
新規人工塩基対及びその利用
【課題】新規人工塩基対に基づく核酸、ならびに当該核酸の調製方法、利用を提供する。
【解決手段】置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドと、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドとが形成する塩基対。置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドを含む核酸を鋳型として転写、逆転写又は複製を行い、前記置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を有するヌクレオチドの相補的な位置に、6位置換された9H−プリン−9−イル基を有するヌクレオチドを組み込む、ことを含む、核酸の調製方法。
【解決手段】置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドと、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドとが形成する塩基対。置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドを含む核酸を鋳型として転写、逆転写又は複製を行い、前記置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を有するヌクレオチドの相補的な位置に、6位置換された9H−プリン−9−イル基を有するヌクレオチドを組み込む、ことを含む、核酸の調製方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、新規人工塩基対に基づく核酸及びその利用に関する。
【背景技術】
【0002】
生体高分子である核酸(DNA,RNA)は、生命活動に必要となる膨大な量の遺伝情報を、僅か4種類の塩基の組合せからなる配列として記録している。また、核酸は自分自身を鋳型としてDNAポリメラーゼにより自己複製し、さらにRNAポリメラーゼによる転写、リボソームによる翻訳というプロセスを介して、DNAからDNAへ、DNAからRNAへ、RNAからタンパク質へと遺伝情報を伝達する。この遺伝情報の複製と伝達を可能としているのは排他的な塩基対形成(A・T/U、G・C)のルールである。また、核酸は、多様な高次構造を形成して様々な機能を発揮する。例えば、in vitroセレクション法によって、アプタマーやリボザイムの機能を有する新規核酸がこれまでに多数見つかってきたこともその一つである。
【0003】
しかし、20種類のアミノ酸からなるタンパク質に比べて、天然の核酸には4種類の塩基(2種類の塩基対)しかないという事実は、核酸の化学的・物理的多様性に限界を与えている。たとえば、生体中のtRNA、rRNA、mRNA等の機能性RNAは自分自身の構造を安定化したりRNA・RNA間、RNA・タンパク質問相互作用を安定化するために、様々な修飾塩基を利用している。従って、新規機能性核酸の開発において、新たな塩基(対)のレパートリーを増やすことは大変有益であると考えられる。
【0004】
核酸のさらなる機能拡張をめざして、非天然型塩基をもつヌクレオシド又はヌクレオチドの創製への取り組みが行われている。核酸に修飾塩基(もしくは非天然型塩基)を導入する手法として、1)化学合成により直接導入する方法、2)核酸合成酵素により導入する方法が考えられる。1)の場合は、アミダイトユニットの安定性や塩基部分の適当な保護基が存在すること等化学合成上の問題の解決が必要である。また、これらの問題が解決されれば様々な非天然型塩基を位置選択的に導入できるが、その核酸の増幅は困難であり、長鎖長の核酸の合成も難しくなる。2)の場合は、もし、基質が酵素に認識され、人工塩基対間で相補的に複製、転写されれば、その核酸の増幅・調製が可能となるが、そのような基質や塩基対(非天然型ヌクレオチド)も開発途中である。
【0005】
非天然型人工塩基対の研究の背景
天然の二本鎖DNA中のAとT及びGとCは、それぞれ特異的な水素結合を介して互いに「排他的」な塩基対を形成している。非天然型塩基対の研究は塩基間の水素結合を利用した組み合わせや塩基の疎水性を利用した組み合わせ等が知られているが、複製、転写、翻訳の工程全てにおいて天然型塩基対に拮抗し得るものは見出されていない。このような状況において複製、転写、翻訳の少なくとも一工程において天然型塩基対に拮抗し得る非天然型塩基対は特異的な有用性を持つものである。
【0006】
最近の非天然型の非水素結合塩基対の研究により、複製における効率的で適合したヌクレオチドの取り込みのために、対合塩基間の水素結合は絶対的に必要なものではなく、その代わり、塩基間の形状の相補性が複製において重要な役割を果たすことが、証明されてきた(非特許文献5−6)。よって、非水素結合塩基対は、新規なバイオテクノロジーを創造するための、遺伝子アルファベットおよびコードの拡張のための潜在的な候補である(非特許文献7−9)。
【0007】
一方、転写に関しては、RNAポリメラーゼによって高い選択性および効率で認識され
る非水素結合塩基対は、まだ報告されていない。具体的には、本発明者は、これまでに2−アミノ−6−(2−チエニル)−9H−プリン−9−イル基(s)と2−オキソ−(1H)ピリジン−3−イル基(y)との組み合わせによる水素結合の相互作用を介した(s−y)塩基対を開発し、試験管内で非天然型アミノ酸を部位特異的に含むタンパク質の合成に成功している(特許文献1、非特許文献14)。
【0008】
さらに、特許文献3は、2−アミノ−6−(2−チアゾリル)プリン−9−イル基(v)と2−オキソ−(1H)ピリジン−3−イル基(y)との組み合わせによる(v−y)塩基対を開示している(特許文献3)。
【0009】
なおsの核酸塩基対としては天然型4塩基、(s−y)の他、(s−z)(zは、2−オキソ−1,3−ジヒドロイミダゾール−1−イル基を意味する)、(s−5位置換y)が、本発明前に報告されていた。これらの(s−y)、(s−z)、(s−5位置換y)の非天然型の塩基対は全て水素結合相互作用を含むものである(特許文献2、非特許文献13−15)。
【0010】
しかしながら、これらの非天然型塩基対のうち(s−y)と(s−5位置換y)は、y
をDNA鋳型としてsをRNAへ挿入する転写工程については、選択性が高くなく、また、(s−z)は、選択性は高いが、必ずしも高収率ではなかった。
【0011】
【化1】

一方、2−ホルミル−1H−ピロール−1−イル基(Pa)は、天然型4塩基のほか9−メチルイミダゾ[(4,5)−b]ピリジン(Q)と非水素結合塩基対を形成しうることが(Pa−Q)で報告されている(非特許文献9)。
【0012】
【化2】
【0013】
しかしながら、上記非水素結合塩基対もDNAポリメラーゼによる翻訳、又は、逆転写酵素によるDNA合成が報告されているのみであり、RNAポリメラーゼによる認識については知られていない。T7様RNAポリメラーゼの単一サブユニットは、構造およびメカニズムの点においてDNAポリメラーゼに類似するが(非特許文献16−17)、T7
RNAポリメラーゼ複合体の最近の構造解析によって、RNAポリメラーゼとDNAポリメラーゼ間の相違が明らかにされた(非特許文献17−18)。転写では、入ってくる基質が鋳型中の塩基と「開いた」コンフォメーションで塩基対を形成し、そして、形成された塩基対は、「開いた」コンフォメーションから「閉じた」コンフォメーションに遷移する間、維持される(非特許文献18)。しかしながら、複製では、「閉じた」コンフォメーションへの遷移の後に塩基対形成が開始される。このことから、転写では複製の場合よりも、対合する塩基間の水素結合が重要であると考えられ、そして、非水素結合塩基対が転写において機能しうるか、という問題が生じる。
【0014】
非天然型塩基によって仲介される特異的な転写が可能になれば、促進された機能性を有する新規なRNA分子を創造し、そして、遺伝子コードが拡張されることとなる(非特許文献1−4)。
【0015】
RNAへの蛍光プローブの導入
最近、RNA医療の関心が高まる中で、RNA中への蛍光プローブの導入は、RNAの蛍光標識化法、そして、RNAの複雑な高次構造を解析するための重要なテクノロジーの1つになっている。後者の構造解析用の蛍光プローブとしては、蛍光性の塩基(2−アミノプリンなど)をもつヌクレオチドを利用する方法が従来から用いられている。この点、上述した2−アミノ−6−(2−チエニル)−9H−プリン−9−イル基(s)は蛍光塩基であり、これを塩基として含むヌクレオチドは強い蛍光特性を有する。従って、sを塩基として含むヌクレオチドをRNA中の任意の部位に導入することができれば、有用な蛍光プローブの調製が可能となる。
【0016】
しかし、これらの蛍光性塩基を有するヌクレオチドを転写によりRNAに導入するための公知の方法は、収率の面で必ずしも十分なものではなかった。従って、sなどの蛍光性塩基を有するヌクレオチドを、転写によりRNA中に部位特異的に導入することは実質的にできなかったため、長鎖RNAの高次構造の解析は難しかった。
【0017】
転写で蛍光プローブをRNA中に導入する方法が開発されれば、溶液中でのRNAの構造のダイナミックスを調べることが可能になる。さらに、この手法は、RNAを利用した医薬品の開発にも役立つ。そこで、不可能であると考えられている長鎖RNA中への蛍光プローブの導入を可能とする方法の確立が重要となる。これにより、生体内の機能性RNAの構造解析、ならびに、他分子との複合体の構造解析に供する新たな手法の開発が可能になる。
【0018】
複製、転写、翻訳の工程全てにおいて天然型塩基対に拮抗し得る非天然型の塩基であって、かつ、蛍光特性を付与する塩基を有するヌクレオチドを核酸へ導入するための、新規な人工塩基対の開発が希求される。
【特許文献1】WO2001/005801
【特許文献2】WO2004/007713
【特許文献3】WO2005/026187
【非特許文献1】Benner, S. A., Burgstaller, P., Battersby, T. R. & Jurczyk, S. in The RNA World (eds Gesteland, R. F., Cech, T. R. & Atkins, J. F.) 163−181 (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1999).
【非特許文献2】Bergstrom, D. E. Orthogonal base pairs continue to evolve. Chem. Biol. 11, 18−20 (2004).
【非特許文献3】Wang, L. & Schultz, P. G. Expanding the genetic code. Chem. Commun. 1−11 (2002).
【非特許文献4】Hendrickson, T. L., de Crecy−Lagard, V. & Schimmel, P. Incorporation of nonnatural amino acids into proteins. Annu. Rev. Biochem. 73, 147−176 (2004).
【非特許文献5】Morales, J. C. & Kool, E. T. Efficient replication between non−hydrogen−bonded nucleoside shape analogs. Nat. Struct. Biol. 5, 950−954 (1998).
【非特許文献6】Kool, E. T. Hydrogen bonding, base stacking, and steric effects in DNA replication. Annu. Rev. Biophys. Biomol. Struct. 30, 1−22 (2001).
【非特許文献7】McMinn, D. L. et al. Efforts toward expansion of the genetic alphabet: DNA polymerase recognition of a highly stable, self−pairing hydrophobic base. J. Am. Chem. Soc. 121, 11585−11586 (1999).
【非特許文献8】Wu,Y. et al. Efforts toward expansion of the genetic alphabet: optimization of interbase hydrophobic interactions. J. Am. Chem. Soc. 122, 7621−7632 (2000).
【非特許文献9】Mitsui, T. et al. An unnatural hydrophobic base pair with shape complementarity between pyrrole−2−carbaldehyde and 9−methylimidazo[(4,5)−b]pyridine. J. Am. Chem. Soc. 125, 5298−5307 (2003).
【非特許文献10】Piccirilli, J. A., Krauch, T., Moroney, S. E. & Benner, S. A. Enzymatic incorporation of a new base pair into DNA and RNA extends the genetic alphabet. Nature 343, 33−37 (1990).
【非特許文献11】Switzer, C. Y., Moroney, S. E. & Benner, S. A. Enzymatic recognition of the base pair between isocytidine and isoguanosine. Biochemistry 32, 10489−10496 (1993).
【非特許文献12】Tor, Y. & Dervan, P. B. Site−specific enzymatic incorporation of an unnatural base, N6−(6−aminohexyl)isoguanosine, into RNA. J. Am. Chem. Soc. 115, 4461−4467 (1993).
【非特許文献13】Ohtsuki, T. et al. Unnatural base pairs for specific transcription. Proc. Natl. Acad. Sci. U.S.A. 98, 4922−4925 (2001).
【非特許文献14】Hirao, I. et al. An unnatural base pair for incorporating amino acid analogs into proteins. Nat. Biotechnol. 20, 177−182 (2002).
【非特許文献15】Hirao, I. et al. A two−unnatural−base−pair system toward the expansion of the genetic code. J. Am. Chem. Soc. 126, 113298−113305 (2004).
【非特許文献16】Cheetham, G. M., Jeruzalmi, D. & Steitz, T. A. Structural basis for initiation of transcription from an RNA polymerase−promoter complex. Nature 399, 80−83 (1999).
【非特許文献17】Yin, Y. W. & Steitz, T. A. The structural mechanism of translocation and helicase activity in T7 RNA polymerase. Cell 116, 393−404 (2004).
【非特許文献18】Temiakov, D. et al. Structural basis for substrate selection by T7 RNA polymerase. Cell 116, 381−391 (2004).
【非特許文献19】Hirao, I., Fujiwara, T., Kimoto, M. & Yokoyama, S. Unnatural base pairs between 2− and 6−substituted purines and 2−oxo(1H)pyridine for expansion of the genetic alphabet. Bioorg. Med. Chem. Lett. 14, 4887−4890 (2004).
【非特許文献20】Jovine, L., Djordjevic, S. & Rhodes, D. The crystal structure of yeast phenylalanine tRNA at 2.0 Å resolution: cleavage by Mg2+ in 15−year old crystals. J. Mol. Biol. 301, 401−414 (2000).
【非特許文献21】Law, S. M., Eritja, R., Goodman, M. F. & Breslauer, K. J. Spectroscopic and calorimetric characterizations of DNA duplexes containing 2−aminopurine. Biochemistry 35, 12329−12337 (1996).
【非特許文献22】Holz, B., Klimasauskas, S., Serva, S. & Weinhold, E. 2−Aminopurine as a fluorescent probe for DNA base flipping by methyltransferases. Nucleic Acids Res. 26, 1076−1083 (1998).
【非特許文献23】Rist, M. J. & Marino, J. P. Association of an RNA kissing complex analyzed using 2−aminopurine fluorescence. Nucleic Acids Res. 29, 2401−2408 (2001).
【非特許文献24】Bain, J. D., Switzer, C., Chamberlin, A. R. & Benner, S. A. Ribosome−mediated incorporation of a non−standard amino acid into a peptide through expansion of the genetic code. Nature 356, 537−539 (1992).
【非特許文献25】Ludwig, J. & Eckstein, F. Rapid and efficient synthesis of nucleoside 5’−O−(1−thiotriphosphates), 5’−triphosphates and 2’,3’−cyclophosphorothioates using 2−chloro−4H−1,3,2−benzodioxaphosphorin−4−one. J. Org. Chem. 54, 631−635 (1989).
【非特許文献26】Frugier, M., Helm, M., Felden, B., Giege, R. & Florentz, C. Sequences outside recognition sets are not neutral for tRNA aminoacylation. J. Biol. Chem. 273, 11605−11610 (1998).
【非特許文献27】Kao, C., Rudisser, S. & Zheng, M. A simple and efficient method to transcribe RNAs with reduced 3’ heterogeneity. Methods 23, 201−205 (2001).
【非特許文献28】Mitsui,T., Kimoto,M., Sato,A., Yokoyama,S. & Hirao, I., Bioorg.Med.Chem.Lett.,13, 4515−4518 (2003)
【非特許文献29】Fujiwara, T., Kimoto, M., Sugiyama, H., Hirao, I. & Yokoyama, S., Bioorg.Med.Chem.Lett.,11, 2221−2223 (2001)
【非特許文献30】Mitsui, T., Kimoto, M., Harada, Y., Yokoyama, S. & Hirao, I., J.Am.Chem.Soc.,127, 8652−8658 (2005)
【発明の開示】
【発明が解決しようとする課題】
【0019】
本発明は、新規人工塩基対に基づく核酸を提供することを目的とする。具体的には、本発明の核酸は、置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドと、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドとが塩基対を形成している。
【0020】
限定されるわけではないが、本発明の核酸は好ましくは、前記置換された若しくは無置
換の2−ホルミル−1H−ピロール−1−イル基が、
A1)2−ホルミル−1H−ピロール−1−イル基;
A2)2−ホルミル−4−(1−プロピン−1−イル)−1H−ピロール−1−イル基;
A3)2−ホルミル−4−メチル−1H−ピロール−1−イル基;及び
A4)2−ホルミル−4−エチニル−1H−ピロール−1−イル基
からなるグループから選択され、そして
前記6位置換された9H−プリン−9−イル基が、
B1)2−アミノ−6−(2−チエニル)−9H−プリン−9−イル基;
B2)6−(2−チエニル)−9H−プリン−9−イル基;
B3)2−アミノ−6−(4−メチル−2−チエニル)−9H−プリン−9−イル基;
B4)6−(4−メチル−2−チエニル)−9H−プリン−9−イル基;
B5)2−アミノ−6−(5−メチル−2−チエニル)−9H−プリン−9−イル基;
B6)6−(5−メチル−2−チエニル)−9H−プリン−9−イル基;
B7)2−アミノ−6−(2−チアゾリル)−9H−プリン−9−イル基;
B8)6−(2−チアゾリル)−9H−プリン−9−イル基;
B9)2−アミノ−6−(4−メチル−2−チアゾリル)−9H−プリン−9−イル基;
B10)6−(4−メチル−2−チアゾリル)−9H−プリン−9−イル基;
B11)2−アミノ−6−(5−メチル−2−チアゾリル)−9H−プリン−9−イル基;及び
B12)6−(5−メチル−2−チアゾリル)−9H−プリン−9−イル基
からなるグループから選択される。
【0021】
本発明の核酸は、好ましくは転写、逆転写、複製又は翻訳の工程で、塩基対を形成している。
本発明はまた、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを含む核酸を調製する方法を提供する。本発明の調製方法は、置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドを含む核酸を鋳型として転写、逆転写又は複製を行い、前記置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を有するヌクレオチドの相補的な位置に、6位置換された9H−プリン−9−イル基を有するヌクレオチドを組み込む、ことを含む。
【0022】
本発明はさらに、上記本発明の方法によって調製された、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを含む核酸を提供することを目的とする。
【0023】
本発明はさらにまた、本発明の核酸における、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを含む局部領域の立体構造の解析方法を提供することを目的とする。本発明の解析方法は、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド中の、塩基に由来する蛍光強度を、温度変化やイオン(例えば、マグネシウム)濃度などの変化等、環境を変えることにより測定することを含む。
【0024】
本発明の解析方法は、一態様において、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド中の、塩基に由来する蛍光強度が、温度の上昇に伴い実質的に上昇する場合には、生体内温度下で、前記ヌクレオチドは立体配座において周囲に存在するヌクレオチドと積み重なっていると判断し、そして、蛍光強度が温度の上昇に伴い実質的に低下又は上昇しない場合には、生体内温度下で、前記ヌクレオチドは外部に暴露していると判断する。
【0025】
本発明はまた、核酸の二本鎖形成の検出方法を提供することを目的とする。本発明の検出方法は、
I) i)本発明の核酸の調製方法によって調製された、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを含む核酸、と、
ii)5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドであって、前記塩基の5位に、5−カルボキシフルオレセイン(5−FAM)、6−カルボキシフルオレセイン(6−FAM)、5−カルボキシテトラメチルローダミン(5−TAMRA)、6−カルボキシテトラメチルローダミン(6−TAMRA)、 5−ジメチルアミノナフタレン−1−スルホン酸(DANSYL)、5−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(5−HEX)、6−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(6−HEX)、5−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(5−TET)、6−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(6−TET)、5−カルボキシ−X−ローダミン(5−ROX)、及び6−カルボキシ−X−ローダミン(6−ROX)からなるグループから選択される蛍光色素が、直接又はリンカーを介して結合している前記ヌクレオチド、を含む核酸、とをハイブリダイズさせ、そして、
II) 蛍光スペクトルの変化を測定する
ことを含む。
【0026】
本発明はさらに、
i) 6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド
並びに、
ii) 5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドであって、前記塩基の5位に、5−カルボキシフルオレセイン(5−FAM)、6−カルボキシフルオレセイン(6−FAM)、5−カルボキシテトラメチルローダミン(5−TAMRA)、6−カルボキシテトラメチルローダミン(6−TAMRA)、 5−ジメチルアミノナフタレン−1−スルホン酸(DANSYL)、5−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(5−HEX)、6−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(6−HEX)、5−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(5−TET)、6−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(6−TET)、5−カルボキシ−X−ローダミン(5−ROX)、及び6−カルボキシ−X−ローダミン(6−ROX)からなるグループから選択される蛍光色素が、直接又はリンカーを介して結合している、前記ヌクレオチド
を、同一の鎖上に含む核酸を調製する方法を提供することを目的とする。本発明の調製方法は、
iii) 置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチド
並びに
iv) 6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド
を含む核酸を鋳型として転写、逆転写又は複製を行い、前記iii)のヌクレオチドの相補的な位置に、前記i)のヌクレオチドを組み込み、そして、前記iv)のヌクレオチドの相補的な位置に、前記ii)のヌクレオチドを組み込む
ことを含む。
【0027】
本発明はさらにまた、上記方法によって調製された核酸も提供する。
本発明はまた、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドと5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドとを、同一の鎖上に含む、本発明の核酸のステム−ループ(ヘアピン)構造形成の検
出方法を提供することを含む。本発明の検出方法は、
1) 本発明の核酸において、i)のヌクレオチドと、ii)のヌクレオチドとをハイブリダイズさせ、そして、
2) 蛍光スペクトルの変化を測定する
ことを含む。
【課題を解決するための手段】
【0028】
本発明者らは、上記問題解決のために、以前に自分たちが開発したいくつかの非天然型の塩基を組み合わせることによって構築される塩基対を研究した。その結果、2−アミノ−6−(2−チエニル)プリン−9−イル(s)(非特許文献13、14)と2−ホルミル−1H−ピロール−1−イル(Pa)(非特許文献9)(図1A)の組み合わせが、転写において高度な選択性および効率を示すことを見出した。即ち、PaヌクレオチドをDNAの鋳型に挿入しRNAへの転写反応を行うと、生成するRNA中の、鋳型DNAのPaの相補的な位置に選択的にかつ高収率でsが導入されることを発見し、本発明を想到した。
【0029】
基質sはT7 RNA ポリメラーゼによって、DNA鋳型から生成されるRNAにおいて、鋳型中のPaの相補的な位置に部位特異的に挿入することができる。プリン塩基であるsは、水素結合を可能とする置換基(1位の水素受容基と2位の水素供与のアミノ基)を有する。しかし、Paは対合表面にそのような親水性の基を有しない。本発明者らは、Pa塩基のホルミル基中の酸素は、二本鎖DNAの小溝(minor groove)に向かい、ポリメラーゼと相互作用していて、塩基対形成における水素受容基としては働いていないことを見出している(非特許文献9)。本発明において、s−Pa対は有意な水素結合相互作用を有しないが、5員環塩基Paはsと形状相補性を共有すると考えられる。
【0030】
本発明の人工塩基対に基づく核酸
よって、本発明はその一態様において、置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドと、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドとが塩基対を形成している、核酸を提供する。
【0031】
本発明における「ヌクレオシド」とは、核酸塩基と糖の還元基とがグリコシド結合によって結合した配糖体化合物を意味する。なお、「核酸塩基」は、アデニン、グアニン、シトシン、チミン、ウラシル、及びこれら塩基の誘導体も含む概念である。「誘導体」の種類は特に限定されるものではないが、具体的には、例えば、置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基に相当する塩基や、6位置換された9H−プリン−9−イル基に相当する塩基、などが挙げられる。「ヌクレオチド」は、前記ヌクレオシドの糖部分が、リン酸とエステルをつくっている化合物をいう。より好ましくは、一、二、又は三リン酸エステルである。ヌクレオシド又はヌクレオチドの糖部分はリボフラノシル、2’−デオキシリボフラノシル、あるいはハロゲンなどの置換基を2’位に有する2’−置換リボフラノシルであってもよく、また、リン酸部分は、チオリン酸であってもよい。つまり、糖部分及びリン酸部分は、公知のヌクレオシド、ヌクレオチド、あるいはこれらの誘導体にみられる構成をとっていればよい。糖部分がリボフラノシルであるリボヌクレオチドはRNAの構成成分となり、デオキシリボフラノシルであるデオキシリボヌクレオチドはDNAの構成成分となる。
【0032】
本発明の「置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基」は、以下の一般式で表される構造を有する。
【0033】
【化3】
【0034】
ここにおいて、ピロール環の4位のRは、水素であるか、あるいは、置換された若しく
は無置換のC1−C3アルキル基、置換された若しくは無置換のC2−C3アルケニル基、又は置換された若しくは無置換のC2−C3アルキニル基から選択される置換基によって置換されていてもよい。上記アルキル基、アルケニル基又はアルキニル基等は、さらに、低級アルキル基、ハロゲン基、ヒドロキシル基、アミノ基、アルキルアミノ基、及び芳香族複素環からなるグループより独立して選択される一つまたはそれより多くの基で置換されていてもよい。本発明の置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を、本明細書において文脈により、「Pa」又は「Pa類似体」と呼称する。
【0035】
限定されるわけではないが、前記置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基は、
A1)2−ホルミル−1H−ピロール−1−イル基(Pa);
A2)2−ホルミル−4−(1−プロピン−1−イル)−1H−ピロール−1−イル基;
A3)2−ホルミル−4−メチル−1H−ピロール−1−イル基;及び
A4)2−ホルミル−4−エチニル−1H−ピロール−1−イル基
からなるグループから選択されてもよい。より好ましくは、2−ホルミル−1H−ピロール−1−イル基である。
【0036】
本発明の「置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基」(「Pa」又は「Pa類似体」)を有するヌクレオシド、ヌクレオチドは、公知の方法を用いて合成することが可能である。その出発原料である例えば、ピロール−2−カルボアルデヒドは、例えば、Aldrich社 [1003−29−8]、Merck社 [807574]から購入することができる。また、Pa誘導体は、基本的にPaから誘導することによって合成できる。例えば、Paの4位にプロピンを導入した誘導体は、Bioorg.Med.Chem.Lett.,13,p.4515−4518(2003)(非特許文献28)に記載されている。
【0037】
本発明の「6位置換された9H−プリン−9−イル基」は、以下の一般式で表される構造を有する。
【0038】
【化4】
【0039】
[ここにおいて、
R1は、水素又はアミノ基であり、
R2は、置換された又は無置換の2−チエニル基又は2−チアゾリル基である。]
R2のチエニル基又はチアゾリル基は、無置換であるかあるいは、4位及び/又は5位が、メチル基、アミノ基、ニトロ基及びヒドロキシ基からなるグループより独立して選択される一つまたはそれより多くの基で置換されていてもよい。本発明の6位置換された9H−プリン−9−イル基のうち、R2が置換された又は無置換の2−チエニル基のものを、本明細書において文脈により、「s」又は「s類似体」と呼称する。本発明の6位置換された9H−プリン−9−イル基のうち、R2が置換された又は無置換の2−チアゾリル基のものを、本明細書において文脈により、「v」又は「v類似体」と呼称する。
【0040】
限定されるわけではないが、前記6位置換された9H−プリン−9−イル基は、
B1)2−アミノ−6−(2−チエニル)−9H−プリン−9−イル基(s);
B2)6−(2−チエニル)−9H−プリン−9−イル基(s’);
B3)2−アミノ−6−(4−メチル−2−チエニル)−9H−プリン−9−イル基;
B4)6−(4−メチル−2−チエニル)−9H−プリン−9−イル基;
B5)2−アミノ−6−(5−メチル−2−チエニル)−9H−プリン−9−イル基;
