
新規ホスホリパーゼC
【課題】 様々なリン脂質を酸性側でも中性付近でも効率よく加水分解する能力を有し、クエン酸緩衝液中でも活性を有するとともにある程度熱的に安定であり、脂質部分を含有しないリン酸エステルを加水分解しない性質を有するホスホリパーゼCを提供すること。
【解決手段】 酸性から中性のpHにおいて活性を示し、かつ、リン脂質以外のリン酸エステルを実質的に加水分解しないホスホリパーゼC。
【解決手段】 酸性から中性のpHにおいて活性を示し、かつ、リン脂質以外のリン酸エステルを実質的に加水分解しないホスホリパーゼC。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ホスホリパーゼC、該ホスホリパーゼCを生産する糸状菌、糸状菌の培養物から該ホスホリパーゼCを分離精製する方法、該ホスホリパーゼCをコードするDNAおよび該ホスホリパーゼCの製造方法等に関する。具体的には、本発明は、食品工業および医薬品工業における使用に特に適するホスホリパーゼC、特に糸状菌アスペルギルス・オリザエ(Aspergillus oryzae)あるいはアスぺルギルス・タマリ(Aspergillus tamarii)が生産するホスホリパーゼC、該ホスホリパーゼCを生産する糸状菌、糸状菌の培養物から該ホスホリパーゼCを分離精製する方法、該ホスホリパーゼCをコードするDNAおよび該ホスホリパーゼCの製造方法等に関する。
【背景技術】
【0002】
〔1〕ホスホリパーゼC
従来、動物および微生物がホスホリパーゼCを生産することが知られている。動物由来の酵素は主としてホスファチジルイノシトール選択的なホスホリパーゼCである。また、微生物のなかでは細菌、放線菌、酵母およびカビ由来のホスホリパーゼCが知られている。細菌、放線菌および酵母が生産するホスホリパーゼCは、ホスファチジルイノシトール選択的あるいはホスファチジルコリン選択的なものがほとんどである。
【0003】
細菌由来のホスホリパーゼCとしては、例えばシュードモーナス・シュルキリエンシス(Psudomonus schuylkilliensis)(例えば、特許文献1参照)、ブルコールデリア・シュードマレイ(Bulkholderia pseudomallei)(例えば、非特許文献1参照)、バチラス・セレウス(Bacillus cereus)(例えば、非特許文献2参照)、スタフィロコッカス・アウレウス(Staphylococcus aureus)(例えば、非特許文献3参照)およびクロストリディウム・ぺルフリンゲンス(Clostridium perfringens)(例えば、非特許文献4参照)等が生産するホスホリパーゼCが知られている。
【0004】
放線菌由来のホスホリパーゼCとしては、例えばストレプトマイセス・ハチジョウエンシス(Streptomyces hachijyoensis)(例えば、特許文献2参照)が生産するホスホリパーゼCが知られている。
【0005】
酵母由来のホスホリパーゼCとしては、例えばキャンディダ・アルビカンス(Candida albicans)(例えば、非特許文献5参照)、サッカロマイセス・セレビシアエ(Saccharomyces cerevisiae)(例えば、非特許文献6参照)等が生産するホスホリパーゼCが知られている。
【0006】
カビ由来のホスホリパーゼCは、従来2例知られている。一つはアスペルギルス・ニガー(Aspergillus niger)(例えば、特許文献3参照)、もうひとつはアスペルギルス・サイトイ(Aspergillus saitoi)(例えば、非特許文献7参照)が生産するホスホリパーゼCである。
【0007】
〔2〕レシチン
レシチンは、動物、植物、菌類に広く分布している代表的なグリセロリン脂質である。グリセロリン脂質とは、1,2−ジアシルグリセロールの3位にホスホリル塩基が共有結合している化合物である。塩基としては、コリン、エタノールアミン、セリン、イノシトールおよびグリセロール等が含まれており、その組成割合は由来により異なる。レシチンはグリセロリン脂質に含有される概念として用いるものとする。
【0008】
レシチンは、界面活性作用、酸化防止作用、生理活性作用等を有し、食品、飼料、医薬品等に利用されている。食品工業においては、卵黄レシチンや大豆レシチンなどに代表される天然レシチンが食品添加物として使用されており、食品の性質を改変するために主に乳化剤等として使用され、豊富に供給されている。
【0009】
〔3〕レシチンの酵素処理
レシチンを酵素により部分的に加水分解し、新たな性質を付与する検討も行われている。この際に用いられる酵素がホスホリパーゼ類であり、ホスホリパーゼA,B,C,Dが知られている。グリセロリン脂質の1位あるいは2位の脂肪酸を選択的に加水分解するのがホスホリパーゼAである。非選択的に加水分解するのがホスホリパーゼB、ジアシルグリセロールとホスホリル塩基に加水分解するのがホスホリパーゼC、ホスファチジン酸と塩基に分解するのがホスホリパーゼDである。
【0010】
食品工業の分野において、現在最も比較的多く使用されているのはホスホリパーゼAである。レシチンは水に難溶性であるが、レシチンにホスホリパーゼAを作用させるとアシル基が部分的に加水分解され、水溶性のリゾレシチンが生成する。リゾレシチンを食品添加物として使用した場合には、得られる食品の物性がこれまでレシチンを用いて得られていた食品の物性とは異なる可能性がある。
【0011】
〔4〕スフィンゴリン脂質
スフィンゴリン脂質は、グリセロリン脂質とともに、リン脂質を構成している。
スフィンゴリン脂質の代表はスフィンゴミエリンであり、セラミドの1級アルコールにコリンリン酸がリン酸ジエステル結合した化合物である。動物の臓器に広く含まれている。母乳にも含有されることから、乳児用粉ミルクに配合されることもある。
スフィンゴミエリンにホスホリパーゼCを作用させるとコリンリン酸がはずれ、セラミドが生成する。セラミドは保湿成分として、化粧品に広く使用されている。また、アトピー性皮膚炎はセラミドが不足することにより惹起されるという報告がある。
【0012】
〔5〕食品中でのホスホリパーゼCの使用
ホスホリパーゼCを用いることにより、下図に示すように水の存在下でレシチンからジアシルグリセロールを生産することができる。
【0013】
【化1】

【0014】
(式中、R1およびR2は、それぞれアルキル基を、R3は、コリン、エタノールアミン、グリセロール、イノシトール等の基を意味する。)
【0015】
食品素材中でこのような反応を起こさせることにより、ホスホリパーゼAとは本質的に異なった性質を付加した商品が提供できる可能性がある。
ここで用いられるホスホリパーゼCには、様々なグリセロリン脂質を加水分解することが求められる。たとえば大豆レシチンにはホスファチジルコリンおよびホスファチジルエタノールアミンを主として、ホスファチジルグリセロールあるいはホスファチジルイノシトール等も含まれていることが知られている。また、卵黄由来レシチンにはホスファチジルコリンおよびホスファチジルエタノールアミンが主として含まれている。したがって、用いられるホスホリパーゼCは、これらを区別無く加水分解することが望ましい。
【0016】
しかし、食品中にはリン脂質以外にもリン酸エステルが多く含有されており、それらを加水分解してリン酸を遊離させることは、当該食品の性質をさらに変質させてしまうことにつながるので、好ましいものではない。したがって、リン脂質以外のリン酸エステルは加水分解しないホスホリパーゼCが望ましく、例えばグリセロホスホリルコリンのように、リン脂質に類似するが脂質部分を含有しないリン酸エステルに対する酵素活性は無いホスホリパーゼCが望ましい。
【0017】
同様の観点から、用いられるホスホリパーゼCは、ホスファターゼ活性を有さないタンパク質であることが望ましい。すなわち、ホスファターゼの基質となるパラニトロフェニルリン酸分解活性を有しないホスホリパーゼCが望ましい。
【0018】
また、食品工業においては、食品の変質を防ぐため中性から弱酸性域でさまざまな処理を行うことが好ましく、ホスホリパーゼ等を含む酵素剤はこのpH域に高い活性を有していることが望まれる。
【0019】
〔6〕食品工業におけるホスホリパーゼCの使用
ホスホリパーゼCの用途には、例えば、パンの冷凍保存生地焼成時に、その表面で発生する老化、梨肌の緩和や食用油精製工程の改善がある。
大豆・菜種等から食用油を製造する際には、着色あるいは食味の劣化の原因となるため、レシチンは除去されるべき物質である。この目的のために、従来ホスホリパーゼAを用いてレシチンを部分的に加水分解し、リゾレシチンとすることで水溶性にして除去する方法が検討されてきた。
【0020】
しかし、ここでホスホリパーゼCを用いてレシチンをジアシルグリセロールとすることにより、トリアシルグリセロールと共に油の一成分にすることができる。すなわち、食用油の製造工程において、歩留まりを向上させる効果が期待される。
【0021】
ここで用いられるホスホリパーゼCには、様々なリン脂質を加水分解することが求められる。たとえば大豆油中には先述した様々なリン脂質が含まれている。また、綿実油あるいは菜種油中にも同様にさまざまなリン脂質が含まれている。用いられるホスホリパーゼCは、これらを区別なく加水分解することが望ましい。
【0022】
また、製油工業においては油以外の不純物を酸性加温条件下で除去するので、酸性域での活性が高く、ある程度温度安定性を有する酵素剤の使用が望まれる。処理される粗油を酸性にするためにクエン酸が使用されることがあるので、用いられるホスホリパーゼCにはクエン酸存在下での活性およびある程度の温度安定性が望まれる。
【0023】
〔7〕既知のホスホリパーゼCの問題点
動物、細菌、放線菌あるいは酵母が生産するホスホリパーゼCは、主としてホスファチジルイノシトールあるいはホスファチジルコリン選択的であり、様々な基質を分解する必要がある食品工業における使用には向かない。さらに、動物起源の酵素剤は、宗教的に受け入れられない国および地域があり、汎用性にも問題がある。さらに、ホスホリパーゼCを生産する細菌は、病原性を示すものがほとんどであり、安全性に問題がある。
【0024】
従来知られている糸状菌由来のホスホリパーゼCは、様々なリン脂質を加水分解する性質を有している。また、従来ホスホリパーゼCの産生に用いられているいずれの糸状菌も食用酵素生産に実績がある特徴を有している。アスペルギルス・ニガーとアスペルギルス・サイトイの両菌株由来の酵素の、温度あるいはpHに関する性質は極めて類似しており、分子量も同一である。いずれも酸性側で強い活性を有しているが、中性付近では全く活性がないという特徴を有している。したがって、酸性側でこれらの酵素を使用する可能性がある製油工業においては、これらの酵素は使用できる可能性がある。しかしクエン酸中での性質については記載がなく、実際に製油工業で使用できるかどうかは不明である(特許文献3および非特許文献7参照)。また、pH6での活性がほぼ0であることが示されており、中性付近で酵素反応を行うことが多いと考えられる食品工業においては、使用しづらい酵素である(特許文献3参照)。
【0025】
さらに、アスペルギルス・ニガーが生産するホスホリパーゼCは、ホスファターゼの基質であるホスファチジン酸の分解活性が非常に高いことが記載されている(特許文献3参照)。したがって、該酵素は、リン酸モノエステルを加水分解する能力を有することから、ホスファターゼ活性をも併せ持つタンパク質であることが推察される。さらに、アスペルギルス・サイトイが生産するホスホリパーゼCは、2−ヘキサデカノイルアミノ−4−ニトロフェニルホスホリルコリンの分解活性が非常に高いことが記載されている(非特許文献7参照)。したがって、該酵素は、リン脂質以外のリン酸ジエステルを加水分解する能力をも併せもつタンパク質であることが推察される。したがって、これらの酵素を用いて加工される食品は、不必要に改質されてしまう可能性がある。
【0026】
このように、従来知られているホスホリパーゼCは、酵素の特性として不十分であるか、あるいは安全性に問題がある等の問題点が挙げられ、ホスホリパーゼCとして市場に流通している酵素剤は現在までに存在しない。望ましい酵素剤の性質としては、既に食品用酵素剤の生産実績がある微生物由来であり、様々なリン脂質を酸性側でも中性付近でも効率よく加水分解する能力を有し、製油工業においても使用可能なようにクエン酸緩衝液中でも活性を有するとともにある程度熱的に安定であることが挙げられる。さらには、グリセロホスホリルコリンに代表されるようなリン脂質以外のリン酸エステルを加水分解しない性質を有するホスホリパーゼCが挙げられる。
【先行技術文献】
【特許文献】
【0027】
【特許文献1】特開昭50−1017183号公報
【特許文献2】特開昭49−55893号公報
【特許文献3】特開2000−166543号公報
【非特許文献】
【0028】
【非特許文献1】コルブスリサテら(Korbsrisate S. et. al.)「ジャーナル・オブ・クリニカル・マイクロバイオロジー」(Jounal of Clinical Microbiology)1999年、37巻、p.3742−3745
【非特許文献2】タンら(Tan C. et. al.)「プロテイン・エクスプレッション・アンド・ピューリフィケイション」(Protein Expression and Purification)1997年、10巻、p.365−372
【非特許文献3】ダウガーティーら(Daugherty S. et. al.)「インフェクション・アンド・イミュニティー」(Infection and Immunity)1993年、61巻、p.5078−5089
【非特許文献4】ティットバールら(Titball R. et. al.)「インフェクション・アンド・イミュニティー」(Infection and Immunity)1989年、57巻、p.367−376
【非特許文献5】アンダルツら(Andaluz E. et. al.)「イースト」(Yeast)2001年、18巻、p.711−721
【非特許文献6】パイネら(Payne W. et. al.)「モレキュラー・アンド・セルラー・バイオロジー」(Molecular and Cellular Biology)1993年、13巻、p.4351−4364
【非特許文献7】マツオカら(Matsuoka S. et. al.)「バイオテクノロジー・アンド・アプライドバイオケミストリー」(Biotechnology and Applied Biochemistry)1987年、9巻、p.401−409
【発明の概要】
【発明が解決しようとする課題】
【0029】
上述のように、望ましいホスホリパーゼCの性質としては、様々なリン脂質を酸性側でも中性付近でも効率よく加水分解する能力を有し、製油工業においても使用可能なようにクエン酸緩衝液中でも活性を有するとともに、ある程度熱的に安定であることが挙げられる。さらには、グリセロホスホリルコリンに代表されるような、リン脂質以外のリン酸エステルを加水分解しない性質を有するホスホリパーゼCであることが挙げられ、さらに好ましくは、既に食品用酵素剤の生産実績がある微生物由来のホスホリパーゼCであることが挙げられる。
このようなホスホリパーゼCを提供することは、この技術分野において非常に関心の高いことであった。
【課題を解決するための手段】
【0030】
本発明者らは、安全性に優れ、様々なリン脂質を酸性側でも中性付近でも効率よく加水分解する能力を有し、クエン酸緩衝液中でも活性を有するとともにある程度熱的に安定であり、リン脂質以外のリン酸エステルを加水分解しないホスホリパーゼCを見出すべく鋭意検討を行ったところ、アスぺルギルス・オリザエFERM ABP−10200株またはNBRC 4190株、あるいはアスぺルギルス・タマリIAM 13907株由来のホスホリパーゼCを精製し、アスぺルギルス・オリザエNBRC 4190株に由来するホスホリパーゼC遺伝子をクローニングし、本発明を完成するに至った。
【0031】
すなわち本発明は、(1) 酸性から中性のpHにおいて活性を示し、かつ、リン脂質以外のリン酸エステルを実質的に加水分解しないホスホリパーゼC。
(2) ホスファチジルイノシトール特異的ではない、(1)記載のホスホリパーゼC。
(3) 至適pHがpH3乃至pH6の範囲にある、(1)または(2)記載のホスホリパーゼC。
(4) pH7における相対活性が20%以上である、(1)〜(3)のいずれか1つ記載のホスホリパーゼC。
【0032】
(5) 糸状菌によって産生される、(1)〜(4)のいずれか1つ記載のホスホリパーゼC。
(6) 糸状菌がアスペルギルス属である、(5)に記載のホスホリパーゼC。
(7) アスぺルギルス・オリザエ(Aspergillus oryzae)あるいはアスぺルギルス・タマリ(Aspergillus tamarii)によって産生される、(6)に記載のホスホリパーゼC。
(8)アスぺルギルス・オリザエFERM ABP−10200株またはNBRC 4190株、あるいはアスぺルギルス・タマリIAM 13907株によって産生される、(7)に記載のホスホリパーゼC。
【0033】
(9) 以下の性質:
1)SDS−PAGE電気泳動法にて分子量約87,000を示す;
2)ホスファチジルコリン、ホスファチジルエタノールアミン、ホスファチジルイノシトールおよびホスファチジルグリセロールを加水分解する;
3)グリセロホスホリルコリンおよびパラニトロフェニルリン酸を実質的に加水分解しない;
4)卵黄由来レシチンをpH3乃至pH9の範囲で加水分解する;
5)0℃乃至80℃の範囲において4)記載の加水分解活性を有する;
6)温度安定性について、pH4.5において45℃以下の温度で安定である;
7)pH安定性について、pH3乃至pH10の範囲で安定である。
を示す(1)〜(8)のいずれか1つに記載のホスホリパーゼC。
【0034】
(10) 下記のa)〜d):
a)配列番号5に記載のアミノ酸配列からなるタンパク質;
b)配列番号4のヌクレオチド配列によりコードされるアミノ酸配列からなるタンパク質;
c)a)またはb)に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ、ホスホリパーゼC活性を有することを特徴とするタンパク質;
d)a)またはb)に記載のアミノ酸配列を含むことからなるタンパク質、
のいずれか一つに記載のタンパク質であるホスホリパーゼC。
【0035】
(11) (1)〜(4)、(9)および(10)のいずれか1つに記載のホスホリパーゼCを生産する能力を有する、単離されたアスぺルギルス・オリザエあるいはアスぺルギルス・タマリに属する糸状菌。ただし、アスぺルギルス・タマリIAM 13907株を除く。
(12)アスぺルギルス・オリザエFERM ABP−10200株またはNBRC 4190株である、(11)に記載の糸状菌。
【0036】
(13) 下記のa)〜d):
a)配列番号4のコード領域(CDS)のヌクレオチド配列からなるDNA;
b)上記a)に記載のDNAと70%以上のヌクレオチド配列相同性を有するヌクレオチド配列からなり、かつ、ホスホリパーゼC活性を有するタンパク質をコードすることを特徴とするDNA;
c)配列番号5に記載のアミノ酸配列からなるタンパク質をコードするDNA;
d)配列番号4のコード領域(CDS)のヌクレオチド配列を含むことからなるDNA、
のいずれか一つに記載のDNA。
(14) (11)に記載のDNAにコードされるタンパク質であるホスホリパーゼC。
【0037】
(15)
1)(9)に記載のアスペルギルス・オリザエあるいはアスぺルギルス・タマリを培養する工程、および、
2)1)の培養産物からホスホリパーゼCを分離・精製する工程、
を含む、ホスホリパーゼCの製造方法。
【0038】
(16) アスペルギルス・オリザエがアスペルギルス・オリザエFERM ABP−10200株またはNBRC 4190株、アスぺルギルス・タマリがアスぺルギルス・タマリIAM 13907株である(15)記載の方法。
に関する。
【図面の簡単な説明】
【0039】
【図1】アスぺルギルス・オリザエFERM ABP−10200株由来の精製ホスホリパーゼCの活性とpHとの関係を表した図である。
