
新規配糖化酵素及びそれをコードするポリヌクレオチド
【課題】酵素の糖供与体特異性、配糖化可能な化合物についての特異性、配糖化効率等の優れた配糖化酵素の提供。
【解決手段】子嚢菌類の糸状菌(好ましくは、Trichoderma属に属する菌、より好ましくは、Trichoderma viride)が生産する、新規配糖化酵素タンパク質、及びそれをコードする特定の塩基配列を有する遺伝子。該酵素タンパク質のうち、特に好ましい例は、Trichoderma viride IAM 5141株の培養上清から得られるものである。該酵素によりフラボノイド類を配糖化することによって、水溶性を向上することができる。
【解決手段】子嚢菌類の糸状菌(好ましくは、Trichoderma属に属する菌、より好ましくは、Trichoderma viride)が生産する、新規配糖化酵素タンパク質、及びそれをコードする特定の塩基配列を有する遺伝子。該酵素タンパク質のうち、特に好ましい例は、Trichoderma viride IAM 5141株の培養上清から得られるものである。該酵素によりフラボノイド類を配糖化することによって、水溶性を向上することができる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、フラボノイド類を配糖化することができる、新規な配糖化酵素及びそれをコードするポリヌクレオチドに関する。本発明により得られるフラボノイド類の配糖体は、食品、医薬品及び化粧品として用いることができる。
【従来技術】
【0002】
プロアントシアニジン(ブドウ種子抽出物)の血管治療薬としての有用性に関する研究が進んだ理由の一つに、対象となる物質が、熱や酸に対し安定で水によく溶け、吸収性が高いことから、生体内での吸収、代謝を追うマーカーとなり得たことが挙げられる。これに対して、カテキン等のポリフェノール化合物は水に溶けにくいものが多く、また生体内で吸収されにくいという問題がある。
【0003】
水に対する溶解度の改善や安定性の向上を目的に、カテキン等を配糖化する技術が検討されてきた。
例えば、特許文献1は、Xanthomonas campestris WU-9701の培養液から回収した分子量約57,000のα−グルコシダーゼを開示する。この酵素は、マルトース等を供与体とし(マルトトリオース、サイクロデキストリン、澱粉等は供与体としない)、特定の受容体にグルコースを転移して配糖体を合成するものである。ここでは、受容体として、メントール、エタノール、1−プロパノール、1−ブタノール、2−ブタノール、イソブチルアルコール、1−アミルアルコール、2−アミルアルコール、5−ノニルアルコール等のアルコール性水酸基を有する化合物、及びカプサイシン、ジヒドロカプサイシン、ノニル酸バニリルアミド、カテキン、エピカテキン、バニリン、ハイドロキノン、カテコール、レゾルシノール、3,4−ジメトキシフェノール等のフェノール性水酸基を有する化合物が挙げられている。また、生成が認められた配糖体はモノグルコシドのみである。
【0004】
特許文献2は、カテキン類とグルコース−1−リン酸又はシュークロースとの混合液にシュークロースホスホリラーゼを作用させ、カテキン類配糖体を製造する方法を開示する。ここでは、シュークロースホスホリラーゼの起源として、Leuconostocmesenteroides、Pseudomonas saccharophila、Pseudomonasputrefaciens、Clostridium pasteurianum、Acetobacter xylinum、Pullulariapullulansが挙げられている。ここでは、受容体として、カテキン類としては、(+)−カテキン、(−)−エピカテキン、(−)−エピカテキン3-O-ガレート、(−)−エピガロカテキン、及び(−)−エピガロカテキン3-O-ガレートが挙げられているが、実施例として記載されているのは、(+)−カテキンを受容体に用いて(+)−カテキン 3'-O-α-D-グルコピラノシドを製造した例のみである。
【0005】
特許文献3は、5、7、3'、4'、5'、3"、4"、5"位の少なくとも1つにグルコース残基又は重合度2〜8のマルトオリゴ糖残基が結合しているエピガロカテキン3-O-ガレート誘導体を開示する。ここで実施例として記載されているのは、特許文献2と同様、(-)-エピガロカテキンガレートとグルコース-1-リン酸又はシュークロースとの混合液にシュークロースホスホリラーゼを作用させ、4'-O-α-D-グルコピラノシル(-)-エピガロカテキンガレート及び4',4”-O-α-D-ジ-グルコピラノシル(-)-エピガロカテキンガレートを製造した例のみである。
【0006】
特許文献4は、ポリフェノール類を配糖化することにより渋みを低減した茶抽出物又は茶飲料を開示する。ここでは、茶抽出物又は茶飲料の渋みを低減する具体的な方法として、茶抽出物又は茶飲料に、デキストリン、サイクロデキストリン、澱粉又はこれらの混合物を添加し、これにサイクロマルトデキストリングルカノトランスフェラーゼを作用させる方法が記載されている。実施例においては、緑茶抽出物とα−サイクロデキストリンにBacillus stearothermophilus由来のサイクロマルトデキストリングルカノトランスフェラーゼを作用させ、得られた反応生成物には渋味が低減されたことが述べられており、またこれはエピガロカテキン3-O-ガレートやエピカテキンといったポリフェノール類が配糖化されていたことを示すものである旨が述べられている。しかしながら、反応生成物の具体的な構造は明らかにされていない。
【0007】
特許文献5は、3'位、3'と5位及び3'と7位が配糖化されたカテキン類の配糖体を開示する。ここでは、そのための具体的な方法として、特許文献4と同様、カテキン類及びデキストリン、サイクロデキストリン、澱粉又はこれらの混合物に、Bacillusstearothermophilus由来のサイクロマルトデキストリングルカノトランスフェラーゼを作用させる方法が記載されている。そして、糖供与体としてデキストリンを用いた上記方法による実施例においては、(-)-エピガロカテキン、(-)-エピガロカテキン3-O-ガレート、(-)-エピカテキン3-O-ガレートの配糖体については、分子吸光係数から、各ポリフェノール1分子当り平均で6〜8個グルコースが結合しているものも得られたと考察している。また、上記方法で得られた配糖体にリゾプス・ニベウス由来のグルコアミラーゼを作用させることで、3',7-ジ-O-α-D-グルコピラノシル(-)-エピガロカテキン、3',5-ジ-O-α-D-グルコピラノシル(-)-エピガロカテキン、3'-O-α-D−グルコピラノシル(-)-エピガロカテキン、3',7-ジ-O-α-D-グルコピラノシル(-)-エピガロカテキン3-O-ガレート、3'-O-α-D-グルコピラノシル(-)-エピガロカテキン3-O-ガレート、3'-O-α-D-グルコピラノシル(-)-ガロカテキン、3'-O-α-D−グルコピラノシル(-)-エピカテキン3-O-ガレートの生成を確認している。
【0008】
また、カテキン配糖体の効果として、非特許文献1には、渋みの低減、水溶性の向上、安定性の改善、チロシナーゼ阻害が、非特許文献2には、変異原性抑制が記載されている。
【特許文献1】特開2001-46096
【特許文献2】特開平5-176786(特許第3024848号)
【特許文献3】特開平7-10897(特許第3071610号)
【特許文献4】特開平8-298930(特許第3579496号)
【特許文献5】特開平9-3089(特許第3712285号)
【非特許文献1】Biosci. Biotech. Biochem., 57 (10), 1666-1669 (1993)
【非特許文献2】Biosci. Biotech. Biochem., 57 (10), 1290-1293 (1993)
【発明の開示】
【発明が解決しようとする課題】
【0009】
これらの配糖化技術は、用いる酵素の糖供与体特異性、配糖化可能な化合物についての特異性、配糖化効率等の観点からは充分なものとはいえず、さらに優れた配糖化酵素の開発が望まれている。
【課題を解決するための手段】
【0010】
本発明者らは、カテキン等を含むフラボノイドの配糖化技術について鋭意検討してきた。その結果、Trichoderma virideの培養上清から新規な配糖化酵素を見いだし、その遺伝子をクローン化することにより本発明を完成した。
【0011】
〔配糖化酵素遺伝子等〕
本発明は、子嚢菌類の糸状菌(好ましくは、Trichoderma属に属する菌、より好ましくは、Trichoderma viride)が生産する配糖化酵素タンパク質をコードするポリヌクレオチド又はそのホモログ、すなわち下記の(A)〜(K)のいずれか一を含むポリヌクレオチド(好ましくは、下記の(A)〜(K)のいずれか一であるポリヌクレオチド)を提供する:
(A)配列番号:6に記載の塩基配列の全部又は一部(例えば、423〜1928番の塩基配列部分の全部又は一部、配列番号:9の塩基配列に対応する部分)からなるポリヌクレオチド;
(B)(A)に記載のポリヌクレオチドの塩基配列と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(C)(A)に記載のポリヌクレオチドの塩基配列において1若しくは複数の塩基が置換、欠失、挿入、及び/又は付加された塩基配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(D)(A)に記載のポリヌクレオチドの塩基配列と少なくとも60%以上の同一性を有し、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(E)配列番号:9に記載の塩基配列からなるポリヌクレオチド;
(F)(E)に記載のポリヌクレオチドの塩基配列と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(G)(E)に記載のポリヌクレオチドの塩基配列において1若しくは複数の塩基が置換、欠失、挿入、及び/又は付加された塩基配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(H)(E)に記載のポリヌクレオチドの塩基配列と少なくとも60%以上の同一性を有し、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(I)配列番号:10に記載のアミノ酸配列からなるタンパク質をコードするポリヌクレオチド;
(J)配列番号:10に記載のアミノ酸配列において1若しくは複数のアミノ酸が置換、欠失、挿入、及び/又は付加したアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(K)配列番号:10に記載のアミノ酸配列と少なくとも60%以上の同一性を有するアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド。
【0012】
配列番号:6にはT. viride IAM5141株から得られた配糖化酵素TRa2のゲノムDNAの塩基配列が、配列番号:9にはそのcDNA配列が、配列番号:10にはTRa2の推定アミノ酸配列が示されている。
【0013】
TRa2の推定アミノ酸配列をSignalPで解析すると、配列番号:10の1〜20番目のアミノ酸配列は、分泌シグナルであると推定される。したがって、配糖化酵素として作用するのに必須の成熟タンパク質は配列番号:10の21番目以降の配列であり、大腸菌など、異種発現系で同酵素を発現させる場合には、シグナル配列を除去して発現させるのが適切であると考えられる。このような成熟タンパク質をコードするポリヌクレオチドもまた、本発明の範囲に含まれる。
【0014】
したがって、本発明はまた、下記の(L)〜(R)のいずれか一を含むポリヌクレオチド(好ましくは、下記の(L)〜(R)のいずれか一であるポリヌクレオチド)を提供する:
(L)配列番号:25に記載の塩基配列からなるポリヌクレオチド;
(M)配列番号:25に記載の塩基配列と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(N)配列番号:25に記載のポリヌクレオチドの塩基配列において1若しくは複数の塩基が置換、欠失、挿入、及び/又は付加された塩基配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(O)配列番号:25に記載の塩基配列と少なくとも60%以上の同一性を有し、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(P)配列番号:26に記載のアミノ酸配列からなるタンパク質をコードするポリヌクレオチド;
(Q)配列番号:26に記載のアミノ酸配列において1若しくは複数のアミノ酸が置換、欠失、挿入、及び/又は付加したアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(R)配列番号:26に記載のアミノ酸配列と少なくとも60%以上の同一性を有するアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド。
【0015】
配列番号:25には成熟タンパク質としてのTRa2をコードするcDNAの塩基配列、すなわち配列番号9の61-1389番目の塩基配列が、配列番号:26には成熟タンパク質としてのTRa2のアミノ酸配列、すなわち配列番号10の21-463番目のアミノ酸配列が示されている。
【0016】
本発明はまた、Trichoderma属由来であり、好ましくはTrichoderma viride、Trichodermareesei、Trichoderma saturnisporum、Trichoderma ghanense、Trichodermakoningii、 Trichoderma hamatum、Trichoderma harzianum又はTrichodermapolysporum由来であり、配列番号:11〜24のいずれか一に記載の塩基配列を含み、フラボノイド類の配糖化活性を有するタンパク質コードするポリヌクレオチドを提供する。このようなポリヌクレオチドの好ましい例は、配列番号:11〜24のいずれか一に記載の塩基配列を含み、配列番号:6、9又は25に記載の塩基配列と高い同一性を有し、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチドである。
【0017】
本明細書で「配糖化活性」というときは、フラボノイド類に糖残基を転移する活性を有することをいう。あるタンパク質が、フラボノイド類に糖残基を転移する活性を有するか否かは、特別な場合を除き、例えば、本明細書の実施例に示されているように、フラボノイド(例えば、カテキン)と適切な糖供与体(例えば、デキストリン)との混合液に、対象酵素を供し、充分な時間反応させ、反応液を高速液体クロマトグラフィー(HPLC)等に供して、分析することにより確認することができる。
【0018】
本発明のポリヌクレオチドによってコードされるタンパク質は、配糖化活性とともに、他の活性、例えばデキストリナーゼ活性を有していてもよい。本明細書で「デキストリナーゼ」というときは、特別な場合を除き、澱粉、デキストリン等、α-グルコシド結合した糖質を加水分解することができる酵素をいう。デキストリナーゼは、アミラーゼの一種である。対象がデキストリナーゼ活性を有するか否かは、適切な条件下で、市販のデキストリン(例えば、澱粉を酸、熱又は酵素により加水分解し、平均分子量を3,500程度としたもの)に対して作用させ、デキストリンが加水分解されているか否かにより、判断することができる。当業者であれば、対象を作用させる反応の適切な条件、及びデキストリンが加水分解されているか否かを判断するための手法を、適宜設計することができる。
【0019】
(フラボノイド類)
本明細書で「フラボノイド類」というときは、特別な場合を除き、フラボノイド及びエスクレチンを含む。
【0020】
本明細書で「フラボノイド」というときは、特別な場合を除き、カテキン類(フラバノール)、フラバノン、フラボン、フラボノール、フラバノノール、イソフラボン、アントシアン及びカルコン、並びにそのメチル化体をいう。フラボノイドには、ナリンゲニン、ケルセチン、ダイゼイン、ゲニステイン、ケンフェロールが含まれる。本発明に用いることのできるフラボノイドは、天然物由来であってもよく、合成されたものであってもよい。
【0021】
本明細書で「カテキン類」というときは、特別な場合を除き、広義のカテキンの意味で用いており、3-オキシフラバンのポリオキシ誘導体をいう。これには、カテキン、ガロカテキン及びそれらの3-ガロイル体が含まれ、またそれらの光学異性体((+)体、(-)体、(+)-epi体、(-)-epi体)及びラセミ体が含まれる。具体的には、カテキン、ガロカテキン(gallocatechin;GC)、カテキンガレート(catechin-3-O-gallate;CG)、ガロカテキンガレート(gallocatechin-3-O-gallate;GCG)、エピカテキン(epicatechin;EC)、エピガロカテキン(epigallocatechin;EGC)、エピカテキンガレート(epicatechin-3-O-gallate;ECG)及びエピガロカテキンガレート(epigallocatechin-3-O-gallate;EGCG)、並びにそれらの光学異性体が含まれる。カテキン類のメチル化体とは、上述したカテキン類において、少なくとも一つのOH基のHがメチルで置き換わったものをいう。カテキン類のメチル化体の例としては、エピカテキン、エピガロカテキン、エピカテキンガレート又はエピガロカテキンガレートにおいて、3'位、4'位、3"位及び4"位のいずれかのOH基のHがメチルで置き換わったものを挙げることができる。本発明に用いることのできるカテキン類及びカテキン類のメチル化体は、天然物由来であってもよく、合成されたものであってもよい。天然物の例としては、茶抽出物、その濃縮物及び精製物(例えば、テアビゴ(DSM ニュートリション ジャパン)、ポリフェノン(三井農林)及びサンフェノン(太陽化学)等の緑茶抽出物)、及びベニフウキ抽出物を挙げることができる。
【0022】
本発明には、フラボノイド類として、単一の化合物を用いることができ、また、2種以上のフラボノイド類の混合物を用いることもできる。
(糖供与体)
本明細書で「糖供与体」というときは、特別な場合を除き、本発明の方法において酵素の基質となり、加水分解されてフラボノイド類に対して糖残基を供給しうる糖質をいう。本発明に用いることができる糖供与体は、マルトトリオース残基を含む糖質であり、これには、マルトトリオース、マルトテトラオース、マルトペンタオース、マルトヘキサオース、マルトヘプタオース、デキストリン、γ-シクロデキストリン、及び可溶性澱粉が含まれる。本明細書で「デキストリン」というときは、特別な場合を除き、澱粉を加水分解したものをいい、「可溶性澱粉」というときは、特別な場合を除き、澱粉の加水分解物であって、熱水に可溶であるものをいう。加水分解は、酸、熱、酵素等のいずれの手法によってもよい。本発明においては、デキストリンとして、例えば平均分子量が約3,500のものを、可溶性澱粉として、例えば平均分子量が約100万のものを用いることができる。
【0023】
本発明でいう「ストリンジェントな条件」とは、特別な場合を除き、6M尿素、0.4% SDS、0.5×SSCの条件又はこれと同等のハイブリダイゼーション条件を指し、さらに必要に応じ、本発明には、よりストリンジェンシーの高い条件、例えば、6M尿素、0.4% SDS、0.1×SSC又はこれと同等のハイブリダイゼーション条件を適用してもよい。それぞれの条件において、温度は約40℃以上とすることができ、よりストリンジェンシーの高い条件が必要であれば、例えば約50℃、さらに約65℃としてもよい。
【0024】
また、明細書において「1若しくは複数の塩基が置換、欠失、挿入、及び/又は付加された塩基配列」というときの置換等されるヌクレオチドの個数は、その塩基配列からなるポリヌクレオチドがコードするタンパク質が所望の機能を有する限り特に限定されないが、1〜9個又は1〜4個程度であるか、同一又は性質の似たアミノ酸配列をコードするような置換等であれば、さらに多くの個数の置換等がありうる。また、本明細書において「1若しくは複数の塩基が置換、欠失、挿入、及び/又は付加されたアミノ酸配列」というときの置換等されるアミノ酸の個数は、そのアミノ酸配列を有するタンパク質が所望の機能を有する限り特に限定されないが、1〜9個又は1〜4個程度であるか、性質の似たアミノ酸への置換であれば、さらに多くの個数の置換等がありうる。このような塩基配列又はアミノ酸配列に係るポリヌクレオチド又はタンパク質を調製するための手段は、当業者にはよく知られている。
