
新規IL−8様ポリペプチド
本発明は、新規IL−8様/TMEM9ポリペプチドをコードするヒトゲノムにおけるオープンリーディングフレーム(ORF)、及びそれに関連する試薬、例えば前記ポリペプチドの変異体、突然変異体及びフラグメント、並びにそれらに対するリガンド及びアンタゴニストを開示する。本発明は、これらの分子の同定方法、生成方法、それらを含む医薬組成物の調製方法、並びに疾患の診断、予防、及び処置におけるそれらの使用方法、を開示する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、新規ポリペプチド、更に具体的にはIL−8様/TMEM9ポリペプチドをコードするものとしてヒトゲノムにおいて同定された核酸配列に関する。
【0002】
本明細書で引用する全ての刊行物、特許及び特許出願は、引用により全体が組み入れられる。
【背景技術】
【0003】
既に多くの新規ポリペプチドが、同じファミリーの公知のポリペプチドに対する厳密な相同性の基準を適用することにより同定されている。しかしIL−8様ポリペプチド(及び任意な他のタンパク質ファミリー)についてのヒトゲノム中のポリペプチド-コード配列中の実際の内容は依然不明であるので、ヒトゲノムに存在するオープンリーディングフレーム(ORF、すなわち、ヌクレオチドの連続的にコードしているトリプレットを含み、終止コドンによって中断されておらず、且つ潜在的にポリペプチドに翻訳可能なDNA配列)全体に、別の、そしてあまり厳密でない相同性/構造の基準を適用することにより、IL−8様ポリペプチド活性を有するポリペプチドをコードするDNA配列を同定することの可能性は尚も存在している。
【0004】
哺乳類の免疫応答は、一連の複雑なネットワーク様の相互作用に基づいており、これらには、細胞活動(移動、増殖、分化等)を調節することができる細胞成分(例えばリンパ球又は顆粒球)及び可溶性タンパク質が関与している。従って、ヒトの障害、特に免疫系に関連するものの診断、予防、及び治療における重大な進歩を提供する目的での、細胞調節因子の単離及び特徴づけに大きな関心が存在している。
【0005】
ケモカインは、それらが細胞の方向付けられた遊走及び活性化に関与しているため、これらの可溶性タンパク質の中に含まれる。この小さい(70〜130アミノ酸)分泌型のヘパリン結合炎症誘発性タンパク質のスーパーファミリーは、これらの細胞の補充を必要とする、血液から組織への白血球の血管外遊走における役割が特に知られている(Fernandez EJ and Lolis E, Annu. Rev. Pharmacol. Toxicol.. 42:469-499. 2002)。
【0006】
ケモカインは、それらが全て、保存されたシステインが分子内結合を形成している中心領域を含んでいるため、機能的に関連しているだけでなく、構造的に関連している。特に、成熟ポリペプチドのN末端配列における保存されたシステイン残基の数及び位置は、一般的に認識されているケモカイン分類の基礎的な基準であり、これらは孤立した又は隣接するシステイン残基、あるいは1〜3個のアミノ酸で分離されたシステイン残基を含むケモカイン間で本質的に分類される。
【0007】
一連の膜受容体は、全て7回膜貫通Gタンパク質共役受容体であって、ケモカインに対して、それらの生物学的活性を標的細胞に発揮させるようにする結合パートナーである。ケモカインの生理学的作用は、同時発生の相互作用の複雑で且つ統合された系に起因する。異なる細胞は、それらの状態及び/又は型に従い、受容体の特定の組み合わせを提示することができる。更に、ケモカイン受容体は、しばしば重複しているリガンド特異性を有し、その結果、単一の受容体は、単一のケモカインが異なる受容体に結合することができるのと同様に、異なるケモカインにより高い親和性で結合することができる。
【0008】
通常ケモカインは、損傷、炎症、又は他の組織変化の部位で産生され、そしてパラ分泌又は自己分泌の様式でそれらの活性を発揮する。しかしながら、炎症及び免疫反応における細胞型特異的な遊走及び活性化はケモカイン単独での活性ではない。他の生理学的活性、例えば造血又は血管形成、及び病理学的症状、例えば転移、移植片拒絶、アルツハイマー病又はアテローム性動脈硬化症は、少なくとも、これらのタンパク質のうちの幾つかによって制御されているようである。これは、ケモカイン及び/又はそれらの受容体は、複数の動物モデル又は臨床試料においてかなりアップレギュレートされ、そして/あるいは活性化されていることが明らかとなっているためである(Haskell CA et al., Curr. Opin. Invest. Drugs, 3: 399-455,2002 ; Lucas AD and Greaves DR, Exp. Rev. Mol. Med. 2001 ; Frederick MJ and Clayman GL, Exp. Rev. Mol. Med. 2001; Godessart N and Kunkel SL, Curr. Opin. Immunol., 13: 670-675, 2001 ; Reape TJ and Groot PH, Atherosclerosis,147 : 213- 25,1999)。
