるそれぞれのヒトテンプレート配列に示される最適なヒトテンプレートを選択するための結果を提供する。
【0148】
この場合、この抗体を設計するために、一種より多いヒトフレームワークテンプレートを使用した。VH鎖に対して選択されたヒトテンプレートは、DP88(アミノ酸残基1〜95)とJH4b(アミノ酸残基103〜113)との組合せであった(図3Bを参照のこと)。VL鎖に対して選択されたヒトテンプレートは、JK4(アミノ酸残基98〜107)と合わせたL16(VKサブグループIII、アミノ酸残基1〜87)の組合せであった(図3Aを参照のこと)。マウス配列とヒトテンプレートとの間のフレームワーク相同性は、VHに関して約70%、VLに関して約74%であった。
【0149】
一旦テンプレートを選択すると、DNA合成、および上に記載され、そして図2に示されるオーバーラッピングPCRによりFabライブラリーを構築した。このライブラリーは、それぞれ選択されたヒトテンプレートであるDP88/JH4bおよびL16/JK4を用いて合成した合成TES−C21 CDRからなった。このライブラリーの複雑さは、4096(=212)であった。部分的なVH配列およびVL配列をコードする重複するヌクレオチドを、18〜21のヌクレオチドの重複を有する約63〜約76のヌクレオチドの範囲で合成した。
【0150】
VL遺伝子およびVH遺伝子のPCR増幅を、フレームワーク領域FR1に対する特異的配列を含むビオチン化フォワードプライマーおよびリーダー配列(遺伝子III)の末端にアニーリングした突出配列および保存された定常領域(CKまたはCH1)由来のリバースプライマーを使用して、標準PCR条件下で行なった。PCR産物を、アガロースゲル電気泳動、または市販のPCR精製キットにより精製し、取り込まれなかったビオチン化プライマーおよび非特異的PCR産物を取り除いた。
【0151】
PCR産物の5’リン酸化を、2μgのPCR産物、1μLのT4ポリヌクレオチドキナーゼ(10ユニット/μL)、2μLの10×PNK緩衝液、1μLの10mM ATPを用いて、ddH2Oで、総体積20μLに調節して行なった。37℃にて45分間インキュベートした後、65℃にて10分間加熱変性し、次の工程のために、ddH2Oを添加して反応体積を200μLに調節した。
【0152】
100μLのストレプトアビジン被覆磁気ビーズを、200μLの2×B&W緩衝液で2度洗浄し、200μLの2×B&W緩衝液に再懸濁した。リン酸化PCR産物を、ビーズと混合し、室温(RT)で穏やかに振盪しながら16分間インキュベートした。
【0153】
このビーズを、沈降させ、200μLの2×B&W緩衝液で2度洗浄した。非ビオチン化ssDNA(マイナス鎖)を、300μLの新鮮に調製した0.15MのNaOHで、室温で穏やかに振盪しながら10分間溶出した。第二のNaOH溶出は、収率をわずかに増加させ得る(任意)。溶離液を遠心分離して、微量のビーズを完全に取り除いた。
【0154】
ssDNAを、1μLグリコゲン(10mg/mL)、1/10体積の3M NaOAc(pH5.2)および2.5体積のEtOHを添加することにより、上清から沈殿させた。次いで、沈殿したssDNAを、70% EtOHで洗浄し、その後、3分間凍結乾燥し、20μLのddH2Oに溶解した。ssDNAを、エチジウムブロマイド(EtBr)アガロースプレート上に、DNA標準と一緒にスポッティングすることにより定量するか、またはOD260を測定することにより定量した。
【0155】
(実施例2)
(VHおよびVLのファージ発現ベクターへのクローニング)
VHおよびVLをハイブリダイゼーション変異誘発により、ファージ発現ベクターにクローニングした。ウリジン化テンプレートを、CJ236 E.coli株(dut「ung」)を、M13ベースのファージで感染させることにより調製した(ファージ発現ベクターTN003)。
【0156】
以下の成分(200ngのウリジン化ファージベクター(8.49kb);92ngのリン酸化一本鎖H鎖(489塩基);100ngのリン酸化一本鎖L鎖(525塩基);1μLの10×アニーリング緩衝液;ddH2Oで体積を10μlに調整)を、85℃の温度で5分間PCR保持(変性)し、次いで、1時間かけて55℃に低下させることによって(ベクターに対して約8倍のモル比の挿入物において)アニーリングさせた。そのサンプルを氷上で冷却した。
【0157】
アニーリングさせた産物に、以下の成分:1.4μLの10×合成緩衝液;0.5μLのT4 DNAリガーゼ(1ユニット/μL);1μL T4 DNAポリメラーゼ(1ユニット/μL)を添加し、その後、氷上で5分間インキュベートし、37℃で1.5時間インキュベートした。次いで、その産物をエタノール沈殿し、10μLのddH2OまたはTEに溶解させた。DNAを、1μL のXbaI(10ユニット/μL)で2時間消化し、65℃で20分間加熱不活性化した。消化したDNAを、エレクトロポレーションにより50μLのエレクトロコンピーテントDH10B細胞にトランスフェクトした。生じたファージを、37℃で一晩、XL−1 Blue細菌ローン(bacterial lawn)上で増殖させることにより、力価を測った。クローンを、組成を確認するために、配列決定した。
【0158】
(実施例3)
(ライブラリースクリーニングのためのディープウェルでの培養)
(A.ファージライブラリーのプレーティング)
ファージライブラリーを、LB培地に希釈し、プレートあたりの所望のプラーク数を達成した。高力価のファージを、200μLのXL−1B細胞培養物と混合した。3mLのLBの表面の寒天を混合し、LBプレートに注ぎ、室温で10分間そのままにしておいた。そのプレートを、37℃にて、一晩インキュベートした。
【0159】
(B.ファージ溶出)
100μLのファージ溶出緩衝液(10mM Tris−Cl、pH7.5、10mM EDTA、100mM NaCl)を滅菌U底96ウェルプレートの各ウェルに添加した。一晩後のライブラリープレート由来の単一のファージプラークを、フィルター処理したピペットチップを用いてウェルに移した。ファージ溶出プレートを、37℃にて1時間インキュベートした。このプレートをインキュベーションに続いて4℃にて保存し得る。
【0160】
(C.ディープウェルプレートでの培養)
50mLの培養物由来のXL1B細胞を、2×YT培地に、1:100希釈で添加した。その細胞を、A600が0.9〜1.2の間になるまで、シェーカー内で37℃にて増殖させた。
【0161】
(C.ディープウェルプレート内でのファージによる感染)
細胞が適切なODに到達したときに、1M IPTG(1:2000)をXL 1B培養物に添加した。IPTGの最終濃度は、0.5mMであった。750μLの細胞培養物を96ウェルのディープウェルプレート(Fisher Scientific)の各ウェルに移した。各ウェルに、25μLの溶出ファージを接種した。このディープウェルプレートをシェーカー(250rpm)内に配置し、37℃にて一晩インキュベートした。
【0162】
(D.ELISAスクリーニングのための上清の調製)
インキュベーション後、このディープウェルプレートを、Beckman JA−5.3プレートローターを使用して、3,250rpmにて20分間遠心した。ELISAのために、各ウェルから50μLの上清を抜き取った。
【0163】
(E.XL−1細胞の15mLの液体培養物の接種)
XL−1を、10μg/mLのテトラサイクリンを含有する2×YT中で、シェーカー内で(250rpm)、37℃にて、A600が0.9〜1.2になるまで増殖させた。IPTGを、0.5mMの最終濃度で添加し、15mLの培養物を50mLの円錐管に、各クローンが特徴付けられるように移した。細胞に、高力価ストック(力価=約1011pfu/mL)由来の10μLのファージを接種し、37℃にて1時間インキュベートした。この細胞を、室温で振盪しながら一晩増殖させた。
【0164】
(F.ペリプラズム由来の可溶性Fabの単離)
この細胞を、4,500rpmでの20分間のIEC遠心分離にて、ペレット化した。培養培地を取り除き、ペレットを、650μLの再懸濁緩衝液(1mMのEDTAおよび500mMのスクロースを含有する、50mM Tris、pH8.0)に再懸濁し、ボルテックスし、その後穏やかに振盪しながら1時間氷上に配置した。細胞の破片を、4℃で、9,000rpmで10分間遠心することにより取り除いた。可溶性Fabを含む上清を、収集し、そして4℃にて保存した。
【0165】
(実施例4)
(フレームワーク改変)
フレームワーク内の上記の潜在的に重要な位置に、12個のマウス/ヒトのゆらぎ残基が存在する。VHの73位は、ヒト化ライブラリーでは、マウス残基スレオニンとして維持されていた。なぜならこの位置は、結合に影響を及ぼすと確定されたからである。しかし、VH73のスレオニンが、ヒト生殖細胞系VHサブグループ1および2の共通のヒト残基であることが示された。
【0166】
TES−C21配列とヒトテンプレートとの間で異なるフレームワーク残基を、上記のように無作為に置換し、次いで、標的結合および抗体フォールディングに対するその潜在的な影響について評価した。結合に影響し得る潜在的なフレームワーク残基を同定した。この場合、それらは、VHの残基12、27、43、48、67、69であり、VLの残基1、3、4、49、60、85であった(Kabat記数法)(図4を参照のこと)。27位および69位のみが、VH領域(クローン番号1136−2C)の結合に顕著に影響することが、後に実証された。
【0167】
使用した一次スクリーニングは、培養培地を用いる一点ELISA(Single Point ELISA)(SPE)であった(以下の記載を参照のこと)。一次スクリーニングは、抗体の標的分子に結合するクローンを選択した。親分子と等しいか、またはそれより良好なシグナルを与えるクローンを、次のラウンドのスクリーニングのために選択した。
【0168】
第2のラウンドのスクリーニングにおいて、個々のファージを、15mlの細菌培養で増殖させ、ペリプラズム調製物を、SPE滴定アッセイおよびELISA滴定アッセイのために使用した。このアッセイにおいて、より高い結合を保持しているクローンをさらに特徴付けした。一旦全ての選択された一次クローンを、加工すると、上から10〜15%のクローンを配列決定し、そのクローンを配列に従って、配置した。各配列群由来の代表を、互いに比較し、最も良いクローンを選択した。これらの選択されたクローンの配列を合わせ、種々の組合せの効果を評価した。
【0169】
構築したライブラリーを、組換えヒトIgE、SE44に対する結合の改善についてELISAスクリーニングに供した。マウスTES−C21Fabより大きい結合親和性を有するクローンを単離し、配列決定した。クローンID番号4、49、72、76および136を、さらに特徴付けした。クローン4、49、72、78および136に対するELIZA滴定曲線を図5Aおよび図5Bに示すが、これは、その親和性が、その親であるTES−C21に類似していることを示唆する。これらのクローンは、ヒトIgEに対する結合について、マウスTES−C21と競合し、このことは、結合するエピトープが、ヒト化プロセスの間に変化しなかったことを示唆する。ヒト化Fabは、FcεRI結合IgEに結合せず、このことは、ヒト化抗体が、二価IgGに構築される場合にヒスタミン放出を引き起こすようにレセプターと架橋するのではなさそうであることを示唆する。ヒト化クローン136は、5個のマウス重鎖フレームワーク残基(=94.3%のヒトVHフレームワークホモロジー)を保持し、親和性成熟により選択された100%のヒト軽鎖フレームワークを有する。IgEのFcεRIへの結合のヒト化Fabによる阻害を実証した(図6)。
【0170】
(実施例5)
(抗IgEをスクリーニングするための一点ELISAプロトコル)
プレートを、炭酸塩被覆緩衝液中の2μg/mLのヒツジ抗ヒトFdで、4℃で一晩被覆した。被覆溶液を取り除き、そして、そのプレートを、ウェルあたり200μLの3% BSA/PBSで、37℃にて1時間ブロックした。プレートを、PBS/0.1% TWEEN(登録商標)(PBST)で4回洗浄した後、50μL/ウェルのFabサンプル(すなわち、高力価ファージおよびDMBブロックから分泌されたFabもしくはペリプラズム調製物、または15mLの調製物を含む上清)を添加した。プレートを室温で1時間インキュベートし、その後、PBSTで4回洗浄した。0.5% BSA/PBS中に0.015μg/mLで希釈した50μL/ウェルのビオチン化SE44および0.05%のTWEEN(登録商標)を添加した。次いで、プレートを室温で2時間インキュベートし、4×PBSTで洗浄した。0.5%のBSA/PBSおよび0.05%のTWEEN(登録商標)中に1:2000で希釈した50μL/ウェルのストレプトアビジンHRPを添加し、プレートを室温で1時間インキュベートした。プレートを、6×PBSTで洗浄した。50μL/ウェルのTMB 基質(Sigma)を添加して展開し、次いで、50μL/ウェルの0.2MのH2SO4を添加することにより停止させた。
【0171】
(実施例6)
(ELISA滴定:抗IgE)
プレートを、炭酸塩被覆緩衝液中の0.25μg/mL(精製したFab 0.1μg/ml)のSE44で4℃で一晩被覆した。被覆溶液を取り除き、このプレートを、200μL/ウェルの3%のBSA/PBSで、37℃にて1時間ブロックした。
【0172】
このプレートを、PBS/0.1%のTWEEN(登録商標)(PBST)で4回洗浄した。50μL/ウェルのFab(15mLのペリプラズム調製物由来)を添加して、1:2の希釈で開始して、0.5%のBSA/PBSおよび0.05%のTWEEN(登録商標)20に連続的に3倍希釈した。プレートを、室温で2時間インキュベートした。
【0173】
このプレートを、PBSTで4回洗浄し、0.5%のBSA/PBSおよび0.05%のTWEEN(登録商標)20中のビオチンヒツジ抗ヒトFdの1:1000希釈(0.8μg/ml)を50μL/ウェルで添加した。このプレートを、再度室温で2時間インキュベートした。
【0174】
PBSTでの4回の洗浄後、0.5%のBSA/PBSおよび0.05%のTWEEN(登録商標)20中のNeutra−アビジン−APの1:2000(0.9μg/ml)を50μl/ウェルで添加し、このプレートを、室温で1時間インキュベートした。
【0175】
このプレートを、PBSTで4回洗浄した。そして、pNPP基質を50μL/ウェルで添加することにより展開した。展開を、50μL/ウェルの3MのNaOHを添加することにより停止させた。各ウェルの吸光度を、405nmまたは410nmで読み取った。
【0176】
(実施例7)
(可溶性Fabを発現するM13ファージの親和性精製のためのプロトコル)
(A.1日目)
10mg/mLのテトラサイクリンを含有する2つの500mL培養物(2×YT)に、5mLの一晩のストックXL1Bを接種し、37℃にてA600が0.9〜1.2になるまで増殖させた。IPTGを、0.5mMの濃度まで添加した。次いで、細胞培養物を、培養あたり200μLのファージで感染させ、振盪しながら37℃にて1時間インキュベートした。感染後、その細胞を25℃にて一晩振盪しながら増殖させた。
【0177】
(B.2日目)
細胞を、250mLの遠心管中で、4℃で、3500×gにて30分間で、ペレット化した。培養培地を吸引し、ペレットを、合計12〜15mLの溶解緩衝液(緩衝液A+プロテアーゼインヒビターの混液)に再懸濁した。
【0178】
【化1】

