
新規生物学的実体およびその使用
本発明は、特異性決定領域と組み合わせたタンパク質足場から生成された人工酵素、その製造、および該人工酵素の、研究、栄養学的ケア、パーソナルケア、および工業目的のための使用を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、疾患に関連する標的基質内の特異的なペプチド結合を加水分解することができる規定された特異性を有するプロテアーゼを含む医薬を適用することによって、疾患を処置するための方法を提供する。そのような特異性を有するプロテアーゼは、さらに関連する治療または診断の目的で使用され得る。
【背景技術】
【0002】
学術的および産業上の研究は、治療剤、研究用の薬剤、診断剤、栄養剤、個人医療用の薬剤、または産業上の薬剤として使用される機能的なタンパク質を継続的に探求する。今日、このような機能的タンパク質は、主に2つのカテゴリー、すなわち、天然タンパク質および人工(engineered)タンパク質に分類され得る。一方で、天然タンパク質は、自然界から、例えば、天然の単離物をスクリーニングすることによって、または多様な種からのゲノムを配列決定することによって発見される。他方、人工タンパク質は、典型的には公知のタンパク質に基づいており、そして改変された機能を獲得するために変異されている。本発明は、出発成分と比較して新規機能を有する人工タンパク質を開示する。そのようなタンパク質は、NBE(新規生物学的実体(New Biologic Entities))と呼ばれる。本願において開示されるNBEは、新たな基質特異性を有する人工酵素またはそのような人工酵素と他の機能性成分との融合タンパク質である。
【0003】
特異性は、酵素機能の必須の要素である。細胞は、多数の異なる高度に反応性の触媒からなる。それでもなお、細胞は、調和された代謝および高度に秩序立った3次元構造を維持し得る。これは部分的に、酵素の特異性、すなわち、それらに対応する基質の選択的な変換に起因する。特異性は定性的かつ定量的な特性である。特定の酵素の特異性は、標的分子の1つの特定の型から特定の化学構造を有する全ての分子型に至るまで広範に変化し得る。本来、生物の酵素の特異性は、その生物の特定の必要性に対して進化してきている。治療、研究、診断、栄養学、または産業上の適用に高価値を有する任意の特異性は、可能な特異性のスペースが大きいために任意の生物の酵素のレパートリーで見出される可能性は低い。このような特異性を得る唯一の現実的な方法は、新たにそれらを作製することである。
【0004】
酵素を結合剤と比較する場合、特異性の枠組みは、大きな分子内において小さな異なる構造として個々のエピトープを認識する抗体によって与えられる。天然に存在する非常に広範な抗体の特異性は、天然の選択と組み合わされた免疫系によって生成される多様性に起因する。いくつかの機構が、抗体特異性の広範なレパートリーに貢献し、免疫応答生成および抗体成熟の異なる段階で生じる(Janeway、C et al.(1999)Immunobiology、Elsevier Science Ltd.、Garland Publishing、New York)。具体的には、抗体は、非常に特異的に抗原と相互作用し、非常に類似したエピトープとの間でさえも区別を可能にする相補性決定領域(CDR)を含む。抗体の軽鎖および重鎖は、各々3つの結合ドメインに対するCDRに寄与する。自然界は、CDRを生成するのに、種々の遺伝子セグメントの組換えを、さらに変異誘発と組み合わせて使用する。結果として、6つのCDRループの配列は、組成および長さにおいて高度に多様であり、これにより抗体の結合特異性の多様性の基礎を形成する。多様な触媒特異性の生成のための同様の原則は、自然界からは知られていない。
【0005】
触媒作用、すなわち、特定の化学反応の速度の増大は、結合すること以外に、最も重要なタンパク質機能である。触媒タンパク質、すなわち、酵素は、それらが触媒する化学反応に従って分類される。
【0006】
トランスフェラーゼは、基、例えば、メチル基またはグリコシル基を、1つの化合物(一般的にはドナーと見なされる)から別の化合物(一般的にはアクセプターと見なされる)へ移動させる酵素である。例えば、グリコシルトランスフェラーゼ(EC 2.4)は、ドナー分子からアクセプター分子へグリコシル残基を移動させる。グリコシルトランスフェラーゼのなかにはまた、加水分解を触媒するものもあり、これらはグリコシル基をドナーから水へ移動させると見なされ得る。サブクラスは、さらにヘキソシルトランスフェラーゼ(EC 2.4.1)、ペントシルトランスフェラーゼ(EC 2.4.2)、および他のグリコシル基(EC 2.4.99、国際生化学および分子生物学連合命名法委員会(Nomenclature Committee of the International Union of Biochemistry and Molecular Biology (NC−IUBMB)))へとさらに分割される。
【0007】
酸化還元酵素は、酸化還元反応を触媒する。酸化される基質は、水素または電子ドナーと見なされる。酸化還元酵素は、脱水素酵素、オキシダーゼ、モノオキシゲナーゼ、およびジオキシゲナーゼとして分類される。脱水素酵素は、水素を水素ドナーから水素アクセプター分子へ移動させる。オキシダーゼは、水素アクセプターとしての分子酸素と反応し、酸化生成物ならびに過酸化水素または水のいずれかを生成する。モノオキシゲナーゼは、分子酸素から酸素原子1個を基質へ移動させ、そして1つは水へと還元される。対照的に、ジオキシゲナーゼは、分子酸素から両方の酸素原子を基質へと挿入させることを触媒する。
【0008】
リアーゼは、排除反応を触媒し、それにより二重結合を生成するか、または逆方向において二重結合での付加を触媒する。イソメラーゼは、分子内再配置を触媒する。リガーゼは、ATP消費の代償として化学結合の形成を触媒する。
【0009】
最終的に、加水分解酵素は、C−OまたはC−Nのような化学結合の加水分解を触媒する酵素である。これらの酵素のE.Cの分類は、一般的に、それらを、加水分解される結合の性質によって、および基質の性質によって分類する。リパーゼおよびプロテアーゼのような加水分解酵素は、生来、および生体触媒の技術的な応用において重要な役割を果たす。プロテアーゼは、オリゴペプチドまたはポリペプチドの状況において、ペプチド結合を加水分解する。触媒機構に依存して、プロテアーゼは、アスパラギン酸、セリン、システイン、メタロ、およびスレオニンプロテアーゼに分類される(Handbook of proteolytic enzymes. (1998) Eds: Barret、A; Rawling、N. ; Woessner、J. ; Academic Press、London)この分類は、触媒作用を担うアミノ酸側鎖に基づいており、これは典型的には、互いに非常に類似して配向した活性部位に示されている。基質の切れやすい結合は、基質のアミノ酸側鎖とプロテアーゼの相補的な領域との間の特異的な相互作用に起因して触媒性残基と一致するようにされる(Perona、J. & Craik、C (1995) Protein Science、4、337−360)。切れやすい結合のN末端側およびC末端側の残基は、通常、P1、P2、P3など、およびP1’、P2’、P3’と呼ばれ、基質に相補的な結合ポケットは、S1、S2、S3およびS1’、S2’、S3’と、それぞれ呼ばれる(Schlechter & Berger、Biochem. Biophys. Res. Commun. 27 (1967) 157−162に基づく命名法)。プロテアーゼの選択性は、実質的に非選択的(例えば、サブチリシンのP位置での厳格な優先傾向;例えば、トリプシンはアルギニンまたはリジン残基のC末端側を選択的に切断する)から、非常に特異的なプロテアーゼまで(例えば、ヒト組織型プラスミノーゲン活性化因子(t−PA)は、配列CPGRVVGにおけるアルギニンのC末端側を切断する(Ding、L etal. (1995) Proc. Natl. Acad. Sci. USA 92、7627−7631; Coombs、Get al. (1996)J.Biol. Chem. 71、4461−4467)非常に様々である。
【0010】
プロテアーゼの特異性、すなわち、特定の基質を優先的に認識し加水分解する能力は、定性的におよび定量的に表現することができる。定性的な特異性は、ペプチド基質の特定の位置でプロテアーゼにより受け入れられるアミノ酸残基の種類のことをいう。例えば、トリプシンおよびt−PAは、定性的な特異性に関して関連している。なぜなら、これらのいずれも、P位置にアルギニンまたは類似の残基を必要とするからである。他方、定量的な特異性は、プロテアーゼにより基質として受け入れられるペプチド基質の相対数、またはより正確には、プロテアーゼによって受け入れられる異なるペプチドに対するそのプロテアーゼの相対的kcat/kMの比のことをいう。全ての可能なペプチドの中のわずかな少量の部分のみを受け入れるプロテアーゼは、高い特異性を有する。ここで、極端な場合として、任意のペプチド基質を切断するプロテアーゼの特異性は、理論的にゼロである。
【0011】
プロテアーゼの一次構造、二次構造、ならびに三次構造の比較により(Fersht、A. 、Enzyme Structure and Mechanism、W. H. Freeman and Company、New York、1995)、高度の保存を示すクラスを同定することが可能である(Rawlings、N. D. & Barrett、A.J. (1997) In : Proteolysis in Cell Functions Eds. Hopsu−Havu、V. K.;Jarvinen、M. ;Kirschke、H、pp. 13−21、OS Press、Amsterdam)。プロテアーゼの分類として広く受け入れられているスキームは、Rawlings & Barrettによって提唱されている(Handbook of proteolytic enzymes. (1998) Eds: Barret、A; Rawling、N.; Woessner、J.; Academic Press、London)。例えば、セリンプロテアーゼファミリーは、キモトリプシン(クラスS1)、サブチリシン(クラスS8)、およびカルボキシペプチダーゼ(クラスSC)の集団を有する構造クラスにさらに分割され得、これらの各々は、非特異的プロテアーゼおよび特異的プロテアーゼを含む(Rawlings、N. D. & Barrett、A. J. (1994) MethodsEnzymol. 244、19−61)。これは、他のプロテアーゼファミリーにも同様に適用される。基質中の切断される結合の相対的な位置によって、さらなる区別がなされ得る。カルボキシペプチダーゼおよびアミノペプチダーゼは、C末端およびN末端から、それぞれアミノ酸を切断する。これに対して、エンドペプチダーゼは、オリゴペプチドに沿ってどこでも切断する。
【0012】
基本的に無限の種類の特異性を有する酵素が入手可能であるならば、多くの適用が考えられる。しかし、このような高い、低い、または任意の規定された特異性を有する酵素の使用は、現在のところ、天然源から単離され得るものに限定されている。このような酵素の適用分野は、治療、研究、診断、栄養学から、パーソナルケアおよび工業的用途まで様々である。
【0013】
洗剤中の酵素添加物は、工業的な酵素市場全体のほぼ3分の1を構成するまでになっている。洗剤酵素には、有機物によるシミを除去するためのプロテアーゼ、油汚れを除去するためのリパーゼ、でんぷんを含む食品の残渣を除去するためのアミラーゼ、および繊維のなめらかな表面を回復させるためのセルラーゼが含まれる。最も良く知られている洗剤酵素は、おそらく、種々のバチルス(Bacillus)種から単離される非特異的プロテアーゼであるサブチリシンである。
【0014】
アミラーゼのような酵素は、食品加工において使用される酵素の大部分を占めている。でんぷん酵素は、布地デザイン、アルコール発酵、紙およびパルプの加工、ならびに洗濯用洗剤添加物のために重要な製品を含むが、最大の適用は、高フルクトースコーンシロップの製造である。工業用酵素を用いたでんぷんからのコーンシロップの製造は、酸加水分解の代わりとして成功していた。
【0015】
でんぷん加工とは別に、酵素は、食品における増大する範囲の適用に使用されている。食品中の酵素は、きめ、外見、および栄養価値を改善することができ、または所望の風味およびアロマを生成することができる。パン屋さんで現在使用されている食品酵素は、アミラーゼ、アミログリコシダーゼ、ペントサンの分解および還元型グルテン産生のためのペントサンアーゼ(pentosanase)、またはパン生地の安定性を増大させるためのグルコースオキシダーゼである。乳製品用において一般的な酵素は、チーズ製造における凝固剤としてのレンネット(プロテアーゼ)、ラクトースの加水分解のためのラクターゼ、乳清タンパク質の加水分解のためのプロテアーゼまたは過酸化水素除去のためのカタラーゼである。醸造工程において使用される酵素は、上記のアミラーゼであるが、セルラーゼまたはタンパク質懸濁物からビールを清澄化するためのプロテアーゼもまた使用される。ワインおよびフルーツジュース中では、濁りは、より一般的にでんぷんおよびペクチンによって生じるため、アミラーゼおよびペクチナーゼは、収量および透明度を増大させる。パパインおよびその他のプロテイナーゼは、肉を柔らかくするために使用される。
【0016】
酵素はまた、飼料を消化することにおいて動物を助けるために開発されてきた。西半球において、コーンは、ウシ、ブタ、および家禽の主要な食料源である。コーンからのリン酸のバイオアベイラビリティーを改善させるために、フィターゼが一般的に添加される(Wyss、M. etal.、Biochemical characterization of fungal phytases (myo−inositolhexakisphosphate phosphohydrolases) ; Catalytic properties. Applied & Environmental Microbiology 65、367−373 (1999))。さらに、フィターゼ加水分解は、タンパク質の消化およびカルシウムのようなミネラルの吸収を改善することが示されている(Bedford、M. R. & Schulze、H.、Exogenous Enzymes for Pigs and Poultry [Review]. Nutrition Research Reviews 11、91−114(1998))。別の主要な飼料用酵素は、キシラナーゼである。この酵素は、その相対的に高い溶解性線維含有量のために、約10%を超える小麦、大麦、もしくはライ麦を含む飼料の補助剤として特に有用である。キシラーゼは、2つの重要な作用:飼料中のゲル様高分子量アラビノキシランを加水分解することによって腸内容物の粘度を低減させる(Murphy、T.C.、Bedford、M.R.&McCracken、K.J. Effect of range of newxylanases on in vitro viscosity and on performance of broiler diets. British Pultry Science 44、S16−S18 (2003));およびタンパク質およびでんぷんのバイオアベイラビリティーを改善する細胞壁中のポリマーの分解。
【0017】
バイオテクの研究開発の研究室は、日常的に、多くの他の試薬とともに少量の特別な酵素を使用する。アルカリホスファターゼ、西洋ワサビペルオキシダーゼ、およびルシフェラーゼのような酵素は、そのうちのいくつかの例に過ぎない。Taqポリメラーゼまたは制限エンドヌクレアーゼのような熱安定性DNAポリメラーゼは、研究室での作業に革命を起こした。治療用酵素は、薬物の特定のクラスであり、FDAによって、他の、特に非生物学的製剤と比較して多くの利点を有する生物学的製剤として分類されている。成功している治療酵素の例は、ヒトの治療に用いる第VIII因子および第IX因子のようなヒト凝固因子である。さらに、ヒトの消化プロセスにおける種々の欠陥のために消化酵素が使用される。他の例は、心血管疾患の処置のためのt−PAおよびストレプトキナーゼ、I型ゴーシェ病の処置のためのベータグルコセレブロシダーゼ、急性リンパ芽球性白血病の処置のためのL−アスパラギナーゼ、および嚢胞性線維症の処置のためのDNAseである。治療剤としてのタンパク質の適用における重要な事項は、それらが免疫原性を有し得ることである。このリスクを軽減するために、ヒト由来の酵素が好まれ、これによって利用可能な酵素のセットが狭められる。新規のテイラーメードの特異性を有する設計された酵素(好ましくは、ヒト由来の)を提供することにより、意図的に標的基質を特異的に改変することが可能となり、他方免疫原性のリスクを最小限にすることができる。治療剤としての高度に特異的な酵素のさらなる利点は、副作用の危険性が低いことである。低分子とタンパク質との間の特異的な相互作用の可能性を限定することにより、標的ではないタンパク質への結合およびそれゆえ副作用が非常に一般的であり、しばしばそれがなければ有用であるリード化合物の使用を中止することになる。他方、特異的酵素は、基質を区別するためのより多くの接触部位および機構を提供し、それゆえより高い特異性が可能であり、それによって副作用の活動が低減される。
【0018】
プロテアーゼは、治療剤の重要なクラスを表す(Drugs of today、33、641−648 (1997))。しかしながら、現在、治療用のプロテアーゼは、通常、身体自体のプロテアーゼの活動が不十分な場合の代替物である。例えば、第VII因子は、血友病者の凝固障害または外科手術の際の特定の症例において投与され得る(Heuer L. ;Blumenberg D、(2002) Anaesthesist 51: 388)。組織型プラスミノーゲン活性化因子(t−PA)は、急性心筋梗塞において適用され、特異的な切断およびプラスミノーゲンの活性化を介してフィブリン凝血塊の溶解を開始する(Verstraete、M. et al. (1995) Drugs、50、29−41)。これまでのところ、テイラーメードの特異性を有するプロテアーゼが、疾患に関連する標的タンパク質を特異的に活性化または不活性化する治療剤を提供するために提供されている。
【0019】
モノクローナル抗体は、治療能を有する物質の別の重要な生物学的クラスである。主要な抗体標的の1つは、サイトカインのファミリーに属する腫瘍壊死因子(TNF)である。TNFは、炎症反応において主要な役割を果たす。ホモ3量体として、それらはほとんど全ての細胞の受容体と結合することができる。それらは、多数の細胞遺伝子、多数のシグナル伝達機構、キナーゼ、および転写因子を活性化する。最も重要なTNFは、TNFアルファおよびTNFベータである。TNFアルファは、マクロファージ、単球、およびその他の細胞によって産生される。TNFアルファは、炎症媒介因子である。それゆえ、過去10年間の研究は、異なる治療的症状(例えば、関節リウマチ、クローン病、または乾癬)の可能性のある治療剤として、モノクローナル抗体のようなTNFアルファ阻害剤に集中してきた(Hamilton et al. (2000) Expert Opin Pharmacother、1 (5): 1041−1052)。モノクローナル抗体の主要な欠点の1つは、それらが高価であることであり、それ故、新たな生物学的代替品が非常に重要である。
【0020】
文献には多数の人工酵素の例が存在する。Fulaniら(Fulani F. etal. (2003) Protein Engineering 16、515−519)は、Cdc25ホスファターゼ酵素の触媒サブユニットに構造的に関連する触媒ドメインを有するAzotobacter vinelandii由来のローダナーゼ(rhodanase)(チオスルファト:シアニドスルフルトランスフェラーゼ)を記載する。触媒機構の差違は、活性化部位の大きさの違いによる。ローダナーゼおよびホスファターゼのいずれも、異なる基質に対して高度に特異的である(硫酸対リン酸)。ローダナーゼの触媒機構は、1残基の挿入によってセリン/スレオニンホスファターゼに向かってシフトし得る。それゆえ、Fulaniらは、異なる酵素クラスからの天然に知られている酵素の構造比較および配列アラインメントによって触媒機構の変化の1つの例を示したが、同じ触媒機構を保ちながらユーザーが任意に定義可能な基質特異性をどのように生成するかについては示していない。
【0021】
Briggsら(WO02/090300 A2)によって記載されたチオレドキシンレダクターゼは、NADHに比べて優先的にNADPHに結合するようにコファクターの特異性を変化させた。このように、開始時の酵素および生成する人工酵素の両方の酵素が、異なる基質に対して高度に特異的である。このような変化した基質特異性を達成する方法は、コンピュータ処理による方法か、または可変残基および保存残基を決定するための関連するタンパク質の配列アラインメントのいずれかである。それらは全て、それらが公知の特異性を有するタンパク質の構造および配列の比較と、続いて同じ骨格から別の骨格への移動に基づいている点で共通している。
【0022】
特異性操作酵素、特に、文献に公開されたプロテアーゼの他の例が存在する。しかしながら、これらの例はいずれも、記載された方法において使用する出発物質の特異性と比べて新規な特異性を生成する手段を提供しない。その方法は、構造指向型単一点変異(Kurth、T. et al. (1998)Biochemistry 37、11434−11440 ; Ballinger、M et al. (1996)、2つの特異的プロテアーゼ間での表面ループの交換(Horrevoets et al. (1993) J. Biol. Chem. 268、779−782)から、インビトロもしくはインビボ選択と組み合わせた位置選択的または全遺伝子にわたったランダム変異誘発(Sices、H. & Kristie、T. (1998) Proc. Natl. Acad.Sci. USA、95、2828−2833)までにわたる。
【0023】
プロテアーゼ特異性の合理的な設計は、非常に少ない例に限られている。このアプローチは、タンパク質におけるフォールディングおよびダイナミクスならびに構造−機能関係を統御する複雑性の理解が不十分なことによって非常に限定されている(Corey、M.J. & Corey、E. (1996) Proc.Natl. Acad. Sci. USA、93: 11428−11434)。したがって、予測しながらプロテアーゼの活性または特異性を変化させるためにその一次アミノ酸配列を変化させることは困難である。1つの成功した例では、Kruthらは、トリプシンが二塩基性の誘因(motives)に対して優先性を示すように操作した(Kurth、T. et al. (1998) Biochemistry、37:11434−11440)。別の例では、Hedstromらが、トリプシンのS1基の特異性をキモトリプシンの特異性に変換した(Hedstrom、L. et al. (1992) Science、255: 1249− 1253)。これは、1つの既知の特性が1つの骨格から別の骨格へと移動した例である。
【0024】
Ballingerら(WO 96/27671)は、組み合わせ変異(N62D/G166D、および必要に応じて Y104D)を有するサブチリシン変異体を記載し、この変異体は、塩基性アミノ酸を基質のP1、P2、およびP4位に有する塩基性アミノ酸を有するペプチドまたはポリペプチド基質に向かっての基質特異性のシフトを有する。変異体サブチリシンの適切な基質は、5つの連続するランダム化残基を含有するファージ粒子(基質ファージ)のライブラリーをソーティングすることによって明らかになった。これらのサブチリシン変異体は、塩基性基質リンカーを有する融合タンパク質を切断するため、および塩基性切断部位を含有するホルモンまたは他のタンパク質(インビトロまたはインビボ)を加工するために有用である。合理的な酵素の設計に関連する問題は、定方向進化によって部分的に克服され得る(PCT/EP03/04864に開示される)。これらの研究は、それらの発現系および選択系によって分類され得る。遺伝子選択は、生物の内部に、結果として生成される生物の増殖挙動を変化させる前駆体タンパク質を切断することができる酵素(例えば、プロテアーゼ)を産生することを意味する。異なるプロテアーゼを有する生物の集団から、変化した増殖挙動を有するものが選択され得る。この原理は、例えば、Davisら(US5258289、WO96/21009)によって報告された。ファージ系の産生は、ファージタンパク質を切断することができるタンパク質分解酵素の存在下でのみ活性化され得るファージタンパク質の切断に依存する。他のアプローチは、遺伝子選択ではなくスクリーニングによる選択を可能とするレポーター系を使用するが、これもまた、酵素の細胞内特徴付けの本来的な不十分さを克服できない。
【0025】
自己分泌酵素を用いた変更された配列特異性を有する酵素を生成する系もまた、報告されている。Duffら(WO 98/11237)は、自己分泌プロテアーゼのための発現系を記載する。実験のデザインの必須の要素は、触媒反応が、膜結合前駆体分子の自己タンパク質分解プロセッシングによってプロテアーゼ自体に作用し、変異したプロテアーゼを細胞膜から細胞外環境に放出することである。それゆえ、標的ペプチド配列が天然の切断部位を自己タンパク質分解のために置換した融合タンパク質が構築されなければならない。このような系の制限は、肯定的に同定されたプロテアーゼが特定のアミノ酸配列を切断する能力を有するが、それらはまた多くの他のペプチド配列も切断し得ることである。それゆえ、高い基質特異性は、達成され得ない。さらに、このような系は、規定されたアミノ酸配列における特定の位置での選択されたプロテアーゼ切断を制御できず、選択されたプロテアーゼの速度定数の正確な特徴付けができない(kcat、KM)。
【0026】
α/β−バレルタンパク質内で新たな触媒活性および特異性の生成を目的とする方法が記載されている(WO 01/42432 ; Fersht et al、Methods of producingnovel enzymes ; Altamirano et al.(2000)Nature 403、617−622)。α/β−バレルタンパク質は、全ての既知の酵素の大きな部分を占めるタンパク質の大きなスーパーファミリーを含む。このタンパク質の構造は、α−ヘリクスによって囲まれたα/β−バレルからなっている。β−鎖およびヘリクスをつないでいるループは、活性部位残基を含む、いわゆる、リッド(lid(ふた))構造を含む。この方法は、触媒リッド構造に基づく2つのクラスへのα/β−バレルタンパク質の分類に基づく。α/β−バレルタンパク質構造の集中的な比較により、執筆者らは、基質結合および特異性は、基本的にバレル構造により規定されるのに対して、化学反応の特異性は、ループ内に存在するという結論に至った。異なる酵素からのバレルおよびリッド構造は、標的化またはランダム化された変異誘発および選択によって、新たな酵素活性を生成し、そして特性を最適化するための出発点を提供するために組み合わせられ得ることが示唆される。
【0027】
まとめると、治療、研究、および診断、工業酵素、食品および食品加工、化粧品、ならびに新規な基質特異性を有する酵素が入手可能になることによって可能になるその他の領域の分野において、多くの適用の可能性が存在することが明らかである。しかし、これまで、わずかに限られた数の特異的酵素が天然源から同定されているにすぎない。配列特異性を修飾し、変更し、または移動させるための合理的な設計方法ならびに上記のランダムアプローチは、利用された出発物質に存在しなかった新規かつユーザーが任意に決定することができる特異性の作成を可能にしなかった。
【0028】
それゆえ、現在利用可能な方法はいずれも、新規かつユーザーにより定義された配列特異性を有する酵素を提供することができない。対照的に、本発明は、このような酵素ならびにそれらを生成する方法を提供する。
【発明の開示】
【0029】
(発明の要旨)
本発明の目的は、人工タンパク質であって、このようなタンパク質を操作するために使用された成分中に存在しない新規な機能を有する人工タンパク質を提供することである。特に、本発明は、ユーザーが任意に定義することができる特異性を有する酵素を提供する。ユーザーが任意に定義することができる特異性とは、酵素が、そのような酵素の操作に使用した成分中に存在しない特異性を有して提供されることを意味する。特異性は、1つ以上の意図された標的基質が優先的に認識され、その酵素によって変換されるように、ユーザーによって選択され得る。さらに、本発明は、ヒトタンパク質と本質的に同一の配列を有するが、異なる特異性を有する酵素を提供する。1つの特定の実施形態では、本発明は、ユーザーが任意に定義することができる特異性を有するプロテアーゼを提供する。
【0030】
さらに、本発明は、1つ以上のさらなる成分と融合された人工酵素に関する。これらのさらなる成分は、好ましくは結合特性を有し、基質結合ドメイン、抗体、受容体、またはこれらのフラグメントからなる群のタンパク質様の成分であり得る。さらに、これらのさらなる成分は、さらなる機能的成分、好ましくは、ポリエチレングリコール、炭水化物、脂質、脂肪酸、核酸、金属、金属キレート剤、およびこれらのフラグメントまたは誘導体からなる群から選択され得る。生じる融合タンパク質は、本発明の範囲内のユーザーが任意に定義することができる特異性を有する酵素として理解される。
【0031】
加えて、本発明は、新規な、ユーザーが任意に定義することができる特異性を有するこのような酵素を、治療、研究、診断、栄養学、パーソナルケア、または工業目的に適用することに関する。さらに、本発明は、ユーザーが任意に定義することができる特異性を有する人工酵素を生成するための方法に関する。特に、本発明は、ヒト酵素と本質的に同一の配列を有するが、異なる特異性を有する酵素を生成することに関する。
【0032】
この課題は、下記の説明および請求の範囲において特定される本発明の実施形態によって解決されている。したがって、本発明は、
(1)下記の成分:(a)スクリーニング1つの基質におけるスクリーニング1つの化学反応を触媒するタンパク質足場、および(b)生じる人工タンパク質が少なくとも1つの標的基質と1つ以上の異なる基質とを区別できるようにする上記タンパク質足場中の部位に存在する1つ以上の特異性決定領域(SDR)であって、本質的に合成ペプチド配列である上記SDR、(c)の組み合わせによって特徴付けられる規定された特異性を有する人工酵素、
(2)治療、研究、診断、栄養学、パーソナルケア、または工業目的のための上記(1)に記載の人工酵素の使用、
(3)標的基質に対する特異性(その特異性は、個々の出発成分には存在しない)を有する上記(2)に記載の人工酵素を作製するための方法であって、少なくとも以下の工程:(a)少なくとも1つの基質上での少なくとも1つの化学反応を触媒するタンパク質足場を提供する工程、(b)工程(a)からの上記タンパク質足場を、完全にまたは部分的に、ランダムペプチド配列と、得られる人工酵素が少なくとも1つの標的基質と1つ以上の異なる基質とを区別できるようなタンパク質足場における部位で組み合わせることによって、人工酵素のライブラリーを作製する工程、ならびに(c)工程(b)で作製された人工酵素の上記ライブラリーから、少なくとも1つの標的基質に対する特異性を有する1つ以上の酵素を選択する工程、を含む方法、
(4)上記(1)に記載の少なくとも1つの人工酵素、ならびに少なくとも1つのさらなる成分、好ましくは、結合特性を有し、より好ましくは、抗体、結合ドメイン、受容体、およびこれらのフラグメントからなる群から選択される少なくとも1つのさらなる成分から構成される融合タンパク質、
(5)上記(1)に記載の1つ以上の人工酵素または上記(4)に記載の融合タンパク質を含む組成物または薬学的組成物であって、上記薬学的組成物は、必要に応じて、許容できる担体、賦形剤、および/もしくは助剤を含み得る、組成物または薬学的組成物、
(6)上記(1)に記載の人工酵素をコードするDNA、
(7)上記(6)に記載のDNAを含有するベクター、
(8)上記(7)に記載のベクターおよび/または上記(6)に記載のDNAを含有するベクターで形質転換/トランスフェクトされた宿主細胞ならびにトランスジェニック生物、ならびに
(9)上記(8)に記載の細胞または生物を培養する工程およびその培養液から酵素を単離する工程を含む人工酵素を製造する方法。
【0033】
(定義)
本発明の枠組みにおいては、以下の用語および定義が使用される。
【0034】
用語「プロテアーゼ」は、ペプチド結合を加水分解し得る任意のタンパク質分子を意味する。これは、天然に存在するタンパク質分解酵素または人工のタンパク質分解酵素、ならびに部位特異的変異誘発またはランダム変異誘発または任意の他のタンパク質操作法によって得られるこれらの変異体、タンパク質分解酵素の活性部位フラグメント、または上述のタンパク質の1つを含む任意の分子複合体もしくは融合タンパク質を含む。「プロテアーゼのキメラ」は、異なる親プロテアーゼに由来する2つ以上のフラグメントの融合タンパク質を意味する。
【0035】
用語「基質」は、酵素による触媒作用によって変換し得る任意の分子を意味する。用語「ペプチド基質」は、任意のペプチド、オリゴペプチド、またはプロテアーゼによって触媒作用により加水分解され得るペプチド結合を含む任意のアミノ酸組成物、配列もしくは長さのタンパク質分子を意味する。加水分解されるペプチド結合は、「切断部位」と呼ばれる。基質中の位置の番号付けは、Schlechter & Bergerによって導入されたシステムにしたがって行った(Biochem. Biophys. Res. Commun. 27 (1967) 157−162)。切断部位のN末端に隣接するアミノ酸残基は、P1、P2、P3などと番号付けされ、切断部位のC末端に隣接する残基は、P1’、P2’、P3’などと番号付けされる。
【0036】
用語「標的基質」は、本発明の酵素によって特異的に認識され、変換されるユーザーによって定義される基質を記載する。用語「標的ペプチド基質」は、ユーザーによって定義されるペプチド基質を記載する。用語「標的特異性」は、標的基質を認識し、変換することができる酵素の定性的および定量的な特異性を記載する。酵素の触媒特性は、ミカエリスおよびメンテンの定義に従う速度論パラメータ「KM」または「ミカエリスメンテン定数」、「kcat」または「触媒速度定数」、「kcat/KM」または「触媒効率」を使用して表される(Fersht、A. 、Enzyme Structure and Mechanism、W. H. Freeman and Company、New York、1995)。用語「触媒活性」は、規定された反応条件下での所定の基質の変換を定量的に記載する。
【0037】
用語「特異性」は、特定の基質を優先的に認識し、変換する酵素の能力を意味する。特異性は、定性的にも定量的にも表現することができる。「定量的特異性」とは、酵素によって認識される基質残基の化学的性質を意味する。「定量的特異性」とは、基質として受け入れられる基質の数のことをいう。定量的特異性は、全ての受け入れられる基質の数を全ての可能な基質の数で除算したものの負の対数として定義される語句「s」によって表され得る。例えば、全ての可能なペプチド基質の小部分を優先的に受け入れるプロテアーゼは、「高い特異性」を有する。ほとんど全てのペプチド基質を受け入れるプロテアーゼは、「低い特異性」を有する。定義は、WO 03/095670(これは本明細書中に参考として組み込まれる)に従ってなされる。非常に低い特異性を有するプロテアーゼはまた「非特異的プロテアーゼ」と呼ばれる。用語「規定された特異性」とは、特定の種類の特異性、すなわち、その他の基質に対して優先的に変換される特定の標的基質または特定の標的基質のセットのことをいう。
【0038】
用語「人工(engineered)」は、用語「酵素」と組み合わされて、異なる成分を含み、個々の成分のみによっては与えられない特徴を有する酵素を記載する。
【0039】
用語「タンパク質足場」または「足場タンパク質」とは、種々の一次、二次、および三次ポリペプチド構造のことをいう。
【0040】
用語「ペプチド配列」とは、タンパク質足場との挿入もしくは置換のため、または組み合わせのために使用される任意のペプチド配列を示す。ペプチド配列は、通常、周知の技術に従って合成され得るか、または天然源から得られ得るDNA配列から発現により得られる。タンパク質足場との挿入、置換、組み合わせは、オリゴヌクレオチドを、タンパク質足場をコードするポリヌクレオチドへ挿入、置換、または組み合わせることによって生成される。用語「合成」は、用語「ペプチド配列」と組み合わせて、そのペプチド配列が挿入もしくは置換されたか、またはそれが組み合わされたタンパク質足場に存在しないペプチド配列のことをいう。
【0041】
用語「成分」は、用語「人工酵素」と組み合わせて、このような酵素の操作において組み合わされるペプチド配列またはポリペプチド配列を意味する。このような成分は、とりわけ、1つ以上のタンパク質足場および1つ以上の合成ペプチド配列を含み得る。用語「人工酵素のライブラリー」は、人工酵素の混合物を記載し、そこではいかなる単一の人工酵素も異なるポリヌクレオチド配列によってコードされる。用語「遺伝子ライブラリー」は、人工酵素のライブラリーをコードするポリヌクレオチドのライブラリーを示す。用語「SDR」または「特異性決定領域」は、得られる酵素が標的基質と1つ以上の例えば、の基質とを区別できるような部位でタンパク質足場と組み合わされたときに規定された特異性を提供する合成ペプチド配列を意味する。このような部位は、「SDR部位」と呼ばれる。
【0042】
用語「の構造と類似した3次構造」および「類似した3次構造」は、用語「酵素」または「タンパク質」と組み合わされて、種類、配列、結合性、およびタンパク質の典型的2次構造要素の相対配位(例えば、アルファへリックス、ベータシート、ベータターン、およびループ)が類似しており、それゆえそのタンパク質が同じ構造的または位相幾何学的なクラスまたは集団にグループ分けされるタンパク質のことをいう。これは、任意の種類の変更された、追加の、または欠失された構造要素を有するが、それ以外は変化していない位相幾何学を有するタンパク質を含む。このような構造クラスの例は、TNFスーパーファミリー、セリンプロテアーゼ内のS1集団もしくはS8集団、GPCR、またはα/βバレル集団である。
【0043】
用語「構造的に対応する位置」は、互いに構造的に対応する類似の3次構造中のアミノ酸を示す。すなわち、それらは通常同じ構造またはその構造の位相幾何学的要素内に位置する。構造要素内で、それらはその構造要素の始めおよび終わりに対して同じ相対的位置を有する。例えば、2つのタンパク質を位相幾何学的に比較することにより、異なる長さの2つの構造的に対応する配列、次いで、例えば、それぞれの領域の長さの20%および40%内のアミノ酸が構造的に違いに対応することが明らかになる。
【0044】
用語「(本発明の)人工酵素のライブラリー」は、混合物または単離形態で存在し得る多数の酵素または酵素変異体のことをいう。
【0045】
アミノ酸残基は、以下の表1に従って、1文字または3文字表記で略称される。
【表1】

【発明を実施するための最良の形態】
【0046】
本発明は、新規の機能を有する人工タンパク質を提供する。特に、本発明は、ユーザーが任意に定義することができる特異性を有する酵素を提供する。特定の実施形態において、本発明は、ユーザーが任意に定義することができる特異性を有するプロテアーゼを提供する。加えて、本発明は、このような新規の、ユーザーが任意に定義することができる特異性を有する酵素の、治療、研究、診断、栄養学、パーソナルケア、または工業用途への適用を提供する。さらに、本発明は、このような酵素を操作するために使用された成分中に存在しない特異性を有する酵素を生成するための方法を提供する。特に、本発明は、哺乳動物、特にヒトの酵素と本質的に同一である配列を有するが、異なる特異性を有する酵素の生成に関する。さらに、本発明は、遺伝子によりコードされた対応する特異性を有する特異的な人工酵素のライブラリー、遺伝子によりコードされた対応する特異性を有する特異的な人工酵素のライブラリーを生成する方法、ならびにこのようなライブラリーの技術、診断、栄養学、パーソナルケア、または研究目的への適用を提供する。
【0047】
本発明の第一の局面は、規定された特異性を有する人工酵素を開示する。これらの人工酵素は、以下の成分によって特徴付けられる:
(a)基質上で少なくとも1つの化学反応を触媒し得るタンパク質足場、および
(b)得られる人工タンパク質が少なくとも1つの標的基質と1つ以上の異なる基質とを区別できるようなタンパク質足場の部位に存在する1つ以上の特異性決定領域(SDR)であって、本質的に合成ペプチド配列である、SDR。
【0048】
好ましくは、人工酵素のこのように規定された特異性は、タンパク質足場によって与えられない。
【0049】
原則として、種々の一次、二次、および三次構造を有し得る。一次構造、すなわち、アミノ酸配列は、人工配列であるか、または任意のウイルス、原核生物、もしくは真核生物起源のものでありえる。しかしながら、ヒトの治療用途には、タンパク質足場は、好ましくは、哺乳動物由来のものであり、より好ましくは、ヒト由来のものである。さらに、タンパク質足場は、1つ以上の反応を触媒し得、そして好ましくは、低い特異性のみを有する。
【0050】
好ましくは、タンパク質足場としての適用可能性のために改善された特性を付与する修飾されたアミノ酸配列を有するタンパク質足場の誘導体が使用される。このような改善された特性は、安定性、発現量もしくは分泌量、折り畳み(特に、タンパク質足場とSDRとの組み合わせ後に)、活性化剤もしくは阻害剤のような調節剤に対する増大もしくは減少した感受性、免疫原性、触媒速度、kMまたは基質親和性が含まれるが、これらに限定されない。
【0051】
人工酵素は、タンパク質足場と組み合わされた合成ペプチド配列から定量的特異性を明らかにする。それゆえ、人工ペプチド配列は、特異性決定領域またはSDRとして作用する。このようなSDRの数、長さ、および位置は、広範に変化し得る。足場内のSDRの数は、少なくとも1個であり、好ましくは、1個以上、より好ましくは、2〜11個、最も好ましくは、2〜6個である。SDRは、1〜50アミノ酸残基の長さを有し、好ましくは、1〜15アミノ酸残基、より好ましくは、1〜6アミノ酸残基の長さを有する。あるいは、SDRは、2〜20アミノ酸残基の長さ、好ましくは、2〜10アミノ酸残基の長さ、より好ましくは、3〜8アミノ酸残基の長さを有する。
【0052】
本発明の人工酵素は、定常領域および可変領域を含むが、非免疫グロブリン骨格を有し、かつ定常領域に活性部位(触媒活性)を有し、そのため活性部位の基質特異性が可変領域によって調節される抗体様分子としてもさらに記載され得る。好ましくは、免疫グロブリン構造においけると同様、可変領域は、標的分子と相互作用する可変の長さおよび組成のループ構造である。
【0053】
本発明の特定の変異体において、人工酵素は、加水分解活性を有する。好ましい変異体において、人工酵素は、タンパク質分解活性を有する。この変異体のための特に好ましいタンパク質足場は、非特異的プロテアーゼもしくは非特異的プロテアーゼ由来の部分であるか、またはそうでなければ非特異的プロテアーゼに由来する。この観点および以下の変異体および実施形態において、「に由来する」または「その誘導体」という表現は、1つ以上のアミノ酸位置で変異し、および/または元々のタンパク質に対して少なくとも70%、好ましくは、90%、より好ましくは、95%、および最も好ましくは、99%の相同性を有し、および/またはタンパク質分解によりプロセスされ、および/または変化したグリコシル化パターンを有し、および/または非タンパク質物質に共有結合しているか、および/またはさらなるタンパク質ドメインに融合し、および/またはC末端および/もしくはN末端切断を有し、および/または特定の挿入、置換、および/もしくは欠失を有するタンパク質の誘導体のことをいう。あるいは、「に由来する」は、2つ以上のタンパク質からの2つ以上のフラグメントの組み合わせまたはキメラである誘導体のことをいうこともある。ここで、各々のフラグメントは、上述のいずれかまたは全ての修飾を必要に応じて含む。タンパク質足場の三次構造は、任意の種類のものであり得る。しかしながら、好ましくは、三次構造は、以下の構造クラスの1つに属する:クラスS1(セリンプロテアーゼファミリーのキモトリプシン集団)、クラスS8(セリンプロテアーゼファミリーのサブチリシン集団)、クラスSC(セリンプロテアーゼファミリーのカルボキシペプチダーゼ集団)、クラスA1(アスパラギン酸プロテアーゼのペプシンA集団)、またはクラスC14(システインプロテアーゼのカスパーゼ−1集団)。ヒト治療薬として使用するための人工タンパク質分解酵素のタンパク質足場として働くプロテアーゼの例は、ヒトトリプシン、ヒトトロンビン、ヒトキモトリプシン、ヒトペプシン、ヒトエンドチアペプシン(human endothiapepsin)、ヒトカスパーゼ1〜14、および/またはヒトフリンであるか、またはこれらに由来する。
【0054】
人工タンパク質分解酵素の規定された特異性は、少なくとも1つの標的ペプチドまたはタンパク質基質と、1つ以上のさらなるペプチドまたはタンパク質基質とを区別する能力の指標である。好ましくは、規定された特異性は、P1部位とは異なる他の位置で異なるペプチドまたはタンパク質基質を区別する能力をいい、より好ましくは、規定された特異性は、P1部位およびP1’部位とは異なる他の位置で異なるペプチドまたはタンパク質基質を区別する能力のことをいう。より好ましくは、人工タンパク質分解酵素は、標的ペプチドまたはタンパク質基質を、必要な限りできるだけ多くの部位で区別して、他のタンパク質に対して優先的に標的基質を加水分解する。例として、ヒトの身体において静脈内に適用される治療的に有用な人工タンパク質分解酵素は、標的基質とヒト血清中の任意の他のタンパク質とを区別するのに十分に特異的であるべきである。好ましくは、このような人工タンパク質分解酵素は、3個以上のアミノ酸位置、より好ましくは、4個以上の位置、そしてさらにより好ましくは、5個以上のアミノ酸位置でペプチド基質を認識し、区別する。これらの位置は、隣接していてもしていなくてもよい。
【0055】
第一の実施形態において、タンパク質足場は、セリンプロテアーゼのS1構造サブクラス(すなわち、キモトリプシン集団)の三次構造もしくは折り畳みと等しいかもしくは類似した三次構造または折り畳みを有し、および/またはセリンプロテアーゼのS1構造サブクラスのタンパク質とアミノ酸レベルで少なくとも70%の同一性を有する。SDRは、ヒトトリプシン1中の領域18−25、38−48、54−63、73−86、122−130、148−156、165−171、および194−204に対して構造的に、またはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域20−23、41−45、57−60、76−83、125−128、150−153、167−169、および197−201(配列番号1に従うアミノ酸配列の番号付け)に構造的に、またはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質足場に挿入されることが好ましい。この種のタンパク質足場と組み合わせられるべきSDRの数は、好ましくは、1〜10、より好ましくは、2〜4である。好ましくは、タンパク質足場は、以下のタンパク質の1つ以上の誘導体またはホモログである:キモトリプシン、グランザイム、カリクレイン、トリプシン、メソトリプシン、好中球エラスターゼ、膵エラスターゼ、エンテロペプチダーゼ、カテプシン、トロンビン、アンクロド、凝固因子IXa、凝固因子VIIa、凝固因子Xa、活性化プロテインC、ウロキナーゼ、組織型プラスミノーゲン活性化因子、プラスミン、Desmodus型プラスミノーゲン活性化因子。より好ましくは、タンパク質足場は、トリプシンもしくはトロンビンであり、またはトリプシンもしくはトロンビン由来の誘導体もしくはホモログである。ヒトの治療のための用途には、トリプシンまたはトロンビン足場は、免疫反応またはアレルギー反応の危険性を最小限にするために、最も好ましくは、ヒト由来である。好ましくは、ヒトトリプシンIに由来する、または類似した三次構造を有するタンパク質に由来する改善された特徴を有する誘導体が使用される。このような誘導体の好ましい例は、ヒトトリプシンI(配列番号1)に由来し、そして1つ以上の以下のアミノ酸置換:E56G;R78W;Y131F;A146T;C183Rを有する。
