
新規シアリダーゼ
本発明は、(a)配列番号3のアミノ酸1〜407と少なくとも60%のアミノ酸配列同一性を有するアミノ酸配列を有するポリペプチド;(b)低度のストリンジェンシー条件下で、(i)60ヌクレオチド超、好ましくは100ヌクレオチド超にわたり少なくとも80%もしくは90%同一であり、より好ましくは、200ヌクレオチド超にわたり少なくとも90%同一である配列番号1または2の核酸配列、あるいは(ii)配列番号1または2の核酸配列に相補的な核酸配列とハイブリダイズするポリヌクレオチドによってコードされるポリペプチドからなる群から選択されるシアリダーゼ活性を有する単離されたポリペプチドに関する。
【発明の詳細な説明】
【発明の詳細な説明】
【0001】
[発明の分野]
本発明は、新規シアリダーゼに関する。
【0002】
[発明の背景]
シアル酸は、9炭素糖のノイラミン酸の約40種類の誘導体のファミリーを含んでなる。それは、約2.2のpKaを伴う強有機酸である。非置換型のノイラミン酸(neuraminic acic)は、天然には存在しない。アミノ基は、通常、アセチル化されて、最も普及した形のシアル酸であるN−アセチルノイラミン酸(N−acetylneuraminc acid)を産出するが、他の形も存在する(トラビング(Traving)ら Cell Mol Life Sci(1998年)54,1330−1349)。シアル酸は、棘皮動物からヒトにまで至る動物界において見出されるが、一方、前口動物系統の下等動物または植物におけるそれらの存在については、示唆されていない。唯一の公知の例外は、昆虫のショウジョウバエ(Drosophila)の幼虫におけるポリシアル酸の存在である。さらに、いくつかの原生動物、ウイルスおよび細菌にもシアル酸が認められる。シアロ糖複合体は、細胞表面ならびに細胞内膜に存在する。高等動物では、それらはまた、血清および粘膜物質の重要な成分でもある。
【0003】
シアル酸は、多様な生物学的機能を有する。それらの負電荷により、シアル酸は、カルシウムイオンのような正に荷電した分子の結合および輸送、ならびに細胞と分子との間の誘引および反発現象に関与する。それらのサイズおよび負電荷に加えて、それらが暴露された炭水化物鎖の末端に位置することは、それらが分子または細胞のサブ末端部の保護シールドとして機能することを可能にする。それらは、例えば、グリコール−タンパク質(glycol−proteins)がプロテアーゼによって分解されるのを防ぐことができ、または呼吸器系の粘膜層を細菌感染から防ぐことができる。興味深い現象は、それらの負の電荷の間に作用する反発力によりシアル酸含有分子に対して及ぼされる延展効果である。これは、酵素または膜(糖)タンパク質の正確なコンホメーションを安定化し、そして粘液性(slimy)の性質ならびに眼の表面および粘膜上皮のような粘膜物質の滑走および保護機能に重要である(トラビング(Traving)ら Cell Mol Life Sci(1998年)54,1330−1349)。明らかに、適切なシアリダーゼによるそのようなシアル酸含有物質の処理は、そのような物質の生物学的特性および物理的特徴に劇的に影響を及ぼすことができる。シアリダーゼによるシアル酸含有タンパク質の処理は、それらを、かなり容易にプロテアーゼによって分解され易くすることができ、シアリダーゼによる粘膜物質の処理は、それらの粘液性特徴を強度に減少または排除し得る。そのような変化は、そのようなタンパク質が、例えば、タンパク質加水分解物のためのプロセシング(例えば、タンパク質分解)の産業的プロセスを必要とする場合、興味深い。
【0004】
シアル酸は、細胞と分子との間の多様な認識プロセスに関与する。それ故、免疫系は、それらのシアル酸パターンに従って、自己構造と非自己構造とを区別することができる。糖は、抗原決定基、例えば血液型物質を表し、そしてホルモンおよびサイトカインのような多くの内因性物質の受容体の必要な成分である。加えて、毒素(例えば、コレラ毒素)のような多くの病原因子、ウイルス(例えば、インフルエンザ)、細菌(例えば、大腸菌(Escherichia coli)、ヘリコバクター・ピロリ(Helicobacter pylori))および原生動物(例えば、トリパノソーマ・クルージ(Trypanosome cruzi)もまた、シアル酸含有受容体を介して宿主細胞に結合する。シアル酸認識分子の別の重要な基は、特定の糖残基に結合する植物、動物および無脊椎動物由来の通常オリゴマー糖タンパク質であるレクチンに属する。例には、コムギ胚芽アグルチニン、アメリカカブトガニ(Limulus polyphemus)アグルチニン(aagglutinin)、セイヨウニワトコ(Sambucus nigra)アグルチニンおよびカライヌエンジュ(Maackia amurensis)アグルチニンがある。これらのレクチンは、植物を、シアル酸含有微生物または草食哺乳動物に対するその防御において援助するようである。レクチンの哺乳動物対応物は、セレクチンおよびシグレックを含み(トラビング(Traving)ら Cell Mol Life Sci(1998年)54,1330−1349)、そして多様な生理学的役割を有する。シアル酸はまた、細胞および分子の遮蔽を補助することができる。赤血球は、シアル酸分子の防御層に覆われ、これは、血液細胞の生活環中に段階的に取り出される。次いで(than)、分解のためのシグナルを表す最後から2番目のガラクトース残基が晒され、次いで(than)、遮蔽されていない血液細胞がマクロファージに結合し、そして貪食される。そのような遮蔽ストラテジーの他のいくらかの例は公知である。遮蔽はまた、有害な影響を有し得るが、これは、対応する組織よりもかなり高い程度でシアル酸付加される腫瘍のいくつかから認められ得る。結果的に、遮蔽された細胞は免疫防御系に晒されず、そして高シアル酸含有量もまた、さらなる細胞増殖の阻害の欠如および拡大に役割を果たし得る。シアル酸の遮蔽効果はまた、寄生体細胞上の抗原部位を隠すのに役立ち、それらを系に晒されないようにする。これは、所定の大腸菌(E coli)株および淋菌(Neisneria gonorrhoeae)のような微生物種の場合に当てはまる。そのような種をシアリダーゼで処理すると、免疫系から隠れるそれらの可能性に影響を及ぼす。
【0005】
シアリダーゼ(ノイラミニダーゼ、EC3.2.1.18)は、糖タンパク質、糖脂質、ガングリオシド、多糖および合成分子における末端の非還元シアル酸結合を加水分解する。トランスシアリダーゼと呼ばれるシアリダーゼはまた、シアル酸残基を1つの分子から別の分子に転移する転移反応を実施することも可能である。シアリダーゼは、新口動物系統(棘皮動物(Echinodermata)から哺乳動物(Mammalia))の動物およびまた動物の共生生物または病原体として最も多く存在する多様な微生物において一般的に認められる。シアリダーゼ、およびそれらのシアリル基質は、植物および他のほとんどの後生動物には存在しないと思われる。細菌の間であっても、シアリダーゼは、関連する種または1つの種の株であってもこの特性が異なるように、不規則に見出される。シアリダーゼはまた、ウイルスおよび原生動物においても見出されている(トラビング(Traving)ら Cell Mol Life Sci(1998年)54,1330−1349)。シアリダーゼを含有する微生物は、しばしば、例えば、寄生体として、宿主としての高等動物と接触しながら生存する。ここで、それらは、栄養機能を有し得、それらの所有者が宿主のシアル酸を炭素源として使用するためにスカベンジすることを可能にする。いくつかの微生物病原体では、シアリダーゼがビルレンス因子として作用すると考えられる。なお、病原性における因子としてのサリダーゼ(salidases)の役割については、議論の余地がある。一方では、それらは、ウェルシュ菌(Clostridium perfringens)のような病原性微生物種の影響を裏付けている。他方では、これらの酵素は、高等動物を含む多くの非病原性種の炭水化物異化に共通の因子である。しかし、それらは、直接的な毒性効果を発揮しない(トラビング(Traving)ら Cell Mol Life Sci(1998年)54,1330−1349)。その代わり、それらの有害な影響は、非生理学的条件下での宿主シアル酸による誘導時に他の毒性因子と共に宿主に放出される大量の酵素に依存する。
【0006】
哺乳動物シアリダーゼは、通常、約40〜45kDのサイズである。哺乳動物シアリダーゼを過剰発現させ、そして産業的に関心のある量にまで産生させる試みについては、報告されていない。ヒトシアリダーゼは、リソソーム酵素、サイトゾル酵素または膜結合酵素であり得る(エイカイウタン(Achyuthan)およびエイカイウタン(Achyuthan)(2001年)Comp.Biochem.Phys.B部、129,29−64)。リソソームシアリダーゼは、グリコシル化酵素である。シアリダーゼは、保存されたモチーフを含有する。最も顕著な保存されたモチーフは、一般式−S−X−D−X−G−X−T−W−[式中、Xは可変の残基を表す]のアミノ酸のストレッチであるいわゆるAspボックスである。このモチーフが僅か1回もしくは2回しか見出されないかまたは存在さえしないウイルスシアリダーゼを例外として、このモチーフは、すべての微生物配列をとおして4〜5回見出される。第3のAspボックスは、Aspボックス2および4より強度に保存されている。連続的な2つのAspボックスの間の空間もまた、異なる一次構造の間で保存される(トラビング(Traving)ら Cell Mol Life Sci(1998年)54,1330−1349)。Aspボックスは、おそらく構造的役割を果たし、そしておそらく触媒に関与しない。Aspボックスとは対照的に、FRIP−モチーフは、アミノ酸配列のN末端部分に局在する。それは、絶対的に保存されたアルギニンおよびプラリネ(praline)残基を伴うアミノ酸−X−R−X−P−を包含する。アルギニンは、基質分子の結合によって触媒に直接関与する。aspボックス3および4の間のグルタミン酸リッチ領域、ならびにさらなる2つのアルギニン残基もまた、触媒作用に重要である(トラビング(Traving)ら Cell Mol Life Sci(1998年)54,1330−1349)。
【0007】
微生物シアリダーゼは、それらのサイズに従って、2つのグループ、すなわち、約42kDaの小さなタンパク質および60〜70kDaの大きなタンパク質に分類することができる。大きなシアリダーゼの一次構造は、N末端と第2のAspボックスとの間、ならびに第5のAspボックスとC末端との間にアミノ酸の追加のストレッチを含有する。それらは大きなシアリダーゼのより広範な基質特異性に寄与すると考えられる。哺乳動物シアリダーゼと同様に、細菌対応物は、F/YRIPモチーフおよびいくらかのAspボックスを含有する。細菌シアリダーゼは、しばしば、粘膜感染およびビルレンス(virulance)に関係している。このため、大きい方の細菌シアリダーゼは、食品または製薬用途における加工助剤としての使用に適切であるとは認識されていない。小さなシアリダーゼ(哺乳動物シアリダーゼと同じサイズ)は、上記に示す細菌において同定されている。即ち、ウェルシュ菌(Clostridium perfringens)は、他の細菌のシアリダーゼに共通の伸長部を伴わない約40kDaのサイズを伴う小さなシアリダーゼを含有する。しかし、このクロストリジウム(Clostridium)シアリダーゼは、細菌によって分泌されず、従ってまた、ビルレンス(virulance)に関与しない(ロッグゲンチン(Roggentin)ら(1995年)Biol Chem Hoppe Seyler 376,569−575)。追加の伸長部を伴う細菌シアリダーゼのみが病原性に関与することは、大変興味深い。大腸菌(E.coli)における細菌シアリダーゼの過剰発現は、一般的に、低い産生率をもたらし;小さなクロストリジウム(Clostridium)シアリダーゼは、大腸菌(E.coli)における細胞内タンパク質として1mg/lまでしか産生され得ない(クルーゼ(Kruse)ら(1996年)Protein Expr Purif.7,415−422)。
【0008】
ウチダ(Uchida)ら(Biochimica et Biophysica Acta,第350巻、第2号、1974年、425〜431頁)は、コロミン酸によって誘導される微生物ノイラミニダーゼのスクリーニングについて記載している。スクリーニングした1000の微生物のうち、スポロトリチウム・シェンキー(Sporotrichium schenckii)、ペニシリウム・ウルチカエ(Penicillium urticae)およびストレプトマイセス(Streptomyces)sp.からノイラミニダーゼが得られた。ペニシリウム・ウルチカエ(Penicillium urticae)は、食物の腐敗に関与する真菌であるため、それは、食品級シアリダーゼに適切な産生生物体ではなく、そしてスポロトリチウム・シェンキー(Sporotrichium schenckii)は病原性の真菌である。細菌ストレプトマイセス(Streptomyces)spのなかでの種名は決定されていない。この論文に記載のシアリダーゼのMWは不明である。イワモリ(Iwamori)ら(J.Biochem.138,327〜334頁)では、細菌アルスロバクター・ウレアファシエンス(Arthrobacter ureafaciens)シアリダーゼが開示されている。しかし、この細菌ノイラミニダーゼは、HIV−1仲介シンシチウム形成およびウイルス結合/進入プロセスに関連することが公知である(サン(Sun)ら、Virology 284,26〜36頁、2001年)。従って、食品および製薬における用途のための良好に産生される小さな非ビルレントなシアリダーゼが明らかに必要である。
【0009】
特に、分泌型酵素は容易に過剰発現され、そして真菌培養物から大量に精製され得るため、分泌型真菌シアリダーゼの所見は有益である。これは、シアリダーゼの産生のための原価を劇的に減少する。
【0010】
[発明の概要]
本発明は、以下からなる群から選択される、シアリダーゼ活性を有する単離されたポリペプチドに関する:
(a)配列番号3のアミノ酸1〜407と少なくとも60%のアミノ酸配列同一性を有するアミノ酸配列を有するポリペプチド;
(b)低度のストリンジェンシー条件で、(i)60ヌクレオチド超、好ましくは100ヌクレオチド超にわたり少なくとも80%もしくは90%同一な、より好ましくは200ヌクレオチド超にわたり少なくとも90%同一な配列番号1または2の核酸配列、あるいは(ii)配列番号1または2の核酸配列に相補的な核酸配列とハイブリダイズするポリヌクレオチドによってコードされるポリペプチド。好ましくは、このポリペプチドは、55kDa未満のMW(分子量)(SDS−page)を有し、より好ましくは、このポリペプチドは52kDa未満のMW(分子量)(SDS−page)を有するか、またはこのポリペプチドは50kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有し、より好ましくは、このポリペプチドは45kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有する。
【0011】
さらに、本発明は、シアリダーゼ活性を有し、そして非ビルレントであるポリペプチドを提供する。また、本発明の一部分は、シアリダーゼ活性を有するポリペプチドであり、前記ポリペプチドは、真菌のシアリダーゼであり、55kDa未満のMW(分子量)(SDS−Page)を有し、より好ましくは、この真菌シアリダーゼは52kDa未満のMW(分子量)(SDS−Page)を有するか、またはこの真菌シアリダーゼは50kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有し、より好ましくは、この真菌シアリダーゼは45kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有する。
【0012】
好ましくは、本発明のポリペプチドは、配列番号3のアミノ酸1〜407と少なくとも65%、好ましくは少なくとも70%、より好ましくは少なくとも80%、さらにより好ましくは少なくとも90%、最も好ましくは少なくとも95%、およびさらに最も好ましくは少なくとも約97%同一性を有するアミノ酸配列を有する。本発明はまた、請求項1に記載のポリペプチドをコードするか、または低度のストリンジェンシー条件下で、より好ましくは、中度のストリンジェンシー条件下で、および最も好ましくは、高度のストリンジェンシー条件下で、配列番号1または2とハイブリダイズする核酸配列を含んでなる単離されたポリヌクレオチドに関する。さらに加えて、本発明は、適切な発現宿主においてポリペプチドの産生を指令する1つもしくはそれ以上の制御配列に作動可能に連結された本発明のポリヌクレオチドを含んでなる核酸構築物を開示する。さらに、本発明は、この核酸構築物を含んでなる組み換え発現ベクター、および前記核酸構築物を含んでなる組み換え宿主細胞を提供する。本発明の別の態様に従えば、上記の株/組み換え宿主細胞を培養して、ポリペプチドを含んでなる上清および/または細胞を生成すること;ならびにポリペプチドを回収することを含んでなる、ポリペプチドの産生させるための方法が開示される。本発明のさらなる態様に従えば、ポリペプチドの産生に適切な条件下でポリペプチドをコードするポリヌクレオチドを含んでなる核酸構築物を含んでなる宿主細胞を培養すること;およびポリペプチドを回収することを含んでなる、本発明のポリペプチドを産生させるための方法が開示される。シアリダーゼ活性を有する本発明のポリペプチドは、有利なことに、非ビルレントである。本発明のポリペプチドは、食品もしくは飼料の調製において、または医薬品もしくは医薬品の一部分として使用することができる。
【0013】
[発明の詳細な説明]
本発明に従えば、好ましくは、55kDa未満のMW(分子量)(SDS−Page)を有する小型のシアリダーゼ(より好ましくは、このポリペプチドは52kDa未満のMW(分子量)(SDS−Page)を有する)、または50kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有するシアリダーゼ(より好ましくは、このポリペプチドは45kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有する)が開示される。有利なことに、本発明のシアリダーゼ、細胞外酵素。さらに、シアリダーゼは、好ましくは真菌由来である。一般に、本発明のポリペプチドは、35kDaを超えるMW(分子量)(SDS−Page)を有し、より好ましくは、このポリペプチドは40kDaを超えるMW(分子量)(SDS−Page)を有するか、またはこのポリペプチドは35kDaを超えるMW(分子量)(アミノ酸配列に基づいて算出される)を有し、より好ましくは、このポリペプチドは40kDaを超えるMW(分子量)(アミノ酸配列に基づいて算出される)を有する。
【0014】
エキソ酵素、または細胞外酵素は、細胞から分泌され、そして細胞の外側で作用する酵素である。それは、分解しなければ細胞に進入し得ない大きな分子を分解するために使用される。生物体が環境に分泌する酵素が、微生物の外側で作用する。エキソ酵素の反意語は、エンド酵素または細胞内酵素と呼ばれる(出典はWikipediaである)。
【0015】
本発明者らの理論では、一般に、大きなシアリダーゼは、粘膜感染およびビルレンスに関連し、従って、食品または製薬用途において適切ではない。それとは対照的に、小さなシアリダーゼは、食品または製薬用途に適切であると考えられる。
【0016】
しかし、本発明は、この理論の正確さについて肯定も否定もしない。本発明は、55kDa未満のMW(分子量)(SDS−Page)を有する細胞外シアリダーゼ(より好ましくは、このシアリダーゼは52kDa未満のMW(分子量)(SDS−Page)を有する)、または50kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)有する細胞外シアリダーゼ(より好ましくは、このシアリダーゼは45kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有する)を提供する。
【0017】
本発明は、真菌ペニシリウム・クリソゲナム(Penicillium chrysogenum)において同定された新たなシアリダーゼに関する。
【0018】
有利なことに、本発明は、多量に産生され得るシアリダーゼの需要を満たす。好ましくは、そのようなシアリダーゼは宿主細胞から分泌される。能動的分泌は、面倒な精製プロセスを経験することなく、ほとんど純粋な形の酵素の回収を可能にするため、それは、経済的な産生プロセスに最も重要である。アスペルギルス(Aspergillus)のような食品級真菌宿主によるそのような能動的に分泌されるシアリダーゼの過剰発現は、食品級酵素および費用効果の高い産生プロセスを生み出し、従って、好適である。本分泌型シアリダーゼは、糸状菌においてはじめて見出されたものである。食品級産生宿主アスペルギルス・ニガー(Aspergillus niger)による大量のシアリダーゼの産生のためのプロセスについて開示する。
【0019】
経済的観点から、哺乳動物および細菌シアリダーゼの低い産生率と比較して、多量かつ比較的純粋な形でシアリダーゼを産生させる改善された方法が明らかに必要である。これを行うための好適な方法は、組み換えDNA技術を使用するそのようなシアリダーゼの過剰産生を介することである。これを行うための特に好適な方法は、真菌由来のシアリダーゼの過剰産生を介することであり、そしてこれを行うための最も好適な方法は、ペニシリウム(Penicillium)由来シアリダーゼの過剰産生を介することである。後者の産生経路を可能にするためには、ペニシリウム(Penicillium)由来シアリダーゼの独特な配列情報が必須である。より好適には、コーディング遺伝子のヌクレオチド配列全体が利用可能でなければならない。
【0020】
多量かつ比較的純粋な形で新たに同定された分泌型シアリダーゼを産生させる改善された手段は、組み換えDNA技術を使用するペニシリウム(Penicillium)によりコードされる酵素の過剰産生を介することである。これを行うための好適な方法は、食品級宿主微生物におけるそのような分泌型シアリダーゼの過剰産生を介することである。周知の食品級微生物として、アスペルギルス(Aspergilli)、トリコデルマ(Trichoderma)、ストレプトマイセス(Streptomyces)、バチルス(Bacilli)ならびにサッカロマイセス(Saccharomyces)およびクリヴェロマイセス(Kluyveromyces)のような酵母が挙げられる。これを行うためのさらにより好適な方法は、アスペルギルス(Aspergillus)のような食品級真菌における分泌型ペニシリウム(Penicillium)由来シアリダーゼの過剰産生を介することである。シアリダーゼをコードする遺伝子のコドン使用頻度が、使用する食品級発現宿主に最適化されている食品級真菌における分泌型シアリダーゼの過剰産生が最も好適である。一般に、後者の最適化経路を可能にするためには、分泌型シアリダーゼの独特な配列情報が望ましい。より好適には、シアリダーゼコーディング遺伝子のヌクレオチド配列全体が利用可能でなければならない。一旦、分泌型シアリダーゼがコードする遺伝子が好適な宿主において形質転換されると、分泌型株を、発酵ブロスからの分泌型シアリダーゼタンパク質の発酵および単離に使用することができる。
【0021】
一旦、新たな酵素が、大量かつ比較的純粋な食物、(チーズのような)食料品において利用可能になれば、改善されたテクスチャーおよび/または免疫学的特性を伴うタンパク質加水分解物を、食品級のおよび経済的な方法で調製することができる。酵素はまた、潜在的に保存剤として使用することもできる。保護的シアル酸残基を細菌壁から取り出すことによって、微生物は、免疫系によって認識され、そして排除される。
【0022】
シアリダーゼ活性を有する本発明のポリペプチドは、単離された形態であってもよい。本明細書において定義されるように、単離されたポリペプチドは、本質的に他の非シアリダーゼポリペプチドを含まず、そして典型的に、SDS−PAGEによって決定されるように、少なくとも約20%純粋、好ましくは少なくとも約40%純粋、より好ましくは少なくとも約60%純粋、さらにより好ましくは少なくとも約80%純粋、なおより好ましくは約90%純粋、および最も好ましくは約95%純粋である、内因的に産生されたまたは組み換えポリペプチドである。ポリペプチドは、遠心分離およびクロマトグラフィー法、または粗溶液から純粋なタンパク質を得るための他の任意の技術によって単離してもよい。ポリペプチドは、ポリペプチドの意図された目的を妨害しないキャリアまたは希釈剤と混合してもよく、それ故、この形のポリペプチドもなお単離されたものと認識されることが理解されよう。それは、一般的に、調製物中のポリペプチドを含んでなり、ここで、本発明のポリペプチドは、調製物中のタンパク質の重量に対し20%を超える、例えば、30%、40%、50%、80%、90%、95%もしくは99%を超える。
【0023】
好ましくは、本発明のポリペプチドは、シアリダーゼ活性を伴う酵素をコードする遺伝子を所有する微生物から入手され得る。より好ましくは、本発明のポリペプチドは、微生物から分泌される。さらにより好ましくは、微生物は真菌であり、そして場合により、糸状菌である。それ故、好適なドナー生物体は、種ペニシリウム・クリソゲナム(Penicillium chrysogenum)のようなペニシリウム(Penicillium)属である。第1の実施形態では、本発明は、配列番号3(即ち、ポリペプチド)のアミノ酸1〜407に対して、少なくとも60%、好ましくは少なくとも70%、より好ましくは少なくとも80%、さらにより好ましくは少なくとも90%、なおより好ましくは少なくとも95%、および最も好ましくは少なくとも97%のアミノ酸配列同一性の程度を有し、かつシアリダーゼ活性を有するアミノ酸配列を有する単離されたポリペプチドを提供する。
【0024】
本発明の目的のために、2つもしくはそれ以上アミノ酸配列間の同一性の程度は、マトリックスBlosum62および10の予想閾値を伴うBLAST Pタンパク質データベースサーチプログラム(アルトシュル(Altschul)ら、1997年、Nucleic Acids Research 25:3389−3402)によって決定される。
【0025】
本発明のポリペプチドは、配列番号3に記載のアミノ酸配列、もしくは実質的に相同な配列、またはシアリダーゼ活性を有するいずれかの配列のフラグメントを含んでなり得る。一般に、配列番号3に示す天然に存在するアミノ酸配列が好適である。
【0026】
本発明の1つの態様は、配列番号3に示されるアミノ酸配列を有するポリペプチドであって、それによって、シグナル配列が取り出される。プレ−もしくはシグナル−配列および/またはプロ−配列を含む配列番号3のアミノ酸配列のような分泌型酵素は、しばしば合成される。これらの配列は、しばしば、分泌プロセス中またはその後のいずれかにタンパク質から取り出される。従って、成熟分泌型タンパク質は、しばしば、もはや、これらのプレ−およびプロ−配列を含有しない。従って、配列番号3のアミノ酸1〜407と少なくとも60%のアミノ酸配列同一性を有するアミノ酸配列を有し、可能なプレ−またはプロ−配列を欠如するポリペプチドは、本発明の一部である。配列番号3のアミノ酸配列における可能なプロセシング部位は、アミノ酸33より後方に存在する。この場合、本発明に従う成熟酵素は、アミノ酸番号34から開始する。アミノ酸の取り出しのようなさらなるプロセシングによる配列番号3のアミノ酸配列からの他の改変も、それらが酵素の活性を妨げない限り、許容される。
【0027】
本発明のポリペプチドもまた、配列番号3のポリペプチドの天然に存在する変異体または種相同体を含んでなり得る。
【0028】
変異体は、例えば、真菌、細菌、酵母または植物細胞において天然に存在するポリペプチドであって、この変異体は、シアリダーゼ活性および配列番号3のタンパク質に実質的に類似の配列を有する。用語「変異体」は、配列番号3のシアリダーゼと同じ本質的特徴または基本的な生物学的機能を有するポリペプチドを指し、そして対立遺伝子変異体を含む。好ましくは、変異ポリペプチドは、配列番号3のポリペプチドと少なくとも同じレベルのシアリダーゼ活性を有する。変異体は、配列番号3のポリペプチドと同じ株または同じ属もしくは種の異なる株のいずれかに由来する対立遺伝子変異体を含む。
【0029】
同様に、本発明のタンパク質の種相同体は、シアリダーゼである類似の配列の等価なタンパク質であり、そして別の種において天然に存在する。
【0030】
変異体および種相同体は、本明細書に記載の手順を使用し、そして適切な細胞供給源、例えば、細菌、酵母、真菌または植物細胞に対してそのような手順を実施して、単離することができる。また、配列番号3のポリペプチドの変異体または種相同体を発現するクローンを入手するために、本発明のプローブを使用して、酵母、細菌、真菌または植物細胞から作製したDNAライブラリーを探索することも可能である。公知の遺伝子の変異体および種相同体を単離するために使用することができる方法は、文献において広範に記載されており、そして当業者に公知である。これらの遺伝子は、従来の技術によって操作して、本発明のポリペプチドを作製することができ、その後、それ自体が公知の組み換えまたは合成技術によって産生され得る。
【0031】
配列番号3のポリペプチドならびに変異体および種相同体の配列を改変して、本発明のポリペプチドを提供することもできる。アミノ酸置換は、例えば、1、2または3〜10、20または30置換から作製され得る。同じ数の欠失および挿入を作製してもよい。これらの変更は、改変されたポリペプチドがそのシアリダーゼ活性を保持するように、ポリペプチドの機能に極めて重要な領域の外側で作製してもよい。
【0032】
本発明のポリペプチドは、上記の全長ポリペプチドおよびその変異体のフラグメントを含み、配列番号3に記載の配列のフラグメントを含む。そのようなフラグメントは、典型的に、シアリダーゼとしての活性を保持する。フラグメントは、少なくとも50、100もしくは200アミノ酸長であってもよく、または配列番号3に示す全長配列より短いこのような数のアミノ酸であってもよい。
【0033】
通常、本発明のポリペプチドは、以下に記載のように組み換え的に作製されるが、必要であれば、それらを、合成手段によって生成することもできる。合成および組み換えポリペプチドは、例えば、それらの同定もしくは精製を補助するためのヒスチジン残基またはT7タグの付加によってか、あるいは細胞からのそれらの分泌を促進するためのシグナル配列の付加によって、改変され得る。
【0034】
それ故、変異配列は、配列番号3のポリペプチドを単離した株以外のペニシリウム(Penicillium)の株から誘導される配列を含んでなり得る。変異体は、シアリダーゼ活性を調べ、そして本明細書に記載のようにクローニングおよび配列決定することによって、他のペニシリウム(Penicillium)株から同定することができる。ペプチドが、配列番号3のシアリダーゼの基本的な生物学的機能を維持する限り、変異体は、タンパク質配列内の単一のアミノ酸もしくはアミノ酸群の欠失、改変または付加を含んでもよい。
【0035】
アミノ酸置換は、例えば、1、2または3〜10、20または30置換から作製され得る。改変されたポリペプチドは、一般的に、シアリダーゼとしての活性を保持する。保存的置換を行ってもよく;そのような置換は、当該技術分野において周知である。
【0036】
