
新規ニューモウィルス組成物及びその使用方法

犬及び猫、潜在的には人間を含む哺乳類に感染しうる、新規に特定されたニューモウィルスが提供される。ウィルスの単離されたポリヌクレオチド及びタンパク質、また、単離されたウィルス自体が提供される。発明は、ウィルスを検出するための組成物及び方法、ウィルスの存在と正の相関関係のある病徴を予防及び/又は治療するための方法及び組成物、及び、ウィルスを含む単離された細胞を含む。完全なウィルス粒子、ウィルス性のタンパク質、及びそれらの断片もまた含まれる。
【発明の詳細な説明】
【技術分野】
【0001】
本願は、その開示内容が引用によりここに援用される、2009年12月21日出願の米国出願番号61/288,401に対して優先権を主張する。
【0002】
本発明は一般的に、ウィルス学の分野、より詳細には、イヌ及びネコを含む哺乳類で見いだされた、パラミクソウィルス科、ニューモウィルス亜科の新規に発見されたウィルスに関する。
【背景技術】
【0003】
シェルター、犬舎又は繁殖施設等に一緒に閉じ込められている家畜犬は、しばしば急性の呼吸器感染症(ケンネルコフ)に感染する。新しい動物が連続的に導入されるにつれて疾患は急速に伝染し、除去が困難となる。病原体のスペクトラムは、診断や処置を困難にする複数で連続的に重複する感染症の複雑な症候群を引き起こしうる。感染した犬は、軽い乾いた咳や鼻汁から肺炎や深刻な場合には死に至る範囲の臨床的な徴候を示す。猫もまた、多様な深刻度の呼吸器の苦痛を引き起こす様々な病原体に感染する。同一又は類似のウィルスはヒトにもまた感染しうる。このように多様な哺乳類に感染する病原体を特定することやこのような感染症のための診断、予防、及び/又は治療において使用するための組成物や方法を発展させることには、継続的かつ未充足のニーズが存在している。本発明はこれらのニーズを満たすものである。
【発明の概要】
【0004】
本発明は、新規なニューモウィルスの我々の発見に基づいている。このウィルスは、イヌ、ネコ及びおそらくヒトを含むがこれらに制限されない、多くの種類の哺乳類に感染しうる。ウィルスの存在は、イヌの急性の呼吸器疾患(ARDC)又はネコの呼吸器疾患、又は、ヒトその他の哺乳類の呼吸器その他の病気と正の相関関係がある。
【0005】
本発明は、ウィルスの単離されたポリヌクレオチド及びタンパク質、また単離されたウィルスそのものを提供する。本発明はウィルスを検出するための組成物及び方法、ウィルスの存在と正の相関関係のある病徴を予防及び/又は治療するための方法及び組成物を含む。またウィルスを含む単離された細胞を提供する。
【0006】
本発明のウィルスはマイナス鎖RNAウィルスである。本発明は、単離されたマイナス鎖、当該マイナス鎖と逆相相補性を有するプラス鎖、RNAポリヌクレオチドのDNA等価物、プラス鎖にコードされているタンパク質、及び、ポリヌクレオチド及びタンパク質の断片を含む。
【0007】
ある実施態様において、単離されたウィルスは配列番号1の配列、又は、配列番号1の配列と少なくとも96%が同一である配列を含むマイナス鎖ポリヌクレオチドを含む。また、配列番号2又は配列番号3の配列を含む単離されたポリヌクレオチド、又は、配列番号2又は3の配列と少なくとも96%が同一である配列を有するポリヌクレオチドが提供される。
【0008】
本発明は、哺乳類において免疫応答を刺激する方法を提供する。刺激された免疫応答は細胞仲介性、体液性、又はそれらの組み合わせでありえ、動物に対して予防及び/又は治療的な利益を提供しうる。免疫応答を刺激する方法は、ここに開示されるポリヌクレオチド及び/又はウィルス、ここに開示されるウィルス性ポリヌクレオチドによってコードされるウィルス性タンパク質、このようなタンパク質の断片、又は、それらの組み合わせを含む組成物を、哺乳類に投与することを含む。ポリヌクレオチド、ウィルス及び/又はタンパク質及び/又はそれらの断片は、いかなる適切なソースから単離されてもよく、また、それらは公知の技術を用いて組み換えによって生産されてもよい。
【0009】
本発明は、哺乳類から取得した生物学的サンプルにおけるウィルスの存在を検出するための方法を提供する。当該方法は、哺乳類から生物学的サンプルを取得すること、及び、当該生物学的サンプルから、ウィルス又はウィルス性のタンパク質又はそれらの断片によって含まれるポリヌクレオチド配列を検出すること、を含む。ポリヌクレオチド、タンパク質、タンパク質の断片、又はそれらの組み合わせの存在は、その哺乳類がウィルスに感染していることを示唆する。
【図面の簡単な説明】
【0010】
【図1】ヒトRSウィルス特異的なモノクローナル抗体(Mab)を用いたA72細胞の免疫蛍光アッセイ。A)感染細胞でのMab 2G122。B)非感染細胞でのMab 2G122。C)感染細胞でのMab 5H5N。D)非感染細胞でのMab 5H5N。一次Mabのストックは製造者から入手したものを1:100希釈して用いられた。赤いバックグラウンドは、エバンスブルーでの対比染色によるものである。
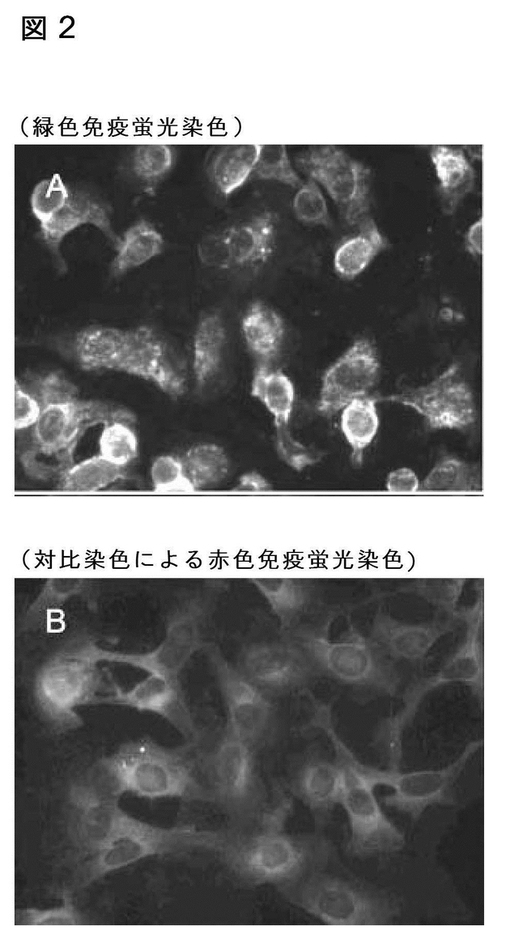
【図2】感染及び非感染のイヌA72細胞に対する、非呼吸器関連のテストのために提出された、ランダムに選択されたイヌ血清の反応性を示す免疫蛍光アッセイである。A)1:320希釈での感染細胞の血清。B)1:320希釈での非感染細胞の血清。赤いバックグラウンドは、エバンスブルーでの対比染色によるものである。
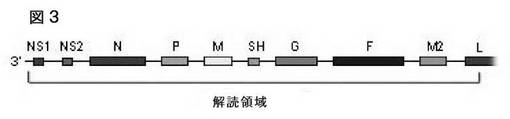
【図3】配列番号1の配列を含むウィルスゲノム組織の図示である。
【発明を実施するための形態】
【0011】
本発明は、新規な単離されたウィルス、単離されたポリヌクレオチド、及びウィルスのタンパク質、ウィルスを検出するための組成物及び方法、及び、哺乳類においてウィルスの存在と正の相関関係のある病徴の予防及び/又は治療のための組成物及び方法を提供する。
【0012】
本発明のウィルスは、ネガティブ(マイナス)鎖RNAウィルスである。つまり、ウィルス粒子に包含されている遺伝子材料は、ポジティブ(プラス)鎖の逆相補物であるRNAの単一鎖である。プラス鎖は、RNA依存性RNAポリメラーゼによってマイナス鎖から転写される。当該プラス鎖によってコードされているウィルス性タンパク質の、少なくとも一部はmRNAとしてのプラス鎖の機能は、感染細胞の細胞質で翻訳され、そして、新しいウィルス粒子の組み立て及びマイナス鎖ゲノムのパッケージングに関与する。
【0013】
特定の実施態様において、本発明は、配列番号1の配列又は配列番号1の配列と少なくとも96%が同一である配列を含むマイナス鎖ポリヌクレオチドを含有する単離されたウィルスを提供する。本発明はまた、当該マイナス鎖に相補的なポリヌクレオチドを含む。特定の実施態様において、これらのポリヌクレオチドは、配列番号4に示される配列又は少なくとも96%が配列番号4と同一である配列を含む。また、配列番号1の配列又は少なくとも96%が配列番号1の配列と同一である配列を含む単離されたポリヌクレオチド、又は、配列番号4の配列又は少なくとも96%が配列番号4の配列と同一である配列を含む単離されたポリヌクレオチドが提供される。また、配列番号2及び/又は配列番号3の配列を含むポリヌクレオチド、又は少なくとも96%が配列番号2,3の配列と同一である配列を有するポリヌクレオチドが提供される。これらの配列の逆相補物、及び、少なくとも96%が逆相補配列と同一性を有する配列もまた提供される。ここに開示されるすべてのポリヌクレオチド及びタンパク質及びそれらの断片の組み合わせもまた、本発明に含まれる。本発明によって提供されるウィルス性のポリヌクレオチドは、生ウィルス(弱毒化ウィルスも含む)において、複製をサポートする働きをすることによって特徴づけられる。
【0014】
また、ウィルスを含有する細胞が提供される。多くの実施態様において、ウィルスを含有する細胞は、単離された細胞、又は、ウィルス弱毒化のために及び/又はワクチンや他の目的のために使用するためのウィルス源として、インビトロで増殖させられる細胞である。このように、ウィルスを含む単離された細胞、ウィルスを含む細胞培養及び/又はセルライン、細胞培養培地に存在するウィルス及び/又はウィルス性生産物、及び、ウィルスを含む単離された組織は全て、本発明の局面である。
【0015】
特定の実施態様において、本発明で開示される各RNA配列のDNA等価物が提供される。つまり、本発明は、RNAの各々のU(ウラシル)がT(チミン)に置換されている、ここに開示される各RNA配列を含む。本発明で開示される各々のウィルス性ポリヌクレオチドは、各ポリヌクレオチドについて指定された配列番号で示される配列と同一であるポリヌクレオチドを含み、さらに、ウィルスの配列番号で示された配列と少なくとも96%が同一であるすべてのポリヌクレオチドをも含む。特定の実施態様では、配列リストで示されたポリヌクレオチド配列と少なくとも96%が同一であるポリヌクレオチドは、全長に渡る配列のうち少なくとも96%がそれらの配列と同一である。いかなる特定の理論に拘束されることを意図するものではないが、ここに示され、また、配列リストで説明されるウィルス性のポリヌクレオチド配列と少なくとも96%の同一性を有する配列を含むポリヌクレオチド配列は、例えばネズミ科の哺乳類に感染することが既に知られているウィルスのような、関連のウィルスによって含まれるポリヌクレオチドとは区別されるものと考えられている。また、これもいかなる特定の理論に拘束されることを意図するものではないが、例えば連続継代によるウィルスの弱毒化は、配列リストに示された配列と少なくとも96%が同一であるポリヌクレオチドを含み、及び/又は転写するウィルスを生じ得る。配列リストに特定して列挙された配列と同一でない配列は、示された配列と96.0%から99.9%同一でありえ、数値には示された数値の間の小数第一位のすべての整数値を含む。96.0%から100.0%の間の配列の同一性の全ての範囲が、包括的に、本発明の実施態様である。
【0016】
ポリヌクレオチドの断片もまた本発明に含まれる。つまり、様々な実施態様において、各々のウィルス性ポリヌクレオチド(その逆相補物及びDNA等価物を含む)は、10ヌクレオチドからポリヌクレオチド全長にわたる範囲、そしてその間のすべての整数値を含む範囲の、ポリヌクレオチド断片を含む。例えば、本発明は、配列番号1の配列を含むポリヌクレオチドを含み、また、長さで10ヌクレオチド以上、8598ヌクレオチドポリヌクレオチドまでの間(また、それらの間の全ての整数値を含む)の全ての配列の断片を含むポリヌクレオチドを含む。本発明で提供されるポリヌクレオチドは、ウィルス粒子に存在するか否かに関わらず、ここに開示されるものよりも長くてもよい。なぜなら、提示される配列は、ウィルスゲノム及び/又はそれらの逆相補物及び/又はDNA等価物の、全体を含むものではないからである。
【0017】
本発明はまた、哺乳類において免疫応答を刺激する方法をも提供する。刺激された免疫応答は、動物に予防的及び/又は治療的利益をもたらしうる免疫応答である。刺激された免疫応答は、体液性の及び/又は細胞仲介性の免疫応答でありえる。刺激された免疫応答は、ウィルスにコードされる、抗原及び/又は免疫原のいずれかに対するものでありえる。つまり、完全なウィルス粒子、各ウィルス性のタンパク質、及び、ウィルス性のタンパク質の断片は、哺乳類において免疫応答を刺激するために使用されうる本発明の実施態様である。多くの実施態様において、免疫応答が刺激される動物はイヌ(Canis familiaris)又はネコ(Felis catus)、すなわち各々家畜化された犬又は猫である。
【0018】
特定の実施態様において、配列番号1の配列又は配列番号1の配列と少なくとも96%が同一の配列、又はそこにコードされるタンパク質又はタンパク質断片、又は配列番号1の逆相補物によってコードされるウィルス粒子、を含むポリヌクレオチドは、イヌにおいて免疫応答を刺激するために用いられる。配列番号2及び/又は配列番号3の配列を含むポリヌクレオチド、又は、配列番号2又は3の配列と少なくとも96%が同一の配列を有するポリヌクレオチド、又は、配列番号2又は3の逆相補物によってコードされるタンパク質又はタンパク質断片又はウィルス性タンパク質、又は、配列番号2又は3の逆相補物と少なくとも96%が同一の配列を有するポリヌクレオチドは、ネコにおいて免疫応答を刺激するために好ましく使用される。イヌ及びネコへの、これらの配列、タンパク質、タンパク質断片、及びウィルス粒子の間の同様の関係が、診断目的のためのこれらの組成物の使用にも適用される。しかしこのことは、ヒトを含む他の哺乳類での本発明の実施についての適用性を、いかなる意味でも減縮するものではない。
【0019】
免疫応答を刺激する方法には、ここに開示されたウィルス、ここに開示されたウィルス性ポリヌクレオチドによってコードされるウィルス性タンパク質、このようなタンパク質の断片又はこれらの組み合わせを含む組成物を、哺乳類に投与することを含む。ウィルス及び/又はタンパク質及び/又はそれらの断片は、いかなる適切なソースから単離されてもよく、また、公知の技術を用いて組み換え的に生産されてもよい。多くの実施態様において、ワクチンにおいて使用されるための本発明によって提供されるウィルス性タンパク質は、NS−1、NS−2、N、P、M、SH、G、F、M2−1、M2−2、Lタンパク質及びそれらの組み合わせを含むが、これらに限定されるものではない。これらのタンパク質をコードする全てのヌクレオチド配列は本発明に含まれ、当業者によって通常の技術を用いて決定されうるものである。
【0020】
本発明の組成物が投与される哺乳類は、ここに開示されるウィルスを含むがそれに制限されない1又は複数のニューモウィルスに対してリスクのある、保菌が疑われる、又はこのウィルスの存在と正の相関関係のある条件で診断された、あらゆる哺乳類でありえる。ある実施態様において、この動物は、ARDCにリスクがあり、疑われ、又は診断されている犬である。他の実施態様において、この哺乳類は、ここに開示されるウィルスと正の関連がありうる呼吸器疾患その他の疾患が疑われ、或いは診断された、リスクのある人間である。この点において、マウス肺炎ウィルスのような、ここに開示される新規に発見されたウィルスと関係するウィルスは、人間にも広範囲の感染を引き起こすことが、当分野では公知である(例えば、Pringle CR, Eglin RP. Murine pneumonia virus: seroepidemiological
evidence of widespread human infection. J Gen Virol. 1986 Jun; 67 (Pt 6):
975-82を参照)。つまり、ここに提供される組成物及び方法が、人間において免疫応答を刺激するために、また、人間においてウィルス感染の診断に用いられるために、有用性を有すると考えることは合理的である。
【0021】
動物への本発明の組成物すなわちワクチンの投与は予防的効果を生じ得る。これはワクチンを受けていない動物よりもワクチンを受けた動物のほうが感染の可能性が低いこと、又は、動物においてウィルスの存在と正の相関関係のある病徴がワクチンを受けていない動物よりもワクチンを受けた動物において深刻度がより低いことを意味している。本発明のワクチンの投与はまた治療的効果を有しうる。これはワクチンを受けていない感染動物に対して、ワクチンを受けた後に感染した動物は感染の度合い(すなわちウィルス負荷又は力価、及び/又は当業者に公知の感染度マーカー)が低いことを意味している。同様に、ワクチンを受けていない動物に対してワクチンを受けた感染動物は、動物においてウィルスの存在と正の相関関係のある病徴の深刻度が低い。
【0022】
ここに開示されるウィルス及び/又はタンパク質及びポリヌクレオチドは単離されうる。これは例えば動物及び/又は動物から得られる生物学的サンプルからウィルスを分離することによって、ウィルスの自然の環境からウィルスが除かれることを意味している。一つの実施態様において、ここに開示されるウィルスを含む単離された細胞は単離されたウィルスを包含すると考えられる。さらに本発明の単離されたウィルス及び/又はタンパク質及び/又はそれらの断片及び/又は単離され及び/又は組み換えられたポリヌクレオチドは、如何なる所望の精製度で精製されてもよい。本発明で提供される組成物はウィルス、ウィルス性タンパク質、それらの断片、及び/又はポリヌクレオチド、ポリヌクレオチドの断片及び/又はそれらの組み合わせを含み、それらからなり、又は、実質的にそれらからなるものであってよい。
【0023】