再懸濁した細胞を、50mLの円錐管に移し、混合物が、小さな塊として一緒に移動する(溶解に起因して)まで、その管を数回逆さにすることによって、100μLの100mg/mLのリゾチームに溶解した。細胞を氷上で超音波処理し、その後、10μLのDNase I(約1000ユニット)を添加し、4℃にて30分間穏やかに振動させた。破片を、50mLの遠心管を用いて、4℃にて、12000×gにて30分間遠心することによりペレット化した。上清を新しい円錐管に移し、4℃にて保存した。
【0179】
Ni−NTアガロース(Qiagen、Valencis、CA)を使用して、製造業者のプロトコルに従って、可溶性Fabを精製した。この溶解物を、Ni−NTAと混合し、カラムにロードした。流れ出たものを、SDS−PAGE分析のために収集した。このカラムを、20mLの緩衝液(50mMのNaH2PO4、300mMのNaCl、15mMのイミダゾール、NaOHでpH8.0に調整)で洗浄し、その後50mM NaH2PO4、300mM NaCl、20mM イミダゾールを20mL用いて洗浄した。Fabを、500μLの溶出緩衝液(50mMのNaH2PO4、300mMのNaCl、450mMのイミダゾール、NaOHでpH8.0に調整)を用いて6回洗浄し、SDS PAGEにより分析した。カラム画分を、4℃にて保存した。カラム画分を、SDS−PAGEにより分析し、Fabの量が最も多い画分を選択し、PBS中に4℃にて透析した。
【0180】
(実施例8)
(可溶性レセプターアッセイ)
ELISAに適した96ウェルアッセイプレートを、0.05mLの0.5μg/mL FcεRIα鎖レセプター被覆緩衝液(50mM 炭酸塩/重炭酸塩、pH9.6)で、4〜8℃にて12時間被覆した。このウェルを吸引し、250μLのブロック緩衝液(PBS、1% BSA、pH7.2)を添加し、37℃にて1時間インキュベートした。別個のアッセイプレートにおいて、サンプルおよび参照のTES−C21 MAbを、200μg/mL〜0.001μg/mLまで、アッセイ緩衝液(0.5% BSAおよび0.05% Tween20、PBS、pH7.2)で1:4希釈して滴定し、等量の100ng/mLのビオチン化IgEを添加して、そのプレートを25℃にて、2〜3時間インキュベートした。FcεRI被覆ウェルを、0.05% TWEEN20を含むPBSを用いて三回洗浄し、サンプルウェルから50μLを移し、25℃にて、30分間攪拌しながらインキュベートした。アッセイ緩衝液に1:2000で希釈した1mg/mLのストレプトアビジン−HRPを50μL/ウェルで、30分間攪拌しながらインキュベートし、次いで、そのプレートを既に述べたように洗浄した。50μL/ウェルのTMB基質を添加し、発色させた。等量の0.2M H2SO4を添加することにより、反応を停止させ、450nmにて吸光度を測定した。
【0181】
(実施例9)
(抗体のIgE負荷FcεRIに対する結合)
FcεRIのαサブユニットと結合したヒトIgEに対する抗体の結合を、10μg/mLのヒトIgEと4℃にて30分間プレインキュベートすることによって、決定した。プレートを三回洗浄し、その後、種々の濃度のマウス抗ヒトIgE MAb E−10−10またはヒト化Fab改変体のどちらかと1時間インキュベートした。Fabの結合を、ビオチン標識した抗ヒトFd抗体を用いて、その後SA−HRPにより検出した。マウスMAb E−10−10を、ヤギ抗マウスIg Fc HRP結合Abにより検出した。
【0182】
(実施例10)
(クローン特徴付け)
各候補を、結合親和性についてアッセイし、ポジティブクローンを配列決定した。結合親和性を増加させる、CDR領域における有益な変異を有する抗体改変体をさらに特徴付けした。アッセイには、Biacore分析;IgEのレセプターに対するIgE結合の阻害;およびレセプターが結合したIgEの架橋が含まれる。
【0183】
改変体のライブラリーを作製した。親和性を改善することが実証された種々のCDRに対するアミノ酸配列を表1に示す。図7は、置換の組合せを有する高親和性の候補を示す。
【0184】
【表1】
19個の重鎖改変体を図9に示し、35個の軽鎖改変体を、図8に示す。さらに3個の候補を、結合親和性について特徴付けし、これらを表2に示す。
【0185】
【表2】
(実施例11)
(抗IgE抗体の発現および精製ならびにHRP結合)
高親和性MAb候補を生成した。インタクトな抗IgE MAbの生成のために、重鎖可変領域および軽鎖可変領域をファージベクターテンプレートからPCR増幅し、CMVプロモーターの発現下で、H鎖発現ベクターおよびL鎖発現ベクターに別個にサブクローニングした。6個の全長抗体クローンを構築した。これらを図10A〜図10Fに示す。適切な重鎖プラスミドおよび適切な軽鎖プラスミドを、当該分野で周知の技術によるエレクトロポレーションを使用して、マウス骨髄腫細胞株NS0に同時トランスフェクトした。例えば、Liouら、J.Immunol.第143巻、第12号:3967〜75(1989)を参照のこと。抗体を、プロテインAセファロース(Pharmacia)を使用して単一の安定な細胞株上清から精製した。抗体の濃度を、280nmでのスペクトロフォトメーターおよびFCAアッセイ(IDEXX)を使用して決定した。
【0186】
ペルオキシダーゼ結合キット(Zymed Labs、San Francisco、CA)を製造業者のプロトコルに従って使用して、精製した抗体に、ホースラディッシュペルオキシダーゼ(HRP)を結合させた。結合した各抗IgE MAbの力価を、モノクローナルヒトIgE(SE44)で被覆したプレートを用いたELISAを使用して決定した。
【0187】
以下の培養物は、American Type Culture Collection、10801 University Boulevard、Manassas Va.20110−2209 USA(ATCC)に寄託された。
【0188】
【表3】
この寄託を、特許手続き上の微生物の寄託の国際的承認に関するブダペスト条約および特許手続き上の微生物の寄託の国際的承認に関するブダペスト条約に基づく規則(ブダペスト条約)の下で行なった。このことは、寄託の日から30年間生存培養の維持を保証する。この生物は、ブダペスト条約の条件の下でATCCにより入手可能にされ、このことは、関連する米国特許の発行の際に、培養の子孫の永久的かつ非制限的な利用可能性を公共に保証する。
【0189】
本出願の譲受人は、寄託されたこの培養が、適切な条件下で培養された場合に死滅するか、失われるか、破壊される場合に、通知に対して、同一の培養の生存可能な試料により、ただちに取り替えられることに同意した。寄託した株の利用可能性は、特許法に従って任意の政府の当局に認可された権利に違反して、本発明の実施に対する実施権として解釈されるべきではない。
【0190】
前述の明細書は、当業者が本発明を実施可能にするのに十分であると考えられる。本発明は、寄託した培養物によってその範囲が限定されるべきではない。なぜならば、寄託された実施形態は、本発明の一局面の例示として意図され、機能的に等価である任意の培養物が、本発明の範囲内であるからである。本明細書中の材料の寄託は、本明細書中に含まれる記載が、その最良の形態を含めて、本発明の任意の局面の実施を可能にするのに不適切であるという承認を構成せず、特許請求の範囲の、本明細書が示す特定の例示への制限として解釈されるべきではない。実際、本明細書中に示され、記載される改変に加えて、本発明の種々の改変が、前述の記載から当業者に明らかになり、それらは、添付された特許請求の範囲の範囲内である。
【0191】
(実施例12)
(ヒトIgEの高親和性結合エピトープのマッピング)
(A.ペプチド合成および抗IgE結合アッセイ)
研究は、IgEが、そのCH3ドメインを介してそのレセプターに結合することを示した。本発明のHA 抗IgE抗体は、非常に効率的にIgEのそのレセプターへの結合をブロックするので、本発明者らは、全長CH3ドメインを含むペプチドを使用してエピトープをマッピングした。第一に、本発明者らは、二つのV5タグ化したペプチドを調製し、その一つは、ヒトIgEの全長定常領域を含み、もう一つは、ヒトIgEのCH2−CH3領域のみを含んでいた。これらの二つのペプチドは、インビトロ転写翻訳によって発現し、HA抗IgE MAb結合を検出するために、ウエスタンブロットアッセイに使用された。CL−2 CMAbおよびCL−5I MAbは両方ともインタクトなヒトIgEに結合し得、両方ともそのペプチドに結合し得る。
【0192】
エピトープをより詳細にマッピングするために、ヒトIgEの残基141〜368のアミノ酸を含み、全長CH3ドメインを含む73個の重複するペプチドを合成した。各ペプチドは、上記ペプチドの3’末端と重複する3アミノ酸を有する12アミノ酸残基からなる。SPOT膜を、セルロース膜上に、フルオレニルメトキシカルボニル(Fmoc)アミノ酸を用いて合成した。その膜をメタノール中でリンスし、次いで、TBS(pH7.5)中で、10分間、3回洗浄した。ブロック溶液(TBS中5%乳汁または3%BSA)中で一晩インキュベートした後、ブロック溶液に希釈したHRP標識抗IgE MAbを、膜と一緒に3時間インキュベートした。TBS−TWEEN(登録商標)中で15分間、3回洗浄した後、SuperSignal HRP基質(Pierce)を使用して、IgE反応性を、所望の時間BioMax MSフィルム(Kodak)の化学発光露光することにより測定した。
【0193】
この実験の結果は、HA抗IgE MAbが、IgEのCH3部分の二つの領域に結合することを示唆し、この二つの領域は、以下の二つのペプチド配列NPRGVSAYLSRP(エピトープ「A」)およびHPHLPRALMRST(エピトープ「B」)により表される(図12を参照のこと)。エピトープAに対する結合は、エピトープBに対する結合より数倍弱かった。
【0194】
(B.アラニンスキャニング変異誘発)
IgG1に見出されるアミノ酸とペプチド内のアミノ酸との置換に伴うアラニンスキャニングを、どのアミノ酸が、HA抗IgE MAbのこれらのペプチドへの結合に決定的に関与しているかを決定するために行なった。HA抗IgE MAb結合に重要であることが決定されたアミノ酸を、IgEのε鎖におけるインビトロ変異誘発方法を使用して置換した。上に記載されるCε2領域およびCε3領域をカバーする別のペプチドをまた、この研究に使用した(図13および図14を参照のこと)。
【0195】
ヒトIgEアミノ酸残基に対してEU番号付けスキームを使用した。ポリメラーゼ連鎖反応(PCR)を使用して、IgEの全長Fc領域および、CH2−CH3ドメインのみを含むIgE Fcの短縮形態を増幅した。このDNA産物を、TOPOクローニング(Invitrogene、Carlsbad、CA)を使用して、pcDNA3発現ベクター(Invitrogene、Carlsbad、CA)に直接クローニングした。
【0196】
IgE−Fcにおける変異誘発を、オーバーラッピングPCR(Hoら、1989)を使用して行った。このDNA産物を、アガロースゲル電気泳動によって精製し、適切な制限酵素を用いて消化し、pcDNA3発現ベクターにサブクローニングした。各改変体構築物について、PCR増幅領域を、DNAの両方の鎖から、ジデオキシヌクレオチド方法を使用して完全に配列決定した。組換えヒトIgE Fcおよびその変異体を、網赤血球溶解物ベースのインビトロ転写とインビトロ翻訳との結合システム(Promega、Madison、WI)を使用して発現させた。
【0197】
このインビトロ転写とインビトロ翻訳との結合システム(10μlの反応混合物)由来の溶解物を、SDS−PAGE(12%)に供し、次いで、ニトロセルロース膜に移した。この膜を、Tris緩衝生理食塩水(TBS)中の5%脱脂粉乳を用いてブロックし、その後一次抗体、抗IgE MAbで染色した。特異的な反応バンドを、ホースラディッシュペルオキシダーゼに結合したヤギ抗ヒトIgG Fc(Jackson Labs、Bar Harbor、Maine)を使用して検出し、その免疫応答性バンドを、SuperSignal ウェスタンブロッティング検出キット(Pierce)により可視化した。抗V5抗体を、これらのペプチドのC末端に導入されたV5タグを検出するポジティブコントロールとして使用した。抗V5抗体を用いたウエスタンブロットは、全てのペプチドが、ほぼ等しいレベルで発現したことを実証した。興味深いことに、HA抗IgE MAbは、エピトープ「A」に変異を保有するペプチドに結合し得たが、それらは、エピトープ「B」に変異を保有するペプチドには結合しなかった。このことは、この第2の部位が、結合に関してより重要であったことを示唆する(図15を参照のこと)。
【0198】
(実施例13)
(エピトープBの免疫原性ペプチドを使用するトランスジェニックマウスの能動免疫)
ヒトIgEを構成的に発現するトランスジェニックマウスを使用して、エピトープBのヒト免疫原性ペプチドに対する抗体の能動的な産生を実証した。二つの融合ペプチド(それぞれが、本発明の免疫原性ペプチドであるシステイン残基およびKLHを含む)を化学的に合成した。ペプチド1の配列は、以下:
(KLH−Cys)−Leu Pro Arg Ala Leu Met Arg Ser Thr
であり、ペプチド2の配列は、以下:
Leu Pro Arg Ala Leu Met Arg Ser Thr−(Cys−KLH)
であった。
【0199】
そのトランスジェニックマウスに、200μLのPBS(pH7.4)中の完全フロイントアジュバント(Difco Laboratories、Detroit、MI)中の20μgの免疫原性ペプチドを皮下に注射した。2週間の間隔で2度、不完全フロイントアジュバント中の20μgのペプチド免疫源を、そのマウスに、皮下に注射した。次いで、二週間後、屠殺の3日前に、そのマウスに再度、腹腔内に、PBS中の20μgの同じ免疫源を注射した。血清を収集し、エピトープBに特異的な抗IgE抗体の存在について試験した。図16に見られるように、このペプチドは、これらのトランスジェニックマウスにおいて、抗IgE抗体を誘発した。
【0200】
当業者は、慣例的にすぎない実験を使用して、本明細書中に記載される本発明の特定の実施形態に対する多くの等価物を認識するか、または確認することができる。このような等価物は、添付の特許請求の範囲によって包含されることが意図される。
【図面の簡単な説明】
【0201】
【図1】図1は、抗体クローニングおよび抗体スクリーニングに使用されるファージベクターの模式的表現である。
【図2】図2は、抗体改変体を生成するために有用なオリゴヌクレオチドの模式的表現である。
【図3】図3Aは、マウス抗IgE抗体TES−C21の軽鎖とL16およびJK4の混合ヒトテンプレートとの比較を示す。図3Bは、TES−C21の重鎖とDP88およびJH4bの混合ヒトテンプレートとの比較を示す。
【図4】図4は、親TES−C21と比較して高親和性を有する、フレームワーク残基改変体の表を提示する。
【図5】図5AおよびBは、TES−C21の親Fabとネガティブコントロール(5D12)と比較した、クローン4、49、72、78および136についてのELISA滴定曲線を示す。
【図6】図6は、親TES−C21およびネガティブコントロール抗体と比較した、クローン2C、5Aおよび5Iの阻害アッセイを示す。
【図7】図7Aは、IgEに対するさらに高い親和性をもたらす有益な変異の組み合わせを有するクローンの配列を示す。
【図8A】図8Aおよび8Bは、クローン136、1、2、4、8、13、15、21、30、31、35、43、44、53、81、90および113についての全軽鎖可変領域のフレームワーク配列を示す。
【図8B】図8Aおよび8Bは、クローン136、1、2、4、8、13、15、21、30、31、35、43、44、53、81、90および113についての全軽鎖可変領域のフレームワーク配列を示す。
【図9A】図9Aおよび9Bは、35個のクローンについての全重鎖可変領域のフレームワーク配列を示す。
【図9B】図9Aおよび9Bは、35個のクローンについての全重鎖可変領域のフレームワーク配列を示す。
【図10A】図10A〜Fは、クローン136、2C、5I、5A、2Bおよび1136〜2Cについての完全重鎖配列および完全軽鎖配列を示す。
【図10B】図10A〜Fは、クローン136、2C、5I、5A、2Bおよび1136〜2Cについての完全重鎖配列および完全軽鎖配列を示す。
【図10C】図10A〜Fは、クローン136、2C、5I、5A、2Bおよび1136〜2Cについての完全重鎖配列および完全軽鎖配列を示す。
【図10D】図10A〜Fは、クローン136、2C、5I、5A、2Bおよび1136〜2Cについての完全重鎖配列および完全軽鎖配列を示す。
【図10E】図10A〜Fは、クローン136、2C、5I、5A、2Bおよび1136〜2Cについての完全重鎖配列および完全軽鎖配列を示す。
【図10F】図10A〜Fは、クローン136、2C、5I、5A、2Bおよび1136〜2Cについての完全重鎖配列および完全軽鎖配列を示す。
【図11】図11は、ヒトIgEのCH3領域アミノ酸配列を示し、そしてエピトープ「A」およびエピトープ「B」を強調表示する。
【図12】図12は、エピトープBを同定するために使用される重複ペプチドを示す。
【図13】図13は、エピトープAの結合領域中の重要な残基の識別を示す。
【図14】図14は、エピトープBの結合領域中の重要な残基の識別を示す。
【図15】図15は、変異体ペプチドの結合するMAbのウエスタンブロット分析を示す。
【図16】図16は、ヒトIgEを発現するトランスジェニック動物における抗IgE抗体の産生を示す。
【技術分野】
【0001】
(相互関連出願)
本出願は、PCT出願番号PCT/US04/02892およびPCT/US04/02894(2004年2月2日出願)の優先権を主張する。上記出願の全ては、本明細書において参考として援用される。
【背景技術】
【0002】
(発明の背景)
アレルギーは、外来の因子(例えば、アレルゲン)に対する過大な免疫応答によって誘導される過敏性状態である。即時(I型)過敏症は、アレルゲンとの接触直後のアレルギー性応答によって特徴付けられ、これは、B細胞を介して媒介され、そして抗原−抗体反応に基づく。遅延性過敏症は、T細胞を介して媒介され、そして細胞免疫の機構に基づく。近年、用語「アレルギー」は、よりI型過敏症と同義になってきている。
【0003】
即時過敏症は、アレルゲンに暴露されて抗体分泌性プラスマ細胞に分化するB細胞による、免疫グロブリンクラスEの抗体(IgE抗体)の産生に基づく反応である。IgEに誘導される反応は、身体へのアレルゲン侵入部位で(すなわち、粘膜表面および/または局所リンパ節において)生じる局所的事象である。局所的に産生されたIgEは、最初に局所性肥満細胞を感作する。すなわち、IgE抗体は、定常領域を用いて、肥満細胞の表面上のFcεレセプターに結合し、次いで、「溢出」IgEが循環に侵入し、全身にわたって循環性好塩基球および組織固定(tissue−fixed)肥満細胞の両方のレセプターに結合する。その結合したIgEが、その後アレルゲンに接触する場合、Fcεレセプターは、アレルゲンと結合することによって交差結合され、細胞を脱顆粒させ、多くのアナフィラキシー性媒介物質(例えば、ヒスタミン、プロスタグランジン、ロイコトリエンなど)を放出させる。即時過敏症に典型的な臨床症状(すなわち、気道または腸の平滑筋の収縮、小血管の拡張、ならびに水および血漿タンパク質に対する透過性の増大、粘液分泌(これらは、例えば、アレルギー性鼻炎、アトピー性湿疹(excema)、および喘息を引き起こす)、そして皮膚内に終わる神経の刺激(これは、かゆみおよび痛みを引き起こす))の原因となるのは、これらの物質の放出である。さらに、アレルゲンとの2回目の接触における反応は、IgEを細胞表面上に発現することによるアレルゲンとの最初の接触の後に、一部のB細胞が表面IgE陽性B細胞(sIgE+B細胞)の「記憶プール」を形成することに起因して、強化される。
【0004】
IgEに対する2つの主要なレセプターが存在する。抗親和性レセプターFcεRIおよび低親和性レセプターFcεRIIである。FcεRIは、肥満細胞および好塩基球の表面上に優先的に発現されるが、ヒトランゲルハンス細胞、樹状細胞、および単球上にもまた、低レベルのFcεRIが見出され、ここで、FcεRIは、IgE媒介性アレルゲン提示において機能する。さらに、FcεRIは、ヒト好酸球および血小板上でも報告されている(非特許文献1)。FcεRIは、B細胞、T細胞または好中球の表面上には見いだされない。ランゲルハンス細胞および皮膚樹状細胞でのFcεRIの発現は、機能的におよび生物学的に、アレルギーの個体におけるIgE結合抗原提示に関して重要である(非特許文献2)。
【0005】
低親和性レセプターFcεRII(CD23)は、細胞質膜からの長いらせん形のコイル状の柄から伸びる頭部構造を有する3つの同一なサブユニットを備えるレクチン様分子である(非特許文献3)。IgEへの結合のとき、FcεRIIは、IgEの合成の調節に関与するB細胞上のCD21と会合する(非特許文献4、非特許文献5)。FccRIIは、アレルゲン提示に関して長く認識されてきた。(非特許文献6)。上皮細胞上のFcεRIIに結合したIgEは、特異的かつ迅速なアレルゲン提示を引き起こす(非特許文献7)。FcεRIIは、数種の細胞型(B細胞、好酸球、血小板、ナチュラルキラー細胞、T細胞、濾胞樹状細胞、およびランゲルハンス細胞が挙げられる)に存在する。
【0006】
FcεRIおよびFcεRIIと相互作用するIgE分子の構造の実体もまた同定されている。突然変異誘発研究は、CH3ドメインが、FcεRI(非特許文献8;非特許文献9)およびFcεRII(非特許文献6;非特許文献10)の両方とのIgEの相互作用を媒介することを示した。高親和性レセプターおよび低親和レセプターの両方に対する結合部位は、2つのCH3ドメインを通る中心回転軸に沿って対称的に位置する。FcεRI結合部位は、CH3ドメインの、CH2ドメインの接合部近くの外側に位置し、一方FcεRII結合部位は、CH3のカルボキシル末端にある。
【0007】
アレルギー処置のための有望な考え方は、IgEアイソタイプ特異的であり、したがってIgEに結合し得る、モノクローナル抗体の適用に関する。このアプローチは、IgE免疫応答(アレルギーの誘導における最も初期の反応であり、そしてアレルギー状態を維持させる)をダウンレギュレートすることによる、アレルギー性反応の阻害に基づく。他の抗体クラスの応答が影響を与えない場合、アレルギー症状に対する即時効果および長期持続効果の両方が達成される。ヒト好塩基球密度の初期の研究は、患者の血漿中のIgEレベルと好塩基球あたりのFcεRIレセプターの数との間の相関を示した(非特許文献11)。彼らは、アレルギーの人物と非アレルギーの人物とにおけるFcεRI密度が、好塩基球あたり104〜106レセプターの範囲に及ぶことを示した。後に、抗IgEによるアレルギー性疾患の処置が、循環性IgEの量を処置前のレベルの1%まで減少させることが示された(非特許文献12)。MacGlashanは、患者の血清中を循環する遊離IgEに結合する、全抗IgE抗体で処置された患者から得られた血清を分析した。彼らは、患者中の循環性IgEのレベルを低下させることは、好塩基球上に存在するレセプターの数を減少させる結果となることを報告した。したがって、彼らは、好塩基球および肥満細胞上のFcεRI密度が循環性IgE抗体のレベルによって直接的もしくは間接的に調節される、という仮説を立てた。
【0008】
より最近では、特許文献1は、FcεRIおよびFcεRIIのIgE結合部位に結合してIgEのレセプターへの結合をブロックする、IgE分子およびフラグメントの使用を開示した。しかし、これらのアレルギー性疾患の管理に対する有害な副作用のない効果的な治療は、限られている。アレルギー性疾患の処置に対する1つの治療アプローチは、ヒト化抗IgE抗体を使用して、アレルギー性鼻炎および喘息を処置することに関する(非特許文献13;非特許文献14;非特許文献15;非特許文献16;非特許文献17)。これらの臨床的データは、IgEのそのレセプターへの結合の阻害が、アレルギー性疾患を処置するための効果的なアプローチであることを実証する。
【0009】
抗アレルギー剤として適切な抗体は、IgE産生性プラスマ細胞へと分化する表面IgE陽性B細胞と反応すべきであり、これによってこれらは、機能的にそれらのB細胞を消失させるために使用され得る。しかし、IgEに対する抗体は、基本的に、Fcεレセプターと交差結合することによって、IgE感作された肥満細胞から放出される媒介物質を誘導し得る。したがって、血清IgEレベルおよびsIgE+B細胞レベルに対して発揮される有利な効果を拮抗する。抗IgE治療を発展させ得ることに伴う潜在的に危険な問題の1つは、既に抗親和性レセプターに結合しているIgEに結合する治療用抗体によって引き起こされ、そしてヒスタミン放出を誘発し、潜在的にアナフィラキシー性の反応を引き起こす、IgE交差結合の可能性である。
【0010】
したがって、アレルギーの治療のために適用可能な抗体は、感作された肥満細胞および好塩基球に結合したIgEと反応してはいけないが、sIgE+B細胞を認識する能力を保持しているべきである。このようなIgEアイソタイプ特異的抗体は、例えば、非特許文献18)によって、特許文献2において、および数件の米国特許(例えば、特許文献3)において、記載されている。
【0011】
抗IgE抗体を産生するために使用されるペプチドはまた、アナフィラキシー性抗体を誘導する危険の可能性を被る。能動的ワクチン接種の間の抗IgE抗体の産生は、免疫の間に産生された抗体が、高親和性IgEレセプターに結合したIgEに結合する場合、または他の機構によって、受動的に投与された抗IgE抗体と同様の方法で、ヒスタミン放出を誘発し得る。
【特許文献1】国際公開第99/62550号パンフレット
【特許文献2】欧州特許第0407392号明細書
【特許文献3】米国特許第5,449,760号明細書
【非特許文献1】Hasegawa,S.ら,Hematopoiesis,1999年,93:2543−2551
【非特許文献2】Klubal R.ら,J.Invest.Dermatol.1997年,108(3):336−42
【非特許文献3】Dierks,A.E.ら,J.Immunol.1993年,150:2372−2382
【非特許文献4】Sanon,A.ら,J.Allergy Clin.Immunol.1990年,86:333−344
【非特許文献5】Bonnefoy,J.ら,Eur.Resp.J.1996年,9:63s−66s
【非特許文献6】SuttonおよびGould,Nature,1993年,366:421−428
【非特許文献7】Yang,P.P.,J.Clin.Invest.,2000年,106:879−886
【非特許文献8】Prestaら,J.Biol.Chem.1994年,269:26368−26373
【非特許文献9】Henry A.J.ら,Biochemistry,1997年,36:15568−15578
【非特許文献10】Shi,J.ら,Biochemistry,1997年,36:2112−2122
【非特許文献11】Malveauxら,J.Clin.Invest.,1978年,62:176
【非特許文献12】MacGlashanら,J.Immunol.,1997年,158:1438−1445
【非特許文献13】Corne,J.ら,J.Clin.Invest.1997年,99:879−887
【非特許文献14】Racine−Poon,A.ら,Clin.Pharmcol.Ther.1997年,62:675−690
【非特許文献15】Fahy,J.V.ら,Am.J.Resp.Crit.Care Med.1997年,155:1824−1834
【非特許文献16】Boulet,L.P.ら,Am.J.Resp.Crit.Care Med.,1997年,155:1835−1840
【非特許文献17】Milgrom,E.ら,N.Engl.J.Med.,1999年,341:1966−1973
【非特許文献18】Changら(Biotechnology 1990年,8,122−126
【発明の開示】
【発明が解決しようとする課題】
【0012】
したがって、IgEに特異的に結合するが、既にその高親和性レセプターに結合したIgEには結合しない、高親和性の非アナフィラキシー性抗体に対する必要性、ならびに、アナフィラキシー性抗体を誘導しない、能動免疫のためのペプチドに対する必要性が存在する。本発明者らは、肥満細胞または好塩基球上のIgEに結合することなく、抗体の高親和性結合を提供する、IgEの特異的エピトープを同定した。これらの特異的エピトープは、次いで、レセプターに結合するIgEの領域に結合するのみである、IgEに対する抗体を産生する能動免疫のための、特異的ペプチドを産生するために使用され得る。このことは、この抗体が、既にレセプターに結合したIgEに交差結合しないこと、したがって非アナフィラキシー性であることを保証する。
【課題を解決するための手段】
【0013】
(発明の要旨)
本発明は、IgEのCH3ドメインに由来する新規のペプチドエピトープに関する。これらのペプチドエピトープは、IgEに特異的に結合する高親和性抗体によって認識される。これらの新規のペプチドは、哺乳動物においてこれらのペプチドを投与して高親和性抗体を産生することによる、哺乳動物の能動免疫のために使用され得る。このペプチドエピトープはまた、非ヒト宿主において、IgEのこれらの領域に特異的に結合する高親和性抗IgE抗体を産生すること、および得られた抗体を哺乳動物の受動免疫のために使用することに使用され得る。
【0014】
本発明の一免疫原(エピトープA、図11)は、以下のアミノ酸配列を含む:
Asn Pro Arg Gly Val Ser Xaa Tyr Xaa Xaa Arg Xaa(配列番号72)
エピトープAの1つの例は、以下である:
Asn Pro Arg Gly Val Ser Ala Tyr Leu Ser Arg Pro(配列番号73)
別の免疫原(エピトープB、図11)は、以下のアミノ酸配列を含む:
Leu Pro Arg Ala Leu Xaa Arg Ser Xaa(配列番号74)
エピトープBの例としては、以下が挙げられる:
Leu Pro Arg Ala Leu Met Arg Ser Thr(配列番号75)
His Pro His Leu Pro Arg Ala Leu Met Arg Ser Thr(配列番号76)
Leu Pro Arg Ala Leu Met Arg Ser Thr Thr Lys Thr(配列番号77)
配列番号72または配列番号74のいずれにおいても、Xaaは、任意のアミノ酸であり得る。
【0015】
これらのペプチドは、上記ペプチドの少なくとも1つ、および生理学的に受容可能なキャリア、希釈剤、安定剤、もしくは賦形剤、ならびに免疫原性キャリアを含む組成物中に含まれ得る。上記免疫原性キャリアは、例えば、BSA、KLH、破傷風トキソイド、およびジフテリアトキソイドであり得る。本発明はまた、配列番号72〜77をコードするポリヌクレオチド、このポリヌクレオチドを含むベクター、およびこのベクターを有する細胞に関する。
【0016】
本発明はまた、エピトープAおよび/またはエピトープBに特異的に結合する抗体に関する。本発明はまた、エピトープAおよび/またはエピトープBに特異的に結合する抗体を作製する方法に関する。
【0017】
本発明は、配列番号72および/または配列番号74を含むペプチドの、IgE媒介性疾患もしくはIgE媒介性状態に罹患する被験体への投与に関する。
【0018】
本発明は、配列番号72および/または配列番号74を含むペプチドを用いて産生された高親和性抗体の、IgE媒介性疾患もしくはIgE媒介性状態に罹患する哺乳動物への投与に関する。この高親和性抗体は、ヒト抗体、ヒト化抗体、またはキメラ抗体であり得る。この抗体は、ポリクローナル抗体またはモノクローナル抗体であり得る。このようなIgE媒介性疾患もしくはIgE媒介性状態としては、例えば、喘息、アトピー性皮膚炎、蕁麻疹、アレルギー性鼻炎、および湿疹が挙げられる。
【0019】
(発明の詳細な説明)
(定義)
本出願を通して使用される用語は、当業者にとって通常かつ典型的な意味で解釈されるべきである。しかし、本出願人らは、以下の用語が以下に定義されるような特定の定義を与えられることを所望する。
【0020】
抗体鎖ポリペプチド配列に関する用語「実質的に同一」は、参照ポリペプチド配列に対して、少なくとも70%、もしくは少なくとも80%、もしくは少なくとも90%、もしくは少なくとも95%の配列同一性を示す抗体鎖として解釈され得る。核酸配列に関するこの用語は、参照核酸配列に対して、少なくとも約85%、もしくは少なくとも約90%、もしくは少なくとも約95%、もしくは少なくとも約97%の配列同一性を示すヌクレオチドの配列として解釈され得る。
【0021】
用語「同一性」または「相同性」は、配列全体に対する最大パーセント同一性を達成することを必要し、そして配列同一性の一部として任意の保存的置換を考慮しない場合、配列をアラインメントさせ、そしてギャップを導入した後に比較される、対応する配列の残基と同一な候補配列中のアミノ酸残基のパーセンテージを意味すると解釈されるべきである。N末端またはC末端の伸張も、それらの挿入も、同一性または相同性の低減として解釈されるべきではない。アラインメントのための方法およびコンピュータプログラムは、当該分野で周知である。配列同一性は、配列分析ソフトウェアを使用して測定され得る。
