
新規真正細菌及びそれを用いた長鎖飽和脂肪酸の製造方法
【課題】遺伝子組み換えを伴わずに遊離の長鎖飽和脂肪酸を生産する新規微生物を提供する。
【解決手段】遊離の長鎖飽和脂肪酸を細胞内に蓄積するErysipelotrichaceae科に帰属する新規細菌、Catenostreptococcus属lactis種GK12株(FERMP-22123)。当該細菌は、絶対嫌気性条件下でグルコース含有培地で培養すると、14〜20の偶数の炭素鎖数を有する直鎖状の飽和脂肪酸を遊離状態で菌体内に蓄積する。さらに、高温条件下(45℃)での培養など培養条件を変化させることで、脂肪酸産生量を増大させ、かつ脂肪酸を構成する炭素鎖数をより長鎖側にシフトさせることができるため、バイオディーゼル生産菌株として有用である。
【解決手段】遊離の長鎖飽和脂肪酸を細胞内に蓄積するErysipelotrichaceae科に帰属する新規細菌、Catenostreptococcus属lactis種GK12株(FERMP-22123)。当該細菌は、絶対嫌気性条件下でグルコース含有培地で培養すると、14〜20の偶数の炭素鎖数を有する直鎖状の飽和脂肪酸を遊離状態で菌体内に蓄積する。さらに、高温条件下(45℃)での培養など培養条件を変化させることで、脂肪酸産生量を増大させ、かつ脂肪酸を構成する炭素鎖数をより長鎖側にシフトさせることができるため、バイオディーゼル生産菌株として有用である。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、長鎖飽和脂肪酸を産生する新規微生物、及び当該微生物を用いた脂肪酸を製造する方法に関する。
【背景技術】
【0002】
近年、地球温暖化対策として石油など化石燃料資源に代わる液体燃料としてのバイオディーゼルが脚光を浴びてきており、実用化も進んでいる。典型的なバイオディーゼルとしては、植物の種子に由来する植物油のアシル基をメタノールなどのアルコールでエステル交換反応して合成された脂肪酸アルキルエステルがある。この方法で得られるバイオディーゼルは、従来の石油由来燃料と比較しても優れた燃焼効率を有しているばかりか、地中に埋蔵されていた化石由来の炭素を燃焼時放出する石油燃料とは異なり、CO2温室効果ガス放射の減少にも寄与できる。しかし、パーム油の場合のように広大なアブラヤシ農園建設のために多くの熱帯雨林が失われるなどの別の環境破壊も問題となっており、大豆などの油糧種子作物の場合は、食糧、飼料源であるため燃料への転用は食糧危機を招くおそれがある。
そこで、最近では、タンクによる大量培養可能な藻類や微生物がバイオディーゼル原料として注目されており、研究開発が進んでいたが、従来の微生物によるバイオディーゼル生産では、生産される油のほとんどがトリグリセリド脂質であるため、メチルエステル化によって、触媒や未変換脂肪酸が混入したグリセロールが副産物として大量に産生してしまい、生産効率の面で問題であった。微生物を用いて遊離の脂肪酸又は脂肪酸エステルを直接的に生産させるための方法として、セルロースを炭素源として遺伝子組換えを施した藻類を培養し、多価不飽和脂肪酸より構成されるトリグリセリドを生産する方法(特許文献1)、組換え微生物を用いて脂肪エステル及び遊離脂肪酸を生産し細胞外に放出させる方法(特許文献2)、組換え藻類あるいは酵母による脂質生産と化学反応による直鎖アルカンの生産方法(特許文献3)などの研究開発が相次いでいる。しかし、いずれも遺伝子組換え操作が必要であって、操作の煩雑性および生産の安定性に不安が残った。また、これら遺伝子組換え微生物を含め、従来微生物によって生産される脂質に含まれる脂肪酸および遊離脂肪酸には、二重結合を1つ以上持つ不飽和脂肪酸を多少に含み酸化しやすいため、燃料として安定的に用いる場合に酸化防止剤の必要性が生じることから、生産コストの面での問題があった。
以上のことから、遊離の長鎖飽和脂肪酸を産生する、遺伝子組み換えを伴わない天然の新規微生物の探索が望まれていた。
【先行技術文献】
【特許文献】
【0003】
【特許文献1】特表2010−538642号公報
【特許文献2】特表2010−538602号公報
【特許文献3】特表2010−528627号公報
【発明の概要】
【発明が解決しようとする課題】
【0004】
本発明は、遊離の長鎖飽和脂肪酸を生産する新規微生物を提供しようとするものである。
【課題を解決するための手段】
【0005】
本発明者らは、微生物資源として未開拓な絶対嫌気性の微生物を分離培養する過程で、遊離の長鎖飽和脂肪酸を細胞内に蓄積する新規の細菌GK12株を見出した。当該細菌GK12株は、Erysipelotrichaceae科に帰属するが、公知のいずれの属、種にも属さない新属の新種細菌であり、本発明者らはCatenostreptococcus属lactis種と命名した。当該細菌は、絶対嫌気性条件下でグルコース含有培地により培養すると、14〜20の偶数の炭素鎖数を有する直鎖状の飽和脂肪酸を遊離状態で菌体内に蓄積する。さらに、本発明微生物は、高温条件下(45℃)で培養した場合の他、培地中にブタノールを添加するなどの培養条件を変化させた場合に、個々の菌体内での脂肪酸生産量が高まり、産生される脂肪酸の総量(乾燥菌体1g当り)が著量に増大する。また、同時に産生される脂肪酸を構成する炭素鎖数が、より長鎖側にシフトした直鎖状の飽和脂肪酸となることを見出した。
以上の知見を得たことで、本発明を完成させた。
【0006】
即ち、本発明は以下の通りのものである。
〔1〕 遊離の炭素鎖数14〜20の直鎖状飽和脂肪酸の産生能を有する、Erysipelotrichaceae科のCatenostreptococcus属lactis種に属する細菌。
〔2〕 細菌が、GK12株(FERM P-22123)又はその変異株である、前記〔1〕に記載の細菌。
〔3〕 遊離の炭素鎖数14〜20の直鎖状飽和脂肪酸の産生能を有し、Erysipelotrichaceae科Catenostreptococcus属lactis種に属する細菌を培地中で培養し、培養物から長鎖飽和脂肪酸を採取することを特徴とする、炭素鎖数14〜20の直鎖状長鎖飽和脂肪酸の製造方法。
〔4〕 細菌が、GK12株(FERM P-22123)又はその変異株である、前記〔3〕に記載の製造方法。
〔5〕 培養温度40〜45℃での温度ストレスに相当するストレス負荷の条件下で培養することを特徴とする、前記〔3〕又は〔4〕に記載の製造方法。
〔6〕 前記ストレス付加の条件が、培地中にブタノールを2.0〜2.5%添加することである、前記〔5〕に記載の製造方法。
〔7〕 前記細菌を、菌体増殖速度が定常期になるまで培養した後に、長鎖飽和脂肪酸を採取することを特徴とする、前記〔3〕〜〔6〕のいずれかに記載の製造方法。
【発明の効果】
【0007】