一つの実施態様において方法は、イヌにおいてウィルスに対する免疫応答が刺激されるように、配列番号1の配列を含むポリヌクレオチド又は配列番号1の配列と少なくとも96%が同一の配列を有するポリヌクレオチドを含む組成物をイヌに投与することを含む。一つの実施態様において、ポリヌクレオチドはDNAワクチンとして投与される。DNAワクチンを含む薬剤組成物を成形すること、及び、DNAワクチンを含む組成物を投与することは当分野で公知である。一つの実施態様において、イヌに投与されるポリヌクレオチドはウィルス粒子中に存在する。ウィルス粒子は単離及び/又は組み換えされうる。多くの実施態様において、イヌに投与されるウィルスはウィルス粒子によって含まれるタンパク質の一部であるアミノ酸配列を含むタンパク質を含んでいる。ウィルスゲノムによってコードされるウィルス粒子によって含まれるウィルスタンパク質の予測されるアミノ酸配列は、イヌウィルス性タンパク質NS1、非構造タンパク質1(配列番号7)、NS2、非構造タンパク質2(配列番号8)、N、核タンパク質(配列番号9)、P、リン酸タンパク質(配列番号10)、M、マトリクスタンパク質(配列番号11)、SH、小疎水性タンパク質(配列番号12)、G、アタッチメントタンパク質(配列番号13)、F、フュージョンタンパク質(配列番号14)、M2、マトリクスタンパク質M2−1(配列番号15)、及びM2−2(配列番号16)、L、一部配列が提供されている(配列番号17)RNA依存性RNAポリメラーゼである。これらのタンパク質のネコウィルス型も本発明によって提供される。
【0024】
他の実施態様においてこの方法は、ネコにおいてこのウィルスに対する免疫応答が刺激されるように配列番号2及び/又は配列番号3の配列を含むポリヌクレオチド、又は、配列番号2又は3の配列と少なくとも96%が同一の配列を有するポリヌクレオチドを含む組成物でウィルスをネコに投与することを含む。一つの実施態様において、ネコに投与されるポリヌクレオチドはウィルス粒子の中に存在する。粒子は単離され及び/又は組み換えられ得る。
【0025】
多くの実施態様において、ネコに投与されるウィルスは配列番号5又は配列番号6から選択されるアミノ酸配列を含むタンパク質を含む。
【0026】
ある実施態様において本発明は、ここに開示されるウィルスによる感染に対する予防的及び/又は治療的効果を提供しうるワクチンを含む。このワクチンは生又は死滅(不活性化)ウィルス又はその断片を含みうる。死滅ウィルスとは複製が不可能であるウィルスである。死滅ウィルスは当業者には周知の多様な技術のいずれを用いて調製されてもよく、熱による不活性化及びウィルス性タンパク質及び/又は核酸の修飾を含みまたこれらに制限されず、その方法はパラホルムアルデヒド、ホルマリン及び紫外光線への暴露を含みまたこれらに制限されない。例えば、ウィルス性タンパク質は例えばウィルス性の細胞侵入及び/又は複製に必要とされる1又は複数のステップを回避するようにウィルス性タンパク質を修飾できうるクロスリンク剤によって共有結合的に修飾されうる。生ウィルスは弱毒化ウィルスを含むと考えられている。弱毒化ウィルスとは弱毒化されていないものに対して毒性が低減されているウィルスである。つまり弱毒化ウィルスは複製可能である、しかしながら、同種の弱毒化されていないものと比して速度は遅く、つまりそれゆえに一般的に疾患を引き起こさない。当業者は、弱毒化されていないもの、弱毒化されたもの、及び、死滅ウィルスの間の他の区別点についても認識しそれらは互いに区別され得るであろう。
【0027】
ウィルスの弱毒化は何らかの標準的な技術を用いて得られうる。一つの実施態様ではウィルスは哺乳類細胞培養における連続継代によって弱毒化される。連続継代は多様な機構を通じて弱毒化を生じ得る。人間の細胞を含みこれに制限されない、ウィルスの複製を支える何らかの細胞はウィルスの増殖や弱毒化の目的のために継代において使用され得る。様々な非制限の例示において継代のための細胞培養は其々Canis familiaris 又はFelis catus のようなイヌ又はネコから得られ又はそれに由来しうる。一つの実施態様において細胞は初代細胞培養である。ある実施態様において、細胞はATCCカタログ番号CRL−1542の番号を有するA−72細胞株由来のイヌ細胞に含まれうる。ウィルスの弱毒化に好適ないかなる継代数も使用することができる。当業者は本開示の有益性を持ったウィルスの弱毒化のための継代パラメータを決定することができるだろう。一般的に5−150(その間の全ての整数値を含む)継代、好ましくは20−50の間の継代が使用され得る。
【0028】
ウィルスを含む組成物はワクチンとして使用するための医薬製剤として提供されうる。ウィルスを含む組成物は懸濁液として又は凍結乾燥形態で提供されることができ、またこのような組成物に通常使用される医薬的に許容されるキャリア及び/又は希釈剤を含有しうる。本発明での使用のために好適なキャリア及び希釈剤のいくつかの例が、The Science and Practice of Pharmacy (2005) 21st edition,
Philadelphia, PA. Lippincott Williams & Wilkins に見出される。特定かつ制限的ではないキャリアの例は安定化剤、防腐剤及びバッファを含む。特定かつ制限的ではない安定化剤の例は、炭水化物(例えばソルビトール、マンニトール、スターチ、スクロース、デキストラン、グルタミン酸、又はグルコースなど)、タンパク質(例えば、乾燥乳清、アルブミン又はカゼインなど)又はそれらの分解生成物を含む。好適な、特定かつ制限的ではないバッファの例はアルカリ金属リン酸塩を含む。好適な、特定かつ制限的ではない希釈剤の例は水、水系バッファ(例えば緩衝生理食塩水)、アルコール及びポリオール(例えばグリセロール)を含む。
【0029】
当業者はここに提供されるワクチン製剤はアジュバント活性を有する1又は複数の組成物を含みうることを認識するであろう。好適なアジュバントはフロイント(Freund)の不完全アジュバント、トレオニルムラミルジペプチド、水酸化アルミニウム、リン酸塩又は酸化物、例えば、鉱物油又は、例えばビタミンE酢酸塩のようなベジタブルオイル及びサポニンに基づく水中油型又は油中水型乳化液を含みまたこれらに制限されない。
【0030】
本発明によって提供されるワクチンはあらゆる好適なルートを通じて投与されうる。多様な実施態様においてワクチンは非経口で、筋肉注射又は皮下に、静脈内投与で、或いは、経鼻を含むがこれに制限されない経口で投与されうる。ワクチンの効果的な投与量は例えば、免疫応答の刺激が所望される動物のタイプ、サイズ、年齢、環境、感染ステージのリスクのような通常の検討を用いて決定され得ることを当業者は認識するであろう。一般的に本発明による生ワクチンは、動物当たり、例えば1mL用量あたり101−106pfu(プラーク形成ユニット)の用量で投与されうる。不活性化されたワクチンは動物当たり106−1010pfuの抗原性等価物を含みうる。
【0031】
本発明のポリヌクレオチドは例えばここに開示されるウィルスだけでなく、他の関連する又は非関連のウィルス及び/又は他の感染性病原体に対して免疫化を提供しうる異種遺伝子(1又は複数)を包含するように修飾されてもよい。同様にここで提供されるワクチン組成物は、ここに開示されるウィルスから区別される感染性病原体に対して免疫応答を刺激することを目的とされる免疫原性の薬剤を含むことができる。つまり本発明は種々の哺乳類において、また、種々の感染性病原体に対して使用するための混合ワクチンを予測するものである。混合ワクチンはここに開示されるウィルス性タンパク質の全部又は一部を含み、また、異なる感染性病原体に対して効果を有する多価ワクチンを提供するために他のタンパク質からのポリペプチド配列を含みうる融合タンパク質を含みうる。本発明はさらに、動物に有益な効果を与え得る他の組成物であってこのような感染のための従来的な治療に先立って、同時に、又は続いて投与される組成物を投与することを含む。
【0032】
本発明はまたここに開示されるウィルスを検出するための方法も提供する。この方法は動物から生物学的サンプルを得ること、及び、サンプル中に又はサンプルから、ウィルス性タンパク質及び/又はウィルス性ポリヌクレオチドの存在を検出することを含む。ウィルス性ポリヌクレオチド及びタンパク質のいずれかがウィルスの存在又は不存在を検出する方法において使用されうる。配列番号1、2又は3の逆相補物又はDNA等価物を検出することは配列番号1、2、又は3の配列、又はこれらの配列と少なくとも96%が同一の配列を含むポリヌクレオチドを検出することと同等であると考えられている。
【0033】
検出は、診断が望まれる哺乳類から得られたいずれの好適な生物学的サンプルを使用して実行されてもよい。生物学的サンプルの好適なソースは、血液、粘膜剥離物、組織生検、又は唾液を含みこれらに制限されない。一つの実施態様においては生物学的サンプルは鼻及び/又は咽頭スワブ、又は気管洗浄液を含む。
【0034】
ウィルスの存在又は不存在は当分野では周知の様々な技術を用いて、DNA、RNA又はタンパク質を検査することによって検出されうる。ある実施態様ではウィルスの存在はウィルスゲノムの全体又は一部を含む、又は、ウィルスゲノムの全部又は一部から増幅されるポリヌクレオチドを検出することによって決定される。この点で本発明はウィルスのマイナス鎖、ウィルスのプラス鎖、又はそれらの組み合わせからのポリヌクレオチド配列を同定することによって、ウィルスの存在(又は不存在)を決定することを含む。本発明はまたウィルスのマイナス鎖、プラス鎖、及びそれらの組み合わせのDNA型を検出することも含む。例えば一つの実施態様において逆転写PCR(ポリメラーゼ連鎖反応)がウィルスのマイナス鎖又はその一部のDNA複製物を生成するために用いられる。またDNA複製物はハイブリダイズされた二本鎖において、プラス及びマイナス鎖又はそれらの一部分のDNA型を含む複数の二本鎖DNA分子を得るためのウィルスゲノムの増幅のためのテンプレートとして用いられうる。当業者は興味の対象である生物学的サンプル中にウィルスがいるか又はいたかどうかの決定において使用するための遺伝子材料の量の増幅のために使用される多様な増幅技術があること、また、この増幅された(又は増幅されない)遺伝子材料はポリヌクレオチド配列を決定するための多くの周知の方法及び試薬を用いて分析されうることを認識するであろう。すなわち本発明は、ウィルス性ポリヌクレオチドの存在又は不存在が確認されうるように生物学的サンプルから核酸を獲得し、また分析するための何らかの及びあらゆる方法を含む。この方法は何らかの公知の技術を用いて、核酸及び/又は他のタイプのプローブをハイブリダイズすることによって、及び、ウィルス性ポリヌクレオチドの配列を決定することによってポリヌクレオチドを検出するための方法を含みこれらに制限されない。配列の全部又は一部分がウィルス性のポリヌクレオチドを同定するために決定され、使用され得る。
【0035】
ある実施態様において本発明は単離されたウィルス性ポリヌクレオチドを含む組成物、及び、核酸ハイブリダイゼーション及び/又は増幅のために使用される要素を提供する。また組成物はDNAポリメラーゼ、逆転写酵素、遊離核酸三リン酸塩、塩、バッファ、及び、核酸をハイブリダイズ及び/又は増幅するのに典型的に用いられる他の試薬を付加的に含むことができる。一つの実施態様において本発明は、単離されたウィルス性ポリヌクレオチド及び/又はウィルス性ポリヌクレオチドから増幅されたポリヌクレオチドであって、rt(real time)核酸増幅技術において用いられる1又は複数の増幅プライマー及び検出プローブにハイブリダイズされたものである単離されたウィルス性ポリヌクレオチド及び/又はウィルス性ポリヌクレオチドから増幅されたポリヌクレオチドを提供する。
【0036】
本発明はまた、単離されたウィルス性ポリヌクレオチド、及び/又は、ポリヌクレオチドがアレイの中に存在する少なくとも一つのプローブに増幅されたものである、ウィルス性ポリヌクレオチドから増幅されたポリヌクレオチドを提供する。このアレイは、例えば複数の特定のポリヌクレオチドの存在又は不存在を決定するために使用されるチップ上に存在しうる。このようなチップは商業的に提供されるものであり、また本質的にあらゆるポリヌクレオチドの存在又は不存在を検出するためにカスタマイズされうる。
【0037】
多様な実施形態において本発明はさらに、哺乳類がここに開示されるウィルスに感染しているか否かの決定を有形的な媒体に固定することを含む。有形的な媒体はどのような有形的な媒体でもよく、例えば、コンピュータ、DVD、CD−ROM又は電子メールメッセージに保存されるデジタルファイルを含みこれらに制限されないデジタル媒体のあらゆるものでありえる。有形的な媒体は感染哺乳類のための治療プロトコルを発展させるようにヘルスケア提供者に提供されうる。
【0038】
当業者には本発明の利益を受けて多様な反応試薬が作られ、ここに開示されるウィルスの検出のための診断方法に使用されうることが認識されるであろう。例えばポリクローナル又はモノクローナル抗体が、ウィルス粒子又はウィルス粒子の一部、又は単離された又は組み換えのウィルス性タンパク質又はその断片を用いて作られ得る。すなわち抗体は、あらゆるウィルス性タンパク質に存在するあらゆる抗原決定要因を特異的に認識するように作られ得る。多様な実施態様において抗体は、NS−1,NS−2,N、P,M、SH、G、F、M2−1、M2−2及びLウィルス性タンパク質、又は、その一部又は組み合わせを認識する。抗体は本発明のウィルスを、MPVのような他の関連するウィルスから識別できるように、作られ得る。
【0039】
抗体は、生物学的サンプルからウィルス性抗原の存在又は不存在が決定されうるようなあらゆる技術において用いられ得る。このような技術はウェスタンブロッティング、免疫蛍光法、免疫組織化学及びビーズ法又は従来のタンパク質アレイのELISAアッセイを含みこれらに制限されない。
【0040】
一つの実施態様において本発明は、抗体との複合体に存在する単離された及び/又は組み換えウィルス又はウィルス性タンパク質を提供する。
続く実施例は本発明の特定の実施形態を描写することを意図しているが、いかなる意味でも本発明を制限するものと解釈されるべきではない。
【実施例】
【0041】
[実施例1]
我々は、雑種犬の鼻及び咽頭スワブからサンプルを採取し、呼吸器ウィルスを単離するために、イヌA72細胞(アメリカ培養細胞系統保存機関(ATCC)、CRL−1542)にそれらを植菌するために用いた。植菌後約21日に対応する、培養の経過の後期段階において、いくつかの培養が僅かに細胞変性の変化を示した。連続する経過の後、培養は培養全体の急速な細胞死に続いて、円形細胞の小巣を現した。このパターンは一般的なARDCに伴うウィルスには非特徴的であると主観的に判断された。13の別個のウィルス単離物が得られ、この実施例において説明される。一般的なイヌ呼吸器病原体に特異的な診断反応試薬のパネルを用いたテストは、公知のウィルスを同定することができなかった。他のウィルスのための追加的な反応試薬でのさらなるテストは、最終的に、ヒト呼吸器合胞体ウィルス(抗HRSV、番号VP−R151、Vector Laboratories、USA、カリフォルニア州バーリンガム)に対するモノクローナル抗体(Mab)プールを用いた免疫蛍光アッセイ(IFA)によって陽性認識を示した。この抗体の調製は、ウシRSV(BRSV)の検出のために我々のラボで一般的に使用されているものである。染色パターンは、RSV感染細胞において観察される典型的なパターンである線維状の膜結合及び遊離のウィルス粒子及び細胞質内封入体を含んでいた。
【0042】
最初のIFA結果に続いて、我々は、ヒト、ウシ、ヒツジのRSV配列のアライメントに基づいてデザインされたPCRプライマーを用いて、ウィルスからヌクレオカプシド遺伝子(N)の断片を増幅することを試みた。これは不成功であった。抗RSVプールの個々のMabのストック及びそれらの特異性は、製造者から入手され、IFA(図1)のために使用された。Mab5H5N(M2タンパク質特異的)での染色は、ウィルス粒子及び封入体の両方を染色した。Mab2G122(Pタンパク質特異的)は、一次的に封入体を染色し、相対的に均一な膜仲介性のシグナルを与えた。Mab1C3(Nタンパク質特異的)又は5A6(Fタンパク質特異的)では染色は得られなかった。4種の個々のMabの全てが、IFAによってBRSVを認識した。4種のMabのうちの2種のみによるイヌウィルスの認識と、RSVの保存領域の増幅が不可能であったことは、RSVに関連性があるが典型的な形態でないことを示唆した。
【0043】
ウィルス配列の解明は、CODEHOPアルゴリズムを用いて、ニューモウィルス亜科の全てのウィルスの複数の配列アライメントのうち高度に保存されたアミノ酸(aa)配列に基づいた変性PCRプライマーをデザインすることによって追跡された。L(ポリメラーゼ)及びN遺伝子の特定領域が目標とされた。反応生成物の配列決定とBLAST分析は、ウィルスが伝統的にマウスのニューモウィルス(又は肺炎ウィルス)として知られているマウスニューモウィルス(MPV)に近いことを明らかにした。2つのL遺伝子PCR産物は、MPVと95%から97%同一であることが見いだされ、N遺伝子断片は約96%が同一であった。
【0044】
新規に同定されたニューモウィルスが、テストされたシェルターのイヌの群に限られるのかどうかという疑問に対処するために、我々はランダムなイヌ血清サンプルをスクリーニングし、27のうち6の動物が感染された培養のウィルスが特異的に認識される抗体を有していたことを見出した。染色パターンは、Mab(図2)で観察されるものと類似であった。BRSVに対する中和抗体を種々の量で含有するウシ血清は、イヌウィルスに感染している細胞を染色しなかった。
【0045】