B6)6−(5−メチル−2−チエニル)−9H−プリン−9−イル基;
B7)2−アミノ−6−(2−チアゾリル)−9H−プリン−9−イル基(v);
B8)6−(2−チアゾリル)−9H−プリン−9−イル基;
B9)2−アミノ−6−(4−メチル−2−チアゾリル)−9H−プリン−9−イル基;
B10)6−(4−メチル−2−チアゾリル)−9H−プリン−9−イル基;
B11)2−アミノ−6−(5−メチル−2−チアゾリル)−9H−プリン−9−イル基;及び
B12)6−(5−メチル−2−チアゾリル)−9H−プリン−9−イル基
からなるグループから選択されてもよい。より好ましくは、2−アミノ−6−(2−チエニル)−9H−プリン−9−イル基である。
【0041】
上記塩基のうち、B1)は、「s」であり、B2)ないしB6)は「s類似体」である。また、B7)は、「v」であり、B8)ないしB12)は「v類似体」である。
さらにより好ましくは、前記置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基が、2−ホルミル−1H−ピロール−1−イル基であり、そして、前記6位置換された9H−プリン−9−イル基が、2−アミノ−6−(2−チエニル)−9H−プリン−9−イル基である。
【0042】
本発明の「6位置換された9H−プリン−9−イル基」、及びこれを含むヌクレオシド、ヌクレオチドは、公知の方法を用いて合成することが可能である。具体的には例えば、本明細書の実施例1には、2−N−フェノキシアセチル−6−(2−チエニル)−9−(2,3−ジ−O−アセチル−1−β−D−リボフラノシル)プリンから、2−アミノ−6−(2−チエニル)−9−(1−β−D−リボフラノシル)プリン 5’−三リン酸(sTP)を合成する方法を開示した。
【0043】
またsの合成は、例えば、Bioorg.Med.Chem.Lett.,11,p.2221−2223(2001)(非特許文献29)の第2頁の第二段落に記載されている。また、vの合成は、例えば特許文献3の段落0026、0027、図5−6等に開示されている。あるいは、J.Am.Chem.Soc.,127,p.8652−8658(2005)(非特許文献30)の第2頁の右側の段落に開示されている。
【0044】
s、v以外の、本発明の「6位置換された9H−プリン−9−イル基」を有するヌクレオシド、ヌクレオチドも、s又はvのいずれかの合成方法と同様の方法を用いて合成することができる。例えば、フランを結合したo は、sと同様の合成法で合成可能である(非
特許文献29)。
【0045】
本発明は、置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドと、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドとが塩基対を形成している、核酸を提供する。本明細書において「核酸」とは、1より多くのヌクレオチドが、5’→3’の方向に結合した核酸鎖の分子を意味する。本発明の核酸は、一本鎖又は二本鎖のRNA又はDNAを含む。二本鎖は、DNA/DNA、RNA/RNA、又はDNA/RNAであってもよい。また、DNAには、RNAを鋳型として逆転写してなるcDNAも含まれる。あるいは、核酸は3本鎖、4本鎖等も形成しうる。
【0046】
本発明者らは、核酸のさらなる機能拡張をめざして、非天然型塩基をもつヌクレオシド又はヌクレオチドの創製へ取り組んだ。新たに開発された人工塩基対の態様として、置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基(例:Pa)として有するヌクレオチドと、6位置換された9H−プリン−9−イル基を塩基(例:s)として有するヌクレオチドとの塩基対が含まれる。(図1)。これにより特に、Paを含むDNA鋳型を用いたT7 RNAポリメラーゼによる転写において、RNAへのsの特異的取り込みが可能となった。
【0047】
sおよびPa間では有意な水素結合相互作用が存在しないにもかかわらず(非水素結合s−Pa対)、転写におけるs−Pa対合の効率および選択性は、天然の塩基対合のそれと同程度に高いものである。このs−Pa対は、先に開発された親水性s−z対よりも高い効率を示す。s及びPaの相補的な形状は互いに適合するが、天然のプリン類及びピリミジン類の形状とは相違する。この特異的な立体化学的適合が、天然塩基との非規範的対合を排除し、その結果、転写の際にs−Paの高い選択性が得られると考えられる。このように、形状−相補性は転写、及び複製における特異的な塩基対合に重要な役割を果たす。
【0048】
本発明の、置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドと、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドとは、異なる2本の核酸上に存在して塩基対により二本鎖を形成しうる。あるいは、両ヌクレオチドは同一の一本の核酸上に存在してもよい。この場合、塩基対により、一本鎖はループ構造を形成してもよい。
【0049】
本発明の置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチド、あるいは6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドは、転写、複製又は逆転写反応により、DNA又はRNA等の核酸に取り込むことが可能である。あるいは、天然型塩基を有するヌクレオシド又はヌクレオチドと同様に、化学合成によってDNA又はRNAに取り込んでもよい。
【0050】
転写、複製又は逆転写反応は公知の方法に従って行うことが可能である。限定されるわけではないが、例えば、転写反応はT7 RNAポリメラーゼ(Takara等)、複製反応は、クレノウフラグメント(KF)、逆転写反応はAMV Reverse Transcriptase XL (AMV−RT)(Life Science社)を使用することが可能である。複製反応は、反応中に本発明のヌクレオチドが除去されてしまうのを防ぐために、例えば、3’→5’エキソヌクレアーゼ活性をもたないTaq DNAポリメラーゼ(Takara TaqTM)を用いて、置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドを含むプライマーによる鋳型DNAのPCR増幅も可能である。
【0051】
限定されるわけではないが、本発明の核酸は一態様において、核酸の転写、逆転写、複製又は翻訳の工程で、塩基対を形成している。本発明の核酸が、転写工程で塩基対を形成している場合、前記置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドがDNAの一部であり、そして、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドがRNAの一部でありうる。
【0052】
核酸の調製方法
本発明はまた、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを含む核酸を調製する方法を提供する。本発明の調製方法は、置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドを含む核酸を鋳型として転写、逆転写又は複製を行い、前記置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を有するヌクレオチドの相補的な位置に、6位置換された9H−プリン−9−イル基を有するヌクレオチドを組み込む、ことを含む。
【0053】
2−ホルミル−1H−ピロール−1−イル基を有するヌクレオチド、並びに6位置換された9H−プリン−9−イル基を有するヌクレオチドの定義については、本明細書の「本発明の人工塩基対に基づく核酸」の項目において前述した通りである。
【0054】
本明細書中の実施例1−2でも明らかにされたように、鋳型中に置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドが2つまたはそれ以上連続して存在する場合でも、転写反応は進行し、相補的な位置に6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドが取り込まれる。よって、本発明の方法により従来不可能であった、非天然型塩基sが2つ又はそれより多く隣接して配置されるDNA及びRNAの作成も可能となった。よって、本発明の調製方法は、一態様において、鋳型において、前記置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドが2個以上存在する。
【0055】
本発明はさらに、上記本発明の方法によって調製された、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを含む核酸を提供することを目的とする。
【0056】
本発明のヌクレオチドが組み込まれた核酸は、tRNA、mRNA、アンチセンスDNA若しくはRNA、リボザイム又はアプタマーとして使用されうる。アンチセンスDNA若しくはRNAとは、ある特定の遺伝子の発現を抑えるDNA又はRNAである。標的とする遺伝子配列(センス鎖)の全長又は部分配列に対して相補的という意味で名付けられた。人為的に遺伝子発現を調節する手法として使用されうる。本発明のヌクレオチドが組み込まれたアンチセンスDNA又はRNAは、非天然型塩基を含むため標的に対する相補性が天然型塩基のみを使用した場合と比較して異なるものを創製しうる。リボザイムは、RNAを構成成分とする触媒の総称である。アプタマーは、in vitroセレクション法によって得られた、タンパク質等の特定の分子に結合する機能を有する核酸である
また、本発明の、ヌクレオチドが組み込まれた核酸(DNA又はRNA)(例えば、mRNA、合成RNA)は、タンパク質、ペプチドの全体又は一部をコードするものであってもよい。本発明の核酸は遺伝子断片やプローブなどとして使用されうる。天然の遺伝子の一部又は全部を本発明の核酸で置換した態様、天然の遺伝子に本発明のヌクレオチドを1個又はそれより多く付加したもの、又はこれらを組み合わせたものも本発明に包含される。
【0057】
さらにまた、本発明の非天然型塩基を有するヌクレオチドが組み込まれた核酸は、RNA干渉(RNA interference、RNAi)においても利用可能である。R
NA干渉は、二本鎖RNA(dsRNA)によってその配列特異的にmRNAが分解され、その結果遺伝子の発現が抑制される現象である。RNA干渉の典型的な例としては、dsRNAは、RNaseIIIファミリーに属するダイサー(Dicer)により、3’末端の側に2塩基程度のオーバーハングを有する約21塩基−23塩基のsiRNA(short interfering RNA)にプロセッシングされる。siRNAはRISCと呼ばれるsiRNA−蛋白質複合体に取り込まれ、配列特異的にmRNAを分解する。RNA干渉は、哺乳動物(ヒト、マウス等)、線虫、植物、ショウジョウバエ、菌類などの広範な生物種間で保存されている現象であることが示されている。本発明の非天然型塩基を有するヌクレオチドが組み込まれた核酸は、RNA干渉におけるsiRNAとして、または分解を受けるmRNAの一部として利用可能である。
【0058】
また、本発明のヌクレオチドを含む新たなコドンを設計することが可能である。前述したように、鋳型中に置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基(例えば、Pa)として有するヌクレオチドが2つまたはそれ以上連続して存在する場合でも、転写反応は進行し、相補的な位置に6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド(例えば、s)が取り込まれる。よって、本発明の方法により従来不可能であった、非天然型塩基sが2つ又はそれより多く隣接して配置されるDNA及びRNAの作成が可能である。よって、sを3つ含むコドン(sss)、sを2つ含むコドン(例えば、ssA、Gss、sGs)、sを1つ含むコドン(例えば、sAG、CsT、AGs)を設計することができる。新たなコドンは、天然型のアミノ酸をコードさせることもできるし、また、非天然型のアミノ酸をコードさせることもできる。このように、本発明は新規な非天然型人工塩基を提供するのみならず、本発明のヌクレオチドを含む新たなコドンの設計による、全く新しい遺伝暗号の設計を可能とするものであり、新たな遺伝暗号の世界を提供するものである。
【0059】
さらに、本発明の新たなコドンに応じたtRNA系を設計することにより、非常に多くのアミノ酸を利用可能とする新たなタンパク質合成システムを設計することができる。利用可能なアミノ酸はリボソームにおけるタンパク質合成酵素系で利用できるものであればよい。従って、本発明は前記本発明のコドンを用いた新たなタンパク質合成システムを提供する。本発明のタンパク質合成システムによれば、所望の位置のコドンの核酸を本発明の核酸によって効率よく置換、又はコドンの所望の位置へ本発明の核酸を導入することにより、所望の非天然型のアミノ酸を含有するタンパク質の製造が可能となる。
【0060】
立体構造の解析方法
6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド(例えば、s)は蛍光塩基であり、これを塩基として含むヌクレオチドは強い蛍光特性を有する。
【0061】
具体的には、6位に芳香族複素環などを結合させて電子共役系を広げたプリン誘導体は蛍光特性がでやすい。さらに、限定されるわけではないが、従来から2−アミノプリンは蛍光特性を有することがわかっている。よって、限定されるわけではないが、特に6位に芳香族複素環が結合しており、かつ2位にアミノ基を有するプリンは強い蛍光特性を示す。特に、s、s’及びvは好ましい蛍光塩基である。従来技術に対して、蛍光色素を結合した第5番目の塩基を人工塩基対を介して、転写により核酸中の特定部位に導入できれば、核酸の標識、そして、それによる核酸の解析、検出が非常に簡便になる。本発明ではPa−sという人工塩基対を用いて、これを可能にした。
【0062】
本発明者らは、蛍光性の人工塩基sをプローブとして、酵母tRNAPheを部位特異的に蛍光標識し、標識部位、温度およびMg2+濃度に応じた、特有の蛍光プロフィールを得た(実施例3)。さらに、蛍光プロフィールを同一のs標識された酵母tRNAPheに対するUVプロフィールと比較した。その結果、以下のことが判明した。
【0063】
第一にsを含む各tRNA転写産物のUV吸収スペクトルの変化から求めた融解温度(Tm値)は、天然のtRNA転写産物のUV吸収スペクトルの変化から求めた融解温度(Tm値)と同じくらいの高さであった。このことはこれらの位置におけるsによる置換は、tRNA構造全体を有意に不安定化しないことを示唆している。
【0064】
第二に、特定の位置にsを含む各tRNAは、局部構造特性を反映する、蛍光強度の特徴的な変化を表示した。すなわちtRNAの高次構造において、sが近隣の塩基と積み重なっている場合は蛍光強度が弱く、積み重なり(stacking)の程度が減少するに従い蛍光強度が増加した。この発見は、RNAの特定の位置にsを導入し、例えば温度変化に伴う蛍光強度の変化を測定することにより、高次構造中におけるそのsの積み重なりの状態を把握することが可能なことを示している。そこで、sを含むRNAの用途として、当該RNAの高次構造の新規な解析方法が提供される。
【0065】
より具体的には、Mg2+(2及び5mM)の存在下における、酵母tRNAPheのs蛍光プロフィールは、明確に2つのグループに分類された。低温度でグループ1(tRNA 16s、17s及び47s)によって放射されるs蛍光の強度は、グループ2(RNA 36s、57s及び59s)によって放射される強度よりも、1.7−3.4倍大きかった。温度が上昇すると、グループ1によって放出される大きな強度は減少し、逆に、グループ2による強度は上昇した。これらの結果は、グループ1のs塩基は生理学的温度で溶液中に露出しており、そして、温度の上昇による折り畳み構造の変性に伴い、s塩基の他の塩基との非特異的な相互作用が高まることを示唆している。対照的に、グループ2の塩基は、低温下では折り畳み構造において近隣の塩基が積み重なっており、そして、温度の上昇によって塩基積み重ねが徐々に変性する。この推論は、tRNAの結晶構造中の各部位における元来の塩基の配座(図4)(非特許文献20)と極めてよく一致する。さらに、グループ1の蛍光強度は、生理学的温度において、Mg2+の添加によって劇的に上昇する。このことは、Mg2+存在下でtRNAが活性型のL字型構造を形成し、これらの位置における塩基が外側に配置されることを示唆している。
【0066】
第三に、蛍光プロフィールから得られるTm値は、UV融解プロフィールから得られるTm値と比較して、tRNA構造中の各局部構造の安定性を反映した。
例えば、tRNA 36sの蛍光プロフィールから得られるTm値は、UVプロフィールから得られるTm値よりも高かった。このことは、全tRNA構造の安定性に比べてアンチコドン ステム−ループがより高い安定性を有することを示唆している。対照的に、tRNA16s及び17sの低い安定性(蛍光プロフィールから得られる低いTm値)は、Dループを含む部分構造は、tRNA構造全体の脆弱な部分の可能性があることを示唆している。よって、各s標識体の蛍光プロフィールから得られるTm値を、UV融解プロフィールから得られるTm値と比較することにより、s標識された局部構造の安定性の高さを判別することができる。
【0067】
このように、s標識を利用した部位特異的な蛍光プローブ化は、三次元RNA分子の局部構造特徴を研究するための有力なツールとなりうる。
よって、本発明は、本発明の核酸における、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを含む局部領域の立体構造の解析方法を提供することを目的とする。本発明の解析方法は、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド中の、塩基に由来する蛍光強度をいろいろな環境下(温度変化、イオン濃度(例えば、マグネシウム)の変化など)で測定することを含む。
【0068】
本発明の解析方法は、一態様において、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド中の、塩基に由来する蛍光強度が、温度の上昇に伴い実
質的に上昇する場合には、生体内温度下で、前記ヌクレオチドは立体配座において周囲に存在するヌクレオチドと積み重なっていると判断し、そして、蛍光強度が温度の上昇に伴い実質的に低下又は上昇しない場合には、生体内温度下で、前記ヌクレオチドは外部に暴露していると判断する。
【0069】
「生体内温度」とは、各生体が自然の条件下で生息している場合の体内の温度を意味し、生体の種類に応じて変化する、例えば、ヒトの場合は約35.0℃ないし約39.0℃であり、ウシの場合は、約36.7℃ないし約39.3℃、マウスの場合は、約35.0℃ないし約39.0℃、イヌの場合は、約37.9℃ないし約39.9℃、ブタの場合は、約38.7℃ないし約39.8℃である。
【0070】
「ヌクレオチドが立体配座において周囲に存在するヌクレオチドと積み重なっている」とは、より具体的には、2つ以上の塩基が好ましくは約5Å以内で積み重なった状態(ス
タッキング)を意味する。
【0071】
「ヌクレオチドが外部に暴露している」とは、より具体的には、塩基部分が他の塩基とスタッキングしていない(好ましくは約5Å以上離れた)状態、あるいは、消光性の塩基と十分にはなれている状態を意味する。
【0072】
蛍光強度は、好ましくは、前記6位置換された9H−プリン−9−イル基の種類に応じた最大の蛍光特性を示す特定の波長で測定する。例えば、B1)2−アミノ−6−(2−チエニル)−9H−プリン−9−イル基(s)は、最大励起波長である352nmで励起すると、434nmを中心とする蛍光波長を示す。同様に、B2)6−(2−チエニル)−9H−プリン−9−イル基(s’)は、325nmで励起すると381nmを中心とする蛍光波長を示し、B7)2−アミノ−6−(2−チアゾリル)−9H−プリン−9−イル基(v)は、363nmで励起すると461nmを中心とする蛍光波長を示す。
【0073】
「蛍光強度が温度の上昇に伴い実質的に上昇する」とは、例えば温度が20℃から90℃まで上昇した場合に、蛍光強度が上昇することが確認できること(例えば1.1倍以上)を意味する。蛍光強度の上昇の割合が大きいほど、生体内温度下で、その蛍光性の塩基が立体配座において周囲に存在する、より多くのヌクレオチドと、及び/又は、より近距離で積み重なっていることを示す。
【0074】
「蛍光強度が温度の上昇に伴い実質的に低下又は上昇しない場合」とは、例えば温度が20℃から90℃まで上昇した場合に蛍光強度が変化しない又は低下することが確認できること(例えば、10%以上減少)を意味する。蛍光強度の低下の割合が大きいほど、生体内温度下で、その蛍光性の塩基が孤立して存在していて、その部分の核酸の構造が特定の構造を取っていないということを示す。
【0075】
核酸の二本鎖形成の検出方法
本発明はさらに、本発明の核酸の調製方法によって調製された、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを含む核酸の別の利用態様の1つである、核酸の二本鎖形成の検出方法を提供する。本発明の検出方法は、
I) i)本発明の核酸の調製方法によって調製された、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを含む核酸、と、
ii)5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドであって、前記塩基の5位に、5−カルボキシフルオレセイン(5−FAM)、6−カルボキシフルオレセイン(6−FAM)、5−カルボキシテトラメチルローダミン(5−TAMRA)、6−カルボキシテトラメチルローダミン(6−TAMRA)、 5−ジメチルアミノナフタレン−1−スルホン酸(DANSYL)、5−カルボキシ−2’,
4,4’,5’,7,7’−ヘキサクロロフルオレセイン(5−HEX)、6−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(6−HEX)、5−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(5−TET)、6−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(6−TET)、5−カルボキシ−X−ローダミン(5−ROX)、及び6−カルボキシ−X−ローダミン(6−ROX)からなるグループから選択される蛍光色素が、直接又はリンカーを介して結合している前記ヌクレオチド、を含む核酸、とをハイブリダイズさせ、そして、
II) 蛍光スペクトルの変化を測定する
ことを含む。
【0076】
本発明の核酸の二本鎖形成の検出方法は、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを含む核酸、と、蛍光色素によって5位が置換された、5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドを含む核酸とのハイブリダイズによる蛍光スペクトルの変化を利用するものである。特許文献1、2、非特許文献30等に記載されているように、6位置換された9H−プリン−9−イル基(例えばs、v)と、2−オキソ(1H)ピリジン−3−イル基(y)は塩基対合を形成しうる。なお、5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオシド又はヌクレオチドの合成方法等については、本明細書中に別途詳述する。
【0077】
6位置換された9H−プリン−9−イル基の蛍光と、これとは別の特性を有する蛍光色素で置換された5位置換−2−オキソ(1H)ピリジン−3−イル基の蛍光が各核酸分子に存在する場合に、核酸の二本鎖が形成され蛍光同士が物理的に近傍に存在すると、蛍光共鳴エネルギー転移(FRET:Fluorescence Resonance Energy Transfer)が生じて蛍光スペクトルが変化する。本発明の検出方法は、核酸の二本鎖の形成をFRETによる蛍光スペクトルの変化の測定によって検出するものである。
【0078】
「蛍光共鳴エネルギー転移(FRET)」は、ある蛍光分子から他の分子へ、共鳴により励起エネルギーが移動する現象を意味する。エネルギーを与える分子はドナー(供与体)、受け取る分子はアクセプター(受容体)と呼ばれる。FRETが生じると、エネルギーを失ったドナーは基底状態に戻り、同時にエネルギーを受け取ったアクセプターは励起状態となる。従って、ドナーの蛍光は弱まり、アクセプターが蛍光分子であればその蛍光が観察される。アクセプターが消光分子であれば、ドナーが単独の場合には観察されていた蛍光がFRETにより観察されなくなる。FRETによるタンパク質の検出、核酸の検出の一般的な方法は公知であり、例えば、RNAアプタマーに2種類のFRETを起こす色素を導入して、標的タンパク質を検出する方法(Jhaveri et al.,2000)や、ヘアピン型核酸を利用した相補鎖核酸の検出法(Tyagi et al.,1996)が知られている。その他、FRETに関しては、Walter et al.
,2001及びKlostermeier et al.,2001に概説されている。
【0079】
FRETが生じるためには、以下の3条件を満たす必要がある。i)ドナーの蛍光スペクトルとアクセプターの吸収スペクトルの重なりがあること。スペクトルの重なり範囲は大きい方が望ましいが、必ずしも完全に重なっている必要はない。ドナーの蛍光スペクトルとアクセプターの吸収スペクトルが30%以上重なっていることが好ましく、50%重なっていることがより好ましい。ii)ドナーとアクセプターが物理的に近距離に存在すること。FRETが50%の確率で生じる距離は3nmないし6nmと考えられており、FRETの効率はこの距離の変化に対して敏感に変化する。例えば、核酸が二本鎖を形成した場合に、蛍光特性を有する6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドに対し、これとは別の特性を有する蛍光色素で置換された5位置換−2
−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドが近接し、両者の距離が好ましくは10nm以内、より好ましくは6nm以内、最も好ましくは3nm以内である場合に蛍光スペクトルの変化が検出可能である。iii)ドナーとアクセプターの相対的な向きが適切であること。
【0080】
本発明の検出方法の一態様において、5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドを含む核酸は、塩基の5位に、5−カルボキシフルオレセイン(5−FAM)又は6−カルボキシフルオレセイン(6−FAM)がリンカーを介して結合している、5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドを含む核酸である。
【0081】
前述したように、例えばsは353nmの最大励起波長、そして、最大励起波長である352nmで励起すると、434nmを中心とする波長を示す。一方、FAMは、FAMの吸収極大波長は493nm、蛍光極大波長は522nmである。353nmの波長で励起した場合、sを有するヌクレオチドは434nmを中心とする波長を示す。これに対し、FAMで置換された5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチド(例えば、FAM−y)がsに近接すると、sからの蛍光を吸収し、蛍光極大波長が522nmの蛍光スペクトルを示すようになる。即ち、353nmの波長で励起した場合に、434nmの蛍光スペクトルが減少し、522nmの蛍光スペクトルが増加するほど、sに対してFAM−yが近接していること、即ち、sを含む核酸とFAM−yを含む核酸とのハイブリダイゼーションにより核酸の二本鎖が形成されていることを検出することができる。
【0082】
5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオシド又はヌクレオチド
5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオシド又はヌクレオチドにおいて、5位に蛍光色素は、直接又はリンカーを介して結合している。これらの標識ヌクレオシド又はヌクレオチドについては、本発明者の先行する特許出願(特願2004−324271、2004年11月8日出願、未公開)に詳述されている。
【0083】
蛍光色素は、公知の蛍光色素を利用可能であるが、好ましくは、5−カルボキシフルオレセイン(5−FAM)、6−カルボキシフルオレセイン(6−FAM)、5−カルボキシテトラメチルローダミン(5−TAMRA)、6−カルボキシテトラメチルローダミン(6−TAMRA)、5−ジメチルアミノナフタレン−1−スルホン酸(DANSYL)、5−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(5−HEX)、6−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(6−HEX)、5−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(5−TET)、6−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(6−TET)、5−カルボキシ−X−ローダミン(5−ROX)、及び6−カルボキシ−X−ローダミン(6−ROX)からなるグループから選択される。フルオレセイン及びローダミンは、一般的に開環型及びスピロ型の二通りで表記される。これらの蛍光色素については、例えば Tuschl et al.,1994;Misra et al
.,2004;Gibson et al.,1996;及び、Chehab et al.,1989;に詳述されている。
【0084】
例えば、FAMの吸収極大波長は493nm、蛍光極大波長は522nmである。また、TAMRAの吸収極大波長は553nm、蛍光極大波長は578nmである。DANSYLの吸収極大波長は335nm、蛍光極大波長は518nmである。HEXの吸収極大波長は535nm、蛍光極大波長は556nmである。TETの吸収極大波長は521n
m、蛍光極大波長は536nmである。5−ROXの吸収極大波長は567nm、蛍光極大波長は591nmである。6−ROXの吸収極大波長は570nm、蛍光極大波長は590nmである。
【0085】
5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオシド又はヌクレオチドは、5位置換基として蛍光分子を有するため、蛍光分子の種類に応じて、核酸の検出を行うことが可能である。よって、標識核酸として当該核酸と相互作用する物質検出のプローブとして使用されうる。また、各蛍光色素によって蛍光色が異なるため、多重染色に使用することも可能である。
【0086】
蛍光色素は、2−オキソ(1H)ピリジン−3−イル基の5位に直接結合させても、あるいはリンカーを介して結合させてもよい。リンカーの種類は特に限定されず、当業者は適宜採用可能である。リンカーは限定されるわけではないが、好ましくは、下記の化学式I−III:
【0087】
【化5】
[式I中、nは1ないし5の整数から選択される]
【0088】
【化6】
[式II中、m及びlは、1ないし5の整数から各々独立に選択される]
及び
【0089】
【化7】
【0090】
からなるグループから選択される
5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオシド
又はヌクレオチドの合成法は、特に限定されることはない。3−(β−D−リボフラノシル)−ピリジン−2(1H)−オンに、先ず5位に置換基を導入し、次いで、三リン酸を導入してもよい。あるいは、3−(β−D−リボフラノシル)−ピリジン−2(1H)−オンに、先ず三リン酸を導入し、次いで、置換基を導入してもよい。特に、大きな基を導入する場合、先ず、5位にアミノプロピニル基等を導入し、これに活性化した置換基を結合させてもよい。
【0091】
あるいは、蛍光色素による、2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドの修飾は、単独のヌクレオチド合成時ではなく、当該ヌクレオチドを含む核酸を転写等によって合成した後に行うことも可能である。
【0092】
5位置換−2−オキソ(1H)ピリジン−3−イル基は、6位置換された9H−プリン−9−イル基と、2箇所で水素結合を生じる。5位置換−2−オキソ(1H)ピリジン−3−イル基は、立体構造上、天然型プリン塩基A(アデニン)及びG(グアニン)とは塩基対を形成できない。そして、6位置換された9H−プリン−9−イル基は立体障害のため、天然型T(チミン)、U(ウラシル)及びC(シトシン)とは塩基対を形成できない。よって、6位置換された9H−プリン−9−イル基(例えばs、v)と、5位置換−2−オキソ(1H)ピリジン−3−イル基は、特異的に塩基対を形成しうる。
【0093】
5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドは、転写、複製又は逆転写反応により、DNA又はRNA等の核酸に取り込むことが可能である。具体的には、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを含む核酸、例えば、sあるいはvを含む鋳型DNAを用いてT7 RNAポリメラーゼによる転写反応を行うと、2−オキソ(1H)ピリジン−3−イル基の基質(例えば、yTP)が部位特異的に、sあるいはvに相補して、RNA中に取り込まれる。yは、その5位を化学的に修飾することができるので、種々の機能性のy誘導体をRNA中の特定の部位に取り込ませることが出来る。
【0094】
あるいは、天然型塩基を有するヌクレオシド又はヌクレオチドと同様に、化学合成によってDNA又はRNAに取り込んでもよい。
同一の鎖上に2種類の人工塩基を含む核酸を調製する方法
本発明はさらに、
i) 6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド
並びに、
ii) 5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドであって、前記塩基の5位に、5−カルボキシフルオレセイン(5−FAM)、6−カルボキシフルオレセイン(6−FAM)、5−カルボキシテトラメチルローダミン(5−TAMRA)、6−カルボキシテトラメチルローダミン(6−TAMRA)、 5−ジメチルアミノナフタレン−1−スルホン酸(DANSYL)、5−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(5−HEX)、6−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(6−HEX)、5−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(5−TET)、6−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(6−TET)、5−カルボキシ−X−ローダミン(5−ROX)、及び6−カルボキシ−X−ローダミン(6−ROX)からなるグループから選択される蛍光色素が、直接又はリンカーを介して結合している、前記ヌクレオチド
を、同一の鎖上に含む核酸を調製する方法を提供することを目的とする。本発明の調製方法は、
iii) 置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチド
並びに
iv) 6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド
を含む核酸を鋳型として転写、逆転写又は複製を行い、前記iii)のヌクレオチドの相補的な位置に、前記i)のヌクレオチドを組み込み、そして、前記iv)のヌクレオチドの相補的な位置に、前記ii)のヌクレオチドを組み込む
