
新規酵素
【課題】新規酵素を提供する。
【解決手段】ルシフェラーゼ活性を有し、かつ、フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス、ホタリア・パロウラ、ピロフォラス・プラギオフタラマス、ラムピリス・ノクチルカ、ピロコエリア・ナヤコ又はフォティナス・ペンシランヴァニカ由来のルシフェラーゼと少なくとも60%の類似性を有するタンパク質であって、前記酵素の配列が対応の野生型配列において現れるアミノ酸とは異なるものであり、かつ、該ルシフェラーゼ酵素が、当該部位に対応の野生型ルシフェラーゼのアミノ酸を有する酵素と比較して増加した熱安定性を有することを特徴とする酵素。
【解決手段】ルシフェラーゼ活性を有し、かつ、フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス、ホタリア・パロウラ、ピロフォラス・プラギオフタラマス、ラムピリス・ノクチルカ、ピロコエリア・ナヤコ又はフォティナス・ペンシランヴァニカ由来のルシフェラーゼと少なくとも60%の類似性を有するタンパク質であって、前記酵素の配列が対応の野生型配列において現れるアミノ酸とは異なるものであり、かつ、該ルシフェラーゼ酵素が、当該部位に対応の野生型ルシフェラーゼのアミノ酸を有する酵素と比較して増加した熱安定性を有することを特徴とする酵素。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、新規酵素、特に対応の野生型酵素と比較して増加した熱安定性を有する変異ルシフェラーゼ酵素、アッセイにおける前記酵素の使用及び前記酵素を含む試験キットに関する。
ホタルのルシフェラーゼは、ATP、Mg2+及び分子酸素の存在下、光の生成を伴うルシフェリンの酸化を触媒する。この反応は約0.88の量子収率を有する。前記ルシフェラーゼの発光特性は、ATPレベルを測定する種々の発光(luminometric)アッセイにおける前記酵素の使用を導く。前記のアッセイの例には、EP-B-680515及びWO96/02665の記載に基づくアッセイが含まれる。
ルシフェラーゼは、昆虫の体、特にホタル又はツチボタル等の甲虫から直接得ることができる。ルシフェラーゼが得られる特定の種には、ニホンゲンジボタル又はケイケ(KEIKE)ボタル、ルシオラ・クルシアタ(Luciola cruciata)及びルシオラ・ラテラリス(Luciola lateralis)、東欧ホタルであるルシオラ・ミングレリカ(Luciola mingrelica)、北米ホタルであるフォティナス・ピラリス(Photinus pyralis)並びにツチボタルであるラムピリス・ノクチルカ(Lampyris noctiluca)が含まれる。ルシフェラーゼを得ることができるその他の種は、Ye ら., Biochimica et Biophysica Acta, 1339 (1997) 39-52に記載されている。Vivianiら, Biochemistry, 38, (1999) 8271-8279に記載される更なる種は、フィリキソシックス(Phrixothrix)(レールロードワーム(railroad-worms))である。
しかしながら、これらの酵素をコードする多くの遺伝子がクローニングされかつ配列決定されているので、組換えDNA技術を使用しても製造することができるだろう。前記酵素をコードする組換えDNA配列を使用して、大腸菌(E.coli)等の微生物を形質転換し、所望の酵素生成物を発現させる。
野生型及び組換え型ルシフェラーゼの熱安定性は、約30℃、特に35℃を超える温度に曝露したときにその活性が急速に消失する程度である。この不安定性は、高い周囲温度下で酵素を保存するか又は、例えば反応速度を高くするために高温反応条件下アッセイを行うときに問題を起こす。
増加した熱安定性を有する変異ルシフェラーゼはEP-A-524448及びW095/25798より既知である。前者は、ニホンボタルのルシフェラーゼにおいてスレオニン残基をイソロイシン残基で置換することにより部位217に変異を有する変異ルシフェラーゼを記載している。後者は、フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼと60%をこえる類似性を有するが、フォティナス・ピラリスの部位354のアミノ酸残基又はルシオラ種の部位356に対応するアミノ酸残基がグルタメート以外になるように変異している変異ルシフェラーゼを記載している。
【発明の概要】
【課題を解決するための手段】
【0002】
本件出願人は、増加した熱安定性をもたらし、かつ当該技術分野において既知の変異体を補完する更なる変異体を見いだした。
本発明は、ルシフェラーゼ活性を有し、かつ、フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス、ホタリア・パロウラ(Hotaria paroula)、ピロフォラス・プラギオフタラマス(Pyrophorus plagiophthalamus)、ラムピリス・ノクチルカ、ピロコエリア・ナヤコ(Pyrocoelia nayako)、フォティナス・ペンシランヴァニカ(Photinus pennsylanvanica)又はフィリキソシックス由来のルシフェラーゼと少なくとも60%の類似性を有するタンパク質であって、前記酵素の配列において、
(a)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基214又はルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基216に対応するアミノ酸残基、
(b)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基232又はルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基234に対応するアミノ酸残基、
(c)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基295又はルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基297に対応するアミノ酸残基、
(d)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基14、ルシオラ・ミングレリカ由来ルシフェラーゼのアミノ酸残基16、又はルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基17に対応するアミノ酸残基、
(e)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基35、ルシオラ・ミングレリカ由来ルシフェラーゼのアミノ酸残基37、又はルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基38に対応するアミノ酸残基、
(f)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基105、ルシオラ・ミングレリカ由来ルシフェラーゼのアミノ酸残基106、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基107、又はルシオラ・ラテラリス遺伝子由来ルシフェラーゼのアミノ酸残基108に対応するアミノ酸残基、
(g)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基234又はルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基236に対応するアミノ酸残基、
(h)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基420又はルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基422に対応するアミノ酸残基、
(i)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基310又はルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリスのアミノ酸残基312に対応するアミノ酸残基、
のなかの少なくとも1つが対応の野生型配列において現れるアミノ酸とは異なるものであり、かつ、前記ルシフェラーゼ酵素は、当該部位に特定種の対応野生型ルシフェラーゼのアミノ酸を有する酵素と比較して増加した熱安定性を有することを特徴とする酵素を提供する。
【0003】
好ましくは、前記タンパク質はルシフェラーゼ活性を有し、かつ、フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス、ホタリア・パロウラ、ピロフォラス・プラギオフタラマス、ラムピリス・ノクチルカ、ピロコエリア・ナヤコ又はフォティナス・ペンシランヴァニカ由来のルシフェラーゼと少なくとも60%の類似性を有している。
特に、前記タンパク質は、ルシフェラーゼ活性を有し、かつ、野生型ルシフェラーゼの配列、例えばフォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス、ホタリア・パロウラ、ピロフォラス・プラギオフタラマス (グリーン−Luc GR)、ピロフォラス・プラギオフタラマス (イエローグリーン Luc YG)、ピロフォラス・プラギオフタラマス (イエロー−Luc YE)、ピロフォラス・プラギオフタラマス (オレンジ−Luc OR)、ラムピリス・ノクチルカ、ピロコエリア・ナヤコ、フォティナス・ペンシランヴァニカ LY、フォティナス・ペンシランヴァニカ KW、フォティナス・ペンシランヴァニカ J19又はフィリキソシックス・グリーン(PvGR)若しくはレッド(PhRE)の配列を実質的に有する。
但し、前記タンパク質は、野生型酵素のアミノ酸とは異なるように操作された1以上、例えば100までのアミノ酸残基、好ましくはせいぜい50、より好ましくはせいぜい30のアミノ酸を含んでいてもよい。
特に、基質D−ルシフェリン(4,5−ジヒドロ−2−[6−ヒドロキシ−2−ベンゾチアゾリル]−4−チアゾール カルボン酸)を使用して光放射を生じることができる種に由来する生物発光酵素は、本発明の変異体酵素の基礎を形成するだろう。
実施例によると、本発明のルシフェラーゼは、フォティナス・ピラリスのルシフェラーゼ配列を実質的に有し、かつ、以下の変化のうちの少なくとも1つを有している。
(a)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基214に対応するアミノ酸残基がスレオニン以外のアミノ酸残基に変化している、
(b)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基232に対応するアミノ酸残基がイソロイシン以外に変化している、
(c)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基295に対応するアミノ酸残基がフェニルアラニン以外に変化している、
(d)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基14に対応するアミノ酸残基がフェニルアラニン以外に変化している、
(e)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基35に対応するアミノ酸残基がロイシン以外に変化している、
(f)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基105に対応するアミノ酸残基がアラニン以外に変化している、
(g)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基234に対応するアミノ酸残基がアスパラギン酸以外に変化している、
(h)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基420に対応するアミノ酸残基がセリン以外に変化している、
(i)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基310に対応するアミノ酸残基がヒスチジン以外に変化している。
【0004】
本発明のタンパク質がルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来酵素の配列を実質的に有する場合、以下の変化のうちの少なくとも1つを有する。
(a)ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基216に対応するアミノ酸残基が、(ルシオラ・ミングレリカを基礎とする配列では)グリシン以外であり、又は、(ルシオラ・クルシアタ又はルシオラ・ラテラリスを基礎とする配列では)アスパラギン以外である。
(b)ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基234に対応するアミノ酸残基がセリン以外である。
(c)ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基297に対応するアミノ酸残基がロイシン以外である。
(d)ルシオラ・ミングレリカ由来ルシフェラーゼのアミノ酸残基16又はルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基17に対応するアミノ酸残基がフェニルアラニン以外である。
(e)ルシオラ・ミングレリカ由来ルシフェラーゼのアミノ酸残基37又はルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基38に対応するアミノ酸残基がリジン以外である。
(f)ルシオラ・ミングレリカ由来ルシフェラーゼのアミノ酸残基106に対応するアミノ酸残基、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基107に対応するアミノ酸残基、又は、ルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基108に対応するアミノ酸残基がグリシン以外である。
(g)ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基236に対応するアミノ酸残基がグリシン以外である。
(h)ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基422に対応するアミノ酸残基がスレオニン以外である。
(i)ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基312に対応するアミノ酸残基が、(ルシオラ・ミングレリカを基本とする配列では)スレオニン以外であり、又は、(ルシオラ・クルシアタ又はルシオラ・ラテラリスを基本とする配列では)バリン以外である。
【0005】
熱安定性を増強させる全ての場合における特定の置換アミノ酸は、以下に説明されるようなルーチンの方法により決定することができる。それぞれの場合において、異なる置換が熱安定性の増強を引き起こすだろう。当業者に理解されるように、置換は、天然又は適切な変異体タンパク質をコードするDNAの部位特異的突然変異誘発により行うことができるだろう。この場合において本発明は熱安定性と関連するタンパク質の同定と関連する。
しかしながら、一般的には、野生型のアミノ酸とは異なる性質を有するアミノ酸で置換することを考慮することが好ましい。したがって、ある場合においては、親水性アミノ酸残基を疎水性アミノ酸残基(逆も又同様)で好ましく置換するだろう。同様に、酸性アミノ酸残基を塩基性アミノ酸残基で置換するだろう。
例えば、本発明のタンパク質は、ルシフェラーゼ活性を有し、かつ、フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来のルシフェラーゼ酵素と少なくとも60%の類似性を有し、その配列内に以下の変化のうち少なくとも1つを有し、野生型ルシフェラーゼ酵素と比較して増加した熱安定性を有している。
(a)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基214に対応するアミノ酸残基及びルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基216に対応するアミノ酸残基が変異しており、かつ、フォティナス・ピラリス由来ルシフェラーゼの場合はスレオニン以外である。
(b)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基232に対応するアミノ酸残基及びルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基234に対応するアミノ酸残基が変異しており、かつ、フォティナス・ピラリス由来ルシフェラーゼの場合はイソロイシン以外である。
(c)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基295に対応するアミノ酸残基及びルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基297に対応するアミノ酸残基が変異しており、かつ、例えば、フォティナス・ピラリス由来ルシフェラーゼの場合、フェニルアラニン以外である。
【0006】
種々のルシフェラーゼの全ての配列は、高度に保存されて、これら酵素間で有意な程度の類似性を有していることを示す。このことは、配列を調査して最も類似している領域を検出することにより酵素配列間の対応領域が容易に決定可能であることを示している。必要ならば、種々の配列間対応領域又は特定のアミノ酸を決定するために市販のソフト(例えば、ウィスコンシン大学遺伝学コンピュータグループの「Bestfit」、Devereuxら(1984) Nucleic Acid Research 12: 387-395を参照のこと)を使用することができる。代替又は追加として、対応の酸を、文献:L. Yeら, Biochim. Biophys Acta 1339 (1997) 39-52により決定することができる。前記文献で使用する番号方式(numbering system)は、本件出願で使用した番号方式の基礎を形成する。
フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基214に対応するアミノ酸残基の可能な変化について、極性アミノ酸スレオニンは、非極性アミノ酸、例えばアラニン、グリシン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン、トリプトファン又はシステインなどにより適切に置換される。フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基214に対応するスレオニン残基の特に好ましい置換はアラニンによるものである。
より好ましい置換はシステインによるものである。しかしながら、この部位における異なる極性残基、例えばアスパラギンも、当該部位にスレオニンを有する対応の酵素の熱安定性を増強するだろう。
野生型ルシフェラーゼ酵素においてこの部位に現れるその他のアミノ酸には、グリシン(ルシオラ・ミングレリカ、ホタリア・パロウラ)、アスパラギン(ピロフォラス・プラギオフタラマス、GR、YC、YE及びOR、ルシオラ・クルシアタ、ルシオラ・ラテラリス、ラムピリス・ノクチルカ、ピロセリア・ナヤコ、フォティナス・ペンシランヴァニカ LY、KW、J19)及びセリン(フィリキソシックス・ルシフェラーゼの部位211)が含まれる。非極性又は異なる非極性側鎖、例えばアラニン及びシステインで置換することは有利であろう。
【0007】
フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基232に対応するアミノ酸残基の可能性のある変化について、非極性アミノ酸であるイソロイシンは、異なる非極性アミノ酸、例えばアラニン、グリシン、バリン、ロイシン、プロリン、フェニルアラニン、メチオニン、トリプトファン又はシステインなどで適切に置換される。野生型の配列においてこの部位に現れるその他のアミノ酸には、セリン及びアスパラギン(それぞれ、フィトシックス・グリーン(Phritothix green)及びレッド(Phritothix red)における対応の229番目の部位におけるバリン又はアラニンと同じ)が含まれる。適切には、これらの極性残基は、非極性残基、例えば前記にて概略を述べたものにより置換される。フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基232に対応する残基及びルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリスの残基234に対応する残基の特に好ましい置換はアラニンによるものである。このことは、野生型配列に対するアミノ酸の変化を示している。
フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基295に対応するアミノ酸残基及びルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基297に対応するアミノ酸残基の変化も、タンパク質の熱安定性に影響するだろう(これは、フィリキソシックス・ルシフェラーゼの残基292に対応する)。一般的に、この部位におけるアミノ酸は非極性アミノ酸であるフェニルアラニン又はロイシンである。これらは異なる非極性アミノ酸で改変される。例えば、フォティナス・ピラリスでは、非極性アミノ酸フェニルアラニンは、異なる非極性アミノ酸、例えばアラニン、ロイシン、グリシン、バリン、イソロイシン、プロリン、メチオニン、トリプトファン又はシステインにより適切に置換される。フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基214に対応するフェニルアラニン残基についての特に好ましい置換はロイシンによるものである。
フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基14に対応するアミノ酸残基又はルシオラ由来ルシフェラーゼのアミノ酸残基16(フィリキソシックス由来ルシフェラーゼではアミノ酸残基13)に対応するアミノ酸残基における変異も可能である。このアミノ酸残基(通常はフェニルアラニンであるが、ロイシン、セリン、アルギニン又はある場合ではチロシンであってもよい)は、異なるアミノ酸、特に異なる非極性アミノ酸、例えばアラニン、バリン、ロイシン、イソロイシン、プロリン、メチオニン又はトリプトファン、好ましくはアラニンへ適切に改変するだろう。
【0008】
フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基35又はルシオラ・ミングレリカ由来ルシフェラーゼのアミノ酸残基37に対応するアミノ酸残基(その他のルシオラ種及びフィリキソシックスにおけるアミノ酸38に対応する)における変異も有効であろう。このアミノ酸は野生型酵素間で変化し、この部位におけるロイシン(フォティナス・ピラリス)だけでなく、リジン、ヒスチジン、グリシン、アラニン、グルタミン及びアスパラギン酸も含まれるだろう。適切には、この部位におけるアミノ酸残基は、非極性アミノ酸残基又は異なる非極性アミノ酸、例えばアラニン、バリン、フェニルアラニン、イソロイシン、プロリン、メチオニン又はトリプトファンにより適切に置換される。この部位における好ましいアミノ酸はアラニンであり、これは野生型酵素とは異なるものである。
フォティナス・ピラリス由来配列の部位14に対応するアミノ酸における変異及び/又はフォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基35に対応するアミノ酸残基における変異は、好ましくはこの酵素における唯一の変異ではない。これらの変異は、前記で定義したその他の変異、特にフォティナス・ピラリス由来ルシフェラーゼの部位214、395又は232に対応する部位における変異を適切に伴う。
フォティナス・ピラリス由来ルシフェラーゼにおける残基105に対応するアミノ酸残基及びルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基106に対応するアミノ酸残基(フィリキソシックスにおける残基102)の変化も、タンパク質の熱安定性に影響するだろう。一般的に、この部位におけるアミノ酸は非極性アミノ酸であるアラニン又はグリシンであり、フィリキソシックスではセリンであろう。これらのアミノ酸は異なる非極性アミノ酸により適切に改変される。例えば、フォティナス・ピラリスにおいて、非極性アミノ酸アラニンは、異なる非極性アミノ酸、例えばフェニルアラニン、ロイシン、グリシン、バリン、イソロイシン、プロリン、メチオニン又はトリプトファン等により適切に置換される。フォティナス・ピラリス由来ルシフェラーゼの残基105に対応するアラニン残基についての特に好ましい置換はバリンによるものである。
【0009】
フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基234に対応するアミノ酸残基及びルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基236(フィリキソシックスにおける残基231)に対応するアミノ酸残基の変化も、タンパク質の熱安定性に影響するだろう。一般的にこの部位におけるアミノ酸はアスパラギン酸又はグリシンであり、ある場合においてはグルタミン又はスレオニンである。これらは、非極性アミノ酸又は異なる非極性アミノ酸により適切に置換される。例えば、フォティナス・ピラリスにおいて、アミノ酸残基アスパラギン酸は、非極性アミノ酸、例えばアラニン、ロイシン、グリシン、バリン、イソロイシン、プロリン、メチオニン又はトリプトファンにより適切に置換される。フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基234に対応するフェニルアラニン残基の置換に特に好ましいのはグリシンによるものである。非極性アミノ酸残基、例えばグリシンが(例えば、ルシオラ由来ルシフェラーゼにおいて)この部位に存在する場合、グリシンは異なる非極性アミノ酸により適切に置換されるだろう。
フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基420に対応するアミノ酸残基及びルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基422(フィリキソシックス・グリーンにおける残基417及びフィリキソシックス・レッドにおける残基418)に対応するアミノ酸残基の変化も、タンパク質の熱安定性に影響するだろう。一般的にこの部位におけるアミノ酸は無電荷極性アミノ酸であるセリン又はスレオリン又はグリシンである。これらは、異なる無電荷極性アミノ酸により適切に置換される。例えば、フォティナス・ピラリスにおいて、セリンは、アスパラギン、グルタミン、スレオニン又はチロシン、特にスレオニンにより適切に置換される。
フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基310に対応するアミノ酸残基及びルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基312に対応するアミノ酸残基の変化も、タンパク質の熱安定性に影響するだろう。この部位におけるアミノ酸は既知のルシフェラーゼタンパク質の間で変化し、フォティナス・ピラリス、ピロセリア・ナヤコ、ラムピリス・ノクチルカ及びある形態のフォティナス・ペンシランヴァニカ由来ルシフェラーゼではヒスチジンであり、ルシオラ・ミングレリカ、ホタリア・パロウラ及びフィリキソシックス(アミノ酸残基307)由来ルシフェラーゼではスレオニンであり、ルシオラ・クルシアタ及びルシオラ・ラテラリスではバリンであり、あるピロフォラス・プラギオフタラマス由来ルシフェラーゼではアスパラギンである。したがって、一般的に、この部位におけるアミノ酸は親水性アミノ酸であり、これを異なるアミノ酸残基に換えて、酵素の熱安定性を増加させてもよい。フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基310に対応するヒスチジン残基に対する特に好ましい置換はアルギニンによるものである。
【0010】
酵素内にその他の変異が存在していてもよい。例えば、好ましい態様では、タンパク質は、フォティナス・ピラリス由来ルシフェラーゼのアミノ酸354に対応する部位(ルシオラ由来ルシフェラーゼにおける部位356及びフィリキソシックス由来ルシフェラーゼの部位351)にグルタミンから、特にグリシン、プロリン又はアスパラギン酸以外のアミノ酸に変化したアミノ酸を有する。適切には、この部位におけるアミノ酸はトリプトファン、バリン、ロイシン、イソロイシン及びアスパラギンであり、特に好ましくはリジン又はアルギニンである。この変異はWO 95/25798に記載されている。
代替の好ましい態様では、EP-A-052448に記載されるように、タンパク質は、ルシオラ由来ルシフェラーゼのアミノ酸残基217に対応する部位が、疎水性アミノ酸、特にイソロイシン、ロイシン又はバリンに換わったアミノ酸を有している。
タンパク質は、当該タンパク質のルシフェラーゼ活性が過度に妥協しないことを条件に配列内に更に変異を含んでいてもよい。その変異は酵素の特性を適切に増強するか、又は、ある場合においては意図する目的により適するようにする。
このことは、更なる変異が熱安定性及び/又はカラーシフト特性及び/又はATPに対する酵素のKmの増強を引き起こすことを意味する。カラーシフトを引き起こす変異の例はWO95/18853に記載されている。Km値に影響する変異は例えばWO96/22376及び国際特許出願第PCT/GB98/01026号に記載されている。これらの文献は参照することにより本明細書に組み込まれる。
本発明のタンパク質は、1つより多い変異、好ましくは前記の変異の3つすべてを適切に有している。
本発明のタンパク質には野生型及び組換え型ルシフェラーゼ酵素の両方が含まれる。本発明のタンパク質は、前記のとおり野生型酵素に存在するアミノ酸の少なくとも60%が本発明のタンパク質に存在するという意味で、フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼの配列又はその他のルシフェラーゼ酵素の配列と少なくとも60%の類似性を有する。本発明のタンパク質は前記の野生型酵素に対し、より高い程度類似性、特に少なくとも70%、より好ましくは少なくとも80%、特に好ましくは少なくとも90%の類似性を有することができる。このタイプの類似のタンパク質には、対立遺伝子変異体、その他の昆虫種由来のタンパク質及び組換えにより生成した酵素が含まれる。
【0011】
本発明のタンパク質は、野生型酵素をコードする配列とストリンジェントなハイブリダイゼーション条件下、好ましくは高ストリンジェントな条件下でハイブリダイズする核酸によりコードされるという点で同定されるだろう。前記の条件は当業者に周知であり、例えばSambrookら(1989) Molecular Cloning, Cold Spring Harbor Laboratory Pressに例示されている。一般的には、低ストリンジェント条件は約周囲温度〜約65℃における3×SSCとして定義され、高ストリンジェント条件は約65℃における0.1×SSCとして定義される。SSCは0.15M NaCl、0.015M クエン酸三ナトリウムからなる緩衝液の名称である。3×SSCは3回の同じ強さのSSCのことである。
特に、本発明の配列と特定の配列との類似性は、Lipman 及び Pearson (Lipman, D.J. & Pearson, W.R. (1985) Rapid and Sensitive Protein Similarity Searches, Science, vol 227, ppl435-1441)に記載される多重整列化(multiple alignment)法を使用して評価されるだろう。「最適化された」百分率のスコアは、Lipman-Pearsonアルゴリズムについての以下のパラメーター:ktup=1、ギャップペナルティー(gap penalty)=4及びギャップペナルティー長さ(gap penalty length)=12を用いて計算されるべきである。類似性を評価する配列は「試験配列」として使用されるべきである。このことは、比較のための塩基配列、例えばフォティナス・ピラリスの配列若しくはYeら、前出に記録された前記のその他の配列又はフィリキソシックスの場合はBiochemistry, 1999, 38, 8271-8279に記載される配列は、参照配列として使用されることを意味する。
本発明の特定の例は、前記の変異を有する野生型ルシフェラーゼ配列である。
このタンパク質は少なくとも1つ、好ましくは1つより多くの前記の変位を有している。
更に本発明は前記のルシフェラーゼをコードする核酸を提供する。適切には、前記核酸は当業者に周知の野生型配列に基づくものである。アミノ酸配列に所望の変異を起こす適切な突然変異は、遺伝子コードの知識に基づき容易に明らかになるだろう。
【0012】
本発明の核酸は、発現ベクター、例えばプロモーター、エンハンサー、ターミネーターなどの制御要素の制御下にあるプラスミドへ適切に組み込まれる。これらのベクターを用いて宿主細胞、例えば原核細胞又は真核細胞、例えば植物又は動物細胞、特に原核細胞、例えば大腸菌(E. coli)を形質転換し、当該細胞に所望のルシフェラーゼ酵素を発現させることができる。当該技術分野で周知の条件を使用して、得られた形質転換細胞の培養にルシフェラーゼ酵素を産生させ、次いで培養培地からルシフェラーゼ酵素を分離することができる。細胞が動物又は植物細胞の場合、当該細胞から植物又は動物を繁殖させてもよい。その後、当該植物からタンパク質を抽出してもよい。トランスジェニック動物の場合、タンパク質を乳汁から回収してもよい。ベクター、形質転換細胞、トランスジェニック植物及び動物並びに前記細胞の培養による酵素の生産方法の全ては本発明の更なる側面を形成する。
フォティナス・ピラリスT214A変異ルシフェラーゼを、以下のランダム突然変異誘発により作成した。T214A単一点変異体は野生型ルシフェラーゼよりも高い熱安定性を有している。
2つの新規な三重変異ルシフェラーゼ:E354K/T214A/A215L 及び E354K/T214A/I232Aを作成したところ、これらも高い熱安定性を示した。
本発明の範囲内にあるフォティナス・ピラリス変異体酵素の特定の例には以下のものが含まれる。
I232A/E354K
T214A/I232A/E354K
A215L/I232A/E354K
T214A/I232A/E354K/A215L
I232A/E354K/T214A/F295L
I232A/E354K/T214A/F295L/F14A/L35A
I232A/E354K/T214A/F295L/Fl4A/L35A/A215L
A105V
T214A
T214C
T214N
T295L
I232A
F14A
L35A
D234G
S420T
H31OR
又はその他の種のルシフェラーゼに由来するときは、それらのすべての等価物。
三重変異体作成のための変異を、プラスミドpET23上のルシフェラーゼ遺伝子へ、部位特異的突然変異(PCR)により導入した。関連する変異を起こすためにPCR反応へ添加したオリゴヌクレオチドは以下の実施例で与えられたものである。
部位354及び215における点変異の効果は相加的であることが既に示されている。本発明はより高い熱安定性を与えるために3又はそれより多い変異を組み合わせる可能性を提供する。
本発明の熱安定性ルシフェラーゼは、シグナル伝達手段としてルシフェラーゼ/ルシフェリン反応を利用する全ての生物発光アッセイにおいて有利に使用されるだろう。文献において非常に多数のアッセイが知られている。それゆえ、本発明のタンパク質は、前記のアッセイの実行を鑑みて調製されるキット中に、適宜ルシフェリン及び特定のアッセイを行うのに必要なその他の試薬と共に含まれるだろう。
本発明は添付図面を参照した実施例により詳述されるだろう。
【図面の簡単な説明】
【0013】
【図1−1】図1は、本発明の変異体の生産に使用するプラスミドを示している。
【図1−2】図1は、本発明の変異体の生産に使用するプラスミドを示している。
【図2】図2は、本発明のルシフェラーゼを含むルシフェラーゼについての熱不活性化研究の結果を示している。
【図3a】図3は、種々のルシフェラーゼ変異体についての熱安定性実験の結果を示している。
【図3b】図3は、種々のルシフェラーゼ変異体についての熱安定性実験の結果を示している。
【図3c】図3は、種々のルシフェラーゼ変異体についての熱安定性実験の結果を示している。
【図3d】図3は、種々のルシフェラーゼ変異体についての熱安定性実験の結果を示している。
【図3e】図3は、種々のルシフェラーゼ変異体についての熱安定性実験の結果を示している。
【図3f】図3は、種々のルシフェラーゼ変異体についての熱安定性実験の結果を示している。
【図3g】図3は、種々のルシフェラーゼ変異体についての熱安定性実験の結果を示している。
【図3h】図3は、種々のルシフェラーゼ変異体についての熱安定性実験の結果を示している。
【図4】図4は、その他のルシフェラーゼ変異体についての熱安定性実験の結果を示している。
【図5】図5は、本発明の変異体酵素の生産に使用したオリゴヌクレオチドを示している。
【0014】
実施例1
熱安定性変異ルシフェラーゼの同定
エラー易発性(error-prone)PCRは、Fromantら, Analytical Biochemistry, 224, 347-353 (1995)により考案されたプロトコルに基づいていた。
本反応におけるdNTP混合物は以下の通りであった。
35mM dTTP
12.5mM dGTP
22.5mM dCTP
14mM dATP
PCR条件は以下の通りであった。
0.5μl(50ng)プラスミドpPW601a J54*
5.0μl 10×KCl反応緩衝液
各1μlのW56及びW57+(60pmoleの各プライマー)
1μl Biotaq(登録商標)ポリメラーゼ(5U)
2μl dNTPs(前記参照)
1.76μl MgCl2(50mMストック)
1μl mNCl2(25mMストック)[反応における終濃度=3.26mM]
36.7μl dH2O
*プラスミドpPW601aJ54はpPW601a(WO95/25798)の変異体バージョンであり、NdeI部位をATG開始コドン前3塩基以内に作成した。これにより、pPW601aからpET23ベクターへの容易なクローニングが許容された。
+プライマー配列:
W56:
5'-AAACAGGGACCCATATGGAAGACGC-3'
W57:
5'-AATTAACTCGAGGAATTTCGTCATCGCTGAATACAG-3')
サイクルのパラメーター配下の通りであった。
94℃で5分、
次いで94℃で30秒
55℃で30秒
72℃で5分
のサイクル×12、
72℃で10分間。
【0015】
PCR産物は、Clontech Advantage(登録商標)PCR−ピュア キット(pure kit)を使用して反応混合物から精製した。精製産物のアリコートを制限酵素NdeI及びXhoIにて消化した。次いで消化したPCR産物を、前記Advantageキットで「クリーンアップ(cleaned up)」し、同一の酵素で消化したベクターpET23aに連結した。
ライゲーション条件:
4μl pET23a(56ng)
5μl PCR産物(200ng)
3μl 5×GibcoBRLリガーゼ反応緩衝液
1μl GibcoBRLリガーゼ(10U)
2μl dH2O
ライゲーションは16℃で一晩行った。
連結したDNAをAdvantage(登録商標)キットを用いて精製し、次いでエレクトロコンピテント(electrocompetent)なHB101細胞へエレクトロポレーションした(1mmキュベット、1.8Kv)。
11回のエレクトロポレーションを行い、ついで細胞を、50μg/mlアンピシリン含有TYブロスの40mlに添加した。次いで細胞を37℃で一晩増殖させた。一晩増殖させた培養物の全50mlを使用してプラスミドDNAを精製した。これをライブラリーとした。
ライブラリーのスクリーニング
プラスミドライブラリーのアリコートを使用して大腸菌BL21 DE3細胞をエレクトロポレーションに付した。これらの細胞を、50μg/mlのアンピシリンを含むLB寒天に撒き、37℃で一晩増殖させた。
翌日、コロニーを選び、LB寒天+ampプレート上のナイロンフィルターにパッチをつけ(patch)、37℃で増殖を一晩続けた。翌日、フィルターをルシフェリン溶液(100mMクエン酸ナトリウム、pH5.0中の500μM)で覆った。次いでパッチを暗室で観察した。更なる分析のために200から1コロニー/パッチを選んだ。
熱安定性変異体の特徴付け
変異体プラスミドを保持する大腸菌クローンを単離した。プラスミドDNAをABI配列決定のために調製した。ルシフェラーゼをコードする完全なオープンリーディングフレームを、4つの異なるオリゴヌクレオチドプライマーを使用して配列決定した。配列決定により、ヌクレオチド640におけるAからGへの単一の点変異が明らかになった。ACT(T)からGCT(A)へのコドン変化は部位214におけるアミノ酸の変化を与えた。
【0016】
実施例2
三重変異酵素の調製
突然変異誘発性オリゴヌクレオチドを使用してpMOD1(A215L/E354K)に同一の変異を作成し、三重変異体pMOD2(A215L/E354K/T214A)を作成した。この変異もpMOD1に唯一のSacI/SstI部位を作成した。
実施例3
更なる三重変異酵素の調製
鋳型としてT214A変異を有するpET23プラスミドを用いて、以下のプライマーを使用した標準PCR反応により三重変異体T214A/I232A/E354Kを作成した。
CTGATTACACCCAAGGGGGATG E354K-センス
CATCCCCCTTGGGTGTAATCAG E354K-アンチセンス
GCAATCAAATCGCTCCGGATACTGC I232A-センス
GCAGTATCCGGAGCGATTTGATTGC I232A-アンチセンス
実施例4
熱安定性295変異体の同定
F295変異体を、Fromantら, Analytical Biochemistry, vol 224, 347-353(1995)に記載のエラー易発性PCR法を用いて作成した。PCR条件は以下の通りであった。
0.5μl(50ng)プラスミドpET23
5.0μl 10×KCl反応緩衝液
1μlプライマー1 60pmolの各プライマー
1μlプライマー2
1μl Biotaq(登録商標)ポリメラーゼ(5U)
2μl dNTPs、混合物中35mM dTTP、12.5mM dGTP、22.5mM dCTP、14mM dATP
1.76μl MgCl2(50mMストック)
1μl MnCl2(25mMストック)[反応中の最終濃度=3.26mM]36.7μl dH2O
プライマー1=5'-AAACAGGGACCCATATGGAAGACGC-3'
プライマー2=5'-AATTAACTCGAGGAATTTCGTCATCGCTGAATACAG-3'
【0017】
サイクルのパラメーターは以下の通りであった。
94℃で5分間、
次いで94℃で30秒
55℃で30秒
72℃で5分
のサイクル×15、
72℃で10分間。
PCR産物は、Clontech Advantage(登録商標)PCR−ピュア キットを使用して反応混合物から精製した。精製産物のアリコートを制限酵素NdeI及びXhoIにて消化した。次いで消化したPCR産物を、前記Advantageキットで「クリーンアップ」し、同一の酵素で消化したベクターpET23aに連結した。
ライゲーション条件は以下の通りであった。
56ng pET23a
200ng PCR産物
3μl 5×GibcoBRLリガーゼ反応緩衝液
1μl GibcoBRLリガーゼ(10U)
dH2Oを用いて、容量を10μlにした。
ライゲーションは16℃で一晩行った。
連結したDNAをAdvantage(登録商標)キットを用いて精製し、次いでエレクトロコンピテントな大腸菌DH5α細胞へエレクトロポレーションした(1mmキュベット、1.8Kv)。1mlのSOCブロスを各エレクトロポレーションに添加し、細胞を回復させ、プラスミドによりコードされる抗生物質耐性遺伝子を発現させた。ライブラリーのアリコートを50μg/mlのアンピシリンを含むLB寒天に撒き、細菌を37℃で一晩増殖させた。ナイロンフィルターディスクで寒天プレートを覆い、コロニーを新鮮なプレートに移した。オリジナルのプレートは室温下に放置し、コロニーを再増殖させた。プレートをナイロンフィルターとともに42℃で2時間インキュベートし、その後、100mM クエン酸緩衝液、pH5.0中の500μMルシフェリンをスプレーし、暗室中で観察した。
42℃で2時間後に増殖したことことを基礎に3つの熱安定性コロニーを選択した。プラスミドDNAをこれらのクローンから単離して、配列決定したところ、それぞれの場合においてF295L変異が明らかになった。
【0018】
実施例5
本発明のその他の変異体を、前記及び下記のオリゴヌクレオチドの適切な組み合わせを用いたPCRにより生産した。
GAAAGGCCCGGCACCAGCCTATCCTCTAGAGG F14A-センス
CCTCTAGCGGATAGGCTGGTGCCGGGCCTTTC F14A-アンチセンス
GAGATACGCCGCGGTTCCTGG L35A-センス
CCAGGAACCGCGGCGTATCTC L35A-アンチセンス
実施例6
ルシフェラーゼの精製及び熱不活性化の研究
組換え変異ルシフェラーゼを発現する細胞を増殖させ、破壊し、WO95/25798に記載の通りに抽出し、ルシフェラーゼの無細胞抽出物を得た。
