
新規GPR101トランスジェニックマウスおよびそれを使用する方法
ゲノムがGPR101遺伝子ノックアウトを含むトランスジェニック非ヒト動物またはそれから単離される細胞と、ゲノムが構成的に活性があるGPR101遺伝子を含むトランスジェニック非ヒト動物またはそれから単離される細胞、ならびに、肥満、代謝症候群、脂質異常症、インスリン耐性症候群、2型糖尿病、拒食症および悪液質を含むエネルギー代謝に関連する疾患の処置、予防または診断を改善するためのモデルとして、トランスジェニック非ヒト動物を使用する方法が記載される。
【発明の詳細な説明】
【技術分野】
【0001】
関連出願
本願は、2008年5月19日に出願された米国仮特許出願第61/128,110号明細書の便益性を主張する。上記出願の教示全体は、参照により本明細書中に援用される。
【背景技術】
【0002】
Gタンパク質受容体101(GPR101)は、公知のリガンドを全く有しないオーファンGタンパク質共役受容体(GPCR)である。GPR101は、特に中枢神経系内に存在し、視床下部および扁桃体、具体的には、前脳領域の弓状核(ARC)、視床下部腹内側部(VMH)、視床下部背内側部(DMH)、視床下部後部(PH)、視床下部傍室核(PVN)、内側視索前野(MPOA)、視交叉上核(SCN)および視床下部前野(AHA)、ならびに、代謝恒常性機能に関与すると考えられる領域である後脳領域内の孤束核(NTS)および外側結合腕傍核(LPB)において豊富に発現される(Nilaweera K.N.ら、「G Protein−Coupled Receptor 101 mRNA Expression in the Mouse Brain:Altered Expression in the Posterior Hypothalamus and Amygdala By Energetic Challenges」、Journal of Neuroendocrinology、19:34−45頁(2006年)(その全体が参照により本明細書中に援用される))。GPR101はまた、前脳における側坐核および中脳におけるセロトニン作動性の核などの動機付けられた挙動の調節に関与する脳の領域内で発現される(Batesら、「Characterization of GPR101 Expression and G−protein Coupling Selectivity」、Brain Research、1087:1−14頁(2006年)(その全体が参照により本明細書中に援用される))。視床下部、扁桃体、LPBおよびNTSがエネルギーバランス調節において重要な役割を発揮することから、これらの領域内でのGPR101 mRNAの高度の発現は、代謝恒常性におけるGPR101の重要な役割を示唆しうる。したがって、GPR101が代謝恒常性における重要な役割を有しうることから、それは代謝性疾患および障害の処置および予防において重要な役割を有しうる。
【発明の概要】
【発明が解決しようとする課題】
【0003】
上記の観点から、受容体調節の考えられる生理的結果を同定し、かつ代謝性疾患の処置または予防における使用にとって潜在的な作用剤または組成物を同定するための信頼性の高いツールが大いに必要とされる。
【課題を解決するための手段】
【0004】
本発明は、GPR101受容体を調節する作用剤をスクリーニングすることを目的とした、新規トランスジェニックマウスモデルまたはそれから単離される細胞を特徴とする。かかるモデルを使用し、エネルギー代謝に関連する疾患の診断を改善するとともに、エネルギー代謝に関連する疾患のより優れた処置および予防用の薬学的組成物を同定し、試験することが可能である。
【0005】
一実施形態では、本発明は、GPR101ノックアウトトランスジェニック非ヒト動物、またはそれから単離される細胞、ならびに肥満、代謝症候群、脂質異常症、インスリン耐性症候群、2型糖尿病、拒食症および悪液質などのエネルギー代謝に関連する疾患におけるモデルとしてのその使用を提供する。
【0006】
別の実施形態では、本発明は、トランスジェニック非ヒト動物、またはそれから単離される細胞、ならびに肥満、代謝症候群、脂質異常症、インスリン耐性症候群、糖尿病、特に2型糖尿病、拒食症および悪液質などのエネルギー代謝に関連する疾患におけるモデルとしてのその使用を提供し、ここでGPR101遺伝子は構成的に活性がある。
【0007】
一実施形態では、本発明は、ゲノムが内因性GPR101遺伝子内に破壊を含むトランスジェニックノックアウト非ヒト哺乳動物に関する。
【0008】
別の実施形態では、前記破壊は、胚幹細胞内で、DNA標的化コンストラクトとの相同組換えによってゲノムに導入されている。
【0009】
一実施形態では、GPR101遺伝子の破壊の結果、前記トランスジェニック非ヒト哺乳動物は、検出可能なレベルのGPR101を生成することができない。
【0010】
別の実施形態では、哺乳動物はマウスである。
【0011】
さらに、本発明は、ゲノムが内因性GPR101遺伝子内に破壊を含むトランスジェニックノックアウト非ヒト哺乳動物に由来する単離細胞に関する。
【0012】
さらに、本発明は、ゲノムが内因性GPR101遺伝子内に破壊を含むノックアウト非ヒト哺乳動物を作製する方法に関する。
【0013】
さらに、本発明は、ノックアウト非ヒト哺乳動物における体重および食物摂取に対する調節能について候補作用剤をスクリーニングするための方法であって、
(a)ゲノムが内因性GPR101遺伝子内での破壊を含むトランスジェニックノックアウト非ヒト哺乳動物を提供するステップと、
(b)前記ノックアウト非ヒト哺乳動物に候補作用剤を投与するステップと、
(c)トランスジェニックノックアウト非ヒト哺乳動物の体重および食物摂取を、野生型対照の体重および食物摂取と比較するステップと、ここで効果における差異は、GPR101の活性の改変によって体重および食物摂取を調節する作用剤を示唆する、
を含む、方法に関する。
【0014】
さらに、本発明は、ノックアウト非ヒト哺乳動物における体重および食物摂取に対する調節能について候補作用剤をスクリーニングするための方法であって、
(a)ゲノムが内因性GPR101遺伝子内での破壊を含むトランスジェニック非ヒト哺乳動物を提供するステップと、
(b)前記トランスジェニック非ヒト哺乳動物に候補作用剤を投与するステップと、
(c)トランスジェニック非ヒト哺乳動物に対する前記候補作用剤の効果を評価するステップと、
を含む、方法に関する。
【0015】
別の実施形態では、本発明は、ゲノムが構成的に活性な内因性GPR101遺伝子を含むトランスジェニック非ヒト哺乳動物に関する。
【0016】
一実施形態では、構成的に活性なGPR101遺伝子は、単一のアミノ酸置換によってゲノムに導入されている。
【0017】
別の実施形態では、アミノ酸置換は、胚幹細胞内で、DNA標的化コンストラクトとの相同組換えによってゲノムに導入されている。
【0018】
さらなる実施形態では、哺乳動物はマウスである。
【0019】
さらに、本発明は、ゲノムが構成的に活性な内因性GPR101遺伝子を含むトランスジェニック非ヒト哺乳動物に由来する単離細胞に関する。
【0020】
さらに、本発明は、ゲノムが構成的に活性な内因性GPR101遺伝子を含むトランスジェニック非ヒト哺乳動物を作製する方法に関する。
【0021】
さらに、本発明は、ゲノムが構成的に活性な内因性GPR101遺伝子を含むトランスジェニック非ヒト哺乳動物における体重および食物摂取に対する調節能について候補作用剤をスクリーニングするための方法であって、
(a)ゲノムが構成的に活性な内因性GPR101遺伝子を含むトランスジェニック非ヒト哺乳動物を提供するステップと、
(b)前記トランスジェニック非ヒト哺乳動物に候補作用剤を投与するステップと、
(c)トランスジェニック非ヒト哺乳動物に対する前記候補作用剤の効果を評価するステップと、
を含む、方法に関する。
【図面の簡単な説明】
【0022】
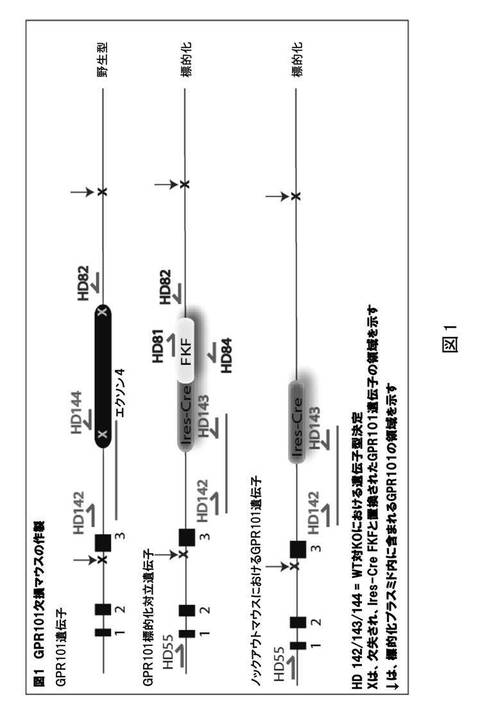
【図1】遺伝子型マウスに対する3つのプライマーによる多重ポリメラーゼ連鎖反応(PCR)法を示す。
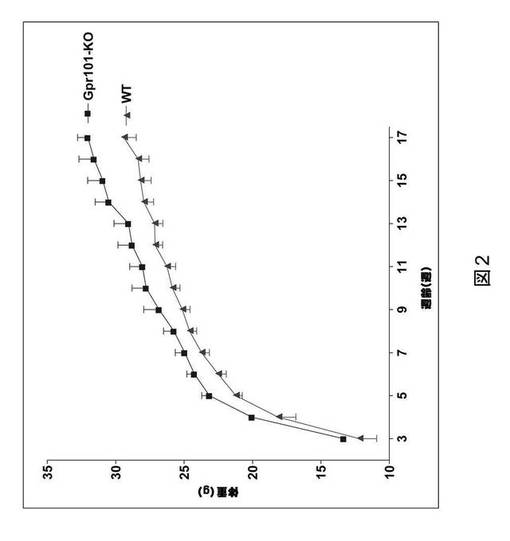
【図2】GPR101ノックアウトマウス(GPR101−KO)および野生型同腹子対照(WT)における体重を示すグラフである。Y軸はグラム(g)での体重を表し、X軸は週齢(wks)を表す。
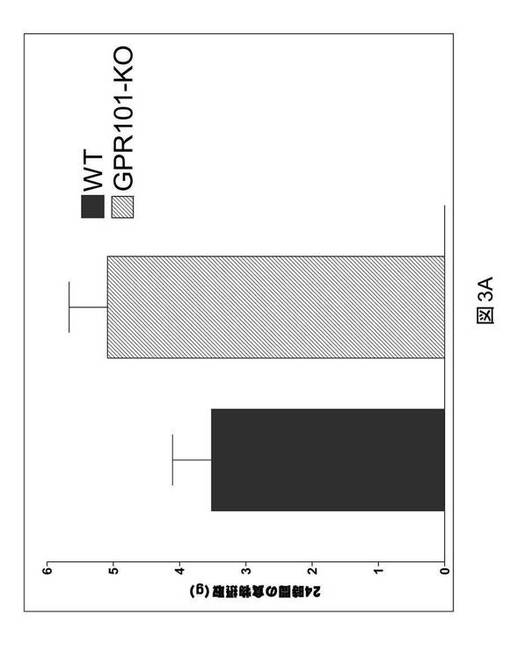
【図3A】GPR101ノックアウトマウス(GPR101−KO)および野生型同腹子対照(WT)における、24時間の期間中のグラム(g)での食物摂取の棒グラフである。
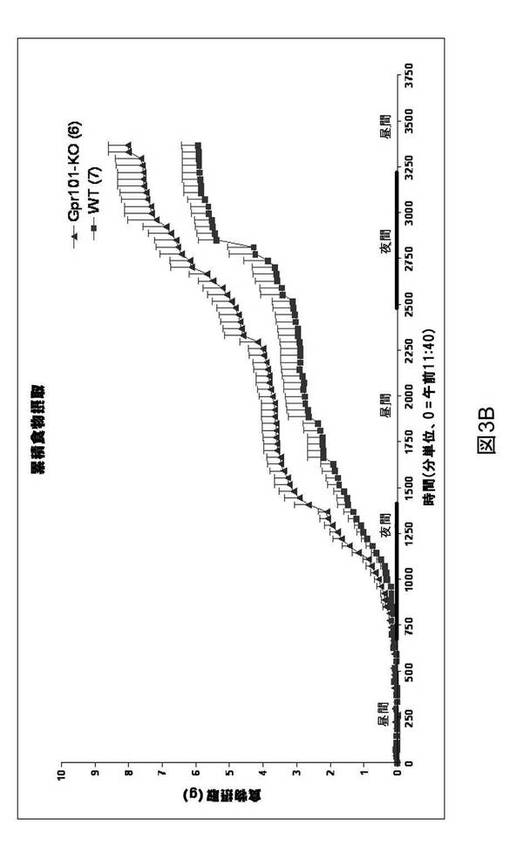
【図3B】GPR101ノックアウトマウス(GPR101−KO)および野生型同腹子対照(WT)における累積食物摂取を示すグラフである。Y軸はグラム(g)での食物摂取を表し、X軸は分単位での時間を表す。
【図4】野生型対照(WT)における食物消費の変化を示すグラフである。Y軸はグラム(g)での食物摂取を表し、X軸は分単位での時間を表す。
【図5】GPR101ノックアウトマウス(GPR101KO)における食物消費の変化を示すグラフである。Y軸はグラム(g)での食物摂取を表し、X軸は分単位での時間を表す。
【図6】食餌(chow diet)に対するGPR101ノックアウトマウス(GPR101KO)および野生型同腹子対照(WT)の体重を示すグラフである。Y軸はグラム(g)での体重を表し、X軸は週齢(wks)を表す。
【図7】高脂肪食に対するGPR101ノックアウトマウス(GPR101KO)および野生型同腹子対照(WT)の体重を示すグラフである。Y軸はグラム(g)での体重を表し、X軸は週齢(wks)を表す。
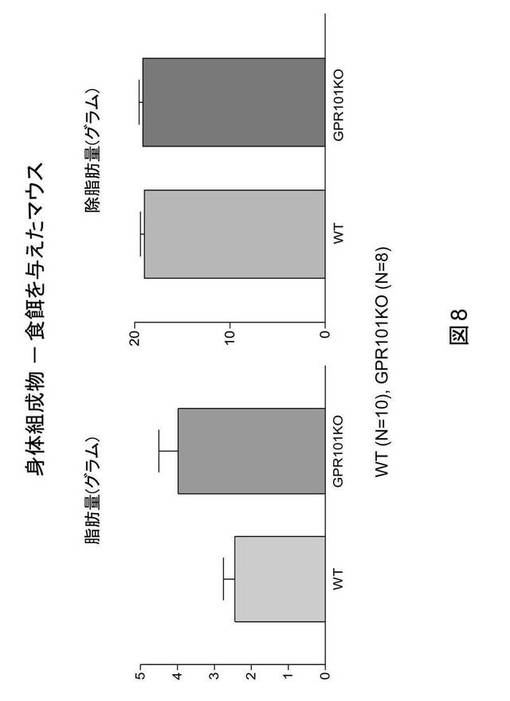
【図8】食餌に対するマウス(野生型(WT)およびGPR101ノックアウトマウス(GPR101KO))における身体組成物(グラムでの脂肪量および除脂肪量)の棒グラフである。
【0023】
前述は、同様の参照符合が異なる図面全体を通じて同じ部分を示す添付の図面中に図示されるように、本発明の例示的な実施形態に関する以下のより詳細な説明から自明であろう。図面は、必ずしも縮尺通りに描かれることはなく、本発明の実施形態を例示することに重点が置かれている。
【発明を実施するための形態】
【0024】
本発明は、トランスジェニック非ヒトGPR101ノックアウト動物、ならびに、エネルギー代謝に関連する疾患、例えば肥満、代謝症候群、脂質異常症、インスリン耐性症候群、糖尿病、特に2型糖尿病、拒食症および悪液質の処置または予防を目的とした薬剤の開発のために動物を使用する方法に関する。本発明はまた、GPR101遺伝子が構成的に活性があるトランスジェニック非ヒト動物、ならびに、エネルギー代謝に関連する疾患、例えば肥満、代謝症候群、脂質異常症、インスリン耐性症候群、糖尿病、特に2型糖尿病、拒食症および悪液質の処置または予防を目的とした薬剤の開発のために動物を使用する方法に関する。特定の機序によって縛られることなく、GPR101遺伝子の量または活性の調節は、かかるエネルギー代謝疾患の処置において有益でありうる。
【0025】
代謝性疾患
代謝性疾患は、エネルギー代謝に関連する疾患、例えば限定はされないが、肥満、代謝症候群、脂質異常症、インスリン耐性症候群、糖尿病、特に2型糖尿病、拒食症および悪液質を含む。
【0026】
定義
本明細書で使用される「トランスジェニック非ヒト動物」は、初代トランスジェニック非ヒト動物および初代動物の子孫、ならびに、動物の1つ以上の細胞が1つ以上のトランス遺伝子を含むかかる動物由来の細胞、細胞系および組織を含む。トランスジェニック非ヒト動物は、ブタ、ヤギ、ヒツジ、ウシ、ウマ、およびウサギなどの家畜動物、ラット、モルモット、およびマウスなどの齧歯動物、ならびにヒヒ、サル、およびチンパンジーなどの非ヒト霊長類であってもよい。トランスジェニックマウスは特に有用である。本明細書で使用されるトランス遺伝子は、トランスジェニック動物がそこから発生する細胞のゲノムに融合され、かつ成熟動物のゲノム内に残る外因性DNAであり、それにより、トランスジェニック動物の1つ以上の細胞種または組織においてコード化遺伝子産物の発現が誘導される。
【0027】
