
新規酸化酵素及びそれを用いたグリオキサールの製造方法
【課題】グリコールアルデヒドをグリオキサールに変換する酸化酵素又は微生物を提供し、グリオキサールを効率的に製造する。
【解決手段】酸素存在下、グリコールアルデヒドに作用し、グリオキサールと過酸化水素を生成し、基質特異性は、グリコールアルデヒド、及び、グリセルアルデヒドに対して活性を示し、メタノール、エタノール、1,2−プロパンジオール、1,3−プロパンジオール、及び、グリオキサールには実質的に活性を示さないものである酸化酵素。
【解決手段】酸素存在下、グリコールアルデヒドに作用し、グリオキサールと過酸化水素を生成し、基質特異性は、グリコールアルデヒド、及び、グリセルアルデヒドに対して活性を示し、メタノール、エタノール、1,2−プロパンジオール、1,3−プロパンジオール、及び、グリオキサールには実質的に活性を示さないものである酸化酵素。
【発明の詳細な説明】
【技術分野】
【0001】
本発明はグリコールアルデヒドをグリオキサールへ変換する能力を有する新規酸化酵素、および当該酵素を用いてグリオキサールを製造する方法に関する。グリオキサールは繊維加工剤、紙加工剤、土壌硬化剤、農薬、グリオキシル酸などの医薬品の原料として有用な化合物である。
【背景技術】
【0002】
現在グリオキサールは化学的な方法で製造されているが、その製造方法は煩雑で、多量の副産物が生成されるため、グリオキサールの収率が低い欠点を有する。この化学的な製造方法の欠点を解決するために、微生物や酵素を用いる方法が研究され、酵母由来のアルコール酸化酵素やアスペルギルス・ジャパニカス(Aspergillus japanicus)由来のグリセロール酸化酵素がエチレングリコールをグリコールアルデヒド経由でグリオキサールに変換できることが明らかにされた(特許文献1、非特許文献1、2、3)。一方、リゾビアレス(Rhizobiales)目およびアースロバクター(Arthrobacter)属に属する微生物より、グリコールアルデヒドをグリオキサールへ変換する酵素が見出されており、本酵素により、グリオキサールが合成できることが報告されている(特許文献2)。
【0003】
従来の酵母由来のアルコール酸化酵素やアスペルギルス・ジャパニカス(Aspergillus japanicus)由来のグリセロール酸化酵素はエチレングリコール高濃度存在下では、グリコールアルデヒドが蓄積され、エチレングリコールをグリオキサールにまで変換することは困難であり、グリコールアルデヒドを効率的にグリオキサールへ変換し、熱安定性が高く、広い範囲のpHで安定な酵素を産生する微生物、および酵素の取得が望まれていた。
【特許文献1】特開平8−84592号公報
【特許文献2】特開2005−245235号公報
【非特許文献1】Biosci.Biotech.Biochem.58,170−173(1994)
【非特許文献2】Biosci.Biotech.Biochem.59,576−581(1995)
【非特許文献3】J.Mol.Catal.B:Enzymatic,1,37−43(1995)
【発明の開示】
【発明が解決しようとする課題】
【0004】
したがって、本発明の目的は、グリコールアルデヒドをグリオキサールに変換する新規な酸化酵素、当該酸化酵素を産生する微生物、当該酸化酵素を製造する方法、及び、当該酸化酵素又は上記微生物を用いてグリオキサールを効率的に製造する方法を提供することにある。
【課題を解決するための手段】
【0005】
本発明者は、上述の問題点を解決すべく鋭意研究を重ねた結果、グリコールアルデヒドをグリオキサールに変換する新規な酸化酵素をパエニバシルス(Paenibacillus)属細菌に見出した。そして、当該微生物から本酸化酵素を単離、精製し、本発明を完成した。
即ち、本発明は、以下の(1)及び(2)の性質を有する新規な酸化酵素を提供するものである。
(1)作用:
酸素存在下、グリコールアルデヒドに作用し、グリオキサールと過酸化水素を生成する。
(2)基質特異性:
グリコールアルデヒド、及び、グリセルアルデヒドに対して活性を示し、メタノール、エタノール、1,2−プロパンジオール、1,3−プロパンジオール、及び、グリオキサールには実質的に活性を示さない。(なお本願明細書において、「実質的に活性を示さない」とは、その化合物に対する活性が、グリコールアルデヒドに対する活性の5%未満であることをいう。)
好ましくは、上記性質に加え、エチレングリコールに対する活性が、グリコールアルデヒドに対する活性の10%以下という性質を有する酸化酵素である。
好ましくは、上記性質に加え、下記(3)及び(4)の性質を有する酸化酵素である。
(3)作用最適pH:6.0−7.0。
(4)作用最適温度:45−55℃。
より好ましくは、上記性質に加え、下記(5)及び(6)の性質を有する酸化酵素である。
(5)熱安定性:pH6.5で40℃、1時間加温する前処理を行った後、85%以上の活性が残存する。
(6)pH安定性:pH5.0〜pH8.5で40℃、1時間加温する前処理を行った後、80%以上の活性が残存する。
さらに好ましくは、上記性質に加え、下記(7)及び(8)の性質を有する酸化酵素である。
(7)分子量:ゲル濾過分析で4.9×104、SDS−ポリアクリルアミド電気泳動分析において2.4×104。
(8)阻害剤:フェニルヒドラジン、ヒドラジン、セミカルバジド、α,α′−ジピリジール、8−オキシキノリン、モノヨード酢酸、、NiCl2、及び、CoCl2により阻害される。
【0006】
また、本発明は、上記酸化酵素を産生する微生物を提供する。さらに、本発明は、上記酸化酵素の製造法も提供する。
一方、本発明は、上記酸化酵素又は上記微生物をグリコールアルデヒドに作用させ、グリコールアルデヒドに存在するアルコール水酸基を酸化して、グリオキサールに変換することを特徴とするグリオキサールの製造方法も提供する。
更には、エチレングリコールに、エチレングリコールをグリコールアルデヒドへ変換する能力を有する酸化酵素又は当該酸化酵素を産生する微生物を作用させてグリコールアルデヒドを生成させて、生じたグリコールアルデヒドに、本発明の酸化酵素又は本発明の微生物を作用させてグリオキサールへ変換することを特徴とするグリオキサールの製造方法にも関する。
以下に本発明を詳述する。
【0007】
本発明の新規な酸化酵素は、グリコールアルデヒドをグリオキサールに酸化するが、反応生成物であるグリオキサールには全く作用しない特異性の高い酸化酵素であり、本発明の酸化酵素を使用すれば、グリオキサールを効率的に製造することができる。
【0008】
本発明は、従来報告されていないグリコールアルデヒドに作用してグリオキサールに作用しない新規な酸化酵素を産生する微生物を自然界から見出し、当該酵素の諸性質を明らかにし、グリオキサールの製造に有効であることを明らかにすることにより完成された。このような微生物は、土壌サンプルをエチレングリコール、グリコールアルデヒド、グリコール酸などを炭素源とした培地に添加して培養を行った後、生育してきた微生物について、グリコールアルデヒドとグリオキサールに対する酸化活性を調べる事により得ることが出来る。
【0009】
本発明の酵素の起源となる微生物は特に限定されないが、細菌などが好適であり、好ましくはPaenibacillus属に属する微生物が挙げられ、特に好ましくは北海道網走市能取湖畔の土壌から分離されたパエニバシルス・スピーシーズ(Paenibacillus sp.)AIU AL311株が挙げられる。当該AIU AL311株は独立法人産業技術総合研究所特許微生物寄託センター(〒305−8566 日本国茨城県つくば市東1丁目1番地1 中央第6)にFERM P−20786として寄託されている。AIU AL311株の菌学的諸性質を表1に示す。
【0010】
【表1】

【0011】
また、AIU AL311株の同定には、該菌株の16SリボゾーマルRNA遺伝子(16SrDNA)のうち5′末端側約500bpの領域をPCRで増幅して、塩基配列を決定し、国際塩基配列データベース(GenBank/DDBJ/EMBL)に対し相同性検索を行い、分子系統樹を作製する方法で行った。
【0012】
本発明の酸化酵素は、該酵素を産生する微生物より、下記のようにして取得することができる。例えば、本活性を有する微生物を好適な条件で培養し、培養終了後に培養液から遠心分離などにより菌体を集め、超音波破砕、ガラスビーズを用いた破砕などの方法により菌体を破砕し、粗酵素液を得る。さらにこの粗酵素液から塩析、各種クロマトグラフィーなどの方法により精製し、本発明の酵素を得ることができる。
【0013】
本発明の酸化酵素、例えばパエニバシルス(Paenibacillus)属に属する細菌、その代表的なものとして、パエニバシルス・スピーシーズ(Paenibacillus sp.)AIU AL311(FERM P−20786)が産生する酸化酵素は、グリコールアルデヒド、グリセルアルデヒドのようなアルデヒドアルコールには良く作用するが、脂肪族アルデヒド類やアルコール類にはほとんど作用せず、従来報告されているアルコールオキシダーゼやアルデヒドオキシダーゼとは基質特異性において明らかに異なる。また、本発明の酸化酵素の安定性は高く、pH6.5で、30℃、1時間の加熱後、ほぼ100%の活性を保持しており、40℃、1時間の加熱でも、約90%の活性が残存している。また、本発明の酸化酵素は、pH5.0から8.5において、40℃、1時間の加熱後、80%の活性を保持するなど、高いpH安定性も有する。
【0014】
本発明の酸化酵素の作用最適pHまたは作用最適温度は、反応条件のpHまたは温度を変えて活性を測定することにより決定される。
