
新規リグニン分解菌とその利用
【課題】植物材料、とりわけイネ科植物材料を効率よく脱リグニンしうる新規菌株、ならびにそれらを用いた植物材料、とりわけイネ科植物材料の脱リグニン方法を提供する。
【解決手段】木材腐朽菌またはその抽出物をイネ科植物材料と接触させることを特徴とする、イネ科植物材料の脱リグニン方法。イネ科植物はいずれのものであってもよく、特に限定はないが、イネ、タケ、ササ、オオムギ、コムギ、ライムギ、カラスムギ、ハトムギなどのムギ類、アワ、ヒエ、トウモロコシ、サトウキビ、カヤ、ヨシ、シバ、エノコログサ、ススキ、アシなどが例示されるが、とくに余剰植物資源としてのタケを有効利用することができる。
【解決手段】木材腐朽菌またはその抽出物をイネ科植物材料と接触させることを特徴とする、イネ科植物材料の脱リグニン方法。イネ科植物はいずれのものであってもよく、特に限定はないが、イネ、タケ、ササ、オオムギ、コムギ、ライムギ、カラスムギ、ハトムギなどのムギ類、アワ、ヒエ、トウモロコシ、サトウキビ、カヤ、ヨシ、シバ、エノコログサ、ススキ、アシなどが例示されるが、とくに余剰植物資源としてのタケを有効利用することができる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、新規リグニン分解菌、ならびにそれを用いた植物材料、特に、イネ科植物材料の脱リグニン方法に関する。
【背景技術】
【0002】
現在、世界各地で余剰の植物資源の処理が問題となっている。例えば、間伐材、廃材、わらくず、作物の残りカス、放置されたタケ林などが大量に存在しており、それらを処理するために、バイオエタノールの製造などの様々な取り組みがなされている。
【0003】
タケは日本では昔から建築材、楽器、食料、すだれなどの生活に密着した道具の材料として用いられてきた。しかし、1960年代以降、中国などの諸外国より安価なタケノコやタケ材が輸入されるようになり、国内産の需要は減少傾向をたどっている。その結果、全国においてタケ林が次第に利用されなくなり放置されたタケ林の拡大および荒廃が進んでいる(徳永・荒木、2007)。そのため現在、このタケの有効利用法が求められている。
【0004】
このような植物資源中には、様々な有用物質の生産の原料となる糖類が多く含まれている。木質中に含まれているリグノセルロースには、多糖類であるブドウ糖から構成されるセルロースとマンノース、アラビノースおよびガラクツロン酸などの六単糖およびキシロースなどの五単糖からなるヘミセルロースが含まれており、これらの低分子化合物は化学的・生物学的プロセスによりさまざまな有用物質の生産の原料として重要である。しかしながら、これらのセルロースは強靭なリグニンによって被覆されている。このリグニンは、分子量が5万以上あり、セルロースやヘミセルロースとともに高等植物の主成分である。草本類であるイネ科に属するタケにおいては、3種のモノリグノール(コニフェニリルアルコール、シナピルアルコール、p−クマリルアルコール)が脱水素架橋重合したもので、三次元網目構造を持つことが知られている。したがって、例えばタケを原料としてバイオエタノールを製造する場合には、リグニンを除去してから、セルロース、ヘミセルロースを利用することが必要である。すなわち、脱リグニンは、植物資源からの有用物質生産の前段階として非常に重要な工程である。
【0005】
リグニン除去には化学的処理・物理的処理などの方法がある。例えば、薬剤を用いて高湿度で脱リグニンをおこなう蒸解法、アルカリ性の薬剤(水酸化ナトリウムおよび亜硫酸水素カルシウム)を用いるクラフト法、酸性の薬剤(亜硫酸および亜硫酸水素カルシウム)を用いるサルフィト法、リグノセルロース原料を高温・高圧の水蒸気にさらし、圧力を急激に常圧に戻し植物組織を破壊させ、ヘミセルロースを加水分解によって低分子化させる爆砕法などが報告されている(非特許文献1参照)。
【0006】
しかし、これらのプロセスには大量のエネルギーが必要であり高コストかつ薬品などによる環境負荷が高いため、近年では菌類による生物学的処理が注目されている。菌類の中でも、担子菌類に所属する木材腐朽菌は、木質を分解する生物として一般的に認識されており、これまでに、白色腐朽菌を用いたリグニン分解が検討されている(非特許文献1〜3参照)。これらの菌は一般にはセルロースやヘミセルロースをエネルギー源および生育源として利用し、この過程でリグニンを分解していると考えられている。そのため、これらの菌はリグニン分解と同時にセルロース類の資化をしている。これらの菌を用いて脱リグニンを行った原料をバイオエタノールの製造に使用すると、エタノールの収量が減少してしまう。セルロースやヘミセルロースよりもリグニンを優先的に除去する菌株を用いた脱リグニンについても研究されているが(非特許文献4および5参照)、タケを原料として脱リグニンを行った例はない。
【先行技術文献】
【非特許文献】
【0007】
【非特許文献1】桑原正章 バイオマスエネルギー利用技術(2001)pp.147-157シーエムシー出版
【非特許文献2】Yu Fukasawa, Takashi Osono, Hiroshi TakedaDecomposition of Japanese beech wood by diverse fungi isolated from a cool temperate deciduous forest. Mycoscience (2005) 46:97-101.
【非特許文献3】Xiaoyu Zhang, Hongbo Yu, Huiyan Huang, Youxun LiuCollege of Life Science and Technology, Huazhong University of Science and Technology, 430074 Wuhan, P.R. China, Evaluation of biological pretreatment with white rot fungi for the enzymatic hydrolysis of bamboo culms, International Biodeterioration & Biodegradation 60 (2007) 159-164.
【非特許文献4】Karin Fackler , Cornelia Gradinger , Barbara Hinterstoisser ,Kurt Messner, Manfred Schwanninger, Lignin degradation by white rot fungi on spruce wood shavings during short-time solid-state fermentations monitored by near infrared spectroscopy, Enzyme and Microbial Technology 39 (2006) 1476-1483.
【非特許文献5】Yasunori Ohashi, Yoshihiko Kan, Takahito Watanabe, Yoichi Honda, Takashi Watanabe, Redox silencing of the Fenton reaction system by an alkylitaconic acid, ceriporic acid B produced by a selective lignin-degrading fungus, Ceriporiopsis subvermispora, Organic & Biomolecular Chemistry 5(2007)840-847.
