
新規糖トランスポーター
SWEET、GLUEまたはGlueと称するトランスポータータンパク質の新規クラスを開示する。これらのトランスポーターは、細胞内および細胞の内と外の間で膜を通過する糖輸送の新規システムを提供する。かかるトランスポーターは、生物の特定の臓器および細胞の特定の細胞小器官内の糖濃度を理解し、変化するために有用である。これらのトランスポーターはまた、病原体侵襲からの植物の保護にも有用である。
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、細胞の膜を通過する新規糖トランスポーターに関する。
【0002】
関連出願
本願は米国仮出願第61/175,267号(2009年5月4日出願)の優先権を主張し、その全体について、参照により本明細書に組み込む。
【背景技術】
【0003】
発明の背景
多くの生化学経路における糖の必要および使用は、数世紀にわたって広範に研究され、報告されている。単糖類、二糖類および多糖類または糖は、多くの生物にとって糖の主たる食事性供給源である。
【0004】
グルコースは、最も容易に利用可能な糖の一つであり、その構造は、多くの生物の生化学系によって容易に作用される。6個の炭素原子からなるグルコースは、アルドヘキソース(aldehexose)のカテゴリーに属する。アルドヘキソースは4個のキラル中心を有し、これらは16個の立体異性体を導く。アルドヘキソースの2個の立体異性体がグルコースと見なされ、主たるものがD−グルコースである。全ての主たる食事性炭水化物が、デンプンおよびグリコーゲンにおけるような唯一の構造ブロックとして、あるいはショ糖およびラクトースにおけるような別の単糖類とともに、グルコースを含む。この炭水化物の代謝は、アデノシン3リン酸(ATP)のようなエネルギーに変換する。グルコースの他の代謝経路は、エネルギー貯蔵を導く。グルコース分子は環の開いた(非環式)形態または環の(環状)形態で存在しうる。
【0005】
グルコースは、デンプン、セルロースおよびグリコーゲンのようないくつかの重要な物質の合成の前駆体として使用できる。牛乳中の糖であるラクトースは、グルコース−ガラクトース二糖である。別の二糖であるショ糖は、グルコースとフルクトースの結合体である。グルコースは後生動物における糖の主要な輸送形態であるが、ショ糖およびその誘導体は、植物における主要な輸送形態として利用される。
【0006】
グルコースは植物およびいくつかの原核生物における光合成の下流産物の一つである。動物および真菌のような真核生物において、グルコースは、グリコーゲン分解と称されるプロセスを介して、グリコーゲンの分解の結果として生産されうる。植物において得られる分解基質はデンプンである。グルコースはまた、植物における主要な輸送糖であるショ糖に対するインベルターゼの作用にも由来しうる(細胞壁、細胞基質または液胞において、各々特異的なアイソフォームによる)。
【0007】
動物では、グルコースはピルビン酸およびグリセロールのような非炭水化物中間体から、糖新生と称されるプロセスを介して肝臓および腎臓において合成されうる。グルコースはまた、例えばデンプンの酵素加水分解によって、合成されることもある。商業的には、市販の農作物、例えばトウモロコシ、コメ、コムギ、ジャガイモ、キャッサバ、サゴおよびクズウコンがデンプンの供給源として使用されうる。
【0008】
グルコースは好気的または嫌気的呼吸のいずれかに使用されうる。炭水化物は生物の重要なエネルギー源である。好気的呼吸は1グラムあたり約3.75 kcalのエネルギーを提供できる。デンプンのような炭水化物の分解は、単糖類および二糖類を生じる。解糖プロセスおよびクエン酸回路(またはクレブス回路)の反応により、グルコースは酸化され、分解されて最終的に二酸化炭素と水を形成し、エネルギー源(主としてATP)を得る。インスリン応答および他のメカニズムは、血中グルコース濃度を制御しうる。
【0009】
脳のような神経中枢エネルギーの必要は、グルコースと心理的プロセスを直接的に関連づける。グルコースは脳の第一次的なエネルギー供給源であり、したがってその利用能は心理的プロセスに影響する。グルコースが少ない場合、精神的な努力を必要とする心理的プロセスは損なわれうる。好気的および嫌気的呼吸のいずれも、解糖代謝経路の初期工程で開始し、最初のステップは、エネルギーを得るための後の分解を準備するための、ヘキソキナーゼによるグルコースのリン酸化である。次いで、ヘキソキナーゼによるグルコースの速やかなリン酸化は、細胞外への拡散を防止しうる。リン酸化の作用は荷電されたリン酸基を加え、それによってグルコース−6−リン酸が容易に細胞膜を通過することを防止する。グルコースはまた、タンパク質の産生および脂質代謝プロセスにおいても重要である。グルコースはまた、アスコルビン酸またはビタミンCの前駆体分子としても利用できる。
【0010】
したがって、グルコースおよびショ糖のような糖の細胞内および生物中の組織の細胞間での取り込み、吸収、処理、代謝、交換および輸送は、細胞または細胞を含む生物の生育に最も重要である。細胞膜を通過し、細胞の小器官間でのグルコースまたはショ糖輸送の機能不全は、破局的でありうる。グルコースの輸送を効率的に制御する方法を開発する必要がある。
【発明の概要】
【0011】
発明の概要
本発明は、細胞内で糖を輸送するためのトランスポータータンパク質の新規クラスを提供する。GLUEと称される(GlueまたはSWEETとも知られる)トランスポーターは、植物中に存在しおり、そして糖(例えばペントース、グルコース、マンノース、要するに単糖類)またはショ糖およびマルトース(要するに二糖類またはオリゴ糖)のトランスポーターをコードする、下記受託番号と少なくとも30%、40%、50%、60%、70%、80%、85%、90%、95%または99%配列同一性を有する核酸によってコードされうる: AT4G15920、AT3G16690、AT5G13170、AtSAG29、AT4G25010、AT5G50800、AT5G23660、AT3G48740、AT5G50790、AT2G39060、AT5G40260、AtRPG1、AT4G10850、AT1G66770、AT1G21460、AT5G62850、AtVEX1、AT3G28007、AT3G14770、AT1G21460、AT5G53190、NEC1、AC202585、AC147714、MtC60432 GC、MtC11004 GC、CT963079、MtD03138 GC、TC 125536、AC146866、AC189276、TC129646、CAA69976 MtNod3 AC2456、TC115479、AC146747、MtC10424 GC、CT954252、CU302340、AC202585、AC147714、MtC60432 GC、MtC11004 GC、CT963079、Os08g42350 (Os8N3) Os08g0535200、Os12g29220 Os03g0347500、Os05g51090 Os05g0588500、Os12g07860、Os09g08440、Os09g08490、Os09g08270、Os09g08030 Os09g0254600、Os01g42090.1 Os01g0605700、Os01g42110.1 Os01g060600、Os02g19820 Os02g0301100、Os05g35140 Os05g0426000、Os01g65880 Os01g0881300、Os01g50460 Os01g0700100、Os01g36070.1 Os01g0541800、Os01g12130.1、Os05g12320 Os05g0214300およびOs01g21230(いずれもその全体について参照により本明細書に組み込む)。核酸はベクター中および/または細胞、例えば植物細胞または動物細胞中に存在していてよい。本発明はまた、GLUEを含むトランスジェニック植物も提供する。該核酸は、下記動物受託番号と少なくとも30%、40%、50%、60%、70%、80%、85%、90%、95%または99%配列同一性を有する、グルコースまたはショ糖トランスポーターをコードする核酸によってコードされうる:(例えば線虫(C. elegans)由来)R10D12.9、K11D12.5、およびK06A4.4、K02D7.5、C54F6.4、C06G8.1、Y39A1A.8、ci-rga、RAG1AP1(例えばショウジョウバエ、ヒト、マウス、ドブネズミ(Rattus norvegicus)およびアフリカツメガエル(Xenopus))(いずれもその全体について参照により本明細書に組み込む)。
【0012】
本発明はさらに、成熟したGLUEタンパク質を提供する。GLUEによる糖の移動が野生型と比較して改善されるように、GLUEは、変異されていてもよい。移動速度の増加または輸送能力の増加によりより多くの糖を輸送できるように、変異がGLUEの機能を改善していてよい。変異は、野生型と比較して、GLUEによる糖の移動を防止または遅延させうる。変異は、野生型配列のアミノ酸または複数のアミノ酸の欠失または置換であってよい。変異はGLUEの切断であってもよい。
【0013】
本発明は、GLUEタンパク質を含む融合タンパク質を提供する。GLUEはタグ、例えばエピトープと融合されてよい。GLUEは膜タンパク質のキメラメンバーの一部、例えば既知の下流カスケードでの他の7回膜貫通タンパク質であってよい。キメラタンパク質はGLUEの第3細胞内ループおよび/または細胞質尾部が別の7回膜貫通タンパク質由来の対応するドメインでの置換を含みうる。GLUEは、細胞内の特定の小器官または領域とGLUEの発現および位置を指定するために、標的配列と結合していてもよい。GLUEは変異GLUEタンパク質であってよい。
【0014】
本発明は、コントロール植物と比較して増加したレベルの炭素を産生する植物を作成する方法であって、植物細胞に単糖、二糖またはオリゴ糖トランスポーターをコードする核酸を導入し、GLUEをコードする核酸を発現する植物に植物細胞を成長させることを含む方法を提供する。
【0015】
本発明は、植物の根への糖輸送を増加させる方法であって、植物の細胞にGLUEをコードする核酸を導入することを含む方法を提供する。植物へのGLUEの導入は、植物の根圏への糖分泌を調節する方法および植物の葉圏への糖輸送または発生種子、花等への糖の送達を調節する方法を提供できる。
【0016】
本発明は、植物における糖のレベルを変化させる方法であって、細胞にGLUEをコードする核酸を導入することを含む方法を提供する。本方法は、細胞内の糖レベルを低下させてもよい。本方法は、植物または動物のある領域、臓器または小器官に蓄積させるために糖濃度を指定しうる。本ほう法は、植物または動物のある領域、臓器または小器官における糖レベルの低下を引き起こしてもよい。本方法は、糖の取り込みを増加させてもよい。本方法は、糖の取り込みを減少させてもよい。本方法は、糖の排出を増加させてもよい。本方法は、糖の排出を減少させてもよい。GLUEはコファクター、例えば別の細胞内タンパク質またはトランスポーター、例えばコトランスポーターによって細胞内で発現される。
【0017】
本発明は、植物に有益微生物を誘引する方法であって、GLUEの導入によって糖濃度を変化させることを含む方法を提供する。本発明はさらに、GLUEの導入によって病原体から植物を保護する方法を提供する。植物侵襲病原体は、植物細胞機構を利用して植物の糖排出を変化させることがある。さらに異なるプロモーターの制御下に存在しうる異種GLUEを植物に導入することにより、病原体の糖排出を変化させる能力が限定または変化されうる。
【0018】
本発明はまた、細胞または生物中でGLUEがどのように作用しているかを決定する方法を提供する。糖検出分子、例えば蛍光ドナードメインと蛍光アクセプタードメイン間で挟まれた糖(例えばグルコースまたはショ糖)結合ドメインのような、糖検出分子と共に、異種GLUEは細胞内で共発現されてよい。糖濃度は蛍光共鳴エネルギー移動によって経時的に測定およびモニターできる。
【0019】
本発明は、細胞中のグルコーストランスポーター促進(GLUT)タンパク質の発現に影響し、および/またはそれを変化させる方法を提供する。
【0020】
本発明は、細胞によって分泌される液体、例えば花蜜または樹液中の糖レベルを変化させる方法を提供する。本発明は、生物の細胞にGLUEを導入することによって生物の発生を変化させる方法を提供する。GLUEは変異されていてもよい。本発明は、生物の細胞中のGLUEを変異することによって生物の発生を変化させる方法を提供する。生物中のGLUEの上昇または低下した機能は、細胞中または生物中の糖濃度および/または糖分布を変化させ、それによって発生に影響を与えうる。
【図面の簡単な説明】
【0021】
【図1】多様な植物種におけるGLUEスーパーファミリーcDNAの系統樹を示す。図1AはGLUEスーパーファミリーの系統樹を示す。距離はネイバージョイニング法を用いた複数配列アラインメント(ClustalW)から計算し、ツリーはブーストラップ値を示す(1000のパーセンテージ)。SWEETファミリーは4つのクレードに分割できる。全ての配列はNCBIまたはAramemnonデータベースから得た。コメ、シロイヌナズナ、ウマゴヤシおよびペチュニアのツリーを示す。示したスケールは示した長さあたり50塩基の変化を意味する。
【0022】
【図2】植物の細胞内、器官および小器官で糖が輸送され、移動する多様な手段の概要を示す。
【0023】
【図3】図3Aおよび3Bは細胞中で発現されたグルコースFRETセンサーによる応答を示す。図3AはGLUEの共発現なしでのグルコースの多様な濃度に対するセンサーの応答を示す。図3BはグルコースFRETセンサーを有する細胞にGLUE1を導入することが、糖の濃度が変化した場合にセンサーに顕著な変化をもたらすことを示す。
【0024】
【図4】図4A〜4Dは発現したGLUEタンパク質の変化による変化したグルコース濃度に対するグルコースFRETセンサーの応答を示す。図4AはGLUE1による応答を示す。図4BはGLUE12による応答を示す。図4CはGLUE8による応答を示す。図4DはGLUE13による応答を示す。
【0025】
【図5】ベクターのみ(EBY-pDRF1)を有するグルコース取り込み欠損株EBY4000がD−グルコースを取り込めないことを示す。放射性標識化14C−グルコース取り込みを経時的に測定した。反対に、GLUE1は酵母株EBY4000(EBY-GLUE1)にグルコース取り込みを可能とさせる。。グルコース取り込みコンピテント野生型株CENPK(CEN)はポジティブコントロールとして使用し、グルコースを取り込める。
【0026】
【図6】試験したGLUEの15種の遺伝子のうち4つが酵母変異をレスキューできる。酵母は異なるpHを有する2%グルコースで増殖した。
【0027】
【図7】授乳中のRAG1AP1(HsSWEET1)の誘導を示す。マイクロアレイデータは、授乳中に推定糖トランスポーターRAG1AP1がアップレギュレートされることを示唆している。
【0028】
【図8】肺胞細胞からのラクトース合成および分泌を示す。グルコースはGLUT/SGLTによって基底膜を介して取り込まれ、次いで未知のトランスポーターによりER(○1)またはゴルジ体(○2)に取り込まれる。ラクトース合成はゴルジ体において生じ、ラクトースは乳管に面した頂側で排出されると推測される。未知のトランスポーターは、頂側の膜を介してグルコースを排出するか、または乳由来グルコースの回復に関与する(○3)。
【0029】
【図9】SWEET(GLUE)トランスポーターの同定および特徴付けを示す。図9AはHEK293T細胞における細胞基質FRETグルコースセンサーFLIPglu600μΔ13Vとの共発現による、SWEET1についてのグルコース輸送活性の同定を示す。個々の細胞はCFPおよびVenus放出の定量比イメージングによって分析した(収集間隔5秒;Fc/Dは正規化した放出強度比に対応する)。HEK293T/FLIPglu600μΔ13V細胞を増加したグルコース濃度の中間パルス、次いで方形パルスでかん流した。センサーのみを発現する細胞は、FRET比変化(オレンジの線)がないことによって示されるように、細胞質中の顕著な量のグルコースを蓄積しなかった。センサーおよびSWEET1を共発現する細胞は、増加した外部グルコース供給と相関する振幅でのネガティブFRET比変化によって示されるとおり、グルコースを蓄積した(青色線)。データ点は平均±SD(n>10)である。図9Bは、細胞質からERへのグルコースの「流出」のFRETイメージングを示す(図9C参照)。センサーFLIPglu600μΔ13VERはERの膜を標的とした(図9Aと同様に分析を行った。収集間隔10秒)。センサーのみを発現する細胞はERにおける顕著な量のグルコースを蓄積しなかった。センサーおよびSWEET1を共発現する細胞は、グルコースの流出によって誘導されるネガティブ比変化によって示されるとおり、ER中にグルコースを蓄積した。データ点は平均±SD(n>10)である。図9CはPMを通過するSWEET1流入および細胞質からERへの流出の模式図を示す。細胞基質センサーFLIPglu600μΔ13Vは、細胞外表面で開始されるグルコースの輸送を同定する(細胞外N末端によって示される)。FLIPglu600μΔ13VERは、細胞内側で開始される輸送を測定する(細胞基質C末端)。図9DはSWEET1、SWEET8または酵母HXT5による18種のヘキソーストランスポーター遺伝子全てを欠く酵母株EBY4000の相補性を示す;コントロール:空のベクター。図9Eは、0、20および100 mMのグルコース添加前後のSWEET1およびFLII12Pglu700μΔ6を共発現するEBY4000におけるグルコースの蓄積を示す。グルコース添加前に2つのサイクルを実行した。データは平均±SD、n=3である。図9FはEBY4000中のSWEET1による14C−グルコース蓄積の動力学を示す。データは平均±SD、n=3である。図9Gは安定的に形質転換されたシロイヌナズナの葉の葉におけるSWEET1−YFPの共焦点イメージングを示す。図21Hは疎水性プロットに基づくSWEETの構造モデルを示す。各タンパク質は推定の細胞外N末端、およびリンカーとしてTMH4で区分された3個のTMH(TMH1〜3および5〜7、赤および青の三角形で強調した)の重複に由来する推定の平行方向の2個の「サブユニット」を有する7個のTMHを含む。図9IはSGLT1によって仲介されるが、OsSWEET11によって仲介されないアフリカツメガエル卵母細胞への[14C]−グルコースの取り込みを示す。SGLT1とOsSWEET11の共発現は、卵母細胞におけるグルコース蓄積を減少させる。データは平均±SD、n=7である。挿入図はSGLT1によるグルコースの集中的取り込みおよびOsSWEET11によって引き起こされるグルコース流出(「リーク」)を示す。図9KはSWEET1またはOsSWEET11を発現する卵母細胞からの直接流出測定を示す。10 mMの放射性標識化グルコース(0.18μCi/μl)50 nlを注射して、放射性トレーサー流出を経時的に測定した。データは平均±SD(n>10細胞)である。
【0030】
【図10】生体栄養細菌または真菌が異なるSWEET遺伝子のmRNAレベルを誘導することを示す。図10Aは細菌性シュードモナス・シリンゲ(Pseudomonas syringae)病原型トマトDC3000(2 x 108 cfu/ml、接種後8時間、qPCRで測定、MgCl2バッファー処理で正規化)、ウドンコ病菌、G. cichoracearum(〜25〜35 分生子 mm-2、接種後48時間、qPCRで測定;0時間値に正規化)、またはシロイヌナズナの葉におけるボトリチス・シネレア菌(Botrytis cinerea)による、SWEET mRNAの誘導を示す。図10BはP.s. DC3000によるSWEET4、5および15の誘導が機能的タイプIII分泌系(T3S)に基づくことを示す。サンプルは2 x 108 cfu/mlのDC3000またはT3S変異体DC3000ΔhrcUでの浸潤後6、12および24時間で採取した。図10CはG. cichoracearumによる感染がSWEET11およびSWEET12の誘導を導くが、SWEET15のダウンレギュレーションを導くことを示す。サンプルは接種後0、8、12、24および72時間後に採取した。
【0031】
【図11】病原体の栄養摂取におけるSWEETの役割についてのスキームモデルを示す。図11Aは病原菌キサントモナス・オリゼ(Xanthomonas oryzae)病原型オリゼ株PXO99 A(X.o. PXO99A)がタイプIII分泌系を介してコメ細胞にTALエフェクターPthXo1を注入することを示す。この転写アクチベーターは直接または間接的に、グルコースの分泌を導くコメOsSWEET11/Os8N3グルコース流出トランスポーター遺伝子の誘導を惹起する。細菌は内因性取り込み系を介してグルコースを取り込み、増殖できる。図11BはPthXo1が変異されている場合(ME)、OsSWEET11/Os8N3の誘導は低下または停止されて、細菌の飢餓を導くことを示す(細菌細胞の減少したサイズ、平均少細胞数として示される)。図11CはOsSWEET11/Os8N3の変異も細菌の飢餓を導くことを示す。図11Dは別のエフェクターAvrXa7を発現する病原体が、SWEETファミリーの別のメンバーを誘導する(または別の炭素源へのアクセスを誘導する)場合、増殖できることを示す。
【0032】
【図12】HEK293T細胞におけるSWEET仲介グルコース輸送の証拠を示す。図12AはGLUT1とコトランスフェクトされたFLIPglu600μΔ13Vセンサーを用いて分析した、20 μMのサイトカラシンBによるGLUT1活性の阻害を示す。図12BはSWEET1とコトランスフェクトされたFLIPglu600μΔ13Vを用いて分析した、20 μMのサイトカラシンBに対するSWEET1活性の非感受性を示す。図12CはHepG2細胞、HEK293T細胞ならびにFLIPglu600μΔ13Vおよび/またはSWEET1を共発現するHEK293T細胞における、SLC2(GLUT)およびSLC5(SGLT)グルコーストランスポーターの発現レベルを示す。
【0033】
【図13】SWEET1の生化学的特徴を示す。図13AはSWEET1の最適pHを示す。放射性トレーサー取り込みを異なるpHで測定した。取り込みの至適pHは約pH8.5である。データは平均±SDである。図13Bは異なる糖による酵母株EBY4000におけるSWEET1によって仲介されるグルコース取り込み(5 mM D−グルコース;0.1 μCi [14C]−D−グルコース)の阻害を示す。コンペティターを10倍過剰(最終濃度50 mM)で加えた。相対活性はD−グルコース取り込み速度(100%)に正規化した。データは平均±SDである。
【0034】
【図14】シロイヌナズナにおけるSWEET1およびSWEET8遺伝子の組織特異的発現パターンを示す。分析はArabadopsis eFP Browser(http://bar.utoronto.ca/efp/cgi-bin/efpWeb.cgi)のマイクロアレイ試験に基づく。
【0035】
【図15】異種系におけるSWEET8の機能的分析を示す。図15Aは細胞基質FRETグルコースセンサーFLIPglu600μΔ13VおよびSWEET8を共発現する細胞が、HEK293T細胞におけるネガティブな細胞基質FRET比変化によって示されるとおり、細胞基質中にグルコースを蓄積したことを示す(図9A参照)。図15BはHEK293T細胞においてER FRETグルコースセンサーFLIPglu-600μΔ13VERおよびSWEET8を共発現する場合、細胞が細胞基質からERへとグルコースを流出することを示す(図9B参照)。データ点は平均±SD(n>10)である。図15Cは酵母グルコース輸送欠損変異体EBY4000におけるSWEET1、SWEETおよびベクターコントロールの相対的取り込み速度を示す(2分;10mM D−グルコース;0.1μCi [14C]−D−グルコース)。値をSWEET1(100%)に正規化する。データは平均±SDである。図15Dは安定的に形質転換されたシロイヌナズナを用いた実生(T1世代)におけるSWEET8局在の共焦点イメージングを示す。
【0036】
【図16】アフリカツメガエル卵母細胞におけるSWEET仲介グルコース取り込みを示す。卵母細胞における相対的グルコース取り込み速度は、コントロールとして水、SWEET1、OsSWEET11、SGLT1またはSGLT1とSWEET1を注射した(時間=1時間;1mM D−グルコース; 4μCi/ml [14C]−D−グルコース)。データは平均±SD、n>9である。
【0037】
【図17】HEK293T細胞におけるグルコースの実時間取り込みを示す。GLUE1はHEK293T細胞において細胞基質またはER FRETセンサーFLIPglu600μD6と共発現された。CFPおよびVFPの正規化した放出比をY軸に示す。ネガティブなFRET比の変化は、GLUE1がグルコーストランスポーターならびにエフラクサー(effluxer)として機能することを示している。
【0038】
【図18】GLUE8、12および13がHEK293T細胞において細胞基質FRETセンサーFLIPglu600μD6と共発現されたことを示している。CFPおよびVFPの正規化した放出比をY軸に示す。細胞基質におけるネガティブなFRET比の変化は、GLUE8、12および13が異なる活性を有するグルコーストランスポーターとして機能することを示している。
【0039】
【図19】アフリカツメガエル卵母細胞におけるGLUEの機能的発現を示す。図19AはSGLT1、GLUE1によって仲介されるがOsGLUE11によって仲介されないアフリカツメガエル卵母細胞への[14C]−グルコースの取り込みを示す。OsGLUE11とSGLT1の共発現はグルコース蓄積を減少させる。図19BはCeSWEET1、3、4、5、7およびRAG1AP1スプライス変異体1および2ならびに変異体(Y216A、L218A、L219A; RAG1AP1-3aa)によって仲介される、アフリカツメガエル卵母細胞への14C−グルコースの取り込みを示す。SGLT1をコントロールとして用いた。図19Cはアフリカツメガエル卵母細胞におけるSGLT1によるグルコース蓄積に対するCeSWEETおよびRAG1AP1変異体の共発現の影響を示す。全実験は独立して少なくとも6回反復した。エラーバーは平均±SDである。
【0040】
【図20】17種のシロイヌナズナGLUE遺伝子による18種のヘキソーストランスポーター遺伝子全てを欠く酵母株YSL2-1の相補性を示す。酵母ホモログHXT5または哺乳類ホモログGLUT1を発現する細胞をコントロールとして用いた。Os8N3およびRAG1AP1は、コメおよび哺乳動物由来ホモログである。酵母細胞を初期対数期に2%マルトースを含むSD-Ura液体培地中で増殖し、2%のマルトース、フルクトースまたはマンノースを含むYPMまたはSD-URAを含む培地に、5m の連続希釈をスポットした。GLUE4およびGLUE7はフルクトースおよびマンノースを輸送する。GLUE5および8はマンノースを輸送する。
【0041】
【図21】17種のシロイヌナズナGLUE遺伝子による18種のヘキソーストランスポーター遺伝子全てを欠く酵母株YSL2-1の相補性を示す。哺乳類ホモログGLUT1を発現する細胞をコントロールとして用いた。Os8N3およびRAG1AP1は、コメおよび哺乳動物由来ホモログである。酵母細胞を初期対数期に2%マルトースを含むSD-Ura液体培地中で増殖し、多様な濃度のガラクトースを含むSD-URAを含む培地に、5m の連続希釈をスポットした。GLUE1、4、5および7を除き、他の全ては5%ガラクトースに感受性であり、これはガラクトース輸送を仲介する可能性を示している。
【0042】
【図22】17種のシロイヌナズナGLUE遺伝子による18種のヘキソーストランスポーター遺伝子全てを欠く酵母株YSL2-1の相補性を示す。酵母ホモログHXT5または哺乳類ホモログGLUT1を発現する細胞をコントロールとして用いた。Os8N3およびRAG1AP1は、コメおよび哺乳動物由来ホモログである。酵母細胞を初期対数期に2%マルトースを含むSD-Ura液体培地中で増殖し、2%のグルコースを含むYPMまたはSD-URAを含む培地に、5m の連続希釈をスポットした。GLUE1、4、5および7はグルコースを輸送する(GLUE8も同様。ここでは示さず)。
【0043】
【図23】17種のシロイヌナズナGLUE遺伝子による18種のヘキソーストランスポーター遺伝子全てを欠く酵母株YSL2-1の相補性を示す。酵母ホモログHXT5または哺乳類ホモログGLUT1を発現する細胞をコントロールとして用いた。Os8N3およびRAG1AP1は、コメおよび哺乳動物由来ホモログである。酵母細胞を初期対数期に2%マルトースを含むSD-Ura液体培地中で増殖し、多様な2−デオキシ−グルコースレベルを含むYPMまたはSD-URAを含む培地に、5m の連続希釈をスポットした。GLUE1、3、4、5、7、8、14、16および17は、糖アナログにより感受性であるため、2−デオキシグルコースを輸送する。
【0044】
【図24】植物体におけるシロイヌナズナGLUEタンパク質の細胞内局在を示す。GLUE−GFP融合タンパク質は、タバコの葉において一時的に発現した場合、細胞膜の近傍または細胞膜に局在する。
【0045】
【図25】根におけるGLUE発現を示す。シロイヌナズナの根におけるGLUE遺伝子発現のqPCR分析。転写産物は10日齢シロイヌナズナ実生から単離し、テンプレートとしてcDNAを作成した。比較Ct法を用いて相対発現レベルを計算した(1000*1/(2^(CtGlU-CtActin8)。ここに示されていないファミリーのメンバーは、顕著な発現レベルを示さなかった(Guo et al.、未公表の結果)。4回の独立した実験からのデータ。
【0046】
【図26】はシロイヌナズナGLUEの発現パターンの組織学的分析を示す。GLUE2、GLUE16およびGLUE17−GUS融合タンパク質を担持するトランスジェニックシロイヌナズナにおけるGUS活性をX-glucで染色して分析した。10日齢シロイヌナズナ実生由来植物全体のイメージを示す。
【0047】
【図27】FRETグルコースセンサーによるCIT3細胞における糖流量分析を示す。細胞基質FRETグルコースセンサーFLIPglu-30μΔ13VとRAG1AP1-mCherryの共発現の非存在(A)および存在(B)下における、ヒト乳腺細胞系としてCIT3細胞におけるFRET分析。細胞を異なる外部グルコース濃度でかん流した(5、25および40 mM)。FRETイメージを得て、データを分析した。データは平均±SD(n=7〜9)である。
【0048】
【図28】RAG1AP1mCherryを発現するCIT3におけるグルコースレベルに対するサイトカラシンBの影響を示す。細胞基質FRETグルコースセンサーFLIPglu-30μΔ13VとRAG1AP1-mCherryの共発現の存在下における、ヒト乳腺細胞系としてCIT3細胞におけるFRET分析。細胞を外部40mMグルコースで、20μMのサイトカラシンBの存在下または非存在下でかん流した。FRETイメージを得て、データを分析した。データは平均±SD(n=5)である。
【0049】
【図29】RAG1AP1mCherryを発現するCIT3細胞におけるグルコースレベルに対する分化の影響を示す。細胞基質FRETグルコースセンサーFLIPglu-30μΔ13VとRAG1AP1-mCherryの共発現の存在下における、ヒト乳腺細胞系としてCIT3細胞におけるFRET分析。10 μg/mL インスリン、3 μg/mL プロラクチンおよび3μg/mL ヒドロコルチゾン(分泌培地)で細胞を分化した。10 μg/mL インスリンおよび5 ng/mL EGFを含むDMEM/F12(増殖培地)中で細胞を培養した。細胞を異なる外部グルコース濃度でかん流した(5、25および40 mM)。FRETイメージを得て、データを分析した。データは平均±SD(n=11〜13)である。
【0050】
【図30】図30AはCIT3細胞におけるRAG1AP1−GFP融合タンパク質の局在を示す。イメージは共焦点顕微鏡で撮影した。図30BはHepG2細胞およびHEK293T細胞由来RNAのRT−PCR分析を示す。RAG1AP1、GLUT1またはβアクチンを逆転写し、PCRで増幅した。未分化または分化細胞を10 μg/mL インスリンおよび5 ng/mL EGFを含むDMEM/F12のGM(増殖培地)または10 μg/mL インスリン、3 μg/mL プロラクチンおよび3μg/mL ヒドロコルチゾンを含むDMEM/F12であるSM(分泌培地)中で培養した。
【0051】
【図31】RAG1AP1およびRAG1AP1mCherryのウェスタンブロットを示す。SDS−PAGE(12.5%ゲル)で酵母の全細胞溶解物、RAG1AP1またはRAG1AP1-mCherryを過剰発現するCIT3を分離した。抗原領域はEQDRNYWLLQTであり、ヒトRAG1AP1のC末端アミノ酸211〜221に対応する(Abcam)。
【0052】
【図32】RAG1AP1を過剰発現するMDCK細胞におけるRAG1AP1の免疫蛍光局在化を示す。RAG1AP1はヒトRAG1AP1(Abcam)のC末端ペプチドに対する抗体およびAlexa 594標識化ロバ抗ヤギIgGで染色した。2,6−シアリルトランスフェラーゼ−GFP(ゴルジ体マーカー)をRAG1AP1と混合した。
【0053】
【図33】図33AはMDCK細胞におけるRAG1AP1-GFP融合タンパク質の局在化を示す。イメージは共焦点顕微鏡で撮影した。図45BはFRETセンサーによるHEK293T細胞の糖流量分析を示す。RAG1AP1を発現し、細胞基質FRETグルコースセンサーFLIPglu-600μΔ13VとRAG1AP1-mCherryを共発現するHEK293T細胞におけるFRET分析。細胞を異なる外部グルコース濃度でかん流した(2.5、5、25および40 mM)。FRETイメージを得て、データを分析した。データは平均±SD(n=)である。
【0054】
【図34】ヒト肝臓断片におけるRAG1AP1の免疫蛍光局在を示す。RAG1AP1はヒトRAG1AP1(Abcam)のC末端ペプチドに対する抗体およびAlexa fluor 594標識化ロバ抗ヤギIgGで染色した。Golgin-97をゴルジ体マーカーとして用い、これをgolgin-97に対するモノクローナル抗体(Invitrogen)およびAlexa fluor 488標識化ロバ抗マウスIgGで染色した。
【0055】
【図35】ヒト肝臓断片におけるRAG1AP1の免疫蛍光局在を示す。RAG1AP1はヒトRAG1AP1(Abcam)のC末端ペプチドに対する抗体およびAlexa fluor 594標識化ロバ抗ヤギIgGで染色した。Golgin-97をゴルジ体マーカーとして用い、これをgolgin-97に対するモノクローナル抗体(Invitrogen)およびAlexa fluor 488標識化ロバ抗マウスIgGで染色した。
【0056】
【図36】Golgi標的化FLIPglu-600μΔ13Vを示す。β−1,4−ガラクトシルトランスフェラーゼ1(galT)のペプチド(14〜44)およびステムを用いてFRETグルコースセンサーをゴルジ体に標的化した(Schaub et al, Mol Biol Cell, 17: 5153-5162, 2006)。
【0057】
【図37】Golgi標的化FLIPglu-600μΔ13Vを示す。β−1,4−ガラクトシルトランスフェラーゼ1(galT)のペプチド(14〜44)およびステムを用いてFRETグルコースセンサーをゴルジ体に標的化した(Schaub et al, Mol Biol Cell, 17: 5153-5162, 2006)。
【0058】
【図38】FRETグルコースセンサーによるMDCK細胞の細胞基質およびゴルジ体中の糖流量分析を示す。RAG1AP1発現の存在下(A,B)または非存在下(C,D)および細胞基質(A,C)またはゴルジ体(B,D)標的化FRETグルコースセンサーFLIPglu-600μΔ13Vを共発現する、MDCK細胞におけるFRET分析。細胞を異なる外部グルコース濃度でかん流した(1、2.5、5、10および40 mM)。FRETイメージを得て、データを分析した。データは平均±SD(n=4〜15)である。
【0059】
【図39】FRET分析センサーによるHela細胞の細胞基質およびゴルジ体における糖流量分析を示す。細胞基質(A)またはゴルジ体(B)標的化FRETグルコースセンサーFLIPglu-600μΔ13Vを発現するHela細胞におけるFRET分析。細胞を異なる外部グルコース濃度(5、10および40 mM)およびガラクトース濃度(5および40mM)でかん流した。FRETイメージを得て、データを分析した。データは平均±SD(n=7〜8)である。
【0060】
【図40】FRETグルコースセンサーによる線虫(C. elegans)GLUEファミリーメンバーを発現するHEK293T細胞における糖流量分析を示す。FRETグルコースセンサーFLIPglu-600μΔ13Vを共発現する、線虫(C. elegans)GLUEメンバーを発現するHEK293T細胞におけるFRET分析。細胞を異なる外部グルコース濃度(2.5、5、10および40 mM)およびガラクトース濃度(5および40mM)でかん流した。FRETイメージを得て、データを分析した。データは平均±SD(n=12〜24)である。
【0061】
【図41】ヒトRAG1AP1ホモログが糖流出トランスポーターであることを示す。ヒトSGLT1ナトリウムグルコースコトランスポーター(「S」)は、二次活性グルコースインポーターであることが既に示されている。SGLT1はアフリカツメガエル卵母細胞において内因的に発現され、14C標識化グルコースの卵母細胞への取り込みを仲介できる。RAG1AP1(「R])が共発現している場合、卵母細胞においてより低い取り込みが見られる。これはSGLT1発現細胞におけるグルコースの高い蓄積を防止するRAG1AP1活性による、グルコース「漏出」と一致する。
【0062】
【図42】5種の線虫(C. elegans)ホモログについての14Cグルコース取り込みデータを示す。少なくとも3、4および5は活性である。(C elegans遺伝子名のコード:Ce1: C06G8.1; Ce2: K06A4.4; Ce3: Y39A1A.8; Ce4: K11D12.5; Ce5: K02D7.5;(Ce5のワーム変異体: K02D7.5は増加した脂肪蓄積を示し、これはこれらの細胞からのグルコースの低下した流出と一致する)
【0063】
【図43−1】図43A〜43Hは、HEK293細胞におけるSWEETのショ糖排出機能を示す(ポジティブコントロール(ポテトショ糖トランスポーターStSUT1; Riesmeier et al. 1993 Plant Cell)は43Aであり、ネガティブコントロール(空のベクター43B))。ショ糖の取り込みはFRETショ糖センサーFLIPsuc90μΔ1を用いて測定した(Chaudhuri et al., 2008 Plant Journal)。SWEET10(図43C)、SWEET11(図43D)、SWEET12(図43E)、SWEET13(図43F)、SWEET14(図43G)およびOsSWEET11/Os8N3(図43H)は、哺乳類細胞へのショ糖取り込みを示すポジティブコントロールStSUT1のものと同様に、ネガティブなFRET応答を示した(ネガティブ比変化は細胞基質中の糖含量の増加に対応する)。この取り込みは、単輸送(促進された分散)、プロトンシンポートまたはプロトンアンチポートによって仲介されている可能性がある。
【図43−2】図43A〜43Hは、HEK293細胞におけるSWEETのショ糖排出機能を示す(ポジティブコントロール(ポテトショ糖トランスポーターStSUT1; Riesmeier et al. 1993 Plant Cell)は43Aであり、ネガティブコントロール(空のベクター43B))。ショ糖の取り込みはFRETショ糖センサーFLIPsuc90μΔ1を用いて測定した(Chaudhuri et al., 2008 Plant Journal)。SWEET10(図43C)、SWEET11(図43D)、SWEET12(図43E)、SWEET13(図43F)、SWEET14(図43G)およびOsSWEET11/Os8N3(図43H)は、哺乳類細胞へのショ糖取り込みを示すポジティブコントロールStSUT1のものと同様に、ネガティブなFRET応答を示した(ネガティブ比変化は細胞基質中の糖含量の増加に対応する)。この取り込みは、単輸送(促進された分散)、プロトンシンポートまたはプロトンアンチポートによって仲介されている可能性がある。
【0064】
【図44】線虫(C. elegans)、マウス、ラット、ヒト、シロイヌナズナ、コメ、ウマゴヤシおよびペチュニアにおける多様なGLUEタンパク質のアミノ酸配列を示す。
【発明を実施するための形態】
【0065】
詳細な説明
下記説明は、本発明の理解に有用であり得る情報を含む。本明細書において提供するあらゆる情報は、先行技術または明らかに特許請求の範囲に記載の発明と関連していること、あるいはいずれかの文献が具体的または暗示的に先行技術であることを認めるものではない。
【0066】
本発明の他の目的、利点および特徴は、本明細書において提供される明細書および図面を検討することによって、当業者に明らかとなる。したがって、本発明のさらなる目的および利点は、以下の説明から明らかである。
【0067】
植物は種子および花粉発生支援、花蜜産生ならびに根圏における有益微生物養育のために糖流出トランスポーターを必要とする。しかし、これらの流出トランスポーターの同一性は、不明なままであった。光学的糖センサーを用いて、植物から糖トランスポーターの新規クラス(GLUE、Glue、またはSWEET)が同定された。シロイヌナズナおよびコメGLUEは、糖のインポーターおよびエクスポーターとして機能する。OsGluE11/Os8N3およびAtGluE8/RPG1は花粉生存に必要とされる。花蜜および根粒におけるGLUEホモログの発現は、受粉媒介者および共生生物の給餌における役割を示唆している。真菌および細菌病原体は、異なるGLUEメンバーのmRNAレベルを調節して、単糖、二糖またはオリゴ糖トランスポーター活性をともに選択している。OsGLUE11/Os8N3は斑点細菌病の宿主感受性因子として機能し、植物および病原体両方の栄養摂取とGLUEを結びつける。
【0068】
ヒトゲノムは少なくとも2つのクラスのグルコーストランスポーター、SLC2およびSLC5を含む。GLUTと呼ばれるSLC2は単輸送体であり、すなわちそれは濃度勾配に沿ってグルコースを輸送する。逆にSGLTは、ナトリウム勾配によって駆動されてグルコースを能動的に輸送できる、Na+共役コトランスポーターである。これらのトランスポーターは、ヒトにおいてみられる取込活動のほとんどを説明することができ、例えばGLUT2ノックアウトマウス変異体は、劇的に低下した取り込み能を示すが、驚くべきことに細胞流出はそうではない。しかし、バイオインフォマティック分析により、動物およびヒトゲノムが溶質輸送体ファミリーSLC50として登録されているSWEETのホモログを含むことが示された。線虫(C. elegans)ゲノムは糖流出トランスポーターの新規クラス(SLC50)の7種のホモログを含むが、ヒトゲノムはRAG1AP1と呼ばれる1種のホモログを有する。
【0069】
糖流出は多細胞生物において、および生物間の相互作用において、炭素骨格とエネルギーの細胞性交換に必要とされる必須プロセスである。タペータムまたは花柱の輸送管からの糖流出は、花粉発生および後の花粉管成長を刺激する。花は受粉媒介者を誘引するため花蜜産生のために糖を分泌し、そして植物は、潜在的には有益微生物に給餌するために根圏に炭水化物を分泌する(T. Bisseling et al., Science 324, 691 (2009))。糖流出輸送体は、葉肉および種皮を含む多くの他の部位で必要とされる(Y. Zhou et al., Plant J. 49, 750 (2007))。流出トランスポーターの分子の性質は知られていない(S. Lalonde et al., Annu. Rev. Plant Biol. 55, 341 (2004))。植物由来の糖はまた、病原体のための基質を提供する。病原体の第一次的ゴールは、効率的に繁殖するために宿主植物から栄養分を摂取することである。シュードモナス(Pseudomonas)属およびザントモナス(Xanthomonas)属の植物病原性細菌は、植物組織の細胞外空間(アポプラズム)に生息でき、ここでそれらはエネルギー源および炭素骨格源として炭水化物を獲得する。成功した病原体は、かかるメカニズムを共に選択して栄養素流動を変化させる可能性がある(J. W. Patrick, Aust. J. Plant Physiol. 16, 53 (1989))。結果として、病原体と植物は、植物が栄養素への病原体のアクセスを制限しようとし、また防御戦略を開始する一方で、病原体が栄養素へのアクセスを得ようとし、宿主免疫を抑制する戦略を考え出す進化的主導権争いに突入する。植物の防御を変化させるための病原体によって用いられるメカニズムに対する洞察が現在出現しつつあるが;どのようにして病原体が、病原体の増殖を支援するために宿主の生理機能、特に糖排出を変化させるのかについて、わずかしか知られていない。本発明は、小胞または細胞膜で糖を分泌するトランスポーターの存在を突き止めた。本発明はさらに、これらの植物流出トランスポーターが病原体によって栄養素の要求を支給するために「共に選択」されることを同定した(J. W. Patrick, Aust. J. Plant Physiol. 16, 53 (1989))。少なくともコムギウドンコ病の場合、グルコースは植物宿主から病原体に移送される主な糖である(J. Aked, J. L. Hall, New Phytol. 123, 271 (1993); P. N. Sutton et al., Planta 208, 426 (1999);およびP. N. Sutton et al., Physiol. Plant. 129, 787 (2007))。それぞれの病原体グルコース/H+コトランスポーターが同定された(R. T. Voegele et al., Proc. Natl. Acad. Sci. USA 98, 8133 (2001));反対に、植物糖流出メカニズムはもとより不明のままであった。
【0070】
多くの代謝経路において、トランスポーターは、基質を酵素に供給し、酵素の存在から各々の産物を除去する具体的な経路のいずれかまたは両方の終点で機能しうる。輸送は受動輸送、能動輸送、分散または浸透作用によりうる。トランスポーターは、酵素での基質の存否に直接または間接的に関与しうる。トランスポーターは細胞膜内またはその近くに局在してよく、あるいはそれらは細胞基質またはその近くまたは他の細胞小器官、例えば小胞体、ミトコンドリア、葉緑体、ペルオキシソーム、ゴルジ装置、ベシクル、核膜または液胞、リソソームまたは細胞膜中に局在していてよい。
【0071】
トランスポーターは固定されていてもよく、それによるかまたはそれを通過する基質の通過を可能とするか、あるいは基質と結合して基質を特定の細胞内目的地に物理的にシャトルする。トランスポーターは、別のタイプの分子の通過または輸送を可能とする一つのタイプの分子と結合できる。トランスポーターは独立して、またはプロテインキナーゼもしくはATP切断ドメインのような他のタンパク質の作用を介して移動してよい。
【0072】
トランスポーターは、細胞または生物内へのまたは外への物質の取り込みまたは放出を決定し、そしてトランスポーターは、細胞間の物質の輸送および分散を制御する。トランスポーターはまた、細胞小器官間、例えば核の中と外での輸送のように、細胞間でも機能しうる。トランスポーターはしばしば代謝経路の始点および終点に存在するため、それらは基本的により高次の制御機能を担当している。トランスポーターは、放出された低分子、例えばモノアミンまたは神経伝達物質の再取り込みに関与している可能性がある。
【0073】
いくつかのトランスポーターは、特定の基質を輸送するためにエネルギーを必要とする。ある例では、エネルギーはATPを介して供給され、得られたトランスポーターのリン酸化は、輸送を進めることができるコンホメーション変化を引き起こす。別の場合では、輸送体と第二基質のカップリング、例えばPまたはVタイプATPaseによって作出されるプロトンまたはナトリウム/カリウム勾配を介して間接的に、エネルギーは供給される。他の例では、トランスポーターと他のタンパク質または分子の相互作用が、同じタンパク質もしくは分子または異なるタンパク質もしくは分子の輸送を進めることができるコンホメーション変化を引き起こす。さらなる例では、トランスポーターと制御タンパク質間の分離が、輸送を進めることができるトランスポーターのコンホメーション変化を引き起こす。トランスポーターは直接結合を介して基質または産物と相互作用しうる。輸送の触媒において、トランスポーターはコンホメーション変化を引き起こしうる。これはまた、PタイプまたはVタイプATPaseのような酵素およびトランスポーターとして使用されるハイブリッドタンパク質を含む。
【0074】
本明細書において使用するとき、用語「コンホメーション変化」は、センサーにおいて生じるいずれかの立体配置の変化に対する影響、例えばタンパク質中の原子および原子団の三次元位置、移動可能な原子および側鎖の平均位置、タンパク質の表面特性の変化、シングルまたは相対位置の位置/平均位置またはコンホメーションに影響するドメインのドメインフォールディング/アンフォールディングの移動、フルオロフォアの平均位置が変化して、当該検出部分によって放出されるエネルギーの変化をがもたらされることを意味する。用語「相対位置」は、2つの検出部分が互いに有しうる空間的関係、例えば距離および方向のいずれかの可能性のある種類を意味する。例えばコンホメーションは、1または複数個の原子、側鎖またはドメインの回転、酵素の折り畳み、外側へのドメインの一方または両方のねじれ、あるいはこれらの移動のいずれかの組合せによって変化してよい。検出部分間の方向または距離が変化されているコンホメーション変化、またはレポーター要素のコンホメーションに対する効果を示す変化が有用である。あるいは、酵素部分におけるコンホメーション変化は、シングル結合フルオロフォアの特性に作用する。この場合、RETシグナルを得るために第二フルオロフォアが使用されうる。かかる場合において、結合の前または結合により、少なくとも最大の半分のエネルギー移動が生じるように検出部分が向けられていることが有利である。
【0075】
本明細書において使用するとき、「リガンド」は、ペリプラズム結合タンパク質のようなタンパク質と結合してそのタンパク質との複合体を形成できる分子または物質を意味する。リガンドとタンパク質の結合は、タンパク質の形状、特に3次元および4次元構造を歪めまたは変化しうる。リガンドは基質であってもよい。基質は遊離体または酵素の支援触媒作用を介して産物に変換される物質を含みうる。リガンドは内因性リガンドのアナログまたは誘導体であってよい。リガンドは結合部位について内因性リガンドと競合しうる。リガンドは例えば、低分子、化学物質、一本鎖オリゴヌクレオチド、二本鎖オリゴヌクレオチド、DNA、RNAまたはポリペプチドであってよい。リガンドはタンパク質と結合するいずれかの化学物質、例えばマグネシウムのようなイオンまたはアロステリック因子または別のタンパク質を含む。リガンドはグルコースのような糖であってよい。
【0076】
本明細書において使用するとき、「蛍光インジケーター」は、PBPと結合した蛍光ドメインまたは化合物を意味する。PBPの形状の変化は、蛍光ドメインまたは化合物の蛍光の変化をもたらし、それによって酵素の形状の変化が示される。ドメインは蛍光タンパク質であってよい。蛍光ドメインは2個のサブドメイン、例えばドナーおよびアクセプターフルオロフォアを含んでいてよい。いくつかの例において、PBPはドナーとアクセプターフルオロフォア間で共有結合されている。蛍光インジケーターの別の使用は、発光またはリン光性分子、ならびにNMR、偏極検出器のような他の手段によって検出されうる化合物である。
【0077】
タンパク質およびポリペプチドに関して本明細書において使用するとき、用語「組換え」は、遺伝子操作によって、例えば非天然核酸の細胞での翻訳によって作成されまたは由来するか、あるいは人工手段またはメカニズムによって組み立てられるタンパク質および/またはポリペプチドおよび/またはペプチドを含みうる。
【0078】
本明細書において使用するとき、「融合」は、本発明に従ってそれらがおかれる順序または文脈において、互いに関連した天然に見られない配列を含む核酸およびポリペプチドを意味しうる。融合核酸またはポリペプチドはその全体において核酸またはポリペプチドの天然配列を必ずしも含んでいる必要がない。融合タンパク質は通常のペプチド結合を介して一体に結合した2個以上のセグメントを有する。融合核酸は通常のホスホジエステル結合を介して一体に結合した2個以上のセグメントを有する。
【0079】
本明細書において使用するとき、用語「生物学的サンプル」は、細胞または細胞内物質の集合体を意味する。サンプルは生物または生物の成分(例えば細胞)から得られる。サンプルはいずれかの生物学的組織または液体から得られる。サンプルは対象由来のサンプルであり得る。対象は患者であってよい。サンプルは患者または患者の成分から得られる。対象は動物であってもよい。動物は哺乳類、例えばヒトまたはヒト患者であってよい。かかるサンプルは、唾液、血液、血球細胞(例えば白血球および赤血球)、組織もしくは生検サンプル(例えば腫瘍生検)、尿、腹腔液および胸膜液またはそれら由来の細胞を含むが、これらに限定されない。生物学的サンプルはまた、組織学的目的で採取した凍結切片のような組織の切片を含んでもよい。生物学的サンプルはまた、インビトロ細胞培養物を含んでもよい。細胞培養物は不死化細胞系または一次細胞系であってもよい。細胞培養物は異なる細胞タイプを含んでいてもよい。
【0080】
本明細書において使用するとき、用語「dsRNA」は二本鎖RNAを意味し、ここでdsRNAは2本の別個のストランドまたは一本鎖ヘアピンによる二本鎖であってよい。dsRNAは標的遺伝子のヌクレオチドと類似するヌクレオチド配列を含んでいてよい。dsRNAは、ヘアピンRNAのような自己相補的dsRNAまたは別個の相補的RNAストランドによって形成されるdsRNAを形成する転写産物および/またはインビボでsiRNAを産生できる転写産物を産生可能な発現ベクター(RNAi発現ベクターとも称される)によって作成できる。ベクターは(1)遺伝子発現において制御の役割を有する遺伝子因子(複数の因子)、例えばプロモーター、オペレーターまたはエンハンサー、それと作動可能に連結した、(2)転写されて二本鎖RNA(細胞内でアニールしてsiRNAを形成する2個のRNA部分、またはsiRNAに処理されうる一本のヘアピンRNA)を産生する「コード」配列、ならびに(3)適切な転写開始および終止配列を含んでなる転写ユニットを含みうる。プロモーターおよび他の制御要素の選択は、一般に、意図する宿主細胞に従って変化する。一般に、組換えDNA技術において有用な発現ベクターは、しばしばベクター形態が染色体と結合しない環状二本鎖DNAループを意味する「プラスミド」の形態で存在する。
【0081】
本明細書において使用するとき、用語「単離された」は、巨大分子の天然源に存在する他の細胞/組織構成物(例えばDNAまたはRNA)から分離された分子を意味する。用語「単離された」は、本明細書において使用するとき、組換えDNA技術で産生された場合には細胞性物質、ウイルス性物質および培養培地を実質的に含まない、あるいは化学的に合成された場合には化学的前駆体または他の化学物質を実質的に含まない核酸またはタンパク質を意味する。さらに、「単離された核酸」は、フラグメントとして天然には生じない、天然状態では見られない核酸フラグメントを含んでいてもよい。
【0082】
本明細書において使用するとき、用語「多量体」は、2個以上の異なる分子間の多量体複合体の形成を意味する。多量体複合体は、例えば同じタンパク質の2個以上の分子(例えばホモ二量体、三量体、四量体、二量体の二量体または高次多量体)または2個以上の異なる(すなわち非同一)タンパク質の混合物(例えばヘテロ二量体、異なる二量体の二量体、三量体、四量体または高次多量体)を含んでいてよい。例えば多量体抗体は、同じ抗体または2個以上の異なる抗体を含んでいてよく、これらはそれぞれ、2種以上の機能または活性を有している(例えば2種以上のエピトープと結合する)。
【0083】
本明細書において使用するとき、「対象」は、本発明のレシピエントを含みうる。対象は植物または植物の構成要素、例えば植物の器官または小器官であってよい。対象は脊椎動物を含むいずれかの動物であってよい。対象は多くの場合、好ましくはヒトであるが、家畜、実験動物(例えばラットまたはマウスのようなげっ歯類を含むがこれらに限定されない)またはペットであってもよい。
【0084】
本明細書において使用するとき、用語「変異体」は、BLAST分析によって天然分子と少なくとも約70%、より好ましくは少なくとも75%同一性、例えば少なくとも80%、90%、95%、96%、97%、98%、99%またはそれ以上の同一性を有するポリペプチドを意味する。多くのかかる変異体は当該技術分野において既知であるか、あるいは天然蛍光分子のランダムまたは特異的突然変異誘発によって容易に製造できる(例えばFradkov et al., FEBS Lett. 479:127-130 (2000)参照)。
【0085】
本明細書において使用するとき、用語「プラスミド」および「ベクター」は、プラスミドが最も一般的に使用されるベクターの形態であるから、相互に交換可能なように使用される。しかし、本発明は同等の機能を有し、この後当該技術分野において既知となるかかる他の形態の発現ベクターを含むことを意図する。ベクターは、異なる遺伝子環境間での輸送または宿主細胞における発現のために制限およびライゲーションによって所望の配列が挿入されていてよい任意の数の核酸であってよい。ベクターは典型的にはDNAから成るが、RNAベクターも利用可能である。ベクターはプラスミドおよびファージミドを含むがこれらに限定されない。クローニングベクターは、宿主細胞で複製でき、決定可能なようにベクターを切断できて、新たな組換えベクターが宿主細胞で複製できる能力を保持するように所望のDNA配列をライゲーションできる1個以上のエンドヌクレアーゼ制限部位によってさらに特徴付けられるものである。プラスミドの場合、プラスミドが宿主細菌内でコピー数を増加するように多くの回数、あるい宿主が有糸分裂によって増殖する前に宿主当たり丁度1回、所望の配列の複製が行われうる。ファージの場合、複製は溶解期に能動的にまたは溶原期に受動的に行われうる。
【0086】
ベクターはさらに、プロモーター配列を含んでいてもよい。プロモーターは、核酸の転写を開始する部位を含むコード領域の通常上流に位置する非翻訳核酸配列を含みうる。プロモーター領域はまた、遺伝子発現の制御因子として作用する他の要素も含んでいてよい。本発明のさらなる態様において、発現ベクターは、組み込まれた発現ベクターが有する細胞の選択に利用するためのさらなる領域を含む。プロモーター配列は、転写開始部位によって3’末端でしばしば結合されており(包括的に)、バックグラウンド以上の検出可能なレベルで転写を開始するために必要な最小数の塩基または要素を含むように上流(5’方向)に伸長する。プロモーター配列内で転写開始部位およびRNAポリメラーゼの結合に関与するタンパク質結合ドメインが見られる。真核プロモーターはしばしば、常ではないが、「TATA」ボックスおよび「CAT」ボックスを含む。プロモーターの活性化は、例えばある組織においてのみ発現される転写因子によってある細胞または組織に特異的であってよく、あるいはプロモーターは偏在してほとんどの細胞または組織で発現可能であってよい。
【0087】
ベクターはさらに、ベクターで形質転換またはトランスフェクトされた細胞の同定および選択に使用するのに好適な1種以上のマーカー配列を含んでいてよい。マーカーは例えば、抗生物質または他の化合物に対する耐性または感受性を増減するタンパク質をコードする遺伝子、活性が当該技術分野において既知の標準的なアッセイによって検出可能な酵素をコードする遺伝子(例えばβガラクトシダーゼまたはアルカリホスファターゼ)ならびに形質転換またはトランスフェクトされた細胞、宿主、コロニーまたはプラークの表現型に視覚的に影響を与える遺伝子を含む。ベクターは、それが作動可能に結合しているDNAセグメントに存在する構造遺伝子産物の自己複製および発現が可能なものであってもよい。
【0088】
発現ベクターは、制御配列と作動可能に連結または作動可能に結合されてRNA転写産物として発現されうるように所望の核酸配列を制限およびライゲーションによって挿入できるものである。発現は、細胞での内因性遺伝子、トランスジーンまたはコード領域の転写および/または翻訳を意味する。
【0089】
コード配列および制御配列は、コード配列の発現または転写が制御配列の影響または制御下におかれているようにそれらが共有結合している場合に、作動可能に連結されている。コード配列が機能的タンパク質に翻訳されることが望まれる場合、5’制御配列におけるプロモーターの誘導がコード配列の転写をもたらすとき、そして2個のDNA配列間の結合の性質が(1)フレームシフト変異の導入をもたらさない、(2)コード配列の転写を指定するプロモーター領域の能力と干渉しない、あるいは(3)タンパク質へと翻訳される対応するRNA転写産物の能力と干渉しないとき、2個のDNA配列は作動可能に結合されていると言われる。したがって、得られる転写産物が所望のタンパク質またはポリペプチドに翻訳されうるようにプロモーター領域がDNA配列の転写に影響できる場合、プロモーター領域はコード配列と作動可能に結合されている。
【0090】
本発明のいくつかの局面は、核酸の形質転換および/またはトランスフェクションを含む。形質転換は、原核細胞の内部への外因性または異種性核酸の導入である。トランスフェクションは、真核細胞の内部への外因性または異種性核酸の導入である。形質転換またはトランスフェクトする核酸は、細胞のゲノムを構成する染色体DNAに組み込まれ(共有結合)ても組み込まれなくてもよい。原核生物において、例えば、形質転換する核酸は、プラスミドまたはウイルス性ベクターのようなエピソーム要素で維持されてよい。真核細胞について、安定的にトランスフェクトされた細胞は、染色体複製を介して娘細胞によって受け継がれるように染色体にトランスフェクトする核酸が組み込まれているものである。この安定性は、トランスフェクトする核酸を含む娘細胞の集団からなる細胞系またはクローンを確立する真核細胞の能力によって示される。
【0091】
本明細書において使用するとき、用語「融合タンパク質」または「キメラタンパク質」は、直接またはスペーサーアミノ酸を用いて一体に融合している少なくとも2個のポリペプチドを含むポリペプチドを意味するものとして使用される。融合ポリペプチドは、融合タンパク質の全体的な機能において協同的または反対の役割を有しうる。
【0092】
本明細書において使用するとき、抗体の「フラグメント」は、Fc、Fab、Fab’、F(ab’)2および一本鎖免疫グロブリンを含むがこれらに限定されない。
【0093】
本明細書において使用するとき、免疫学的有効量は、単回投与または一連の一部のいずれかにおいて疾患または障害の処置に有効である対象への投与量を意味する。この量は処置する対象の健康および身体状態、処置する対象の種(例えば非ヒト哺乳類、霊長類など)、抗体を合成する対象の免疫系の能力、望まれる保護の程度、ワクチンの形成および他の関連する要因に依存して変化する。この量は日常的な試験を通じて決定されうる相対的に広い範囲に含まれると予想される。
【0094】
本明細書において使用するとき、「医薬組成物」または「製剤」は、薬学的に許容される担体または希釈剤と共に薬物または化合物を含む組成物を意味する。薬学的に許容される担体は、生理食塩水、リンゲル液、リン酸緩衝化食塩水および当該技術分野において既知の他の担体を含むが、これらに限定されない。医薬組成物はまた、安定化剤、抗酸化剤、着色剤および希釈剤を含んでいてもよい。薬学的に許容される担体および添加剤は、医薬製剤からもたらされる副作用が最小限となり、処置が不奏効となる程度に薬物のパフォーマンスを没却または阻害しないように、選択される。
【0095】
本明細書において使用するとき、「治療上有効量」は、それを必要とする対象に投与したとき処置を達成するために必要な薬物または化合物の量を意味する。「治療上有効量」を構成する架橋Aβオリゴマー反応性抗体のような抗体の量は、状態または疾患の重症度ならびに処置する対象の年齢および体重に依存して変化するが、当業者によってその知識および本開示に基づいて日常的に決定されうる。
【0096】
「コファクター」は、タンパク質と相互作用して、その生理的な機能の実行において当該タンパク質を補助する要素を意味する。コファクターは反応を触媒しうる。コファクターは、強い相互作用または緩いつながりによってタンパク質、例えばトランスポーターまたは酵素と結合しうる。コファクターは「コエンザイム」または「補欠分子族」であってもよい。「コエンザイム」は、酵素間で化学団をシャトルまたは輸送する有機分子を意味する。「補欠分子族」は酵素と結合して酵素の一部となるコファクターを意味する。コファクターは、金属イオン、例えばカルシウム、マグネシウム、マンガン、鉄、カリウム、ナトリウム、アルミニウム、銅、ニッケル、セレン、モリブデンまたは亜鉛であってもよい。コファクターの制限された供給は、速度制限因子でありうる。
【0097】
用語「生物学的液体」は、循環タンパク質を含む体液、例えば血漿、血清および全血、唾液、脳脊髄液、羊水、滑液、水性体液、胆汁、耳垢、カウパー腺液、乳び、女性の射精および膣潤滑液、間質液、リンパ液、月経液、粘液、胸水、膿、皮脂、精液、汗、涙液、嘔吐物、尿、乳汁および他の分泌物を含む。生物学的液体のタンパク質含有抽出物は、生物学的液体から回収または分離されるいずれかの調製物、例えば免疫グロブリン分画である。本発明に使用できる血液、血清または血漿は、個体から新たに得ることができ、あるいは血液バンクまたは他の血液回収組織から得た貯蔵された血液または血漿調製物のような供給源から得てもよい。本発明の目的のために、血液、血清または血漿は、期限切れの回収物または血液バンクまたは血液回収組織によって基準を満たさないとされた他のもの由来であってもよい。本発明の同一のプロセスは、動物血液に適用してもよく、獣医学的医薬に関する目的のために同様の動物抗体を得ることができる。液体はそれを得たそのままの状態で使用してよく、あるいは例えば沈降させるか遠心分離してさらに処理してもよい。液体は植物由来であってもよく、例えば樹液、師部樹液、木部樹液、花蜜、樹脂、ラテックスまたはオイルであってよい。液体は上清、回収した沈降物または外部重力、例えば遠心分離またはろ過によって得たペレットであってよい。
【0098】
本発明は、部分的に、糖トランスポータータンパク質の新規クラスの発見に基づく。糖は単糖、二糖またはオリゴ糖であってよく、例えばグルコース、フルクトース、リボース、ラクトース、ガラクトース、アラビノース、マルトース、アミロース、セルロースまたはショ糖であってよい。トランスポーターは細胞の膜を通過して糖を輸送できる。膜は細胞膜または細胞壁であってよい。膜は細胞の細胞基質を取り巻きうる。膜は細胞小器官、例えばミトコンドリア、小胞体、ゴルジ装置、核、エンドソームまたは液胞を取り巻いてもよい。トランスポーターはある細胞/細胞小器官の膜と別の細胞/細胞小器官の膜の間で糖を輸送できる。
【0099】
本発明は、臓器内または対象の臓器間で糖を輸送する方法を提供できる。臓器は炭水化物、例えば糖およびグルコースの処理、取込または排出に関与しうる。臓器は消化系に関与していてもよい。例えば臓器は、腸(大腸または小腸)、胃または肝臓であってよい。本発明のトランスポーターは、臓器で細胞の内部でのまたは外部への糖の輸送を補助できる。トランスポーターは糖の移動に作用することが知られている他のタンパク質、例えばSGLT、GLUT1およびGLUT2と共同で働きうる。トランスポーターは細胞小器官の外へと糖を流出しうる。トランスポーターは小胞の形成を介して受動的に糖を流出しうる。例えば、ゴルジ装置におけるトランスポーターの発現は、形成された小胞に糖を流出させ、次いでその内容物を細胞外へ移動する。
【0100】
最初はグルコース単輸送体GLUT2が肝臓および腸におけるグルコースの取込および流出の両方に関与していると考えられた。しかし、肝細胞およびトランスジェニックマウスにおけるGLUT2のノックダウンは、GLUT2がグルコース取込に必須であるが、グルコース流出にはそうではないことを示した。GLUT2ノックアウトマウスの経口グルコース負荷は、血中に現れるグルコースの正常な速度をもたらした(Thorens et al.. J. Biol. Chem. 275, 23751-23758, (2000))。同様に、GLUT2対立遺伝子の両方の不活性化によって引き起こされる症候群であるFanconi-Bickel症候群(Santer et al. Nat. Genet. 17, 324-326, (1997))に罹患したヒトは、腸細胞からの流出を必要とするプロセス(Manz, F. et al. Pediatr Nephrol 1, 509-518, (1987))である異常な炭水化物摂取を導かなかった。その機能に基づいて、HsSWEET1/RAG1AP1は肝臓からグルコースのような糖の流出を補助しうる。
【0101】
トランスポーター、例えばGLUEトランスポーターは膜に位置しうる。トランスポーターは膜を貫通しうる。トランスポーターは膜を貫通する孔を含みうる。糖輸送はトランスポーターによって作り出される膜の孔を介しうる。孔は刺激に応答して幅を変えて、それによって糖が孔を通過できるように変化しうる。トランスポーターはゲートであってよい。ゲート機構は刺激に応答してトランスポーターを糖が通過できる。ゲート機構は結合するリガンドを必要とするか、あるいは電圧センサーを含みうる。トランスポーターは受動拡散によって糖を通過させうる。トランスポーターは糖を輸送するためにアデノシン三リン酸のようなエネルギーを必要としうる。トランスポーターは平衡に達するように高濃度から低濃度へと糖を輸送しうる。トランスポーターは低濃度の領域から高濃度の領域へと糖を輸送し、それによって勾配を増加させうる。トランスポーターは単輸送体であってもよい。単輸送体は、本明細書において使用するとき、促進因子として機能するトランスポーターを意味し、トランスポーターの向きは輸送される基質の膜内外の勾配または濃度差によって決定される。単輸送体は、共輸送分子または活性化分子のようなコファクターに頼ることなく自己充足的に作動できるトランスポーターを意味する。単輸送体は典型的には、濃度勾配の方向に、すなわち高濃度の糖を有する側から低濃度の糖を有する側へと、糖の流出を可能とする。
【0102】
トランスポーター、例えばGLUEトランスポーターは、膜の片側での糖濃度を上昇させうる。トランスポーターは細胞内の糖レベルを上昇させうる。これは、イオンまたは他の代謝産物、例えばプロトン、ヒドロキシアニオン、ナトリウムまたはカリウムでありうる第二化合物の輸送とその輸送の共役を介して達成されうる。共役は、共輸送もしくはアンチポートによるか、またはピンポン機構によるものでありうる。トランスポーターは細胞内の糖濃度を低下させうる。トランスポーターは細胞から糖を排出しうる。トランスポーターは細胞に糖を取り込みうる。
【0103】
トランスポーターはさらに、キナーゼ、第二メッセンジャー、アニオン、カチオンまたはリガンドのようなシグナルに影響されることがある。トランスポーターはコファクターに影響されることがある。コファクターは、マグネシウム、亜鉛、鉄、銅、ヨウ素、塩素、ナトリウム、カリウム、カルシウム、マグネシウム、硫酸塩、サルフェート、アンモニウム、硝酸塩、亜硝酸塩、カルボネート、カルボン酸またはリン酸塩のイオン化形態であってよい。コファクターは、トランスポーターまたは第二メッセンジャーと基質またはリガンドの結合を補助するために必要であり得る。コファクターはトランスポーターの活性に必要であり得る。いくつかの例において、コファクターの添加がな場合には、トランスポーターの機能喪失または低下が生じる。他の例において、コファクターの存在はトランスポーターの活性をダウンレギュレートする。トランスポーターはクラスリン内在化機構を介するような内在化を介してダウンレギュレートされうる。
【0104】
トランスポーターは細胞または細胞から得られた抽出物中に存在しうる。細胞は原核細胞であってよい。細胞は真核細胞であってもよい。
【0105】
細胞は動物またはその一部(rt)に存在していてよい。トランスポーターは動物細胞の小胞体、ゴルジ装置、ベシクルまたは細胞膜におけるグルコース輸送に関与しうる。細胞はグルコース輸送または分泌に関与する動物細胞であってよい。細胞は腺細胞であってよい。腺細胞は乳腺の肺胞細胞であってもよい。
【0106】
細胞は植物またはその一部、例えば根、幹、葉、種子、花、果実、葯、蜜腺、子房、花弁、タペータム、木部または師部に存在していてよい。例示として、植物は、有胚植物、コケ植物、種子植物(spermatophye)、ネマトファイト(nematophyte)、管束植物、ダイズ、コメ、トマト、アルファルファ、ジャガイモ、エンドウ、イネ科植物、ハーブ、樹木、藻類、蘚類、菌類、つる植物、シダ類、低木、オオムギ、コムギ、ホップ、トウモロコシ、レタス、オレンジ、モモ、柑橘類、レモン、ライム、ココナッツ、ヤシ、マツ、オーク、シーダー、マンゴー、パイナップル、ダイオウ、イチゴ、ブラックベリー、ブラックカラント、ブルーベリー、ラズベリー、キウイ、ブドウ、ルタバガ、パースニップ、サツマイモ、カブ、マッシュルーム(菌類)、コショウ、シラントロ、タマネギ、ニラネギ、フェンネル、クローブ、アボカドまたはキュウリを含む。またススキ属(Miscanthus)またはスイッチグラス、ポプラ、モロコシ属(Sorghum)およびヤマカモジグサ属(Brachypodium)のようなバイオ燃料作物も含む。
【0107】
本発明のトランスポーター、例えばGLUEトランスポーターは、植物または動物のような生物内の糖、特に単糖類、二糖類またはオリゴ糖、例えばグルコースまたはショ糖を輸送できる。植物において、トランスポーターは、花蜜の産生のために糖を輸送しうる。トランスポーターは、植物の蜜腺におよび/またはそれから糖を輸送しうる。本発明のトランスポーターは、蜜腺に局在化されうる。本明細書において使用するとき、「蜜腺」は、花蜜を産生する分泌構造を意味する。花蜜はグルコースおよび/またはフルクトースおよびまたは他の糖類を含む、受粉媒介者への報償として利用されうる組成物である。
【0108】
トランスポーターは植物の葯におよび/またはそれから糖を輸送しうる。葯は、雄ずいおよびフィラメントからなる植物の生殖器官を意味する。本発明のトランスポーターは、葯に局在化されていてもよい。本発明のトランスポーターは、葯の雄ずいおよび/またはフィラメントに局在化されていてもよい。それはタペータムまたは花粉それ自体に局在しうる。花粉において、それは栄養細胞または雄原細胞に局在しうる。本発明のトランスポーターの存在は、葯の機能に影響を与えうる。タペータムにおけるトランスポーターの機能は、花粉の栄養に関与しうる。花粉における機能は、雄原細胞への栄養補給を補助しうる。葯における機能は、デンプンの突然の加水分解を引き起こし、これは浸透能の増加を導き、次いで周囲組織からの水の吸い戻しを導き、葯の脱水および開裂を促進しうる。この機能はまた、花粉への糖の取込または輸送管もしくは精巣上体管における糖の放出に寄与しうる。
【0109】
本発明のトランスポーターは、微小胞子嚢または巨大胞子嚢のような植物の胞子嚢におよび/またはそれから糖を輸送しうる。本発明のトランスポーターは、胞子嚢または生殖腺を放出する胞子に局在化されていてもよい。トランスポーターは胞子嚢の機能に影響を与えうる。
【0110】
本発明のトランスポーターは、輸送管におよび/またはそれから糖を輸送して、伸長花粉管に栄養およびエネルギーを供給できる。
【0111】
本発明のトランスポーターは、細胞膜を通過する取込およびERへの「流出」を仲介しうる。本発明のトランスポーターは、輸送の方向が膜内外のグルコース勾配のみに依存するグルコース単輸送体として機能する。
【0112】
トランスポーター、例えばGLUEトランスポーターは、師部細胞の発生を誘導しうる。トランスポーターは師部に局在化されていてもよい。トランスポーターは師部細胞の機能に影響しうる。師部は、細胞における栄養、例えば樹液の輸送に関与する組織を意味する。師部はある領域、例えばねまたは植物の糖供給源から植物の別の領域、例えば葉または植物の糖貯蔵部へと栄養素を輸送しうる。師部に沿った輸送は、多方向性または一方向性でありうる。師部は柔組織細胞、師細胞、葉肉細胞および伴細胞、例えば常伴細胞、転送細胞および中間細胞を含みうる。師部はさらに、タンパク細胞、繊維および厚膜細胞を含みうる。
【0113】
本発明のトランスポーターは、植物または動物のような生物の疾患感受性に影響を与えうる。本発明のトランスポーターは、ウイルスまたは細菌または昆虫のような病原体に対する植物の感受性に影響を与えうる。病原体は宿主細胞の遺伝子転写に影響を与えうることが知られている。病原体は糖トランスポーターの遺伝子転写に影響を与えうる。本発明は、宿主細胞を保護する新規方法を提供する。それは地上および地下の共生生物および病原体両方の栄養に影響を与えうる。それは微生物を誘引しうる。それは土壌への糖の分泌に関与しうる。したがって、根の周辺の微生物相および植物の生産性に影響を与えうる。それは受粉媒介者との相互作用に影響を与えうる。それは外部の供給に依存する植物の細胞、例えば表皮、孔辺細胞、種子への糖の供給に影響を与えうる。
【0114】
トランスポータータンパク質
【0115】
本発明は、膜を通過する糖の移動において集合的に機能するポリペプチドの連続配列を提供する。トランスポーターは、膜に埋め込まれるかまたは完全に膜を貫通していてよい。トランスポーターは複数回、例えば2、3、4、5、6、7またはそれ以上膜を貫通していてよい。当業者は、膜を通過するトランスポーターの位置が、膜の外側(いずれかの側、例えば細胞外または細胞内)に位置するトランスポーターの部分と比較して疎水性および親水性で変化すると理解する。トランスポーターは少なくとも2個のサブユニット、例えば2個の膜貫通タンパク質、例えばホモまたはヘテロトリマーを含んでいてよく、ここで用語「トリマー」はタンパク質が膜を貫通する回数を意味する。さらなる例において、サブユニットはリンカーペプチド、例えばさらなる細胞内ドメインまたはさらなる膜貫通ドメインによって結合されていてもよい。
【0116】
トランスポーターはポアを形成しうる。ポアはトランスポーターの膜貫通ドメインまたはそのサブドメインの球状配置によって形成されうる。ポアはそれを介した糖の通過を可能としうる。ポアは糖の移動についてのみ選択的でありうる。ポアは、特定のサイズまたは特定の形状の分子の移動を制限する選択点または複数の点を有していてもよい。ポアを通る移動は、濃度勾配のみに基づきうる。ポアはさらに、コファクターの活性、例えば相互作用タンパク質の活性またはイオンの結合もしくは電荷の存在、例えば負または正の電荷に基づいて、開閉しうる。
【0117】
本発明はまた、キメラトランスポーターも提供する。キメラは、2種以上の異なるタンパク質に由来する機能的ドメインの組合せである。キメラトランスポーターは他の膜関連タンパク質、例えばチロシンキナーゼ、Gタンパク共役受容体、アクアポリンまたは他のトランスポーター由来のドメインと相互作用/採用する第二メッセンジャーと、トランスポーターのポアを融合しうる。キメラトランスポーターは、本明細書に記載の2種以上のトランスポーターの融合体であってもよい。例として、トランスポーターは、グルコーストランスポーター、グルタミン酸トランスポーター(ナトリウム依存的および小胞)、アクアポリン、Na/K ATPase、セロトニントランスポーター(SERT)、ドーパミントランスポーター(DAT)、ノルエピネフリントランスポーター(NET)、アンモニウムトランスポーターおよびカリウムチャネルを含みうる。
【0118】
本発明はまた、トランスポーターの融合タンパク質を提供する。例えば、既知のエピトープまたはタグをトランスポーターと融合してよい。タグは蛍光タグ、例えば緑色蛍光タンパク質、赤色蛍光タンパク質、オレンジ色蛍光タンパク質、黄色蛍光タンパク質、シアン色蛍光タンパク質または青色蛍光タンパク質であってよい。融合タンパク質を製造する方法は当該技術分野において既知である。
【0119】
本発明は、糖の輸送を提供しうるトランスポーターを提供する。輸送は膜を通過しうる。輸送は細胞または細胞の小器官から糖を排出しうる。輸送は細胞または細胞の小器官に糖を取り込みうる。糖は単糖、二糖もしくはオリゴ糖またはその誘導体であってよい。例えば、糖は、リボース、アラビノース、ペントース、例えばグルコース、フルクトース、ガラクトースおよびマンノース、ヘキソース、例えばマルトースおよびショ糖を含みうる。二糖類は、ラフィノースおよびスタキオースを含みうる。誘導体は、アミノ酸およびホルモンのグリコシル誘導体を含みうる。
【0120】
本発明はまた、本発明のトランスポーター、例えばGLUEトランスポーターをコードする拡散も提供する。本発明は、本発明のトランスポーターをコードする多用なcDNAを開示する。ウマゴヤシ由来のタンパク質MtN3およびそのホモログは、GLUEとして機能しうる。シロイヌナズナにおいて、下記受託番号がトランスポーターをコードする:AT4G15920、AT3G16690、AT5G13170、AtSAG29、AT4G25010、AT5G50800、AT5G23660、AT3G48740、AT5G50790、AT2G39060、AT5G40260、AtRPG1、AT4G10850、AT1G66770、AT5G62850、AtVEX1、AT3G28007、AT3G14770、AT1G21460およびAT5G53190(全て全体において参照により本明細書に組み込む)。ペチュニア植物において、NEC1は糖トランスポーターの例である(その全体において参照により本明細書に組み込む)。ウマゴヤシ植物において、本発明のトランスポーターをコードする下記cDNA受託番号が同定されている:AC202585、AC147714、MtC60432 GC、MtC11004 GC、CT963079、MtD03138 GC、TC 125536、AC146866、AC189276、TC129646、CAA69976 MtNod3 AC2456、TC115479、AC146747、MtC10424 GC、CT954252、CU302340、AC202585、AC147714、MtC60432 GC、MtC11004 GCおよびCT963079(全て全体において参照により本明細書に組み込む)。コメ植物において、本発明のトランスポーターをコードする下記cDNA受託番号が同定されている:Os08g42350 (Os8N3) Os08g0535200、Os12g29220 Os03g0347500、Os05g51090 Os05g0588500、Os12g07860、Os09g08440、Os09g08490、Os09g08270、Os09g08030 Os09g0254600、Os01g42090.1 Os01g0605700、Os01g42110.1 Os01g060600、Os02g19820 Os02g0301100、Os05g35140 Os05g0426000、Os01g65880 Os01g0881300、Os01g50460 Os01g0700100、)s01g36070.1 Os01g0541800、Os01g12130.1、Os05g12320 Os05g0214300およびOs01g21230(全て全体において参照により本明細書に組み込む)。
【0121】
本発明はまた、動物ホモログRAG1AP1およびこのファミリーの細菌ホモログ、例えば下記受託番号によってコードされるものを含む:A1BJ76、A1VHH8、A3IH65、A4AVY5、A5ERR3、A5FEJ3、A5G4U0、A5IEV6、A8AYJ9、B0SHL1、B0SR19、B1MYL5、B1MZF9、B1WTC6、B3EHG6、B5EHF6、B5YGD6、B6IU72、Q11VQ0、Q21M9、Q300V9、Q39VX0、Q3B6J0、Q5WTV4、Q5X228、Q72RB5、Q72FY5、Q89G85、Q8F4F7(全て全体において参照により本明細書に組み込む)。GLUEは原核生物、例えばレジオネラ属(Legionella)、デスルホビブリオ属(Desulfovibrio)、ブラディリゾビウム属(Bradyrhizobium)、レプトスピラ属(Leptospira)、ロドシュードモナス属(Rhodopseudomonas)、連鎖球菌、ジオバクター属(Geobacter)、ペロディクチオン属(Pelodictyon)、サイトファーガ属(Cytophaga)、ロドスピリラム属(Rhodospirillum)、サーモデスルフォビブリオ属(Thermodesulfovibrio)、クロロビウム属(Chlorobium)、ウォルバキア属(Wolbachia)、シアノセイス(Cyanothece、ロイコノストック属(Leuconostoc)およびフラボバクテリウム属(Flavobacterium)から得られる。
【0122】
糖の輸送は必須である。例えばグルコース流出は生体の多くの時点で、例えば花粉の発生または精子細胞発生のための精巣上体栄養供給の役割において必要とされる。GLUEはある生理的プロセス、例えば授乳においてアップレギュレートされ、乳管にラクトースを分泌する腺細胞に局在化されうる。同様に、肝臓のような臓器は血中グルコースレベルを一定に保つためにグルコースを流出する必要がある。GLUEは小胞流出経路のためのベシクルまたはゴルジ体へのグルコース負荷に関与しうる。
【0123】
トランスポーターファミリーのメンバーは実質的な同一性を共有する。GLUE1はそのパラログGLUE8と41%同一であり、4種のシロイヌナズナGLUEクレードの第2に属する。GLUE8/RPG1の変異は、雄不妊を導くことが示されている。哺乳類細胞におけるGLUE88/RPG1とグルコースのFRETセンサーの共発現は、いくつかのGLUE、例えばGLUE8も単輸送体として機能することを示す。さらに、GLUE8/RPG1は酵母グルコース輸送変異を補完する。GLUE8/RPG1はタペータムにおいて発現されてよく、これは花粉栄養供給における関与を示す。
【0124】
GLUE1およびGLUE8はコメタンパク質OsGLUE11/Os8N3(系統発生学に基づいてOsGLUE11と命名)と34%アミノ酸配列同一性を共有する。最も近いシロイヌナズナホモログは、OsGLUE11/Os8N3と40%同一性を共有し、第3GLUEクレードに属する。RNA阻害による発現の現象が花粉における低下したデンプン濃度ならびに花粉不妊を導くため、GLUE8と同様に、OsGLUE11/Os8N3は花粉栄養供給に機能しうる。クレード3の他のGLUEホモログであるPetunia Neclも、雄不妊を導きうる。Nec1は蜜腺で発現され、その発生制御は蜜腺のデンプン含量と逆相関しており、これは蜜腺の糖分泌におけるNec1の第二の役割を示す。
【0125】
本発明は、他の生物のトランスポーターを提供する。例えば、線虫(C. elegans)遺伝子は糖流出トランスポーターの新規クラス(SLC50)の7種のホモログを含むが、ヒトゲノムは1種のホモログ、すなわちRAG1AP1(またはHsGLUE1)を有する。シロイヌナズナGLUE1同様、線虫(C. elegans)CeGLUE1はグルコース取込を仲介しうる。CeGLUE1およびヒトRAG1AP1は、Na+/グルコースコトランスポーターSGLT1によって仲介される第二活性グルコース蓄積を相殺しうる。CeGLUE1およびヒトRAG1AP1の変異は、脂質の蓄積を導く細胞性グルコース流出の欠損と一致して、脂肪蓄積を導きうる。
【0126】
本発明は糖トランスポーター、例えばGLUEをコードする核酸を提供する。本発明はまた、保存的アミノ酸置換を有するポリペプチドをコードする核酸を提供する。本発明の核酸は、糖を輸送するポリペプチドをコードしうる。単離された核酸は上記同定した配列と少なくとも約30%、40%、50%、60%、70%、80%、85%、90%、95%または99%配列同一性を有しうる。単離された核酸は、上記同定された受託番号によってコードされるアミノ酸配列と少なくとも約30%、40%、50%、60%、70%、80%、85%、90%、95%または99%配列同一性を有するアミノ酸配列を有するポリペプチドをコードしうる。トランスポーターをコードする単離された核酸は、上記同定された核酸配列とハイブリダイズしうる。
【0127】
GLUEのタンパク質は配列およびサブドメインホモロジーを共有する。下記表1は、GLUEの選択における配列保存量の図を提供し、下記表2は、配列によるGLUEの比較を提供する。
【表1】