【0022】
用語「抗体」は、最も広い意味で使用され、そして具体的には、モノクローナル抗体(全長モノクローナル抗体が挙げられる)、ポリクローナル抗体、および多重特異性抗体(例えば、二重特異性抗体)を網羅する。抗体(Ab)および免疫グロブリン(Ig)は、同じ構造特性を有する、糖タンパク質である。抗体が特定の標的に対する結合特異性を示す一方で、免疫グロブリンは、抗体および標的特異性を欠く他の抗体様分子の両方を含む。ネイティブな抗体および免疫グロブリンは、通常、約150,000ダルトンのヘテロテトラマー糖タンパク質であり、2つの同一な軽(L)鎖と2つの同一な重(H)鎖から構成される。各重鎖は、一端に可変ドメイン(VH)と、それに続く多数の定常ドメインを有する。各軽鎖は、一方の端に可変ドメイン(VL)を有し、そして他方の端に定常ドメインを有する。「高親和性」抗体とは、少なくとも10−10、好ましくは10−12の結合親和性を有するこれらの抗体をいう。
【0023】
本明細書で使用される場合、「抗ヒトIgE抗体」とは、このようなIgEの高親和性レセプターFcεRIへの結合を、阻害または実質的に低下させるような様式で、ヒトIgEを結合する抗体を意味する。
【0024】
抗体の可変ドメインに関連する用語「可変」とは、可変ドメインの特定の部分が、抗体間で配列がかなり異なり、そして各特定の抗体の、その特定の標的に対する結合および特異性において使用されるという事実を意味する。しかし、可変性は、抗体の可変ドメインを通して均一に分布していない。これは、軽鎖可変ドメインと重鎖可変ドメインとの両方において相補性決定領域(CDR)と呼ばれる(または超可変領域としても知られる)3つのセグメントに集中されている。可変ドメインのより高度に保存された部分は、フレームワーク(FR)と呼ばれる。ネイティブな重鎖および軽鎖の可変ドメインは、各々、4つのFR領域を含み、その多くはβ−シート構造を採用し、3つのCDRによって結合され、これらはループを形成し、上記β−シート構造を結合し、いくつかの場合はその一部を形成する。各鎖のCDRは、FR領域によって、他の鎖からのCDRと一緒に近接して保持され、抗体の標的結合部位の形成に寄与する(Kabatらを参照のこと)。本明細書において使用される場合、免疫グロブリンアミノ酸残基の番号付けは、他に示されない限り、Kabatら(Sequences of Proteins of Immunological Interest,National Institute of Health,Bethesda,Md.1987)の免疫グロブリンアミノ酸残基の番号付けシステムに従って行われる。
【0025】
用語「抗体フラグメント」とは、全長抗体の一部(一般的には、標的結合領域または可変領域)をいう。抗体フラグメントの例としては、Fab、Fab’、F(ab’)2およびFvフラグメントが挙げられる。語句、抗体の「機能的フラグメントまたはアナログ」は、全長抗体に共通の定性的生物学的活性を有する化合物である。例えば、抗IgE抗体の機能的フラグメントまたはアナログは、IgE免疫グロブリンに対して、このような分子の能力を、高親和性レセプターFcεRIに結合する能力を有することから防ぐかまたは実質的に低減させる様式で、結合し得るものをいう。本明細書において使用される場合、抗体に関して、「機能的フラグメント」とは、Fv、F(ab)およびF(ab’)2フラグメントをいう。「Fv」フラグメントは、完全な標的認識部位および完全な標的結合部位を含む、最小抗体フラグメントである。この領域は、1つの重鎖可変領域と1つの軽鎖可変領域との強い、非共有結合性結合のダイマー(VH−VLダイマー)からなる。これは、各可変ドメインの3つのCDRがVH−VLダイマーの表面における標的結合部位を規定するために相互作用する構造にある。集合的に、6つのCDRが、抗体に対する標的結合特異性を与える。しかし、単一の可変ドメイン(または、標的に関して特異的な3つのCDRのみを含む、Fvの半分)でさえ、全結合部位よりも低い親和性ではあるが、標的に認識および結合する能力を有する。「単鎖Fv」抗体フラグメントまたは「sFv」抗体フラグメントは、抗体のVHドメインおよびVLドメインを含み、これらのドメインは、単一のポリペプチド鎖中に存在する。一般的に、Fvポリペプチドはさらに、VHドメインとVLドメインとの間に、標的結合に関する所望の構造を形成するsFvを形成し得るポリペプチドリンカーを含む。
【0026】
Fabフラグメントは、軽鎖の定常ドメインおよび重鎖の第一定常ドメイン(CH1)を含む。Fab’フラグメントは、重鎖CH1ドメインのカルボキシル末端における数個の残基(抗体ヒンジ領域からの1つ以上のシステインを含む)の付加によってFabフラグメントと異なる。F(ab’)フラグメントは、F(ab’)2ペプシン消化産物のヒンジシステインにおけるジスルフィド結合の切断によって生成される。抗体フラグメントのさらなる化学結合は、当業者に公知である。
【0027】
用語「モノクローナル抗体」は、本明細書において使用される場合、実質的に均一な抗体の集団から得られた抗体をいう(すなわち、この集団を構成する個々の抗体は、少数の量で存在し得る、可能性のある天然に存在する変異を除いて、同一である)。モノクローナル抗体は、高度に特異的であり、単一の標的部位に対して指向する。さらに、種々の決定基(エピトープ)に対して指向する種々の抗体を代表的に含む従来の(ポリクローナル)抗体調製物に対して、各モノクローナル抗体は、標的上の単一の決定基に対して指向する。それらの特異性に加えて、モノクローナル抗体は、ハイブリドーマ培養によって合成され得、他の免疫グロブリンによって汚染されない可能性があるという点で有利である。修飾語「モノクローナル」は、実質的に均一の抗体集団から得られるような抗体の特徴を示し、そして任意の特定の方法による抗体の産生を必要とすると解釈されるべきではない。例えば、本発明で使用するためのモノクローナル抗体は、周知の技術を使用して、ファージ抗体ライブラリーから単離され得る。本発明によって使用されるべき親モノクローナル抗体は、KohlerおよびMilstein,Nature 256,495(1975)によって最初に記載されたハイブリドーマ法によって作製され得るか、または組換え法によって作製され得る。
【0028】
非ヒト(例えば、マウス)抗体の「ヒト化」形態は、非ヒト免疫グロブリンに由来する最低の配列を含む、キメラ免疫グロブリン、キメラ免疫グロブリン鎖、またはそれらのフラグメント(例えば、Fv、Fab、Fab’、F(ab’)2または他の抗体の標的結合配列)である。一般に、ヒト化抗体は、少なくとも1つ、そして代表的には2つの可変ドメインの実質的に全てを含む。ここで、CDR領域の全てまたは実質的に全ては、非ヒト免疫グロブリンに対応し、そしてFR領域の全てまたは実質的に全ては、ヒト免疫グロブリンコンセンサス配列のそれらの領域である。非ヒト抗体はまた、免疫グロブリン定常領域(Fc)(代表的に、選択されたヒト免疫グロブリンテンプレートの免疫グロブリン定常領域)の少なくとも一部を含み得る。
【0029】
用語「細胞」、「細胞株」および「細胞培養物」は、子孫を含む。全ての子孫は、意図的な変異または不慮の変異に起因して、DNA内容物において正確には同一でない可能性があることが、また理解される。最初に形質転換された細胞についてスクリーニングされる場合、同じ機能特性または生物学的特性を有する改変体子孫が含まれる。本発明において使用される「宿主細胞」は、一般的に、原核生物宿主または真核生物宿主である。
【0030】
DNAによる細胞生物体の「形質転換」とは、生物体にDNAを導入し、そのDNAが、染色体外エレメントとして、または染色体組み込みによって複製されることを意味する。DNAによる細胞生物体の「トランスフェクション」とは、任意のコード配列が実際に発現されていてもいなくとも、細胞または生物体によるDNA(例えば、発現ベクター)の取り込みをいう。用語「トランスフェクトされた宿主細胞」および「形質転換された」とは、DNAが導入された細胞をいう。この細胞は、「宿主細胞」と称され、原核生物または真核生物のいずれかであり得る。代表的な原核生物宿主細胞としては、E.coliの種々の系統が挙げられる。代表的な真核生物宿主細胞としては、哺乳動物(例えば、チャイニーズハムスター卵巣またはヒト起原の細胞)が挙げられる。導入されるDNA配列は、宿主細胞と同種由来、または(of)宿主細胞と異なる種であり得るか、あるいは、一部の外来DNAおよび一部の同種DNAを含む、ハイブリッドDNA配列であり得る。
【0031】
用語「ベクター」とは、適切な宿主中におけるDNAの発現を生じ得る適切な制御配列に作動可能に連結されたDNA配列を含む、DNA構築物を意味する。このような制御配列としては、転写を引き起こすプロモーター、このような転写を制御する任意のオペレーター配列、適切なmRNAリボゾーム結合部位をコードする配列、ならびに転写および翻訳の終止を制御する配列が挙げられる。ベクターは、プラスミド、ファージ粒子、または、単に潜在的ゲノムインサートであり得る。一旦適切な宿主中へと形質転換された場合、ベクターは、複製して宿主ゲノムと独立して機能し得るか、またはいくつかの例では、ゲノムそのものへと一体化し得る。本明細書において、「プラスミド」および「ベクター」は、時として、交換可能に使用される。なぜなら、プラスミドは最も一般的に使用されるベクターの形態だからである。しかし、本発明は、当該分野で公知であるか、公知となる機能と等価な機能を果たすような他のベクターの形態を包含することを意図する。
【0032】
発現「制御配列」とは、特定の宿主生物において作動可能に連結されたコード配列の発現に必要なDNA配列をいう。原核生物に適切な制御配列としては、例えば、プロモーター、必要に応じて、オペレーター配列、およびリボゾーム結合部位が挙げられる。真核生物細胞は、プロモーター、ポリアデニル化シグナル、およびエンハンサーを利用することが知られている。前配列(presequence)または分泌リーダーのためのDNAは、それがポリペプチドの分泌に関わる前タンパク質(preprotein)として発現される場合、そのポリペプチドのためのDNAと作動可能に連結されている可能性がある;プロモーターまたはエンハンサーは、それがコード配列の転写に影響する場合、そのコード配列に作動可能に連結されている;あるいは、リボゾーム結合部位は、それが翻訳を促進するように配置されている場合、コード配列に作動可能に連結されている。一般的に、「操作可能に連結された」とは、連結されたDNA配列が、隣接しており、そして分泌リーダーの場合、隣接していてリーディングフェーズにあることを意味する。しかし、エンハンサーは、連続している必要はない。
【0033】
処置目的のための「哺乳動物」とは、哺乳動物として分類される任意の動物をいい、ヒト、家畜動物(domestic animal)および家畜動物(farm animal)、非ヒト霊長類、および動物園の動物、競技用動物、またはペット用動物(例えば、イヌ、ウマ、ネコ、ウシなど)が挙げられる。
【0034】
用語「エピトープタグ化」とは、ポリペプチドに関して本明細書において使用される場合、「エピトープタグ」に融合されたポリペプチドをいう。エピトープタグポリペプチドは、そのエピトープに対して抗体が産生され得るエピトープを提供するのに十分な残基を有するが、ポリペプチドの活性を干渉しないように十分短い。エピトープタグはまた、好ましくは、かなり独特であり、したがって抗体は、他のエピトープと実質的に交差反応しない。適切なタグポリペプチドは、一般的に、少なくとも6アミノ酸残基を有し、通常、約8〜50アミノ酸残基(好ましくは、約9〜30残基)の間である。例としては、以下が挙げられる:インフルエンザHAタグポリペプチドおよびその抗体12CA5(Fieldら,Mol Cell.Biol.8:2159−2165(1988));c−mycタグならびにそれに対する8F9抗体、3C7抗体、6E10抗体、G4抗体、B7抗体および9E10抗体(Evanら,Mol Cell.Biol.5(12):3610−3616(1985));そして単純ヘルペスウイルス糖タンパク質D(gD)タグおよびその抗体(Paborskyら,Protein Engineering 3(6):547−553(1990))。特定の実施形態において、エピトープタグは、IgG分子(例えば、IgG1、IgG2、IgG3またはIgG4)のインビボ血清半減期の延長を担う、IgG分子のFc領域のエピトープであり得る。
【0035】
語「標識」とは、本明細書において使用される場合、分子またはタンパク質(例えば、抗体)と直接的または間接的に結合され得る検出可能な化合物または組成物をいう。標識は、それ自身検出可能(例えば、放射性同位体標識または蛍光標識)であり得るか、または酵素標識の場合、検出可能な基質化合物または組成物の化学的変化を触媒し得る。
【0036】
本明細書において使用される場合、「固相」とは、本発明の抗体が接着し得る非水性マトリックスを意味する。本明細書に包含される固相の例としては、部分的もしくは全体的に、ガラス(例えば、制御された多孔質ガラス)、多糖類(例えば、アガロース)、ポリアクリルアミド、ポリスチレン、ポリビニルアルコール、およびシリコーンから形成されたものが挙げられる。特定の実施形態において、状況に依存して、固相は、アッセイプレートのウェルを包含し得;他方、それは精製カラム(例えば、アフィニティークロマトグラフィーカラム)である。
【0037】
本明細書中で使用される場合、用語「IgE媒介性障害」とは、免疫グロブリンIgEの過剰生成および/または免疫グロブリンIgEに対する過剰な感受性によって特徴付けられる状態または疾患を意味する。具体的には、アナフィラキシー性過敏症ならびにアトピー性アレルギー(例えば、喘息、アレルギー性鼻炎、およびアレルギー性結膜炎(花粉症)、湿疹、蕁麻疹、アトピー性皮膚炎、および食物アレルギーが挙げられる)に関する状態を包含すると解釈される。例えば、蜂に刺されること、蛇にかまれること、食物、または医薬によって引き起こされるアナフィラキシーショックの重篤な生理学的状態もまた、本用語の範囲下に包含される。
【0038】
(抗体の生成)
出発抗体または「親」抗体は、このような抗体を生成するために当該分野で利用可能な技術を用いて調製され得る。これらの技術は、当該分野で周知である。出発抗体を生成するための例示的方法は、以下の節においてより詳細に記載される。これらの記載は、親抗体を作製または選択するための考えられる代替案であり、このような分子が生成され得る方法を決して限定しない。
【0039】
抗体の結合親和性は、本発明の高親和性抗体を生成する前に決定される。また、この抗体は、他の生物学的に活性なアッセイに(例えば、治療剤としての有効性を評価するために)供され得る。このようなアッセイは、当該分野で公知であり、そして抗体に対する標的および意図される用途に依存する。
【0040】
特定のエピトープに結合する抗体(例えば、IgEの、その高親和性レセプターへの結合をブロックするもの)についてスクリーニングするために、慣習的な交差ブロックアッセイ(例えば、Antibodies:A Laboratory Manual」(Cold Spring Harbor Laboratory,Ed HarlowおよびDavid Lane(1988))に記載されるようなアッセイ)が実施される。あるいは、エピトープマッピングは、抗体が目的のエピトープにどこで結合するかを決定するために実施され得る。必要に応じて、抗体を生成するために使用される標的のホモログに対する抗体の結合親和性(ここで、このホモログは、異なる種に由来する)は、当該分野で公知の技術を用いて評価され得る。一実施形態において、他の種は、前臨床研究において抗体が投与される非ヒト動物である。したがって、この種は、アカゲザル、カニクイザル、ヒヒ、チンパンジー、およびマカクのような非ヒト動物であり得る。他の実施形態において、この種は、例えば、げっ歯類、ネコ、またはイヌであり得る。
【0041】
親抗体は、標的に対して親抗体よりもより高いかもしくはより強い結合親和性を有する抗体を生成するように、本発明に従って変化される。抗体特異性は、抗体とその標的との間に形成される独特の界面から生じる;この表面は、独特のはめ合いを生じるように互いに補完する(Jones,S.およびThornton,J.M.(1996)Proc.Natl.Acad.Sci.USA 93:13−20)。その界面に沿った接触をさらに改善することによって、結合パートナーの会合を助けるために必要とされるエネルギーコストがより低いことの結果として、全体的な親和性が増加し得る。
【0042】
抗体の結合表面は、一般的に、6つの相補性決定領域(CDR)から構成される。これらは、コアから外に伸びるループである。CDRは、特異的標的への結合のために独特の配列を有するアミノ酸から構成される。抗体の、その抗原に対する親和性を増加させるために、これらのアミノ酸の周辺の環境は、種々の非共有結合力を導入または改善し、最終的に、相互作用のエネルギー特性を低下させ、その結果としてより高い親和性をもたらすことによって、より都合よくならねばならない。
【0043】
ファンデルワールス力は、2つの電気的に中性な分子の間で生じる非共有結合性相互作用である(Voet,D.およびVoet,J.G.(1990)Biochemistry John Wiley and Sons,NY,NY)。会合は、永久双極子または誘起双極子から生じる静電気的相互作用に由来して、2つの表面の間で起こり得る。これらの双極子は、αヘリックスの端に沿って存在し得るか、または極性アミノ酸の近くに存在し得る。結合界面に沿ったファンデルワールス力の数の増加によって、より都合よい会合が生じる。
【0044】
水素結合の導入もまた、抗体とその抗原との間の相互作用の特異性を増加させる。水素結合に関与する一般的なドナーおよびアクセプターは、主にアミノ酸を構成する窒素原子、酸素原子および硫黄原子である(上述のVoetらを参照のこと)。水素結合は、短い距離のみにわたる傾向にあり(通常2.7〜3.1Å)、そのため、その結合パートナーは、これらの相互作用が生じるためには密接しなければならない。したがって、親和性を改善し得る1つの様式は、潜在的ドナー分子と潜在的アクセプター分子とを密接して接触させて、水素結合を確立させることである。
【0045】
最後に、疎水性相互作用の改善もまた、2つの結合パートナーの間の有利なエネルギー特性を増加させる。結合表面に密接して位置する非極性残基は、他の非極性残基によって囲まれるべきであり、それによって有利な環境に存在する。非極性側鎖の囲み(the burial of nonpolar side chains)を満足することによって、相互作用のエネルギー特性は、強い結合界面のために有利となる。
【0046】
タンパク質−タンパク質界面を安定化させる相互作用は、これらの接触を維持するエネルギーコストを低下させ、したがって、全体的親和性を増加させる。結合界面近くの個々のアミノ酸の周りの環境を改善することによって、より有利な環境が生成され、結果としてより高い親和性が得られる。したがって、有利な接触を導入し、そしてさらなる相補性を介して界面を改善することによって、抗体と抗原との間の全体的な結合相互作用は大きく改善される。
【0047】
得られた抗親和性抗体は、好ましくは、親抗体の標的に対する結合親和性よりも少なくとも約10倍高い、または少なくとも約20倍高い、または少なくとも約500倍高い、または1000倍〜5000倍高くあり得る、標的に対する結合親和性を有する。必要な、もしくは所望される、結合親和性における増強の程度は、親抗体の最初の結合親和性に依存する。
【0048】
一般的に、親抗体由来の高親和性抗体を作製するための方法は、以下の工程を包含する。
【0049】
1.目的の標的に結合する、軽鎖可変ドメインおよび重鎖可変ドメインを含む親抗体を得るかまたか選択する工程。これは、伝統的なハイブリドーマ技術、ファージディスプレイ技術、または標的特異的抗体を生成する任意の他の方法によって行われ得る。
【0050】
2.配列が親フレームワーク配列(好ましくはヒトテンプレート配列)に近いフレームワーク配列を選択する工程。このテンプレートは、例えば、その比較される全体の長さ、CDRのサイズ、フレームワークとCDRとの間の接合部に位置するアミノ酸残基、全体的な相同性などに基づいて選択され得る。選択されるテンプレートは、1より多くの配列の混合物であり得るか、またはコンセンサステンプレートであり得る。
【0051】
3.考えられるCDR位置の各々および全てにおいてランダムアミノ酸置換を作製することによってクローンのライブラリーを生成する工程。これは、例えば、CDRに隣接するか、または全ての考えられるアミノ酸と結合するか、それをフォールディングすることに影響を及ぼすヒトフレームワークテンプレート中のアミノ酸をランダムに置換し得、フレームワーク置換のライブラリーを生成する。これらのフレームワーク置換は、標的結合および抗体フォールディングに対するそれらの潜在的効果について評価され得る。フレームワーク中のアミノ酸の置換は、CDR中のアミノ酸の置換と同時にか、または順次にかのいずれかで行われ得る。改変体のライブラリーを生成するための1つの方法は、オリゴヌクレオチド合成による。
【0052】
4.工程(3)において生成された重鎖および/または軽鎖改変体を含む発現ベクターを構築する工程。上記改変体は、以下の式を含み得る:FRH1−CDRH1−FRH2−CDRH2−FRH3−CDRH3−FRH4(I)およびFRL1−CDRL1−FRL2−CDRL2−FRL3−CDRL3−FRL4(II)、ここで、FRL1、FRL2、FRL3、FRL4、FRH1、FRH2、FRH3およびFRH4は、工程3において選択されたフレームワークテンプレートの軽鎖配列および重鎖配列の改変体を表し、そしてCDRは、親抗体CDRの改変体CDRを表す。このような重鎖配列および軽鎖配列を含むベクターの例は、図1に示される。
【0053】
5.特異的標的に対するクローンのライブラリーをスクリーニングする工程。標的に結合するこれらのクローンは、改善された結合親和性についてスクリーニングされる。親分子より大きな親和性で結合するこれらのクローンが選択され得る。最適な高親和性候補は、親抗体と比較して、(好ましくは、20倍より大きい、100倍より大きい、1000倍より大きい、または5000倍より大きい)最大の結合親和性の可能性を有する。選択された改変体が、望ましくない特定のアミノ酸(例えば、導入されたグリコシル化部位または潜在的に免疫原性の部位)を含む場合、それらのアミノ酸は、より有利なアミノ酸残基で置き換えられ得、そして結合親和性が再評価され得る。
【0054】
また、この方法を使用して、完全ヒト親抗体から、CDR領域のみをランダムに置換することによって、ヒトフレームワーク領域をインタクトなままにして、高親和性抗体を生成し得る。
【0055】
改善されたハイスループットスクリーニング技術および図1に記載されたようなベクターによって、当業者は、所与のCDR中および/またはフレームワーク領域中の全ての部位における置換の包括的なライブラリーを、迅速かつ効果的にスクリーニングし得る。全てのアミノ酸を全ての位置において同時に、ランダムに置換することによって、個々の置換によっては予測または同定できない(例えば、相乗作用に起因する)親和性を有意に増加させる可能性のある組合せをスクリーニングすることができる。
【0056】
(親抗体の調製)
(標的の調製)
可溶性の標的またはそれらのフラグメントは、抗体を生成するための免疫原として使用され得る。この抗体は、目的の標的に対して指向する。好ましくは、この標的は、生物学的に重要なポリペプチドであり、疾患または障害に罹患する哺乳動物へのこの抗体の投与は、哺乳動物に治療的利益をもたらし得る。しかし、抗体は、非ポリペプチド標的に対して指向し得る。標的がポリペプチドである場合、それは、膜貫通分子(例えば、レセプター)またはリガンド(例えば、成長因子)であり得る。本発明の1つの標的は、IgEである。全細胞が抗体を作製するための免疫原として使用され得る。標的は、組み換え的に使用され得るか、または合成法を用いて使用され得る。標的はまた、天然の供給源から単離され得る。
【0057】
本発明の抗体を生成するために使用される抗原は、本発明のポリペプチドおよびポリペプチドフラグメント(エピトープAおよび/またはエピトープBが挙げられる)を包含し得る。動物を免疫するために使用されるポリペプチドは、標準的な組み換え法、化学合成法、または精製法によって得ることができる。当該分野で周知であるように、免疫原性を増加させるため、抗原は、キャリアタンパク質に結合され得る。一般的に使用されるキャリアとしては、キーホールリンペットヘモシアニン(KLH)、サイログロブリン、ウシ血清アルブミン(BSA)および破傷風トキソイドが挙げられるが、これらに限定されない。次いで、結合されたペプチドは、動物(例えば、マウス、ラット、もしくはウサギ)を免疫するために使用される。このようなキャリアに加えて、周知のアジュバントが抗原とともに投与されて、強力な免疫応答の誘導を促進し得る。
【0058】
(ポリクローナル抗体)
ポリクローナル抗体は、通常、非ヒト哺乳動物において、アジュバントと組み合わせての関連する標的の複数回の皮下(sc)注射または腹腔内(ip)注射によって生成される。免疫応答を誘起し得る多くの因子は当該分野において周知である。
【0059】
動物は、(各々、ウサギまたはマウスについての)タンパク質または結合体をフロイント完全アジュバントと組み合わせて、そしてその溶液を皮内に注射することによって、標的、免疫原性結合体、または誘導体に対して免疫される。一ヵ月後、その動物は、フロイント不完全アジュバント中の最初の量の1/5〜1/10のペプチドまたは結合体を用いて、複数部位への皮下注射によって、追加免疫される。7日〜14日後、この動物は、採血されて、その血清が抗体力価についてアッセイされる。動物は、力価がプラトーに達するまで追加免疫される。
【0060】
選択された哺乳動物抗体は、通常、標的に対して十分に強力な結合親和性を有する。例えば、この抗体は、ヒト抗IgE標的と、約1×10−8Mの結合親和性(Kd)の値で結合し得る。抗体親和性は、飽和結合;酵素結合イムノソルベントアッセイ(ELISA);および競合アッセイ(例えば、ラジオイムノアッセイ)によって決定され得る。
【0061】
目的の標的に結合する抗体をスクリーニングするため、慣習的な交差結合アッセイ(例えば、Antibodies,A Laboratory Manual,Cold Spring Harbor Laboratory,Ed HarlowおよびDavid Lane(1988)に記載されるようなアッセイ)が行われ得る。あるいは、エピトープマッピング(例えば、Champeら,J.Biol.Chem.270:1388−1394(1995)に記載される)が行われて、結合を決定し得る。
【0062】
(モノクローナル抗体)
モノクローナル抗体は、単一の抗原部位を認識する抗体である。これらの均一な特異性は、モノクローナル抗体を、種々の異なる抗原部位を認識する抗体を通常含むポリクローナル抗体よりもずっと有用にしている。モノクローナル抗体は、Kohlerら,Nature,256:495(1975)によって最初に記載されたハイブリドーマ法を用いて作製され得るか、または組み換えDNA法によって作製され得る。
【0063】
ハイブリドーマ法において、マウスまたは他の適切な宿主動物(例えば、げっ歯類)は、本明細書において上記されるように免疫されて、免疫のために使用されたタンパク質に特異的に結合する抗体を産生するか、または産生し得るリンパ球を誘起される。あるいは、リンパ球は、インビトロで免疫され得る。次いで、リンパ球は、ハイブリドーマ細胞を形成するために適切な融合因子(例えば、ポリエチレングリコール)を用いて、骨髄腫細胞と融合される(Goding,Monoclonal Antibodies:Principals and Practice,pp.590−103(Academic Press,1986))。
【0064】
このようにして調製されたハイブリドーマ細胞は、好ましくは非融合の親骨髄腫細胞の成長または生存を阻害する1つ以上の物質を含む適切な培養培地中に播種され、そして増殖する。例えば、親骨髄腫細胞が酵素ヒポキサンチングアニンホスホリボシルトランスフェラーゼ(HGPRTまたはHPRT)を欠く場合、ハイブリドーマのための培養培地は、代表的に、ヒポキサンチン、アミノプテリンおよびチミジン(HGPRT欠損細胞の成長を妨害する物質)を含む(HAT培地)。好ましい骨髄腫細胞は、効果的に融合し、選択された抗体産生細胞による抗体の安定な高レベル産生を支持し、そしてHAT培地のような培地に対する感受性のある細胞である。ヒト骨髄腫細胞株およびマウス−ヒトヘテロ骨髄腫細胞株は、ヒトモノクローナル抗体の産生に関して記載されている(Kozbar,J.Immunol.133:3001(1984);Brodeurら,Monoclonal Antibody Production Techniques and Applications,pp.51−63(Marcel Dekker,Inc.,New York,1987))。
【0065】
所望の特異性、親和性、および/または活性の抗体を産生するハイブリドーマ細胞が同定された後、クローンが、限界希釈手順によってサブクローニングされ得、そして標準的方法によって増殖され得る(Goding,Monoclonal Antibodies:Principals and Practice,pp.59−103,Academic Press,1986)。この目的のための適切な培養培地は含む。サブクローンによって分泌されるモノクローナル抗体は、培養培地から従来の免疫グロブリン精製手順(例えば、プロテインAセファロース、ヒドロキシアパタイトクロマトグラフィー、ゲル電気泳動、透析、またはアフィニティークロマトグラフィー)によって適切に分離される。
【0066】
モノクローナル抗体をコードするDNAは、従来の手順を用いて(例えば、そのモノクローナル抗体の重鎖および軽鎖をコードする遺伝子に特異的に結合し得るオリゴヌクレオチドプローブを用いることによって)容易に単離され、配列決定される。ハイブリドーマ細胞は、このようなDNAの供給源として役立つ。一旦単離された場合、このDNAは、発現ベクター中に配置され得る。次いでこの発現ベクターは、宿主細胞(例えば、E.coli細胞、NS0細胞、チャイニーズハムスター卵巣(CHO)細胞、または骨髄腫細胞)に移送されて、この組み換え宿主細胞におけるモノクローナル抗体の合成を得る。このDNAはまた、例えば、相同なマウス配列の代わりに、ヒト重鎖および軽鎖の定常ドメインについてのコード配列を置換することによって(米国特許第4,816,567号;Morrisonら,Proc.Natl Acad.Sci.USA 81:6851(1984))、または免疫グロブリンポリペプチドに共有結合性に結合することによって、改変され得る。
【0067】
(ヒト化抗体)
ヒト化は、キメラ抗体を作製するための技術であり、キメラ抗体においては、インタクトなヒト可変ドメインよりもずっとわずかが、対応する非ヒト種由来の配列によって置換されている。ヒト化抗体は、非ヒトである供給源からその中に導入された1つ以上のアミノ酸残基を有する。これらの非ヒトアミノ酸残基は、多くの場合、「移入(import)」残基と称され、これは代表的に、「移入」可変ドメインから取られる。ヒト化は、本質的に、Winterおよび共同研究者(Jonesら,Nature 321:522−525(1986);Riechmanら,Nature 332:323−327(1988);Verhoeyensら,Science 239:1534−1536(1988))の方法に従って、非ヒトCDRまたはヒト抗体中の対応する配列についてのCDR配列で置換することによって行われる(例えば、米国特許第4,816,567号を参照のこと)。本発明において実施される場合、ヒト化抗体は、マウス抗体における類似部位由来の残基によって置換された、いくつかのCDR残基およびいくつかのFR残基を有し得る。
【0068】
ヒト化抗体を作製するのに使用されるべきヒト可変ドメイン(軽鎖および重鎖の両方)の選択は、抗原性を低減させるために非常に重要である。いわゆる「ベストフィット(best fit)」法に従って、非ヒト抗体の可変ドメインの配列は、公知のヒト可変ドメイン配列のライブラリーと比較される。非ヒト親抗体の配列に最も近いヒト配列が、ヒト化抗体についてのヒトフレームワークとして受容される(Simsら,J.Immunol.151:2296(1993);Chothiaら,J.Mol.Biol.196:901(1987))。別の方法は、軽鎖または重鎖の特定のサブグループの全てのヒト抗体のコンセンサス配列に由来する、特定のフレームワークを使用する。同じフレームワークが、数種の異なるヒト化抗体に対して使用され得る(Carterら,Proc.Natl.Acad.Sci.USA,89:4285(1992);Prestaら,J.Immunol.151:2623(1993))。
【0069】
(抗体フラグメント)