【0056】
2つのSDRのうちの少なくとも1つが、ヒトトリプシンIまたはその誘導体に、残基42と43との間(SDR1)、および残基123と124との間(SDR2)にそれぞれ挿入されていることが好ましい(配列番号1に従う番号付け)。さらに、SDR1は、6という好ましい長さを、SDR2は、5という好ましい長さを、それぞれ有する。この実施形態の好ましい変異体において、SDR1およびSDR2の配列は、表2に列挙するアミノ酸配列の1つを含む。このような人工タンパク質分解酵素は、実施例IVに例示されるように標的基質Bに対する特異性を有する。
【0057】
さらなる実施形態において、タンパク質足場は、セリンプロテアーゼのS8構造サブクラスに属するか、および/または枯草菌由来のサブチリシンEに類似する三次構造を有するか、および/またはセリンプロテアーゼのS8構造サブクラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有する。好ましくは、足場は、サブチリシンファミリーまたはヒトプロタンパク質コンバターゼに属する。SDRは、枯草菌由来のサブチリシンE中の領域6−17、25−29、47−55、59−69、101−111、117−125、129−137、139−154、158−169、185−195、および204−225に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域59−69、101−111、129−137、158−169、および204−225に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号7に従う番号付け)。タンパク質足場は、1つ以上の以下のタンパク質に等しいかまたはそれらの誘導体またはホモログであることが好ましい:サブチリシンカールスバーグ、枯草菌サブチリシンE、サブチリシンBPN’、リケニホルミス菌、B.lentusサブチリシン、Bacillus alcalophilusアルカリホスファターゼ、プロテインキナーゼK、ケクシン(kexin)、ヒトプロタンパク質コンバターゼ、ヒトフリン。好ましい変異体において、サブチリシンBPN’またはタンパク質SPC1〜7の1つが、タンパク質足場として使用される。
【0058】
さらなる実施形態において、タンパク質足場は、アスパラギン酸プロテアーゼのファミリーに属するか、および/またはヒトペプチドに類似する三次構造を有する。好ましくは、足場は、プロテアーゼのA1クラスに属するか、および/またはプロテアーゼのA1クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有する。SDRは、枯草菌由来のサブチリシンE中の領域6−18、49−55、74−83、91−97、112−120、126−137、159−164、184−194、242−247、262−267、および277−300に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域10−15、75−80、114−118、130−134、186−191、および280−296に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号11に従う番号付け)。タンパク質足場は、1つ以上の以下のタンパク質に等しいかまたはそれらの誘導体またはホモログであることが好ましい:ペプシン、キモシン、レニン、カテプシン、ヤプシン。好ましくは、ペプチドもしくはエンドチオペプシンまたはこれらの誘導体もしくはホモログがタンパク質足場として使用される。
【0059】
さらなる実施形態において、タンパク質足場は、システインプロテアーゼのファミリーに属するか、および/またはヒトカスパーゼ7に類似する三次構造を有する。好ましくは、足場は、システインプロテアーゼのC14クラスに属するか、またはシステインプロテアーゼのC14クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有する。SDRは、ヒトカスパーゼ7中の領域78−91、144−160、186−198、226−243、および271−291に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域80−86、149−157、190−194、および233−238に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号14に従う番号付け)。タンパク質足場は、タンパク質足場は、カスパーゼ1〜9の1つに等しいか、またはその誘導体もしくはホモログであることが好ましい。
【0060】
さらなる実施形態において、タンパク質足場は、セリンプロテアーゼのS11クラスに属するか、またはセリンプロテアーゼのS11クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはStreptomyces種K15由来のD−アラニン−D−アラニントランスペプチダーゼに類似した三次構造を有する。SDRは、Streptomyces種K15由来のD−アラニン−D−アラニントランスペプチダーゼ中の領域67−79、137−150、191−206、212−222、および241−251に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域70−75、141−147、195−202、および216−220に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号15に従う番号付け)。Streptomyces種K15由来のD−アラニン−D−アラニントランスペプチダーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0061】
さらなる実施形態において、タンパク質足場は、セリンプロテアーゼのS21クラスに属するか、またはセリンプロテアーゼのS21クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはヒトサイトメガロウイルス由来のアセンブリン(assemblin)に類似した三次構造を有する。SDRは、ヒトサイトメガロウイルス由来のアセンブリン中の領域25−33、64−69、134−155、162−169、および217−244に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域27−31、164−168、および222−239に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号16に従う番号付け)。ヒトサイトメガロウイルス由来のアセンブリンまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0062】
さらなる実施形態において、タンパク質足場は、セリンプロテアーゼのS26クラスに属するか、またはセリンプロテアーゼのS26クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/または大腸菌由来のシグナルペプチダーゼに類似した三次構造を有する。SDRは、大腸菌由来のシグナルペプチダーゼ中の領域8−14、57−68、125−134、239−254、200−211、および228−239に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域9−13、60−67、127−132、および203−209に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号17に従う番号付け)。大腸菌由来のシグナルペプチダーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0063】
さらなる実施形態において、タンパク質足場は、セリンプロテアーゼのS33クラスに属するか、またはセリンプロテアーゼのS33クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/または大腸菌由来のシグナルペプチダーゼに類似した三次構造を有する。SDRは、セラチア・マルセッセンス由来のプロピルアミノペプチダーゼ中の領域47−54、152−160、203−212、および297−302に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域50−53、154−158、および206−210に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号18に従う番号付け)。セラチア・マルセッセンス由来のプロピルアミノペプチダーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0064】
さらなる実施形態において、タンパク質足場は、セリンプロテアーゼのS51クラスに属するか、またはセリンプロテアーゼのS51クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/または大腸菌由来のアスパルチルジペプチダーゼに類似した三次構造を有する。SDRは、大腸菌由来のアスパルチルジペプチダーゼ中の領域8−16、38−46、85−92、132−140、159−170、および205−211に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域10−14、87−90、134−138、および160−165に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号19に従う番号付け)。大腸菌由来のアスパルチルジペプチダーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0065】
さらなる実施形態において、タンパク質足場は、アスパラギン酸プロテアーゼのA2クラスに属するか、またはアスパラギン酸プロテアーゼのA2クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはヒト免疫不全ウイルス由来のプロテアーゼに類似した三次構造を有する。SDRは、ヒト免疫不全ウイルス由来のプロテアーゼ中の領域5−12、17−23、27−30、33−38、および77−83に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域7−10、18−21、34−37、および79−82に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号20に従う番号付け)。ヒト免疫不全ウイルス、好ましくは、HIV−1プロテアーゼ由来のプロテアーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0066】
さらなる実施形態において、タンパク質足場は、アスパラギン酸プロテアーゼのA26クラスに属するか、またはアスパラギン酸プロテアーゼのA26クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/または大腸菌由来のオンプチン(omptin)に類似した三次構造を有する。SDRは、大腸菌由来のオンプチン中の領域28−40、86−98、150−168、213−219、および267−278に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域33−38、161−168、および273−277に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号21に従う番号付け)。大腸菌由来のオンプチン由来のプロテアーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0067】
さらなる実施形態において、タンパク質足場は、システインプロテアーゼのC1クラスに属するか、またはシステインプロテアーゼのC1クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはカリーカ・パパイヤ由来のパパインに類似した三次構造を有する。SDRは、カリーカ・パパイヤ由来のパパイン中の領域17−24、61−68、88−95、135−142、153−158、および176−184に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域63−66、136−139、および177−181に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号22に従う番号付け)。カリーカ・パパイヤ由来のパパイン由来のプロテアーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0068】
さらなる実施形態において、タンパク質足場は、システインプロテアーゼのC2クラスに属するか、またはシステインプロテアーゼのC2クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはヒトカルパイン−2に類似した三次構造を有する。SDRは、ヒトカルパイン−2中の領域90−103、160−172、193−199、243−260、286−294、および316−322に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域92−101、245−250、および287−291に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号23に従う番号付け)。ヒトカルパイン−2由来のプロテアーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0069】
さらなる実施形態において、タンパク質足場は、システインプロテアーゼのC4クラスに属するか、またはシステインプロテアーゼのC4クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはタバコエッチウイルス(tobacco etch virus)由来のNIaプロテアーゼに類似した三次構造を有する。SDRは、タバコエッチウイルス由来のNIaプロテアーゼ中の領域23−31、112−120、144−150、168−176、および205−218に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域145−149、169−174、および212−218に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号24に従う番号付け)。タバコエッチウイルス由来のNIaプロテアーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0070】
さらなる実施形態において、タンパク質足場は、システインプロテアーゼのC10クラスに属するか、またはシステインプロテアーゼのC10クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/または化膿性連鎖球菌由来のストレプトパイン(streptopain)に類似した三次構造を有する。SDRは、化膿性連鎖球菌由来のストレプトパイン中の領域81−90、133−140、150−164、191−199、219−229、246−256、306−312、および330−337に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域82−87、134−138、250−254、および331−335に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号25に従う番号付け)。化膿性連鎖球菌由来のストレプトパインまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0071】
さらなる実施形態において、タンパク質足場は、システインプロテアーゼのC19クラスに属するか、またはシステインプロテアーゼのC19クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはヒトユビキチン特異的プロテアーゼ7に類似した三次構造を有する。SDRは、ヒトユビキチン特異的プロテアーゼ7中の領域3−15、63−70、80−86、248−256、272−283、および292−304に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域10−15、251−255、277−281、および298−304に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号26に従う番号付け)。ヒトユビキチン特異的プロテアーゼ7またはその誘導体もしくはホモログが足場として使用されることが、好ましい
【0072】
さらなる実施形態において、タンパク質足場は、システインプロテアーゼのC47クラスに属するか、またはシステインプロテアーゼのC47クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/または黄色ブドウ球菌由来のスタフォパイン(staphopain)に類似した三次構造を有する。SDRは、黄色ブドウ球菌由来のスタフォパイン中の領域15−23、57−66、108−119、142−149、および157−164に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域17−22、111−117、143−147、および159−163に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号27に従う番号付け)。黄色ブドウ球菌由来のスタフォパインまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0073】
さらなる実施形態において、タンパク質足場は、システインプロテアーゼのC48クラスに属するか、またはシステインプロテアーゼのC48クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/または出芽酵母(Saccharomyces cerevisiae)由来のUlp1エンドペプチダーゼに類似した三次構造を有する。SDRは、出芽酵母由来のUlp1エンドペプチダーゼ中の領域40−51、108−115、132−141、173−179、および597−605に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域43−49、110−113、133−137、および175−178に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号28に従う番号付け)。出芽酵母由来のUlp1エンドペプチダーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0074】
さらなる実施形態において、タンパク質足場は、システインプロテアーゼのC56クラスに属するか、またはシステインプロテアーゼのC56クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはPyrococcus horikoshii由来のPfplエンドペプチダーゼに類似した三次構造を有する。SDRは、Pyrococcus horikoshii由来のPfplエンドペプチダーゼ中の領域8−16、40−47、66−73、118−125、および147−153に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域9−14、68−71、120−123、および148−151に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号29に従う番号付け)。Pyrococcus horikoshii由来のPfplエンドペプチダーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0075】
さらなる実施形態において、タンパク質足場は、メタロプロテアーゼのM4クラスに属するか、またはメタロプロテアーゼのM4クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはBacillus thermoproteolyticus由来のサーモリシンに類似した三次構造を有する。SDRは、Bacillus thermoproteolyticus由来のサーモリシン中の領域106−118、125−130、152−160、197−204、210−213、および221−229に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域108−115、126−129、199−203、および223−227に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号30に従う番号付け)。Bacillus thermoproteolyticus由来のサーモリシンまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0076】
さらなる実施形態において、タンパク質足場は、メタロプロテアーゼのM10クラスに属するか、またはメタロプロテアーゼのM10クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはヒトコラゲナーゼに類似した三次構造を有する。SDRは、ヒトコラゲナーゼ中の領域2−7、68−79、85−90、107−111、および135−141に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域3−6、71−78、および136−140に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号31に従う番号付け)。ヒトコラゲナーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0077】
人工酵素は、グリコシダーゼ活性を有することがさらに好ましい。この変異体について特に適したタンパク質足場は、グリコシダーゼであるか、またはグリコシダーゼに由来するものである。好ましくは、三次構造は、以下の構造クラスの1つに属する:クラスGH13、GH7、GH12、GH11、GH10、GH28、GH26、およびGH18(ベータ/アルファ)8バレル。
【0078】
第一の実施形態において、タンパク質足場は、グリコシラーゼのGH13クラスに属するか、またはグリコシラーゼのGH13クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはヒト膵アルファアミラーゼに類似した三次構造を有する。SDRは、ヒト膵アルファアミラーゼ中の領域50−60、100−110、148−167、235−244、302−310、および346−359に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域51−58、148−155、および303−309に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号32に従う番号付け)。ヒト膵アルファアミラーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0079】
さらなる実施形態において、タンパク質足場は、グリコシラーゼのGH7クラスに属するか、またはグリコシラーゼのGH7クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはTrichoderma reesei由来のセルラーゼに類似した三次構造を有する。SDRは、Trichoderma reesei由来のセルラーゼ中の領域47−56、93−104、173−182、215−223、229−236、および322−334に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域175−180、218−222、および324−332に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号33に従う番号付け)。Trichoderma reesei由来のセルラーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0080】
さらなる実施形態において、タンパク質足場は、グリコシラーゼのGH12クラスに属するか、またはグリコシラーゼのGH12クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはクロカビ(Aspergillus niger)由来のセルラーゼに類似した三次構造を有する。SDRは、Aspergillus niger由来のセルラーゼ中の領域18−28、55−60、106−113、126−132、および149−159に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域20−26、56−59、108−112、および151−156に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号34に従う番号付け)。Aspergillus niger由来のセルラーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0081】
さらなる実施形態において、タンパク質足場は、グリコシラーゼのGH11クラスに属するか、またはグリコシラーゼのGH11クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはAspergillus niger由来のトキシラナーゼ(toxylanase)に類似した三次構造を有する。SDRは、Aspergillus niger由来のトキシラナーゼ中の領域7−14、33−39、88−97、114−126、および158−167に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域20−26、56−59、108−112、および151−156に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号35に従う番号付け)。Aspergillus niger由来のトキシラナーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0082】
さらなる実施形態において、タンパク質足場は、グリコシラーゼのGH10クラスに属するか、またはグリコシラーゼのGH10クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはStreptomyces lividans由来のトキシラナーゼ(toxylanase)に類似した三次構造を有する。SDRは、Streptomyces lividans由来のトキシラナーゼ中の領域21−29、42−50、84−92、130−136、206−217、および269−278に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域43−49、86−90、208−213、および271−276に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号36に従う番号付け)。Streptomyces lividans由来のトキシラナーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0083】
さらなる実施形態において、タンパク質足場は、グリコシラーゼのGH28クラスに属するか、またはグリコシラーゼのGH28クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはAspergillus niger由来のペクチナーゼに類似した三次構造を有する。SDRは、Aspergillus niger由来のペクチナーゼ中の領域82−88、118−126、171−178、228−236、256−264、および289−299に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域116−124、174−178、および291−296に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号37に従う番号付け)。Aspergillus niger由来のペクチナーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0084】
さらなる実施形態において、タンパク質足場は、グリコシラーゼのGH26クラスに属するか、またはグリコシラーゼのGH26クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはPseudomonas cellulosa由来のマンナナーゼに類似した三次構造を有する。SDRは、Pseudomonas cellulosa由来のマンナナーゼ中の領域75−83、113−125、174−182、217−224、247−254、324−332、および325−340に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域115−123、176−180、286−291、および328−337に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号38に従う番号付け)。Pseudomonas cellulosa由来のマンナナーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0085】
さらなる実施形態において、タンパク質足場は、グリコシラーゼのGH18(ベータ/アルファ)8クラスに属するか、またはグリコシラーゼのGH18(ベータ/アルファ)8クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはBacillus circulans由来のキチナーゼに類似した三次構造を有する。SDRは、Bacillus circulans由来のキチナーゼ中の領域21−29、57−65、130−136、176−183、221−229、249−257、および327−337に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域59−63、178−181、250−254、および330−336に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号39に従う番号付け)。Bacillus circulans由来のキチナーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0086】
人工酵素がエステル加水分解酵素活性を有することがさらに好ましい。好ましくは、この変異体についてのタンパク質足場は、リパーゼ、ホスファターゼ、フィターゼ、またはホスホジエステラーゼ活性を有する。
【0087】
第一の実施形態において、タンパク質足場は、エステラーゼのGXクラスに属するか、またはエステラーゼのGXクラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはCandida antarctica由来のリパーゼBに類似した三次構造を有する。好ましくは、足場はリパーゼ活性を有する。SDRは、Candida antarctica由来のリパーゼB中の領域139−148、188−195、216−224、256−266、272−287に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域141−146、218−222、259−263、および275−283に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号40に従う番号付け)。Candida antarctica由来のリパーゼBまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0088】
さらなる実施形態において、タンパク質足場は、エステラーゼのGXクラスに属するか、またはエステラーゼのGXクラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはモルモット由来の膵リパーゼに類似した三次構造を有する。好ましくは、足場はリパーゼ活性を有する。SDRは、モルモット由来の膵リパーゼ中の領域78−90、91−100、112−120、179−186、207−218、238−247、および248−260に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域80−87、114−118、209−215、および239−246に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号41に従う番号付け)。モルモット由来の膵リパーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0089】
さらなる実施形態において、タンパク質足場は、大腸菌由来のアルカリホスファターゼの構造に類似した三次構造を有するか、または大腸菌由来のアルカリホスファターゼの構造に類似した三次構造を有するタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有する。好ましくは、足場はホスファターゼ活性を有する。SDRは、大腸菌由来のアルカリホスファターゼ中の領域110−122、187−142、170−175、186−193、280−287、および425−435に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域171−174、187−191、282−286、および426−433に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号42に従う番号付け)。大腸菌由来のアルカリホスファターゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0090】
さらなる実施形態において、タンパク質足場は、ウシ膵臓デスオキシリボヌレアーゼIの構造に類似した三次構造を有するか、またはウシ膵臓デスオキシリボヌレアーゼIの構造に類似した三次構造を有するタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有する。好ましくは、足場はホスホジエステラーゼ活性を有する。より好ましくは、ヌクレアーゼ、そして最も好ましくは、非特異的エンドヌクレアーゼまたはその誘導体が、足場として使用される。SDRは、ウシ膵臓デスオキシリボヌレアーゼI中の領域14−21、41−47、72−77、97−111、135−143、171−178、202−209、および242−251に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域16−19、42−46、136−141、および172−176に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号43に従う番号付け)。ウシ膵臓デスオキシリボヌレアーゼIもしくはヒトデスオキシリボヌレアーゼ1またはその誘導体もしくはホモログが足場として使用されることが、好ましい。人工酵素がトランスフェラーゼ活性を有することがさらに好ましい。この変異体について特に適したタンパク質足場は、グリコシルトランスフェラーゼ、ホスホトランスフェラーゼまたはメチルトランスフェラーゼであり、またはその誘導体である。この変異体について特に好ましいタンパク質足場は、グリコシルトランスフェラーゼであり、またはグリコシルトランスフェラーゼに由来する。タンパク質足場の三次構造は、任意の種類のものであり得る。しかしながら、好ましくは、三次構造は、以下の構造クラスの1つに属する:GH13、およびGT1。
【0091】
第一の実施形態において、タンパク質足場は、トランスフェラーゼのGH13クラスに属するか、またはトランスフェラーゼのGH13クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはBacillus circulans由来のシクロマルトデキストリン グルカノトランスフェラーゼの構造に類似した三次構造を有する。好ましくは、足場はトランスフェラーゼ活性を有し、より好ましくは、グリコシルトランスフェラーゼが足場として使用される。SDRは、Bacillus circulans由来のシクロマルトデキストリン グルカノトランスフェラーゼ中の領域38−48、85−94、142−154、178−186、259−266、331−340、および367−377に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域87−92、180−185、261−264、および269−275に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号44に従う番号付け)。Bacillus circulans由来のシクロマルトデキストリン グルカノトランスフェラーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0092】
さらなる実施形態において、タンパク質足場は、トランスフェラーゼのGT1クラスに属するか、またはトランスフェラーゼのGT1クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはAmycolatopsis orientalis A82846由来のグリコシルトランスフェラーゼの構造に類似した三次構造を有する。好ましくは、足場はトランスフェラーゼ活性を有し、より好ましくは、グリコシルトランスフェラーゼ活性を有する。SDRは、Amycolatopsis orientalis A82846由来のグリコシルトランスフェラーゼ中の領域58−74、130−138、185−193、228−236、および314−323に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域61−71、230−234、および316−321に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号45に従う番号付け)。Amycolatopsis orientalis A82846由来のグリコシルトランスフェラーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0093】
人工酵素は、酸化還元酵素活性を有することがさらに好ましい。この変異体に特に適したタンパク質足場は、モノオキシゲナーゼ、ジオキシゲナーゼ、もしくはアルコールデヒドロゲナーゼ、またはこれらの誘導体である。タンパク質足場の三次構造は、任意の種類のものであり得る。
【0094】
第一の実施形態において、タンパク質足場は、Pseudomonas種由来の2、3−ジヒドロキシビフェニル ジオキシゲナーゼ(2、3−diphydroxybiphenyl dioxygenase)の構造に類似した三次構造を有するか、またはPseudomonas種由来の2、3−ジヒドロキシビフェニル ジオキシゲナーゼの構造に類似した三次構造を有するタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有する。好ましくは、足場はジヒドロゲナーゼ活性を有する。SDRは、Pseudomonas種由来の2、3−ジヒドロキシビフェニル ジオキシゲナーゼ中の領域172−185、198−206、231−237、250−259、および282−287に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域175−182、200−204、252−257、および284−287に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号46に従う番号付け)。Pseudomonas種由来の2、3−ジヒドロキシビフェニル ジオキシゲナーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0095】
さらなる実施形態において、タンパク質足場は、Acinetobacter種由来のカテコールジオキシゲナーゼの構造に類似した三次構造を有するか、またはAcinetobacter種由来のカテコールジオキシゲナーゼの構造に類似した三次構造を有するタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有する。好ましくは、足場はジオキシゲナーゼ活性を有し、より好ましくは、カテコールジオキシゲナーゼ活性を有する。SDRは、Acinetobacter種由来のカテコールジオキシゲナーゼ中の領域66−72、105−112、156−171、および198−207に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域107−110、161−171、および201−205に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号47に従う番号付け)。Acinetobacter種由来のカテコールジオキシゲナーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0096】
さらなる実施形態において、タンパク質足場は、Pseudomonas putida由来のカンファー−5−モノオキシゲナーゼの構造に類似した三次構造を有するか、またはPseudomonas putida由来のカンファー−5−モノオキシゲナーゼの構造に類似した三次構造を有するタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有する。好ましくは、足場はモノオキシゲナーゼ活性を有し、より好ましくは、カンファーモノオキシゲナーゼ活性を有する。SDRは、Pseudomonas putida由来のカンファー−5−モノオキシゲナーゼ中の領域26−31、57−63、84−98、182−191、242−256、292−299、および392−399に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域85−96、183−188、244−253、293−298、および393−398に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号48に従う番号付け)。Pseudomonas putida由来のカンファー−5−モノオキシゲナーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0097】
さらなる実施形態において、タンパク質足場は、Equus callabus由来のアルコールデヒドロゲナーゼの構造に類似した三次構造を有するか、またはEquus callabus由来のアルコールデヒドロゲナーゼの構造に類似した三次構造を有するタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有する。好ましくは、足場はアルコールデヒドロゲナーゼ活性を有する。SDRは、Equus callabus由来のアルコールデヒドロゲナーゼ中の領域49−63、111−112、294−301、および361−369に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域51−61、および295−299に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号48に従う番号付け)。Equus callabus由来のアルコールデヒドロゲナーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0098】
人工酵素は、リアーゼ活性を有することがさらに好ましい。この変異体に特に適したタンパク質足場は、オキソ酸リアーゼ、またはその誘導体である。この変異体について特に好ましいタンパク質足場は、アルドラーゼもしくはシンターゼ、またはこれらの誘導体である。タンパク質足場の三次構造は、任意の種類のものであり得るが、(ベータ/アルファ)8バレル構造が好ましい。
【0099】
第一の実施形態において、タンパク質足場は、Escherichia coli由来のN−アセチル−d−ニューラミン酸アルドラーゼ(N−acetyl−d−neuramic acid aldolase)の構造に類似した三次構造を有するか、またはEscherichia coli由来のN−アセチル−d−ニューラミン酸アルドラーゼの構造に類似した三次構造を有するタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有する。好ましくは、足場はアルドラーゼ活性を有する。SDRは、Escherichia coli由来のN−アセチル−d−ニューラミン酸アルドラーゼ中の領域45−55、78−87、105−113、137−146、164−171、187−193、205−210、244−255、および269−276に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域45−52、138−144、189−192、247−253、および271−275に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号50に従う番号付け)。Escherichia coli由来のN−アセチル−d−ニューラミン酸アルドラーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0100】
さらなる実施形態において、タンパク質足場は、Salmonella typhimurium由来のトリプトファンシンターゼの構造に類似した三次構造を有するか、またはSalmonella typhimurium由来のトリプトファンシンターゼの構造に類似した三次構造を有するタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有する。好ましくは、足場はシンターゼ活性を有する。SDRは、Salmonella typhimurium由来のトリプトファンシンターゼ中の領域56−63、127−134、154−161、175−193、209−216、および230−240に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域57−62、155−160、178−190、および210−215に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号51に従う番号付け)。Salmonella typhimurium由来のトリプトファンシンターゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0101】
人工酵素は、イソメラーゼ活性を有することがさらに好ましい。この変異体に特に適したタンパク質足場は、変換アルドラーゼ(converting aldose)もしくは変換ケトース(converting ketose)、またはこれらの誘導体である。
【0102】