本明細書で使用される遺伝子の「ノックアウト」は、好ましくは標的遺伝子の発現が検出できないかまたは有意でない程度に標的遺伝子の機能の低下をもたらす、遺伝子の配列内での改変または破壊を意味する。内因性遺伝子のノックアウトは、遺伝子の機能が、発現が検出できないかまたは有意でないレベルでしか認められない程度に実質的に低下していることを意味する。本明細書で使用される用語「破壊」および「改変」は、GPR101遺伝子の発現および/または機能における部分的または完全な低下を示唆する。GPR101遺伝子の改変または破壊は、当業者に公知の種々の方法によって行ってもよい。例えば、相同組換え、突然変異誘発(例えば点突然変異)およびアンチセンス技術を用いる遺伝子標的化を用いて、GPR101遺伝子を破壊してもよい。
【0028】
本明細書で使用される用語「遺伝子標的化」は、哺乳類細胞(例えばES細胞)への標的化コンストラクト(例えばベクター)の導入の結果として生じる相同組換えのタイプを示し、それは改変(例えば破壊)の標的とされる遺伝子座の核酸配列の対応部分を探索し、それとの組換えを行い、それによって計画された改変を付与可能な外因性組換え核酸配列を内因性遺伝子に導入するように設計される。したがって、相同組換えは、特定のDNA配列が外因性の遺伝子操作された配列によって置換されることが可能なプロセス(例えば方法)である。より詳細には、破壊の標的とされる遺伝子の内因性ヌクレオチド配列に対して相同(例えば相補的)であるように遺伝子操作されている標的化ベクターの領域が、内因性遺伝子の対応位置に、標的化ベクターのヌクレオチド配列が組み込まれる(例えばそれと融合される)ように互いに整列されるかまたは組換えられる。
【0029】
本明細書で使用される「コンストラクト」は、組換え核酸、一般に組換えDNAを意味し、それは特定のヌクレオチド配列の発現を目的として作成されているか、または他の組換えヌクレオチド配列の作成において使用されることになっているものである。
【0030】
本明細書で使用される用語「遺伝子型」は、GPR101染色体座に関連した動物の遺伝子組成を示す。より詳細には、遺伝子型という用語は、動物のGPR101対立遺伝子の状態を示す。GPR101は、X染色体上に位置する。雌マウスが2つのX染色体を有することから、マウスは、次の3つの遺伝子型、すなわち野生型マウス(XGPR101+、XGPR101+)、ヘテロ接合体マウス(XGPR101+、XGPR101−null)、またはホモ接合体ヌルマウス(XGPR101−null、XGPR101−null)でありうる。雌マウスにおいては、1つのX染色体のみが任意の所与の細胞内で活性がある。このX染色体の不活性化は、細胞間でランダムであり、「ライオニゼーション(lyonization)」と称されることが多い。したがって、ヘテロ接合体である雌マウスにおいては、生体における細胞の約半分が、GPR101を全く発現しないことになる(野生型対立遺伝子を担持するX染色体(XGPR101+)が不活性化されているため)一方、その生体における細胞の残り半分は、正常なレベルのGPR101を発現することになる(「ヌル」対立遺伝子を担持するX染色体(XGPR101−null)が不活性化され、ひいては野生型対立遺伝子を担持するX染色体(XGPR101+)が活性を保持するため)。したがって、雌GPR101ヘテロ接合体は複雑であり、その細胞の半分が通常量のGPR101を有する一方、その細胞の残り半分はGPR101を全く有しないことになる。野生型雌がすべての細胞内に通常量のGPR101を常に有する一方、ホモ接合体ヌル雌マウスは、どの細胞内にも常にGPR101を全く有しないことになる。雄マウスは、(GPR101遺伝子が欠損する)1つのY染色体および(GPR101遺伝子を有する)1つのX染色体を有する。したがって、雄マウスは、単にどちらか1つの遺伝子型を有し、すなわちすべての細胞内に通常量のGPR101を有する野生型(Y、XGPR101+)マウス、およびどの細胞内にもGPR101が全く欠損したノックアウト(Y、XGPR101−null)マウスでありうる。
【0031】
GPR101遺伝子の改変または破壊の結果として、本発明のGPR101ノックアウト哺乳動物は特定の表現型を呈しうる。本明細書で使用される用語「表現型」は、特定の遺伝子型に起因する、生じた生化学または生理的結果を示す。一実施形態では、GPR101ノックアウト哺乳動物は、代謝恒常性を改変している。
【0032】
「ノックアウト」トランスジェニックは、遺伝子のヘテロ接合体ノックアウトまたは遺伝子のホモ接合体ノックアウトを有するトランスジェニック動物であってもよい。「ノックアウト」はまた、標的遺伝子の改変が、例えば、標的遺伝子の改変を促進する物質への動物の暴露、標的遺伝子部位で組換えを促進する酵素の導入(例えばCre−lox系におけるCre)、または生後に標的遺伝子の改変を誘導するための他の方法によって生じうる場合での条件的ノックアウトを含む。
【0033】
リコンビニアリング(Recombineering)(組換えを介する遺伝子工学)は、ベクター、例えばBACの一部である標的配列への突然変異の導入に使用可能な相同組換えに基づく高効率の遺伝子工学システムである。リコンビニアリングの方法は当業者に公知である(例えば、Zhangら、Nature Biotech.18:1314−7頁、2000年;Zhangら、Nature Genetics 20:123−8頁(1998年);ならびにDatsenkoおよびWanner、Proc.Natl.Acad.Sci.USA、97:6640−5頁(2000年)を参照)。リコンビニアリングのレビューは、Courtら、Annu.Rev.Genet.36:361−88頁(2002年)およびCopelandら、Nature Rev.Genet.、2:769−779頁、2001年において見出すことが可能である(いずれもそれら全体が参照により本明細書中に援用される)。
【0034】
本明細書で使用される「構成的に活性な受容体」は、受容体のそのリガンドまたはその化学的等価物への結合を介する場合以外の手段によって活性状態で安定化された受容体を意味するものとする。構成的に活性な受容体は、内因性または非内因性のいずれであってもよい。
【0035】
「構成的に活性化された受容体」は、構成的に活性があるように修飾されているか、またはより構成的に活性があるように修飾されている内因性受容体を意味するものとする。
【0036】
「構成的受容体の活性化」は、受容体の、そのリガンドまたはその化学的等価物への結合の不在下での活性化を意味するものとする。
【0037】
本明細書で使用される用語「ES細胞」は、多能性胚幹細胞と、限定はされないが、胚細胞の発生段階における細胞を含む、胚発生の非常に早い段階でのかかる多能性細胞を示す。
【0038】
「部位特異的突然変異誘発(site specific mutagenesis)」または「部位特異的突然変異誘発(site directed mutagenesis)」は、DNA配列内で特異的な所定の変化をもたらすことである。部位特異的突然変異誘発のための方法は、Sambrookら、「Molecular Cloning:A Laboratory Manual」、CSH Press 1989年、15.3−15.108頁;Weinerら、Gene、126:35−41頁(1993年);Sayersら、Biotechniques、13:592−6頁(1992年);JonesおよびWinistorfer、Biotechniques、12:528−30頁(1992年);Bartonら、Nucleic Acids Res、18:7349−55頁(1990年);MarottiおよびTomich、Gene Anal Tech、6:67−70頁(1989年);ならびにZhu、Anal Biochem、177:1204頁(1989年)(すべてはそれら全体が参照により本明細書中に援用される)において見出すことが可能である。
【0039】
トランスジェニックマウスを作製する方法
一般的態様では、トランスジェニック動物は、トランス遺伝子の発現を可能にする方法での、所与のトランス遺伝子のゲノムへの組込みによって作製される。トランスジェニック動物を作製するための方法は、一般に、WagnerおよびHoppe(米国特許第4,873,191号明細書);Brinsterら、Proc.Natl.Acad.Sci.USA82:4438−4442頁(1985年);ならびに「Manipulating the mouse Embryo;A Laboratory Manual」 第2版(Hogan、Beddington、CostantimiおよびLong編、Cold Spring Harbor Laboratory Press、1994年)によって説明されている(すべてはそれら全体が参照により本明細書中に援用される)。典型的には、ゲノム配列によって隣接される遺伝子は、マイクロインジェクションによって受精卵に移される。マイクロインジェクトした卵は宿主雌に移植され、子孫はトランス遺伝子の発現についてスクリーニングされる。トランスジェニック動物は、限定はされないが、爬虫類、両生類、鳥類、哺乳類、および魚類を含む多数の動物に由来する受精卵から作製することが可能である。
【0040】
あるいは、トランスジェニック動物は、トランス遺伝子を作製するために胚幹(ES)細胞を使用することによって得てもよい。トランス遺伝子は胚幹細胞に導入され、形質移入された幹細胞は胚を形成するために利用される。ES細胞は、適切な条件下で、インビトロで着床前胚を培養することによって得られる(Evansら、Nature 292:154−156頁(1981年);Bradleyら、Nature 309:255−256頁(1984年);Gosslerら、Proc.Acad.Sci.USA 83:9065−9069頁(1986年);およびRobertsonら、Nature 323:445−448頁(1986年)(それら全体が参照により本明細書中に援用される))。子孫は、尾の組織からゲノムDNAを単離することにより、トランス遺伝子の融合について分析し、遺伝子をコードする断片は、従来のDNA−ハイブリダイゼーション技術によって同定することが可能である(Southern J.Mol.Biol.98:503−517頁(1975年)(その全体が参照により本明細書中に援用される))。
【0041】
本発明の一態様は、本発明の非ヒトトランスジェニック動物から細胞または細胞系を単離することと、細胞を培地中で成長させることに関する。本発明の別の態様は、宿主細胞のゲノムの特定の部位への相同組換えが可能になる配列あるいは宿主細胞のゲノムへのランダムまたは準ランダム組換えが可能になる配列を有する本発明の組換え発現ベクターが導入されている宿主細胞に関する。かかる細胞が特定の対象細胞だけでなく、かかる細胞の子孫または潜在的子孫を示すことは理解されている。突然変異または環境の影響のいずれかが原因で後世において特定の修飾が生じうることから、かかる子孫は、実際には親細胞と同一でない場合があるが、本明細書で使用される用語の範囲内に含まれる。
【0042】
本発明の別の態様は、ベクターに関する。本明細書で使用される用語「ベクター」は、核酸分子であって、それが連結されている別の核酸を輸送可能なものを示す。ベクターの1つのタイプは「プラスミド」であり、それは追加的なDNAセグメントのライゲートが可能な環状二本鎖DNAループを示す。ベクターの別のタイプはウイルスベクターであり、ここでは追加的なDNAセグメントがウイルスゲノムにライゲート可能である。特定のベクターは、それが導入される宿主細胞内で自律複製可能である(例えば、細菌起源の複製を有する細菌ベクターおよびエピソーム哺乳類ベクター)。他のベクター(例えば非エピソーム哺乳類ベクター)は、宿主細胞への導入時、宿主細胞のゲノムに融合され、それによって宿主ゲノムとともに複製される。さらに、特定のベクターは、それが作動可能に連結される遺伝子の発現を誘導可能である。かかるベクターは、本明細書中で「発現ベクター」と称される。一般に、組換えDNA技術において有用な発現ベクターは、プラスミドの形態であることが多い。本明細書における「プラスミド」および「ベクター」は、プラスミドがベクターの最も一般的に使用される形態であることから同義的に使用してもよい。しかし、本発明は、等価な機能を果たす、かかる他の形態の発現ベクター、例えばウイルスベクター(例えば、複製欠損性レトロウイルス、アデノウイルスおよびアデノ関連ウイルス)を含むように意図されている。
【0043】
