
新規ポリペプチドおよびそのポリペプチドをコードするポリヌクレオチド、並びにその利用

【課題】高マンノース型の糖鎖と高い親和性を有する新規ポリペプチド、およびそのポリペプチドをコードするポリヌクレオチド、並びにそれらの代表的利用を提供する。
【解決手段】食用紅藻であるトサカノリ(Meristothecapapulosa)由来の高マンノース型糖鎖と結合するポリペプチドであって、特定な配列からなるアミノ酸配列;または(b)特定な配列からなるアミノ酸配列において、1個もしくは数個のアミノ酸が置換、欠失、挿入、もしくは付加されたアミノ酸配列、からなる坑ウイルス剤。
【解決手段】食用紅藻であるトサカノリ(Meristothecapapulosa)由来の高マンノース型糖鎖と結合するポリペプチドであって、特定な配列からなるアミノ酸配列;または(b)特定な配列からなるアミノ酸配列において、1個もしくは数個のアミノ酸が置換、欠失、挿入、もしくは付加されたアミノ酸配列、からなる坑ウイルス剤。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、高マンノース型糖鎖と高い親和性を有する新規ポリペプチド、およびそのポリペプチドをコードするポリヌクレオチド、並びにそれらの代表的利用に関するものである。
【背景技術】
【0002】
ヒト免疫不全ウイルス(以下「HIV」)、インフルエンザウイルス、C型肝炎ウイルス(以下「HCV」)、エボラウイルス、ヒトヘルペスウイルス6(以下「HHV−6」)、重症急性呼吸症候群(以下「SARS」)ウイルスなどのウイルスは、しばしば人体に対して重篤な症状を惹起し、その感染拡大が社会問題ともなっていることから、効果的に感染を阻害する薬剤の開発が望まれている。これらのウイルスは、表面に高マンノース型糖鎖を有し、これが宿主への感染に重要な役割を果たすことが知られている(例えば、非特許文献1)。そのため、これらのウイルスが備える高マンノース型糖鎖への結合能を有する物質は、上記ウイルスに対する感染阻害剤として機能することが期待できる。
【0003】
これまでに、海藻類または藻類(淡水産藍藻)から多くの種類のレクチンが単離され、その生化学的性質が明らかにされている。上記レクチンの一部は、HIV、インフルエンザウイルス等の上述のウイルスに特異的に結合することが知られている(非特許文献2〜12)。
【0004】
他にも、レクチンとHIVとの結合に関する発明として、例えば、ConAなどのレクチンが結合した疎水性基材からなる微粒子担体にHIVまたはその一部のタンパクもしくはペプチドを捕捉してなる微粒子を含有するエイズワクチン(特許文献1)、GNAなどのレクチンを固定した多孔性中空糸膜に血液を通過させ、ウイルスに感染した個体の血液中のウイルス粒子及びそのレクチン結合性フラグメントを減じる方法(特許文献2)、癌患者における感染症の予防および/または処置用の医薬の製造において、マンナン結合レクチンサブユニット等を含むマンナン結合レクチンオリゴマーを含有する組成物を使用する発明(特許文献3)、スクシニル−コンカナバリンAなどのレクチンを結合成分として用い、ウイルス粒子を含んでなるサンプルのウイルス力価を高める方法(特許文献4)などが知られている。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開2001−335510号公報(2001年12月4日公開)
【特許文献2】特表2007−525232号公報(2007年9月6日公表)
【特許文献3】特表2008−535872号公報(2008年9月4日公表)
【特許文献4】特表2004−500831号公報(2004年1月15日公表)
【非特許文献】
【0006】
【非特許文献1】New England Journal of Medicine, 348, 2228-2238, 2003.
【非特許文献2】Boyd, M. R. et al., Antimicrob. Agents Chemother.41, 1521-1530, 1997.
【非特許文献3】O’Keefe, B. R. et al., Antimicrob. Agents Chemother. 47, 2518-2525, 2003.
【非特許文献4】Helle, F., .et al., J. Biol. Chem. 281, 25177-25183, 2006.
【非特許文献5】Barrientos, L. G., et al., Antiviral. Res. 58, 47-56, 2003.
【非特許文献6】Dey, B., et al., J. Virol. 74, 4562-4569, 2000.
【非特許文献7】O’Keefe, B. R. et al.,J. Virol. 84, 2511-2521, 2010.
【非特許文献8】Hori,K. et al., Glycobiology, 17, 479-491, 2007.
【非特許文献9】Sato,Y., Okuyama, S., and Hori, K., J. Biol. Chem. 282, 11021-11029, 2007.
【非特許文献10】Sato,Y., Morimoto,K., Hirayama, M., and Hori, K. Biochem. Biophys. Res. commun. 405, 291-296, 2011.
【非特許文献11】佐藤雄一郎、平山 真、藤原佳史、森本金治郎、堀 貫治 (2010) 第13回マリンバイオテクノロジー学会大会講演要旨 (2010. 5.29発表)
【非特許文献12】Sato,Y., Hirayama, M., Morimoto,K., Yamamoto, N., Okuyama, S., and Hori, K. J. Biol. Chem. 286, No.22, 19446-19458, 2011.
【発明の概要】
【発明が解決しようとする課題】
【0007】
しかしながら、現在のところHIVウイルスやインフルエンザウイルス等が表面に有する高マンノース型糖鎖に特異的に結合し、上記ウイルスの感染を効果的に阻害する物質はまだ十分に知られているとは言いがたく、その数は限られているため、上記物質が十分に供給できる状況にはなっていない。したがって、新規な上記物質がさらに多く見出され、その特性が明らかにされることが必要である。
【0008】
本発明は、このような問題点に鑑みてなされたものであり、その目的は、高マンノース型糖鎖と高い親和性を有する新規ポリペプチド、およびそのポリペプチドをコードするポリヌクレオチド、並びにそれらの代表的利用を提供することにある。
【課題を解決するための手段】
【0009】
本発明者は、上記課題を解決すべく、レクチン、特にトサカノリ(Meristotheca papulosa (Montagne) J. Agardh)由来のレクチンに着目し、HIVウイルス等が表面に備える高マンノース型糖鎖と特異的に結合しうるレクチンの探索を行った。その結果、従来公知のレクチンとアミノ酸配列の相同性が非常に低い、本発明にかかるポリペプチドを見出すに至った。
【0010】
すなわち、本発明にかかるポリペプチドは、上記課題を解決すべく、高マンノース型糖鎖と結合するポリペプチドであって、(a)配列番号2に示されるアミノ酸配列;または(b)配列番号2に示されるアミノ酸配列において、1個もしくは数個のアミノ酸が置換、欠失、挿入、もしくは付加されたアミノ酸配列、からなることを特徴としている。
【0011】
本発明にかかるポリヌクレオチドは、上記課題を解決すべく、上記本発明にかかるポリペプチドをコードすることを特徴としている。
【0012】
また、本発明にかかるポリヌクレオチドは、上記課題を解決すべく、下記の(a)または(b)のいずれかであるポリヌクレオチド:(a)配列番号1に示される塩基配列からなるポリヌクレオチド;または(b)以下の(i)もしくは(ii)のいずれかとストリンジェントな条件下でハイブリダイズするポリヌクレオチド:(i)配列番号1に示される塩基配列からなるポリヌクレオチド;もしくは(ii)配列番号1に示される塩基配列と相補的な塩基配列からなるポリヌクレオチド、であってもよい。
【0013】
本発明にかかる抗ウイルス剤は、上記課題を解決すべく、上記本発明にかかるポリペプチドを含有し、高マンノース型糖鎖を備えるウイルスを標的とすることを特徴としている。
【0014】
上記本発明にかかる抗ウイルス剤は、上記課題を解決すべく、上記抗ウイルス剤の標的であるウイルスが、エイズウイルス、インフルエンザウイルス、C型肝炎ウイルス、エボラウイルス、ヒトヘルペスウイルス6および重症急性呼吸症候群ウイルスからなる群より選ばれるウイルスであってもよい。
【0015】
本発明にかかるウイルス除去膜は、上記課題を解決すべく、上記本発明にかかるポリペプチドが固定されてなることを特徴としている。
【0016】
本発明にかかるベクターは、上記課題を解決すべく、上記本発明にかかるポリヌクレオチドを含むことを特徴としている。
【0017】
本発明にかかるポリペプチドを生産する方法は、上記課題を解決すべく、上記本発明にかかるベクターを用いることを特徴としている。
【0018】
本発明にかかる形質転換体は、上記課題を解決すべく、上記本発明にかかるポリヌクレオチドが導入されていることを特徴としている。
【0019】
本発明にかかるポリペプチドを生産する方法は、上記課題を解決すべく、上記本発明にかかる形質転換体を用いることを特徴としている。
【発明の効果】
【0020】
以上のように、本発明にかかるポリペプチドは、高マンノース型糖鎖と結合するポリペプチドであって、(a)配列番号2に示されるアミノ酸配列;または(b)配列番号2に示されるアミノ酸配列において、1個もしくは数個のアミノ酸が置換、欠失、挿入、もしくは付加されたアミノ酸配列、からなる構成である。上記アミノ酸配列は従来公知のレクチンと非常に相同性の低いものである。
【0021】
それゆえ、HIVやインフルエンザウイルス等が表面に備える高マンノース型糖鎖と効率的に結合することができ、これらのウイルスの感染を効果的に阻害することができるという効果を奏する。
【0022】
本発明にかかるポリヌクレオチドは、上記本発明にかかるポリペプチドをコードしている。したがって、本発明にかかるポリヌクレオチド、当該ポリヌクレオチドを含むベクター、当該ポリヌクレオチドが導入された形質転換体を用いることによって、上記本発明にかかるポリペプチドを簡便かつ大量に調製することが可能となる。その結果、上記ウイルスの感染を効果的に阻害することができるという効果を奏する。
【図面の簡単な説明】
【0023】
【図1】MPLのcDNAの全長塩基配列、およびその演繹アミノ酸配列を示す図である。
【図2】図2のAは固定化したgp120とMPLとの相互作用のセンサーグラムであり、図2のBはgp120とMPLとの親和定数を示す図である。
【図3】MPLをSDS−PAGE(10%ゲル)に供した結果を示す図である。
【図4】20〜60%飽和硫安塩析沈殿画分を、HiPrep QXL 16/10カラムを用いた陰イオン交換クロマトグラフィー(ステップワイズ溶出)に供した結果を示す図である。
【図5】粗レクチン画分を、HiPrep QXL 16/10カラムを用いた陰イオン交換クロマトグラフィー(グラジエント溶出)に供した結果を示す図である。
【図6】粗レクチン画分を、HiPrep QXL 16/10カラムを用いた陰イオン交換クロマトグラフィー(グラジエント溶出)に供した結果得られた溶出液の画分をSDS−PAGE(10%ゲル)に供した結果を示す図である。
【図7】MPLをTSKgel DEAE-5PWカラムを用いた陰イオン交換クロマトグラフィーに供した結果を示す図である。
【図8】MPLをTSKgel DEAE-5PWカラムによる陰イオン交換クロマトグラフィーに供して得られた4つのピークを、SDS−PAGE(10%ゲル)に供した結果を示す図である。
【図9】MPL−1およびMPL−2の分子量測定結果を示す図である。
【図10】MPL画分をブロッティングし、CBB染色したPVDF膜を示す図である。
【図11】遠心限外ろ過−HPLC法に供試したピリジルアミノ化糖鎖27種の構造と、当該糖鎖へのMPL−1の結合活性を示すものである。
【発明を実施するための形態】
【0024】
以下、本発明の実施の一形態について説明すれば以下のとおりである。なお、本発明はこれに限定されるものではない。本明細書において、範囲を示す「A〜B」は、A以上B以下であることを表す。また、本明細書中に記載された特許文献および非特許文献は、本明細書中において参考として援用される。
【0025】
(1)ポリペプチド
本発明者は、食用紅藻であるトサカノリ(Meristotheca papulosa (Montagne) J. Agardh)から単離した新規ポリペプチド(以下「MPL」という)が、高マンノース型糖鎖への結合特異性を有することを見出し、本発明を完成するに至った。上記MPLのアミノ酸配列は、後述するように、バナナMusa acuminataレクチンおよび紅藻Griffithsia sp.由来レクチンGriffithsin(GRFT)等の従来公知のレクチンとの相同性が最大33%と、非常に低いものであった。
【0026】
本明細書中で使用される場合、用語「ポリペプチド」は、「ペプチド」または「タンパク質」と交換可能に使用される。本発明にかかるポリペプチドはまた、天然供給源より単離されても、化学合成されてもよい。
【0027】
用語「単離された」ポリペプチドまたはタンパク質とは、その天然の環境から取り出されたポリペプチドまたはタンパク質が意図される。例えば、宿主細胞中で発現された組換え産生されたポリペプチドおよびタンパク質は、任意の適切な技術によって実質的に精製されている天然または組換えのポリペプチドおよびタンパク質と同様に、単離されていると考えられる。
【0028】
本発明にかかるポリペプチドは、天然の精製産物、化学合成手順の産物、および原核生物宿主または真核生物宿主(例えば、細菌細胞、酵母細胞、高等植物細胞、昆虫細胞、および哺乳動物細胞を含む)から組換え技術によって産生された産物を含む。組換え産生手順において用いられる宿主に依存して、本発明にかかるポリペプチドは、グリコシル化され得るか、または非グリコシル化され得る。さらに、本発明にかかるポリペプチドはまた、いくつかの場合、宿主媒介プロセスの結果として、開始の改変メチオニン残基を含み得る。
【0029】
本発明は、本発明にかかるポリペプチドを提供する。一実施形態において、本発明にかかるポリペプチドは、配列番号2に示されるアミノ酸配列からなるポリペプチド、または配列番号2に示されるアミノ酸配列からなるポリペプチドの変異体でありかつ本発明にかかるポリペプチドである。上記ポリペプチドは、従来公知のレクチンのアミノ酸配列との相同性が非常に低い新規ポリペプチドであるという特徴を有する。
【0030】
変異体としては、欠失、挿入、逆転、反復、およびタイプ置換(例えば、親水性の残基の別の残基への置換、しかし通常は強く親水性の残基を強く疎水性の残基には置換しない)を含む変異体が挙げられる。特に、ポリペプチドにおける「中性」アミノ酸置換は、一般的にそのポリペプチドの活性にほとんど影響しない。
【0031】
ポリペプチドのアミノ酸配列中のいくつかのアミノ酸が、このポリペプチドの構造または機能に有意に影響することなく容易に改変され得ることは、当該分野において周知である。さらに、人為的に改変させるだけではく、天然のタンパク質において、当該タンパク質の構造または機能を有意に変化させない変異体が存在することもまた周知である。
【0032】
当業者は、周知技術を使用してポリペプチドのアミノ酸配列において1または数個のアミノ酸を容易に変異させることができる。例えば、公知の点変異導入法に従えば、ポリペプチドをコードするポリヌクレオチドの任意の塩基を変異させることができる。また、ポリペプチドをコードするポリヌクレオチドの任意の部位に対応するプライマーを設計して欠失変異体または付加変異体を作製することができる。さらに、本明細書中に記載される方法を用いれば、作製した変異体が、本発明にかかる所望の変異体であるか否かを容易に決定し得る。
【0033】
本実施形態にかかるポリペプチドは、高マンノース型の糖鎖と結合するポリペプチドであって、
(a)配列番号2に示されるアミノ酸配列;または
(b)配列番号2に示されるアミノ酸配列において、1個もしくは数個のアミノ酸が置換、欠失、挿入、もしくは付加されたアミノ酸配列、
からなることが好ましい。
【0034】
上記「1個もしくはそれ以上のアミノ酸が置換、欠失、挿入、もしくは付加された」とは、部位特異的突然変異誘発法等の公知の変異ポリペプチド作製法により置換、欠失、挿入、もしくは付加できる程度の数(好ましくは1から10個、より好ましくは1から7個、さらに好ましくは1個から5個、特に好ましくは1個から3個)のアミノ酸が置換、欠失、挿入もしくは付加されていることを意味する。このような変異ポリペプチドは、上述したように、公知の変異ポリペプチド作製法により人為的に導入された変異を有するポリペプチドに限定されるものではなく、天然に存在するポリペプチドを単離精製したものであってもよい。
【0035】
なお、本発明にかかるポリペプチドは、アミノ酸がペプチド結合しているポリペプチドであればよいが、これに限定されるものではなく、ポリペプチド以外の構造を含む複合ポリペプチドであってもよい。本明細書中で使用される場合、「ポリペプチド以外の構造」としては、糖鎖やイソプレノイド基等を挙げることができるが、特に限定されるものではない。
【0036】
また、本発明にかかるポリペプチドは、付加的なポリペプチドを含むものであってもよい。付加的なポリペプチドとしては、例えば、HisやMyc、Flag等のエピトープ標識ポリペプチドが挙げられる。
【0037】
また、本発明にかかるポリペプチドは、後述する本発明にかかるポリヌクレオチド(本発明にかかるポリペプチドをコードする遺伝子)を宿主細胞に導入して、そのポリペプチドを細胞内発現させた状態であってもよいし、細胞、組織などから単離精製された場合であってもよい。また、本発明にかかるポリペプチドは、化学合成されたものであってもよい。
【0038】
他の実施形態において、本発明にかかるポリペプチドは、融合タンパク質のような改変された形態で組換え発現され得る。例えば、本発明にかかるポリペプチドの付加的なアミノ酸、特に荷電性アミノ酸の領域が、宿主細胞内での、精製の間または引き続く操作および保存の間の安定性および持続性を改善するために、ポリペプチドのN末端に付加され得る。
【0039】
別の実施形態において、本発明にかかるポリペプチドは、下記で詳述されるように組換え生成されても、化学合成されてもよい。
【0040】
組換え生成は、当該分野において周知の方法を使用して行なうことができ、例えば、以下に詳述されるようなベクターおよび細胞を用いて行なうことができる。
【0041】
合成ペプチドは化学合成の公知の方法を使用して合成され得る。例えばHoughtenは、4週間未満で調製され、そして特徴付けられたHA1ポリペプチドセグメントの単一アミノ酸改変体を示す10〜20mgの248の異なる13残基ペプチドのような、多数のペプチドの合成のための簡単な方法を記載している(Houghten,R.A.,Proc.Natl.Acad.Sci.USA, 82:5131-5135(1985))。
【0042】
この「Simultaneous Multiple Peptide Synthesis(SMPS)」プロセスは、さらにHoughtenら(1986)の米国特許第4,631,211号に記載される。この手順において、種々のペプチドの固相合成のための個々の樹脂は、別々の溶媒透過性パケットに含まれ、固相法に関連する多くの同一の反復工程の最適な使用を可能にする。完全なマニュアル手順は、500〜1000以上の合成が同時に行われるのを可能にする(Houghtenら、前出、5134)。これらの文献は、本明細書中に参考として援用される。
【0043】
本発明者は、上記本発明にかかるポリペプチドが高マンノース型糖鎖に特異的に結合することを見出した。ここで、「糖鎖」とは、直鎖または分岐したオリゴ糖または多糖を意味する。また上記糖鎖は、タンパク質との結合様式によって、アスパラギンと結合するN−グリコシド結合糖鎖(以下、「N型糖鎖」という)およびセリン、スレオニンなどと結合するO−グリコシド結合糖鎖(以下、「O型糖鎖」という)に大別され、N型糖鎖には高マンノース型糖鎖、複合型糖鎖、混成型糖鎖がある。
【0044】
なおオリゴ糖とは、単糖または単糖の置換誘導体が2〜10個脱水結合して生じたものをいう。さらに多数の単糖が結合している糖質を多糖という。多糖は、構成糖の種類によって異なるが、ウロン酸やエステル硫酸を多く含む糖質を酸性多糖、中性糖のみのものを中性多糖という。多糖のうち、ムコ多糖とよばれる一群の多糖は、ほとんどがタンパク質と結合しており、プロテオグリカンという。単糖とは、糖鎖の構成単位となるもので、加水分解によってそれ以上簡単な分子にならない基本的物質である。
【0045】
さらに単糖は、カルボキシル基などの酸性側鎖を有する酸性糖、ヒドロキシル基がアミノ基で置換されたアミノ糖、それ以外の中性糖の3つに大別される。生体内に存在する単糖としては、酸性糖はN−アセチルノイラミン酸やN−グリコリルノイラミン酸(以下、「Neu5Gc」という)などのシアル酸や、ウロン酸などがあり、アミノ糖としてはN−アセチルグルコサミン(以下、「GlcNAc」という)やN−アセチルガラクトサミンなどがあり、中性糖としてはグルコース、マンノース、ガラクトース、フコースなどがあげられる。
【0046】
N型糖鎖は全て、「トリマンノシルコア」と呼ばれる〔Manα1-6(Manα1-3)Manβ1-4GlcNAcβ1-4GlcNAc〕からなる共通母核構造を持っている。高マンノース型糖鎖は、トリマンノシルコアに加え、分岐構造部分にα−マンノース残基のみを含む。この糖鎖には〔Manα1-6(Manα1-3)Manα1-6(Manα1-3 )Manβ1-4GlcNAcβ1-4GlcNAc〕という七糖が共通の母核として含まれている。また混成型糖鎖は、複合型と高マンノース型の両方の特徴を併せ持っていることからそう呼ばれている。1つまたは2つのα−マンノシル基が、高マンノース型の場合と同様に、トリマンノシルコアのManα1-6腕と結合し、複合型糖鎖の側鎖と同じものがコアのManα1-3腕に結合している。
【0047】
トリマンノシルコアの還元末端に位置するGlcNAc のC-6位へのフコースの結合の有無、またβ-マンノシル残基のC-4位へのβ-GlcNAcの結合(バィセクテイングGlcNAc と呼ばれる)の有無は、複合型や混成型糖鎖の構造の多様性に寄与している。3つのN型糖鎖の間で、複合型が最も多様な構造を含んでいる。
【0048】
この多様性は、主に2つの要素で作り出され、トリマンノシルコアに1個から5個の側鎖がそれぞれ異なる結合位置で結合しており、一、二、三、四、または五本側鎖糖鎖を形成している。三本側鎖の複合型糖鎖には、〔GlcNAcβ1-4(GlcNAcβ1-2)Manα1-3〕あるいは〔GlcNAcβ1-6(GlcNAcβ1-2)Manα1-6〕のどちらかを含む2つの異性体が見つかっている。
【0049】
なお上記トリマンノシルコアにおいて、アスパラギンと結合する糖鎖の末端、すなわちGlcNAc側の末端を還元末端、その反対側、すなわちMan側の末端を非還元末端という。
【0050】
ポリペプチドが糖鎖と結合するか否かは、例えば標的となる糖鎖、または糖鎖が結合した糖タンパク質等を固定化したカラムに、試験対象であるポリペプチドを通し、当該カラムにポリペプチドが結合したか否かをその通過液に含まれるポリペプチドの量、または特異的溶出剤でカラムから溶出したポリペプチドの量により評価することができる。また標的となる糖鎖が結合した糖タンパク質をメンブレン等に固定化し、ビオチン、フルオレセインイソチオシアネート、ペルオキシダーゼ等で標識したポリペプチドを用いて検出するウエスタンブロット法(法医学の実際と研究、37, 155, 1994 参照)、ドットブロット法(Analytical Biochemistry, 204(1), 198, 1992 参照)を用いて評価することができる。
【0051】
また標的となる糖鎖、または糖鎖が結合した糖タンパク質等を固定化したチップと、試験対象であるポリペプチドとの親和性を表面プラズモン共鳴法(SPR法)を用いて測定すればよい。上記方法によれば、その親和性の有無のみならず、その強度まで測定できるために好ましい方法であるといえる。このとき得られる結合定数(親和定数)(KA)が、10(M-1)以上、より好ましくは103(M-1)以上、最も好ましくは104(M-1)以上であればポリペプチドと糖鎖とが結合していると判断できる。
【0052】
本発明にかかるポリペプチドには上記(a)または(b)に示すアミノ酸配列と、特定の機能(例えば、タグ)を有する任意のアミノ酸配列とからなるポリペプチドも含まれることに留意すべきである。また、上記(a)または(b)に示すアミノ酸配列と、当該任意のアミノ酸配列とは、それぞれの機能を阻害しないように適切なリンカーペプチドで連結されていてもよい。
【0053】
つまり、本発明の目的は、本発明にかかるポリペプチドを提供することにあるのであって、本明細書中に具体的に記載したポリペプチド作製方法等に存するのではない。したがって、上記各方法以外によって取得される本発明にかかるポリペプチドも本発明の技術的範囲に属することに留意しなければならない。
