
新規酸性プロテアーゼ及びその用途

【課題】担子菌系酵母であるクリプトコッカス属菌Cryptococcus sp. S-2が生産するプロテアーゼを同定し、これを産業上利用可能にする手段を提供すること。
【解決手段】S-2株が菌体外に分泌するプロテアーゼを培養上清より精製し、さらにこれをクローニングすることに成功した。この新規アスパラギン酸プロテアーゼCAP1は、公知のプロテアーゼと最大でも37%の相同性しか有さず、また、微生物由来のアスパラギン酸プロテアーゼよりも哺乳類が胃内に分泌するペプシンに相同性があり、非常に珍しい配列を有する。CAP1は凝乳活性の比率が高く、凝乳酵素として利用可能である。
【解決手段】S-2株が菌体外に分泌するプロテアーゼを培養上清より精製し、さらにこれをクローニングすることに成功した。この新規アスパラギン酸プロテアーゼCAP1は、公知のプロテアーゼと最大でも37%の相同性しか有さず、また、微生物由来のアスパラギン酸プロテアーゼよりも哺乳類が胃内に分泌するペプシンに相同性があり、非常に珍しい配列を有する。CAP1は凝乳活性の比率が高く、凝乳酵素として利用可能である。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、担子菌系酵母Cryptococcus sp. S-2から単離された新規酸性プロテアーゼ及びその用途に関する。
【背景技術】
【0002】
担子菌系酵母Cryptococcus sp. S-2 (S-2 株)は、膨大な微生物からのスクリーニングにより取得された、優れた油脂分解能を有する公知の菌株である。本発明者らは、このS-2株の培養物から生分解性プラスチックの分解に優れ有機溶媒中でも高い油脂分解能を発揮する新規リパーゼCS2を製造する技術の開発、並びにその遺伝子の取得に成功している(特許文献1及び2)。また、この株の培養物から酸性キシラナーゼやα-アミラーゼが精製され、クローニングされている(非特許文献1及び2)。
【0003】
しかしながら、S-2株が分泌するプロテアーゼに関する報告は未だ存在しない。
【先行技術文献】
【特許文献】
【0004】
【特許文献1】特許第3507860号
【特許文献2】特許第4085157号
【非特許文献】
【0005】
【非特許文献1】Iefuji H et al., Biosci. Biotechnol. Biochem., 1996, vol. 60: 1331-1338
【非特許文献2】Iefuji H et al., Biochem. J., 1996, vol. 318: 989-996
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明は、担子菌系酵母であるクリプトコッカス属菌Cryptococcus sp. S-2が生産するプロテアーゼを同定し、これを産業上利用可能にする手段を提供することを目的とする。
【課題を解決するための手段】
【0007】
本願発明者らは、鋭意研究の結果、S-2株が菌体外に分泌するプロテアーゼを培養上清より精製し、さらにこれをクローニングすることに成功した。このプロテアーゼの機能解析及び配列解析の結果、該プロテアーゼがアスパラギン酸プロテアーゼであること、公知のプロテアーゼとは最大でも37%の相同性しか有さず、また、微生物由来のアスパラギン酸プロテアーゼよりも哺乳類が胃内に分泌するペプシンに相同性があり、非常に珍しい配列を有する新規アスパラギン酸プロテアーゼであること、さらに、凝乳活性の比率が高く、凝乳酵素として利用可能であることを見出し、本願発明を完成した。
【0008】
すなわち、本発明は、以下の(1)〜(3)のいずれかであるポリペプチドを提供する。
(1) 配列番号4に示すアミノ酸配列から成るポリペプチド
(2) 配列番号4に示すアミノ酸配列と70%以上の同一性を有するアミノ酸配列から成り、酸性プロテアーゼ活性を有するポリペプチド
(3) (1)又は(2)のポリペプチドのアミノ酸配列を含み、酸性プロテアーゼ活性を有するポリペプチド
【0009】
また、本発明は、上記本発明のポリペプチドをコードするポリヌクレオチドを提供する。さらに、本発明は、配列表の配列番号1若しくは3に示す塩基配列から成るポリヌクレオチド、又は該塩基配列と70%以上の相同性を有し、酸性プロテアーゼ活性を有するポリペプチドをコードするポリヌクレオチドを提供する。さらに、本発明は、上記本発明のポリヌクレオチドを含む組換えベクターを提供する。さらに、本発明は、該組換えベクターで形質転換された形質転換体を提供する。さらに、本発明は、上記本発明のポリペプチドを含有する凝乳酵素組成物を提供する。さらに、本発明は、上記本発明のポリペプチドを乳タンパク質と接触させることを含む凝乳方法を提供する。
【発明の効果】
【0010】
本発明により新規な酸性プロテアーゼが提供された。本発明のプロテアーゼはアスパラギン酸プロテアーゼであり、公知のプロテアーゼとは最大でも37%の相同性しか有さず、また、微生物由来のアスパラギン酸プロテアーゼよりも哺乳類が胃内に分泌するペプシンに相同性があるという非常に珍しい配列を有する。該プロテアーゼは、タンパク質分解活性に対して凝乳活性の比率が高いため、凝乳酵素としてチーズの製造等に利用することができる。また、インスリンB鎖の切断パターンが示す通り、本プロテアーゼによる切断パターンはヒトペプシンによる切断パターンと非常に類似しているため、本プロテアーゼはヒトの消化補助剤やヒトにとって消化性の良いペプチドの作成等にも有用である。
【図面の簡単な説明】
【0011】
【図1】実施例で同定されたCap1遺伝子の塩基配列及び推定アミノ酸配列を示す図である。
【図2】実施例で精製したCap1の凝乳活性を示す図である。
【図3】Cap1で切断されたヒトInsulin B鎖のクロマトグラムである。
【図4】Cap1及び公知の各種ペプシンによるInsulin B鎖の切断パターンを示す図である。疎水性アミノ酸をグレーのボックスで示す。
【発明を実施するための形態】
【0012】
本発明が提供するポリペプチドは酸性プロテアーゼ活性を有するポリペプチドである(以下、単に「酸性プロテアーゼ」と呼ぶことがある)。具体的には、以下の(1)〜(3)のいずれかから選択されるポリペプチドである。
(1) 配列番号4に示すアミノ酸配列から成るポリペプチド
(2) 配列番号4に示すアミノ酸配列と70%以上の同一性を有するアミノ酸配列から成り、酸性プロテアーゼ活性を有するポリペプチド
(3) (1)又は(2)のポリペプチドのアミノ酸配列を含み、酸性プロテアーゼ活性を有するポリペプチド
【0013】
配列表の配列番号4に示されるアミノ酸配列は、下記実施例において、担子菌系酵母であるクリプトコッカス属菌Cryptococcus sp. S-2(S-2株)の培養上清から精製されたプロテアーゼの配列である。本願発明者らは、S-2株が産生するこのプロテアーゼをCryptococcus sp. S-2 asparatic protease(Cap1)と命名した。
【0014】
S-2株が有するCap1遺伝子の塩基配列及び該遺伝子の推定アミノ酸配列を配列番号1及び2に示す。公知のシグナル領域予測プログラムSOSUIを用いた解析によると、配列番号2中、aa1-18がシグナルペプチド領域、aa19-85がプロ領域、aa86-417が成熟タンパク質となる領域である(図1参照)。S-2株は、aa86-417の領域を成熟タンパク質として菌体外に分泌する。従って、実際にプロテアーゼとして酵素活性を発揮する領域はaa86-417である。配列番号3及び4には、この成熟タンパク質をコードする塩基配列及びそのアミノ酸配列を示す。
【0015】
