
新規純粋コハク酸生成変異微生物及びそれを用いたコハク酸の製造方法
本発明は純粋コハク酸生成新規ルーメンバクテリア変異菌株及び前記ルーメンバクテリア変異菌株を用いた純粋なコハク酸の製造方法に関して、より詳しくは、コハク酸を生成する微生物でピルビン酸ギ酸リアーゼをコードする遺伝子(pfl)を含み、乳酸脱水素酵素をコードする遺伝子(ldhA)、ホスホトランスアセチラーゼをコードする遺伝子(pta)及び酢酸キナーゼをコードする遺伝子(ackA)が欠失しており、嫌気条件でコハク酸以外の有機酸はほとんど生成せず、コハク酸のみを高濃度で生成する特性を有するルーメンバクテリア変異菌株及び前記ルーメンバクテリア変異菌株を用いたコハク酸の製造方法に関する。本発明によるルーメンバクテリア変異菌株は従来のコハク酸生成微生物より菌株の生長速度及びコハク酸生産性が高く、他の有機酸を生成せずに高濃度でコハク酸を生成するから、コハク酸の産業的生産菌株として有用である。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は純粋コハク酸生成新規ルーメンバクテリア変異菌株及び前記ルーメンバクテリア変異菌株を用いた純粋なコハク酸の製造方法に関し、より詳しくは、コハク酸を生成する微生物でピルビン酸ギ酸リアーゼをコードする遺伝子(以下、「pfl」という)は欠失されず、乳酸脱水素酵素をコードする遺伝子(以下、「ldhA」という)、ホスホトランスアセチラーゼをコードする遺伝子(以下、「pta」という)及び酢酸キナーゼをコードする遺伝子(以下、「ackA」という)が欠失しており、嫌気条件でコハク酸以外の有機酸はほとんど生成せず、コハク酸のみを高濃度で生成する特性を有するルーメンバクテリア変異菌株及び前記ルーメンバクテリア変異菌株を用いたコハク酸の製造方法に関する。
【背景技術】
【0002】
コハク酸(HOOCCH2CH2COOH)は4個の炭素からなるジカルボン酸で、医薬用、食品用、化粧品及び他の産業で使用される化学製品の前具体として広く使用される産業的、経済的な利用価値が高い有機酸である(Zeikus et al., Appl. Microbiol. Biotechnol., 51:545, 1999; Song et al., Enzyme Microbial Technol., 39:352, 2006)。特に最近急激な原油価格の上昇と伴に生分解性高分子の主な原料物質として、のコハク酸の使用可能性が証明され、急激な需要増大が予想されている(Willke et al., Appl. Microbiol. Biotechnol., 66:131, 2004)。
【0003】
コハク酸は化学合成と醗酵により生産できるが、現在まで産業的に使用される大部分のコハク酸はBASF,DuPont,BPケミカルなど巨大な化学会社と中国及び日本の化学会社によって原油から由来したn−ブタンとアセチレンを原料物質に使用して化学合成法によって生産されており、医薬品などの特殊な用途に使用される少量のコハク酸だけが古典的な微生物醗酵法により生産されている。このようなコハク酸の化学合成法は製造過程から多量の有害廃棄物、廃溶液及び廃ガス(一酸化炭素含み)を排出するという問題点を有しており、特に、資源枯渇の可能性の高い化石原料を其礎物質として使用しており、従って、これの代わりに使用できるコハク酸の生産方法の開発が緊急課題である。
【0004】
このようなコハク酸の化学合成工程による問題点を解決するため、微生物醗酵を通じて再生可能な原料からコハク酸を生産する研究が多くの研究者らによって集中的に行われていた。醗酵によって、コハク酸の生産に使用される微生物は非常に多様だが、大きく組換え大腸菌とルーメンバクテリア(ruminal bacteria: Actinobacillus, Anaerobiospirillum, Bacteroides, Mannheimia, Succinimonas, Succinivibrio等)に分けることができる(Song et al., Enzyme Microbial Technol., 39:352, 2006)。
【0005】
組換え大腸菌を利用したコハク酸生産に関する研究を調べてみると、先ずアメリカのシカゴ大学の研究チームが大腸菌の乳酸及びギ酸の生産に関与する遺伝子ら(ldh及びpfl)を除去すると同時にブドウ糖伝達システム遺伝子(ptsG)を操作した変異菌株であるAFP111(ATCC202021)を製作してコハク酸の生産増加を試みした(米国特許5,770,435号明細書)。
【0006】
本発明者らはコハク酸生産に関与するリンゴ酸酵素遺伝子(sfcA)をldh遺伝子及びpfl遺伝子が除去された組換え大腸菌であるNZN111菌株に増幅させ、NZN111菌株の醗酵過程から蓄積されるピルビン酸を抑制することによってコハク酸の生産を増大したことがあり(Hong et al., Biotechnol. Bioeng., 74:89, 2001)、アメリカのジョージア大学の研究チームはRhizobium etli菌株のピルビン酸カルボキシラーゼ遺伝子(pyc)を前記AFP111菌株に発現させることによりAFP111/pTrc99A−pyc菌株を製作し、これをコハク酸生産に利用した(Vemuri et al., J. Ind. Microbiol. Biotechnol., 28:325, 2001)。最近、好気条件でコハク酸生産を誘導するために、アメリカのライス大学研究チームが解糖過程、TCA経路及びグリオキシレート経路に関与する遺伝子を操作した組換え大腸菌株を製作して発表した(Lin et al., Metabol. Eng., 7:116, 2005; Lin et al., Biotechnol. Bioeng., 90:775, 2005)。
【0007】
コハク酸の生産においてはルーメンバクテリアの一種であるActinobacillus,Anaerobiospirillum及びMannheimia菌株が最も優れていることが知られており、これら菌株に対する研究が多く行われてきた。アメリカのMBI(Michgan Biotechnology Institute)研究チームはActinobacillus succiniciproducens130Z菌株(ATCC55681)を発見し、コハク酸生産方法を開発し(米国特許5,504,004号明細書)、そして古典的化学突然変異方法を用いてActinobacillus succiniciproducensの多様な変異菌株を開発して、これをコハク酸生産及び精製工程の開発に利用した(米国特許5,521,075号明細書;米国特許5,168,055号明細書;米国特許5,143,834号明細書)。
【0008】
しかし、現在まで開発された微生物の醗酵を通じたコハク酸生産方法は生産性が2g/L/h以下で非常に低く、特に、醗酵時にコハク酸と伴に副産物として各種有機酸とエタノールを同時に生産することにより、莫大な分離及び精製費用が要求されている。たとえ前記結果が一部組換え菌株において、副産物である乳酸、ギ酸、酢酸及びエタノールの生成をある程度減少させる効果を見せたとしても、これを完全に除去することはできなかった。また一部の組換え変異菌株では菌株の生長速度が非常に低下して、全体的なコハク酸の生産性増加が、できない場合もあった。したがって、高いコハク酸生産性を持ちながら、同時に副産物の生産も遮断できる、新しいコハク酸生産菌株システムの開発が要求されている(Hong et al., Biotechnol. Lett., 22:871, 2000)。
【0009】
このような要求に応える新しいコハク酸生産菌株を開発するためには優秀なコハク酸生産能力を有する菌株の同定及び菌株の遺伝情報の確保、菌株の代謝特性の把握及び遺伝子組み換え菌株製作に必要な遺伝子操作技術の確保が先行されるべきである。現在まで優秀なコハク酸生産能力を表すルーメンバクテリアの場合、Mannheimia succiniciproducens MBEL55E菌株だけが遺伝情報が確保されているが、ActinobacillusとAnaerobiospirillumなどのルーメンバクテリアは完全な遺伝情報が確保できない実情である。A.succinogenesとA.succiniciproducensのホスホエノールピルビン酸カルボキシキナーぜ(pckA)遺伝子を大腸菌に増幅させることによるコハク酸生産の試みが報告されているが(Kim et al., Appl. Environ. Microbiol., 70:1238, 2004; Laivenieks et al., Appl. Environ. Microbiol., 63:2273, 1997)、ゲノム配列を基にした遺伝子組み換えコハク酸生産菌株の開発は、試みられていない。
【0010】
本発明者らは韓牛からコハク酸を高効率に生産するM. succiniciproducens MBEL55E(KCTC 0769BP)菌株を同定して、前記菌株のゲノム配列及び代謝特性を把握して発表した(Hong et al., Nature Biotechnol., 22:1275, 2004)。また、本発明者らはルーメンバクテリアの一種であるM. succiniciproducens MBEL55Eから乳酸とギ酸の生成を抑制するため乳酸脱水素酵素遺伝子(ldhA)とピルビン酸ギ酸リアーゼをコードする遺伝子(pfl)を欠失した変異菌株であるM. succiniciproducensLPK(KCTC 10558BP)を製作し、酢酸の生成を阻害するため、前記変異菌株M. succiniciproducens LPKからホスホトランスアセチラーゼ遺伝子(pta)と酢酸キナーゼ遺伝子(ackA)を欠失させた変異菌株であるM. succiniciproducens LPK7(KCTC 10626BP)を製作して、これらを嫌気条件で培養してコハク酸を製造した(WO05/052135A1;Lee et al., Appl. Environ. Microbiol.72:1939, 2006)。しかし、前記菌株の場合、副産物であるギ酸と酢酸の生成をある程度抑制することができたが、醗酵時副産物として多量のピルビン酸の蓄積をもたらし、なによりも野生菌株と比較して菌株の生長速度が顕著に低くなって全体的に非常に優れたコハク酸生産性の向上を得ることができなかった。
【0011】
一方、最近ピルビン酸ギ酸リアーゼをコードする遺伝子(pfl)がピルビン酸からアセチル補助酵素A(acetyl−CoA)への変換過程に関与することにより細胞生長とピルビン酸の再分配に大きい影響を及ぼすことが発表された(Wolfe, Microbial. Mol. Biol. Rev., 69:12, 2005)。
【0012】
そこで、本発明者らは、微生物の生長速度の低下を最小化して、且つピルビン酸を含む多様な副産物らの生成を完全に遮断して高収率で純粋なコハク酸が製造できる変異微生物及び醗酵方法を開発するため、鋭意努力した結果、ルーメンバクテリアの一種であるM. succiniciproducens MBEL55Eからピルビン酸ギ酸リアーゼ遺伝子(pfl)は欠失せず、乳酸脱水素酵素を遺伝子(ldhA)、ホスホトランスアセチラーゼ遺伝子(pta)及び酢酸キナーゼ遺伝子(ackA)を欠失させた変異菌株M. succiniciproducens PALK(KCTC10973BP)を製作した後、前記菌株を嫌気条件でブドウ糖またはグリセロールを炭素原として使用して醗酵させた結果、高収率と高生産性で純粋なコハク酸が製造できることを確認して、本発明を完成することに至った。
【発明の詳細な説明】
【0013】
《技術的課題》
従って、本発明の目的は、生長速度及びコハク酸の生産性が優れて、嫌気条件で培養時、他の有機酸は生成せず、コハク酸のみを高濃度で生産する変異微生物を提供することにある。
【0014】
本発明の他の目的は、前記変異微生物を嫌気条件下でブドウ糖またはグリセロールを炭素原として使用して他の有機酸は生成せず、純粋なコハク酸を製造する方法を提供することにある。
【技術的解決方法】
【0015】