【表2】
【0128】
GLUEタンパク質をコードする核酸は、一般に、発現のための発現制御配列と遺伝的に融合されていてよい。好適な発現制御配列は、標的宿主生物に適用可能なプロモーターを含む。かかるプロモーターは、原核生物および真核生物からの多様な宿主について、当業者に周知であり、文献に記載されている。例えばかかるプロモーターは、天然由来の遺伝子から単離されてよく、あるいは合成またはキメラプロモーターであってもよい。同様に、プロモーターは標的ゲノム中に既に存在していてよく、例えば相同組換えのような当該技術分野において既知の好適な技術によって核酸分子と連結されてもよい。
【0129】
本発明はまた、GLUEをコードする核酸を標的核酸分子、例えばベクターまたはゲノムDNAに挿入するための発現カセットを提供する。この目的で、発現カセットは、特異的配列位置など、例えば制限酵素認識部位または例えばリコンビナーゼによって触媒される場合の相同組換えの標的部位からの除去またはそれへの挿入を促進するための、5’または3’隣接ヌクレオチド配列を提供する。
【0130】
本発明はまた、遺伝子操作において通常使用される、本発明の核酸分子および発現カセットを含むベクター、特にプラスミド、コスミド、ウイルスおよびバクテリオファージにも関する。
【0131】
本発明の好ましい態様において、本発明のベクターは、真菌細胞、植物細胞、微生物の細胞(すなわち細菌、原生生物、酵母、藻類等)または動物細胞、特に哺乳動物細胞の形質転換に好適である。好ましくは、かかるベクターはヒト細胞の形質転換に好適である。当業者に周知の方法を用いて、組換えベクターを構築できる:例えば、Sambrook and Russell, Molecular Cloning: A Laboratory Manual, CSH Press, 2001および Ausubel, Current Protocols in Molecular Biology, Green Publishing Associates and Wiley Interscience, N.Y., 1989に記載の技術を参照。あるいは、ベクターは、本発明の核酸分子または発現カセットが標的細胞への送達のために再構成されてよいリポソームであってもよい。同様に、用語「ベクター」は、細胞への遺伝子トランスファーを促進することが知られている化合物、例えばポリカチオン、カチオン性ペプチド等をさらに含む、核酸分子または発現カセットを含む複合体を意味する。
【0132】
本発明の核酸分子または発現カセットに加えて、ベクターは、好適な宿主細胞において好適な条件下でベクターの選択が可能な遺伝子、例えばマーカー遺伝子をさらに含んでいてよい。一般に、ベクターは1個以上の複製起点も含む。ベクターはまた、本発明のトランスポーターをコードする核酸を超えた転写の長さを制限する終止配列を含んでいてもよい。
【0133】
有利には、ベクターに含まれる核酸分子は、原核細胞または真核細胞において、発現を可能とする、すなわち翻訳可能なRNAの転写および合成を保証する発現制御配列と作動可能に連結されている。
【0134】
遺伝子操作について、例えば原核細胞において、本発明の核酸分子およびそれらの分子の一部は、DNA配列の組換えによる突然変異または配列修飾を許容するプラスミドに導入されていてよい。標準的な方法(Sambrook and Russell, Molecular Cloning: A Laboratory Manual, CSH Press, 2001参照)は、ベース交換の実施または天然もしくは合成配列の添加を可能とする。DNAフラグメントは、該フラグメントにアダプターまたはリンカーを適用することにより互いに結合していてよい。さらにまた、好適な制限部位を提供するかまたは余剰DNAもしくは制限部位を除去する操作手法が使用できる。これらの場合、挿入、欠失または置換が可能であり、インビトロ突然変異、「プライマー修復」、制限またはライゲーションが使用できる。配列分析、制限分析または生化学および分子生物学の他の方法が分析法として実施される。
【0135】
本発明はまた、トランスポーターをコードする核酸の指定された発現を提供する。遺伝子の発現はコードヌクレオチド配列の上流(5’)の特定のプロモーターによって制御されうることが当該技術分野において知られている。動物の特定の組織での発現を指示する組織特異的プロモーターは、当該技術分野において既知である。例えば、データベースがこれらのプロモーターを収集し、共有している(Chen et al., Nucleic Acids Res. 34: D104-D107, 2006)。植物では、根、種子または果実での発現を指示するプロモーターが知られている。
【0136】
本発明はさらに、さらなるポリペプチドと融合したトランスポーターを含む単離されたポリペプチドを提供する。さらなるポリペプチドは、大型ポリペプチドのフラグメントであってもよい。一つの態様において、トランスポーターと融合した1、2、3、4またはそれ以上のさらなるポリペプチドが存在する。いくつかの態様において、さらなるポリペプチドは、トランスポーターのアミノ末端に対して融合している。他の態様において、さらなるポリペプチドはトランスポーターのカルボキシル末端に対して融合している。さらなる態様において、さらなるポリペプチドはトランスポーターと隣接している。いくつかの態様において、核酸分子は、トランスポーターをコードする核酸と融合した核酸を含む融合タンパク質をコードする。融合核酸は、トランスポーターのコドンリーディングフレームを移動させることなく、精製および/または免疫原性および/または安定性に役立ちうるポリペプチドをコードできる。いくつかの態様において、融合核酸は、トランスポーターの精製に役立つポリペプチドをコードする。いくつかの態様において、融合核酸は、エピトープおよび/または親和性タグをコードする。他の態様において、融合核酸は、部位特異的または誤りがちな切断に関するポリペプチドをコードする。好ましい態様において、融合核酸は、酵素的切断の部位であるポリペプチドをコードする。さらなる態様において、酵素的切断は、トランスポーターの単離に役立つ。
【0137】
他の態様において、複数の核酸がトランスポーターをコードする核酸と融合される。融合核酸は、精製および/または酵素的切断および/または安定性に役立つポリペプチドをコードしうる。さらなる態様において、融合核酸は発現されるポリペプチドを顕著に伸長しない。
【0138】
いくつかの態様において、さらなるポリペプチドはエピトープを含んでもよい。他の態様において、さらなるポリペプチドは親和性タグを含んでもよい。例として、エピトープおよび/または親和性タグを含むポリペプチドとトランスポーターの融合体は、ポリペプチドの精製および/または同定に役立ちうる。例として、ポリペプチドセグメントはHisタグ、mycタグ、Sペプチドタグ、MBPタグ(マルトース結合タンパク質)、GSTタグ(グルタチオンSトランスフェラーゼ)、FLAGタグ、チオレドキシンタグ、GFPタグ(緑色蛍光タンパク質)、BCCP(ビオチンカルボキシル担体タンパク質)、カルモジュリンタグ、Strepタグ、HSVエピトープタグ、V5エピトープタグおよびCBPタグであってよい。かかるエピトープおよび親和性タグの使用は、当業者に既知である。
【0139】
さらなる態様において、さらなるポリペプチドは、ポリペプチドの切断のための部位を含む融合タンパク質を提供できる。切断部位は、例えばターゲティングポリペプチドで融合ポリペプチドからトランスポーターを後で切り出すために有用である。例として、ポリペプチドはペプチド結合の加水分解によって切断されうる。いくつかの態様において、切断は酵素によって実施される。いくつかの態様において、切断は細胞において生じる。他の態様において、切断は人工操作および/または切断酵素の人工導入によって生じる。例として、切断酵素はペプシン、トリプシン、キモトリプシンおよび/またはXa因子を含みうる。
【0140】
融合ポリペプチドは、同じ有機的に合成されたペプチドとは共有されないさらなる構造修飾、例えばアデニル化、カルボキシル化、グリコシル化、ヒドロキシル化、メチル化、リン酸化またはミリスチル化をさらに有していてよい。これらのさらなる構造修飾は、組換え発現系の適切な選択によってさらに選択または選好されうる。他方、融合ポリペプチドは、有機合成の原理および実施によって伸長された配列を有していてもよい。
【0141】
一般に、本発明の融合タンパク質は、先行技術に記載の技術に従って作成できる。例えば、これらの技術は、Sambrook and Russell, Molecular Cloning: A Laboratory Manual, CSH Press, 2001またはAusubel, Current Protocols in Molecular BiologyのVolumes1および2, Current Protocols, 1994に記載のとおりに実施できる組換え技術を含む。したがって、融合タンパク質の個々の部分は、宿主生物またはインビトロで組み合わされ、次いで発現される、それらをコードする核酸分子の形態で提供できる。あるいは、融合タンパク質またはその一部の提供は、化学合成または天然供給源からのかかる部分の単離を含んでもよく、それによって組換え技術によって部分的に作成されうる要素が好適な方法、例えば化学的架橋、例えばWO 94/04686に記載のとおりに、タンパク質レベルで融合されてよい。さらにまた、適切である場合、融合タンパク質は、それぞれの目的のためにその特性を改善するため、例えば溶解性を向上し、pH耐性を向上し、宿主生物にいてより良好な耐性であり、インビボもしくはインビトロで特定の基質に接着させる(後者は固相への融合タンパク質の不動化に潜在的に有用である)等のために、翻訳後修飾されてよい。当業者はかかる修飾およびその有用性を周知している。例示的説明は、1アミノ酸側鎖の修飾(例えばグリコシル化、ミリスチル化、リン酸化、カルボエトキシル化またはアミド化による)、ポリエチレングリコール、炭水化物等のようなポリマーまたはタンパク質部分、例えば抗体もしくはその一部または他の酵素等とのカップリングを含む。
【0142】
本発明の他の態様において、融合タンパク質はさらに、ターゲティングシグナル配列を含む。葉緑体、液胞、ペルオキシソーム、グリオキシソーム、細胞壁またはミトコンドリアのような細胞内区画へのタンパク質の輸送またはアポプラストへの分泌は、目的のタンパク質をコードする遺伝子の5’および/または3’領域と、シグナル配列をコードするヌクレオチド配列を作動可能に連結することによって達成される。構造遺伝子の5’および/または3’末端のターゲティング配列は、コードされるタンパク質が最終的に区分されるタンパク質合成およびプロセッシングにおいて決定されうる。
【0143】
シグナル配列の存在は、細胞内小器官または細胞内区画にポリペプチドを指向させるか、あるいはアポプラストに分泌させる。多くのシグナル配列が当該技術分野において既知である。例えばBecker et al., Plant Mol. Biol. 20:49 (1992); Close, P. S., Master's Thesis, Iowa State University (1993); Knox, C., et al., Plant Mol. Biol. 9:3-17 (1987); Lerner et al., Plant Physiol. 91:124-129 (1989); Frontes et al., Plant Cell 3:483-496 (1991); Matsuoka et al., Proc. Natl. Acad. Sci. 88:834 (1991); Gould et al., J. Cell. Biol. 108:1657 (1989); Creissen et al., Plant J. 2:129 (1991); Kalderon, et al., Cell 39:499-509 (1984); Steifel, et al., Plant Cell 2:785-793 (1990)参照。
【0144】
用語「ターゲティングシグナル配列」は、発現されたタンパク質におけるその存在のためにそれが特定の細胞内局在を標的とするアミノ酸配列を意味する。例えば、対応するターゲティングシグナルは、精製を単純化するために例えば細菌宿主から、発現されたトランスポーターの分泌を導きうる。好ましくは、トランスポーターのターゲティングは、特定の細胞内または細胞外区画における糖濃度に影響を与えるために使用できる。異なる生物群に有用な適切なターゲティングシグナル配列は、当業者に既知であり、文献または配列データベースから取り出すことができる。
【0145】
本発明のトランスポーターは、細胞基質、細胞表面または細胞内小器官、例えば核、ベシクル、ER、液胞等の細胞の任意の場所で発現されてよい。異なる細胞区画にタンパク質の発現をターゲティングする方法およびベクター成分は、バイオセンサーが発現される具体的な細胞または生物に基づく選択と共に、当該技術分野において周知である。例えばOkumoto et al. PNAS 102: 8740-8745, 2005; Fehr et al. J Fluoresc 14: 603-609, 2005を参照されたい(その全体について参照により本明細書に組み込む)。葉緑体、液胞、ペルオキシソーム、グリオキシソーム、細胞壁またはミトコンドリアのような細胞内区画へのタンパク質の輸送またはアポプラストへの分泌は、トランスポーターをコードする遺伝子の5’および/または3’領域と、シグナル配列をコードするヌクレオチド配列を作動可能に連結することによって達成されうる。構造遺伝子の5’および/または3’末端のターゲティング配列は、コードされるタンパク質が最終的に区分されるタンパク質合成およびプロセッシングにおいて決定されうる。
【0146】
植物細胞の色素体へのターゲティングが望まれる場合、例えば次のターゲティングシグナルペプチドが使用できる:シロイヌナズナ色素体RNAポリメラーゼ(AtRpoT 3)のアミノ酸残基1〜124(Plant Journal 17: 557-561, 1999);ホウレンソウの色素体Ferredoxin:NADP+オキシドレダクターゼ(FNR)のターゲティングシグナルペプチド(Jansen et al., Current Genetics 13: 517-522, 1988)、特にそれに開示のcDNA配列のヌクレオチド−171〜165によってコードされるアミノ酸配列;成熟ロウタンパク質の最初の34アミノ酸残基を含むかまたは含まないトウモロコシのロウタンパク質の輸送ペプチド(Klosgen et al., Mol. Gen. Genet. 217: 155-161, 1989);リブロースビスホスフェートカルボキシラーゼ小サブユニットのシグナルペプチド(Wolter et al., PNAS 85: 846-850, 1988; Nawrath et al., PNAS 91: 12760-12764, 1994)、NADPリンゴ酸デヒドロゲナーゼのシグナルペプチド(Gallardo et al., Planta 197: 324-332, 1995)、グルタチオンレダクターゼのシグナルペプチド(Creissen et al., Plant J. 8: 167-175, 1995)またはR1タンパク質のシグナルペプチド(Lorberth et al., Nature Biotechnology 16: 473-477, 1998)。
【0147】
植物細胞のミトコンドリアへのターゲティングは、次のターゲティングシグナルペプチドを用いて達成できる:シロイヌナズナミトコンドリアRNAポリメラーゼのアミノ酸残基1〜131(AtRpoT 1)(Plant Journal 17: 557-561, 1999)またはBraunに記載の輸送ペプチド(EMBO J. 11: 3219-3227、1992)。
【0148】
植物細胞の液胞へのターゲティングは、次のターゲティングシグナルペプチドを用いて達成できる:パタチンタンパク質のN末端配列(146アミノ酸)(Sonnewald et al., Plant J. 1: 95-106, 1991)またはMatsuoka and Neuhaus(Journal of Exp. Botany 50: 165-174, 1999);Chrispeels and Raikhel(Cell 68: 613-616, 1992); Matsuoka and Nakamura(PNAS 88: 834-838, 1991); Bednarek and Raikhel(Plant Cell 3: 1195-1206, 1991)またはNakamura and Matsuoka(Plant Phys. 101: 1-5, 1993)に記載のシグナル配列。
【0149】
植物細胞のERへのターゲティングは、例えばC末端伸長HDEL(Haselhoff, PNAS 94: 2122-2127, 1997)と組み合わせたERターゲティングペプチドHKTMLPLPLIPSLLLSLSSAEFを用いて達成できる。植物細胞の核へのターゲティングは、例えばタバコC2ポリペプチドQPSLKRMKIQPSSQPの核局在シグナル(NLS)を用いて達成できる。
【0150】
細胞外空隙へのターゲティングは、例えば下記輸送ペプチドの一つを用いて達成できる:プロテイナーゼ阻害剤II遺伝子のシグナル配列(Keil et al., Nucleic Acid Res. 14: 5641-5650, 1986; von Schaewen et al., EMBO J. 9: 30-33, 1990)、火傷病菌由来レバンスクラーゼ遺伝子のシグナル配列(Geier and Geider, Phys. Mol. Plant Pathol. 42: 387-404, 1993)、最初の33アミノ酸をコードするジャガイモ由来パタチン遺伝子B33のフラグメントのシグナル配列(Rosahl et al., Mol Gen. Genet. 203: 214-220, 1986)またはOshima et al.に記載のもののシグナル配列(Nucleic Acids Res. 18: 181, 1990)。
【0151】
さらにまた、膜へのターゲティングは、ウサギスクラーゼ−イソマルターゼのN末端シグナルアンカーを用いて達成できる(Hegner et al., J. Biol. Chem. 276: 16928-16933, 1992)。
【0152】
哺乳類細胞の膜へのターゲティングは、N末端ミリスチン酸結合配列can be accomplished by using the N-terminal myristate attachment sequence MGSSKSKまたはC末端プレニル化配列CaaXを用いて達成できる(Garabet, Methods Enzymol. 332: 77-87, 2001)。ここで「a」は脂肪族アミノ酸(すなわちVal、Leu またはIle)であり「X」は任意のアミノ酸である。
【0153】
細胞膜または植物細胞へのさらなるターゲティングは、トランスポーター、好ましくはショ糖トランスポーターSUT1との融合によって達成できる(Riesmeier, EMBO J. 11: 4705-4713, 1992)。多様な細胞内膜へのターゲティングは、空胞水チャネル(γTIP)のような特定の区画における膜タンパク質(Karlsson、Plant J. 21: 83-90、2000)、ミトコンドリアのMCFタンパク質(Kuan, Crit. Rev. Biochem. Mol. Biol. 28: 209-233, 1993)、色素体の内包膜におけるトリオースリン酸トランスロケーター(Flugge, EMBO J. 8: 39-46, 1989)およびチラコイドの光化学系との融合によって達成できる。
【0154】
ゴルジ装置へのターゲティングは、C末端認識配列K(X)KXXを用いて達成できる(Garabet, Methods Enzymol. 332: 77-87, 2001)。ここで「X」は任意のアミノ酸である。
【0155】
ペルオキシソームへのターゲティングは、ペルオキシソームターゲティング配列PTS IまたはPTS IIを用いて達成できる(Garabet, Methods Enzymol. 332: 77-87, 2001)。
【0156】
哺乳動物細胞の核へのターゲティングは、SV-40大型T抗原核局在化配列PKKKRKVを用いて達成できる(Garabet, Methods Enzymol. 332: 77-87, 2001)。
【0157】
哺乳動物細胞のミトコンドリアへのターゲティングは、N末端ターゲティング配列MSVLTPLLLRGLTGSARRLPVPRAKISLを用いて達成できる(Garabet, Methods Enzymol. 332: 77-87, 2001)。
【0158】
いくつかの態様において、トランスポーターの発現は、特定の組織または細胞タイプに標的化されうる。例えば、特定のプロモーターを使用して、トランスポーターをコードする核酸の転写を駆動できる。プロモーターは核酸の転写を指示する核酸コントロール配列のアレイである。プロモーターは転写開始部位、例えばポリメラーゼII型プロモーターの場合にはTATA要素の近くの必要な核酸配列を含む。プロモーターはまた、所望により、転写開始部位から数千塩基対に位置しうる離れたエンハンサーまたはリプレッサーを含む。恒常プロモーターは、ほとんどの環境および発生条件下で活性なプロモーターである。誘導性プロモーターは、環境または発生制御下で活性なプロモーターである。いずれかの誘導性プロモーターが使用でき、例えばWard et al., Plant Mol. Biol. 22:361-366, 1993を参照されたい。誘導性プロモーターの例は、ACEI系由来のもの(銅に応答性)(Meft et al., Proc. Natl. Acad. Sci. USA 90:4567-4571, 1993);トウモロコシ由来のIn2遺伝子(ベンゼンスルホンアミド除草剤薬害軽減剤に応答性)(Hershey et al., Mol. Gen. Genetics 227:229-237, 1991およびGatz et al., Mol. Gen. Genetics 243:32-38, 1994)またはTn10由来のTetリプレッサー(Gatz et al., Mol. Gen. Genetics 227:229-237, 1991)を含むがこれらに限定されない。誘導性プロモーターは、宿主細胞の外来物質に応答しうる。例えばSchena et al., PNAS 88: 10421-10425, 1991参照。
【0159】
プロモーターは恒常プロモーターであってもよい。恒常プロモーターは発現のために遺伝子と作動可能に連結されているか、または発現のために遺伝子と作動可能に連結しているシグナル配列をコードするヌクレオチド配列と作動可能に連結している。例えば植物細胞において、恒常プロモーターは、CaMV由来の35Sプロモーターのような植物ウイルス由来のプロモーター(Odell et al., Nature 313: 810-812, 1985)およびコメアクチンのような遺伝子由来のプロモーター(McElroy et al., Plant Cell 2: 163-171, 1990);ユビキチン(Christensen et al., Plant Mol. Biol. 12:619-632, 1989およびChristensen et al., Plant Mol. Biol. 18: 675-689, 1992);pEMU(Last et al., Theor. Appl. Genet. 81:581-588, 1991);MAS(Velten et al., EMBO J. 3:2723-2730, 1984)およびトウモロコシH3ヒストン(Lepetit et al., Mol. Gen. Genetics 231: 276-285, 1992およびAtanassova et al., Plant Journal 2(3): 291-300, 1992)を含むが、これらに限定されない。原核生物プロモーター要素は、大腸菌におけるRNAポリメラーゼによる転写に最適な−35〜−10(Pribnowボックス)を担持するものを含む。いくつかの原核生物プロモーター要素は、制御リプレッサーの重複結合部位を含みうる(例えばLacおよびTACプロモーター、これらはlacリプレッサーの重複結合部位を含み、それによって基質ホモログIPTGによる誘導性を与える)。好適なプロモーター配列が得られうる原核生物遺伝子の例は、大腸菌lac、araおよびtrpを含む。本発明の原核生物ウイルス性プロモーター要素は、λファージプロモーター(例えばPRMおよびPR)、T7ファージプロモーター要素およびSP6プロモーター要素を含む。本発明の真核生物プロモーターベクター要素は、酵母(例えばGAL1、GAL10、CYC1)および哺乳類(例えばグロビン遺伝子およびインターフェロン遺伝子のプロモーター)の両方を含む。さらなる真核生物プロモーターベクター要素は、ウイルス性遺伝子プロモーター、例えばSV40プロモーター、CMVプロモーター、単純ヘルペスチミジンキナーゼプロモーターならびに多様なレトロウイルスLTRプロモーター要素(例えばMMTV LTR)のものを含む。他の真核生物の例は、hMTIIaプロモーター(例えば米国特許5,457,034)、HSV−1 4/5 プロモーター(例えば米国特許5,501,979)および早期中間体HCMVプロモーター(WO 92/17581)を含む。
【0160】
プロモーターは組織特異的または組織選好プロモーターであってもよい。組織特異的プロモーターは、特定の組織で排他的または優先的なトランスポーターの産生を補助する。あらゆる組織特異的または組織選好プロモーターが利用できる。植物細胞において、例えば組織特異的または組織選好プロモーターは、根選好プロモーター、例えばファゼオリン遺伝子に由来するもの(Murai et al., Science 23: 476-482, 1983およびSengupta-Gopalan et al., PNAS 82: 3320-3324, 1985);葉特異的および光誘導性プロモーター、例えばcabまたはrubisco由来のもの(Simpson et al., EMBO J. 4(11): 2723-2729, 1985および Timko et al., Nature 318: 579-582, 1985);葯特異的プロモーター、例えばLAT52由来のもの(Twell et al., Mol. Gen. Genetics 217: 240-245, 1989);花粉特異的プロモーター、例えばZm13由来のもの(Guerrero et al., Mol. Gen. Genetics 244: 161-168, 1993)または小胞子選好プロモーター、例えばapg由来のもの(Twell et al., Sex. Plant Reprod. 6: 217-224, 1993)を含むが、これらに限定されない。
【0161】
さらに本発明は、上記本発明の核酸分子を含み、原核細胞または真核細胞において発現可能なように制御配列と作動可能に連結している発現カセットに関する。
【0162】
さらなる態様において、本発明は、本発明のトランスポーターを発現できる細胞を作成する方法であって、上記本発明の核酸分子、発現カセットまたはベクターで細胞を遺伝的に操作することを含む方法に関する。
【0163】
本発明の別の態様は、上記本発明の核酸分子、発現カセットまたはベクターで遺伝的に操作された宿主細胞、特に原核細胞または真核細胞、ならびにかかる形質転換された細胞の子孫細胞であって、本発明の核酸分子、発現カセットまたはベクターを含む細胞、ならびにそれを作成する上記方法によって得られた細胞に関する。
【0164】
宿主細胞緒は細菌、真菌、昆虫、植物または動物宿主細胞であってよい。一つの態様において、宿主細胞は、導入されてゲノムに安定に組み込まれた核酸分子を含むように遺伝的に操作されている。他の態様において、核酸分子は、本発明の融合タンパク質の産生を導くように発現されてよい。
【0165】
多様な発現系の概略は、例えばBitter et al.のMethods in Enzymology 153: 385-516, 1987 (Methods in Enzymology 153: 516-544, 1987)およびSawers et al.(Applied Microbiology and Biotechnology 46: 1-9, 1996)、Billman-Jacobe(Current Opinion in Biotechnology 7: 500-4, 1996)、Hockney(Trends in Biotechnology 12: 456-463, 1994)およびGriffiths et al.(Methods in Molecular Biology 75: 427-440, 1997)に含まれる。酵母発現系の概略は、例えばHensing et al.(Antoine von Leuwenhoek 67: 261-279, 1995)、Bussineau(Developments in Biological Standardization 83: 13-19, 1994)、Gellissen et al.(Antoine van Leuwenhoek 62: 79-93, 1992)、Fleer(Current Opinion in Biotechnology 3: 486-496, 1992)、Vedvick(Current Opinion in Biotechnology 2: 742-745, 1991)およびBuckholz(Bio/Technology 9: 1067-1072, 1991)で得られる。
【0166】
発現ベクターは文献に広く記載されている。一般に、それは選択マーカー遺伝子および選択した宿主における複製を保証する複製起点だけでなく、細菌性またはウイルス性プロモーターおよび多くの場合転写の終止シグナルを含む。一般に、プロモーターと終止シグナルの間に、コードヌクレオチド配列の挿入が可能な少なくとも1個の制限部位またはポリリンカーが存在する。遺伝子発現の意図的な制御を可能とする、遺伝子および誘導性プロモーターの恒常的発現を保証するプロモーターの使用が可能である。これらの特徴を有する細菌性またはウイルス性プロモーター配列は、文献に詳細に説明されている。微生物(例えば大腸菌、出芽酵母)における発現のための制御配列は、十分に文献に記載されている。下流配列の特に高い発現を可能とするプロモーターは、例えばT7プロモーター(Studier et al., Methods in Enzymology 185: 60-89、1990)、lacUV5、trp、trp-lacUV5(DeBoer et al., in Rodriguez and Chamberlin (Eds)、Promoters、Structure and Function; Praeger、New York、1982、p. 462-481; DeBoer et al., PNAS 80: 21-25, 1983)、Ip1、rac(Boros et al., Gene 42: 97-100, 1986)である。誘導性プロモーターは、タンパク質の合成に使用できる。これらのプロモーターはしばしば恒常プロモーターよりも高いタンパク質収量を導く。最適な量のタンパク質を得るために、二段階法がしばしば用いられる。第一に、宿主細胞を比較的高い細胞密度まで最適条件下で培養する。第二段階で、使用するプロモーターのタイプに依存して転写を誘導する。これに関して、ラクトースまたはIPTG(イソプロピル−β−D−チオガラクトピラノシド)によって誘導可能なtacプロモーターが特に好適である(DeBoer et al., PNAS 80: 21-25, 1983)。哺乳動物細胞における適用に有用なSV40-ポリA部位またはtk-ポリA部位のような転写の終止シグナルも文献に記載されている。好適な発現ベクターは当該技術分野において既知であり、例えばOkayama-Berg cDNA発現ベクターpcDV1(Pharmacia)、pCDM8、pRc/CMV、pcDNA1、pcDNA3(In-vitrogene)、pSPORT1 (GIBCO BRL)またはpCI(Promega)である。
【0167】
本発明はまた、本発明のトランスポーターをコードするベクターまたは発現ベクターでトランスフェクトされた宿主細胞、例えば原核細胞、例えば大腸菌もしくは他の細菌または真核細胞、例えば酵母細胞もしくは動物細胞も含む。生細胞培養は、原核細胞または真核細胞を含んでもよい。原核細胞源の例は、細菌または巨細筋を含むがこれらに限定されない。真核細胞源の例は、酵母、真菌、原生生物、哺乳類、節足動物、ヒト、動物、軟体動物、環形動物、線形動物、甲殻類、扁形動物、単孔類、魚類、有袋類、爬虫類、両生類、鳥類、げっ歯類、昆虫および植物を含むが、これらに限定されない。
【0168】
本発明の核酸分子またはベクターによる宿主細胞の形質転換は、標準的な方法、例えばSambrook and Russell, Molecular Cloning: A Laboratory Manual, CSH Press, 2001; Methods in Yeast Genetics, A Laboratory Course Manual, Cold Spring Harbor Laboratory Press, 1990に記載のとおりに実施できる。例えば、塩酸カルシウムトランスフェクションは、一般に原核細胞に利用され、一方例えばリン酸カルシウムもしくはDEAEデキストラン介在トランスフェクションまたはエレクトロポレーションは、他の細胞宿主に使用できる。宿主細胞は使用する具体的な宿主細胞の、特にpH値、温度、塩濃度、通気、抗生物質、ビタミン、微量元素等に関する要求に合う栄養培地中で培養する。本発明のトランスポーターは、硫酸アンモニウムもしくはエタノール沈殿、酸抽出、アニオンもしくはカチオン交換クロマトグラフィー、ホスホセルロースクロマトグラフィー、疎水性相互作用クロマトグラフィー、アフィニティークロマトグラフィー、ヒドロキシルアパタイトクロマトグラフィーおよびレクチンクロマトグラフィーを含む方法によって、組換え細胞から回収し、精製できる。トランスポーターのリガンドまたは基質、例えばグルコースはアフィニティー精製に使用できるか、あるいはトランスポーターの融合タンパク質は、親和性タグのような融合タンパク質が結合する基質またはリガンドによるアフィニティークロマトグラフィーを適用して精製できる。タンパク質再フォールディング段階は、必要により、タンパク質の立体配置を完成させるために使用できる。最後に、高速液体クロマトグラフィー(HPLC)を最終生成段階に使用できる。
【0169】
したがって、本発明のさらなる態様は、本発明のトランスポーターを製造する方法であって、トランスポーターの発現が可能な条件下で前記宿主細胞を培養し、培養物からトランスポーターを回収することを含む方法に関する。発現タンパク質が宿主細胞中に局在するか細胞から分泌されるかに従って、タンパク質は、培養細胞および/または培地の上清から回収できる。
【0170】
あるいは、トランスポーターはマイクロインジェクション、微粒子銃、封入センサーの導入または細胞へのバイオセンサーの取り込みを導くペプチド配列の融合によって、細胞に送達されうる。
【0171】
さらにまた、本発明は、上記製造方法によって得られるトランスポーターに関する。
【0172】
本発明のトランスポーターは、例えば化学的合成法の産物であるか、あるいは原核または真核宿主(例えば、培養の細菌、酵母、高等植物、昆虫または哺乳類細胞による)から組換え技術により産生されてよい。組換え製造法において使用される宿主に依存して、発現されるトランスポーターは、グリコシル化されているかまたは非グリコシル化されていてよい。本発明のトランスポーターはまた、初期メチオニンアミノ酸残基を含んでいてもよい。本発明のトランスポーターはさらに、通常タンパク質の一部ではないさらなる化学部分を含むように修飾されていてもよい。これらの誘導体化部分は、例えばタンパク質の安定性、溶解性、生物学的半減期またはタンパク質の吸着を改善しうる。当該部分はまた、タンパク質の望ましくないいずれかの副作用などを低減または削除しうる。これらの部分の概要は、例えばRemington's Pharmaceutical Sciences, Mack Publishing Co., Easton, Paに見られる。
【0173】
トランスジェニック
【0174】
本発明は、本発明のトランスポーター、例えばGLUEをコードする核酸分子を含むトランスジェニック植物および非ヒトトランスジェニック生物、すなわち多細胞生物、または少なくとも当該生物または組織もしくは臓器のようなその一部の細胞のサブセットにおいて、そのゲノムに安定的に組み込まれた上記発現カセットまたはベクターを含む、生物を提供する。
【0175】
本発明は、トランスジェニック植物細胞、すなわちゲノムに安定的に組み込まれた上記本発明の核酸分子、発現カセットまたはベクターを含む、トランスジェニック植物または植物組織を提供する。本発明はまた、以下に説明する製造方法によって得られたトランスジェニック植物、植物細胞または植物組織を提供する。
【0176】
一つの態様において、本発明は、トランスジェニック植物、植物組織または植物細胞を製造する方法であって、本発明の核酸分子、発現カセットまたはベクターを植物細胞に導入し、所望によりそれからトランスジェニック植物または植物組織を再生することを含む方法を提供する。トランスポーターを発現するトランスジェニック植物は、生物の器官を介するおよびそれの間の、例えば土壌へのまたは土壌からの、糖の輸送に影響を与えるために使用できる。本発明のトランスポーターを発現するトランスジェニック植物は、例えば有機化合物の代謝または輸送プロセスを、先行技術では得られなかった時間および空間分解能で調査するために使用できる。
【0177】
外来核酸分子の植物への導入方法は、当該技術分野において周知である。例えば植物形質転換は、アグロバクテリウム(Agrobacterium)介在遺伝子導入、マイクロインジェクション、エレクトロポレーションまたは微粒子銃法を用いて、例えばPotrykus and Spangenberg (Eds.), Gene Transfer to Plants. Springer Verlag, Berlin, New York, 1995に記載のとおりに実施できる。これにおよび多数の他の先行技術文献において、有用な植物形質転換ベクター、形質転換された細胞および組織の選択方法ならびに再生技術が記載されており、それらは当業者に既知であり、本発明の目的のために適用されうる。
【0178】
別の局面において、本発明は、上記トランスジェニック植物細胞を含む、本発明のトランスジェニック植物の収穫可能な部分および伝播物質を提供する。収穫可能な部分は、基本的に、植物のあらゆる有用な部分、例えば葉、幹、果実、種子、根などであってよい。伝播物質は、例えば種子、果実、挿し木、実生、塊茎、根茎等を含む。
【0179】
ある局面において、本発明は、トランスポーターの発現をコードする核酸配列の発現によって特徴付けられる表現型を有するトランスジェニック非ヒト動物を提供する。表現型は動物の体細胞および生殖細胞に含まれるトランスジーンによって与えられ、これは(a)動物の接合体にトランスポーターをコードするDNA構築物を含むトランスジーンを導入し;(b)接合体を偽妊娠動物に移植し;(c)接合体を発生、出産させ;そして(d)トランスジーンを含む少なくとも1体のトランスジェニック子孫を同定することによって製造できる。胚にトランスジーンを導入する工程は、胚にトランスジーンを含む胚性幹細胞を導入するか、またはトランスジーンを含むレトロウイルスに胚を感染させることを含んでいてよい。好ましいトランスジェニック動物は、コードされたトランスポーターを発現する。本発明のトランスジェニック動物は、トランスジェニック出芽酵母(S. cerevisae)、線虫(C. elegans)、ショウジョウバエ、特に、キイロショウジョウバエおよびトランスジェニックマウスおよび他の動物を含む。
【0180】
本発明はまた、トランスポーターをコードする少なくとも1個の核酸分子、ゲノムに安定的に組み込まれうる核酸を含む発現カセットまたはベクターを含む、トランスジェニック非ヒト動物を提供する。
【0181】
本発明はまた、トランスジェニック非ヒト動物の作成方法であって、本発明の核酸分子、発現カセットまたはベクターを、生殖細胞、胚性細胞、肝細胞または卵またはそれらに由来する細胞に導入することを含む方法を包含する。本発明のトランスポーターを発現するかかるトランスジェニック動物または接合体から始まるその任意の発生段階が、組織の完全性を破壊することなく実時間である化合物(融合タンパク質に存在する酵素に依存する)の分布を決定できるモデル生物として使用できる。これらのモデル生物は、栄養学的もしくは薬理学的研究または薬物スクリーニングに特に有用でありうる。トランスジェニック胚の作成およびそのスクリーニングは、例えばA. L. Joyner (Ed.), Gene Targeting, A Practical Approach, Oxford University Press, 1993に記載のとおりに実施できる。胚のDNAは、例えば適切なプローブによるサザンブロットまたはPCR技術に基づいて分析できる。
【0182】
本発明によるトランスジェニック非ヒト動物は、本発明の核酸分子、発現カセットまたはベクターを含む、好ましくはゲノムに安定的に組み込まれている、あるいは上記方法によって作成された、例えばトランスジェニックマウス、ラット、ハムスター、有袋類、単孔類、イヌ、サル、ウサギ、翼手類、ブタ、カエル、線形動物、例えば線虫(Caenorhabditis elegans)、ショウジョウバエ、例えばキイロショウジョウバエ、または魚類、例えばシビレエイ目(torpediniforms)、例えばシビレエイ、フグ目、カラシン目、ネズミザメ目もしくはコイ目、例えばゼブラフィッシュであってよい。かかるトランスジェニック非ヒト動物は、1個または複数個の同一または異なる本発明の核酸分子のコピーを含んでいてよい。かかるトランスジェニック非ヒト動物中の本発明の核酸分子、発現カセットまたはベクターの存在は、本発明のトランスポーターの発現を導く。本発明のトランスジェニック非ヒト動物は、研究モデルを含む、多様な有用性を有する。したがって、この場合、哺乳類は好ましくは実験動物、例えばチンパンジー、マウスまたはラットである。
【0183】
したがって、一つの態様において、本発明のトランスジェニック非ヒト動物は、マウス、ラット、イヌ、例えばビーグルまたはゼブラフィッシュである。多くの報告により当該動物が薬物代謝およびその欠点または発がん性の研究のためのモデル生物として特に十分に好適であることが示された。有利には、糖輸送、特にグルコース輸送を研究するための当該技術分野において周知の多様な好適な技術の利用可能性のため、当該モデル生物を用いてトランスジェニック動物は容易に作成できる。
【0184】
抗体
【0185】
本発明の別の局面は、本発明のトランスポーターと結合する抗体の作製に関する。本発明に包含される抗体の例は、特許請求の範囲に記載の発明のトランスポーターに特異的な抗体および中和抗体を含むが、これらに限定されない。本発明の抗体は、当該技術分野において周知の方法を用いて特徴付けてよい。
【0186】
本発明に有用な抗体は、モノクローナル抗体、ポリクローナル抗体、抗体フラグメント(例えばFab、Fab’、F(ab’)2、Fv、Fc等)、キメラ抗体、二重特異性抗体、ヘテロコンジュゲート抗体、一本鎖(ScFv)、その変異体、抗体の一部を含む融合タンパク質、ヒト化抗体、ならびに抗体のグリコシル化変異体、抗体のアミノ酸配列変異体および共有修飾された抗体を含む、必要な特異性抗原認識部位を含む、任意の他の修飾された立体配置の免疫グロブリン分子を含みうる。抗体はマウス、ラット、ヒト、霊長類または他の起源(キメラおよびヒト化抗体を含む)に由来しうる。
【0187】
一つの態様において、抗体はポリクローナルまたはモノクローナル抗体であってよい。モノクローナルおよびポリクローナル抗体の製造方法は当該技術分野において周知である。
【0188】
用語「モノクローナル抗体」は、本明細書において使用するとき、実質的に均一な抗体の集団から得られる抗体を意味し、すなわち個々の抗体を含む集団が微量で存在しうる可能性のある天然由来の変異体を除き同一であり、本明細書に定義の抗体フラグメントを含む。モノクローナル抗体は、極めて特異的であり、単一の抗原部位に対する。さらに、異なる決定基(エピトープ)に対する異なる抗体を含むポリクローナル抗体調製物とは異なって、各モノクローナル抗体は、抗原の単一の決定基に対する。特異性に加えて、モノクローナル抗体は、他の抗体で汚染されることなく合成できる点で有利である。修飾語「モノクローナル」は、いずれかの具体的な方法で抗体が製造されることを要求するものではない。例えば、本発明に有用なモノクローナル抗体は、Kohler et al. (1975) Nature 256, 495で初めて記載されたハイブリドーマ法によって製造してよく、あるいは細菌、真核動物または植物細胞における組換えDNA法を用いて製造してもよい(米国特許4,816,567参照)。「モノクローナル抗体」は、例えばClackson et al. (1991) Nature 352:624-628およびMarks et al. (1991) J. Mol. Biol. 222, 581-597に記載の技術を用いたファージ抗体ライブラリーから単離してもよい。「ポリクローナル」抗体は、特定のタンパク質またはそのフラグメントに対する抗体の選択を意味し、ここで該抗体は、異なるエピトープと結合しうる。
【0189】
他の態様において、抗体は当該技術分野において既知の方法によってヒト化してもよい。ヒト化抗体は非ヒト免疫グロブリン由来の最小配列を含む免疫グロブリン分子である。さらに別の態様において、完全ヒト抗体は、特定のヒト免疫グロブリンタンパク質を発現するように操作されている商業的に入手可能なマウスを用いて得られる。他の態様において、抗体はキメラである。キメラ抗体は2種の異なる抗体由来の特徴を組み合わせた抗体である。キメラ抗体の作製方法は当該技術分野において既知である。
【0190】
他の態様において、抗体をコードするヌクレオチド配列を得て、次いで発現または増殖のためにベクターにクローン化する。他の態様において、抗体は組換え的に作製され、当該技術分野において既知の方法を用いて発現される。例として、トランスポーターまたはそのフラグメントは、これらの技術によって組換え抗体を単離する目的で抗原として使用できる。トランスポーターの抗原モチーフは、当該技術分野において既知の方法、例えばJameson-Wolf法(CABIOS, 4: 181-186, 1988)によって容易に決定できる。抗体は、宿主細胞において抗体を組換え的に発現する遺伝子配列を用いて、組換え的に作製できる。抗体の誘導体および組換え抗体を作製する方法は当該技術分野において既知である。
【0191】
他の態様において、例えば天然トランスポーターの単離もしくは精製または生物学的サンプルまたは標本中の天然トランスポーターの検出に使用するために、当該技術分野において常套の方法によって、抗体は単体と結合されている。
【0192】
用語「抗体またはそのフラグメント」は、本明細書において使用するとき、糖トランスポーターまたはそのフラグメントと特異的に結合し、他の非トランスポーターと特異的に結合しない抗体またはそのフラグメントを意味する。トランスポーターまたはそのフラグメントと免疫特異的に結合する抗体またはフラグメントは、他の抗原と非特異的に交差反応しない(例えば、適切なアッセイにおいて非トランスポーター、例えばBSAと結合が競合しない)。トランスポーターと免疫特異的に結合する抗体またはフラグメントは、例えばイムノアッセイまたは当業者に既知の他の技術によって同定できる。本発明の抗体は、合成抗体、モノクローナル抗体、重鎖のみの抗体、組換え的に作製した抗体、細胞内抗体、二重特異性抗体、多重特異性抗体(二重特異性抗体を含む)、ヒト抗体、ヒト化抗体、キメラ抗体、一本鎖Fv(scFv)(二重特異性scfvを含む)、一本鎖抗体、Fab’フラグメント、F(ab’)2フラグメント、ジスルフィド結合Fvs(sdFv)および抗イディオタイプ(抗−Id)抗体、ならびに上記のいずれかのエピトープ結合フラグメントを含むが、それらに限定されない。特に本発明の抗体は、免疫グロブリン分子および免疫グロブリン分子の免疫学的に活性な部分、すなわちトランスポーターと免疫特異的に結合する抗原結合部位(例えば抗トランスポーター抗体の1個以上の相補性決定領域(CDR))を含む分子を含む。
【0193】
本明細書において使用するとき、「インタクトな」抗体は、抗原結合部位ならびにCLおよび少なくとも重鎖定常ドメインCH1およびCH2およびCH3を含むものである。定常ドメインは天然配列定常ドメイン(たとえがヒト天然配列定常ドメイン)またはそのアミノ酸配列変異体であってよい。好ましくは、インタクトな抗体は1個以上のエフェクター機能を有する。
【0194】
「抗体フラグメント」は、インタクトな抗体の一部、好ましくはインタクトな抗体の抗原結合CDRまたは可変領域を含む。抗体フラグメントの例は、Fab、Fv、Fab’およびF(ab’)2 フラグメント;二重特異性抗体;線状抗体(米国特許5,641,870およびZapata et al. (1995) Protein Eng. 8, 1057-1062参照);一本鎖抗体分子;および抗体フラグメントから形成された多重特異性抗体を含む。
【0195】
抗体のパパイン消化は、「Fab」フラグメントと呼ばれる、2個の同一の抗原結合フラグメントと、容易に結晶化できる能力を反映して名付けられた残りの1個の「Fc」フラグメントとを産生する。FabフラグメントはH鎖の可変領域ドメイン(VH)と共にL鎖全体および1個の重鎖の最初の定常ドメイン(CH1)からなる。各Fabフラグメントは抗原結合について一価であり、すなわちそれは単一の抗原結合部位を有する。抗体のペプシン処理によって、1個の大きなF(ab’)2フラグメントが得られ、これは二価の抗原結合活性を有する2個のジスルフィド結合したFabフラグメントにほぼ対応しており、抗原と交差結合できる。Fab’フラグメントは、抗体ヒンジ領域由来の1個以上のシステインを含むCH1ドメインのカルボキシ末端でさらに数個の残基を有する点で、Fabフラグメントと異なっている。Fab’−SHは、本明細書において、定常ドメインのシステイン残基が遊離チオール基を有するFab’を意味する。F(ab’)2抗体フラグメントは、それらの間にヒンジシステインを有するFab’フラグメントの対として本来作製された。抗体フラグメントの他の化学結合も知られている。
【0196】
Fcフラグメントは、ジスルフィドで一体に固定された両方のH鎖のカルボキシ末端部分を含む。抗体のエフェクター機能は、あるタイプの細胞で見られるFcレセプター(FcR)によって認識される部分でもあるFc領域の配列によって決定される。
【0197】
本明細書において使用するとき、「Fv」は、完全な抗原認識部位および結合部位を含む最小抗体フラグメントである。このフラグメントは、密接な非共有結合での1個の重鎖可変領域および1個の軽鎖可変領域の二量体からなる。これらの2個のドメインの折り畳みから、6個の超可変ループ(H鎖およびL鎖それぞれから3個のループ)が生じ、これらは抗原結合のためのアミノ酸残基の一因であり、抗体に抗原結合特異性を与える。しかし、1個の可変ドメイン(または抗原に特異的な3個のCDRのみを含むFvの半分)さえも抗原を認識し、結合する能力を有するが、全結合部位よりも親和性は低い。
【0198】
本明細書において使用するとき、「sFv」または「scFv」とも略される「一本鎖Fv」は、1個のポリペプチド鎖に結合されたVHおよびVL抗体ドメインを含む抗体フラグメントである。scFvポリペプチドはさらに、sFvが所望の抗原結合構造を形成できるようにVHとVLドメイン間のポリペプチドリンカーを含んでいてもよい(Rosenburg et al. (1994) The Pharmacology of Monoclonal Antibodies, Springer-Verlag, pp. 269-315参照)。
【0199】
本明細書において使用するとき、用語「二重特異性抗体」は、鎖間でVドメインが対となるが鎖内ではならないようにVHおよびVLドメイン間の短いリンカー(約5〜約10残基)を有するsFvフラグメント(上段参照)を構築することによって作製される小さな抗体フラグメントを意味し、二価フラグメント、すなわち2個の抗原結合部位を有するフラグメントである。二重特異性二重特異性抗体は、2個の抗体のVHおよびVLドメインが異なるポリペプチド鎖上に存在する2個の「クロスオーバー」sFvフラグメントのヘテロ二量体である。二重特異性抗体は、例えばWO 93/11161およびHollinger et al. (1993) Proc. Natl. Acad. Sci. USA 90, 6444-6448により十分に記載されている。
【0200】
「単離された抗体」は、その天然環境の成分から同定および分離および/または回収されているものである。その天然環境の混入成分は、抗体の診断または治療的使用を妨げる物質であり、酵素、ホルモンおよび他のタンパク質性または非タンパク質性成分である。好ましい態様において、抗体は抗体の95重量%以上、最も好ましくは99重量%以上に精製されている。単離された抗体は、少なくとも1種の抗体の天然環境の成分が存在しないため、組換え細胞内のインサイチュの抗体を含む。しかし通常、単離された抗体は、少なくとも1回の精製段階によって製造される。
【0201】
本発明の一つの態様において、コンジュゲートした抗体は、がん細胞に特異的なタンパク質の細胞質ドメイン上のエピトープ(すなわちがん細胞マーカー)と結合する。他の態様において、コンジュゲートした抗体は、sFの細胞質ドメイン上のエピトープと結合する抗体を含むが、これに限定されない。
【0202】
医薬組成物
本発明の別の局面は、医薬組成物の一部としてのトランスポーターの使用に関する。本発明の抗体および核酸はまた、医薬組成物の一部として使用できる。本発明の方法に使用される組成物は一般に、例として、有効量の本発明の核酸またはポリペプチド(例えば免疫応答を誘導するために必要な量)または本発明の抗体(例えば、感染の緩和、感染の症状の緩解および/または感染の予防に必要な中和抗体の量)を含むが、これに限らない。本発明の核酸、ポリペプチドおよび抗体は、さらに、当該技術分野において既知の薬学的に許容される担体、賦形剤または安定化剤を含む(一般に、Remington, (2005) The Science and Practice of Pharmacy, Lippincott, Williams and Wilkins参照)。
【0203】
本発明の核酸、ポリペプチドおよび抗体は、凍結乾燥製剤または水溶液の形態であってよい。許容される担体、賦形剤または安定化剤は、投与する量および濃度で受容者に非毒性でありうる。担体、賦形剤または安定化剤はさらにバッファーを含んでいてもよい。バッファーの例は、炭水化物(例えば単糖類および二糖類)、糖(例えばショ糖、マンニトールおよびソルビトール)、リン酸、クエン酸、抗酸化剤(例えばアスコルビン酸およびメチオニン)、保存剤(例えばフェノール、ブタノール、ベンザノール; アルキルパラベン、カテコール、オクタデシルジメチルベンジル塩化アンモニウム、塩化ヘキサメトニウム、レゾルシノール、シクロヘキサノール、3−ペンタノール、塩化ベンザルコニウム、塩化ベンズエトニウム、およびm−クレゾール)、低分子量ポリペプチド、タンパク質(例えば血清アルブミンまたは免疫グロブリン)、親水性ポリマーアミノ酸、キレート剤(例えばEDTA)、塩形成対イオン、金属錯体(例えばZnタンパク質錯体)および非イオン性界面活性剤(例えばTWEEN(商標)およびポリエチレングリコール)を含むがこれらに限定されない。
【0204】
本発明の医薬組成物は、さらに、所望の効果を向上および/または補足するために使用される添加剤を含んでいてもよい。例として、本発明のトランスポーターの免疫原性を向上させるため、医薬組成物はさらに、アジュバントを含んでいてよい。アジュバントはアルミニウム塩(alum)、完全フロイントアジュバント(CFA)、不完全フロイントアジュバント(IFA)、ムラミルジペプチド(MDP)、MDPの合成アナログである、N−アセチルムラミル−L−アラニル−D−イソグルタミル−L−アラニン−2−[1,2−ジパルミトイル−s−グリセロ−3−(ヒドロキシホスホリルオキシ)]エチルアミド(MTP−PE)ならびに代謝可能な油と乳化剤を含む組成物(油と乳化剤は実質的に全てが直径1ミクロン未満の油滴を有する水中油型エマルジョンの形態で存在している(例えばEP 0399843参照))を含む。
【0205】
いくつかの態様において、アジュバントは、サポニンと組み合わせたToll様受容体(TLR)4リガンドを含む。Toll様受容体(TLR)4リガンドは例えば、リピドA誘導体、特にモノホスホリルリピドAまたはより具体的には3−デアシル化モノホスホリルリピドA(3D−MPL)であってよい。3D−MPLは商標名MPL(登録商標)としてCorixa Corporationから販売されており、IFN−g(Th1)表現型を有するCD4+T細胞応答を第一に促進する。それはGB 2220211Aに記載の方法に従って製造できる。化学的には、それは3−デアシル化モノホスホリルリピドAと3、4、5または6アシル化鎖の混合物である。一つの態様において、本発明の組成物において、小粒子3D−MPLを用いる。小粒子3D−MPLは、0.22 μmのフィルターで滅菌ろ過できるような粒子サイズを有する。かかる調製物は、PCT特許出願WO 94/21292に記載されている。
【0206】
アジュバントはまた、 may also comprise one or more synthetic 誘導体s of lipid A which are known to be TLR4アゴニストであると知られている1種以上のリピドAの合成誘導体、例えば:PCT特許出願WO 95/14026に記載のOM174(2−デオキシ−6−o−[2−デオキシ−2−[(R)−3−ドデカノイルオキシテトラ−デカノイルアミノ]−4−o−ホスホノ−β−D−グルコピラノシル]−2−[(R)−3−ヒドロキシテトラデカノイルアミノ]−α−D−グルコピラノシルジヒドロゲンホスフェート);WO 9964301およびWO 00/0462に記載のOM 294 DP(3S,9R)−3−[(R)−ドデカノイルオキシテトラデカノイルアミノ]−4−オキソ−5−アザ−9(R)−[(R)−3−ヒドロキシテトラデカノイルアミノ]デカン−1,10−ジオール,1,10−ビス(ジヒドロゲノホスフェート);ならびにOM 197 MP−Ac DP(3S−、9R)−3−[(R)−ドデカノイルオキシテトラデカノイルアミノ]−4−オキソ−5−アザ−9−[(R)−3−ヒドロキシテトラデカノイルアミノ]デカン−1,10−ジオール,1−ジヒドロゲノホスフェート10−(6−アミノヘキサノエート)(WO 01/46127)も含むが、これらに限定されない。
【0207】
他の使用できるTLR4リガンドは、アルキルグルコサミニドホスフェート(AGP)、例えばWO 98/50399または米国特許6,303,347(AGPの製造法も記載されている)に記載のもの、または米国特許6,764,840に記載のAGPの薬学的に許容される塩を含むが、これらに限定されない。いくつかのAGPはTLR4アゴニストであり、いくつかはTLR4アンタゴニストである。いずれも本発明の組成物の1種以上のアジュバントとして使用できる。
【0208】
本発明に使用するためのサポニン担体は、Quil Aおよびその誘導体である。Quil AはSouth American tree Quilaja Saponaria Molinaから単離されたサポニン調製物であり、Dalsgaard et al. (1974) Saponin adjuvants, Archiv. fuer die gesamte Virusforschung, Vol. 44, Springer Verlag, pp. 243-254で初めてアジュバント活性を有することが記載された。Quil A に関連した毒性がなく、アジュバント活性を保持しているQuil Aの精製されたフラグメントはHPLCによって単離されており(EP 0 362 278)例えばQS7およびQS21(QA7およびQA21とも知られる)である。QS21は、CD8+細胞傷害性T細胞(CTL)、Th1細胞および主なIgG2a抗体応答を誘導するQuillaja saponaria Molinaの樹皮由来の天然サポニンであり、本発明の文脈で好ましいサポニンである。
【0209】
QS21の特定の製剤が記載されており、これは特に好ましく、これらの製剤はさらにステロールを含む(WO 96/33739)。本発明のサポニン形成部分は、ミセル、混合ミセルの形態で分離されていてよく(好ましくは、しかし排他的ではなく胆汁酸塩で)、あるいはISCOMマトリックス(EP 0109942 B1)、リポソームまたは関連するコロイド構造、例えばワーム状またはリング状マルチマー複合体または脂質/層状構造ならびにコレステロールおよび脂質で製剤した場合ラメラ、または水中油エマルジョン(例えばWO 95/17210のとおり)の形態であってよい。サポニンは酸化アルミニウムまたはリン酸アルミニウムのような金属塩と結合していてよい(WO 98/15287)。いくつかの態様において、サポニンはリポソーム、ISCOMまたは水中油エマルジョンの形態で存在する。
【0210】
いくつかの態様において、アジュバントは3D−MPLとQS21(EP 0671948 B1)の組合せ、ならびに3D−MPLおよびQS21を含む水中油エマルジョン(WO 95/17210、WO 98/56414)である。
【0211】
糖トランスポーターの使用法
【0212】
本発明は、糖トランスポーター、例えばGLUEの使用法を提供する。方法は、細胞、例えばインビトロまたは生物の細胞に糖トランスポーターを導入することを含む。細胞に導入された糖トランスポーターは、野生型または変異型トランスポーターであってよい。トランスポーターは、トランスポーターをコードする核酸またはアミノ酸ポリペプチドとして導入できる。トランスポーターの機能する能力は、所望の結果に基づいて当業者に理解される。例えば、恒常的活性型または野生型トランスポーターを用いて、糖輸送不全を克服できる。変異型トランスポーターを用いて、糖輸送に関する問題を克服できる。糖輸送の変異体は、病原体の所望の遺伝子発現を変化させるために導入してもよい。
【0213】
本発明の方法は、生物発生の変化を提供する。生物は成体または胚であってよい。生物細胞への導入は、生物の発生に影響を与えうる。植物において、糖トランスポーターの導入は、葉発生、茎発生、花蜜発生、根発生、葯発生、木部発生、生殖系発生、幹発生および果実発生に影響を与えうる。動物において、本発明の糖トランスポーターの導入は、臓器、例えば脳、心臓、肺、循環系、皮膚、肝臓、腎臓、脳、脊髄、骨、筋肉(平滑および骨格)、肢、耳、脾臓、腸、膵臓、副腎、胆嚢、睾丸、卵巣、前立腺、膀胱、胃、甲状腺、副甲状腺、海馬、松果体、リンパ節、乳腺、免疫系または管系の発生に影響を与えうる。
【0214】
本発明の方法は、生物の臓器、例えば循環系、神経系(交感神経および副交感神経系を含む)、呼吸器系、消化系、排泄系および生殖系の間の機能的系に影響を与えうる。植物において、糖トランスポーターの導入は、根および幹および葉の間の糖輸送に影響を与えうる。動物において、糖トランスポーターの導入は、乳産生、生殖腺の発生および機能、排卵、酸素と二酸化炭素の交換、食物の消化および栄養素の吸収に影響を与えうる。
【0215】
本発明の方法は、病原体侵襲のような生物への侵襲の感受性に影響する新規メカニズムを提供する。病原体は原核生物または真核生物でありうる。病原体は、細菌、ウイルス、真菌、蠕虫または昆虫であってよい。病原体は、例えば宿主の感受性遺伝子を介して、栄養素または滋養を病原体に与えるように宿主生物の遺伝子制御に影響しうる。本発明のトランスポーターは宿主生物遺伝子転写に影響を与える病原体の能力を変化させうる。宿主細胞に導入される本発明の糖トランスポーターは、病原体侵襲から宿主細胞を保護できる。
【0216】
当業者は、発生中の花粉、発芽中の花粉、発生中の胚およびアポプラスト経路(細胞壁を介した細胞間)を介して細胞が炭素を交換する全ての他の場合を補給するために同様の流出段階が必要とされることを理解する。GLUEホモログであるRPG1は、タペータムに局在化され、RPG1の変異は花粉の生存不能を導く(Guan YF, Huang XY, Zhu J, Gao JF, Zhang HX, Yang ZN. 2008 Plant Physiol. 147:852-63)。したがって、トランスポーターの操作は、割り当てに影響を与えうる。
【0217】
本発明のトランスポーターは、植物の根圏への糖、例えばグルコースの分泌を調節できる。例えば、植物は植物期間あたり1.5t/haの炭素を土壌に分泌する。本発明の方法は、糖分泌の操作が可能であり、したがって植物生産性に影響を与えうる。例示として、糖分泌の増加は、多くの微生物を細胞に誘引し、より多くの炭素を土壌に預けうる(例えば大気中CO2の隔離)。糖分泌の減少は、植物中バイオマスの増加を導く。糖分泌の減少は、植物の増加したバイオマスを導きうる。根におけるかかるトランスポーターの一般的概念は、Chaudhuri et al. 2008, Plant Journalに部分的に記載されている。
【0218】
本発明のトランスポーターは、植物の葉圏への糖、例えばグルコースの分泌を調節できる。例えば、葉圏における糖分泌の調節は、より多くの有益微生物を誘引でき、あるいは病原体に食餌を与える。操作(農薬として特定の化学的阻害剤の発達も介する)。
【0219】
本発明のトランスポーターは糖、例えばグルコースの分泌を調節して、植物の受粉パターンに影響を与えうる。糖分泌の操作は、受粉パターンに影響を与えうる。例えば、変化した糖分泌は、異なる受粉媒介者は異なる花蜜組成物を必要とするため、異なる受粉媒介者を誘引しうる(Ge et al. Plant J. 24:725-734, 2000)。
【0220】
本発明のトランスポーターは、糖、例えばグルコースの分泌を調節でき、植物の葉および師部の発生に影響を与えうる。例えば、Ge et al.は、関連タンパク質の過剰発現が師部発生の刺激を導き、葉の対称性を変化させることを開示している。植物の構造はまた、本明細書に記載の方法で操作してもよい。
【0221】
本発明の別の局面は、植物および植物細胞に対する病原体の活性を調節する方法に関する。病原体(共生生物を含む)は餌を得るためにあるトランスポーターを採用する(Yang et al. 2006 PNAS 103:10503-10508)。Yang et al.は、その遺伝子は、病原型特異的にタイプIII分泌系依存メカニズムによって誘導される、感受性因子であると考えている。マイクロアレイデータを分析することにより、異なる病原体が異なるGLUEファミリーのメンバーを採用することが示される。病原体がトランスポーターを採用できる一つの手段が、細胞に存在するトランスポーター数またはその活性を増加するために、トランスポーターをコードする核酸のプロモーター領域に影響を与えることである。異なるトランスポーターの導入またはトランスポーターの上流への異なるプロモーターの導入のような操作は、病原体感染を予防でき、共生生物を改善または移動できる。トランスポーターを遮断して病原体感染を予防する化学的阻害剤が同定されうる。葉からの糖の排出は、師部負荷のためのプロトンショ糖コトランスポーターだけでなく、葉肉または師部細胞から排出する細胞エフラクサーも必要とする。
【0222】
乳は新生児、小児および成人にとって重要な栄養源である。米国では、一人当たりの乳消費量は80リットルを超える(www.foodsci.uoguelph.ca/dairyedu/intro.html)。乳はバター、ヨーグルトおよびチーズを製造するためにも使用される。動物では、乳は新生児の一次栄養源である。母乳および牛乳は、多くの重要な栄養素および抗体を新生児に与える。タンパク質および脂質に加えて、乳は、グルコースおよび二糖ラクトースのような可溶性の糖も含む。ヒトの乳のラクトース含量は〜7%(200mM)であり、牛乳は〜4.5%(140 mM)である。ラクトースは、乳腺の乳管の内側を覆う肺胞細胞において産生される。特に、ラクトース合成はゴルジ体で生じ、β−1,4−ガラクトシルトランスフェラーゼサブユニットおよびラクトアルブミンからなるヘテロマー酵素ラクトースシンターゼによって仲介され、授乳中に高度に誘導される。