Cap1のアミノ酸配列と既知のタンパク質配列データベースとの相同性をBLASTを用いて解析した結果、豚やキクガシラコウモリといった哺乳類のアスパラギン酸プロテアーゼであるペプシノーゲン(ペプシン)と35%程度の同一性が確認されたのみであり、公知のタンパク質との同一性は非常に低かった。なお、この同一性が算出されたオーバーラップ領域は、配列番号2のうちのaa101-413の領域である。微生物由来のアスパラギン酸プロテアーゼが、他の微生物が産生するアスパラギン酸プロテアーゼではなく、哺乳類の胃において分泌されるペプシンと相同性を有するということは、従来のプロテアーゼでは報告がない。Cap1は非常に珍しい配列を有するプロテアーゼである。
【0016】
Cap1の機能解析の結果、以下の機能を有することが判明した。
(a) アスパラギン酸プロテアーゼである
ペプスタチンAによってプロテアーゼ活性が顕著に阻害されることから、Cap1はアスパラギン酸プロテアーゼである。
(b) 反応最適pHが5.0、安定pH範囲が2.0〜7.0
タンパク質分解反応の最適pHは5.0である。また、プロテアーゼ活性が50%以上保たれる安定pH範囲は2.0〜7.0である。
(c) 反応最適温度が30℃であり、55℃以上で速やかに失活する
(d) カルシウムイオンで活性が上昇する
1 mMのCaイオンで、プロテアーゼ活性が1.4倍程度活性化される。
(e) タンパク質分解活性に対して凝乳活性の比率が高い
乳タンパク質と接触させると乳タンパク質の凝固沈殿が観察される。チーズの製造工程で用いられるレンネットのように、乳タンパク質のミセル構造を破壊して凝固・沈殿させる活性がある。
【0017】
本発明のポリペプチドが有する酸性プロテアーゼ活性とは、ポリペプチド鎖の中間からペプチド結合を切断するエンドペプチダーゼ活性であって、そのタンパク質分解反応の最適pHが酸性側、例えば4.5〜5.5程度である活性をいう。ポリペプチドが酸性プロテアーゼ活性を有するか否かは、当業者であれば常法により容易に測定することができる。例えば、下記実施例に記載されるように、Freeman SA et. al., Biochem J. 1993 Oct 15;295 (Pt 2):463-9(PMID: 8240244)記載の方法に従い、アゾカゼインを基質として活性を求めることができる。
【0018】
好ましくは、本発明のポリペプチドはさらに凝乳活性を有する。すなわち、タンパク質分解活性に対して凝乳活性の比率が高い。この分野で周知の通り、乳タンパク質は、主にカゼインからなり、αs1、αs2、β、κの4種類に分けられる。これらのカゼインタンパク質は、液体中では、親水性のκカゼインが、結合・沈殿しやすい他のカゼインタンパク質の外側を取り囲んだミセルの状態で存在する。凝乳活性とは、κカゼインのペプチド結合(通常は105-106番間の結合)を切断し、κカゼインに守られていた他のカゼイン同士の凝集沈殿を生じさせる活性である。ポリペプチドが凝乳活性を有するか否かは、当業者であれば常法により容易に確認することができる。例えば、下記実施例に具体的に記載されるように10%程度の脱脂乳等の乳タンパク質溶液にポリペプチドを添加して30〜35℃で1時間程度反応させ、乳タンパク質の凝固沈殿の有無を調べればよい。
【0019】
本発明のポリペプチドの1つ目の例は、配列番号4に示すアミノ酸配列から成るポリペプチドである。該ポリペプチドは、例えば、S-2株の培養上清から精製される天然のCap1成熟体であってもよいし、また、周知の遺伝子工学的手法により製造された組換えポリペプチドであってもよい。天然のCap1成熟体が酸性プロテアーゼ活性及び凝乳活性を有することは上記した通りであり、下記実施例にも具体的に示されている。
【0020】
本発明のポリペプチドの2つ目の例は、配列番号4に示すアミノ酸配列のうちの少数のアミノ酸残基が置換され、欠失され又は挿入されたアミノ酸配列から成り、酸性プロテアーゼ活性を有するポリペプチドである。一般に、タンパク質において、そのアミノ酸配列のうちの少数のアミノ酸残基が置換され、欠失され又は挿入された場合であっても、もとのタンパク質とほぼ同じ機能を有している場合があることは、当業者において広く知られている。従って、配列番号4に示すアミノ酸配列のうちの少数(例えば好ましくは1個又は数個)のアミノ酸残基が置換され、欠失され又は挿入されたアミノ酸配列から成り、配列番号4のアミノ酸配列との同一性が70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、さらに好ましくは98%以上であるポリペプチドも、酸性プロテアーゼ活性を有する蓋然性が高く、本発明の範囲に包含される。
【0021】
ここで、アミノ酸配列の「同一性」とは、比較すべき2つのアミノ酸配列のアミノ酸残基ができるだけ多く一致するように両アミノ酸配列を整列させ、一致したアミノ酸残基数を、全アミノ酸残基数で除したものを百分率で表したものである。上記整列の際には、必要に応じ、比較する2つの配列の一方又は双方に適宜ギャップを挿入する。このような配列の整列化は、例えばBLAST、FASTA、CLUSTAL W等の周知のプログラムを用いて行なうことができる。ギャップが挿入される場合、上記全アミノ酸残基数は、1つのギャップを1つのアミノ酸残基として数えた残基数となる。このようにして数えた全アミノ酸残基数が、比較する2つの配列間で異なる場合には、同一性(%)は、長い方の配列の全アミノ酸残基数で、一致したアミノ酸残基数を除して算出される。塩基配列の相同性についても同様である。
【0022】
なお、天然のタンパク質を構成する20種類のアミノ酸は、低極性側鎖を有する中性アミノ酸(Gly, Ile, Val, Leu, Ala, Met, Pro)、親水性側鎖を有する中性アミノ酸(Asn, Gln, Thr, Ser, Tyr, Cys)、酸性アミノ酸(Asp, Glu)、塩基性アミノ酸(Arg, Lys, His)、芳香族アミノ酸(Phe, Tyr, Trp)のように類似の性質を有するものにグループ分けでき、これらの間での置換であればポリペプチドの性質が変化しないことが多いことが知られている。従って、配列番号4に示すアミノ酸配列中のアミノ酸残基を置換する場合には、これらの各グループの間で置換することにより、酸性プロテアーゼ活性が維持される可能性が高くなる。
【0023】
また、配列番号4に示すアミノ酸配列から成るポリペプチドがアスパラギン酸プロテアーゼであるという事実から、アスパラギン酸残基以外の残基が置換又は欠失しているものでは、酸性プロテアーゼ活性を維持している可能性が高い。とりわけ、配列番号4中の38番目と216番目の2つのアスパラギン酸残基が酸性プロテアーゼ活性に重要であると考えられるため、これらのアスパラギン酸残基が維持されたアミノ酸配列から成るものが好ましい。
【0024】
本発明のポリペプチドの3つ目の例は、配列番号4に示すアミノ酸配列((1)のポリペプチドのアミノ酸配列)及び該アミノ酸配列と70%以上の同一性を有するアミノ酸配列((2)のポリペプチドのアミノ酸配列)のいずれかを含むポリペプチドであって、酸性プロテアーゼ活性を有するポリペプチドである。すなわち、(1)又は(2)のポリペプチドの片末端又は両末端に1個以上のアミノ酸残基が付加されたポリペプチドであって、酸性プロテアーゼ活性を有するポリペプチドである。一般に、遺伝子組み換え技術によりポリペプチドを作製する場合には、細胞内での発現、細胞外への分泌、又は精製の便宜等のために各種のシグナル領域やタグ等を付加することがある。例えば、天然のCap1遺伝子がコードするCap1タンパク質(配列番号2)にも、シグナルペプチド領域及びプロ領域が含まれ、これらが切断されて成熟タンパク質(配列番号4)として菌体外に分泌されている。あるいは、使用目的に応じて、蛍光タンパク質や他の酵素等の異なるタンパク質との融合タンパク質として作製することもある。本発明のポリペプチドも、作製の便宜のために各種のシグナル領域やタグ等を付加してもよいし、他のタンパク質との融合タンパク質として作製してもよい。上記(1)又は(2)のポリペプチドの領域が酸性プロテアーゼ活性を発揮する限り、任意の付加領域を有するポリペプチドも本発明の範囲に包含される。このような(3)のポリペプチドの具体例としては、例えば配列番号2に示すアミノ酸配列から成るポリペプチドが挙げられるが、これに限定されない。