無細胞抽出物を含むエッペンドルフチューブを特に述べない限りは通常40℃でインキュベートした。野生型ルシフェラーゼの精製調節物(比較目的)を、10%飽和硫酸アンモニウム、1mMジチオスレイトール及び0.2%ウシ血清アルブミン(BSA)を含む50mMリン酸カリウム緩衝液(pH7.8)を含む熱安定性緩衝液中でインキュベートした。設定時間に、チューブを取り出し、氷/水浴中で冷却し、その後、初期活性又は相対生物発光の百分率として計算される残存するアッセイされる活性を測定した。
結果を図2及び図3に示す。図2から、本発明のルシフェラーゼ変異体は既知の変異体と比べて改善された熱安定性を有していることが理解される。
野生型ルシフェラーゼ(RWT)に対して劇的に増加した安定性は図3より明らかである。
【0019】
実施例7
214の変異体の活性についての研究
214変異体からなるライブラリーを、カセットオリゴ(cassette oligos)を用いた部位特異的突然変異により調製し(図5)、熱安定性変異体を選択し、実施例1と同様にして試験した。3つの特定の熱安定性変異体を、実施例1と同様にして配列決定し、T214A、T214C及びT214Nとして特徴づけた。
T214、T214A、T214C及びT214Nをコードするプラスミドを保持する大腸菌XL1−ブルーのO/N培養を、Promega溶菌緩衝液を使用して溶解した。50μlの液体抽出物を37℃及び40℃下、種々の時間で熱不活性化した。10μlの加熱抽出物のアリコートを、Promegaライブアッセイ(live assay)緩衝液中で試験した(100μl)。
結果を以下の表に示す。
【0020】

【0021】
【0022】
3つの変異体について40℃で実験を繰り返した。
【0023】
【0024】
これらの結果は、T214Cがr−wt(野生型)又はT214A若しくはNのいずれかよりも有意に熱安定性が高いことを示している。存在するシステイン残基が多くなる程、熱安定性が悪くなると予想していたので、特性におけるこの変化は予想外のものであった。
実施例8
その他の点変異体の研究
単一の点変異を有するその他のフォティナス・ピラリス変異体のシリーズを、ランダムエラー易発性PCRを使用して調製した(図5)。生成した変異体のスクリーニング及び配列決定の後、その配列決定を、部位特異的突然変異誘発、続く更なる配列決定を用いてチェックした。D234G、A105V及びF295Lが存在した。これらの変異体の熱安定性は、試験した組換え野生型フォティナス・ピラリス由来ルシフェラーゼと同様であった。Promega溶菌緩衝液中タンパク質サンプルを37℃で10分間インキュベートし、その活性を2、5及び10分後にアッセイした。図4に示す結果は、各変異体が野生型よりも増強された熱安定性を生じることを示している。
【0025】
本発明は以下に関するものである。
1.ルシフェラーゼ活性を有し、かつ、フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス、ホタリア・パロウラ、ピロフォラス・プラギオフタラマス、ラムピリス・ノクチルカ、ピロコエリア・ナヤコ又はフォティナス・ペンシランヴァニカ由来のルシフェラーゼと少なくとも60%の類似性を有するタンパク質であって、
前記酵素の配列において、
(a)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基214又はルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基216に対応するアミノ酸残基、
(b)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基232又はルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基234に対応するアミノ酸残基、
(c)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基295又はルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基297に対応するアミノ酸残基、
(d)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基14、ルシオラ・ミングレリカ由来ルシフェラーゼのアミノ酸残基16、又はルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基17に対応するアミノ酸残基、
(e)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基35、ルシオラ・ミングレリカ由来ルシフェラーゼのアミノ酸残基37、又はルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基38に対応するアミノ酸残基、
(f)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基105、ルシオラ・ミングレリカ由来ルシフェラーゼのアミノ酸残基106、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基107、又はルシオラ・ラテラリス遺伝子由来ルシフェラーゼのアミノ酸残基108に対応するアミノ酸残基、
(g)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基234又はルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基236に対応するアミノ酸残基、
(h)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基420又はルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基422に対応するアミノ酸残基、
(i)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基310又はルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリスのアミノ酸残基312に対応するアミノ酸残基、
のなかの少なくとも1つが対応の野生型配列において現れるアミノ酸とは異なるものであり、かつ、該ルシフェラーゼ酵素が、当該部位に対応の野生型ルシフェラーゼのアミノ酸を有する酵素と比較して増加した熱安定性を有することを特徴とする酵素。
2.野生型ルシフェラーゼの配列を有し、該配列中の1より多いアミノ酸残基が該野生型酵素のアミノ酸残基とは異なる、前記1に記載のタンパク質。
3.50までのアミノ酸が野生型酵素のアミノ酸と異なっている、前記2に記載のタンパク質。
4.ルシフェラーゼが、フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼから修飾されたものである、前記1〜3のいずれかに記載のタンパク質。
5.前記1〜4のいずれかに記載のタンパク質であって、フォティナス・ピラリス由来ルシフェラーゼの配列において、以下の変化のうちの少なくとも1つを有しているタンパク質。
(a)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基214に対応するアミノ酸残基がスレオニン以外のアミノ酸残基である、
(b)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基232に対応するアミノ酸残基がイソロイシン以外のアミノ酸残基である、
(c)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基295に対応するアミノ酸残基がフェニルアラニン以外のアミノ酸残基である、
(d)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基14に対応するアミノ酸残基がフェニルアラニン以外のアミノ酸残基である、
(e)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基35に対応するアミノ酸残基がロイシン以外のアミノ酸残基である、
(f)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基105に対応するアミノ酸残基がアラニン以外のアミノ酸残基である、
(g)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基234に対応するアミノ酸残基がアスパラギン酸以外のアミノ酸残基である、
(h)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基420に対応するアミノ酸残基がセリン以外のアミノ酸残基である、
(i)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基310に対応するアミノ酸残基がヒスチジン以外のアミノ酸残基である。
6.前記1〜4のいずれかに記載のタンパク質であって、該タンパク質がルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来酵素の配列を実質的に有し、かつ、以下の変化のうちの少なくとも1つを有しているタンパク質。
(a)ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基216に対応するアミノ酸残基が、(ルシオラ・ミングレリカを基礎とする配列では)グリシン以外のアミノ酸残基であり、又は、(ルシオラ・クルシアタ又はルシオラ・ラテラリスを基礎とする配列では)アスパラギン以外のアミノ酸残基である、
(b)ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基234に対応するアミノ酸残基がセリン以外のアミノ酸残基である、
(c)ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基297に対応するアミノ酸残基がロイシン以外のアミノ酸残基である、
(d)ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基16に対応するアミノ酸残基がフェニルアラニン以外のアミノ酸残基である、
(e)ルシオラ・ミングレリカ由来ルシフェラーゼのアミノ酸残基37又はルシオラ・クルシアタ及びルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基38に対応するアミノ酸残基がリジン以外のアミノ酸残基である、
(f)ルシオラ・ミングレリカ由来ルシフェラーゼのアミノ酸残基106に対応するアミノ酸残基、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基107に対応するアミノ酸残基、又は、ルシオラ・ラテラリス遺伝子由来ルシフェラーゼのアミノ酸残基108に対応するアミノ酸残基がグリシン以外のアミノ酸残基である、
(g)ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基236に対応するアミノ酸残基がグリシン以外のアミノ酸残基である、
(h)ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基422に対応するアミノ酸残基がスレオニン以外のアミノ酸残基である、
(i)ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基312に対応するアミノ酸残基が、(ルシオラ・ミングレリカを基本とする配列では)スレオニン以外のアミノ酸残基であり、又は、(ルシオラ・クルシアタ又はルシオラ・ラテラリスを基本とする配列では)バリン以外のアミノ酸残基である。
7.前記1〜6のいずれかに記載のタンパク質であって、ルシフェラーゼ活性を有し、フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来のルシフェラーゼ酵素と少なくとも60%の類似性を有し、該タンパク質の配列内に以下の変化のうち少なくとも1つを有し、かつ、野生型ルシフェラーゼ酵素と比較して増加した熱安定性を有するタンパク質。
(a)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基214に対応するアミノ酸残基及びルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基216に対応するアミノ酸残基が変異しており、かつ、フォティナス・ピラリス由来ルシフェラーゼの場合はスレオニン以外である、
(b)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基232に対応するアミノ酸残基及びルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基234に対応するアミノ酸残基が変異しており、かつ、フォティナス・ピラリス由来ルシフェラーゼの場合はイソロイシン以外である、又は、
(c)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基295に対応するアミノ酸残基及びルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基297に対応するアミノ酸残基が変異しており、かつ、例えば、フォティナス・ピラリス由来ルシフェラーゼの場合、フェニルアラニン以外である。
8.フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基214に対応するアミノ酸残基及びルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基216に対応するアミノ酸残基がアラニンである、前記1に記載のタンパク質。
9.フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基232に対応するアミノ酸残基及びルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基234に対応するアミノ酸残基がアラニンである、前記1〜8のいずれかに記載のタンパク質。
10.変異したフォティナス・ピラリス由来ルシフェラーゼであり、フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基295に対応するアミノ酸残基がロイシンである、前記1〜9のいずれかに記載のタンパク質。
11.フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基14に対応するアミノ酸残基又はルシオラ由来ルシフェラーゼのアミノ酸残基16に対応するアミノ酸残基がアラニンである、前記1〜10のいずれかに記載のタンパク質。
12.ルシフェラーゼが変異したフォティナス・ピラリス又はルシオラ種由来ルシフェラーゼであり、フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基35に対応するアミノ酸残基、ルシオラ・ミングレリカ由来ルシフェラーゼのアミノ酸残基37に対応するアミノ酸残基、又はルシオラ・ラテラリス若しくはルシオラ・クルシアタ由来ルシフェラーゼのアミノ酸残基38に対応するアミノ酸残基がアラニンである、前記1〜11のいずれかに記載のタンパク質。
13.フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基105に対応するアミノ酸残基、ルシオラ・ミングレリカ由来ルシフェラーゼのアミノ酸残基106に対応するアミノ酸残基、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基107に対応するアミノ酸残基、又はルシオラ・ラテラリス遺伝子由来ルシフェラーゼのアミノ酸残基108に対応するアミノ酸残基がバリンである、前記1〜12のいずれかに記載のタンパク質。
14.変異したフォティナス・ピラリス由来ルシフェラーゼを含み、フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基234に対応するアミノ酸残基がグリシンである、前記1〜13のいずれかに記載のタンパク質。
15.変異したフォティナス・ピラリス由来ルシフェラーゼを含み、フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基420に対応するアミノ酸残基がスレオニンである、前記1〜14のいずれかに記載のタンパク質。
16.変異したフォティナス・ピラリス由来ルシフェラーゼを含み、フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基310に対応するアミノ酸残基がアルギニンである、前記1〜15のいずれかに記載のタンパク質。
17.フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基354(ルシオラ由来ルシフェラーゼの部位356)に対応する部位のアミノ酸がグルタメート以外のアミノ酸である、前記1〜16のいずれかに記載のタンパク質。
18.フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基354(ルシオラ由来ルシフェラーゼの部位356)に対応する部位のアミノ酸がリジン又はアルギニンである、前記17に記載のタンパク質。
19.ルシオラ由来ルシフェラーゼのアミノ酸残基217(フォティナス・ピラリス由来ルシフェラーゼの部位215)に対応する部位のアミノ酸が異なる疎水性アミノ酸である、前記1〜18のいずれかに記載のタンパク質。
20.ルシオラ由来ルシフェラーゼのアミノ酸残基217(フォティナス・ピラリス由来ルシフェラーゼの部位215)に対応する部位のアミノ酸がイソロイシン、ロイシン又はバリンである、前記19に記載のタンパク質。
21.前記1〜20のいずれかに記載のルシフェラーゼをコードする核酸。
22.前記21に記載の核酸を含むベクター。
23.前記22に記載のベクターで形質転換した細胞。
24.原核細胞である、前記23に記載の細胞。
25.植物細胞である、前記23に記載の細胞。
26.前記25に記載の細胞を含む植物。
27.前記1〜20のいずれかに記載のタンパク質を生産する方法であって、前記23に記載の細胞を培養する工程又は前記26に記載の植物を増殖させる工程を含むことを特徴とする方法。
28.前記1〜20のいずれかに記載のタンパク質の生物発光アッセイにおける使用。
29.前記1〜20のいずれかに記載のタンパク質を含むキット。
30.更にルシフェリンを含む、前記29に記載のキット。
【0026】
SEQUENCE LISTING
<110> The Secretary of State for Defence in Her Britannic Majesty's
Government of the United Kingdom of Great Britain and Northern Ireland
<120> Novel enzyme
<130> IPD/P1206/WOD
<140> PCT/GB99/03538
<141> 1999-10-26
<150> GB 9823468.5
<151> 1998-10-28
<160> 35
<170> PatentIn Ver. 2.1
<210> 1
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 1
cgccggtgag ctccccgccg ccg 23
<210> 2
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 2
cggcggcggg gagctcaccg gcg 23
<210> 3
<211> 51
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 3
cgaacacttc ttcatcgttg accgccttaa gtctttaatt aaatacaaag g 51
<210> 4
<211> 51
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 4
cctttgtatt taattaaaga cttaaggcgg tcaactatga agaagtgttc g 51
<210> 5
<211> 32
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 5
gaaaggcccg gcaccagcct atcctctaga gg 32
<210> 6
<211> 32
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 6
cctctagcgg ataggctggt gccgggcctt tc 32
<210> 7
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 7
ccataaattt accgaattcg tcgacttcga tcgagg 36
<210> 8
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 8
gtgtggaatt gtgagcgg 18
<210> 9
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 9
gagatacgcc gcggttcctg g 21
<210> 10
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 10
ccaggaaccg cggcgtatct c 21
<210> 11
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 11
ccctattttc attcctggcc aaaagcactc 30
<210> 12
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 12
gagtgctttt ggccaggaat gaaaataggg 30
<210> 13
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 13
ccgcatagag ctctctgcgt cagattc 27
<210> 14
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 14
gaatctgacg cagagagctc tatgcgg 27
<210> 15
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 15
gttgaccgct tgggatcctt aattaaatac 30