種々の技術が、抗体フラグメントの生成のために開発されている。伝統的には、これらのフラグメントは、インタクトな抗体のタンパク質分解性消化を介して得られた(例えば、Morimotoら,Journal of Biochemical and Biophysical Methods 24:107−117(1992)およびBrennanら,Science 229:81(1985))。しかし、これらのフラグメントは、現在、組み換え宿主細胞によって直接的に生成され得る。例えば、抗体フラグメントは、抗体ファージライブラリーから単離され得る。あるいは、F(ab’)2−SHフラグメントが、E.coliから直接的に回収され得、化学的に結合されてF(ab’)2フラグメントを形成し得る(Carterら,Bio/Technology 10:163−167(1992))。別のアプローチに従うと、F(ab’)2フラグメントは、組み換え宿主細胞培養物から直接的に単離され得る。抗体フラグメントを生成するための他の技術は、当業者に明らかである。別の実施形態において、最適な抗体は、単鎖Fvフラグメントである(scFv)(PCT特許出願 国際公開番号93/16185)。
【0070】
(高親和性抗体の調製)
一旦親抗体が同定され、単離された場合、親抗体の可変領域の1つ以上において、1つ以上のアミノ酸残基が変化され得る。あるいは、または加えて、フレームワーク残基の1つ以上の置換が、親抗体中に導入され得、これらは、抗体の(例えば、ヒトIgEに対する)結合親和性に改善をもたらす。改変するフレームワーク領域の残基の例としては、以下が挙げられる:直接的に標的に非共有結合性に結合するもの(Amitら,Science 233:747−753(1986));CDRの高次構造と相互作用するもの/CDRの高次構造に影響するもの(Chothiaら,J.Mol.Biol.196:901−917(1987));および/またはVL−VH界面に(EP 239 400 B1)関与するもの。特定の実施形態において、このようなフレームワーク領域の残基の1つ以上の改変は、目的の標的に対する抗体の結合親和性の増強をもたらす。
【0071】
抗体の生物学的特性の改変は、例えば、以下を維持することに対するそれらの影響において有意に異なる置換を選択することによって達成され得る:(a)置換の範囲におけるポリペプチド骨格の構造(例えば、シート構造もしくはヘリックス構造);(b)標的部位における分子の電荷もしくは疎水性、または(c)側鎖の大きさ(bulk)。非保存的置換は、これらの分類の1つのメンバーと別の分類との交換を伴う。
【0072】
アミノ酸配列改変体をコードする核酸分子は、当該分野で公知の種々の方法によって調製される。これらの方法としては、より早期に調製された、種依存性抗体の改変体または非改変バージョンの、オリゴヌクレオチド媒介性(または部位指向性)突然変異誘発、PCR突然変異誘発、およびカセット突然変異誘発が挙げられるが、これらに限定されない。改変体を生成するための好ましい方法は、オリゴヌクレオチド媒介性合成である。特定の実施形態において、抗体改変体は、例えば、約2〜約15の超可変領域置換を置換した、単一の超可変領域残基を有するのみである。
【0073】
改変体のライブラリーを生成するための1つの方法は、図2に示されるスキームに従うオリゴヌクレオチド媒介性合成による方法である。各々約100ヌクレオチドの3つのオリゴヌクレオチドが、全軽鎖可変領域または全重鎖可変領域にわたって合成され得る。各オリゴヌクレオチドは、以下を含む:(1)トリプレット(NNK)20によって生成された60アミノ酸伸長(ここで、Nは任意のヌクレオチドであり、KはGまたはTである)、および(2)隣のオリゴか、または各端のベクター配列とのおよそ15〜30ヌクレオチドの重複。PCR反応におけるこれら3つのオリゴヌクレオチドのアニーリングのとき、ポリメラーゼは、反対側の鎖を充填し、完全二重鎖化された重鎖可変領域配列または軽鎖可変領域配列を生成する。トリプレットの数は、任意の反復長に調整され得、そして、オリゴヌクレオチド中のそれらの位置は、所与のCDRまたはフレームワーク領域におけるアミノ酸のみを置換するように選択され得る。(NNK)を用いることによって、コードされた改変体中の各々の位置において、全20アミノ酸が考えられる。5〜10アミノ酸(15〜30ヌクレオチド)の重複配列は、置換されないが、これはフレームワークの積み重なり(stacking)領域内に含まれるように選択され得るか、または合成の別のラウンドまたはその後のラウンドで置換され得る。オリゴヌクレオチドを合成するための方法は、当該分野で周知であり、そしてまた市販されている。これらのオリゴヌクレオチドから抗体改変体を生成するための方法もまた、当該分野で周知である(例えば、PCR)。
【0074】
重鎖改変体および軽鎖改変体のライブラリーは、それらの配列のランダムな位置において異なり、任意の発現ベクター(例えば、バクテリオファージ、具体的には、図1のベクター)中で構築され得る。これらのベクターの各々は、特定の重鎖改変体および軽鎖改変体をコードするDNAを含む。
【0075】
抗体改変体の生成に続いて、親抗体に比較した改変体の生物学的活性が決定される。上記のように、これは、改変体の標的に対する結合親和性を決定する工程を包含する。多くのハイスループット法が、抗体改変体を、目的の標的と結合するそれらの能力に関して迅速にスクリーニングするために存在する。
【0076】
次いで、この最初のスクリーニングから選択された1つ以上の抗体改変体が、親抗体に対して増強された結合親和性についてスクリーニングされ得る。結合親和性を決定するための1つの一般的な方法は、BIAcoreTM表面プラズモン共鳴システム(BIAcore,Inc.)を用いて会合速度定数および解離速度定数を評価することによる方法である。バイオセンサーチップは、製造元(BIAcore)の指示に従って、標的の共有結合のために活性化される。次いで、標的は、希釈され、そしてチップ上に注入されて、固定材料の反応ユニット(RU)におけるシグナルを得る。RUのシグナルは、固定材料の質量に比例するので、このシグナルは、マトリックス上の固定された標的密度の範囲を示す。解離データは、一部位モデル(one−site model)に適合されて、koff +/− s.d.(測定の標準偏差)を得る。偽第一次速度定数(ks)は、各会合曲線について計算されて、タンパク質濃度の関数としてプロットされてkon +/− s.e.(適合の標準誤差)を得る。結合の平衡解離定数KDは、koff/konとしてSPR測定から計算される。平衡解離定数KDは、koffに反比例するので、親和性改善の推定は、会合速度(kon)が全改変体について一定であると仮定して行われ得る。
【0077】
得られた高親和性を有する候補は、必要に応じて、1つ以上のさらなる生物学的活性アッセイに供されて、増強された結合親和性を有する抗体改変体が、さらに所望される治療特性を保有することを確認し得る。例えば、抗IgE抗体の場合、IgEのレセプターへの結合をブロックし、ヒスタミンの放出を阻害するものについてスクリーニングし得る。最適な抗体改変体は、親抗体よりも有意に高い結合親和性で標的に結合する能力を保有する。
【0078】
そのようにして選択された抗体改変体は、多くの場合抗体の意図された用途に依存して、さらなる改変に供され得る。このような改変は、アミノ酸配列のさらなる変化、異種ポリペプチドとの融合、および/または以下に詳述されるような共有結合性の改変を含む。例えば、抗体の適切な高次構造の維持に関与しない任意のシステイン残基が、一般的にセリンによって置換されて、分子の酸化的安定性を改善し得、そして異常な交差結合を防止し得る。逆に、システイン結合が抗体に付加されて、(特に、この抗体が、Fvフラグメントのような抗体フラグメントである場合)その安定性を改善し得る。
【0079】
(ベクター)
本発明はまた、本明細書において開示されるような抗体改変体をコードする単離された核酸、この核酸を含むベクターおよび宿主細胞、ならびに上記抗体改変体の生成のための組み換え技術を提供する。抗体改変体の組み換え生成のため、それをコードする核酸が単離され、そしてさらなるクローニング(DNAの増幅)または発現のため、複製可能なベクターに挿入される。抗体改変体をコードするDNAは、容易に単離され、従来の手順を用いて(例えば、この抗体改変体の重鎖および軽鎖をコードする遺伝子に対して特異的に結合し得るオリゴヌクレオチドプローブを用いることによって)配列決定される。
【0080】
多くのベクターが利用可能である。ベクター構成要素としては、一般的に以下の1つ以上が挙げられるが、これらに限定されない:シグナル配列、複製起点、1つ以上のマーカー遺伝子、エンハンサーエレメント、プロモーター、および転写終止配列。
【0081】
図1に示されるファージ発現ベクターは、一般的に使用されるM13ベクターと、迅速な分泌のためおよび適切な結合特異性および最低の親和性基準に関して改変体Fabをスクリーニングするため、M13の自身の遺伝子IIIウイルス分泌シグナルとから構成される。このベクターは、全遺伝子III配列を使用せず、したがって細菌細胞の表面上に表示が存在しないが、Fabはペリプラズム空間に分泌される。あるいは、Fabは、細胞質中で発現され得、そして単離され得る。重鎖および軽鎖の各々は、それら自身の分泌シグナルを有するが、単一の強力な誘導プロモーターに依存して発現される。
【0082】
図1におけるベクターはまた、容易な精製および検出のために、Hisタグおよびmycタグを提供する。当業者は、Fabが、別個のプロモーターから独立して発現され得るか、または分泌シグナルが選択されたウイルス配列である必要はないが、選択された宿主細胞からの抗体フラグメントの分泌に適切な原核生物もしくは真核生物のシグナル配列であり得ることを認識する、重鎖および軽鎖が異なるベクター上に位置していてもよいことが、また認識されるべきである。
【0083】
(A.シグナル配列構成要素)
本発明の抗体改変体は、組み換え的に生成され得る。この改変体はまた、異種ポリペプチド(好ましくは、シグナル配列、または成熟タンパク質または成熟ポリペプチドのN末端に特異的切断部位を有する他のポリペプチド)と融合された融合ポリペプチドとして発現され得る。好んで選択される異種シグナル配列は、宿主細胞によって認識およびプロセシングされる(すなわち、シグナルペプチダーゼによって切断される)配列である。ネイティブ抗体シグナル配列を認識およびプロセシングしない原核生物宿主細胞に対しては、このシグナル配列は、例えば、アルカリホスファターゼ、ペニシリナーゼ、Ippまたは熱安定性エンテロトキシンIIリーダーの群から選択された原核生物シグナル配列によって置換され得る。あるいは、図1のベクターの場合、選択されたシグナル配列は、遺伝子III由来のウイルスシグナル配列であった。酵母分泌については、ネイティブシグナル配列は、例えば、酵母インベルターゼリーダー、α因子リーダー(SaccharomycesおよびKluyveromyces α因子リーダーが挙げられる)もしくは酸性ホスファターゼリーダー、C.albicansグルコアミラーゼリーダー、または、例えば国際公開番号90/13646において記載されるシグナルによって置換され得る。哺乳動物細胞発現においては、哺乳動物シグナル配列およびウイルス分泌リーダー(例えば、単純ヘルペスgDシグナル)が利用可能である。このような前駆領域のためのDNAは、リーディングフレームにおいて、抗体改変体をコードするDNAに連結される。
【0084】
(B.複製起点の構成要素)
ベクターは、通常、ベクターが1種以上の選択された宿主細胞において複製することを可能にする核酸配列を含む。一般的に、この配列は、ベクターが宿主の染色体DNAと独立に複製することを可能にする配列であり、そして複製起点または自立的に複製する配列を含む。このような配列は、種々の細菌、酵母、およびウイルスに関して周知である。プラスミドpBR322由来の複製起点は、ほとんどのグラム陰性細菌に対して適切であり、2μプラスミド起点は、酵母に対して適切であり、そして種々のウイルス起点(SV40、ポリオーマ、アデノウイルス、VSVもしくはBPV)は、哺乳動物細胞中のベクターに対して有用である。一般的に、複製起点の構成要素は、哺乳動物発現ベクターにとって必要ではない(SV40起点は、初期プロモーターを含むので、代表的に唯一使用され得る)。
【0085】
(C.選択遺伝子構成要素)
ベクターは、選択遺伝子(選択マーカーとも称される)を含み得る。代表的な選択遺伝子は、(a)抗生物質もしくは他の毒素(例えば、アンピシリン、ネオマイシン、メトトレキサート、もしくはテトラサイクリン)に対する耐性を与えるか、(b)栄養素要求性欠損を補完するか、または(c)複合培地から得られない重要な栄養素を供給する、タンパク質をコードする(例えば、Bacillus属にとってのD−アラニンラセマーゼをコードする遺伝子)。
【0086】
選択スキームの1つの例は、宿主細胞の増殖を阻止する薬物を利用する。異種遺伝子によって首尾よく形質転換されたこれらの細胞は、薬物耐性を与えるタンパク質を産生し、したがって選択レジメンを生き延びる。このような優性選択の例は、薬物(ネオマイシン、ミコフェノール酸およびハイグロマイシン)を使用する。
【0087】
哺乳動物細胞のための適切な選択マーカーの別の例は、抗体核酸(例えば、DHFR、チミジンキナーゼ、メタロチオネイン−Iおよび−II、好ましくは、霊長類メタロチオネイン遺伝子、アデノシンデアミナーゼ、オルニチンデカルボキシラーゼなど)を取り込む能力のある細胞の同定を可能にするものである。
【0088】
例えば、DHFR選択遺伝子によって形質転換された細胞は、全ての形質転換体をメトトレキサート(Mtx)(DHFRの競合的アンタゴニスト)を含む培養培地中で培養することによって、最初に同定される。野生型DHFRが利用される場合、適切な宿主細胞は、DHFR活性が欠損したチャイニーズハムスター卵巣(CHO)細胞株である。
【0089】
あるいは、抗体、野生型DHFRタンパク質およびアミノグリコシド3’−ホスホトランスフェラーゼ(APH)のような別の選択マーカーをコードするDNA配列によって形質転換または同時形質転換された宿主細胞(特に、内因性DHFRを含む野生型宿主)は、上記選択マーカーに対する選択因子(例えば、アミノグリコシド系抗生物質(例えば、カナマイシン、ネオマイシンまたはG418(米国特許第4,965,199号)))を含む培地中での細胞増殖によって選択され得る。
【0090】
酵母中での使用のための適切な選択遺伝子は、酵母プラスミドYrp7に存在するtrp1遺伝子である(Stinchcombら,Nature 282:39(1979))。trp1遺伝子は、トリプトファン中で増殖する能力を欠く酵母の改変体系統(例えば、ATCC No.44076もしくはPEP4−1)に対する選択マーカーを提供する(Jones,Genetics 85:12(1977))。次いで、酵母宿主細胞ゲノムにおけるtrp1障害の存在は、トリプトファンの非存在下での増殖によって形質転換を検出するための有効な環境を提供する。同様に、Leu2−欠損性酵母系統(ATCC 20,622もしくは38,626)は、Leu2遺伝子を保有する公知のプラスミドによって補完される。
【0091】
(D.プロモーター構成要素)
発現ベクターおよびクローニングベクターは、通常、宿主生物体によって認識され、かつ抗体核酸に作動可能に連結されたプロモーターを含む。原核生物宿主とともに使用するために適切なプロモーターとしては、phoAプロモーター系、β−ラクタマーゼプロモーター系およびラクトースプロモーター系、アルカリホスファターゼプロモーター系、トリプトファン(trp)プロモーター系、ならびにハイブリッドプロモーター(例えば、tacプロモーター)が挙げられる。しかし、他の公知の細菌プロモーターが適切である。細菌系において使用するためのプロモーターはまた、抗体をコードするDNAに作動可能に連結された、シャイン−ダルガーノ(S.D.)配列を含む。
【0092】
プロモーター配列は、真核生物に関して知られている。実質的に全ての真核生物遺伝子は、転写が開始される部位から約25〜30塩基上流に位置するATリッチな領域を有する。多くの遺伝子の転写の開始部から70〜80塩基上流に見出される別の配列は、CNCAAT領域である(ここで、Nは、任意のヌクレオチドであり得る)。ほとんどの真核生物遺伝子の3’末端には、コード配列の3’末端へのポリAテイルの付加のためのシグナルであり得るAATAAA配列がある。これらの配列の全ては、真核生物発現ベクター中に適切に挿入される。
【0093】
酵母宿主とともに使用するための適切なプロモーター配列の例としては、3−ホスホグリセリン酸キナーゼまたは他の糖分解酵素(例えば、エノラーゼ、グリセルアルデヒド3−リン酸デヒドロゲナーゼ、ヘキソキナーゼ、ピルビン酸デカルボキシラーゼ、ホスホフルクトキナーゼ、グルコース6−リン酸イソメラーゼ、3−ホスホグリセリン酸ムターゼ、ピルビン酸キナーゼ、トリオースリン酸イソメラーゼ、グルコースリン酸イソメラーゼおよびグルコキナーゼ)に対するプロモーターが挙げられる。
【0094】
他の酵母プロモーター(増殖条件によって制御される転写というさらなる利点を有する誘導プロモーター)は、アルコールデヒドロゲナーゼ2、イソシトクロムC、酸性ホスファターゼ、窒素代謝に関連する分解酵素、メタロチオネイン、グルセルアルデヒド3−リン酸デヒドロゲナーゼ、ならびにマルトース利用およびガラクトース利用を担う酵素に対するプロモーター領域である。酵母発現において使用するための適切なベクターおよびプロモーターは、EP 73,657にさらに記載されている。酵母エンハンサーもまた、酵母プロモーターとともに有利に使用される。
【0095】
哺乳動物宿主細胞におけるベクターからの抗体転写は、例えば、ウイルス(例えば、ポリオーマウイルス、トリポックスウイルス、アデノウイルス2のようなアデノウイルス、ウシパピローマウイルス、トリ肉腫ウイルス、サイトメガロウイルス、レトロウイルス、B型肝炎ウイルス、および最も好ましくは、シミアンウイルス40(SV40))のゲノムから得られたプロモーター、異種哺乳動物プロモーター(例えば、アクチンプロモーターまたは免疫グロブリンプロモーター)、熱ショックプロモーターによって制御される(ただし、このようなプロモーターが宿主細胞系と適合することを条件とする)。
【0096】
SV40ウイルスの初期プロモーターおよび後期プロモーターは、SV40ウイルス複製起点もまた含むSV40制限フラグメントとして、都合よく得られる。ヒトサイトメガロウイルスの極初期プロモーターは、HindIII E制限フラグメントとして都合よく得られる。ウシパピローマウイルスをベクターとして使用して、哺乳動物宿主においてDNAを発現するための系は、米国特許第4,419,446号に開示されている。この系の改変形態は、米国特許第4,601,978号に記載されている。あるいは、ヒトβ−インターフェロンcDNAは、マウス細胞において、単純ヘルペスウイルス由来のチミジンキナーゼプロモーターの制御下で発現された。あるいは、ラウス肉腫ウイルスの長い末端反復(long terminal repeat)が、プロモーターとして使用され得る。
【0097】
(E.エンハンサーエレメント構成要素)
高等真核生物による本発明の抗体をコードするDNAの転写は、多くの場合、ベクター中にエンハンサー配列を挿入することによって増大する。多くのエンハンサー配列が、現在、哺乳動物遺伝子から知られている(グロブリン、エラスターゼ、アルブミン、α−フェトプロテイン、およびインシュリン)。しかし代表的には、真核生物細胞ウイルス由来のエンハンサーを使用する。例としては、複製起点の後期側におけるSV40エンハンサー(bp100〜270)、サイトメガロウイルス初期プロモーターエンハンサー、複製起点の後期側におけるポリオーマエンハンサー、およびアデノウイルスエンハンサーが挙げられる。真核生物プロモーターの活性化のためのエンハンサーエレメントに関する、Yaniv,Nature 297:17−18(1982)もまた参照のこと。エンハンサーは、抗体をコードする配列に対して5’位置または3’位置において、ベクター中にスプライシングされ得るが、好ましくはプロモーターの5’部位に位置する。
【0098】
(F.転写終止構成要素)
真核生物宿主細胞(酵母、真菌、昆虫、植物、動物、ヒト、もしくは他の多細胞生物由来の有核細胞)において使用される発現ベクターはまた、転写の終止のためおよびmRNAを安定化するために必要な配列を含み得る。このような配列は、一般的に、真核生物またはウイルスのDNAもしくはcDNAの、5’非翻訳領域から、そしてときには3’非翻訳領域から得られる。これらの領域は、抗体をコードするmRNAの非翻訳部分においてポリアデニル化フラグメントとして転写される、ヌクレオチドセグメントを含む。1つの有用な転写終止構成要素は、ウシ成長ホルモンポリアデニル化領域である。例えば、国際公開番号94/11026を参照のこと。
【0099】
(宿主細胞の選択および形質転換)
本明細書中に記載のベクターにDNAをクローニングまたは発現させるのに適した宿主細胞は、原核生物、酵母または高等真核生物の細胞である。この目的に適した原核生物としては、グラム陰性生物およびグラム陽性生物(例えば、E.coli、Enterobacter属、エルウィニア属、Klebsiella属、Proteus属、サルモネラ属、Serratia属およびShigella属などの腸内細菌、ならびにバチルス属、シュードモナス属およびストレプトミセス属)の両方が挙げられる。一つの好ましいE.coliクローニング宿主は、E.coli294(ATCC 31,446)であるが、E.coli B、E.coli X1776(ATCC 31,537)およびE.coli W3110(ATCC 27,325)のような他の菌株も適している。これらの例は、限定的ではなく、むしろ例示的である。
【0100】
原核生物に加えて真核生物微生物(例えば、糸状菌または酵母)は、抗体をコードするベクターのための適したクローニング宿主または発現宿主である。Saccharomyces cerevisiaeは、下等真核宿主微生物の内で最も一般的に使用される。しかし、例えば、Schizosaccharomyces pombe;Kluyveromyces属;カンジダ属;Trichoderma属;Neurospora crassaおよび糸状菌(例えば、アカパンカビ属、ペニシリウム属、イトクズグサ属およびコウジカビ属(例えば、A.nidulansおよびA.niger)の宿主などの他の多数の属、種および菌株が、本明細書中で一般的に利用可能であり、そして有用である。
【0101】
グリコシル化抗体の発現のために適した宿主細胞は、多細胞生物に由来する。第一に、脊椎動物培養または無脊椎動物培養に由来しようと、任意の高等真核細胞培養が機能し得る。無脊椎動物細胞の例としては、植物細胞および昆虫細胞が挙げられる(Luckowら、Bio/Technology 6、47〜55(1988);Millerら、Genetic Engineering、Setlowら編、第8巻、p.277−279(Plenam publishing 1986);Msedaら、Nature 第315巻、592〜594(1985))。多数のバキュロウイルス株および改変体ならびに宿主由来の対応する許容される昆虫宿主細胞(例えば、Spodoptera frugiperda(毛虫)、ヤブカ属(蚊)、Drosophila melanogaster(ショウジョウバエ)およびBombyx mori)が同定されている。トランスフェクションのための種々のウイルス株(例えば、Autographa californica NPVのL−1改変体、Bombyx mori NPVのBm−5株)が、公けに利用可能であり、このようなウイルスが、本発明による本明細書中のウイルスとして、特にSpodoptera frugiperda細胞のトランスフェクションに使用され得る。さらに、綿、トウモロコシ、ジャガイモ、大豆、ペチュニア、トマトおよびタバコの植物細胞培養物もまた宿主として利用される。
【0102】
脊椎動物細胞および培養(組織培養)における脊椎動物細胞の増殖は、慣例的になっている。Tissue Culture、Academic Press、KruseおよびPatterson編(1973)を参照のこと。有用な哺乳動物宿主細胞系統の例は、サル腎臓系統、ヒト胚腎臓系統、生まれたばかりのハムスターの腎臓細胞;チャイニーズハムスターの卵巣細胞/−DHFR(CHO、Urlaubら、Proc.Natl.Acad.Sci.USA 77:4216(1980);マウスセルトリ細胞;ヒト子宮頸部癌細胞(HELA);イヌ腎臓細胞;ヒト肺細胞;ヒト肝細胞;マウス乳腺癌;およびNS0細胞である。
【0103】
宿主細胞は、抗体産生のために上記ベクターで形質転換され、プロモーターを含むため、形質転換体を選択するため、または所望の配列をコードする遺伝子を増幅するために適切なものとして改変された通常の栄養培地で培養される。
【0104】
本発明の抗体改変体を産生するために使用される宿主細胞は、種々の培地で培養され得る。例えば、Ham’s F10(Sigma)、最小必須培地(MEM、Sigma)、RPMI−1640(Sigma)およびダルベッコ改変イーグル培地(DMEM、Sigma)などの市販の培地が、宿主細胞の培養に適している。さらに、Hamら、Meth.Enzymol.58:44(1979)、Barnesら、Anal.Biochem.102:255(1980)、米国特許第4,767,704号、同第4,657,866号;同第4,560,655号;同第5,122,469号;同第5,712,163号または同第6,048,728号に記載される任意の培地が、宿主細胞のための培養培地として使用され得る。これらの培地のいずれもが、必要な場合、ホルモンおよび/または他の増殖因子(例えば、インスリン、トランスフェリン、または表皮増殖因子)、塩(例えば、X塩化物、ここで、Xは、ナトリウム、カルシウム、マグネシウムである;およびリン酸塩)、緩衝液(例えば、HEPES)、ヌクレオチド(例えば、アデノシンおよびチミジン)、抗生物質(例えば、GENTAMYCIN.TM.薬物)、痕跡元素(通常マイクロモル範囲の最終濃度で存在する無機化合物として定義される)、およびグルコースまたは同等のエネルギー源を補充され得る。任意の他の必要な補充物はまた、当業者に公知である適切な濃度で含まれ得る。培養条件(例えば、温度、pHなど)は、発現のために選択された宿主細胞にこれまで用いられてきたものであり、当業者に明らかである。
【0105】
(抗体精製)
組換え技術を使用する場合、抗体改変体は、細胞内で産生され得るか、ペリプラズム空間で産生され得るか、または培地に直接分泌され得る。抗体改変体が、細胞内で産生される場合、第一の工程として、粒状の破片である、宿主細胞の断片または溶解した断片のいずれかは、例えば、遠心分離または限外濾過により取り除かれ得る。Carterら、Bio/Technology 10:163〜167(1992)は、E.coliのペリプラズム空間に分泌される抗体を単離するための手順を記述する。手短かには、細胞ペーストを、酢酸ナトリウム(pH3.5)、EDTAおよびフェニルメチルスルホニルフルオリド(PMSF)の存在下で約30分間にわたって溶解する。細胞の破片は、遠心分離によって取り除かれ得る。抗体改変体が、培地中に分泌される場合、そのような発現系由来の上清は、一般的には最初に市販のタンパク質濃縮フィルター(例えば、Amicon限外濾過ユニットまたはMillipore Pellicon限外濾過ユニット)を使用して濃縮される。タンパク質分解を阻害するために、前の工程のいずれかに例えば、PMSFのようなプロテアーゼインヒビターが含まれてもよく、外来性の夾雑物の増殖を防止するために抗生物質が含まれてもよい。
【0106】
細胞から調製された抗体組成物は、例えば、ヒドロキシアパタイトクロマトグラフィー、ゲル電気泳動、透析およびアフィニティークロマトグラフィーを使用して精製され得、アフィニティークロマトグラフィーが好ましい精製技術である。親和性リガンドとしてのプロテインAの適合性は、抗体改変体に存在するいずれかの免疫グロブリンFcドメインの種およびアイソタイプに依存する。プロテインAは、ヒトIgG1重鎖、ヒトIgG2重鎖またはヒトIgG4重鎖に基づく抗体を精製するために使用され得る(Lindmarkら、J.Immunol Meth.62:1〜13(1983))。プロテインGは、全てのマウスアイソタイプおよびヒトIgG3に対して推奨される(Gussら、EMBO J.5:1567〜1575(1986))。親和性リガンドが取り付けられるマトリクスは、多くの場合アガロースであるが、他のマトリクスも利用可能である。機械的に安定なマトリクス(例えば、調節された多孔ガラスまたはポリ(スチレンジビニル)ベンゼン)は、アガロースを用いて達成され得るより早い流速および短いプロセス時間を可能にする。抗体改変体が、CH3ドメインを含む場合、Bakerbond ABXTM樹脂(J.T.Baker、Phillipsburg,N.J.)が、精製に有用である。イオン交換カラムによる分画、エタノール沈殿、逆相HPLC、シリカによるクロマトグラフィー、アニオン交換樹脂もしくはカチオン交換樹脂(例えば、ポリアスパラギン酸カラム)上のヘパリンSEPHAROSETMクロマトグラフィーによるクロマトグラフィー、等電点電気泳動、SDS−PAGEおよび硫酸アンモニウム沈殿などのタンパク質精製のための技術がまた回収されるべき抗体改変体によって利用可能である。
【0107】
任意の予備精製工程の後、目的の抗体改変体および夾雑物を含む混合物は、約2.5〜4.5の間のpHで、好ましくは低塩濃度(例えば、約0〜0.25Mの塩)で、溶出緩衝液を使用して、低pHの疎水性相互作用クロマトグラフィーに供され得る。
【0108】
(薬学的処方物)
ポリペプチドまたは抗体の治療処方物は、凍結乾燥処方物または水溶液として保存するために、所望の程度の精製度を有するポリペプチドを、当該分野で代表的に利用される任意の「薬学的に受容可能な」キャリア、賦形剤または安定化剤(これらは全て「賦形剤」と称される)と混合することにより調製され得る。例えば、緩衝剤、安定化剤、保存剤、等張化剤、非イオン性洗浄剤、抗酸化剤および他の種々の添加剤である(Remington’s Pharmaceutical Sciences、第16版、A.Osol編(1980)を参照のこと)。このような添加剤は、用いられる投薬量および濃度では、レシピエントに対して無毒性でなければならない。
【0109】
緩衝剤は、生理学的条件に近い範囲にpHを維持するのを補助する。それらは、好ましくは約2mM〜約50mMの範囲の濃度で存在する。本発明に使用するための適した緩衝剤としては、有機酸とその塩および無機酸とその塩の両方が挙げられ、例えば、クエン酸緩衝液(例えば、クエン酸一ナトリウム−クエン酸二ナトリウムの混合物、クエン酸三ナトリウムの混合物、クエン酸−クエン酸一ナトリウムの混合物など)、コハク酸緩衝液(例えば、コハク酸−コハク酸一ナトリウムの混合物、コハク酸−水酸化ナトリウムの混合物、コハク酸−コハク酸二ナトリウムの混合物など)、酒石酸緩衝液(例えば、酒石酸−酒石酸ナトリウムの混合物、酒石酸−酒石酸カリウムの混合物、酒石酸−水酸化ナトリウムの混合物など)、フマル酸緩衝液(例えば、フマル酸−フマル酸一ナトリウムの混合物など)、フマル酸緩衝液(例えば、フマル酸−フマル酸一ナトリウムの混合物、フマル酸−フマル酸二ナトリウムの混合物、フマル酸一ナトリウム−フマル酸二ナトリウムの混合物など)、グルコン酸緩衝液(例えば、グルコン酸−グルコン酸ナトリウム混合物、グルコン酸−水酸化ナトリウム混合物、グルコン酸−グルコン酸カリウム混合物など)、シュウ酸緩衝液(例えば、シュウ酸−シュウ酸ナトリウム混合物、シュウ酸−水酸化ナトリウム混合物、シュウ酸−シュウ酸カリウム混合物など)、乳酸緩衝液(例えば、乳酸−乳酸ナトリウム混合物、乳酸−水酸化ナトリウム混合物、乳酸−乳酸カリウム混合物など)および酢酸緩衝液(例えば、酢酸−酢酸ナトリウム混合物、酢酸−水酸化ナトリウム混合物など)が挙げられる。さらに、リン酸緩衝液、ヒスチジン緩衝液およびトリメチルアミン塩(例えば、Tris)が言及され得る。
【0110】
保存剤は、微生物の増殖を妨げるために添加され得、0.2%〜1%(w/v)の範囲の量で添加され得る。本発明に使用するための適切な保存剤としては、フェノール、ベンジルアルコール、メタ−クレゾール、メチルパラベン、プロピルパラベン、オクタデシルジメチルベンジルアンモニウムクロライド、ベンザルコニウムハロゲン化物(例えば、塩化物、臭化物、ヨウ化物)、塩化ヘキサメトニウム、アルキルパラベン(例えば、メチルパラベンまたはプロピルパラベン)、カテコール、レゾルシノール、シクロヘキサノールおよび3−ペンタノールが挙げられる。
【0111】
「安定化剤」として公知であることもある等張化剤は、本発明の液体組成物の等張性を確実にするために添加され得、多価糖アルコール、好ましくは三価アルコールまたはそれより多価の糖アルコール(例えば、グリセリン、エリスリトール、アラビトール、キシリトール、ソルビトールおよびマンニトール)が挙げられる。
【0112】