本発明微生物は、天然的に飽和長鎖脂肪酸を生産する新規微生物であり、遺伝子組換え操作をすることなく、培養工程のみで遊離の長鎖脂肪酸を簡便に取得できる。しかも、本発明により得られる長鎖脂肪酸は不飽和脂肪酸を含まないので酸化しないために扱い易く、簡単なエステル化反応によりバイオディーゼル燃料の長鎖飽和脂肪酸エステルを提供することができる。そして、その際にグリセロールという副産物が産生されない上に、炭素鎖長が14から20の長鎖脂肪酸のみを選択的に生産するため、燃料効率が高いメリットもある。
本発明微生物は、培養温度を上げるだけで増殖速度及び遊離脂肪酸生産量が顕著に増加するだけでなく、より炭素鎖数の多い脂肪酸の割合が高まるために、さらに得られるバイオディーゼル燃料の燃料効率アップが期待できる。また、その際、高温下での培養を菌株の増殖速度が定常期に至るまでの間続けることで、飛躍的な遊離脂肪酸量の増大が達成できる。
また、遺伝子組換え技術を適用することで、細胞内に蓄積する脂肪酸を細胞外に放出することなどにより、さらなる長鎖飽和脂肪酸生産量の増加の可能性も高い。
【図面の簡単な説明】
【0008】
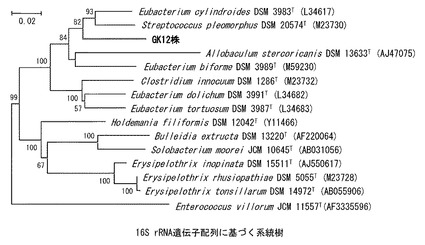
【図1】16S rRNA遺伝子配列に基づく系統樹。 Erysipelotrichaceae科基準種およびGK12株の配列を近隣結合法を用いて作成した。分岐上の数字はブーツストラップ値を、バーの長さは100塩基中に2塩基置換する割合を示す。
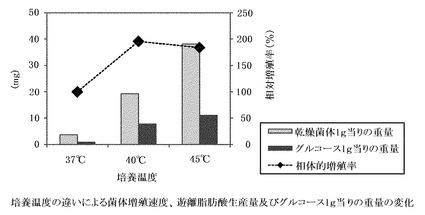
【図2】培養温度の違いによる菌体増殖速度、遊離脂肪酸生産量、及び、グルコース1gあたりの生産量の変化。菌体増殖速度はコントロール(37℃)を100%とした場合の相対値を示す。
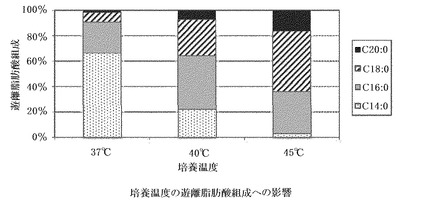
【図3】培養温度の遊離脂肪酸組成への影響。全遊離脂肪酸中の割合をそれぞれ示す。
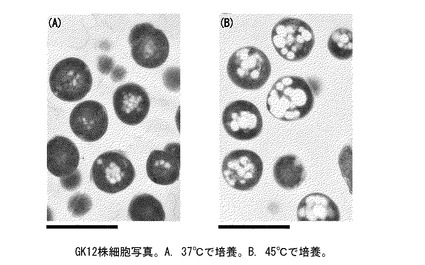
【図4】走査型電子顕微鏡によるGK12細胞写真。バーの長さは1μm。 (A)37℃で培養した菌体。 (B)45℃で培養した菌体。
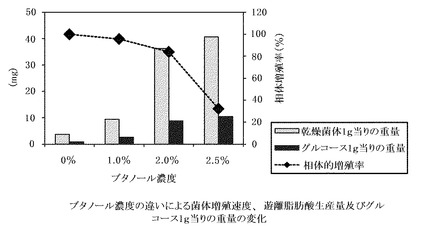
【図5】ブタノール濃度の違いによる菌体増殖速度、遊離脂肪酸生産量、及び、グルコース1gあたりの生産量の変化。菌体増殖速度はコントロール(0% ブタノール)を100%とした場合の相対値を示す。
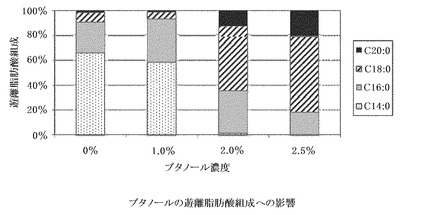
【図6】ブタノールの遊離脂肪酸組成への影響。全遊離脂肪酸中の割合をそれぞれ示す。
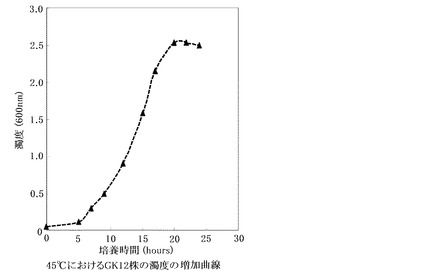
【図7】45℃培養条件下での培地の濁度の増加曲線
【発明を実施するための形態】
【0009】
1.遊離飽和脂肪酸生産菌GK12株の単離方法
北海道深川市、北空知衛生センターの高温メタン発酵槽から採取した内容物を生理食塩水に懸濁した。これを2%(v/v)のブタノールを添加した液体培地(表1〜3)に10%(v/v)の割合で加え、37℃で嫌気培養を行った。なお、培養瓶の液相及び気相は、混合ガス(N2:CO2=80:20)で置換し、ブチルゴム栓及びアルミシールで密閉した。2週間毎に継代培養を計5回行った後、ロールチューブ法にてコロニーの単離を行った。
菌体内に多数の顆粒状の遊離飽和脂肪酸を蓄積し、かつ高増殖性を示す菌株を単離し、GK12株と命名した。なお、下記に述べるようにGK12株は、その菌学的性質から新属新種であることが判明したため、GK12株と同様の菌学的性質を示す細菌の属種をCatenostreptococcus属lactis種と命名したので、Catenostreptococcus属lactis種GK12株ともいう。
以上の通りに単離したGK12株は、2011年6月8日付で、独立行政法人産業技術総合研究所特許生物寄託センターに、受託番号FERM P-22123として寄託されている。
【表1】

【表2】
【表3】
【0010】
2.菌学的性質(菌株の特徴)
本発明のGK12株の同定を行い、下記の結果を得た。
(a)形態的性質
球菌細胞、直径0.5〜0.6μm、細胞の多形性及び運動性ならびに胞子は無し。
(b)培養的性質
表1〜表3に示した培地を用いたロールチューブ法による寒天培養では、白色の楕円の平たいコロニーを寒天中に形成する。表1〜表3に示した培地を用いた液体培養では、表面発育は無し、白色に混濁する。
(c)生理学的性質
硝酸塩の還元、脱窒反応、デンプンの加水分解、及び、クエン酸の利用は無し。30℃から50℃で増殖し、増殖に至適な温度は40℃から45℃であった。絶対嫌気性。D-グルコース、D-フルクトース、D-マンノース、シュークロースから酸を生産する。L-アラビノース、D-キシロース、D-ガラクトース、マルトース、ラクトースから酸を産生しない。
(e)化学分類学的性質
細胞壁ペプチドグリカンのアミノ酸としてジアミノピメリン酸を持たない。リン脂質として、フォスファチジルグリセロール及びフォスファチジルイノシトールを持つ。細胞膜脂質由来脂肪酸として、ミリスチン酸(C14:0)、パルミチン酸(C16:0)、ステアリン酸(C18:0)、アラキジン酸(C20:0)を持つ。その他に細胞脂質成分として、炭素鎖数14、16及び18のジメチルアセタールを持つ。
(f)16S rRNA遺伝子配列の系統解析
16S rRNA遺伝子配列の系統解析の結果、Firmicutes門、Erysipelotrichaceae科に帰属し、最も近縁の公知の属・種はEubacterium cylindroides及びStreptococcus
pleomorphusであり、これらとそれぞれ91%および90%の相同性を示しており、系統樹上で異なる分岐を示す(図1)。
【0011】
以上の結果を得たことで、本発明のGK12株は、Erysipelotrichaceae科に帰属する新属新種細菌であることが判明したため、本発明者らは、その属種をCatenostreptococcus属lactis種と命名した。発明において「Catenostreptococcus属lactis種細菌」というとき、上記(a)〜(f)の菌学的性質を有する微生物を指す。GK12株の突然変異株、形質転換株などのGK12株から誘導される変異株も含まれる。
即ち、本発明のCatenostreptococcus属lactis種GK12株又はその変異株は、絶対嫌気性、従属栄養性の細菌であり、無芽胞、運動性無しの連鎖球菌で、グルコースを発酵して主に乳酸を生産する。30℃から50℃で増殖し、増殖に至適な温度は40℃から45℃である。