【図2】アスぺルギルス・オリザエFERM ABP−10200株由来の精製ホスホリパーゼCの活性と温度との関係を表した図である。
【図3】アスぺルギルス・オリザエFERM ABP−10200株由来の精製ホスホリパーゼCの温度安定性を表した図である。
【図4】アスぺルギルス・オリザエFERM ABP−10200株由来の精製ホスホリパーゼCのpH安定性を表した図である。
【図5】アスぺルギルス・タマリIAM 13907株由来の精製ホスホリパーゼCの活性とpHとの関係を表した図である。
【図6】アスぺルギルス・タマリIAM 13907株由来の精製ホスホリパーゼCの活性と温度との関係を表した図である。
【図7】アスぺルギルス・タマリIAM 13907株由来の精製ホスホリパーゼCの温度安定性を表した図である。
【図8】アスぺルギルス・タマリIAM 13907株由来の精製ホスホリパーゼCのpH安定性を表した図である。
【図9】アスぺルギルス・オリザエNBRC 4190株由来の精製ホスホリパーゼCの活性とpHとの関係を表した図である。
【発明を実施するための形態】
【0040】
以下、本発明を詳細に説明する。なお、特に指定しない限り、以下に示す各特性の測定方法は、後述する実施例または試験例に記載の測定方法による。
本発明は、酸性から中性のpHにおいて活性を示し、かつ、リン脂質以外のリン酸エステルを実質的に加水分解しないホスホリパーゼCであるか、あるいは酸性から中性のpHにおいて活性を示し、かつ、ホスファターゼ活性を有しないホスホリパーゼCである。
【0041】
「酸性から中性のpHにおいて活性を示す」とは、pH約3乃至約7の範囲において、相対活性が20%以上であることをいう。(酵素活性の測定法は「試験例1−1)pH活性」の記載に従う。)
【0042】
また、「リン脂質以外のリン酸エステルを実質的に加水分解しない」とは、好ましくは、ホスファチジルコリン(卵黄由来)に対する加水分解活性を100%とした場合における加水分解活性が、ホスファチジン酸に対しては30%以下、さらに好ましくは25%以下であること、および/またはグリセロホスホリルコリンに対しては15%以下、さらに好ましくは10%以下であること、および/またはパラニトロフェニルリン酸に対しては10%以下、さらに好ましくは5%以下であることをいう。「グリセロホスホリルコリンおよびパラニトロフェニルリン酸を実質的に加水分解しない」についても同様である。
ここで、ホスファチジン酸は、中性脂肪およびグリセロリン脂質の合成中間体であると認識されるため、本願の明細書等においてはグリセロリン脂質(リン脂質)には含まれないものとする。
【0043】
「ホスファターゼ活性を有しない」とは、ホスファチジン酸またはパラニトロフェニルリン酸の分解活性がホスファチジルコリンの分解活性の50%以下であることをいい、ホスファチジン酸の分解活性については、好ましくはホスファチジルコリンの分解活性の40%以下、さらに好ましくは30%以下、最も好ましくは25%以下である。また、パラニトロフェニルリン酸の分解活性については、好ましくはホスファチジルコリンの分解活性の30%以下、さらに好ましくは20%以下、最も好ましくは10%以下である。
【0044】
本発明のホスホリパーゼCは、スフィンゴミエリンに対する分解活性を有し、好ましくはその分解活性が、ホスファチジルコリンに対する分解活性と同等かそれ以上である。具体的には、ホスファチジルコリン(卵黄由来)に対する分解活性を100%とした場合に、ホスファチジルコリンに対する分解活性が、好ましくは90%以上、さらに好ましくは105%以上、最も好ましくは115%以上である。
【0045】
また、本発明のホスホリパーゼCは、ホスファチジルエタノールアミンに対する分解活性を有し、好ましくはその分解活性が、ホスファチジルコリンに対する分解活性と同等である。具体的には、ホスファチジルコリン(卵黄由来)に対する分解活性を100%とした場合にホスファチジルエタノールアミンに対する分解活性が、好ましくは80%以上、さらに好ましくは85%以上、最も好ましくは90%以上であり、150%以下である。
【0046】
ホスホリパーゼCが、「ホスファチジルイノシトール特異的ではない」とは、ホスファチジルイノシトールの分解活性(相対活性)に比べて、ホスファチジルイノシトール以外の基質の分解活性の方が高いことを意味する。ここで、ホスファチジルイノシトール以外の基質としては、好ましくはホスファチジルコリン、ホスファチジルエタノールアミン、ホスファチジルイノシトールまたはホスファチジルグリセロールである。
【0047】
本発明のホスホリパーゼCの至適pHは、酸性から中性のpHの範囲にあることが好ましく、さらにpH3乃至pH6の範囲にあることが好ましく、さらに好ましくはpH3乃至pH5の範囲であり、最も好ましくはpH4乃至pH5の範囲である。
【0048】
また、「pH7における相対活性」とは、最も活性が高いpHでの加水分解活性を100%とし、pH7における酵素の加水分解活性を相対値として百分率(%)で表したものをいう。本発明のホスホリパーゼCは、pH7における相対活性が、好ましくは20%以上であり、さらに好ましくは40%以上、最も好ましくは50%以上である。
【0049】
温度活性について、本発明のホスホリパーゼCは、至適温度が、45℃乃至70℃の範囲にあることが好ましく、さらには55℃乃至70℃の範囲にあることが好ましく、最も好ましくは60℃乃至70℃の範囲である。また、本発明のホスホリパーゼCは、55℃乃至70℃の範囲において相対活性が50%以上であることが好ましく、さらに好ましくは45℃乃至70℃の範囲において相対活性が50%以上であり、最も好ましくは35℃乃至70℃の範囲において相対活性が50%である。また、55℃乃至65℃の範囲においては、相対活性が80%以上であることが好ましい。
【0050】
また、温度安定性について、「安定である」とは、40%以上の残存加水分解活性を有していることを意味し、本発明のホスホリパーゼCは、45℃以下の処理温度において、残存活性が好ましくは50%以上、さらに好ましくは70%以上、最も好ましくは80%以上である。また、本発明のホスホリパーゼCは、処理温度が、好ましくは45℃、さらに好ましくは50℃、最も好ましくは60℃において、残存加水分解活性が40%以上である。
【0051】
また、pH安定性について、「安定である」とは、保存処理後において、5%以上の残存加水分解活性を有していることを意味し、本発明のホスホリパーゼCは、処理pHがpH5乃至pH9の範囲において、好ましくは残存加水分解活性が40%以上であり、さらに好ましくは50%以上であり、最も好ましくは60%以上である。また、処理pHがpH6乃至pH8の範囲において、好ましくは残存加水分解活性が60%以上であり、さらに好ましくは70%以上であり、最も好ましくは80%以上である。
【0052】
また、本発明のホスホリパーゼCの別の例としては、配列番号1,2および3に示されるアミノ酸配列のいずれか1つ、好ましくはいずれか2つ、最も好ましくは3つ全てを含み、且つ、ホスホリパーゼC活性を有するタンパク質が挙げられる。該タンパク質は、これら3つのうちの2つ以上のアミノ酸配列を含む場合には、各々のアミノ酸配列の順序はいかなる順序であってもよい。
【0053】
また、配列番号5のアミノ酸配列からなるタンパク質において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ、ホスホリパーゼC活性を有することを特徴とするタンパク質も本発明に含まれる。置換したアミノ酸配列を有するタンパク質が、天然型タンパク質と同等の活性を有する例として、例えば、インターロイキン2(IL−2)遺伝子のシステインに相当するヌクレオチド配列をセリンに相当するヌクレオチド配列に置換して得られたタンパク質が、IL−2活性を保持することが知られている(Wang, A. et al. (1984) Science 224, 1431-1433)。
【0054】
また、本発明のホスホリパーゼCの別の例としては、配列番号5に記載のアミノ酸配列からなるタンパク質を挙げる事ができる。また、配列番号5に記載のアミノ酸配列を含むことからなるタンパク質であっても、ホスホリパーゼC活性を有する限り本発明に含まれる。
【0055】
また、本発明のホスホリパーゼCの別の例としては配列番号5に記載のアミノ酸配列からなり、糖鎖によって修飾されているタンパク質が挙げられる。また、このようなタンパク質を含むことからなるタンパク質も、ホスホリパーゼC活性を有する限り本発明に含まれる。
【0056】
本発明のホスホリパーゼCとして好適なものは、アスペルギルス・オリザエFERM ABP−10200株またはNBRC 4190株あるいはアスぺルギルス・タマリIAM 13907株由来のホスホリパーゼC、配列番号5に記載のアミノ酸配列からなるタンパク質および配列番号5に記載のアミノ酸配列からなり、糖鎖によって修飾されているタンパク質が挙げられるが、より好適なものは、アスペルギルス・オリザエFERM ABP−10200株またはNBRC 4190株あるいはアスぺルギルス・タマリIAM 13907株由来のホスホリパーゼCおよび配列番号5に記載のアミノ酸配列からなり、且つ糖鎖によって修飾されているタンパク質である。
【0057】
本発明において、「本発明のDNA」とは、本発明のホスホリパーゼCをコードするDNAをいう。DNAとしては、cDNA、ゲノムDNA、人工的に改変されたDNA、化学的に合成されたDNAなど、現在知られる限りどのような形態であっても良い。
【0058】
本発明のDNAの例としては、配列番号4のコード領域(CDS)のヌクレオチド配列であり、且つ、ホスホリパーゼC活性を有するタンパク質をコードするDNAが挙げられる。
【0059】
本発明のDNAの別の例としては、配列番号4のコード領域(CDS)のヌクレオチド配列と70%以上のヌクレオチド配列相同性を有するDNAを挙げることができる。このようなDNAとしては、自然界で発見される変異型DNA、人為的に改変した変異型DNA、異種生物由来の相同DNAなどが含まれる。
【0060】
また、本発明のヌクレオチドのさらに他の例としては、配列番号4に記載のアミノ酸配列からなるタンパク質をコードするDNAが挙げられる。なお、所望のアミノ酸に対応するコドンは、その選択も任意でよく、例えば利用する宿主のコドン使用頻度を考慮して常法に従い決定できる。(Grantham, R. et al. (1981) Nucleic Acids Res. 9, 143-174)。さらに、これらヌクレオチド配列のコドンの一部改変は、常法に従い、所望の改変をコードする合成オリゴヌクレオチドからなるプライマーを利用した、部位特異的変異導入法(site specific mutagenesis/Mark, D. F. et al. (1984) Proc. Natl. Acad. Sci. USA 81, 5662-5666)などに従うことができる。
【0061】
本発明のDNAのまた別の例としては、配列番号4のコード領域(CDS)のヌクレオチド配列からなるDNAが挙げられる。また、配列番号4のコード領域(CDS)のヌクレオチド配列を含むDNAも、ホスホリパーゼC活性を有するタンパク質をコードしている限り、本発明に含まれるものである。
【0062】
また、本発明のホスホリパーゼCには、本発明のDNAによりコードされるアミノ酸配列からなるタンパク質を挙げることができる。また、本発明のホスホリパーゼCにおいて、任意の一つもしくは二つ以上のアミノ酸を欠失させた改変体を作製するためには、エキソヌクレアーゼBal31等を用いてDNAを末端から削る方法(岸本 利光ら“続生化学実験講座1・遺伝子研究法II”335-354)、カセット変異法(岸本 利光、“新生化学実験講座2・核酸III 組換えDNA技術”242-251)などに従うことができる。このように、本発明のDNAを元に遺伝子工学的手法により得られるタンパク質であっても、ホスホリパーゼC活性を有する限り本発明に含まれる。このようなホスホリパーゼCは、必ずしも配列番号5に記載のアミノ酸配列の全てを有するものである必要はなく、例えばその部分配列からなるタンパク質であっても、該タンパク質がホスホリパーゼC活性を示す限り本発明のホスホリパーゼCに包含される。また、このようなホスホリパーゼCをコードするDNAも本発明に含まれる。
【0063】
本発明に用いるホスホリパーゼCはホスホリパーゼC生産菌から精製したもの、粗精製したもの、菌体の破砕液の他、菌体の培養上清をそのまま用いたものでも良い。ホスホリパーゼC生産菌を培養する際には、培地中に炭素源、窒素源の他に界面活性剤を添加して培養するのが好ましい。あるいは魚粉、スリゴマ、綿実粕等天然素材の培地で培養するのが好ましい。界面活性剤としては、トライトン、トゥイーン、ショ糖脂肪酸エステル、コール酸ナトリウム、デオキシコール酸ナトリウムおよびサポニン等を挙げることができる。
【0064】
ホスホリパーゼC生産菌の培養は通常の培養装置、培地を用いて行なうことができる。培養は液体培養、固体培養等の方法を適宜選択することができる。液体培養の場合はフラスコ培養や発酵槽を用いた培養を行なうことができ、培養開始後は培地の追加のないバッチ培養法や培養中に適宜培地を添加していく流加培養法を用いることができる。培地には炭素源、窒素源を添加し、必要に応じてビタミン、微量金属等を添加することができる。炭素源としては、グルコース、マンノース、ガラクトース、フルクトース等の単糖類、マルトース、セロビオース、イソマルトース、ラクトース、スクロース等の二糖類、デンプン等の多糖類、マルトエクストラクト等を挙げることができる。窒素源としてはアンモニア、硫酸アンモニウム、硝酸アンモニウム等の無機窒素、イーストエクストラクト、マルトエクストラクト、コーンスティープリカー、ペプトン等の有機窒素を用いることができる。これらの培地中の組成物量は適宜選択することができる。培養温度、pH、通気攪拌量はホスホリパーゼC生産に適するように適宜選択することができる。
【0065】
ホスホリパーゼC生産菌の培養終了後に遠心分離を行ない、菌体を除いた培養上清をそのまま粗酵素液として用いることができる。また、粗酵素液をイオン交換クロマトグラフィー等によって粗精製したり、精製したりしたものを用いることもできる。
【0066】
本発明にいうアスペルギルス・オリザエFERM ABP−10200株またはNBRC 4190株あるいはアスぺルギルス・タマリIAM 13907株は、本発明のホスホリパーゼCを産生することができるものである限り、その全ての変異株を包含する。また、これらの変異株の中には、遺伝的方法、たとえば組み換え、形質導入、形質転換等によりえられたものも含有される。即ち、本発明のホスホリパーゼCを生産するアスペルギルス・オリザエFERM ABP−10200またはNBRC 4190株あるいはアスぺルギルス・タマリIAM 13907株、それらの変異株およびそれらと明確に区別されない菌株は全てアスペルギルス・オリザエFERM ABP−10200株またはNBRC 4190株あるいはアスぺルギルス・タマリIAM 13907株に包含される。
【0067】
また、本発明のホスホリパーゼCは、ベクターに本発明のDNAが挿入された組換えプラスミドで宿主細胞を形質転換し、該形質転換された細胞の培養産物から得る事もできる。このように適当なベクターに本発明のDNAが挿入された組換えプラスミドも本発明に含まれる。このような目的に用いるベクターとしては、一般に知られているさまざまなベクターを用いることができる。好適なものとしては、原核細胞用ベクター、真核細胞用ベクター、哺乳動物由来の細胞用ベクターなどが挙げられるが、これに限定されない。このような組換えプラスミドにより、他の原核生物、または真核生物の宿主細胞を形質転換させることができる。さらに、適当なプロモーター配列および/または形質発現に関わる配列を有するベクターを用いるか、もしくはそのような配列を導入することにより、発現ベクターとすることで、それぞれの宿主において遺伝子を発現させることが可能である。このような発現ベクターは、本発明の組換えプラスミドの好適な態様である。
【0068】
本発明の組換えプラスミドを、各種細胞に導入することにより、宿主細胞を得ることができる。このような細胞は、プラスミドを導入することができる細胞であれば原核細胞であっても真核細胞であってもよい。
【0069】
原核細胞の宿主としては、例えば、大腸菌(Escherichia coli)や枯草菌(Bacillus subtilis)などが挙げられる。目的の遺伝子をこれらの宿主細胞内で形質転換させるには、宿主と適合し得る種由来のレプリコンすなわち複製起点と、調節配列を含んでいるプラスミドベクターで宿主細胞を形質転換させる。また、ベクターとしては、形質転換細胞に表現形質(表現型)の選択性を付与することができる配列を有するものが好ましい。
【0070】
例えば、大腸菌としてはK12株などがよく用いられ、ベクターとしては、一般にpBR322やpUC系のプラスミドが用いられるが、これらに限定されず、公知の各種菌株、およびベクターを使用することができる。
【0071】
プロモーターとしては、大腸菌においては、トリプトファン(trp)プロモーター、ラクトース(lac)プロモーター、トリプトファン・ラクトース(tac)プロモーター、リポプロテイン(lpp)プロモーター、ポリペプチド鎖伸張因子Tu(tufB)プロモーター等が挙げられ、どのプロモーターも本発明のホスホリパーゼCの産生に使用することができる。
【0072】
枯草菌としては、例えば207−25株が好ましく、ベクターとしてはpTUB228(Ohmura, K. et al. (1984) J. Biochem. 95, 87-93)などが用いられるが、これに限定されるものではない。
【0073】
プロモーターとしては、枯草菌のα−アミラーゼのシグナルペプチド配列をコードするDNA配列を連結することにより、菌体外での分泌発現も可能となる。
【0074】
真核細胞の宿主細胞には、脊椎動物、昆虫、酵母などの細胞が含まれ、脊椎動物細胞としては、哺乳動物由来の細胞、例えば、サルの細胞であるCOS細胞(Gluzman, Y. (1981) Cell 23, 175-182、ATCC CRL−1650)やチャイニーズ・ハムスター卵巣細胞(CHO細胞、ATCC CCL−61)のジヒドロ葉酸還元酵素欠損株(Urlaub, G. and Chasin, L. A. (1980) Proc. Natl. Acad. Sci. USA 77, 4126-4220)等を用いることができる。
【0075】
脊椎動物細胞の発現プロモーターとしては、通常発現しようとする遺伝子の上流に位置するプロモーター、RNAのスプライス部位、ポリアデニル化部位、および転写終結配列等を有するものを使用することができ、さらにこれは必要により複製起点を有していてもよい。該発現ベクターの例としては、SV40の初期プロモーターを有するpSV2dhfr(Subramani, S. et al. (1981) Mol. Cell. Biol. 1, 854-864)等を挙げることができるが、これに限定されない。
【0076】
宿主細胞として、COS細胞を用いる場合を例に挙げると、発現ベクターとしては、SV40複製起点を有し、COS細胞において自立増殖が可能であり、さらに、転写プロモーター、転写終結シグナル、およびRNAスプライス部位を具えたものを用いることができる。該発現ベクターは、ジエチルアミノエチル(DEAE)−デキストラン法(Luthman, H. and Magnusson, G. (1983) Nucleic Acids Res, 11, 1295-1308)、リン酸カルシウム−DNA共沈殿法(Graham, F. L. and van der Eb, A. J. (1973) Virology 52, 456-457)、および電気パルス穿孔法(Neumann, E. et al. (1982) EMBO J. 1, 841-845)などによりCOS細胞に取り込ませることができ、かくして所望の形質転換細胞を得ることができる。また、宿主細胞としてCHO細胞を用いる場合には、発現ベクターと共に、抗生物質G418耐性マーカーとして機能するneo遺伝子を発現し得るベクター、例えばpRSVneo(Sambrook, J. et al. (1989) : “Molecular Cloning A Laboratory Manual“ Cold Spring Harbor Laboratory, NY)やpSV2−neo(Southern, P. J. and Berg, P. (1982) J. Mol. Appl. Genet. 1, 327-341)などをコ・トランスフェクトし、G418耐性のコロニーを選択することにより、本発明のホスホリパーゼCを安定に産生する形質転換細胞を得ることができる。