ことを含む。
【0095】
本発明の上記調製方法は、置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドを鋳型とした場合の、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドの取り込みと、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを鋳型とした場合の、5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドの取り込みを組み合わせて利用したものである。
【0096】
本発明はさらにまた、上記方法によって調製された
i) 6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド
並びに、
ii) 5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドであって、前記塩基の5位に、5−カルボキシフルオレセイン(5−FAM)、6−カルボキシフルオレセイン(6−FAM)、5−カルボキシテトラメチルローダミン(5−TAMRA)、6−カルボキシテトラメチルローダミン(6−TAMRA)、 5−ジメチルアミノナフタレン−1−スルホン酸(DANSYL)、5−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(5−HEX)、6−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(6−HEX)、5−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(5−TET)、6−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(6−TET)、5−カルボキシ−X−ローダミン(5−ROX)、及び6−カルボキシ−X−ローダミン(6−ROX)からなるグループから選択される蛍光色素が、直接又はリンカーを介して結合している、前記ヌクレオチド
を、同一の鎖上に含む核酸も提供する。
【0097】
本発明の上記核酸は、tRNA、mRNA、アンチセンスDNA若しくはRNA、リボザイム又はアプタマーとして利用可能である。さらに、RNA干渉(RNA interference、RNAi)においても利用可能である。
【0098】
本明細書中の実施例4では、Pa及びvを含むDNAを鋳型として転写を行い、sとy、あるいは、sとFAM−yを同一鎖上に含むRNAを作成した。
核酸のステム−ループ(ヘアピン)構造形成の検出方法
本発明はまた、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドと5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドとを同一の鎖上に含む、本発明の核酸のステム−ループ(ヘアピン)構造形成の検出方法を提供することを含む。本発明の検出方法は、
1) 本発明の核酸において、i)のヌクレオチドと、ii)のヌクレオチドとをハイブリダイズさせ、そして、
2) 蛍光スペクトルの変化を測定する
ことを含む。
【0099】
本発明の核酸のステム−ループ(ヘアピン)構造形成の検出方法は、「核酸の二本鎖形成の検出方法」の項目で上述したFERTを利用するものである、即ち、6位置換された9H−プリン−9−イル基の蛍光と、これとは別の特性を有する蛍光色素で置換された5
位置換−2−オキソ(1H)ピリジン−3−イル基の蛍光が、「同一分子内に」存在する場合に、ステム−ループ(ヘアピン)構造が形成され蛍光同士が物理的に近傍に存在すると、FRETが生じて蛍光スペクトルが変化する。本発明の検出方法は、核酸のステム−ループ(ヘアピン)構造の形成をFRETによる蛍光スペクトルの変化の測定によって検出するものである。
【0100】
核酸のステム−ループ(ヘアピン)構造の形成をFRETによる蛍光スペクトルの変化は、別個の二本の核酸同士で二本鎖が形成される場合の蛍光スペクトルの変化と同様に観察することができる。
【0101】
本発明の好ましい一態様において、核酸のステム−ループ(ヘアピン)構造は、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドと5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドとを同一の鎖上に含む、本発明の核酸が、分子内でsとFAM−yを含む自己相補の配列がステムを作ること
により形成される。このように、核酸分子内のsとFAM−yをプローブにして両者間のFRETの効率を測定することにより、ステム−ループ構造などの核酸の高次構造を知ることが出来る。
【0102】
さらに、別の標的分子が結合した場合のみにステム−ループ(ヘアピン)構造を形成するような核酸分子を設計すれば、蛍光スペクトルの変化を利用して、その標的分子を検出するシステムを作ることが出来る。したがって、本発明は、リアルタイム分子間相互作用検出システム、リアルタイム分子内構造変化の検出システム、モレキュラービーコン、FRETの効率を利用した分子間距離の推定等に利用可能である。標的分子としては、DNA,RNA、タンパク質等が含まれる。
【0103】
本明細書中の実施例4では、sとFAM−yを同一鎖上に含むRNAについて、sとFAM−y間のFRETを調べた。本明細書中の実施例4では、350nmでsを励起した場合に、ステム−ループ構造を形成させてsとFAM−yの間で塩基対が作られた場合と、相補鎖RNAを加えて分子間での二本鎖構造を形成させてsとFAM−yが離れた場合の、いずれにおいてもFRETが観測されて、FAM−y由来の521nm付近の蛍光が現れた。しかしながら、sとFAM−yを導入した断片のsの蛍光強度(図12bおよび
cの実線のスペクトルの440nm付近の強度)とその対照のsとyを導入した断片のsの蛍光強度(図12bおよびcの破線のスペクトルの440nm付近の強度)を比較すると、ステム−ループ構造を形成している場合のほうが、顕著にsの蛍光強度が減少した。ステム−ループ構造中のsとFAM−yの距離は8−10Åであり、二本鎖構造中のsとFAM−yの距離は65−67Åに広がる。したがって、sとFAM−yの距離が近づいた場合に、両者間のFRETの効率が上がっている。
【0104】
6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを、2個以上隣接して含む核酸
6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを2個以上隣接して含む核酸もまた、本発明の範囲内である。
【0105】
前記核酸において、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドが2個以上同一の鎖上に隣接することにより、隣接した人工塩基の間で自己消光が生じるため、9H−プリン−9−イル基の蛍光強度が、9H−プリン−9−イル基を塩基として有するヌクレオチドが隣接しない場合と比較して、有意に減少する(図13cおよびdを参照)。一方、9H−プリン−9−イル基の間で自己消光が生じていても、蛍光色素によって5位が置換された、5位置換−2−オキソ(1H)ピリジン−3−イル基とのハイブリダイズにより、9H−プリン−9−イル基と蛍光色素との間のFRETを観察
することができる(図14cおよびdを参照)。さらに、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを2個以上隣接して含む核酸においては、9H−プリン−9−イル基の蛍光強度が減少している、すなわちバックグラウンド蛍光が低いため、蛍光色素との間のFRETを効率よく観察することができる(図15を参照)。すなわち、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを2個以上隣接して含む自己消光プローブを使用する本発明のFRET系は、蛍光ドナーのバックグラウンドを低く抑えることができるため、大量のドナープローブの存在下においても蛍光アクセプターを含有する相補鎖の検出が可能である点で特に有用である。
【0106】
6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを2個以上隣接して含む核酸は、上述した核酸の二本鎖形成の検出方法や核酸のステム−ループまたはヘアピン構造形成の検出方法に使用することができる。
【0107】
6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを2個以上隣接して含む核酸において、隣接して含まれる6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドの数は、2個以上であれば特に限定されないが、好ましくは、2ないし10個、2ないし5個、2ないし3個、最も好ましくは2個である。
【0108】
6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを2個以上隣接して含む核酸は、本発明の核酸の調製方法、または公知の化学合成によって製造することが可能であるが、これらに限定されない。
【発明の効果】
【0109】
本発明によって、非天然型塩基の新たな塩基対合Pa−sが見出されたことから、転写においても、対合する塩基間の形状の相補性が重要であることが示唆された(非特許文献5−6)。これにより、従来困難と考えられていた転写においても、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドの取り込みが可能となった。
【0110】
本発明の方法は、所望の位置にsを有すRNAを得る方法として従来知られている、(1)化学的RNA合成法、(2)yヌクレオチドを有する鋳型DNAの転写反応、(3)2−オキソ−1,3−ジヒドロ−イミダゾール−1−イル基を有するヌクレオチド(z)を有する鋳型DNAの転写反応よりも有意に高収率である。
【0111】
従来の2−アミノプリンヌクレオチド類(非特許文献21−23)のような慣用された蛍光プローブは、転写によってRNAに部位特異的に取り込まれることができないため、特に、長鎖RNA分子で使用することが困難であった。即ち、従来法では、2−アミノプリンのヌクレオチド誘導体を化学合成によりRNA中に導入していたが、RNAの化学合成法では、長鎖のRNA(50ヌクレオチド以上)を合成することは難しく、その操作も煩雑で時間がかかっていた。その結果、従来法は、限定的な利用に限られていた。
【0112】
これに対し、本発明では、蛍光特性を有する6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドの、RNAへの部位特異的取り込みが可能となった。本発明では、鋳型となるDNAを化学合成する必要があるが、DNAの化学合成は、RNAよりも長鎖長(150ヌクレオチド程度)の合成が可能であり、また、化学合成したDNAを酵素を用いて結合させることも出来る。これにより特に、従来の蛍光プローブ法では不可能であった長鎖RNAの分析が可能になった。これは、大きなRNA分子内における特定された位置における局部配座変化の動態の研究のための、強力な道具となりうる。本発明の実施例3では、実際にtRNA内の局部の高次構造(スタッキング)の解析が行われた。
【0113】
また、現在、RNAは、RNA干渉など、生体内の新たな機能が数多く見つかり、これらの現象を利用した医薬品の開発も進んでいる。しかし、それらのRNA分子の生体内での挙動を調べる手法は、それほど多くは無い。したがって、本蛍光プローブ法は、生体内のRNAの解析、ならびに、RNA医薬品の生体内での働きを調べる良い方法になる。
【0114】
さらに、本発明で新たに明らかにされたs−Pa塩基対は、翻訳によってタンパク質にアミノ酸類似体を部位特異的に取り込ませることを可能にするため、遺伝子コードの拡張に実用的に使用しうる(非特許文献15)。
【実施例】
【0115】
以下、実施例によって本発明を具体的に説明するが、これらは本発明の技術的範囲を限定するためのものではない。当業者は本明細書の記載に基づいて容易に本発明に修飾・変更を加えることができ、それらは本発明の技術的範囲に含まれる。
【0116】
実施例1 s−Pa塩基対合を利用したT7転写
本実施例では、s−Pa塩基対合を利用してT7転写を行った。
1)試薬、溶媒等
試薬及び溶媒は、標準的な供給業者から購入し、さらに精製することなく使用した。エレクトロスプレイ−イオン化マススペクトル(ESI−MS)は、Waters ZMD
4000 LC/MSシステム上に記録した。DNA鋳型は、Pa(非特許文献9)及び天然塩基のホスホルアミド類を用いて、自動DNA合成機(model392,PerkinElmer Applied Biosystems,Foster City,CA)によって化学合成した。ゲル電気泳動によってオリゴヌクレオチドを精製した。
【0117】
2)2−アミノ−6−(2−チエニル)−9−(1−β−D−リボフラノシル)プリン
5’−三リン酸(sTP)の合成
2−N−フェノキシアセチル−6−(2−チエニル)−9−(2,3−ジ−O−アセチル−1−β−D−リボフラノシル)プリン(57mg、0.1mmol)(非特許文献29)をピリジン中に溶解し、そして真空下で溶媒を留去し、試料中の水分を共沸除去した。残渣をピリジン(100μl)及びジオキサン(300μl)中に溶解した。次いで、2−クロロ−4H−1,2,3−ベンゾジオキサホスフォリン−4−オン(0.11mm
ol)の1Mジオキサン溶液(100μl)を添加した(非特許文献25)。10分後、
この溶液に0.5M ビス(トリブチルアンモニウム)ピロリン酸(0.15mmol)のDMF溶液(300μl)とトリ−n−ブチルアミン(100μl)をすばやく加えた。その反応混合物を室温で10分間かき混ぜた。次いで、1%ヨウ素溶液(ピリジン/水(98:2、v/v)溶液、2ml)を加えた。15分後、5%NaHSO3水溶液(150μl)、次いで、5mlの水を反応溶液に加えた。
【0118】
溶液を室温で30分間攪拌し、次いで、濃アンモニア水(20ml)を加えた。55℃で12時間、アンモノリシスを行った。溶液を真空下で濃縮し、そして産物をDEAEセファデックス(A−25)カラムクロマトグラフィー(50mMないし1M TEABの直線勾配によって溶出した)によって、そして次いで、C18−HPLC(100mM 酢酸トリエチルアンモニウム中の0%ないし30%CH3CNの勾配によって溶出した)(Eichorom Technologies社のSynchropak RPPを伴うGilson HPLC精製システム)によって精製し、sTPを得た。
【0119】
【化8】
【0120】
3)T7転写のための鋳型の調製
化学合成されたDNA鋳型(17−mer RNA合成のための、10μMの35−merのコード鎖及び21−merの非コード鎖)を、10mM NaClを含む10mM
Tris−HCl(pH7.6)緩衝溶液中で、95℃で加熱し、その後、4℃まで徐冷して、アニーリングした。
【0121】
4)T7転写(17−merRNA)
24mM MgCl2、2 mM スペルミジン、5 mM DTT、0.01% Triton X−100を含む40mM Tris−HCl(pH 8.0)緩衝溶液(2
0μl)中、1mM天然NTPs、若しくは1mM sTP、2μCi[γ−32P]GTP、2μM鋳型及び50単位のT7 RNAポリメラーゼ(Takara、Kyoto)の存在下で転写を行った。[γ−32P]GTPの使用により、転写産物は5’末端のみ標識され、これにより収率を求めた。37℃での3時間のインキュベーションの後、10M尿素及び0.05%BPBを含む色素溶液(20μl)を加えることにより反応を止めた。混合物を75℃で3時間加熱し、20%ポリアクリルアミド−7M尿素ゲル上で産物を分析した。
【0122】
5)結果
本実施例では、基質としてs(sTP)、および、非天然塩基が転写産物中の13−15番目に相当する相補的な部位に位置する、1つ若しくは2つのPa塩基を含むDNA鋳型を用いて、転写を行った。図2Aに実験スキームを示す。図2Aにおいて、17−mer RNA合成のための10μMの35−merのコード鎖及び21−merの非コード鎖の配列、並びに17−mer RNA転写産物の塩基配列は下記の通りである。
【0123】
鋳型相補鎖 (配列番号1)
5’−ataatacgactcactataggg−3’
鋳型鎖 (配列番号2)
3’−tattatgctgagtgatatccctcgaagggannntc−5’
17−merRNA転写産物 (配列番号3)
5’−gggagcuucccunnnag−3’
本発明者らはまた、対照として、イミダゾリン−2−オン(z)(図1B)を含む別の
DNA鋳型も調べた(図1B);zは、T7転写によるRNAへのsの部位特異的挿入のためのよい鋳型塩基であるが、転写効率は、天然の転写と比較して効率が低い。s−z対は塩基間に2つの水素結合が形成される可能性があるが、その形はs−Pa対に類似する。
【0124】
これらの鋳型を用いて3時間転写を行い、[γ−32P]GTPで5’末端を標識した転写産物をゲル上で分析した結果を、図2Bに示す。具体的には、図2Bは、1つまたは2つのPa又はz塩基を含む鋳型を用いて、天然型NTP(1mM、N=A,G,C,U)とsTP(1mM)を加えて行った転写の産物のゲル電気泳動の結果である。転写産物は、[γ−32P]GTPでその5’末端を標識されている。各転写産物の相対収率は、天然型塩基のみからなる鋳型からの転写産物を100%として、それとの比較によって決定した。各収率は3−4データセットからの平均をとった。
【0125】
図2Bに示すように、Paを1つ含む鋳型DNAからの全長(17−mer)の転写産物(1つのs塩基を含む)の相対収率は92%であり(図2B、レーン1、N1=Pa)、これはzを1つ含む鋳型DNAからの転写の収率(35%)(図2B、レーン4)よりも高く、そして、天然型のみの鋳型(N1=C)とNTPを用いた転写の収率(図2B、レーン8)と同じくらい高かった。2つのPa塩基を含む鋳型を用いた転写では、その相対収率は、天然型の転写(図2B、レーン8)と比較して低いものの、全長の産物が得られた(図2B、レーン2及び3)。対照的に、2つのzを含む鋳型を用いた転写では、全長の転写物が得られなかった(図2B、レーン5及び6)。
【0126】
実施例2 T7転写産物(17−mer RNA)中のヌクレオチドの組成分析
本実施例では、転写におけるs−Pa塩基対の選択性を調べるために、Pa、z及び天然の鋳型からの転写で得られる全長(17−mer)の転写物中のヌクレオチドの組成分析を行った。転写物を[α−32P]UTP、[α−32P]ATP又は[α−32P]GTPのどれかで内部標識し、この転写産物をRNase T2でヌクレオシド3’リン酸体にまで完全に分解した。得られた標識ヌクレオチドを2D−TLCで分析し(図3)、そして各ヌクレオシド3’−一リン酸のスポットを定量した(表1)。
【0127】
具体的には、24mM MgCl2、2mM スペルミジン、5mM DTT、0.01% Triton X−100を含む40mM Tris−HCl(pH8.0)緩衝液(20μl)中、10mM GMP、1mM 天然型NTP(1mM、N=A,G,C,U)、0、1若しくは3mM sTP、2μCi[γ−32P]UTP、[γ−32P]ATP若しくは[γ−32P]GTP(アマーシャム)、2μM鋳型DNA、及び50単位のT7 RNAポリメラーゼ(Takara)(非特許文献14、15)の存在下で転写を行った。3時間、37℃でインキュベーション後、色素溶液を加えて転写を終了させた。
【0128】
この混合物を75℃で3時間加熱し、そして次いで、15%ポリアクリルアミド−7M尿素ゲルで電気泳動した。全長の産物をゲルから溶出し、そして、これに0.05A260ユニットの大腸菌由来のtRNAを加えて、産物をエタノール沈殿させた。転写産物を、15mM 酢酸ナトリウム緩衝液(pH4.5)中、0.075U/μlのRNase
T2で、37℃、2時間消化した。この処理により、転写産物はRNase T2でヌクレオシド3’−一リン酸にまで完全に分解された。消化産物を、Merck HPTLCプレート(100×100mm)(Merck,Darmstadt,Germany)を用いた2D−TLCによって分析した。使用した展開溶剤は、1次元目がイソ酪酸/NH4OH/H2O(66:1:33 v/v/v)、そして、2次元目がイソプロピルアルコール/HCl/H2O(70:15:15 v/v/v)であった。TLCプレート上の産物をバイオイメージングアナライザーで分析した。各スポットの定量は3−9の
データセットから平均化した。
【0129】
結果を図3に示す。さらに、各ヌクレオシド3’−一リン酸を定量した結果を表1にまとめた。
表1 T7転写産物のヌクレオチド組成分析
【0130】
【表1】
【0131】
a: 図2に示した転写産物のU(エントリー1−8)、A(エントリー9−12)、又はG(エントリー13−16)の5’側に取り込まれたヌクレオチドの組成
b: 値は、下記の式を用いて決定された:
(各ヌクレオチドの放射活性)/[全てのヌクレオチド3’−一リン酸]×([α−32P]NTPの5’側のヌクレオチドの総数]
c: 各ヌクレオチドの理論的な数は、角型括弧書きで示す。
【0132】
d: 標準偏差は、括弧書きで示す。
e: 検出されず
図3及び表1に示した結果より、1つのPa塩基を含む鋳型を用いた場合では、高い選択性でRNA中へのsの取り込みが確認された。図3では、Paを含む鋳型を用いて[γ−32P]UTPで標識した場合にのみ、標識されたs−ヌクレオシド3’−一リン酸が2D−TLC上で観察された。さらに、表1に示されたように、鋳型DNA中のPaに相
補してRNA中に取り込まれたsの選択性(97%)(表1、エントリー1)は、s−z塩基対での場合の選択性(表1、エントリー2)、又は天然型塩基対のみでの転写の選択性と同じくらい高く(表1、エントリー3及び5)、そして、鋳型中の天然型の塩基に相補した間違ったsTPの取り込みは観察されなかった(表1、エントリー4及び6)。2つのPa塩基を含む鋳型を用いた転写の場合、1番目の位置におけるsの取り込みの選択性は十分に高かったが(95%)、2番目の位置におけるsの取り込みの選択性は84−87%程度であった。しかし、sTPの濃度を上げる(3mM)ことによって、この選択性はある程度(89−93%)改善された。従って、RNA中へのsの部位特異的取り込みに用いる鋳型塩基として、Paはzよりも優れており、そして、この結果は、非水素結合性の塩基対も転写において機能しうることを示唆している。
【0133】
実施例3 16s、17s、36s、47s、57s及び59s tRNAの調製及びその蛍光スペクトル及びUV融解曲線
1)16s、17s、36s、47s、57s及び59s tRNAの調製
蛍光プローブとしてのs塩基の有用性を示すために、本発明者らは、sを酵母由来のtRNAPheの特定の部位に取り込ませ、そしてtRNAの各部位におけるsの蛍光特性をいろいろな条件下で分析することにより、tRNAの高次構造を調べた。s塩基の蛍光は、近隣の塩基による塩基積み重ね(stacking)によって消光されるため、RNA分子中のsの周囲の局部構造についての情報が得られる。
【0134】
本発明者らは、tRNA中の6箇所のそれぞれにsを導入した:具体的には、D−ループ中の16番目又は17番目(tRNA 16s又は17s)、アンチコドン中の36番目(tRNA 36s)、エキストラループ中の47番目(tRNA 47s)、並びにTΨCループ中の57又は59番目(tRNA 57s又は59s)である。それぞれの部位のsは、tRNA中の他の塩基と塩基対を形成しない部位に導入されている(図4)。
【0135】
sを含む酵母tRNAPhe分子を調製するための各鋳型の配列(RNA合成のための5μMの94−merの鋳型鎖DNA及びその相補鎖のDNA)は、以下の表2の通りである(配列番号4−11)。
【0136】
【表2】
【0137】
RNA合成のための5μMの94−merの鋳型DNA及びその相補鎖DNAの配列
この配列では、C2−G71塩基対はG2−C71(非特許文献26)で置換されている。鋳型鎖DNAの5’末端の2つのヌクレオシド(G及びT)は、2'−O−メチルリボ
ヌクレオシドで置換することにより、目的とする産物よりも長くなってしまう鋳型非依存の転写産物の生成を抑えている(非特許文献27)。
【0138】
RNA合成のための94−merの鋳型DNA及び鋳型の相補鎖DNAの調製は、実施例1の「3)T7転写のための鋳型の調製」の項目で記載したのと同様に行った。
図4−6に、酵母由来のtRNAPhe中のsの導入部位及びtRNAの構造(非特許文献20)を示す。図4は、オリジナルなtRNAの2次構造である。sで置換された塩
基は○で囲んである。波線は塩基−塩基相互作用を示す。ボックスで囲んだG−C塩基対はオリジナルのtRNA中ではC−G塩基対であるが、この置換ではtRNAの高次構造(非特許文献26)は有意に変わっていない。
【0139】
図5は天然型のtRNAの3次構造であり、図6A−Cが、sが取り込まれる各部位の拡大図を示す。濃い色の塩基(群)はsと置換されていて、これは薄い色の塩基(群)と積み重なっている。そして、球(黄色)は、tRNA中のMg2+をあらわす。図中の塩
基は天然型のtRNAにおいて修飾されている塩基を示すが、括弧内に記載された塩基は転写産物における塩基を示している。
【0140】
それぞれの部位にsが導入されたtRNA転写産物は、Paを含む鋳型DNAを用いて、1mM sTPと天然型NTPsの存在下で、T7 RNAポリメラーゼによって調製された。具体的には先ず、24mM MgCl2、2mM スペルミジン、5mM DTT、0.01% Triton X−100を含む40mM Tris−HCl(pH8.0)緩衝液中、10mM GMP、1mM 天然型NTPs、1mM sTP、0.5μM鋳型DNA、及び2.5U/μl T7 RNAポリメラーゼの存在下で、転写を行った。37℃で6時間インキュベーションし、1.75倍量の色素溶液を加えて転写を終了させた。この溶液を75℃で3分加熱し、そして、10%ポリアクリルアミド−7M 尿素ゲルで電気泳動した。全長の産物をゲルから溶出し、そして、エタノールで沈殿した。450μlの10mM EDTA(pH8)中に産物を溶かし、75℃で5分間インキュベートした。
【0141】
次いで、Microcon YM−10 フィルター(Amicon)を用いて、緩衝溶液をTm測定用の緩衝液(50mM カコジル酸ナトリウム(pH7.2)及び50mM KClを含む)に交換した。260nmの吸光度によってtRNAの量を決定した。そして、蛍光及びUV融解測定のために、0.1mM EDTA、2mM MgCl2又は5mM MgCl2を含むTm緩衝液中に1μM tRNAを含むように溶液を調製した。
【0142】
2)蛍光スペクトル及びUV融解曲線
sを特定の部位に含む各tRNAの蛍光スペクトル及びUV溶解曲線を、20から90℃までの範囲で、0.5℃/分の加熱速度で、各々、FP−6500分光蛍光計(JASCO)、及びUV−2450分光光度計(SHIMADZU)を用いて記録した。
【0143】
s塩基は2つの励起最大(299及び352nm)を有する、図7Aは、sのリボヌクレオシド(1μM)の励起及び蛍光スペクトルを示す。図7Aから明らかなように、sは、434nmを中心とする蛍光スペクトルを示し、その蛍光の量子収率はpH7.0で0.41であった。図7B及びCは、352nmでの励起による(B)tRNA 47s(1μM)の蛍光スペクトル;及び(C)tRNA 57s(1μM)の蛍光スペクトルを示す。蛍光スペクトルは、50mM カコジル酸ナトリウム(pH7.2)、50mM KCl、及び0.1mM EDTA中、20℃において測定した。
【0144】
本実施例では、50mM カコジル酸ナトリウム、50mM KCl、及び0.1mM
EDTA又は、EDTAを除いて2mM若しくは5mM MgCl2を加えた溶液中、(352nmを中心に3nmのスペクトルバンド幅で励起された)sの434nmでの蛍光強度及び260nmでのUV吸収を測定して、各tRNA転写産物(1μM)の熱力学的融解プロフィールを調べた。
【0145】
溶媒分子との衝突によるsの蛍光の消光は、温度が高くなるほど生じやすくなる(図8A)。そこで、各tRNA転写産物の蛍光強度の温度依存変化を、sのヌクレオシドモノ
マーによって規格化した。蛍光強度の温度依存変化によって求められるtRNAの融解温度は、IGOR Proソフトウェア(WaveMetrics.Inc.)を用いて計算した。具体的には、図8は、(A)sのリボヌクレオシドの蛍光強度の温度依存による変化、及び(B)sのリボヌクレオシドの蛍光強度の温度依存による変化を補正したtRNA 36sの蛍光強度の温度依存性を示す。tRNA転写産物の蛍光強度は、等式:Yct=Yt/(Rt/R20)によって補正された。ここにおいて、Yctは、t℃での各tRNAの補正された強度であり、Ytは、t℃での各tRNAの実測された蛍光強度であり、Rtは、t℃でのsのリボヌクレオシドの蛍光強度であり、そして、R20は、20℃でのsのリボヌクレオシドの蛍光強度である。
【0146】
3)結果
16s、17s、36s、47s、57s及び59s tRNAの、434nmでの蛍光強度の温度依存変化によって得られる融解曲線(実線)及び260nmにおけるUV吸収の変化によって得られる融解曲線(波線)を図9に示す。
【0147】
先ず、第一にsを含む各tRNA転写産物のUV融解温度(2mM MgCl2中、64.6−66.4℃)は天然のtRNA転写産物の融解温度(2mM MgCl2中、65.5℃)と同じくらいの高さであった。このことは、これらの部位をsで置換しても、tRNAの構造は有意に不安定化されないことを示唆している。
【0148】
第二に、特定の位置にsを含む各tRNAは、局部構造の特性を反映する特徴的な蛍光強度の変化を表示した。Mg2+(2及び5mM)の存在下におけるsの蛍光特性は、明確に2つのグループに分類される:1つは、tRNA 16s、17s及び47sを含み(グループ1)、そしてもう一方は、tRNA 36s、57s及び59sを含む(グループ2)。2mM又は5mM Mg2+の存在下では、低温度でのグループ1のsの蛍光強度は、グループ2のsの蛍光強度よりも、1.7−3.4倍大きかった。グループ1の大きなsの蛍光強度は、温度が上昇すると減少するが、グループ2によるsの蛍光強度は温度の上昇とともに大きくなった。
【0149】
これらの結果は、16、17又は47番目のs塩基は生理学的温度で溶液中に露出しており、そして、温度の上昇によるtRNAの変性に伴い、s塩基と他の塩基との非特異的な相互作用が高まることを示唆している。対照的に、36、57又は59番目のs塩基は、低温下ではL字型構造を形成したtRNA中で近隣の塩基と積み重なっており、そして、温度の上昇によって徐々にtRNAが変性し、sと隣接する塩基との積み重なり構造が壊れる。これらの推測は、X線結晶構造解析によるtRNAのL字型構造中の各部位における元来の塩基の配置(図4)(非特許文献20)と極めてよく一致する。tRNA 16s、17s及び47sの蛍光強度は、生理学的温度において、Mg2+の添加によって劇的に上昇する。このことは、Mg2+のtRNAへの結合によって、tRNAがL字型構造を形成し、その結果、これらの位置における塩基が溶液中に露出される位置に配置されることが示唆している。
【0150】
第三に、蛍光プロフィールから得られるTm値は、UV融解プロフィールから得られるTm値と比較して、tRNA構造中の各局部構造の安定性を反映している。表3に、特定部位にsを含む酵母由来のtRNAPheの蛍光及びUVプロフィールから得られた融解温度(Tm)を示す。
【0151】
表3 特定部位にsを含む酵母由来のtRNAPheの蛍光及びUVプロフィールから得られた融解温度(Tm)
【0152】
【表3】
【0153】
蛍光及びUVプロフィールは、50mMカコジル酸ナトリウム(pH7.2)及び50mM KCl中、MgCl2共存下あるいは非共存下で、各tRNA転写産物(1μM)を用いて得られた。各融解温度は、IgorProソフトウェア(WaveMetrics. Inc.)を用いることによって計算された。n.c.計算されず。
【0154】
例えば、tRNA 36sの蛍光プロフィールから得られるTm値(2mM MgCl2中、69.5℃)は、UVプロフィールから得られるTm値(2mM MgCl2中、65.2℃)よりも高かった。このことは、tRNAの全体構造の安定性に比べてアンチコドン ステム−ループがより高い安定性を有することを示唆している。対照的に、tRNA16s(2mM MgCl2中、58.0℃)及び17s(2mM MgCl2中、59.6℃)の低い安定性は、Dループを含む部分構造は、tRNA構造全体の脆弱な部分であることを示唆している。
【0155】
実施例4 人工塩基sとFAM−yを含むRNA鎖のFRET実験
s−Pa塩基対を転写に用いることにより、蛍光性の人工塩基sをRNA中に効率よく導入することが出来るようになった。このs−Pa塩基対を、本発明者らが既に開発しているv−y塩基対と組み合わせることにより、1本のRNA中にsとy、あるいは、sと5位を修飾したyを、それぞれ任意の位置に導入することができる。特に、FAMを導入したyを利用することにより、sとFAM−yの間でFRETを観察することができる。
【0156】
本実施例では、いろいろな部位にPaとvを導入した転写用の鋳型DNAを化学合成し、これらの鋳型とsとy、あるいはFAM−yのそれぞれの基質(sTP、yTP、FAM−yTP)を用いて転写を行った。
【0157】
T7 RNA ポリメラーゼによる転写反応
(sとyを部位特異的に含むRNA 39−merの解析).
二本鎖の鋳型DNA(5μM)のアニーリング操作は、10mM NaClを含む10mM Tris−HCl緩衝液(pH7.6)中で行われた(温度条件:95℃で3分間加熱後、−0.1℃/秒の勾配で4℃まで徐冷)。T7転写反応は、40mM Tris−HCl緩衝液(pH8.0)、8mM MgCl2、2mM スペルミジン、5mM DTT、0.01% TritonX−100、2μCi[γ−32P]GTP、1mM
NTPs、0−1mM sTP、0−1mM yTP、0.5μM 鋳型DNA、および50unitsのT7 RNA polymerase(Takara)を含む反応溶液(全量 20μl)で行った。37℃、3時間反応を行った後、10M 尿素を含む電気泳動用色素溶液を等量(20μl)加えて反応を停止させた。この溶液を75℃で3分
間加熱後、15% ポリアクリルアミド−7M 尿素ゲルで電気泳動を行い、転写物をバイオイメージアナライザーで解析した(図10)。
【0158】
T7 RNA ポリメラーゼによる転写反応生成物の塩基組成分析
T7 RNAポリメラーゼによる転写反応を、40mM Tris−HCl緩衝液(pH8.0)、8mM MgCl2、2mM スペルミジン、5mM DTT、0.01%
TritonX−100、2μCi[α−32P]ATPまたは2μCi[α−32P]GTPまたは2μCi[α−32P]CTPまたは2μCi[α−32P]UTP、1mM NTPs、0−1mM sTP、0−1mM yTP、0.5μM 鋳型DNA、および50unitsのT7 RNA polymerase(Takara)を含む反応溶液で行った。転写物を、15% ポリアクリルアミド−7M 尿素ゲルによる電気泳動により精製し、ゲルからの溶出を行った後、tRNA(0.05 OD)を加えてエタノール沈殿を行い回収した。このサンプルを滅菌水10μlに溶解し、溶液(8.5μl)を用いてRNaseT2による完全分解反応を行った。溶液(8.5μl)にRNase T2(1.5μl,0.5U/μl)を加えて、37℃で2時間反応を行った。この溶液の一部をTLC(Merck,10cmx10cm)にスポットし、二次元展開を行った。展開液として、一次元目はイソ酪酸:濃アンモニア水:水=66:1:33(v/v/v)、二次元目は2−プロパノール:塩酸:水=65:10:25(v/v/v)を用いた。展開後のスポットをバイオイメージアナライザーで検出し、解析を行った(図11、表4)。
【0159】
T7 RNA ポリメラーゼによる転写反応
(sとy、もしくはsとFAM−yを部位特異的に含むRNA 39−merの調製).