細胞壁ペプチドグリカンのアミノ酸として、ジアミノピメリン酸を持たず、細胞内単糖として、ガラクトース、ラムノース及びリボースを持ち、細胞膜リン脂質として、フォスファチジルグリセロール及びフォスファチジルイノシトールを持っており、細胞膜脂肪酸として、ミリスチン酸、パルミチン酸、ステアリン酸、アラキジン酸及び炭素鎖数14、16及び18のジメチルアセタールを持っている点が、上記の近縁種と異なる特徴的な点である。
そして、下記に述べるように、細胞膜脂肪酸を構成するミリスチン酸(C14:0)、パルミチン酸(C16:0)、ステアリン酸(C18:0)、アラキジン酸(C20:0)を遊離脂肪酸の状態で菌体内に蓄積する(図4)。また、培養条件を調整して、菌体が増殖可能な程度の適度なストレスをかけることで、脂肪酸の産生量を著量に増大させることができ、その際の産生する脂肪酸の炭素鎖数を、より長鎖側にシフトさせることができる。
【0012】
3.GK12株からの飽和脂肪酸の回収方法
GK12株を表1〜表3に示した培地で37℃で嫌気的に培養後、培養液から遠心分離によって菌体を回収し、細胞破砕装置を用いて破砕する。次いで、細胞内容物から全脂質を抽出する。本実施例では、ブライダイヤー法(Bligh,E.G.and Dyer,W.J.1959.A rapid method for total lipid extraction and purification. Can.J.Biochem.Physiol.,37:911-917.)を用いて細胞内容物から全脂質を抽出した。
実際にバイオ燃料として用いる場合には、この全脂質に対してエステル化反応を施せばよいが、本実施例では、以下のようにして、遊離脂肪酸を分離し、かつ各々の脂肪酸を単離してそれぞれの組成割合を決定した。
すなわち、薄層クロマトグラフィーを用いて、遊離脂肪酸を分離後、遊離脂肪酸のスポットをかきとった後、メチルエステル化し、ヘキサンにより抽出処理した。ヘキサン層をガスクロマトグラフ質量分析計にて脂肪酸メチルエステルを分析して、得られた遊離脂肪酸の組成を決定した。
37℃の培養条件下でのGK12株は、ミリスチン酸:66.4質量%、パルミチン酸:24.3質量%、ステアリン酸:7.9質量%、アラキジン酸:1.4質量%であった。総重量は乾燥菌体1g当り3.7mgで、グルコース1g当りの遊離脂肪酸量は、約1mgであった。
【0013】
4.脂肪酸生産量を向上させるための培養条件の検討
GK12株が菌体内に蓄積する脂肪酸は、細胞膜脂肪酸を構成するミリスチン酸(C14:0)、パルミチン酸(C16:0)、ステアリン酸(C18:0)、アラキジン酸(C20:0)のみであることから、細胞膜に大きく影響するストレスをかけることで、その応答として産生脂肪酸量が増大する可能性があると考えて、生産量が向上する適度なストレスとなる培養条件の検討を行った。
【0014】
(1)温度条件でのストレス付加
GK12株を、37℃(コントロール)、40℃、45℃、50℃の温度下で培養し、菌体増殖速度を観察し、対数増殖後期の菌体から遊離脂肪酸を抽出し、生産量、収率と共に脂肪酸組成をそれぞれ分析した。温度変化に対応する菌体増殖速度と、菌体1gあたりの脂肪酸生産量、及びグルコース1gあたりの生産量を、まとめて図2に示す。この図から、温度ストレスにより菌体増殖速度は、約40〜45℃がピークで、50℃ではほとんど増殖しないが、菌体1gあたりの脂肪酸生産量は、45℃付近がピークとなる。グルコース1gあたりの生産量、すなわち、培地中で消費されるグルコースがどれだけ脂肪酸に変換されるかの変換効率は、生産量と同じく約45℃付近にピークが来る。また、温度によって脂肪酸組成も大きく異なる(図3)。このことから、ストレスが高まると、産生脂肪酸の炭素鎖数がより長鎖側にシフトすることがわかる。なお、GK12株を45℃で培養すると、乾燥菌体1g当り38.1mg、グルコース収率約1.2%の、パルミチン酸及びステアリン酸を主成分として構成する遊離脂肪酸が生産された。また、45℃で培養した菌体において、細胞内に遊離脂肪酸の顆粒が顕著に発達している様子が観察された(図4)。
【0015】
(2)培地内への有機溶媒によるストレス
(2−1)ブタノールによるストレス
培地中に、ブタノールを1%、1.5%、2%、2.5%まで添加し、上記(1)と同様に、菌体増殖速度、対数増殖後期に抽出した遊離脂肪酸の生産量、収率、脂肪酸組成を分析した。
その結果、ブタノール2.5%の添加が最も生産量を増加させるものであったが、菌体増殖速度は顕著に減少した(図5)。また、添加量の変化により、脂肪酸組成も大きく変化し、温度ストレスと同様に、添加量が増えてストレスが高まると、産生脂肪酸の炭素鎖数がより長鎖側にシフトしている(図6)。
(2−2)他の有機溶媒によるストレス
他の有機溶媒として、エタノール、トルエン及びヘキサンを試したところ、エタノール及びヘキサンについては、顕著な生産量の増加及び脂肪酸組成の変化は見られなかった(図示せず)。トルエンの場合では、生産量が増加し、脂肪酸組成の長鎖へのシフトが見られたものの、増殖速度は顕著に減少した。
【0016】
(3)浸透圧変化によるストレス
培地中に塩化ナトリウムを50、100、200mol/Lまで添加し、浸透圧変化を与えたところ、塩濃度に依存して増殖は低下し、200mol/Lにおいては増殖しなかったが、遊離脂肪酸の増加は認められなかった(図示せず)。
【0017】
(4)脂肪酸生産量をさらに増大させるための培養時間の検討
さらに、本発明のストレス負荷条件下での最適な培養時間を検討した。
上記の検討において最も脂肪酸生産量が向上する培養条件である45℃で、GK12株を複数回培養し、一定時間ごとに培地の濁度を分光光度計で測定したところ、約22時間から24時間程度で濁度の増加曲線の傾きが横ばいになり定常期に至ることがわかる(図7)。
そこで、GK12株の45℃でのストレス条件下での培養を、遊離脂肪酸が十分に生産する定常期(約1日程度)にまで培養した後でのステアリン酸など遊離飽和脂肪酸総量を測定すると、対数増殖後期の菌体の約7倍もの生産量(乾燥菌体1g当り255.1mg)が達成できていることを確認した。
【0018】
(5)脂肪酸生産量を最大限にするための培養条件
上記(1)〜(3)の各種ストレス負荷実験の結果からみて、本発明のGK12株による遊離飽和長鎖脂肪酸生産量を増大させるためには適度なストレス負荷をかけることが好ましく、その際の培養温度40℃〜45℃、好ましくは45℃という温度条件によるストレスが、その適度なストレスの典型例といえる。すなわち、本発明は40℃〜45℃の温度条件に対応するストレス条件下で培養することによる遊離飽和長鎖脂肪酸の培養方法にも特徴を有するものでもある。