【0015】
本発明の酸化酵素の分子量は例えばTSK−G3000SW(7.8mm×30cm)(東ソー株式会社製)カラムを用いたゲル濾過分析により、標準タンパク質との相対溶出時間から算出することができ、サブユニット分子量はSDS−ポリアクリルアミド電気泳動により、標準タンパク質との相対移動度から算出することができる。
【0016】
また、N末端アミノ酸配列は、上記方法により精製した酵素、またはさらに逆相HPLCにより精製したものをプロテインシークエンサーにかけて決定することができる。
【0017】
本発明において、グリコールアルデヒドをグリオキサールへ変換する酸化酵素を産生する微生物を培養するための培地は、その微生物が増殖し得るものであれば特に限定されない。例えば、炭素源として、グルコース、シュークロースなどの糖類、エタノール、グリセリン、エチレングリコール、プロピレングリコールなどのアルコール類、グリコール酸などの有機酸類、オレイン酸、ステアリン酸などの脂肪酸ならびにそのエステル類、菜種油および大豆油などの油類;窒素源として、硫酸アンモニウム、硝酸アンモニウム、ペプトン、カザミノ酸、酵母エキス、肉エキスおよびコーンスチープリカーなど;無機塩類として、硫酸マグネシウム、塩化ナトリウム、炭酸カルシウム、リン酸水素カリウム、リン酸水素ナトリウムなど;その他に麦芽エキス、肉エキスなどを有する通常の液体培地が使用され得る。
【0018】
本発明によるグリオキサールの製造方法は、本発明の酸化酵素、又は、当該酸化酵素を産生する本発明の微生物をグリコールアルデヒドに作用させ、グリオキサールへと変換蓄積せしめることを特徴とする。
【0019】
更に本発明によるグリオキサールの製造方法は、エチレングリコールに、エチレングリコールをグリコールアルデヒドへ変換する能力を有する酸化酵素、又は、当該酸化酵素を産生する微生物を作用させてグリコールアルデヒドを生成させて、生じたグリコールアルデヒドに、本発明の酸化酵素又は本発明の微生物を作用させてグリオキサールへ変換することを特徴とする。
【0020】
エチレングリコールをグリコールアルデヒドへ変換する能力を有する酸化酵素としては、エチレングリコールをグリコールアルデヒドへ変換する能力を有するものであれば特に限定されないが、例えば、アルコールオキシダーゼ、グリセロールオキシダーゼが挙げられる。アルコールオキシダーゼの起源として特に限定されないが、酵母、カビ、細菌が産生するアルコールオキシダーゼが挙げられ、例えばピキア(Pichia)属、カンジダ(Candida)属、ハンセヌラ(Hansenula)属、トルロプシス(Toluropsis)属、オガタエア(Ogataea)属に属する酵母が産生するアルコールオキシダーゼが挙げられる。グリセロールオキシダーゼとしては、酵母、カビ、細菌が産生するグリセロールオキシダーゼが挙げられ、例えばアスペルギルス(Aspergillus)属産生するグリセロールオキシダーゼが挙げられる。上記以外の酵素では、アスペルギルス属に属する微生物、例えばアスペルギルス・オクラゼウス(Aspergillus ochraceus)、その一例としてアスペルギルス・オクラゼウス(Aspergillus ochraceus)AIU 031株(FERM P−20785)が産生するエチレングリコールをグリコールアルデヒドへ変換する能力を有する酸化酵素が挙げられる。同該AIU 031株は本発明者らにより土壌から分離・同定され、前記寄託番号にて独立法人産業技術総合研究所特許微生物寄託センター(〒305−8566 茨城県つくば市東1丁目1番地 中央第6)に寄託されている。
【0021】
本発明で使用する酸化酵素としては、単一または部分的に精製された酵素であってもよい。本発明で使用する微生物としては、当該微生物の培養物またはその処理物を使用することも可能である。ここで、「微生物の培養物」とは、菌体を含む培養液あるいは培養菌体を意味し、「その処理物」とは、例えば粗酵素液、凍結乾燥菌体、アセトン乾燥菌体、あるいはそれらの破砕物、これらの混合物などを意味する。更に上記酸化酵素又は上記微生物は、公知の手段(例えば、架橋法、物理的吸着法、包括法など)で固定化されて使用できる。本発明の微生物又はエチレングリコールをグリコールアルデヒドへ変換する能力を有する酸化酵素を産生する微生物としては、上記酸化酵素の産生能を有している限り、野生株または変異株、あるいは上記酸化酵素をコードするDNAをベクターに組込み、これを宿主内に導入してなる形質転換体(組替え体)であってもよい。また、本発明で使用する酸化酵素の産生能を有する形質転換体は、酸化酵素をコードするDNAを宿主のゲノムに安定的に組み込むことによっても製造できる。
【0022】
本発明の酸化酵素をグリコールアルデヒドに反応させる際の条件又はエチレングリコールをグリコールアルデヒドへ変換する能力を有する酸化酵素をエチレングリコールに反応させる際の条件は使用する酵素により異なるが、温度は5℃〜80℃、好ましくは5〜60℃の範囲、pHは4〜12、好ましくはpH5〜9の範囲である。エチレングリコールをグリコールアルデヒドへ変換する能力を有する酸化酵素、又は、当該酸化酵素を産生する微生物を作用させてグリコールアルデヒドを生成させて、生じたグリコールアルデヒドに本発明の酸化酵素又は本発明の微生物を作用させてグリオキサールへ変換する場合、エチレングリコールの濃度は、エチレングリコールをグリコールアルデヒドへ変換する酵素を効率的に働かせるために、高く設定した方がよく、100mM以上、更に1M以上が好ましい。また、グリコールアルデヒドは酵素を失活させる性質を有するため、反応時に高濃度に蓄積することは好ましくない。したがって、エチレングリコールをグリコールアルデヒドへ変換する能力を有する酵素の使用量とグリコールアルデヒドをグリオキサールへ変換する能力を有する酵素の使用量を調節することによりグリコールアルデヒドの蓄積量をコントロールすることは有効である。一方、グリコールアルデヒドを初発基質として、本発明の酵素によりグリオキサールへ変換する場合、反応系にグリコ−ルアルデヒドを高濃度添加することは酵素の安定性の観点から好ましくなく、その添加量としては、好ましくは1M以下、さらに好ましくは100mM以下が望ましい。尚、この場合、反応の進行と共にグリコールアルデヒドを断続的または連続的に添加することは、生産性の向上に繋がり好ましい。反応は、酸素条件下で行うことが好ましい。また酸素の反応液への溶解を促進するため、反応は振とう、攪拌条件下で行なわれることが好ましい。さらに大気圧以上の加圧下で反応を行うことにより、反応液への酸素の溶解度が向上し、反応がより進む場合もある。
【0023】
尚、本発明の酸化酵素又はエチレングリコールをグリコールアルデヒドへ変換する能力を有する酸化酵素による酸化反応により、過酸化水素が生成するが、この過酸化水素は酵素を失活させる場合がある。しかし、反応系にカタラーゼを添加することにより、生成した過酸化水素を分解、除去することが可能である。使用するカタラーゼは、過酸化酵素を速やかに分解、除去するという観点から、使用する酸化酵素の活性の10倍以上、好ましくは100倍以上、さらに好ましくは1000倍以上の活性量を使用する事が望ましい。また、上述した形質転換体に酸化酵素と共にカタラーゼを組換え発現させることにより、効率良く過酸化水素を分解することが可能である。一方、形質転換体の宿主として、もともとカタラーゼを産生する能力を有する微生物を使用することが好ましい。
【発明の効果】
【0024】
本発明の新規な酸化酵素は、グリコールアルデヒドをグリオキサールに酸化するが、反応生成物であるグリオキサールやエチレングリコールには全く作用しない特異性の高い酸化酵素であり、本発明の酸化酵素を使用すれば、グリオキサールを効率的に製造することができる。
【発明を実施するための最良の形態】
【0025】
以下に実施例を掲げて本発明を更に詳しく説明するが、本発明はこれら実施例のみに限定されるものではない。
(実施例1)グリコールアルデヒドに作用してグリオキサールに作用しない酸化酵素を産生する能力を持つ微生物の分離
スクリーニングのための培地は、表2に示す1%グリコール酸含有培地を120℃で15分間加圧殺菌し、このグリコール酸含有培地5mlを分注した試験管に少量の土壌を添加し、30℃で1〜7日間振盪培養した。微生物の生育が確認された培養液について、その培養液0.1mlを上記と同一組成の培地に植菌し、同様に30℃で1〜7日間振盪培養した。この操作をもう一度繰り返した後、培養液を上記液体培地と同一組成に2%寒天を加えて調製した寒天平板培地にプレートアウトし、30℃で培養した。そして微生物の生育が確認された後、この寒天平板培地にグリコールアルデヒドと過酸化水素の生成を確認するためのグリコールアルデヒドを50mMになるように添加した表3に示す発色液を摘下して、30℃で反応させた。この方法で、過酸化水素の生成が認められた微生物を、この平板培地と同一組成の斜面培地に植菌し、30℃で培養して保存した。次に、斜面培地に分離した微生物を、表2に示す5%エチレングリコール含有液体培地で振盪培養した。
【0026】