T7転写反応は、40mM Tris−HCl緩衝液(pH8.0)、8mM MgCl2、2mM スペルミジン、5mM DTT、0.01% TritonX−100、1mM NTPs、1mM sTP、1mM yTPまたは1mM FAM−yTP、0.5μM 鋳型DNA、および125unitsのT7 RNA polymerase(Takara)を含む反応溶液(全量50μl)で行った。37℃、6時間反応を行っ
た後、DNase I溶液を加えて、37℃で15分間保温することで鋳型DNAを分解した。10M 尿素を含む電気泳動用色素溶液を等量(50μl)加えて反応を停止させ
た後、反応溶液を75℃で3分間加熱してから、15% ポリアクリルアミド−7M 尿素ゲルで電気泳動を行い、目的の転写物39−merを精製した。ゲルから滅菌水で溶出した後、エタノール沈澱を行い、転写物を回収し、UV吸収から目的の転写物RNA 39−merの濃度を決定した。
【0160】
蛍光スペクトル測定
sとy、もしくはsとFAM−yを部位特異的に含むRNA 39−mer(0.2μM)を用いて、10mM リン酸ナトリウム緩衝液(pH7.0)、100mM NaCl、0.1mM EDTA中での蛍光スペクトルを20℃で測定した。測定前のRNAサンプルのアニーリング操作では、RNA 39−mer(0.2μM)を含む溶液、およびRNA 39−merとその39−merに相補的な配列を持つ相補性RNA 42−mer(0.4μM)を含む溶液を75℃で3分間加熱した後、−0.1℃/3秒の勾配で4℃まで徐冷を行った。アニーリング操作を行ったサンプルは、4℃で保管され、測定時は、サンプルを測定セルに移してから20℃に2分以上放置した後にスペクトル走査(360−650nm)を開始した。蛍光測定(励起波長350nm)には、3x3mmセルを用い、レスポンス 0.5s、走査速度 2000nm/分、感度はマニュアル設定でPMT 500Vに固定、スリットバンド幅は5nmに固定してスペクトル測定を行った(図12)。
【0161】
結果
図10は、v−y塩基対とs−Pa塩基対を用いた転写によるRNAへのyとsの部位特異的導入の戦略と転写産物のポリアクリルアミド電気泳動の結果を示す。
【0162】
詳細には、図10aに鋳型DNAの配列と転写反応の条件を示した(配列中でPaは、Pで示している)。転写物(Bhp2−1,Bhp2−2,Bhp2−3)をポリアクリルアミドゲル電気泳動で調べると(図10b)、sTPとyTPをどちらも加えた転写で、転写物の量が多くなった。そこで、sとyが選択的にRNA中に取り込まれているかどうかを調べるために、α位に放射性のリンを導入した[α−32P]ATP,[α−32P]GTP,[α−32P]CTP,[α−32P]UTPのそれぞれの基質を別々に加えて転写を行い、放射性標識された転写物をRNase T2でヌクレオシド 3’−一リン酸に加水分解し、リン酸が放射性標識されているヌクレオシド 3’−一リン酸を2次元のTLCで分析し、転写物のヌクレオチド組成を調べた(図11)。
【0163】
この方法では、放射性の基質が取り込まれると、その5’側のヌクレオチドが2次元のTLC上で検出できる。たとえば、図11bには、Bhp2−1の2次元TLCの結果を示した。Bhp2−1では、yがAの5’側に、また、sがGの5’側にそれぞれあるので、もし、鋳型上のvに相補してyが、また、Paに相補してsが、それぞれ選択的にRNA中に取り込まれていれば、[α−32P]ATPで標識した転写物の2次元TLCにはyp(yのヌクレオシド 3’−一リン酸)のスポットが現れ、また、[α−32P]GTP,で標識した転写物では、sp(sのヌクレオシド 3’−一リン酸)のスポットが現れることになる。実際に、図11bでは、それぞれのTLC上に期待されるスポットが現れている。また、[α−32P]CTP,あるいは、[α−32P]UTPで標識した転写物では、ypとspのどちらのスポットも認められないことから、鋳型上の天然型塩基に相補してそれぞれの人工塩基が間違って取り込まれていないこともわかった。さらに、2次元TLC上のそれぞれのスポットを定量し、その結果を表4に示した。
【0164】
【表4】
【0165】
a: 図11に示された、A(エントリー1,2及び9−11)、G(エントリー3,4及び12−14)、C(エントリー5及び6)又はU(エントリー7及び8)の5’隣りに取り込まれたヌクレオチドの組成
b: 値は、下記の式を用いて決定された:
(各ヌクレオチドの放射活性)/[全てのヌクレオチド(3’−一リン酸)]×([α−32P]NTPの5’隣りのヌクレオチドの総数]
c: 各ヌクレオチドの理論的な数は、角型括弧書きで示す。
【0166】
d: 標準偏差は、括弧書きで示す。
e: 検出されず
転写では、yTPの量を0.5mMと1mMで行った結果を示してある。エントリー1−8の結果は、全て、理論値(カッコ内の数値)と良く一致している。エントリー9−14では、天然型塩基のみの鋳型DNAを用いた場合についての結果であるが、エントリー10において、1mMのyTPを用いた場合にわずかにypが天然型の鋳型塩基に相補して取り込まれていることがわかった。したがって、転写におけるyTPの量は、0.5mMのほうが良い。以上の結果から、s−Pa塩基対とv−y塩基対を組み合わせて転写を行うことにより、1本のRNA中にsとyをそれぞれ選択的に導入できることがわかった。
【0167】
そこで、yの基質に代えて、yの5位にFAMを結合した基質(FAM−yTP)とsTPを用いて、Bhp2−1の転写を行い、転写物をゲル電気泳動で精製し、その蛍光特
性を調べた。Bhp2−1は、単独ではヘアピン型の構造を形成し、sとFAM−yが塩基対を形成する位置に配置されるので、両者でFRETが観測される。そして、このRNAの相補鎖のRNAを加えると二本鎖構造を形成し、sとFAM−yが離れ、FRETの効率が低下する。この二本鎖構造のRNAの蛍光スペクトルを図12bに、また、ヘアピン型のRNAの蛍光スペクトルを図12cに示した(実線)。それぞれの蛍光スペクトルは、sとyを導入したRNAのスペクトルと比較している(破線:このRNAでは、FRETが起こらず、sの蛍光スペクトルを示すコントロールとなる)。どちらも、350nmでsを励起しているが、FRETが観測されて、FAM−yの521nm付近の蛍光が現れている。しかし、ヘアピン型のRNAのほうが、FAM−yの蛍光強度が強く、また、コントロール(破線)と比較したsの消光も顕著である。
【0168】
この結果から、sとFAM−yの両者の距離に相関してFRETが観測されることがわかった。したがって、sとFAM−yをRNA中のいろいろな部位に導入し、両者のFRETの効率を測定することにより、両者間の距離を見積もることができ、RNAの構造解析に役立たせることができる。従来のX線結晶構造解析やNMRによる構造解析では、静的なRNAの構造に関する情報しか得ることが出来なかったが、本手法で、溶液中のRNAの構造変化のダイナミクスを知ることが可能になる。
【0169】
実施例5 核酸二重鎖におけるsまたはvとFAM−yの人工塩基対のFRET実験
DNA/RNA二本鎖において、蛍光性の人工塩基である2−アミノ−6−(2−チエニル)プリン−9−イル基または2−アミノ−6−(チアゾリル)プリン−9−イル基(以下、本実施例においてそれぞれsまたはvと称する)、およびフルオロフォアを連結した2−オキソ−(1H)ピリジン−3−イル基(以下、本実施例においてyと称する)の間でFRET実験を行った。sまたはvの発光スペクトルは、FAMが連結した5−(3−アミノ−1−プロピニル)−y(FAM−yTP)またはFAMが連結した5−[3−(6−アミノヘキサンアミド)−1−プロピニル]−y(FAM−hx−yTP)であるヌクレオチド誘導体の吸収スペクトルと重なった(図13a)。よって、DNAフラグメント中のsまたはvは蛍光ドナーとして作用し、そして相補的なRNAフラグメント中のFAM−yまたはFAM−hx−yは蛍光アクセプターとして作用する。
【0170】
人工塩基sまたはvを含むDNAの合成
人工塩基sまたはvを含むDNAは、例えば、国際公開WO 01/05801号または国際公開WO 2005/026187号に記載の方法に従って、人工塩基sまたはvを含むヌクレオチドのシアノエチルホスホロアミダイト誘導体を用いて、当該技術分野に周知のDNA化学合成法により合成した。化学的に合成された35merのDNAの配列は、5’−CAN1N2N3CTCGGGATTCCCTATAGTGAGTCGTATTAT−3’(配列番号20;ここで、N1N2N3が;CTsまたはCTvのDNAをそれぞれDNA35s1またはDNA35v1;CssまたはCvvのDNAをそれぞれDNA35s2aまたはDNA35v2a;sTsまたはvTvのDNAをそれぞれDNA35s2bまたはDNA35v2b;と称する)とした。得られたDNAはゲル電気泳動によって精製した。
【0171】
FAM−yまたはFAM−hx−yを含むRNAの調製
FAM−yまたはFAM−hx−yをN’の位置で含有する17merのRNA(5’−GGGAAUCCCGAGN’AGUG−3’:配列番号21)を、T7 RNAポリメラーゼおよびFAM−yまたはFAM−hx−yの基質(FAM−yTPまたはFAM−hx−yTP)を用いてT7転写により調製した。鋳型(コード鎖としてDNA35v1、非コード鎖として5’−ATAATACGACTCACTATAGGG−3’(配列番号22)をそれぞれ10μM)を、10mM Tris−HCl(pH7.6)および10mM NaClを含む緩衝溶液中で、95℃で加熱し、その後、4℃まで徐冷して、
アニーリングした。転写反応は、40mM Tris−HCl(pH8.0)、24mM
MgCl2、2mM スペルミジン、5mM DTT、および0.01% Triton X−100を含む緩衝溶液中、1mM 天然NTPs、1mM FAM−yTPまたはFAM−hx−yTP、2μM 鋳型、および2.5ユニット/μL T7 RNAポリメラーゼ(Takara)の存在下で行った。37℃で3時間インキュベーションした後、10M 尿素および0.05% BPBを含有する色素溶液を等量加えることにより反応を止めた。反応混合物は75℃で3分間加熱し、そして17−merの転写物をゲル電気泳動により精製した。
【0172】
蛍光スペクトル測定
蛍光スペクトルは10mM リン酸ナトリウム(pH7.0)、100mM NaCl、および0.1mM EDTAを含む緩衝液中で測定した。発光スペクトルはsについて350nm、vについて360nmの励起波長で記録した。発光のスペクトルバンドパスは5nmであった。
【0173】
FAM−yまたはFAM−hx−yを部位特異的に含む17−mer RNAを、1個または2個のsまたはv塩基を部位特異的に含む35−mer DNA(DNA35s1、35s2a、35s2b、35v1、35v2a、および35v2b)とハイブリダイズさせ、DNA/RNA二本鎖の発光スペクトルを、異なる温度(20および70℃)で測定した。
【0174】
結果
共鳴エネルギー転移は、sまたはvの低蛍光状態からFAM−yまたはFAM−hx−yの高蛍光状態の間で観察された。DNA35s1またはDNA35v1が、FAM−yまたはFAM−hx−yを部位特異的に含む17−mer RNAと20℃で二本鎖を形成したときにFRETが生じ、そして70℃で二本鎖が変性したときにFRETは有意に減少した(図14aおよびb)。加えて、DNA35s2aおよびDNA35v2aと、FAM−yまたはFAM−hx−yを部位特異的に含むRNAとの二本鎖においても、FRETが効率よく生じた(図14cおよびd)。
【0175】
興味深いことに、二つの隣接した人工塩基を含むDNA35s2aまたはDNA35v2aの一本鎖の蛍光強度は有意に減少する一方、1つのTで隔てられた二つの人工塩基を含むDNA35s2bまたはDNA35v2bの一本鎖の蛍光強度の有意な減少は観察さなかった(図13cおよびd)。DNA35s2aまたはDNA35v2aの一本鎖の蛍光強度の減少は、二つの隣接した人工塩基の間での自己消光によって起こるものである。
【0176】
二つの隣接した人工塩基の間での自己消光が起こるDNA35s2aまたはDNA35v2aとの二本鎖形成によっても、17−mer RNAの強いFAM蛍光が観察されたので、自己消光プローブを用いるこのFRET系は、標的の鎖に対して過剰量のプローブの使用を容易にする。17−mer RNAに対して1つの人工塩基を含むDNA35s1およびDNA35v1を10モル等量用いた場合、共鳴エネルギー転移は、それぞれ、DNA35s1およびDNA35v1のバックグラウンド蛍光によって完全に隠れてしまった(図15aおよびb)。対照的に、17−mer RNAに対して隣接する二つの人工塩基を含むDNA35s2aおよびDNA35v2aを10モル等量用いた場合は、それぞれのss−およびvv−含有DNAおよびFAM含有RNAの間のFRETがはっきりと観察された(図15cおよびd)。したがって、隣接する二つの人工塩基を含む自己消光プローブを使用するFRET系は、蛍光ドナーのバックグラウンドを減少させつつ、FRET現象を観察することができるため、大量のドナープローブの存在下においても蛍光アクセプターを含む相補鎖の検出が可能である。また、大量の自己消光DNAプローブは、相補的な蛍光アクセプター鎖との二本鎖形成を容易にするため、FRET効率を増加
させることができる。
【図面の簡単な説明】
【0177】
【図1】図1は、非天然型s−Pa(A)塩基対及びs−z(B)塩基対を示す。
【図2】図2は、s−Pa塩基対によって媒介されるT7転写の実験スキーム及び、転写産物のゲル電気泳動の結果を示す。図2A:実験スキーム 図2B:1つまたは2つのPa又はz塩基を含む鋳型DNAを用いて、天然型NTPs(1mM)とsTP(1mM)を加えた場合の転写産物のゲル電気泳動。転写産物は、[γ−32P]GTPでその5’末端を標識された。各転写産物の相対収率は、全て天然型塩基からなる鋳型からの転写産物の収率との比較によって決定し、各収率は3−4データセットから平均化した。
【図3】図3は、転写産物(17−mer)のヌクレアーゼ消化から得られた標識リボヌクレオシド3’−一リン酸の2D−TLC分析を示す。転写産物を[α−32P]NTPで内部標識し、1mM sTPの存在下で転写された17−mer断片からの二次元TLCである。中パネルは鋳型DNA中のN1N2N3がPaACであり、右パネルはN1N2N3がCACである。
【図4】図4は、酵母由来のtRNAPhe中へのsの導入部位及びtRNAの構造(非特許文献20)を示す。図4はオリジナルのtRNA転写産物の2次構造である。sで置換された場所は○で囲んである。波線は3次構造の塩基−塩基相互作用を示す。ボックスで囲んだG−C対はオリジナルのC−G対から変更されているが、この変位はオリジナルのtRNA構造(非特許文献26)を有意には変えない。
【図5】図5は、酵母由来のtRNAPhe中へのsの導入部位及びtRNAの構造(非特許文献20)を示す。図5は原型のtRNA転写産物の3次構造である。
【図6】図6A−Cは、酵母由来のtRNAPhe中へのsの導入部位を含む領域の拡大図である。濃い色の塩基(群)はsと置換される部位で、この塩基は薄い色の塩基(群)と積み重なっており、そして、球(黄色)はMg2+を表す。図中の塩基は修飾された塩基を示すが、括弧内に記載された塩基はオリジナルな転写産物中の塩基に相当する。
【図7】図7Aは、sのリボヌクレオシド(1μM)の励起及び蛍光スペクトルを示す。黒丸実線は、352nmでの励起による蛍光スペクトルを示す。白丸波線は、434nmでの励起による蛍光スペクトルを示す。 図7B及びCは、352nmでの励起による(B)tRNA 47s(1μM)の蛍光スペクトル;及び(C)tRNA 57s(1μM)の蛍光スペクトルを示す。蛍光スペクトルは、50mM カコジル酸ナトリウム(pH7.2)、50mM KCl、及び0.1mM EDTA中、20℃において測定した。(B)において、黒丸実線、白丸波線、及び×印実線は、tRNA47sの各々0mM Mg2+、2mM Mg2+及び5mM Mg2+存在下における蛍光スペクトルを示す。(C)において、黒丸実線、白丸波線、及び×印実線は、tRNA57sの各々0mM Mg2+、2mM Mg2+及び5mM Mg2+存在下における蛍光スペクトルを示す。
【図8】図8は、(A)sリボヌクレオシドの蛍光強度の温度依存性、及び(B)sのリボヌクレオシドでtRNA 36sの蛍光強度を補正した例を示す。tRNA転写産物の蛍光強度は、等式:Yct=Yt/(Rt/R20)によって補正された。ここにおいて、Yctは、t℃での各tRNAの補正された強度であり、Ytは、t℃での各tRNAの測定された蛍光強度であり、Rtは、t℃でのsのリボヌクレオシドの観察された蛍光強度であり、そして、R20は、20℃でのsのリボヌクレオシドの観察された蛍光強度である。
【図9】図9は、種々の位置においてsを含むtRNAの、434nmでの蛍光強度の変化によって得られる溶解曲線(実線)及び260nmにおけるUV吸収の変化によって得られる溶解曲線(波線)を示す。太い(黒)実線、中くらいの(青)実線、細い実線(赤)の実線は、各々0mM Mg2+、2mM Mg2+及び5mM Mg2+存在下における434nmでの蛍光強度の変化によって得られる融解曲線である。太い(黒)波線、中くらいの(青)波線及び細い実線(赤)の波線は、各々0mM Mg2+、2mM Mg2+及び5mM Mg2+存在下における260nmにおけるUV吸収の変化によって得られる融解曲線である。
【図10】図10は、v−y塩基対とs−Pa塩基対を用いた転写によるRNAへのyとsの部位特異的導入の方法と転写産物のポリアクリルアミド電気泳動の結果を示す。図10aにおいて、配列中のPは、人工塩基Paを示している。
【図11】図11は、v−y塩基対とs−Pa塩基対を用いた転写により調製された転写産物(Bhp2−1)のヌクレオチド組成の分析の結果を示す。図11bにおける、各スポットの定量は、表4にまとめた。
【図12】図12は、人工塩基sとy若しくはFAM−yを導入したRNA(Bhp2−1)の蛍光測定の結果を示す。図12bは、Bhp2−1が相補性RNAと二本鎖を形成している場合の蛍光スペクトルを示す。図12cは、Bhp2−1が1本鎖のままヘアピン構造を形成している場合の結果を示す。図12b及び図12cのいずれの場合も、実線は、位置13にFAM−y、位置36にsを有するBhp2−1の結果である。波線は、位置13にy、位置36にsを有するBhp2−1の結果である
【図13】図13aは、リン酸緩衝液(pH7.0)中におけるsデオキシリボヌクレオシド(細い線)およびFAM−yTP(太線)の蛍光(flu、黒線)および吸収(abs、破線)スペクトルである。図13bは、FRET実験のスキームである。非天然塩基は、sまたvについて励起させたとき(λex)、低蛍光状態を黒丸で、高蛍光状態を白丸で示した。図13cおよびdは、s塩基を含有する(c:DNA35s1、DNA35s2a、およびDNA35s2b)およびv塩基を含有する(c:DNA35v1、DNA35v2a、およびDNA35v2b)一本鎖DNAの定常状態蛍光発光スペクトルである。
【図14】図14は、DNA/RNA二本鎖の定常状態蛍光発光スペクトルを示す。35−mer DNAのそれぞれ(a,DNA35s1;b,DNA35v1;c,DNA35s2a;d,DNA35v2a;e,DNA35s2b;f,DNA35v2b)を、FAM−y(白丸および黒丸)またはFAM−hx−y(白三角または黒三角)を含む17−mer RNAとハイブリダイズさせた。白丸および白三角は20℃、黒丸および黒三角は70℃でのスペクトルである。
【図15】図15は、人工塩基を含むDNAプローブを過剰量用いた場合のDNA/RNA二本鎖の定常状態蛍光発光スペクトルを示す。35−mer DNAのそれぞれ(1μM)(a,DNA35s1;b,DNA35v1;c,DNA35s2a;d,DNA35v2a)を、FAM−y(白丸および黒丸)またはFAM−hx−y(白三角および黒丸)を含む17−mer RNA(100nM)とハイブリダイズさせた。白丸および白三角は20℃、黒丸および黒三角は70℃でのスペクトルである。
【技術分野】
【0001】
本発明は、新規人工塩基対に基づく核酸及びその利用に関する。
【背景技術】
【0002】
生体高分子である核酸(DNA,RNA)は、生命活動に必要となる膨大な量の遺伝情報を、僅か4種類の塩基の組合せからなる配列として記録している。また、核酸は自分自身を鋳型としてDNAポリメラーゼにより自己複製し、さらにRNAポリメラーゼによる転写、リボソームによる翻訳というプロセスを介して、DNAからDNAへ、DNAからRNAへ、RNAからタンパク質へと遺伝情報を伝達する。この遺伝情報の複製と伝達を可能としているのは排他的な塩基対形成(A・T/U、G・C)のルールである。また、核酸は、多様な高次構造を形成して様々な機能を発揮する。例えば、in vitroセレクション法によって、アプタマーやリボザイムの機能を有する新規核酸がこれまでに多数見つかってきたこともその一つである。
【0003】
しかし、20種類のアミノ酸からなるタンパク質に比べて、天然の核酸には4種類の塩基(2種類の塩基対)しかないという事実は、核酸の化学的・物理的多様性に限界を与えている。たとえば、生体中のtRNA、rRNA、mRNA等の機能性RNAは自分自身の構造を安定化したりRNA・RNA間、RNA・タンパク質問相互作用を安定化するために、様々な修飾塩基を利用している。従って、新規機能性核酸の開発において、新たな塩基(対)のレパートリーを増やすことは大変有益であると考えられる。
【0004】
核酸のさらなる機能拡張をめざして、非天然型塩基をもつヌクレオシド又はヌクレオチドの創製への取り組みが行われている。核酸に修飾塩基(もしくは非天然型塩基)を導入する手法として、1)化学合成により直接導入する方法、2)核酸合成酵素により導入する方法が考えられる。1)の場合は、アミダイトユニットの安定性や塩基部分の適当な保護基が存在すること等化学合成上の問題の解決が必要である。また、これらの問題が解決されれば様々な非天然型塩基を位置選択的に導入できるが、その核酸の増幅は困難であり、長鎖長の核酸の合成も難しくなる。2)の場合は、もし、基質が酵素に認識され、人工塩基対間で相補的に複製、転写されれば、その核酸の増幅・調製が可能となるが、そのような基質や塩基対(非天然型ヌクレオチド)も開発途中である。
【0005】
非天然型人工塩基対の研究の背景
天然の二本鎖DNA中のAとT及びGとCは、それぞれ特異的な水素結合を介して互いに「排他的」な塩基対を形成している。非天然型塩基対の研究は塩基間の水素結合を利用した組み合わせや塩基の疎水性を利用した組み合わせ等が知られているが、複製、転写、翻訳の工程全てにおいて天然型塩基対に拮抗し得るものは見出されていない。このような状況において複製、転写、翻訳の少なくとも一工程において天然型塩基対に拮抗し得る非天然型塩基対は特異的な有用性を持つものである。
【0006】
最近の非天然型の非水素結合塩基対の研究により、複製における効率的で適合したヌクレオチドの取り込みのために、対合塩基間の水素結合は絶対的に必要なものではなく、その代わり、塩基間の形状の相補性が複製において重要な役割を果たすことが、証明されてきた(非特許文献5−6)。よって、非水素結合塩基対は、新規なバイオテクノロジーを創造するための、遺伝子アルファベットおよびコードの拡張のための潜在的な候補である(非特許文献7−9)。
【0007】
一方、転写に関しては、RNAポリメラーゼによって高い選択性および効率で認識され
る非水素結合塩基対は、まだ報告されていない。具体的には、本発明者は、これまでに2−アミノ−6−(2−チエニル)−9H−プリン−9−イル基(s)と2−オキソ−(1H)ピリジン−3−イル基(y)との組み合わせによる水素結合の相互作用を介した(s−y)塩基対を開発し、試験管内で非天然型アミノ酸を部位特異的に含むタンパク質の合成に成功している(特許文献1、非特許文献14)。
【0008】
さらに、特許文献3は、2−アミノ−6−(2−チアゾリル)プリン−9−イル基(v)と2−オキソ−(1H)ピリジン−3−イル基(y)との組み合わせによる(v−y)塩基対を開示している(特許文献3)。
【0009】
なおsの核酸塩基対としては天然型4塩基、(s−y)の他、(s−z)(zは、2−オキソ−1,3−ジヒドロイミダゾール−1−イル基を意味する)、(s−5位置換y)が、本発明前に報告されていた。これらの(s−y)、(s−z)、(s−5位置換y)の非天然型の塩基対は全て水素結合相互作用を含むものである(特許文献2、非特許文献13−15)。
【0010】
しかしながら、これらの非天然型塩基対のうち(s−y)と(s−5位置換y)は、y
をDNA鋳型としてsをRNAへ挿入する転写工程については、選択性が高くなく、また、(s−z)は、選択性は高いが、必ずしも高収率ではなかった。
【0011】
【化1】
一方、2−ホルミル−1H−ピロール−1−イル基(Pa)は、天然型4塩基のほか9−メチルイミダゾ[(4,5)−b]ピリジン(Q)と非水素結合塩基対を形成しうることが(Pa−Q)で報告されている(非特許文献9)。
【0012】
【化2】
【0013】
しかしながら、上記非水素結合塩基対もDNAポリメラーゼによる翻訳、又は、逆転写酵素によるDNA合成が報告されているのみであり、RNAポリメラーゼによる認識については知られていない。T7様RNAポリメラーゼの単一サブユニットは、構造およびメカニズムの点においてDNAポリメラーゼに類似するが(非特許文献16−17)、T7
RNAポリメラーゼ複合体の最近の構造解析によって、RNAポリメラーゼとDNAポリメラーゼ間の相違が明らかにされた(非特許文献17−18)。転写では、入ってくる基質が鋳型中の塩基と「開いた」コンフォメーションで塩基対を形成し、そして、形成された塩基対は、「開いた」コンフォメーションから「閉じた」コンフォメーションに遷移する間、維持される(非特許文献18)。しかしながら、複製では、「閉じた」コンフォメーションへの遷移の後に塩基対形成が開始される。このことから、転写では複製の場合よりも、対合する塩基間の水素結合が重要であると考えられ、そして、非水素結合塩基対が転写において機能しうるか、という問題が生じる。
【0014】
非天然型塩基によって仲介される特異的な転写が可能になれば、促進された機能性を有する新規なRNA分子を創造し、そして、遺伝子コードが拡張されることとなる(非特許文献1−4)。
【0015】
RNAへの蛍光プローブの導入
最近、RNA医療の関心が高まる中で、RNA中への蛍光プローブの導入は、RNAの蛍光標識化法、そして、RNAの複雑な高次構造を解析するための重要なテクノロジーの1つになっている。後者の構造解析用の蛍光プローブとしては、蛍光性の塩基(2−アミノプリンなど)をもつヌクレオチドを利用する方法が従来から用いられている。この点、上述した2−アミノ−6−(2−チエニル)−9H−プリン−9−イル基(s)は蛍光塩基であり、これを塩基として含むヌクレオチドは強い蛍光特性を有する。従って、sを塩基として含むヌクレオチドをRNA中の任意の部位に導入することができれば、有用な蛍光プローブの調製が可能となる。
【0016】
しかし、これらの蛍光性塩基を有するヌクレオチドを転写によりRNAに導入するための公知の方法は、収率の面で必ずしも十分なものではなかった。従って、sなどの蛍光性塩基を有するヌクレオチドを、転写によりRNA中に部位特異的に導入することは実質的にできなかったため、長鎖RNAの高次構造の解析は難しかった。
【0017】
転写で蛍光プローブをRNA中に導入する方法が開発されれば、溶液中でのRNAの構造のダイナミックスを調べることが可能になる。さらに、この手法は、RNAを利用した医薬品の開発にも役立つ。そこで、不可能であると考えられている長鎖RNA中への蛍光プローブの導入を可能とする方法の確立が重要となる。これにより、生体内の機能性RNAの構造解析、ならびに、他分子との複合体の構造解析に供する新たな手法の開発が可能になる。
【0018】
複製、転写、翻訳の工程全てにおいて天然型塩基対に拮抗し得る非天然型の塩基であって、かつ、蛍光特性を付与する塩基を有するヌクレオチドを核酸へ導入するための、新規な人工塩基対の開発が希求される。
【特許文献1】WO2001/005801
【特許文献2】WO2004/007713
【特許文献3】WO2005/026187
【非特許文献1】Benner, S. A., Burgstaller, P., Battersby, T. R. & Jurczyk, S. in The RNA World (eds Gesteland, R. F., Cech, T. R. & Atkins, J. F.) 163−181 (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1999).
【非特許文献2】Bergstrom, D. E. Orthogonal base pairs continue to evolve. Chem. Biol. 11, 18−20 (2004).
【非特許文献3】Wang, L. & Schultz, P. G. Expanding the genetic code. Chem. Commun. 1−11 (2002).
【非特許文献4】Hendrickson, T. L., de Crecy−Lagard, V. & Schimmel, P. Incorporation of nonnatural amino acids into proteins. Annu. Rev. Biochem. 73, 147−176 (2004).
【非特許文献5】Morales, J. C. & Kool, E. T. Efficient replication between non−hydrogen−bonded nucleoside shape analogs. Nat. Struct. Biol. 5, 950−954 (1998).
【非特許文献6】Kool, E. T. Hydrogen bonding, base stacking, and steric effects in DNA replication. Annu. Rev. Biophys. Biomol. Struct. 30, 1−22 (2001).
【非特許文献7】McMinn, D. L. et al. Efforts toward expansion of the genetic alphabet: DNA polymerase recognition of a highly stable, self−pairing hydrophobic base. J. Am. Chem. Soc. 121, 11585−11586 (1999).
【非特許文献8】Wu,Y. et al. Efforts toward expansion of the genetic alphabet: optimization of interbase hydrophobic interactions. J. Am. Chem. Soc. 122, 7621−7632 (2000).
【非特許文献9】Mitsui, T. et al. An unnatural hydrophobic base pair with shape complementarity between pyrrole−2−carbaldehyde and 9−methylimidazo[(4,5)−b]pyridine. J. Am. Chem. Soc. 125, 5298−5307 (2003).
【非特許文献10】Piccirilli, J. A., Krauch, T., Moroney, S. E. & Benner, S. A. Enzymatic incorporation of a new base pair into DNA and RNA extends the genetic alphabet. Nature 343, 33−37 (1990).
【非特許文献11】Switzer, C. Y., Moroney, S. E. & Benner, S. A. Enzymatic recognition of the base pair between isocytidine and isoguanosine. Biochemistry 32, 10489−10496 (1993).
【非特許文献12】Tor, Y. & Dervan, P. B. Site−specific enzymatic incorporation of an unnatural base, N6−(6−aminohexyl)isoguanosine, into RNA. J. Am. Chem. Soc. 115, 4461−4467 (1993).
【非特許文献13】Ohtsuki, T. et al. Unnatural base pairs for specific transcription. Proc. Natl. Acad. Sci. U.S.A. 98, 4922−4925 (2001).
【非特許文献14】Hirao, I. et al. An unnatural base pair for incorporating amino acid analogs into proteins. Nat. Biotechnol. 20, 177−182 (2002).
【非特許文献15】Hirao, I. et al. A two−unnatural−base−pair system toward the expansion of the genetic code. J. Am. Chem. Soc. 126, 113298−113305 (2004).
【非特許文献16】Cheetham, G. M., Jeruzalmi, D. & Steitz, T. A. Structural basis for initiation of transcription from an RNA polymerase−promoter complex. Nature 399, 80−83 (1999).
【非特許文献17】Yin, Y. W. & Steitz, T. A. The structural mechanism of translocation and helicase activity in T7 RNA polymerase. Cell 116, 393−404 (2004).
【非特許文献18】Temiakov, D. et al. Structural basis for substrate selection by T7 RNA polymerase. Cell 116, 381−391 (2004).
【非特許文献19】Hirao, I., Fujiwara, T., Kimoto, M. & Yokoyama, S. Unnatural base pairs between 2− and 6−substituted purines and 2−oxo(1H)pyridine for expansion of the genetic alphabet. Bioorg. Med. Chem. Lett. 14, 4887−4890 (2004).
【非特許文献20】Jovine, L., Djordjevic, S. & Rhodes, D. The crystal structure of yeast phenylalanine tRNA at 2.0 Å resolution: cleavage by Mg2+ in 15−year old crystals. J. Mol. Biol. 301, 401−414 (2000).
【非特許文献21】Law, S. M., Eritja, R., Goodman, M. F. & Breslauer, K. J. Spectroscopic and calorimetric characterizations of DNA duplexes containing 2−aminopurine. Biochemistry 35, 12329−12337 (1996).
【非特許文献22】Holz, B., Klimasauskas, S., Serva, S. & Weinhold, E. 2−Aminopurine as a fluorescent probe for DNA base flipping by methyltransferases. Nucleic Acids Res. 26, 1076−1083 (1998).
【非特許文献23】Rist, M. J. & Marino, J. P. Association of an RNA kissing complex analyzed using 2−aminopurine fluorescence. Nucleic Acids Res. 29, 2401−2408 (2001).
【非特許文献24】Bain, J. D., Switzer, C., Chamberlin, A. R. & Benner, S. A. Ribosome−mediated incorporation of a non−standard amino acid into a peptide through expansion of the genetic code. Nature 356, 537−539 (1992).
【非特許文献25】Ludwig, J. & Eckstein, F. Rapid and efficient synthesis of nucleoside 5’−O−(1−thiotriphosphates), 5’−triphosphates and 2’,3’−cyclophosphorothioates using 2−chloro−4H−1,3,2−benzodioxaphosphorin−4−one. J. Org. Chem. 54, 631−635 (1989).
【非特許文献26】Frugier, M., Helm, M., Felden, B., Giege, R. & Florentz, C. Sequences outside recognition sets are not neutral for tRNA aminoacylation. J. Biol. Chem. 273, 11605−11610 (1998).
【非特許文献27】Kao, C., Rudisser, S. & Zheng, M. A simple and efficient method to transcribe RNAs with reduced 3’ heterogeneity. Methods 23, 201−205 (2001).