【発明の概要】
【発明が解決しようとする課題】
【0008】
植物材料中のリグニンに対してさらに高い選択的分解能を有する菌株を提供することが本発明の解決課題であった。特に、近年我が国で問題となっている余剰のタケの脱リグニンに有効な菌株を提供することも本発明の解決課題であった。
【課題を解決するための手段】
【0009】
本発明者らは、上記課題に鑑みて鋭意研究を重ね、植物材料、とりわけタケのリグリンに対して非常に高い選択的分解能を有する菌株を見出し、本発明を完成させるに至った。
【0010】
すなわち、本発明は下記のものを提供する:
(1)木材腐朽菌またはその抽出物をイネ科植物材料と接触させることを特徴とする、イネ科植物材料の脱リグニン方法;
(2)イネ科植物がタケである(1)記載の方法;
(3)木材腐朽菌が、Punctularia属の菌株である(1)または(2)記載の方法;
(4)Punctularia属の菌株が、受領番号NITE AP−970として独立行政法人製品評価技術基盤機構特許微生物寄託センターに受領された菌株である(3)記載の方法;
(5)木材腐朽菌が、受領番号NITE AP−971として独立行政法人製品評価技術基盤機構特許微生物寄託センターに受領された菌株である(1)または(2)記載の方法;
(6)受領番号NITE AP−970として独立行政法人製品評価技術基盤機構特許微生物寄託センターに受領された菌株;
(7)受領番号NITE AP−971として独立行政法人製品評価技術基盤機構特許微生物寄託センターに受領された菌株。
【発明の効果】
【0011】
本発明により、植物材料、特に、イネ科植物材料を効率よく脱リグニンしうる新規菌株、ならびにそれらを用いた植物材料、特に、イネ科植物の脱リグニン方法が提供される。本発明によれば、余剰植物資源、とりわけ、近年我が国で問題となっている余剰のタケの脱リグニンを効果的に行うことができ、バイエタノール等の有用物質の製造に資することができる。
【図面の簡単な説明】
【0012】
【図1】図1は2年堆積地の様子を示す。
【図2】図2はグァイアコール呈色反応の一例を示す。
【図3】図3はタケ粉末スラント培地を用いた脱色試験の結果を示す。上3本がPunctularia sp.(TUFC20083)処理区、一番下は無処理区である。
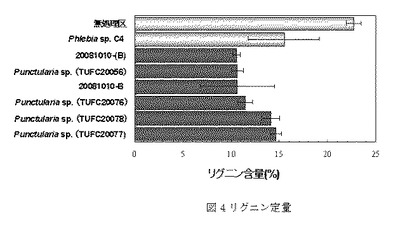
【図4】図4は腐朽実験後のリグニン残存量を示す。一番上が無処理区、上から2番目がポジティブコントロール(C4)、下6つが供試菌のなかでリグニン含量が少なかった菌株での結果である。
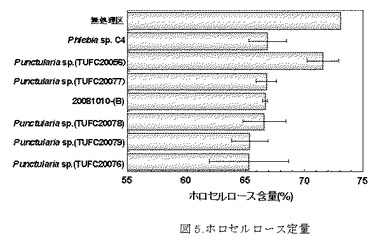
【図5】図5は腐朽実験後のホロセルロース残存量を示す。一番上が無処理区、上から2番目がポジティブコントロール(C4)、下6つが供試菌のなかでホロセルロース含量が多かった菌株での結果である。
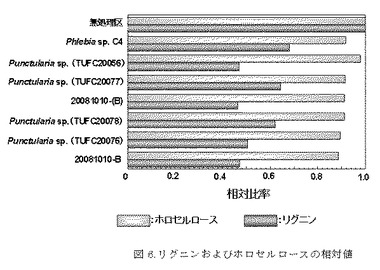
【図6】図6は腐朽実験後のリグニンおよびホロセルロースの相対値を示す。一番上が無処理区、上から2番目がポジティブコントロール(C4)、下6つは図4、図5に示した菌株と同じである。
【発明を実施するための形態】
【0013】
本発明は、1の態様において、木材腐朽菌またはその抽出物をイネ科植物材料と接触させることを特徴とする、イネ科植物材料の脱リグニン方法を提供する。本発明により脱リグニンされるイネ科植物はいずれのものであってもよく、特に限定はないが、イネ、タケ、ササ、オオムギ、コムギ、ライムギ、カラスムギ、ハトムギなどのムギ類、アワ、ヒエ、トウモロコシ、サトウキビ、カヤ、ヨシ、シバ、エノコログサ、ススキ、アシなどが例示される。このうち、タケについては放置されたタケ林などが大量に存在しており問題となっているので、本発明の方法を適用することにより余剰植物資源としてのタケを有効利用することができる。もちろん、他のイネ科植物であっても、タケと同様に本発明の方法にて脱リグニンすることができる。
【0014】
イネ科植物材料は、生育しているイネ科植物体のみならず、収穫、伐採等により得られたイネ科植物体の部分であってもよい。
【0015】
本発明の脱リグニン方法に使用できる菌類は木材腐朽菌であって、リグニン分解活性の高いものであればいずれの種類の菌であってもよい。このような木材腐朽菌の例としては、Coprinellus属、Coprinopsis 属、Collybia属、Ganoderma 属、Gymnopilus属、Lycoperdon属、Psathyrella属、Punctularia属、Pleurotus属、Lentinula属、Panus属、Trametes属、Flammulina属、Pholiota属、Kuehneromyces属、Stereum属、Mycoleptodonoides属、Polyporellus属、Porostereum属、Hericium属、Grifora属の菌株が挙げられるがこれらに限定されない。菌を自然界から得てもよく、保存機関のストック菌株を用いてもよい。本発明において好ましいのは白色木材腐朽菌である。白色木材腐朽菌の例としては、Ceriporiopsis属、Ceripora属、Coriolus属、Hyphoderma属、Hyphodontia属、Phlebia属、Punctularia属、Phanerochaete属、Trametes属、Phlebia属、Phlebia属、Bjerkandera属、Pycnoporus属、Polyporus属、Sphaerobolus属、Crucibulum属、Cyathus属、Pseudocolus属、Phallus属、Peniophora属、Resinicium属の菌株が挙げられるがこれらに限定されない。白色木材腐朽菌のなかでも、Punctularia属の菌がより好ましく本発明に用いられる。特に、独立行政法人製品評価技術基盤機構特許微生物寄託センターに寄託され受領番号AP−970を付与されたPunctularia属の菌(TUFC20056)および独立行政法人製品評価技術基盤機構特許微生物寄託センターに寄託され受領番号AP−971を付与された白色木材腐朽菌(TUFC20057/図表中では20081010−(B)、20081010−Bとして記載)は本発明において非常に好ましい菌株であり、イネ科植物材料のリグニンに対して非常に高い選択的分解能を有し、セルロースやヘミセルロースをほとんど分解しないという特徴を有する。これらの菌株は、自然界から採取され、スクリーニングされたものである。なお、「TUFC」は鳥取大学附属菌類きのこ遺伝資源研究センターの保存株であることを表す。
【0016】
リグニン分解活性は公知の方法にて測定することができる。リグニン分解酵素として、リグニンペルオキシダーゼ、マンガンペルオキシダーゼ、ラッカーゼおよび/またはポリフェノールオキシダーゼなどが知られており、これらの酵素活性を公知方法により測定することにより、菌のリグニン分解活性を調べることができる。
【0017】
本発明の脱リグニン方法において、イネ科植物材料に木材腐朽菌の菌糸または胞子を接種することにより、脱リグニンを開始させることができる。その際に、イネ科植物材料を公知の手段・方法にて細断あるいは粉砕等して、チップ、ペレット、粉末などの形態にしたものに、木材腐朽菌の菌糸または胞子を接種することができる。接種する菌糸は、固体培地または液体培地にて数日〜数週間培養されたものであってもよい。固体培養は、例えば寒天平板培地で行ってもよく、液体培養は、フラスコ等において振盪しながら行ってもよい。培養の際に、脱リグニンすべきイネ科植物材料を培地に添加して(例えば、タケ粉を培地に混合して)、菌体の脱リグニン酵素活性を誘導してもよい。木材腐朽菌に使用する培地の種類や培養温度、pH、通気条件、培養時間などの培養条件は当業者が選択、決定することができる。さらに、イネ科植物材料に木材腐朽菌を接種してリグニン分解を行う際の諸条件、例えば、材料の形状(チップ、粉末など)、材料への菌の接種量、脱リグニン反応時の温度、時間、湿度、通気条件、材料の水分量などは、材料の種類、形状、サイズ、量、および木材腐朽菌の種類などの諸因子を考慮して、当業者が適宜選択、決定できるものである。例えば、タケチップを上記菌株(TUFC20056またはTUFC20057)にて脱リグニンする場合には、約15℃〜約30℃で約1ヶ月〜約6ヶ月行ってもよい。実用的には、サイロのような施設内,もしくは大型のコンテナのような容器のなかで、タケチップと培養菌株を混合し、適宜攪拌しながら、好ましくは温度および湿度をコントロールしながら処理を行なってもよい。
【0018】
また、本発明において、木材腐朽菌の抽出液をイネ科植物材料と接触させることにより脱リグニンを行ってもよい。例えば、木材腐朽菌を液体培養した場合には、培養液を濾過し、硫安やエタノールなどを用いて濃縮された抽出液を得てもよい。また、木材腐朽菌を固体培養した場合には、菌糸が生育した培養物を適当な抽出溶媒、例えば水、あるいは水とアルコール(メタノール、エタノールなど)またはグリセリンなどの混合物を用いて抽出してもよい。得られた抽出液を公知の方法にてさらに濃縮、精製してもよい。木材腐朽菌の抽出液を得る場合において、脱リグニンすべきイネ科植物材料を培地に添加して培養し、菌体の脱リグニン酵素活性を誘導しておいてもよい。