【0025】
本発明のポリペプチドには、翻訳後修飾されたポリペプチドも包含される。翻訳修飾の具体例としては、N末端メチオニンの脱離、N末端アセチル化、糖鎖付加、細胞内プロテアーゼによる限定分解、ミリストイル化、イソプレニル化、リン酸化などが例示できる。
【0026】
上記した本発明のポリペプチドのうちでは、配列番号4に示すアミノ酸配列から成るポリペプチド及び配列番号4に示すアミノ酸配列を含むポリペプチドが好ましく、中でも、配列番号4に示すアミノ酸配列から成るポリペプチドが最も好ましい。
【0027】
本発明のポリペプチドは、例えば、Cryptococcus sp. S-2の培養上清から精製して得ることができる。S-2株は、特許文献1及び2にも記載される公知の菌株であり、工業技術院生命工学工業研究所(現・産業技術総合研究所 特許微生物寄託センター)にFERM P-15155として寄託されているため、入手は容易である。精製方法の具体例は下記実施例に記載される通りである。簡潔に記載すると、2日間程度前培養したS-2株を1日間程度培養後、培養上清を回収し、上清を限外濾過して濾液を回収する。この濾液を酢酸ナトリウム緩衝液(20 mM、pH5.5程度)にて透析後、DEAEカラムを通して酵素を吸着させる。カラムを洗浄後、0〜0.5 MのNaCl直線濃度勾配をかけて酵素を溶出させる。プロテアーゼ活性を示す画分が目的のポリペプチドを含む画分である。この画分を再度酢酸ナトリウム緩衝液にて透析し、適当なカラムに吸着させ、NaCl勾配により溶出させることで、精製されたポリペプチドを得ることができる。
【0028】
また、所定のアミノ酸配列から成る上記した(1)〜(3)のポリペプチドは、例えばFmoc法(フルオレニルメチルオキシカルボニル法)、tBoc法(t-ブチルオキシカルボニル法)等の化学合成法により合成することもできる。また、各種の市販のペプチド合成機を利用して常法により合成することもできる。さらにまた、公知の遺伝子工学的手法を用いて、上記ポリペプチドをコードするポリヌクレオチドを調製し、該ポリヌクレオチドを発現ベクターに組み込んで宿主細胞に導入し、該宿主細胞中でポリペプチドを生産させることにより、目的とするポリペプチドを得ることができる。
【0029】
上記ポリペプチドをコードするポリヌクレオチドは、公知の遺伝子工学的手法や市販の核酸合成機を用いた常法により、容易に調製することができる。例えば、Cryptococcus sp. S-2の菌体からmRNAを抽出し、RT-PCRによりcDNAを合成することで、配列番号1又は3に示す塩基配列から成るDNAを得ることができる。cDNAの合成に使用するプライマーは、配列番号1又は3に記載されたCap1の塩基配列に基づいて容易に設計し作成することができる。また、各アミノ酸をコードするコドンは公知であるから、特定のアミノ酸配列をコードするポリヌクレオチドの塩基配列は容易に特定することができる。従って、(2)や(3)のポリペプチドをコードするポリヌクレオチドの塩基配列も容易に特定することができ、それらのポリヌクレオチドは、市販の核酸合成機を用いて、又は配列番号1若しくは3に示す塩基配列から成るDNAに常法により適宜変異を導入することで、容易に合成することができる。所定のポリペプチドをコードするDNAを適当な発現ベクターに組み込んで適当な宿主細胞内に導入し、該細胞内でポリペプチドを発現させ、これを回収することで、目的とするポリペプチドを得ることができる。mRNAの抽出、RT-PCR、ベクターへのDNAの組み込み、宿主細胞への導入等の手法自体は周知の常法である。用いるベクターや宿主細胞も周知であり、種々の市販品が存在するため、当業者であれば適宜選択して使用することができる。また、各種タグや他のタンパク質を融合させた融合タンパク質の調製方法も周知の常法である。
【0030】
本発明のポリペプチドをコードするポリヌクレオチドの具体例としては、配列番号1又は3に示す塩基配列から成るポリヌクレオチド、及び配列番号1又は3の塩基配列と70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、さらに好ましくは98%以上の相同性を有する塩基配列から成るポリヌクレオチドが挙げられる。中でも配列番号1又は3に示す塩基配列から成るポリヌクレオチドが好ましい。
【0031】
本発明のポリペプチドは、上記の通り、好ましくは凝乳活性を有する。そのため、乳タンパク質と本発明のポリペプチドを接触させ、30〜35℃程度で30分間〜数時間程度反応させれば、乳タンパク質を凝固・沈殿させることができる。本発明の凝乳方法は、チーズの製造工程等に採用することができる。本発明の凝乳酵素組成物は、凝乳活性をさらに有する本発明のポリペプチド、例えば配列番号4に示すアミノ酸配列から成るポリペプチドを含有する。該組成物は、凝乳活性を有する本発明のポリペプチドのみを含むものであってよいし、また、凝乳活性を有する他の酵素をさらに含んでいてもよい。本発明のポリペプチド及び任意の他の凝乳酵素の安定化等に有用な保存剤等の他の成分を含んでいてもよい。
【実施例】
【0032】
以下、本発明を実施例に基づきより具体的に説明する。もっとも、本発明は下記実施例に限定されるものではない。
【0033】
実施例1 S-2株が産生するプロテアーゼの精製
YM ブロスにより、25℃, 48 時間培養されたCryptococcus sp. S-2株を、OD660 = 0.1 となるように YG (1% Yeast extract, 2% Glucose) 液体培地に接種して、25℃, 120 rpm で振盪培養を行った。24 時間培養後、遠心 (7,000 rpm, 20min, 4℃) を行い、上清を回収した。得られた上清を Millipore lab scale TFF システム(ミリポア社)を用いて限外濾過を行い、得られた濾液を粗酵素液とした。
【0034】
以後の精製過程は全て 4℃で行った。粗酵素液を 20 mM 酢酸ナトリウム緩衝液 (pH 5.5) にて透析し、同緩衝液で平衡化した DEAE-FF カラム(GE ヘルスケア社)に供し、酵素を吸着させた。充分量の同緩衝液を流してカラムを洗浄した後、これに0 M〜0.5 MのNaClの直線濃度勾配をかけて酵素を溶出させた。酵素画分は、Freeman SA et. al., Biochem J. 1993 Oct 15;295 (Pt 2):463-9(PMID: 8240244)に記載の方法に従い、アゾカゼインを基質としてプロテアーゼの活性測定を行うことで、目的画分を取得した。得られた目的画分を再度、20 mM 酢酸ナトリウム緩衝液 (pH 5.5) にて透析し、同緩衝液で平衡化した Q HP カラム(GE ヘルスケア社)に供し、酵素を吸着させた。充分量の同緩衝液を流してカラムを洗浄した後、これに0 M〜0.5 MのNaClの直線濃度勾配をかけて酵素を溶出させることで、目的のプロテアーゼの精製を行った。活性は、12.5 倍、収量は11.8 %であった。SDS-PAGE 上で単バンドとなり、分子量は SDS-PAGE により36 kDである事が判明した。
【0035】
比活性
アゾカゼインを基質として本酵素の比活性を求めた。活性の測定は上記Freemanらの方法に従って行なった。また、タンパク質量は、Bradford法を用いて測定した。以上から、精製酵素の比活性は、0.050 U/μgである事が判明した。
【0036】
最適pH及び安定pH範囲
反応最適pH及び50%以上活性が安定に保たれるpH範囲を調べた結果は以下の通りであった。