【0025】
本明細書においてアミノ酸配列に関し、高い同一性というときは、少なくとも60%以上、好ましくは70%以上、より好ましくは80%以上、さらに好ましくは90%以上、最も好ましくは95%以上の配列の同一性を指す。
【0026】
ポリヌクレオチド配列又はアミノ酸配列の同一性に関する検索・解析は、当業者には周知のアルゴリズム又はプログラム(例えば、BLASTN、BLASTP、BLASTX、ClustalW)により行うことができる。プログラムを用いる場合のパラメーターは、当業者であれば適切に設定することができ、また各プログラムのデフォルトパラメーターを用いてもよい。これらの解析方法の具体的な手法もまた、当業者には周知である。
【0027】
本発明のポリヌクレオチドは、天然物からハイブリダイゼーション技術、ポリメラーゼ連鎖反応(PCR)技術等を利用して得ることができる。
本発明のポリヌクレオチドは、好ましくは、Trichoderma属由来、より好ましくは、Trichoderma viride由来である。
【0028】
本発明はまた、本発明のポリヌクレオチドを含む組換えベクター、その組換えベクターにより形質転換された、形質転換体(例えば形質転換大腸菌、形質転換酵母、形質転換昆虫細胞)も提供する。本発明はさらに、本発明のポリヌクレオチドを用いて宿主(例えば大腸菌、酵母、昆虫細胞)を形質転換する工程(例えば、本発明の組換えベクターにより形質転換する工程)を含む、形質転換方法も提供する。
【0029】
本発明のポリヌクレオチドが挿入されるベクターは、宿主内で挿入物を発現させることが可能なものであれば特に制限はなく、ベクターは、通常、プロモーター配列、ターミネーター配列、外的な刺激により誘導的に挿入物を発現させるための配列、目的遺伝子を挿入するための制限酵素により認識される配列、及び形質転換体を選択するためのマーカーをコードする配列を有する。組換えベクターの作成、組換えベクターによる形質転換方法は、当業者に周知の方法を適用することができる。
【0030】
〔配糖化酵素タンパク質〕
本発明はまた、新規配糖化酵素タンパク質及びそのホモログ、すなわち下記の(i)、(j)又は(k)を含むタンパク質(好ましくは、下記の(i)、(j)又は(k)であるタンパク質)を提供する:
(i)配列番号:10に記載のアミノ酸配列からなるタンパク質;
(j)配列番号:10に記載のアミノ酸配列において1若しくは複数のアミノ酸が置換、欠失、挿入、及び/又は付加したアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質;
(k)配列番号:10に記載のアミノ酸配列と少なくとも60%以上の同一性を有するアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質。
【0031】
本発明はまた、上述の新規配糖化酵素タンパク質において推定分泌シグナル配列部分を除いた成熟タンパク質及びそのホモログ、すなわち下記の(p)、(q)又は(r)を含むタンパク質(好ましくは、下記の(p)、(q)又は(r)であるタンパク質)も提供する:
(p)配列番号:26に記載のアミノ酸配列からなるタンパク質;
(q)配列番号:26に記載のアミノ酸配列において1若しくは複数のアミノ酸が置換、欠失、挿入、及び/又は付加したアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質;
(r)配列番号:26に記載のアミノ酸配列と少なくとも60%以上の同一性を有するアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質。
【0032】
本発明はまた、Trichoderma属由来であり、好ましくはTrichoderma viride、Trichodermareesei、Trichoderma saturnisporum、Trichoderma ghanense、Trichodermakoningii、 Trichoderma hamatum、Trichoderma harzianum又はTrichodermapolysporum由来であり、配列番号:11〜24のいずれか一に記載の塩基配列を含むポリヌクレオチドによってコードされる、フラボノイド類の配糖化活性を有するタンパク質を提供する。このようなタンパク質の好ましい例は、配列番号:11〜24のいずれか一に記載の塩基配列を含み、配列番号:6、9又は25に記載の塩基配列と高い同一性を有するポリヌクレオチドによってコードされる、フラボノイド類の配糖化活性を有するタンパク質である。
【0033】
本発明のタンパク質は、Trichoderma属に属する菌、例えばTrichoderma viride、Trichodermareesei、Trichoderma saturnisporum、Trichoderma ghanense、Trichodermakoningii、 Trichoderma hamatum、Trichoderma harzianum又はTrichodermapolysporumを公知の培養液で培養して得られる培養上清から単離・精製することができる。また、本発明のポリヌクレオチドを挿入した組換えベクターにより形質転換された形質転換体(例えば、形質転換酵母や形質転換大腸菌)により、組換えタンパク質として得ることができる。
【0034】
〔酵素TRa2の酵素学的性質〕
本発明によって提供される新規な酵素タンパク質のうち、特に好ましい例は、Trichoderma viride IAM 5141株の培養上清から得られる、配列番号:10に記載のアミノ酸配列からなる配糖化酵素(本明細書では、「TRa2」ということもある。)である。この酵素は、フラボノイドと糖供与体との反応において、以下のような酵素学的特徴を有する。
【0035】
糖供与体選択性:
本酵素は、実施例に示した条件では、マルトトリオース、マルトテトラオース、マルトペンタオース、マルトヘキサオース、マルトヘプタオース、可溶性澱粉、デキストリン、γ-シクロデキストリン等を糖供与体とするが、セロビオース、デキストラン、マルトース一水和物、カルボキシメチルセルロースナトリウム、イソマルトオリゴ糖、α-シクロデキストリン、β-シクロデキストリン等は糖供与体としない。また、グルコース1分子又は2分子からなる糖のみならず、グルコース3分子(G3)以上の糖鎖長の配糖体を産生することができる配糖化酵素である。
【0036】
基質特異性:
本酵素は、カテキン、エピガロカテキンガレート、ナリンゲニン、ケルセチン、ダイゼイン、ゲニステイン、ケンフェロール等の主要なフラボノイドやエスクレチンなどを含むポリフェノールに対し、広く作用し、配糖化することができる。
【0037】
反応最適温度、pH(反応温度依存性、pH依存性):
本酵素は、加水分解反応においては、実施例に示した条件では、約20〜約60℃、特に約35〜約55℃でよく反応し、またpH約4.0〜約7.0、特にpH約4.5〜約5.5でよく反応する。
【0038】
また、本酵素は、糖転移反応においては、実施例に示した条件では、約20〜約60℃、特に約45〜約55℃でよく反応し、またpH約4.0〜約9.0、特にpH約4.5〜約7.0でよく反応する。
【0039】
温度安定性、pH安定性:
本酵素は、実施例に示した条件では、約20〜約45℃の1hr処理によっては活性を失わず、またpH約5.0〜約9.0での室温、16hr処理によっても活性を失わない。
【0040】
〔フラボノイド類の配糖体の製造方法〕
本発明はまた、フラボノイド類と糖供与体(好ましくは、デキストリン、可溶性澱粉、γ-シクロデキストリン)との混合液に、本発明の新規配糖化酵素を作用させることを特徴とする、フラボノイド類の配糖体の製造方法を提供する。
【0041】
本発明によって提供されるフラボノイド類の配糖体は、下式で表されるものである:
【0042】
【化1】

【0043】
式(I)
式中:
R1〜R5は、少なくとも一つが糖残基であり、他はOH若しくはOCH3であるか、又は
R1〜R4は、少なくとも一つが糖残基であり、他はOH若しくはOCH3であり、R5はHであり;そして
Xは、H、CH3、ガロイル基又はメチル化されたガロイル基である。
【0044】
これには、さらに、下式で表されるフラボノイド類の配糖体が含まれる:
5-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(+)-カテキン;
5-O-α-D-グルコピラノシル-(+)-カテキン;
4'-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(+)-カテキン;
4'-O-α-D-グルコピラノシル-(+)-カテキン;
3'-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(+)-カテキン;
3'-O-α-D-グルコピラノシル-(+)-カテキン;
7-O-α-D-グルコピラノシル-(-)-エピガロカテキン-3-O-ガレート;
7-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(-)-エピガロカテキン-3-O-ガレート;
3'-O-α-D-グルコピラノシル-(-)-エピガロカテキン-3-O-ガレート;及び
3'-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(-)-エピガロカテキン-3-O-ガレート及びそれらの光学異性体。
【0045】
〔配糖体の用途〕
本発明により得られた配糖体は、食品用組成物、医薬用組成物又は化粧用組成物として用いることができる。より詳細には、例えばカテキン類配糖体を配合したものは、カテキンと同様に、抗アレルギー、抗酸化、抗癌、抗炎症、抗菌・抗う歯、抗ウイルス、解毒、腸内フローラ改善、消臭、血漿コレステロール上昇抑制、血圧上昇抑制、血糖上昇抑制、血小板凝集抑制、痴呆予防、体脂肪燃焼、体脂肪蓄積抑制、持久力向上、抗疲労、腎機能改善のための剤として、あるいは食品用組成物、医薬用組成物又は化粧用組成物として用いることができる。
【0046】
食品用組成物は、栄養補助食品、健康食品、食事療法用食品、総合健康食品、サプリメント及び飲料を含む。飲料は、茶飲料、ジュース、清涼飲料、ドリンク剤を含む。医薬品組成物は、医薬品又は医薬部外品とすることができ、経口製剤又は皮膚用外用剤であることが好ましく、液剤、錠剤、顆粒剤、丸剤、シロップ剤、又はローション剤、スプレー剤、硬膏剤、軟膏剤の形態とすることができる。化粧用組成物は、クリーム、液状ローション、乳液状ローション、スプレーの形態とすることができる。
【0047】
本発明の食品用組成物、医薬用組成物又は化粧用組成物中における配糖体の配合量は、特に限定されないが、当業者であれば、対応するフラボノイド類の好ましい一日摂取量を参考に、溶解性、呈味等を考慮して、適宜設定することができる。例えば、組成物中における本発明の配糖体の配合量を0.01〜99.9重量%とすることができ、また本発明の配糖体を一日当たり100 mg〜20 gを1〜数回(例えば3回)に分けて摂取することができるように、組成物中の配合量を決定することができる。
【0048】
本発明の食品用組成物、医薬用組成物又は化粧用組成物には、食品、医薬又は化粧品として許容できる種々の成分を添加することができる。これらの添加剤及び/又は成分の例として、ビタミン類、糖類、賦形剤、崩壊剤、結合剤、潤沢剤、乳化剤、緊張化剤(等張化剤)、緩衝剤、溶解補助剤、防腐剤、安定化剤、抗酸化剤、着色剤、矯味剤、香料、凝固剤、pH調整剤、増粘剤、茶エキス、生薬エキス、無機塩等が挙げられる。
【0049】
〔他の態様〕
本発明において、配糖化酵素タンパク質は、適当な担体に固定化し、固定化酵素として用いることができる。担体としては、同様の目的で使用される従来の樹脂、例えば、塩基性樹脂(例えば、MARATHON WBA(Dow Chemical社製)、SAシリーズ、WAシリーズ又はFPシリーズ(三菱化学)、及びアンバーライトIRA904(オルガノ))、疎水性樹脂(例えば、ダイアイオンFPHA13(三菱化学)、HPシリーズ(三菱化学)、アンバーライトXAD7(オルガノ))等を用いることができる。他に、エキスプレスイオンD(Whatman)、DEAE-トヨパール650M(東ソー)、DEAE-セファロースCL4B(Amersham Biosciences)等も好適に用いうる。固定化の方法としては、従来の、物理的吸着、イオン結合又は共有結合を介して固定化する結合法、二価性官能基をもつ試薬で架橋固定化する架橋法、網目構造のゲル又は半透膜の中に酵素を包埋する包括法のいずれも採りうる。例えば、まず各樹脂5mlに対し、蒸留水に溶解した酵素20〜2,000 mg、例えば50〜400 mgを吸着させた後、上清を除去し、乾燥することによる。
【0050】
本発明はまた、配列番号:6、配列番号:9、配列番号:10、配列番号:11〜26のいずれか一に記載の塩基配列又はアミノ酸配列の全部若しくは一部を用いることを特徴とする、フラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチドの塩基配列又はフラボノイド類の配糖化活性を有するタンパク質のアミノ酸配列を決定する方法も提供する。
【発明の効果】
【0051】
本発明により、フラボノイドを効率的に配糖化することができる新規酵素及びその遺伝子が提供される。
本発明の新規酵素によりフラボノイド類を配糖化することによって、水溶性を向上することができる。そのため、本発明により、フラボノイド類の経口吸収性を高めることができると考えられる。また、水溶性の向上は、水への溶解速度の向上、ひいては生体内での吸収速度の向上に資するものである。したがって、本発明により、フラボノイド類の有用な活性、例えば抗酸化活性を、生体内で効率よく発揮させることが可能となる。
【0052】
また、本発明によりフラボノイド類を配糖化することによって、フラボノイド類の呈味を改変することができる。特にフラボノイド類としてカテキン類のように苦味、渋味を呈するものを本発明により配糖化する場合には、そのような呈味を低減することができる。
【0053】
また、本発明によりフラボノイドを配糖化することによって、熱安定性を向上することができる。
【実施例】
【0054】
〔実施例1:Trichoderma培養液のカテキン配糖化活性〕
Trichoderma viride IAM 5141株のスラントから、酵母エキス(Difco)1%、ポリペプトン(日本製薬)1%、デキストリン(ナカライテスク)2%の液体培地 10mlに植菌し、30℃で1日間振とう培養を行い、前培養液とした。さらに、上記液体培地900mlに前培養液を全量植菌して30℃で3日間培養を行い、フィルターろ過により培養上清液を調製した。培養上清690mlに対して、硫酸アンモニウム387g(80%飽和)を添加し攪拌した後、遠心分離により沈殿を回収した。得られた沈殿に10mlの0.1M酢酸バッファー(pH5.0)を加え、粗酵素液とした。
【0055】
粗酵素液100μlに対して、カテキン3mg、デキストリン10mgを添加し、50℃で24時間攪拌して酵素反応を行った。反応液を0.1%トリフルオロ酢酸(TFA)で10倍希釈し、10μlを高速液体クロマトグラフィー(HPLC)で分析を行った。
【0056】
分析条件
カラム:Develosil C30-UG-5(4.6x150mm)
グラジエント条件: 5%B液 → 50%B液 / 20分間
溶離液A:0.1%TFA / 蒸留水
溶離液B:90%アセトニトリル/ 0.08%TFA
流量:1ml/min
検出波長:280nm
図1に示したとおり、上記反応によるカテキン配糖体の生成を確認できた。また、糖供与体としてγ-サイクロデキストリンを用いた場合にも同様の配糖体の生成を確認できた。このことから、T. viride IAM5141株はデキストリン又はγサイクロデキストリンを糖供与体として、カテキンを配糖化する酵素を分泌生産していることが示唆された。
【0057】
〔実施例2:PCRによるα−アミラーゼホモログの部分配列のクローニング〕
デキストリン及びγ‐サイクロデキストリンは、グルコースがα-1,4結合で結合したポリマーであり、目的とする酵素はこれを分解する活性を有していることから、α−アミラーゼファミリー様の酵素である可能性が示唆された。
【0058】
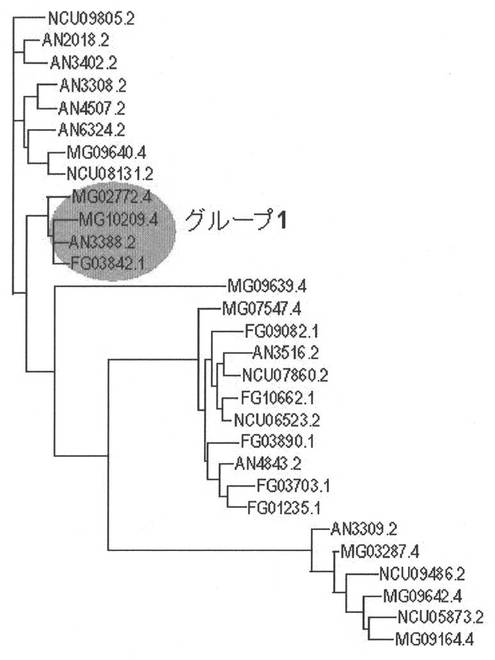
そこで、トリコデルマと同じ子嚢菌類の糸状菌でゲノム配列が明らかになっている微生物のうち、Aspergillus nidulans、Neurosporacrassa、Magnaporthe grisea、Fusarium graminearum のゲノム情報データベースから、プロテインファミリーデータベース(PFAM)のalpha-amylase, catalytic domain(accession No. PF00128)のモチーフを持つ推定ORFのアミノ酸配列をそれぞれ9個、6個、8個、6個抽出した。これらを配列相同性検索プログラムであるClustalWによりアライメントを作成し、系統樹作成プログラムであるTree viewにより系統樹を作成し、相同性をもとにグループ分けを行った。図2のグループ1の4つのアミノ酸配列(括弧内はGenebank アクセッションNo.)MG02772.4(EAA47529)、MG10209.4(EAA48146)、AN3388.2(EAA63356)、FG03842.1(EAA71544)をアライメントし、保存性の高い部分(図3、下線部)のアミノ酸配列に対応するオリゴDNAを合成した。
【0059】
AMY-12f:
5'-TAYTGYGGNGGNACNTTYAARGGNYT-3'(配列番号:1)
AMY-15r:
5'-TTYTCNACRTGYTTNACNGTRTCDAT-3'(配列番号:2)
AMY-17r:
5'-GGTNAYRTCYTCNCKRTTNGCNGGRTC-3'(配列番号:3)
先に培養したT. viride IAM5141菌体約1gの湿菌体よりDNeasy plant Maxi Kit(QIAGEN)により、ゲノムDNAを抽出した。このゲノムDNA50ngを鋳型として、プライマーAMY-12fとプライマーAMY-15r、又はプライマーAMY-12fとプライマーAMY-17rを用いてPCR反応を行った。すなわち、ExTaq(タカラバイオ社製)を用いて、94℃ 2分、(94℃ 1分、50℃1分、72℃ 1分)を30サイクル、72℃ 10分の反応を行った。PCR産物をアガロースゲル電気泳動で分析したところ、プライマーAMY-12fとプライマーAMY-15rの組み合わせで約0.6kbp、プライマーAMY-12fとプライマーAMY-17rの組み合わせで約1.0kbpの断片が確認できた。そこで、これらのDNA断片をアガロースゲルから切り出し、GFX PCR DNA and Gel Band Purifiction Kit(アマシャムバイオサイエンス)によりDNAを精製した。これをTOPO-TA cloning kit(インビトロジェン)にてクローニングし、塩基配列の解析をABI3100Avant(アプライドバイオシステムズ社製)にて行った。前者で得られた塩基配列は後者で得られた塩基配列に含まれていた。この塩基配列をGenBankに登録されているアミノ酸配列に対してBlastxにより相同性検索を行ったところ、MG10209.4(EAA48146)と最も高い相同性を示した。
【0060】
〔実施例3:α−アミラーゼホモログのゲノム配列の決定〕
得られた約1.0kbpの塩基配列をもとに、以下のプライマーを設計し、Inverse PCR(逆PCR)を行った。
【0061】
TRa2-2:
5'-CCAACCTGGTATCTACATAC-3'(配列番号:4)
TRa2-3:
5'-AGATGGCATCAAATCCCAT-3'(配列番号:5)
まず、T. viride IAM5141より調製したゲノムDNAをHindIIIあるいはPstIで完全消化し、ligation high(東洋紡)により、16℃で一晩反応させ、セルフライゲーションにより閉環させた。これらのDNA 0.1μgを鋳型として、上記プライマーTRa2-2及びTRa2-3を用いてPCR反応を行った。PCRは、LA Taq(タカラバイオ)を使用して、94℃ 2分、(95℃ 30秒、66℃ 15分)を30サイクル、72℃ 10分の反応を行った。得られたPCR産物をアガロースゲル電気泳動で分析したところ、HindIIIで消化したゲノムDNAを鋳型としたものでは約2kb、PstIで消化したゲノムを鋳型としたものでは約4.5kbのDNA断片が確認できた。これらをそれぞれアガロースゲルから切り出し、前述と同様にクローニングした。挿入された断片の両端からそれぞれ塩基配列を決定した。HindIIIで消化したゲノム由来のものとPstIで消化したゲノム由来のものは、当該制限酵素サイトまでの塩基配列が一致していた。こうして得られた塩基配列を、先に得られた部分配列と連結した。この塩基配列を図4(TRa2-gDNA)及び配列番号:6に示す。図2グループ1の4つの配列との比較及び、開始コドンや終始コドンの出現などからα−アミラーゼホモログのコード領域を推定した。開始コドンは塩基番号423-425のATG、終始コドンは1926-1928のTAAであると考えられた。