【0054】
(2)ポリヌクレオチド
本発明は、上述した本発明にかかるポリペプチドをコードするポリヌクレオチドを提供する。本明細書中で使用される場合、用語「ポリヌクレオチド」は「核酸」または「核酸分子」と交換可能に使用され、ヌクレオチドの重合体が意図される。本明細書中で使用される場合、用語「塩基配列」は、「核酸配列」または「ヌクレオチド配列」と交換可能に使用され、デオキシリボヌクレオチド(A、G、CおよびTと省略される)の配列として示される。
【0055】
本発明にかかるポリヌクレオチドは、RNA(例えば、mRNA)の形態、またはDNAの形態(例えば、cDNAまたはゲノムDNA)で存在し得る。DNAは、二本鎖または一本鎖であり得る。一本鎖DNAまたはRNAは、コード鎖(センス鎖としても知られる)であり得、または、非コード鎖(アンチセンス鎖としても知られる)であり得る。
【0056】
本明細書中で使用される場合、用語「オリゴヌクレオチド」は、ヌクレオチドが数個ないし数十個結合したものが意図され、「ポリヌクレオチド」と交換可能に使用される。オリゴヌクレオチドは、短いものはジヌクレオチド(二量体)、トリヌクレオチド(三量体)といわれ、長いものは30マーまたは100マーというように重合しているヌクレオチドの数で表される。オリゴヌクレオチドは、より長いポリヌクレオチドのフラグメントとして生成されても、化学合成されてもよい。
【0057】
本発明にかかるポリヌクレオチドのフラグメントは、少なくとも12nt(ヌクレオチド)、好ましくは約15nt、そしてより好ましくは少なくとも約20nt、なおより好ましくは少なくとも約30nt、そしてさらにより好ましくは少なくとも約40ntの長さのフラグメントが意図される。少なくとも20ntの長さのフラグメントによって、例えば、配列番号1に示される塩基配列からの20以上の連続した塩基を含むフラグメントが意図される。本明細書を参照すれば配列番号1に示される塩基配列が提供されるので、当業者は、配列番号1に基づくDNAフラグメントを容易に作製することができる。
【0058】
例えば、制限エンドヌクレアーゼ切断または超音波による剪断は、種々のサイズのフラグメントを作製するために容易に使用され得る。あるいは、このようなフラグメントは、合成的に作製され得る。適切なフラグメント(オリゴヌクレオチド)が、Applied Biosystems Incorporated(ABI,850 Lincoln Center Dr., Foster City, CA 94404)392型シンセサイザーなどによって合成される。
【0059】
合成的に作製した上記フラグメントとしては、例えば宿主のコドンに対して配列を最適化した合成ポリヌクレオチドを挙げることができる。このような合成ポリヌクレオチドは、各種の発現系を用いる場合に利用可能である。
【0060】
また本発明にかかるポリヌクレオチドは、その5’側または3’側で上述のタグ標識(タグ配列またはマーカー配列)をコードするポリヌクレオチドに融合され得る。
【0061】
本発明はさらに、本発明にかかるポリペプチドをコードするポリヌクレオチドの変異体に関する。変異体は、天然の対立遺伝子変異体のように、天然に生じ得る。「対立遺伝子変異体」によって、生物の染色体上の所定の遺伝子座を占める遺伝子のいくつかの交換可能な形態の1つが意図される。天然に存在しない変異体は、例えば当該分野で周知の変異誘発技術を用いて生成され得る。
【0062】
このような変異体としては、本発明にかかるポリペプチドをコードするポリヌクレオチドの塩基配列において1または数個の塩基が欠失、置換、または付加した変異体が挙げられる。変異体は、コードもしくは非コード領域、またはその両方において変異され得る。コード領域における変異は、保存的もしくは非保存的なアミノ酸欠失、置換、または付加を生成し得る。
【0063】
本発明はさらに、ストリンジェントなハイブリダイゼーション条件下で、本発明にかかるポリペプチドをコードするポリヌクレオチドまたは当該ポリヌクレオチドにハイブリダイズするポリヌクレオチドを含む、単離したポリヌクレオチドを提供する。
【0064】
一実施形態において、本発明にかかるポリヌクレオチドは、本発明にかかるポリペプチドをコードするポリヌクレオチドであり、かつ、
(a)配列番号2に示されるアミノ酸配列;または
(b)配列番号2に示されるアミノ酸配列において、1個もしくは数個のアミノ酸が置換、欠失、挿入、もしくは付加されたアミノ酸配列、からなるポリペプチドをコードするポリヌクレオチド、
のいずれかであることが好ましい。
【0065】
他の実施形態において、本発明にかかるポリヌクレオチドは、本発明にかかるポリペプチドをコードするポリヌクレオチドであって、以下の(a)または(b):
(a)配列番号1に示される塩基配列からなるポリヌクレオチド;または
(b)以下の(i)もしくは(ii)のいずれかとストリンジェントな条件下でハイブリダイズするポリヌクレオチド:
(i)配列番号1に示される塩基配列からなるポリヌクレオチド;もしくは
(ii)配列番号1に示される塩基配列と相補的な塩基配列からなるポリヌクレオチド、
のいずれかであることが好ましい。
【0066】
なお、上記「ストリンジェントな条件」とは、少なくとも90%以上の同一性、好ましくは少なくとも95%以上の同一性、最も好ましくは97%以上の同一性が配列間に存在する時にのみハイブリダイゼーションが起こることを意味する。
【0067】
上記ハイブリダイゼーションは、Sambrookら、Molecular Cloning,A Laboratory Manual,2d Ed.,Cold Spring Harbor Laboratory(1989)に記載されている方法のような周知の方法で行なうことができる。通常、温度が高いほど、塩濃度が低いほどストリンジェンシーは高くなり(ハイブリダイズし難くなる)、より相同なポリヌクレオチドを取得することができる。
【0068】
ハイブリダイゼーションの条件としては、従来公知の条件を好適に用いることができ、特に限定しないが、例えば、42℃、6×SSPE、50%ホルムアミド、1%SDS、100μg/ml サケ精子DNA、5×デンハルト液(ただし、1×SSPE;0.18M 塩化ナトリウム、10mMリン酸ナトリウム、pH7.7、1mM EDTA。5×デンハルト液;0.1%牛血清アルブミン、0.1%フィコール、0.1%ポリビニルピロリドン)が挙げられる。
【0069】
本発明にかかるポリヌクレオチドまたはオリゴヌクレオチドは、2本鎖DNAのみならず、それを構成するセンス鎖およびアンチセンス鎖といった各1本鎖DNAやRNAを包含する。またDNAには例えばクローニングや化学合成技術またはそれらの組み合わせで得られるようなcDNAやゲノムDNAなどが含まれる。さらに、本発明にかかるポリヌクレオチドまたはオリゴヌクレオチドは、非翻訳領域(UTR)の配列やベクター配列(発現ベクター配列を含む)などの配列を含むものであってもよい。
【0070】
本発明にかかるポリヌクレオチドまたはオリゴヌクレオチドを取得する方法として、公知の技術により、本発明にかかるポリヌクレオチドまたはオリゴヌクレオチドを含むDNA断片を単離し、クローニングする方法が挙げられる。例えば、本発明におけるポリヌクレオチドの塩基配列の一部と特異的にハイブリダイズするプローブを調製し、ゲノムDNAライブラリーやcDNAライブラリーをスクリーニングすればよい。このようなプローブとしては、本発明にかかるポリヌクレオチドの塩基配列またはその相補配列の少なくとも一部に特異的にハイブリダイズするプローブであれば、いずれの配列および/または長さのものを用いてもよい。
【0071】
あるいは、本発明にかかるポリヌクレオチドを取得する方法として、PCR等の増幅手段を用いる方法を挙げることができる。例えば、本発明におけるポリヌクレオチドのcDNAのうち、5’側および3’側の配列(またはその相補配列)の中からそれぞれプライマーを調製し、これらプライマーを用いてゲノムDNA(またはcDNA)等を鋳型にしてPCR等を行い、両プライマー間に挟まれるDNA領域を増幅することで、本発明にかかるポリヌクレオチドを含むDNA断片を大量に取得できる。
【0072】
本発明にかかるポリヌクレオチドを取得するための供給源としては、特に限定されないが、所望のポリヌクレオチドを含む生物材料であることが好ましい。特に、本発明にかかるポリペプチドの起源であるトサカノリ(Meristotheca papulosa (Montagne) J. Agardh)が好ましい。ただし、これに限定されるものではない。
【0073】
なお本発明の目的は、本発明にかかるポリペプチドをコードするポリヌクレオチド、および当該ポリヌクレオチドとハイブリダイズするオリゴヌクレオチドを提供することにあるのであって、本明細書中に具体的に記載したポリヌクレオチドおよびオリゴヌクレオチドの作製方法等に存するのではない。したがって、上記各方法以外によって取得される本発明にかかるポリペプチドをコードするポリヌクレオチドもまた本発明の技術的範囲に属することに留意しなければならない。
【0074】
(3)本発明にかかるポリペプチドおよび/またはポリヌクレオチドの利用
(3−1)抗ウイルス剤
本発明にかかる抗ウイルス剤は、上記本発明にかかるポリペプチドを含有し、高マンノース型糖鎖を備えるウイルスを標的とする。上記ウイルスは、高マンノース型糖鎖を備えるウイルスであれば特に限定されるものではなく、HIV、インフルエンザウイルス、HCV、エボラウイルス、HHV−6、SARSウイルス等を挙げることができる。
【0075】
上記ウイルスには、表面に備える高マンノース型糖鎖を宿主細胞の表面レセプターに結合させることによって宿主に感染するものが存在する。例えばHIVは、表面にgp120という糖タンパク質を備えており、gp120が有する高マンノース型糖鎖が宿主リンパ細胞表面レセプターであるCD4を認識することによって宿主に感染する。
【0076】
一方、上記抗ウイルス剤に含まれる本発明にかかるポリペプチドは、高マンノース型糖鎖と特異的に結合することができる。それゆえ、上記抗ウイルス剤を用いれば上記ウイルスの感染を効果的に阻害することができる。
【0077】
また、ウイルスの宿主への感染が、gp120とCD4との関係のような、ウイルス表面糖タンパク質の高マンノース型糖鎖と、宿主側の高マンノース型糖鎖のレセプターとの直接結合によるものではない場合もある。例えば、高マンノース型糖鎖を有するものの、高マンノース型糖鎖は宿主のレセプターと直接結合せず、糖タンパク質によって上記レセプターに結合するタイプのウイルスを挙げることができる。このようなウイルスとしては例えばインフルエンザウイルスを挙げることができる。
【0078】
上記抗ウイルス剤に含まれる本発明にかかるポリペプチドは、高マンノース型糖鎖と特異的に結合することができる。これにより糖タンパク質に対するアロステリック効果が得られ、高マンノース型糖鎖がレセプターとの結合に直接関与しない場合でもウイルスの感染を防ぐことができると考えられる。よって、上記抗ウイルス剤は、インフルエンザウイルス等の感染も有効に阻害することができる。
【0079】
なお、ウイルスの型は特に限定されるものではない。例えばHIVは1型でも2型でもよく、インフルエンザウイルスはA型、B型、C型のいずれであってもよい。
【0080】
「高マンノース型糖鎖を備える」とは、高マンノース型糖鎖がウイルスの表面に存在することを言う。例えば、高マンノース型糖鎖がキャプシドの外部へ露出した状態や、ウイルスの表面糖タンパク質の構成成分として存在する場合を挙げることができる。
【0081】
高マンノース型糖鎖は、上述のように、宿主細胞のレセプターへ直接結合するものであってもよいし、宿主細胞のレセプターへ直接結合しないものであってもよい。上述のように、本発明にかかる抗ウイルス剤は、いずれの高マンノース型糖鎖を有するウイルスであっても有効に感染を阻害することが可能である。
【0082】
本発明にかかる抗ウイルス剤は経口製剤、非経口製剤のいずれであってもよい。また、剤型は特に限定されるものではなく、常法に従い、錠剤、顆粒剤、散剤、カプセル剤、エリキシル剤、シロップ剤、マイクロカプセル剤あるいは懸濁液剤等に製剤化して用いることができる。
【0083】
非経口的に投与する場合には、例えば、本発明にかかるポリペプチドを含有する溶液を点鼻噴霧することや、注射剤として投与することができる。経口的に投与する場合には、食前、食後、食間のいずれに投与してもよい。
【0084】
本発明にかかる抗ウイルス剤は、必要に応じて、担体、賦形剤、結合剤、膨化剤、潤滑剤、甘味剤、香味剤、防腐剤、安定剤、被覆剤等の材料を含有することができる。
【0085】
本発明にかかる抗ウイルス剤において、例えば錠剤、カプセル剤等に含有することができる具体的な成分としては、トラガント、アラビアゴム、コーンスターチおよびゼラチンのような結合剤; 微晶性セルロース、結晶セルロースのような賦形剤; コーンスターチ、前ゼラチン化デンプン、アルギン酸、デキストリンのような膨化剤; ステアリン酸マグネシウムのような潤滑剤; 微粒二酸化ケイ素のような流動性改善剤; グリセリン脂肪酸エステルのような滑沢剤; ショ糖、乳糖およびアスパルテームのような甘味剤; ペパーミント、ワニラ香料およびチェリーのような香味剤等を挙げることができる。
【0086】
調剤単位形態がカプセル剤である場合には上記のタイプの材料にさらに油脂のような液
状担体を含有することができる。
【0087】
また、種々の他の材料を、被覆剤としてまたは調剤単位の物理的形態を変化させるために含有させることができる。錠剤の被覆剤としては、例えば、シェラック、砂糖またはその両方が挙げられる。シロップ剤またはエリキシル剤は、例えば、甘味剤としてショ糖、防腐剤としてメチルパラベンおよびプロピルパラベン、色素およびチェリーまたはオレンジ香味等を含有することができる。その他、各種ビタミン類、各種アミノ酸類を含有しても良い。
【0088】
本発明にかかる抗ウイルス剤の適用対象は、ヒト、ヒト以外の動物〔例えば、ヒト以外の哺乳類(ブタ、ウシ、ウマ、イヌ等の家畜) 、鳥類(ニワトリ等の家禽)等〕のいずれであってもよい。家畜や家禽等へ適用することにより、それらの経済的損失を軽減することができる。また、それらを食用とする場合も多いため、ヒトへの感染予防という点からも重要である。
【0089】
本発明にかかる抗ウイルス剤の投与量は、適用対象が必要とする量を確保できるように設定すればよく、製剤化して用いたり、飲食品に配合して上記抗ウイルス剤を用いることができる。
【0090】
(3−2)ウイルス除去膜
本発明にかかるウイルス除去膜は、本発明にかかる上記ポリペプチドが固定されてなるものである。上記ウイルス除去膜は、例えば従来公知の限外ろ過膜に上記ポリペプチドを固定することによって作製することができる。
【0091】
上記限外ろ過膜としては、従来公知のポリマー、例えば、ポリスルホン、ポリエーテルスルホン、ポリアミド類、ポリイミド類、酢酸セルロース、ポリアクリルアミド等によって形成された膜を挙げることができ、平膜型であっても多孔性中空糸膜型であってもよい。
【0092】
上記限外ろ過膜に上記ポリペプチドを結合するためには、従来公知のプロセスを用いて、限外ろ過膜のポリマーをまず賦活化し、タンパク質と化学的に結合しやすい状態にする。上記ポリマーとしては複数の異なるポリマーを用いてもよい。
【0093】
ポリマーを賦活化させた後、本発明にかかる上記ポリペプチドを直接あるいはリンカーを用いて付着させ、アフィニティーマトリクスを形成する。好ましいリンカーとして、アビジン、ストレプトアビジン、ビオチン、プロテインA、プロテインG等が挙げられるが、これらに限定されない。
【0094】
上記ポリペプチドは、二価性試薬のようなカップリング剤を用いて、限外ろ過膜のポリマーに直接結合されてもよく、あるいは間接的に結合されてもよい。アフィニティーマトリクスを形成するためには、アガロースに共有結合的に連結されたMPLを使用することが好ましいが、これに限定されるものではない。
【0095】
HIVのような、高マンノース型糖鎖を備えるウイルスの粒子および/またはそのフラグメントを含有する血液等を上記ウイルス除去膜にアプライすると、上記ウイルス除去膜に固定された本発明にかかるポリペプチドが、ウイルスの粒子および/またはそのフラグメントが有する高マンノース型糖鎖と結合するため、血液は孔を通過し、ウイルスの粒子および/またはそのフラグメントは捕捉されるため孔を通過することができない。それゆえ、高マンノース型糖鎖を備えるウイルスを血液等から除去することができる。
【0096】
(3−3)ベクター
本発明は、本発明にかかるポリペプチドを生成するために使用されるベクターを提供する。本発明にかかるベクターは、インビトロ翻訳に用いるベクターであっても組換え発現に用いるベクターであってもよい。
【0097】
本発明にかかるベクターは、上述した本発明にかかるポリヌクレオチドを含むものであれば、特に限定されない。例えば、本発明にかかるポリペプチドをコードするポリヌクレオチドのcDNAが挿入された組換え発現ベクターなどが挙げられる。組換え発現ベクターの作製方法としては、プラスミド、ファージ、またはコスミドなどを用いる方法が挙げられるが特に限定されない。
【0098】
ベクターの具体的な種類は特に限定されず、宿主細胞中で発現可能なベクターを適宜選択すればよい。すなわち、宿主細胞の種類に応じて、確実に本発明にかかるポリヌクレオチドを発現させるために適宜プロモーター配列を選択し、これと本発明にかかるポリヌクレオチドを各種プラスミド等に組み込んだベクターを発現ベクターとして用いればよい。
【0099】
発現ベクターは、好ましくは少なくとも1つの選択マーカーを含む。このようなマーカーとしては、真核生物細胞培養についてはジヒドロ葉酸レダクターゼまたはネオマイシン耐性、およびE.coliおよび他の細菌における培養についてはテトラサイクリン耐性遺伝子またはアンピシリン耐性遺伝子が挙げられる。
【0100】
上記選択マーカーを用いれば、本発明にかかるポリヌクレオチドが宿主細胞に導入されたか否か、さらには宿主細胞中で確実に発現しているか否かを確認することができる。あるいは、本発明にかかるポリペプチドを融合ポリペプチドとして発現させてもよく、例えば、オワンクラゲ由来の緑色蛍光ポリペプチドGFP(Green Fluorescent Protein)をマーカーとして用い、本発明にかかるポリペプチドをGFP融合ポリペプチドとして発現させてもよい。
【0101】
上記の宿主細胞は、特に限定されるものではなく、従来公知の各種細胞を好適に用いることができる。具体的には、例えば、大腸菌(Escherichia coli)等の細菌、酵母(出芽酵母Saccharomyces cerevisiae、分裂酵母Schizosaccharomyces pombe)、線虫(Caenorhabditis elegans)、アフリカツメガエル(Xenopus laevis)の卵母細胞等を挙げることができるが、特に限定されるものではない。上記の宿主細胞のための適切な培養培地および条件は当分野で周知である。
【0102】
上記発現ベクターを宿主細胞に導入する方法、すなわち形質転換法も特に限定されるものではなく、電気穿孔法、リン酸カルシウム法、リポソーム法、DEAEデキストラン法等の従来公知の方法を好適に用いることができる。また、例えば、本発明にかかるポリペプチドを昆虫で転移発現させる場合には、バキュロウイルスを用いた発現系を用いればよい。
【0103】
このように、本発明にかかるベクターは、少なくとも、本発明にかかるポリペプチドをコードするポリヌクレオチドを含めばよいといえる。すなわち、発現ベクター以外のベクターも、本発明の技術的範囲に含まれる点に留意すべきである。
【0104】
つまり、本発明の目的は、本発明にかかるポリペプチドをコードするポリヌクレオチドを含有するベクターを提供することにあるのであって、本明細書中に具体的に記載した個々のベクター種および細胞種、ならびにベクター作製方法および細胞導入方法に存するのではない。したがって、上記以外のベクター種およびベクター作製方法を用いて取得したベクターも本発明の技術的範囲に属することに留意しなければならない。
【0105】
(3−4)形質転換体または細胞
本発明は、上述した本発明にかかるポリペプチドをコードするポリヌクレオチドが導入された形質転換体または細胞を提供する。ここで「形質転換体」とは、組織または器官だけでなく、生物個体を含むことを意味する。
【0106】
形質転換体または細胞の作製方法(生産方法)は特に限定されるものではないが、例えば、上述した組換えベクターを宿主に導入して形質転換する方法を挙げることができる。また、形質転換の対象となる生物も特に限定されるものではなく、上記宿主細胞で例示した各種微生物、植物または動物を挙げることができる。
【0107】
本発明にかかる形質転換体または細胞は、海藻類もしくはその子孫、またはこれら由来の組織であることが好ましく、トサカノリ(Meristotheca papulosa (Montagne) J. Agardh)であることが特に好ましい。
【0108】
本発明にかかるポリペプチドをコードするポリヌクレオチドを含む形質転換体は、当該ポリヌクレオチドを含む組換えベクターを、当該遺伝子が発現し得るように宿主細胞中に導入することにより得ることができる。
【0109】
このように、本発明にかかる形質転換体または細胞は、少なくとも、本発明にかかるポリペプチドをコードするポリヌクレオチドが導入されていればよいといえる。すなわち、組換え発現ベクター以外の手段によって生成された形質転換体または細胞も、本発明の技術的範囲に含まれる点に留意すべきである。
【0110】
本発明の目的は、本発明にかかるポリペプチドをコードするポリヌクレオチドが導入されていることを特徴とする形質転換体または細胞を提供することにあるのであって、本明細書中に具体的に記載した個々のベクター種および導入方法に存するのではない。したがって、上記以外のベクター種および細胞種、ならびにベクター作製方法および細胞導入方法を用いて取得した形質転換体または細胞も本発明の技術的範囲に属することに留意しなければならない。
【0111】
(3−5)ポリペプチドの生産方法
本発明は、本発明にかかるポリペプチドを生産する方法を提供する。
【0112】
一実施形態において、本発明にかかるポリペプチドの生産方法は、本発明にかかるポリペプチドをコードするポリヌクレオチドを含むベクターを用いることを特徴とする。
【0113】
本実施形態の1つの局面において、本実施形態にかかるポリペプチドの生産方法は、上記ベクターを無細胞タンパク質合成系に用いることが好ましい。無細胞タンパク質合成系を用いる場合、種々の市販のキットを用いればよい。好ましくは、本実施形態にかかるポリペプチドの生産方法は、上記ベクターと無細胞タンパク質合成液とをインキュベートする工程を包含する。
【0114】
本実施形態の他の局面において、本実施形態にかかるポリペプチドの生産方法は、組換え発現系を用いることが好ましい。組換え発現系を用いる場合、本発明にかかるポリヌクレオチドを組換え発現ベクターに組み込んだ後、公知の方法により発現可能な宿主に導入し、宿主内で翻訳されて得られる上記ポリペプチドを精製するという方法などを採用することができる。組換え発現ベクターは、プラスミドであってもなくてもよく、宿主に目的ポリヌクレオチドを導入することができればよい。好ましくは、本実施形態にかかるポリペプチドの生産方法は、上記ベクターを宿主に導入する工程を包含する。
【0115】
このように宿主に外来ポリヌクレオチドを導入する場合、発現ベクターは、外来ポリヌクレオチドを発現するように宿主内で機能するプロモーターを組み込んであることが好ましい。組換え的に産生されたポリペプチドを精製する方法は、用いた宿主、ポリペプチドの性質によって異なるが、タグの利用等によって比較的容易に目的のポリペプチドを精製することが可能である。
【0116】
本実施形態にかかるポリペプチドの生産方法は、本発明にかかるポリペプチドを含む細胞または組織の抽出液から当該ポリペプチドを精製する工程をさらに包含することが好ましい。
【0117】
ポリペプチドを精製する工程は、周知の方法(例えば、細胞または組織を破壊した後に遠心分離して可溶性画分を回収する方法)で細胞や組織から細胞抽出液を調製した後、この細胞抽出液から周知の方法(例えば、硫安沈殿またはエタノール沈殿、酸抽出、陰イオンまたは陽イオン交換クロマトグラフィー、ホスホセルロースクロマトグラフィー、疎水性相互作用クロマトグラフィー、アフィニティークロマトグラフィー、ヒドロキシアパタイトクロマトグラフィー、およびレクチンクロマトグラフィー)によって精製する工程が好ましいが、これらに限定されない。最も好ましくは、高速液体クロマトグラフィー(「HPLC」)が精製のために用いられる。
【0118】
別の実施形態において、本発明にかかるポリペプチドの生産方法は、本発明にかかるポリペプチドを天然に発現する細胞または組織から当該ポリペプチドを精製することを特徴とする。本実施形態にかかるポリペプチドの生産方法は、上述したオリゴヌクレオチドを用いて、本発明にかかるポリペプチドを天然に発現する細胞または組織を同定する工程を包含することが好ましい。また、本実施形態にかかるポリペプチドの生産方法は、上述したポリペプチドを精製する工程をさらに包含することが好ましい。
【0119】