本発明の組換え発現ベクターは、本発明の核酸を、宿主細胞内での核酸の発現に適した形態で含み、それは、組換え発現ベクターが、発現されるべき核酸配列に作動可能に連結される、発現のために使用されるべき宿主細胞に基づいて選択される1つ以上の調節配列を含むことを意味する。かかる調節配列は、例えば、Goeddel、「Gene Expression Technology:Methods in Enzymology」、第185巻、Academic Press(San Diego,Calif.)(1990年)において記載されている(その全体が参照により本明細書中に援用される)。調節配列は、多種の宿主細胞内でのヌクレオチド配列の構成的発現を誘導するものと、特定の宿主細胞内に限ってヌクレオチド配列の発現を誘導するものを含む。発現ベクターの設計が、形質転換されるべき宿主細胞の選択、所望されるタンパク質の発現のレベルなどの要素に依存しうることは、当業者によって理解されるであろう。本発明の発現ベクターは、宿主細胞内に導入されることで、本明細書中に記載される核酸によってコードされる、融合タンパク質または融合ペプチドを含むタンパク質またはペプチドを生成することが可能である。
【0044】
本発明の組換え発現ベクターは、原核または真核細胞内での標的遺伝子の発現のために設計してもよい。例えば、標的遺伝子または断片は、大腸菌(E.coli)、昆虫細胞(バキュロウイルス発現ベクターを使用)、酵母細胞または哺乳類細胞などの細菌細胞内で発現させてもよい。好適な宿主細胞は、Goeddel、「Gene Expression Technology:Methods in Enzymology」、第185巻、Academic Press(San Diego,Calif.)(1990年)(その全体が参照により本明細書中に援用される)においてさらに考察されている。あるいは、組換え発現ベクターは、インビトロで転写または翻訳してもよい。
【0045】
標的遺伝子はまた、哺乳類発現ベクターを使用して、哺乳類細胞内で発現させてもよい。哺乳類発現ベクターの例として、pCDM8(Seed B.、Nature、329:840頁(1987年))およびpMT2PC(Kaufmanら、EMBO J.、6:187頁(1987年))が挙げられる。発現ベクターの制御機能は、哺乳類細胞内で使用される場合、ウイルス調節因子によって提供されることが多い。例えば、一般に使用されるプロモーターは、ポリオーマ、アデノウイルス2型、サイトメガロウイルスおよびサルウイルス40に由来する。原核および真核細胞の双方における他の好適な発現系については、Sambrook J.、Fritsh E.F.およびManiatis T.「Molecular Cloning:A Laboratory Manual」、第2版、Cold Spring Harbor Laboratory、Cold Spring Harbor Laboratory Press(Cold Spring Harbor,N.Y.)、1989年の第16章および第17章を参照のこと。
【0046】
宿主細胞は、任意の原核または真核細胞であってもよい。例えば、宿主細胞は、大腸菌(E.coli)などの細菌細胞、昆虫細胞、酵母、アフリカツメガエル細胞、または哺乳類細胞(例えば、チャイニーズハムスター卵巣細胞(CHO)、アフリカミドリザル腎細胞(COS)、またはヒト胎児細胞(293T))であってもよい。他の好適な宿主細胞は、当業者に公知である。本発明の一態様では、宿主細胞は、本明細書中に記載のトランスジェニック非ヒト動物に由来する。
【0047】
ベクターDNAは、従来の形質転換または形質移入技術を介して、原核または真核細胞に導入してもよい。本明細書で使用される用語「形質転換」および「形質移入」は、リン酸カルシウムまたは塩化カルシウム共沈、DEAE−デキストラン媒介性形質移入、リポフェクション、またはエレクトロポレーションを含む、外来核酸(例えばDNA)を宿主細胞に導入するための種々の当該技術分野で公知の技術を示すように意図されている。宿主細胞を形質転換または形質移入するための好適な方法については、Sambrookら(「Molecular Cloning:A Laboratory Manual」、第2版、Cold Spring Harbor Laboratory、Cold Spring Harbor Laboratory Press(Cold Spring Harbor,N.Y.)、1989年)および他の実験マニュアルにおいて見出すことが可能である。
【0048】
トランスジェニック非ヒト動物を使用する方法
本発明は、エネルギー代謝に関連する疾患、例えば肥満、代謝症候群、脂質異常症、インスリン耐性症候群、2型糖尿病、拒食症および悪液質に対して作用する(すなわち調節する、阻害する、低下させる、予防するまたは改善する)調節因子、すなわち候補もしくは試験化合物または作用物質(例えばペプチド、環状ペプチド、ペプチドミメティック、小分子、小有機分子、または他の薬剤)を同定するための方法(本明細書中で「スクリーニングアッセイ」とも称される)を提供する。本明細書で使用される「調節する(modulating)」または「調節因子(modulator)」または「調節する(modulate)」は、GPR101受容体に対して作動または拮抗させることを示す。特に、本発明のトランスジェニック非ヒト動物を使用し、エネルギー代謝に関連する疾患の処置または予防にとって有効な化合物または組成物を同定することが可能である。化合物または組成物は、試験化合物または組成物を本発明のトランスジェニック非ヒト動物に投与するか、あるいは試験化合物または組成物を、トランスジェニック非ヒト動物に由来する器官、組織(例えば骨格筋)または細胞(例えば神経細胞または筋肉細胞)と接触させることにより、同定することが可能である。トランスジェニック非ヒト動物、器官、組織または細胞におけるエネルギー代謝に対する試験化合物または組成物の効果が評価される。例えば、候補作用剤は、トランスジェニック非ヒト動物において評価してもよい。エネルギー代謝を改変する試験化合物または組成物は、エネルギー代謝に関連する疾患の処置または予防にとって有効でありうる。
【0049】
さらに、本発明は、上記のスクリーニングアッセイによって同定される新規作用剤に関する。したがって、本明細書中に記載の適切な動物モデルにおいて、本明細書中に記載のように同定される試験化合物または組成物をさらに使用することは、本発明の範囲内に含まれる。例えば、本明細書中に記載のように同定される試験化合物または組成物は、動物モデル(例えばトランスジェニックGPR101ノックアウト非ヒト動物)において使用し、かかる試験化合物または組成物での処置の有効性、毒性、または副作用を判定することが可能である。あるいは、本明細書中に記載のように同定される試験化合物または組成物は、動物モデルにおいて使用し、かかる試験化合物または組成物の作用機序を判定することが可能である。
【0050】
試験化合物は、医薬的に許容できる非毒性賦形剤またはキャリアとの混合によって薬学的組成物に調合し、任意の投与経路によって本発明のトランスジェニック非ヒト動物に投与してもよい。例えば、皮下、筋肉内、血管内、皮内、鼻腔内、吸入、髄腔内、または腹腔内投与などの非経口経路、および舌下、経口、または直腸投与などの経腸経路を使用してもよい。
【0051】
試験化合物または組成物は、例えば試験化合物または組成物を野生型マウスおよびGPR101ノックアウトマウスに投与することにより、GPR101受容体に対する生理作用について試験してもよい。試験化合物または組成物が野生型マウスにおいて効果をもたらすが、GPR101ノックアウトマウスにおいてもたらさない場合、その試験化合物または組成物は、GPR101の活性を変化させることによって効果をもたらすものである。試験化合物または組成物が野生型マウスにおいて体重の減少、体脂肪の減少、および/または食物摂取の低下をもたらすが、GPR101ノックアウトマウスにおいてもたらさない場合、その試験化合物または組成物はGPR101の活性化因子である可能性が高い。それに対し、試験化合物または組成物が体重の増加、体脂肪の増加、および/または食物摂取の増加をもたらす場合、その試験化合物または組成物はGPR101の阻害剤である可能性が高い。試験化合物または組成物が野生型マウスとGPR101ノックアウトマウスの双方において効果をもたらす場合、その試験化合物または組成物は、GRP101の活性を変化させることによってその効果を発揮するものではない。
【0052】
試験化合物または組成物の特異性は、GPR101ノックアウトマウスにおいて試験してもよい。例えば、野生型マウスにおいて見られる効果がGPR101ノックアウトマウスにおいても見られる場合、その試験化合物または組成物はGPR101に対して特異的でない。
【0053】
GPR101ノックアウトマウスを使用し、試験化合物または組成物によってもたらされる毒性が、GPR101の活性の改変によるもの(的確な毒性)か、または他の何らかの機序の改変によるもの(的外れの毒性)かを判定してもよい。例えば、試験化合物または組成物が野生型マウスにおいて毒性または望ましくない効果をもたらすが、GPR101ノックアウトマウスにおいてもたらさない場合、その毒性効果は、GPR101の活性の改変による副産物(的確な効果)である。試験化合物または組成物が野生型マウスとGPR101ノックアウトマウスの双方において毒性または望ましくない効果をもたらす場合、その試験化合物または組成物の毒性効果は、GPR101の活性の変化に何も関連性がない(的外れの効果)。
【0054】
GPR101の構成的に活性なマウスは、例えばGPR101の活性を低下させかねない試験化合物または組成物を試験するのに特に有用でありうる。かかる試験化合物または組成物であれば、拒食症または悪液質を有する患者における食物摂取を刺激するのに有用でありうる。これらのマウスにおいて受容体が活性化されることから、それらはGPR101の活性の潜在的な阻害剤に対して感受性を示すはずである。
【0055】
本発明の例示的な実施形態の説明は以下の通りである。
【実施例】
【0056】
実験手順
GPR101ノックアウトマウスの作製
胚幹(ES)細胞における遺伝子標的化を用いて、GPR101欠損マウスを作製した。置換標的化コンストラクトを、約100kb上流への、および約40kb下流へのGPR101配列を有する細菌人工染色体(BAC)ゲノムクローンを使用して調製した。BACクローンを、開始コドンを含むコードエクソンに及ぶGPR101配列の2216bp部分(NCBI登録番号NC_000086)が除去され、Ires−Cre−Frt−カナマイシン−Frt(Ires−Cre FKF)カセットと置換されるように設計した。GPR101におけるゲノムDNA配列は、NC_000086内にあり、アセンブリ上の54756894から54749845の位置に由来する。欠失の設計に使用されるプライマーは、(配列番号1)HD73フォワード5’−TCC TCT GCA AGG CAC TAA CCC TAG CCA CAT GTT TCT CTC GTC CTC AAT CTA GTG ATG TAA TTC CGC CCC TCT CCC T−3’および(配列番号2)HD74リバース5’−CCT AGC TCC TCA TTT CAG GCT TGC CCT TTT CTG GAT CCC TTT TGA AGA CCT AAA CAA AAT ATT AAC GCT TAC A−3であった。欠失を有するBACの11.4kb部分を、Zeocinを有するpCR−Bluntに挿入した。次いで、この標的化プラスミドを線状化し、胚幹細胞にエレクトロポレートした。標的化クローンを、標的化遺伝子座の3’および5’末端に及ぶプライマーを使用するPCR分析によって同定した(図1)。標的化ESクローンから拡張された細胞をC57BL6胚盤胞に注入し、生殖細胞系を伝達するキメラ動物を得て、次いでFLPe−リコンビナーゼマウスと交配させた。GPR101がX染色体上に位置することから、得られたヘテロ接合性の子らを交配させ、野生型(+/+)およびホモ接合性(−/−)試験対象を作製した。すべての試験を雄マウスにおいて実施した。3つのプライマーによる多重PCR法を、図1に示される遺伝子型マウスに対して用いた。以下は、図1からのプライマー配列である。
(配列番号3)HD55 TTC TTT GCT CCC TCT TCA TTC TCA
(配列番号4)HD81 CGC ATC GCC TTC TAT CG
(配列番号5)HD82 ACC TAC TTC ATG TTT ATTT ACG
(配列番号6)HD84 CGT CAA GAA GGC GAT AGA
(配列番号7)HD142 TGAGACCCCCAAGAATTAGAAAAA