【非特許文献28】Mitsui,T., Kimoto,M., Sato,A., Yokoyama,S. & Hirao, I., Bioorg.Med.Chem.Lett.,13, 4515−4518 (2003)
【非特許文献29】Fujiwara, T., Kimoto, M., Sugiyama, H., Hirao, I. & Yokoyama, S., Bioorg.Med.Chem.Lett.,11, 2221−2223 (2001)
【非特許文献30】Mitsui, T., Kimoto, M., Harada, Y., Yokoyama, S. & Hirao, I., J.Am.Chem.Soc.,127, 8652−8658 (2005)
【発明の開示】
【発明が解決しようとする課題】
【0019】
本発明は、新規人工塩基対に基づく核酸を提供することを目的とする。具体的には、本発明の核酸は、置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドと、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドとが塩基対を形成している。
【0020】
限定されるわけではないが、本発明の核酸は好ましくは、前記置換された若しくは無置
換の2−ホルミル−1H−ピロール−1−イル基が、
A1)2−ホルミル−1H−ピロール−1−イル基;
A2)2−ホルミル−4−(1−プロピン−1−イル)−1H−ピロール−1−イル基;
A3)2−ホルミル−4−メチル−1H−ピロール−1−イル基;及び
A4)2−ホルミル−4−エチニル−1H−ピロール−1−イル基
からなるグループから選択され、そして
前記6位置換された9H−プリン−9−イル基が、
B1)2−アミノ−6−(2−チエニル)−9H−プリン−9−イル基;
B2)6−(2−チエニル)−9H−プリン−9−イル基;
B3)2−アミノ−6−(4−メチル−2−チエニル)−9H−プリン−9−イル基;
B4)6−(4−メチル−2−チエニル)−9H−プリン−9−イル基;
B5)2−アミノ−6−(5−メチル−2−チエニル)−9H−プリン−9−イル基;
B6)6−(5−メチル−2−チエニル)−9H−プリン−9−イル基;
B7)2−アミノ−6−(2−チアゾリル)−9H−プリン−9−イル基;
B8)6−(2−チアゾリル)−9H−プリン−9−イル基;
B9)2−アミノ−6−(4−メチル−2−チアゾリル)−9H−プリン−9−イル基;
B10)6−(4−メチル−2−チアゾリル)−9H−プリン−9−イル基;
B11)2−アミノ−6−(5−メチル−2−チアゾリル)−9H−プリン−9−イル基;及び
B12)6−(5−メチル−2−チアゾリル)−9H−プリン−9−イル基
からなるグループから選択される。
【0021】
本発明の核酸は、好ましくは転写、逆転写、複製又は翻訳の工程で、塩基対を形成している。
本発明はまた、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを含む核酸を調製する方法を提供する。本発明の調製方法は、置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドを含む核酸を鋳型として転写、逆転写又は複製を行い、前記置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を有するヌクレオチドの相補的な位置に、6位置換された9H−プリン−9−イル基を有するヌクレオチドを組み込む、ことを含む。
【0022】
本発明はさらに、上記本発明の方法によって調製された、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを含む核酸を提供することを目的とする。
【0023】
本発明はさらにまた、本発明の核酸における、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを含む局部領域の立体構造の解析方法を提供することを目的とする。本発明の解析方法は、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド中の、塩基に由来する蛍光強度を、温度変化やイオン(例えば、マグネシウム)濃度などの変化等、環境を変えることにより測定することを含む。
【0024】
本発明の解析方法は、一態様において、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド中の、塩基に由来する蛍光強度が、温度の上昇に伴い実質的に上昇する場合には、生体内温度下で、前記ヌクレオチドは立体配座において周囲に存在するヌクレオチドと積み重なっていると判断し、そして、蛍光強度が温度の上昇に伴い実質的に低下又は上昇しない場合には、生体内温度下で、前記ヌクレオチドは外部に暴露していると判断する。
【0025】
本発明はまた、核酸の二本鎖形成の検出方法を提供することを目的とする。本発明の検出方法は、
I) i)本発明の核酸の調製方法によって調製された、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを含む核酸、と、
ii)5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドであって、前記塩基の5位に、5−カルボキシフルオレセイン(5−FAM)、6−カルボキシフルオレセイン(6−FAM)、5−カルボキシテトラメチルローダミン(5−TAMRA)、6−カルボキシテトラメチルローダミン(6−TAMRA)、 5−ジメチルアミノナフタレン−1−スルホン酸(DANSYL)、5−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(5−HEX)、6−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(6−HEX)、5−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(5−TET)、6−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(6−TET)、5−カルボキシ−X−ローダミン(5−ROX)、及び6−カルボキシ−X−ローダミン(6−ROX)からなるグループから選択される蛍光色素が、直接又はリンカーを介して結合している前記ヌクレオチド、を含む核酸、とをハイブリダイズさせ、そして、
II) 蛍光スペクトルの変化を測定する
ことを含む。
【0026】
本発明はさらに、
i) 6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド
並びに、
ii) 5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドであって、前記塩基の5位に、5−カルボキシフルオレセイン(5−FAM)、6−カルボキシフルオレセイン(6−FAM)、5−カルボキシテトラメチルローダミン(5−TAMRA)、6−カルボキシテトラメチルローダミン(6−TAMRA)、 5−ジメチルアミノナフタレン−1−スルホン酸(DANSYL)、5−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(5−HEX)、6−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(6−HEX)、5−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(5−TET)、6−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(6−TET)、5−カルボキシ−X−ローダミン(5−ROX)、及び6−カルボキシ−X−ローダミン(6−ROX)からなるグループから選択される蛍光色素が、直接又はリンカーを介して結合している、前記ヌクレオチド
を、同一の鎖上に含む核酸を調製する方法を提供することを目的とする。本発明の調製方法は、
iii) 置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチド
並びに
iv) 6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド
を含む核酸を鋳型として転写、逆転写又は複製を行い、前記iii)のヌクレオチドの相補的な位置に、前記i)のヌクレオチドを組み込み、そして、前記iv)のヌクレオチドの相補的な位置に、前記ii)のヌクレオチドを組み込む
ことを含む。
【0027】
本発明はさらにまた、上記方法によって調製された核酸も提供する。
本発明はまた、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドと5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドとを、同一の鎖上に含む、本発明の核酸のステム−ループ(ヘアピン)構造形成の検
出方法を提供することを含む。本発明の検出方法は、
1) 本発明の核酸において、i)のヌクレオチドと、ii)のヌクレオチドとをハイブリダイズさせ、そして、
2) 蛍光スペクトルの変化を測定する
ことを含む。
【課題を解決するための手段】
【0028】
本発明者らは、上記問題解決のために、以前に自分たちが開発したいくつかの非天然型の塩基を組み合わせることによって構築される塩基対を研究した。その結果、2−アミノ−6−(2−チエニル)プリン−9−イル(s)(非特許文献13、14)と2−ホルミル−1H−ピロール−1−イル(Pa)(非特許文献9)(図1A)の組み合わせが、転写において高度な選択性および効率を示すことを見出した。即ち、PaヌクレオチドをDNAの鋳型に挿入しRNAへの転写反応を行うと、生成するRNA中の、鋳型DNAのPaの相補的な位置に選択的にかつ高収率でsが導入されることを発見し、本発明を想到した。
【0029】
基質sはT7 RNA ポリメラーゼによって、DNA鋳型から生成されるRNAにおいて、鋳型中のPaの相補的な位置に部位特異的に挿入することができる。プリン塩基であるsは、水素結合を可能とする置換基(1位の水素受容基と2位の水素供与のアミノ基)を有する。しかし、Paは対合表面にそのような親水性の基を有しない。本発明者らは、Pa塩基のホルミル基中の酸素は、二本鎖DNAの小溝(minor groove)に向かい、ポリメラーゼと相互作用していて、塩基対形成における水素受容基としては働いていないことを見出している(非特許文献9)。本発明において、s−Pa対は有意な水素結合相互作用を有しないが、5員環塩基Paはsと形状相補性を共有すると考えられる。
【0030】
本発明の人工塩基対に基づく核酸
よって、本発明はその一態様において、置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドと、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドとが塩基対を形成している、核酸を提供する。
【0031】
本発明における「ヌクレオシド」とは、核酸塩基と糖の還元基とがグリコシド結合によって結合した配糖体化合物を意味する。なお、「核酸塩基」は、アデニン、グアニン、シトシン、チミン、ウラシル、及びこれら塩基の誘導体も含む概念である。「誘導体」の種類は特に限定されるものではないが、具体的には、例えば、置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基に相当する塩基や、6位置換された9H−プリン−9−イル基に相当する塩基、などが挙げられる。「ヌクレオチド」は、前記ヌクレオシドの糖部分が、リン酸とエステルをつくっている化合物をいう。より好ましくは、一、二、又は三リン酸エステルである。ヌクレオシド又はヌクレオチドの糖部分はリボフラノシル、2’−デオキシリボフラノシル、あるいはハロゲンなどの置換基を2’位に有する2’−置換リボフラノシルであってもよく、また、リン酸部分は、チオリン酸であってもよい。つまり、糖部分及びリン酸部分は、公知のヌクレオシド、ヌクレオチド、あるいはこれらの誘導体にみられる構成をとっていればよい。糖部分がリボフラノシルであるリボヌクレオチドはRNAの構成成分となり、デオキシリボフラノシルであるデオキシリボヌクレオチドはDNAの構成成分となる。
【0032】
本発明の「置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基」は、以下の一般式で表される構造を有する。
【0033】
【化3】
【0034】
ここにおいて、ピロール環の4位のRは、水素であるか、あるいは、置換された若しく
は無置換のC1−C3アルキル基、置換された若しくは無置換のC2−C3アルケニル基、又は置換された若しくは無置換のC2−C3アルキニル基から選択される置換基によって置換されていてもよい。上記アルキル基、アルケニル基又はアルキニル基等は、さらに、低級アルキル基、ハロゲン基、ヒドロキシル基、アミノ基、アルキルアミノ基、及び芳香族複素環からなるグループより独立して選択される一つまたはそれより多くの基で置換されていてもよい。本発明の置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を、本明細書において文脈により、「Pa」又は「Pa類似体」と呼称する。
【0035】
限定されるわけではないが、前記置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基は、
A1)2−ホルミル−1H−ピロール−1−イル基(Pa);
A2)2−ホルミル−4−(1−プロピン−1−イル)−1H−ピロール−1−イル基;
A3)2−ホルミル−4−メチル−1H−ピロール−1−イル基;及び
A4)2−ホルミル−4−エチニル−1H−ピロール−1−イル基
からなるグループから選択されてもよい。より好ましくは、2−ホルミル−1H−ピロール−1−イル基である。
【0036】
本発明の「置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基」(「Pa」又は「Pa類似体」)を有するヌクレオシド、ヌクレオチドは、公知の方法を用いて合成することが可能である。その出発原料である例えば、ピロール−2−カルボアルデヒドは、例えば、Aldrich社 [1003−29−8]、Merck社 [807574]から購入することができる。また、Pa誘導体は、基本的にPaから誘導することによって合成できる。例えば、Paの4位にプロピンを導入した誘導体は、Bioorg.Med.Chem.Lett.,13,p.4515−4518(2003)(非特許文献28)に記載されている。
【0037】
本発明の「6位置換された9H−プリン−9−イル基」は、以下の一般式で表される構造を有する。
【0038】
【化4】
【0039】
[ここにおいて、
R1は、水素又はアミノ基であり、
R2は、置換された又は無置換の2−チエニル基又は2−チアゾリル基である。]
R2のチエニル基又はチアゾリル基は、無置換であるかあるいは、4位及び/又は5位が、メチル基、アミノ基、ニトロ基及びヒドロキシ基からなるグループより独立して選択される一つまたはそれより多くの基で置換されていてもよい。本発明の6位置換された9H−プリン−9−イル基のうち、R2が置換された又は無置換の2−チエニル基のものを、本明細書において文脈により、「s」又は「s類似体」と呼称する。本発明の6位置換された9H−プリン−9−イル基のうち、R2が置換された又は無置換の2−チアゾリル基のものを、本明細書において文脈により、「v」又は「v類似体」と呼称する。
【0040】
限定されるわけではないが、前記6位置換された9H−プリン−9−イル基は、
B1)2−アミノ−6−(2−チエニル)−9H−プリン−9−イル基(s);
B2)6−(2−チエニル)−9H−プリン−9−イル基(s’);
B3)2−アミノ−6−(4−メチル−2−チエニル)−9H−プリン−9−イル基;
B4)6−(4−メチル−2−チエニル)−9H−プリン−9−イル基;
B5)2−アミノ−6−(5−メチル−2−チエニル)−9H−プリン−9−イル基;
B6)6−(5−メチル−2−チエニル)−9H−プリン−9−イル基;
B7)2−アミノ−6−(2−チアゾリル)−9H−プリン−9−イル基(v);
B8)6−(2−チアゾリル)−9H−プリン−9−イル基;
B9)2−アミノ−6−(4−メチル−2−チアゾリル)−9H−プリン−9−イル基;
B10)6−(4−メチル−2−チアゾリル)−9H−プリン−9−イル基;
B11)2−アミノ−6−(5−メチル−2−チアゾリル)−9H−プリン−9−イル基;及び
B12)6−(5−メチル−2−チアゾリル)−9H−プリン−9−イル基
からなるグループから選択されてもよい。より好ましくは、2−アミノ−6−(2−チエニル)−9H−プリン−9−イル基である。
【0041】
上記塩基のうち、B1)は、「s」であり、B2)ないしB6)は「s類似体」である。また、B7)は、「v」であり、B8)ないしB12)は「v類似体」である。
さらにより好ましくは、前記置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基が、2−ホルミル−1H−ピロール−1−イル基であり、そして、前記6位置換された9H−プリン−9−イル基が、2−アミノ−6−(2−チエニル)−9H−プリン−9−イル基である。
【0042】
本発明の「6位置換された9H−プリン−9−イル基」、及びこれを含むヌクレオシド、ヌクレオチドは、公知の方法を用いて合成することが可能である。具体的には例えば、本明細書の実施例1には、2−N−フェノキシアセチル−6−(2−チエニル)−9−(2,3−ジ−O−アセチル−1−β−D−リボフラノシル)プリンから、2−アミノ−6−(2−チエニル)−9−(1−β−D−リボフラノシル)プリン 5’−三リン酸(sTP)を合成する方法を開示した。
【0043】
またsの合成は、例えば、Bioorg.Med.Chem.Lett.,11,p.2221−2223(2001)(非特許文献29)の第2頁の第二段落に記載されている。また、vの合成は、例えば特許文献3の段落0026、0027、図5−6等に開示されている。あるいは、J.Am.Chem.Soc.,127,p.8652−8658(2005)(非特許文献30)の第2頁の右側の段落に開示されている。
【0044】
s、v以外の、本発明の「6位置換された9H−プリン−9−イル基」を有するヌクレオシド、ヌクレオチドも、s又はvのいずれかの合成方法と同様の方法を用いて合成することができる。例えば、フランを結合したo は、sと同様の合成法で合成可能である(非
特許文献29)。
【0045】
本発明は、置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドと、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドとが塩基対を形成している、核酸を提供する。本明細書において「核酸」とは、1より多くのヌクレオチドが、5’→3’の方向に結合した核酸鎖の分子を意味する。本発明の核酸は、一本鎖又は二本鎖のRNA又はDNAを含む。二本鎖は、DNA/DNA、RNA/RNA、又はDNA/RNAであってもよい。また、DNAには、RNAを鋳型として逆転写してなるcDNAも含まれる。あるいは、核酸は3本鎖、4本鎖等も形成しうる。
【0046】
本発明者らは、核酸のさらなる機能拡張をめざして、非天然型塩基をもつヌクレオシド又はヌクレオチドの創製へ取り組んだ。新たに開発された人工塩基対の態様として、置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基(例:Pa)として有するヌクレオチドと、6位置換された9H−プリン−9−イル基を塩基(例:s)として有するヌクレオチドとの塩基対が含まれる。(図1)。これにより特に、Paを含むDNA鋳型を用いたT7 RNAポリメラーゼによる転写において、RNAへのsの特異的取り込みが可能となった。
【0047】
sおよびPa間では有意な水素結合相互作用が存在しないにもかかわらず(非水素結合s−Pa対)、転写におけるs−Pa対合の効率および選択性は、天然の塩基対合のそれと同程度に高いものである。このs−Pa対は、先に開発された親水性s−z対よりも高い効率を示す。s及びPaの相補的な形状は互いに適合するが、天然のプリン類及びピリミジン類の形状とは相違する。この特異的な立体化学的適合が、天然塩基との非規範的対合を排除し、その結果、転写の際にs−Paの高い選択性が得られると考えられる。このように、形状−相補性は転写、及び複製における特異的な塩基対合に重要な役割を果たす。
【0048】
本発明の、置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドと、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドとは、異なる2本の核酸上に存在して塩基対により二本鎖を形成しうる。あるいは、両ヌクレオチドは同一の一本の核酸上に存在してもよい。この場合、塩基対により、一本鎖はループ構造を形成してもよい。
【0049】
本発明の置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチド、あるいは6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドは、転写、複製又は逆転写反応により、DNA又はRNA等の核酸に取り込むことが可能である。あるいは、天然型塩基を有するヌクレオシド又はヌクレオチドと同様に、化学合成によってDNA又はRNAに取り込んでもよい。
【0050】
転写、複製又は逆転写反応は公知の方法に従って行うことが可能である。限定されるわけではないが、例えば、転写反応はT7 RNAポリメラーゼ(Takara等)、複製反応は、クレノウフラグメント(KF)、逆転写反応はAMV Reverse Transcriptase XL (AMV−RT)(Life Science社)を使用することが可能である。複製反応は、反応中に本発明のヌクレオチドが除去されてしまうのを防ぐために、例えば、3’→5’エキソヌクレアーゼ活性をもたないTaq DNAポリメラーゼ(Takara TaqTM)を用いて、置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドを含むプライマーによる鋳型DNAのPCR増幅も可能である。
【0051】
限定されるわけではないが、本発明の核酸は一態様において、核酸の転写、逆転写、複製又は翻訳の工程で、塩基対を形成している。本発明の核酸が、転写工程で塩基対を形成している場合、前記置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドがDNAの一部であり、そして、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドがRNAの一部でありうる。
【0052】
核酸の調製方法
本発明はまた、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを含む核酸を調製する方法を提供する。本発明の調製方法は、置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドを含む核酸を鋳型として転写、逆転写又は複製を行い、前記置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を有するヌクレオチドの相補的な位置に、6位置換された9H−プリン−9−イル基を有するヌクレオチドを組み込む、ことを含む。
【0053】
2−ホルミル−1H−ピロール−1−イル基を有するヌクレオチド、並びに6位置換された9H−プリン−9−イル基を有するヌクレオチドの定義については、本明細書の「本発明の人工塩基対に基づく核酸」の項目において前述した通りである。
【0054】
本明細書中の実施例1−2でも明らかにされたように、鋳型中に置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドが2つまたはそれ以上連続して存在する場合でも、転写反応は進行し、相補的な位置に6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドが取り込まれる。よって、本発明の方法により従来不可能であった、非天然型塩基sが2つ又はそれより多く隣接して配置されるDNA及びRNAの作成も可能となった。よって、本発明の調製方法は、一態様において、鋳型において、前記置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドが2個以上存在する。
【0055】
本発明はさらに、上記本発明の方法によって調製された、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを含む核酸を提供することを目的とする。
【0056】
本発明のヌクレオチドが組み込まれた核酸は、tRNA、mRNA、アンチセンスDNA若しくはRNA、リボザイム又はアプタマーとして使用されうる。アンチセンスDNA若しくはRNAとは、ある特定の遺伝子の発現を抑えるDNA又はRNAである。標的とする遺伝子配列(センス鎖)の全長又は部分配列に対して相補的という意味で名付けられた。人為的に遺伝子発現を調節する手法として使用されうる。本発明のヌクレオチドが組み込まれたアンチセンスDNA又はRNAは、非天然型塩基を含むため標的に対する相補性が天然型塩基のみを使用した場合と比較して異なるものを創製しうる。リボザイムは、RNAを構成成分とする触媒の総称である。アプタマーは、in vitroセレクション法によって得られた、タンパク質等の特定の分子に結合する機能を有する核酸である
また、本発明の、ヌクレオチドが組み込まれた核酸(DNA又はRNA)(例えば、mRNA、合成RNA)は、タンパク質、ペプチドの全体又は一部をコードするものであってもよい。本発明の核酸は遺伝子断片やプローブなどとして使用されうる。天然の遺伝子の一部又は全部を本発明の核酸で置換した態様、天然の遺伝子に本発明のヌクレオチドを1個又はそれより多く付加したもの、又はこれらを組み合わせたものも本発明に包含される。
【0057】
さらにまた、本発明の非天然型塩基を有するヌクレオチドが組み込まれた核酸は、RNA干渉(RNA interference、RNAi)においても利用可能である。R
NA干渉は、二本鎖RNA(dsRNA)によってその配列特異的にmRNAが分解され、その結果遺伝子の発現が抑制される現象である。RNA干渉の典型的な例としては、dsRNAは、RNaseIIIファミリーに属するダイサー(Dicer)により、3’末端の側に2塩基程度のオーバーハングを有する約21塩基−23塩基のsiRNA(short interfering RNA)にプロセッシングされる。siRNAはRISCと呼ばれるsiRNA−蛋白質複合体に取り込まれ、配列特異的にmRNAを分解する。RNA干渉は、哺乳動物(ヒト、マウス等)、線虫、植物、ショウジョウバエ、菌類などの広範な生物種間で保存されている現象であることが示されている。本発明の非天然型塩基を有するヌクレオチドが組み込まれた核酸は、RNA干渉におけるsiRNAとして、または分解を受けるmRNAの一部として利用可能である。
【0058】
また、本発明のヌクレオチドを含む新たなコドンを設計することが可能である。前述したように、鋳型中に置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基(例えば、Pa)として有するヌクレオチドが2つまたはそれ以上連続して存在する場合でも、転写反応は進行し、相補的な位置に6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド(例えば、s)が取り込まれる。よって、本発明の方法により従来不可能であった、非天然型塩基sが2つ又はそれより多く隣接して配置されるDNA及びRNAの作成が可能である。よって、sを3つ含むコドン(sss)、sを2つ含むコドン(例えば、ssA、Gss、sGs)、sを1つ含むコドン(例えば、sAG、CsT、AGs)を設計することができる。新たなコドンは、天然型のアミノ酸をコードさせることもできるし、また、非天然型のアミノ酸をコードさせることもできる。このように、本発明は新規な非天然型人工塩基を提供するのみならず、本発明のヌクレオチドを含む新たなコドンの設計による、全く新しい遺伝暗号の設計を可能とするものであり、新たな遺伝暗号の世界を提供するものである。
【0059】
さらに、本発明の新たなコドンに応じたtRNA系を設計することにより、非常に多くのアミノ酸を利用可能とする新たなタンパク質合成システムを設計することができる。利用可能なアミノ酸はリボソームにおけるタンパク質合成酵素系で利用できるものであればよい。従って、本発明は前記本発明のコドンを用いた新たなタンパク質合成システムを提供する。本発明のタンパク質合成システムによれば、所望の位置のコドンの核酸を本発明の核酸によって効率よく置換、又はコドンの所望の位置へ本発明の核酸を導入することにより、所望の非天然型のアミノ酸を含有するタンパク質の製造が可能となる。
【0060】
立体構造の解析方法
6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド(例えば、s)は蛍光塩基であり、これを塩基として含むヌクレオチドは強い蛍光特性を有する。
【0061】
具体的には、6位に芳香族複素環などを結合させて電子共役系を広げたプリン誘導体は蛍光特性がでやすい。さらに、限定されるわけではないが、従来から2−アミノプリンは蛍光特性を有することがわかっている。よって、限定されるわけではないが、特に6位に芳香族複素環が結合しており、かつ2位にアミノ基を有するプリンは強い蛍光特性を示す。特に、s、s’及びvは好ましい蛍光塩基である。従来技術に対して、蛍光色素を結合した第5番目の塩基を人工塩基対を介して、転写により核酸中の特定部位に導入できれば、核酸の標識、そして、それによる核酸の解析、検出が非常に簡便になる。本発明ではPa−sという人工塩基対を用いて、これを可能にした。
【0062】
本発明者らは、蛍光性の人工塩基sをプローブとして、酵母tRNAPheを部位特異的に蛍光標識し、標識部位、温度およびMg2+濃度に応じた、特有の蛍光プロフィールを得た(実施例3)。さらに、蛍光プロフィールを同一のs標識された酵母tRNAPheに対するUVプロフィールと比較した。その結果、以下のことが判明した。
【0063】
第一にsを含む各tRNA転写産物のUV吸収スペクトルの変化から求めた融解温度(Tm値)は、天然のtRNA転写産物のUV吸収スペクトルの変化から求めた融解温度(Tm値)と同じくらいの高さであった。このことはこれらの位置におけるsによる置換は、tRNA構造全体を有意に不安定化しないことを示唆している。
【0064】
第二に、特定の位置にsを含む各tRNAは、局部構造特性を反映する、蛍光強度の特徴的な変化を表示した。すなわちtRNAの高次構造において、sが近隣の塩基と積み重なっている場合は蛍光強度が弱く、積み重なり(stacking)の程度が減少するに従い蛍光強度が増加した。この発見は、RNAの特定の位置にsを導入し、例えば温度変化に伴う蛍光強度の変化を測定することにより、高次構造中におけるそのsの積み重なりの状態を把握することが可能なことを示している。そこで、sを含むRNAの用途として、当該RNAの高次構造の新規な解析方法が提供される。
【0065】
より具体的には、Mg2+(2及び5mM)の存在下における、酵母tRNAPheのs蛍光プロフィールは、明確に2つのグループに分類された。低温度でグループ1(tRNA 16s、17s及び47s)によって放射されるs蛍光の強度は、グループ2(RNA 36s、57s及び59s)によって放射される強度よりも、1.7−3.4倍大きかった。温度が上昇すると、グループ1によって放出される大きな強度は減少し、逆に、グループ2による強度は上昇した。これらの結果は、グループ1のs塩基は生理学的温度で溶液中に露出しており、そして、温度の上昇による折り畳み構造の変性に伴い、s塩基の他の塩基との非特異的な相互作用が高まることを示唆している。対照的に、グループ2の塩基は、低温下では折り畳み構造において近隣の塩基が積み重なっており、そして、温度の上昇によって塩基積み重ねが徐々に変性する。この推論は、tRNAの結晶構造中の各部位における元来の塩基の配座(図4)(非特許文献20)と極めてよく一致する。さらに、グループ1の蛍光強度は、生理学的温度において、Mg2+の添加によって劇的に上昇する。このことは、Mg2+存在下でtRNAが活性型のL字型構造を形成し、これらの位置における塩基が外側に配置されることを示唆している。
【0066】
第三に、蛍光プロフィールから得られるTm値は、UV融解プロフィールから得られるTm値と比較して、tRNA構造中の各局部構造の安定性を反映した。
例えば、tRNA 36sの蛍光プロフィールから得られるTm値は、UVプロフィールから得られるTm値よりも高かった。このことは、全tRNA構造の安定性に比べてアンチコドン ステム−ループがより高い安定性を有することを示唆している。対照的に、tRNA16s及び17sの低い安定性(蛍光プロフィールから得られる低いTm値)は、Dループを含む部分構造は、tRNA構造全体の脆弱な部分の可能性があることを示唆している。よって、各s標識体の蛍光プロフィールから得られるTm値を、UV融解プロフィールから得られるTm値と比較することにより、s標識された局部構造の安定性の高さを判別することができる。
【0067】
このように、s標識を利用した部位特異的な蛍光プローブ化は、三次元RNA分子の局部構造特徴を研究するための有力なツールとなりうる。
よって、本発明は、本発明の核酸における、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを含む局部領域の立体構造の解析方法を提供することを目的とする。本発明の解析方法は、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド中の、塩基に由来する蛍光強度をいろいろな環境下(温度変化、イオン濃度(例えば、マグネシウム)の変化など)で測定することを含む。
【0068】
本発明の解析方法は、一態様において、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド中の、塩基に由来する蛍光強度が、温度の上昇に伴い実
質的に上昇する場合には、生体内温度下で、前記ヌクレオチドは立体配座において周囲に存在するヌクレオチドと積み重なっていると判断し、そして、蛍光強度が温度の上昇に伴い実質的に低下又は上昇しない場合には、生体内温度下で、前記ヌクレオチドは外部に暴露していると判断する。
【0069】
「生体内温度」とは、各生体が自然の条件下で生息している場合の体内の温度を意味し、生体の種類に応じて変化する、例えば、ヒトの場合は約35.0℃ないし約39.0℃であり、ウシの場合は、約36.7℃ないし約39.3℃、マウスの場合は、約35.0℃ないし約39.0℃、イヌの場合は、約37.9℃ないし約39.9℃、ブタの場合は、約38.7℃ないし約39.8℃である。
【0070】
「ヌクレオチドが立体配座において周囲に存在するヌクレオチドと積み重なっている」とは、より具体的には、2つ以上の塩基が好ましくは約5Å以内で積み重なった状態(ス
タッキング)を意味する。
【0071】
「ヌクレオチドが外部に暴露している」とは、より具体的には、塩基部分が他の塩基とスタッキングしていない(好ましくは約5Å以上離れた)状態、あるいは、消光性の塩基と十分にはなれている状態を意味する。
【0072】
蛍光強度は、好ましくは、前記6位置換された9H−プリン−9−イル基の種類に応じた最大の蛍光特性を示す特定の波長で測定する。例えば、B1)2−アミノ−6−(2−チエニル)−9H−プリン−9−イル基(s)は、最大励起波長である352nmで励起すると、434nmを中心とする蛍光波長を示す。同様に、B2)6−(2−チエニル)−9H−プリン−9−イル基(s’)は、325nmで励起すると381nmを中心とする蛍光波長を示し、B7)2−アミノ−6−(2−チアゾリル)−9H−プリン−9−イル基(v)は、363nmで励起すると461nmを中心とする蛍光波長を示す。
【0073】
「蛍光強度が温度の上昇に伴い実質的に上昇する」とは、例えば温度が20℃から90℃まで上昇した場合に、蛍光強度が上昇することが確認できること(例えば1.1倍以上)を意味する。蛍光強度の上昇の割合が大きいほど、生体内温度下で、その蛍光性の塩基が立体配座において周囲に存在する、より多くのヌクレオチドと、及び/又は、より近距離で積み重なっていることを示す。
【0074】
「蛍光強度が温度の上昇に伴い実質的に低下又は上昇しない場合」とは、例えば温度が20℃から90℃まで上昇した場合に蛍光強度が変化しない又は低下することが確認できること(例えば、10%以上減少)を意味する。蛍光強度の低下の割合が大きいほど、生体内温度下で、その蛍光性の塩基が孤立して存在していて、その部分の核酸の構造が特定の構造を取っていないということを示す。
【0075】
核酸の二本鎖形成の検出方法
本発明はさらに、本発明の核酸の調製方法によって調製された、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを含む核酸の別の利用態様の1つである、核酸の二本鎖形成の検出方法を提供する。本発明の検出方法は、
I) i)本発明の核酸の調製方法によって調製された、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを含む核酸、と、
ii)5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドであって、前記塩基の5位に、5−カルボキシフルオレセイン(5−FAM)、6−カルボキシフルオレセイン(6−FAM)、5−カルボキシテトラメチルローダミン(5−TAMRA)、6−カルボキシテトラメチルローダミン(6−TAMRA)、 5−ジメチルアミノナフタレン−1−スルホン酸(DANSYL)、5−カルボキシ−2’,
4,4’,5’,7,7’−ヘキサクロロフルオレセイン(5−HEX)、6−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(6−HEX)、5−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(5−TET)、6−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(6−TET)、5−カルボキシ−X−ローダミン(5−ROX)、及び6−カルボキシ−X−ローダミン(6−ROX)からなるグループから選択される蛍光色素が、直接又はリンカーを介して結合している前記ヌクレオチド、を含む核酸、とをハイブリダイズさせ、そして、
II) 蛍光スペクトルの変化を測定する
ことを含む。
【0076】
本発明の核酸の二本鎖形成の検出方法は、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを含む核酸、と、蛍光色素によって5位が置換された、5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドを含む核酸とのハイブリダイズによる蛍光スペクトルの変化を利用するものである。特許文献1、2、非特許文献30等に記載されているように、6位置換された9H−プリン−9−イル基(例えばs、v)と、2−オキソ(1H)ピリジン−3−イル基(y)は塩基対合を形成しうる。なお、5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオシド又はヌクレオチドの合成方法等については、本明細書中に別途詳述する。
【0077】
6位置換された9H−プリン−9−イル基の蛍光と、これとは別の特性を有する蛍光色素で置換された5位置換−2−オキソ(1H)ピリジン−3−イル基の蛍光が各核酸分子に存在する場合に、核酸の二本鎖が形成され蛍光同士が物理的に近傍に存在すると、蛍光共鳴エネルギー転移(FRET:Fluorescence Resonance Energy Transfer)が生じて蛍光スペクトルが変化する。本発明の検出方法は、核酸の二本鎖の形成をFRETによる蛍光スペクトルの変化の測定によって検出するものである。
【0078】
「蛍光共鳴エネルギー転移(FRET)」は、ある蛍光分子から他の分子へ、共鳴により励起エネルギーが移動する現象を意味する。エネルギーを与える分子はドナー(供与体)、受け取る分子はアクセプター(受容体)と呼ばれる。FRETが生じると、エネルギーを失ったドナーは基底状態に戻り、同時にエネルギーを受け取ったアクセプターは励起状態となる。従って、ドナーの蛍光は弱まり、アクセプターが蛍光分子であればその蛍光が観察される。アクセプターが消光分子であれば、ドナーが単独の場合には観察されていた蛍光がFRETにより観察されなくなる。FRETによるタンパク質の検出、核酸の検出の一般的な方法は公知であり、例えば、RNAアプタマーに2種類のFRETを起こす色素を導入して、標的タンパク質を検出する方法(Jhaveri et al.,2000)や、ヘアピン型核酸を利用した相補鎖核酸の検出法(Tyagi et al.,1996)が知られている。その他、FRETに関しては、Walter et al.