この例は、ARDCの13頭のイヌから単離されたウィルスはMPVに非常に近い関係にあるとみられることを示し、ゆえに我々は、それらはイヌニューモウィルス(CnPnV)であると考える。マウスニューモウィルスは、パラミクソウィルス科のニューモウィルス亜科の中でニューモウィルス属として分類されている3種のうちの1種である。ヒトRSVはタイプ種であり、BRSVと非常に近縁関係であるがMPVはより遠い関係である。例えば、ヒト及びウシRSVのNタンパク質ヌクレオチド配列は約94%が互いに同一であるが、MPVに対しては60%の同一性を有するにとどまる。MPVの完全に配列決定された株は2種のみで、‘Strain 15’及びJ3666であるが、それらはヌクレオチドのレベルでは99.7%の同一性を有する。
【0046】
ARDCでのCnPnVの連関は、いくらかの場合、特に複合的な病因論に関与する時には病原性でありえる。類推では、MPVは一般に齧歯類の飼育株に感染することが知られ、また、血清学的な証拠は幾らかの野生型齧歯類での感染を指摘するが、自然的な生態学については多くは知られていない。実際、齧歯類がMPVの唯一の自然的なホストであること、又は、近縁関係のウィルスが他種に拡散するのかどうかは、明確ではない。呼吸器症状を伴うヒトの感染の証拠が報告されている(Pringle CRら、J.Gen.Virol. 1986;67:975-82)。自然界では齧歯類の感染は無症状で潜在的であるかもしれない。飼育マウスでの臨床症状は無症状から高い罹患率及び死亡率を伴う肺浮腫への発展まで幅広い。J3666とStrain15の両方を含む病原株は、深刻な肺炎を引き起こし、少量で植菌された時には6−10日で死に至りうる。病原性、またはその欠如はウィルス及びマウス株依存性であるのだろう。
【0047】
複数の微生物とウィルス病原体がイヌの呼吸器疾患に関与しうる。すなわちCnPnVは、より深刻な疾患を導く上部呼吸器胴の防御機構を損傷する一連の事象を開始する能力のある他の病原体であると考えられる。
【0048】
[実施例2]
この実施例の材料及び方法は、残りの実施例で説明される結果を得るために使用された。
【0049】
[ウィルスの単離]
鼻及び咽頭検査サンプルは、湿潤スワブを用いて呼吸器疾患の症状のある雑種犬から集められ、コーネル大学動物健康診断センターにおいて受領から24時間以内に処置された。植菌の前に、スワブはアール塩(Earle’s salt)(Gibco 10370, Invitrogen,
Carlsbad, CA, USA)、0.5%ウシ血清アルブミン、ペニシリン(200 U/ml)、ストレプトマイシン(200 μg/ml)、及びファンギゾン(2.5 μg/ml)を含む最少基礎培地(MEM培地)3mLに30分間浸漬し、続いて10秒間機械的に撹拌した。それぞれ0.5mLの鼻及び咽頭抽出物の分割単位は集められ、セミコンフルエントのA72細胞(Binnら、1980)(CRL-1542,
American Type Culture Collection, Manassas, VA, USA)のシングルT25フラスコに、培地を追加することなく植菌するのに用いられた。抽出物は、1−3時間モノレイヤーの上に残置され、その後リン酸緩衝生理食塩水(PBS)でリンスオフされた。細胞は6mLの培養培地(ライボビッツのL−15培地、10% 熱非活性化ウシ胎児血清(FBS)、200 U/ml ペニシリン、200 μg/ml ストレプトマイシン、50 μg/mlゲンタマイシン)にて37℃で維持され、6−8日毎に継代培養された。植菌されていないコントロールのA−72培養が単離プロセスを通じて並行して行われた。
【0050】
[免疫蛍光アッセイ]
細胞はガラススライドに接着させられ、その後PBSでリンスされ、冷アセトン10mLで固定化された。スライドは空気乾燥され、染色の前に−20℃で貯蔵された。PBSで希釈された一次抗体及び二次抗体がスライドに滴下され、37℃で30分間インキュベートされた。各インキュベーションの後、スライドはPBS中で15分間リンスされた。染色は蛍光顕微鏡で可視化された。HRSVに対するMabのプールが希釈率1:400にて当初の同定において使用された(VP−R151、ベクターラボラトリー、バーリンガム、カリフォルニア州、USA)。1倍濃度で供給されたマウス抗−PVM(CL-603IFA, Charles River Laboratories, ウィルミントン、マサチューセッツ州、USA)が、希釈率1:20にて使用された。二次抗体FITC標識ヤギ抗マウスIgG(02−18−06、KPL、ガイザースバーグ、メリーランド州、USA)は、希釈率1:40(12.5 μg/mL)で使用された。
【0051】
[RNA精製]
全細胞性RNAがA72細胞から精製された(74106、Qiagen、バレンシア、カリフォルニア州、USA)。約50−75%の細胞変性効果(CPE)を示す一つの25cm2フラスコからの細胞が、ペレットにされ、各カラムを通して精製された。細胞フリーの上清及びスワブ溶出物を含む培地はサンプル140μLを用いて精製された(52906、Qiagen)。精製されたRNAsはRNaseフリーの水40μLで再懸濁された。
【0052】
[PCR]
ゲノム断片を得るために当初使用された縮重プライマー配列は、ニューモウィルス科ウィルスのアライメントに基づき、CODEHOPアルゴリズム(blocks.fhcrc.org/codehop.html)(Roseら、1998)を用いて設計された。逆転写PCR(RT−PCR)は製造者の指示に従い25μLボリュームで1段階反応(210212、Qiagen)で行われた。1:5〜1:10で希釈された全RNAの1μL及び各プライマー10pmolが各反応で使用された。RTステップの反応条件は、50℃/30分、続いて95℃/15分であった。サイクル条件は、95℃/60秒、54−56℃/30秒及び72℃/60−90秒を35サイクル行った。プライマーは次の通りである:N遺伝子:N276F,tccgtgcaggccgaratggarcarg/P1R,(配列番号35)ggaactcgggggcgaayttytccat;配列番号36);L遺伝子 no 1:L428F, ccggatcttcggccayccnatggt/(配列番号37)L538R, ttcttaggaggggagatggcyttrtcrtt;(配列番号38)L遺伝子 no. 2:L698F,
catcaccgacctgtccaagttyaaycargc/(配列番号39)L894R,
ttgaagtcgtccaggatggtrttdatcca配列番号40。様々な地理的位置からのスワブサンプルと用いられた2つのPCRプライマーセットは、CnPnV、MPV J3666及びstrain15の配列のアライメントに基づいて設計された。Gタンパク質:G715F, ggcttctgtttcttcctttctgg/(配列番号41)G1062R,
ccgtggtggtgcctgtg(配列番号42);SH1タンパク質:SH1F,atggatcctaacatgacctcayac(配列番号43)/SH187R,
gattgggatgaacygtgcattg(配列番号44)。低継代Brne単離物を増幅するのに使用されたプライマー:G84F,tgtaaaagtgaaccaaatgtgta(配列番号45)/G404R,aaatcttcaggtaaatcaggtc(配列番号46);G849F,ttttaacaacaagaatcagtc(配列番号47)/G1048R,
ctcctaggtgcgggggttgg(配列番号48)。G及びSH単位複製配列のための反応条件は、RTが50℃/30分、不活性化/変性が95℃/15分、及び、PCRは95℃/60秒、54℃/30秒及び72℃/90秒を35サイクルであった。
【0053】
[ゲノム分析]
PCR生成物は、Applied Biosystems Automated 3730 DNA Analyzerを用いてコーネル大学Sequencing and Genotyping Core Laboratoryにて配列決定された。完全配列を決定するのに使用されたプライマーはMPV配列又は先に決定されたCnPnV配列に基づくものであった。重複するPCR生成物はギャップとプライマー結合領域をカバーするように作り出された。すべての領域は両鎖分析された。CnPnV単離物は、Lasergene(DNASTAR,マディソン, ウィスコンシン州, USA)を用いて、MPV Strain15(GenBank AY729016)及びJ3666(GenBank NC006579)配列と比較された。
【0054】
[実施例3]
[単離と同定]
鼻及び咽頭スワブ溶出液が、A72細胞に植菌するように用いられた。相対的に長期間の培養の後、通常は3継代又は4継代の後に出現する、限定的なCPEが観察された。初期のCPEは典型的に円形細胞の分散性の小巣を含み、いくらかは小合胞体や空胞化を伴っていた。このCPEは、静置培養において数日間以上ゆっくりと進行した。継代培養はしばしばこの低レベルCPEが維持されるモノレイヤーを導いたが、いくらかの培養では24−48時間以内に迅速な進行及びモノレイヤーの崩壊が生じた。このパターンはいくつかのヘルペスウィルス感染と同様であるが、イヌヘルペスウィルスには試験陽性ではなく、同様の方法を用いて一般に単離されている犬呼吸器ウィルスのいずれでも典型的なものではないようであった。新鮮な細胞への上清の移送に続くCPEは僅かであるか又は生じず、初期継代の単離物の幾らかは培養における複製が少なかった。しかしながら継代の連続につれていくつかの培養は、より安定的な移送可能CPEを発展させ、それらはさらなる分析のために選択された。初期のテストでは、いくつかの無関連のイヌからの血清はCPEの培養においてウィルス性抗原を認識するように見え(データは不図示)たが、しかし、一般的なイヌ呼吸器ウィルスに対するテストはネガティブであった。実施例1で報告されたように、HRSV特異的なMabはウィルス性抗原を強く認識した。抗PMVポリクローナル血清でのポジティブIFA染色が確認のために使用された。抗PMVマウス血清はIFAによってBRSVを弱く認識するのみであった。
【0055】
実施例1で説明されたCnPnVの元の13の単離物は、同じ組織によって管理されている2つの動物シェルターに由来するものであった。ウィルスがこれらの2施設に制限されるのかどうか調査するために、他の地理的位置からの病気のからの鼻又は咽頭スワブ試料がPCRによって試験された。USAの8つの州(コロラド、ジョージア、フロリダ、インディアナ、ミズーリ、ネバダ、サウスカロライナ、バージニア)からの19頭のイヌが、G及びSH遺伝子のプライマーセットを用いてテストされた。19サンプルのうち、ネバダからの1例、サウスカロライナからの9頭のグループのうち5例がG及びSHの両方のPCRに陽性を示し、配列決定により確認された。他の13頭からはPCR陽性の結果は得られなかった。A72細胞でのウィルス単離が、イヌ呼吸器診断のためのニューヨーク及びペンシルヴァニアからの鼻スワブの6サンプルについてさらに行われた。これらのうち、2つのさらなる単離物はニューヨーク市の獣医学病院からのものであり、他は、ペンシルヴァニア州フィラデルフィアのシェルター由来のものであった。これらの単離物は、抗RSV及び抗PVM抗体でIFAによって、続いてRT−PCRで、CnPnVと確認された。これらのRT−PCRとウィルス単離結果は正確な有病率の評価を提供するものではないが、それらは、CnPnV感染が広汎であることを示すものではなかった。
【0056】
[配列分析]
以前の研究は自然的な準種の存在、培養における継代中の変異やPCR産物に起因しうるウィルス性単離物の間の差異を指摘していた。CnPnV単離物の最も正確なコンセンサス配列を決定するために、テンプレートとして全細胞性RNAを用いる複数の重複するRT−PCR反応が使われた。2頭の犬からのウィルス性単離物が配列決定されたが、一つは4継代の組織培養(Ane4)、一つは17継代(Brne17)であり、J3666(Thorpe及びEaston、2005)及びStrain15(Kremplら、2005)の配列と比較された。NS−1に近接する3´リーダーの中から開始し、Lコード領域中へと小距離延在し、ゲノム中の予測された10の遺伝子のうち9をカバーしている、8598ヌクレオチドの配列(配列番号1)がCnPnV-Ane4から得られた。遺伝子の順番、NS1−NS2−N−P−M−SH−G−F−M2−Lが確認された。MPV株と比較した、CnPnV-Ane4のこの領域の全体的なヌクレオチド配列の同一性は、J3666に対して95.0%でありStrain15に対して94.7%であった。MPV株は互いに99.7%同一であった。CnPnV-Ane4とMPV株を比較したときにコード領域中のギャップは見いだされなかった。セクション3.4及び3.5で説明したようにGとSHの長さの違いを除いて、MPVとCnPnV-Ane4のコード領域の全てが同じ長さのタンパク質をコードしていることが見いだされた。
【0057】
[非翻訳領域の比較分析]
CnPnVのGS及びGE配列の境界が、J3666及びStrain15の配列のアライメントによって決定された(Thorpe, L.C.,
Easton, A.J., 2005. J. Gen. Virol. 86, 159-169、 Kremplら 2005. Virus Genes 30, 237-249)。MPVのGS配列は、Chambersら(1991)によって説明され、より最近では (Dibben及びEaston, 2007) 、Collinら(1986)によって決定されたRSV GS配列に基づく長さ9ヌクレオチドのものとして説明されている。Kuoら(1997)は、10番目のヌクレオチドもまたRSVのGSにおいて重要であり、これは長さ10ヌクレオチドであるMPV GS(Kremplら、2005)と同等であることの証拠を提供している。表1において、GSは、両方の解釈を含む10ヌクレオチドとして示されている。表1は、MPVstrain15、J3666と比較したCnPnV-Ane4のコード領域間のGS、GE及びIGRsを提供する。
【0058】
【表1】

【0059】
表1では2つのMPV株の間でNTR長の違いはない。RNA配列の向きは5’から3’方向である。*:J3666及びstrain15から少なくとも1ヌクレオチドの変化がある。ヌクレオチドの差異は太字・下線で示されている。GE:遺伝子末端;IGR:遺伝子間領域;GS:遺伝子開始、無印;長さの差異がない。aM−SHについてはNTR全長において差異がない;一つのuがIGRから消失し、他のuがNTRの上流に追加されている。b3ヌクレオチドIGR及びM2もGSはDibbon及びEastonによる分析に基づいている(Dibbon,O., Easton, A.J.,
2007. Mutational Analysis of the Gene Start Sequences of Pneumonia Virus of
Mice. Virus Res. 130, 303-309)。表1に示された10又はそれ以上のヌクレオチドを有する配列は次の配列番号を有している:“GE”カラムの配列:NS1
uaguuaauuaaaa(配列番号18);NS2 uaguuauagaaaaa(配列番号19);N uauuuaauuaaaaa(配列番号20);P(配列番号18);M
uaguuaaauaaaa (配列番号21);SH uaguuaacaaaaaa (配列番号22);
G uaguuaaugaaaaa (配列番号23); F
uaguuaauuaaaaaa (配列番号33);及びM2 uaguuauauaaaaa (配列番号34)。“IGR”カラムの配列:N
cuggaaaauau (配列番号24); 及び G uaagcuaugauauaau
(配列番号25)。“GS”カラムの配列;NS1 aggacaagug (配列番号26); NS2 aggacaaauc (配列番号27);
N aggauaaaua (配列番号28);P (配列番号28);M
aggacaaaua (配列番号29);SH aggauaagua (配列番号30);
G (配列番号30); M2aggauaagug (配列番号31);F (配列番号29)及びL aggaucaaua(配列番号32)。
【0060】
わずかな長さのバリエーション及びヌクレオチドの相違が、CnPnV-Ane4 NTRで同定され、大部分はIGRの中で見いだされた。IGRにおける2つのMPV株の間には差異がみられなかった。Chambersら(Chambers, P., Matthews, D.A.,
Pringle, C.R., Easton, A.J., 1991. Virus Res. 18, 263-270)はcDNAクローンからのM2のGSを決定したが、56ヌクレオチドのIGR中に2つの付加的な潜在的なGS配列(GS1、GS2)の存在も指摘している。変異分析は後に、6番位置のAの代わりであるGによって推定されるGS配列は非機能的であることを決定した。続いて彼らは、GS1及びGS2の両方の配列が転写開始を指示する能力を有し、第一の配列(GS1)が最も重要であることを示した。彼らは、元々同定されたGS配列はcDNAクローニングの間の自然的な削除によって誤って同定されたものだと結論づけた。CnPnVにおいて、FのためのGE配列のすぐ後ろに続くM2のためのGS1配列は、10番目のヌクレオチドを除いてSH及びG遺伝子の開始配列と同じである。GS1は一次的に使用されるものと考えられており、表1に示されている。M2のためのGS2AGGACAGGGは(Dibben, O., Easton, A.J., 2007.