より短いまたはより長いポリペプチド配列も本発明の範囲内にある。例えば、少なくとも50アミノ酸または100、150、200、300、400、500、600、700もしくは800アミノ酸長までのペプチドが、それが配列番号3のシアリダーゼの基本的な生物学的機能を実証する限り、本発明の範囲内に当てはまると考えられる。特に、但し排他的ではないが、本発明のこの態様は、タンパク質が完全なタンパク質配列のフラグメントである状況を包含する。
【0037】
本発明はまた、シアリダーゼ活性を有するポリペプチドをコードするポリヌクレオチドに関し、前記ポリヌクレオチドは、アミノ酸配列番号3をコードするポリヌクレオチド配列を含んでなる。
【0038】
本発明では、目的のタンパク質が、能動的に増殖培地に分泌されることが特に関連する。分泌型タンパク質は、通常、プレ−タンパク質として最初は合成され、続いてプレ−配列(シグナル配列)が、分泌プロセス中に取り出される。分泌プロセスは、原核細胞および真核生物において基本的に類似する:能動的に分泌されるプレ−タンパク質は膜を通り抜け、シグナル配列が特異的シグナルペプチダーゼによって取り出され、そして成熟タンパク質が(リ)フォールディングされる。また、シグナル配列について、一般構造を認識することができる。分泌のためのシグナル配列は、プレ−タンパク質のアミノ末端に局在し、そして一般的に、15〜35アミノ酸長である。アミノ末端は、好ましくは、正に荷電したアミノ酸を含有し、そして好ましくは、酸性アミノ酸を含有しない。この正に荷電した領域は、膜のリン脂質の負に荷電したヘッド基と相互作用すると考えられる。この領域の後に、疎水性の膜貫通コア領域が続く。この領域は、一般的に、10〜20アミノ酸長であり、そして主に、疎水性アミノ酸よりなる。荷電したアミノ酸は、通常、この領域には存在しない。膜貫通領域の後には、シグナルペプチダーゼの認識部位が続く。認識部位は、小−X−小を選好するアミノ酸よりなる。小さなアミノ酸は、アラニン、グリシン、セリンまたはシステインであり得る。Xは、任意のアミノ酸であり得る。そのような規則を使用して、真核生物および原核細胞由来のそのようなシグナル配列を認識することが可能なアルゴリズムが作成されている(ベントセン(Bendtsen)、ニールセン(Nielsen)、フォン・ヘイネ(von Heijne)およびブルナック(Brunak).(2004年)J.Mol.Biol.,340:783−795)。タンパク質のシグナル配列を算出し、そして認識するためのSignalPプログラムが、一般的に利用可能である(http://www.cbs.dtu.dk/services/SignalP/)。
【0039】
シグナル配列が配列決定した遺伝子の推測されたタンパク質配列から認識され得ることは、本発明に関連する。遺伝子がSignalPプログラムを使用してシグナル配列が予測されるタンパク質をコードする場合、このタンパク質が分泌される可能性は高い。
【0040】
第2の実施形態では、本発明は、シアリダーゼ活性を有し、そして低度のストリンジェンシー条件下で、より好ましくは、中度のストリンジェンシー条件下で、および最も好ましくは、高度のストリンジェンシー条件下で、(i)配列番号1の核酸配列、または(ii)配列番号1の少なくとも一部を含んでなる核酸フラグメント、もしくは(iii)配列番号1の塩基とは異なる塩基を有する核酸フラグメントと;あるいは(iv)配列番号1に相補的な核酸鎖とハイブリダイズするかまたはハイブリダイズすることが可能であるポリヌクレオチドによってコードされる単離されたポリペプチドを提供する。
【0041】
用語「ハイブリダイズすることが可能」は、本発明の標的ポリヌクレオチドが、プローブとして使用される核酸(例えば、配列番号1のヌクレオチド配列、もしくはそのフラグメント、または配列番号1の相補体もしくはそのフラグメント)と、バックグランドよりも有意に高いレベルでハイブリダイズすることができることを意味する。本発明はまた、本発明のシアリダーゼをコードするポリヌクレオチド、ならびにそれに相補的であるヌクレオチド配列を含む。ヌクレオチド配列は、RNA、またはゲノムDNA、合成DNAもしくはcDNAを含むDNAであってもよい。好ましくは、ヌクレオチド配列は、DNAおよび最も好ましくは、ゲノムDNA配列である。典型的に、本発明のポリヌクレオチドは、選択的条件下で、配列番号1のコーディング配列またはコーディング配列の相補体にハイブリダイズすることが可能であるヌクレオチドの連続配列を含んでなる。そのようなヌクレオチドは、当該技術分野において周知の方法に従って合成することができる。
【0042】
本発明のポリヌクレオチドは、配列番号2のコーディング配列またはコーディング配列の相補体に、バックグランドよりも有意に高いレベルでハイブリダイズすることができる。例えば、cDNAライブラリーに存在する他のcDNAのため、バックグランドのハイブリダイゼーションが生じ得る。本発明のポリヌクレオチドと配列番号2のコーディング配列またはコーディング配列の相補体との間の相互作用によって作製されるシグナルレベルは、典型的に、他のポリヌクレオチドと配列番号2のコーディング配列との間の相互作用の少なくとも10倍、好ましくは少なくとも20倍、より好ましくは少なくとも50倍、およびさらにより好ましくは少なくとも100倍の強度である。相互作用の強度は、例えば、プローブを、例えば、32Pで放射性標識することによって、測定することができる。選択的ハイブリダイゼーションは、典型的に、低いストリンジェンシー(0.3M塩化ナトリウムおよび0.03Mクエン酸ナトリウム、約40℃)、中等度のストリンジェンシー(例えば、0.3M塩化ナトリウムおよび0.03Mクエン酸ナトリウム、約50℃)または高いストリンジェンシー(例えば、0.3M塩化ナトリウムおよび0.03Mクエン酸ナトリウム、約60℃)の条件を使用して、達成することができる。
【0043】
本発明のポリヌクレオチドはまた、配列番号3のポリペプチドまたはその変異体をコードすることができる合成遺伝子を含む。時々、遺伝子のコドン使用頻度を産生宿主の好適なバイアスに適応することも好適である。合成遺伝子を設計および構築するための技術は、一般的に利用可能である(即ち、http://www.dnatwopointo.com/)。
【0044】
[改変]
本発明のポリヌクレオチドは、DNAまたはRNAを含んでなり得る。それらは、一本鎖であってもまたは二本鎖であってもよい。それらはまた、それらの内部にペプチド核酸を含む合成または改変されたヌクレオチドを含むポリヌクレオチドであってもよい。ポリヌクレオチドに対する異なる多くのタイプの改変が、当該技術分野において公知である。これらは、メチルホスホネートおよびホスホロチオエート骨格、ならびに分子の3’および/または5’末端におけるアクリジン鎖またはポリリジン鎖の付加を含む。本発明の目的のために、本明細書に記載のポリヌクレオチドは、当該技術分野において利用可能な任意の方法によって改変され得ることが理解されるべきである。
【0045】
当業者は、日常的な技術を使用して、本発明のポリヌクレオチドによってコードされるポリペプチド配列に影響を及ぼさないヌクレオチド置換を行って、本発明のポリペプチドを発現させようとする任意の特定の宿主生物体のコドン使用頻度に反映させ得ることが理解されるべきである。
【0046】
配列番号2のコーディング配列は、ヌクレオチド置換、例えば、1、2もしくは3〜10、25、50、100、もしくはそれ以上の置換によって、改変してもよい。配列番号2のポリヌクレオチドは、1つもしくはそれ以上の挿入および/または欠失によって、ならびに/あるいはいずれか一方もしくは両方の末端における伸長によって代替的または付加的に改変され得る。改変されたポリヌクレオチドは、一般的に、シアリダーゼ活性を有するポリペプチドをコードする。例えば、後に、ポリペプチドに関して考察するように、改変された配列が翻訳される場合、縮重置換を行ってもよく、および/または保存的アミノ酸置換を生じる置換を行ってもよい。
【0047】
[相同体]
配列番号2のDNAコーディング配列の相補体に選択的にハイブリダイズすることが可能であるヌクレオチド配列は、本発明に含まれ、そして一般的に、配列番号2の少なくとも60、好ましくは少なくとも100、より好ましくは少なくとも200連続ヌクレオチドの領域にわたって、または最も好ましくは全長にわたって配列番号2のコーディング配列と少なくとも50%もしくは60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%または少なくとも99%の配列同一性を有する。同様に、活性なシアリダーゼをコードし、そして配列番号2のDNAコーディング配列の相補体のフラグメントと選択的にハイブリダイズすることが可能であるヌクレオチドもまた、本発明に包含される。
【0048】
上記の程度の同一性および最小サイズの任意の組み合わせを使用して、本発明のポリヌクレオチドを定義してもよく、よりストリンジェントな組み合わせ(即ち、より長い長さにわたって同一性がより高い)が好適である。それ故、例えば、60、好ましくは、100ヌクレオチドにわたって少なくとも80%または90%同一であるポリヌクレオチドは、200ヌクレオチドにわたって少なくとも90%同一であるポリヌクレオチドと同様に、本発明の1つの態様を形成する。
【0049】
BLAST Nアルゴリズムを使用して、配列同一性を算出するかまたは(例えば、それらのデフォルト設定値に対する均等もしくは対応する配列を同定するような)配列を列挙することができる。
【0050】
BLAST解析を実施するためのソフトウェアは、National Center for Biotechnology Information(http://www.ncbi.nlm.nih.gov/)を介して公的に入手することができる。このアルゴリズムは、データベース配列において同じ長さのワードと共に整列される場合、いくつかのポジティブ評価された閾値スコアTに一致するか、または満たすかのいずれかである問い合わせ配列における長さWの短いワードを同定することによって、ハイスコアリング配列対(high scoring sequence pair)(HSP)を第1に同定することに関与する。Tは隣接ワードスコア閾値と称される。これらの初期の隣接ワードのヒットは、それらを含有するHSPを見出すための検索を初期化するためのシードとして作用する。ワードヒットは、累積アラインメントスコアが増加し得る限り、各配列に沿って両方向において拡張される。累積アラインメントスコアがその最大到達値から量Xだけ低下する場合:1つもしくはそれ以上のネガティブスコアリング残基アラインメントの累積により累積スコアが0もしくはそれ以下となる場合;またはいずれかの配列の末端に到達する場合、各方向におけるワードヒットの拡張は停止する。BLASTアルゴリズムパラメータW、TおよびXは、アラインメントの感度および速度を決定する。BLASTプラグラムは、デフォルトとして、11のワード長(W)、50のBLOSUM62スコアリングマトリックスアラインメント(B)、10の期待値(E)、M=5、N=4、および両鎖の比較を使用する。
【0051】
BLASTアルゴリズムは、2つの配列間の類似性の統計解析を実施する。BLASTアルゴリズムによって提供される類似性の1つの測定は、2つのヌクレオチドまたはアミノ酸配列間の一致が偶然に生じる確率の表示を提供する最小和確率(P(N))である。例えば、配列は、第1の配列と第2の配列との比較における最小和確率が約1未満、好ましくは約0.1未満、より好ましくは約0.01未満、および最も好ましくは約0.001未満である場合、もう1つの配列に類似であるとみなされる。
【0052】
[プライマーおよびプローブ]
本発明のポリヌクレオチドは、プライマー、例えば、ポリメラーゼ連鎖反応(PCR)プライマー、代替的増幅反応のためのプライマー、または例えば、放射性もしくは非放射性標識を使用する従来の手段によって明示用標識(revealing label)で標識されたプローブを含み、そして上記プライマーまたはプローブとして使用してもよく、あるいはポリヌクレオチドをベクターにクローニングしてもよい。そのようなプライマー、プローブおよび他のフラグメントは、少なくとも15、例えば、少なくとも20、25、30もしくは40ヌクレオチド長である。それらは、典型的に、40、50、60、70、100、150、200もしくは300ヌクレオチド長までであるか、またはなお、配列番号2のコーディング配列より数ヌクレオチド(例えば、5もしくは10ヌクレオチド)短い長さまでである。
【0053】
一般に、プライマーは、1回に1ヌクレオチドの所望される核酸配列の段階的な製造を含めた合成手段によって生成される。これを達成するための技術およびプロトコルは、当該技術分野において容易に利用可能である。より長いポリヌクレオチドは、一般的に、組み換え手段、例えば、PCRクローニング技術を使用して、生成される。これは、(典型的に、約15〜30ヌクレオチドの)プライマーの対を作製して、クローニングしようとするシアリダーゼの所望される領域を増幅し、プライマーと、酵母、細菌、植物、原核細胞または真菌細胞、好ましくは、ペニシリウム(Penicillium)株から得られるmRNA、cDNAもしくはゲノムDNAとを接触させ、所望される領域の増幅に適切な条件下でポリメラーゼ連鎖反応を実施し、(例えば、アガロースゲル上で反応混合物を精製することによって)増幅したフラグメントを単離し、そして増幅したDNAを回収することを含む。実施例1に記載のように、増幅したDNAを適切なクローニングベクターにクローニングすることができるように、適切な制限酵素認識部位を含有するようにプライマーを設計してもよい。
【0054】
あるいは、分泌型シアリダーゼまたはその変異体のコーディング領域を包含する合成遺伝子を構築することができる。これらの技術を使用して、多くの位置で変更されているが、なお同じタンパク質をコードするポリヌクレオチドを簡便に設計し、そして構築することができる。これは、コドン使用頻度を好適な発現宿主に適応することができ、そのようにしてこの宿主におけるタンパク質の産生率を改善することができることを利点として有する。また、遺伝子のポリヌクレオチド配列を変更して、mRNA安定性または減少したターンオーバーを改善することができる。これは、所望されるタンパク質またはその変異体の改善された発現をもたらすことができる。さらに、分泌効率、安定性、タンパク質分解に対する脆弱性、至適温度、比活性またはタンパク質の産業的産生もしくは用途のための他の関連特性に対してポジティブな効果を有する変異がタンパク質配列において作製されるように、合成遺伝子においてポリヌクレオチド配列を変更することができる。一般的に、合成遺伝子を構築し、そしてコドン使用頻度を最適化する業務を提供する企業が利用可能である。
【0055】
そのような技術を使用して、本明細書に記載のシアリダーゼ配列をコードするポリヌクレオチドのすべてまたは一部分を入手してもよい。イントロン、プロモーターおよびトレーラー領域は、本発明の範囲内にあり、そしてまた、真菌、酵母、細菌、植物または原核細胞由来のゲノムDNAから開始して、類似の様式で(例えば、組み換え手段、PCRもしくはクローニング技術によって)入手してもよい。
【0056】
ポリヌクレオチドまたはプライマーは、明示用標識を担持してもよい。適切な標識として、放射性同位元素、例えば、32Pまたは35S、蛍光標識、酵素標識または他のタンパク質標識、例えば、ビオチンが挙げられる。そのような標識は、本発明のポリヌクレオチドまたはプライマーに付加し得、そして当業者に公知の技術を使用して検出し得る。
【0057】
標識されたまたは標識されていないポリヌクレオチドまたはプライマー(もしくはそのフラグメント)を、真菌サンプルにおけるシアリダーゼまたはその変異体を検出または配列決定するための核酸に基づく試験に使用してもよい。そのような検出試験は、一般的に、目的のDNAを含有することが疑わしい真菌サンプルと、本発明のポリヌクレオチドまたはプライマーを含んでなるプローブとを、ハイブリダイジング条件下で接触させ、そしてプローブとサンプル中の核酸との間に形成される任意の二重鎖を検出することを含んでなる。検出は、PCRのような技術を使用するか、またはプローブを固相支持体上に固定化し、プローブにハイブリダイズしないサンプル中の任意の核酸を取り出し、次いで、プローブにハイブリダイズする任意の核酸を検出することによって、達成され得る。あるいは、サンプル核酸を固相支持体上に固定化し、プローブをハイブリダイズさせ、そして、任意の非結合プローブの取り出し後、そのような支持体に結合したプローブの量を検出してもよい。
【0058】
本発明のプローブを、適切な容器に試験キットの形で簡便に包装してもよい。そのようなキットでは、プローブを固相支持体に結合させてもよく、ここで、キットが設計されるアッセイ形式は、そのような結合を必要とする。キットはまた、探索しようとするサンプルを処理し、プローブをサンプル中の核酸にハイブリダイズさせるための適切な試薬、コントロール試薬、取扱説明書などを含有してもよい。本発明のプローブおよびポリヌクレオチドはまた、マイクロアッセイに使用してもよい。
【0059】
好ましくは、本発明のポリヌクレオチドは、真菌、特に、アスペルギルス(Aspergillus)属の真菌のようなポリペプチドと同じ生物体から入手可能である。
【0060】
[ポリヌクレオチドの生成]
配列番号2と100%の同一性を有さないが、本発明の範囲内に当てはまるポリヌクレオチドは、多くの方法で入手することができる。それ故、本明細書に記載のシアリダーゼ配列の変異体は、例えば、本発明のポリペプチドの供給源として考察したような生物体の範囲から作製されるゲノムDNAライブラリーを探索することによって、入手してもよい。加えて、シアリダーゼの他の真菌、植物または原核生物相同体が入手され得、そしてそのような相同体およびそのフラグメントは、一般に、配列番号1にハイブリダイズすることが可能である。そのような配列は、他の種由来のcDNAライブラリーまたはゲノムDNAライブラリーを探索すること、およびそのようなライブラリーを、(先に記載のように)低度、中度〜高度のストリンジェンシーの条件下、配列番号1のすべてまたは一部分を含んでなるプローブで探索することによって、入手され得る。配列番号1のすべてまたは一部分を含んでなる核酸プローブを使用して、本発明のポリペプチドの供給源として記載したような他の種由来のcDNAまたはゲノムライブラリーを探索してもよい。
【0061】
種相同体はまた、保存されたアミノ酸配列をコードする変異体および相同体内の配列を標的化するように設計されたプライマーを使用する縮重PCRを使用して、入手してもよい。プライマーは、1つもしくはそれ以上の縮重位置を含有することができ、そして既知の配列に対する単一の配列プライマーを伴う配列をクローニングするために使用される条件より低いストリンジェンシー条件で使用される。
【0062】
あるいは、そのようなポリヌクレオチドは、シアリダーゼ配列またはその変異体の部位特異的変異によって入手され得る。これは、例えば、配列に対するサイレントコドンの変化が、ポリヌクレオチド配列が発現される特定の宿主細胞についてコドン選択を最適化するために必要とされる場合、有用であり得る。制限酵素認識部位を導入するため、またはポリヌクレオチドによってコードされるポリペプチドの特性または機能を改変するために、他の配列変化を作製してもよい。
【0063】
本発明は、本発明のポリヌクレオチドおよびその相補体を含んでなる二本鎖ポリヌクレオチドを含む。
【0064】
本発明はまた、上記の本発明のポリペプチドをコードするポリヌクレオチドを提供する。そのようなポリヌクレオチドは、本発明のポリペプチドの組み換え産生のための配列に有用であるため、これは一般的に望ましいが、それらが、配列番号1または配列番号2の配列にハイブリダイズすることが可能である必要はない。そうでなければ、所望であれば、そのようなポリヌクレオチドを、上記のように標識、使用、および作製してもよい。
【0065】
[組み換えポリヌクレオチド]
本発明はまた、クローニングベクターおよび発現ベクターを含む本発明のポリヌクレオチドを含んでなるベクター、ならびに別の態様において、増殖、形質転換、または例えば、本発明のポリペプチド、もしくは本発明の配列によってコードされるポリペプチドの発現が生じる条件下でそのようなベクターを適切な宿主細胞にトランスフェクトする方法を提供する。本発明のポリヌクレオチドまたはベクターを含んでなる宿主細胞もまた提供され、ここで、ポリヌクレオチドは、宿主細胞のゲノムに対して異種である。通常、宿主細胞に関する用語「異種」は、ポリヌクレオチドが宿主細胞のゲノムにおいて天然には存在しないか、またはポリペプチドが細胞によって天然には産生されないことを意味する。好ましくは、宿主細胞は、酵母細胞、例えば、クリヴェロマイセス(Kluyveromyces)、ピキア(Pichia)、ハンセヌラ(Hansenula)もしくはサッカロマイセス(Saccharomyces)属の酵母細胞または例えば、アスペルギルス(Aspergillus)、ペニシリウム(Penicillium)、トリコデルマ(Trichoderma)もしくはフザリウム(Fusarium)属の糸状菌細胞である。
【0066】
[ベクター]
本発明の発現カセットが挿入されるベクターは、組み換えDNA手順に簡便に供され得る任意のベクターであり得、そしてベクターの選択は、しばしば、それが導入されるべき宿主細胞に依存する。それ故、ベクターは、自律的に複製するベクター、即ち、プラスミドのように染色体外の実体として存在し、その複製は、染色体の複製に依存しないベクターであってもよい。あるいは、ベクターは、宿主細胞に導入される場合、宿主細胞のゲノムに組込まれ、それが組込まれている染色体と共に複製するものであってもよい。
【0067】
好ましくは、本発明のポリヌクレオチドがベクター中にある場合、それは、宿主細胞によるコーディング配列の発現を提供することが可能である調節配列に作動可能に連結される(即ち、ベクターは発現ベクターである)。用語「作動可能に連結された」は並置を指し、ここで、記載の成分は、それらが、それらの意図された様式で機能することを可能にする関係にある。プロモーター、エンハンサーまたはコーディング配列に「作動可能に連結された」他の発現調節シグナルのような調節配列は、コーディング配列の発現が、産生条件下で達成されるような方法で配置される。
【0068】
ベクターは、例えば、プラスミド、コスミド、ウイルスもしくはファージベクターの場合、複製開始点、場合により、ポリヌクレオチドの発現のためのプロモーター、ならびに場合により、プロモーターのエンハンサーおよび/またはレギュレーターを伴って提供され得る。ターミネーター配列が存在してもよく、それはポリアデニル化配列であってもよい。ベクターは、1つもしくはそれ以上の選択マーカー遺伝子、例えば、細菌プラスミドの場合にはアンピシリン耐性遺伝子または哺乳動物ベクターではネオマイシン耐性遺伝子を含有してもよい。ベクターは、例えば、RNAの産生のためにインビトロで使用してもよく、または宿主細胞をトランスフェクトもしくは形質転換するために使用することができる。
【0069】
ポリペプチドをコードするDNA配列は、好ましくは、DNA配列が、宿主細胞におけるDNA配列の発現を指令することが可能である発現シグナルに作動可能に連結される発現構築物の一部分として適切な宿主に導入される。発現構築物による適切な宿主の形質転換のために、当業者に周知である形質転換手順が利用可能である。発現構築物は、選択マーカーを担持するベクターの一部分として、宿主の形質転換に使用することができ、または発現構築物は、選択マーカーを担持するベクターと共に個別の分子として同時形質転換される。ベクターは、1つもしくはそれ以上の選択マーカー遺伝子を含有してもよい。
【0070】
好適な選択マーカーとして、宿主細胞における欠損を補うか、または薬物に対する耐性を付与するものが挙げられるが、これらに限定されない。それらは、例えば、アセトアミダーゼ遺伝子またはcDNA(amdS、niaD、facA遺伝子またはA.ニダランス(A.nidulans)、麹菌(A.oryzae)、もしくはA.ニガー(A.niger)由来のcDNA)のようなほとんどの糸状菌および酵母の形質転換に使用することができる多目的マーカー遺伝子、あるいはG418、ハイグロマイシン、ブレオマイシン、カナマイシン、フレオマイシンまたはベノミル耐性(benA)のような抗生物質に対する耐性を提供する遺伝子を含む。あるいは、対応する変異宿主株を必要とする栄養要求性マーカーのような特定の選択マーカー:例えば、URA3(S.セレビシエ(S.cerevisiae)由来もしくは他の酵母由来の類似の遺伝子)、pyrGまたはpyrA(A.ニダランス(A.nidulans)もしくはA.ニガー(A.niger)由来)、argB(A.ニダランス(A.nidulans)もしくはA.ニガー(A.niger)由来)あるいはtrpCを使用することができる。好適な実施形態では、発現構築物の導入後、選択マーカー遺伝子を含まないポリペプチドを産生することが可能な形質転換された宿主細胞が得られるように、選択マーカーを、形質転換された宿主細胞から欠失する。
【0071】
他のマーカーとして、ATPシンテターゼサブユニット9(oliC)、オロチジン−5’−リン酸−デカルボキシラーゼ(pvrA)、細菌G418耐性遺伝子(酵母において有用であるが、糸状菌では有用ではない)、アンピシリン耐性遺伝子(大腸菌(E.coli))、ネオマイシン耐性遺伝子(バチルス(Bacillus))およびグルクロニダーゼ(GUS)をコードする大腸菌(E.coli)uidA遺伝子が挙げられる。ベクターは、例えば、RNAの産生のために、または宿主細胞をトランスフェクトもしくは形質転換するためにインビトロで使用してもよい。
【0072】
ほとんどの糸状菌および酵母では、発現構築物は、好ましくは、安定な形質転換体を得るために、宿主細胞のゲノムに組み込まれる。しかし、所定の酵母では、発現構築物を、安定かつ高いレベルの発現のために組み入れることができる適切なエピソームベクター系もまた、入手可能である。それらの例として、それぞれ、サッカロマイセス(Saccharomyces)およびクリヴェロマイセス(Kluyveromyces)の2μm、CENおよびpKD1プラスミドから誘導されるベクター、またはAMA配列(例えば、アスペルギルス(Aspergillus)由来のAMA1)を含有するベクターが挙げられる。発現構築物を宿主細胞ゲノムに組み込む場合、構築物は、ゲノムの無作為の座位か、または相同組み換えを使用して、予め決定された標的座位(この場合、標的座位は、好ましくは、高度に発現される遺伝子を含んでなる)のいずれかに組み込まれる。高度に発現される遺伝子は、例えば、誘導条件下で、そのmRNAが少なくとも0.01%(w/w)の全細胞mRNAを構成し得る遺伝子、あるいは、その遺伝子産物が少なくとも0.2%(w/w)の全細胞性タンパク質を構成し得るか、または、分泌される遺伝子産物の場合、少なくとも0.05g/lのレベルまで分泌され得る遺伝子である。
【0073】
所定の宿主細胞のための発現構築物は、通常、第1の態様のポリペプチドをコードする配列のコーディング鎖に対して5’末端から3’末端への連続した順番で相互に作動可能に連結した次のエレメントを含有する:(1)所与の宿主細胞においてポリペプチドをコードするDNA配列の転写を指令することが可能なプロモーター配列、(2)好ましくは、5’非翻訳領域(リーダー)、(3)場合により、所与の宿主細胞由来のポリペプチドの培養培地への分泌を指令することが可能なシグナル配列、(4)成熟かつ好ましくは活性型のポリペプチドをコードするDNA配列、および好ましくはまた、(5)ポリペプチドをコードするDNA配列の下流で転写を終結することが可能な転写終結領域(ターミネーター)。
【0074】
ポリペプチドをコードするDNA配列の下流の発現構築物は、好ましくは、ターミネーターとも称される1つもしくはそれ以上の転写終結部位を含有する3’非翻訳領域を含有する。ターミネーターの起源はそれほど重要ではない。ターミネーターは、例えば、ポリペプチドをコードするDNA配列に由来し得る。しかし、好ましくは、細菌ターミネーターが、細菌宿主細胞において使用され、酵母ターミネーターが、酵母宿主細胞において使用され、そして糸状菌ターミネーターが、糸状菌宿主細胞において使用される。より好ましくは、ターミネーターは、ポリペプチドをコードするDNA配列が発現される宿主細胞に内因性である。
【0075】
本発明のポリペプチドをコードするポリヌクレオチドの増強された発現はまた、選択された発現宿主からの目的のタンパク質の発現、および所望であれば、分泌レベルを増加し、および/または本発明のポリペプチドの発現の誘導可能な制御を提供する役割を果たす異種の調節領域、例えば、プロモーター、シグナル配列およびターミネーター領域の選択によっても達成され得る。
【0076】
本発明のポリペプチドをコードする遺伝子に由来するプロモーターに加えて、他のプロモーターを使用して、本発明のポリペプチドの発現を指令してもよい。プロモーターは、所望される発現宿主において本発明のポリペプチドの発現を指令するその効率について、選択してもよい。
【0077】
プロモーター/エンハンサーおよび他の発現調節シグナルは、発現ベクターが設計される宿主細胞に適合可能であるように選択することができる。例えば、原核プロモーター、特に、大腸菌(E.coli)株に使用するのに適切なプロモーターを使用してもよい。本発明のポリペプチドの発現が哺乳動物細胞において行われる場合、哺乳動物のプロモーターを使用してもよい。組織特異的プロモーター、例えば、肝細胞特異的プロモーターを使用してもよい。ウイルスプロモーター、例えば、モロニーマウス白血病ウイルス長末端反復(MMLV LTR)、ラウス肉腫ウイルス(RSV)LTRプロモーター、SV40プロモーター、ヒトサイトメガロウイルス(CMV)IEプロモーター、単純ヘルペスウイルスプロモーターまたはアデノウイルスプロモーターを使用することもできる。
【0078】
適切な酵母プロモーターとして、S.セレビシエ(S.cerevisiae)GAL4およびADHプロモーターならびにS.ポンベ(S.pombe)nmt1およびadhプロモーターが挙げられる。哺乳動物のプロモーターとしては、重金属、例えば、カドミウムに応答して誘導され得るメタロチオネインプロモーターが挙げられる。ウイルスプロモーター、例えば、SV40ラージT抗原プロモーターまたはアデノウイルスプロモーターを使用してもよい。これらのすべてのプロモーターは、当該技術分野において容易に利用可能である。
【0079】
哺乳動物のプロモーター、例えば、β−アクチンプロモーターを使用してもよい。組織特異的プロモーター、特に、内皮または神経細胞特異的プロモーター(例えば、DDAHIおよびDDAHIIプロモーター)が特に好適である。ウイルスプロモーター、例えば、モロニーマウス白血病ウイルス長末端反復(MMLV LTR)、ラウス肉腫ウイルス(RSV)LTRプロモーター、SV40プロモーター、ヒトサイトメガロウイルス(CMV)IEプロモーター、アデノウイルス、HSVプロモーター(例えば、HSV IEプロモーター)、またはHPVプロモーター、特に、HPV上流調節領域(URR)を使用することもできる。ウイルスプロモーターは、当該技術分野において容易に利用可能である。
【0080】