前記目的を達成するために、本発明は、コハク酸を生成する微生物において、ピルビン酸ギ酸リアーゼをコードする遺伝子(pfl)を含み、乳酸脱水素酵素をコードする遺伝子(ldhA)、ホスホトランスアセチラーゼをコードする遺伝子(pta)及び酢酸キナーゼをコードする遺伝子(ackA)が欠失しており、嫌気条件でコハク酸以外の有機酸はほとんど生成せず、コハク酸のみを高濃度で生成する特性を有するルーメンバクテリア変異菌株を提供する。
【0016】
また、本発明は、Mannheimia succiniciproducensにおいて、ピルビン酸ギ酸リアーゼをコードする遺伝子(pfl)を含み、乳酸脱水素酵素をコードする遺伝子(ldhA)、ホスホトランスアセチラーゼをコードする遺伝子(pta)及び酢酸キナーゼをコードする遺伝子(ackA)が欠失しており、嫌気条件でコハク酸以外の有機酸はほとんど生成せず、コハク酸のみを高濃度で生成する特性を有するルーメンバクテリアMannheimia succiniciproducens PALK(KCTC10973BP)を提供する。
【0017】
また、本発明は、(a)コハク酸の生成能を有するルーメンバクテリアのゲノムから相同組換え方法で乳酸脱水素酵素をコードする遺伝子(ldhA)を欠失させ、乳酸脱水素酵素をコードする遺伝子(ldhA)が欠失したルーメンバクテリア変異菌株を収得するステップ、及び(b)前記乳酸脱水素酵素をコードする遺伝子(ldhA)が欠失したルーメンバクテリア変異菌株のゲノムから相同組換え方法でホスホトランスアセチラーゼをコードする遺伝子(pta)及び酢酸キナーゼをコードする遺伝子(ackA)を欠失させ、乳酸脱水素酵素をコードする遺伝子(ldhA)、ホスホトランスアセチラーゼをコードする遺伝子(pta)及び酢酸キナーゼ遺伝子(ackA)が欠失したルーメンバクテリア変異菌株を収得するステップを含む変異微生物の製造方法を提供する。
【0018】
また、本発明は、前記ルーメンバクテリア変異菌株を嫌気条件で培養するステップ、及び前記培養液からコハク酸を回収するステップを含むコハク酸の製造方法を提供する。
【0019】
本発明の他の特徴及び実施態様は、下記の詳細な説明及び特許請求の範囲からなお一層明らかになる。
【図面の簡単な説明】
【0020】
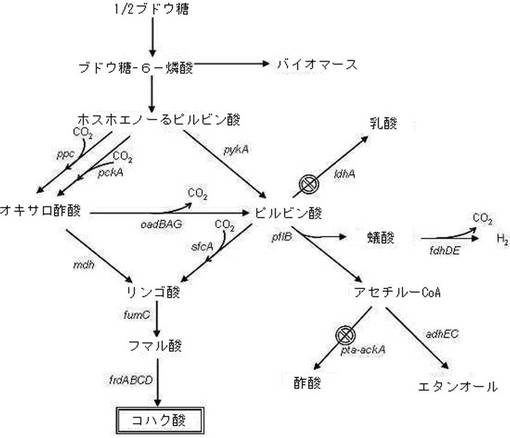
【図1】本発明による変異微生物からコハク酸が合成される経路を示す図である。
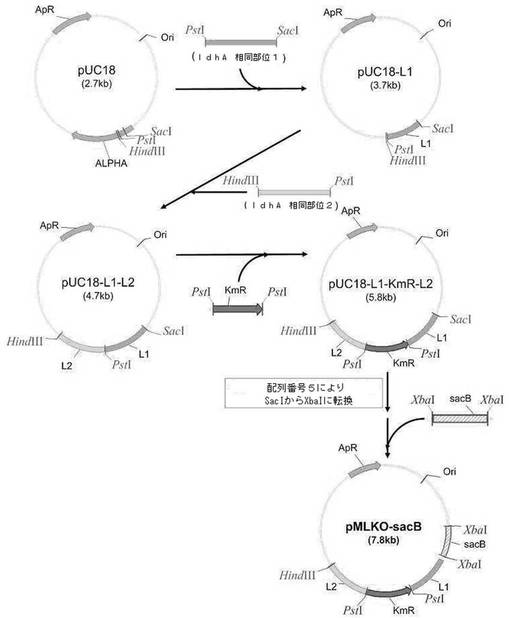
【図2】相同組換えによるldhA欠失用置換ベクター(pMLKO−sacB)の作製過程を示す図である。
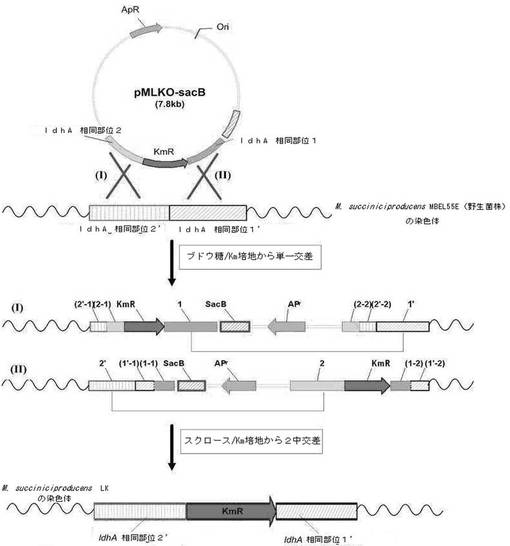
【図3】相同組換え方法でMannheimia succiniciproducens MBEL55EからldhAを欠失させた変異菌株(M. succiniciproducensLK)を作製する過程を示す図である。
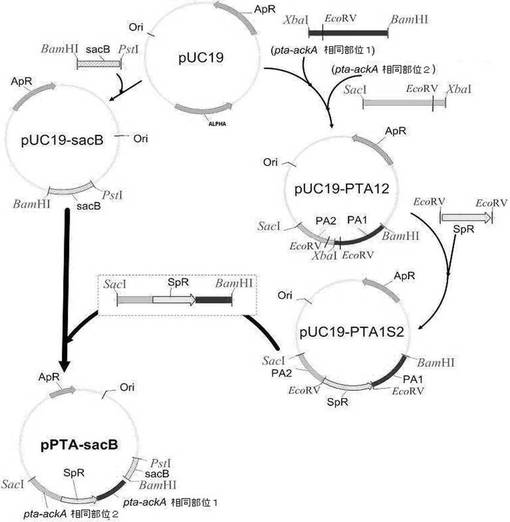
【図4】相同組換えによるpta及びackAの欠失用ベクター(pPTA−sacB)の作製過程を示す図である。
【図5】M. succiniciproducens LKからldhA及びpta―ackAを欠失させた変異菌株(M. succiniciproducens PALK)を作製する過程を示す図である。
【図6】M. succiniciproducens PALKからpta―ackAの欠失を確認した電気泳動写真である[レーン1〜6:PAU1プライマ−及びSP1プライマーを用いたPCR断片(1.1kb);レーンA〜F:SP2プライマー及びPAD2プライマーを用いたPCR断片(1.5kb)]。
【図7】CO2で飽和された嫌気条件でMannheimiasp. PALKの培養特性を示すグラフである。
【発明を実施するための最良の形態】
【0021】
本発明は一つの観点において、コハク酸を生成する微生物において、ピルビン酸ギ酸リアーゼをコードする遺伝子(pfl)を含み、乳酸脱水素酵素をコードする遺伝子(ldhA)、ホスホトランスアセチラーゼをコードする遺伝子(pta)及び酢酸キナーゼをコードする遺伝子(以下、ackA)が欠失しており、嫌気条件でコハク酸以外の有機酸はほとんど生成せず、コハク酸のみを高濃度で生成する特性を有するルーメンバクテリア変異菌株に関する。
【0022】
本発明において、コハク酸生成微生物はエタノール、又は他の有機酸に比べ、コハク酸を過量に生産する微生物で、醗酵によるコハク酸合成に産業的に使用できる微生物を意味する。
【0023】
代表的コハク酸生成微生物としてはルーメンバクテリアをあげることができ、ルーメンバクテリアの一種であるActinobacillus succinogenesとAnaerobiospirillum succiniciproducens及び現在まで知られているコハク酸を生産する多様なルーメンバクテリアの部分的な遺伝情報(16srRNA)、酵素分析及び醗酵結果を見ると、ルーメンバクテリアのコハク酸生産のための炭素原の主な生合成経路がルーメンバクテリアの一種であるマンヘミア属のコハク酸生産の生合成経路と殆ど同一であることが分かる(表1;Van der Werf et al., Arch Microbiol., 167:332, 1997; Laivenieks et al., Appl. Environ. Microbiol., 63:2273, 1997; Samuelov et al., App. Environ. Microbiol., 65:2260, 1999; Kim et al., Appl. Environ. Microbiol., 70: 1238, 2004)。特に、コハク酸生成に関する全てのルーメンバクテリアはコハク酸生成の時、二酸化炭素固定化酵素を利用してC3化合物であるホスホエノ−ルピルビン酸とピルビン酸をC4化合物であるオキサロ酢酸とリンゴ酸に転換した後、最終的にコハク酸を生産する。また、ルーメンバクテリアと嫌気条件下で醗酵副産物として酢酸、ギ酸及び乳酸を生成する。従って、マンヘミア属を含む全てのルーメンバクテリアが同一なコハク酸生合成経路を持つといえる。
【0024】
【表1】
【0025】
本発明ではコハク酸生産微生物であるルーメンバクテリアのゲノムを変形させ、菌株の成長速度が高く、且つ他の有機酸を生成せずに、高濃度でコハク酸を生成するコハク酸生成変異微生物を製作した。即ち、Mannheimia succiniciproducens MBEL55E(KCTC 0769BP)のゲノムDNAから乳酸脱水素酵素をコードする遺伝子(ldhA)、ホスホトランスアセチラーゼ遺伝子(pta)及び酢酸キナーゼ遺伝子(ackA)を欠失させ,菌株の生長速度が高く、他の有機酸を生成せずに高濃度でコハク酸を生成する変異菌株であるMannheimia succiniciproducens PALK(KCTC10973BP)を製作した。
【0026】
本発明には前記各遺伝子の欠失は相同組換えを利用して遺伝子を置換して不活性化させる方法を使用したが、該当遺伝子によってコードされる酵素が生成されないように該当遺伝子を修飾または削除することができる遺伝子操作方法であれば、制限なく使用できる。
【0027】
本発明において、前記ルーメンバクテリアはマンヘミア(Mannheimia)属、アクチノバチルス(Actinobacillus)属及びアネロビオスピリルム(Anaerobiospirillum)属からなる群より選択されることを特徴とすることができる。
【0028】
本発明において、前記変異微生物は副産物として他の有機酸は生成せず、コハク酸のみを生成する同型発酵菌株であることを特徴とすることができ、前記他の有機酸の生成量がコハク酸の1重量%以下であることを特徴とすることができ、前記他の有機酸は乳酸、酢酸、ギ酸及びピルビン酸からなる群より選択される一種以上であることを特徴とすることができる。
【0029】
本発明は他の観点において、Mannheimia succiniciproducensでピルビン酸ギ酸リアーゼをコードする遺伝子(pfl)を含み、乳酸脱水素酵素をコードする遺伝子(ldhA)、ホスホトランスアセチラーゼをコードする遺伝子(pta)及び酢酸キナーゼをコードする遺伝子(ackA)が欠失しており、嫌気条件でコハク酸以外の有機酸はほとんど生成せず、コハク酸のみを高濃度で生成する特性を有するルーメンバクテリアMannheimia succiniciproducens PALK(KCTC10973BP)に関する。
【0030】
本発明において、Mannheimia succiniciproducens PALK(KCTC10973BP)のコハク酸生成能と成長速度を既存の変異菌株と比較するため使用した変異菌株であるM. succiniciproducens LPK7(KCTC10626BP)はマンヘミア属菌株のゲノムから乳酸脱水素酵素をコードする遺伝子(ldhA)、ピルビン酸ギ酸リアーゼをコードする遺伝子(pfl)、ホスホトランスアセチラーゼをコードする遺伝子(pta)及び酢酸キナーゼをコードする遺伝子(ackA)が欠失されているした変異微生物で、本発明による変異微生物であるM. succiniciproducens PALK(KCTC10973BP)からピルビン酸ギ酸リアーゼをコードする遺伝子(pfl)が更に欠失した変異菌株である(WO2005/052135; Lee et al., Appl. Environ. Microbiol. 72:1939, 2006)。
【0031】
本発明によるコハク酸生成変異微生物M. succiniciproducens PALK(KCTC10973BP)はコハク酸の生成に用いられる野生形のMannheimia succiniciproducens MBEL55E(KCTC0769BP)及び既に確立された変異微生物であるM. succiniciproducens LPK7(KCTC10626BP)と比較してコハク酸生産能が優れており、乳酸、ピルビン酸、酢酸、ギ酸などの副産物を殆ど生産しないから、コハク酸生産菌株として優れた特徴をもち、既に確立された変異微生物であるM. succiniciproducens LPK7(KCTC10626BP)と比較して、高い菌株成長速度とコハク酸生産性及びピルビン酸の蓄積を抑えて純粋なコハク酸のみを生産して、高い収率でコハク酸が生産できる。
【0032】
本発明はまた他の観点において、(a)コハク酸の生成能を有するルーメンバクテリアのゲノムから相同組換え方法で乳酸脱水素酵素をコードする遺伝子(ldhA)を欠失させ、乳酸脱水素酵素をコードする遺伝子(ldhA)が欠失したルーメンバクテリア変異菌株を収得するステップ、及び(b)前記乳酸脱水素酵素をコードする遺伝子(ldhA)が欠失したルーメンバクテリア変異菌株のゲノムから相同組換え方法でホスホトランスアセチラーゼをコードする遺伝子(pta)及び酢酸キナーゼをコードする遺伝子(ackA)を欠失させ、乳酸脱水素酵素をコードする遺伝子(ldhA)、ホスホトランスアセチラーゼをコードする遺伝子(pta)及び酢酸キナーゼをコードする遺伝子(ackA)が欠失したルーメンバクテリア変異菌株を収得するステップを含む変異微生物の製造方法に関する。
【0033】
本発明において、前記(a)ステップの相同組換えは、欠失されたldhAを含む遺伝子交換ベクターを使用して行われることを特徴とすることができるし、前記(b)ステップの相同組換えは、欠失されたpta−ackAを含む遺伝子交換ベクターを使用して行われることを特徴とすることができる。
【0034】
本発明において、欠失されたldhAを含む遺伝子交換ベクターはpMLKO−sacBであることを特徴とすることができ、欠失されたpta−ackAを含む遺伝子交換ベクターはpMPKO−sacBであることを特徴とすることができる。
【0035】
また、本発明は前記変異微生物を嫌気条件で培養するステップ、及び前記培養液からコハク酸を回収するステップを含むコハク酸の製造方法を提供する。
【0036】