第一の実施形態において、タンパク質足場は、Actinoplanes missouriensis由来のキシロースイソメラーゼの構造に類似した三次構造を有するか、またはActinoplanes missouriensis由来のキシロースイソメラーゼの構造に類似した三次構造を有するタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有する。SDRは、Actinoplanes missouriensis由来のキシロースイソメラーゼ中の領域18−31、92−103、136−147、178−188、および250−257に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域20−27、92−99、および180−186に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号52に従う番号付け)。Actinoplanes missouriensis由来のキシロースイソメラーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0103】
人工酵素は、リガーゼ活性を有することがさらに好ましい。この変異体に特に適したタンパク質足場は、DNAリガーゼまたはその誘導体である。
【0104】
第一の実施形態において、タンパク質足場は、Bacteriophage T7由来のDNAリガーゼの構造に類似した三次構造を有するか、またはBacteriophage T7由来のDNAリガーゼの構造に類似した三次構造を有するタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有する。SDRは、Bacteriophage T7由来のDNAリガーゼ中の領域52−60、94−108、119−131、241−248、255−263、および302−318に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域96−106、121−129、256−262、および304−316に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号53に従う番号付け)。Bacteriophage T7由来のDNAリガーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0105】
本発明の第二の局面は、治療、研究、診断、栄養学、パーソナルケア、工業用途のための特異性を有する人工酵素の適用に関する。この適用は、少なくとも以下の工程を含む:
(a)その加水分解が意図された目的、例えば、疾患の治癒、疾患の診断、ヒトもしくは動物栄養学のための成分の加工、またはその他の技術的プロセスに関して正の効果を有する標的ペプチド基質の同定;
(b)工程(a)において同定された標的ペプチドに特異的な酵素である、人工酵素の提供;ならびに
(c)意図された目的のための、工程(b)において提供された酵素の使用。
【0106】
本発明の第一の局面の第一の変異体において、人工酵素は、疾患関連標的基質を不活性化させるための治療手段として使用される。この適用は、少なくとも以下の工程を包含する:
(a)疾患に関連し、その不活性化が疾患に関して正の効果を有する標的基質の同定、および標的基質内の標的部位の決定であって、その標的部位での修飾がその標的基質の不活性化を導くという事実によって特徴付けられる標的部位の決定、
(b)人工酵素の提供であって、その酵素が工程(a)で同定した標的部位に対して特異的である、酵素の提供、ならびに
(c)ヒトの体内または体外での上記標的基質の不活性化のための上記酵素の使用。
【0107】
好ましい実施形態において、工程(c)に提供される人工酵素の足場は、ヒトの身体への上記酵素の適用に伴う免疫原性またはアレルギー性の効果を避けるかまたは低減するために、ヒト由来のものである。この変異体のより好ましい実施形態において、足場は、ヒトプロテアーゼであり、加水分解はそのペプチドまたはタンパク質標的の活性化または不活性化を導く。潜在的なペプチドまたはタンパク質標的には、サイトカイン、成長因子、ペプチドホルモン、インターロイキン、インターフェロン、凝固カスケード由来の酵素、セルピン、免疫グロブリン、可溶性もしくは膜結合レセプター、細胞もしくはウイルス表面タンパク質、ペプチドドラッグ、タンパク質ドラッグが含まれる。
【0108】
特に好ましい実施形態は、人工酵素がヒト腫瘍壊死因子アルファ(TNF−α)を切断し得るという知見に基づいている。したがって、人工酵素または融合タンパク質は、炎症性疾患(ならびにTNF−αに関連する他の疾患)の処置のための医薬の調製のために使用され得る。好ましくは、上記人工酵素または上記融合タンパク質は、ヒト腫瘍壊死因子アルファ(hTNF−α)を特異的に不活性化し得、より好ましくは、上記人工酵素または上記融合タンパク質は、hTNF−α(配列番号96)中の31/32、32/33、44/45、87/88、128/129、および/または141/142位間(最も好ましくは、31/32および32/33位間)のペプチド結合を加水分解し得る。
【0109】
さらなる実施形態において、標的基質は、人工酵素によって活性化されるプロドラッグである。この変異体の特定の実施形態において、人工酵素は、タンパク質分解活性を有し、標的基質は、タンパク質分解的に活性化されているタンパク質標的である。このようなプロドラッグの例は、凝固因子の不活化形態のようなプロタンパク質である。別の特定の変異体において、人工酵素は、酸化還元酵素であり、標的基質は、酸化により活性化され得る化学物質である。
【0110】
本発明のこの局面の第二の変異体において、人工酵素は、工業的にまたは栄養学的に重要な反応を規定された特異性でもって触媒するための技術的手段として使用される。この変異体の特定の実施形態において、人工酵素はタンパク質分解活性を有し、触媒される反応は、タンパク質分解プロセスであり、人工酵素は、1つ以上の工業的または栄養学的に重要なタンパク質を特異的に加水分解する。この変異体の好ましい実施形態において、人工酵素は、1つ以上の工業的または栄養学的に重要なタンパク質基質を、特定の部位で加水分解し、それによって工業的または栄養学的に所望の製品特性(例えば、きめ、風味、または沈殿特性)を導く。この変異体のさらなる特定の実施形態において、人工酵素は、グリコシド結合の加水分解を触媒する(グリコシダーゼまたはグリコシラーゼ活性)。次いで、好ましくは、触媒された反応は、ポリサッカライドプロセスであり、人工酵素は、工業的に、技術的に、または栄養学的に重要なポリサッカライド基質の1つ以上を特異的に加水分解する。この変異体のさらに特定の実施形態において、人工酵素は、トリグリセリドエステルまたは脂質の加水分解を触媒する(リパーゼ活性)。
【0111】
次いで、好ましくは、触媒された反応は、脂質プロセス工程であり、人工酵素は、1つ以上の工業的、技術的、または栄養学的に重要な脂質基質を特異的に加水分解する。この実施形態のさらに特定の変異体において、人工酵素は、基質の酸化または還元を触媒する(酸化還元活性)。次いで、好ましくは、人工酵素は、1つ以上の工業的、技術的、または栄養学的に重要な化学物質基質を特異的に酸化または還元する。
【0112】
本発明の第三の局面は、定性的におよび/または定量的に新規な特異性を有する人工酵素を、タンパク質足場と組み合わせて作製する方法に関する。本発明の方法は、少なくとも以下の工程を包含する:
(a)少なくとも1つの標的基質における少なくとも1つの化学反応を触媒することができるタンパク質足場を提供する工程、
(b)工程(a)からの前記タンパク質足場と、1つ以上の完全または部分的にランダムなペプチド配列の変異体とを、生じる酵素が少なくとも1つの標的基質と1つ以上の異なる基質とを区別できるようにする前記タンパク質足場の部位において組み合わせることによって、酵素または単離された酵素のライブラリーを生成する工程、ならびに
(c)工程(b)で生成した前記酵素のライブラリーから、少なくとも1つの標的基質に対する規定された特異性を有する1つ以上の酵素を選択する工程。
【0113】
本発明のこの局面の第一の変異体において、本発明の方法は、少なくとも以下の工程を包含する:
(a)少なくとも1つの標的基質における少なくとも1つの化学反応を触媒することができるタンパク質足場を提供する工程、
(b)工程(a)からの前記タンパク質足場に、1つ以上の完全または部分的にランダムなペプチド配列を、生じる酵素が少なくとも1つの標的基質と1つ以上の異なる基質とを区別できるようにする前記タンパク質足場の部位に挿入することによって、酵素または単離された酵素のライブラリーを生成する工程、ならびに
(c)工程(b)で生成した前記酵素のライブラリーから、少なくとも1つの標的基質に対する規定された特異性を有する1つ以上の酵素を選択する工程。
【0114】
好ましくは、1つ以上の完全にまたは部分的にランダム化されたペプチド配列がタンパク質足場と組み合わされるかまたはそれに挿入される位置は、その組み合わせまたは挿入の前に同定されている。
【0115】
完全にまたは部分的にランダムなペプチド配列の挿入の数または他の組み合わせ、ならびにそれらの長さは、広範に変化し得る。その数は、少なくとも1個、好ましくは、1個以上、より好ましくは、2〜11個、最も好ましくは、2〜6個である。このような完全または部分的にランダム化されたペプチド配列の長さは、通常、50アミノ酸残基よりも短い。好ましくは、その長さは、1〜15アミノ酸残基、より好ましくは、1〜6アミノ酸残基である。あるいは、その長さは、2〜20アミノ酸残基、好ましくは、2〜10アミノ酸残基、より好ましくは、3〜8アミノ酸残基である。
【0116】
好ましくは、このような挿入または他の組み合わせは、このようなタンパク質足場をコードするポリヌクレオチド、およびこのような完全にまたは部分的にランダム化したペプチド配列をコードするポリヌクレオチドもしくはオリゴヌクレオチドを使用してDNAレベルで行われる。
【0117】
必要に応じて、工程(a)〜(c)は、循環的に繰り返され、それにより工程(c)で選択された酵素が、将来のサイクルの工程(a)におけるタンパク質足場として働き、そしてランダム化したペプチド配列が挿入されるか、あるいは以前のサイクルで挿入されたペプチド配列で置換される。それにより、挿入されたペプチド配列の数は、サイクルにわたって一定であるか、または増大する。意図される特異性を有する1つ以上の酵素が生成されるまで、サイクルが繰り返される
【0118】
さらに、工程(a)〜(c)の1ラウンド以上の間またはその後に、足場は1つ以上の位置で変異されて、SDR配列との組み合わせのためにより受け入れ可能な足場が作製され得るか、および/または特定のpHおよび温度での触媒活性を増大し得るか、および/またはグリコシル化のパターンを変化させ得るか、および/または酵素阻害剤に対する感受性を減少させ得るか、および/または酵素の安定性を変化させ得る。
【0119】
本発明のこの局面の第二の変異体において、本発明の方法は、少なくとも以下の工程を包含する:
(a)第一のタンパク質足場フラグメントを提供する工程、
(b)上記タンパク質足場フラグメントをペプチド結合を介して第一のSDRと連結する工程、および、必要に応じて
(c)上記工程(b)の生成物をペプチド結合を介してさらなるSDRペプチドまたはさらなるタンパク質足場フラグメントと連結する工程、および、必要に応じて、
(d)十分に特異的な酵素が生成されるために必要なサイクル工程(c)を繰り返す工程、ならびに
(e)工程(a)〜(d)において生成された集団から、1つ以上の標的基質に対する所望の特異性を有する1つ以上の酵素を選択する工程。
【0120】
タンパク質足場フラグメントは、タンパク質足場の配列の一部を意味する。タンパク質足場は、少なくとも2つのタンパク質足場フラグメントから構成される。
【0121】
本発明のこの局面の第三の変異体において、タンパク質足場、SDR、および人工酵素は、DNA配列によってコードされ、発現系がタンパク質を生成するために使用される。代替的な変異体において、タンパク質足場、SDR、および/または人工酵素は、ペプチドビルディングブロックから化学合成される。
【0122】
本発明のこの局面の第四の変異体において、本発明の方法は、少なくとも以下の工程を包含する:
(a)1つ以上の標的基質における1つ以上の化学反応を触媒し得るタンパク質足場をコードするポリヌクレオチドを提供する工程、
(b)1つ以上の完全にまたは部分的にランダム化されたオリゴヌクレオチド配列を、タンパク質足場をコードするポリヌクレオチドと組み合わせる工程であって、上記完全にまたは部分的にランダム化されたオリゴヌクレオチド配列が、コードされる人工酵素が1つ以上の標的基質と1つ以上の他の基質とを区別できるようなポリヌクレオチド内の部位に位置する、工程、ならびに
(c)工程(b)において生成された集団から、1つ以上の基質に対する規定された特異性を有する酵素をコードする1つ以上のポリヌクレオチドを選択する工程。
【0123】
任意の酵素が工程(a)におけるタンパク質足場として働き得る。それは、天然に存在する酵素、その変異体もしくは短縮型誘導体、または人工酵素であり得る。ヒトの治療用途には、タンパク質足場は、好ましくは、哺乳動物の酵素であり、より好ましくは、ヒトの酵素である。その局面において、本発明は、哺乳動物ゲノムまたはヒトゲノムにおいてそれぞれコードされるいかなる酵素の特異性とも異なる特異性を有する本質的に哺乳動物の、特に本質的にヒトの酵素の作製方法に関する。
【0124】
本発明に従って、この局面の工程(a)において提供されるタンパク質足場は、標的基質上で1つ以上の化学反応を触媒し得ることが要求される。それゆえ、タンパク質足場は、標的基質に対するその活性によって可能性のあるタンパク質足場の群から選択される。
【0125】
本発明のこの局面の好ましい変異体において、加水分解活性を有するタンパク質足場が使用される。好ましくは、タンパク質分解活性を有するタンパク質足場が使用され、より好ましくは、標的基質上で塩基性の活性を有する非常に低い特異性を有するプロテアーゼが、タンパク質足場として使用される。基質特異性の低い異なる構造クラス由来のプロテアーゼの例は、パパイン、トリプシン、サブチリシン、SET(Streptomyces erythraeus由来のトリプシン様セリンプロテアーゼ)、エラスターゼ、カテプシンG、またはキマーゼである。タンパク質足場として使用される前に、プロテアーゼのアミノ酸配列は、特異性以外のタンパク質特性、例えば、触媒活性、安定性、阻害剤感受性、または発現量を、本質的にWO 92/18645に記載されるように、または特異性を、本質的にEP 02020576.3およびPCT/EP03/04864に記載されるように変化させるために、修飾され得る。
【0126】
ふさわしいタンパク質足場についての別の選択肢は、リパーゼである。肝性リパーゼ、リポタンパク質リパーゼ、および膵リパーゼが、「リポタンパク質リパーゼスーパーファミリー」に属し、これは言い換えるとリパーゼのGXクラスの例である(M. Fischer、J. Pleiss (2003)、Nucl. Acid. Res. 、31、319−321)。リパーゼの基質特異性は、荷電したヘッド基を有する脂肪酸およびリン脂質のトリグリセロールエステルに対するそれらの相対活性によって特徴付けられる。あるいは、エステラーゼ、グリコシラーゼ、アミダーゼ、またはニトリラーゼのような他の加水分解酵素が、足場として使用され得る。トラスフェラーゼはまた、ふさわしいタンパク質足場である。グリコシルトランスフェラーゼは、種々のドナーおよびアクセプターを含む多くの生物学的合成に関わっている。あるいは、タンパク質足場は、リガーゼ、リアーゼ、酸化還元酵素、またはイソメラーゼ活性を有し得る。
【0127】
第一の実施形態において、1つ以上の完全または部分的にランダム化されたペプチド配列が、タンパク質足場の特定の部位に挿入される。これらの挿入部位は、挿入されたペプチド配列が、異なる基質間の識別子、すなわち、特異性決定領域(SDR)として作用し得るという事実によって特徴付けられる。このような挿入部位は、いくつかのアプローチによって同定され得る。好ましくは、挿入部位は、タンパク質足場の3次元構造の分析によって、タンパク質足場の一次構造の、異なる定量的特異性を有する他の酵素との比較分析により、またはアラニンスキャニング、ランダム変異誘発、もしくはランダム欠失、または任意のその組み合わせのような技術によって実験的に、同定される。
【0128】
SDRのための挿入部位を同定するための第一のアプローチは、X線結晶学、または核磁気共鳴研究によって得ることができるタンパク質足場の3次元構造に基づいている。同じ構造クラスであるが異なる定量的特異性を有する他の酵素との比較におけるタンパク質足場の構造アラインメントは、高い構造類似性の領域および低い構造類似性の領域を明らかにする。例えば、このような分析は、PBDビューアのような公のソフトウェア(Guex、N. and Peitsch、M. C. (1997)Electrophoresis 18、2714−2723))を使用してなされ得る。低い構造類似性の領域は、好ましいSDR挿入部位である。SDRのための挿入部位を同定する第二のアプローチにおいて、競合阻害剤または基質アナログと複合体を形成した足場タンパク質の3次元構造が分析される。競合阻害剤の結合部位は、基質の結合部位と有意に重なり合うことが仮定される。その場合、阻害剤の特定の原子間距離内にあるタンパク質の原子は、基質に対しても類似する距離にある傾向がある。例えば、5Å未満の短い距離を選択することによって、基質との密接に施蝕しているタンパク質原子の組み合わせを生じる。これらの残基は、第一のシェルコンタクト(shell contacts)を構成し、それゆえSDRにとって好ましい挿入部位である。一旦第一のシェルコンタクトが同定されると、第二のシェルコンタクトが、第一のシェル原子から開始する距離分析を繰り返すことによって見出され得る。本発明のなお別の代替において、上記の距離分析は、活性部位残基から開始して行われる。
【0129】
SDRに対する挿入部位を同定する第三のアプローチにおいて、足場タンパク質の一次配列は、アラインメントアルゴリズムを使用することによって、同じ構造クラスであるが異なる定量的特異性を有する他の酵素とアラインメントされる。このようなアラインメントアルゴリズムの例は公開されている(Altschul、S. F. 、Gish、W.、Miller、W. 、Myers、E. W. & Lipman、D. J. (1990) J. Mol. Biol. 215: 403−410; 「Statistical methods in Bioinformatics: an introduction」 by Ewens、W. & Grant、G.R. 2001、Springer、New York)。このようなアラインメントは、種々の配列相同性、特に足場タンパク質と比較して1つ以上の酵素におけるさらなる配列エレメントを有する、保存された領域または保存されていない領域を明らかにし得る。保存された領域は、異なるタンパク質の間で共有されている表現型に寄与する(例えば、三次元構造の折り畳みを安定化する)可能性が高い。保存されていない領域、および特に、定量的に高い特異性を有する酵素におけるさらなる配列(Turner、R. etal. (2002) J. Biol.Chem.、277、33068−33074)は、SDRのための好ましい挿入部位である。
【0130】
プロテアーゼとして、現在5つのファミリー、すなわち、アスパラギン酸プロテアーゼ、システインプロテアーゼ、セリンプロテアーゼ、メタロプロテアーゼ、およびスレオニンプロテアーゼが知られている。各々のファミリーには、類似の集団を共有するプロテアーゼのグループが含まれる。これらのグループのメンバーの結晶構造は解析されており、公のデータベース(例えば、Brookhavenタンパク質データベース)を通じてアクセス可能である(H. M. Berman et al. Nucleic Acids Research、28 pp. 235−242(2000))。このようなデータベースはまた、他の酵素クラスおよび各クラスの非酵素的に活性なタンパク質における構造ホモログを含んでいる。いくつかのツールが、構造ホモログについて公のデータベースを検索するために利用可能である:SCOP−配列および構造の調査のためのタンパク質データベースの構造分類(Murzin A. G. etal. (1995) J. Mol. Biol. 247、536−540);CATH−クラス、アーキテクチャ、トポロジー、および相同スーパーファミリー:タンパク質ドメイン構造の階層型分類(Orengo et al. (1997) Structure 5 (8) 1093−1108);FSSP−タンパク質の構造間アラインメントに基づく折り畳み分類(Holm and Sander (1998) Nucl. Acids Res. 26 316−319);またはVAST−ベクターアラインメントサーチツール(Gibrat、Madej and Bryant (1996) Current Opinion in Structural Biology 6、377−385)。
【0131】
上記のアプローチにおいて、構造クラスのメンバーは、SDRの挿入部位を同定するために比較される。
【0132】
これらのアプローチの好ましい変形において、構造クラスS1のセリンプロテアーゼが、互いに比較される。トリプシンは、低い基質特異性を有するメンバーである。なぜならそれは、R位置にアルギニンまたはリジン残基のみを必要とする。他方、トロンビン、組織型プラスミノーゲン活性化因子、またはエンテロキナーゼは、全て、それらの基質配列(すなわち、それぞれ、(L/I/V/F)XPR^NA、CPGR^VVGG、およびDDDK^)に対して高い特異性を有する(Perona、J. & Craik、C. (1997) J.Biol.Chem.、272、29987−29990 ; Perona、J. & Craik、C (1995) Protein Science、4、337−360)。これらのプロテアーゼのアミノ酸配列のアラインメントは、実施例1(図2)に同定したSDRとともに記載されている。
【0133】
セリンプロテアーゼのファミリー内のさらなる例は、構造クラスS8(サブチリシン折り畳み)のメンバーによって与えられる。サブチリシンは、このクラスの型プロテアーゼであり、非特異的プロテアーゼである(Ottesen、M. & Svendsen、A. (1998) Methods Enzymol 19、199−215)。フリン、PC1、およびPC5は、プロペプチドのプロセッシングに関与する同じ構造クラスのプロテアーゼであり、高い基質特異性を有する(Seidah、N. & Chretien、M. (1997) Curr. Opin. Biotech. 、8: 602−607; Bergeron、F. etal. (2000) J. Mol.ER7docrin.、24: 1−22)。アプローチの好ましい変形において、一次アミノ酸配列のアラインメント(図4)が、サブチリシンと比較して特異的なプロがさらに有し、それゆえ潜在的な特異性決定領域である3アミノ酸より長い11個の配列ストレッチを同定するために使用される。アプローチのさらなる変形において、サブチリシンの3次元構造からの情報が、選択をさらに狭めるために使用され得る(図3)。11の挿入配列ストレッチのうち、3つ、すなわち、ストレッチ番号7、8、および11が、特に活性部位残基に近く、これらはそれぞれ、PC5、PC1、および全ての3つの特異的プロテアーゼにおける挿入である(図3)。好ましい変形において、可変長および可変組成の1または数個のアミノ酸ストレッチが、11個の部位のうちの1または数個の位置でサブチリシン配列に挿入され得る。アプローチのより好ましい変形において、挿入が領域7、8、もしくは11、またはこれらの任意の組み合わせで行われる。アプローチの別の好ましい変形において、構造クラスS8由来のサブチリシン以外のタンパク質足場が使用される。
【0134】
このアプローチのさらに好ましい変形において、構造クラスA1のアスパラギン酸プロテアーゼが分析される(Rawlings、N. D. & Barrett、A. J. (1995). Methods Enzymol. 248、105−120 ; Chitpinityol、S. & Crabbe、MJ. (1998)、Food Chemistry、61、395−418)。アスパラギン酸プロテアーゼのA1構造クラスの例は、低い基質特異性を有するペプチド、ならびに比較的高い基質特異性を有するベータセクレターゼ(Grninger−Leitch、F. 、et al. (2002) JBiol. Chem. 277、4687−4693)、およびレニンである。レトロウイルスプロテアーゼもまたこのクラスに属するが、活性酵素は、2つの同一のサブユニットのダイマーである。ウイルスプロテアーゼは、ポリタンパク質前駆体の正確なプロセッシングと、各場合において高い基質特異性を必要とする機能的タンパク質の生成にとって必須である(Wu、J. et al. (1998) Biochemistry、37、4518−4526 ; Pettit、S. etal. (1991) J.Biol. Chem. 、266、14539−14547)。ペプシンはこのクラスの型プロテアーゼであり、非特異的プロテアーゼを代表する(Kageyama、T. (2002) Cell. Mol.Life Sci. 59、288−306)。B−セクレターゼおよびカテプシンD(Aguilar、C.F. et al. (1995) Adv. Exp. Med.Bio/. 362、155−166)は、同じクラスのプロテアーゼであり、高い基質特異性を有する。アプローチの好ましい変形において、一次アミノ酸配列のアラインメント(図6)が、ペプチドと比較して特異的プロテアーゼに挿入され、それゆえ潜在的な特異性決定領域である、3つのアミノ酸よりも長い6つの配列ストレッチを同定するために使用される。アプローチのさらなる変形において、b−セクレターゼの3次元構造からの情報が、選択をさらに狭めるために使用され得る。6個の挿入配列のなかから、3つ、すなわち、それぞれストレッチ番号1、3、および4(これらはカテプシンDおよびベータセクレターゼにおける挿入出る)が活性部位残基に特に近い(図5)。アプローチの好ましい変形において、可変数および可変組成の1または数個のアミノ酸ストレッチが、6個の位置のうち1または数個でペプシン配列に挿入され得る。本発明のより好ましい実施形態において、挿入が、位置1、3.もしくは4、またはこれらの任意の組み合わせで行われる。本発明の別の好ましい実施形態において、ペプシン以外のタンパク質足場が使用される。
【0135】
特定の構造クラスが、低い特異性および高い特異性の公知のメンバーを含まない場合がある。これは、システインプロテアーゼファミリーに属し(Rawlings、N. D. & Barrett、A. J. (1994) Methods Enzymol. 244、461−486)、P4〜P1位置に対して全て高い特異性を示す、カスパーゼのC14クラスによって例示される。例えば、カスパーゼ1、カスパーゼ3、およびカスパーゼ9は、配列YVDA^、DEVD^、またはLEHD^をそれぞれ認識する。カスパーゼ間で異なる領域の同定は、基質特異性においける差違を担う領域が含まれる(図7および8)。
【0136】
最後に、酵素足場と同じ折り畳みの非酵素タンパク質もまた、SDRの挿入部位の同定に起用する。例えば、ハプトグロブリン(Arcoleo、J. & Greer、J.; (1982)d Biol. Chem. 257、10063−10068)およびアズロシジン(azurocidin)(Almeida、R. et al. (1991) Biochem. Biophys. Res. Commun. 177、688−695)は、全てのS1プロテアーゼと同じキモトリプシン様の折り畳みを共有する。活性部位残基での置換により、これらのタンパク質は、如何なるタンパク質分解機能も有しないが、活性プロテアーゼとの高い相同性を示す。これらのタンパク質と特定のプロテアーゼとの間の差違は、SDRのための挿入部位として働き得る領域を含む。
【0137】
第四のアプローチにおいて、SDRの挿入部位は、アラニンスキャニング、ランダム変異誘発、ランダム挿入、またはランダム欠失のような技術によって実験的に同定される。上記のアプローチとは対照的に、このアプローチは、足場タンパク質の三次元構造についての詳細な知識を必要としない。このアプローチの1つの好ましい変形において、タンパク質足場と同じ構造クラス由来の比較的高い特異性を有する酵素のランダム変異誘発および特異性の消失または変化についてのスクリーニングを使用して、タンパク質足場におけるSDRの挿入部位を同定し得る。ランダム変異誘発、アラニンスキャニング、ランダム挿入、またはランダム欠失は全て、酵素をコードするポリヌクレオチドのレベルに対して行われる。文献において公知の種々のプロトコルが存在する(例えば、Sambrook、J. F; Fritsch、E. F.; Maniatis、T.;Gold Spring Harbor Laboratory Press、Second Edition、1989、New York)。例えば、ランダム変異誘発は、特許WO 9218645に記載されるポリメラーゼを使用することによって達成され得る。この特許によれば、1つ以上のプロテアーゼをコードする1つ以上の遺伝子が、高いエラー率または誤取り込みの率を増大する条件下でDNAポリメラーゼを使用することによって増幅される。例えば、CadwellおよびJoyceの方法が使用され得る(Cadwell、R. C. and Joyce、G. F. 、PCR methods.Appl. 2 (1992)28−33)。突然変異誘発株、化学突然変異誘発物質、またはUV放射線の使用のような、しかしこれらに限定されないランダム変異誘発の他の方法もまた使用され得る。
【0138】
あるいは、オリゴヌクレオチドが、ランダムに分布したアミノ酸残基をアラニンと置換する変異誘発のために使用され得る。この方法は、一般的に、アラニンスキャニング範囲誘発と呼ばれる(Fersht、A. R. Biochemistry (1989) 8031−8036)。さらなる代替方法として、2項変異誘発のようなアラニンスキャニングの改変(Gregoret、L. M. and Sauer、R. T. PNAS (1993) 4246−4250)またはコンビナトリアルアラニンスキャニング(Weiss etal.、PNAS (2000) 8950−8954)が使用され得る。
【0139】
人工酵素を発現するために、このような人工タンパク質をコードするDNAが、表群的な分子クローニング技術によって適切な発現ベクターに連結される(例えば、Sambrook、J. F; Fritsch、E. F.; Maniatis、T.; Cold Spring Harbor Laboratory Press、Second Edition、1989、New York)。ベクターは、適切な発現宿主細胞に導入され、これは、対応する人工酵素変異体を発現する。特に適切な発言宿主は、大腸菌もしくは枯草菌(Bacillus subtilis)のような細菌性発現宿主、またはSaccharomyces cerevisaeもしくはPichia pastorisのような酵母発現宿主、またはチャイニーズハムスター卵巣(CHO)細胞株もしくはベビーハムスター腎臓(BHK)細胞株のような哺乳動物発現宿主、またはバクテリオファージ(例:M13もしくはラムダ)もしくはバキュロウイルス発現系のようなウイルスのようなウイルス発現宿主である。さらなる代替として、インビトロタンパク質発現のためのシステムが使用され得る。代表的には、DNAが、発現ベクター中、適切なシグナル配列の後ろに挿入される。シグナル配列は、酵素変異体が細胞外空間に分泌されることを導き、それによって細胞上清中でのプロテアーゼ活性の直接的な検出を可能にする。それぞれ、大腸菌にとって特に適切なシグナル配列は、HlyAであり、枯草菌にとっては、AprE、NprE、Mpr、AmyA、AmyE、Blac、SacBであり、S. cerevisiaeにとっては、Barl、Suc2、Mata、Inu1A、Ggpipである。あるいは、酵素変異体は細胞内で発現され、そして基質もまた細胞内で発現される。好ましくは、これは本質的に特許出願WO 0212543に記載されるように、基質アミノ酸配列と連結された2つの自己蛍光タンパク質を含む融合ペプチド基質を使用して行われる。さらなる代替として、酵素変異体の細胞内発現またはペリプラズム空間への分泌の後に、大腸菌についてのDsbA、PhoA、PelB、OmpA、OmpT、またはgIIIのようなシグナル配列を使用して、透過化または溶解工程により、酵素変異体を上清中へ放出する。膜バリアの破壊は、超音波、フレンチプレスのような機械的手段、またはリゾチームのような膜消化酵素の使用により強制され得る。別のさらなる代替として、酵素変異体をコードする遺伝子が、細胞を含まずに、適切な細胞を含まない発現系を使用して発現される。例えば、大腸菌細胞由来のS30抽出物を、Leslyらに記載のように、この目的のために使用し得る(Methods in Molecular Biology 37 (1995) 265−278)。任意の上記の方法によって生成され発現される遺伝子変異体の集団は、例えば、特許出願PCT/EP03/04864に詳細に記載されるように、適切なアッセイおよびスクリーニング方法によってそれらの親和性、基質特異性、または活性が分析される。元々の酵素と比較して減少した特異性を有する触媒活性変異体由来の遺伝子は、配列決定によって分析される。変異および/または挿入および/または欠失が生じる部位は、SDRが部位特異的に挿入され得る好ましい挿入部位である。
【0140】
第二の実施形態において、1つ以上の完全または部分的にランダムなペプチド配列が、タンパク質足場中のランダム部位に挿入される。この修飾は、通常、ポリヌクレオチドレベルで、すなわち、タンパク質足場をコードする遺伝子にヌクレオチド配列を挿入することによって行われる。ヌクレオチド配列のランダム挿入を可能にするいくつかの方法が利用可能である。ランダム挿入に使用され得るシステムは、例えば、ライゲーションに基づくシステム(Murakami etal、Nature Biotechnology 20 (2002) 76−81)、DNA重合反応に基づくシステムおよびトランスポゾンに基づくシステム(e.g、GPS−MTM mutagenesis system、NEBBiolabs ; MGTM mutation generation system、Finnzymes)である。トランスポゾンに基づく方法は、トランスポーゼースの認識配列をその末端に、ならびにまれな切断制限エンドヌクレアーゼ(rare cutting restriction endonuclease)を含有する選択可マーカー遺伝子のトランスポーゼース仲介挿入を利用する。後者のエンドヌクレアーゼを使用して、通常選択マーカーを放出し、ライゲーションの後、挿入断片を得る。再ライゲーションを行う代わりに、代替的に、1つまたは2つの外側切断制限エンドヌクレアーゼについての末端認識配列を有するフラグメントならびに選択マーカーを挿入し得る。ライゲーションの後、このフラグメントを1つまたは2つの外側切断エンドヌクレアーゼを使用して放出する。標準的な方法によって平滑末端を作製した後、平滑末端ランダムフラグメントを遺伝子にランダムな位置に挿入する。
【0141】
さらに好ましい実施形態において、インビトロ相同組換えのための方法が、上記方法によって導入される変異を組み合わせて、酵素集団を生成するために使用される。適用され得る方法の例は、特許出願WO0134835による組換え連鎖反応(RCR)、特許出願WO 9522625によるDNAシャッフリング方法、特許出願WO9842728によるねじれ伸長法(Staggered Extension method)、または特許出願WO9842728によるランダムプライミング組換えである。さらに、イッチー法(Itchy method)のような非相同組換えのための方法もまた、適用され得る(Ostermeier、M. et al. Nature Biotechnology 17 (1999) 1205−1209)。
【0142】
ヌクレオチド配列のタンパク質足場へのランダム挿入に際して、酵素変異体をコードする異なる遺伝子のライブラリーを得る。ポリヌクレオチドライブラリーは、引き続き、適切な発現ベクターに移される。適切な宿主における発現の際、またはインビトロ発現系の使用により、ランダムに挿入されたアミノ酸のストレッチを含有する酵素のライブラリーが得られる。
【0143】
本発明のこの局面の工程(b)にしたがって、1つ以上の完全または部分的ランダムペプチド配列が、タンパク質足場に挿入される。このような挿入されたSDRの実際の数が、意図された特異性が高いほど、SDRがより多く挿入されるという関係に従って、意図された定量的特異性によって決定される。単一のSDRが中程度に特異的な酵素の生成を可能にするのに対して、2つのSDRは、有意に特異的な酵素の生成をすでに可能にする。しかしながら、6個までまたはそれ以上のSDRがタンパク質足場に挿入され得る。類似の関係がSDRの長さについて有効である:意図される特異性が高いほど、挿入されるSDRが長い。SDRは、4アミノ酸残基程度の短さであり得る。しかしながら、それらはまた、50アミノ酸残基程度まで長くてもよい。有意な特性がすでに、4〜6アミノ酸残基の長さのSDRの使用によって生成され得る。
【0144】
挿入されるペプチド配列は、完全にまたは部分的にランダムであり得る。この状況において、完全にランダムとは、各位置または全ての位置において互いに異なる配列を含む一組の配列が並行して挿入されることを意味する。部分的にランダムとは、少なくとも1つの位置で違いに異なる配列を含む一組の配列が並行して挿入されることを意味する。この違いは、ペアごとまたは単一の配列に関してであり得る。例えば、アミノ酸の長さの挿入に関しての場合、部分的ランダムは、(i)AGGG、GVGG、GGLG、GGGIを含む、または(ii)AGGG、VGGG、LGGG、およびIGGGを含むセットであり得る。あるいは、ランダム配列はまた、長さが互いに異なる配列を含む。ペプチド配列のランダム化は、それぞれの部位で遺伝子に挿入されたヌクレオチド配列のランダム化によって達成される。それにより、ランダム化は、オリゴヌクレオチドの化学合成の間に核酸塩基の混合物をモノマーとして利用することによって達成され得る。ストップコドンの可能性をさらに最小化する完全にランダムなコドンのためのモノマーの特に好ましい混合物は、NN(GTC)である。あるいは、ランダムオリゴヌクレオチドが、それぞれの部位で遺伝子に挿入される短いフラグメントへのDNAのフラグメント化によって得られ得る。フラグメント化されるDNAの供給源は、合成オリゴヌクレオチドであり得るが、あるいは、クローン化された遺伝子、cDNA、またはゲノムDNAに由来するものでもよい。好ましくは、DNAは酵素をコードする遺伝子である。フラグメント化は、例えば、DNAのランダムエンドヌクレアーゼ消化によって達成され得る。好ましくは、DNAseI(例えば、ウシ膵臓由来の)のような非特異的エンドヌクレアーゼは、エンドヌクレアーゼ消化のために利用される。
【0145】
本発明の方法の工程(a)〜(c)が循環的に繰り返される場合、連続するラウンドにおいて挿入されるランダムペプチド配列を得るための異なる代替が存在する。好ましくは、酵素の増大した特異性を導くものとして1つのラウンドにおいて同定されたSDRは、次のラウンドにおいて挿入されるランダムペプチド配列のための鋳型として使用される。
【0146】
好ましい代替において、1つのラウンドで選択された配列が分析され、そしてランダム化されたオリゴヌクレオチドがこれらの配列に基づいて生成される。これは、例えば、元々のヌクレオチドに加えて、他の3つのヌクレオチドモノマーの特定の割合(%)の混合物をオリゴヌクレオチド合成において各位置で使用することによって達成され得る。例えば、第一ラウンドにおいて、アミノ酸配列ARLT(例えば、ヌクレオチド配列GCG CGC CTT ACCによってコードされる)を有するSDRが同定された場合、このSDR部位に挿入されるランダムペプチド配列は、70%G、10%A、10%T、および10%Cを第一の位置に、70%C、10%G、10%T、および10%Aを第二の位置に、など、有するオリゴヌクレオチドによってコードされる。これは、各位置に、およそ3回に1回の割合で、鋳型アミノ酸を、3回に2回の割合で別のアミノ酸を導く。
【0147】
別の好ましい代替において、1つのラウンドにおいて選択される配列が分析され、そしてコンセンサスライブラリーがこれらの配列に基づいて生成される。これは、例えば、オリゴヌクレオチド合成における各位置においてヌクレオチドの規定された混合物を、前回のラウンドで選択されたSDRの各位置で同定されたアミノ酸残基の混合物を導くように、使用することによって達成され得る。例えば、第一のラウンドにおいて、アミノ酸配列ARLTおよびVPGSを有する2つのSDRが同定された場合、次のラウンドでこのSDRに挿入されるコンセンサスライブラリーは、配列G(C/T)GC(G/C)C(G/T)(G/T)G(A/T)CCを有するオリゴヌクレオチドによってコードされ得るこれは、ランダムペプチド配列(A/V)(R/P)(L/G/V/W)(T/S)に対応し、それにより、第一のラウンドで同定された全てのアミノ酸残基の組み合わせを可能にし、そして遺伝コードの縮重により、いくつかの位置でより低い程度に代替のアミノ酸を許容する。
【0148】
別の好ましい代替において、1つのラウンドで選択された配列は、以前の分析なしで、ポリヌクレオチドのインビトロ組換えのための方法(例えば、WO01/34835に記載される方法)を使用して組換えされる。(以下は、また、本発明の第8および第9の局面の詳細を提供する。)
部分的にまたは完全にランダムな配列を足場タンパク質をコードする遺伝子に挿入し、最終的に標準的な分子クローニング技術(Sambrook、J. F; Fritsch、E.F. ; Maniatis、T.; Cold Spring Harbor Laboratory Press、Second Edition、1989、New York)を使用して、得られた遺伝子を適切な発現ベクターにライゲーションした後、そのベクターは、対応する酵素変異体を発現する適切な発現宿主細胞へ導入される。特に適切な発現宿主は、大腸菌もしくは枯草菌のような細菌発現宿主、またはSaccharomyces cerevisae もしくはPichia pastorisのような酵母宿主、またはチャイニーズハムスター卵巣(CHO)細胞株またはベビーハムスター腎臓(BHK)細胞株のような哺乳動物宿主、またはバクテリオファージ(例:M13、T7ファージ、ラムダ)のようなウイルス発現系、またはバキュウロウイルス発現系のようなウイルスである。さらなる代替として、インビトロタンパク質発現のためのシステムが使用され得る。代表的には、DNAが、細胞外空間に酵素変異体を分泌させる適切なシグナル配列の後に、発現ベクターにライゲーションされ、それにより細胞上清中の酵素活性の直接検出を可能にする。大腸菌について特に適切なシグナル配列は、ompA、pelB、HlyAであり、枯草菌についてはAprE、NprB、Mpr、AmyA、AmyE、Blac、SacBであり、S.CerevisiaeについてはBar1、Suc2、Mata、I nu1A、Ggplpである。あるいは、酵素変異体が細胞内で発現され、基質もまた細胞内で発現される。プロテアーゼ変異体に従って、これは、本質的に特許出願WO 0212543に記載されるように、基質アミノ酸配列によって連結された2つの自己蛍光タンパク質を含む融合ペプチド基質を使用して、行われる。さらなる代替として、酵素変異体の細胞内発現、または大腸菌のDsbA、PhoA、PelB、OmpA、OmpT orgi 11のようなシグナル配列を使用したペリプラズム空間への分泌の後、透過化または溶解工程によいr、酵素辺が上清へ放出される。膜バリアの破壊は、超音波、フレンチプレスのような機械的手段を使用して、またはリボチームのような膜消化酵素を使用して、強制され得る。別のさらなる代替として、酵素変異体をコードする遺伝子が、細胞を含まないで、適切な細胞を含まない発現系を使用して発現される。例えば、大腸菌細胞由来のS30抽出物が、Leslyらによって記載されるようにこの目的のために使用される(Methods in Molecular Biology 37 (1995) 265−278)。
【0149】
ベクターの宿主細胞への導入の後、これらの細胞を、意図された標的基質に対する特異性を有する酵素の発現についてスクリーニングする。このようなスクリーニングは、典型的には、遺伝子型と表現型とを相関付けるために細胞を互いに分離し、そして増殖および発現期間の後に各細胞クローンの活性をアッセイすることによって行われる。このような分離は、例えば、細胞を、例えば、WO01/24933に記載されるようにサンプルキャリアの区画へ分布させることによって行われ得る。あるいは、細胞を、アガロースプレート上でストリーキングすることにより、アガロースのようなポリマー中で包むことにより、キャピラリー中に充填することにより、または同様の方法により分離される。
【0150】
意図される特異性を有する変異体の分離は、異なるアプローチによってなされ得る。プロテアーゼの場合、好ましくは、本質的にPCT/EP03/04864に記載されるペプチド基質を用いるアッセイが使用される。
【0151】
発現のフォーマットにかかわらず、酵素変異体を選択することにより、好ましくは、標的配列を認識し、変換する酵素の同定が可能になる。第一の代替として、標的配列を認識し、変換する酵素は、好ましくは、標的基質配列に対する高い親和性を有する酵素をスクリーニングことによって同定される。高い親和性は、実質的に第一の酵素のKMより低い標的基質濃度でスクリーニングすることによって選択される低いKMに対応する。好ましくは、使用される基質は、10μM未満、好ましくは、1μM未満、より好ましくは、100nM未満、そして最も好ましくは、10nM未満の濃度で基質の修飾を検出することを可能にする1つ以上のフルオロフォアに連結される。
【0152】
第二の代替として、好ましく標的基質を認識し変換する酵素が、2つ以上の基質をアッセイにおいて使用し、そしてこれらの2つ以上の基質の活性を比較してスクリーニングすることによって同定される。好ましくは、使用される2つ以上の基質は、異なるマーカー分子に連結され、それにより、2つ以上の基質の修飾を連続的にまたは並行して検出することが可能になる。プロテアーゼの場合、特に好ましくは、2つのペプチド基質が使用される。一方のペプチド基質は、任意に選択されたかまたは部分的もしくは完全にランダムなアミノ酸配列を有し、それにより、任意の基質上での活性がモニターでき、そして他方のペプチド基質は、意図された標的基質配列に同一かまたは類似するアミノ酸配列を有し、それにより標的基質上での活性をモニターできる。特に好ましくは、これらの2つのペプチド基質が蛍光マーカー分子に連結され、そして連続的にまたは並行して測定された場合に両方の活性を区別するために、2つのペプチド基質の蛍光特性は十分に異なる。例えば、特許出願WO 0212543による第一の蛍光タンパク質、ペプチド、および第二の蛍光タンパク質を含む融合タンパク質が、この目的に使用され得る。あるいは、ローダミンのような蛍光色素がペプチド基質に化学的に連結される。
【0153】
第三の代替として、標的酵素を好ましく認識し変換する酵素が、標的基質に類似する1つ以上の基質を、高過剰の競合基質とともに使用することによって同定される。次いで、標的基質に類似する基質上の活性に関してのスクリーニングが、競合基質の存在下で行われる。標的特異性に定性的に対応する特異性を有するが、低い定量的特異性のみを有する酵素は、このようなスクリーニングにおいて負のサンプルとして同定される。これに対して、定量的におよび定性的に標的特異性に対応する特異性を有する酵素は、正に同定される。好ましくは、標的基質に類似する1つ以上の基質はマーカー分子に連結され、それによりそれらの修飾の検出が可能となり、一方、競合基質はマーカー分子を有しない。競合基質は、任意に選択されたかまたはランダムアミノ酸配列を有し、それによりマーカー保有基質の加水分解に対する競合阻害剤として作用する。例えば、タンパク質加水分解酵素(例:トリプトン)は、本発明の人工タンパク質分解酵素に対する競合基質として作用し得る。
【0154】
第四の代替として、標的基質を認識し変換する酵素が同定され、増幅結合または増殖結合選択工程によって同定および選択される。さらに、活性が細胞内で測定され得、そして選択がセルソーター(例えば、蛍光活性化セルソーター)によってなされ得る。
