
新規微生物および該微生物由来の酵素、ならびにこれらを用いた糖化液の製造方法
【課題】リグノセルロース系バイオマスから、高効率で糖化液を得るための手段を提供することを目的とする。
【解決手段】上記課題は、リグノセルロース系バイオマス分解能を有するペニシリウム属(Penicillium sp.)に属する微生物、好ましくは、S11D6608−M1(受託番号:FERM BP−11083)および/またはS11D6608−M2(受託番号:FERM BP−11084)により解決される。
【解決手段】上記課題は、リグノセルロース系バイオマス分解能を有するペニシリウム属(Penicillium sp.)に属する微生物、好ましくは、S11D6608−M1(受託番号:FERM BP−11083)および/またはS11D6608−M2(受託番号:FERM BP−11084)により解決される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、新規微生物および該微生物由来の酵素、ならびにこれらを用いた糖化液の製造方法に関する。詳しくは、本発明は、リグノセルロース系バイオマスから効率よく糖化液を製造することができる新規微生物および該微生物由来の酵素、ならびにこれらを用いた糖化液の製造方法に関する。
【背景技術】
【0002】
産業革命以来、エネルギー資源として、石油、石炭、および天然ガスなどの化石燃料が大量に使用されてきた。その結果、今日、二酸化炭素などの温室効果ガスによる地球温暖化は、もはや一刻の猶予もないほど深刻化している。さらに、これまでの主要なエネルギー資源であった石油の可採年数が数十年との予測がされており、これらの化石燃料に取って代わる、地球に優しい新たなエネルギー資源の確保が急務となっている。
【0003】
これらの問題の一つの解決策として、バイオマス(再生可能な、生物由来の有機性資源で化石資源を除いたもの)から燃料や、生分解性ポリマーなどの原料となる化成品を生産する、バイオリファイナリーの研究・開発が世界各地で進められている。
【0004】
こうしたバイオリファイナリーの中でも、特に、バイオマスエタノール(以下、「バイオエタノール」とも称する)の生産は既に実用化されており、トウモロコシやサトウキビなどの可食部に含まれるデンプン質または糖質を用いたエタノール生産が、米国やブラジルなどで大規模に行われている。しかしながら、これらの食糧資源をバイオエタノールの原料とすることで、食糧価格の高騰や、発展途上国での深刻な食糧不足を引き起こすことが懸念されている。本発明者は、こうした状況を鑑みて、作物の非可食部である葉や茎など(ソフトバイオマス)、あるいは木材など(ハードバイオマス)に含まれるリグノセルロース系バイオマスを原料とするバイオリファイナリー技術の開発に取り組んできた。
【0005】
一般に、バイオリファイナリーのプロセスは、(i)バイオマスに含まれる多糖類を糖化(加水分解)することによって単糖類を含む糖化液を得る工程と、(ii)得られた糖化液に含まれる単糖類に、微生物または酵素を作用させることによって各種の有用化合物に変換する工程との2つの工程を含む。しかしながら、作物の葉や茎または木材のようなリグノセルロース系バイオマスは、構造性多糖であるセルロースに芳香族化合物の重合体であるリグニンおよびヘミセルロースが強固に結合する構造を有しており、デンプン質や糖質を主成分とするデンプン系バイオマスや糖質系バイオマスと比較して、加水分解反応の効率が著しく低いことが知られている。そこで、リグノセルロース系バイオマスを高効率で糖化するための前処理または加水分解反応に関する技術開発が進められている。
【0006】
リグノセルロース系バイオマスの加水分解法は、主に酸または塩基を用いる化学的加水分解法と、微生物等が生産する酵素(セルラーゼ)を用いる生物学的加水分解法の2種に大別される。後者の生物学的加水分解法によると、温和な条件で、発酵阻害物質が少ない高濃度の糖化液を得ることができる。そこで、より高活性なセルラーゼを産生する微生物の探索が進められている。
【0007】
例えば、特許文献1ではリグノセルロース系バイオマスに含まれる結晶性セルロースに対して高い加水分解活性を有するトリコデルマ・リーゼイ(Trichoderma reesei)および該微生物の製造方法が開示されている。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】特開2006−166848号公報
【発明の概要】
【発明が解決しようとする課題】
【0009】
しかしながら、上記特許文献1に記載のトリコデルマ・リーゼイを用いても、糖化効率が未だ不十分であり、さらなる改良が望まれているのが現状である。
【0010】
そこで本発明は、高効率で糖化液を得るための手段を提供することを目的とする。
【課題を解決するための手段】
【0011】
本発明者は、上記の問題を解決すべく、鋭意研究を行った。その結果、リグノセルロース系バイオマス分解能を有するペニシリウム属(Penicillium sp.)に属する微生物を用いることによってリグノセルロース系バイオマスから高効率で糖化液を製造できることを見出し、本発明を完成させた。
【0012】
すなわち、本発明は、リグノセルロース系バイオマス分解能を有するペニシリウム属(Penicillium sp.)に属する微生物である。
【発明の効果】
【0013】
本発明によれば、リグノセルロース系バイオマスから高効率で糖化液を製造することができる。
【図面の簡単な説明】
【0014】
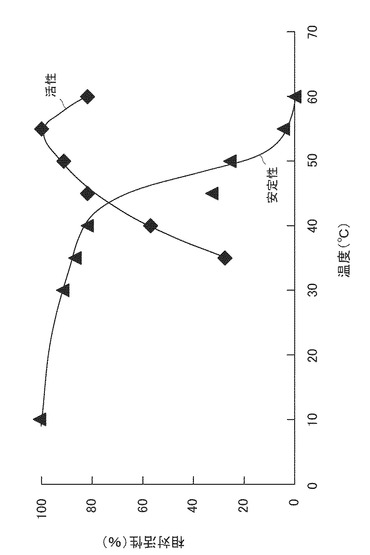
【図1】酵素活性および安定性に対する温度の影響を表すグラフである。
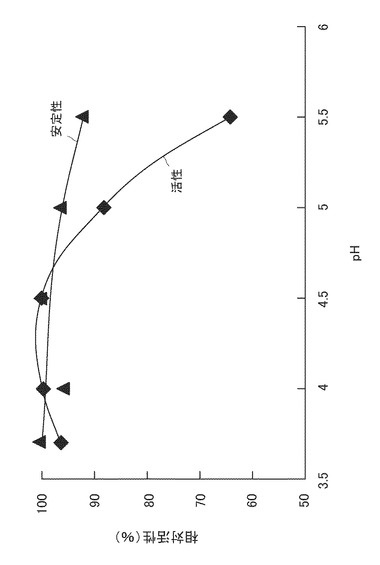
【図2】酵素活性および安定性に対するpHの影響を表すグラフである。
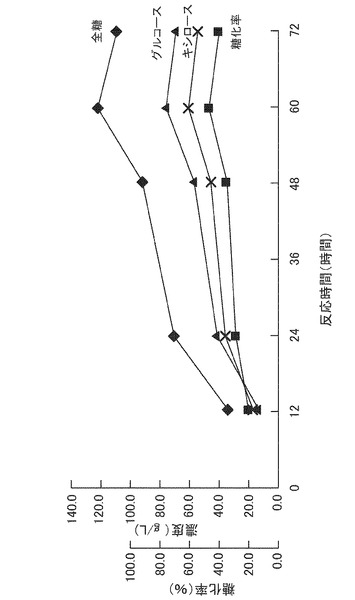
【図3】稲わらの糖化反応における反応溶液中の全糖量、グルコース量、キシロース量、および糖化率の経時変化を表すグラフである。
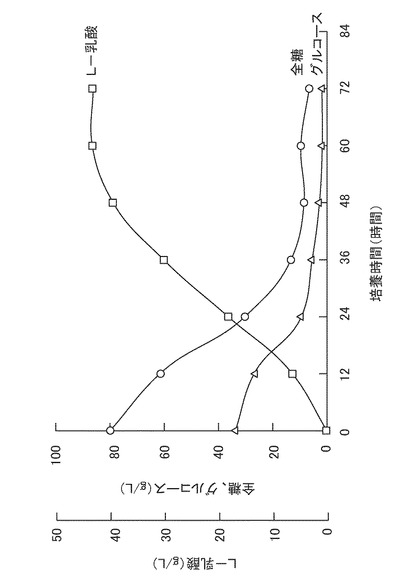
【図4】稲わら糖化液中でのリゾプス エスピー MK96−1156(受託番号:FERM BP−6777)の培養における、全糖量、グルコース量、およびL−乳酸量の経時変化を表すグラフである。
【図5】コーンコブの糖化反応における反応溶液中の全糖量およびグルコース量の経時変化を表すグラフである。
【図6】コーンコブ糖化液中でのリゾプス エスピー MK96−1156(受託番号:FERM BP−6777)の培養における、全糖量、グルコース量、およびL−乳酸量の経時変化を表すグラフである。
【発明を実施するための形態】
【0015】
以下、本発明の好ましい形態を説明する。なお、本発明の技術的範囲は特許請求の範囲の記載に基づいて定められるべきであり、以下の形態のみに制限されない。
【0016】
<微生物>
本形態の微生物は、ペニシリウム属(Penicillium sp.)に属し、リグノセルロース系バイオマス分解能を有する微生物である。該微生物はリグノセルロース系バイオマスの分解活性に優れるため、リグノセルロース系バイオマスから高効率で糖化液を製造することができる。このようなペニシリウム属に属する微生物のうち、本発明者によって新たに見出されたペニシリウム S11D6608−M1(受託番号:FERM BP−11083)および/またはペニシリウム S11D6608−M2(受託番号:FERM BP−11084)であることが好ましい。なお、本明細書において、ペニシリウム S11D6608−M1を略して「菌株M1」と、ペニシリウム S11D6608−M2を略して「菌株M2」と、それぞれ称する場合がある。以下で、菌株M1および菌株M2について説明する。
【0017】
[1次スクリーニング]
中国東北部で生産されたトウモロコシのコーンコブを入手した。該コーンコブの表面の一部にはカビが観察された。該コーンコブを室温で約2ヵ月間風乾し、金切ばさみで1〜5cm程度に切断した。そして、これを家庭用ミキサーに5分間かけることにより粗粉砕した。該粗粉砕物をさらにボールミル(ANS−60S、日陶科学社製)を用いて微粉化した。得られたコーンコブ粉砕物の粒径分布を下記表1に示す。なお、本明細書において「粒径」とは、粒子の輪郭線上の任意の2点間の距離のうち、最大の距離を意味する。また、粒径の値は、試験用ふるいによって測定した値を採用する。下記表1の各粒径における粉砕物の割合の求め方は以下のとおりである。まず、3種類の試験用ふるい(呼び寸法0.5mm、1.0mm、1.7mm;SANPO社製)を呼び寸法が小さいものが下になるように順次重ね、最上部のふるいに上記粉砕物5gを入れた。これを間欠的に5分間ふるい、それぞれのふるいに残存した粉砕物の質量を測定し、それぞれの割合を求めた。
【0018】
【表1】

【0019】
該コーンコブ粉砕物のうち、粒径が1.0mm未満の粉砕物1gを試験管に取り、滅菌水道水10mlを加えて懸濁し、恒温振盪培養装置を用いて32℃、回転数220ppmで7日間振盪培養した。培養後の各試験管中の培養液を一部サンプリングし、培養液中に含まれる全糖量をフェノール硫酸法により分析して、全糖量が特に多かった培養液を選定した。全糖量が多かった培養液では、コーンコブに付着した微生物が、コーンコブに含まれるリグノセルロースを効率よく糖化したものと考えられた。なお、本明細書において、フェノール硫酸法による分析は、中村道徳編、貝沼圭二編、「生物化学実験法(25)澱粉・関連糖質酵素実験法」、学会出版センター版、1989年10月発行、206頁に記載の方法に準じて行った。
【0020】
[1次スクリーニングした菌の単胞子分離]
上記で選定した培養液を滅菌水で適当に希釈し、下記表2に示す組成のPDY寒天培地(オートクレーブにて121℃、20分間滅菌)に塗布し、32℃のインキュベーター中でプレート培養した。培養開始から14日後に培地上にコロニーの形成が認められた。このうち、明らかにカビ以外の微生物であると思われるコロニーを除外し、カビ状でかつ形状が大きなコロニーを選別した。
【0021】
【表2】
【0022】
[2次スクリーニング]
(前培養)
200ml三角フラスコに下記表3に示す組成のA培地20mlを入れ、オートクレーブで121℃、20分間滅菌した。該A培地に上記1次スクリーニングで選定した菌株を1白金耳接種した。これを、恒温振盪培養装置を用いて32℃、回転数220ppmで24時間振盪培養した。
【0023】
【表3】
【0024】
次に、200ml三角フラスコに下記表4に示す組成のB培地20mlを入れ、オートクレーブで121℃、20分間滅菌した。該B培地に、上記で培養した前培養液を5(v/v)%植菌した。これを、恒温振盪培養装置を用いて32℃、回転数220ppmで6日間振盪培養した。