【0223】
前駆体グルコースは、GLUTおよびSGLTファミリーに属するグルコーストランスポーターによって腺細胞への基底膜を介して取り込まれる。グルコースおよびUDP−ガラクトーストランスポーターはいずれも、ゴルジ体でのラクトース合成のための前駆体の取り込みを必要とするが、ゴルジ体グルコースインポーターは同定されてない。ラクトースはエキソサイトーシスによって生じると推測される(2)。ゴルジ体ベシクルの膜は二糖類を透過しないと思われるため、高い浸透力が水取込を引きつける。エキソサイトーシスにおいて水は排出され、これは乳の水含量の主な割合に寄与する。ラクトース合成に寄与する細胞メカニズムの理解は、したがって、乳産生および組成の複数の局面にとって重要である。さらに、牛乳におけるラクトース含量は、腸内でラクターゼによってラクトースを効率的に代謝する能力がないために、集団の大部分にとって健康問題を意味する。ラクターゼ欠損は先天的(腸でのラクターゼ活性に影響する稀な変異)または後天的(二次的ラクターゼ欠損)でありうる。ラクターゼ欠損の最も一般的な原因は、小児の後に生じて成人まで持続する、成人型低ラクターゼ症と呼ばれるラクターゼ量の減少である。ほぼ100%のアジア人が低ラクターゼ症に罹患し、ラクトースを含まない食品を食べる必要がある。したがって本発明は、対象における乳の産生を変化させるために利用できる。本発明は、ラクターゼを取り込むまたは排出する細胞の能力を変化させうる。
【実施例】
【0224】
実施例
【0225】
栄養取込および代謝、ならびに植物での生物相互作用における栄養素交換、例えば受粉媒介者を誘引するための花における花蜜産生、微生物に給餌するための根圏への植物の根による糖分泌および病原体によるこれらのシステムの乗っ取りを制御するシグナル伝達カスケード。生殖細胞、蜜腺、根圏および病原体への糖の供給に関与する新規トランスポーターが最近同定された。次いで、この植物トランスポーターが動物、特に哺乳動物においてホモログを有すること、このタンパク質は乳腺における糖分泌に関与する多くの特徴を示すことが見出された。FRETナノセンサーは細胞内分解による定量的流量分析が可能なユニークなツールを提供する(Okumoto et al. New Phytol. 180:271-295, 2008)。これらのナノセンサーは、レポーター要素として緑色蛍光タンパク質(GFP)のスペクトル変異体の対とアロステリックに結合した、認識要素として使用される最近ペリプラズム結合タンパク質からなる(Fehr et al. Proc. Natl. Acad. Sci. USA 99:9846-9851, 2002;Fehr et al. J. Biol. Chem. 278:19127-19133, 2003; Deuschle et al. Protein Sci 14:2304-2314, 2005)。
【0226】
認識要素とのリガンド結合によって誘導されるコンホメーション変化は、結合したシアンと黄色蛍光タンパク質部分の間での蛍光共鳴エネルギー転移(FRET)の変化に翻訳される。これらのセンサーは、生体細胞に遺伝的に導入でき、生体細胞および組織における分析物レベルの非侵襲的測定が可能である(Fehr et al. J. Biol. Chem. 278:19127-19133, 2003)。これらのセンサーにより、インタクトなシロイヌナズナの根におけるグルコース流量(Chaudhuri et al. Plant J. 56:948-962, 2008)、海馬ニューロンからのグルタミン酸放出(Okumoto et al. Proc Natl Acad Sci U S A 102:8740-8745, 2005)およびがん細胞におけるトリプトファン交換(Kaper et al. PLoS Biol 5:e257, 2007)が測定されている。
【0227】
実施例1:ER膜を通過するグルコース流量の分析
【0228】
小器官内の分析物レベルを測定するため、これらのFRETナノセンサーを各細胞内区画に標的化した(Fehr et al.J. Fluoresc. 14:603-609, 2004)。ER膜を通過するグルコース流量を直接モニターするために、FRETグルコースセンサーをERシグナル配列およびKDEL保留シグナルと隣接させて、ER内腔に標的化した。このアプローチは、ER膜を通過する高グルコース流量の同定が可能であり、HepG2細胞におけるグルコースの速やかな双方向性高性能輸送活性の存在を示唆した(Fehr et al. Mol Cell Biol 25:11102-11112, 2005)。
【0229】
実施例2:植物の根における新規糖輸送機能の同定
【0230】
土壌は微量のみの溶解性炭水化物を含むが、植物の根は、人工培地に供給された場合、効率的にグルコースおよびショ糖を吸い上げる。土壌中に見られる溶解性炭水化物および小さな代謝産物は、部分的には植物の根からの浸出に由来する。取込および浸出に関するトランスポーター分子の性質は知られていない。FRETグルコースおよびショ糖センサーを展開して、シロイヌナズナの根端におけるグルコースおよびショ糖の蓄積および排出を特徴付けた(Chaudhuri et al. Plant J. 56:948-962, 2008)。グルコースおよびショ糖蓄積は、プロトノフォアに非感受性であり、pH5.8、6.8および7.8でも同様であった。これは流入および流出の両方がプロトンから独立した新規クラスの輸送系によって仲介されうることを示唆している。さらに、全ての既知の植物グルコーストランスポーターとは異なって、この新たな根輸送系は、グルコースアナログである3−O−メチルグルコースの輸送を仲介しなかった。
【0231】
実施例3:グルコーストランスポーターの発現系としてのHEK293T細胞
【0232】
細胞膜およびER膜を通過するグルコース輸送をよりよく特徴付けられるように、低内因性糖取込能を有する細胞をアッセイした。HEK293T細胞が極めて低い内因性取込を有し、FRETグルコースセンサーの補助の下でGLUTおよびSGLT糖トランスポーターの特性を定義する発現系として使用できることが見出された。
【0233】
実施例4:花蜜産生および根圏分泌に関与するグルコーストランスポーターの同定
【0234】
上記HEK293T細胞系を用いて、植物における花蜜産生に関与し、根端に記載のグルコース輸送活性の特徴を有するシロイヌナズナ糖トランスポーターの新規クラスを同定した。次いで、HEK293T細胞発現系においてシロイヌナズナ由来の候補トランスポーター遺伝子がFRETグルコースセンサーと共発現されるスクリーニングアッセイを開始した。膜タンパク質の未知のクラス(GLUE1と命名)がグルコース濃度依存的FRET応答を誘導することを見出した(例えば図3および4参照)。
【0235】
次いで、GLUE1がHEK293T細胞において発現されているとき既知の哺乳類GLUTまたはSGLTトランスポーター遺伝子のいずれもが誘導されないことを確認し、そしてGLUE1介在性取込がGLUT阻害剤のサイトカラシンBに非感受性であることを示した。さらに、GLUE1が3−O−メチルグルコースを輸送できないという結果が示された。小胞体内腔内で発現されるセンサーの使用により、GLUEが細胞基質への取込を仲介するだけでなく、細胞基質の外へのグルコースの排出もできることが示された。同様の結果がこの遺伝子ファミリーの複数のシロイヌナズナパラログについて得られた。
【0236】
GLUE1が内因性シグナル伝達カスケードと相互作用している可能性を排除するため、グルコース取込欠損酵母株EBY4000においてGLUE1を発現した。結果は、GLUE1がグルコースの取込を10mMのKMで、エネルギー独立的に仲介することを示した。GLUE1は、水および溶質輸送アクアポリンと同様の、7個の膜貫通ドメインを含む小さなタンパク質をコードする。GLUE1−GFP融合体は細胞膜に局在化する(例えば図9参照)。
【0237】
併せて考えると、これらのデータは、GLUE1がシロイヌナズナの根における根輸送系と同一の特性を有する新規クラスの糖単輸送体をコードすることを示している(Chaudhuri et al. Plant J. 56:948-962, 2008)。マイクロアレイデータの分析は、このファミリーのメンバーが根において実際に発現されていることを示している。
【0238】
このタンパク質ファミリーのメンバーは、例えば蜜腺(Ge et al. Plant J. 24:725-734, 2000)およびタペータム(Guan et al. Plant Physiol. 147:852-863, 2008)において発現される。その糖を流出する能力に鑑みて、GLUEは、花において花蜜を産せ宇するグルコースの分泌に関与していてよく、花粉を発生し、有益微生物を誘引し給餌するために根圏にグルコースを分泌するために、タペータムからグルコースを排出する(Chaudhuri et al. Plant J. 56:948-962, 2008)。
【0239】
実施例5:植物GLUEトランスポーターのヒトホモログ
【0240】
同様の調査により、RAG1AP1と命名したマウスおよびヒトゲノムにおけるGLUEホモログを同定した(Tagoh et al. Biochem Biophys Res Commun 221:744-749, 1996)。RAG1AP1は植物GLUEと顕著な相同性を有し、また7個の推定膜貫通ドメインを有するタンパク質をコードする。シロイヌナズナとは異なって、マウスおよびヒトゲノムは各々、1個のメンバーのみを含む。RAG1AP1活性を欠く変異リンパ球細胞系は、抗体成熟に関与する遺伝子の発現を制御することが示された(Tagoh et al. Biochem Biophys Res Commun 221:744-749, 1996)。これはグルコースまたはグルコースアナログを分泌できないことによって引き起こされる間接的な影響である可能性がある。RAG1AP1は抗体変動に関与する遺伝子の発現の制御に間接的に関与している可能性がある。マイクロアレイ試験によって、この遺伝子が授乳中に高度に誘導されることが示される(図9)。
【0241】
植物GLUEが植物におけるグルコースの分泌に関与する機能的証拠と共に、RAG1AP1は、乳腺の肺胞細胞におけるラクトース分泌および/またはグルコース輸送の役割において機能しうる。このことは、大型プロテオミクスプログラム、Human Protein Atlasのデータによって支持され、これはRAG1AP1(RAG1活性化タンパク質1)が乳房の腺細胞において特異的に発現されることを示唆している(www.proteinatlas.org/normal_unit.php?antibody_id=18095&mainannotation_id=1747078)。さらに、該タンパク質は人体の他の腺細胞、例えば潜在的に精子細胞に給餌する精巣上体においても局在化していると思われる。
【0242】
実施例6:病原体の栄養のための植物糖流出トランスポーター
【0243】
材料および方法
【0244】
qPCRおよびRT−PCR分析。RNeasy MINIキット(QIAGEN, Hilden)を用いて全RNAをHepG2およびHEK293T細胞から抽出し、第一ストランドcDNAを作製し(New England Biolabs)、既に公開されているGLUTおよびSGLTプライマーのセットを用いたRT−PCRによって推定長のフラグメントを得た。サンプルを2%アガロースゲル上で分離した。Pst DC3000によって播種したサンプルについて、Trizol試薬(Invitrogen)を用いて葉から全RNAを抽出した。HotStart-IT SYBR Green qPCR Master Mix (USB)を用いて7300 PCRシステム(Applied Biosystems)の製造業者の指示書に従って実時間定量PCR(qPCR)を実施した。アクチン(ACT8)発現を用いて各サンプルの発現レベルを正規化した;相対発現値は、比較2−ΔΔCt法を用いて各時点で1 mMのMgCl2で浸潤したサンプルの値に対して決定した。G.cichoracearumによって感染したサンプルについて、LightCyclerO 480(Roche)を用いてqPCRアッセイを行った。定量について、各遺伝子の相対的転写レベルを2-ΔΔCt法に従ってACT8に正規化した。変動比は未処理サンプルに対して計算した。分析を独立して2回反復した。マイクロアレイデータによって誘導を確認する(Genevestigator)。
【0245】
構築物。pOO2におけるSGLT1 ORFのクローニングを説明した。シロイヌナズナおよびコメそれぞれ由来の特定のプライマー(補足表1)を用いたRT−PCRによって、SWEET1、SWEET8およびOsSWEET11 ORFを増幅した。コメ由来の第一ストランスcDNAはPamela Ronald, UC Davisが快く提供下さった。Gateway BPクロナーゼ反応によってORFをpDONR221(Invitrogen)にクローン化し、Gateway LR組換え反応によって酵母発現ベクターpDRf1-GWに固定化した。pDRf1-GWにおけるFLII12Pglu700mD6との酵母コトランスフォーメーションのためにSWEET1をp112-A1NE-GWにクローン化した。p112-A1NEのSmaI制限部位にGatewayカセットを挿入してp112-A1NE-GWを作製した。放射性トレーサー実験のために、SWEET1、SWEET8およびOsSWEET11の停止コドンを有するORFを、Gateway LR組換え反応により卵母細胞発現ベクターpOO2-GW(D. Loque、未公表の結果)にクローン化した。
【0246】
FRET分析、細胞培養、トランスフェクション、イメージ獲得およびFRET分析を上記の通りに実施した。
【0247】
細胞基質インベルターゼ(YSL2-1)を担持する酵母株EBY4000の修飾型(hxt1 through -17Δ::loxP gal2Δ::loxP stl1Δ::loxP agt1Δ::loxP ydl247wΔ::loxP yjr160cΔ::loxP)をSWEETおよびHXT5で形質転換して、2%マルトースおよび必要な栄養要求マーカーを補ったSD(合成欠乏)培地で増殖した。補完増殖アッセイのために、細胞を液体最少培地でOD600 〜0.6に一夜増殖させ、次いで水を用いてOD600 〜0.2に希釈した。連続希釈(1x、5x、25xおよび125x)を2%マルトース(コントロールとして)または2%グルコースおよび関連した栄養マーカーを含むSD培地に播種した。30℃で2〜5日後、プレートをスキャンして(CanoScan, Canon)増殖を実証した。
【0248】
酵母取込。2%グルコースおよび栄養マーカを補ったSD培地中で、酵母細胞を増殖した。OD600 0.5〜0.7で、細胞を遠心分離で回収し、氷冷蒸留水で2回洗浄した。上清を除去した後、細胞ペレットを秤量した。40 mM リン酸カリウムバッファー、pH 6.0中に細胞を5〜10%(w/v)で再懸濁した。細胞をリン酸カリウムバッファー中、30℃で5分間プレインキュベートした。各反応について、20mM グルコース(0.55μCi D-[U-14C] グルコース; 590 KBq/μmol、Amersham Pharmacia Biotech Inc.)を含む予め温めたバッファー330 μlを等体積の細胞に加えた。120 μlのアリコートを取り除き、氷冷水に写した。ガラスファイバーフィルター(GF/C、Whatman)での真空ろ過によって細胞を回収し、10 mlの氷冷水で2回洗浄した。5 mlのUltima Gold XR Scintillator液(Perkin Elmer)を含むシンチレーションバイアルにフィルターを写した。液体シンチレーション分光法によって、細胞が取り込んだ放射活性を測定した。D−グルコースと比較したSWEET1の基質特異性を決定するため、10倍過剰の競合糖種を用いた。SWEET1活性のpH依存性を決定するため、特定のpHの40 mMのリン酸カリウム取込バッファーを用いた。3つの独立の形質転換体を取込実験に用いた。
【0249】
アフリカツメガエル卵母細胞単離およびRNAインジェクション。MluIでのpOO2プラスミドの直線化の後、キャップしたcRNAをインビトロで、mMESSAGE mMACHINEキット(Ambion, Inc., Austin, TX)を用いたSP6 RNAポリメラーゼによって合成した。アフリカツメガエル卵母細胞単離はM. Goodman(Stanford University)が快く提供下さった。Ballatori et al.に記載のとおりにマイクロインジェクションを実施した。25 ng〜50 ngの各cRNAを健康な外見の卵母細胞にインジェクションした(RNAseを含まない水をコントロールとして用いた)。100μM ゲンタマイシン、100 U/ml ペニシリンおよび100μM ストレプトマイシン溶液を含む修飾バース食塩水(MBS:(単位mM) 88 NaCl、1 KCl、2.4 NaHCO3、0.82 MgSO4、0.33 Ca(NO3)2、0.41 CaC12および20 HEPES-Tris、pH 7.5)中、18℃で2〜3日間、インジェクションした卵母細胞を維持した。インキュベーションバッファーを1日1回交換した。
【0250】
アフリカツメガエル卵母細胞におけるトレーサー取込。Detaille et al.に記載のとおりに修飾してアッセイを実施した。インジェクションの2日後、200μLのMBSおよび1 mMのD−グルコース(4 μCi/ml D-[14C(U)]-グルコース; 319 mCi/mmol、PerkinElmer)を含むチューブに7〜16卵母細胞のグループを移した。20℃で1時間インキュベーションした後、1 mlの氷冷MBSを加えて細胞を洗浄した。氷冷MBSバッファーを加えてインキュベーションを停止した。卵母細胞は氷冷MBSバッファーで3回洗浄した。細胞を100μlの1% (w/v)SDSで可溶化し、個々に測定した。
【0251】
アフリカツメガエル卵母細胞におけるトレーサー流出アッセイ。流出は本質的には上記の通りに測定した。cRNAインジェクションの3日後、0.18 μCi/μl D-[14C(U)]-グルコースを有する10 mMのD−グルコースを含む50 nlの溶液で卵母細胞をインジェクションした。細胞を速やかにMBSで1回洗浄した。所定の時点で、シンチレーションカウンティングのために反応バッファー(950 μl)を除去し、新鮮な培地と交換した。最後に、卵母細胞を1% SDSで可溶化し、放射活性を分析した。
【0252】
FRETセンサーによる酵母細胞におけるグルコース蓄積の分析。酵母細胞におけるFRET測定を上記の通り実施した。
【0253】
植物成長および病原体感染。シロイヌナズナCol-0植物を8時間明/14時間暗下、22℃の成長チャンバーで成長させた。1 mMのMgCl2バッファー、1 mMのMgCl2中の2x108cfu/mlのPst DC3000またはPst DC3000 ΔhrcU懸濁液で、無針注射器を用いて5週齢の葉を浸潤した。光下で6、12および24時間インキュベーション後、葉サンプルを回収した。G.cichoracearum接種を上記の通りに実施した。植物を「移動タワー」(段ボール)中に置き、感染したカボチャの葉を移動タワーの上にかぶせ、カボチャの葉から胞子が飛んでシロイヌナズナに移動するように圧縮空気(散布カン)を用いて、シロイヌナズナ植物にG. cichoracearum胞子を接種した。接種密度は〜25-35 分生子s/mm-2であった。接種後、植物を1時間、暗露チャンバーでインキュベートし、次いで日長16時間、相対湿度70%の成長チャンバーに移した。
【0254】
アラインメントおよび系統発生解析。デフォルトパラメーターを用いたCLUSTALWで、SWEETアミノ酸配列の複数アラインメントを実施し、ソフトウェアMega V3.1. Bootstrappingを用いて系統発生解析を1000回実施して、各ブランチのサポート値を得た。対比較のために、Vector NTIアドバンス11.0を用いて完全アミノ酸配列の複数アラインメントを実施した。
【0255】
共焦点顕微鏡。YFP::SWEET1およびYFP::SWEET8を発現する植物のイメージングをLeica TCS SP5顕微鏡で実施した。アルゴンレーザーで514 nmでの励起および放出につき525〜560 nmのスペクトル検出セットによってYFPを可視化した。40/0.75-1.25NA HCX PL APO CS対物レンズによって標本を観察した。
【0256】
結果および考察
【0257】
糖流出は多細胞生物において、および生物間の相互作用において、炭素骨格とエネルギーの細胞性交換に必要とされる必須プロセスである。タペータムまたは花柱の輸送管からの糖流出は、花粉発生および後の花粉管成長を刺激する。花は受粉媒介者を誘引するため花蜜産生のために糖を分泌し、そして植物は、潜在的には有益微生物に給餌するために根圏に炭水化物を分泌する(T. Bisseling et al., Science 324, 691 (2009))。糖流出輸送体は、葉肉および種皮を含む多くの他の部位で必要とされる(Y. Zhou et al., Plant J. 49, 750 (2007))。流出トランスポーターの分子の性質は知られていない(S. Lalonde et al., Annu. Rev. Plant Biol. 55, 341 (2004))。植物由来の糖はまた、病原体のための基質を提供する。病原体の第一次的ゴールは、効率的に繁殖するために宿主植物から栄養分を摂取することである。シュードモナス(Pseudomonas)属およびザントモナス(Xanthomonas)属の植物病原性細菌は、植物組織の細胞外空間(アポプラズム)に生息でき、ここでそれらはエネルギー源および炭素骨格源として炭水化物を獲得する。成功した病原体は、かかるメカニズムを共に選択して栄養素流動を変化させる可能性がある(J. W. Patrick, Aust. J. Plant Physiol. 16, 53 (1989))。結果として、病原体と植物は、植物が栄養素への病原体のアクセスを制限しようとし、また防御戦略を開始する一方で、病原体が栄養素へのアクセスを得ようとし、宿主免疫を抑制する戦略を考え出す進化的主導権争いに突入する。植物の防御を変化させるための病原体によって用いられるメカニズムに対する洞察が現在出現しつつあるが;どのようにして病原体が、病原体の増殖を支援するために宿主の生理機能、特に糖排出を変化させるのかについて、わずかしか知られていない。したがって我々は、小胞または細胞膜で糖を分泌するトランスポーターの存在を突き止めた。我々はまた、これらの植物流出トランスポーターが病原体によって栄養素の要求を支給するために「共に選択」されると仮定した。少なくともコムギウドンコ病の場合、グルコースは植物宿主から病原体に移送される主な糖である。それぞれの病原体グルコース/H+コトランスポーターを同定した;反対に、植物糖流出メカニズムはもとより不明のままであった。
【0258】
参照植物シロイヌナズナから新規なグルコーストランスポーターを同定するため、哺乳動物発現系を用いて、植物タンパク質データベースAramemnon (http://aramemnon.botanik.uni-koeln.de)の特徴付けられていないポリトピックな膜タンパク質をコードする遺伝子をスクリーニングした。低内因性グルコース取込活性によって特徴付けられるヒト胚性腎臓HEK293T細胞において、候補遺伝子を高感受性FRETグルコースセンサー(すなわちFLIPglu600μΔ13V)と共発現した。グルコース依存ネガティブFRET比変化によって検出したとき、試験した遺伝子のうち、SWEET1(AT1G21460)発現によってHEK293T細胞の糖蓄積が可能となった;輸送機能と一致した(図9A)。SWEET1が細胞基質からの流出も仲介できるかを決定するため、小胞体の内腔にFRETグルコースセンサーFLIPglu600μΔ13VERを発現した(ER;図9B)。形態的に、細胞膜(PM)を通過する取込は運搬体の細胞外側から開始されるが、一方でERへの「排出」は、トランスポーターの細胞質側から開始される(図9C)。ERセンサーのグルコース依存的応答は、SWEET1がPMを通過した取り込みおよびERへの「流出」を仲介できることを示す。したがってSWEET1はグルコース単輸送体として機能でき、輸送の方向は、膜内外のグルコース勾配のみに依存する。HEK293T細胞の内因性グルコーストランスポーター(GLUT)は、GLUT阻害剤であるサイトカラシンBがSWEET1誘導性グルコース取り込みに影響しなかったため、グルコース取り込みに関与しなかった(図12A)。さらに、GLUTおよびSGLTファミリーの既知のヒトグルコーストランスポーターのmRNAレベルは、SWEET1を発現するHEK293T細胞において誘導されなかった(図12B)。SWEET1活性がグルコース取り込みに必要であることを独立して示すため、18種のヘキソーストランスポーター全てを欠く酵母変異体においてSWEET1を発現した。SWEET1は、グルコースでの増殖(図9D)およびFRETグルコースセンサーFLII12Pglu700μ∂6を用いて測定したとき細胞内グルコース蓄積(図9E)を酵母変異体に可能とした。さらなる特徴付けにより、SWEET1が酵母における低親和性トランスポーターとして、グルコースについて9 mMのKmで機能することが示された(図9F)。単輸送メカニズムと一致して、取り込みは通電によって刺激されず、またほぼpH非依存的であった(図13)。シロイヌナズナの根において記載したグルコース輸送活性と同様に、SWEET1介在性取込は、グルコースアナログ3−O−メチルグルコースによって僅かに阻害された(図13B)。細胞取込および流出における役割の一助として、恒常的に発現されるSWEET1-YFP融合体は、シロイヌナズナの葉のPMに局在している(図9G)。マイクロアレイ試験に基づくと、SWEET1は根において弱くしか発現されないが、シロイヌナズナの花では高度に発現され、このことは配偶体または蜜腺に栄養素を供給する役割を示唆している(図14)。推定糖トランスポーターの根系における生化学的特性の顕著な類似性にもかかわらず、根におけるSWEET1発現は低く、このことは根からのグルコース流出にあまり関与していないことを示唆している。SWEET1と関連している可能性がある他のタンパク質は、根における糖輸送に関与している可能性がある。
【0259】
SWEET1は、シロイヌナズナにおいて17種のメンバーおよびコメにおいて19種のメンバーを有する新たなトランスポーターファミリー(図1)の最初に特徴付けられたメンバーである。シロイヌナズナSWEETは多様であり、4つのサブクレードに属し(図1A)、28〜86%の範囲の同一性を有する(表1および2)。輸送における機能と一致して、SWEETは、7個の膜貫通ヘリックス(TMH)によって構築される疎水性ポアを形成すると予測される小さな疎水性タンパク質である。コンピューター解析では、SWEETの7個のTMHはTMH4を介して融合した3個のTMHドメインポリペプチド(1−3および5−7)の古い重複によってもたらされることが示唆される(図9H)。
【0260】
このファミリーのいずれのメンバーも機能的に特徴付けられていなかったが、複数のSWEET変異体の表現型が記載されている。SWEET1はそのパラログであるSWEET8と41%同一であり、4つのシロイヌナズナSWEETクレードの第二に属する。SWEET8/RPG1の変異は、雄不妊を導くことが示された。HEK293T細胞におけるFRETセンサーFLIPglu600μΔ13VまたはFLIPglu600μΔ13VERとSWEET8/RPG1の共発現は、SWEET8も単輸送体として機能することを示唆する(図15A:図9C)。さらにSWEET8/RPG1は、酵母グルコース輸送変異体を補足する(図9D)。SWEET8/RPG1はタペータムにおいて発現され、このことは花粉の栄養補給に必要なグルコースエフラクサーとしての役割を強く示唆している。
【0261】
SWEET1およびSWEET8はコメタンパク質OsSWEET11/Os8N3(系統樹に基づいてOsSWEET11と命名、図1)と34%のアミノ酸配列同一性を有する。最も近いシロイヌナズナホモログは、OsSWEET11/Os8N3と40%同一性を有し、第三SWEETクレードに属する(図1)。SWEET8と同様に、OsSWEET11/Os8N3は、RNA阻害によるその発現の低下が花粉において低下したデンプン含量および花粉の不稔性を導くため、花粉の栄養補給において機能すると思われる。クレード3のSWEETの別のホモログ(図1)であるペチュニアNec1のサイレンシングはまた、雄不稔を導いた。Nec1は蜜腺において発現され、その発生制御は蜜腺のデンプン含量と逆相関し、このことは蜜腺における糖分泌におけるNec1の第二の役割を示唆した。併せて考えると、これらのデータは単子葉および双子葉植物両方でSWEETSが重要な生殖目的に炭水化物を供給する重要な役割を有することを強く示唆する。
【0262】
病原体は宿主植物の光合成能を用いて、増殖複製のためのエネルギーおよび栄養素を得る。広範な病原体がそれらの宿主からのグルコースを必要とすることは十分に確立されている。様々な病原体が配偶体への給餌のような植物発生のために捧げられる宿主糖流出系を乗っ取ると仮定した。したがって、次にシロイヌナズナSWEETファミリーメンバーのmRNAレベルが細菌または真菌感染によって変化するかを試験した(図10)。シュードモナス・シリンゲ(Pseudomonas syringae)病原型トマト株DC3000感染は、シロイヌナズナの葉においてSWEET4、5、7、8および15 mRNAレベルを高度に誘導した。反対に、宿主にタイプIIIエフェクタータンパク質を注入できず、したがって病原性において危険にさらされているDC3000タイプIII分泌変異体(ΔhrcU)は5つの遺伝子の4つを誘導せず、このことはSWEET mRNA存在量がタイプIII依存的に調節されていることを示している。次に他の病原体が同じまたは異なるファミリーメンバーを標的とするかを試験した。ウドンコ病菌Golovinomyces cichoracearumは異なるセットのSWEET mRNA、主としてSWEET12を誘導した(図10A、C)。マイクロアレイデータは、真菌病原体灰色カビ病菌(Botrytis cinerea)が再び異なるSWEETのセット(すなわちSWEET4、15、17)を標的とすることを示した。併せて考えると、SWEET mRNAレベルの病原体特異的調節は、感染部位での糖輸送を変化させ、病原体増殖および植物免疫に影響を与える可能性がある。
【0263】
この仮説と一致して、花粉の栄養素供給に重要なコメ遺伝子OsSWEET11/Os8N3が、病原体感受性因子として機能する。コメossweet11/os8n3変異体は、細菌病原体 mutant was found to be resistant to the bacterial pathogenイネ白葉枯病菌(Xanthomonas oryzae)病原型オリゼ(Xoo)株PXO99Aに抵抗性であることが見出され、このことはOsSWEET11/Os8N3が感染において病原体に糖を供給することを強く示唆している(図11A)。したがって、OsSWEET11/Os8N3もグルコーストランスポーターとして機能するかを試験した。SWEET1およびSWEET8とは異なって、OsSWEET11/Os8N3は、HEK293T細胞におけるグルコース取込を仲介せず、酵素ヘキソース輸送変異体を補足しなかった(データは示さず)が、このことはグルコース取込においてそれが機能しないことを示している。しかし、OsSWEET11/Os8N3がグルコースエフラクサーとして機能することは考えられた。この仮説を検証するため、流出試験に受け入れられる系であるOsSWEET11/Os8N3をアフリカツメガエル卵母細胞において発現させた。OsSWEET11は、SWEET1および哺乳類Na+依存的グルコーストランスポーターSGLT1とは異なって、卵母細胞における[14C]−グルコース取込も仲介できなかった(図9I;図16)。しかし、SGLT1およびOsSWEET11/Os8N3の共発現は、卵母細胞における低下した[14C]−グルコース蓄積を導き、OsSWEET11/Os8N3の流出(すなわち「漏出」)活性と適合する発見であった(図9I、K)。The hypothesis that OsSWEET11/Os8N3がグルコースを排出できるという仮説は、直接流出測定によって実証される。植物タンパク質を発現する卵母細胞に[14C]−グルコースを注入して、グルコース流出を測定した。SWEET1およびOsSWEET11/Os8N3はいずれも[14C]−グルコースを流出でき(図9G)、これはSWEET1は促進因子として機能するが、OsSWEET11/Os8N3はエフラクサー(潜在的にはH+/グルコースアンチポーター)であることを示唆している。さらに、OsSWEET11(Os8N3)はショ糖を輸送できる(図43G)。したがって、OsSWEET11/Os8N3は、生殖のためにグルコースおよびショ糖を得るために病原体によって採用されると思われる。
【0264】
OsSWEET11/Os8N3が糖エフラクサーとして機能するという発見は、どのように病原体が植物のエネルギー源へのアクセスを得るために基本的植物機能を共に選択するかのモデルを提供する(図11)。Xoo株PXO99Aは、コメの感染についてタイプIIIエフェクター遺伝子に依存する。PthXo1はTAL(転写アクチベーター様)エフェクターであり、これはDNAと直接相互作用して標的遺伝子の転写を促進する。Xoo PXO99Aによって分泌されるPthXo1は、OsSWEET11/Os8N3の転写を特に活性化して、おそらくアポプラスト細菌に給餌するために糖流出を誘導する(図11A)。PthXo1が変異されると(ME)、OsSWEET11/Os8N3の転写および病原性が低下し、これは病原体の飢餓と一致する(図11B)。変異またはRNA阻害によりOsSWEET11/Os8N3が利用できなくなった場合、糖(例えばグルコース、glc)は十分な量で排出されず、病原体は飢餓となるであろう(図11C)。実際、ossweet11/os8n3変異体はXoo PXO99Aに抵抗性である。別のTALエフェクター(AvrXa7)を担持するPXO99A細菌はossweet11/os8n3変異体においてさえ病原性であり(図11D)、これは他のSWEETが病原体増殖を支援するために病原体によって共に選択されるという最も簡単な仮説と一致している(図11D)。実際、PthXo1によって標的とされる推定DNA配列は、TRCA・CT・CCATTACTRTAAAA・N・(OsSWEET11/Os8N3の上流プロモーターにおいて見られる)であり、一方でAvrXa7によって標的とされるものはTA・AANCRCCCN・・CCNNRRATRA・Nである。この配列は潜在的標的を同定するのに十分ではなかった。これらの知見は、免疫におけるそれらの役割に加えて、タイプIIIエフェクターはまた、宿主植物の栄養源へのアクセスをえることにも関与しているという考えを支持する。どのように真菌病原体がこれらのトランスポーターのプロモーターを標的とするかは未だ理解されていないが、トランスポーター遺伝子は、真菌増殖を支える制御ネットワークを解明するための好適な診断ツールでありうる。明らかに、この病原ターベースの選択圧に拘わらず進化において維持されるために、SWEETトランスポーターは植物において必須機能を有するはずである;変異体の分析は、少なくともそれらの1個が配偶体への炭水化物の供給に関与していることを示唆している。したがって、他のパラログの活性も、他の植物機能に重要であり得る。特に疾患感受性についての残りのSWEETパラログの特徴付けおよび変異体の分析は、次の段階で重要である。
【0265】
病原体エフェクター分子の全スペクトルおよびどのようにそれが病原体増殖を支持するために植物代謝を阻害するかについての知識は、宿主−病原体相互作用の我々の理解を改善し、年間10%以上もの農作物損失を導くグローバル規模での病原体感染と闘う新たな戦略を導くことができる。さらに、SWEETファミリーの他の遺伝子の分析は、花粉の栄養、花蜜産生および植物の根からの炭素隔離の謎のいくつかを解決する助けとなりうる。興味深いことに、動物ゲノムは糖輸送にも関与するSWEETホモログを含む。
【0266】
実施例7:線虫(C. elegans)およびヒトにおけるグルコーストランスポーターの第三ファミリー
【0267】
材料および方法
【0268】
SGLT1のORF(Invitrogen, Carlsbad, CA)をPCRで増幅し、pCR2.1-TOPO(Invitrogen)にクローン化した。SGLT1 ORFをEcoRI/XhoIで切り取り、p002の対応する部位にクローン化した。pDNR-LIBのスプライス変異体RAG1AP1-1(Clone ID: 4076256、Open Biosystems, Huntsville, AL)をEcoRI/XhoIで制限し、pOO2にクローン化した。インビトロLR組換え(Gateway)によってpCMV-SPORT6のRAG1AP1-2(Clone ID: 3896154、Open Biosystems, Huntsville, AL)をpOO2-GW(D. Loque、未公表)に移した。部位特異的突然変異誘発によってRAG1AP1-3aaをY216A、L218A、L219A(推定内在化モチーフ)に変異させて、インビトロBP組換えによってpDONR221-f1にクローン化し、さらにLR反応によってpOO2-GWに移動させた。LR反応を用いてCeSWEET1(K02D7.5)、CeSWEET3(C06G8.1)、CeSWEET4(Y39A1A.8)、CeSWEET5(K06A4.4)およびCeSWEET7(K11D12.5)(Open Biosystems, Huntsville, AL)をpOO2-GWにクローン化した。次いで、部位特異的突然変異誘発によって開始および終止コドンを加えた。卵母細胞発現および輸送試験は、全取込実験卵母細胞をMBS中1 mMのグルコース中で一夜プレインキュベートする以外は、Chen et alに記載のとおりに実施した。
【0269】
結果および考察
【0270】
線虫(C. elegans)ゲノムは糖流出トランスポーター(SLC50)の新規クラスの7種のホモログを含むが、ヒトゲノムはRAG1AP1と命名された1種のホモログを有する。シロイヌナズナSWEET1と同様に、線虫(C. elegans)CeSWEET1はアフリカツメガエル卵母細胞で発現された場合グルコース取込を仲介する。CeSWEET1およびヒトRAG1AP1は、Na+/グルコースコトランスポーターSGLT1によって仲介される卵母細胞の第二活性グルコース蓄積を相殺する。CeSWEET1の変異は、脂質の蓄積を導く細胞グルコース流出の欠損と両立して、脂肪蓄積を導いた。これらの知見は、糖輸送におけるヒトホモログRAG1AP1の役割に新たな光を投げかけうる。
【0271】
ヒトゲノムはグルコーストランスポーターの少なくとも2つのクラス、SLC2およびSLC5を含む。GLUTと名付けられたSLC2は単輸送体であり、すなわちそれらは濃度勾配に沿ってグルコースを輸送する。反対にSGLTは、ナトリウム勾配によって駆動されてグルコースを能動的に取り込むことができるNa+共役コトランスポーターである。これらのトランスポーターは、ヒトにおいて見られるほとんどの取込活性を説明でき、例えばGLUT2ノックアウト変異マウスは、劇的に低下した取り込み能を示す。しかし驚くべきことに、グルコースクリアランスは正常であり、このことは別の流出経路の存在を示唆している。FRETセンサーを利用した発現クローニング系の使用の使用は、植物におけるグルコーストランスポーターの新規クラスの同定を導く(msに伴う)。シロイヌナズナSWEET1および8は単輸送体として機能するが、コメOsSWEET11はグルコースおよびショ糖を流出させると思われる。バイオインフォマティック分析によって、動物およびヒトゲノムが溶質輸送体ファミリーSLC50として登録されている(www.bioparadigms.org/slc/intro.htm)ホモログを含むことが示された(図1)。ここで我々は、アフリカツメガエル卵母細胞において発現されたとき、線虫(C. elegans)CeSWEET1が弱い、しかし重要なグルコース取込を仲介することを示す(図19B)。活性はSGLT1または植物ホモログSWEET1のものよりも顕著に弱い。比較として、線虫(C. elegans)ホモログCeSWEET3、4、5および7は、卵母細胞において検出可能なグルコース取込を仲介しなかった(図19B)。グルコースおよびショ糖を流出させる植物ホモログOsSWEET11は、両方のタンパク質が共発現された場合、卵母細胞におけるSGLT1介在性グルコース蓄積を相殺することが示された。作業仮説は、OsSWEET11介在性流出が「漏出」として作用し、卵母細胞におけるグルコース蓄積を予防することである(3)。線虫(C. elegans)ホモログがエフラクサーとして潜在的に機能しうるかを検証するため、我々は、SGLT1とそれらを共発現させた(図19C)。試験した5種のホモログ全てが、SGLT1のみと比較して低下したグルコース蓄積を導いた。[14C]−グルコースを注入した卵母細胞からの直接流出測定は、RAP1AP1が卵母細胞からのグルコースの流出を誘導することを示す(2分以内に放出されたグルコースの%: RAG1AP1: 5.27 ± 0.29; コントロール: 0.91 ± 0.23; S.E.; n>7)。線虫(C. elegans)ホモログの一つの変異体(CeSWEET1; K02D7.5)は、糖を流出する能力の欠如と一致して、脂肪蓄積を導く。CeSWEET3、4、5および7と同様に、ヒトホモログRAG1AP1(HsSWEET1と命名)はまた、卵母細胞においてグルコース蓄積を仲介することはできなかったが、SGLT1介在性グルコース蓄積を相殺した。これは2種のスプライス変異体および推定内在化モチーフを形成する3個の残基における3つの変異を担持する変異型についても妥当した(図19C)。ホヤCiona由来のホモログCi-Rga(CiSWEET1)における変異は初期発生以上を導き、後生生物にとってのこれらの遺伝子の重要性を裏付けている。哺乳類において、肝臓からの少なくとも1つのグルコース流出経路が不明のままである。
【0272】
RAG1AP1は、細胞系における欠損が組換えに影響を与えるため、組換え活性化遺伝子1活性化タンパク質1(Recombination Activating Gene 1 Activating Protein 1)から名付けられた。さらに、該遺伝子はRGAと名付けられており、TRPVイオンチャネルのターゲティングを意味している。RAG1AP1が肝臓におけるグルコース流出に寄与しうるという仮説を検証することが興味深い。
【0273】
実施例8:HEK293細胞におけるSWEETショ糖排出
【0274】
SWEET1〜4および6〜11および14〜16をFRETショ糖センサーと共にHEK293において発現させた。SWEETファミリーの複数のメンバーがショ糖を排出した。この能力を示したSWEETメンバーは全て、同じクレードに属する(図1参照)。したがって、このクレードはショ糖を排出する強い能力を示すと思われる。OsSWEET11はこのクレードに属し、従ってグルコースを排出することに加えて、これらのタンパク質はショ糖も排出する。これはショ糖を取り込むUstilagoのような病原体のターゲティングにおいて顕著であり得る。植物病原体トウモロコシ黒穂病菌(Ustilago maydis)の病原性のために、新たな高親和性ショ糖トランスポーターが必要とされることは報告されている。PLoS Biol. 8(2):e1000303)または次いで細胞壁インベルターゼがショ糖を切断し、病原体がグルコースを取り込む。
【0275】
ショ糖エフラクサーが葉における細胞間輸送に必要とされることは長らく知られていたが、これらのタンパク質の同一性は不明なままであったため、SWEETのショ糖を排出する示された能力は重要である。SUTは葉のどこかで細胞壁からショ糖を取り込み、葉肉細胞で産生されたショ糖を細胞壁に流出する。したがって、ショ糖を排出するSWEETの能力は、どのように植物がこれを達成するかを理解する上で重要なデータを提供する。さらに、植物の母系組織は植物における主な輸送される糖として発生中の種子にショ糖を供給するためにショ糖を排出する必要がある。したがって、ショ糖エクスポーターとしてのSWEETの役割は、発生中の種子において重要であり得る。
【0276】
本明細書において全ての文献および特許出願は、各個々の文献または特許出願が具体的および個々に記載されているのと同程度に、参照により本明細書に組み込まれる。本明細書に記載の文献は、本願の出願日前のそれらの開示について単に提供される。本明細書のあらゆる事項は、本願発明が先行発明の理由でかかる文献に劣後すると認めるものではない。
【技術分野】
【0001】
発明の分野
本発明は、細胞の膜を通過する新規糖トランスポーターに関する。
【0002】
関連出願
本願は米国仮出願第61/175,267号(2009年5月4日出願)の優先権を主張し、その全体について、参照により本明細書に組み込む。
【背景技術】
【0003】
発明の背景
多くの生化学経路における糖の必要および使用は、数世紀にわたって広範に研究され、報告されている。単糖類、二糖類および多糖類または糖は、多くの生物にとって糖の主たる食事性供給源である。
【0004】
グルコースは、最も容易に利用可能な糖の一つであり、その構造は、多くの生物の生化学系によって容易に作用される。6個の炭素原子からなるグルコースは、アルドヘキソース(aldehexose)のカテゴリーに属する。アルドヘキソースは4個のキラル中心を有し、これらは16個の立体異性体を導く。アルドヘキソースの2個の立体異性体がグルコースと見なされ、主たるものがD−グルコースである。全ての主たる食事性炭水化物が、デンプンおよびグリコーゲンにおけるような唯一の構造ブロックとして、あるいはショ糖およびラクトースにおけるような別の単糖類とともに、グルコースを含む。この炭水化物の代謝は、アデノシン3リン酸(ATP)のようなエネルギーに変換する。グルコースの他の代謝経路は、エネルギー貯蔵を導く。グルコース分子は環の開いた(非環式)形態または環の(環状)形態で存在しうる。
【0005】
グルコースは、デンプン、セルロースおよびグリコーゲンのようないくつかの重要な物質の合成の前駆体として使用できる。牛乳中の糖であるラクトースは、グルコース−ガラクトース二糖である。別の二糖であるショ糖は、グルコースとフルクトースの結合体である。グルコースは後生動物における糖の主要な輸送形態であるが、ショ糖およびその誘導体は、植物における主要な輸送形態として利用される。
【0006】
グルコースは植物およびいくつかの原核生物における光合成の下流産物の一つである。動物および真菌のような真核生物において、グルコースは、グリコーゲン分解と称されるプロセスを介して、グリコーゲンの分解の結果として生産されうる。植物において得られる分解基質はデンプンである。グルコースはまた、植物における主要な輸送糖であるショ糖に対するインベルターゼの作用にも由来しうる(細胞壁、細胞基質または液胞において、各々特異的なアイソフォームによる)。
【0007】
動物では、グルコースはピルビン酸およびグリセロールのような非炭水化物中間体から、糖新生と称されるプロセスを介して肝臓および腎臓において合成されうる。グルコースはまた、例えばデンプンの酵素加水分解によって、合成されることもある。商業的には、市販の農作物、例えばトウモロコシ、コメ、コムギ、ジャガイモ、キャッサバ、サゴおよびクズウコンがデンプンの供給源として使用されうる。
【0008】
グルコースは好気的または嫌気的呼吸のいずれかに使用されうる。炭水化物は生物の重要なエネルギー源である。好気的呼吸は1グラムあたり約3.75 kcalのエネルギーを提供できる。デンプンのような炭水化物の分解は、単糖類および二糖類を生じる。解糖プロセスおよびクエン酸回路(またはクレブス回路)の反応により、グルコースは酸化され、分解されて最終的に二酸化炭素と水を形成し、エネルギー源(主としてATP)を得る。インスリン応答および他のメカニズムは、血中グルコース濃度を制御しうる。
【0009】
脳のような神経中枢エネルギーの必要は、グルコースと心理的プロセスを直接的に関連づける。グルコースは脳の第一次的なエネルギー供給源であり、したがってその利用能は心理的プロセスに影響する。グルコースが少ない場合、精神的な努力を必要とする心理的プロセスは損なわれうる。好気的および嫌気的呼吸のいずれも、解糖代謝経路の初期工程で開始し、最初のステップは、エネルギーを得るための後の分解を準備するための、ヘキソキナーゼによるグルコースのリン酸化である。次いで、ヘキソキナーゼによるグルコースの速やかなリン酸化は、細胞外への拡散を防止しうる。リン酸化の作用は荷電されたリン酸基を加え、それによってグルコース−6−リン酸が容易に細胞膜を通過することを防止する。グルコースはまた、タンパク質の産生および脂質代謝プロセスにおいても重要である。グルコースはまた、アスコルビン酸またはビタミンCの前駆体分子としても利用できる。
【0010】
したがって、グルコースおよびショ糖のような糖の細胞内および生物中の組織の細胞間での取り込み、吸収、処理、代謝、交換および輸送は、細胞または細胞を含む生物の生育に最も重要である。細胞膜を通過し、細胞の小器官間でのグルコースまたはショ糖輸送の機能不全は、破局的でありうる。グルコースの輸送を効率的に制御する方法を開発する必要がある。
【発明の概要】
【0011】
発明の概要
本発明は、細胞内で糖を輸送するためのトランスポータータンパク質の新規クラスを提供する。GLUEと称される(GlueまたはSWEETとも知られる)トランスポーターは、植物中に存在しおり、そして糖(例えばペントース、グルコース、マンノース、要するに単糖類)またはショ糖およびマルトース(要するに二糖類またはオリゴ糖)のトランスポーターをコードする、下記受託番号と少なくとも30%、40%、50%、60%、70%、80%、85%、90%、95%または99%配列同一性を有する核酸によってコードされうる: AT4G15920、AT3G16690、AT5G13170、AtSAG29、AT4G25010、AT5G50800、AT5G23660、AT3G48740、AT5G50790、AT2G39060、AT5G40260、AtRPG1、AT4G10850、AT1G66770、AT1G21460、AT5G62850、AtVEX1、AT3G28007、AT3G14770、AT1G21460、AT5G53190、NEC1、AC202585、AC147714、MtC60432 GC、MtC11004 GC、CT963079、MtD03138 GC、TC 125536、AC146866、AC189276、TC129646、CAA69976 MtNod3 AC2456、TC115479、AC146747、MtC10424 GC、CT954252、CU302340、AC202585、AC147714、MtC60432 GC、MtC11004 GC、CT963079、Os08g42350 (Os8N3) Os08g0535200、Os12g29220 Os03g0347500、Os05g51090 Os05g0588500、Os12g07860、Os09g08440、Os09g08490、Os09g08270、Os09g08030 Os09g0254600、Os01g42090.1 Os01g0605700、Os01g42110.1 Os01g060600、Os02g19820 Os02g0301100、Os05g35140 Os05g0426000、Os01g65880 Os01g0881300、Os01g50460 Os01g0700100、Os01g36070.1 Os01g0541800、Os01g12130.1、Os05g12320 Os05g0214300およびOs01g21230(いずれもその全体について参照により本明細書に組み込む)。核酸はベクター中および/または細胞、例えば植物細胞または動物細胞中に存在していてよい。本発明はまた、GLUEを含むトランスジェニック植物も提供する。該核酸は、下記動物受託番号と少なくとも30%、40%、50%、60%、70%、80%、85%、90%、95%または99%配列同一性を有する、グルコースまたはショ糖トランスポーターをコードする核酸によってコードされうる:(例えば線虫(C. elegans)由来)R10D12.9、K11D12.5、およびK06A4.4、K02D7.5、C54F6.4、C06G8.1、Y39A1A.8、ci-rga、RAG1AP1(例えばショウジョウバエ、ヒト、マウス、ドブネズミ(Rattus norvegicus)およびアフリカツメガエル(Xenopus))(いずれもその全体について参照により本明細書に組み込む)。
【0012】
本発明はさらに、成熟したGLUEタンパク質を提供する。GLUEによる糖の移動が野生型と比較して改善されるように、GLUEは、変異されていてもよい。移動速度の増加または輸送能力の増加によりより多くの糖を輸送できるように、変異がGLUEの機能を改善していてよい。変異は、野生型と比較して、GLUEによる糖の移動を防止または遅延させうる。変異は、野生型配列のアミノ酸または複数のアミノ酸の欠失または置換であってよい。変異はGLUEの切断であってもよい。
【0013】
本発明は、GLUEタンパク質を含む融合タンパク質を提供する。GLUEはタグ、例えばエピトープと融合されてよい。GLUEは膜タンパク質のキメラメンバーの一部、例えば既知の下流カスケードでの他の7回膜貫通タンパク質であってよい。キメラタンパク質はGLUEの第3細胞内ループおよび/または細胞質尾部が別の7回膜貫通タンパク質由来の対応するドメインでの置換を含みうる。GLUEは、細胞内の特定の小器官または領域とGLUEの発現および位置を指定するために、標的配列と結合していてもよい。GLUEは変異GLUEタンパク質であってよい。
【0014】
本発明は、コントロール植物と比較して増加したレベルの炭素を産生する植物を作成する方法であって、植物細胞に単糖、二糖またはオリゴ糖トランスポーターをコードする核酸を導入し、GLUEをコードする核酸を発現する植物に植物細胞を成長させることを含む方法を提供する。
【0015】
本発明は、植物の根への糖輸送を増加させる方法であって、植物の細胞にGLUEをコードする核酸を導入することを含む方法を提供する。植物へのGLUEの導入は、植物の根圏への糖分泌を調節する方法および植物の葉圏への糖輸送または発生種子、花等への糖の送達を調節する方法を提供できる。
【0016】
本発明は、植物における糖のレベルを変化させる方法であって、細胞にGLUEをコードする核酸を導入することを含む方法を提供する。本方法は、細胞内の糖レベルを低下させてもよい。本方法は、植物または動物のある領域、臓器または小器官に蓄積させるために糖濃度を指定しうる。本ほう法は、植物または動物のある領域、臓器または小器官における糖レベルの低下を引き起こしてもよい。本方法は、糖の取り込みを増加させてもよい。本方法は、糖の取り込みを減少させてもよい。本方法は、糖の排出を増加させてもよい。本方法は、糖の排出を減少させてもよい。GLUEはコファクター、例えば別の細胞内タンパク質またはトランスポーター、例えばコトランスポーターによって細胞内で発現される。
【0017】
本発明は、植物に有益微生物を誘引する方法であって、GLUEの導入によって糖濃度を変化させることを含む方法を提供する。本発明はさらに、GLUEの導入によって病原体から植物を保護する方法を提供する。植物侵襲病原体は、植物細胞機構を利用して植物の糖排出を変化させることがある。さらに異なるプロモーターの制御下に存在しうる異種GLUEを植物に導入することにより、病原体の糖排出を変化させる能力が限定または変化されうる。
【0018】
本発明はまた、細胞または生物中でGLUEがどのように作用しているかを決定する方法を提供する。糖検出分子、例えば蛍光ドナードメインと蛍光アクセプタードメイン間で挟まれた糖(例えばグルコースまたはショ糖)結合ドメインのような、糖検出分子と共に、異種GLUEは細胞内で共発現されてよい。糖濃度は蛍光共鳴エネルギー移動によって経時的に測定およびモニターできる。
【0019】
本発明は、細胞中のグルコーストランスポーター促進(GLUT)タンパク質の発現に影響し、および/またはそれを変化させる方法を提供する。
【0020】
本発明は、細胞によって分泌される液体、例えば花蜜または樹液中の糖レベルを変化させる方法を提供する。本発明は、生物の細胞にGLUEを導入することによって生物の発生を変化させる方法を提供する。GLUEは変異されていてもよい。本発明は、生物の細胞中のGLUEを変異することによって生物の発生を変化させる方法を提供する。生物中のGLUEの上昇または低下した機能は、細胞中または生物中の糖濃度および/または糖分布を変化させ、それによって発生に影響を与えうる。
【図面の簡単な説明】
【0021】
【図1】多様な植物種におけるGLUEスーパーファミリーcDNAの系統樹を示す。図1AはGLUEスーパーファミリーの系統樹を示す。距離はネイバージョイニング法を用いた複数配列アラインメント(ClustalW)から計算し、ツリーはブーストラップ値を示す(1000のパーセンテージ)。SWEETファミリーは4つのクレードに分割できる。全ての配列はNCBIまたはAramemnonデータベースから得た。コメ、シロイヌナズナ、ウマゴヤシおよびペチュニアのツリーを示す。示したスケールは示した長さあたり50塩基の変化を意味する。
【0022】
【図2】植物の細胞内、器官および小器官で糖が輸送され、移動する多様な手段の概要を示す。
【0023】
【図3】図3Aおよび3Bは細胞中で発現されたグルコースFRETセンサーによる応答を示す。図3AはGLUEの共発現なしでのグルコースの多様な濃度に対するセンサーの応答を示す。図3BはグルコースFRETセンサーを有する細胞にGLUE1を導入することが、糖の濃度が変化した場合にセンサーに顕著な変化をもたらすことを示す。
【0024】
【図4】図4A〜4Dは発現したGLUEタンパク質の変化による変化したグルコース濃度に対するグルコースFRETセンサーの応答を示す。図4AはGLUE1による応答を示す。図4BはGLUE12による応答を示す。図4CはGLUE8による応答を示す。図4DはGLUE13による応答を示す。
【0025】
【図5】ベクターのみ(EBY-pDRF1)を有するグルコース取り込み欠損株EBY4000がD−グルコースを取り込めないことを示す。放射性標識化14C−グルコース取り込みを経時的に測定した。反対に、GLUE1は酵母株EBY4000(EBY-GLUE1)にグルコース取り込みを可能とさせる。。グルコース取り込みコンピテント野生型株CENPK(CEN)はポジティブコントロールとして使用し、グルコースを取り込める。
【0026】
【図6】試験したGLUEの15種の遺伝子のうち4つが酵母変異をレスキューできる。酵母は異なるpHを有する2%グルコースで増殖した。
【0027】
【図7】授乳中のRAG1AP1(HsSWEET1)の誘導を示す。マイクロアレイデータは、授乳中に推定糖トランスポーターRAG1AP1がアップレギュレートされることを示唆している。
【0028】
【図8】肺胞細胞からのラクトース合成および分泌を示す。グルコースはGLUT/SGLTによって基底膜を介して取り込まれ、次いで未知のトランスポーターによりER(○1)またはゴルジ体(○2)に取り込まれる。ラクトース合成はゴルジ体において生じ、ラクトースは乳管に面した頂側で排出されると推測される。未知のトランスポーターは、頂側の膜を介してグルコースを排出するか、または乳由来グルコースの回復に関与する(○3)。
【0029】
【図9】SWEET(GLUE)トランスポーターの同定および特徴付けを示す。図9AはHEK293T細胞における細胞基質FRETグルコースセンサーFLIPglu600μΔ13Vとの共発現による、SWEET1についてのグルコース輸送活性の同定を示す。個々の細胞はCFPおよびVenus放出の定量比イメージングによって分析した(収集間隔5秒;Fc/Dは正規化した放出強度比に対応する)。HEK293T/FLIPglu600μΔ13V細胞を増加したグルコース濃度の中間パルス、次いで方形パルスでかん流した。センサーのみを発現する細胞は、FRET比変化(オレンジの線)がないことによって示されるように、細胞質中の顕著な量のグルコースを蓄積しなかった。センサーおよびSWEET1を共発現する細胞は、増加した外部グルコース供給と相関する振幅でのネガティブFRET比変化によって示されるとおり、グルコースを蓄積した(青色線)。データ点は平均±SD(n>10)である。図9Bは、細胞質からERへのグルコースの「流出」のFRETイメージングを示す(図9C参照)。センサーFLIPglu600μΔ13VERはERの膜を標的とした(図9Aと同様に分析を行った。収集間隔10秒)。センサーのみを発現する細胞はERにおける顕著な量のグルコースを蓄積しなかった。センサーおよびSWEET1を共発現する細胞は、グルコースの流出によって誘導されるネガティブ比変化によって示されるとおり、ER中にグルコースを蓄積した。データ点は平均±SD(n>10)である。図9CはPMを通過するSWEET1流入および細胞質からERへの流出の模式図を示す。細胞基質センサーFLIPglu600μΔ13Vは、細胞外表面で開始されるグルコースの輸送を同定する(細胞外N末端によって示される)。FLIPglu600μΔ13VERは、細胞内側で開始される輸送を測定する(細胞基質C末端)。図9DはSWEET1、SWEET8または酵母HXT5による18種のヘキソーストランスポーター遺伝子全てを欠く酵母株EBY4000の相補性を示す;コントロール:空のベクター。図9Eは、0、20および100 mMのグルコース添加前後のSWEET1およびFLII12Pglu700μΔ6を共発現するEBY4000におけるグルコースの蓄積を示す。グルコース添加前に2つのサイクルを実行した。データは平均±SD、n=3である。図9FはEBY4000中のSWEET1による14C−グルコース蓄積の動力学を示す。データは平均±SD、n=3である。図9Gは安定的に形質転換されたシロイヌナズナの葉の葉におけるSWEET1−YFPの共焦点イメージングを示す。図21Hは疎水性プロットに基づくSWEETの構造モデルを示す。各タンパク質は推定の細胞外N末端、およびリンカーとしてTMH4で区分された3個のTMH(TMH1〜3および5〜7、赤および青の三角形で強調した)の重複に由来する推定の平行方向の2個の「サブユニット」を有する7個のTMHを含む。図9IはSGLT1によって仲介されるが、OsSWEET11によって仲介されないアフリカツメガエル卵母細胞への[14C]−グルコースの取り込みを示す。SGLT1とOsSWEET11の共発現は、卵母細胞におけるグルコース蓄積を減少させる。データは平均±SD、n=7である。挿入図はSGLT1によるグルコースの集中的取り込みおよびOsSWEET11によって引き起こされるグルコース流出(「リーク」)を示す。図9KはSWEET1またはOsSWEET11を発現する卵母細胞からの直接流出測定を示す。10 mMの放射性標識化グルコース(0.18μCi/μl)50 nlを注射して、放射性トレーサー流出を経時的に測定した。データは平均±SD(n>10細胞)である。
【0030】
【図10】生体栄養細菌または真菌が異なるSWEET遺伝子のmRNAレベルを誘導することを示す。図10Aは細菌性シュードモナス・シリンゲ(Pseudomonas syringae)病原型トマトDC3000(2 x 108 cfu/ml、接種後8時間、qPCRで測定、MgCl2バッファー処理で正規化)、ウドンコ病菌、G. cichoracearum(〜25〜35 分生子 mm-2、接種後48時間、qPCRで測定;0時間値に正規化)、またはシロイヌナズナの葉におけるボトリチス・シネレア菌(Botrytis cinerea)による、SWEET mRNAの誘導を示す。図10BはP.s. DC3000によるSWEET4、5および15の誘導が機能的タイプIII分泌系(T3S)に基づくことを示す。サンプルは2 x 108 cfu/mlのDC3000またはT3S変異体DC3000ΔhrcUでの浸潤後6、12および24時間で採取した。図10CはG. cichoracearumによる感染がSWEET11およびSWEET12の誘導を導くが、SWEET15のダウンレギュレーションを導くことを示す。サンプルは接種後0、8、12、24および72時間後に採取した。
【0031】
【図11】病原体の栄養摂取におけるSWEETの役割についてのスキームモデルを示す。図11Aは病原菌キサントモナス・オリゼ(Xanthomonas oryzae)病原型オリゼ株PXO99 A(X.o. PXO99A)がタイプIII分泌系を介してコメ細胞にTALエフェクターPthXo1を注入することを示す。この転写アクチベーターは直接または間接的に、グルコースの分泌を導くコメOsSWEET11/Os8N3グルコース流出トランスポーター遺伝子の誘導を惹起する。細菌は内因性取り込み系を介してグルコースを取り込み、増殖できる。図11BはPthXo1が変異されている場合(ME)、OsSWEET11/Os8N3の誘導は低下または停止されて、細菌の飢餓を導くことを示す(細菌細胞の減少したサイズ、平均少細胞数として示される)。図11CはOsSWEET11/Os8N3の変異も細菌の飢餓を導くことを示す。図11Dは別のエフェクターAvrXa7を発現する病原体が、SWEETファミリーの別のメンバーを誘導する(または別の炭素源へのアクセスを誘導する)場合、増殖できることを示す。
【0032】
【図12】HEK293T細胞におけるSWEET仲介グルコース輸送の証拠を示す。図12AはGLUT1とコトランスフェクトされたFLIPglu600μΔ13Vセンサーを用いて分析した、20 μMのサイトカラシンBによるGLUT1活性の阻害を示す。図12BはSWEET1とコトランスフェクトされたFLIPglu600μΔ13Vを用いて分析した、20 μMのサイトカラシンBに対するSWEET1活性の非感受性を示す。図12CはHepG2細胞、HEK293T細胞ならびにFLIPglu600μΔ13Vおよび/またはSWEET1を共発現するHEK293T細胞における、SLC2(GLUT)およびSLC5(SGLT)グルコーストランスポーターの発現レベルを示す。
【0033】
【図13】SWEET1の生化学的特徴を示す。図13AはSWEET1の最適pHを示す。放射性トレーサー取り込みを異なるpHで測定した。取り込みの至適pHは約pH8.5である。データは平均±SDである。図13Bは異なる糖による酵母株EBY4000におけるSWEET1によって仲介されるグルコース取り込み(5 mM D−グルコース;0.1 μCi [14C]−D−グルコース)の阻害を示す。コンペティターを10倍過剰(最終濃度50 mM)で加えた。相対活性はD−グルコース取り込み速度(100%)に正規化した。データは平均±SDである。
【0034】
【図14】シロイヌナズナにおけるSWEET1およびSWEET8遺伝子の組織特異的発現パターンを示す。分析はArabadopsis eFP Browser(http://bar.utoronto.ca/efp/cgi-bin/efpWeb.cgi)のマイクロアレイ試験に基づく。
【0035】
【図15】異種系におけるSWEET8の機能的分析を示す。図15Aは細胞基質FRETグルコースセンサーFLIPglu600μΔ13VおよびSWEET8を共発現する細胞が、HEK293T細胞におけるネガティブな細胞基質FRET比変化によって示されるとおり、細胞基質中にグルコースを蓄積したことを示す(図9A参照)。図15BはHEK293T細胞においてER FRETグルコースセンサーFLIPglu-600μΔ13VERおよびSWEET8を共発現する場合、細胞が細胞基質からERへとグルコースを流出することを示す(図9B参照)。データ点は平均±SD(n>10)である。図15Cは酵母グルコース輸送欠損変異体EBY4000におけるSWEET1、SWEETおよびベクターコントロールの相対的取り込み速度を示す(2分;10mM D−グルコース;0.1μCi [14C]−D−グルコース)。値をSWEET1(100%)に正規化する。データは平均±SDである。図15Dは安定的に形質転換されたシロイヌナズナを用いた実生(T1世代)におけるSWEET8局在の共焦点イメージングを示す。