【0077】
昆虫細胞を宿主細胞として用いる場合には、鱗翅類ヤガ科のSpodoptera frugiperdaの卵巣細胞由来株化細胞(Sf−9またはSf−21)やTrichoplusia niの卵細胞由来High Five細胞(Wickham, T. J. et al, (1992) Biotechnol. Prog. I: 391-396)などが宿主細胞としてよく用いられ、バキュロウイルストランスファーベクターとしてはオートグラファ核多角体ウイルス(AcNPV)のポリヘドリンタンパク質のプロモーターを利用したpVL1392/1393がよく用いられる(Kidd, I. M. and V.C. Emery (1993) The use of baculoviruses as expression vectors. Applied Biochemistry and Biotechnology 42, 137-159)。この他にも、バキュロウイルスのP10や同塩基性タンパク質のプロモーターを利用したベクターも使用できる。さらに、AcNPVのエンベロープ表面タンパク質GP67の分泌シグナル配列を目的タンパク質のN末端側に繋げることにより、組換えタンパク質を分泌タンパク質として発現させることも可能である(Zhe-mei Wang, et al. (1998) Biol. Chem., 379, 167-174)。
【0078】
真核微生物を宿主細胞とした発現系としては、酵母が一般によく知られており、その中でもサッカロミセス属酵母、例えばパン酵母Saccharomyces cerevisiaeや石油酵母Pichia pastorisが好ましい。該酵母などの真核微生物の発現ベクターとしては、例えば、アルコール脱水素酵素遺伝子のプロモーター(Bennetzen, J. L. and Hall, B. D. (1982) J. Biol. Chem. 257, 3018-3025)や酸性フォスファターゼ遺伝子のプロモーター(Miyanohara, A. et al. (1983) Proc. Natl. Acad. Sci. USA 80, 1-5)などを好ましく利用できる。また、分泌型タンパク質として発現させる場合には、分泌シグナル配列と宿主細胞の持つ内在性プロテアーゼあるいは既知のプロテアーゼの切断部位をN末端側に持つ組換え体として発現することも可能である。例えば、トリプシン型セリンプロテアーゼのヒトマスト細胞トリプターゼを石油酵母で発現させた系では、N末端側に酵母のαファクターの分泌シグナル配列と石油酵母の持つKEX2プロテアーゼの切断部位をつなぎ発現させることにより、活性型トリプターゼが培地中に分泌されることが知られている(Andrew, L. Niles,et al. (1998) Biotechnol.Appl. Biochem. 28, 125-131)。
【0079】
上記のようにして得られる形質転換体は、常法に従い培養することができ、該培養により細胞内、または細胞外に本発明のホスホリパーゼCが産生される。該培養に用いられる培地としては、採用した宿主細胞に応じて慣用される各種のものを適宜選択でき、例えば、上記COS細胞であれば、RPMI1640培地やダルベッコ改変イーグル培地(以下「DMEM」という)などの培地に、必要に応じウシ胎児血清などの血清成分を添加したものを使用できる。培養条件としては、CO2濃度は0乃至50%の範囲であればよく、好適には1乃至10%でありより好適には5%である。培養温度は0乃至99℃であればよく、好適には20乃至50℃であり、より好適には35乃至40℃である。
【0080】
上記培養により形質転換体の細胞内または細胞外に組換えタンパク質として産生される本発明のホスホリパーゼCは、培養産物中から、そのタンパク質の物理化学的性質、化学的性質、生化学的性質(酵素活性など)等を利用した各種の分離操作(「生化学データブックII」、1175-1259項、第1版第1刷、1980年6月23日株式会社東京化学同人発行;Biochemistry, vol. 25, No.25, p8274-8277 (1986); Eur. J. Biochem., 163, p313-321 (1987)等参照)により分離、精製することができる。該方法としては、具体的には例えば通常の再構成処理、タンパク質沈殿剤による処理(塩析法)、遠心分離、浸透圧ショック法、凍結融解法、超音波破砕、限外ろ過、ゲル濾過、吸着クロマトグラフィー、イオン交換クロマトグラフィー、アフィニティークロマトグラフィー、高速液体クロマトグラフィー(HPLC)等の各種液体クロマトグラフィー、透析法、それらの組み合わせ等を例示することができる。上記により、高収率で所望の組換えタンパク質を工業的規模で製造することができる。また、発現させる組換えタンパク質に6残基からなるヒスチジンを繋げることにより、ニッケルアフィニティーカラムで効率的に精製することができる。上記方法を組み合わせることにより容易に高収率、高純度で本発明のホスホリパーゼCを大量に製造することができる。
【0081】
以上のような方法により製造されたホスホリパーゼCも本発明の好適な例としてあげることができる。
【0082】
ホスホリパーゼC生産菌とは、実質的にホスホリパーゼC生産能を有する微生物をいい、ホスホリパーゼCを菌体内に蓄積する微生物や、菌体外に分泌する微生物等を含む。ホスホリパーゼC生産菌の培養上清または培養上清から精製したホスホリパーゼCを用いる場合には菌体外にホスホリパーゼCを分泌する菌を用いることができる。
【0083】
本発明に用いるホスホリパーゼCとしては、アスペルギルス・オリザエまたはアスぺルギルス・タマリ由来のホスホリパーゼCを用いることができ、より好適なものは、アスペルギルス・オリザエFERM ABP−10200株またはNBRC 4190株あるいはアスぺルギルス・タマリIAM 13907株由来のホスホリパーゼCを用いることができる。ホスホリパーゼCは、これらホスホリパーゼC生産菌自身が生産するものでもよいし、その変異体または修飾体が生産するものであってもよく、更に、これらホスホリパーゼC生産菌のホスホリパーゼCをコードする遺伝子を宿主に導入して得られた形質転換体から生産される組換えタンパク質であってもよい。
【0084】
ホスホリパーゼC生産菌の入手
アスペルギルス・オリザエNBRC 4190株は独立行政法人製品評価技術基盤機構 バイオテクノロジー本部 生物遺伝資源部門(NBRC: NITE Biological Resource Center;〒292−0818 日本国千葉県木更津市かずさ鎌足2−5−8、ホームページアドレス <http://www.nite.go.jp/>)より入手することができる。
アスペルギルス・タマリIAM 13907(=IAM 13907)株は東京大学分子細胞生物学研究所(IAM: Institute of Molecular and Cellular Biosciences, The University of Tokyo ;〒113−0032 東京都文京区弥生1−1−1、ホームページアドレス <http://www.iam.u-tokyo.ac.jp/indexe.html>)より入手することができる。
【0085】
アスペルギルス・オリザエFERM ABP−10200株の菌学的性質を以下に示す。
アスペルギルス・オリザエFERM ABP−10200株をクリックの文献(Klich, M. A. (2002) Identification of common Aspergillus Centraalbureau voor Schimmelcultures, Utrecht, The Netherlands)に従い、4種類の培地(CYA培地, CY20S培地, CZ培地, MEA培地)に接種して、菌学的性状を観察した。
【0086】
色調の表示は「メチューン・ハンドブック・オブ・カラー」(Kornerup, A. and Wanscher, J. H. (1978) Methuen handbook of colour (3rd. edition). Erye Methuen, London.)に従った。
【0087】
4種類の培地(CYA培地, CY20S培地, CZ培地, MEA培地)の組成は以下の通りである。
【0088】
CYA培地[ザペック イースト アガー(Czapek Yeast Extract Agar)培地]
(K2HPO4 1.0g,*ザペック濃縮液 10ml,イーストエキス 5g,シュークロース 30g,寒天 15g,蒸留水 1000ml)
*ザペック濃縮液(NaNO3 30g, KCl 5g,MgSO4・7H2O 5g,FeSO4・7H2O 0.1g,ZnSO4・7H2O 0.1g,CuSO4・5H2O 0.05g,蒸留水100ml)
【0089】
CY20S培地[20% シュークロース ザペック イーストアガー(Czapek Yeast ExtractAgar with 20% Sucrose)培地]
(K2HPO4 1.0g,*ザペック濃縮液 10ml,イーストエキス 5g,シュークロース 200g,寒天 15g,蒸留水 1000ml)
【0090】
CZ培地[ザペック ドックス アガー (Czapek Dox Agar)培地]
(K2HPO4 1.0 g, *ザペック濃縮液 10 ml, シュークロース 30 g, 寒天 17.5 g, 蒸留水 1000 ml)
【0091】
MEA培地[モルト エキス アガー(Malt Extract Agar)培地]
(モルトエキス 20g,ペプトン 1g,グルコース 20g,寒天 20g,蒸留水 1000ml)
【0092】
1)菌学的性状
アスペルギルス・オリザエFERM ABP−10200株の菌学的性状
CYA培地でのコロニーは、25℃、1週間の培養で直径36−40mmである。コロニーはやや厚く、羊毛状で、中心部では綿毛状となり、中心部から放射状の溝を形成する。菌糸は白色である。分生子は中心部に疎に形成され、灰黄色(4B4)から黄白色(4A2)を呈する。浸出液、菌核は観察されない。裏面は淡黄色(2A4)から白色(2A1)で、中心部から放射状の溝を形成する。可溶性色素は観察されない。
CYA培地でのコロニーは、37℃、1週間の培養で直径54−58mmである。コロニーは厚く、羊毛状である。菌糸は白色である。分生子は、中心部に形成され、灰黄色(4B4)から黄白色(4A2)を呈する。浸出液、菌核は観察されない。裏面は薄橙色(4A4)から白色(4A2)で、中心部から放射状の溝を形成する。可溶性色素は観察されない。
【0093】
CY20S培地でのコロニーは、25℃、1週間の培養で直径35−41mmである。コロニー性状はCYA培地と同様となるが、中心部の綿毛状の菌糸がやや多い。
【0094】
CZ培地でのコロニーは、25℃、1週間の培養で直径17−21mmである。コロニー性状はCYA培地と同様となるが、コロニーは小さく、分生子の形成が疎となる。
【0095】
MEA培地でのコロニーは、25℃、1週間の培養で直径37−41mmである。コロニーは薄く、綿毛状である。菌糸は白色である。分生子は中心部に疎に形成され、暗緑色(26D4)から灰緑色(26D6)を呈する。浸出液、菌核は観察されない。裏面は灰黄色(4B3)から黄白色(4A2)で、可溶性色素は観察されない。
14℃から42℃まで生育し、18℃から38℃まで分生子形成が観察された。
【0096】
分生子頭は放射状から緩い円柱状である。分生子柄は幅6.7−13.6μm、長さ302.2−1398.0μm、無色、粗面である。頂嚢は亜球形からフラスコ形、幅は14.6−29.3μmである。アスペルギラ(aspergilla)は主に単列(uniseriate)、まれに2列(biseriate)である。メトレまたはフィアライドは頂嚢の上部半分から形成される。メトレは11.3−28.3×5.2−9.9μmである。フィアライドはフラスコ形で、8.4−21.3×3.8−7.7μmである。分生子は滑面、亜球形から卵形、直径4.2−6.3μmである。
【0097】
以上の菌学的性状より、本菌に該当する菌を検索したところ、クリックの文献に記載されているアスペルギルス オリザエ(アールブルク)コーン(Aspergillus oryzae(Ahlburg)Chon)の性状とほぼ一致した。よって、FERM ABP−10200株をアスペルギルス・オリザエ(アールブルク)コーン(Aspergillus oryzae (Ahlburg)Chon)と同定し、独立行政法人産業技術総合研究所特許生物寄託センターに寄託した。
【0098】
ホスホリパーゼC生産菌から得られたホスホリパーゼCの具体的な性質について以下に示すが、本発明のホスホリパーゼCの有する性質は必ずしもこれらに限定されるものではない。
【0099】
アスペルギルス・オリザエFERM ABP−10200株またはNBRC 4190株あるいはアスぺルギルス・タマリIAM 13907株により生産され、精製されたホスホリパーゼCは、以下の性質を有する。
1)SDS−PAGE電気泳動法にて分子量約87,000を示す。
2)卵黄由来レシチン(ナカライ・テスク(株)製)をpH3乃至pH9の範囲で加水分解する。
3)0℃乃至80℃の範囲で4)記載の加水分解活性を有する。
4)pH4.5において45℃以下の温度で安定である。
5)pH3乃至pH10の範囲で安定である。
6)2)記載の加水分解活性の最適pHはpH4.5である。
7)3)記載の加水分解活性の最適温度はpH4.5では65℃である。
8)ホスファチジルコリン、ホスファチジルエタノールアミン、ホスファチジルイノシトールおよびホスファチジルグリセロールを加水分解するが、グリセロホスホリルコリンおよびパラニトロフェニルリン酸を実質的に加水分解しない。
【0100】
一例として、卵黄由来ホスファチジルコリンに対する活性を100%としたときの相対活性を表1に示した。なお、表中卵黄由来基質および大豆由来基質はその旨を記載した。また、卵黄由来レシチンはナカライテスク(株)、大豆由来レシチンは辻製油(株)から購入し、それ以外の基質はシグマアルドリッチジャパン(株)から購入した。
【0101】
【表1】
【0102】
9)下記に示す部分アミノ酸配列を有する。配列はN末端から記す。
Thr−Ala−Asp−Ser−Ala−Thr−Ala−Ile−Gly−
Tyr−Val−Thr−Pro−Ser−Met(配列番号1)。
【0103】
Glu−Ala−Tyr−Gly−Ser−Leu−Leu−Thr−Pro−
Pro(配列番号2)。
【0104】
Val−Pro−Pro−Ser−His−Asn−Pro−Gln−Trp−
Ala(配列番号3)。
【0105】
10)タンパク質が糖鎖で修飾されている。
【0106】
以上のことから、本発明のホスホリパーゼCの有する性質としては以下のようなものが挙げられるが、これに限定されるものではない。
1)SDS−PAGE電気泳動法にて分子量約87,000を示す。
2)卵黄由来レシチン(ナカライテスク(株))をpH3乃至pH9の範囲で加水分解する。
3)0℃乃至80℃の範囲で4)記載の加水分解活性を有する。
4)pH4.5において45℃以下の温度で安定である。
5)pH3乃至pH10の範囲で安定である。
【0107】
また、本発明のホスホリパーゼCを製造する方法も本発明に含まれる。
アスペルギルス・オリザエFERM ABP−10200株またはNBRC 4190株あるいはアスぺルギルス・タマリIAM 13907株を初めとするホスホリパーゼC産生微生物を培地で培養することにより、ホスホリパーゼCを生産することができる。例えば、0.1乃至5.0%ポリペプトン(和光純薬工業(株))、0.1乃至1.0%イーストエクストラクト(日本ベクトン・ディッキンソン(株))に0.05乃至1.0%デオキシコール酸ナトリウムを添加した培地または0.1乃至4.0%ファーマメディア(TRANDERS PROTEIN(株))に0.05から1.0%トライトンX−100(シグマアルドリッチジャパン(株))もしくは0.05乃至0.3%卵黄レシチンを添加した培地あるいは1乃至10%魚粉(池口喜一郎商店)の培地で、16乃至45℃で1乃至15日間、100乃至250rpmで振とう培養する。
【実施例】
【0108】
以下に、実施例および試験例を挙げるが、本発明の範囲はこれらに限定されるものではない。
【0109】
実施例1 アスペルギルス・オリザエFERM ABP−10200株からのホスホリパーゼCの精製
1)粗酵素液の調製
滅菌した表2の組成の培地100mlが入っている500ml容の三角フラスコ(種フラスコ)にフィルター滅菌した5%デオキシコール酸ナトリウム溶液1mlおよびアスペルギルス・オリザエFERM ABP−10200株の菌体を接種し、26℃にて7日間、170rpmの振とう培養を行った。
【0110】
【表2】
【0111】
培養終了後、4℃、10,000×Gにて10分間の遠心分離を行った。得られた上清を粗酵素液とした。
【0112】
2)酵素活性測定法
ホスホリパーゼCの加水分解活性は以下のようにして測定した。
<1>レシチンの加水分解反応
卵黄レシチン(ナカライテスク(株))1.5gを4%(wt/v)TritonX−100 50mlに溶解した基質溶液60μlに200mM酢酸緩衝液(pH5.5)60μlを加えて37℃で保温した。この混合液に酵素液60μlを加え撹拌して均一にし、37℃で保温して3時間酵素反応を行った。
【0113】
<2>遊離ホスホリル塩基の加水分解反応
酵素反応の結果生じるホスホリル塩基をアルカリホスファターゼで加水分解した。<1>で調整した酵素反応液50μlに200mMトリス・塩酸緩衝液(pH8.0)50μlおよびアルカリホスファターゼ(シグマアルドリッチジャパン(株))1μlを加えて37℃で40分反応を行った。なおこのときブランクとしてアルカリホスファターゼを加えないサンプルも同時に調整した。
【0114】
<3>無機リン酸の定量
<2>の結果生じた無機リン酸をホスファCテストワコー(和光純薬工業(株))で定量した。<2>で得られた反応液100μlにホスファCテストワコーのA液およびB液を各1ml添加し、37℃で20分間反応した。この混合液の750nmにおける吸光度を測定した。ブランクとの差がホスホリパーゼC活性になる。酵素反応1分間当たり1μmolのホスホリル塩基を生成する酵素活性を1単位とした。
【0115】
3)精製酵素液の調製
1)で得られた粗酵素液1,200mlを10mMトリス・塩酸緩衝液(pH7.5)8,000mlに対して12時間ずつ8回透析した。これを、予め10mMトリス・塩酸緩衝液(pH7.5)で平衡化したDEAEトヨパール(東ソー(株))カラム(直径2.2cm×長さ20cm)に添加し、吸着させた。10mMトリス・塩酸緩衝液(pH7.5)で該カラムを十分洗浄した後、10mMトリス・塩酸緩衝液(pH7.5)600ml中に0乃至0.2Mの塩化ナトリウムの直線的濃度勾配を作製して該カラムに吸着した成分を溶出させた。卵黄由来レシチン分解活性は塩化ナトリウム濃度が0.05M乃至0.08Mの画分(90ml)に溶出された。これを粗精製酵素画分とした。
【0116】
得られた活性画分90mlを20mMトリス・塩酸緩衝液(pH7.5)4,000mlに対して12時間ずつ3回透析した後、予め20mMトリス・塩酸緩衝液(pH7.5)で平衡化したMonoQ(アマシャムバイオサイエンス(株))カラム(直径10mm×長さ10cm)に添加し、吸着させた。該カラムを20mMトリス・塩酸緩衝液(pH7.5)で十分洗浄した後、20mMトリス・塩酸緩衝液(pH7.5)250ml中に0乃至0.2Mの塩化ナトリウムの直線的濃度勾配を作製して該カラムに吸着した成分を溶出させた。卵黄由来レシチン分解活性は塩化ナトリウム濃度が0.1M乃至0.12Mの画分(25ml)に溶出された。