(配列番号8)HD143 TTGGCGAGAGGGGAAAGAC
(配列番号9)HD144 GGGGCCACGAGAGCAACCT
【0057】
GPR101が構成的に活性なマウスの作製
GPCRの特定の残基の突然変異の結果、オープン構造ひいては構成的活性(リガンドの不在下での活性)を有する受容体が得られる。受容体の潜在的な構成的活性に関与することになるGPR101内の残基を同定するため、インビトロ試験を実施した。マウスGPR101をコードする完全長cDNAを、マウス脳RNAから増幅し、それを発現プラスミドpCDNA3.1に挿入した。部位特異的突然変異誘発を用いて、活性の増大につながるGPR101の膜貫通ループにおける突然変異を同定したが、ここでは標的としてGPR101タンパク質のアラニン397を同定し、リジンに突然変異される場合、GPR101の活性における2倍の増大をもたらす(データは示さず)。単一のアミノ酸置換、すなわちA397→K397を伴う新規マウスモデルを作製した。
【0058】
A397→K397を伴う構成的に活性化型のGPR101を生成するための標的化コンストラクトを、リコンビニアリング(recombineering)を含む標準技術を用いて作成した。galK陽性および対抗選択のスキームを用いて、GPR101のBACにおいて点突然変異を生じさせた(Warming S.ら、「Simple and Highly Efficient BAC Recombineering Using galK Selection」、Nucleic Acids Research、33:1−12頁(2005年)(その全体が参照により本明細書中に援用される))。次いで、点突然変異を有するマウスを作製するため、修飾BACをプラスミドに挿入し、胚盤胞に注入した。
【0059】
結果
GPR101ノックアウトマウスにおける体重
体重調節におけるGPR101の機能的重要性を判定するため、GPR101ノックアウトマウスを作製した。GPR101ノックアウトマウスに標準の齧歯類用食餌を与え、3週齢以降の雄GPR101ノックアウトマウス(n=8)の体重を、野生型同腹子(n=12)のそれと比較した。
【0060】
図2に見られるように、雄GPR101ノックアウトマウスは、野生型同腹子対照と比較して体重増加傾向で推移した。17週齢のGPR101ノックアウトマウスは、体重において、野生型マウスに見られる場合よりも約10%の増加を有した。これらの結果は、中枢神経系におけるGPR101が正常な体重恒常性にとって必要とされることを示す。
【0061】
GPR101ノックアウトマウスにおける食物摂取
全食物摂取を、GPR101ノックアウトマウスにおいて評価した。GPR101ノックアウトマウスに標準の齧歯類用食餌を与えた。6週齢目、食物摂取を24時間記録した(図3A)。結果は、GPR101ノックアウトマウスにおける24時間内での食物摂取が、その野生型同腹子に対して増加したことを示す(5.1±0.58、GPR101ノックアウト対3.52±0.58野生型、n=6〜7/群)。8週齢目、全食物摂取を包括的な実験動物モニタリングシステムにおいて記録し、GPR101ノックアウトマウスによって消費される全食物の、その野生型対照に対する有意な増加が見られた(図3B)。結果は、GPR101が食餌サイズの調節における役割を果たすことを示唆する。
【0062】
GPR101ノックアウトマウスにおける食餌パターン
上で考察した24時間の食物摂取における増加に加え、8週齢のGPR101ノックアウトマウスはまた、図5に示されるように、食物消費の全体パターンにおける変化を示す。GPR101ノックアウトマウスは、図4および5に示されるように、野生型対照に対して食餌サイズにおける変化を示した。GPR101ノックアウトマウスは、暗期中により大量の食餌を消費した。
【0063】
データは、GPR101ノックアウトマウスが体重および食物摂取の増加を有し、それによりGPR101ノックアウトマウスが代謝性疾患の処置に使用可能なGPR101のアゴニストおよびアンタゴニストの同定において有用でありうることを示す。
【0064】
GPR101ノックアウトマウスにおける高脂肪食誘発性肥満
野生型およびGPR101遺伝子ノックアウト雄マウスを、離乳(3週齢)時に、食餌(Forumulab、食餌#5008)(WTにおいてn=10およびGPR101−KOにおいてn=8)または高脂肪食(カロリーの45%が脂肪由来、Research Diets、D12451)(WTにおいてn=7およびGPR101−KOにおいてn=7)を与え、体重を毎週モニタリングした。先に認められたように、食餌を与えたGPR101KOマウスの体重は、野生型対照の場合より重かった(図6)。注目すべきことに、野生型とGPR101KOマウスの間での体重における差異は、マウスに高脂肪食を与える場合に大きくなる(図7)。食餌を与えたマウスの身体組成物は、12週齢時でのEcho−MRIによる評価によると、体重の増加が脂肪量の増加に起因することを示す(図8)。これらの実験結果は、GPR101の遺伝子ノックアウトが肥満を誘発することを示す。さらに、GPR101の遺伝子ノックアウトは、高脂肪食誘発性肥満に対する感受性の増大の原因になる。まとめると、これらの実験結果は、GPR101アゴニストが、抗肥満効果を有する可能性が高く、故に肥満の処置にとって有用でありうることを示す。
【0065】
本明細書中に引用されるすべての特許、公開された出願および参考文献の教示内容は、それら全体が参照により援用される。
【0066】
本発明は、特にその例示的な実施形態を参照して示され、説明されている一方、形態におけるさまざまな変化および詳細化が、添付の特許請求の範囲によって包含される本発明の範囲から逸脱することなく、本明細書中でなされうることは当業者によって理解されるであろう。
【技術分野】
【0001】
関連出願
本願は、2008年5月19日に出願された米国仮特許出願第61/128,110号明細書の便益性を主張する。上記出願の教示全体は、参照により本明細書中に援用される。
【背景技術】
【0002】
Gタンパク質受容体101(GPR101)は、公知のリガンドを全く有しないオーファンGタンパク質共役受容体(GPCR)である。GPR101は、特に中枢神経系内に存在し、視床下部および扁桃体、具体的には、前脳領域の弓状核(ARC)、視床下部腹内側部(VMH)、視床下部背内側部(DMH)、視床下部後部(PH)、視床下部傍室核(PVN)、内側視索前野(MPOA)、視交叉上核(SCN)および視床下部前野(AHA)、ならびに、代謝恒常性機能に関与すると考えられる領域である後脳領域内の孤束核(NTS)および外側結合腕傍核(LPB)において豊富に発現される(Nilaweera K.N.ら、「G Protein−Coupled Receptor 101 mRNA Expression in the Mouse Brain:Altered Expression in the Posterior Hypothalamus and Amygdala By Energetic Challenges」、Journal of Neuroendocrinology、19:34−45頁(2006年)(その全体が参照により本明細書中に援用される))。GPR101はまた、前脳における側坐核および中脳におけるセロトニン作動性の核などの動機付けられた挙動の調節に関与する脳の領域内で発現される(Batesら、「Characterization of GPR101 Expression and G−protein Coupling Selectivity」、Brain Research、1087:1−14頁(2006年)(その全体が参照により本明細書中に援用される))。視床下部、扁桃体、LPBおよびNTSがエネルギーバランス調節において重要な役割を発揮することから、これらの領域内でのGPR101 mRNAの高度の発現は、代謝恒常性におけるGPR101の重要な役割を示唆しうる。したがって、GPR101が代謝恒常性における重要な役割を有しうることから、それは代謝性疾患および障害の処置および予防において重要な役割を有しうる。
【発明の概要】
【発明が解決しようとする課題】
【0003】
上記の観点から、受容体調節の考えられる生理的結果を同定し、かつ代謝性疾患の処置または予防における使用にとって潜在的な作用剤または組成物を同定するための信頼性の高いツールが大いに必要とされる。
【課題を解決するための手段】
【0004】
本発明は、GPR101受容体を調節する作用剤をスクリーニングすることを目的とした、新規トランスジェニックマウスモデルまたはそれから単離される細胞を特徴とする。かかるモデルを使用し、エネルギー代謝に関連する疾患の診断を改善するとともに、エネルギー代謝に関連する疾患のより優れた処置および予防用の薬学的組成物を同定し、試験することが可能である。
【0005】
一実施形態では、本発明は、GPR101ノックアウトトランスジェニック非ヒト動物、またはそれから単離される細胞、ならびに肥満、代謝症候群、脂質異常症、インスリン耐性症候群、2型糖尿病、拒食症および悪液質などのエネルギー代謝に関連する疾患におけるモデルとしてのその使用を提供する。
【0006】
別の実施形態では、本発明は、トランスジェニック非ヒト動物、またはそれから単離される細胞、ならびに肥満、代謝症候群、脂質異常症、インスリン耐性症候群、糖尿病、特に2型糖尿病、拒食症および悪液質などのエネルギー代謝に関連する疾患におけるモデルとしてのその使用を提供し、ここでGPR101遺伝子は構成的に活性がある。
【0007】
一実施形態では、本発明は、ゲノムが内因性GPR101遺伝子内に破壊を含むトランスジェニックノックアウト非ヒト哺乳動物に関する。
【0008】
別の実施形態では、前記破壊は、胚幹細胞内で、DNA標的化コンストラクトとの相同組換えによってゲノムに導入されている。
【0009】
一実施形態では、GPR101遺伝子の破壊の結果、前記トランスジェニック非ヒト哺乳動物は、検出可能なレベルのGPR101を生成することができない。
【0010】
別の実施形態では、哺乳動物はマウスである。
【0011】
さらに、本発明は、ゲノムが内因性GPR101遺伝子内に破壊を含むトランスジェニックノックアウト非ヒト哺乳動物に由来する単離細胞に関する。
【0012】
さらに、本発明は、ゲノムが内因性GPR101遺伝子内に破壊を含むノックアウト非ヒト哺乳動物を作製する方法に関する。
【0013】
さらに、本発明は、ノックアウト非ヒト哺乳動物における体重および食物摂取に対する調節能について候補作用剤をスクリーニングするための方法であって、
(a)ゲノムが内因性GPR101遺伝子内での破壊を含むトランスジェニックノックアウト非ヒト哺乳動物を提供するステップと、
(b)前記ノックアウト非ヒト哺乳動物に候補作用剤を投与するステップと、
(c)トランスジェニックノックアウト非ヒト哺乳動物の体重および食物摂取を、野生型対照の体重および食物摂取と比較するステップと、ここで効果における差異は、GPR101の活性の改変によって体重および食物摂取を調節する作用剤を示唆する、
を含む、方法に関する。
【0014】
さらに、本発明は、ノックアウト非ヒト哺乳動物における体重および食物摂取に対する調節能について候補作用剤をスクリーニングするための方法であって、
(a)ゲノムが内因性GPR101遺伝子内での破壊を含むトランスジェニック非ヒト哺乳動物を提供するステップと、
(b)前記トランスジェニック非ヒト哺乳動物に候補作用剤を投与するステップと、
(c)トランスジェニック非ヒト哺乳動物に対する前記候補作用剤の効果を評価するステップと、
を含む、方法に関する。
【0015】
別の実施形態では、本発明は、ゲノムが構成的に活性な内因性GPR101遺伝子を含むトランスジェニック非ヒト哺乳動物に関する。
【0016】
一実施形態では、構成的に活性なGPR101遺伝子は、単一のアミノ酸置換によってゲノムに導入されている。
【0017】
別の実施形態では、アミノ酸置換は、胚幹細胞内で、DNA標的化コンストラクトとの相同組換えによってゲノムに導入されている。
【0018】
さらなる実施形態では、哺乳動物はマウスである。
【0019】
さらに、本発明は、ゲノムが構成的に活性な内因性GPR101遺伝子を含むトランスジェニック非ヒト哺乳動物に由来する単離細胞に関する。
【0020】
さらに、本発明は、ゲノムが構成的に活性な内因性GPR101遺伝子を含むトランスジェニック非ヒト哺乳動物を作製する方法に関する。
【0021】