【0062】
〔実施例4:α−アミラーゼホモログのcDNAのクローニング〕
先に培養したT. viride IAM5141株の菌体約0.1gより、RNeasy plant mini kitにより全RNAを抽出した。1μgの全RNAをスーパースクリプトファーストストランドシステム for RT-PCR(インビトロジェン)でランダムヘキサマーを用いて、cDNAを合成した。
【0063】
先に得られたゲノム配列をもとに、以下のプライマーを設計した。
TRa2EcoRI-f2:
5'-GGAATTCATGAAGCTTCGATCCGCCGTCCC-3'(配列番号:7)
TRa2XhoI-r2:
5'-CCGCTCGAGTTATGAAGACAGCAGCACAAT-3'(配列番号:8)
合成されたcDNAを鋳型として、上記プライマーTRa2EcoRI-f2及びTRa2XhoI-r2を用いて、PCR反応を行った。PCRは、Ex Taq(タカラバイオ)を使用し、94℃ 2分、(94℃ 1分、55℃ 1分、72℃ 2分)を30サイクル、72℃ 10分の反応を行った。得られたPCR産物をアガロースゲル電気泳動で分析したところ、約1.5kbのDNA断片が確認できた。これをアガロースゲルより切り出し、GFXにて精製した。得られたDNA断片をTOPO-TA cloning kit(インビトロジェン)にてクローニングして、プラスミドpCRTRa2-cDNAを構築し、cDNAの塩基配列を決定した(図4、図5及び配列番号:9)。先に得られたゲノムDNA配列とcDNA配列を比較したところ、ゲノム配列には2つのイントロンが含まれていた(図4)。cDNA配列には、1392bpのORFがあり、463アミノ酸残基からなるタンパク質をコードしていた(図5及び配列番号:10)。この遺伝子をTRa2と命名した。本遺伝子によりコードされる推定アミノ酸配列をSignalP(Nielsen H. et.al., Protein Eng., 10, 1-6, 1997 )で解析した結果、N末端から20アミノ酸残基は分泌シグナル配列であると考えられた。さらに、TRa2によりコードされる推定アミノ酸配列を前述の方法に従って相同性検索を行ったところ、AN3388.2(EAA63356)ともっとも高い相同性を示した。TRa2タンパク質の推定アミノ酸配列を、既知のα-アミラーゼであるタカアミラーゼのアミノ酸配列と比較した。その結果、α-アミラーゼファミリー酵素の4つの保存領域が本酵素においても保存されており(図6、2重下線)、活性中心とされるアスパラギン酸残基、グルタミン酸残基、アスパラギン酸残基がすべて保存されていた(図6、*印アミノ酸残基)。
【0064】
〔実施例5:TRa2タンパク質の酵母での分泌発現系の構築〕
プラスミドpCRTRa2-cDNAを制限酵素EcoRI及びXhoIで消化して得られた約1.5kbの断片を、プラスミドpYE22m(Biosci. Biotech. Biochem., 59(7), 1221-1228, 1995)を制限酵素EcoRI及びSalIで消化した断片とligation high(東洋紡)を用いて連結し、プラスミドpYETRa2を得た。
【0065】
プラスミドpYETRa2により、酵母S. cerevisiae EH1315株を酢酸リチウム法により、形質転換した。得られた形質転換株をTRa2-1株とした。TRa2-1株をYPD(Difco)液体培地 10mlに一白金耳植菌し、30℃で2日間振とう培養した。TRa2タンパク質はN末に20アミノ酸残基からなる分泌シグナル配列が存在するので、培養液中に分泌されると考えられた。そこで遠心分離により、菌体を沈殿させ培養上清を回収した。
【0066】
〔実施例6:TRa2の糖分解活性の測定〕
培養上清500μlをMicrocon YM-30(アミコン)を用いて、約5倍に濃縮した。上記濃縮液10μlを、0.5%のマルトース、マルトトリオース、マルトテトラオース、デキストリン、α-サイクロデキストリン、β-サイクロデキストリン、又は γ-サイクロデキストリンを含む20mM 酢酸バッファー(pH5.0)100μlに添加し、50℃で1時間反応させた。
【0067】
反応終了後、以下のとおりTLCにて分析した。プレートは、シリカゲルG-60プレート(メルク)を使用し、展開液として、2−プロパノール:アセトン:0.5M乳酸=2:2:1を使用した。検出は、硫酸:エタノール=1:9を噴霧して風乾したのち、ホットプレートで熱することにより行った。その結果、ベクターpYE22mにて形質転換したコントロール株(C-1株)の培養液では、いずれの糖の分解も確認できなかった。一方、TRa2-1株の培養液ではマルトトリオース、マルトテトラオース、デキストリン及びγ-サイクロデキストリンが分解されて主にマルトース及びグルコースが生成することが確認できたが、マルトース、α-サイクロデキストリン及びβ-サイクロデキストリンの分解は確認できなかった。
【0068】
〔実施例7:TRa2の糖転移活性の測定〕
培養上清の原液、又はVIVASPIN 10,000MWCO/PES製(VIVASCIENCE社)にて約5倍に濃縮した培養上清濃縮液100μlに(+)-カテキン又は(-)-エピガロカテキン-3-O-ガレート3mgと、デキストリン10mgを加えて、50℃にて1日間攪拌しながら反応させた。反応終了後、反応液を0.1%トリフルオロ酢酸溶液にて10倍希釈し、高速液体クロマトグラフィー(HPLC)にて、実施例1と同様の条件で分析した。その結果、ベクターpYE22mにて形質転換したコントロール株(C-1株)の培養上清と反応させた反応液には、反応生成物が確認できなかったのに対し、TRa2-1株では、カテキン配糖体及びエピガロカテキン-3-O-ガレート配糖体の生成が確認できた(図7)。
【0069】
〔実施例8:TRa2によるカテキン配糖体の製造〕
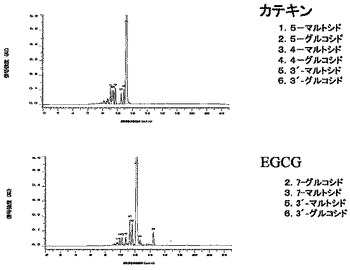
TRa2-1株をYPD液体培地200mlに植菌し30℃で3日間振とう培養した。遠心分離により菌体を集め、培養上清を得た。この培養上清100mlを限外ろ過ディスク:NMWL30000/再生セルロースで0.1M 酢酸バッファー(pH5) 100mlを加えながら50mlになるまで濃縮し、TRa2酵素液とした。上記TRa2酵素液50mlに(+)-カテキン1.5gとデキストリン 5gを混合し、45℃,18hr撹拌した。反応液を遠心分離し、上清をLH20(Amersham Biosciences)樹脂60ml/Ф2.5×20cmカラムに吸着させた。蒸留水120ml、10%エタノール 240mlで溶出し、配糖体画分を回収・凍結乾燥し、凍乾末530mgを得た。その50mgをとって蒸留水 5mlに溶かし、DevelosilC30-UG-5カラム20×250mm、A:0.1%TFA/蒸留水,B:90%メタノール/0.1%TFA,30%B、3ml/min、280nmで分取した。HPLCの保持時間の早い順にピーク1〜6を回収・凍結乾燥した。MS及びNMR解析から、ピーク1は5-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(+)-カテキン、ピーク2は5-O-α-D-グルコピラノシル-(+)-カテキン、ピーク3は4'-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(+)-カテキン、ピーク4は4'-O-α-D-グルコピラノシル-(+)-カテキン、ピーク5は3'-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(+)-カテキン、ピーク6は3'-O-α-D-グルコピラノシル-(+)-カテキンであることが示唆された。
【0070】
5-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(+)-カテキン:m/z 615.2、NMRδppm (D2O);2.71 (1H, dd), 2.85 (1H, dd), 3.42 (1H, t), 3.56-3.85 (9H, m), 4.19 (1H, t), 4.26 (1H, dd), 4.87 (1H, d), 5.70 (1H, d), 6.19 (1H, d), 6.39 (1H, d), 6.83 (1H, dd), 6.90-6.93 (2H, m) 。
【0071】
5-O-α-D-グルコピラノシル-(+)-カテキン:m/z 450.8、NMRδppm (D2O);2.62 (1H, dd), 2.81 (1H, dd), 3.43 (1H, t), 3.45-3.55 (1H, m), 3.6-3.7 (3H, m), 3.83 (1H, t), 4.18 (1H, dd), 4.76 (1H, d), 5.61 (1H, d), 6.09 (1H, d), 6.31 (1H, d), 6.77 (1H, d), 6.8-6.9 (2H, m) 。
【0072】
4'-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(+)-カテキン:m/z 615.2、NMRδppm (D2O);2.54 (1H, dd), 2.81 (1H, dd), 3.43 (1H, t), 3.60 (1H, dd), 3.68-3.94 (9H, m), 4.19-4.28 (2H, m), 4.82 (1H, d), 5.44 (1H, d), 5.62 (1H, d), 6.04 (1H, d), 6.11 (1H, d), 6.91 (1H, dd), 7.00 (1H, d), 7.22 (1H, d)。
【0073】
4'-O-α-D-グルコピラノシル-(+)-カテキン:m/z 453.2、NMRδppm (D2O);2.45 (1H, dd), 2.73 (1H, dd), 3.45 (1H, t), 3.65-3.75 (4H, m), 4.11 (1H, dd), 4.7-4.75 (2H, m), 5.53 (1H, d), 5.95 (1H, d), 6.02 (1H, d), 6.83 (1H, dd), 6.91 (1H, d), 7.15 (1H, d)。
【0074】
3'-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(+)-カテキン:m/z 615.2、NMRδppm (D2O);2.54 (1H, dd), 2.80 (1H, dd), 3.44 (1H, t), 3.59 ( 1H, dd), 3.67-3.90 (9H, m), 4.17-4.24 (2H, m), 4.83 (1H, d), 5.41 (1H, d), 5.55 (1H, d), 6.03 (1H, d), 6.10 (1H, d), 7.10 (1H, d), 7.06 (1H, d), 7.26 (1H, d)。
【0075】
3'-O-α-D-グルコピラノシル-(+)-カテキン:m/z 453.2、NMRδppm (D2O);2.43 (1H, dd), 2.73 (1H, dd), 3.27 (1H, s), 3.44 (1H, t), 3.6-3.7 (4H, m), 3.88 (1H, t), 4.10 (1H, dd), 4.69 (1H, d), 5.46 (1H, d), 5.93 (1H, s), 6.01 (1H, s), 6.89 (1H, d), 6.94 (1H, dd), 7.18 (1H, d)。
【0076】
〔実施例9:TRa2によるエピガロカテキン-3-O-ガレート配糖体の製造〕
TRa2-1株をYPD液体培地100mlに植菌し30℃で3日間振とう培養した。遠心分離により菌体を集め、培養上清を得た。この培養上清45mlを限外ろ過ディスク:NMWL30000/再生セルロースで0.1M 酢酸バッファー(pH5) 50mlを加えながら20mlになるまで濃縮し、TRa2酵素液とした。TRa酵素液 20mlに(-)-エピガロカテキン-3-O-ガレート600mgとデキストリン 2gを混合し、50℃、1日撹拌した。反応液を遠心し、上清をLH20樹脂25ml/Ф1.5×30cmカラムに吸着させた。蒸留水100ml、10%エタノール 100ml、20%エタノール 100ml、30%エタノール 200mlで溶出し、30%エタノール画分を回収・凍結乾燥した。凍乾末120mgを蒸留水12mlに溶かし、DevelosilC30-UG-5カラム20×250mm、A:0.1%TFA/蒸留水,B:90%メタノール/0.1%TFA,40%B、3ml/min、280nmで分取した。MS及びNMR解析から、ピーク2は7-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(-)-エピガロカテキン-3-O-ガレート、ピーク5は3'-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(-)-エピガロカテキン-3-O-ガレート、ピーク6は3'-O-α-D-グルコピラノシル-(-)-エピガロカテキン-3-O-ガレートであることが示唆された。一方、ピーク3はMS(m/z 621.2、1107.3)から、グルコシドとマルトテトラオシドの混合物であることが示唆され、保持時間からグルコシドは7-O-α-D-グルコピラノシル-(-)-エピガロカテキン-3-O-ガレートであると考えられた。
【0077】
7-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(-)-エピガロカテキン-3-O-ガレート:m/z 783.2、NMRδppm (CD3OD); 2.88 (1H, dd), 2.01 (1H, dd), 3.26 (1H, t), 3.46 (1H, dd), 3.6-3.9 (9H, m), 4.08 (1H, t), 5.00 (1H, s), 5.20 (1H, d), 5.43 (1H, d), 5.54 (1H, s), 6.27 (1H, d), 6.34 (1H, d), 6.51 (2H, d), 6.94 (2H, d)。
【0078】
3'-O-α-D-グルコピラノシル-(-)-エピガロカテキン-3-O-ガレート:m/z 621.1, δppm (CD3OD);2.88 (1H, dd), 2.99 (1H, dd), 3.42 (1H, dd), 3.51 (1H, t), 3.69 (1H, m), 3.8-3.9 (3H, m), 4.88 (1H, d), 4.98 (1H, s), 5.49 (1H, broad s), 5.95 (1H, d), 5.96 (1H, d), 6.65 (1H, d), 7.01 (2H, s), 7.11 (1H, d)。
【0079】
3'-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(-)-エピガロカテキン-3-O-ガレート:m/z 783.2, δppm (CD3OD);2.87 (1H, broad d), 2.99 (1H, dd), 3.27 (1H, t), 3.44-3.48 (2H, m), 3.6-3.8 (4H, m), 3.85 (2H, d), 3.98 (1H, dd), 4.06 (H, t), 4.85 (1H, d), 4.99 (1H, s), 5.28 (1H, d), 5.49 (1H, broad s), 5.94 (1H, d), 5.96 (1H, d), 6.64 (1H, d), 7.01 (2H, s), 7.09 (1H, d)。
【0080】
〔実施例10:TRa2タンパク質の酵母表層発現系の構築〕
TRa2タンパク質を酵母の表層に呈示発現させるために、以下のベクターを構築した。
プラスミドpCRTRa2-cDNAを鋳型として、プライマーTraEcoRI-f2とプライマーTRa2XhoI-r3を用いて、PCR反応を行った。PCRは、Ex Taq(タカラバイオ)を使用し、94℃ 2分、(94℃ 1分、58℃ 1分、72℃ 2分)を25サイクル、72℃ 10分の反応を行った。得られたPCR産物をアガロースゲル電気泳動で分析したところ、約1.5kbのDNA断片が確認できた。これをアガロースゲルより切り出し、GFXにて精製した。得られたDNA断片をTOPO-TA cloning kit(インビトロジェン)にてクローニングして、塩基配列を確認しプラスミドpCRTRa2-cDNA2とした。プラスミドpCRTRa2-cDNA-2をEcoRIとXhoIで消化して得られた約1.5kbのDNA断片とプラスミドpGA11(Appl.Environ.Microbiol.,63,1362-1366(1997)をEcoRIとXhoIで消化して得られた約9.2kbのDNA断片を連結し、プラスミドpCAS-TRa2を得た。
【0081】
〔実施例11:TRa2タンパク質表層発現酵母の活性確認〕
プラスミドpCAS-TRa2にて酵母S. cerevisiaeEH1315株を形質転換した。得られた形質転換株をTRa2-2株とした。TRa2-2株及び、プラスミドpGA11のグルコアミラーゼ遺伝子の分泌シグナルとα-アグルチニンの3'側半分をコードする遺伝子の間にマルチクローニングサイトを挿入したプラスミドpCAS1にて酵母S. cerevisiae EH1315株を形質転換したコントロール株(C-2株)をYPD液体培地10mlに1白金耳植菌し、30℃で2日間培養した。遠心分離により菌体を回収し、PBSにて洗浄後、1ml PBSに懸濁した。この懸濁液を用いて実施例6及び実施例7と同様に、糖分解反応及び糖転移反応を行い反応生成物の分析を行った。その結果、TRa2-2株はデキストリン分解活性及びデキストリンを糖供与体とするカテキンへの糖転移活性を有するのに対し、C-2株ではこれらの活性がないことが確認できた。すなわち、TRa2-2株ではTRa2が活性を有する形で酵母表層に呈示されたことが示された。
【0082】
〔実施例12:Trichoderma属の各種菌株からのTRa2ホモログの取得〕
Trichoderma viride IFO31137株、Trichodermaviride SAM1427株、Trichoderma viride IFO31327株、Trichodermaviride IFO30498株、Trichoderma viride IFO5720株、Trichodermareesei IFO31329株、Trichoderma reesei IFO31328株、Trichodermareesei IFO31326株、 Trichoderma saturnisporum IAM12535株、Trichoderma ghanenseIAM13109株、Trichoderma koningii IAM12534株、 Trichoderma hamatumIAM12505株、 Trichoderma harzianum IFO31292株、Trichoderma polysporumSAM357株をそれぞれ、酵母エキス(Difco)1%、ポリペプトン(日本製薬)1%、デキストリン(ナカライテスク)2%の液体培地 10mlに植菌し、30℃で4日間振とう培養した。フィルターろ過により菌体を回収し、DNeasy plant mini Kit(QIAGEN)により、ゲノムDNAを抽出した。このゲノムDNA50ngを鋳型として、プライマーAMY-12fとプライマーAMY-15rを用いてPCR反応をおこなった。すなわち、ExTaq(タカラバイオ社製)を用いて、94℃ 2分、(94℃ 1分、50℃1分、72℃ 1分)を30サイクル、72℃ 10分の反応を行った。PCR産物をアガロースゲル電気泳動で分析したところ、約0.6kbpの断片が確認できた。これらのDNA断片をアガロースゲルから切り出し、GFX PCR DNA and Gel Band Purifiction Kit(アマシャムバイオサイエンス)によりDNAを精製した。これをTOPO-TA cloning kit(インビトロジェン)にてクローニングし、塩基配列の解析をABI3100Avant(アプライドバイオシステムズ社製)にて行った。その結果、配列番号:11〜24の配列が得られた。アライメント結果を図8に示す。TRa2遺伝子のこれらに相当する領域及びこれらの互いの相同性は63〜99.5%であった。