さらに他の実施形態において、本発明にかかるポリペプチドの生産方法は、本発明にかかるポリペプチドを化学合成することを特徴とする。当業者は、本明細書中に記載される本発明にかかるポリペプチドのアミノ酸配列に基づいて周知の化学合成技術を適用すれば、本発明にかかるポリペプチドを化学合成できることを、容易に理解する。
【0120】
以上のように、本発明にかかるポリペプチドを生産する方法によって取得されるポリペプチドは、天然に存在する変異ポリペプチドであっても、人為的に作製された変異ポリペプチドであってもよい。
【0121】
変異ポリペプチドを作製する方法についても、特に限定されるものではない。例えば、部位特異的変異誘発法(例えば、Hashimoto-Gotoh, Gene, 152,271-275(1995)参照)、PCR法を利用して塩基配列に点変異を導入し変異ポリペプチドを作製する方法、またはトランスポゾンの挿入による突然変異株作製法などの周知の変異ポリペプチド作製法を用いることによって、変異ポリペプチドを作製することができる。変異ポリペプチドの作製には市販のキットを利用してもよい。
【0122】
このように、本発明にかかるポリペプチドの生産方法は、少なくとも、本発明にかかるポリペプチドのアミノ酸配列、または本発明にかかるポリペプチドをコードするポリヌクレオチドの塩基配列に基づいて公知慣用技術を用いればよいといえる。
【0123】
つまり、本発明の目的は、本発明にかかるポリペプチドの生産方法を提供することにあるのであって、上述した種々の工程以外の工程を包含する生産方法も本発明の技術的範囲に属することに留意しなければならない。
【実施例】
【0124】
以下、実施例に基づいて本発明をより詳細に説明するが、本発明は以下の実施例に限定されるものではない。
【0125】
〔実施例1:トサカノリからのポリペプチド(MPL)の単離〕
(抽出液の調製および硫安沈殿)
九州西岸にて採集したトサカノリ(Meristotheca papulosa (Montagne) J. Agardh)の凍結藻体500gを液体窒素下で粉末とした。これに500mlの20mM PBSA(Phosphate-buffered saline sodium azide;0.02%アジ化ナトリウム入り20mM PBS;pH7.0)を加え、4℃で一晩撹拌した後、遠心分離を行なって一次抽出液を得た。残渣を700mlの20mM PBSAで同様に抽出し、二次抽出液を得た。両抽出液を合一し、UV280nmの吸収および凝集活性を測定した。
【0126】
上記抽出液に硫安粉末を20%飽和となるように少しずつ撹拌しながら加え、この混合液を4℃で一晩静置した。これを遠心分離(8500rpm、30分間)して得られた沈殿を、PBSAに溶解後、同溶媒に対し十分透析した。透析終了後、内液を遠心分離し、得られた上清を20%飽和硫安塩析沈殿画分とした。一方、20%飽和硫安塩析処理で得られた上清に、硫安粉末を60%飽和となるように加え、同様に処理して、20〜60%飽和硫安塩析沈殿画分を得た。当該画分につき、UV280nmの吸収および凝集活性を測定した。
【0127】
(疎水クロマトグラフィー)
上記20〜60%飽和硫安塩析沈殿画分106mlのうち、10mlについて、20mMトリス−塩酸緩衝液(pH7.0)に対して透析を行ない、内液6mlを得た。得られた内液を遠心分離(10,000 rpm、10分間)して上清を回収した。得られた上清5mlを同緩衝液で平衡化したHiPrep Phenyl FFカラム(16×100mm, Vt=20 ml)に注入し、20mMトリス−塩酸緩衝液,pH7.0(溶媒A)と、蒸留水(溶媒B)の2液を用いる濃度勾配法により溶出した。濃度勾配[溶媒B100%(10分)、溶媒:B0%−溶媒A100%(10−60分)、溶媒A100%(60−110分)]は、グラジェントプログラマー(CCPコントローラー、東ソー社製)を用いて設定し、流速は4ml/分とした。溶出液は5mlずつ分取し、UV280nmの吸光度を測定するとともに凝集活性を測定した。
【0128】
(ゲルろ過)
疎水クロマトグラフィーで分取した活性画分を合一し、限外濾過により5mlに濃縮した。そのうち1mlを、0.3 M NaCl を含む20mM PB (pH 7.0)で平衡化したTSKgel G3SWxLゲルろ過カラム(東ソー社製、7.8 x 300 mm、Vt=14.7 ml)に供した。20mMトリス−塩酸緩衝液(pH 7.0)を用いて流速1ml/minで溶出した。溶出液は1mlずつ分取し、各フラクションのUV280nmの吸光度および凝集活性を測定した。
【0129】
(凝集活性の検討)
赤血球凝集活性は、マイクロタイター法を用いて測定した。生理食塩水にて調製した各精製画分溶液の2倍段階希釈液、各25μlをマイクロタイタープレート上に作製した。各希釈液に2%赤血球浮遊液25μlを加えて軽く撹拌し、室温で1.5時間静置後、凝集能を観察した。凝集能は肉眼で判定し、赤血球の50%以上が凝集している場合を陽性とした。凝集活性は、赤血球凝集活性(力価)、すなわち凝集活性を示す最大希釈液の希釈倍数の逆数、および凝集素価(力価)、すなわち凝集活性を示す最大希釈液のタンパク質の濃度(最小凝集濃度)で示した。
【0130】
本実施例においては、赤血球としてトリプシン処理ウサギ赤血球(TRBC)を用いた。なお、赤血球浮遊液の調製は次のように行なった。まず、実験室で飼育中のウサギの耳から血液2mlを採取し、これを約50mlの生理食塩水で3回洗浄後、50mlの生理食塩水を加えて2%のウサギ赤血球浮遊液を調製した。これに1/10容の0.5%トリプシン−生理食塩水を加え、37℃で1.5時間静置した。このトリプシン処理赤血球を生理食塩水で3回洗浄後、45mlの生理食塩水を加え、トリプシン処理2%ウサギ赤血球浮遊液(TRBC)とした。
【0131】
トリプシン処理ウサギ赤血球(TRBC)に対する凝集活性成分を検討したところ、表1に示すように、上記抽出液、上記20〜60%飽和硫安塩析沈殿画分、上記疎水クロマトグラフィーによって得られた画分、および上記ゲルろ過によって得られた画分に強い赤血球凝集活性が検出された。
【0132】
【表1】

【0133】
表1中、「タンパク質濃度」は、「a」を付したものについてはUV280nmの吸収(A280)を測定し、A280(1mg/ml)=1.0より算出した。「b」を付したものについてはUV280nmの吸収(A280)を測定し、本発明にかかるポリペプチドのアミノ酸配列から求めた分子吸光係数を用いてA280(1mg/ml)=0.97より算出した。
【0134】
(SDS−PAGE)
上記ゲルろ過で得られた凝集活性を有する画分(精製画分)をSDS−PAGE(10%ゲル)に供した。結果を図3に示す。レーン1は分子量マーカー、レーン2は20〜60%飽和硫安塩析沈殿画分(非還元下)、レーン3は20〜60%飽和硫安塩析沈殿画分(還元下)、レーン4は上記精製画分(非還元下)、レーン5は上記精製画分(還元下)を示す。なお、タンパク染色はCBB(Coomassie brilliant blue R-250)染色を行なった。
【0135】
SDS−PAGEの結果、精製画分は、還元下で14kDaの、非還元下で26kDaの単一バンドを与えるものであることが確認されたことから、同一サブユニット(14kDa)がジスルフィド結合(S−S結合)した2量体からなると推定される。以上のように、上記抽出液を硫安塩析、疎水クロマトグラフィーおよびゲルろ過に順次供した精製過程により最終的に0.4mgの精製画分が得られた。表1に示す収量6mgは藻体500g(湿重量)当たりに換算した値である。本発明者らは、当該精製画分を「MPL」と命名した。
【0136】
なお、本発明者は、上記MPLは、上記20〜60%飽和硫安塩析沈殿画分を陰イオン交換クロマトグラフィーに供することによって効果的に調製できることも確認した。陰イオン交換クロマトグラフィーによって、トサカノリに含まれる他のレクチン(MPAと称する)と上記MPLとを効果的に分離することができる。
【0137】
上記陰イオン交換クロマトグラフィーの条件を以下に示す。
【0138】
MPAとMPLの両レクチンを同時に精製することを意図して、先に調製した20〜60%飽和硫安塩析沈殿画分を、HiPrep QXL 16/10カラム(φ1.6×10 cm、Vt = 20 ml、GE Healthcare社)を用いる陰イオン交換クロマトグラフィーに供した。
【0139】
すなわち、上記カラムを0.02M トリス−塩酸緩衝液(pH 8.0)で平衡化し、20〜60%飽和硫安塩析沈殿画分5mlを添加した。カラムを上記緩衝液で十分洗浄した後、上記緩衝液中0M〜1M NaClの濃度勾配で溶出した。流速は5ml/minとした。洗浄液は10mlずつ、濃度勾配溶出液は5mlずつ分取した。
【0140】
各フラクションにつき、A280吸光度および赤血球凝集活性を測定した。赤血球凝集活性が見られた画分は、還元下SDS-PAGEに付した。その結果より、各フラクションを5つの画分に分けた。各画分につき、Folin-Lowry法によるタンパク質量の測定および赤血球凝集活性試験に供した。
【0141】
その結果、両レクチンはカラムに吸着し、MPAは0.37M NaCl付近、MPLは0.57M NaCl付近にそれぞれ活性ピークとして溶出した。このように、陰イオンクロトマトグラフィーを用いることにより、20〜60%飽和硫安塩析沈殿画分に混在する2種類のレクチン、MPA(29kDa)とMPL(26kDa)とを分離することができる。
【0142】
〔実施例2:MPLのcDNAのクローニング〕
以下に示すように、実施例1で得られた精製画分「MPL」の14kDaサブユニットの23N末端アミノ酸配列を、プロテインシーケンサー(Procise 492HT, Applied Biosystems)を用いてGVVDQLIVTYSDGTRVSHGQPGS(配列番号12)と決定した。当該配列の情報を参考に縮重プライマーを設計した。別途調製したトサカノリ由来1st strand cDNA溶液を鋳型にRapid Amplification of cDNA Ends(RACE)法を行うことにより、MPLのcDNAの全長塩基配列 949bpを明らかにした。
【0143】
まず、MPLの上記23N末端のアミノ酸配列を参考にして、MPL_d_R1およびMPL_d_R2の各プライマーを作製した。
【0144】
MPL_d_R1の塩基配列は、CCRTGISWIACICKIGTICCRTC(配列番号3)であり、MPL_d_R2の塩基配列は、TAIGTIACDATIARYTGRTC(配列番号4)である。ここで、DはG、AまたはTを、Iはイノシンを、KはGまたはTを、RはAまたはGを、YはCまたはTを、WはAまたはTを示すIUBコードである。
【0145】
RNAlater 中に−20℃で保存したトサカノリ藻体より Plant RNA Isolation Reagent(Life Technologies Corp.)を用いて全RNAを抽出後、NucleoTrap mRNA(Macherey-Nagel)によりmRNAを精製し、さらに GeneRacer Kit(Life Technologies Corp.)により完全長cDNAを調製した。
【0146】
次に、上記完全長cDNAを鋳型に GeneRacer_5'_Primer および MPL_d_R1のプライマーペアを用いてPCRを行った。GeneRacer_5'_Primer の塩基配列はCGACTGGAGCACGAGGACACTGA(配列番号5)である。
【0147】
上記PCRにより得られた産物の100倍希釈溶液を鋳型に、GeneRacer_5’_Nested_PrimerおよびMPL_d_R2のプライマーペアを用いてNested PCRを行った。GeneRacer_5’_Nested_Primerの塩基配列は、GGACACTGACATGGACTGAAGGAGTA(配列番号6)である。
【0148】
得られたNested PCRの産物を低融点アガロースにより精製後、pGEM-T Easy Vector System(PROMEGA製)を用いてサブクローニングを行い、得られたクローンから精製プラスミドを回収して、ダイデオキシ法により塩基配列の決定を行った(以上5’RACE)。
【0149】
得られた塩基配列に基づいて新たにプライマーMPL_F1を作製した。MPL_F1の塩基配列はTGTAGCAGCCATGTCTTTGC(配列番号7)である。
【0150】
上記完全長cDNAを鋳型として、MPL_F1およびGeneRacer_3’_Primerのプライマーペアを用いてPCRを行った。得られた増幅産物を低融点アガロースにより精製後、pGEM-T Easy Vector System(PROMEGA製)を用いてサブクローニングを行い、得られたクローンから精製プラスミドを回収して、ダイデオキシ法により塩基配列の決定を行った(以上、3’RACE)。
【0151】
GeneRacer_3’_Primerの塩基配列はGCTGTCAACGATACGCTACGTAACG(配列番号8)である。
【0152】
上記5’RACEおよび3’RACEから明らかとなったMPL cDNAの5’および3’末端配列から作製したMPL_5'_End_FおよびMPL_3'_End_Rのプライマーペア、ならびに高正確性DNAポリメラーゼを用いてPCRを行い、増幅産物につき塩基配列を決定することで、MPL cDNAの全長塩基配列を確認した。
【0153】
MPL_5'_End_Fプライマーの塩基配列はAATCCACATTCAACTGCACTG(配列番号9)であり、MPL_3'_End_Rプライマーの塩基配列はAAATCGACGCACACAGAAGTC(配列番号10)である。
【0154】
MPLのcDNAの全長塩基配列、およびその演繹アミノ酸配列を図1に示す。図1において、星印は終止コドン、破線はシグナルペプチド(SignalP 3.0を用いて予測)、実線は実施例1で得られた精製画分「MPL」の14kDaサブユニットの23N末端配列を示す。
【0155】
演繹アミノ酸配列の解析から、図1に示すcDNAは、シグナルペプチド領域22残基(1−22)を含め、計198残基をコードすることがわかった。一方、演繹アミノ酸配列の解析から、図1に示すcDNAは、プロペプチド領域74残基(23−96)および成熟タンパク質領域102残基(97−198)をコードすると考えられたが、後述する実施例6においてさらに詳細に検討したところ、成熟タンパク質領域は53番目−198番目の146アミノ酸残基であることが分かった。
【0156】
〔実施例3:MPLの糖結合特異性〕
MPLの糖結合特異性を赤血球凝集阻止試験により明らかにした。本赤血球凝集阻止試験には、実施例1で調製した20〜60%飽和硫安塩析沈殿画分および実施例1で調製したMPLを供試し、これらの赤血球凝集活性(力価4)を阻止する糖化合物を検索した。
【0157】
赤血球凝集阻止試験は以下のようにして行った。まず生理食塩水にて調製した糖溶液の2倍段階希釈液、各25μlをマイクロタイタープレート上に作製した。なお、使用した糖類の原液の濃度は単糖類および二糖類の場合は100mM、糖タンパク質の場合は2mg/mlとした。これに、それぞれ凝集素価(力価)4に調整した上記飽和硫安塩析沈殿画分の溶液およびMPL溶液、各25μlを加えて軽く撹拌後、室温で1.5時間静置した。これに25μlのTRBCを加え、室温で2時間静置後、凝集阻止能を観察した。
【0158】
凝集阻止能の有無は肉眼で判定し、赤血球の約100%が凝集していない場合を陽性とした。凝集阻止能(凝集阻止活性)は、最小阻止濃度すなわち凝集阻止能を示す最小濃度(mMまたはμg/ml)で表示した。結果を表2に示した。表2において、「最小凝集阻止濃度」は、上述の力価4の検液の赤血球凝集活性を阻止する糖化合物の最小濃度を表し、「NT」は未測定であることを表す。
【0159】
【表2】
【0160】
表2に示すように、本赤血球凝集阻止試験には、単糖類および二糖類としてD−グルコース(表中Glcと表示。以下、糖の名称に続く括弧内には、表2に示した略称を記載する。)、D−マンノース(Man)、D−ガラクトース(Gal)、N−アセチル−D−グルコサミン(GlcNAc)、N−アセチル−D−ガラクトサミン(GalNAc)、N−アセチルノイラミン酸(NeuAc)、L−フコース(Fuc)、D−キシロース(Xyl)、L−ラムノース(Rha)、ラクトースを用いた。
【0161】
糖タンパク質としては、複合型糖鎖を含有するものとして、トランスフェリン、アシアロトランスフェリンを用いた。また、高マンノース型糖鎖を含有する糖タンパク質として、イーストマンナンを用い、複合型糖鎖および高マンノース型糖鎖を含有する糖タンパク質として、ブタチログロブリン、アシアロブタチログロブリンを用いた。
【0162】
表2から明らかなように、MPLの赤血球凝集活性は供試した単糖類および二糖類では阻止されず、高マンノース型糖鎖を含む糖タンパク質、並びに、複合型糖鎖および高マンノース型糖鎖を含む糖タンパク質によってのみ阻止された。このように、MPLは従来公知のレクチンとの配列相同性が非常に低く、かつ、高マンノース型糖鎖に高い親和性を有することが示された。
〔実施例4:エイズウイルスの表面糖タンパク質・gp120とMPLとの相互作用〕
HIVの宿主への感染は、HIVの表面糖タンパク質であるgp120と宿主リンパ細胞の表面レセプターCD4との相互作用により成立する。該相互作用にはgp120の高マンノース型糖鎖が不可欠である。そのため、高マンノース型糖鎖に特異的に結合するレクチンはHIV感染の有力な阻害剤となりうる。
【0163】
そこで、MPLと市販品のリコンビナント糖付加HIV-1 IIIB gp120 (バキュロウイルス;1分子あたり15本の高マンノース型糖鎖を有する。ImmunoDiagnostics製)との相互作用を表面プラズモン共鳴(SPR)法を用いて定量解析した。SPR法にはBIAcore2000(GE Healthcare社製)を用いて、リコンビナント糖付加HIV-1 IIIB gp120をCM5センサーチップ(GE Healthcare社製)上に固定化し、MPLの溶液をアナライトとして用いて、マニュアルに従って測定、解析した。
【0164】
上記固定化はマニュアルに従って行った。具体的には、アミンカップリング法を用いて固定化を行った。なお、固定化量は300〜400RUの範囲になるよう、マニュアルインジェクション法によって調整した。
【0165】
SPR法では、生体分子を標識することなく、生体分子間の特異的な相互作用を微量かつ短時間で定量的に測定できる。本法では、リガンドをセンサーチップ表面上に固定化し、これに作用する物質(アナライト)を含む溶液を添加すると、分子の結合・解離により生ずる微量の質量変化がSPRシグナルの変化として検出される。
【0166】
質量変化はレゾナンスユニット(RU)で表され、1000RUは共鳴による反射角度0.1°の変化に相当し、アナライトがリガンドに 1 ng/mm2 結合したことを意味する。センサーチップ表面の金薄膜上にはデキストランがコーティングされており、主としてこのデキストラン内に導入されたカルボキシル基を介してリガンドを固定化する。
【0167】
MPLと上記リコンビナントgp120とのSPR法による親和性解析に先立ち、予備実験で解析法を検討し、非線形最小二乗法によるカイネティクス解析が適当と判断した。そこで、得られたセンサーグラムからおおよそのKD値を算出し、0.1〜10KD[M]を濃度の目安として、2倍希釈列で5段階以上のアナライト(MPL)溶液を調製した。
【0168】
分析プログラムの作成には、マニュアルに従って“Customaized Application”を用い、センサーチップ内の4つのフローセルのうち、何も固定化していないフローセル1をコントロールとして、リコンビナントgp120を固定化したフローセルからの差し引き機能を使用した。本分析プログラム下で、アナライト(MPL)溶液をそれぞれセンサーチップ上に流速30μl/minで3分間流した後、バッファーを3分間流し、レクチンの結合・解離量を測定した。なお、アナライト添加開始後5秒から添加終了5秒前までのRUの増加量を結合量、バッファー添加開始後10秒から添加終了10秒前までのRU減少量を解離量とした。
【0169】
次に、100mM HClおよび100mM NaOHを用いて、センサーチップを洗浄して再生した。得られたセンサーグラムについて、結合相と解離相を同時にカーブフィッティングさせ、結合速度定数ka、解離速度定数kd、親和定数KA、および解離定数KDを算出した。
【0170】
結果を図2に示す。図2のAは固定化したgp120とMPLとの相互作用のセンサーグラムであり、図2のBはgp120とMPLとの親和定数を示す。図2のBには、結合速度定数Ka(M-1s-1)、解離速度定数Kd(s-1)、親和定数KA(M-1)、および解離定数KD(M)を示した。なお親和定数は、その値が大きくなればなるほど、親和性が高い(結合力が強い)ことを意味する。
【0171】
図2に示すように、MPLはgp120に強い親和性(KD= 4.50×10-9 M)を持つことが認められた。このことから、MPLは、従来公知のレクチンとの配列相同性が非常に低い上に、HIVのgp120と宿主リンパ細胞のCD4との相互作用を十分に阻害できるといえる。それゆえ、MPLは新たな抗HIV剤として有望である。
【0172】
また、実施例3に示したように、MPLは高マンノース型糖鎖に高い親和性を有する。そのため、MPLは抗HIV剤として有望である。また、高マンノース型糖鎖を有する糖タンパク質が宿主のレセプターと結合することによって感染するタイプのウイルスに対しても有力な感染阻害剤となりうる。
【0173】
〔実施例5:MPLのさらなる精製〕
(実施例5−1:陰イオン交換クロマトグラフィーによるトサカノリレクチンの精製)
実施例1で述べたように、MPAとMPLとの分離に陰イオン交換クロマトグラフィーが有用であることが判明したので、陰イオン交換クロマトグラフィーによる精製についてさらなる検討を行い、精製方法を改良した。
【0174】
実施例1で調製した20〜60%飽和硫安塩析沈殿画分68mlを、20mMのトリス塩酸緩衝液(pH8.0)(以下、当該緩衝液を「TB」と称する)で平衡化したHiPrep QXL 16/10カラム(φ1.6×10 cm、Vt = 20 ml、GE Healthcare社)に添加し、カラムをTBで十分洗浄後、TB中0.2Mおよび1M NaClで順次、段階的に溶出した(ステップワイズ溶出)。その結果、活性成分は0.2M NaCl溶出画分に認められた。
【0175】
図4は、20〜60%飽和硫安塩析沈殿画分を、HiPrep QXL 16/10カラムを用いた陰イオン交換クロマトグラフィー(ステップワイズ溶出)に供した結果を示す図である。
【0176】
図4中、横軸はフラクション番号であり、縦軸は左側が280nmの吸光度を表し、右側が赤血球凝集活性(力価)を表す。図中、当該吸光度測定結果を丸印で示し、赤血球凝集活性(力価)の測定結果を三角印で示した。
【0177】
括弧書き数字1は、上記カラムに20〜60%飽和硫安塩析沈殿画分を添加し、非吸着成分を洗浄後、TB中0.2M NaClにより吸着成分の溶出を開始した位置を示している。括弧書き数字2は、上記溶出後、TB中1M NaClにより他の吸着成分の洗浄を開始した位置を示している。
【0178】
図4に示すように、赤血球凝集活性成分は0.2M NaCl溶出画分に認められた。この赤血球凝集活性成分(粗レクチン画分)70mlを、TBで平衡化したHiPrep QXL 16/10カラムに添加し、カラムをTBで洗浄後、TB中NaClの濃度勾配(0〜0.2M)で溶出させた(グラジエント溶出法)。
【0179】
溶出液は280nmの吸光度をモニターするとともに、2mlずつ分取し、各画分の赤血球凝集活性を測定した。図5は、上記粗レクチン画分を、HiPrep QXL 16/10カラムを用いた陰イオン交換クロマトグラフィー(グラジエント溶出)に供した結果を示す図である。図5に実線で示すように、上記溶出液は、凝集活性を示す3つのタンパク質のピークに分離した。
【0180】
図5において、横軸はフラクション番号である。左側の縦軸は280nmの吸光度を表し、吸光度の測定結果は実線で示されている。右側の縦軸では、「HA」が赤血球凝集活性(力価)を表し、赤血球凝集活性(力価)の測定結果は三角印で示されている。右側の縦軸のうち、”NaCl in TB”は、NaCl(TB中)の濃度勾配を示し、図中、濃度勾配は点線で示されている。
【0181】
上記溶出液を、図5に示す4つの画分(I〜IV)に分画し、各画分とも、20μgのタンパク質に相当する量をSDS−PAGE(10%ゲル)に供した。結果を図6に示す。なお、タンパク染色はCBB(Coomassie brilliant blue R-250)染色を行なった。
【0182】
図6のAは非還元下における泳動結果を示し、図6のBは還元下における泳動結果を示す。Mは分子量マーカー、レーン1は20〜60%飽和硫安塩析沈殿画分、レーン2はHiPrep QXL 16/10カラム(ステップワイズ溶出)で得られた赤血球凝集活性成分(0.2M NaCl溶出画分)、レーン3は画分I、レーン4は画分II、レーン5は画分III、レーン6は画分IVを示す。なお、上記赤血球凝集活性成分は、「粗レクチン画分」とも称される。
【0183】
画分IおよびIIは、図6のBに示すように、還元下で29kDaのほぼ単一のバンドを与えることが確認された。そこで、画分IをMPA−1、画分IIをMPA−2と命名し、最終精製レクチンとした。