【0009】
治療物質としてケモカインを用いることには潜在的な欠点(特に、凝集する傾向及び無差別な結合)があるが、ケモカインに対してアンタゴニスト的な特性を有する分子は、過剰なケモカイン活性に関連する障害における治療的介入の価値のある機会を提供すると広く考えられる。特異的なケモカイン及びそれらの受容体の阻害は、不所望な又は制御されない細胞過程、例えば補充又は活性化を防ぐ解決策とみなされている(Baggiolini M, J. Intern. Med., 250: 91-104,2001 ; Proudfoot A, et al. Immunol. Rev., 177: 246-256,2000 ; Rossi D and Zlotnik A, Annu. Rev. Immunol., 18: 217-42,2000)。
【0010】
現在利用可能なヒトゲノム及び生理学に対する技術及び情報も新規で且つ有用な治療分子及び治療標的を提供するかもしれない新規なケモカイン及び受容体を発見するために使用された。最初に、ケモカイン遺伝子は、ヒトゲノムのジーンリッチ領域の、第4及び第17染色体上に規則正しくマッピングされたが(Nomiyama H et al., Genes Immun, 2: 110-113,2001)、この文献は、それぞれ特徴的なパターンでリンパ様組織及び他の組織において発現しており、且つ伝統的な遺伝子クラスターと異なる染色体の座位にマッピングされている転写物のバイオインフォマティクス解析を利用することにより新規なケモカインを特徴付けるための異なるアプローチの多数の例を提供している。ケモカインは、通常リンパ様組織及び他の組織において発現しているが、新規ケモカインは特異的な発現パターンを有することがあり、そして伝統的な遺伝子クラスターと異なる染色体の座位にマッピングされうる(WO 02/70706; Wells TN. and Peitsch MC. Methods Mol. Biol., 138: 65-73,2000 ; Chantry DF et al., J Leukoc Biol, 64: 49-54, 1998; Rossi D et al., J. Immunol, 158: 1033-1036,1997)。
【0011】
インターロイキン−8(IL−8)は、好中球活性化ペプチド−1又はSCYB8とも称される、炎症性刺激に応じて複数の細胞型によって分泌される組織由来のペプチドである。
【0012】
インターロイキン−8は、13のヒトCXCケモカインから成るファミリーのうちの1つである。これらの小さい塩基性のヘパリン結合タンパク質は炎症誘発性であり、そして主に好中球の活性化及び末梢血から組織への好中球の遊走を媒介する。IL−8は、呼吸器合胞体ウイルス(RSV)によって引き起こされるウイルス性の下気道感染気管支炎の発病に関与している。この疾患は毎年主要な伝染病に関与しており、何千人もの乳児が入院治療を必要としている。高レベルのIL−8がRSV気管支炎を有する乳児の鼻汁及び気管の吸引物で見られ、そしてIL−8のレベルは疾患の重症度と関連しているようである(Smyth, R. L. et al., Arch. Dis. Child. 82 (Suppl. 1) : A4-A5, 2000)。
【0013】
IL−8は造血前駆細胞(HPC)の迅速な動員を誘導する。動員は、マウスにおいて、ベータ−2−インテグリンLfa1(CD11A)に対する中和抗体による前処理によって完全に防止されうる。尚、マウスHPCはLfa1を発現せず、これは動員が補助細胞集団を必要とすることを示唆している。
【0014】
Pruijt, J. F. Mらは(Proc. Nat. Acad. Sci. 99: 6228-6233,2002)、多形核細胞(PMN)がIL−8誘導型のHPC動員における主要な制御因子としての役割を果たすことを示した。PMNの役割は、単回投与の抗好中球抗体の投与により好中球減少性となったマウスにおいて研究された。完全な好中球減少症が最大3〜5日で観察され、但し、7日目にリバウンドして好中球が増大する。IL−8誘導型の動員能力は、好中球減少期の間に有意に低下し、PMNの循環と共に再び現れ、そして好中球期(neutrophilic phase)の間に比例して増大した。このデータは、HPCのIL−8誘導型の動員が循環しているPMNのin vivoでの活性化を必要とすることを示した。
【0015】
多数の新規ケモカインが既知のケモカインに対し厳しいホモロジー基準を適用することによって同定されている。しかしながら、ケモカインについてのヒトゲノムにおけるポリペプチドコード配列の実際の内容(及びあらゆるほかのタンパク質ファミリーに関してのもの)は未だに知られておらず、オープンリーディングフレーム(ORF、すなわち、ヌクレオチドの連続的にコードしているトリプレットを含み、終止コドンによって中断されておらず、且つ潜在的にポリペプチドに翻訳可能なDNA配列)の解析において別の基準を適用することにより、アミノ酸をコードするDNA配列を同定することの可能性は尚も存在している。
【0016】