最適pH:5.0
安定pH範囲:2.0〜7.0
【0037】
最適温度及び失活温度
反応最適温度及び温度による失活の条件について調べた結果は以下の通りであった。
最適温度:30℃
温度による失活の条件:55℃以上で速やかに失活する
【0038】
金属イオンによる影響
Ca, Rb, Mn, Mg, Zn, Cu, Cd, Co, Fe, Hgイオンが本酵素の活性に及ぼす影響を調べたところ、1 mMのCaイオンで、1.44倍活性化される事が明らかになった。
【0039】
阻害剤
ペプスタチンAによって顕著に阻害された。これにより本酵素はアスパラギン酸プロテアーゼであることが判明した。
【0040】
実施例2 アスパラギン酸プロテアーゼのクローニング
アスパラギン酸プロテアーゼ遺伝子をクローニングするために、精製したアスパラギン酸プロテアーゼのN末端アミノ酸配列を決定した。また、内部アミノ酸配列については、タンパク質分解酵素(リジルエンドペプチダーゼ)で処理し断片化した後、そのいくつかの断片につきそのアミノ酸配列を決定した。
N末端配列: GSSATIGL(配列番号5)
内部配列: TPVTSRTYYEVGTNGMTV, FPAAIDTGT, NFGSNYVGFTPAHNY(配列番号6〜8)
【0041】
アスパラギン酸プロテアーゼ遺伝子の内部塩基配列の決定のため、得られた部分配列をもとにDNAプライマー(混合プライマー、配列番号10及び11)を設計した。
【0042】
続いて、液体培養した Cryptococcus sp. S-2の菌体よりmRNAを調製し、逆転写によりcDNAの合成を行った。上記のプライマーを用いてアスパラギン酸プロテアーゼ遺伝子の内部配列(約400塩基)を決定した。決定された内部配列を利用して新たなプライマーを設計し(配列番号12〜18)、それらを用いて3'-及び5'-RACE法によりアスパラギン酸プロテアーゼ遺伝子のcDNA配列を決定した。決定したアスパラギン酸プロテアーゼ遺伝子の全塩基配列を配列番号1に示し、この塩基配列から推定されるタンパク質のアミノ酸配列を配列番号2に示す。
【0043】
推定されたアミノ酸配列は、上記で決定したN末端配列及び内部配列を全て有していた。また、公知のシグナル領域予測プログラムSOSUIを用いた解析により、本酵素はシグナルペプチド領域、プロ領域を有していることが判明し、417アミノ酸残基(成熟体は332アミノ酸残基)であることが明らかとなった(図1)。以上から本酵素を Cryptococcus sp. S-2 aspartic protease 1 (Cap1) と命名した。
【0044】
本酵素の成熟体の推定アミノ酸配列から計算される分子量は、本精製酵素のSDS-PAGE解析による結果と矛盾しないものであった。本酵素のアミノ酸配列と既知のタンパク質配列データベースとの相同性をBLASTを用いて解析した結果、豚やキクガシラコウモリといった哺乳類のアスパラギン酸プロテアーゼであるペプシノーゲン(ペプシン)と35%程度の相同性が確認された(表1)。この結果より、本酵素はアミノ酸配列からもアスパラギン酸プロテアーゼである事が確認された。また、微生物より菌体外に分泌されるアスパラギン酸プロテアーゼが、他の微生物のアスパラギン酸プロテアーゼではなく、哺乳類の胃において分泌されるペプシンと相同性を有していたことは、従来知られておらず、全く新規である事も判明した。
【0045】
【表1】

【0046】
実施例3 凝乳活性(レンネットとしての作用)
1.10% (W/V) 脱脂乳の調整
10g脱脂乳粉末に20mM酢酸ナトリウム緩衝液pH5.5を加え、脱脂乳溶液のpHを5.5になるまで酢酸で調整し、100mlにメスアップした。
【0047】
2.凝乳活性の確認
10% (W/V) 脱脂乳を基質として用い、35℃に保温した基質5mlに酵素液0.5mlを添加し、1 時間反応させた。コントロールとして酵素液の代わりに 20 mM 酢酸ナトリウム緩衝液 (pH 5.5) を用いた。その結果、脱脂乳と酵素液の混合液のみで乳タンパク質の凝固沈殿が観察された(図2)。分解活性より乳タンパク質を凝固させる活性の比率が高く、凝乳酵素としての機能を有していることが明らかになった。
【0048】
実施例4 Insulin B鎖を基質としたCap1の基質特異性
実施例1で精製したCap1によるヒトInsulin B鎖の切断パターンを調べることにより、Cap1の基質特異性を調べた。実施例1で精製したCap1を以下の組成で混合して反応液を調製した。
【0049】
【表2】
【0050】
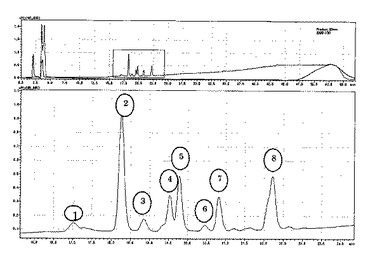
上記反応液を30℃で12時間インキュベートした後、100℃、5 minで反応を停止させた。下記の条件でHPLCに供し、220 nmにおける吸収で得られたクロマトグラム(図3)のピーク毎に分取し、各フラクションをペプチドシークエンサーに供し配列を決定した。
<HPLC 条件>
カラム : μBondasphere (ウォーターズ社製) C18、5μm (3.9μmφ x 150 mm)
移動相流速: 1.0 ml / min
検出 : Absorption of 220 nm
移動相 : A ; 10% acetonitrile / 0.1% TFA, B ; 100% acetonitrile / 0.1% TFA
B=0%→100%(直線勾配)
【0051】
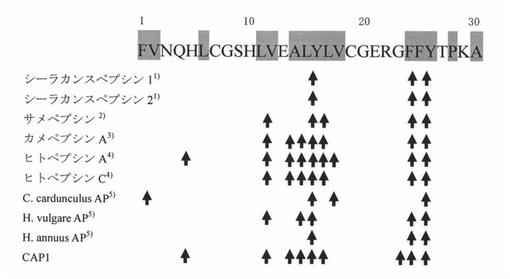
Insulin B鎖全長のアミノ酸配列はFVNQHLCGSHLVEALYLVCGERGFFYTPKA(配列番号9)である。得られた8本のピークのアミノ酸配列は、1: ALY, 2: FYTPKA, 3: LVCGERGF, 4: FVNQHLCGSHLVE, 5: FVNQHLCGSHLVEA, 6: FFYTPKA, YLVCGERG, 7: YLVCGERGF, 8: LYLVCGERGFF, ALYLVCGERGF, FVNQHLCGSHLVEA であった。得られた配列をもとに、Insulin B鎖に対する本酵素の特異性と他の明らかにされているペプシンの特異性(参考文献:Tanji M et al., 2007. Comparative Biochemistry and Physiology, Part B 146 412-420.及びPark, H. et al., 2000. Biosci. Biotechnol. Biochem., 64 (5), 931-939.)とを合わせて図4に示した。なお、図4中に記載の公知のペプシンはそれぞれ以下の論文に記載のものである。
1) シーラカンスペプシン1及び2
Tanji M et al., 2007. Comparative Biochemistry and Physiology, Part B 146 412-420.
2) サメペプシン
Nguyen, A.D. et al., 1998. J. Biochem. 124, 287-293.
3) カメペプシンA(accession No. AAB35842)
Hirasawa, A. et al., 1996. J. Biochem.120, 407-414.
4) ヒトペプシンA(accession No. AAA60061)、ヒトペプシンC(accession No. AAA60063)
Athauda, S.B.P. et al., 2002. Protein, Peptide Lett. 9, 289-294.
5) 各種植物アスパラギン酸プロテアーゼ(Cc AP: CAL07969, Hv AP: CAA39602, Ha AP: BAA76870)
Park, H. et al., 2000. Biosci. Biotechnol. Biochem., 64 (5), 931-939.