【0184】
MPA−1およびMPA−2は、詳細は示さないが、分子量測定、N末端アミノ酸配列解析およびcDNAクローニングの結果から、実施例1で述べたMPA(29kDa)のアイソフォームであることが分かった。
【0185】
実施例1で述べたMPLは画分IVに溶出した。画分IVは、非還元下では26kDaおよび55kDaの2つのバンド成分を与え(図6のA)、還元下では14kDaの主要バンド成分と、8kDaおよび5kDaの微量バンド成分を与えた。なお、画分IIIは還元下で29kDaおよび14kDaのバンド成分を与えることから、MPLとMPAとを混有すると考えられる。
【0186】
(実施例5−2:陰イオン交換クロマトグラフィーによるMPL−1およびMPL−2の単離)
上記MPL画分(画分IV)5mlを、TBで平衡化したTSKgel DEAE-5PWカラム(7.5×75 mm、東ソー社)に添加し、TB中NaClの濃度勾配(0−0.2M)によるグラジエント溶出を行った。操作中、280nmの吸光度をモニターしながらピークをそれぞれ分取した。
【0187】
図7は、MPLのTSKgel DEAE-5PWカラムを用いた陰イオン交換クロマトグラフィーによる精製結果を示す図である。図中、横軸は溶出時間を表し、左側の縦軸は280nmの吸光度、右側の縦軸はNaClの濃度を示す。また、実線は280nmの吸光度の測定結果を示し、点線はNaClの濃度を示す。(1)〜(4)はピーク番号である。
【0188】
TSKgel DEAE-5PWカラムによる陰イオン交換クロマトグラフィーの結果、図7に示すように、MPLは4つのピーク(ピーク(1)〜(4))に分離された。そこで次に、ピーク(1)〜(4)をそれぞれ分取後、20μgのタンパク質に相当する量をSDS−PAGE(10%ゲル)に供した。結果を図8に示す。
【0189】
図中、「非還元」は非還元下における泳動結果を示し、「還元」は還元下における泳動結果を示す。Mは分子量マーカーを示し、SはMPL、レーン1〜4はそれぞれピーク(1)〜(4)を泳動させた結果を示す。
【0190】
還元下において、ピーク(1)および(3)で14kDaのほぼ単一のバンドが認められた。ピーク(1)および(3)は、非還元下において26kDa付近のタンパク質性バンドを与えた。
【0191】
ピーク(2)および(4)については、非還元下で55kDa、還元下では14kDaの他に8kDaおよび5kDaのペプチドの存在が確認された。全ピークに共通して非還元下で26kDaおよび55kDaにタンパク質性バンドの存在が認められたが、ピーク(1)および(3)では26kDa成分、ピーク(2)および(4)では55kDa成分を主成分としていることがわかった。
【0192】
そこで、ピーク(1)をMPL−1、ピーク(3)をMPL−2と命名し、最終精製標品とした。MPL−1およびMPL−2につき、LTQ Orbitrap XL(Thermo Fisher Scientific)を用いる ESI-MSにより分子量の測定を行った。図9はMPL−1およびMPL−2の分子量測定結果を示すものである。
【0193】
図9のA,Bより、MPL−1およびMPL−2の分子量はそれぞれ31,104.1 Daおよび31,132.0 Daであることが分かった。
【0194】
MPL−1およびMPL−2について、実施例3と同様の方法により、赤血球凝集阻止試験を行った。結果を表3に示す。
【0195】
【表3】
【0196】
表3では「単糖類および二糖類」と記載しているが、これは、実施例3と同様に、単糖類としてD−グルコース(Glc)、D−マンノース(Man)、D−ガラクトース(Gal)、N−アセチル−D−グルコサミン(GlcNAc)、N−アセチル−D−ガラクトサミン(GalNAc)、N−アセチルノイラミン酸(NeuAc)、L−フコース(Fuc)、D−キシロース(Xyl)またはL−ラムノース(Rha)を各100mMで用い、二糖類としてラクトース(Lac)を100mM用いたことを示す。
【0197】
複合型糖鎖を含有する糖タンパク質、高マンノース型糖鎖を含有する糖タンパク質、並びに、複合型糖鎖および高マンノース型糖鎖を含有する糖タンパク質としては、実施例3と同じものを用いた。加えて、N型糖鎖(複合型)およびO型糖鎖を含有する糖タンパク質としてフェツイン、アシアロフェツインを用い、O型糖鎖を含有する糖タンパク質としてウシ顎下線ムチン、アシアロウシ顎下線ムチンを用いた。
【0198】
表3から明らかなように、MPL−1およびMPL−2の赤血球凝集活性は、実施例3に示したMPLと同様に、高マンノース型糖鎖を含む糖タンパク質、並びに、複合型糖鎖および高マンノース型糖鎖を含む糖タンパク質によってのみ阻止された。このように、MPL−1およびMPL−2は、MPLと同様に、高マンノース型糖鎖に高い親和性を有することが示された。
【0199】
〔実施例6:MPLの構造解析〕
(実施例6−1:MPL画分のブロッティング)
実施例1で得られたMPL画分(26kDa)を非還元下および還元下(2-メルカプトエタノール添加)で12%トリス-トリシン系SDS−PAGEに付し、Polyvinylidene difluoride(PVDF)膜(Immobilon-P(登録商標)、Millipore)へ転写した。
【0200】
PVDF膜への転写は、セミドライブロッティング装置(AE-6677:ATTO CORPORATION)を用いてブロッティング緩衝液(CAPS buffer:メタノール:超純水= 1:1:8)中、144mAの定電流で1時間通電することによって行った。
【0201】
CAPS bufferはCAPS(3-Cyclohexylaminopropane sulfonic acid、和光純薬工業)11.1gを450mlの超純水に溶解し、NaOHを用いてpH11.0に調整した後、超純水で550 mlにメスアップしたものを使用した。転写後のPVDF膜はCBB染色を行い、一晩風乾後、目的とする14kDa成分および8kDaの成分を切り出して、実施例2同様に、Procise(登録商標)492HTを用いて、N末端アミノ酸配列を解析した。
【0202】
図10は、MPL画分をブロッティングし、CBB染色したPVDF膜を示す図である。図10において、Mは分子量マーカーを示し、「非還元」は非還元下において得られたバンド、「還元」は還元下において得られたバンドを示す。
【0203】
(実施例6−2:N末端アミノ酸配列の解析およびMPL−1のアミノ酸配列の決定)
図10に示す8kDaの成分のN末端アミノ酸配列はGVVDQLI(配列番号14)と決定された。この配列は、実施例2で決定された配列番号12に示すアミノ酸配列のN末端と一致した。
【0204】
一方、14kDaの成分については、配列が解析できず、N末端がブロッキングされていると推測された。そこで、ピログルタミル基によるN末端ブロッキングと予測し、脱ピログルタミル化を試みた。
【0205】
すなわち、14kDa成分を転写したPVDF膜を60%メタノール1mlで2回洗浄後、90%メタノール1mlで1回洗浄し、これを0.5%(w/v)ポリビニルピロリドン(PVP)-55含有100mM 酢酸500μlに浸漬し、37℃で30分間静置した。次に、蒸留水で膜を10回以上洗浄し、膜をメタノールで湿らせ、酵素消化を行った。
【0206】
酵素消化は、Pfu Pyroglutamate Aminopeptitase(TAKARA)2mUを、酵素反応緩衝液(10mM ジチオトレイトール(DTT)、1mM EDTAおよび10mM PB(pH 7.0))200μl中に溶解し、本溶液中に膜を浸漬後、50℃で5時間反応させることによって行った。本反応によりピログルタミル基を除去後、膜を蒸留水で3回洗浄し、Procise(登録商標)492 HTを用いて分析した。
【0207】
その結果、2残基以降の配列がTGSCNTFQRS(配列番号15)と決定された。これらの結果から、14kDaの成分のN末端はグルタミンがピログルタミル化することによりブロッキングされ、そのN末端アミノ酸配列は、QTGSCNTFQRS(配列番号16)であることがわかった。
【0208】
本配列は、図1に示すように、実施例2においてcDNAクローニングにより明らかとされたMPL cDNAの演繹アミノ酸配列中、実施例2でプロペプチドと予想された領域(23−96)の53番目の残基以降に認められた。図1において、今回決定した14 kDa成分のN末端アミノ酸配列をボックスで示す。
【0209】
MPL cDNAの演繹アミノ酸配列うち、53−198残基からなる領域の算出分子量は15,553.3 Daであるが、ESI−MSによるMPL−1およびMPL−2の測定分子量は、実施例5−2で求めたように、それぞれ31,104.1 Daおよび31,132.0 Daである。SDS−PAGEの結果(図6、8)から、MPLはS−S結合によるホモダイマーであると考えられることから、53−198残基からなる成分が1つのS−S結合により二量体を形成していると仮定すると、その算出分子量は31,104.6となり、MPL−1の測定分子量と一致する。
【0210】
これらの結果から、実施例2で得られたcDNAはMPL−1をコードするものであり、MPL−1は15.6kDaサブユニットのジスルフィド結合による二量体であることがわかった。
【0211】
すなわち、MPL−1のサブユニット(上記15.6kDaサブユニット)のcDNAの全長塩基配列は配列番号11に示される配列であり、当該塩基配列の演繹アミノ酸配列は配列番号13に示される配列である。また、MPL−1のサブユニットの成熟タンパク質をコードする塩基配列を配列番号1に示し、その演繹アミノ酸配列を配列番号2に示した。
【0212】
このことから、MPL−1は、高マンノース型糖鎖と結合するポリペプチドであって、(a)配列番号2に示されるアミノ酸配列;または(b)配列番号2に示されるアミノ酸配列において、1個もしくは数個のアミノ酸が置換、欠失、挿入、もしくは付加されたアミノ酸配列、からなるポリペプチドの二量体であるということができる。
【0213】
そして、当該二量体を含有する抗ウイルス剤も、高マンノース型糖鎖を備えるウイルスを標的とする抗ウイルス剤として用いることができる。また、当該二量体を例えば限外ろ過膜などに固定されてなる膜も、ウイルス除去膜として用いることができる。
【0214】
加えて、実施例2で得られたMPLのN末端アミノ酸配列(配列番号12)は8kDa成分のものであることがわかった。この8kDa成分は15.6kDa成分に由来することが明らかであるが、15.6kDa成分の分解産物であるか、MPLの構成成分として機能しているかどうかについてはさらなる解析が必要である。
【0215】
なお、ESI−MSによる測定分子量とSDS−PAGE上の推定分子量に差異がみられたが、SDS−PAGE によるタンパク質の移動度は、成分の性質(電荷、疎水性、構造等)により実際の分子量と数kDaのズレが生じることがある。
【0216】
そのため、SDS−PAGE上の成分について、例えば、SDS−PAGE上の推定分子量である14kDaと、ESI−MSによる測定分子量である15.6kDa成分とにずれがあることは特に問題はないと考えられる。
【0217】
Pfamプログラムによる分子内ドメイン検索にMPL−1を供したところ、MPL−1はJacalin-like lectinドメインを含み、紅藻 Griffithsia sp.由来レクチン Griffithsin(GRFT)およびバナナ Musa acuminataレクチンとの配列相同性(同一率)はそれぞれ33%および27%と非常に低いものであった。
【0218】
成熟タンパク質領域につきBasic Local Alignment Search Tool(BLAST)を用いて相同配列検索を行ったところ、上記2種のレクチンを含むJacalin-like lectinドメインを有するものが見られたが、その多くはhypothetical proteinであり、MPL−1と最も高い同一率を示したものは細菌Brevibacillus laterosporus由来hypothetical protein(35%)であった。
【0219】
このように、MPL−1には、部分的には既知レクチンのモチーフと類似するモチーフが存在することが示されたが、従来公知のレクチンとの配列相同性は非常に低く、全体配列から新規タンパク質であることが分かった。
【0220】
〔実施例7:MPL−1の糖鎖結合特異性〕
(実施例7−1:遠心限外ろ過−HPLC法)
上述のように精製されたMPL−1につき、糖鎖結合特異性を調べた。すなわち、まず、50mM トリス-塩酸緩衝液(pH7.0)中の500nM MPL−1溶液90μl(45pmol)と300nM ピリジルアミノ化(PA化)糖鎖水溶液10μl(3pmol)を軽く混合後、室温で60分間保温した。
【0221】
この反応液を微量遠心限外ろ過器(Nanosep 10K Omega、PALL)を用いて遠心ろ過(10,000×g、30秒)し、ろ液の20μlをHPLCに供し、溶出するPA化糖鎖の量を測定し遊離糖鎖量とした。
【0222】
次に、50mMトリス−塩酸緩衝液(pH7.0)の90mlとPA化糖鎖水溶液10μlを混合後、同様に処理し、そのろ液の20μlをHPLCに供し、溶出するPA化糖鎖の量を測定して添加糖鎖量とした。
【0223】
結合糖鎖量は、添加糖鎖量から反応液の遊離糖鎖量を差し引いた値として算出した。レクチンの糖鎖結合活性は結合率、すなわち添加糖鎖量に対する結合糖鎖量の割合(%)で表示した。なお、結合試験は各糖鎖につき2回行い、結合率は平均値として算出した。
【0224】
PA化糖鎖の分離、定量は逆相系HPLCを用いて行った。すなわち、検液を40℃のカラムオーブン中、TSKgel ODS-80TM カラム(4.6×150 mm)に注入し、0.1M 酢酸アンモニウム−15%メタノールで溶出した。流速は1.0 ml/minとし、溶出液は励起波長320nm、蛍光波長400nmでモニターし、各糖鎖のピーク面積をEZChrom Elite(アジレントテクノロジー)で解析、定量した。
【0225】
供試糖鎖としてPA化糖鎖27種:N−グリコシド型糖鎖の複合型6種(PA-Sugar Chain 001、002、004、009、010、023、TAKARA)、高マンノース型11種(PA-Sugar Chain 017、018、019、020、051、052、053、054、055、056、058、TAKARA)、共通コア構造1種(PA-Sugar Chain 016、TAKARA)、共通コア関連糖鎖1種(PA-042、増田化学工業)、糖脂質系糖鎖3種(PA-Sugar Chain 027、028、038、TAKARA)、およびオリゴマンノース5種を用いた。このうち、PA−オリゴマンノースを除き、市販品(TAKARA)を用いた。
【0226】
PA−オリゴマンノースは、PA化装置(PALSTATION、TAKARA)を用いて以下のように調製した。
【0227】
すなわち、各50nmolのManα1-2Man、Manα1-3Man、Manα1-6Man、Manα1-6(Manα1-3)Man、Manα1-6(Manα1-3)Manα1-6(Manα1-3)Man(DextraLaboratories、Funakoshi)を凍結乾燥後、カップリング試薬(2-アミノピリジン/酢酸溶液、Pyridylamination Reagent Kit、TAKARA)20μlを加え、90℃で60分間反応させた。これに還元試薬(ボラン-ジメチルアミン/酢酸溶液)20μlを加え、80℃で60分間反応させた。この反応液にトリエチルアミン-メタノール20μlを加えてよく撹拌した後、トルエン40 μlを加えて撹拌し、窒素気流化で60℃、10分間減圧乾固した。
【0228】
これにメタノール20μlとトルエン40μlとを加えて撹拌し、同様に減圧乾固した後、トルエン50μlを加えて再び減圧乾固した。過剰の試薬は順相系HPLCにより除去した。
【0229】
すなわち、上記PA化物を25μlの超純水に溶解した後、40℃のカラムオーブン中、50mM 酢酸−トリエチルアミン(pH7.3)/アセトニトリル(25/75)で平衡化したTSKgel NH2-60カラム(4.6×250 mm)に注入し、50mM酢酸−トリエチルアミン(pH7.3)/アセトニトリル(25/75)(溶媒A)と50mM酢酸−トリエチルアミン(pH7.3)/アセトニトリル(50/50)(溶媒B)の2液による濃度勾配[溶媒A 100%(0−5分)、溶媒A 0%−溶媒B 100%(5−55分)]を用いて、流速1.0ml/minで溶出した。溶出液は励起波長310nm、蛍光波長380nmでモニターし、PA化糖鎖(オリゴマンノース)ピークを分取した。
【0230】
得られたPA−オリゴマンノースの定量は以下のようにして行った。すなわち、上記の各オリゴマンノース画分の50μlを減圧乾固後、4N HCl/4M TFA(1/1(v/v))中100℃で4時間、気相酸加水分解した。
【0231】
この加水分解物を50μlの超純水に溶解し、その10μlを40℃のカラムオーブン中、TSKgel ODS-80TMカラム(4.6×150 mm)に注入し、0.1 M 酢酸アンモニウム−10%メタノールで溶出した。流速は1.0ml/minとし、溶出液は励起波長320nm、蛍光波長400mmでモニターした。
【0232】
これにより、PA−オリゴマンノースから加水分解により遊離したPA−マンノースのピーク面積を測定した。このPA−マンノースの定量は、10pmolの標準PA−マンノース(TAKARA)の同カラムからの溶出ピーク面積との比較により行い、PA−オリゴマンノース量とした。
【0233】
(実施例7−2:糖鎖結合特異性試験結果)
遠心限外ろ過−HPLC法により27種類のピリジルアミノ化(PA化)糖鎖に対するMPL−1の結合性を精査した。図11は、PA化糖鎖27種の構造と、当該糖鎖へのMPL−1の結合活性を示すものである。図中、結合活性(単位%、結合糖鎖量/添加糖鎖量×100)が10%以下ものはハイフンで示した。
【0234】
図11に示すように、MPL−1は供試した糖鎖中、N−グリコシド型糖鎖のうち高マンノース型糖鎖とのみ特異的に結合することがわかった。複合型(図11に示す1−6)、共通コア構造(図11に示す23−24)とは結合しなかったことから、本レクチンは高マンノース型糖鎖構造のオリゴマンノースで構成される分岐糖鎖部分を認識することが判明した。さらに興味深いことに、高マンノース型糖鎖(図11に示す7−17)に対する結合性について、分岐糖鎖部分のオリゴマンノース構造の違いにより結合活性に明瞭な差異が認められた。
【0235】
ここで、高マンノース型糖鎖の非還元末端は、トリマンノースコアのMan(α1-3)アーム(D1アーム)、同コアのMan(α1-6)アームから分岐したMan(α1-3)アーム(D2アーム)および同Man(α1-6)アームから分岐したMan(α1-6)アーム(D3アーム)の3つのアームからなる。
【0236】
MPL−1はD3アームの非還元末端にα1-2Manを有するもの(図11に示す9、10、13、15)とのみ高い結合活性が認められ、その結合率は96%以上であった。一方、同Man残基を含まない高マンノース型糖鎖(図11に示す7、8、11、12、14、16、17)およびオリゴマンノース(図11に示す18−22)に対するMPL−1の結合活性は、図11に示す糖鎖12を除いて10%以下であった。糖鎖12はD3アームにα1-2Manを含まず、D1アームの非還元末端にMan(α1-2)Man(α1-2)を有するM7糖鎖であるが、同様の構造を有するM8糖鎖(図11に示す14)では結合がみられないことから、MPL−1の主要認識糖鎖構造ではないと考えられる。
【0237】
これらの結果から、MPL−1はD3アームの非還元末端にα1-2Man残基を有する高マンノース型糖鎖とのみ結合すると考えられる。単糖類(マンノースを含む)に結合せず、このような厳密な高マンノース型糖鎖認識をもつものは既知レクチン中に見出されていない。糖化合物による赤血球凝集阻止試験でMPL−1と同様のプロファイル(表3)を示したイソレクチンMPL−2においても同様の糖鎖結合特異性を有すると予測される。
【0238】
本発明は上述した各実施形態に限定されるものではなく、請求項に示した範囲で種々の変更が可能であり、異なる実施形態にそれぞれ開示された技術的手段を適宜組み合わせて得られる実施形態についても本発明の技術的範囲に含まれる。
【産業上の利用可能性】
【0239】
本発明にかかるポリペプチドは、HIVなどのウイルスが備える高マンノース型糖鎖と特異的に結合することができる新規ポリペプチドであり、抗ウイルス剤、ウイルス除去膜等への適用が可能である。それゆえ、医療産業、薬品産業等において広く利用することが可能である。
【技術分野】
【0001】
本発明は、高マンノース型糖鎖と高い親和性を有する新規ポリペプチド、およびそのポリペプチドをコードするポリヌクレオチド、並びにそれらの代表的利用に関するものである。
【背景技術】
【0002】
ヒト免疫不全ウイルス(以下「HIV」)、インフルエンザウイルス、C型肝炎ウイルス(以下「HCV」)、エボラウイルス、ヒトヘルペスウイルス6(以下「HHV−6」)、重症急性呼吸症候群(以下「SARS」)ウイルスなどのウイルスは、しばしば人体に対して重篤な症状を惹起し、その感染拡大が社会問題ともなっていることから、効果的に感染を阻害する薬剤の開発が望まれている。これらのウイルスは、表面に高マンノース型糖鎖を有し、これが宿主への感染に重要な役割を果たすことが知られている(例えば、非特許文献1)。そのため、これらのウイルスが備える高マンノース型糖鎖への結合能を有する物質は、上記ウイルスに対する感染阻害剤として機能することが期待できる。
【0003】
これまでに、海藻類または藻類(淡水産藍藻)から多くの種類のレクチンが単離され、その生化学的性質が明らかにされている。上記レクチンの一部は、HIV、インフルエンザウイルス等の上述のウイルスに特異的に結合することが知られている(非特許文献2〜12)。
【0004】
他にも、レクチンとHIVとの結合に関する発明として、例えば、ConAなどのレクチンが結合した疎水性基材からなる微粒子担体にHIVまたはその一部のタンパクもしくはペプチドを捕捉してなる微粒子を含有するエイズワクチン(特許文献1)、GNAなどのレクチンを固定した多孔性中空糸膜に血液を通過させ、ウイルスに感染した個体の血液中のウイルス粒子及びそのレクチン結合性フラグメントを減じる方法(特許文献2)、癌患者における感染症の予防および/または処置用の医薬の製造において、マンナン結合レクチンサブユニット等を含むマンナン結合レクチンオリゴマーを含有する組成物を使用する発明(特許文献3)、スクシニル−コンカナバリンAなどのレクチンを結合成分として用い、ウイルス粒子を含んでなるサンプルのウイルス力価を高める方法(特許文献4)などが知られている。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開2001−335510号公報(2001年12月4日公開)
【特許文献2】特表2007−525232号公報(2007年9月6日公表)
【特許文献3】特表2008−535872号公報(2008年9月4日公表)
【特許文献4】特表2004−500831号公報(2004年1月15日公表)
【非特許文献】
【0006】
【非特許文献1】New England Journal of Medicine, 348, 2228-2238, 2003.
【非特許文献2】Boyd, M. R. et al., Antimicrob. Agents Chemother.41, 1521-1530, 1997.
【非特許文献3】O’Keefe, B. R. et al., Antimicrob. Agents Chemother. 47, 2518-2525, 2003.
【非特許文献4】Helle, F., .et al., J. Biol. Chem. 281, 25177-25183, 2006.
【非特許文献5】Barrientos, L. G., et al., Antiviral. Res. 58, 47-56, 2003.
【非特許文献6】Dey, B., et al., J. Virol. 74, 4562-4569, 2000.
【非特許文献7】O’Keefe, B. R. et al.,J. Virol. 84, 2511-2521, 2010.
【非特許文献8】Hori,K. et al., Glycobiology, 17, 479-491, 2007.
【非特許文献9】Sato,Y., Okuyama, S., and Hori, K., J. Biol. Chem. 282, 11021-11029, 2007.
【非特許文献10】Sato,Y., Morimoto,K., Hirayama, M., and Hori, K. Biochem. Biophys. Res. commun. 405, 291-296, 2011.
【非特許文献11】佐藤雄一郎、平山 真、藤原佳史、森本金治郎、堀 貫治 (2010) 第13回マリンバイオテクノロジー学会大会講演要旨 (2010. 5.29発表)
【非特許文献12】Sato,Y., Hirayama, M., Morimoto,K., Yamamoto, N., Okuyama, S., and Hori, K. J. Biol. Chem. 286, No.22, 19446-19458, 2011.