IL−8用タンパク質が疾患において有する役割は現在研究中である。しかしながら、新規IL−8様タンパク質の同定は、これらのタンパク質が関与しているとある病状に至る基礎的な経路の理解を深め、且つ、これらの障害を処置するのにより有効な遺伝子又は薬物治療を開発するのにかなり重要であるのは明らかである。
【発明の開示】
【発明が解決しようとする課題】
【0017】
本発明は、新規IL−8様/TMEM9ポリペプチドをコードするヒトゲノムのオープンリーディングフレーム(ORF)の同定に基づいている。このポリペプチドは、本明細書ではSCS0010ポリペプチドと称する。
【課題を解決するための手段】
【0018】
従って、本発明は、配列番号2で示すアミノ酸配列を有する、単離されたSCS0010ポリペプチド、並びに、IL−8様/TMEM9ポリペプチドの活性を有するポリペプチドとしての、それらの成熟型、それらのヒスチジンタグ付き型、変異体、及びフラグメント、を提供する。本発明はまた、それらをコードする核酸、当該核酸を含むベクター、及びこれらのベクター又は核酸を含む細胞、並びに他の関連する試薬、例えば融合タンパク質、リガンド、及びアンタゴニスト、を含む。
【0019】
本発明は、これらの分子を同定し、且つ生成するための方法、それらを含む医薬組成物を調製するための方法、並びに、疾患の診断、予防及び処置においてそれらを使用するための方法、を提供する。
【0020】
本発明の詳細な説明
本発明の第一の側面に従い、IL−8様/TMEM9活性を有する単離されたポリペプチドであって:
a)配列番号2に記載のアミノ酸配列;
b)配列番号2に記載の配列のポリペプチドの成熟型(配列番号4又は配列番号7);
c)配列番号4に記載の配列のポリペプチドのヒスチジンタグ付き型(配列番号5)
d)配列番号2に記載のアミノ酸配列の変異体であって、選択した配列において特定されている任意のアミノ酸が非保存的に置換されており、但し、当該配列中の15%未満のアミノ酸残基がそのように変化している、変異体;
e) a)〜c)に示すアミノ酸配列の活性フラグメント、前駆体、塩、又は誘導体、
から成る群から選択されるポリペプチド、が提供される。
【0021】
本明細書に記載の新規ポリペプチドは、クエリー配列としてIL−8様タンパク質を用いて同定され、そして最終的なアノテーションは、アミノ酸配列のホモロジーに基づいて行った。新規ポリペプチドはまた、内在性膜貫通タンパク質9のスプライス変異体としても同定された(TMEM9;実施例を参照のこと)。
【0022】
ヒトゲノムにおける既知のORF配列を翻訳することによって得られるアミノ酸配列の全体が、この共通配列を用いて挑戦され、そしてポジティブなヒットは、予測される構造的且つ機能的な「サイン」(N末端のシグナル配列及びC末端のアルファヘリックス)の存在について更にスクリーニングされ、そして最終的に、配列の特徴を既知のIL−8様ポリペプチドと比較することによって選択される。従って、本発明の新規ポリペプチドはIL−8様活性及びTMEM9活性を有することが予想されうる。
【0023】
用語「活性な」及び「活性」は、本願において配列番号2に提示するアミノ酸配列のIL−8様/TMEM9ポリペプチドについて予測されるIL−8様/TMEM9の特性を意味する。IL−8及び関連タンパク質は、炎症細胞、線維芽細胞及びケラチノサイトを創傷部位に引き付ける走化性活性を有する;それらはマイトジェンとして働いて細胞増殖を刺激する;サイトカインは血管形成、すなわち新規血管が創傷内部に増殖することを刺激することができる;それらはECMの産生及び分解に対しての著しい作用を有する;それらはまた、隣接細胞による他のサイトカイン及び増殖因子の合成に影響を及ぼす。IL−8様ケモカインとして機能する能力は、70pg/ml程度の低さのIL−8濃度を検出することができるヒトIL−8ELISAのようなアッセイキット(IBL, Hamburg)を用いて測定されうる。TMEM9は実施例で説明する。
【0024】
第二の側面において、本発明は、本発明の第一の側面のポリペプチドをコードしている精製された核酸分子を提供する。好ましくは、精製された核酸分子は、配列番号1に記載の核酸配列(配列番号2に記載のアミノ酸配列のIL−8様/TMEM9ポリペプチドをコードしている)又は配列番号3に記載の核酸配列(前記ポリペプチドの成熟型であって、アミノ酸配列が配列番号4に記載のものをコードしている)又は配列番号6に記載の核酸配列(前記ポリペプチドの成熟型であって、アミノ酸配列が配列番号7に記載のものをコードしている)を有する。好ましい成熟ポリヌクレオチド配列は、配列番号4に記載の好ましい成熟ポリペプチド型をコードしている配列番号3である。
【0025】
第三の側面において、本発明は、高ストリンジェントな条件下で、本発明の第二の側面の核酸分子とハイブリダイズする、精製された核酸分子を提供する。
【0026】
第四の側面において、本発明は、本発明の第二又は第三の側面の核酸分子を含む、発現ベクターなどの、ベクターを提供する。
【0027】
第五の側面において、本発明は、本発明の第四の側面のベクターにより形質転換された宿主細胞を提供する。
【0028】