,2001及びKlostermeier et al.,2001に概説されている。
【0079】
FRETが生じるためには、以下の3条件を満たす必要がある。i)ドナーの蛍光スペクトルとアクセプターの吸収スペクトルの重なりがあること。スペクトルの重なり範囲は大きい方が望ましいが、必ずしも完全に重なっている必要はない。ドナーの蛍光スペクトルとアクセプターの吸収スペクトルが30%以上重なっていることが好ましく、50%重なっていることがより好ましい。ii)ドナーとアクセプターが物理的に近距離に存在すること。FRETが50%の確率で生じる距離は3nmないし6nmと考えられており、FRETの効率はこの距離の変化に対して敏感に変化する。例えば、核酸が二本鎖を形成した場合に、蛍光特性を有する6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドに対し、これとは別の特性を有する蛍光色素で置換された5位置換−2
−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドが近接し、両者の距離が好ましくは10nm以内、より好ましくは6nm以内、最も好ましくは3nm以内である場合に蛍光スペクトルの変化が検出可能である。iii)ドナーとアクセプターの相対的な向きが適切であること。
【0080】
本発明の検出方法の一態様において、5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドを含む核酸は、塩基の5位に、5−カルボキシフルオレセイン(5−FAM)又は6−カルボキシフルオレセイン(6−FAM)がリンカーを介して結合している、5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドを含む核酸である。
【0081】
前述したように、例えばsは353nmの最大励起波長、そして、最大励起波長である352nmで励起すると、434nmを中心とする波長を示す。一方、FAMは、FAMの吸収極大波長は493nm、蛍光極大波長は522nmである。353nmの波長で励起した場合、sを有するヌクレオチドは434nmを中心とする波長を示す。これに対し、FAMで置換された5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチド(例えば、FAM−y)がsに近接すると、sからの蛍光を吸収し、蛍光極大波長が522nmの蛍光スペクトルを示すようになる。即ち、353nmの波長で励起した場合に、434nmの蛍光スペクトルが減少し、522nmの蛍光スペクトルが増加するほど、sに対してFAM−yが近接していること、即ち、sを含む核酸とFAM−yを含む核酸とのハイブリダイゼーションにより核酸の二本鎖が形成されていることを検出することができる。
【0082】
5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオシド又はヌクレオチド
5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオシド又はヌクレオチドにおいて、5位に蛍光色素は、直接又はリンカーを介して結合している。これらの標識ヌクレオシド又はヌクレオチドについては、本発明者の先行する特許出願(特願2004−324271、2004年11月8日出願、未公開)に詳述されている。
【0083】
蛍光色素は、公知の蛍光色素を利用可能であるが、好ましくは、5−カルボキシフルオレセイン(5−FAM)、6−カルボキシフルオレセイン(6−FAM)、5−カルボキシテトラメチルローダミン(5−TAMRA)、6−カルボキシテトラメチルローダミン(6−TAMRA)、5−ジメチルアミノナフタレン−1−スルホン酸(DANSYL)、5−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(5−HEX)、6−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(6−HEX)、5−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(5−TET)、6−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(6−TET)、5−カルボキシ−X−ローダミン(5−ROX)、及び6−カルボキシ−X−ローダミン(6−ROX)からなるグループから選択される。フルオレセイン及びローダミンは、一般的に開環型及びスピロ型の二通りで表記される。これらの蛍光色素については、例えば Tuschl et al.,1994;Misra et al
.,2004;Gibson et al.,1996;及び、Chehab et al.,1989;に詳述されている。
【0084】
例えば、FAMの吸収極大波長は493nm、蛍光極大波長は522nmである。また、TAMRAの吸収極大波長は553nm、蛍光極大波長は578nmである。DANSYLの吸収極大波長は335nm、蛍光極大波長は518nmである。HEXの吸収極大波長は535nm、蛍光極大波長は556nmである。TETの吸収極大波長は521n
m、蛍光極大波長は536nmである。5−ROXの吸収極大波長は567nm、蛍光極大波長は591nmである。6−ROXの吸収極大波長は570nm、蛍光極大波長は590nmである。
【0085】
5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオシド又はヌクレオチドは、5位置換基として蛍光分子を有するため、蛍光分子の種類に応じて、核酸の検出を行うことが可能である。よって、標識核酸として当該核酸と相互作用する物質検出のプローブとして使用されうる。また、各蛍光色素によって蛍光色が異なるため、多重染色に使用することも可能である。
【0086】
蛍光色素は、2−オキソ(1H)ピリジン−3−イル基の5位に直接結合させても、あるいはリンカーを介して結合させてもよい。リンカーの種類は特に限定されず、当業者は適宜採用可能である。リンカーは限定されるわけではないが、好ましくは、下記の化学式I−III:
【0087】
【化5】
[式I中、nは1ないし5の整数から選択される]
【0088】
【化6】
[式II中、m及びlは、1ないし5の整数から各々独立に選択される]
及び
【0089】
【化7】
【0090】
からなるグループから選択される
5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオシド
又はヌクレオチドの合成法は、特に限定されることはない。3−(β−D−リボフラノシル)−ピリジン−2(1H)−オンに、先ず5位に置換基を導入し、次いで、三リン酸を導入してもよい。あるいは、3−(β−D−リボフラノシル)−ピリジン−2(1H)−オンに、先ず三リン酸を導入し、次いで、置換基を導入してもよい。特に、大きな基を導入する場合、先ず、5位にアミノプロピニル基等を導入し、これに活性化した置換基を結合させてもよい。
【0091】
あるいは、蛍光色素による、2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドの修飾は、単独のヌクレオチド合成時ではなく、当該ヌクレオチドを含む核酸を転写等によって合成した後に行うことも可能である。
【0092】
5位置換−2−オキソ(1H)ピリジン−3−イル基は、6位置換された9H−プリン−9−イル基と、2箇所で水素結合を生じる。5位置換−2−オキソ(1H)ピリジン−3−イル基は、立体構造上、天然型プリン塩基A(アデニン)及びG(グアニン)とは塩基対を形成できない。そして、6位置換された9H−プリン−9−イル基は立体障害のため、天然型T(チミン)、U(ウラシル)及びC(シトシン)とは塩基対を形成できない。よって、6位置換された9H−プリン−9−イル基(例えばs、v)と、5位置換−2−オキソ(1H)ピリジン−3−イル基は、特異的に塩基対を形成しうる。
【0093】
5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドは、転写、複製又は逆転写反応により、DNA又はRNA等の核酸に取り込むことが可能である。具体的には、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを含む核酸、例えば、sあるいはvを含む鋳型DNAを用いてT7 RNAポリメラーゼによる転写反応を行うと、2−オキソ(1H)ピリジン−3−イル基の基質(例えば、yTP)が部位特異的に、sあるいはvに相補して、RNA中に取り込まれる。yは、その5位を化学的に修飾することができるので、種々の機能性のy誘導体をRNA中の特定の部位に取り込ませることが出来る。
【0094】
あるいは、天然型塩基を有するヌクレオシド又はヌクレオチドと同様に、化学合成によってDNA又はRNAに取り込んでもよい。
同一の鎖上に2種類の人工塩基を含む核酸を調製する方法
本発明はさらに、
i) 6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド
並びに、
ii) 5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドであって、前記塩基の5位に、5−カルボキシフルオレセイン(5−FAM)、6−カルボキシフルオレセイン(6−FAM)、5−カルボキシテトラメチルローダミン(5−TAMRA)、6−カルボキシテトラメチルローダミン(6−TAMRA)、 5−ジメチルアミノナフタレン−1−スルホン酸(DANSYL)、5−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(5−HEX)、6−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(6−HEX)、5−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(5−TET)、6−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(6−TET)、5−カルボキシ−X−ローダミン(5−ROX)、及び6−カルボキシ−X−ローダミン(6−ROX)からなるグループから選択される蛍光色素が、直接又はリンカーを介して結合している、前記ヌクレオチド
を、同一の鎖上に含む核酸を調製する方法を提供することを目的とする。本発明の調製方法は、
iii) 置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチド
並びに
iv) 6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド
を含む核酸を鋳型として転写、逆転写又は複製を行い、前記iii)のヌクレオチドの相補的な位置に、前記i)のヌクレオチドを組み込み、そして、前記iv)のヌクレオチドの相補的な位置に、前記ii)のヌクレオチドを組み込む
ことを含む。
【0095】
本発明の上記調製方法は、置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドを鋳型とした場合の、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドの取り込みと、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを鋳型とした場合の、5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドの取り込みを組み合わせて利用したものである。
【0096】
本発明はさらにまた、上記方法によって調製された
i) 6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド
並びに、
ii) 5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドであって、前記塩基の5位に、5−カルボキシフルオレセイン(5−FAM)、6−カルボキシフルオレセイン(6−FAM)、5−カルボキシテトラメチルローダミン(5−TAMRA)、6−カルボキシテトラメチルローダミン(6−TAMRA)、 5−ジメチルアミノナフタレン−1−スルホン酸(DANSYL)、5−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(5−HEX)、6−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(6−HEX)、5−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(5−TET)、6−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(6−TET)、5−カルボキシ−X−ローダミン(5−ROX)、及び6−カルボキシ−X−ローダミン(6−ROX)からなるグループから選択される蛍光色素が、直接又はリンカーを介して結合している、前記ヌクレオチド
を、同一の鎖上に含む核酸も提供する。
【0097】
本発明の上記核酸は、tRNA、mRNA、アンチセンスDNA若しくはRNA、リボザイム又はアプタマーとして利用可能である。さらに、RNA干渉(RNA interference、RNAi)においても利用可能である。
【0098】
本明細書中の実施例4では、Pa及びvを含むDNAを鋳型として転写を行い、sとy、あるいは、sとFAM−yを同一鎖上に含むRNAを作成した。
核酸のステム−ループ(ヘアピン)構造形成の検出方法
本発明はまた、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドと5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドとを同一の鎖上に含む、本発明の核酸のステム−ループ(ヘアピン)構造形成の検出方法を提供することを含む。本発明の検出方法は、
1) 本発明の核酸において、i)のヌクレオチドと、ii)のヌクレオチドとをハイブリダイズさせ、そして、
2) 蛍光スペクトルの変化を測定する
ことを含む。
【0099】
本発明の核酸のステム−ループ(ヘアピン)構造形成の検出方法は、「核酸の二本鎖形成の検出方法」の項目で上述したFERTを利用するものである、即ち、6位置換された9H−プリン−9−イル基の蛍光と、これとは別の特性を有する蛍光色素で置換された5
位置換−2−オキソ(1H)ピリジン−3−イル基の蛍光が、「同一分子内に」存在する場合に、ステム−ループ(ヘアピン)構造が形成され蛍光同士が物理的に近傍に存在すると、FRETが生じて蛍光スペクトルが変化する。本発明の検出方法は、核酸のステム−ループ(ヘアピン)構造の形成をFRETによる蛍光スペクトルの変化の測定によって検出するものである。
【0100】
核酸のステム−ループ(ヘアピン)構造の形成をFRETによる蛍光スペクトルの変化は、別個の二本の核酸同士で二本鎖が形成される場合の蛍光スペクトルの変化と同様に観察することができる。
【0101】
本発明の好ましい一態様において、核酸のステム−ループ(ヘアピン)構造は、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドと5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドとを同一の鎖上に含む、本発明の核酸が、分子内でsとFAM−yを含む自己相補の配列がステムを作ること
により形成される。このように、核酸分子内のsとFAM−yをプローブにして両者間のFRETの効率を測定することにより、ステム−ループ構造などの核酸の高次構造を知ることが出来る。
【0102】
さらに、別の標的分子が結合した場合のみにステム−ループ(ヘアピン)構造を形成するような核酸分子を設計すれば、蛍光スペクトルの変化を利用して、その標的分子を検出するシステムを作ることが出来る。したがって、本発明は、リアルタイム分子間相互作用検出システム、リアルタイム分子内構造変化の検出システム、モレキュラービーコン、FRETの効率を利用した分子間距離の推定等に利用可能である。標的分子としては、DNA,RNA、タンパク質等が含まれる。
【0103】
本明細書中の実施例4では、sとFAM−yを同一鎖上に含むRNAについて、sとFAM−y間のFRETを調べた。本明細書中の実施例4では、350nmでsを励起した場合に、ステム−ループ構造を形成させてsとFAM−yの間で塩基対が作られた場合と、相補鎖RNAを加えて分子間での二本鎖構造を形成させてsとFAM−yが離れた場合の、いずれにおいてもFRETが観測されて、FAM−y由来の521nm付近の蛍光が現れた。しかしながら、sとFAM−yを導入した断片のsの蛍光強度(図12bおよび
cの実線のスペクトルの440nm付近の強度)とその対照のsとyを導入した断片のsの蛍光強度(図12bおよびcの破線のスペクトルの440nm付近の強度)を比較すると、ステム−ループ構造を形成している場合のほうが、顕著にsの蛍光強度が減少した。ステム−ループ構造中のsとFAM−yの距離は8−10Åであり、二本鎖構造中のsとFAM−yの距離は65−67Åに広がる。したがって、sとFAM−yの距離が近づいた場合に、両者間のFRETの効率が上がっている。
【0104】
6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを、2個以上隣接して含む核酸
6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを2個以上隣接して含む核酸もまた、本発明の範囲内である。
【0105】
前記核酸において、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドが2個以上同一の鎖上に隣接することにより、隣接した人工塩基の間で自己消光が生じるため、9H−プリン−9−イル基の蛍光強度が、9H−プリン−9−イル基を塩基として有するヌクレオチドが隣接しない場合と比較して、有意に減少する(図13cおよびdを参照)。一方、9H−プリン−9−イル基の間で自己消光が生じていても、蛍光色素によって5位が置換された、5位置換−2−オキソ(1H)ピリジン−3−イル基とのハイブリダイズにより、9H−プリン−9−イル基と蛍光色素との間のFRETを観察
することができる(図14cおよびdを参照)。さらに、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを2個以上隣接して含む核酸においては、9H−プリン−9−イル基の蛍光強度が減少している、すなわちバックグラウンド蛍光が低いため、蛍光色素との間のFRETを効率よく観察することができる(図15を参照)。すなわち、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを2個以上隣接して含む自己消光プローブを使用する本発明のFRET系は、蛍光ドナーのバックグラウンドを低く抑えることができるため、大量のドナープローブの存在下においても蛍光アクセプターを含有する相補鎖の検出が可能である点で特に有用である。
【0106】
6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを2個以上隣接して含む核酸は、上述した核酸の二本鎖形成の検出方法や核酸のステム−ループまたはヘアピン構造形成の検出方法に使用することができる。
【0107】
6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを2個以上隣接して含む核酸において、隣接して含まれる6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドの数は、2個以上であれば特に限定されないが、好ましくは、2ないし10個、2ないし5個、2ないし3個、最も好ましくは2個である。
【0108】
6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを2個以上隣接して含む核酸は、本発明の核酸の調製方法、または公知の化学合成によって製造することが可能であるが、これらに限定されない。
【発明の効果】
【0109】
本発明によって、非天然型塩基の新たな塩基対合Pa−sが見出されたことから、転写においても、対合する塩基間の形状の相補性が重要であることが示唆された(非特許文献5−6)。これにより、従来困難と考えられていた転写においても、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドの取り込みが可能となった。
【0110】
本発明の方法は、所望の位置にsを有すRNAを得る方法として従来知られている、(1)化学的RNA合成法、(2)yヌクレオチドを有する鋳型DNAの転写反応、(3)2−オキソ−1,3−ジヒドロ−イミダゾール−1−イル基を有するヌクレオチド(z)を有する鋳型DNAの転写反応よりも有意に高収率である。
【0111】
従来の2−アミノプリンヌクレオチド類(非特許文献21−23)のような慣用された蛍光プローブは、転写によってRNAに部位特異的に取り込まれることができないため、特に、長鎖RNA分子で使用することが困難であった。即ち、従来法では、2−アミノプリンのヌクレオチド誘導体を化学合成によりRNA中に導入していたが、RNAの化学合成法では、長鎖のRNA(50ヌクレオチド以上)を合成することは難しく、その操作も煩雑で時間がかかっていた。その結果、従来法は、限定的な利用に限られていた。
【0112】
これに対し、本発明では、蛍光特性を有する6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドの、RNAへの部位特異的取り込みが可能となった。本発明では、鋳型となるDNAを化学合成する必要があるが、DNAの化学合成は、RNAよりも長鎖長(150ヌクレオチド程度)の合成が可能であり、また、化学合成したDNAを酵素を用いて結合させることも出来る。これにより特に、従来の蛍光プローブ法では不可能であった長鎖RNAの分析が可能になった。これは、大きなRNA分子内における特定された位置における局部配座変化の動態の研究のための、強力な道具となりうる。本発明の実施例3では、実際にtRNA内の局部の高次構造(スタッキング)の解析が行われた。
【0113】
また、現在、RNAは、RNA干渉など、生体内の新たな機能が数多く見つかり、これらの現象を利用した医薬品の開発も進んでいる。しかし、それらのRNA分子の生体内での挙動を調べる手法は、それほど多くは無い。したがって、本蛍光プローブ法は、生体内のRNAの解析、ならびに、RNA医薬品の生体内での働きを調べる良い方法になる。
【0114】
さらに、本発明で新たに明らかにされたs−Pa塩基対は、翻訳によってタンパク質にアミノ酸類似体を部位特異的に取り込ませることを可能にするため、遺伝子コードの拡張に実用的に使用しうる(非特許文献15)。
【実施例】
【0115】
以下、実施例によって本発明を具体的に説明するが、これらは本発明の技術的範囲を限定するためのものではない。当業者は本明細書の記載に基づいて容易に本発明に修飾・変更を加えることができ、それらは本発明の技術的範囲に含まれる。
【0116】
実施例1 s−Pa塩基対合を利用したT7転写
本実施例では、s−Pa塩基対合を利用してT7転写を行った。
1)試薬、溶媒等
試薬及び溶媒は、標準的な供給業者から購入し、さらに精製することなく使用した。エレクトロスプレイ−イオン化マススペクトル(ESI−MS)は、Waters ZMD
4000 LC/MSシステム上に記録した。DNA鋳型は、Pa(非特許文献9)及び天然塩基のホスホルアミド類を用いて、自動DNA合成機(model392,PerkinElmer Applied Biosystems,Foster City,CA)によって化学合成した。ゲル電気泳動によってオリゴヌクレオチドを精製した。
【0117】
2)2−アミノ−6−(2−チエニル)−9−(1−β−D−リボフラノシル)プリン
5’−三リン酸(sTP)の合成
2−N−フェノキシアセチル−6−(2−チエニル)−9−(2,3−ジ−O−アセチル−1−β−D−リボフラノシル)プリン(57mg、0.1mmol)(非特許文献29)をピリジン中に溶解し、そして真空下で溶媒を留去し、試料中の水分を共沸除去した。残渣をピリジン(100μl)及びジオキサン(300μl)中に溶解した。次いで、2−クロロ−4H−1,2,3−ベンゾジオキサホスフォリン−4−オン(0.11mm
ol)の1Mジオキサン溶液(100μl)を添加した(非特許文献25)。10分後、
この溶液に0.5M ビス(トリブチルアンモニウム)ピロリン酸(0.15mmol)のDMF溶液(300μl)とトリ−n−ブチルアミン(100μl)をすばやく加えた。その反応混合物を室温で10分間かき混ぜた。次いで、1%ヨウ素溶液(ピリジン/水(98:2、v/v)溶液、2ml)を加えた。15分後、5%NaHSO3水溶液(150μl)、次いで、5mlの水を反応溶液に加えた。
【0118】
溶液を室温で30分間攪拌し、次いで、濃アンモニア水(20ml)を加えた。55℃で12時間、アンモノリシスを行った。溶液を真空下で濃縮し、そして産物をDEAEセファデックス(A−25)カラムクロマトグラフィー(50mMないし1M TEABの直線勾配によって溶出した)によって、そして次いで、C18−HPLC(100mM 酢酸トリエチルアンモニウム中の0%ないし30%CH3CNの勾配によって溶出した)(Eichorom Technologies社のSynchropak RPPを伴うGilson HPLC精製システム)によって精製し、sTPを得た。
【0119】
【化8】
【0120】
3)T7転写のための鋳型の調製
化学合成されたDNA鋳型(17−mer RNA合成のための、10μMの35−merのコード鎖及び21−merの非コード鎖)を、10mM NaClを含む10mM
Tris−HCl(pH7.6)緩衝溶液中で、95℃で加熱し、その後、4℃まで徐冷して、アニーリングした。
【0121】
4)T7転写(17−merRNA)
24mM MgCl2、2 mM スペルミジン、5 mM DTT、0.01% Triton X−100を含む40mM Tris−HCl(pH 8.0)緩衝溶液(2
0μl)中、1mM天然NTPs、若しくは1mM sTP、2μCi[γ−32P]GTP、2μM鋳型及び50単位のT7 RNAポリメラーゼ(Takara、Kyoto)の存在下で転写を行った。[γ−32P]GTPの使用により、転写産物は5’末端のみ標識され、これにより収率を求めた。37℃での3時間のインキュベーションの後、10M尿素及び0.05%BPBを含む色素溶液(20μl)を加えることにより反応を止めた。混合物を75℃で3時間加熱し、20%ポリアクリルアミド−7M尿素ゲル上で産物を分析した。
【0122】
5)結果
本実施例では、基質としてs(sTP)、および、非天然塩基が転写産物中の13−15番目に相当する相補的な部位に位置する、1つ若しくは2つのPa塩基を含むDNA鋳型を用いて、転写を行った。図2Aに実験スキームを示す。図2Aにおいて、17−mer RNA合成のための10μMの35−merのコード鎖及び21−merの非コード鎖の配列、並びに17−mer RNA転写産物の塩基配列は下記の通りである。
【0123】
鋳型相補鎖 (配列番号1)
5’−ataatacgactcactataggg−3’
鋳型鎖 (配列番号2)
3’−tattatgctgagtgatatccctcgaagggannntc−5’
17−merRNA転写産物 (配列番号3)
5’−gggagcuucccunnnag−3’
本発明者らはまた、対照として、イミダゾリン−2−オン(z)(図1B)を含む別の
DNA鋳型も調べた(図1B);zは、T7転写によるRNAへのsの部位特異的挿入のためのよい鋳型塩基であるが、転写効率は、天然の転写と比較して効率が低い。s−z対は塩基間に2つの水素結合が形成される可能性があるが、その形はs−Pa対に類似する。
【0124】
これらの鋳型を用いて3時間転写を行い、[γ−32P]GTPで5’末端を標識した転写産物をゲル上で分析した結果を、図2Bに示す。具体的には、図2Bは、1つまたは2つのPa又はz塩基を含む鋳型を用いて、天然型NTP(1mM、N=A,G,C,U)とsTP(1mM)を加えて行った転写の産物のゲル電気泳動の結果である。転写産物は、[γ−32P]GTPでその5’末端を標識されている。各転写産物の相対収率は、天然型塩基のみからなる鋳型からの転写産物を100%として、それとの比較によって決定した。各収率は3−4データセットからの平均をとった。
【0125】
図2Bに示すように、Paを1つ含む鋳型DNAからの全長(17−mer)の転写産物(1つのs塩基を含む)の相対収率は92%であり(図2B、レーン1、N1=Pa)、これはzを1つ含む鋳型DNAからの転写の収率(35%)(図2B、レーン4)よりも高く、そして、天然型のみの鋳型(N1=C)とNTPを用いた転写の収率(図2B、レーン8)と同じくらい高かった。2つのPa塩基を含む鋳型を用いた転写では、その相対収率は、天然型の転写(図2B、レーン8)と比較して低いものの、全長の産物が得られた(図2B、レーン2及び3)。対照的に、2つのzを含む鋳型を用いた転写では、全長の転写物が得られなかった(図2B、レーン5及び6)。
【0126】
実施例2 T7転写産物(17−mer RNA)中のヌクレオチドの組成分析
本実施例では、転写におけるs−Pa塩基対の選択性を調べるために、Pa、z及び天然の鋳型からの転写で得られる全長(17−mer)の転写物中のヌクレオチドの組成分析を行った。転写物を[α−32P]UTP、[α−32P]ATP又は[α−32P]GTPのどれかで内部標識し、この転写産物をRNase T2でヌクレオシド3’リン酸体にまで完全に分解した。得られた標識ヌクレオチドを2D−TLCで分析し(図3)、そして各ヌクレオシド3’−一リン酸のスポットを定量した(表1)。
【0127】
具体的には、24mM MgCl2、2mM スペルミジン、5mM DTT、0.01% Triton X−100を含む40mM Tris−HCl(pH8.0)緩衝液(20μl)中、10mM GMP、1mM 天然型NTP(1mM、N=A,G,C,U)、0、1若しくは3mM sTP、2μCi[γ−32P]UTP、[γ−32P]ATP若しくは[γ−32P]GTP(アマーシャム)、2μM鋳型DNA、及び50単位のT7 RNAポリメラーゼ(Takara)(非特許文献14、15)の存在下で転写を行った。3時間、37℃でインキュベーション後、色素溶液を加えて転写を終了させた。
【0128】
この混合物を75℃で3時間加熱し、そして次いで、15%ポリアクリルアミド−7M尿素ゲルで電気泳動した。全長の産物をゲルから溶出し、そして、これに0.05A260ユニットの大腸菌由来のtRNAを加えて、産物をエタノール沈殿させた。転写産物を、15mM 酢酸ナトリウム緩衝液(pH4.5)中、0.075U/μlのRNase
T2で、37℃、2時間消化した。この処理により、転写産物はRNase T2でヌクレオシド3’−一リン酸にまで完全に分解された。消化産物を、Merck HPTLCプレート(100×100mm)(Merck,Darmstadt,Germany)を用いた2D−TLCによって分析した。使用した展開溶剤は、1次元目がイソ酪酸/NH4OH/H2O(66:1:33 v/v/v)、そして、2次元目がイソプロピルアルコール/HCl/H2O(70:15:15 v/v/v)であった。TLCプレート上の産物をバイオイメージングアナライザーで分析した。各スポットの定量は3−9の
データセットから平均化した。
【0129】
結果を図3に示す。さらに、各ヌクレオシド3’−一リン酸を定量した結果を表1にまとめた。
表1 T7転写産物のヌクレオチド組成分析
【0130】
【表1】
【0131】
a: 図2に示した転写産物のU(エントリー1−8)、A(エントリー9−12)、又はG(エントリー13−16)の5’側に取り込まれたヌクレオチドの組成
b: 値は、下記の式を用いて決定された:
(各ヌクレオチドの放射活性)/[全てのヌクレオチド3’−一リン酸]×([α−32P]NTPの5’側のヌクレオチドの総数]
c: 各ヌクレオチドの理論的な数は、角型括弧書きで示す。
【0132】
d: 標準偏差は、括弧書きで示す。
e: 検出されず
図3及び表1に示した結果より、1つのPa塩基を含む鋳型を用いた場合では、高い選択性でRNA中へのsの取り込みが確認された。図3では、Paを含む鋳型を用いて[γ−32P]UTPで標識した場合にのみ、標識されたs−ヌクレオシド3’−一リン酸が2D−TLC上で観察された。さらに、表1に示されたように、鋳型DNA中のPaに相
補してRNA中に取り込まれたsの選択性(97%)(表1、エントリー1)は、s−z塩基対での場合の選択性(表1、エントリー2)、又は天然型塩基対のみでの転写の選択性と同じくらい高く(表1、エントリー3及び5)、そして、鋳型中の天然型の塩基に相補した間違ったsTPの取り込みは観察されなかった(表1、エントリー4及び6)。2つのPa塩基を含む鋳型を用いた転写の場合、1番目の位置におけるsの取り込みの選択性は十分に高かったが(95%)、2番目の位置におけるsの取り込みの選択性は84−87%程度であった。しかし、sTPの濃度を上げる(3mM)ことによって、この選択性はある程度(89−93%)改善された。従って、RNA中へのsの部位特異的取り込みに用いる鋳型塩基として、Paはzよりも優れており、そして、この結果は、非水素結合性の塩基対も転写において機能しうることを示唆している。
【0133】
実施例3 16s、17s、36s、47s、57s及び59s tRNAの調製及びその蛍光スペクトル及びUV融解曲線
1)16s、17s、36s、47s、57s及び59s tRNAの調製
蛍光プローブとしてのs塩基の有用性を示すために、本発明者らは、sを酵母由来のtRNAPheの特定の部位に取り込ませ、そしてtRNAの各部位におけるsの蛍光特性をいろいろな条件下で分析することにより、tRNAの高次構造を調べた。s塩基の蛍光は、近隣の塩基による塩基積み重ね(stacking)によって消光されるため、RNA分子中のsの周囲の局部構造についての情報が得られる。
【0134】
本発明者らは、tRNA中の6箇所のそれぞれにsを導入した:具体的には、D−ループ中の16番目又は17番目(tRNA 16s又は17s)、アンチコドン中の36番目(tRNA 36s)、エキストラループ中の47番目(tRNA 47s)、並びにTΨCループ中の57又は59番目(tRNA 57s又は59s)である。それぞれの部位のsは、tRNA中の他の塩基と塩基対を形成しない部位に導入されている(図4)。
【0135】
sを含む酵母tRNAPhe分子を調製するための各鋳型の配列(RNA合成のための5μMの94−merの鋳型鎖DNA及びその相補鎖のDNA)は、以下の表2の通りである(配列番号4−11)。
【0136】
【表2】
【0137】
RNA合成のための5μMの94−merの鋳型DNA及びその相補鎖DNAの配列
この配列では、C2−G71塩基対はG2−C71(非特許文献26)で置換されている。鋳型鎖DNAの5’末端の2つのヌクレオシド(G及びT)は、2'−O−メチルリボ
ヌクレオシドで置換することにより、目的とする産物よりも長くなってしまう鋳型非依存の転写産物の生成を抑えている(非特許文献27)。
【0138】
RNA合成のための94−merの鋳型DNA及び鋳型の相補鎖DNAの調製は、実施例1の「3)T7転写のための鋳型の調製」の項目で記載したのと同様に行った。
図4−6に、酵母由来のtRNAPhe中のsの導入部位及びtRNAの構造(非特許文献20)を示す。図4は、オリジナルなtRNAの2次構造である。sで置換された塩
基は○で囲んである。波線は塩基−塩基相互作用を示す。ボックスで囲んだG−C塩基対はオリジナルのtRNA中ではC−G塩基対であるが、この置換ではtRNAの高次構造(非特許文献26)は有意に変わっていない。
【0139】
図5は天然型のtRNAの3次構造であり、図6A−Cが、sが取り込まれる各部位の拡大図を示す。濃い色の塩基(群)はsと置換されていて、これは薄い色の塩基(群)と積み重なっている。そして、球(黄色)は、tRNA中のMg2+をあらわす。図中の塩
基は天然型のtRNAにおいて修飾されている塩基を示すが、括弧内に記載された塩基は転写産物における塩基を示している。
【0140】
それぞれの部位にsが導入されたtRNA転写産物は、Paを含む鋳型DNAを用いて、1mM sTPと天然型NTPsの存在下で、T7 RNAポリメラーゼによって調製された。具体的には先ず、24mM MgCl2、2mM スペルミジン、5mM DTT、0.01% Triton X−100を含む40mM Tris−HCl(pH8.0)緩衝液中、10mM GMP、1mM 天然型NTPs、1mM sTP、0.5μM鋳型DNA、及び2.5U/μl T7 RNAポリメラーゼの存在下で、転写を行った。37℃で6時間インキュベーションし、1.75倍量の色素溶液を加えて転写を終了させた。この溶液を75℃で3分加熱し、そして、10%ポリアクリルアミド−7M 尿素ゲルで電気泳動した。全長の産物をゲルから溶出し、そして、エタノールで沈殿した。450μlの10mM EDTA(pH8)中に産物を溶かし、75℃で5分間インキュベートした。
【0141】
次いで、Microcon YM−10 フィルター(Amicon)を用いて、緩衝溶液をTm測定用の緩衝液(50mM カコジル酸ナトリウム(pH7.2)及び50mM KClを含む)に交換した。260nmの吸光度によってtRNAの量を決定した。そして、蛍光及びUV融解測定のために、0.1mM EDTA、2mM MgCl2又は5mM MgCl2を含むTm緩衝液中に1μM tRNAを含むように溶液を調製した。
【0142】
2)蛍光スペクトル及びUV融解曲線
sを特定の部位に含む各tRNAの蛍光スペクトル及びUV溶解曲線を、20から90℃までの範囲で、0.5℃/分の加熱速度で、各々、FP−6500分光蛍光計(JASCO)、及びUV−2450分光光度計(SHIMADZU)を用いて記録した。
【0143】
s塩基は2つの励起最大(299及び352nm)を有する、図7Aは、sのリボヌクレオシド(1μM)の励起及び蛍光スペクトルを示す。図7Aから明らかなように、sは、434nmを中心とする蛍光スペクトルを示し、その蛍光の量子収率はpH7.0で0.41であった。図7B及びCは、352nmでの励起による(B)tRNA 47s(1μM)の蛍光スペクトル;及び(C)tRNA 57s(1μM)の蛍光スペクトルを示す。蛍光スペクトルは、50mM カコジル酸ナトリウム(pH7.2)、50mM KCl、及び0.1mM EDTA中、20℃において測定した。
【0144】
本実施例では、50mM カコジル酸ナトリウム、50mM KCl、及び0.1mM
EDTA又は、EDTAを除いて2mM若しくは5mM MgCl2を加えた溶液中、(352nmを中心に3nmのスペクトルバンド幅で励起された)sの434nmでの蛍光強度及び260nmでのUV吸収を測定して、各tRNA転写産物(1μM)の熱力学的融解プロフィールを調べた。
【0145】
溶媒分子との衝突によるsの蛍光の消光は、温度が高くなるほど生じやすくなる(図8A)。そこで、各tRNA転写産物の蛍光強度の温度依存変化を、sのヌクレオシドモノ
マーによって規格化した。蛍光強度の温度依存変化によって求められるtRNAの融解温度は、IGOR Proソフトウェア(WaveMetrics.Inc.)を用いて計算した。具体的には、図8は、(A)sのリボヌクレオシドの蛍光強度の温度依存による変化、及び(B)sのリボヌクレオシドの蛍光強度の温度依存による変化を補正したtRNA 36sの蛍光強度の温度依存性を示す。tRNA転写産物の蛍光強度は、等式:Yct=Yt/(Rt/R20)によって補正された。ここにおいて、Yctは、t℃での各tRNAの補正された強度であり、Ytは、t℃での各tRNAの実測された蛍光強度であり、Rtは、t℃でのsのリボヌクレオシドの蛍光強度であり、そして、R20は、20℃でのsのリボヌクレオシドの蛍光強度である。
【0146】
3)結果
16s、17s、36s、47s、57s及び59s tRNAの、434nmでの蛍光強度の温度依存変化によって得られる融解曲線(実線)及び260nmにおけるUV吸収の変化によって得られる融解曲線(波線)を図9に示す。
【0147】
先ず、第一にsを含む各tRNA転写産物のUV融解温度(2mM MgCl2中、64.6−66.4℃)は天然のtRNA転写産物の融解温度(2mM MgCl2中、65.5℃)と同じくらいの高さであった。このことは、これらの部位をsで置換しても、tRNAの構造は有意に不安定化されないことを示唆している。
【0148】
第二に、特定の位置にsを含む各tRNAは、局部構造の特性を反映する特徴的な蛍光強度の変化を表示した。Mg2+(2及び5mM)の存在下におけるsの蛍光特性は、明確に2つのグループに分類される:1つは、tRNA 16s、17s及び47sを含み(グループ1)、そしてもう一方は、tRNA 36s、57s及び59sを含む(グループ2)。2mM又は5mM Mg2+の存在下では、低温度でのグループ1のsの蛍光強度は、グループ2のsの蛍光強度よりも、1.7−3.4倍大きかった。グループ1の大きなsの蛍光強度は、温度が上昇すると減少するが、グループ2によるsの蛍光強度は温度の上昇とともに大きくなった。
【0149】
これらの結果は、16、17又は47番目のs塩基は生理学的温度で溶液中に露出しており、そして、温度の上昇によるtRNAの変性に伴い、s塩基と他の塩基との非特異的な相互作用が高まることを示唆している。対照的に、36、57又は59番目のs塩基は、低温下ではL字型構造を形成したtRNA中で近隣の塩基と積み重なっており、そして、温度の上昇によって徐々にtRNAが変性し、sと隣接する塩基との積み重なり構造が壊れる。これらの推測は、X線結晶構造解析によるtRNAのL字型構造中の各部位における元来の塩基の配置(図4)(非特許文献20)と極めてよく一致する。tRNA 16s、17s及び47sの蛍光強度は、生理学的温度において、Mg2+の添加によって劇的に上昇する。このことは、Mg2+のtRNAへの結合によって、tRNAがL字型構造を形成し、その結果、これらの位置における塩基が溶液中に露出される位置に配置されることが示唆している。
【0150】
第三に、蛍光プロフィールから得られるTm値は、UV融解プロフィールから得られるTm値と比較して、tRNA構造中の各局部構造の安定性を反映している。表3に、特定部位にsを含む酵母由来のtRNAPheの蛍光及びUVプロフィールから得られた融解温度(Tm)を示す。
【0151】
表3 特定部位にsを含む酵母由来のtRNAPheの蛍光及びUVプロフィールから得られた融解温度(Tm)
【0152】
【表3】
【0153】
蛍光及びUVプロフィールは、50mMカコジル酸ナトリウム(pH7.2)及び50mM KCl中、MgCl2共存下あるいは非共存下で、各tRNA転写産物(1μM)を用いて得られた。各融解温度は、IgorProソフトウェア(WaveMetrics. Inc.)を用いることによって計算された。n.c.計算されず。
【0154】
例えば、tRNA 36sの蛍光プロフィールから得られるTm値(2mM MgCl2中、69.5℃)は、UVプロフィールから得られるTm値(2mM MgCl2中、65.2℃)よりも高かった。このことは、tRNAの全体構造の安定性に比べてアンチコドン ステム−ループがより高い安定性を有することを示唆している。対照的に、tRNA16s(2mM MgCl2中、58.0℃)及び17s(2mM MgCl2中、59.6℃)の低い安定性は、Dループを含む部分構造は、tRNA構造全体の脆弱な部分であることを示唆している。
【0155】
実施例4 人工塩基sとFAM−yを含むRNA鎖のFRET実験
s−Pa塩基対を転写に用いることにより、蛍光性の人工塩基sをRNA中に効率よく導入することが出来るようになった。このs−Pa塩基対を、本発明者らが既に開発しているv−y塩基対と組み合わせることにより、1本のRNA中にsとy、あるいは、sと5位を修飾したyを、それぞれ任意の位置に導入することができる。特に、FAMを導入したyを利用することにより、sとFAM−yの間でFRETを観察することができる。
【0156】
本実施例では、いろいろな部位にPaとvを導入した転写用の鋳型DNAを化学合成し、これらの鋳型とsとy、あるいはFAM−yのそれぞれの基質(sTP、yTP、FAM−yTP)を用いて転写を行った。
【0157】
T7 RNA ポリメラーゼによる転写反応
(sとyを部位特異的に含むRNA 39−merの解析).