【発明の概要】
【発明が解決しようとする課題】
【0007】
しかしながら、現在のところHIVウイルスやインフルエンザウイルス等が表面に有する高マンノース型糖鎖に特異的に結合し、上記ウイルスの感染を効果的に阻害する物質はまだ十分に知られているとは言いがたく、その数は限られているため、上記物質が十分に供給できる状況にはなっていない。したがって、新規な上記物質がさらに多く見出され、その特性が明らかにされることが必要である。
【0008】
本発明は、このような問題点に鑑みてなされたものであり、その目的は、高マンノース型糖鎖と高い親和性を有する新規ポリペプチド、およびそのポリペプチドをコードするポリヌクレオチド、並びにそれらの代表的利用を提供することにある。
【課題を解決するための手段】
【0009】
本発明者は、上記課題を解決すべく、レクチン、特にトサカノリ(Meristotheca papulosa (Montagne) J. Agardh)由来のレクチンに着目し、HIVウイルス等が表面に備える高マンノース型糖鎖と特異的に結合しうるレクチンの探索を行った。その結果、従来公知のレクチンとアミノ酸配列の相同性が非常に低い、本発明にかかるポリペプチドを見出すに至った。
【0010】
すなわち、本発明にかかるポリペプチドは、上記課題を解決すべく、高マンノース型糖鎖と結合するポリペプチドであって、(a)配列番号2に示されるアミノ酸配列;または(b)配列番号2に示されるアミノ酸配列において、1個もしくは数個のアミノ酸が置換、欠失、挿入、もしくは付加されたアミノ酸配列、からなることを特徴としている。
【0011】
本発明にかかるポリヌクレオチドは、上記課題を解決すべく、上記本発明にかかるポリペプチドをコードすることを特徴としている。
【0012】
また、本発明にかかるポリヌクレオチドは、上記課題を解決すべく、下記の(a)または(b)のいずれかであるポリヌクレオチド:(a)配列番号1に示される塩基配列からなるポリヌクレオチド;または(b)以下の(i)もしくは(ii)のいずれかとストリンジェントな条件下でハイブリダイズするポリヌクレオチド:(i)配列番号1に示される塩基配列からなるポリヌクレオチド;もしくは(ii)配列番号1に示される塩基配列と相補的な塩基配列からなるポリヌクレオチド、であってもよい。
【0013】
本発明にかかる抗ウイルス剤は、上記課題を解決すべく、上記本発明にかかるポリペプチドを含有し、高マンノース型糖鎖を備えるウイルスを標的とすることを特徴としている。
【0014】
上記本発明にかかる抗ウイルス剤は、上記課題を解決すべく、上記抗ウイルス剤の標的であるウイルスが、エイズウイルス、インフルエンザウイルス、C型肝炎ウイルス、エボラウイルス、ヒトヘルペスウイルス6および重症急性呼吸症候群ウイルスからなる群より選ばれるウイルスであってもよい。
【0015】
本発明にかかるウイルス除去膜は、上記課題を解決すべく、上記本発明にかかるポリペプチドが固定されてなることを特徴としている。
【0016】
本発明にかかるベクターは、上記課題を解決すべく、上記本発明にかかるポリヌクレオチドを含むことを特徴としている。
【0017】
本発明にかかるポリペプチドを生産する方法は、上記課題を解決すべく、上記本発明にかかるベクターを用いることを特徴としている。
【0018】
本発明にかかる形質転換体は、上記課題を解決すべく、上記本発明にかかるポリヌクレオチドが導入されていることを特徴としている。
【0019】
本発明にかかるポリペプチドを生産する方法は、上記課題を解決すべく、上記本発明にかかる形質転換体を用いることを特徴としている。
【発明の効果】
【0020】
以上のように、本発明にかかるポリペプチドは、高マンノース型糖鎖と結合するポリペプチドであって、(a)配列番号2に示されるアミノ酸配列;または(b)配列番号2に示されるアミノ酸配列において、1個もしくは数個のアミノ酸が置換、欠失、挿入、もしくは付加されたアミノ酸配列、からなる構成である。上記アミノ酸配列は従来公知のレクチンと非常に相同性の低いものである。
【0021】
それゆえ、HIVやインフルエンザウイルス等が表面に備える高マンノース型糖鎖と効率的に結合することができ、これらのウイルスの感染を効果的に阻害することができるという効果を奏する。
【0022】
本発明にかかるポリヌクレオチドは、上記本発明にかかるポリペプチドをコードしている。したがって、本発明にかかるポリヌクレオチド、当該ポリヌクレオチドを含むベクター、当該ポリヌクレオチドが導入された形質転換体を用いることによって、上記本発明にかかるポリペプチドを簡便かつ大量に調製することが可能となる。その結果、上記ウイルスの感染を効果的に阻害することができるという効果を奏する。
【図面の簡単な説明】
【0023】
【図1】MPLのcDNAの全長塩基配列、およびその演繹アミノ酸配列を示す図である。
【図2】図2のAは固定化したgp120とMPLとの相互作用のセンサーグラムであり、図2のBはgp120とMPLとの親和定数を示す図である。
【図3】MPLをSDS−PAGE(10%ゲル)に供した結果を示す図である。
【図4】20〜60%飽和硫安塩析沈殿画分を、HiPrep QXL 16/10カラムを用いた陰イオン交換クロマトグラフィー(ステップワイズ溶出)に供した結果を示す図である。
【図5】粗レクチン画分を、HiPrep QXL 16/10カラムを用いた陰イオン交換クロマトグラフィー(グラジエント溶出)に供した結果を示す図である。
【図6】粗レクチン画分を、HiPrep QXL 16/10カラムを用いた陰イオン交換クロマトグラフィー(グラジエント溶出)に供した結果得られた溶出液の画分をSDS−PAGE(10%ゲル)に供した結果を示す図である。
【図7】MPLをTSKgel DEAE-5PWカラムを用いた陰イオン交換クロマトグラフィーに供した結果を示す図である。
【図8】MPLをTSKgel DEAE-5PWカラムによる陰イオン交換クロマトグラフィーに供して得られた4つのピークを、SDS−PAGE(10%ゲル)に供した結果を示す図である。
【図9】MPL−1およびMPL−2の分子量測定結果を示す図である。
【図10】MPL画分をブロッティングし、CBB染色したPVDF膜を示す図である。
【図11】遠心限外ろ過−HPLC法に供試したピリジルアミノ化糖鎖27種の構造と、当該糖鎖へのMPL−1の結合活性を示すものである。
【発明を実施するための形態】
【0024】
以下、本発明の実施の一形態について説明すれば以下のとおりである。なお、本発明はこれに限定されるものではない。本明細書において、範囲を示す「A〜B」は、A以上B以下であることを表す。また、本明細書中に記載された特許文献および非特許文献は、本明細書中において参考として援用される。
【0025】
(1)ポリペプチド
本発明者は、食用紅藻であるトサカノリ(Meristotheca papulosa (Montagne) J. Agardh)から単離した新規ポリペプチド(以下「MPL」という)が、高マンノース型糖鎖への結合特異性を有することを見出し、本発明を完成するに至った。上記MPLのアミノ酸配列は、後述するように、バナナMusa acuminataレクチンおよび紅藻Griffithsia sp.由来レクチンGriffithsin(GRFT)等の従来公知のレクチンとの相同性が最大33%と、非常に低いものであった。
【0026】
本明細書中で使用される場合、用語「ポリペプチド」は、「ペプチド」または「タンパク質」と交換可能に使用される。本発明にかかるポリペプチドはまた、天然供給源より単離されても、化学合成されてもよい。
【0027】
用語「単離された」ポリペプチドまたはタンパク質とは、その天然の環境から取り出されたポリペプチドまたはタンパク質が意図される。例えば、宿主細胞中で発現された組換え産生されたポリペプチドおよびタンパク質は、任意の適切な技術によって実質的に精製されている天然または組換えのポリペプチドおよびタンパク質と同様に、単離されていると考えられる。
【0028】
本発明にかかるポリペプチドは、天然の精製産物、化学合成手順の産物、および原核生物宿主または真核生物宿主(例えば、細菌細胞、酵母細胞、高等植物細胞、昆虫細胞、および哺乳動物細胞を含む)から組換え技術によって産生された産物を含む。組換え産生手順において用いられる宿主に依存して、本発明にかかるポリペプチドは、グリコシル化され得るか、または非グリコシル化され得る。さらに、本発明にかかるポリペプチドはまた、いくつかの場合、宿主媒介プロセスの結果として、開始の改変メチオニン残基を含み得る。
【0029】
本発明は、本発明にかかるポリペプチドを提供する。一実施形態において、本発明にかかるポリペプチドは、配列番号2に示されるアミノ酸配列からなるポリペプチド、または配列番号2に示されるアミノ酸配列からなるポリペプチドの変異体でありかつ本発明にかかるポリペプチドである。上記ポリペプチドは、従来公知のレクチンのアミノ酸配列との相同性が非常に低い新規ポリペプチドであるという特徴を有する。
【0030】
変異体としては、欠失、挿入、逆転、反復、およびタイプ置換(例えば、親水性の残基の別の残基への置換、しかし通常は強く親水性の残基を強く疎水性の残基には置換しない)を含む変異体が挙げられる。特に、ポリペプチドにおける「中性」アミノ酸置換は、一般的にそのポリペプチドの活性にほとんど影響しない。
【0031】
ポリペプチドのアミノ酸配列中のいくつかのアミノ酸が、このポリペプチドの構造または機能に有意に影響することなく容易に改変され得ることは、当該分野において周知である。さらに、人為的に改変させるだけではく、天然のタンパク質において、当該タンパク質の構造または機能を有意に変化させない変異体が存在することもまた周知である。
【0032】
当業者は、周知技術を使用してポリペプチドのアミノ酸配列において1または数個のアミノ酸を容易に変異させることができる。例えば、公知の点変異導入法に従えば、ポリペプチドをコードするポリヌクレオチドの任意の塩基を変異させることができる。また、ポリペプチドをコードするポリヌクレオチドの任意の部位に対応するプライマーを設計して欠失変異体または付加変異体を作製することができる。さらに、本明細書中に記載される方法を用いれば、作製した変異体が、本発明にかかる所望の変異体であるか否かを容易に決定し得る。
【0033】
本実施形態にかかるポリペプチドは、高マンノース型の糖鎖と結合するポリペプチドであって、
(a)配列番号2に示されるアミノ酸配列;または
(b)配列番号2に示されるアミノ酸配列において、1個もしくは数個のアミノ酸が置換、欠失、挿入、もしくは付加されたアミノ酸配列、
からなることが好ましい。
【0034】
上記「1個もしくはそれ以上のアミノ酸が置換、欠失、挿入、もしくは付加された」とは、部位特異的突然変異誘発法等の公知の変異ポリペプチド作製法により置換、欠失、挿入、もしくは付加できる程度の数(好ましくは1から10個、より好ましくは1から7個、さらに好ましくは1個から5個、特に好ましくは1個から3個)のアミノ酸が置換、欠失、挿入もしくは付加されていることを意味する。このような変異ポリペプチドは、上述したように、公知の変異ポリペプチド作製法により人為的に導入された変異を有するポリペプチドに限定されるものではなく、天然に存在するポリペプチドを単離精製したものであってもよい。
【0035】
なお、本発明にかかるポリペプチドは、アミノ酸がペプチド結合しているポリペプチドであればよいが、これに限定されるものではなく、ポリペプチド以外の構造を含む複合ポリペプチドであってもよい。本明細書中で使用される場合、「ポリペプチド以外の構造」としては、糖鎖やイソプレノイド基等を挙げることができるが、特に限定されるものではない。
【0036】
また、本発明にかかるポリペプチドは、付加的なポリペプチドを含むものであってもよい。付加的なポリペプチドとしては、例えば、HisやMyc、Flag等のエピトープ標識ポリペプチドが挙げられる。
【0037】
また、本発明にかかるポリペプチドは、後述する本発明にかかるポリヌクレオチド(本発明にかかるポリペプチドをコードする遺伝子)を宿主細胞に導入して、そのポリペプチドを細胞内発現させた状態であってもよいし、細胞、組織などから単離精製された場合であってもよい。また、本発明にかかるポリペプチドは、化学合成されたものであってもよい。
【0038】
他の実施形態において、本発明にかかるポリペプチドは、融合タンパク質のような改変された形態で組換え発現され得る。例えば、本発明にかかるポリペプチドの付加的なアミノ酸、特に荷電性アミノ酸の領域が、宿主細胞内での、精製の間または引き続く操作および保存の間の安定性および持続性を改善するために、ポリペプチドのN末端に付加され得る。
【0039】
別の実施形態において、本発明にかかるポリペプチドは、下記で詳述されるように組換え生成されても、化学合成されてもよい。
【0040】
組換え生成は、当該分野において周知の方法を使用して行なうことができ、例えば、以下に詳述されるようなベクターおよび細胞を用いて行なうことができる。
【0041】
合成ペプチドは化学合成の公知の方法を使用して合成され得る。例えばHoughtenは、4週間未満で調製され、そして特徴付けられたHA1ポリペプチドセグメントの単一アミノ酸改変体を示す10〜20mgの248の異なる13残基ペプチドのような、多数のペプチドの合成のための簡単な方法を記載している(Houghten,R.A.,Proc.Natl.Acad.Sci.USA, 82:5131-5135(1985))。
【0042】
この「Simultaneous Multiple Peptide Synthesis(SMPS)」プロセスは、さらにHoughtenら(1986)の米国特許第4,631,211号に記載される。この手順において、種々のペプチドの固相合成のための個々の樹脂は、別々の溶媒透過性パケットに含まれ、固相法に関連する多くの同一の反復工程の最適な使用を可能にする。完全なマニュアル手順は、500〜1000以上の合成が同時に行われるのを可能にする(Houghtenら、前出、5134)。これらの文献は、本明細書中に参考として援用される。
【0043】
本発明者は、上記本発明にかかるポリペプチドが高マンノース型糖鎖に特異的に結合することを見出した。ここで、「糖鎖」とは、直鎖または分岐したオリゴ糖または多糖を意味する。また上記糖鎖は、タンパク質との結合様式によって、アスパラギンと結合するN−グリコシド結合糖鎖(以下、「N型糖鎖」という)およびセリン、スレオニンなどと結合するO−グリコシド結合糖鎖(以下、「O型糖鎖」という)に大別され、N型糖鎖には高マンノース型糖鎖、複合型糖鎖、混成型糖鎖がある。
【0044】
なおオリゴ糖とは、単糖または単糖の置換誘導体が2〜10個脱水結合して生じたものをいう。さらに多数の単糖が結合している糖質を多糖という。多糖は、構成糖の種類によって異なるが、ウロン酸やエステル硫酸を多く含む糖質を酸性多糖、中性糖のみのものを中性多糖という。多糖のうち、ムコ多糖とよばれる一群の多糖は、ほとんどがタンパク質と結合しており、プロテオグリカンという。単糖とは、糖鎖の構成単位となるもので、加水分解によってそれ以上簡単な分子にならない基本的物質である。
【0045】
さらに単糖は、カルボキシル基などの酸性側鎖を有する酸性糖、ヒドロキシル基がアミノ基で置換されたアミノ糖、それ以外の中性糖の3つに大別される。生体内に存在する単糖としては、酸性糖はN−アセチルノイラミン酸やN−グリコリルノイラミン酸(以下、「Neu5Gc」という)などのシアル酸や、ウロン酸などがあり、アミノ糖としてはN−アセチルグルコサミン(以下、「GlcNAc」という)やN−アセチルガラクトサミンなどがあり、中性糖としてはグルコース、マンノース、ガラクトース、フコースなどがあげられる。
【0046】
N型糖鎖は全て、「トリマンノシルコア」と呼ばれる〔Manα1-6(Manα1-3)Manβ1-4GlcNAcβ1-4GlcNAc〕からなる共通母核構造を持っている。高マンノース型糖鎖は、トリマンノシルコアに加え、分岐構造部分にα−マンノース残基のみを含む。この糖鎖には〔Manα1-6(Manα1-3)Manα1-6(Manα1-3 )Manβ1-4GlcNAcβ1-4GlcNAc〕という七糖が共通の母核として含まれている。また混成型糖鎖は、複合型と高マンノース型の両方の特徴を併せ持っていることからそう呼ばれている。1つまたは2つのα−マンノシル基が、高マンノース型の場合と同様に、トリマンノシルコアのManα1-6腕と結合し、複合型糖鎖の側鎖と同じものがコアのManα1-3腕に結合している。
【0047】
トリマンノシルコアの還元末端に位置するGlcNAc のC-6位へのフコースの結合の有無、またβ-マンノシル残基のC-4位へのβ-GlcNAcの結合(バィセクテイングGlcNAc と呼ばれる)の有無は、複合型や混成型糖鎖の構造の多様性に寄与している。3つのN型糖鎖の間で、複合型が最も多様な構造を含んでいる。
【0048】
この多様性は、主に2つの要素で作り出され、トリマンノシルコアに1個から5個の側鎖がそれぞれ異なる結合位置で結合しており、一、二、三、四、または五本側鎖糖鎖を形成している。三本側鎖の複合型糖鎖には、〔GlcNAcβ1-4(GlcNAcβ1-2)Manα1-3〕あるいは〔GlcNAcβ1-6(GlcNAcβ1-2)Manα1-6〕のどちらかを含む2つの異性体が見つかっている。
【0049】
なお上記トリマンノシルコアにおいて、アスパラギンと結合する糖鎖の末端、すなわちGlcNAc側の末端を還元末端、その反対側、すなわちMan側の末端を非還元末端という。
【0050】
ポリペプチドが糖鎖と結合するか否かは、例えば標的となる糖鎖、または糖鎖が結合した糖タンパク質等を固定化したカラムに、試験対象であるポリペプチドを通し、当該カラムにポリペプチドが結合したか否かをその通過液に含まれるポリペプチドの量、または特異的溶出剤でカラムから溶出したポリペプチドの量により評価することができる。また標的となる糖鎖が結合した糖タンパク質をメンブレン等に固定化し、ビオチン、フルオレセインイソチオシアネート、ペルオキシダーゼ等で標識したポリペプチドを用いて検出するウエスタンブロット法(法医学の実際と研究、37, 155, 1994 参照)、ドットブロット法(Analytical Biochemistry, 204(1), 198, 1992 参照)を用いて評価することができる。
【0051】
また標的となる糖鎖、または糖鎖が結合した糖タンパク質等を固定化したチップと、試験対象であるポリペプチドとの親和性を表面プラズモン共鳴法(SPR法)を用いて測定すればよい。上記方法によれば、その親和性の有無のみならず、その強度まで測定できるために好ましい方法であるといえる。このとき得られる結合定数(親和定数)(KA)が、10(M-1)以上、より好ましくは103(M-1)以上、最も好ましくは104(M-1)以上であればポリペプチドと糖鎖とが結合していると判断できる。
【0052】
本発明にかかるポリペプチドには上記(a)または(b)に示すアミノ酸配列と、特定の機能(例えば、タグ)を有する任意のアミノ酸配列とからなるポリペプチドも含まれることに留意すべきである。また、上記(a)または(b)に示すアミノ酸配列と、当該任意のアミノ酸配列とは、それぞれの機能を阻害しないように適切なリンカーペプチドで連結されていてもよい。
【0053】
つまり、本発明の目的は、本発明にかかるポリペプチドを提供することにあるのであって、本明細書中に具体的に記載したポリペプチド作製方法等に存するのではない。したがって、上記各方法以外によって取得される本発明にかかるポリペプチドも本発明の技術的範囲に属することに留意しなければならない。
【0054】
(2)ポリヌクレオチド
本発明は、上述した本発明にかかるポリペプチドをコードするポリヌクレオチドを提供する。本明細書中で使用される場合、用語「ポリヌクレオチド」は「核酸」または「核酸分子」と交換可能に使用され、ヌクレオチドの重合体が意図される。本明細書中で使用される場合、用語「塩基配列」は、「核酸配列」または「ヌクレオチド配列」と交換可能に使用され、デオキシリボヌクレオチド(A、G、CおよびTと省略される)の配列として示される。
【0055】
本発明にかかるポリヌクレオチドは、RNA(例えば、mRNA)の形態、またはDNAの形態(例えば、cDNAまたはゲノムDNA)で存在し得る。DNAは、二本鎖または一本鎖であり得る。一本鎖DNAまたはRNAは、コード鎖(センス鎖としても知られる)であり得、または、非コード鎖(アンチセンス鎖としても知られる)であり得る。
【0056】
本明細書中で使用される場合、用語「オリゴヌクレオチド」は、ヌクレオチドが数個ないし数十個結合したものが意図され、「ポリヌクレオチド」と交換可能に使用される。オリゴヌクレオチドは、短いものはジヌクレオチド(二量体)、トリヌクレオチド(三量体)といわれ、長いものは30マーまたは100マーというように重合しているヌクレオチドの数で表される。オリゴヌクレオチドは、より長いポリヌクレオチドのフラグメントとして生成されても、化学合成されてもよい。
【0057】
本発明にかかるポリヌクレオチドのフラグメントは、少なくとも12nt(ヌクレオチド)、好ましくは約15nt、そしてより好ましくは少なくとも約20nt、なおより好ましくは少なくとも約30nt、そしてさらにより好ましくは少なくとも約40ntの長さのフラグメントが意図される。少なくとも20ntの長さのフラグメントによって、例えば、配列番号1に示される塩基配列からの20以上の連続した塩基を含むフラグメントが意図される。本明細書を参照すれば配列番号1に示される塩基配列が提供されるので、当業者は、配列番号1に基づくDNAフラグメントを容易に作製することができる。
【0058】
例えば、制限エンドヌクレアーゼ切断または超音波による剪断は、種々のサイズのフラグメントを作製するために容易に使用され得る。あるいは、このようなフラグメントは、合成的に作製され得る。適切なフラグメント(オリゴヌクレオチド)が、Applied Biosystems Incorporated(ABI,850 Lincoln Center Dr., Foster City, CA 94404)392型シンセサイザーなどによって合成される。
【0059】
合成的に作製した上記フラグメントとしては、例えば宿主のコドンに対して配列を最適化した合成ポリヌクレオチドを挙げることができる。このような合成ポリヌクレオチドは、各種の発現系を用いる場合に利用可能である。
【0060】
また本発明にかかるポリヌクレオチドは、その5’側または3’側で上述のタグ標識(タグ配列またはマーカー配列)をコードするポリヌクレオチドに融合され得る。
【0061】
本発明はさらに、本発明にかかるポリペプチドをコードするポリヌクレオチドの変異体に関する。変異体は、天然の対立遺伝子変異体のように、天然に生じ得る。「対立遺伝子変異体」によって、生物の染色体上の所定の遺伝子座を占める遺伝子のいくつかの交換可能な形態の1つが意図される。天然に存在しない変異体は、例えば当該分野で周知の変異誘発技術を用いて生成され得る。
【0062】
このような変異体としては、本発明にかかるポリペプチドをコードするポリヌクレオチドの塩基配列において1または数個の塩基が欠失、置換、または付加した変異体が挙げられる。変異体は、コードもしくは非コード領域、またはその両方において変異され得る。コード領域における変異は、保存的もしくは非保存的なアミノ酸欠失、置換、または付加を生成し得る。
【0063】
本発明はさらに、ストリンジェントなハイブリダイゼーション条件下で、本発明にかかるポリペプチドをコードするポリヌクレオチドまたは当該ポリヌクレオチドにハイブリダイズするポリヌクレオチドを含む、単離したポリヌクレオチドを提供する。
【0064】
一実施形態において、本発明にかかるポリヌクレオチドは、本発明にかかるポリペプチドをコードするポリヌクレオチドであり、かつ、
(a)配列番号2に示されるアミノ酸配列;または
(b)配列番号2に示されるアミノ酸配列において、1個もしくは数個のアミノ酸が置換、欠失、挿入、もしくは付加されたアミノ酸配列、からなるポリペプチドをコードするポリヌクレオチド、
のいずれかであることが好ましい。
【0065】
他の実施形態において、本発明にかかるポリヌクレオチドは、本発明にかかるポリペプチドをコードするポリヌクレオチドであって、以下の(a)または(b):
(a)配列番号1に示される塩基配列からなるポリヌクレオチド;または