そして、その際の培養時間をGK12株の増殖が定常期に至るまでの間(具体的には22〜24時間)、40℃〜45℃の温度条件に対応するストレス条件下に曝して培養することで、遊離飽和長鎖脂肪酸生産量を最大限に増大させることが可能となる。温度的ストレスのみならず、他のストレス条件下でも同様である。定常期に至るまでの一般的な時間は22〜24時間であるため、培養時間を22〜24時間とすることもできるが、与えるストレスの種類及び強さ、又は培地成分などその他の条件によっては、定常期に至るまでの時間に幅があるため、培地中の濁度を観察し、濁度の増加が止まったことを指標として、定常期に至ったことを判断する方が好ましい。
【実施例】
【0019】
以下、実施例を挙げて本発明を説明するが、本発明はそれに限られるものではない。
(実施例1)分離方法
北海道深川市、北空知衛生センターの高温メタン発酵槽から採取した内容物を生理食塩水に懸濁した。これを2%(v/v)のブタノールを添加した液体培地(表1〜3)に10%(v/v)の割合で加え、37℃で嫌気培養を行った。なお、培養瓶の液相及び気相は、混合ガス(N2:CO2=80:20)で置換し、ブチルゴム栓及びアルミシールで密閉した。2週間毎に継代培養を計5回行った後、ロールチューブ法にてコロニーの単離を行った。
【0020】
(実施例2)菌株の特徴
GK12株の同定を行い、以下の結果を得た。絶対嫌気性、従属栄養性の細菌。無芽胞、運動性無しの連鎖球菌で、グルコースを発酵して主に乳酸を生産した。30℃から50℃で増殖し、増殖に至適な温度は40℃から45℃であった。16S rRNA遺伝子配列(大腸菌における同遺伝子の28−1491の位置に相当する)の系統解析の結果、Firmicutes門、Erysipelotrichaceae科に帰属する公知のEubacterium cylindroidesおよびStreptococcus pleomorphusとそれぞれ91%および90%の相同性を示し、系統樹上で異なる分岐を示した(図1)。細胞壁ペプチドグリカンのアミノ酸として、ジアミノピメリン酸を持たず、ガラクトース、ラムノース及びリボースを細胞内単糖として、フォスファチジルグリセロール及びフォスファチジルイノシトールを細胞膜リン脂質として、ミリスチン酸、パルミチン酸、ステアリン酸及び炭素鎖数14、16及び18のジメチルアセタールを細胞膜脂肪酸として持ち、これらは上記の近縁種と異なっていた。これらの結果から、GK12株はErysipelotrichaceae科に帰属する新属新種細菌であるものと決定し、Catenostreptococcus属lactis種と命名した。
【0021】
(実施例3)遊離脂肪酸の分離と分析(37℃)
GK12株を表1に示した培地で37℃で嫌気的に培養した。18時間後の対数増殖後期の培養液から遠心分離によって回収した菌体を、細胞破砕装置を用いて破砕した。ブライダイヤー法を用いて細胞内容物から全脂質を抽出した。薄層クロマトグラフィー(固定層:シリカゲル、展開溶媒:ヘキサン/ジエチルエーテル/酢酸(容量比=70/30/20)、呈色:プリムリン(UV照射下))を用いて、遊離脂肪酸を分離した。遊離脂肪酸のスポットをかきとった後、10%塩酸メタノール溶液を加え100℃で3時間反応させた(メチルエステル化)。ヘキサンを用いて抽出処理し、ヘキサン層をガスクロマトグラフ質量分析計にて脂肪酸メチルエステルを分析した。得られた遊離脂肪酸の組成は、ミリスチン酸:66.4質量%、パルミチン酸:24.3質量%、ステアリン酸:7.9質量%、アラキジン酸:1.4質量%であった。総重量は乾燥菌体1g当り3.7mgで、グルコース1g当りの遊離脂肪酸量は、約1mgであった。
【0022】
(実施例4)遊離脂肪酸を生産する培養条件の検討
生産量が向上する培養条件の検討を行った。GK12株を40℃、あるいは45℃、50℃、1%、2%、2.5%ブタノール存在下、5%エタノール存在下、0.2%トルエン存在下、0.6%ヘキサン存在下、50mol/L、100mol/L、200mol/L塩化ナトリウム存在下で対数増殖後期まで培養した菌体から遊離脂肪酸を抽出し、分析を行った。なお、37℃で培養した菌体をコントロールとした。条件の違いによる生産量を表4に、遊離脂肪酸組成を表5に示した。
【表4】
【表5】
【0023】
菌体増殖速度、生産量、収率、ならびに、脂肪酸組成を比較検討した結果、遊離脂肪酸生産の最適条件は45℃での培養と結論付けられた。なお、GK12株を45℃で培養すると、乾燥菌体1g当り38.1mg、グルコース収率約1.2%の、パルミチン酸及びステアリン酸を主成分として構成する遊離脂肪酸が生産された。45℃で培養した菌体において、細胞内に遊離脂肪酸の顆粒が顕著に発達している様子が観察された(図4)。
【0024】
(実施例5)遊離脂肪酸の最大生産条件の検討
(実施例4)において、菌体増殖速度、生産量、収率、及び脂肪酸組成のいずれの観点から見ても、本発明のGK12株を用いた場合の遊離脂肪酸生産の最適な培養条件は45℃での培養であることが結論付けられた。
そこで、このGK12株を45℃の温度条件で培養し、22時間後に濁度などから定常期に至ったことを確認し、菌体を採取して遊離脂肪酸を抽出し、分析を行った。
その結果、グルコース収率は約9.8%であったが、ステアリン酸を主成分として構成する遊離脂肪酸の総生産量が乾燥菌体1g当り255.1mgであって、(実施例4)での遊離脂肪酸総生産量が乾燥菌体1g当り38.1mgであったものを約7倍近くにまで生産量を増大させることができた。この条件の遊離脂肪酸組成および生産量を表6に示した。
この結果からみて、GK12株において長鎖飽和脂肪酸の生産性を最大限発揮させるためには、45℃の温度条件下で、遊離脂肪酸を十分に生産させる定常期まで、すなわち培養開始後22時間まで培養することが好ましいことがわかる。
【表6】
【技術分野】
【0001】
本発明は、長鎖飽和脂肪酸を産生する新規微生物、及び当該微生物を用いた脂肪酸を製造する方法に関する。
【背景技術】
【0002】
近年、地球温暖化対策として石油など化石燃料資源に代わる液体燃料としてのバイオディーゼルが脚光を浴びてきており、実用化も進んでいる。典型的なバイオディーゼルとしては、植物の種子に由来する植物油のアシル基をメタノールなどのアルコールでエステル交換反応して合成された脂肪酸アルキルエステルがある。この方法で得られるバイオディーゼルは、従来の石油由来燃料と比較しても優れた燃焼効率を有しているばかりか、地中に埋蔵されていた化石由来の炭素を燃焼時放出する石油燃料とは異なり、CO2温室効果ガス放射の減少にも寄与できる。しかし、パーム油の場合のように広大なアブラヤシ農園建設のために多くの熱帯雨林が失われるなどの別の環境破壊も問題となっており、大豆などの油糧種子作物の場合は、食糧、飼料源であるため燃料への転用は食糧危機を招くおそれがある。