<210> 16
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 16
gtatagattt gaaaaagagc tg 22
<210> 17
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 17
cagctctttt tcaaatctat ac 22
<210> 18
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 18
ggctacatac tggagacata gc 22
<210> 19
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 19
gctatgtctc cagtatgtag cc 22
<210> 20
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 20
gcagttgcgc ccgtgaacga c 21
<210> 21
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 21
gtcgttcacg ggcgcaactg c 21
<210> 22
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 22
caaatcattc cgggtactgc gattttaag 29
<210> 23
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 23
cttaaaatcg cagtacccgg aatgatttg 29
<210> 24
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 24
ccgcatagaa ctctctgcgt cagattc 27
<210> 25
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 25
gaatctgacg cagagagttc tatgcgc 27
<210> 26
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 26
ctgattacac ccaaggggga tg 22
<210> 27
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 27
catccccctt gggtgtaatc ag 22
<210> 28
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<220>
<221> misc_feature
<222> (15)..(17)
<223> n=a or g or c or t
<400> 28
cccttccgca tagannngcc tgcgtcagt 29
<210> 29
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<220>
<221> misc_feature
<222> (13)..(15)
<223> n=a or g or c or t
<400> 29
actgacgcag gcnnntctat gcggaaggg 29
<210> 30
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 30
gcaatcaaat cgctccggat actgc 25
<210> 31
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 31
gcagtatccg gagcgatttg attgc 25
<210> 32
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 32
ccattccatc aaggttttgg 20
<210> 33
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 33
ccaaaacctt gatggaatgg 20
<210> 34
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Primer
<400> 34
aaacagggac ccatatggaa gacgc 25
<210> 35
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Primer
<400> 35
aattaactcg aggaatttcg tcatcgctga atacag 36
【技術分野】
【0001】
本発明は、新規酵素、特に対応の野生型酵素と比較して増加した熱安定性を有する変異ルシフェラーゼ酵素、アッセイにおける前記酵素の使用及び前記酵素を含む試験キットに関する。
ホタルのルシフェラーゼは、ATP、Mg2+及び分子酸素の存在下、光の生成を伴うルシフェリンの酸化を触媒する。この反応は約0.88の量子収率を有する。前記ルシフェラーゼの発光特性は、ATPレベルを測定する種々の発光(luminometric)アッセイにおける前記酵素の使用を導く。前記のアッセイの例には、EP-B-680515及びWO96/02665の記載に基づくアッセイが含まれる。
ルシフェラーゼは、昆虫の体、特にホタル又はツチボタル等の甲虫から直接得ることができる。ルシフェラーゼが得られる特定の種には、ニホンゲンジボタル又はケイケ(KEIKE)ボタル、ルシオラ・クルシアタ(Luciola cruciata)及びルシオラ・ラテラリス(Luciola lateralis)、東欧ホタルであるルシオラ・ミングレリカ(Luciola mingrelica)、北米ホタルであるフォティナス・ピラリス(Photinus pyralis)並びにツチボタルであるラムピリス・ノクチルカ(Lampyris noctiluca)が含まれる。ルシフェラーゼを得ることができるその他の種は、Ye ら., Biochimica et Biophysica Acta, 1339 (1997) 39-52に記載されている。Vivianiら, Biochemistry, 38, (1999) 8271-8279に記載される更なる種は、フィリキソシックス(Phrixothrix)(レールロードワーム(railroad-worms))である。
しかしながら、これらの酵素をコードする多くの遺伝子がクローニングされかつ配列決定されているので、組換えDNA技術を使用しても製造することができるだろう。前記酵素をコードする組換えDNA配列を使用して、大腸菌(E.coli)等の微生物を形質転換し、所望の酵素生成物を発現させる。
野生型及び組換え型ルシフェラーゼの熱安定性は、約30℃、特に35℃を超える温度に曝露したときにその活性が急速に消失する程度である。この不安定性は、高い周囲温度下で酵素を保存するか又は、例えば反応速度を高くするために高温反応条件下アッセイを行うときに問題を起こす。
増加した熱安定性を有する変異ルシフェラーゼはEP-A-524448及びW095/25798より既知である。前者は、ニホンボタルのルシフェラーゼにおいてスレオニン残基をイソロイシン残基で置換することにより部位217に変異を有する変異ルシフェラーゼを記載している。後者は、フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼと60%をこえる類似性を有するが、フォティナス・ピラリスの部位354のアミノ酸残基又はルシオラ種の部位356に対応するアミノ酸残基がグルタメート以外になるように変異している変異ルシフェラーゼを記載している。
【発明の概要】
【課題を解決するための手段】
【0002】
本件出願人は、増加した熱安定性をもたらし、かつ当該技術分野において既知の変異体を補完する更なる変異体を見いだした。
本発明は、ルシフェラーゼ活性を有し、かつ、フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス、ホタリア・パロウラ(Hotaria paroula)、ピロフォラス・プラギオフタラマス(Pyrophorus plagiophthalamus)、ラムピリス・ノクチルカ、ピロコエリア・ナヤコ(Pyrocoelia nayako)、フォティナス・ペンシランヴァニカ(Photinus pennsylanvanica)又はフィリキソシックス由来のルシフェラーゼと少なくとも60%の類似性を有するタンパク質であって、前記酵素の配列において、
(a)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基214又はルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基216に対応するアミノ酸残基、
(b)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基232又はルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基234に対応するアミノ酸残基、
(c)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基295又はルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基297に対応するアミノ酸残基、
(d)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基14、ルシオラ・ミングレリカ由来ルシフェラーゼのアミノ酸残基16、又はルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基17に対応するアミノ酸残基、
(e)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基35、ルシオラ・ミングレリカ由来ルシフェラーゼのアミノ酸残基37、又はルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基38に対応するアミノ酸残基、
(f)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基105、ルシオラ・ミングレリカ由来ルシフェラーゼのアミノ酸残基106、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基107、又はルシオラ・ラテラリス遺伝子由来ルシフェラーゼのアミノ酸残基108に対応するアミノ酸残基、
(g)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基234又はルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基236に対応するアミノ酸残基、
(h)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基420又はルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基422に対応するアミノ酸残基、
(i)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基310又はルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリスのアミノ酸残基312に対応するアミノ酸残基、
のなかの少なくとも1つが対応の野生型配列において現れるアミノ酸とは異なるものであり、かつ、前記ルシフェラーゼ酵素は、当該部位に特定種の対応野生型ルシフェラーゼのアミノ酸を有する酵素と比較して増加した熱安定性を有することを特徴とする酵素を提供する。
【0003】
好ましくは、前記タンパク質はルシフェラーゼ活性を有し、かつ、フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス、ホタリア・パロウラ、ピロフォラス・プラギオフタラマス、ラムピリス・ノクチルカ、ピロコエリア・ナヤコ又はフォティナス・ペンシランヴァニカ由来のルシフェラーゼと少なくとも60%の類似性を有している。
特に、前記タンパク質は、ルシフェラーゼ活性を有し、かつ、野生型ルシフェラーゼの配列、例えばフォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス、ホタリア・パロウラ、ピロフォラス・プラギオフタラマス (グリーン−Luc GR)、ピロフォラス・プラギオフタラマス (イエローグリーン Luc YG)、ピロフォラス・プラギオフタラマス (イエロー−Luc YE)、ピロフォラス・プラギオフタラマス (オレンジ−Luc OR)、ラムピリス・ノクチルカ、ピロコエリア・ナヤコ、フォティナス・ペンシランヴァニカ LY、フォティナス・ペンシランヴァニカ KW、フォティナス・ペンシランヴァニカ J19又はフィリキソシックス・グリーン(PvGR)若しくはレッド(PhRE)の配列を実質的に有する。
但し、前記タンパク質は、野生型酵素のアミノ酸とは異なるように操作された1以上、例えば100までのアミノ酸残基、好ましくはせいぜい50、より好ましくはせいぜい30のアミノ酸を含んでいてもよい。
特に、基質D−ルシフェリン(4,5−ジヒドロ−2−[6−ヒドロキシ−2−ベンゾチアゾリル]−4−チアゾール カルボン酸)を使用して光放射を生じることができる種に由来する生物発光酵素は、本発明の変異体酵素の基礎を形成するだろう。
実施例によると、本発明のルシフェラーゼは、フォティナス・ピラリスのルシフェラーゼ配列を実質的に有し、かつ、以下の変化のうちの少なくとも1つを有している。
(a)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基214に対応するアミノ酸残基がスレオニン以外のアミノ酸残基に変化している、
(b)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基232に対応するアミノ酸残基がイソロイシン以外に変化している、
(c)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基295に対応するアミノ酸残基がフェニルアラニン以外に変化している、
(d)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基14に対応するアミノ酸残基がフェニルアラニン以外に変化している、
(e)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基35に対応するアミノ酸残基がロイシン以外に変化している、
(f)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基105に対応するアミノ酸残基がアラニン以外に変化している、
(g)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基234に対応するアミノ酸残基がアスパラギン酸以外に変化している、
(h)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基420に対応するアミノ酸残基がセリン以外に変化している、
(i)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基310に対応するアミノ酸残基がヒスチジン以外に変化している。
【0004】
本発明のタンパク質がルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来酵素の配列を実質的に有する場合、以下の変化のうちの少なくとも1つを有する。
(a)ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基216に対応するアミノ酸残基が、(ルシオラ・ミングレリカを基礎とする配列では)グリシン以外であり、又は、(ルシオラ・クルシアタ又はルシオラ・ラテラリスを基礎とする配列では)アスパラギン以外である。
(b)ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基234に対応するアミノ酸残基がセリン以外である。
(c)ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基297に対応するアミノ酸残基がロイシン以外である。
(d)ルシオラ・ミングレリカ由来ルシフェラーゼのアミノ酸残基16又はルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基17に対応するアミノ酸残基がフェニルアラニン以外である。
(e)ルシオラ・ミングレリカ由来ルシフェラーゼのアミノ酸残基37又はルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基38に対応するアミノ酸残基がリジン以外である。
(f)ルシオラ・ミングレリカ由来ルシフェラーゼのアミノ酸残基106に対応するアミノ酸残基、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基107に対応するアミノ酸残基、又は、ルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基108に対応するアミノ酸残基がグリシン以外である。
(g)ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基236に対応するアミノ酸残基がグリシン以外である。
(h)ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基422に対応するアミノ酸残基がスレオニン以外である。
(i)ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基312に対応するアミノ酸残基が、(ルシオラ・ミングレリカを基本とする配列では)スレオニン以外であり、又は、(ルシオラ・クルシアタ又はルシオラ・ラテラリスを基本とする配列では)バリン以外である。
【0005】
熱安定性を増強させる全ての場合における特定の置換アミノ酸は、以下に説明されるようなルーチンの方法により決定することができる。それぞれの場合において、異なる置換が熱安定性の増強を引き起こすだろう。当業者に理解されるように、置換は、天然又は適切な変異体タンパク質をコードするDNAの部位特異的突然変異誘発により行うことができるだろう。この場合において本発明は熱安定性と関連するタンパク質の同定と関連する。
しかしながら、一般的には、野生型のアミノ酸とは異なる性質を有するアミノ酸で置換することを考慮することが好ましい。したがって、ある場合においては、親水性アミノ酸残基を疎水性アミノ酸残基(逆も又同様)で好ましく置換するだろう。同様に、酸性アミノ酸残基を塩基性アミノ酸残基で置換するだろう。
例えば、本発明のタンパク質は、ルシフェラーゼ活性を有し、かつ、フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来のルシフェラーゼ酵素と少なくとも60%の類似性を有し、その配列内に以下の変化のうち少なくとも1つを有し、野生型ルシフェラーゼ酵素と比較して増加した熱安定性を有している。
(a)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基214に対応するアミノ酸残基及びルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基216に対応するアミノ酸残基が変異しており、かつ、フォティナス・ピラリス由来ルシフェラーゼの場合はスレオニン以外である。
(b)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基232に対応するアミノ酸残基及びルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基234に対応するアミノ酸残基が変異しており、かつ、フォティナス・ピラリス由来ルシフェラーゼの場合はイソロイシン以外である。
(c)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基295に対応するアミノ酸残基及びルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基297に対応するアミノ酸残基が変異しており、かつ、例えば、フォティナス・ピラリス由来ルシフェラーゼの場合、フェニルアラニン以外である。
【0006】
種々のルシフェラーゼの全ての配列は、高度に保存されて、これら酵素間で有意な程度の類似性を有していることを示す。このことは、配列を調査して最も類似している領域を検出することにより酵素配列間の対応領域が容易に決定可能であることを示している。必要ならば、種々の配列間対応領域又は特定のアミノ酸を決定するために市販のソフト(例えば、ウィスコンシン大学遺伝学コンピュータグループの「Bestfit」、Devereuxら(1984) Nucleic Acid Research 12: 387-395を参照のこと)を使用することができる。代替又は追加として、対応の酸を、文献:L. Yeら, Biochim. Biophys Acta 1339 (1997) 39-52により決定することができる。前記文献で使用する番号方式(numbering system)は、本件出願で使用した番号方式の基礎を形成する。
フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基214に対応するアミノ酸残基の可能な変化について、極性アミノ酸スレオニンは、非極性アミノ酸、例えばアラニン、グリシン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン、トリプトファン又はシステインなどにより適切に置換される。フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基214に対応するスレオニン残基の特に好ましい置換はアラニンによるものである。
より好ましい置換はシステインによるものである。しかしながら、この部位における異なる極性残基、例えばアスパラギンも、当該部位にスレオニンを有する対応の酵素の熱安定性を増強するだろう。
野生型ルシフェラーゼ酵素においてこの部位に現れるその他のアミノ酸には、グリシン(ルシオラ・ミングレリカ、ホタリア・パロウラ)、アスパラギン(ピロフォラス・プラギオフタラマス、GR、YC、YE及びOR、ルシオラ・クルシアタ、ルシオラ・ラテラリス、ラムピリス・ノクチルカ、ピロセリア・ナヤコ、フォティナス・ペンシランヴァニカ LY、KW、J19)及びセリン(フィリキソシックス・ルシフェラーゼの部位211)が含まれる。非極性又は異なる非極性側鎖、例えばアラニン及びシステインで置換することは有利であろう。