(b)以下の(i)もしくは(ii)のいずれかとストリンジェントな条件下でハイブリダイズするポリヌクレオチド:
(i)配列番号1に示される塩基配列からなるポリヌクレオチド;もしくは
(ii)配列番号1に示される塩基配列と相補的な塩基配列からなるポリヌクレオチド、
のいずれかであることが好ましい。
【0066】
なお、上記「ストリンジェントな条件」とは、少なくとも90%以上の同一性、好ましくは少なくとも95%以上の同一性、最も好ましくは97%以上の同一性が配列間に存在する時にのみハイブリダイゼーションが起こることを意味する。
【0067】
上記ハイブリダイゼーションは、Sambrookら、Molecular Cloning,A Laboratory Manual,2d Ed.,Cold Spring Harbor Laboratory(1989)に記載されている方法のような周知の方法で行なうことができる。通常、温度が高いほど、塩濃度が低いほどストリンジェンシーは高くなり(ハイブリダイズし難くなる)、より相同なポリヌクレオチドを取得することができる。
【0068】
ハイブリダイゼーションの条件としては、従来公知の条件を好適に用いることができ、特に限定しないが、例えば、42℃、6×SSPE、50%ホルムアミド、1%SDS、100μg/ml サケ精子DNA、5×デンハルト液(ただし、1×SSPE;0.18M 塩化ナトリウム、10mMリン酸ナトリウム、pH7.7、1mM EDTA。5×デンハルト液;0.1%牛血清アルブミン、0.1%フィコール、0.1%ポリビニルピロリドン)が挙げられる。
【0069】
本発明にかかるポリヌクレオチドまたはオリゴヌクレオチドは、2本鎖DNAのみならず、それを構成するセンス鎖およびアンチセンス鎖といった各1本鎖DNAやRNAを包含する。またDNAには例えばクローニングや化学合成技術またはそれらの組み合わせで得られるようなcDNAやゲノムDNAなどが含まれる。さらに、本発明にかかるポリヌクレオチドまたはオリゴヌクレオチドは、非翻訳領域(UTR)の配列やベクター配列(発現ベクター配列を含む)などの配列を含むものであってもよい。
【0070】
本発明にかかるポリヌクレオチドまたはオリゴヌクレオチドを取得する方法として、公知の技術により、本発明にかかるポリヌクレオチドまたはオリゴヌクレオチドを含むDNA断片を単離し、クローニングする方法が挙げられる。例えば、本発明におけるポリヌクレオチドの塩基配列の一部と特異的にハイブリダイズするプローブを調製し、ゲノムDNAライブラリーやcDNAライブラリーをスクリーニングすればよい。このようなプローブとしては、本発明にかかるポリヌクレオチドの塩基配列またはその相補配列の少なくとも一部に特異的にハイブリダイズするプローブであれば、いずれの配列および/または長さのものを用いてもよい。
【0071】
あるいは、本発明にかかるポリヌクレオチドを取得する方法として、PCR等の増幅手段を用いる方法を挙げることができる。例えば、本発明におけるポリヌクレオチドのcDNAのうち、5’側および3’側の配列(またはその相補配列)の中からそれぞれプライマーを調製し、これらプライマーを用いてゲノムDNA(またはcDNA)等を鋳型にしてPCR等を行い、両プライマー間に挟まれるDNA領域を増幅することで、本発明にかかるポリヌクレオチドを含むDNA断片を大量に取得できる。
【0072】
本発明にかかるポリヌクレオチドを取得するための供給源としては、特に限定されないが、所望のポリヌクレオチドを含む生物材料であることが好ましい。特に、本発明にかかるポリペプチドの起源であるトサカノリ(Meristotheca papulosa (Montagne) J. Agardh)が好ましい。ただし、これに限定されるものではない。
【0073】
なお本発明の目的は、本発明にかかるポリペプチドをコードするポリヌクレオチド、および当該ポリヌクレオチドとハイブリダイズするオリゴヌクレオチドを提供することにあるのであって、本明細書中に具体的に記載したポリヌクレオチドおよびオリゴヌクレオチドの作製方法等に存するのではない。したがって、上記各方法以外によって取得される本発明にかかるポリペプチドをコードするポリヌクレオチドもまた本発明の技術的範囲に属することに留意しなければならない。
【0074】
(3)本発明にかかるポリペプチドおよび/またはポリヌクレオチドの利用
(3−1)抗ウイルス剤
本発明にかかる抗ウイルス剤は、上記本発明にかかるポリペプチドを含有し、高マンノース型糖鎖を備えるウイルスを標的とする。上記ウイルスは、高マンノース型糖鎖を備えるウイルスであれば特に限定されるものではなく、HIV、インフルエンザウイルス、HCV、エボラウイルス、HHV−6、SARSウイルス等を挙げることができる。
【0075】
上記ウイルスには、表面に備える高マンノース型糖鎖を宿主細胞の表面レセプターに結合させることによって宿主に感染するものが存在する。例えばHIVは、表面にgp120という糖タンパク質を備えており、gp120が有する高マンノース型糖鎖が宿主リンパ細胞表面レセプターであるCD4を認識することによって宿主に感染する。
【0076】
一方、上記抗ウイルス剤に含まれる本発明にかかるポリペプチドは、高マンノース型糖鎖と特異的に結合することができる。それゆえ、上記抗ウイルス剤を用いれば上記ウイルスの感染を効果的に阻害することができる。
【0077】
また、ウイルスの宿主への感染が、gp120とCD4との関係のような、ウイルス表面糖タンパク質の高マンノース型糖鎖と、宿主側の高マンノース型糖鎖のレセプターとの直接結合によるものではない場合もある。例えば、高マンノース型糖鎖を有するものの、高マンノース型糖鎖は宿主のレセプターと直接結合せず、糖タンパク質によって上記レセプターに結合するタイプのウイルスを挙げることができる。このようなウイルスとしては例えばインフルエンザウイルスを挙げることができる。
【0078】
上記抗ウイルス剤に含まれる本発明にかかるポリペプチドは、高マンノース型糖鎖と特異的に結合することができる。これにより糖タンパク質に対するアロステリック効果が得られ、高マンノース型糖鎖がレセプターとの結合に直接関与しない場合でもウイルスの感染を防ぐことができると考えられる。よって、上記抗ウイルス剤は、インフルエンザウイルス等の感染も有効に阻害することができる。
【0079】
なお、ウイルスの型は特に限定されるものではない。例えばHIVは1型でも2型でもよく、インフルエンザウイルスはA型、B型、C型のいずれであってもよい。
【0080】
「高マンノース型糖鎖を備える」とは、高マンノース型糖鎖がウイルスの表面に存在することを言う。例えば、高マンノース型糖鎖がキャプシドの外部へ露出した状態や、ウイルスの表面糖タンパク質の構成成分として存在する場合を挙げることができる。
【0081】
高マンノース型糖鎖は、上述のように、宿主細胞のレセプターへ直接結合するものであってもよいし、宿主細胞のレセプターへ直接結合しないものであってもよい。上述のように、本発明にかかる抗ウイルス剤は、いずれの高マンノース型糖鎖を有するウイルスであっても有効に感染を阻害することが可能である。
【0082】
本発明にかかる抗ウイルス剤は経口製剤、非経口製剤のいずれであってもよい。また、剤型は特に限定されるものではなく、常法に従い、錠剤、顆粒剤、散剤、カプセル剤、エリキシル剤、シロップ剤、マイクロカプセル剤あるいは懸濁液剤等に製剤化して用いることができる。
【0083】
非経口的に投与する場合には、例えば、本発明にかかるポリペプチドを含有する溶液を点鼻噴霧することや、注射剤として投与することができる。経口的に投与する場合には、食前、食後、食間のいずれに投与してもよい。
【0084】
本発明にかかる抗ウイルス剤は、必要に応じて、担体、賦形剤、結合剤、膨化剤、潤滑剤、甘味剤、香味剤、防腐剤、安定剤、被覆剤等の材料を含有することができる。
【0085】
本発明にかかる抗ウイルス剤において、例えば錠剤、カプセル剤等に含有することができる具体的な成分としては、トラガント、アラビアゴム、コーンスターチおよびゼラチンのような結合剤; 微晶性セルロース、結晶セルロースのような賦形剤; コーンスターチ、前ゼラチン化デンプン、アルギン酸、デキストリンのような膨化剤; ステアリン酸マグネシウムのような潤滑剤; 微粒二酸化ケイ素のような流動性改善剤; グリセリン脂肪酸エステルのような滑沢剤; ショ糖、乳糖およびアスパルテームのような甘味剤; ペパーミント、ワニラ香料およびチェリーのような香味剤等を挙げることができる。
【0086】
調剤単位形態がカプセル剤である場合には上記のタイプの材料にさらに油脂のような液
状担体を含有することができる。
【0087】
また、種々の他の材料を、被覆剤としてまたは調剤単位の物理的形態を変化させるために含有させることができる。錠剤の被覆剤としては、例えば、シェラック、砂糖またはその両方が挙げられる。シロップ剤またはエリキシル剤は、例えば、甘味剤としてショ糖、防腐剤としてメチルパラベンおよびプロピルパラベン、色素およびチェリーまたはオレンジ香味等を含有することができる。その他、各種ビタミン類、各種アミノ酸類を含有しても良い。
【0088】
本発明にかかる抗ウイルス剤の適用対象は、ヒト、ヒト以外の動物〔例えば、ヒト以外の哺乳類(ブタ、ウシ、ウマ、イヌ等の家畜) 、鳥類(ニワトリ等の家禽)等〕のいずれであってもよい。家畜や家禽等へ適用することにより、それらの経済的損失を軽減することができる。また、それらを食用とする場合も多いため、ヒトへの感染予防という点からも重要である。
【0089】
本発明にかかる抗ウイルス剤の投与量は、適用対象が必要とする量を確保できるように設定すればよく、製剤化して用いたり、飲食品に配合して上記抗ウイルス剤を用いることができる。
【0090】
(3−2)ウイルス除去膜
本発明にかかるウイルス除去膜は、本発明にかかる上記ポリペプチドが固定されてなるものである。上記ウイルス除去膜は、例えば従来公知の限外ろ過膜に上記ポリペプチドを固定することによって作製することができる。
【0091】
上記限外ろ過膜としては、従来公知のポリマー、例えば、ポリスルホン、ポリエーテルスルホン、ポリアミド類、ポリイミド類、酢酸セルロース、ポリアクリルアミド等によって形成された膜を挙げることができ、平膜型であっても多孔性中空糸膜型であってもよい。
【0092】
上記限外ろ過膜に上記ポリペプチドを結合するためには、従来公知のプロセスを用いて、限外ろ過膜のポリマーをまず賦活化し、タンパク質と化学的に結合しやすい状態にする。上記ポリマーとしては複数の異なるポリマーを用いてもよい。
【0093】
ポリマーを賦活化させた後、本発明にかかる上記ポリペプチドを直接あるいはリンカーを用いて付着させ、アフィニティーマトリクスを形成する。好ましいリンカーとして、アビジン、ストレプトアビジン、ビオチン、プロテインA、プロテインG等が挙げられるが、これらに限定されない。
【0094】
上記ポリペプチドは、二価性試薬のようなカップリング剤を用いて、限外ろ過膜のポリマーに直接結合されてもよく、あるいは間接的に結合されてもよい。アフィニティーマトリクスを形成するためには、アガロースに共有結合的に連結されたMPLを使用することが好ましいが、これに限定されるものではない。
【0095】
HIVのような、高マンノース型糖鎖を備えるウイルスの粒子および/またはそのフラグメントを含有する血液等を上記ウイルス除去膜にアプライすると、上記ウイルス除去膜に固定された本発明にかかるポリペプチドが、ウイルスの粒子および/またはそのフラグメントが有する高マンノース型糖鎖と結合するため、血液は孔を通過し、ウイルスの粒子および/またはそのフラグメントは捕捉されるため孔を通過することができない。それゆえ、高マンノース型糖鎖を備えるウイルスを血液等から除去することができる。
【0096】
(3−3)ベクター
本発明は、本発明にかかるポリペプチドを生成するために使用されるベクターを提供する。本発明にかかるベクターは、インビトロ翻訳に用いるベクターであっても組換え発現に用いるベクターであってもよい。
【0097】
本発明にかかるベクターは、上述した本発明にかかるポリヌクレオチドを含むものであれば、特に限定されない。例えば、本発明にかかるポリペプチドをコードするポリヌクレオチドのcDNAが挿入された組換え発現ベクターなどが挙げられる。組換え発現ベクターの作製方法としては、プラスミド、ファージ、またはコスミドなどを用いる方法が挙げられるが特に限定されない。
【0098】
ベクターの具体的な種類は特に限定されず、宿主細胞中で発現可能なベクターを適宜選択すればよい。すなわち、宿主細胞の種類に応じて、確実に本発明にかかるポリヌクレオチドを発現させるために適宜プロモーター配列を選択し、これと本発明にかかるポリヌクレオチドを各種プラスミド等に組み込んだベクターを発現ベクターとして用いればよい。
【0099】
発現ベクターは、好ましくは少なくとも1つの選択マーカーを含む。このようなマーカーとしては、真核生物細胞培養についてはジヒドロ葉酸レダクターゼまたはネオマイシン耐性、およびE.coliおよび他の細菌における培養についてはテトラサイクリン耐性遺伝子またはアンピシリン耐性遺伝子が挙げられる。
【0100】
上記選択マーカーを用いれば、本発明にかかるポリヌクレオチドが宿主細胞に導入されたか否か、さらには宿主細胞中で確実に発現しているか否かを確認することができる。あるいは、本発明にかかるポリペプチドを融合ポリペプチドとして発現させてもよく、例えば、オワンクラゲ由来の緑色蛍光ポリペプチドGFP(Green Fluorescent Protein)をマーカーとして用い、本発明にかかるポリペプチドをGFP融合ポリペプチドとして発現させてもよい。
【0101】
上記の宿主細胞は、特に限定されるものではなく、従来公知の各種細胞を好適に用いることができる。具体的には、例えば、大腸菌(Escherichia coli)等の細菌、酵母(出芽酵母Saccharomyces cerevisiae、分裂酵母Schizosaccharomyces pombe)、線虫(Caenorhabditis elegans)、アフリカツメガエル(Xenopus laevis)の卵母細胞等を挙げることができるが、特に限定されるものではない。上記の宿主細胞のための適切な培養培地および条件は当分野で周知である。
【0102】
上記発現ベクターを宿主細胞に導入する方法、すなわち形質転換法も特に限定されるものではなく、電気穿孔法、リン酸カルシウム法、リポソーム法、DEAEデキストラン法等の従来公知の方法を好適に用いることができる。また、例えば、本発明にかかるポリペプチドを昆虫で転移発現させる場合には、バキュロウイルスを用いた発現系を用いればよい。
【0103】
このように、本発明にかかるベクターは、少なくとも、本発明にかかるポリペプチドをコードするポリヌクレオチドを含めばよいといえる。すなわち、発現ベクター以外のベクターも、本発明の技術的範囲に含まれる点に留意すべきである。
【0104】
つまり、本発明の目的は、本発明にかかるポリペプチドをコードするポリヌクレオチドを含有するベクターを提供することにあるのであって、本明細書中に具体的に記載した個々のベクター種および細胞種、ならびにベクター作製方法および細胞導入方法に存するのではない。したがって、上記以外のベクター種およびベクター作製方法を用いて取得したベクターも本発明の技術的範囲に属することに留意しなければならない。
【0105】
(3−4)形質転換体または細胞
本発明は、上述した本発明にかかるポリペプチドをコードするポリヌクレオチドが導入された形質転換体または細胞を提供する。ここで「形質転換体」とは、組織または器官だけでなく、生物個体を含むことを意味する。
【0106】
形質転換体または細胞の作製方法(生産方法)は特に限定されるものではないが、例えば、上述した組換えベクターを宿主に導入して形質転換する方法を挙げることができる。また、形質転換の対象となる生物も特に限定されるものではなく、上記宿主細胞で例示した各種微生物、植物または動物を挙げることができる。
【0107】
本発明にかかる形質転換体または細胞は、海藻類もしくはその子孫、またはこれら由来の組織であることが好ましく、トサカノリ(Meristotheca papulosa (Montagne) J. Agardh)であることが特に好ましい。
【0108】
本発明にかかるポリペプチドをコードするポリヌクレオチドを含む形質転換体は、当該ポリヌクレオチドを含む組換えベクターを、当該遺伝子が発現し得るように宿主細胞中に導入することにより得ることができる。
【0109】
このように、本発明にかかる形質転換体または細胞は、少なくとも、本発明にかかるポリペプチドをコードするポリヌクレオチドが導入されていればよいといえる。すなわち、組換え発現ベクター以外の手段によって生成された形質転換体または細胞も、本発明の技術的範囲に含まれる点に留意すべきである。
【0110】
本発明の目的は、本発明にかかるポリペプチドをコードするポリヌクレオチドが導入されていることを特徴とする形質転換体または細胞を提供することにあるのであって、本明細書中に具体的に記載した個々のベクター種および導入方法に存するのではない。したがって、上記以外のベクター種および細胞種、ならびにベクター作製方法および細胞導入方法を用いて取得した形質転換体または細胞も本発明の技術的範囲に属することに留意しなければならない。
【0111】
(3−5)ポリペプチドの生産方法
本発明は、本発明にかかるポリペプチドを生産する方法を提供する。
【0112】
一実施形態において、本発明にかかるポリペプチドの生産方法は、本発明にかかるポリペプチドをコードするポリヌクレオチドを含むベクターを用いることを特徴とする。
【0113】
本実施形態の1つの局面において、本実施形態にかかるポリペプチドの生産方法は、上記ベクターを無細胞タンパク質合成系に用いることが好ましい。無細胞タンパク質合成系を用いる場合、種々の市販のキットを用いればよい。好ましくは、本実施形態にかかるポリペプチドの生産方法は、上記ベクターと無細胞タンパク質合成液とをインキュベートする工程を包含する。
【0114】
本実施形態の他の局面において、本実施形態にかかるポリペプチドの生産方法は、組換え発現系を用いることが好ましい。組換え発現系を用いる場合、本発明にかかるポリヌクレオチドを組換え発現ベクターに組み込んだ後、公知の方法により発現可能な宿主に導入し、宿主内で翻訳されて得られる上記ポリペプチドを精製するという方法などを採用することができる。組換え発現ベクターは、プラスミドであってもなくてもよく、宿主に目的ポリヌクレオチドを導入することができればよい。好ましくは、本実施形態にかかるポリペプチドの生産方法は、上記ベクターを宿主に導入する工程を包含する。
【0115】
このように宿主に外来ポリヌクレオチドを導入する場合、発現ベクターは、外来ポリヌクレオチドを発現するように宿主内で機能するプロモーターを組み込んであることが好ましい。組換え的に産生されたポリペプチドを精製する方法は、用いた宿主、ポリペプチドの性質によって異なるが、タグの利用等によって比較的容易に目的のポリペプチドを精製することが可能である。
【0116】
本実施形態にかかるポリペプチドの生産方法は、本発明にかかるポリペプチドを含む細胞または組織の抽出液から当該ポリペプチドを精製する工程をさらに包含することが好ましい。
【0117】
ポリペプチドを精製する工程は、周知の方法(例えば、細胞または組織を破壊した後に遠心分離して可溶性画分を回収する方法)で細胞や組織から細胞抽出液を調製した後、この細胞抽出液から周知の方法(例えば、硫安沈殿またはエタノール沈殿、酸抽出、陰イオンまたは陽イオン交換クロマトグラフィー、ホスホセルロースクロマトグラフィー、疎水性相互作用クロマトグラフィー、アフィニティークロマトグラフィー、ヒドロキシアパタイトクロマトグラフィー、およびレクチンクロマトグラフィー)によって精製する工程が好ましいが、これらに限定されない。最も好ましくは、高速液体クロマトグラフィー(「HPLC」)が精製のために用いられる。
【0118】
別の実施形態において、本発明にかかるポリペプチドの生産方法は、本発明にかかるポリペプチドを天然に発現する細胞または組織から当該ポリペプチドを精製することを特徴とする。本実施形態にかかるポリペプチドの生産方法は、上述したオリゴヌクレオチドを用いて、本発明にかかるポリペプチドを天然に発現する細胞または組織を同定する工程を包含することが好ましい。また、本実施形態にかかるポリペプチドの生産方法は、上述したポリペプチドを精製する工程をさらに包含することが好ましい。
【0119】
さらに他の実施形態において、本発明にかかるポリペプチドの生産方法は、本発明にかかるポリペプチドを化学合成することを特徴とする。当業者は、本明細書中に記載される本発明にかかるポリペプチドのアミノ酸配列に基づいて周知の化学合成技術を適用すれば、本発明にかかるポリペプチドを化学合成できることを、容易に理解する。
【0120】
以上のように、本発明にかかるポリペプチドを生産する方法によって取得されるポリペプチドは、天然に存在する変異ポリペプチドであっても、人為的に作製された変異ポリペプチドであってもよい。
【0121】
変異ポリペプチドを作製する方法についても、特に限定されるものではない。例えば、部位特異的変異誘発法(例えば、Hashimoto-Gotoh, Gene, 152,271-275(1995)参照)、PCR法を利用して塩基配列に点変異を導入し変異ポリペプチドを作製する方法、またはトランスポゾンの挿入による突然変異株作製法などの周知の変異ポリペプチド作製法を用いることによって、変異ポリペプチドを作製することができる。変異ポリペプチドの作製には市販のキットを利用してもよい。
【0122】
このように、本発明にかかるポリペプチドの生産方法は、少なくとも、本発明にかかるポリペプチドのアミノ酸配列、または本発明にかかるポリペプチドをコードするポリヌクレオチドの塩基配列に基づいて公知慣用技術を用いればよいといえる。
【0123】
つまり、本発明の目的は、本発明にかかるポリペプチドの生産方法を提供することにあるのであって、上述した種々の工程以外の工程を包含する生産方法も本発明の技術的範囲に属することに留意しなければならない。
【実施例】
【0124】
以下、実施例に基づいて本発明をより詳細に説明するが、本発明は以下の実施例に限定されるものではない。
【0125】
〔実施例1:トサカノリからのポリペプチド(MPL)の単離〕
(抽出液の調製および硫安沈殿)
九州西岸にて採集したトサカノリ(Meristotheca papulosa (Montagne) J. Agardh)の凍結藻体500gを液体窒素下で粉末とした。これに500mlの20mM PBSA(Phosphate-buffered saline sodium azide;0.02%アジ化ナトリウム入り20mM PBS;pH7.0)を加え、4℃で一晩撹拌した後、遠心分離を行なって一次抽出液を得た。残渣を700mlの20mM PBSAで同様に抽出し、二次抽出液を得た。両抽出液を合一し、UV280nmの吸収および凝集活性を測定した。
【0126】
上記抽出液に硫安粉末を20%飽和となるように少しずつ撹拌しながら加え、この混合液を4℃で一晩静置した。これを遠心分離(8500rpm、30分間)して得られた沈殿を、PBSAに溶解後、同溶媒に対し十分透析した。透析終了後、内液を遠心分離し、得られた上清を20%飽和硫安塩析沈殿画分とした。一方、20%飽和硫安塩析処理で得られた上清に、硫安粉末を60%飽和となるように加え、同様に処理して、20〜60%飽和硫安塩析沈殿画分を得た。当該画分につき、UV280nmの吸収および凝集活性を測定した。
【0127】
(疎水クロマトグラフィー)
上記20〜60%飽和硫安塩析沈殿画分106mlのうち、10mlについて、20mMトリス−塩酸緩衝液(pH7.0)に対して透析を行ない、内液6mlを得た。得られた内液を遠心分離(10,000 rpm、10分間)して上清を回収した。得られた上清5mlを同緩衝液で平衡化したHiPrep Phenyl FFカラム(16×100mm, Vt=20 ml)に注入し、20mMトリス−塩酸緩衝液,pH7.0(溶媒A)と、蒸留水(溶媒B)の2液を用いる濃度勾配法により溶出した。濃度勾配[溶媒B100%(10分)、溶媒:B0%−溶媒A100%(10−60分)、溶媒A100%(60−110分)]は、グラジェントプログラマー(CCPコントローラー、東ソー社製)を用いて設定し、流速は4ml/分とした。溶出液は5mlずつ分取し、UV280nmの吸光度を測定するとともに凝集活性を測定した。