【0036】
【図16】アフリカツメガエル卵母細胞におけるSWEET仲介グルコース取り込みを示す。卵母細胞における相対的グルコース取り込み速度は、コントロールとして水、SWEET1、OsSWEET11、SGLT1またはSGLT1とSWEET1を注射した(時間=1時間;1mM D−グルコース; 4μCi/ml [14C]−D−グルコース)。データは平均±SD、n>9である。
【0037】
【図17】HEK293T細胞におけるグルコースの実時間取り込みを示す。GLUE1はHEK293T細胞において細胞基質またはER FRETセンサーFLIPglu600μD6と共発現された。CFPおよびVFPの正規化した放出比をY軸に示す。ネガティブなFRET比の変化は、GLUE1がグルコーストランスポーターならびにエフラクサー(effluxer)として機能することを示している。
【0038】
【図18】GLUE8、12および13がHEK293T細胞において細胞基質FRETセンサーFLIPglu600μD6と共発現されたことを示している。CFPおよびVFPの正規化した放出比をY軸に示す。細胞基質におけるネガティブなFRET比の変化は、GLUE8、12および13が異なる活性を有するグルコーストランスポーターとして機能することを示している。
【0039】
【図19】アフリカツメガエル卵母細胞におけるGLUEの機能的発現を示す。図19AはSGLT1、GLUE1によって仲介されるがOsGLUE11によって仲介されないアフリカツメガエル卵母細胞への[14C]−グルコースの取り込みを示す。OsGLUE11とSGLT1の共発現はグルコース蓄積を減少させる。図19BはCeSWEET1、3、4、5、7およびRAG1AP1スプライス変異体1および2ならびに変異体(Y216A、L218A、L219A; RAG1AP1-3aa)によって仲介される、アフリカツメガエル卵母細胞への14C−グルコースの取り込みを示す。SGLT1をコントロールとして用いた。図19Cはアフリカツメガエル卵母細胞におけるSGLT1によるグルコース蓄積に対するCeSWEETおよびRAG1AP1変異体の共発現の影響を示す。全実験は独立して少なくとも6回反復した。エラーバーは平均±SDである。
【0040】
【図20】17種のシロイヌナズナGLUE遺伝子による18種のヘキソーストランスポーター遺伝子全てを欠く酵母株YSL2-1の相補性を示す。酵母ホモログHXT5または哺乳類ホモログGLUT1を発現する細胞をコントロールとして用いた。Os8N3およびRAG1AP1は、コメおよび哺乳動物由来ホモログである。酵母細胞を初期対数期に2%マルトースを含むSD-Ura液体培地中で増殖し、2%のマルトース、フルクトースまたはマンノースを含むYPMまたはSD-URAを含む培地に、5m の連続希釈をスポットした。GLUE4およびGLUE7はフルクトースおよびマンノースを輸送する。GLUE5および8はマンノースを輸送する。
【0041】
【図21】17種のシロイヌナズナGLUE遺伝子による18種のヘキソーストランスポーター遺伝子全てを欠く酵母株YSL2-1の相補性を示す。哺乳類ホモログGLUT1を発現する細胞をコントロールとして用いた。Os8N3およびRAG1AP1は、コメおよび哺乳動物由来ホモログである。酵母細胞を初期対数期に2%マルトースを含むSD-Ura液体培地中で増殖し、多様な濃度のガラクトースを含むSD-URAを含む培地に、5m の連続希釈をスポットした。GLUE1、4、5および7を除き、他の全ては5%ガラクトースに感受性であり、これはガラクトース輸送を仲介する可能性を示している。
【0042】
【図22】17種のシロイヌナズナGLUE遺伝子による18種のヘキソーストランスポーター遺伝子全てを欠く酵母株YSL2-1の相補性を示す。酵母ホモログHXT5または哺乳類ホモログGLUT1を発現する細胞をコントロールとして用いた。Os8N3およびRAG1AP1は、コメおよび哺乳動物由来ホモログである。酵母細胞を初期対数期に2%マルトースを含むSD-Ura液体培地中で増殖し、2%のグルコースを含むYPMまたはSD-URAを含む培地に、5m の連続希釈をスポットした。GLUE1、4、5および7はグルコースを輸送する(GLUE8も同様。ここでは示さず)。
【0043】
【図23】17種のシロイヌナズナGLUE遺伝子による18種のヘキソーストランスポーター遺伝子全てを欠く酵母株YSL2-1の相補性を示す。酵母ホモログHXT5または哺乳類ホモログGLUT1を発現する細胞をコントロールとして用いた。Os8N3およびRAG1AP1は、コメおよび哺乳動物由来ホモログである。酵母細胞を初期対数期に2%マルトースを含むSD-Ura液体培地中で増殖し、多様な2−デオキシ−グルコースレベルを含むYPMまたはSD-URAを含む培地に、5m の連続希釈をスポットした。GLUE1、3、4、5、7、8、14、16および17は、糖アナログにより感受性であるため、2−デオキシグルコースを輸送する。
【0044】
【図24】植物体におけるシロイヌナズナGLUEタンパク質の細胞内局在を示す。GLUE−GFP融合タンパク質は、タバコの葉において一時的に発現した場合、細胞膜の近傍または細胞膜に局在する。
【0045】
【図25】根におけるGLUE発現を示す。シロイヌナズナの根におけるGLUE遺伝子発現のqPCR分析。転写産物は10日齢シロイヌナズナ実生から単離し、テンプレートとしてcDNAを作成した。比較Ct法を用いて相対発現レベルを計算した(1000*1/(2^(CtGlU-CtActin8)。ここに示されていないファミリーのメンバーは、顕著な発現レベルを示さなかった(Guo et al.、未公表の結果)。4回の独立した実験からのデータ。
【0046】
【図26】はシロイヌナズナGLUEの発現パターンの組織学的分析を示す。GLUE2、GLUE16およびGLUE17−GUS融合タンパク質を担持するトランスジェニックシロイヌナズナにおけるGUS活性をX-glucで染色して分析した。10日齢シロイヌナズナ実生由来植物全体のイメージを示す。
【0047】
【図27】FRETグルコースセンサーによるCIT3細胞における糖流量分析を示す。細胞基質FRETグルコースセンサーFLIPglu-30μΔ13VとRAG1AP1-mCherryの共発現の非存在(A)および存在(B)下における、ヒト乳腺細胞系としてCIT3細胞におけるFRET分析。細胞を異なる外部グルコース濃度でかん流した(5、25および40 mM)。FRETイメージを得て、データを分析した。データは平均±SD(n=7〜9)である。
【0048】
【図28】RAG1AP1mCherryを発現するCIT3におけるグルコースレベルに対するサイトカラシンBの影響を示す。細胞基質FRETグルコースセンサーFLIPglu-30μΔ13VとRAG1AP1-mCherryの共発現の存在下における、ヒト乳腺細胞系としてCIT3細胞におけるFRET分析。細胞を外部40mMグルコースで、20μMのサイトカラシンBの存在下または非存在下でかん流した。FRETイメージを得て、データを分析した。データは平均±SD(n=5)である。
【0049】
【図29】RAG1AP1mCherryを発現するCIT3細胞におけるグルコースレベルに対する分化の影響を示す。細胞基質FRETグルコースセンサーFLIPglu-30μΔ13VとRAG1AP1-mCherryの共発現の存在下における、ヒト乳腺細胞系としてCIT3細胞におけるFRET分析。10 μg/mL インスリン、3 μg/mL プロラクチンおよび3μg/mL ヒドロコルチゾン(分泌培地)で細胞を分化した。10 μg/mL インスリンおよび5 ng/mL EGFを含むDMEM/F12(増殖培地)中で細胞を培養した。細胞を異なる外部グルコース濃度でかん流した(5、25および40 mM)。FRETイメージを得て、データを分析した。データは平均±SD(n=11〜13)である。
【0050】
【図30】図30AはCIT3細胞におけるRAG1AP1−GFP融合タンパク質の局在を示す。イメージは共焦点顕微鏡で撮影した。図30BはHepG2細胞およびHEK293T細胞由来RNAのRT−PCR分析を示す。RAG1AP1、GLUT1またはβアクチンを逆転写し、PCRで増幅した。未分化または分化細胞を10 μg/mL インスリンおよび5 ng/mL EGFを含むDMEM/F12のGM(増殖培地)または10 μg/mL インスリン、3 μg/mL プロラクチンおよび3μg/mL ヒドロコルチゾンを含むDMEM/F12であるSM(分泌培地)中で培養した。
【0051】
【図31】RAG1AP1およびRAG1AP1mCherryのウェスタンブロットを示す。SDS−PAGE(12.5%ゲル)で酵母の全細胞溶解物、RAG1AP1またはRAG1AP1-mCherryを過剰発現するCIT3を分離した。抗原領域はEQDRNYWLLQTであり、ヒトRAG1AP1のC末端アミノ酸211〜221に対応する(Abcam)。
【0052】
【図32】RAG1AP1を過剰発現するMDCK細胞におけるRAG1AP1の免疫蛍光局在化を示す。RAG1AP1はヒトRAG1AP1(Abcam)のC末端ペプチドに対する抗体およびAlexa 594標識化ロバ抗ヤギIgGで染色した。2,6−シアリルトランスフェラーゼ−GFP(ゴルジ体マーカー)をRAG1AP1と混合した。
【0053】
【図33】図33AはMDCK細胞におけるRAG1AP1-GFP融合タンパク質の局在化を示す。イメージは共焦点顕微鏡で撮影した。図45BはFRETセンサーによるHEK293T細胞の糖流量分析を示す。RAG1AP1を発現し、細胞基質FRETグルコースセンサーFLIPglu-600μΔ13VとRAG1AP1-mCherryを共発現するHEK293T細胞におけるFRET分析。細胞を異なる外部グルコース濃度でかん流した(2.5、5、25および40 mM)。FRETイメージを得て、データを分析した。データは平均±SD(n=)である。
【0054】
【図34】ヒト肝臓断片におけるRAG1AP1の免疫蛍光局在を示す。RAG1AP1はヒトRAG1AP1(Abcam)のC末端ペプチドに対する抗体およびAlexa fluor 594標識化ロバ抗ヤギIgGで染色した。Golgin-97をゴルジ体マーカーとして用い、これをgolgin-97に対するモノクローナル抗体(Invitrogen)およびAlexa fluor 488標識化ロバ抗マウスIgGで染色した。
【0055】
【図35】ヒト肝臓断片におけるRAG1AP1の免疫蛍光局在を示す。RAG1AP1はヒトRAG1AP1(Abcam)のC末端ペプチドに対する抗体およびAlexa fluor 594標識化ロバ抗ヤギIgGで染色した。Golgin-97をゴルジ体マーカーとして用い、これをgolgin-97に対するモノクローナル抗体(Invitrogen)およびAlexa fluor 488標識化ロバ抗マウスIgGで染色した。
【0056】
【図36】Golgi標的化FLIPglu-600μΔ13Vを示す。β−1,4−ガラクトシルトランスフェラーゼ1(galT)のペプチド(14〜44)およびステムを用いてFRETグルコースセンサーをゴルジ体に標的化した(Schaub et al, Mol Biol Cell, 17: 5153-5162, 2006)。
【0057】
【図37】Golgi標的化FLIPglu-600μΔ13Vを示す。β−1,4−ガラクトシルトランスフェラーゼ1(galT)のペプチド(14〜44)およびステムを用いてFRETグルコースセンサーをゴルジ体に標的化した(Schaub et al, Mol Biol Cell, 17: 5153-5162, 2006)。
【0058】
【図38】FRETグルコースセンサーによるMDCK細胞の細胞基質およびゴルジ体中の糖流量分析を示す。RAG1AP1発現の存在下(A,B)または非存在下(C,D)および細胞基質(A,C)またはゴルジ体(B,D)標的化FRETグルコースセンサーFLIPglu-600μΔ13Vを共発現する、MDCK細胞におけるFRET分析。細胞を異なる外部グルコース濃度でかん流した(1、2.5、5、10および40 mM)。FRETイメージを得て、データを分析した。データは平均±SD(n=4〜15)である。
【0059】
【図39】FRET分析センサーによるHela細胞の細胞基質およびゴルジ体における糖流量分析を示す。細胞基質(A)またはゴルジ体(B)標的化FRETグルコースセンサーFLIPglu-600μΔ13Vを発現するHela細胞におけるFRET分析。細胞を異なる外部グルコース濃度(5、10および40 mM)およびガラクトース濃度(5および40mM)でかん流した。FRETイメージを得て、データを分析した。データは平均±SD(n=7〜8)である。
【0060】
【図40】FRETグルコースセンサーによる線虫(C. elegans)GLUEファミリーメンバーを発現するHEK293T細胞における糖流量分析を示す。FRETグルコースセンサーFLIPglu-600μΔ13Vを共発現する、線虫(C. elegans)GLUEメンバーを発現するHEK293T細胞におけるFRET分析。細胞を異なる外部グルコース濃度(2.5、5、10および40 mM)およびガラクトース濃度(5および40mM)でかん流した。FRETイメージを得て、データを分析した。データは平均±SD(n=12〜24)である。
【0061】
【図41】ヒトRAG1AP1ホモログが糖流出トランスポーターであることを示す。ヒトSGLT1ナトリウムグルコースコトランスポーター(「S」)は、二次活性グルコースインポーターであることが既に示されている。SGLT1はアフリカツメガエル卵母細胞において内因的に発現され、14C標識化グルコースの卵母細胞への取り込みを仲介できる。RAG1AP1(「R])が共発現している場合、卵母細胞においてより低い取り込みが見られる。これはSGLT1発現細胞におけるグルコースの高い蓄積を防止するRAG1AP1活性による、グルコース「漏出」と一致する。
【0062】
【図42】5種の線虫(C. elegans)ホモログについての14Cグルコース取り込みデータを示す。少なくとも3、4および5は活性である。(C elegans遺伝子名のコード:Ce1: C06G8.1; Ce2: K06A4.4; Ce3: Y39A1A.8; Ce4: K11D12.5; Ce5: K02D7.5;(Ce5のワーム変異体: K02D7.5は増加した脂肪蓄積を示し、これはこれらの細胞からのグルコースの低下した流出と一致する)
【0063】
【図43−1】図43A〜43Hは、HEK293細胞におけるSWEETのショ糖排出機能を示す(ポジティブコントロール(ポテトショ糖トランスポーターStSUT1; Riesmeier et al. 1993 Plant Cell)は43Aであり、ネガティブコントロール(空のベクター43B))。ショ糖の取り込みはFRETショ糖センサーFLIPsuc90μΔ1を用いて測定した(Chaudhuri et al., 2008 Plant Journal)。SWEET10(図43C)、SWEET11(図43D)、SWEET12(図43E)、SWEET13(図43F)、SWEET14(図43G)およびOsSWEET11/Os8N3(図43H)は、哺乳類細胞へのショ糖取り込みを示すポジティブコントロールStSUT1のものと同様に、ネガティブなFRET応答を示した(ネガティブ比変化は細胞基質中の糖含量の増加に対応する)。この取り込みは、単輸送(促進された分散)、プロトンシンポートまたはプロトンアンチポートによって仲介されている可能性がある。
【図43−2】図43A〜43Hは、HEK293細胞におけるSWEETのショ糖排出機能を示す(ポジティブコントロール(ポテトショ糖トランスポーターStSUT1; Riesmeier et al. 1993 Plant Cell)は43Aであり、ネガティブコントロール(空のベクター43B))。ショ糖の取り込みはFRETショ糖センサーFLIPsuc90μΔ1を用いて測定した(Chaudhuri et al., 2008 Plant Journal)。SWEET10(図43C)、SWEET11(図43D)、SWEET12(図43E)、SWEET13(図43F)、SWEET14(図43G)およびOsSWEET11/Os8N3(図43H)は、哺乳類細胞へのショ糖取り込みを示すポジティブコントロールStSUT1のものと同様に、ネガティブなFRET応答を示した(ネガティブ比変化は細胞基質中の糖含量の増加に対応する)。この取り込みは、単輸送(促進された分散)、プロトンシンポートまたはプロトンアンチポートによって仲介されている可能性がある。
【0064】
【図44】線虫(C. elegans)、マウス、ラット、ヒト、シロイヌナズナ、コメ、ウマゴヤシおよびペチュニアにおける多様なGLUEタンパク質のアミノ酸配列を示す。
【発明を実施するための形態】
【0065】
詳細な説明
下記説明は、本発明の理解に有用であり得る情報を含む。本明細書において提供するあらゆる情報は、先行技術または明らかに特許請求の範囲に記載の発明と関連していること、あるいはいずれかの文献が具体的または暗示的に先行技術であることを認めるものではない。
【0066】
本発明の他の目的、利点および特徴は、本明細書において提供される明細書および図面を検討することによって、当業者に明らかとなる。したがって、本発明のさらなる目的および利点は、以下の説明から明らかである。
【0067】
植物は種子および花粉発生支援、花蜜産生ならびに根圏における有益微生物養育のために糖流出トランスポーターを必要とする。しかし、これらの流出トランスポーターの同一性は、不明なままであった。光学的糖センサーを用いて、植物から糖トランスポーターの新規クラス(GLUE、Glue、またはSWEET)が同定された。シロイヌナズナおよびコメGLUEは、糖のインポーターおよびエクスポーターとして機能する。OsGluE11/Os8N3およびAtGluE8/RPG1は花粉生存に必要とされる。花蜜および根粒におけるGLUEホモログの発現は、受粉媒介者および共生生物の給餌における役割を示唆している。真菌および細菌病原体は、異なるGLUEメンバーのmRNAレベルを調節して、単糖、二糖またはオリゴ糖トランスポーター活性をともに選択している。OsGLUE11/Os8N3は斑点細菌病の宿主感受性因子として機能し、植物および病原体両方の栄養摂取とGLUEを結びつける。
【0068】
ヒトゲノムは少なくとも2つのクラスのグルコーストランスポーター、SLC2およびSLC5を含む。GLUTと呼ばれるSLC2は単輸送体であり、すなわちそれは濃度勾配に沿ってグルコースを輸送する。逆にSGLTは、ナトリウム勾配によって駆動されてグルコースを能動的に輸送できる、Na+共役コトランスポーターである。これらのトランスポーターは、ヒトにおいてみられる取込活動のほとんどを説明することができ、例えばGLUT2ノックアウトマウス変異体は、劇的に低下した取り込み能を示すが、驚くべきことに細胞流出はそうではない。しかし、バイオインフォマティック分析により、動物およびヒトゲノムが溶質輸送体ファミリーSLC50として登録されているSWEETのホモログを含むことが示された。線虫(C. elegans)ゲノムは糖流出トランスポーターの新規クラス(SLC50)の7種のホモログを含むが、ヒトゲノムはRAG1AP1と呼ばれる1種のホモログを有する。
【0069】
糖流出は多細胞生物において、および生物間の相互作用において、炭素骨格とエネルギーの細胞性交換に必要とされる必須プロセスである。タペータムまたは花柱の輸送管からの糖流出は、花粉発生および後の花粉管成長を刺激する。花は受粉媒介者を誘引するため花蜜産生のために糖を分泌し、そして植物は、潜在的には有益微生物に給餌するために根圏に炭水化物を分泌する(T. Bisseling et al., Science 324, 691 (2009))。糖流出輸送体は、葉肉および種皮を含む多くの他の部位で必要とされる(Y. Zhou et al., Plant J. 49, 750 (2007))。流出トランスポーターの分子の性質は知られていない(S. Lalonde et al., Annu. Rev. Plant Biol. 55, 341 (2004))。植物由来の糖はまた、病原体のための基質を提供する。病原体の第一次的ゴールは、効率的に繁殖するために宿主植物から栄養分を摂取することである。シュードモナス(Pseudomonas)属およびザントモナス(Xanthomonas)属の植物病原性細菌は、植物組織の細胞外空間(アポプラズム)に生息でき、ここでそれらはエネルギー源および炭素骨格源として炭水化物を獲得する。成功した病原体は、かかるメカニズムを共に選択して栄養素流動を変化させる可能性がある(J. W. Patrick, Aust. J. Plant Physiol. 16, 53 (1989))。結果として、病原体と植物は、植物が栄養素への病原体のアクセスを制限しようとし、また防御戦略を開始する一方で、病原体が栄養素へのアクセスを得ようとし、宿主免疫を抑制する戦略を考え出す進化的主導権争いに突入する。植物の防御を変化させるための病原体によって用いられるメカニズムに対する洞察が現在出現しつつあるが;どのようにして病原体が、病原体の増殖を支援するために宿主の生理機能、特に糖排出を変化させるのかについて、わずかしか知られていない。本発明は、小胞または細胞膜で糖を分泌するトランスポーターの存在を突き止めた。本発明はさらに、これらの植物流出トランスポーターが病原体によって栄養素の要求を支給するために「共に選択」されることを同定した(J. W. Patrick, Aust. J. Plant Physiol. 16, 53 (1989))。少なくともコムギウドンコ病の場合、グルコースは植物宿主から病原体に移送される主な糖である(J. Aked, J. L. Hall, New Phytol. 123, 271 (1993); P. N. Sutton et al., Planta 208, 426 (1999);およびP. N. Sutton et al., Physiol. Plant. 129, 787 (2007))。それぞれの病原体グルコース/H+コトランスポーターが同定された(R. T. Voegele et al., Proc. Natl. Acad. Sci. USA 98, 8133 (2001));反対に、植物糖流出メカニズムはもとより不明のままであった。
【0070】
多くの代謝経路において、トランスポーターは、基質を酵素に供給し、酵素の存在から各々の産物を除去する具体的な経路のいずれかまたは両方の終点で機能しうる。輸送は受動輸送、能動輸送、分散または浸透作用によりうる。トランスポーターは、酵素での基質の存否に直接または間接的に関与しうる。トランスポーターは細胞膜内またはその近くに局在してよく、あるいはそれらは細胞基質またはその近くまたは他の細胞小器官、例えば小胞体、ミトコンドリア、葉緑体、ペルオキシソーム、ゴルジ装置、ベシクル、核膜または液胞、リソソームまたは細胞膜中に局在していてよい。
【0071】
トランスポーターは固定されていてもよく、それによるかまたはそれを通過する基質の通過を可能とするか、あるいは基質と結合して基質を特定の細胞内目的地に物理的にシャトルする。トランスポーターは、別のタイプの分子の通過または輸送を可能とする一つのタイプの分子と結合できる。トランスポーターは独立して、またはプロテインキナーゼもしくはATP切断ドメインのような他のタンパク質の作用を介して移動してよい。
【0072】
トランスポーターは、細胞または生物内へのまたは外への物質の取り込みまたは放出を決定し、そしてトランスポーターは、細胞間の物質の輸送および分散を制御する。トランスポーターはまた、細胞小器官間、例えば核の中と外での輸送のように、細胞間でも機能しうる。トランスポーターはしばしば代謝経路の始点および終点に存在するため、それらは基本的により高次の制御機能を担当している。トランスポーターは、放出された低分子、例えばモノアミンまたは神経伝達物質の再取り込みに関与している可能性がある。
【0073】
いくつかのトランスポーターは、特定の基質を輸送するためにエネルギーを必要とする。ある例では、エネルギーはATPを介して供給され、得られたトランスポーターのリン酸化は、輸送を進めることができるコンホメーション変化を引き起こす。別の場合では、輸送体と第二基質のカップリング、例えばPまたはVタイプATPaseによって作出されるプロトンまたはナトリウム/カリウム勾配を介して間接的に、エネルギーは供給される。他の例では、トランスポーターと他のタンパク質または分子の相互作用が、同じタンパク質もしくは分子または異なるタンパク質もしくは分子の輸送を進めることができるコンホメーション変化を引き起こす。さらなる例では、トランスポーターと制御タンパク質間の分離が、輸送を進めることができるトランスポーターのコンホメーション変化を引き起こす。トランスポーターは直接結合を介して基質または産物と相互作用しうる。輸送の触媒において、トランスポーターはコンホメーション変化を引き起こしうる。これはまた、PタイプまたはVタイプATPaseのような酵素およびトランスポーターとして使用されるハイブリッドタンパク質を含む。
【0074】
本明細書において使用するとき、用語「コンホメーション変化」は、センサーにおいて生じるいずれかの立体配置の変化に対する影響、例えばタンパク質中の原子および原子団の三次元位置、移動可能な原子および側鎖の平均位置、タンパク質の表面特性の変化、シングルまたは相対位置の位置/平均位置またはコンホメーションに影響するドメインのドメインフォールディング/アンフォールディングの移動、フルオロフォアの平均位置が変化して、当該検出部分によって放出されるエネルギーの変化をがもたらされることを意味する。用語「相対位置」は、2つの検出部分が互いに有しうる空間的関係、例えば距離および方向のいずれかの可能性のある種類を意味する。例えばコンホメーションは、1または複数個の原子、側鎖またはドメインの回転、酵素の折り畳み、外側へのドメインの一方または両方のねじれ、あるいはこれらの移動のいずれかの組合せによって変化してよい。検出部分間の方向または距離が変化されているコンホメーション変化、またはレポーター要素のコンホメーションに対する効果を示す変化が有用である。あるいは、酵素部分におけるコンホメーション変化は、シングル結合フルオロフォアの特性に作用する。この場合、RETシグナルを得るために第二フルオロフォアが使用されうる。かかる場合において、結合の前または結合により、少なくとも最大の半分のエネルギー移動が生じるように検出部分が向けられていることが有利である。
【0075】
本明細書において使用するとき、「リガンド」は、ペリプラズム結合タンパク質のようなタンパク質と結合してそのタンパク質との複合体を形成できる分子または物質を意味する。リガンドとタンパク質の結合は、タンパク質の形状、特に3次元および4次元構造を歪めまたは変化しうる。リガンドは基質であってもよい。基質は遊離体または酵素の支援触媒作用を介して産物に変換される物質を含みうる。リガンドは内因性リガンドのアナログまたは誘導体であってよい。リガンドは結合部位について内因性リガンドと競合しうる。リガンドは例えば、低分子、化学物質、一本鎖オリゴヌクレオチド、二本鎖オリゴヌクレオチド、DNA、RNAまたはポリペプチドであってよい。リガンドはタンパク質と結合するいずれかの化学物質、例えばマグネシウムのようなイオンまたはアロステリック因子または別のタンパク質を含む。リガンドはグルコースのような糖であってよい。
【0076】
本明細書において使用するとき、「蛍光インジケーター」は、PBPと結合した蛍光ドメインまたは化合物を意味する。PBPの形状の変化は、蛍光ドメインまたは化合物の蛍光の変化をもたらし、それによって酵素の形状の変化が示される。ドメインは蛍光タンパク質であってよい。蛍光ドメインは2個のサブドメイン、例えばドナーおよびアクセプターフルオロフォアを含んでいてよい。いくつかの例において、PBPはドナーとアクセプターフルオロフォア間で共有結合されている。蛍光インジケーターの別の使用は、発光またはリン光性分子、ならびにNMR、偏極検出器のような他の手段によって検出されうる化合物である。
【0077】
タンパク質およびポリペプチドに関して本明細書において使用するとき、用語「組換え」は、遺伝子操作によって、例えば非天然核酸の細胞での翻訳によって作成されまたは由来するか、あるいは人工手段またはメカニズムによって組み立てられるタンパク質および/またはポリペプチドおよび/またはペプチドを含みうる。
【0078】
本明細書において使用するとき、「融合」は、本発明に従ってそれらがおかれる順序または文脈において、互いに関連した天然に見られない配列を含む核酸およびポリペプチドを意味しうる。融合核酸またはポリペプチドはその全体において核酸またはポリペプチドの天然配列を必ずしも含んでいる必要がない。融合タンパク質は通常のペプチド結合を介して一体に結合した2個以上のセグメントを有する。融合核酸は通常のホスホジエステル結合を介して一体に結合した2個以上のセグメントを有する。
【0079】
本明細書において使用するとき、用語「生物学的サンプル」は、細胞または細胞内物質の集合体を意味する。サンプルは生物または生物の成分(例えば細胞)から得られる。サンプルはいずれかの生物学的組織または液体から得られる。サンプルは対象由来のサンプルであり得る。対象は患者であってよい。サンプルは患者または患者の成分から得られる。対象は動物であってもよい。動物は哺乳類、例えばヒトまたはヒト患者であってよい。かかるサンプルは、唾液、血液、血球細胞(例えば白血球および赤血球)、組織もしくは生検サンプル(例えば腫瘍生検)、尿、腹腔液および胸膜液またはそれら由来の細胞を含むが、これらに限定されない。生物学的サンプルはまた、組織学的目的で採取した凍結切片のような組織の切片を含んでもよい。生物学的サンプルはまた、インビトロ細胞培養物を含んでもよい。細胞培養物は不死化細胞系または一次細胞系であってもよい。細胞培養物は異なる細胞タイプを含んでいてもよい。
【0080】
本明細書において使用するとき、用語「dsRNA」は二本鎖RNAを意味し、ここでdsRNAは2本の別個のストランドまたは一本鎖ヘアピンによる二本鎖であってよい。dsRNAは標的遺伝子のヌクレオチドと類似するヌクレオチド配列を含んでいてよい。dsRNAは、ヘアピンRNAのような自己相補的dsRNAまたは別個の相補的RNAストランドによって形成されるdsRNAを形成する転写産物および/またはインビボでsiRNAを産生できる転写産物を産生可能な発現ベクター(RNAi発現ベクターとも称される)によって作成できる。ベクターは(1)遺伝子発現において制御の役割を有する遺伝子因子(複数の因子)、例えばプロモーター、オペレーターまたはエンハンサー、それと作動可能に連結した、(2)転写されて二本鎖RNA(細胞内でアニールしてsiRNAを形成する2個のRNA部分、またはsiRNAに処理されうる一本のヘアピンRNA)を産生する「コード」配列、ならびに(3)適切な転写開始および終止配列を含んでなる転写ユニットを含みうる。プロモーターおよび他の制御要素の選択は、一般に、意図する宿主細胞に従って変化する。一般に、組換えDNA技術において有用な発現ベクターは、しばしばベクター形態が染色体と結合しない環状二本鎖DNAループを意味する「プラスミド」の形態で存在する。
【0081】
本明細書において使用するとき、用語「単離された」は、巨大分子の天然源に存在する他の細胞/組織構成物(例えばDNAまたはRNA)から分離された分子を意味する。用語「単離された」は、本明細書において使用するとき、組換えDNA技術で産生された場合には細胞性物質、ウイルス性物質および培養培地を実質的に含まない、あるいは化学的に合成された場合には化学的前駆体または他の化学物質を実質的に含まない核酸またはタンパク質を意味する。さらに、「単離された核酸」は、フラグメントとして天然には生じない、天然状態では見られない核酸フラグメントを含んでいてもよい。
【0082】
本明細書において使用するとき、用語「多量体」は、2個以上の異なる分子間の多量体複合体の形成を意味する。多量体複合体は、例えば同じタンパク質の2個以上の分子(例えばホモ二量体、三量体、四量体、二量体の二量体または高次多量体)または2個以上の異なる(すなわち非同一)タンパク質の混合物(例えばヘテロ二量体、異なる二量体の二量体、三量体、四量体または高次多量体)を含んでいてよい。例えば多量体抗体は、同じ抗体または2個以上の異なる抗体を含んでいてよく、これらはそれぞれ、2種以上の機能または活性を有している(例えば2種以上のエピトープと結合する)。
【0083】
本明細書において使用するとき、「対象」は、本発明のレシピエントを含みうる。対象は植物または植物の構成要素、例えば植物の器官または小器官であってよい。対象は脊椎動物を含むいずれかの動物であってよい。対象は多くの場合、好ましくはヒトであるが、家畜、実験動物(例えばラットまたはマウスのようなげっ歯類を含むがこれらに限定されない)またはペットであってもよい。
【0084】
本明細書において使用するとき、用語「変異体」は、BLAST分析によって天然分子と少なくとも約70%、より好ましくは少なくとも75%同一性、例えば少なくとも80%、90%、95%、96%、97%、98%、99%またはそれ以上の同一性を有するポリペプチドを意味する。多くのかかる変異体は当該技術分野において既知であるか、あるいは天然蛍光分子のランダムまたは特異的突然変異誘発によって容易に製造できる(例えばFradkov et al., FEBS Lett. 479:127-130 (2000)参照)。
【0085】
本明細書において使用するとき、用語「プラスミド」および「ベクター」は、プラスミドが最も一般的に使用されるベクターの形態であるから、相互に交換可能なように使用される。しかし、本発明は同等の機能を有し、この後当該技術分野において既知となるかかる他の形態の発現ベクターを含むことを意図する。ベクターは、異なる遺伝子環境間での輸送または宿主細胞における発現のために制限およびライゲーションによって所望の配列が挿入されていてよい任意の数の核酸であってよい。ベクターは典型的にはDNAから成るが、RNAベクターも利用可能である。ベクターはプラスミドおよびファージミドを含むがこれらに限定されない。クローニングベクターは、宿主細胞で複製でき、決定可能なようにベクターを切断できて、新たな組換えベクターが宿主細胞で複製できる能力を保持するように所望のDNA配列をライゲーションできる1個以上のエンドヌクレアーゼ制限部位によってさらに特徴付けられるものである。プラスミドの場合、プラスミドが宿主細菌内でコピー数を増加するように多くの回数、あるい宿主が有糸分裂によって増殖する前に宿主当たり丁度1回、所望の配列の複製が行われうる。ファージの場合、複製は溶解期に能動的にまたは溶原期に受動的に行われうる。
【0086】
ベクターはさらに、プロモーター配列を含んでいてもよい。プロモーターは、核酸の転写を開始する部位を含むコード領域の通常上流に位置する非翻訳核酸配列を含みうる。プロモーター領域はまた、遺伝子発現の制御因子として作用する他の要素も含んでいてよい。本発明のさらなる態様において、発現ベクターは、組み込まれた発現ベクターが有する細胞の選択に利用するためのさらなる領域を含む。プロモーター配列は、転写開始部位によって3’末端でしばしば結合されており(包括的に)、バックグラウンド以上の検出可能なレベルで転写を開始するために必要な最小数の塩基または要素を含むように上流(5’方向)に伸長する。プロモーター配列内で転写開始部位およびRNAポリメラーゼの結合に関与するタンパク質結合ドメインが見られる。真核プロモーターはしばしば、常ではないが、「TATA」ボックスおよび「CAT」ボックスを含む。プロモーターの活性化は、例えばある組織においてのみ発現される転写因子によってある細胞または組織に特異的であってよく、あるいはプロモーターは偏在してほとんどの細胞または組織で発現可能であってよい。
【0087】
ベクターはさらに、ベクターで形質転換またはトランスフェクトされた細胞の同定および選択に使用するのに好適な1種以上のマーカー配列を含んでいてよい。マーカーは例えば、抗生物質または他の化合物に対する耐性または感受性を増減するタンパク質をコードする遺伝子、活性が当該技術分野において既知の標準的なアッセイによって検出可能な酵素をコードする遺伝子(例えばβガラクトシダーゼまたはアルカリホスファターゼ)ならびに形質転換またはトランスフェクトされた細胞、宿主、コロニーまたはプラークの表現型に視覚的に影響を与える遺伝子を含む。ベクターは、それが作動可能に結合しているDNAセグメントに存在する構造遺伝子産物の自己複製および発現が可能なものであってもよい。
【0088】
発現ベクターは、制御配列と作動可能に連結または作動可能に結合されてRNA転写産物として発現されうるように所望の核酸配列を制限およびライゲーションによって挿入できるものである。発現は、細胞での内因性遺伝子、トランスジーンまたはコード領域の転写および/または翻訳を意味する。
【0089】
コード配列および制御配列は、コード配列の発現または転写が制御配列の影響または制御下におかれているようにそれらが共有結合している場合に、作動可能に連結されている。コード配列が機能的タンパク質に翻訳されることが望まれる場合、5’制御配列におけるプロモーターの誘導がコード配列の転写をもたらすとき、そして2個のDNA配列間の結合の性質が(1)フレームシフト変異の導入をもたらさない、(2)コード配列の転写を指定するプロモーター領域の能力と干渉しない、あるいは(3)タンパク質へと翻訳される対応するRNA転写産物の能力と干渉しないとき、2個のDNA配列は作動可能に結合されていると言われる。したがって、得られる転写産物が所望のタンパク質またはポリペプチドに翻訳されうるようにプロモーター領域がDNA配列の転写に影響できる場合、プロモーター領域はコード配列と作動可能に結合されている。
【0090】
本発明のいくつかの局面は、核酸の形質転換および/またはトランスフェクションを含む。形質転換は、原核細胞の内部への外因性または異種性核酸の導入である。トランスフェクションは、真核細胞の内部への外因性または異種性核酸の導入である。形質転換またはトランスフェクトする核酸は、細胞のゲノムを構成する染色体DNAに組み込まれ(共有結合)ても組み込まれなくてもよい。原核生物において、例えば、形質転換する核酸は、プラスミドまたはウイルス性ベクターのようなエピソーム要素で維持されてよい。真核細胞について、安定的にトランスフェクトされた細胞は、染色体複製を介して娘細胞によって受け継がれるように染色体にトランスフェクトする核酸が組み込まれているものである。この安定性は、トランスフェクトする核酸を含む娘細胞の集団からなる細胞系またはクローンを確立する真核細胞の能力によって示される。
【0091】
本明細書において使用するとき、用語「融合タンパク質」または「キメラタンパク質」は、直接またはスペーサーアミノ酸を用いて一体に融合している少なくとも2個のポリペプチドを含むポリペプチドを意味するものとして使用される。融合ポリペプチドは、融合タンパク質の全体的な機能において協同的または反対の役割を有しうる。
【0092】
本明細書において使用するとき、抗体の「フラグメント」は、Fc、Fab、Fab’、F(ab’)2および一本鎖免疫グロブリンを含むがこれらに限定されない。
【0093】
本明細書において使用するとき、免疫学的有効量は、単回投与または一連の一部のいずれかにおいて疾患または障害の処置に有効である対象への投与量を意味する。この量は処置する対象の健康および身体状態、処置する対象の種(例えば非ヒト哺乳類、霊長類など)、抗体を合成する対象の免疫系の能力、望まれる保護の程度、ワクチンの形成および他の関連する要因に依存して変化する。この量は日常的な試験を通じて決定されうる相対的に広い範囲に含まれると予想される。
【0094】
本明細書において使用するとき、「医薬組成物」または「製剤」は、薬学的に許容される担体または希釈剤と共に薬物または化合物を含む組成物を意味する。薬学的に許容される担体は、生理食塩水、リンゲル液、リン酸緩衝化食塩水および当該技術分野において既知の他の担体を含むが、これらに限定されない。医薬組成物はまた、安定化剤、抗酸化剤、着色剤および希釈剤を含んでいてもよい。薬学的に許容される担体および添加剤は、医薬製剤からもたらされる副作用が最小限となり、処置が不奏効となる程度に薬物のパフォーマンスを没却または阻害しないように、選択される。
【0095】
本明細書において使用するとき、「治療上有効量」は、それを必要とする対象に投与したとき処置を達成するために必要な薬物または化合物の量を意味する。「治療上有効量」を構成する架橋Aβオリゴマー反応性抗体のような抗体の量は、状態または疾患の重症度ならびに処置する対象の年齢および体重に依存して変化するが、当業者によってその知識および本開示に基づいて日常的に決定されうる。
【0096】
「コファクター」は、タンパク質と相互作用して、その生理的な機能の実行において当該タンパク質を補助する要素を意味する。コファクターは反応を触媒しうる。コファクターは、強い相互作用または緩いつながりによってタンパク質、例えばトランスポーターまたは酵素と結合しうる。コファクターは「コエンザイム」または「補欠分子族」であってもよい。「コエンザイム」は、酵素間で化学団をシャトルまたは輸送する有機分子を意味する。「補欠分子族」は酵素と結合して酵素の一部となるコファクターを意味する。コファクターは、金属イオン、例えばカルシウム、マグネシウム、マンガン、鉄、カリウム、ナトリウム、アルミニウム、銅、ニッケル、セレン、モリブデンまたは亜鉛であってもよい。コファクターの制限された供給は、速度制限因子でありうる。
【0097】
用語「生物学的液体」は、循環タンパク質を含む体液、例えば血漿、血清および全血、唾液、脳脊髄液、羊水、滑液、水性体液、胆汁、耳垢、カウパー腺液、乳び、女性の射精および膣潤滑液、間質液、リンパ液、月経液、粘液、胸水、膿、皮脂、精液、汗、涙液、嘔吐物、尿、乳汁および他の分泌物を含む。生物学的液体のタンパク質含有抽出物は、生物学的液体から回収または分離されるいずれかの調製物、例えば免疫グロブリン分画である。本発明に使用できる血液、血清または血漿は、個体から新たに得ることができ、あるいは血液バンクまたは他の血液回収組織から得た貯蔵された血液または血漿調製物のような供給源から得てもよい。本発明の目的のために、血液、血清または血漿は、期限切れの回収物または血液バンクまたは血液回収組織によって基準を満たさないとされた他のもの由来であってもよい。本発明の同一のプロセスは、動物血液に適用してもよく、獣医学的医薬に関する目的のために同様の動物抗体を得ることができる。液体はそれを得たそのままの状態で使用してよく、あるいは例えば沈降させるか遠心分離してさらに処理してもよい。液体は植物由来であってもよく、例えば樹液、師部樹液、木部樹液、花蜜、樹脂、ラテックスまたはオイルであってよい。液体は上清、回収した沈降物または外部重力、例えば遠心分離またはろ過によって得たペレットであってよい。
【0098】
本発明は、部分的に、糖トランスポータータンパク質の新規クラスの発見に基づく。糖は単糖、二糖またはオリゴ糖であってよく、例えばグルコース、フルクトース、リボース、ラクトース、ガラクトース、アラビノース、マルトース、アミロース、セルロースまたはショ糖であってよい。トランスポーターは細胞の膜を通過して糖を輸送できる。膜は細胞膜または細胞壁であってよい。膜は細胞の細胞基質を取り巻きうる。膜は細胞小器官、例えばミトコンドリア、小胞体、ゴルジ装置、核、エンドソームまたは液胞を取り巻いてもよい。トランスポーターはある細胞/細胞小器官の膜と別の細胞/細胞小器官の膜の間で糖を輸送できる。
【0099】
本発明は、臓器内または対象の臓器間で糖を輸送する方法を提供できる。臓器は炭水化物、例えば糖およびグルコースの処理、取込または排出に関与しうる。臓器は消化系に関与していてもよい。例えば臓器は、腸(大腸または小腸)、胃または肝臓であってよい。本発明のトランスポーターは、臓器で細胞の内部でのまたは外部への糖の輸送を補助できる。トランスポーターは糖の移動に作用することが知られている他のタンパク質、例えばSGLT、GLUT1およびGLUT2と共同で働きうる。トランスポーターは細胞小器官の外へと糖を流出しうる。トランスポーターは小胞の形成を介して受動的に糖を流出しうる。例えば、ゴルジ装置におけるトランスポーターの発現は、形成された小胞に糖を流出させ、次いでその内容物を細胞外へ移動する。
【0100】
最初はグルコース単輸送体GLUT2が肝臓および腸におけるグルコースの取込および流出の両方に関与していると考えられた。しかし、肝細胞およびトランスジェニックマウスにおけるGLUT2のノックダウンは、GLUT2がグルコース取込に必須であるが、グルコース流出にはそうではないことを示した。GLUT2ノックアウトマウスの経口グルコース負荷は、血中に現れるグルコースの正常な速度をもたらした(Thorens et al.. J. Biol. Chem. 275, 23751-23758, (2000))。同様に、GLUT2対立遺伝子の両方の不活性化によって引き起こされる症候群であるFanconi-Bickel症候群(Santer et al. Nat. Genet. 17, 324-326, (1997))に罹患したヒトは、腸細胞からの流出を必要とするプロセス(Manz, F. et al. Pediatr Nephrol 1, 509-518, (1987))である異常な炭水化物摂取を導かなかった。その機能に基づいて、HsSWEET1/RAG1AP1は肝臓からグルコースのような糖の流出を補助しうる。
【0101】
トランスポーター、例えばGLUEトランスポーターは膜に位置しうる。トランスポーターは膜を貫通しうる。トランスポーターは膜を貫通する孔を含みうる。糖輸送はトランスポーターによって作り出される膜の孔を介しうる。孔は刺激に応答して幅を変えて、それによって糖が孔を通過できるように変化しうる。トランスポーターはゲートであってよい。ゲート機構は刺激に応答してトランスポーターを糖が通過できる。ゲート機構は結合するリガンドを必要とするか、あるいは電圧センサーを含みうる。トランスポーターは受動拡散によって糖を通過させうる。トランスポーターは糖を輸送するためにアデノシン三リン酸のようなエネルギーを必要としうる。トランスポーターは平衡に達するように高濃度から低濃度へと糖を輸送しうる。トランスポーターは低濃度の領域から高濃度の領域へと糖を輸送し、それによって勾配を増加させうる。トランスポーターは単輸送体であってもよい。単輸送体は、本明細書において使用するとき、促進因子として機能するトランスポーターを意味し、トランスポーターの向きは輸送される基質の膜内外の勾配または濃度差によって決定される。単輸送体は、共輸送分子または活性化分子のようなコファクターに頼ることなく自己充足的に作動できるトランスポーターを意味する。単輸送体は典型的には、濃度勾配の方向に、すなわち高濃度の糖を有する側から低濃度の糖を有する側へと、糖の流出を可能とする。
【0102】
トランスポーター、例えばGLUEトランスポーターは、膜の片側での糖濃度を上昇させうる。トランスポーターは細胞内の糖レベルを上昇させうる。これは、イオンまたは他の代謝産物、例えばプロトン、ヒドロキシアニオン、ナトリウムまたはカリウムでありうる第二化合物の輸送とその輸送の共役を介して達成されうる。共役は、共輸送もしくはアンチポートによるか、またはピンポン機構によるものでありうる。トランスポーターは細胞内の糖濃度を低下させうる。トランスポーターは細胞から糖を排出しうる。トランスポーターは細胞に糖を取り込みうる。
【0103】
トランスポーターはさらに、キナーゼ、第二メッセンジャー、アニオン、カチオンまたはリガンドのようなシグナルに影響されることがある。トランスポーターはコファクターに影響されることがある。コファクターは、マグネシウム、亜鉛、鉄、銅、ヨウ素、塩素、ナトリウム、カリウム、カルシウム、マグネシウム、硫酸塩、サルフェート、アンモニウム、硝酸塩、亜硝酸塩、カルボネート、カルボン酸またはリン酸塩のイオン化形態であってよい。コファクターは、トランスポーターまたは第二メッセンジャーと基質またはリガンドの結合を補助するために必要であり得る。コファクターはトランスポーターの活性に必要であり得る。いくつかの例において、コファクターの添加がな場合には、トランスポーターの機能喪失または低下が生じる。他の例において、コファクターの存在はトランスポーターの活性をダウンレギュレートする。トランスポーターはクラスリン内在化機構を介するような内在化を介してダウンレギュレートされうる。
【0104】
トランスポーターは細胞または細胞から得られた抽出物中に存在しうる。細胞は原核細胞であってよい。細胞は真核細胞であってもよい。
【0105】
細胞は動物またはその一部(rt)に存在していてよい。トランスポーターは動物細胞の小胞体、ゴルジ装置、ベシクルまたは細胞膜におけるグルコース輸送に関与しうる。細胞はグルコース輸送または分泌に関与する動物細胞であってよい。細胞は腺細胞であってよい。腺細胞は乳腺の肺胞細胞であってもよい。
【0106】
細胞は植物またはその一部、例えば根、幹、葉、種子、花、果実、葯、蜜腺、子房、花弁、タペータム、木部または師部に存在していてよい。例示として、植物は、有胚植物、コケ植物、種子植物(spermatophye)、ネマトファイト(nematophyte)、管束植物、ダイズ、コメ、トマト、アルファルファ、ジャガイモ、エンドウ、イネ科植物、ハーブ、樹木、藻類、蘚類、菌類、つる植物、シダ類、低木、オオムギ、コムギ、ホップ、トウモロコシ、レタス、オレンジ、モモ、柑橘類、レモン、ライム、ココナッツ、ヤシ、マツ、オーク、シーダー、マンゴー、パイナップル、ダイオウ、イチゴ、ブラックベリー、ブラックカラント、ブルーベリー、ラズベリー、キウイ、ブドウ、ルタバガ、パースニップ、サツマイモ、カブ、マッシュルーム(菌類)、コショウ、シラントロ、タマネギ、ニラネギ、フェンネル、クローブ、アボカドまたはキュウリを含む。またススキ属(Miscanthus)またはスイッチグラス、ポプラ、モロコシ属(Sorghum)およびヤマカモジグサ属(Brachypodium)のようなバイオ燃料作物も含む。
【0107】
本発明のトランスポーター、例えばGLUEトランスポーターは、植物または動物のような生物内の糖、特に単糖類、二糖類またはオリゴ糖、例えばグルコースまたはショ糖を輸送できる。植物において、トランスポーターは、花蜜の産生のために糖を輸送しうる。トランスポーターは、植物の蜜腺におよび/またはそれから糖を輸送しうる。本発明のトランスポーターは、蜜腺に局在化されうる。本明細書において使用するとき、「蜜腺」は、花蜜を産生する分泌構造を意味する。花蜜はグルコースおよび/またはフルクトースおよびまたは他の糖類を含む、受粉媒介者への報償として利用されうる組成物である。
【0108】
トランスポーターは植物の葯におよび/またはそれから糖を輸送しうる。葯は、雄ずいおよびフィラメントからなる植物の生殖器官を意味する。本発明のトランスポーターは、葯に局在化されていてもよい。本発明のトランスポーターは、葯の雄ずいおよび/またはフィラメントに局在化されていてもよい。それはタペータムまたは花粉それ自体に局在しうる。花粉において、それは栄養細胞または雄原細胞に局在しうる。本発明のトランスポーターの存在は、葯の機能に影響を与えうる。タペータムにおけるトランスポーターの機能は、花粉の栄養に関与しうる。花粉における機能は、雄原細胞への栄養補給を補助しうる。葯における機能は、デンプンの突然の加水分解を引き起こし、これは浸透能の増加を導き、次いで周囲組織からの水の吸い戻しを導き、葯の脱水および開裂を促進しうる。この機能はまた、花粉への糖の取込または輸送管もしくは精巣上体管における糖の放出に寄与しうる。
【0109】
本発明のトランスポーターは、微小胞子嚢または巨大胞子嚢のような植物の胞子嚢におよび/またはそれから糖を輸送しうる。本発明のトランスポーターは、胞子嚢または生殖腺を放出する胞子に局在化されていてもよい。トランスポーターは胞子嚢の機能に影響を与えうる。
【0110】
本発明のトランスポーターは、輸送管におよび/またはそれから糖を輸送して、伸長花粉管に栄養およびエネルギーを供給できる。
【0111】
本発明のトランスポーターは、細胞膜を通過する取込およびERへの「流出」を仲介しうる。本発明のトランスポーターは、輸送の方向が膜内外のグルコース勾配のみに依存するグルコース単輸送体として機能する。
【0112】
トランスポーター、例えばGLUEトランスポーターは、師部細胞の発生を誘導しうる。トランスポーターは師部に局在化されていてもよい。トランスポーターは師部細胞の機能に影響しうる。師部は、細胞における栄養、例えば樹液の輸送に関与する組織を意味する。師部はある領域、例えばねまたは植物の糖供給源から植物の別の領域、例えば葉または植物の糖貯蔵部へと栄養素を輸送しうる。師部に沿った輸送は、多方向性または一方向性でありうる。師部は柔組織細胞、師細胞、葉肉細胞および伴細胞、例えば常伴細胞、転送細胞および中間細胞を含みうる。師部はさらに、タンパク細胞、繊維および厚膜細胞を含みうる。
【0113】
本発明のトランスポーターは、植物または動物のような生物の疾患感受性に影響を与えうる。本発明のトランスポーターは、ウイルスまたは細菌または昆虫のような病原体に対する植物の感受性に影響を与えうる。病原体は宿主細胞の遺伝子転写に影響を与えうることが知られている。病原体は糖トランスポーターの遺伝子転写に影響を与えうる。本発明は、宿主細胞を保護する新規方法を提供する。それは地上および地下の共生生物および病原体両方の栄養に影響を与えうる。それは微生物を誘引しうる。それは土壌への糖の分泌に関与しうる。したがって、根の周辺の微生物相および植物の生産性に影響を与えうる。それは受粉媒介者との相互作用に影響を与えうる。それは外部の供給に依存する植物の細胞、例えば表皮、孔辺細胞、種子への糖の供給に影響を与えうる。
【0114】
トランスポータータンパク質
【0115】
本発明は、膜を通過する糖の移動において集合的に機能するポリペプチドの連続配列を提供する。トランスポーターは、膜に埋め込まれるかまたは完全に膜を貫通していてよい。トランスポーターは複数回、例えば2、3、4、5、6、7またはそれ以上膜を貫通していてよい。当業者は、膜を通過するトランスポーターの位置が、膜の外側(いずれかの側、例えば細胞外または細胞内)に位置するトランスポーターの部分と比較して疎水性および親水性で変化すると理解する。トランスポーターは少なくとも2個のサブユニット、例えば2個の膜貫通タンパク質、例えばホモまたはヘテロトリマーを含んでいてよく、ここで用語「トリマー」はタンパク質が膜を貫通する回数を意味する。さらなる例において、サブユニットはリンカーペプチド、例えばさらなる細胞内ドメインまたはさらなる膜貫通ドメインによって結合されていてもよい。
【0116】
トランスポーターはポアを形成しうる。ポアはトランスポーターの膜貫通ドメインまたはそのサブドメインの球状配置によって形成されうる。ポアはそれを介した糖の通過を可能としうる。ポアは糖の移動についてのみ選択的でありうる。ポアは、特定のサイズまたは特定の形状の分子の移動を制限する選択点または複数の点を有していてもよい。ポアを通る移動は、濃度勾配のみに基づきうる。ポアはさらに、コファクターの活性、例えば相互作用タンパク質の活性またはイオンの結合もしくは電荷の存在、例えば負または正の電荷に基づいて、開閉しうる。
【0117】
本発明はまた、キメラトランスポーターも提供する。キメラは、2種以上の異なるタンパク質に由来する機能的ドメインの組合せである。キメラトランスポーターは他の膜関連タンパク質、例えばチロシンキナーゼ、Gタンパク共役受容体、アクアポリンまたは他のトランスポーター由来のドメインと相互作用/採用する第二メッセンジャーと、トランスポーターのポアを融合しうる。キメラトランスポーターは、本明細書に記載の2種以上のトランスポーターの融合体であってもよい。例として、トランスポーターは、グルコーストランスポーター、グルタミン酸トランスポーター(ナトリウム依存的および小胞)、アクアポリン、Na/K ATPase、セロトニントランスポーター(SERT)、ドーパミントランスポーター(DAT)、ノルエピネフリントランスポーター(NET)、アンモニウムトランスポーターおよびカリウムチャネルを含みうる。
【0118】
本発明はまた、トランスポーターの融合タンパク質を提供する。例えば、既知のエピトープまたはタグをトランスポーターと融合してよい。タグは蛍光タグ、例えば緑色蛍光タンパク質、赤色蛍光タンパク質、オレンジ色蛍光タンパク質、黄色蛍光タンパク質、シアン色蛍光タンパク質または青色蛍光タンパク質であってよい。融合タンパク質を製造する方法は当該技術分野において既知である。
【0119】
本発明は、糖の輸送を提供しうるトランスポーターを提供する。輸送は膜を通過しうる。輸送は細胞または細胞の小器官から糖を排出しうる。輸送は細胞または細胞の小器官に糖を取り込みうる。糖は単糖、二糖もしくはオリゴ糖またはその誘導体であってよい。例えば、糖は、リボース、アラビノース、ペントース、例えばグルコース、フルクトース、ガラクトースおよびマンノース、ヘキソース、例えばマルトースおよびショ糖を含みうる。二糖類は、ラフィノースおよびスタキオースを含みうる。誘導体は、アミノ酸およびホルモンのグリコシル誘導体を含みうる。
【0120】
本発明はまた、本発明のトランスポーター、例えばGLUEトランスポーターをコードする拡散も提供する。本発明は、本発明のトランスポーターをコードする多用なcDNAを開示する。ウマゴヤシ由来のタンパク質MtN3およびそのホモログは、GLUEとして機能しうる。シロイヌナズナにおいて、下記受託番号がトランスポーターをコードする:AT4G15920、AT3G16690、AT5G13170、AtSAG29、AT4G25010、AT5G50800、AT5G23660、AT3G48740、AT5G50790、AT2G39060、AT5G40260、AtRPG1、AT4G10850、AT1G66770、AT5G62850、AtVEX1、AT3G28007、AT3G14770、AT1G21460およびAT5G53190(全て全体において参照により本明細書に組み込む)。ペチュニア植物において、NEC1は糖トランスポーターの例である(その全体において参照により本明細書に組み込む)。ウマゴヤシ植物において、本発明のトランスポーターをコードする下記cDNA受託番号が同定されている:AC202585、AC147714、MtC60432 GC、MtC11004 GC、CT963079、MtD03138 GC、TC 125536、AC146866、AC189276、TC129646、CAA69976 MtNod3 AC2456、TC115479、AC146747、MtC10424 GC、CT954252、CU302340、AC202585、AC147714、MtC60432 GC、MtC11004 GCおよびCT963079(全て全体において参照により本明細書に組み込む)。コメ植物において、本発明のトランスポーターをコードする下記cDNA受託番号が同定されている:Os08g42350 (Os8N3) Os08g0535200、Os12g29220 Os03g0347500、Os05g51090 Os05g0588500、Os12g07860、Os09g08440、Os09g08490、Os09g08270、Os09g08030 Os09g0254600、Os01g42090.1 Os01g0605700、Os01g42110.1 Os01g060600、Os02g19820 Os02g0301100、Os05g35140 Os05g0426000、Os01g65880 Os01g0881300、Os01g50460 Os01g0700100、)s01g36070.1 Os01g0541800、Os01g12130.1、Os05g12320 Os05g0214300およびOs01g21230(全て全体において参照により本明細書に組み込む)。
【0121】
本発明はまた、動物ホモログRAG1AP1およびこのファミリーの細菌ホモログ、例えば下記受託番号によってコードされるものを含む:A1BJ76、A1VHH8、A3IH65、A4AVY5、A5ERR3、A5FEJ3、A5G4U0、A5IEV6、A8AYJ9、B0SHL1、B0SR19、B1MYL5、B1MZF9、B1WTC6、B3EHG6、B5EHF6、B5YGD6、B6IU72、Q11VQ0、Q21M9、Q300V9、Q39VX0、Q3B6J0、Q5WTV4、Q5X228、Q72RB5、Q72FY5、Q89G85、Q8F4F7(全て全体において参照により本明細書に組み込む)。GLUEは原核生物、例えばレジオネラ属(Legionella)、デスルホビブリオ属(Desulfovibrio)、ブラディリゾビウム属(Bradyrhizobium)、レプトスピラ属(Leptospira)、ロドシュードモナス属(Rhodopseudomonas)、連鎖球菌、ジオバクター属(Geobacter)、ペロディクチオン属(Pelodictyon)、サイトファーガ属(Cytophaga)、ロドスピリラム属(Rhodospirillum)、サーモデスルフォビブリオ属(Thermodesulfovibrio)、クロロビウム属(Chlorobium)、ウォルバキア属(Wolbachia)、シアノセイス(Cyanothece、ロイコノストック属(Leuconostoc)およびフラボバクテリウム属(Flavobacterium)から得られる。
【0122】
糖の輸送は必須である。例えばグルコース流出は生体の多くの時点で、例えば花粉の発生または精子細胞発生のための精巣上体栄養供給の役割において必要とされる。GLUEはある生理的プロセス、例えば授乳においてアップレギュレートされ、乳管にラクトースを分泌する腺細胞に局在化されうる。同様に、肝臓のような臓器は血中グルコースレベルを一定に保つためにグルコースを流出する必要がある。GLUEは小胞流出経路のためのベシクルまたはゴルジ体へのグルコース負荷に関与しうる。
【0123】
トランスポーターファミリーのメンバーは実質的な同一性を共有する。GLUE1はそのパラログGLUE8と41%同一であり、4種のシロイヌナズナGLUEクレードの第2に属する。GLUE8/RPG1の変異は、雄不妊を導くことが示されている。哺乳類細胞におけるGLUE88/RPG1とグルコースのFRETセンサーの共発現は、いくつかのGLUE、例えばGLUE8も単輸送体として機能することを示す。さらに、GLUE8/RPG1は酵母グルコース輸送変異を補完する。GLUE8/RPG1はタペータムにおいて発現されてよく、これは花粉栄養供給における関与を示す。
【0124】
GLUE1およびGLUE8はコメタンパク質OsGLUE11/Os8N3(系統発生学に基づいてOsGLUE11と命名)と34%アミノ酸配列同一性を共有する。最も近いシロイヌナズナホモログは、OsGLUE11/Os8N3と40%同一性を共有し、第3GLUEクレードに属する。RNA阻害による発現の現象が花粉における低下したデンプン濃度ならびに花粉不妊を導くため、GLUE8と同様に、OsGLUE11/Os8N3は花粉栄養供給に機能しうる。クレード3の他のGLUEホモログであるPetunia Neclも、雄不妊を導きうる。Nec1は蜜腺で発現され、その発生制御は蜜腺のデンプン含量と逆相関しており、これは蜜腺の糖分泌におけるNec1の第二の役割を示す。
【0125】
本発明は、他の生物のトランスポーターを提供する。例えば、線虫(C. elegans)遺伝子は糖流出トランスポーターの新規クラス(SLC50)の7種のホモログを含むが、ヒトゲノムは1種のホモログ、すなわちRAG1AP1(またはHsGLUE1)を有する。シロイヌナズナGLUE1同様、線虫(C. elegans)CeGLUE1はグルコース取込を仲介しうる。CeGLUE1およびヒトRAG1AP1は、Na+/グルコースコトランスポーターSGLT1によって仲介される第二活性グルコース蓄積を相殺しうる。CeGLUE1およびヒトRAG1AP1の変異は、脂質の蓄積を導く細胞性グルコース流出の欠損と一致して、脂肪蓄積を導きうる。
【0126】
本発明は糖トランスポーター、例えばGLUEをコードする核酸を提供する。本発明はまた、保存的アミノ酸置換を有するポリペプチドをコードする核酸を提供する。本発明の核酸は、糖を輸送するポリペプチドをコードしうる。単離された核酸は上記同定した配列と少なくとも約30%、40%、50%、60%、70%、80%、85%、90%、95%または99%配列同一性を有しうる。単離された核酸は、上記同定された受託番号によってコードされるアミノ酸配列と少なくとも約30%、40%、50%、60%、70%、80%、85%、90%、95%または99%配列同一性を有するアミノ酸配列を有するポリペプチドをコードしうる。トランスポーターをコードする単離された核酸は、上記同定された核酸配列とハイブリダイズしうる。
【0127】
GLUEのタンパク質は配列およびサブドメインホモロジーを共有する。下記表1は、GLUEの選択における配列保存量の図を提供し、下記表2は、配列によるGLUEの比較を提供する。
【表1】
【表2】
【0128】
GLUEタンパク質をコードする核酸は、一般に、発現のための発現制御配列と遺伝的に融合されていてよい。好適な発現制御配列は、標的宿主生物に適用可能なプロモーターを含む。かかるプロモーターは、原核生物および真核生物からの多様な宿主について、当業者に周知であり、文献に記載されている。例えばかかるプロモーターは、天然由来の遺伝子から単離されてよく、あるいは合成またはキメラプロモーターであってもよい。同様に、プロモーターは標的ゲノム中に既に存在していてよく、例えば相同組換えのような当該技術分野において既知の好適な技術によって核酸分子と連結されてもよい。
【0129】
本発明はまた、GLUEをコードする核酸を標的核酸分子、例えばベクターまたはゲノムDNAに挿入するための発現カセットを提供する。この目的で、発現カセットは、特異的配列位置など、例えば制限酵素認識部位または例えばリコンビナーゼによって触媒される場合の相同組換えの標的部位からの除去またはそれへの挿入を促進するための、5’または3’隣接ヌクレオチド配列を提供する。
【0130】
本発明はまた、遺伝子操作において通常使用される、本発明の核酸分子および発現カセットを含むベクター、特にプラスミド、コスミド、ウイルスおよびバクテリオファージにも関する。
【0131】
本発明の好ましい態様において、本発明のベクターは、真菌細胞、植物細胞、微生物の細胞(すなわち細菌、原生生物、酵母、藻類等)または動物細胞、特に哺乳動物細胞の形質転換に好適である。好ましくは、かかるベクターはヒト細胞の形質転換に好適である。当業者に周知の方法を用いて、組換えベクターを構築できる:例えば、Sambrook and Russell, Molecular Cloning: A Laboratory Manual, CSH Press, 2001および Ausubel, Current Protocols in Molecular Biology, Green Publishing Associates and Wiley Interscience, N.Y., 1989に記載の技術を参照。あるいは、ベクターは、本発明の核酸分子または発現カセットが標的細胞への送達のために再構成されてよいリポソームであってもよい。同様に、用語「ベクター」は、細胞への遺伝子トランスファーを促進することが知られている化合物、例えばポリカチオン、カチオン性ペプチド等をさらに含む、核酸分子または発現カセットを含む複合体を意味する。
【0132】
本発明の核酸分子または発現カセットに加えて、ベクターは、好適な宿主細胞において好適な条件下でベクターの選択が可能な遺伝子、例えばマーカー遺伝子をさらに含んでいてよい。一般に、ベクターは1個以上の複製起点も含む。ベクターはまた、本発明のトランスポーターをコードする核酸を超えた転写の長さを制限する終止配列を含んでいてもよい。
【0133】
有利には、ベクターに含まれる核酸分子は、原核細胞または真核細胞において、発現を可能とする、すなわち翻訳可能なRNAの転写および合成を保証する発現制御配列と作動可能に連結されている。
【0134】