Virus Res. 130, 303-309)で報告されたコンセンサス配列から2箇所のみが異なるものである、しかしながら、7番目に保存されたAを有していないので活性は低いかもしれない。これは、Aが保存されているJ3666及びStrain15からは相違点である。CnPnV-Ane4での第三の潜在的なM2 GS(GS3)は、J3666及びStrain15と同一であり、それゆえ6番目のGによって貧弱な機能であることが予測されるであろう。
【0061】
[非構造タンパク質の分析]
非構造の小疎水性タンパク質は92アミノ酸を共有し、92.4%のアミノ酸同一性のみを有する2つのMPV株の間で最も不一致のタンパク質であるように、大幅な変動性を許容するようである。CnPnV-Ane4において、SHタンパク質はMPVstrain15と比較した時にすべての試験したタンパク質のうちでもっとも不一致であった。CnPnV-Ane4 SHは、J3666と比較して4アミノ酸の相違(95.7%)、strain15と比較して9アミノ酸の相違(90.2%)を有し、すなわちMPV株の相互の相同性よりもJ3666により類似している(表2)。他の特徴的な相違は、異なる単離物におけるSH ORFの長さの相違である。CnPnV-Ane4及びstrain15の両方が279ヌクレオチド(92アミノ酸)のSH ORFを有しているが、公開されたJ3666配列は345ヌクレオチド(114アミノ酸)である。しかしながら、J3666の独立に増幅された配列は、92、96又は114アミノ酸というバリエーションをもってコードされており、これらは抗体回避変異体を表していると示唆されている。表2は、CnPnV-Ane4遺伝子及びタンパク質のニューモウィルス亜科におけるウィルスに対する同一性(%)のまとめを提供している。
【0062】
【表2】
【0063】
表2ではGenBank配列に言及している:HRSV、NC_001781;BRSV、NC_001989;HMPV、NC_004148.2;AMPV、NC_007652。これらは、ヒトRSV、ウシRSV、ヒトメタニューモウィルス及び鳥メタニューモウィルスの配列全長に対応している。nt:ヌクレオチド;aa:アミノ酸;NA:適用無し、をそれぞれ表す。aの付された数字は各遺伝子におけるヌクレオチド、アミノ酸の相違の数を示す。CnPnV-Ane4Pタンパク質は、J3666に対して10アミノ酸の差異、strain15に対して15アミノ酸の差異を有することが見いだされた。MPV及びRSVPのアライメントは明確な類似性を有さず、アライメント中に6から7ギャップを含むアミノ酸22−124のアミノ末端近傍領域に隣接する、高い相同性を有する2つの領域を示す。CnPnV-Ane4がMPV株の両方と比較されたときに、中程度の相違である2つの領域が、Pにおいて相同性の低いこの領域の中で同定された。7アミノ酸の相違がアミノ酸58−74の領域に集まっており、他の4つの変異はアミノ酸93−102に見出された。確率プロットは、アミノ酸58−74の相違の第一領域は相対的に表面への露出しやすさ(surface probability)及び抗原指数(antigenic index)が高いことを予測する。第二の相違領域アミノ酸93−102は疎水性で表面への露出しやすさが低い(データは示さず)。137アミノ酸のタンパク質をコードしうるものであるPにおける第二のオーバーラップする内部開始ORFは、MPVにおいて同定された。合成ミニゲノム構築物が、第二のORFから作られるタンパク質を調査するために使用され、それは転写インヒビターとして機能することが見いだされた。CnPnV-Ane4は、同位置から始まるが終止位置は早い、第二のオーバーラップP ORFを有しており、ヌクレオチド285のTがAに変異してTAGコドンになることによって、半分以下のサイズである54アミノ酸のタンパク質を産生する(Dibben, O.ら、(2008) Virus Res. 131, 47-53)。このことは、もし生産されるならばより短いP−2タンパク質が同じ機能を有するかどうかという疑問を残している。
【0064】
残る非構造タンパク質はNS1及びNS2、及びM1−1及びM1−2を含んでおり、これらはM2遺伝子のARF(alternate
reading frame)から翻訳されるものである。strain15とJ3666におけるこれらのタンパク質の間には差異がない。しかしながら、CnPnVは、これらのタンパク質それぞれのMPV単離物と差異を有している。CnPnV-Ane4 NS−1は2つのMPV株と比較して7アミノ酸の差異を有しており、4つはC末端近傍に位置している。NS2では、配列全体にわたって比較的均等に分散している9アミノ酸の差異がある。CnPnV-Ane4は6アミノ酸の差異でM2−1と96.6%同一であり、4アミノ酸の差異でM2−2と95.9%同一である。
【0065】
[構造タンパク質の分析]
ニューモウィルスのG付着タンパク質は典型的に最大の相違があり、抗体応答の無効化のための第一目標となるため特に興味深い。Gタンパク質は2つのMPV単離物の間で396アミノ酸のうち相違は3つのみであり(99.2%)、比較的高い保存性を有している。CnPnVにおけるGタンパク質は、J3666との比較ではもっとも保存されておらず、strain15ではSHに続いて2番目に保存されていないタンパク質である。J3666、strain15とそれぞれ32アミノ酸、33アミノ酸の差異がある。アミノ酸の相違は、113−124領域に数個の差異が集まっていること、及び、331と367アミノ酸(Ane4ORFに基づく数字)の間に11アミノ酸の差異があることを除いて、一般的にランダムに分散している。一つの注目すべき観察は、CnPnV-Ane4のG ORFは、比較対象のJ3666及びstrain15に比べて、54ヌクレオチド長いことである。GSから+29の位置でのACGからAUGへの変異は、J3666にはあるがstrain15にはない代替的な開始コドンを作り出している。+65位置の、第二のUAGからAAGへの変異はインフレーム終止コドンを抹消するものである。G ORFは53アミノ酸の細胞質末端があり、公知のJ3666とstrain15の配列よりも18アミノ酸長いタンパク質をコードしている。J3666の配列はこの位置に終止コドンを有しているがstrain15にはない。すなわち、J3666とstrain15の両方において、単一点変異がより長いG変異体を産生しうる。これはJ3666に感染したマウス肺から増幅されたクローンでないRT−PCR産物の配列も+65位置でUからAへの変異を有していたように、前例の無いことではない。すなわちより長いGの変異体はCnPnVに特異的な特徴ではないように見える。本開示の利益を受けて、いずれの変異体が自然感染において優位であるかを決定するためのイヌ及びマウスのRT−PCR産物からの、さらなる配列データが当分野の通常の技術で決定されうる。G細胞質尾の長さにはいくらかの自然的な柔軟性があるようであり、また、尾自体が病原性の決定要因である。G細胞質尾を欠く組み換えMPV変異体はBalbCマウスで弱毒化されたが、複製レベルは野生型ウィルスと区別できなかった(Krempl, C.D.ら、2007.. J. Virol. 81, 9490-9501)。RSVにおいて細胞質尾の最初の6アミノ酸はMタンパク質との相互作用に本質的である。GのN末端近傍アミノ酸がMPVにおけるMへの結合のために本質的であるかどうかは知られていない。しかしそうであれば、細胞質尾の2つの変異体は、Mとの相互作用能力に差がありえる。
【0066】
CnPnV-Ane4のFタンパク質は、Gで見いだされたのと類似のハイレベルな多様性を有しておらず、実際それはMPVとの比較において、より保存されたタンパク質の一つである。J3666と14アミノ酸、strain15と16アミノ酸の差異があり、97%もしくはそれ以上が保存されている。アミノ酸の差異の約半分が、COOH末端の最後55アミノ酸の中、膜貫通ドメインの中もしくは近傍にあり、ほとんどの置換アミノ酸は類似の機能グループのタイプである。全体として、他のニューモウィルスとの関係においてNが最も高度に保存されたタンパク質である。CnPnV-Ane4のアミノ酸245−333のタンパク質の中央領域は、HRSV及びBRSVと92から93%の同一性に達している。CnPnV-Ane4 Nは、J3666と9アミノ酸の差異、strain15と8アミノ酸の差異を有しておりそれらは比較的均等に分散している。ニューモウィルスで同定された不変位置、又はRNAゲノムと相互作用する残基に対応するアミノ酸の差異はない。これらの差異のいずれもタンパク質の構造又は機能に大きな影響を有するものとは思われない。Mタンパク質はMPV単離物と98.1%という、最も高いレベルの保存性を有している。5アミノ酸の差異は配列中に均等に分散されており、どのケースでも機能グループは保存されている。アミノ酸18におけるVからIへのただ1つの変異のみが、ニューモウィルス及びメタニューモウィルスのMタンパク質において通常は不変とみられる残基に関する。
【0067】
[低継代及び高継代培養における分析]
我々は、培養によく適合され、また、一貫して複製されナイーブ培養に感染させるために用いられるときにはCPEが産生される一つの単離物を同定した。この単離物は17継代(Brne17)であったが、配列決定され、続いて、より低い継代数のAne4単離物と比較された。配列決定された8423ヌクレオチドのうち、Brne17では7ヌクレオチドの変異が同定された(表3、CnPnV-Ane4及び2種の継代数及びex vivoのBrneの間のヌクレオチド及びアミノ酸の差異を提供している)。表3について、a:各遺伝子の開始位置からのヌクレオチド位置、b:各ORFでの位置に基づくアミノ酸番号、ND:未決定、nt:ヌクレオチド、aa:アミノ酸。
【0068】
【表3】
【0069】
Nでの単一のヌクレオチドの差異、Fでの2つの差異は同義の置換体である。Brne17のSH GE配列におけるA残基の付加もまた非コードである。2つのヌクレオチドの差異は、Mにおいてアミノ酸178をQからRに変異させ、また、Gにおいてアミノ酸342をLからPに変異させる非同義の置換である。Brne17で見いだされた最も顕著な差異は、G ORFでの364ヌクレオチドのUからAへの置換であり、これはアミノ酸位置122においてK残基を終止コドンに置換するものである。この位置は、MPV株において、ヌクレオチド310(アミノ酸104)に相当する。この位置での終止は、Brne17に53アミノ酸の細胞質尾、24アミノ酸の膜貫通ドメイン及び44アミノ酸のみの細胞外ドメインのある切り取られたGを与えるであろう。Brne17における短縮されたGの発見は、3継代のストック及びオリジナルのスワブ溶出物(Brne3及びBrneSW)からの増幅産物のさらなる配列決定を促した。興味深いことにこのことは短縮された変異体が早くも3継代目において既に形成されていることを示した。しかしながら、BrneSWではヌクレオチド364にはAが存在し、これはイヌにおける優位なウィルス種は、短縮されたGを有していないことを意味した。RNA抽出物及び増幅物の第二の独立したセットは、1継代及び2継代からのサンプルを含み、1継代のもののみにヌクレオチド364がAであるマイナーな亜集団を示唆する電気泳動図の潜在的なピークが見られた。つまり培養において短縮された形態は迅速に選択されることを表している。同様の変異が継代されたstrain15(Warwick)の培養において同定されたが、そこではフレームシフトを生じる1つのヌクレオチド挿入がペプチドの中途終止を引き起こした。代替的な下流サイトでの翻訳の開始は33アミノ酸短く、細胞質尾を欠くGタンパク質を産生しうるものである。切り取られたGは少なくとも、strain15の低減された病原性についての部分的な説明を示唆した(Warwick)。
【0070】
RSVにおいて、低温継代されたcp−52変異体はG及びSHの産生を抹消する大きな欠失を有するが、培養において複製回避はされないことが見いだされた。この状況はBHK−21細胞培養では完全な組み換えウィルスと同じくG遺伝子全体を欠く組み換え体MPVが複製されたが、マウスにおいては検出可能なレベルに複製されなかったことと相同である。このことは、Gは常にin vivoでは本質的であることを示唆しており、すなわちBrneSwでは短縮されていない変異体のみの検出が予測され得る。なぜ2継代の終わりまでに短縮された変異体が完全に選択されるのかは明らかではない。もしも選択が単に複製効率の向上のみによるならば、Gの不存在は複製のレートや力価を向上させると予測される。しかし、これが真実だと支持する証拠は無い(Kremplら、2007、supra)。G欠損変異体の選択に影響を与える要素は、A72細胞に特異的でありえる。
【0071】
図3は配列番号1のゲノム組織の図示を提供する。当業者は図3の図は3´から5´方向に示されていることを認識するであろう。なぜならこれは、配列番号1は従来の方向としてマイナス鎖を左から右に読んで5´から3´に表している一方で、マイナス鎖ウィルスであるからである。つまり図3において、L ORFは、プラス鎖の3´末端にある。表4は配列番号1に示された配列の注釈を示している。略号は次の通りである。GS:遺伝子開始シグナル配列、ORF:オープンリーディングフレーム、GE:遺伝子終止シグナル配列、IGR:非翻訳インテグリン領域。リストされていないヌクレオチドは翻訳されないと考えられている。
【0072】
【表4】
【0073】
[実施例4]
我々はまた、ここで同定されるイヌのウィルスと関係がありえるウィルスの存在のためネコから得たサンプルを分析した。配列は2つの単離物から得られ、それらは29ケンタッキー(29KY)及び77ケンタッキー(77KY)と称呼される。29KY及び77KYからの部分的なG遺伝子配列は上記で説明されたイヌニューモウィルス(CnPnV)Ane4単離物の726−1078ヌクレオチドに対応する、353ヌクレオチドを提供する。29KYは、CnPnV-Ane4の2−208ヌクレオチドに対応する207ヌクレオチドを有する。
【0074】
両方の単離物の部分的なGヌクレオチド配列はマウスニューモウィルス(MPV)単離物J3666と94.7%が同一であり、Ane4と98.0%が同一であった。117アミノ酸(aa)配列はJ3666と90.6%が同一、Ane4と97.4%が同一であった。つまり、ネコ29KY/77KY G配列はJ3666よりもAne4により関連している。
【0075】
29KYの部分SHヌクレオチド配列はJ3666と94.7%が同一であり、Ane4と97.1%が同一である。69アミノ酸配列はJ3666と98.5%が同一であり、Ane4と97.1%が同一である。つまり、ネコ29KY配列はAne4よりもJ3666にわずかにより関連している。しかしながら、これらの配列は新規に発見されたネコニューモウィルスを表すと考えられている。つまり、上記で説明されたイヌニューモウィルスについて開示された発明の組成物及び方法及びすべての実施態様の開示は、ネコウィルスにも同様に適用され、それらの開示はすなわちネコウィルスについても反復される。
【0076】
当業者には前記から、我々は、CnPnV感染はそれが元々単離されたシェルターの犬に留まらないことを示したことを理解するであろう。それは、急性の呼吸器疾患の犬からのサンプルにおいて容易に同定される。疾患における病原性の役割は明確に証明されていないが、本発明はウィルスの存在を同定し、防御する組成物と方法を提供するものであって、それらは哺乳類においてウィルスの存在と正の相関関係のあるイヌやネコ両方の疾患を含みまたこれらに限定されない呼吸器疾患をコントロールするのに重要であろうと予測されるものである。
【技術分野】
【0001】
本願は、その開示内容が引用によりここに援用される、2009年12月21日出願の米国出願番号61/288,401に対して優先権を主張する。
【0002】
本発明は一般的に、ウィルス学の分野、より詳細には、イヌ及びネコを含む哺乳類で見いだされた、パラミクソウィルス科、ニューモウィルス亜科の新規に発見されたウィルスに関する。
【背景技術】
【0003】
シェルター、犬舎又は繁殖施設等に一緒に閉じ込められている家畜犬は、しばしば急性の呼吸器感染症(ケンネルコフ)に感染する。新しい動物が連続的に導入されるにつれて疾患は急速に伝染し、除去が困難となる。病原体のスペクトラムは、診断や処置を困難にする複数で連続的に重複する感染症の複雑な症候群を引き起こしうる。感染した犬は、軽い乾いた咳や鼻汁から肺炎や深刻な場合には死に至る範囲の臨床的な徴候を示す。猫もまた、多様な深刻度の呼吸器の苦痛を引き起こす様々な病原体に感染する。同一又は類似のウィルスはヒトにもまた感染しうる。このように多様な哺乳類に感染する病原体を特定することやこのような感染症のための診断、予防、及び/又は治療において使用するための組成物や方法を発展させることには、継続的かつ未充足のニーズが存在している。本発明はこれらのニーズを満たすものである。