さらに、本発明は、ゲノムが構成的に活性な内因性GPR101遺伝子を含むトランスジェニック非ヒト哺乳動物における体重および食物摂取に対する調節能について候補作用剤をスクリーニングするための方法であって、
(a)ゲノムが構成的に活性な内因性GPR101遺伝子を含むトランスジェニック非ヒト哺乳動物を提供するステップと、
(b)前記トランスジェニック非ヒト哺乳動物に候補作用剤を投与するステップと、
(c)トランスジェニック非ヒト哺乳動物に対する前記候補作用剤の効果を評価するステップと、
を含む、方法に関する。
【図面の簡単な説明】
【0022】
【図1】遺伝子型マウスに対する3つのプライマーによる多重ポリメラーゼ連鎖反応(PCR)法を示す。
【図2】GPR101ノックアウトマウス(GPR101−KO)および野生型同腹子対照(WT)における体重を示すグラフである。Y軸はグラム(g)での体重を表し、X軸は週齢(wks)を表す。
【図3A】GPR101ノックアウトマウス(GPR101−KO)および野生型同腹子対照(WT)における、24時間の期間中のグラム(g)での食物摂取の棒グラフである。
【図3B】GPR101ノックアウトマウス(GPR101−KO)および野生型同腹子対照(WT)における累積食物摂取を示すグラフである。Y軸はグラム(g)での食物摂取を表し、X軸は分単位での時間を表す。
【図4】野生型対照(WT)における食物消費の変化を示すグラフである。Y軸はグラム(g)での食物摂取を表し、X軸は分単位での時間を表す。
【図5】GPR101ノックアウトマウス(GPR101KO)における食物消費の変化を示すグラフである。Y軸はグラム(g)での食物摂取を表し、X軸は分単位での時間を表す。
【図6】食餌(chow diet)に対するGPR101ノックアウトマウス(GPR101KO)および野生型同腹子対照(WT)の体重を示すグラフである。Y軸はグラム(g)での体重を表し、X軸は週齢(wks)を表す。
【図7】高脂肪食に対するGPR101ノックアウトマウス(GPR101KO)および野生型同腹子対照(WT)の体重を示すグラフである。Y軸はグラム(g)での体重を表し、X軸は週齢(wks)を表す。
【図8】食餌に対するマウス(野生型(WT)およびGPR101ノックアウトマウス(GPR101KO))における身体組成物(グラムでの脂肪量および除脂肪量)の棒グラフである。
【0023】
前述は、同様の参照符合が異なる図面全体を通じて同じ部分を示す添付の図面中に図示されるように、本発明の例示的な実施形態に関する以下のより詳細な説明から自明であろう。図面は、必ずしも縮尺通りに描かれることはなく、本発明の実施形態を例示することに重点が置かれている。
【発明を実施するための形態】
【0024】
本発明は、トランスジェニック非ヒトGPR101ノックアウト動物、ならびに、エネルギー代謝に関連する疾患、例えば肥満、代謝症候群、脂質異常症、インスリン耐性症候群、糖尿病、特に2型糖尿病、拒食症および悪液質の処置または予防を目的とした薬剤の開発のために動物を使用する方法に関する。本発明はまた、GPR101遺伝子が構成的に活性があるトランスジェニック非ヒト動物、ならびに、エネルギー代謝に関連する疾患、例えば肥満、代謝症候群、脂質異常症、インスリン耐性症候群、糖尿病、特に2型糖尿病、拒食症および悪液質の処置または予防を目的とした薬剤の開発のために動物を使用する方法に関する。特定の機序によって縛られることなく、GPR101遺伝子の量または活性の調節は、かかるエネルギー代謝疾患の処置において有益でありうる。
【0025】
代謝性疾患
代謝性疾患は、エネルギー代謝に関連する疾患、例えば限定はされないが、肥満、代謝症候群、脂質異常症、インスリン耐性症候群、糖尿病、特に2型糖尿病、拒食症および悪液質を含む。
【0026】
定義
本明細書で使用される「トランスジェニック非ヒト動物」は、初代トランスジェニック非ヒト動物および初代動物の子孫、ならびに、動物の1つ以上の細胞が1つ以上のトランス遺伝子を含むかかる動物由来の細胞、細胞系および組織を含む。トランスジェニック非ヒト動物は、ブタ、ヤギ、ヒツジ、ウシ、ウマ、およびウサギなどの家畜動物、ラット、モルモット、およびマウスなどの齧歯動物、ならびにヒヒ、サル、およびチンパンジーなどの非ヒト霊長類であってもよい。トランスジェニックマウスは特に有用である。本明細書で使用されるトランス遺伝子は、トランスジェニック動物がそこから発生する細胞のゲノムに融合され、かつ成熟動物のゲノム内に残る外因性DNAであり、それにより、トランスジェニック動物の1つ以上の細胞種または組織においてコード化遺伝子産物の発現が誘導される。
【0027】
本明細書で使用される遺伝子の「ノックアウト」は、好ましくは標的遺伝子の発現が検出できないかまたは有意でない程度に標的遺伝子の機能の低下をもたらす、遺伝子の配列内での改変または破壊を意味する。内因性遺伝子のノックアウトは、遺伝子の機能が、発現が検出できないかまたは有意でないレベルでしか認められない程度に実質的に低下していることを意味する。本明細書で使用される用語「破壊」および「改変」は、GPR101遺伝子の発現および/または機能における部分的または完全な低下を示唆する。GPR101遺伝子の改変または破壊は、当業者に公知の種々の方法によって行ってもよい。例えば、相同組換え、突然変異誘発(例えば点突然変異)およびアンチセンス技術を用いる遺伝子標的化を用いて、GPR101遺伝子を破壊してもよい。
【0028】
本明細書で使用される用語「遺伝子標的化」は、哺乳類細胞(例えばES細胞)への標的化コンストラクト(例えばベクター)の導入の結果として生じる相同組換えのタイプを示し、それは改変(例えば破壊)の標的とされる遺伝子座の核酸配列の対応部分を探索し、それとの組換えを行い、それによって計画された改変を付与可能な外因性組換え核酸配列を内因性遺伝子に導入するように設計される。したがって、相同組換えは、特定のDNA配列が外因性の遺伝子操作された配列によって置換されることが可能なプロセス(例えば方法)である。より詳細には、破壊の標的とされる遺伝子の内因性ヌクレオチド配列に対して相同(例えば相補的)であるように遺伝子操作されている標的化ベクターの領域が、内因性遺伝子の対応位置に、標的化ベクターのヌクレオチド配列が組み込まれる(例えばそれと融合される)ように互いに整列されるかまたは組換えられる。
【0029】
本明細書で使用される「コンストラクト」は、組換え核酸、一般に組換えDNAを意味し、それは特定のヌクレオチド配列の発現を目的として作成されているか、または他の組換えヌクレオチド配列の作成において使用されることになっているものである。
【0030】
本明細書で使用される用語「遺伝子型」は、GPR101染色体座に関連した動物の遺伝子組成を示す。より詳細には、遺伝子型という用語は、動物のGPR101対立遺伝子の状態を示す。GPR101は、X染色体上に位置する。雌マウスが2つのX染色体を有することから、マウスは、次の3つの遺伝子型、すなわち野生型マウス(XGPR101+、XGPR101+)、ヘテロ接合体マウス(XGPR101+、XGPR101−null)、またはホモ接合体ヌルマウス(XGPR101−null、XGPR101−null)でありうる。雌マウスにおいては、1つのX染色体のみが任意の所与の細胞内で活性がある。このX染色体の不活性化は、細胞間でランダムであり、「ライオニゼーション(lyonization)」と称されることが多い。したがって、ヘテロ接合体である雌マウスにおいては、生体における細胞の約半分が、GPR101を全く発現しないことになる(野生型対立遺伝子を担持するX染色体(XGPR101+)が不活性化されているため)一方、その生体における細胞の残り半分は、正常なレベルのGPR101を発現することになる(「ヌル」対立遺伝子を担持するX染色体(XGPR101−null)が不活性化され、ひいては野生型対立遺伝子を担持するX染色体(XGPR101+)が活性を保持するため)。したがって、雌GPR101ヘテロ接合体は複雑であり、その細胞の半分が通常量のGPR101を有する一方、その細胞の残り半分はGPR101を全く有しないことになる。野生型雌がすべての細胞内に通常量のGPR101を常に有する一方、ホモ接合体ヌル雌マウスは、どの細胞内にも常にGPR101を全く有しないことになる。雄マウスは、(GPR101遺伝子が欠損する)1つのY染色体および(GPR101遺伝子を有する)1つのX染色体を有する。したがって、雄マウスは、単にどちらか1つの遺伝子型を有し、すなわちすべての細胞内に通常量のGPR101を有する野生型(Y、XGPR101+)マウス、およびどの細胞内にもGPR101が全く欠損したノックアウト(Y、XGPR101−null)マウスでありうる。
【0031】
GPR101遺伝子の改変または破壊の結果として、本発明のGPR101ノックアウト哺乳動物は特定の表現型を呈しうる。本明細書で使用される用語「表現型」は、特定の遺伝子型に起因する、生じた生化学または生理的結果を示す。一実施形態では、GPR101ノックアウト哺乳動物は、代謝恒常性を改変している。
【0032】
「ノックアウト」トランスジェニックは、遺伝子のヘテロ接合体ノックアウトまたは遺伝子のホモ接合体ノックアウトを有するトランスジェニック動物であってもよい。「ノックアウト」はまた、標的遺伝子の改変が、例えば、標的遺伝子の改変を促進する物質への動物の暴露、標的遺伝子部位で組換えを促進する酵素の導入(例えばCre−lox系におけるCre)、または生後に標的遺伝子の改変を誘導するための他の方法によって生じうる場合での条件的ノックアウトを含む。
【0033】
リコンビニアリング(Recombineering)(組換えを介する遺伝子工学)は、ベクター、例えばBACの一部である標的配列への突然変異の導入に使用可能な相同組換えに基づく高効率の遺伝子工学システムである。リコンビニアリングの方法は当業者に公知である(例えば、Zhangら、Nature Biotech.18:1314−7頁、2000年;Zhangら、Nature Genetics 20:123−8頁(1998年);ならびにDatsenkoおよびWanner、Proc.Natl.Acad.Sci.USA、97:6640−5頁(2000年)を参照)。リコンビニアリングのレビューは、Courtら、Annu.Rev.Genet.36:361−88頁(2002年)およびCopelandら、Nature Rev.Genet.、2:769−779頁、2001年において見出すことが可能である(いずれもそれら全体が参照により本明細書中に援用される)。
【0034】
本明細書で使用される「構成的に活性な受容体」は、受容体のそのリガンドまたはその化学的等価物への結合を介する場合以外の手段によって活性状態で安定化された受容体を意味するものとする。構成的に活性な受容体は、内因性または非内因性のいずれであってもよい。
【0035】
「構成的に活性化された受容体」は、構成的に活性があるように修飾されているか、またはより構成的に活性があるように修飾されている内因性受容体を意味するものとする。
【0036】
「構成的受容体の活性化」は、受容体の、そのリガンドまたはその化学的等価物への結合の不在下での活性化を意味するものとする。
【0037】
本明細書で使用される用語「ES細胞」は、多能性胚幹細胞と、限定はされないが、胚細胞の発生段階における細胞を含む、胚発生の非常に早い段階でのかかる多能性細胞を示す。
【0038】
「部位特異的突然変異誘発(site specific mutagenesis)」または「部位特異的突然変異誘発(site directed mutagenesis)」は、DNA配列内で特異的な所定の変化をもたらすことである。部位特異的突然変異誘発のための方法は、Sambrookら、「Molecular Cloning:A Laboratory Manual」、CSH Press 1989年、15.3−15.108頁;Weinerら、Gene、126:35−41頁(1993年);Sayersら、Biotechniques、13:592−6頁(1992年);JonesおよびWinistorfer、Biotechniques、12:528−30頁(1992年);Bartonら、Nucleic Acids Res、18:7349−55頁(1990年);MarottiおよびTomich、Gene Anal Tech、6:67−70頁(1989年);ならびにZhu、Anal Biochem、177:1204頁(1989年)(すべてはそれら全体が参照により本明細書中に援用される)において見出すことが可能である。
【0039】
トランスジェニックマウスを作製する方法