【0019】
木材腐朽菌またはその抽出物をイネ科植物材料と「接触」させるとは、木材腐朽菌の菌糸またはその抽出液に由来するリグニン分解酵素がイネ科植物材料に直接的に接触して作用し、イネ科植物材料中のリグニンが分解されうる状況にすることをいう。例えば、チップ状にしたイネ科植物材料に木材腐朽菌の菌糸または胞子をまんべんなく接種して、十分にリグニンが分解されるまで、あるいは目安として材料が十分に白化するまで培養することにより、脱リグニンを行うことができる。菌糸または胞子の接種量、培養温度や培養時間などの培養条件は、イネ科植物材料の種類やサイズ、木材腐朽菌の種類などの諸条件に応じて、当業者が選択、決定することができる。
【0020】
イネ科植物材料の脱リグニンの確認は、例えば、材料の白化を確認すること、材料中のリグニンを定量すること等によって行うことができる。材料の白化は肉眼で確認することができる。リグニンは、硫酸(クラーソン)法(木質科学実験マニュアル、日本木材学会編、文永堂出版、2000年、ISBN4-8300-4094-7)により定量することができる。これらの方法は当業者に公知である。
【0021】
さらに本発明は、リグニンに対して高い選択性を有し、ヘミセルロース、セルロースなどのホロセルロースに対して低い選択性を有するリグニン分解菌であって、イネ科植物材料の脱リグニンに好適な木材腐朽菌を提供する。これらの菌株なかで好ましいものは、独立行政法人製品評価技術基盤機構特許微生物寄託センターに寄託され受領番号AP−970を付与されたPunctularia属の菌(TUFC20056)および独立行政法人製品評価技術基盤機構特許微生物寄託センターに寄託され受領番号AP−971を付与された白色木材腐朽菌(TUFC20057)である。これらの菌株を用いることにより、イネ科植物材料の脱リグニンを効率的に行うことができ、しかも、セルロース、ヘミセルロースなどのホロセルロース成分の分解が少ないので、バイオエタノールなどの有用物質の製造等に資することができる。
【0022】
以下に実施例を示して本発明をさらに詳細かつ具体的に説明するが、実施例は単なる例示説明であり、本発明を限定するものと解釈してはならない。
【実施例1】
【0023】
(サンプリングおよび分離)
0年、半年、2年間(09年4月時点)堆積(W 2m×D 2m ×H 20cm、図1)した腐朽段階の異なるPhyllostachys pubescens(モウソウチク)のタケチップから担子菌類の子実体の発生が見られるものを中心にそれぞれ5サンプルずつ採取し、分離株の単離を行った。分離方法は、子実体が発生していたものは、マルトエキス寒天培地[MA培地、1.5%(w/v)麦芽エキス、1.5%(w/v)寒天]に多胞子分離により分離を行った。子実体の発生が見られないものは、材の腐朽部から組織分離[0.1%(v/v)のビオコートを含むMA培地]を行い、その後MA培地に移植した。このようにして単離した菌株63株を以下の実験に用いた。そして、鳥取大学附属菌類きのこ遺伝資源研究センター保有のタケを基質とする菌として分離された菌株4株、および行木(2008)の検討でスギ材に対して高い選択的リグニン分解能を示した菌株1株を供試菌とした。ポジティブコントロールとしてPhanerochaete sordida (TMIC31227)、Phlebia sp.(C4)、Trametes versicolor(TUFC10537)を用いた(表1)。以下に説明する一次スクリーニングに用いた供試菌は、ポジティブコントロールを含めて合計68株であった。
【実施例2】
【0024】
(一次スクリーニング(グァイアコール試験))
0.02%(v/v)グァイアコールを含むMA平板培地に植菌し、24℃、暗所下で2週間培養後に菌糸の直径と変色部の長さを測定した。選抜基準は呈色領域半径1cm以上を示す菌株とした。一次スクリーニングを行った結果、67菌株のうち40菌株において酸化酵素活性の指標となるグァイアコールの呈色反応が認められた。グァイアコールの呈色反応の一例を図2に示す。そのうち呈色半径が1.0cm以上を優良菌株とし36菌株を選抜した(表2、ポジティブコントロールとして用いたPhlebia sp.(C4)および20050710−03を含む)。
【0025】
【表1】

【0026】
【表2】
【0027】
一番活性が強いものはTUFC20077で3.7cmの値を示した。呈色反応が見られなかった菌株は27株あった。選抜した36菌株および、ポジティブコントロールである20050710−03、Phlebia sp.(C4)、Trametes versicolor(TUFC10537)、Phanerochaete sordida(TUFC31227)の4菌株を合わせた40菌株を二次スクリーニングに用いた。
【実施例3】
【0028】
(二次スクリーニング(脱色度試験))
タケに対する腐朽力を検定するために、タケ粉末培地[0.3%(w/v)麦芽溶液1400mLとタケ粉末1000gを混合]を口径18mmの試験管に3分の1程度詰め、タケ粉末が斜面状になるように薬さじで成形した。その後、シリコン栓で蓋をしてオートクレーブ(121℃、15分)で滅菌し、タケ粉末培地を作成した。タケ粉末スラント培地にグァイアコール呈色反応の見られた菌株を、MA培地にあらかじめ培養した各供試菌の菌糸体を接種し3ヶ月間の培養を行った。接種源にはMA平板培地に生育させた菌糸体の先端付近を同心円状に孔径5mmのコルクボーラーで打ち抜き、菌糸体ディスク2個をランダムに選んで用いた。1ヶ月ごとに目視で培地の脱色度を観察し、腐朽の観測を行った。3ヶ月培養後、強い脱色が確認された26菌株を選抜した。目視評価はコントロールと比較して、(−)〜(++++)の5段階で評価した(表2)。
【0029】
【表3】
【実施例4】
【0030】
(リグニン定量、ホロセルロース定量、および重量減少率の測定)
二次スクリーニングの結果、脱色度が一番高かったものはPunctularia sp.であり(図3に結果を例示する)、処理前のタケ粉末培地と比較して、顕著な脱色を示した。二次スクリーニングで脱色度の高かった26菌株について、リグニン定量、ホロセルロース定量、および重量減少率の測定を行った(表3に結果をまとめた)。
【0031】
【表4】
【0032】
(重量減少率の測定)
培養ビンに20mLのMA培地を入れ、供試菌を植菌し2週間培養後、あらかじめ秤量したタケ片(約1.2g)をそれぞれ1つずつビンに入れ21℃、暗所下で2ヶ月間培養した。2ヶ月後、タケのみを取り出して、タケに付着した菌糸を取り除き105℃で3日間乾燥させ、タケの重量減少率を測定した。重量減少率は次式のとおりである。
【化1】
【0033】
重量減少率は、菌株により低い(3.4%)もの(TUFC20083)から約21%の菌株(TUFC20065)まで、様々な値であった。
【0034】
(リグニン、ホロセルロース定量)
A.アルベン抽出
脱脂はアルベン抽出法を用いた。3ヶ月培養したタケチップ培地を2日間風乾させ、100mLの三角フラスコに移した。エタノール(99.5%、Wako)とn−へキサン(96.0%、Wako)の混合溶液(容量比、1:2)75mLを加え38℃、30分超音波処理し、ろ過を行った。この操作を8回繰り返し、2日間風乾させ、リグニン定量、ホロセルロース定量に用いた。
【0035】
B. リグニン定量
アルベン抽出後、風乾したタケ粉末を0.3g秤量し、72%硫酸4.5mLを3回に分け加えて100rpm、2.5時間、38℃で振とう後、170mLの純水を加えオートクレーブ(120℃、30分)処理し、あらかじめ秤量済みのガラス繊維ろ紙(GA200、size 47mm;Toyo Roshi)で吸引ろ過した。沈殿物は熱水400mL、および冷水400mLで洗浄し、105℃に設定した滅菌乾燥機に一晩かけ、デシケーターで半日放冷した。その後秤量し、増加した重量を酸不溶性リグニン量とした。ろ液は酸可溶性リグニン量を求めるため少量残し、ろ液の210nmの吸光度が0.3〜0.7の範囲になるように3%硫酸水溶液で希釈して210nmの波長で吸光度測定を行い、次式により酸可溶性リグニン量を算出した。
【化2】
【0036】
リグニン定量に関する操作は、木質科学実験マニュアル(黒田、94頁(2001))にしたがって行った。
【0037】
リグニン定量の結果を表3、図4に示す。無処理区のリグニン量は22.7%であった。ポジティブコントロールで用いたPhlebia sp.C4のリグニン残存量(15.5%)と比較して、TUFC20056(10.6%)、TUFC20076(11.5%)、TUFC20077(14.6%)、TUFC20078(14.1%)、20081010−(B)(10.5%)および20081010−B(10.7%)の6菌株(括弧内はリグニン残存量)がリグニンをより多く分解した。また、特にリグニン減少が顕著に見られた3菌株(TUFC20056および20081010−(B)、20081010−B)は、無処理区のリグニン含量と比較して50%以上の減少が確認できた。ただし、20081010−(B)、20081010−Bは再現性試験のための同一の菌株をもちいており、現在はTUFC20057を付与している。
【0038】
C. ホロセルロース定量