本発明の宿主細胞において転写を指令することが可能である多様なプロモーターを使用することができる。好ましくは、プロモーター配列は、先に定義したように、高度に発現される遺伝子から誘導される。好ましくは、プロモーターが誘導され、および/または発現構築物の組み込みのために、好適な予め決定された標的座位に含まれる好適な高度に発現される遺伝子の例として、トリオース−リン酸イソメラーゼ(TPI)、グリセルアルデヒド−リン酸デヒドロゲナーゼ(GAPDH)、ホスホグリセリン酸キナーゼ(PGK)、ピルビン酸キナーゼ(PYK)、アルコールデヒドロゲナーゼ(ADH)のような解糖系酵素をコードする遺伝子、ならびにアミラーゼ、グルコアミラーゼ、プロテアーゼ、キシラナーゼ、セロビオヒドロラーゼ、β−ガラクトシダーゼ、アルコール(メタノール)オキシダーゼ、伸長因子およびリボソームタンパク質をコードする遺伝子が挙げられるが、これらに限定されない。適切な高度に発現される遺伝子の具体例として、例えば、クリヴェロマイセス(Kluyveromyces)sp.由来のLAC4遺伝子、ハンセヌラ(Hansenula)およびピキア(Pichia)由来のそれぞれメタノールオキシダーゼ遺伝子(AOXおよびMOX)、A.ニガー(A.niger)およびA.アワモリ(A.awamori)由来のグルコアミラーゼ(glaA)遺伝子、麹菌(A.oryzae)TAKA−アミラーゼ遺伝子、A.ニダランス(A.nidulans)gpdA遺伝子およびT.リーセイ(T.reesei)セロビオヒドロラーゼ遺伝子が挙げられる。
【0081】
真菌発現宿主に使用するために好適である強力な構成性および/または誘導性プロモーターの例には、キシラナーゼ(xlnA)、フィターゼ、ATP−シンテターゼサブユニット9(oliC)、トリオースリン酸イソメラーゼ(tpi)、アルコールデヒドロゲナーゼ(AdhA)、アミラーゼ(amy)、アミログルコシダーゼ(glaA遺伝子由来のAG−)、アセトアミダーゼ(amdS)およびグリセルアルデヒド−3−リン酸デヒドロゲナーゼ(gpd)プロモーターのための真菌から入手可能なプロモーターがある。
【0082】
使用することができる強力な酵母プロモーターの例として、アルコールデヒドロゲナーゼ、グリセルアルデヒド−3−リン酸デヒドロゲナーゼ、ラクターゼ、3−ホスホグリセリン酸キナーゼ、形質膜ATPase(PMA1)およびトリオースリン酸イソメラーゼの遺伝子から入手可能なプロモーターが挙げられる。
【0083】
使用することができる強力な細菌プロモーターの例として、アミラーゼおよびSPo2プロモーター、ならびに細胞外プロテアーゼ遺伝子由来のプロモーターが挙げられる。
【0084】
使用することができる植物細胞に適切なプロモーターとして、ナパリンシンターゼ(nos)、オクトピンシンターゼ(ocs)、マンノピンシンターゼ(mas)、リブロース小サブユニット(rubisco ssu)、ヒストン、イネアクチン、ファゼオリン、カリフラワーモザイクウイルス(CMV)35Sおよび19Sならびにサーコウイルスプロモーターが挙げられる。
【0085】
ベクターは、真核生物のゲノム配列、好ましくは、真菌のゲノム配列、または酵母ゲノム配列に相同な配列を含んでなるRNAを生じるポリヌクレオチドにフランキングする配列をさらに含んでもよい。これは、相同組換えによる本発明のポリヌクレオチドの真菌または酵母のゲノムへの導入を可能にする。特に、真菌配列によってフランキングされる発現カセットを含んでなるプラスミドベクターを使用して、本発明のポリヌクレオチドを真菌細胞に送達するのに適切なベクターを調製することができる。これらの真菌ベクターを使用する形質転換技術は当業者に公知である。
【0086】
ベクターは、アンチセンスRNAの産生を提供するためにアンチセンス方向に配向された本発明のポリヌクレオチドを含有してもよい。所望であれば、これを使用して、ポリペプチドの発現のレベルを減少してもよい。
【0087】
[宿主細胞および発現]
さらなる態様において、本発明は、ポリペプチドをコードするコーディング配列のベクターによる発現に適切な条件下で、上記の発現ベクターで形質転換またはトランスフェクトされた宿主細胞を培養し、そして発現されたポリペプチドを回収することを含んでなる本発明のポリペプチドを調製するための方法を提供する。本発明のポリヌクレオチドは、複製可能な組み換えベクター、例えば、発現ベクターに組み入れることができる。ベクターは、適合可能な宿主細胞において核酸を複製するために使用され得る。それ故、さらなる実施形態では、本発明は、本発明のポリヌクレオチドを複製可能なベクターに導入し、ベクターを適合可能な宿主細胞に導入し、そしてベクターの複製を生じる条件下で宿主細胞を培養することによって、本発明のポリヌクレオチドを作製する方法を提供する。適切な宿主細胞として、大腸菌(E.coli)のような細菌、酵母、哺乳動物細胞系統、および他の真核細胞系統、例えば、Sf9細胞のような昆虫細胞、および(例えば、糸状)真菌細胞が挙げられる。
【0088】
好ましくは、発現構築物においてポリペプチドの成熟形態をコードするDNA配列が、シグナル配列をコードするDNA配列に作動可能に結合される場合、ポリペプチドは、分泌型タンパク質として産生される。分泌型タンパク質をコードする遺伝子が野生型株内にシグナル配列を有する場合、好ましくは、使用されるシグナル配列は、ポリペプチドをコードするDNA配列に由来する(それと同種である)。あるいは、シグナル配列は、ポリペプチドをコードするDNA配列に外来性(異種)であり、その場合、シグナル配列は、DNA配列が発現される宿主細胞に好ましくは内因性である。酵母宿主細胞に適切なシグナル配列の例には、酵母MFα遺伝子由来のシグナル配列がある。同様に、糸状菌宿主細胞に適切なシグナル配列には、例えば、糸状菌のアミログルコシダーゼ(AG)遺伝子、例えば、A.ニガー(A.niger)glaA遺伝子由来のシグナル配列がある。このシグナル配列は、アミログルコシダーゼ((グルコ)アミラーゼとも呼ばれる)プロモーター自体と組み合わせて、ならびに他のプロモーターと組み合わせて使用してもよい。ハイブリッドシグナル配列もまた、本発明に関して使用され得る。
【0089】
好適な異種分泌リーダー配列は、真菌アミログルコシダーゼ(AG)遺伝子(glaA−両方とも、例えば、アスペルギルス(Aspergillus)由来の18および24アミノ酸バージョン)、MFα遺伝子(酵母、例えば、サッカロマイセス(Saccharomyces)、ピキア(Pichia)ならびにクリヴェロマイセス(Kluyveromyces))またはα−アミラーゼ遺伝子(バチルス(Bacillus))由来のそれらである。
【0090】
ベクターは、本発明のポリペプチドの発現を提供するために、上記の適切な宿主細胞に形質転換またはトランスフェクトしてもよい。このプロセスは、ポリペプチドの発現に適切な条件下で上記のように発現ベクターで形質転換された宿主細胞を培養し、そして場合により、発現されたポリペプチドを回収することを含んでなり得る。
【0091】
それ故、本発明のさらなる態様は、本発明のポリヌクレオチドもしくはベクターで形質転換またはトランスフェクトされたか、あるいは本発明のポリヌクレオチドもしくはベクターを含んでなる宿主細胞を提供する。好ましくは、ポリヌクレオチドは、ポリヌクレオチドの複製および発現を可能にするベクターにおいて担持される。細胞は、前記ベクターに適合可能であるように選択され、そして例えば、原核生物(例えば、細菌)、または真核生物の真菌、酵母もしくは植物細胞であってもよい。
【0092】
本発明は、ポリヌクレオチドをコードするDNA配列の組み換え発現による本発明のポリペプチドの生産のためのプロセスを包含する。この目的のために、本発明のDNA配列は、適切な同種または異種の宿主細胞におけるポリペプチドの経済的な産生を可能にするために、遺伝子増幅および/または発現シグナル、例えば、プロモーター、分泌シグナル配列の交換のために使用され得る。同種の宿主細胞は、同じ種であるか、またはDNA配列が由来する種と同じ種内の変異体である宿主細胞として、本明細書において定義される。
【0093】
適切な宿主細胞は、好ましくは、細菌のような原核微生物、あるいはより好ましくは、真核生物体、例えば、酵母もしくは糸状菌のような真菌、または植物細胞である。一般に、酵母細胞の方が操作が容易であるため、酵母細胞は、糸状菌細胞より好適である。しかし、いくつかのタンパク質は、酵母からの分泌が不十分であるか、またはある場合に適切に処理されない(例えば、酵母における過グリコシル化)かのいずれかである。これらの場合、糸状菌宿主生物体が選択されるべきである。
【0094】
バチルス(Bacillus)属由来の細菌は、培養培地にタンパク質を分泌するそれらの能力のため、異種の宿主として極めて適切である。宿主として適切な他の細菌は、ストレプトマイセス(Streptomyces)およびシュードモナス(Pseudomonas)属由来の宿主である。ポリペプチドをコードするDNA配列の発現のための好適な酵母宿主細胞は、サッカロマイセス(Saccharomyces)、クリヴェロマイセス(Kluyveromyces)、ハンセヌラ(Hansenula)、ピキア(Pichia)、ヤロウイア(Yarrowia)、またはシゾサッカロマイセス(Schizosaccharomyces)属の1つである。より好ましくは、酵母宿主細胞は、サッカロマイセス・セレビシエ(Saccharomyces cerevisiae)、クリュイベロマイセス・ラクティス(Kluyveromyces lactis)(クリヴェロマイセス・マルキサナス・バー・ラクティス(Kluyveromyces marxianus var.lactis)としても公知である)、ハンセヌラ・ポリモルファ(Hansenula polymorpha)、ピキア・パストリス(Pichia pastoris)、ヤロウイア・リポリティカ(Yarrowia lipolytica)、およびシゾサッカロマイセス・ポンベ(Schizosaccharomyces pombe)の種からなる群から選択される。
【0095】
しかし、糸状菌宿主細胞が、ポリペプチドをコードするDNA配列の発現に最も好適である。好適な糸状菌宿主細胞は、アスペルギルス(Aspergillus)、トリコデルマ(Trichoderma)、フザリウム(Fusarium)、ディスポロトリクム(Disporotrichum)、ペニシリウム(Penicillium)、アクレモニウム(Acremonium)、ニューロスポラ(Neurospora)、サーモアスカス(Thermoascus)、ミセリオフトラ(Myceliophtora)、スポロトリクム(Sporotrichum)、シーラビア(Thielavia)、およびタラロマイセス(Talaromyces)属からなる群から選択される。より好ましくは、糸状菌宿主細胞は、アスペルギルス・オイザエ(Aspergillus oyzae)、アスペルギルス・ソーヤ(Aspergillus sojae)もしくはアスペルギルス・ニダランス(Aspergillus nidulans)の種またはアスペルギルス・ニガー(Aspergillus niger)群由来の種(これらは、レイパー(Raper)およびフェンネル(Fennell)、The Genus Aspergillus,The Williams & Wilkins Company,Baltimore,293〜344頁、1965年によって定義される)である。これらには、アスペルギルス・ニガー(Aspergillus niger)、アスペルギルス・アワモリ(Aspergillus awamori)、アスペルギルス・ツビゲンシス(Aspergillus tubigensis)、アスペルギルス・アクレアータス(Aspergillus aculeatus)、アスペルギルス・フォエティダス(Aspergillus foetidus)、アスペルギルス・ニダランス(Aspergillus nidulans)、アスペルギルス・ジャポニクス(Aspergillus japonicus)、麹菌(Aspergillus oryzae)およびアスペルギルス・フィクウム(Aspergillus ficuum)、ならびにまた、トリコデルマ・リーセイ(Trichoderma reesei)、フザリウム・グラミネアラム(Fusarium graminearum)、ペニシリウム・クリソゲナム(Penicillium chrysogenum)、アクレモニウム・アラバメンセ(Acremonium alabamense)、アカパンカビ(Neurospora crassa)、ミセリオフィトラ・サーモフィルム(Myceliophtora thermophilum)、スポロトリクム・セルロフィルム(Sporotrichum cellulophilum)、 ディスポロトリカム・ディモルフォスポラム(Disporotrichum dimorphosporum)およびシーラビア・テレストリス(Thielavia terrestris)種のものが挙げられるが、これらに限定されない。
【0096】
本発明の範囲内の好適な発現宿主の例には、真菌、例えば、アスペルギルス(Aspergillus)種(特に、欧州特許出願公開第A−184,438号明細書、および欧州特許出願公開第A−284,603号明細書に記載のもの)ならびにトリコデルマ(Trichoderma)種;細菌、例えば、バチルス(Bacillus)種(特に、欧州特許出願公開第A−134,048号明細書および欧州特許出願公開第A−253,455号明細書に記載のもの)、特に、枯草菌(Bacillus subtilis)、バチルス・リケニフォルミス(Bacillus licheniformis)、バチルス・アミロリケファシエンス(Bacillus amyloliquefaciens)、シュードモナス(Pseudomonas)種、ならびに酵母、例えば、クリヴェロマイセス(Kluyveromyces)種(特に、欧州特許出願公開第A−096,430号明細書、例えば、クリュイベロマイセス・ラクティス(Kluyveromyces lactis)、および欧州特許出願公開第A−301,670号明細書に記載のもの)およびサッカロマイセス(Saccharomyces)種、例えば、サッカロマイセス・セレビシエ(Saccharomyces cerevisiae)がある。
【0097】
本発明に従う宿主細胞は植物細胞を含み、従って、本発明はトランスジェニック生物体、例えば、植物体およびそれらの一部分に及び、それらは本発明の1つもしくはそれ以上の細胞を含有する。細胞は、本発明のポリペプチドを異種として発現してもよく、または1つもしくはそれ以上の本発明のポリヌクレオチドを異種として含有してもよい。従って、トランスジェニック(または遺伝子的に改変された)植物体は、本発明のポリペプチドをコードする配列をそのゲノムに(典型的には安定に)挿入し得る。植物細胞の形質転換は、公知の技術を使用して、例えば、アグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)由来のTiまたはRiプラスミドを使用して、実施することができる。それ故、プラスミド(もしくはベクター)は、植物に感染するために必要な配列を含んでもよく、そしてTiおよび/またはRiプラスミドの誘導体を用いてもよい。
【0098】
宿主細胞は、ポリペプチドを過剰発現してもよく、そして過剰発現を操作するための技術は周知であり、かつ本発明において使用することができる。それ故、宿主は、2コピーもしくはそれ以上のポリヌクレオチドを有してもよい。
【0099】
あるいは、葉、根または茎のような植物体の一部の直接感染を行うことができる。この技術では、感染させようとする植物体は、例えば、カミソリで植物体を切断するか、針で植物体に穴を開けるか、または研磨剤で植物体を擦ることによって損傷され得る。次いで、損傷部にアグロバクテリウム(Agrobacterium)が播種される。次いで、植物体または植物部分を、適切な培養培地上で増殖させ、そして成熟植物体に生育させることができる。形質転換された細胞の遺伝子的に改変された植物体への再生は、公知の技術、例えば、抗生物質を使用して、形質転換されたシュートを選択することによって、および適切な栄養素、植物ホルモンなどを含有する培地上でシュートを継代培養することによって達成することができる。
【0100】
[宿主細胞の培養および組み換え産生]
本発明はまた、シアリダーゼまたはその変異体を発現するために改変されている細胞も含む。そのような細胞として、一過的、または好ましくは安定に改変された高等真核細胞系統、例えば、哺乳類細胞もしくは昆虫細胞、下等真核細胞、例えば、酵母および糸状菌細胞、または原核細胞、例えば、細菌細胞が挙げられる。
【0101】
また、本発明のポリペプチドを、細胞系統、または例えば、バキュロウイルス発現系のような膜上で一過的に発現させることも可能である。本発明に従うタンパク質を発現するために適応されるそのような系もまた、本発明の範囲内に含まれる。
【0102】
本発明に従えば、本発明のポリペプチドの産生は、微生物の発現宿主を培養することによって行うことができ、それは、従来の栄養発酵培地において、本発明の1つもしくはそれ以上のポリヌクレオチドで形質転換されている。
【0103】
本発明に従う組み換え宿主細胞は、当業者に公知の手順を使用して培養することができる。プロモーターおよび宿主細胞の各組み合わせにとって、ポリペプチドをコードするDNA配列の発現につながる培養条件が利用可能である。所望の細胞密度またはポリペプチドの力価に達した後、培養は中止され、そしてポリペプチドは公知の手順を使用して回収される。
【0104】
発酵培地は、炭素源(例えば、グルコース、マルトース、モラセスなど)、窒素源(例えば、硫酸アンモニウム、硝酸アンモニウム、塩化アンモニウムなど)、有機窒素源(例えば、酵母抽出物、麦芽抽出物、ペプトンなど)および無機栄養源(例えば、リン酸、マグネシウム、カリウム、亜鉛、鉄など)を含有する公知の培養培地を含んでなり得る。場合により、(使用される発現構築物に依存する)誘導因子が含まれていてもよく、または後で添加してもよい。
【0105】
適切な培地の選択は、発現宿主の選択に基づいてもよく、および/または発現構築物の調節要件に基づいてもよい。適切な培地は、当業者に周知である。培地は、所望であれば、他の潜在的に混入する微生物より形質転換された発現宿主の方を選好するさらなる成分を含有してもよい。
【0106】
発酵は、0.5〜30日間の期間にわたって実施し得る。発酵は、0℃〜45℃の間の範囲の温度、および例えば、2から10までのpHにおけるバッチ、継続または流加培養プロセスであり得る。好適な発酵条件は、20℃〜37℃の間の範囲の温度および/または3〜9の間のpHを含む。適切な条件は、通常、発現宿主および発現させようとするタンパク質の選択に基づいて選択される。
【0107】
発酵後、必要であれば、遠心分離またはろ過によって、発酵ブロスから細胞を取り出すことができる。発酵が停止した後または細胞の取り出し後、次いで、本発明のポリペプチドを回収し、そして所望であれば、従来の手段によって精製および単離してもよい。本発明のシアリダーゼは、真菌の菌糸体から、または培養真菌細胞によってシアリダーゼが放出される培養ブロスから、精製することができる。
【0108】
好適な実施形態では、ポリペプチドは、真菌、より好ましくはアスペルギルス(Aspergillus)、最も好ましくはアスペルギルス・ニガー(Aspergillus niger)から産生される。
【0109】
[修飾]
本発明のポリペプチドは、化学的に修飾されていてもよく、例えば、翻訳後修飾される。例えば、それらは、グリコシル化(1回もしくはそれ以上の回数)されるか、または修飾アミノ酸残基を含んでなり得る。それらはまた、それらの精製を補助するためにヒスチジン残基の添加によってか、または細胞からの分泌を促進するためのシグナル配列の添加によって、修飾してもよい。ポリペプチドは、アミノ末端メチオニン残基、約20〜25残基までの小さなリンカーペプチドのようなアミノもしくはカルボキシル末端伸長、またはポリヒスチジントラクト、抗原エピトープもしくは結合ドメインのような精製を容易にする小さな伸長を有してもよい。
【0110】
本発明のポリペプチドは、明示用標識で標識されていてもよい。明示用標識は、ポリペプチドが検出されることを可能にする任意の適切な標識であり得る。適切な標識として、放射性同位元素、例えば、125I、35S、酵素、抗体、ポリヌクレオチドおよびビオチンのようなリンカーが挙げられる。
【0111】
ポリペプチドは、天然に存在しないアミノ酸を含むか、またはポリペプチドの安定性を増加するように修飾してもよい。タンパク質またはペプチドが合成手段によって生成される場合、生成中にそのようなアミノ酸を導入してもよい。タンパク質またはペプチドはまた、合成または組み換え産生後に修飾してもよい。
【0112】
本発明のポリペプチドはまた、D−アミノ酸を使用して生成してもよい。そのような場合、アミノ酸は、CからNへの配向で、逆向きの配列で連結される。これは、そのようなタンパク質またはペプチドを生成するための当該技術分野において慣用的である。
【0113】
多くの側鎖修飾が当該術分野において公知であり、そして本発明のタンパク質またはペプチドの側鎖に対して行うことができる。そのような修飾として、例えば、還元的アルキル化、アルデヒドとの反応、それに続く、NaBH4による還元、アセトイミド酸メチルによるアミジン化または無水酢酸によるアシル化によるアミノ酸の修飾が挙げられる。
【0114】
本発明によって提供される配列はまた、「第2世代」の酵素の構築のための出発物質として使用してもよい。「第2世代」のシアリダーゼは、野生型シアリダーゼまたは本発明によって産生されるような組み換えシアリダーゼとは異なる特性を有する変異技術(例えば、部位特異的変異誘発または遺伝子シャッフリング技術)によって改変されたシアリダーゼである。特定のプロセスにおける使用により良好に適合されるように、例えば、それらの温度または至適pH値、比活性、基質親和性または耐熱性を変更してもよい。
【0115】
本発明のシアリダーゼの活性に必須であり、従って、好ましくは置換に供されるアミノ酸は、当業者に公知の手順、例えば、部位特異的変異誘発またはアラニン−スキャンニング変異誘発に従って、同定してもよい。後者の技術では、変異は、分子の全ての残基で導入され、そして得られる変異が、分子の活性に極めて重要であるアミノ酸残基を同定するために、生物活性(例えば、シアリダーゼ活性)について試験される。酵素−基質相互作用の部位はまた、核磁気共鳴、結晶学または光親和性標識のような技術によって決定されるように、結晶構造の分析によっても決定することができる。
【0116】
遺伝子シャッフリング技術は、ポリヌクレオチド配列に変異を導入するための無作為な方法を提供する。発現後、最良の特性を伴う単離体は、再単離され、組み合わされ、そして再度シャッフリングされて、遺伝的多様性を増加する。この手順を多くの回数反復することによって、迅速に改善されたタンパク質をコードする遺伝子を単離することができる。好ましくは、遺伝子シャッフリング手順は、類似の機能を伴うタンパク質をコードする遺伝子のファミリーから開始される。本発明によって提供されるポリヌクレオチド配列のファミリーは、分泌型シアリダーゼの特性を改善するための遺伝子シャッフリングに良好に適合される。
【0117】
あるいは、NTG処理またはUV変異誘発のような古典的な無作為変異技術および選択を使用して、タンパク質の特性を改善することができる。変異誘発は、単離されたDNA、または目的のDNAで形質転換された細胞に対して直接実施することができる。あるいは、変異は、当業者に公知である多くの技術によって、単離されたDNAに導入することができる。これらの方法の例には、エラープローンPCR、修復欠損宿主細胞におけるプラスミドDNAの増幅などがある。
【0118】
酵母および糸状菌宿主細胞の使用は、翻訳後修飾(例えば、タンパク質分解プロセシング、ミリスチル化(myristilation)、グリコシル化、トランケーション、およびチロシン、セリンまたはスレオニンリン酸化)を提供することが予想され、これは、本発明の組み換え発現産物に対して至適生物活性を付与するのに必要であり得る。
【0119】
[調製]
本発明のポリペプチドは、単離された形であり得る。ポリペプチドは、ポリペプチドの意図された目的を妨害しないキャリアまたは希釈剤と混合してもよく、そしてなお単離されたものと認識されることが理解されよう。本発明のポリペプチドはまた、実質的に精製された形であり得、その場合、それは、一般的に、本発明のポリペプチドである調製物中のタンパク質の70%を超える、例えば、80%、90%、95%、98%もしくは99%を超える調製物中のポリペプチドを含んでなる。
【0120】
本発明のポリペプチドは、それらがそれらの天然の細胞環境外に存在するような形で提供され得る。それ故、それらは、上記のように、実質的に単離もしくは精製され得るか、またはそれらが天然では存在しない細胞、例えば、他の真菌種、動物、植物もしくは細菌細胞に存在し得る。
【0121】
[シアリダーゼ活性の取り出しまたは減少]
本発明はまた、親細胞の変異細胞を生成するための方法であって、ポリペプチドをコードする内因性核酸配列もしくはその制御配列を破壊または欠失することを含んでなり、親細胞より少なくポリペプチドを産生する変異細胞を生じる方法に関する。
【0122】
減少したシアリダーゼ活性を有する株の構築は、細胞におけるシアリダーゼの発現に必要な核酸配列の改変または不活化によって簡便に達成され得る。改変または不活化しようとする核酸配列は、例えば、シアリダーゼ活性を示すのに必須のポリペプチドもしくはその一部をコードする核酸配列であってもよく、または核酸配列は、核酸配列のコーディング配列由来のポリペプチドの発現に必要な調節機能を有してもよい。そのような調節または制御配列の例は、プロモーター配列またはその機能的部分、即ち、ポリペプチドの発現に影響を及ぼすのに十分な部分であり得る。可能な改変のための他の制御配列として、リーダー配列、ポリアデニル化配列、プロペプチド配列、シグナル配列、および終結配列が挙げられるが、これらに限定されない。
【0123】
核酸配列の改変または不活化は、細胞を変異誘発に供し、そしてシアリダーゼ産生能が減少または排除されている細胞を選択することによって、達成され得る。特異的であってもまたは無作為であってもよい変異誘発は、例えば、適切な物理的または化学的変異誘発因子の使用によって、適切なオリゴヌクレオチドの使用によって、またはDNA配列をPCR変異誘発に供することによって、実施され得る。さらに加えて、変異誘発は、これらの変異誘発因子の任意の組み合わせの使用によって、実施され得る。
【0124】
本目的に適切な物理的または化学的変異誘発因子の例として、紫外線(UV)照射、ヒドロキシルアミン、N−メチル−N’−ニトロ−N−ニトロソグアニジン(NTG)、O−メチルヒドロキシルアミン、亜硝酸、メタンスルホン酸エチル(EMS)、亜硫酸水素ナトリウム、ギ酸、およびヌクレオチドアナログが挙げられる。
【0125】
そのような因子を使用する場合、変異誘発は、典型的に、変異誘発しようとする細胞を、適切な条件下で選好される変異誘発因子の存在下でインキュベートし、そしてシアリダーゼ活性の発現の減少を示すか、または発現を示さない細胞を選択することによって、実施される。
【0126】
本発明のポリペプチドの産生の改変または不活化は、ポリペプチドをコードする核酸配列、またはその転写もしくは翻訳に必要な調節エレメントにおける1つもしくはそれ以上のヌクレオチドの導入、置換、あるいは取出しによって、達成され得る。例えば、ヌクレオチドは、終止コドンの導入、開始コドンの取り出し、もしくはオープンリーディングフレームの変更を生じるように、挿入してもよく、または取り出してもよい。そのような改変または不活化は、当該技術分野において公知の方法に従って、部位特異的変異誘発またはPCR変異誘発によって達成され得る。
【0127】
原則として、改変は、インビボで、即ち、改変しようとする核酸配列を発現する細胞に対して直接実施してもよいが、改変は、以下に例示するようにインビトロで実施するのが好ましい。
【0128】
選好される宿主細胞によるシアリダーゼの産生を不活化または減少するための簡便な方法の例は、遺伝子置換または遺伝子中断(gene interruption)の技術に基づく。例えば、遺伝子中断方法では、内因性遺伝子または目的の遺伝子フラグメントに対応する核酸配列をインビトロで変異誘発して、欠損核酸配列を生成し、次いで、宿主細胞に形質転換して、欠損遺伝子を生成する。相同組み換えによって、欠損核酸配列が、内因性遺伝子または遺伝子フラグメントに取って代わる。好ましくは、欠損遺伝子または遺伝子フラグメントはまた、ポリペプチドをコードする遺伝子が改変または破壊されている形質転換体を選択するために使用され得るマーカーをコードする。
【0129】
あるいは、本発明のポリペプチドをコードする核酸配列の改変または不活化は、ポリペプチドコーディング配列に相補的なヌクレオチド配列を使用する確立されたアンチセンス技術によって、達成され得る。より具体的には、細胞によるポリペプチドの産生は、ポリペプチドをコードする核酸配列に相補的なヌクレオチド配列を導入することによって、減少または排除され得る。次いで、アンチセンスポリヌクレオチドは、典型的に、細胞において転写され、そしてシアリダーゼをコードするmRNAにハイブリダイズすることが可能である。相補的なアンチセンスヌクレオチド配列をmRNAにハイブリダイズさせる条件下で、細胞において産生されるシアリダーゼの量は、減少または排除される。
【0130】
本発明の方法に従って改変しようとする細胞は、微生物由来、例えば、細胞に同種または異種のいずれかである所望されるタンパク質産物の産生に適切である真菌株由来であることが好ましい。
【0131】
本発明は、さらに、ポリペプチドをコードする内因性核酸配列もしくはその制御配列の破壊または欠失を含んでなり、親細胞より少なくポリペプチドを産生する変異細胞を生じる、親細胞の変異細胞に関する。
【0132】
そのようにして作製されるポリペプチド欠損変異細胞は、同種および/または異種ポリペプチドの発現のための宿主細胞として特に有用である。従って、本発明は、さらに、(a)ポリペプチドの産生につながる条件下で変異細胞を培養すること;および(b)ポリペプチドを回収することを含んでなる、同種または異種ポリペプチドを産生させるための方法に関する。これに関して、用語「異種ポリペプチド」は、本明細書において、宿主細胞に由来しないポリペプチド、天然配列を変更するために改変が施されている天然タンパク質、または発現が、組み換えDNA技術による宿主細胞の操作の結果として量的に変更される天然タンパク質として定義される。
【0133】
なおさらなる態様では、本発明は、本発明のシアリダーゼポリペプチドならびに目的のタンパク質産物の両方を産生する細胞の発酵によって、シアリダーゼ活性を本質的に含まないタンパク質産物を産生するための方法を提供する。本方法は、発酵中、または発酵が完了した後のいずれかに、シアリダーゼ活性を阻害することが可能な有効量の薬剤を発酵ブロスに添加し、発酵ブロスから目的の産物を回収し、そして場合により、回収した産物をさらなる精製に供することを含んでなる。あるいは、培養後、得られた培養ブロスを、シアリダーゼ活性を実質的に減少するようにpHまたは温度処理に供し、そして培養ブロスからの産物の回収を可能にすることができる。組み合わされたpHまたは温度処理は、培養ブロスから回収したタンパク質調製物に対して実施し得る。
【0134】
本質的にシアリダーゼを含まない産物を産生させるための本発明の方法は、真核生物ポリペプチドの産生、特に、酵素のような真菌タンパク質の産生において得に興味深い。また、シアリダーゼ欠損細胞を使用して、食品産業、または製薬目的のために目的の異種タンパク質を発現させてもよい。
【0135】
[チーズ作製および乳汁凝固]
凝固は、牛乳のような酪農組成物からのチーズの伝統的な生産における必須の工程である。凝固は、酸性化および/またはキモシンのような酵素(凝固剤)の添加によって開始され得る。凝固後、乳汁はカードおよび乳清に分離される。カードはさらにチーズに処理される。