第六の側面において、本発明は、本発明の第一の側面のポリペプチドと特異的に結合し、且つ好ましくは本発明の第一の側面のポリペプチドのIL−8様/TMEM9活性を阻害するリガンドを提供する。本発明のポリペプチドに対するリガンドは、天然の又は修飾された基質、酵素、受容体、最大2000Da、好ましくは800Da又はそれ未満の小型の天然又は合成の有機分子のような小型有機分子、ペプチドミメティクス、無機分子、ペプチド、ポリペプチド、抗体、前述のものの構造的又は機能的ミメティクスを含む、様々な形であってよい。
【0029】
第七の側面において、本発明は、本発明の第一の側面のポリペプチドをコードしている天然の遺伝子の発現を変更するか、又は本発明の第一の側面のポリペプチドの活性を制御するのに有効である化合物を提供する。
【0030】
本発明の第七の側面の化合物は、遺伝子の発現のレベル又はポリペプチドの活性を増加する(アゴニスト作用)又は減少する(アンタゴニスト作用)のいずれかであってもよい。重要なことは、本発明のIL−8様/TMEM9ポリペプチドの機能の同定は、疾患の治療及び/又は診断に有効である化合物を同定することが可能であるスクリーニング法のデザインを可能にする。
【0031】
第八の側面において、本発明は、治療又は診断において使用するための、本発明の第一の側面のポリペプチド、又は本発明の第二もしくは第三の側面の核酸分子、又は本発明の第四の側面のベクター、又は本発明の第五の側面の宿主細胞、又は本発明の第六の側面のリガンド、又は本発明の第七の側面の化合物を提供する。これらの分子は、IL−8様ポリペプチドが関与している疾患及び症状、例えば、免疫障害、転移、移植片拒絶、血管形成、アルツハイマー病、アテローム性動脈硬化症、亜致死性の内毒素血症、敗血症ショック、羊膜腔の微生物感染、回帰熱のヤーリッシュ・ヘルクスハイマー反応、中枢神経系の感染病、急性膵炎、潰瘍性大腸炎、膿胸、溶血性尿毒症症候群、髄膜炎菌性の疾患、胃の感染症、腹膜炎、乾癬、関節リウマチ、敗血症、喘息及び糸球体腎炎、の予防及び処置のための薬物の製造においても使用されうる。
【0032】
第九の側面において、本発明は、患者の疾患を診断する方法であって、当該患者の組織において、本発明の第一の側面のポリペプチドをコードしている天然の遺伝子の発現レベル又は本発明の第一の側面のポリペプチドの活性を評価すること、及び当該発現のレベル又は活性を対照レベルと比較することを含む方法を提供し、ここで当該対照レベルとは異なるレベルは、疾患の指標である。このような方法は、in vitroで実行されることが好ましいであろう。同様の方法を用い、患者における疾患の治療的処置をモニタリングすることができ、ここで経時的に対照レベルへと向かうポリペプチド又は核酸分子の発現のレベル又は活性の変化は、疾患の回復の指標である。
【0033】
本発明の第一の側面のポリペプチドを検出する好ましい方法は:(a)本発明の第六の側面のリガンド、例えば抗体を、生物学的試料と、リガンド−ポリペプチド複合体を形成するのに適した条件下で接触する工程;及び(b)当該複合体を検出する工程、を含んで成る。
【0034】
本発明の第九の観点に従う多数の異なるそのような方法が存在するのは当業者にとって自明であり、例えば、短いプローブによる核酸ハイブリダイゼーション法、点変異解析、ポリメラーゼ連鎖反応(PCR)増幅、及び、抗体を用いて異常なタンパク質レベルを検出する方法である。同様の方法を短期又は長期的に用いて、疾患の治療的処置が患者においてモニタリングされることが可能となりうる。本発明はまた、これらの疾患診断方法に有用なキットも提供する。
【0035】
第十の側面において、IL−8様/TMEM9タンパク質として本発明の第一の側面のポリペプチドの使用を提供する。適当な使用には、細胞増殖、代謝又は分化の制御因子としての使用、受容体/リガンド対の一部としての使用及び上記リストから選択される生理学的又は病理学的症状の診断用マーカーとしての使用が含まれる。
【0036】
第十一の側面において、本発明は、本発明の第一の側面のポリペプチド、又は本発明の第二もしくは第三の側面の核酸分子、又は本発明の第四の側面のベクター、又は本発明の第五の側面の宿主細胞、又は本発明の第六の側面のリガンド、あるいは本発明の第七の側面の化合物を、医薬として許容される担体と組合わせて含んで成る医薬組成物を提供する。
【0037】
第十二の側面において、本発明は、IL−8様ポリペプチドが関与している疾患及び症状、例えば、免疫障害、転移、移植片拒絶、血管形成、アルツハイマー病、アテローム性動脈硬化症、亜致死性の内毒素血症、敗血症ショック、羊膜腔の微生物感染、回帰熱のヤーリッシュ・ヘルクスハイマー反応、中枢神経系の感染病、急性膵炎、潰瘍性大腸炎、膿胸、溶血性尿毒症症候群、髄膜炎菌性の疾患、胃の感染症、腹膜炎、乾癬、関節リウマチ、敗血症、喘息及び糸球体腎炎、の診断又は処置のための薬物の製造における使用のための、本発明の第一の側面のポリペプチド、又は本発明の第二もしくは第三の側面の核酸分子、又は本発明の第四の側面のベクター、又は本発明の第五の側面の宿主細胞、又は本発明の第六の側面のリガンド、あるいは本発明の第七の側面の化合物を提供する。
【0038】