脱脂を終えたタケ粉末0.75gを300mL容三角フラスコに移し、蒸留水150mL、亜塩素酸ナトリウム(Wako)1.0gおよび酢酸(Wako)0.2mLを加え、100mL容三角フラスコでゆるく蓋をして、70〜80℃のウォーターバス内で1時間反応させた。実験はドラフト中で行った。次いで、亜塩素酸ナトリウム1.0gおよび酢酸0.2mLを加える操作を、内容物が白く変色するまで1時間おきに計4回繰り返した。この際、内容物を混和させるために軽く振った。内容物が白く変色した後、あらかじめ秤量済みのガラス繊維ろ紙を用いて白色内容物を吸引ろ過し、アセトンおよび冷水で洗浄後、105℃の乾燥機中で乾燥させ、デシケーター中で半日放冷後秤量し、増加した重量をホロセルロース量とした。
【0039】
ホロセルロース定量に関する操作は、木質科学実験マニュアル(黒田、94頁(2000))にしたがって行った。
【0040】
ホロセルロース定量の結果を表3、図5に示した。無処理区のホロセルロース量は73.11%であった。Phlebia sp.(C4)のホロセルロース残存量(66.9%)と比較して、TUFC20056(71.59%)が高い値を示した。次いで、20081010−(B)(66.7%)が高い値を示した。優れた白色腐朽菌とされるカワラタケTUFC10537では、59.5%と低い値を示した。他の菌株ではTUFC20072(57.8%)を除いて、62−66%の値を示した。
【0041】
リグニン定量およびホロセルロース定量の結果、タケ材のリグニン含有量22.7%およびホロセルロース含有量73.1%に対して、TUFC20056がリグニン残存量で10.6%、ホロセルロース残存量で71.6%の高い選択的リグニン分解能を示した。ポジティブコントロールとして用いたPhlebia sp.(C4)のリグニン残存量(15.5%)およびホロセルロース量(66.9%)と比較しても優れた選択的リグニン分解能を有していた。今回、リグニン含量が10.5%−14.6%の高いリグニン分解能を示した菌株は6株あり、そのうち4株は形態学的、遺伝子学的同定の結果、Punctularia sp.であることが分かった(図4参照)。これら6株を作用させた後のリグニンおよびホロセルロースの相対値を図6に示す。
【実施例5】
【0042】
(酵素活性測定)
リグニン定量およびホロセルロース定量試験の結果が良かった上位10菌株(C4、TUFC20056、20081010−B、20081010−(B)、TUFC20076、TUFC20077、TUFC20083、TUFC20079、20090803−12、20091013−08)を液体培地で2週間培養し、各リグニン分解酵素の活性を測定した(表4)。TUFC20079およびTUFC20083の2菌株は液体培地では生育しなかった。
【0043】
A.供試菌の培養
100mL容三角フラスコに10mLのタケ粉末液体培地[1.5%(w/v)麦芽エキス、0.5%(w/v)タケ粉末]を分注し、通気性のあるシリコンキャップで蓋をし、オートクレーブ(121℃、20分)で滅菌した。その後、あらかじめMA平板培地に培養した供試菌の菌糸体を接種した。接種源にはMA平板培地に生育している菌糸体の先端付近を同心円状に口径5mmのコルクボーラーで打ち抜き、菌糸体ディスク3個をランダムに選んで用いた。クリーンベンチ内で接種後、シリコンキャップでふたをし、25℃、暗所下で2週間静置培養を行った。その後、菌糸体を吸引ろ過により除去した。ろ過にはガラス繊維ろ紙(GA−200、サイズ47mm;Toyo Roshi)およびメンブランフィルター(PTFE、サイズ47mm; 孔径3.0μm、0.45μm、0.2μm;Toyo Roshi)を用いて4段階ろ過した。回収したろ液を粗酵素溶液とし酵素活性測定に用いた。
【0044】
B.リグニンペルオキシダーゼ(LiP)活性測定
71.43mmol/Lコハク酸緩衝液(pH3.0)700μL、10mmol/Lベラトリルアルコール溶液50μL、粗酵素液200μLをキュベットにそれぞれ入れ、最後に2mmol/L過酸化水素水50μLを添加し、素早く混合した。対照区には、コハク酸緩衝液700μL、ベラトリルアルコール溶液50μL、粗酵素液200μLをキュベットに入れ混合し、310nmでの吸光度を分光光度計(日本分光、東京)により180秒間測定した。吸光度が直線的に増加したことを確認し、直線部の傾きから1分間あたりの吸光度変化を求めた後、酵素活性を算出した。ネガティブコントロールは過酸化水素を加えてないものを用いた。酵素活性1Uは1分間に1μmolの反応生成物を生じる酵素量とした。その結果、ほとんどの菌においてLiP活性を確認することができなかった。ほとんどの菌が0に近い値であったが、20081010−B(0.3U/L)にわずかな活性が見られた。
【0045】
C.マンガンペルオキシダーゼ(MnP)活性測定
71.43mmol/Lマロン酸緩衝液(pH4.5)700μL、20mmol/L硫酸マンガン溶液50μL、粗酵素液200μLをキュベットにそれぞれ入れ、最後に2mmol/L過酸化水素水50μLを添加し、素早く混合した。対照区には、マロン酸緩衝液700μL、20mmol/L硫酸マンガン溶液50μL、粗酵素液200μLをキュベットに入れ混合し、270nmにおける吸光度を分光光度計により180秒間測定した。吸光度が直線的に増加したことを確認し、直線部の傾きから1分間あたりの吸光度変化を求めた後、酵素活性を算出した。酵素活性1Uは1分間に1μmolの反応生成物を生じる酵素量とした。その結果、MnP活性に関して最も活性が高い菌株は、20081010−Bで72.5U/Lの値を示した。Phlebia sp.(C4)(28.5U/L)と比較してTUFC20077(10.4U/L)以外の全ての菌がPhlebia sp.(C4)よりも高い値を示した。また、TUFC20056(68.2U/L)、20081010−B(72.5U/L)、20081010−(B)(69.9U/L)およびTUFC20076(66.4U/L)においては、C4の2倍以上の活性が確認できた。
【0046】
D.ラッカーゼ(Lac)活性測定
71.43mmol/L酒石酸緩衝液(pH3.0)700μL、0.5mmol/L 2,2’−アジノ−ビス(3−エチルベンゾチアゾリン−6−スルホネート)(ABTS)50μL、粗酵素液200μLをキュベットにそれぞれ入れ、最後に2mmol/L過酸化水素水50μLを添加し、素早く混合した。対照区には、71.43mmol/L酒石酸酸緩衝液(pH3.0)700μL、0.5mmol/L ABTS 50μL、蒸留水200μLをキュベットに入れ混合し、436nmにおける吸光度を分光光度計により3分間モニターした。吸光度が直線的に増加したことを確認し、直線部の傾きから1分間あたりの吸光度変化を求めた後、酵素活性を算出した。酵素活性1Uは1分間に1μmolの反応生成物を生じる酵素量とした。その結果、Lac活性において最も活性が高い菌株はTUFC20056であり、36.2U/Lの値を示した。C4の活性は15.0U/Lであり、全ての菌株の中で最も低い値を示した。C4と最も活性が高かったTUFC20056との差は2倍以上であった。
【0047】
E.ポリフェノールオキシダーゼ(PPO)活性測定
71.43mmol/Lコハク酸緩衝液(pH3.0)700μL、50mmol/l 2,6−ジメトキシ−フェノール溶液50μL、粗酵素液200 μL、最後に2mmol/L過酸化水素水50 μLを添加し素早く混合した。対照区には、71.43mmol/Lコハク酸緩衝液700μL、50mmol/L 2,6−ジメトキシ−フェノール溶液50μL、蒸留水200μLを入れ混合し、270nmにおける吸光度を分光光度計により180秒間測定した。吸光度が直線的に増加したことを確認し、直線部の傾きから1分間あたりの吸光度変化を求めた後、酵素活性を算出した。酵素活性1Uは1分間に1μmolの反応生成物を生じる酵素量とした。その結果、PPO活性に関して最も活性が高い菌株はTUFC20077で34.2U/Lの値を示した。C4の値(16.7U/L)と比較して、2倍以上の値であった。次いで、20081010−(B)、TUFC20056および20081010−BのPPO活性が高かった。
【0048】
【表5】
【0049】
上記結果を総合して、リグニンに対する選択性、分解能が高く、ヘミセルロース、セルロースなどのホロセルロースに対する選択性、分解能が低い木材腐朽菌として、TUFC20056、20081010−Bおよび20081010−(B)を選択した。さらなる検討により、20081010−Bおよび20081010−(B)は同一菌株であることが判明し、TUFC20057と命名した。TUFC20056株を2010年7月22日に独立行政法人製品評価技術基盤機構特許微生物寄託センターに寄託し、2010年7月29日に受領番号NITE AP−970を付与された。TUFC20057株を2010年7月22日に独立行政法人製品評価技術基盤機構特許微生物寄託センターに寄託し、2010年7月29日に受領番号NITE AP−971を付与された。
【産業上の利用可能性】
【0050】
本発明は、林業、製材業、製紙業、エネルギー分野、環境分野、製薬業などにおいて利用可能である。
【受託番号】
【0051】