【0136】
様々な乳汁供給源からのチーズ製造プロセスは、かなり以前より公知であり、そして異なる多くのタイプのチーズ変異体について詳細に記載されている。(例えば、Cheese:Chemistry,Physics and Microbiology、第1および2巻、1999年、フォックス(Fox)編、Aspen Publications,Gaithersburg、メリーランド州;Encyclopedia of Dairy Sciences、第1〜4巻、2003年、Academic Press,Londonを参照のこと)。チーズ製造において極めて重要な点は凝固のプロセスであって、ここで、カゼインミセルおよびサブミセルの溶解度が減少する。酵素誘導性凝固は、極めて一般的に使用される。仔ウシキモシンのような酵素、キモシンの微生物等価物および他の供給源由来の他の酵素についても記載されており、そしていくつかは、様々な商品名で利用可能である。それらのすべてを使用して、凝固プロセスを開始することができる。凝固の一次工程は、κ−カゼインにおけるPhe105−Met106結合の切断である。これは、κ−カゼインのC末端部:グリコマクロペプチド(GMP)の取り出しをもたらす。GMPの取り出しは、カゼインミセルの会合、即ち、カゼイン凝固をもたらす。カゼイン凝固はゲル形成をもたらし、そして特定の酪農組成物においてゲル化を得るのに必要な時間は、凝固剤の活性に直接関連する。
【0137】
凝固剤の添加と初期のカゼイン凝集物の出現との間を経過する時間は、凝固時間として定義される。チーズ用乳汁においてゲルが形成される速度およびゲルの密集度は、添加される酵素の量、カルシウムイオンの濃度、リン、温度およびpHに密接に依存する。初期の凝固後、ゲルが形成し、そしてミセル間結合の増加に従ってゲルの粘稠度が増加する。ミセルは共に移動し、そして凝塊が収縮し、それによって乳清を放出する。この現象は離液として公知であり、そしてカードを切断することによって加速され、温度を増加し、そして発育する乳酸菌によって生じる酸性度を増加する。
【0138】
凝固剤の添加と離液を開始するためのゲルの切断との間の時間を、本明細書において以後、凝固時間と称する。例えば、乳汁または凝固剤の質の変化の結果として、少しのばらつき(5〜10%まで)が生じ得るが、凝固時間は、特定のチーズ製造工場において特定のチーズの種類について一定である。多くのチーズ製造工場では、凝固時間は、プロセスにおいて時間を制限する工程である。凝固時間が減少する場合、工場は、より多くの乳汁を処理してカードにし、次いで(than)、さらにチーズに処理することができる。従って、凝固時間の減少は、経済的に興味深く、そしてそのような凝固時間の減少が必要とされている。
【0139】
添加する凝固剤の増加、乳汁のpHの降下または添加する塩化カルシウムのレベルの増加のようないくらかの方法で、凝固時間を減少することができる。しかし、それらの解決策は、チーズの品質にネガティブに影響を及ぼすため、チーズ作製産業において広範には使用されていない。凝固剤のレベルの増加は、しばしば、不均衡なプロテアーゼ活性の結果として苦味の形成をもたらす。乳汁pHの減少は、粗悪な品質のカードをもたらし、そしてしばしば、収量の損失をもたらす。塩化カルシウム濃度の増加は、味にネガティブな影響を及ぼし、そしてしばしば法的に制限されている。ネガティブな副作用を伴わずに凝固時間を減少するための現在利用可能な方法は存在しない。
【0140】
凝集に対するカゼインミセルの安定性は、凝集プロセスにおいて極めて重要である。κ−カゼインは、カゼインミセル安定化において極めて重要な役割を果たす。ミセル表面に局在するκ−カゼイン鎖の親水性かつ負に荷電したC末端部は、立体構造上の障害および静電反発の原因となり、ミセル状カゼインの凝固を妨害する(ミンキーウィックズ(Minkiewicz)ら、Pol J Food Nutr Sci(1993年)243,39−48)。κ−カゼインは、既知の位置においてシアル酸残基を含有する糖タンパク質である(ケーシズ(Cases)ら、J Food Sci(2003年)68,2406−2410;フォルネ(Fournet)ら、Biochim Biophys Acta(1979年)576,339−346)。グリコシル化の差異およびシアル酸含有量に基づき、κ−カゼインの集団においてミクロ的不均質が存在することが公知である(ロビテイル(Robitaille)ら、Food Res Int(1995年)28,17−21;ロビテイル(Robitaille)ら、J Dairy Res(1991年)58,107−114)。いくつかの研究は、ウェルシュ菌(Clostridium perfringens)由来のサイリダーゼ(sailidase)を使用するシアル酸残基の取り出しの効果について説明している。調べたパラメータは、加熱およびキモシン分解に対するカゼインミセルの安定性である。ギボンス(Gibbons)ら(Biochim Biophys Acta(1962年)56,354−356)は、精製されたκ−カゼインの溶液を使用し、そしてシアル酸残基の取出しが、κ−カゼインに対するキモシンの作用に影響を及ぼさないことを示した。ヴリーマン(Vreeman)ら(Biochem J(1986年)240,87−97)は、グリコシル化を伴うまたは伴わない精製されたκ−カゼイン画分に対するキモシンの動力学的挙動を比較し、そして脱グリコシル化κ−カゼインがキモシンのより良好な基質であることを見出した。また、ミンキーウィックズ(Minkiewicz)ら(Pol J Food Nutr Sci(1993年),243,39−48)は、人工的に再構成されたカゼインミセル系を使用して、シアル酸残基が、カゼインミセルの耐熱性に寄与することを示し、これは後にロビテイル(Robitaille)ら(Food Res Int(1995年)28,17−21)によって確認された。個別の研究において、ロビテイル(Robitaille)ら(Food Res Int(1993年)26,365−369)は、κ−カゼインからのシアル酸残基の取り出しは、凝固時間に有意な影響を及ぼさないが、カードの堅固性が減少したことを示した。凝固時間の減少を伴うが、チーズの特性に影響を及ぼさないチーズ作製プロセスについての記載は認められておらず、そしてこれは、産業上必要とされている。
【0141】
最近の特許出願(欧州特許第1370146号明細書)は、N−アセチルノイラミニダーゼを含む単一のグリコシダーゼを使用して、乳汁を凝固するプロセスについて記載している。記載された構成では、酵素凝固剤は使用されていない。凝固剤の省略は、代替的チーズ作製プロセスのための方法への道を開くが、(タンパク質分解性)凝固剤は、チーズにおけるタンパク質分解系において重要な部分であり、特徴的な風味化合物の形成をもたらすため、伝統的なチーズ作製プロセスには魅力的なプロセスではない(例えば、チーズ風味形成におけるキモシンの役割に関する参考文献について、Cheese:Chemistry,Physics and Microbiology、第1および2巻、1999年、フォックス(Fox)編、Aspen Publications,Gaithersburg,Maryland;Encyclopedia of Dairy Sciences 第1〜4巻、2003年、Academic Press,Londonを参照のこと)。
【0142】
これに関して、用語「チーズ」は、例えば、ナチュラルチーズ、チーズ類似物およびプロセスチーズのような任意の種類のチーズを指す。チーズは、例えば、レンネットを伴う酪農組成物の酵素的凝固、または食品級酸もしくは乳酸菌増殖によって産生される酸による酪農組成物の酸性凝固によるような当該技術分野において公知の任意の適切なプロセスによって入手され得る。一実施形態では、チーズは、レンネット−カードチーズである。酪農組成物は、従来のチーズ作製プロセスに供され得る。プロセスチーズは、好ましくは、チーズを調理ならびに例えば、乳化塩(例えば、リン酸塩およびクエン酸塩)で乳化することによって、ナチュラルチーズまたはチーズ類似物から製造される。プロセスは、香辛料/調味料の添加をさらに含んでもよい。
【0143】
用語「チーズ類似物」は、組成物の一部分として(例えば、乳脂肪(例えば、クリーム)のような脂肪を含有し、そして一組成物の部分として、例えば、植物油のような非乳汁構成分をさらに含有するチーズ様製品を指す。チーズは、軟質チーズ、半硬質チーズおよび硬質チーズのようなすべての種類のチーズを含んでなり得る。チーズ製造では、酪農組成物の凝固は、好ましくは、それぞれ、レンネット−カードおよび酸−カードチーズを生じるレンネットまたは酸性化のいずれか単独によって、実施される。フレッシュ酸−カードチーズとは、酸性化もしくは酸および加熱の組み合わせを介して乳汁、クリームまたは乳清の凝固によって生成され、そして製造が完了したら成熟を伴わなくとも消費される状態にある種類のチーズを指す。高い温度を使用する場合、カゼインの等電点、即ち、例えば、pH4.6もしくはそれ以上の値付近で、通常、凝固が生じる(例えば、リコッタ(Ricotta)では、典型的に約6.0のpH、および典型的に約80℃の温度)点で、フレッシュ酸−カードチーズは、一般的に、レンネット−カードチーズの種類(例えば、カマンベール(Camembert)、チェダー(Cheddar)、エメンタール(Emmenthal))とは異なり、ここで、凝固は、通常、pH値6.4〜6.6でレンネットの作用により誘導される。好適な実施形態では、チーズは、レンネットカードチーズのクラスに属する。
【0144】
酪農組成物は、牛乳構成分を含んでなる任意の組成物であり得る。乳構成分は、乳脂肪、乳タンパク質、カゼイン、乳清タンパク質およびラクトースのような乳汁の任意の構成分であり得る。乳汁画分は、例えば、スキムミルク、バターミルク、乳清、クリーム、粉乳、全粉乳、脱脂粉乳のような乳汁の任意の画分であり得る。好適な実施形態では、酪農組成物は、乳汁、スキムミルク、バターミルク、全乳、乳清、クリーム、またはそれらの任意の組み合わせを含んでなる。より好適な実施形態では、酪農組成物は、スキムミルク、全乳、クリームまたはそれらの任意の組み合わせのような乳汁からなる。さらなる実施形態では、酪農組成物は、例えば、全粉乳、脱脂粉乳、カゼイン、カゼイン塩、総乳タンパク質もしくは加糖粉乳、またはそれらの任意の組み合わせのような乾燥乳汁画分から全体的または部分的に調製される。酪農組成物は、牛乳およびまたは1つもしくはそれ以上の牛乳画分を含んでなる。牛乳画分は、ウシの任意の品種(ウシ(Bos taurus taurus)、コブウシ(Bos indicus taurus)およびこれらの交雑種由来であり得る。一実施形態では酪農組成物は、ウシの2つもしくはそれ以上の品種由来の牛乳および/または牛乳画分を含んでなる。酪農組成物はまた、ヤギ、スイギュウまたはラクダ由来の乳汁のようなチーズ調製物に使用される他の哺乳動物由来の乳汁を含んでなる。チーズの調製のための酪農組成物は、生乳汁成分のすべてもしくは一部の取り出しによって、および/またはそれにさらなる量のそのような成分を添加することによって、所望される組成物に標準化され得る。これは、例えば、乳製品製造所到着時の乳汁のクリームと乳汁とへの分離によって行われ得る。それ故、酪農組成物は、乳汁を分画し、そして酪農組成物の所望される最終組成物が得られるように、画分を再度組み合わせることによって、簡便に行われるように、調製され得る。分離は、連続遠心分離において行ってもよく、極めて低い脂肪含有量(即ち<0.5%)を伴うスキムミルク画分および例えば、>35%脂肪を伴うクリームがもたらされる。酪農組成物は、クリームおよびスキムミルクを混合することによって調製してもよい。別の実施形態では、タンパク質および/またはカゼイン含有量は、限外ろ過の使用によって標準化され得る。酪農組成物は、本発明のプロセスによって生成されるべきチーズに適切であることが認められる任意の全脂肪含有量を有し得る。
【0145】
カルシウムは、スターターカルチャーの添加の前、同時、または後のようなチーズ作製前および/または最中に任意の適切な段階で酪農組成物に添加され得る。好適な実施形態では、カルシウムは、加熱処置の前および後の両方に添加される。カルシウムは、適切ないずれの形で添加してもよい。好適な実施形態では、カルシウムは、カルシウム塩、例えば、CaCl2として添加される。適切な任意の量のカルシウムが、酪農組成物に添加され得る。添加されるカルシウムの濃度は、通常、1〜3mMの間のような0.1〜5.0mMの範囲である。CaCl2を酪農組成物に添加する場合、量は、通常、100リットルの酪農組成物あたり1〜50gの範囲、例えば、1000リットル酪農組成物あたり5〜30gの範囲、好ましくは、100リットル酪農組成物あたり10〜20gの範囲にある。
【0146】
スキムミルクの細菌含有量は、従来の好適によって低減され得る。酪農組成物は、デンマークブルーチーズ(Danish Blue Cheese)の生成におけるように、チーズの生成前に、均質化プロセスに供してもよい。
【図面の簡単な説明】
【0147】
【図1】図1は、ZJW発現ベクターの構築を示す。
【0148】
以下の実施例によって、本発明を例示する。
【0149】
[実施例]
[実施例1]
[シアリダーゼ遺伝子ZJWのクローニングおよび発現]
ペニシリウム・クリソゲナム(Penicillium chrysogenum)株Wisconsin54−1255(ATCC28089)を、3日間、30℃、PDB(ポテトデキストロースブロス、Difco)中で増殖させ、そして染色体DNAを、供給者の取扱説明書を使用し、Q−Biogeneキット(カタログ番号6540−600;Omnilabo International BV,Breda、蘭国)を使用して、菌糸体から単離した。染色体DNAを、PCRを使用するシアリダーゼ遺伝子のコーディング配列の増幅に使用した。
【0150】
ペニシリウム・クリソゲナム(Penicillium chrysogenum)株Wisconsin54−1255(ATCC28089)の染色体DNAからシアリダーゼ遺伝子ZJWを特異的に増幅するために、2つのPCRプライマーを設計した。プライマー配列を、ペニシリウム・クリソゲナム(Penicillium chrysogenum)Wisconsin54−1255(ATCC28089)のゲノムDNAにおいて見出された配列から部分的に入手したが、これを配列番号1に示す。本発明者らは、この配列は、アクチノマイセス(Actinomyces)およびアルスロバクター(Arthrobacter)のシアリダーゼ配列と相同性を有することを見出した。しかし、相同な真菌シアリダーゼについての記載はまだ認められていない。従って、本発明者らが、真菌から分泌型シアリダーゼをコードする遺伝子を見出すことができたことは、意外である。本発明者らは、本明細書において、分泌型真菌シアリダーゼの効率的な発現および特徴付けについてはじめて説明する。潜在的なプレ−およびプロ−配列を含んでなる完全なシアリダーゼタンパク質のタンパク質配列を配列番号3に示す。細菌相同体と比較した真菌酵素の利点は、真菌酵素が容易に過剰発現され、そして食物産業におけるアプリケーションに関連する量で分泌され得ることである。
【化1】

【0151】
第1の直接PCRプライマー(ZJW−dir)は、ATG開始コドンから開始する23ヌクレオチドZJWコーディング配列、およびそれより前方のPacI制限部位を含む23ヌクレオチド配列を含有する(配列番号4)。第2の逆方向プライマー(ZJW−rev)は、ZJWコーディング配列の下流の領域の逆方向鎖に相補的なヌクレオチド、およびそれより前方のAscI制限部位を含有する(配列番号5)。これらのプライマーを使用して、本発明者らは、テンプレートとしてペニシリウム・クリソゲナム(Penicillium chrysogenum)株Wisconsin54−1255(ATCC28089)由来の染色体DNAを伴う1.4kbのサイズのフラグメントを増幅することができた。このようにして得られた1.4kbサイズのフラグメントを単離し、PacIおよびAscIで消化し、そして精製した。ZJWコーディング配列を含んでなるPacI/AscIフラグメントを、pGBFIN−5(国際公開第99/32617号パンフレット)由来のPacI/AscI phyAフラグメントと交換した。得られたプラスミドは、pGBFINZJWと命名したZJW発現ベクターである(図1を参照のこと)。発現ベクターpGBFINZJWを、NotIによる消化によって線状化した(これは、発現ベクターからすべての大腸菌(E.coli)由来の配列を取り出す)。消化したDNAを、フェノール:クロロホルム:イソアミルアルコール(24:23:1)抽出を使用して精製し、そしてエタノールで沈殿させた。これらのベクターを使用して、アスペルギルス・ニガー(Aspergillus niger)CBS513.88を形質転換した。アスペルギルス・ニガー(Aspergillus niger)形質転換手順ついては、国際公開第98/46772号パンフレットに広範に記載されている。アセトアミドを含有する寒天プレート上で形質転換体を選択し、そして標的化されたマルチコピー組込み体を選択する仕方についても記載されている。好ましくは、複数コピーの発現カセットを含有するA.ニガー(A.niger)形質転換体を、サンプル材料のさらなる作製のために選択する。pGBFINZJW発現ベクターについて、まず、個々の形質転換体を選択倍地上にプレート化し、続いて、単一のコロニーを、PDA(ポテトデキストロース寒天:PDB+1.5%寒天)プレートにプレート化することによって、30個のA.ニガー(A.niger)形質転換体を精製した。1週間、30℃での増殖後、個々の形質転換体の胞子を回収した。胞子を冷蔵庫に貯蔵し、そして液体培地の播種のために使用した。
【0152】
複数コピーの発現カセットを含有するA.ニガー(A.niger)株を、振盪フラスコ培養における株の培養によってサンプル材料の作製に使用した。A.ニガー(A.niger)株の培養および培養ブロスからの菌糸体の分離のための有用な方法についても、国際公開第98/46772号パンフレットに記載されている。培養培地は、CSM−MES(1リットル培地あたり150gマルトース、60gのSoytone(Difco)、15gの(NH4)2SO4、1gのNaH2PO4・H2O、1gのMgSO4・7H2O、1gのL−アルギニン、80mgのTween−80、20gのMES、pH6.2)であった。発酵の4〜8日目に5mlサンプルを採取し、10分間、5000rpm、Hereaus labofuge RF中で遠心分離し、そしてさらなる分析まで、上清を−20℃で貯蔵した。
【0153】
SDS−Page分子量(MW)決定:NuPAGE MES−SDS泳動用緩衝液を使用するNuPAGE Novex Bis−Tris4〜12%勾配系(Invitrogen(Breda−蘭国))上でのSDS−PAGEによって、サンプルを分析する。供給者のプロトコルに従い、NuPAGE還元剤を使用して、充填前にサンプルを還元する。産生されたタンパク質の分子量を見積もるために、マーカータンパク質(SeeBlue Plus2Pre−Stained Standard(Invitrogen))を使用する。ゲルを泳動させ、そして供給者の取扱説明書に従って、Simply Blue Protein Strain(Invitrogen)を使用して染色し、そして産生されたタンパク質の分子量を、標準タンパク質との比較によって見積もった。本実施例では、形質転換体の発酵ブロスの13マイクロリットルの上清を分析した。
【0154】
SDS−PAGEで分析する場合、pGBFINZJWベクターを含有する形質転換体は、約50kDaの見かけの分子量のタンパク質を分泌したことが明らかになった。これは、タンパク質配列から推定される分子量よりわずかに大きいため、本発明者らは、シグナル配列の取り出し後、ペニシリウム・クリソゲナム(Penicillium chrysogenum)シアリダーゼZJWがアスペルギルス・ニガー(Aspergillus niger)から分泌される場合、いくつかのグリコシル化が生じると推定している。
【0155】
発酵および下流のプロセシングが大規模化される場合、選択された株は、より大量の真菌シアリダーゼの単離および精製のために使用することができる。次いで、この酵素を、さらなる分析、および多様な個々のアプリケーションにおける使用のために使用することができる。
【0156】
[実施例2]
[シアリダーゼの精製および特徴付け]
シアリダーゼを、実施例1に記載のような発酵を介して産生させた。酵素活性を、Amplex Redノイラミニダーゼアッセイキット(Invitrogenから入手した)を使用して、測定した。培養ろ過物(100ml)を、milliQ−水で、4.8mS/cmの導電率に希釈し、そしてBiomax−10膜(Milliporeから入手した)を使用する限外ろ過によって、70mlまで濃縮した。pHを、NaOHを使用して6.0に調整し、そしてサンプルを、5mlのHiTrapQイオン交換カラム(Amershamから入手した、5ml/分)上に充填し、20mMクエン酸ナトリウム(pH6.0)において平衡化した。シアリダーゼを含有するカラムのフロースルーを回収し、そして25mMのTris,HCl(pH7.0)に対して透析し、そして5mlのHiTrap Q FF(5ml/分)上に充填し、同じ緩衝液において平衡化した。シアリダーゼはフロースルー画分に存在し、そして回収した。次いで、酵素溶液を30mMクエン酸ナトリウム(pH4.0、緩衝液A)に対して透析し、そして5mlのHiTrap SPカラム(Amershamから入手した、5ml/分)上に適用し、緩衝液Aにおいて平衡化した。酵素を充填した後、カラムを3カラム容積の緩衝液Aで洗浄し、そして酵素を、直線勾配で20カラム容積の緩衝液Aから緩衝液B(緩衝液B:1MのNaClを含有する30mMナトリウム−クエン酸、pH4.0)により溶出させた。シアリダーゼ含有画分を同定し、そしてプールした。タンパク質濃度を、対照タンパク質としてウシ血清アルブミンを使用するBradford試薬(Sigmaから入手した)によって決定した。タンパク質は、ナトリウム−ドデシルポリアクリルアミドゲル電気泳動上で夾雑するバンドが認められなかったことから、>95%純粋であると判断した。シアリダーゼは、47kDの見かけの分子量まで泳動し、これは、推定されたアミノ酸配列に基づいて算出される42.7kDの分子量より僅かに大きい。酵素調製物は、一連の基質ZAAXpNA(Z=ベンゾイル基、A=アラニン、X=任意のアミノ酸残基、pNA=パラ−パラニトロアニリド)に対してタンパク質分解活性を示さず、エンド−プロテアーゼ活性の不在を示す。
【0157】
[実施例3]
[ミニチュアチーズの調製方法]
ミニチュアチーズを、シャキール−Ur−レーマン(Shakeel−Ur−Rehman)ら(Protocol for the manufacture of miniature cheeses、Lait,78,(1998年),607−620)により記載の通りに、生成した。低温殺菌された全脂均質化牛乳を使用したが、生の牛乳または還元牛乳を使用することもできる。乳汁を広口プラスチック製ボトル(ボトルあたり200mL)に移し、31℃まで冷却した。続いて、1.8単位のスターターカルチャーDS 5LT1(DSM Gist B.V.、Delft、蘭国)を、遠心ボトル中の200mlの低温殺菌した乳汁のそれぞれに添加し、乳汁を20分間、成熟させた。次いで、CaCl2(200mL発酵乳あたり132μLの1mol.L−1溶液)を添加した。最後に、凝固剤(1mlあたり0.04IMCU)を添加した。シアリダーゼを使用した場合、これを、凝固剤と共に添加した。凝塊が形成されるまで、乳汁溶液を40〜50分間、31℃で保持した。凝塊を、フレーム上に1cm間隔で張り渡されたワイヤのカッターで、手動で切断した。2分間回復させ、続いて、10分間、緩徐に撹拌した。その後、カード/乳清混合物の連続撹拌下、30分間、温度を徐々に39℃に上昇させた。6.2のpHに到達したら、カード/乳清混合物を、室温で60分間、1,700gで遠心分離した。乳清を排出させ、カードを水浴中、36℃に保持した。pHが5.2〜5.3に低下するまで、チーズを15分間ごとに反転させ、次いで、室温で20分間、1,700gで遠心分離した。製造後、チーズを秤量した。
【0158】
[実施例4]
[チーズ凝固を決定するための方法]
Kubarsepら(Acta Agric ScandセクションA(2005年)55,145−148)に記載のように、Optigraph(Alliance Instruments、仏国)を使用して、チーズ凝固を追跡した。凝乳酵素溶液の添加後、標準的な手順(IDF157 A[2])に従って、標準的な乳汁基質溶液の凝固時間を決定した。11グラムの低加熱脱脂粉乳(Nilac、NIZO、蘭国から入手した)を、4.5mMのCaCl2を含有する100mlのmilliQ−水に添加することによって、乳汁基質溶液を調製した。乳汁溶液を、磁気スターラーで、30分間、撹拌した。乳汁を安定にするために、溶液を、室温で30分間、暗所に置いた。乳汁のpHは約6.5である。8〜15分間の間のレンネット凝固時間(r)を達成するために、凝乳酵素をmilli−Q水で希釈した。10ml乳汁溶液を、Optigraphのサンプルキュベットに移した。乳汁を、32℃の反応温度に調整した。200μlの希釈酵素サンプルを多スプーン装置(multiple spoon apparatus)にピペッティングし、そして1回で乳汁に添加した。この操作が、凝固試験の開始時点である。酵素および乳汁をスプーンで混合し、この目的のためにOptigraphで供給された。凝固を45分間、モニターした。
【発明の詳細な説明】
【0001】
[発明の分野]
本発明は、新規シアリダーゼに関する。
【0002】
[発明の背景]
シアル酸は、9炭素糖のノイラミン酸の約40種類の誘導体のファミリーを含んでなる。それは、約2.2のpKaを伴う強有機酸である。非置換型のノイラミン酸(neuraminic acic)は、天然には存在しない。アミノ基は、通常、アセチル化されて、最も普及した形のシアル酸であるN−アセチルノイラミン酸(N−acetylneuraminc acid)を産出するが、他の形も存在する(トラビング(Traving)ら Cell Mol Life Sci(1998年)54,1330−1349)。シアル酸は、棘皮動物からヒトにまで至る動物界において見出されるが、一方、前口動物系統の下等動物または植物におけるそれらの存在については、示唆されていない。唯一の公知の例外は、昆虫のショウジョウバエ(Drosophila)の幼虫におけるポリシアル酸の存在である。さらに、いくつかの原生動物、ウイルスおよび細菌にもシアル酸が認められる。シアロ糖複合体は、細胞表面ならびに細胞内膜に存在する。高等動物では、それらはまた、血清および粘膜物質の重要な成分でもある。
【0003】
シアル酸は、多様な生物学的機能を有する。それらの負電荷により、シアル酸は、カルシウムイオンのような正に荷電した分子の結合および輸送、ならびに細胞と分子との間の誘引および反発現象に関与する。それらのサイズおよび負電荷に加えて、それらが暴露された炭水化物鎖の末端に位置することは、それらが分子または細胞のサブ末端部の保護シールドとして機能することを可能にする。それらは、例えば、グリコール−タンパク質(glycol−proteins)がプロテアーゼによって分解されるのを防ぐことができ、または呼吸器系の粘膜層を細菌感染から防ぐことができる。興味深い現象は、それらの負の電荷の間に作用する反発力によりシアル酸含有分子に対して及ぼされる延展効果である。これは、酵素または膜(糖)タンパク質の正確なコンホメーションを安定化し、そして粘液性(slimy)の性質ならびに眼の表面および粘膜上皮のような粘膜物質の滑走および保護機能に重要である(トラビング(Traving)ら Cell Mol Life Sci(1998年)54,1330−1349)。明らかに、適切なシアリダーゼによるそのようなシアル酸含有物質の処理は、そのような物質の生物学的特性および物理的特徴に劇的に影響を及ぼすことができる。シアリダーゼによるシアル酸含有タンパク質の処理は、それらを、かなり容易にプロテアーゼによって分解され易くすることができ、シアリダーゼによる粘膜物質の処理は、それらの粘液性特徴を強度に減少または排除し得る。そのような変化は、そのようなタンパク質が、例えば、タンパク質加水分解物のためのプロセシング(例えば、タンパク質分解)の産業的プロセスを必要とする場合、興味深い。
【0004】
シアル酸は、細胞と分子との間の多様な認識プロセスに関与する。それ故、免疫系は、それらのシアル酸パターンに従って、自己構造と非自己構造とを区別することができる。糖は、抗原決定基、例えば血液型物質を表し、そしてホルモンおよびサイトカインのような多くの内因性物質の受容体の必要な成分である。加えて、毒素(例えば、コレラ毒素)のような多くの病原因子、ウイルス(例えば、インフルエンザ)、細菌(例えば、大腸菌(Escherichia coli)、ヘリコバクター・ピロリ(Helicobacter pylori))および原生動物(例えば、トリパノソーマ・クルージ(Trypanosome cruzi)もまた、シアル酸含有受容体を介して宿主細胞に結合する。シアル酸認識分子の別の重要な基は、特定の糖残基に結合する植物、動物および無脊椎動物由来の通常オリゴマー糖タンパク質であるレクチンに属する。例には、コムギ胚芽アグルチニン、アメリカカブトガニ(Limulus polyphemus)アグルチニン(aagglutinin)、セイヨウニワトコ(Sambucus nigra)アグルチニンおよびカライヌエンジュ(Maackia amurensis)アグルチニンがある。これらのレクチンは、植物を、シアル酸含有微生物または草食哺乳動物に対するその防御において援助するようである。レクチンの哺乳動物対応物は、セレクチンおよびシグレックを含み(トラビング(Traving)ら Cell Mol Life Sci(1998年)54,1330−1349)、そして多様な生理学的役割を有する。シアル酸はまた、細胞および分子の遮蔽を補助することができる。赤血球は、シアル酸分子の防御層に覆われ、これは、血液細胞の生活環中に段階的に取り出される。次いで(than)、分解のためのシグナルを表す最後から2番目のガラクトース残基が晒され、次いで(than)、遮蔽されていない血液細胞がマクロファージに結合し、そして貪食される。そのような遮蔽ストラテジーの他のいくらかの例は公知である。遮蔽はまた、有害な影響を有し得るが、これは、対応する組織よりもかなり高い程度でシアル酸付加される腫瘍のいくつかから認められ得る。結果的に、遮蔽された細胞は免疫防御系に晒されず、そして高シアル酸含有量もまた、さらなる細胞増殖の阻害の欠如および拡大に役割を果たし得る。シアル酸の遮蔽効果はまた、寄生体細胞上の抗原部位を隠すのに役立ち、それらを系に晒されないようにする。これは、所定の大腸菌(E coli)株および淋菌(Neisneria gonorrhoeae)のような微生物種の場合に当てはまる。そのような種をシアリダーゼで処理すると、免疫系から隠れるそれらの可能性に影響を及ぼす。
【0005】