安定化剤は、治療剤を溶解させるか、または変性もしくは容器の壁への付着を防止するのを補助する充填剤から添加剤の機能の範囲であり得る広い範疇の賦形剤をいう。代表的な安定化剤は、多価糖アルコール(上記で列挙した);アミノ酸(例えば、アルギニン、リジン、グリシン、グルタミン、アスパラギン、ヒスチジン、アラニン、オルニチン、L−ロイシン、2−フェニルアラニン、グルタミン酸、スレオニンなど)、有機糖または糖アルコール(例えば、ラクトース、トレハロース、スタキオース、マンニトール、ソルビトール、キシリトール、リビトール、ミオイノシトール、ガラクティトール、グリセロールなど、イノシトールなどのシクリトールを含む);ポリエチレングリコール;アミノ酸ポリマー;硫黄含有還元剤(例えば、尿素、グルタチオン、チオクト酸、チオグリコール酸ナトリウム、チオグリセロール、α−モノチオグリセロールおよびチオ硫酸ナトリウム);低分子量ポリペプチド(すなわち、10残基未満);タンパク質(例えば、ヒト血清アルブミン、ウシ血清アルブミン、ゼラチンまたは免疫グロブリン)、親水性ポリマー(例えば、ポリビニルピロリドン)、単糖(例えば、キシロース、マンノース、フルクトース、グルコース);二糖(例えば、ラクトース、マルトース、スクロース)および三糖(例えば、ラフィノース);多糖(例えば、デキストラン)であり得る。安定化剤は、活性タンパク質の重量部あたり0.1〜10,000の重量の範囲で存在し得る。
【0113】
非イオン性界面活性剤または洗浄剤(「湿潤剤」としても公知である)は、治療剤の可溶化を補助するため、および治療タンパク質を攪拌に誘導される凝集に対して保護するために添加され得、また処方物が、タンパク質の変性を引き起こすことなく、圧力を加えた剪断表面に露出されることを可能にする。適切な非イオン性界面活性剤としては、ポリソルベート(20、80など)、ポロキサマー(184、188など)、Pluronic(登録商標)ポリオール、ポリオキシエチレンソルビタンモノエーテル(TWEEN(登録商標)−20、TWEEN(登録商標)−80など)が挙げられる。非イオン性界面活性剤は、約0.05mg/ml〜約1.0mg/mlの範囲、好ましくは約0.07mg/ml〜約0.2mg/mlの範囲で存在し得る。
【0114】
さらなる種々の賦形剤としては、充填剤(例えば、デンプン)、キレート剤(例えば、EDTA)、抗酸化剤(例えば、アスコルビン酸、メチオニン、ビタミンE)および共溶媒が挙げられる。本明細書中の処方物はまた、必要な場合、処置される特定の適応症に必要な一種より多い活性化合物、好ましくは互いに不利に影響しない相補的活性を有する化合物を含有し得る。例えば、さらに免疫抑制剤を提供することが望ましくあり得る。このような分子は、意図される目的に対して有効である量で、組合せて適切に存在する。活性成分はまた、例えば、コアセルベーション技術により、または例えばヒドロキシメチルセルロースまたはゼラチンマイクロカプセルおよびポリ(メチルメタクリレート)マイクロカプセルそれぞれのコロイド状薬物送達系(例えば、リポソーム、アルブミンマイクロフェア、マイクロエマルジョン、ナノ粒子およびナノカプセル)またはマクロエマルジョン)中での界面のポリマー化により調製されたマイクロカプセルに閉じ込められ得る。このような技術は、Remington’s Pharmaeutical Sciences、第16版、A.Osal編(1980)に開示される。
【0115】
インビボ投与のために使用される処方物は、無菌性でなければならない。このことは、例えば、滅菌濾過膜を通す濾過によって容易に達成される。徐放調製物が調製され得る。徐放調製物の適切な例としては、抗体改変体を含む固体疎水性ポリマーの半透性マトリクスが挙げられ、このマトリクスは、成形した物品(例えば、フィルムまたはマイクロカプセル)の形態である。徐放マトリクスの例としては、ポリエステル、ヒドロゲル(例えば、ポリ(2−ヒドロキシエチルメタクリレート)またはポリ(ビニルアルコール)、ポリラクチド(米国特許第3,773,919号)、L−グルタミン酸とエチル−L−グルタメートのコポリマー、非分解酢酸ビニルエチレン、分解可能乳酸−グリコール酸コポリマー(例えば、LUPRON DEPOTTM(乳酸−グリコール酸のコポリマーと酢酸ロイプロリドからなる注射可能マイクロスフェア)およびポリ−D−(−)−3−ヒドロキシ酪酸が挙げられる。例えば、酢酸ビニルエチレンと乳酸−グリコール酸ポリマーは、100日間に亘って分子の放出を可能にし、特定のヒドロゲルは、より短い期間タンパク質を放出する。封じ込められた抗体が、体内に長い期間保持される場合、それらは、37℃にて湿気に曝露された結果として変性し得るか、または凝集し得、生物学的活性の喪失および免疫原性における可能な変化を生じる。合理的な方法は、関係する機構に依存して安定化のために、工夫され得る。例えば、凝集機構が、チオ−ジスルフィド交換を介した細胞間S−S結合形成であることが発見される場合、安定化は、スルフヒドリル残基を改変することによって、酸性溶液から凍結乾燥することによって、含水量を制御することによって、適切な添加剤を使用することによって、そして、特定のポリマーマトリクス組成物を開発することによって達成され得る。
【0116】
特定の障害または状態の処置に有効である治療ポリペプチド、抗体またはそのフラグメントの量は、障害もしくは状態の性質に依存し、そして、標準的な臨床技術により決定され得る。可能な場合、用量−応答曲線を決定し、ヒトで試験する前に、最初にインビトロで、次いで有用な動物モデル系で本発明の薬学的組成物を決定することが望ましい。
【0117】
好ましい実施形態において、治療ポリペプチド、抗体またはそのフラグメントの水溶液は、皮下注射によって投与される。各用量は、体重1kgあたり約0.5μg〜約50μgの範囲であり得るか、またはより好ましくは、体重1kgあたり約3μg〜約30μgの範囲であり得る。
【0118】
皮下投与のための投薬計画は、多数の臨床的因子(疾患の型、疾患の重篤度、および治療剤に対する患者の感受性)に依存して一ヶ月に一回から一日一回に変動し得る。
【0119】
(抗体改変体の使用)
本発明の抗体改変体は、親和性精製因子として使用され得る。このプロセスにおいて、抗体は、当該分野で周知の方法を使用して、固相(例えば、SEPHADEXTM樹脂、またはろ紙)上に固定される。固定された抗体改変体は、精製されるべき標的を含むサンプルと接触され、その後、その支持体は、精製されるべき標的を除いてサンプル中の実質的に全ての物質を取り除く適した溶媒で洗浄される。その支持体は、固定された抗体改変体結合される。最後に、その支持体は、標的を抗体改変体から放出する別の適切な溶媒(例えば、グリシン緩衝液)で洗浄される。
【0120】
改変抗体はまた、診断アッセイ(例えば、特定の細胞、組織または血清における目的の標的の発現の検出)に有用であり得る。診断適用について、抗体改変体は、代表的に検出可能部分で標識される。多数の標識が、蛍光の変化を定量するために利用可能な技術であり、上に記した。化学発光基質は、化学反応により電気的に励起され、次いで、放射光は、(例えば、化学発光計を使用して)測定され得るか、または蛍光受容体にエネルギーを供与し得る。酵素標識の例としては、ルシフェラーゼ(例えば、ホタルルシフェラーゼおよび細菌ルシフェラーゼ;米国特許第4,737,456号)、ルシフェリン、2,3−ジヒドロフタラジンジオン、リンゴ酸デヒドロゲナーゼ、ウレアーゼ、ペルオキシダーゼ(例えば、ホースラディッシュペルオキシダーゼ(HPRO)、アルカリホスファターゼ、β−ガラクトシダーゼ、グルコアミラーゼ、リゾチーム、糖類オキシダーゼ(例えば、グルコースオキシダーゼ、ガラクトースオキシダーゼおよびグルコース6リン酸デヒドロゲナーゼ)、複素環式オキシダーゼ(例えば、ウリカーゼおよびキサンチンオキシダーゼ)、ラクトペルオキシダーゼ、ミクロペルオキシダーゼなどが挙げられる。酵素を抗体に結合するための技術は、Methods in Enzym.(J.LangoneおよびH.Van Vunakis編)、Academic press、New York、73:147〜166(1981)中のO’Sullivanら、Methods for the Preparation of Enzyme−Antibody Conjugates for Use in Enzyme Immunoassayに記載されている。
【0121】
標識は、抗体改変体と間接的に結合されることもある。当業者は、このことを達成するための種々の技術を知っている。例えば、抗体改変体は、ビオチンと結合され得、上記の3種の広範な範疇の標識のいずれかが、アビジンと結合され得、逆もまた同様である。ビオチンは、アビジンに選択的に結合し、従って標識は、この間接的な様式で抗体改変体と結合され得る。あるいは、標識の抗体改変体との間接的な結合を達成するために、抗体改変体は、小型のハプテン(例えば、ジグロキシン(digloxin))と結合され、上記の異なる型の標識の一つが、抗ハプテン抗体改変体(例えば、抗ジグロキシン抗体)と結合される。従って、標識の抗体改変体との間接的な結合は、達成され得る。
【0122】
本発明の別の実施形態において、抗体改変体は、標識される必要はなく、その存在は、抗体改変体に結合する標識された抗体を使用して検出され得る。
【0123】
本発明の抗体は、任意の公知のアッセイ方法(例えば、競合結合アッセイ、直接サンドイッチアッセイおよび間接サンドイッチアッセイならびに免疫沈降アッセイ)で使用され得る。Zola、Monoclonal Antibodies:A Manual of Techniques、pp.147〜158(CRC Press Inc.1987)。
【0124】
競合結合アッセイは、標識された標準が、限られた量の抗体改変体との結合について試験サンプルと競合する能力に依存する。試験サンプル中の標的の量は、抗体に結する標準の量に反比例する。結合する標準の量の決定を容易にするために、抗体は、一般的に、競合の前か後に、不溶化される。結果として、抗体に結合する標準サンプルおよび試験サンプルは、結合しないままである標準サンプルおよび試験サンプルから好都合に分離され得る。
【0125】
サンドイッチアッセイは、二つの抗体の使用を伴い、それぞれは、検出されるべき、異なる免疫原性部分またはエピトープまたはタンパク質に結合し得る。サンドイッチアッセイにおいて、分析されるべき試験サンプルは、固体支持体上に固定された一次抗体によって結合され、その後、二次抗体は、試験サンプルに結合し、それによって不溶性の3つの部分の複合体を形成する。例えば、米国特許第4,376,110号を参照のこと。二次抗体はそれ自体、検出可能な部分で標識され得る(直接サンドイッチアッセイ)か、または検出可能な部分で標識される抗免疫グロブリン抗体を使用して測定され得る(間接サンドイッチアッセイ)。例えば、一つの型のサンドイッチアッセイは、ELISAアッセイであり、その場合、検出可能な部分は、酵素である。
【0126】
免疫組織化学に関して、腫瘍サンプルは、新鮮であっても凍結されていてもよく、パラフィンに包埋されていても保存剤(例えばホルマリン)で固定されていてもよい。
【0127】
その抗体はまたインビボ診断アッセイに使用され得る。一般的に、抗体改変体は、放射性ヌクレオチド(例えば、111In、99Tc、14C、131I、3H、32Pまたは35S)で標識されて、その結果腫瘍は、免疫シンチグラフィを使用して局在化され得る。例えば、本発明の高親和性抗IgE抗体は、例えば、喘息患者の肺に存在するIgEの量を検出するために使用され得る。
【0128】
本発明の抗体は、キット、すなわち、診断アッセイを行なうための指示書を備える、所定量の試薬のパッケージングした組合せで提供され得る。抗体改変体が、酵素で標識される場合、そのキットは、酵素に必要とされる基質および補因子(例えば、検出可能な発色団またはフルオロフォアを提供する基質前駆体)を備え得る。さらに、他の添加剤(例えば、安定化剤、緩衝液(例えば、ブロック緩衝液または溶解緩衝液)など)が含まれ得る。種々の試薬の相対量は、アッセイの感度を実質的に最適化する試薬の溶液中の濃度を提供するために、大きく変動し得る。特に、試薬は、溶解の際に、適切な濃度を有する試薬溶液を提供する賦形剤を含む、乾燥粉末、通常は、凍結乾燥した粉末として提供され得る。
【0129】
(抗体のインビボ使用)
本発明の抗体は、哺乳動物を処置するために使用され得ることが企図される。一実施形態において、この抗体は、例えば、前臨床データを得る目的で、非ヒト哺乳動物に投与される。処置されるべき例示的な非ヒト哺乳動物としては、非ヒト霊長類、イヌ、ネコ、げっ歯類、および前臨床研究が行われる他の哺乳動物が挙げられる。このような哺乳動物は、この抗体を用いて処置されるべき疾患に対する確立された動物モデルであり得るか、または、目的の抗体の毒性を研究するために使用され得る。これらの実施形態のそれぞれにおいて、用量増大研究が、哺乳動物に対して行なわれ得る。この抗体またはポリペプチドは、任意の適切な手段で投与され、その手段としては、非経口投与、皮下投与、腹腔内投与、肺内投与、鼻腔内投与が挙げられ、局所的な免疫抑制処置について所望の場合、病巣内投与が挙げられる。非経口注入としては、筋肉内投与、静脈内投与、動脈内投与、腹腔内投与または皮下投与が挙げられる。さらに、抗体改変体は、パルス注入、特に低下用量の抗体改変体を用いたパルス注入により適切に投与される。好ましくは、この用量は、注射によって与えられ、最も好ましくは、投与が、短期であるか、長期であるかに一部依存して、静脈内注射または皮下注射によって与えられる。
【0130】
疾患の予防または処置のために、適切な投薬量の抗体またはポリペプチドは、処置されるべき疾患の型、疾患の重篤度および過程、抗体改変体が、予防目的または治療目的で投与されたか否か、これまでの治療、患者の臨床履歴および抗体改変体に対する応答、および主治医の裁量に依存する。本発明の非常に高い親和性の抗ヒトIgE抗体は、患者に、一時に、または一連の処置にわたって適切に投与され得る。疾患の型および重篤度に依存して、約0.1mg/kg〜150mg/kg(例えば、0.1〜20mg/kg)の抗体が、患者に投与するための初期候補投薬量であり、この量は例えば、一回以上の別個の投与であっても、連続注入であってもよい。代表的な毎日の投薬量は、上記の因子に依存して、約1mg/kg〜100mg/kgまたはそれ以上の範囲であり得る。数日以上に亘る繰り返しの投与については、その条件に依存して、その処置は、疾患症状の所望の抑制が生じるまで維持される。しかし、他の投薬レジメンも有用であり得る。この治療の進行は、従来の技術およびアッセイにより容易にモニタリングされる。抗LFA−1抗体または抗ICAM−1抗体についての例示的な用量レジメンは、国際公開番号94/04188に開示される。抗体改変体組成物は、良好な医療行為と一致した様式で処方され、投薬され、投与される。この文脈において、考慮のための因子としては、処置される特定の障害、処置される特定の哺乳動物、個々の患者の臨床状態、障害の原因、薬剤の送達の部位、投与の方法、投与のスケジュール、および医師に公知の他の因子が挙げられる。投与されるべき抗体改変体の「治療有効量」は、このような考慮すべき事項によって決定され、疾患または障害を予防するか、改善するか、または処置するために必要である最少量である。この抗体改変体は、必ずしもではないが、必要に応じて、問題の障害を予防または処置するために現在使用されている一種以上の薬剤と一緒に処方される。このような他の薬剤の有効量は、処方物中に存在する抗体の量、障害または処置の型、および上で考察した他の因子に依存する。これらは、一般的に同じ投薬量で、上で使用した投与経路で、またはこれまでに用いた投薬量の約1〜99%で使用される。
【0131】
IgEをその標的として認識する本発明の抗体は、「IgE媒介性障害」を処置するために、使用され得る。これらとしては、例えば、ぜん息、アレルギー性鼻炎およびアレルギー性結膜炎(枯草熱)、湿疹、じんま疹、アトピー性皮膚炎および食物アレルギーのような疾患が挙げられる。例えば、蜂に刺されること、蛇にかまれること、食物または薬物によって引き起こされるアナフィラキシーショックの深刻な生理学的状態もまた本発明の範囲に包含される。
【0132】
(抗体エピトープマッピング)
用語「エピトープ」とは、B細胞および/またはT細胞が抗原に対して応答する部位をいう。B細胞エピトープは、連続するアミノ酸またはタンパク質の三次元のフォールディングによって並列する連続しないアミノ酸の両方から形成され得る。連続するアミノ酸から形成されたエピトープは、代表的に、変性溶媒に曝露されて保持されるのに対して、三次元フォールディングによって形成されるエピトープは、代表的に変性溶媒を用いた処理に対して失われる。エピトープは、代表的に、少なくとも3個、より通常は、少なくとも5個または8〜10個のアミノ酸を、固有の空間コンフォメーションに含む。同一のエピトープを認識する抗体は、ある抗体の、別の抗体の標的抗原への結合をブロックする能力を示す単純な免疫学的検定において同定され得る。
【0133】
本発明の高親和性抗体IgEに対する結合部位のエピトープマッピングは、結合についてのウエスタンブロット分析、IgEのCH3ドメインのペプチドスキャン、結合を示した領域のアラニン走査、IgG1の対応する領域からのアミノ酸置換および特定部位の変異誘発を含む。
【0134】
IgEの全CH3ドメインのペプチドスキャンは、73個の重複するペプチドを必要とした。各ペプチドを、本発明の標識した抗IgE抗体による結合に供し、IgEの、その高親和性レセプターへの結合をブロックするIgEの特異的なエピトープを決定した。ペプチドスキャンは、二つのペプチドを、IgE上の潜在的な抗IgE MAb接触部位と同定し、エピトープ「A」およびエピトープ「B」と名付けた(図11を参照のこと)。エピトープ「A」配列およびエピトープ「B」配列は、直鎖状配列で約80アミノ酸離れているが、IgEの三次元構造では互いに近くに位置している。両方とも表面に露出されていて、それらは、IgEのFcεRI結合部位に重複し、両方ともペプチドであり、Argの正に荷電した残基およびProの疎水性残基が存在する。図12は、ペプチドスキャンを使用してELISAにより決定したエピトープBの結合領域を図示する。
【0135】
どのアミノ酸残基が、これらのエピトープ内での高親和性抗体の結合に重要であるかの決定を、アラニンスキャニング変異誘発によって行なった(Cunninghamら、「High−Resolution Epitope Mapping of hGH−Receptor Interactions by Alanine−Scanning Mutagenesis」、Science、第244巻、p.1081〜1085)。エピトープAおよびエピトープBの各残基の代わりに、アラニンを用い、高親和性モノクローナル抗体の結合を決定した(実施例12の下ならびに図13および図14を参照のこと)。
【0136】
(能動免疫および受動免疫)
本発明はまた、薬学的組成物(例えば、配列番号72および/または配列番号74を含む本発明のペプチド免疫源分子、ならびに希釈剤、賦形剤、アジュバントまたはキャリアを含有するワクチン)に関する。本発明は、さらに、本発明の少なくとも一つのペプチドを、そのペプチドに対する免疫応答を誘発し得る部分と共有結合させる工程を包含する、本発明の免疫源の調製のためのプロセスに関する。
【0137】
本発明はまた、例えば、IgE媒介性疾患またはIgE媒介性状態(例えば、アレルギーおよびアトピー性皮膚炎)の処置における医薬として使用するための上に定義された免疫原性ペプチドに関する。
【0138】
本発明はさらに、上に定義する免疫原性ペプチドの治療有効量を、そのような処置を必要とする患者に投与する工程を包含する、IgE媒介性疾患またはIgE媒介性状態(例えば、アレルギーおよびアトピー性皮膚炎)に対して哺乳動物を免疫状態にする方法に関する。
【0139】
本発明の免疫原性ペプチドは、非細胞溶解性のヒスタミン放出を実質的に媒介し得ないが、エピトープAおよび/またはエピトープBの標的アミノ酸配列との血清学的な強い交差反応性を抗体に誘発し得る。
【0140】
ペプチドの初期用量(例えば、約0.2mg〜約5mg)が、例えば、筋肉内に投与され得、その後14日後〜28日後に、同じ用量で繰り返し投与される(追加抗原)。用量は、当然のことながらある程度は、患者の年齢、体重、全体的な健康に依存する。免疫処置は、「能動的」または「受動的」であり得る。「能動」免疫において、被験体は、本発明の免疫原性ペプチドを受容し、抗IgE応答が、被験体の免疫系によって能動的に誘導される。
【0141】
「能動」免疫、これは、ヒト使用について好ましいが、他の哺乳動物種(例えば、イヌ)も同様に処置され得る。用語「免疫原性キャリア」は、本明細書中では、宿主動物内で免疫原性応答を独立に誘発する特性を有し、ポリペプチドおよび免疫原性キャリア物質の対応する基の中の遊離のカルボキシル基、アミノ基またはヒドロキシル基の間のペプチド結合またはエステル結合の形成を介して、または、従来の二官能性連結基を介した結合、または融合タンパク質として直接ポリペプチドに共有結合し得る物質を包含する。
【0142】
このような免疫原性キャリアの例としては、以下:アルブミン(例えば、BSA);グロブリン;サイログロブリン;ヘモグロブリン;ヘモシアニン(特に、キーホールリンペットヘモシアニン(KLH));回虫属から抽出したタンパク質(例えば、J.Immunol.、111巻、(1973)260〜268、J.Immunol.、122巻、(1979)302〜308、J.Immun.、98巻、(1967)893〜900、Am.J.Physiol.、199巻、(1960)575〜578に記載される回虫属抽出物またはその精製産物);ポリリシン;ポリグルタミン酸;リシン−グルタミン酸コポリマー;リシンまたはオルニチンなどを含むコポリマーなどが挙げられる。ワクチンは、ジフテリアトキソイドまたは破傷風トキソイドを免疫原性キャリア物質として使用して産生され(Lepow M.L.ら、J.of Infectious Diseases、第150巻、(1984)402〜406;Coen Beuvery,E.ら、Infection and Immunity、第40巻、(1983)39〜45)、これらのトキソイド物質はまた、本発明に使用され得る。ツベルクリンの精製されたタンパク質誘導体(PPD)は、(1)それ自体ではT細胞応答を誘導せず(すなわち、それは、事実上「T細胞ハプテン」である)、なお十分にプロセスされた抗原として挙動し、従ってT細胞により認識される;(2)連結した認識様式では、最も強力なハプテン「キャリア」の一つであることが公知である;そして(3)さらなる試験を行なわずに、ヒトにおいて使用され得るので、「能動」免疫スキームにおける利用に特に好ましい。
【0143】
本発明はまた、本発明のペプチドをコードするポリヌクレオチド、このポリヌクレオチドを含むベクターおよびこのベクターを有する細胞に関する。さらに、能動免疫は、本発明のペプチドをコードするポリヌクレオチドを投与することにより達成され得る。このような治療に適したベクターは、当該分野で公知であり、例えば、アデノウイルスベクターが挙げられる。
【0144】
「受動」免疫は、本発明の抗IgE抗体を、IgE媒介性疾患またはIgE媒介性状態を罹患する患者に投与することによって達成される。これらの抗体は、本発明の免疫原性ペプチドを非ヒト哺乳動物に投与し、生じた抗血清を収集することにより調製され得る。改善された力価は、長期間にわたる繰り返しの注射によって獲得され得る。抗体を誘発するために使用され得る哺乳動物の種に特に制限はなく、一般的には、ウサギまたはモルモットを使用することが好ましいが、ウマ、ネコ、イヌ、ヤギ、ブタ、ラット、ウシ、ヒツジなども使用され得る。抗体は、最後の投与の後、1〜2週間の経過後、免疫状態にした動物から血液を収集し、血液を遠心分離し、血清を血液から単離することにより回収される。モノクローナル抗体は、例えば、ヒトまたはマウスのものであり得る。
【0145】
被験体を免疫状態にする場合、本発明の抗体は、哺乳動物に、例えば、筋肉内注射により導入され得る。しかし、抗体投与の任意の形態が使用され得る。被験体に許容され、有害な副作用を有さない任意の従来の液体または固体のビヒクルが、使用され得る。生理学的pH(例えば、pH約6.8〜7.2、好ましくはpH約7.0)のリン酸緩衝生理食塩水(PBS)が、それのみでビヒクルとして、または例えば、水酸化アルミニウムベースのアジュバントなどの適切なアジュバントと一緒にビヒクルとして使用され得る。
【0146】
以下の実施例は、限定の目的で提供されるのではなく、例示の目的で提供される。
【実施例】
【0147】
(実施例1)
(抗IgEマウスMAb TES−C21のヒト化)
マウスmAb TES−C21の重鎖可変領域(VH)および軽鎖可変領域(VL)の配列を、公開されたデータベースで利用可能なヒト抗体生殖細胞系配列と比較した。上記工程1に記載されるテンプレートを決定する場合、いくつかの基準を用い、これらの基準としては、全長、フレームワーク内の類似したCDR位置、全体の相同性、CDRの大きさなどが挙げられる。これらの基準は全てひとまとめにして、TES−C21MAb重鎖配列とTES−C21MAb軽鎖配列との間の配列アラインメント、ならびに図3Aおよび図3Bに示されるそれぞれのヒトテンプレート配列に示される最適なヒトテンプレートを選択するための結果を提供する。
【0148】
この場合、この抗体を設計するために、一種より多いヒトフレームワークテンプレートを使用した。VH鎖に対して選択されたヒトテンプレートは、DP88(アミノ酸残基1〜95)とJH4b(アミノ酸残基103〜113)との組合せであった(図3Bを参照のこと)。VL鎖に対して選択されたヒトテンプレートは、JK4(アミノ酸残基98〜107)と合わせたL16(VKサブグループIII、アミノ酸残基1〜87)の組合せであった(図3Aを参照のこと)。マウス配列とヒトテンプレートとの間のフレームワーク相同性は、VHに関して約70%、VLに関して約74%であった。
【0149】
一旦テンプレートを選択すると、DNA合成、および上に記載され、そして図2に示されるオーバーラッピングPCRによりFabライブラリーを構築した。このライブラリーは、それぞれ選択されたヒトテンプレートであるDP88/JH4bおよびL16/JK4を用いて合成した合成TES−C21 CDRからなった。このライブラリーの複雑さは、4096(=212)であった。部分的なVH配列およびVL配列をコードする重複するヌクレオチドを、18〜21のヌクレオチドの重複を有する約63〜約76のヌクレオチドの範囲で合成した。
【0150】
VL遺伝子およびVH遺伝子のPCR増幅を、フレームワーク領域FR1に対する特異的配列を含むビオチン化フォワードプライマーおよびリーダー配列(遺伝子III)の末端にアニーリングした突出配列および保存された定常領域(CKまたはCH1)由来のリバースプライマーを使用して、標準PCR条件下で行なった。PCR産物を、アガロースゲル電気泳動、または市販のPCR精製キットにより精製し、取り込まれなかったビオチン化プライマーおよび非特異的PCR産物を取り除いた。
【0151】
PCR産物の5’リン酸化を、2μgのPCR産物、1μLのT4ポリヌクレオチドキナーゼ(10ユニット/μL)、2μLの10×PNK緩衝液、1μLの10mM ATPを用いて、ddH2Oで、総体積20μLに調節して行なった。37℃にて45分間インキュベートした後、65℃にて10分間加熱変性し、次の工程のために、ddH2Oを添加して反応体積を200μLに調節した。
【0152】
100μLのストレプトアビジン被覆磁気ビーズを、200μLの2×B&W緩衝液で2度洗浄し、200μLの2×B&W緩衝液に再懸濁した。リン酸化PCR産物を、ビーズと混合し、室温(RT)で穏やかに振盪しながら16分間インキュベートした。
【0153】
このビーズを、沈降させ、200μLの2×B&W緩衝液で2度洗浄した。非ビオチン化ssDNA(マイナス鎖)を、300μLの新鮮に調製した0.15MのNaOHで、室温で穏やかに振盪しながら10分間溶出した。第二のNaOH溶出は、収率をわずかに増加させ得る(任意)。溶離液を遠心分離して、微量のビーズを完全に取り除いた。
【0154】
ssDNAを、1μLグリコゲン(10mg/mL)、1/10体積の3M NaOAc(pH5.2)および2.5体積のEtOHを添加することにより、上清から沈殿させた。次いで、沈殿したssDNAを、70% EtOHで洗浄し、その後、3分間凍結乾燥し、20μLのddH2Oに溶解した。ssDNAを、エチジウムブロマイド(EtBr)アガロースプレート上に、DNA標準と一緒にスポッティングすることにより定量するか、またはOD260を測定することにより定量した。
【0155】
(実施例2)
(VHおよびVLのファージ発現ベクターへのクローニング)
VHおよびVLをハイブリダイゼーション変異誘発により、ファージ発現ベクターにクローニングした。ウリジン化テンプレートを、CJ236 E.coli株(dut「ung」)を、M13ベースのファージで感染させることにより調製した(ファージ発現ベクターTN003)。
【0156】
以下の成分(200ngのウリジン化ファージベクター(8.49kb);92ngのリン酸化一本鎖H鎖(489塩基);100ngのリン酸化一本鎖L鎖(525塩基);1μLの10×アニーリング緩衝液;ddH2Oで体積を10μlに調整)を、85℃の温度で5分間PCR保持(変性)し、次いで、1時間かけて55℃に低下させることによって(ベクターに対して約8倍のモル比の挿入物において)アニーリングさせた。そのサンプルを氷上で冷却した。
【0157】
アニーリングさせた産物に、以下の成分:1.4μLの10×合成緩衝液;0.5μLのT4 DNAリガーゼ(1ユニット/μL);1μL T4 DNAポリメラーゼ(1ユニット/μL)を添加し、その後、氷上で5分間インキュベートし、37℃で1.5時間インキュベートした。次いで、その産物をエタノール沈殿し、10μLのddH2OまたはTEに溶解させた。DNAを、1μL のXbaI(10ユニット/μL)で2時間消化し、65℃で20分間加熱不活性化した。消化したDNAを、エレクトロポレーションにより50μLのエレクトロコンピーテントDH10B細胞にトランスフェクトした。生じたファージを、37℃で一晩、XL−1 Blue細菌ローン(bacterial lawn)上で増殖させることにより、力価を測った。クローンを、組成を確認するために、配列決定した。
【0158】
(実施例3)
(ライブラリースクリーニングのためのディープウェルでの培養)
(A.ファージライブラリーのプレーティング)
ファージライブラリーを、LB培地に希釈し、プレートあたりの所望のプラーク数を達成した。高力価のファージを、200μLのXL−1B細胞培養物と混合した。3mLのLBの表面の寒天を混合し、LBプレートに注ぎ、室温で10分間そのままにしておいた。そのプレートを、37℃にて、一晩インキュベートした。
【0159】
(B.ファージ溶出)
100μLのファージ溶出緩衝液(10mM Tris−Cl、pH7.5、10mM EDTA、100mM NaCl)を滅菌U底96ウェルプレートの各ウェルに添加した。一晩後のライブラリープレート由来の単一のファージプラークを、フィルター処理したピペットチップを用いてウェルに移した。ファージ溶出プレートを、37℃にて1時間インキュベートした。このプレートをインキュベーションに続いて4℃にて保存し得る。
【0160】
(C.ディープウェルプレートでの培養)
50mLの培養物由来のXL1B細胞を、2×YT培地に、1:100希釈で添加した。その細胞を、A600が0.9〜1.2の間になるまで、シェーカー内で37℃にて増殖させた。
【0161】
(C.ディープウェルプレート内でのファージによる感染)
細胞が適切なODに到達したときに、1M IPTG(1:2000)をXL 1B培養物に添加した。IPTGの最終濃度は、0.5mMであった。750μLの細胞培養物を96ウェルのディープウェルプレート(Fisher Scientific)の各ウェルに移した。各ウェルに、25μLの溶出ファージを接種した。このディープウェルプレートをシェーカー(250rpm)内に配置し、37℃にて一晩インキュベートした。
【0162】
(D.ELISAスクリーニングのための上清の調製)
インキュベーション後、このディープウェルプレートを、Beckman JA−5.3プレートローターを使用して、3,250rpmにて20分間遠心した。ELISAのために、各ウェルから50μLの上清を抜き取った。
【0163】
(E.XL−1細胞の15mLの液体培養物の接種)
XL−1を、10μg/mLのテトラサイクリンを含有する2×YT中で、シェーカー内で(250rpm)、37℃にて、A600が0.9〜1.2になるまで増殖させた。IPTGを、0.5mMの最終濃度で添加し、15mLの培養物を50mLの円錐管に、各クローンが特徴付けられるように移した。細胞に、高力価ストック(力価=約1011pfu/mL)由来の10μLのファージを接種し、37℃にて1時間インキュベートした。この細胞を、室温で振盪しながら一晩増殖させた。
【0164】
(F.ペリプラズム由来の可溶性Fabの単離)
この細胞を、4,500rpmでの20分間のIEC遠心分離にて、ペレット化した。培養培地を取り除き、ペレットを、650μLの再懸濁緩衝液(1mMのEDTAおよび500mMのスクロースを含有する、50mM Tris、pH8.0)に再懸濁し、ボルテックスし、その後穏やかに振盪しながら1時間氷上に配置した。細胞の破片を、4℃で、9,000rpmで10分間遠心することにより取り除いた。可溶性Fabを含む上清を、収集し、そして4℃にて保存した。