独立行政法人製品評価技術基盤機構特許微生物寄託センター受領番号NITE AP−970
独立行政法人製品評価技術基盤機構特許微生物寄託センター受領番号NITE AP−971
【技術分野】
【0001】
本発明は、新規リグニン分解菌、ならびにそれを用いた植物材料、特に、イネ科植物材料の脱リグニン方法に関する。
【背景技術】
【0002】
現在、世界各地で余剰の植物資源の処理が問題となっている。例えば、間伐材、廃材、わらくず、作物の残りカス、放置されたタケ林などが大量に存在しており、それらを処理するために、バイオエタノールの製造などの様々な取り組みがなされている。
【0003】
タケは日本では昔から建築材、楽器、食料、すだれなどの生活に密着した道具の材料として用いられてきた。しかし、1960年代以降、中国などの諸外国より安価なタケノコやタケ材が輸入されるようになり、国内産の需要は減少傾向をたどっている。その結果、全国においてタケ林が次第に利用されなくなり放置されたタケ林の拡大および荒廃が進んでいる(徳永・荒木、2007)。そのため現在、このタケの有効利用法が求められている。
【0004】
このような植物資源中には、様々な有用物質の生産の原料となる糖類が多く含まれている。木質中に含まれているリグノセルロースには、多糖類であるブドウ糖から構成されるセルロースとマンノース、アラビノースおよびガラクツロン酸などの六単糖およびキシロースなどの五単糖からなるヘミセルロースが含まれており、これらの低分子化合物は化学的・生物学的プロセスによりさまざまな有用物質の生産の原料として重要である。しかしながら、これらのセルロースは強靭なリグニンによって被覆されている。このリグニンは、分子量が5万以上あり、セルロースやヘミセルロースとともに高等植物の主成分である。草本類であるイネ科に属するタケにおいては、3種のモノリグノール(コニフェニリルアルコール、シナピルアルコール、p−クマリルアルコール)が脱水素架橋重合したもので、三次元網目構造を持つことが知られている。したがって、例えばタケを原料としてバイオエタノールを製造する場合には、リグニンを除去してから、セルロース、ヘミセルロースを利用することが必要である。すなわち、脱リグニンは、植物資源からの有用物質生産の前段階として非常に重要な工程である。
【0005】
リグニン除去には化学的処理・物理的処理などの方法がある。例えば、薬剤を用いて高湿度で脱リグニンをおこなう蒸解法、アルカリ性の薬剤(水酸化ナトリウムおよび亜硫酸水素カルシウム)を用いるクラフト法、酸性の薬剤(亜硫酸および亜硫酸水素カルシウム)を用いるサルフィト法、リグノセルロース原料を高温・高圧の水蒸気にさらし、圧力を急激に常圧に戻し植物組織を破壊させ、ヘミセルロースを加水分解によって低分子化させる爆砕法などが報告されている(非特許文献1参照)。
【0006】
しかし、これらのプロセスには大量のエネルギーが必要であり高コストかつ薬品などによる環境負荷が高いため、近年では菌類による生物学的処理が注目されている。菌類の中でも、担子菌類に所属する木材腐朽菌は、木質を分解する生物として一般的に認識されており、これまでに、白色腐朽菌を用いたリグニン分解が検討されている(非特許文献1〜3参照)。これらの菌は一般にはセルロースやヘミセルロースをエネルギー源および生育源として利用し、この過程でリグニンを分解していると考えられている。そのため、これらの菌はリグニン分解と同時にセルロース類の資化をしている。これらの菌を用いて脱リグニンを行った原料をバイオエタノールの製造に使用すると、エタノールの収量が減少してしまう。セルロースやヘミセルロースよりもリグニンを優先的に除去する菌株を用いた脱リグニンについても研究されているが(非特許文献4および5参照)、タケを原料として脱リグニンを行った例はない。
【先行技術文献】
【非特許文献】
【0007】
【非特許文献1】桑原正章 バイオマスエネルギー利用技術(2001)pp.147-157シーエムシー出版
【非特許文献2】Yu Fukasawa, Takashi Osono, Hiroshi TakedaDecomposition of Japanese beech wood by diverse fungi isolated from a cool temperate deciduous forest. Mycoscience (2005) 46:97-101.
【非特許文献3】Xiaoyu Zhang, Hongbo Yu, Huiyan Huang, Youxun LiuCollege of Life Science and Technology, Huazhong University of Science and Technology, 430074 Wuhan, P.R. China, Evaluation of biological pretreatment with white rot fungi for the enzymatic hydrolysis of bamboo culms, International Biodeterioration & Biodegradation 60 (2007) 159-164.
【非特許文献4】Karin Fackler , Cornelia Gradinger , Barbara Hinterstoisser ,Kurt Messner, Manfred Schwanninger, Lignin degradation by white rot fungi on spruce wood shavings during short-time solid-state fermentations monitored by near infrared spectroscopy, Enzyme and Microbial Technology 39 (2006) 1476-1483.
【非特許文献5】Yasunori Ohashi, Yoshihiko Kan, Takahito Watanabe, Yoichi Honda, Takashi Watanabe, Redox silencing of the Fenton reaction system by an alkylitaconic acid, ceriporic acid B produced by a selective lignin-degrading fungus, Ceriporiopsis subvermispora, Organic & Biomolecular Chemistry 5(2007)840-847.
【発明の概要】
【発明が解決しようとする課題】
【0008】
植物材料中のリグニンに対してさらに高い選択的分解能を有する菌株を提供することが本発明の解決課題であった。特に、近年我が国で問題となっている余剰のタケの脱リグニンに有効な菌株を提供することも本発明の解決課題であった。
【課題を解決するための手段】
【0009】
本発明者らは、上記課題に鑑みて鋭意研究を重ね、植物材料、とりわけタケのリグリンに対して非常に高い選択的分解能を有する菌株を見出し、本発明を完成させるに至った。
【0010】
すなわち、本発明は下記のものを提供する:
(1)木材腐朽菌またはその抽出物をイネ科植物材料と接触させることを特徴とする、イネ科植物材料の脱リグニン方法;
(2)イネ科植物がタケである(1)記載の方法;
(3)木材腐朽菌が、Punctularia属の菌株である(1)または(2)記載の方法;
(4)Punctularia属の菌株が、受領番号NITE AP−970として独立行政法人製品評価技術基盤機構特許微生物寄託センターに受領された菌株である(3)記載の方法;
(5)木材腐朽菌が、受領番号NITE AP−971として独立行政法人製品評価技術基盤機構特許微生物寄託センターに受領された菌株である(1)または(2)記載の方法;
(6)受領番号NITE AP−970として独立行政法人製品評価技術基盤機構特許微生物寄託センターに受領された菌株;
(7)受領番号NITE AP−971として独立行政法人製品評価技術基盤機構特許微生物寄託センターに受領された菌株。
【発明の効果】
【0011】
本発明により、植物材料、特に、イネ科植物材料を効率よく脱リグニンしうる新規菌株、ならびにそれらを用いた植物材料、特に、イネ科植物の脱リグニン方法が提供される。本発明によれば、余剰植物資源、とりわけ、近年我が国で問題となっている余剰のタケの脱リグニンを効果的に行うことができ、バイエタノール等の有用物質の製造に資することができる。
【図面の簡単な説明】
【0012】
【図1】図1は2年堆積地の様子を示す。
【図2】図2はグァイアコール呈色反応の一例を示す。
【図3】図3はタケ粉末スラント培地を用いた脱色試験の結果を示す。上3本がPunctularia sp.(TUFC20083)処理区、一番下は無処理区である。
【図4】図4は腐朽実験後のリグニン残存量を示す。一番上が無処理区、上から2番目がポジティブコントロール(C4)、下6つが供試菌のなかでリグニン含量が少なかった菌株での結果である。
【図5】図5は腐朽実験後のホロセルロース残存量を示す。一番上が無処理区、上から2番目がポジティブコントロール(C4)、下6つが供試菌のなかでホロセルロース含量が多かった菌株での結果である。