シアリダーゼ(ノイラミニダーゼ、EC3.2.1.18)は、糖タンパク質、糖脂質、ガングリオシド、多糖および合成分子における末端の非還元シアル酸結合を加水分解する。トランスシアリダーゼと呼ばれるシアリダーゼはまた、シアル酸残基を1つの分子から別の分子に転移する転移反応を実施することも可能である。シアリダーゼは、新口動物系統(棘皮動物(Echinodermata)から哺乳動物(Mammalia))の動物およびまた動物の共生生物または病原体として最も多く存在する多様な微生物において一般的に認められる。シアリダーゼ、およびそれらのシアリル基質は、植物および他のほとんどの後生動物には存在しないと思われる。細菌の間であっても、シアリダーゼは、関連する種または1つの種の株であってもこの特性が異なるように、不規則に見出される。シアリダーゼはまた、ウイルスおよび原生動物においても見出されている(トラビング(Traving)ら Cell Mol Life Sci(1998年)54,1330−1349)。シアリダーゼを含有する微生物は、しばしば、例えば、寄生体として、宿主としての高等動物と接触しながら生存する。ここで、それらは、栄養機能を有し得、それらの所有者が宿主のシアル酸を炭素源として使用するためにスカベンジすることを可能にする。いくつかの微生物病原体では、シアリダーゼがビルレンス因子として作用すると考えられる。なお、病原性における因子としてのサリダーゼ(salidases)の役割については、議論の余地がある。一方では、それらは、ウェルシュ菌(Clostridium perfringens)のような病原性微生物種の影響を裏付けている。他方では、これらの酵素は、高等動物を含む多くの非病原性種の炭水化物異化に共通の因子である。しかし、それらは、直接的な毒性効果を発揮しない(トラビング(Traving)ら Cell Mol Life Sci(1998年)54,1330−1349)。その代わり、それらの有害な影響は、非生理学的条件下での宿主シアル酸による誘導時に他の毒性因子と共に宿主に放出される大量の酵素に依存する。
【0006】
哺乳動物シアリダーゼは、通常、約40〜45kDのサイズである。哺乳動物シアリダーゼを過剰発現させ、そして産業的に関心のある量にまで産生させる試みについては、報告されていない。ヒトシアリダーゼは、リソソーム酵素、サイトゾル酵素または膜結合酵素であり得る(エイカイウタン(Achyuthan)およびエイカイウタン(Achyuthan)(2001年)Comp.Biochem.Phys.B部、129,29−64)。リソソームシアリダーゼは、グリコシル化酵素である。シアリダーゼは、保存されたモチーフを含有する。最も顕著な保存されたモチーフは、一般式−S−X−D−X−G−X−T−W−[式中、Xは可変の残基を表す]のアミノ酸のストレッチであるいわゆるAspボックスである。このモチーフが僅か1回もしくは2回しか見出されないかまたは存在さえしないウイルスシアリダーゼを例外として、このモチーフは、すべての微生物配列をとおして4〜5回見出される。第3のAspボックスは、Aspボックス2および4より強度に保存されている。連続的な2つのAspボックスの間の空間もまた、異なる一次構造の間で保存される(トラビング(Traving)ら Cell Mol Life Sci(1998年)54,1330−1349)。Aspボックスは、おそらく構造的役割を果たし、そしておそらく触媒に関与しない。Aspボックスとは対照的に、FRIP−モチーフは、アミノ酸配列のN末端部分に局在する。それは、絶対的に保存されたアルギニンおよびプラリネ(praline)残基を伴うアミノ酸−X−R−X−P−を包含する。アルギニンは、基質分子の結合によって触媒に直接関与する。aspボックス3および4の間のグルタミン酸リッチ領域、ならびにさらなる2つのアルギニン残基もまた、触媒作用に重要である(トラビング(Traving)ら Cell Mol Life Sci(1998年)54,1330−1349)。
【0007】
微生物シアリダーゼは、それらのサイズに従って、2つのグループ、すなわち、約42kDaの小さなタンパク質および60〜70kDaの大きなタンパク質に分類することができる。大きなシアリダーゼの一次構造は、N末端と第2のAspボックスとの間、ならびに第5のAspボックスとC末端との間にアミノ酸の追加のストレッチを含有する。それらは大きなシアリダーゼのより広範な基質特異性に寄与すると考えられる。哺乳動物シアリダーゼと同様に、細菌対応物は、F/YRIPモチーフおよびいくらかのAspボックスを含有する。細菌シアリダーゼは、しばしば、粘膜感染およびビルレンス(virulance)に関係している。このため、大きい方の細菌シアリダーゼは、食品または製薬用途における加工助剤としての使用に適切であるとは認識されていない。小さなシアリダーゼ(哺乳動物シアリダーゼと同じサイズ)は、上記に示す細菌において同定されている。即ち、ウェルシュ菌(Clostridium perfringens)は、他の細菌のシアリダーゼに共通の伸長部を伴わない約40kDaのサイズを伴う小さなシアリダーゼを含有する。しかし、このクロストリジウム(Clostridium)シアリダーゼは、細菌によって分泌されず、従ってまた、ビルレンス(virulance)に関与しない(ロッグゲンチン(Roggentin)ら(1995年)Biol Chem Hoppe Seyler 376,569−575)。追加の伸長部を伴う細菌シアリダーゼのみが病原性に関与することは、大変興味深い。大腸菌(E.coli)における細菌シアリダーゼの過剰発現は、一般的に、低い産生率をもたらし;小さなクロストリジウム(Clostridium)シアリダーゼは、大腸菌(E.coli)における細胞内タンパク質として1mg/lまでしか産生され得ない(クルーゼ(Kruse)ら(1996年)Protein Expr Purif.7,415−422)。
【0008】
ウチダ(Uchida)ら(Biochimica et Biophysica Acta,第350巻、第2号、1974年、425〜431頁)は、コロミン酸によって誘導される微生物ノイラミニダーゼのスクリーニングについて記載している。スクリーニングした1000の微生物のうち、スポロトリチウム・シェンキー(Sporotrichium schenckii)、ペニシリウム・ウルチカエ(Penicillium urticae)およびストレプトマイセス(Streptomyces)sp.からノイラミニダーゼが得られた。ペニシリウム・ウルチカエ(Penicillium urticae)は、食物の腐敗に関与する真菌であるため、それは、食品級シアリダーゼに適切な産生生物体ではなく、そしてスポロトリチウム・シェンキー(Sporotrichium schenckii)は病原性の真菌である。細菌ストレプトマイセス(Streptomyces)spのなかでの種名は決定されていない。この論文に記載のシアリダーゼのMWは不明である。イワモリ(Iwamori)ら(J.Biochem.138,327〜334頁)では、細菌アルスロバクター・ウレアファシエンス(Arthrobacter ureafaciens)シアリダーゼが開示されている。しかし、この細菌ノイラミニダーゼは、HIV−1仲介シンシチウム形成およびウイルス結合/進入プロセスに関連することが公知である(サン(Sun)ら、Virology 284,26〜36頁、2001年)。従って、食品および製薬における用途のための良好に産生される小さな非ビルレントなシアリダーゼが明らかに必要である。
【0009】
特に、分泌型酵素は容易に過剰発現され、そして真菌培養物から大量に精製され得るため、分泌型真菌シアリダーゼの所見は有益である。これは、シアリダーゼの産生のための原価を劇的に減少する。
【0010】
[発明の概要]
本発明は、以下からなる群から選択される、シアリダーゼ活性を有する単離されたポリペプチドに関する:
(a)配列番号3のアミノ酸1〜407と少なくとも60%のアミノ酸配列同一性を有するアミノ酸配列を有するポリペプチド;
(b)低度のストリンジェンシー条件で、(i)60ヌクレオチド超、好ましくは100ヌクレオチド超にわたり少なくとも80%もしくは90%同一な、より好ましくは200ヌクレオチド超にわたり少なくとも90%同一な配列番号1または2の核酸配列、あるいは(ii)配列番号1または2の核酸配列に相補的な核酸配列とハイブリダイズするポリヌクレオチドによってコードされるポリペプチド。好ましくは、このポリペプチドは、55kDa未満のMW(分子量)(SDS−page)を有し、より好ましくは、このポリペプチドは52kDa未満のMW(分子量)(SDS−page)を有するか、またはこのポリペプチドは50kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有し、より好ましくは、このポリペプチドは45kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有する。
【0011】
さらに、本発明は、シアリダーゼ活性を有し、そして非ビルレントであるポリペプチドを提供する。また、本発明の一部分は、シアリダーゼ活性を有するポリペプチドであり、前記ポリペプチドは、真菌のシアリダーゼであり、55kDa未満のMW(分子量)(SDS−Page)を有し、より好ましくは、この真菌シアリダーゼは52kDa未満のMW(分子量)(SDS−Page)を有するか、またはこの真菌シアリダーゼは50kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有し、より好ましくは、この真菌シアリダーゼは45kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有する。
【0012】
好ましくは、本発明のポリペプチドは、配列番号3のアミノ酸1〜407と少なくとも65%、好ましくは少なくとも70%、より好ましくは少なくとも80%、さらにより好ましくは少なくとも90%、最も好ましくは少なくとも95%、およびさらに最も好ましくは少なくとも約97%同一性を有するアミノ酸配列を有する。本発明はまた、請求項1に記載のポリペプチドをコードするか、または低度のストリンジェンシー条件下で、より好ましくは、中度のストリンジェンシー条件下で、および最も好ましくは、高度のストリンジェンシー条件下で、配列番号1または2とハイブリダイズする核酸配列を含んでなる単離されたポリヌクレオチドに関する。さらに加えて、本発明は、適切な発現宿主においてポリペプチドの産生を指令する1つもしくはそれ以上の制御配列に作動可能に連結された本発明のポリヌクレオチドを含んでなる核酸構築物を開示する。さらに、本発明は、この核酸構築物を含んでなる組み換え発現ベクター、および前記核酸構築物を含んでなる組み換え宿主細胞を提供する。本発明の別の態様に従えば、上記の株/組み換え宿主細胞を培養して、ポリペプチドを含んでなる上清および/または細胞を生成すること;ならびにポリペプチドを回収することを含んでなる、ポリペプチドの産生させるための方法が開示される。本発明のさらなる態様に従えば、ポリペプチドの産生に適切な条件下でポリペプチドをコードするポリヌクレオチドを含んでなる核酸構築物を含んでなる宿主細胞を培養すること;およびポリペプチドを回収することを含んでなる、本発明のポリペプチドを産生させるための方法が開示される。シアリダーゼ活性を有する本発明のポリペプチドは、有利なことに、非ビルレントである。本発明のポリペプチドは、食品もしくは飼料の調製において、または医薬品もしくは医薬品の一部分として使用することができる。
【0013】
[発明の詳細な説明]
本発明に従えば、好ましくは、55kDa未満のMW(分子量)(SDS−Page)を有する小型のシアリダーゼ(より好ましくは、このポリペプチドは52kDa未満のMW(分子量)(SDS−Page)を有する)、または50kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有するシアリダーゼ(より好ましくは、このポリペプチドは45kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有する)が開示される。有利なことに、本発明のシアリダーゼ、細胞外酵素。さらに、シアリダーゼは、好ましくは真菌由来である。一般に、本発明のポリペプチドは、35kDaを超えるMW(分子量)(SDS−Page)を有し、より好ましくは、このポリペプチドは40kDaを超えるMW(分子量)(SDS−Page)を有するか、またはこのポリペプチドは35kDaを超えるMW(分子量)(アミノ酸配列に基づいて算出される)を有し、より好ましくは、このポリペプチドは40kDaを超えるMW(分子量)(アミノ酸配列に基づいて算出される)を有する。
【0014】
エキソ酵素、または細胞外酵素は、細胞から分泌され、そして細胞の外側で作用する酵素である。それは、分解しなければ細胞に進入し得ない大きな分子を分解するために使用される。生物体が環境に分泌する酵素が、微生物の外側で作用する。エキソ酵素の反意語は、エンド酵素または細胞内酵素と呼ばれる(出典はWikipediaである)。
【0015】
本発明者らの理論では、一般に、大きなシアリダーゼは、粘膜感染およびビルレンスに関連し、従って、食品または製薬用途において適切ではない。それとは対照的に、小さなシアリダーゼは、食品または製薬用途に適切であると考えられる。
【0016】
しかし、本発明は、この理論の正確さについて肯定も否定もしない。本発明は、55kDa未満のMW(分子量)(SDS−Page)を有する細胞外シアリダーゼ(より好ましくは、このシアリダーゼは52kDa未満のMW(分子量)(SDS−Page)を有する)、または50kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)有する細胞外シアリダーゼ(より好ましくは、このシアリダーゼは45kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有する)を提供する。
【0017】
本発明は、真菌ペニシリウム・クリソゲナム(Penicillium chrysogenum)において同定された新たなシアリダーゼに関する。
【0018】
有利なことに、本発明は、多量に産生され得るシアリダーゼの需要を満たす。好ましくは、そのようなシアリダーゼは宿主細胞から分泌される。能動的分泌は、面倒な精製プロセスを経験することなく、ほとんど純粋な形の酵素の回収を可能にするため、それは、経済的な産生プロセスに最も重要である。アスペルギルス(Aspergillus)のような食品級真菌宿主によるそのような能動的に分泌されるシアリダーゼの過剰発現は、食品級酵素および費用効果の高い産生プロセスを生み出し、従って、好適である。本分泌型シアリダーゼは、糸状菌においてはじめて見出されたものである。食品級産生宿主アスペルギルス・ニガー(Aspergillus niger)による大量のシアリダーゼの産生のためのプロセスについて開示する。
【0019】
経済的観点から、哺乳動物および細菌シアリダーゼの低い産生率と比較して、多量かつ比較的純粋な形でシアリダーゼを産生させる改善された方法が明らかに必要である。これを行うための好適な方法は、組み換えDNA技術を使用するそのようなシアリダーゼの過剰産生を介することである。これを行うための特に好適な方法は、真菌由来のシアリダーゼの過剰産生を介することであり、そしてこれを行うための最も好適な方法は、ペニシリウム(Penicillium)由来シアリダーゼの過剰産生を介することである。後者の産生経路を可能にするためには、ペニシリウム(Penicillium)由来シアリダーゼの独特な配列情報が必須である。より好適には、コーディング遺伝子のヌクレオチド配列全体が利用可能でなければならない。
【0020】
多量かつ比較的純粋な形で新たに同定された分泌型シアリダーゼを産生させる改善された手段は、組み換えDNA技術を使用するペニシリウム(Penicillium)によりコードされる酵素の過剰産生を介することである。これを行うための好適な方法は、食品級宿主微生物におけるそのような分泌型シアリダーゼの過剰産生を介することである。周知の食品級微生物として、アスペルギルス(Aspergilli)、トリコデルマ(Trichoderma)、ストレプトマイセス(Streptomyces)、バチルス(Bacilli)ならびにサッカロマイセス(Saccharomyces)およびクリヴェロマイセス(Kluyveromyces)のような酵母が挙げられる。これを行うためのさらにより好適な方法は、アスペルギルス(Aspergillus)のような食品級真菌における分泌型ペニシリウム(Penicillium)由来シアリダーゼの過剰産生を介することである。シアリダーゼをコードする遺伝子のコドン使用頻度が、使用する食品級発現宿主に最適化されている食品級真菌における分泌型シアリダーゼの過剰産生が最も好適である。一般に、後者の最適化経路を可能にするためには、分泌型シアリダーゼの独特な配列情報が望ましい。より好適には、シアリダーゼコーディング遺伝子のヌクレオチド配列全体が利用可能でなければならない。一旦、分泌型シアリダーゼがコードする遺伝子が好適な宿主において形質転換されると、分泌型株を、発酵ブロスからの分泌型シアリダーゼタンパク質の発酵および単離に使用することができる。
【0021】
一旦、新たな酵素が、大量かつ比較的純粋な食物、(チーズのような)食料品において利用可能になれば、改善されたテクスチャーおよび/または免疫学的特性を伴うタンパク質加水分解物を、食品級のおよび経済的な方法で調製することができる。酵素はまた、潜在的に保存剤として使用することもできる。保護的シアル酸残基を細菌壁から取り出すことによって、微生物は、免疫系によって認識され、そして排除される。
【0022】
シアリダーゼ活性を有する本発明のポリペプチドは、単離された形態であってもよい。本明細書において定義されるように、単離されたポリペプチドは、本質的に他の非シアリダーゼポリペプチドを含まず、そして典型的に、SDS−PAGEによって決定されるように、少なくとも約20%純粋、好ましくは少なくとも約40%純粋、より好ましくは少なくとも約60%純粋、さらにより好ましくは少なくとも約80%純粋、なおより好ましくは約90%純粋、および最も好ましくは約95%純粋である、内因的に産生されたまたは組み換えポリペプチドである。ポリペプチドは、遠心分離およびクロマトグラフィー法、または粗溶液から純粋なタンパク質を得るための他の任意の技術によって単離してもよい。ポリペプチドは、ポリペプチドの意図された目的を妨害しないキャリアまたは希釈剤と混合してもよく、それ故、この形のポリペプチドもなお単離されたものと認識されることが理解されよう。それは、一般的に、調製物中のポリペプチドを含んでなり、ここで、本発明のポリペプチドは、調製物中のタンパク質の重量に対し20%を超える、例えば、30%、40%、50%、80%、90%、95%もしくは99%を超える。
【0023】
好ましくは、本発明のポリペプチドは、シアリダーゼ活性を伴う酵素をコードする遺伝子を所有する微生物から入手され得る。より好ましくは、本発明のポリペプチドは、微生物から分泌される。さらにより好ましくは、微生物は真菌であり、そして場合により、糸状菌である。それ故、好適なドナー生物体は、種ペニシリウム・クリソゲナム(Penicillium chrysogenum)のようなペニシリウム(Penicillium)属である。第1の実施形態では、本発明は、配列番号3(即ち、ポリペプチド)のアミノ酸1〜407に対して、少なくとも60%、好ましくは少なくとも70%、より好ましくは少なくとも80%、さらにより好ましくは少なくとも90%、なおより好ましくは少なくとも95%、および最も好ましくは少なくとも97%のアミノ酸配列同一性の程度を有し、かつシアリダーゼ活性を有するアミノ酸配列を有する単離されたポリペプチドを提供する。
【0024】
本発明の目的のために、2つもしくはそれ以上アミノ酸配列間の同一性の程度は、マトリックスBlosum62および10の予想閾値を伴うBLAST Pタンパク質データベースサーチプログラム(アルトシュル(Altschul)ら、1997年、Nucleic Acids Research 25:3389−3402)によって決定される。
【0025】
本発明のポリペプチドは、配列番号3に記載のアミノ酸配列、もしくは実質的に相同な配列、またはシアリダーゼ活性を有するいずれかの配列のフラグメントを含んでなり得る。一般に、配列番号3に示す天然に存在するアミノ酸配列が好適である。
【0026】
本発明の1つの態様は、配列番号3に示されるアミノ酸配列を有するポリペプチドであって、それによって、シグナル配列が取り出される。プレ−もしくはシグナル−配列および/またはプロ−配列を含む配列番号3のアミノ酸配列のような分泌型酵素は、しばしば合成される。これらの配列は、しばしば、分泌プロセス中またはその後のいずれかにタンパク質から取り出される。従って、成熟分泌型タンパク質は、しばしば、もはや、これらのプレ−およびプロ−配列を含有しない。従って、配列番号3のアミノ酸1〜407と少なくとも60%のアミノ酸配列同一性を有するアミノ酸配列を有し、可能なプレ−またはプロ−配列を欠如するポリペプチドは、本発明の一部である。配列番号3のアミノ酸配列における可能なプロセシング部位は、アミノ酸33より後方に存在する。この場合、本発明に従う成熟酵素は、アミノ酸番号34から開始する。アミノ酸の取り出しのようなさらなるプロセシングによる配列番号3のアミノ酸配列からの他の改変も、それらが酵素の活性を妨げない限り、許容される。
【0027】
本発明のポリペプチドもまた、配列番号3のポリペプチドの天然に存在する変異体または種相同体を含んでなり得る。
【0028】
変異体は、例えば、真菌、細菌、酵母または植物細胞において天然に存在するポリペプチドであって、この変異体は、シアリダーゼ活性および配列番号3のタンパク質に実質的に類似の配列を有する。用語「変異体」は、配列番号3のシアリダーゼと同じ本質的特徴または基本的な生物学的機能を有するポリペプチドを指し、そして対立遺伝子変異体を含む。好ましくは、変異ポリペプチドは、配列番号3のポリペプチドと少なくとも同じレベルのシアリダーゼ活性を有する。変異体は、配列番号3のポリペプチドと同じ株または同じ属もしくは種の異なる株のいずれかに由来する対立遺伝子変異体を含む。
【0029】
同様に、本発明のタンパク質の種相同体は、シアリダーゼである類似の配列の等価なタンパク質であり、そして別の種において天然に存在する。
【0030】
変異体および種相同体は、本明細書に記載の手順を使用し、そして適切な細胞供給源、例えば、細菌、酵母、真菌または植物細胞に対してそのような手順を実施して、単離することができる。また、配列番号3のポリペプチドの変異体または種相同体を発現するクローンを入手するために、本発明のプローブを使用して、酵母、細菌、真菌または植物細胞から作製したDNAライブラリーを探索することも可能である。公知の遺伝子の変異体および種相同体を単離するために使用することができる方法は、文献において広範に記載されており、そして当業者に公知である。これらの遺伝子は、従来の技術によって操作して、本発明のポリペプチドを作製することができ、その後、それ自体が公知の組み換えまたは合成技術によって産生され得る。
【0031】
配列番号3のポリペプチドならびに変異体および種相同体の配列を改変して、本発明のポリペプチドを提供することもできる。アミノ酸置換は、例えば、1、2または3〜10、20または30置換から作製され得る。同じ数の欠失および挿入を作製してもよい。これらの変更は、改変されたポリペプチドがそのシアリダーゼ活性を保持するように、ポリペプチドの機能に極めて重要な領域の外側で作製してもよい。
【0032】
本発明のポリペプチドは、上記の全長ポリペプチドおよびその変異体のフラグメントを含み、配列番号3に記載の配列のフラグメントを含む。そのようなフラグメントは、典型的に、シアリダーゼとしての活性を保持する。フラグメントは、少なくとも50、100もしくは200アミノ酸長であってもよく、または配列番号3に示す全長配列より短いこのような数のアミノ酸であってもよい。
【0033】
通常、本発明のポリペプチドは、以下に記載のように組み換え的に作製されるが、必要であれば、それらを、合成手段によって生成することもできる。合成および組み換えポリペプチドは、例えば、それらの同定もしくは精製を補助するためのヒスチジン残基またはT7タグの付加によってか、あるいは細胞からのそれらの分泌を促進するためのシグナル配列の付加によって、改変され得る。
【0034】
それ故、変異配列は、配列番号3のポリペプチドを単離した株以外のペニシリウム(Penicillium)の株から誘導される配列を含んでなり得る。変異体は、シアリダーゼ活性を調べ、そして本明細書に記載のようにクローニングおよび配列決定することによって、他のペニシリウム(Penicillium)株から同定することができる。ペプチドが、配列番号3のシアリダーゼの基本的な生物学的機能を維持する限り、変異体は、タンパク質配列内の単一のアミノ酸もしくはアミノ酸群の欠失、改変または付加を含んでもよい。
【0035】
アミノ酸置換は、例えば、1、2または3〜10、20または30置換から作製され得る。改変されたポリペプチドは、一般的に、シアリダーゼとしての活性を保持する。保存的置換を行ってもよく;そのような置換は、当該技術分野において周知である。
【0036】
より短いまたはより長いポリペプチド配列も本発明の範囲内にある。例えば、少なくとも50アミノ酸または100、150、200、300、400、500、600、700もしくは800アミノ酸長までのペプチドが、それが配列番号3のシアリダーゼの基本的な生物学的機能を実証する限り、本発明の範囲内に当てはまると考えられる。特に、但し排他的ではないが、本発明のこの態様は、タンパク質が完全なタンパク質配列のフラグメントである状況を包含する。
【0037】
本発明はまた、シアリダーゼ活性を有するポリペプチドをコードするポリヌクレオチドに関し、前記ポリヌクレオチドは、アミノ酸配列番号3をコードするポリヌクレオチド配列を含んでなる。
【0038】
本発明では、目的のタンパク質が、能動的に増殖培地に分泌されることが特に関連する。分泌型タンパク質は、通常、プレ−タンパク質として最初は合成され、続いてプレ−配列(シグナル配列)が、分泌プロセス中に取り出される。分泌プロセスは、原核細胞および真核生物において基本的に類似する:能動的に分泌されるプレ−タンパク質は膜を通り抜け、シグナル配列が特異的シグナルペプチダーゼによって取り出され、そして成熟タンパク質が(リ)フォールディングされる。また、シグナル配列について、一般構造を認識することができる。分泌のためのシグナル配列は、プレ−タンパク質のアミノ末端に局在し、そして一般的に、15〜35アミノ酸長である。アミノ末端は、好ましくは、正に荷電したアミノ酸を含有し、そして好ましくは、酸性アミノ酸を含有しない。この正に荷電した領域は、膜のリン脂質の負に荷電したヘッド基と相互作用すると考えられる。この領域の後に、疎水性の膜貫通コア領域が続く。この領域は、一般的に、10〜20アミノ酸長であり、そして主に、疎水性アミノ酸よりなる。荷電したアミノ酸は、通常、この領域には存在しない。膜貫通領域の後には、シグナルペプチダーゼの認識部位が続く。認識部位は、小−X−小を選好するアミノ酸よりなる。小さなアミノ酸は、アラニン、グリシン、セリンまたはシステインであり得る。Xは、任意のアミノ酸であり得る。そのような規則を使用して、真核生物および原核細胞由来のそのようなシグナル配列を認識することが可能なアルゴリズムが作成されている(ベントセン(Bendtsen)、ニールセン(Nielsen)、フォン・ヘイネ(von Heijne)およびブルナック(Brunak).(2004年)J.Mol.Biol.,340:783−795)。タンパク質のシグナル配列を算出し、そして認識するためのSignalPプログラムが、一般的に利用可能である(http://www.cbs.dtu.dk/services/SignalP/)。
【0039】
シグナル配列が配列決定した遺伝子の推測されたタンパク質配列から認識され得ることは、本発明に関連する。遺伝子がSignalPプログラムを使用してシグナル配列が予測されるタンパク質をコードする場合、このタンパク質が分泌される可能性は高い。
【0040】
第2の実施形態では、本発明は、シアリダーゼ活性を有し、そして低度のストリンジェンシー条件下で、より好ましくは、中度のストリンジェンシー条件下で、および最も好ましくは、高度のストリンジェンシー条件下で、(i)配列番号1の核酸配列、または(ii)配列番号1の少なくとも一部を含んでなる核酸フラグメント、もしくは(iii)配列番号1の塩基とは異なる塩基を有する核酸フラグメントと;あるいは(iv)配列番号1に相補的な核酸鎖とハイブリダイズするかまたはハイブリダイズすることが可能であるポリヌクレオチドによってコードされる単離されたポリペプチドを提供する。
【0041】
用語「ハイブリダイズすることが可能」は、本発明の標的ポリヌクレオチドが、プローブとして使用される核酸(例えば、配列番号1のヌクレオチド配列、もしくはそのフラグメント、または配列番号1の相補体もしくはそのフラグメント)と、バックグランドよりも有意に高いレベルでハイブリダイズすることができることを意味する。本発明はまた、本発明のシアリダーゼをコードするポリヌクレオチド、ならびにそれに相補的であるヌクレオチド配列を含む。ヌクレオチド配列は、RNA、またはゲノムDNA、合成DNAもしくはcDNAを含むDNAであってもよい。好ましくは、ヌクレオチド配列は、DNAおよび最も好ましくは、ゲノムDNA配列である。典型的に、本発明のポリヌクレオチドは、選択的条件下で、配列番号1のコーディング配列またはコーディング配列の相補体にハイブリダイズすることが可能であるヌクレオチドの連続配列を含んでなる。そのようなヌクレオチドは、当該技術分野において周知の方法に従って合成することができる。
【0042】
本発明のポリヌクレオチドは、配列番号2のコーディング配列またはコーディング配列の相補体に、バックグランドよりも有意に高いレベルでハイブリダイズすることができる。例えば、cDNAライブラリーに存在する他のcDNAのため、バックグランドのハイブリダイゼーションが生じ得る。本発明のポリヌクレオチドと配列番号2のコーディング配列またはコーディング配列の相補体との間の相互作用によって作製されるシグナルレベルは、典型的に、他のポリヌクレオチドと配列番号2のコーディング配列との間の相互作用の少なくとも10倍、好ましくは少なくとも20倍、より好ましくは少なくとも50倍、およびさらにより好ましくは少なくとも100倍の強度である。相互作用の強度は、例えば、プローブを、例えば、32Pで放射性標識することによって、測定することができる。選択的ハイブリダイゼーションは、典型的に、低いストリンジェンシー(0.3M塩化ナトリウムおよび0.03Mクエン酸ナトリウム、約40℃)、中等度のストリンジェンシー(例えば、0.3M塩化ナトリウムおよび0.03Mクエン酸ナトリウム、約50℃)または高いストリンジェンシー(例えば、0.3M塩化ナトリウムおよび0.03Mクエン酸ナトリウム、約60℃)の条件を使用して、達成することができる。
【0043】
本発明のポリヌクレオチドはまた、配列番号3のポリペプチドまたはその変異体をコードすることができる合成遺伝子を含む。時々、遺伝子のコドン使用頻度を産生宿主の好適なバイアスに適応することも好適である。合成遺伝子を設計および構築するための技術は、一般的に利用可能である(即ち、http://www.dnatwopointo.com/)。
【0044】
[改変]