【0155】
さらなる代替として、標的基質を認識し変換する酵素は、第一に、標的基質に優先的に結合する酵素を選択し、第二に、酵素変異体のこのサブグループから標的基質を変換する酵素を選択することによって同定される。標的基質に優先的に結合する酵素の選択は、標的基質に対するバインダーの選択か、または他の基質へ結合する酵素の対抗選択のいずれかによって行われる。バインダーの選択または非バインダーの対抗選択のための方法は当該分野で公知である。このような方法は、典型的には、表面ディスプレイ発現法を使用することによって解決され得る表現型−遺伝子型共役を必要とする。このような方法には、例えば、ファージもしくはウイルスディスプレイ、細胞表面ディスプレイ、およびインビトロディスプレイが含まれる。ファージもしくはウイルスディスプレイは、典型的には、目的のタンパク質のウイルス/ファージタンパク質への融合を含む。細胞表面ディスプレイ(すなわち、細菌もしくは真核生物細胞ディスプレイのいずれか)は、典型的には、目的のタンパク質の細胞表面に存在するペプチドもしくはタンパク質への融合を含む。インビトロディスプレイにおいて、タンパク質は、典型的には、インビトロで作製され、それらに直接または間接的に、そのタンパク質をコードするRNAを結合させる(DE 19646372)。
【0156】
本発明はまた、本明細書中上記で規定した本発明の第一の局面に従う1つ以上の人口酵素を含有する組成物または薬学的組成物を提供する。その組成物は、必要に応じて、受容可能な担体、賦形剤、および/または助剤を含んでいても良い。本明細書中で規定される非薬学的組成物は、研究用組成物、栄養学組成物、洗浄組成物、消毒組成物、化粧用組成物、またはパーソナルケア用組成物である。さらに、本明細書中に規定する人工酵素をコードするDNA配列およびこのDNAを含有するベクターもまた、提供される。最後に、このようなDNA配列および/もしくはベクターを含有する形質転換された宿主細胞(原核生物もしくは真核生物)またはトランスジェニック生物、ならびにこのような宿主細胞もしくはトランスジェニック動物を本発明の第一の局面の人工酵素を作製するために使用する方法もまた、意図される。
【0157】
(図面の詳細な説明)
図1:「球棒」表示で示された活性部位残基を有するヒトトリプシンIの3次元構造。印を付けた領域は、潜在的SDR挿入部位を示す。
【0158】
図2:ヒトプロテアーゼトリプシン1、アルファトロンビンおよびエンテロペプチダーゼ(これらの全てがセリンプロテアーゼファミリーの構造クラスS1に属する)の一次アミノ酸配列のアラインメント。トリプシンは、この構造クラスの非特異的プロテアーゼであるのに対して、アルファトロンビンおよびエンテロペプチダーゼは、高い基質特異性を有するプロテアーゼである。トリプシンと比較して、トロンビンおよびエンテロキナーゼの一次配列中に3以上のアミノ酸の挿入の領域がいくつか見られる。(−1−)と記された領域および(−3−)と記された領域は、SDR挿入部位である。
【0159】
アルファトロンビンの三次構造において、両方の領域が基質結合部位の近傍にある。これらの領域は、それゆえ、SDRの候補として選択されるための2つの基準:第一に、それらが非特異的プロテアーゼと比較して特異的プロテアーゼ中にある挿入であること、および第二に、それらが基質結合部位の近くにあること、を満たす。三次元構造の表示が図3に与えられる。
【0160】
図3:「球棒」表示で示された活性部位残基を有するサブチリシンの三次元構造。番号を付された領域は、潜在的SDR挿入部位を示す。
【0161】
図4:サブチリシンE、フリン、PC1、およびPC5(これらの全てが、セリンプロテアーゼファミリーの構造クラスS8に属する)の一次アミノ酸配列のアラインメント。サブチリシンEは、この構造クラスの非特異的プロテアーゼである。一方、フリン、PC1、およびPC5は、高い基質特異性を有するプロテアーゼである。サブチリシンと比較して、フリン、PC1、およびPC5の一次配列への3つ以上のアミノ酸の挿入のいくつかの領域が見られる。(−4−)、(−5−)、(−7−)、(−9−)、および(−11−)と記された領域は、好ましいSDR挿入部位である。これらの領域ストレッチは、SDRの候補として選択されるための2つの基準を満たす。第一に、これらは、非特異的なプロテアーゼと比較して特異的なプロテアーゼにおける挿入である。第二に、それらは活性部位残基に近い。
【0162】
図5:「球棒」表示で示された活性部位残基を有するベータセクレターゼの3次元構造。番号を付された領域は、潜在的SDR挿入部位を示す。
【0163】
図6:ペプシン、b−セクレターゼ、およびカテプシンD(これらの全てが、アスパラギン酸プロテアーゼファミリーの構造クラスA1に属する)の一次アミノ酸配列のアラインメント。ペプシンは、この構造クラスの非特異的プロテアーゼである。一方、b−セクレターゼおよびカテプシンDは、高い特異性を有するプロテアーゼである。ペプチドに比べて、b−セクレターゼおよびカテプシンDの一次配列中への3以上のアミノ酸の挿入のいくつかの領域が見られる。−1−〜−11−で記された領域は、可能性のあるSDR組み合わせ部位に対応し、そして図5にも記されている。
【0164】
図7:「球棒」表示で示された活性部位残基を有するカスパーゼ7の3次元構造を示す。番号を付した領域は、潜在的SDR挿入部位を示す。
【0165】
図8:システインプロテアーゼクラスC14ファミリーのメンバーとしてのカスパーゼ7の一次アミノ酸配列を示す(配列番号14もまた参照)。
【0166】
図9:本発明の第三の局面による方法の模式図である。
【0167】
図10:トリプシン発現のウエスタンブロット分析である。トリプシンの変異体を発現する細胞培養の上清が、ネガティブコントロールと比較されている。レーン1:分子量標準:レーン2:ネガティブコントロール;レーン3:変異体aの上清;レーン4:ネガティブコントロール;レーン5:変異体bの上清;発現された担に対して特異的な一次抗体およびシグナルの生成のための二次抗体を使用した。
【0168】
図11:標的基質のタンパク質分解切断の時間経過。ヒトトリプシンの遺伝子を有するベクターを含有する細胞の上清、および遺伝子を含まないベクターを含有する細胞の上清を、本文中に記載したペプチド基質とインキュベートした。ペプチドの切断により、読み出し値が低下した。タンパク質分解活性は、陽性クローンについて確認される。
【0169】
図12:異なるペプチド基質に対するヒトトリプシンIと比較した3つの人工タンパク質分解酵素の相対活性。2つの基質のタンパク質分解消化の時間経過を行い、評価した。基質Bをスクリーニングに使用し、基質Aは密接に関連した配列である。相対活性;3つの変異体の相対活性を、ヒトトリプシンIに対して活性に対して正規化した。変異体1および変異体2は、標的基質に対して明らかに増大した特異性を示す。一方、変異体3は、ヒトトリプシンIと類似した活性を有するネガティブコントロールとして働く。
【0170】
図13:それぞれ、トリプシン、および1つまたは2つのSDRを有する人工タンパク質分解酵素の変異体の相対活性。プロテアーゼの活性を、競合基質の存在下(すなわち、10mg/mlの濃度のペプトン)および非存在下で決定した。タンパク質分解切断の時間経過を記録紙、時間定数kを決定した。競合剤ありとなしとの間の時間定数の比が形成され、プロテアーゼの特異性の定量的尺度を表す。比をトリプシンに対して正規化した。2つのSDRを含有する変異体の特異性は、SDR2のみを有する変異体の特異性よりも2.5倍高かった。
【0171】
図14:競合基質の非存在下および存在下でのプロテアーゼ変異体の相対的特異性を示す。異なる配列を有する2つのインサートを含有するプロテアーゼ変異体はおよび非修飾足場ヒトトリプシンIを適切な宿主のおいて発現した。プロテアーゼ変異体の活性を、競合基質の非存在下および存在下でのTNFアルファの所望の標的配列を有するペプチドの切断速度として決定した。特異性を、競合剤の存在下および非存在下での切断速度の比として表した。
【0172】
図15:図は、ヒトTNFアルファに対して特異的な本発明の人工タンパク質分解酵素を発現する培養物由来の濃縮された上清とともにヒトTNFあるいはフラグメントをインキュベートした時のヒトTNFアルファによって誘導される細胞傷害性の減少を示す。これは、本発明の人工タンパク質分解酵素の効率を示す。
【0173】
図16:図は、ヒトTNFアルファに対して特異的な異なる濃度の精製した本発明の人工タンパク質分解酵素とともにヒトTNFアルファをインキュベートした時のヒトTNFアルファによって誘導される細胞傷害性の減少を示す。変異体gは、SDR1として配列番号72およびSDR2として配列番号73を含む。これは、本発明の人工タンパク質分解酵素の効率を示す。
【0174】
図17:図は、2つのタンパク質基質(a)ヒトTNFアルファ、(b)ヒト血清タンパク質の混合物において、ヒトTNFアルファに対して特異的な本発明の人工タンパク質分解酵素の活性を、ヒトトリプシンIの活性と比較する。これは、本発明の人工タンパク質分解酵素の安全性を示す。変異体xは、配列番号89(SDR1)および配列番号95(SDR2)に従うSDRを含有する配列番号75に対応する。変異体xiおよびxiiは、同じSDR配列を含有するその誘導体に対応する。
【0175】
図18:ヒトトリプシン由来の人工タンパク質分解酵素によるヒトVEGFの特異的加水分解。
【実施例】
【0176】
以下の実施例では、本発明の方法により得られた酵素の触媒特性の決定を含む本発明の材料および方法が提供される。これらの実施例は、例示の目的のためのみであり、いかなる様式においても本発明を限定すると解釈されるものではないことは理解されるべきである。ここに引用される全ての文献、特許、および特許出願は、その全体が、本明細書中で参考として援用される。
【0177】
以下に記載される実験例では、種々の文献(例えば、Sambrook et al. (1989)、Molecular Cloning : A Laboratory Manual、Cold Spring Harbor Laboratory、or Ausubel et al. (1987)、Current Protocols in Molecular Biology 1987−1988、Wiley Interscience)に記載された組換えDNA技術の標準技術を使用した。特に示されない限り、製造業者の指示書に従って、制限酵素、ポリメラーゼ、および他の酵素、ならびにDNA精製キットを使用した。
【0178】
(実施例I:ヒトトリプシンにおけるSDR部位の同定)
SDRの挿入部位は、セリンプロテアーゼヒトトリプシンI(構造クラスS1)において、より高い配列特異性を有する同じ構造クラスのメンバーとの比較によって、同定されている。トリプシンは、P1位置にアルギニンまたはリジン残基のみを必要とするので、低い基質特異性を有するメンバーである。他方、トロンビン、組織型プラスミノーゲン活性化因子、またはエンテロキナーゼは全て、それらの基質配列、すなわち、(L/I/V/F)XPR^NA、CPGR^VVGG、およびDDDK^それぞれに対して高い特異性を有する。これらおよびさらいS1セリンプロテアーゼの一次配列および三次構造は、配列および構造の低いおよび高い相同性の領域、特により特異的なプロテアーゼの配列の挿入に対応する領域を決定するためにアラインメントされている(図2)。潜在的SDR部位を表す3アミノ酸と同じかまたはそれよりも長い挿入のいくつかの領域は、図1に示されるように同定されている。これらの領域を、以下の実施例においてSDRの挿入のための標的部位として選択した。例えば、それぞれ、6の長さを有するSDR1(図2における領域1、配列番号1のアミノ酸42の後)および5アミノ酸の長さを有するSDR2(図2の領域3、配列番号1のアミノ酸23の後)。
【0179】
(実施例II:足場タンパク質として使用されるヒトトリプシンI遺伝子の分子クローニングおよびB.subtilisにおける成熟プロテアーゼの発現)
非特異的プロテアーゼヒトトリプシノーゲンIをコードする遺伝子を、ベクターpUC18にクローニングした。クローニングを以下のとおり行った。タンパク質のコード配列を、KpnI部位を5’末端に、そしてBamHI部位を3’末端に導入したプライマーを使用してPCRによって増幅した。このPCRフラグメントを、ベクターpUC18の適切な部位にクローニングした。同一性を配列決定により確認した。配列決定した後、成熟タンパク質のコード配列を、異なるBglI部位を5’末端および3’末端に導入したPCRによって増幅した。
【0180】
このフラグメントを、大腸菌B.subtilisシャトルベクターの適切な部位にクローニングした。このベクターは、大腸菌中のpMB1増幅起点、大腸菌中での選択のためのネオマイシン耐性マーカー、ならびにB.subtilisにおける構成的発現のためのP43プロモーターを含有する。B.subtiilsのsacB遺伝子由来のシグナルペプチドをコードするリーダー配列を含有する87bpのフラグメントを、P43プロモーターの後ろに導入した。異なるBglI制限部位は、異種遺伝子が発現するための挿入部位として作用する。
【0181】
ヒトトリプシンIの発現を、遺伝子を有するベクターを含有する細胞の上清中でタンパク質分解活性を、ネガティブコントロールと比較して測定することにより確認した。アルギニン切断部位を含むペプチドを基質として選択した。ペプチドは、N末端ビオチン化および蛍光色素によりC末端を標識された。そのペプチドを培養上清とともにインキュベートした後、ストレプトアビジンを添加した。切断されていないペプチドはストレプトアビジンと結合し、高い読み出し値を示すが、切断されたものは低い読み出し値を生じる。図11は、トリプシンI遺伝子を含まないベクターを含有するB.subtilis細胞(ネガティブコントロール)と比較して、トリプシンI遺伝子を含有するB.subtilis細胞のタンパク質分解消化の時間経過を示す。プロテアーゼの発現のさらなる確認として、遺伝子を有するベクターを含有する細胞およびコントロール細胞の上清を、ポリアクリルアミドゲル電気泳動、およびその後の標的プロテアーゼに対する抗体を使用したウエスタンブロットによって分析した。手順は、標準的な方法にしたがって行った(Sambrook、J. F; Fritsch、E. F.; Maniatis、T.; Cold Spring Harbor Laboratory Press、Second Edition、1989、New York)。図8は、トリプシンの遺伝子を有するベクターを有する細胞中のみのタンパク質の発現を確認する。
【0182】
(実施例III:足場タンパク質の提供)
この実施例では、ヒトトリプシンIを足場タンパク質として使用した。遺伝子を、その天然の形態で使用したか、または触媒活性が増大し、もしくはさらに改善した特性を有する足場タンパク質を生じるように修飾した。
【0183】
修飾は、遺伝子のランダム修飾、次いでその酵素の発現、および続いて活性の増大したものの選択によって行った。第一に、遺伝子を、本質的にCadwell、R.C.およびJoyce、G.F.(PCR MethodsAppl. 2 (1992) 28−33)によって記載されるように変異性(error−prone)条件下でPCR増幅した。変異性PCRを、30pmolの各プライマー、20mmolのGTPおよびdATP、100nmolのCTPおよびdTTP、20fmolのテンプレート、および5UのTaqDNAポリメラーゼを、10mM TrisHCl(pH7.6)、50mM KCl、7mM MgCl2、0.5mM MnCl2、0.01% ゼラチン中で、94℃で1分、65℃で1分、および72℃で1分の20サイクルで行った。得られるDNAライブラリーを、Qiaquick PCR精製キットを供給業者の指示書に従って使用して精製した。PCR産物を制限酵素BglIを用いて消化し、精製した。その後、PCR産物を、上記のように大腸菌B.subtilisシャトルベクター中へライゲーションし、これをBglIで消化して、脱リン酸化した。ライゲーション産物を大腸菌へ形質転換し、LB中で増幅し、そしてプラスミドをQiagenプラスミド精製キットを供給業者の指示書にしたがって使用して精製した。得られるプラスミドをB.subtilis細胞に形質転換した。
【0184】
あるいは、またはランダム変異誘発に加えて、遺伝子の変異体を、本質的にWO 0134835に記載されるように、組換え連鎖反応を使用して相同位置で統計学的に組み換えた。プロテアーゼ変異体をコードする遺伝子のPCR産物を、QlAquick PCR精製キットを供給業者の指示書にしたがって使用して精製し、アガロースゲル電気泳動によって正確なサイズを供給業者の指示書にしたがってチェックし、そして等モル量で混合した。150mM TrisHCl(pH7.6)、6.6mM MgCl2中のこのPCR混合物の80μgを、94℃で5分間加熱し、続いて1秒ごとに0.05℃で37℃まで冷却し、鎖を再アニーリングし、それによりヘテロ二重鎖を確率論的な様式で生成した。次いで、1ΜGのDNA当たり2.5U エンドヌクレアーゼIIIを添加し、20、40、または60分間、37℃でインキュベートして、ヘテロ二重鎖の両方の3’末端から異なる長さを消化した。部分的に消化したPCR産物を、供給業者の指示書にしたがって0.17mM dNTPおよびPfuポリメラーゼ緩衝液中で72℃で15分間インキュベートすることによって、1μg DNA当たり0.6U Pfuのポリメラーゼで再充填した。一回のPCRサイクルを行った後、得られたDNAをQIAquick PCR精製キットを供給業者の指示書にしたがって使用して精製し、BglIで消化し、直線化したベクターにライゲーションした。ライゲーション産物を大腸菌中へ形質転換し、アンピシリンをマーカーとして含有するLB中で増幅し、そのプラスミドをQiagenプラスミド精製キットを供給業者の指示書に従って使用して精製した。得られたプラスミドを、B.subtilis細胞中へ形質転換した。
【0185】
(実施例IV:ヒトトリプシンIのタンパク質足場へのSDRの挿入および配列KKWLGRVPGGPVを有するペプチド基質に対する特異性を有する人工タンパク質分解酵素の生成)
ヒトトリプシンI中にSDRのための挿入部位を作製するために、異なる制限部位の2つのペアを、アミノ酸配列を変更せずに潜在的SDR部位として同定された部位(上記実施例Iを参照)に遺伝子を導入した。制限部位の挿入は、オーバーラップPCRによって行われた。プライマーrestrIおよびrestr2をSacIIおよびBamHI制限部位の導入のために使用し、restr3およびrestr4を、KpnIおよびNheI制限部位の導入のために使用した。プライマーの配列は以下の通りである:
【0186】
restr1およびrestr2についての結合部位および対応するアミノ酸配列(配列番号54):
【化1】
フォワードプライマーrestr1(配列番号56):
【化2】
リバースプライマーrestr2(配列番号57):
【化3】
restr3およびrestr4についての結合部位および対応するアミノ酸配列(配列番号58):
【化4】
フォワードプライマーrestr3(配列番号60):
【化5】
リバースプライマーrestr4(配列番号61):
【化6】
【0187】
第一のオーバーラップ伸長PCRにおいて、SacII/BamHI部位を導入し、SDR1を挿入できるようにし、第二のオーバーラップ伸長PCRにおいて、KpnI/NheI部位を導入し、SDR2を挿入できるようにした。オーバーラップ伸長PCRの産物を、プライマーpUCフォワードおよびpUCリバースを用いて増幅した。pUCフォワードおよびpUCリバースの配列は、以下の通りである:
pUCフォワード(配列番号62):
【0188】
5’−GGGGTACCCCACCACCATGAATCCACTCCT−3’
pUCリバース(配列番号63):
【0189】
5’−CGGGATCCGGTATAGAGACTGAAGAGATAC−3’
【0190】
続いて、それにより生成された制限酵素部位を使用して、規定されたまたはランダムオリゴヌクレオチドをSDR1およびSDR2挿入部位に、標準的な制限およびライゲーション法によって挿入した。典型的には、2つの相補性合成5’リン酸化オリゴヌクレオチドをアニーリングし、それぞれの制限酵素で切断した修飾ヒトトリプシンI遺伝子を有するベクターにライゲーションした。SDR1をコードするオリゴヌクレオチドを、SacII/BamHI部位を介して挿入し、一方、SDR2をコードするオリゴヌクレオチドを、KpnI/NheI部位を介して挿入した。各挿入について、以下の一般的な配列に従うオリゴヌクレオチドペアを使用した([P]は、5’リン酸化を示す。NおよびXは、任意のヌクレオチドまたはアミノ酸残基をそれぞれ示す。):
oligox−SDR1f(配列番号64):
【化7】
oligox−SDR1r(配列番号66):
【化8】
oligox−SDR2f(配列番号67):
【化9】
oligox−SDR2r(配列番号69):
【化10】
【0191】
上記方法の代替方法として、修飾されたヒトトリプシンI中のSDR1およびSDR2挿入部位へのランダム配列の組み込みのためにPCRに基づく方法が使用される。各SDRについて、1つのプライマーが使用され、ここでSDR領域は、完全にランダム化されている。プライマーの配列は以下の通りである(N=A/C/G/t、B=C/G/T、V=A/C/G):
プライマーSDR1−mutnnbフォワード(配列番号70):
【化11】
プライマーSDR2−mutnnbリバース(配列番号71):
【化12】
【0192】
さらなる代替方法として、特異性を増大させるSDRを同定した後、これらのSDRをさらなるランダム化のための鋳型として使用した。それにより、各位置で部分的にランダム化したランダム化したランダムペプチド配列、および元々の配列に対して各位置で同一部分的に同一であるランダムペプチド配列を挿入した。
【0193】
例として、3つの場合のなかの約1において鋳型アミノ酸残基を有し、3つの場合のうちの約2において任意の他のアミノ酸残基を各位置で有するランダムペプチド配列を、修飾ヒトトリプシンLの2つのSDR挿入部位に挿入した。この目的のために、SDRの各ヌクレオチド位置で鋳型塩基の約70%および3つの他の塩基の混合物を30%を含有するプライマーを使用した。各プライマーペアを用いて、標準的な条件下でヒトトリプシンI遺伝子を鋳型として用いてPCRを行った。得られたDNAをQIAquickPCR精製キットを、供給業者の指示書に従って使用して精製し、SacIIおよびNheIで消化した。消化後、DNAを精製し、SacIIおよびNheI消化され脱リン酸化されたベクターにライゲーションした。ライゲーション産物を、大腸菌に形質転換し、それぞれのマーカーを含有するLB中で増幅し、そのプラスミドをQiagenプラスミド精製キットを供給業者の指示書にしたがって用いて精製した。得られたプラスミドをB.subtilis細胞中へ形質転換した。次いで、これらの細胞を、単一の細胞へ分離し、クローンへと増殖させ、プロテアーゼ遺伝子の発現後、タンパク質分解活性についてスクリーニングした。
【0194】
以下の基質を、タンパク質分解活性のスクリーニングのために使用した(配列番号76および77)
【数1】
【0195】
プロテアーゼ変異体を、基質Bについて、共焦点蛍光顕微鏡によって106変異体の複雑性(complexities)でスクリーニングした。基質は、N末端でビオチン化され、C末端で蛍光標識されていた。ペプチドをプロテアーゼの異なる変異体を発現する細胞の上清とともにインキュベーションした後、ストレプトアビジンを添加し、そのサンプルを共焦点蛍光定量法によって分析した。ペプチドの低い濃度(20nM)は、高いkcat/KM値を有するプロテアーゼ(すなわち、標的配列に対する高い特異性を有するプロテアーゼ)による優先的な切断を導いた。
【0196】
スクリーニング手順において選択された変異体を、さらにそれらの基質Bおよび密接に関連する基質Aに対する特異性について、タンパク質分解酵素消化の時間経過を測定することにより、およびkcat/KM値に比例する速度定数を決定することにより、評価した。明らかに、足場タンパク質として使用されたヒトトリプシンと比較して、変異体1および2の特異的活性は、基質Bのほうへシフトしている(それぞれ、配列番号2および3)。他方、変異体3(配列番号4)は、ヒトトリプシン1と同様の活性を有するネガティブコントロールとして作用する。3つの変異体の遺伝子の配列決定により、SDR中の以下のアミノ酸配列が明らかになった。
表2:基質Bの特異的加水分解について選択された3つの異なる変異体中の2つのSDRの配列(配列番号78〜83)
【表2】
【0197】
さらなる実験において、1遺伝子当たり異なる数のSDRを含有する変異体のプールを、規定された基質および競合基質としてのペプトンの混合物を使用して、増大した特異性に関してスクリーニングした。1遺伝子当たりのSDRをさらに分析した。特異性の指標として、ペプチド切断アッセイにおける活性を、競合基質の存在下および非存在下で比較した。競合基質の濃度は10mg/mlであった。これらの条件下で、非特異的プロテアーゼは、特異的プロテアーゼと比較して、競合剤濃度の増加につれて(0〜100mg/mlの範囲)より強い活性の減少を示す。基質ありとなしのタンパク質分解活性の比は、プロテアーゼの特異性の定量的指標である。図9は、競合基質ありとなしとの相対的活性を示す。足場タンパク質として使用されたヒトトリプシンIおよび2つの変異体(1つはSDR2のみを含み、1つは両方のSDRを含む)を比較した。両方のSDRを有する変異体の特異性は、SDR2のみを有する変異体の特異性よりも2.5倍高い。これは、SDRの数と得られる人工タンパク質分解酵素の定量的特異性との間に直接的関連性があることを確証する。
【0198】
(実施例V:ヒトTNFアルファを特異的に不活性化する人工タンパク質分解酵素の生成)
ヒトトリプシンアルファI、または1つ以上の下記のアミノ酸置換E56G;R78W;Y131F;A146T;C183Rを含む誘導体を、ヒトTNFアルファに対する高い特異性を有する人工タンパク質分解酵素の生成のためのタンパク質足場として使用した。ヒトトリプシンIまたはその誘導体におけるSDR部位の同定を、上記のように行った。足場内の2つの挿入部位を、SDRについて選択した。異なる配列を有する2つの挿入断片を含有するプロテアーゼ変異体および挿入物を有さないヒトトリプシンI自体は、枯草菌細胞内で発現された。変異体プロテアーゼ細胞を、単一細胞クローンに分離し、プロテアーゼ発現変異体を、TNFアルファの所望の標的配列を有するペプチドにおけるタンパク質分解活性についてスクリーニングした。プロテアーゼ変異体の活性を、競合基質の存在下または非存在下でのTNFアルファの所望の標的配列を有するペプチドの切断速度として決定した。特異性を、競合剤の存在下および非存在下における切断速度の比として表した(図14)。
【0199】
表3:競合基質の存在下または非存在下での異なるSDR配列を有する人工タンパク質分解酵素の変異体の相対活性(配列番号84−95)。
【表3】
【0200】
3つの本発明のプロテアーゼ変異体のヒトTNFアルファに対する拮抗性効果を、図15に示す。変異体の使用により、アポトーシスの誘導は、ほとんど完全に排除された。これはTNFアルファの分解を開始する本発明のプロテアーゼの抗炎症性効果を示す。TNFアルファを、変異体i〜iiiを発現する培養由来の濃縮した上清とともに2時間インキュベートした。得られたTNFアルファを、非修飾細胞とともに4時間インキュベートした。残りのTNFアルファ活性の効果を、アポトーシス細胞のマーカーとしての活性化カスパーゼ3を検出することによって、アポトーシスの誘導の程度として決定した。コントロールとして、それぞれ、ヒトTNFアルファ(死細胞)にプロテアーゼを加えないか、またはヒトTNFアルファ(生細胞)の代わりに緩衝液を使用した。精製した変異体xiiiを使用した類似の実験を図16に示した。TNFアルファを、異なる濃度の精製した本発明のプロテアーゼ変異体とともにインキュベートした。
【0201】
本発明のプロテアーゼ変異体の特異性を実証するために、ヒト血清由来のタンパク質または精製したヒトTNFアルファを、ヒトトリプシンIまたは本発明の人工タンパク質分解酵素変異体とともにそれぞれインキュベートした。ここで、変異体xは、配列番号75に対応し、変異体fと同じSDR、すなわち、配列番号89(SDR1)および配列番号95(SDR2)のSDRを含む。変異体xiおよびxiiは、同じSDR配列を含むその誘導体に対応する。残りのインタクトなタンパク質を時間の関数として決定した。変異体ならびにヒトトリプシンIがヒトTNFアルファを消化するのに対して、トリプシンのみが血清タンパク質に対して活性を示す(図17aおよびb)。これは、本発明のタンパク質分解酵素の高いTNFアルファ特異性を示し、それらの安全性、したがって、それらの治療用途のための副作用の低さを示す。
【0202】
(実施例VI:ヒトVEGFを特異的に加水分解する人工タンパク質分解酵素の生成)
ヒトトリプシンIを、ヒトVEGFに対して高い特異性を有する人工タンパク質分解酵素の生成のためにタンパク質足場として使用した。ヒトトリプシンI内のSDR部位の同定は、上記のように行った。足場内の2つの挿入部位をSDRのために選択した。異なる配列を有する2つの挿入断片を含有するプロテアーゼ変異体を、枯草菌細胞中で発現させた。変異体プロテアーゼ細胞を、単一細胞クローンへと分離し、そのプロテアーゼ発現変異体を上記のようにスクリーニングした。プロテアーゼ変異体の活性をVEGF切断の速度として決定した。4μgの組換えヒトVEGF165を、室温でPBS/pH7.4中にて0.18μgの精製プロテアーゼとともにインキュベートした。示された時間にアリコートをとり、ポリアクリルアミドゲル上で分析した。切断の程度を、バンドの濃度測定分析によって定量した。活性をインキュベーション時間に対して図18にプロットした。特異的切断を、さらなるSDSポリアクリルアミドゲル分析によって制御した。
【0203】
以下の図面は、詳細な説明を補充し、本発明をさらに説明するために提供される。
【図面の簡単な説明】
【0204】
【図1】「球棒」表示で示される活性化部位残基を有するヒトトリプシンIの3次元構造を示す。印をつけた領域は潜在的SDR挿入部位を示す。
【図2】セリンプロテアーゼクラスS1ファミリーである3つのメンバー:ヒトトリプシン1、ヒトアルファトロンビン、およびヒトエンテロペプチダーゼの一次アミノ酸配列のアラインメントを示す(配列番号1、5、および6もまた参照)。
【図3】「球棒」表示で示された活性部位残基を有するサブチリシンの3次元構造を示す。番号を付された領域は、潜在的SDR挿入部位を示す。
【図4】セリンプロテアーゼS8ファミリーの4つのメンバー:サブチリシンE、フリン、PC1、およびPC5の一次アミノ酸配列のアラインメントを示す(配列番号7〜10もまた参照)。
【図5】「球棒」表示で示された活性部位残基を有するペプシンの3次元構造を示す。番号を付された領域は、潜在的SDR挿入部位を示す。
【図6】A1アスパラギン酸プロテアーゼファミリーの3つのメンバー:ペプシン、β−セクレターゼ、およびカテプシンDの一次アミノ酸配列のアラインメントを示す(配列番号11〜13もまた参照)。
【図7】「球棒」表示で示された活性部位残基を有するカスパーゼ7の3次元構造を示す。番号を付した領域は、潜在的SDR挿入部位を示す。
【図8】システインプロテアーゼクラスC14ファミリーのメンバーとしてのカスパーゼ7の一次アミノ酸配列を示す(配列番号14もまた参照)。
【図9】本発明の第3の局面を模式的に示す。
【図10】ネガティブコントロールと比較して、SDR1およびSDR2を有するヒトトリプシンIの変異体を発現する細胞の培養上清のウエスタンブロットを示す。
【図11】ヒトトリプシンIによる標的基質のタンパク質分解による切断の時間経過を示す。
【図12】本発明の人工タンパク質分解酵素の3つの変異体の、2つの異なるペプチド基質上でのヒトトリプシンIとの比較における相対的活性を示す。
【図13】ヒトトリプシンIおよび1つ以上のSDRを有する本発明の人工タンパク質分解酵素の相対的特異性をそれぞれ示す。
【図14】ヒトトリプシンIおよび、ヒトTNF−アルファの標的基質を有するペプチド上の足場を有数するヒトTNF−アルファに対して特異的な本発明の人工タンパク質分解酵素の相対的特異性を示す。
【図15】TNFアルファを、ヒトTNF−アルファに対して特異的な本発明の人工タンパク質分解酵素を発現する培養物からの濃縮された上清と組み合わせたときにTNFアルファによって誘導される細胞傷害性の減少を示す。
【図16】TNF−アルファを、ヒトTNF−アルファに対して特異的である精製された本発明の人工タンパク質分解酵素とインキュベートしたときの、TNF−アルファによって誘導される細胞傷害性の減少を示す。
【図17】ヒトTNF−アルファに対して特異的である本発明の人工タンパク質分解酵素の活性を、ヒトトリプシンIIの活性と、2つのタンパク質基質:(a)ヒトTNF−アルファ;(b)ヒトセリンタンパク質の混合物において比較する。
【図18】ヒトVEGFに対する特異性を有する本発明の人工タンパク質分解酵素の特異的活性を示す。
【技術分野】
【0001】
本発明は、疾患に関連する標的基質内の特異的なペプチド結合を加水分解することができる規定された特異性を有するプロテアーゼを含む医薬を適用することによって、疾患を処置するための方法を提供する。そのような特異性を有するプロテアーゼは、さらに関連する治療または診断の目的で使用され得る。
【背景技術】
【0002】
学術的および産業上の研究は、治療剤、研究用の薬剤、診断剤、栄養剤、個人医療用の薬剤、または産業上の薬剤として使用される機能的なタンパク質を継続的に探求する。今日、このような機能的タンパク質は、主に2つのカテゴリー、すなわち、天然タンパク質および人工(engineered)タンパク質に分類され得る。一方で、天然タンパク質は、自然界から、例えば、天然の単離物をスクリーニングすることによって、または多様な種からのゲノムを配列決定することによって発見される。他方、人工タンパク質は、典型的には公知のタンパク質に基づいており、そして改変された機能を獲得するために変異されている。本発明は、出発成分と比較して新規機能を有する人工タンパク質を開示する。そのようなタンパク質は、NBE(新規生物学的実体(New Biologic Entities))と呼ばれる。本願において開示されるNBEは、新たな基質特異性を有する人工酵素またはそのような人工酵素と他の機能性成分との融合タンパク質である。
【0003】
特異性は、酵素機能の必須の要素である。細胞は、多数の異なる高度に反応性の触媒からなる。それでもなお、細胞は、調和された代謝および高度に秩序立った3次元構造を維持し得る。これは部分的に、酵素の特異性、すなわち、それらに対応する基質の選択的な変換に起因する。特異性は定性的かつ定量的な特性である。特定の酵素の特異性は、標的分子の1つの特定の型から特定の化学構造を有する全ての分子型に至るまで広範に変化し得る。本来、生物の酵素の特異性は、その生物の特定の必要性に対して進化してきている。治療、研究、診断、栄養学、または産業上の適用に高価値を有する任意の特異性は、可能な特異性のスペースが大きいために任意の生物の酵素のレパートリーで見出される可能性は低い。このような特異性を得る唯一の現実的な方法は、新たにそれらを作製することである。
【0004】
酵素を結合剤と比較する場合、特異性の枠組みは、大きな分子内において小さな異なる構造として個々のエピトープを認識する抗体によって与えられる。天然に存在する非常に広範な抗体の特異性は、天然の選択と組み合わされた免疫系によって生成される多様性に起因する。いくつかの機構が、抗体特異性の広範なレパートリーに貢献し、免疫応答生成および抗体成熟の異なる段階で生じる(Janeway、C et al.(1999)Immunobiology、Elsevier Science Ltd.、Garland Publishing、New York)。具体的には、抗体は、非常に特異的に抗原と相互作用し、非常に類似したエピトープとの間でさえも区別を可能にする相補性決定領域(CDR)を含む。抗体の軽鎖および重鎖は、各々3つの結合ドメインに対するCDRに寄与する。自然界は、CDRを生成するのに、種々の遺伝子セグメントの組換えを、さらに変異誘発と組み合わせて使用する。結果として、6つのCDRループの配列は、組成および長さにおいて高度に多様であり、これにより抗体の結合特異性の多様性の基礎を形成する。多様な触媒特異性の生成のための同様の原則は、自然界からは知られていない。
【0005】
触媒作用、すなわち、特定の化学反応の速度の増大は、結合すること以外に、最も重要なタンパク質機能である。触媒タンパク質、すなわち、酵素は、それらが触媒する化学反応に従って分類される。
【0006】
トランスフェラーゼは、基、例えば、メチル基またはグリコシル基を、1つの化合物(一般的にはドナーと見なされる)から別の化合物(一般的にはアクセプターと見なされる)へ移動させる酵素である。例えば、グリコシルトランスフェラーゼ(EC 2.4)は、ドナー分子からアクセプター分子へグリコシル残基を移動させる。グリコシルトランスフェラーゼのなかにはまた、加水分解を触媒するものもあり、これらはグリコシル基をドナーから水へ移動させると見なされ得る。サブクラスは、さらにヘキソシルトランスフェラーゼ(EC 2.4.1)、ペントシルトランスフェラーゼ(EC 2.4.2)、および他のグリコシル基(EC 2.4.99、国際生化学および分子生物学連合命名法委員会(Nomenclature Committee of the International Union of Biochemistry and Molecular Biology (NC−IUBMB)))へとさらに分割される。
【0007】
酸化還元酵素は、酸化還元反応を触媒する。酸化される基質は、水素または電子ドナーと見なされる。酸化還元酵素は、脱水素酵素、オキシダーゼ、モノオキシゲナーゼ、およびジオキシゲナーゼとして分類される。脱水素酵素は、水素を水素ドナーから水素アクセプター分子へ移動させる。オキシダーゼは、水素アクセプターとしての分子酸素と反応し、酸化生成物ならびに過酸化水素または水のいずれかを生成する。モノオキシゲナーゼは、分子酸素から酸素原子1個を基質へ移動させ、そして1つは水へと還元される。対照的に、ジオキシゲナーゼは、分子酸素から両方の酸素原子を基質へと挿入させることを触媒する。
【0008】
リアーゼは、排除反応を触媒し、それにより二重結合を生成するか、または逆方向において二重結合での付加を触媒する。イソメラーゼは、分子内再配置を触媒する。リガーゼは、ATP消費の代償として化学結合の形成を触媒する。
【0009】
最終的に、加水分解酵素は、C−OまたはC−Nのような化学結合の加水分解を触媒する酵素である。これらの酵素のE.Cの分類は、一般的に、それらを、加水分解される結合の性質によって、および基質の性質によって分類する。リパーゼおよびプロテアーゼのような加水分解酵素は、生来、および生体触媒の技術的な応用において重要な役割を果たす。プロテアーゼは、オリゴペプチドまたはポリペプチドの状況において、ペプチド結合を加水分解する。触媒機構に依存して、プロテアーゼは、アスパラギン酸、セリン、システイン、メタロ、およびスレオニンプロテアーゼに分類される(Handbook of proteolytic enzymes. (1998) Eds: Barret、A; Rawling、N. ; Woessner、J. ; Academic Press、London)この分類は、触媒作用を担うアミノ酸側鎖に基づいており、これは典型的には、互いに非常に類似して配向した活性部位に示されている。基質の切れやすい結合は、基質のアミノ酸側鎖とプロテアーゼの相補的な領域との間の特異的な相互作用に起因して触媒性残基と一致するようにされる(Perona、J. & Craik、C (1995) Protein Science、4、337−360)。切れやすい結合のN末端側およびC末端側の残基は、通常、P1、P2、P3など、およびP1’、P2’、P3’と呼ばれ、基質に相補的な結合ポケットは、S1、S2、S3およびS1’、S2’、S3’と、それぞれ呼ばれる(Schlechter & Berger、Biochem. Biophys. Res. Commun. 27 (1967) 157−162に基づく命名法)。プロテアーゼの選択性は、実質的に非選択的(例えば、サブチリシンのP位置での厳格な優先傾向;例えば、トリプシンはアルギニンまたはリジン残基のC末端側を選択的に切断する)から、非常に特異的なプロテアーゼまで(例えば、ヒト組織型プラスミノーゲン活性化因子(t−PA)は、配列CPGRVVGにおけるアルギニンのC末端側を切断する(Ding、L etal. (1995) Proc. Natl. Acad. Sci. USA 92、7627−7631; Coombs、Get al. (1996)J.Biol. Chem. 71、4461−4467)非常に様々である。
【0010】
プロテアーゼの特異性、すなわち、特定の基質を優先的に認識し加水分解する能力は、定性的におよび定量的に表現することができる。定性的な特異性は、ペプチド基質の特定の位置でプロテアーゼにより受け入れられるアミノ酸残基の種類のことをいう。例えば、トリプシンおよびt−PAは、定性的な特異性に関して関連している。なぜなら、これらのいずれも、P位置にアルギニンまたは類似の残基を必要とするからである。他方、定量的な特異性は、プロテアーゼにより基質として受け入れられるペプチド基質の相対数、またはより正確には、プロテアーゼによって受け入れられる異なるペプチドに対するそのプロテアーゼの相対的kcat/kMの比のことをいう。全ての可能なペプチドの中のわずかな少量の部分のみを受け入れるプロテアーゼは、高い特異性を有する。ここで、極端な場合として、任意のペプチド基質を切断するプロテアーゼの特異性は、理論的にゼロである。
【0011】
プロテアーゼの一次構造、二次構造、ならびに三次構造の比較により(Fersht、A. 、Enzyme Structure and Mechanism、W. H. Freeman and Company、New York、1995)、高度の保存を示すクラスを同定することが可能である(Rawlings、N. D. & Barrett、A.J. (1997) In : Proteolysis in Cell Functions Eds. Hopsu−Havu、V. K.;Jarvinen、M. ;Kirschke、H、pp. 13−21、OS Press、Amsterdam)。プロテアーゼの分類として広く受け入れられているスキームは、Rawlings & Barrettによって提唱されている(Handbook of proteolytic enzymes. (1998) Eds: Barret、A; Rawling、N.; Woessner、J.; Academic Press、London)。例えば、セリンプロテアーゼファミリーは、キモトリプシン(クラスS1)、サブチリシン(クラスS8)、およびカルボキシペプチダーゼ(クラスSC)の集団を有する構造クラスにさらに分割され得、これらの各々は、非特異的プロテアーゼおよび特異的プロテアーゼを含む(Rawlings、N. D. & Barrett、A. J. (1994) MethodsEnzymol. 244、19−61)。これは、他のプロテアーゼファミリーにも同様に適用される。基質中の切断される結合の相対的な位置によって、さらなる区別がなされ得る。カルボキシペプチダーゼおよびアミノペプチダーゼは、C末端およびN末端から、それぞれアミノ酸を切断する。これに対して、エンドペプチダーゼは、オリゴペプチドに沿ってどこでも切断する。
【0012】
基本的に無限の種類の特異性を有する酵素が入手可能であるならば、多くの適用が考えられる。しかし、このような高い、低い、または任意の規定された特異性を有する酵素の使用は、現在のところ、天然源から単離され得るものに限定されている。このような酵素の適用分野は、治療、研究、診断、栄養学から、パーソナルケアおよび工業的用途まで様々である。
【0013】
洗剤中の酵素添加物は、工業的な酵素市場全体のほぼ3分の1を構成するまでになっている。洗剤酵素には、有機物によるシミを除去するためのプロテアーゼ、油汚れを除去するためのリパーゼ、でんぷんを含む食品の残渣を除去するためのアミラーゼ、および繊維のなめらかな表面を回復させるためのセルラーゼが含まれる。最も良く知られている洗剤酵素は、おそらく、種々のバチルス(Bacillus)種から単離される非特異的プロテアーゼであるサブチリシンである。
【0014】
アミラーゼのような酵素は、食品加工において使用される酵素の大部分を占めている。でんぷん酵素は、布地デザイン、アルコール発酵、紙およびパルプの加工、ならびに洗濯用洗剤添加物のために重要な製品を含むが、最大の適用は、高フルクトースコーンシロップの製造である。工業用酵素を用いたでんぷんからのコーンシロップの製造は、酸加水分解の代わりとして成功していた。
【0015】
でんぷん加工とは別に、酵素は、食品における増大する範囲の適用に使用されている。食品中の酵素は、きめ、外見、および栄養価値を改善することができ、または所望の風味およびアロマを生成することができる。パン屋さんで現在使用されている食品酵素は、アミラーゼ、アミログリコシダーゼ、ペントサンの分解および還元型グルテン産生のためのペントサンアーゼ(pentosanase)、またはパン生地の安定性を増大させるためのグルコースオキシダーゼである。乳製品用において一般的な酵素は、チーズ製造における凝固剤としてのレンネット(プロテアーゼ)、ラクトースの加水分解のためのラクターゼ、乳清タンパク質の加水分解のためのプロテアーゼまたは過酸化水素除去のためのカタラーゼである。醸造工程において使用される酵素は、上記のアミラーゼであるが、セルラーゼまたはタンパク質懸濁物からビールを清澄化するためのプロテアーゼもまた使用される。ワインおよびフルーツジュース中では、濁りは、より一般的にでんぷんおよびペクチンによって生じるため、アミラーゼおよびペクチナーゼは、収量および透明度を増大させる。パパインおよびその他のプロテイナーゼは、肉を柔らかくするために使用される。
【0016】
酵素はまた、飼料を消化することにおいて動物を助けるために開発されてきた。西半球において、コーンは、ウシ、ブタ、および家禽の主要な食料源である。コーンからのリン酸のバイオアベイラビリティーを改善させるために、フィターゼが一般的に添加される(Wyss、M. etal.、Biochemical characterization of fungal phytases (myo−inositolhexakisphosphate phosphohydrolases) ; Catalytic properties. Applied & Environmental Microbiology 65、367−373 (1999))。さらに、フィターゼ加水分解は、タンパク質の消化およびカルシウムのようなミネラルの吸収を改善することが示されている(Bedford、M. R. & Schulze、H.、Exogenous Enzymes for Pigs and Poultry [Review]. Nutrition Research Reviews 11、91−114(1998))。別の主要な飼料用酵素は、キシラナーゼである。この酵素は、その相対的に高い溶解性線維含有量のために、約10%を超える小麦、大麦、もしくはライ麦を含む飼料の補助剤として特に有用である。キシラーゼは、2つの重要な作用:飼料中のゲル様高分子量アラビノキシランを加水分解することによって腸内容物の粘度を低減させる(Murphy、T.C.、Bedford、M.R.&McCracken、K.J. Effect of range of newxylanases on in vitro viscosity and on performance of broiler diets. British Pultry Science 44、S16−S18 (2003));およびタンパク質およびでんぷんのバイオアベイラビリティーを改善する細胞壁中のポリマーの分解。
【0017】