そして培養液から遠心分離で集菌し、集めた菌体を0.1Mリン酸緩衝液(pH7.0)に懸濁し、グリコールアルデヒドまたはグリオキサールを50mMになるように添加した表4に示す酵素活性測定用発色液に添加し、反応を行った。この反応で、グリコールアルデヒド存在下で反応液が紫色に着色し(過酸化水素が生成した)、グリオキサール存在下では、反応液の色がほとんど変化しない(過酸化水素が生成しない)微生物を選抜した。続いて、選抜した微生物を、表2に示す5%エチレングリコールを含む上記組成の液体培地150mlを入れた坂口フラスコを用いて30℃で振盪培養した。この培養で得られた900ml分の菌体を遠心分離で集め、0.1Mリン酸緩衝液(pH7.0)に懸濁してガラスビーズを用いて菌体を破砕した。そして、菌体破砕残渣を遠心分離で除去し、得られた上清液をDEAE−Toyopearlカラクムクロマトグラフィーに供した。即ち、菌体破砕残渣を除去して集めた上清液を10mMリン酸緩衝液と同程度の塩濃度まで脱塩した後、10mMリン酸緩衝液(pH7.0)で平衡化した20mlのDEAE−Toyopearlカラムに通液し、このカラムを10mMリン酸緩衝液(pH7.0)で洗浄した。吸着した酵素は、10mMリン酸緩衝液(pH7.0)と0.5MNaClを含む10mMリン酸緩衝液(pH7.0)を用いた直線濃度勾配法で溶出し、全ての画分についてグリコールアルデヒドおよびグリオキサールに対するオキシダーゼ活性を調べた。そして、グリコールアルデヒドに良く作用してグリオキサールに対するオキシダーゼ活性を示さない酵素を産生する微生物を選抜した。
【0027】
このような方法で得られた菌株は、上記の菌学的性質を示す微生物であり、Paenibacillus sp.AIU AL311と命名した。
尚、寒天平板培地上でのグリコールアルデヒドに対するオキシダーゼ活性は表3に示す発色液を用い、溶液でのグリコールアルデヒドに対するオキシダーゼ活性は表4に示す発色液を用いた。
【0028】
【表2】
【0029】
【表3】
【0030】
【表4】
【0031】
(実施例2)グリコールアルデヒドに作用してグリオキサールに作用しない酸化酵素の製造
実施例1で分離した微生物を、表2に示す5%エチレングリコールを含む液体培地を用いて、試験管で30℃、2日間振盪培養して種培養液を調製した。次に、この種培養液1.5mlを、種培養と同一組成の液体培地150mlを入れた500ml容フラスコに植菌し、30℃で振盪培養して、培養時間と酵素生産量との関係を調べた。その結果、グリコールアルデヒドに作用する酸化酵素の生産量は1日培養で最も高い値を示し、当該酸化酵素の活性は培養6日付近でも高い値を維持した。
【0032】
(実施例3)グリコールアルデヒドに作用してグリオキサールに作用しない酸化酵素の精製
実施例2の結果を基に、表2に示す5%エチレングリコールを含む液体培地を用いて試験管で30℃、2日間振盪培養して一次種培養液を調製した。この一次種培養液1.5mlを同一組成の液体培地を150ml分注した500ml容フラスコに植菌し、30℃で2日間振盪培養して二次種培養液を調製した。続いて、この二次種培養液20mlを一次種培養と同一組成の液体培地2Lを入れた3L容フラスコに植菌し、30℃で1日間振盪培養した。このようにして培養した9Lの培養液から菌体を集めて、以下の方法で酵素を精製した。尚、酵素の精製にはpH7.0のリン酸緩衝液を用いた。
【0033】
1)粗酵素液の調製
9L培養液から集菌した菌体(湿重量:7.5g)を10mMリン酸緩衝液に懸濁し、10℃以下の温度で0.5mmガラスビーズを用いて12分間(2分x6回)細胞破砕した。この細胞破砕液を遠心分離して菌体残渣を除去し、上清画分を粗酵素液として酵素の精製に用いた。
2)DEAE−トヨパールカラムクロマトグラフィー
粗酵素酵素液を限外濾過膜で2ms/cmまで脱塩濃縮した後、10mMリン酸緩衝液で平衡化したDEAE−トヨパールカラム(2.2×17.5cm)に吸着させた。本カラムを10mMリン酸緩衝液650mlで洗浄した後、酵素を10mMリン酸緩衝液(340ml)と0.5M NaClを含む10mMリン酸緩衝液(340ml)を用いて直線濃度勾配法で溶出した。
3)ヒドロキシアパタイトカラムクロマトグラフィー
DEAE−トヨパールカラムからの溶出液を限外濾過膜で脱塩濃縮した後、5mMリン酸緩衝液で緩衝化済みのヒドロキシアパタイトカラム(1×6cm)に通液し、続いて本カラムを5mMリン酸緩衝液100mlで洗浄し、目的とする酵素を溶出した。
以上の方法で得られた上記3)の酵素標品をNativeおよびSDS−ポリアクリルアミド電気泳動法で分析した結果、電気泳動的に単一であった。尚、本精製法による酵素の精製収率は、表5の通りであった。
【0034】
【表5】
【0035】
(実施例4)グリコールアルデヒドに作用してグリオキサールに作用しない酸化酵素の性質
実施例3の方法で精製した酵素標品は、NativeおよびSDS−ポリアクリルアミド電気泳動法を用いて分析した結果、いずれの分析でも蛋白的に単一であった。よって、本精製酵素標品を用いて諸性質を検討した。
(作用)
グリコールアルデヒドを21mMになるように添加した表4に示す酵素活性測定用発色液0.95mlに精製酵素液0.05mlを添加し、30℃で反応を行い、555nmの吸光度の増加を経時的に測定した。その結果、時間の経過と共に555nmの吸光度が増大し、エチレングリコールと本酵素の反応によって、過酸化水素が生成されることが明らかになった。
【0036】
また、反応生成物をIsobe and Nishiseの方法[Biosci.Biotech.Biochem.,58,170−173,(1994)]に従ってN−methyl−2−benzothiazolinone hydrazone(MBTH)と反応させ、その吸収スペクトルおよびC18の逆相カラムからの溶出時間を分析した。まず、反応生成物を0.2Mグリシン−HCl緩衝液(pH4.0)0.75mlに溶解し、それに1.0%(w/v)MBTH液を0.3ml添加した(反応1)。その後、反応1で得られた反応液について吸収スペクトルを測定した。その結果、グリオキサールの反応1で得られた反応液の吸収スペクトルと同様に、上記反応生成物の反応1の反応液では、410nm付近に極大吸収を持つスペクトルが得られた。そして反応1の反応液をC18の逆相カラムで分析した場合も、グリオキサールの反応1で得られる生成物と同じ溶出位置にピークが得られた。よって、本酵素は、下記の式に従ってグリコールアルデヒドをグリオキサールに酸化することが明らかになった。
OHCCHO+O2+H2O→OHCCOOH+H2O2
【0037】
次に、グリオキサールを21mMになるように添加した表4に示す酵素活性測定用発色液0.95mlに精製酵素液0.05mlを添加し、30℃で反応を行い555nmの吸光度の増加を経時的に測定した。その結果、555nmの吸光度の増大は認められなかった。よって、本酵素は、グリコールアルデヒドをグリオキサールに酸化するが、反応生成物であるグリオキサールには作用しないことが明らかになった。
【0038】
(基質特異性)
表6に示すアルデヒド類、有機酸類又はアルコール類を53mMになるように添加した表4に示す酵素活性測定用発色液0.95mlに精製酵素液0.05mlを添加し、30℃で反応を行い、555nmの吸光度の増加を分光光度計にて連続して測定し、時間あたりの555nmの増加量を算出し、各化合物に対する活性を調べた。
その結果、表6に示すように、本酵素は、グリコールアルデヒドとグリセルアルデヒドに良く作用し、ホルムアルデヒド、アセトアルデヒド、エチレングリコール、プロピオンアルデヒドにもわずかに作用を示すが、イソブチルアルデヒド、バレルアルデヒド、イソバレルアルデヒド、グリオキサール、グリオキシル酸には実質的に作用しなかった。また、メタノール、エタノール、1−プロパノール、1−ブタノールなどの一級アルコールや1,2−プロパンジオール、1,3−プロパンジオールなどにも作用しなかった。なお、表6中の各数値は、グリコールアルデヒドに対する活性を100とした場合の相対値である。
【0039】
【表6】
【0040】
(グリコールアルデヒドに対するKm値)
グリコールアルデヒドを10mMから45mMの濃度になるように添加した表4に示す酵素活性測定用発色液0.95mlに精製酵素液0.05mlを添加し、30℃で反応を行い、555nmの吸光度の増加を分光光度計にて連続して測定し、時間あたりの555nmの増加量を算出し、各濃度のグリコールアルデヒドに対する活性を測定した。更に、得られた活性値と基質濃度より作成したLineweaver−Burk plotにより、Km値を算出した。その結果、グリコールアルデヒドに対するKm値は約13mMであった。
【0041】
(分子量)
精製酵素を高速液体クロマトグラフィーによるTSK−G3000SW(7.8mm×30cm)(東ソー株式会社製)カラムを用いたゲル濾過、および1%の2−メルカプトエタノール存在下、10%SDS−ポリアクリルアミド電気泳動に供し、標準タンパクとの相対移動度より、その分子量を推定した。その結果、分子量は、ゲル濾過法で約49,000、SDS−ポリアクリルアミド電気泳動法で約24,000であった
[ゲル濾過法による分子量測定条件]
カラム:TSK−G3000SW(7.8mm×30cm)(東ソー株式会社製)
溶離液:0.1Mリン酸緩衝液+0.3M塩化ナトリウム(pH7.0)
流 速:1.0ml/min
温 度:室温
検 出:220nm
【0042】
(各種化合物の影響)
グリコールアルデヒドを53mM、更に表7に示す化合物を1.05mM濃度になるように添加した表4に示す酵素活性測定用発色液0.95mlに精製酵素液0.05mlを添加し、30℃で反応を行い、555nmの吸光度の増加を分光光度計にて連続して測定し、時間あたりの555nmの増加量を算出し、活性を調べた。
その結果、表7に示すように、本酵素は、フェニルヒドラジン、ヒドラジン、セミカルバジド、α,α′−ジピリジール、8−オキシキノリン、モノヨード酢酸、NiCl2、CoCl2で阻害され、MnCl2で活性化されることが明らかになった。
【0043】
【表7】
【0044】
(最適pH)