【発明の概要】
【0004】
本発明は、新規なニューモウィルスの我々の発見に基づいている。このウィルスは、イヌ、ネコ及びおそらくヒトを含むがこれらに制限されない、多くの種類の哺乳類に感染しうる。ウィルスの存在は、イヌの急性の呼吸器疾患(ARDC)又はネコの呼吸器疾患、又は、ヒトその他の哺乳類の呼吸器その他の病気と正の相関関係がある。
【0005】
本発明は、ウィルスの単離されたポリヌクレオチド及びタンパク質、また単離されたウィルスそのものを提供する。本発明はウィルスを検出するための組成物及び方法、ウィルスの存在と正の相関関係のある病徴を予防及び/又は治療するための方法及び組成物を含む。またウィルスを含む単離された細胞を提供する。
【0006】
本発明のウィルスはマイナス鎖RNAウィルスである。本発明は、単離されたマイナス鎖、当該マイナス鎖と逆相相補性を有するプラス鎖、RNAポリヌクレオチドのDNA等価物、プラス鎖にコードされているタンパク質、及び、ポリヌクレオチド及びタンパク質の断片を含む。
【0007】
ある実施態様において、単離されたウィルスは配列番号1の配列、又は、配列番号1の配列と少なくとも96%が同一である配列を含むマイナス鎖ポリヌクレオチドを含む。また、配列番号2又は配列番号3の配列を含む単離されたポリヌクレオチド、又は、配列番号2又は3の配列と少なくとも96%が同一である配列を有するポリヌクレオチドが提供される。
【0008】
本発明は、哺乳類において免疫応答を刺激する方法を提供する。刺激された免疫応答は細胞仲介性、体液性、又はそれらの組み合わせでありえ、動物に対して予防及び/又は治療的な利益を提供しうる。免疫応答を刺激する方法は、ここに開示されるポリヌクレオチド及び/又はウィルス、ここに開示されるウィルス性ポリヌクレオチドによってコードされるウィルス性タンパク質、このようなタンパク質の断片、又は、それらの組み合わせを含む組成物を、哺乳類に投与することを含む。ポリヌクレオチド、ウィルス及び/又はタンパク質及び/又はそれらの断片は、いかなる適切なソースから単離されてもよく、また、それらは公知の技術を用いて組み換えによって生産されてもよい。
【0009】
本発明は、哺乳類から取得した生物学的サンプルにおけるウィルスの存在を検出するための方法を提供する。当該方法は、哺乳類から生物学的サンプルを取得すること、及び、当該生物学的サンプルから、ウィルス又はウィルス性のタンパク質又はそれらの断片によって含まれるポリヌクレオチド配列を検出すること、を含む。ポリヌクレオチド、タンパク質、タンパク質の断片、又はそれらの組み合わせの存在は、その哺乳類がウィルスに感染していることを示唆する。
【図面の簡単な説明】
【0010】
【図1】ヒトRSウィルス特異的なモノクローナル抗体(Mab)を用いたA72細胞の免疫蛍光アッセイ。A)感染細胞でのMab 2G122。B)非感染細胞でのMab 2G122。C)感染細胞でのMab 5H5N。D)非感染細胞でのMab 5H5N。一次Mabのストックは製造者から入手したものを1:100希釈して用いられた。赤いバックグラウンドは、エバンスブルーでの対比染色によるものである。
【図2】感染及び非感染のイヌA72細胞に対する、非呼吸器関連のテストのために提出された、ランダムに選択されたイヌ血清の反応性を示す免疫蛍光アッセイである。A)1:320希釈での感染細胞の血清。B)1:320希釈での非感染細胞の血清。赤いバックグラウンドは、エバンスブルーでの対比染色によるものである。
【図3】配列番号1の配列を含むウィルスゲノム組織の図示である。
【発明を実施するための形態】
【0011】
本発明は、新規な単離されたウィルス、単離されたポリヌクレオチド、及びウィルスのタンパク質、ウィルスを検出するための組成物及び方法、及び、哺乳類においてウィルスの存在と正の相関関係のある病徴の予防及び/又は治療のための組成物及び方法を提供する。
【0012】
本発明のウィルスは、ネガティブ(マイナス)鎖RNAウィルスである。つまり、ウィルス粒子に包含されている遺伝子材料は、ポジティブ(プラス)鎖の逆相補物であるRNAの単一鎖である。プラス鎖は、RNA依存性RNAポリメラーゼによってマイナス鎖から転写される。当該プラス鎖によってコードされているウィルス性タンパク質の、少なくとも一部はmRNAとしてのプラス鎖の機能は、感染細胞の細胞質で翻訳され、そして、新しいウィルス粒子の組み立て及びマイナス鎖ゲノムのパッケージングに関与する。
【0013】
特定の実施態様において、本発明は、配列番号1の配列又は配列番号1の配列と少なくとも96%が同一である配列を含むマイナス鎖ポリヌクレオチドを含有する単離されたウィルスを提供する。本発明はまた、当該マイナス鎖に相補的なポリヌクレオチドを含む。特定の実施態様において、これらのポリヌクレオチドは、配列番号4に示される配列又は少なくとも96%が配列番号4と同一である配列を含む。また、配列番号1の配列又は少なくとも96%が配列番号1の配列と同一である配列を含む単離されたポリヌクレオチド、又は、配列番号4の配列又は少なくとも96%が配列番号4の配列と同一である配列を含む単離されたポリヌクレオチドが提供される。また、配列番号2及び/又は配列番号3の配列を含むポリヌクレオチド、又は少なくとも96%が配列番号2,3の配列と同一である配列を有するポリヌクレオチドが提供される。これらの配列の逆相補物、及び、少なくとも96%が逆相補配列と同一性を有する配列もまた提供される。ここに開示されるすべてのポリヌクレオチド及びタンパク質及びそれらの断片の組み合わせもまた、本発明に含まれる。本発明によって提供されるウィルス性のポリヌクレオチドは、生ウィルス(弱毒化ウィルスも含む)において、複製をサポートする働きをすることによって特徴づけられる。
【0014】
また、ウィルスを含有する細胞が提供される。多くの実施態様において、ウィルスを含有する細胞は、単離された細胞、又は、ウィルス弱毒化のために及び/又はワクチンや他の目的のために使用するためのウィルス源として、インビトロで増殖させられる細胞である。このように、ウィルスを含む単離された細胞、ウィルスを含む細胞培養及び/又はセルライン、細胞培養培地に存在するウィルス及び/又はウィルス性生産物、及び、ウィルスを含む単離された組織は全て、本発明の局面である。
【0015】
特定の実施態様において、本発明で開示される各RNA配列のDNA等価物が提供される。つまり、本発明は、RNAの各々のU(ウラシル)がT(チミン)に置換されている、ここに開示される各RNA配列を含む。本発明で開示される各々のウィルス性ポリヌクレオチドは、各ポリヌクレオチドについて指定された配列番号で示される配列と同一であるポリヌクレオチドを含み、さらに、ウィルスの配列番号で示された配列と少なくとも96%が同一であるすべてのポリヌクレオチドをも含む。特定の実施態様では、配列リストで示されたポリヌクレオチド配列と少なくとも96%が同一であるポリヌクレオチドは、全長に渡る配列のうち少なくとも96%がそれらの配列と同一である。いかなる特定の理論に拘束されることを意図するものではないが、ここに示され、また、配列リストで説明されるウィルス性のポリヌクレオチド配列と少なくとも96%の同一性を有する配列を含むポリヌクレオチド配列は、例えばネズミ科の哺乳類に感染することが既に知られているウィルスのような、関連のウィルスによって含まれるポリヌクレオチドとは区別されるものと考えられている。また、これもいかなる特定の理論に拘束されることを意図するものではないが、例えば連続継代によるウィルスの弱毒化は、配列リストに示された配列と少なくとも96%が同一であるポリヌクレオチドを含み、及び/又は転写するウィルスを生じ得る。配列リストに特定して列挙された配列と同一でない配列は、示された配列と96.0%から99.9%同一でありえ、数値には示された数値の間の小数第一位のすべての整数値を含む。96.0%から100.0%の間の配列の同一性の全ての範囲が、包括的に、本発明の実施態様である。
【0016】
ポリヌクレオチドの断片もまた本発明に含まれる。つまり、様々な実施態様において、各々のウィルス性ポリヌクレオチド(その逆相補物及びDNA等価物を含む)は、10ヌクレオチドからポリヌクレオチド全長にわたる範囲、そしてその間のすべての整数値を含む範囲の、ポリヌクレオチド断片を含む。例えば、本発明は、配列番号1の配列を含むポリヌクレオチドを含み、また、長さで10ヌクレオチド以上、8598ヌクレオチドポリヌクレオチドまでの間(また、それらの間の全ての整数値を含む)の全ての配列の断片を含むポリヌクレオチドを含む。本発明で提供されるポリヌクレオチドは、ウィルス粒子に存在するか否かに関わらず、ここに開示されるものよりも長くてもよい。なぜなら、提示される配列は、ウィルスゲノム及び/又はそれらの逆相補物及び/又はDNA等価物の、全体を含むものではないからである。
【0017】
本発明はまた、哺乳類において免疫応答を刺激する方法をも提供する。刺激された免疫応答は、動物に予防的及び/又は治療的利益をもたらしうる免疫応答である。刺激された免疫応答は、体液性の及び/又は細胞仲介性の免疫応答でありえる。刺激された免疫応答は、ウィルスにコードされる、抗原及び/又は免疫原のいずれかに対するものでありえる。つまり、完全なウィルス粒子、各ウィルス性のタンパク質、及び、ウィルス性のタンパク質の断片は、哺乳類において免疫応答を刺激するために使用されうる本発明の実施態様である。多くの実施態様において、免疫応答が刺激される動物はイヌ(Canis familiaris)又はネコ(Felis catus)、すなわち各々家畜化された犬又は猫である。
【0018】
特定の実施態様において、配列番号1の配列又は配列番号1の配列と少なくとも96%が同一の配列、又はそこにコードされるタンパク質又はタンパク質断片、又は配列番号1の逆相補物によってコードされるウィルス粒子、を含むポリヌクレオチドは、イヌにおいて免疫応答を刺激するために用いられる。配列番号2及び/又は配列番号3の配列を含むポリヌクレオチド、又は、配列番号2又は3の配列と少なくとも96%が同一の配列を有するポリヌクレオチド、又は、配列番号2又は3の逆相補物によってコードされるタンパク質又はタンパク質断片又はウィルス性タンパク質、又は、配列番号2又は3の逆相補物と少なくとも96%が同一の配列を有するポリヌクレオチドは、ネコにおいて免疫応答を刺激するために好ましく使用される。イヌ及びネコへの、これらの配列、タンパク質、タンパク質断片、及びウィルス粒子の間の同様の関係が、診断目的のためのこれらの組成物の使用にも適用される。しかしこのことは、ヒトを含む他の哺乳類での本発明の実施についての適用性を、いかなる意味でも減縮するものではない。
【0019】
免疫応答を刺激する方法には、ここに開示されたウィルス、ここに開示されたウィルス性ポリヌクレオチドによってコードされるウィルス性タンパク質、このようなタンパク質の断片又はこれらの組み合わせを含む組成物を、哺乳類に投与することを含む。ウィルス及び/又はタンパク質及び/又はそれらの断片は、いかなる適切なソースから単離されてもよく、また、公知の技術を用いて組み換え的に生産されてもよい。多くの実施態様において、ワクチンにおいて使用されるための本発明によって提供されるウィルス性タンパク質は、NS−1、NS−2、N、P、M、SH、G、F、M2−1、M2−2、Lタンパク質及びそれらの組み合わせを含むが、これらに限定されるものではない。これらのタンパク質をコードする全てのヌクレオチド配列は本発明に含まれ、当業者によって通常の技術を用いて決定されうるものである。
【0020】
本発明の組成物が投与される哺乳類は、ここに開示されるウィルスを含むがそれに制限されない1又は複数のニューモウィルスに対してリスクのある、保菌が疑われる、又はこのウィルスの存在と正の相関関係のある条件で診断された、あらゆる哺乳類でありえる。ある実施態様において、この動物は、ARDCにリスクがあり、疑われ、又は診断されている犬である。他の実施態様において、この哺乳類は、ここに開示されるウィルスと正の関連がありうる呼吸器疾患その他の疾患が疑われ、或いは診断された、リスクのある人間である。この点において、マウス肺炎ウィルスのような、ここに開示される新規に発見されたウィルスと関係するウィルスは、人間にも広範囲の感染を引き起こすことが、当分野では公知である(例えば、Pringle CR, Eglin RP. Murine pneumonia virus: seroepidemiological
evidence of widespread human infection. J Gen Virol. 1986 Jun; 67 (Pt 6):
975-82を参照)。つまり、ここに提供される組成物及び方法が、人間において免疫応答を刺激するために、また、人間においてウィルス感染の診断に用いられるために、有用性を有すると考えることは合理的である。
【0021】
動物への本発明の組成物すなわちワクチンの投与は予防的効果を生じ得る。これはワクチンを受けていない動物よりもワクチンを受けた動物のほうが感染の可能性が低いこと、又は、動物においてウィルスの存在と正の相関関係のある病徴がワクチンを受けていない動物よりもワクチンを受けた動物において深刻度がより低いことを意味している。本発明のワクチンの投与はまた治療的効果を有しうる。これはワクチンを受けていない感染動物に対して、ワクチンを受けた後に感染した動物は感染の度合い(すなわちウィルス負荷又は力価、及び/又は当業者に公知の感染度マーカー)が低いことを意味している。同様に、ワクチンを受けていない動物に対してワクチンを受けた感染動物は、動物においてウィルスの存在と正の相関関係のある病徴の深刻度が低い。
【0022】
ここに開示されるウィルス及び/又はタンパク質及びポリヌクレオチドは単離されうる。これは例えば動物及び/又は動物から得られる生物学的サンプルからウィルスを分離することによって、ウィルスの自然の環境からウィルスが除かれることを意味している。一つの実施態様において、ここに開示されるウィルスを含む単離された細胞は単離されたウィルスを包含すると考えられる。さらに本発明の単離されたウィルス及び/又はタンパク質及び/又はそれらの断片及び/又は単離され及び/又は組み換えられたポリヌクレオチドは、如何なる所望の精製度で精製されてもよい。本発明で提供される組成物はウィルス、ウィルス性タンパク質、それらの断片、及び/又はポリヌクレオチド、ポリヌクレオチドの断片及び/又はそれらの組み合わせを含み、それらからなり、又は、実質的にそれらからなるものであってよい。
【0023】
一つの実施態様において方法は、イヌにおいてウィルスに対する免疫応答が刺激されるように、配列番号1の配列を含むポリヌクレオチド又は配列番号1の配列と少なくとも96%が同一の配列を有するポリヌクレオチドを含む組成物をイヌに投与することを含む。一つの実施態様において、ポリヌクレオチドはDNAワクチンとして投与される。DNAワクチンを含む薬剤組成物を成形すること、及び、DNAワクチンを含む組成物を投与することは当分野で公知である。一つの実施態様において、イヌに投与されるポリヌクレオチドはウィルス粒子中に存在する。ウィルス粒子は単離及び/又は組み換えされうる。多くの実施態様において、イヌに投与されるウィルスはウィルス粒子によって含まれるタンパク質の一部であるアミノ酸配列を含むタンパク質を含んでいる。ウィルスゲノムによってコードされるウィルス粒子によって含まれるウィルスタンパク質の予測されるアミノ酸配列は、イヌウィルス性タンパク質NS1、非構造タンパク質1(配列番号7)、NS2、非構造タンパク質2(配列番号8)、N、核タンパク質(配列番号9)、P、リン酸タンパク質(配列番号10)、M、マトリクスタンパク質(配列番号11)、SH、小疎水性タンパク質(配列番号12)、G、アタッチメントタンパク質(配列番号13)、F、フュージョンタンパク質(配列番号14)、M2、マトリクスタンパク質M2−1(配列番号15)、及びM2−2(配列番号16)、L、一部配列が提供されている(配列番号17)RNA依存性RNAポリメラーゼである。これらのタンパク質のネコウィルス型も本発明によって提供される。
【0024】
他の実施態様においてこの方法は、ネコにおいてこのウィルスに対する免疫応答が刺激されるように配列番号2及び/又は配列番号3の配列を含むポリヌクレオチド、又は、配列番号2又は3の配列と少なくとも96%が同一の配列を有するポリヌクレオチドを含む組成物でウィルスをネコに投与することを含む。一つの実施態様において、ネコに投与されるポリヌクレオチドはウィルス粒子の中に存在する。粒子は単離され及び/又は組み換えられ得る。