本発明において、前記培養はブドウ糖またはグリセロールを炭素原として使用することを特徴とすることができ、副産物としての他の有機酸の生成量はコハク酸の1重量%以下であることを特徴とすることができる。
【0037】
本発明はまた他の観点において、前記ルーメンバクテリア変異菌朱を嫌気条件で培養するステップ、及び前記培養液からコハク酸を回収するステップを含むコハク酸の製造方法に関する。
【0038】
本発明によるコハク酸生成変異微生物の培養及びコハク酸の収得過程は従来醗酵工程で通常的に知られている培養方法及びコハク酸分離精製方法を用いて行われる。
【0039】
本発明において、前記培養はブドウ糖またはグリセロールを炭素原として使用することを特徴とすることができるし、副産物としての他の有機酸の生成量はコハク酸の1重量%以下であることを特徴とすることができる。
【実施例】
【0040】
以下、実施例を挙げて本発明をより詳細に説明する。但し、これらの実施例は単に本発明を一層詳しく説明するためのものであり、本発明の範囲がこれらの実施例に限定されるものではないということは、当業界における通常の知識を有する者にとって明らかである。
【0041】
特に、下記実施例では、本発明による遺伝子を除去するために宿主細胞としてコハク酸生成微生物であるマンヘミア属微生物だけを例示したが、他の種類のコハク酸生成微生物を使用しても純粋コハク酸生成変異微生物が収得できることは、当業界における通常の知識を持つ者にとっては明らかであろう。
【0042】
《実施例1:ldhA欠失ベクター(pMLKO−sacB)の作製》
コハク酸生成微生物のゲノムからldhAを相同組換え方法で除去するために、遺伝子交換ベクターを次のように作製した。まず、M. succiniciproducens MBEL55E(KCTC 0769BP)のゲノムDNAを鋳型とし、下記配列番号1及び2のプライマーを用いてPCRを行った後、得られたldhA相同部位1(L1)を含むPCR断片をSacI及びPstIで切断し、これをpUC18(New England Biolabs, Inc.,USA)に導入してpUC18−L1を作製した。
配列番号1:5’−CAGTGAAGGAGCTCCGTAACGCATCCGCCG
配列番号2:5’−CTTTATCGAATCTGCAGGCGGTTTCCAAAA
【0043】
その後、M. succiniciproducens MBEL55E(KCTC 0769BP)のゲノムDNAを鋳型とし、下記配列番号3及び4のプライマーを用いてPCRを行った後、得られたldhA相同部位2(L2)を含むPCR断片をPstI及びHindIIIで切断し、これを前記pUC18−L1に導入してpUC18−L1−L2を作製した。
配列番号3:5’−GTACTGTAAACTGCAGCTTTCATAGTTAGC
配列番号4:5’−GCCGAAAGTCAAGCTTGCCGTCGTTTAGTG
【0044】
前記pUC18−L1−L2に選択マーカーとしてカナマイシン耐性遺伝子(KmR)を挿入するため、pUC4K(Pharmacia,Freiburg,Germany)をPstIで切断して、得られたカナマイシン耐性遺伝子を、PstIで切断したpUC18−L1−L2と融合させてpUC18−L1−KmR−L2を作製した。SacIで切断したpUC18−L1−KmR−L2に下記配列番号5のリンカーを挿入して、新たなXbaI切断部位を作製した。
配列番号5:5’−TCTAGAAGCT
【0045】
前記XbaI切断部位が挿入されたpUC18−L1−KmR−L2ベクターにsacB遺伝子を挿入するため、pKmobsacB(Schafer et al.,Gene,145:69,1994)を鋳型とし、下記配列番号6及び7のプライマーを用いてPCRを行った後、得られたsacB遺伝子を含むPCR断片をXbaIで切断し、これを前記XbaI制限酵素サイトが挿入されたpUC18−L1−KmR−L2ベクターの新たなXbaI制限酵素サイトに挿入してldhA遺伝子交換ベクターpMLKO−sacBを作製した(図2)。
配列番号6:5’−GCTCTAGACCTTCTATCGCCTTCTTGACG
配列番号7:5’−GCTCTAGAGGCTACAAAATCACGGGCGTC
【0046】
《実施例2:M. succiniciproducens LK菌株の作製》
実施例1から製作されたldhA遺伝子除去用交換ベクターpMLKO−sacBを利用してMannheimia succiniciproducens MBEL55E(KCTC 0769BP)のゲノムからldhAを欠失させて変異菌株を作製した(図3)。
【0047】
即ち、Mannheimia succiniciproducens MBEL55E(KCTC 0769BP)を10g/Lのグルコースを含有するLB−グルコース培地に塗抹して、37℃で36時間培養した後、コロニーをLB−グルコース液体培地10mLに接種して12時間培養した。培養が終わった細菌培養液をLB−グルコース液体培地100mLに1%接種して、200rpm、37℃の振盪培養器で培養した。
【0048】
約4〜5時間後に細菌培養液のOD600が約0.3〜0.4程度になり、前記細菌培養液を4℃、20分間、4500rpmの条件で遠心分離して細胞を得た後、4℃、10%のグリセロール溶液200mLで細胞を再懸濁した。懸濁された細胞溶液を4℃、5500rpmで20分間、再び遠心分離して細胞を得た。再懸濁時、使用されるグリセロール溶液を半分に減らして前記過程を2回繰り返した後、細胞とグリセロールとの体積比が1:1になるように再懸濁して細胞濃縮液を得た。
【0049】
このようにして得られた細胞濃縮液と、実施例1で作製された遺伝子交換ベクターpMLKO−sacBを混合した後、1.8kV、25μF,200ohmsの条件でエレクトロポレーション方法でpMLKO−sacBをMannheimia succiniciproducens MBEL55E(KCTC 0769BP)に導入した。pMLKO−sacBが導入された細菌にLB−グルコース液体培地1mLを加えて、200rpm、37℃の振盪培養器で1時間前培養した後、培養液を抗生剤カナマイシン(最終濃度25μg/mL)を含むLB−グルコース固体培地に塗抹して、37℃で48時間以上培養した。二重交差(double crossover)が起こされたコロニーのみを選び出すために、形成されたコロニーをカナマイシン(25μg/mL)と100g/Lのスクロースを含有したLB−スクロース培地にストリークした後、24時間培養した後、形成されたコロニーを同じ固体培地に再びストリークした。
【0050】
前記培地で形成されたコロニー(変異株)を、抗生剤が含まれたLB−グルコース液体培地で培養し、ロッシェルなどの方法(Rochelle et al.,FEMS Microbiol.Lett.,100:59,1992)を応用して、培養菌株のゲノムDNAを分離した。前記分離された変異株のゲノムDNAを鋳型としてPCRを行った後、得られたPCR産物を電気泳動することによってゲノムDNAのldhA遺伝子の欠失の可否を確認した。
【0051】
ldhAの欠失の可否は下記のように、PCRを2回行っテ確認した。まず、前記変異株のゲノムDNAを鋳型とし、下記配列番号8及び9のプライマーを用いてPCRを行った。
配列番号8:5’−GACGTTTCCCGTTGAATATGGC(KM1)
配列番号9:5’−CATTGAGGCGTATTATCAGGAAAC(LU1)
【0052】
次いで、前記変異株のゲノムDNAを鋳型とし、下記配列番号10及び11のプライマーを用いてPCRを行った。
配列番号10:5’−GCAGTTTCATTTGATGCTCGATG(KM2)
配列番号11:5’−CCTCTTACGATGACGCATCTTTCC(LD2)
【0053】
前記2回のPCR反応で得られたPCR断片を電気泳動して、PCR断片のサイズを確認してldhAの欠失の可否を確認した。ldhAの欠失されたゲノムDNAのPCR断片は配列番号8(KM1)及び9(LU1)を用いたPCR産物が1.5kbで得られ、同時に配列番号10(KM2)及び11(LD2)を用いたPCR産物が1.7kbに得られることで確認できた。前記各プライマーの位置は図3に表した。
【0054】
前記のような方法でM. succiniciproducens MBEL55EのゲノムからldhAを欠失させた変異菌株M. succiniciproducens LKを製作した。
【0055】
《実施例3:pta及びackA欠失ベクター(pPTA−sacB)の作製》
M. succiniciproducens LK菌株のゲノムからpta及びackAを相同組換え方法で欠失させるために、遺伝子交換ベクターを次のように作製した。sacB遺伝子を含有するpKmobsacBを鋳型とし、下記配列番号12及び13プライマーを用いてPCRを行った後、得られたsacB産物をPstI及びBamHIで切断し、これをpUC19(Stratagene Cloning Systems.USA)に導入してpUC19−sacBを作製した。
配列番号12:5’−AGCGGATCCCCTTCTATCGCCTTCTTGACG
配列番号13:5’−GTCCTGCAGGGCTACAAAATCACGGGCGTC
【0056】
一方、M. succiniciproducens LKのゲノムDNAを鋳型とし、下記配列番号14及び15のプライマーを用いてPCRを行った後、得られたpta−ackA相同部位1を含むPCR断片をXbaI及びBamHIで切断し、下記配列番号16及び17のプライマーを用いてPCRを行って得られたpta−ackA相同部位2を含むPCR断片をXbaI及びSacIで切断した後、この断片らをpUC19のBamHIとSacI部位に導入してpUC19−PTA12を作製した。
配列番号14:5’−GCTCTAGATATCCGCAGTATCACTTTCTGCGC
配列番号15:5’−TCCGCAGTCGGATCCGGGTTAACCGCACAG
配列番号16:5’−GGGGAGCTCGCTAACTTAGCTTCTAAAGGCCATGTTTCC
配列番号17:5’−GCTCTAGATATCCGGGTCAATATCGCCGCAAC
【0057】
前記pUC19−PTA12の選択マーカーとしてスペクチノマイシン耐性遺伝子を挿入するため、スペクチノマイシン耐性遺伝子(GenBankX02588;SpR)を含有するプラスミドpIC156(Steinmetz et al.,Gene,142:79,1994)を鋳型とし、下記配列番号18及び19のプライマーを用いてPCRを行った後、得られたSpR遺伝子を含むPCR断片をEcoRVで切断し、これを前記製作されたpUC19−PTA12に導入してスペクチノマイシン耐性遺伝子を有するpUC19−PTA1S2を作製した。前記作製されたpUC19−PTA1S2をSacI及びBamHIで切断した後、前、製作したpUC19−SacBに導入してpta及びackA遺伝子除去用交換ベクターpPTA−sacBベクターを作製した(図4)。
配列番号18:5’−GAATTCGAGCTCGCCCGGGGATCGATCCTC
配列番号19:5’−CCCGGGCCGACAGGCTTTGAAGCATGCAAATGTCAC
【0058】
《実施例4:M. succiniciproducens PALK菌株の作製》
実施例3で製作されたホスホトランスアセチラーゼ遺伝子(pta)及び酢酸キナーゼ遺伝子(ackA)除去用交換ベクターpPTA−sacBを利用してM. succiniciproducens LKのゲノムからpta及びackA遺伝子を欠失させて変異菌株を製作した(図4)。
【0059】
即ち、実施例2で製作されたM. succiniciproducens LKを、10g/Lのグルコースを含有したLB−グルコース培地に塗抹して、37℃で36時間培養した後、コロニーをLB−グルコース液体培地10mLに接種して12時間培養した。充分に成長した培養液をLB−グルコース液体培地100mLに1%接種して、37℃の振盪培養器で培養した。
【0060】