【0052】
その結果、本酵素は疎水性アミノ酸を好んで切断する傾向が明らかとなった。さらに、魚類や植物ペプシンの Insulin B鎖に対する基質特異性と比較して、本酵素は、よりヒトペプシンの基質特異性に類似している事が判明した。
【技術分野】
【0001】
本発明は、担子菌系酵母Cryptococcus sp. S-2から単離された新規酸性プロテアーゼ及びその用途に関する。
【背景技術】
【0002】
担子菌系酵母Cryptococcus sp. S-2 (S-2 株)は、膨大な微生物からのスクリーニングにより取得された、優れた油脂分解能を有する公知の菌株である。本発明者らは、このS-2株の培養物から生分解性プラスチックの分解に優れ有機溶媒中でも高い油脂分解能を発揮する新規リパーゼCS2を製造する技術の開発、並びにその遺伝子の取得に成功している(特許文献1及び2)。また、この株の培養物から酸性キシラナーゼやα-アミラーゼが精製され、クローニングされている(非特許文献1及び2)。
【0003】
しかしながら、S-2株が分泌するプロテアーゼに関する報告は未だ存在しない。
【先行技術文献】
【特許文献】
【0004】
【特許文献1】特許第3507860号
【特許文献2】特許第4085157号
【非特許文献】
【0005】
【非特許文献1】Iefuji H et al., Biosci. Biotechnol. Biochem., 1996, vol. 60: 1331-1338
【非特許文献2】Iefuji H et al., Biochem. J., 1996, vol. 318: 989-996
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明は、担子菌系酵母であるクリプトコッカス属菌Cryptococcus sp. S-2が生産するプロテアーゼを同定し、これを産業上利用可能にする手段を提供することを目的とする。
【課題を解決するための手段】
【0007】
本願発明者らは、鋭意研究の結果、S-2株が菌体外に分泌するプロテアーゼを培養上清より精製し、さらにこれをクローニングすることに成功した。このプロテアーゼの機能解析及び配列解析の結果、該プロテアーゼがアスパラギン酸プロテアーゼであること、公知のプロテアーゼとは最大でも37%の相同性しか有さず、また、微生物由来のアスパラギン酸プロテアーゼよりも哺乳類が胃内に分泌するペプシンに相同性があり、非常に珍しい配列を有する新規アスパラギン酸プロテアーゼであること、さらに、凝乳活性の比率が高く、凝乳酵素として利用可能であることを見出し、本願発明を完成した。
【0008】
すなわち、本発明は、以下の(1)〜(3)のいずれかであるポリペプチドを提供する。
(1) 配列番号4に示すアミノ酸配列から成るポリペプチド
(2) 配列番号4に示すアミノ酸配列と70%以上の同一性を有するアミノ酸配列から成り、酸性プロテアーゼ活性を有するポリペプチド
(3) (1)又は(2)のポリペプチドのアミノ酸配列を含み、酸性プロテアーゼ活性を有するポリペプチド
【0009】
また、本発明は、上記本発明のポリペプチドをコードするポリヌクレオチドを提供する。さらに、本発明は、配列表の配列番号1若しくは3に示す塩基配列から成るポリヌクレオチド、又は該塩基配列と70%以上の相同性を有し、酸性プロテアーゼ活性を有するポリペプチドをコードするポリヌクレオチドを提供する。さらに、本発明は、上記本発明のポリヌクレオチドを含む組換えベクターを提供する。さらに、本発明は、該組換えベクターで形質転換された形質転換体を提供する。さらに、本発明は、上記本発明のポリペプチドを含有する凝乳酵素組成物を提供する。さらに、本発明は、上記本発明のポリペプチドを乳タンパク質と接触させることを含む凝乳方法を提供する。
【発明の効果】
【0010】
本発明により新規な酸性プロテアーゼが提供された。本発明のプロテアーゼはアスパラギン酸プロテアーゼであり、公知のプロテアーゼとは最大でも37%の相同性しか有さず、また、微生物由来のアスパラギン酸プロテアーゼよりも哺乳類が胃内に分泌するペプシンに相同性があるという非常に珍しい配列を有する。該プロテアーゼは、タンパク質分解活性に対して凝乳活性の比率が高いため、凝乳酵素としてチーズの製造等に利用することができる。また、インスリンB鎖の切断パターンが示す通り、本プロテアーゼによる切断パターンはヒトペプシンによる切断パターンと非常に類似しているため、本プロテアーゼはヒトの消化補助剤やヒトにとって消化性の良いペプチドの作成等にも有用である。
【図面の簡単な説明】
【0011】
【図1】実施例で同定されたCap1遺伝子の塩基配列及び推定アミノ酸配列を示す図である。
【図2】実施例で精製したCap1の凝乳活性を示す図である。
【図3】Cap1で切断されたヒトInsulin B鎖のクロマトグラムである。
【図4】Cap1及び公知の各種ペプシンによるInsulin B鎖の切断パターンを示す図である。疎水性アミノ酸をグレーのボックスで示す。
【発明を実施するための形態】
【0012】
本発明が提供するポリペプチドは酸性プロテアーゼ活性を有するポリペプチドである(以下、単に「酸性プロテアーゼ」と呼ぶことがある)。具体的には、以下の(1)〜(3)のいずれかから選択されるポリペプチドである。
(1) 配列番号4に示すアミノ酸配列から成るポリペプチド
(2) 配列番号4に示すアミノ酸配列と70%以上の同一性を有するアミノ酸配列から成り、酸性プロテアーゼ活性を有するポリペプチド
(3) (1)又は(2)のポリペプチドのアミノ酸配列を含み、酸性プロテアーゼ活性を有するポリペプチド
【0013】
配列表の配列番号4に示されるアミノ酸配列は、下記実施例において、担子菌系酵母であるクリプトコッカス属菌Cryptococcus sp. S-2(S-2株)の培養上清から精製されたプロテアーゼの配列である。本願発明者らは、S-2株が産生するこのプロテアーゼをCryptococcus sp. S-2 asparatic protease(Cap1)と命名した。
【0014】
S-2株が有するCap1遺伝子の塩基配列及び該遺伝子の推定アミノ酸配列を配列番号1及び2に示す。公知のシグナル領域予測プログラムSOSUIを用いた解析によると、配列番号2中、aa1-18がシグナルペプチド領域、aa19-85がプロ領域、aa86-417が成熟タンパク質となる領域である(図1参照)。S-2株は、aa86-417の領域を成熟タンパク質として菌体外に分泌する。従って、実際にプロテアーゼとして酵素活性を発揮する領域はaa86-417である。配列番号3及び4には、この成熟タンパク質をコードする塩基配列及びそのアミノ酸配列を示す。
【0015】
Cap1のアミノ酸配列と既知のタンパク質配列データベースとの相同性をBLASTを用いて解析した結果、豚やキクガシラコウモリといった哺乳類のアスパラギン酸プロテアーゼであるペプシノーゲン(ペプシン)と35%程度の同一性が確認されたのみであり、公知のタンパク質との同一性は非常に低かった。なお、この同一性が算出されたオーバーラップ領域は、配列番号2のうちのaa101-413の領域である。微生物由来のアスパラギン酸プロテアーゼが、他の微生物が産生するアスパラギン酸プロテアーゼではなく、哺乳類の胃において分泌されるペプシンと相同性を有するということは、従来のプロテアーゼでは報告がない。Cap1は非常に珍しい配列を有するプロテアーゼである。
【0016】
Cap1の機能解析の結果、以下の機能を有することが判明した。
(a) アスパラギン酸プロテアーゼである
ペプスタチンAによってプロテアーゼ活性が顕著に阻害されることから、Cap1はアスパラギン酸プロテアーゼである。
(b) 反応最適pHが5.0、安定pH範囲が2.0〜7.0
タンパク質分解反応の最適pHは5.0である。また、プロテアーゼ活性が50%以上保たれる安定pH範囲は2.0〜7.0である。
(c) 反応最適温度が30℃であり、55℃以上で速やかに失活する
(d) カルシウムイオンで活性が上昇する
1 mMのCaイオンで、プロテアーゼ活性が1.4倍程度活性化される。
(e) タンパク質分解活性に対して凝乳活性の比率が高い
乳タンパク質と接触させると乳タンパク質の凝固沈殿が観察される。チーズの製造工程で用いられるレンネットのように、乳タンパク質のミセル構造を破壊して凝固・沈殿させる活性がある。
【0017】
本発明のポリペプチドが有する酸性プロテアーゼ活性とは、ポリペプチド鎖の中間からペプチド結合を切断するエンドペプチダーゼ活性であって、そのタンパク質分解反応の最適pHが酸性側、例えば4.5〜5.5程度である活性をいう。ポリペプチドが酸性プロテアーゼ活性を有するか否かは、当業者であれば常法により容易に測定することができる。例えば、下記実施例に記載されるように、Freeman SA et. al., Biochem J. 1993 Oct 15;295 (Pt 2):463-9(PMID: 8240244)記載の方法に従い、アゾカゼインを基質として活性を求めることができる。