【0165】
(実施例4)
(フレームワーク改変)
フレームワーク内の上記の潜在的に重要な位置に、12個のマウス/ヒトのゆらぎ残基が存在する。VHの73位は、ヒト化ライブラリーでは、マウス残基スレオニンとして維持されていた。なぜならこの位置は、結合に影響を及ぼすと確定されたからである。しかし、VH73のスレオニンが、ヒト生殖細胞系VHサブグループ1および2の共通のヒト残基であることが示された。
【0166】
TES−C21配列とヒトテンプレートとの間で異なるフレームワーク残基を、上記のように無作為に置換し、次いで、標的結合および抗体フォールディングに対するその潜在的な影響について評価した。結合に影響し得る潜在的なフレームワーク残基を同定した。この場合、それらは、VHの残基12、27、43、48、67、69であり、VLの残基1、3、4、49、60、85であった(Kabat記数法)(図4を参照のこと)。27位および69位のみが、VH領域(クローン番号1136−2C)の結合に顕著に影響することが、後に実証された。
【0167】
使用した一次スクリーニングは、培養培地を用いる一点ELISA(Single Point ELISA)(SPE)であった(以下の記載を参照のこと)。一次スクリーニングは、抗体の標的分子に結合するクローンを選択した。親分子と等しいか、またはそれより良好なシグナルを与えるクローンを、次のラウンドのスクリーニングのために選択した。
【0168】
第2のラウンドのスクリーニングにおいて、個々のファージを、15mlの細菌培養で増殖させ、ペリプラズム調製物を、SPE滴定アッセイおよびELISA滴定アッセイのために使用した。このアッセイにおいて、より高い結合を保持しているクローンをさらに特徴付けした。一旦全ての選択された一次クローンを、加工すると、上から10〜15%のクローンを配列決定し、そのクローンを配列に従って、配置した。各配列群由来の代表を、互いに比較し、最も良いクローンを選択した。これらの選択されたクローンの配列を合わせ、種々の組合せの効果を評価した。
【0169】
構築したライブラリーを、組換えヒトIgE、SE44に対する結合の改善についてELISAスクリーニングに供した。マウスTES−C21Fabより大きい結合親和性を有するクローンを単離し、配列決定した。クローンID番号4、49、72、76および136を、さらに特徴付けした。クローン4、49、72、78および136に対するELIZA滴定曲線を図5Aおよび図5Bに示すが、これは、その親和性が、その親であるTES−C21に類似していることを示唆する。これらのクローンは、ヒトIgEに対する結合について、マウスTES−C21と競合し、このことは、結合するエピトープが、ヒト化プロセスの間に変化しなかったことを示唆する。ヒト化Fabは、FcεRI結合IgEに結合せず、このことは、ヒト化抗体が、二価IgGに構築される場合にヒスタミン放出を引き起こすようにレセプターと架橋するのではなさそうであることを示唆する。ヒト化クローン136は、5個のマウス重鎖フレームワーク残基(=94.3%のヒトVHフレームワークホモロジー)を保持し、親和性成熟により選択された100%のヒト軽鎖フレームワークを有する。IgEのFcεRIへの結合のヒト化Fabによる阻害を実証した(図6)。
【0170】
(実施例5)
(抗IgEをスクリーニングするための一点ELISAプロトコル)
プレートを、炭酸塩被覆緩衝液中の2μg/mLのヒツジ抗ヒトFdで、4℃で一晩被覆した。被覆溶液を取り除き、そして、そのプレートを、ウェルあたり200μLの3% BSA/PBSで、37℃にて1時間ブロックした。プレートを、PBS/0.1% TWEEN(登録商標)(PBST)で4回洗浄した後、50μL/ウェルのFabサンプル(すなわち、高力価ファージおよびDMBブロックから分泌されたFabもしくはペリプラズム調製物、または15mLの調製物を含む上清)を添加した。プレートを室温で1時間インキュベートし、その後、PBSTで4回洗浄した。0.5% BSA/PBS中に0.015μg/mLで希釈した50μL/ウェルのビオチン化SE44および0.05%のTWEEN(登録商標)を添加した。次いで、プレートを室温で2時間インキュベートし、4×PBSTで洗浄した。0.5%のBSA/PBSおよび0.05%のTWEEN(登録商標)中に1:2000で希釈した50μL/ウェルのストレプトアビジンHRPを添加し、プレートを室温で1時間インキュベートした。プレートを、6×PBSTで洗浄した。50μL/ウェルのTMB 基質(Sigma)を添加して展開し、次いで、50μL/ウェルの0.2MのH2SO4を添加することにより停止させた。
【0171】
(実施例6)
(ELISA滴定:抗IgE)
プレートを、炭酸塩被覆緩衝液中の0.25μg/mL(精製したFab 0.1μg/ml)のSE44で4℃で一晩被覆した。被覆溶液を取り除き、このプレートを、200μL/ウェルの3%のBSA/PBSで、37℃にて1時間ブロックした。
【0172】
このプレートを、PBS/0.1%のTWEEN(登録商標)(PBST)で4回洗浄した。50μL/ウェルのFab(15mLのペリプラズム調製物由来)を添加して、1:2の希釈で開始して、0.5%のBSA/PBSおよび0.05%のTWEEN(登録商標)20に連続的に3倍希釈した。プレートを、室温で2時間インキュベートした。
【0173】
このプレートを、PBSTで4回洗浄し、0.5%のBSA/PBSおよび0.05%のTWEEN(登録商標)20中のビオチンヒツジ抗ヒトFdの1:1000希釈(0.8μg/ml)を50μL/ウェルで添加した。このプレートを、再度室温で2時間インキュベートした。
【0174】
PBSTでの4回の洗浄後、0.5%のBSA/PBSおよび0.05%のTWEEN(登録商標)20中のNeutra−アビジン−APの1:2000(0.9μg/ml)を50μl/ウェルで添加し、このプレートを、室温で1時間インキュベートした。
【0175】
このプレートを、PBSTで4回洗浄した。そして、pNPP基質を50μL/ウェルで添加することにより展開した。展開を、50μL/ウェルの3MのNaOHを添加することにより停止させた。各ウェルの吸光度を、405nmまたは410nmで読み取った。
【0176】
(実施例7)
(可溶性Fabを発現するM13ファージの親和性精製のためのプロトコル)
(A.1日目)
10mg/mLのテトラサイクリンを含有する2つの500mL培養物(2×YT)に、5mLの一晩のストックXL1Bを接種し、37℃にてA600が0.9〜1.2になるまで増殖させた。IPTGを、0.5mMの濃度まで添加した。次いで、細胞培養物を、培養あたり200μLのファージで感染させ、振盪しながら37℃にて1時間インキュベートした。感染後、その細胞を25℃にて一晩振盪しながら増殖させた。
【0177】
(B.2日目)
細胞を、250mLの遠心管中で、4℃で、3500×gにて30分間で、ペレット化した。培養培地を吸引し、ペレットを、合計12〜15mLの溶解緩衝液(緩衝液A+プロテアーゼインヒビターの混液)に再懸濁した。
【0178】
【化1】
再懸濁した細胞を、50mLの円錐管に移し、混合物が、小さな塊として一緒に移動する(溶解に起因して)まで、その管を数回逆さにすることによって、100μLの100mg/mLのリゾチームに溶解した。細胞を氷上で超音波処理し、その後、10μLのDNase I(約1000ユニット)を添加し、4℃にて30分間穏やかに振動させた。破片を、50mLの遠心管を用いて、4℃にて、12000×gにて30分間遠心することによりペレット化した。上清を新しい円錐管に移し、4℃にて保存した。
【0179】
Ni−NTアガロース(Qiagen、Valencis、CA)を使用して、製造業者のプロトコルに従って、可溶性Fabを精製した。この溶解物を、Ni−NTAと混合し、カラムにロードした。流れ出たものを、SDS−PAGE分析のために収集した。このカラムを、20mLの緩衝液(50mMのNaH2PO4、300mMのNaCl、15mMのイミダゾール、NaOHでpH8.0に調整)で洗浄し、その後50mM NaH2PO4、300mM NaCl、20mM イミダゾールを20mL用いて洗浄した。Fabを、500μLの溶出緩衝液(50mMのNaH2PO4、300mMのNaCl、450mMのイミダゾール、NaOHでpH8.0に調整)を用いて6回洗浄し、SDS PAGEにより分析した。カラム画分を、4℃にて保存した。カラム画分を、SDS−PAGEにより分析し、Fabの量が最も多い画分を選択し、PBS中に4℃にて透析した。
【0180】
(実施例8)
(可溶性レセプターアッセイ)
ELISAに適した96ウェルアッセイプレートを、0.05mLの0.5μg/mL FcεRIα鎖レセプター被覆緩衝液(50mM 炭酸塩/重炭酸塩、pH9.6)で、4〜8℃にて12時間被覆した。このウェルを吸引し、250μLのブロック緩衝液(PBS、1% BSA、pH7.2)を添加し、37℃にて1時間インキュベートした。別個のアッセイプレートにおいて、サンプルおよび参照のTES−C21 MAbを、200μg/mL〜0.001μg/mLまで、アッセイ緩衝液(0.5% BSAおよび0.05% Tween20、PBS、pH7.2)で1:4希釈して滴定し、等量の100ng/mLのビオチン化IgEを添加して、そのプレートを25℃にて、2〜3時間インキュベートした。FcεRI被覆ウェルを、0.05% TWEEN20を含むPBSを用いて三回洗浄し、サンプルウェルから50μLを移し、25℃にて、30分間攪拌しながらインキュベートした。アッセイ緩衝液に1:2000で希釈した1mg/mLのストレプトアビジン−HRPを50μL/ウェルで、30分間攪拌しながらインキュベートし、次いで、そのプレートを既に述べたように洗浄した。50μL/ウェルのTMB基質を添加し、発色させた。等量の0.2M H2SO4を添加することにより、反応を停止させ、450nmにて吸光度を測定した。
【0181】
(実施例9)
(抗体のIgE負荷FcεRIに対する結合)
FcεRIのαサブユニットと結合したヒトIgEに対する抗体の結合を、10μg/mLのヒトIgEと4℃にて30分間プレインキュベートすることによって、決定した。プレートを三回洗浄し、その後、種々の濃度のマウス抗ヒトIgE MAb E−10−10またはヒト化Fab改変体のどちらかと1時間インキュベートした。Fabの結合を、ビオチン標識した抗ヒトFd抗体を用いて、その後SA−HRPにより検出した。マウスMAb E−10−10を、ヤギ抗マウスIg Fc HRP結合Abにより検出した。
【0182】
(実施例10)
(クローン特徴付け)
各候補を、結合親和性についてアッセイし、ポジティブクローンを配列決定した。結合親和性を増加させる、CDR領域における有益な変異を有する抗体改変体をさらに特徴付けした。アッセイには、Biacore分析;IgEのレセプターに対するIgE結合の阻害;およびレセプターが結合したIgEの架橋が含まれる。
【0183】
改変体のライブラリーを作製した。親和性を改善することが実証された種々のCDRに対するアミノ酸配列を表1に示す。図7は、置換の組合せを有する高親和性の候補を示す。
【0184】
【表1】
19個の重鎖改変体を図9に示し、35個の軽鎖改変体を、図8に示す。さらに3個の候補を、結合親和性について特徴付けし、これらを表2に示す。
【0185】
【表2】
(実施例11)
(抗IgE抗体の発現および精製ならびにHRP結合)
高親和性MAb候補を生成した。インタクトな抗IgE MAbの生成のために、重鎖可変領域および軽鎖可変領域をファージベクターテンプレートからPCR増幅し、CMVプロモーターの発現下で、H鎖発現ベクターおよびL鎖発現ベクターに別個にサブクローニングした。6個の全長抗体クローンを構築した。これらを図10A〜図10Fに示す。適切な重鎖プラスミドおよび適切な軽鎖プラスミドを、当該分野で周知の技術によるエレクトロポレーションを使用して、マウス骨髄腫細胞株NS0に同時トランスフェクトした。例えば、Liouら、J.Immunol.第143巻、第12号:3967〜75(1989)を参照のこと。抗体を、プロテインAセファロース(Pharmacia)を使用して単一の安定な細胞株上清から精製した。抗体の濃度を、280nmでのスペクトロフォトメーターおよびFCAアッセイ(IDEXX)を使用して決定した。
【0186】
ペルオキシダーゼ結合キット(Zymed Labs、San Francisco、CA)を製造業者のプロトコルに従って使用して、精製した抗体に、ホースラディッシュペルオキシダーゼ(HRP)を結合させた。結合した各抗IgE MAbの力価を、モノクローナルヒトIgE(SE44)で被覆したプレートを用いたELISAを使用して決定した。
【0187】
以下の培養物は、American Type Culture Collection、10801 University Boulevard、Manassas Va.20110−2209 USA(ATCC)に寄託された。
【0188】
【表3】
この寄託を、特許手続き上の微生物の寄託の国際的承認に関するブダペスト条約および特許手続き上の微生物の寄託の国際的承認に関するブダペスト条約に基づく規則(ブダペスト条約)の下で行なった。このことは、寄託の日から30年間生存培養の維持を保証する。この生物は、ブダペスト条約の条件の下でATCCにより入手可能にされ、このことは、関連する米国特許の発行の際に、培養の子孫の永久的かつ非制限的な利用可能性を公共に保証する。
【0189】
本出願の譲受人は、寄託されたこの培養が、適切な条件下で培養された場合に死滅するか、失われるか、破壊される場合に、通知に対して、同一の培養の生存可能な試料により、ただちに取り替えられることに同意した。寄託した株の利用可能性は、特許法に従って任意の政府の当局に認可された権利に違反して、本発明の実施に対する実施権として解釈されるべきではない。
【0190】
前述の明細書は、当業者が本発明を実施可能にするのに十分であると考えられる。本発明は、寄託した培養物によってその範囲が限定されるべきではない。なぜならば、寄託された実施形態は、本発明の一局面の例示として意図され、機能的に等価である任意の培養物が、本発明の範囲内であるからである。本明細書中の材料の寄託は、本明細書中に含まれる記載が、その最良の形態を含めて、本発明の任意の局面の実施を可能にするのに不適切であるという承認を構成せず、特許請求の範囲の、本明細書が示す特定の例示への制限として解釈されるべきではない。実際、本明細書中に示され、記載される改変に加えて、本発明の種々の改変が、前述の記載から当業者に明らかになり、それらは、添付された特許請求の範囲の範囲内である。
【0191】
(実施例12)
(ヒトIgEの高親和性結合エピトープのマッピング)
(A.ペプチド合成および抗IgE結合アッセイ)
研究は、IgEが、そのCH3ドメインを介してそのレセプターに結合することを示した。本発明のHA 抗IgE抗体は、非常に効率的にIgEのそのレセプターへの結合をブロックするので、本発明者らは、全長CH3ドメインを含むペプチドを使用してエピトープをマッピングした。第一に、本発明者らは、二つのV5タグ化したペプチドを調製し、その一つは、ヒトIgEの全長定常領域を含み、もう一つは、ヒトIgEのCH2−CH3領域のみを含んでいた。これらの二つのペプチドは、インビトロ転写翻訳によって発現し、HA抗IgE MAb結合を検出するために、ウエスタンブロットアッセイに使用された。CL−2 CMAbおよびCL−5I MAbは両方ともインタクトなヒトIgEに結合し得、両方ともそのペプチドに結合し得る。
【0192】
エピトープをより詳細にマッピングするために、ヒトIgEの残基141〜368のアミノ酸を含み、全長CH3ドメインを含む73個の重複するペプチドを合成した。各ペプチドは、上記ペプチドの3’末端と重複する3アミノ酸を有する12アミノ酸残基からなる。SPOT膜を、セルロース膜上に、フルオレニルメトキシカルボニル(Fmoc)アミノ酸を用いて合成した。その膜をメタノール中でリンスし、次いで、TBS(pH7.5)中で、10分間、3回洗浄した。ブロック溶液(TBS中5%乳汁または3%BSA)中で一晩インキュベートした後、ブロック溶液に希釈したHRP標識抗IgE MAbを、膜と一緒に3時間インキュベートした。TBS−TWEEN(登録商標)中で15分間、3回洗浄した後、SuperSignal HRP基質(Pierce)を使用して、IgE反応性を、所望の時間BioMax MSフィルム(Kodak)の化学発光露光することにより測定した。
【0193】
この実験の結果は、HA抗IgE MAbが、IgEのCH3部分の二つの領域に結合することを示唆し、この二つの領域は、以下の二つのペプチド配列NPRGVSAYLSRP(エピトープ「A」)およびHPHLPRALMRST(エピトープ「B」)により表される(図12を参照のこと)。エピトープAに対する結合は、エピトープBに対する結合より数倍弱かった。
【0194】
(B.アラニンスキャニング変異誘発)
IgG1に見出されるアミノ酸とペプチド内のアミノ酸との置換に伴うアラニンスキャニングを、どのアミノ酸が、HA抗IgE MAbのこれらのペプチドへの結合に決定的に関与しているかを決定するために行なった。HA抗IgE MAb結合に重要であることが決定されたアミノ酸を、IgEのε鎖におけるインビトロ変異誘発方法を使用して置換した。上に記載されるCε2領域およびCε3領域をカバーする別のペプチドをまた、この研究に使用した(図13および図14を参照のこと)。
【0195】
ヒトIgEアミノ酸残基に対してEU番号付けスキームを使用した。ポリメラーゼ連鎖反応(PCR)を使用して、IgEの全長Fc領域および、CH2−CH3ドメインのみを含むIgE Fcの短縮形態を増幅した。このDNA産物を、TOPOクローニング(Invitrogene、Carlsbad、CA)を使用して、pcDNA3発現ベクター(Invitrogene、Carlsbad、CA)に直接クローニングした。
【0196】
IgE−Fcにおける変異誘発を、オーバーラッピングPCR(Hoら、1989)を使用して行った。このDNA産物を、アガロースゲル電気泳動によって精製し、適切な制限酵素を用いて消化し、pcDNA3発現ベクターにサブクローニングした。各改変体構築物について、PCR増幅領域を、DNAの両方の鎖から、ジデオキシヌクレオチド方法を使用して完全に配列決定した。組換えヒトIgE Fcおよびその変異体を、網赤血球溶解物ベースのインビトロ転写とインビトロ翻訳との結合システム(Promega、Madison、WI)を使用して発現させた。
【0197】
このインビトロ転写とインビトロ翻訳との結合システム(10μlの反応混合物)由来の溶解物を、SDS−PAGE(12%)に供し、次いで、ニトロセルロース膜に移した。この膜を、Tris緩衝生理食塩水(TBS)中の5%脱脂粉乳を用いてブロックし、その後一次抗体、抗IgE MAbで染色した。特異的な反応バンドを、ホースラディッシュペルオキシダーゼに結合したヤギ抗ヒトIgG Fc(Jackson Labs、Bar Harbor、Maine)を使用して検出し、その免疫応答性バンドを、SuperSignal ウェスタンブロッティング検出キット(Pierce)により可視化した。抗V5抗体を、これらのペプチドのC末端に導入されたV5タグを検出するポジティブコントロールとして使用した。抗V5抗体を用いたウエスタンブロットは、全てのペプチドが、ほぼ等しいレベルで発現したことを実証した。興味深いことに、HA抗IgE MAbは、エピトープ「A」に変異を保有するペプチドに結合し得たが、それらは、エピトープ「B」に変異を保有するペプチドには結合しなかった。このことは、この第2の部位が、結合に関してより重要であったことを示唆する(図15を参照のこと)。
【0198】
(実施例13)
(エピトープBの免疫原性ペプチドを使用するトランスジェニックマウスの能動免疫)
ヒトIgEを構成的に発現するトランスジェニックマウスを使用して、エピトープBのヒト免疫原性ペプチドに対する抗体の能動的な産生を実証した。二つの融合ペプチド(それぞれが、本発明の免疫原性ペプチドであるシステイン残基およびKLHを含む)を化学的に合成した。ペプチド1の配列は、以下:
(KLH−Cys)−Leu Pro Arg Ala Leu Met Arg Ser Thr
であり、ペプチド2の配列は、以下:
Leu Pro Arg Ala Leu Met Arg Ser Thr−(Cys−KLH)
であった。
【0199】
そのトランスジェニックマウスに、200μLのPBS(pH7.4)中の完全フロイントアジュバント(Difco Laboratories、Detroit、MI)中の20μgの免疫原性ペプチドを皮下に注射した。2週間の間隔で2度、不完全フロイントアジュバント中の20μgのペプチド免疫源を、そのマウスに、皮下に注射した。次いで、二週間後、屠殺の3日前に、そのマウスに再度、腹腔内に、PBS中の20μgの同じ免疫源を注射した。血清を収集し、エピトープBに特異的な抗IgE抗体の存在について試験した。図16に見られるように、このペプチドは、これらのトランスジェニックマウスにおいて、抗IgE抗体を誘発した。
【0200】
当業者は、慣例的にすぎない実験を使用して、本明細書中に記載される本発明の特定の実施形態に対する多くの等価物を認識するか、または確認することができる。このような等価物は、添付の特許請求の範囲によって包含されることが意図される。
【図面の簡単な説明】
【0201】
【図1】図1は、抗体クローニングおよび抗体スクリーニングに使用されるファージベクターの模式的表現である。
【図2】図2は、抗体改変体を生成するために有用なオリゴヌクレオチドの模式的表現である。
【図3】図3Aは、マウス抗IgE抗体TES−C21の軽鎖とL16およびJK4の混合ヒトテンプレートとの比較を示す。図3Bは、TES−C21の重鎖とDP88およびJH4bの混合ヒトテンプレートとの比較を示す。
【図4】図4は、親TES−C21と比較して高親和性を有する、フレームワーク残基改変体の表を提示する。
【図5】図5AおよびBは、TES−C21の親Fabとネガティブコントロール(5D12)と比較した、クローン4、49、72、78および136についてのELISA滴定曲線を示す。
【図6】図6は、親TES−C21およびネガティブコントロール抗体と比較した、クローン2C、5Aおよび5Iの阻害アッセイを示す。
【図7】図7Aは、IgEに対するさらに高い親和性をもたらす有益な変異の組み合わせを有するクローンの配列を示す。
【図8A】図8Aおよび8Bは、クローン136、1、2、4、8、13、15、21、30、31、35、43、44、53、81、90および113についての全軽鎖可変領域のフレームワーク配列を示す。
【図8B】図8Aおよび8Bは、クローン136、1、2、4、8、13、15、21、30、31、35、43、44、53、81、90および113についての全軽鎖可変領域のフレームワーク配列を示す。
【図9A】図9Aおよび9Bは、35個のクローンについての全重鎖可変領域のフレームワーク配列を示す。
【図9B】図9Aおよび9Bは、35個のクローンについての全重鎖可変領域のフレームワーク配列を示す。
【図10A】図10A〜Fは、クローン136、2C、5I、5A、2Bおよび1136〜2Cについての完全重鎖配列および完全軽鎖配列を示す。
【図10B】図10A〜Fは、クローン136、2C、5I、5A、2Bおよび1136〜2Cについての完全重鎖配列および完全軽鎖配列を示す。
【図10C】図10A〜Fは、クローン136、2C、5I、5A、2Bおよび1136〜2Cについての完全重鎖配列および完全軽鎖配列を示す。
【図10D】図10A〜Fは、クローン136、2C、5I、5A、2Bおよび1136〜2Cについての完全重鎖配列および完全軽鎖配列を示す。
【図10E】図10A〜Fは、クローン136、2C、5I、5A、2Bおよび1136〜2Cについての完全重鎖配列および完全軽鎖配列を示す。
【図10F】図10A〜Fは、クローン136、2C、5I、5A、2Bおよび1136〜2Cについての完全重鎖配列および完全軽鎖配列を示す。
【図11】図11は、ヒトIgEのCH3領域アミノ酸配列を示し、そしてエピトープ「A」およびエピトープ「B」を強調表示する。
【図12】図12は、エピトープBを同定するために使用される重複ペプチドを示す。
【図13】図13は、エピトープAの結合領域中の重要な残基の識別を示す。
【図14】図14は、エピトープBの結合領域中の重要な残基の識別を示す。
【図15】図15は、変異体ペプチドの結合するMAbのウエスタンブロット分析を示す。
【図16】図16は、ヒトIgEを発現するトランスジェニック動物における抗IgE抗体の産生を示す。
【特許請求の範囲】
【請求項1】
以下:Asn Pro Arg Gly Val Ser Xaa Tyr Xaa Xaa Arg Xaa(配列番号72)のアミノ酸配列から実質的に構成される、単離されたペプチド。
【請求項2】
前記アミノ酸配列が、Asn Pro Arg Gly Val Ser Ala Tyr Leu Ser Arg Pro(配列番号73)である、請求項1に記載のアミノ酸。
【請求項3】
以下:Leu Pro Arg Ala Leu Xaa Arg Ser Xaa(配列番号74)のアミノ酸配列から実質的に構成される、単離されたペプチド。
【請求項4】
前記アミノ酸配列が、以下:
a)Leu Pro Arg Ala Leu Met Arg Ser Thr(配列番号75);
b)Hi Pro His Leu Pro Arg Ala Leu Met Arg Ser Thr(配列番号76);または
c)Leu Pro Arg Ala Leu Met Arg Ser Thr Thr Lys Thr(配列番号77)
である、請求項3に記載のペプチド。
【請求項5】
請求項1または3に記載のペプチドと、生理学的に受容可能なキャリア、希釈剤、安定剤、または賦形剤とを含む、組成物。
【請求項6】
請求項5に記載の組成物であって、免疫原性キャリアをさらに含む、組成物。
【請求項7】
請求項6に記載の組成物であって、前記免疫原性キャリアが、BSA、KLH、破傷風トキソイド、およびジフテリアトキソイドからなる群より選択される、組成物。
【請求項8】
請求項1〜4のいずれか1項に記載のペプチドに特異的に結合する、単離された抗体。
【請求項9】
標識をさらに含む、請求項8に記載の抗体。
【請求項10】
請求項8に記載の抗体であって、該抗体は、以下:
a)キメラ抗体、
b)単鎖抗体、
c)Fabフラグメント、
d)F(ab’)、フラグメント、
e)ヒト抗体、または
f)ヒト化抗体
である、抗体。
【請求項11】
請求項8に記載の抗体と、受容可能なキャリアとを含む、組成物。
【請求項12】
ポリクローナル抗体を調製する方法であって、該方法は、以下:
a)抗体応答を誘発する条件下において、配列番号72または配列番号74のアミノ酸配列から構成されるポリペプチドで、動物を免疫する工程、
b)該動物から抗体を単離する工程、および
c)該単離された抗体を前記ポリペプチドによってスクリーニングし、該スクリーニングによって、配列番号72または配列番号74のアミノ酸配列を含むポリペプチドに対して、高親和性に特異的に結合するポリクローナル抗体を同定する工程、
を包含する、方法。
【請求項13】
請求項12に記載の方法によって生成されるポリクローナル抗体。
【請求項14】
請求項13に記載のポリクローナル抗体と適切なキャリアとを含む、組成物。
【請求項15】
モノクローナル抗体を作製する方法であって、該方法は、以下:
a)抗体応答を誘発する条件下において、配列番号72または配列番号74のアミノ酸配列から実質的に構成されるポリペプチドで、動物を免疫する工程、
b)該動物から抗体産生細胞を単離する工程、
c)該抗体産生細胞と不死化細胞とを融合させて、モノクローナル抗体産生ハイブリドーマ細胞を形成する工程、
d)該ハイブリドーマ細胞を培養する工程、および
e)該培養から、配列番号72または配列番号74のアミノ酸配列を含むポリペプチドに対して高親和性に特異的に結合するモノクローナル抗体を単離する工程、
を包含する、方法。
【請求項16】
請求項15に記載の方法によって生成されるモノクローナル抗体。
【請求項17】
請求項16に記載のモノクローナル抗体と適切なキャリアとを含む、組成物。
【請求項18】
前記抗体が、Fab発現ライブラリーをスクリーニングすることによって生成される、請求項8に記載の抗体。
【請求項19】
前記抗体がコンビナトリアル免疫グロブリンライブラリをスクリーニングすることによって生成される、請求項8に記載の抗体。
【請求項20】
請求項8〜19のいずれか1項に記載の抗体を含む、キット。
【請求項21】
哺乳動物においてIgEに対する免疫応答を誘導する方法であって、該哺乳動物に、応答を誘導するのに十分な量の、請求項1〜7のいずれか1項に記載のペプチドまたは組成物を投与する工程を包含する、方法。
【請求項1】
以下:Asn Pro Arg Gly Val Ser Xaa Tyr Xaa Xaa Arg Xaa(配列番号72)のアミノ酸配列から実質的に構成される、単離されたペプチド。
【請求項2】
前記アミノ酸配列が、Asn Pro Arg Gly Val Ser Ala Tyr Leu Ser Arg Pro(配列番号73)である、請求項1に記載のアミノ酸。
【請求項3】
以下:Leu Pro Arg Ala Leu Xaa Arg Ser Xaa(配列番号74)のアミノ酸配列から実質的に構成される、単離されたペプチド。
【請求項4】
前記アミノ酸配列が、以下:
a)Leu Pro Arg Ala Leu Met Arg Ser Thr(配列番号75);
b)Hi Pro His Leu Pro Arg Ala Leu Met Arg Ser Thr(配列番号76);または
c)Leu Pro Arg Ala Leu Met Arg Ser Thr Thr Lys Thr(配列番号77)
である、請求項3に記載のペプチド。
【請求項5】
請求項1または3に記載のペプチドと、生理学的に受容可能なキャリア、希釈剤、安定剤、または賦形剤とを含む、組成物。
【請求項6】
請求項5に記載の組成物であって、免疫原性キャリアをさらに含む、組成物。
【請求項7】
請求項6に記載の組成物であって、前記免疫原性キャリアが、BSA、KLH、破傷風トキソイド、およびジフテリアトキソイドからなる群より選択される、組成物。
【請求項8】
請求項1〜4のいずれか1項に記載のペプチドに特異的に結合する、単離された抗体。
【請求項9】
標識をさらに含む、請求項8に記載の抗体。
【請求項10】
請求項8に記載の抗体であって、該抗体は、以下:
a)キメラ抗体、
b)単鎖抗体、
c)Fabフラグメント、
d)F(ab’)、フラグメント、
e)ヒト抗体、または
f)ヒト化抗体
である、抗体。
【請求項11】
請求項8に記載の抗体と、受容可能なキャリアとを含む、組成物。
【請求項12】
ポリクローナル抗体を調製する方法であって、該方法は、以下:
a)抗体応答を誘発する条件下において、配列番号72または配列番号74のアミノ酸配列から構成されるポリペプチドで、動物を免疫する工程、
b)該動物から抗体を単離する工程、および
c)該単離された抗体を前記ポリペプチドによってスクリーニングし、該スクリーニングによって、配列番号72または配列番号74のアミノ酸配列を含むポリペプチドに対して、高親和性に特異的に結合するポリクローナル抗体を同定する工程、
を包含する、方法。
【請求項13】
請求項12に記載の方法によって生成されるポリクローナル抗体。
【請求項14】
請求項13に記載のポリクローナル抗体と適切なキャリアとを含む、組成物。
【請求項15】
モノクローナル抗体を作製する方法であって、該方法は、以下:
a)抗体応答を誘発する条件下において、配列番号72または配列番号74のアミノ酸配列から実質的に構成されるポリペプチドで、動物を免疫する工程、
b)該動物から抗体産生細胞を単離する工程、
c)該抗体産生細胞と不死化細胞とを融合させて、モノクローナル抗体産生ハイブリドーマ細胞を形成する工程、
d)該ハイブリドーマ細胞を培養する工程、および
e)該培養から、配列番号72または配列番号74のアミノ酸配列を含むポリペプチドに対して高親和性に特異的に結合するモノクローナル抗体を単離する工程、
を包含する、方法。
【請求項16】
請求項15に記載の方法によって生成されるモノクローナル抗体。
【請求項17】
請求項16に記載のモノクローナル抗体と適切なキャリアとを含む、組成物。
【請求項18】
前記抗体が、Fab発現ライブラリーをスクリーニングすることによって生成される、請求項8に記載の抗体。
【請求項19】
前記抗体がコンビナトリアル免疫グロブリンライブラリをスクリーニングすることによって生成される、請求項8に記載の抗体。
【請求項20】
請求項8〜19のいずれか1項に記載の抗体を含む、キット。
【請求項21】
哺乳動物においてIgEに対する免疫応答を誘導する方法であって、該哺乳動物に、応答を誘導するのに十分な量の、請求項1〜7のいずれか1項に記載のペプチドまたは組成物を投与する工程を包含する、方法。
【図1】