【0025】
多くの実施態様において、ネコに投与されるウィルスは配列番号5又は配列番号6から選択されるアミノ酸配列を含むタンパク質を含む。
【0026】
ある実施態様において本発明は、ここに開示されるウィルスによる感染に対する予防的及び/又は治療的効果を提供しうるワクチンを含む。このワクチンは生又は死滅(不活性化)ウィルス又はその断片を含みうる。死滅ウィルスとは複製が不可能であるウィルスである。死滅ウィルスは当業者には周知の多様な技術のいずれを用いて調製されてもよく、熱による不活性化及びウィルス性タンパク質及び/又は核酸の修飾を含みまたこれらに制限されず、その方法はパラホルムアルデヒド、ホルマリン及び紫外光線への暴露を含みまたこれらに制限されない。例えば、ウィルス性タンパク質は例えばウィルス性の細胞侵入及び/又は複製に必要とされる1又は複数のステップを回避するようにウィルス性タンパク質を修飾できうるクロスリンク剤によって共有結合的に修飾されうる。生ウィルスは弱毒化ウィルスを含むと考えられている。弱毒化ウィルスとは弱毒化されていないものに対して毒性が低減されているウィルスである。つまり弱毒化ウィルスは複製可能である、しかしながら、同種の弱毒化されていないものと比して速度は遅く、つまりそれゆえに一般的に疾患を引き起こさない。当業者は、弱毒化されていないもの、弱毒化されたもの、及び、死滅ウィルスの間の他の区別点についても認識しそれらは互いに区別され得るであろう。
【0027】
ウィルスの弱毒化は何らかの標準的な技術を用いて得られうる。一つの実施態様ではウィルスは哺乳類細胞培養における連続継代によって弱毒化される。連続継代は多様な機構を通じて弱毒化を生じ得る。人間の細胞を含みこれに制限されない、ウィルスの複製を支える何らかの細胞はウィルスの増殖や弱毒化の目的のために継代において使用され得る。様々な非制限の例示において継代のための細胞培養は其々Canis familiaris 又はFelis catus のようなイヌ又はネコから得られ又はそれに由来しうる。一つの実施態様において細胞は初代細胞培養である。ある実施態様において、細胞はATCCカタログ番号CRL−1542の番号を有するA−72細胞株由来のイヌ細胞に含まれうる。ウィルスの弱毒化に好適ないかなる継代数も使用することができる。当業者は本開示の有益性を持ったウィルスの弱毒化のための継代パラメータを決定することができるだろう。一般的に5−150(その間の全ての整数値を含む)継代、好ましくは20−50の間の継代が使用され得る。
【0028】
ウィルスを含む組成物はワクチンとして使用するための医薬製剤として提供されうる。ウィルスを含む組成物は懸濁液として又は凍結乾燥形態で提供されることができ、またこのような組成物に通常使用される医薬的に許容されるキャリア及び/又は希釈剤を含有しうる。本発明での使用のために好適なキャリア及び希釈剤のいくつかの例が、The Science and Practice of Pharmacy (2005) 21st edition,
Philadelphia, PA. Lippincott Williams & Wilkins に見出される。特定かつ制限的ではないキャリアの例は安定化剤、防腐剤及びバッファを含む。特定かつ制限的ではない安定化剤の例は、炭水化物(例えばソルビトール、マンニトール、スターチ、スクロース、デキストラン、グルタミン酸、又はグルコースなど)、タンパク質(例えば、乾燥乳清、アルブミン又はカゼインなど)又はそれらの分解生成物を含む。好適な、特定かつ制限的ではないバッファの例はアルカリ金属リン酸塩を含む。好適な、特定かつ制限的ではない希釈剤の例は水、水系バッファ(例えば緩衝生理食塩水)、アルコール及びポリオール(例えばグリセロール)を含む。
【0029】
当業者はここに提供されるワクチン製剤はアジュバント活性を有する1又は複数の組成物を含みうることを認識するであろう。好適なアジュバントはフロイント(Freund)の不完全アジュバント、トレオニルムラミルジペプチド、水酸化アルミニウム、リン酸塩又は酸化物、例えば、鉱物油又は、例えばビタミンE酢酸塩のようなベジタブルオイル及びサポニンに基づく水中油型又は油中水型乳化液を含みまたこれらに制限されない。
【0030】
本発明によって提供されるワクチンはあらゆる好適なルートを通じて投与されうる。多様な実施態様においてワクチンは非経口で、筋肉注射又は皮下に、静脈内投与で、或いは、経鼻を含むがこれに制限されない経口で投与されうる。ワクチンの効果的な投与量は例えば、免疫応答の刺激が所望される動物のタイプ、サイズ、年齢、環境、感染ステージのリスクのような通常の検討を用いて決定され得ることを当業者は認識するであろう。一般的に本発明による生ワクチンは、動物当たり、例えば1mL用量あたり101−106pfu(プラーク形成ユニット)の用量で投与されうる。不活性化されたワクチンは動物当たり106−1010pfuの抗原性等価物を含みうる。
【0031】
本発明のポリヌクレオチドは例えばここに開示されるウィルスだけでなく、他の関連する又は非関連のウィルス及び/又は他の感染性病原体に対して免疫化を提供しうる異種遺伝子(1又は複数)を包含するように修飾されてもよい。同様にここで提供されるワクチン組成物は、ここに開示されるウィルスから区別される感染性病原体に対して免疫応答を刺激することを目的とされる免疫原性の薬剤を含むことができる。つまり本発明は種々の哺乳類において、また、種々の感染性病原体に対して使用するための混合ワクチンを予測するものである。混合ワクチンはここに開示されるウィルス性タンパク質の全部又は一部を含み、また、異なる感染性病原体に対して効果を有する多価ワクチンを提供するために他のタンパク質からのポリペプチド配列を含みうる融合タンパク質を含みうる。本発明はさらに、動物に有益な効果を与え得る他の組成物であってこのような感染のための従来的な治療に先立って、同時に、又は続いて投与される組成物を投与することを含む。
【0032】
本発明はまたここに開示されるウィルスを検出するための方法も提供する。この方法は動物から生物学的サンプルを得ること、及び、サンプル中に又はサンプルから、ウィルス性タンパク質及び/又はウィルス性ポリヌクレオチドの存在を検出することを含む。ウィルス性ポリヌクレオチド及びタンパク質のいずれかがウィルスの存在又は不存在を検出する方法において使用されうる。配列番号1、2又は3の逆相補物又はDNA等価物を検出することは配列番号1、2、又は3の配列、又はこれらの配列と少なくとも96%が同一の配列を含むポリヌクレオチドを検出することと同等であると考えられている。
【0033】
検出は、診断が望まれる哺乳類から得られたいずれの好適な生物学的サンプルを使用して実行されてもよい。生物学的サンプルの好適なソースは、血液、粘膜剥離物、組織生検、又は唾液を含みこれらに制限されない。一つの実施態様においては生物学的サンプルは鼻及び/又は咽頭スワブ、又は気管洗浄液を含む。
【0034】
ウィルスの存在又は不存在は当分野では周知の様々な技術を用いて、DNA、RNA又はタンパク質を検査することによって検出されうる。ある実施態様ではウィルスの存在はウィルスゲノムの全体又は一部を含む、又は、ウィルスゲノムの全部又は一部から増幅されるポリヌクレオチドを検出することによって決定される。この点で本発明はウィルスのマイナス鎖、ウィルスのプラス鎖、又はそれらの組み合わせからのポリヌクレオチド配列を同定することによって、ウィルスの存在(又は不存在)を決定することを含む。本発明はまたウィルスのマイナス鎖、プラス鎖、及びそれらの組み合わせのDNA型を検出することも含む。例えば一つの実施態様において逆転写PCR(ポリメラーゼ連鎖反応)がウィルスのマイナス鎖又はその一部のDNA複製物を生成するために用いられる。またDNA複製物はハイブリダイズされた二本鎖において、プラス及びマイナス鎖又はそれらの一部分のDNA型を含む複数の二本鎖DNA分子を得るためのウィルスゲノムの増幅のためのテンプレートとして用いられうる。当業者は興味の対象である生物学的サンプル中にウィルスがいるか又はいたかどうかの決定において使用するための遺伝子材料の量の増幅のために使用される多様な増幅技術があること、また、この増幅された(又は増幅されない)遺伝子材料はポリヌクレオチド配列を決定するための多くの周知の方法及び試薬を用いて分析されうることを認識するであろう。すなわち本発明は、ウィルス性ポリヌクレオチドの存在又は不存在が確認されうるように生物学的サンプルから核酸を獲得し、また分析するための何らかの及びあらゆる方法を含む。この方法は何らかの公知の技術を用いて、核酸及び/又は他のタイプのプローブをハイブリダイズすることによって、及び、ウィルス性ポリヌクレオチドの配列を決定することによってポリヌクレオチドを検出するための方法を含みこれらに制限されない。配列の全部又は一部分がウィルス性のポリヌクレオチドを同定するために決定され、使用され得る。
【0035】
ある実施態様において本発明は単離されたウィルス性ポリヌクレオチドを含む組成物、及び、核酸ハイブリダイゼーション及び/又は増幅のために使用される要素を提供する。また組成物はDNAポリメラーゼ、逆転写酵素、遊離核酸三リン酸塩、塩、バッファ、及び、核酸をハイブリダイズ及び/又は増幅するのに典型的に用いられる他の試薬を付加的に含むことができる。一つの実施態様において本発明は、単離されたウィルス性ポリヌクレオチド及び/又はウィルス性ポリヌクレオチドから増幅されたポリヌクレオチドであって、rt(real time)核酸増幅技術において用いられる1又は複数の増幅プライマー及び検出プローブにハイブリダイズされたものである単離されたウィルス性ポリヌクレオチド及び/又はウィルス性ポリヌクレオチドから増幅されたポリヌクレオチドを提供する。
【0036】
本発明はまた、単離されたウィルス性ポリヌクレオチド、及び/又は、ポリヌクレオチドがアレイの中に存在する少なくとも一つのプローブに増幅されたものである、ウィルス性ポリヌクレオチドから増幅されたポリヌクレオチドを提供する。このアレイは、例えば複数の特定のポリヌクレオチドの存在又は不存在を決定するために使用されるチップ上に存在しうる。このようなチップは商業的に提供されるものであり、また本質的にあらゆるポリヌクレオチドの存在又は不存在を検出するためにカスタマイズされうる。
【0037】
多様な実施形態において本発明はさらに、哺乳類がここに開示されるウィルスに感染しているか否かの決定を有形的な媒体に固定することを含む。有形的な媒体はどのような有形的な媒体でもよく、例えば、コンピュータ、DVD、CD−ROM又は電子メールメッセージに保存されるデジタルファイルを含みこれらに制限されないデジタル媒体のあらゆるものでありえる。有形的な媒体は感染哺乳類のための治療プロトコルを発展させるようにヘルスケア提供者に提供されうる。
【0038】
当業者には本発明の利益を受けて多様な反応試薬が作られ、ここに開示されるウィルスの検出のための診断方法に使用されうることが認識されるであろう。例えばポリクローナル又はモノクローナル抗体が、ウィルス粒子又はウィルス粒子の一部、又は単離された又は組み換えのウィルス性タンパク質又はその断片を用いて作られ得る。すなわち抗体は、あらゆるウィルス性タンパク質に存在するあらゆる抗原決定要因を特異的に認識するように作られ得る。多様な実施態様において抗体は、NS−1,NS−2,N、P,M、SH、G、F、M2−1、M2−2及びLウィルス性タンパク質、又は、その一部又は組み合わせを認識する。抗体は本発明のウィルスを、MPVのような他の関連するウィルスから識別できるように、作られ得る。
【0039】
抗体は、生物学的サンプルからウィルス性抗原の存在又は不存在が決定されうるようなあらゆる技術において用いられ得る。このような技術はウェスタンブロッティング、免疫蛍光法、免疫組織化学及びビーズ法又は従来のタンパク質アレイのELISAアッセイを含みこれらに制限されない。
【0040】
一つの実施態様において本発明は、抗体との複合体に存在する単離された及び/又は組み換えウィルス又はウィルス性タンパク質を提供する。
続く実施例は本発明の特定の実施形態を描写することを意図しているが、いかなる意味でも本発明を制限するものと解釈されるべきではない。
【実施例】
【0041】
[実施例1]
我々は、雑種犬の鼻及び咽頭スワブからサンプルを採取し、呼吸器ウィルスを単離するために、イヌA72細胞(アメリカ培養細胞系統保存機関(ATCC)、CRL−1542)にそれらを植菌するために用いた。植菌後約21日に対応する、培養の経過の後期段階において、いくつかの培養が僅かに細胞変性の変化を示した。連続する経過の後、培養は培養全体の急速な細胞死に続いて、円形細胞の小巣を現した。このパターンは一般的なARDCに伴うウィルスには非特徴的であると主観的に判断された。13の別個のウィルス単離物が得られ、この実施例において説明される。一般的なイヌ呼吸器病原体に特異的な診断反応試薬のパネルを用いたテストは、公知のウィルスを同定することができなかった。他のウィルスのための追加的な反応試薬でのさらなるテストは、最終的に、ヒト呼吸器合胞体ウィルス(抗HRSV、番号VP−R151、Vector Laboratories、USA、カリフォルニア州バーリンガム)に対するモノクローナル抗体(Mab)プールを用いた免疫蛍光アッセイ(IFA)によって陽性認識を示した。この抗体の調製は、ウシRSV(BRSV)の検出のために我々のラボで一般的に使用されているものである。染色パターンは、RSV感染細胞において観察される典型的なパターンである線維状の膜結合及び遊離のウィルス粒子及び細胞質内封入体を含んでいた。
【0042】
最初のIFA結果に続いて、我々は、ヒト、ウシ、ヒツジのRSV配列のアライメントに基づいてデザインされたPCRプライマーを用いて、ウィルスからヌクレオカプシド遺伝子(N)の断片を増幅することを試みた。これは不成功であった。抗RSVプールの個々のMabのストック及びそれらの特異性は、製造者から入手され、IFA(図1)のために使用された。Mab5H5N(M2タンパク質特異的)での染色は、ウィルス粒子及び封入体の両方を染色した。Mab2G122(Pタンパク質特異的)は、一次的に封入体を染色し、相対的に均一な膜仲介性のシグナルを与えた。Mab1C3(Nタンパク質特異的)又は5A6(Fタンパク質特異的)では染色は得られなかった。4種の個々のMabの全てが、IFAによってBRSVを認識した。4種のMabのうちの2種のみによるイヌウィルスの認識と、RSVの保存領域の増幅が不可能であったことは、RSVに関連性があるが典型的な形態でないことを示唆した。
【0043】
ウィルス配列の解明は、CODEHOPアルゴリズムを用いて、ニューモウィルス亜科の全てのウィルスの複数の配列アライメントのうち高度に保存されたアミノ酸(aa)配列に基づいた変性PCRプライマーをデザインすることによって追跡された。L(ポリメラーゼ)及びN遺伝子の特定領域が目標とされた。反応生成物の配列決定とBLAST分析は、ウィルスが伝統的にマウスのニューモウィルス(又は肺炎ウィルス)として知られているマウスニューモウィルス(MPV)に近いことを明らかにした。2つのL遺伝子PCR産物は、MPVと95%から97%同一であることが見いだされ、N遺伝子断片は約96%が同一であった。
【0044】
新規に同定されたニューモウィルスが、テストされたシェルターのイヌの群に限られるのかどうかという疑問に対処するために、我々はランダムなイヌ血清サンプルをスクリーニングし、27のうち6の動物が感染された培養のウィルスが特異的に認識される抗体を有していたことを見出した。染色パターンは、Mab(図2)で観察されるものと類似であった。BRSVに対する中和抗体を種々の量で含有するウシ血清は、イヌウィルスに感染している細胞を染色しなかった。
【0045】
この例は、ARDCの13頭のイヌから単離されたウィルスはMPVに非常に近い関係にあるとみられることを示し、ゆえに我々は、それらはイヌニューモウィルス(CnPnV)であると考える。マウスニューモウィルスは、パラミクソウィルス科のニューモウィルス亜科の中でニューモウィルス属として分類されている3種のうちの1種である。ヒトRSVはタイプ種であり、BRSVと非常に近縁関係であるがMPVはより遠い関係である。例えば、ヒト及びウシRSVのNタンパク質ヌクレオチド配列は約94%が互いに同一であるが、MPVに対しては60%の同一性を有するにとどまる。MPVの完全に配列決定された株は2種のみで、‘Strain 15’及びJ3666であるが、それらはヌクレオチドのレベルでは99.7%の同一性を有する。
【0046】