そこで、最近では、タンクによる大量培養可能な藻類や微生物がバイオディーゼル原料として注目されており、研究開発が進んでいたが、従来の微生物によるバイオディーゼル生産では、生産される油のほとんどがトリグリセリド脂質であるため、メチルエステル化によって、触媒や未変換脂肪酸が混入したグリセロールが副産物として大量に産生してしまい、生産効率の面で問題であった。微生物を用いて遊離の脂肪酸又は脂肪酸エステルを直接的に生産させるための方法として、セルロースを炭素源として遺伝子組換えを施した藻類を培養し、多価不飽和脂肪酸より構成されるトリグリセリドを生産する方法(特許文献1)、組換え微生物を用いて脂肪エステル及び遊離脂肪酸を生産し細胞外に放出させる方法(特許文献2)、組換え藻類あるいは酵母による脂質生産と化学反応による直鎖アルカンの生産方法(特許文献3)などの研究開発が相次いでいる。しかし、いずれも遺伝子組換え操作が必要であって、操作の煩雑性および生産の安定性に不安が残った。また、これら遺伝子組換え微生物を含め、従来微生物によって生産される脂質に含まれる脂肪酸および遊離脂肪酸には、二重結合を1つ以上持つ不飽和脂肪酸を多少に含み酸化しやすいため、燃料として安定的に用いる場合に酸化防止剤の必要性が生じることから、生産コストの面での問題があった。
以上のことから、遊離の長鎖飽和脂肪酸を産生する、遺伝子組み換えを伴わない天然の新規微生物の探索が望まれていた。
【先行技術文献】
【特許文献】
【0003】
【特許文献1】特表2010−538642号公報
【特許文献2】特表2010−538602号公報
【特許文献3】特表2010−528627号公報
【発明の概要】
【発明が解決しようとする課題】
【0004】
本発明は、遊離の長鎖飽和脂肪酸を生産する新規微生物を提供しようとするものである。
【課題を解決するための手段】
【0005】
本発明者らは、微生物資源として未開拓な絶対嫌気性の微生物を分離培養する過程で、遊離の長鎖飽和脂肪酸を細胞内に蓄積する新規の細菌GK12株を見出した。当該細菌GK12株は、Erysipelotrichaceae科に帰属するが、公知のいずれの属、種にも属さない新属の新種細菌であり、本発明者らはCatenostreptococcus属lactis種と命名した。当該細菌は、絶対嫌気性条件下でグルコース含有培地により培養すると、14〜20の偶数の炭素鎖数を有する直鎖状の飽和脂肪酸を遊離状態で菌体内に蓄積する。さらに、本発明微生物は、高温条件下(45℃)で培養した場合の他、培地中にブタノールを添加するなどの培養条件を変化させた場合に、個々の菌体内での脂肪酸生産量が高まり、産生される脂肪酸の総量(乾燥菌体1g当り)が著量に増大する。また、同時に産生される脂肪酸を構成する炭素鎖数が、より長鎖側にシフトした直鎖状の飽和脂肪酸となることを見出した。
以上の知見を得たことで、本発明を完成させた。
【0006】
即ち、本発明は以下の通りのものである。
〔1〕 遊離の炭素鎖数14〜20の直鎖状飽和脂肪酸の産生能を有する、Erysipelotrichaceae科のCatenostreptococcus属lactis種に属する細菌。
〔2〕 細菌が、GK12株(FERM P-22123)又はその変異株である、前記〔1〕に記載の細菌。
〔3〕 遊離の炭素鎖数14〜20の直鎖状飽和脂肪酸の産生能を有し、Erysipelotrichaceae科Catenostreptococcus属lactis種に属する細菌を培地中で培養し、培養物から長鎖飽和脂肪酸を採取することを特徴とする、炭素鎖数14〜20の直鎖状長鎖飽和脂肪酸の製造方法。
〔4〕 細菌が、GK12株(FERM P-22123)又はその変異株である、前記〔3〕に記載の製造方法。
〔5〕 培養温度40〜45℃での温度ストレスに相当するストレス負荷の条件下で培養することを特徴とする、前記〔3〕又は〔4〕に記載の製造方法。
〔6〕 前記ストレス付加の条件が、培地中にブタノールを2.0〜2.5%添加することである、前記〔5〕に記載の製造方法。
〔7〕 前記細菌を、菌体増殖速度が定常期になるまで培養した後に、長鎖飽和脂肪酸を採取することを特徴とする、前記〔3〕〜〔6〕のいずれかに記載の製造方法。
【発明の効果】
【0007】
本発明微生物は、天然的に飽和長鎖脂肪酸を生産する新規微生物であり、遺伝子組換え操作をすることなく、培養工程のみで遊離の長鎖脂肪酸を簡便に取得できる。しかも、本発明により得られる長鎖脂肪酸は不飽和脂肪酸を含まないので酸化しないために扱い易く、簡単なエステル化反応によりバイオディーゼル燃料の長鎖飽和脂肪酸エステルを提供することができる。そして、その際にグリセロールという副産物が産生されない上に、炭素鎖長が14から20の長鎖脂肪酸のみを選択的に生産するため、燃料効率が高いメリットもある。
本発明微生物は、培養温度を上げるだけで増殖速度及び遊離脂肪酸生産量が顕著に増加するだけでなく、より炭素鎖数の多い脂肪酸の割合が高まるために、さらに得られるバイオディーゼル燃料の燃料効率アップが期待できる。また、その際、高温下での培養を菌株の増殖速度が定常期に至るまでの間続けることで、飛躍的な遊離脂肪酸量の増大が達成できる。
また、遺伝子組換え技術を適用することで、細胞内に蓄積する脂肪酸を細胞外に放出することなどにより、さらなる長鎖飽和脂肪酸生産量の増加の可能性も高い。
【図面の簡単な説明】
【0008】
【図1】16S rRNA遺伝子配列に基づく系統樹。 Erysipelotrichaceae科基準種およびGK12株の配列を近隣結合法を用いて作成した。分岐上の数字はブーツストラップ値を、バーの長さは100塩基中に2塩基置換する割合を示す。
【図2】培養温度の違いによる菌体増殖速度、遊離脂肪酸生産量、及び、グルコース1gあたりの生産量の変化。菌体増殖速度はコントロール(37℃)を100%とした場合の相対値を示す。
【図3】培養温度の遊離脂肪酸組成への影響。全遊離脂肪酸中の割合をそれぞれ示す。
【図4】走査型電子顕微鏡によるGK12細胞写真。バーの長さは1μm。 (A)37℃で培養した菌体。 (B)45℃で培養した菌体。
【図5】ブタノール濃度の違いによる菌体増殖速度、遊離脂肪酸生産量、及び、グルコース1gあたりの生産量の変化。菌体増殖速度はコントロール(0% ブタノール)を100%とした場合の相対値を示す。
【図6】ブタノールの遊離脂肪酸組成への影響。全遊離脂肪酸中の割合をそれぞれ示す。
【図7】45℃培養条件下での培地の濁度の増加曲線
【発明を実施するための形態】
【0009】
1.遊離飽和脂肪酸生産菌GK12株の単離方法