【0007】
フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基232に対応するアミノ酸残基の可能性のある変化について、非極性アミノ酸であるイソロイシンは、異なる非極性アミノ酸、例えばアラニン、グリシン、バリン、ロイシン、プロリン、フェニルアラニン、メチオニン、トリプトファン又はシステインなどで適切に置換される。野生型の配列においてこの部位に現れるその他のアミノ酸には、セリン及びアスパラギン(それぞれ、フィトシックス・グリーン(Phritothix green)及びレッド(Phritothix red)における対応の229番目の部位におけるバリン又はアラニンと同じ)が含まれる。適切には、これらの極性残基は、非極性残基、例えば前記にて概略を述べたものにより置換される。フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基232に対応する残基及びルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリスの残基234に対応する残基の特に好ましい置換はアラニンによるものである。このことは、野生型配列に対するアミノ酸の変化を示している。
フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基295に対応するアミノ酸残基及びルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基297に対応するアミノ酸残基の変化も、タンパク質の熱安定性に影響するだろう(これは、フィリキソシックス・ルシフェラーゼの残基292に対応する)。一般的に、この部位におけるアミノ酸は非極性アミノ酸であるフェニルアラニン又はロイシンである。これらは異なる非極性アミノ酸で改変される。例えば、フォティナス・ピラリスでは、非極性アミノ酸フェニルアラニンは、異なる非極性アミノ酸、例えばアラニン、ロイシン、グリシン、バリン、イソロイシン、プロリン、メチオニン、トリプトファン又はシステインにより適切に置換される。フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基214に対応するフェニルアラニン残基についての特に好ましい置換はロイシンによるものである。
フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基14に対応するアミノ酸残基又はルシオラ由来ルシフェラーゼのアミノ酸残基16(フィリキソシックス由来ルシフェラーゼではアミノ酸残基13)に対応するアミノ酸残基における変異も可能である。このアミノ酸残基(通常はフェニルアラニンであるが、ロイシン、セリン、アルギニン又はある場合ではチロシンであってもよい)は、異なるアミノ酸、特に異なる非極性アミノ酸、例えばアラニン、バリン、ロイシン、イソロイシン、プロリン、メチオニン又はトリプトファン、好ましくはアラニンへ適切に改変するだろう。
【0008】
フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基35又はルシオラ・ミングレリカ由来ルシフェラーゼのアミノ酸残基37に対応するアミノ酸残基(その他のルシオラ種及びフィリキソシックスにおけるアミノ酸38に対応する)における変異も有効であろう。このアミノ酸は野生型酵素間で変化し、この部位におけるロイシン(フォティナス・ピラリス)だけでなく、リジン、ヒスチジン、グリシン、アラニン、グルタミン及びアスパラギン酸も含まれるだろう。適切には、この部位におけるアミノ酸残基は、非極性アミノ酸残基又は異なる非極性アミノ酸、例えばアラニン、バリン、フェニルアラニン、イソロイシン、プロリン、メチオニン又はトリプトファンにより適切に置換される。この部位における好ましいアミノ酸はアラニンであり、これは野生型酵素とは異なるものである。
フォティナス・ピラリス由来配列の部位14に対応するアミノ酸における変異及び/又はフォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基35に対応するアミノ酸残基における変異は、好ましくはこの酵素における唯一の変異ではない。これらの変異は、前記で定義したその他の変異、特にフォティナス・ピラリス由来ルシフェラーゼの部位214、395又は232に対応する部位における変異を適切に伴う。
フォティナス・ピラリス由来ルシフェラーゼにおける残基105に対応するアミノ酸残基及びルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基106に対応するアミノ酸残基(フィリキソシックスにおける残基102)の変化も、タンパク質の熱安定性に影響するだろう。一般的に、この部位におけるアミノ酸は非極性アミノ酸であるアラニン又はグリシンであり、フィリキソシックスではセリンであろう。これらのアミノ酸は異なる非極性アミノ酸により適切に改変される。例えば、フォティナス・ピラリスにおいて、非極性アミノ酸アラニンは、異なる非極性アミノ酸、例えばフェニルアラニン、ロイシン、グリシン、バリン、イソロイシン、プロリン、メチオニン又はトリプトファン等により適切に置換される。フォティナス・ピラリス由来ルシフェラーゼの残基105に対応するアラニン残基についての特に好ましい置換はバリンによるものである。
【0009】
フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基234に対応するアミノ酸残基及びルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基236(フィリキソシックスにおける残基231)に対応するアミノ酸残基の変化も、タンパク質の熱安定性に影響するだろう。一般的にこの部位におけるアミノ酸はアスパラギン酸又はグリシンであり、ある場合においてはグルタミン又はスレオニンである。これらは、非極性アミノ酸又は異なる非極性アミノ酸により適切に置換される。例えば、フォティナス・ピラリスにおいて、アミノ酸残基アスパラギン酸は、非極性アミノ酸、例えばアラニン、ロイシン、グリシン、バリン、イソロイシン、プロリン、メチオニン又はトリプトファンにより適切に置換される。フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基234に対応するフェニルアラニン残基の置換に特に好ましいのはグリシンによるものである。非極性アミノ酸残基、例えばグリシンが(例えば、ルシオラ由来ルシフェラーゼにおいて)この部位に存在する場合、グリシンは異なる非極性アミノ酸により適切に置換されるだろう。
フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基420に対応するアミノ酸残基及びルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基422(フィリキソシックス・グリーンにおける残基417及びフィリキソシックス・レッドにおける残基418)に対応するアミノ酸残基の変化も、タンパク質の熱安定性に影響するだろう。一般的にこの部位におけるアミノ酸は無電荷極性アミノ酸であるセリン又はスレオリン又はグリシンである。これらは、異なる無電荷極性アミノ酸により適切に置換される。例えば、フォティナス・ピラリスにおいて、セリンは、アスパラギン、グルタミン、スレオニン又はチロシン、特にスレオニンにより適切に置換される。
フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基310に対応するアミノ酸残基及びルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基312に対応するアミノ酸残基の変化も、タンパク質の熱安定性に影響するだろう。この部位におけるアミノ酸は既知のルシフェラーゼタンパク質の間で変化し、フォティナス・ピラリス、ピロセリア・ナヤコ、ラムピリス・ノクチルカ及びある形態のフォティナス・ペンシランヴァニカ由来ルシフェラーゼではヒスチジンであり、ルシオラ・ミングレリカ、ホタリア・パロウラ及びフィリキソシックス(アミノ酸残基307)由来ルシフェラーゼではスレオニンであり、ルシオラ・クルシアタ及びルシオラ・ラテラリスではバリンであり、あるピロフォラス・プラギオフタラマス由来ルシフェラーゼではアスパラギンである。したがって、一般的に、この部位におけるアミノ酸は親水性アミノ酸であり、これを異なるアミノ酸残基に換えて、酵素の熱安定性を増加させてもよい。フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基310に対応するヒスチジン残基に対する特に好ましい置換はアルギニンによるものである。
【0010】
酵素内にその他の変異が存在していてもよい。例えば、好ましい態様では、タンパク質は、フォティナス・ピラリス由来ルシフェラーゼのアミノ酸354に対応する部位(ルシオラ由来ルシフェラーゼにおける部位356及びフィリキソシックス由来ルシフェラーゼの部位351)にグルタミンから、特にグリシン、プロリン又はアスパラギン酸以外のアミノ酸に変化したアミノ酸を有する。適切には、この部位におけるアミノ酸はトリプトファン、バリン、ロイシン、イソロイシン及びアスパラギンであり、特に好ましくはリジン又はアルギニンである。この変異はWO 95/25798に記載されている。
代替の好ましい態様では、EP-A-052448に記載されるように、タンパク質は、ルシオラ由来ルシフェラーゼのアミノ酸残基217に対応する部位が、疎水性アミノ酸、特にイソロイシン、ロイシン又はバリンに換わったアミノ酸を有している。
タンパク質は、当該タンパク質のルシフェラーゼ活性が過度に妥協しないことを条件に配列内に更に変異を含んでいてもよい。その変異は酵素の特性を適切に増強するか、又は、ある場合においては意図する目的により適するようにする。
このことは、更なる変異が熱安定性及び/又はカラーシフト特性及び/又はATPに対する酵素のKmの増強を引き起こすことを意味する。カラーシフトを引き起こす変異の例はWO95/18853に記載されている。Km値に影響する変異は例えばWO96/22376及び国際特許出願第PCT/GB98/01026号に記載されている。これらの文献は参照することにより本明細書に組み込まれる。
本発明のタンパク質は、1つより多い変異、好ましくは前記の変異の3つすべてを適切に有している。
本発明のタンパク質には野生型及び組換え型ルシフェラーゼ酵素の両方が含まれる。本発明のタンパク質は、前記のとおり野生型酵素に存在するアミノ酸の少なくとも60%が本発明のタンパク質に存在するという意味で、フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼの配列又はその他のルシフェラーゼ酵素の配列と少なくとも60%の類似性を有する。本発明のタンパク質は前記の野生型酵素に対し、より高い程度類似性、特に少なくとも70%、より好ましくは少なくとも80%、特に好ましくは少なくとも90%の類似性を有することができる。このタイプの類似のタンパク質には、対立遺伝子変異体、その他の昆虫種由来のタンパク質及び組換えにより生成した酵素が含まれる。
【0011】
本発明のタンパク質は、野生型酵素をコードする配列とストリンジェントなハイブリダイゼーション条件下、好ましくは高ストリンジェントな条件下でハイブリダイズする核酸によりコードされるという点で同定されるだろう。前記の条件は当業者に周知であり、例えばSambrookら(1989) Molecular Cloning, Cold Spring Harbor Laboratory Pressに例示されている。一般的には、低ストリンジェント条件は約周囲温度〜約65℃における3×SSCとして定義され、高ストリンジェント条件は約65℃における0.1×SSCとして定義される。SSCは0.15M NaCl、0.015M クエン酸三ナトリウムからなる緩衝液の名称である。3×SSCは3回の同じ強さのSSCのことである。
特に、本発明の配列と特定の配列との類似性は、Lipman 及び Pearson (Lipman, D.J. & Pearson, W.R. (1985) Rapid and Sensitive Protein Similarity Searches, Science, vol 227, ppl435-1441)に記載される多重整列化(multiple alignment)法を使用して評価されるだろう。「最適化された」百分率のスコアは、Lipman-Pearsonアルゴリズムについての以下のパラメーター:ktup=1、ギャップペナルティー(gap penalty)=4及びギャップペナルティー長さ(gap penalty length)=12を用いて計算されるべきである。類似性を評価する配列は「試験配列」として使用されるべきである。このことは、比較のための塩基配列、例えばフォティナス・ピラリスの配列若しくはYeら、前出に記録された前記のその他の配列又はフィリキソシックスの場合はBiochemistry, 1999, 38, 8271-8279に記載される配列は、参照配列として使用されることを意味する。
本発明の特定の例は、前記の変異を有する野生型ルシフェラーゼ配列である。
このタンパク質は少なくとも1つ、好ましくは1つより多くの前記の変位を有している。
更に本発明は前記のルシフェラーゼをコードする核酸を提供する。適切には、前記核酸は当業者に周知の野生型配列に基づくものである。アミノ酸配列に所望の変異を起こす適切な突然変異は、遺伝子コードの知識に基づき容易に明らかになるだろう。
【0012】
本発明の核酸は、発現ベクター、例えばプロモーター、エンハンサー、ターミネーターなどの制御要素の制御下にあるプラスミドへ適切に組み込まれる。これらのベクターを用いて宿主細胞、例えば原核細胞又は真核細胞、例えば植物又は動物細胞、特に原核細胞、例えば大腸菌(E. coli)を形質転換し、当該細胞に所望のルシフェラーゼ酵素を発現させることができる。当該技術分野で周知の条件を使用して、得られた形質転換細胞の培養にルシフェラーゼ酵素を産生させ、次いで培養培地からルシフェラーゼ酵素を分離することができる。細胞が動物又は植物細胞の場合、当該細胞から植物又は動物を繁殖させてもよい。その後、当該植物からタンパク質を抽出してもよい。トランスジェニック動物の場合、タンパク質を乳汁から回収してもよい。ベクター、形質転換細胞、トランスジェニック植物及び動物並びに前記細胞の培養による酵素の生産方法の全ては本発明の更なる側面を形成する。
フォティナス・ピラリスT214A変異ルシフェラーゼを、以下のランダム突然変異誘発により作成した。T214A単一点変異体は野生型ルシフェラーゼよりも高い熱安定性を有している。
2つの新規な三重変異ルシフェラーゼ:E354K/T214A/A215L 及び E354K/T214A/I232Aを作成したところ、これらも高い熱安定性を示した。
本発明の範囲内にあるフォティナス・ピラリス変異体酵素の特定の例には以下のものが含まれる。
I232A/E354K
T214A/I232A/E354K
A215L/I232A/E354K
T214A/I232A/E354K/A215L
I232A/E354K/T214A/F295L
I232A/E354K/T214A/F295L/F14A/L35A
I232A/E354K/T214A/F295L/Fl4A/L35A/A215L
A105V
T214A
T214C
T214N
T295L
I232A
F14A
L35A
D234G
S420T
H31OR
又はその他の種のルシフェラーゼに由来するときは、それらのすべての等価物。
三重変異体作成のための変異を、プラスミドpET23上のルシフェラーゼ遺伝子へ、部位特異的突然変異(PCR)により導入した。関連する変異を起こすためにPCR反応へ添加したオリゴヌクレオチドは以下の実施例で与えられたものである。
部位354及び215における点変異の効果は相加的であることが既に示されている。本発明はより高い熱安定性を与えるために3又はそれより多い変異を組み合わせる可能性を提供する。
本発明の熱安定性ルシフェラーゼは、シグナル伝達手段としてルシフェラーゼ/ルシフェリン反応を利用する全ての生物発光アッセイにおいて有利に使用されるだろう。文献において非常に多数のアッセイが知られている。それゆえ、本発明のタンパク質は、前記のアッセイの実行を鑑みて調製されるキット中に、適宜ルシフェリン及び特定のアッセイを行うのに必要なその他の試薬と共に含まれるだろう。
本発明は添付図面を参照した実施例により詳述されるだろう。
【図面の簡単な説明】
【0013】
【図1−1】図1は、本発明の変異体の生産に使用するプラスミドを示している。
【図1−2】図1は、本発明の変異体の生産に使用するプラスミドを示している。
【図2】図2は、本発明のルシフェラーゼを含むルシフェラーゼについての熱不活性化研究の結果を示している。
【図3a】図3は、種々のルシフェラーゼ変異体についての熱安定性実験の結果を示している。
【図3b】図3は、種々のルシフェラーゼ変異体についての熱安定性実験の結果を示している。
【図3c】図3は、種々のルシフェラーゼ変異体についての熱安定性実験の結果を示している。
【図3d】図3は、種々のルシフェラーゼ変異体についての熱安定性実験の結果を示している。
【図3e】図3は、種々のルシフェラーゼ変異体についての熱安定性実験の結果を示している。
【図3f】図3は、種々のルシフェラーゼ変異体についての熱安定性実験の結果を示している。
【図3g】図3は、種々のルシフェラーゼ変異体についての熱安定性実験の結果を示している。
【図3h】図3は、種々のルシフェラーゼ変異体についての熱安定性実験の結果を示している。
【図4】図4は、その他のルシフェラーゼ変異体についての熱安定性実験の結果を示している。
【図5】図5は、本発明の変異体酵素の生産に使用したオリゴヌクレオチドを示している。
【0014】
実施例1
熱安定性変異ルシフェラーゼの同定
エラー易発性(error-prone)PCRは、Fromantら, Analytical Biochemistry, 224, 347-353 (1995)により考案されたプロトコルに基づいていた。
本反応におけるdNTP混合物は以下の通りであった。
35mM dTTP
12.5mM dGTP
22.5mM dCTP
14mM dATP
PCR条件は以下の通りであった。
0.5μl(50ng)プラスミドpPW601a J54*
5.0μl 10×KCl反応緩衝液
各1μlのW56及びW57+(60pmoleの各プライマー)
1μl Biotaq(登録商標)ポリメラーゼ(5U)
2μl dNTPs(前記参照)
1.76μl MgCl2(50mMストック)
1μl mNCl2(25mMストック)[反応における終濃度=3.26mM]
36.7μl dH2O
*プラスミドpPW601aJ54はpPW601a(WO95/25798)の変異体バージョンであり、NdeI部位をATG開始コドン前3塩基以内に作成した。これにより、pPW601aからpET23ベクターへの容易なクローニングが許容された。
+プライマー配列:
W56:
5'-AAACAGGGACCCATATGGAAGACGC-3'
W57:
5'-AATTAACTCGAGGAATTTCGTCATCGCTGAATACAG-3')
サイクルのパラメーター配下の通りであった。
94℃で5分、
次いで94℃で30秒
55℃で30秒
72℃で5分
のサイクル×12、
72℃で10分間。
【0015】
PCR産物は、Clontech Advantage(登録商標)PCR−ピュア キット(pure kit)を使用して反応混合物から精製した。精製産物のアリコートを制限酵素NdeI及びXhoIにて消化した。次いで消化したPCR産物を、前記Advantageキットで「クリーンアップ(cleaned up)」し、同一の酵素で消化したベクターpET23aに連結した。
ライゲーション条件:
4μl pET23a(56ng)
5μl PCR産物(200ng)
3μl 5×GibcoBRLリガーゼ反応緩衝液
1μl GibcoBRLリガーゼ(10U)
2μl dH2O
ライゲーションは16℃で一晩行った。
連結したDNAをAdvantage(登録商標)キットを用いて精製し、次いでエレクトロコンピテント(electrocompetent)なHB101細胞へエレクトロポレーションした(1mmキュベット、1.8Kv)。
11回のエレクトロポレーションを行い、ついで細胞を、50μg/mlアンピシリン含有TYブロスの40mlに添加した。次いで細胞を37℃で一晩増殖させた。一晩増殖させた培養物の全50mlを使用してプラスミドDNAを精製した。これをライブラリーとした。
ライブラリーのスクリーニング
プラスミドライブラリーのアリコートを使用して大腸菌BL21 DE3細胞をエレクトロポレーションに付した。これらの細胞を、50μg/mlのアンピシリンを含むLB寒天に撒き、37℃で一晩増殖させた。
翌日、コロニーを選び、LB寒天+ampプレート上のナイロンフィルターにパッチをつけ(patch)、37℃で増殖を一晩続けた。翌日、フィルターをルシフェリン溶液(100mMクエン酸ナトリウム、pH5.0中の500μM)で覆った。次いでパッチを暗室で観察した。更なる分析のために200から1コロニー/パッチを選んだ。
熱安定性変異体の特徴付け
変異体プラスミドを保持する大腸菌クローンを単離した。プラスミドDNAをABI配列決定のために調製した。ルシフェラーゼをコードする完全なオープンリーディングフレームを、4つの異なるオリゴヌクレオチドプライマーを使用して配列決定した。配列決定により、ヌクレオチド640におけるAからGへの単一の点変異が明らかになった。ACT(T)からGCT(A)へのコドン変化は部位214におけるアミノ酸の変化を与えた。
【0016】
実施例2
三重変異酵素の調製
突然変異誘発性オリゴヌクレオチドを使用してpMOD1(A215L/E354K)に同一の変異を作成し、三重変異体pMOD2(A215L/E354K/T214A)を作成した。この変異もpMOD1に唯一のSacI/SstI部位を作成した。
実施例3
更なる三重変異酵素の調製
鋳型としてT214A変異を有するpET23プラスミドを用いて、以下のプライマーを使用した標準PCR反応により三重変異体T214A/I232A/E354Kを作成した。
CTGATTACACCCAAGGGGGATG E354K-センス
CATCCCCCTTGGGTGTAATCAG E354K-アンチセンス
GCAATCAAATCGCTCCGGATACTGC I232A-センス
GCAGTATCCGGAGCGATTTGATTGC I232A-アンチセンス
実施例4
熱安定性295変異体の同定
F295変異体を、Fromantら, Analytical Biochemistry, vol 224, 347-353(1995)に記載のエラー易発性PCR法を用いて作成した。PCR条件は以下の通りであった。