前記培養液から、実施例2と同じ方法で細胞濃縮液を得た後、これを、実施例3で作製された遺伝子交換ベクターpPTA−sacBと混合した後、2.5kV、50μF,200ohmsの条件でエレクトロポレーションを行って、pPTA−sacBをM. succiniciproducens LKに導入させた。pPTA−sacBが導入された細菌にLB−グルコース液体培地800mLを加えて、37℃の恒温槽で1時間30分間、前培養した後、二重交差(double crossover)を誘導するため、培養液を、スペクチノマイシン(最終濃度50μg/mL)を含有したTSB−スクロース固体培地[100g/Lのスクロースを含有したTryptic Soy Broth(Becton,Dickinson and Company)固体培地]に塗抹して、37℃で48時間以上培養した。コロニーが形成され、二重交差のみが起こったものを選び出すために、各コロニーを、スペクチノマイシン50μg/mLを含有したTSB−スクロース培地とアンピシリン50μg/mLを含有したTSB培地にストリークした。37℃で12時間程度培養した後、スペクチノマイシン50μg/mLを含有したTSB培地では生長するが、アンピシリン50μg/mLを含有したTSB培地には生長しないコロニーを選別してスペクチノマイシン50μg/mLを含有したTSB−スクロース培地にストリークした。ここでに形成されたコロニーをスペクチノマイシンとアンピシリンが合有されている培地で再び選別して、望ましい結果を表したコロニーを最終選択した。前記選択された菌株のゲノムを分離して鋳型として使用し、PCRを通じてpta−ackAの欠失の可否を確認した。
【0061】
pta−ackAの欠失の可否は、下記のようにPCRを2回行って確認した。まず、前記変異株のゲノムDNAを鋳型とし、下記配列番号20及び21のプライマーを用いてPCRを行った。次に、前記変異株のゲノムDNAを鋳型とし、下記配列番号22及び23のプライマーを用いてPCRを行った。
配列番号20:5’−CCTGCAGGCATGCAAGCTTGGGCTGCAGGTCGACTC(SP1)
配列番号21:5’−GCTGCCAAACAACCGAAAATACCGCAATAAACGGC(PAU1)
配列番号22:5’−GCATGTAACTTTACTGGATATAGCTAGAAAAGGCATCGGGGAG(SP2)
配列番号23:5’−GCAACGCGAGGGTCAATACCGAAGGATTTCGCCG(PAD2)
【0062】
前記2回のPCR反応で得られた産物をゲル電気泳動して、そのサイズによってpta−ackAの欠失の可否を確認した(図6)。pta−ackAの欠失は配列番号20及び21のプライマー(SP1とPAU1)を使用したPCR産物が1.1kbに得られ、同時に、配列番号22及び23のプライマー(SP2及びPAD2)を用いたPCRの産物が1.5kbに得られることを確認した。前記プライマーの位置は図5に示した。
【0063】
前記方法で作製されたM. succiniciproducens LKのゲノムからpta−ackAが欠失された菌株、即ち、M. succiniciproducensからldhA、pta及びackAが欠失された変異菌株をM. succiniciproducens PALKと命名して、2006年7月26日付に国際寄託機関であるKCTC(韓国生命工学研究院内生物資源センター;韓国デジョンユスングヲウンドン52)に寄託番号‘KCTC 10973BP’として寄託した。
【0064】
《実施例5:M. succiniciproducens PALK菌株を利用した純粋コハク酸の生産》
前記実施例4で作製されたM. succiniciproducens PALK(KCTC 10973BP)菌株を5g/Lのブドウ糖を含む10mLの合成培地で接種し、嫌気条件で39℃で8時間前培養した後、これを再び5g/Lのブドウ糖を含む250mLの合成培地に移して39℃で培養した。この場合、抗生剤として50μg/mLのスペクチノマイシンを添加した。醗酵は前記250mLのM. succiniciproducens PALK培養液を2.25Lの合成培地(1g/LのNaCl,2g/Lの(NH4)2HPO4,0.02g/LのCaCl2・2H2O,0.2g/LのMgCl26H2O,8.709g/LのK2HPO4,0.5g/Lのシステイン,0.5g/Lのメチオニン,0.5g/Lのアラニン,0.5g/Lのアスパラギン,0.5g/Lのアスパラギン酸,0.5g/Lのプロリン,0.5g/Lのセリン,0.005g/Lのニコチン酸,0.005g/LのCa−パントセネート(pantothenate),0.005g/LのピリドキシンHCl(pyridoxine・HCl),0.005g/Lのチアミン,0.005g/Lのアスコルビン酸及び0.005g/Lのビオチン)を合有する微生物反応器に接種して行って、醗酵条件は18.2g/L(100mM)の初期ブドウ糖の濃度9.2g/L(100mM)の初期グリセロール濃度、温度39℃で醗酵した。醗酵中pHはアンモニア水を使用して6.5に調整し、抗生剤として使用したスペクチノマイシンは50μg/mLの濃度で添加した。高濃度のコハク酸生産のためにブドウ糖を完全に使い果たした場合、濃縮されたブドウ糖溶液を添加して培養液のブドウ糖の濃度が約18.2g/L(100mM)に成るように調整した。
【0065】
前記と同じ方法でM. succiniciproducens MBEL55E(KCTC 0769BP)及びM. succiniciproducens LPK7(KCTC 10626BP)を培養してコハク酸を醗酵させた。
培養液内の細胞濃度を分光光度計を用いて測定して、既に測定した分光光度計の吸光度と乾燥細胞重量の検量線を利用して細胞濃度を計算した。醗酵過程中の生物反応器から周期的にサンプルを採取して、採取された試料は13、000rpmで10分間遠心分離した後、上層液の各種代謝産物及びコハク酸濃度、ブドウ糖とグリセロール濃度を液体クロマトグラフ(HPLC)で分析した。
【0066】
その結果、図7及び表2に表すように、本発明のM. succiniciproducens PALK(KCTC 10973BP)はM. succiniciproducens MBEL55E(KCTC 0769BP)及びM. succiniciproducens LPK7(KCTC 10626B)と比較すると、より優れたコハク酸生産性を表すだけではなく、副産物としてほとんど有機酸及びエタノールが生成されないことがわかる。0.45g/Lの酢酸と0.24g/Lのピルビン酸が検出されたけれども、これは菌株の生長のため生成される最小量で生産されたコハク酸に比べ、1重量%以下の極微な量で無視することができるのは当業者に自明な事実である。
【0067】
【表2】
【産業上の利用可能性】
【0068】
以上詳細に説明したように、本発明は生長速度が速く、且つ嫌気的条件で培養する場合、コハク酸を除き、他の有機酸を殆ど生成せずにコハク酸だけを高濃度で生産するコハク酸生成変異微生物及び前記変異微生物を利用したコハク酸の製造方法を提供する効果がある。本発明による変異微生物は既存のコハク酸生成微生物より菌株の生長速度及びコハク酸生産性が高く、他の有機酸を生成せずに高濃度でコハク酸を生成するから、コハク酸の産業的生産菌株として有用である。
【0069】
以上、本発明の内容の特定の部分を詳述したが、当業界における通常の知識を有する者にとって、このような具体的な記述は単なる好適な実施様態に過ぎず、これにより本発明の範囲が制限されることはないという点は明らかであろう。よって、本発明の実質的な範囲は特許請求の範囲とこれらの等価物により定まると言えるであろう。
【技術分野】
【0001】
本発明は純粋コハク酸生成新規ルーメンバクテリア変異菌株及び前記ルーメンバクテリア変異菌株を用いた純粋なコハク酸の製造方法に関し、より詳しくは、コハク酸を生成する微生物でピルビン酸ギ酸リアーゼをコードする遺伝子(以下、「pfl」という)は欠失されず、乳酸脱水素酵素をコードする遺伝子(以下、「ldhA」という)、ホスホトランスアセチラーゼをコードする遺伝子(以下、「pta」という)及び酢酸キナーゼをコードする遺伝子(以下、「ackA」という)が欠失しており、嫌気条件でコハク酸以外の有機酸はほとんど生成せず、コハク酸のみを高濃度で生成する特性を有するルーメンバクテリア変異菌株及び前記ルーメンバクテリア変異菌株を用いたコハク酸の製造方法に関する。
【背景技術】
【0002】
コハク酸(HOOCCH2CH2COOH)は4個の炭素からなるジカルボン酸で、医薬用、食品用、化粧品及び他の産業で使用される化学製品の前具体として広く使用される産業的、経済的な利用価値が高い有機酸である(Zeikus et al., Appl. Microbiol. Biotechnol., 51:545, 1999; Song et al., Enzyme Microbial Technol., 39:352, 2006)。特に最近急激な原油価格の上昇と伴に生分解性高分子の主な原料物質として、のコハク酸の使用可能性が証明され、急激な需要増大が予想されている(Willke et al., Appl. Microbiol. Biotechnol., 66:131, 2004)。
【0003】
コハク酸は化学合成と醗酵により生産できるが、現在まで産業的に使用される大部分のコハク酸はBASF,DuPont,BPケミカルなど巨大な化学会社と中国及び日本の化学会社によって原油から由来したn−ブタンとアセチレンを原料物質に使用して化学合成法によって生産されており、医薬品などの特殊な用途に使用される少量のコハク酸だけが古典的な微生物醗酵法により生産されている。このようなコハク酸の化学合成法は製造過程から多量の有害廃棄物、廃溶液及び廃ガス(一酸化炭素含み)を排出するという問題点を有しており、特に、資源枯渇の可能性の高い化石原料を其礎物質として使用しており、従って、これの代わりに使用できるコハク酸の生産方法の開発が緊急課題である。
【0004】
このようなコハク酸の化学合成工程による問題点を解決するため、微生物醗酵を通じて再生可能な原料からコハク酸を生産する研究が多くの研究者らによって集中的に行われていた。醗酵によって、コハク酸の生産に使用される微生物は非常に多様だが、大きく組換え大腸菌とルーメンバクテリア(ruminal bacteria: Actinobacillus, Anaerobiospirillum, Bacteroides, Mannheimia, Succinimonas, Succinivibrio等)に分けることができる(Song et al., Enzyme Microbial Technol., 39:352, 2006)。
【0005】
組換え大腸菌を利用したコハク酸生産に関する研究を調べてみると、先ずアメリカのシカゴ大学の研究チームが大腸菌の乳酸及びギ酸の生産に関与する遺伝子ら(ldh及びpfl)を除去すると同時にブドウ糖伝達システム遺伝子(ptsG)を操作した変異菌株であるAFP111(ATCC202021)を製作してコハク酸の生産増加を試みした(米国特許5,770,435号明細書)。
【0006】
本発明者らはコハク酸生産に関与するリンゴ酸酵素遺伝子(sfcA)をldh遺伝子及びpfl遺伝子が除去された組換え大腸菌であるNZN111菌株に増幅させ、NZN111菌株の醗酵過程から蓄積されるピルビン酸を抑制することによってコハク酸の生産を増大したことがあり(Hong et al., Biotechnol. Bioeng., 74:89, 2001)、アメリカのジョージア大学の研究チームはRhizobium etli菌株のピルビン酸カルボキシラーゼ遺伝子(pyc)を前記AFP111菌株に発現させることによりAFP111/pTrc99A−pyc菌株を製作し、これをコハク酸生産に利用した(Vemuri et al., J. Ind. Microbiol. Biotechnol., 28:325, 2001)。最近、好気条件でコハク酸生産を誘導するために、アメリカのライス大学研究チームが解糖過程、TCA経路及びグリオキシレート経路に関与する遺伝子を操作した組換え大腸菌株を製作して発表した(Lin et al., Metabol. Eng., 7:116, 2005; Lin et al., Biotechnol. Bioeng., 90:775, 2005)。