【0128】
(ゲルろ過)
疎水クロマトグラフィーで分取した活性画分を合一し、限外濾過により5mlに濃縮した。そのうち1mlを、0.3 M NaCl を含む20mM PB (pH 7.0)で平衡化したTSKgel G3SWxLゲルろ過カラム(東ソー社製、7.8 x 300 mm、Vt=14.7 ml)に供した。20mMトリス−塩酸緩衝液(pH 7.0)を用いて流速1ml/minで溶出した。溶出液は1mlずつ分取し、各フラクションのUV280nmの吸光度および凝集活性を測定した。
【0129】
(凝集活性の検討)
赤血球凝集活性は、マイクロタイター法を用いて測定した。生理食塩水にて調製した各精製画分溶液の2倍段階希釈液、各25μlをマイクロタイタープレート上に作製した。各希釈液に2%赤血球浮遊液25μlを加えて軽く撹拌し、室温で1.5時間静置後、凝集能を観察した。凝集能は肉眼で判定し、赤血球の50%以上が凝集している場合を陽性とした。凝集活性は、赤血球凝集活性(力価)、すなわち凝集活性を示す最大希釈液の希釈倍数の逆数、および凝集素価(力価)、すなわち凝集活性を示す最大希釈液のタンパク質の濃度(最小凝集濃度)で示した。
【0130】
本実施例においては、赤血球としてトリプシン処理ウサギ赤血球(TRBC)を用いた。なお、赤血球浮遊液の調製は次のように行なった。まず、実験室で飼育中のウサギの耳から血液2mlを採取し、これを約50mlの生理食塩水で3回洗浄後、50mlの生理食塩水を加えて2%のウサギ赤血球浮遊液を調製した。これに1/10容の0.5%トリプシン−生理食塩水を加え、37℃で1.5時間静置した。このトリプシン処理赤血球を生理食塩水で3回洗浄後、45mlの生理食塩水を加え、トリプシン処理2%ウサギ赤血球浮遊液(TRBC)とした。
【0131】
トリプシン処理ウサギ赤血球(TRBC)に対する凝集活性成分を検討したところ、表1に示すように、上記抽出液、上記20〜60%飽和硫安塩析沈殿画分、上記疎水クロマトグラフィーによって得られた画分、および上記ゲルろ過によって得られた画分に強い赤血球凝集活性が検出された。
【0132】
【表1】
【0133】
表1中、「タンパク質濃度」は、「a」を付したものについてはUV280nmの吸収(A280)を測定し、A280(1mg/ml)=1.0より算出した。「b」を付したものについてはUV280nmの吸収(A280)を測定し、本発明にかかるポリペプチドのアミノ酸配列から求めた分子吸光係数を用いてA280(1mg/ml)=0.97より算出した。
【0134】
(SDS−PAGE)
上記ゲルろ過で得られた凝集活性を有する画分(精製画分)をSDS−PAGE(10%ゲル)に供した。結果を図3に示す。レーン1は分子量マーカー、レーン2は20〜60%飽和硫安塩析沈殿画分(非還元下)、レーン3は20〜60%飽和硫安塩析沈殿画分(還元下)、レーン4は上記精製画分(非還元下)、レーン5は上記精製画分(還元下)を示す。なお、タンパク染色はCBB(Coomassie brilliant blue R-250)染色を行なった。
【0135】
SDS−PAGEの結果、精製画分は、還元下で14kDaの、非還元下で26kDaの単一バンドを与えるものであることが確認されたことから、同一サブユニット(14kDa)がジスルフィド結合(S−S結合)した2量体からなると推定される。以上のように、上記抽出液を硫安塩析、疎水クロマトグラフィーおよびゲルろ過に順次供した精製過程により最終的に0.4mgの精製画分が得られた。表1に示す収量6mgは藻体500g(湿重量)当たりに換算した値である。本発明者らは、当該精製画分を「MPL」と命名した。
【0136】
なお、本発明者は、上記MPLは、上記20〜60%飽和硫安塩析沈殿画分を陰イオン交換クロマトグラフィーに供することによって効果的に調製できることも確認した。陰イオン交換クロマトグラフィーによって、トサカノリに含まれる他のレクチン(MPAと称する)と上記MPLとを効果的に分離することができる。
【0137】
上記陰イオン交換クロマトグラフィーの条件を以下に示す。
【0138】
MPAとMPLの両レクチンを同時に精製することを意図して、先に調製した20〜60%飽和硫安塩析沈殿画分を、HiPrep QXL 16/10カラム(φ1.6×10 cm、Vt = 20 ml、GE Healthcare社)を用いる陰イオン交換クロマトグラフィーに供した。
【0139】
すなわち、上記カラムを0.02M トリス−塩酸緩衝液(pH 8.0)で平衡化し、20〜60%飽和硫安塩析沈殿画分5mlを添加した。カラムを上記緩衝液で十分洗浄した後、上記緩衝液中0M〜1M NaClの濃度勾配で溶出した。流速は5ml/minとした。洗浄液は10mlずつ、濃度勾配溶出液は5mlずつ分取した。
【0140】
各フラクションにつき、A280吸光度および赤血球凝集活性を測定した。赤血球凝集活性が見られた画分は、還元下SDS-PAGEに付した。その結果より、各フラクションを5つの画分に分けた。各画分につき、Folin-Lowry法によるタンパク質量の測定および赤血球凝集活性試験に供した。
【0141】
その結果、両レクチンはカラムに吸着し、MPAは0.37M NaCl付近、MPLは0.57M NaCl付近にそれぞれ活性ピークとして溶出した。このように、陰イオンクロトマトグラフィーを用いることにより、20〜60%飽和硫安塩析沈殿画分に混在する2種類のレクチン、MPA(29kDa)とMPL(26kDa)とを分離することができる。
【0142】
〔実施例2:MPLのcDNAのクローニング〕
以下に示すように、実施例1で得られた精製画分「MPL」の14kDaサブユニットの23N末端アミノ酸配列を、プロテインシーケンサー(Procise 492HT, Applied Biosystems)を用いてGVVDQLIVTYSDGTRVSHGQPGS(配列番号12)と決定した。当該配列の情報を参考に縮重プライマーを設計した。別途調製したトサカノリ由来1st strand cDNA溶液を鋳型にRapid Amplification of cDNA Ends(RACE)法を行うことにより、MPLのcDNAの全長塩基配列 949bpを明らかにした。
【0143】
まず、MPLの上記23N末端のアミノ酸配列を参考にして、MPL_d_R1およびMPL_d_R2の各プライマーを作製した。
【0144】
MPL_d_R1の塩基配列は、CCRTGISWIACICKIGTICCRTC(配列番号3)であり、MPL_d_R2の塩基配列は、TAIGTIACDATIARYTGRTC(配列番号4)である。ここで、DはG、AまたはTを、Iはイノシンを、KはGまたはTを、RはAまたはGを、YはCまたはTを、WはAまたはTを示すIUBコードである。
【0145】
RNAlater 中に−20℃で保存したトサカノリ藻体より Plant RNA Isolation Reagent(Life Technologies Corp.)を用いて全RNAを抽出後、NucleoTrap mRNA(Macherey-Nagel)によりmRNAを精製し、さらに GeneRacer Kit(Life Technologies Corp.)により完全長cDNAを調製した。
【0146】
次に、上記完全長cDNAを鋳型に GeneRacer_5'_Primer および MPL_d_R1のプライマーペアを用いてPCRを行った。GeneRacer_5'_Primer の塩基配列はCGACTGGAGCACGAGGACACTGA(配列番号5)である。
【0147】
上記PCRにより得られた産物の100倍希釈溶液を鋳型に、GeneRacer_5’_Nested_PrimerおよびMPL_d_R2のプライマーペアを用いてNested PCRを行った。GeneRacer_5’_Nested_Primerの塩基配列は、GGACACTGACATGGACTGAAGGAGTA(配列番号6)である。
【0148】
得られたNested PCRの産物を低融点アガロースにより精製後、pGEM-T Easy Vector System(PROMEGA製)を用いてサブクローニングを行い、得られたクローンから精製プラスミドを回収して、ダイデオキシ法により塩基配列の決定を行った(以上5’RACE)。
【0149】
得られた塩基配列に基づいて新たにプライマーMPL_F1を作製した。MPL_F1の塩基配列はTGTAGCAGCCATGTCTTTGC(配列番号7)である。
【0150】
上記完全長cDNAを鋳型として、MPL_F1およびGeneRacer_3’_Primerのプライマーペアを用いてPCRを行った。得られた増幅産物を低融点アガロースにより精製後、pGEM-T Easy Vector System(PROMEGA製)を用いてサブクローニングを行い、得られたクローンから精製プラスミドを回収して、ダイデオキシ法により塩基配列の決定を行った(以上、3’RACE)。
【0151】
GeneRacer_3’_Primerの塩基配列はGCTGTCAACGATACGCTACGTAACG(配列番号8)である。
【0152】
上記5’RACEおよび3’RACEから明らかとなったMPL cDNAの5’および3’末端配列から作製したMPL_5'_End_FおよびMPL_3'_End_Rのプライマーペア、ならびに高正確性DNAポリメラーゼを用いてPCRを行い、増幅産物につき塩基配列を決定することで、MPL cDNAの全長塩基配列を確認した。
【0153】
MPL_5'_End_Fプライマーの塩基配列はAATCCACATTCAACTGCACTG(配列番号9)であり、MPL_3'_End_Rプライマーの塩基配列はAAATCGACGCACACAGAAGTC(配列番号10)である。
【0154】
MPLのcDNAの全長塩基配列、およびその演繹アミノ酸配列を図1に示す。図1において、星印は終止コドン、破線はシグナルペプチド(SignalP 3.0を用いて予測)、実線は実施例1で得られた精製画分「MPL」の14kDaサブユニットの23N末端配列を示す。
【0155】
演繹アミノ酸配列の解析から、図1に示すcDNAは、シグナルペプチド領域22残基(1−22)を含め、計198残基をコードすることがわかった。一方、演繹アミノ酸配列の解析から、図1に示すcDNAは、プロペプチド領域74残基(23−96)および成熟タンパク質領域102残基(97−198)をコードすると考えられたが、後述する実施例6においてさらに詳細に検討したところ、成熟タンパク質領域は53番目−198番目の146アミノ酸残基であることが分かった。
【0156】
〔実施例3:MPLの糖結合特異性〕
MPLの糖結合特異性を赤血球凝集阻止試験により明らかにした。本赤血球凝集阻止試験には、実施例1で調製した20〜60%飽和硫安塩析沈殿画分および実施例1で調製したMPLを供試し、これらの赤血球凝集活性(力価4)を阻止する糖化合物を検索した。
【0157】
赤血球凝集阻止試験は以下のようにして行った。まず生理食塩水にて調製した糖溶液の2倍段階希釈液、各25μlをマイクロタイタープレート上に作製した。なお、使用した糖類の原液の濃度は単糖類および二糖類の場合は100mM、糖タンパク質の場合は2mg/mlとした。これに、それぞれ凝集素価(力価)4に調整した上記飽和硫安塩析沈殿画分の溶液およびMPL溶液、各25μlを加えて軽く撹拌後、室温で1.5時間静置した。これに25μlのTRBCを加え、室温で2時間静置後、凝集阻止能を観察した。
【0158】
凝集阻止能の有無は肉眼で判定し、赤血球の約100%が凝集していない場合を陽性とした。凝集阻止能(凝集阻止活性)は、最小阻止濃度すなわち凝集阻止能を示す最小濃度(mMまたはμg/ml)で表示した。結果を表2に示した。表2において、「最小凝集阻止濃度」は、上述の力価4の検液の赤血球凝集活性を阻止する糖化合物の最小濃度を表し、「NT」は未測定であることを表す。
【0159】
【表2】
【0160】
表2に示すように、本赤血球凝集阻止試験には、単糖類および二糖類としてD−グルコース(表中Glcと表示。以下、糖の名称に続く括弧内には、表2に示した略称を記載する。)、D−マンノース(Man)、D−ガラクトース(Gal)、N−アセチル−D−グルコサミン(GlcNAc)、N−アセチル−D−ガラクトサミン(GalNAc)、N−アセチルノイラミン酸(NeuAc)、L−フコース(Fuc)、D−キシロース(Xyl)、L−ラムノース(Rha)、ラクトースを用いた。
【0161】
糖タンパク質としては、複合型糖鎖を含有するものとして、トランスフェリン、アシアロトランスフェリンを用いた。また、高マンノース型糖鎖を含有する糖タンパク質として、イーストマンナンを用い、複合型糖鎖および高マンノース型糖鎖を含有する糖タンパク質として、ブタチログロブリン、アシアロブタチログロブリンを用いた。
【0162】
表2から明らかなように、MPLの赤血球凝集活性は供試した単糖類および二糖類では阻止されず、高マンノース型糖鎖を含む糖タンパク質、並びに、複合型糖鎖および高マンノース型糖鎖を含む糖タンパク質によってのみ阻止された。このように、MPLは従来公知のレクチンとの配列相同性が非常に低く、かつ、高マンノース型糖鎖に高い親和性を有することが示された。
〔実施例4:エイズウイルスの表面糖タンパク質・gp120とMPLとの相互作用〕
HIVの宿主への感染は、HIVの表面糖タンパク質であるgp120と宿主リンパ細胞の表面レセプターCD4との相互作用により成立する。該相互作用にはgp120の高マンノース型糖鎖が不可欠である。そのため、高マンノース型糖鎖に特異的に結合するレクチンはHIV感染の有力な阻害剤となりうる。
【0163】
そこで、MPLと市販品のリコンビナント糖付加HIV-1 IIIB gp120 (バキュロウイルス;1分子あたり15本の高マンノース型糖鎖を有する。ImmunoDiagnostics製)との相互作用を表面プラズモン共鳴(SPR)法を用いて定量解析した。SPR法にはBIAcore2000(GE Healthcare社製)を用いて、リコンビナント糖付加HIV-1 IIIB gp120をCM5センサーチップ(GE Healthcare社製)上に固定化し、MPLの溶液をアナライトとして用いて、マニュアルに従って測定、解析した。
【0164】
上記固定化はマニュアルに従って行った。具体的には、アミンカップリング法を用いて固定化を行った。なお、固定化量は300〜400RUの範囲になるよう、マニュアルインジェクション法によって調整した。
【0165】
SPR法では、生体分子を標識することなく、生体分子間の特異的な相互作用を微量かつ短時間で定量的に測定できる。本法では、リガンドをセンサーチップ表面上に固定化し、これに作用する物質(アナライト)を含む溶液を添加すると、分子の結合・解離により生ずる微量の質量変化がSPRシグナルの変化として検出される。
【0166】
質量変化はレゾナンスユニット(RU)で表され、1000RUは共鳴による反射角度0.1°の変化に相当し、アナライトがリガンドに 1 ng/mm2 結合したことを意味する。センサーチップ表面の金薄膜上にはデキストランがコーティングされており、主としてこのデキストラン内に導入されたカルボキシル基を介してリガンドを固定化する。
【0167】
MPLと上記リコンビナントgp120とのSPR法による親和性解析に先立ち、予備実験で解析法を検討し、非線形最小二乗法によるカイネティクス解析が適当と判断した。そこで、得られたセンサーグラムからおおよそのKD値を算出し、0.1〜10KD[M]を濃度の目安として、2倍希釈列で5段階以上のアナライト(MPL)溶液を調製した。
【0168】
分析プログラムの作成には、マニュアルに従って“Customaized Application”を用い、センサーチップ内の4つのフローセルのうち、何も固定化していないフローセル1をコントロールとして、リコンビナントgp120を固定化したフローセルからの差し引き機能を使用した。本分析プログラム下で、アナライト(MPL)溶液をそれぞれセンサーチップ上に流速30μl/minで3分間流した後、バッファーを3分間流し、レクチンの結合・解離量を測定した。なお、アナライト添加開始後5秒から添加終了5秒前までのRUの増加量を結合量、バッファー添加開始後10秒から添加終了10秒前までのRU減少量を解離量とした。
【0169】
次に、100mM HClおよび100mM NaOHを用いて、センサーチップを洗浄して再生した。得られたセンサーグラムについて、結合相と解離相を同時にカーブフィッティングさせ、結合速度定数ka、解離速度定数kd、親和定数KA、および解離定数KDを算出した。
【0170】
結果を図2に示す。図2のAは固定化したgp120とMPLとの相互作用のセンサーグラムであり、図2のBはgp120とMPLとの親和定数を示す。図2のBには、結合速度定数Ka(M-1s-1)、解離速度定数Kd(s-1)、親和定数KA(M-1)、および解離定数KD(M)を示した。なお親和定数は、その値が大きくなればなるほど、親和性が高い(結合力が強い)ことを意味する。
【0171】
図2に示すように、MPLはgp120に強い親和性(KD= 4.50×10-9 M)を持つことが認められた。このことから、MPLは、従来公知のレクチンとの配列相同性が非常に低い上に、HIVのgp120と宿主リンパ細胞のCD4との相互作用を十分に阻害できるといえる。それゆえ、MPLは新たな抗HIV剤として有望である。
【0172】
また、実施例3に示したように、MPLは高マンノース型糖鎖に高い親和性を有する。そのため、MPLは抗HIV剤として有望である。また、高マンノース型糖鎖を有する糖タンパク質が宿主のレセプターと結合することによって感染するタイプのウイルスに対しても有力な感染阻害剤となりうる。
【0173】
〔実施例5:MPLのさらなる精製〕
(実施例5−1:陰イオン交換クロマトグラフィーによるトサカノリレクチンの精製)
実施例1で述べたように、MPAとMPLとの分離に陰イオン交換クロマトグラフィーが有用であることが判明したので、陰イオン交換クロマトグラフィーによる精製についてさらなる検討を行い、精製方法を改良した。
【0174】
実施例1で調製した20〜60%飽和硫安塩析沈殿画分68mlを、20mMのトリス塩酸緩衝液(pH8.0)(以下、当該緩衝液を「TB」と称する)で平衡化したHiPrep QXL 16/10カラム(φ1.6×10 cm、Vt = 20 ml、GE Healthcare社)に添加し、カラムをTBで十分洗浄後、TB中0.2Mおよび1M NaClで順次、段階的に溶出した(ステップワイズ溶出)。その結果、活性成分は0.2M NaCl溶出画分に認められた。
【0175】
図4は、20〜60%飽和硫安塩析沈殿画分を、HiPrep QXL 16/10カラムを用いた陰イオン交換クロマトグラフィー(ステップワイズ溶出)に供した結果を示す図である。
【0176】
図4中、横軸はフラクション番号であり、縦軸は左側が280nmの吸光度を表し、右側が赤血球凝集活性(力価)を表す。図中、当該吸光度測定結果を丸印で示し、赤血球凝集活性(力価)の測定結果を三角印で示した。
【0177】
括弧書き数字1は、上記カラムに20〜60%飽和硫安塩析沈殿画分を添加し、非吸着成分を洗浄後、TB中0.2M NaClにより吸着成分の溶出を開始した位置を示している。括弧書き数字2は、上記溶出後、TB中1M NaClにより他の吸着成分の洗浄を開始した位置を示している。
【0178】
図4に示すように、赤血球凝集活性成分は0.2M NaCl溶出画分に認められた。この赤血球凝集活性成分(粗レクチン画分)70mlを、TBで平衡化したHiPrep QXL 16/10カラムに添加し、カラムをTBで洗浄後、TB中NaClの濃度勾配(0〜0.2M)で溶出させた(グラジエント溶出法)。
【0179】
溶出液は280nmの吸光度をモニターするとともに、2mlずつ分取し、各画分の赤血球凝集活性を測定した。図5は、上記粗レクチン画分を、HiPrep QXL 16/10カラムを用いた陰イオン交換クロマトグラフィー(グラジエント溶出)に供した結果を示す図である。図5に実線で示すように、上記溶出液は、凝集活性を示す3つのタンパク質のピークに分離した。
【0180】
図5において、横軸はフラクション番号である。左側の縦軸は280nmの吸光度を表し、吸光度の測定結果は実線で示されている。右側の縦軸では、「HA」が赤血球凝集活性(力価)を表し、赤血球凝集活性(力価)の測定結果は三角印で示されている。右側の縦軸のうち、”NaCl in TB”は、NaCl(TB中)の濃度勾配を示し、図中、濃度勾配は点線で示されている。
【0181】
上記溶出液を、図5に示す4つの画分(I〜IV)に分画し、各画分とも、20μgのタンパク質に相当する量をSDS−PAGE(10%ゲル)に供した。結果を図6に示す。なお、タンパク染色はCBB(Coomassie brilliant blue R-250)染色を行なった。
【0182】
図6のAは非還元下における泳動結果を示し、図6のBは還元下における泳動結果を示す。Mは分子量マーカー、レーン1は20〜60%飽和硫安塩析沈殿画分、レーン2はHiPrep QXL 16/10カラム(ステップワイズ溶出)で得られた赤血球凝集活性成分(0.2M NaCl溶出画分)、レーン3は画分I、レーン4は画分II、レーン5は画分III、レーン6は画分IVを示す。なお、上記赤血球凝集活性成分は、「粗レクチン画分」とも称される。
【0183】
画分IおよびIIは、図6のBに示すように、還元下で29kDaのほぼ単一のバンドを与えることが確認された。そこで、画分IをMPA−1、画分IIをMPA−2と命名し、最終精製レクチンとした。
【0184】
MPA−1およびMPA−2は、詳細は示さないが、分子量測定、N末端アミノ酸配列解析およびcDNAクローニングの結果から、実施例1で述べたMPA(29kDa)のアイソフォームであることが分かった。
【0185】
実施例1で述べたMPLは画分IVに溶出した。画分IVは、非還元下では26kDaおよび55kDaの2つのバンド成分を与え(図6のA)、還元下では14kDaの主要バンド成分と、8kDaおよび5kDaの微量バンド成分を与えた。なお、画分IIIは還元下で29kDaおよび14kDaのバンド成分を与えることから、MPLとMPAとを混有すると考えられる。
【0186】
(実施例5−2:陰イオン交換クロマトグラフィーによるMPL−1およびMPL−2の単離)
上記MPL画分(画分IV)5mlを、TBで平衡化したTSKgel DEAE-5PWカラム(7.5×75 mm、東ソー社)に添加し、TB中NaClの濃度勾配(0−0.2M)によるグラジエント溶出を行った。操作中、280nmの吸光度をモニターしながらピークをそれぞれ分取した。
【0187】
図7は、MPLのTSKgel DEAE-5PWカラムを用いた陰イオン交換クロマトグラフィーによる精製結果を示す図である。図中、横軸は溶出時間を表し、左側の縦軸は280nmの吸光度、右側の縦軸はNaClの濃度を示す。また、実線は280nmの吸光度の測定結果を示し、点線はNaClの濃度を示す。(1)〜(4)はピーク番号である。
【0188】
TSKgel DEAE-5PWカラムによる陰イオン交換クロマトグラフィーの結果、図7に示すように、MPLは4つのピーク(ピーク(1)〜(4))に分離された。そこで次に、ピーク(1)〜(4)をそれぞれ分取後、20μgのタンパク質に相当する量をSDS−PAGE(10%ゲル)に供した。結果を図8に示す。
【0189】
図中、「非還元」は非還元下における泳動結果を示し、「還元」は還元下における泳動結果を示す。Mは分子量マーカーを示し、SはMPL、レーン1〜4はそれぞれピーク(1)〜(4)を泳動させた結果を示す。
【0190】
還元下において、ピーク(1)および(3)で14kDaのほぼ単一のバンドが認められた。ピーク(1)および(3)は、非還元下において26kDa付近のタンパク質性バンドを与えた。
【0191】
ピーク(2)および(4)については、非還元下で55kDa、還元下では14kDaの他に8kDaおよび5kDaのペプチドの存在が確認された。全ピークに共通して非還元下で26kDaおよび55kDaにタンパク質性バンドの存在が認められたが、ピーク(1)および(3)では26kDa成分、ピーク(2)および(4)では55kDa成分を主成分としていることがわかった。
【0192】