【0083】
これらの配列をもとに、T. viride IAM5141のTRa2遺伝子、cDNA全長を取得したときと同様の方法を用いれば、TRa2と同様な活性を持つTRa2ホモログを取得できる。
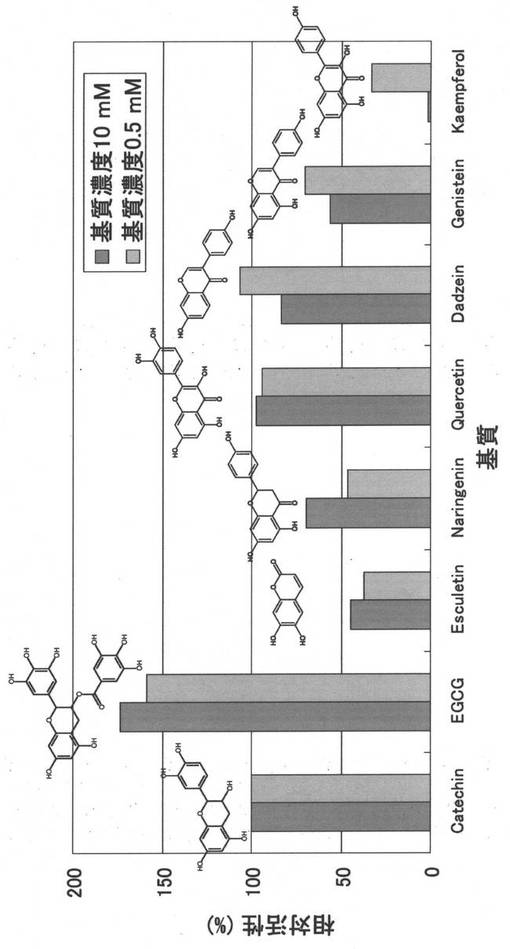
〔実施例13:TRa2の基質特異性解析〕
TRa2-1株をYPD培地10 mlにて、30℃で一晩振とう培養した。静止期に達した培養液を同培地に接種し(2%(v/v))、30℃で3日間振とう培養した。培養後、遠心分離にて上清を回収し、5倍に濃縮してTRa2の粗酵素溶液を得た。酵素反応溶液(0.5 mM 及び 10 mM 糖受容体基質((+)-カテキン、(-)-エピガロカテキン-3-O-ガレート、エスクレチン、ナリンゲニン、ケルセチン、ダイゼイン、ゲニステイン、ケンフェロール)、10 mg デキストリン、100 mM 酢酸バッファー (pH 5.2)、粗酵素溶液)100μlにて45℃、24 hr反応させ、HPLCにて分析した。結果を図9に示した。
【0084】
受容体基質と配糖体生成物の面積比(%)は、(+)-カテキン:10%、(-)-エピガロカテキン-3-O-ガレート:17.7%、エスクレチン:3.5%、ナリンゲニン:4.4%、ケルセチン:9.4%、ダイゼイン:10.7%、ゲニステイン:6.8%、ケンフェロール:3.1%であった。
【0085】
〔実施例14:TRa2キャラクタリゼーション〕
His-tagを付加したTRa2タンパク質(TRa2-His)の発現:
TRa2-His発現用プラスミドの構築と形質転換酵母の取得
TRa2タンパク質のC末端にHis-tagを付加して酵母で発現させるために、以下のプライマーを設計した。
TRa2HisXhoI-r2: Gctcgagttagtggtggtggtggtggtgtgaagacagcagcaa(配列番号:28)
プラスミドpCRTRa2-cDNAを鋳型として、プライマーTraEcoRI-f2とプライマーTRa2HisXhoI-r2を用いて、PCR反応を行った。PCRは、Ex Taq(タカラバイオ)を使用し、94℃ 2分、(94℃ 1分、58℃ 1分、72℃ 2分)を25サイクル、72℃ 10分の反応を行った。得られたPCR産物をアガロースゲル電気泳動で分析したところ、約1.5kbのDNA断片が確認できた。これをアガロースゲルより切り出し、GFXにて精製した。得られたDNA断片をTOPO-TA cloning kit(インビトロジェン)にてクローニングして、塩基配列を確認しプラスミドpCRTRa2-cDNA-Hisとした。pCRTRa2-cDNA-HisをEcoRIとXhoIで消化して約1.5kbのDNA断片を、プラスミドpYE22mを制限酵素EcoRI及びSalIで消化した断片とligation high(東洋紡)を用いて連結し、プラスミドpYE-TRa2-Hisを得た。
プラスミドpYE-TRa2-Hisにて酵母S. cerevisiae EH1315株を形質転換した。得られた形質転換株をTRa2-3株とした。
【0086】
培養
TRa2-3株をSD(-Trp) 20 mlにて30℃、16 hr培養した。前培養液をSD(-Trp) + 100 mM KH2PO4-KOH (pH 6.0) 1 Lに植菌し、30℃、3 days培養した。遠心分離により培養液上清を回収した。
【0087】
精製
培養液上清をBuffer S1[20 mM NaH2PO4-NaOH (pH 7.4)、10 mM イミダゾール、0.5 M NaCl、15 mM 2-メルカプトエタノール]にて平衡化したN2+キレート済みのChelating Sepharose Fast Flow (5 ml、Pharmacia Biotech)カラムにアプライし、同バッファー(40 ml)で洗浄した。続いてBuffer E1[20 mM NaH2PO4-NaOH (pH 7.4)、200 mM イミダゾール、0.5 M NaCl、15 mM 2-メルカプトエタノール]によりカラムに結合したタンパク質を溶出した。活性画分を集め、VIVASPIN(30,000 MWCO、VIVASCIENCE)を用いて脱塩、濃縮した。
【0088】
続いて、酵素溶液をBuffer S2[20 mM KH2PO4-KOH (pH 7.4)、15 mM 2-メルカプトエタノール、0.1% CHAPS]にて平衡化したResource Q (1 ml、Pharmacia Biotech)カラムにアプライ(1.5 ml/min)し、同バッファー(10 ml)で洗浄した。続いてBuffer E2[20 mM KH2PO4-KOH (pH 7.4)、0.6 M NaCl、15 mM 2-メルカプトエタノール、0.1% CHAPS]の0-100%直線濃度勾配(60 ml)によりカラムに結合したタンパク質を溶出した。活性画分を集め、VIVASPIN(30,000 MWCO、VIVASCIENCE)を用いて脱塩、濃縮した。
【0089】
再度、同手順でResource Qカラムクロマトグラフィーを行い、活性を示しかつSDS-PAGE上で単一バンドの画分を集め、VIVASPIN(30,000 MWCO、VIVASCIENCE)を用いて脱塩、濃縮した。
【0090】
酵素活性測定:
糖転移活性
反応液(10 mM カテキン、10 mg デキストリン、100 mM Acetate-NaOH(pH 5.3)、酵素溶液)100 μlを45℃、24 hr攪拌し、0.5% TFA 100 μlを添加することで反応を停止した。反応停止後のサンプルを遠心分離し、上清を回収した。生成物を以下のような条件でHPLCにて分析した。HPLC条件:溶離液A、0.1% TFA;溶離液B、90% アセトニトリル、0.08% TFA;分析カラム、Devolosil C30-UG-5 (4.6 x 150 mm、NOMURA CHEMICAL);流速、1 ml/min;分離モード、0 min-5%B、20 min-50%B、20.5 min-5%B、25 min-5%B
加水分解活性
反応液(5 mM マルトテトラオース、20 mM KH2PO4-KOH(pH 6.0)、酵素溶液)100 μlを35℃、15min反応させ、熱処理(100℃、5 min)することで反応を停止した。反応停止後のサンプルを遠心分離し、上清を回収した。生成物を以下のような条件でHPLCにて分析した。HPLC条件:溶離液、68% アセトニトリル;分析カラム、SUPELCOSIL LC-NH2 (5 μm、4.6 x 250 mm、SUPELCO);流速、1 ml/min;分離モード、isocratic
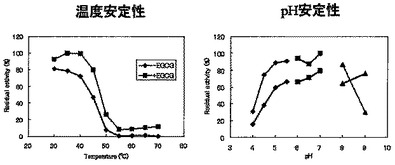
温度安定性解析:
酵素溶液(±10 mMエピガロカテキンガレート)を20〜60℃の各温度で、1 hr処理後、4℃に冷却した。冷却後のサンプルに、10mMエピガロカテキンガレート、10 mg デキストリンを添加して反応(各温度,pH 5.3,24 hr)した。0.5% TFA添加で反応停止後、逆相HPLCにて分析した。
【0091】
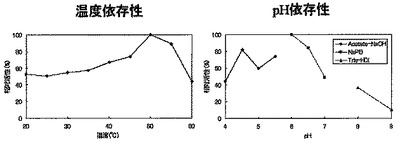
結果を図10(左)に示した。
pH安定性解析:
酵素溶液(±10 mMエピガロカテキンガレート)に各種pHの終濃度167 mM 緩衝液(pH 4、4.5、5、Acetate-NaOH;pH 5.5、6、6.5、7、NaH2PO4-NaOH;pH 8、9、Tris-HCl)を添加し、4℃、16 hr処理後、終濃度400 mM Acetate-NaOH(pH 5.3)を添加した。処理後のサンプルに、10 mMエピガロカテキンガレート、10 mg デキストリンを添加して反応(45℃,各pH,24 hr)した。0.5% TFA添加で反応停止後、逆相HPLCにて分析した。活性測定時の反応液のAcetate-NaOH濃度は200 mMとなる。
【0092】
結果を図10(右)に示した。
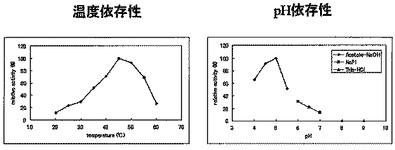
反応温度依存性解析:
5 mM マルトテトラオースに酵素を添加して、加水分解反応(20〜60℃の各温度,pH 6.0、15 min)を行った。100℃、5 minで反応停止後、順相HPLCにて分析した。結果を図11(左)に示した。
【0093】
また、10 mM カテキン,10 mgデキストリンに酵素を添加して、糖転移反応(20〜60℃の各温度,pH 5.3,24 hr)を行った。TFA添加にて反応停止後、逆相HPLCにて分析した。結果を図12(左)に示した。
【0094】
反応pH依存性解析:
5 mM マルトテトラオースに酵素を添加して、各種pHの緩衝液中で加水分解反応(35℃、pH 4、4.5、5、Acetate-NaOH;pH 5.5、6、6.5、7、NaH2PO4-NaOH;pH 8、9、Tris-HCl、15 min)を行った。100℃、5 minで反応停止後、順相HPLCにて分析した。結果を図11(右)に示した。
【0095】
また、10 mM カテキン,10 mgデキストリンに酵素を添加して、糖転移反応(45℃、pH 4、4.5、5、Acetate-NaOH;pH 5.5、6、6.5、7、NaH2PO4-NaOH;pH 8、9、Tris-HCl、24 hr)を行った。TFA添加にて反応停止後、逆相HPLCにて分析した。結果を図12(右)に示した。
【0096】
〔実施例15:配糖体のキャラクタリゼーション〕
熱安定性:
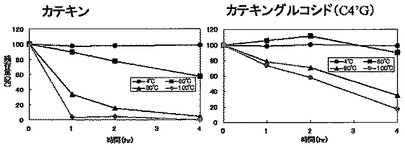
100μMの(+)-カテキン、又は4'-O-α-D-グルコピラノシル-(+)-カテキンを含む10mM リン酸カリウムバッファー(pH7.0) 30μlを4〜100℃の各温度で0〜4時間処理した後、氷中に移し、続いて0.1% TFA 60μlを加えて実施例1と同様にHPLCで分析した。各温度で処理したときの(+)-カテキン又は4'-O-α-D-グルコピラノシル-(+)-カテキンの残存量を図13に示した。カテキンに比べ4'-O-α-D-グルコピラノシル-(+)-カテキンは熱に対して安定であった。
【0097】
溶解性:
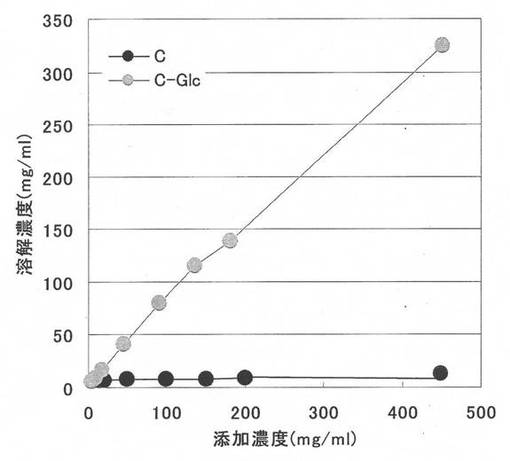
(+)-カテキン又は5-O-α-D-グルコピラノシル-(+)-カテキンを10〜450mg/mlの各濃度になるよう水に添加し、激しく攪拌することにより溶解させた。その後、遠心分離により沈殿物を除去し上清をHPLCにより分析し、(+)-カテキン及び5-O-α-D-グルコピラノシル-(+)-カテキンの量を定量した。その結果を図14に示す。(+)-カテキンはほとんど水に溶けないのに対し、5-O-α-D-グルコピラノシル-(+)-カテキンは少なくとも40倍以上の溶解性を示した。
【産業上の利用可能性】
【0098】
本発明により得られる新規酵素遺伝子又は本酵素遺伝子を改変した遺伝子、それを導入した形質転換体、及び新規酵素タンパク質又は本酵素タンパク質を改変したタンパク質は、フラボノイド類の配糖体又はそれらを用いる食品、医薬品、化粧品を生産する上で有用である。
【図面の簡単な説明】
【0099】
【図1】図1は、Trichoderma viride IAM 5141 粗酵素液でカテキンを処理した際の、HPLC分析チャートである。
【図2】図2は、Aspergillus nidulans、Neurospora crassa、Magnaporthe grisea、Fusariumgraminearum のゲノム情報データベースから抽出した、alpha-amylase, catalytic domain(accession No. PF00128)のモチーフを持つ推定ORFのアミノ酸配列について、系統樹作成プログラムであるTree viewにより作成した系統樹を示した図である。
【図3】図3は、図2のグループ1の4つのアミノ酸配列をアライメントし、保存性の高い部分(下線部)を示した図である。
【図4】図4は、TRa2のゲノムDNA配列(配列番号6)とcDNA配列(配列番号9)を比較した図である。
【図5】図5は、TRa2のcDNA塩基配列、及びそれに対応する推定アミノ酸配列を示した図である。2重下線部は推定分泌シグナル配列。
【図6】図6は、TRa2の推定アミノ酸配列とタカアミラーゼ前駆体アミノ酸配列(GBNo.BAA00336)の一次構造を比較した図である。下線部:TRa2の推定分泌シグナル、破線下線部:タカアミラーゼの分泌シグナル、2重下線部:α‐アミラーゼファミリー酵素で高度に保存されている4つの領域、*印アミノ酸残基:触媒部位に位置するアミノ酸残基。
【図7】図7は、T. viride IAM5141の培養上清の原液、又はその濃縮液に(+)-カテキン又は(-)-エピガロカテキン-3-O-ガレートと、デキストリンを加えて反応させた際の、反応液のHPLC分析チャートである。
【図8】図8は、Trichoderma属の各種菌株から調製されたゲノムDNAより得られた、TRa2ホモログの部分塩基配列のアライメント結果を示した図である。
【図9】図9は、形質転換体(TRa2-1株)の培養上清から調製したTRa2の粗酵素溶液に、各糖受容体基質((+)-カテキン、(-)-エピガロカテキン-3-O-ガレート、エスクレチン、ナリンゲニン、ケルセチン、ダイゼイン、ゲニステイン、ケンフェロール)、デキストリンを加えて反応させた際の、配糖化活性を示したグラフである。
【図10】図10は、TRa2の温度安定性及びpH安定性を示したグラフである。
【図11】図11は、TRa2による糖質加水分解反応の、温度依存性及びpH依存性を示したグラフである。
【図12】図12は、TRa2による糖転移反応の、温度依存性及びpH依存性を示したグラフである。
【図13】図13は、(+)-カテキン又は4'-O-α-D-グルコピラノシル-(+)-カテキンを含む溶液を4〜100℃の各温度で0〜4時間処理した後の、それぞれの残存量を示したグラフである。
【図14】図14は、(+)-カテキン及び5-O-α-D-グルコピラノシル-(+)-カテキンの水への溶解性を示したグラフである。
【技術分野】
【0001】
本発明は、フラボノイド類を配糖化することができる、新規な配糖化酵素及びそれをコードするポリヌクレオチドに関する。本発明により得られるフラボノイド類の配糖体は、食品、医薬品及び化粧品として用いることができる。
【従来技術】
【0002】
プロアントシアニジン(ブドウ種子抽出物)の血管治療薬としての有用性に関する研究が進んだ理由の一つに、対象となる物質が、熱や酸に対し安定で水によく溶け、吸収性が高いことから、生体内での吸収、代謝を追うマーカーとなり得たことが挙げられる。これに対して、カテキン等のポリフェノール化合物は水に溶けにくいものが多く、また生体内で吸収されにくいという問題がある。
【0003】
水に対する溶解度の改善や安定性の向上を目的に、カテキン等を配糖化する技術が検討されてきた。
例えば、特許文献1は、Xanthomonas campestris WU-9701の培養液から回収した分子量約57,000のα−グルコシダーゼを開示する。この酵素は、マルトース等を供与体とし(マルトトリオース、サイクロデキストリン、澱粉等は供与体としない)、特定の受容体にグルコースを転移して配糖体を合成するものである。ここでは、受容体として、メントール、エタノール、1−プロパノール、1−ブタノール、2−ブタノール、イソブチルアルコール、1−アミルアルコール、2−アミルアルコール、5−ノニルアルコール等のアルコール性水酸基を有する化合物、及びカプサイシン、ジヒドロカプサイシン、ノニル酸バニリルアミド、カテキン、エピカテキン、バニリン、ハイドロキノン、カテコール、レゾルシノール、3,4−ジメトキシフェノール等のフェノール性水酸基を有する化合物が挙げられている。また、生成が認められた配糖体はモノグルコシドのみである。
【0004】
特許文献2は、カテキン類とグルコース−1−リン酸又はシュークロースとの混合液にシュークロースホスホリラーゼを作用させ、カテキン類配糖体を製造する方法を開示する。ここでは、シュークロースホスホリラーゼの起源として、Leuconostocmesenteroides、Pseudomonas saccharophila、Pseudomonasputrefaciens、Clostridium pasteurianum、Acetobacter xylinum、Pullulariapullulansが挙げられている。ここでは、受容体として、カテキン類としては、(+)−カテキン、(−)−エピカテキン、(−)−エピカテキン3-O-ガレート、(−)−エピガロカテキン、及び(−)−エピガロカテキン3-O-ガレートが挙げられているが、実施例として記載されているのは、(+)−カテキンを受容体に用いて(+)−カテキン 3'-O-α-D-グルコピラノシドを製造した例のみである。
【0005】
特許文献3は、5、7、3'、4'、5'、3"、4"、5"位の少なくとも1つにグルコース残基又は重合度2〜8のマルトオリゴ糖残基が結合しているエピガロカテキン3-O-ガレート誘導体を開示する。ここで実施例として記載されているのは、特許文献2と同様、(-)-エピガロカテキンガレートとグルコース-1-リン酸又はシュークロースとの混合液にシュークロースホスホリラーゼを作用させ、4'-O-α-D-グルコピラノシル(-)-エピガロカテキンガレート及び4',4”-O-α-D-ジ-グルコピラノシル(-)-エピガロカテキンガレートを製造した例のみである。
【0006】
特許文献4は、ポリフェノール類を配糖化することにより渋みを低減した茶抽出物又は茶飲料を開示する。ここでは、茶抽出物又は茶飲料の渋みを低減する具体的な方法として、茶抽出物又は茶飲料に、デキストリン、サイクロデキストリン、澱粉又はこれらの混合物を添加し、これにサイクロマルトデキストリングルカノトランスフェラーゼを作用させる方法が記載されている。実施例においては、緑茶抽出物とα−サイクロデキストリンにBacillus stearothermophilus由来のサイクロマルトデキストリングルカノトランスフェラーゼを作用させ、得られた反応生成物には渋味が低減されたことが述べられており、またこれはエピガロカテキン3-O-ガレートやエピカテキンといったポリフェノール類が配糖化されていたことを示すものである旨が述べられている。しかしながら、反応生成物の具体的な構造は明らかにされていない。
【0007】
特許文献5は、3'位、3'と5位及び3'と7位が配糖化されたカテキン類の配糖体を開示する。ここでは、そのための具体的な方法として、特許文献4と同様、カテキン類及びデキストリン、サイクロデキストリン、澱粉又はこれらの混合物に、Bacillusstearothermophilus由来のサイクロマルトデキストリングルカノトランスフェラーゼを作用させる方法が記載されている。そして、糖供与体としてデキストリンを用いた上記方法による実施例においては、(-)-エピガロカテキン、(-)-エピガロカテキン3-O-ガレート、(-)-エピカテキン3-O-ガレートの配糖体については、分子吸光係数から、各ポリフェノール1分子当り平均で6〜8個グルコースが結合しているものも得られたと考察している。また、上記方法で得られた配糖体にリゾプス・ニベウス由来のグルコアミラーゼを作用させることで、3',7-ジ-O-α-D-グルコピラノシル(-)-エピガロカテキン、3',5-ジ-O-α-D-グルコピラノシル(-)-エピガロカテキン、3'-O-α-D−グルコピラノシル(-)-エピガロカテキン、3',7-ジ-O-α-D-グルコピラノシル(-)-エピガロカテキン3-O-ガレート、3'-O-α-D-グルコピラノシル(-)-エピガロカテキン3-O-ガレート、3'-O-α-D-グルコピラノシル(-)-ガロカテキン、3'-O-α-D−グルコピラノシル(-)-エピカテキン3-O-ガレートの生成を確認している。
【0008】
また、カテキン配糖体の効果として、非特許文献1には、渋みの低減、水溶性の向上、安定性の改善、チロシナーゼ阻害が、非特許文献2には、変異原性抑制が記載されている。
【特許文献1】特開2001-46096
【特許文献2】特開平5-176786(特許第3024848号)
【特許文献3】特開平7-10897(特許第3071610号)
【特許文献4】特開平8-298930(特許第3579496号)
【特許文献5】特開平9-3089(特許第3712285号)
【非特許文献1】Biosci. Biotech. Biochem., 57 (10), 1666-1669 (1993)