0.5μl(50ng)プラスミドpET23
5.0μl 10×KCl反応緩衝液
1μlプライマー1 60pmolの各プライマー
1μlプライマー2
1μl Biotaq(登録商標)ポリメラーゼ(5U)
2μl dNTPs、混合物中35mM dTTP、12.5mM dGTP、22.5mM dCTP、14mM dATP
1.76μl MgCl2(50mMストック)
1μl MnCl2(25mMストック)[反応中の最終濃度=3.26mM]36.7μl dH2O
プライマー1=5'-AAACAGGGACCCATATGGAAGACGC-3'
プライマー2=5'-AATTAACTCGAGGAATTTCGTCATCGCTGAATACAG-3'
【0017】
サイクルのパラメーターは以下の通りであった。
94℃で5分間、
次いで94℃で30秒
55℃で30秒
72℃で5分
のサイクル×15、
72℃で10分間。
PCR産物は、Clontech Advantage(登録商標)PCR−ピュア キットを使用して反応混合物から精製した。精製産物のアリコートを制限酵素NdeI及びXhoIにて消化した。次いで消化したPCR産物を、前記Advantageキットで「クリーンアップ」し、同一の酵素で消化したベクターpET23aに連結した。
ライゲーション条件は以下の通りであった。
56ng pET23a
200ng PCR産物
3μl 5×GibcoBRLリガーゼ反応緩衝液
1μl GibcoBRLリガーゼ(10U)
dH2Oを用いて、容量を10μlにした。
ライゲーションは16℃で一晩行った。
連結したDNAをAdvantage(登録商標)キットを用いて精製し、次いでエレクトロコンピテントな大腸菌DH5α細胞へエレクトロポレーションした(1mmキュベット、1.8Kv)。1mlのSOCブロスを各エレクトロポレーションに添加し、細胞を回復させ、プラスミドによりコードされる抗生物質耐性遺伝子を発現させた。ライブラリーのアリコートを50μg/mlのアンピシリンを含むLB寒天に撒き、細菌を37℃で一晩増殖させた。ナイロンフィルターディスクで寒天プレートを覆い、コロニーを新鮮なプレートに移した。オリジナルのプレートは室温下に放置し、コロニーを再増殖させた。プレートをナイロンフィルターとともに42℃で2時間インキュベートし、その後、100mM クエン酸緩衝液、pH5.0中の500μMルシフェリンをスプレーし、暗室中で観察した。
42℃で2時間後に増殖したことことを基礎に3つの熱安定性コロニーを選択した。プラスミドDNAをこれらのクローンから単離して、配列決定したところ、それぞれの場合においてF295L変異が明らかになった。
【0018】
実施例5
本発明のその他の変異体を、前記及び下記のオリゴヌクレオチドの適切な組み合わせを用いたPCRにより生産した。
GAAAGGCCCGGCACCAGCCTATCCTCTAGAGG F14A-センス
CCTCTAGCGGATAGGCTGGTGCCGGGCCTTTC F14A-アンチセンス
GAGATACGCCGCGGTTCCTGG L35A-センス
CCAGGAACCGCGGCGTATCTC L35A-アンチセンス
実施例6
ルシフェラーゼの精製及び熱不活性化の研究
組換え変異ルシフェラーゼを発現する細胞を増殖させ、破壊し、WO95/25798に記載の通りに抽出し、ルシフェラーゼの無細胞抽出物を得た。
無細胞抽出物を含むエッペンドルフチューブを特に述べない限りは通常40℃でインキュベートした。野生型ルシフェラーゼの精製調節物(比較目的)を、10%飽和硫酸アンモニウム、1mMジチオスレイトール及び0.2%ウシ血清アルブミン(BSA)を含む50mMリン酸カリウム緩衝液(pH7.8)を含む熱安定性緩衝液中でインキュベートした。設定時間に、チューブを取り出し、氷/水浴中で冷却し、その後、初期活性又は相対生物発光の百分率として計算される残存するアッセイされる活性を測定した。
結果を図2及び図3に示す。図2から、本発明のルシフェラーゼ変異体は既知の変異体と比べて改善された熱安定性を有していることが理解される。
野生型ルシフェラーゼ(RWT)に対して劇的に増加した安定性は図3より明らかである。
【0019】
実施例7
214の変異体の活性についての研究
214変異体からなるライブラリーを、カセットオリゴ(cassette oligos)を用いた部位特異的突然変異により調製し(図5)、熱安定性変異体を選択し、実施例1と同様にして試験した。3つの特定の熱安定性変異体を、実施例1と同様にして配列決定し、T214A、T214C及びT214Nとして特徴づけた。
T214、T214A、T214C及びT214Nをコードするプラスミドを保持する大腸菌XL1−ブルーのO/N培養を、Promega溶菌緩衝液を使用して溶解した。50μlの液体抽出物を37℃及び40℃下、種々の時間で熱不活性化した。10μlの加熱抽出物のアリコートを、Promegaライブアッセイ(live assay)緩衝液中で試験した(100μl)。
結果を以下の表に示す。
【0020】
【0021】
【0022】
3つの変異体について40℃で実験を繰り返した。
【0023】
【0024】
これらの結果は、T214Cがr−wt(野生型)又はT214A若しくはNのいずれかよりも有意に熱安定性が高いことを示している。存在するシステイン残基が多くなる程、熱安定性が悪くなると予想していたので、特性におけるこの変化は予想外のものであった。
実施例8
その他の点変異体の研究
単一の点変異を有するその他のフォティナス・ピラリス変異体のシリーズを、ランダムエラー易発性PCRを使用して調製した(図5)。生成した変異体のスクリーニング及び配列決定の後、その配列決定を、部位特異的突然変異誘発、続く更なる配列決定を用いてチェックした。D234G、A105V及びF295Lが存在した。これらの変異体の熱安定性は、試験した組換え野生型フォティナス・ピラリス由来ルシフェラーゼと同様であった。Promega溶菌緩衝液中タンパク質サンプルを37℃で10分間インキュベートし、その活性を2、5及び10分後にアッセイした。図4に示す結果は、各変異体が野生型よりも増強された熱安定性を生じることを示している。
【0025】
本発明は以下に関するものである。
1.ルシフェラーゼ活性を有し、かつ、フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス、ホタリア・パロウラ、ピロフォラス・プラギオフタラマス、ラムピリス・ノクチルカ、ピロコエリア・ナヤコ又はフォティナス・ペンシランヴァニカ由来のルシフェラーゼと少なくとも60%の類似性を有するタンパク質であって、
前記酵素の配列において、
(a)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基214又はルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基216に対応するアミノ酸残基、
(b)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基232又はルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基234に対応するアミノ酸残基、
(c)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基295又はルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基297に対応するアミノ酸残基、
(d)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基14、ルシオラ・ミングレリカ由来ルシフェラーゼのアミノ酸残基16、又はルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基17に対応するアミノ酸残基、
(e)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基35、ルシオラ・ミングレリカ由来ルシフェラーゼのアミノ酸残基37、又はルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基38に対応するアミノ酸残基、
(f)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基105、ルシオラ・ミングレリカ由来ルシフェラーゼのアミノ酸残基106、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基107、又はルシオラ・ラテラリス遺伝子由来ルシフェラーゼのアミノ酸残基108に対応するアミノ酸残基、
(g)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基234又はルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基236に対応するアミノ酸残基、
(h)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基420又はルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基422に対応するアミノ酸残基、
(i)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基310又はルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリスのアミノ酸残基312に対応するアミノ酸残基、
のなかの少なくとも1つが対応の野生型配列において現れるアミノ酸とは異なるものであり、かつ、該ルシフェラーゼ酵素が、当該部位に対応の野生型ルシフェラーゼのアミノ酸を有する酵素と比較して増加した熱安定性を有することを特徴とする酵素。
2.野生型ルシフェラーゼの配列を有し、該配列中の1より多いアミノ酸残基が該野生型酵素のアミノ酸残基とは異なる、前記1に記載のタンパク質。
3.50までのアミノ酸が野生型酵素のアミノ酸と異なっている、前記2に記載のタンパク質。
4.ルシフェラーゼが、フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼから修飾されたものである、前記1〜3のいずれかに記載のタンパク質。
5.前記1〜4のいずれかに記載のタンパク質であって、フォティナス・ピラリス由来ルシフェラーゼの配列において、以下の変化のうちの少なくとも1つを有しているタンパク質。
(a)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基214に対応するアミノ酸残基がスレオニン以外のアミノ酸残基である、
(b)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基232に対応するアミノ酸残基がイソロイシン以外のアミノ酸残基である、
(c)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基295に対応するアミノ酸残基がフェニルアラニン以外のアミノ酸残基である、
(d)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基14に対応するアミノ酸残基がフェニルアラニン以外のアミノ酸残基である、
(e)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基35に対応するアミノ酸残基がロイシン以外のアミノ酸残基である、
(f)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基105に対応するアミノ酸残基がアラニン以外のアミノ酸残基である、
(g)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基234に対応するアミノ酸残基がアスパラギン酸以外のアミノ酸残基である、
(h)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基420に対応するアミノ酸残基がセリン以外のアミノ酸残基である、
(i)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基310に対応するアミノ酸残基がヒスチジン以外のアミノ酸残基である。
6.前記1〜4のいずれかに記載のタンパク質であって、該タンパク質がルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来酵素の配列を実質的に有し、かつ、以下の変化のうちの少なくとも1つを有しているタンパク質。
(a)ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基216に対応するアミノ酸残基が、(ルシオラ・ミングレリカを基礎とする配列では)グリシン以外のアミノ酸残基であり、又は、(ルシオラ・クルシアタ又はルシオラ・ラテラリスを基礎とする配列では)アスパラギン以外のアミノ酸残基である、
(b)ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基234に対応するアミノ酸残基がセリン以外のアミノ酸残基である、
(c)ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基297に対応するアミノ酸残基がロイシン以外のアミノ酸残基である、
(d)ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基16に対応するアミノ酸残基がフェニルアラニン以外のアミノ酸残基である、
(e)ルシオラ・ミングレリカ由来ルシフェラーゼのアミノ酸残基37又はルシオラ・クルシアタ及びルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基38に対応するアミノ酸残基がリジン以外のアミノ酸残基である、
(f)ルシオラ・ミングレリカ由来ルシフェラーゼのアミノ酸残基106に対応するアミノ酸残基、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基107に対応するアミノ酸残基、又は、ルシオラ・ラテラリス遺伝子由来ルシフェラーゼのアミノ酸残基108に対応するアミノ酸残基がグリシン以外のアミノ酸残基である、
(g)ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基236に対応するアミノ酸残基がグリシン以外のアミノ酸残基である、
(h)ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基422に対応するアミノ酸残基がスレオニン以外のアミノ酸残基である、
(i)ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基312に対応するアミノ酸残基が、(ルシオラ・ミングレリカを基本とする配列では)スレオニン以外のアミノ酸残基であり、又は、(ルシオラ・クルシアタ又はルシオラ・ラテラリスを基本とする配列では)バリン以外のアミノ酸残基である。
7.前記1〜6のいずれかに記載のタンパク質であって、ルシフェラーゼ活性を有し、フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来のルシフェラーゼ酵素と少なくとも60%の類似性を有し、該タンパク質の配列内に以下の変化のうち少なくとも1つを有し、かつ、野生型ルシフェラーゼ酵素と比較して増加した熱安定性を有するタンパク質。
(a)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基214に対応するアミノ酸残基及びルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基216に対応するアミノ酸残基が変異しており、かつ、フォティナス・ピラリス由来ルシフェラーゼの場合はスレオニン以外である、
(b)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基232に対応するアミノ酸残基及びルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基234に対応するアミノ酸残基が変異しており、かつ、フォティナス・ピラリス由来ルシフェラーゼの場合はイソロイシン以外である、又は、
(c)フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基295に対応するアミノ酸残基及びルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基297に対応するアミノ酸残基が変異しており、かつ、例えば、フォティナス・ピラリス由来ルシフェラーゼの場合、フェニルアラニン以外である。
8.フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基214に対応するアミノ酸残基及びルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基216に対応するアミノ酸残基がアラニンである、前記1に記載のタンパク質。
9.フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基232に対応するアミノ酸残基及びルシオラ・ミングレリカ、ルシオラ・クルシアタ又はルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基234に対応するアミノ酸残基がアラニンである、前記1〜8のいずれかに記載のタンパク質。
10.変異したフォティナス・ピラリス由来ルシフェラーゼであり、フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基295に対応するアミノ酸残基がロイシンである、前記1〜9のいずれかに記載のタンパク質。
11.フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基14に対応するアミノ酸残基又はルシオラ由来ルシフェラーゼのアミノ酸残基16に対応するアミノ酸残基がアラニンである、前記1〜10のいずれかに記載のタンパク質。
12.ルシフェラーゼが変異したフォティナス・ピラリス又はルシオラ種由来ルシフェラーゼであり、フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基35に対応するアミノ酸残基、ルシオラ・ミングレリカ由来ルシフェラーゼのアミノ酸残基37に対応するアミノ酸残基、又はルシオラ・ラテラリス若しくはルシオラ・クルシアタ由来ルシフェラーゼのアミノ酸残基38に対応するアミノ酸残基がアラニンである、前記1〜11のいずれかに記載のタンパク質。
13.フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基105に対応するアミノ酸残基、ルシオラ・ミングレリカ由来ルシフェラーゼのアミノ酸残基106に対応するアミノ酸残基、ルシオラ・クルシアタ若しくはルシオラ・ラテラリス由来ルシフェラーゼのアミノ酸残基107に対応するアミノ酸残基、又はルシオラ・ラテラリス遺伝子由来ルシフェラーゼのアミノ酸残基108に対応するアミノ酸残基がバリンである、前記1〜12のいずれかに記載のタンパク質。
14.変異したフォティナス・ピラリス由来ルシフェラーゼを含み、フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基234に対応するアミノ酸残基がグリシンである、前記1〜13のいずれかに記載のタンパク質。
15.変異したフォティナス・ピラリス由来ルシフェラーゼを含み、フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基420に対応するアミノ酸残基がスレオニンである、前記1〜14のいずれかに記載のタンパク質。
16.変異したフォティナス・ピラリス由来ルシフェラーゼを含み、フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基310に対応するアミノ酸残基がアルギニンである、前記1〜15のいずれかに記載のタンパク質。
17.フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基354(ルシオラ由来ルシフェラーゼの部位356)に対応する部位のアミノ酸がグルタメート以外のアミノ酸である、前記1〜16のいずれかに記載のタンパク質。
18.フォティナス・ピラリス由来ルシフェラーゼのアミノ酸残基354(ルシオラ由来ルシフェラーゼの部位356)に対応する部位のアミノ酸がリジン又はアルギニンである、前記17に記載のタンパク質。
19.ルシオラ由来ルシフェラーゼのアミノ酸残基217(フォティナス・ピラリス由来ルシフェラーゼの部位215)に対応する部位のアミノ酸が異なる疎水性アミノ酸である、前記1〜18のいずれかに記載のタンパク質。
20.ルシオラ由来ルシフェラーゼのアミノ酸残基217(フォティナス・ピラリス由来ルシフェラーゼの部位215)に対応する部位のアミノ酸がイソロイシン、ロイシン又はバリンである、前記19に記載のタンパク質。
21.前記1〜20のいずれかに記載のルシフェラーゼをコードする核酸。
22.前記21に記載の核酸を含むベクター。
23.前記22に記載のベクターで形質転換した細胞。
24.原核細胞である、前記23に記載の細胞。
25.植物細胞である、前記23に記載の細胞。
26.前記25に記載の細胞を含む植物。
27.前記1〜20のいずれかに記載のタンパク質を生産する方法であって、前記23に記載の細胞を培養する工程又は前記26に記載の植物を増殖させる工程を含むことを特徴とする方法。
28.前記1〜20のいずれかに記載のタンパク質の生物発光アッセイにおける使用。
29.前記1〜20のいずれかに記載のタンパク質を含むキット。
30.更にルシフェリンを含む、前記29に記載のキット。
【0026】
SEQUENCE LISTING
<110> The Secretary of State for Defence in Her Britannic Majesty's
Government of the United Kingdom of Great Britain and Northern Ireland
<120> Novel enzyme
<130> IPD/P1206/WOD
<140> PCT/GB99/03538
<141> 1999-10-26
<150> GB 9823468.5
<151> 1998-10-28
<160> 35
<170> PatentIn Ver. 2.1
<210> 1
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 1
cgccggtgag ctccccgccg ccg 23
<210> 2