遺伝子操作について、例えば原核細胞において、本発明の核酸分子およびそれらの分子の一部は、DNA配列の組換えによる突然変異または配列修飾を許容するプラスミドに導入されていてよい。標準的な方法(Sambrook and Russell, Molecular Cloning: A Laboratory Manual, CSH Press, 2001参照)は、ベース交換の実施または天然もしくは合成配列の添加を可能とする。DNAフラグメントは、該フラグメントにアダプターまたはリンカーを適用することにより互いに結合していてよい。さらにまた、好適な制限部位を提供するかまたは余剰DNAもしくは制限部位を除去する操作手法が使用できる。これらの場合、挿入、欠失または置換が可能であり、インビトロ突然変異、「プライマー修復」、制限またはライゲーションが使用できる。配列分析、制限分析または生化学および分子生物学の他の方法が分析法として実施される。
【0135】
本発明はまた、トランスポーターをコードする核酸の指定された発現を提供する。遺伝子の発現はコードヌクレオチド配列の上流(5’)の特定のプロモーターによって制御されうることが当該技術分野において知られている。動物の特定の組織での発現を指示する組織特異的プロモーターは、当該技術分野において既知である。例えば、データベースがこれらのプロモーターを収集し、共有している(Chen et al., Nucleic Acids Res. 34: D104-D107, 2006)。植物では、根、種子または果実での発現を指示するプロモーターが知られている。
【0136】
本発明はさらに、さらなるポリペプチドと融合したトランスポーターを含む単離されたポリペプチドを提供する。さらなるポリペプチドは、大型ポリペプチドのフラグメントであってもよい。一つの態様において、トランスポーターと融合した1、2、3、4またはそれ以上のさらなるポリペプチドが存在する。いくつかの態様において、さらなるポリペプチドは、トランスポーターのアミノ末端に対して融合している。他の態様において、さらなるポリペプチドはトランスポーターのカルボキシル末端に対して融合している。さらなる態様において、さらなるポリペプチドはトランスポーターと隣接している。いくつかの態様において、核酸分子は、トランスポーターをコードする核酸と融合した核酸を含む融合タンパク質をコードする。融合核酸は、トランスポーターのコドンリーディングフレームを移動させることなく、精製および/または免疫原性および/または安定性に役立ちうるポリペプチドをコードできる。いくつかの態様において、融合核酸は、トランスポーターの精製に役立つポリペプチドをコードする。いくつかの態様において、融合核酸は、エピトープおよび/または親和性タグをコードする。他の態様において、融合核酸は、部位特異的または誤りがちな切断に関するポリペプチドをコードする。好ましい態様において、融合核酸は、酵素的切断の部位であるポリペプチドをコードする。さらなる態様において、酵素的切断は、トランスポーターの単離に役立つ。
【0137】
他の態様において、複数の核酸がトランスポーターをコードする核酸と融合される。融合核酸は、精製および/または酵素的切断および/または安定性に役立つポリペプチドをコードしうる。さらなる態様において、融合核酸は発現されるポリペプチドを顕著に伸長しない。
【0138】
いくつかの態様において、さらなるポリペプチドはエピトープを含んでもよい。他の態様において、さらなるポリペプチドは親和性タグを含んでもよい。例として、エピトープおよび/または親和性タグを含むポリペプチドとトランスポーターの融合体は、ポリペプチドの精製および/または同定に役立ちうる。例として、ポリペプチドセグメントはHisタグ、mycタグ、Sペプチドタグ、MBPタグ(マルトース結合タンパク質)、GSTタグ(グルタチオンSトランスフェラーゼ)、FLAGタグ、チオレドキシンタグ、GFPタグ(緑色蛍光タンパク質)、BCCP(ビオチンカルボキシル担体タンパク質)、カルモジュリンタグ、Strepタグ、HSVエピトープタグ、V5エピトープタグおよびCBPタグであってよい。かかるエピトープおよび親和性タグの使用は、当業者に既知である。
【0139】
さらなる態様において、さらなるポリペプチドは、ポリペプチドの切断のための部位を含む融合タンパク質を提供できる。切断部位は、例えばターゲティングポリペプチドで融合ポリペプチドからトランスポーターを後で切り出すために有用である。例として、ポリペプチドはペプチド結合の加水分解によって切断されうる。いくつかの態様において、切断は酵素によって実施される。いくつかの態様において、切断は細胞において生じる。他の態様において、切断は人工操作および/または切断酵素の人工導入によって生じる。例として、切断酵素はペプシン、トリプシン、キモトリプシンおよび/またはXa因子を含みうる。
【0140】
融合ポリペプチドは、同じ有機的に合成されたペプチドとは共有されないさらなる構造修飾、例えばアデニル化、カルボキシル化、グリコシル化、ヒドロキシル化、メチル化、リン酸化またはミリスチル化をさらに有していてよい。これらのさらなる構造修飾は、組換え発現系の適切な選択によってさらに選択または選好されうる。他方、融合ポリペプチドは、有機合成の原理および実施によって伸長された配列を有していてもよい。
【0141】
一般に、本発明の融合タンパク質は、先行技術に記載の技術に従って作成できる。例えば、これらの技術は、Sambrook and Russell, Molecular Cloning: A Laboratory Manual, CSH Press, 2001またはAusubel, Current Protocols in Molecular BiologyのVolumes1および2, Current Protocols, 1994に記載のとおりに実施できる組換え技術を含む。したがって、融合タンパク質の個々の部分は、宿主生物またはインビトロで組み合わされ、次いで発現される、それらをコードする核酸分子の形態で提供できる。あるいは、融合タンパク質またはその一部の提供は、化学合成または天然供給源からのかかる部分の単離を含んでもよく、それによって組換え技術によって部分的に作成されうる要素が好適な方法、例えば化学的架橋、例えばWO 94/04686に記載のとおりに、タンパク質レベルで融合されてよい。さらにまた、適切である場合、融合タンパク質は、それぞれの目的のためにその特性を改善するため、例えば溶解性を向上し、pH耐性を向上し、宿主生物にいてより良好な耐性であり、インビボもしくはインビトロで特定の基質に接着させる(後者は固相への融合タンパク質の不動化に潜在的に有用である)等のために、翻訳後修飾されてよい。当業者はかかる修飾およびその有用性を周知している。例示的説明は、1アミノ酸側鎖の修飾(例えばグリコシル化、ミリスチル化、リン酸化、カルボエトキシル化またはアミド化による)、ポリエチレングリコール、炭水化物等のようなポリマーまたはタンパク質部分、例えば抗体もしくはその一部または他の酵素等とのカップリングを含む。
【0142】
本発明の他の態様において、融合タンパク質はさらに、ターゲティングシグナル配列を含む。葉緑体、液胞、ペルオキシソーム、グリオキシソーム、細胞壁またはミトコンドリアのような細胞内区画へのタンパク質の輸送またはアポプラストへの分泌は、目的のタンパク質をコードする遺伝子の5’および/または3’領域と、シグナル配列をコードするヌクレオチド配列を作動可能に連結することによって達成される。構造遺伝子の5’および/または3’末端のターゲティング配列は、コードされるタンパク質が最終的に区分されるタンパク質合成およびプロセッシングにおいて決定されうる。
【0143】
シグナル配列の存在は、細胞内小器官または細胞内区画にポリペプチドを指向させるか、あるいはアポプラストに分泌させる。多くのシグナル配列が当該技術分野において既知である。例えばBecker et al., Plant Mol. Biol. 20:49 (1992); Close, P. S., Master's Thesis, Iowa State University (1993); Knox, C., et al., Plant Mol. Biol. 9:3-17 (1987); Lerner et al., Plant Physiol. 91:124-129 (1989); Frontes et al., Plant Cell 3:483-496 (1991); Matsuoka et al., Proc. Natl. Acad. Sci. 88:834 (1991); Gould et al., J. Cell. Biol. 108:1657 (1989); Creissen et al., Plant J. 2:129 (1991); Kalderon, et al., Cell 39:499-509 (1984); Steifel, et al., Plant Cell 2:785-793 (1990)参照。
【0144】
用語「ターゲティングシグナル配列」は、発現されたタンパク質におけるその存在のためにそれが特定の細胞内局在を標的とするアミノ酸配列を意味する。例えば、対応するターゲティングシグナルは、精製を単純化するために例えば細菌宿主から、発現されたトランスポーターの分泌を導きうる。好ましくは、トランスポーターのターゲティングは、特定の細胞内または細胞外区画における糖濃度に影響を与えるために使用できる。異なる生物群に有用な適切なターゲティングシグナル配列は、当業者に既知であり、文献または配列データベースから取り出すことができる。
【0145】
本発明のトランスポーターは、細胞基質、細胞表面または細胞内小器官、例えば核、ベシクル、ER、液胞等の細胞の任意の場所で発現されてよい。異なる細胞区画にタンパク質の発現をターゲティングする方法およびベクター成分は、バイオセンサーが発現される具体的な細胞または生物に基づく選択と共に、当該技術分野において周知である。例えばOkumoto et al. PNAS 102: 8740-8745, 2005; Fehr et al. J Fluoresc 14: 603-609, 2005を参照されたい(その全体について参照により本明細書に組み込む)。葉緑体、液胞、ペルオキシソーム、グリオキシソーム、細胞壁またはミトコンドリアのような細胞内区画へのタンパク質の輸送またはアポプラストへの分泌は、トランスポーターをコードする遺伝子の5’および/または3’領域と、シグナル配列をコードするヌクレオチド配列を作動可能に連結することによって達成されうる。構造遺伝子の5’および/または3’末端のターゲティング配列は、コードされるタンパク質が最終的に区分されるタンパク質合成およびプロセッシングにおいて決定されうる。
【0146】
植物細胞の色素体へのターゲティングが望まれる場合、例えば次のターゲティングシグナルペプチドが使用できる:シロイヌナズナ色素体RNAポリメラーゼ(AtRpoT 3)のアミノ酸残基1〜124(Plant Journal 17: 557-561, 1999);ホウレンソウの色素体Ferredoxin:NADP+オキシドレダクターゼ(FNR)のターゲティングシグナルペプチド(Jansen et al., Current Genetics 13: 517-522, 1988)、特にそれに開示のcDNA配列のヌクレオチド−171〜165によってコードされるアミノ酸配列;成熟ロウタンパク質の最初の34アミノ酸残基を含むかまたは含まないトウモロコシのロウタンパク質の輸送ペプチド(Klosgen et al., Mol. Gen. Genet. 217: 155-161, 1989);リブロースビスホスフェートカルボキシラーゼ小サブユニットのシグナルペプチド(Wolter et al., PNAS 85: 846-850, 1988; Nawrath et al., PNAS 91: 12760-12764, 1994)、NADPリンゴ酸デヒドロゲナーゼのシグナルペプチド(Gallardo et al., Planta 197: 324-332, 1995)、グルタチオンレダクターゼのシグナルペプチド(Creissen et al., Plant J. 8: 167-175, 1995)またはR1タンパク質のシグナルペプチド(Lorberth et al., Nature Biotechnology 16: 473-477, 1998)。
【0147】
植物細胞のミトコンドリアへのターゲティングは、次のターゲティングシグナルペプチドを用いて達成できる:シロイヌナズナミトコンドリアRNAポリメラーゼのアミノ酸残基1〜131(AtRpoT 1)(Plant Journal 17: 557-561, 1999)またはBraunに記載の輸送ペプチド(EMBO J. 11: 3219-3227、1992)。
【0148】
植物細胞の液胞へのターゲティングは、次のターゲティングシグナルペプチドを用いて達成できる:パタチンタンパク質のN末端配列(146アミノ酸)(Sonnewald et al., Plant J. 1: 95-106, 1991)またはMatsuoka and Neuhaus(Journal of Exp. Botany 50: 165-174, 1999);Chrispeels and Raikhel(Cell 68: 613-616, 1992); Matsuoka and Nakamura(PNAS 88: 834-838, 1991); Bednarek and Raikhel(Plant Cell 3: 1195-1206, 1991)またはNakamura and Matsuoka(Plant Phys. 101: 1-5, 1993)に記載のシグナル配列。
【0149】
植物細胞のERへのターゲティングは、例えばC末端伸長HDEL(Haselhoff, PNAS 94: 2122-2127, 1997)と組み合わせたERターゲティングペプチドHKTMLPLPLIPSLLLSLSSAEFを用いて達成できる。植物細胞の核へのターゲティングは、例えばタバコC2ポリペプチドQPSLKRMKIQPSSQPの核局在シグナル(NLS)を用いて達成できる。
【0150】
細胞外空隙へのターゲティングは、例えば下記輸送ペプチドの一つを用いて達成できる:プロテイナーゼ阻害剤II遺伝子のシグナル配列(Keil et al., Nucleic Acid Res. 14: 5641-5650, 1986; von Schaewen et al., EMBO J. 9: 30-33, 1990)、火傷病菌由来レバンスクラーゼ遺伝子のシグナル配列(Geier and Geider, Phys. Mol. Plant Pathol. 42: 387-404, 1993)、最初の33アミノ酸をコードするジャガイモ由来パタチン遺伝子B33のフラグメントのシグナル配列(Rosahl et al., Mol Gen. Genet. 203: 214-220, 1986)またはOshima et al.に記載のもののシグナル配列(Nucleic Acids Res. 18: 181, 1990)。
【0151】
さらにまた、膜へのターゲティングは、ウサギスクラーゼ−イソマルターゼのN末端シグナルアンカーを用いて達成できる(Hegner et al., J. Biol. Chem. 276: 16928-16933, 1992)。
【0152】
哺乳類細胞の膜へのターゲティングは、N末端ミリスチン酸結合配列can be accomplished by using the N-terminal myristate attachment sequence MGSSKSKまたはC末端プレニル化配列CaaXを用いて達成できる(Garabet, Methods Enzymol. 332: 77-87, 2001)。ここで「a」は脂肪族アミノ酸(すなわちVal、Leu またはIle)であり「X」は任意のアミノ酸である。
【0153】
細胞膜または植物細胞へのさらなるターゲティングは、トランスポーター、好ましくはショ糖トランスポーターSUT1との融合によって達成できる(Riesmeier, EMBO J. 11: 4705-4713, 1992)。多様な細胞内膜へのターゲティングは、空胞水チャネル(γTIP)のような特定の区画における膜タンパク質(Karlsson、Plant J. 21: 83-90、2000)、ミトコンドリアのMCFタンパク質(Kuan, Crit. Rev. Biochem. Mol. Biol. 28: 209-233, 1993)、色素体の内包膜におけるトリオースリン酸トランスロケーター(Flugge, EMBO J. 8: 39-46, 1989)およびチラコイドの光化学系との融合によって達成できる。
【0154】
ゴルジ装置へのターゲティングは、C末端認識配列K(X)KXXを用いて達成できる(Garabet, Methods Enzymol. 332: 77-87, 2001)。ここで「X」は任意のアミノ酸である。
【0155】
ペルオキシソームへのターゲティングは、ペルオキシソームターゲティング配列PTS IまたはPTS IIを用いて達成できる(Garabet, Methods Enzymol. 332: 77-87, 2001)。
【0156】
哺乳動物細胞の核へのターゲティングは、SV-40大型T抗原核局在化配列PKKKRKVを用いて達成できる(Garabet, Methods Enzymol. 332: 77-87, 2001)。
【0157】
哺乳動物細胞のミトコンドリアへのターゲティングは、N末端ターゲティング配列MSVLTPLLLRGLTGSARRLPVPRAKISLを用いて達成できる(Garabet, Methods Enzymol. 332: 77-87, 2001)。
【0158】
いくつかの態様において、トランスポーターの発現は、特定の組織または細胞タイプに標的化されうる。例えば、特定のプロモーターを使用して、トランスポーターをコードする核酸の転写を駆動できる。プロモーターは核酸の転写を指示する核酸コントロール配列のアレイである。プロモーターは転写開始部位、例えばポリメラーゼII型プロモーターの場合にはTATA要素の近くの必要な核酸配列を含む。プロモーターはまた、所望により、転写開始部位から数千塩基対に位置しうる離れたエンハンサーまたはリプレッサーを含む。恒常プロモーターは、ほとんどの環境および発生条件下で活性なプロモーターである。誘導性プロモーターは、環境または発生制御下で活性なプロモーターである。いずれかの誘導性プロモーターが使用でき、例えばWard et al., Plant Mol. Biol. 22:361-366, 1993を参照されたい。誘導性プロモーターの例は、ACEI系由来のもの(銅に応答性)(Meft et al., Proc. Natl. Acad. Sci. USA 90:4567-4571, 1993);トウモロコシ由来のIn2遺伝子(ベンゼンスルホンアミド除草剤薬害軽減剤に応答性)(Hershey et al., Mol. Gen. Genetics 227:229-237, 1991およびGatz et al., Mol. Gen. Genetics 243:32-38, 1994)またはTn10由来のTetリプレッサー(Gatz et al., Mol. Gen. Genetics 227:229-237, 1991)を含むがこれらに限定されない。誘導性プロモーターは、宿主細胞の外来物質に応答しうる。例えばSchena et al., PNAS 88: 10421-10425, 1991参照。
【0159】
プロモーターは恒常プロモーターであってもよい。恒常プロモーターは発現のために遺伝子と作動可能に連結されているか、または発現のために遺伝子と作動可能に連結しているシグナル配列をコードするヌクレオチド配列と作動可能に連結している。例えば植物細胞において、恒常プロモーターは、CaMV由来の35Sプロモーターのような植物ウイルス由来のプロモーター(Odell et al., Nature 313: 810-812, 1985)およびコメアクチンのような遺伝子由来のプロモーター(McElroy et al., Plant Cell 2: 163-171, 1990);ユビキチン(Christensen et al., Plant Mol. Biol. 12:619-632, 1989およびChristensen et al., Plant Mol. Biol. 18: 675-689, 1992);pEMU(Last et al., Theor. Appl. Genet. 81:581-588, 1991);MAS(Velten et al., EMBO J. 3:2723-2730, 1984)およびトウモロコシH3ヒストン(Lepetit et al., Mol. Gen. Genetics 231: 276-285, 1992およびAtanassova et al., Plant Journal 2(3): 291-300, 1992)を含むが、これらに限定されない。原核生物プロモーター要素は、大腸菌におけるRNAポリメラーゼによる転写に最適な−35〜−10(Pribnowボックス)を担持するものを含む。いくつかの原核生物プロモーター要素は、制御リプレッサーの重複結合部位を含みうる(例えばLacおよびTACプロモーター、これらはlacリプレッサーの重複結合部位を含み、それによって基質ホモログIPTGによる誘導性を与える)。好適なプロモーター配列が得られうる原核生物遺伝子の例は、大腸菌lac、araおよびtrpを含む。本発明の原核生物ウイルス性プロモーター要素は、λファージプロモーター(例えばPRMおよびPR)、T7ファージプロモーター要素およびSP6プロモーター要素を含む。本発明の真核生物プロモーターベクター要素は、酵母(例えばGAL1、GAL10、CYC1)および哺乳類(例えばグロビン遺伝子およびインターフェロン遺伝子のプロモーター)の両方を含む。さらなる真核生物プロモーターベクター要素は、ウイルス性遺伝子プロモーター、例えばSV40プロモーター、CMVプロモーター、単純ヘルペスチミジンキナーゼプロモーターならびに多様なレトロウイルスLTRプロモーター要素(例えばMMTV LTR)のものを含む。他の真核生物の例は、hMTIIaプロモーター(例えば米国特許5,457,034)、HSV−1 4/5 プロモーター(例えば米国特許5,501,979)および早期中間体HCMVプロモーター(WO 92/17581)を含む。
【0160】
プロモーターは組織特異的または組織選好プロモーターであってもよい。組織特異的プロモーターは、特定の組織で排他的または優先的なトランスポーターの産生を補助する。あらゆる組織特異的または組織選好プロモーターが利用できる。植物細胞において、例えば組織特異的または組織選好プロモーターは、根選好プロモーター、例えばファゼオリン遺伝子に由来するもの(Murai et al., Science 23: 476-482, 1983およびSengupta-Gopalan et al., PNAS 82: 3320-3324, 1985);葉特異的および光誘導性プロモーター、例えばcabまたはrubisco由来のもの(Simpson et al., EMBO J. 4(11): 2723-2729, 1985および Timko et al., Nature 318: 579-582, 1985);葯特異的プロモーター、例えばLAT52由来のもの(Twell et al., Mol. Gen. Genetics 217: 240-245, 1989);花粉特異的プロモーター、例えばZm13由来のもの(Guerrero et al., Mol. Gen. Genetics 244: 161-168, 1993)または小胞子選好プロモーター、例えばapg由来のもの(Twell et al., Sex. Plant Reprod. 6: 217-224, 1993)を含むが、これらに限定されない。
【0161】
さらに本発明は、上記本発明の核酸分子を含み、原核細胞または真核細胞において発現可能なように制御配列と作動可能に連結している発現カセットに関する。
【0162】
さらなる態様において、本発明は、本発明のトランスポーターを発現できる細胞を作成する方法であって、上記本発明の核酸分子、発現カセットまたはベクターで細胞を遺伝的に操作することを含む方法に関する。
【0163】
本発明の別の態様は、上記本発明の核酸分子、発現カセットまたはベクターで遺伝的に操作された宿主細胞、特に原核細胞または真核細胞、ならびにかかる形質転換された細胞の子孫細胞であって、本発明の核酸分子、発現カセットまたはベクターを含む細胞、ならびにそれを作成する上記方法によって得られた細胞に関する。
【0164】
宿主細胞緒は細菌、真菌、昆虫、植物または動物宿主細胞であってよい。一つの態様において、宿主細胞は、導入されてゲノムに安定に組み込まれた核酸分子を含むように遺伝的に操作されている。他の態様において、核酸分子は、本発明の融合タンパク質の産生を導くように発現されてよい。
【0165】
多様な発現系の概略は、例えばBitter et al.のMethods in Enzymology 153: 385-516, 1987 (Methods in Enzymology 153: 516-544, 1987)およびSawers et al.(Applied Microbiology and Biotechnology 46: 1-9, 1996)、Billman-Jacobe(Current Opinion in Biotechnology 7: 500-4, 1996)、Hockney(Trends in Biotechnology 12: 456-463, 1994)およびGriffiths et al.(Methods in Molecular Biology 75: 427-440, 1997)に含まれる。酵母発現系の概略は、例えばHensing et al.(Antoine von Leuwenhoek 67: 261-279, 1995)、Bussineau(Developments in Biological Standardization 83: 13-19, 1994)、Gellissen et al.(Antoine van Leuwenhoek 62: 79-93, 1992)、Fleer(Current Opinion in Biotechnology 3: 486-496, 1992)、Vedvick(Current Opinion in Biotechnology 2: 742-745, 1991)およびBuckholz(Bio/Technology 9: 1067-1072, 1991)で得られる。
【0166】
発現ベクターは文献に広く記載されている。一般に、それは選択マーカー遺伝子および選択した宿主における複製を保証する複製起点だけでなく、細菌性またはウイルス性プロモーターおよび多くの場合転写の終止シグナルを含む。一般に、プロモーターと終止シグナルの間に、コードヌクレオチド配列の挿入が可能な少なくとも1個の制限部位またはポリリンカーが存在する。遺伝子発現の意図的な制御を可能とする、遺伝子および誘導性プロモーターの恒常的発現を保証するプロモーターの使用が可能である。これらの特徴を有する細菌性またはウイルス性プロモーター配列は、文献に詳細に説明されている。微生物(例えば大腸菌、出芽酵母)における発現のための制御配列は、十分に文献に記載されている。下流配列の特に高い発現を可能とするプロモーターは、例えばT7プロモーター(Studier et al., Methods in Enzymology 185: 60-89、1990)、lacUV5、trp、trp-lacUV5(DeBoer et al., in Rodriguez and Chamberlin (Eds)、Promoters、Structure and Function; Praeger、New York、1982、p. 462-481; DeBoer et al., PNAS 80: 21-25, 1983)、Ip1、rac(Boros et al., Gene 42: 97-100, 1986)である。誘導性プロモーターは、タンパク質の合成に使用できる。これらのプロモーターはしばしば恒常プロモーターよりも高いタンパク質収量を導く。最適な量のタンパク質を得るために、二段階法がしばしば用いられる。第一に、宿主細胞を比較的高い細胞密度まで最適条件下で培養する。第二段階で、使用するプロモーターのタイプに依存して転写を誘導する。これに関して、ラクトースまたはIPTG(イソプロピル−β−D−チオガラクトピラノシド)によって誘導可能なtacプロモーターが特に好適である(DeBoer et al., PNAS 80: 21-25, 1983)。哺乳動物細胞における適用に有用なSV40-ポリA部位またはtk-ポリA部位のような転写の終止シグナルも文献に記載されている。好適な発現ベクターは当該技術分野において既知であり、例えばOkayama-Berg cDNA発現ベクターpcDV1(Pharmacia)、pCDM8、pRc/CMV、pcDNA1、pcDNA3(In-vitrogene)、pSPORT1 (GIBCO BRL)またはpCI(Promega)である。
【0167】
本発明はまた、本発明のトランスポーターをコードするベクターまたは発現ベクターでトランスフェクトされた宿主細胞、例えば原核細胞、例えば大腸菌もしくは他の細菌または真核細胞、例えば酵母細胞もしくは動物細胞も含む。生細胞培養は、原核細胞または真核細胞を含んでもよい。原核細胞源の例は、細菌または巨細筋を含むがこれらに限定されない。真核細胞源の例は、酵母、真菌、原生生物、哺乳類、節足動物、ヒト、動物、軟体動物、環形動物、線形動物、甲殻類、扁形動物、単孔類、魚類、有袋類、爬虫類、両生類、鳥類、げっ歯類、昆虫および植物を含むが、これらに限定されない。
【0168】
本発明の核酸分子またはベクターによる宿主細胞の形質転換は、標準的な方法、例えばSambrook and Russell, Molecular Cloning: A Laboratory Manual, CSH Press, 2001; Methods in Yeast Genetics, A Laboratory Course Manual, Cold Spring Harbor Laboratory Press, 1990に記載のとおりに実施できる。例えば、塩酸カルシウムトランスフェクションは、一般に原核細胞に利用され、一方例えばリン酸カルシウムもしくはDEAEデキストラン介在トランスフェクションまたはエレクトロポレーションは、他の細胞宿主に使用できる。宿主細胞は使用する具体的な宿主細胞の、特にpH値、温度、塩濃度、通気、抗生物質、ビタミン、微量元素等に関する要求に合う栄養培地中で培養する。本発明のトランスポーターは、硫酸アンモニウムもしくはエタノール沈殿、酸抽出、アニオンもしくはカチオン交換クロマトグラフィー、ホスホセルロースクロマトグラフィー、疎水性相互作用クロマトグラフィー、アフィニティークロマトグラフィー、ヒドロキシルアパタイトクロマトグラフィーおよびレクチンクロマトグラフィーを含む方法によって、組換え細胞から回収し、精製できる。トランスポーターのリガンドまたは基質、例えばグルコースはアフィニティー精製に使用できるか、あるいはトランスポーターの融合タンパク質は、親和性タグのような融合タンパク質が結合する基質またはリガンドによるアフィニティークロマトグラフィーを適用して精製できる。タンパク質再フォールディング段階は、必要により、タンパク質の立体配置を完成させるために使用できる。最後に、高速液体クロマトグラフィー(HPLC)を最終生成段階に使用できる。
【0169】
したがって、本発明のさらなる態様は、本発明のトランスポーターを製造する方法であって、トランスポーターの発現が可能な条件下で前記宿主細胞を培養し、培養物からトランスポーターを回収することを含む方法に関する。発現タンパク質が宿主細胞中に局在するか細胞から分泌されるかに従って、タンパク質は、培養細胞および/または培地の上清から回収できる。
【0170】
あるいは、トランスポーターはマイクロインジェクション、微粒子銃、封入センサーの導入または細胞へのバイオセンサーの取り込みを導くペプチド配列の融合によって、細胞に送達されうる。
【0171】
さらにまた、本発明は、上記製造方法によって得られるトランスポーターに関する。
【0172】
本発明のトランスポーターは、例えば化学的合成法の産物であるか、あるいは原核または真核宿主(例えば、培養の細菌、酵母、高等植物、昆虫または哺乳類細胞による)から組換え技術により産生されてよい。組換え製造法において使用される宿主に依存して、発現されるトランスポーターは、グリコシル化されているかまたは非グリコシル化されていてよい。本発明のトランスポーターはまた、初期メチオニンアミノ酸残基を含んでいてもよい。本発明のトランスポーターはさらに、通常タンパク質の一部ではないさらなる化学部分を含むように修飾されていてもよい。これらの誘導体化部分は、例えばタンパク質の安定性、溶解性、生物学的半減期またはタンパク質の吸着を改善しうる。当該部分はまた、タンパク質の望ましくないいずれかの副作用などを低減または削除しうる。これらの部分の概要は、例えばRemington's Pharmaceutical Sciences, Mack Publishing Co., Easton, Paに見られる。
【0173】
トランスジェニック
【0174】
本発明は、本発明のトランスポーター、例えばGLUEをコードする核酸分子を含むトランスジェニック植物および非ヒトトランスジェニック生物、すなわち多細胞生物、または少なくとも当該生物または組織もしくは臓器のようなその一部の細胞のサブセットにおいて、そのゲノムに安定的に組み込まれた上記発現カセットまたはベクターを含む、生物を提供する。
【0175】
本発明は、トランスジェニック植物細胞、すなわちゲノムに安定的に組み込まれた上記本発明の核酸分子、発現カセットまたはベクターを含む、トランスジェニック植物または植物組織を提供する。本発明はまた、以下に説明する製造方法によって得られたトランスジェニック植物、植物細胞または植物組織を提供する。
【0176】
一つの態様において、本発明は、トランスジェニック植物、植物組織または植物細胞を製造する方法であって、本発明の核酸分子、発現カセットまたはベクターを植物細胞に導入し、所望によりそれからトランスジェニック植物または植物組織を再生することを含む方法を提供する。トランスポーターを発現するトランスジェニック植物は、生物の器官を介するおよびそれの間の、例えば土壌へのまたは土壌からの、糖の輸送に影響を与えるために使用できる。本発明のトランスポーターを発現するトランスジェニック植物は、例えば有機化合物の代謝または輸送プロセスを、先行技術では得られなかった時間および空間分解能で調査するために使用できる。
【0177】
外来核酸分子の植物への導入方法は、当該技術分野において周知である。例えば植物形質転換は、アグロバクテリウム(Agrobacterium)介在遺伝子導入、マイクロインジェクション、エレクトロポレーションまたは微粒子銃法を用いて、例えばPotrykus and Spangenberg (Eds.), Gene Transfer to Plants. Springer Verlag, Berlin, New York, 1995に記載のとおりに実施できる。これにおよび多数の他の先行技術文献において、有用な植物形質転換ベクター、形質転換された細胞および組織の選択方法ならびに再生技術が記載されており、それらは当業者に既知であり、本発明の目的のために適用されうる。
【0178】
別の局面において、本発明は、上記トランスジェニック植物細胞を含む、本発明のトランスジェニック植物の収穫可能な部分および伝播物質を提供する。収穫可能な部分は、基本的に、植物のあらゆる有用な部分、例えば葉、幹、果実、種子、根などであってよい。伝播物質は、例えば種子、果実、挿し木、実生、塊茎、根茎等を含む。
【0179】
ある局面において、本発明は、トランスポーターの発現をコードする核酸配列の発現によって特徴付けられる表現型を有するトランスジェニック非ヒト動物を提供する。表現型は動物の体細胞および生殖細胞に含まれるトランスジーンによって与えられ、これは(a)動物の接合体にトランスポーターをコードするDNA構築物を含むトランスジーンを導入し;(b)接合体を偽妊娠動物に移植し;(c)接合体を発生、出産させ;そして(d)トランスジーンを含む少なくとも1体のトランスジェニック子孫を同定することによって製造できる。胚にトランスジーンを導入する工程は、胚にトランスジーンを含む胚性幹細胞を導入するか、またはトランスジーンを含むレトロウイルスに胚を感染させることを含んでいてよい。好ましいトランスジェニック動物は、コードされたトランスポーターを発現する。本発明のトランスジェニック動物は、トランスジェニック出芽酵母(S. cerevisae)、線虫(C. elegans)、ショウジョウバエ、特に、キイロショウジョウバエおよびトランスジェニックマウスおよび他の動物を含む。
【0180】
本発明はまた、トランスポーターをコードする少なくとも1個の核酸分子、ゲノムに安定的に組み込まれうる核酸を含む発現カセットまたはベクターを含む、トランスジェニック非ヒト動物を提供する。
【0181】
本発明はまた、トランスジェニック非ヒト動物の作成方法であって、本発明の核酸分子、発現カセットまたはベクターを、生殖細胞、胚性細胞、肝細胞または卵またはそれらに由来する細胞に導入することを含む方法を包含する。本発明のトランスポーターを発現するかかるトランスジェニック動物または接合体から始まるその任意の発生段階が、組織の完全性を破壊することなく実時間である化合物(融合タンパク質に存在する酵素に依存する)の分布を決定できるモデル生物として使用できる。これらのモデル生物は、栄養学的もしくは薬理学的研究または薬物スクリーニングに特に有用でありうる。トランスジェニック胚の作成およびそのスクリーニングは、例えばA. L. Joyner (Ed.), Gene Targeting, A Practical Approach, Oxford University Press, 1993に記載のとおりに実施できる。胚のDNAは、例えば適切なプローブによるサザンブロットまたはPCR技術に基づいて分析できる。
【0182】
本発明によるトランスジェニック非ヒト動物は、本発明の核酸分子、発現カセットまたはベクターを含む、好ましくはゲノムに安定的に組み込まれている、あるいは上記方法によって作成された、例えばトランスジェニックマウス、ラット、ハムスター、有袋類、単孔類、イヌ、サル、ウサギ、翼手類、ブタ、カエル、線形動物、例えば線虫(Caenorhabditis elegans)、ショウジョウバエ、例えばキイロショウジョウバエ、または魚類、例えばシビレエイ目(torpediniforms)、例えばシビレエイ、フグ目、カラシン目、ネズミザメ目もしくはコイ目、例えばゼブラフィッシュであってよい。かかるトランスジェニック非ヒト動物は、1個または複数個の同一または異なる本発明の核酸分子のコピーを含んでいてよい。かかるトランスジェニック非ヒト動物中の本発明の核酸分子、発現カセットまたはベクターの存在は、本発明のトランスポーターの発現を導く。本発明のトランスジェニック非ヒト動物は、研究モデルを含む、多様な有用性を有する。したがって、この場合、哺乳類は好ましくは実験動物、例えばチンパンジー、マウスまたはラットである。
【0183】
したがって、一つの態様において、本発明のトランスジェニック非ヒト動物は、マウス、ラット、イヌ、例えばビーグルまたはゼブラフィッシュである。多くの報告により当該動物が薬物代謝およびその欠点または発がん性の研究のためのモデル生物として特に十分に好適であることが示された。有利には、糖輸送、特にグルコース輸送を研究するための当該技術分野において周知の多様な好適な技術の利用可能性のため、当該モデル生物を用いてトランスジェニック動物は容易に作成できる。
【0184】
抗体
【0185】
本発明の別の局面は、本発明のトランスポーターと結合する抗体の作製に関する。本発明に包含される抗体の例は、特許請求の範囲に記載の発明のトランスポーターに特異的な抗体および中和抗体を含むが、これらに限定されない。本発明の抗体は、当該技術分野において周知の方法を用いて特徴付けてよい。
【0186】
本発明に有用な抗体は、モノクローナル抗体、ポリクローナル抗体、抗体フラグメント(例えばFab、Fab’、F(ab’)2、Fv、Fc等)、キメラ抗体、二重特異性抗体、ヘテロコンジュゲート抗体、一本鎖(ScFv)、その変異体、抗体の一部を含む融合タンパク質、ヒト化抗体、ならびに抗体のグリコシル化変異体、抗体のアミノ酸配列変異体および共有修飾された抗体を含む、必要な特異性抗原認識部位を含む、任意の他の修飾された立体配置の免疫グロブリン分子を含みうる。抗体はマウス、ラット、ヒト、霊長類または他の起源(キメラおよびヒト化抗体を含む)に由来しうる。
【0187】
一つの態様において、抗体はポリクローナルまたはモノクローナル抗体であってよい。モノクローナルおよびポリクローナル抗体の製造方法は当該技術分野において周知である。
【0188】
用語「モノクローナル抗体」は、本明細書において使用するとき、実質的に均一な抗体の集団から得られる抗体を意味し、すなわち個々の抗体を含む集団が微量で存在しうる可能性のある天然由来の変異体を除き同一であり、本明細書に定義の抗体フラグメントを含む。モノクローナル抗体は、極めて特異的であり、単一の抗原部位に対する。さらに、異なる決定基(エピトープ)に対する異なる抗体を含むポリクローナル抗体調製物とは異なって、各モノクローナル抗体は、抗原の単一の決定基に対する。特異性に加えて、モノクローナル抗体は、他の抗体で汚染されることなく合成できる点で有利である。修飾語「モノクローナル」は、いずれかの具体的な方法で抗体が製造されることを要求するものではない。例えば、本発明に有用なモノクローナル抗体は、Kohler et al. (1975) Nature 256, 495で初めて記載されたハイブリドーマ法によって製造してよく、あるいは細菌、真核動物または植物細胞における組換えDNA法を用いて製造してもよい(米国特許4,816,567参照)。「モノクローナル抗体」は、例えばClackson et al. (1991) Nature 352:624-628およびMarks et al. (1991) J. Mol. Biol. 222, 581-597に記載の技術を用いたファージ抗体ライブラリーから単離してもよい。「ポリクローナル」抗体は、特定のタンパク質またはそのフラグメントに対する抗体の選択を意味し、ここで該抗体は、異なるエピトープと結合しうる。
【0189】
他の態様において、抗体は当該技術分野において既知の方法によってヒト化してもよい。ヒト化抗体は非ヒト免疫グロブリン由来の最小配列を含む免疫グロブリン分子である。さらに別の態様において、完全ヒト抗体は、特定のヒト免疫グロブリンタンパク質を発現するように操作されている商業的に入手可能なマウスを用いて得られる。他の態様において、抗体はキメラである。キメラ抗体は2種の異なる抗体由来の特徴を組み合わせた抗体である。キメラ抗体の作製方法は当該技術分野において既知である。
【0190】
他の態様において、抗体をコードするヌクレオチド配列を得て、次いで発現または増殖のためにベクターにクローン化する。他の態様において、抗体は組換え的に作製され、当該技術分野において既知の方法を用いて発現される。例として、トランスポーターまたはそのフラグメントは、これらの技術によって組換え抗体を単離する目的で抗原として使用できる。トランスポーターの抗原モチーフは、当該技術分野において既知の方法、例えばJameson-Wolf法(CABIOS, 4: 181-186, 1988)によって容易に決定できる。抗体は、宿主細胞において抗体を組換え的に発現する遺伝子配列を用いて、組換え的に作製できる。抗体の誘導体および組換え抗体を作製する方法は当該技術分野において既知である。
【0191】
他の態様において、例えば天然トランスポーターの単離もしくは精製または生物学的サンプルまたは標本中の天然トランスポーターの検出に使用するために、当該技術分野において常套の方法によって、抗体は単体と結合されている。
【0192】
用語「抗体またはそのフラグメント」は、本明細書において使用するとき、糖トランスポーターまたはそのフラグメントと特異的に結合し、他の非トランスポーターと特異的に結合しない抗体またはそのフラグメントを意味する。トランスポーターまたはそのフラグメントと免疫特異的に結合する抗体またはフラグメントは、他の抗原と非特異的に交差反応しない(例えば、適切なアッセイにおいて非トランスポーター、例えばBSAと結合が競合しない)。トランスポーターと免疫特異的に結合する抗体またはフラグメントは、例えばイムノアッセイまたは当業者に既知の他の技術によって同定できる。本発明の抗体は、合成抗体、モノクローナル抗体、重鎖のみの抗体、組換え的に作製した抗体、細胞内抗体、二重特異性抗体、多重特異性抗体(二重特異性抗体を含む)、ヒト抗体、ヒト化抗体、キメラ抗体、一本鎖Fv(scFv)(二重特異性scfvを含む)、一本鎖抗体、Fab’フラグメント、F(ab’)2フラグメント、ジスルフィド結合Fvs(sdFv)および抗イディオタイプ(抗−Id)抗体、ならびに上記のいずれかのエピトープ結合フラグメントを含むが、それらに限定されない。特に本発明の抗体は、免疫グロブリン分子および免疫グロブリン分子の免疫学的に活性な部分、すなわちトランスポーターと免疫特異的に結合する抗原結合部位(例えば抗トランスポーター抗体の1個以上の相補性決定領域(CDR))を含む分子を含む。
【0193】
本明細書において使用するとき、「インタクトな」抗体は、抗原結合部位ならびにCLおよび少なくとも重鎖定常ドメインCH1およびCH2およびCH3を含むものである。定常ドメインは天然配列定常ドメイン(たとえがヒト天然配列定常ドメイン)またはそのアミノ酸配列変異体であってよい。好ましくは、インタクトな抗体は1個以上のエフェクター機能を有する。
【0194】
「抗体フラグメント」は、インタクトな抗体の一部、好ましくはインタクトな抗体の抗原結合CDRまたは可変領域を含む。抗体フラグメントの例は、Fab、Fv、Fab’およびF(ab’)2 フラグメント;二重特異性抗体;線状抗体(米国特許5,641,870およびZapata et al. (1995) Protein Eng. 8, 1057-1062参照);一本鎖抗体分子;および抗体フラグメントから形成された多重特異性抗体を含む。
【0195】
抗体のパパイン消化は、「Fab」フラグメントと呼ばれる、2個の同一の抗原結合フラグメントと、容易に結晶化できる能力を反映して名付けられた残りの1個の「Fc」フラグメントとを産生する。FabフラグメントはH鎖の可変領域ドメイン(VH)と共にL鎖全体および1個の重鎖の最初の定常ドメイン(CH1)からなる。各Fabフラグメントは抗原結合について一価であり、すなわちそれは単一の抗原結合部位を有する。抗体のペプシン処理によって、1個の大きなF(ab’)2フラグメントが得られ、これは二価の抗原結合活性を有する2個のジスルフィド結合したFabフラグメントにほぼ対応しており、抗原と交差結合できる。Fab’フラグメントは、抗体ヒンジ領域由来の1個以上のシステインを含むCH1ドメインのカルボキシ末端でさらに数個の残基を有する点で、Fabフラグメントと異なっている。Fab’−SHは、本明細書において、定常ドメインのシステイン残基が遊離チオール基を有するFab’を意味する。F(ab’)2抗体フラグメントは、それらの間にヒンジシステインを有するFab’フラグメントの対として本来作製された。抗体フラグメントの他の化学結合も知られている。
【0196】
Fcフラグメントは、ジスルフィドで一体に固定された両方のH鎖のカルボキシ末端部分を含む。抗体のエフェクター機能は、あるタイプの細胞で見られるFcレセプター(FcR)によって認識される部分でもあるFc領域の配列によって決定される。
【0197】
本明細書において使用するとき、「Fv」は、完全な抗原認識部位および結合部位を含む最小抗体フラグメントである。このフラグメントは、密接な非共有結合での1個の重鎖可変領域および1個の軽鎖可変領域の二量体からなる。これらの2個のドメインの折り畳みから、6個の超可変ループ(H鎖およびL鎖それぞれから3個のループ)が生じ、これらは抗原結合のためのアミノ酸残基の一因であり、抗体に抗原結合特異性を与える。しかし、1個の可変ドメイン(または抗原に特異的な3個のCDRのみを含むFvの半分)さえも抗原を認識し、結合する能力を有するが、全結合部位よりも親和性は低い。
【0198】
本明細書において使用するとき、「sFv」または「scFv」とも略される「一本鎖Fv」は、1個のポリペプチド鎖に結合されたVHおよびVL抗体ドメインを含む抗体フラグメントである。scFvポリペプチドはさらに、sFvが所望の抗原結合構造を形成できるようにVHとVLドメイン間のポリペプチドリンカーを含んでいてもよい(Rosenburg et al. (1994) The Pharmacology of Monoclonal Antibodies, Springer-Verlag, pp. 269-315参照)。
【0199】
本明細書において使用するとき、用語「二重特異性抗体」は、鎖間でVドメインが対となるが鎖内ではならないようにVHおよびVLドメイン間の短いリンカー(約5〜約10残基)を有するsFvフラグメント(上段参照)を構築することによって作製される小さな抗体フラグメントを意味し、二価フラグメント、すなわち2個の抗原結合部位を有するフラグメントである。二重特異性二重特異性抗体は、2個の抗体のVHおよびVLドメインが異なるポリペプチド鎖上に存在する2個の「クロスオーバー」sFvフラグメントのヘテロ二量体である。二重特異性抗体は、例えばWO 93/11161およびHollinger et al. (1993) Proc. Natl. Acad. Sci. USA 90, 6444-6448により十分に記載されている。
【0200】
「単離された抗体」は、その天然環境の成分から同定および分離および/または回収されているものである。その天然環境の混入成分は、抗体の診断または治療的使用を妨げる物質であり、酵素、ホルモンおよび他のタンパク質性または非タンパク質性成分である。好ましい態様において、抗体は抗体の95重量%以上、最も好ましくは99重量%以上に精製されている。単離された抗体は、少なくとも1種の抗体の天然環境の成分が存在しないため、組換え細胞内のインサイチュの抗体を含む。しかし通常、単離された抗体は、少なくとも1回の精製段階によって製造される。
【0201】
本発明の一つの態様において、コンジュゲートした抗体は、がん細胞に特異的なタンパク質の細胞質ドメイン上のエピトープ(すなわちがん細胞マーカー)と結合する。他の態様において、コンジュゲートした抗体は、sFの細胞質ドメイン上のエピトープと結合する抗体を含むが、これに限定されない。
【0202】
医薬組成物
本発明の別の局面は、医薬組成物の一部としてのトランスポーターの使用に関する。本発明の抗体および核酸はまた、医薬組成物の一部として使用できる。本発明の方法に使用される組成物は一般に、例として、有効量の本発明の核酸またはポリペプチド(例えば免疫応答を誘導するために必要な量)または本発明の抗体(例えば、感染の緩和、感染の症状の緩解および/または感染の予防に必要な中和抗体の量)を含むが、これに限らない。本発明の核酸、ポリペプチドおよび抗体は、さらに、当該技術分野において既知の薬学的に許容される担体、賦形剤または安定化剤を含む(一般に、Remington, (2005) The Science and Practice of Pharmacy, Lippincott, Williams and Wilkins参照)。
【0203】
本発明の核酸、ポリペプチドおよび抗体は、凍結乾燥製剤または水溶液の形態であってよい。許容される担体、賦形剤または安定化剤は、投与する量および濃度で受容者に非毒性でありうる。担体、賦形剤または安定化剤はさらにバッファーを含んでいてもよい。バッファーの例は、炭水化物(例えば単糖類および二糖類)、糖(例えばショ糖、マンニトールおよびソルビトール)、リン酸、クエン酸、抗酸化剤(例えばアスコルビン酸およびメチオニン)、保存剤(例えばフェノール、ブタノール、ベンザノール; アルキルパラベン、カテコール、オクタデシルジメチルベンジル塩化アンモニウム、塩化ヘキサメトニウム、レゾルシノール、シクロヘキサノール、3−ペンタノール、塩化ベンザルコニウム、塩化ベンズエトニウム、およびm−クレゾール)、低分子量ポリペプチド、タンパク質(例えば血清アルブミンまたは免疫グロブリン)、親水性ポリマーアミノ酸、キレート剤(例えばEDTA)、塩形成対イオン、金属錯体(例えばZnタンパク質錯体)および非イオン性界面活性剤(例えばTWEEN(商標)およびポリエチレングリコール)を含むがこれらに限定されない。
【0204】
本発明の医薬組成物は、さらに、所望の効果を向上および/または補足するために使用される添加剤を含んでいてもよい。例として、本発明のトランスポーターの免疫原性を向上させるため、医薬組成物はさらに、アジュバントを含んでいてよい。アジュバントはアルミニウム塩(alum)、完全フロイントアジュバント(CFA)、不完全フロイントアジュバント(IFA)、ムラミルジペプチド(MDP)、MDPの合成アナログである、N−アセチルムラミル−L−アラニル−D−イソグルタミル−L−アラニン−2−[1,2−ジパルミトイル−s−グリセロ−3−(ヒドロキシホスホリルオキシ)]エチルアミド(MTP−PE)ならびに代謝可能な油と乳化剤を含む組成物(油と乳化剤は実質的に全てが直径1ミクロン未満の油滴を有する水中油型エマルジョンの形態で存在している(例えばEP 0399843参照))を含む。
【0205】
いくつかの態様において、アジュバントは、サポニンと組み合わせたToll様受容体(TLR)4リガンドを含む。Toll様受容体(TLR)4リガンドは例えば、リピドA誘導体、特にモノホスホリルリピドAまたはより具体的には3−デアシル化モノホスホリルリピドA(3D−MPL)であってよい。3D−MPLは商標名MPL(登録商標)としてCorixa Corporationから販売されており、IFN−g(Th1)表現型を有するCD4+T細胞応答を第一に促進する。それはGB 2220211Aに記載の方法に従って製造できる。化学的には、それは3−デアシル化モノホスホリルリピドAと3、4、5または6アシル化鎖の混合物である。一つの態様において、本発明の組成物において、小粒子3D−MPLを用いる。小粒子3D−MPLは、0.22 μmのフィルターで滅菌ろ過できるような粒子サイズを有する。かかる調製物は、PCT特許出願WO 94/21292に記載されている。
【0206】
アジュバントはまた、 may also comprise one or more synthetic 誘導体s of lipid A which are known to be TLR4アゴニストであると知られている1種以上のリピドAの合成誘導体、例えば:PCT特許出願WO 95/14026に記載のOM174(2−デオキシ−6−o−[2−デオキシ−2−[(R)−3−ドデカノイルオキシテトラ−デカノイルアミノ]−4−o−ホスホノ−β−D−グルコピラノシル]−2−[(R)−3−ヒドロキシテトラデカノイルアミノ]−α−D−グルコピラノシルジヒドロゲンホスフェート);WO 9964301およびWO 00/0462に記載のOM 294 DP(3S,9R)−3−[(R)−ドデカノイルオキシテトラデカノイルアミノ]−4−オキソ−5−アザ−9(R)−[(R)−3−ヒドロキシテトラデカノイルアミノ]デカン−1,10−ジオール,1,10−ビス(ジヒドロゲノホスフェート);ならびにOM 197 MP−Ac DP(3S−、9R)−3−[(R)−ドデカノイルオキシテトラデカノイルアミノ]−4−オキソ−5−アザ−9−[(R)−3−ヒドロキシテトラデカノイルアミノ]デカン−1,10−ジオール,1−ジヒドロゲノホスフェート10−(6−アミノヘキサノエート)(WO 01/46127)も含むが、これらに限定されない。
【0207】
他の使用できるTLR4リガンドは、アルキルグルコサミニドホスフェート(AGP)、例えばWO 98/50399または米国特許6,303,347(AGPの製造法も記載されている)に記載のもの、または米国特許6,764,840に記載のAGPの薬学的に許容される塩を含むが、これらに限定されない。いくつかのAGPはTLR4アゴニストであり、いくつかはTLR4アンタゴニストである。いずれも本発明の組成物の1種以上のアジュバントとして使用できる。
【0208】
本発明に使用するためのサポニン担体は、Quil Aおよびその誘導体である。Quil AはSouth American tree Quilaja Saponaria Molinaから単離されたサポニン調製物であり、Dalsgaard et al. (1974) Saponin adjuvants, Archiv. fuer die gesamte Virusforschung, Vol. 44, Springer Verlag, pp. 243-254で初めてアジュバント活性を有することが記載された。Quil A に関連した毒性がなく、アジュバント活性を保持しているQuil Aの精製されたフラグメントはHPLCによって単離されており(EP 0 362 278)例えばQS7およびQS21(QA7およびQA21とも知られる)である。QS21は、CD8+細胞傷害性T細胞(CTL)、Th1細胞および主なIgG2a抗体応答を誘導するQuillaja saponaria Molinaの樹皮由来の天然サポニンであり、本発明の文脈で好ましいサポニンである。
【0209】
QS21の特定の製剤が記載されており、これは特に好ましく、これらの製剤はさらにステロールを含む(WO 96/33739)。本発明のサポニン形成部分は、ミセル、混合ミセルの形態で分離されていてよく(好ましくは、しかし排他的ではなく胆汁酸塩で)、あるいはISCOMマトリックス(EP 0109942 B1)、リポソームまたは関連するコロイド構造、例えばワーム状またはリング状マルチマー複合体または脂質/層状構造ならびにコレステロールおよび脂質で製剤した場合ラメラ、または水中油エマルジョン(例えばWO 95/17210のとおり)の形態であってよい。サポニンは酸化アルミニウムまたはリン酸アルミニウムのような金属塩と結合していてよい(WO 98/15287)。いくつかの態様において、サポニンはリポソーム、ISCOMまたは水中油エマルジョンの形態で存在する。
【0210】
いくつかの態様において、アジュバントは3D−MPLとQS21(EP 0671948 B1)の組合せ、ならびに3D−MPLおよびQS21を含む水中油エマルジョン(WO 95/17210、WO 98/56414)である。
【0211】
糖トランスポーターの使用法
【0212】
本発明は、糖トランスポーター、例えばGLUEの使用法を提供する。方法は、細胞、例えばインビトロまたは生物の細胞に糖トランスポーターを導入することを含む。細胞に導入された糖トランスポーターは、野生型または変異型トランスポーターであってよい。トランスポーターは、トランスポーターをコードする核酸またはアミノ酸ポリペプチドとして導入できる。トランスポーターの機能する能力は、所望の結果に基づいて当業者に理解される。例えば、恒常的活性型または野生型トランスポーターを用いて、糖輸送不全を克服できる。変異型トランスポーターを用いて、糖輸送に関する問題を克服できる。糖輸送の変異体は、病原体の所望の遺伝子発現を変化させるために導入してもよい。
【0213】
本発明の方法は、生物発生の変化を提供する。生物は成体または胚であってよい。生物細胞への導入は、生物の発生に影響を与えうる。植物において、糖トランスポーターの導入は、葉発生、茎発生、花蜜発生、根発生、葯発生、木部発生、生殖系発生、幹発生および果実発生に影響を与えうる。動物において、本発明の糖トランスポーターの導入は、臓器、例えば脳、心臓、肺、循環系、皮膚、肝臓、腎臓、脳、脊髄、骨、筋肉(平滑および骨格)、肢、耳、脾臓、腸、膵臓、副腎、胆嚢、睾丸、卵巣、前立腺、膀胱、胃、甲状腺、副甲状腺、海馬、松果体、リンパ節、乳腺、免疫系または管系の発生に影響を与えうる。
【0214】
本発明の方法は、生物の臓器、例えば循環系、神経系(交感神経および副交感神経系を含む)、呼吸器系、消化系、排泄系および生殖系の間の機能的系に影響を与えうる。植物において、糖トランスポーターの導入は、根および幹および葉の間の糖輸送に影響を与えうる。動物において、糖トランスポーターの導入は、乳産生、生殖腺の発生および機能、排卵、酸素と二酸化炭素の交換、食物の消化および栄養素の吸収に影響を与えうる。
【0215】
本発明の方法は、病原体侵襲のような生物への侵襲の感受性に影響する新規メカニズムを提供する。病原体は原核生物または真核生物でありうる。病原体は、細菌、ウイルス、真菌、蠕虫または昆虫であってよい。病原体は、例えば宿主の感受性遺伝子を介して、栄養素または滋養を病原体に与えるように宿主生物の遺伝子制御に影響しうる。本発明のトランスポーターは宿主生物遺伝子転写に影響を与える病原体の能力を変化させうる。宿主細胞に導入される本発明の糖トランスポーターは、病原体侵襲から宿主細胞を保護できる。
【0216】
当業者は、発生中の花粉、発芽中の花粉、発生中の胚およびアポプラスト経路(細胞壁を介した細胞間)を介して細胞が炭素を交換する全ての他の場合を補給するために同様の流出段階が必要とされることを理解する。GLUEホモログであるRPG1は、タペータムに局在化され、RPG1の変異は花粉の生存不能を導く(Guan YF, Huang XY, Zhu J, Gao JF, Zhang HX, Yang ZN. 2008 Plant Physiol. 147:852-63)。したがって、トランスポーターの操作は、割り当てに影響を与えうる。
【0217】
本発明のトランスポーターは、植物の根圏への糖、例えばグルコースの分泌を調節できる。例えば、植物は植物期間あたり1.5t/haの炭素を土壌に分泌する。本発明の方法は、糖分泌の操作が可能であり、したがって植物生産性に影響を与えうる。例示として、糖分泌の増加は、多くの微生物を細胞に誘引し、より多くの炭素を土壌に預けうる(例えば大気中CO2の隔離)。糖分泌の減少は、植物中バイオマスの増加を導く。糖分泌の減少は、植物の増加したバイオマスを導きうる。根におけるかかるトランスポーターの一般的概念は、Chaudhuri et al. 2008, Plant Journalに部分的に記載されている。
【0218】
本発明のトランスポーターは、植物の葉圏への糖、例えばグルコースの分泌を調節できる。例えば、葉圏における糖分泌の調節は、より多くの有益微生物を誘引でき、あるいは病原体に食餌を与える。操作(農薬として特定の化学的阻害剤の発達も介する)。
【0219】
本発明のトランスポーターは糖、例えばグルコースの分泌を調節して、植物の受粉パターンに影響を与えうる。糖分泌の操作は、受粉パターンに影響を与えうる。例えば、変化した糖分泌は、異なる受粉媒介者は異なる花蜜組成物を必要とするため、異なる受粉媒介者を誘引しうる(Ge et al. Plant J. 24:725-734, 2000)。
【0220】
本発明のトランスポーターは、糖、例えばグルコースの分泌を調節でき、植物の葉および師部の発生に影響を与えうる。例えば、Ge et al.は、関連タンパク質の過剰発現が師部発生の刺激を導き、葉の対称性を変化させることを開示している。植物の構造はまた、本明細書に記載の方法で操作してもよい。
【0221】
本発明の別の局面は、植物および植物細胞に対する病原体の活性を調節する方法に関する。病原体(共生生物を含む)は餌を得るためにあるトランスポーターを採用する(Yang et al. 2006 PNAS 103:10503-10508)。Yang et al.は、その遺伝子は、病原型特異的にタイプIII分泌系依存メカニズムによって誘導される、感受性因子であると考えている。マイクロアレイデータを分析することにより、異なる病原体が異なるGLUEファミリーのメンバーを採用することが示される。病原体がトランスポーターを採用できる一つの手段が、細胞に存在するトランスポーター数またはその活性を増加するために、トランスポーターをコードする核酸のプロモーター領域に影響を与えることである。異なるトランスポーターの導入またはトランスポーターの上流への異なるプロモーターの導入のような操作は、病原体感染を予防でき、共生生物を改善または移動できる。トランスポーターを遮断して病原体感染を予防する化学的阻害剤が同定されうる。葉からの糖の排出は、師部負荷のためのプロトンショ糖コトランスポーターだけでなく、葉肉または師部細胞から排出する細胞エフラクサーも必要とする。
【0222】
乳は新生児、小児および成人にとって重要な栄養源である。米国では、一人当たりの乳消費量は80リットルを超える(www.foodsci.uoguelph.ca/dairyedu/intro.html)。乳はバター、ヨーグルトおよびチーズを製造するためにも使用される。動物では、乳は新生児の一次栄養源である。母乳および牛乳は、多くの重要な栄養素および抗体を新生児に与える。タンパク質および脂質に加えて、乳は、グルコースおよび二糖ラクトースのような可溶性の糖も含む。ヒトの乳のラクトース含量は〜7%(200mM)であり、牛乳は〜4.5%(140 mM)である。ラクトースは、乳腺の乳管の内側を覆う肺胞細胞において産生される。特に、ラクトース合成はゴルジ体で生じ、β−1,4−ガラクトシルトランスフェラーゼサブユニットおよびラクトアルブミンからなるヘテロマー酵素ラクトースシンターゼによって仲介され、授乳中に高度に誘導される。
【0223】
前駆体グルコースは、GLUTおよびSGLTファミリーに属するグルコーストランスポーターによって腺細胞への基底膜を介して取り込まれる。グルコースおよびUDP−ガラクトーストランスポーターはいずれも、ゴルジ体でのラクトース合成のための前駆体の取り込みを必要とするが、ゴルジ体グルコースインポーターは同定されてない。ラクトースはエキソサイトーシスによって生じると推測される(2)。ゴルジ体ベシクルの膜は二糖類を透過しないと思われるため、高い浸透力が水取込を引きつける。エキソサイトーシスにおいて水は排出され、これは乳の水含量の主な割合に寄与する。ラクトース合成に寄与する細胞メカニズムの理解は、したがって、乳産生および組成の複数の局面にとって重要である。さらに、牛乳におけるラクトース含量は、腸内でラクターゼによってラクトースを効率的に代謝する能力がないために、集団の大部分にとって健康問題を意味する。ラクターゼ欠損は先天的(腸でのラクターゼ活性に影響する稀な変異)または後天的(二次的ラクターゼ欠損)でありうる。ラクターゼ欠損の最も一般的な原因は、小児の後に生じて成人まで持続する、成人型低ラクターゼ症と呼ばれるラクターゼ量の減少である。ほぼ100%のアジア人が低ラクターゼ症に罹患し、ラクトースを含まない食品を食べる必要がある。したがって本発明は、対象における乳の産生を変化させるために利用できる。本発明は、ラクターゼを取り込むまたは排出する細胞の能力を変化させうる。
【実施例】
【0224】
実施例
【0225】
栄養取込および代謝、ならびに植物での生物相互作用における栄養素交換、例えば受粉媒介者を誘引するための花における花蜜産生、微生物に給餌するための根圏への植物の根による糖分泌および病原体によるこれらのシステムの乗っ取りを制御するシグナル伝達カスケード。生殖細胞、蜜腺、根圏および病原体への糖の供給に関与する新規トランスポーターが最近同定された。次いで、この植物トランスポーターが動物、特に哺乳動物においてホモログを有すること、このタンパク質は乳腺における糖分泌に関与する多くの特徴を示すことが見出された。FRETナノセンサーは細胞内分解による定量的流量分析が可能なユニークなツールを提供する(Okumoto et al. New Phytol. 180:271-295, 2008)。これらのナノセンサーは、レポーター要素として緑色蛍光タンパク質(GFP)のスペクトル変異体の対とアロステリックに結合した、認識要素として使用される最近ペリプラズム結合タンパク質からなる(Fehr et al. Proc. Natl. Acad. Sci. USA 99:9846-9851, 2002;Fehr et al. J. Biol. Chem. 278:19127-19133, 2003; Deuschle et al. Protein Sci 14:2304-2314, 2005)。
【0226】