【0025】
【表4】
【0026】
(本培養)
上記1次スクリーニングで調製した粒径が1.0mm未満のコーンコブ粉砕物0.2gを、上記培養液のろ液を適当に水道水で希釈した希釈培養液20mlに懸濁し、50℃で3日間糖化を行った。得られた糖化液中に含まれる全糖量をフェノール硫酸法により分析し、全糖量が多かった糖化液に対応した2菌株を、リグノセルロース系バイオマスの糖化力に優れている菌株として選定した。これらをそれぞれ菌株Iおよび菌株IIと称した。
【0027】
[分類学的性質]
上記でスクリーニングした菌株Iおよび菌株IIの分類学的性質を以下で述べる。
【0028】
(1)菌株I
(a)形態的性質
以下の(i)〜(iv)培地において、25℃で3週間培養後のコロニーの形態を観察した。なお、以下の表5〜8において、色調の括弧内の英数文字は「Kornerup and Wanscher(1978)」で用いられている色のコード ナンバーを表す。
【0029】
(i)ポテトデキストロース寒天培地(商品名:ダイゴ、日本製薬社製)
【0030】
【表5】
【0031】
(ii)2% 麦芽寒天培地(Malt Agar)
【0032】
【表6】
【0033】
(iii)バクトオートミール寒天培地(Bacto Oatmeal Agar、ベクトン ディッキンソン社製)
【0034】
【表7】
【0035】
(iv)LCA(三浦培地)
【0036】
【表8】
【0037】
その他の微視的観察結果を表9に示す。
【0038】
【表9】
【0039】
(b)同定
アポロンDB−FUに対するBLAST(Altschul et al.,1997)相同性検索の結果、菌株Iの28S rDNA−D1/D2塩基配列は、子嚢菌類の一種であるPenicillium aculeatumの塩基配列と99.5%の相同率を示した。GenBank/DDBJ/EMBLなどの国際塩基配列データベースに対する相同性検索の結果においては、菌株Iの28S rDNA−D1/D2塩基配列は子嚢菌類の一種であるPenicillium cf. verruculosum RS7PF(この菌種名の「cf.」は、P. verruculosumに類似する菌株であることを表す)、P. aculeatum、P. verruculosumの塩基配列と高い相同率を示した。アポロンDB−FUおよび国際塩基配列データベースに対する相同性検索で得られた上位の塩基配列をもとに作成した系統樹において、菌株IはPenicillim cf. verruculosum RS7PFと同一の系統枝を形成した。
【0040】
菌株IのITS−5.8S rDNA塩基配列は、アポロンDB−FUに対するBLAST相同性検索の結果、子嚢菌類の一種である、P. verruculosum の塩基配列と98.8%の相同率を示した。また、GenBank/DDBJ/EMBLなどの国際塩基配列データベースに対する相同性検索の結果においては、菌株IのITS−5.8S rDNA塩基配列は、P. pinophilumの塩基配列と100%の相同率を示した。国際塩基配列データベースおよびアポロンDB−FUに対する相同性検索で得られた上位の塩基配列をもとに作成した系統樹において、菌株IはP. pinophilumと同一の系統枝を形成した。
【0041】
したがって、28S rDNA−D1/D2塩基配列の解析結果から、菌株IはPenicillim cf. verruculosumに近縁であるものの、ITS−5.8S rDNA塩基配列の解析で本検体と同一系統枝を形成した28S rDNA−D1/D2塩基配列のデータが登録されてないことから、本検体の種レベルの推定は困難であると思われた。よって、菌株IはP. pinophilumに近縁なPenicillium属の一種であると考えられた。なお、P. verruculosumとP. pinophilumとは、分類学的に非常に近縁であることが知られている。
【0042】
コロニー性状および形態観察の結果、菌株Iは赤褐色〜茶褐色系のコロニーを形成し、培地中に顕著な赤色系の可溶性色素を産出した。また、有隔壁の栄養菌糸のみの形成が観察された。今回の観察結果では、胞子などの生殖器官の形成が観察されなかったことから、分類群の推定はできなかった。なお、ITS−5.8S rDNA塩基配列解析結果より菌株Iと近縁を推定されるP. pinophilumはまれに赤色系の色素を産出することが知られている。
【0043】
以上の28S rDNA−D1/D2およびITS−5.8S rDNA塩基配列解析、ならびにコロニー性状および形態観察の結果から総合的に鑑みて、菌株IはP. pinophilumに近縁なPenicillium属に帰属する新規な菌であると判断し、ペニシリウム S11D6608−M1(Penicillium sp. S11D6608−M1)と命名した。
【0044】
なお、本菌株は、2008年12月24日付けで独立行政法人産業技術総合研究所特許生物寄託センターに受託番号FERM BP−11083として、S11D6608−M1の名称で寄託されている。
【0045】
(2)菌株II
(a)形態的性質
以下の(i)〜(iv)培地において、25℃で3週間培養後のコロニーの形態を観察した。なお、以下の表10〜13において、色調の括弧内の英数文字は「Kornerup and Wanscher(1978)」で用いられている色のコード ナンバーを表す。
【0046】
(i)ポテトデキストロース寒天培地(商品名:ダイゴ、日本製薬社製)
【0047】
【表10】
【0048】
(ii)2% 麦芽寒天培地(Malt Agar)
【0049】
【表11】
【0050】
(iii)バクトオートミール寒天培地(Bacto Oatmeal Agar、ベクトン ディッキンソン社製)
【0051】
【表12】
【0052】
(iv)LCA(三浦培地)
【0053】
【表13】
【0054】
その他の微視的観察結果を表14に示す。
【0055】
【表14】
【0056】
(b)同定
アポロンDB−FUに対するBLAST(Altschul et al.,1997)相同性検索の結果、菌株IIの28S rDNA−D1/D2塩基配列は、子嚢菌類の一種であるPenicillium aculeatumの塩基配列と99.5%の相同率を示した。GenBank/DDBJ/EMBLなどの国際塩基配列データベースに対する相同性検索の結果においては、菌株IIの28S rDNA−D1/D2塩基配列は子嚢菌類の一種であるPenicillium cf. verruculosum RS7PF(この菌種名の「cf.」は、P. verruculosumに類似する菌株であることを表す)、P. aculeatum、P. verruculosumの塩基配列と高い相同率を示した。アポロンDB−FUおよび国際塩基配列データベースに対する相同性検索で得られた上位の塩基配列をもとに作成した系統樹において、菌株IIはPenicillim cf. verruculosum RS7PFと同一の系統枝を形成した。
【0057】
菌株IIのITS−5.8S rDNA塩基配列は、アポロンDB−FUに対するBLAST相同性検索の結果、子嚢菌類の一種である、P. verruculosum の塩基配列と98.8%の相同率を示した。また、GenBank/DDBJ/EMBLなどの国際塩基配列データベースに対する相同性検索の結果においては、菌株IIのITS−5.8S rDNA塩基配列は、P. pinophilumの塩基配列と100%の相同率を示した。国際塩基配列データベースおよびアポロンDB−FUに対する相同性検索で得られた上位の塩基配列をもとに作成した系統樹において、菌株IIはP. pinophilumと同一の系統枝を形成した。
【0058】
したがって、28S rDNA−D1/D2塩基配列の解析結果から、菌株IIはPenicillim cf. verruculosumに近縁であるものの、ITS−5.8S rDNA塩基配列の解析で本検体と同一系統枝を形成した28S rDNA−D1/D2塩基配列のデータが登録されてないことから、本検体の種レベルの推定は困難であると思われた。よって、菌株IIはP. pinophilumに近縁なPenicillium属の一種であると考えられた。なお、P. verruculosumとP. pinophilumとは、分類学的に非常に近縁であることが知られている。
【0059】
コロニー性状および形態観察の結果、菌株IIは白色〜赤黄色系のコロニーを形成し、有隔壁の栄養菌糸のみの形成が観察された。今回の観察結果では、胞子などの生殖器官の形成が観察されなかったことから、分類群の推定はできなかった。上述の菌株Iのコロニー性状とは顕著な違いが認められたが、菌糸の形状において差異は認められなかった。
【0060】
以上の28S rDNA−D1/D2およびITS−5.8S rDNA塩基配列解析、ならびにコロニー性状および形態観察の結果から総合的に鑑みて、菌株IはP. pinophilumに近縁なPenicillium属に帰属する新規な菌であると判断し、ペニシリウム S11D6608−M2(Penicillium sp. S11D6608−M2)と命名した。
【0061】
なお、本菌株は、2008年12月24日付けで独立行政法人産業技術総合研究所特許生物寄託センターに受託番号FERM BP−11084として、S11D6608−M2の名称で寄託されている。
【0062】
また、本発明の微生物として、より好ましくは、より高効率で糖化液を得られるよう、菌株M1または菌株M2に公知の変異処理を施すことによって得られた変異株であることが好ましい。変異処理の方法は、特に制限はないが、例えば、紫外線照射やニトロソグアニジンなどの変異誘発剤を使用することによって変異誘導処理する方法が挙げられる。そして、変異処理を施した菌株から、リグノセルロース系バイオマスの分解活性に優れる菌株を選定する。
【0063】
<酵素>
本発明の他の形態としては、上述の微生物に由来する酵素が挙げられる。該酵素は、リグノセルロース系バイオマスに対して優れた加水分解活性能を有するので、該酵素をリグノセルロース系バイオマスに作用させることにより高効率で糖化液を製造することができる。
【0064】
本形態の酵素は、上述の微生物の培養物から、従来公知の酵素採取手段および精製手段を用いて調製することができる。酵素採取法としては、例えば、微生物を含む培養物を遠心分離またはろ過することによって、菌体とろ液とを分離する。そして、該菌体およびろ液から、塩析法、等電点沈殿法、溶媒沈殿法(メタノール、エタノール、アセトン、およびイソプロパノールなど)によりタンパク質である酵素を沈殿させたり、限外ろ過により濃縮したりする方法が挙げられる。