グリコールアルデヒドを53mMになるように添加し、緩衝液の種類を変更してpH5.5からpH8.5で調製した表4に示す酵素活性測定用発色液0.95mlに精製酵素液0.05mlを添加し、30℃で反応を行い、555nmの吸光度の増加を分光光度計にて連続して測定し、時間あたりの555nmの増加量を算出し、活性を調べた。
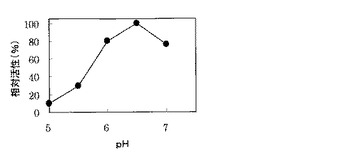
その結果、図1に示すように、本酵素は酸性領域から弱アルカリ性領域の広い範囲で活性を示し、反応の最適pHは6.0−7.0であった。
【0045】
(最適温度)
グリコールアルデヒドを53mMになるように添加した表4に示す酵素活性測定用発色液0.95mlに精製酵素液0.05mlを添加し、30℃から60℃の範囲で反応を行い、555nmの吸光度の増加を分光光度計にて連続して測定し、時間あたりの555nmの増加量を算出し、活性を調べた。
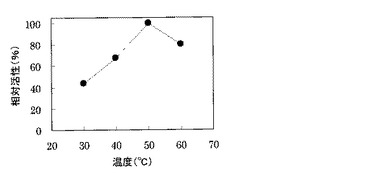
その結果、図2に示すように、本酵素は測定したいずれの温度でも活性を示し、反応の最適温度は45−55℃であった。
【0046】
(pH安定性)
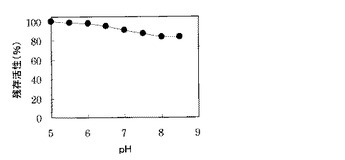
pH5.0からpH8.5で酵素液を40℃、1時間加温する前処理を行った。その後、グリコールアルデヒドを53mMになるように添加した表4に示す酵素活性測定用発色液0.95mlに上記前処理後の酵素液0.05mlを添加し、30℃で反応を行い、555nmの吸光度の増加を分光光度計にて連続して測定し、時間あたりの555nmの増加量を算出し、残存する酵素活性を調べた。各処理後の残存活性(前処理前の活性に対する各処理後の相対活性:前処理前の活性を100とした)を図3に示す。
その結果、図3に示すように、酵素活性は、酸性領域からアルカリ性領域の広い範囲で残存しており、特にpH5.0からpH8.5の範囲では、80%以上の活性が残存した。よって、本酵素は、酸性領域からアルカリ性の領域領域の広い範囲で安定であることが明らかになった。
【0047】
(熱安定性)
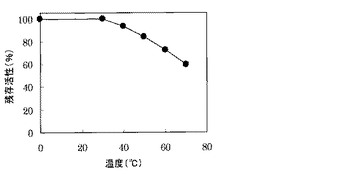
pH6.5で酵素液を0℃から70℃、1時間加温する前処理を行った後、上記pH安定性と同様な方法で、残存酵素活性を測定した。その結果、図4に示すように、酵素活性は、70℃で1時間加温しても60%以上の活性が残存した。よって、本酵素は、熱に安定であることが明らかになった。
【0048】
(N末端アミノ酸配列)
精製酵素を用いて、本酵素のN末端アミノ酸配列を調べた。本精製酵素のN末端を配列番号1に示す。
【0049】
(実施例5)粗酵素液を用いたグリオキサールの合成
実施例2記載の方法で得たAIU AL311株培養液250mlから集菌した菌体を10mMリン酸緩衝液5mlに懸濁し、10℃以下の温度で0.5mmガラスビーズを用いて10分間(2分×5回)細胞破砕した。この細胞破砕液を遠心分離して菌体残渣を除去し、上清分画を粗酵素液として得た。得られた粗酵素液0.7mlに250mMグリコールアルデヒド水溶液0.2ml、50,000U/mlカタラーゼ(Bovive Liver由来;ナカライ社製)溶液0.1mlを添加し、試験管中で28℃、10時間、振盪し、反応を行った。その後、得られた反応液を高速液体クロマトグラフィー(HPLC)により分析した。その結果、反応液中に13mMのグリオキサールが生成していた。
【0050】
[HPLC分析条件]
カラム:バイオラッド社製アミネックスHPX−87H(7.8mm×300mm)
検出:示差屈折率(RI)
カラム温度:25℃
溶離液:5mM H2SO4水溶液
流速:0.4ml/分
溶離時間:グリオキサール−16分、
【0051】
(実施例6)精製酵素を用いたグリオキサールの合成
実施例3で得た精製酵素0.1U、グリコールアルデヒド50mM、カタラーゼ(Bovive Liver由来;ナカライ社製)500Uを含む100mMリン酸緩衝液(pH6.5)1mlを試験管に加え、30℃、24時間、振盪し、反応を行った。その後、得られた反応液をHPLCにより分析した。その結果、反応液中に20mMのグリオキサールが生成していた。
【0052】
(実施例7)エチレングリコールからのグリオキサールの合成
1Mエチレングリコール、2U/ml(メタノール酸化活性)のピキア属(Pichia pastris)由来アルコールオキシダーゼ(シグマ社製)、0.2U/ml(グリコールアルデヒド酸化活性)の実施例3で得た精製酸化酵素、10000U/mlのカタラーゼ(Bovive Liver由来;ナカライ社製)を含む300mMリン酸緩衝液(pH8.0)0.5mlを試験管中で、28℃、4時間振盪しながら反応を行った。その後、得られた反応液をHPLCにより分析した。その結果、5mMのグリコールアルデヒドおよび18mMのグリオキサールが生成していた。
[HPLC分析条件]
カラム:Shodex Rspak KC−811(昭和電工社製)を二本連結
検出:示差屈折率(RI)
カラム温度:40℃
溶離液:50mM過塩素酸水溶液
流速:1ml/min
溶離時間:グリコールアルデヒド;17.9分、グリオキサール;15.7分
【図面の簡単な説明】
【0053】
【図1】実施例3で取得した酸化酵素の作用最適pHを示すグラフ
【図2】実施例3で取得した酸化酵素の作用最適温度を示すグラフ
【図3】実施例3で取得した酸化酵素のpH安定性を示すグラフ
【図4】実施例3で取得した酸化酵素の熱安定性を示すグラフ
【技術分野】
【0001】
本発明はグリコールアルデヒドをグリオキサールへ変換する能力を有する新規酸化酵素、および当該酵素を用いてグリオキサールを製造する方法に関する。グリオキサールは繊維加工剤、紙加工剤、土壌硬化剤、農薬、グリオキシル酸などの医薬品の原料として有用な化合物である。
【背景技術】
【0002】
現在グリオキサールは化学的な方法で製造されているが、その製造方法は煩雑で、多量の副産物が生成されるため、グリオキサールの収率が低い欠点を有する。この化学的な製造方法の欠点を解決するために、微生物や酵素を用いる方法が研究され、酵母由来のアルコール酸化酵素やアスペルギルス・ジャパニカス(Aspergillus japanicus)由来のグリセロール酸化酵素がエチレングリコールをグリコールアルデヒド経由でグリオキサールに変換できることが明らかにされた(特許文献1、非特許文献1、2、3)。一方、リゾビアレス(Rhizobiales)目およびアースロバクター(Arthrobacter)属に属する微生物より、グリコールアルデヒドをグリオキサールへ変換する酵素が見出されており、本酵素により、グリオキサールが合成できることが報告されている(特許文献2)。
【0003】
従来の酵母由来のアルコール酸化酵素やアスペルギルス・ジャパニカス(Aspergillus japanicus)由来のグリセロール酸化酵素はエチレングリコール高濃度存在下では、グリコールアルデヒドが蓄積され、エチレングリコールをグリオキサールにまで変換することは困難であり、グリコールアルデヒドを効率的にグリオキサールへ変換し、熱安定性が高く、広い範囲のpHで安定な酵素を産生する微生物、および酵素の取得が望まれていた。
【特許文献1】特開平8−84592号公報
【特許文献2】特開2005−245235号公報
【非特許文献1】Biosci.Biotech.Biochem.58,170−173(1994)
【非特許文献2】Biosci.Biotech.Biochem.59,576−581(1995)
【非特許文献3】J.Mol.Catal.B:Enzymatic,1,37−43(1995)
【発明の開示】
【発明が解決しようとする課題】
【0004】
したがって、本発明の目的は、グリコールアルデヒドをグリオキサールに変換する新規な酸化酵素、当該酸化酵素を産生する微生物、当該酸化酵素を製造する方法、及び、当該酸化酵素又は上記微生物を用いてグリオキサールを効率的に製造する方法を提供することにある。
【課題を解決するための手段】
【0005】
本発明者は、上述の問題点を解決すべく鋭意研究を重ねた結果、グリコールアルデヒドをグリオキサールに変換する新規な酸化酵素をパエニバシルス(Paenibacillus)属細菌に見出した。そして、当該微生物から本酸化酵素を単離、精製し、本発明を完成した。
即ち、本発明は、以下の(1)及び(2)の性質を有する新規な酸化酵素を提供するものである。
(1)作用:
酸素存在下、グリコールアルデヒドに作用し、グリオキサールと過酸化水素を生成する。
(2)基質特異性:
グリコールアルデヒド、及び、グリセルアルデヒドに対して活性を示し、メタノール、エタノール、1,2−プロパンジオール、1,3−プロパンジオール、及び、グリオキサールには実質的に活性を示さない。(なお本願明細書において、「実質的に活性を示さない」とは、その化合物に対する活性が、グリコールアルデヒドに対する活性の5%未満であることをいう。)
好ましくは、上記性質に加え、エチレングリコールに対する活性が、グリコールアルデヒドに対する活性の10%以下という性質を有する酸化酵素である。
好ましくは、上記性質に加え、下記(3)及び(4)の性質を有する酸化酵素である。
(3)作用最適pH:6.0−7.0。
(4)作用最適温度:45−55℃。
より好ましくは、上記性質に加え、下記(5)及び(6)の性質を有する酸化酵素である。
(5)熱安定性:pH6.5で40℃、1時間加温する前処理を行った後、85%以上の活性が残存する。
(6)pH安定性:pH5.0〜pH8.5で40℃、1時間加温する前処理を行った後、80%以上の活性が残存する。