認識要素とのリガンド結合によって誘導されるコンホメーション変化は、結合したシアンと黄色蛍光タンパク質部分の間での蛍光共鳴エネルギー転移(FRET)の変化に翻訳される。これらのセンサーは、生体細胞に遺伝的に導入でき、生体細胞および組織における分析物レベルの非侵襲的測定が可能である(Fehr et al. J. Biol. Chem. 278:19127-19133, 2003)。これらのセンサーにより、インタクトなシロイヌナズナの根におけるグルコース流量(Chaudhuri et al. Plant J. 56:948-962, 2008)、海馬ニューロンからのグルタミン酸放出(Okumoto et al. Proc Natl Acad Sci U S A 102:8740-8745, 2005)およびがん細胞におけるトリプトファン交換(Kaper et al. PLoS Biol 5:e257, 2007)が測定されている。
【0227】
実施例1:ER膜を通過するグルコース流量の分析
【0228】
小器官内の分析物レベルを測定するため、これらのFRETナノセンサーを各細胞内区画に標的化した(Fehr et al.J. Fluoresc. 14:603-609, 2004)。ER膜を通過するグルコース流量を直接モニターするために、FRETグルコースセンサーをERシグナル配列およびKDEL保留シグナルと隣接させて、ER内腔に標的化した。このアプローチは、ER膜を通過する高グルコース流量の同定が可能であり、HepG2細胞におけるグルコースの速やかな双方向性高性能輸送活性の存在を示唆した(Fehr et al. Mol Cell Biol 25:11102-11112, 2005)。
【0229】
実施例2:植物の根における新規糖輸送機能の同定
【0230】
土壌は微量のみの溶解性炭水化物を含むが、植物の根は、人工培地に供給された場合、効率的にグルコースおよびショ糖を吸い上げる。土壌中に見られる溶解性炭水化物および小さな代謝産物は、部分的には植物の根からの浸出に由来する。取込および浸出に関するトランスポーター分子の性質は知られていない。FRETグルコースおよびショ糖センサーを展開して、シロイヌナズナの根端におけるグルコースおよびショ糖の蓄積および排出を特徴付けた(Chaudhuri et al. Plant J. 56:948-962, 2008)。グルコースおよびショ糖蓄積は、プロトノフォアに非感受性であり、pH5.8、6.8および7.8でも同様であった。これは流入および流出の両方がプロトンから独立した新規クラスの輸送系によって仲介されうることを示唆している。さらに、全ての既知の植物グルコーストランスポーターとは異なって、この新たな根輸送系は、グルコースアナログである3−O−メチルグルコースの輸送を仲介しなかった。
【0231】
実施例3:グルコーストランスポーターの発現系としてのHEK293T細胞
【0232】
細胞膜およびER膜を通過するグルコース輸送をよりよく特徴付けられるように、低内因性糖取込能を有する細胞をアッセイした。HEK293T細胞が極めて低い内因性取込を有し、FRETグルコースセンサーの補助の下でGLUTおよびSGLT糖トランスポーターの特性を定義する発現系として使用できることが見出された。
【0233】
実施例4:花蜜産生および根圏分泌に関与するグルコーストランスポーターの同定
【0234】
上記HEK293T細胞系を用いて、植物における花蜜産生に関与し、根端に記載のグルコース輸送活性の特徴を有するシロイヌナズナ糖トランスポーターの新規クラスを同定した。次いで、HEK293T細胞発現系においてシロイヌナズナ由来の候補トランスポーター遺伝子がFRETグルコースセンサーと共発現されるスクリーニングアッセイを開始した。膜タンパク質の未知のクラス(GLUE1と命名)がグルコース濃度依存的FRET応答を誘導することを見出した(例えば図3および4参照)。
【0235】
次いで、GLUE1がHEK293T細胞において発現されているとき既知の哺乳類GLUTまたはSGLTトランスポーター遺伝子のいずれもが誘導されないことを確認し、そしてGLUE1介在性取込がGLUT阻害剤のサイトカラシンBに非感受性であることを示した。さらに、GLUE1が3−O−メチルグルコースを輸送できないという結果が示された。小胞体内腔内で発現されるセンサーの使用により、GLUEが細胞基質への取込を仲介するだけでなく、細胞基質の外へのグルコースの排出もできることが示された。同様の結果がこの遺伝子ファミリーの複数のシロイヌナズナパラログについて得られた。
【0236】
GLUE1が内因性シグナル伝達カスケードと相互作用している可能性を排除するため、グルコース取込欠損酵母株EBY4000においてGLUE1を発現した。結果は、GLUE1がグルコースの取込を10mMのKMで、エネルギー独立的に仲介することを示した。GLUE1は、水および溶質輸送アクアポリンと同様の、7個の膜貫通ドメインを含む小さなタンパク質をコードする。GLUE1−GFP融合体は細胞膜に局在化する(例えば図9参照)。
【0237】
併せて考えると、これらのデータは、GLUE1がシロイヌナズナの根における根輸送系と同一の特性を有する新規クラスの糖単輸送体をコードすることを示している(Chaudhuri et al. Plant J. 56:948-962, 2008)。マイクロアレイデータの分析は、このファミリーのメンバーが根において実際に発現されていることを示している。
【0238】
このタンパク質ファミリーのメンバーは、例えば蜜腺(Ge et al. Plant J. 24:725-734, 2000)およびタペータム(Guan et al. Plant Physiol. 147:852-863, 2008)において発現される。その糖を流出する能力に鑑みて、GLUEは、花において花蜜を産せ宇するグルコースの分泌に関与していてよく、花粉を発生し、有益微生物を誘引し給餌するために根圏にグルコースを分泌するために、タペータムからグルコースを排出する(Chaudhuri et al. Plant J. 56:948-962, 2008)。
【0239】
実施例5:植物GLUEトランスポーターのヒトホモログ
【0240】
同様の調査により、RAG1AP1と命名したマウスおよびヒトゲノムにおけるGLUEホモログを同定した(Tagoh et al. Biochem Biophys Res Commun 221:744-749, 1996)。RAG1AP1は植物GLUEと顕著な相同性を有し、また7個の推定膜貫通ドメインを有するタンパク質をコードする。シロイヌナズナとは異なって、マウスおよびヒトゲノムは各々、1個のメンバーのみを含む。RAG1AP1活性を欠く変異リンパ球細胞系は、抗体成熟に関与する遺伝子の発現を制御することが示された(Tagoh et al. Biochem Biophys Res Commun 221:744-749, 1996)。これはグルコースまたはグルコースアナログを分泌できないことによって引き起こされる間接的な影響である可能性がある。RAG1AP1は抗体変動に関与する遺伝子の発現の制御に間接的に関与している可能性がある。マイクロアレイ試験によって、この遺伝子が授乳中に高度に誘導されることが示される(図9)。
【0241】
植物GLUEが植物におけるグルコースの分泌に関与する機能的証拠と共に、RAG1AP1は、乳腺の肺胞細胞におけるラクトース分泌および/またはグルコース輸送の役割において機能しうる。このことは、大型プロテオミクスプログラム、Human Protein Atlasのデータによって支持され、これはRAG1AP1(RAG1活性化タンパク質1)が乳房の腺細胞において特異的に発現されることを示唆している(www.proteinatlas.org/normal_unit.php?antibody_id=18095&mainannotation_id=1747078)。さらに、該タンパク質は人体の他の腺細胞、例えば潜在的に精子細胞に給餌する精巣上体においても局在化していると思われる。
【0242】
実施例6:病原体の栄養のための植物糖流出トランスポーター
【0243】
材料および方法
【0244】
qPCRおよびRT−PCR分析。RNeasy MINIキット(QIAGEN, Hilden)を用いて全RNAをHepG2およびHEK293T細胞から抽出し、第一ストランドcDNAを作製し(New England Biolabs)、既に公開されているGLUTおよびSGLTプライマーのセットを用いたRT−PCRによって推定長のフラグメントを得た。サンプルを2%アガロースゲル上で分離した。Pst DC3000によって播種したサンプルについて、Trizol試薬(Invitrogen)を用いて葉から全RNAを抽出した。HotStart-IT SYBR Green qPCR Master Mix (USB)を用いて7300 PCRシステム(Applied Biosystems)の製造業者の指示書に従って実時間定量PCR(qPCR)を実施した。アクチン(ACT8)発現を用いて各サンプルの発現レベルを正規化した;相対発現値は、比較2−ΔΔCt法を用いて各時点で1 mMのMgCl2で浸潤したサンプルの値に対して決定した。G.cichoracearumによって感染したサンプルについて、LightCyclerO 480(Roche)を用いてqPCRアッセイを行った。定量について、各遺伝子の相対的転写レベルを2-ΔΔCt法に従ってACT8に正規化した。変動比は未処理サンプルに対して計算した。分析を独立して2回反復した。マイクロアレイデータによって誘導を確認する(Genevestigator)。
【0245】
構築物。pOO2におけるSGLT1 ORFのクローニングを説明した。シロイヌナズナおよびコメそれぞれ由来の特定のプライマー(補足表1)を用いたRT−PCRによって、SWEET1、SWEET8およびOsSWEET11 ORFを増幅した。コメ由来の第一ストランスcDNAはPamela Ronald, UC Davisが快く提供下さった。Gateway BPクロナーゼ反応によってORFをpDONR221(Invitrogen)にクローン化し、Gateway LR組換え反応によって酵母発現ベクターpDRf1-GWに固定化した。pDRf1-GWにおけるFLII12Pglu700mD6との酵母コトランスフォーメーションのためにSWEET1をp112-A1NE-GWにクローン化した。p112-A1NEのSmaI制限部位にGatewayカセットを挿入してp112-A1NE-GWを作製した。放射性トレーサー実験のために、SWEET1、SWEET8およびOsSWEET11の停止コドンを有するORFを、Gateway LR組換え反応により卵母細胞発現ベクターpOO2-GW(D. Loque、未公表の結果)にクローン化した。
【0246】
FRET分析、細胞培養、トランスフェクション、イメージ獲得およびFRET分析を上記の通りに実施した。
【0247】
細胞基質インベルターゼ(YSL2-1)を担持する酵母株EBY4000の修飾型(hxt1 through -17Δ::loxP gal2Δ::loxP stl1Δ::loxP agt1Δ::loxP ydl247wΔ::loxP yjr160cΔ::loxP)をSWEETおよびHXT5で形質転換して、2%マルトースおよび必要な栄養要求マーカーを補ったSD(合成欠乏)培地で増殖した。補完増殖アッセイのために、細胞を液体最少培地でOD600 〜0.6に一夜増殖させ、次いで水を用いてOD600 〜0.2に希釈した。連続希釈(1x、5x、25xおよび125x)を2%マルトース(コントロールとして)または2%グルコースおよび関連した栄養マーカーを含むSD培地に播種した。30℃で2〜5日後、プレートをスキャンして(CanoScan, Canon)増殖を実証した。
【0248】
酵母取込。2%グルコースおよび栄養マーカを補ったSD培地中で、酵母細胞を増殖した。OD600 0.5〜0.7で、細胞を遠心分離で回収し、氷冷蒸留水で2回洗浄した。上清を除去した後、細胞ペレットを秤量した。40 mM リン酸カリウムバッファー、pH 6.0中に細胞を5〜10%(w/v)で再懸濁した。細胞をリン酸カリウムバッファー中、30℃で5分間プレインキュベートした。各反応について、20mM グルコース(0.55μCi D-[U-14C] グルコース; 590 KBq/μmol、Amersham Pharmacia Biotech Inc.)を含む予め温めたバッファー330 μlを等体積の細胞に加えた。120 μlのアリコートを取り除き、氷冷水に写した。ガラスファイバーフィルター(GF/C、Whatman)での真空ろ過によって細胞を回収し、10 mlの氷冷水で2回洗浄した。5 mlのUltima Gold XR Scintillator液(Perkin Elmer)を含むシンチレーションバイアルにフィルターを写した。液体シンチレーション分光法によって、細胞が取り込んだ放射活性を測定した。D−グルコースと比較したSWEET1の基質特異性を決定するため、10倍過剰の競合糖種を用いた。SWEET1活性のpH依存性を決定するため、特定のpHの40 mMのリン酸カリウム取込バッファーを用いた。3つの独立の形質転換体を取込実験に用いた。
【0249】
アフリカツメガエル卵母細胞単離およびRNAインジェクション。MluIでのpOO2プラスミドの直線化の後、キャップしたcRNAをインビトロで、mMESSAGE mMACHINEキット(Ambion, Inc., Austin, TX)を用いたSP6 RNAポリメラーゼによって合成した。アフリカツメガエル卵母細胞単離はM. Goodman(Stanford University)が快く提供下さった。Ballatori et al.に記載のとおりにマイクロインジェクションを実施した。25 ng〜50 ngの各cRNAを健康な外見の卵母細胞にインジェクションした(RNAseを含まない水をコントロールとして用いた)。100μM ゲンタマイシン、100 U/ml ペニシリンおよび100μM ストレプトマイシン溶液を含む修飾バース食塩水(MBS:(単位mM) 88 NaCl、1 KCl、2.4 NaHCO3、0.82 MgSO4、0.33 Ca(NO3)2、0.41 CaC12および20 HEPES-Tris、pH 7.5)中、18℃で2〜3日間、インジェクションした卵母細胞を維持した。インキュベーションバッファーを1日1回交換した。
【0250】
アフリカツメガエル卵母細胞におけるトレーサー取込。Detaille et al.に記載のとおりに修飾してアッセイを実施した。インジェクションの2日後、200μLのMBSおよび1 mMのD−グルコース(4 μCi/ml D-[14C(U)]-グルコース; 319 mCi/mmol、PerkinElmer)を含むチューブに7〜16卵母細胞のグループを移した。20℃で1時間インキュベーションした後、1 mlの氷冷MBSを加えて細胞を洗浄した。氷冷MBSバッファーを加えてインキュベーションを停止した。卵母細胞は氷冷MBSバッファーで3回洗浄した。細胞を100μlの1% (w/v)SDSで可溶化し、個々に測定した。
【0251】
アフリカツメガエル卵母細胞におけるトレーサー流出アッセイ。流出は本質的には上記の通りに測定した。cRNAインジェクションの3日後、0.18 μCi/μl D-[14C(U)]-グルコースを有する10 mMのD−グルコースを含む50 nlの溶液で卵母細胞をインジェクションした。細胞を速やかにMBSで1回洗浄した。所定の時点で、シンチレーションカウンティングのために反応バッファー(950 μl)を除去し、新鮮な培地と交換した。最後に、卵母細胞を1% SDSで可溶化し、放射活性を分析した。
【0252】
FRETセンサーによる酵母細胞におけるグルコース蓄積の分析。酵母細胞におけるFRET測定を上記の通り実施した。
【0253】
植物成長および病原体感染。シロイヌナズナCol-0植物を8時間明/14時間暗下、22℃の成長チャンバーで成長させた。1 mMのMgCl2バッファー、1 mMのMgCl2中の2x108cfu/mlのPst DC3000またはPst DC3000 ΔhrcU懸濁液で、無針注射器を用いて5週齢の葉を浸潤した。光下で6、12および24時間インキュベーション後、葉サンプルを回収した。G.cichoracearum接種を上記の通りに実施した。植物を「移動タワー」(段ボール)中に置き、感染したカボチャの葉を移動タワーの上にかぶせ、カボチャの葉から胞子が飛んでシロイヌナズナに移動するように圧縮空気(散布カン)を用いて、シロイヌナズナ植物にG. cichoracearum胞子を接種した。接種密度は〜25-35 分生子s/mm-2であった。接種後、植物を1時間、暗露チャンバーでインキュベートし、次いで日長16時間、相対湿度70%の成長チャンバーに移した。
【0254】
アラインメントおよび系統発生解析。デフォルトパラメーターを用いたCLUSTALWで、SWEETアミノ酸配列の複数アラインメントを実施し、ソフトウェアMega V3.1. Bootstrappingを用いて系統発生解析を1000回実施して、各ブランチのサポート値を得た。対比較のために、Vector NTIアドバンス11.0を用いて完全アミノ酸配列の複数アラインメントを実施した。
【0255】
共焦点顕微鏡。YFP::SWEET1およびYFP::SWEET8を発現する植物のイメージングをLeica TCS SP5顕微鏡で実施した。アルゴンレーザーで514 nmでの励起および放出につき525〜560 nmのスペクトル検出セットによってYFPを可視化した。40/0.75-1.25NA HCX PL APO CS対物レンズによって標本を観察した。
【0256】
結果および考察
【0257】
糖流出は多細胞生物において、および生物間の相互作用において、炭素骨格とエネルギーの細胞性交換に必要とされる必須プロセスである。タペータムまたは花柱の輸送管からの糖流出は、花粉発生および後の花粉管成長を刺激する。花は受粉媒介者を誘引するため花蜜産生のために糖を分泌し、そして植物は、潜在的には有益微生物に給餌するために根圏に炭水化物を分泌する(T. Bisseling et al., Science 324, 691 (2009))。糖流出輸送体は、葉肉および種皮を含む多くの他の部位で必要とされる(Y. Zhou et al., Plant J. 49, 750 (2007))。流出トランスポーターの分子の性質は知られていない(S. Lalonde et al., Annu. Rev. Plant Biol. 55, 341 (2004))。植物由来の糖はまた、病原体のための基質を提供する。病原体の第一次的ゴールは、効率的に繁殖するために宿主植物から栄養分を摂取することである。シュードモナス(Pseudomonas)属およびザントモナス(Xanthomonas)属の植物病原性細菌は、植物組織の細胞外空間(アポプラズム)に生息でき、ここでそれらはエネルギー源および炭素骨格源として炭水化物を獲得する。成功した病原体は、かかるメカニズムを共に選択して栄養素流動を変化させる可能性がある(J. W. Patrick, Aust. J. Plant Physiol. 16, 53 (1989))。結果として、病原体と植物は、植物が栄養素への病原体のアクセスを制限しようとし、また防御戦略を開始する一方で、病原体が栄養素へのアクセスを得ようとし、宿主免疫を抑制する戦略を考え出す進化的主導権争いに突入する。植物の防御を変化させるための病原体によって用いられるメカニズムに対する洞察が現在出現しつつあるが;どのようにして病原体が、病原体の増殖を支援するために宿主の生理機能、特に糖排出を変化させるのかについて、わずかしか知られていない。したがって我々は、小胞または細胞膜で糖を分泌するトランスポーターの存在を突き止めた。我々はまた、これらの植物流出トランスポーターが病原体によって栄養素の要求を支給するために「共に選択」されると仮定した。少なくともコムギウドンコ病の場合、グルコースは植物宿主から病原体に移送される主な糖である。それぞれの病原体グルコース/H+コトランスポーターを同定した;反対に、植物糖流出メカニズムはもとより不明のままであった。
【0258】
参照植物シロイヌナズナから新規なグルコーストランスポーターを同定するため、哺乳動物発現系を用いて、植物タンパク質データベースAramemnon (http://aramemnon.botanik.uni-koeln.de)の特徴付けられていないポリトピックな膜タンパク質をコードする遺伝子をスクリーニングした。低内因性グルコース取込活性によって特徴付けられるヒト胚性腎臓HEK293T細胞において、候補遺伝子を高感受性FRETグルコースセンサー(すなわちFLIPglu600μΔ13V)と共発現した。グルコース依存ネガティブFRET比変化によって検出したとき、試験した遺伝子のうち、SWEET1(AT1G21460)発現によってHEK293T細胞の糖蓄積が可能となった;輸送機能と一致した(図9A)。SWEET1が細胞基質からの流出も仲介できるかを決定するため、小胞体の内腔にFRETグルコースセンサーFLIPglu600μΔ13VERを発現した(ER;図9B)。形態的に、細胞膜(PM)を通過する取込は運搬体の細胞外側から開始されるが、一方でERへの「排出」は、トランスポーターの細胞質側から開始される(図9C)。ERセンサーのグルコース依存的応答は、SWEET1がPMを通過した取り込みおよびERへの「流出」を仲介できることを示す。したがってSWEET1はグルコース単輸送体として機能でき、輸送の方向は、膜内外のグルコース勾配のみに依存する。HEK293T細胞の内因性グルコーストランスポーター(GLUT)は、GLUT阻害剤であるサイトカラシンBがSWEET1誘導性グルコース取り込みに影響しなかったため、グルコース取り込みに関与しなかった(図12A)。さらに、GLUTおよびSGLTファミリーの既知のヒトグルコーストランスポーターのmRNAレベルは、SWEET1を発現するHEK293T細胞において誘導されなかった(図12B)。SWEET1活性がグルコース取り込みに必要であることを独立して示すため、18種のヘキソーストランスポーター全てを欠く酵母変異体においてSWEET1を発現した。SWEET1は、グルコースでの増殖(図9D)およびFRETグルコースセンサーFLII12Pglu700μ∂6を用いて測定したとき細胞内グルコース蓄積(図9E)を酵母変異体に可能とした。さらなる特徴付けにより、SWEET1が酵母における低親和性トランスポーターとして、グルコースについて9 mMのKmで機能することが示された(図9F)。単輸送メカニズムと一致して、取り込みは通電によって刺激されず、またほぼpH非依存的であった(図13)。シロイヌナズナの根において記載したグルコース輸送活性と同様に、SWEET1介在性取込は、グルコースアナログ3−O−メチルグルコースによって僅かに阻害された(図13B)。細胞取込および流出における役割の一助として、恒常的に発現されるSWEET1-YFP融合体は、シロイヌナズナの葉のPMに局在している(図9G)。マイクロアレイ試験に基づくと、SWEET1は根において弱くしか発現されないが、シロイヌナズナの花では高度に発現され、このことは配偶体または蜜腺に栄養素を供給する役割を示唆している(図14)。推定糖トランスポーターの根系における生化学的特性の顕著な類似性にもかかわらず、根におけるSWEET1発現は低く、このことは根からのグルコース流出にあまり関与していないことを示唆している。SWEET1と関連している可能性がある他のタンパク質は、根における糖輸送に関与している可能性がある。
【0259】
SWEET1は、シロイヌナズナにおいて17種のメンバーおよびコメにおいて19種のメンバーを有する新たなトランスポーターファミリー(図1)の最初に特徴付けられたメンバーである。シロイヌナズナSWEETは多様であり、4つのサブクレードに属し(図1A)、28〜86%の範囲の同一性を有する(表1および2)。輸送における機能と一致して、SWEETは、7個の膜貫通ヘリックス(TMH)によって構築される疎水性ポアを形成すると予測される小さな疎水性タンパク質である。コンピューター解析では、SWEETの7個のTMHはTMH4を介して融合した3個のTMHドメインポリペプチド(1−3および5−7)の古い重複によってもたらされることが示唆される(図9H)。
【0260】
このファミリーのいずれのメンバーも機能的に特徴付けられていなかったが、複数のSWEET変異体の表現型が記載されている。SWEET1はそのパラログであるSWEET8と41%同一であり、4つのシロイヌナズナSWEETクレードの第二に属する。SWEET8/RPG1の変異は、雄不妊を導くことが示された。HEK293T細胞におけるFRETセンサーFLIPglu600μΔ13VまたはFLIPglu600μΔ13VERとSWEET8/RPG1の共発現は、SWEET8も単輸送体として機能することを示唆する(図15A:図9C)。さらにSWEET8/RPG1は、酵母グルコース輸送変異体を補足する(図9D)。SWEET8/RPG1はタペータムにおいて発現され、このことは花粉の栄養補給に必要なグルコースエフラクサーとしての役割を強く示唆している。
【0261】
SWEET1およびSWEET8はコメタンパク質OsSWEET11/Os8N3(系統樹に基づいてOsSWEET11と命名、図1)と34%のアミノ酸配列同一性を有する。最も近いシロイヌナズナホモログは、OsSWEET11/Os8N3と40%同一性を有し、第三SWEETクレードに属する(図1)。SWEET8と同様に、OsSWEET11/Os8N3は、RNA阻害によるその発現の低下が花粉において低下したデンプン含量および花粉の不稔性を導くため、花粉の栄養補給において機能すると思われる。クレード3のSWEETの別のホモログ(図1)であるペチュニアNec1のサイレンシングはまた、雄不稔を導いた。Nec1は蜜腺において発現され、その発生制御は蜜腺のデンプン含量と逆相関し、このことは蜜腺における糖分泌におけるNec1の第二の役割を示唆した。併せて考えると、これらのデータは単子葉および双子葉植物両方でSWEETSが重要な生殖目的に炭水化物を供給する重要な役割を有することを強く示唆する。
【0262】
病原体は宿主植物の光合成能を用いて、増殖複製のためのエネルギーおよび栄養素を得る。広範な病原体がそれらの宿主からのグルコースを必要とすることは十分に確立されている。様々な病原体が配偶体への給餌のような植物発生のために捧げられる宿主糖流出系を乗っ取ると仮定した。したがって、次にシロイヌナズナSWEETファミリーメンバーのmRNAレベルが細菌または真菌感染によって変化するかを試験した(図10)。シュードモナス・シリンゲ(Pseudomonas syringae)病原型トマト株DC3000感染は、シロイヌナズナの葉においてSWEET4、5、7、8および15 mRNAレベルを高度に誘導した。反対に、宿主にタイプIIIエフェクタータンパク質を注入できず、したがって病原性において危険にさらされているDC3000タイプIII分泌変異体(ΔhrcU)は5つの遺伝子の4つを誘導せず、このことはSWEET mRNA存在量がタイプIII依存的に調節されていることを示している。次に他の病原体が同じまたは異なるファミリーメンバーを標的とするかを試験した。ウドンコ病菌Golovinomyces cichoracearumは異なるセットのSWEET mRNA、主としてSWEET12を誘導した(図10A、C)。マイクロアレイデータは、真菌病原体灰色カビ病菌(Botrytis cinerea)が再び異なるSWEETのセット(すなわちSWEET4、15、17)を標的とすることを示した。併せて考えると、SWEET mRNAレベルの病原体特異的調節は、感染部位での糖輸送を変化させ、病原体増殖および植物免疫に影響を与える可能性がある。
【0263】
この仮説と一致して、花粉の栄養素供給に重要なコメ遺伝子OsSWEET11/Os8N3が、病原体感受性因子として機能する。コメossweet11/os8n3変異体は、細菌病原体 mutant was found to be resistant to the bacterial pathogenイネ白葉枯病菌(Xanthomonas oryzae)病原型オリゼ(Xoo)株PXO99Aに抵抗性であることが見出され、このことはOsSWEET11/Os8N3が感染において病原体に糖を供給することを強く示唆している(図11A)。したがって、OsSWEET11/Os8N3もグルコーストランスポーターとして機能するかを試験した。SWEET1およびSWEET8とは異なって、OsSWEET11/Os8N3は、HEK293T細胞におけるグルコース取込を仲介せず、酵素ヘキソース輸送変異体を補足しなかった(データは示さず)が、このことはグルコース取込においてそれが機能しないことを示している。しかし、OsSWEET11/Os8N3がグルコースエフラクサーとして機能することは考えられた。この仮説を検証するため、流出試験に受け入れられる系であるOsSWEET11/Os8N3をアフリカツメガエル卵母細胞において発現させた。OsSWEET11は、SWEET1および哺乳類Na+依存的グルコーストランスポーターSGLT1とは異なって、卵母細胞における[14C]−グルコース取込も仲介できなかった(図9I;図16)。しかし、SGLT1およびOsSWEET11/Os8N3の共発現は、卵母細胞における低下した[14C]−グルコース蓄積を導き、OsSWEET11/Os8N3の流出(すなわち「漏出」)活性と適合する発見であった(図9I、K)。The hypothesis that OsSWEET11/Os8N3がグルコースを排出できるという仮説は、直接流出測定によって実証される。植物タンパク質を発現する卵母細胞に[14C]−グルコースを注入して、グルコース流出を測定した。SWEET1およびOsSWEET11/Os8N3はいずれも[14C]−グルコースを流出でき(図9G)、これはSWEET1は促進因子として機能するが、OsSWEET11/Os8N3はエフラクサー(潜在的にはH+/グルコースアンチポーター)であることを示唆している。さらに、OsSWEET11(Os8N3)はショ糖を輸送できる(図43G)。したがって、OsSWEET11/Os8N3は、生殖のためにグルコースおよびショ糖を得るために病原体によって採用されると思われる。
【0264】
OsSWEET11/Os8N3が糖エフラクサーとして機能するという発見は、どのように病原体が植物のエネルギー源へのアクセスを得るために基本的植物機能を共に選択するかのモデルを提供する(図11)。Xoo株PXO99Aは、コメの感染についてタイプIIIエフェクター遺伝子に依存する。PthXo1はTAL(転写アクチベーター様)エフェクターであり、これはDNAと直接相互作用して標的遺伝子の転写を促進する。Xoo PXO99Aによって分泌されるPthXo1は、OsSWEET11/Os8N3の転写を特に活性化して、おそらくアポプラスト細菌に給餌するために糖流出を誘導する(図11A)。PthXo1が変異されると(ME)、OsSWEET11/Os8N3の転写および病原性が低下し、これは病原体の飢餓と一致する(図11B)。変異またはRNA阻害によりOsSWEET11/Os8N3が利用できなくなった場合、糖(例えばグルコース、glc)は十分な量で排出されず、病原体は飢餓となるであろう(図11C)。実際、ossweet11/os8n3変異体はXoo PXO99Aに抵抗性である。別のTALエフェクター(AvrXa7)を担持するPXO99A細菌はossweet11/os8n3変異体においてさえ病原性であり(図11D)、これは他のSWEETが病原体増殖を支援するために病原体によって共に選択されるという最も簡単な仮説と一致している(図11D)。実際、PthXo1によって標的とされる推定DNA配列は、TRCA・CT・CCATTACTRTAAAA・N・(OsSWEET11/Os8N3の上流プロモーターにおいて見られる)であり、一方でAvrXa7によって標的とされるものはTA・AANCRCCCN・・CCNNRRATRA・Nである。この配列は潜在的標的を同定するのに十分ではなかった。これらの知見は、免疫におけるそれらの役割に加えて、タイプIIIエフェクターはまた、宿主植物の栄養源へのアクセスをえることにも関与しているという考えを支持する。どのように真菌病原体がこれらのトランスポーターのプロモーターを標的とするかは未だ理解されていないが、トランスポーター遺伝子は、真菌増殖を支える制御ネットワークを解明するための好適な診断ツールでありうる。明らかに、この病原ターベースの選択圧に拘わらず進化において維持されるために、SWEETトランスポーターは植物において必須機能を有するはずである;変異体の分析は、少なくともそれらの1個が配偶体への炭水化物の供給に関与していることを示唆している。したがって、他のパラログの活性も、他の植物機能に重要であり得る。特に疾患感受性についての残りのSWEETパラログの特徴付けおよび変異体の分析は、次の段階で重要である。
【0265】
病原体エフェクター分子の全スペクトルおよびどのようにそれが病原体増殖を支持するために植物代謝を阻害するかについての知識は、宿主−病原体相互作用の我々の理解を改善し、年間10%以上もの農作物損失を導くグローバル規模での病原体感染と闘う新たな戦略を導くことができる。さらに、SWEETファミリーの他の遺伝子の分析は、花粉の栄養、花蜜産生および植物の根からの炭素隔離の謎のいくつかを解決する助けとなりうる。興味深いことに、動物ゲノムは糖輸送にも関与するSWEETホモログを含む。
【0266】
実施例7:線虫(C. elegans)およびヒトにおけるグルコーストランスポーターの第三ファミリー
【0267】
材料および方法
【0268】
SGLT1のORF(Invitrogen, Carlsbad, CA)をPCRで増幅し、pCR2.1-TOPO(Invitrogen)にクローン化した。SGLT1 ORFをEcoRI/XhoIで切り取り、p002の対応する部位にクローン化した。pDNR-LIBのスプライス変異体RAG1AP1-1(Clone ID: 4076256、Open Biosystems, Huntsville, AL)をEcoRI/XhoIで制限し、pOO2にクローン化した。インビトロLR組換え(Gateway)によってpCMV-SPORT6のRAG1AP1-2(Clone ID: 3896154、Open Biosystems, Huntsville, AL)をpOO2-GW(D. Loque、未公表)に移した。部位特異的突然変異誘発によってRAG1AP1-3aaをY216A、L218A、L219A(推定内在化モチーフ)に変異させて、インビトロBP組換えによってpDONR221-f1にクローン化し、さらにLR反応によってpOO2-GWに移動させた。LR反応を用いてCeSWEET1(K02D7.5)、CeSWEET3(C06G8.1)、CeSWEET4(Y39A1A.8)、CeSWEET5(K06A4.4)およびCeSWEET7(K11D12.5)(Open Biosystems, Huntsville, AL)をpOO2-GWにクローン化した。次いで、部位特異的突然変異誘発によって開始および終止コドンを加えた。卵母細胞発現および輸送試験は、全取込実験卵母細胞をMBS中1 mMのグルコース中で一夜プレインキュベートする以外は、Chen et alに記載のとおりに実施した。
【0269】
結果および考察
【0270】
線虫(C. elegans)ゲノムは糖流出トランスポーター(SLC50)の新規クラスの7種のホモログを含むが、ヒトゲノムはRAG1AP1と命名された1種のホモログを有する。シロイヌナズナSWEET1と同様に、線虫(C. elegans)CeSWEET1はアフリカツメガエル卵母細胞で発現された場合グルコース取込を仲介する。CeSWEET1およびヒトRAG1AP1は、Na+/グルコースコトランスポーターSGLT1によって仲介される卵母細胞の第二活性グルコース蓄積を相殺する。CeSWEET1の変異は、脂質の蓄積を導く細胞グルコース流出の欠損と両立して、脂肪蓄積を導いた。これらの知見は、糖輸送におけるヒトホモログRAG1AP1の役割に新たな光を投げかけうる。
【0271】
ヒトゲノムはグルコーストランスポーターの少なくとも2つのクラス、SLC2およびSLC5を含む。GLUTと名付けられたSLC2は単輸送体であり、すなわちそれらは濃度勾配に沿ってグルコースを輸送する。反対にSGLTは、ナトリウム勾配によって駆動されてグルコースを能動的に取り込むことができるNa+共役コトランスポーターである。これらのトランスポーターは、ヒトにおいて見られるほとんどの取込活性を説明でき、例えばGLUT2ノックアウト変異マウスは、劇的に低下した取り込み能を示す。しかし驚くべきことに、グルコースクリアランスは正常であり、このことは別の流出経路の存在を示唆している。FRETセンサーを利用した発現クローニング系の使用の使用は、植物におけるグルコーストランスポーターの新規クラスの同定を導く(msに伴う)。シロイヌナズナSWEET1および8は単輸送体として機能するが、コメOsSWEET11はグルコースおよびショ糖を流出させると思われる。バイオインフォマティック分析によって、動物およびヒトゲノムが溶質輸送体ファミリーSLC50として登録されている(www.bioparadigms.org/slc/intro.htm)ホモログを含むことが示された(図1)。ここで我々は、アフリカツメガエル卵母細胞において発現されたとき、線虫(C. elegans)CeSWEET1が弱い、しかし重要なグルコース取込を仲介することを示す(図19B)。活性はSGLT1または植物ホモログSWEET1のものよりも顕著に弱い。比較として、線虫(C. elegans)ホモログCeSWEET3、4、5および7は、卵母細胞において検出可能なグルコース取込を仲介しなかった(図19B)。グルコースおよびショ糖を流出させる植物ホモログOsSWEET11は、両方のタンパク質が共発現された場合、卵母細胞におけるSGLT1介在性グルコース蓄積を相殺することが示された。作業仮説は、OsSWEET11介在性流出が「漏出」として作用し、卵母細胞におけるグルコース蓄積を予防することである(3)。線虫(C. elegans)ホモログがエフラクサーとして潜在的に機能しうるかを検証するため、我々は、SGLT1とそれらを共発現させた(図19C)。試験した5種のホモログ全てが、SGLT1のみと比較して低下したグルコース蓄積を導いた。[14C]−グルコースを注入した卵母細胞からの直接流出測定は、RAP1AP1が卵母細胞からのグルコースの流出を誘導することを示す(2分以内に放出されたグルコースの%: RAG1AP1: 5.27 ± 0.29; コントロール: 0.91 ± 0.23; S.E.; n>7)。線虫(C. elegans)ホモログの一つの変異体(CeSWEET1; K02D7.5)は、糖を流出する能力の欠如と一致して、脂肪蓄積を導く。CeSWEET3、4、5および7と同様に、ヒトホモログRAG1AP1(HsSWEET1と命名)はまた、卵母細胞においてグルコース蓄積を仲介することはできなかったが、SGLT1介在性グルコース蓄積を相殺した。これは2種のスプライス変異体および推定内在化モチーフを形成する3個の残基における3つの変異を担持する変異型についても妥当した(図19C)。ホヤCiona由来のホモログCi-Rga(CiSWEET1)における変異は初期発生以上を導き、後生生物にとってのこれらの遺伝子の重要性を裏付けている。哺乳類において、肝臓からの少なくとも1つのグルコース流出経路が不明のままである。
【0272】
RAG1AP1は、細胞系における欠損が組換えに影響を与えるため、組換え活性化遺伝子1活性化タンパク質1(Recombination Activating Gene 1 Activating Protein 1)から名付けられた。さらに、該遺伝子はRGAと名付けられており、TRPVイオンチャネルのターゲティングを意味している。RAG1AP1が肝臓におけるグルコース流出に寄与しうるという仮説を検証することが興味深い。
【0273】
実施例8:HEK293細胞におけるSWEETショ糖排出
【0274】
SWEET1〜4および6〜11および14〜16をFRETショ糖センサーと共にHEK293において発現させた。SWEETファミリーの複数のメンバーがショ糖を排出した。この能力を示したSWEETメンバーは全て、同じクレードに属する(図1参照)。したがって、このクレードはショ糖を排出する強い能力を示すと思われる。OsSWEET11はこのクレードに属し、従ってグルコースを排出することに加えて、これらのタンパク質はショ糖も排出する。これはショ糖を取り込むUstilagoのような病原体のターゲティングにおいて顕著であり得る。植物病原体トウモロコシ黒穂病菌(Ustilago maydis)の病原性のために、新たな高親和性ショ糖トランスポーターが必要とされることは報告されている。PLoS Biol. 8(2):e1000303)または次いで細胞壁インベルターゼがショ糖を切断し、病原体がグルコースを取り込む。
【0275】
ショ糖エフラクサーが葉における細胞間輸送に必要とされることは長らく知られていたが、これらのタンパク質の同一性は不明なままであったため、SWEETのショ糖を排出する示された能力は重要である。SUTは葉のどこかで細胞壁からショ糖を取り込み、葉肉細胞で産生されたショ糖を細胞壁に流出する。したがって、ショ糖を排出するSWEETの能力は、どのように植物がこれを達成するかを理解する上で重要なデータを提供する。さらに、植物の母系組織は植物における主な輸送される糖として発生中の種子にショ糖を供給するためにショ糖を排出する必要がある。したがって、ショ糖エクスポーターとしてのSWEETの役割は、発生中の種子において重要であり得る。
【0276】
本明細書において全ての文献および特許出願は、各個々の文献または特許出願が具体的および個々に記載されているのと同程度に、参照により本明細書に組み込まれる。本明細書に記載の文献は、本願の出願日前のそれらの開示について単に提供される。本明細書のあらゆる事項は、本願発明が先行発明の理由でかかる文献に劣後すると認めるものではない。
【特許請求の範囲】
【請求項1】
AT4G15920、AT3G16690、AT5G13170、AtSAG29、AT4G25010、AT5G50800、AT5G23660、AT3G48740、AT5G50790、AT2G39060、AT5G40260、AtRPG1、AT4G10850、AT1G66770、AT5G62850、AtVEX1、AT3G28007、AT3G14770、AT1G21460、AT5G53190、NEC1、AC202585、AC147714、MtC60432 GC、MtC11004 GC、CT963079、MtD03138 GC、TC 125536、AC146866、AC189276、TC129646、CAA69976 MtNod3 AC2456、TC115479、AC146747、MtC10424 GC、CT954252、CU302340、AC202585、AC147714、MtC60432 GC、MtC11004 GC、CT963079、Os08g42350 (Os8N3) Os08g0535200、Os12g29220 Os03g0347500、Os05g51090 Os05g0588500、Os12g07860、Os09g08440、Os09g08490、Os09g08270、Os09g08030 Os09g0254600、Os01g42090.1 Os01g0605700、Os01g42110.1 Os01g060600、Os02g19820 Os02g0301100、Os05g35140 Os05g0426000、Os01g65880 Os01g0881300、Os01g50460 Os01g0700100、Os01g36070.1 Os01g0541800、Os01g12130.1、Os05g12320 Os05g0214300およびOs01g21230ならびに線虫(C. elegans)(R10D12.9、K11D12.5、K06A4.4、K02D7.5、C54F6.4、C06G8.1、Y39A1A.8)Ciona遺伝子Ci-RgaおよびマウスまたはヒトRAG1AP1遺伝子からなる群から選択される配列と少なくとも85%配列同一性を有する、糖トランスポーターをコードする単離された核酸。
【請求項2】
請求項1の核酸を含むベクター。
【請求項3】
請求項1の核酸を含む細胞。
【請求項4】
植物細胞である、請求項3の細胞。
【請求項5】
動物細胞である、請求項3の細胞。
【請求項6】
糖トランスポーターをコードする核酸を含むトランスジェニック植物であって、核酸がAT4G15920、AT3G16690、AT5G13170、AtSAG29、AT4G25010、AT5G50800、AT5G23660、AT3G48740、AT5G50790、AT2G39060、AT5G40260、AtRPG1、AT4G10850、AT1G66770、AT5G62850、AtVEX1、AT3G28007、AT3G14770、AT1G21460、AT5G53190、NEC1、AC202585、AC147714、MtC60432 GC、MtC11004 GC、CT963079、MtD03138 GC、TC 125536、AC146866、AC189276、TC129646、CAA69976 MtNod3 AC2456、TC115479、AC146747、MtC10424 GC、CT954252、CU302340、AC202585、AC147714、MtC60432 GC、MtC11004 GC、CT963079、Os08g42350 (Os8N3) Os08g0535200、Os12g29220 Os03g0347500、Os05g51090 Os05g0588500、Os12g07860、Os09g08440、Os09g08490、Os09g08270、Os09g08030 Os09g0254600、Os01g42090.1 Os01g0605700、Os01g42110.1 Os01g060600、Os02g19820 Os02g0301100、Os05g35140 Os05g0426000、Os01g65880 Os01g0881300、Os01g50460 Os01g0700100、Os01g36070.1 Os01g0541800、Os01g12130.1、Os05g12320 Os05g0214300およびOs01g21230ならびに線虫(C. elegans)(R10D12.9、K11D12.5、K06A4.4、K02D7.5、C54F6.4、C06G8.1、Y39A1A.8)Ciona遺伝子Ci-RgaおよびマウスまたはヒトRAG1AP1遺伝子からなる群から選択される配列と少なくとも85%配列同一性を有する、トランスジェニック植物。
【請求項7】
コントロール植物と比較して増加したレベルの炭素を産生する植物を作成する方法であって、植物細胞に糖トランスポーターをコードする核酸を導入し、植物細胞を該核酸を発現する植物へと成長させることを含み、ここでグルコーストランスポーターをコードする前記核酸は、AT4G15920、AT3G16690、AT5G13170、AtSAG29、AT4G25010、AT5G50800、AT5G23660、AT3G48740、AT5G50790、AT2G39060、AT5G40260、AtRPG1、AT4G10850、AT1G66770、AT5G62850、AtVEX1、AT3G28007、AT3G14770、AT1G21460、AT5G53190、NEC1、AC202585、AC147714、MtC60432 GC、MtC11004 GC、CT963079、MtD03138 GC、TC 125536、AC146866、AC189276、TC129646、CAA69976 MtNod3 AC2456、TC115479、AC146747、MtC10424 GC、CT954252、CU302340、AC202585、AC147714、MtC60432 GC、MtC11004 GC、CT963079、Os08g42350 (Os8N3) Os08g0535200、Os12g29220 Os03g0347500、Os05g51090 Os05g0588500、Os12g07860、Os09g08440、Os09g08490、Os09g08270、Os09g08030 Os09g0254600、Os01g42090.1 Os01g0605700、Os01g42110.1 Os01g060600、Os02g19820 Os02g0301100、Os05g35140 Os05g0426000、Os01g65880 Os01g0881300、Os01g50460 Os01g0700100、Os01g36070.1 Os01g0541800、Os01g12130.1、Os05g12320 Os05g0214300およびOs01g21230ならびに(C. elegans)(R10D12.9、K11D12.5、K06A4.4、K02D7.5、C54F6.4、C06G8.1、Y39A1A.8)Ciona遺伝子Ci-RgaおよびマウスまたはヒトRAG1AP1遺伝子からなる群から選択される配列と85%配列同一性を有する、方法。
【請求項8】
植物の根への糖輸送を増加させる方法であって、植物の細胞に請求項1の核酸を導入することを含む方法。
【請求項9】
植物の根圏への糖分泌を調節する方法であって、植物の細胞に請求項1の核酸を導入することを含む方法。
【請求項10】
植物の葉圏への糖輸送を調節する方法であって、植物の細胞に請求項1の核酸を導入することを含む方法。
【請求項11】
葉圏における糖含量の増加が存在する、請求項10の方法。
【請求項12】
糖含量の増加が有益微生物を誘引する、請求項11の方法。.
【請求項13】
病原体による侵襲を予防する糖含量の減少が存在する、請求項10の方法。
【請求項14】
細胞膜を通過して糖を輸送する単離されたポリペプチドであって、請求項1の核酸によってコードされる、単離されたポリペプチド。
【請求項15】
請求項1の核酸によってコードされる、単離されたポリペプチド。
【請求項16】
請求項1のポリペプチドをコードする、単離された核酸。
【請求項17】
細胞が植物細胞である、請求項16のポリペプチド。
【請求項18】
細胞が動物細胞である、請求項16のポリペプチド。
【請求項19】
細胞内の糖を調節する方法であって、細胞にグルコースエクスポーター(GLUE)ポリペプチドまたはその機能的均等物を導入することを含む方法。
【請求項20】
糖濃度がFRETセンサーによってモニターされる、請求項19の方法。
【請求項21】
細胞がインビトロに存在する、請求項19の方法。
【請求項22】
細胞がインビボに存在する、請求項19の方法。
【請求項23】
細胞が動物細胞である、請求項19の方法。
【請求項24】
細胞が植物細胞である、請求項19の方法。
【請求項25】
調節が増加したグルコース排出である、請求項19の方法。
【請求項26】
調節が増加したグルコース取込である、請求項19の方法。
【請求項27】
調節が減少したグルコース排出である、請求項19の方法。
【請求項28】
調節が減少したグルコース取込である、請求項19の方法。
【請求項29】
細胞内のグルコースを変化させる方法であって、細胞に変異を含むGLUEポリペプチドを導入することを含む方法。
【請求項30】
変異が欠失である、請求項29の方法。
【請求項31】
欠失が一アミノ酸である、請求項29の方法。
【請求項32】
変異がアミノ産地間である、請求項29の方法。
【請求項33】
変異が野生型アミノ酸配列のトランケーションである、請求項29の方法。
【請求項34】
植物の葉からの糖排出を制御する方法であって、植物にGLUEポリペプチドを導入することを含む方法。
【請求項35】
さらにコファクターを含む、請求項34の方法。
【請求項36】
コファクターがコトランスポーターである、請求項35の方法。
【請求項37】
生物の発生を変化させる方法であって、生物の細胞にGLUEポリペプチドを導入することを含む方法。
【請求項38】
GLUEをコードする核酸の導入によってGLUEポリペプチドを導入する、請求項37の方法。
【請求項39】
生物が植物である、請求項37の方法。
【請求項40】
GLUEが生物の組織に局在して発現する、請求項37の方法。
【請求項41】
組織における発現が生物の他の組織へのグルコース移動に影響する、請求項37の方法。
【請求項42】
組織が蜜腺またはタペータムである、請求項41の方法。
【請求項43】
細胞におけるGLUTまたはSGLTの発現を抑制する方法であって、細胞にGLUEポリペプチドを導入することを含む方法。
【請求項44】
細胞によって分泌される液体中の糖濃度を変化させる方法であって、細胞にGLUEポリペプチドを導入することを含む方法。
【請求項45】
液体が花蜜である、請求項44の方法。
【請求項46】
液体が樹液である、請求項44の方法。
【請求項47】
GLUEが野生型である、請求項44の方法。
【請求項48】
GLUEが変異型である、請求項44の方法。
【請求項49】
生物の発生を変化させる方法であって、生物の細胞にGLUEトランスポーターを導入することを含み、ここでGLUEが細胞内の糖輸送を制御する、方法。
【請求項50】
生物が植物または動物である、請求項49の方法。
【請求項51】
GLUEが野生型または変異型である、請求項49の方法。
【請求項52】
生物が動物である、請求項49の方法。
【請求項53】
動物がショウジョウバエである、請求項49の方法。
【請求項54】
細胞が神経細胞である、請求項49の方法。
【請求項55】
細胞が生殖細胞である、請求項49の方法。
【請求項56】
胚盤胞/胚にGLUEを導入する、請求項49の方法。.
【請求項57】
生物が成人である、請求項49の方法。
【請求項58】
GLUEが神経発生を制御する、請求項49の方法。
【請求項59】
生物からの侵襲に対する細胞の感受性を調節する方法であって、生物にGLUEトランスポーターを導入することを含む方法。
【請求項60】
生物の細胞にGLUEを導入する、請求項59の方法。
【請求項61】
GLUEをコードする核酸としてGLUEを導入する、請求項59の方法。
【請求項62】
生物が病原体によって侵襲される、請求項59の方法。
【請求項63】
病原体が生物の遺伝子発現を誘導する、請求項59の方法。
【請求項64】
GLUEが遺伝子発現に影響する病原体の能力を変化させる、請求項59の方法。
【請求項65】
GLUEが病原体によって吸収される、請求項59の方法。
【請求項66】
GLUEが病原体における糖輸送に影響する、請求項59の方法。
【請求項67】
病原体が生物におけるGLUE遺伝子発現に影響する、請求項59の方法。
【請求項68】
細胞内のグルコース濃度を変化させる方法であって、GLUEタンパク質に影響する薬物と細胞を接触させることを含む方法。
【請求項69】
細胞内のグルコースを調節する方法であって、細胞にグルコースエクスポータータンパク質をコードする核酸を導入することを含む方法。
【請求項70】
植物生産性に影響を与える方法であって、植物の細胞にGLUEポリペプチドを導入することを含む方法。
【請求項71】
請求項1のポリペプチドを含む免疫原性組成物。
【請求項72】
GLUEを検出する方法であって、請求項71の免疫原性組成物と動物を接触させて産生した抗体と、細胞またはその抽出物を接触させることを含む方法。
【請求項73】
GLUEトランスポーターを調節する物質を同定する方法。
【請求項1】
AT4G15920、AT3G16690、AT5G13170、AtSAG29、AT4G25010、AT5G50800、AT5G23660、AT3G48740、AT5G50790、AT2G39060、AT5G40260、AtRPG1、AT4G10850、AT1G66770、AT5G62850、AtVEX1、AT3G28007、AT3G14770、AT1G21460、AT5G53190、NEC1、AC202585、AC147714、MtC60432 GC、MtC11004 GC、CT963079、MtD03138 GC、TC 125536、AC146866、AC189276、TC129646、CAA69976 MtNod3 AC2456、TC115479、AC146747、MtC10424 GC、CT954252、CU302340、AC202585、AC147714、MtC60432 GC、MtC11004 GC、CT963079、Os08g42350 (Os8N3) Os08g0535200、Os12g29220 Os03g0347500、Os05g51090 Os05g0588500、Os12g07860、Os09g08440、Os09g08490、Os09g08270、Os09g08030 Os09g0254600、Os01g42090.1 Os01g0605700、Os01g42110.1 Os01g060600、Os02g19820 Os02g0301100、Os05g35140 Os05g0426000、Os01g65880 Os01g0881300、Os01g50460 Os01g0700100、Os01g36070.1 Os01g0541800、Os01g12130.1、Os05g12320 Os05g0214300およびOs01g21230ならびに線虫(C. elegans)(R10D12.9、K11D12.5、K06A4.4、K02D7.5、C54F6.4、C06G8.1、Y39A1A.8)Ciona遺伝子Ci-RgaおよびマウスまたはヒトRAG1AP1遺伝子からなる群から選択される配列と少なくとも85%配列同一性を有する、糖トランスポーターをコードする単離された核酸。
【請求項2】
請求項1の核酸を含むベクター。
【請求項3】
請求項1の核酸を含む細胞。
【請求項4】
植物細胞である、請求項3の細胞。
【請求項5】
動物細胞である、請求項3の細胞。
【請求項6】
糖トランスポーターをコードする核酸を含むトランスジェニック植物であって、核酸がAT4G15920、AT3G16690、AT5G13170、AtSAG29、AT4G25010、AT5G50800、AT5G23660、AT3G48740、AT5G50790、AT2G39060、AT5G40260、AtRPG1、AT4G10850、AT1G66770、AT5G62850、AtVEX1、AT3G28007、AT3G14770、AT1G21460、AT5G53190、NEC1、AC202585、AC147714、MtC60432 GC、MtC11004 GC、CT963079、MtD03138 GC、TC 125536、AC146866、AC189276、TC129646、CAA69976 MtNod3 AC2456、TC115479、AC146747、MtC10424 GC、CT954252、CU302340、AC202585、AC147714、MtC60432 GC、MtC11004 GC、CT963079、Os08g42350 (Os8N3) Os08g0535200、Os12g29220 Os03g0347500、Os05g51090 Os05g0588500、Os12g07860、Os09g08440、Os09g08490、Os09g08270、Os09g08030 Os09g0254600、Os01g42090.1 Os01g0605700、Os01g42110.1 Os01g060600、Os02g19820 Os02g0301100、Os05g35140 Os05g0426000、Os01g65880 Os01g0881300、Os01g50460 Os01g0700100、Os01g36070.1 Os01g0541800、Os01g12130.1、Os05g12320 Os05g0214300およびOs01g21230ならびに線虫(C. elegans)(R10D12.9、K11D12.5、K06A4.4、K02D7.5、C54F6.4、C06G8.1、Y39A1A.8)Ciona遺伝子Ci-RgaおよびマウスまたはヒトRAG1AP1遺伝子からなる群から選択される配列と少なくとも85%配列同一性を有する、トランスジェニック植物。
【請求項7】
コントロール植物と比較して増加したレベルの炭素を産生する植物を作成する方法であって、植物細胞に糖トランスポーターをコードする核酸を導入し、植物細胞を該核酸を発現する植物へと成長させることを含み、ここでグルコーストランスポーターをコードする前記核酸は、AT4G15920、AT3G16690、AT5G13170、AtSAG29、AT4G25010、AT5G50800、AT5G23660、AT3G48740、AT5G50790、AT2G39060、AT5G40260、AtRPG1、AT4G10850、AT1G66770、AT5G62850、AtVEX1、AT3G28007、AT3G14770、AT1G21460、AT5G53190、NEC1、AC202585、AC147714、MtC60432 GC、MtC11004 GC、CT963079、MtD03138 GC、TC 125536、AC146866、AC189276、TC129646、CAA69976 MtNod3 AC2456、TC115479、AC146747、MtC10424 GC、CT954252、CU302340、AC202585、AC147714、MtC60432 GC、MtC11004 GC、CT963079、Os08g42350 (Os8N3) Os08g0535200、Os12g29220 Os03g0347500、Os05g51090 Os05g0588500、Os12g07860、Os09g08440、Os09g08490、Os09g08270、Os09g08030 Os09g0254600、Os01g42090.1 Os01g0605700、Os01g42110.1 Os01g060600、Os02g19820 Os02g0301100、Os05g35140 Os05g0426000、Os01g65880 Os01g0881300、Os01g50460 Os01g0700100、Os01g36070.1 Os01g0541800、Os01g12130.1、Os05g12320 Os05g0214300およびOs01g21230ならびに(C. elegans)(R10D12.9、K11D12.5、K06A4.4、K02D7.5、C54F6.4、C06G8.1、Y39A1A.8)Ciona遺伝子Ci-RgaおよびマウスまたはヒトRAG1AP1遺伝子からなる群から選択される配列と85%配列同一性を有する、方法。
【請求項8】
植物の根への糖輸送を増加させる方法であって、植物の細胞に請求項1の核酸を導入することを含む方法。
【請求項9】
植物の根圏への糖分泌を調節する方法であって、植物の細胞に請求項1の核酸を導入することを含む方法。
【請求項10】
植物の葉圏への糖輸送を調節する方法であって、植物の細胞に請求項1の核酸を導入することを含む方法。
【請求項11】
葉圏における糖含量の増加が存在する、請求項10の方法。
【請求項12】
糖含量の増加が有益微生物を誘引する、請求項11の方法。.
【請求項13】
病原体による侵襲を予防する糖含量の減少が存在する、請求項10の方法。
【請求項14】
細胞膜を通過して糖を輸送する単離されたポリペプチドであって、請求項1の核酸によってコードされる、単離されたポリペプチド。
【請求項15】
請求項1の核酸によってコードされる、単離されたポリペプチド。
【請求項16】
請求項1のポリペプチドをコードする、単離された核酸。
【請求項17】
細胞が植物細胞である、請求項16のポリペプチド。
【請求項18】
細胞が動物細胞である、請求項16のポリペプチド。
【請求項19】
細胞内の糖を調節する方法であって、細胞にグルコースエクスポーター(GLUE)ポリペプチドまたはその機能的均等物を導入することを含む方法。
【請求項20】
糖濃度がFRETセンサーによってモニターされる、請求項19の方法。
【請求項21】
細胞がインビトロに存在する、請求項19の方法。
【請求項22】
細胞がインビボに存在する、請求項19の方法。
【請求項23】
細胞が動物細胞である、請求項19の方法。
【請求項24】
細胞が植物細胞である、請求項19の方法。
【請求項25】
調節が増加したグルコース排出である、請求項19の方法。
【請求項26】
調節が増加したグルコース取込である、請求項19の方法。
【請求項27】
調節が減少したグルコース排出である、請求項19の方法。
【請求項28】
調節が減少したグルコース取込である、請求項19の方法。
【請求項29】
細胞内のグルコースを変化させる方法であって、細胞に変異を含むGLUEポリペプチドを導入することを含む方法。
【請求項30】
変異が欠失である、請求項29の方法。
【請求項31】
欠失が一アミノ酸である、請求項29の方法。
【請求項32】
変異がアミノ産地間である、請求項29の方法。
【請求項33】
変異が野生型アミノ酸配列のトランケーションである、請求項29の方法。
【請求項34】
植物の葉からの糖排出を制御する方法であって、植物にGLUEポリペプチドを導入することを含む方法。
【請求項35】
さらにコファクターを含む、請求項34の方法。
【請求項36】
コファクターがコトランスポーターである、請求項35の方法。
【請求項37】
生物の発生を変化させる方法であって、生物の細胞にGLUEポリペプチドを導入することを含む方法。
【請求項38】
GLUEをコードする核酸の導入によってGLUEポリペプチドを導入する、請求項37の方法。
【請求項39】
生物が植物である、請求項37の方法。
【請求項40】
GLUEが生物の組織に局在して発現する、請求項37の方法。
【請求項41】
組織における発現が生物の他の組織へのグルコース移動に影響する、請求項37の方法。
【請求項42】
組織が蜜腺またはタペータムである、請求項41の方法。
【請求項43】
細胞におけるGLUTまたはSGLTの発現を抑制する方法であって、細胞にGLUEポリペプチドを導入することを含む方法。
【請求項44】
細胞によって分泌される液体中の糖濃度を変化させる方法であって、細胞にGLUEポリペプチドを導入することを含む方法。
【請求項45】
液体が花蜜である、請求項44の方法。
【請求項46】
液体が樹液である、請求項44の方法。
【請求項47】
GLUEが野生型である、請求項44の方法。
【請求項48】
GLUEが変異型である、請求項44の方法。
【請求項49】
生物の発生を変化させる方法であって、生物の細胞にGLUEトランスポーターを導入することを含み、ここでGLUEが細胞内の糖輸送を制御する、方法。
【請求項50】
生物が植物または動物である、請求項49の方法。
【請求項51】
GLUEが野生型または変異型である、請求項49の方法。
【請求項52】
生物が動物である、請求項49の方法。
【請求項53】
動物がショウジョウバエである、請求項49の方法。
【請求項54】
細胞が神経細胞である、請求項49の方法。
【請求項55】
細胞が生殖細胞である、請求項49の方法。
【請求項56】
胚盤胞/胚にGLUEを導入する、請求項49の方法。.
【請求項57】
生物が成人である、請求項49の方法。
【請求項58】
GLUEが神経発生を制御する、請求項49の方法。
【請求項59】
生物からの侵襲に対する細胞の感受性を調節する方法であって、生物にGLUEトランスポーターを導入することを含む方法。
【請求項60】
生物の細胞にGLUEを導入する、請求項59の方法。
【請求項61】
GLUEをコードする核酸としてGLUEを導入する、請求項59の方法。
【請求項62】
生物が病原体によって侵襲される、請求項59の方法。
【請求項63】
病原体が生物の遺伝子発現を誘導する、請求項59の方法。
【請求項64】
GLUEが遺伝子発現に影響する病原体の能力を変化させる、請求項59の方法。
【請求項65】
GLUEが病原体によって吸収される、請求項59の方法。
【請求項66】
GLUEが病原体における糖輸送に影響する、請求項59の方法。
【請求項67】
病原体が生物におけるGLUE遺伝子発現に影響する、請求項59の方法。
【請求項68】
細胞内のグルコース濃度を変化させる方法であって、GLUEタンパク質に影響する薬物と細胞を接触させることを含む方法。
【請求項69】
細胞内のグルコースを調節する方法であって、細胞にグルコースエクスポータータンパク質をコードする核酸を導入することを含む方法。
【請求項70】
植物生産性に影響を与える方法であって、植物の細胞にGLUEポリペプチドを導入することを含む方法。
【請求項71】
請求項1のポリペプチドを含む免疫原性組成物。
【請求項72】
GLUEを検出する方法であって、請求項71の免疫原性組成物と動物を接触させて産生した抗体と、細胞またはその抽出物を接触させることを含む方法。
【請求項73】
GLUEトランスポーターを調節する物質を同定する方法。
【図1】