【図6】図6は腐朽実験後のリグニンおよびホロセルロースの相対値を示す。一番上が無処理区、上から2番目がポジティブコントロール(C4)、下6つは図4、図5に示した菌株と同じである。
【発明を実施するための形態】
【0013】
本発明は、1の態様において、木材腐朽菌またはその抽出物をイネ科植物材料と接触させることを特徴とする、イネ科植物材料の脱リグニン方法を提供する。本発明により脱リグニンされるイネ科植物はいずれのものであってもよく、特に限定はないが、イネ、タケ、ササ、オオムギ、コムギ、ライムギ、カラスムギ、ハトムギなどのムギ類、アワ、ヒエ、トウモロコシ、サトウキビ、カヤ、ヨシ、シバ、エノコログサ、ススキ、アシなどが例示される。このうち、タケについては放置されたタケ林などが大量に存在しており問題となっているので、本発明の方法を適用することにより余剰植物資源としてのタケを有効利用することができる。もちろん、他のイネ科植物であっても、タケと同様に本発明の方法にて脱リグニンすることができる。
【0014】
イネ科植物材料は、生育しているイネ科植物体のみならず、収穫、伐採等により得られたイネ科植物体の部分であってもよい。
【0015】
本発明の脱リグニン方法に使用できる菌類は木材腐朽菌であって、リグニン分解活性の高いものであればいずれの種類の菌であってもよい。このような木材腐朽菌の例としては、Coprinellus属、Coprinopsis 属、Collybia属、Ganoderma 属、Gymnopilus属、Lycoperdon属、Psathyrella属、Punctularia属、Pleurotus属、Lentinula属、Panus属、Trametes属、Flammulina属、Pholiota属、Kuehneromyces属、Stereum属、Mycoleptodonoides属、Polyporellus属、Porostereum属、Hericium属、Grifora属の菌株が挙げられるがこれらに限定されない。菌を自然界から得てもよく、保存機関のストック菌株を用いてもよい。本発明において好ましいのは白色木材腐朽菌である。白色木材腐朽菌の例としては、Ceriporiopsis属、Ceripora属、Coriolus属、Hyphoderma属、Hyphodontia属、Phlebia属、Punctularia属、Phanerochaete属、Trametes属、Phlebia属、Phlebia属、Bjerkandera属、Pycnoporus属、Polyporus属、Sphaerobolus属、Crucibulum属、Cyathus属、Pseudocolus属、Phallus属、Peniophora属、Resinicium属の菌株が挙げられるがこれらに限定されない。白色木材腐朽菌のなかでも、Punctularia属の菌がより好ましく本発明に用いられる。特に、独立行政法人製品評価技術基盤機構特許微生物寄託センターに寄託され受領番号AP−970を付与されたPunctularia属の菌(TUFC20056)および独立行政法人製品評価技術基盤機構特許微生物寄託センターに寄託され受領番号AP−971を付与された白色木材腐朽菌(TUFC20057/図表中では20081010−(B)、20081010−Bとして記載)は本発明において非常に好ましい菌株であり、イネ科植物材料のリグニンに対して非常に高い選択的分解能を有し、セルロースやヘミセルロースをほとんど分解しないという特徴を有する。これらの菌株は、自然界から採取され、スクリーニングされたものである。なお、「TUFC」は鳥取大学附属菌類きのこ遺伝資源研究センターの保存株であることを表す。
【0016】
リグニン分解活性は公知の方法にて測定することができる。リグニン分解酵素として、リグニンペルオキシダーゼ、マンガンペルオキシダーゼ、ラッカーゼおよび/またはポリフェノールオキシダーゼなどが知られており、これらの酵素活性を公知方法により測定することにより、菌のリグニン分解活性を調べることができる。
【0017】
本発明の脱リグニン方法において、イネ科植物材料に木材腐朽菌の菌糸または胞子を接種することにより、脱リグニンを開始させることができる。その際に、イネ科植物材料を公知の手段・方法にて細断あるいは粉砕等して、チップ、ペレット、粉末などの形態にしたものに、木材腐朽菌の菌糸または胞子を接種することができる。接種する菌糸は、固体培地または液体培地にて数日〜数週間培養されたものであってもよい。固体培養は、例えば寒天平板培地で行ってもよく、液体培養は、フラスコ等において振盪しながら行ってもよい。培養の際に、脱リグニンすべきイネ科植物材料を培地に添加して(例えば、タケ粉を培地に混合して)、菌体の脱リグニン酵素活性を誘導してもよい。木材腐朽菌に使用する培地の種類や培養温度、pH、通気条件、培養時間などの培養条件は当業者が選択、決定することができる。さらに、イネ科植物材料に木材腐朽菌を接種してリグニン分解を行う際の諸条件、例えば、材料の形状(チップ、粉末など)、材料への菌の接種量、脱リグニン反応時の温度、時間、湿度、通気条件、材料の水分量などは、材料の種類、形状、サイズ、量、および木材腐朽菌の種類などの諸因子を考慮して、当業者が適宜選択、決定できるものである。例えば、タケチップを上記菌株(TUFC20056またはTUFC20057)にて脱リグニンする場合には、約15℃〜約30℃で約1ヶ月〜約6ヶ月行ってもよい。実用的には、サイロのような施設内,もしくは大型のコンテナのような容器のなかで、タケチップと培養菌株を混合し、適宜攪拌しながら、好ましくは温度および湿度をコントロールしながら処理を行なってもよい。
【0018】
また、本発明において、木材腐朽菌の抽出液をイネ科植物材料と接触させることにより脱リグニンを行ってもよい。例えば、木材腐朽菌を液体培養した場合には、培養液を濾過し、硫安やエタノールなどを用いて濃縮された抽出液を得てもよい。また、木材腐朽菌を固体培養した場合には、菌糸が生育した培養物を適当な抽出溶媒、例えば水、あるいは水とアルコール(メタノール、エタノールなど)またはグリセリンなどの混合物を用いて抽出してもよい。得られた抽出液を公知の方法にてさらに濃縮、精製してもよい。木材腐朽菌の抽出液を得る場合において、脱リグニンすべきイネ科植物材料を培地に添加して培養し、菌体の脱リグニン酵素活性を誘導しておいてもよい。
【0019】
木材腐朽菌またはその抽出物をイネ科植物材料と「接触」させるとは、木材腐朽菌の菌糸またはその抽出液に由来するリグニン分解酵素がイネ科植物材料に直接的に接触して作用し、イネ科植物材料中のリグニンが分解されうる状況にすることをいう。例えば、チップ状にしたイネ科植物材料に木材腐朽菌の菌糸または胞子をまんべんなく接種して、十分にリグニンが分解されるまで、あるいは目安として材料が十分に白化するまで培養することにより、脱リグニンを行うことができる。菌糸または胞子の接種量、培養温度や培養時間などの培養条件は、イネ科植物材料の種類やサイズ、木材腐朽菌の種類などの諸条件に応じて、当業者が選択、決定することができる。
【0020】
イネ科植物材料の脱リグニンの確認は、例えば、材料の白化を確認すること、材料中のリグニンを定量すること等によって行うことができる。材料の白化は肉眼で確認することができる。リグニンは、硫酸(クラーソン)法(木質科学実験マニュアル、日本木材学会編、文永堂出版、2000年、ISBN4-8300-4094-7)により定量することができる。これらの方法は当業者に公知である。
【0021】
さらに本発明は、リグニンに対して高い選択性を有し、ヘミセルロース、セルロースなどのホロセルロースに対して低い選択性を有するリグニン分解菌であって、イネ科植物材料の脱リグニンに好適な木材腐朽菌を提供する。これらの菌株なかで好ましいものは、独立行政法人製品評価技術基盤機構特許微生物寄託センターに寄託され受領番号AP−970を付与されたPunctularia属の菌(TUFC20056)および独立行政法人製品評価技術基盤機構特許微生物寄託センターに寄託され受領番号AP−971を付与された白色木材腐朽菌(TUFC20057)である。これらの菌株を用いることにより、イネ科植物材料の脱リグニンを効率的に行うことができ、しかも、セルロース、ヘミセルロースなどのホロセルロース成分の分解が少ないので、バイオエタノールなどの有用物質の製造等に資することができる。
【0022】
以下に実施例を示して本発明をさらに詳細かつ具体的に説明するが、実施例は単なる例示説明であり、本発明を限定するものと解釈してはならない。
【実施例1】
【0023】
(サンプリングおよび分離)
0年、半年、2年間(09年4月時点)堆積(W 2m×D 2m ×H 20cm、図1)した腐朽段階の異なるPhyllostachys pubescens(モウソウチク)のタケチップから担子菌類の子実体の発生が見られるものを中心にそれぞれ5サンプルずつ採取し、分離株の単離を行った。分離方法は、子実体が発生していたものは、マルトエキス寒天培地[MA培地、1.5%(w/v)麦芽エキス、1.5%(w/v)寒天]に多胞子分離により分離を行った。子実体の発生が見られないものは、材の腐朽部から組織分離[0.1%(v/v)のビオコートを含むMA培地]を行い、その後MA培地に移植した。このようにして単離した菌株63株を以下の実験に用いた。そして、鳥取大学附属菌類きのこ遺伝資源研究センター保有のタケを基質とする菌として分離された菌株4株、および行木(2008)の検討でスギ材に対して高い選択的リグニン分解能を示した菌株1株を供試菌とした。ポジティブコントロールとしてPhanerochaete sordida (TMIC31227)、Phlebia sp.(C4)、Trametes versicolor(TUFC10537)を用いた(表1)。以下に説明する一次スクリーニングに用いた供試菌は、ポジティブコントロールを含めて合計68株であった。