【0007】
コハク酸の生産においてはルーメンバクテリアの一種であるActinobacillus,Anaerobiospirillum及びMannheimia菌株が最も優れていることが知られており、これら菌株に対する研究が多く行われてきた。アメリカのMBI(Michgan Biotechnology Institute)研究チームはActinobacillus succiniciproducens130Z菌株(ATCC55681)を発見し、コハク酸生産方法を開発し(米国特許5,504,004号明細書)、そして古典的化学突然変異方法を用いてActinobacillus succiniciproducensの多様な変異菌株を開発して、これをコハク酸生産及び精製工程の開発に利用した(米国特許5,521,075号明細書;米国特許5,168,055号明細書;米国特許5,143,834号明細書)。
【0008】
しかし、現在まで開発された微生物の醗酵を通じたコハク酸生産方法は生産性が2g/L/h以下で非常に低く、特に、醗酵時にコハク酸と伴に副産物として各種有機酸とエタノールを同時に生産することにより、莫大な分離及び精製費用が要求されている。たとえ前記結果が一部組換え菌株において、副産物である乳酸、ギ酸、酢酸及びエタノールの生成をある程度減少させる効果を見せたとしても、これを完全に除去することはできなかった。また一部の組換え変異菌株では菌株の生長速度が非常に低下して、全体的なコハク酸の生産性増加が、できない場合もあった。したがって、高いコハク酸生産性を持ちながら、同時に副産物の生産も遮断できる、新しいコハク酸生産菌株システムの開発が要求されている(Hong et al., Biotechnol. Lett., 22:871, 2000)。
【0009】
このような要求に応える新しいコハク酸生産菌株を開発するためには優秀なコハク酸生産能力を有する菌株の同定及び菌株の遺伝情報の確保、菌株の代謝特性の把握及び遺伝子組み換え菌株製作に必要な遺伝子操作技術の確保が先行されるべきである。現在まで優秀なコハク酸生産能力を表すルーメンバクテリアの場合、Mannheimia succiniciproducens MBEL55E菌株だけが遺伝情報が確保されているが、ActinobacillusとAnaerobiospirillumなどのルーメンバクテリアは完全な遺伝情報が確保できない実情である。A.succinogenesとA.succiniciproducensのホスホエノールピルビン酸カルボキシキナーぜ(pckA)遺伝子を大腸菌に増幅させることによるコハク酸生産の試みが報告されているが(Kim et al., Appl. Environ. Microbiol., 70:1238, 2004; Laivenieks et al., Appl. Environ. Microbiol., 63:2273, 1997)、ゲノム配列を基にした遺伝子組み換えコハク酸生産菌株の開発は、試みられていない。
【0010】
本発明者らは韓牛からコハク酸を高効率に生産するM. succiniciproducens MBEL55E(KCTC 0769BP)菌株を同定して、前記菌株のゲノム配列及び代謝特性を把握して発表した(Hong et al., Nature Biotechnol., 22:1275, 2004)。また、本発明者らはルーメンバクテリアの一種であるM. succiniciproducens MBEL55Eから乳酸とギ酸の生成を抑制するため乳酸脱水素酵素遺伝子(ldhA)とピルビン酸ギ酸リアーゼをコードする遺伝子(pfl)を欠失した変異菌株であるM. succiniciproducensLPK(KCTC 10558BP)を製作し、酢酸の生成を阻害するため、前記変異菌株M. succiniciproducens LPKからホスホトランスアセチラーゼ遺伝子(pta)と酢酸キナーゼ遺伝子(ackA)を欠失させた変異菌株であるM. succiniciproducens LPK7(KCTC 10626BP)を製作して、これらを嫌気条件で培養してコハク酸を製造した(WO05/052135A1;Lee et al., Appl. Environ. Microbiol.72:1939, 2006)。しかし、前記菌株の場合、副産物であるギ酸と酢酸の生成をある程度抑制することができたが、醗酵時副産物として多量のピルビン酸の蓄積をもたらし、なによりも野生菌株と比較して菌株の生長速度が顕著に低くなって全体的に非常に優れたコハク酸生産性の向上を得ることができなかった。
【0011】
一方、最近ピルビン酸ギ酸リアーゼをコードする遺伝子(pfl)がピルビン酸からアセチル補助酵素A(acetyl−CoA)への変換過程に関与することにより細胞生長とピルビン酸の再分配に大きい影響を及ぼすことが発表された(Wolfe, Microbial. Mol. Biol. Rev., 69:12, 2005)。
【0012】
そこで、本発明者らは、微生物の生長速度の低下を最小化して、且つピルビン酸を含む多様な副産物らの生成を完全に遮断して高収率で純粋なコハク酸が製造できる変異微生物及び醗酵方法を開発するため、鋭意努力した結果、ルーメンバクテリアの一種であるM. succiniciproducens MBEL55Eからピルビン酸ギ酸リアーゼ遺伝子(pfl)は欠失せず、乳酸脱水素酵素を遺伝子(ldhA)、ホスホトランスアセチラーゼ遺伝子(pta)及び酢酸キナーゼ遺伝子(ackA)を欠失させた変異菌株M. succiniciproducens PALK(KCTC10973BP)を製作した後、前記菌株を嫌気条件でブドウ糖またはグリセロールを炭素原として使用して醗酵させた結果、高収率と高生産性で純粋なコハク酸が製造できることを確認して、本発明を完成することに至った。
【発明の詳細な説明】
【0013】
《技術的課題》
従って、本発明の目的は、生長速度及びコハク酸の生産性が優れて、嫌気条件で培養時、他の有機酸は生成せず、コハク酸のみを高濃度で生産する変異微生物を提供することにある。
【0014】
本発明の他の目的は、前記変異微生物を嫌気条件下でブドウ糖またはグリセロールを炭素原として使用して他の有機酸は生成せず、純粋なコハク酸を製造する方法を提供することにある。
【技術的解決方法】
【0015】
前記目的を達成するために、本発明は、コハク酸を生成する微生物において、ピルビン酸ギ酸リアーゼをコードする遺伝子(pfl)を含み、乳酸脱水素酵素をコードする遺伝子(ldhA)、ホスホトランスアセチラーゼをコードする遺伝子(pta)及び酢酸キナーゼをコードする遺伝子(ackA)が欠失しており、嫌気条件でコハク酸以外の有機酸はほとんど生成せず、コハク酸のみを高濃度で生成する特性を有するルーメンバクテリア変異菌株を提供する。
【0016】
また、本発明は、Mannheimia succiniciproducensにおいて、ピルビン酸ギ酸リアーゼをコードする遺伝子(pfl)を含み、乳酸脱水素酵素をコードする遺伝子(ldhA)、ホスホトランスアセチラーゼをコードする遺伝子(pta)及び酢酸キナーゼをコードする遺伝子(ackA)が欠失しており、嫌気条件でコハク酸以外の有機酸はほとんど生成せず、コハク酸のみを高濃度で生成する特性を有するルーメンバクテリアMannheimia succiniciproducens PALK(KCTC10973BP)を提供する。
【0017】
また、本発明は、(a)コハク酸の生成能を有するルーメンバクテリアのゲノムから相同組換え方法で乳酸脱水素酵素をコードする遺伝子(ldhA)を欠失させ、乳酸脱水素酵素をコードする遺伝子(ldhA)が欠失したルーメンバクテリア変異菌株を収得するステップ、及び(b)前記乳酸脱水素酵素をコードする遺伝子(ldhA)が欠失したルーメンバクテリア変異菌株のゲノムから相同組換え方法でホスホトランスアセチラーゼをコードする遺伝子(pta)及び酢酸キナーゼをコードする遺伝子(ackA)を欠失させ、乳酸脱水素酵素をコードする遺伝子(ldhA)、ホスホトランスアセチラーゼをコードする遺伝子(pta)及び酢酸キナーゼ遺伝子(ackA)が欠失したルーメンバクテリア変異菌株を収得するステップを含む変異微生物の製造方法を提供する。
【0018】
また、本発明は、前記ルーメンバクテリア変異菌株を嫌気条件で培養するステップ、及び前記培養液からコハク酸を回収するステップを含むコハク酸の製造方法を提供する。
【0019】
本発明の他の特徴及び実施態様は、下記の詳細な説明及び特許請求の範囲からなお一層明らかになる。
【図面の簡単な説明】
【0020】
【図1】本発明による変異微生物からコハク酸が合成される経路を示す図である。
【図2】相同組換えによるldhA欠失用置換ベクター(pMLKO−sacB)の作製過程を示す図である。
【図3】相同組換え方法でMannheimia succiniciproducens MBEL55EからldhAを欠失させた変異菌株(M. succiniciproducensLK)を作製する過程を示す図である。
【図4】相同組換えによるpta及びackAの欠失用ベクター(pPTA−sacB)の作製過程を示す図である。
【図5】M. succiniciproducens LKからldhA及びpta―ackAを欠失させた変異菌株(M. succiniciproducens PALK)を作製する過程を示す図である。
【図6】M. succiniciproducens PALKからpta―ackAの欠失を確認した電気泳動写真である[レーン1〜6:PAU1プライマ−及びSP1プライマーを用いたPCR断片(1.1kb);レーンA〜F:SP2プライマー及びPAD2プライマーを用いたPCR断片(1.5kb)]。
【図7】CO2で飽和された嫌気条件でMannheimiasp. PALKの培養特性を示すグラフである。
【発明を実施するための最良の形態】
【0021】
本発明は一つの観点において、コハク酸を生成する微生物において、ピルビン酸ギ酸リアーゼをコードする遺伝子(pfl)を含み、乳酸脱水素酵素をコードする遺伝子(ldhA)、ホスホトランスアセチラーゼをコードする遺伝子(pta)及び酢酸キナーゼをコードする遺伝子(以下、ackA)が欠失しており、嫌気条件でコハク酸以外の有機酸はほとんど生成せず、コハク酸のみを高濃度で生成する特性を有するルーメンバクテリア変異菌株に関する。
【0022】
本発明において、コハク酸生成微生物はエタノール、又は他の有機酸に比べ、コハク酸を過量に生産する微生物で、醗酵によるコハク酸合成に産業的に使用できる微生物を意味する。
【0023】
代表的コハク酸生成微生物としてはルーメンバクテリアをあげることができ、ルーメンバクテリアの一種であるActinobacillus succinogenesとAnaerobiospirillum succiniciproducens及び現在まで知られているコハク酸を生産する多様なルーメンバクテリアの部分的な遺伝情報(16srRNA)、酵素分析及び醗酵結果を見ると、ルーメンバクテリアのコハク酸生産のための炭素原の主な生合成経路がルーメンバクテリアの一種であるマンヘミア属のコハク酸生産の生合成経路と殆ど同一であることが分かる(表1;Van der Werf et al., Arch Microbiol., 167:332, 1997; Laivenieks et al., Appl. Environ. Microbiol., 63:2273, 1997; Samuelov et al., App. Environ. Microbiol., 65:2260, 1999; Kim et al., Appl. Environ. Microbiol., 70: 1238, 2004)。特に、コハク酸生成に関する全てのルーメンバクテリアはコハク酸生成の時、二酸化炭素固定化酵素を利用してC3化合物であるホスホエノ−ルピルビン酸とピルビン酸をC4化合物であるオキサロ酢酸とリンゴ酸に転換した後、最終的にコハク酸を生産する。また、ルーメンバクテリアと嫌気条件下で醗酵副産物として酢酸、ギ酸及び乳酸を生成する。従って、マンヘミア属を含む全てのルーメンバクテリアが同一なコハク酸生合成経路を持つといえる。