【0117】
得られた活性画分25mlを濃縮し、予め0.15M塩化ナトリウムを含む10mMトリス・塩酸緩衝液(pH7.5)で平衡化したHiLoad Sephadex200pg(アマシャムバイオサイエンス(株))カラム(直径16mm×60cm)に添加した後、0.15M塩化ナトリウムを含む10mMトリス・塩酸緩衝液(pH7.5)にて溶出させた。卵黄由来レシチン分解活性は、溶出量が60ml乃至70mlの画分に溶出された。
この画分を精製酵素溶液とした。
【0118】
4)精製酵素の分子量測定
12.5%ポリアクリルアミドゲルを用いたSDS−PAGE電気泳動法(Laemmli, U.K., Nature, 227, 680(1970)参照)により、精製酵素の分子量を求めた。標準タンパク質として次のものを用いた:a.ホスホリラーゼ(phosphorylase)、分子量94,000:b.アルブミン(albumin)、分子量67,000:c.オバルブミン(ovalbumin)、分子量43,000:d.カルボニック・アンヒドラーゼ(carbonic anhydrase)、分子量30,000:e.トリプシン・インヒビター(trypsin inhibitor)、分子量20,100:f.α−ラクタルブミン(α-lactalbumin)、分子量14,400。
精製酵素は分子量約87,000の単一バンドを示した。
【0119】
実施例2 アスぺルギルス・タマリIAM 13907株からのホスホリパーゼCの精製
1)粗酵素液の調製
表2に示した組成の培地100mlが入っている500ml容の三角フラスコ(種フラスコ)にフィルター滅菌した5%デオキシコール酸ナトリウム溶液1mlおよびアスぺルギルス・タマリIAM 13907株の菌体を接種し、26℃にて5日間、170rpmの振とう培養を行った。培養終了後、4℃、10,000×Gにて10分間の遠心分離を行った。得られた上清を粗酵素液とした。
【0120】
2)精製酵素液の調製
実施例1. 3)と同様に精製を行い、精製酵素溶液を得た。
【0121】
3)精製酵素の分子量測定
実施例1. 4)と同じ方法で測定した。
精製酵素は分子量約87,000の単一バンドを示した。
【0122】
実施例3 アスぺルギルス・オリザエNBRC 4190株からのホスホリパーゼCの精製
1)粗酵素液の調製
滅菌した表3の組成の培地100mlが入っている500ml容の三角フラスコ(種フラスコ)にアスペルギルス・オリザエNBRC 4190株の菌体を接種し、26℃にて7日間、170rpmの振とう培養を行った。
【0123】
【表3】
【0124】
培養終了後、4℃、10,000×Gにて10分間の遠心分離を行った。得られた上清を粗酵素液とした。
【0125】
2)精製酵素液の調製
1)で得られた粗酵素液600mlを10mMトリス・塩酸緩衝液(pH7.5)8,000mlに対して12時間ずつ5回透析した。これを、予め10mMトリス・塩酸緩衝液(pH7.5)で平衡化したDEAEトヨパール(東ソー(株))カラム(直径2.2cm×長さ20cm)に添加し、吸着させた。10mMトリス・塩酸緩衝液(pH7.5)で該カラムを十分洗浄した後、10mMトリス・塩酸緩衝液(pH7.5)600ml中に0乃至0.6Mの塩化ナトリウムの直線的濃度勾配を作製して該カラムに吸着した成分を溶出させた。卵黄由来レシチン分解活性は塩化ナトリウム濃度が0.30M乃至0.35Mの画分(50ml)に溶出された。これを粗精製酵素画分とした。
【0126】
得られた活性画分50mlを20mMトリス・塩酸緩衝液(pH7.5)4,000mlに対して12時間ずつ3回透析した後、予め20mMトリス・塩酸緩衝液(pH7.5)で平衡化したMonoQ(アマシャムバイオサイエンス(株))カラム(直径10mm×長さ10cm)に添加し、吸着させた。該カラムを20mMトリス・塩酸緩衝液(pH7.5)で十分洗浄した後、20mMトリス・塩酸緩衝液(pH7.5)250ml中に0乃至0.6Mの塩化ナトリウムの直線的濃度勾配を作製して該カラムに吸着した成分を溶出させた。卵黄由来レシチン分解活性は塩化ナトリウム濃度が0.12M乃至0.20Mの画分(35ml)に溶出された。
【0127】
得られた活性画分35mlを濃縮し、予め0.15M塩化ナトリウムを含む10mMトリス・塩酸緩衝液(pH7.5)で平衡化したHiLoad Sephadex200pg(アマシャムバイオサイエンス(株))カラム(直径16mm×60cm)に添加した後、0.15M塩化ナトリウムを含む10mMトリス・塩酸緩衝液(pH7.5)にて溶出させた。卵黄由来レシチン分解活性は、溶出量が60ml乃至70mlの画分に溶出された。
この画分を精製酵素溶液とした。
【0128】
3)精製酵素の分子量測定
実施例1の4)と同じ方法で測定した。
精製酵素は分子量約87,000の単一バンドを示した。
【0129】
実施例4 アスぺルギルス・オリザエNBRC 4190株からのホスホリパーゼCの部分アミノ酸配列の決定
精製した酵素溶液を約1mg/ml程度まで限外濾過膜(ザルトリウス(株)VIVASPIN2、分子量分画1万)を用いて濃縮した。濃縮酵素液150μlにDenature buffer(6M塩酸グアニジン、10mM EDTA、0.1M炭酸水素アンモニウムpH7.8)150μlおよび50mM ジチオスレイトール6μlを加え、95℃で10分間反応させた。室温まで冷却後、反応液にDenature buffer溶液に溶解した50mMヨードアセトアミド30μlを加え、暗所室温で1時間反応させた。この溶液を20mM炭酸水素アンモニウム(pH8.0)で予め平衡化しておいたHitrap Desalting(アマシャムバイオサイエンス(株))カラムに添加した後、20mM炭酸水素アンモニウム(pH8.0)にて溶出させた。溶出量が1.5ml乃至2.5mlの画分に溶出した溶液を凍結乾燥し、20mM炭酸水素アンモニウム(pH8.0)100μlに溶解した。得られた溶液にトリプシン(Modified、プロメガ(株))60ユニットを加え、37℃で18時間酵素反応させた。反応液を高速液体クロマトグラフィー(日立製作所(株))にかけて分解アミノ酸のピークを分離した。分離条件を以下に示す。
【0130】
カラム:東ソー(株) TSKgel ODS-120T (4.6mm×150mm)
A buffer:0.1%TFA/Water
B buffer:0.1%TFA/Acetonitril
グラジエント:10→70%B 2%/ml
Flow:1ml/分
【0131】
分離した分解アミノ酸のうち、3本のピーク(B buffer濃度 30から38%程度)を分取し、これについてアミノ酸配列解析装置(Procise cLC、アプライドバイオシステムズジャパン(株))でアミノ酸配列を解析した。その結果得られた部分アミノ酸配列をアミノ末端側から記す。
【0132】
Thr−Ala−Asp−Ser−Ala−Thr−Ala−Ile−Gly−
Tyr−Val−Thr−Pro−Ser−Met(配列番号1)。
【0133】
Glu−Ala−Tyr−Gly−Ser−Leu−Leu−Thr−Pro−
Pro(配列番号2)。
【0134】
Val−Pro−Pro−Ser−His−Asn−Pro−Gln−Trp−
Ala(配列番号3)。
【0135】
実施例5 アスぺルギルス・オリザエNBRC 4190株由来のホスホリパーゼCをコードするDNAの同定
1)全RNAの精製
アスぺルギルス・オリザエNBRC 4190株を液体培地(2% ポリペプトン、0.5% イースト エクストラクト、0.02% リン酸水素2カリウム、0.05% 硫酸マグネシウム)20mlで26℃、1日間前培養した。その後、液体培地(5%魚粉)に1%植菌し、26℃で4日間培養した。培養した菌体を吸引集菌し、−80℃で冷やした乳鉢(オートクレーブ滅菌済)に移した。液体窒素を加えながら、乳棒で菌体を破砕し、粉末状にした。完全に粉末状になった菌体をRNeasy Plant Mini Kit(キアゲン(株))を用いて全RNAの精製を行った。905ng/μlの濃度の溶液が、50μl得られた。
【0136】
2)ホスホリパーゼC遺伝子の解読
5'RACE法および3'RACE法にて遺伝子配列の解読をおこなった。具体的には、5'RACE Systemおよび3'RACE System(いずれもインビトロジェン(株))を使用し、ポリメラーゼとしてEx TaqTM(タカラバイオ(株))を用いてPCRをおこなった。このときに使用したPCRプライマーは、5'側の遺伝子配列増幅用に5'-GGCCACGCGTCGACTAGTAC-3'および5'-GACAGTGTAGTCGAGCACAGCGAA-3'、3'側の遺伝子配列増幅用に5'-GACTCTGCCACCGCAATCGGCTA-3'および5'-GGCCACGCGTCGACTAGTAC-3'を用いた。PCRサイクルは、94℃・5分、(94℃・30秒、55℃・30秒、72℃・2分30秒)×30、72℃・10分、4℃で増幅した。5'側の遺伝子配列約1200b.p.、3'側の遺伝子配列約800b.p.の長さのDNAが増幅された。
【0137】
各々のPCR産物をアガロースゲル電気泳動をおこなった後、QIAquick Gel Extarction Kit(キアゲン(株))で精製した。精製物をTOPOTMTAクローニングキット(インビトロジェン(株))を用いてベクターに連結し、形質転換をおこなった。形質転換した大腸菌を37℃、一晩寒天培地(LB/Agar(和光純薬工業(株)))上で培養した後、生育したコロニーを37℃、一晩液体培地(LBbroth(和光純薬工業(株)))で培養した。増殖した大腸菌からプラスミドをQIAprep Spin Miniprep Kit(キアゲン(株))を用いて精製し、DNA配列解析をおこなった。DNA配列解析の結果を配列番号4に示した。また、DNA配列から推定されるアミノ酸配列を配列番号5に示した。
【0138】
実施例6 アスぺルギルス・オリザエNBRC 4190株由来ホスホリパーゼCのグリコペプチダーゼ処理
1)酵素反応
精製した酵素溶液を限外濾過膜(ザルトリウス(株)VIVASPIN2、分子量分画1万)を用いて濃縮した。濃縮酵素液20μlに、蒸留水15μl、0.25mMリン酸緩衝液(pH7.5)10μlおよび1M 2-メルカプトエタノール/2%ドデシル硫酸ナトリウム溶液2.5μlを加え、95℃にて5分間反応した。急冷後、15%Triton X-100(シグマアルドリッチジャパン(株))2.5μlを加え、グリコペプチダーゼF(シグマアルドリッチジャパン(株))10ユニットを加えて、37℃にて20時間反応した。
【0139】
2)反応後の分子量測定
実施例1の4)と同じ方法で測定した。
酵素反応後の分子量約63,000の単一バンドを示した。
【0140】
試験例1 アスペルギルス・オリザエFERM ABP−10200株由来ホスホリパーゼCの精製酵素液の諸性質
実施例1の3)で得られた精製酵素液について、活性測定を行なった。
【0141】
1)pH活性
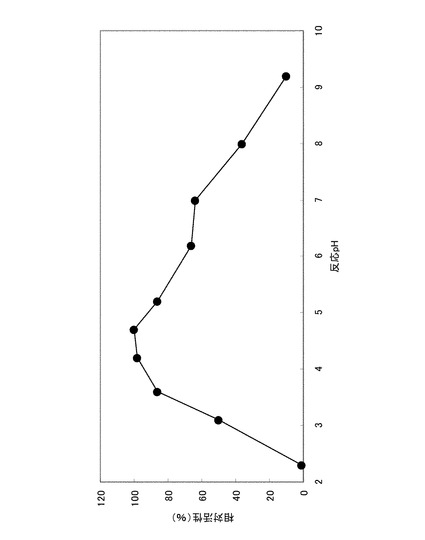
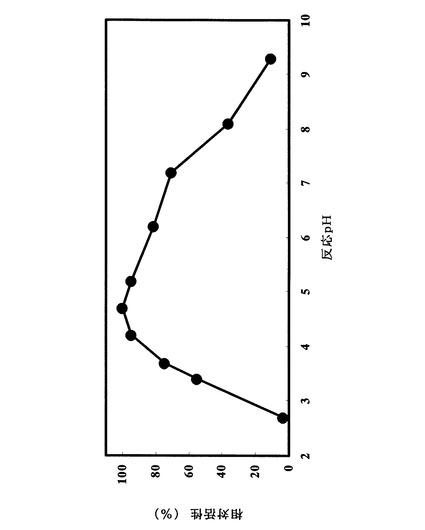
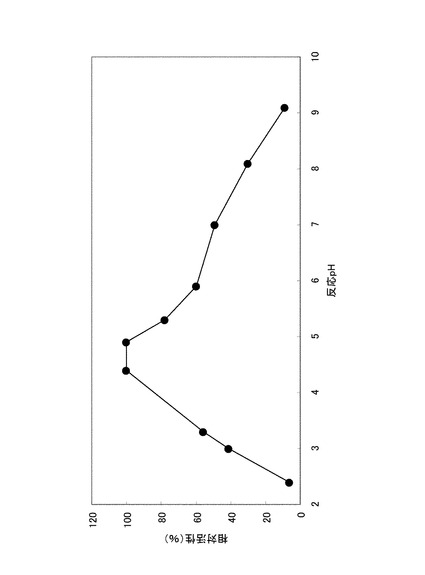
実施例1の2)に示した方法に拠った。ただし酵素反応時間は37℃、10分間とした。また、緩衝液は次のものを用いた:pH2.3乃至pH3.7の場合、グリシン−塩酸緩衝液:pH3.3乃至pH6.2の場合、クエン酸−クエン酸ナトリウム緩衝液:pH6.1乃至pH8.0の場合、MOPS緩衝液:pH8.2乃至pH9.2の場合、Atkins−Pantin緩衝液。最も活性が高かったpH条件での加水分解活性を100%とし、各pHにおける酵素の加水分解活性を相対値として図1に記載した。至適pHはクエン酸緩衝液中pH4.5付近であった。
【0142】
2)温度活性
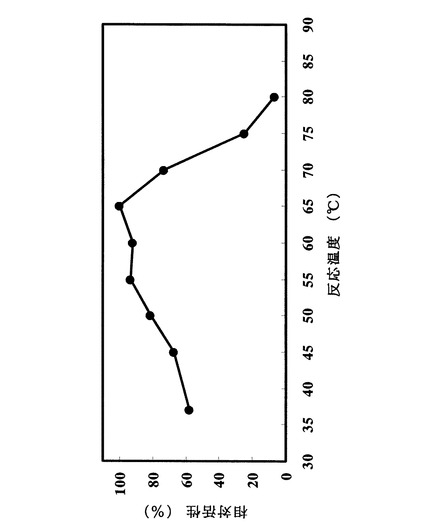
クエン酸緩衝液pH4.5における温度活性を測定した。測定法は実施例1の2)に示した方法に拠った。ただし酵素反応時間は37℃、20分間とした。最も活性が高かった温度条件での加水分解活性を100%とし、各温度における酵素の加水分解活性を相対値として図2に記載した。至適温度は65℃付近であった。
【0143】
3)温度安定性
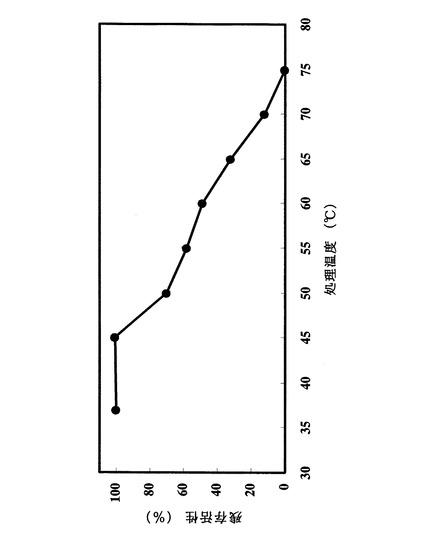
精製酵素溶液を種々の温度で30分間処理した後、その残存加水分解活性を測定した。あらかじめ処理温度に保持した25mMクエン酸緩衝液(pH4.5)90μlに、精製酵素液10μlを加え撹拌して均一にし、30分間保温した。実施例1で作製した卵黄レシチン溶液60μlに200mMクエン酸緩衝液(pH4.5)60μlを加え37℃に保持し、加温処理した酵素液60μlを加え、37℃で30分間酵素反応を行った。遊離ホスホリル塩基の定量は実施例1の2)にしたがった。最も高い残存加水分解活性を100%とし、各温度における加水分解活性を相対値として図3にまとめた。pH4.5において少なくとも60℃以下の温度で安定であった。
【0144】
4)pH安定性
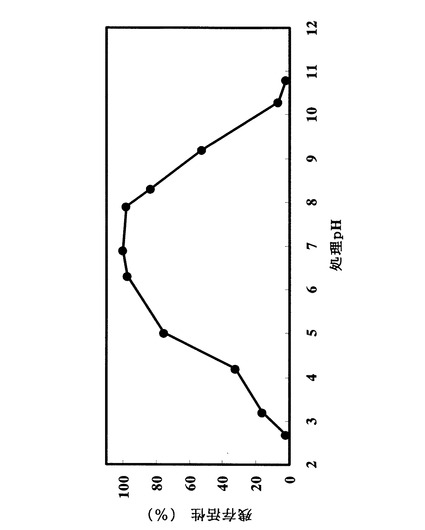
精製酵素液30μlに、以下に述べる各pHの200mM緩衝液30μlを添加し、37℃にて30分間保温した。緩衝液は次のものを用いた:pH2.7乃至pH3.2の場合、グリシン−塩酸緩衝液:pH3.5乃至pH6.1の場合、酢酸−酢酸ナトリウム緩衝液:pH6.3乃至pH7.9の場合、MOPS緩衝液:pH8.3乃至pH10.8の場合、Atkins−Pantin緩衝液。200mMクエン酸緩衝液(pH4.5)60μl、および実施例1の2)記載の卵黄レシチン溶液60μlの混合液に、加温した酵素混合液60μlに水60μlを加えた溶液のうち60μlを加え撹拌して均一にし、37℃で10分間酵素反応を行った。遊離ホスホリル塩基の定量は実施例1の2)にしたがった。最も高い残存加水分解活性を100%とし、各pHにおける加水分解活性を相対値として図4にまとめた。pH3乃至pH10の範囲で安定であった。
【0145】
5)精製酵素の基質選択性
次に、基質選択性を測定した。実施例1の3)で調製した精製酵素液を用いた。測定方法は実施例1の2)に拠った。ただし、酵素反応は200mMクエン酸緩衝液pH4.5中で37℃で保温して10分間行った。卵黄由来ホスファチジルコリンに対する加水分解活性を100%としたときの、相対活性を表4に示す。
【0146】
【表4】
【0147】
試験例2.アスペルギルス・タマリIAM 13907株由来ホスホリパーゼCの精製酵素液の諸性質
実施例2の2)で得られた精製酵素液について、活性測定を行なった。
【0148】
1)pH活性
試験例1.に示した方法に拠った。最も活性が高かったpH条件での加水分解活性を100%とし、各pHにおける酵素の加水分解活性を相対値として図5に記載した。至適pHはpH4.5付近であった。
【0149】
2)温度活性
試験例1.に示した方法に拠った。最も活性が高かった温度条件での加水分解活性を100%とし、各温度における酵素の加水分解活性を相対値として図6に記載した。至適温度は65℃付近であった。
【0150】
3)温度安定性
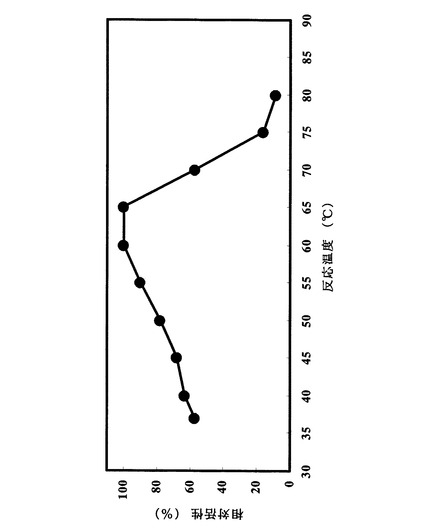
試験例1.に示した方法に拠った。最も高い残存加水分解活性を100%とし、各温度における加水分解活性を相対値として図7にまとめた。pH4.5において45℃以下の温度で安定であった。
【0151】
4)pH安定性
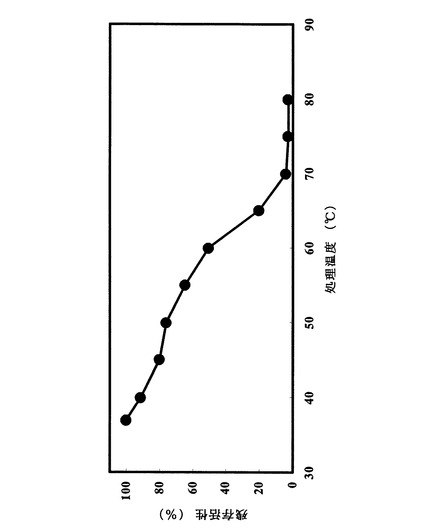
試験例1.に示した方法に拠った。最も高い残存加水分解活性を100%とし、各pHにおける加水分解活性を相対値として図8にまとめた。pH3乃至pH10の範囲で安定であった。
【0152】
2)精製酵素の基質選択性
試験例1.に示した方法に拠った。卵黄由来ホスファチジルコリンに対する加水分解活性を100%としたときの、相対活性を表5に示す。
【0153】
【表5】
【0154】
試験例3.アスペルギルス・オリザエNBRC4190株由来ホスホリパーゼCの精製酵素液の諸性質
実施例3の2)で得られた精製酵素液について、活性測定を行なった。
【0155】
1)pH活性
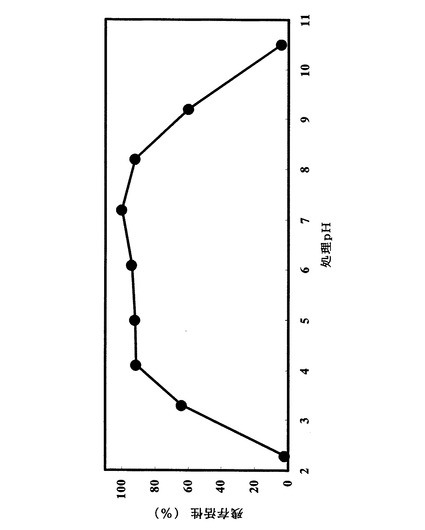
試験例1.に示した方法に拠った。最も活性が高かったpH条件での加水分解活性を100%とし、各pHにおける酵素の加水分解活性を相対値として図9に記載した。至適pHはpH4.5付近であった。
【0156】
2)精製酵素の基質選択性
試験例1.に示した方法に拠った。卵黄由来ホスファチジルコリンに対する加水分解活性を100%としたときの、相対活性を表6に示す。
【0157】
【表6】
【0158】
発明の効果
以上述べたように、本発明のホスホリパーゼCはアスぺルギルス・オリザエFERM ABP−10200株またはNBRC 4190株あるいはアスぺルギルス・タマリIAM 13907株由来であり安全性に優れ、様々なグリセロリン脂質を酸性側でも中性付近でも効率よく加水分解する能力を有し、クエン酸緩衝液中でも活性を有するとともにある程度熱的に安定であり、リン脂質以外のリン酸エステルを加水分解しない酵素であり、食品工業および製油工業いずれの分野においても優れた効果をあげられる酵素である。
【技術分野】
【0001】
本発明は、ホスホリパーゼC、該ホスホリパーゼCを生産する糸状菌、糸状菌の培養物から該ホスホリパーゼCを分離精製する方法、該ホスホリパーゼCをコードするDNAおよび該ホスホリパーゼCの製造方法等に関する。具体的には、本発明は、食品工業および医薬品工業における使用に特に適するホスホリパーゼC、特に糸状菌アスペルギルス・オリザエ(Aspergillus oryzae)あるいはアスぺルギルス・タマリ(Aspergillus tamarii)が生産するホスホリパーゼC、該ホスホリパーゼCを生産する糸状菌、糸状菌の培養物から該ホスホリパーゼCを分離精製する方法、該ホスホリパーゼCをコードするDNAおよび該ホスホリパーゼCの製造方法等に関する。
【背景技術】
【0002】
〔1〕ホスホリパーゼC
従来、動物および微生物がホスホリパーゼCを生産することが知られている。動物由来の酵素は主としてホスファチジルイノシトール選択的なホスホリパーゼCである。また、微生物のなかでは細菌、放線菌、酵母およびカビ由来のホスホリパーゼCが知られている。細菌、放線菌および酵母が生産するホスホリパーゼCは、ホスファチジルイノシトール選択的あるいはホスファチジルコリン選択的なものがほとんどである。
【0003】
細菌由来のホスホリパーゼCとしては、例えばシュードモーナス・シュルキリエンシス(Psudomonus schuylkilliensis)(例えば、特許文献1参照)、ブルコールデリア・シュードマレイ(Bulkholderia pseudomallei)(例えば、非特許文献1参照)、バチラス・セレウス(Bacillus cereus)(例えば、非特許文献2参照)、スタフィロコッカス・アウレウス(Staphylococcus aureus)(例えば、非特許文献3参照)およびクロストリディウム・ぺルフリンゲンス(Clostridium perfringens)(例えば、非特許文献4参照)等が生産するホスホリパーゼCが知られている。
【0004】
放線菌由来のホスホリパーゼCとしては、例えばストレプトマイセス・ハチジョウエンシス(Streptomyces hachijyoensis)(例えば、特許文献2参照)が生産するホスホリパーゼCが知られている。
【0005】
酵母由来のホスホリパーゼCとしては、例えばキャンディダ・アルビカンス(Candida albicans)(例えば、非特許文献5参照)、サッカロマイセス・セレビシアエ(Saccharomyces cerevisiae)(例えば、非特許文献6参照)等が生産するホスホリパーゼCが知られている。