さらに好ましくは、上記性質に加え、下記(7)及び(8)の性質を有する酸化酵素である。
(7)分子量:ゲル濾過分析で4.9×104、SDS−ポリアクリルアミド電気泳動分析において2.4×104。
(8)阻害剤:フェニルヒドラジン、ヒドラジン、セミカルバジド、α,α′−ジピリジール、8−オキシキノリン、モノヨード酢酸、、NiCl2、及び、CoCl2により阻害される。
【0006】
また、本発明は、上記酸化酵素を産生する微生物を提供する。さらに、本発明は、上記酸化酵素の製造法も提供する。
一方、本発明は、上記酸化酵素又は上記微生物をグリコールアルデヒドに作用させ、グリコールアルデヒドに存在するアルコール水酸基を酸化して、グリオキサールに変換することを特徴とするグリオキサールの製造方法も提供する。
更には、エチレングリコールに、エチレングリコールをグリコールアルデヒドへ変換する能力を有する酸化酵素又は当該酸化酵素を産生する微生物を作用させてグリコールアルデヒドを生成させて、生じたグリコールアルデヒドに、本発明の酸化酵素又は本発明の微生物を作用させてグリオキサールへ変換することを特徴とするグリオキサールの製造方法にも関する。
以下に本発明を詳述する。
【0007】
本発明の新規な酸化酵素は、グリコールアルデヒドをグリオキサールに酸化するが、反応生成物であるグリオキサールには全く作用しない特異性の高い酸化酵素であり、本発明の酸化酵素を使用すれば、グリオキサールを効率的に製造することができる。
【0008】
本発明は、従来報告されていないグリコールアルデヒドに作用してグリオキサールに作用しない新規な酸化酵素を産生する微生物を自然界から見出し、当該酵素の諸性質を明らかにし、グリオキサールの製造に有効であることを明らかにすることにより完成された。このような微生物は、土壌サンプルをエチレングリコール、グリコールアルデヒド、グリコール酸などを炭素源とした培地に添加して培養を行った後、生育してきた微生物について、グリコールアルデヒドとグリオキサールに対する酸化活性を調べる事により得ることが出来る。
【0009】
本発明の酵素の起源となる微生物は特に限定されないが、細菌などが好適であり、好ましくはPaenibacillus属に属する微生物が挙げられ、特に好ましくは北海道網走市能取湖畔の土壌から分離されたパエニバシルス・スピーシーズ(Paenibacillus sp.)AIU AL311株が挙げられる。当該AIU AL311株は独立法人産業技術総合研究所特許微生物寄託センター(〒305−8566 日本国茨城県つくば市東1丁目1番地1 中央第6)にFERM P−20786として寄託されている。AIU AL311株の菌学的諸性質を表1に示す。
【0010】
【表1】
【0011】
また、AIU AL311株の同定には、該菌株の16SリボゾーマルRNA遺伝子(16SrDNA)のうち5′末端側約500bpの領域をPCRで増幅して、塩基配列を決定し、国際塩基配列データベース(GenBank/DDBJ/EMBL)に対し相同性検索を行い、分子系統樹を作製する方法で行った。
【0012】
本発明の酸化酵素は、該酵素を産生する微生物より、下記のようにして取得することができる。例えば、本活性を有する微生物を好適な条件で培養し、培養終了後に培養液から遠心分離などにより菌体を集め、超音波破砕、ガラスビーズを用いた破砕などの方法により菌体を破砕し、粗酵素液を得る。さらにこの粗酵素液から塩析、各種クロマトグラフィーなどの方法により精製し、本発明の酵素を得ることができる。
【0013】
本発明の酸化酵素、例えばパエニバシルス(Paenibacillus)属に属する細菌、その代表的なものとして、パエニバシルス・スピーシーズ(Paenibacillus sp.)AIU AL311(FERM P−20786)が産生する酸化酵素は、グリコールアルデヒド、グリセルアルデヒドのようなアルデヒドアルコールには良く作用するが、脂肪族アルデヒド類やアルコール類にはほとんど作用せず、従来報告されているアルコールオキシダーゼやアルデヒドオキシダーゼとは基質特異性において明らかに異なる。また、本発明の酸化酵素の安定性は高く、pH6.5で、30℃、1時間の加熱後、ほぼ100%の活性を保持しており、40℃、1時間の加熱でも、約90%の活性が残存している。また、本発明の酸化酵素は、pH5.0から8.5において、40℃、1時間の加熱後、80%の活性を保持するなど、高いpH安定性も有する。
【0014】
本発明の酸化酵素の作用最適pHまたは作用最適温度は、反応条件のpHまたは温度を変えて活性を測定することにより決定される。
【0015】
本発明の酸化酵素の分子量は例えばTSK−G3000SW(7.8mm×30cm)(東ソー株式会社製)カラムを用いたゲル濾過分析により、標準タンパク質との相対溶出時間から算出することができ、サブユニット分子量はSDS−ポリアクリルアミド電気泳動により、標準タンパク質との相対移動度から算出することができる。
【0016】
また、N末端アミノ酸配列は、上記方法により精製した酵素、またはさらに逆相HPLCにより精製したものをプロテインシークエンサーにかけて決定することができる。
【0017】
本発明において、グリコールアルデヒドをグリオキサールへ変換する酸化酵素を産生する微生物を培養するための培地は、その微生物が増殖し得るものであれば特に限定されない。例えば、炭素源として、グルコース、シュークロースなどの糖類、エタノール、グリセリン、エチレングリコール、プロピレングリコールなどのアルコール類、グリコール酸などの有機酸類、オレイン酸、ステアリン酸などの脂肪酸ならびにそのエステル類、菜種油および大豆油などの油類;窒素源として、硫酸アンモニウム、硝酸アンモニウム、ペプトン、カザミノ酸、酵母エキス、肉エキスおよびコーンスチープリカーなど;無機塩類として、硫酸マグネシウム、塩化ナトリウム、炭酸カルシウム、リン酸水素カリウム、リン酸水素ナトリウムなど;その他に麦芽エキス、肉エキスなどを有する通常の液体培地が使用され得る。
【0018】
本発明によるグリオキサールの製造方法は、本発明の酸化酵素、又は、当該酸化酵素を産生する本発明の微生物をグリコールアルデヒドに作用させ、グリオキサールへと変換蓄積せしめることを特徴とする。
【0019】
更に本発明によるグリオキサールの製造方法は、エチレングリコールに、エチレングリコールをグリコールアルデヒドへ変換する能力を有する酸化酵素、又は、当該酸化酵素を産生する微生物を作用させてグリコールアルデヒドを生成させて、生じたグリコールアルデヒドに、本発明の酸化酵素又は本発明の微生物を作用させてグリオキサールへ変換することを特徴とする。
【0020】
エチレングリコールをグリコールアルデヒドへ変換する能力を有する酸化酵素としては、エチレングリコールをグリコールアルデヒドへ変換する能力を有するものであれば特に限定されないが、例えば、アルコールオキシダーゼ、グリセロールオキシダーゼが挙げられる。アルコールオキシダーゼの起源として特に限定されないが、酵母、カビ、細菌が産生するアルコールオキシダーゼが挙げられ、例えばピキア(Pichia)属、カンジダ(Candida)属、ハンセヌラ(Hansenula)属、トルロプシス(Toluropsis)属、オガタエア(Ogataea)属に属する酵母が産生するアルコールオキシダーゼが挙げられる。グリセロールオキシダーゼとしては、酵母、カビ、細菌が産生するグリセロールオキシダーゼが挙げられ、例えばアスペルギルス(Aspergillus)属産生するグリセロールオキシダーゼが挙げられる。上記以外の酵素では、アスペルギルス属に属する微生物、例えばアスペルギルス・オクラゼウス(Aspergillus ochraceus)、その一例としてアスペルギルス・オクラゼウス(Aspergillus ochraceus)AIU 031株(FERM P−20785)が産生するエチレングリコールをグリコールアルデヒドへ変換する能力を有する酸化酵素が挙げられる。同該AIU 031株は本発明者らにより土壌から分離・同定され、前記寄託番号にて独立法人産業技術総合研究所特許微生物寄託センター(〒305−8566 茨城県つくば市東1丁目1番地 中央第6)に寄託されている。
【0021】
本発明で使用する酸化酵素としては、単一または部分的に精製された酵素であってもよい。本発明で使用する微生物としては、当該微生物の培養物またはその処理物を使用することも可能である。ここで、「微生物の培養物」とは、菌体を含む培養液あるいは培養菌体を意味し、「その処理物」とは、例えば粗酵素液、凍結乾燥菌体、アセトン乾燥菌体、あるいはそれらの破砕物、これらの混合物などを意味する。更に上記酸化酵素又は上記微生物は、公知の手段(例えば、架橋法、物理的吸着法、包括法など)で固定化されて使用できる。本発明の微生物又はエチレングリコールをグリコールアルデヒドへ変換する能力を有する酸化酵素を産生する微生物としては、上記酸化酵素の産生能を有している限り、野生株または変異株、あるいは上記酸化酵素をコードするDNAをベクターに組込み、これを宿主内に導入してなる形質転換体(組替え体)であってもよい。また、本発明で使用する酸化酵素の産生能を有する形質転換体は、酸化酵素をコードするDNAを宿主のゲノムに安定的に組み込むことによっても製造できる。
【0022】