【非特許文献2】Biosci. Biotech. Biochem., 57 (10), 1290-1293 (1993)
【発明の開示】
【発明が解決しようとする課題】
【0009】
これらの配糖化技術は、用いる酵素の糖供与体特異性、配糖化可能な化合物についての特異性、配糖化効率等の観点からは充分なものとはいえず、さらに優れた配糖化酵素の開発が望まれている。
【課題を解決するための手段】
【0010】
本発明者らは、カテキン等を含むフラボノイドの配糖化技術について鋭意検討してきた。その結果、Trichoderma virideの培養上清から新規な配糖化酵素を見いだし、その遺伝子をクローン化することにより本発明を完成した。
【0011】
〔配糖化酵素遺伝子等〕
本発明は、子嚢菌類の糸状菌(好ましくは、Trichoderma属に属する菌、より好ましくは、Trichoderma viride)が生産する配糖化酵素タンパク質をコードするポリヌクレオチド又はそのホモログ、すなわち下記の(A)〜(K)のいずれか一を含むポリヌクレオチド(好ましくは、下記の(A)〜(K)のいずれか一であるポリヌクレオチド)を提供する:
(A)配列番号:6に記載の塩基配列の全部又は一部(例えば、423〜1928番の塩基配列部分の全部又は一部、配列番号:9の塩基配列に対応する部分)からなるポリヌクレオチド;
(B)(A)に記載のポリヌクレオチドの塩基配列と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(C)(A)に記載のポリヌクレオチドの塩基配列において1若しくは複数の塩基が置換、欠失、挿入、及び/又は付加された塩基配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(D)(A)に記載のポリヌクレオチドの塩基配列と少なくとも60%以上の同一性を有し、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(E)配列番号:9に記載の塩基配列からなるポリヌクレオチド;
(F)(E)に記載のポリヌクレオチドの塩基配列と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(G)(E)に記載のポリヌクレオチドの塩基配列において1若しくは複数の塩基が置換、欠失、挿入、及び/又は付加された塩基配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(H)(E)に記載のポリヌクレオチドの塩基配列と少なくとも60%以上の同一性を有し、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(I)配列番号:10に記載のアミノ酸配列からなるタンパク質をコードするポリヌクレオチド;
(J)配列番号:10に記載のアミノ酸配列において1若しくは複数のアミノ酸が置換、欠失、挿入、及び/又は付加したアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(K)配列番号:10に記載のアミノ酸配列と少なくとも60%以上の同一性を有するアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド。
【0012】
配列番号:6にはT. viride IAM5141株から得られた配糖化酵素TRa2のゲノムDNAの塩基配列が、配列番号:9にはそのcDNA配列が、配列番号:10にはTRa2の推定アミノ酸配列が示されている。
【0013】
TRa2の推定アミノ酸配列をSignalPで解析すると、配列番号:10の1〜20番目のアミノ酸配列は、分泌シグナルであると推定される。したがって、配糖化酵素として作用するのに必須の成熟タンパク質は配列番号:10の21番目以降の配列であり、大腸菌など、異種発現系で同酵素を発現させる場合には、シグナル配列を除去して発現させるのが適切であると考えられる。このような成熟タンパク質をコードするポリヌクレオチドもまた、本発明の範囲に含まれる。
【0014】
したがって、本発明はまた、下記の(L)〜(R)のいずれか一を含むポリヌクレオチド(好ましくは、下記の(L)〜(R)のいずれか一であるポリヌクレオチド)を提供する:
(L)配列番号:25に記載の塩基配列からなるポリヌクレオチド;
(M)配列番号:25に記載の塩基配列と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(N)配列番号:25に記載のポリヌクレオチドの塩基配列において1若しくは複数の塩基が置換、欠失、挿入、及び/又は付加された塩基配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(O)配列番号:25に記載の塩基配列と少なくとも60%以上の同一性を有し、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(P)配列番号:26に記載のアミノ酸配列からなるタンパク質をコードするポリヌクレオチド;
(Q)配列番号:26に記載のアミノ酸配列において1若しくは複数のアミノ酸が置換、欠失、挿入、及び/又は付加したアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(R)配列番号:26に記載のアミノ酸配列と少なくとも60%以上の同一性を有するアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド。
【0015】
配列番号:25には成熟タンパク質としてのTRa2をコードするcDNAの塩基配列、すなわち配列番号9の61-1389番目の塩基配列が、配列番号:26には成熟タンパク質としてのTRa2のアミノ酸配列、すなわち配列番号10の21-463番目のアミノ酸配列が示されている。
【0016】
本発明はまた、Trichoderma属由来であり、好ましくはTrichoderma viride、Trichodermareesei、Trichoderma saturnisporum、Trichoderma ghanense、Trichodermakoningii、 Trichoderma hamatum、Trichoderma harzianum又はTrichodermapolysporum由来であり、配列番号:11〜24のいずれか一に記載の塩基配列を含み、フラボノイド類の配糖化活性を有するタンパク質コードするポリヌクレオチドを提供する。このようなポリヌクレオチドの好ましい例は、配列番号:11〜24のいずれか一に記載の塩基配列を含み、配列番号:6、9又は25に記載の塩基配列と高い同一性を有し、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチドである。
【0017】
本明細書で「配糖化活性」というときは、フラボノイド類に糖残基を転移する活性を有することをいう。あるタンパク質が、フラボノイド類に糖残基を転移する活性を有するか否かは、特別な場合を除き、例えば、本明細書の実施例に示されているように、フラボノイド(例えば、カテキン)と適切な糖供与体(例えば、デキストリン)との混合液に、対象酵素を供し、充分な時間反応させ、反応液を高速液体クロマトグラフィー(HPLC)等に供して、分析することにより確認することができる。
【0018】
本発明のポリヌクレオチドによってコードされるタンパク質は、配糖化活性とともに、他の活性、例えばデキストリナーゼ活性を有していてもよい。本明細書で「デキストリナーゼ」というときは、特別な場合を除き、澱粉、デキストリン等、α-グルコシド結合した糖質を加水分解することができる酵素をいう。デキストリナーゼは、アミラーゼの一種である。対象がデキストリナーゼ活性を有するか否かは、適切な条件下で、市販のデキストリン(例えば、澱粉を酸、熱又は酵素により加水分解し、平均分子量を3,500程度としたもの)に対して作用させ、デキストリンが加水分解されているか否かにより、判断することができる。当業者であれば、対象を作用させる反応の適切な条件、及びデキストリンが加水分解されているか否かを判断するための手法を、適宜設計することができる。
【0019】
(フラボノイド類)
本明細書で「フラボノイド類」というときは、特別な場合を除き、フラボノイド及びエスクレチンを含む。
【0020】
本明細書で「フラボノイド」というときは、特別な場合を除き、カテキン類(フラバノール)、フラバノン、フラボン、フラボノール、フラバノノール、イソフラボン、アントシアン及びカルコン、並びにそのメチル化体をいう。フラボノイドには、ナリンゲニン、ケルセチン、ダイゼイン、ゲニステイン、ケンフェロールが含まれる。本発明に用いることのできるフラボノイドは、天然物由来であってもよく、合成されたものであってもよい。
【0021】
本明細書で「カテキン類」というときは、特別な場合を除き、広義のカテキンの意味で用いており、3-オキシフラバンのポリオキシ誘導体をいう。これには、カテキン、ガロカテキン及びそれらの3-ガロイル体が含まれ、またそれらの光学異性体((+)体、(-)体、(+)-epi体、(-)-epi体)及びラセミ体が含まれる。具体的には、カテキン、ガロカテキン(gallocatechin;GC)、カテキンガレート(catechin-3-O-gallate;CG)、ガロカテキンガレート(gallocatechin-3-O-gallate;GCG)、エピカテキン(epicatechin;EC)、エピガロカテキン(epigallocatechin;EGC)、エピカテキンガレート(epicatechin-3-O-gallate;ECG)及びエピガロカテキンガレート(epigallocatechin-3-O-gallate;EGCG)、並びにそれらの光学異性体が含まれる。カテキン類のメチル化体とは、上述したカテキン類において、少なくとも一つのOH基のHがメチルで置き換わったものをいう。カテキン類のメチル化体の例としては、エピカテキン、エピガロカテキン、エピカテキンガレート又はエピガロカテキンガレートにおいて、3'位、4'位、3"位及び4"位のいずれかのOH基のHがメチルで置き換わったものを挙げることができる。本発明に用いることのできるカテキン類及びカテキン類のメチル化体は、天然物由来であってもよく、合成されたものであってもよい。天然物の例としては、茶抽出物、その濃縮物及び精製物(例えば、テアビゴ(DSM ニュートリション ジャパン)、ポリフェノン(三井農林)及びサンフェノン(太陽化学)等の緑茶抽出物)、及びベニフウキ抽出物を挙げることができる。
【0022】
本発明には、フラボノイド類として、単一の化合物を用いることができ、また、2種以上のフラボノイド類の混合物を用いることもできる。
(糖供与体)
本明細書で「糖供与体」というときは、特別な場合を除き、本発明の方法において酵素の基質となり、加水分解されてフラボノイド類に対して糖残基を供給しうる糖質をいう。本発明に用いることができる糖供与体は、マルトトリオース残基を含む糖質であり、これには、マルトトリオース、マルトテトラオース、マルトペンタオース、マルトヘキサオース、マルトヘプタオース、デキストリン、γ-シクロデキストリン、及び可溶性澱粉が含まれる。本明細書で「デキストリン」というときは、特別な場合を除き、澱粉を加水分解したものをいい、「可溶性澱粉」というときは、特別な場合を除き、澱粉の加水分解物であって、熱水に可溶であるものをいう。加水分解は、酸、熱、酵素等のいずれの手法によってもよい。本発明においては、デキストリンとして、例えば平均分子量が約3,500のものを、可溶性澱粉として、例えば平均分子量が約100万のものを用いることができる。
【0023】
本発明でいう「ストリンジェントな条件」とは、特別な場合を除き、6M尿素、0.4% SDS、0.5×SSCの条件又はこれと同等のハイブリダイゼーション条件を指し、さらに必要に応じ、本発明には、よりストリンジェンシーの高い条件、例えば、6M尿素、0.4% SDS、0.1×SSC又はこれと同等のハイブリダイゼーション条件を適用してもよい。それぞれの条件において、温度は約40℃以上とすることができ、よりストリンジェンシーの高い条件が必要であれば、例えば約50℃、さらに約65℃としてもよい。
【0024】
また、明細書において「1若しくは複数の塩基が置換、欠失、挿入、及び/又は付加された塩基配列」というときの置換等されるヌクレオチドの個数は、その塩基配列からなるポリヌクレオチドがコードするタンパク質が所望の機能を有する限り特に限定されないが、1〜9個又は1〜4個程度であるか、同一又は性質の似たアミノ酸配列をコードするような置換等であれば、さらに多くの個数の置換等がありうる。また、本明細書において「1若しくは複数の塩基が置換、欠失、挿入、及び/又は付加されたアミノ酸配列」というときの置換等されるアミノ酸の個数は、そのアミノ酸配列を有するタンパク質が所望の機能を有する限り特に限定されないが、1〜9個又は1〜4個程度であるか、性質の似たアミノ酸への置換であれば、さらに多くの個数の置換等がありうる。このような塩基配列又はアミノ酸配列に係るポリヌクレオチド又はタンパク質を調製するための手段は、当業者にはよく知られている。
【0025】
本明細書においてアミノ酸配列に関し、高い同一性というときは、少なくとも60%以上、好ましくは70%以上、より好ましくは80%以上、さらに好ましくは90%以上、最も好ましくは95%以上の配列の同一性を指す。
【0026】
ポリヌクレオチド配列又はアミノ酸配列の同一性に関する検索・解析は、当業者には周知のアルゴリズム又はプログラム(例えば、BLASTN、BLASTP、BLASTX、ClustalW)により行うことができる。プログラムを用いる場合のパラメーターは、当業者であれば適切に設定することができ、また各プログラムのデフォルトパラメーターを用いてもよい。これらの解析方法の具体的な手法もまた、当業者には周知である。
【0027】
本発明のポリヌクレオチドは、天然物からハイブリダイゼーション技術、ポリメラーゼ連鎖反応(PCR)技術等を利用して得ることができる。
本発明のポリヌクレオチドは、好ましくは、Trichoderma属由来、より好ましくは、Trichoderma viride由来である。
【0028】
本発明はまた、本発明のポリヌクレオチドを含む組換えベクター、その組換えベクターにより形質転換された、形質転換体(例えば形質転換大腸菌、形質転換酵母、形質転換昆虫細胞)も提供する。本発明はさらに、本発明のポリヌクレオチドを用いて宿主(例えば大腸菌、酵母、昆虫細胞)を形質転換する工程(例えば、本発明の組換えベクターにより形質転換する工程)を含む、形質転換方法も提供する。
【0029】
本発明のポリヌクレオチドが挿入されるベクターは、宿主内で挿入物を発現させることが可能なものであれば特に制限はなく、ベクターは、通常、プロモーター配列、ターミネーター配列、外的な刺激により誘導的に挿入物を発現させるための配列、目的遺伝子を挿入するための制限酵素により認識される配列、及び形質転換体を選択するためのマーカーをコードする配列を有する。組換えベクターの作成、組換えベクターによる形質転換方法は、当業者に周知の方法を適用することができる。
【0030】
〔配糖化酵素タンパク質〕
本発明はまた、新規配糖化酵素タンパク質及びそのホモログ、すなわち下記の(i)、(j)又は(k)を含むタンパク質(好ましくは、下記の(i)、(j)又は(k)であるタンパク質)を提供する:
(i)配列番号:10に記載のアミノ酸配列からなるタンパク質;
(j)配列番号:10に記載のアミノ酸配列において1若しくは複数のアミノ酸が置換、欠失、挿入、及び/又は付加したアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質;
(k)配列番号:10に記載のアミノ酸配列と少なくとも60%以上の同一性を有するアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質。
【0031】
本発明はまた、上述の新規配糖化酵素タンパク質において推定分泌シグナル配列部分を除いた成熟タンパク質及びそのホモログ、すなわち下記の(p)、(q)又は(r)を含むタンパク質(好ましくは、下記の(p)、(q)又は(r)であるタンパク質)も提供する:
(p)配列番号:26に記載のアミノ酸配列からなるタンパク質;
(q)配列番号:26に記載のアミノ酸配列において1若しくは複数のアミノ酸が置換、欠失、挿入、及び/又は付加したアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質;
(r)配列番号:26に記載のアミノ酸配列と少なくとも60%以上の同一性を有するアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質。
【0032】
本発明はまた、Trichoderma属由来であり、好ましくはTrichoderma viride、Trichodermareesei、Trichoderma saturnisporum、Trichoderma ghanense、Trichodermakoningii、 Trichoderma hamatum、Trichoderma harzianum又はTrichodermapolysporum由来であり、配列番号:11〜24のいずれか一に記載の塩基配列を含むポリヌクレオチドによってコードされる、フラボノイド類の配糖化活性を有するタンパク質を提供する。このようなタンパク質の好ましい例は、配列番号:11〜24のいずれか一に記載の塩基配列を含み、配列番号:6、9又は25に記載の塩基配列と高い同一性を有するポリヌクレオチドによってコードされる、フラボノイド類の配糖化活性を有するタンパク質である。
【0033】
本発明のタンパク質は、Trichoderma属に属する菌、例えばTrichoderma viride、Trichodermareesei、Trichoderma saturnisporum、Trichoderma ghanense、Trichodermakoningii、 Trichoderma hamatum、Trichoderma harzianum又はTrichodermapolysporumを公知の培養液で培養して得られる培養上清から単離・精製することができる。また、本発明のポリヌクレオチドを挿入した組換えベクターにより形質転換された形質転換体(例えば、形質転換酵母や形質転換大腸菌)により、組換えタンパク質として得ることができる。
【0034】
〔酵素TRa2の酵素学的性質〕
本発明によって提供される新規な酵素タンパク質のうち、特に好ましい例は、Trichoderma viride IAM 5141株の培養上清から得られる、配列番号:10に記載のアミノ酸配列からなる配糖化酵素(本明細書では、「TRa2」ということもある。)である。この酵素は、フラボノイドと糖供与体との反応において、以下のような酵素学的特徴を有する。
【0035】
糖供与体選択性:
本酵素は、実施例に示した条件では、マルトトリオース、マルトテトラオース、マルトペンタオース、マルトヘキサオース、マルトヘプタオース、可溶性澱粉、デキストリン、γ-シクロデキストリン等を糖供与体とするが、セロビオース、デキストラン、マルトース一水和物、カルボキシメチルセルロースナトリウム、イソマルトオリゴ糖、α-シクロデキストリン、β-シクロデキストリン等は糖供与体としない。また、グルコース1分子又は2分子からなる糖のみならず、グルコース3分子(G3)以上の糖鎖長の配糖体を産生することができる配糖化酵素である。
【0036】
基質特異性:
本酵素は、カテキン、エピガロカテキンガレート、ナリンゲニン、ケルセチン、ダイゼイン、ゲニステイン、ケンフェロール等の主要なフラボノイドやエスクレチンなどを含むポリフェノールに対し、広く作用し、配糖化することができる。
【0037】
反応最適温度、pH(反応温度依存性、pH依存性):
本酵素は、加水分解反応においては、実施例に示した条件では、約20〜約60℃、特に約35〜約55℃でよく反応し、またpH約4.0〜約7.0、特にpH約4.5〜約5.5でよく反応する。
【0038】
また、本酵素は、糖転移反応においては、実施例に示した条件では、約20〜約60℃、特に約45〜約55℃でよく反応し、またpH約4.0〜約9.0、特にpH約4.5〜約7.0でよく反応する。
【0039】
温度安定性、pH安定性:
本酵素は、実施例に示した条件では、約20〜約45℃の1hr処理によっては活性を失わず、またpH約5.0〜約9.0での室温、16hr処理によっても活性を失わない。
【0040】
〔フラボノイド類の配糖体の製造方法〕
本発明はまた、フラボノイド類と糖供与体(好ましくは、デキストリン、可溶性澱粉、γ-シクロデキストリン)との混合液に、本発明の新規配糖化酵素を作用させることを特徴とする、フラボノイド類の配糖体の製造方法を提供する。