本発明のポリヌクレオチドは、DNAまたはRNAを含んでなり得る。それらは、一本鎖であってもまたは二本鎖であってもよい。それらはまた、それらの内部にペプチド核酸を含む合成または改変されたヌクレオチドを含むポリヌクレオチドであってもよい。ポリヌクレオチドに対する異なる多くのタイプの改変が、当該技術分野において公知である。これらは、メチルホスホネートおよびホスホロチオエート骨格、ならびに分子の3’および/または5’末端におけるアクリジン鎖またはポリリジン鎖の付加を含む。本発明の目的のために、本明細書に記載のポリヌクレオチドは、当該技術分野において利用可能な任意の方法によって改変され得ることが理解されるべきである。
【0045】
当業者は、日常的な技術を使用して、本発明のポリヌクレオチドによってコードされるポリペプチド配列に影響を及ぼさないヌクレオチド置換を行って、本発明のポリペプチドを発現させようとする任意の特定の宿主生物体のコドン使用頻度に反映させ得ることが理解されるべきである。
【0046】
配列番号2のコーディング配列は、ヌクレオチド置換、例えば、1、2もしくは3〜10、25、50、100、もしくはそれ以上の置換によって、改変してもよい。配列番号2のポリヌクレオチドは、1つもしくはそれ以上の挿入および/または欠失によって、ならびに/あるいはいずれか一方もしくは両方の末端における伸長によって代替的または付加的に改変され得る。改変されたポリヌクレオチドは、一般的に、シアリダーゼ活性を有するポリペプチドをコードする。例えば、後に、ポリペプチドに関して考察するように、改変された配列が翻訳される場合、縮重置換を行ってもよく、および/または保存的アミノ酸置換を生じる置換を行ってもよい。
【0047】
[相同体]
配列番号2のDNAコーディング配列の相補体に選択的にハイブリダイズすることが可能であるヌクレオチド配列は、本発明に含まれ、そして一般的に、配列番号2の少なくとも60、好ましくは少なくとも100、より好ましくは少なくとも200連続ヌクレオチドの領域にわたって、または最も好ましくは全長にわたって配列番号2のコーディング配列と少なくとも50%もしくは60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%または少なくとも99%の配列同一性を有する。同様に、活性なシアリダーゼをコードし、そして配列番号2のDNAコーディング配列の相補体のフラグメントと選択的にハイブリダイズすることが可能であるヌクレオチドもまた、本発明に包含される。
【0048】
上記の程度の同一性および最小サイズの任意の組み合わせを使用して、本発明のポリヌクレオチドを定義してもよく、よりストリンジェントな組み合わせ(即ち、より長い長さにわたって同一性がより高い)が好適である。それ故、例えば、60、好ましくは、100ヌクレオチドにわたって少なくとも80%または90%同一であるポリヌクレオチドは、200ヌクレオチドにわたって少なくとも90%同一であるポリヌクレオチドと同様に、本発明の1つの態様を形成する。
【0049】
BLAST Nアルゴリズムを使用して、配列同一性を算出するかまたは(例えば、それらのデフォルト設定値に対する均等もしくは対応する配列を同定するような)配列を列挙することができる。
【0050】
BLAST解析を実施するためのソフトウェアは、National Center for Biotechnology Information(http://www.ncbi.nlm.nih.gov/)を介して公的に入手することができる。このアルゴリズムは、データベース配列において同じ長さのワードと共に整列される場合、いくつかのポジティブ評価された閾値スコアTに一致するか、または満たすかのいずれかである問い合わせ配列における長さWの短いワードを同定することによって、ハイスコアリング配列対(high scoring sequence pair)(HSP)を第1に同定することに関与する。Tは隣接ワードスコア閾値と称される。これらの初期の隣接ワードのヒットは、それらを含有するHSPを見出すための検索を初期化するためのシードとして作用する。ワードヒットは、累積アラインメントスコアが増加し得る限り、各配列に沿って両方向において拡張される。累積アラインメントスコアがその最大到達値から量Xだけ低下する場合:1つもしくはそれ以上のネガティブスコアリング残基アラインメントの累積により累積スコアが0もしくはそれ以下となる場合;またはいずれかの配列の末端に到達する場合、各方向におけるワードヒットの拡張は停止する。BLASTアルゴリズムパラメータW、TおよびXは、アラインメントの感度および速度を決定する。BLASTプラグラムは、デフォルトとして、11のワード長(W)、50のBLOSUM62スコアリングマトリックスアラインメント(B)、10の期待値(E)、M=5、N=4、および両鎖の比較を使用する。
【0051】
BLASTアルゴリズムは、2つの配列間の類似性の統計解析を実施する。BLASTアルゴリズムによって提供される類似性の1つの測定は、2つのヌクレオチドまたはアミノ酸配列間の一致が偶然に生じる確率の表示を提供する最小和確率(P(N))である。例えば、配列は、第1の配列と第2の配列との比較における最小和確率が約1未満、好ましくは約0.1未満、より好ましくは約0.01未満、および最も好ましくは約0.001未満である場合、もう1つの配列に類似であるとみなされる。
【0052】
[プライマーおよびプローブ]
本発明のポリヌクレオチドは、プライマー、例えば、ポリメラーゼ連鎖反応(PCR)プライマー、代替的増幅反応のためのプライマー、または例えば、放射性もしくは非放射性標識を使用する従来の手段によって明示用標識(revealing label)で標識されたプローブを含み、そして上記プライマーまたはプローブとして使用してもよく、あるいはポリヌクレオチドをベクターにクローニングしてもよい。そのようなプライマー、プローブおよび他のフラグメントは、少なくとも15、例えば、少なくとも20、25、30もしくは40ヌクレオチド長である。それらは、典型的に、40、50、60、70、100、150、200もしくは300ヌクレオチド長までであるか、またはなお、配列番号2のコーディング配列より数ヌクレオチド(例えば、5もしくは10ヌクレオチド)短い長さまでである。
【0053】
一般に、プライマーは、1回に1ヌクレオチドの所望される核酸配列の段階的な製造を含めた合成手段によって生成される。これを達成するための技術およびプロトコルは、当該技術分野において容易に利用可能である。より長いポリヌクレオチドは、一般的に、組み換え手段、例えば、PCRクローニング技術を使用して、生成される。これは、(典型的に、約15〜30ヌクレオチドの)プライマーの対を作製して、クローニングしようとするシアリダーゼの所望される領域を増幅し、プライマーと、酵母、細菌、植物、原核細胞または真菌細胞、好ましくは、ペニシリウム(Penicillium)株から得られるmRNA、cDNAもしくはゲノムDNAとを接触させ、所望される領域の増幅に適切な条件下でポリメラーゼ連鎖反応を実施し、(例えば、アガロースゲル上で反応混合物を精製することによって)増幅したフラグメントを単離し、そして増幅したDNAを回収することを含む。実施例1に記載のように、増幅したDNAを適切なクローニングベクターにクローニングすることができるように、適切な制限酵素認識部位を含有するようにプライマーを設計してもよい。
【0054】
あるいは、分泌型シアリダーゼまたはその変異体のコーディング領域を包含する合成遺伝子を構築することができる。これらの技術を使用して、多くの位置で変更されているが、なお同じタンパク質をコードするポリヌクレオチドを簡便に設計し、そして構築することができる。これは、コドン使用頻度を好適な発現宿主に適応することができ、そのようにしてこの宿主におけるタンパク質の産生率を改善することができることを利点として有する。また、遺伝子のポリヌクレオチド配列を変更して、mRNA安定性または減少したターンオーバーを改善することができる。これは、所望されるタンパク質またはその変異体の改善された発現をもたらすことができる。さらに、分泌効率、安定性、タンパク質分解に対する脆弱性、至適温度、比活性またはタンパク質の産業的産生もしくは用途のための他の関連特性に対してポジティブな効果を有する変異がタンパク質配列において作製されるように、合成遺伝子においてポリヌクレオチド配列を変更することができる。一般的に、合成遺伝子を構築し、そしてコドン使用頻度を最適化する業務を提供する企業が利用可能である。
【0055】
そのような技術を使用して、本明細書に記載のシアリダーゼ配列をコードするポリヌクレオチドのすべてまたは一部分を入手してもよい。イントロン、プロモーターおよびトレーラー領域は、本発明の範囲内にあり、そしてまた、真菌、酵母、細菌、植物または原核細胞由来のゲノムDNAから開始して、類似の様式で(例えば、組み換え手段、PCRもしくはクローニング技術によって)入手してもよい。
【0056】
ポリヌクレオチドまたはプライマーは、明示用標識を担持してもよい。適切な標識として、放射性同位元素、例えば、32Pまたは35S、蛍光標識、酵素標識または他のタンパク質標識、例えば、ビオチンが挙げられる。そのような標識は、本発明のポリヌクレオチドまたはプライマーに付加し得、そして当業者に公知の技術を使用して検出し得る。
【0057】
標識されたまたは標識されていないポリヌクレオチドまたはプライマー(もしくはそのフラグメント)を、真菌サンプルにおけるシアリダーゼまたはその変異体を検出または配列決定するための核酸に基づく試験に使用してもよい。そのような検出試験は、一般的に、目的のDNAを含有することが疑わしい真菌サンプルと、本発明のポリヌクレオチドまたはプライマーを含んでなるプローブとを、ハイブリダイジング条件下で接触させ、そしてプローブとサンプル中の核酸との間に形成される任意の二重鎖を検出することを含んでなる。検出は、PCRのような技術を使用するか、またはプローブを固相支持体上に固定化し、プローブにハイブリダイズしないサンプル中の任意の核酸を取り出し、次いで、プローブにハイブリダイズする任意の核酸を検出することによって、達成され得る。あるいは、サンプル核酸を固相支持体上に固定化し、プローブをハイブリダイズさせ、そして、任意の非結合プローブの取り出し後、そのような支持体に結合したプローブの量を検出してもよい。
【0058】
本発明のプローブを、適切な容器に試験キットの形で簡便に包装してもよい。そのようなキットでは、プローブを固相支持体に結合させてもよく、ここで、キットが設計されるアッセイ形式は、そのような結合を必要とする。キットはまた、探索しようとするサンプルを処理し、プローブをサンプル中の核酸にハイブリダイズさせるための適切な試薬、コントロール試薬、取扱説明書などを含有してもよい。本発明のプローブおよびポリヌクレオチドはまた、マイクロアッセイに使用してもよい。
【0059】
好ましくは、本発明のポリヌクレオチドは、真菌、特に、アスペルギルス(Aspergillus)属の真菌のようなポリペプチドと同じ生物体から入手可能である。
【0060】
[ポリヌクレオチドの生成]
配列番号2と100%の同一性を有さないが、本発明の範囲内に当てはまるポリヌクレオチドは、多くの方法で入手することができる。それ故、本明細書に記載のシアリダーゼ配列の変異体は、例えば、本発明のポリペプチドの供給源として考察したような生物体の範囲から作製されるゲノムDNAライブラリーを探索することによって、入手してもよい。加えて、シアリダーゼの他の真菌、植物または原核生物相同体が入手され得、そしてそのような相同体およびそのフラグメントは、一般に、配列番号1にハイブリダイズすることが可能である。そのような配列は、他の種由来のcDNAライブラリーまたはゲノムDNAライブラリーを探索すること、およびそのようなライブラリーを、(先に記載のように)低度、中度〜高度のストリンジェンシーの条件下、配列番号1のすべてまたは一部分を含んでなるプローブで探索することによって、入手され得る。配列番号1のすべてまたは一部分を含んでなる核酸プローブを使用して、本発明のポリペプチドの供給源として記載したような他の種由来のcDNAまたはゲノムライブラリーを探索してもよい。
【0061】
種相同体はまた、保存されたアミノ酸配列をコードする変異体および相同体内の配列を標的化するように設計されたプライマーを使用する縮重PCRを使用して、入手してもよい。プライマーは、1つもしくはそれ以上の縮重位置を含有することができ、そして既知の配列に対する単一の配列プライマーを伴う配列をクローニングするために使用される条件より低いストリンジェンシー条件で使用される。
【0062】
あるいは、そのようなポリヌクレオチドは、シアリダーゼ配列またはその変異体の部位特異的変異によって入手され得る。これは、例えば、配列に対するサイレントコドンの変化が、ポリヌクレオチド配列が発現される特定の宿主細胞についてコドン選択を最適化するために必要とされる場合、有用であり得る。制限酵素認識部位を導入するため、またはポリヌクレオチドによってコードされるポリペプチドの特性または機能を改変するために、他の配列変化を作製してもよい。
【0063】
本発明は、本発明のポリヌクレオチドおよびその相補体を含んでなる二本鎖ポリヌクレオチドを含む。
【0064】
本発明はまた、上記の本発明のポリペプチドをコードするポリヌクレオチドを提供する。そのようなポリヌクレオチドは、本発明のポリペプチドの組み換え産生のための配列に有用であるため、これは一般的に望ましいが、それらが、配列番号1または配列番号2の配列にハイブリダイズすることが可能である必要はない。そうでなければ、所望であれば、そのようなポリヌクレオチドを、上記のように標識、使用、および作製してもよい。
【0065】
[組み換えポリヌクレオチド]
本発明はまた、クローニングベクターおよび発現ベクターを含む本発明のポリヌクレオチドを含んでなるベクター、ならびに別の態様において、増殖、形質転換、または例えば、本発明のポリペプチド、もしくは本発明の配列によってコードされるポリペプチドの発現が生じる条件下でそのようなベクターを適切な宿主細胞にトランスフェクトする方法を提供する。本発明のポリヌクレオチドまたはベクターを含んでなる宿主細胞もまた提供され、ここで、ポリヌクレオチドは、宿主細胞のゲノムに対して異種である。通常、宿主細胞に関する用語「異種」は、ポリヌクレオチドが宿主細胞のゲノムにおいて天然には存在しないか、またはポリペプチドが細胞によって天然には産生されないことを意味する。好ましくは、宿主細胞は、酵母細胞、例えば、クリヴェロマイセス(Kluyveromyces)、ピキア(Pichia)、ハンセヌラ(Hansenula)もしくはサッカロマイセス(Saccharomyces)属の酵母細胞または例えば、アスペルギルス(Aspergillus)、ペニシリウム(Penicillium)、トリコデルマ(Trichoderma)もしくはフザリウム(Fusarium)属の糸状菌細胞である。
【0066】
[ベクター]
本発明の発現カセットが挿入されるベクターは、組み換えDNA手順に簡便に供され得る任意のベクターであり得、そしてベクターの選択は、しばしば、それが導入されるべき宿主細胞に依存する。それ故、ベクターは、自律的に複製するベクター、即ち、プラスミドのように染色体外の実体として存在し、その複製は、染色体の複製に依存しないベクターであってもよい。あるいは、ベクターは、宿主細胞に導入される場合、宿主細胞のゲノムに組込まれ、それが組込まれている染色体と共に複製するものであってもよい。
【0067】
好ましくは、本発明のポリヌクレオチドがベクター中にある場合、それは、宿主細胞によるコーディング配列の発現を提供することが可能である調節配列に作動可能に連結される(即ち、ベクターは発現ベクターである)。用語「作動可能に連結された」は並置を指し、ここで、記載の成分は、それらが、それらの意図された様式で機能することを可能にする関係にある。プロモーター、エンハンサーまたはコーディング配列に「作動可能に連結された」他の発現調節シグナルのような調節配列は、コーディング配列の発現が、産生条件下で達成されるような方法で配置される。
【0068】
ベクターは、例えば、プラスミド、コスミド、ウイルスもしくはファージベクターの場合、複製開始点、場合により、ポリヌクレオチドの発現のためのプロモーター、ならびに場合により、プロモーターのエンハンサーおよび/またはレギュレーターを伴って提供され得る。ターミネーター配列が存在してもよく、それはポリアデニル化配列であってもよい。ベクターは、1つもしくはそれ以上の選択マーカー遺伝子、例えば、細菌プラスミドの場合にはアンピシリン耐性遺伝子または哺乳動物ベクターではネオマイシン耐性遺伝子を含有してもよい。ベクターは、例えば、RNAの産生のためにインビトロで使用してもよく、または宿主細胞をトランスフェクトもしくは形質転換するために使用することができる。
【0069】
ポリペプチドをコードするDNA配列は、好ましくは、DNA配列が、宿主細胞におけるDNA配列の発現を指令することが可能である発現シグナルに作動可能に連結される発現構築物の一部分として適切な宿主に導入される。発現構築物による適切な宿主の形質転換のために、当業者に周知である形質転換手順が利用可能である。発現構築物は、選択マーカーを担持するベクターの一部分として、宿主の形質転換に使用することができ、または発現構築物は、選択マーカーを担持するベクターと共に個別の分子として同時形質転換される。ベクターは、1つもしくはそれ以上の選択マーカー遺伝子を含有してもよい。
【0070】
好適な選択マーカーとして、宿主細胞における欠損を補うか、または薬物に対する耐性を付与するものが挙げられるが、これらに限定されない。それらは、例えば、アセトアミダーゼ遺伝子またはcDNA(amdS、niaD、facA遺伝子またはA.ニダランス(A.nidulans)、麹菌(A.oryzae)、もしくはA.ニガー(A.niger)由来のcDNA)のようなほとんどの糸状菌および酵母の形質転換に使用することができる多目的マーカー遺伝子、あるいはG418、ハイグロマイシン、ブレオマイシン、カナマイシン、フレオマイシンまたはベノミル耐性(benA)のような抗生物質に対する耐性を提供する遺伝子を含む。あるいは、対応する変異宿主株を必要とする栄養要求性マーカーのような特定の選択マーカー:例えば、URA3(S.セレビシエ(S.cerevisiae)由来もしくは他の酵母由来の類似の遺伝子)、pyrGまたはpyrA(A.ニダランス(A.nidulans)もしくはA.ニガー(A.niger)由来)、argB(A.ニダランス(A.nidulans)もしくはA.ニガー(A.niger)由来)あるいはtrpCを使用することができる。好適な実施形態では、発現構築物の導入後、選択マーカー遺伝子を含まないポリペプチドを産生することが可能な形質転換された宿主細胞が得られるように、選択マーカーを、形質転換された宿主細胞から欠失する。
【0071】
他のマーカーとして、ATPシンテターゼサブユニット9(oliC)、オロチジン−5’−リン酸−デカルボキシラーゼ(pvrA)、細菌G418耐性遺伝子(酵母において有用であるが、糸状菌では有用ではない)、アンピシリン耐性遺伝子(大腸菌(E.coli))、ネオマイシン耐性遺伝子(バチルス(Bacillus))およびグルクロニダーゼ(GUS)をコードする大腸菌(E.coli)uidA遺伝子が挙げられる。ベクターは、例えば、RNAの産生のために、または宿主細胞をトランスフェクトもしくは形質転換するためにインビトロで使用してもよい。
【0072】
ほとんどの糸状菌および酵母では、発現構築物は、好ましくは、安定な形質転換体を得るために、宿主細胞のゲノムに組み込まれる。しかし、所定の酵母では、発現構築物を、安定かつ高いレベルの発現のために組み入れることができる適切なエピソームベクター系もまた、入手可能である。それらの例として、それぞれ、サッカロマイセス(Saccharomyces)およびクリヴェロマイセス(Kluyveromyces)の2μm、CENおよびpKD1プラスミドから誘導されるベクター、またはAMA配列(例えば、アスペルギルス(Aspergillus)由来のAMA1)を含有するベクターが挙げられる。発現構築物を宿主細胞ゲノムに組み込む場合、構築物は、ゲノムの無作為の座位か、または相同組み換えを使用して、予め決定された標的座位(この場合、標的座位は、好ましくは、高度に発現される遺伝子を含んでなる)のいずれかに組み込まれる。高度に発現される遺伝子は、例えば、誘導条件下で、そのmRNAが少なくとも0.01%(w/w)の全細胞mRNAを構成し得る遺伝子、あるいは、その遺伝子産物が少なくとも0.2%(w/w)の全細胞性タンパク質を構成し得るか、または、分泌される遺伝子産物の場合、少なくとも0.05g/lのレベルまで分泌され得る遺伝子である。
【0073】
所定の宿主細胞のための発現構築物は、通常、第1の態様のポリペプチドをコードする配列のコーディング鎖に対して5’末端から3’末端への連続した順番で相互に作動可能に連結した次のエレメントを含有する:(1)所与の宿主細胞においてポリペプチドをコードするDNA配列の転写を指令することが可能なプロモーター配列、(2)好ましくは、5’非翻訳領域(リーダー)、(3)場合により、所与の宿主細胞由来のポリペプチドの培養培地への分泌を指令することが可能なシグナル配列、(4)成熟かつ好ましくは活性型のポリペプチドをコードするDNA配列、および好ましくはまた、(5)ポリペプチドをコードするDNA配列の下流で転写を終結することが可能な転写終結領域(ターミネーター)。
【0074】
ポリペプチドをコードするDNA配列の下流の発現構築物は、好ましくは、ターミネーターとも称される1つもしくはそれ以上の転写終結部位を含有する3’非翻訳領域を含有する。ターミネーターの起源はそれほど重要ではない。ターミネーターは、例えば、ポリペプチドをコードするDNA配列に由来し得る。しかし、好ましくは、細菌ターミネーターが、細菌宿主細胞において使用され、酵母ターミネーターが、酵母宿主細胞において使用され、そして糸状菌ターミネーターが、糸状菌宿主細胞において使用される。より好ましくは、ターミネーターは、ポリペプチドをコードするDNA配列が発現される宿主細胞に内因性である。
【0075】
本発明のポリペプチドをコードするポリヌクレオチドの増強された発現はまた、選択された発現宿主からの目的のタンパク質の発現、および所望であれば、分泌レベルを増加し、および/または本発明のポリペプチドの発現の誘導可能な制御を提供する役割を果たす異種の調節領域、例えば、プロモーター、シグナル配列およびターミネーター領域の選択によっても達成され得る。
【0076】
本発明のポリペプチドをコードする遺伝子に由来するプロモーターに加えて、他のプロモーターを使用して、本発明のポリペプチドの発現を指令してもよい。プロモーターは、所望される発現宿主において本発明のポリペプチドの発現を指令するその効率について、選択してもよい。
【0077】
プロモーター/エンハンサーおよび他の発現調節シグナルは、発現ベクターが設計される宿主細胞に適合可能であるように選択することができる。例えば、原核プロモーター、特に、大腸菌(E.coli)株に使用するのに適切なプロモーターを使用してもよい。本発明のポリペプチドの発現が哺乳動物細胞において行われる場合、哺乳動物のプロモーターを使用してもよい。組織特異的プロモーター、例えば、肝細胞特異的プロモーターを使用してもよい。ウイルスプロモーター、例えば、モロニーマウス白血病ウイルス長末端反復(MMLV LTR)、ラウス肉腫ウイルス(RSV)LTRプロモーター、SV40プロモーター、ヒトサイトメガロウイルス(CMV)IEプロモーター、単純ヘルペスウイルスプロモーターまたはアデノウイルスプロモーターを使用することもできる。
【0078】
適切な酵母プロモーターとして、S.セレビシエ(S.cerevisiae)GAL4およびADHプロモーターならびにS.ポンベ(S.pombe)nmt1およびadhプロモーターが挙げられる。哺乳動物のプロモーターとしては、重金属、例えば、カドミウムに応答して誘導され得るメタロチオネインプロモーターが挙げられる。ウイルスプロモーター、例えば、SV40ラージT抗原プロモーターまたはアデノウイルスプロモーターを使用してもよい。これらのすべてのプロモーターは、当該技術分野において容易に利用可能である。
【0079】
哺乳動物のプロモーター、例えば、β−アクチンプロモーターを使用してもよい。組織特異的プロモーター、特に、内皮または神経細胞特異的プロモーター(例えば、DDAHIおよびDDAHIIプロモーター)が特に好適である。ウイルスプロモーター、例えば、モロニーマウス白血病ウイルス長末端反復(MMLV LTR)、ラウス肉腫ウイルス(RSV)LTRプロモーター、SV40プロモーター、ヒトサイトメガロウイルス(CMV)IEプロモーター、アデノウイルス、HSVプロモーター(例えば、HSV IEプロモーター)、またはHPVプロモーター、特に、HPV上流調節領域(URR)を使用することもできる。ウイルスプロモーターは、当該技術分野において容易に利用可能である。
【0080】
本発明の宿主細胞において転写を指令することが可能である多様なプロモーターを使用することができる。好ましくは、プロモーター配列は、先に定義したように、高度に発現される遺伝子から誘導される。好ましくは、プロモーターが誘導され、および/または発現構築物の組み込みのために、好適な予め決定された標的座位に含まれる好適な高度に発現される遺伝子の例として、トリオース−リン酸イソメラーゼ(TPI)、グリセルアルデヒド−リン酸デヒドロゲナーゼ(GAPDH)、ホスホグリセリン酸キナーゼ(PGK)、ピルビン酸キナーゼ(PYK)、アルコールデヒドロゲナーゼ(ADH)のような解糖系酵素をコードする遺伝子、ならびにアミラーゼ、グルコアミラーゼ、プロテアーゼ、キシラナーゼ、セロビオヒドロラーゼ、β−ガラクトシダーゼ、アルコール(メタノール)オキシダーゼ、伸長因子およびリボソームタンパク質をコードする遺伝子が挙げられるが、これらに限定されない。適切な高度に発現される遺伝子の具体例として、例えば、クリヴェロマイセス(Kluyveromyces)sp.由来のLAC4遺伝子、ハンセヌラ(Hansenula)およびピキア(Pichia)由来のそれぞれメタノールオキシダーゼ遺伝子(AOXおよびMOX)、A.ニガー(A.niger)およびA.アワモリ(A.awamori)由来のグルコアミラーゼ(glaA)遺伝子、麹菌(A.oryzae)TAKA−アミラーゼ遺伝子、A.ニダランス(A.nidulans)gpdA遺伝子およびT.リーセイ(T.reesei)セロビオヒドロラーゼ遺伝子が挙げられる。
【0081】
真菌発現宿主に使用するために好適である強力な構成性および/または誘導性プロモーターの例には、キシラナーゼ(xlnA)、フィターゼ、ATP−シンテターゼサブユニット9(oliC)、トリオースリン酸イソメラーゼ(tpi)、アルコールデヒドロゲナーゼ(AdhA)、アミラーゼ(amy)、アミログルコシダーゼ(glaA遺伝子由来のAG−)、アセトアミダーゼ(amdS)およびグリセルアルデヒド−3−リン酸デヒドロゲナーゼ(gpd)プロモーターのための真菌から入手可能なプロモーターがある。
【0082】
使用することができる強力な酵母プロモーターの例として、アルコールデヒドロゲナーゼ、グリセルアルデヒド−3−リン酸デヒドロゲナーゼ、ラクターゼ、3−ホスホグリセリン酸キナーゼ、形質膜ATPase(PMA1)およびトリオースリン酸イソメラーゼの遺伝子から入手可能なプロモーターが挙げられる。
【0083】
使用することができる強力な細菌プロモーターの例として、アミラーゼおよびSPo2プロモーター、ならびに細胞外プロテアーゼ遺伝子由来のプロモーターが挙げられる。
【0084】
使用することができる植物細胞に適切なプロモーターとして、ナパリンシンターゼ(nos)、オクトピンシンターゼ(ocs)、マンノピンシンターゼ(mas)、リブロース小サブユニット(rubisco ssu)、ヒストン、イネアクチン、ファゼオリン、カリフラワーモザイクウイルス(CMV)35Sおよび19Sならびにサーコウイルスプロモーターが挙げられる。
【0085】
ベクターは、真核生物のゲノム配列、好ましくは、真菌のゲノム配列、または酵母ゲノム配列に相同な配列を含んでなるRNAを生じるポリヌクレオチドにフランキングする配列をさらに含んでもよい。これは、相同組換えによる本発明のポリヌクレオチドの真菌または酵母のゲノムへの導入を可能にする。特に、真菌配列によってフランキングされる発現カセットを含んでなるプラスミドベクターを使用して、本発明のポリヌクレオチドを真菌細胞に送達するのに適切なベクターを調製することができる。これらの真菌ベクターを使用する形質転換技術は当業者に公知である。
【0086】
ベクターは、アンチセンスRNAの産生を提供するためにアンチセンス方向に配向された本発明のポリヌクレオチドを含有してもよい。所望であれば、これを使用して、ポリペプチドの発現のレベルを減少してもよい。
【0087】
[宿主細胞および発現]
さらなる態様において、本発明は、ポリペプチドをコードするコーディング配列のベクターによる発現に適切な条件下で、上記の発現ベクターで形質転換またはトランスフェクトされた宿主細胞を培養し、そして発現されたポリペプチドを回収することを含んでなる本発明のポリペプチドを調製するための方法を提供する。本発明のポリヌクレオチドは、複製可能な組み換えベクター、例えば、発現ベクターに組み入れることができる。ベクターは、適合可能な宿主細胞において核酸を複製するために使用され得る。それ故、さらなる実施形態では、本発明は、本発明のポリヌクレオチドを複製可能なベクターに導入し、ベクターを適合可能な宿主細胞に導入し、そしてベクターの複製を生じる条件下で宿主細胞を培養することによって、本発明のポリヌクレオチドを作製する方法を提供する。適切な宿主細胞として、大腸菌(E.coli)のような細菌、酵母、哺乳動物細胞系統、および他の真核細胞系統、例えば、Sf9細胞のような昆虫細胞、および(例えば、糸状)真菌細胞が挙げられる。
【0088】
好ましくは、発現構築物においてポリペプチドの成熟形態をコードするDNA配列が、シグナル配列をコードするDNA配列に作動可能に結合される場合、ポリペプチドは、分泌型タンパク質として産生される。分泌型タンパク質をコードする遺伝子が野生型株内にシグナル配列を有する場合、好ましくは、使用されるシグナル配列は、ポリペプチドをコードするDNA配列に由来する(それと同種である)。あるいは、シグナル配列は、ポリペプチドをコードするDNA配列に外来性(異種)であり、その場合、シグナル配列は、DNA配列が発現される宿主細胞に好ましくは内因性である。酵母宿主細胞に適切なシグナル配列の例には、酵母MFα遺伝子由来のシグナル配列がある。同様に、糸状菌宿主細胞に適切なシグナル配列には、例えば、糸状菌のアミログルコシダーゼ(AG)遺伝子、例えば、A.ニガー(A.niger)glaA遺伝子由来のシグナル配列がある。このシグナル配列は、アミログルコシダーゼ((グルコ)アミラーゼとも呼ばれる)プロモーター自体と組み合わせて、ならびに他のプロモーターと組み合わせて使用してもよい。ハイブリッドシグナル配列もまた、本発明に関して使用され得る。
【0089】
好適な異種分泌リーダー配列は、真菌アミログルコシダーゼ(AG)遺伝子(glaA−両方とも、例えば、アスペルギルス(Aspergillus)由来の18および24アミノ酸バージョン)、MFα遺伝子(酵母、例えば、サッカロマイセス(Saccharomyces)、ピキア(Pichia)ならびにクリヴェロマイセス(Kluyveromyces))またはα−アミラーゼ遺伝子(バチルス(Bacillus))由来のそれらである。
【0090】
ベクターは、本発明のポリペプチドの発現を提供するために、上記の適切な宿主細胞に形質転換またはトランスフェクトしてもよい。このプロセスは、ポリペプチドの発現に適切な条件下で上記のように発現ベクターで形質転換された宿主細胞を培養し、そして場合により、発現されたポリペプチドを回収することを含んでなり得る。