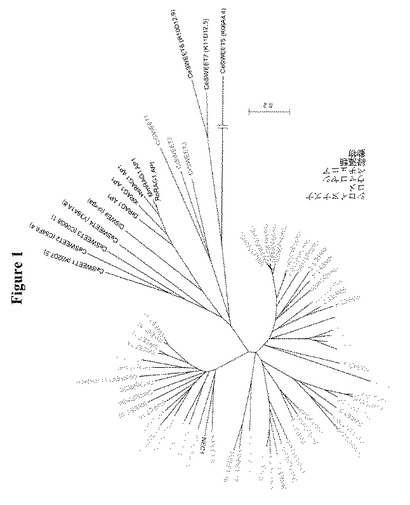
【図2】
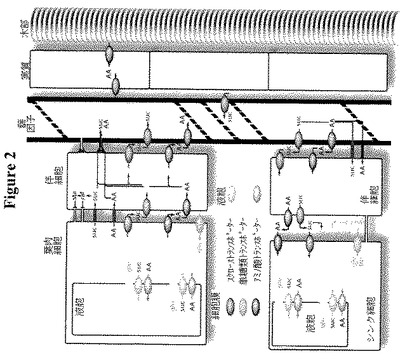
【図3】
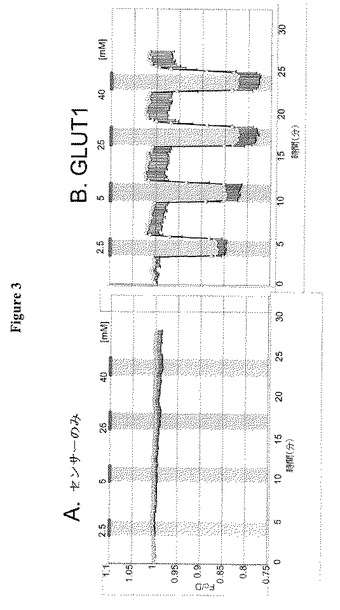
【図4】
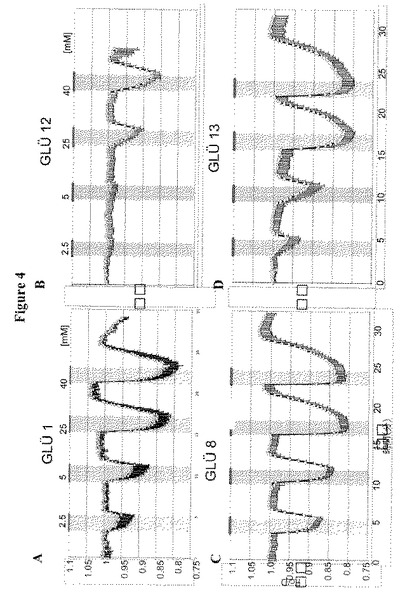
【図5】
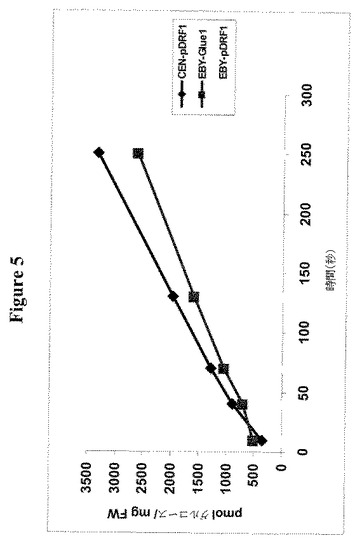
【図6】
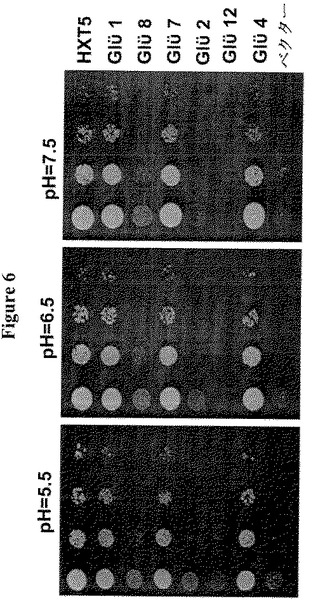
【図7】
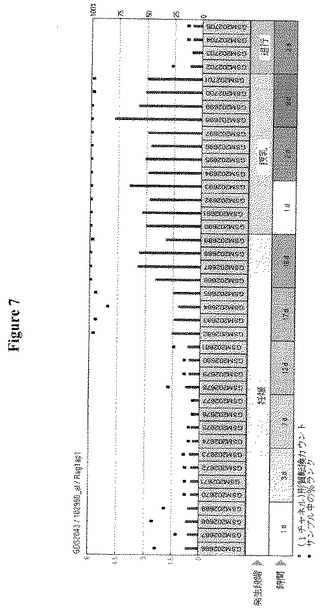
【図8】
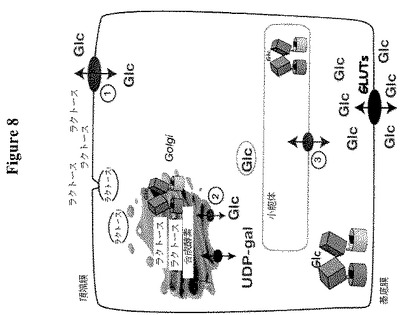
【図9】
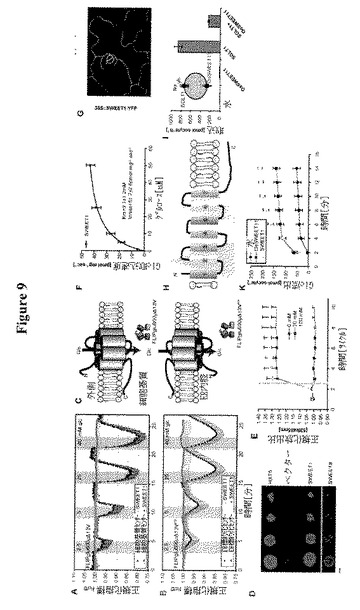
【図10】
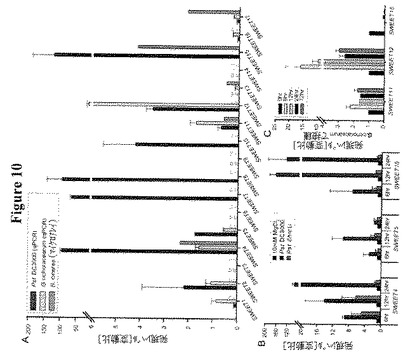
【図11】
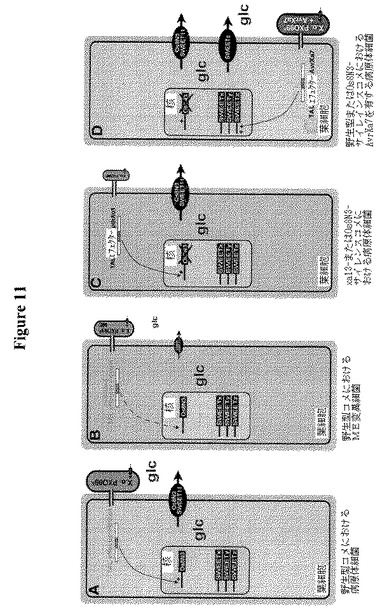
【図12】
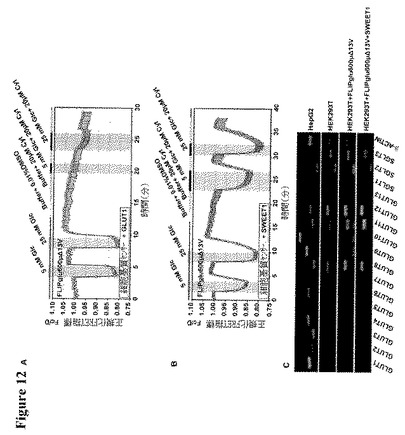
【図13】
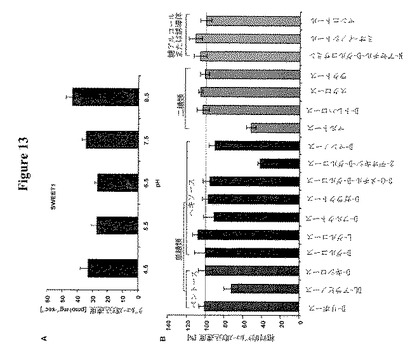
【図14】
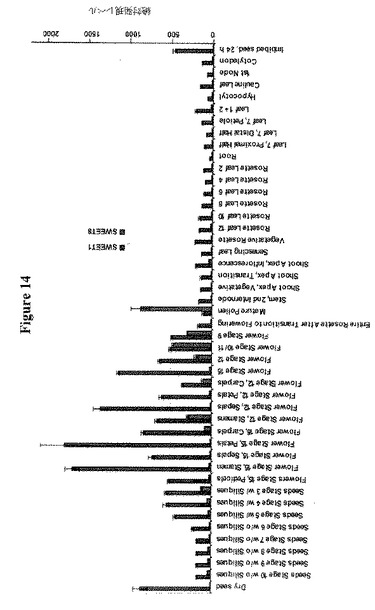
【図15】
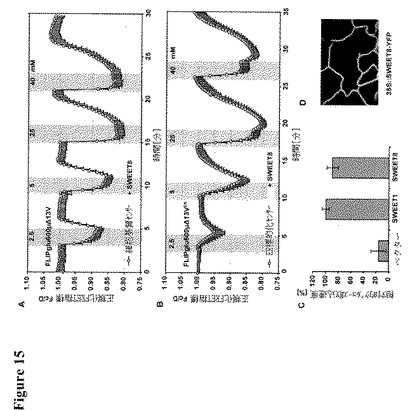
【図16】
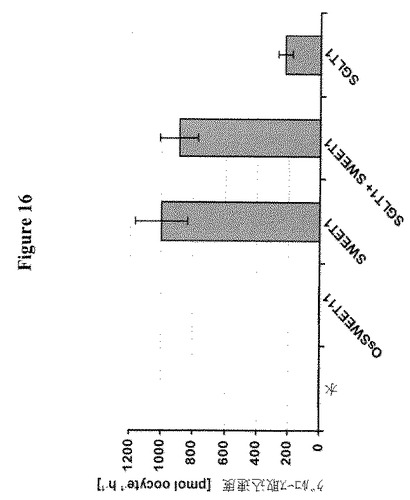
【図17】
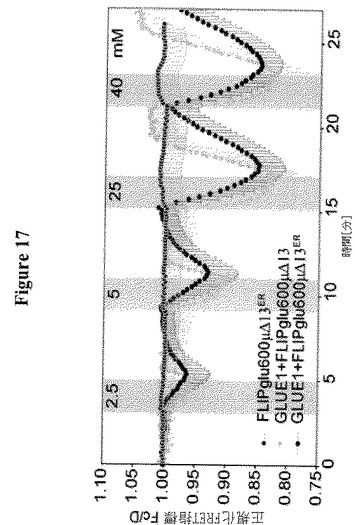
【図18】
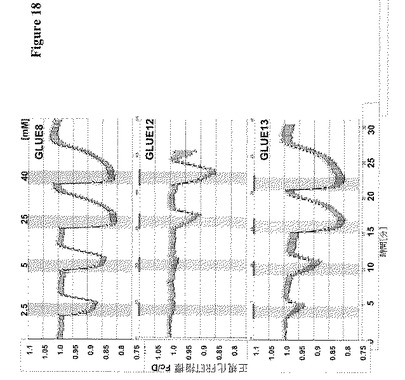
【図19】
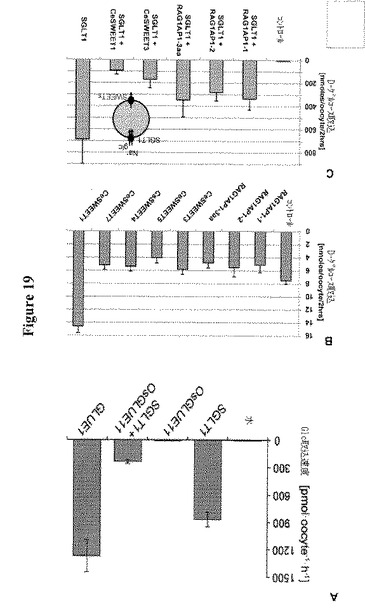
【図20】
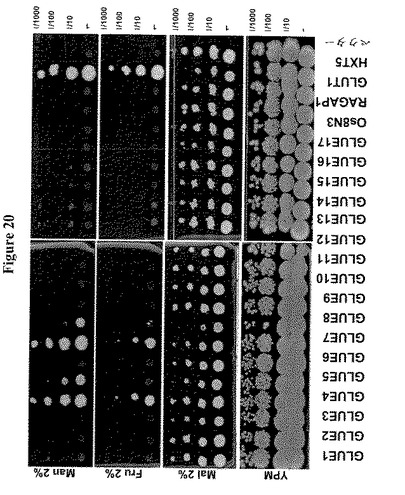
【図21】
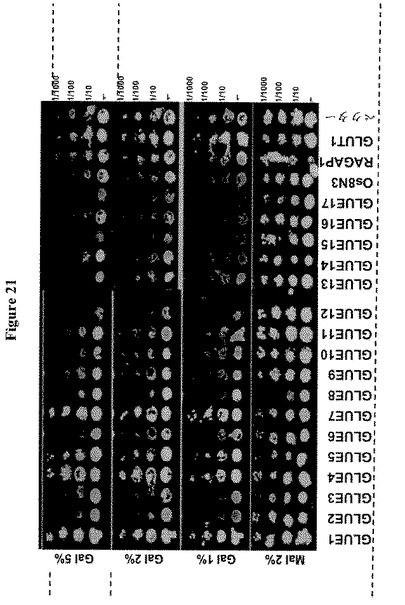
【図22】
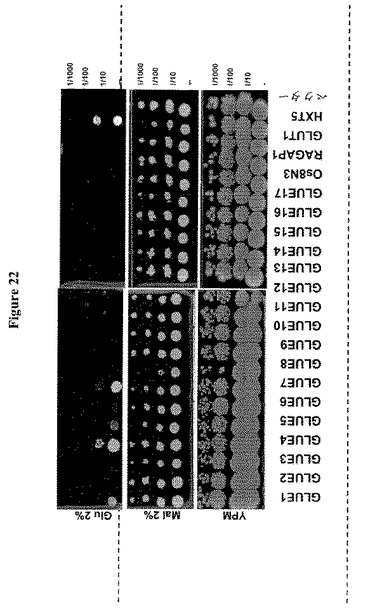
【図23】
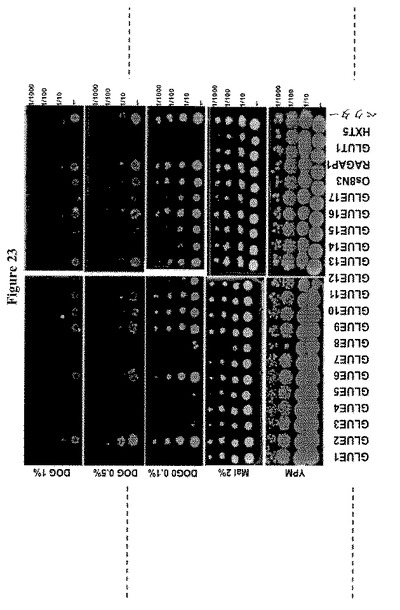
【図24】
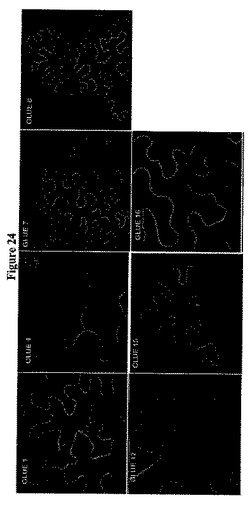
【図25】
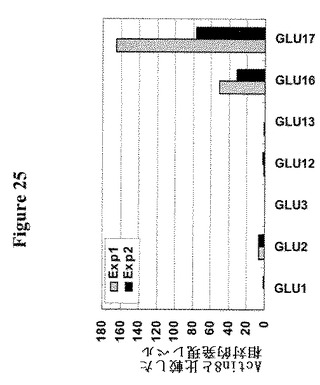
【図26】
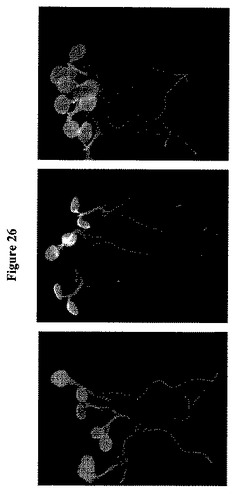
【図27】
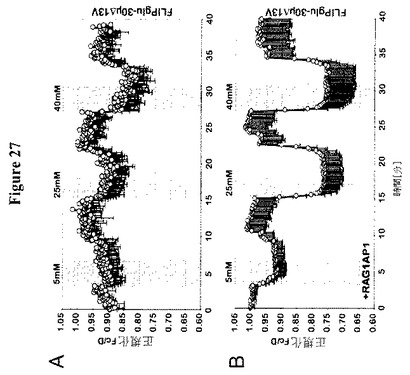
【図28】
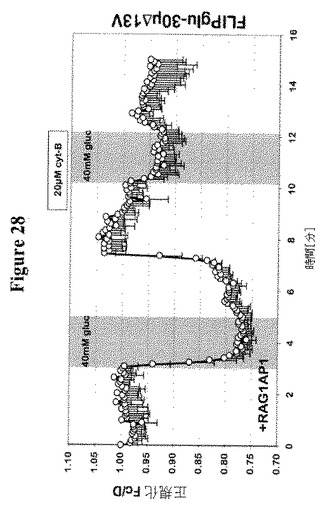
【図29】
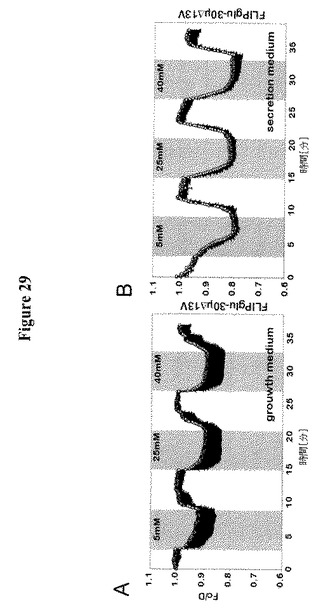
【図30】
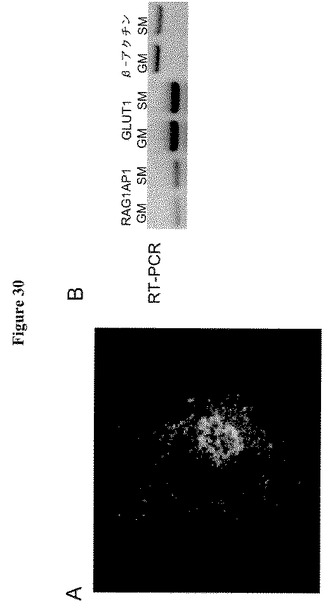
【図31】
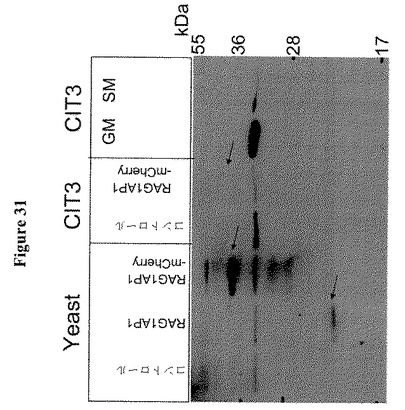
【図32】
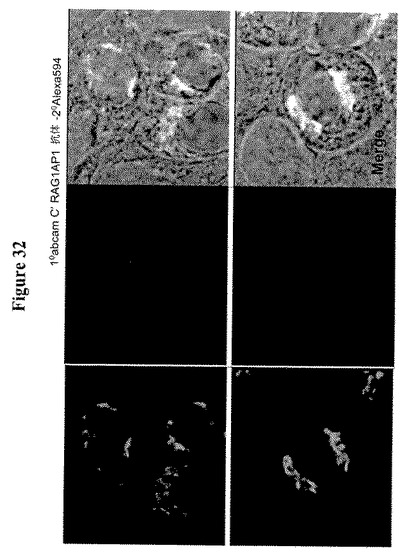
【図33】
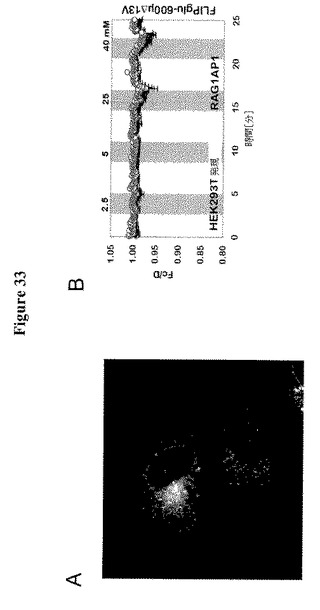
【図34】