北海道深川市、北空知衛生センターの高温メタン発酵槽から採取した内容物を生理食塩水に懸濁した。これを2%(v/v)のブタノールを添加した液体培地(表1〜3)に10%(v/v)の割合で加え、37℃で嫌気培養を行った。なお、培養瓶の液相及び気相は、混合ガス(N2:CO2=80:20)で置換し、ブチルゴム栓及びアルミシールで密閉した。2週間毎に継代培養を計5回行った後、ロールチューブ法にてコロニーの単離を行った。
菌体内に多数の顆粒状の遊離飽和脂肪酸を蓄積し、かつ高増殖性を示す菌株を単離し、GK12株と命名した。なお、下記に述べるようにGK12株は、その菌学的性質から新属新種であることが判明したため、GK12株と同様の菌学的性質を示す細菌の属種をCatenostreptococcus属lactis種と命名したので、Catenostreptococcus属lactis種GK12株ともいう。
以上の通りに単離したGK12株は、2011年6月8日付で、独立行政法人産業技術総合研究所特許生物寄託センターに、受託番号FERM P-22123として寄託されている。
【表1】
【表2】
【表3】
【0010】
2.菌学的性質(菌株の特徴)
本発明のGK12株の同定を行い、下記の結果を得た。
(a)形態的性質
球菌細胞、直径0.5〜0.6μm、細胞の多形性及び運動性ならびに胞子は無し。
(b)培養的性質
表1〜表3に示した培地を用いたロールチューブ法による寒天培養では、白色の楕円の平たいコロニーを寒天中に形成する。表1〜表3に示した培地を用いた液体培養では、表面発育は無し、白色に混濁する。
(c)生理学的性質
硝酸塩の還元、脱窒反応、デンプンの加水分解、及び、クエン酸の利用は無し。30℃から50℃で増殖し、増殖に至適な温度は40℃から45℃であった。絶対嫌気性。D-グルコース、D-フルクトース、D-マンノース、シュークロースから酸を生産する。L-アラビノース、D-キシロース、D-ガラクトース、マルトース、ラクトースから酸を産生しない。
(e)化学分類学的性質
細胞壁ペプチドグリカンのアミノ酸としてジアミノピメリン酸を持たない。リン脂質として、フォスファチジルグリセロール及びフォスファチジルイノシトールを持つ。細胞膜脂質由来脂肪酸として、ミリスチン酸(C14:0)、パルミチン酸(C16:0)、ステアリン酸(C18:0)、アラキジン酸(C20:0)を持つ。その他に細胞脂質成分として、炭素鎖数14、16及び18のジメチルアセタールを持つ。
(f)16S rRNA遺伝子配列の系統解析
16S rRNA遺伝子配列の系統解析の結果、Firmicutes門、Erysipelotrichaceae科に帰属し、最も近縁の公知の属・種はEubacterium cylindroides及びStreptococcus
pleomorphusであり、これらとそれぞれ91%および90%の相同性を示しており、系統樹上で異なる分岐を示す(図1)。
【0011】
以上の結果を得たことで、本発明のGK12株は、Erysipelotrichaceae科に帰属する新属新種細菌であることが判明したため、本発明者らは、その属種をCatenostreptococcus属lactis種と命名した。発明において「Catenostreptococcus属lactis種細菌」というとき、上記(a)〜(f)の菌学的性質を有する微生物を指す。GK12株の突然変異株、形質転換株などのGK12株から誘導される変異株も含まれる。
即ち、本発明のCatenostreptococcus属lactis種GK12株又はその変異株は、絶対嫌気性、従属栄養性の細菌であり、無芽胞、運動性無しの連鎖球菌で、グルコースを発酵して主に乳酸を生産する。30℃から50℃で増殖し、増殖に至適な温度は40℃から45℃である。
細胞壁ペプチドグリカンのアミノ酸として、ジアミノピメリン酸を持たず、細胞内単糖として、ガラクトース、ラムノース及びリボースを持ち、細胞膜リン脂質として、フォスファチジルグリセロール及びフォスファチジルイノシトールを持っており、細胞膜脂肪酸として、ミリスチン酸、パルミチン酸、ステアリン酸、アラキジン酸及び炭素鎖数14、16及び18のジメチルアセタールを持っている点が、上記の近縁種と異なる特徴的な点である。
そして、下記に述べるように、細胞膜脂肪酸を構成するミリスチン酸(C14:0)、パルミチン酸(C16:0)、ステアリン酸(C18:0)、アラキジン酸(C20:0)を遊離脂肪酸の状態で菌体内に蓄積する(図4)。また、培養条件を調整して、菌体が増殖可能な程度の適度なストレスをかけることで、脂肪酸の産生量を著量に増大させることができ、その際の産生する脂肪酸の炭素鎖数を、より長鎖側にシフトさせることができる。
【0012】
3.GK12株からの飽和脂肪酸の回収方法
GK12株を表1〜表3に示した培地で37℃で嫌気的に培養後、培養液から遠心分離によって菌体を回収し、細胞破砕装置を用いて破砕する。次いで、細胞内容物から全脂質を抽出する。本実施例では、ブライダイヤー法(Bligh,E.G.and Dyer,W.J.1959.A rapid method for total lipid extraction and purification. Can.J.Biochem.Physiol.,37:911-917.)を用いて細胞内容物から全脂質を抽出した。
実際にバイオ燃料として用いる場合には、この全脂質に対してエステル化反応を施せばよいが、本実施例では、以下のようにして、遊離脂肪酸を分離し、かつ各々の脂肪酸を単離してそれぞれの組成割合を決定した。
すなわち、薄層クロマトグラフィーを用いて、遊離脂肪酸を分離後、遊離脂肪酸のスポットをかきとった後、メチルエステル化し、ヘキサンにより抽出処理した。ヘキサン層をガスクロマトグラフ質量分析計にて脂肪酸メチルエステルを分析して、得られた遊離脂肪酸の組成を決定した。
37℃の培養条件下でのGK12株は、ミリスチン酸:66.4質量%、パルミチン酸:24.3質量%、ステアリン酸:7.9質量%、アラキジン酸:1.4質量%であった。総重量は乾燥菌体1g当り3.7mgで、グルコース1g当りの遊離脂肪酸量は、約1mgであった。
【0013】
4.脂肪酸生産量を向上させるための培養条件の検討
GK12株が菌体内に蓄積する脂肪酸は、細胞膜脂肪酸を構成するミリスチン酸(C14:0)、パルミチン酸(C16:0)、ステアリン酸(C18:0)、アラキジン酸(C20:0)のみであることから、細胞膜に大きく影響するストレスをかけることで、その応答として産生脂肪酸量が増大する可能性があると考えて、生産量が向上する適度なストレスとなる培養条件の検討を行った。
【0014】
(1)温度条件でのストレス付加