【0091】
それ故、本発明のさらなる態様は、本発明のポリヌクレオチドもしくはベクターで形質転換またはトランスフェクトされたか、あるいは本発明のポリヌクレオチドもしくはベクターを含んでなる宿主細胞を提供する。好ましくは、ポリヌクレオチドは、ポリヌクレオチドの複製および発現を可能にするベクターにおいて担持される。細胞は、前記ベクターに適合可能であるように選択され、そして例えば、原核生物(例えば、細菌)、または真核生物の真菌、酵母もしくは植物細胞であってもよい。
【0092】
本発明は、ポリヌクレオチドをコードするDNA配列の組み換え発現による本発明のポリペプチドの生産のためのプロセスを包含する。この目的のために、本発明のDNA配列は、適切な同種または異種の宿主細胞におけるポリペプチドの経済的な産生を可能にするために、遺伝子増幅および/または発現シグナル、例えば、プロモーター、分泌シグナル配列の交換のために使用され得る。同種の宿主細胞は、同じ種であるか、またはDNA配列が由来する種と同じ種内の変異体である宿主細胞として、本明細書において定義される。
【0093】
適切な宿主細胞は、好ましくは、細菌のような原核微生物、あるいはより好ましくは、真核生物体、例えば、酵母もしくは糸状菌のような真菌、または植物細胞である。一般に、酵母細胞の方が操作が容易であるため、酵母細胞は、糸状菌細胞より好適である。しかし、いくつかのタンパク質は、酵母からの分泌が不十分であるか、またはある場合に適切に処理されない(例えば、酵母における過グリコシル化)かのいずれかである。これらの場合、糸状菌宿主生物体が選択されるべきである。
【0094】
バチルス(Bacillus)属由来の細菌は、培養培地にタンパク質を分泌するそれらの能力のため、異種の宿主として極めて適切である。宿主として適切な他の細菌は、ストレプトマイセス(Streptomyces)およびシュードモナス(Pseudomonas)属由来の宿主である。ポリペプチドをコードするDNA配列の発現のための好適な酵母宿主細胞は、サッカロマイセス(Saccharomyces)、クリヴェロマイセス(Kluyveromyces)、ハンセヌラ(Hansenula)、ピキア(Pichia)、ヤロウイア(Yarrowia)、またはシゾサッカロマイセス(Schizosaccharomyces)属の1つである。より好ましくは、酵母宿主細胞は、サッカロマイセス・セレビシエ(Saccharomyces cerevisiae)、クリュイベロマイセス・ラクティス(Kluyveromyces lactis)(クリヴェロマイセス・マルキサナス・バー・ラクティス(Kluyveromyces marxianus var.lactis)としても公知である)、ハンセヌラ・ポリモルファ(Hansenula polymorpha)、ピキア・パストリス(Pichia pastoris)、ヤロウイア・リポリティカ(Yarrowia lipolytica)、およびシゾサッカロマイセス・ポンベ(Schizosaccharomyces pombe)の種からなる群から選択される。
【0095】
しかし、糸状菌宿主細胞が、ポリペプチドをコードするDNA配列の発現に最も好適である。好適な糸状菌宿主細胞は、アスペルギルス(Aspergillus)、トリコデルマ(Trichoderma)、フザリウム(Fusarium)、ディスポロトリクム(Disporotrichum)、ペニシリウム(Penicillium)、アクレモニウム(Acremonium)、ニューロスポラ(Neurospora)、サーモアスカス(Thermoascus)、ミセリオフトラ(Myceliophtora)、スポロトリクム(Sporotrichum)、シーラビア(Thielavia)、およびタラロマイセス(Talaromyces)属からなる群から選択される。より好ましくは、糸状菌宿主細胞は、アスペルギルス・オイザエ(Aspergillus oyzae)、アスペルギルス・ソーヤ(Aspergillus sojae)もしくはアスペルギルス・ニダランス(Aspergillus nidulans)の種またはアスペルギルス・ニガー(Aspergillus niger)群由来の種(これらは、レイパー(Raper)およびフェンネル(Fennell)、The Genus Aspergillus,The Williams & Wilkins Company,Baltimore,293〜344頁、1965年によって定義される)である。これらには、アスペルギルス・ニガー(Aspergillus niger)、アスペルギルス・アワモリ(Aspergillus awamori)、アスペルギルス・ツビゲンシス(Aspergillus tubigensis)、アスペルギルス・アクレアータス(Aspergillus aculeatus)、アスペルギルス・フォエティダス(Aspergillus foetidus)、アスペルギルス・ニダランス(Aspergillus nidulans)、アスペルギルス・ジャポニクス(Aspergillus japonicus)、麹菌(Aspergillus oryzae)およびアスペルギルス・フィクウム(Aspergillus ficuum)、ならびにまた、トリコデルマ・リーセイ(Trichoderma reesei)、フザリウム・グラミネアラム(Fusarium graminearum)、ペニシリウム・クリソゲナム(Penicillium chrysogenum)、アクレモニウム・アラバメンセ(Acremonium alabamense)、アカパンカビ(Neurospora crassa)、ミセリオフィトラ・サーモフィルム(Myceliophtora thermophilum)、スポロトリクム・セルロフィルム(Sporotrichum cellulophilum)、 ディスポロトリカム・ディモルフォスポラム(Disporotrichum dimorphosporum)およびシーラビア・テレストリス(Thielavia terrestris)種のものが挙げられるが、これらに限定されない。
【0096】
本発明の範囲内の好適な発現宿主の例には、真菌、例えば、アスペルギルス(Aspergillus)種(特に、欧州特許出願公開第A−184,438号明細書、および欧州特許出願公開第A−284,603号明細書に記載のもの)ならびにトリコデルマ(Trichoderma)種;細菌、例えば、バチルス(Bacillus)種(特に、欧州特許出願公開第A−134,048号明細書および欧州特許出願公開第A−253,455号明細書に記載のもの)、特に、枯草菌(Bacillus subtilis)、バチルス・リケニフォルミス(Bacillus licheniformis)、バチルス・アミロリケファシエンス(Bacillus amyloliquefaciens)、シュードモナス(Pseudomonas)種、ならびに酵母、例えば、クリヴェロマイセス(Kluyveromyces)種(特に、欧州特許出願公開第A−096,430号明細書、例えば、クリュイベロマイセス・ラクティス(Kluyveromyces lactis)、および欧州特許出願公開第A−301,670号明細書に記載のもの)およびサッカロマイセス(Saccharomyces)種、例えば、サッカロマイセス・セレビシエ(Saccharomyces cerevisiae)がある。
【0097】
本発明に従う宿主細胞は植物細胞を含み、従って、本発明はトランスジェニック生物体、例えば、植物体およびそれらの一部分に及び、それらは本発明の1つもしくはそれ以上の細胞を含有する。細胞は、本発明のポリペプチドを異種として発現してもよく、または1つもしくはそれ以上の本発明のポリヌクレオチドを異種として含有してもよい。従って、トランスジェニック(または遺伝子的に改変された)植物体は、本発明のポリペプチドをコードする配列をそのゲノムに(典型的には安定に)挿入し得る。植物細胞の形質転換は、公知の技術を使用して、例えば、アグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)由来のTiまたはRiプラスミドを使用して、実施することができる。それ故、プラスミド(もしくはベクター)は、植物に感染するために必要な配列を含んでもよく、そしてTiおよび/またはRiプラスミドの誘導体を用いてもよい。
【0098】
宿主細胞は、ポリペプチドを過剰発現してもよく、そして過剰発現を操作するための技術は周知であり、かつ本発明において使用することができる。それ故、宿主は、2コピーもしくはそれ以上のポリヌクレオチドを有してもよい。
【0099】
あるいは、葉、根または茎のような植物体の一部の直接感染を行うことができる。この技術では、感染させようとする植物体は、例えば、カミソリで植物体を切断するか、針で植物体に穴を開けるか、または研磨剤で植物体を擦ることによって損傷され得る。次いで、損傷部にアグロバクテリウム(Agrobacterium)が播種される。次いで、植物体または植物部分を、適切な培養培地上で増殖させ、そして成熟植物体に生育させることができる。形質転換された細胞の遺伝子的に改変された植物体への再生は、公知の技術、例えば、抗生物質を使用して、形質転換されたシュートを選択することによって、および適切な栄養素、植物ホルモンなどを含有する培地上でシュートを継代培養することによって達成することができる。
【0100】
[宿主細胞の培養および組み換え産生]
本発明はまた、シアリダーゼまたはその変異体を発現するために改変されている細胞も含む。そのような細胞として、一過的、または好ましくは安定に改変された高等真核細胞系統、例えば、哺乳類細胞もしくは昆虫細胞、下等真核細胞、例えば、酵母および糸状菌細胞、または原核細胞、例えば、細菌細胞が挙げられる。
【0101】
また、本発明のポリペプチドを、細胞系統、または例えば、バキュロウイルス発現系のような膜上で一過的に発現させることも可能である。本発明に従うタンパク質を発現するために適応されるそのような系もまた、本発明の範囲内に含まれる。
【0102】
本発明に従えば、本発明のポリペプチドの産生は、微生物の発現宿主を培養することによって行うことができ、それは、従来の栄養発酵培地において、本発明の1つもしくはそれ以上のポリヌクレオチドで形質転換されている。
【0103】
本発明に従う組み換え宿主細胞は、当業者に公知の手順を使用して培養することができる。プロモーターおよび宿主細胞の各組み合わせにとって、ポリペプチドをコードするDNA配列の発現につながる培養条件が利用可能である。所望の細胞密度またはポリペプチドの力価に達した後、培養は中止され、そしてポリペプチドは公知の手順を使用して回収される。
【0104】
発酵培地は、炭素源(例えば、グルコース、マルトース、モラセスなど)、窒素源(例えば、硫酸アンモニウム、硝酸アンモニウム、塩化アンモニウムなど)、有機窒素源(例えば、酵母抽出物、麦芽抽出物、ペプトンなど)および無機栄養源(例えば、リン酸、マグネシウム、カリウム、亜鉛、鉄など)を含有する公知の培養培地を含んでなり得る。場合により、(使用される発現構築物に依存する)誘導因子が含まれていてもよく、または後で添加してもよい。
【0105】
適切な培地の選択は、発現宿主の選択に基づいてもよく、および/または発現構築物の調節要件に基づいてもよい。適切な培地は、当業者に周知である。培地は、所望であれば、他の潜在的に混入する微生物より形質転換された発現宿主の方を選好するさらなる成分を含有してもよい。
【0106】
発酵は、0.5〜30日間の期間にわたって実施し得る。発酵は、0℃〜45℃の間の範囲の温度、および例えば、2から10までのpHにおけるバッチ、継続または流加培養プロセスであり得る。好適な発酵条件は、20℃〜37℃の間の範囲の温度および/または3〜9の間のpHを含む。適切な条件は、通常、発現宿主および発現させようとするタンパク質の選択に基づいて選択される。
【0107】
発酵後、必要であれば、遠心分離またはろ過によって、発酵ブロスから細胞を取り出すことができる。発酵が停止した後または細胞の取り出し後、次いで、本発明のポリペプチドを回収し、そして所望であれば、従来の手段によって精製および単離してもよい。本発明のシアリダーゼは、真菌の菌糸体から、または培養真菌細胞によってシアリダーゼが放出される培養ブロスから、精製することができる。
【0108】
好適な実施形態では、ポリペプチドは、真菌、より好ましくはアスペルギルス(Aspergillus)、最も好ましくはアスペルギルス・ニガー(Aspergillus niger)から産生される。
【0109】
[修飾]
本発明のポリペプチドは、化学的に修飾されていてもよく、例えば、翻訳後修飾される。例えば、それらは、グリコシル化(1回もしくはそれ以上の回数)されるか、または修飾アミノ酸残基を含んでなり得る。それらはまた、それらの精製を補助するためにヒスチジン残基の添加によってか、または細胞からの分泌を促進するためのシグナル配列の添加によって、修飾してもよい。ポリペプチドは、アミノ末端メチオニン残基、約20〜25残基までの小さなリンカーペプチドのようなアミノもしくはカルボキシル末端伸長、またはポリヒスチジントラクト、抗原エピトープもしくは結合ドメインのような精製を容易にする小さな伸長を有してもよい。
【0110】
本発明のポリペプチドは、明示用標識で標識されていてもよい。明示用標識は、ポリペプチドが検出されることを可能にする任意の適切な標識であり得る。適切な標識として、放射性同位元素、例えば、125I、35S、酵素、抗体、ポリヌクレオチドおよびビオチンのようなリンカーが挙げられる。
【0111】
ポリペプチドは、天然に存在しないアミノ酸を含むか、またはポリペプチドの安定性を増加するように修飾してもよい。タンパク質またはペプチドが合成手段によって生成される場合、生成中にそのようなアミノ酸を導入してもよい。タンパク質またはペプチドはまた、合成または組み換え産生後に修飾してもよい。
【0112】
本発明のポリペプチドはまた、D−アミノ酸を使用して生成してもよい。そのような場合、アミノ酸は、CからNへの配向で、逆向きの配列で連結される。これは、そのようなタンパク質またはペプチドを生成するための当該技術分野において慣用的である。
【0113】
多くの側鎖修飾が当該術分野において公知であり、そして本発明のタンパク質またはペプチドの側鎖に対して行うことができる。そのような修飾として、例えば、還元的アルキル化、アルデヒドとの反応、それに続く、NaBH4による還元、アセトイミド酸メチルによるアミジン化または無水酢酸によるアシル化によるアミノ酸の修飾が挙げられる。
【0114】
本発明によって提供される配列はまた、「第2世代」の酵素の構築のための出発物質として使用してもよい。「第2世代」のシアリダーゼは、野生型シアリダーゼまたは本発明によって産生されるような組み換えシアリダーゼとは異なる特性を有する変異技術(例えば、部位特異的変異誘発または遺伝子シャッフリング技術)によって改変されたシアリダーゼである。特定のプロセスにおける使用により良好に適合されるように、例えば、それらの温度または至適pH値、比活性、基質親和性または耐熱性を変更してもよい。
【0115】
本発明のシアリダーゼの活性に必須であり、従って、好ましくは置換に供されるアミノ酸は、当業者に公知の手順、例えば、部位特異的変異誘発またはアラニン−スキャンニング変異誘発に従って、同定してもよい。後者の技術では、変異は、分子の全ての残基で導入され、そして得られる変異が、分子の活性に極めて重要であるアミノ酸残基を同定するために、生物活性(例えば、シアリダーゼ活性)について試験される。酵素−基質相互作用の部位はまた、核磁気共鳴、結晶学または光親和性標識のような技術によって決定されるように、結晶構造の分析によっても決定することができる。
【0116】
遺伝子シャッフリング技術は、ポリヌクレオチド配列に変異を導入するための無作為な方法を提供する。発現後、最良の特性を伴う単離体は、再単離され、組み合わされ、そして再度シャッフリングされて、遺伝的多様性を増加する。この手順を多くの回数反復することによって、迅速に改善されたタンパク質をコードする遺伝子を単離することができる。好ましくは、遺伝子シャッフリング手順は、類似の機能を伴うタンパク質をコードする遺伝子のファミリーから開始される。本発明によって提供されるポリヌクレオチド配列のファミリーは、分泌型シアリダーゼの特性を改善するための遺伝子シャッフリングに良好に適合される。
【0117】
あるいは、NTG処理またはUV変異誘発のような古典的な無作為変異技術および選択を使用して、タンパク質の特性を改善することができる。変異誘発は、単離されたDNA、または目的のDNAで形質転換された細胞に対して直接実施することができる。あるいは、変異は、当業者に公知である多くの技術によって、単離されたDNAに導入することができる。これらの方法の例には、エラープローンPCR、修復欠損宿主細胞におけるプラスミドDNAの増幅などがある。
【0118】
酵母および糸状菌宿主細胞の使用は、翻訳後修飾(例えば、タンパク質分解プロセシング、ミリスチル化(myristilation)、グリコシル化、トランケーション、およびチロシン、セリンまたはスレオニンリン酸化)を提供することが予想され、これは、本発明の組み換え発現産物に対して至適生物活性を付与するのに必要であり得る。
【0119】
[調製]
本発明のポリペプチドは、単離された形であり得る。ポリペプチドは、ポリペプチドの意図された目的を妨害しないキャリアまたは希釈剤と混合してもよく、そしてなお単離されたものと認識されることが理解されよう。本発明のポリペプチドはまた、実質的に精製された形であり得、その場合、それは、一般的に、本発明のポリペプチドである調製物中のタンパク質の70%を超える、例えば、80%、90%、95%、98%もしくは99%を超える調製物中のポリペプチドを含んでなる。
【0120】
本発明のポリペプチドは、それらがそれらの天然の細胞環境外に存在するような形で提供され得る。それ故、それらは、上記のように、実質的に単離もしくは精製され得るか、またはそれらが天然では存在しない細胞、例えば、他の真菌種、動物、植物もしくは細菌細胞に存在し得る。
【0121】
[シアリダーゼ活性の取り出しまたは減少]
本発明はまた、親細胞の変異細胞を生成するための方法であって、ポリペプチドをコードする内因性核酸配列もしくはその制御配列を破壊または欠失することを含んでなり、親細胞より少なくポリペプチドを産生する変異細胞を生じる方法に関する。
【0122】
減少したシアリダーゼ活性を有する株の構築は、細胞におけるシアリダーゼの発現に必要な核酸配列の改変または不活化によって簡便に達成され得る。改変または不活化しようとする核酸配列は、例えば、シアリダーゼ活性を示すのに必須のポリペプチドもしくはその一部をコードする核酸配列であってもよく、または核酸配列は、核酸配列のコーディング配列由来のポリペプチドの発現に必要な調節機能を有してもよい。そのような調節または制御配列の例は、プロモーター配列またはその機能的部分、即ち、ポリペプチドの発現に影響を及ぼすのに十分な部分であり得る。可能な改変のための他の制御配列として、リーダー配列、ポリアデニル化配列、プロペプチド配列、シグナル配列、および終結配列が挙げられるが、これらに限定されない。
【0123】
核酸配列の改変または不活化は、細胞を変異誘発に供し、そしてシアリダーゼ産生能が減少または排除されている細胞を選択することによって、達成され得る。特異的であってもまたは無作為であってもよい変異誘発は、例えば、適切な物理的または化学的変異誘発因子の使用によって、適切なオリゴヌクレオチドの使用によって、またはDNA配列をPCR変異誘発に供することによって、実施され得る。さらに加えて、変異誘発は、これらの変異誘発因子の任意の組み合わせの使用によって、実施され得る。
【0124】
本目的に適切な物理的または化学的変異誘発因子の例として、紫外線(UV)照射、ヒドロキシルアミン、N−メチル−N’−ニトロ−N−ニトロソグアニジン(NTG)、O−メチルヒドロキシルアミン、亜硝酸、メタンスルホン酸エチル(EMS)、亜硫酸水素ナトリウム、ギ酸、およびヌクレオチドアナログが挙げられる。
【0125】
そのような因子を使用する場合、変異誘発は、典型的に、変異誘発しようとする細胞を、適切な条件下で選好される変異誘発因子の存在下でインキュベートし、そしてシアリダーゼ活性の発現の減少を示すか、または発現を示さない細胞を選択することによって、実施される。
【0126】
本発明のポリペプチドの産生の改変または不活化は、ポリペプチドをコードする核酸配列、またはその転写もしくは翻訳に必要な調節エレメントにおける1つもしくはそれ以上のヌクレオチドの導入、置換、あるいは取出しによって、達成され得る。例えば、ヌクレオチドは、終止コドンの導入、開始コドンの取り出し、もしくはオープンリーディングフレームの変更を生じるように、挿入してもよく、または取り出してもよい。そのような改変または不活化は、当該技術分野において公知の方法に従って、部位特異的変異誘発またはPCR変異誘発によって達成され得る。
【0127】
原則として、改変は、インビボで、即ち、改変しようとする核酸配列を発現する細胞に対して直接実施してもよいが、改変は、以下に例示するようにインビトロで実施するのが好ましい。
【0128】
選好される宿主細胞によるシアリダーゼの産生を不活化または減少するための簡便な方法の例は、遺伝子置換または遺伝子中断(gene interruption)の技術に基づく。例えば、遺伝子中断方法では、内因性遺伝子または目的の遺伝子フラグメントに対応する核酸配列をインビトロで変異誘発して、欠損核酸配列を生成し、次いで、宿主細胞に形質転換して、欠損遺伝子を生成する。相同組み換えによって、欠損核酸配列が、内因性遺伝子または遺伝子フラグメントに取って代わる。好ましくは、欠損遺伝子または遺伝子フラグメントはまた、ポリペプチドをコードする遺伝子が改変または破壊されている形質転換体を選択するために使用され得るマーカーをコードする。
【0129】
あるいは、本発明のポリペプチドをコードする核酸配列の改変または不活化は、ポリペプチドコーディング配列に相補的なヌクレオチド配列を使用する確立されたアンチセンス技術によって、達成され得る。より具体的には、細胞によるポリペプチドの産生は、ポリペプチドをコードする核酸配列に相補的なヌクレオチド配列を導入することによって、減少または排除され得る。次いで、アンチセンスポリヌクレオチドは、典型的に、細胞において転写され、そしてシアリダーゼをコードするmRNAにハイブリダイズすることが可能である。相補的なアンチセンスヌクレオチド配列をmRNAにハイブリダイズさせる条件下で、細胞において産生されるシアリダーゼの量は、減少または排除される。
【0130】
本発明の方法に従って改変しようとする細胞は、微生物由来、例えば、細胞に同種または異種のいずれかである所望されるタンパク質産物の産生に適切である真菌株由来であることが好ましい。
【0131】
本発明は、さらに、ポリペプチドをコードする内因性核酸配列もしくはその制御配列の破壊または欠失を含んでなり、親細胞より少なくポリペプチドを産生する変異細胞を生じる、親細胞の変異細胞に関する。
【0132】
そのようにして作製されるポリペプチド欠損変異細胞は、同種および/または異種ポリペプチドの発現のための宿主細胞として特に有用である。従って、本発明は、さらに、(a)ポリペプチドの産生につながる条件下で変異細胞を培養すること;および(b)ポリペプチドを回収することを含んでなる、同種または異種ポリペプチドを産生させるための方法に関する。これに関して、用語「異種ポリペプチド」は、本明細書において、宿主細胞に由来しないポリペプチド、天然配列を変更するために改変が施されている天然タンパク質、または発現が、組み換えDNA技術による宿主細胞の操作の結果として量的に変更される天然タンパク質として定義される。
【0133】
なおさらなる態様では、本発明は、本発明のシアリダーゼポリペプチドならびに目的のタンパク質産物の両方を産生する細胞の発酵によって、シアリダーゼ活性を本質的に含まないタンパク質産物を産生するための方法を提供する。本方法は、発酵中、または発酵が完了した後のいずれかに、シアリダーゼ活性を阻害することが可能な有効量の薬剤を発酵ブロスに添加し、発酵ブロスから目的の産物を回収し、そして場合により、回収した産物をさらなる精製に供することを含んでなる。あるいは、培養後、得られた培養ブロスを、シアリダーゼ活性を実質的に減少するようにpHまたは温度処理に供し、そして培養ブロスからの産物の回収を可能にすることができる。組み合わされたpHまたは温度処理は、培養ブロスから回収したタンパク質調製物に対して実施し得る。
【0134】
本質的にシアリダーゼを含まない産物を産生させるための本発明の方法は、真核生物ポリペプチドの産生、特に、酵素のような真菌タンパク質の産生において得に興味深い。また、シアリダーゼ欠損細胞を使用して、食品産業、または製薬目的のために目的の異種タンパク質を発現させてもよい。
【0135】
[チーズ作製および乳汁凝固]
凝固は、牛乳のような酪農組成物からのチーズの伝統的な生産における必須の工程である。凝固は、酸性化および/またはキモシンのような酵素(凝固剤)の添加によって開始され得る。凝固後、乳汁はカードおよび乳清に分離される。カードはさらにチーズに処理される。
【0136】
様々な乳汁供給源からのチーズ製造プロセスは、かなり以前より公知であり、そして異なる多くのタイプのチーズ変異体について詳細に記載されている。(例えば、Cheese:Chemistry,Physics and Microbiology、第1および2巻、1999年、フォックス(Fox)編、Aspen Publications,Gaithersburg、メリーランド州;Encyclopedia of Dairy Sciences、第1〜4巻、2003年、Academic Press,Londonを参照のこと)。チーズ製造において極めて重要な点は凝固のプロセスであって、ここで、カゼインミセルおよびサブミセルの溶解度が減少する。酵素誘導性凝固は、極めて一般的に使用される。仔ウシキモシンのような酵素、キモシンの微生物等価物および他の供給源由来の他の酵素についても記載されており、そしていくつかは、様々な商品名で利用可能である。それらのすべてを使用して、凝固プロセスを開始することができる。凝固の一次工程は、κ−カゼインにおけるPhe105−Met106結合の切断である。これは、κ−カゼインのC末端部:グリコマクロペプチド(GMP)の取り出しをもたらす。GMPの取り出しは、カゼインミセルの会合、即ち、カゼイン凝固をもたらす。カゼイン凝固はゲル形成をもたらし、そして特定の酪農組成物においてゲル化を得るのに必要な時間は、凝固剤の活性に直接関連する。
【0137】
凝固剤の添加と初期のカゼイン凝集物の出現との間を経過する時間は、凝固時間として定義される。チーズ用乳汁においてゲルが形成される速度およびゲルの密集度は、添加される酵素の量、カルシウムイオンの濃度、リン、温度およびpHに密接に依存する。初期の凝固後、ゲルが形成し、そしてミセル間結合の増加に従ってゲルの粘稠度が増加する。ミセルは共に移動し、そして凝塊が収縮し、それによって乳清を放出する。この現象は離液として公知であり、そしてカードを切断することによって加速され、温度を増加し、そして発育する乳酸菌によって生じる酸性度を増加する。
【0138】
凝固剤の添加と離液を開始するためのゲルの切断との間の時間を、本明細書において以後、凝固時間と称する。例えば、乳汁または凝固剤の質の変化の結果として、少しのばらつき(5〜10%まで)が生じ得るが、凝固時間は、特定のチーズ製造工場において特定のチーズの種類について一定である。多くのチーズ製造工場では、凝固時間は、プロセスにおいて時間を制限する工程である。凝固時間が減少する場合、工場は、より多くの乳汁を処理してカードにし、次いで(than)、さらにチーズに処理することができる。従って、凝固時間の減少は、経済的に興味深く、そしてそのような凝固時間の減少が必要とされている。
【0139】
添加する凝固剤の増加、乳汁のpHの降下または添加する塩化カルシウムのレベルの増加のようないくらかの方法で、凝固時間を減少することができる。しかし、それらの解決策は、チーズの品質にネガティブに影響を及ぼすため、チーズ作製産業において広範には使用されていない。凝固剤のレベルの増加は、しばしば、不均衡なプロテアーゼ活性の結果として苦味の形成をもたらす。乳汁pHの減少は、粗悪な品質のカードをもたらし、そしてしばしば、収量の損失をもたらす。塩化カルシウム濃度の増加は、味にネガティブな影響を及ぼし、そしてしばしば法的に制限されている。ネガティブな副作用を伴わずに凝固時間を減少するための現在利用可能な方法は存在しない。
【0140】
凝集に対するカゼインミセルの安定性は、凝集プロセスにおいて極めて重要である。κ−カゼインは、カゼインミセル安定化において極めて重要な役割を果たす。ミセル表面に局在するκ−カゼイン鎖の親水性かつ負に荷電したC末端部は、立体構造上の障害および静電反発の原因となり、ミセル状カゼインの凝固を妨害する(ミンキーウィックズ(Minkiewicz)ら、Pol J Food Nutr Sci(1993年)243,39−48)。κ−カゼインは、既知の位置においてシアル酸残基を含有する糖タンパク質である(ケーシズ(Cases)ら、J Food Sci(2003年)68,2406−2410;フォルネ(Fournet)ら、Biochim Biophys Acta(1979年)576,339−346)。グリコシル化の差異およびシアル酸含有量に基づき、κ−カゼインの集団においてミクロ的不均質が存在することが公知である(ロビテイル(Robitaille)ら、Food Res Int(1995年)28,17−21;ロビテイル(Robitaille)ら、J Dairy Res(1991年)58,107−114)。いくつかの研究は、ウェルシュ菌(Clostridium perfringens)由来のサイリダーゼ(sailidase)を使用するシアル酸残基の取り出しの効果について説明している。調べたパラメータは、加熱およびキモシン分解に対するカゼインミセルの安定性である。ギボンス(Gibbons)ら(Biochim Biophys Acta(1962年)56,354−356)は、精製されたκ−カゼインの溶液を使用し、そしてシアル酸残基の取出しが、κ−カゼインに対するキモシンの作用に影響を及ぼさないことを示した。ヴリーマン(Vreeman)ら(Biochem J(1986年)240,87−97)は、グリコシル化を伴うまたは伴わない精製されたκ−カゼイン画分に対するキモシンの動力学的挙動を比較し、そして脱グリコシル化κ−カゼインがキモシンのより良好な基質であることを見出した。また、ミンキーウィックズ(Minkiewicz)ら(Pol J Food Nutr Sci(1993年),243,39−48)は、人工的に再構成されたカゼインミセル系を使用して、シアル酸残基が、カゼインミセルの耐熱性に寄与することを示し、これは後にロビテイル(Robitaille)ら(Food Res Int(1995年)28,17−21)によって確認された。個別の研究において、ロビテイル(Robitaille)ら(Food Res Int(1993年)26,365−369)は、κ−カゼインからのシアル酸残基の取り出しは、凝固時間に有意な影響を及ぼさないが、カードの堅固性が減少したことを示した。凝固時間の減少を伴うが、チーズの特性に影響を及ぼさないチーズ作製プロセスについての記載は認められておらず、そしてこれは、産業上必要とされている。
【0141】