第十三の側面において、本発明は、患者の疾患を処置する方法であって、当該患者へ、本発明の第一の側面のポリペプチド、又は本発明の第二もしくは第三の側面の核酸分子、又は本発明の第四の側面のベクター、又は本発明の第五の側面の宿主細胞、又は本発明の第六の側面のリガンド、あるいは本発明の第七の側面の化合物を投与することを含んで成る方法を提供する。
【0039】
本発明の第一の側面のポリペプチドをコードする天然の遺伝子の発現、又は本発明の第一の側面のポリペプチドの活性が、健常な対象者の発現のレベル又は活性と比較したとき罹患患者で低下する疾患については、この患者に投与される前記ポリペプチド、核酸分子、リガンド又は化合物はアゴニストであるはずである。逆に、前記天然の遺伝子の発現又はポリペプチドの活性が、健常な対象者の発現のレベル又は活性と比較したとき罹患患者で上昇する疾患については、この患者に投与される前記ポリペプチド、核酸分子、リガンド又は化合物はアンタゴニストであるはずである。このようなアンタゴニストの例にはアンチセンス核酸分子、リボザイム及びリガンド、例えば抗体が含まれる。
【0040】
第十四の側面において、本発明は、本発明の第一の側面のポリペプチドを高レベル、低レベルで発現、又は発現しないように形質転換したトランスジェニック又はノックアウト非ヒト動物を提供する。このようなトランスジェニック動物は、疾患の研究用モデルとして非常に有用であり、さらに前記疾患の治療又は診断に有効な化合物の同定を目的とするスクリーニング方法で用いることもできる。
【0041】
本発明を利用するために用いることができる標準的な技術及び手法の要約は後述する。本発明は、記載される特定の方法論、プロトコール、細胞系、ベクター及び試薬に限定されないことは理解されよう。本明細書で用いられる専門用語は単に特定の態様を説明するためのものであり、この用語によって本発明の範囲を限定しようとするものではないこともまた理解されよう。本発明の範囲は特許請求の範囲の用語によってのみ限定される。
【0042】
本明細書では、ヌクレオチド及びアミノ酸についての標準的な略語が用いられる。
【0043】
本発明の実施では別に指示がなければ、分子生物学、微生物学、組換えDNA技術及び免疫学の通常の技術が用いられ、これらの技術は当業者の技術範囲内である。
【0044】
このような技術は、文献で十分に説明されている。特に適切な解説書の例には以下が含まれる:Sambrook Molecular Cloning; A Laboratory Manual, Second Edition (1989);DNA Cloning, Vol.I及びII (D.N. Glover編集、1985);Oligonucleotide Synthesis (M.J. Gait編集、1984);Nucleic Acid Hybridization (B.D. Hames及びS.J. Higgins編集、1984);Transcription and Translation (B.D. Hames及びS.J. Higgins編集、1984);Animal Cell Culture (R.I. Freshney編集、1986);Immobilized Cells and Enzymes (IRL Press, 1986);B. Perbal, A Practical Guide to Molecular Cloning (1984);the Methods in Enzymology series (Academic Press, Inc.)特にVol. 154及び155;Gene Transfer Vectors for Mammalian Cells (J.H. Miller and M.P. Calos編集、1987, Cold Spring Harbor Laboratory);Immunochemical Methods in Cell and Molecular Biology (Mayer及びWalker編集、1987, Academic Press, London);Scopes, (1987) Protein Purification: Principles and Practice, Second Edition (Springer Verlag, NY);及びHandbook of Experimental Immunology, Vols. I-IV (D.M. Weir and C.C. Blackwell編集、1986)。
【0045】
本発明の第一の側面は、配列番号2又は配列番号4又は配列番号5又は配列番号6に記載のアミノ酸配列の変異体を含み、ここで、選択された配列で特定された任意のアミノ酸は、非保存的に置換され、但し当該配列中の15%未満のアミノ酸残基がそのように変化している。指定された数の非保存的置換を有するタンパク質配列は、通常利用可能なバイオインフォマティクスツールを用いて同定することができる(Mulder NJ及びApweiler R, 2002; Rehm BH, 2001)。
【0046】