本発明の酸化酵素をグリコールアルデヒドに反応させる際の条件又はエチレングリコールをグリコールアルデヒドへ変換する能力を有する酸化酵素をエチレングリコールに反応させる際の条件は使用する酵素により異なるが、温度は5℃〜80℃、好ましくは5〜60℃の範囲、pHは4〜12、好ましくはpH5〜9の範囲である。エチレングリコールをグリコールアルデヒドへ変換する能力を有する酸化酵素、又は、当該酸化酵素を産生する微生物を作用させてグリコールアルデヒドを生成させて、生じたグリコールアルデヒドに本発明の酸化酵素又は本発明の微生物を作用させてグリオキサールへ変換する場合、エチレングリコールの濃度は、エチレングリコールをグリコールアルデヒドへ変換する酵素を効率的に働かせるために、高く設定した方がよく、100mM以上、更に1M以上が好ましい。また、グリコールアルデヒドは酵素を失活させる性質を有するため、反応時に高濃度に蓄積することは好ましくない。したがって、エチレングリコールをグリコールアルデヒドへ変換する能力を有する酵素の使用量とグリコールアルデヒドをグリオキサールへ変換する能力を有する酵素の使用量を調節することによりグリコールアルデヒドの蓄積量をコントロールすることは有効である。一方、グリコールアルデヒドを初発基質として、本発明の酵素によりグリオキサールへ変換する場合、反応系にグリコ−ルアルデヒドを高濃度添加することは酵素の安定性の観点から好ましくなく、その添加量としては、好ましくは1M以下、さらに好ましくは100mM以下が望ましい。尚、この場合、反応の進行と共にグリコールアルデヒドを断続的または連続的に添加することは、生産性の向上に繋がり好ましい。反応は、酸素条件下で行うことが好ましい。また酸素の反応液への溶解を促進するため、反応は振とう、攪拌条件下で行なわれることが好ましい。さらに大気圧以上の加圧下で反応を行うことにより、反応液への酸素の溶解度が向上し、反応がより進む場合もある。
【0023】
尚、本発明の酸化酵素又はエチレングリコールをグリコールアルデヒドへ変換する能力を有する酸化酵素による酸化反応により、過酸化水素が生成するが、この過酸化水素は酵素を失活させる場合がある。しかし、反応系にカタラーゼを添加することにより、生成した過酸化水素を分解、除去することが可能である。使用するカタラーゼは、過酸化酵素を速やかに分解、除去するという観点から、使用する酸化酵素の活性の10倍以上、好ましくは100倍以上、さらに好ましくは1000倍以上の活性量を使用する事が望ましい。また、上述した形質転換体に酸化酵素と共にカタラーゼを組換え発現させることにより、効率良く過酸化水素を分解することが可能である。一方、形質転換体の宿主として、もともとカタラーゼを産生する能力を有する微生物を使用することが好ましい。
【発明の効果】
【0024】
本発明の新規な酸化酵素は、グリコールアルデヒドをグリオキサールに酸化するが、反応生成物であるグリオキサールやエチレングリコールには全く作用しない特異性の高い酸化酵素であり、本発明の酸化酵素を使用すれば、グリオキサールを効率的に製造することができる。
【発明を実施するための最良の形態】
【0025】
以下に実施例を掲げて本発明を更に詳しく説明するが、本発明はこれら実施例のみに限定されるものではない。
(実施例1)グリコールアルデヒドに作用してグリオキサールに作用しない酸化酵素を産生する能力を持つ微生物の分離
スクリーニングのための培地は、表2に示す1%グリコール酸含有培地を120℃で15分間加圧殺菌し、このグリコール酸含有培地5mlを分注した試験管に少量の土壌を添加し、30℃で1〜7日間振盪培養した。微生物の生育が確認された培養液について、その培養液0.1mlを上記と同一組成の培地に植菌し、同様に30℃で1〜7日間振盪培養した。この操作をもう一度繰り返した後、培養液を上記液体培地と同一組成に2%寒天を加えて調製した寒天平板培地にプレートアウトし、30℃で培養した。そして微生物の生育が確認された後、この寒天平板培地にグリコールアルデヒドと過酸化水素の生成を確認するためのグリコールアルデヒドを50mMになるように添加した表3に示す発色液を摘下して、30℃で反応させた。この方法で、過酸化水素の生成が認められた微生物を、この平板培地と同一組成の斜面培地に植菌し、30℃で培養して保存した。次に、斜面培地に分離した微生物を、表2に示す5%エチレングリコール含有液体培地で振盪培養した。
【0026】
そして培養液から遠心分離で集菌し、集めた菌体を0.1Mリン酸緩衝液(pH7.0)に懸濁し、グリコールアルデヒドまたはグリオキサールを50mMになるように添加した表4に示す酵素活性測定用発色液に添加し、反応を行った。この反応で、グリコールアルデヒド存在下で反応液が紫色に着色し(過酸化水素が生成した)、グリオキサール存在下では、反応液の色がほとんど変化しない(過酸化水素が生成しない)微生物を選抜した。続いて、選抜した微生物を、表2に示す5%エチレングリコールを含む上記組成の液体培地150mlを入れた坂口フラスコを用いて30℃で振盪培養した。この培養で得られた900ml分の菌体を遠心分離で集め、0.1Mリン酸緩衝液(pH7.0)に懸濁してガラスビーズを用いて菌体を破砕した。そして、菌体破砕残渣を遠心分離で除去し、得られた上清液をDEAE−Toyopearlカラクムクロマトグラフィーに供した。即ち、菌体破砕残渣を除去して集めた上清液を10mMリン酸緩衝液と同程度の塩濃度まで脱塩した後、10mMリン酸緩衝液(pH7.0)で平衡化した20mlのDEAE−Toyopearlカラムに通液し、このカラムを10mMリン酸緩衝液(pH7.0)で洗浄した。吸着した酵素は、10mMリン酸緩衝液(pH7.0)と0.5MNaClを含む10mMリン酸緩衝液(pH7.0)を用いた直線濃度勾配法で溶出し、全ての画分についてグリコールアルデヒドおよびグリオキサールに対するオキシダーゼ活性を調べた。そして、グリコールアルデヒドに良く作用してグリオキサールに対するオキシダーゼ活性を示さない酵素を産生する微生物を選抜した。
【0027】
このような方法で得られた菌株は、上記の菌学的性質を示す微生物であり、Paenibacillus sp.AIU AL311と命名した。
尚、寒天平板培地上でのグリコールアルデヒドに対するオキシダーゼ活性は表3に示す発色液を用い、溶液でのグリコールアルデヒドに対するオキシダーゼ活性は表4に示す発色液を用いた。
【0028】
【表2】
【0029】
【表3】
【0030】
【表4】
【0031】
(実施例2)グリコールアルデヒドに作用してグリオキサールに作用しない酸化酵素の製造
実施例1で分離した微生物を、表2に示す5%エチレングリコールを含む液体培地を用いて、試験管で30℃、2日間振盪培養して種培養液を調製した。次に、この種培養液1.5mlを、種培養と同一組成の液体培地150mlを入れた500ml容フラスコに植菌し、30℃で振盪培養して、培養時間と酵素生産量との関係を調べた。その結果、グリコールアルデヒドに作用する酸化酵素の生産量は1日培養で最も高い値を示し、当該酸化酵素の活性は培養6日付近でも高い値を維持した。
【0032】
(実施例3)グリコールアルデヒドに作用してグリオキサールに作用しない酸化酵素の精製
実施例2の結果を基に、表2に示す5%エチレングリコールを含む液体培地を用いて試験管で30℃、2日間振盪培養して一次種培養液を調製した。この一次種培養液1.5mlを同一組成の液体培地を150ml分注した500ml容フラスコに植菌し、30℃で2日間振盪培養して二次種培養液を調製した。続いて、この二次種培養液20mlを一次種培養と同一組成の液体培地2Lを入れた3L容フラスコに植菌し、30℃で1日間振盪培養した。このようにして培養した9Lの培養液から菌体を集めて、以下の方法で酵素を精製した。尚、酵素の精製にはpH7.0のリン酸緩衝液を用いた。
【0033】
1)粗酵素液の調製
9L培養液から集菌した菌体(湿重量:7.5g)を10mMリン酸緩衝液に懸濁し、10℃以下の温度で0.5mmガラスビーズを用いて12分間(2分x6回)細胞破砕した。この細胞破砕液を遠心分離して菌体残渣を除去し、上清画分を粗酵素液として酵素の精製に用いた。
2)DEAE−トヨパールカラムクロマトグラフィー
粗酵素酵素液を限外濾過膜で2ms/cmまで脱塩濃縮した後、10mMリン酸緩衝液で平衡化したDEAE−トヨパールカラム(2.2×17.5cm)に吸着させた。本カラムを10mMリン酸緩衝液650mlで洗浄した後、酵素を10mMリン酸緩衝液(340ml)と0.5M NaClを含む10mMリン酸緩衝液(340ml)を用いて直線濃度勾配法で溶出した。
3)ヒドロキシアパタイトカラムクロマトグラフィー
DEAE−トヨパールカラムからの溶出液を限外濾過膜で脱塩濃縮した後、5mMリン酸緩衝液で緩衝化済みのヒドロキシアパタイトカラム(1×6cm)に通液し、続いて本カラムを5mMリン酸緩衝液100mlで洗浄し、目的とする酵素を溶出した。
以上の方法で得られた上記3)の酵素標品をNativeおよびSDS−ポリアクリルアミド電気泳動法で分析した結果、電気泳動的に単一であった。尚、本精製法による酵素の精製収率は、表5の通りであった。
【0034】
【表5】
【0035】
(実施例4)グリコールアルデヒドに作用してグリオキサールに作用しない酸化酵素の性質
実施例3の方法で精製した酵素標品は、NativeおよびSDS−ポリアクリルアミド電気泳動法を用いて分析した結果、いずれの分析でも蛋白的に単一であった。よって、本精製酵素標品を用いて諸性質を検討した。
(作用)
グリコールアルデヒドを21mMになるように添加した表4に示す酵素活性測定用発色液0.95mlに精製酵素液0.05mlを添加し、30℃で反応を行い、555nmの吸光度の増加を経時的に測定した。その結果、時間の経過と共に555nmの吸光度が増大し、エチレングリコールと本酵素の反応によって、過酸化水素が生成されることが明らかになった。
【0036】