【0065】
リグノセルロース系バイオマスの加水分解には、上記の方法によって得られる粗酵素を用いてもよいが、必要によりさらに精製した酵素を用いることもできる。酵素の精製法としては、例えば、吸着クロマトグラフィー、イオン交換クロマトグラフィー、およびゲルクロマトグラフィーなどのクロマトグラフィーを単独で、または2種以上を組み合わせた精製法が挙げられる。
【0066】
<糖化液の製造方法>
本発明の他の形態にかかる糖化液の製造方法は、上述の本発明の微生物をリグノセルロース系バイオマスを含む培地中で培養する工程、または上述の本発明の酵素をリグノセルロース系バイオマスに作用させることにより加水分解する工程を含む。
【0067】
本形態の糖化液の製造方法で用いるリグノセルロース系バイオマスは、セルロースおよびヘミセルロース、ならびにリグニンを含む有機資源であれば植物性でも動物性でも特に制限はなく、農作物、草木類、および海藻類、ならびにこれらの処理物または廃棄物などが含まれる。より具体的には、稲わら、籾殻、麦わら、ふすま、バガス、ヤシ殻、コーンコブ、コットン、ラミー、ケナフ、ビート、大豆粕、コーヒー粕、雑草、木材、竹、バクテリアセルロース、パルプ、紙、セルロース粉末、結晶セルロース、レーヨン、アルカリセルロース、リン酸膨潤セルロース、およびカルボキシメチルセルロースなどが挙げられる。なお、本発明の微生物または酵素は、上記リグノセルロース系バイオマスの糖化能に優れるだけでなく、でんぷん系バイオマスまたは糖質系バイオマスに対しても優れた糖化能を示す。
【0068】
上記リグノセルロース系バイオマスは、微生物の培養または酵素による加水分解に供する前に、糖化効率を高めるための前処理を行うことが好ましい。前処理としては、例えば、脱水処理または乾燥処理、粉砕処理、および蒸煮処理などの従来公知の技術を適宜採用することができる。特に、稲わら、麦わら、とうもろこしの茎葉、コーンコブなどの農作物の非可食部などのリグノセルロース系バイオマスを用いる場合は、まず、風乾によりバイオマス中に含まれる水分を除く。その後、切断装置や粉砕装置を用いて、好ましくは1.0mm以下であり、より好ましくは0.01〜1.0mmであり、さらに好ましくは0.1〜1.0mmの粉末状とすることが好ましい。
【0069】
上記リグノセルロース系バイオマスは、微生物の培養または酵素による加水分解に供するために、水性媒体に懸濁して用いる。水性媒体は微生物の増殖または酵素反応の進行が可能な範囲においては特に制限はないが、水、緩衝液、酸性水溶液、またはアルカリ性水溶液を用いることが好ましい。懸濁液中に含まれるリグノセルロース系バイオマスの粉砕物の濃度は、水性媒体に対して、通常、50〜300g/Lであり、好ましくは100〜200g/Lである。
【0070】
上述の本発明の微生物を用いて糖化液を製造する場合、本発明の微生物は、上記懸濁液のみを培地とした場合であっても、十分な増殖能力を有するが、必要によっては炭素源、窒素源、または無機物などを懸濁液に補充したものを培地として用いることも可能である。これらの栄養源は、微生物の数を増やすための前培養の培地に添加してもよいし、本培養の培地に添加しても構わない。
【0071】
炭素源としては、例えば、グルコース、ガラクトース、フルクトース、およびキシロースなどの単糖、マルトース、ラクトース、およびスクロースなどの二糖、グリセリンおよびキシリトールなどの糖アルコールといった糖類が挙げられ、これらを単独であるいは2種以上を組み合わせて使用できる。特に、前培養の培地の培地にグルコースを添加することにより、本培養のときの微生物のリグノセルロース系バイオマスの分解活性をより向上させることができる。
【0072】
窒素源としては、例えば、アンモニア、塩化アンモニウム、硫酸アンモニウム、硝酸アンモニウムなどのアンモニウム塩、尿素、L−グルタミン酸などのアミノ酸類、あるいは尿酸などの無機あるいは有機の窒素化合物が使用できる。さらに、窒素源としては、ペプトン、ポリペプトン、肉エキス、酵母エキス、大豆加水分解物、大豆粉末、ミルクカゼイン、カザミノ酸、コーンスティープリカー(CSL)などの窒素含有天然物を使用してもよい。これらのうち、塩化アンモニウム、硫酸アンモニウム、尿素、L−グルタミン酸などのアミノ酸類、尿酸などの無機あるいは有機窒素化合物、ペプトン、肉エキス、酵母エキスなの窒素含有天然物が好ましい。これらの窒素源は、単独であるいは2種以上を組み合わせて使用できる。これらの窒素源のうち、大豆粉末またはコーンスティープリカーを使用することが好ましい。
【0073】
無機物としては、例えば、塩化ナトリウム、塩化カリウム、塩化カルシウム、リン酸カリウム、リン酸ナトリウム、硫酸マグネシウム、硫酸アンモニウムなどの、マグネシウム、マンガン、カルシウム、ナトリウム、カリウム、銅、鉄および亜鉛などのリン酸塩、塩酸塩、硫酸塩および酢酸塩などが用いられる。そのほか、チアミン、ビオチンなどのビタミン類、さらに必要に応じて、アデニン、ウラシルなどの核酸関連物質が使用されてもよい。これらの無機物は、単独であるいは2種以上を組み合わせて使用できる。
【0074】
微生物の培養方法は、本発明の微生物の増殖が実質的に阻害されず、リグノセルロース系バイオマスから単糖類を生産可能な範囲であれば、特に制限なく当業者によって適宜調整されうるが、培養温度は、通常20〜50℃であり、好ましくは28〜37℃である。また、培地のpHは、通常3〜9であり、好ましくは4〜7である。培養前または培養中にpHが4未満となる場合は、アンモニアなどを用いてpH制御を行うことが好ましい。本発明の微生物を培養する場合は、一定量の酸素を供給することが必要である。供給方法は、特に制限はないが、フラスコ培養の場合は回転振盪による攪拌、バイオリアクターを用いる培養の場合は通気攪拌をすることによって培地に酸素を供給する。
【0075】
一方、本発明の酵素を用いた加水分解反応は、リグノセルロース系バイオマスを含む懸濁液中に酵素を添加し、懸濁液を攪拌下、インキュベートすることによって行われうる。懸濁液に添加する酵素量は、懸濁するリグノセルロース系バイオマスの粉砕物の質量に対して、好ましくは0.1〜3質量%、より好ましくは1〜2質量%である。かような量の酵素を用いることによって、通常12〜144時間、好ましくは24〜72時間で加水分解反応が完了しうる。また、加水分解条件は、用いる酵素の至適条件を用いることが好ましい。pHは通常3〜6であり、好ましくは3.5〜5である。また、反応温度は通常30〜60℃であり、好ましくは40〜60℃である。
【0076】
微生物の培養、または酵素による加水分解により得られた糖化液は、糖化液中の不溶物を適宜ろ過してもよいし、そのままの状態で、微生物代謝産物(エタノール、乳酸などの有用化合物)を産生する他の微生物の培養に供しても構わない。撹拌効率および収率向上の観点の観点からは、糖化液中の不溶物を予めろ過することが好ましい。
【0077】
他の微生物としては、特に制限はないが、例えば、乳酸を産生するRizopus属、イタコン酸を生産するAspergillus terreus、エタノールを産生するPachysolen tannophilus、フマル酸を産生するRhizopus oryzae、リンゴ酸を産生するAspergillus flavus、ピルビン酸を産生するTorulopsis glabrata、キシリトールを産生するCandida tropicalisが挙げられる。本発明によって得られる糖化液を含む倍中でこれらの微生物を培養することによって、燃料や化成品を工業的規模で製造するが可能である。
【実施例】
【0078】
本発明の作用効果を、以下の実施例および比較例を用いて説明する。ただし、本発明の技術的範囲が以下の実施例のみに制限されるわけではない。
【0079】
<菌株M1および菌株M2の稲わら分解活性の評価>
[実施例1]
1.生産菌の培養
(前培養)
200ml三角フラスコに下記表15に示す組成のC培地20mlを入れ、オートクレーブで121℃、20分間滅菌した。該C培地に菌株M1および菌株M2をそれぞれ1白金耳ずつ接種し、これを恒温振盪培養装置を用いて32℃、回転数220ppmで24時間振盪培養した(混合培養)。
【0080】
【表15】
【0081】
(本培養)
次に、下記表16の組成を有する各培地を調製した。下記表16において、対象なる成分が含まれている場合を「○」であらわし、含まれていない場合を「×」で表す。
【0082】
【表16】
【0083】
なお、上記表16における稲わらは、粒径が1.0mm未満の粉砕物を用いた。稲わらの粉砕方法は以下のとおりである。稲わらを室温で約2ヵ月間風乾し、金切ばさみで1〜5cm程度に切断した。そして、これを家庭用ミキサーに5分間かけることにより粗粉砕した。該粗粉砕物をさらにボールミル(ANS−60S、日陶科学社製)を用いて微粉化し、稲わら粉砕物を得た。
【0084】
次に、上記の培地1〜6を200ml三角フラスコにそれぞれ20mlずつ入れ、オートクレーブで121℃、20分間滅菌した。該培地1〜6に、上記前培養液を7(v/v)%植菌した。これを恒温振盪培養装置を用いて32℃、回転数220ppmで6日間振盪培養した。
【0085】
2.稲わら分解活性の評価(糖化)
上述の方法で得た粒径が1.0mm未満の稲わら粉砕物2gを水道水9mlに懸濁した。該懸濁液に、上記培地1〜6で培養した培養液、またはそのろ液を希釈した酵素液1mlを加え、50℃で24時間糖化した。この後、それぞれの糖化液をろ過し、得られたろ液中に含まれる全糖量をフェノール硫酸法により測定した。そして、稲わら分解活性を下記式(1)より算出した。なお、酵素活性は1[U]は、(1/60)[μkat]に相当する。
【0086】
【数1】
【0087】
[比較例1]
菌株M1および菌株M2に変えて、トリコデルマ・リーゼイ(Tricoderma reesei)NBRC31329を用いたことを除いては、実施例1と同様の方法で稲わら分解活性を求めた。
【0088】
結果を下記表17に示す。
【0089】
【表17】
【0090】
表17の結果より、本発明の菌株M1およびM2は、従来優れたセルラーゼ産生菌として知られるトリコデルマ・リーゼイ NBRC31329よりも顕著な稲わら分解活性能を有することが示された。
【0091】
また、前培養の培地に、窒素源として大豆粉を、炭素源としてグルコースを添加することにより、本培養のときの微生物の増殖を促進し、微生物のリグノセルロース系バイオマスの分解活性をより向上させることができることが示された。
【0092】
<酵素活性の評価>
[実施例2]
1.酵素の調製
(前培養)