GK12株を、37℃(コントロール)、40℃、45℃、50℃の温度下で培養し、菌体増殖速度を観察し、対数増殖後期の菌体から遊離脂肪酸を抽出し、生産量、収率と共に脂肪酸組成をそれぞれ分析した。温度変化に対応する菌体増殖速度と、菌体1gあたりの脂肪酸生産量、及びグルコース1gあたりの生産量を、まとめて図2に示す。この図から、温度ストレスにより菌体増殖速度は、約40〜45℃がピークで、50℃ではほとんど増殖しないが、菌体1gあたりの脂肪酸生産量は、45℃付近がピークとなる。グルコース1gあたりの生産量、すなわち、培地中で消費されるグルコースがどれだけ脂肪酸に変換されるかの変換効率は、生産量と同じく約45℃付近にピークが来る。また、温度によって脂肪酸組成も大きく異なる(図3)。このことから、ストレスが高まると、産生脂肪酸の炭素鎖数がより長鎖側にシフトすることがわかる。なお、GK12株を45℃で培養すると、乾燥菌体1g当り38.1mg、グルコース収率約1.2%の、パルミチン酸及びステアリン酸を主成分として構成する遊離脂肪酸が生産された。また、45℃で培養した菌体において、細胞内に遊離脂肪酸の顆粒が顕著に発達している様子が観察された(図4)。
【0015】
(2)培地内への有機溶媒によるストレス
(2−1)ブタノールによるストレス
培地中に、ブタノールを1%、1.5%、2%、2.5%まで添加し、上記(1)と同様に、菌体増殖速度、対数増殖後期に抽出した遊離脂肪酸の生産量、収率、脂肪酸組成を分析した。
その結果、ブタノール2.5%の添加が最も生産量を増加させるものであったが、菌体増殖速度は顕著に減少した(図5)。また、添加量の変化により、脂肪酸組成も大きく変化し、温度ストレスと同様に、添加量が増えてストレスが高まると、産生脂肪酸の炭素鎖数がより長鎖側にシフトしている(図6)。
(2−2)他の有機溶媒によるストレス
他の有機溶媒として、エタノール、トルエン及びヘキサンを試したところ、エタノール及びヘキサンについては、顕著な生産量の増加及び脂肪酸組成の変化は見られなかった(図示せず)。トルエンの場合では、生産量が増加し、脂肪酸組成の長鎖へのシフトが見られたものの、増殖速度は顕著に減少した。
【0016】
(3)浸透圧変化によるストレス
培地中に塩化ナトリウムを50、100、200mol/Lまで添加し、浸透圧変化を与えたところ、塩濃度に依存して増殖は低下し、200mol/Lにおいては増殖しなかったが、遊離脂肪酸の増加は認められなかった(図示せず)。
【0017】
(4)脂肪酸生産量をさらに増大させるための培養時間の検討
さらに、本発明のストレス負荷条件下での最適な培養時間を検討した。
上記の検討において最も脂肪酸生産量が向上する培養条件である45℃で、GK12株を複数回培養し、一定時間ごとに培地の濁度を分光光度計で測定したところ、約22時間から24時間程度で濁度の増加曲線の傾きが横ばいになり定常期に至ることがわかる(図7)。
そこで、GK12株の45℃でのストレス条件下での培養を、遊離脂肪酸が十分に生産する定常期(約1日程度)にまで培養した後でのステアリン酸など遊離飽和脂肪酸総量を測定すると、対数増殖後期の菌体の約7倍もの生産量(乾燥菌体1g当り255.1mg)が達成できていることを確認した。
【0018】
(5)脂肪酸生産量を最大限にするための培養条件
上記(1)〜(3)の各種ストレス負荷実験の結果からみて、本発明のGK12株による遊離飽和長鎖脂肪酸生産量を増大させるためには適度なストレス負荷をかけることが好ましく、その際の培養温度40℃〜45℃、好ましくは45℃という温度条件によるストレスが、その適度なストレスの典型例といえる。すなわち、本発明は40℃〜45℃の温度条件に対応するストレス条件下で培養することによる遊離飽和長鎖脂肪酸の培養方法にも特徴を有するものでもある。
そして、その際の培養時間をGK12株の増殖が定常期に至るまでの間(具体的には22〜24時間)、40℃〜45℃の温度条件に対応するストレス条件下に曝して培養することで、遊離飽和長鎖脂肪酸生産量を最大限に増大させることが可能となる。温度的ストレスのみならず、他のストレス条件下でも同様である。定常期に至るまでの一般的な時間は22〜24時間であるため、培養時間を22〜24時間とすることもできるが、与えるストレスの種類及び強さ、又は培地成分などその他の条件によっては、定常期に至るまでの時間に幅があるため、培地中の濁度を観察し、濁度の増加が止まったことを指標として、定常期に至ったことを判断する方が好ましい。
【実施例】
【0019】
以下、実施例を挙げて本発明を説明するが、本発明はそれに限られるものではない。
(実施例1)分離方法
北海道深川市、北空知衛生センターの高温メタン発酵槽から採取した内容物を生理食塩水に懸濁した。これを2%(v/v)のブタノールを添加した液体培地(表1〜3)に10%(v/v)の割合で加え、37℃で嫌気培養を行った。なお、培養瓶の液相及び気相は、混合ガス(N2:CO2=80:20)で置換し、ブチルゴム栓及びアルミシールで密閉した。2週間毎に継代培養を計5回行った後、ロールチューブ法にてコロニーの単離を行った。
【0020】
(実施例2)菌株の特徴
GK12株の同定を行い、以下の結果を得た。絶対嫌気性、従属栄養性の細菌。無芽胞、運動性無しの連鎖球菌で、グルコースを発酵して主に乳酸を生産した。30℃から50℃で増殖し、増殖に至適な温度は40℃から45℃であった。16S rRNA遺伝子配列(大腸菌における同遺伝子の28−1491の位置に相当する)の系統解析の結果、Firmicutes門、Erysipelotrichaceae科に帰属する公知のEubacterium cylindroidesおよびStreptococcus pleomorphusとそれぞれ91%および90%の相同性を示し、系統樹上で異なる分岐を示した(図1)。細胞壁ペプチドグリカンのアミノ酸として、ジアミノピメリン酸を持たず、ガラクトース、ラムノース及びリボースを細胞内単糖として、フォスファチジルグリセロール及びフォスファチジルイノシトールを細胞膜リン脂質として、ミリスチン酸、パルミチン酸、ステアリン酸及び炭素鎖数14、16及び18のジメチルアセタールを細胞膜脂肪酸として持ち、これらは上記の近縁種と異なっていた。これらの結果から、GK12株はErysipelotrichaceae科に帰属する新属新種細菌であるものと決定し、Catenostreptococcus属lactis種と命名した。
【0021】
(実施例3)遊離脂肪酸の分離と分析(37℃)
GK12株を表1に示した培地で37℃で嫌気的に培養した。18時間後の対数増殖後期の培養液から遠心分離によって回収した菌体を、細胞破砕装置を用いて破砕した。ブライダイヤー法を用いて細胞内容物から全脂質を抽出した。薄層クロマトグラフィー(固定層:シリカゲル、展開溶媒:ヘキサン/ジエチルエーテル/酢酸(容量比=70/30/20)、呈色:プリムリン(UV照射下))を用いて、遊離脂肪酸を分離した。遊離脂肪酸のスポットをかきとった後、10%塩酸メタノール溶液を加え100℃で3時間反応させた(メチルエステル化)。ヘキサンを用いて抽出処理し、ヘキサン層をガスクロマトグラフ質量分析計にて脂肪酸メチルエステルを分析した。得られた遊離脂肪酸の組成は、ミリスチン酸:66.4質量%、パルミチン酸:24.3質量%、ステアリン酸:7.9質量%、アラキジン酸:1.4質量%であった。総重量は乾燥菌体1g当り3.7mgで、グルコース1g当りの遊離脂肪酸量は、約1mgであった。