また、反応生成物をIsobe and Nishiseの方法[Biosci.Biotech.Biochem.,58,170−173,(1994)]に従ってN−methyl−2−benzothiazolinone hydrazone(MBTH)と反応させ、その吸収スペクトルおよびC18の逆相カラムからの溶出時間を分析した。まず、反応生成物を0.2Mグリシン−HCl緩衝液(pH4.0)0.75mlに溶解し、それに1.0%(w/v)MBTH液を0.3ml添加した(反応1)。その後、反応1で得られた反応液について吸収スペクトルを測定した。その結果、グリオキサールの反応1で得られた反応液の吸収スペクトルと同様に、上記反応生成物の反応1の反応液では、410nm付近に極大吸収を持つスペクトルが得られた。そして反応1の反応液をC18の逆相カラムで分析した場合も、グリオキサールの反応1で得られる生成物と同じ溶出位置にピークが得られた。よって、本酵素は、下記の式に従ってグリコールアルデヒドをグリオキサールに酸化することが明らかになった。
OHCCHO+O2+H2O→OHCCOOH+H2O2
【0037】
次に、グリオキサールを21mMになるように添加した表4に示す酵素活性測定用発色液0.95mlに精製酵素液0.05mlを添加し、30℃で反応を行い555nmの吸光度の増加を経時的に測定した。その結果、555nmの吸光度の増大は認められなかった。よって、本酵素は、グリコールアルデヒドをグリオキサールに酸化するが、反応生成物であるグリオキサールには作用しないことが明らかになった。
【0038】
(基質特異性)
表6に示すアルデヒド類、有機酸類又はアルコール類を53mMになるように添加した表4に示す酵素活性測定用発色液0.95mlに精製酵素液0.05mlを添加し、30℃で反応を行い、555nmの吸光度の増加を分光光度計にて連続して測定し、時間あたりの555nmの増加量を算出し、各化合物に対する活性を調べた。
その結果、表6に示すように、本酵素は、グリコールアルデヒドとグリセルアルデヒドに良く作用し、ホルムアルデヒド、アセトアルデヒド、エチレングリコール、プロピオンアルデヒドにもわずかに作用を示すが、イソブチルアルデヒド、バレルアルデヒド、イソバレルアルデヒド、グリオキサール、グリオキシル酸には実質的に作用しなかった。また、メタノール、エタノール、1−プロパノール、1−ブタノールなどの一級アルコールや1,2−プロパンジオール、1,3−プロパンジオールなどにも作用しなかった。なお、表6中の各数値は、グリコールアルデヒドに対する活性を100とした場合の相対値である。
【0039】
【表6】
【0040】
(グリコールアルデヒドに対するKm値)
グリコールアルデヒドを10mMから45mMの濃度になるように添加した表4に示す酵素活性測定用発色液0.95mlに精製酵素液0.05mlを添加し、30℃で反応を行い、555nmの吸光度の増加を分光光度計にて連続して測定し、時間あたりの555nmの増加量を算出し、各濃度のグリコールアルデヒドに対する活性を測定した。更に、得られた活性値と基質濃度より作成したLineweaver−Burk plotにより、Km値を算出した。その結果、グリコールアルデヒドに対するKm値は約13mMであった。
【0041】
(分子量)
精製酵素を高速液体クロマトグラフィーによるTSK−G3000SW(7.8mm×30cm)(東ソー株式会社製)カラムを用いたゲル濾過、および1%の2−メルカプトエタノール存在下、10%SDS−ポリアクリルアミド電気泳動に供し、標準タンパクとの相対移動度より、その分子量を推定した。その結果、分子量は、ゲル濾過法で約49,000、SDS−ポリアクリルアミド電気泳動法で約24,000であった
[ゲル濾過法による分子量測定条件]
カラム:TSK−G3000SW(7.8mm×30cm)(東ソー株式会社製)
溶離液:0.1Mリン酸緩衝液+0.3M塩化ナトリウム(pH7.0)
流 速:1.0ml/min
温 度:室温
検 出:220nm
【0042】
(各種化合物の影響)
グリコールアルデヒドを53mM、更に表7に示す化合物を1.05mM濃度になるように添加した表4に示す酵素活性測定用発色液0.95mlに精製酵素液0.05mlを添加し、30℃で反応を行い、555nmの吸光度の増加を分光光度計にて連続して測定し、時間あたりの555nmの増加量を算出し、活性を調べた。
その結果、表7に示すように、本酵素は、フェニルヒドラジン、ヒドラジン、セミカルバジド、α,α′−ジピリジール、8−オキシキノリン、モノヨード酢酸、NiCl2、CoCl2で阻害され、MnCl2で活性化されることが明らかになった。
【0043】
【表7】
【0044】
(最適pH)
グリコールアルデヒドを53mMになるように添加し、緩衝液の種類を変更してpH5.5からpH8.5で調製した表4に示す酵素活性測定用発色液0.95mlに精製酵素液0.05mlを添加し、30℃で反応を行い、555nmの吸光度の増加を分光光度計にて連続して測定し、時間あたりの555nmの増加量を算出し、活性を調べた。
その結果、図1に示すように、本酵素は酸性領域から弱アルカリ性領域の広い範囲で活性を示し、反応の最適pHは6.0−7.0であった。
【0045】
(最適温度)
グリコールアルデヒドを53mMになるように添加した表4に示す酵素活性測定用発色液0.95mlに精製酵素液0.05mlを添加し、30℃から60℃の範囲で反応を行い、555nmの吸光度の増加を分光光度計にて連続して測定し、時間あたりの555nmの増加量を算出し、活性を調べた。
その結果、図2に示すように、本酵素は測定したいずれの温度でも活性を示し、反応の最適温度は45−55℃であった。
【0046】
(pH安定性)
pH5.0からpH8.5で酵素液を40℃、1時間加温する前処理を行った。その後、グリコールアルデヒドを53mMになるように添加した表4に示す酵素活性測定用発色液0.95mlに上記前処理後の酵素液0.05mlを添加し、30℃で反応を行い、555nmの吸光度の増加を分光光度計にて連続して測定し、時間あたりの555nmの増加量を算出し、残存する酵素活性を調べた。各処理後の残存活性(前処理前の活性に対する各処理後の相対活性:前処理前の活性を100とした)を図3に示す。
その結果、図3に示すように、酵素活性は、酸性領域からアルカリ性領域の広い範囲で残存しており、特にpH5.0からpH8.5の範囲では、80%以上の活性が残存した。よって、本酵素は、酸性領域からアルカリ性の領域領域の広い範囲で安定であることが明らかになった。
【0047】
(熱安定性)
pH6.5で酵素液を0℃から70℃、1時間加温する前処理を行った後、上記pH安定性と同様な方法で、残存酵素活性を測定した。その結果、図4に示すように、酵素活性は、70℃で1時間加温しても60%以上の活性が残存した。よって、本酵素は、熱に安定であることが明らかになった。
【0048】
(N末端アミノ酸配列)
精製酵素を用いて、本酵素のN末端アミノ酸配列を調べた。本精製酵素のN末端を配列番号1に示す。
【0049】
(実施例5)粗酵素液を用いたグリオキサールの合成
実施例2記載の方法で得たAIU AL311株培養液250mlから集菌した菌体を10mMリン酸緩衝液5mlに懸濁し、10℃以下の温度で0.5mmガラスビーズを用いて10分間(2分×5回)細胞破砕した。この細胞破砕液を遠心分離して菌体残渣を除去し、上清分画を粗酵素液として得た。得られた粗酵素液0.7mlに250mMグリコールアルデヒド水溶液0.2ml、50,000U/mlカタラーゼ(Bovive Liver由来;ナカライ社製)溶液0.1mlを添加し、試験管中で28℃、10時間、振盪し、反応を行った。その後、得られた反応液を高速液体クロマトグラフィー(HPLC)により分析した。その結果、反応液中に13mMのグリオキサールが生成していた。
【0050】
[HPLC分析条件]
カラム:バイオラッド社製アミネックスHPX−87H(7.8mm×300mm)
検出:示差屈折率(RI)
カラム温度:25℃
溶離液:5mM H2SO4水溶液
流速:0.4ml/分
溶離時間:グリオキサール−16分、
【0051】
(実施例6)精製酵素を用いたグリオキサールの合成
実施例3で得た精製酵素0.1U、グリコールアルデヒド50mM、カタラーゼ(Bovive Liver由来;ナカライ社製)500Uを含む100mMリン酸緩衝液(pH6.5)1mlを試験管に加え、30℃、24時間、振盪し、反応を行った。その後、得られた反応液をHPLCにより分析した。その結果、反応液中に20mMのグリオキサールが生成していた。
【0052】
(実施例7)エチレングリコールからのグリオキサールの合成
1Mエチレングリコール、2U/ml(メタノール酸化活性)のピキア属(Pichia pastris)由来アルコールオキシダーゼ(シグマ社製)、0.2U/ml(グリコールアルデヒド酸化活性)の実施例3で得た精製酸化酵素、10000U/mlのカタラーゼ(Bovive Liver由来;ナカライ社製)を含む300mMリン酸緩衝液(pH8.0)0.5mlを試験管中で、28℃、4時間振盪しながら反応を行った。その後、得られた反応液をHPLCにより分析した。その結果、5mMのグリコールアルデヒドおよび18mMのグリオキサールが生成していた。
[HPLC分析条件]
カラム:Shodex Rspak KC−811(昭和電工社製)を二本連結
検出:示差屈折率(RI)
カラム温度:40℃
溶離液:50mM過塩素酸水溶液
流速:1ml/min
溶離時間:グリコールアルデヒド;17.9分、グリオキサール;15.7分
【図面の簡単な説明】
【0053】
【図1】実施例3で取得した酸化酵素の作用最適pHを示すグラフ
【図2】実施例3で取得した酸化酵素の作用最適温度を示すグラフ