【0006】
カビ由来のホスホリパーゼCは、従来2例知られている。一つはアスペルギルス・ニガー(Aspergillus niger)(例えば、特許文献3参照)、もうひとつはアスペルギルス・サイトイ(Aspergillus saitoi)(例えば、非特許文献7参照)が生産するホスホリパーゼCである。
【0007】
〔2〕レシチン
レシチンは、動物、植物、菌類に広く分布している代表的なグリセロリン脂質である。グリセロリン脂質とは、1,2−ジアシルグリセロールの3位にホスホリル塩基が共有結合している化合物である。塩基としては、コリン、エタノールアミン、セリン、イノシトールおよびグリセロール等が含まれており、その組成割合は由来により異なる。レシチンはグリセロリン脂質に含有される概念として用いるものとする。
【0008】
レシチンは、界面活性作用、酸化防止作用、生理活性作用等を有し、食品、飼料、医薬品等に利用されている。食品工業においては、卵黄レシチンや大豆レシチンなどに代表される天然レシチンが食品添加物として使用されており、食品の性質を改変するために主に乳化剤等として使用され、豊富に供給されている。
【0009】
〔3〕レシチンの酵素処理
レシチンを酵素により部分的に加水分解し、新たな性質を付与する検討も行われている。この際に用いられる酵素がホスホリパーゼ類であり、ホスホリパーゼA,B,C,Dが知られている。グリセロリン脂質の1位あるいは2位の脂肪酸を選択的に加水分解するのがホスホリパーゼAである。非選択的に加水分解するのがホスホリパーゼB、ジアシルグリセロールとホスホリル塩基に加水分解するのがホスホリパーゼC、ホスファチジン酸と塩基に分解するのがホスホリパーゼDである。
【0010】
食品工業の分野において、現在最も比較的多く使用されているのはホスホリパーゼAである。レシチンは水に難溶性であるが、レシチンにホスホリパーゼAを作用させるとアシル基が部分的に加水分解され、水溶性のリゾレシチンが生成する。リゾレシチンを食品添加物として使用した場合には、得られる食品の物性がこれまでレシチンを用いて得られていた食品の物性とは異なる可能性がある。
【0011】
〔4〕スフィンゴリン脂質
スフィンゴリン脂質は、グリセロリン脂質とともに、リン脂質を構成している。
スフィンゴリン脂質の代表はスフィンゴミエリンであり、セラミドの1級アルコールにコリンリン酸がリン酸ジエステル結合した化合物である。動物の臓器に広く含まれている。母乳にも含有されることから、乳児用粉ミルクに配合されることもある。
スフィンゴミエリンにホスホリパーゼCを作用させるとコリンリン酸がはずれ、セラミドが生成する。セラミドは保湿成分として、化粧品に広く使用されている。また、アトピー性皮膚炎はセラミドが不足することにより惹起されるという報告がある。
【0012】
〔5〕食品中でのホスホリパーゼCの使用
ホスホリパーゼCを用いることにより、下図に示すように水の存在下でレシチンからジアシルグリセロールを生産することができる。
【0013】
【化1】
【0014】
(式中、R1およびR2は、それぞれアルキル基を、R3は、コリン、エタノールアミン、グリセロール、イノシトール等の基を意味する。)
【0015】
食品素材中でこのような反応を起こさせることにより、ホスホリパーゼAとは本質的に異なった性質を付加した商品が提供できる可能性がある。
ここで用いられるホスホリパーゼCには、様々なグリセロリン脂質を加水分解することが求められる。たとえば大豆レシチンにはホスファチジルコリンおよびホスファチジルエタノールアミンを主として、ホスファチジルグリセロールあるいはホスファチジルイノシトール等も含まれていることが知られている。また、卵黄由来レシチンにはホスファチジルコリンおよびホスファチジルエタノールアミンが主として含まれている。したがって、用いられるホスホリパーゼCは、これらを区別無く加水分解することが望ましい。
【0016】
しかし、食品中にはリン脂質以外にもリン酸エステルが多く含有されており、それらを加水分解してリン酸を遊離させることは、当該食品の性質をさらに変質させてしまうことにつながるので、好ましいものではない。したがって、リン脂質以外のリン酸エステルは加水分解しないホスホリパーゼCが望ましく、例えばグリセロホスホリルコリンのように、リン脂質に類似するが脂質部分を含有しないリン酸エステルに対する酵素活性は無いホスホリパーゼCが望ましい。
【0017】
同様の観点から、用いられるホスホリパーゼCは、ホスファターゼ活性を有さないタンパク質であることが望ましい。すなわち、ホスファターゼの基質となるパラニトロフェニルリン酸分解活性を有しないホスホリパーゼCが望ましい。
【0018】
また、食品工業においては、食品の変質を防ぐため中性から弱酸性域でさまざまな処理を行うことが好ましく、ホスホリパーゼ等を含む酵素剤はこのpH域に高い活性を有していることが望まれる。
【0019】
〔6〕食品工業におけるホスホリパーゼCの使用
ホスホリパーゼCの用途には、例えば、パンの冷凍保存生地焼成時に、その表面で発生する老化、梨肌の緩和や食用油精製工程の改善がある。
大豆・菜種等から食用油を製造する際には、着色あるいは食味の劣化の原因となるため、レシチンは除去されるべき物質である。この目的のために、従来ホスホリパーゼAを用いてレシチンを部分的に加水分解し、リゾレシチンとすることで水溶性にして除去する方法が検討されてきた。
【0020】
しかし、ここでホスホリパーゼCを用いてレシチンをジアシルグリセロールとすることにより、トリアシルグリセロールと共に油の一成分にすることができる。すなわち、食用油の製造工程において、歩留まりを向上させる効果が期待される。
【0021】
ここで用いられるホスホリパーゼCには、様々なリン脂質を加水分解することが求められる。たとえば大豆油中には先述した様々なリン脂質が含まれている。また、綿実油あるいは菜種油中にも同様にさまざまなリン脂質が含まれている。用いられるホスホリパーゼCは、これらを区別なく加水分解することが望ましい。
【0022】
また、製油工業においては油以外の不純物を酸性加温条件下で除去するので、酸性域での活性が高く、ある程度温度安定性を有する酵素剤の使用が望まれる。処理される粗油を酸性にするためにクエン酸が使用されることがあるので、用いられるホスホリパーゼCにはクエン酸存在下での活性およびある程度の温度安定性が望まれる。
【0023】
〔7〕既知のホスホリパーゼCの問題点
動物、細菌、放線菌あるいは酵母が生産するホスホリパーゼCは、主としてホスファチジルイノシトールあるいはホスファチジルコリン選択的であり、様々な基質を分解する必要がある食品工業における使用には向かない。さらに、動物起源の酵素剤は、宗教的に受け入れられない国および地域があり、汎用性にも問題がある。さらに、ホスホリパーゼCを生産する細菌は、病原性を示すものがほとんどであり、安全性に問題がある。
【0024】
従来知られている糸状菌由来のホスホリパーゼCは、様々なリン脂質を加水分解する性質を有している。また、従来ホスホリパーゼCの産生に用いられているいずれの糸状菌も食用酵素生産に実績がある特徴を有している。アスペルギルス・ニガーとアスペルギルス・サイトイの両菌株由来の酵素の、温度あるいはpHに関する性質は極めて類似しており、分子量も同一である。いずれも酸性側で強い活性を有しているが、中性付近では全く活性がないという特徴を有している。したがって、酸性側でこれらの酵素を使用する可能性がある製油工業においては、これらの酵素は使用できる可能性がある。しかしクエン酸中での性質については記載がなく、実際に製油工業で使用できるかどうかは不明である(特許文献3および非特許文献7参照)。また、pH6での活性がほぼ0であることが示されており、中性付近で酵素反応を行うことが多いと考えられる食品工業においては、使用しづらい酵素である(特許文献3参照)。
【0025】
さらに、アスペルギルス・ニガーが生産するホスホリパーゼCは、ホスファターゼの基質であるホスファチジン酸の分解活性が非常に高いことが記載されている(特許文献3参照)。したがって、該酵素は、リン酸モノエステルを加水分解する能力を有することから、ホスファターゼ活性をも併せ持つタンパク質であることが推察される。さらに、アスペルギルス・サイトイが生産するホスホリパーゼCは、2−ヘキサデカノイルアミノ−4−ニトロフェニルホスホリルコリンの分解活性が非常に高いことが記載されている(非特許文献7参照)。したがって、該酵素は、リン脂質以外のリン酸ジエステルを加水分解する能力をも併せもつタンパク質であることが推察される。したがって、これらの酵素を用いて加工される食品は、不必要に改質されてしまう可能性がある。
【0026】
このように、従来知られているホスホリパーゼCは、酵素の特性として不十分であるか、あるいは安全性に問題がある等の問題点が挙げられ、ホスホリパーゼCとして市場に流通している酵素剤は現在までに存在しない。望ましい酵素剤の性質としては、既に食品用酵素剤の生産実績がある微生物由来であり、様々なリン脂質を酸性側でも中性付近でも効率よく加水分解する能力を有し、製油工業においても使用可能なようにクエン酸緩衝液中でも活性を有するとともにある程度熱的に安定であることが挙げられる。さらには、グリセロホスホリルコリンに代表されるようなリン脂質以外のリン酸エステルを加水分解しない性質を有するホスホリパーゼCが挙げられる。
【先行技術文献】
【特許文献】
【0027】
【特許文献1】特開昭50−1017183号公報
【特許文献2】特開昭49−55893号公報
【特許文献3】特開2000−166543号公報
【非特許文献】
【0028】
【非特許文献1】コルブスリサテら(Korbsrisate S. et. al.)「ジャーナル・オブ・クリニカル・マイクロバイオロジー」(Jounal of Clinical Microbiology)1999年、37巻、p.3742−3745
【非特許文献2】タンら(Tan C. et. al.)「プロテイン・エクスプレッション・アンド・ピューリフィケイション」(Protein Expression and Purification)1997年、10巻、p.365−372
【非特許文献3】ダウガーティーら(Daugherty S. et. al.)「インフェクション・アンド・イミュニティー」(Infection and Immunity)1993年、61巻、p.5078−5089
【非特許文献4】ティットバールら(Titball R. et. al.)「インフェクション・アンド・イミュニティー」(Infection and Immunity)1989年、57巻、p.367−376
【非特許文献5】アンダルツら(Andaluz E. et. al.)「イースト」(Yeast)2001年、18巻、p.711−721
【非特許文献6】パイネら(Payne W. et. al.)「モレキュラー・アンド・セルラー・バイオロジー」(Molecular and Cellular Biology)1993年、13巻、p.4351−4364
【非特許文献7】マツオカら(Matsuoka S. et. al.)「バイオテクノロジー・アンド・アプライドバイオケミストリー」(Biotechnology and Applied Biochemistry)1987年、9巻、p.401−409
【発明の概要】
【発明が解決しようとする課題】
【0029】
上述のように、望ましいホスホリパーゼCの性質としては、様々なリン脂質を酸性側でも中性付近でも効率よく加水分解する能力を有し、製油工業においても使用可能なようにクエン酸緩衝液中でも活性を有するとともに、ある程度熱的に安定であることが挙げられる。さらには、グリセロホスホリルコリンに代表されるような、リン脂質以外のリン酸エステルを加水分解しない性質を有するホスホリパーゼCであることが挙げられ、さらに好ましくは、既に食品用酵素剤の生産実績がある微生物由来のホスホリパーゼCであることが挙げられる。
このようなホスホリパーゼCを提供することは、この技術分野において非常に関心の高いことであった。
【課題を解決するための手段】
【0030】
本発明者らは、安全性に優れ、様々なリン脂質を酸性側でも中性付近でも効率よく加水分解する能力を有し、クエン酸緩衝液中でも活性を有するとともにある程度熱的に安定であり、リン脂質以外のリン酸エステルを加水分解しないホスホリパーゼCを見出すべく鋭意検討を行ったところ、アスぺルギルス・オリザエFERM ABP−10200株またはNBRC 4190株、あるいはアスぺルギルス・タマリIAM 13907株由来のホスホリパーゼCを精製し、アスぺルギルス・オリザエNBRC 4190株に由来するホスホリパーゼC遺伝子をクローニングし、本発明を完成するに至った。
【0031】
すなわち本発明は、(1) 酸性から中性のpHにおいて活性を示し、かつ、リン脂質以外のリン酸エステルを実質的に加水分解しないホスホリパーゼC。
(2) ホスファチジルイノシトール特異的ではない、(1)記載のホスホリパーゼC。
(3) 至適pHがpH3乃至pH6の範囲にある、(1)または(2)記載のホスホリパーゼC。
(4) pH7における相対活性が20%以上である、(1)〜(3)のいずれか1つ記載のホスホリパーゼC。
【0032】
(5) 糸状菌によって産生される、(1)〜(4)のいずれか1つ記載のホスホリパーゼC。
(6) 糸状菌がアスペルギルス属である、(5)に記載のホスホリパーゼC。
(7) アスぺルギルス・オリザエ(Aspergillus oryzae)あるいはアスぺルギルス・タマリ(Aspergillus tamarii)によって産生される、(6)に記載のホスホリパーゼC。
(8)アスぺルギルス・オリザエFERM ABP−10200株またはNBRC 4190株、あるいはアスぺルギルス・タマリIAM 13907株によって産生される、(7)に記載のホスホリパーゼC。
【0033】
(9) 以下の性質:
1)SDS−PAGE電気泳動法にて分子量約87,000を示す;
2)ホスファチジルコリン、ホスファチジルエタノールアミン、ホスファチジルイノシトールおよびホスファチジルグリセロールを加水分解する;
3)グリセロホスホリルコリンおよびパラニトロフェニルリン酸を実質的に加水分解しない;
4)卵黄由来レシチンをpH3乃至pH9の範囲で加水分解する;
5)0℃乃至80℃の範囲において4)記載の加水分解活性を有する;
6)温度安定性について、pH4.5において45℃以下の温度で安定である;
7)pH安定性について、pH3乃至pH10の範囲で安定である。
を示す(1)〜(8)のいずれか1つに記載のホスホリパーゼC。
【0034】
(10) 下記のa)〜d):
a)配列番号5に記載のアミノ酸配列からなるタンパク質;
b)配列番号4のヌクレオチド配列によりコードされるアミノ酸配列からなるタンパク質;
c)a)またはb)に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ、ホスホリパーゼC活性を有することを特徴とするタンパク質;
d)a)またはb)に記載のアミノ酸配列を含むことからなるタンパク質、
のいずれか一つに記載のタンパク質であるホスホリパーゼC。
【0035】
(11) (1)〜(4)、(9)および(10)のいずれか1つに記載のホスホリパーゼCを生産する能力を有する、単離されたアスぺルギルス・オリザエあるいはアスぺルギルス・タマリに属する糸状菌。ただし、アスぺルギルス・タマリIAM 13907株を除く。
(12)アスぺルギルス・オリザエFERM ABP−10200株またはNBRC 4190株である、(11)に記載の糸状菌。
【0036】
(13) 下記のa)〜d):
a)配列番号4のコード領域(CDS)のヌクレオチド配列からなるDNA;
b)上記a)に記載のDNAと70%以上のヌクレオチド配列相同性を有するヌクレオチド配列からなり、かつ、ホスホリパーゼC活性を有するタンパク質をコードすることを特徴とするDNA;
c)配列番号5に記載のアミノ酸配列からなるタンパク質をコードするDNA;
d)配列番号4のコード領域(CDS)のヌクレオチド配列を含むことからなるDNA、
のいずれか一つに記載のDNA。
(14) (11)に記載のDNAにコードされるタンパク質であるホスホリパーゼC。
【0037】
(15)
1)(9)に記載のアスペルギルス・オリザエあるいはアスぺルギルス・タマリを培養する工程、および、
2)1)の培養産物からホスホリパーゼCを分離・精製する工程、
を含む、ホスホリパーゼCの製造方法。
【0038】
(16) アスペルギルス・オリザエがアスペルギルス・オリザエFERM ABP−10200株またはNBRC 4190株、アスぺルギルス・タマリがアスぺルギルス・タマリIAM 13907株である(15)記載の方法。
に関する。
【図面の簡単な説明】
【0039】
【図1】アスぺルギルス・オリザエFERM ABP−10200株由来の精製ホスホリパーゼCの活性とpHとの関係を表した図である。
【図2】アスぺルギルス・オリザエFERM ABP−10200株由来の精製ホスホリパーゼCの活性と温度との関係を表した図である。
【図3】アスぺルギルス・オリザエFERM ABP−10200株由来の精製ホスホリパーゼCの温度安定性を表した図である。
【図4】アスぺルギルス・オリザエFERM ABP−10200株由来の精製ホスホリパーゼCのpH安定性を表した図である。
【図5】アスぺルギルス・タマリIAM 13907株由来の精製ホスホリパーゼCの活性とpHとの関係を表した図である。
【図6】アスぺルギルス・タマリIAM 13907株由来の精製ホスホリパーゼCの活性と温度との関係を表した図である。
【図7】アスぺルギルス・タマリIAM 13907株由来の精製ホスホリパーゼCの温度安定性を表した図である。
【図8】アスぺルギルス・タマリIAM 13907株由来の精製ホスホリパーゼCのpH安定性を表した図である。
【図9】アスぺルギルス・オリザエNBRC 4190株由来の精製ホスホリパーゼCの活性とpHとの関係を表した図である。
【発明を実施するための形態】
【0040】
以下、本発明を詳細に説明する。なお、特に指定しない限り、以下に示す各特性の測定方法は、後述する実施例または試験例に記載の測定方法による。
本発明は、酸性から中性のpHにおいて活性を示し、かつ、リン脂質以外のリン酸エステルを実質的に加水分解しないホスホリパーゼCであるか、あるいは酸性から中性のpHにおいて活性を示し、かつ、ホスファターゼ活性を有しないホスホリパーゼCである。
【0041】
「酸性から中性のpHにおいて活性を示す」とは、pH約3乃至約7の範囲において、相対活性が20%以上であることをいう。(酵素活性の測定法は「試験例1−1)pH活性」の記載に従う。)
【0042】
また、「リン脂質以外のリン酸エステルを実質的に加水分解しない」とは、好ましくは、ホスファチジルコリン(卵黄由来)に対する加水分解活性を100%とした場合における加水分解活性が、ホスファチジン酸に対しては30%以下、さらに好ましくは25%以下であること、および/またはグリセロホスホリルコリンに対しては15%以下、さらに好ましくは10%以下であること、および/またはパラニトロフェニルリン酸に対しては10%以下、さらに好ましくは5%以下であることをいう。「グリセロホスホリルコリンおよびパラニトロフェニルリン酸を実質的に加水分解しない」についても同様である。
ここで、ホスファチジン酸は、中性脂肪およびグリセロリン脂質の合成中間体であると認識されるため、本願の明細書等においてはグリセロリン脂質(リン脂質)には含まれないものとする。
【0043】
「ホスファターゼ活性を有しない」とは、ホスファチジン酸またはパラニトロフェニルリン酸の分解活性がホスファチジルコリンの分解活性の50%以下であることをいい、ホスファチジン酸の分解活性については、好ましくはホスファチジルコリンの分解活性の40%以下、さらに好ましくは30%以下、最も好ましくは25%以下である。また、パラニトロフェニルリン酸の分解活性については、好ましくはホスファチジルコリンの分解活性の30%以下、さらに好ましくは20%以下、最も好ましくは10%以下である。
【0044】