【実施例2】
【0024】
(一次スクリーニング(グァイアコール試験))
0.02%(v/v)グァイアコールを含むMA平板培地に植菌し、24℃、暗所下で2週間培養後に菌糸の直径と変色部の長さを測定した。選抜基準は呈色領域半径1cm以上を示す菌株とした。一次スクリーニングを行った結果、67菌株のうち40菌株において酸化酵素活性の指標となるグァイアコールの呈色反応が認められた。グァイアコールの呈色反応の一例を図2に示す。そのうち呈色半径が1.0cm以上を優良菌株とし36菌株を選抜した(表2、ポジティブコントロールとして用いたPhlebia sp.(C4)および20050710−03を含む)。
【0025】
【表1】
【0026】
【表2】
【0027】
一番活性が強いものはTUFC20077で3.7cmの値を示した。呈色反応が見られなかった菌株は27株あった。選抜した36菌株および、ポジティブコントロールである20050710−03、Phlebia sp.(C4)、Trametes versicolor(TUFC10537)、Phanerochaete sordida(TUFC31227)の4菌株を合わせた40菌株を二次スクリーニングに用いた。
【実施例3】
【0028】
(二次スクリーニング(脱色度試験))
タケに対する腐朽力を検定するために、タケ粉末培地[0.3%(w/v)麦芽溶液1400mLとタケ粉末1000gを混合]を口径18mmの試験管に3分の1程度詰め、タケ粉末が斜面状になるように薬さじで成形した。その後、シリコン栓で蓋をしてオートクレーブ(121℃、15分)で滅菌し、タケ粉末培地を作成した。タケ粉末スラント培地にグァイアコール呈色反応の見られた菌株を、MA培地にあらかじめ培養した各供試菌の菌糸体を接種し3ヶ月間の培養を行った。接種源にはMA平板培地に生育させた菌糸体の先端付近を同心円状に孔径5mmのコルクボーラーで打ち抜き、菌糸体ディスク2個をランダムに選んで用いた。1ヶ月ごとに目視で培地の脱色度を観察し、腐朽の観測を行った。3ヶ月培養後、強い脱色が確認された26菌株を選抜した。目視評価はコントロールと比較して、(−)〜(++++)の5段階で評価した(表2)。
【0029】
【表3】
【実施例4】
【0030】
(リグニン定量、ホロセルロース定量、および重量減少率の測定)
二次スクリーニングの結果、脱色度が一番高かったものはPunctularia sp.であり(図3に結果を例示する)、処理前のタケ粉末培地と比較して、顕著な脱色を示した。二次スクリーニングで脱色度の高かった26菌株について、リグニン定量、ホロセルロース定量、および重量減少率の測定を行った(表3に結果をまとめた)。
【0031】
【表4】
【0032】
(重量減少率の測定)
培養ビンに20mLのMA培地を入れ、供試菌を植菌し2週間培養後、あらかじめ秤量したタケ片(約1.2g)をそれぞれ1つずつビンに入れ21℃、暗所下で2ヶ月間培養した。2ヶ月後、タケのみを取り出して、タケに付着した菌糸を取り除き105℃で3日間乾燥させ、タケの重量減少率を測定した。重量減少率は次式のとおりである。
【化1】
【0033】
重量減少率は、菌株により低い(3.4%)もの(TUFC20083)から約21%の菌株(TUFC20065)まで、様々な値であった。
【0034】
(リグニン、ホロセルロース定量)
A.アルベン抽出
脱脂はアルベン抽出法を用いた。3ヶ月培養したタケチップ培地を2日間風乾させ、100mLの三角フラスコに移した。エタノール(99.5%、Wako)とn−へキサン(96.0%、Wako)の混合溶液(容量比、1:2)75mLを加え38℃、30分超音波処理し、ろ過を行った。この操作を8回繰り返し、2日間風乾させ、リグニン定量、ホロセルロース定量に用いた。
【0035】
B. リグニン定量
アルベン抽出後、風乾したタケ粉末を0.3g秤量し、72%硫酸4.5mLを3回に分け加えて100rpm、2.5時間、38℃で振とう後、170mLの純水を加えオートクレーブ(120℃、30分)処理し、あらかじめ秤量済みのガラス繊維ろ紙(GA200、size 47mm;Toyo Roshi)で吸引ろ過した。沈殿物は熱水400mL、および冷水400mLで洗浄し、105℃に設定した滅菌乾燥機に一晩かけ、デシケーターで半日放冷した。その後秤量し、増加した重量を酸不溶性リグニン量とした。ろ液は酸可溶性リグニン量を求めるため少量残し、ろ液の210nmの吸光度が0.3〜0.7の範囲になるように3%硫酸水溶液で希釈して210nmの波長で吸光度測定を行い、次式により酸可溶性リグニン量を算出した。
【化2】
【0036】
リグニン定量に関する操作は、木質科学実験マニュアル(黒田、94頁(2001))にしたがって行った。
【0037】
リグニン定量の結果を表3、図4に示す。無処理区のリグニン量は22.7%であった。ポジティブコントロールで用いたPhlebia sp.C4のリグニン残存量(15.5%)と比較して、TUFC20056(10.6%)、TUFC20076(11.5%)、TUFC20077(14.6%)、TUFC20078(14.1%)、20081010−(B)(10.5%)および20081010−B(10.7%)の6菌株(括弧内はリグニン残存量)がリグニンをより多く分解した。また、特にリグニン減少が顕著に見られた3菌株(TUFC20056および20081010−(B)、20081010−B)は、無処理区のリグニン含量と比較して50%以上の減少が確認できた。ただし、20081010−(B)、20081010−Bは再現性試験のための同一の菌株をもちいており、現在はTUFC20057を付与している。
【0038】
C. ホロセルロース定量
脱脂を終えたタケ粉末0.75gを300mL容三角フラスコに移し、蒸留水150mL、亜塩素酸ナトリウム(Wako)1.0gおよび酢酸(Wako)0.2mLを加え、100mL容三角フラスコでゆるく蓋をして、70〜80℃のウォーターバス内で1時間反応させた。実験はドラフト中で行った。次いで、亜塩素酸ナトリウム1.0gおよび酢酸0.2mLを加える操作を、内容物が白く変色するまで1時間おきに計4回繰り返した。この際、内容物を混和させるために軽く振った。内容物が白く変色した後、あらかじめ秤量済みのガラス繊維ろ紙を用いて白色内容物を吸引ろ過し、アセトンおよび冷水で洗浄後、105℃の乾燥機中で乾燥させ、デシケーター中で半日放冷後秤量し、増加した重量をホロセルロース量とした。
【0039】
ホロセルロース定量に関する操作は、木質科学実験マニュアル(黒田、94頁(2000))にしたがって行った。
【0040】
ホロセルロース定量の結果を表3、図5に示した。無処理区のホロセルロース量は73.11%であった。Phlebia sp.(C4)のホロセルロース残存量(66.9%)と比較して、TUFC20056(71.59%)が高い値を示した。次いで、20081010−(B)(66.7%)が高い値を示した。優れた白色腐朽菌とされるカワラタケTUFC10537では、59.5%と低い値を示した。他の菌株ではTUFC20072(57.8%)を除いて、62−66%の値を示した。
【0041】
リグニン定量およびホロセルロース定量の結果、タケ材のリグニン含有量22.7%およびホロセルロース含有量73.1%に対して、TUFC20056がリグニン残存量で10.6%、ホロセルロース残存量で71.6%の高い選択的リグニン分解能を示した。ポジティブコントロールとして用いたPhlebia sp.(C4)のリグニン残存量(15.5%)およびホロセルロース量(66.9%)と比較しても優れた選択的リグニン分解能を有していた。今回、リグニン含量が10.5%−14.6%の高いリグニン分解能を示した菌株は6株あり、そのうち4株は形態学的、遺伝子学的同定の結果、Punctularia sp.であることが分かった(図4参照)。これら6株を作用させた後のリグニンおよびホロセルロースの相対値を図6に示す。
【実施例5】
【0042】
(酵素活性測定)
リグニン定量およびホロセルロース定量試験の結果が良かった上位10菌株(C4、TUFC20056、20081010−B、20081010−(B)、TUFC20076、TUFC20077、TUFC20083、TUFC20079、20090803−12、20091013−08)を液体培地で2週間培養し、各リグニン分解酵素の活性を測定した(表4)。TUFC20079およびTUFC20083の2菌株は液体培地では生育しなかった。
【0043】
A.供試菌の培養