ARDCでのCnPnVの連関は、いくらかの場合、特に複合的な病因論に関与する時には病原性でありえる。類推では、MPVは一般に齧歯類の飼育株に感染することが知られ、また、血清学的な証拠は幾らかの野生型齧歯類での感染を指摘するが、自然的な生態学については多くは知られていない。実際、齧歯類がMPVの唯一の自然的なホストであること、又は、近縁関係のウィルスが他種に拡散するのかどうかは、明確ではない。呼吸器症状を伴うヒトの感染の証拠が報告されている(Pringle CRら、J.Gen.Virol. 1986;67:975-82)。自然界では齧歯類の感染は無症状で潜在的であるかもしれない。飼育マウスでの臨床症状は無症状から高い罹患率及び死亡率を伴う肺浮腫への発展まで幅広い。J3666とStrain15の両方を含む病原株は、深刻な肺炎を引き起こし、少量で植菌された時には6−10日で死に至りうる。病原性、またはその欠如はウィルス及びマウス株依存性であるのだろう。
【0047】
複数の微生物とウィルス病原体がイヌの呼吸器疾患に関与しうる。すなわちCnPnVは、より深刻な疾患を導く上部呼吸器胴の防御機構を損傷する一連の事象を開始する能力のある他の病原体であると考えられる。
【0048】
[実施例2]
この実施例の材料及び方法は、残りの実施例で説明される結果を得るために使用された。
【0049】
[ウィルスの単離]
鼻及び咽頭検査サンプルは、湿潤スワブを用いて呼吸器疾患の症状のある雑種犬から集められ、コーネル大学動物健康診断センターにおいて受領から24時間以内に処置された。植菌の前に、スワブはアール塩(Earle’s salt)(Gibco 10370, Invitrogen,
Carlsbad, CA, USA)、0.5%ウシ血清アルブミン、ペニシリン(200 U/ml)、ストレプトマイシン(200 μg/ml)、及びファンギゾン(2.5 μg/ml)を含む最少基礎培地(MEM培地)3mLに30分間浸漬し、続いて10秒間機械的に撹拌した。それぞれ0.5mLの鼻及び咽頭抽出物の分割単位は集められ、セミコンフルエントのA72細胞(Binnら、1980)(CRL-1542,
American Type Culture Collection, Manassas, VA, USA)のシングルT25フラスコに、培地を追加することなく植菌するのに用いられた。抽出物は、1−3時間モノレイヤーの上に残置され、その後リン酸緩衝生理食塩水(PBS)でリンスオフされた。細胞は6mLの培養培地(ライボビッツのL−15培地、10% 熱非活性化ウシ胎児血清(FBS)、200 U/ml ペニシリン、200 μg/ml ストレプトマイシン、50 μg/mlゲンタマイシン)にて37℃で維持され、6−8日毎に継代培養された。植菌されていないコントロールのA−72培養が単離プロセスを通じて並行して行われた。
【0050】
[免疫蛍光アッセイ]
細胞はガラススライドに接着させられ、その後PBSでリンスされ、冷アセトン10mLで固定化された。スライドは空気乾燥され、染色の前に−20℃で貯蔵された。PBSで希釈された一次抗体及び二次抗体がスライドに滴下され、37℃で30分間インキュベートされた。各インキュベーションの後、スライドはPBS中で15分間リンスされた。染色は蛍光顕微鏡で可視化された。HRSVに対するMabのプールが希釈率1:400にて当初の同定において使用された(VP−R151、ベクターラボラトリー、バーリンガム、カリフォルニア州、USA)。1倍濃度で供給されたマウス抗−PVM(CL-603IFA, Charles River Laboratories, ウィルミントン、マサチューセッツ州、USA)が、希釈率1:20にて使用された。二次抗体FITC標識ヤギ抗マウスIgG(02−18−06、KPL、ガイザースバーグ、メリーランド州、USA)は、希釈率1:40(12.5 μg/mL)で使用された。
【0051】
[RNA精製]
全細胞性RNAがA72細胞から精製された(74106、Qiagen、バレンシア、カリフォルニア州、USA)。約50−75%の細胞変性効果(CPE)を示す一つの25cm2フラスコからの細胞が、ペレットにされ、各カラムを通して精製された。細胞フリーの上清及びスワブ溶出物を含む培地はサンプル140μLを用いて精製された(52906、Qiagen)。精製されたRNAsはRNaseフリーの水40μLで再懸濁された。
【0052】
[PCR]
ゲノム断片を得るために当初使用された縮重プライマー配列は、ニューモウィルス科ウィルスのアライメントに基づき、CODEHOPアルゴリズム(blocks.fhcrc.org/codehop.html)(Roseら、1998)を用いて設計された。逆転写PCR(RT−PCR)は製造者の指示に従い25μLボリュームで1段階反応(210212、Qiagen)で行われた。1:5〜1:10で希釈された全RNAの1μL及び各プライマー10pmolが各反応で使用された。RTステップの反応条件は、50℃/30分、続いて95℃/15分であった。サイクル条件は、95℃/60秒、54−56℃/30秒及び72℃/60−90秒を35サイクル行った。プライマーは次の通りである:N遺伝子:N276F,tccgtgcaggccgaratggarcarg/P1R,(配列番号35)ggaactcgggggcgaayttytccat;配列番号36);L遺伝子 no 1:L428F, ccggatcttcggccayccnatggt/(配列番号37)L538R, ttcttaggaggggagatggcyttrtcrtt;(配列番号38)L遺伝子 no. 2:L698F,
catcaccgacctgtccaagttyaaycargc/(配列番号39)L894R,
ttgaagtcgtccaggatggtrttdatcca配列番号40。様々な地理的位置からのスワブサンプルと用いられた2つのPCRプライマーセットは、CnPnV、MPV J3666及びstrain15の配列のアライメントに基づいて設計された。Gタンパク質:G715F, ggcttctgtttcttcctttctgg/(配列番号41)G1062R,
ccgtggtggtgcctgtg(配列番号42);SH1タンパク質:SH1F,atggatcctaacatgacctcayac(配列番号43)/SH187R,
gattgggatgaacygtgcattg(配列番号44)。低継代Brne単離物を増幅するのに使用されたプライマー:G84F,tgtaaaagtgaaccaaatgtgta(配列番号45)/G404R,aaatcttcaggtaaatcaggtc(配列番号46);G849F,ttttaacaacaagaatcagtc(配列番号47)/G1048R,
ctcctaggtgcgggggttgg(配列番号48)。G及びSH単位複製配列のための反応条件は、RTが50℃/30分、不活性化/変性が95℃/15分、及び、PCRは95℃/60秒、54℃/30秒及び72℃/90秒を35サイクルであった。
【0053】
[ゲノム分析]
PCR生成物は、Applied Biosystems Automated 3730 DNA Analyzerを用いてコーネル大学Sequencing and Genotyping Core Laboratoryにて配列決定された。完全配列を決定するのに使用されたプライマーはMPV配列又は先に決定されたCnPnV配列に基づくものであった。重複するPCR生成物はギャップとプライマー結合領域をカバーするように作り出された。すべての領域は両鎖分析された。CnPnV単離物は、Lasergene(DNASTAR,マディソン, ウィスコンシン州, USA)を用いて、MPV Strain15(GenBank AY729016)及びJ3666(GenBank NC006579)配列と比較された。
【0054】
[実施例3]
[単離と同定]
鼻及び咽頭スワブ溶出液が、A72細胞に植菌するように用いられた。相対的に長期間の培養の後、通常は3継代又は4継代の後に出現する、限定的なCPEが観察された。初期のCPEは典型的に円形細胞の分散性の小巣を含み、いくらかは小合胞体や空胞化を伴っていた。このCPEは、静置培養において数日間以上ゆっくりと進行した。継代培養はしばしばこの低レベルCPEが維持されるモノレイヤーを導いたが、いくらかの培養では24−48時間以内に迅速な進行及びモノレイヤーの崩壊が生じた。このパターンはいくつかのヘルペスウィルス感染と同様であるが、イヌヘルペスウィルスには試験陽性ではなく、同様の方法を用いて一般に単離されている犬呼吸器ウィルスのいずれでも典型的なものではないようであった。新鮮な細胞への上清の移送に続くCPEは僅かであるか又は生じず、初期継代の単離物の幾らかは培養における複製が少なかった。しかしながら継代の連続につれていくつかの培養は、より安定的な移送可能CPEを発展させ、それらはさらなる分析のために選択された。初期のテストでは、いくつかの無関連のイヌからの血清はCPEの培養においてウィルス性抗原を認識するように見え(データは不図示)たが、しかし、一般的なイヌ呼吸器ウィルスに対するテストはネガティブであった。実施例1で報告されたように、HRSV特異的なMabはウィルス性抗原を強く認識した。抗PMVポリクローナル血清でのポジティブIFA染色が確認のために使用された。抗PMVマウス血清はIFAによってBRSVを弱く認識するのみであった。
【0055】
実施例1で説明されたCnPnVの元の13の単離物は、同じ組織によって管理されている2つの動物シェルターに由来するものであった。ウィルスがこれらの2施設に制限されるのかどうか調査するために、他の地理的位置からの病気のからの鼻又は咽頭スワブ試料がPCRによって試験された。USAの8つの州(コロラド、ジョージア、フロリダ、インディアナ、ミズーリ、ネバダ、サウスカロライナ、バージニア)からの19頭のイヌが、G及びSH遺伝子のプライマーセットを用いてテストされた。19サンプルのうち、ネバダからの1例、サウスカロライナからの9頭のグループのうち5例がG及びSHの両方のPCRに陽性を示し、配列決定により確認された。他の13頭からはPCR陽性の結果は得られなかった。A72細胞でのウィルス単離が、イヌ呼吸器診断のためのニューヨーク及びペンシルヴァニアからの鼻スワブの6サンプルについてさらに行われた。これらのうち、2つのさらなる単離物はニューヨーク市の獣医学病院からのものであり、他は、ペンシルヴァニア州フィラデルフィアのシェルター由来のものであった。これらの単離物は、抗RSV及び抗PVM抗体でIFAによって、続いてRT−PCRで、CnPnVと確認された。これらのRT−PCRとウィルス単離結果は正確な有病率の評価を提供するものではないが、それらは、CnPnV感染が広汎であることを示すものではなかった。
【0056】
[配列分析]
以前の研究は自然的な準種の存在、培養における継代中の変異やPCR産物に起因しうるウィルス性単離物の間の差異を指摘していた。CnPnV単離物の最も正確なコンセンサス配列を決定するために、テンプレートとして全細胞性RNAを用いる複数の重複するRT−PCR反応が使われた。2頭の犬からのウィルス性単離物が配列決定されたが、一つは4継代の組織培養(Ane4)、一つは17継代(Brne17)であり、J3666(Thorpe及びEaston、2005)及びStrain15(Kremplら、2005)の配列と比較された。NS−1に近接する3´リーダーの中から開始し、Lコード領域中へと小距離延在し、ゲノム中の予測された10の遺伝子のうち9をカバーしている、8598ヌクレオチドの配列(配列番号1)がCnPnV-Ane4から得られた。遺伝子の順番、NS1−NS2−N−P−M−SH−G−F−M2−Lが確認された。MPV株と比較した、CnPnV-Ane4のこの領域の全体的なヌクレオチド配列の同一性は、J3666に対して95.0%でありStrain15に対して94.7%であった。MPV株は互いに99.7%同一であった。CnPnV-Ane4とMPV株を比較したときにコード領域中のギャップは見いだされなかった。セクション3.4及び3.5で説明したようにGとSHの長さの違いを除いて、MPVとCnPnV-Ane4のコード領域の全てが同じ長さのタンパク質をコードしていることが見いだされた。
【0057】
[非翻訳領域の比較分析]
CnPnVのGS及びGE配列の境界が、J3666及びStrain15の配列のアライメントによって決定された(Thorpe, L.C.,
Easton, A.J., 2005. J. Gen. Virol. 86, 159-169、 Kremplら 2005. Virus Genes 30, 237-249)。MPVのGS配列は、Chambersら(1991)によって説明され、より最近では (Dibben及びEaston, 2007) 、Collinら(1986)によって決定されたRSV GS配列に基づく長さ9ヌクレオチドのものとして説明されている。Kuoら(1997)は、10番目のヌクレオチドもまたRSVのGSにおいて重要であり、これは長さ10ヌクレオチドであるMPV GS(Kremplら、2005)と同等であることの証拠を提供している。表1において、GSは、両方の解釈を含む10ヌクレオチドとして示されている。表1は、MPVstrain15、J3666と比較したCnPnV-Ane4のコード領域間のGS、GE及びIGRsを提供する。
【0058】
【表1】
【0059】
表1では2つのMPV株の間でNTR長の違いはない。RNA配列の向きは5’から3’方向である。*:J3666及びstrain15から少なくとも1ヌクレオチドの変化がある。ヌクレオチドの差異は太字・下線で示されている。GE:遺伝子末端;IGR:遺伝子間領域;GS:遺伝子開始、無印;長さの差異がない。aM−SHについてはNTR全長において差異がない;一つのuがIGRから消失し、他のuがNTRの上流に追加されている。b3ヌクレオチドIGR及びM2もGSはDibbon及びEastonによる分析に基づいている(Dibbon,O., Easton, A.J.,
2007. Mutational Analysis of the Gene Start Sequences of Pneumonia Virus of
Mice. Virus Res. 130, 303-309)。表1に示された10又はそれ以上のヌクレオチドを有する配列は次の配列番号を有している:“GE”カラムの配列:NS1
uaguuaauuaaaa(配列番号18);NS2 uaguuauagaaaaa(配列番号19);N uauuuaauuaaaaa(配列番号20);P(配列番号18);M
uaguuaaauaaaa (配列番号21);SH uaguuaacaaaaaa (配列番号22);
G uaguuaaugaaaaa (配列番号23); F
uaguuaauuaaaaaa (配列番号33);及びM2 uaguuauauaaaaa (配列番号34)。“IGR”カラムの配列:N
cuggaaaauau (配列番号24); 及び G uaagcuaugauauaau
(配列番号25)。“GS”カラムの配列;NS1 aggacaagug (配列番号26); NS2 aggacaaauc (配列番号27);
N aggauaaaua (配列番号28);P (配列番号28);M
aggacaaaua (配列番号29);SH aggauaagua (配列番号30);
G (配列番号30); M2aggauaagug (配列番号31);F (配列番号29)及びL aggaucaaua(配列番号32)。
【0060】
わずかな長さのバリエーション及びヌクレオチドの相違が、CnPnV-Ane4 NTRで同定され、大部分はIGRの中で見いだされた。IGRにおける2つのMPV株の間には差異がみられなかった。Chambersら(Chambers, P., Matthews, D.A.,
Pringle, C.R., Easton, A.J., 1991. Virus Res. 18, 263-270)はcDNAクローンからのM2のGSを決定したが、56ヌクレオチドのIGR中に2つの付加的な潜在的なGS配列(GS1、GS2)の存在も指摘している。変異分析は後に、6番位置のAの代わりであるGによって推定されるGS配列は非機能的であることを決定した。続いて彼らは、GS1及びGS2の両方の配列が転写開始を指示する能力を有し、第一の配列(GS1)が最も重要であることを示した。彼らは、元々同定されたGS配列はcDNAクローニングの間の自然的な削除によって誤って同定されたものだと結論づけた。CnPnVにおいて、FのためのGE配列のすぐ後ろに続くM2のためのGS1配列は、10番目のヌクレオチドを除いてSH及びG遺伝子の開始配列と同じである。GS1は一次的に使用されるものと考えられており、表1に示されている。M2のためのGS2AGGACAGGGは(Dibben, O., Easton, A.J., 2007.