200ml三角フラスコに下記表18に示す組成のD培地20mlを入れ、オートクレーブで121℃、20分間滅菌した。該D培地に菌株M1および菌株M2をそれぞれ1白金耳ずつ接種し、これを恒温振盪培養装置を用いて32℃、回転数220ppmで24時間振盪培養した(混合培養)。
【0093】
【表18】
【0094】
(本培養)
次に、3Lジャーファーメンター(3MDL型、丸菱バイオエンジ社製)に下記表19に示す組成のE培地1.5Lを入れ、オートクレーブで121℃、20分間滅菌した。該B培地に、上記で培養したD培地を7(v/v)%植菌した。これを、恒温振盪培養装置を用いて32℃、回転数600ppm、通気量2vvmで6日間培養した。
【0095】
【表19】
【0096】
上記の6日間の培養中、培養開始から0(培養開始直後)、24、48、72、および84時間後の培養液を1ml取り出し、上記実施例1の3.稲わら分解活性の評価と同様の方法で、各時間の培養液の稲わら分解活性を評価した。結果を下記表20に示す。
【0097】
【表20】
【0098】
6日間培養後の培養液をろ過し、ケーキ(残渣)を水洗した。ろ液とケーキの水洗液とをあわせて濃縮乾固した。次に、この濃縮乾固物を5℃の冷水200mlに溶解し、不溶分をろ別した後、これにアセトンを1000ml加えて、アセトン沈殿物を得た。そして、これを凍結乾燥して酵素(「酵素M1M2」と称する)を得た。なお、上記の各工程で得られた生成物の収量、酵素活性または比活性、総活性、および収率を下記表21に示す。なお、酵素活性は上記実施例1の分解活性の評価と同様の方法を用いて測定した。
【0099】
【表21】
【0100】
2.酵素化学的性質の評価
(安定性に対する温度の影響)
0.05M酢酸バッファー(pH5.5)9mlに、上記で得た酵素M1M21gを溶解したものを酵素溶液とした。ふた付試験管に該酵素溶液1mlと、上記酢酸バッファー9mlとを入れ、それぞれ所定の温度で10時間振盪した後、試験管を5℃に冷却した。ついで、この試験管に上記実施例1で調製した稲わら粉末(粒径1.0mm未満のもの)を1g加え、50℃で24時間糖化反応を行った。この後、それぞれの糖化液をろ過し、得られたろ液中に含まれる全糖量をフェノール硫酸法により測定した。そして、分解活性(糖化活性)を上記式(1)より算出した。得られた値のうち最も高い安定性を与えた10℃における分解活性に対する、それぞれの温度における活性を百分率で表した。結果を図1に示す。
【0101】
(糖化活性に対する温度の影響)
ふた付試験管に、上記酵素溶液1mlと、上記酢酸バッファー9mlとを入れ、上記実施例1で調製した稲わら粉末(粒径1.0mm未満のもの)を1g加え、所定の温度で24時間糖化反応を行った。この後、それぞれの糖化液をろ過し、得られたろ液中に含まれる全糖量をフェノール硫酸法により測定した。そして、分解活性(糖化活性)を上記式(1)より算出した。得られた値のうち最も高い安定性を与えた55℃における分解活性に対する、それぞれの温度における活性を百分率で表した。結果を図1に示す。
【0102】
(安定性に対するpHの影響)
ふた付試験管に上記酵素溶液1mlと、それぞれ所定のpHに調整した0.05M酢酸バッファー9mlを入れ、50℃で10時間振盪した後、試験管を5℃に冷却した。ついで、この試験管に上記実施例1で調製した稲わら粉末(粒径1.0mm未満のもの)を1g加え、50℃で24時間糖化反応を行った。この後、それぞれの糖化液をろ過し、得られたろ液中に含まれる全糖量をフェノール硫酸法により測定した。そして、分解活性(糖化活性)を上記式(1)より算出した。得られた値のうち最も高い安定性を与えたpH3.7における分解活性に対する、それぞれのpHにおける活性を百分率で表した。結果を図2に示す。
【0103】
(糖化活性に対する温度の影響)
ふた付試験管に、上記酵素溶液1mlと、それぞれ所定のpHに調整した0.05M酢酸バッファー9mlを入れ、上記実施例1で調製した稲わら粉末(粒径1.0mm未満のもの)を1g加え、50℃で24時間糖化反応を行った。この後、それぞれの糖化液をろ過し、得られたろ液中に含まれる全糖量をフェノール硫酸法により測定した。そして、分解活性(糖化活性)を上記式(1)より算出した。得られた値のうち最も高い安定性を与えたpH4.5における分解活性に対する、それぞれのpHにおける活性を百分率で表した。結果を図2に示す。
【0104】
[実施例3]
D培地に接種した微生物を菌株M1のみとしたことを除いては、実施例2と同様の方法で酵素を調製した。得られた酵素を「酵素M1」と称する。そして、実施例2と同様の方法で酵素M1の比活性を測定した。
【0105】
[実施例4]
D培地に接種した微生物を菌株M2のみとしたことを除いては、実施例2と同様の方法で酵素を調製した。得られた酵素を「酵素M2」と称する。そして、実施例2と同様の方法で酵素M2の比活性を測定した。
【0106】
[比較例2]
アクレモニウムセルラーゼ(明治製菓社製)を用いて、実施例2と同様の方法で比活性を測定した。
【0107】
[比較例3]
メイセラーゼ(明治製菓社製)を用いて、実施例2と同様の方法で比活性を測定した。
【0108】
[比較例4]
スミチーム(新日本化学工業社製)を用いて、実施例2と同様の方法で比活性を測定した。
【0109】
酵素の比活性の結果を下記表22に示す。
【0110】
【表22】
【0111】
表22の結果より、本発明の酵素M1M2、酵素M1、および酵素M2は、従来のセルラーゼよりも優れた稲わら分解活性を有することが示された。
【0112】
<酵素M1M2を用いた稲わら糖化液の製造、および該糖化液を用いたL−乳酸の製造>
[実施例5]
3Lジャーファーメンター(MDL−3型、丸菱バイオエンジ社製)に水道水1.0Lを入れ、上記実施例1で調製した稲わら粉末(粒径1.0mm未満のもの)200gを懸濁し、さらに水道水を加えて液量を1.5Lとした。該懸濁液のpHを、25%塩酸を用いて4.5に調整した。そして、実施例2で得た酵素M1M2を20U/mlとなるように添加し、50℃で攪拌(200rpm)下、72時間糖化反応を行った。反応液中の全糖量、グルコース量、キシロース量、および糖化率の経時変化を図3に示す。
【0113】
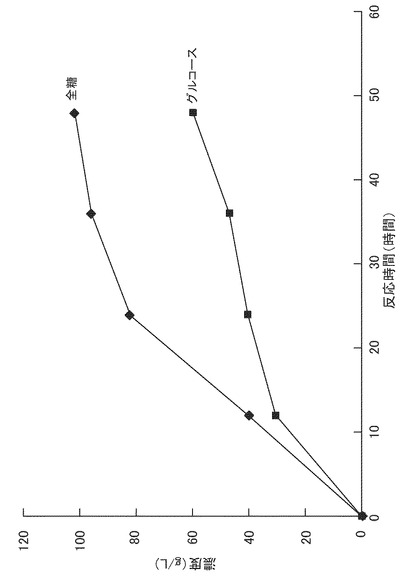
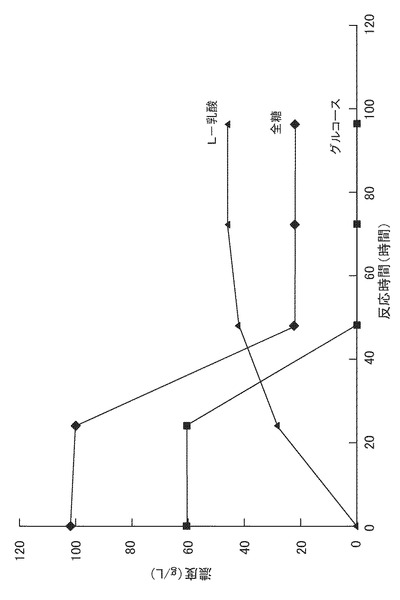
得られた糖化液を3Lジャーファーメンターに仕込み、これにリン酸一カリウムを0.3(g/L)、硫酸マグネシウムを0.25(g/L)となるように添加した。該糖化液をオートクレーブを用いて121℃で10分間滅菌した。次に、予め300ml三角フラスコで前培養したリゾプス エスピー(Rizopus sp.)MK96−1156(受託番号:FERM BP−6777)の培養液100mlを添加し、回転数300rpm、通器量0.5vvm、培養温度30℃で72時間培養した。培養中、25%アンモニア水を用いて培養液のpHを6.0に制御した。全糖量、グルコース量、およびL−乳酸量の経時変化を図4に示す。リゾプス エスピー MK96−1156(受託番号:FERM BP−6777)の培養により、全糖90g/Lから42g/LのL−乳酸が得られた。また、稲わら粉末からのL−乳酸の収率は21(w/w)%であった。
【0114】
<酵素M1M2を用いたコーンコブ糖化液の製造、および該糖化液を用いたL−乳酸の製造>
[実施例6]
稲わら粉末に変えて、上述の[1次スクリーニング]で調製したコーンコブ粉末(粒径1.0mm未満のもの)を用い、糖化時間を48時間、Rizopus sp. MK96−1156(受託番号:FERM BP−6777)の培養時間を96時間としたことを除いては、実施例5と同様の方法で、L−乳酸を生産した。糖化反応液中の全糖量、グルコース量の経時変化を図5に示す。また、Rizopus sp. MK96−1156(受託番号:FERM BP−6777)の培養液中の全糖量、グルコース量、およびL−乳酸量の経時変化を図6に示す。Rizopus sp. MK96−1156(受託番号:FERM BP−6777)の培養により、全糖102g/Lから46g/LのL−乳酸が得られた。また、コーンコブ粉末からのL−乳酸の収率は21(w/w)%であった。
【0115】
<稲わら糖化液を用いたエタノールの製造>
硫酸玉つきガラス管を備えた100ml三角フラスコに、実施例5と同様の方法で製造した稲わら糖化液および下記表23に示す添加物を加えた培養液20mlを準備した。これにパン酵母(オリエンタルドライイースト、オリエンタル酵母工業社製)0.5gを加え、30℃で静置培養を行った。経時的にフラスコ全体の重量を測定し、その重量の減少量を下記式(2)にあてはめ、各培養時間後のエタノール量を算出した。下記式(2)において、フラスコの重量の減少量はエタノール発酵による炭酸ガス発生量(g)を意味する。アルコール発酵により、グルコース1モルからエタノール2モルと炭酸ガス(CO2
)2モルが生産されることから、炭酸ガス発生量(g)に1.047(46.068/44.0095)を乗じて得られる値が、エタノール量(g)となる。そして、培養開始前の糖化液中のグルコース量に対するエタノール量を百分率で表し、各培養時間におけるエタノール収率(%)とした。結果を下記表23に示す。
【0116】
【数2】
【0117】
【表23】
【0118】
表23の結果より、本発明の酵素M1M2を用いて製造された糖化液は、酵母エキス、乾燥酵母、尿素、硫安などの窒素源を補填することのなく、46%の高収率でエタノールを製造できることが示された。
【技術分野】
【0001】
本発明は、新規微生物および該微生物由来の酵素、ならびにこれらを用いた糖化液の製造方法に関する。詳しくは、本発明は、リグノセルロース系バイオマスから効率よく糖化液を製造することができる新規微生物および該微生物由来の酵素、ならびにこれらを用いた糖化液の製造方法に関する。
【背景技術】
【0002】
産業革命以来、エネルギー資源として、石油、石炭、および天然ガスなどの化石燃料が大量に使用されてきた。その結果、今日、二酸化炭素などの温室効果ガスによる地球温暖化は、もはや一刻の猶予もないほど深刻化している。さらに、これまでの主要なエネルギー資源であった石油の可採年数が数十年との予測がされており、これらの化石燃料に取って代わる、地球に優しい新たなエネルギー資源の確保が急務となっている。
【0003】
これらの問題の一つの解決策として、バイオマス(再生可能な、生物由来の有機性資源で化石資源を除いたもの)から燃料や、生分解性ポリマーなどの原料となる化成品を生産する、バイオリファイナリーの研究・開発が世界各地で進められている。
【0004】