【0041】
本発明によって提供されるフラボノイド類の配糖体は、下式で表されるものである:
【0042】
【化1】
【0043】
式(I)
式中:
R1〜R5は、少なくとも一つが糖残基であり、他はOH若しくはOCH3であるか、又は
R1〜R4は、少なくとも一つが糖残基であり、他はOH若しくはOCH3であり、R5はHであり;そして
Xは、H、CH3、ガロイル基又はメチル化されたガロイル基である。
【0044】
これには、さらに、下式で表されるフラボノイド類の配糖体が含まれる:
5-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(+)-カテキン;
5-O-α-D-グルコピラノシル-(+)-カテキン;
4'-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(+)-カテキン;
4'-O-α-D-グルコピラノシル-(+)-カテキン;
3'-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(+)-カテキン;
3'-O-α-D-グルコピラノシル-(+)-カテキン;
7-O-α-D-グルコピラノシル-(-)-エピガロカテキン-3-O-ガレート;
7-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(-)-エピガロカテキン-3-O-ガレート;
3'-O-α-D-グルコピラノシル-(-)-エピガロカテキン-3-O-ガレート;及び
3'-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(-)-エピガロカテキン-3-O-ガレート及びそれらの光学異性体。
【0045】
〔配糖体の用途〕
本発明により得られた配糖体は、食品用組成物、医薬用組成物又は化粧用組成物として用いることができる。より詳細には、例えばカテキン類配糖体を配合したものは、カテキンと同様に、抗アレルギー、抗酸化、抗癌、抗炎症、抗菌・抗う歯、抗ウイルス、解毒、腸内フローラ改善、消臭、血漿コレステロール上昇抑制、血圧上昇抑制、血糖上昇抑制、血小板凝集抑制、痴呆予防、体脂肪燃焼、体脂肪蓄積抑制、持久力向上、抗疲労、腎機能改善のための剤として、あるいは食品用組成物、医薬用組成物又は化粧用組成物として用いることができる。
【0046】
食品用組成物は、栄養補助食品、健康食品、食事療法用食品、総合健康食品、サプリメント及び飲料を含む。飲料は、茶飲料、ジュース、清涼飲料、ドリンク剤を含む。医薬品組成物は、医薬品又は医薬部外品とすることができ、経口製剤又は皮膚用外用剤であることが好ましく、液剤、錠剤、顆粒剤、丸剤、シロップ剤、又はローション剤、スプレー剤、硬膏剤、軟膏剤の形態とすることができる。化粧用組成物は、クリーム、液状ローション、乳液状ローション、スプレーの形態とすることができる。
【0047】
本発明の食品用組成物、医薬用組成物又は化粧用組成物中における配糖体の配合量は、特に限定されないが、当業者であれば、対応するフラボノイド類の好ましい一日摂取量を参考に、溶解性、呈味等を考慮して、適宜設定することができる。例えば、組成物中における本発明の配糖体の配合量を0.01〜99.9重量%とすることができ、また本発明の配糖体を一日当たり100 mg〜20 gを1〜数回(例えば3回)に分けて摂取することができるように、組成物中の配合量を決定することができる。
【0048】
本発明の食品用組成物、医薬用組成物又は化粧用組成物には、食品、医薬又は化粧品として許容できる種々の成分を添加することができる。これらの添加剤及び/又は成分の例として、ビタミン類、糖類、賦形剤、崩壊剤、結合剤、潤沢剤、乳化剤、緊張化剤(等張化剤)、緩衝剤、溶解補助剤、防腐剤、安定化剤、抗酸化剤、着色剤、矯味剤、香料、凝固剤、pH調整剤、増粘剤、茶エキス、生薬エキス、無機塩等が挙げられる。
【0049】
〔他の態様〕
本発明において、配糖化酵素タンパク質は、適当な担体に固定化し、固定化酵素として用いることができる。担体としては、同様の目的で使用される従来の樹脂、例えば、塩基性樹脂(例えば、MARATHON WBA(Dow Chemical社製)、SAシリーズ、WAシリーズ又はFPシリーズ(三菱化学)、及びアンバーライトIRA904(オルガノ))、疎水性樹脂(例えば、ダイアイオンFPHA13(三菱化学)、HPシリーズ(三菱化学)、アンバーライトXAD7(オルガノ))等を用いることができる。他に、エキスプレスイオンD(Whatman)、DEAE-トヨパール650M(東ソー)、DEAE-セファロースCL4B(Amersham Biosciences)等も好適に用いうる。固定化の方法としては、従来の、物理的吸着、イオン結合又は共有結合を介して固定化する結合法、二価性官能基をもつ試薬で架橋固定化する架橋法、網目構造のゲル又は半透膜の中に酵素を包埋する包括法のいずれも採りうる。例えば、まず各樹脂5mlに対し、蒸留水に溶解した酵素20〜2,000 mg、例えば50〜400 mgを吸着させた後、上清を除去し、乾燥することによる。
【0050】
本発明はまた、配列番号:6、配列番号:9、配列番号:10、配列番号:11〜26のいずれか一に記載の塩基配列又はアミノ酸配列の全部若しくは一部を用いることを特徴とする、フラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチドの塩基配列又はフラボノイド類の配糖化活性を有するタンパク質のアミノ酸配列を決定する方法も提供する。
【発明の効果】
【0051】
本発明により、フラボノイドを効率的に配糖化することができる新規酵素及びその遺伝子が提供される。
本発明の新規酵素によりフラボノイド類を配糖化することによって、水溶性を向上することができる。そのため、本発明により、フラボノイド類の経口吸収性を高めることができると考えられる。また、水溶性の向上は、水への溶解速度の向上、ひいては生体内での吸収速度の向上に資するものである。したがって、本発明により、フラボノイド類の有用な活性、例えば抗酸化活性を、生体内で効率よく発揮させることが可能となる。
【0052】
また、本発明によりフラボノイド類を配糖化することによって、フラボノイド類の呈味を改変することができる。特にフラボノイド類としてカテキン類のように苦味、渋味を呈するものを本発明により配糖化する場合には、そのような呈味を低減することができる。
【0053】
また、本発明によりフラボノイドを配糖化することによって、熱安定性を向上することができる。
【実施例】
【0054】
〔実施例1:Trichoderma培養液のカテキン配糖化活性〕
Trichoderma viride IAM 5141株のスラントから、酵母エキス(Difco)1%、ポリペプトン(日本製薬)1%、デキストリン(ナカライテスク)2%の液体培地 10mlに植菌し、30℃で1日間振とう培養を行い、前培養液とした。さらに、上記液体培地900mlに前培養液を全量植菌して30℃で3日間培養を行い、フィルターろ過により培養上清液を調製した。培養上清690mlに対して、硫酸アンモニウム387g(80%飽和)を添加し攪拌した後、遠心分離により沈殿を回収した。得られた沈殿に10mlの0.1M酢酸バッファー(pH5.0)を加え、粗酵素液とした。
【0055】
粗酵素液100μlに対して、カテキン3mg、デキストリン10mgを添加し、50℃で24時間攪拌して酵素反応を行った。反応液を0.1%トリフルオロ酢酸(TFA)で10倍希釈し、10μlを高速液体クロマトグラフィー(HPLC)で分析を行った。
【0056】
分析条件
カラム:Develosil C30-UG-5(4.6x150mm)
グラジエント条件: 5%B液 → 50%B液 / 20分間
溶離液A:0.1%TFA / 蒸留水
溶離液B:90%アセトニトリル/ 0.08%TFA
流量:1ml/min
検出波長:280nm
図1に示したとおり、上記反応によるカテキン配糖体の生成を確認できた。また、糖供与体としてγ-サイクロデキストリンを用いた場合にも同様の配糖体の生成を確認できた。このことから、T. viride IAM5141株はデキストリン又はγサイクロデキストリンを糖供与体として、カテキンを配糖化する酵素を分泌生産していることが示唆された。
【0057】
〔実施例2:PCRによるα−アミラーゼホモログの部分配列のクローニング〕
デキストリン及びγ‐サイクロデキストリンは、グルコースがα-1,4結合で結合したポリマーであり、目的とする酵素はこれを分解する活性を有していることから、α−アミラーゼファミリー様の酵素である可能性が示唆された。
【0058】
そこで、トリコデルマと同じ子嚢菌類の糸状菌でゲノム配列が明らかになっている微生物のうち、Aspergillus nidulans、Neurosporacrassa、Magnaporthe grisea、Fusarium graminearum のゲノム情報データベースから、プロテインファミリーデータベース(PFAM)のalpha-amylase, catalytic domain(accession No. PF00128)のモチーフを持つ推定ORFのアミノ酸配列をそれぞれ9個、6個、8個、6個抽出した。これらを配列相同性検索プログラムであるClustalWによりアライメントを作成し、系統樹作成プログラムであるTree viewにより系統樹を作成し、相同性をもとにグループ分けを行った。図2のグループ1の4つのアミノ酸配列(括弧内はGenebank アクセッションNo.)MG02772.4(EAA47529)、MG10209.4(EAA48146)、AN3388.2(EAA63356)、FG03842.1(EAA71544)をアライメントし、保存性の高い部分(図3、下線部)のアミノ酸配列に対応するオリゴDNAを合成した。
【0059】
AMY-12f:
5'-TAYTGYGGNGGNACNTTYAARGGNYT-3'(配列番号:1)
AMY-15r:
5'-TTYTCNACRTGYTTNACNGTRTCDAT-3'(配列番号:2)
AMY-17r:
5'-GGTNAYRTCYTCNCKRTTNGCNGGRTC-3'(配列番号:3)
先に培養したT. viride IAM5141菌体約1gの湿菌体よりDNeasy plant Maxi Kit(QIAGEN)により、ゲノムDNAを抽出した。このゲノムDNA50ngを鋳型として、プライマーAMY-12fとプライマーAMY-15r、又はプライマーAMY-12fとプライマーAMY-17rを用いてPCR反応を行った。すなわち、ExTaq(タカラバイオ社製)を用いて、94℃ 2分、(94℃ 1分、50℃1分、72℃ 1分)を30サイクル、72℃ 10分の反応を行った。PCR産物をアガロースゲル電気泳動で分析したところ、プライマーAMY-12fとプライマーAMY-15rの組み合わせで約0.6kbp、プライマーAMY-12fとプライマーAMY-17rの組み合わせで約1.0kbpの断片が確認できた。そこで、これらのDNA断片をアガロースゲルから切り出し、GFX PCR DNA and Gel Band Purifiction Kit(アマシャムバイオサイエンス)によりDNAを精製した。これをTOPO-TA cloning kit(インビトロジェン)にてクローニングし、塩基配列の解析をABI3100Avant(アプライドバイオシステムズ社製)にて行った。前者で得られた塩基配列は後者で得られた塩基配列に含まれていた。この塩基配列をGenBankに登録されているアミノ酸配列に対してBlastxにより相同性検索を行ったところ、MG10209.4(EAA48146)と最も高い相同性を示した。
【0060】
〔実施例3:α−アミラーゼホモログのゲノム配列の決定〕
得られた約1.0kbpの塩基配列をもとに、以下のプライマーを設計し、Inverse PCR(逆PCR)を行った。
【0061】
TRa2-2:
5'-CCAACCTGGTATCTACATAC-3'(配列番号:4)
TRa2-3:
5'-AGATGGCATCAAATCCCAT-3'(配列番号:5)
まず、T. viride IAM5141より調製したゲノムDNAをHindIIIあるいはPstIで完全消化し、ligation high(東洋紡)により、16℃で一晩反応させ、セルフライゲーションにより閉環させた。これらのDNA 0.1μgを鋳型として、上記プライマーTRa2-2及びTRa2-3を用いてPCR反応を行った。PCRは、LA Taq(タカラバイオ)を使用して、94℃ 2分、(95℃ 30秒、66℃ 15分)を30サイクル、72℃ 10分の反応を行った。得られたPCR産物をアガロースゲル電気泳動で分析したところ、HindIIIで消化したゲノムDNAを鋳型としたものでは約2kb、PstIで消化したゲノムを鋳型としたものでは約4.5kbのDNA断片が確認できた。これらをそれぞれアガロースゲルから切り出し、前述と同様にクローニングした。挿入された断片の両端からそれぞれ塩基配列を決定した。HindIIIで消化したゲノム由来のものとPstIで消化したゲノム由来のものは、当該制限酵素サイトまでの塩基配列が一致していた。こうして得られた塩基配列を、先に得られた部分配列と連結した。この塩基配列を図4(TRa2-gDNA)及び配列番号:6に示す。図2グループ1の4つの配列との比較及び、開始コドンや終始コドンの出現などからα−アミラーゼホモログのコード領域を推定した。開始コドンは塩基番号423-425のATG、終始コドンは1926-1928のTAAであると考えられた。
【0062】
〔実施例4:α−アミラーゼホモログのcDNAのクローニング〕
先に培養したT. viride IAM5141株の菌体約0.1gより、RNeasy plant mini kitにより全RNAを抽出した。1μgの全RNAをスーパースクリプトファーストストランドシステム for RT-PCR(インビトロジェン)でランダムヘキサマーを用いて、cDNAを合成した。
【0063】
先に得られたゲノム配列をもとに、以下のプライマーを設計した。
TRa2EcoRI-f2:
5'-GGAATTCATGAAGCTTCGATCCGCCGTCCC-3'(配列番号:7)
TRa2XhoI-r2:
5'-CCGCTCGAGTTATGAAGACAGCAGCACAAT-3'(配列番号:8)
合成されたcDNAを鋳型として、上記プライマーTRa2EcoRI-f2及びTRa2XhoI-r2を用いて、PCR反応を行った。PCRは、Ex Taq(タカラバイオ)を使用し、94℃ 2分、(94℃ 1分、55℃ 1分、72℃ 2分)を30サイクル、72℃ 10分の反応を行った。得られたPCR産物をアガロースゲル電気泳動で分析したところ、約1.5kbのDNA断片が確認できた。これをアガロースゲルより切り出し、GFXにて精製した。得られたDNA断片をTOPO-TA cloning kit(インビトロジェン)にてクローニングして、プラスミドpCRTRa2-cDNAを構築し、cDNAの塩基配列を決定した(図4、図5及び配列番号:9)。先に得られたゲノムDNA配列とcDNA配列を比較したところ、ゲノム配列には2つのイントロンが含まれていた(図4)。cDNA配列には、1392bpのORFがあり、463アミノ酸残基からなるタンパク質をコードしていた(図5及び配列番号:10)。この遺伝子をTRa2と命名した。本遺伝子によりコードされる推定アミノ酸配列をSignalP(Nielsen H. et.al., Protein Eng., 10, 1-6, 1997 )で解析した結果、N末端から20アミノ酸残基は分泌シグナル配列であると考えられた。さらに、TRa2によりコードされる推定アミノ酸配列を前述の方法に従って相同性検索を行ったところ、AN3388.2(EAA63356)ともっとも高い相同性を示した。TRa2タンパク質の推定アミノ酸配列を、既知のα-アミラーゼであるタカアミラーゼのアミノ酸配列と比較した。その結果、α-アミラーゼファミリー酵素の4つの保存領域が本酵素においても保存されており(図6、2重下線)、活性中心とされるアスパラギン酸残基、グルタミン酸残基、アスパラギン酸残基がすべて保存されていた(図6、*印アミノ酸残基)。
【0064】
〔実施例5:TRa2タンパク質の酵母での分泌発現系の構築〕
プラスミドpCRTRa2-cDNAを制限酵素EcoRI及びXhoIで消化して得られた約1.5kbの断片を、プラスミドpYE22m(Biosci. Biotech. Biochem., 59(7), 1221-1228, 1995)を制限酵素EcoRI及びSalIで消化した断片とligation high(東洋紡)を用いて連結し、プラスミドpYETRa2を得た。
【0065】
プラスミドpYETRa2により、酵母S. cerevisiae EH1315株を酢酸リチウム法により、形質転換した。得られた形質転換株をTRa2-1株とした。TRa2-1株をYPD(Difco)液体培地 10mlに一白金耳植菌し、30℃で2日間振とう培養した。TRa2タンパク質はN末に20アミノ酸残基からなる分泌シグナル配列が存在するので、培養液中に分泌されると考えられた。そこで遠心分離により、菌体を沈殿させ培養上清を回収した。
【0066】
〔実施例6:TRa2の糖分解活性の測定〕
培養上清500μlをMicrocon YM-30(アミコン)を用いて、約5倍に濃縮した。上記濃縮液10μlを、0.5%のマルトース、マルトトリオース、マルトテトラオース、デキストリン、α-サイクロデキストリン、β-サイクロデキストリン、又は γ-サイクロデキストリンを含む20mM 酢酸バッファー(pH5.0)100μlに添加し、50℃で1時間反応させた。
【0067】
反応終了後、以下のとおりTLCにて分析した。プレートは、シリカゲルG-60プレート(メルク)を使用し、展開液として、2−プロパノール:アセトン:0.5M乳酸=2:2:1を使用した。検出は、硫酸:エタノール=1:9を噴霧して風乾したのち、ホットプレートで熱することにより行った。その結果、ベクターpYE22mにて形質転換したコントロール株(C-1株)の培養液では、いずれの糖の分解も確認できなかった。一方、TRa2-1株の培養液ではマルトトリオース、マルトテトラオース、デキストリン及びγ-サイクロデキストリンが分解されて主にマルトース及びグルコースが生成することが確認できたが、マルトース、α-サイクロデキストリン及びβ-サイクロデキストリンの分解は確認できなかった。
【0068】
〔実施例7:TRa2の糖転移活性の測定〕
培養上清の原液、又はVIVASPIN 10,000MWCO/PES製(VIVASCIENCE社)にて約5倍に濃縮した培養上清濃縮液100μlに(+)-カテキン又は(-)-エピガロカテキン-3-O-ガレート3mgと、デキストリン10mgを加えて、50℃にて1日間攪拌しながら反応させた。反応終了後、反応液を0.1%トリフルオロ酢酸溶液にて10倍希釈し、高速液体クロマトグラフィー(HPLC)にて、実施例1と同様の条件で分析した。その結果、ベクターpYE22mにて形質転換したコントロール株(C-1株)の培養上清と反応させた反応液には、反応生成物が確認できなかったのに対し、TRa2-1株では、カテキン配糖体及びエピガロカテキン-3-O-ガレート配糖体の生成が確認できた(図7)。
【0069】
〔実施例8:TRa2によるカテキン配糖体の製造〕
TRa2-1株をYPD液体培地200mlに植菌し30℃で3日間振とう培養した。遠心分離により菌体を集め、培養上清を得た。この培養上清100mlを限外ろ過ディスク:NMWL30000/再生セルロースで0.1M 酢酸バッファー(pH5) 100mlを加えながら50mlになるまで濃縮し、TRa2酵素液とした。上記TRa2酵素液50mlに(+)-カテキン1.5gとデキストリン 5gを混合し、45℃,18hr撹拌した。反応液を遠心分離し、上清をLH20(Amersham Biosciences)樹脂60ml/Ф2.5×20cmカラムに吸着させた。蒸留水120ml、10%エタノール 240mlで溶出し、配糖体画分を回収・凍結乾燥し、凍乾末530mgを得た。その50mgをとって蒸留水 5mlに溶かし、DevelosilC30-UG-5カラム20×250mm、A:0.1%TFA/蒸留水,B:90%メタノール/0.1%TFA,30%B、3ml/min、280nmで分取した。HPLCの保持時間の早い順にピーク1〜6を回収・凍結乾燥した。MS及びNMR解析から、ピーク1は5-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(+)-カテキン、ピーク2は5-O-α-D-グルコピラノシル-(+)-カテキン、ピーク3は4'-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(+)-カテキン、ピーク4は4'-O-α-D-グルコピラノシル-(+)-カテキン、ピーク5は3'-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(+)-カテキン、ピーク6は3'-O-α-D-グルコピラノシル-(+)-カテキンであることが示唆された。