【図35】

【図36】
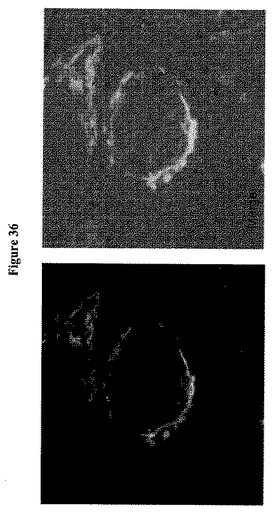
【図37】
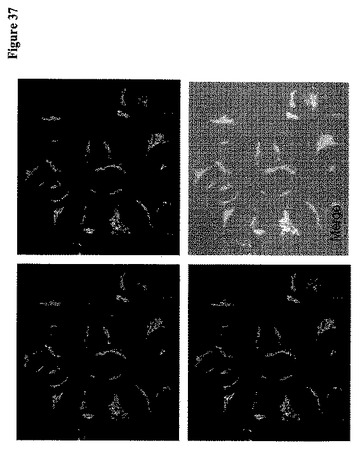
【図38】
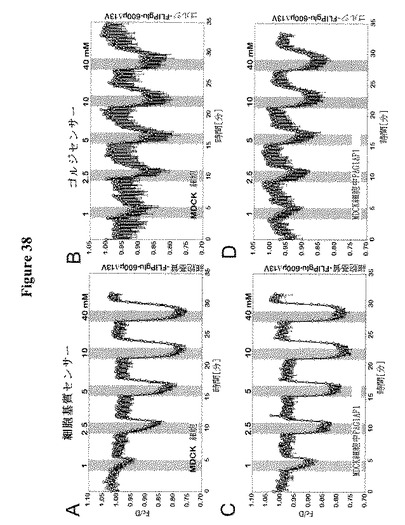
【図39】
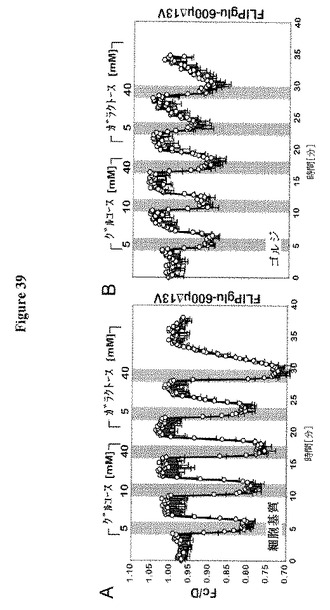
【図40】
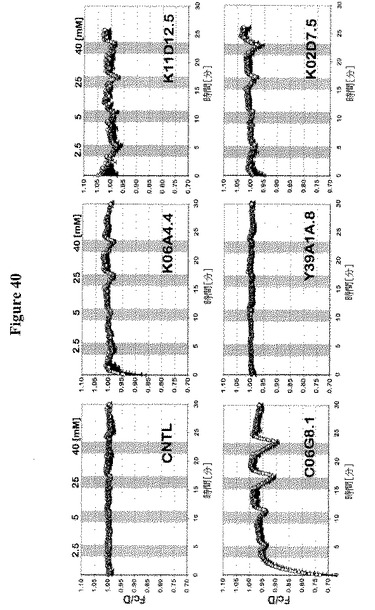
【図41】
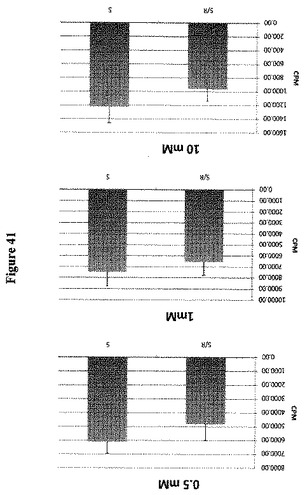
【図42】
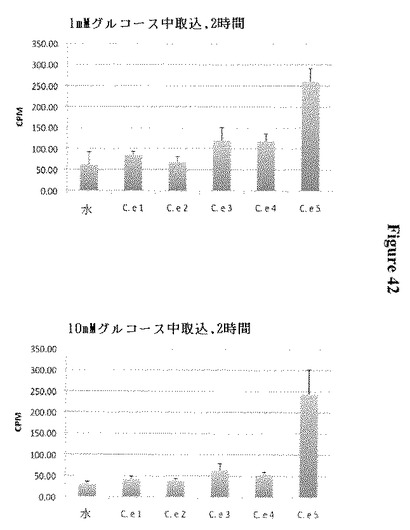
【図43−1】
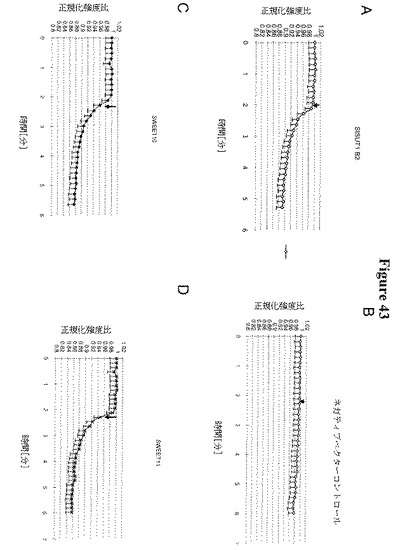
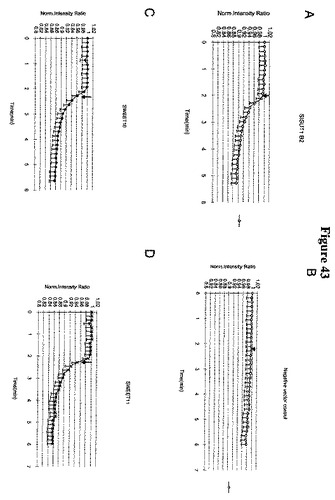
【図43−2】
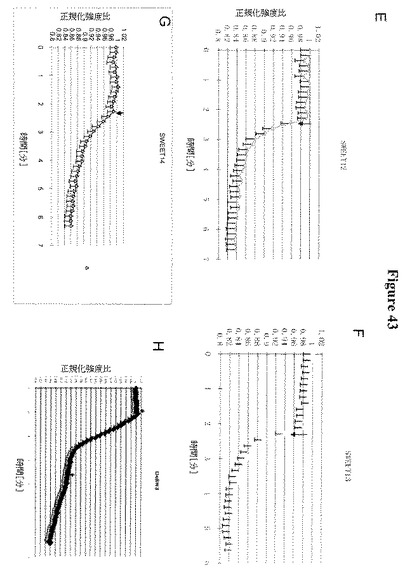
【図44−1】

【図44−2】

【図44−3】

【図44−4】

【図44−5】

【図44−6】

【図44−7】

【図44−8】

【図44−9】

【図44−10】

【図44−11】

【図44−12】

【図44−13】

【図44−14】

【図44−15】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【図30】
【図31】
【図32】
【図33】
【図34】
【図35】
【図36】
【図37】
【図38】
【図39】
【図40】
【図41】
【図42】
【図43−1】
【図43−2】
【図44−1】
【図44−2】
【図44−3】
【図44−4】
【図44−5】
【図44−6】
【図44−7】
【図44−8】
【図44−9】
【図44−10】
【図44−11】
【図44−12】
【図44−13】
【図44−14】
【図44−15】
【公表番号】特表2012−525845(P2012−525845A)
【公表日】平成24年10月25日(2012.10.25)
【国際特許分類】
【出願番号】特願2012−509899(P2012−509899)
【出願日】平成22年5月4日(2010.5.4)
【国際出願番号】PCT/US2010/033535
【国際公開番号】WO2010/129540
【国際公開日】平成22年11月11日(2010.11.11)
【出願人】(500026234)カーネギー インスチチューション オブ ワシントン (25)
【Fターム(参考)】
【公表日】平成24年10月25日(2012.10.25)
【国際特許分類】
【出願日】平成22年5月4日(2010.5.4)
【国際出願番号】PCT/US2010/033535
【国際公開番号】WO2010/129540
【国際公開日】平成22年11月11日(2010.11.11)
【出願人】(500026234)カーネギー インスチチューション オブ ワシントン (25)
【Fターム(参考)】
[ Back to top ]