一般的態様では、トランスジェニック動物は、トランス遺伝子の発現を可能にする方法での、所与のトランス遺伝子のゲノムへの組込みによって作製される。トランスジェニック動物を作製するための方法は、一般に、WagnerおよびHoppe(米国特許第4,873,191号明細書);Brinsterら、Proc.Natl.Acad.Sci.USA82:4438−4442頁(1985年);ならびに「Manipulating the mouse Embryo;A Laboratory Manual」 第2版(Hogan、Beddington、CostantimiおよびLong編、Cold Spring Harbor Laboratory Press、1994年)によって説明されている(すべてはそれら全体が参照により本明細書中に援用される)。典型的には、ゲノム配列によって隣接される遺伝子は、マイクロインジェクションによって受精卵に移される。マイクロインジェクトした卵は宿主雌に移植され、子孫はトランス遺伝子の発現についてスクリーニングされる。トランスジェニック動物は、限定はされないが、爬虫類、両生類、鳥類、哺乳類、および魚類を含む多数の動物に由来する受精卵から作製することが可能である。
【0040】
あるいは、トランスジェニック動物は、トランス遺伝子を作製するために胚幹(ES)細胞を使用することによって得てもよい。トランス遺伝子は胚幹細胞に導入され、形質移入された幹細胞は胚を形成するために利用される。ES細胞は、適切な条件下で、インビトロで着床前胚を培養することによって得られる(Evansら、Nature 292:154−156頁(1981年);Bradleyら、Nature 309:255−256頁(1984年);Gosslerら、Proc.Acad.Sci.USA 83:9065−9069頁(1986年);およびRobertsonら、Nature 323:445−448頁(1986年)(それら全体が参照により本明細書中に援用される))。子孫は、尾の組織からゲノムDNAを単離することにより、トランス遺伝子の融合について分析し、遺伝子をコードする断片は、従来のDNA−ハイブリダイゼーション技術によって同定することが可能である(Southern J.Mol.Biol.98:503−517頁(1975年)(その全体が参照により本明細書中に援用される))。
【0041】
本発明の一態様は、本発明の非ヒトトランスジェニック動物から細胞または細胞系を単離することと、細胞を培地中で成長させることに関する。本発明の別の態様は、宿主細胞のゲノムの特定の部位への相同組換えが可能になる配列あるいは宿主細胞のゲノムへのランダムまたは準ランダム組換えが可能になる配列を有する本発明の組換え発現ベクターが導入されている宿主細胞に関する。かかる細胞が特定の対象細胞だけでなく、かかる細胞の子孫または潜在的子孫を示すことは理解されている。突然変異または環境の影響のいずれかが原因で後世において特定の修飾が生じうることから、かかる子孫は、実際には親細胞と同一でない場合があるが、本明細書で使用される用語の範囲内に含まれる。
【0042】
本発明の別の態様は、ベクターに関する。本明細書で使用される用語「ベクター」は、核酸分子であって、それが連結されている別の核酸を輸送可能なものを示す。ベクターの1つのタイプは「プラスミド」であり、それは追加的なDNAセグメントのライゲートが可能な環状二本鎖DNAループを示す。ベクターの別のタイプはウイルスベクターであり、ここでは追加的なDNAセグメントがウイルスゲノムにライゲート可能である。特定のベクターは、それが導入される宿主細胞内で自律複製可能である(例えば、細菌起源の複製を有する細菌ベクターおよびエピソーム哺乳類ベクター)。他のベクター(例えば非エピソーム哺乳類ベクター)は、宿主細胞への導入時、宿主細胞のゲノムに融合され、それによって宿主ゲノムとともに複製される。さらに、特定のベクターは、それが作動可能に連結される遺伝子の発現を誘導可能である。かかるベクターは、本明細書中で「発現ベクター」と称される。一般に、組換えDNA技術において有用な発現ベクターは、プラスミドの形態であることが多い。本明細書における「プラスミド」および「ベクター」は、プラスミドがベクターの最も一般的に使用される形態であることから同義的に使用してもよい。しかし、本発明は、等価な機能を果たす、かかる他の形態の発現ベクター、例えばウイルスベクター(例えば、複製欠損性レトロウイルス、アデノウイルスおよびアデノ関連ウイルス)を含むように意図されている。
【0043】
本発明の組換え発現ベクターは、本発明の核酸を、宿主細胞内での核酸の発現に適した形態で含み、それは、組換え発現ベクターが、発現されるべき核酸配列に作動可能に連結される、発現のために使用されるべき宿主細胞に基づいて選択される1つ以上の調節配列を含むことを意味する。かかる調節配列は、例えば、Goeddel、「Gene Expression Technology:Methods in Enzymology」、第185巻、Academic Press(San Diego,Calif.)(1990年)において記載されている(その全体が参照により本明細書中に援用される)。調節配列は、多種の宿主細胞内でのヌクレオチド配列の構成的発現を誘導するものと、特定の宿主細胞内に限ってヌクレオチド配列の発現を誘導するものを含む。発現ベクターの設計が、形質転換されるべき宿主細胞の選択、所望されるタンパク質の発現のレベルなどの要素に依存しうることは、当業者によって理解されるであろう。本発明の発現ベクターは、宿主細胞内に導入されることで、本明細書中に記載される核酸によってコードされる、融合タンパク質または融合ペプチドを含むタンパク質またはペプチドを生成することが可能である。
【0044】
本発明の組換え発現ベクターは、原核または真核細胞内での標的遺伝子の発現のために設計してもよい。例えば、標的遺伝子または断片は、大腸菌(E.coli)、昆虫細胞(バキュロウイルス発現ベクターを使用)、酵母細胞または哺乳類細胞などの細菌細胞内で発現させてもよい。好適な宿主細胞は、Goeddel、「Gene Expression Technology:Methods in Enzymology」、第185巻、Academic Press(San Diego,Calif.)(1990年)(その全体が参照により本明細書中に援用される)においてさらに考察されている。あるいは、組換え発現ベクターは、インビトロで転写または翻訳してもよい。
【0045】
標的遺伝子はまた、哺乳類発現ベクターを使用して、哺乳類細胞内で発現させてもよい。哺乳類発現ベクターの例として、pCDM8(Seed B.、Nature、329:840頁(1987年))およびpMT2PC(Kaufmanら、EMBO J.、6:187頁(1987年))が挙げられる。発現ベクターの制御機能は、哺乳類細胞内で使用される場合、ウイルス調節因子によって提供されることが多い。例えば、一般に使用されるプロモーターは、ポリオーマ、アデノウイルス2型、サイトメガロウイルスおよびサルウイルス40に由来する。原核および真核細胞の双方における他の好適な発現系については、Sambrook J.、Fritsh E.F.およびManiatis T.「Molecular Cloning:A Laboratory Manual」、第2版、Cold Spring Harbor Laboratory、Cold Spring Harbor Laboratory Press(Cold Spring Harbor,N.Y.)、1989年の第16章および第17章を参照のこと。
【0046】
宿主細胞は、任意の原核または真核細胞であってもよい。例えば、宿主細胞は、大腸菌(E.coli)などの細菌細胞、昆虫細胞、酵母、アフリカツメガエル細胞、または哺乳類細胞(例えば、チャイニーズハムスター卵巣細胞(CHO)、アフリカミドリザル腎細胞(COS)、またはヒト胎児細胞(293T))であってもよい。他の好適な宿主細胞は、当業者に公知である。本発明の一態様では、宿主細胞は、本明細書中に記載のトランスジェニック非ヒト動物に由来する。
【0047】
ベクターDNAは、従来の形質転換または形質移入技術を介して、原核または真核細胞に導入してもよい。本明細書で使用される用語「形質転換」および「形質移入」は、リン酸カルシウムまたは塩化カルシウム共沈、DEAE−デキストラン媒介性形質移入、リポフェクション、またはエレクトロポレーションを含む、外来核酸(例えばDNA)を宿主細胞に導入するための種々の当該技術分野で公知の技術を示すように意図されている。宿主細胞を形質転換または形質移入するための好適な方法については、Sambrookら(「Molecular Cloning:A Laboratory Manual」、第2版、Cold Spring Harbor Laboratory、Cold Spring Harbor Laboratory Press(Cold Spring Harbor,N.Y.)、1989年)および他の実験マニュアルにおいて見出すことが可能である。
【0048】
トランスジェニック非ヒト動物を使用する方法
本発明は、エネルギー代謝に関連する疾患、例えば肥満、代謝症候群、脂質異常症、インスリン耐性症候群、2型糖尿病、拒食症および悪液質に対して作用する(すなわち調節する、阻害する、低下させる、予防するまたは改善する)調節因子、すなわち候補もしくは試験化合物または作用物質(例えばペプチド、環状ペプチド、ペプチドミメティック、小分子、小有機分子、または他の薬剤)を同定するための方法(本明細書中で「スクリーニングアッセイ」とも称される)を提供する。本明細書で使用される「調節する(modulating)」または「調節因子(modulator)」または「調節する(modulate)」は、GPR101受容体に対して作動または拮抗させることを示す。特に、本発明のトランスジェニック非ヒト動物を使用し、エネルギー代謝に関連する疾患の処置または予防にとって有効な化合物または組成物を同定することが可能である。化合物または組成物は、試験化合物または組成物を本発明のトランスジェニック非ヒト動物に投与するか、あるいは試験化合物または組成物を、トランスジェニック非ヒト動物に由来する器官、組織(例えば骨格筋)または細胞(例えば神経細胞または筋肉細胞)と接触させることにより、同定することが可能である。トランスジェニック非ヒト動物、器官、組織または細胞におけるエネルギー代謝に対する試験化合物または組成物の効果が評価される。例えば、候補作用剤は、トランスジェニック非ヒト動物において評価してもよい。エネルギー代謝を改変する試験化合物または組成物は、エネルギー代謝に関連する疾患の処置または予防にとって有効でありうる。
【0049】
さらに、本発明は、上記のスクリーニングアッセイによって同定される新規作用剤に関する。したがって、本明細書中に記載の適切な動物モデルにおいて、本明細書中に記載のように同定される試験化合物または組成物をさらに使用することは、本発明の範囲内に含まれる。例えば、本明細書中に記載のように同定される試験化合物または組成物は、動物モデル(例えばトランスジェニックGPR101ノックアウト非ヒト動物)において使用し、かかる試験化合物または組成物での処置の有効性、毒性、または副作用を判定することが可能である。あるいは、本明細書中に記載のように同定される試験化合物または組成物は、動物モデルにおいて使用し、かかる試験化合物または組成物の作用機序を判定することが可能である。
【0050】
試験化合物は、医薬的に許容できる非毒性賦形剤またはキャリアとの混合によって薬学的組成物に調合し、任意の投与経路によって本発明のトランスジェニック非ヒト動物に投与してもよい。例えば、皮下、筋肉内、血管内、皮内、鼻腔内、吸入、髄腔内、または腹腔内投与などの非経口経路、および舌下、経口、または直腸投与などの経腸経路を使用してもよい。
【0051】
試験化合物または組成物は、例えば試験化合物または組成物を野生型マウスおよびGPR101ノックアウトマウスに投与することにより、GPR101受容体に対する生理作用について試験してもよい。試験化合物または組成物が野生型マウスにおいて効果をもたらすが、GPR101ノックアウトマウスにおいてもたらさない場合、その試験化合物または組成物は、GPR101の活性を変化させることによって効果をもたらすものである。試験化合物または組成物が野生型マウスにおいて体重の減少、体脂肪の減少、および/または食物摂取の低下をもたらすが、GPR101ノックアウトマウスにおいてもたらさない場合、その試験化合物または組成物はGPR101の活性化因子である可能性が高い。それに対し、試験化合物または組成物が体重の増加、体脂肪の増加、および/または食物摂取の増加をもたらす場合、その試験化合物または組成物はGPR101の阻害剤である可能性が高い。試験化合物または組成物が野生型マウスとGPR101ノックアウトマウスの双方において効果をもたらす場合、その試験化合物または組成物は、GRP101の活性を変化させることによってその効果を発揮するものではない。