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 2
cggcggcggg gagctcaccg gcg 23
<210> 3
<211> 51
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 3
cgaacacttc ttcatcgttg accgccttaa gtctttaatt aaatacaaag g 51
<210> 4
<211> 51
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 4
cctttgtatt taattaaaga cttaaggcgg tcaactatga agaagtgttc g 51
<210> 5
<211> 32
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 5
gaaaggcccg gcaccagcct atcctctaga gg 32
<210> 6
<211> 32
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 6
cctctagcgg ataggctggt gccgggcctt tc 32
<210> 7
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 7
ccataaattt accgaattcg tcgacttcga tcgagg 36
<210> 8
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 8
gtgtggaatt gtgagcgg 18
<210> 9
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 9
gagatacgcc gcggttcctg g 21
<210> 10
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 10
ccaggaaccg cggcgtatct c 21
<210> 11
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 11
ccctattttc attcctggcc aaaagcactc 30
<210> 12
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 12
gagtgctttt ggccaggaat gaaaataggg 30
<210> 13
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 13
ccgcatagag ctctctgcgt cagattc 27
<210> 14
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 14
gaatctgacg cagagagctc tatgcgg 27
<210> 15
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 15
gttgaccgct tgggatcctt aattaaatac 30
<210> 16
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 16
gtatagattt gaaaaagagc tg 22
<210> 17
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 17
cagctctttt tcaaatctat ac 22
<210> 18
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 18
ggctacatac tggagacata gc 22
<210> 19
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 19
gctatgtctc cagtatgtag cc 22
<210> 20
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 20
gcagttgcgc ccgtgaacga c 21
<210> 21
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 21
gtcgttcacg ggcgcaactg c 21
<210> 22
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 22
caaatcattc cgggtactgc gattttaag 29
<210> 23
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 23
cttaaaatcg cagtacccgg aatgatttg 29
<210> 24
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 24
ccgcatagaa ctctctgcgt cagattc 27
<210> 25
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 25
gaatctgacg cagagagttc tatgcgc 27
<210> 26
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 26
ctgattacac ccaaggggga tg 22
<210> 27
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 27
catccccctt gggtgtaatc ag 22
<210> 28
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<220>
<221> misc_feature
<222> (15)..(17)
<223> n=a or g or c or t
<400> 28
cccttccgca tagannngcc tgcgtcagt 29
<210> 29
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<220>
<221> misc_feature
<222> (13)..(15)
<223> n=a or g or c or t
<400> 29
actgacgcag gcnnntctat gcggaaggg 29
<210> 30
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 30
gcaatcaaat cgctccggat actgc 25
<210> 31
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 31
gcagtatccg gagcgatttg attgc 25
<210> 32
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 32
ccattccatc aaggttttgg 20
<210> 33
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence:
Oligonucleotide
<400> 33
ccaaaacctt gatggaatgg 20
<210> 34
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Primer
<400> 34
aaacagggac ccatatggaa gacgc 25
<210> 35
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Primer
<400> 35
aattaactcg aggaatttcg tcatcgctga atacag 36
【特許請求の範囲】
【請求項1】
以下の(a)又は(b)のタンパク質。
(a)フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ、ルシオラ・ラテラリス、ホタリア・パロウラ、ピロフォラス・プラギオフタラマス、ラムピリス・ノクチルカ又はフォティナス・ペンシランヴァニカの1つに由来する野生型ルシフェラーゼに由来するタンパク質であって、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの232位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの234位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの234位又は
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの234位
のアミノ酸に対応するアミノ酸が変異している、タンパク質、
(b)(a)のタンパク質のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ、ルシフェラーゼ活性及び各野生型ルシフェラーゼと比較して増加した熱安定性を有するタンパク質、但し、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの232位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの234位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの234位又は
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの234位
のアミノ酸に対応するアミノ酸が変異している、タンパク質。
【請求項2】
以下の(a)又は(b)のタンパク質。
(a)フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ、ルシオラ・ラテラリス、ホタリア・パロウラ、ピロフォラス・プラギオフタラマス、ラムピリス・ノクチルカ又はフォティナス・ペンシランヴァニカの1つに由来する野生型ルシフェラーゼに由来するタンパク質であって、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの295位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの297位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの297位又は
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの297位
のアミノ酸に対応するアミノ酸が変異している、タンパク質、
(b)(a)のタンパク質のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ、ルシフェラーゼ活性及び各野生型ルシフェラーゼと比較して増加した熱安定性を有するタンパク質、但し、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの295位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの297位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの297位又は
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの297位
のアミノ酸に対応するアミノ酸が変異している、タンパク質。
【請求項3】
以下の(a)又は(b)のタンパク質。
(a)フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ、ルシオラ・ラテラリス、ホタリア・パロウラ、ピロフォラス・プラギオフタラマス、ラムピリス・ノクチルカ又はフォティナス・ペンシランヴァニカの1つに由来する野生型ルシフェラーゼに由来するタンパク質であって、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの14位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの16位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの17位又は
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの17位
のアミノ酸に対応するアミノ酸が変異している、タンパク質、
(b)(a)のタンパク質のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ、ルシフェラーゼ活性及び各野生型ルシフェラーゼと比較して増加した熱安定性を有するタンパク質、但し、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの14位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの16位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの17位又は
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの17位
のアミノ酸に対応するアミノ酸が変異している、タンパク質。
【請求項4】
以下の(a)又は(b)のタンパク質。
(a)フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ、ルシオラ・ラテラリス、ホタリア・パロウラ、ピロフォラス・プラギオフタラマス、ラムピリス・ノクチルカ又はフォティナス・ペンシランヴァニカの1つに由来する野生型ルシフェラーゼに由来するタンパク質であって、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの35位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの37位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの38位又は
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの38位
のアミノ酸に対応するアミノ酸が変異している、タンパク質、
(b)(a)のタンパク質のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ、ルシフェラーゼ活性及び各野生型ルシフェラーゼと比較して増加した熱安定性を有するタンパク質、但し、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの35位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの37位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの38位又は
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの38位
のアミノ酸に対応するアミノ酸が変異している、タンパク質。
【請求項5】
以下の(a)又は(b)のタンパク質。
(a)フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ、ルシオラ・ラテラリス、ホタリア・パロウラ、ピロフォラス・プラギオフタラマス、ラムピリス・ノクチルカ又はフォティナス・ペンシランヴァニカの1つに由来する野生型ルシフェラーゼに由来するタンパク質であって、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの105位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの106位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの107位、
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの107位又は
ルシオラ・ラテラリス遺伝子に由来する野生型ルシフェラーゼの108位
のアミノ酸に対応するアミノ酸が変異している、タンパク質、
(b)(a)のタンパク質のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ、ルシフェラーゼ活性及び各野生型ルシフェラーゼと比較して増加した熱安定性を有するタンパク質、但し、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの105位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの106位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの107位、
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの107位又は
ルシオラ・ラテラリス遺伝子に由来する野生型ルシフェラーゼの108位
のアミノ酸に対応するアミノ酸が変異している、タンパク質。
【請求項6】
以下の(a)又は(b)のタンパク質。
(a)フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ、ルシオラ・ラテラリス、ホタリア・パロウラ、ピロフォラス・プラギオフタラマス、ラムピリス・ノクチルカ又はフォティナス・ペンシランヴァニカの1つに由来する野生型ルシフェラーゼに由来するタンパク質であって、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの234位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの236位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの236位又は
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの236位
のアミノ酸に対応するアミノ酸が変異している、タンパク質、
(b)(a)のタンパク質のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ、ルシフェラーゼ活性及び各野生型ルシフェラーゼと比較して増加した熱安定性を有するタンパク質、但し、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの234位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの236位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの236位又は
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの236位
のアミノ酸に対応するアミノ酸が変異している、タンパク質。
【請求項7】
以下の(a)又は(b)のタンパク質。
(a)フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ、ルシオラ・ラテラリス、ホタリア・パロウラ、ピロフォラス・プラギオフタラマス、ラムピリス・ノクチルカ又はフォティナス・ペンシランヴァニカの1つに由来する野生型ルシフェラーゼに由来するタンパク質であって、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの420位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの422位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの422位又は
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの422位
のアミノ酸に対応するアミノ酸が変異している、タンパク質、
(b)(a)のタンパク質のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ、ルシフェラーゼ活性及び各野生型ルシフェラーゼと比較して増加した熱安定性を有するタンパク質、但し、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの420位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの422位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの422位又は
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの422位
のアミノ酸に対応するアミノ酸が変異している、タンパク質。
【請求項8】
以下の(a)又は(b)のタンパク質。
(a)フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ、ルシオラ・ラテラリス、ホタリア・パロウラ、ピロフォラス・プラギオフタラマス、ラムピリス・ノクチルカ又はフォティナス・ペンシランヴァニカの1つに由来する野生型ルシフェラーゼに由来するタンパク質であって、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの310位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの312位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの312位又は
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの312位
のアミノ酸に対応するアミノ酸が変異している、タンパク質、
(b)(a)のタンパク質のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ、ルシフェラーゼ活性及び各野生型ルシフェラーゼと比較して増加した熱安定性を有するタンパク質、但し、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの310位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの312位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの312位又は