【図3】実施例3で取得した酸化酵素のpH安定性を示すグラフ
【図4】実施例3で取得した酸化酵素の熱安定性を示すグラフ
【特許請求の範囲】
【請求項1】
下記(1)及び(2)の性質を有する酸化酵素。
(1)作用:
酸素存在下、グリコールアルデヒドに作用し、グリオキサールと過酸化水素を生成する。
(2)基質特異性:
グリコールアルデヒド、及び、グリセルアルデヒドに対して活性を示し、メタノール、エタノール、1,2−プロパンジオール、1,3−プロパンジオール、及び、グリオキサールには実質的に活性を示さない。
【請求項2】
エチレングリコールに対する活性が、グリコールアルデヒドに対する活性の10%以下である請求項1記載の酸化酵素。
【請求項3】
さらに、下記(3)及び(4)の性質を有する請求項1又は2記載の酸化酵素。
(3)作用最適pH:6.0−7.0。
(4)作用最適温度:45−55℃。
【請求項4】
さらに、下記(5)及び(6)の性質を有する請求項1〜3のいずれか記載の酸化酵素。
(5)熱安定性:pH6.5で40℃、1時間加温する前処理を行った後、85%以上の活性が残存する。
(6)pH安定性:pH5.0〜8.5で40℃、1時間加温する前処理を行った後、80%以上の活性が残存する。
【請求項5】
更に、下記(7)及び(8)の性質を有する請求項1〜4のいずれか記載の酸化酵素。
(7)分子量:ゲル濾過分析で4.9×104、SDS−ポリアクリルアミド電気泳動分析において2.4×104。
(8)阻害剤:フェニルヒドラジン、ヒドラジン、セミカルバジド、α,α′−ジピリジール、8−オキシキノリン、モノヨード酢酸、NiCl2、及び、CoCl2により阻害される。
【請求項6】
配列表の配列番号1で表されるアミノ酸配列をN末端に有する請求項1〜5いずれか記載の酸化酵素。
【請求項7】
パエニバシルス(Paenibacillus)属細菌が産生する請求項1〜6いずれか記載の酸化酵素。
【請求項8】
パエニバシルス(Paenibacillus)属細菌がパエニバシルス・スピーシーズ(Paenibacillus sp.)AIU AL311株(FERM P−20786)である請求項7記載の酸化酵素。
【請求項9】
請求項1〜8いずれか記載の酸化酵素を産生する微生物。
【請求項10】
請求項9記載の微生物を培養して請求項1〜8いずれか記載の酸化酵素を製造する方法。
【請求項11】
請求項1〜8のいずれか記載の酸化酵素、又は、請求項9記載の微生物の培養物若しくはその処理物をグリコールアルデヒドに接触させてグリオキサールへと変換することを特徴とするグリオキサールの製造方法。
【請求項12】
エチレングリコールに、エチレングリコールをグリコールアルデヒドへ変換する能力を有する酸化酵素、又は、当該酸化酵素を産生する微生物の培養物若しくはその処理物を作用させてグリコールアルデヒドを生成させて、生じたグリコールアルデヒドに、請求項1〜8のいずれか記載の酸化酵素、又は、請求項9記載の微生物の培養物若しくはその処理物を作用させてグリオキサールへ変換することを特徴とするグリオキサールの製造方法。
【請求項13】
エチレングリコールをグリコールアルデヒドへ変換する能力を有する酸化酵素が、アルコールオキシダーゼ、グリセロールオキシダーゼ、及び、アスペルギルス・オクラゼウス(Aspergillus ochraceus)から得られる酸化酵素からなる群から選ばれる少なくとも1つの酸化酵素である請求項12記載のグリオキサールの製造方法。
【請求項14】
前記アルコールオキシダーゼが、ピキア(Pichia)属、カンジダ(Candida)属、ハンセヌラ(Hansenula)属、トルロプシス(Toluropsis)属、及び、オガタエア(Ogataea)属からなる群から選ばれる少なくとも1つの微生物から得られるアルコールオキシダーゼであり、グリセロールオキシダーゼがAspergillus属に属する微生物から得られるグリセロールオキシダーゼである請求項13記載のグリオキサールの製造方法。
【請求項15】
反応時にカタラーゼを共存させることを特徴とする請求項11〜14いずれか記載のグリオキサールの製造方法。
【請求項16】
酸素存在下で反応を行うことを特徴とする請求項11〜15記載のグリオキサールの製造方法。
【請求項1】
下記(1)及び(2)の性質を有する酸化酵素。
(1)作用:
酸素存在下、グリコールアルデヒドに作用し、グリオキサールと過酸化水素を生成する。
(2)基質特異性:
グリコールアルデヒド、及び、グリセルアルデヒドに対して活性を示し、メタノール、エタノール、1,2−プロパンジオール、1,3−プロパンジオール、及び、グリオキサールには実質的に活性を示さない。
【請求項2】
エチレングリコールに対する活性が、グリコールアルデヒドに対する活性の10%以下である請求項1記載の酸化酵素。
【請求項3】
さらに、下記(3)及び(4)の性質を有する請求項1又は2記載の酸化酵素。
(3)作用最適pH:6.0−7.0。
(4)作用最適温度:45−55℃。
【請求項4】
さらに、下記(5)及び(6)の性質を有する請求項1〜3のいずれか記載の酸化酵素。
(5)熱安定性:pH6.5で40℃、1時間加温する前処理を行った後、85%以上の活性が残存する。
(6)pH安定性:pH5.0〜8.5で40℃、1時間加温する前処理を行った後、80%以上の活性が残存する。
【請求項5】
更に、下記(7)及び(8)の性質を有する請求項1〜4のいずれか記載の酸化酵素。
(7)分子量:ゲル濾過分析で4.9×104、SDS−ポリアクリルアミド電気泳動分析において2.4×104。
(8)阻害剤:フェニルヒドラジン、ヒドラジン、セミカルバジド、α,α′−ジピリジール、8−オキシキノリン、モノヨード酢酸、NiCl2、及び、CoCl2により阻害される。
【請求項6】
配列表の配列番号1で表されるアミノ酸配列をN末端に有する請求項1〜5いずれか記載の酸化酵素。
【請求項7】
パエニバシルス(Paenibacillus)属細菌が産生する請求項1〜6いずれか記載の酸化酵素。
【請求項8】
パエニバシルス(Paenibacillus)属細菌がパエニバシルス・スピーシーズ(Paenibacillus sp.)AIU AL311株(FERM P−20786)である請求項7記載の酸化酵素。
【請求項9】
請求項1〜8いずれか記載の酸化酵素を産生する微生物。
【請求項10】
請求項9記載の微生物を培養して請求項1〜8いずれか記載の酸化酵素を製造する方法。
【請求項11】
請求項1〜8のいずれか記載の酸化酵素、又は、請求項9記載の微生物の培養物若しくはその処理物をグリコールアルデヒドに接触させてグリオキサールへと変換することを特徴とするグリオキサールの製造方法。
【請求項12】
エチレングリコールに、エチレングリコールをグリコールアルデヒドへ変換する能力を有する酸化酵素、又は、当該酸化酵素を産生する微生物の培養物若しくはその処理物を作用させてグリコールアルデヒドを生成させて、生じたグリコールアルデヒドに、請求項1〜8のいずれか記載の酸化酵素、又は、請求項9記載の微生物の培養物若しくはその処理物を作用させてグリオキサールへ変換することを特徴とするグリオキサールの製造方法。
【請求項13】
エチレングリコールをグリコールアルデヒドへ変換する能力を有する酸化酵素が、アルコールオキシダーゼ、グリセロールオキシダーゼ、及び、アスペルギルス・オクラゼウス(Aspergillus ochraceus)から得られる酸化酵素からなる群から選ばれる少なくとも1つの酸化酵素である請求項12記載のグリオキサールの製造方法。
【請求項14】
前記アルコールオキシダーゼが、ピキア(Pichia)属、カンジダ(Candida)属、ハンセヌラ(Hansenula)属、トルロプシス(Toluropsis)属、及び、オガタエア(Ogataea)属からなる群から選ばれる少なくとも1つの微生物から得られるアルコールオキシダーゼであり、グリセロールオキシダーゼがAspergillus属に属する微生物から得られるグリセロールオキシダーゼである請求項13記載のグリオキサールの製造方法。
【請求項15】
反応時にカタラーゼを共存させることを特徴とする請求項11〜14いずれか記載のグリオキサールの製造方法。
【請求項16】
酸素存在下で反応を行うことを特徴とする請求項11〜15記載のグリオキサールの製造方法。
【図1】

【図2】
【図3】
【図4】
【図2】
【図3】
【図4】
【公開番号】特開2007−228925(P2007−228925A)
【公開日】平成19年9月13日(2007.9.13)
【国際特許分類】
【出願番号】特願2006−56943(P2006−56943)
【出願日】平成18年3月2日(2006.3.2)
【国等の委託研究の成果に係る記載事項】(出願人による申告)国等の委託研究の成果に係る特許出願(平成17年度新エネルギー・産業技術総合開発機構「生物機能を活用した生産プロセスの基盤技術開発事業の委託研究」、産業活力再生特別措置法30条の適用を受けるもの)
【出願人】(000000941)株式会社カネカ (3,932)
【Fターム(参考)】
【公開日】平成19年9月13日(2007.9.13)
【国際特許分類】
【出願日】平成18年3月2日(2006.3.2)
【国等の委託研究の成果に係る記載事項】(出願人による申告)国等の委託研究の成果に係る特許出願(平成17年度新エネルギー・産業技術総合開発機構「生物機能を活用した生産プロセスの基盤技術開発事業の委託研究」、産業活力再生特別措置法30条の適用を受けるもの)
【出願人】(000000941)株式会社カネカ (3,932)
【Fターム(参考)】
[ Back to top ]