【0018】
好ましくは、本発明のポリペプチドはさらに凝乳活性を有する。すなわち、タンパク質分解活性に対して凝乳活性の比率が高い。この分野で周知の通り、乳タンパク質は、主にカゼインからなり、αs1、αs2、β、κの4種類に分けられる。これらのカゼインタンパク質は、液体中では、親水性のκカゼインが、結合・沈殿しやすい他のカゼインタンパク質の外側を取り囲んだミセルの状態で存在する。凝乳活性とは、κカゼインのペプチド結合(通常は105-106番間の結合)を切断し、κカゼインに守られていた他のカゼイン同士の凝集沈殿を生じさせる活性である。ポリペプチドが凝乳活性を有するか否かは、当業者であれば常法により容易に確認することができる。例えば、下記実施例に具体的に記載されるように10%程度の脱脂乳等の乳タンパク質溶液にポリペプチドを添加して30〜35℃で1時間程度反応させ、乳タンパク質の凝固沈殿の有無を調べればよい。
【0019】
本発明のポリペプチドの1つ目の例は、配列番号4に示すアミノ酸配列から成るポリペプチドである。該ポリペプチドは、例えば、S-2株の培養上清から精製される天然のCap1成熟体であってもよいし、また、周知の遺伝子工学的手法により製造された組換えポリペプチドであってもよい。天然のCap1成熟体が酸性プロテアーゼ活性及び凝乳活性を有することは上記した通りであり、下記実施例にも具体的に示されている。
【0020】
本発明のポリペプチドの2つ目の例は、配列番号4に示すアミノ酸配列のうちの少数のアミノ酸残基が置換され、欠失され又は挿入されたアミノ酸配列から成り、酸性プロテアーゼ活性を有するポリペプチドである。一般に、タンパク質において、そのアミノ酸配列のうちの少数のアミノ酸残基が置換され、欠失され又は挿入された場合であっても、もとのタンパク質とほぼ同じ機能を有している場合があることは、当業者において広く知られている。従って、配列番号4に示すアミノ酸配列のうちの少数(例えば好ましくは1個又は数個)のアミノ酸残基が置換され、欠失され又は挿入されたアミノ酸配列から成り、配列番号4のアミノ酸配列との同一性が70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、さらに好ましくは98%以上であるポリペプチドも、酸性プロテアーゼ活性を有する蓋然性が高く、本発明の範囲に包含される。
【0021】
ここで、アミノ酸配列の「同一性」とは、比較すべき2つのアミノ酸配列のアミノ酸残基ができるだけ多く一致するように両アミノ酸配列を整列させ、一致したアミノ酸残基数を、全アミノ酸残基数で除したものを百分率で表したものである。上記整列の際には、必要に応じ、比較する2つの配列の一方又は双方に適宜ギャップを挿入する。このような配列の整列化は、例えばBLAST、FASTA、CLUSTAL W等の周知のプログラムを用いて行なうことができる。ギャップが挿入される場合、上記全アミノ酸残基数は、1つのギャップを1つのアミノ酸残基として数えた残基数となる。このようにして数えた全アミノ酸残基数が、比較する2つの配列間で異なる場合には、同一性(%)は、長い方の配列の全アミノ酸残基数で、一致したアミノ酸残基数を除して算出される。塩基配列の相同性についても同様である。
【0022】
なお、天然のタンパク質を構成する20種類のアミノ酸は、低極性側鎖を有する中性アミノ酸(Gly, Ile, Val, Leu, Ala, Met, Pro)、親水性側鎖を有する中性アミノ酸(Asn, Gln, Thr, Ser, Tyr, Cys)、酸性アミノ酸(Asp, Glu)、塩基性アミノ酸(Arg, Lys, His)、芳香族アミノ酸(Phe, Tyr, Trp)のように類似の性質を有するものにグループ分けでき、これらの間での置換であればポリペプチドの性質が変化しないことが多いことが知られている。従って、配列番号4に示すアミノ酸配列中のアミノ酸残基を置換する場合には、これらの各グループの間で置換することにより、酸性プロテアーゼ活性が維持される可能性が高くなる。
【0023】
また、配列番号4に示すアミノ酸配列から成るポリペプチドがアスパラギン酸プロテアーゼであるという事実から、アスパラギン酸残基以外の残基が置換又は欠失しているものでは、酸性プロテアーゼ活性を維持している可能性が高い。とりわけ、配列番号4中の38番目と216番目の2つのアスパラギン酸残基が酸性プロテアーゼ活性に重要であると考えられるため、これらのアスパラギン酸残基が維持されたアミノ酸配列から成るものが好ましい。
【0024】
本発明のポリペプチドの3つ目の例は、配列番号4に示すアミノ酸配列((1)のポリペプチドのアミノ酸配列)及び該アミノ酸配列と70%以上の同一性を有するアミノ酸配列((2)のポリペプチドのアミノ酸配列)のいずれかを含むポリペプチドであって、酸性プロテアーゼ活性を有するポリペプチドである。すなわち、(1)又は(2)のポリペプチドの片末端又は両末端に1個以上のアミノ酸残基が付加されたポリペプチドであって、酸性プロテアーゼ活性を有するポリペプチドである。一般に、遺伝子組み換え技術によりポリペプチドを作製する場合には、細胞内での発現、細胞外への分泌、又は精製の便宜等のために各種のシグナル領域やタグ等を付加することがある。例えば、天然のCap1遺伝子がコードするCap1タンパク質(配列番号2)にも、シグナルペプチド領域及びプロ領域が含まれ、これらが切断されて成熟タンパク質(配列番号4)として菌体外に分泌されている。あるいは、使用目的に応じて、蛍光タンパク質や他の酵素等の異なるタンパク質との融合タンパク質として作製することもある。本発明のポリペプチドも、作製の便宜のために各種のシグナル領域やタグ等を付加してもよいし、他のタンパク質との融合タンパク質として作製してもよい。上記(1)又は(2)のポリペプチドの領域が酸性プロテアーゼ活性を発揮する限り、任意の付加領域を有するポリペプチドも本発明の範囲に包含される。このような(3)のポリペプチドの具体例としては、例えば配列番号2に示すアミノ酸配列から成るポリペプチドが挙げられるが、これに限定されない。
【0025】
本発明のポリペプチドには、翻訳後修飾されたポリペプチドも包含される。翻訳修飾の具体例としては、N末端メチオニンの脱離、N末端アセチル化、糖鎖付加、細胞内プロテアーゼによる限定分解、ミリストイル化、イソプレニル化、リン酸化などが例示できる。
【0026】
上記した本発明のポリペプチドのうちでは、配列番号4に示すアミノ酸配列から成るポリペプチド及び配列番号4に示すアミノ酸配列を含むポリペプチドが好ましく、中でも、配列番号4に示すアミノ酸配列から成るポリペプチドが最も好ましい。
【0027】
本発明のポリペプチドは、例えば、Cryptococcus sp. S-2の培養上清から精製して得ることができる。S-2株は、特許文献1及び2にも記載される公知の菌株であり、工業技術院生命工学工業研究所(現・産業技術総合研究所 特許微生物寄託センター)にFERM P-15155として寄託されているため、入手は容易である。精製方法の具体例は下記実施例に記載される通りである。簡潔に記載すると、2日間程度前培養したS-2株を1日間程度培養後、培養上清を回収し、上清を限外濾過して濾液を回収する。この濾液を酢酸ナトリウム緩衝液(20 mM、pH5.5程度)にて透析後、DEAEカラムを通して酵素を吸着させる。カラムを洗浄後、0〜0.5 MのNaCl直線濃度勾配をかけて酵素を溶出させる。プロテアーゼ活性を示す画分が目的のポリペプチドを含む画分である。この画分を再度酢酸ナトリウム緩衝液にて透析し、適当なカラムに吸着させ、NaCl勾配により溶出させることで、精製されたポリペプチドを得ることができる。
【0028】
また、所定のアミノ酸配列から成る上記した(1)〜(3)のポリペプチドは、例えばFmoc法(フルオレニルメチルオキシカルボニル法)、tBoc法(t-ブチルオキシカルボニル法)等の化学合成法により合成することもできる。また、各種の市販のペプチド合成機を利用して常法により合成することもできる。さらにまた、公知の遺伝子工学的手法を用いて、上記ポリペプチドをコードするポリヌクレオチドを調製し、該ポリヌクレオチドを発現ベクターに組み込んで宿主細胞に導入し、該宿主細胞中でポリペプチドを生産させることにより、目的とするポリペプチドを得ることができる。
【0029】