バイオテクの研究開発の研究室は、日常的に、多くの他の試薬とともに少量の特別な酵素を使用する。アルカリホスファターゼ、西洋ワサビペルオキシダーゼ、およびルシフェラーゼのような酵素は、そのうちのいくつかの例に過ぎない。Taqポリメラーゼまたは制限エンドヌクレアーゼのような熱安定性DNAポリメラーゼは、研究室での作業に革命を起こした。治療用酵素は、薬物の特定のクラスであり、FDAによって、他の、特に非生物学的製剤と比較して多くの利点を有する生物学的製剤として分類されている。成功している治療酵素の例は、ヒトの治療に用いる第VIII因子および第IX因子のようなヒト凝固因子である。さらに、ヒトの消化プロセスにおける種々の欠陥のために消化酵素が使用される。他の例は、心血管疾患の処置のためのt−PAおよびストレプトキナーゼ、I型ゴーシェ病の処置のためのベータグルコセレブロシダーゼ、急性リンパ芽球性白血病の処置のためのL−アスパラギナーゼ、および嚢胞性線維症の処置のためのDNAseである。治療剤としてのタンパク質の適用における重要な事項は、それらが免疫原性を有し得ることである。このリスクを軽減するために、ヒト由来の酵素が好まれ、これによって利用可能な酵素のセットが狭められる。新規のテイラーメードの特異性を有する設計された酵素(好ましくは、ヒト由来の)を提供することにより、意図的に標的基質を特異的に改変することが可能となり、他方免疫原性のリスクを最小限にすることができる。治療剤としての高度に特異的な酵素のさらなる利点は、副作用の危険性が低いことである。低分子とタンパク質との間の特異的な相互作用の可能性を限定することにより、標的ではないタンパク質への結合およびそれゆえ副作用が非常に一般的であり、しばしばそれがなければ有用であるリード化合物の使用を中止することになる。他方、特異的酵素は、基質を区別するためのより多くの接触部位および機構を提供し、それゆえより高い特異性が可能であり、それによって副作用の活動が低減される。
【0018】
プロテアーゼは、治療剤の重要なクラスを表す(Drugs of today、33、641−648 (1997))。しかしながら、現在、治療用のプロテアーゼは、通常、身体自体のプロテアーゼの活動が不十分な場合の代替物である。例えば、第VII因子は、血友病者の凝固障害または外科手術の際の特定の症例において投与され得る(Heuer L. ;Blumenberg D、(2002) Anaesthesist 51: 388)。組織型プラスミノーゲン活性化因子(t−PA)は、急性心筋梗塞において適用され、特異的な切断およびプラスミノーゲンの活性化を介してフィブリン凝血塊の溶解を開始する(Verstraete、M. et al. (1995) Drugs、50、29−41)。これまでのところ、テイラーメードの特異性を有するプロテアーゼが、疾患に関連する標的タンパク質を特異的に活性化または不活性化する治療剤を提供するために提供されている。
【0019】
モノクローナル抗体は、治療能を有する物質の別の重要な生物学的クラスである。主要な抗体標的の1つは、サイトカインのファミリーに属する腫瘍壊死因子(TNF)である。TNFは、炎症反応において主要な役割を果たす。ホモ3量体として、それらはほとんど全ての細胞の受容体と結合することができる。それらは、多数の細胞遺伝子、多数のシグナル伝達機構、キナーゼ、および転写因子を活性化する。最も重要なTNFは、TNFアルファおよびTNFベータである。TNFアルファは、マクロファージ、単球、およびその他の細胞によって産生される。TNFアルファは、炎症媒介因子である。それゆえ、過去10年間の研究は、異なる治療的症状(例えば、関節リウマチ、クローン病、または乾癬)の可能性のある治療剤として、モノクローナル抗体のようなTNFアルファ阻害剤に集中してきた(Hamilton et al. (2000) Expert Opin Pharmacother、1 (5): 1041−1052)。モノクローナル抗体の主要な欠点の1つは、それらが高価であることであり、それ故、新たな生物学的代替品が非常に重要である。
【0020】
文献には多数の人工酵素の例が存在する。Fulaniら(Fulani F. etal. (2003) Protein Engineering 16、515−519)は、Cdc25ホスファターゼ酵素の触媒サブユニットに構造的に関連する触媒ドメインを有するAzotobacter vinelandii由来のローダナーゼ(rhodanase)(チオスルファト:シアニドスルフルトランスフェラーゼ)を記載する。触媒機構の差違は、活性化部位の大きさの違いによる。ローダナーゼおよびホスファターゼのいずれも、異なる基質に対して高度に特異的である(硫酸対リン酸)。ローダナーゼの触媒機構は、1残基の挿入によってセリン/スレオニンホスファターゼに向かってシフトし得る。それゆえ、Fulaniらは、異なる酵素クラスからの天然に知られている酵素の構造比較および配列アラインメントによって触媒機構の変化の1つの例を示したが、同じ触媒機構を保ちながらユーザーが任意に定義可能な基質特異性をどのように生成するかについては示していない。
【0021】
Briggsら(WO02/090300 A2)によって記載されたチオレドキシンレダクターゼは、NADHに比べて優先的にNADPHに結合するようにコファクターの特異性を変化させた。このように、開始時の酵素および生成する人工酵素の両方の酵素が、異なる基質に対して高度に特異的である。このような変化した基質特異性を達成する方法は、コンピュータ処理による方法か、または可変残基および保存残基を決定するための関連するタンパク質の配列アラインメントのいずれかである。それらは全て、それらが公知の特異性を有するタンパク質の構造および配列の比較と、続いて同じ骨格から別の骨格への移動に基づいている点で共通している。
【0022】
特異性操作酵素、特に、文献に公開されたプロテアーゼの他の例が存在する。しかしながら、これらの例はいずれも、記載された方法において使用する出発物質の特異性と比べて新規な特異性を生成する手段を提供しない。その方法は、構造指向型単一点変異(Kurth、T. et al. (1998)Biochemistry 37、11434−11440 ; Ballinger、M et al. (1996)、2つの特異的プロテアーゼ間での表面ループの交換(Horrevoets et al. (1993) J. Biol. Chem. 268、779−782)から、インビトロもしくはインビボ選択と組み合わせた位置選択的または全遺伝子にわたったランダム変異誘発(Sices、H. & Kristie、T. (1998) Proc. Natl. Acad.Sci. USA、95、2828−2833)までにわたる。
【0023】
プロテアーゼ特異性の合理的な設計は、非常に少ない例に限られている。このアプローチは、タンパク質におけるフォールディングおよびダイナミクスならびに構造−機能関係を統御する複雑性の理解が不十分なことによって非常に限定されている(Corey、M.J. & Corey、E. (1996) Proc.Natl. Acad. Sci. USA、93: 11428−11434)。したがって、予測しながらプロテアーゼの活性または特異性を変化させるためにその一次アミノ酸配列を変化させることは困難である。1つの成功した例では、Kruthらは、トリプシンが二塩基性の誘因(motives)に対して優先性を示すように操作した(Kurth、T. et al. (1998) Biochemistry、37:11434−11440)。別の例では、Hedstromらが、トリプシンのS1基の特異性をキモトリプシンの特異性に変換した(Hedstrom、L. et al. (1992) Science、255: 1249− 1253)。これは、1つの既知の特性が1つの骨格から別の骨格へと移動した例である。
【0024】
Ballingerら(WO 96/27671)は、組み合わせ変異(N62D/G166D、および必要に応じて Y104D)を有するサブチリシン変異体を記載し、この変異体は、塩基性アミノ酸を基質のP1、P2、およびP4位に有する塩基性アミノ酸を有するペプチドまたはポリペプチド基質に向かっての基質特異性のシフトを有する。変異体サブチリシンの適切な基質は、5つの連続するランダム化残基を含有するファージ粒子(基質ファージ)のライブラリーをソーティングすることによって明らかになった。これらのサブチリシン変異体は、塩基性基質リンカーを有する融合タンパク質を切断するため、および塩基性切断部位を含有するホルモンまたは他のタンパク質(インビトロまたはインビボ)を加工するために有用である。合理的な酵素の設計に関連する問題は、定方向進化によって部分的に克服され得る(PCT/EP03/04864に開示される)。これらの研究は、それらの発現系および選択系によって分類され得る。遺伝子選択は、生物の内部に、結果として生成される生物の増殖挙動を変化させる前駆体タンパク質を切断することができる酵素(例えば、プロテアーゼ)を産生することを意味する。異なるプロテアーゼを有する生物の集団から、変化した増殖挙動を有するものが選択され得る。この原理は、例えば、Davisら(US5258289、WO96/21009)によって報告された。ファージ系の産生は、ファージタンパク質を切断することができるタンパク質分解酵素の存在下でのみ活性化され得るファージタンパク質の切断に依存する。他のアプローチは、遺伝子選択ではなくスクリーニングによる選択を可能とするレポーター系を使用するが、これもまた、酵素の細胞内特徴付けの本来的な不十分さを克服できない。
【0025】
自己分泌酵素を用いた変更された配列特異性を有する酵素を生成する系もまた、報告されている。Duffら(WO 98/11237)は、自己分泌プロテアーゼのための発現系を記載する。実験のデザインの必須の要素は、触媒反応が、膜結合前駆体分子の自己タンパク質分解プロセッシングによってプロテアーゼ自体に作用し、変異したプロテアーゼを細胞膜から細胞外環境に放出することである。それゆえ、標的ペプチド配列が天然の切断部位を自己タンパク質分解のために置換した融合タンパク質が構築されなければならない。このような系の制限は、肯定的に同定されたプロテアーゼが特定のアミノ酸配列を切断する能力を有するが、それらはまた多くの他のペプチド配列も切断し得ることである。それゆえ、高い基質特異性は、達成され得ない。さらに、このような系は、規定されたアミノ酸配列における特定の位置での選択されたプロテアーゼ切断を制御できず、選択されたプロテアーゼの速度定数の正確な特徴付けができない(kcat、KM)。
【0026】
α/β−バレルタンパク質内で新たな触媒活性および特異性の生成を目的とする方法が記載されている(WO 01/42432 ; Fersht et al、Methods of producingnovel enzymes ; Altamirano et al.(2000)Nature 403、617−622)。α/β−バレルタンパク質は、全ての既知の酵素の大きな部分を占めるタンパク質の大きなスーパーファミリーを含む。このタンパク質の構造は、α−ヘリクスによって囲まれたα/β−バレルからなっている。β−鎖およびヘリクスをつないでいるループは、活性部位残基を含む、いわゆる、リッド(lid(ふた))構造を含む。この方法は、触媒リッド構造に基づく2つのクラスへのα/β−バレルタンパク質の分類に基づく。α/β−バレルタンパク質構造の集中的な比較により、執筆者らは、基質結合および特異性は、基本的にバレル構造により規定されるのに対して、化学反応の特異性は、ループ内に存在するという結論に至った。異なる酵素からのバレルおよびリッド構造は、標的化またはランダム化された変異誘発および選択によって、新たな酵素活性を生成し、そして特性を最適化するための出発点を提供するために組み合わせられ得ることが示唆される。
【0027】
まとめると、治療、研究、および診断、工業酵素、食品および食品加工、化粧品、ならびに新規な基質特異性を有する酵素が入手可能になることによって可能になるその他の領域の分野において、多くの適用の可能性が存在することが明らかである。しかし、これまで、わずかに限られた数の特異的酵素が天然源から同定されているにすぎない。配列特異性を修飾し、変更し、または移動させるための合理的な設計方法ならびに上記のランダムアプローチは、利用された出発物質に存在しなかった新規かつユーザーが任意に決定することができる特異性の作成を可能にしなかった。
【0028】
それゆえ、現在利用可能な方法はいずれも、新規かつユーザーにより定義された配列特異性を有する酵素を提供することができない。対照的に、本発明は、このような酵素ならびにそれらを生成する方法を提供する。
【発明の開示】
【0029】
(発明の要旨)
本発明の目的は、人工タンパク質であって、このようなタンパク質を操作するために使用された成分中に存在しない新規な機能を有する人工タンパク質を提供することである。特に、本発明は、ユーザーが任意に定義することができる特異性を有する酵素を提供する。ユーザーが任意に定義することができる特異性とは、酵素が、そのような酵素の操作に使用した成分中に存在しない特異性を有して提供されることを意味する。特異性は、1つ以上の意図された標的基質が優先的に認識され、その酵素によって変換されるように、ユーザーによって選択され得る。さらに、本発明は、ヒトタンパク質と本質的に同一の配列を有するが、異なる特異性を有する酵素を提供する。1つの特定の実施形態では、本発明は、ユーザーが任意に定義することができる特異性を有するプロテアーゼを提供する。
【0030】
さらに、本発明は、1つ以上のさらなる成分と融合された人工酵素に関する。これらのさらなる成分は、好ましくは結合特性を有し、基質結合ドメイン、抗体、受容体、またはこれらのフラグメントからなる群のタンパク質様の成分であり得る。さらに、これらのさらなる成分は、さらなる機能的成分、好ましくは、ポリエチレングリコール、炭水化物、脂質、脂肪酸、核酸、金属、金属キレート剤、およびこれらのフラグメントまたは誘導体からなる群から選択され得る。生じる融合タンパク質は、本発明の範囲内のユーザーが任意に定義することができる特異性を有する酵素として理解される。
【0031】
加えて、本発明は、新規な、ユーザーが任意に定義することができる特異性を有するこのような酵素を、治療、研究、診断、栄養学、パーソナルケア、または工業目的に適用することに関する。さらに、本発明は、ユーザーが任意に定義することができる特異性を有する人工酵素を生成するための方法に関する。特に、本発明は、ヒト酵素と本質的に同一の配列を有するが、異なる特異性を有する酵素を生成することに関する。
【0032】
この課題は、下記の説明および請求の範囲において特定される本発明の実施形態によって解決されている。したがって、本発明は、
(1)下記の成分:(a)スクリーニング1つの基質におけるスクリーニング1つの化学反応を触媒するタンパク質足場、および(b)生じる人工タンパク質が少なくとも1つの標的基質と1つ以上の異なる基質とを区別できるようにする上記タンパク質足場中の部位に存在する1つ以上の特異性決定領域(SDR)であって、本質的に合成ペプチド配列である上記SDR、(c)の組み合わせによって特徴付けられる規定された特異性を有する人工酵素、
(2)治療、研究、診断、栄養学、パーソナルケア、または工業目的のための上記(1)に記載の人工酵素の使用、
(3)標的基質に対する特異性(その特異性は、個々の出発成分には存在しない)を有する上記(2)に記載の人工酵素を作製するための方法であって、少なくとも以下の工程:(a)少なくとも1つの基質上での少なくとも1つの化学反応を触媒するタンパク質足場を提供する工程、(b)工程(a)からの上記タンパク質足場を、完全にまたは部分的に、ランダムペプチド配列と、得られる人工酵素が少なくとも1つの標的基質と1つ以上の異なる基質とを区別できるようなタンパク質足場における部位で組み合わせることによって、人工酵素のライブラリーを作製する工程、ならびに(c)工程(b)で作製された人工酵素の上記ライブラリーから、少なくとも1つの標的基質に対する特異性を有する1つ以上の酵素を選択する工程、を含む方法、
(4)上記(1)に記載の少なくとも1つの人工酵素、ならびに少なくとも1つのさらなる成分、好ましくは、結合特性を有し、より好ましくは、抗体、結合ドメイン、受容体、およびこれらのフラグメントからなる群から選択される少なくとも1つのさらなる成分から構成される融合タンパク質、
(5)上記(1)に記載の1つ以上の人工酵素または上記(4)に記載の融合タンパク質を含む組成物または薬学的組成物であって、上記薬学的組成物は、必要に応じて、許容できる担体、賦形剤、および/もしくは助剤を含み得る、組成物または薬学的組成物、
(6)上記(1)に記載の人工酵素をコードするDNA、
(7)上記(6)に記載のDNAを含有するベクター、
(8)上記(7)に記載のベクターおよび/または上記(6)に記載のDNAを含有するベクターで形質転換/トランスフェクトされた宿主細胞ならびにトランスジェニック生物、ならびに
(9)上記(8)に記載の細胞または生物を培養する工程およびその培養液から酵素を単離する工程を含む人工酵素を製造する方法。
【0033】
(定義)
本発明の枠組みにおいては、以下の用語および定義が使用される。
【0034】
用語「プロテアーゼ」は、ペプチド結合を加水分解し得る任意のタンパク質分子を意味する。これは、天然に存在するタンパク質分解酵素または人工のタンパク質分解酵素、ならびに部位特異的変異誘発またはランダム変異誘発または任意の他のタンパク質操作法によって得られるこれらの変異体、タンパク質分解酵素の活性部位フラグメント、または上述のタンパク質の1つを含む任意の分子複合体もしくは融合タンパク質を含む。「プロテアーゼのキメラ」は、異なる親プロテアーゼに由来する2つ以上のフラグメントの融合タンパク質を意味する。
【0035】
用語「基質」は、酵素による触媒作用によって変換し得る任意の分子を意味する。用語「ペプチド基質」は、任意のペプチド、オリゴペプチド、またはプロテアーゼによって触媒作用により加水分解され得るペプチド結合を含む任意のアミノ酸組成物、配列もしくは長さのタンパク質分子を意味する。加水分解されるペプチド結合は、「切断部位」と呼ばれる。基質中の位置の番号付けは、Schlechter & Bergerによって導入されたシステムにしたがって行った(Biochem. Biophys. Res. Commun. 27 (1967) 157−162)。切断部位のN末端に隣接するアミノ酸残基は、P1、P2、P3などと番号付けされ、切断部位のC末端に隣接する残基は、P1’、P2’、P3’などと番号付けされる。
【0036】
用語「標的基質」は、本発明の酵素によって特異的に認識され、変換されるユーザーによって定義される基質を記載する。用語「標的ペプチド基質」は、ユーザーによって定義されるペプチド基質を記載する。用語「標的特異性」は、標的基質を認識し、変換することができる酵素の定性的および定量的な特異性を記載する。酵素の触媒特性は、ミカエリスおよびメンテンの定義に従う速度論パラメータ「KM」または「ミカエリスメンテン定数」、「kcat」または「触媒速度定数」、「kcat/KM」または「触媒効率」を使用して表される(Fersht、A. 、Enzyme Structure and Mechanism、W. H. Freeman and Company、New York、1995)。用語「触媒活性」は、規定された反応条件下での所定の基質の変換を定量的に記載する。
【0037】
用語「特異性」は、特定の基質を優先的に認識し、変換する酵素の能力を意味する。特異性は、定性的にも定量的にも表現することができる。「定量的特異性」とは、酵素によって認識される基質残基の化学的性質を意味する。「定量的特異性」とは、基質として受け入れられる基質の数のことをいう。定量的特異性は、全ての受け入れられる基質の数を全ての可能な基質の数で除算したものの負の対数として定義される語句「s」によって表され得る。例えば、全ての可能なペプチド基質の小部分を優先的に受け入れるプロテアーゼは、「高い特異性」を有する。ほとんど全てのペプチド基質を受け入れるプロテアーゼは、「低い特異性」を有する。定義は、WO 03/095670(これは本明細書中に参考として組み込まれる)に従ってなされる。非常に低い特異性を有するプロテアーゼはまた「非特異的プロテアーゼ」と呼ばれる。用語「規定された特異性」とは、特定の種類の特異性、すなわち、その他の基質に対して優先的に変換される特定の標的基質または特定の標的基質のセットのことをいう。
【0038】
用語「人工(engineered)」は、用語「酵素」と組み合わされて、異なる成分を含み、個々の成分のみによっては与えられない特徴を有する酵素を記載する。
【0039】
用語「タンパク質足場」または「足場タンパク質」とは、種々の一次、二次、および三次ポリペプチド構造のことをいう。
【0040】
用語「ペプチド配列」とは、タンパク質足場との挿入もしくは置換のため、または組み合わせのために使用される任意のペプチド配列を示す。ペプチド配列は、通常、周知の技術に従って合成され得るか、または天然源から得られ得るDNA配列から発現により得られる。タンパク質足場との挿入、置換、組み合わせは、オリゴヌクレオチドを、タンパク質足場をコードするポリヌクレオチドへ挿入、置換、または組み合わせることによって生成される。用語「合成」は、用語「ペプチド配列」と組み合わせて、そのペプチド配列が挿入もしくは置換されたか、またはそれが組み合わされたタンパク質足場に存在しないペプチド配列のことをいう。
【0041】
用語「成分」は、用語「人工酵素」と組み合わせて、このような酵素の操作において組み合わされるペプチド配列またはポリペプチド配列を意味する。このような成分は、とりわけ、1つ以上のタンパク質足場および1つ以上の合成ペプチド配列を含み得る。用語「人工酵素のライブラリー」は、人工酵素の混合物を記載し、そこではいかなる単一の人工酵素も異なるポリヌクレオチド配列によってコードされる。用語「遺伝子ライブラリー」は、人工酵素のライブラリーをコードするポリヌクレオチドのライブラリーを示す。用語「SDR」または「特異性決定領域」は、得られる酵素が標的基質と1つ以上の例えば、の基質とを区別できるような部位でタンパク質足場と組み合わされたときに規定された特異性を提供する合成ペプチド配列を意味する。このような部位は、「SDR部位」と呼ばれる。
【0042】
用語「の構造と類似した3次構造」および「類似した3次構造」は、用語「酵素」または「タンパク質」と組み合わされて、種類、配列、結合性、およびタンパク質の典型的2次構造要素の相対配位(例えば、アルファへリックス、ベータシート、ベータターン、およびループ)が類似しており、それゆえそのタンパク質が同じ構造的または位相幾何学的なクラスまたは集団にグループ分けされるタンパク質のことをいう。これは、任意の種類の変更された、追加の、または欠失された構造要素を有するが、それ以外は変化していない位相幾何学を有するタンパク質を含む。このような構造クラスの例は、TNFスーパーファミリー、セリンプロテアーゼ内のS1集団もしくはS8集団、GPCR、またはα/βバレル集団である。
【0043】
用語「構造的に対応する位置」は、互いに構造的に対応する類似の3次構造中のアミノ酸を示す。すなわち、それらは通常同じ構造またはその構造の位相幾何学的要素内に位置する。構造要素内で、それらはその構造要素の始めおよび終わりに対して同じ相対的位置を有する。例えば、2つのタンパク質を位相幾何学的に比較することにより、異なる長さの2つの構造的に対応する配列、次いで、例えば、それぞれの領域の長さの20%および40%内のアミノ酸が構造的に違いに対応することが明らかになる。
【0044】
用語「(本発明の)人工酵素のライブラリー」は、混合物または単離形態で存在し得る多数の酵素または酵素変異体のことをいう。
【0045】
アミノ酸残基は、以下の表1に従って、1文字または3文字表記で略称される。
【表1】
【発明を実施するための最良の形態】
【0046】
本発明は、新規の機能を有する人工タンパク質を提供する。特に、本発明は、ユーザーが任意に定義することができる特異性を有する酵素を提供する。特定の実施形態において、本発明は、ユーザーが任意に定義することができる特異性を有するプロテアーゼを提供する。加えて、本発明は、このような新規の、ユーザーが任意に定義することができる特異性を有する酵素の、治療、研究、診断、栄養学、パーソナルケア、または工業用途への適用を提供する。さらに、本発明は、このような酵素を操作するために使用された成分中に存在しない特異性を有する酵素を生成するための方法を提供する。特に、本発明は、哺乳動物、特にヒトの酵素と本質的に同一である配列を有するが、異なる特異性を有する酵素の生成に関する。さらに、本発明は、遺伝子によりコードされた対応する特異性を有する特異的な人工酵素のライブラリー、遺伝子によりコードされた対応する特異性を有する特異的な人工酵素のライブラリーを生成する方法、ならびにこのようなライブラリーの技術、診断、栄養学、パーソナルケア、または研究目的への適用を提供する。
【0047】
本発明の第一の局面は、規定された特異性を有する人工酵素を開示する。これらの人工酵素は、以下の成分によって特徴付けられる:
(a)基質上で少なくとも1つの化学反応を触媒し得るタンパク質足場、および
(b)得られる人工タンパク質が少なくとも1つの標的基質と1つ以上の異なる基質とを区別できるようなタンパク質足場の部位に存在する1つ以上の特異性決定領域(SDR)であって、本質的に合成ペプチド配列である、SDR。
【0048】
好ましくは、人工酵素のこのように規定された特異性は、タンパク質足場によって与えられない。
【0049】
原則として、種々の一次、二次、および三次構造を有し得る。一次構造、すなわち、アミノ酸配列は、人工配列であるか、または任意のウイルス、原核生物、もしくは真核生物起源のものでありえる。しかしながら、ヒトの治療用途には、タンパク質足場は、好ましくは、哺乳動物由来のものであり、より好ましくは、ヒト由来のものである。さらに、タンパク質足場は、1つ以上の反応を触媒し得、そして好ましくは、低い特異性のみを有する。
【0050】
好ましくは、タンパク質足場としての適用可能性のために改善された特性を付与する修飾されたアミノ酸配列を有するタンパク質足場の誘導体が使用される。このような改善された特性は、安定性、発現量もしくは分泌量、折り畳み(特に、タンパク質足場とSDRとの組み合わせ後に)、活性化剤もしくは阻害剤のような調節剤に対する増大もしくは減少した感受性、免疫原性、触媒速度、kMまたは基質親和性が含まれるが、これらに限定されない。
【0051】
人工酵素は、タンパク質足場と組み合わされた合成ペプチド配列から定量的特異性を明らかにする。それゆえ、人工ペプチド配列は、特異性決定領域またはSDRとして作用する。このようなSDRの数、長さ、および位置は、広範に変化し得る。足場内のSDRの数は、少なくとも1個であり、好ましくは、1個以上、より好ましくは、2〜11個、最も好ましくは、2〜6個である。SDRは、1〜50アミノ酸残基の長さを有し、好ましくは、1〜15アミノ酸残基、より好ましくは、1〜6アミノ酸残基の長さを有する。あるいは、SDRは、2〜20アミノ酸残基の長さ、好ましくは、2〜10アミノ酸残基の長さ、より好ましくは、3〜8アミノ酸残基の長さを有する。
【0052】
本発明の人工酵素は、定常領域および可変領域を含むが、非免疫グロブリン骨格を有し、かつ定常領域に活性部位(触媒活性)を有し、そのため活性部位の基質特異性が可変領域によって調節される抗体様分子としてもさらに記載され得る。好ましくは、免疫グロブリン構造においけると同様、可変領域は、標的分子と相互作用する可変の長さおよび組成のループ構造である。
【0053】
本発明の特定の変異体において、人工酵素は、加水分解活性を有する。好ましい変異体において、人工酵素は、タンパク質分解活性を有する。この変異体のための特に好ましいタンパク質足場は、非特異的プロテアーゼもしくは非特異的プロテアーゼ由来の部分であるか、またはそうでなければ非特異的プロテアーゼに由来する。この観点および以下の変異体および実施形態において、「に由来する」または「その誘導体」という表現は、1つ以上のアミノ酸位置で変異し、および/または元々のタンパク質に対して少なくとも70%、好ましくは、90%、より好ましくは、95%、および最も好ましくは、99%の相同性を有し、および/またはタンパク質分解によりプロセスされ、および/または変化したグリコシル化パターンを有し、および/または非タンパク質物質に共有結合しているか、および/またはさらなるタンパク質ドメインに融合し、および/またはC末端および/もしくはN末端切断を有し、および/または特定の挿入、置換、および/もしくは欠失を有するタンパク質の誘導体のことをいう。あるいは、「に由来する」は、2つ以上のタンパク質からの2つ以上のフラグメントの組み合わせまたはキメラである誘導体のことをいうこともある。ここで、各々のフラグメントは、上述のいずれかまたは全ての修飾を必要に応じて含む。タンパク質足場の三次構造は、任意の種類のものであり得る。しかしながら、好ましくは、三次構造は、以下の構造クラスの1つに属する:クラスS1(セリンプロテアーゼファミリーのキモトリプシン集団)、クラスS8(セリンプロテアーゼファミリーのサブチリシン集団)、クラスSC(セリンプロテアーゼファミリーのカルボキシペプチダーゼ集団)、クラスA1(アスパラギン酸プロテアーゼのペプシンA集団)、またはクラスC14(システインプロテアーゼのカスパーゼ−1集団)。ヒト治療薬として使用するための人工タンパク質分解酵素のタンパク質足場として働くプロテアーゼの例は、ヒトトリプシン、ヒトトロンビン、ヒトキモトリプシン、ヒトペプシン、ヒトエンドチアペプシン(human endothiapepsin)、ヒトカスパーゼ1〜14、および/またはヒトフリンであるか、またはこれらに由来する。
【0054】
人工タンパク質分解酵素の規定された特異性は、少なくとも1つの標的ペプチドまたはタンパク質基質と、1つ以上のさらなるペプチドまたはタンパク質基質とを区別する能力の指標である。好ましくは、規定された特異性は、P1部位とは異なる他の位置で異なるペプチドまたはタンパク質基質を区別する能力をいい、より好ましくは、規定された特異性は、P1部位およびP1’部位とは異なる他の位置で異なるペプチドまたはタンパク質基質を区別する能力のことをいう。より好ましくは、人工タンパク質分解酵素は、標的ペプチドまたはタンパク質基質を、必要な限りできるだけ多くの部位で区別して、他のタンパク質に対して優先的に標的基質を加水分解する。例として、ヒトの身体において静脈内に適用される治療的に有用な人工タンパク質分解酵素は、標的基質とヒト血清中の任意の他のタンパク質とを区別するのに十分に特異的であるべきである。好ましくは、このような人工タンパク質分解酵素は、3個以上のアミノ酸位置、より好ましくは、4個以上の位置、そしてさらにより好ましくは、5個以上のアミノ酸位置でペプチド基質を認識し、区別する。これらの位置は、隣接していてもしていなくてもよい。
【0055】
第一の実施形態において、タンパク質足場は、セリンプロテアーゼのS1構造サブクラス(すなわち、キモトリプシン集団)の三次構造もしくは折り畳みと等しいかもしくは類似した三次構造または折り畳みを有し、および/またはセリンプロテアーゼのS1構造サブクラスのタンパク質とアミノ酸レベルで少なくとも70%の同一性を有する。SDRは、ヒトトリプシン1中の領域18−25、38−48、54−63、73−86、122−130、148−156、165−171、および194−204に対して構造的に、またはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域20−23、41−45、57−60、76−83、125−128、150−153、167−169、および197−201(配列番号1に従うアミノ酸配列の番号付け)に構造的に、またはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質足場に挿入されることが好ましい。この種のタンパク質足場と組み合わせられるべきSDRの数は、好ましくは、1〜10、より好ましくは、2〜4である。好ましくは、タンパク質足場は、以下のタンパク質の1つ以上の誘導体またはホモログである:キモトリプシン、グランザイム、カリクレイン、トリプシン、メソトリプシン、好中球エラスターゼ、膵エラスターゼ、エンテロペプチダーゼ、カテプシン、トロンビン、アンクロド、凝固因子IXa、凝固因子VIIa、凝固因子Xa、活性化プロテインC、ウロキナーゼ、組織型プラスミノーゲン活性化因子、プラスミン、Desmodus型プラスミノーゲン活性化因子。より好ましくは、タンパク質足場は、トリプシンもしくはトロンビンであり、またはトリプシンもしくはトロンビン由来の誘導体もしくはホモログである。ヒトの治療のための用途には、トリプシンまたはトロンビン足場は、免疫反応またはアレルギー反応の危険性を最小限にするために、最も好ましくは、ヒト由来である。好ましくは、ヒトトリプシンIに由来する、または類似した三次構造を有するタンパク質に由来する改善された特徴を有する誘導体が使用される。このような誘導体の好ましい例は、ヒトトリプシンI(配列番号1)に由来し、そして1つ以上の以下のアミノ酸置換:E56G;R78W;Y131F;A146T;C183Rを有する。
【0056】
2つのSDRのうちの少なくとも1つが、ヒトトリプシンIまたはその誘導体に、残基42と43との間(SDR1)、および残基123と124との間(SDR2)にそれぞれ挿入されていることが好ましい(配列番号1に従う番号付け)。さらに、SDR1は、6という好ましい長さを、SDR2は、5という好ましい長さを、それぞれ有する。この実施形態の好ましい変異体において、SDR1およびSDR2の配列は、表2に列挙するアミノ酸配列の1つを含む。このような人工タンパク質分解酵素は、実施例IVに例示されるように標的基質Bに対する特異性を有する。
【0057】
さらなる実施形態において、タンパク質足場は、セリンプロテアーゼのS8構造サブクラスに属するか、および/または枯草菌由来のサブチリシンEに類似する三次構造を有するか、および/またはセリンプロテアーゼのS8構造サブクラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有する。好ましくは、足場は、サブチリシンファミリーまたはヒトプロタンパク質コンバターゼに属する。SDRは、枯草菌由来のサブチリシンE中の領域6−17、25−29、47−55、59−69、101−111、117−125、129−137、139−154、158−169、185−195、および204−225に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域59−69、101−111、129−137、158−169、および204−225に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号7に従う番号付け)。タンパク質足場は、1つ以上の以下のタンパク質に等しいかまたはそれらの誘導体またはホモログであることが好ましい:サブチリシンカールスバーグ、枯草菌サブチリシンE、サブチリシンBPN’、リケニホルミス菌、B.lentusサブチリシン、Bacillus alcalophilusアルカリホスファターゼ、プロテインキナーゼK、ケクシン(kexin)、ヒトプロタンパク質コンバターゼ、ヒトフリン。好ましい変異体において、サブチリシンBPN’またはタンパク質SPC1〜7の1つが、タンパク質足場として使用される。
【0058】
さらなる実施形態において、タンパク質足場は、アスパラギン酸プロテアーゼのファミリーに属するか、および/またはヒトペプチドに類似する三次構造を有する。好ましくは、足場は、プロテアーゼのA1クラスに属するか、および/またはプロテアーゼのA1クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有する。SDRは、枯草菌由来のサブチリシンE中の領域6−18、49−55、74−83、91−97、112−120、126−137、159−164、184−194、242−247、262−267、および277−300に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域10−15、75−80、114−118、130−134、186−191、および280−296に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号11に従う番号付け)。タンパク質足場は、1つ以上の以下のタンパク質に等しいかまたはそれらの誘導体またはホモログであることが好ましい:ペプシン、キモシン、レニン、カテプシン、ヤプシン。好ましくは、ペプチドもしくはエンドチオペプシンまたはこれらの誘導体もしくはホモログがタンパク質足場として使用される。
【0059】
さらなる実施形態において、タンパク質足場は、システインプロテアーゼのファミリーに属するか、および/またはヒトカスパーゼ7に類似する三次構造を有する。好ましくは、足場は、システインプロテアーゼのC14クラスに属するか、またはシステインプロテアーゼのC14クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有する。SDRは、ヒトカスパーゼ7中の領域78−91、144−160、186−198、226−243、および271−291に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域80−86、149−157、190−194、および233−238に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号14に従う番号付け)。タンパク質足場は、タンパク質足場は、カスパーゼ1〜9の1つに等しいか、またはその誘導体もしくはホモログであることが好ましい。
【0060】
さらなる実施形態において、タンパク質足場は、セリンプロテアーゼのS11クラスに属するか、またはセリンプロテアーゼのS11クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはStreptomyces種K15由来のD−アラニン−D−アラニントランスペプチダーゼに類似した三次構造を有する。SDRは、Streptomyces種K15由来のD−アラニン−D−アラニントランスペプチダーゼ中の領域67−79、137−150、191−206、212−222、および241−251に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域70−75、141−147、195−202、および216−220に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号15に従う番号付け)。Streptomyces種K15由来のD−アラニン−D−アラニントランスペプチダーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0061】
さらなる実施形態において、タンパク質足場は、セリンプロテアーゼのS21クラスに属するか、またはセリンプロテアーゼのS21クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはヒトサイトメガロウイルス由来のアセンブリン(assemblin)に類似した三次構造を有する。SDRは、ヒトサイトメガロウイルス由来のアセンブリン中の領域25−33、64−69、134−155、162−169、および217−244に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域27−31、164−168、および222−239に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号16に従う番号付け)。ヒトサイトメガロウイルス由来のアセンブリンまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0062】
さらなる実施形態において、タンパク質足場は、セリンプロテアーゼのS26クラスに属するか、またはセリンプロテアーゼのS26クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/または大腸菌由来のシグナルペプチダーゼに類似した三次構造を有する。SDRは、大腸菌由来のシグナルペプチダーゼ中の領域8−14、57−68、125−134、239−254、200−211、および228−239に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域9−13、60−67、127−132、および203−209に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号17に従う番号付け)。大腸菌由来のシグナルペプチダーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0063】
さらなる実施形態において、タンパク質足場は、セリンプロテアーゼのS33クラスに属するか、またはセリンプロテアーゼのS33クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/または大腸菌由来のシグナルペプチダーゼに類似した三次構造を有する。SDRは、セラチア・マルセッセンス由来のプロピルアミノペプチダーゼ中の領域47−54、152−160、203−212、および297−302に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域50−53、154−158、および206−210に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号18に従う番号付け)。セラチア・マルセッセンス由来のプロピルアミノペプチダーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0064】
さらなる実施形態において、タンパク質足場は、セリンプロテアーゼのS51クラスに属するか、またはセリンプロテアーゼのS51クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/または大腸菌由来のアスパルチルジペプチダーゼに類似した三次構造を有する。SDRは、大腸菌由来のアスパルチルジペプチダーゼ中の領域8−16、38−46、85−92、132−140、159−170、および205−211に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域10−14、87−90、134−138、および160−165に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号19に従う番号付け)。大腸菌由来のアスパルチルジペプチダーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0065】
さらなる実施形態において、タンパク質足場は、アスパラギン酸プロテアーゼのA2クラスに属するか、またはアスパラギン酸プロテアーゼのA2クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはヒト免疫不全ウイルス由来のプロテアーゼに類似した三次構造を有する。SDRは、ヒト免疫不全ウイルス由来のプロテアーゼ中の領域5−12、17−23、27−30、33−38、および77−83に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域7−10、18−21、34−37、および79−82に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号20に従う番号付け)。ヒト免疫不全ウイルス、好ましくは、HIV−1プロテアーゼ由来のプロテアーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0066】
さらなる実施形態において、タンパク質足場は、アスパラギン酸プロテアーゼのA26クラスに属するか、またはアスパラギン酸プロテアーゼのA26クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/または大腸菌由来のオンプチン(omptin)に類似した三次構造を有する。SDRは、大腸菌由来のオンプチン中の領域28−40、86−98、150−168、213−219、および267−278に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域33−38、161−168、および273−277に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号21に従う番号付け)。大腸菌由来のオンプチン由来のプロテアーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0067】
さらなる実施形態において、タンパク質足場は、システインプロテアーゼのC1クラスに属するか、またはシステインプロテアーゼのC1クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはカリーカ・パパイヤ由来のパパインに類似した三次構造を有する。SDRは、カリーカ・パパイヤ由来のパパイン中の領域17−24、61−68、88−95、135−142、153−158、および176−184に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域63−66、136−139、および177−181に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号22に従う番号付け)。カリーカ・パパイヤ由来のパパイン由来のプロテアーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0068】
さらなる実施形態において、タンパク質足場は、システインプロテアーゼのC2クラスに属するか、またはシステインプロテアーゼのC2クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはヒトカルパイン−2に類似した三次構造を有する。SDRは、ヒトカルパイン−2中の領域90−103、160−172、193−199、243−260、286−294、および316−322に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域92−101、245−250、および287−291に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号23に従う番号付け)。ヒトカルパイン−2由来のプロテアーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0069】
さらなる実施形態において、タンパク質足場は、システインプロテアーゼのC4クラスに属するか、またはシステインプロテアーゼのC4クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはタバコエッチウイルス(tobacco etch virus)由来のNIaプロテアーゼに類似した三次構造を有する。SDRは、タバコエッチウイルス由来のNIaプロテアーゼ中の領域23−31、112−120、144−150、168−176、および205−218に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域145−149、169−174、および212−218に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号24に従う番号付け)。タバコエッチウイルス由来のNIaプロテアーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0070】