このような配列に加え、一連のポリペプチドは、本発明の開示の一部を形成する。IL−8様ポリペプチドはN末端配列のタンパク質分解性の除去(シグナルペプチダーゼ及び他のタンパク質分解酵素による)を含む成熟過程を進行することが分かっているので、本願は、配列番号2に記載の配列のポリペプチドの成熟型も特許請求する。これらのポリペプチド配列は配列番号4又は配列番号7に記載されている。好ましい成熟型は配列番号4に記載されている。成熟型は、IL−8様/TMEM9活性を示し、且つin vivo(発現している細胞又は動物によるもの)又はin vitro(精製されたポリペプチドの特異的酵素による修飾によるもの)での翻訳後成熟過程から生じるあらゆるポリペプチドを含むことが意図されている。他の選択的(alternative)成熟型も、糖又はリン酸のような化学基の付加から生じ得る。
【0047】
本願はまた、配列番号4に記載の配列のポリペプチドのヒスチジンタグ付き型も特許請求する。このポリペプチド配列は配列番号5に記載されている。
【0048】
特許請求する他のポリペプチドは、配列番号2、配列番号4又は配列番号5又は配列番号7で示すアミノ酸配列の活性変異体であり、ここで、選択した配列において特定されている任意のアミノ酸が非保存的に置換されており、但し、当該配列中の15%未満、好ましくは10%、5%、3%、又は1%未満のアミノ酸残基がそのように変化している。指定のパーセンテージは、開示した新規アミノ酸配列全体について測定されされなければならない。
【0049】
本発明によると、あらゆる置換が、分子の構造及び生物学的機能を保存するために、好ましくは「保存的」置換又は「安全」置換であるべきであり、これは、十分に類似の化学的特性を有するアミノ酸を導入する、一般的に定義されている置換のことである(例えば、塩基性正電荷アミノ酸は別の塩基性正電荷アミノ酸によって置換されるべきである)。
【0050】
文献は、保存的アミノ酸置換の選択が、タンパク質の配列及び/又は構造に対する統計的及び/又は物理化学的研究に基づいて実施されうる多数のモデルを提供している(Rogov Si and Nekrasov AN, 2001)。タンパク質デザイン実験は、アミノ酸の特異的なサブセットの使用が、折りたたみ可能で且つ活性なタンパク質を生成することができ、その結果、タンパク質構造内に容易に適合され且つ、機能的且つ構造的ホモログ及びパラログを検出するために使用されうるアミノ酸の「同義」置換の分類に役立つ(Murphy LR et al., 2000)。同義アミノ酸群及び更に好ましい同義アミノ酸群を表1に示す。
【0051】
相当のIL−8様/TMEM9ポリペプチドに対し、同等の、又は改善された活性を有する活性変異体は、コードDNAの常用の突然変異誘発技術、コードDNA配列のレベルでのコンビナトリアル技術(例えば、DNAシャッフリング、ファージディスプレー/セレクション)から、あるいはコンピューター支援設計研究、続く従来技術において説明されているような所望の活性についてのバリデーション、から生じることがある。
【0052】
特異的な非保存的突然変異も、異なる目的で本発明のポリペプチド内に導入されることがある。IL−8様/TMEM9ポリペプチドの親和性を低下する突然変異は、その再使用能及び再循環能を高め、これはその治療効能を高める可能性がある(Robinson CR, 2002)。最終的には本発明のポリペプチド中に存在する免疫原性エピトープが、ワクチン開発のために活用されるか(Stevanovic S, 2002)、もしくはタンパク質安定性を増大する変異を選択し、それらを補正するための公知の方法に従いそれらの配列を修飾することにより排除され得る(van den Burg B及びEijsink V, 2002;国際公開公報第02/05146号、第00/34317号、第98/52976号)。
【0053】
本発明の更に別のポリペプチドは、上述の配列のアミノ酸配列の活性なフラグメント、前駆体、塩、又は機能的に等価の誘導体である。
【0054】
フラグメントは、それらの機能を変化することなく末端又は内部のアミノ酸の欠失を提示すべきであり、そして、前記タンパク質の機能的な高次構造に必須のアミノ酸を除去又は置換することなく、一般的に数個のアミノ酸、例えば10個以下、好ましくは3個以下のものを包含すべきである。小さいフラグメントは、抗原決定基を形成することがある。
【0055】
「前駆体」は、細胞又は身体への投与前又は後に代謝的で且つ酵素的なプロセシングにより本発明の化合物へと変換されうる化合物である。
【0056】
用語「塩」は、本明細書では、本発明のポリペプチドのカルボキシル基の塩及びアミノ基の酸付加塩の両方を意味する。カルボキシル基の塩は、当業界で知られている手段によって形成することができ、そして無機塩、例えばナトリウム、カルシウム、アンモニウム、第二鉄又は亜鉛の塩など、及び、例えばアミンで形成されるような有機性塩、例えばトリエタノールアミン、アルギニン又はリジン、ピペリジン、プロカインなどを含む。酸付加塩には、例えば、鉱酸、例えば塩酸又は硫酸との塩、及び有機酸、例えば酢酸又はシュウ酸との塩、が含まれる。そのような塩のいずれも、本発明のペプチド及びポリペプチド又はそれらの類似体と実質的に同様の活性を有するはずである。
【0057】