Virus Res. 130, 303-309)で報告されたコンセンサス配列から2箇所のみが異なるものである、しかしながら、7番目に保存されたAを有していないので活性は低いかもしれない。これは、Aが保存されているJ3666及びStrain15からは相違点である。CnPnV-Ane4での第三の潜在的なM2 GS(GS3)は、J3666及びStrain15と同一であり、それゆえ6番目のGによって貧弱な機能であることが予測されるであろう。
【0061】
[非構造タンパク質の分析]
非構造の小疎水性タンパク質は92アミノ酸を共有し、92.4%のアミノ酸同一性のみを有する2つのMPV株の間で最も不一致のタンパク質であるように、大幅な変動性を許容するようである。CnPnV-Ane4において、SHタンパク質はMPVstrain15と比較した時にすべての試験したタンパク質のうちでもっとも不一致であった。CnPnV-Ane4 SHは、J3666と比較して4アミノ酸の相違(95.7%)、strain15と比較して9アミノ酸の相違(90.2%)を有し、すなわちMPV株の相互の相同性よりもJ3666により類似している(表2)。他の特徴的な相違は、異なる単離物におけるSH ORFの長さの相違である。CnPnV-Ane4及びstrain15の両方が279ヌクレオチド(92アミノ酸)のSH ORFを有しているが、公開されたJ3666配列は345ヌクレオチド(114アミノ酸)である。しかしながら、J3666の独立に増幅された配列は、92、96又は114アミノ酸というバリエーションをもってコードされており、これらは抗体回避変異体を表していると示唆されている。表2は、CnPnV-Ane4遺伝子及びタンパク質のニューモウィルス亜科におけるウィルスに対する同一性(%)のまとめを提供している。
【0062】
【表2】
【0063】
表2ではGenBank配列に言及している:HRSV、NC_001781;BRSV、NC_001989;HMPV、NC_004148.2;AMPV、NC_007652。これらは、ヒトRSV、ウシRSV、ヒトメタニューモウィルス及び鳥メタニューモウィルスの配列全長に対応している。nt:ヌクレオチド;aa:アミノ酸;NA:適用無し、をそれぞれ表す。aの付された数字は各遺伝子におけるヌクレオチド、アミノ酸の相違の数を示す。CnPnV-Ane4Pタンパク質は、J3666に対して10アミノ酸の差異、strain15に対して15アミノ酸の差異を有することが見いだされた。MPV及びRSVPのアライメントは明確な類似性を有さず、アライメント中に6から7ギャップを含むアミノ酸22−124のアミノ末端近傍領域に隣接する、高い相同性を有する2つの領域を示す。CnPnV-Ane4がMPV株の両方と比較されたときに、中程度の相違である2つの領域が、Pにおいて相同性の低いこの領域の中で同定された。7アミノ酸の相違がアミノ酸58−74の領域に集まっており、他の4つの変異はアミノ酸93−102に見出された。確率プロットは、アミノ酸58−74の相違の第一領域は相対的に表面への露出しやすさ(surface probability)及び抗原指数(antigenic index)が高いことを予測する。第二の相違領域アミノ酸93−102は疎水性で表面への露出しやすさが低い(データは示さず)。137アミノ酸のタンパク質をコードしうるものであるPにおける第二のオーバーラップする内部開始ORFは、MPVにおいて同定された。合成ミニゲノム構築物が、第二のORFから作られるタンパク質を調査するために使用され、それは転写インヒビターとして機能することが見いだされた。CnPnV-Ane4は、同位置から始まるが終止位置は早い、第二のオーバーラップP ORFを有しており、ヌクレオチド285のTがAに変異してTAGコドンになることによって、半分以下のサイズである54アミノ酸のタンパク質を産生する(Dibben, O.ら、(2008) Virus Res. 131, 47-53)。このことは、もし生産されるならばより短いP−2タンパク質が同じ機能を有するかどうかという疑問を残している。
【0064】
残る非構造タンパク質はNS1及びNS2、及びM1−1及びM1−2を含んでおり、これらはM2遺伝子のARF(alternate
reading frame)から翻訳されるものである。strain15とJ3666におけるこれらのタンパク質の間には差異がない。しかしながら、CnPnVは、これらのタンパク質それぞれのMPV単離物と差異を有している。CnPnV-Ane4 NS−1は2つのMPV株と比較して7アミノ酸の差異を有しており、4つはC末端近傍に位置している。NS2では、配列全体にわたって比較的均等に分散している9アミノ酸の差異がある。CnPnV-Ane4は6アミノ酸の差異でM2−1と96.6%同一であり、4アミノ酸の差異でM2−2と95.9%同一である。
【0065】
[構造タンパク質の分析]
ニューモウィルスのG付着タンパク質は典型的に最大の相違があり、抗体応答の無効化のための第一目標となるため特に興味深い。Gタンパク質は2つのMPV単離物の間で396アミノ酸のうち相違は3つのみであり(99.2%)、比較的高い保存性を有している。CnPnVにおけるGタンパク質は、J3666との比較ではもっとも保存されておらず、strain15ではSHに続いて2番目に保存されていないタンパク質である。J3666、strain15とそれぞれ32アミノ酸、33アミノ酸の差異がある。アミノ酸の相違は、113−124領域に数個の差異が集まっていること、及び、331と367アミノ酸(Ane4ORFに基づく数字)の間に11アミノ酸の差異があることを除いて、一般的にランダムに分散している。一つの注目すべき観察は、CnPnV-Ane4のG ORFは、比較対象のJ3666及びstrain15に比べて、54ヌクレオチド長いことである。GSから+29の位置でのACGからAUGへの変異は、J3666にはあるがstrain15にはない代替的な開始コドンを作り出している。+65位置の、第二のUAGからAAGへの変異はインフレーム終止コドンを抹消するものである。G ORFは53アミノ酸の細胞質末端があり、公知のJ3666とstrain15の配列よりも18アミノ酸長いタンパク質をコードしている。J3666の配列はこの位置に終止コドンを有しているがstrain15にはない。すなわち、J3666とstrain15の両方において、単一点変異がより長いG変異体を産生しうる。これはJ3666に感染したマウス肺から増幅されたクローンでないRT−PCR産物の配列も+65位置でUからAへの変異を有していたように、前例の無いことではない。すなわちより長いGの変異体はCnPnVに特異的な特徴ではないように見える。本開示の利益を受けて、いずれの変異体が自然感染において優位であるかを決定するためのイヌ及びマウスのRT−PCR産物からの、さらなる配列データが当分野の通常の技術で決定されうる。G細胞質尾の長さにはいくらかの自然的な柔軟性があるようであり、また、尾自体が病原性の決定要因である。G細胞質尾を欠く組み換えMPV変異体はBalbCマウスで弱毒化されたが、複製レベルは野生型ウィルスと区別できなかった(Krempl, C.D.ら、2007.. J. Virol. 81, 9490-9501)。RSVにおいて細胞質尾の最初の6アミノ酸はMタンパク質との相互作用に本質的である。GのN末端近傍アミノ酸がMPVにおけるMへの結合のために本質的であるかどうかは知られていない。しかしそうであれば、細胞質尾の2つの変異体は、Mとの相互作用能力に差がありえる。
【0066】
CnPnV-Ane4のFタンパク質は、Gで見いだされたのと類似のハイレベルな多様性を有しておらず、実際それはMPVとの比較において、より保存されたタンパク質の一つである。J3666と14アミノ酸、strain15と16アミノ酸の差異があり、97%もしくはそれ以上が保存されている。アミノ酸の差異の約半分が、COOH末端の最後55アミノ酸の中、膜貫通ドメインの中もしくは近傍にあり、ほとんどの置換アミノ酸は類似の機能グループのタイプである。全体として、他のニューモウィルスとの関係においてNが最も高度に保存されたタンパク質である。CnPnV-Ane4のアミノ酸245−333のタンパク質の中央領域は、HRSV及びBRSVと92から93%の同一性に達している。CnPnV-Ane4 Nは、J3666と9アミノ酸の差異、strain15と8アミノ酸の差異を有しておりそれらは比較的均等に分散している。ニューモウィルスで同定された不変位置、又はRNAゲノムと相互作用する残基に対応するアミノ酸の差異はない。これらの差異のいずれもタンパク質の構造又は機能に大きな影響を有するものとは思われない。Mタンパク質はMPV単離物と98.1%という、最も高いレベルの保存性を有している。5アミノ酸の差異は配列中に均等に分散されており、どのケースでも機能グループは保存されている。アミノ酸18におけるVからIへのただ1つの変異のみが、ニューモウィルス及びメタニューモウィルスのMタンパク質において通常は不変とみられる残基に関する。
【0067】
[低継代及び高継代培養における分析]
我々は、培養によく適合され、また、一貫して複製されナイーブ培養に感染させるために用いられるときにはCPEが産生される一つの単離物を同定した。この単離物は17継代(Brne17)であったが、配列決定され、続いて、より低い継代数のAne4単離物と比較された。配列決定された8423ヌクレオチドのうち、Brne17では7ヌクレオチドの変異が同定された(表3、CnPnV-Ane4及び2種の継代数及びex vivoのBrneの間のヌクレオチド及びアミノ酸の差異を提供している)。表3について、a:各遺伝子の開始位置からのヌクレオチド位置、b:各ORFでの位置に基づくアミノ酸番号、ND:未決定、nt:ヌクレオチド、aa:アミノ酸。
【0068】
【表3】
【0069】
Nでの単一のヌクレオチドの差異、Fでの2つの差異は同義の置換体である。Brne17のSH GE配列におけるA残基の付加もまた非コードである。2つのヌクレオチドの差異は、Mにおいてアミノ酸178をQからRに変異させ、また、Gにおいてアミノ酸342をLからPに変異させる非同義の置換である。Brne17で見いだされた最も顕著な差異は、G ORFでの364ヌクレオチドのUからAへの置換であり、これはアミノ酸位置122においてK残基を終止コドンに置換するものである。この位置は、MPV株において、ヌクレオチド310(アミノ酸104)に相当する。この位置での終止は、Brne17に53アミノ酸の細胞質尾、24アミノ酸の膜貫通ドメイン及び44アミノ酸のみの細胞外ドメインのある切り取られたGを与えるであろう。Brne17における短縮されたGの発見は、3継代のストック及びオリジナルのスワブ溶出物(Brne3及びBrneSW)からの増幅産物のさらなる配列決定を促した。興味深いことにこのことは短縮された変異体が早くも3継代目において既に形成されていることを示した。しかしながら、BrneSWではヌクレオチド364にはAが存在し、これはイヌにおける優位なウィルス種は、短縮されたGを有していないことを意味した。RNA抽出物及び増幅物の第二の独立したセットは、1継代及び2継代からのサンプルを含み、1継代のもののみにヌクレオチド364がAであるマイナーな亜集団を示唆する電気泳動図の潜在的なピークが見られた。つまり培養において短縮された形態は迅速に選択されることを表している。同様の変異が継代されたstrain15(Warwick)の培養において同定されたが、そこではフレームシフトを生じる1つのヌクレオチド挿入がペプチドの中途終止を引き起こした。代替的な下流サイトでの翻訳の開始は33アミノ酸短く、細胞質尾を欠くGタンパク質を産生しうるものである。切り取られたGは少なくとも、strain15の低減された病原性についての部分的な説明を示唆した(Warwick)。
【0070】
RSVにおいて、低温継代されたcp−52変異体はG及びSHの産生を抹消する大きな欠失を有するが、培養において複製回避はされないことが見いだされた。この状況はBHK−21細胞培養では完全な組み換えウィルスと同じくG遺伝子全体を欠く組み換え体MPVが複製されたが、マウスにおいては検出可能なレベルに複製されなかったことと相同である。このことは、Gは常にin vivoでは本質的であることを示唆しており、すなわちBrneSwでは短縮されていない変異体のみの検出が予測され得る。なぜ2継代の終わりまでに短縮された変異体が完全に選択されるのかは明らかではない。もしも選択が単に複製効率の向上のみによるならば、Gの不存在は複製のレートや力価を向上させると予測される。しかし、これが真実だと支持する証拠は無い(Kremplら、2007、supra)。G欠損変異体の選択に影響を与える要素は、A72細胞に特異的でありえる。
【0071】
図3は配列番号1のゲノム組織の図示を提供する。当業者は図3の図は3´から5´方向に示されていることを認識するであろう。なぜならこれは、配列番号1は従来の方向としてマイナス鎖を左から右に読んで5´から3´に表している一方で、マイナス鎖ウィルスであるからである。つまり図3において、L ORFは、プラス鎖の3´末端にある。表4は配列番号1に示された配列の注釈を示している。略号は次の通りである。GS:遺伝子開始シグナル配列、ORF:オープンリーディングフレーム、GE:遺伝子終止シグナル配列、IGR:非翻訳インテグリン領域。リストされていないヌクレオチドは翻訳されないと考えられている。
【0072】
【表4】
【0073】
[実施例4]
我々はまた、ここで同定されるイヌのウィルスと関係がありえるウィルスの存在のためネコから得たサンプルを分析した。配列は2つの単離物から得られ、それらは29ケンタッキー(29KY)及び77ケンタッキー(77KY)と称呼される。29KY及び77KYからの部分的なG遺伝子配列は上記で説明されたイヌニューモウィルス(CnPnV)Ane4単離物の726−1078ヌクレオチドに対応する、353ヌクレオチドを提供する。29KYは、CnPnV-Ane4の2−208ヌクレオチドに対応する207ヌクレオチドを有する。
【0074】
両方の単離物の部分的なGヌクレオチド配列はマウスニューモウィルス(MPV)単離物J3666と94.7%が同一であり、Ane4と98.0%が同一であった。117アミノ酸(aa)配列はJ3666と90.6%が同一、Ane4と97.4%が同一であった。つまり、ネコ29KY/77KY G配列はJ3666よりもAne4により関連している。
【0075】
29KYの部分SHヌクレオチド配列はJ3666と94.7%が同一であり、Ane4と97.1%が同一である。69アミノ酸配列はJ3666と98.5%が同一であり、Ane4と97.1%が同一である。つまり、ネコ29KY配列はAne4よりもJ3666にわずかにより関連している。しかしながら、これらの配列は新規に発見されたネコニューモウィルスを表すと考えられている。つまり、上記で説明されたイヌニューモウィルスについて開示された発明の組成物及び方法及びすべての実施態様の開示は、ネコウィルスにも同様に適用され、それらの開示はすなわちネコウィルスについても反復される。
【0076】
当業者には前記から、我々は、CnPnV感染はそれが元々単離されたシェルターの犬に留まらないことを示したことを理解するであろう。それは、急性の呼吸器疾患の犬からのサンプルにおいて容易に同定される。疾患における病原性の役割は明確に証明されていないが、本発明はウィルスの存在を同定し、防御する組成物と方法を提供するものであって、それらは哺乳類においてウィルスの存在と正の相関関係のあるイヌやネコ両方の疾患を含みまたこれらに限定されない呼吸器疾患をコントロールするのに重要であろうと予測されるものである。
【特許請求の範囲】
【請求項1】
配列番号1、2又は3の配列と少なくとも96%が同一である単離されたポリヌクレオチドを含む組成物。
【請求項2】
前記ポリヌクレオチドが、配列番号1の配列と少なくとも96%が同一であるポリヌクレオチドである、請求項1に記載の組成物。
【請求項3】
前記ポリヌクレオチドが、配列番号2の配列と少なくとも96%が同一であるポリヌクレオチドである、請求項1に記載の組成物。
【請求項4】
前記ポリヌクレオチドが、配列番号3の配列と少なくとも96%が同一であるポリヌクレオチドである、請求項1に記載の組成物。
【請求項5】
配列番号1、2又は3の配列と少なくとも96%が同一であるポリヌクレオチドを含む単離されたウィルス。
【請求項6】
前記単離されたウィルスが弱毒化されている、請求項5に記載の単離されたウィルス。
【請求項7】
前記単離されたウィルスが、配列番号1の配列と少なくとも96%が同一であるポリヌクレオチドを含む、請求項5に記載の単離されたウィルス。
【請求項8】
前記単離されたウィルスが、配列番号2の配列と少なくとも96%が同一であるポリヌクレオチドを含む、請求項6に記載の単離されたウィルス。
【請求項9】
前記単離されたウィルスが、配列番号3の配列と少なくとも96%が同一であるポリヌクレオチドを含む、請求項7に記載の単離されたウィルス。
【請求項10】
配列番号1、2又は3の配列と少なくとも96%が同一であるポリヌクレオチドを含むウィルスを含む、単離された細胞又はインビトロの細胞培養。
【請求項11】
請求項5の単離されたウィルス、又は、前記単離されたウィルスの一部である単離されたタンパク質を含む組成物を哺乳類に投与することを含む方法であって、投与後に、動物において免疫応答が刺激されることを特徴とする、哺乳類において免疫応答を刺激する方法。
【請求項12】
前記請求項5の単離されたウィルスが、配列番号1の配列と少なくとも96%が同一であるポリヌクレオチドを含み、前記組成物がイヌに投与されることを特徴とする、請求項11に記載の方法。
【請求項13】
前記請求項5の単離されたウィルスが、配列番号2又は配列番号3の配列と少なくとも96%が同一であるポリヌクレオチドを含み、前記組成物がネコに投与されることを特徴とする、請求項11に記載の方法。
【請求項14】
哺乳類から生物学的サンプルを得ること、配列番号1、2又は3の配列と少なくとも96%が同一であるポリヌクレオチドの存在を決定すること、又は、配列番号1、2又は3の逆相相補配列によってコードされるタンパク質の存在を検出することによる方法であって、前記ポリヌクレオチド又は前記タンパク質の存在は、哺乳類がニューモウィルスに感染していることを示すことを特徴とする、哺乳類がニューモウィルスに感染しているか否かを決定する方法。
【請求項15】
前記哺乳類がイヌ又はネコである、請求項14に記載の方法。
【請求項1】
配列番号1、2又は3の配列と少なくとも96%が同一である単離されたポリヌクレオチドを含む組成物。
【請求項2】
前記ポリヌクレオチドが、配列番号1の配列と少なくとも96%が同一であるポリヌクレオチドである、請求項1に記載の組成物。
【請求項3】
前記ポリヌクレオチドが、配列番号2の配列と少なくとも96%が同一であるポリヌクレオチドである、請求項1に記載の組成物。
【請求項4】
前記ポリヌクレオチドが、配列番号3の配列と少なくとも96%が同一であるポリヌクレオチドである、請求項1に記載の組成物。
【請求項5】
配列番号1、2又は3の配列と少なくとも96%が同一であるポリヌクレオチドを含む単離されたウィルス。
【請求項6】
前記単離されたウィルスが弱毒化されている、請求項5に記載の単離されたウィルス。
【請求項7】
前記単離されたウィルスが、配列番号1の配列と少なくとも96%が同一であるポリヌクレオチドを含む、請求項5に記載の単離されたウィルス。
【請求項8】
前記単離されたウィルスが、配列番号2の配列と少なくとも96%が同一であるポリヌクレオチドを含む、請求項6に記載の単離されたウィルス。
【請求項9】
前記単離されたウィルスが、配列番号3の配列と少なくとも96%が同一であるポリヌクレオチドを含む、請求項7に記載の単離されたウィルス。
【請求項10】
配列番号1、2又は3の配列と少なくとも96%が同一であるポリヌクレオチドを含むウィルスを含む、単離された細胞又はインビトロの細胞培養。
【請求項11】
請求項5の単離されたウィルス、又は、前記単離されたウィルスの一部である単離されたタンパク質を含む組成物を哺乳類に投与することを含む方法であって、投与後に、動物において免疫応答が刺激されることを特徴とする、哺乳類において免疫応答を刺激する方法。
【請求項12】
前記請求項5の単離されたウィルスが、配列番号1の配列と少なくとも96%が同一であるポリヌクレオチドを含み、前記組成物がイヌに投与されることを特徴とする、請求項11に記載の方法。
【請求項13】
前記請求項5の単離されたウィルスが、配列番号2又は配列番号3の配列と少なくとも96%が同一であるポリヌクレオチドを含み、前記組成物がネコに投与されることを特徴とする、請求項11に記載の方法。
【請求項14】
哺乳類から生物学的サンプルを得ること、配列番号1、2又は3の配列と少なくとも96%が同一であるポリヌクレオチドの存在を決定すること、又は、配列番号1、2又は3の逆相相補配列によってコードされるタンパク質の存在を検出することによる方法であって、前記ポリヌクレオチド又は前記タンパク質の存在は、哺乳類がニューモウィルスに感染していることを示すことを特徴とする、哺乳類がニューモウィルスに感染しているか否かを決定する方法。
【請求項15】
前記哺乳類がイヌ又はネコである、請求項14に記載の方法。
【図1】


【図2】
【図3】
【図2】
【図3】
【公表番号】特表2013−514813(P2013−514813A)
【公表日】平成25年5月2日(2013.5.2)
【国際特許分類】
【出願番号】特願2012−546141(P2012−546141)
【出願日】平成22年12月21日(2010.12.21)
【国際出願番号】PCT/US2010/061510
【国際公開番号】WO2011/084783
【国際公開日】平成23年7月14日(2011.7.14)
【出願人】(511040838)コーネル ユニバーシティ (4)
【氏名又は名称原語表記】CORNELL UNIVERSITY
【Fターム(参考)】
【公表日】平成25年5月2日(2013.5.2)
【国際特許分類】
【出願日】平成22年12月21日(2010.12.21)
【国際出願番号】PCT/US2010/061510
【国際公開番号】WO2011/084783
【国際公開日】平成23年7月14日(2011.7.14)
【出願人】(511040838)コーネル ユニバーシティ (4)
【氏名又は名称原語表記】CORNELL UNIVERSITY
【Fターム(参考)】
[ Back to top ]