さらなる実施形態において、タンパク質足場は、システインプロテアーゼのC10クラスに属するか、またはシステインプロテアーゼのC10クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/または化膿性連鎖球菌由来のストレプトパイン(streptopain)に類似した三次構造を有する。SDRは、化膿性連鎖球菌由来のストレプトパイン中の領域81−90、133−140、150−164、191−199、219−229、246−256、306−312、および330−337に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域82−87、134−138、250−254、および331−335に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号25に従う番号付け)。化膿性連鎖球菌由来のストレプトパインまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0071】
さらなる実施形態において、タンパク質足場は、システインプロテアーゼのC19クラスに属するか、またはシステインプロテアーゼのC19クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはヒトユビキチン特異的プロテアーゼ7に類似した三次構造を有する。SDRは、ヒトユビキチン特異的プロテアーゼ7中の領域3−15、63−70、80−86、248−256、272−283、および292−304に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域10−15、251−255、277−281、および298−304に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号26に従う番号付け)。ヒトユビキチン特異的プロテアーゼ7またはその誘導体もしくはホモログが足場として使用されることが、好ましい
【0072】
さらなる実施形態において、タンパク質足場は、システインプロテアーゼのC47クラスに属するか、またはシステインプロテアーゼのC47クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/または黄色ブドウ球菌由来のスタフォパイン(staphopain)に類似した三次構造を有する。SDRは、黄色ブドウ球菌由来のスタフォパイン中の領域15−23、57−66、108−119、142−149、および157−164に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域17−22、111−117、143−147、および159−163に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号27に従う番号付け)。黄色ブドウ球菌由来のスタフォパインまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0073】
さらなる実施形態において、タンパク質足場は、システインプロテアーゼのC48クラスに属するか、またはシステインプロテアーゼのC48クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/または出芽酵母(Saccharomyces cerevisiae)由来のUlp1エンドペプチダーゼに類似した三次構造を有する。SDRは、出芽酵母由来のUlp1エンドペプチダーゼ中の領域40−51、108−115、132−141、173−179、および597−605に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域43−49、110−113、133−137、および175−178に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号28に従う番号付け)。出芽酵母由来のUlp1エンドペプチダーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0074】
さらなる実施形態において、タンパク質足場は、システインプロテアーゼのC56クラスに属するか、またはシステインプロテアーゼのC56クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはPyrococcus horikoshii由来のPfplエンドペプチダーゼに類似した三次構造を有する。SDRは、Pyrococcus horikoshii由来のPfplエンドペプチダーゼ中の領域8−16、40−47、66−73、118−125、および147−153に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域9−14、68−71、120−123、および148−151に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号29に従う番号付け)。Pyrococcus horikoshii由来のPfplエンドペプチダーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0075】
さらなる実施形態において、タンパク質足場は、メタロプロテアーゼのM4クラスに属するか、またはメタロプロテアーゼのM4クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはBacillus thermoproteolyticus由来のサーモリシンに類似した三次構造を有する。SDRは、Bacillus thermoproteolyticus由来のサーモリシン中の領域106−118、125−130、152−160、197−204、210−213、および221−229に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域108−115、126−129、199−203、および223−227に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号30に従う番号付け)。Bacillus thermoproteolyticus由来のサーモリシンまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0076】
さらなる実施形態において、タンパク質足場は、メタロプロテアーゼのM10クラスに属するか、またはメタロプロテアーゼのM10クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはヒトコラゲナーゼに類似した三次構造を有する。SDRは、ヒトコラゲナーゼ中の領域2−7、68−79、85−90、107−111、および135−141に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域3−6、71−78、および136−140に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号31に従う番号付け)。ヒトコラゲナーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0077】
人工酵素は、グリコシダーゼ活性を有することがさらに好ましい。この変異体について特に適したタンパク質足場は、グリコシダーゼであるか、またはグリコシダーゼに由来するものである。好ましくは、三次構造は、以下の構造クラスの1つに属する:クラスGH13、GH7、GH12、GH11、GH10、GH28、GH26、およびGH18(ベータ/アルファ)8バレル。
【0078】
第一の実施形態において、タンパク質足場は、グリコシラーゼのGH13クラスに属するか、またはグリコシラーゼのGH13クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはヒト膵アルファアミラーゼに類似した三次構造を有する。SDRは、ヒト膵アルファアミラーゼ中の領域50−60、100−110、148−167、235−244、302−310、および346−359に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域51−58、148−155、および303−309に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号32に従う番号付け)。ヒト膵アルファアミラーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0079】
さらなる実施形態において、タンパク質足場は、グリコシラーゼのGH7クラスに属するか、またはグリコシラーゼのGH7クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはTrichoderma reesei由来のセルラーゼに類似した三次構造を有する。SDRは、Trichoderma reesei由来のセルラーゼ中の領域47−56、93−104、173−182、215−223、229−236、および322−334に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域175−180、218−222、および324−332に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号33に従う番号付け)。Trichoderma reesei由来のセルラーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0080】
さらなる実施形態において、タンパク質足場は、グリコシラーゼのGH12クラスに属するか、またはグリコシラーゼのGH12クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはクロカビ(Aspergillus niger)由来のセルラーゼに類似した三次構造を有する。SDRは、Aspergillus niger由来のセルラーゼ中の領域18−28、55−60、106−113、126−132、および149−159に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域20−26、56−59、108−112、および151−156に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号34に従う番号付け)。Aspergillus niger由来のセルラーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0081】
さらなる実施形態において、タンパク質足場は、グリコシラーゼのGH11クラスに属するか、またはグリコシラーゼのGH11クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはAspergillus niger由来のトキシラナーゼ(toxylanase)に類似した三次構造を有する。SDRは、Aspergillus niger由来のトキシラナーゼ中の領域7−14、33−39、88−97、114−126、および158−167に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域20−26、56−59、108−112、および151−156に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号35に従う番号付け)。Aspergillus niger由来のトキシラナーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0082】
さらなる実施形態において、タンパク質足場は、グリコシラーゼのGH10クラスに属するか、またはグリコシラーゼのGH10クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはStreptomyces lividans由来のトキシラナーゼ(toxylanase)に類似した三次構造を有する。SDRは、Streptomyces lividans由来のトキシラナーゼ中の領域21−29、42−50、84−92、130−136、206−217、および269−278に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域43−49、86−90、208−213、および271−276に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号36に従う番号付け)。Streptomyces lividans由来のトキシラナーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0083】
さらなる実施形態において、タンパク質足場は、グリコシラーゼのGH28クラスに属するか、またはグリコシラーゼのGH28クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはAspergillus niger由来のペクチナーゼに類似した三次構造を有する。SDRは、Aspergillus niger由来のペクチナーゼ中の領域82−88、118−126、171−178、228−236、256−264、および289−299に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域116−124、174−178、および291−296に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号37に従う番号付け)。Aspergillus niger由来のペクチナーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0084】
さらなる実施形態において、タンパク質足場は、グリコシラーゼのGH26クラスに属するか、またはグリコシラーゼのGH26クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはPseudomonas cellulosa由来のマンナナーゼに類似した三次構造を有する。SDRは、Pseudomonas cellulosa由来のマンナナーゼ中の領域75−83、113−125、174−182、217−224、247−254、324−332、および325−340に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域115−123、176−180、286−291、および328−337に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号38に従う番号付け)。Pseudomonas cellulosa由来のマンナナーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0085】
さらなる実施形態において、タンパク質足場は、グリコシラーゼのGH18(ベータ/アルファ)8クラスに属するか、またはグリコシラーゼのGH18(ベータ/アルファ)8クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはBacillus circulans由来のキチナーゼに類似した三次構造を有する。SDRは、Bacillus circulans由来のキチナーゼ中の領域21−29、57−65、130−136、176−183、221−229、249−257、および327−337に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域59−63、178−181、250−254、および330−336に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号39に従う番号付け)。Bacillus circulans由来のキチナーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0086】
人工酵素がエステル加水分解酵素活性を有することがさらに好ましい。好ましくは、この変異体についてのタンパク質足場は、リパーゼ、ホスファターゼ、フィターゼ、またはホスホジエステラーゼ活性を有する。
【0087】
第一の実施形態において、タンパク質足場は、エステラーゼのGXクラスに属するか、またはエステラーゼのGXクラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはCandida antarctica由来のリパーゼBに類似した三次構造を有する。好ましくは、足場はリパーゼ活性を有する。SDRは、Candida antarctica由来のリパーゼB中の領域139−148、188−195、216−224、256−266、272−287に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域141−146、218−222、259−263、および275−283に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号40に従う番号付け)。Candida antarctica由来のリパーゼBまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0088】
さらなる実施形態において、タンパク質足場は、エステラーゼのGXクラスに属するか、またはエステラーゼのGXクラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはモルモット由来の膵リパーゼに類似した三次構造を有する。好ましくは、足場はリパーゼ活性を有する。SDRは、モルモット由来の膵リパーゼ中の領域78−90、91−100、112−120、179−186、207−218、238−247、および248−260に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域80−87、114−118、209−215、および239−246に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号41に従う番号付け)。モルモット由来の膵リパーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0089】
さらなる実施形態において、タンパク質足場は、大腸菌由来のアルカリホスファターゼの構造に類似した三次構造を有するか、または大腸菌由来のアルカリホスファターゼの構造に類似した三次構造を有するタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有する。好ましくは、足場はホスファターゼ活性を有する。SDRは、大腸菌由来のアルカリホスファターゼ中の領域110−122、187−142、170−175、186−193、280−287、および425−435に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域171−174、187−191、282−286、および426−433に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号42に従う番号付け)。大腸菌由来のアルカリホスファターゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0090】
さらなる実施形態において、タンパク質足場は、ウシ膵臓デスオキシリボヌレアーゼIの構造に類似した三次構造を有するか、またはウシ膵臓デスオキシリボヌレアーゼIの構造に類似した三次構造を有するタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有する。好ましくは、足場はホスホジエステラーゼ活性を有する。より好ましくは、ヌクレアーゼ、そして最も好ましくは、非特異的エンドヌクレアーゼまたはその誘導体が、足場として使用される。SDRは、ウシ膵臓デスオキシリボヌレアーゼI中の領域14−21、41−47、72−77、97−111、135−143、171−178、202−209、および242−251に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域16−19、42−46、136−141、および172−176に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号43に従う番号付け)。ウシ膵臓デスオキシリボヌレアーゼIもしくはヒトデスオキシリボヌレアーゼ1またはその誘導体もしくはホモログが足場として使用されることが、好ましい。人工酵素がトランスフェラーゼ活性を有することがさらに好ましい。この変異体について特に適したタンパク質足場は、グリコシルトランスフェラーゼ、ホスホトランスフェラーゼまたはメチルトランスフェラーゼであり、またはその誘導体である。この変異体について特に好ましいタンパク質足場は、グリコシルトランスフェラーゼであり、またはグリコシルトランスフェラーゼに由来する。タンパク質足場の三次構造は、任意の種類のものであり得る。しかしながら、好ましくは、三次構造は、以下の構造クラスの1つに属する:GH13、およびGT1。
【0091】
第一の実施形態において、タンパク質足場は、トランスフェラーゼのGH13クラスに属するか、またはトランスフェラーゼのGH13クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはBacillus circulans由来のシクロマルトデキストリン グルカノトランスフェラーゼの構造に類似した三次構造を有する。好ましくは、足場はトランスフェラーゼ活性を有し、より好ましくは、グリコシルトランスフェラーゼが足場として使用される。SDRは、Bacillus circulans由来のシクロマルトデキストリン グルカノトランスフェラーゼ中の領域38−48、85−94、142−154、178−186、259−266、331−340、および367−377に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域87−92、180−185、261−264、および269−275に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号44に従う番号付け)。Bacillus circulans由来のシクロマルトデキストリン グルカノトランスフェラーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0092】
さらなる実施形態において、タンパク質足場は、トランスフェラーゼのGT1クラスに属するか、またはトランスフェラーゼのGT1クラスのタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有するか、および/またはAmycolatopsis orientalis A82846由来のグリコシルトランスフェラーゼの構造に類似した三次構造を有する。好ましくは、足場はトランスフェラーゼ活性を有し、より好ましくは、グリコシルトランスフェラーゼ活性を有する。SDRは、Amycolatopsis orientalis A82846由来のグリコシルトランスフェラーゼ中の領域58−74、130−138、185−193、228−236、および314−323に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域61−71、230−234、および316−321に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号45に従う番号付け)。Amycolatopsis orientalis A82846由来のグリコシルトランスフェラーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0093】
人工酵素は、酸化還元酵素活性を有することがさらに好ましい。この変異体に特に適したタンパク質足場は、モノオキシゲナーゼ、ジオキシゲナーゼ、もしくはアルコールデヒドロゲナーゼ、またはこれらの誘導体である。タンパク質足場の三次構造は、任意の種類のものであり得る。
【0094】
第一の実施形態において、タンパク質足場は、Pseudomonas種由来の2、3−ジヒドロキシビフェニル ジオキシゲナーゼ(2、3−diphydroxybiphenyl dioxygenase)の構造に類似した三次構造を有するか、またはPseudomonas種由来の2、3−ジヒドロキシビフェニル ジオキシゲナーゼの構造に類似した三次構造を有するタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有する。好ましくは、足場はジヒドロゲナーゼ活性を有する。SDRは、Pseudomonas種由来の2、3−ジヒドロキシビフェニル ジオキシゲナーゼ中の領域172−185、198−206、231−237、250−259、および282−287に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域175−182、200−204、252−257、および284−287に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号46に従う番号付け)。Pseudomonas種由来の2、3−ジヒドロキシビフェニル ジオキシゲナーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0095】
さらなる実施形態において、タンパク質足場は、Acinetobacter種由来のカテコールジオキシゲナーゼの構造に類似した三次構造を有するか、またはAcinetobacter種由来のカテコールジオキシゲナーゼの構造に類似した三次構造を有するタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有する。好ましくは、足場はジオキシゲナーゼ活性を有し、より好ましくは、カテコールジオキシゲナーゼ活性を有する。SDRは、Acinetobacter種由来のカテコールジオキシゲナーゼ中の領域66−72、105−112、156−171、および198−207に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域107−110、161−171、および201−205に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号47に従う番号付け)。Acinetobacter種由来のカテコールジオキシゲナーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0096】
さらなる実施形態において、タンパク質足場は、Pseudomonas putida由来のカンファー−5−モノオキシゲナーゼの構造に類似した三次構造を有するか、またはPseudomonas putida由来のカンファー−5−モノオキシゲナーゼの構造に類似した三次構造を有するタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有する。好ましくは、足場はモノオキシゲナーゼ活性を有し、より好ましくは、カンファーモノオキシゲナーゼ活性を有する。SDRは、Pseudomonas putida由来のカンファー−5−モノオキシゲナーゼ中の領域26−31、57−63、84−98、182−191、242−256、292−299、および392−399に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域85−96、183−188、244−253、293−298、および393−398に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号48に従う番号付け)。Pseudomonas putida由来のカンファー−5−モノオキシゲナーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0097】
さらなる実施形態において、タンパク質足場は、Equus callabus由来のアルコールデヒドロゲナーゼの構造に類似した三次構造を有するか、またはEquus callabus由来のアルコールデヒドロゲナーゼの構造に類似した三次構造を有するタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有する。好ましくは、足場はアルコールデヒドロゲナーゼ活性を有する。SDRは、Equus callabus由来のアルコールデヒドロゲナーゼ中の領域49−63、111−112、294−301、および361−369に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域51−61、および295−299に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号48に従う番号付け)。Equus callabus由来のアルコールデヒドロゲナーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0098】
人工酵素は、リアーゼ活性を有することがさらに好ましい。この変異体に特に適したタンパク質足場は、オキソ酸リアーゼ、またはその誘導体である。この変異体について特に好ましいタンパク質足場は、アルドラーゼもしくはシンターゼ、またはこれらの誘導体である。タンパク質足場の三次構造は、任意の種類のものであり得るが、(ベータ/アルファ)8バレル構造が好ましい。
【0099】
第一の実施形態において、タンパク質足場は、Escherichia coli由来のN−アセチル−d−ニューラミン酸アルドラーゼ(N−acetyl−d−neuramic acid aldolase)の構造に類似した三次構造を有するか、またはEscherichia coli由来のN−アセチル−d−ニューラミン酸アルドラーゼの構造に類似した三次構造を有するタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有する。好ましくは、足場はアルドラーゼ活性を有する。SDRは、Escherichia coli由来のN−アセチル−d−ニューラミン酸アルドラーゼ中の領域45−55、78−87、105−113、137−146、164−171、187−193、205−210、244−255、および269−276に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域45−52、138−144、189−192、247−253、および271−275に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号50に従う番号付け)。Escherichia coli由来のN−アセチル−d−ニューラミン酸アルドラーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0100】
さらなる実施形態において、タンパク質足場は、Salmonella typhimurium由来のトリプトファンシンターゼの構造に類似した三次構造を有するか、またはSalmonella typhimurium由来のトリプトファンシンターゼの構造に類似した三次構造を有するタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有する。好ましくは、足場はシンターゼ活性を有する。SDRは、Salmonella typhimurium由来のトリプトファンシンターゼ中の領域56−63、127−134、154−161、175−193、209−216、および230−240に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域57−62、155−160、178−190、および210−215に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号51に従う番号付け)。Salmonella typhimurium由来のトリプトファンシンターゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0101】
人工酵素は、イソメラーゼ活性を有することがさらに好ましい。この変異体に特に適したタンパク質足場は、変換アルドラーゼ(converting aldose)もしくは変換ケトース(converting ketose)、またはこれらの誘導体である。
【0102】
第一の実施形態において、タンパク質足場は、Actinoplanes missouriensis由来のキシロースイソメラーゼの構造に類似した三次構造を有するか、またはActinoplanes missouriensis由来のキシロースイソメラーゼの構造に類似した三次構造を有するタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有する。SDRは、Actinoplanes missouriensis由来のキシロースイソメラーゼ中の領域18−31、92−103、136−147、178−188、および250−257に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域20−27、92−99、および180−186に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号52に従う番号付け)。Actinoplanes missouriensis由来のキシロースイソメラーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0103】
人工酵素は、リガーゼ活性を有することがさらに好ましい。この変異体に特に適したタンパク質足場は、DNAリガーゼまたはその誘導体である。
【0104】
第一の実施形態において、タンパク質足場は、Bacteriophage T7由来のDNAリガーゼの構造に類似した三次構造を有するか、またはBacteriophage T7由来のDNAリガーゼの構造に類似した三次構造を有するタンパク質に対してアミノ酸レベルで少なくとも70%の同一性を有する。SDRは、Bacteriophage T7由来のDNAリガーゼ中の領域52−60、94−108、119−131、241−248、255−263、および302−318に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、より好ましくは、領域96−106、121−129、256−262、および304−316に構造的にまたはアミノ酸配列相同性により対応する位置の群からの1つ以上の位置で、タンパク質に挿入される(配列番号53に従う番号付け)。Bacteriophage T7由来のDNAリガーゼまたはその誘導体もしくはホモログが足場として使用されることが、好ましい。
【0105】
本発明の第二の局面は、治療、研究、診断、栄養学、パーソナルケア、工業用途のための特異性を有する人工酵素の適用に関する。この適用は、少なくとも以下の工程を含む:
(a)その加水分解が意図された目的、例えば、疾患の治癒、疾患の診断、ヒトもしくは動物栄養学のための成分の加工、またはその他の技術的プロセスに関して正の効果を有する標的ペプチド基質の同定;
(b)工程(a)において同定された標的ペプチドに特異的な酵素である、人工酵素の提供;ならびに
(c)意図された目的のための、工程(b)において提供された酵素の使用。
【0106】
本発明の第一の局面の第一の変異体において、人工酵素は、疾患関連標的基質を不活性化させるための治療手段として使用される。この適用は、少なくとも以下の工程を包含する:
(a)疾患に関連し、その不活性化が疾患に関して正の効果を有する標的基質の同定、および標的基質内の標的部位の決定であって、その標的部位での修飾がその標的基質の不活性化を導くという事実によって特徴付けられる標的部位の決定、
(b)人工酵素の提供であって、その酵素が工程(a)で同定した標的部位に対して特異的である、酵素の提供、ならびに
(c)ヒトの体内または体外での上記標的基質の不活性化のための上記酵素の使用。
【0107】
好ましい実施形態において、工程(c)に提供される人工酵素の足場は、ヒトの身体への上記酵素の適用に伴う免疫原性またはアレルギー性の効果を避けるかまたは低減するために、ヒト由来のものである。この変異体のより好ましい実施形態において、足場は、ヒトプロテアーゼであり、加水分解はそのペプチドまたはタンパク質標的の活性化または不活性化を導く。潜在的なペプチドまたはタンパク質標的には、サイトカイン、成長因子、ペプチドホルモン、インターロイキン、インターフェロン、凝固カスケード由来の酵素、セルピン、免疫グロブリン、可溶性もしくは膜結合レセプター、細胞もしくはウイルス表面タンパク質、ペプチドドラッグ、タンパク質ドラッグが含まれる。
【0108】
特に好ましい実施形態は、人工酵素がヒト腫瘍壊死因子アルファ(TNF−α)を切断し得るという知見に基づいている。したがって、人工酵素または融合タンパク質は、炎症性疾患(ならびにTNF−αに関連する他の疾患)の処置のための医薬の調製のために使用され得る。好ましくは、上記人工酵素または上記融合タンパク質は、ヒト腫瘍壊死因子アルファ(hTNF−α)を特異的に不活性化し得、より好ましくは、上記人工酵素または上記融合タンパク質は、hTNF−α(配列番号96)中の31/32、32/33、44/45、87/88、128/129、および/または141/142位間(最も好ましくは、31/32および32/33位間)のペプチド結合を加水分解し得る。
【0109】
さらなる実施形態において、標的基質は、人工酵素によって活性化されるプロドラッグである。この変異体の特定の実施形態において、人工酵素は、タンパク質分解活性を有し、標的基質は、タンパク質分解的に活性化されているタンパク質標的である。このようなプロドラッグの例は、凝固因子の不活化形態のようなプロタンパク質である。別の特定の変異体において、人工酵素は、酸化還元酵素であり、標的基質は、酸化により活性化され得る化学物質である。
【0110】
本発明のこの局面の第二の変異体において、人工酵素は、工業的にまたは栄養学的に重要な反応を規定された特異性でもって触媒するための技術的手段として使用される。この変異体の特定の実施形態において、人工酵素はタンパク質分解活性を有し、触媒される反応は、タンパク質分解プロセスであり、人工酵素は、1つ以上の工業的または栄養学的に重要なタンパク質を特異的に加水分解する。この変異体の好ましい実施形態において、人工酵素は、1つ以上の工業的または栄養学的に重要なタンパク質基質を、特定の部位で加水分解し、それによって工業的または栄養学的に所望の製品特性(例えば、きめ、風味、または沈殿特性)を導く。この変異体のさらなる特定の実施形態において、人工酵素は、グリコシド結合の加水分解を触媒する(グリコシダーゼまたはグリコシラーゼ活性)。次いで、好ましくは、触媒された反応は、ポリサッカライドプロセスであり、人工酵素は、工業的に、技術的に、または栄養学的に重要なポリサッカライド基質の1つ以上を特異的に加水分解する。この変異体のさらに特定の実施形態において、人工酵素は、トリグリセリドエステルまたは脂質の加水分解を触媒する(リパーゼ活性)。
【0111】
次いで、好ましくは、触媒された反応は、脂質プロセス工程であり、人工酵素は、1つ以上の工業的、技術的、または栄養学的に重要な脂質基質を特異的に加水分解する。この実施形態のさらに特定の変異体において、人工酵素は、基質の酸化または還元を触媒する(酸化還元活性)。次いで、好ましくは、人工酵素は、1つ以上の工業的、技術的、または栄養学的に重要な化学物質基質を特異的に酸化または還元する。
【0112】
本発明の第三の局面は、定性的におよび/または定量的に新規な特異性を有する人工酵素を、タンパク質足場と組み合わせて作製する方法に関する。本発明の方法は、少なくとも以下の工程を包含する:
(a)少なくとも1つの標的基質における少なくとも1つの化学反応を触媒することができるタンパク質足場を提供する工程、
(b)工程(a)からの前記タンパク質足場と、1つ以上の完全または部分的にランダムなペプチド配列の変異体とを、生じる酵素が少なくとも1つの標的基質と1つ以上の異なる基質とを区別できるようにする前記タンパク質足場の部位において組み合わせることによって、酵素または単離された酵素のライブラリーを生成する工程、ならびに
(c)工程(b)で生成した前記酵素のライブラリーから、少なくとも1つの標的基質に対する規定された特異性を有する1つ以上の酵素を選択する工程。
【0113】
本発明のこの局面の第一の変異体において、本発明の方法は、少なくとも以下の工程を包含する:
(a)少なくとも1つの標的基質における少なくとも1つの化学反応を触媒することができるタンパク質足場を提供する工程、
(b)工程(a)からの前記タンパク質足場に、1つ以上の完全または部分的にランダムなペプチド配列を、生じる酵素が少なくとも1つの標的基質と1つ以上の異なる基質とを区別できるようにする前記タンパク質足場の部位に挿入することによって、酵素または単離された酵素のライブラリーを生成する工程、ならびに
(c)工程(b)で生成した前記酵素のライブラリーから、少なくとも1つの標的基質に対する規定された特異性を有する1つ以上の酵素を選択する工程。
【0114】
好ましくは、1つ以上の完全にまたは部分的にランダム化されたペプチド配列がタンパク質足場と組み合わされるかまたはそれに挿入される位置は、その組み合わせまたは挿入の前に同定されている。
【0115】
完全にまたは部分的にランダムなペプチド配列の挿入の数または他の組み合わせ、ならびにそれらの長さは、広範に変化し得る。その数は、少なくとも1個、好ましくは、1個以上、より好ましくは、2〜11個、最も好ましくは、2〜6個である。このような完全または部分的にランダム化されたペプチド配列の長さは、通常、50アミノ酸残基よりも短い。好ましくは、その長さは、1〜15アミノ酸残基、より好ましくは、1〜6アミノ酸残基である。あるいは、その長さは、2〜20アミノ酸残基、好ましくは、2〜10アミノ酸残基、より好ましくは、3〜8アミノ酸残基である。
【0116】
好ましくは、このような挿入または他の組み合わせは、このようなタンパク質足場をコードするポリヌクレオチド、およびこのような完全にまたは部分的にランダム化したペプチド配列をコードするポリヌクレオチドもしくはオリゴヌクレオチドを使用してDNAレベルで行われる。
【0117】
必要に応じて、工程(a)〜(c)は、循環的に繰り返され、それにより工程(c)で選択された酵素が、将来のサイクルの工程(a)におけるタンパク質足場として働き、そしてランダム化したペプチド配列が挿入されるか、あるいは以前のサイクルで挿入されたペプチド配列で置換される。それにより、挿入されたペプチド配列の数は、サイクルにわたって一定であるか、または増大する。意図される特異性を有する1つ以上の酵素が生成されるまで、サイクルが繰り返される
【0118】
さらに、工程(a)〜(c)の1ラウンド以上の間またはその後に、足場は1つ以上の位置で変異されて、SDR配列との組み合わせのためにより受け入れ可能な足場が作製され得るか、および/または特定のpHおよび温度での触媒活性を増大し得るか、および/またはグリコシル化のパターンを変化させ得るか、および/または酵素阻害剤に対する感受性を減少させ得るか、および/または酵素の安定性を変化させ得る。
【0119】
本発明のこの局面の第二の変異体において、本発明の方法は、少なくとも以下の工程を包含する:
(a)第一のタンパク質足場フラグメントを提供する工程、
(b)上記タンパク質足場フラグメントをペプチド結合を介して第一のSDRと連結する工程、および、必要に応じて
(c)上記工程(b)の生成物をペプチド結合を介してさらなるSDRペプチドまたはさらなるタンパク質足場フラグメントと連結する工程、および、必要に応じて、
(d)十分に特異的な酵素が生成されるために必要なサイクル工程(c)を繰り返す工程、ならびに
(e)工程(a)〜(d)において生成された集団から、1つ以上の標的基質に対する所望の特異性を有する1つ以上の酵素を選択する工程。
【0120】
タンパク質足場フラグメントは、タンパク質足場の配列の一部を意味する。タンパク質足場は、少なくとも2つのタンパク質足場フラグメントから構成される。
【0121】
本発明のこの局面の第三の変異体において、タンパク質足場、SDR、および人工酵素は、DNA配列によってコードされ、発現系がタンパク質を生成するために使用される。代替的な変異体において、タンパク質足場、SDR、および/または人工酵素は、ペプチドビルディングブロックから化学合成される。
【0122】
本発明のこの局面の第四の変異体において、本発明の方法は、少なくとも以下の工程を包含する:
(a)1つ以上の標的基質における1つ以上の化学反応を触媒し得るタンパク質足場をコードするポリヌクレオチドを提供する工程、
(b)1つ以上の完全にまたは部分的にランダム化されたオリゴヌクレオチド配列を、タンパク質足場をコードするポリヌクレオチドと組み合わせる工程であって、上記完全にまたは部分的にランダム化されたオリゴヌクレオチド配列が、コードされる人工酵素が1つ以上の標的基質と1つ以上の他の基質とを区別できるようなポリヌクレオチド内の部位に位置する、工程、ならびに
(c)工程(b)において生成された集団から、1つ以上の基質に対する規定された特異性を有する酵素をコードする1つ以上のポリヌクレオチドを選択する工程。
【0123】
任意の酵素が工程(a)におけるタンパク質足場として働き得る。それは、天然に存在する酵素、その変異体もしくは短縮型誘導体、または人工酵素であり得る。ヒトの治療用途には、タンパク質足場は、好ましくは、哺乳動物の酵素であり、より好ましくは、ヒトの酵素である。その局面において、本発明は、哺乳動物ゲノムまたはヒトゲノムにおいてそれぞれコードされるいかなる酵素の特異性とも異なる特異性を有する本質的に哺乳動物の、特に本質的にヒトの酵素の作製方法に関する。
【0124】
本発明に従って、この局面の工程(a)において提供されるタンパク質足場は、標的基質上で1つ以上の化学反応を触媒し得ることが要求される。それゆえ、タンパク質足場は、標的基質に対するその活性によって可能性のあるタンパク質足場の群から選択される。
【0125】
本発明のこの局面の好ましい変異体において、加水分解活性を有するタンパク質足場が使用される。好ましくは、タンパク質分解活性を有するタンパク質足場が使用され、より好ましくは、標的基質上で塩基性の活性を有する非常に低い特異性を有するプロテアーゼが、タンパク質足場として使用される。基質特異性の低い異なる構造クラス由来のプロテアーゼの例は、パパイン、トリプシン、サブチリシン、SET(Streptomyces erythraeus由来のトリプシン様セリンプロテアーゼ)、エラスターゼ、カテプシンG、またはキマーゼである。タンパク質足場として使用される前に、プロテアーゼのアミノ酸配列は、特異性以外のタンパク質特性、例えば、触媒活性、安定性、阻害剤感受性、または発現量を、本質的にWO 92/18645に記載されるように、または特異性を、本質的にEP 02020576.3およびPCT/EP03/04864に記載されるように変化させるために、修飾され得る。
【0126】
ふさわしいタンパク質足場についての別の選択肢は、リパーゼである。肝性リパーゼ、リポタンパク質リパーゼ、および膵リパーゼが、「リポタンパク質リパーゼスーパーファミリー」に属し、これは言い換えるとリパーゼのGXクラスの例である(M. Fischer、J. Pleiss (2003)、Nucl. Acid. Res. 、31、319−321)。リパーゼの基質特異性は、荷電したヘッド基を有する脂肪酸およびリン脂質のトリグリセロールエステルに対するそれらの相対活性によって特徴付けられる。あるいは、エステラーゼ、グリコシラーゼ、アミダーゼ、またはニトリラーゼのような他の加水分解酵素が、足場として使用され得る。トラスフェラーゼはまた、ふさわしいタンパク質足場である。グリコシルトランスフェラーゼは、種々のドナーおよびアクセプターを含む多くの生物学的合成に関わっている。あるいは、タンパク質足場は、リガーゼ、リアーゼ、酸化還元酵素、またはイソメラーゼ活性を有し得る。
【0127】