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの312位
のアミノ酸に対応するアミノ酸が変異している、タンパク質。
【請求項9】
フォティナス・ピラリス由来の野生型ルシフェラーゼに由来する、請求項1〜8のいずれか1項に記載のタンパク質。
【請求項10】
請求項9に記載のタンパク質であって、
フォティナス・ピラリス由来の野生型ルシフェラーゼと少なくとも60%の類似性を有する、タンパク質。
【請求項11】
請求項9に記載のタンパク質であって、
フォティナス・ピラリス由来の野生型ルシフェラーゼと少なくとも70%の類似性を有する、タンパク質。
【請求項12】
請求項9に記載のタンパク質であって、
フォティナス・ピラリス由来の野生型ルシフェラーゼと少なくとも80%の類似性を有する、タンパク質。
【請求項13】
請求項9に記載のタンパク質であって、
フォティナス・ピラリス由来の野生型ルシフェラーゼと少なくとも90%の類似性を有する、タンパク質。
【請求項14】
フォティナス・ピラリス由来の野生型ルシフェラーゼの354位、又は
ルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリスに由来する野生型ルシフェラーゼの356位
のアミノ酸に対応するアミノ酸がグルタメート以外のアミノ酸に変異している、請求項1〜8のいずれかに記載のタンパク質。
【請求項15】
フォティナス・ピラリス由来の野生型ルシフェラーゼの354位、又は
ルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリスに由来する野生型ルシフェラーゼの356位
のアミノ酸に対応するアミノ酸がリジン又はアルギニンに変異している、請求項14に記載のタンパク質。
【請求項16】
フォティナス・ピラリス由来の野生型ルシフェラーゼの215位、又は
ルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリスに由来する野生型ルシフェラーゼの217位
のアミノ酸に対応するアミノ酸が疎水性アミノ酸に変異している、請求項1〜8のいずれかに記載のタンパク質。
【請求項17】
フォティナス・ピラリス由来の野生型ルシフェラーゼの215位、又は
ルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリスに由来する野生型ルシフェラーゼの217位
のアミノ酸に対応するアミノ酸がイソロイシン、ロイシン又はバリンに変異している、請求項16に記載のタンパク質。
【請求項18】
請求項1〜17のいずれかに記載のルシフェラーゼをコードする核酸。
【請求項19】
請求項18に記載の核酸を含むベクター。
【請求項20】
請求項19に記載のベクターで形質転換した細胞。
【請求項21】
原核細胞である、請求項20に記載の細胞。
【請求項22】
請求項1〜17のいずれかに記載のタンパク質の生物発光アッセイにおける使用。
【請求項23】
請求項1〜17のいずれかに記載のタンパク質を含むキット。
【請求項1】
以下の(a)又は(b)のタンパク質。
(a)フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ、ルシオラ・ラテラリス、ホタリア・パロウラ、ピロフォラス・プラギオフタラマス、ラムピリス・ノクチルカ又はフォティナス・ペンシランヴァニカの1つに由来する野生型ルシフェラーゼに由来するタンパク質であって、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの232位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの234位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの234位又は
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの234位
のアミノ酸に対応するアミノ酸が変異している、タンパク質、
(b)(a)のタンパク質のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ、ルシフェラーゼ活性及び各野生型ルシフェラーゼと比較して増加した熱安定性を有するタンパク質、但し、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの232位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの234位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの234位又は
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの234位
のアミノ酸に対応するアミノ酸が変異している、タンパク質。
【請求項2】
以下の(a)又は(b)のタンパク質。
(a)フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ、ルシオラ・ラテラリス、ホタリア・パロウラ、ピロフォラス・プラギオフタラマス、ラムピリス・ノクチルカ又はフォティナス・ペンシランヴァニカの1つに由来する野生型ルシフェラーゼに由来するタンパク質であって、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの295位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの297位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの297位又は
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの297位
のアミノ酸に対応するアミノ酸が変異している、タンパク質、
(b)(a)のタンパク質のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ、ルシフェラーゼ活性及び各野生型ルシフェラーゼと比較して増加した熱安定性を有するタンパク質、但し、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの295位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの297位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの297位又は
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの297位
のアミノ酸に対応するアミノ酸が変異している、タンパク質。
【請求項3】
以下の(a)又は(b)のタンパク質。
(a)フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ、ルシオラ・ラテラリス、ホタリア・パロウラ、ピロフォラス・プラギオフタラマス、ラムピリス・ノクチルカ又はフォティナス・ペンシランヴァニカの1つに由来する野生型ルシフェラーゼに由来するタンパク質であって、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの14位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの16位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの17位又は
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの17位
のアミノ酸に対応するアミノ酸が変異している、タンパク質、
(b)(a)のタンパク質のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ、ルシフェラーゼ活性及び各野生型ルシフェラーゼと比較して増加した熱安定性を有するタンパク質、但し、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの14位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの16位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの17位又は
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの17位
のアミノ酸に対応するアミノ酸が変異している、タンパク質。
【請求項4】
以下の(a)又は(b)のタンパク質。
(a)フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ、ルシオラ・ラテラリス、ホタリア・パロウラ、ピロフォラス・プラギオフタラマス、ラムピリス・ノクチルカ又はフォティナス・ペンシランヴァニカの1つに由来する野生型ルシフェラーゼに由来するタンパク質であって、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの35位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの37位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの38位又は
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの38位
のアミノ酸に対応するアミノ酸が変異している、タンパク質、
(b)(a)のタンパク質のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ、ルシフェラーゼ活性及び各野生型ルシフェラーゼと比較して増加した熱安定性を有するタンパク質、但し、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの35位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの37位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの38位又は
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの38位
のアミノ酸に対応するアミノ酸が変異している、タンパク質。
【請求項5】
以下の(a)又は(b)のタンパク質。
(a)フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ、ルシオラ・ラテラリス、ホタリア・パロウラ、ピロフォラス・プラギオフタラマス、ラムピリス・ノクチルカ又はフォティナス・ペンシランヴァニカの1つに由来する野生型ルシフェラーゼに由来するタンパク質であって、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの105位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの106位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの107位、
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの107位又は
ルシオラ・ラテラリス遺伝子に由来する野生型ルシフェラーゼの108位
のアミノ酸に対応するアミノ酸が変異している、タンパク質、
(b)(a)のタンパク質のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ、ルシフェラーゼ活性及び各野生型ルシフェラーゼと比較して増加した熱安定性を有するタンパク質、但し、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの105位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの106位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの107位、
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの107位又は
ルシオラ・ラテラリス遺伝子に由来する野生型ルシフェラーゼの108位
のアミノ酸に対応するアミノ酸が変異している、タンパク質。
【請求項6】
以下の(a)又は(b)のタンパク質。
(a)フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ、ルシオラ・ラテラリス、ホタリア・パロウラ、ピロフォラス・プラギオフタラマス、ラムピリス・ノクチルカ又はフォティナス・ペンシランヴァニカの1つに由来する野生型ルシフェラーゼに由来するタンパク質であって、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの234位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの236位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの236位又は
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの236位
のアミノ酸に対応するアミノ酸が変異している、タンパク質、
(b)(a)のタンパク質のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ、ルシフェラーゼ活性及び各野生型ルシフェラーゼと比較して増加した熱安定性を有するタンパク質、但し、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの234位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの236位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの236位又は
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの236位
のアミノ酸に対応するアミノ酸が変異している、タンパク質。
【請求項7】
以下の(a)又は(b)のタンパク質。
(a)フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ、ルシオラ・ラテラリス、ホタリア・パロウラ、ピロフォラス・プラギオフタラマス、ラムピリス・ノクチルカ又はフォティナス・ペンシランヴァニカの1つに由来する野生型ルシフェラーゼに由来するタンパク質であって、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの420位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの422位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの422位又は
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの422位
のアミノ酸に対応するアミノ酸が変異している、タンパク質、
(b)(a)のタンパク質のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ、ルシフェラーゼ活性及び各野生型ルシフェラーゼと比較して増加した熱安定性を有するタンパク質、但し、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの420位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの422位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの422位又は
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの422位
のアミノ酸に対応するアミノ酸が変異している、タンパク質。
【請求項8】
以下の(a)又は(b)のタンパク質。
(a)フォティナス・ピラリス、ルシオラ・ミングレリカ、ルシオラ・クルシアタ、ルシオラ・ラテラリス、ホタリア・パロウラ、ピロフォラス・プラギオフタラマス、ラムピリス・ノクチルカ又はフォティナス・ペンシランヴァニカの1つに由来する野生型ルシフェラーゼに由来するタンパク質であって、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの310位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの312位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの312位又は
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの312位
のアミノ酸に対応するアミノ酸が変異している、タンパク質、
(b)(a)のタンパク質のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ、ルシフェラーゼ活性及び各野生型ルシフェラーゼと比較して増加した熱安定性を有するタンパク質、但し、
フォティナス・ピラリスに由来する野生型ルシフェラーゼの310位、
ルシオラ・ミングレリカに由来する野生型ルシフェラーゼの312位、
ルシオラ・クルシアタに由来する野生型ルシフェラーゼの312位又は
ルシオラ・ラテラリスに由来する野生型ルシフェラーゼの312位
のアミノ酸に対応するアミノ酸が変異している、タンパク質。
【請求項9】
フォティナス・ピラリス由来の野生型ルシフェラーゼに由来する、請求項1〜8のいずれか1項に記載のタンパク質。
【請求項10】
請求項9に記載のタンパク質であって、
フォティナス・ピラリス由来の野生型ルシフェラーゼと少なくとも60%の類似性を有する、タンパク質。
【請求項11】
請求項9に記載のタンパク質であって、
フォティナス・ピラリス由来の野生型ルシフェラーゼと少なくとも70%の類似性を有する、タンパク質。
【請求項12】
請求項9に記載のタンパク質であって、
フォティナス・ピラリス由来の野生型ルシフェラーゼと少なくとも80%の類似性を有する、タンパク質。
【請求項13】
請求項9に記載のタンパク質であって、
フォティナス・ピラリス由来の野生型ルシフェラーゼと少なくとも90%の類似性を有する、タンパク質。
【請求項14】
フォティナス・ピラリス由来の野生型ルシフェラーゼの354位、又は
ルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリスに由来する野生型ルシフェラーゼの356位
のアミノ酸に対応するアミノ酸がグルタメート以外のアミノ酸に変異している、請求項1〜8のいずれかに記載のタンパク質。
【請求項15】
フォティナス・ピラリス由来の野生型ルシフェラーゼの354位、又は
ルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリスに由来する野生型ルシフェラーゼの356位
のアミノ酸に対応するアミノ酸がリジン又はアルギニンに変異している、請求項14に記載のタンパク質。
【請求項16】
フォティナス・ピラリス由来の野生型ルシフェラーゼの215位、又は
ルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリスに由来する野生型ルシフェラーゼの217位
のアミノ酸に対応するアミノ酸が疎水性アミノ酸に変異している、請求項1〜8のいずれかに記載のタンパク質。
【請求項17】
フォティナス・ピラリス由来の野生型ルシフェラーゼの215位、又は
ルシオラ・ミングレリカ、ルシオラ・クルシアタ若しくはルシオラ・ラテラリスに由来する野生型ルシフェラーゼの217位
のアミノ酸に対応するアミノ酸がイソロイシン、ロイシン又はバリンに変異している、請求項16に記載のタンパク質。
【請求項18】
請求項1〜17のいずれかに記載のルシフェラーゼをコードする核酸。
【請求項19】
請求項18に記載の核酸を含むベクター。
【請求項20】
請求項19に記載のベクターで形質転換した細胞。
【請求項21】
原核細胞である、請求項20に記載の細胞。
【請求項22】
請求項1〜17のいずれかに記載のタンパク質の生物発光アッセイにおける使用。
【請求項23】
請求項1〜17のいずれかに記載のタンパク質を含むキット。
【図1−1】

【図1−2】
【図2】
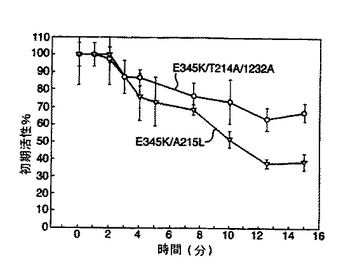
【図3a】
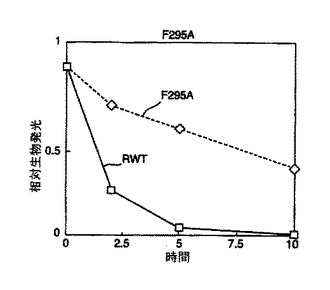
【図3b】
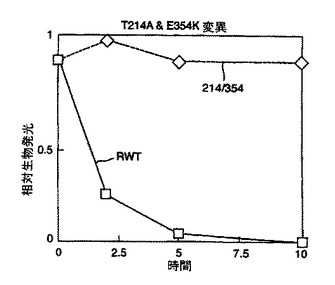
【図3c】
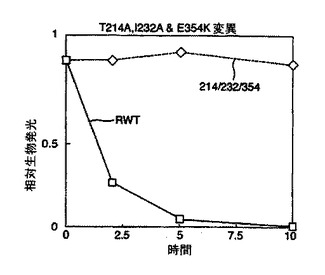
【図3d】
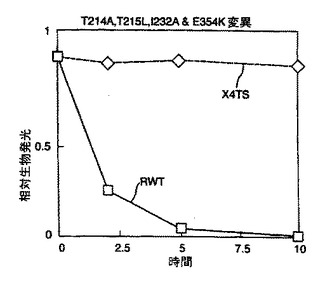
【図3e】
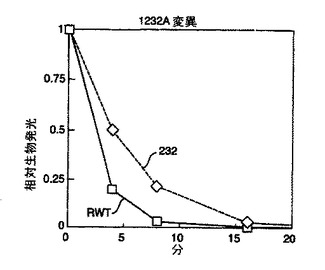
【図3f】
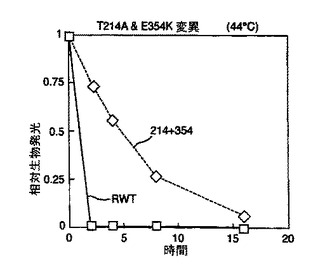
【図3g】
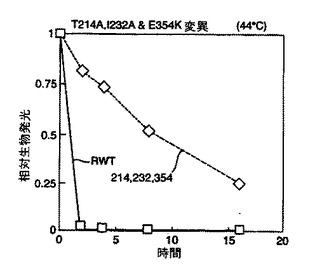
【図3h】
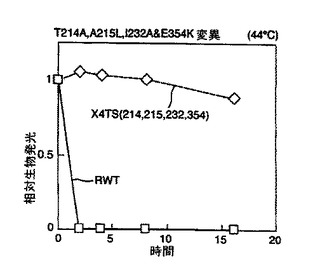
【図4】
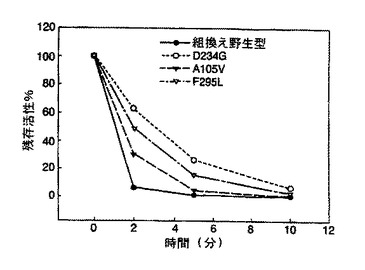
【図5】

【図1−2】
【図2】
【図3a】
【図3b】
【図3c】
【図3d】
【図3e】
【図3f】
【図3g】
【図3h】
【図4】
【図5】
【公開番号】特開2011−120605(P2011−120605A)
【公開日】平成23年6月23日(2011.6.23)
【国際特許分類】
【出願番号】特願2011−31071(P2011−31071)
【出願日】平成23年2月16日(2011.2.16)
【分割の表示】特願2000−578432(P2000−578432)の分割
【原出願日】平成11年10月26日(1999.10.26)
【出願人】(505005049)スリーエム イノベイティブ プロパティズ カンパニー (2,080)
【Fターム(参考)】
【公開日】平成23年6月23日(2011.6.23)
【国際特許分類】
【出願日】平成23年2月16日(2011.2.16)
【分割の表示】特願2000−578432(P2000−578432)の分割
【原出願日】平成11年10月26日(1999.10.26)
【出願人】(505005049)スリーエム イノベイティブ プロパティズ カンパニー (2,080)
【Fターム(参考)】
[ Back to top ]