こうしたバイオリファイナリーの中でも、特に、バイオマスエタノール(以下、「バイオエタノール」とも称する)の生産は既に実用化されており、トウモロコシやサトウキビなどの可食部に含まれるデンプン質または糖質を用いたエタノール生産が、米国やブラジルなどで大規模に行われている。しかしながら、これらの食糧資源をバイオエタノールの原料とすることで、食糧価格の高騰や、発展途上国での深刻な食糧不足を引き起こすことが懸念されている。本発明者は、こうした状況を鑑みて、作物の非可食部である葉や茎など(ソフトバイオマス)、あるいは木材など(ハードバイオマス)に含まれるリグノセルロース系バイオマスを原料とするバイオリファイナリー技術の開発に取り組んできた。
【0005】
一般に、バイオリファイナリーのプロセスは、(i)バイオマスに含まれる多糖類を糖化(加水分解)することによって単糖類を含む糖化液を得る工程と、(ii)得られた糖化液に含まれる単糖類に、微生物または酵素を作用させることによって各種の有用化合物に変換する工程との2つの工程を含む。しかしながら、作物の葉や茎または木材のようなリグノセルロース系バイオマスは、構造性多糖であるセルロースに芳香族化合物の重合体であるリグニンおよびヘミセルロースが強固に結合する構造を有しており、デンプン質や糖質を主成分とするデンプン系バイオマスや糖質系バイオマスと比較して、加水分解反応の効率が著しく低いことが知られている。そこで、リグノセルロース系バイオマスを高効率で糖化するための前処理または加水分解反応に関する技術開発が進められている。
【0006】
リグノセルロース系バイオマスの加水分解法は、主に酸または塩基を用いる化学的加水分解法と、微生物等が生産する酵素(セルラーゼ)を用いる生物学的加水分解法の2種に大別される。後者の生物学的加水分解法によると、温和な条件で、発酵阻害物質が少ない高濃度の糖化液を得ることができる。そこで、より高活性なセルラーゼを産生する微生物の探索が進められている。
【0007】
例えば、特許文献1ではリグノセルロース系バイオマスに含まれる結晶性セルロースに対して高い加水分解活性を有するトリコデルマ・リーゼイ(Trichoderma reesei)および該微生物の製造方法が開示されている。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】特開2006−166848号公報
【発明の概要】
【発明が解決しようとする課題】
【0009】
しかしながら、上記特許文献1に記載のトリコデルマ・リーゼイを用いても、糖化効率が未だ不十分であり、さらなる改良が望まれているのが現状である。
【0010】
そこで本発明は、高効率で糖化液を得るための手段を提供することを目的とする。
【課題を解決するための手段】
【0011】
本発明者は、上記の問題を解決すべく、鋭意研究を行った。その結果、リグノセルロース系バイオマス分解能を有するペニシリウム属(Penicillium sp.)に属する微生物を用いることによってリグノセルロース系バイオマスから高効率で糖化液を製造できることを見出し、本発明を完成させた。
【0012】
すなわち、本発明は、リグノセルロース系バイオマス分解能を有するペニシリウム属(Penicillium sp.)に属する微生物である。
【発明の効果】
【0013】
本発明によれば、リグノセルロース系バイオマスから高効率で糖化液を製造することができる。
【図面の簡単な説明】
【0014】
【図1】酵素活性および安定性に対する温度の影響を表すグラフである。
【図2】酵素活性および安定性に対するpHの影響を表すグラフである。
【図3】稲わらの糖化反応における反応溶液中の全糖量、グルコース量、キシロース量、および糖化率の経時変化を表すグラフである。
【図4】稲わら糖化液中でのリゾプス エスピー MK96−1156(受託番号:FERM BP−6777)の培養における、全糖量、グルコース量、およびL−乳酸量の経時変化を表すグラフである。
【図5】コーンコブの糖化反応における反応溶液中の全糖量およびグルコース量の経時変化を表すグラフである。
【図6】コーンコブ糖化液中でのリゾプス エスピー MK96−1156(受託番号:FERM BP−6777)の培養における、全糖量、グルコース量、およびL−乳酸量の経時変化を表すグラフである。
【発明を実施するための形態】
【0015】
以下、本発明の好ましい形態を説明する。なお、本発明の技術的範囲は特許請求の範囲の記載に基づいて定められるべきであり、以下の形態のみに制限されない。
【0016】
<微生物>
本形態の微生物は、ペニシリウム属(Penicillium sp.)に属し、リグノセルロース系バイオマス分解能を有する微生物である。該微生物はリグノセルロース系バイオマスの分解活性に優れるため、リグノセルロース系バイオマスから高効率で糖化液を製造することができる。このようなペニシリウム属に属する微生物のうち、本発明者によって新たに見出されたペニシリウム S11D6608−M1(受託番号:FERM BP−11083)および/またはペニシリウム S11D6608−M2(受託番号:FERM BP−11084)であることが好ましい。なお、本明細書において、ペニシリウム S11D6608−M1を略して「菌株M1」と、ペニシリウム S11D6608−M2を略して「菌株M2」と、それぞれ称する場合がある。以下で、菌株M1および菌株M2について説明する。
【0017】
[1次スクリーニング]
中国東北部で生産されたトウモロコシのコーンコブを入手した。該コーンコブの表面の一部にはカビが観察された。該コーンコブを室温で約2ヵ月間風乾し、金切ばさみで1〜5cm程度に切断した。そして、これを家庭用ミキサーに5分間かけることにより粗粉砕した。該粗粉砕物をさらにボールミル(ANS−60S、日陶科学社製)を用いて微粉化した。得られたコーンコブ粉砕物の粒径分布を下記表1に示す。なお、本明細書において「粒径」とは、粒子の輪郭線上の任意の2点間の距離のうち、最大の距離を意味する。また、粒径の値は、試験用ふるいによって測定した値を採用する。下記表1の各粒径における粉砕物の割合の求め方は以下のとおりである。まず、3種類の試験用ふるい(呼び寸法0.5mm、1.0mm、1.7mm;SANPO社製)を呼び寸法が小さいものが下になるように順次重ね、最上部のふるいに上記粉砕物5gを入れた。これを間欠的に5分間ふるい、それぞれのふるいに残存した粉砕物の質量を測定し、それぞれの割合を求めた。
【0018】
【表1】
【0019】
該コーンコブ粉砕物のうち、粒径が1.0mm未満の粉砕物1gを試験管に取り、滅菌水道水10mlを加えて懸濁し、恒温振盪培養装置を用いて32℃、回転数220ppmで7日間振盪培養した。培養後の各試験管中の培養液を一部サンプリングし、培養液中に含まれる全糖量をフェノール硫酸法により分析して、全糖量が特に多かった培養液を選定した。全糖量が多かった培養液では、コーンコブに付着した微生物が、コーンコブに含まれるリグノセルロースを効率よく糖化したものと考えられた。なお、本明細書において、フェノール硫酸法による分析は、中村道徳編、貝沼圭二編、「生物化学実験法(25)澱粉・関連糖質酵素実験法」、学会出版センター版、1989年10月発行、206頁に記載の方法に準じて行った。
【0020】
[1次スクリーニングした菌の単胞子分離]
上記で選定した培養液を滅菌水で適当に希釈し、下記表2に示す組成のPDY寒天培地(オートクレーブにて121℃、20分間滅菌)に塗布し、32℃のインキュベーター中でプレート培養した。培養開始から14日後に培地上にコロニーの形成が認められた。このうち、明らかにカビ以外の微生物であると思われるコロニーを除外し、カビ状でかつ形状が大きなコロニーを選別した。
【0021】
【表2】
【0022】
[2次スクリーニング]
(前培養)
200ml三角フラスコに下記表3に示す組成のA培地20mlを入れ、オートクレーブで121℃、20分間滅菌した。該A培地に上記1次スクリーニングで選定した菌株を1白金耳接種した。これを、恒温振盪培養装置を用いて32℃、回転数220ppmで24時間振盪培養した。
【0023】
【表3】
【0024】
次に、200ml三角フラスコに下記表4に示す組成のB培地20mlを入れ、オートクレーブで121℃、20分間滅菌した。該B培地に、上記で培養した前培養液を5(v/v)%植菌した。これを、恒温振盪培養装置を用いて32℃、回転数220ppmで6日間振盪培養した。
【0025】
【表4】
【0026】
(本培養)
上記1次スクリーニングで調製した粒径が1.0mm未満のコーンコブ粉砕物0.2gを、上記培養液のろ液を適当に水道水で希釈した希釈培養液20mlに懸濁し、50℃で3日間糖化を行った。得られた糖化液中に含まれる全糖量をフェノール硫酸法により分析し、全糖量が多かった糖化液に対応した2菌株を、リグノセルロース系バイオマスの糖化力に優れている菌株として選定した。これらをそれぞれ菌株Iおよび菌株IIと称した。
【0027】
[分類学的性質]
上記でスクリーニングした菌株Iおよび菌株IIの分類学的性質を以下で述べる。
【0028】
(1)菌株I
(a)形態的性質
以下の(i)〜(iv)培地において、25℃で3週間培養後のコロニーの形態を観察した。なお、以下の表5〜8において、色調の括弧内の英数文字は「Kornerup and Wanscher(1978)」で用いられている色のコード ナンバーを表す。
【0029】
(i)ポテトデキストロース寒天培地(商品名:ダイゴ、日本製薬社製)
【0030】
【表5】
【0031】
(ii)2% 麦芽寒天培地(Malt Agar)
【0032】
【表6】
【0033】
(iii)バクトオートミール寒天培地(Bacto Oatmeal Agar、ベクトン ディッキンソン社製)
【0034】
【表7】
【0035】
(iv)LCA(三浦培地)
【0036】
【表8】
【0037】
その他の微視的観察結果を表9に示す。
【0038】
【表9】
【0039】
(b)同定
アポロンDB−FUに対するBLAST(Altschul et al.,1997)相同性検索の結果、菌株Iの28S rDNA−D1/D2塩基配列は、子嚢菌類の一種であるPenicillium aculeatumの塩基配列と99.5%の相同率を示した。GenBank/DDBJ/EMBLなどの国際塩基配列データベースに対する相同性検索の結果においては、菌株Iの28S rDNA−D1/D2塩基配列は子嚢菌類の一種であるPenicillium cf. verruculosum RS7PF(この菌種名の「cf.」は、P. verruculosumに類似する菌株であることを表す)、P. aculeatum、P. verruculosumの塩基配列と高い相同率を示した。アポロンDB−FUおよび国際塩基配列データベースに対する相同性検索で得られた上位の塩基配列をもとに作成した系統樹において、菌株IはPenicillim cf. verruculosum RS7PFと同一の系統枝を形成した。