第一の実施形態において、1つ以上の完全または部分的にランダム化されたペプチド配列が、タンパク質足場の特定の部位に挿入される。これらの挿入部位は、挿入されたペプチド配列が、異なる基質間の識別子、すなわち、特異性決定領域(SDR)として作用し得るという事実によって特徴付けられる。このような挿入部位は、いくつかのアプローチによって同定され得る。好ましくは、挿入部位は、タンパク質足場の3次元構造の分析によって、タンパク質足場の一次構造の、異なる定量的特異性を有する他の酵素との比較分析により、またはアラニンスキャニング、ランダム変異誘発、もしくはランダム欠失、または任意のその組み合わせのような技術によって実験的に、同定される。
【0128】
SDRのための挿入部位を同定するための第一のアプローチは、X線結晶学、または核磁気共鳴研究によって得ることができるタンパク質足場の3次元構造に基づいている。同じ構造クラスであるが異なる定量的特異性を有する他の酵素との比較におけるタンパク質足場の構造アラインメントは、高い構造類似性の領域および低い構造類似性の領域を明らかにする。例えば、このような分析は、PBDビューアのような公のソフトウェア(Guex、N. and Peitsch、M. C. (1997)Electrophoresis 18、2714−2723))を使用してなされ得る。低い構造類似性の領域は、好ましいSDR挿入部位である。SDRのための挿入部位を同定する第二のアプローチにおいて、競合阻害剤または基質アナログと複合体を形成した足場タンパク質の3次元構造が分析される。競合阻害剤の結合部位は、基質の結合部位と有意に重なり合うことが仮定される。その場合、阻害剤の特定の原子間距離内にあるタンパク質の原子は、基質に対しても類似する距離にある傾向がある。例えば、5Å未満の短い距離を選択することによって、基質との密接に施蝕しているタンパク質原子の組み合わせを生じる。これらの残基は、第一のシェルコンタクト(shell contacts)を構成し、それゆえSDRにとって好ましい挿入部位である。一旦第一のシェルコンタクトが同定されると、第二のシェルコンタクトが、第一のシェル原子から開始する距離分析を繰り返すことによって見出され得る。本発明のなお別の代替において、上記の距離分析は、活性部位残基から開始して行われる。
【0129】
SDRに対する挿入部位を同定する第三のアプローチにおいて、足場タンパク質の一次配列は、アラインメントアルゴリズムを使用することによって、同じ構造クラスであるが異なる定量的特異性を有する他の酵素とアラインメントされる。このようなアラインメントアルゴリズムの例は公開されている(Altschul、S. F. 、Gish、W.、Miller、W. 、Myers、E. W. & Lipman、D. J. (1990) J. Mol. Biol. 215: 403−410; 「Statistical methods in Bioinformatics: an introduction」 by Ewens、W. & Grant、G.R. 2001、Springer、New York)。このようなアラインメントは、種々の配列相同性、特に足場タンパク質と比較して1つ以上の酵素におけるさらなる配列エレメントを有する、保存された領域または保存されていない領域を明らかにし得る。保存された領域は、異なるタンパク質の間で共有されている表現型に寄与する(例えば、三次元構造の折り畳みを安定化する)可能性が高い。保存されていない領域、および特に、定量的に高い特異性を有する酵素におけるさらなる配列(Turner、R. etal. (2002) J. Biol.Chem.、277、33068−33074)は、SDRのための好ましい挿入部位である。
【0130】
プロテアーゼとして、現在5つのファミリー、すなわち、アスパラギン酸プロテアーゼ、システインプロテアーゼ、セリンプロテアーゼ、メタロプロテアーゼ、およびスレオニンプロテアーゼが知られている。各々のファミリーには、類似の集団を共有するプロテアーゼのグループが含まれる。これらのグループのメンバーの結晶構造は解析されており、公のデータベース(例えば、Brookhavenタンパク質データベース)を通じてアクセス可能である(H. M. Berman et al. Nucleic Acids Research、28 pp. 235−242(2000))。このようなデータベースはまた、他の酵素クラスおよび各クラスの非酵素的に活性なタンパク質における構造ホモログを含んでいる。いくつかのツールが、構造ホモログについて公のデータベースを検索するために利用可能である:SCOP−配列および構造の調査のためのタンパク質データベースの構造分類(Murzin A. G. etal. (1995) J. Mol. Biol. 247、536−540);CATH−クラス、アーキテクチャ、トポロジー、および相同スーパーファミリー:タンパク質ドメイン構造の階層型分類(Orengo et al. (1997) Structure 5 (8) 1093−1108);FSSP−タンパク質の構造間アラインメントに基づく折り畳み分類(Holm and Sander (1998) Nucl. Acids Res. 26 316−319);またはVAST−ベクターアラインメントサーチツール(Gibrat、Madej and Bryant (1996) Current Opinion in Structural Biology 6、377−385)。
【0131】
上記のアプローチにおいて、構造クラスのメンバーは、SDRの挿入部位を同定するために比較される。
【0132】
これらのアプローチの好ましい変形において、構造クラスS1のセリンプロテアーゼが、互いに比較される。トリプシンは、低い基質特異性を有するメンバーである。なぜならそれは、R位置にアルギニンまたはリジン残基のみを必要とする。他方、トロンビン、組織型プラスミノーゲン活性化因子、またはエンテロキナーゼは、全て、それらの基質配列(すなわち、それぞれ、(L/I/V/F)XPR^NA、CPGR^VVGG、およびDDDK^)に対して高い特異性を有する(Perona、J. & Craik、C. (1997) J.Biol.Chem.、272、29987−29990 ; Perona、J. & Craik、C (1995) Protein Science、4、337−360)。これらのプロテアーゼのアミノ酸配列のアラインメントは、実施例1(図2)に同定したSDRとともに記載されている。
【0133】
セリンプロテアーゼのファミリー内のさらなる例は、構造クラスS8(サブチリシン折り畳み)のメンバーによって与えられる。サブチリシンは、このクラスの型プロテアーゼであり、非特異的プロテアーゼである(Ottesen、M. & Svendsen、A. (1998) Methods Enzymol 19、199−215)。フリン、PC1、およびPC5は、プロペプチドのプロセッシングに関与する同じ構造クラスのプロテアーゼであり、高い基質特異性を有する(Seidah、N. & Chretien、M. (1997) Curr. Opin. Biotech. 、8: 602−607; Bergeron、F. etal. (2000) J. Mol.ER7docrin.、24: 1−22)。アプローチの好ましい変形において、一次アミノ酸配列のアラインメント(図4)が、サブチリシンと比較して特異的なプロがさらに有し、それゆえ潜在的な特異性決定領域である3アミノ酸より長い11個の配列ストレッチを同定するために使用される。アプローチのさらなる変形において、サブチリシンの3次元構造からの情報が、選択をさらに狭めるために使用され得る(図3)。11の挿入配列ストレッチのうち、3つ、すなわち、ストレッチ番号7、8、および11が、特に活性部位残基に近く、これらはそれぞれ、PC5、PC1、および全ての3つの特異的プロテアーゼにおける挿入である(図3)。好ましい変形において、可変長および可変組成の1または数個のアミノ酸ストレッチが、11個の部位のうちの1または数個の位置でサブチリシン配列に挿入され得る。アプローチのより好ましい変形において、挿入が領域7、8、もしくは11、またはこれらの任意の組み合わせで行われる。アプローチの別の好ましい変形において、構造クラスS8由来のサブチリシン以外のタンパク質足場が使用される。
【0134】
このアプローチのさらに好ましい変形において、構造クラスA1のアスパラギン酸プロテアーゼが分析される(Rawlings、N. D. & Barrett、A. J. (1995). Methods Enzymol. 248、105−120 ; Chitpinityol、S. & Crabbe、MJ. (1998)、Food Chemistry、61、395−418)。アスパラギン酸プロテアーゼのA1構造クラスの例は、低い基質特異性を有するペプチド、ならびに比較的高い基質特異性を有するベータセクレターゼ(Grninger−Leitch、F. 、et al. (2002) JBiol. Chem. 277、4687−4693)、およびレニンである。レトロウイルスプロテアーゼもまたこのクラスに属するが、活性酵素は、2つの同一のサブユニットのダイマーである。ウイルスプロテアーゼは、ポリタンパク質前駆体の正確なプロセッシングと、各場合において高い基質特異性を必要とする機能的タンパク質の生成にとって必須である(Wu、J. et al. (1998) Biochemistry、37、4518−4526 ; Pettit、S. etal. (1991) J.Biol. Chem. 、266、14539−14547)。ペプシンはこのクラスの型プロテアーゼであり、非特異的プロテアーゼを代表する(Kageyama、T. (2002) Cell. Mol.Life Sci. 59、288−306)。B−セクレターゼおよびカテプシンD(Aguilar、C.F. et al. (1995) Adv. Exp. Med.Bio/. 362、155−166)は、同じクラスのプロテアーゼであり、高い基質特異性を有する。アプローチの好ましい変形において、一次アミノ酸配列のアラインメント(図6)が、ペプチドと比較して特異的プロテアーゼに挿入され、それゆえ潜在的な特異性決定領域である、3つのアミノ酸よりも長い6つの配列ストレッチを同定するために使用される。アプローチのさらなる変形において、b−セクレターゼの3次元構造からの情報が、選択をさらに狭めるために使用され得る。6個の挿入配列のなかから、3つ、すなわち、それぞれストレッチ番号1、3、および4(これらはカテプシンDおよびベータセクレターゼにおける挿入出る)が活性部位残基に特に近い(図5)。アプローチの好ましい変形において、可変数および可変組成の1または数個のアミノ酸ストレッチが、6個の位置のうち1または数個でペプシン配列に挿入され得る。本発明のより好ましい実施形態において、挿入が、位置1、3.もしくは4、またはこれらの任意の組み合わせで行われる。本発明の別の好ましい実施形態において、ペプシン以外のタンパク質足場が使用される。
【0135】
特定の構造クラスが、低い特異性および高い特異性の公知のメンバーを含まない場合がある。これは、システインプロテアーゼファミリーに属し(Rawlings、N. D. & Barrett、A. J. (1994) Methods Enzymol. 244、461−486)、P4〜P1位置に対して全て高い特異性を示す、カスパーゼのC14クラスによって例示される。例えば、カスパーゼ1、カスパーゼ3、およびカスパーゼ9は、配列YVDA^、DEVD^、またはLEHD^をそれぞれ認識する。カスパーゼ間で異なる領域の同定は、基質特異性においける差違を担う領域が含まれる(図7および8)。
【0136】
最後に、酵素足場と同じ折り畳みの非酵素タンパク質もまた、SDRの挿入部位の同定に起用する。例えば、ハプトグロブリン(Arcoleo、J. & Greer、J.; (1982)d Biol. Chem. 257、10063−10068)およびアズロシジン(azurocidin)(Almeida、R. et al. (1991) Biochem. Biophys. Res. Commun. 177、688−695)は、全てのS1プロテアーゼと同じキモトリプシン様の折り畳みを共有する。活性部位残基での置換により、これらのタンパク質は、如何なるタンパク質分解機能も有しないが、活性プロテアーゼとの高い相同性を示す。これらのタンパク質と特定のプロテアーゼとの間の差違は、SDRのための挿入部位として働き得る領域を含む。
【0137】
第四のアプローチにおいて、SDRの挿入部位は、アラニンスキャニング、ランダム変異誘発、ランダム挿入、またはランダム欠失のような技術によって実験的に同定される。上記のアプローチとは対照的に、このアプローチは、足場タンパク質の三次元構造についての詳細な知識を必要としない。このアプローチの1つの好ましい変形において、タンパク質足場と同じ構造クラス由来の比較的高い特異性を有する酵素のランダム変異誘発および特異性の消失または変化についてのスクリーニングを使用して、タンパク質足場におけるSDRの挿入部位を同定し得る。ランダム変異誘発、アラニンスキャニング、ランダム挿入、またはランダム欠失は全て、酵素をコードするポリヌクレオチドのレベルに対して行われる。文献において公知の種々のプロトコルが存在する(例えば、Sambrook、J. F; Fritsch、E. F.; Maniatis、T.;Gold Spring Harbor Laboratory Press、Second Edition、1989、New York)。例えば、ランダム変異誘発は、特許WO 9218645に記載されるポリメラーゼを使用することによって達成され得る。この特許によれば、1つ以上のプロテアーゼをコードする1つ以上の遺伝子が、高いエラー率または誤取り込みの率を増大する条件下でDNAポリメラーゼを使用することによって増幅される。例えば、CadwellおよびJoyceの方法が使用され得る(Cadwell、R. C. and Joyce、G. F. 、PCR methods.Appl. 2 (1992)28−33)。突然変異誘発株、化学突然変異誘発物質、またはUV放射線の使用のような、しかしこれらに限定されないランダム変異誘発の他の方法もまた使用され得る。
【0138】
あるいは、オリゴヌクレオチドが、ランダムに分布したアミノ酸残基をアラニンと置換する変異誘発のために使用され得る。この方法は、一般的に、アラニンスキャニング範囲誘発と呼ばれる(Fersht、A. R. Biochemistry (1989) 8031−8036)。さらなる代替方法として、2項変異誘発のようなアラニンスキャニングの改変(Gregoret、L. M. and Sauer、R. T. PNAS (1993) 4246−4250)またはコンビナトリアルアラニンスキャニング(Weiss etal.、PNAS (2000) 8950−8954)が使用され得る。
【0139】
人工酵素を発現するために、このような人工タンパク質をコードするDNAが、表群的な分子クローニング技術によって適切な発現ベクターに連結される(例えば、Sambrook、J. F; Fritsch、E. F.; Maniatis、T.; Cold Spring Harbor Laboratory Press、Second Edition、1989、New York)。ベクターは、適切な発現宿主細胞に導入され、これは、対応する人工酵素変異体を発現する。特に適切な発言宿主は、大腸菌もしくは枯草菌(Bacillus subtilis)のような細菌性発現宿主、またはSaccharomyces cerevisaeもしくはPichia pastorisのような酵母発現宿主、またはチャイニーズハムスター卵巣(CHO)細胞株もしくはベビーハムスター腎臓(BHK)細胞株のような哺乳動物発現宿主、またはバクテリオファージ(例:M13もしくはラムダ)もしくはバキュロウイルス発現系のようなウイルスのようなウイルス発現宿主である。さらなる代替として、インビトロタンパク質発現のためのシステムが使用され得る。代表的には、DNAが、発現ベクター中、適切なシグナル配列の後ろに挿入される。シグナル配列は、酵素変異体が細胞外空間に分泌されることを導き、それによって細胞上清中でのプロテアーゼ活性の直接的な検出を可能にする。それぞれ、大腸菌にとって特に適切なシグナル配列は、HlyAであり、枯草菌にとっては、AprE、NprE、Mpr、AmyA、AmyE、Blac、SacBであり、S. cerevisiaeにとっては、Barl、Suc2、Mata、Inu1A、Ggpipである。あるいは、酵素変異体は細胞内で発現され、そして基質もまた細胞内で発現される。好ましくは、これは本質的に特許出願WO 0212543に記載されるように、基質アミノ酸配列と連結された2つの自己蛍光タンパク質を含む融合ペプチド基質を使用して行われる。さらなる代替として、酵素変異体の細胞内発現またはペリプラズム空間への分泌の後に、大腸菌についてのDsbA、PhoA、PelB、OmpA、OmpT、またはgIIIのようなシグナル配列を使用して、透過化または溶解工程により、酵素変異体を上清中へ放出する。膜バリアの破壊は、超音波、フレンチプレスのような機械的手段、またはリゾチームのような膜消化酵素の使用により強制され得る。別のさらなる代替として、酵素変異体をコードする遺伝子が、細胞を含まずに、適切な細胞を含まない発現系を使用して発現される。例えば、大腸菌細胞由来のS30抽出物を、Leslyらに記載のように、この目的のために使用し得る(Methods in Molecular Biology 37 (1995) 265−278)。任意の上記の方法によって生成され発現される遺伝子変異体の集団は、例えば、特許出願PCT/EP03/04864に詳細に記載されるように、適切なアッセイおよびスクリーニング方法によってそれらの親和性、基質特異性、または活性が分析される。元々の酵素と比較して減少した特異性を有する触媒活性変異体由来の遺伝子は、配列決定によって分析される。変異および/または挿入および/または欠失が生じる部位は、SDRが部位特異的に挿入され得る好ましい挿入部位である。
【0140】
第二の実施形態において、1つ以上の完全または部分的にランダムなペプチド配列が、タンパク質足場中のランダム部位に挿入される。この修飾は、通常、ポリヌクレオチドレベルで、すなわち、タンパク質足場をコードする遺伝子にヌクレオチド配列を挿入することによって行われる。ヌクレオチド配列のランダム挿入を可能にするいくつかの方法が利用可能である。ランダム挿入に使用され得るシステムは、例えば、ライゲーションに基づくシステム(Murakami etal、Nature Biotechnology 20 (2002) 76−81)、DNA重合反応に基づくシステムおよびトランスポゾンに基づくシステム(e.g、GPS−MTM mutagenesis system、NEBBiolabs ; MGTM mutation generation system、Finnzymes)である。トランスポゾンに基づく方法は、トランスポーゼースの認識配列をその末端に、ならびにまれな切断制限エンドヌクレアーゼ(rare cutting restriction endonuclease)を含有する選択可マーカー遺伝子のトランスポーゼース仲介挿入を利用する。後者のエンドヌクレアーゼを使用して、通常選択マーカーを放出し、ライゲーションの後、挿入断片を得る。再ライゲーションを行う代わりに、代替的に、1つまたは2つの外側切断制限エンドヌクレアーゼについての末端認識配列を有するフラグメントならびに選択マーカーを挿入し得る。ライゲーションの後、このフラグメントを1つまたは2つの外側切断エンドヌクレアーゼを使用して放出する。標準的な方法によって平滑末端を作製した後、平滑末端ランダムフラグメントを遺伝子にランダムな位置に挿入する。
【0141】
さらに好ましい実施形態において、インビトロ相同組換えのための方法が、上記方法によって導入される変異を組み合わせて、酵素集団を生成するために使用される。適用され得る方法の例は、特許出願WO0134835による組換え連鎖反応(RCR)、特許出願WO 9522625によるDNAシャッフリング方法、特許出願WO9842728によるねじれ伸長法(Staggered Extension method)、または特許出願WO9842728によるランダムプライミング組換えである。さらに、イッチー法(Itchy method)のような非相同組換えのための方法もまた、適用され得る(Ostermeier、M. et al. Nature Biotechnology 17 (1999) 1205−1209)。
【0142】
ヌクレオチド配列のタンパク質足場へのランダム挿入に際して、酵素変異体をコードする異なる遺伝子のライブラリーを得る。ポリヌクレオチドライブラリーは、引き続き、適切な発現ベクターに移される。適切な宿主における発現の際、またはインビトロ発現系の使用により、ランダムに挿入されたアミノ酸のストレッチを含有する酵素のライブラリーが得られる。
【0143】
本発明のこの局面の工程(b)にしたがって、1つ以上の完全または部分的ランダムペプチド配列が、タンパク質足場に挿入される。このような挿入されたSDRの実際の数が、意図された特異性が高いほど、SDRがより多く挿入されるという関係に従って、意図された定量的特異性によって決定される。単一のSDRが中程度に特異的な酵素の生成を可能にするのに対して、2つのSDRは、有意に特異的な酵素の生成をすでに可能にする。しかしながら、6個までまたはそれ以上のSDRがタンパク質足場に挿入され得る。類似の関係がSDRの長さについて有効である:意図される特異性が高いほど、挿入されるSDRが長い。SDRは、4アミノ酸残基程度の短さであり得る。しかしながら、それらはまた、50アミノ酸残基程度まで長くてもよい。有意な特性がすでに、4〜6アミノ酸残基の長さのSDRの使用によって生成され得る。
【0144】
挿入されるペプチド配列は、完全にまたは部分的にランダムであり得る。この状況において、完全にランダムとは、各位置または全ての位置において互いに異なる配列を含む一組の配列が並行して挿入されることを意味する。部分的にランダムとは、少なくとも1つの位置で違いに異なる配列を含む一組の配列が並行して挿入されることを意味する。この違いは、ペアごとまたは単一の配列に関してであり得る。例えば、アミノ酸の長さの挿入に関しての場合、部分的ランダムは、(i)AGGG、GVGG、GGLG、GGGIを含む、または(ii)AGGG、VGGG、LGGG、およびIGGGを含むセットであり得る。あるいは、ランダム配列はまた、長さが互いに異なる配列を含む。ペプチド配列のランダム化は、それぞれの部位で遺伝子に挿入されたヌクレオチド配列のランダム化によって達成される。それにより、ランダム化は、オリゴヌクレオチドの化学合成の間に核酸塩基の混合物をモノマーとして利用することによって達成され得る。ストップコドンの可能性をさらに最小化する完全にランダムなコドンのためのモノマーの特に好ましい混合物は、NN(GTC)である。あるいは、ランダムオリゴヌクレオチドが、それぞれの部位で遺伝子に挿入される短いフラグメントへのDNAのフラグメント化によって得られ得る。フラグメント化されるDNAの供給源は、合成オリゴヌクレオチドであり得るが、あるいは、クローン化された遺伝子、cDNA、またはゲノムDNAに由来するものでもよい。好ましくは、DNAは酵素をコードする遺伝子である。フラグメント化は、例えば、DNAのランダムエンドヌクレアーゼ消化によって達成され得る。好ましくは、DNAseI(例えば、ウシ膵臓由来の)のような非特異的エンドヌクレアーゼは、エンドヌクレアーゼ消化のために利用される。
【0145】
本発明の方法の工程(a)〜(c)が循環的に繰り返される場合、連続するラウンドにおいて挿入されるランダムペプチド配列を得るための異なる代替が存在する。好ましくは、酵素の増大した特異性を導くものとして1つのラウンドにおいて同定されたSDRは、次のラウンドにおいて挿入されるランダムペプチド配列のための鋳型として使用される。
【0146】
好ましい代替において、1つのラウンドで選択された配列が分析され、そしてランダム化されたオリゴヌクレオチドがこれらの配列に基づいて生成される。これは、例えば、元々のヌクレオチドに加えて、他の3つのヌクレオチドモノマーの特定の割合(%)の混合物をオリゴヌクレオチド合成において各位置で使用することによって達成され得る。例えば、第一ラウンドにおいて、アミノ酸配列ARLT(例えば、ヌクレオチド配列GCG CGC CTT ACCによってコードされる)を有するSDRが同定された場合、このSDR部位に挿入されるランダムペプチド配列は、70%G、10%A、10%T、および10%Cを第一の位置に、70%C、10%G、10%T、および10%Aを第二の位置に、など、有するオリゴヌクレオチドによってコードされる。これは、各位置に、およそ3回に1回の割合で、鋳型アミノ酸を、3回に2回の割合で別のアミノ酸を導く。
【0147】
別の好ましい代替において、1つのラウンドにおいて選択される配列が分析され、そしてコンセンサスライブラリーがこれらの配列に基づいて生成される。これは、例えば、オリゴヌクレオチド合成における各位置においてヌクレオチドの規定された混合物を、前回のラウンドで選択されたSDRの各位置で同定されたアミノ酸残基の混合物を導くように、使用することによって達成され得る。例えば、第一のラウンドにおいて、アミノ酸配列ARLTおよびVPGSを有する2つのSDRが同定された場合、次のラウンドでこのSDRに挿入されるコンセンサスライブラリーは、配列G(C/T)GC(G/C)C(G/T)(G/T)G(A/T)CCを有するオリゴヌクレオチドによってコードされ得るこれは、ランダムペプチド配列(A/V)(R/P)(L/G/V/W)(T/S)に対応し、それにより、第一のラウンドで同定された全てのアミノ酸残基の組み合わせを可能にし、そして遺伝コードの縮重により、いくつかの位置でより低い程度に代替のアミノ酸を許容する。
【0148】
別の好ましい代替において、1つのラウンドで選択された配列は、以前の分析なしで、ポリヌクレオチドのインビトロ組換えのための方法(例えば、WO01/34835に記載される方法)を使用して組換えされる。(以下は、また、本発明の第8および第9の局面の詳細を提供する。)
部分的にまたは完全にランダムな配列を足場タンパク質をコードする遺伝子に挿入し、最終的に標準的な分子クローニング技術(Sambrook、J. F; Fritsch、E.F. ; Maniatis、T.; Cold Spring Harbor Laboratory Press、Second Edition、1989、New York)を使用して、得られた遺伝子を適切な発現ベクターにライゲーションした後、そのベクターは、対応する酵素変異体を発現する適切な発現宿主細胞へ導入される。特に適切な発現宿主は、大腸菌もしくは枯草菌のような細菌発現宿主、またはSaccharomyces cerevisae もしくはPichia pastorisのような酵母宿主、またはチャイニーズハムスター卵巣(CHO)細胞株またはベビーハムスター腎臓(BHK)細胞株のような哺乳動物宿主、またはバクテリオファージ(例:M13、T7ファージ、ラムダ)のようなウイルス発現系、またはバキュウロウイルス発現系のようなウイルスである。さらなる代替として、インビトロタンパク質発現のためのシステムが使用され得る。代表的には、DNAが、細胞外空間に酵素変異体を分泌させる適切なシグナル配列の後に、発現ベクターにライゲーションされ、それにより細胞上清中の酵素活性の直接検出を可能にする。大腸菌について特に適切なシグナル配列は、ompA、pelB、HlyAであり、枯草菌についてはAprE、NprB、Mpr、AmyA、AmyE、Blac、SacBであり、S.CerevisiaeについてはBar1、Suc2、Mata、I nu1A、Ggplpである。あるいは、酵素変異体が細胞内で発現され、基質もまた細胞内で発現される。プロテアーゼ変異体に従って、これは、本質的に特許出願WO 0212543に記載されるように、基質アミノ酸配列によって連結された2つの自己蛍光タンパク質を含む融合ペプチド基質を使用して、行われる。さらなる代替として、酵素変異体の細胞内発現、または大腸菌のDsbA、PhoA、PelB、OmpA、OmpT orgi 11のようなシグナル配列を使用したペリプラズム空間への分泌の後、透過化または溶解工程によいr、酵素辺が上清へ放出される。膜バリアの破壊は、超音波、フレンチプレスのような機械的手段を使用して、またはリボチームのような膜消化酵素を使用して、強制され得る。別のさらなる代替として、酵素変異体をコードする遺伝子が、細胞を含まないで、適切な細胞を含まない発現系を使用して発現される。例えば、大腸菌細胞由来のS30抽出物が、Leslyらによって記載されるようにこの目的のために使用される(Methods in Molecular Biology 37 (1995) 265−278)。
【0149】
ベクターの宿主細胞への導入の後、これらの細胞を、意図された標的基質に対する特異性を有する酵素の発現についてスクリーニングする。このようなスクリーニングは、典型的には、遺伝子型と表現型とを相関付けるために細胞を互いに分離し、そして増殖および発現期間の後に各細胞クローンの活性をアッセイすることによって行われる。このような分離は、例えば、細胞を、例えば、WO01/24933に記載されるようにサンプルキャリアの区画へ分布させることによって行われ得る。あるいは、細胞を、アガロースプレート上でストリーキングすることにより、アガロースのようなポリマー中で包むことにより、キャピラリー中に充填することにより、または同様の方法により分離される。
【0150】
意図される特異性を有する変異体の分離は、異なるアプローチによってなされ得る。プロテアーゼの場合、好ましくは、本質的にPCT/EP03/04864に記載されるペプチド基質を用いるアッセイが使用される。
【0151】
発現のフォーマットにかかわらず、酵素変異体を選択することにより、好ましくは、標的配列を認識し、変換する酵素の同定が可能になる。第一の代替として、標的配列を認識し、変換する酵素は、好ましくは、標的基質配列に対する高い親和性を有する酵素をスクリーニングことによって同定される。高い親和性は、実質的に第一の酵素のKMより低い標的基質濃度でスクリーニングすることによって選択される低いKMに対応する。好ましくは、使用される基質は、10μM未満、好ましくは、1μM未満、より好ましくは、100nM未満、そして最も好ましくは、10nM未満の濃度で基質の修飾を検出することを可能にする1つ以上のフルオロフォアに連結される。
【0152】
第二の代替として、好ましく標的基質を認識し変換する酵素が、2つ以上の基質をアッセイにおいて使用し、そしてこれらの2つ以上の基質の活性を比較してスクリーニングすることによって同定される。好ましくは、使用される2つ以上の基質は、異なるマーカー分子に連結され、それにより、2つ以上の基質の修飾を連続的にまたは並行して検出することが可能になる。プロテアーゼの場合、特に好ましくは、2つのペプチド基質が使用される。一方のペプチド基質は、任意に選択されたかまたは部分的もしくは完全にランダムなアミノ酸配列を有し、それにより、任意の基質上での活性がモニターでき、そして他方のペプチド基質は、意図された標的基質配列に同一かまたは類似するアミノ酸配列を有し、それにより標的基質上での活性をモニターできる。特に好ましくは、これらの2つのペプチド基質が蛍光マーカー分子に連結され、そして連続的にまたは並行して測定された場合に両方の活性を区別するために、2つのペプチド基質の蛍光特性は十分に異なる。例えば、特許出願WO 0212543による第一の蛍光タンパク質、ペプチド、および第二の蛍光タンパク質を含む融合タンパク質が、この目的に使用され得る。あるいは、ローダミンのような蛍光色素がペプチド基質に化学的に連結される。
【0153】
第三の代替として、標的酵素を好ましく認識し変換する酵素が、標的基質に類似する1つ以上の基質を、高過剰の競合基質とともに使用することによって同定される。次いで、標的基質に類似する基質上の活性に関してのスクリーニングが、競合基質の存在下で行われる。標的特異性に定性的に対応する特異性を有するが、低い定量的特異性のみを有する酵素は、このようなスクリーニングにおいて負のサンプルとして同定される。これに対して、定量的におよび定性的に標的特異性に対応する特異性を有する酵素は、正に同定される。好ましくは、標的基質に類似する1つ以上の基質はマーカー分子に連結され、それによりそれらの修飾の検出が可能となり、一方、競合基質はマーカー分子を有しない。競合基質は、任意に選択されたかまたはランダムアミノ酸配列を有し、それによりマーカー保有基質の加水分解に対する競合阻害剤として作用する。例えば、タンパク質加水分解酵素(例:トリプトン)は、本発明の人工タンパク質分解酵素に対する競合基質として作用し得る。
【0154】
第四の代替として、標的基質を認識し変換する酵素が同定され、増幅結合または増殖結合選択工程によって同定および選択される。さらに、活性が細胞内で測定され得、そして選択がセルソーター(例えば、蛍光活性化セルソーター)によってなされ得る。
【0155】
さらなる代替として、標的基質を認識し変換する酵素は、第一に、標的基質に優先的に結合する酵素を選択し、第二に、酵素変異体のこのサブグループから標的基質を変換する酵素を選択することによって同定される。標的基質に優先的に結合する酵素の選択は、標的基質に対するバインダーの選択か、または他の基質へ結合する酵素の対抗選択のいずれかによって行われる。バインダーの選択または非バインダーの対抗選択のための方法は当該分野で公知である。このような方法は、典型的には、表面ディスプレイ発現法を使用することによって解決され得る表現型−遺伝子型共役を必要とする。このような方法には、例えば、ファージもしくはウイルスディスプレイ、細胞表面ディスプレイ、およびインビトロディスプレイが含まれる。ファージもしくはウイルスディスプレイは、典型的には、目的のタンパク質のウイルス/ファージタンパク質への融合を含む。細胞表面ディスプレイ(すなわち、細菌もしくは真核生物細胞ディスプレイのいずれか)は、典型的には、目的のタンパク質の細胞表面に存在するペプチドもしくはタンパク質への融合を含む。インビトロディスプレイにおいて、タンパク質は、典型的には、インビトロで作製され、それらに直接または間接的に、そのタンパク質をコードするRNAを結合させる(DE 19646372)。
【0156】
本発明はまた、本明細書中上記で規定した本発明の第一の局面に従う1つ以上の人口酵素を含有する組成物または薬学的組成物を提供する。その組成物は、必要に応じて、受容可能な担体、賦形剤、および/または助剤を含んでいても良い。本明細書中で規定される非薬学的組成物は、研究用組成物、栄養学組成物、洗浄組成物、消毒組成物、化粧用組成物、またはパーソナルケア用組成物である。さらに、本明細書中に規定する人工酵素をコードするDNA配列およびこのDNAを含有するベクターもまた、提供される。最後に、このようなDNA配列および/もしくはベクターを含有する形質転換された宿主細胞(原核生物もしくは真核生物)またはトランスジェニック生物、ならびにこのような宿主細胞もしくはトランスジェニック動物を本発明の第一の局面の人工酵素を作製するために使用する方法もまた、意図される。
【0157】
(図面の詳細な説明)
図1:「球棒」表示で示された活性部位残基を有するヒトトリプシンIの3次元構造。印を付けた領域は、潜在的SDR挿入部位を示す。
【0158】
図2:ヒトプロテアーゼトリプシン1、アルファトロンビンおよびエンテロペプチダーゼ(これらの全てがセリンプロテアーゼファミリーの構造クラスS1に属する)の一次アミノ酸配列のアラインメント。トリプシンは、この構造クラスの非特異的プロテアーゼであるのに対して、アルファトロンビンおよびエンテロペプチダーゼは、高い基質特異性を有するプロテアーゼである。トリプシンと比較して、トロンビンおよびエンテロキナーゼの一次配列中に3以上のアミノ酸の挿入の領域がいくつか見られる。(−1−)と記された領域および(−3−)と記された領域は、SDR挿入部位である。
【0159】
アルファトロンビンの三次構造において、両方の領域が基質結合部位の近傍にある。これらの領域は、それゆえ、SDRの候補として選択されるための2つの基準:第一に、それらが非特異的プロテアーゼと比較して特異的プロテアーゼ中にある挿入であること、および第二に、それらが基質結合部位の近くにあること、を満たす。三次元構造の表示が図3に与えられる。
【0160】
図3:「球棒」表示で示された活性部位残基を有するサブチリシンの三次元構造。番号を付された領域は、潜在的SDR挿入部位を示す。
【0161】
図4:サブチリシンE、フリン、PC1、およびPC5(これらの全てが、セリンプロテアーゼファミリーの構造クラスS8に属する)の一次アミノ酸配列のアラインメント。サブチリシンEは、この構造クラスの非特異的プロテアーゼである。一方、フリン、PC1、およびPC5は、高い基質特異性を有するプロテアーゼである。サブチリシンと比較して、フリン、PC1、およびPC5の一次配列への3つ以上のアミノ酸の挿入のいくつかの領域が見られる。(−4−)、(−5−)、(−7−)、(−9−)、および(−11−)と記された領域は、好ましいSDR挿入部位である。これらの領域ストレッチは、SDRの候補として選択されるための2つの基準を満たす。第一に、これらは、非特異的なプロテアーゼと比較して特異的なプロテアーゼにおける挿入である。第二に、それらは活性部位残基に近い。
【0162】
図5:「球棒」表示で示された活性部位残基を有するベータセクレターゼの3次元構造。番号を付された領域は、潜在的SDR挿入部位を示す。
【0163】
図6:ペプシン、b−セクレターゼ、およびカテプシンD(これらの全てが、アスパラギン酸プロテアーゼファミリーの構造クラスA1に属する)の一次アミノ酸配列のアラインメント。ペプシンは、この構造クラスの非特異的プロテアーゼである。一方、b−セクレターゼおよびカテプシンDは、高い特異性を有するプロテアーゼである。ペプチドに比べて、b−セクレターゼおよびカテプシンDの一次配列中への3以上のアミノ酸の挿入のいくつかの領域が見られる。−1−〜−11−で記された領域は、可能性のあるSDR組み合わせ部位に対応し、そして図5にも記されている。
【0164】
図7:「球棒」表示で示された活性部位残基を有するカスパーゼ7の3次元構造を示す。番号を付した領域は、潜在的SDR挿入部位を示す。
【0165】
図8:システインプロテアーゼクラスC14ファミリーのメンバーとしてのカスパーゼ7の一次アミノ酸配列を示す(配列番号14もまた参照)。
【0166】
図9:本発明の第三の局面による方法の模式図である。
【0167】
図10:トリプシン発現のウエスタンブロット分析である。トリプシンの変異体を発現する細胞培養の上清が、ネガティブコントロールと比較されている。レーン1:分子量標準:レーン2:ネガティブコントロール;レーン3:変異体aの上清;レーン4:ネガティブコントロール;レーン5:変異体bの上清;発現された担に対して特異的な一次抗体およびシグナルの生成のための二次抗体を使用した。
【0168】
図11:標的基質のタンパク質分解切断の時間経過。ヒトトリプシンの遺伝子を有するベクターを含有する細胞の上清、および遺伝子を含まないベクターを含有する細胞の上清を、本文中に記載したペプチド基質とインキュベートした。ペプチドの切断により、読み出し値が低下した。タンパク質分解活性は、陽性クローンについて確認される。
【0169】
図12:異なるペプチド基質に対するヒトトリプシンIと比較した3つの人工タンパク質分解酵素の相対活性。2つの基質のタンパク質分解消化の時間経過を行い、評価した。基質Bをスクリーニングに使用し、基質Aは密接に関連した配列である。相対活性;3つの変異体の相対活性を、ヒトトリプシンIに対して活性に対して正規化した。変異体1および変異体2は、標的基質に対して明らかに増大した特異性を示す。一方、変異体3は、ヒトトリプシンIと類似した活性を有するネガティブコントロールとして働く。
【0170】
図13:それぞれ、トリプシン、および1つまたは2つのSDRを有する人工タンパク質分解酵素の変異体の相対活性。プロテアーゼの活性を、競合基質の存在下(すなわち、10mg/mlの濃度のペプトン)および非存在下で決定した。タンパク質分解切断の時間経過を記録紙、時間定数kを決定した。競合剤ありとなしとの間の時間定数の比が形成され、プロテアーゼの特異性の定量的尺度を表す。比をトリプシンに対して正規化した。2つのSDRを含有する変異体の特異性は、SDR2のみを有する変異体の特異性よりも2.5倍高かった。
【0171】
図14:競合基質の非存在下および存在下でのプロテアーゼ変異体の相対的特異性を示す。異なる配列を有する2つのインサートを含有するプロテアーゼ変異体はおよび非修飾足場ヒトトリプシンIを適切な宿主のおいて発現した。プロテアーゼ変異体の活性を、競合基質の非存在下および存在下でのTNFアルファの所望の標的配列を有するペプチドの切断速度として決定した。特異性を、競合剤の存在下および非存在下での切断速度の比として表した。
【0172】
図15:図は、ヒトTNFアルファに対して特異的な本発明の人工タンパク質分解酵素を発現する培養物由来の濃縮された上清とともにヒトTNFあるいはフラグメントをインキュベートした時のヒトTNFアルファによって誘導される細胞傷害性の減少を示す。これは、本発明の人工タンパク質分解酵素の効率を示す。
【0173】
図16:図は、ヒトTNFアルファに対して特異的な異なる濃度の精製した本発明の人工タンパク質分解酵素とともにヒトTNFアルファをインキュベートした時のヒトTNFアルファによって誘導される細胞傷害性の減少を示す。変異体gは、SDR1として配列番号72およびSDR2として配列番号73を含む。これは、本発明の人工タンパク質分解酵素の効率を示す。
【0174】
図17:図は、2つのタンパク質基質(a)ヒトTNFアルファ、(b)ヒト血清タンパク質の混合物において、ヒトTNFアルファに対して特異的な本発明の人工タンパク質分解酵素の活性を、ヒトトリプシンIの活性と比較する。これは、本発明の人工タンパク質分解酵素の安全性を示す。変異体xは、配列番号89(SDR1)および配列番号95(SDR2)に従うSDRを含有する配列番号75に対応する。変異体xiおよびxiiは、同じSDR配列を含有するその誘導体に対応する。
【0175】
図18:ヒトトリプシン由来の人工タンパク質分解酵素によるヒトVEGFの特異的加水分解。
【実施例】
【0176】
以下の実施例では、本発明の方法により得られた酵素の触媒特性の決定を含む本発明の材料および方法が提供される。これらの実施例は、例示の目的のためのみであり、いかなる様式においても本発明を限定すると解釈されるものではないことは理解されるべきである。ここに引用される全ての文献、特許、および特許出願は、その全体が、本明細書中で参考として援用される。
【0177】
以下に記載される実験例では、種々の文献(例えば、Sambrook et al. (1989)、Molecular Cloning : A Laboratory Manual、Cold Spring Harbor Laboratory、or Ausubel et al. (1987)、Current Protocols in Molecular Biology 1987−1988、Wiley Interscience)に記載された組換えDNA技術の標準技術を使用した。特に示されない限り、製造業者の指示書に従って、制限酵素、ポリメラーゼ、および他の酵素、ならびにDNA精製キットを使用した。
【0178】
(実施例I:ヒトトリプシンにおけるSDR部位の同定)
SDRの挿入部位は、セリンプロテアーゼヒトトリプシンI(構造クラスS1)において、より高い配列特異性を有する同じ構造クラスのメンバーとの比較によって、同定されている。トリプシンは、P1位置にアルギニンまたはリジン残基のみを必要とするので、低い基質特異性を有するメンバーである。他方、トロンビン、組織型プラスミノーゲン活性化因子、またはエンテロキナーゼは全て、それらの基質配列、すなわち、(L/I/V/F)XPR^NA、CPGR^VVGG、およびDDDK^それぞれに対して高い特異性を有する。これらおよびさらいS1セリンプロテアーゼの一次配列および三次構造は、配列および構造の低いおよび高い相同性の領域、特により特異的なプロテアーゼの配列の挿入に対応する領域を決定するためにアラインメントされている(図2)。潜在的SDR部位を表す3アミノ酸と同じかまたはそれよりも長い挿入のいくつかの領域は、図1に示されるように同定されている。これらの領域を、以下の実施例においてSDRの挿入のための標的部位として選択した。例えば、それぞれ、6の長さを有するSDR1(図2における領域1、配列番号1のアミノ酸42の後)および5アミノ酸の長さを有するSDR2(図2の領域3、配列番号1のアミノ酸23の後)。
【0179】
(実施例II:足場タンパク質として使用されるヒトトリプシンI遺伝子の分子クローニングおよびB.subtilisにおける成熟プロテアーゼの発現)
非特異的プロテアーゼヒトトリプシノーゲンIをコードする遺伝子を、ベクターpUC18にクローニングした。クローニングを以下のとおり行った。タンパク質のコード配列を、KpnI部位を5’末端に、そしてBamHI部位を3’末端に導入したプライマーを使用してPCRによって増幅した。このPCRフラグメントを、ベクターpUC18の適切な部位にクローニングした。同一性を配列決定により確認した。配列決定した後、成熟タンパク質のコード配列を、異なるBglI部位を5’末端および3’末端に導入したPCRによって増幅した。
【0180】
このフラグメントを、大腸菌B.subtilisシャトルベクターの適切な部位にクローニングした。このベクターは、大腸菌中のpMB1増幅起点、大腸菌中での選択のためのネオマイシン耐性マーカー、ならびにB.subtilisにおける構成的発現のためのP43プロモーターを含有する。B.subtiilsのsacB遺伝子由来のシグナルペプチドをコードするリーダー配列を含有する87bpのフラグメントを、P43プロモーターの後ろに導入した。異なるBglI制限部位は、異種遺伝子が発現するための挿入部位として作用する。
【0181】
ヒトトリプシンIの発現を、遺伝子を有するベクターを含有する細胞の上清中でタンパク質分解活性を、ネガティブコントロールと比較して測定することにより確認した。アルギニン切断部位を含むペプチドを基質として選択した。ペプチドは、N末端ビオチン化および蛍光色素によりC末端を標識された。そのペプチドを培養上清とともにインキュベートした後、ストレプトアビジンを添加した。切断されていないペプチドはストレプトアビジンと結合し、高い読み出し値を示すが、切断されたものは低い読み出し値を生じる。図11は、トリプシンI遺伝子を含まないベクターを含有するB.subtilis細胞(ネガティブコントロール)と比較して、トリプシンI遺伝子を含有するB.subtilis細胞のタンパク質分解消化の時間経過を示す。プロテアーゼの発現のさらなる確認として、遺伝子を有するベクターを含有する細胞およびコントロール細胞の上清を、ポリアクリルアミドゲル電気泳動、およびその後の標的プロテアーゼに対する抗体を使用したウエスタンブロットによって分析した。手順は、標準的な方法にしたがって行った(Sambrook、J. F; Fritsch、E. F.; Maniatis、T.; Cold Spring Harbor Laboratory Press、Second Edition、1989、New York)。図8は、トリプシンの遺伝子を有するベクターを有する細胞中のみのタンパク質の発現を確認する。
【0182】
(実施例III:足場タンパク質の提供)
この実施例では、ヒトトリプシンIを足場タンパク質として使用した。遺伝子を、その天然の形態で使用したか、または触媒活性が増大し、もしくはさらに改善した特性を有する足場タンパク質を生じるように修飾した。
【0183】