最近の特許出願(欧州特許第1370146号明細書)は、N−アセチルノイラミニダーゼを含む単一のグリコシダーゼを使用して、乳汁を凝固するプロセスについて記載している。記載された構成では、酵素凝固剤は使用されていない。凝固剤の省略は、代替的チーズ作製プロセスのための方法への道を開くが、(タンパク質分解性)凝固剤は、チーズにおけるタンパク質分解系において重要な部分であり、特徴的な風味化合物の形成をもたらすため、伝統的なチーズ作製プロセスには魅力的なプロセスではない(例えば、チーズ風味形成におけるキモシンの役割に関する参考文献について、Cheese:Chemistry,Physics and Microbiology、第1および2巻、1999年、フォックス(Fox)編、Aspen Publications,Gaithersburg,Maryland;Encyclopedia of Dairy Sciences 第1〜4巻、2003年、Academic Press,Londonを参照のこと)。
【0142】
これに関して、用語「チーズ」は、例えば、ナチュラルチーズ、チーズ類似物およびプロセスチーズのような任意の種類のチーズを指す。チーズは、例えば、レンネットを伴う酪農組成物の酵素的凝固、または食品級酸もしくは乳酸菌増殖によって産生される酸による酪農組成物の酸性凝固によるような当該技術分野において公知の任意の適切なプロセスによって入手され得る。一実施形態では、チーズは、レンネット−カードチーズである。酪農組成物は、従来のチーズ作製プロセスに供され得る。プロセスチーズは、好ましくは、チーズを調理ならびに例えば、乳化塩(例えば、リン酸塩およびクエン酸塩)で乳化することによって、ナチュラルチーズまたはチーズ類似物から製造される。プロセスは、香辛料/調味料の添加をさらに含んでもよい。
【0143】
用語「チーズ類似物」は、組成物の一部分として(例えば、乳脂肪(例えば、クリーム)のような脂肪を含有し、そして一組成物の部分として、例えば、植物油のような非乳汁構成分をさらに含有するチーズ様製品を指す。チーズは、軟質チーズ、半硬質チーズおよび硬質チーズのようなすべての種類のチーズを含んでなり得る。チーズ製造では、酪農組成物の凝固は、好ましくは、それぞれ、レンネット−カードおよび酸−カードチーズを生じるレンネットまたは酸性化のいずれか単独によって、実施される。フレッシュ酸−カードチーズとは、酸性化もしくは酸および加熱の組み合わせを介して乳汁、クリームまたは乳清の凝固によって生成され、そして製造が完了したら成熟を伴わなくとも消費される状態にある種類のチーズを指す。高い温度を使用する場合、カゼインの等電点、即ち、例えば、pH4.6もしくはそれ以上の値付近で、通常、凝固が生じる(例えば、リコッタ(Ricotta)では、典型的に約6.0のpH、および典型的に約80℃の温度)点で、フレッシュ酸−カードチーズは、一般的に、レンネット−カードチーズの種類(例えば、カマンベール(Camembert)、チェダー(Cheddar)、エメンタール(Emmenthal))とは異なり、ここで、凝固は、通常、pH値6.4〜6.6でレンネットの作用により誘導される。好適な実施形態では、チーズは、レンネットカードチーズのクラスに属する。
【0144】
酪農組成物は、牛乳構成分を含んでなる任意の組成物であり得る。乳構成分は、乳脂肪、乳タンパク質、カゼイン、乳清タンパク質およびラクトースのような乳汁の任意の構成分であり得る。乳汁画分は、例えば、スキムミルク、バターミルク、乳清、クリーム、粉乳、全粉乳、脱脂粉乳のような乳汁の任意の画分であり得る。好適な実施形態では、酪農組成物は、乳汁、スキムミルク、バターミルク、全乳、乳清、クリーム、またはそれらの任意の組み合わせを含んでなる。より好適な実施形態では、酪農組成物は、スキムミルク、全乳、クリームまたはそれらの任意の組み合わせのような乳汁からなる。さらなる実施形態では、酪農組成物は、例えば、全粉乳、脱脂粉乳、カゼイン、カゼイン塩、総乳タンパク質もしくは加糖粉乳、またはそれらの任意の組み合わせのような乾燥乳汁画分から全体的または部分的に調製される。酪農組成物は、牛乳およびまたは1つもしくはそれ以上の牛乳画分を含んでなる。牛乳画分は、ウシの任意の品種(ウシ(Bos taurus taurus)、コブウシ(Bos indicus taurus)およびこれらの交雑種由来であり得る。一実施形態では酪農組成物は、ウシの2つもしくはそれ以上の品種由来の牛乳および/または牛乳画分を含んでなる。酪農組成物はまた、ヤギ、スイギュウまたはラクダ由来の乳汁のようなチーズ調製物に使用される他の哺乳動物由来の乳汁を含んでなる。チーズの調製のための酪農組成物は、生乳汁成分のすべてもしくは一部の取り出しによって、および/またはそれにさらなる量のそのような成分を添加することによって、所望される組成物に標準化され得る。これは、例えば、乳製品製造所到着時の乳汁のクリームと乳汁とへの分離によって行われ得る。それ故、酪農組成物は、乳汁を分画し、そして酪農組成物の所望される最終組成物が得られるように、画分を再度組み合わせることによって、簡便に行われるように、調製され得る。分離は、連続遠心分離において行ってもよく、極めて低い脂肪含有量(即ち<0.5%)を伴うスキムミルク画分および例えば、>35%脂肪を伴うクリームがもたらされる。酪農組成物は、クリームおよびスキムミルクを混合することによって調製してもよい。別の実施形態では、タンパク質および/またはカゼイン含有量は、限外ろ過の使用によって標準化され得る。酪農組成物は、本発明のプロセスによって生成されるべきチーズに適切であることが認められる任意の全脂肪含有量を有し得る。
【0145】
カルシウムは、スターターカルチャーの添加の前、同時、または後のようなチーズ作製前および/または最中に任意の適切な段階で酪農組成物に添加され得る。好適な実施形態では、カルシウムは、加熱処置の前および後の両方に添加される。カルシウムは、適切ないずれの形で添加してもよい。好適な実施形態では、カルシウムは、カルシウム塩、例えば、CaCl2として添加される。適切な任意の量のカルシウムが、酪農組成物に添加され得る。添加されるカルシウムの濃度は、通常、1〜3mMの間のような0.1〜5.0mMの範囲である。CaCl2を酪農組成物に添加する場合、量は、通常、100リットルの酪農組成物あたり1〜50gの範囲、例えば、1000リットル酪農組成物あたり5〜30gの範囲、好ましくは、100リットル酪農組成物あたり10〜20gの範囲にある。
【0146】
スキムミルクの細菌含有量は、従来の好適によって低減され得る。酪農組成物は、デンマークブルーチーズ(Danish Blue Cheese)の生成におけるように、チーズの生成前に、均質化プロセスに供してもよい。
【図面の簡単な説明】
【0147】
【図1】図1は、ZJW発現ベクターの構築を示す。
【0148】
以下の実施例によって、本発明を例示する。
【0149】
[実施例]
[実施例1]
[シアリダーゼ遺伝子ZJWのクローニングおよび発現]
ペニシリウム・クリソゲナム(Penicillium chrysogenum)株Wisconsin54−1255(ATCC28089)を、3日間、30℃、PDB(ポテトデキストロースブロス、Difco)中で増殖させ、そして染色体DNAを、供給者の取扱説明書を使用し、Q−Biogeneキット(カタログ番号6540−600;Omnilabo International BV,Breda、蘭国)を使用して、菌糸体から単離した。染色体DNAを、PCRを使用するシアリダーゼ遺伝子のコーディング配列の増幅に使用した。
【0150】
ペニシリウム・クリソゲナム(Penicillium chrysogenum)株Wisconsin54−1255(ATCC28089)の染色体DNAからシアリダーゼ遺伝子ZJWを特異的に増幅するために、2つのPCRプライマーを設計した。プライマー配列を、ペニシリウム・クリソゲナム(Penicillium chrysogenum)Wisconsin54−1255(ATCC28089)のゲノムDNAにおいて見出された配列から部分的に入手したが、これを配列番号1に示す。本発明者らは、この配列は、アクチノマイセス(Actinomyces)およびアルスロバクター(Arthrobacter)のシアリダーゼ配列と相同性を有することを見出した。しかし、相同な真菌シアリダーゼについての記載はまだ認められていない。従って、本発明者らが、真菌から分泌型シアリダーゼをコードする遺伝子を見出すことができたことは、意外である。本発明者らは、本明細書において、分泌型真菌シアリダーゼの効率的な発現および特徴付けについてはじめて説明する。潜在的なプレ−およびプロ−配列を含んでなる完全なシアリダーゼタンパク質のタンパク質配列を配列番号3に示す。細菌相同体と比較した真菌酵素の利点は、真菌酵素が容易に過剰発現され、そして食物産業におけるアプリケーションに関連する量で分泌され得ることである。
【化1】
【0151】
第1の直接PCRプライマー(ZJW−dir)は、ATG開始コドンから開始する23ヌクレオチドZJWコーディング配列、およびそれより前方のPacI制限部位を含む23ヌクレオチド配列を含有する(配列番号4)。第2の逆方向プライマー(ZJW−rev)は、ZJWコーディング配列の下流の領域の逆方向鎖に相補的なヌクレオチド、およびそれより前方のAscI制限部位を含有する(配列番号5)。これらのプライマーを使用して、本発明者らは、テンプレートとしてペニシリウム・クリソゲナム(Penicillium chrysogenum)株Wisconsin54−1255(ATCC28089)由来の染色体DNAを伴う1.4kbのサイズのフラグメントを増幅することができた。このようにして得られた1.4kbサイズのフラグメントを単離し、PacIおよびAscIで消化し、そして精製した。ZJWコーディング配列を含んでなるPacI/AscIフラグメントを、pGBFIN−5(国際公開第99/32617号パンフレット)由来のPacI/AscI phyAフラグメントと交換した。得られたプラスミドは、pGBFINZJWと命名したZJW発現ベクターである(図1を参照のこと)。発現ベクターpGBFINZJWを、NotIによる消化によって線状化した(これは、発現ベクターからすべての大腸菌(E.coli)由来の配列を取り出す)。消化したDNAを、フェノール:クロロホルム:イソアミルアルコール(24:23:1)抽出を使用して精製し、そしてエタノールで沈殿させた。これらのベクターを使用して、アスペルギルス・ニガー(Aspergillus niger)CBS513.88を形質転換した。アスペルギルス・ニガー(Aspergillus niger)形質転換手順ついては、国際公開第98/46772号パンフレットに広範に記載されている。アセトアミドを含有する寒天プレート上で形質転換体を選択し、そして標的化されたマルチコピー組込み体を選択する仕方についても記載されている。好ましくは、複数コピーの発現カセットを含有するA.ニガー(A.niger)形質転換体を、サンプル材料のさらなる作製のために選択する。pGBFINZJW発現ベクターについて、まず、個々の形質転換体を選択倍地上にプレート化し、続いて、単一のコロニーを、PDA(ポテトデキストロース寒天:PDB+1.5%寒天)プレートにプレート化することによって、30個のA.ニガー(A.niger)形質転換体を精製した。1週間、30℃での増殖後、個々の形質転換体の胞子を回収した。胞子を冷蔵庫に貯蔵し、そして液体培地の播種のために使用した。
【0152】
複数コピーの発現カセットを含有するA.ニガー(A.niger)株を、振盪フラスコ培養における株の培養によってサンプル材料の作製に使用した。A.ニガー(A.niger)株の培養および培養ブロスからの菌糸体の分離のための有用な方法についても、国際公開第98/46772号パンフレットに記載されている。培養培地は、CSM−MES(1リットル培地あたり150gマルトース、60gのSoytone(Difco)、15gの(NH4)2SO4、1gのNaH2PO4・H2O、1gのMgSO4・7H2O、1gのL−アルギニン、80mgのTween−80、20gのMES、pH6.2)であった。発酵の4〜8日目に5mlサンプルを採取し、10分間、5000rpm、Hereaus labofuge RF中で遠心分離し、そしてさらなる分析まで、上清を−20℃で貯蔵した。
【0153】
SDS−Page分子量(MW)決定:NuPAGE MES−SDS泳動用緩衝液を使用するNuPAGE Novex Bis−Tris4〜12%勾配系(Invitrogen(Breda−蘭国))上でのSDS−PAGEによって、サンプルを分析する。供給者のプロトコルに従い、NuPAGE還元剤を使用して、充填前にサンプルを還元する。産生されたタンパク質の分子量を見積もるために、マーカータンパク質(SeeBlue Plus2Pre−Stained Standard(Invitrogen))を使用する。ゲルを泳動させ、そして供給者の取扱説明書に従って、Simply Blue Protein Strain(Invitrogen)を使用して染色し、そして産生されたタンパク質の分子量を、標準タンパク質との比較によって見積もった。本実施例では、形質転換体の発酵ブロスの13マイクロリットルの上清を分析した。
【0154】
SDS−PAGEで分析する場合、pGBFINZJWベクターを含有する形質転換体は、約50kDaの見かけの分子量のタンパク質を分泌したことが明らかになった。これは、タンパク質配列から推定される分子量よりわずかに大きいため、本発明者らは、シグナル配列の取り出し後、ペニシリウム・クリソゲナム(Penicillium chrysogenum)シアリダーゼZJWがアスペルギルス・ニガー(Aspergillus niger)から分泌される場合、いくつかのグリコシル化が生じると推定している。
【0155】
発酵および下流のプロセシングが大規模化される場合、選択された株は、より大量の真菌シアリダーゼの単離および精製のために使用することができる。次いで、この酵素を、さらなる分析、および多様な個々のアプリケーションにおける使用のために使用することができる。
【0156】
[実施例2]
[シアリダーゼの精製および特徴付け]
シアリダーゼを、実施例1に記載のような発酵を介して産生させた。酵素活性を、Amplex Redノイラミニダーゼアッセイキット(Invitrogenから入手した)を使用して、測定した。培養ろ過物(100ml)を、milliQ−水で、4.8mS/cmの導電率に希釈し、そしてBiomax−10膜(Milliporeから入手した)を使用する限外ろ過によって、70mlまで濃縮した。pHを、NaOHを使用して6.0に調整し、そしてサンプルを、5mlのHiTrapQイオン交換カラム(Amershamから入手した、5ml/分)上に充填し、20mMクエン酸ナトリウム(pH6.0)において平衡化した。シアリダーゼを含有するカラムのフロースルーを回収し、そして25mMのTris,HCl(pH7.0)に対して透析し、そして5mlのHiTrap Q FF(5ml/分)上に充填し、同じ緩衝液において平衡化した。シアリダーゼはフロースルー画分に存在し、そして回収した。次いで、酵素溶液を30mMクエン酸ナトリウム(pH4.0、緩衝液A)に対して透析し、そして5mlのHiTrap SPカラム(Amershamから入手した、5ml/分)上に適用し、緩衝液Aにおいて平衡化した。酵素を充填した後、カラムを3カラム容積の緩衝液Aで洗浄し、そして酵素を、直線勾配で20カラム容積の緩衝液Aから緩衝液B(緩衝液B:1MのNaClを含有する30mMナトリウム−クエン酸、pH4.0)により溶出させた。シアリダーゼ含有画分を同定し、そしてプールした。タンパク質濃度を、対照タンパク質としてウシ血清アルブミンを使用するBradford試薬(Sigmaから入手した)によって決定した。タンパク質は、ナトリウム−ドデシルポリアクリルアミドゲル電気泳動上で夾雑するバンドが認められなかったことから、>95%純粋であると判断した。シアリダーゼは、47kDの見かけの分子量まで泳動し、これは、推定されたアミノ酸配列に基づいて算出される42.7kDの分子量より僅かに大きい。酵素調製物は、一連の基質ZAAXpNA(Z=ベンゾイル基、A=アラニン、X=任意のアミノ酸残基、pNA=パラ−パラニトロアニリド)に対してタンパク質分解活性を示さず、エンド−プロテアーゼ活性の不在を示す。
【0157】
[実施例3]
[ミニチュアチーズの調製方法]
ミニチュアチーズを、シャキール−Ur−レーマン(Shakeel−Ur−Rehman)ら(Protocol for the manufacture of miniature cheeses、Lait,78,(1998年),607−620)により記載の通りに、生成した。低温殺菌された全脂均質化牛乳を使用したが、生の牛乳または還元牛乳を使用することもできる。乳汁を広口プラスチック製ボトル(ボトルあたり200mL)に移し、31℃まで冷却した。続いて、1.8単位のスターターカルチャーDS 5LT1(DSM Gist B.V.、Delft、蘭国)を、遠心ボトル中の200mlの低温殺菌した乳汁のそれぞれに添加し、乳汁を20分間、成熟させた。次いで、CaCl2(200mL発酵乳あたり132μLの1mol.L−1溶液)を添加した。最後に、凝固剤(1mlあたり0.04IMCU)を添加した。シアリダーゼを使用した場合、これを、凝固剤と共に添加した。凝塊が形成されるまで、乳汁溶液を40〜50分間、31℃で保持した。凝塊を、フレーム上に1cm間隔で張り渡されたワイヤのカッターで、手動で切断した。2分間回復させ、続いて、10分間、緩徐に撹拌した。その後、カード/乳清混合物の連続撹拌下、30分間、温度を徐々に39℃に上昇させた。6.2のpHに到達したら、カード/乳清混合物を、室温で60分間、1,700gで遠心分離した。乳清を排出させ、カードを水浴中、36℃に保持した。pHが5.2〜5.3に低下するまで、チーズを15分間ごとに反転させ、次いで、室温で20分間、1,700gで遠心分離した。製造後、チーズを秤量した。
【0158】
[実施例4]
[チーズ凝固を決定するための方法]
Kubarsepら(Acta Agric ScandセクションA(2005年)55,145−148)に記載のように、Optigraph(Alliance Instruments、仏国)を使用して、チーズ凝固を追跡した。凝乳酵素溶液の添加後、標準的な手順(IDF157 A[2])に従って、標準的な乳汁基質溶液の凝固時間を決定した。11グラムの低加熱脱脂粉乳(Nilac、NIZO、蘭国から入手した)を、4.5mMのCaCl2を含有する100mlのmilliQ−水に添加することによって、乳汁基質溶液を調製した。乳汁溶液を、磁気スターラーで、30分間、撹拌した。乳汁を安定にするために、溶液を、室温で30分間、暗所に置いた。乳汁のpHは約6.5である。8〜15分間の間のレンネット凝固時間(r)を達成するために、凝乳酵素をmilli−Q水で希釈した。10ml乳汁溶液を、Optigraphのサンプルキュベットに移した。乳汁を、32℃の反応温度に調整した。200μlの希釈酵素サンプルを多スプーン装置(multiple spoon apparatus)にピペッティングし、そして1回で乳汁に添加した。この操作が、凝固試験の開始時点である。酵素および乳汁をスプーンで混合し、この目的のためにOptigraphで供給された。凝固を45分間、モニターした。
【特許請求の範囲】
【請求項1】
シアリダーゼ活性を有し、そして配列番号3のアミノ酸1〜407と少なくとも60%のアミノ酸配列同一性を有するアミノ酸配列を有する、単離されたポリペプチド。
【請求項2】
ポリペプチドは55kDa未満のMW(分子量)(SDS−Page)を有し、より好ましくは、前記ポリペプチドは52kDa未満のMW(分子量)(SDS−Page)を有するか、または前記ポリペプチドは50kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有し、より好ましくは、前記ポリペプチドは45kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有する、請求項1に記載のポリペプチド。
【請求項3】
55kDa未満のMW(分子量)(SDS−Page)を有する真菌ポリペプチドであって、より好ましくは、前記ポリペプチドは52kDa未満のMW(分子量)(SDS−Page)を有するか、または前記ポリペプチドは50kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有し、より好ましくは、前記ポリペプチドは45kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有する、真菌ポリペプチド。
【請求項4】
細胞外酵素である、請求項1または3に記載のポリペプチド。
【請求項5】
配列番号3のアミノ酸1〜407と少なくとも65%、好ましくは少なくとも70%、より好ましくは少なくとも80%、さらにより好ましくは少なくとも90%、最も好ましくは少なくとも95%、およびさらに最も好ましくは少なくとも約97%同一性を有するアミノ酸配列を有する、請求項1〜4のいずれか一項に記載のポリペプチド。
【請求項6】
配列番号3のアミノ酸配列を含んでなる、請求項1に記載のポリペプチド。
【請求項7】
真菌、好ましくは、ペニシリウム(Penicillium)、より好ましくは、ペニシリウム・クリソゲナム(Penicilium chrysogenum)から得られる、請求項1〜6のいずれか一項に記載のポリペプチド。
【請求項8】
請求項1〜7のいずれか一項に記載のポリペプチドをコードする核酸配列を含んでなる、単離されたポリヌクレオチド。
【請求項9】
適切な発現宿主においてポリペプチドの産生を指令する1つもしくはそれ以上の制御配列に作動可能に連結された請求項8に記載のポリヌクレオチドを含んでなる、核酸構築物。
【請求項10】
請求項9に記載の核酸構築物を含んでなる、組み換え発現ベクター。
【請求項11】
請求項10に記載の核酸構築物または請求項9に記載の核酸構築物を含んでなる、組み換え宿主細胞。
【請求項12】
請求項9に記載の株/組み換え宿主細胞を培養して、ポリペプチドを含んでなる上清および/または細胞を生成すること;ならびにポリペプチドを回収することを含んでなる、請求項1〜8のいずれか一項に記載のポリペプチドを産生させるための方法。
【請求項13】
請求項12に記載の方法によって入手可能であるか、または55kDa未満のMW(分子量)(SDS−page)を有する分泌型シアリダーゼであるポリペプチドであって、より好ましくは、前記ポリペプチドは52kDa未満のMW(分子量)(SDS−page)を有する、ポリペプチド。
【請求項14】
請求項12に記載の方法によって入手可能であるか、または50kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有する分泌型シアリダーゼであるポリペプチドであって、より好ましくは、前記ポリペプチドは45kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有する、ポリペプチド。
【請求項15】
請求項1〜8のいずれか一項に記載のポリペプチドを産生させるための方法であって、ポリペプチドの産生に適切な条件下でポリペプチドをコードするポリヌクレオチドを含んでなる核酸構築物を含んでなる宿主細胞を培養すること;およびポリペプチドを回収することを含んでなる、方法。
【請求項16】
請求項15の方法によって産生される、ポリペプチド。
【請求項17】
シアリダーゼ活性を有し、そして非ビルレントであるポリペプチドか、または真菌シアリダーゼであり、そして55kDa未満のMW(分子量)(SDS−Page)を有する、シアリダーゼ活性を有するポリペプチドであって、より好ましくは、前記シアリダーゼは52kDa未満のMW(分子量)(SDS−Page)を有するか、または前記シアリダーゼは50kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有し、より好ましくは、前記シアリダーゼは45kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有する、ポリペプチド。
【請求項18】
請求項1〜8、13または14のいずれか一項に記載のポリペプチド、あるいは55kDa未満(SDS−page)のMW(分子量)を有する細胞外シアリダーゼであって、好ましくは、前記ポリペプチドは52kDa未満のMW(分子量)(SDS−page)を有するか、または50kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有する細胞外シアリダーゼであって、好ましくは、前記ポリペプチドは45kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有するポリペプチドの、食物または飼料の調製における、あるいは医薬品としてもしくは医薬品の成分としてまたは医薬品調製における使用。
【請求項1】
シアリダーゼ活性を有し、そして配列番号3のアミノ酸1〜407と少なくとも60%のアミノ酸配列同一性を有するアミノ酸配列を有する、単離されたポリペプチド。
【請求項2】
ポリペプチドは55kDa未満のMW(分子量)(SDS−Page)を有し、より好ましくは、前記ポリペプチドは52kDa未満のMW(分子量)(SDS−Page)を有するか、または前記ポリペプチドは50kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有し、より好ましくは、前記ポリペプチドは45kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有する、請求項1に記載のポリペプチド。
【請求項3】
55kDa未満のMW(分子量)(SDS−Page)を有する真菌ポリペプチドであって、より好ましくは、前記ポリペプチドは52kDa未満のMW(分子量)(SDS−Page)を有するか、または前記ポリペプチドは50kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有し、より好ましくは、前記ポリペプチドは45kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有する、真菌ポリペプチド。
【請求項4】
細胞外酵素である、請求項1または3に記載のポリペプチド。
【請求項5】
配列番号3のアミノ酸1〜407と少なくとも65%、好ましくは少なくとも70%、より好ましくは少なくとも80%、さらにより好ましくは少なくとも90%、最も好ましくは少なくとも95%、およびさらに最も好ましくは少なくとも約97%同一性を有するアミノ酸配列を有する、請求項1〜4のいずれか一項に記載のポリペプチド。
【請求項6】
配列番号3のアミノ酸配列を含んでなる、請求項1に記載のポリペプチド。
【請求項7】
真菌、好ましくは、ペニシリウム(Penicillium)、より好ましくは、ペニシリウム・クリソゲナム(Penicilium chrysogenum)から得られる、請求項1〜6のいずれか一項に記載のポリペプチド。
【請求項8】
請求項1〜7のいずれか一項に記載のポリペプチドをコードする核酸配列を含んでなる、単離されたポリヌクレオチド。
【請求項9】
適切な発現宿主においてポリペプチドの産生を指令する1つもしくはそれ以上の制御配列に作動可能に連結された請求項8に記載のポリヌクレオチドを含んでなる、核酸構築物。
【請求項10】
請求項9に記載の核酸構築物を含んでなる、組み換え発現ベクター。
【請求項11】
請求項10に記載の核酸構築物または請求項9に記載の核酸構築物を含んでなる、組み換え宿主細胞。
【請求項12】
請求項9に記載の株/組み換え宿主細胞を培養して、ポリペプチドを含んでなる上清および/または細胞を生成すること;ならびにポリペプチドを回収することを含んでなる、請求項1〜8のいずれか一項に記載のポリペプチドを産生させるための方法。
【請求項13】
請求項12に記載の方法によって入手可能であるか、または55kDa未満のMW(分子量)(SDS−page)を有する分泌型シアリダーゼであるポリペプチドであって、より好ましくは、前記ポリペプチドは52kDa未満のMW(分子量)(SDS−page)を有する、ポリペプチド。
【請求項14】
請求項12に記載の方法によって入手可能であるか、または50kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有する分泌型シアリダーゼであるポリペプチドであって、より好ましくは、前記ポリペプチドは45kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有する、ポリペプチド。
【請求項15】
請求項1〜8のいずれか一項に記載のポリペプチドを産生させるための方法であって、ポリペプチドの産生に適切な条件下でポリペプチドをコードするポリヌクレオチドを含んでなる核酸構築物を含んでなる宿主細胞を培養すること;およびポリペプチドを回収することを含んでなる、方法。
【請求項16】
請求項15の方法によって産生される、ポリペプチド。
【請求項17】
シアリダーゼ活性を有し、そして非ビルレントであるポリペプチドか、または真菌シアリダーゼであり、そして55kDa未満のMW(分子量)(SDS−Page)を有する、シアリダーゼ活性を有するポリペプチドであって、より好ましくは、前記シアリダーゼは52kDa未満のMW(分子量)(SDS−Page)を有するか、または前記シアリダーゼは50kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有し、より好ましくは、前記シアリダーゼは45kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有する、ポリペプチド。
【請求項18】
請求項1〜8、13または14のいずれか一項に記載のポリペプチド、あるいは55kDa未満(SDS−page)のMW(分子量)を有する細胞外シアリダーゼであって、好ましくは、前記ポリペプチドは52kDa未満のMW(分子量)(SDS−page)を有するか、または50kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有する細胞外シアリダーゼであって、好ましくは、前記ポリペプチドは45kDa未満のMW(分子量)(アミノ酸配列に基づいて算出される)を有するポリペプチドの、食物または飼料の調製における、あるいは医薬品としてもしくは医薬品の成分としてまたは医薬品調製における使用。
【図1】

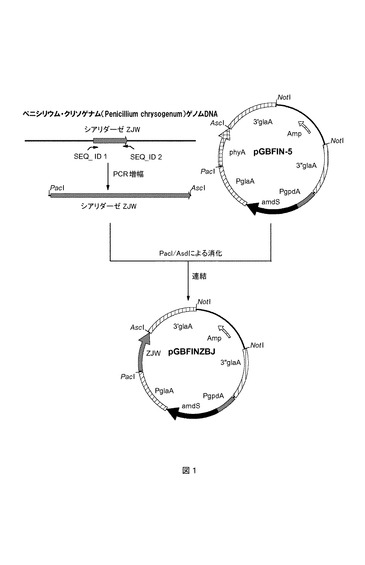
【公表番号】特表2010−520746(P2010−520746A)
【公表日】平成22年6月17日(2010.6.17)
【国際特許分類】
【出願番号】特願2009−549428(P2009−549428)
【出願日】平成20年2月18日(2008.2.18)
【国際出願番号】PCT/EP2008/051930
【国際公開番号】WO2008/101893
【国際公開日】平成20年8月28日(2008.8.28)
【出願人】(503220392)ディーエスエム アイピー アセッツ ビー.ブイ. (873)
【Fターム(参考)】
【公表日】平成22年6月17日(2010.6.17)
【国際特許分類】
【出願日】平成20年2月18日(2008.2.18)
【国際出願番号】PCT/EP2008/051930
【国際公開番号】WO2008/101893
【国際公開日】平成20年8月28日(2008.8.28)
【出願人】(503220392)ディーエスエム アイピー アセッツ ビー.ブイ. (873)
【Fターム(参考)】
[ Back to top ]