【0052】
試験化合物または組成物の特異性は、GPR101ノックアウトマウスにおいて試験してもよい。例えば、野生型マウスにおいて見られる効果がGPR101ノックアウトマウスにおいても見られる場合、その試験化合物または組成物はGPR101に対して特異的でない。
【0053】
GPR101ノックアウトマウスを使用し、試験化合物または組成物によってもたらされる毒性が、GPR101の活性の改変によるもの(的確な毒性)か、または他の何らかの機序の改変によるもの(的外れの毒性)かを判定してもよい。例えば、試験化合物または組成物が野生型マウスにおいて毒性または望ましくない効果をもたらすが、GPR101ノックアウトマウスにおいてもたらさない場合、その毒性効果は、GPR101の活性の改変による副産物(的確な効果)である。試験化合物または組成物が野生型マウスとGPR101ノックアウトマウスの双方において毒性または望ましくない効果をもたらす場合、その試験化合物または組成物の毒性効果は、GPR101の活性の変化に何も関連性がない(的外れの効果)。
【0054】
GPR101の構成的に活性なマウスは、例えばGPR101の活性を低下させかねない試験化合物または組成物を試験するのに特に有用でありうる。かかる試験化合物または組成物であれば、拒食症または悪液質を有する患者における食物摂取を刺激するのに有用でありうる。これらのマウスにおいて受容体が活性化されることから、それらはGPR101の活性の潜在的な阻害剤に対して感受性を示すはずである。
【0055】
本発明の例示的な実施形態の説明は以下の通りである。
【実施例】
【0056】
実験手順
GPR101ノックアウトマウスの作製
胚幹(ES)細胞における遺伝子標的化を用いて、GPR101欠損マウスを作製した。置換標的化コンストラクトを、約100kb上流への、および約40kb下流へのGPR101配列を有する細菌人工染色体(BAC)ゲノムクローンを使用して調製した。BACクローンを、開始コドンを含むコードエクソンに及ぶGPR101配列の2216bp部分(NCBI登録番号NC_000086)が除去され、Ires−Cre−Frt−カナマイシン−Frt(Ires−Cre FKF)カセットと置換されるように設計した。GPR101におけるゲノムDNA配列は、NC_000086内にあり、アセンブリ上の54756894から54749845の位置に由来する。欠失の設計に使用されるプライマーは、(配列番号1)HD73フォワード5’−TCC TCT GCA AGG CAC TAA CCC TAG CCA CAT GTT TCT CTC GTC CTC AAT CTA GTG ATG TAA TTC CGC CCC TCT CCC T−3’および(配列番号2)HD74リバース5’−CCT AGC TCC TCA TTT CAG GCT TGC CCT TTT CTG GAT CCC TTT TGA AGA CCT AAA CAA AAT ATT AAC GCT TAC A−3であった。欠失を有するBACの11.4kb部分を、Zeocinを有するpCR−Bluntに挿入した。次いで、この標的化プラスミドを線状化し、胚幹細胞にエレクトロポレートした。標的化クローンを、標的化遺伝子座の3’および5’末端に及ぶプライマーを使用するPCR分析によって同定した(図1)。標的化ESクローンから拡張された細胞をC57BL6胚盤胞に注入し、生殖細胞系を伝達するキメラ動物を得て、次いでFLPe−リコンビナーゼマウスと交配させた。GPR101がX染色体上に位置することから、得られたヘテロ接合性の子らを交配させ、野生型(+/+)およびホモ接合性(−/−)試験対象を作製した。すべての試験を雄マウスにおいて実施した。3つのプライマーによる多重PCR法を、図1に示される遺伝子型マウスに対して用いた。以下は、図1からのプライマー配列である。
(配列番号3)HD55 TTC TTT GCT CCC TCT TCA TTC TCA
(配列番号4)HD81 CGC ATC GCC TTC TAT CG
(配列番号5)HD82 ACC TAC TTC ATG TTT ATTT ACG
(配列番号6)HD84 CGT CAA GAA GGC GAT AGA
(配列番号7)HD142 TGAGACCCCCAAGAATTAGAAAAA
(配列番号8)HD143 TTGGCGAGAGGGGAAAGAC
(配列番号9)HD144 GGGGCCACGAGAGCAACCT
【0057】
GPR101が構成的に活性なマウスの作製
GPCRの特定の残基の突然変異の結果、オープン構造ひいては構成的活性(リガンドの不在下での活性)を有する受容体が得られる。受容体の潜在的な構成的活性に関与することになるGPR101内の残基を同定するため、インビトロ試験を実施した。マウスGPR101をコードする完全長cDNAを、マウス脳RNAから増幅し、それを発現プラスミドpCDNA3.1に挿入した。部位特異的突然変異誘発を用いて、活性の増大につながるGPR101の膜貫通ループにおける突然変異を同定したが、ここでは標的としてGPR101タンパク質のアラニン397を同定し、リジンに突然変異される場合、GPR101の活性における2倍の増大をもたらす(データは示さず)。単一のアミノ酸置換、すなわちA397→K397を伴う新規マウスモデルを作製した。
【0058】
A397→K397を伴う構成的に活性化型のGPR101を生成するための標的化コンストラクトを、リコンビニアリング(recombineering)を含む標準技術を用いて作成した。galK陽性および対抗選択のスキームを用いて、GPR101のBACにおいて点突然変異を生じさせた(Warming S.ら、「Simple and Highly Efficient BAC Recombineering Using galK Selection」、Nucleic Acids Research、33:1−12頁(2005年)(その全体が参照により本明細書中に援用される))。次いで、点突然変異を有するマウスを作製するため、修飾BACをプラスミドに挿入し、胚盤胞に注入した。
【0059】
結果
GPR101ノックアウトマウスにおける体重
体重調節におけるGPR101の機能的重要性を判定するため、GPR101ノックアウトマウスを作製した。GPR101ノックアウトマウスに標準の齧歯類用食餌を与え、3週齢以降の雄GPR101ノックアウトマウス(n=8)の体重を、野生型同腹子(n=12)のそれと比較した。
【0060】
図2に見られるように、雄GPR101ノックアウトマウスは、野生型同腹子対照と比較して体重増加傾向で推移した。17週齢のGPR101ノックアウトマウスは、体重において、野生型マウスに見られる場合よりも約10%の増加を有した。これらの結果は、中枢神経系におけるGPR101が正常な体重恒常性にとって必要とされることを示す。
【0061】
GPR101ノックアウトマウスにおける食物摂取
全食物摂取を、GPR101ノックアウトマウスにおいて評価した。GPR101ノックアウトマウスに標準の齧歯類用食餌を与えた。6週齢目、食物摂取を24時間記録した(図3A)。結果は、GPR101ノックアウトマウスにおける24時間内での食物摂取が、その野生型同腹子に対して増加したことを示す(5.1±0.58、GPR101ノックアウト対3.52±0.58野生型、n=6〜7/群)。8週齢目、全食物摂取を包括的な実験動物モニタリングシステムにおいて記録し、GPR101ノックアウトマウスによって消費される全食物の、その野生型対照に対する有意な増加が見られた(図3B)。結果は、GPR101が食餌サイズの調節における役割を果たすことを示唆する。
【0062】
GPR101ノックアウトマウスにおける食餌パターン
上で考察した24時間の食物摂取における増加に加え、8週齢のGPR101ノックアウトマウスはまた、図5に示されるように、食物消費の全体パターンにおける変化を示す。GPR101ノックアウトマウスは、図4および5に示されるように、野生型対照に対して食餌サイズにおける変化を示した。GPR101ノックアウトマウスは、暗期中により大量の食餌を消費した。
【0063】
データは、GPR101ノックアウトマウスが体重および食物摂取の増加を有し、それによりGPR101ノックアウトマウスが代謝性疾患の処置に使用可能なGPR101のアゴニストおよびアンタゴニストの同定において有用でありうることを示す。
【0064】
GPR101ノックアウトマウスにおける高脂肪食誘発性肥満
野生型およびGPR101遺伝子ノックアウト雄マウスを、離乳(3週齢)時に、食餌(Forumulab、食餌#5008)(WTにおいてn=10およびGPR101−KOにおいてn=8)または高脂肪食(カロリーの45%が脂肪由来、Research Diets、D12451)(WTにおいてn=7およびGPR101−KOにおいてn=7)を与え、体重を毎週モニタリングした。先に認められたように、食餌を与えたGPR101KOマウスの体重は、野生型対照の場合より重かった(図6)。注目すべきことに、野生型とGPR101KOマウスの間での体重における差異は、マウスに高脂肪食を与える場合に大きくなる(図7)。食餌を与えたマウスの身体組成物は、12週齢時でのEcho−MRIによる評価によると、体重の増加が脂肪量の増加に起因することを示す(図8)。これらの実験結果は、GPR101の遺伝子ノックアウトが肥満を誘発することを示す。さらに、GPR101の遺伝子ノックアウトは、高脂肪食誘発性肥満に対する感受性の増大の原因になる。まとめると、これらの実験結果は、GPR101アゴニストが、抗肥満効果を有する可能性が高く、故に肥満の処置にとって有用でありうることを示す。
【0065】
本明細書中に引用されるすべての特許、公開された出願および参考文献の教示内容は、それら全体が参照により援用される。
【0066】
本発明は、特にその例示的な実施形態を参照して示され、説明されている一方、形態におけるさまざまな変化および詳細化が、添付の特許請求の範囲によって包含される本発明の範囲から逸脱することなく、本明細書中でなされうることは当業者によって理解されるであろう。
【特許請求の範囲】
【請求項1】
ゲノムが内因性GPR101遺伝子内に破壊を含むトランスジェニックノックアウト非ヒト哺乳動物。
【請求項2】
前記破壊が、胚幹細胞内で、DNA標的化コンストラクトとの相同組換えによって前記ゲノムに導入されている、請求項1に記載のトランスジェニック非ヒト哺乳動物。
【請求項3】
前記GPR101遺伝子の破壊の結果、前記トランスジェニック非ヒト哺乳動物は、検出可能なレベルのGPR101を生成することができない、請求項1に記載のトランスジェニック非ヒト哺乳動物。
【請求項4】
前記哺乳動物はマウスである、請求項1に記載のトランスジェニック非ヒト哺乳動物。
【請求項5】
ゲノムが、前記内因性GPR101遺伝子内に破壊を含む、トランスジェニックノックアウト非ヒト哺乳動物に由来する単離細胞。
【請求項6】
前記破壊が、胚幹細胞内で、DNA標的化コンストラクトとの相同組換えによって前記ゲノムに導入されている、請求項5に記載の単離細胞。
【請求項7】
前記GPR101遺伝子の破壊の結果、前記マウスは、検出可能なレベルのGPR101を生成することができない、請求項5に記載の単離細胞。
【請求項8】
前記哺乳動物はマウスである、請求項5に記載の単離細胞。
【請求項9】
ゲノムが、前記内因性GPR101遺伝子内に破壊を含む、ノックアウト非ヒト哺乳動物を作製する方法。
【請求項10】
前記破壊が、胚幹細胞内で、DNA標的化コンストラクトとの相同組換えによって前記ゲノムに導入されている、請求項9に記載の方法。
【請求項11】
前記GPR101遺伝子の破壊の結果、前記マウスは、検出可能なレベルのGPR101を生成することができない、請求項9に記載の方法。
【請求項12】
前記哺乳動物はマウスである、請求項9に記載の方法。
【請求項13】
ノックアウト非ヒト哺乳動物における体重および食物摂取に対する調節能について候補作用剤をスクリーニングするための方法であって、
(a)ゲノムが前記内因性GPR101遺伝子内で破壊を含むトランスジェニックノックアウト非ヒト哺乳動物を提供するステップと、
(b)前記ノックアウト非ヒト哺乳動物に候補作用剤を投与するステップと、