【0040】
菌株IのITS−5.8S rDNA塩基配列は、アポロンDB−FUに対するBLAST相同性検索の結果、子嚢菌類の一種である、P. verruculosum の塩基配列と98.8%の相同率を示した。また、GenBank/DDBJ/EMBLなどの国際塩基配列データベースに対する相同性検索の結果においては、菌株IのITS−5.8S rDNA塩基配列は、P. pinophilumの塩基配列と100%の相同率を示した。国際塩基配列データベースおよびアポロンDB−FUに対する相同性検索で得られた上位の塩基配列をもとに作成した系統樹において、菌株IはP. pinophilumと同一の系統枝を形成した。
【0041】
したがって、28S rDNA−D1/D2塩基配列の解析結果から、菌株IはPenicillim cf. verruculosumに近縁であるものの、ITS−5.8S rDNA塩基配列の解析で本検体と同一系統枝を形成した28S rDNA−D1/D2塩基配列のデータが登録されてないことから、本検体の種レベルの推定は困難であると思われた。よって、菌株IはP. pinophilumに近縁なPenicillium属の一種であると考えられた。なお、P. verruculosumとP. pinophilumとは、分類学的に非常に近縁であることが知られている。
【0042】
コロニー性状および形態観察の結果、菌株Iは赤褐色〜茶褐色系のコロニーを形成し、培地中に顕著な赤色系の可溶性色素を産出した。また、有隔壁の栄養菌糸のみの形成が観察された。今回の観察結果では、胞子などの生殖器官の形成が観察されなかったことから、分類群の推定はできなかった。なお、ITS−5.8S rDNA塩基配列解析結果より菌株Iと近縁を推定されるP. pinophilumはまれに赤色系の色素を産出することが知られている。
【0043】
以上の28S rDNA−D1/D2およびITS−5.8S rDNA塩基配列解析、ならびにコロニー性状および形態観察の結果から総合的に鑑みて、菌株IはP. pinophilumに近縁なPenicillium属に帰属する新規な菌であると判断し、ペニシリウム S11D6608−M1(Penicillium sp. S11D6608−M1)と命名した。
【0044】
なお、本菌株は、2008年12月24日付けで独立行政法人産業技術総合研究所特許生物寄託センターに受託番号FERM BP−11083として、S11D6608−M1の名称で寄託されている。
【0045】
(2)菌株II
(a)形態的性質
以下の(i)〜(iv)培地において、25℃で3週間培養後のコロニーの形態を観察した。なお、以下の表10〜13において、色調の括弧内の英数文字は「Kornerup and Wanscher(1978)」で用いられている色のコード ナンバーを表す。
【0046】
(i)ポテトデキストロース寒天培地(商品名:ダイゴ、日本製薬社製)
【0047】
【表10】
【0048】
(ii)2% 麦芽寒天培地(Malt Agar)
【0049】
【表11】
【0050】
(iii)バクトオートミール寒天培地(Bacto Oatmeal Agar、ベクトン ディッキンソン社製)
【0051】
【表12】
【0052】
(iv)LCA(三浦培地)
【0053】
【表13】
【0054】
その他の微視的観察結果を表14に示す。
【0055】
【表14】
【0056】
(b)同定
アポロンDB−FUに対するBLAST(Altschul et al.,1997)相同性検索の結果、菌株IIの28S rDNA−D1/D2塩基配列は、子嚢菌類の一種であるPenicillium aculeatumの塩基配列と99.5%の相同率を示した。GenBank/DDBJ/EMBLなどの国際塩基配列データベースに対する相同性検索の結果においては、菌株IIの28S rDNA−D1/D2塩基配列は子嚢菌類の一種であるPenicillium cf. verruculosum RS7PF(この菌種名の「cf.」は、P. verruculosumに類似する菌株であることを表す)、P. aculeatum、P. verruculosumの塩基配列と高い相同率を示した。アポロンDB−FUおよび国際塩基配列データベースに対する相同性検索で得られた上位の塩基配列をもとに作成した系統樹において、菌株IIはPenicillim cf. verruculosum RS7PFと同一の系統枝を形成した。
【0057】
菌株IIのITS−5.8S rDNA塩基配列は、アポロンDB−FUに対するBLAST相同性検索の結果、子嚢菌類の一種である、P. verruculosum の塩基配列と98.8%の相同率を示した。また、GenBank/DDBJ/EMBLなどの国際塩基配列データベースに対する相同性検索の結果においては、菌株IIのITS−5.8S rDNA塩基配列は、P. pinophilumの塩基配列と100%の相同率を示した。国際塩基配列データベースおよびアポロンDB−FUに対する相同性検索で得られた上位の塩基配列をもとに作成した系統樹において、菌株IIはP. pinophilumと同一の系統枝を形成した。
【0058】
したがって、28S rDNA−D1/D2塩基配列の解析結果から、菌株IIはPenicillim cf. verruculosumに近縁であるものの、ITS−5.8S rDNA塩基配列の解析で本検体と同一系統枝を形成した28S rDNA−D1/D2塩基配列のデータが登録されてないことから、本検体の種レベルの推定は困難であると思われた。よって、菌株IIはP. pinophilumに近縁なPenicillium属の一種であると考えられた。なお、P. verruculosumとP. pinophilumとは、分類学的に非常に近縁であることが知られている。
【0059】
コロニー性状および形態観察の結果、菌株IIは白色〜赤黄色系のコロニーを形成し、有隔壁の栄養菌糸のみの形成が観察された。今回の観察結果では、胞子などの生殖器官の形成が観察されなかったことから、分類群の推定はできなかった。上述の菌株Iのコロニー性状とは顕著な違いが認められたが、菌糸の形状において差異は認められなかった。
【0060】
以上の28S rDNA−D1/D2およびITS−5.8S rDNA塩基配列解析、ならびにコロニー性状および形態観察の結果から総合的に鑑みて、菌株IはP. pinophilumに近縁なPenicillium属に帰属する新規な菌であると判断し、ペニシリウム S11D6608−M2(Penicillium sp. S11D6608−M2)と命名した。
【0061】
なお、本菌株は、2008年12月24日付けで独立行政法人産業技術総合研究所特許生物寄託センターに受託番号FERM BP−11084として、S11D6608−M2の名称で寄託されている。
【0062】
また、本発明の微生物として、より好ましくは、より高効率で糖化液を得られるよう、菌株M1または菌株M2に公知の変異処理を施すことによって得られた変異株であることが好ましい。変異処理の方法は、特に制限はないが、例えば、紫外線照射やニトロソグアニジンなどの変異誘発剤を使用することによって変異誘導処理する方法が挙げられる。そして、変異処理を施した菌株から、リグノセルロース系バイオマスの分解活性に優れる菌株を選定する。
【0063】
<酵素>
本発明の他の形態としては、上述の微生物に由来する酵素が挙げられる。該酵素は、リグノセルロース系バイオマスに対して優れた加水分解活性能を有するので、該酵素をリグノセルロース系バイオマスに作用させることにより高効率で糖化液を製造することができる。
【0064】
本形態の酵素は、上述の微生物の培養物から、従来公知の酵素採取手段および精製手段を用いて調製することができる。酵素採取法としては、例えば、微生物を含む培養物を遠心分離またはろ過することによって、菌体とろ液とを分離する。そして、該菌体およびろ液から、塩析法、等電点沈殿法、溶媒沈殿法(メタノール、エタノール、アセトン、およびイソプロパノールなど)によりタンパク質である酵素を沈殿させたり、限外ろ過により濃縮したりする方法が挙げられる。
【0065】
リグノセルロース系バイオマスの加水分解には、上記の方法によって得られる粗酵素を用いてもよいが、必要によりさらに精製した酵素を用いることもできる。酵素の精製法としては、例えば、吸着クロマトグラフィー、イオン交換クロマトグラフィー、およびゲルクロマトグラフィーなどのクロマトグラフィーを単独で、または2種以上を組み合わせた精製法が挙げられる。
【0066】
<糖化液の製造方法>
本発明の他の形態にかかる糖化液の製造方法は、上述の本発明の微生物をリグノセルロース系バイオマスを含む培地中で培養する工程、または上述の本発明の酵素をリグノセルロース系バイオマスに作用させることにより加水分解する工程を含む。
【0067】
本形態の糖化液の製造方法で用いるリグノセルロース系バイオマスは、セルロースおよびヘミセルロース、ならびにリグニンを含む有機資源であれば植物性でも動物性でも特に制限はなく、農作物、草木類、および海藻類、ならびにこれらの処理物または廃棄物などが含まれる。より具体的には、稲わら、籾殻、麦わら、ふすま、バガス、ヤシ殻、コーンコブ、コットン、ラミー、ケナフ、ビート、大豆粕、コーヒー粕、雑草、木材、竹、バクテリアセルロース、パルプ、紙、セルロース粉末、結晶セルロース、レーヨン、アルカリセルロース、リン酸膨潤セルロース、およびカルボキシメチルセルロースなどが挙げられる。なお、本発明の微生物または酵素は、上記リグノセルロース系バイオマスの糖化能に優れるだけでなく、でんぷん系バイオマスまたは糖質系バイオマスに対しても優れた糖化能を示す。
【0068】
上記リグノセルロース系バイオマスは、微生物の培養または酵素による加水分解に供する前に、糖化効率を高めるための前処理を行うことが好ましい。前処理としては、例えば、脱水処理または乾燥処理、粉砕処理、および蒸煮処理などの従来公知の技術を適宜採用することができる。特に、稲わら、麦わら、とうもろこしの茎葉、コーンコブなどの農作物の非可食部などのリグノセルロース系バイオマスを用いる場合は、まず、風乾によりバイオマス中に含まれる水分を除く。その後、切断装置や粉砕装置を用いて、好ましくは1.0mm以下であり、より好ましくは0.01〜1.0mmであり、さらに好ましくは0.1〜1.0mmの粉末状とすることが好ましい。
【0069】