そこで、ピーク(1)をMPL−1、ピーク(3)をMPL−2と命名し、最終精製標品とした。MPL−1およびMPL−2につき、LTQ Orbitrap XL(Thermo Fisher Scientific)を用いる ESI-MSにより分子量の測定を行った。図9はMPL−1およびMPL−2の分子量測定結果を示すものである。
【0193】
図9のA,Bより、MPL−1およびMPL−2の分子量はそれぞれ31,104.1 Daおよび31,132.0 Daであることが分かった。
【0194】
MPL−1およびMPL−2について、実施例3と同様の方法により、赤血球凝集阻止試験を行った。結果を表3に示す。
【0195】
【表3】
【0196】
表3では「単糖類および二糖類」と記載しているが、これは、実施例3と同様に、単糖類としてD−グルコース(Glc)、D−マンノース(Man)、D−ガラクトース(Gal)、N−アセチル−D−グルコサミン(GlcNAc)、N−アセチル−D−ガラクトサミン(GalNAc)、N−アセチルノイラミン酸(NeuAc)、L−フコース(Fuc)、D−キシロース(Xyl)またはL−ラムノース(Rha)を各100mMで用い、二糖類としてラクトース(Lac)を100mM用いたことを示す。
【0197】
複合型糖鎖を含有する糖タンパク質、高マンノース型糖鎖を含有する糖タンパク質、並びに、複合型糖鎖および高マンノース型糖鎖を含有する糖タンパク質としては、実施例3と同じものを用いた。加えて、N型糖鎖(複合型)およびO型糖鎖を含有する糖タンパク質としてフェツイン、アシアロフェツインを用い、O型糖鎖を含有する糖タンパク質としてウシ顎下線ムチン、アシアロウシ顎下線ムチンを用いた。
【0198】
表3から明らかなように、MPL−1およびMPL−2の赤血球凝集活性は、実施例3に示したMPLと同様に、高マンノース型糖鎖を含む糖タンパク質、並びに、複合型糖鎖および高マンノース型糖鎖を含む糖タンパク質によってのみ阻止された。このように、MPL−1およびMPL−2は、MPLと同様に、高マンノース型糖鎖に高い親和性を有することが示された。
【0199】
〔実施例6:MPLの構造解析〕
(実施例6−1:MPL画分のブロッティング)
実施例1で得られたMPL画分(26kDa)を非還元下および還元下(2-メルカプトエタノール添加)で12%トリス-トリシン系SDS−PAGEに付し、Polyvinylidene difluoride(PVDF)膜(Immobilon-P(登録商標)、Millipore)へ転写した。
【0200】
PVDF膜への転写は、セミドライブロッティング装置(AE-6677:ATTO CORPORATION)を用いてブロッティング緩衝液(CAPS buffer:メタノール:超純水= 1:1:8)中、144mAの定電流で1時間通電することによって行った。
【0201】
CAPS bufferはCAPS(3-Cyclohexylaminopropane sulfonic acid、和光純薬工業)11.1gを450mlの超純水に溶解し、NaOHを用いてpH11.0に調整した後、超純水で550 mlにメスアップしたものを使用した。転写後のPVDF膜はCBB染色を行い、一晩風乾後、目的とする14kDa成分および8kDaの成分を切り出して、実施例2同様に、Procise(登録商標)492HTを用いて、N末端アミノ酸配列を解析した。
【0202】
図10は、MPL画分をブロッティングし、CBB染色したPVDF膜を示す図である。図10において、Mは分子量マーカーを示し、「非還元」は非還元下において得られたバンド、「還元」は還元下において得られたバンドを示す。
【0203】
(実施例6−2:N末端アミノ酸配列の解析およびMPL−1のアミノ酸配列の決定)
図10に示す8kDaの成分のN末端アミノ酸配列はGVVDQLI(配列番号14)と決定された。この配列は、実施例2で決定された配列番号12に示すアミノ酸配列のN末端と一致した。
【0204】
一方、14kDaの成分については、配列が解析できず、N末端がブロッキングされていると推測された。そこで、ピログルタミル基によるN末端ブロッキングと予測し、脱ピログルタミル化を試みた。
【0205】
すなわち、14kDa成分を転写したPVDF膜を60%メタノール1mlで2回洗浄後、90%メタノール1mlで1回洗浄し、これを0.5%(w/v)ポリビニルピロリドン(PVP)-55含有100mM 酢酸500μlに浸漬し、37℃で30分間静置した。次に、蒸留水で膜を10回以上洗浄し、膜をメタノールで湿らせ、酵素消化を行った。
【0206】
酵素消化は、Pfu Pyroglutamate Aminopeptitase(TAKARA)2mUを、酵素反応緩衝液(10mM ジチオトレイトール(DTT)、1mM EDTAおよび10mM PB(pH 7.0))200μl中に溶解し、本溶液中に膜を浸漬後、50℃で5時間反応させることによって行った。本反応によりピログルタミル基を除去後、膜を蒸留水で3回洗浄し、Procise(登録商標)492 HTを用いて分析した。
【0207】
その結果、2残基以降の配列がTGSCNTFQRS(配列番号15)と決定された。これらの結果から、14kDaの成分のN末端はグルタミンがピログルタミル化することによりブロッキングされ、そのN末端アミノ酸配列は、QTGSCNTFQRS(配列番号16)であることがわかった。
【0208】
本配列は、図1に示すように、実施例2においてcDNAクローニングにより明らかとされたMPL cDNAの演繹アミノ酸配列中、実施例2でプロペプチドと予想された領域(23−96)の53番目の残基以降に認められた。図1において、今回決定した14 kDa成分のN末端アミノ酸配列をボックスで示す。
【0209】
MPL cDNAの演繹アミノ酸配列うち、53−198残基からなる領域の算出分子量は15,553.3 Daであるが、ESI−MSによるMPL−1およびMPL−2の測定分子量は、実施例5−2で求めたように、それぞれ31,104.1 Daおよび31,132.0 Daである。SDS−PAGEの結果(図6、8)から、MPLはS−S結合によるホモダイマーであると考えられることから、53−198残基からなる成分が1つのS−S結合により二量体を形成していると仮定すると、その算出分子量は31,104.6となり、MPL−1の測定分子量と一致する。
【0210】
これらの結果から、実施例2で得られたcDNAはMPL−1をコードするものであり、MPL−1は15.6kDaサブユニットのジスルフィド結合による二量体であることがわかった。
【0211】
すなわち、MPL−1のサブユニット(上記15.6kDaサブユニット)のcDNAの全長塩基配列は配列番号11に示される配列であり、当該塩基配列の演繹アミノ酸配列は配列番号13に示される配列である。また、MPL−1のサブユニットの成熟タンパク質をコードする塩基配列を配列番号1に示し、その演繹アミノ酸配列を配列番号2に示した。
【0212】
このことから、MPL−1は、高マンノース型糖鎖と結合するポリペプチドであって、(a)配列番号2に示されるアミノ酸配列;または(b)配列番号2に示されるアミノ酸配列において、1個もしくは数個のアミノ酸が置換、欠失、挿入、もしくは付加されたアミノ酸配列、からなるポリペプチドの二量体であるということができる。
【0213】
そして、当該二量体を含有する抗ウイルス剤も、高マンノース型糖鎖を備えるウイルスを標的とする抗ウイルス剤として用いることができる。また、当該二量体を例えば限外ろ過膜などに固定されてなる膜も、ウイルス除去膜として用いることができる。
【0214】
加えて、実施例2で得られたMPLのN末端アミノ酸配列(配列番号12)は8kDa成分のものであることがわかった。この8kDa成分は15.6kDa成分に由来することが明らかであるが、15.6kDa成分の分解産物であるか、MPLの構成成分として機能しているかどうかについてはさらなる解析が必要である。
【0215】
なお、ESI−MSによる測定分子量とSDS−PAGE上の推定分子量に差異がみられたが、SDS−PAGE によるタンパク質の移動度は、成分の性質(電荷、疎水性、構造等)により実際の分子量と数kDaのズレが生じることがある。
【0216】
そのため、SDS−PAGE上の成分について、例えば、SDS−PAGE上の推定分子量である14kDaと、ESI−MSによる測定分子量である15.6kDa成分とにずれがあることは特に問題はないと考えられる。
【0217】
Pfamプログラムによる分子内ドメイン検索にMPL−1を供したところ、MPL−1はJacalin-like lectinドメインを含み、紅藻 Griffithsia sp.由来レクチン Griffithsin(GRFT)およびバナナ Musa acuminataレクチンとの配列相同性(同一率)はそれぞれ33%および27%と非常に低いものであった。
【0218】
成熟タンパク質領域につきBasic Local Alignment Search Tool(BLAST)を用いて相同配列検索を行ったところ、上記2種のレクチンを含むJacalin-like lectinドメインを有するものが見られたが、その多くはhypothetical proteinであり、MPL−1と最も高い同一率を示したものは細菌Brevibacillus laterosporus由来hypothetical protein(35%)であった。
【0219】
このように、MPL−1には、部分的には既知レクチンのモチーフと類似するモチーフが存在することが示されたが、従来公知のレクチンとの配列相同性は非常に低く、全体配列から新規タンパク質であることが分かった。
【0220】
〔実施例7:MPL−1の糖鎖結合特異性〕
(実施例7−1:遠心限外ろ過−HPLC法)
上述のように精製されたMPL−1につき、糖鎖結合特異性を調べた。すなわち、まず、50mM トリス-塩酸緩衝液(pH7.0)中の500nM MPL−1溶液90μl(45pmol)と300nM ピリジルアミノ化(PA化)糖鎖水溶液10μl(3pmol)を軽く混合後、室温で60分間保温した。
【0221】
この反応液を微量遠心限外ろ過器(Nanosep 10K Omega、PALL)を用いて遠心ろ過(10,000×g、30秒)し、ろ液の20μlをHPLCに供し、溶出するPA化糖鎖の量を測定し遊離糖鎖量とした。
【0222】
次に、50mMトリス−塩酸緩衝液(pH7.0)の90mlとPA化糖鎖水溶液10μlを混合後、同様に処理し、そのろ液の20μlをHPLCに供し、溶出するPA化糖鎖の量を測定して添加糖鎖量とした。
【0223】
結合糖鎖量は、添加糖鎖量から反応液の遊離糖鎖量を差し引いた値として算出した。レクチンの糖鎖結合活性は結合率、すなわち添加糖鎖量に対する結合糖鎖量の割合(%)で表示した。なお、結合試験は各糖鎖につき2回行い、結合率は平均値として算出した。
【0224】
PA化糖鎖の分離、定量は逆相系HPLCを用いて行った。すなわち、検液を40℃のカラムオーブン中、TSKgel ODS-80TM カラム(4.6×150 mm)に注入し、0.1M 酢酸アンモニウム−15%メタノールで溶出した。流速は1.0 ml/minとし、溶出液は励起波長320nm、蛍光波長400nmでモニターし、各糖鎖のピーク面積をEZChrom Elite(アジレントテクノロジー)で解析、定量した。
【0225】
供試糖鎖としてPA化糖鎖27種:N−グリコシド型糖鎖の複合型6種(PA-Sugar Chain 001、002、004、009、010、023、TAKARA)、高マンノース型11種(PA-Sugar Chain 017、018、019、020、051、052、053、054、055、056、058、TAKARA)、共通コア構造1種(PA-Sugar Chain 016、TAKARA)、共通コア関連糖鎖1種(PA-042、増田化学工業)、糖脂質系糖鎖3種(PA-Sugar Chain 027、028、038、TAKARA)、およびオリゴマンノース5種を用いた。このうち、PA−オリゴマンノースを除き、市販品(TAKARA)を用いた。
【0226】
PA−オリゴマンノースは、PA化装置(PALSTATION、TAKARA)を用いて以下のように調製した。
【0227】
すなわち、各50nmolのManα1-2Man、Manα1-3Man、Manα1-6Man、Manα1-6(Manα1-3)Man、Manα1-6(Manα1-3)Manα1-6(Manα1-3)Man(DextraLaboratories、Funakoshi)を凍結乾燥後、カップリング試薬(2-アミノピリジン/酢酸溶液、Pyridylamination Reagent Kit、TAKARA)20μlを加え、90℃で60分間反応させた。これに還元試薬(ボラン-ジメチルアミン/酢酸溶液)20μlを加え、80℃で60分間反応させた。この反応液にトリエチルアミン-メタノール20μlを加えてよく撹拌した後、トルエン40 μlを加えて撹拌し、窒素気流化で60℃、10分間減圧乾固した。
【0228】
これにメタノール20μlとトルエン40μlとを加えて撹拌し、同様に減圧乾固した後、トルエン50μlを加えて再び減圧乾固した。過剰の試薬は順相系HPLCにより除去した。
【0229】
すなわち、上記PA化物を25μlの超純水に溶解した後、40℃のカラムオーブン中、50mM 酢酸−トリエチルアミン(pH7.3)/アセトニトリル(25/75)で平衡化したTSKgel NH2-60カラム(4.6×250 mm)に注入し、50mM酢酸−トリエチルアミン(pH7.3)/アセトニトリル(25/75)(溶媒A)と50mM酢酸−トリエチルアミン(pH7.3)/アセトニトリル(50/50)(溶媒B)の2液による濃度勾配[溶媒A 100%(0−5分)、溶媒A 0%−溶媒B 100%(5−55分)]を用いて、流速1.0ml/minで溶出した。溶出液は励起波長310nm、蛍光波長380nmでモニターし、PA化糖鎖(オリゴマンノース)ピークを分取した。
【0230】
得られたPA−オリゴマンノースの定量は以下のようにして行った。すなわち、上記の各オリゴマンノース画分の50μlを減圧乾固後、4N HCl/4M TFA(1/1(v/v))中100℃で4時間、気相酸加水分解した。
【0231】
この加水分解物を50μlの超純水に溶解し、その10μlを40℃のカラムオーブン中、TSKgel ODS-80TMカラム(4.6×150 mm)に注入し、0.1 M 酢酸アンモニウム−10%メタノールで溶出した。流速は1.0ml/minとし、溶出液は励起波長320nm、蛍光波長400mmでモニターした。
【0232】
これにより、PA−オリゴマンノースから加水分解により遊離したPA−マンノースのピーク面積を測定した。このPA−マンノースの定量は、10pmolの標準PA−マンノース(TAKARA)の同カラムからの溶出ピーク面積との比較により行い、PA−オリゴマンノース量とした。
【0233】
(実施例7−2:糖鎖結合特異性試験結果)
遠心限外ろ過−HPLC法により27種類のピリジルアミノ化(PA化)糖鎖に対するMPL−1の結合性を精査した。図11は、PA化糖鎖27種の構造と、当該糖鎖へのMPL−1の結合活性を示すものである。図中、結合活性(単位%、結合糖鎖量/添加糖鎖量×100)が10%以下ものはハイフンで示した。
【0234】
図11に示すように、MPL−1は供試した糖鎖中、N−グリコシド型糖鎖のうち高マンノース型糖鎖とのみ特異的に結合することがわかった。複合型(図11に示す1−6)、共通コア構造(図11に示す23−24)とは結合しなかったことから、本レクチンは高マンノース型糖鎖構造のオリゴマンノースで構成される分岐糖鎖部分を認識することが判明した。さらに興味深いことに、高マンノース型糖鎖(図11に示す7−17)に対する結合性について、分岐糖鎖部分のオリゴマンノース構造の違いにより結合活性に明瞭な差異が認められた。
【0235】
ここで、高マンノース型糖鎖の非還元末端は、トリマンノースコアのMan(α1-3)アーム(D1アーム)、同コアのMan(α1-6)アームから分岐したMan(α1-3)アーム(D2アーム)および同Man(α1-6)アームから分岐したMan(α1-6)アーム(D3アーム)の3つのアームからなる。
【0236】
MPL−1はD3アームの非還元末端にα1-2Manを有するもの(図11に示す9、10、13、15)とのみ高い結合活性が認められ、その結合率は96%以上であった。一方、同Man残基を含まない高マンノース型糖鎖(図11に示す7、8、11、12、14、16、17)およびオリゴマンノース(図11に示す18−22)に対するMPL−1の結合活性は、図11に示す糖鎖12を除いて10%以下であった。糖鎖12はD3アームにα1-2Manを含まず、D1アームの非還元末端にMan(α1-2)Man(α1-2)を有するM7糖鎖であるが、同様の構造を有するM8糖鎖(図11に示す14)では結合がみられないことから、MPL−1の主要認識糖鎖構造ではないと考えられる。
【0237】
これらの結果から、MPL−1はD3アームの非還元末端にα1-2Man残基を有する高マンノース型糖鎖とのみ結合すると考えられる。単糖類(マンノースを含む)に結合せず、このような厳密な高マンノース型糖鎖認識をもつものは既知レクチン中に見出されていない。糖化合物による赤血球凝集阻止試験でMPL−1と同様のプロファイル(表3)を示したイソレクチンMPL−2においても同様の糖鎖結合特異性を有すると予測される。
【0238】
本発明は上述した各実施形態に限定されるものではなく、請求項に示した範囲で種々の変更が可能であり、異なる実施形態にそれぞれ開示された技術的手段を適宜組み合わせて得られる実施形態についても本発明の技術的範囲に含まれる。
【産業上の利用可能性】
【0239】
本発明にかかるポリペプチドは、HIVなどのウイルスが備える高マンノース型糖鎖と特異的に結合することができる新規ポリペプチドであり、抗ウイルス剤、ウイルス除去膜等への適用が可能である。それゆえ、医療産業、薬品産業等において広く利用することが可能である。
【特許請求の範囲】
【請求項1】
高マンノース型糖鎖と結合するポリペプチドであって、
(a)配列番号2に示されるアミノ酸配列;または
(b)配列番号2に示されるアミノ酸配列において、1個もしくは数個のアミノ酸が置換、欠失、挿入、もしくは付加されたアミノ酸配列、
からなることを特徴とするポリペプチド。
【請求項2】
請求項1に記載のポリペプチドをコードすることを特徴とするポリヌクレオチド。
【請求項3】
下記の(a)または(b)のいずれかであることを特徴とする請求項2に記載のポリヌクレオチド:
(a)配列番号1に示される塩基配列からなるポリヌクレオチド;または
(b)以下の(i)もしくは(ii)のいずれかとストリンジェントな条件下でハイブリダイズするポリヌクレオチド:
(i)配列番号1に示される塩基配列からなるポリヌクレオチド;もしくは
(ii)配列番号1に示される塩基配列と相補的な塩基配列からなるポリヌクレオチド。
【請求項4】
請求項1に記載のポリペプチドを含有し、高マンノース型糖鎖を備えるウイルスを標的とすることを特徴とする抗ウイルス剤。
【請求項5】
上記ウイルスは、エイズウイルス、インフルエンザウイルス、C型肝炎ウイルス、エボラウイルス、ヒトヘルペスウイルス6および重症急性呼吸症候群ウイルスからなる群より選ばれるウイルスであることを特徴とする請求項4に記載の抗ウイルス剤。
【請求項6】
請求項1に記載のポリペプチドが固定されてなることを特徴とするウイルス除去膜。
【請求項7】
請求項2または3に記載のポリヌクレオチドを含むことを特徴とするベクター。
【請求項8】
請求項7に記載のベクターを用いることを特徴とする、請求項1に記載のポリペプチドを生産する方法。
【請求項9】
請求項2または3に記載のポリヌクレオチドが導入されていることを特徴とする形質転換体。
【請求項10】
請求項9に記載の形質転換体を用いることを特徴とする、請求項1に記載のポリペプチドを生産する方法。
【請求項1】
高マンノース型糖鎖と結合するポリペプチドであって、
(a)配列番号2に示されるアミノ酸配列;または
(b)配列番号2に示されるアミノ酸配列において、1個もしくは数個のアミノ酸が置換、欠失、挿入、もしくは付加されたアミノ酸配列、
からなることを特徴とするポリペプチド。
【請求項2】
請求項1に記載のポリペプチドをコードすることを特徴とするポリヌクレオチド。
【請求項3】
下記の(a)または(b)のいずれかであることを特徴とする請求項2に記載のポリヌクレオチド:
(a)配列番号1に示される塩基配列からなるポリヌクレオチド;または
(b)以下の(i)もしくは(ii)のいずれかとストリンジェントな条件下でハイブリダイズするポリヌクレオチド:
(i)配列番号1に示される塩基配列からなるポリヌクレオチド;もしくは
(ii)配列番号1に示される塩基配列と相補的な塩基配列からなるポリヌクレオチド。
【請求項4】
請求項1に記載のポリペプチドを含有し、高マンノース型糖鎖を備えるウイルスを標的とすることを特徴とする抗ウイルス剤。
【請求項5】
上記ウイルスは、エイズウイルス、インフルエンザウイルス、C型肝炎ウイルス、エボラウイルス、ヒトヘルペスウイルス6および重症急性呼吸症候群ウイルスからなる群より選ばれるウイルスであることを特徴とする請求項4に記載の抗ウイルス剤。
【請求項6】
請求項1に記載のポリペプチドが固定されてなることを特徴とするウイルス除去膜。
【請求項7】
請求項2または3に記載のポリヌクレオチドを含むことを特徴とするベクター。
【請求項8】
請求項7に記載のベクターを用いることを特徴とする、請求項1に記載のポリペプチドを生産する方法。
【請求項9】
請求項2または3に記載のポリヌクレオチドが導入されていることを特徴とする形質転換体。
【請求項10】
請求項9に記載の形質転換体を用いることを特徴とする、請求項1に記載のポリペプチドを生産する方法。
【図1】


【図2】
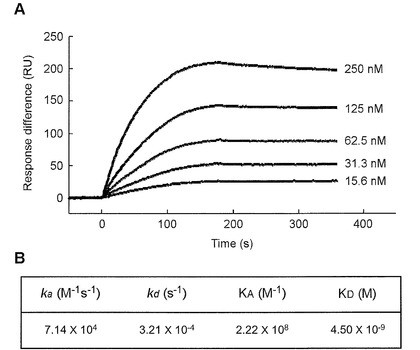
【図4】
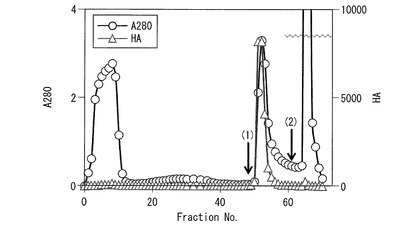
【図5】
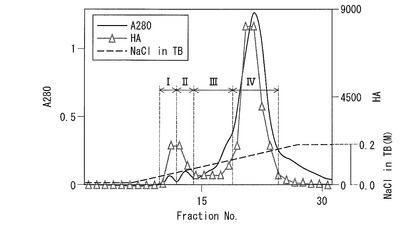
【図7】
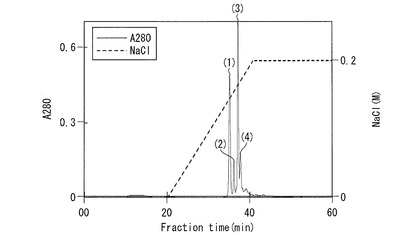
【図9】
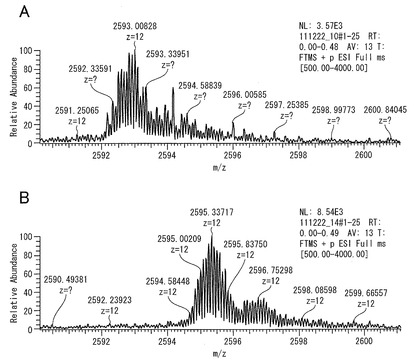
【図11】

【図3】
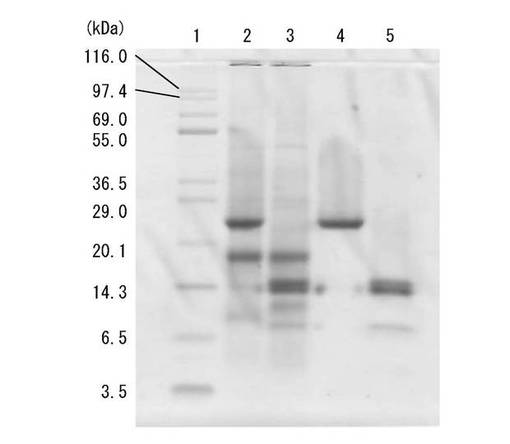
【図6】
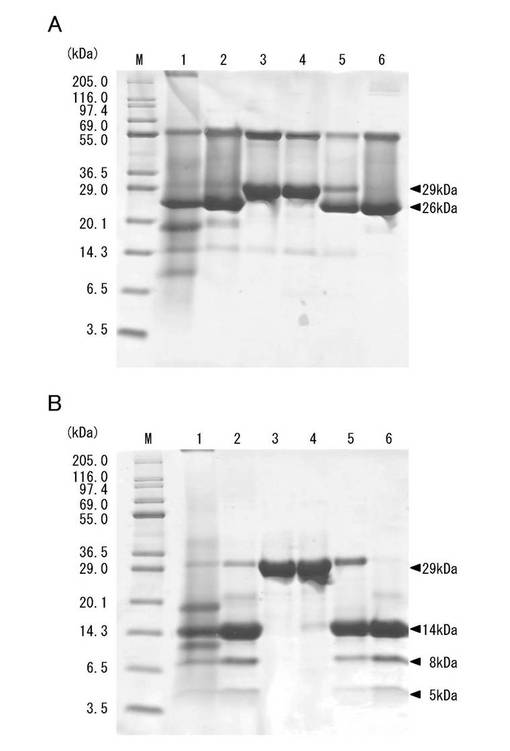
【図8】
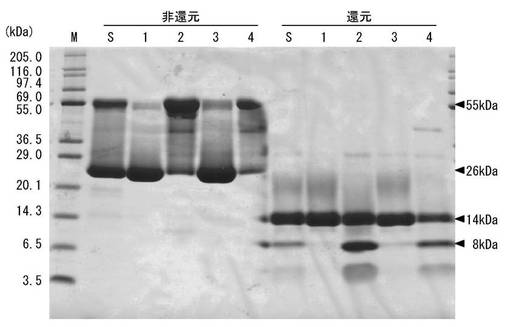
【図10】
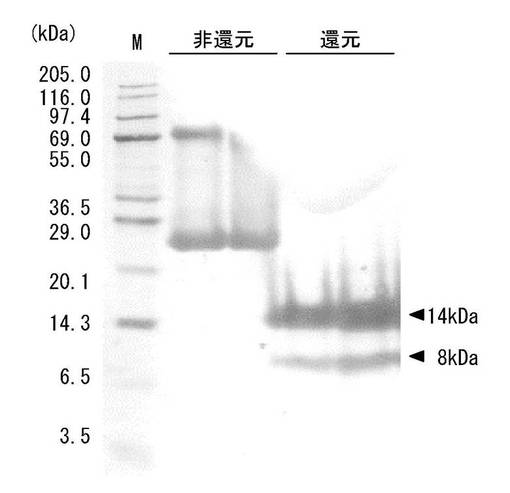
【図2】
【図4】
【図5】
【図7】
【図9】
【図11】
【図3】
【図6】
【図8】
【図10】
【公開番号】特開2012−213382(P2012−213382A)
【公開日】平成24年11月8日(2012.11.8)
【国際特許分類】
【出願番号】特願2012−32205(P2012−32205)
【出願日】平成24年2月16日(2012.2.16)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り ・刊行物名 平成23年度日本水産学会秋季大会(日本農学大会水産部会)講演要旨集 発行日 平成23年9月28日 発行所 平成23年度日本水産学会秋季大会 ・研究集会名 平成23年度日本水産学会秋季大会(日本農学大会水産部会) 主催者名 公益社団法人 日本水産学会 開催日 平成23年9月28日〜10月2日
【出願人】(504136568)国立大学法人広島大学 (924)
【Fターム(参考)】
【公開日】平成24年11月8日(2012.11.8)
【国際特許分類】
【出願日】平成24年2月16日(2012.2.16)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り ・刊行物名 平成23年度日本水産学会秋季大会(日本農学大会水産部会)講演要旨集 発行日 平成23年9月28日 発行所 平成23年度日本水産学会秋季大会 ・研究集会名 平成23年度日本水産学会秋季大会(日本農学大会水産部会) 主催者名 公益社団法人 日本水産学会 開催日 平成23年9月28日〜10月2日
【出願人】(504136568)国立大学法人広島大学 (924)
【Fターム(参考)】
[ Back to top ]