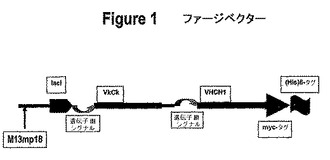
【図2】
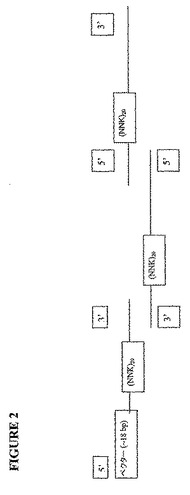
【図3】

【図4】
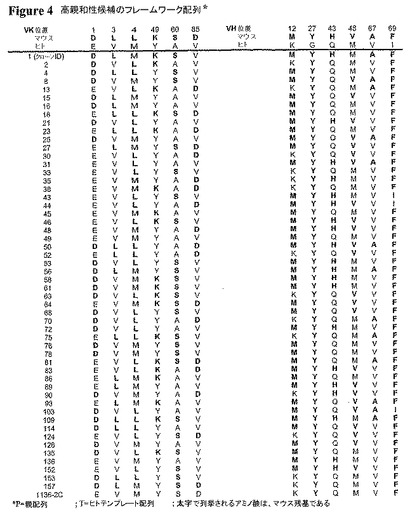
【図5】
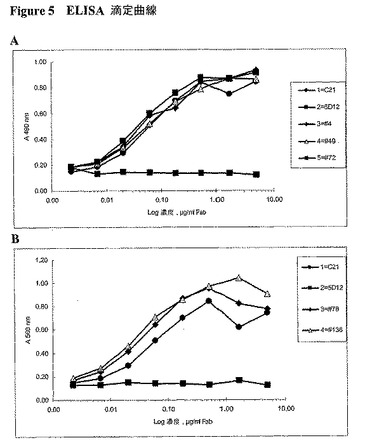
【図6】
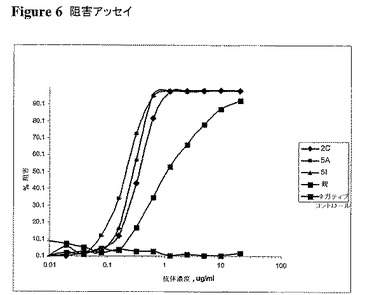
【図7】
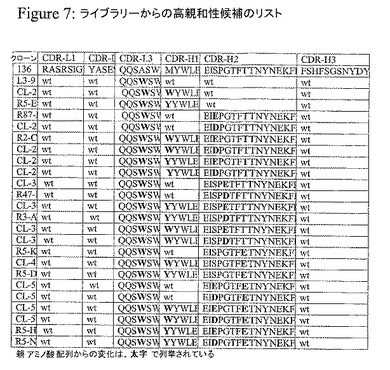
【図8A】