上記リグノセルロース系バイオマスは、微生物の培養または酵素による加水分解に供するために、水性媒体に懸濁して用いる。水性媒体は微生物の増殖または酵素反応の進行が可能な範囲においては特に制限はないが、水、緩衝液、酸性水溶液、またはアルカリ性水溶液を用いることが好ましい。懸濁液中に含まれるリグノセルロース系バイオマスの粉砕物の濃度は、水性媒体に対して、通常、50〜300g/Lであり、好ましくは100〜200g/Lである。
【0070】
上述の本発明の微生物を用いて糖化液を製造する場合、本発明の微生物は、上記懸濁液のみを培地とした場合であっても、十分な増殖能力を有するが、必要によっては炭素源、窒素源、または無機物などを懸濁液に補充したものを培地として用いることも可能である。これらの栄養源は、微生物の数を増やすための前培養の培地に添加してもよいし、本培養の培地に添加しても構わない。
【0071】
炭素源としては、例えば、グルコース、ガラクトース、フルクトース、およびキシロースなどの単糖、マルトース、ラクトース、およびスクロースなどの二糖、グリセリンおよびキシリトールなどの糖アルコールといった糖類が挙げられ、これらを単独であるいは2種以上を組み合わせて使用できる。特に、前培養の培地の培地にグルコースを添加することにより、本培養のときの微生物のリグノセルロース系バイオマスの分解活性をより向上させることができる。
【0072】
窒素源としては、例えば、アンモニア、塩化アンモニウム、硫酸アンモニウム、硝酸アンモニウムなどのアンモニウム塩、尿素、L−グルタミン酸などのアミノ酸類、あるいは尿酸などの無機あるいは有機の窒素化合物が使用できる。さらに、窒素源としては、ペプトン、ポリペプトン、肉エキス、酵母エキス、大豆加水分解物、大豆粉末、ミルクカゼイン、カザミノ酸、コーンスティープリカー(CSL)などの窒素含有天然物を使用してもよい。これらのうち、塩化アンモニウム、硫酸アンモニウム、尿素、L−グルタミン酸などのアミノ酸類、尿酸などの無機あるいは有機窒素化合物、ペプトン、肉エキス、酵母エキスなの窒素含有天然物が好ましい。これらの窒素源は、単独であるいは2種以上を組み合わせて使用できる。これらの窒素源のうち、大豆粉末またはコーンスティープリカーを使用することが好ましい。
【0073】
無機物としては、例えば、塩化ナトリウム、塩化カリウム、塩化カルシウム、リン酸カリウム、リン酸ナトリウム、硫酸マグネシウム、硫酸アンモニウムなどの、マグネシウム、マンガン、カルシウム、ナトリウム、カリウム、銅、鉄および亜鉛などのリン酸塩、塩酸塩、硫酸塩および酢酸塩などが用いられる。そのほか、チアミン、ビオチンなどのビタミン類、さらに必要に応じて、アデニン、ウラシルなどの核酸関連物質が使用されてもよい。これらの無機物は、単独であるいは2種以上を組み合わせて使用できる。
【0074】
微生物の培養方法は、本発明の微生物の増殖が実質的に阻害されず、リグノセルロース系バイオマスから単糖類を生産可能な範囲であれば、特に制限なく当業者によって適宜調整されうるが、培養温度は、通常20〜50℃であり、好ましくは28〜37℃である。また、培地のpHは、通常3〜9であり、好ましくは4〜7である。培養前または培養中にpHが4未満となる場合は、アンモニアなどを用いてpH制御を行うことが好ましい。本発明の微生物を培養する場合は、一定量の酸素を供給することが必要である。供給方法は、特に制限はないが、フラスコ培養の場合は回転振盪による攪拌、バイオリアクターを用いる培養の場合は通気攪拌をすることによって培地に酸素を供給する。
【0075】
一方、本発明の酵素を用いた加水分解反応は、リグノセルロース系バイオマスを含む懸濁液中に酵素を添加し、懸濁液を攪拌下、インキュベートすることによって行われうる。懸濁液に添加する酵素量は、懸濁するリグノセルロース系バイオマスの粉砕物の質量に対して、好ましくは0.1〜3質量%、より好ましくは1〜2質量%である。かような量の酵素を用いることによって、通常12〜144時間、好ましくは24〜72時間で加水分解反応が完了しうる。また、加水分解条件は、用いる酵素の至適条件を用いることが好ましい。pHは通常3〜6であり、好ましくは3.5〜5である。また、反応温度は通常30〜60℃であり、好ましくは40〜60℃である。
【0076】
微生物の培養、または酵素による加水分解により得られた糖化液は、糖化液中の不溶物を適宜ろ過してもよいし、そのままの状態で、微生物代謝産物(エタノール、乳酸などの有用化合物)を産生する他の微生物の培養に供しても構わない。撹拌効率および収率向上の観点の観点からは、糖化液中の不溶物を予めろ過することが好ましい。
【0077】
他の微生物としては、特に制限はないが、例えば、乳酸を産生するRizopus属、イタコン酸を生産するAspergillus terreus、エタノールを産生するPachysolen tannophilus、フマル酸を産生するRhizopus oryzae、リンゴ酸を産生するAspergillus flavus、ピルビン酸を産生するTorulopsis glabrata、キシリトールを産生するCandida tropicalisが挙げられる。本発明によって得られる糖化液を含む倍中でこれらの微生物を培養することによって、燃料や化成品を工業的規模で製造するが可能である。
【実施例】
【0078】
本発明の作用効果を、以下の実施例および比較例を用いて説明する。ただし、本発明の技術的範囲が以下の実施例のみに制限されるわけではない。
【0079】
<菌株M1および菌株M2の稲わら分解活性の評価>
[実施例1]
1.生産菌の培養
(前培養)
200ml三角フラスコに下記表15に示す組成のC培地20mlを入れ、オートクレーブで121℃、20分間滅菌した。該C培地に菌株M1および菌株M2をそれぞれ1白金耳ずつ接種し、これを恒温振盪培養装置を用いて32℃、回転数220ppmで24時間振盪培養した(混合培養)。
【0080】
【表15】
【0081】
(本培養)
次に、下記表16の組成を有する各培地を調製した。下記表16において、対象なる成分が含まれている場合を「○」であらわし、含まれていない場合を「×」で表す。
【0082】
【表16】
【0083】
なお、上記表16における稲わらは、粒径が1.0mm未満の粉砕物を用いた。稲わらの粉砕方法は以下のとおりである。稲わらを室温で約2ヵ月間風乾し、金切ばさみで1〜5cm程度に切断した。そして、これを家庭用ミキサーに5分間かけることにより粗粉砕した。該粗粉砕物をさらにボールミル(ANS−60S、日陶科学社製)を用いて微粉化し、稲わら粉砕物を得た。
【0084】
次に、上記の培地1〜6を200ml三角フラスコにそれぞれ20mlずつ入れ、オートクレーブで121℃、20分間滅菌した。該培地1〜6に、上記前培養液を7(v/v)%植菌した。これを恒温振盪培養装置を用いて32℃、回転数220ppmで6日間振盪培養した。
【0085】
2.稲わら分解活性の評価(糖化)
上述の方法で得た粒径が1.0mm未満の稲わら粉砕物2gを水道水9mlに懸濁した。該懸濁液に、上記培地1〜6で培養した培養液、またはそのろ液を希釈した酵素液1mlを加え、50℃で24時間糖化した。この後、それぞれの糖化液をろ過し、得られたろ液中に含まれる全糖量をフェノール硫酸法により測定した。そして、稲わら分解活性を下記式(1)より算出した。なお、酵素活性は1[U]は、(1/60)[μkat]に相当する。
【0086】
【数1】
【0087】
[比較例1]
菌株M1および菌株M2に変えて、トリコデルマ・リーゼイ(Tricoderma reesei)NBRC31329を用いたことを除いては、実施例1と同様の方法で稲わら分解活性を求めた。
【0088】
結果を下記表17に示す。
【0089】
【表17】
【0090】
表17の結果より、本発明の菌株M1およびM2は、従来優れたセルラーゼ産生菌として知られるトリコデルマ・リーゼイ NBRC31329よりも顕著な稲わら分解活性能を有することが示された。
【0091】
また、前培養の培地に、窒素源として大豆粉を、炭素源としてグルコースを添加することにより、本培養のときの微生物の増殖を促進し、微生物のリグノセルロース系バイオマスの分解活性をより向上させることができることが示された。
【0092】
<酵素活性の評価>
[実施例2]
1.酵素の調製
(前培養)
200ml三角フラスコに下記表18に示す組成のD培地20mlを入れ、オートクレーブで121℃、20分間滅菌した。該D培地に菌株M1および菌株M2をそれぞれ1白金耳ずつ接種し、これを恒温振盪培養装置を用いて32℃、回転数220ppmで24時間振盪培養した(混合培養)。
【0093】
【表18】
【0094】
(本培養)
次に、3Lジャーファーメンター(3MDL型、丸菱バイオエンジ社製)に下記表19に示す組成のE培地1.5Lを入れ、オートクレーブで121℃、20分間滅菌した。該B培地に、上記で培養したD培地を7(v/v)%植菌した。これを、恒温振盪培養装置を用いて32℃、回転数600ppm、通気量2vvmで6日間培養した。
【0095】
【表19】
【0096】
上記の6日間の培養中、培養開始から0(培養開始直後)、24、48、72、および84時間後の培養液を1ml取り出し、上記実施例1の3.稲わら分解活性の評価と同様の方法で、各時間の培養液の稲わら分解活性を評価した。結果を下記表20に示す。
【0097】
【表20】
【0098】
6日間培養後の培養液をろ過し、ケーキ(残渣)を水洗した。ろ液とケーキの水洗液とをあわせて濃縮乾固した。次に、この濃縮乾固物を5℃の冷水200mlに溶解し、不溶分をろ別した後、これにアセトンを1000ml加えて、アセトン沈殿物を得た。そして、これを凍結乾燥して酵素(「酵素M1M2」と称する)を得た。なお、上記の各工程で得られた生成物の収量、酵素活性または比活性、総活性、および収率を下記表21に示す。なお、酵素活性は上記実施例1の分解活性の評価と同様の方法を用いて測定した。
【0099】
【表21】
【0100】
2.酵素化学的性質の評価
(安定性に対する温度の影響)
0.05M酢酸バッファー(pH5.5)9mlに、上記で得た酵素M1M21gを溶解したものを酵素溶液とした。ふた付試験管に該酵素溶液1mlと、上記酢酸バッファー9mlとを入れ、それぞれ所定の温度で10時間振盪した後、試験管を5℃に冷却した。ついで、この試験管に上記実施例1で調製した稲わら粉末(粒径1.0mm未満のもの)を1g加え、50℃で24時間糖化反応を行った。この後、それぞれの糖化液をろ過し、得られたろ液中に含まれる全糖量をフェノール硫酸法により測定した。そして、分解活性(糖化活性)を上記式(1)より算出した。得られた値のうち最も高い安定性を与えた10℃における分解活性に対する、それぞれの温度における活性を百分率で表した。結果を図1に示す。
【0101】
(糖化活性に対する温度の影響)