上記ポリペプチドをコードするポリヌクレオチドは、公知の遺伝子工学的手法や市販の核酸合成機を用いた常法により、容易に調製することができる。例えば、Cryptococcus sp. S-2の菌体からmRNAを抽出し、RT-PCRによりcDNAを合成することで、配列番号1又は3に示す塩基配列から成るDNAを得ることができる。cDNAの合成に使用するプライマーは、配列番号1又は3に記載されたCap1の塩基配列に基づいて容易に設計し作成することができる。また、各アミノ酸をコードするコドンは公知であるから、特定のアミノ酸配列をコードするポリヌクレオチドの塩基配列は容易に特定することができる。従って、(2)や(3)のポリペプチドをコードするポリヌクレオチドの塩基配列も容易に特定することができ、それらのポリヌクレオチドは、市販の核酸合成機を用いて、又は配列番号1若しくは3に示す塩基配列から成るDNAに常法により適宜変異を導入することで、容易に合成することができる。所定のポリペプチドをコードするDNAを適当な発現ベクターに組み込んで適当な宿主細胞内に導入し、該細胞内でポリペプチドを発現させ、これを回収することで、目的とするポリペプチドを得ることができる。mRNAの抽出、RT-PCR、ベクターへのDNAの組み込み、宿主細胞への導入等の手法自体は周知の常法である。用いるベクターや宿主細胞も周知であり、種々の市販品が存在するため、当業者であれば適宜選択して使用することができる。また、各種タグや他のタンパク質を融合させた融合タンパク質の調製方法も周知の常法である。
【0030】
本発明のポリペプチドをコードするポリヌクレオチドの具体例としては、配列番号1又は3に示す塩基配列から成るポリヌクレオチド、及び配列番号1又は3の塩基配列と70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、さらに好ましくは98%以上の相同性を有する塩基配列から成るポリヌクレオチドが挙げられる。中でも配列番号1又は3に示す塩基配列から成るポリヌクレオチドが好ましい。
【0031】
本発明のポリペプチドは、上記の通り、好ましくは凝乳活性を有する。そのため、乳タンパク質と本発明のポリペプチドを接触させ、30〜35℃程度で30分間〜数時間程度反応させれば、乳タンパク質を凝固・沈殿させることができる。本発明の凝乳方法は、チーズの製造工程等に採用することができる。本発明の凝乳酵素組成物は、凝乳活性をさらに有する本発明のポリペプチド、例えば配列番号4に示すアミノ酸配列から成るポリペプチドを含有する。該組成物は、凝乳活性を有する本発明のポリペプチドのみを含むものであってよいし、また、凝乳活性を有する他の酵素をさらに含んでいてもよい。本発明のポリペプチド及び任意の他の凝乳酵素の安定化等に有用な保存剤等の他の成分を含んでいてもよい。
【実施例】
【0032】
以下、本発明を実施例に基づきより具体的に説明する。もっとも、本発明は下記実施例に限定されるものではない。
【0033】
実施例1 S-2株が産生するプロテアーゼの精製
YM ブロスにより、25℃, 48 時間培養されたCryptococcus sp. S-2株を、OD660 = 0.1 となるように YG (1% Yeast extract, 2% Glucose) 液体培地に接種して、25℃, 120 rpm で振盪培養を行った。24 時間培養後、遠心 (7,000 rpm, 20min, 4℃) を行い、上清を回収した。得られた上清を Millipore lab scale TFF システム(ミリポア社)を用いて限外濾過を行い、得られた濾液を粗酵素液とした。
【0034】
以後の精製過程は全て 4℃で行った。粗酵素液を 20 mM 酢酸ナトリウム緩衝液 (pH 5.5) にて透析し、同緩衝液で平衡化した DEAE-FF カラム(GE ヘルスケア社)に供し、酵素を吸着させた。充分量の同緩衝液を流してカラムを洗浄した後、これに0 M〜0.5 MのNaClの直線濃度勾配をかけて酵素を溶出させた。酵素画分は、Freeman SA et. al., Biochem J. 1993 Oct 15;295 (Pt 2):463-9(PMID: 8240244)に記載の方法に従い、アゾカゼインを基質としてプロテアーゼの活性測定を行うことで、目的画分を取得した。得られた目的画分を再度、20 mM 酢酸ナトリウム緩衝液 (pH 5.5) にて透析し、同緩衝液で平衡化した Q HP カラム(GE ヘルスケア社)に供し、酵素を吸着させた。充分量の同緩衝液を流してカラムを洗浄した後、これに0 M〜0.5 MのNaClの直線濃度勾配をかけて酵素を溶出させることで、目的のプロテアーゼの精製を行った。活性は、12.5 倍、収量は11.8 %であった。SDS-PAGE 上で単バンドとなり、分子量は SDS-PAGE により36 kDである事が判明した。
【0035】
比活性
アゾカゼインを基質として本酵素の比活性を求めた。活性の測定は上記Freemanらの方法に従って行なった。また、タンパク質量は、Bradford法を用いて測定した。以上から、精製酵素の比活性は、0.050 U/μgである事が判明した。
【0036】
最適pH及び安定pH範囲
反応最適pH及び50%以上活性が安定に保たれるpH範囲を調べた結果は以下の通りであった。
最適pH:5.0
安定pH範囲:2.0〜7.0
【0037】
最適温度及び失活温度
反応最適温度及び温度による失活の条件について調べた結果は以下の通りであった。
最適温度:30℃
温度による失活の条件:55℃以上で速やかに失活する
【0038】
金属イオンによる影響
Ca, Rb, Mn, Mg, Zn, Cu, Cd, Co, Fe, Hgイオンが本酵素の活性に及ぼす影響を調べたところ、1 mMのCaイオンで、1.44倍活性化される事が明らかになった。
【0039】
阻害剤
ペプスタチンAによって顕著に阻害された。これにより本酵素はアスパラギン酸プロテアーゼであることが判明した。
【0040】
実施例2 アスパラギン酸プロテアーゼのクローニング
アスパラギン酸プロテアーゼ遺伝子をクローニングするために、精製したアスパラギン酸プロテアーゼのN末端アミノ酸配列を決定した。また、内部アミノ酸配列については、タンパク質分解酵素(リジルエンドペプチダーゼ)で処理し断片化した後、そのいくつかの断片につきそのアミノ酸配列を決定した。
N末端配列: GSSATIGL(配列番号5)
内部配列: TPVTSRTYYEVGTNGMTV, FPAAIDTGT, NFGSNYVGFTPAHNY(配列番号6〜8)
【0041】
アスパラギン酸プロテアーゼ遺伝子の内部塩基配列の決定のため、得られた部分配列をもとにDNAプライマー(混合プライマー、配列番号10及び11)を設計した。
【0042】
続いて、液体培養した Cryptococcus sp. S-2の菌体よりmRNAを調製し、逆転写によりcDNAの合成を行った。上記のプライマーを用いてアスパラギン酸プロテアーゼ遺伝子の内部配列(約400塩基)を決定した。決定された内部配列を利用して新たなプライマーを設計し(配列番号12〜18)、それらを用いて3'-及び5'-RACE法によりアスパラギン酸プロテアーゼ遺伝子のcDNA配列を決定した。決定したアスパラギン酸プロテアーゼ遺伝子の全塩基配列を配列番号1に示し、この塩基配列から推定されるタンパク質のアミノ酸配列を配列番号2に示す。
【0043】
推定されたアミノ酸配列は、上記で決定したN末端配列及び内部配列を全て有していた。また、公知のシグナル領域予測プログラムSOSUIを用いた解析により、本酵素はシグナルペプチド領域、プロ領域を有していることが判明し、417アミノ酸残基(成熟体は332アミノ酸残基)であることが明らかとなった(図1)。以上から本酵素を Cryptococcus sp. S-2 aspartic protease 1 (Cap1) と命名した。
【0044】
本酵素の成熟体の推定アミノ酸配列から計算される分子量は、本精製酵素のSDS-PAGE解析による結果と矛盾しないものであった。本酵素のアミノ酸配列と既知のタンパク質配列データベースとの相同性をBLASTを用いて解析した結果、豚やキクガシラコウモリといった哺乳類のアスパラギン酸プロテアーゼであるペプシノーゲン(ペプシン)と35%程度の相同性が確認された(表1)。この結果より、本酵素はアミノ酸配列からもアスパラギン酸プロテアーゼである事が確認された。また、微生物より菌体外に分泌されるアスパラギン酸プロテアーゼが、他の微生物のアスパラギン酸プロテアーゼではなく、哺乳類の胃において分泌されるペプシンと相同性を有していたことは、従来知られておらず、全く新規である事も判明した。
【0045】
【表1】
【0046】
実施例3 凝乳活性(レンネットとしての作用)
1.10% (W/V) 脱脂乳の調整
10g脱脂乳粉末に20mM酢酸ナトリウム緩衝液pH5.5を加え、脱脂乳溶液のpHを5.5になるまで酢酸で調整し、100mlにメスアップした。
【0047】
2.凝乳活性の確認