用語「誘導体」は、本明細書で使用する場合、アミノ酸部分の側鎖又はアミノ末端若しくはカルボキシ末端の基の上に存在する官能基から、既知の方法に従い調製されうる誘導体を意味する。そのような分子は、通常一次配列を変化させない他の修飾、例えば、ポリペプチドのin vivo又はin vitroでの化学的誘導体化(アセチル化又はカルボキシル化)であって、ペプチドの合成及びプロセシングの間に又は更なるプロセシング段階において、当該ペプチドのリン酸化(ホスホチロシン、ホスホセリン、又はホスホスレオニン残基の導入)又はグリコシル化(哺乳類のグリコシル化酵素にポリペプチドを曝露することによるもの)のパターンを修飾することによって生成されるもの、からも生じることがある。あるいは、誘導体は、前記カルボキシル基のエステル又は脂肪族アミド及び遊離アミノ基のN−アシル誘導体又は遊離ヒドロキシル基のO−アシル誘導体を含み、且つ、例えばアルカノイル基又はアリール基として、アシル基により形成される。
【0058】
前記誘導体の生成は、内部又は末端の位置において、適切な残基の部位指定の修飾を包含してもよい。付着に使用する残基は、ポリマーの付着が可能な側鎖(すなわち、官能基を有する側鎖、例えばリジン、アスパラギン酸、グルタミン酸、システイン、ヒスチジン等)を有するはずである。あるいは、ポリマーの付着が可能な側鎖を有する残基は、前記ポリペプチドのアミノ酸を置換することができ、あるいは前記ポリペプチドの内部又は末端の位置で付加することができる。また、遺伝学的にコードされているアミノ酸の側鎖は、ポリマー付着のために化学的に修飾されることがあり、あるいは適切な側鎖の官能基を有する非天然アミノ酸も利用されることがある。好ましい付着方法は、ペプチド合成と化学的ライゲーションの組み合わせを利用する。有利には、水溶性ポリマーの付着が、生分解性リンカーを介して、特にタンパク質のアミノ末端領域でなされる。そのような修飾は、前記リンカーの分解時にポリマーの修飾無しに前記タンパク質を放出する前駆体(又は「プロドラッグ」)型のタンパク質を提供するように働く。
【0059】
ポリマー付着は、アンタゴニストの特定の位置で自然に発生するアミノ酸の側鎖又はアンタゴニストの特定の位置で自然に発生するアミノ酸を置換する天然若しくは非天然の側鎖に対してだけでなく、標的の位置でアミノ酸の側鎖に付着する炭水化物又は他の部分に対してもなされうる。希少な又は非天然のアミノ酸も、特異的に操作された菌種においてタンパク質を発現することによって導入されうる(Bock A, 2001)。
【0060】
上文で示した変異体は全て、自然に発生するものでも、ヒト以外の生物において同定されるものでも、あるいは、化学合成、部位指定突然変異誘発技術、又は任意な他の適当な既知の技術により調製される人工的なもの、であってもよく、これは当業界で提示されている技術を用いて当業者によってルーチンに得ることができ、そして試験することができる、実質的に相当の突然変異型又は短縮型のペプチドの有限集合を提供する。
【0061】
本願で開示されている新規アミノ酸配列は、異なる種類の試薬及び分子を提供するために使用されうる。これらの化合物の例は、それらの完全配列又は特異的フラグメント、例えば抗原決定基を用いて同定されうる結合タンパク質又は抗体である。ペプチドライブラリーは、特許請求の範囲のアミノ酸配列に結合する抗体又は他のタンパク質をスクリーニングし、そして特徴付けるための、そして同様の結合特性を有する前記ポリペプチドの別の型を同定するための既知の方法(Tribbick G, 2002)において使用されうる。
【0062】
本願は、上述のポリペプチドのいずれかを含む融合タンパク質も開示している。これらのポリペプチドは、ポリペプチドのIL−8様/TMEM9活性を著しく損なうことも、追加の特性を提供する可能性もなく、本願で開示されたタンパク質配列とは異種のタンパク質配列を含むべきである。このような特性の例は、より容易な精製手法、より長い体液中の半減期の維持、追加の結合部分、末端タンパク質分解性の消化による成熟、又は細胞外局在化がある。この後者の特徴は、先の定義に含まれる融合タンパク質又はキメラタンパク質の特定群を定義するために特に重要であり、その理由は、特許請求する分子が、これらのポリペプチドの単離及び精製が促進される空間のみではなく、一般にIL−8様/TMEM9ポリペプチド及びそれらの受容体が相互作用する空間にも、局在化することを可能にするからである。
【0063】
前記部分、リガンド及びリンカーの設計は、融合タンパク質の構築、精製、検出及び使用のための方法及び戦略同様、文献において開示されている(Nilsson J et al., 1997; Methods Enzymol, Vol. 326-328, Academic Press, 2000)。本発明の融合タンパク質に含まれうる好ましいタンパク質配列は、これらのタンパク質配列:膜結合タンパク質、イムノグロブリンの定常領域、多量体化ドメイン、細胞外タンパク質、シグナルペプチド含有タンパク質、エクスポートシグナル含有タンパク質、に属する。これらの配列の特徴及びそれらの特異的な使用は詳細に開示されており、例えばアルブミン融合タンパク質(WO01/77137)、多量体化ドメインを含む融合タンパク質(WO01/02240、WO00/24782)、免疫複合体(Garnett MC, 2001)、又はアフィニティークロマトグラフィーによる組み換え産物の精製を可能にする配列を含む融合タンパク質(Constans A, 2002; Burgess RR and Thompson NE, 2002; Lowe CR et al., 2001; J. Bioch. Biophy. Meth., vol. 49 (1-3),2001; Sheibani N, 1999)である。