【図8B】

【図9A】
【図9B】
【図10A】

【図10B】

【図10C】

【図10D】

【図10E】

【図10F】

【図11】

【図12】

【図13】

【図14】

【図15】
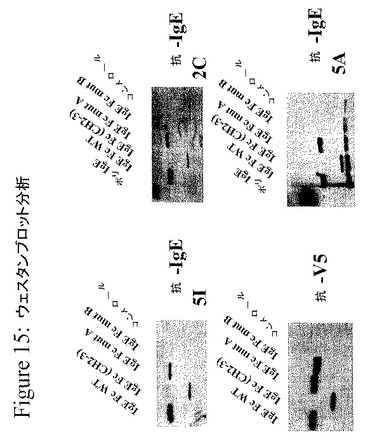
【図16】
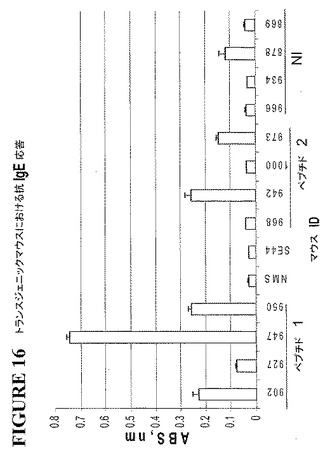
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8A】
【図8B】
【図9A】
【図9B】
【図10A】
【図10B】
【図10C】
【図10D】
【図10E】
【図10F】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【公表番号】特表2008−507474(P2008−507474A)
【公表日】平成20年3月13日(2008.3.13)
【国際特許分類】
【出願番号】特願2006−551035(P2006−551035)
【出願日】平成16年7月29日(2004.7.29)
【国際出願番号】PCT/US2004/024360
【国際公開番号】WO2005/075504
【国際公開日】平成17年8月18日(2005.8.18)
【出願人】(500352579)タノックス インコーポレイテッド (10)
【Fターム(参考)】
【公表日】平成20年3月13日(2008.3.13)
【国際特許分類】
【出願日】平成16年7月29日(2004.7.29)
【国際出願番号】PCT/US2004/024360
【国際公開番号】WO2005/075504
【国際公開日】平成17年8月18日(2005.8.18)
【出願人】(500352579)タノックス インコーポレイテッド (10)
【Fターム(参考)】
[ Back to top ]