【0070】
5-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(+)-カテキン:m/z 615.2、NMRδppm (D2O);2.71 (1H, dd), 2.85 (1H, dd), 3.42 (1H, t), 3.56-3.85 (9H, m), 4.19 (1H, t), 4.26 (1H, dd), 4.87 (1H, d), 5.70 (1H, d), 6.19 (1H, d), 6.39 (1H, d), 6.83 (1H, dd), 6.90-6.93 (2H, m) 。
【0071】
5-O-α-D-グルコピラノシル-(+)-カテキン:m/z 450.8、NMRδppm (D2O);2.62 (1H, dd), 2.81 (1H, dd), 3.43 (1H, t), 3.45-3.55 (1H, m), 3.6-3.7 (3H, m), 3.83 (1H, t), 4.18 (1H, dd), 4.76 (1H, d), 5.61 (1H, d), 6.09 (1H, d), 6.31 (1H, d), 6.77 (1H, d), 6.8-6.9 (2H, m) 。
【0072】
4'-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(+)-カテキン:m/z 615.2、NMRδppm (D2O);2.54 (1H, dd), 2.81 (1H, dd), 3.43 (1H, t), 3.60 (1H, dd), 3.68-3.94 (9H, m), 4.19-4.28 (2H, m), 4.82 (1H, d), 5.44 (1H, d), 5.62 (1H, d), 6.04 (1H, d), 6.11 (1H, d), 6.91 (1H, dd), 7.00 (1H, d), 7.22 (1H, d)。
【0073】
4'-O-α-D-グルコピラノシル-(+)-カテキン:m/z 453.2、NMRδppm (D2O);2.45 (1H, dd), 2.73 (1H, dd), 3.45 (1H, t), 3.65-3.75 (4H, m), 4.11 (1H, dd), 4.7-4.75 (2H, m), 5.53 (1H, d), 5.95 (1H, d), 6.02 (1H, d), 6.83 (1H, dd), 6.91 (1H, d), 7.15 (1H, d)。
【0074】
3'-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(+)-カテキン:m/z 615.2、NMRδppm (D2O);2.54 (1H, dd), 2.80 (1H, dd), 3.44 (1H, t), 3.59 ( 1H, dd), 3.67-3.90 (9H, m), 4.17-4.24 (2H, m), 4.83 (1H, d), 5.41 (1H, d), 5.55 (1H, d), 6.03 (1H, d), 6.10 (1H, d), 7.10 (1H, d), 7.06 (1H, d), 7.26 (1H, d)。
【0075】
3'-O-α-D-グルコピラノシル-(+)-カテキン:m/z 453.2、NMRδppm (D2O);2.43 (1H, dd), 2.73 (1H, dd), 3.27 (1H, s), 3.44 (1H, t), 3.6-3.7 (4H, m), 3.88 (1H, t), 4.10 (1H, dd), 4.69 (1H, d), 5.46 (1H, d), 5.93 (1H, s), 6.01 (1H, s), 6.89 (1H, d), 6.94 (1H, dd), 7.18 (1H, d)。
【0076】
〔実施例9:TRa2によるエピガロカテキン-3-O-ガレート配糖体の製造〕
TRa2-1株をYPD液体培地100mlに植菌し30℃で3日間振とう培養した。遠心分離により菌体を集め、培養上清を得た。この培養上清45mlを限外ろ過ディスク:NMWL30000/再生セルロースで0.1M 酢酸バッファー(pH5) 50mlを加えながら20mlになるまで濃縮し、TRa2酵素液とした。TRa酵素液 20mlに(-)-エピガロカテキン-3-O-ガレート600mgとデキストリン 2gを混合し、50℃、1日撹拌した。反応液を遠心し、上清をLH20樹脂25ml/Ф1.5×30cmカラムに吸着させた。蒸留水100ml、10%エタノール 100ml、20%エタノール 100ml、30%エタノール 200mlで溶出し、30%エタノール画分を回収・凍結乾燥した。凍乾末120mgを蒸留水12mlに溶かし、DevelosilC30-UG-5カラム20×250mm、A:0.1%TFA/蒸留水,B:90%メタノール/0.1%TFA,40%B、3ml/min、280nmで分取した。MS及びNMR解析から、ピーク2は7-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(-)-エピガロカテキン-3-O-ガレート、ピーク5は3'-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(-)-エピガロカテキン-3-O-ガレート、ピーク6は3'-O-α-D-グルコピラノシル-(-)-エピガロカテキン-3-O-ガレートであることが示唆された。一方、ピーク3はMS(m/z 621.2、1107.3)から、グルコシドとマルトテトラオシドの混合物であることが示唆され、保持時間からグルコシドは7-O-α-D-グルコピラノシル-(-)-エピガロカテキン-3-O-ガレートであると考えられた。
【0077】
7-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(-)-エピガロカテキン-3-O-ガレート:m/z 783.2、NMRδppm (CD3OD); 2.88 (1H, dd), 2.01 (1H, dd), 3.26 (1H, t), 3.46 (1H, dd), 3.6-3.9 (9H, m), 4.08 (1H, t), 5.00 (1H, s), 5.20 (1H, d), 5.43 (1H, d), 5.54 (1H, s), 6.27 (1H, d), 6.34 (1H, d), 6.51 (2H, d), 6.94 (2H, d)。
【0078】
3'-O-α-D-グルコピラノシル-(-)-エピガロカテキン-3-O-ガレート:m/z 621.1, δppm (CD3OD);2.88 (1H, dd), 2.99 (1H, dd), 3.42 (1H, dd), 3.51 (1H, t), 3.69 (1H, m), 3.8-3.9 (3H, m), 4.88 (1H, d), 4.98 (1H, s), 5.49 (1H, broad s), 5.95 (1H, d), 5.96 (1H, d), 6.65 (1H, d), 7.01 (2H, s), 7.11 (1H, d)。
【0079】
3'-O-(4-O-α-D-グルコピラノシル-α-D-グルコピラノシル)-(-)-エピガロカテキン-3-O-ガレート:m/z 783.2, δppm (CD3OD);2.87 (1H, broad d), 2.99 (1H, dd), 3.27 (1H, t), 3.44-3.48 (2H, m), 3.6-3.8 (4H, m), 3.85 (2H, d), 3.98 (1H, dd), 4.06 (H, t), 4.85 (1H, d), 4.99 (1H, s), 5.28 (1H, d), 5.49 (1H, broad s), 5.94 (1H, d), 5.96 (1H, d), 6.64 (1H, d), 7.01 (2H, s), 7.09 (1H, d)。
【0080】
〔実施例10:TRa2タンパク質の酵母表層発現系の構築〕
TRa2タンパク質を酵母の表層に呈示発現させるために、以下のベクターを構築した。
プラスミドpCRTRa2-cDNAを鋳型として、プライマーTraEcoRI-f2とプライマーTRa2XhoI-r3を用いて、PCR反応を行った。PCRは、Ex Taq(タカラバイオ)を使用し、94℃ 2分、(94℃ 1分、58℃ 1分、72℃ 2分)を25サイクル、72℃ 10分の反応を行った。得られたPCR産物をアガロースゲル電気泳動で分析したところ、約1.5kbのDNA断片が確認できた。これをアガロースゲルより切り出し、GFXにて精製した。得られたDNA断片をTOPO-TA cloning kit(インビトロジェン)にてクローニングして、塩基配列を確認しプラスミドpCRTRa2-cDNA2とした。プラスミドpCRTRa2-cDNA-2をEcoRIとXhoIで消化して得られた約1.5kbのDNA断片とプラスミドpGA11(Appl.Environ.Microbiol.,63,1362-1366(1997)をEcoRIとXhoIで消化して得られた約9.2kbのDNA断片を連結し、プラスミドpCAS-TRa2を得た。
【0081】
〔実施例11:TRa2タンパク質表層発現酵母の活性確認〕
プラスミドpCAS-TRa2にて酵母S. cerevisiaeEH1315株を形質転換した。得られた形質転換株をTRa2-2株とした。TRa2-2株及び、プラスミドpGA11のグルコアミラーゼ遺伝子の分泌シグナルとα-アグルチニンの3'側半分をコードする遺伝子の間にマルチクローニングサイトを挿入したプラスミドpCAS1にて酵母S. cerevisiae EH1315株を形質転換したコントロール株(C-2株)をYPD液体培地10mlに1白金耳植菌し、30℃で2日間培養した。遠心分離により菌体を回収し、PBSにて洗浄後、1ml PBSに懸濁した。この懸濁液を用いて実施例6及び実施例7と同様に、糖分解反応及び糖転移反応を行い反応生成物の分析を行った。その結果、TRa2-2株はデキストリン分解活性及びデキストリンを糖供与体とするカテキンへの糖転移活性を有するのに対し、C-2株ではこれらの活性がないことが確認できた。すなわち、TRa2-2株ではTRa2が活性を有する形で酵母表層に呈示されたことが示された。
【0082】
〔実施例12:Trichoderma属の各種菌株からのTRa2ホモログの取得〕
Trichoderma viride IFO31137株、Trichodermaviride SAM1427株、Trichoderma viride IFO31327株、Trichodermaviride IFO30498株、Trichoderma viride IFO5720株、Trichodermareesei IFO31329株、Trichoderma reesei IFO31328株、Trichodermareesei IFO31326株、 Trichoderma saturnisporum IAM12535株、Trichoderma ghanenseIAM13109株、Trichoderma koningii IAM12534株、 Trichoderma hamatumIAM12505株、 Trichoderma harzianum IFO31292株、Trichoderma polysporumSAM357株をそれぞれ、酵母エキス(Difco)1%、ポリペプトン(日本製薬)1%、デキストリン(ナカライテスク)2%の液体培地 10mlに植菌し、30℃で4日間振とう培養した。フィルターろ過により菌体を回収し、DNeasy plant mini Kit(QIAGEN)により、ゲノムDNAを抽出した。このゲノムDNA50ngを鋳型として、プライマーAMY-12fとプライマーAMY-15rを用いてPCR反応をおこなった。すなわち、ExTaq(タカラバイオ社製)を用いて、94℃ 2分、(94℃ 1分、50℃1分、72℃ 1分)を30サイクル、72℃ 10分の反応を行った。PCR産物をアガロースゲル電気泳動で分析したところ、約0.6kbpの断片が確認できた。これらのDNA断片をアガロースゲルから切り出し、GFX PCR DNA and Gel Band Purifiction Kit(アマシャムバイオサイエンス)によりDNAを精製した。これをTOPO-TA cloning kit(インビトロジェン)にてクローニングし、塩基配列の解析をABI3100Avant(アプライドバイオシステムズ社製)にて行った。その結果、配列番号:11〜24の配列が得られた。アライメント結果を図8に示す。TRa2遺伝子のこれらに相当する領域及びこれらの互いの相同性は63〜99.5%であった。
【0083】
これらの配列をもとに、T. viride IAM5141のTRa2遺伝子、cDNA全長を取得したときと同様の方法を用いれば、TRa2と同様な活性を持つTRa2ホモログを取得できる。
〔実施例13:TRa2の基質特異性解析〕
TRa2-1株をYPD培地10 mlにて、30℃で一晩振とう培養した。静止期に達した培養液を同培地に接種し(2%(v/v))、30℃で3日間振とう培養した。培養後、遠心分離にて上清を回収し、5倍に濃縮してTRa2の粗酵素溶液を得た。酵素反応溶液(0.5 mM 及び 10 mM 糖受容体基質((+)-カテキン、(-)-エピガロカテキン-3-O-ガレート、エスクレチン、ナリンゲニン、ケルセチン、ダイゼイン、ゲニステイン、ケンフェロール)、10 mg デキストリン、100 mM 酢酸バッファー (pH 5.2)、粗酵素溶液)100μlにて45℃、24 hr反応させ、HPLCにて分析した。結果を図9に示した。
【0084】
受容体基質と配糖体生成物の面積比(%)は、(+)-カテキン:10%、(-)-エピガロカテキン-3-O-ガレート:17.7%、エスクレチン:3.5%、ナリンゲニン:4.4%、ケルセチン:9.4%、ダイゼイン:10.7%、ゲニステイン:6.8%、ケンフェロール:3.1%であった。
【0085】
〔実施例14:TRa2キャラクタリゼーション〕
His-tagを付加したTRa2タンパク質(TRa2-His)の発現:
TRa2-His発現用プラスミドの構築と形質転換酵母の取得
TRa2タンパク質のC末端にHis-tagを付加して酵母で発現させるために、以下のプライマーを設計した。
TRa2HisXhoI-r2: Gctcgagttagtggtggtggtggtggtgtgaagacagcagcaa(配列番号:28)
プラスミドpCRTRa2-cDNAを鋳型として、プライマーTraEcoRI-f2とプライマーTRa2HisXhoI-r2を用いて、PCR反応を行った。PCRは、Ex Taq(タカラバイオ)を使用し、94℃ 2分、(94℃ 1分、58℃ 1分、72℃ 2分)を25サイクル、72℃ 10分の反応を行った。得られたPCR産物をアガロースゲル電気泳動で分析したところ、約1.5kbのDNA断片が確認できた。これをアガロースゲルより切り出し、GFXにて精製した。得られたDNA断片をTOPO-TA cloning kit(インビトロジェン)にてクローニングして、塩基配列を確認しプラスミドpCRTRa2-cDNA-Hisとした。pCRTRa2-cDNA-HisをEcoRIとXhoIで消化して約1.5kbのDNA断片を、プラスミドpYE22mを制限酵素EcoRI及びSalIで消化した断片とligation high(東洋紡)を用いて連結し、プラスミドpYE-TRa2-Hisを得た。
プラスミドpYE-TRa2-Hisにて酵母S. cerevisiae EH1315株を形質転換した。得られた形質転換株をTRa2-3株とした。
【0086】
培養
TRa2-3株をSD(-Trp) 20 mlにて30℃、16 hr培養した。前培養液をSD(-Trp) + 100 mM KH2PO4-KOH (pH 6.0) 1 Lに植菌し、30℃、3 days培養した。遠心分離により培養液上清を回収した。
【0087】
精製
培養液上清をBuffer S1[20 mM NaH2PO4-NaOH (pH 7.4)、10 mM イミダゾール、0.5 M NaCl、15 mM 2-メルカプトエタノール]にて平衡化したN2+キレート済みのChelating Sepharose Fast Flow (5 ml、Pharmacia Biotech)カラムにアプライし、同バッファー(40 ml)で洗浄した。続いてBuffer E1[20 mM NaH2PO4-NaOH (pH 7.4)、200 mM イミダゾール、0.5 M NaCl、15 mM 2-メルカプトエタノール]によりカラムに結合したタンパク質を溶出した。活性画分を集め、VIVASPIN(30,000 MWCO、VIVASCIENCE)を用いて脱塩、濃縮した。
【0088】
続いて、酵素溶液をBuffer S2[20 mM KH2PO4-KOH (pH 7.4)、15 mM 2-メルカプトエタノール、0.1% CHAPS]にて平衡化したResource Q (1 ml、Pharmacia Biotech)カラムにアプライ(1.5 ml/min)し、同バッファー(10 ml)で洗浄した。続いてBuffer E2[20 mM KH2PO4-KOH (pH 7.4)、0.6 M NaCl、15 mM 2-メルカプトエタノール、0.1% CHAPS]の0-100%直線濃度勾配(60 ml)によりカラムに結合したタンパク質を溶出した。活性画分を集め、VIVASPIN(30,000 MWCO、VIVASCIENCE)を用いて脱塩、濃縮した。
【0089】
再度、同手順でResource Qカラムクロマトグラフィーを行い、活性を示しかつSDS-PAGE上で単一バンドの画分を集め、VIVASPIN(30,000 MWCO、VIVASCIENCE)を用いて脱塩、濃縮した。
【0090】
酵素活性測定:
糖転移活性
反応液(10 mM カテキン、10 mg デキストリン、100 mM Acetate-NaOH(pH 5.3)、酵素溶液)100 μlを45℃、24 hr攪拌し、0.5% TFA 100 μlを添加することで反応を停止した。反応停止後のサンプルを遠心分離し、上清を回収した。生成物を以下のような条件でHPLCにて分析した。HPLC条件:溶離液A、0.1% TFA;溶離液B、90% アセトニトリル、0.08% TFA;分析カラム、Devolosil C30-UG-5 (4.6 x 150 mm、NOMURA CHEMICAL);流速、1 ml/min;分離モード、0 min-5%B、20 min-50%B、20.5 min-5%B、25 min-5%B
加水分解活性
反応液(5 mM マルトテトラオース、20 mM KH2PO4-KOH(pH 6.0)、酵素溶液)100 μlを35℃、15min反応させ、熱処理(100℃、5 min)することで反応を停止した。反応停止後のサンプルを遠心分離し、上清を回収した。生成物を以下のような条件でHPLCにて分析した。HPLC条件:溶離液、68% アセトニトリル;分析カラム、SUPELCOSIL LC-NH2 (5 μm、4.6 x 250 mm、SUPELCO);流速、1 ml/min;分離モード、isocratic
温度安定性解析:
酵素溶液(±10 mMエピガロカテキンガレート)を20〜60℃の各温度で、1 hr処理後、4℃に冷却した。冷却後のサンプルに、10mMエピガロカテキンガレート、10 mg デキストリンを添加して反応(各温度,pH 5.3,24 hr)した。0.5% TFA添加で反応停止後、逆相HPLCにて分析した。
【0091】
結果を図10(左)に示した。
pH安定性解析:
酵素溶液(±10 mMエピガロカテキンガレート)に各種pHの終濃度167 mM 緩衝液(pH 4、4.5、5、Acetate-NaOH;pH 5.5、6、6.5、7、NaH2PO4-NaOH;pH 8、9、Tris-HCl)を添加し、4℃、16 hr処理後、終濃度400 mM Acetate-NaOH(pH 5.3)を添加した。処理後のサンプルに、10 mMエピガロカテキンガレート、10 mg デキストリンを添加して反応(45℃,各pH,24 hr)した。0.5% TFA添加で反応停止後、逆相HPLCにて分析した。活性測定時の反応液のAcetate-NaOH濃度は200 mMとなる。
【0092】
結果を図10(右)に示した。
反応温度依存性解析:
5 mM マルトテトラオースに酵素を添加して、加水分解反応(20〜60℃の各温度,pH 6.0、15 min)を行った。100℃、5 minで反応停止後、順相HPLCにて分析した。結果を図11(左)に示した。
【0093】
また、10 mM カテキン,10 mgデキストリンに酵素を添加して、糖転移反応(20〜60℃の各温度,pH 5.3,24 hr)を行った。TFA添加にて反応停止後、逆相HPLCにて分析した。結果を図12(左)に示した。
【0094】
反応pH依存性解析:
5 mM マルトテトラオースに酵素を添加して、各種pHの緩衝液中で加水分解反応(35℃、pH 4、4.5、5、Acetate-NaOH;pH 5.5、6、6.5、7、NaH2PO4-NaOH;pH 8、9、Tris-HCl、15 min)を行った。100℃、5 minで反応停止後、順相HPLCにて分析した。結果を図11(右)に示した。
【0095】
また、10 mM カテキン,10 mgデキストリンに酵素を添加して、糖転移反応(45℃、pH 4、4.5、5、Acetate-NaOH;pH 5.5、6、6.5、7、NaH2PO4-NaOH;pH 8、9、Tris-HCl、24 hr)を行った。TFA添加にて反応停止後、逆相HPLCにて分析した。結果を図12(右)に示した。
【0096】
〔実施例15:配糖体のキャラクタリゼーション〕
熱安定性:
100μMの(+)-カテキン、又は4'-O-α-D-グルコピラノシル-(+)-カテキンを含む10mM リン酸カリウムバッファー(pH7.0) 30μlを4〜100℃の各温度で0〜4時間処理した後、氷中に移し、続いて0.1% TFA 60μlを加えて実施例1と同様にHPLCで分析した。各温度で処理したときの(+)-カテキン又は4'-O-α-D-グルコピラノシル-(+)-カテキンの残存量を図13に示した。カテキンに比べ4'-O-α-D-グルコピラノシル-(+)-カテキンは熱に対して安定であった。
【0097】
溶解性:
(+)-カテキン又は5-O-α-D-グルコピラノシル-(+)-カテキンを10〜450mg/mlの各濃度になるよう水に添加し、激しく攪拌することにより溶解させた。その後、遠心分離により沈殿物を除去し上清をHPLCにより分析し、(+)-カテキン及び5-O-α-D-グルコピラノシル-(+)-カテキンの量を定量した。その結果を図14に示す。(+)-カテキンはほとんど水に溶けないのに対し、5-O-α-D-グルコピラノシル-(+)-カテキンは少なくとも40倍以上の溶解性を示した。
【産業上の利用可能性】
【0098】
本発明により得られる新規酵素遺伝子又は本酵素遺伝子を改変した遺伝子、それを導入した形質転換体、及び新規酵素タンパク質又は本酵素タンパク質を改変したタンパク質は、フラボノイド類の配糖体又はそれらを用いる食品、医薬品、化粧品を生産する上で有用である。
【図面の簡単な説明】
【0099】
【図1】図1は、Trichoderma viride IAM 5141 粗酵素液でカテキンを処理した際の、HPLC分析チャートである。
【図2】図2は、Aspergillus nidulans、Neurospora crassa、Magnaporthe grisea、Fusariumgraminearum のゲノム情報データベースから抽出した、alpha-amylase, catalytic domain(accession No. PF00128)のモチーフを持つ推定ORFのアミノ酸配列について、系統樹作成プログラムであるTree viewにより作成した系統樹を示した図である。
【図3】図3は、図2のグループ1の4つのアミノ酸配列をアライメントし、保存性の高い部分(下線部)を示した図である。
【図4】図4は、TRa2のゲノムDNA配列(配列番号6)とcDNA配列(配列番号9)を比較した図である。
【図5】図5は、TRa2のcDNA塩基配列、及びそれに対応する推定アミノ酸配列を示した図である。2重下線部は推定分泌シグナル配列。
【図6】図6は、TRa2の推定アミノ酸配列とタカアミラーゼ前駆体アミノ酸配列(GBNo.BAA00336)の一次構造を比較した図である。下線部:TRa2の推定分泌シグナル、破線下線部:タカアミラーゼの分泌シグナル、2重下線部:α‐アミラーゼファミリー酵素で高度に保存されている4つの領域、*印アミノ酸残基:触媒部位に位置するアミノ酸残基。
【図7】図7は、T. viride IAM5141の培養上清の原液、又はその濃縮液に(+)-カテキン又は(-)-エピガロカテキン-3-O-ガレートと、デキストリンを加えて反応させた際の、反応液のHPLC分析チャートである。
【図8】図8は、Trichoderma属の各種菌株から調製されたゲノムDNAより得られた、TRa2ホモログの部分塩基配列のアライメント結果を示した図である。
【図9】図9は、形質転換体(TRa2-1株)の培養上清から調製したTRa2の粗酵素溶液に、各糖受容体基質((+)-カテキン、(-)-エピガロカテキン-3-O-ガレート、エスクレチン、ナリンゲニン、ケルセチン、ダイゼイン、ゲニステイン、ケンフェロール)、デキストリンを加えて反応させた際の、配糖化活性を示したグラフである。
【図10】図10は、TRa2の温度安定性及びpH安定性を示したグラフである。
【図11】図11は、TRa2による糖質加水分解反応の、温度依存性及びpH依存性を示したグラフである。
【図12】図12は、TRa2による糖転移反応の、温度依存性及びpH依存性を示したグラフである。
【図13】図13は、(+)-カテキン又は4'-O-α-D-グルコピラノシル-(+)-カテキンを含む溶液を4〜100℃の各温度で0〜4時間処理した後の、それぞれの残存量を示したグラフである。
【図14】図14は、(+)-カテキン及び5-O-α-D-グルコピラノシル-(+)-カテキンの水への溶解性を示したグラフである。
【特許請求の範囲】
【請求項1】
下記の(A)〜(K)のいずれか一を含むポリヌクレオチド:
(A)配列番号:6に記載の塩基配列の全部又は一部からなるポリヌクレオチド;
(B)(A)に記載のポリヌクレオチドの塩基配列と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(C)(A)に記載のポリヌクレオチドの塩基配列において1若しくは複数の塩基が置換、欠失、挿入、及び/又は付加された塩基配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(D)(A)に記載のポリヌクレオチドの塩基配列と少なくとも60%以上の同一性を有し、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(E)配列番号:9に記載の塩基配列からなるポリヌクレオチド;
(F)(E)に記載のポリヌクレオチドの塩基配列と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(G)(E)に記載のポリヌクレオチドの塩基配列において1若しくは複数の塩基が置換、欠失、挿入、及び/又は付加された塩基配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(H)(E)に記載のポリヌクレオチドの塩基配列と少なくとも60%以上の同一性を有し、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(I)配列番号:10に記載のアミノ酸配列からなるタンパク質をコードするポリヌクレオチド;
(J)配列番号:10に記載のアミノ酸配列において1若しくは複数のアミノ酸が置換、欠失、挿入、及び/又は付加したアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(K)配列番号:10に記載のアミノ酸配列と少なくとも60%以上の同一性を有するアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド。
【請求項2】
下記の(L)〜(R)のいずれか一を含むポリヌクレオチド:
(L)配列番号:25に記載の塩基配列からなるポリヌクレオチド;
(M)配列番号:25に記載の塩基配列と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(N)配列番号:25に記載のポリヌクレオチドの塩基配列において1若しくは複数の塩基が置換、欠失、挿入、及び/又は付加された塩基配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(O)配列番号:25に記載の塩基配列と少なくとも60%以上の同一性を有し、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(P)配列番号:26に記載のアミノ酸配列からなるタンパク質をコードするポリヌクレオチド;
(Q)配列番号:26に記載のアミノ酸配列において1若しくは複数のアミノ酸が置換、欠失、挿入、及び/又は付加したアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(R)配列番号:26に記載のアミノ酸配列と少なくとも60%以上の同一性を有するアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド。
【請求項3】
子嚢菌類の糸状菌由来である(好ましくは、Trichoderma属由来、より好ましくは、Trichoderma viride由来である)、請求項1又は2に記載のポリヌクレオチド。
【請求項4】
請求項1、2又は3に記載のポリヌクレオチドを含むベクター。
【請求項5】
請求項4に記載のベクターにより形質転換された、形質転換体。
【請求項6】
下記の(i)、(j)又は(k)を含むタンパク質:
(i)配列番号:10に記載のアミノ酸配列からなるタンパク質;
(j)配列番号:10に記載のアミノ酸配列において1若しくは複数のアミノ酸が置換、欠失、挿入、及び/又は付加したアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質;
(k)配列番号:10に記載のアミノ酸配列と少なくとも60%以上の同一性を有するアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質。
【請求項7】
下記の(p)、(q)又は(r)を含むタンパク質:
(p)配列番号:26に記載のアミノ酸配列からなるタンパク質;
(q)配列番号:26に記載のアミノ酸配列において1若しくは複数のアミノ酸が置換、欠失、挿入、及び/又は付加したアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質;
(r)配列番号:26に記載のアミノ酸配列と少なくとも60%以上の同一性を有するアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質。
【請求項8】
樹脂に固定化されている、請求項6又は7に記載のタンパク質。
【請求項9】
Trichoderma属由来であり(好ましくはTrichoderma viride、Trichoderma reesei、Trichodermasaturnisporum、Trichoderma ghanense、Trichoderma koningii、 Trichoderma hamatum、Trichoderma harzianum又はTrichodermapolysporum由来であり)、配列番号:11〜24のいずれか一に記載の塩基配列を含み、フラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド。
【請求項10】
Trichoderma属由来であり(好ましくはTrichoderma viride、Trichoderma reesei、Trichodermasaturnisporum、Trichoderma ghanense、Trichoderma koningii、 Trichoderma hamatum、Trichoderma harzianum又はTrichodermapolysporum由来であり)、配列番号:11〜24のいずれか一に記載の塩基配列を含むポリヌクレオチドによってコードされる、フラボノイド類の配糖化活性を有するタンパク質。
【請求項11】
樹脂に固定化されている、請求項10に記載のタンパク質。
【請求項12】
配列番号:6、配列番号:9、配列番号:10、配列番号:11〜26のいずれか一に記載の塩基配列又はアミノ酸配列の全部若しくは一部を用いることを特徴とする、フラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチドの塩基配列又はフラボノイド類の配糖化活性を有するタンパク質のアミノ酸配列を決定する方法。
【請求項1】
下記の(A)〜(K)のいずれか一を含むポリヌクレオチド:
(A)配列番号:6に記載の塩基配列の全部又は一部からなるポリヌクレオチド;
(B)(A)に記載のポリヌクレオチドの塩基配列と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(C)(A)に記載のポリヌクレオチドの塩基配列において1若しくは複数の塩基が置換、欠失、挿入、及び/又は付加された塩基配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(D)(A)に記載のポリヌクレオチドの塩基配列と少なくとも60%以上の同一性を有し、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(E)配列番号:9に記載の塩基配列からなるポリヌクレオチド;
(F)(E)に記載のポリヌクレオチドの塩基配列と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(G)(E)に記載のポリヌクレオチドの塩基配列において1若しくは複数の塩基が置換、欠失、挿入、及び/又は付加された塩基配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(H)(E)に記載のポリヌクレオチドの塩基配列と少なくとも60%以上の同一性を有し、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(I)配列番号:10に記載のアミノ酸配列からなるタンパク質をコードするポリヌクレオチド;
(J)配列番号:10に記載のアミノ酸配列において1若しくは複数のアミノ酸が置換、欠失、挿入、及び/又は付加したアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(K)配列番号:10に記載のアミノ酸配列と少なくとも60%以上の同一性を有するアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド。
【請求項2】
下記の(L)〜(R)のいずれか一を含むポリヌクレオチド:
(L)配列番号:25に記載の塩基配列からなるポリヌクレオチド;
(M)配列番号:25に記載の塩基配列と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(N)配列番号:25に記載のポリヌクレオチドの塩基配列において1若しくは複数の塩基が置換、欠失、挿入、及び/又は付加された塩基配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(O)配列番号:25に記載の塩基配列と少なくとも60%以上の同一性を有し、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(P)配列番号:26に記載のアミノ酸配列からなるタンパク質をコードするポリヌクレオチド;
(Q)配列番号:26に記載のアミノ酸配列において1若しくは複数のアミノ酸が置換、欠失、挿入、及び/又は付加したアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド;
(R)配列番号:26に記載のアミノ酸配列と少なくとも60%以上の同一性を有するアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド。
【請求項3】
子嚢菌類の糸状菌由来である(好ましくは、Trichoderma属由来、より好ましくは、Trichoderma viride由来である)、請求項1又は2に記載のポリヌクレオチド。
【請求項4】
請求項1、2又は3に記載のポリヌクレオチドを含むベクター。
【請求項5】
請求項4に記載のベクターにより形質転換された、形質転換体。
【請求項6】
下記の(i)、(j)又は(k)を含むタンパク質:
(i)配列番号:10に記載のアミノ酸配列からなるタンパク質;
(j)配列番号:10に記載のアミノ酸配列において1若しくは複数のアミノ酸が置換、欠失、挿入、及び/又は付加したアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質;
(k)配列番号:10に記載のアミノ酸配列と少なくとも60%以上の同一性を有するアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質。
【請求項7】
下記の(p)、(q)又は(r)を含むタンパク質:
(p)配列番号:26に記載のアミノ酸配列からなるタンパク質;
(q)配列番号:26に記載のアミノ酸配列において1若しくは複数のアミノ酸が置換、欠失、挿入、及び/又は付加したアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質;
(r)配列番号:26に記載のアミノ酸配列と少なくとも60%以上の同一性を有するアミノ酸配列からなり、かつフラボノイド類の配糖化活性を有するタンパク質。
【請求項8】
樹脂に固定化されている、請求項6又は7に記載のタンパク質。
【請求項9】
Trichoderma属由来であり(好ましくはTrichoderma viride、Trichoderma reesei、Trichodermasaturnisporum、Trichoderma ghanense、Trichoderma koningii、 Trichoderma hamatum、Trichoderma harzianum又はTrichodermapolysporum由来であり)、配列番号:11〜24のいずれか一に記載の塩基配列を含み、フラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチド。
【請求項10】
Trichoderma属由来であり(好ましくはTrichoderma viride、Trichoderma reesei、Trichodermasaturnisporum、Trichoderma ghanense、Trichoderma koningii、 Trichoderma hamatum、Trichoderma harzianum又はTrichodermapolysporum由来であり)、配列番号:11〜24のいずれか一に記載の塩基配列を含むポリヌクレオチドによってコードされる、フラボノイド類の配糖化活性を有するタンパク質。
【請求項11】
樹脂に固定化されている、請求項10に記載のタンパク質。
【請求項12】
配列番号:6、配列番号:9、配列番号:10、配列番号:11〜26のいずれか一に記載の塩基配列又はアミノ酸配列の全部若しくは一部を用いることを特徴とする、フラボノイド類の配糖化活性を有するタンパク質をコードするポリヌクレオチドの塩基配列又はフラボノイド類の配糖化活性を有するタンパク質のアミノ酸配列を決定する方法。
【図1】

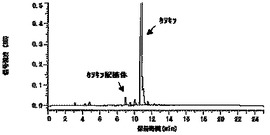
【図7】
【図10】
【図11】
【図12】
【図13】
【図2】
【図3】

【図4】

【図5】

【図6】

【図8】

【図9】
【図14】
【図7】
【図10】
【図11】
【図12】
【図13】
【図2】
【図3】
【図4】
【図5】
【図6】
【図8】
【図9】
【図14】
【公開番号】特開2008−173075(P2008−173075A)
【公開日】平成20年7月31日(2008.7.31)
【国際特許分類】
【出願番号】特願2007−10759(P2007−10759)
【出願日】平成19年1月19日(2007.1.19)
【出願人】(000001904)サントリー株式会社 (319)
【Fターム(参考)】
【公開日】平成20年7月31日(2008.7.31)
【国際特許分類】
【出願日】平成19年1月19日(2007.1.19)
【出願人】(000001904)サントリー株式会社 (319)
【Fターム(参考)】
[ Back to top ]