10% (W/V) 脱脂乳を基質として用い、35℃に保温した基質5mlに酵素液0.5mlを添加し、1 時間反応させた。コントロールとして酵素液の代わりに 20 mM 酢酸ナトリウム緩衝液 (pH 5.5) を用いた。その結果、脱脂乳と酵素液の混合液のみで乳タンパク質の凝固沈殿が観察された(図2)。分解活性より乳タンパク質を凝固させる活性の比率が高く、凝乳酵素としての機能を有していることが明らかになった。
【0048】
実施例4 Insulin B鎖を基質としたCap1の基質特異性
実施例1で精製したCap1によるヒトInsulin B鎖の切断パターンを調べることにより、Cap1の基質特異性を調べた。実施例1で精製したCap1を以下の組成で混合して反応液を調製した。
【0049】
【表2】
【0050】
上記反応液を30℃で12時間インキュベートした後、100℃、5 minで反応を停止させた。下記の条件でHPLCに供し、220 nmにおける吸収で得られたクロマトグラム(図3)のピーク毎に分取し、各フラクションをペプチドシークエンサーに供し配列を決定した。
<HPLC 条件>
カラム : μBondasphere (ウォーターズ社製) C18、5μm (3.9μmφ x 150 mm)
移動相流速: 1.0 ml / min
検出 : Absorption of 220 nm
移動相 : A ; 10% acetonitrile / 0.1% TFA, B ; 100% acetonitrile / 0.1% TFA
B=0%→100%(直線勾配)
【0051】
Insulin B鎖全長のアミノ酸配列はFVNQHLCGSHLVEALYLVCGERGFFYTPKA(配列番号9)である。得られた8本のピークのアミノ酸配列は、1: ALY, 2: FYTPKA, 3: LVCGERGF, 4: FVNQHLCGSHLVE, 5: FVNQHLCGSHLVEA, 6: FFYTPKA, YLVCGERG, 7: YLVCGERGF, 8: LYLVCGERGFF, ALYLVCGERGF, FVNQHLCGSHLVEA であった。得られた配列をもとに、Insulin B鎖に対する本酵素の特異性と他の明らかにされているペプシンの特異性(参考文献:Tanji M et al., 2007. Comparative Biochemistry and Physiology, Part B 146 412-420.及びPark, H. et al., 2000. Biosci. Biotechnol. Biochem., 64 (5), 931-939.)とを合わせて図4に示した。なお、図4中に記載の公知のペプシンはそれぞれ以下の論文に記載のものである。
1) シーラカンスペプシン1及び2
Tanji M et al., 2007. Comparative Biochemistry and Physiology, Part B 146 412-420.
2) サメペプシン
Nguyen, A.D. et al., 1998. J. Biochem. 124, 287-293.
3) カメペプシンA(accession No. AAB35842)
Hirasawa, A. et al., 1996. J. Biochem.120, 407-414.
4) ヒトペプシンA(accession No. AAA60061)、ヒトペプシンC(accession No. AAA60063)
Athauda, S.B.P. et al., 2002. Protein, Peptide Lett. 9, 289-294.
5) 各種植物アスパラギン酸プロテアーゼ(Cc AP: CAL07969, Hv AP: CAA39602, Ha AP: BAA76870)
Park, H. et al., 2000. Biosci. Biotechnol. Biochem., 64 (5), 931-939.
【0052】
その結果、本酵素は疎水性アミノ酸を好んで切断する傾向が明らかとなった。さらに、魚類や植物ペプシンの Insulin B鎖に対する基質特異性と比較して、本酵素は、よりヒトペプシンの基質特異性に類似している事が判明した。
【特許請求の範囲】
【請求項1】
以下の(1)〜(3)のいずれかであるポリペプチド。
(1) 配列番号4に示すアミノ酸配列から成るポリペプチド
(2) 配列番号4に示すアミノ酸配列と70%以上の同一性を有するアミノ酸配列から成り、酸性プロテアーゼ活性を有するポリペプチド
(3) (1)又は(2)のポリペプチドのアミノ酸配列を含み、酸性プロテアーゼ活性を有するポリペプチド
【請求項2】
前記(2)のポリペプチドが、配列番号4に示すアミノ酸配列と80%以上の同一性を有するアミノ酸配列から成る請求項1記載のポリペプチド。
【請求項3】
前記(2)のポリペプチドが、配列番号4に示すアミノ酸配列と90%以上の同一性を有するアミノ酸配列から成る請求項2記載のポリペプチド。
【請求項4】
配列番号4に示すアミノ酸配列又は該アミノ酸配列を含むアミノ酸配列から成る請求項1記載のポリペプチド。
【請求項5】
配列番号4に示すアミノ酸配列から成る請求項4記載のポリペプチド。
【請求項6】
凝乳活性をさらに有する請求項1ないし5のいずれか1項に記載のポリペプチド。
【請求項7】
請求項1ないし6のいずれか1項に記載のポリペプチドをコードするポリヌクレオチド。
【請求項8】
配列表の配列番号1若しくは3に示す塩基配列から成るポリヌクレオチド、又は該塩基配列と70%以上の相同性を有し、酸性プロテアーゼ活性を有するポリペプチドをコードするポリヌクレオチド。
【請求項9】
請求項7又は8記載のポリヌクレオチドを含む組換えベクター。
【請求項10】
請求項9記載の組換えベクターで形質転換された形質転換体。
【請求項11】
請求項6記載のポリペプチドを含有する凝乳酵素組成物。
【請求項12】
請求項6記載のポリペプチドを乳タンパク質と接触させることを含む凝乳方法。
【請求項1】
以下の(1)〜(3)のいずれかであるポリペプチド。
(1) 配列番号4に示すアミノ酸配列から成るポリペプチド
(2) 配列番号4に示すアミノ酸配列と70%以上の同一性を有するアミノ酸配列から成り、酸性プロテアーゼ活性を有するポリペプチド
(3) (1)又は(2)のポリペプチドのアミノ酸配列を含み、酸性プロテアーゼ活性を有するポリペプチド
【請求項2】
前記(2)のポリペプチドが、配列番号4に示すアミノ酸配列と80%以上の同一性を有するアミノ酸配列から成る請求項1記載のポリペプチド。
【請求項3】
前記(2)のポリペプチドが、配列番号4に示すアミノ酸配列と90%以上の同一性を有するアミノ酸配列から成る請求項2記載のポリペプチド。
【請求項4】
配列番号4に示すアミノ酸配列又は該アミノ酸配列を含むアミノ酸配列から成る請求項1記載のポリペプチド。
【請求項5】
配列番号4に示すアミノ酸配列から成る請求項4記載のポリペプチド。
【請求項6】
凝乳活性をさらに有する請求項1ないし5のいずれか1項に記載のポリペプチド。
【請求項7】
請求項1ないし6のいずれか1項に記載のポリペプチドをコードするポリヌクレオチド。
【請求項8】
配列表の配列番号1若しくは3に示す塩基配列から成るポリヌクレオチド、又は該塩基配列と70%以上の相同性を有し、酸性プロテアーゼ活性を有するポリペプチドをコードするポリヌクレオチド。
【請求項9】
請求項7又は8記載のポリヌクレオチドを含む組換えベクター。
【請求項10】
請求項9記載の組換えベクターで形質転換された形質転換体。
【請求項11】
請求項6記載のポリペプチドを含有する凝乳酵素組成物。
【請求項12】
請求項6記載のポリペプチドを乳タンパク質と接触させることを含む凝乳方法。
【図1】


【図3】
【図2】

【図4】
【図3】
【図2】
【図4】
【公開番号】特開2011−193870(P2011−193870A)
【公開日】平成23年10月6日(2011.10.6)
【国際特許分類】
【出願番号】特願2010−189422(P2010−189422)
【出願日】平成22年8月26日(2010.8.26)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 日本農芸化学会2010年度(平成22年度)大会講演要旨集、第131頁、2010年3月5日発行
【出願人】(301025634)独立行政法人酒類総合研究所 (55)
【Fターム(参考)】
【公開日】平成23年10月6日(2011.10.6)
【国際特許分類】
【出願日】平成22年8月26日(2010.8.26)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 日本農芸化学会2010年度(平成22年度)大会講演要旨集、第131頁、2010年3月5日発行
【出願人】(301025634)独立行政法人酒類総合研究所 (55)
【Fターム(参考)】
[ Back to top ]