本発明のホスホリパーゼCは、スフィンゴミエリンに対する分解活性を有し、好ましくはその分解活性が、ホスファチジルコリンに対する分解活性と同等かそれ以上である。具体的には、ホスファチジルコリン(卵黄由来)に対する分解活性を100%とした場合に、ホスファチジルコリンに対する分解活性が、好ましくは90%以上、さらに好ましくは105%以上、最も好ましくは115%以上である。
【0045】
また、本発明のホスホリパーゼCは、ホスファチジルエタノールアミンに対する分解活性を有し、好ましくはその分解活性が、ホスファチジルコリンに対する分解活性と同等である。具体的には、ホスファチジルコリン(卵黄由来)に対する分解活性を100%とした場合にホスファチジルエタノールアミンに対する分解活性が、好ましくは80%以上、さらに好ましくは85%以上、最も好ましくは90%以上であり、150%以下である。
【0046】
ホスホリパーゼCが、「ホスファチジルイノシトール特異的ではない」とは、ホスファチジルイノシトールの分解活性(相対活性)に比べて、ホスファチジルイノシトール以外の基質の分解活性の方が高いことを意味する。ここで、ホスファチジルイノシトール以外の基質としては、好ましくはホスファチジルコリン、ホスファチジルエタノールアミン、ホスファチジルイノシトールまたはホスファチジルグリセロールである。
【0047】
本発明のホスホリパーゼCの至適pHは、酸性から中性のpHの範囲にあることが好ましく、さらにpH3乃至pH6の範囲にあることが好ましく、さらに好ましくはpH3乃至pH5の範囲であり、最も好ましくはpH4乃至pH5の範囲である。
【0048】
また、「pH7における相対活性」とは、最も活性が高いpHでの加水分解活性を100%とし、pH7における酵素の加水分解活性を相対値として百分率(%)で表したものをいう。本発明のホスホリパーゼCは、pH7における相対活性が、好ましくは20%以上であり、さらに好ましくは40%以上、最も好ましくは50%以上である。
【0049】
温度活性について、本発明のホスホリパーゼCは、至適温度が、45℃乃至70℃の範囲にあることが好ましく、さらには55℃乃至70℃の範囲にあることが好ましく、最も好ましくは60℃乃至70℃の範囲である。また、本発明のホスホリパーゼCは、55℃乃至70℃の範囲において相対活性が50%以上であることが好ましく、さらに好ましくは45℃乃至70℃の範囲において相対活性が50%以上であり、最も好ましくは35℃乃至70℃の範囲において相対活性が50%である。また、55℃乃至65℃の範囲においては、相対活性が80%以上であることが好ましい。
【0050】
また、温度安定性について、「安定である」とは、40%以上の残存加水分解活性を有していることを意味し、本発明のホスホリパーゼCは、45℃以下の処理温度において、残存活性が好ましくは50%以上、さらに好ましくは70%以上、最も好ましくは80%以上である。また、本発明のホスホリパーゼCは、処理温度が、好ましくは45℃、さらに好ましくは50℃、最も好ましくは60℃において、残存加水分解活性が40%以上である。
【0051】
また、pH安定性について、「安定である」とは、保存処理後において、5%以上の残存加水分解活性を有していることを意味し、本発明のホスホリパーゼCは、処理pHがpH5乃至pH9の範囲において、好ましくは残存加水分解活性が40%以上であり、さらに好ましくは50%以上であり、最も好ましくは60%以上である。また、処理pHがpH6乃至pH8の範囲において、好ましくは残存加水分解活性が60%以上であり、さらに好ましくは70%以上であり、最も好ましくは80%以上である。
【0052】
また、本発明のホスホリパーゼCの別の例としては、配列番号1,2および3に示されるアミノ酸配列のいずれか1つ、好ましくはいずれか2つ、最も好ましくは3つ全てを含み、且つ、ホスホリパーゼC活性を有するタンパク質が挙げられる。該タンパク質は、これら3つのうちの2つ以上のアミノ酸配列を含む場合には、各々のアミノ酸配列の順序はいかなる順序であってもよい。
【0053】
また、配列番号5のアミノ酸配列からなるタンパク質において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ、ホスホリパーゼC活性を有することを特徴とするタンパク質も本発明に含まれる。置換したアミノ酸配列を有するタンパク質が、天然型タンパク質と同等の活性を有する例として、例えば、インターロイキン2(IL−2)遺伝子のシステインに相当するヌクレオチド配列をセリンに相当するヌクレオチド配列に置換して得られたタンパク質が、IL−2活性を保持することが知られている(Wang, A. et al. (1984) Science 224, 1431-1433)。
【0054】
また、本発明のホスホリパーゼCの別の例としては、配列番号5に記載のアミノ酸配列からなるタンパク質を挙げる事ができる。また、配列番号5に記載のアミノ酸配列を含むことからなるタンパク質であっても、ホスホリパーゼC活性を有する限り本発明に含まれる。
【0055】
また、本発明のホスホリパーゼCの別の例としては配列番号5に記載のアミノ酸配列からなり、糖鎖によって修飾されているタンパク質が挙げられる。また、このようなタンパク質を含むことからなるタンパク質も、ホスホリパーゼC活性を有する限り本発明に含まれる。
【0056】
本発明のホスホリパーゼCとして好適なものは、アスペルギルス・オリザエFERM ABP−10200株またはNBRC 4190株あるいはアスぺルギルス・タマリIAM 13907株由来のホスホリパーゼC、配列番号5に記載のアミノ酸配列からなるタンパク質および配列番号5に記載のアミノ酸配列からなり、糖鎖によって修飾されているタンパク質が挙げられるが、より好適なものは、アスペルギルス・オリザエFERM ABP−10200株またはNBRC 4190株あるいはアスぺルギルス・タマリIAM 13907株由来のホスホリパーゼCおよび配列番号5に記載のアミノ酸配列からなり、且つ糖鎖によって修飾されているタンパク質である。
【0057】
本発明において、「本発明のDNA」とは、本発明のホスホリパーゼCをコードするDNAをいう。DNAとしては、cDNA、ゲノムDNA、人工的に改変されたDNA、化学的に合成されたDNAなど、現在知られる限りどのような形態であっても良い。
【0058】
本発明のDNAの例としては、配列番号4のコード領域(CDS)のヌクレオチド配列であり、且つ、ホスホリパーゼC活性を有するタンパク質をコードするDNAが挙げられる。
【0059】
本発明のDNAの別の例としては、配列番号4のコード領域(CDS)のヌクレオチド配列と70%以上のヌクレオチド配列相同性を有するDNAを挙げることができる。このようなDNAとしては、自然界で発見される変異型DNA、人為的に改変した変異型DNA、異種生物由来の相同DNAなどが含まれる。
【0060】
また、本発明のヌクレオチドのさらに他の例としては、配列番号4に記載のアミノ酸配列からなるタンパク質をコードするDNAが挙げられる。なお、所望のアミノ酸に対応するコドンは、その選択も任意でよく、例えば利用する宿主のコドン使用頻度を考慮して常法に従い決定できる。(Grantham, R. et al. (1981) Nucleic Acids Res. 9, 143-174)。さらに、これらヌクレオチド配列のコドンの一部改変は、常法に従い、所望の改変をコードする合成オリゴヌクレオチドからなるプライマーを利用した、部位特異的変異導入法(site specific mutagenesis/Mark, D. F. et al. (1984) Proc. Natl. Acad. Sci. USA 81, 5662-5666)などに従うことができる。
【0061】
本発明のDNAのまた別の例としては、配列番号4のコード領域(CDS)のヌクレオチド配列からなるDNAが挙げられる。また、配列番号4のコード領域(CDS)のヌクレオチド配列を含むDNAも、ホスホリパーゼC活性を有するタンパク質をコードしている限り、本発明に含まれるものである。
【0062】
また、本発明のホスホリパーゼCには、本発明のDNAによりコードされるアミノ酸配列からなるタンパク質を挙げることができる。また、本発明のホスホリパーゼCにおいて、任意の一つもしくは二つ以上のアミノ酸を欠失させた改変体を作製するためには、エキソヌクレアーゼBal31等を用いてDNAを末端から削る方法(岸本 利光ら“続生化学実験講座1・遺伝子研究法II”335-354)、カセット変異法(岸本 利光、“新生化学実験講座2・核酸III 組換えDNA技術”242-251)などに従うことができる。このように、本発明のDNAを元に遺伝子工学的手法により得られるタンパク質であっても、ホスホリパーゼC活性を有する限り本発明に含まれる。このようなホスホリパーゼCは、必ずしも配列番号5に記載のアミノ酸配列の全てを有するものである必要はなく、例えばその部分配列からなるタンパク質であっても、該タンパク質がホスホリパーゼC活性を示す限り本発明のホスホリパーゼCに包含される。また、このようなホスホリパーゼCをコードするDNAも本発明に含まれる。
【0063】
本発明に用いるホスホリパーゼCはホスホリパーゼC生産菌から精製したもの、粗精製したもの、菌体の破砕液の他、菌体の培養上清をそのまま用いたものでも良い。ホスホリパーゼC生産菌を培養する際には、培地中に炭素源、窒素源の他に界面活性剤を添加して培養するのが好ましい。あるいは魚粉、スリゴマ、綿実粕等天然素材の培地で培養するのが好ましい。界面活性剤としては、トライトン、トゥイーン、ショ糖脂肪酸エステル、コール酸ナトリウム、デオキシコール酸ナトリウムおよびサポニン等を挙げることができる。
【0064】
ホスホリパーゼC生産菌の培養は通常の培養装置、培地を用いて行なうことができる。培養は液体培養、固体培養等の方法を適宜選択することができる。液体培養の場合はフラスコ培養や発酵槽を用いた培養を行なうことができ、培養開始後は培地の追加のないバッチ培養法や培養中に適宜培地を添加していく流加培養法を用いることができる。培地には炭素源、窒素源を添加し、必要に応じてビタミン、微量金属等を添加することができる。炭素源としては、グルコース、マンノース、ガラクトース、フルクトース等の単糖類、マルトース、セロビオース、イソマルトース、ラクトース、スクロース等の二糖類、デンプン等の多糖類、マルトエクストラクト等を挙げることができる。窒素源としてはアンモニア、硫酸アンモニウム、硝酸アンモニウム等の無機窒素、イーストエクストラクト、マルトエクストラクト、コーンスティープリカー、ペプトン等の有機窒素を用いることができる。これらの培地中の組成物量は適宜選択することができる。培養温度、pH、通気攪拌量はホスホリパーゼC生産に適するように適宜選択することができる。
【0065】
ホスホリパーゼC生産菌の培養終了後に遠心分離を行ない、菌体を除いた培養上清をそのまま粗酵素液として用いることができる。また、粗酵素液をイオン交換クロマトグラフィー等によって粗精製したり、精製したりしたものを用いることもできる。
【0066】
本発明にいうアスペルギルス・オリザエFERM ABP−10200株またはNBRC 4190株あるいはアスぺルギルス・タマリIAM 13907株は、本発明のホスホリパーゼCを産生することができるものである限り、その全ての変異株を包含する。また、これらの変異株の中には、遺伝的方法、たとえば組み換え、形質導入、形質転換等によりえられたものも含有される。即ち、本発明のホスホリパーゼCを生産するアスペルギルス・オリザエFERM ABP−10200またはNBRC 4190株あるいはアスぺルギルス・タマリIAM 13907株、それらの変異株およびそれらと明確に区別されない菌株は全てアスペルギルス・オリザエFERM ABP−10200株またはNBRC 4190株あるいはアスぺルギルス・タマリIAM 13907株に包含される。
【0067】
また、本発明のホスホリパーゼCは、ベクターに本発明のDNAが挿入された組換えプラスミドで宿主細胞を形質転換し、該形質転換された細胞の培養産物から得る事もできる。このように適当なベクターに本発明のDNAが挿入された組換えプラスミドも本発明に含まれる。このような目的に用いるベクターとしては、一般に知られているさまざまなベクターを用いることができる。好適なものとしては、原核細胞用ベクター、真核細胞用ベクター、哺乳動物由来の細胞用ベクターなどが挙げられるが、これに限定されない。このような組換えプラスミドにより、他の原核生物、または真核生物の宿主細胞を形質転換させることができる。さらに、適当なプロモーター配列および/または形質発現に関わる配列を有するベクターを用いるか、もしくはそのような配列を導入することにより、発現ベクターとすることで、それぞれの宿主において遺伝子を発現させることが可能である。このような発現ベクターは、本発明の組換えプラスミドの好適な態様である。
【0068】
本発明の組換えプラスミドを、各種細胞に導入することにより、宿主細胞を得ることができる。このような細胞は、プラスミドを導入することができる細胞であれば原核細胞であっても真核細胞であってもよい。
【0069】
原核細胞の宿主としては、例えば、大腸菌(Escherichia coli)や枯草菌(Bacillus subtilis)などが挙げられる。目的の遺伝子をこれらの宿主細胞内で形質転換させるには、宿主と適合し得る種由来のレプリコンすなわち複製起点と、調節配列を含んでいるプラスミドベクターで宿主細胞を形質転換させる。また、ベクターとしては、形質転換細胞に表現形質(表現型)の選択性を付与することができる配列を有するものが好ましい。
【0070】
例えば、大腸菌としてはK12株などがよく用いられ、ベクターとしては、一般にpBR322やpUC系のプラスミドが用いられるが、これらに限定されず、公知の各種菌株、およびベクターを使用することができる。
【0071】
プロモーターとしては、大腸菌においては、トリプトファン(trp)プロモーター、ラクトース(lac)プロモーター、トリプトファン・ラクトース(tac)プロモーター、リポプロテイン(lpp)プロモーター、ポリペプチド鎖伸張因子Tu(tufB)プロモーター等が挙げられ、どのプロモーターも本発明のホスホリパーゼCの産生に使用することができる。
【0072】
枯草菌としては、例えば207−25株が好ましく、ベクターとしてはpTUB228(Ohmura, K. et al. (1984) J. Biochem. 95, 87-93)などが用いられるが、これに限定されるものではない。
【0073】
プロモーターとしては、枯草菌のα−アミラーゼのシグナルペプチド配列をコードするDNA配列を連結することにより、菌体外での分泌発現も可能となる。
【0074】
真核細胞の宿主細胞には、脊椎動物、昆虫、酵母などの細胞が含まれ、脊椎動物細胞としては、哺乳動物由来の細胞、例えば、サルの細胞であるCOS細胞(Gluzman, Y. (1981) Cell 23, 175-182、ATCC CRL−1650)やチャイニーズ・ハムスター卵巣細胞(CHO細胞、ATCC CCL−61)のジヒドロ葉酸還元酵素欠損株(Urlaub, G. and Chasin, L. A. (1980) Proc. Natl. Acad. Sci. USA 77, 4126-4220)等を用いることができる。
【0075】
脊椎動物細胞の発現プロモーターとしては、通常発現しようとする遺伝子の上流に位置するプロモーター、RNAのスプライス部位、ポリアデニル化部位、および転写終結配列等を有するものを使用することができ、さらにこれは必要により複製起点を有していてもよい。該発現ベクターの例としては、SV40の初期プロモーターを有するpSV2dhfr(Subramani, S. et al. (1981) Mol. Cell. Biol. 1, 854-864)等を挙げることができるが、これに限定されない。
【0076】
宿主細胞として、COS細胞を用いる場合を例に挙げると、発現ベクターとしては、SV40複製起点を有し、COS細胞において自立増殖が可能であり、さらに、転写プロモーター、転写終結シグナル、およびRNAスプライス部位を具えたものを用いることができる。該発現ベクターは、ジエチルアミノエチル(DEAE)−デキストラン法(Luthman, H. and Magnusson, G. (1983) Nucleic Acids Res, 11, 1295-1308)、リン酸カルシウム−DNA共沈殿法(Graham, F. L. and van der Eb, A. J. (1973) Virology 52, 456-457)、および電気パルス穿孔法(Neumann, E. et al. (1982) EMBO J. 1, 841-845)などによりCOS細胞に取り込ませることができ、かくして所望の形質転換細胞を得ることができる。また、宿主細胞としてCHO細胞を用いる場合には、発現ベクターと共に、抗生物質G418耐性マーカーとして機能するneo遺伝子を発現し得るベクター、例えばpRSVneo(Sambrook, J. et al. (1989) : “Molecular Cloning A Laboratory Manual“ Cold Spring Harbor Laboratory, NY)やpSV2−neo(Southern, P. J. and Berg, P. (1982) J. Mol. Appl. Genet. 1, 327-341)などをコ・トランスフェクトし、G418耐性のコロニーを選択することにより、本発明のホスホリパーゼCを安定に産生する形質転換細胞を得ることができる。
【0077】
昆虫細胞を宿主細胞として用いる場合には、鱗翅類ヤガ科のSpodoptera frugiperdaの卵巣細胞由来株化細胞(Sf−9またはSf−21)やTrichoplusia niの卵細胞由来High Five細胞(Wickham, T. J. et al, (1992) Biotechnol. Prog. I: 391-396)などが宿主細胞としてよく用いられ、バキュロウイルストランスファーベクターとしてはオートグラファ核多角体ウイルス(AcNPV)のポリヘドリンタンパク質のプロモーターを利用したpVL1392/1393がよく用いられる(Kidd, I. M. and V.C. Emery (1993) The use of baculoviruses as expression vectors. Applied Biochemistry and Biotechnology 42, 137-159)。この他にも、バキュロウイルスのP10や同塩基性タンパク質のプロモーターを利用したベクターも使用できる。さらに、AcNPVのエンベロープ表面タンパク質GP67の分泌シグナル配列を目的タンパク質のN末端側に繋げることにより、組換えタンパク質を分泌タンパク質として発現させることも可能である(Zhe-mei Wang, et al. (1998) Biol. Chem., 379, 167-174)。
【0078】
真核微生物を宿主細胞とした発現系としては、酵母が一般によく知られており、その中でもサッカロミセス属酵母、例えばパン酵母Saccharomyces cerevisiaeや石油酵母Pichia pastorisが好ましい。該酵母などの真核微生物の発現ベクターとしては、例えば、アルコール脱水素酵素遺伝子のプロモーター(Bennetzen, J. L. and Hall, B. D. (1982) J. Biol. Chem. 257, 3018-3025)や酸性フォスファターゼ遺伝子のプロモーター(Miyanohara, A. et al. (1983) Proc. Natl. Acad. Sci. USA 80, 1-5)などを好ましく利用できる。また、分泌型タンパク質として発現させる場合には、分泌シグナル配列と宿主細胞の持つ内在性プロテアーゼあるいは既知のプロテアーゼの切断部位をN末端側に持つ組換え体として発現することも可能である。例えば、トリプシン型セリンプロテアーゼのヒトマスト細胞トリプターゼを石油酵母で発現させた系では、N末端側に酵母のαファクターの分泌シグナル配列と石油酵母の持つKEX2プロテアーゼの切断部位をつなぎ発現させることにより、活性型トリプターゼが培地中に分泌されることが知られている(Andrew, L. Niles,et al. (1998) Biotechnol.Appl. Biochem. 28, 125-131)。