【0024】
【表1】
【0025】
本発明ではコハク酸生産微生物であるルーメンバクテリアのゲノムを変形させ、菌株の成長速度が高く、且つ他の有機酸を生成せずに、高濃度でコハク酸を生成するコハク酸生成変異微生物を製作した。即ち、Mannheimia succiniciproducens MBEL55E(KCTC 0769BP)のゲノムDNAから乳酸脱水素酵素をコードする遺伝子(ldhA)、ホスホトランスアセチラーゼ遺伝子(pta)及び酢酸キナーゼ遺伝子(ackA)を欠失させ,菌株の生長速度が高く、他の有機酸を生成せずに高濃度でコハク酸を生成する変異菌株であるMannheimia succiniciproducens PALK(KCTC10973BP)を製作した。
【0026】
本発明には前記各遺伝子の欠失は相同組換えを利用して遺伝子を置換して不活性化させる方法を使用したが、該当遺伝子によってコードされる酵素が生成されないように該当遺伝子を修飾または削除することができる遺伝子操作方法であれば、制限なく使用できる。
【0027】
本発明において、前記ルーメンバクテリアはマンヘミア(Mannheimia)属、アクチノバチルス(Actinobacillus)属及びアネロビオスピリルム(Anaerobiospirillum)属からなる群より選択されることを特徴とすることができる。
【0028】
本発明において、前記変異微生物は副産物として他の有機酸は生成せず、コハク酸のみを生成する同型発酵菌株であることを特徴とすることができ、前記他の有機酸の生成量がコハク酸の1重量%以下であることを特徴とすることができ、前記他の有機酸は乳酸、酢酸、ギ酸及びピルビン酸からなる群より選択される一種以上であることを特徴とすることができる。
【0029】
本発明は他の観点において、Mannheimia succiniciproducensでピルビン酸ギ酸リアーゼをコードする遺伝子(pfl)を含み、乳酸脱水素酵素をコードする遺伝子(ldhA)、ホスホトランスアセチラーゼをコードする遺伝子(pta)及び酢酸キナーゼをコードする遺伝子(ackA)が欠失しており、嫌気条件でコハク酸以外の有機酸はほとんど生成せず、コハク酸のみを高濃度で生成する特性を有するルーメンバクテリアMannheimia succiniciproducens PALK(KCTC10973BP)に関する。
【0030】
本発明において、Mannheimia succiniciproducens PALK(KCTC10973BP)のコハク酸生成能と成長速度を既存の変異菌株と比較するため使用した変異菌株であるM. succiniciproducens LPK7(KCTC10626BP)はマンヘミア属菌株のゲノムから乳酸脱水素酵素をコードする遺伝子(ldhA)、ピルビン酸ギ酸リアーゼをコードする遺伝子(pfl)、ホスホトランスアセチラーゼをコードする遺伝子(pta)及び酢酸キナーゼをコードする遺伝子(ackA)が欠失されているした変異微生物で、本発明による変異微生物であるM. succiniciproducens PALK(KCTC10973BP)からピルビン酸ギ酸リアーゼをコードする遺伝子(pfl)が更に欠失した変異菌株である(WO2005/052135; Lee et al., Appl. Environ. Microbiol. 72:1939, 2006)。
【0031】
本発明によるコハク酸生成変異微生物M. succiniciproducens PALK(KCTC10973BP)はコハク酸の生成に用いられる野生形のMannheimia succiniciproducens MBEL55E(KCTC0769BP)及び既に確立された変異微生物であるM. succiniciproducens LPK7(KCTC10626BP)と比較してコハク酸生産能が優れており、乳酸、ピルビン酸、酢酸、ギ酸などの副産物を殆ど生産しないから、コハク酸生産菌株として優れた特徴をもち、既に確立された変異微生物であるM. succiniciproducens LPK7(KCTC10626BP)と比較して、高い菌株成長速度とコハク酸生産性及びピルビン酸の蓄積を抑えて純粋なコハク酸のみを生産して、高い収率でコハク酸が生産できる。
【0032】
本発明はまた他の観点において、(a)コハク酸の生成能を有するルーメンバクテリアのゲノムから相同組換え方法で乳酸脱水素酵素をコードする遺伝子(ldhA)を欠失させ、乳酸脱水素酵素をコードする遺伝子(ldhA)が欠失したルーメンバクテリア変異菌株を収得するステップ、及び(b)前記乳酸脱水素酵素をコードする遺伝子(ldhA)が欠失したルーメンバクテリア変異菌株のゲノムから相同組換え方法でホスホトランスアセチラーゼをコードする遺伝子(pta)及び酢酸キナーゼをコードする遺伝子(ackA)を欠失させ、乳酸脱水素酵素をコードする遺伝子(ldhA)、ホスホトランスアセチラーゼをコードする遺伝子(pta)及び酢酸キナーゼをコードする遺伝子(ackA)が欠失したルーメンバクテリア変異菌株を収得するステップを含む変異微生物の製造方法に関する。
【0033】
本発明において、前記(a)ステップの相同組換えは、欠失されたldhAを含む遺伝子交換ベクターを使用して行われることを特徴とすることができるし、前記(b)ステップの相同組換えは、欠失されたpta−ackAを含む遺伝子交換ベクターを使用して行われることを特徴とすることができる。
【0034】
本発明において、欠失されたldhAを含む遺伝子交換ベクターはpMLKO−sacBであることを特徴とすることができ、欠失されたpta−ackAを含む遺伝子交換ベクターはpMPKO−sacBであることを特徴とすることができる。
【0035】
また、本発明は前記変異微生物を嫌気条件で培養するステップ、及び前記培養液からコハク酸を回収するステップを含むコハク酸の製造方法を提供する。
【0036】
本発明において、前記培養はブドウ糖またはグリセロールを炭素原として使用することを特徴とすることができ、副産物としての他の有機酸の生成量はコハク酸の1重量%以下であることを特徴とすることができる。
【0037】
本発明はまた他の観点において、前記ルーメンバクテリア変異菌朱を嫌気条件で培養するステップ、及び前記培養液からコハク酸を回収するステップを含むコハク酸の製造方法に関する。
【0038】
本発明によるコハク酸生成変異微生物の培養及びコハク酸の収得過程は従来醗酵工程で通常的に知られている培養方法及びコハク酸分離精製方法を用いて行われる。
【0039】
本発明において、前記培養はブドウ糖またはグリセロールを炭素原として使用することを特徴とすることができるし、副産物としての他の有機酸の生成量はコハク酸の1重量%以下であることを特徴とすることができる。
【実施例】
【0040】
以下、実施例を挙げて本発明をより詳細に説明する。但し、これらの実施例は単に本発明を一層詳しく説明するためのものであり、本発明の範囲がこれらの実施例に限定されるものではないということは、当業界における通常の知識を有する者にとって明らかである。
【0041】
特に、下記実施例では、本発明による遺伝子を除去するために宿主細胞としてコハク酸生成微生物であるマンヘミア属微生物だけを例示したが、他の種類のコハク酸生成微生物を使用しても純粋コハク酸生成変異微生物が収得できることは、当業界における通常の知識を持つ者にとっては明らかであろう。
【0042】
《実施例1:ldhA欠失ベクター(pMLKO−sacB)の作製》
コハク酸生成微生物のゲノムからldhAを相同組換え方法で除去するために、遺伝子交換ベクターを次のように作製した。まず、M. succiniciproducens MBEL55E(KCTC 0769BP)のゲノムDNAを鋳型とし、下記配列番号1及び2のプライマーを用いてPCRを行った後、得られたldhA相同部位1(L1)を含むPCR断片をSacI及びPstIで切断し、これをpUC18(New England Biolabs, Inc.,USA)に導入してpUC18−L1を作製した。
配列番号1:5’−CAGTGAAGGAGCTCCGTAACGCATCCGCCG
配列番号2:5’−CTTTATCGAATCTGCAGGCGGTTTCCAAAA
【0043】
その後、M. succiniciproducens MBEL55E(KCTC 0769BP)のゲノムDNAを鋳型とし、下記配列番号3及び4のプライマーを用いてPCRを行った後、得られたldhA相同部位2(L2)を含むPCR断片をPstI及びHindIIIで切断し、これを前記pUC18−L1に導入してpUC18−L1−L2を作製した。
配列番号3:5’−GTACTGTAAACTGCAGCTTTCATAGTTAGC
配列番号4:5’−GCCGAAAGTCAAGCTTGCCGTCGTTTAGTG
【0044】
前記pUC18−L1−L2に選択マーカーとしてカナマイシン耐性遺伝子(KmR)を挿入するため、pUC4K(Pharmacia,Freiburg,Germany)をPstIで切断して、得られたカナマイシン耐性遺伝子を、PstIで切断したpUC18−L1−L2と融合させてpUC18−L1−KmR−L2を作製した。SacIで切断したpUC18−L1−KmR−L2に下記配列番号5のリンカーを挿入して、新たなXbaI切断部位を作製した。
配列番号5:5’−TCTAGAAGCT
【0045】
前記XbaI切断部位が挿入されたpUC18−L1−KmR−L2ベクターにsacB遺伝子を挿入するため、pKmobsacB(Schafer et al.,Gene,145:69,1994)を鋳型とし、下記配列番号6及び7のプライマーを用いてPCRを行った後、得られたsacB遺伝子を含むPCR断片をXbaIで切断し、これを前記XbaI制限酵素サイトが挿入されたpUC18−L1−KmR−L2ベクターの新たなXbaI制限酵素サイトに挿入してldhA遺伝子交換ベクターpMLKO−sacBを作製した(図2)。
配列番号6:5’−GCTCTAGACCTTCTATCGCCTTCTTGACG
配列番号7:5’−GCTCTAGAGGCTACAAAATCACGGGCGTC
【0046】
《実施例2:M. succiniciproducens LK菌株の作製》
実施例1から製作されたldhA遺伝子除去用交換ベクターpMLKO−sacBを利用してMannheimia succiniciproducens MBEL55E(KCTC 0769BP)のゲノムからldhAを欠失させて変異菌株を作製した(図3)。
【0047】
即ち、Mannheimia succiniciproducens MBEL55E(KCTC 0769BP)を10g/Lのグルコースを含有するLB−グルコース培地に塗抹して、37℃で36時間培養した後、コロニーをLB−グルコース液体培地10mLに接種して12時間培養した。培養が終わった細菌培養液をLB−グルコース液体培地100mLに1%接種して、200rpm、37℃の振盪培養器で培養した。
【0048】
約4〜5時間後に細菌培養液のOD600が約0.3〜0.4程度になり、前記細菌培養液を4℃、20分間、4500rpmの条件で遠心分離して細胞を得た後、4℃、10%のグリセロール溶液200mLで細胞を再懸濁した。懸濁された細胞溶液を4℃、5500rpmで20分間、再び遠心分離して細胞を得た。再懸濁時、使用されるグリセロール溶液を半分に減らして前記過程を2回繰り返した後、細胞とグリセロールとの体積比が1:1になるように再懸濁して細胞濃縮液を得た。
【0049】