(c)トランスジェニックノックアウト非ヒト哺乳動物の体重および食物摂取を野生型対照の体重および食物摂取と比較するステップであって、ここで効果における差異は、GPR101の活性の改変によって体重および食物摂取を調節する作用剤を示唆するステップと、
を含む、方法。
【請求項14】
前記破壊は、ES細胞内で、DNA標的化コンストラクトとの相同組換えによって前記ゲノムに導入されている、請求項13に記載の方法。
【請求項15】
前記GPR101遺伝子の前記破壊の結果、前記マウスは、検出可能なレベルのGPR101を生成することができない、請求項13に記載の方法。
【請求項16】
前記哺乳動物はマウスである、請求項13に記載の方法。
【請求項17】
候補作用剤を、ノックアウト非ヒト哺乳動物における体重および食物摂取に対する調節能についてスクリーニングするための方法であって、
(a)ゲノムが内因性GPR101遺伝子内での破壊を含むトランスジェニック非ヒト哺乳動物を提供するステップと、
(b)前記トランスジェニック非ヒト哺乳動物に候補作用剤を投与するステップと、
(c)前記トランスジェニック非ヒト哺乳動物に対する前記候補作用剤の効果を評価するステップと、
を含む、方法。
【請求項18】
前記破壊が、ES細胞内で、DNA標的化コンストラクトとの相同組換えによって前記ゲノムに導入されている、請求項17に記載の方法。
【請求項19】
前記GPR101遺伝子の前記破壊の結果、前記マウスは、検出可能なレベルのGPR101を生成することができない、請求項17に記載の方法。
【請求項20】
前記哺乳動物はマウスである、請求項17に記載の方法。
【請求項21】
ゲノムが構成的に活性な内因性GPR101遺伝子を含むトランスジェニック非ヒト哺乳動物。
【請求項22】
前記構成的に活性なGPR101遺伝子が、単一のアミノ酸置換によって前記ゲノムに導入されている、請求項21に記載のトランスジェニック非ヒト哺乳動物。
【請求項23】
前記アミノ酸置換が、ES細胞内で、DNA標的化コンストラクトとの相同組換えによって前記ゲノムに導入されている、請求項22に記載のトランスジェニック非ヒト哺乳動物。
【請求項24】
前記哺乳動物はマウスである、請求項21に記載のトランスジェニック非ヒト哺乳動物。
【請求項25】
ゲノムが構成的に活性な内因性GPR101遺伝子を含むトランスジェニック非ヒト哺乳動物に由来する単離細胞。
【請求項26】
前記構成的に活性なGPR101遺伝子が、単一のアミノ酸置換によって前記ゲノムに導入されている、請求項25に記載の単離細胞。
【請求項27】
前記アミノ酸置換は、ES細胞内で、DNA標的化コンストラクトとの相同組換えによって前記ゲノムに導入されている、請求項26に記載の単離細胞。
【請求項28】
前記哺乳動物はマウスである、請求項25に記載の単離細胞。
【請求項29】
ゲノムが構成的に活性な内因性GPR101遺伝子を含むトランスジェニック非ヒト哺乳動物を作製するための方法。
【請求項30】
前記構成的に活性なGPR101遺伝子は、単一のアミノ酸置換によって前記ゲノムに導入されている、請求項29に記載の方法。
【請求項31】
前記アミノ酸置換が、ES細胞内で、DNA標的化コンストラクトとの相同組換えによって前記ゲノムに導入されている、請求項30に記載の方法。
【請求項32】
前記哺乳動物はマウスである、請求項29に記載の方法。
【請求項33】
候補作用剤を、ゲノムが構成的に活性な内因性GPR101遺伝子を含むトランスジェニック非ヒト哺乳動物における体重および食物摂取に対する調節能についてスクリーニングするための方法であって、
(a)ゲノムが構成的に活性な内因性GPR101遺伝子を含むトランスジェニック非ヒト哺乳動物を提供するステップと、
(b)前記トランスジェニック非ヒト哺乳動物に候補作用剤を投与するステップと、
(c)前記トランスジェニック非ヒト哺乳動物に対する前記候補作用剤の効果を評価するステップと、
を含む、方法。
【請求項34】
前記構成的に活性なGPR101遺伝子は、単一のアミノ酸置換によって前記ゲノムに導入されている、請求項33に記載の方法。
【請求項35】
前記アミノ酸置換は、ES細胞内で、DNA標的化コンストラクトとの相同組換えによって前記ゲノムに導入されている、請求項34に記載の方法。
【請求項36】
前記哺乳動物はマウスである、請求項33に記載の方法。
【請求項1】
ゲノムが内因性GPR101遺伝子内に破壊を含むトランスジェニックノックアウト非ヒト哺乳動物。
【請求項2】
前記破壊が、胚幹細胞内で、DNA標的化コンストラクトとの相同組換えによって前記ゲノムに導入されている、請求項1に記載のトランスジェニック非ヒト哺乳動物。
【請求項3】
前記GPR101遺伝子の破壊の結果、前記トランスジェニック非ヒト哺乳動物は、検出可能なレベルのGPR101を生成することができない、請求項1に記載のトランスジェニック非ヒト哺乳動物。
【請求項4】
前記哺乳動物はマウスである、請求項1に記載のトランスジェニック非ヒト哺乳動物。
【請求項5】
ゲノムが、前記内因性GPR101遺伝子内に破壊を含む、トランスジェニックノックアウト非ヒト哺乳動物に由来する単離細胞。
【請求項6】
前記破壊が、胚幹細胞内で、DNA標的化コンストラクトとの相同組換えによって前記ゲノムに導入されている、請求項5に記載の単離細胞。
【請求項7】
前記GPR101遺伝子の破壊の結果、前記マウスは、検出可能なレベルのGPR101を生成することができない、請求項5に記載の単離細胞。
【請求項8】
前記哺乳動物はマウスである、請求項5に記載の単離細胞。
【請求項9】
ゲノムが、前記内因性GPR101遺伝子内に破壊を含む、ノックアウト非ヒト哺乳動物を作製する方法。
【請求項10】
前記破壊が、胚幹細胞内で、DNA標的化コンストラクトとの相同組換えによって前記ゲノムに導入されている、請求項9に記載の方法。
【請求項11】
前記GPR101遺伝子の破壊の結果、前記マウスは、検出可能なレベルのGPR101を生成することができない、請求項9に記載の方法。
【請求項12】
前記哺乳動物はマウスである、請求項9に記載の方法。
【請求項13】
ノックアウト非ヒト哺乳動物における体重および食物摂取に対する調節能について候補作用剤をスクリーニングするための方法であって、
(a)ゲノムが前記内因性GPR101遺伝子内で破壊を含むトランスジェニックノックアウト非ヒト哺乳動物を提供するステップと、
(b)前記ノックアウト非ヒト哺乳動物に候補作用剤を投与するステップと、
(c)トランスジェニックノックアウト非ヒト哺乳動物の体重および食物摂取を野生型対照の体重および食物摂取と比較するステップであって、ここで効果における差異は、GPR101の活性の改変によって体重および食物摂取を調節する作用剤を示唆するステップと、
を含む、方法。
【請求項14】
前記破壊は、ES細胞内で、DNA標的化コンストラクトとの相同組換えによって前記ゲノムに導入されている、請求項13に記載の方法。
【請求項15】
前記GPR101遺伝子の前記破壊の結果、前記マウスは、検出可能なレベルのGPR101を生成することができない、請求項13に記載の方法。
【請求項16】
前記哺乳動物はマウスである、請求項13に記載の方法。
【請求項17】
候補作用剤を、ノックアウト非ヒト哺乳動物における体重および食物摂取に対する調節能についてスクリーニングするための方法であって、
(a)ゲノムが内因性GPR101遺伝子内での破壊を含むトランスジェニック非ヒト哺乳動物を提供するステップと、
(b)前記トランスジェニック非ヒト哺乳動物に候補作用剤を投与するステップと、
(c)前記トランスジェニック非ヒト哺乳動物に対する前記候補作用剤の効果を評価するステップと、
を含む、方法。
【請求項18】
前記破壊が、ES細胞内で、DNA標的化コンストラクトとの相同組換えによって前記ゲノムに導入されている、請求項17に記載の方法。
【請求項19】
前記GPR101遺伝子の前記破壊の結果、前記マウスは、検出可能なレベルのGPR101を生成することができない、請求項17に記載の方法。
【請求項20】
前記哺乳動物はマウスである、請求項17に記載の方法。
【請求項21】
ゲノムが構成的に活性な内因性GPR101遺伝子を含むトランスジェニック非ヒト哺乳動物。
【請求項22】
前記構成的に活性なGPR101遺伝子が、単一のアミノ酸置換によって前記ゲノムに導入されている、請求項21に記載のトランスジェニック非ヒト哺乳動物。
【請求項23】
前記アミノ酸置換が、ES細胞内で、DNA標的化コンストラクトとの相同組換えによって前記ゲノムに導入されている、請求項22に記載のトランスジェニック非ヒト哺乳動物。
【請求項24】
前記哺乳動物はマウスである、請求項21に記載のトランスジェニック非ヒト哺乳動物。
【請求項25】
ゲノムが構成的に活性な内因性GPR101遺伝子を含むトランスジェニック非ヒト哺乳動物に由来する単離細胞。
【請求項26】
前記構成的に活性なGPR101遺伝子が、単一のアミノ酸置換によって前記ゲノムに導入されている、請求項25に記載の単離細胞。
【請求項27】
前記アミノ酸置換は、ES細胞内で、DNA標的化コンストラクトとの相同組換えによって前記ゲノムに導入されている、請求項26に記載の単離細胞。
【請求項28】
前記哺乳動物はマウスである、請求項25に記載の単離細胞。
【請求項29】
ゲノムが構成的に活性な内因性GPR101遺伝子を含むトランスジェニック非ヒト哺乳動物を作製するための方法。
【請求項30】
前記構成的に活性なGPR101遺伝子は、単一のアミノ酸置換によって前記ゲノムに導入されている、請求項29に記載の方法。
【請求項31】
前記アミノ酸置換が、ES細胞内で、DNA標的化コンストラクトとの相同組換えによって前記ゲノムに導入されている、請求項30に記載の方法。
【請求項32】
前記哺乳動物はマウスである、請求項29に記載の方法。
【請求項33】
候補作用剤を、ゲノムが構成的に活性な内因性GPR101遺伝子を含むトランスジェニック非ヒト哺乳動物における体重および食物摂取に対する調節能についてスクリーニングするための方法であって、
(a)ゲノムが構成的に活性な内因性GPR101遺伝子を含むトランスジェニック非ヒト哺乳動物を提供するステップと、
(b)前記トランスジェニック非ヒト哺乳動物に候補作用剤を投与するステップと、
(c)前記トランスジェニック非ヒト哺乳動物に対する前記候補作用剤の効果を評価するステップと、
を含む、方法。
【請求項34】
前記構成的に活性なGPR101遺伝子は、単一のアミノ酸置換によって前記ゲノムに導入されている、請求項33に記載の方法。
【請求項35】
前記アミノ酸置換は、ES細胞内で、DNA標的化コンストラクトとの相同組換えによって前記ゲノムに導入されている、請求項34に記載の方法。
【請求項36】
前記哺乳動物はマウスである、請求項33に記載の方法。
【図1】


【図2】
【図3A】
【図3B】
【図4】
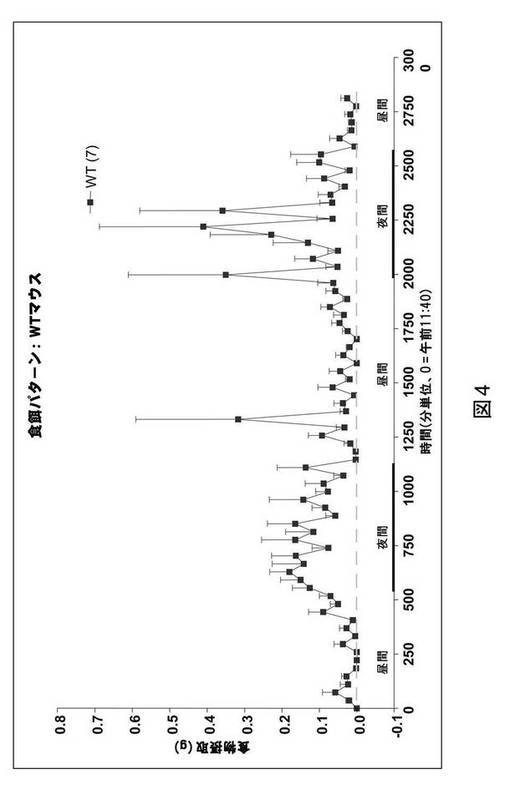
【図5】
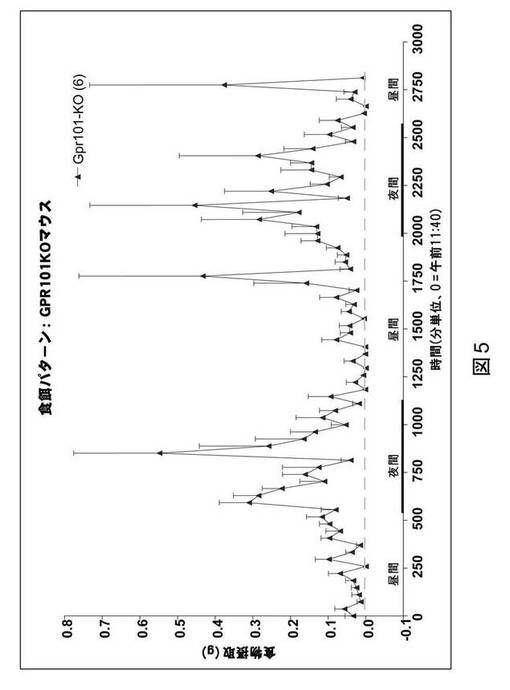
【図6】
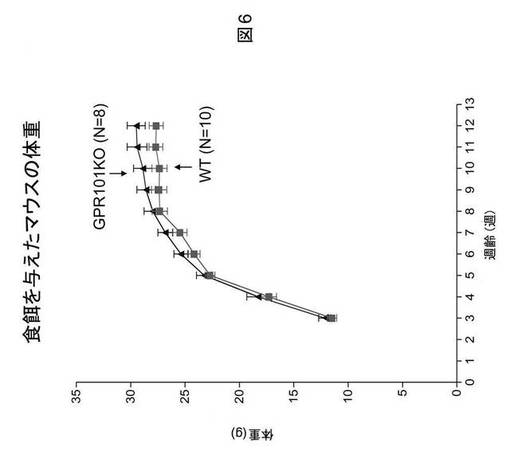
【図7】
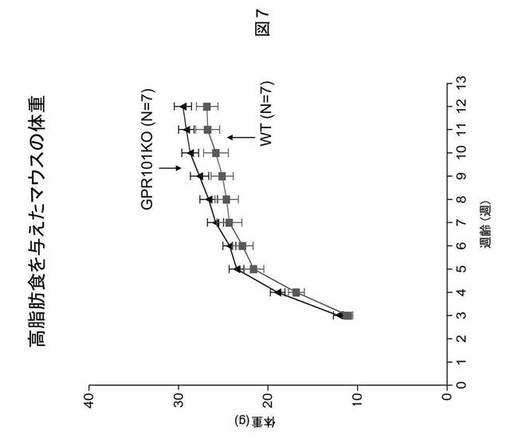
【図8】
【図2】
【図3A】
【図3B】
【図4】
【図5】
【図6】
【図7】
【図8】
【公表番号】特表2011−520465(P2011−520465A)
【公表日】平成23年7月21日(2011.7.21)
【国際特許分類】
【出願番号】特願2011−510505(P2011−510505)
【出願日】平成21年5月18日(2009.5.18)
【国際出願番号】PCT/US2009/003094
【国際公開番号】WO2009/142724
【国際公開日】平成21年11月26日(2009.11.26)
【出願人】(301033259)ベス・イスラエル・ディーコネス・メディカル・センター,インコーポレイテッド (11)
【Fターム(参考)】
【公表日】平成23年7月21日(2011.7.21)
【国際特許分類】
【出願日】平成21年5月18日(2009.5.18)
【国際出願番号】PCT/US2009/003094
【国際公開番号】WO2009/142724
【国際公開日】平成21年11月26日(2009.11.26)
【出願人】(301033259)ベス・イスラエル・ディーコネス・メディカル・センター,インコーポレイテッド (11)
【Fターム(参考)】
[ Back to top ]