【0079】
上記のようにして得られる形質転換体は、常法に従い培養することができ、該培養により細胞内、または細胞外に本発明のホスホリパーゼCが産生される。該培養に用いられる培地としては、採用した宿主細胞に応じて慣用される各種のものを適宜選択でき、例えば、上記COS細胞であれば、RPMI1640培地やダルベッコ改変イーグル培地(以下「DMEM」という)などの培地に、必要に応じウシ胎児血清などの血清成分を添加したものを使用できる。培養条件としては、CO2濃度は0乃至50%の範囲であればよく、好適には1乃至10%でありより好適には5%である。培養温度は0乃至99℃であればよく、好適には20乃至50℃であり、より好適には35乃至40℃である。
【0080】
上記培養により形質転換体の細胞内または細胞外に組換えタンパク質として産生される本発明のホスホリパーゼCは、培養産物中から、そのタンパク質の物理化学的性質、化学的性質、生化学的性質(酵素活性など)等を利用した各種の分離操作(「生化学データブックII」、1175-1259項、第1版第1刷、1980年6月23日株式会社東京化学同人発行;Biochemistry, vol. 25, No.25, p8274-8277 (1986); Eur. J. Biochem., 163, p313-321 (1987)等参照)により分離、精製することができる。該方法としては、具体的には例えば通常の再構成処理、タンパク質沈殿剤による処理(塩析法)、遠心分離、浸透圧ショック法、凍結融解法、超音波破砕、限外ろ過、ゲル濾過、吸着クロマトグラフィー、イオン交換クロマトグラフィー、アフィニティークロマトグラフィー、高速液体クロマトグラフィー(HPLC)等の各種液体クロマトグラフィー、透析法、それらの組み合わせ等を例示することができる。上記により、高収率で所望の組換えタンパク質を工業的規模で製造することができる。また、発現させる組換えタンパク質に6残基からなるヒスチジンを繋げることにより、ニッケルアフィニティーカラムで効率的に精製することができる。上記方法を組み合わせることにより容易に高収率、高純度で本発明のホスホリパーゼCを大量に製造することができる。
【0081】
以上のような方法により製造されたホスホリパーゼCも本発明の好適な例としてあげることができる。
【0082】
ホスホリパーゼC生産菌とは、実質的にホスホリパーゼC生産能を有する微生物をいい、ホスホリパーゼCを菌体内に蓄積する微生物や、菌体外に分泌する微生物等を含む。ホスホリパーゼC生産菌の培養上清または培養上清から精製したホスホリパーゼCを用いる場合には菌体外にホスホリパーゼCを分泌する菌を用いることができる。
【0083】
本発明に用いるホスホリパーゼCとしては、アスペルギルス・オリザエまたはアスぺルギルス・タマリ由来のホスホリパーゼCを用いることができ、より好適なものは、アスペルギルス・オリザエFERM ABP−10200株またはNBRC 4190株あるいはアスぺルギルス・タマリIAM 13907株由来のホスホリパーゼCを用いることができる。ホスホリパーゼCは、これらホスホリパーゼC生産菌自身が生産するものでもよいし、その変異体または修飾体が生産するものであってもよく、更に、これらホスホリパーゼC生産菌のホスホリパーゼCをコードする遺伝子を宿主に導入して得られた形質転換体から生産される組換えタンパク質であってもよい。
【0084】
ホスホリパーゼC生産菌の入手
アスペルギルス・オリザエNBRC 4190株は独立行政法人製品評価技術基盤機構 バイオテクノロジー本部 生物遺伝資源部門(NBRC: NITE Biological Resource Center;〒292−0818 日本国千葉県木更津市かずさ鎌足2−5−8、ホームページアドレス <http://www.nite.go.jp/>)より入手することができる。
アスペルギルス・タマリIAM 13907(=IAM 13907)株は東京大学分子細胞生物学研究所(IAM: Institute of Molecular and Cellular Biosciences, The University of Tokyo ;〒113−0032 東京都文京区弥生1−1−1、ホームページアドレス <http://www.iam.u-tokyo.ac.jp/indexe.html>)より入手することができる。
【0085】
アスペルギルス・オリザエFERM ABP−10200株の菌学的性質を以下に示す。
アスペルギルス・オリザエFERM ABP−10200株をクリックの文献(Klich, M. A. (2002) Identification of common Aspergillus Centraalbureau voor Schimmelcultures, Utrecht, The Netherlands)に従い、4種類の培地(CYA培地, CY20S培地, CZ培地, MEA培地)に接種して、菌学的性状を観察した。
【0086】
色調の表示は「メチューン・ハンドブック・オブ・カラー」(Kornerup, A. and Wanscher, J. H. (1978) Methuen handbook of colour (3rd. edition). Erye Methuen, London.)に従った。
【0087】
4種類の培地(CYA培地, CY20S培地, CZ培地, MEA培地)の組成は以下の通りである。
【0088】
CYA培地[ザペック イースト アガー(Czapek Yeast Extract Agar)培地]
(K2HPO4 1.0g,*ザペック濃縮液 10ml,イーストエキス 5g,シュークロース 30g,寒天 15g,蒸留水 1000ml)
*ザペック濃縮液(NaNO3 30g, KCl 5g,MgSO4・7H2O 5g,FeSO4・7H2O 0.1g,ZnSO4・7H2O 0.1g,CuSO4・5H2O 0.05g,蒸留水100ml)
【0089】
CY20S培地[20% シュークロース ザペック イーストアガー(Czapek Yeast ExtractAgar with 20% Sucrose)培地]
(K2HPO4 1.0g,*ザペック濃縮液 10ml,イーストエキス 5g,シュークロース 200g,寒天 15g,蒸留水 1000ml)
【0090】
CZ培地[ザペック ドックス アガー (Czapek Dox Agar)培地]
(K2HPO4 1.0 g, *ザペック濃縮液 10 ml, シュークロース 30 g, 寒天 17.5 g, 蒸留水 1000 ml)
【0091】
MEA培地[モルト エキス アガー(Malt Extract Agar)培地]
(モルトエキス 20g,ペプトン 1g,グルコース 20g,寒天 20g,蒸留水 1000ml)
【0092】
1)菌学的性状
アスペルギルス・オリザエFERM ABP−10200株の菌学的性状
CYA培地でのコロニーは、25℃、1週間の培養で直径36−40mmである。コロニーはやや厚く、羊毛状で、中心部では綿毛状となり、中心部から放射状の溝を形成する。菌糸は白色である。分生子は中心部に疎に形成され、灰黄色(4B4)から黄白色(4A2)を呈する。浸出液、菌核は観察されない。裏面は淡黄色(2A4)から白色(2A1)で、中心部から放射状の溝を形成する。可溶性色素は観察されない。
CYA培地でのコロニーは、37℃、1週間の培養で直径54−58mmである。コロニーは厚く、羊毛状である。菌糸は白色である。分生子は、中心部に形成され、灰黄色(4B4)から黄白色(4A2)を呈する。浸出液、菌核は観察されない。裏面は薄橙色(4A4)から白色(4A2)で、中心部から放射状の溝を形成する。可溶性色素は観察されない。
【0093】
CY20S培地でのコロニーは、25℃、1週間の培養で直径35−41mmである。コロニー性状はCYA培地と同様となるが、中心部の綿毛状の菌糸がやや多い。
【0094】
CZ培地でのコロニーは、25℃、1週間の培養で直径17−21mmである。コロニー性状はCYA培地と同様となるが、コロニーは小さく、分生子の形成が疎となる。
【0095】
MEA培地でのコロニーは、25℃、1週間の培養で直径37−41mmである。コロニーは薄く、綿毛状である。菌糸は白色である。分生子は中心部に疎に形成され、暗緑色(26D4)から灰緑色(26D6)を呈する。浸出液、菌核は観察されない。裏面は灰黄色(4B3)から黄白色(4A2)で、可溶性色素は観察されない。
14℃から42℃まで生育し、18℃から38℃まで分生子形成が観察された。
【0096】
分生子頭は放射状から緩い円柱状である。分生子柄は幅6.7−13.6μm、長さ302.2−1398.0μm、無色、粗面である。頂嚢は亜球形からフラスコ形、幅は14.6−29.3μmである。アスペルギラ(aspergilla)は主に単列(uniseriate)、まれに2列(biseriate)である。メトレまたはフィアライドは頂嚢の上部半分から形成される。メトレは11.3−28.3×5.2−9.9μmである。フィアライドはフラスコ形で、8.4−21.3×3.8−7.7μmである。分生子は滑面、亜球形から卵形、直径4.2−6.3μmである。
【0097】
以上の菌学的性状より、本菌に該当する菌を検索したところ、クリックの文献に記載されているアスペルギルス オリザエ(アールブルク)コーン(Aspergillus oryzae(Ahlburg)Chon)の性状とほぼ一致した。よって、FERM ABP−10200株をアスペルギルス・オリザエ(アールブルク)コーン(Aspergillus oryzae (Ahlburg)Chon)と同定し、独立行政法人産業技術総合研究所特許生物寄託センターに寄託した。
【0098】
ホスホリパーゼC生産菌から得られたホスホリパーゼCの具体的な性質について以下に示すが、本発明のホスホリパーゼCの有する性質は必ずしもこれらに限定されるものではない。
【0099】
アスペルギルス・オリザエFERM ABP−10200株またはNBRC 4190株あるいはアスぺルギルス・タマリIAM 13907株により生産され、精製されたホスホリパーゼCは、以下の性質を有する。
1)SDS−PAGE電気泳動法にて分子量約87,000を示す。
2)卵黄由来レシチン(ナカライ・テスク(株)製)をpH3乃至pH9の範囲で加水分解する。
3)0℃乃至80℃の範囲で4)記載の加水分解活性を有する。
4)pH4.5において45℃以下の温度で安定である。
5)pH3乃至pH10の範囲で安定である。
6)2)記載の加水分解活性の最適pHはpH4.5である。
7)3)記載の加水分解活性の最適温度はpH4.5では65℃である。
8)ホスファチジルコリン、ホスファチジルエタノールアミン、ホスファチジルイノシトールおよびホスファチジルグリセロールを加水分解するが、グリセロホスホリルコリンおよびパラニトロフェニルリン酸を実質的に加水分解しない。
【0100】
一例として、卵黄由来ホスファチジルコリンに対する活性を100%としたときの相対活性を表1に示した。なお、表中卵黄由来基質および大豆由来基質はその旨を記載した。また、卵黄由来レシチンはナカライテスク(株)、大豆由来レシチンは辻製油(株)から購入し、それ以外の基質はシグマアルドリッチジャパン(株)から購入した。
【0101】
【表1】
【0102】
9)下記に示す部分アミノ酸配列を有する。配列はN末端から記す。
Thr−Ala−Asp−Ser−Ala−Thr−Ala−Ile−Gly−
Tyr−Val−Thr−Pro−Ser−Met(配列番号1)。
【0103】
Glu−Ala−Tyr−Gly−Ser−Leu−Leu−Thr−Pro−
Pro(配列番号2)。
【0104】
Val−Pro−Pro−Ser−His−Asn−Pro−Gln−Trp−
Ala(配列番号3)。
【0105】
10)タンパク質が糖鎖で修飾されている。
【0106】
以上のことから、本発明のホスホリパーゼCの有する性質としては以下のようなものが挙げられるが、これに限定されるものではない。
1)SDS−PAGE電気泳動法にて分子量約87,000を示す。
2)卵黄由来レシチン(ナカライテスク(株))をpH3乃至pH9の範囲で加水分解する。
3)0℃乃至80℃の範囲で4)記載の加水分解活性を有する。
4)pH4.5において45℃以下の温度で安定である。
5)pH3乃至pH10の範囲で安定である。
【0107】
また、本発明のホスホリパーゼCを製造する方法も本発明に含まれる。
アスペルギルス・オリザエFERM ABP−10200株またはNBRC 4190株あるいはアスぺルギルス・タマリIAM 13907株を初めとするホスホリパーゼC産生微生物を培地で培養することにより、ホスホリパーゼCを生産することができる。例えば、0.1乃至5.0%ポリペプトン(和光純薬工業(株))、0.1乃至1.0%イーストエクストラクト(日本ベクトン・ディッキンソン(株))に0.05乃至1.0%デオキシコール酸ナトリウムを添加した培地または0.1乃至4.0%ファーマメディア(TRANDERS PROTEIN(株))に0.05から1.0%トライトンX−100(シグマアルドリッチジャパン(株))もしくは0.05乃至0.3%卵黄レシチンを添加した培地あるいは1乃至10%魚粉(池口喜一郎商店)の培地で、16乃至45℃で1乃至15日間、100乃至250rpmで振とう培養する。
【実施例】
【0108】
以下に、実施例および試験例を挙げるが、本発明の範囲はこれらに限定されるものではない。
【0109】
実施例1 アスペルギルス・オリザエFERM ABP−10200株からのホスホリパーゼCの精製
1)粗酵素液の調製
滅菌した表2の組成の培地100mlが入っている500ml容の三角フラスコ(種フラスコ)にフィルター滅菌した5%デオキシコール酸ナトリウム溶液1mlおよびアスペルギルス・オリザエFERM ABP−10200株の菌体を接種し、26℃にて7日間、170rpmの振とう培養を行った。
【0110】
【表2】
【0111】
培養終了後、4℃、10,000×Gにて10分間の遠心分離を行った。得られた上清を粗酵素液とした。
【0112】
2)酵素活性測定法
ホスホリパーゼCの加水分解活性は以下のようにして測定した。
<1>レシチンの加水分解反応
卵黄レシチン(ナカライテスク(株))1.5gを4%(wt/v)TritonX−100 50mlに溶解した基質溶液60μlに200mM酢酸緩衝液(pH5.5)60μlを加えて37℃で保温した。この混合液に酵素液60μlを加え撹拌して均一にし、37℃で保温して3時間酵素反応を行った。
【0113】
<2>遊離ホスホリル塩基の加水分解反応
酵素反応の結果生じるホスホリル塩基をアルカリホスファターゼで加水分解した。<1>で調整した酵素反応液50μlに200mMトリス・塩酸緩衝液(pH8.0)50μlおよびアルカリホスファターゼ(シグマアルドリッチジャパン(株))1μlを加えて37℃で40分反応を行った。なおこのときブランクとしてアルカリホスファターゼを加えないサンプルも同時に調整した。
【0114】
<3>無機リン酸の定量
<2>の結果生じた無機リン酸をホスファCテストワコー(和光純薬工業(株))で定量した。<2>で得られた反応液100μlにホスファCテストワコーのA液およびB液を各1ml添加し、37℃で20分間反応した。この混合液の750nmにおける吸光度を測定した。ブランクとの差がホスホリパーゼC活性になる。酵素反応1分間当たり1μmolのホスホリル塩基を生成する酵素活性を1単位とした。
【0115】
3)精製酵素液の調製
1)で得られた粗酵素液1,200mlを10mMトリス・塩酸緩衝液(pH7.5)8,000mlに対して12時間ずつ8回透析した。これを、予め10mMトリス・塩酸緩衝液(pH7.5)で平衡化したDEAEトヨパール(東ソー(株))カラム(直径2.2cm×長さ20cm)に添加し、吸着させた。10mMトリス・塩酸緩衝液(pH7.5)で該カラムを十分洗浄した後、10mMトリス・塩酸緩衝液(pH7.5)600ml中に0乃至0.2Mの塩化ナトリウムの直線的濃度勾配を作製して該カラムに吸着した成分を溶出させた。卵黄由来レシチン分解活性は塩化ナトリウム濃度が0.05M乃至0.08Mの画分(90ml)に溶出された。これを粗精製酵素画分とした。
【0116】
得られた活性画分90mlを20mMトリス・塩酸緩衝液(pH7.5)4,000mlに対して12時間ずつ3回透析した後、予め20mMトリス・塩酸緩衝液(pH7.5)で平衡化したMonoQ(アマシャムバイオサイエンス(株))カラム(直径10mm×長さ10cm)に添加し、吸着させた。該カラムを20mMトリス・塩酸緩衝液(pH7.5)で十分洗浄した後、20mMトリス・塩酸緩衝液(pH7.5)250ml中に0乃至0.2Mの塩化ナトリウムの直線的濃度勾配を作製して該カラムに吸着した成分を溶出させた。卵黄由来レシチン分解活性は塩化ナトリウム濃度が0.1M乃至0.12Mの画分(25ml)に溶出された。
【0117】
得られた活性画分25mlを濃縮し、予め0.15M塩化ナトリウムを含む10mMトリス・塩酸緩衝液(pH7.5)で平衡化したHiLoad Sephadex200pg(アマシャムバイオサイエンス(株))カラム(直径16mm×60cm)に添加した後、0.15M塩化ナトリウムを含む10mMトリス・塩酸緩衝液(pH7.5)にて溶出させた。卵黄由来レシチン分解活性は、溶出量が60ml乃至70mlの画分に溶出された。
この画分を精製酵素溶液とした。
【0118】
4)精製酵素の分子量測定
12.5%ポリアクリルアミドゲルを用いたSDS−PAGE電気泳動法(Laemmli, U.K., Nature, 227, 680(1970)参照)により、精製酵素の分子量を求めた。標準タンパク質として次のものを用いた:a.ホスホリラーゼ(phosphorylase)、分子量94,000:b.アルブミン(albumin)、分子量67,000:c.オバルブミン(ovalbumin)、分子量43,000:d.カルボニック・アンヒドラーゼ(carbonic anhydrase)、分子量30,000:e.トリプシン・インヒビター(trypsin inhibitor)、分子量20,100:f.α−ラクタルブミン(α-lactalbumin)、分子量14,400。
精製酵素は分子量約87,000の単一バンドを示した。
【0119】
実施例2 アスぺルギルス・タマリIAM 13907株からのホスホリパーゼCの精製
1)粗酵素液の調製
表2に示した組成の培地100mlが入っている500ml容の三角フラスコ(種フラスコ)にフィルター滅菌した5%デオキシコール酸ナトリウム溶液1mlおよびアスぺルギルス・タマリIAM 13907株の菌体を接種し、26℃にて5日間、170rpmの振とう培養を行った。培養終了後、4℃、10,000×Gにて10分間の遠心分離を行った。得られた上清を粗酵素液とした。
【0120】
2)精製酵素液の調製
実施例1. 3)と同様に精製を行い、精製酵素溶液を得た。
【0121】
3)精製酵素の分子量測定
実施例1. 4)と同じ方法で測定した。
精製酵素は分子量約87,000の単一バンドを示した。
【0122】
実施例3 アスぺルギルス・オリザエNBRC 4190株からのホスホリパーゼCの精製
1)粗酵素液の調製
滅菌した表3の組成の培地100mlが入っている500ml容の三角フラスコ(種フラスコ)にアスペルギルス・オリザエNBRC 4190株の菌体を接種し、26℃にて7日間、170rpmの振とう培養を行った。
【0123】
【表3】
【0124】
培養終了後、4℃、10,000×Gにて10分間の遠心分離を行った。得られた上清を粗酵素液とした。
【0125】
2)精製酵素液の調製