【0022】
(実施例4)遊離脂肪酸を生産する培養条件の検討
生産量が向上する培養条件の検討を行った。GK12株を40℃、あるいは45℃、50℃、1%、2%、2.5%ブタノール存在下、5%エタノール存在下、0.2%トルエン存在下、0.6%ヘキサン存在下、50mol/L、100mol/L、200mol/L塩化ナトリウム存在下で対数増殖後期まで培養した菌体から遊離脂肪酸を抽出し、分析を行った。なお、37℃で培養した菌体をコントロールとした。条件の違いによる生産量を表4に、遊離脂肪酸組成を表5に示した。
【表4】
【表5】
【0023】
菌体増殖速度、生産量、収率、ならびに、脂肪酸組成を比較検討した結果、遊離脂肪酸生産の最適条件は45℃での培養と結論付けられた。なお、GK12株を45℃で培養すると、乾燥菌体1g当り38.1mg、グルコース収率約1.2%の、パルミチン酸及びステアリン酸を主成分として構成する遊離脂肪酸が生産された。45℃で培養した菌体において、細胞内に遊離脂肪酸の顆粒が顕著に発達している様子が観察された(図4)。
【0024】
(実施例5)遊離脂肪酸の最大生産条件の検討
(実施例4)において、菌体増殖速度、生産量、収率、及び脂肪酸組成のいずれの観点から見ても、本発明のGK12株を用いた場合の遊離脂肪酸生産の最適な培養条件は45℃での培養であることが結論付けられた。
そこで、このGK12株を45℃の温度条件で培養し、22時間後に濁度などから定常期に至ったことを確認し、菌体を採取して遊離脂肪酸を抽出し、分析を行った。
その結果、グルコース収率は約9.8%であったが、ステアリン酸を主成分として構成する遊離脂肪酸の総生産量が乾燥菌体1g当り255.1mgであって、(実施例4)での遊離脂肪酸総生産量が乾燥菌体1g当り38.1mgであったものを約7倍近くにまで生産量を増大させることができた。この条件の遊離脂肪酸組成および生産量を表6に示した。
この結果からみて、GK12株において長鎖飽和脂肪酸の生産性を最大限発揮させるためには、45℃の温度条件下で、遊離脂肪酸を十分に生産させる定常期まで、すなわち培養開始後22時間まで培養することが好ましいことがわかる。
【表6】
【特許請求の範囲】
【請求項1】
遊離の炭素鎖数14〜20の直鎖状飽和脂肪酸の産生能を有する、Erysipelotrichaceae科のCatenostreptococcus属lactis種に属する細菌。
【請求項2】
細菌が、GK12株(FERM P-22123)又はその変異株である、請求項1に記載の細菌。
【請求項3】
遊離の炭素鎖数14〜20の直鎖状飽和脂肪酸の産生能を有し、Erysipelotrichaceae科Catenostreptococcus属lactis種に属する細菌を培地中で培養し、培養物から長鎖飽和脂肪酸を採取することを特徴とする、炭素鎖数14〜20の直鎖状長鎖飽和脂肪酸の製造方法。
【請求項4】
細菌が、GK12株(FERM P-22123)又はその変異株である、請求項3に記載の製造方法。
【請求項5】
培養温度40〜45℃での温度ストレスに相当するストレス負荷の条件下で培養することを特徴とする、請求項3又は4に記載の製造方法。
【請求項6】
前記ストレス付加の条件が、培地中にブタノールを2.0〜2.5%添加することである、請求項5に記載の製造方法。
【請求項7】
前記細菌を、菌体増殖速度が定常期になるまで培養した後に、長鎖飽和脂肪酸を採取することを特徴とする、請求項3〜6のいずれかに記載の製造方法。
【請求項1】
遊離の炭素鎖数14〜20の直鎖状飽和脂肪酸の産生能を有する、Erysipelotrichaceae科のCatenostreptococcus属lactis種に属する細菌。
【請求項2】
細菌が、GK12株(FERM P-22123)又はその変異株である、請求項1に記載の細菌。
【請求項3】
遊離の炭素鎖数14〜20の直鎖状飽和脂肪酸の産生能を有し、Erysipelotrichaceae科Catenostreptococcus属lactis種に属する細菌を培地中で培養し、培養物から長鎖飽和脂肪酸を採取することを特徴とする、炭素鎖数14〜20の直鎖状長鎖飽和脂肪酸の製造方法。
【請求項4】
細菌が、GK12株(FERM P-22123)又はその変異株である、請求項3に記載の製造方法。
【請求項5】
培養温度40〜45℃での温度ストレスに相当するストレス負荷の条件下で培養することを特徴とする、請求項3又は4に記載の製造方法。
【請求項6】
前記ストレス付加の条件が、培地中にブタノールを2.0〜2.5%添加することである、請求項5に記載の製造方法。
【請求項7】
前記細菌を、菌体増殖速度が定常期になるまで培養した後に、長鎖飽和脂肪酸を採取することを特徴とする、請求項3〜6のいずれかに記載の製造方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【公開番号】特開2013−17474(P2013−17474A)
【公開日】平成25年1月31日(2013.1.31)
【国際特許分類】
【出願番号】特願2012−123173(P2012−123173)
【出願日】平成24年5月30日(2012.5.30)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成22年度独立行政法人新エネルギー・産業技術総合開発機構委託研究「新エネルギー技術研究開発/バイオマスエネルギー等高効率転換技術開発(先導技術開発)/酵素糖化・効率的発酵に関する基盤研究」産業技術力強化法第19条の適用を受ける特許出願
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
【公開日】平成25年1月31日(2013.1.31)
【国際特許分類】
【出願日】平成24年5月30日(2012.5.30)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成22年度独立行政法人新エネルギー・産業技術総合開発機構委託研究「新エネルギー技術研究開発/バイオマスエネルギー等高効率転換技術開発(先導技術開発)/酵素糖化・効率的発酵に関する基盤研究」産業技術力強化法第19条の適用を受ける特許出願
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
[ Back to top ]