ふた付試験管に、上記酵素溶液1mlと、上記酢酸バッファー9mlとを入れ、上記実施例1で調製した稲わら粉末(粒径1.0mm未満のもの)を1g加え、所定の温度で24時間糖化反応を行った。この後、それぞれの糖化液をろ過し、得られたろ液中に含まれる全糖量をフェノール硫酸法により測定した。そして、分解活性(糖化活性)を上記式(1)より算出した。得られた値のうち最も高い安定性を与えた55℃における分解活性に対する、それぞれの温度における活性を百分率で表した。結果を図1に示す。
【0102】
(安定性に対するpHの影響)
ふた付試験管に上記酵素溶液1mlと、それぞれ所定のpHに調整した0.05M酢酸バッファー9mlを入れ、50℃で10時間振盪した後、試験管を5℃に冷却した。ついで、この試験管に上記実施例1で調製した稲わら粉末(粒径1.0mm未満のもの)を1g加え、50℃で24時間糖化反応を行った。この後、それぞれの糖化液をろ過し、得られたろ液中に含まれる全糖量をフェノール硫酸法により測定した。そして、分解活性(糖化活性)を上記式(1)より算出した。得られた値のうち最も高い安定性を与えたpH3.7における分解活性に対する、それぞれのpHにおける活性を百分率で表した。結果を図2に示す。
【0103】
(糖化活性に対する温度の影響)
ふた付試験管に、上記酵素溶液1mlと、それぞれ所定のpHに調整した0.05M酢酸バッファー9mlを入れ、上記実施例1で調製した稲わら粉末(粒径1.0mm未満のもの)を1g加え、50℃で24時間糖化反応を行った。この後、それぞれの糖化液をろ過し、得られたろ液中に含まれる全糖量をフェノール硫酸法により測定した。そして、分解活性(糖化活性)を上記式(1)より算出した。得られた値のうち最も高い安定性を与えたpH4.5における分解活性に対する、それぞれのpHにおける活性を百分率で表した。結果を図2に示す。
【0104】
[実施例3]
D培地に接種した微生物を菌株M1のみとしたことを除いては、実施例2と同様の方法で酵素を調製した。得られた酵素を「酵素M1」と称する。そして、実施例2と同様の方法で酵素M1の比活性を測定した。
【0105】
[実施例4]
D培地に接種した微生物を菌株M2のみとしたことを除いては、実施例2と同様の方法で酵素を調製した。得られた酵素を「酵素M2」と称する。そして、実施例2と同様の方法で酵素M2の比活性を測定した。
【0106】
[比較例2]
アクレモニウムセルラーゼ(明治製菓社製)を用いて、実施例2と同様の方法で比活性を測定した。
【0107】
[比較例3]
メイセラーゼ(明治製菓社製)を用いて、実施例2と同様の方法で比活性を測定した。
【0108】
[比較例4]
スミチーム(新日本化学工業社製)を用いて、実施例2と同様の方法で比活性を測定した。
【0109】
酵素の比活性の結果を下記表22に示す。
【0110】
【表22】
【0111】
表22の結果より、本発明の酵素M1M2、酵素M1、および酵素M2は、従来のセルラーゼよりも優れた稲わら分解活性を有することが示された。
【0112】
<酵素M1M2を用いた稲わら糖化液の製造、および該糖化液を用いたL−乳酸の製造>
[実施例5]
3Lジャーファーメンター(MDL−3型、丸菱バイオエンジ社製)に水道水1.0Lを入れ、上記実施例1で調製した稲わら粉末(粒径1.0mm未満のもの)200gを懸濁し、さらに水道水を加えて液量を1.5Lとした。該懸濁液のpHを、25%塩酸を用いて4.5に調整した。そして、実施例2で得た酵素M1M2を20U/mlとなるように添加し、50℃で攪拌(200rpm)下、72時間糖化反応を行った。反応液中の全糖量、グルコース量、キシロース量、および糖化率の経時変化を図3に示す。
【0113】
得られた糖化液を3Lジャーファーメンターに仕込み、これにリン酸一カリウムを0.3(g/L)、硫酸マグネシウムを0.25(g/L)となるように添加した。該糖化液をオートクレーブを用いて121℃で10分間滅菌した。次に、予め300ml三角フラスコで前培養したリゾプス エスピー(Rizopus sp.)MK96−1156(受託番号:FERM BP−6777)の培養液100mlを添加し、回転数300rpm、通器量0.5vvm、培養温度30℃で72時間培養した。培養中、25%アンモニア水を用いて培養液のpHを6.0に制御した。全糖量、グルコース量、およびL−乳酸量の経時変化を図4に示す。リゾプス エスピー MK96−1156(受託番号:FERM BP−6777)の培養により、全糖90g/Lから42g/LのL−乳酸が得られた。また、稲わら粉末からのL−乳酸の収率は21(w/w)%であった。
【0114】
<酵素M1M2を用いたコーンコブ糖化液の製造、および該糖化液を用いたL−乳酸の製造>
[実施例6]
稲わら粉末に変えて、上述の[1次スクリーニング]で調製したコーンコブ粉末(粒径1.0mm未満のもの)を用い、糖化時間を48時間、Rizopus sp. MK96−1156(受託番号:FERM BP−6777)の培養時間を96時間としたことを除いては、実施例5と同様の方法で、L−乳酸を生産した。糖化反応液中の全糖量、グルコース量の経時変化を図5に示す。また、Rizopus sp. MK96−1156(受託番号:FERM BP−6777)の培養液中の全糖量、グルコース量、およびL−乳酸量の経時変化を図6に示す。Rizopus sp. MK96−1156(受託番号:FERM BP−6777)の培養により、全糖102g/Lから46g/LのL−乳酸が得られた。また、コーンコブ粉末からのL−乳酸の収率は21(w/w)%であった。
【0115】
<稲わら糖化液を用いたエタノールの製造>
硫酸玉つきガラス管を備えた100ml三角フラスコに、実施例5と同様の方法で製造した稲わら糖化液および下記表23に示す添加物を加えた培養液20mlを準備した。これにパン酵母(オリエンタルドライイースト、オリエンタル酵母工業社製)0.5gを加え、30℃で静置培養を行った。経時的にフラスコ全体の重量を測定し、その重量の減少量を下記式(2)にあてはめ、各培養時間後のエタノール量を算出した。下記式(2)において、フラスコの重量の減少量はエタノール発酵による炭酸ガス発生量(g)を意味する。アルコール発酵により、グルコース1モルからエタノール2モルと炭酸ガス(CO2
)2モルが生産されることから、炭酸ガス発生量(g)に1.047(46.068/44.0095)を乗じて得られる値が、エタノール量(g)となる。そして、培養開始前の糖化液中のグルコース量に対するエタノール量を百分率で表し、各培養時間におけるエタノール収率(%)とした。結果を下記表23に示す。
【0116】
【数2】
【0117】
【表23】
【0118】
表23の結果より、本発明の酵素M1M2を用いて製造された糖化液は、酵母エキス、乾燥酵母、尿素、硫安などの窒素源を補填することのなく、46%の高収率でエタノールを製造できることが示された。
【特許請求の範囲】
【請求項1】
リグノセルロース系バイオマス分解能を有するペニシリウム属(Penicillium sp.)に属する微生物。
【請求項2】
S11D6608−M1(受託番号:FERM BP−11083)および/またはS11D6608−M2(受託番号:FERM BP−11084)である、請求項1に記載の微生物。
【請求項3】
請求項1または2に記載の微生物由来の酵素。
【請求項4】
リグノセルロース系バイオマスを含む培地中で請求項1もしくは2のいずれか1項に記載の微生物を培養する培養工程を含む、糖化液の製造方法。
【請求項5】
前記培養工程の前に、リグノセルロース系バイオマスおよびグルコースを含む培地中で、前記微生物を前培養する工程をさらに含む、請求項4に記載の糖化液の製造方法。
【請求項6】
リグノセルロース系バイオマスを請求項3に記載の酵素により加水分解する、糖化液の製造方法。
【請求項7】
前記加水分解において、反応温度が30〜60℃であり、反応溶液のpHが3〜6である、請求項6に記載の糖化液の製造方法。
【請求項8】
前記リグノセルロース系バイオマスが、稲わら、籾殻、麦わら、ふすま、バガス、ヤシ殻、コーンコブ、コットン、ラミー、ケナフ、ビート、大豆粕、コーヒー粕、雑草、木材、竹、バクテリアセルロース、パルプ、紙、セルロース粉末、結晶セルロース、レーヨン、アルカリセルロース、リン酸膨潤セルロース、およびカルボキシメチルセルロースからなる群から選択される少なくとも1種を含む、請求項4〜7のいずれか1項に記載の糖化液の製造方法。
【請求項1】
リグノセルロース系バイオマス分解能を有するペニシリウム属(Penicillium sp.)に属する微生物。
【請求項2】
S11D6608−M1(受託番号:FERM BP−11083)および/またはS11D6608−M2(受託番号:FERM BP−11084)である、請求項1に記載の微生物。
【請求項3】
請求項1または2に記載の微生物由来の酵素。
【請求項4】
リグノセルロース系バイオマスを含む培地中で請求項1もしくは2のいずれか1項に記載の微生物を培養する培養工程を含む、糖化液の製造方法。
【請求項5】
前記培養工程の前に、リグノセルロース系バイオマスおよびグルコースを含む培地中で、前記微生物を前培養する工程をさらに含む、請求項4に記載の糖化液の製造方法。
【請求項6】
リグノセルロース系バイオマスを請求項3に記載の酵素により加水分解する、糖化液の製造方法。
【請求項7】
前記加水分解において、反応温度が30〜60℃であり、反応溶液のpHが3〜6である、請求項6に記載の糖化液の製造方法。
【請求項8】
前記リグノセルロース系バイオマスが、稲わら、籾殻、麦わら、ふすま、バガス、ヤシ殻、コーンコブ、コットン、ラミー、ケナフ、ビート、大豆粕、コーヒー粕、雑草、木材、竹、バクテリアセルロース、パルプ、紙、セルロース粉末、結晶セルロース、レーヨン、アルカリセルロース、リン酸膨潤セルロース、およびカルボキシメチルセルロースからなる群から選択される少なくとも1種を含む、請求項4〜7のいずれか1項に記載の糖化液の製造方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図2】
【図3】
【図4】
【図5】
【図6】
【公開番号】特開2011−50359(P2011−50359A)
【公開日】平成23年3月17日(2011.3.17)
【国際特許分類】
【出願番号】特願2009−204940(P2009−204940)
【出願日】平成21年9月4日(2009.9.4)
【出願人】(390022301)株式会社武蔵野化学研究所 (63)
【Fターム(参考)】
【公開日】平成23年3月17日(2011.3.17)
【国際特許分類】
【出願日】平成21年9月4日(2009.9.4)
【出願人】(390022301)株式会社武蔵野化学研究所 (63)
【Fターム(参考)】
[ Back to top ]