100mL容三角フラスコに10mLのタケ粉末液体培地[1.5%(w/v)麦芽エキス、0.5%(w/v)タケ粉末]を分注し、通気性のあるシリコンキャップで蓋をし、オートクレーブ(121℃、20分)で滅菌した。その後、あらかじめMA平板培地に培養した供試菌の菌糸体を接種した。接種源にはMA平板培地に生育している菌糸体の先端付近を同心円状に口径5mmのコルクボーラーで打ち抜き、菌糸体ディスク3個をランダムに選んで用いた。クリーンベンチ内で接種後、シリコンキャップでふたをし、25℃、暗所下で2週間静置培養を行った。その後、菌糸体を吸引ろ過により除去した。ろ過にはガラス繊維ろ紙(GA−200、サイズ47mm;Toyo Roshi)およびメンブランフィルター(PTFE、サイズ47mm; 孔径3.0μm、0.45μm、0.2μm;Toyo Roshi)を用いて4段階ろ過した。回収したろ液を粗酵素溶液とし酵素活性測定に用いた。
【0044】
B.リグニンペルオキシダーゼ(LiP)活性測定
71.43mmol/Lコハク酸緩衝液(pH3.0)700μL、10mmol/Lベラトリルアルコール溶液50μL、粗酵素液200μLをキュベットにそれぞれ入れ、最後に2mmol/L過酸化水素水50μLを添加し、素早く混合した。対照区には、コハク酸緩衝液700μL、ベラトリルアルコール溶液50μL、粗酵素液200μLをキュベットに入れ混合し、310nmでの吸光度を分光光度計(日本分光、東京)により180秒間測定した。吸光度が直線的に増加したことを確認し、直線部の傾きから1分間あたりの吸光度変化を求めた後、酵素活性を算出した。ネガティブコントロールは過酸化水素を加えてないものを用いた。酵素活性1Uは1分間に1μmolの反応生成物を生じる酵素量とした。その結果、ほとんどの菌においてLiP活性を確認することができなかった。ほとんどの菌が0に近い値であったが、20081010−B(0.3U/L)にわずかな活性が見られた。
【0045】
C.マンガンペルオキシダーゼ(MnP)活性測定
71.43mmol/Lマロン酸緩衝液(pH4.5)700μL、20mmol/L硫酸マンガン溶液50μL、粗酵素液200μLをキュベットにそれぞれ入れ、最後に2mmol/L過酸化水素水50μLを添加し、素早く混合した。対照区には、マロン酸緩衝液700μL、20mmol/L硫酸マンガン溶液50μL、粗酵素液200μLをキュベットに入れ混合し、270nmにおける吸光度を分光光度計により180秒間測定した。吸光度が直線的に増加したことを確認し、直線部の傾きから1分間あたりの吸光度変化を求めた後、酵素活性を算出した。酵素活性1Uは1分間に1μmolの反応生成物を生じる酵素量とした。その結果、MnP活性に関して最も活性が高い菌株は、20081010−Bで72.5U/Lの値を示した。Phlebia sp.(C4)(28.5U/L)と比較してTUFC20077(10.4U/L)以外の全ての菌がPhlebia sp.(C4)よりも高い値を示した。また、TUFC20056(68.2U/L)、20081010−B(72.5U/L)、20081010−(B)(69.9U/L)およびTUFC20076(66.4U/L)においては、C4の2倍以上の活性が確認できた。
【0046】
D.ラッカーゼ(Lac)活性測定
71.43mmol/L酒石酸緩衝液(pH3.0)700μL、0.5mmol/L 2,2’−アジノ−ビス(3−エチルベンゾチアゾリン−6−スルホネート)(ABTS)50μL、粗酵素液200μLをキュベットにそれぞれ入れ、最後に2mmol/L過酸化水素水50μLを添加し、素早く混合した。対照区には、71.43mmol/L酒石酸酸緩衝液(pH3.0)700μL、0.5mmol/L ABTS 50μL、蒸留水200μLをキュベットに入れ混合し、436nmにおける吸光度を分光光度計により3分間モニターした。吸光度が直線的に増加したことを確認し、直線部の傾きから1分間あたりの吸光度変化を求めた後、酵素活性を算出した。酵素活性1Uは1分間に1μmolの反応生成物を生じる酵素量とした。その結果、Lac活性において最も活性が高い菌株はTUFC20056であり、36.2U/Lの値を示した。C4の活性は15.0U/Lであり、全ての菌株の中で最も低い値を示した。C4と最も活性が高かったTUFC20056との差は2倍以上であった。
【0047】
E.ポリフェノールオキシダーゼ(PPO)活性測定
71.43mmol/Lコハク酸緩衝液(pH3.0)700μL、50mmol/l 2,6−ジメトキシ−フェノール溶液50μL、粗酵素液200 μL、最後に2mmol/L過酸化水素水50 μLを添加し素早く混合した。対照区には、71.43mmol/Lコハク酸緩衝液700μL、50mmol/L 2,6−ジメトキシ−フェノール溶液50μL、蒸留水200μLを入れ混合し、270nmにおける吸光度を分光光度計により180秒間測定した。吸光度が直線的に増加したことを確認し、直線部の傾きから1分間あたりの吸光度変化を求めた後、酵素活性を算出した。酵素活性1Uは1分間に1μmolの反応生成物を生じる酵素量とした。その結果、PPO活性に関して最も活性が高い菌株はTUFC20077で34.2U/Lの値を示した。C4の値(16.7U/L)と比較して、2倍以上の値であった。次いで、20081010−(B)、TUFC20056および20081010−BのPPO活性が高かった。
【0048】
【表5】
【0049】
上記結果を総合して、リグニンに対する選択性、分解能が高く、ヘミセルロース、セルロースなどのホロセルロースに対する選択性、分解能が低い木材腐朽菌として、TUFC20056、20081010−Bおよび20081010−(B)を選択した。さらなる検討により、20081010−Bおよび20081010−(B)は同一菌株であることが判明し、TUFC20057と命名した。TUFC20056株を2010年7月22日に独立行政法人製品評価技術基盤機構特許微生物寄託センターに寄託し、2010年7月29日に受領番号NITE AP−970を付与された。TUFC20057株を2010年7月22日に独立行政法人製品評価技術基盤機構特許微生物寄託センターに寄託し、2010年7月29日に受領番号NITE AP−971を付与された。
【産業上の利用可能性】
【0050】
本発明は、林業、製材業、製紙業、エネルギー分野、環境分野、製薬業などにおいて利用可能である。
【受託番号】
【0051】
独立行政法人製品評価技術基盤機構特許微生物寄託センター受領番号NITE AP−970
独立行政法人製品評価技術基盤機構特許微生物寄託センター受領番号NITE AP−971
【特許請求の範囲】
【請求項1】
木材腐朽菌またはその抽出物をイネ科植物材料と接触させることを特徴とする、イネ科植物材料の脱リグニン方法。
【請求項2】
イネ科植物がタケである請求項1記載の方法。
【請求項3】
木材腐朽菌が、Punctularia属の菌株である請求項1または2記載の方法。
【請求項4】
Punctularia属の菌株が、受領番号NITE AP−970として独立行政法人製品評価技術基盤機構特許微生物寄託センターに受領された菌株である請求項3記載の方法。
【請求項5】
木材腐朽菌が、受領番号NITE AP−971として独立行政法人製品評価技術基盤機構特許微生物寄託センターに受領された菌株である請求項1または2記載の方法。
【請求項6】
受領番号NITE AP−970として独立行政法人製品評価技術基盤機構特許微生物寄託センターに受領された菌株。
【請求項7】
受領番号NITE AP−971として独立行政法人製品評価技術基盤機構特許微生物寄託センターに受領された菌株。
【請求項1】
木材腐朽菌またはその抽出物をイネ科植物材料と接触させることを特徴とする、イネ科植物材料の脱リグニン方法。
【請求項2】
イネ科植物がタケである請求項1記載の方法。
【請求項3】
木材腐朽菌が、Punctularia属の菌株である請求項1または2記載の方法。
【請求項4】
Punctularia属の菌株が、受領番号NITE AP−970として独立行政法人製品評価技術基盤機構特許微生物寄託センターに受領された菌株である請求項3記載の方法。
【請求項5】
木材腐朽菌が、受領番号NITE AP−971として独立行政法人製品評価技術基盤機構特許微生物寄託センターに受領された菌株である請求項1または2記載の方法。
【請求項6】
受領番号NITE AP−970として独立行政法人製品評価技術基盤機構特許微生物寄託センターに受領された菌株。
【請求項7】
受領番号NITE AP−971として独立行政法人製品評価技術基盤機構特許微生物寄託センターに受領された菌株。
【図1】


【図2】

【図3】

【図4】
【図5】
【図6】
【図2】
【図3】
【図4】
【図5】
【図6】
【公開番号】特開2012−39904(P2012−39904A)
【公開日】平成24年3月1日(2012.3.1)
【国際特許分類】
【出願番号】特願2010−182279(P2010−182279)
【出願日】平成22年8月17日(2010.8.17)
【出願人】(504150461)国立大学法人鳥取大学 (271)
【Fターム(参考)】
【公開日】平成24年3月1日(2012.3.1)
【国際特許分類】
【出願日】平成22年8月17日(2010.8.17)
【出願人】(504150461)国立大学法人鳥取大学 (271)
【Fターム(参考)】
[ Back to top ]