このようにして得られた細胞濃縮液と、実施例1で作製された遺伝子交換ベクターpMLKO−sacBを混合した後、1.8kV、25μF,200ohmsの条件でエレクトロポレーション方法でpMLKO−sacBをMannheimia succiniciproducens MBEL55E(KCTC 0769BP)に導入した。pMLKO−sacBが導入された細菌にLB−グルコース液体培地1mLを加えて、200rpm、37℃の振盪培養器で1時間前培養した後、培養液を抗生剤カナマイシン(最終濃度25μg/mL)を含むLB−グルコース固体培地に塗抹して、37℃で48時間以上培養した。二重交差(double crossover)が起こされたコロニーのみを選び出すために、形成されたコロニーをカナマイシン(25μg/mL)と100g/Lのスクロースを含有したLB−スクロース培地にストリークした後、24時間培養した後、形成されたコロニーを同じ固体培地に再びストリークした。
【0050】
前記培地で形成されたコロニー(変異株)を、抗生剤が含まれたLB−グルコース液体培地で培養し、ロッシェルなどの方法(Rochelle et al.,FEMS Microbiol.Lett.,100:59,1992)を応用して、培養菌株のゲノムDNAを分離した。前記分離された変異株のゲノムDNAを鋳型としてPCRを行った後、得られたPCR産物を電気泳動することによってゲノムDNAのldhA遺伝子の欠失の可否を確認した。
【0051】
ldhAの欠失の可否は下記のように、PCRを2回行っテ確認した。まず、前記変異株のゲノムDNAを鋳型とし、下記配列番号8及び9のプライマーを用いてPCRを行った。
配列番号8:5’−GACGTTTCCCGTTGAATATGGC(KM1)
配列番号9:5’−CATTGAGGCGTATTATCAGGAAAC(LU1)
【0052】
次いで、前記変異株のゲノムDNAを鋳型とし、下記配列番号10及び11のプライマーを用いてPCRを行った。
配列番号10:5’−GCAGTTTCATTTGATGCTCGATG(KM2)
配列番号11:5’−CCTCTTACGATGACGCATCTTTCC(LD2)
【0053】
前記2回のPCR反応で得られたPCR断片を電気泳動して、PCR断片のサイズを確認してldhAの欠失の可否を確認した。ldhAの欠失されたゲノムDNAのPCR断片は配列番号8(KM1)及び9(LU1)を用いたPCR産物が1.5kbで得られ、同時に配列番号10(KM2)及び11(LD2)を用いたPCR産物が1.7kbに得られることで確認できた。前記各プライマーの位置は図3に表した。
【0054】
前記のような方法でM. succiniciproducens MBEL55EのゲノムからldhAを欠失させた変異菌株M. succiniciproducens LKを製作した。
【0055】
《実施例3:pta及びackA欠失ベクター(pPTA−sacB)の作製》
M. succiniciproducens LK菌株のゲノムからpta及びackAを相同組換え方法で欠失させるために、遺伝子交換ベクターを次のように作製した。sacB遺伝子を含有するpKmobsacBを鋳型とし、下記配列番号12及び13プライマーを用いてPCRを行った後、得られたsacB産物をPstI及びBamHIで切断し、これをpUC19(Stratagene Cloning Systems.USA)に導入してpUC19−sacBを作製した。
配列番号12:5’−AGCGGATCCCCTTCTATCGCCTTCTTGACG
配列番号13:5’−GTCCTGCAGGGCTACAAAATCACGGGCGTC
【0056】
一方、M. succiniciproducens LKのゲノムDNAを鋳型とし、下記配列番号14及び15のプライマーを用いてPCRを行った後、得られたpta−ackA相同部位1を含むPCR断片をXbaI及びBamHIで切断し、下記配列番号16及び17のプライマーを用いてPCRを行って得られたpta−ackA相同部位2を含むPCR断片をXbaI及びSacIで切断した後、この断片らをpUC19のBamHIとSacI部位に導入してpUC19−PTA12を作製した。
配列番号14:5’−GCTCTAGATATCCGCAGTATCACTTTCTGCGC
配列番号15:5’−TCCGCAGTCGGATCCGGGTTAACCGCACAG
配列番号16:5’−GGGGAGCTCGCTAACTTAGCTTCTAAAGGCCATGTTTCC
配列番号17:5’−GCTCTAGATATCCGGGTCAATATCGCCGCAAC
【0057】
前記pUC19−PTA12の選択マーカーとしてスペクチノマイシン耐性遺伝子を挿入するため、スペクチノマイシン耐性遺伝子(GenBankX02588;SpR)を含有するプラスミドpIC156(Steinmetz et al.,Gene,142:79,1994)を鋳型とし、下記配列番号18及び19のプライマーを用いてPCRを行った後、得られたSpR遺伝子を含むPCR断片をEcoRVで切断し、これを前記製作されたpUC19−PTA12に導入してスペクチノマイシン耐性遺伝子を有するpUC19−PTA1S2を作製した。前記作製されたpUC19−PTA1S2をSacI及びBamHIで切断した後、前、製作したpUC19−SacBに導入してpta及びackA遺伝子除去用交換ベクターpPTA−sacBベクターを作製した(図4)。
配列番号18:5’−GAATTCGAGCTCGCCCGGGGATCGATCCTC
配列番号19:5’−CCCGGGCCGACAGGCTTTGAAGCATGCAAATGTCAC
【0058】
《実施例4:M. succiniciproducens PALK菌株の作製》
実施例3で製作されたホスホトランスアセチラーゼ遺伝子(pta)及び酢酸キナーゼ遺伝子(ackA)除去用交換ベクターpPTA−sacBを利用してM. succiniciproducens LKのゲノムからpta及びackA遺伝子を欠失させて変異菌株を製作した(図4)。
【0059】
即ち、実施例2で製作されたM. succiniciproducens LKを、10g/Lのグルコースを含有したLB−グルコース培地に塗抹して、37℃で36時間培養した後、コロニーをLB−グルコース液体培地10mLに接種して12時間培養した。充分に成長した培養液をLB−グルコース液体培地100mLに1%接種して、37℃の振盪培養器で培養した。
【0060】
前記培養液から、実施例2と同じ方法で細胞濃縮液を得た後、これを、実施例3で作製された遺伝子交換ベクターpPTA−sacBと混合した後、2.5kV、50μF,200ohmsの条件でエレクトロポレーションを行って、pPTA−sacBをM. succiniciproducens LKに導入させた。pPTA−sacBが導入された細菌にLB−グルコース液体培地800mLを加えて、37℃の恒温槽で1時間30分間、前培養した後、二重交差(double crossover)を誘導するため、培養液を、スペクチノマイシン(最終濃度50μg/mL)を含有したTSB−スクロース固体培地[100g/Lのスクロースを含有したTryptic Soy Broth(Becton,Dickinson and Company)固体培地]に塗抹して、37℃で48時間以上培養した。コロニーが形成され、二重交差のみが起こったものを選び出すために、各コロニーを、スペクチノマイシン50μg/mLを含有したTSB−スクロース培地とアンピシリン50μg/mLを含有したTSB培地にストリークした。37℃で12時間程度培養した後、スペクチノマイシン50μg/mLを含有したTSB培地では生長するが、アンピシリン50μg/mLを含有したTSB培地には生長しないコロニーを選別してスペクチノマイシン50μg/mLを含有したTSB−スクロース培地にストリークした。ここでに形成されたコロニーをスペクチノマイシンとアンピシリンが合有されている培地で再び選別して、望ましい結果を表したコロニーを最終選択した。前記選択された菌株のゲノムを分離して鋳型として使用し、PCRを通じてpta−ackAの欠失の可否を確認した。
【0061】
pta−ackAの欠失の可否は、下記のようにPCRを2回行って確認した。まず、前記変異株のゲノムDNAを鋳型とし、下記配列番号20及び21のプライマーを用いてPCRを行った。次に、前記変異株のゲノムDNAを鋳型とし、下記配列番号22及び23のプライマーを用いてPCRを行った。
配列番号20:5’−CCTGCAGGCATGCAAGCTTGGGCTGCAGGTCGACTC(SP1)
配列番号21:5’−GCTGCCAAACAACCGAAAATACCGCAATAAACGGC(PAU1)
配列番号22:5’−GCATGTAACTTTACTGGATATAGCTAGAAAAGGCATCGGGGAG(SP2)
配列番号23:5’−GCAACGCGAGGGTCAATACCGAAGGATTTCGCCG(PAD2)
【0062】
前記2回のPCR反応で得られた産物をゲル電気泳動して、そのサイズによってpta−ackAの欠失の可否を確認した(図6)。pta−ackAの欠失は配列番号20及び21のプライマー(SP1とPAU1)を使用したPCR産物が1.1kbに得られ、同時に、配列番号22及び23のプライマー(SP2及びPAD2)を用いたPCRの産物が1.5kbに得られることを確認した。前記プライマーの位置は図5に示した。
【0063】
前記方法で作製されたM. succiniciproducens LKのゲノムからpta−ackAが欠失された菌株、即ち、M. succiniciproducensからldhA、pta及びackAが欠失された変異菌株をM. succiniciproducens PALKと命名して、2006年7月26日付に国際寄託機関であるKCTC(韓国生命工学研究院内生物資源センター;韓国デジョンユスングヲウンドン52)に寄託番号‘KCTC 10973BP’として寄託した。
【0064】
《実施例5:M. succiniciproducens PALK菌株を利用した純粋コハク酸の生産》
前記実施例4で作製されたM. succiniciproducens PALK(KCTC 10973BP)菌株を5g/Lのブドウ糖を含む10mLの合成培地で接種し、嫌気条件で39℃で8時間前培養した後、これを再び5g/Lのブドウ糖を含む250mLの合成培地に移して39℃で培養した。この場合、抗生剤として50μg/mLのスペクチノマイシンを添加した。醗酵は前記250mLのM. succiniciproducens PALK培養液を2.25Lの合成培地(1g/LのNaCl,2g/Lの(NH4)2HPO4,0.02g/LのCaCl2・2H2O,0.2g/LのMgCl26H2O,8.709g/LのK2HPO4,0.5g/Lのシステイン,0.5g/Lのメチオニン,0.5g/Lのアラニン,0.5g/Lのアスパラギン,0.5g/Lのアスパラギン酸,0.5g/Lのプロリン,0.5g/Lのセリン,0.005g/Lのニコチン酸,0.005g/LのCa−パントセネート(pantothenate),0.005g/LのピリドキシンHCl(pyridoxine・HCl),0.005g/Lのチアミン,0.005g/Lのアスコルビン酸及び0.005g/Lのビオチン)を合有する微生物反応器に接種して行って、醗酵条件は18.2g/L(100mM)の初期ブドウ糖の濃度9.2g/L(100mM)の初期グリセロール濃度、温度39℃で醗酵した。醗酵中pHはアンモニア水を使用して6.5に調整し、抗生剤として使用したスペクチノマイシンは50μg/mLの濃度で添加した。高濃度のコハク酸生産のためにブドウ糖を完全に使い果たした場合、濃縮されたブドウ糖溶液を添加して培養液のブドウ糖の濃度が約18.2g/L(100mM)に成るように調整した。
【0065】
前記と同じ方法でM. succiniciproducens MBEL55E(KCTC 0769BP)及びM. succiniciproducens LPK7(KCTC 10626BP)を培養してコハク酸を醗酵させた。
培養液内の細胞濃度を分光光度計を用いて測定して、既に測定した分光光度計の吸光度と乾燥細胞重量の検量線を利用して細胞濃度を計算した。醗酵過程中の生物反応器から周期的にサンプルを採取して、採取された試料は13、000rpmで10分間遠心分離した後、上層液の各種代謝産物及びコハク酸濃度、ブドウ糖とグリセロール濃度を液体クロマトグラフ(HPLC)で分析した。