二本鎖の鋳型DNA(5μM)のアニーリング操作は、10mM NaClを含む10mM Tris−HCl緩衝液(pH7.6)中で行われた(温度条件:95℃で3分間加熱後、−0.1℃/秒の勾配で4℃まで徐冷)。T7転写反応は、40mM Tris−HCl緩衝液(pH8.0)、8mM MgCl2、2mM スペルミジン、5mM DTT、0.01% TritonX−100、2μCi[γ−32P]GTP、1mM
NTPs、0−1mM sTP、0−1mM yTP、0.5μM 鋳型DNA、および50unitsのT7 RNA polymerase(Takara)を含む反応溶液(全量 20μl)で行った。37℃、3時間反応を行った後、10M 尿素を含む電気泳動用色素溶液を等量(20μl)加えて反応を停止させた。この溶液を75℃で3分
間加熱後、15% ポリアクリルアミド−7M 尿素ゲルで電気泳動を行い、転写物をバイオイメージアナライザーで解析した(図10)。
【0158】
T7 RNA ポリメラーゼによる転写反応生成物の塩基組成分析
T7 RNAポリメラーゼによる転写反応を、40mM Tris−HCl緩衝液(pH8.0)、8mM MgCl2、2mM スペルミジン、5mM DTT、0.01%
TritonX−100、2μCi[α−32P]ATPまたは2μCi[α−32P]GTPまたは2μCi[α−32P]CTPまたは2μCi[α−32P]UTP、1mM NTPs、0−1mM sTP、0−1mM yTP、0.5μM 鋳型DNA、および50unitsのT7 RNA polymerase(Takara)を含む反応溶液で行った。転写物を、15% ポリアクリルアミド−7M 尿素ゲルによる電気泳動により精製し、ゲルからの溶出を行った後、tRNA(0.05 OD)を加えてエタノール沈殿を行い回収した。このサンプルを滅菌水10μlに溶解し、溶液(8.5μl)を用いてRNaseT2による完全分解反応を行った。溶液(8.5μl)にRNase T2(1.5μl,0.5U/μl)を加えて、37℃で2時間反応を行った。この溶液の一部をTLC(Merck,10cmx10cm)にスポットし、二次元展開を行った。展開液として、一次元目はイソ酪酸:濃アンモニア水:水=66:1:33(v/v/v)、二次元目は2−プロパノール:塩酸:水=65:10:25(v/v/v)を用いた。展開後のスポットをバイオイメージアナライザーで検出し、解析を行った(図11、表4)。
【0159】
T7 RNA ポリメラーゼによる転写反応
(sとy、もしくはsとFAM−yを部位特異的に含むRNA 39−merの調製).
T7転写反応は、40mM Tris−HCl緩衝液(pH8.0)、8mM MgCl2、2mM スペルミジン、5mM DTT、0.01% TritonX−100、1mM NTPs、1mM sTP、1mM yTPまたは1mM FAM−yTP、0.5μM 鋳型DNA、および125unitsのT7 RNA polymerase(Takara)を含む反応溶液(全量50μl)で行った。37℃、6時間反応を行っ
た後、DNase I溶液を加えて、37℃で15分間保温することで鋳型DNAを分解した。10M 尿素を含む電気泳動用色素溶液を等量(50μl)加えて反応を停止させ
た後、反応溶液を75℃で3分間加熱してから、15% ポリアクリルアミド−7M 尿素ゲルで電気泳動を行い、目的の転写物39−merを精製した。ゲルから滅菌水で溶出した後、エタノール沈澱を行い、転写物を回収し、UV吸収から目的の転写物RNA 39−merの濃度を決定した。
【0160】
蛍光スペクトル測定
sとy、もしくはsとFAM−yを部位特異的に含むRNA 39−mer(0.2μM)を用いて、10mM リン酸ナトリウム緩衝液(pH7.0)、100mM NaCl、0.1mM EDTA中での蛍光スペクトルを20℃で測定した。測定前のRNAサンプルのアニーリング操作では、RNA 39−mer(0.2μM)を含む溶液、およびRNA 39−merとその39−merに相補的な配列を持つ相補性RNA 42−mer(0.4μM)を含む溶液を75℃で3分間加熱した後、−0.1℃/3秒の勾配で4℃まで徐冷を行った。アニーリング操作を行ったサンプルは、4℃で保管され、測定時は、サンプルを測定セルに移してから20℃に2分以上放置した後にスペクトル走査(360−650nm)を開始した。蛍光測定(励起波長350nm)には、3x3mmセルを用い、レスポンス 0.5s、走査速度 2000nm/分、感度はマニュアル設定でPMT 500Vに固定、スリットバンド幅は5nmに固定してスペクトル測定を行った(図12)。
【0161】
結果
図10は、v−y塩基対とs−Pa塩基対を用いた転写によるRNAへのyとsの部位特異的導入の戦略と転写産物のポリアクリルアミド電気泳動の結果を示す。
【0162】
詳細には、図10aに鋳型DNAの配列と転写反応の条件を示した(配列中でPaは、Pで示している)。転写物(Bhp2−1,Bhp2−2,Bhp2−3)をポリアクリルアミドゲル電気泳動で調べると(図10b)、sTPとyTPをどちらも加えた転写で、転写物の量が多くなった。そこで、sとyが選択的にRNA中に取り込まれているかどうかを調べるために、α位に放射性のリンを導入した[α−32P]ATP,[α−32P]GTP,[α−32P]CTP,[α−32P]UTPのそれぞれの基質を別々に加えて転写を行い、放射性標識された転写物をRNase T2でヌクレオシド 3’−一リン酸に加水分解し、リン酸が放射性標識されているヌクレオシド 3’−一リン酸を2次元のTLCで分析し、転写物のヌクレオチド組成を調べた(図11)。
【0163】
この方法では、放射性の基質が取り込まれると、その5’側のヌクレオチドが2次元のTLC上で検出できる。たとえば、図11bには、Bhp2−1の2次元TLCの結果を示した。Bhp2−1では、yがAの5’側に、また、sがGの5’側にそれぞれあるので、もし、鋳型上のvに相補してyが、また、Paに相補してsが、それぞれ選択的にRNA中に取り込まれていれば、[α−32P]ATPで標識した転写物の2次元TLCにはyp(yのヌクレオシド 3’−一リン酸)のスポットが現れ、また、[α−32P]GTP,で標識した転写物では、sp(sのヌクレオシド 3’−一リン酸)のスポットが現れることになる。実際に、図11bでは、それぞれのTLC上に期待されるスポットが現れている。また、[α−32P]CTP,あるいは、[α−32P]UTPで標識した転写物では、ypとspのどちらのスポットも認められないことから、鋳型上の天然型塩基に相補してそれぞれの人工塩基が間違って取り込まれていないこともわかった。さらに、2次元TLC上のそれぞれのスポットを定量し、その結果を表4に示した。
【0164】
【表4】
【0165】
a: 図11に示された、A(エントリー1,2及び9−11)、G(エントリー3,4及び12−14)、C(エントリー5及び6)又はU(エントリー7及び8)の5’隣りに取り込まれたヌクレオチドの組成
b: 値は、下記の式を用いて決定された:
(各ヌクレオチドの放射活性)/[全てのヌクレオチド(3’−一リン酸)]×([α−32P]NTPの5’隣りのヌクレオチドの総数]
c: 各ヌクレオチドの理論的な数は、角型括弧書きで示す。
【0166】
d: 標準偏差は、括弧書きで示す。
e: 検出されず
転写では、yTPの量を0.5mMと1mMで行った結果を示してある。エントリー1−8の結果は、全て、理論値(カッコ内の数値)と良く一致している。エントリー9−14では、天然型塩基のみの鋳型DNAを用いた場合についての結果であるが、エントリー10において、1mMのyTPを用いた場合にわずかにypが天然型の鋳型塩基に相補して取り込まれていることがわかった。したがって、転写におけるyTPの量は、0.5mMのほうが良い。以上の結果から、s−Pa塩基対とv−y塩基対を組み合わせて転写を行うことにより、1本のRNA中にsとyをそれぞれ選択的に導入できることがわかった。
【0167】
そこで、yの基質に代えて、yの5位にFAMを結合した基質(FAM−yTP)とsTPを用いて、Bhp2−1の転写を行い、転写物をゲル電気泳動で精製し、その蛍光特
性を調べた。Bhp2−1は、単独ではヘアピン型の構造を形成し、sとFAM−yが塩基対を形成する位置に配置されるので、両者でFRETが観測される。そして、このRNAの相補鎖のRNAを加えると二本鎖構造を形成し、sとFAM−yが離れ、FRETの効率が低下する。この二本鎖構造のRNAの蛍光スペクトルを図12bに、また、ヘアピン型のRNAの蛍光スペクトルを図12cに示した(実線)。それぞれの蛍光スペクトルは、sとyを導入したRNAのスペクトルと比較している(破線:このRNAでは、FRETが起こらず、sの蛍光スペクトルを示すコントロールとなる)。どちらも、350nmでsを励起しているが、FRETが観測されて、FAM−yの521nm付近の蛍光が現れている。しかし、ヘアピン型のRNAのほうが、FAM−yの蛍光強度が強く、また、コントロール(破線)と比較したsの消光も顕著である。
【0168】
この結果から、sとFAM−yの両者の距離に相関してFRETが観測されることがわかった。したがって、sとFAM−yをRNA中のいろいろな部位に導入し、両者のFRETの効率を測定することにより、両者間の距離を見積もることができ、RNAの構造解析に役立たせることができる。従来のX線結晶構造解析やNMRによる構造解析では、静的なRNAの構造に関する情報しか得ることが出来なかったが、本手法で、溶液中のRNAの構造変化のダイナミクスを知ることが可能になる。
【0169】
実施例5 核酸二重鎖におけるsまたはvとFAM−yの人工塩基対のFRET実験
DNA/RNA二本鎖において、蛍光性の人工塩基である2−アミノ−6−(2−チエニル)プリン−9−イル基または2−アミノ−6−(チアゾリル)プリン−9−イル基(以下、本実施例においてそれぞれsまたはvと称する)、およびフルオロフォアを連結した2−オキソ−(1H)ピリジン−3−イル基(以下、本実施例においてyと称する)の間でFRET実験を行った。sまたはvの発光スペクトルは、FAMが連結した5−(3−アミノ−1−プロピニル)−y(FAM−yTP)またはFAMが連結した5−[3−(6−アミノヘキサンアミド)−1−プロピニル]−y(FAM−hx−yTP)であるヌクレオチド誘導体の吸収スペクトルと重なった(図13a)。よって、DNAフラグメント中のsまたはvは蛍光ドナーとして作用し、そして相補的なRNAフラグメント中のFAM−yまたはFAM−hx−yは蛍光アクセプターとして作用する。
【0170】
人工塩基sまたはvを含むDNAの合成
人工塩基sまたはvを含むDNAは、例えば、国際公開WO 01/05801号または国際公開WO 2005/026187号に記載の方法に従って、人工塩基sまたはvを含むヌクレオチドのシアノエチルホスホロアミダイト誘導体を用いて、当該技術分野に周知のDNA化学合成法により合成した。化学的に合成された35merのDNAの配列は、5’−CAN1N2N3CTCGGGATTCCCTATAGTGAGTCGTATTAT−3’(配列番号20;ここで、N1N2N3が;CTsまたはCTvのDNAをそれぞれDNA35s1またはDNA35v1;CssまたはCvvのDNAをそれぞれDNA35s2aまたはDNA35v2a;sTsまたはvTvのDNAをそれぞれDNA35s2bまたはDNA35v2b;と称する)とした。得られたDNAはゲル電気泳動によって精製した。
【0171】
FAM−yまたはFAM−hx−yを含むRNAの調製
FAM−yまたはFAM−hx−yをN’の位置で含有する17merのRNA(5’−GGGAAUCCCGAGN’AGUG−3’:配列番号21)を、T7 RNAポリメラーゼおよびFAM−yまたはFAM−hx−yの基質(FAM−yTPまたはFAM−hx−yTP)を用いてT7転写により調製した。鋳型(コード鎖としてDNA35v1、非コード鎖として5’−ATAATACGACTCACTATAGGG−3’(配列番号22)をそれぞれ10μM)を、10mM Tris−HCl(pH7.6)および10mM NaClを含む緩衝溶液中で、95℃で加熱し、その後、4℃まで徐冷して、
アニーリングした。転写反応は、40mM Tris−HCl(pH8.0)、24mM
MgCl2、2mM スペルミジン、5mM DTT、および0.01% Triton X−100を含む緩衝溶液中、1mM 天然NTPs、1mM FAM−yTPまたはFAM−hx−yTP、2μM 鋳型、および2.5ユニット/μL T7 RNAポリメラーゼ(Takara)の存在下で行った。37℃で3時間インキュベーションした後、10M 尿素および0.05% BPBを含有する色素溶液を等量加えることにより反応を止めた。反応混合物は75℃で3分間加熱し、そして17−merの転写物をゲル電気泳動により精製した。
【0172】
蛍光スペクトル測定
蛍光スペクトルは10mM リン酸ナトリウム(pH7.0)、100mM NaCl、および0.1mM EDTAを含む緩衝液中で測定した。発光スペクトルはsについて350nm、vについて360nmの励起波長で記録した。発光のスペクトルバンドパスは5nmであった。
【0173】
FAM−yまたはFAM−hx−yを部位特異的に含む17−mer RNAを、1個または2個のsまたはv塩基を部位特異的に含む35−mer DNA(DNA35s1、35s2a、35s2b、35v1、35v2a、および35v2b)とハイブリダイズさせ、DNA/RNA二本鎖の発光スペクトルを、異なる温度(20および70℃)で測定した。
【0174】
結果
共鳴エネルギー転移は、sまたはvの低蛍光状態からFAM−yまたはFAM−hx−yの高蛍光状態の間で観察された。DNA35s1またはDNA35v1が、FAM−yまたはFAM−hx−yを部位特異的に含む17−mer RNAと20℃で二本鎖を形成したときにFRETが生じ、そして70℃で二本鎖が変性したときにFRETは有意に減少した(図14aおよびb)。加えて、DNA35s2aおよびDNA35v2aと、FAM−yまたはFAM−hx−yを部位特異的に含むRNAとの二本鎖においても、FRETが効率よく生じた(図14cおよびd)。
【0175】
興味深いことに、二つの隣接した人工塩基を含むDNA35s2aまたはDNA35v2aの一本鎖の蛍光強度は有意に減少する一方、1つのTで隔てられた二つの人工塩基を含むDNA35s2bまたはDNA35v2bの一本鎖の蛍光強度の有意な減少は観察さなかった(図13cおよびd)。DNA35s2aまたはDNA35v2aの一本鎖の蛍光強度の減少は、二つの隣接した人工塩基の間での自己消光によって起こるものである。
【0176】
二つの隣接した人工塩基の間での自己消光が起こるDNA35s2aまたはDNA35v2aとの二本鎖形成によっても、17−mer RNAの強いFAM蛍光が観察されたので、自己消光プローブを用いるこのFRET系は、標的の鎖に対して過剰量のプローブの使用を容易にする。17−mer RNAに対して1つの人工塩基を含むDNA35s1およびDNA35v1を10モル等量用いた場合、共鳴エネルギー転移は、それぞれ、DNA35s1およびDNA35v1のバックグラウンド蛍光によって完全に隠れてしまった(図15aおよびb)。対照的に、17−mer RNAに対して隣接する二つの人工塩基を含むDNA35s2aおよびDNA35v2aを10モル等量用いた場合は、それぞれのss−およびvv−含有DNAおよびFAM含有RNAの間のFRETがはっきりと観察された(図15cおよびd)。したがって、隣接する二つの人工塩基を含む自己消光プローブを使用するFRET系は、蛍光ドナーのバックグラウンドを減少させつつ、FRET現象を観察することができるため、大量のドナープローブの存在下においても蛍光アクセプターを含む相補鎖の検出が可能である。また、大量の自己消光DNAプローブは、相補的な蛍光アクセプター鎖との二本鎖形成を容易にするため、FRET効率を増加
させることができる。
【図面の簡単な説明】
【0177】
【図1】図1は、非天然型s−Pa(A)塩基対及びs−z(B)塩基対を示す。
【図2】図2は、s−Pa塩基対によって媒介されるT7転写の実験スキーム及び、転写産物のゲル電気泳動の結果を示す。図2A:実験スキーム 図2B:1つまたは2つのPa又はz塩基を含む鋳型DNAを用いて、天然型NTPs(1mM)とsTP(1mM)を加えた場合の転写産物のゲル電気泳動。転写産物は、[γ−32P]GTPでその5’末端を標識された。各転写産物の相対収率は、全て天然型塩基からなる鋳型からの転写産物の収率との比較によって決定し、各収率は3−4データセットから平均化した。
【図3】図3は、転写産物(17−mer)のヌクレアーゼ消化から得られた標識リボヌクレオシド3’−一リン酸の2D−TLC分析を示す。転写産物を[α−32P]NTPで内部標識し、1mM sTPの存在下で転写された17−mer断片からの二次元TLCである。中パネルは鋳型DNA中のN1N2N3がPaACであり、右パネルはN1N2N3がCACである。
【図4】図4は、酵母由来のtRNAPhe中へのsの導入部位及びtRNAの構造(非特許文献20)を示す。図4はオリジナルのtRNA転写産物の2次構造である。sで置換された場所は○で囲んである。波線は3次構造の塩基−塩基相互作用を示す。ボックスで囲んだG−C対はオリジナルのC−G対から変更されているが、この変位はオリジナルのtRNA構造(非特許文献26)を有意には変えない。
【図5】図5は、酵母由来のtRNAPhe中へのsの導入部位及びtRNAの構造(非特許文献20)を示す。図5は原型のtRNA転写産物の3次構造である。
【図6】図6A−Cは、酵母由来のtRNAPhe中へのsの導入部位を含む領域の拡大図である。濃い色の塩基(群)はsと置換される部位で、この塩基は薄い色の塩基(群)と積み重なっており、そして、球(黄色)はMg2+を表す。図中の塩基は修飾された塩基を示すが、括弧内に記載された塩基はオリジナルな転写産物中の塩基に相当する。
【図7】図7Aは、sのリボヌクレオシド(1μM)の励起及び蛍光スペクトルを示す。黒丸実線は、352nmでの励起による蛍光スペクトルを示す。白丸波線は、434nmでの励起による蛍光スペクトルを示す。 図7B及びCは、352nmでの励起による(B)tRNA 47s(1μM)の蛍光スペクトル;及び(C)tRNA 57s(1μM)の蛍光スペクトルを示す。蛍光スペクトルは、50mM カコジル酸ナトリウム(pH7.2)、50mM KCl、及び0.1mM EDTA中、20℃において測定した。(B)において、黒丸実線、白丸波線、及び×印実線は、tRNA47sの各々0mM Mg2+、2mM Mg2+及び5mM Mg2+存在下における蛍光スペクトルを示す。(C)において、黒丸実線、白丸波線、及び×印実線は、tRNA57sの各々0mM Mg2+、2mM Mg2+及び5mM Mg2+存在下における蛍光スペクトルを示す。
【図8】図8は、(A)sリボヌクレオシドの蛍光強度の温度依存性、及び(B)sのリボヌクレオシドでtRNA 36sの蛍光強度を補正した例を示す。tRNA転写産物の蛍光強度は、等式:Yct=Yt/(Rt/R20)によって補正された。ここにおいて、Yctは、t℃での各tRNAの補正された強度であり、Ytは、t℃での各tRNAの測定された蛍光強度であり、Rtは、t℃でのsのリボヌクレオシドの観察された蛍光強度であり、そして、R20は、20℃でのsのリボヌクレオシドの観察された蛍光強度である。
【図9】図9は、種々の位置においてsを含むtRNAの、434nmでの蛍光強度の変化によって得られる溶解曲線(実線)及び260nmにおけるUV吸収の変化によって得られる溶解曲線(波線)を示す。太い(黒)実線、中くらいの(青)実線、細い実線(赤)の実線は、各々0mM Mg2+、2mM Mg2+及び5mM Mg2+存在下における434nmでの蛍光強度の変化によって得られる融解曲線である。太い(黒)波線、中くらいの(青)波線及び細い実線(赤)の波線は、各々0mM Mg2+、2mM Mg2+及び5mM Mg2+存在下における260nmにおけるUV吸収の変化によって得られる融解曲線である。
【図10】図10は、v−y塩基対とs−Pa塩基対を用いた転写によるRNAへのyとsの部位特異的導入の方法と転写産物のポリアクリルアミド電気泳動の結果を示す。図10aにおいて、配列中のPは、人工塩基Paを示している。
【図11】図11は、v−y塩基対とs−Pa塩基対を用いた転写により調製された転写産物(Bhp2−1)のヌクレオチド組成の分析の結果を示す。図11bにおける、各スポットの定量は、表4にまとめた。
【図12】図12は、人工塩基sとy若しくはFAM−yを導入したRNA(Bhp2−1)の蛍光測定の結果を示す。図12bは、Bhp2−1が相補性RNAと二本鎖を形成している場合の蛍光スペクトルを示す。図12cは、Bhp2−1が1本鎖のままヘアピン構造を形成している場合の結果を示す。図12b及び図12cのいずれの場合も、実線は、位置13にFAM−y、位置36にsを有するBhp2−1の結果である。波線は、位置13にy、位置36にsを有するBhp2−1の結果である
【図13】図13aは、リン酸緩衝液(pH7.0)中におけるsデオキシリボヌクレオシド(細い線)およびFAM−yTP(太線)の蛍光(flu、黒線)および吸収(abs、破線)スペクトルである。図13bは、FRET実験のスキームである。非天然塩基は、sまたvについて励起させたとき(λex)、低蛍光状態を黒丸で、高蛍光状態を白丸で示した。図13cおよびdは、s塩基を含有する(c:DNA35s1、DNA35s2a、およびDNA35s2b)およびv塩基を含有する(c:DNA35v1、DNA35v2a、およびDNA35v2b)一本鎖DNAの定常状態蛍光発光スペクトルである。
【図14】図14は、DNA/RNA二本鎖の定常状態蛍光発光スペクトルを示す。35−mer DNAのそれぞれ(a,DNA35s1;b,DNA35v1;c,DNA35s2a;d,DNA35v2a;e,DNA35s2b;f,DNA35v2b)を、FAM−y(白丸および黒丸)またはFAM−hx−y(白三角または黒三角)を含む17−mer RNAとハイブリダイズさせた。白丸および白三角は20℃、黒丸および黒三角は70℃でのスペクトルである。
【図15】図15は、人工塩基を含むDNAプローブを過剰量用いた場合のDNA/RNA二本鎖の定常状態蛍光発光スペクトルを示す。35−mer DNAのそれぞれ(1μM)(a,DNA35s1;b,DNA35v1;c,DNA35s2a;d,DNA35v2a)を、FAM−y(白丸および黒丸)またはFAM−hx−y(白三角および黒丸)を含む17−mer RNA(100nM)とハイブリダイズさせた。白丸および白三角は20℃、黒丸および黒三角は70℃でのスペクトルである。
【特許請求の範囲】
【請求項1】
置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドと、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドとが塩基対を形成している、核酸。
【請求項2】
前記置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基が、
A1)2−ホルミル−1H−ピロール−1−イル基;
A2)2−ホルミル−4−(1−プロピン−1−イル)−1H−ピロール−1−イル基;
A3)2−ホルミル−4−メチル−1H−ピロール−1−イル基;及び
A4)2−ホルミル−4−エチニル−1H−ピロール−1−イル基
からなるグループから選択され、そして
前記6位置換された9H−プリン−9−イル基が、
B1)2−アミノ−6−(2−チエニル)−9H−プリン−9−イル基;
B2)6−(2−チエニル)−9H−プリン−9−イル基;
B3)2−アミノ−6−(4−メチル−2−チエニル)−9H−プリン−9−イル基;
B4)6−(4−メチル−2−チエニル)−9H−プリン−9−イル基;
B5)2−アミノ−6−(5−メチル−2−チエニル)−9H−プリン−9−イル基;
B6)6−(5−メチル−2−チエニル)−9H−プリン−9−イル基;
B7)2−アミノ−6−(2−チアゾリル)−9H−プリン−9−イル基;
B8)6−(2−チアゾリル)−9H−プリン−9−イル基;
B9)2−アミノ−6−(4−メチル−2−チアゾリル)−9H−プリン−9−イル基;
B10)6−(4−メチル−2−チアゾリル)−9H−プリン−9−イル基;
B11)2−アミノ−6−(5−メチル−2−チアゾリル)−9H−プリン−9−イル基;及び
B12)6−(5−メチル−2−チアゾリル)−9H−プリン−9−イル基
からなるグループから選択される、
請求項1に記載の核酸。
【請求項3】
前記置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基が、2−ホルミル−1H−ピロール−1−イル基であり、そして、前記6位置換された9H−プリン−9−イル基が、2−アミノ−6−(2−チエニル)−9H−プリン−9−イル基である、請求項1又は2に記載の核酸。
【請求項4】
転写、逆転写、複製又は翻訳の工程で、塩基対を形成している、請求項1ないし3のいずれか1項に記載の核酸。
【請求項5】
前記置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドがDNAの一部であり、そして、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドがRNAの一部である、請求項1ないし4のいずれか1項に記載の核酸。
【請求項6】
6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを含む核酸を調製する方法であって、
置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドを含む核酸を鋳型として転写、逆転写又は複製を行い、前記置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を有するヌクレオチドの相補的な位置に、6位置換された9H−プリン−9−イル基を有するヌクレオチドを組み
込む
ことを含む、前記調製方法。
【請求項7】
前記置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基が、
A1)2−ホルミル−1H−ピロール−1−イル基;
A2)2−ホルミル−4−(1−プロピン−1−イル)−1H−ピロール−1−イル基;
A3)2−ホルミル−4−メチル−1H−ピロール−1−イル基;及び
A4)2−ホルミル−4−エチニル−1H−ピロール−1−イル基
からなるグループから選択され、そして
前記6位置換された9H−プリン−9−イル基が、
B1)2−アミノ−6−(2−チエニル)−9H−プリン−9−イル基;
B2)6−(2−チエニル)−9H−プリン−9−イル基;
B3)2−アミノ−6−(4−メチル−2−チエニル)−9H−プリン−9−イル基;
B4)6−(4−メチル−2−チエニル)−9H−プリン−9−イル基;
B5)2−アミノ−6−(5−メチル−2−チエニル)−9H−プリン−9−イル基;
B6)6−(5−メチル−2−チエニル)−9H−プリン−9−イル基;
B7)2−アミノ−6−(2−チアゾリル)−9H−プリン−9−イル基;
B8)6−(2−チアゾリル)−9H−プリン−9−イル基;
B9)2−アミノ−6−(4−メチル−2−チアゾリル)−9H−プリン−9−イル基;
B10)6−(4−メチル−2−チアゾリル)−9H−プリン−9−イル基;
B11)2−アミノ−6−(5−メチル−2−チアゾリル)−9H−プリン−9−イル基;及び
B12)6−(5−メチル−2−チアゾリル)−9H−プリン−9−イル基
からなるグループから選択される、
請求項6に記載の調製方法。
【請求項8】
鋳型において、前記置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドが2個以上存在する、請求項6又は7に記載の調製方法。
【請求項9】
請求項6ないし8に記載の方法によって調製された、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを含む核酸。
【請求項10】
tRNA、mRNA、アンチセンスDNA若しくはRNA、リボザイム、アプタマー又はsiRNAである、請求項9に記載の核酸。
【請求項11】
前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド中の、塩基に由来する蛍光強度の温度変化および/またはイオン濃度変化を測定する、ことを含む、請求項9又は10に記載の核酸における、前記ヌクレオチドを含む局部領域の立体構造の解析方法。
【請求項12】
前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド中の、塩基に由来する蛍光強度が、温度の上昇に伴い実質的に上昇する場合には、生体内温度下で、前記ヌクレオチドは立体配座において周囲に存在するヌクレオチドと積み重なっていると判断し、そして、蛍光強度が温度の上昇に伴い実質的に低下又は上昇しない場合には、生体内温度下で、前記ヌクレオチドは外部に暴露していると判断する、請求項11に記載の解析方法。
【請求項13】
I) i)請求項6ないし8に記載の方法によって調製された、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを含む核酸、と、
ii)5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドであって、前記塩基の5位に、5−カルボキシフルオレセイン(5−FAM)、6−カルボキシフルオレセイン(6−FAM)、5−カルボキシテトラメチルローダミン(5−TAMRA)、6−カルボキシテトラメチルローダミン(6−TAMRA)、 5−ジメチルアミノナフタレン−1−スルホン酸(DANSYL)、5−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(5−HEX)、6−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(6−HEX)、5−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(5−TET)、6−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(6−TET)、5−カルボキシ−X−ローダミン(5−ROX)、及び6−カルボキシ−X−ローダミン(6−ROX)からなるグループから選択される蛍光色素が、直接又はリンカーを介して結合している前記ヌクレオチド、を含む核酸、とをハイブリダイズさせ、そして、
II) 蛍光スペクトルの変化を測定する
ことを含む、核酸の二本鎖形成の検出方法。
【請求項14】
I)i)の6位置換された9H−プリン−9−イル基が、
B1)2−アミノ−6−(2−チエニル)−9H−プリン−9−イル基;
B2)6−(2−チエニル)−9H−プリン−9−イル基;
B3)2−アミノ−6−(4−メチル−2−チエニル)−9H−プリン−9−イル基;
B4)6−(4−メチル−2−チエニル)−9H−プリン−9−イル基;
B5)2−アミノ−6−(5−メチル−2−チエニル)−9H−プリン−9−イル基;
B6)6−(5−メチル−2−チエニル)−9H−プリン−9−イル基;
B7)2−アミノ−6−(2−チアゾリル)−9H−プリン−9−イル基;
B8)6−(2−チアゾリル)−9H−プリン−9−イル基;
B9)2−アミノ−6−(4−メチル−2−チアゾリル)−9H−プリン−9−イル基;
B10)6−(4−メチル−2−チアゾリル)−9H−プリン−9−イル基;
B11)2−アミノ−6−(5−メチル−2−チアゾリル)−9H−プリン−9−イル基;及び
B12)6−(5−メチル−2−チアゾリル)−9H−プリン−9−イル基
からなるグループから選択される、請求項13に記載の検出方法。
【請求項15】