修飾は、遺伝子のランダム修飾、次いでその酵素の発現、および続いて活性の増大したものの選択によって行った。第一に、遺伝子を、本質的にCadwell、R.C.およびJoyce、G.F.(PCR MethodsAppl. 2 (1992) 28−33)によって記載されるように変異性(error−prone)条件下でPCR増幅した。変異性PCRを、30pmolの各プライマー、20mmolのGTPおよびdATP、100nmolのCTPおよびdTTP、20fmolのテンプレート、および5UのTaqDNAポリメラーゼを、10mM TrisHCl(pH7.6)、50mM KCl、7mM MgCl2、0.5mM MnCl2、0.01% ゼラチン中で、94℃で1分、65℃で1分、および72℃で1分の20サイクルで行った。得られるDNAライブラリーを、Qiaquick PCR精製キットを供給業者の指示書に従って使用して精製した。PCR産物を制限酵素BglIを用いて消化し、精製した。その後、PCR産物を、上記のように大腸菌B.subtilisシャトルベクター中へライゲーションし、これをBglIで消化して、脱リン酸化した。ライゲーション産物を大腸菌へ形質転換し、LB中で増幅し、そしてプラスミドをQiagenプラスミド精製キットを供給業者の指示書にしたがって使用して精製した。得られるプラスミドをB.subtilis細胞に形質転換した。
【0184】
あるいは、またはランダム変異誘発に加えて、遺伝子の変異体を、本質的にWO 0134835に記載されるように、組換え連鎖反応を使用して相同位置で統計学的に組み換えた。プロテアーゼ変異体をコードする遺伝子のPCR産物を、QlAquick PCR精製キットを供給業者の指示書にしたがって使用して精製し、アガロースゲル電気泳動によって正確なサイズを供給業者の指示書にしたがってチェックし、そして等モル量で混合した。150mM TrisHCl(pH7.6)、6.6mM MgCl2中のこのPCR混合物の80μgを、94℃で5分間加熱し、続いて1秒ごとに0.05℃で37℃まで冷却し、鎖を再アニーリングし、それによりヘテロ二重鎖を確率論的な様式で生成した。次いで、1ΜGのDNA当たり2.5U エンドヌクレアーゼIIIを添加し、20、40、または60分間、37℃でインキュベートして、ヘテロ二重鎖の両方の3’末端から異なる長さを消化した。部分的に消化したPCR産物を、供給業者の指示書にしたがって0.17mM dNTPおよびPfuポリメラーゼ緩衝液中で72℃で15分間インキュベートすることによって、1μg DNA当たり0.6U Pfuのポリメラーゼで再充填した。一回のPCRサイクルを行った後、得られたDNAをQIAquick PCR精製キットを供給業者の指示書にしたがって使用して精製し、BglIで消化し、直線化したベクターにライゲーションした。ライゲーション産物を大腸菌中へ形質転換し、アンピシリンをマーカーとして含有するLB中で増幅し、そのプラスミドをQiagenプラスミド精製キットを供給業者の指示書に従って使用して精製した。得られたプラスミドを、B.subtilis細胞中へ形質転換した。
【0185】
(実施例IV:ヒトトリプシンIのタンパク質足場へのSDRの挿入および配列KKWLGRVPGGPVを有するペプチド基質に対する特異性を有する人工タンパク質分解酵素の生成)
ヒトトリプシンI中にSDRのための挿入部位を作製するために、異なる制限部位の2つのペアを、アミノ酸配列を変更せずに潜在的SDR部位として同定された部位(上記実施例Iを参照)に遺伝子を導入した。制限部位の挿入は、オーバーラップPCRによって行われた。プライマーrestrIおよびrestr2をSacIIおよびBamHI制限部位の導入のために使用し、restr3およびrestr4を、KpnIおよびNheI制限部位の導入のために使用した。プライマーの配列は以下の通りである:
【0186】
restr1およびrestr2についての結合部位および対応するアミノ酸配列(配列番号54):
【化1】
フォワードプライマーrestr1(配列番号56):
【化2】
リバースプライマーrestr2(配列番号57):
【化3】
restr3およびrestr4についての結合部位および対応するアミノ酸配列(配列番号58):
【化4】
フォワードプライマーrestr3(配列番号60):
【化5】
リバースプライマーrestr4(配列番号61):
【化6】
【0187】
第一のオーバーラップ伸長PCRにおいて、SacII/BamHI部位を導入し、SDR1を挿入できるようにし、第二のオーバーラップ伸長PCRにおいて、KpnI/NheI部位を導入し、SDR2を挿入できるようにした。オーバーラップ伸長PCRの産物を、プライマーpUCフォワードおよびpUCリバースを用いて増幅した。pUCフォワードおよびpUCリバースの配列は、以下の通りである:
pUCフォワード(配列番号62):
【0188】
5’−GGGGTACCCCACCACCATGAATCCACTCCT−3’
pUCリバース(配列番号63):
【0189】
5’−CGGGATCCGGTATAGAGACTGAAGAGATAC−3’
【0190】
続いて、それにより生成された制限酵素部位を使用して、規定されたまたはランダムオリゴヌクレオチドをSDR1およびSDR2挿入部位に、標準的な制限およびライゲーション法によって挿入した。典型的には、2つの相補性合成5’リン酸化オリゴヌクレオチドをアニーリングし、それぞれの制限酵素で切断した修飾ヒトトリプシンI遺伝子を有するベクターにライゲーションした。SDR1をコードするオリゴヌクレオチドを、SacII/BamHI部位を介して挿入し、一方、SDR2をコードするオリゴヌクレオチドを、KpnI/NheI部位を介して挿入した。各挿入について、以下の一般的な配列に従うオリゴヌクレオチドペアを使用した([P]は、5’リン酸化を示す。NおよびXは、任意のヌクレオチドまたはアミノ酸残基をそれぞれ示す。):
oligox−SDR1f(配列番号64):
【化7】
oligox−SDR1r(配列番号66):
【化8】
oligox−SDR2f(配列番号67):
【化9】
oligox−SDR2r(配列番号69):
【化10】
【0191】
上記方法の代替方法として、修飾されたヒトトリプシンI中のSDR1およびSDR2挿入部位へのランダム配列の組み込みのためにPCRに基づく方法が使用される。各SDRについて、1つのプライマーが使用され、ここでSDR領域は、完全にランダム化されている。プライマーの配列は以下の通りである(N=A/C/G/t、B=C/G/T、V=A/C/G):
プライマーSDR1−mutnnbフォワード(配列番号70):
【化11】
プライマーSDR2−mutnnbリバース(配列番号71):
【化12】
【0192】
さらなる代替方法として、特異性を増大させるSDRを同定した後、これらのSDRをさらなるランダム化のための鋳型として使用した。それにより、各位置で部分的にランダム化したランダム化したランダムペプチド配列、および元々の配列に対して各位置で同一部分的に同一であるランダムペプチド配列を挿入した。
【0193】
例として、3つの場合のなかの約1において鋳型アミノ酸残基を有し、3つの場合のうちの約2において任意の他のアミノ酸残基を各位置で有するランダムペプチド配列を、修飾ヒトトリプシンLの2つのSDR挿入部位に挿入した。この目的のために、SDRの各ヌクレオチド位置で鋳型塩基の約70%および3つの他の塩基の混合物を30%を含有するプライマーを使用した。各プライマーペアを用いて、標準的な条件下でヒトトリプシンI遺伝子を鋳型として用いてPCRを行った。得られたDNAをQIAquickPCR精製キットを、供給業者の指示書に従って使用して精製し、SacIIおよびNheIで消化した。消化後、DNAを精製し、SacIIおよびNheI消化され脱リン酸化されたベクターにライゲーションした。ライゲーション産物を、大腸菌に形質転換し、それぞれのマーカーを含有するLB中で増幅し、そのプラスミドをQiagenプラスミド精製キットを供給業者の指示書にしたがって用いて精製した。得られたプラスミドをB.subtilis細胞中へ形質転換した。次いで、これらの細胞を、単一の細胞へ分離し、クローンへと増殖させ、プロテアーゼ遺伝子の発現後、タンパク質分解活性についてスクリーニングした。
【0194】
以下の基質を、タンパク質分解活性のスクリーニングのために使用した(配列番号76および77)
【数1】
【0195】
プロテアーゼ変異体を、基質Bについて、共焦点蛍光顕微鏡によって106変異体の複雑性(complexities)でスクリーニングした。基質は、N末端でビオチン化され、C末端で蛍光標識されていた。ペプチドをプロテアーゼの異なる変異体を発現する細胞の上清とともにインキュベーションした後、ストレプトアビジンを添加し、そのサンプルを共焦点蛍光定量法によって分析した。ペプチドの低い濃度(20nM)は、高いkcat/KM値を有するプロテアーゼ(すなわち、標的配列に対する高い特異性を有するプロテアーゼ)による優先的な切断を導いた。
【0196】
スクリーニング手順において選択された変異体を、さらにそれらの基質Bおよび密接に関連する基質Aに対する特異性について、タンパク質分解酵素消化の時間経過を測定することにより、およびkcat/KM値に比例する速度定数を決定することにより、評価した。明らかに、足場タンパク質として使用されたヒトトリプシンと比較して、変異体1および2の特異的活性は、基質Bのほうへシフトしている(それぞれ、配列番号2および3)。他方、変異体3(配列番号4)は、ヒトトリプシン1と同様の活性を有するネガティブコントロールとして作用する。3つの変異体の遺伝子の配列決定により、SDR中の以下のアミノ酸配列が明らかになった。
表2:基質Bの特異的加水分解について選択された3つの異なる変異体中の2つのSDRの配列(配列番号78〜83)
【表2】
【0197】
さらなる実験において、1遺伝子当たり異なる数のSDRを含有する変異体のプールを、規定された基質および競合基質としてのペプトンの混合物を使用して、増大した特異性に関してスクリーニングした。1遺伝子当たりのSDRをさらに分析した。特異性の指標として、ペプチド切断アッセイにおける活性を、競合基質の存在下および非存在下で比較した。競合基質の濃度は10mg/mlであった。これらの条件下で、非特異的プロテアーゼは、特異的プロテアーゼと比較して、競合剤濃度の増加につれて(0〜100mg/mlの範囲)より強い活性の減少を示す。基質ありとなしのタンパク質分解活性の比は、プロテアーゼの特異性の定量的指標である。図9は、競合基質ありとなしとの相対的活性を示す。足場タンパク質として使用されたヒトトリプシンIおよび2つの変異体(1つはSDR2のみを含み、1つは両方のSDRを含む)を比較した。両方のSDRを有する変異体の特異性は、SDR2のみを有する変異体の特異性よりも2.5倍高い。これは、SDRの数と得られる人工タンパク質分解酵素の定量的特異性との間に直接的関連性があることを確証する。
【0198】
(実施例V:ヒトTNFアルファを特異的に不活性化する人工タンパク質分解酵素の生成)
ヒトトリプシンアルファI、または1つ以上の下記のアミノ酸置換E56G;R78W;Y131F;A146T;C183Rを含む誘導体を、ヒトTNFアルファに対する高い特異性を有する人工タンパク質分解酵素の生成のためのタンパク質足場として使用した。ヒトトリプシンIまたはその誘導体におけるSDR部位の同定を、上記のように行った。足場内の2つの挿入部位を、SDRについて選択した。異なる配列を有する2つの挿入断片を含有するプロテアーゼ変異体および挿入物を有さないヒトトリプシンI自体は、枯草菌細胞内で発現された。変異体プロテアーゼ細胞を、単一細胞クローンに分離し、プロテアーゼ発現変異体を、TNFアルファの所望の標的配列を有するペプチドにおけるタンパク質分解活性についてスクリーニングした。プロテアーゼ変異体の活性を、競合基質の存在下または非存在下でのTNFアルファの所望の標的配列を有するペプチドの切断速度として決定した。特異性を、競合剤の存在下および非存在下における切断速度の比として表した(図14)。
【0199】
表3:競合基質の存在下または非存在下での異なるSDR配列を有する人工タンパク質分解酵素の変異体の相対活性(配列番号84−95)。
【表3】
【0200】
3つの本発明のプロテアーゼ変異体のヒトTNFアルファに対する拮抗性効果を、図15に示す。変異体の使用により、アポトーシスの誘導は、ほとんど完全に排除された。これはTNFアルファの分解を開始する本発明のプロテアーゼの抗炎症性効果を示す。TNFアルファを、変異体i〜iiiを発現する培養由来の濃縮した上清とともに2時間インキュベートした。得られたTNFアルファを、非修飾細胞とともに4時間インキュベートした。残りのTNFアルファ活性の効果を、アポトーシス細胞のマーカーとしての活性化カスパーゼ3を検出することによって、アポトーシスの誘導の程度として決定した。コントロールとして、それぞれ、ヒトTNFアルファ(死細胞)にプロテアーゼを加えないか、またはヒトTNFアルファ(生細胞)の代わりに緩衝液を使用した。精製した変異体xiiiを使用した類似の実験を図16に示した。TNFアルファを、異なる濃度の精製した本発明のプロテアーゼ変異体とともにインキュベートした。
【0201】
本発明のプロテアーゼ変異体の特異性を実証するために、ヒト血清由来のタンパク質または精製したヒトTNFアルファを、ヒトトリプシンIまたは本発明の人工タンパク質分解酵素変異体とともにそれぞれインキュベートした。ここで、変異体xは、配列番号75に対応し、変異体fと同じSDR、すなわち、配列番号89(SDR1)および配列番号95(SDR2)のSDRを含む。変異体xiおよびxiiは、同じSDR配列を含むその誘導体に対応する。残りのインタクトなタンパク質を時間の関数として決定した。変異体ならびにヒトトリプシンIがヒトTNFアルファを消化するのに対して、トリプシンのみが血清タンパク質に対して活性を示す(図17aおよびb)。これは、本発明のタンパク質分解酵素の高いTNFアルファ特異性を示し、それらの安全性、したがって、それらの治療用途のための副作用の低さを示す。
【0202】
(実施例VI:ヒトVEGFを特異的に加水分解する人工タンパク質分解酵素の生成)
ヒトトリプシンIを、ヒトVEGFに対して高い特異性を有する人工タンパク質分解酵素の生成のためにタンパク質足場として使用した。ヒトトリプシンI内のSDR部位の同定は、上記のように行った。足場内の2つの挿入部位をSDRのために選択した。異なる配列を有する2つの挿入断片を含有するプロテアーゼ変異体を、枯草菌細胞中で発現させた。変異体プロテアーゼ細胞を、単一細胞クローンへと分離し、そのプロテアーゼ発現変異体を上記のようにスクリーニングした。プロテアーゼ変異体の活性をVEGF切断の速度として決定した。4μgの組換えヒトVEGF165を、室温でPBS/pH7.4中にて0.18μgの精製プロテアーゼとともにインキュベートした。示された時間にアリコートをとり、ポリアクリルアミドゲル上で分析した。切断の程度を、バンドの濃度測定分析によって定量した。活性をインキュベーション時間に対して図18にプロットした。特異的切断を、さらなるSDSポリアクリルアミドゲル分析によって制御した。
【0203】
以下の図面は、詳細な説明を補充し、本発明をさらに説明するために提供される。
【図面の簡単な説明】
【0204】
【図1】「球棒」表示で示される活性化部位残基を有するヒトトリプシンIの3次元構造を示す。印をつけた領域は潜在的SDR挿入部位を示す。
【図2】セリンプロテアーゼクラスS1ファミリーである3つのメンバー:ヒトトリプシン1、ヒトアルファトロンビン、およびヒトエンテロペプチダーゼの一次アミノ酸配列のアラインメントを示す(配列番号1、5、および6もまた参照)。
【図3】「球棒」表示で示された活性部位残基を有するサブチリシンの3次元構造を示す。番号を付された領域は、潜在的SDR挿入部位を示す。
【図4】セリンプロテアーゼS8ファミリーの4つのメンバー:サブチリシンE、フリン、PC1、およびPC5の一次アミノ酸配列のアラインメントを示す(配列番号7〜10もまた参照)。
【図5】「球棒」表示で示された活性部位残基を有するペプシンの3次元構造を示す。番号を付された領域は、潜在的SDR挿入部位を示す。
【図6】A1アスパラギン酸プロテアーゼファミリーの3つのメンバー:ペプシン、β−セクレターゼ、およびカテプシンDの一次アミノ酸配列のアラインメントを示す(配列番号11〜13もまた参照)。
【図7】「球棒」表示で示された活性部位残基を有するカスパーゼ7の3次元構造を示す。番号を付した領域は、潜在的SDR挿入部位を示す。
【図8】システインプロテアーゼクラスC14ファミリーのメンバーとしてのカスパーゼ7の一次アミノ酸配列を示す(配列番号14もまた参照)。
【図9】本発明の第3の局面を模式的に示す。
【図10】ネガティブコントロールと比較して、SDR1およびSDR2を有するヒトトリプシンIの変異体を発現する細胞の培養上清のウエスタンブロットを示す。
【図11】ヒトトリプシンIによる標的基質のタンパク質分解による切断の時間経過を示す。
【図12】本発明の人工タンパク質分解酵素の3つの変異体の、2つの異なるペプチド基質上でのヒトトリプシンIとの比較における相対的活性を示す。
【図13】ヒトトリプシンIおよび1つ以上のSDRを有する本発明の人工タンパク質分解酵素の相対的特異性をそれぞれ示す。
【図14】ヒトトリプシンIおよび、ヒトTNF−アルファの標的基質を有するペプチド上の足場を有数するヒトTNF−アルファに対して特異的な本発明の人工タンパク質分解酵素の相対的特異性を示す。
【図15】TNFアルファを、ヒトTNF−アルファに対して特異的な本発明の人工タンパク質分解酵素を発現する培養物からの濃縮された上清と組み合わせたときにTNFアルファによって誘導される細胞傷害性の減少を示す。
【図16】TNF−アルファを、ヒトTNF−アルファに対して特異的である精製された本発明の人工タンパク質分解酵素とインキュベートしたときの、TNF−アルファによって誘導される細胞傷害性の減少を示す。
【図17】ヒトTNF−アルファに対して特異的である本発明の人工タンパク質分解酵素の活性を、ヒトトリプシンIIの活性と、2つのタンパク質基質:(a)ヒトTNF−アルファ;(b)ヒトセリンタンパク質の混合物において比較する。
【図18】ヒトVEGFに対する特異性を有する本発明の人工タンパク質分解酵素の特異的活性を示す。
【特許請求の範囲】
【請求項1】
以下の成分:
(a)少なくとも1つの標的基質における少なくとも1つの化学反応を触媒し得るタンパク質足場、および
(b)得られる人工タンパク質が、少なくとも1つの標的基質と、1つ以上の異なる基とを区別することを可能にするタンパク質足場中の部位に位置する一つ以上の特異性決定領域(SDR)、
の組み合わせによって特徴付けられる、規定された特異性の触媒活性を有する人工酵素。
【請求項2】
(I)前記SDR(b)が、50アミノ酸残基未満の長さを有し、好ましくは、2〜20アミノ酸残基の長さを有し、より好ましくは、2〜10アミノ酸残基の長さを有し、さらにより好ましくは、3〜8アミノ酸残基の長さを有し、ここで、SDRの数は、少なくとも1個、好ましくは、1個より多く、より好ましくは、2〜11個、最も好ましくは、2〜6個であり、および/または
(II)前記タンパク質足場(a)は、同じかまたは異なる(i)ウイルス、原核生物もしくは真核生物起源の遺伝子によってコードされるタンパク質、および/または(ii)天然の酵素、もしくはその変異誘導体もしくは切断型誘導体、および/または(iii)哺乳動物酵素、好ましくは、ヒト酵素に由来する1以上のポリペプチドから構成される、
請求項1に記載の人工酵素。
【請求項3】
前記タンパク質足場(a)が、加水分解酵素(好ましくは、プロテアーゼ);リパーゼ;グリコシラーゼ;トランスフェラーゼ(好ましくは、グリコシルトランスフェラーゼ);酸化還元酵素(好ましくは、モノオキシゲナーゼおよびジオキシゲナーゼ);リアーゼ;イソメラーゼ、およびリガーゼからなる群から選択される酵素に由来し、
より好ましくは、前記タンパク質足場(a)が、アスパラギン酸プロテアーゼ、システインプロテアーゼ、セリンプロテアーゼ、メタロプロテアーゼ、およびスレオニンプロテアーゼから選択されるプロテアーゼに由来し、
さらにより好ましくは、前記タンパク質足場(a)は、構造クラスS1、S8、S11、S21、S26、S33、またはS51(最も好ましくは、クラスS1もしくはS8)のセリンプロテアーゼ、構造クラスC1、C2、C4、C10、C14、C19、C47、C48、もしくはC56(最も好ましくは、クラスC14)のシステインプロテアーゼ、または構造クラスA1、A2、もしくはA26(最も好ましくは、クラスA1)のアスパラギン酸プロテアーゼ、または構造クラスM4もしくはM10のメタロプロテアーゼに由来する、請求項1または2に記載の人工酵素。
【請求項4】
(i)前記タンパク質足場(a)が、構造クラスS1のセリンプロテアーゼに由来し、および/または
(ii)前記SDRが、配列番号1に示されるアミノ酸配列を有するヒトトリプシンI中の領域18−25、38−48、54−63、73−86、122−130、148−156、165−171、および194−204に、構造的にもしくはアミノ酸配列相同性により対応する位置の群からの一つ以上の位置に存在し、より好ましくは、ヒトトリプシンI中の領域20−23、41−45、57−60、76−83、125−128、150−153、167−169、および197−201に構造的にまたはアミノ酸配列相同性により対応する位置の群から選択される一つ以上の位置に存在する、請求項3に記載の人工酵素。
【請求項5】
(i)タンパク質足場(a)が、セリンプロテアーゼであるトリプシン、好ましくは、配列番号1に示されるアミノ酸配列を有するヒトトリプシンIもしくはその誘導体であるか、または配列番号1のアミノ酸配列に以下のアミノ酸置換:E56G、R78W、Y131F、A146T、およびC183Rを有するものであり、
(ii)2つのSDRのうちの少なくとも1つが、足場中に位置し、第一のSDRが6アミノ酸までの長さを有し、残基42と43との間に挿入されており、第二のSDRが5アミノ酸までの長さを有し、残基123と124との間に挿入されている(配列番号1に示されるアミノ酸配列を有するヒトトリプシンIに対しての番号付け)、請求項4に記載の人工酵素。
【請求項6】
残基42と43との間に第一のSDRとして挿入された以下の群:配列番号72、78、79、80、84、85、86、87、88、および89のペプチド配列の1つ、および/または
残基123と124との間に第二のSDRとして挿入された以下の群:配列番号73、81、82、83、90、91、92、93、94、および95のペプチド配列の1つ、を含み、
前記人工酵素が、配列番号74または配列番号75に示されるアミノ酸配列を含む、請求項5に記載の人工酵素。
【請求項7】
(i)前記タンパク質足場(a)が、構造クラスS8のセリンプロテアーゼに由来し、および/または
(ii)前記SDRが、配列番号7に示されるアミノ酸配列を有する枯草菌由来のサブチリシン中の領域6−17、25−29、47−55、59−69、101−111、117−125、129−137、139−154、158−169、185−195、および204−225に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの一つ以上の位置、好ましくは、枯草菌由来のサブチリシン中の領域59−69、101−111、129−137、158−169、および204−225に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの一つ以上の位置に存在する、請求項3に記載の人工酵素。
【請求項8】
(i)前記タンパク質足場(a)が、構造クラスA1のアスパラギン酸プロテアーゼに由来し、および/または
(ii)前記SDRが、配列番号11に示されるアミノ酸配列を有するヒトペプシン中の領域6−18、49−55、74−83、91−97、112−120、126−137、159−164、184−194、242−247、262−267、および277−300に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの一つ以上の位置、より好ましくは、ヒトペプシン中の領域10−15、75−80、114−118、130−134、186−191、および280−296に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの一つ以上の位置に存在する、請求項3に記載の人工酵素。
【請求項9】
(i)前記タンパク質足場(a)が、構造クラスC14のシステインプロテアーゼに由来し、および/または
(ii)前記SDRが、配列番号14に示されるアミノ酸配列を有するヒトカスパーゼ7中の領域78−91、144−160、186−198、226−243、および271−291に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの一つ以上の位置、より好ましくは、ヒトカスパーゼ7中の領域80−86、149−157、190−194、および233−238に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの一つ以上の位置に存在する、請求項3に記載の人工酵素。
【請求項10】
請求項1〜7のいずれかに記載の少なくとも1つの人工酵素、ならびに
(i)少なくとも1つのさらなるタンパク質様成分であって、好ましくは、結合ドメイン、受容体、抗体、調節ドメイン、プロ配列、およびこれらのフラグメントからなる群から選択されるタンパク質様成分、および/または
(ii)少なくとも1つのさらなる機能的成分であって、好ましくは、ポリエチレングリコール、炭水化物、脂質、脂肪酸、核酸、金属、金属キレート剤、およびこれらのフラグメントもしくは誘導体からなる群から選択される機能的成分、
から構成される融合タンパク質。
【請求項11】
請求項1〜9のいずれかに記載の酵素または請求項10に記載の融合タンパク質をコードする核酸配列を含む核酸分子。
【請求項12】
請求項11に記載の核酸を含むベクター。
【請求項13】
請求項12に記載のベクターまたは請求項11に記載の核酸分子を含む宿主細胞。
【請求項14】
大腸菌、枯草菌、出芽酵母、Pichia pastonis、CHO、およびBHKからなる群から選択される、請求項13に記載の宿主細胞。
【請求項15】
請求項1に記載の酵素または請求項10に記載の融合タンパク質を製造する方法であって、請求項13または14に記載の宿主細胞を培養する工程を包含する、方法。
【請求項16】
少なくとも1つの標的基質に対する規定された特異性を有する請求項1〜9のいずれかに記載の人工酵素を作製する方法であって、
少なくとも1つの以下の工程:
(a)少なくとも1つの基質における少なくとも1つの化学反応を触媒するタンパク質足場を提供する工程、
(b)工程(a)からの前記タンパク質足場をコードするポリヌクレオチドと、合成ペプチド配列をコードする一つ以上の完全または部分的ランダム合成オリゴヌクレオチド配列とを、得られるコードされた人工酵素が少なくとも1つの標的基質と一つ以上の異なる基質とを区別できるような部位で、組み合わせ、該酵素を発現させることにより、人工酵素または単離された人工酵素のライブラリーを生成する工程、ならびに
(c)工程(b)において生成された人工酵素のライブラリーから、少なくとも1つの標的基質に対する規定された特異性を有する一つ以上の酵素を選択する工程、
を包含する、方法。
【請求項17】
(I)工程(b)の組み合わせが行われる部位が、タンパク質足場の特定の部位であり、組み合わせ部位として適した部位が、活性部位に近い領域を同定することにより、好ましくは、該タンパク質足場と競合阻害剤もしくは基質アナログとの複合体の構造分析により、および/または異なる定量的もしくは定性的特異性を有する同じ構造クラスの異なる酵素の構造アラインメントおよび異種領域の同定により、および/または異なる定性的もしくは定量的特異性を有する同じ構造クラスの酵素からのアミノ酸配列の競合アッセイおよび異種領域の同定により、および/またはアラニンスキャニング、ランダム変異誘発、ランダム挿入、もしくはランダム欠失のような変異誘発技術を含む実験分析およびその後の特異性に必須または感受性のタンパク質足場中の領域を同定することにより、同定され、ならびに/または
(ii)工程(b)の組み合わせ部位が、前記タンパク質足場にわたってランダムに分布している、請求項16に記載の方法。
【請求項18】
工程(b)において組み合わされる前記ペプチド配列が、完全または部分的にランダムであり、および/もしくは長さのバリエーションを有し;ならびに/または
工程(c)における選択が、
(i)低い標的基質濃度で、もしくは
(ii)標的基質および少なくとも1つのさらなる基質を比較して使用することによって、もしくは
(iii)前記標的基質よりも他の基質を過剰に添加し、該添加された基質を競合剤として使用することによって、もしくは
(iv)酵素阻害剤を添加することにより、もしくは
(v)標的基質に優先的に結合する酵素を選択し、このサブグループから基質を変換する酵素を選択することにより、もしくは
(vi)これらの任意の組み合わせにより、
酵素活性および/もしくは酵素親和性に関してスクリーニングすることによって達成される、請求項16または17に記載の方法。
【請求項19】
(i)工程(a)〜(c)が、少なくとももう1サイクル繰り返され、および1つのサイクルの工程(c)で選択される前記SDRが、将来のサイクルの工程(b)において挿入されるタンパク質配列のランダム化のための鋳型として働き、および/または
(ii)工程(a)〜(c)の一つ以上のラウンドの間またはその後に、前記足場が、SDR配列との組み合わせにより受け入れ可能な足場を作製するために、および/もしくは特定のpHおよび温度で触媒活性を増大させるために、および/もしくはグリコシル化パターンを変化させるために、および/もしくは酵素阻害剤に対する感受性を減少させるために、および/もしくは酵素安定性を変化させるために、一つ以上の位置で変異される、
請求項16〜18のいずれかに記載の方法。
【請求項20】
(I)少なくとも以下の工程:
(a)第一のタンパク質足場フラグメントを提供する工程、
(b)前記タンパク質足場フラグメントを、第一のSDRと、ペプチド結合を介して結合する工程、および必要に応じて、
(c)前記工程(b)の生成物を、さらなるSDRペプチドまたはさらなる融合タンパク質足場フラグメントと、ペプチド結合を介して結合する工程、および必要に応じて、
(d)十分に特異的な酵素を生成するために必要なサイクル、工程(c)を繰り返す工程、
(e)工程(a)〜(d)において生成された集団から、一つ以上の標的基質に対する所望の特異性を有する一つ以上の酵素を選択する工程、を包含するか、または
(ii)少なくとも以下の工程:
(a)1つ以上の標的基質における1つ以上の化学反応を触媒し得るタンパク質足場をコードするポリヌクレオチドを提供する工程、
(b)1つ以上の完全または部分的ランダム合成オリゴヌクレオチド配列を、タンパク質足場をコードするポリヌクレオチドと組み合わせる工程であって、該完全または部分的ランダム合成オリゴヌクレオチド配列が、コードされる人工酵素が1つ以上の標的基質と1つ以上の他の基質とを区別し得るような前記ポリヌクレオチド中の部位に位置している、工程、
(c)工程(b)において生成された集団から、1つ以上の標的基質に対する所望の特異性を有する酵素をコードする1つ以上のポリヌクレオチドを選択する工程、
を包含する、請求項16に記載の方法。
【請求項21】
請求項1〜9のいずれかに記載の1つ以上の人工酵素または請求項10に記載の融合タンパク質を含む組成物であって、
該組成物が、好ましくは、研究用組成物、栄養学組成物、食物添加物組成物、洗浄用組成物、脱感染(desinfection)組成物、化粧用組成物、もしくはパーソナルケア用組成物であり、および/または
前記組成物が、必要に応じて、許容できる担体および/もしくは助剤を含有する、
組成物。
【請求項22】
請求項1〜9のいずれかに記載の人工酵素、または請求項10に記載の融合タンパク質の、研究、栄養学、パーソナルケア、または工業目的のための、使用。
【請求項1】
以下の成分:
(a)少なくとも1つの標的基質における少なくとも1つの化学反応を触媒し得るタンパク質足場、および
(b)得られる人工タンパク質が、少なくとも1つの標的基質と、1つ以上の異なる基とを区別することを可能にするタンパク質足場中の部位に位置する一つ以上の特異性決定領域(SDR)、
の組み合わせによって特徴付けられる、規定された特異性の触媒活性を有する人工酵素。
【請求項2】
(I)前記SDR(b)が、50アミノ酸残基未満の長さを有し、好ましくは、2〜20アミノ酸残基の長さを有し、より好ましくは、2〜10アミノ酸残基の長さを有し、さらにより好ましくは、3〜8アミノ酸残基の長さを有し、ここで、SDRの数は、少なくとも1個、好ましくは、1個より多く、より好ましくは、2〜11個、最も好ましくは、2〜6個であり、および/または
(II)前記タンパク質足場(a)は、同じかまたは異なる(i)ウイルス、原核生物もしくは真核生物起源の遺伝子によってコードされるタンパク質、および/または(ii)天然の酵素、もしくはその変異誘導体もしくは切断型誘導体、および/または(iii)哺乳動物酵素、好ましくは、ヒト酵素に由来する1以上のポリペプチドから構成される、
請求項1に記載の人工酵素。
【請求項3】
前記タンパク質足場(a)が、加水分解酵素(好ましくは、プロテアーゼ);リパーゼ;グリコシラーゼ;トランスフェラーゼ(好ましくは、グリコシルトランスフェラーゼ);酸化還元酵素(好ましくは、モノオキシゲナーゼおよびジオキシゲナーゼ);リアーゼ;イソメラーゼ、およびリガーゼからなる群から選択される酵素に由来し、
より好ましくは、前記タンパク質足場(a)が、アスパラギン酸プロテアーゼ、システインプロテアーゼ、セリンプロテアーゼ、メタロプロテアーゼ、およびスレオニンプロテアーゼから選択されるプロテアーゼに由来し、
さらにより好ましくは、前記タンパク質足場(a)は、構造クラスS1、S8、S11、S21、S26、S33、またはS51(最も好ましくは、クラスS1もしくはS8)のセリンプロテアーゼ、構造クラスC1、C2、C4、C10、C14、C19、C47、C48、もしくはC56(最も好ましくは、クラスC14)のシステインプロテアーゼ、または構造クラスA1、A2、もしくはA26(最も好ましくは、クラスA1)のアスパラギン酸プロテアーゼ、または構造クラスM4もしくはM10のメタロプロテアーゼに由来する、請求項1または2に記載の人工酵素。
【請求項4】
(i)前記タンパク質足場(a)が、構造クラスS1のセリンプロテアーゼに由来し、および/または
(ii)前記SDRが、配列番号1に示されるアミノ酸配列を有するヒトトリプシンI中の領域18−25、38−48、54−63、73−86、122−130、148−156、165−171、および194−204に、構造的にもしくはアミノ酸配列相同性により対応する位置の群からの一つ以上の位置に存在し、より好ましくは、ヒトトリプシンI中の領域20−23、41−45、57−60、76−83、125−128、150−153、167−169、および197−201に構造的にまたはアミノ酸配列相同性により対応する位置の群から選択される一つ以上の位置に存在する、請求項3に記載の人工酵素。
【請求項5】
(i)タンパク質足場(a)が、セリンプロテアーゼであるトリプシン、好ましくは、配列番号1に示されるアミノ酸配列を有するヒトトリプシンIもしくはその誘導体であるか、または配列番号1のアミノ酸配列に以下のアミノ酸置換:E56G、R78W、Y131F、A146T、およびC183Rを有するものであり、
(ii)2つのSDRのうちの少なくとも1つが、足場中に位置し、第一のSDRが6アミノ酸までの長さを有し、残基42と43との間に挿入されており、第二のSDRが5アミノ酸までの長さを有し、残基123と124との間に挿入されている(配列番号1に示されるアミノ酸配列を有するヒトトリプシンIに対しての番号付け)、請求項4に記載の人工酵素。
【請求項6】
残基42と43との間に第一のSDRとして挿入された以下の群:配列番号72、78、79、80、84、85、86、87、88、および89のペプチド配列の1つ、および/または
残基123と124との間に第二のSDRとして挿入された以下の群:配列番号73、81、82、83、90、91、92、93、94、および95のペプチド配列の1つ、を含み、
前記人工酵素が、配列番号74または配列番号75に示されるアミノ酸配列を含む、請求項5に記載の人工酵素。
【請求項7】
(i)前記タンパク質足場(a)が、構造クラスS8のセリンプロテアーゼに由来し、および/または
(ii)前記SDRが、配列番号7に示されるアミノ酸配列を有する枯草菌由来のサブチリシン中の領域6−17、25−29、47−55、59−69、101−111、117−125、129−137、139−154、158−169、185−195、および204−225に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの一つ以上の位置、好ましくは、枯草菌由来のサブチリシン中の領域59−69、101−111、129−137、158−169、および204−225に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの一つ以上の位置に存在する、請求項3に記載の人工酵素。
【請求項8】
(i)前記タンパク質足場(a)が、構造クラスA1のアスパラギン酸プロテアーゼに由来し、および/または
(ii)前記SDRが、配列番号11に示されるアミノ酸配列を有するヒトペプシン中の領域6−18、49−55、74−83、91−97、112−120、126−137、159−164、184−194、242−247、262−267、および277−300に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの一つ以上の位置、より好ましくは、ヒトペプシン中の領域10−15、75−80、114−118、130−134、186−191、および280−296に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの一つ以上の位置に存在する、請求項3に記載の人工酵素。
【請求項9】
(i)前記タンパク質足場(a)が、構造クラスC14のシステインプロテアーゼに由来し、および/または
(ii)前記SDRが、配列番号14に示されるアミノ酸配列を有するヒトカスパーゼ7中の領域78−91、144−160、186−198、226−243、および271−291に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの一つ以上の位置、より好ましくは、ヒトカスパーゼ7中の領域80−86、149−157、190−194、および233−238に対して構造的にまたはアミノ酸配列相同性により対応する位置の群からの一つ以上の位置に存在する、請求項3に記載の人工酵素。
【請求項10】
請求項1〜7のいずれかに記載の少なくとも1つの人工酵素、ならびに
(i)少なくとも1つのさらなるタンパク質様成分であって、好ましくは、結合ドメイン、受容体、抗体、調節ドメイン、プロ配列、およびこれらのフラグメントからなる群から選択されるタンパク質様成分、および/または
(ii)少なくとも1つのさらなる機能的成分であって、好ましくは、ポリエチレングリコール、炭水化物、脂質、脂肪酸、核酸、金属、金属キレート剤、およびこれらのフラグメントもしくは誘導体からなる群から選択される機能的成分、
から構成される融合タンパク質。
【請求項11】
請求項1〜9のいずれかに記載の酵素または請求項10に記載の融合タンパク質をコードする核酸配列を含む核酸分子。
【請求項12】
請求項11に記載の核酸を含むベクター。
【請求項13】
請求項12に記載のベクターまたは請求項11に記載の核酸分子を含む宿主細胞。
【請求項14】
大腸菌、枯草菌、出芽酵母、Pichia pastonis、CHO、およびBHKからなる群から選択される、請求項13に記載の宿主細胞。
【請求項15】
請求項1に記載の酵素または請求項10に記載の融合タンパク質を製造する方法であって、請求項13または14に記載の宿主細胞を培養する工程を包含する、方法。
【請求項16】
少なくとも1つの標的基質に対する規定された特異性を有する請求項1〜9のいずれかに記載の人工酵素を作製する方法であって、
少なくとも1つの以下の工程:
(a)少なくとも1つの基質における少なくとも1つの化学反応を触媒するタンパク質足場を提供する工程、
(b)工程(a)からの前記タンパク質足場をコードするポリヌクレオチドと、合成ペプチド配列をコードする一つ以上の完全または部分的ランダム合成オリゴヌクレオチド配列とを、得られるコードされた人工酵素が少なくとも1つの標的基質と一つ以上の異なる基質とを区別できるような部位で、組み合わせ、該酵素を発現させることにより、人工酵素または単離された人工酵素のライブラリーを生成する工程、ならびに
(c)工程(b)において生成された人工酵素のライブラリーから、少なくとも1つの標的基質に対する規定された特異性を有する一つ以上の酵素を選択する工程、
を包含する、方法。
【請求項17】
(I)工程(b)の組み合わせが行われる部位が、タンパク質足場の特定の部位であり、組み合わせ部位として適した部位が、活性部位に近い領域を同定することにより、好ましくは、該タンパク質足場と競合阻害剤もしくは基質アナログとの複合体の構造分析により、および/または異なる定量的もしくは定性的特異性を有する同じ構造クラスの異なる酵素の構造アラインメントおよび異種領域の同定により、および/または異なる定性的もしくは定量的特異性を有する同じ構造クラスの酵素からのアミノ酸配列の競合アッセイおよび異種領域の同定により、および/またはアラニンスキャニング、ランダム変異誘発、ランダム挿入、もしくはランダム欠失のような変異誘発技術を含む実験分析およびその後の特異性に必須または感受性のタンパク質足場中の領域を同定することにより、同定され、ならびに/または
(ii)工程(b)の組み合わせ部位が、前記タンパク質足場にわたってランダムに分布している、請求項16に記載の方法。
【請求項18】
工程(b)において組み合わされる前記ペプチド配列が、完全または部分的にランダムであり、および/もしくは長さのバリエーションを有し;ならびに/または
工程(c)における選択が、
(i)低い標的基質濃度で、もしくは
(ii)標的基質および少なくとも1つのさらなる基質を比較して使用することによって、もしくは
(iii)前記標的基質よりも他の基質を過剰に添加し、該添加された基質を競合剤として使用することによって、もしくは
(iv)酵素阻害剤を添加することにより、もしくは
(v)標的基質に優先的に結合する酵素を選択し、このサブグループから基質を変換する酵素を選択することにより、もしくは
(vi)これらの任意の組み合わせにより、
酵素活性および/もしくは酵素親和性に関してスクリーニングすることによって達成される、請求項16または17に記載の方法。
【請求項19】
(i)工程(a)〜(c)が、少なくとももう1サイクル繰り返され、および1つのサイクルの工程(c)で選択される前記SDRが、将来のサイクルの工程(b)において挿入されるタンパク質配列のランダム化のための鋳型として働き、および/または
(ii)工程(a)〜(c)の一つ以上のラウンドの間またはその後に、前記足場が、SDR配列との組み合わせにより受け入れ可能な足場を作製するために、および/もしくは特定のpHおよび温度で触媒活性を増大させるために、および/もしくはグリコシル化パターンを変化させるために、および/もしくは酵素阻害剤に対する感受性を減少させるために、および/もしくは酵素安定性を変化させるために、一つ以上の位置で変異される、
請求項16〜18のいずれかに記載の方法。
【請求項20】
(I)少なくとも以下の工程:
(a)第一のタンパク質足場フラグメントを提供する工程、
(b)前記タンパク質足場フラグメントを、第一のSDRと、ペプチド結合を介して結合する工程、および必要に応じて、
(c)前記工程(b)の生成物を、さらなるSDRペプチドまたはさらなる融合タンパク質足場フラグメントと、ペプチド結合を介して結合する工程、および必要に応じて、
(d)十分に特異的な酵素を生成するために必要なサイクル、工程(c)を繰り返す工程、
(e)工程(a)〜(d)において生成された集団から、一つ以上の標的基質に対する所望の特異性を有する一つ以上の酵素を選択する工程、を包含するか、または
(ii)少なくとも以下の工程:
(a)1つ以上の標的基質における1つ以上の化学反応を触媒し得るタンパク質足場をコードするポリヌクレオチドを提供する工程、
(b)1つ以上の完全または部分的ランダム合成オリゴヌクレオチド配列を、タンパク質足場をコードするポリヌクレオチドと組み合わせる工程であって、該完全または部分的ランダム合成オリゴヌクレオチド配列が、コードされる人工酵素が1つ以上の標的基質と1つ以上の他の基質とを区別し得るような前記ポリヌクレオチド中の部位に位置している、工程、
(c)工程(b)において生成された集団から、1つ以上の標的基質に対する所望の特異性を有する酵素をコードする1つ以上のポリヌクレオチドを選択する工程、
を包含する、請求項16に記載の方法。
【請求項21】
請求項1〜9のいずれかに記載の1つ以上の人工酵素または請求項10に記載の融合タンパク質を含む組成物であって、
該組成物が、好ましくは、研究用組成物、栄養学組成物、食物添加物組成物、洗浄用組成物、脱感染(desinfection)組成物、化粧用組成物、もしくはパーソナルケア用組成物であり、および/または
前記組成物が、必要に応じて、許容できる担体および/もしくは助剤を含有する、
組成物。
【請求項22】
請求項1〜9のいずれかに記載の人工酵素、または請求項10に記載の融合タンパク質の、研究、栄養学、パーソナルケア、または工業目的のための、使用。
【図1】

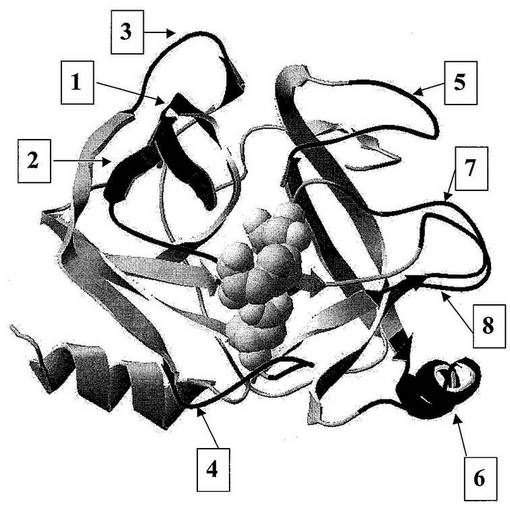
【図2】

【図3】
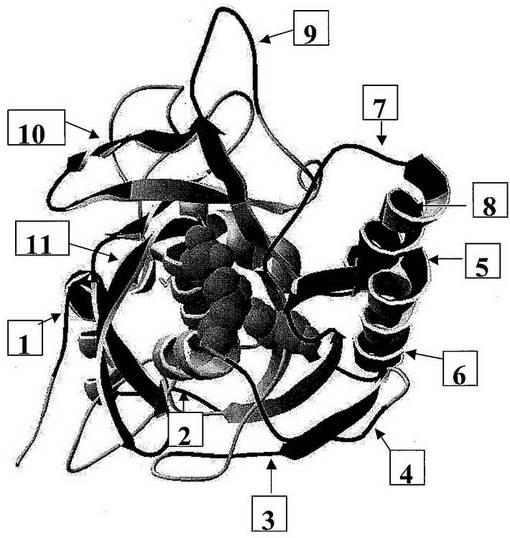
【図4】
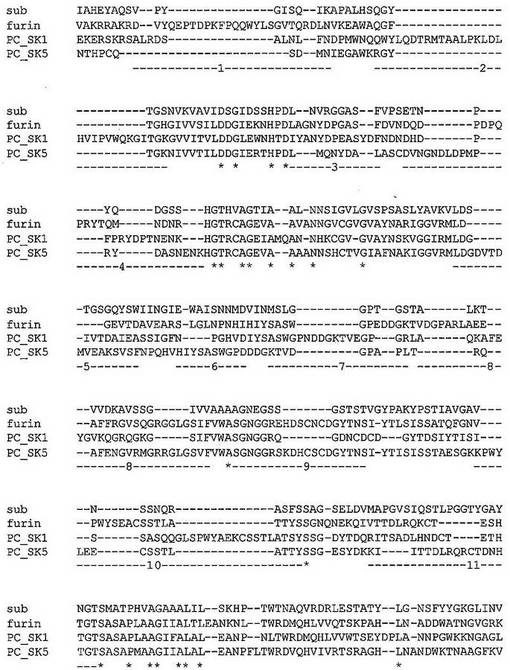
【図5】
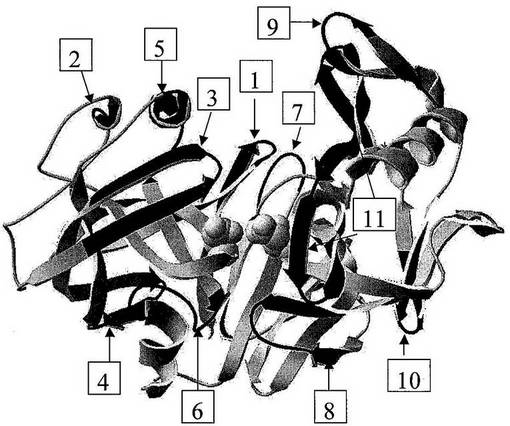
【図6】

【図7】
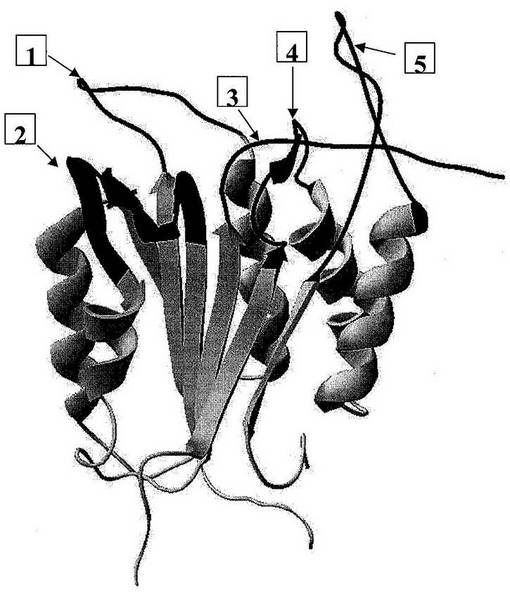
【図8】
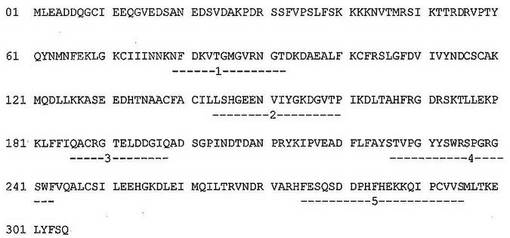
【図9】
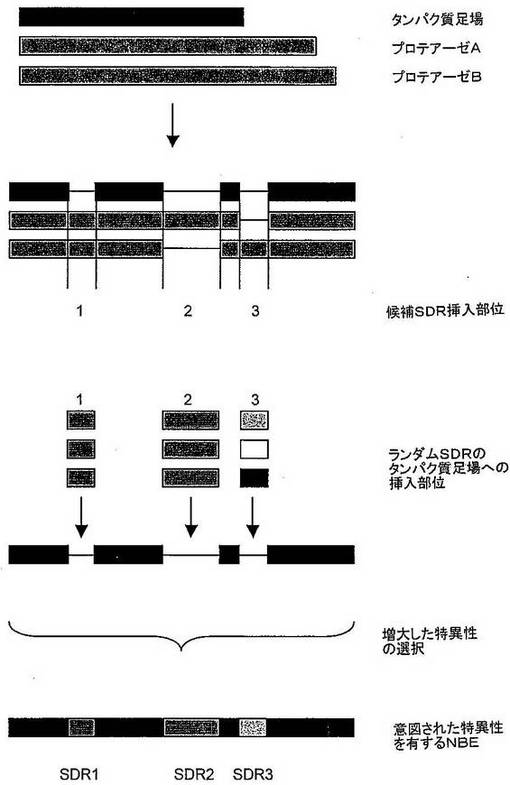
【図10】
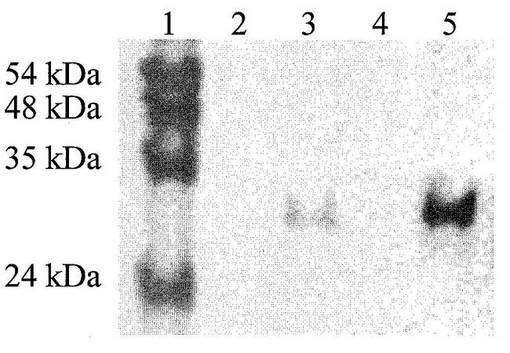
【図11】
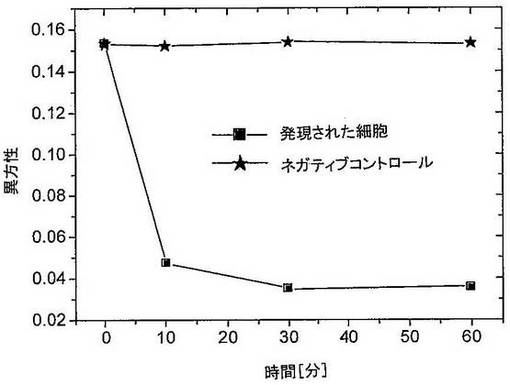
【図12】
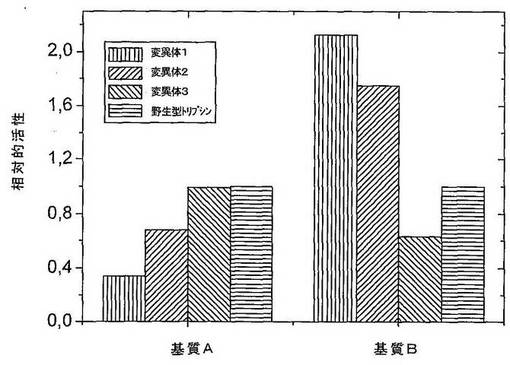
【図13】
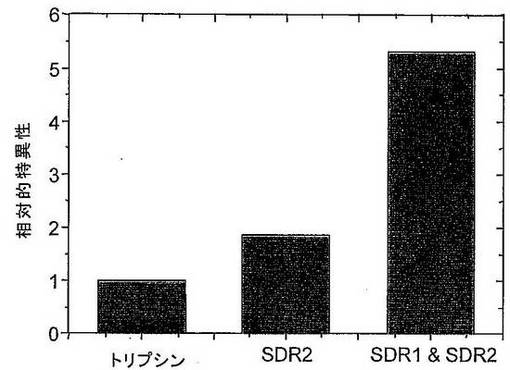
【図14】
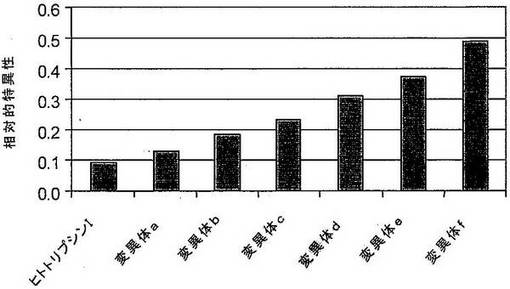
【図15】
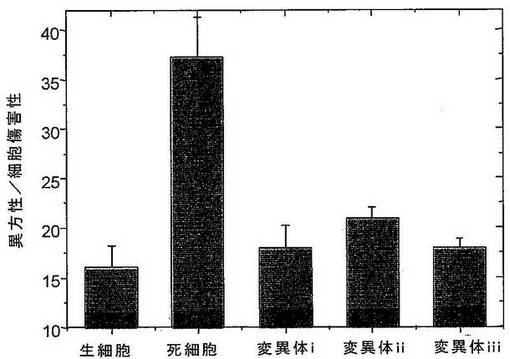
【図16】
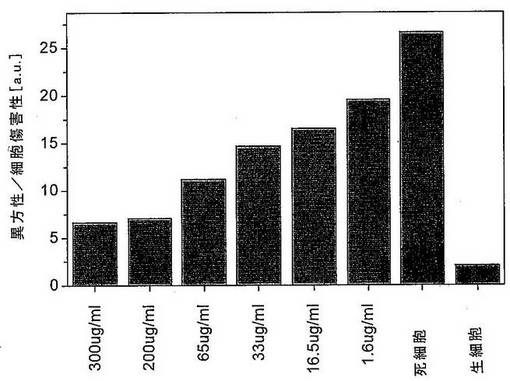
【図17】
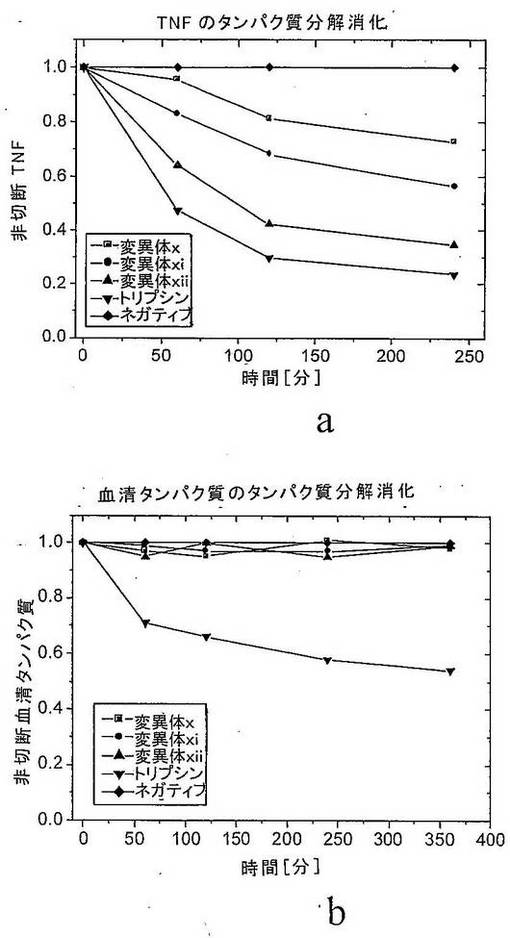
【図18】
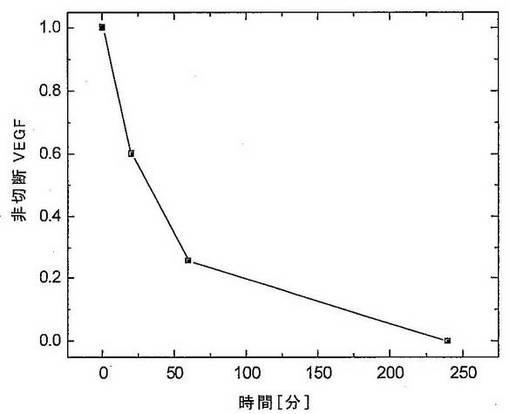
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【公表番号】特表2006−527590(P2006−527590A)
【公表日】平成18年12月7日(2006.12.7)
【国際特許分類】
【出願番号】特願2006−516170(P2006−516170)
【出願日】平成16年6月18日(2004.6.18)
【国際出願番号】PCT/EP2004/051172
【国際公開番号】WO2004/113521
【国際公開日】平成16年12月29日(2004.12.29)
【出願人】(500587403)ディレボ・ビオテク・アーゲー (2)
【氏名又は名称原語表記】DIREVO BioTech AG
【Fターム(参考)】
【公表日】平成18年12月7日(2006.12.7)
【国際特許分類】
【出願日】平成16年6月18日(2004.6.18)
【国際出願番号】PCT/EP2004/051172
【国際公開番号】WO2004/113521
【国際公開日】平成16年12月29日(2004.12.29)
【出願人】(500587403)ディレボ・ビオテク・アーゲー (2)
【氏名又は名称原語表記】DIREVO BioTech AG
【Fターム(参考)】
[ Back to top ]