【0066】
その結果、図7及び表2に表すように、本発明のM. succiniciproducens PALK(KCTC 10973BP)はM. succiniciproducens MBEL55E(KCTC 0769BP)及びM. succiniciproducens LPK7(KCTC 10626B)と比較すると、より優れたコハク酸生産性を表すだけではなく、副産物としてほとんど有機酸及びエタノールが生成されないことがわかる。0.45g/Lの酢酸と0.24g/Lのピルビン酸が検出されたけれども、これは菌株の生長のため生成される最小量で生産されたコハク酸に比べ、1重量%以下の極微な量で無視することができるのは当業者に自明な事実である。
【0067】
【表2】
【産業上の利用可能性】
【0068】
以上詳細に説明したように、本発明は生長速度が速く、且つ嫌気的条件で培養する場合、コハク酸を除き、他の有機酸を殆ど生成せずにコハク酸だけを高濃度で生産するコハク酸生成変異微生物及び前記変異微生物を利用したコハク酸の製造方法を提供する効果がある。本発明による変異微生物は既存のコハク酸生成微生物より菌株の生長速度及びコハク酸生産性が高く、他の有機酸を生成せずに高濃度でコハク酸を生成するから、コハク酸の産業的生産菌株として有用である。
【0069】
以上、本発明の内容の特定の部分を詳述したが、当業界における通常の知識を有する者にとって、このような具体的な記述は単なる好適な実施様態に過ぎず、これにより本発明の範囲が制限されることはないという点は明らかであろう。よって、本発明の実質的な範囲は特許請求の範囲とこれらの等価物により定まると言えるであろう。
【特許請求の範囲】
【請求項1】
コハク酸を生成する微生物において、ピルビン酸ギ酸リアーゼをコードする遺伝子(pfl)を含み、乳酸脱水素酵素をコードする遺伝子(ldhA)、ホスホトランスアセチラーゼ遺伝子(pta)及び酢酸キナーゼ遺伝子(ackA)が欠失しており、嫌気条件でコハク酸以外の有機酸はほとんど生成せず、コハク酸のみを高濃度で生成する特性を有するルーメンバクテリア変異菌株。
【請求項2】
前記ルーメンバクテリアは、マンヘミア(Mannheimia)属、アクチノバチルス(Actinobacillus)属及びアネロビオスピリルム(Anaerobiospirillum)属からなる群より選択されることを特徴とする請求項1に記載のルーメンバクテリア変異菌株。
【請求項3】
前記ルーメンバクテリアは、副産物として他の有機酸は生成せず、コハク酸のみを生成する同型発酵菌株であることを特徴とする請求項1に記載のルーメンバクテリア変異菌株。
【請求項4】
他の有機酸の生成量が、コハク酸の1重量%以下であることを特徴とする請求項1に記載のルーメンバクテリア変異菌株。
【請求項5】
他の有機酸は、乳酸、酢酸、ギ酸及びピルビン酸からなる群より選択される一種以上であることを特徴とする請求項1に記載のルーメンバクテリア変異菌株。
【請求項6】
Mannheimia succiniciproducensにおいてピルビン酸ギ酸リアーゼをコードする遺伝子(pfl)を含み、乳酸脱水素酵素をコードする遺伝子(ldhA)、ホスホトランスアセチラーゼをコードする遺伝子(pta)及び酢酸キナーゼをコードする遺伝子(ackA)が欠失しており、嫌気条件でコハク酸以外の有機酸はほとんど生成せず、コハク酸のみを高濃度で生成する特性を有するルーメンバクテリアMannheimia succiniciproducens PALK(KCTC10973BP)。
【請求項7】
以下のステップを含む請求項1に記載のルーメンバクテリア変異菌株の製造方法:
(a)コハク酸の生成能を有するルーメンバクテリアのゲノムから相同組換え方法で乳酸脱水素酵素をコードする遺伝子(ldhA)を欠失させ、乳酸脱水素酵素をコードする遺伝子(ldhA)が欠失したルーメンバクテリア変異菌株を収得するステップ、及び
(b)前記乳酸脱水素酵素をコードする遺伝子(ldhA)が欠失されているしたルーメンバクテリア変異菌株のゲノムから相同組換え方法でホスホトランスアセチラーゼをコードする遺伝子(pta)及び酢酸キナーゼをコードする遺伝子(ackA)を欠失させ、乳酸脱水素酵素をコードする遺伝子(ldhA)、ホスホトランスアセチラーゼをコードする遺伝子(pta)及び酢酸キナーゼをコードする遺伝子(ackA)が欠失したルーメンバクテリア変異菌株を収得するステップ。
【請求項8】
前記(a)ステップの相同組換えは、欠失されたldhAを含む遺伝子交換ベクターを使用して行われることを特徴とする請求項7に記載の方法。
【請求項9】
前記(b)ステップの相同組換えは、欠失されたpta−ackAを含む遺伝子交換ベクターを使用して行われることを特徴とする請求項7に記載の方法。
【請求項10】
前記欠失されたldhAを含む遺伝子交換ベクターは、pMLKO−sacBであることを特徴とする請求項8に記載の方法。
【請求項11】
前記欠失されたpta−ackAを含む遺伝子交換ベクターは、pMPKO−sacBであることを特徴とする請求項9に記載の方法。
【請求項12】
請求項1〜5の中いずれか一項に記載のルーメンバクテリア変異菌株を嫌気条件で培養するステップ、及び前記培養液からコハク酸を回収するステップを含むコハク酸の製造方法。
【請求項13】
前記培養は、ブドウ糖またはグリセロールを炭素原として使用することを特徴とする請求項12に記載の方法。
【請求項14】
副産物としての他の有機酸の生成量は、コハク酸の1重量%以下であることを特徴とする請求項12に記載の方法。
【請求項15】
ルーメンバクテリア変異菌朱Mannheimia succiniciproducens PALK(KCTC10973BP)を嫌気条件で培養するステップ、及び前記培養液からコハク酸を回収するステップを含むコハク酸の製造方法。
【請求項16】
前記培養は、ブドウ糖またはグリセロールを炭素原として使用することを特徴とする請求項15に記載の方法。
【請求項17】
副産物としての他の有機酸の生成量は、コハク酸の1重量%以下であることを特徴とする請求項15に記載の方法。
【請求項1】
コハク酸を生成する微生物において、ピルビン酸ギ酸リアーゼをコードする遺伝子(pfl)を含み、乳酸脱水素酵素をコードする遺伝子(ldhA)、ホスホトランスアセチラーゼ遺伝子(pta)及び酢酸キナーゼ遺伝子(ackA)が欠失しており、嫌気条件でコハク酸以外の有機酸はほとんど生成せず、コハク酸のみを高濃度で生成する特性を有するルーメンバクテリア変異菌株。
【請求項2】
前記ルーメンバクテリアは、マンヘミア(Mannheimia)属、アクチノバチルス(Actinobacillus)属及びアネロビオスピリルム(Anaerobiospirillum)属からなる群より選択されることを特徴とする請求項1に記載のルーメンバクテリア変異菌株。
【請求項3】
前記ルーメンバクテリアは、副産物として他の有機酸は生成せず、コハク酸のみを生成する同型発酵菌株であることを特徴とする請求項1に記載のルーメンバクテリア変異菌株。
【請求項4】
他の有機酸の生成量が、コハク酸の1重量%以下であることを特徴とする請求項1に記載のルーメンバクテリア変異菌株。
【請求項5】
他の有機酸は、乳酸、酢酸、ギ酸及びピルビン酸からなる群より選択される一種以上であることを特徴とする請求項1に記載のルーメンバクテリア変異菌株。
【請求項6】
Mannheimia succiniciproducensにおいてピルビン酸ギ酸リアーゼをコードする遺伝子(pfl)を含み、乳酸脱水素酵素をコードする遺伝子(ldhA)、ホスホトランスアセチラーゼをコードする遺伝子(pta)及び酢酸キナーゼをコードする遺伝子(ackA)が欠失しており、嫌気条件でコハク酸以外の有機酸はほとんど生成せず、コハク酸のみを高濃度で生成する特性を有するルーメンバクテリアMannheimia succiniciproducens PALK(KCTC10973BP)。
【請求項7】
以下のステップを含む請求項1に記載のルーメンバクテリア変異菌株の製造方法:
(a)コハク酸の生成能を有するルーメンバクテリアのゲノムから相同組換え方法で乳酸脱水素酵素をコードする遺伝子(ldhA)を欠失させ、乳酸脱水素酵素をコードする遺伝子(ldhA)が欠失したルーメンバクテリア変異菌株を収得するステップ、及び
(b)前記乳酸脱水素酵素をコードする遺伝子(ldhA)が欠失されているしたルーメンバクテリア変異菌株のゲノムから相同組換え方法でホスホトランスアセチラーゼをコードする遺伝子(pta)及び酢酸キナーゼをコードする遺伝子(ackA)を欠失させ、乳酸脱水素酵素をコードする遺伝子(ldhA)、ホスホトランスアセチラーゼをコードする遺伝子(pta)及び酢酸キナーゼをコードする遺伝子(ackA)が欠失したルーメンバクテリア変異菌株を収得するステップ。
【請求項8】
前記(a)ステップの相同組換えは、欠失されたldhAを含む遺伝子交換ベクターを使用して行われることを特徴とする請求項7に記載の方法。
【請求項9】
前記(b)ステップの相同組換えは、欠失されたpta−ackAを含む遺伝子交換ベクターを使用して行われることを特徴とする請求項7に記載の方法。
【請求項10】
前記欠失されたldhAを含む遺伝子交換ベクターは、pMLKO−sacBであることを特徴とする請求項8に記載の方法。
【請求項11】
前記欠失されたpta−ackAを含む遺伝子交換ベクターは、pMPKO−sacBであることを特徴とする請求項9に記載の方法。
【請求項12】
請求項1〜5の中いずれか一項に記載のルーメンバクテリア変異菌株を嫌気条件で培養するステップ、及び前記培養液からコハク酸を回収するステップを含むコハク酸の製造方法。
【請求項13】
前記培養は、ブドウ糖またはグリセロールを炭素原として使用することを特徴とする請求項12に記載の方法。
【請求項14】
副産物としての他の有機酸の生成量は、コハク酸の1重量%以下であることを特徴とする請求項12に記載の方法。
【請求項15】
ルーメンバクテリア変異菌朱Mannheimia succiniciproducens PALK(KCTC10973BP)を嫌気条件で培養するステップ、及び前記培養液からコハク酸を回収するステップを含むコハク酸の製造方法。
【請求項16】
前記培養は、ブドウ糖またはグリセロールを炭素原として使用することを特徴とする請求項15に記載の方法。
【請求項17】
副産物としての他の有機酸の生成量は、コハク酸の1重量%以下であることを特徴とする請求項15に記載の方法。
【図1】

【図2】
【図3】
【図4】
【図5】
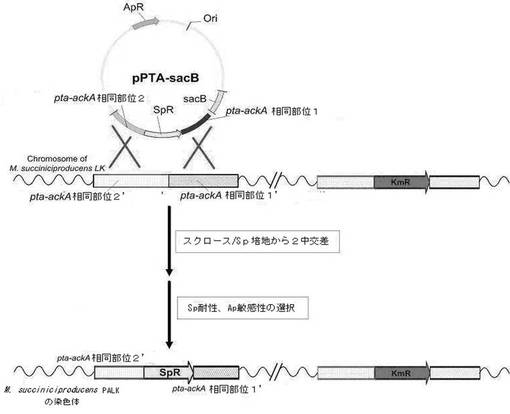
【図6】
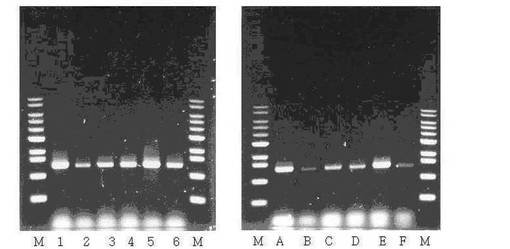
【図7】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【公表番号】特表2009−544335(P2009−544335A)
【公表日】平成21年12月17日(2009.12.17)
【国際特許分類】
【出願番号】特願2009−522705(P2009−522705)
【出願日】平成19年7月25日(2007.7.25)
【国際出願番号】PCT/KR2007/003574
【国際公開番号】WO2008/013405
【国際公開日】平成20年1月31日(2008.1.31)
【出願人】(502318478)コリア アドバンスド インスティチュート オブ サイエンス アンド テクノロジィ (27)
【Fターム(参考)】
【公表日】平成21年12月17日(2009.12.17)
【国際特許分類】
【出願日】平成19年7月25日(2007.7.25)
【国際出願番号】PCT/KR2007/003574
【国際公開番号】WO2008/013405
【国際公開日】平成20年1月31日(2008.1.31)
【出願人】(502318478)コリア アドバンスド インスティチュート オブ サイエンス アンド テクノロジィ (27)
【Fターム(参考)】
[ Back to top ]