I)ii)の核酸が、塩基の5位に、5−カルボキシフルオレセイン(5−FAM)又は6−カルボキシフルオレセイン(6−FAM)が直接又はリンカーを介して-結合して
いる、5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドを含む核酸である、請求項13又は14に記載の検出方法。
【請求項16】
i) 6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド
並びに、
ii) 5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドであって、前記塩基の5位に、5−カルボキシフルオレセイン(5−FAM)、6−カルボキシフルオレセイン(6−FAM)、5−カルボキシテトラメチルローダミン(5−TAMRA)、6−カルボキシテトラメチルローダミン(6−TAMRA)、 5−ジメチルアミノナフタレン−1−スルホン酸(DANSYL)、5−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(5−HEX)、6−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(6−HEX)、5−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(5−TET)、6−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(6−TET)、5−
カルボキシ−X−ローダミン(5−ROX)、及び6−カルボキシ−X−ローダミン(6−ROX)からなるグループから選択される蛍光色素が、直接又はリンカーを介して結合している、前記ヌクレオチド
を、同一の鎖上に含む核酸を調製する方法であって、
iii) 置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチド
並びに
iv) 6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド
を含む核酸を鋳型として転写、逆転写又は複製を行い、前記iii)のヌクレオチドの相補的な位置に、前記i)のヌクレオチドを組み込み、そして、前記iv)のヌクレオチドの相補的な位置に、前記ii)のヌクレオチドを組み込む
ことを含む、前記調製方法。
【請求項17】
請求項16に記載の方法によって調製された、
i) 6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド
並びに、
ii) 5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドであって、前記塩基の5位に、5−カルボキシフルオレセイン(5−FAM)、6−カルボキシフルオレセイン(6−FAM)、5−カルボキシテトラメチルローダミン(5−TAMRA)、6−カルボキシテトラメチルローダミン(6−TAMRA)、 5−ジメチルアミノナフタレン−1−スルホン酸(DANSYL)、5−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(5−HEX)、6−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(6−HEX)、5−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(5−TET)、6−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(6−TET)、5−カルボキシ−X−ローダミン(5−ROX)、及び6−カルボキシ−X−ローダミン(6−ROX)からなるグループから選択される蛍光色素が、直接又はリンカーを介して結合している、前記ヌクレオチド
を、同一の鎖上に含む核酸。
【請求項18】
tRNA、mRNA、アンチセンスDNA若しくはRNA、リボザイム、アプタマー又はsiRNAである、請求項17に記載の核酸。
【請求項19】
核酸のステム−ループ又はヘアピン構造形成の検出方法であって、
1) 請求項17又は18に記載の核酸において、i)のヌクレオチドと、ii)のヌクレオチドとをハイブリダイズさせ、そして、
2) 蛍光スペクトルの変化を測定する
ことを含む、核酸のステム−ループ又はヘアピン構造形成の検出方法。
【請求項20】
核酸のステム−ループ又はヘアピン構造が、請求項17又は18に記載の核酸が、標的分子と結合することによって形成される、請求項19の検出方法。
【請求項21】
6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを、2個以上隣接して含む核酸。
【請求項22】
tRNA、mRNA、アンチセンスDNA若しくはRNA、リボザイム、アプタマー又はsiRNAである、請求項21に記載の核酸。
【請求項23】
I) i)6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを、2個以上隣接して含む核酸、と、
ii)5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドであって、前記塩基の5位に、5−カルボキシフルオレセイン(5−FAM)、6−カルボキシフルオレセイン(6−FAM)、5−カルボキシテトラメチルローダミン(5−TAMRA)、6−カルボキシテトラメチルローダミン(6−TAMRA)、 5−ジメチルアミノナフタレン−1−スルホン酸(DANSYL)、5−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(5−HEX)、6−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(6−HEX)、5−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(5−TET)、6−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(6−TET)、5−カルボキシ−X−ローダミン(5−ROX)、及び6−カルボキシ−X−ローダミン(6−ROX)からなるグループから選択される蛍光色素が、直接又はリンカーを介して結合している前記ヌクレオチド、を含む核酸、とをハイブリダイズさせ、そして、
II) 蛍光スペクトルの変化を測定する
ことを含む、核酸の二本鎖形成の検出方法。
【請求項24】
I)i)の6位置換された9H−プリン−9−イル基が、
B1)2−アミノ−6−(2−チエニル)−9H−プリン−9−イル基;
B2)6−(2−チエニル)−9H−プリン−9−イル基;
B3)2−アミノ−6−(4−メチル−2−チエニル)−9H−プリン−9−イル基;
B4)6−(4−メチル−2−チエニル)−9H−プリン−9−イル基;
B5)2−アミノ−6−(5−メチル−2−チエニル)−9H−プリン−9−イル基;
B6)6−(5−メチル−2−チエニル)−9H−プリン−9−イル基;
B7)2−アミノ−6−(2−チアゾリル)−9H−プリン−9−イル基;
B8)6−(2−チアゾリル)−9H−プリン−9−イル基;
B9)2−アミノ−6−(4−メチル−2−チアゾリル)−9H−プリン−9−イル基;
B10)6−(4−メチル−2−チアゾリル)−9H−プリン−9−イル基;
B11)2−アミノ−6−(5−メチル−2−チアゾリル)−9H−プリン−9−イル基;及び
B12)6−(5−メチル−2−チアゾリル)−9H−プリン−9−イル基
からなるグループから選択される、請求項23に記載の検出方法。
【請求項25】
I)ii)の核酸が、塩基の5位に、5−カルボキシフルオレセイン(5−FAM)又は6−カルボキシフルオレセイン(6−FAM)が直接又はリンカーを介して結合している、5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドを含む核酸である、請求項23又は24に記載の検出方法。
【請求項26】
核酸のステム−ループ又はヘアピン構造形成の検出方法であって、
1) i)6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド、並びに、
ii)5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドであって、前記塩基の5位に、5−カルボキシフルオレセイン(5−FAM)、6−カルボキシフルオレセイン(6−FAM)、5−カルボキシテトラメチルローダミン(5−TAMRA)、6−カルボキシテトラメチルローダミン(6−TAMRA)、 5−ジメチルアミノナフタレン−1−スルホン酸(DANSYL)、5−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(5−HEX)、6−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(6−HEX)、5−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(5−TET)、6−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(6−TET)、5−カルボキシ−X−ローダミン(5−ROX)、及び6−カルボキシ−X−ローダミン(6−
ROX)からなるグループから選択される蛍光色素が、直接又はリンカーを介して結合している前記ヌクレオチド、を同一鎖上に含む核酸であって、i)のヌクレオチドを隣接して2個以上含む前記核酸、において、i)のヌクレオチドと、ii)のヌクレオチドをハイブリダイズさせ、そして
2)蛍光スペクトルの変化を測定する
ことを含む、前記方法。
【請求項1】
置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドと、6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドとが塩基対を形成している、核酸。
【請求項2】
前記置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基が、
A1)2−ホルミル−1H−ピロール−1−イル基;
A2)2−ホルミル−4−(1−プロピン−1−イル)−1H−ピロール−1−イル基;
A3)2−ホルミル−4−メチル−1H−ピロール−1−イル基;及び
A4)2−ホルミル−4−エチニル−1H−ピロール−1−イル基
からなるグループから選択され、そして
前記6位置換された9H−プリン−9−イル基が、
B1)2−アミノ−6−(2−チエニル)−9H−プリン−9−イル基;
B2)6−(2−チエニル)−9H−プリン−9−イル基;
B3)2−アミノ−6−(4−メチル−2−チエニル)−9H−プリン−9−イル基;
B4)6−(4−メチル−2−チエニル)−9H−プリン−9−イル基;
B5)2−アミノ−6−(5−メチル−2−チエニル)−9H−プリン−9−イル基;
B6)6−(5−メチル−2−チエニル)−9H−プリン−9−イル基;
B7)2−アミノ−6−(2−チアゾリル)−9H−プリン−9−イル基;
B8)6−(2−チアゾリル)−9H−プリン−9−イル基;
B9)2−アミノ−6−(4−メチル−2−チアゾリル)−9H−プリン−9−イル基;
B10)6−(4−メチル−2−チアゾリル)−9H−プリン−9−イル基;
B11)2−アミノ−6−(5−メチル−2−チアゾリル)−9H−プリン−9−イル基;及び
B12)6−(5−メチル−2−チアゾリル)−9H−プリン−9−イル基
からなるグループから選択される、
請求項1に記載の核酸。
【請求項3】
前記置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基が、2−ホルミル−1H−ピロール−1−イル基であり、そして、前記6位置換された9H−プリン−9−イル基が、2−アミノ−6−(2−チエニル)−9H−プリン−9−イル基である、請求項1又は2に記載の核酸。
【請求項4】
転写、逆転写、複製又は翻訳の工程で、塩基対を形成している、請求項1ないし3のいずれか1項に記載の核酸。
【請求項5】
前記置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドがDNAの一部であり、そして、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドがRNAの一部である、請求項1ないし4のいずれか1項に記載の核酸。
【請求項6】
6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを含む核酸を調製する方法であって、
置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドを含む核酸を鋳型として転写、逆転写又は複製を行い、前記置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を有するヌクレオチドの相補的な位置に、6位置換された9H−プリン−9−イル基を有するヌクレオチドを組み
込む
ことを含む、前記調製方法。
【請求項7】
前記置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基が、
A1)2−ホルミル−1H−ピロール−1−イル基;
A2)2−ホルミル−4−(1−プロピン−1−イル)−1H−ピロール−1−イル基;
A3)2−ホルミル−4−メチル−1H−ピロール−1−イル基;及び
A4)2−ホルミル−4−エチニル−1H−ピロール−1−イル基
からなるグループから選択され、そして
前記6位置換された9H−プリン−9−イル基が、
B1)2−アミノ−6−(2−チエニル)−9H−プリン−9−イル基;
B2)6−(2−チエニル)−9H−プリン−9−イル基;
B3)2−アミノ−6−(4−メチル−2−チエニル)−9H−プリン−9−イル基;
B4)6−(4−メチル−2−チエニル)−9H−プリン−9−イル基;
B5)2−アミノ−6−(5−メチル−2−チエニル)−9H−プリン−9−イル基;
B6)6−(5−メチル−2−チエニル)−9H−プリン−9−イル基;
B7)2−アミノ−6−(2−チアゾリル)−9H−プリン−9−イル基;
B8)6−(2−チアゾリル)−9H−プリン−9−イル基;
B9)2−アミノ−6−(4−メチル−2−チアゾリル)−9H−プリン−9−イル基;
B10)6−(4−メチル−2−チアゾリル)−9H−プリン−9−イル基;
B11)2−アミノ−6−(5−メチル−2−チアゾリル)−9H−プリン−9−イル基;及び
B12)6−(5−メチル−2−チアゾリル)−9H−プリン−9−イル基
からなるグループから選択される、
請求項6に記載の調製方法。
【請求項8】
鋳型において、前記置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチドが2個以上存在する、請求項6又は7に記載の調製方法。
【請求項9】
請求項6ないし8に記載の方法によって調製された、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを含む核酸。
【請求項10】
tRNA、mRNA、アンチセンスDNA若しくはRNA、リボザイム、アプタマー又はsiRNAである、請求項9に記載の核酸。
【請求項11】
前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド中の、塩基に由来する蛍光強度の温度変化および/またはイオン濃度変化を測定する、ことを含む、請求項9又は10に記載の核酸における、前記ヌクレオチドを含む局部領域の立体構造の解析方法。
【請求項12】
前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド中の、塩基に由来する蛍光強度が、温度の上昇に伴い実質的に上昇する場合には、生体内温度下で、前記ヌクレオチドは立体配座において周囲に存在するヌクレオチドと積み重なっていると判断し、そして、蛍光強度が温度の上昇に伴い実質的に低下又は上昇しない場合には、生体内温度下で、前記ヌクレオチドは外部に暴露していると判断する、請求項11に記載の解析方法。
【請求項13】
I) i)請求項6ないし8に記載の方法によって調製された、前記6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを含む核酸、と、
ii)5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドであって、前記塩基の5位に、5−カルボキシフルオレセイン(5−FAM)、6−カルボキシフルオレセイン(6−FAM)、5−カルボキシテトラメチルローダミン(5−TAMRA)、6−カルボキシテトラメチルローダミン(6−TAMRA)、 5−ジメチルアミノナフタレン−1−スルホン酸(DANSYL)、5−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(5−HEX)、6−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(6−HEX)、5−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(5−TET)、6−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(6−TET)、5−カルボキシ−X−ローダミン(5−ROX)、及び6−カルボキシ−X−ローダミン(6−ROX)からなるグループから選択される蛍光色素が、直接又はリンカーを介して結合している前記ヌクレオチド、を含む核酸、とをハイブリダイズさせ、そして、
II) 蛍光スペクトルの変化を測定する
ことを含む、核酸の二本鎖形成の検出方法。
【請求項14】
I)i)の6位置換された9H−プリン−9−イル基が、
B1)2−アミノ−6−(2−チエニル)−9H−プリン−9−イル基;
B2)6−(2−チエニル)−9H−プリン−9−イル基;
B3)2−アミノ−6−(4−メチル−2−チエニル)−9H−プリン−9−イル基;
B4)6−(4−メチル−2−チエニル)−9H−プリン−9−イル基;
B5)2−アミノ−6−(5−メチル−2−チエニル)−9H−プリン−9−イル基;
B6)6−(5−メチル−2−チエニル)−9H−プリン−9−イル基;
B7)2−アミノ−6−(2−チアゾリル)−9H−プリン−9−イル基;
B8)6−(2−チアゾリル)−9H−プリン−9−イル基;
B9)2−アミノ−6−(4−メチル−2−チアゾリル)−9H−プリン−9−イル基;
B10)6−(4−メチル−2−チアゾリル)−9H−プリン−9−イル基;
B11)2−アミノ−6−(5−メチル−2−チアゾリル)−9H−プリン−9−イル基;及び
B12)6−(5−メチル−2−チアゾリル)−9H−プリン−9−イル基
からなるグループから選択される、請求項13に記載の検出方法。
【請求項15】
I)ii)の核酸が、塩基の5位に、5−カルボキシフルオレセイン(5−FAM)又は6−カルボキシフルオレセイン(6−FAM)が直接又はリンカーを介して-結合して
いる、5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドを含む核酸である、請求項13又は14に記載の検出方法。
【請求項16】
i) 6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド
並びに、
ii) 5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドであって、前記塩基の5位に、5−カルボキシフルオレセイン(5−FAM)、6−カルボキシフルオレセイン(6−FAM)、5−カルボキシテトラメチルローダミン(5−TAMRA)、6−カルボキシテトラメチルローダミン(6−TAMRA)、 5−ジメチルアミノナフタレン−1−スルホン酸(DANSYL)、5−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(5−HEX)、6−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(6−HEX)、5−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(5−TET)、6−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(6−TET)、5−
カルボキシ−X−ローダミン(5−ROX)、及び6−カルボキシ−X−ローダミン(6−ROX)からなるグループから選択される蛍光色素が、直接又はリンカーを介して結合している、前記ヌクレオチド
を、同一の鎖上に含む核酸を調製する方法であって、
iii) 置換された若しくは無置換の2−ホルミル−1H−ピロール−1−イル基を塩基として有するヌクレオチド
並びに
iv) 6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド
を含む核酸を鋳型として転写、逆転写又は複製を行い、前記iii)のヌクレオチドの相補的な位置に、前記i)のヌクレオチドを組み込み、そして、前記iv)のヌクレオチドの相補的な位置に、前記ii)のヌクレオチドを組み込む
ことを含む、前記調製方法。
【請求項17】
請求項16に記載の方法によって調製された、
i) 6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド
並びに、
ii) 5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドであって、前記塩基の5位に、5−カルボキシフルオレセイン(5−FAM)、6−カルボキシフルオレセイン(6−FAM)、5−カルボキシテトラメチルローダミン(5−TAMRA)、6−カルボキシテトラメチルローダミン(6−TAMRA)、 5−ジメチルアミノナフタレン−1−スルホン酸(DANSYL)、5−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(5−HEX)、6−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(6−HEX)、5−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(5−TET)、6−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(6−TET)、5−カルボキシ−X−ローダミン(5−ROX)、及び6−カルボキシ−X−ローダミン(6−ROX)からなるグループから選択される蛍光色素が、直接又はリンカーを介して結合している、前記ヌクレオチド
を、同一の鎖上に含む核酸。
【請求項18】
tRNA、mRNA、アンチセンスDNA若しくはRNA、リボザイム、アプタマー又はsiRNAである、請求項17に記載の核酸。
【請求項19】
核酸のステム−ループ又はヘアピン構造形成の検出方法であって、
1) 請求項17又は18に記載の核酸において、i)のヌクレオチドと、ii)のヌクレオチドとをハイブリダイズさせ、そして、
2) 蛍光スペクトルの変化を測定する
ことを含む、核酸のステム−ループ又はヘアピン構造形成の検出方法。
【請求項20】
核酸のステム−ループ又はヘアピン構造が、請求項17又は18に記載の核酸が、標的分子と結合することによって形成される、請求項19の検出方法。
【請求項21】
6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを、2個以上隣接して含む核酸。
【請求項22】
tRNA、mRNA、アンチセンスDNA若しくはRNA、リボザイム、アプタマー又はsiRNAである、請求項21に記載の核酸。
【請求項23】
I) i)6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチドを、2個以上隣接して含む核酸、と、
ii)5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドであって、前記塩基の5位に、5−カルボキシフルオレセイン(5−FAM)、6−カルボキシフルオレセイン(6−FAM)、5−カルボキシテトラメチルローダミン(5−TAMRA)、6−カルボキシテトラメチルローダミン(6−TAMRA)、 5−ジメチルアミノナフタレン−1−スルホン酸(DANSYL)、5−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(5−HEX)、6−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(6−HEX)、5−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(5−TET)、6−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(6−TET)、5−カルボキシ−X−ローダミン(5−ROX)、及び6−カルボキシ−X−ローダミン(6−ROX)からなるグループから選択される蛍光色素が、直接又はリンカーを介して結合している前記ヌクレオチド、を含む核酸、とをハイブリダイズさせ、そして、
II) 蛍光スペクトルの変化を測定する
ことを含む、核酸の二本鎖形成の検出方法。
【請求項24】
I)i)の6位置換された9H−プリン−9−イル基が、
B1)2−アミノ−6−(2−チエニル)−9H−プリン−9−イル基;
B2)6−(2−チエニル)−9H−プリン−9−イル基;
B3)2−アミノ−6−(4−メチル−2−チエニル)−9H−プリン−9−イル基;
B4)6−(4−メチル−2−チエニル)−9H−プリン−9−イル基;
B5)2−アミノ−6−(5−メチル−2−チエニル)−9H−プリン−9−イル基;
B6)6−(5−メチル−2−チエニル)−9H−プリン−9−イル基;
B7)2−アミノ−6−(2−チアゾリル)−9H−プリン−9−イル基;
B8)6−(2−チアゾリル)−9H−プリン−9−イル基;
B9)2−アミノ−6−(4−メチル−2−チアゾリル)−9H−プリン−9−イル基;
B10)6−(4−メチル−2−チアゾリル)−9H−プリン−9−イル基;
B11)2−アミノ−6−(5−メチル−2−チアゾリル)−9H−プリン−9−イル基;及び
B12)6−(5−メチル−2−チアゾリル)−9H−プリン−9−イル基
からなるグループから選択される、請求項23に記載の検出方法。
【請求項25】
I)ii)の核酸が、塩基の5位に、5−カルボキシフルオレセイン(5−FAM)又は6−カルボキシフルオレセイン(6−FAM)が直接又はリンカーを介して結合している、5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドを含む核酸である、請求項23又は24に記載の検出方法。
【請求項26】
核酸のステム−ループ又はヘアピン構造形成の検出方法であって、
1) i)6位置換された9H−プリン−9−イル基を塩基として有するヌクレオチド、並びに、
ii)5位置換−2−オキソ(1H)ピリジン−3−イル基を塩基として有するヌクレオチドであって、前記塩基の5位に、5−カルボキシフルオレセイン(5−FAM)、6−カルボキシフルオレセイン(6−FAM)、5−カルボキシテトラメチルローダミン(5−TAMRA)、6−カルボキシテトラメチルローダミン(6−TAMRA)、 5−ジメチルアミノナフタレン−1−スルホン酸(DANSYL)、5−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(5−HEX)、6−カルボキシ−2’,4,4’,5’,7,7’−ヘキサクロロフルオレセイン(6−HEX)、5−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(5−TET)、6−カルボキシ−2’,4,7,7’−テトラクロロフルオレセイン(6−TET)、5−カルボキシ−X−ローダミン(5−ROX)、及び6−カルボキシ−X−ローダミン(6−
ROX)からなるグループから選択される蛍光色素が、直接又はリンカーを介して結合している前記ヌクレオチド、を同一鎖上に含む核酸であって、i)のヌクレオチドを隣接して2個以上含む前記核酸、において、i)のヌクレオチドと、ii)のヌクレオチドをハイブリダイズさせ、そして
2)蛍光スペクトルの変化を測定する
ことを含む、前記方法。
【図1】

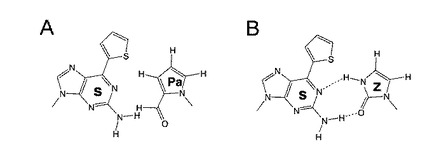
【図2】
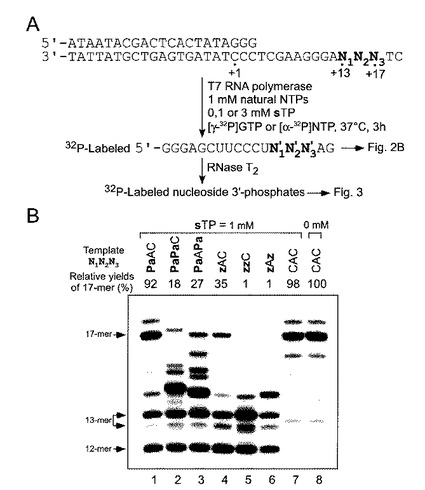
【図3】
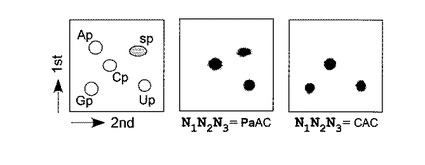
【図4】
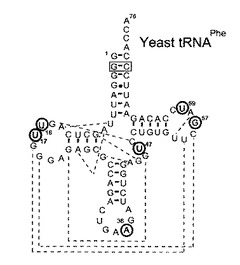
【図5】
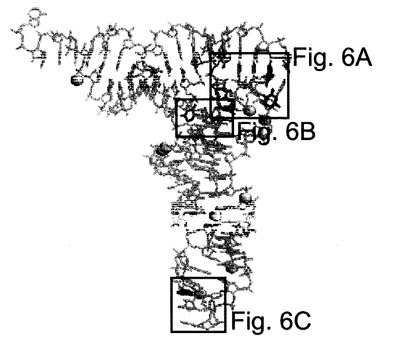
【図6】
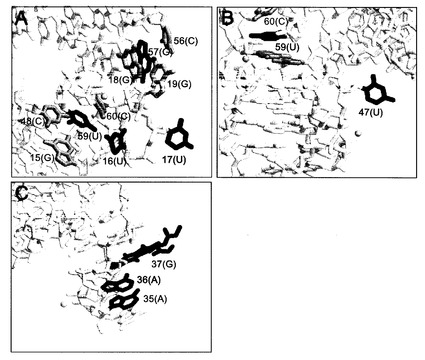
【図7】
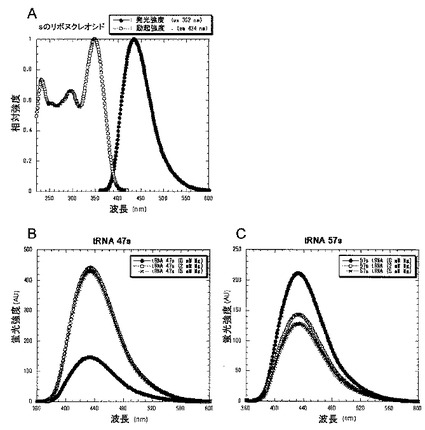
【図8】
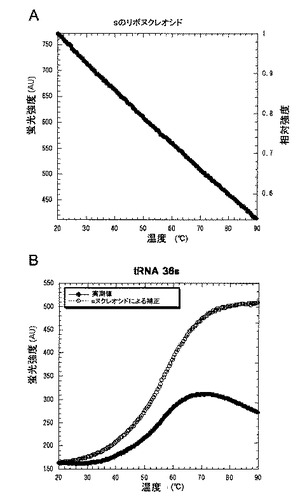
【図9】
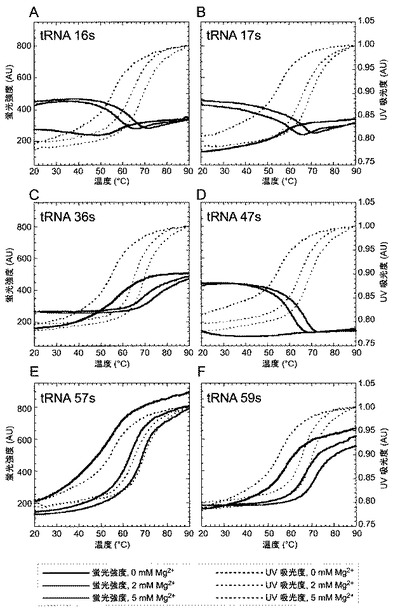
【図10】
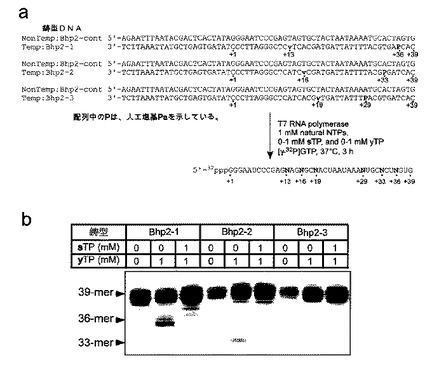
【図11】
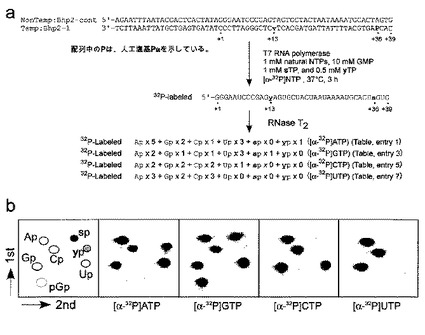
【図12】
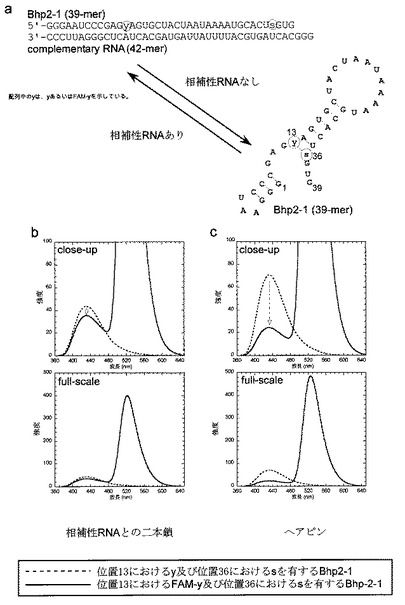
【図13】
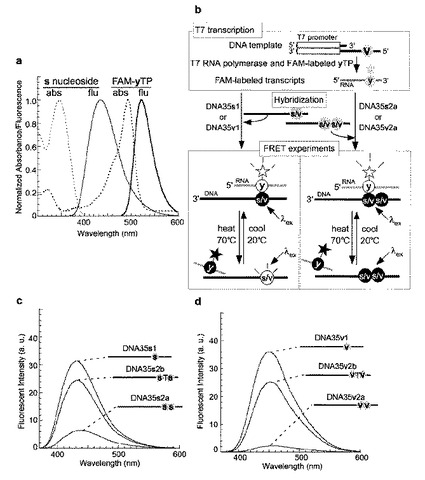
【図14】
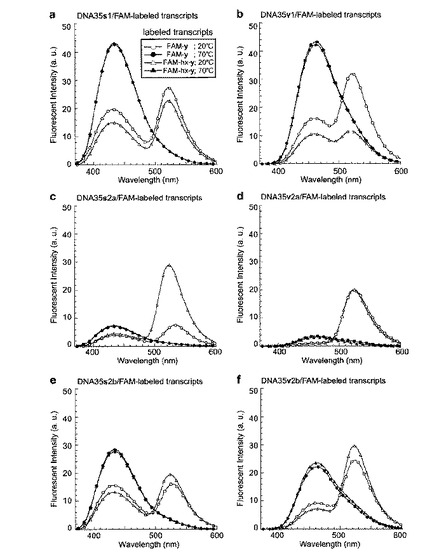
【図15】
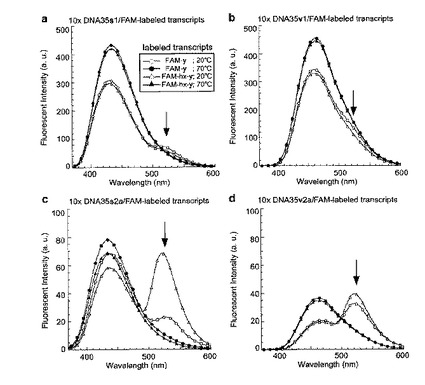
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【公開番号】特開2012−110332(P2012−110332A)
【公開日】平成24年6月14日(2012.6.14)
【国際特許分類】
【出願番号】特願2012−274(P2012−274)
【出願日】平成24年1月4日(2012.1.4)
【分割の表示】特願2006−182062(P2006−182062)の分割
【原出願日】平成18年6月30日(2006.6.30)
【国等の委託研究の成果に係る記載事項】(出願人による申告)国等の委託研究の成果に係る特許出願(平成18度、文部科学省、タンパク3000委託研究「タンパク質基本構造の網羅的解析プログラム」、産業活力再生特別措置法第30条の適用を受けるもの)
【出願人】(508098800)タグシクス・バイオ株式会社 (3)
【出願人】(503359821)独立行政法人理化学研究所 (1,056)
【Fターム(参考)】
【公開日】平成24年6月14日(2012.6.14)
【国際特許分類】
【出願日】平成24年1月4日(2012.1.4)
【分割の表示】特願2006−182062(P2006−182062)の分割
【原出願日】平成18年6月30日(2006.6.30)
【国等の委託研究の成果に係る記載事項】(出願人による申告)国等の委託研究の成果に係る特許出願(平成18度、文部科学省、タンパク3000委託研究「タンパク質基本構造の網羅的解析プログラム」、産業活力再生特別措置法第30条の適用を受けるもの)
【出願人】(508098800)タグシクス・バイオ株式会社 (3)
【出願人】(503359821)独立行政法人理化学研究所 (1,056)
【Fターム(参考)】
[ Back to top ]