1)で得られた粗酵素液600mlを10mMトリス・塩酸緩衝液(pH7.5)8,000mlに対して12時間ずつ5回透析した。これを、予め10mMトリス・塩酸緩衝液(pH7.5)で平衡化したDEAEトヨパール(東ソー(株))カラム(直径2.2cm×長さ20cm)に添加し、吸着させた。10mMトリス・塩酸緩衝液(pH7.5)で該カラムを十分洗浄した後、10mMトリス・塩酸緩衝液(pH7.5)600ml中に0乃至0.6Mの塩化ナトリウムの直線的濃度勾配を作製して該カラムに吸着した成分を溶出させた。卵黄由来レシチン分解活性は塩化ナトリウム濃度が0.30M乃至0.35Mの画分(50ml)に溶出された。これを粗精製酵素画分とした。
【0126】
得られた活性画分50mlを20mMトリス・塩酸緩衝液(pH7.5)4,000mlに対して12時間ずつ3回透析した後、予め20mMトリス・塩酸緩衝液(pH7.5)で平衡化したMonoQ(アマシャムバイオサイエンス(株))カラム(直径10mm×長さ10cm)に添加し、吸着させた。該カラムを20mMトリス・塩酸緩衝液(pH7.5)で十分洗浄した後、20mMトリス・塩酸緩衝液(pH7.5)250ml中に0乃至0.6Mの塩化ナトリウムの直線的濃度勾配を作製して該カラムに吸着した成分を溶出させた。卵黄由来レシチン分解活性は塩化ナトリウム濃度が0.12M乃至0.20Mの画分(35ml)に溶出された。
【0127】
得られた活性画分35mlを濃縮し、予め0.15M塩化ナトリウムを含む10mMトリス・塩酸緩衝液(pH7.5)で平衡化したHiLoad Sephadex200pg(アマシャムバイオサイエンス(株))カラム(直径16mm×60cm)に添加した後、0.15M塩化ナトリウムを含む10mMトリス・塩酸緩衝液(pH7.5)にて溶出させた。卵黄由来レシチン分解活性は、溶出量が60ml乃至70mlの画分に溶出された。
この画分を精製酵素溶液とした。
【0128】
3)精製酵素の分子量測定
実施例1の4)と同じ方法で測定した。
精製酵素は分子量約87,000の単一バンドを示した。
【0129】
実施例4 アスぺルギルス・オリザエNBRC 4190株からのホスホリパーゼCの部分アミノ酸配列の決定
精製した酵素溶液を約1mg/ml程度まで限外濾過膜(ザルトリウス(株)VIVASPIN2、分子量分画1万)を用いて濃縮した。濃縮酵素液150μlにDenature buffer(6M塩酸グアニジン、10mM EDTA、0.1M炭酸水素アンモニウムpH7.8)150μlおよび50mM ジチオスレイトール6μlを加え、95℃で10分間反応させた。室温まで冷却後、反応液にDenature buffer溶液に溶解した50mMヨードアセトアミド30μlを加え、暗所室温で1時間反応させた。この溶液を20mM炭酸水素アンモニウム(pH8.0)で予め平衡化しておいたHitrap Desalting(アマシャムバイオサイエンス(株))カラムに添加した後、20mM炭酸水素アンモニウム(pH8.0)にて溶出させた。溶出量が1.5ml乃至2.5mlの画分に溶出した溶液を凍結乾燥し、20mM炭酸水素アンモニウム(pH8.0)100μlに溶解した。得られた溶液にトリプシン(Modified、プロメガ(株))60ユニットを加え、37℃で18時間酵素反応させた。反応液を高速液体クロマトグラフィー(日立製作所(株))にかけて分解アミノ酸のピークを分離した。分離条件を以下に示す。
【0130】
カラム:東ソー(株) TSKgel ODS-120T (4.6mm×150mm)
A buffer:0.1%TFA/Water
B buffer:0.1%TFA/Acetonitril
グラジエント:10→70%B 2%/ml
Flow:1ml/分
【0131】
分離した分解アミノ酸のうち、3本のピーク(B buffer濃度 30から38%程度)を分取し、これについてアミノ酸配列解析装置(Procise cLC、アプライドバイオシステムズジャパン(株))でアミノ酸配列を解析した。その結果得られた部分アミノ酸配列をアミノ末端側から記す。
【0132】
Thr−Ala−Asp−Ser−Ala−Thr−Ala−Ile−Gly−
Tyr−Val−Thr−Pro−Ser−Met(配列番号1)。
【0133】
Glu−Ala−Tyr−Gly−Ser−Leu−Leu−Thr−Pro−
Pro(配列番号2)。
【0134】
Val−Pro−Pro−Ser−His−Asn−Pro−Gln−Trp−
Ala(配列番号3)。
【0135】
実施例5 アスぺルギルス・オリザエNBRC 4190株由来のホスホリパーゼCをコードするDNAの同定
1)全RNAの精製
アスぺルギルス・オリザエNBRC 4190株を液体培地(2% ポリペプトン、0.5% イースト エクストラクト、0.02% リン酸水素2カリウム、0.05% 硫酸マグネシウム)20mlで26℃、1日間前培養した。その後、液体培地(5%魚粉)に1%植菌し、26℃で4日間培養した。培養した菌体を吸引集菌し、−80℃で冷やした乳鉢(オートクレーブ滅菌済)に移した。液体窒素を加えながら、乳棒で菌体を破砕し、粉末状にした。完全に粉末状になった菌体をRNeasy Plant Mini Kit(キアゲン(株))を用いて全RNAの精製を行った。905ng/μlの濃度の溶液が、50μl得られた。
【0136】
2)ホスホリパーゼC遺伝子の解読
5'RACE法および3'RACE法にて遺伝子配列の解読をおこなった。具体的には、5'RACE Systemおよび3'RACE System(いずれもインビトロジェン(株))を使用し、ポリメラーゼとしてEx TaqTM(タカラバイオ(株))を用いてPCRをおこなった。このときに使用したPCRプライマーは、5'側の遺伝子配列増幅用に5'-GGCCACGCGTCGACTAGTAC-3'および5'-GACAGTGTAGTCGAGCACAGCGAA-3'、3'側の遺伝子配列増幅用に5'-GACTCTGCCACCGCAATCGGCTA-3'および5'-GGCCACGCGTCGACTAGTAC-3'を用いた。PCRサイクルは、94℃・5分、(94℃・30秒、55℃・30秒、72℃・2分30秒)×30、72℃・10分、4℃で増幅した。5'側の遺伝子配列約1200b.p.、3'側の遺伝子配列約800b.p.の長さのDNAが増幅された。
【0137】
各々のPCR産物をアガロースゲル電気泳動をおこなった後、QIAquick Gel Extarction Kit(キアゲン(株))で精製した。精製物をTOPOTMTAクローニングキット(インビトロジェン(株))を用いてベクターに連結し、形質転換をおこなった。形質転換した大腸菌を37℃、一晩寒天培地(LB/Agar(和光純薬工業(株)))上で培養した後、生育したコロニーを37℃、一晩液体培地(LBbroth(和光純薬工業(株)))で培養した。増殖した大腸菌からプラスミドをQIAprep Spin Miniprep Kit(キアゲン(株))を用いて精製し、DNA配列解析をおこなった。DNA配列解析の結果を配列番号4に示した。また、DNA配列から推定されるアミノ酸配列を配列番号5に示した。
【0138】
実施例6 アスぺルギルス・オリザエNBRC 4190株由来ホスホリパーゼCのグリコペプチダーゼ処理
1)酵素反応
精製した酵素溶液を限外濾過膜(ザルトリウス(株)VIVASPIN2、分子量分画1万)を用いて濃縮した。濃縮酵素液20μlに、蒸留水15μl、0.25mMリン酸緩衝液(pH7.5)10μlおよび1M 2-メルカプトエタノール/2%ドデシル硫酸ナトリウム溶液2.5μlを加え、95℃にて5分間反応した。急冷後、15%Triton X-100(シグマアルドリッチジャパン(株))2.5μlを加え、グリコペプチダーゼF(シグマアルドリッチジャパン(株))10ユニットを加えて、37℃にて20時間反応した。
【0139】
2)反応後の分子量測定
実施例1の4)と同じ方法で測定した。
酵素反応後の分子量約63,000の単一バンドを示した。
【0140】
試験例1 アスペルギルス・オリザエFERM ABP−10200株由来ホスホリパーゼCの精製酵素液の諸性質
実施例1の3)で得られた精製酵素液について、活性測定を行なった。
【0141】
1)pH活性
実施例1の2)に示した方法に拠った。ただし酵素反応時間は37℃、10分間とした。また、緩衝液は次のものを用いた:pH2.3乃至pH3.7の場合、グリシン−塩酸緩衝液:pH3.3乃至pH6.2の場合、クエン酸−クエン酸ナトリウム緩衝液:pH6.1乃至pH8.0の場合、MOPS緩衝液:pH8.2乃至pH9.2の場合、Atkins−Pantin緩衝液。最も活性が高かったpH条件での加水分解活性を100%とし、各pHにおける酵素の加水分解活性を相対値として図1に記載した。至適pHはクエン酸緩衝液中pH4.5付近であった。
【0142】
2)温度活性
クエン酸緩衝液pH4.5における温度活性を測定した。測定法は実施例1の2)に示した方法に拠った。ただし酵素反応時間は37℃、20分間とした。最も活性が高かった温度条件での加水分解活性を100%とし、各温度における酵素の加水分解活性を相対値として図2に記載した。至適温度は65℃付近であった。
【0143】
3)温度安定性
精製酵素溶液を種々の温度で30分間処理した後、その残存加水分解活性を測定した。あらかじめ処理温度に保持した25mMクエン酸緩衝液(pH4.5)90μlに、精製酵素液10μlを加え撹拌して均一にし、30分間保温した。実施例1で作製した卵黄レシチン溶液60μlに200mMクエン酸緩衝液(pH4.5)60μlを加え37℃に保持し、加温処理した酵素液60μlを加え、37℃で30分間酵素反応を行った。遊離ホスホリル塩基の定量は実施例1の2)にしたがった。最も高い残存加水分解活性を100%とし、各温度における加水分解活性を相対値として図3にまとめた。pH4.5において少なくとも60℃以下の温度で安定であった。
【0144】
4)pH安定性
精製酵素液30μlに、以下に述べる各pHの200mM緩衝液30μlを添加し、37℃にて30分間保温した。緩衝液は次のものを用いた:pH2.7乃至pH3.2の場合、グリシン−塩酸緩衝液:pH3.5乃至pH6.1の場合、酢酸−酢酸ナトリウム緩衝液:pH6.3乃至pH7.9の場合、MOPS緩衝液:pH8.3乃至pH10.8の場合、Atkins−Pantin緩衝液。200mMクエン酸緩衝液(pH4.5)60μl、および実施例1の2)記載の卵黄レシチン溶液60μlの混合液に、加温した酵素混合液60μlに水60μlを加えた溶液のうち60μlを加え撹拌して均一にし、37℃で10分間酵素反応を行った。遊離ホスホリル塩基の定量は実施例1の2)にしたがった。最も高い残存加水分解活性を100%とし、各pHにおける加水分解活性を相対値として図4にまとめた。pH3乃至pH10の範囲で安定であった。
【0145】
5)精製酵素の基質選択性
次に、基質選択性を測定した。実施例1の3)で調製した精製酵素液を用いた。測定方法は実施例1の2)に拠った。ただし、酵素反応は200mMクエン酸緩衝液pH4.5中で37℃で保温して10分間行った。卵黄由来ホスファチジルコリンに対する加水分解活性を100%としたときの、相対活性を表4に示す。
【0146】
【表4】
【0147】
試験例2.アスペルギルス・タマリIAM 13907株由来ホスホリパーゼCの精製酵素液の諸性質
実施例2の2)で得られた精製酵素液について、活性測定を行なった。
【0148】
1)pH活性
試験例1.に示した方法に拠った。最も活性が高かったpH条件での加水分解活性を100%とし、各pHにおける酵素の加水分解活性を相対値として図5に記載した。至適pHはpH4.5付近であった。
【0149】
2)温度活性
試験例1.に示した方法に拠った。最も活性が高かった温度条件での加水分解活性を100%とし、各温度における酵素の加水分解活性を相対値として図6に記載した。至適温度は65℃付近であった。
【0150】
3)温度安定性
試験例1.に示した方法に拠った。最も高い残存加水分解活性を100%とし、各温度における加水分解活性を相対値として図7にまとめた。pH4.5において45℃以下の温度で安定であった。
【0151】
4)pH安定性
試験例1.に示した方法に拠った。最も高い残存加水分解活性を100%とし、各pHにおける加水分解活性を相対値として図8にまとめた。pH3乃至pH10の範囲で安定であった。
【0152】
2)精製酵素の基質選択性
試験例1.に示した方法に拠った。卵黄由来ホスファチジルコリンに対する加水分解活性を100%としたときの、相対活性を表5に示す。
【0153】
【表5】
【0154】
試験例3.アスペルギルス・オリザエNBRC4190株由来ホスホリパーゼCの精製酵素液の諸性質
実施例3の2)で得られた精製酵素液について、活性測定を行なった。
【0155】
1)pH活性
試験例1.に示した方法に拠った。最も活性が高かったpH条件での加水分解活性を100%とし、各pHにおける酵素の加水分解活性を相対値として図9に記載した。至適pHはpH4.5付近であった。
【0156】
2)精製酵素の基質選択性
試験例1.に示した方法に拠った。卵黄由来ホスファチジルコリンに対する加水分解活性を100%としたときの、相対活性を表6に示す。
【0157】
【表6】
【0158】
発明の効果
以上述べたように、本発明のホスホリパーゼCはアスぺルギルス・オリザエFERM ABP−10200株またはNBRC 4190株あるいはアスぺルギルス・タマリIAM 13907株由来であり安全性に優れ、様々なグリセロリン脂質を酸性側でも中性付近でも効率よく加水分解する能力を有し、クエン酸緩衝液中でも活性を有するとともにある程度熱的に安定であり、リン脂質以外のリン酸エステルを加水分解しない酵素であり、食品工業および製油工業いずれの分野においても優れた効果をあげられる酵素である。
【特許請求の範囲】
【請求項1】
酸性から中性のpHにおいて活性を示し、かつ、リン脂質以外のリン酸エステルを実質的に加水分解しないホスホリパーゼC。
【請求項2】
ホスファチジルイノシトール特異的ではない、請求項1記載のホスホリパーゼC。
【請求項3】
至適pHがpH3乃至pH6の範囲にある、請求項1または2記載のホスホリパーゼC。
【請求項4】
pH7における相対活性が20%以上である、請求項1〜3のいずれか1項記載のホスホリパーゼC。
【請求項5】
糸状菌によって産生される、請求項1〜4のいずれか1項記載のホスホリパーゼC。
【請求項6】
糸状菌がアスペルギルス属である、請求項5記載のホスホリパーゼC。
【請求項7】
アスぺルギルス・オリザエ(Aspergillus oryzae)あるいはアスぺルギルス・タマリ(Aspergillus tamarii)によって産生される、請求項6記載のホスホリパーゼC。
【請求項8】
アスぺルギルス・オリザエFERM ABP−10200株またはNBRC 4190株、あるいはアスぺルギルス・タマリIAM 13907株によって産生される、請求項7に記載のホスホリパーゼC。
【請求項9】
以下の性質:
1)SDS−PAGE電気泳動法にて分子量約87,000を示す;
2)ホスファチジルコリン、ホスファチジルエタノールアミン、ホスファチジルイノシトールおよびホスファチジルグリセロールを加水分解する;
3)グリセロホスホリルコリンおよびパラニトロフェニルリン酸を実質的に加水分解しない;
4)卵黄由来レシチンをpH3乃至pH9の範囲で加水分解する;
5)0℃乃至80℃の範囲において4)記載の加水分解活性を有する;
6)温度安定性について、pH4.5において45℃以下の温度で安定である;
7)pH安定性について、pH3乃至pH10の範囲で安定である、
を示す請求項1〜8のいずれか1項記載のホスホリパーゼC。
【請求項10】
請求項1〜4および9のいずれか1項記載のホスホリパーゼCを生産する能力を有する、単離されたアスぺルギルス・オリザエあるいはアスぺルギルス・タマリに属する糸状菌、ただし、アスぺルギルス・タマリIAM 13907株を除く。
【請求項11】
配列番号4のコード領域(CDS)のヌクレオチド配列からなるDNAと70%以上のヌクレオチド配列相同性を有するヌクレオチド配列からなり、かつ、ホスホリパーゼC活性を有するタンパク質をコードすることを特徴とするDNA。
【請求項12】
請求項11に記載のDNAにコードされるタンパク質であるホスホリパーゼC。
【請求項13】
1)請求項8に記載のアスペルギルス・オリザエあるいはアスぺルギルス・タマリを培養する工程、および、
2)1)の培養産物からホスホリパーゼCを分離・精製する工程、
を含む、ホスホリパーゼCの製造方法。
【請求項1】
酸性から中性のpHにおいて活性を示し、かつ、リン脂質以外のリン酸エステルを実質的に加水分解しないホスホリパーゼC。
【請求項2】
ホスファチジルイノシトール特異的ではない、請求項1記載のホスホリパーゼC。
【請求項3】
至適pHがpH3乃至pH6の範囲にある、請求項1または2記載のホスホリパーゼC。
【請求項4】
pH7における相対活性が20%以上である、請求項1〜3のいずれか1項記載のホスホリパーゼC。
【請求項5】
糸状菌によって産生される、請求項1〜4のいずれか1項記載のホスホリパーゼC。
【請求項6】
糸状菌がアスペルギルス属である、請求項5記載のホスホリパーゼC。
【請求項7】
アスぺルギルス・オリザエ(Aspergillus oryzae)あるいはアスぺルギルス・タマリ(Aspergillus tamarii)によって産生される、請求項6記載のホスホリパーゼC。
【請求項8】
アスぺルギルス・オリザエFERM ABP−10200株またはNBRC 4190株、あるいはアスぺルギルス・タマリIAM 13907株によって産生される、請求項7に記載のホスホリパーゼC。
【請求項9】
以下の性質:
1)SDS−PAGE電気泳動法にて分子量約87,000を示す;
2)ホスファチジルコリン、ホスファチジルエタノールアミン、ホスファチジルイノシトールおよびホスファチジルグリセロールを加水分解する;
3)グリセロホスホリルコリンおよびパラニトロフェニルリン酸を実質的に加水分解しない;
4)卵黄由来レシチンをpH3乃至pH9の範囲で加水分解する;
5)0℃乃至80℃の範囲において4)記載の加水分解活性を有する;
6)温度安定性について、pH4.5において45℃以下の温度で安定である;
7)pH安定性について、pH3乃至pH10の範囲で安定である、
を示す請求項1〜8のいずれか1項記載のホスホリパーゼC。
【請求項10】
請求項1〜4および9のいずれか1項記載のホスホリパーゼCを生産する能力を有する、単離されたアスぺルギルス・オリザエあるいはアスぺルギルス・タマリに属する糸状菌、ただし、アスぺルギルス・タマリIAM 13907株を除く。
【請求項11】
配列番号4のコード領域(CDS)のヌクレオチド配列からなるDNAと70%以上のヌクレオチド配列相同性を有するヌクレオチド配列からなり、かつ、ホスホリパーゼC活性を有するタンパク質をコードすることを特徴とするDNA。
【請求項12】
請求項11に記載のDNAにコードされるタンパク質であるホスホリパーゼC。
【請求項13】
1)請求項8に記載のアスペルギルス・オリザエあるいはアスぺルギルス・タマリを培養する工程、および、
2)1)の培養産物からホスホリパーゼCを分離・精製する工程、
を含む、ホスホリパーゼCの製造方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公開番号】特開2010−252815(P2010−252815A)
【公開日】平成22年11月11日(2010.11.11)
【国際特許分類】
【出願番号】特願2010−184963(P2010−184963)
【出願日】平成22年8月20日(2010.8.20)
【分割の表示】特願2005−37829(P2005−37829)の分割
【原出願日】平成17年2月15日(2005.2.15)
【出願人】(593204214)三菱化学フーズ株式会社 (45)
【Fターム(参考)】
【公開日】平成22年11月11日(2010.11.11)
【国際特許分類】
【出願日】平成22年8月20日(2010.8.20)
【分割の表示】特願2005−37829(P2005−37829)の分割
【原出願日】平成17年2月15日(2005.2.15)
【出願人】(593204214)三菱化学フーズ株式会社 (45)
【Fターム(参考)】
[ Back to top ]