【0064】
本発明のポリペプチドは、それらに特異的に結合するリガンドを生成し、そして特徴付けるために使用されうる。これらの分子は、天然又は人工の、化学的観点(結合タンパク質、抗体、分子インプリンティングポリマー)から非常に異なるものであってもよく、そして当業界の技術(WO 02/74938 ; Kuroiwa Y et al., 2002; Haupt K, 2002; van Dijk MA and van de Winkel JG, 2001; Gavilondo JV and Larrick JW, 2000)を適用することによって生成されてもよい。そのようなリガンドは、それらを生成せしめたポリペプチドのIL−8様活性を拮抗化し、又は阻害することができる。特に、共通で且つ効率的なリガンドは、膜結合タンパク質又は抗体の細胞外ドメインで表され、これは、モノクローナル抗体、ポリクローナル抗体、ヒト化抗体、又は抗原結合フラグメントの形態であってもよい。
【0065】
上述のポリペプチド及びポリペプチドベースの誘導体化された試薬は、所望の使用方法及び/又は製造方法に従い、別の型、例えば、放射性標識、蛍光標識、ビオチン、又は細胞毒性物質の中から選択される分子との活性のある接合体又は複合体であってもよい。
【0066】
特定の分子、例えばペプチドミメティクス(peptide mimetic)も、本発明のポリペプチドの配列及び/又は構造を基に設計されることがある。ペプチドミメティクス(peptidomimeticとも称される)は、アミノ酸の側鎖の、アミノ酸のキラリティーの、及び/又はペプチド主鎖のレベルで化学的に修飾されたペプチドである。これらの変更は、改善された調製、性能及び/又は薬物動態の特徴を有する本発明のポリペプチドのアゴニスト又はアンタゴニストを提供することが意図される。
【0067】
例えば、前記ペプチドが、問題のある対象へのインジェクションの後にペプチダーゼによる開裂を受けやすい場合、非開裂性のペプチドミメティクスによる特に感受性のあるペプチド結合の置換は、より安定で且つその結果治療薬として更に有用なペプチドを提供することができる。同様に、L−アミノ酸残基の置換は、タンパク質分解に対してほとんど感受性がないペプチドであって、最終的にペプチド以外の有機化合物により類似のペプチドを賦与する標準的な方法である。アミノ末端のブロッキング基、例えばt−ブチルオキシカルボニル、アセチル、テイル(theyl)、スクシニル、メトキシスクシニル、スベリル(suberyl)、アジピル、アゼライル(azelayl)、ダンシル、ベンジルオキシカルボニル、フルオレニルメトキシカルボニル、メトキシアゼライル、メトキシアジピル、メトキシスベリル、及び2,4−ジニトロフェニルも有用である。増大した性能、延長された活性、精製の容易さ、及び/又は増大した半減期を提供する多くの他の修飾が従来技術で開示されている(WO 02/10195; Villain M et al., 2001)。
【0068】
ペプチドミメティクスに含まれるアミノ酸誘導体にとって好ましい別の同義群は、表IIに規定されているものである。アミノ酸誘導体の非限定的なリストには、アミノイソ酪酸(Aib)、ヒドロキシプロリン(Hyp)、1,2,3,4−テトラヒドロ−イソキノリン−3−COOH、インドリン−2カルボン酸、4−ジフルオロ−プロリン、L−チアゾリジン−4−カルボン酸、L−ホモプロリン、3,4−デヒドロプロリン、3,4−ジヒドロキシ−フェニルアラニン、シクロヘキシル−グリシン、及びフェニルグリシンも含まれる。
【0069】
「アミノ酸誘導体」とは、20個の遺伝子でコードされる天然のアミノ酸とは異なるアミノ酸又はアミノ酸様の化学的存在物を意図する。特に、アミノ酸誘導体は、置換型又は非置換型の直鎖若しくは分枝、又は環状のアルキル部分を含むことがあり、且つ1又は複数のヘテロ原子を含むことがある。アミノ酸誘導体は、新規に生成されてもよく、又は商業的供給源(Calbiochem-Novabiochem AG, Switzerland; Bachem, USA)から入手してもよい。
【0070】
タンパク質の構造及び機能をプローブし、そして/あるいは改善させるために、in vitro及びin vivo両方の翻訳系を用いて非天然アミノ酸誘導体をタンパク質に組み込む種々の方法論が文献において開示されている(Dougherty DA, 2000)。非ペプチドミメティクスと同様、ペプチドミメティクスの合成及び開発のための技術も当業界で周知である(Golebiowski A et al., 2001; Hruby VJ and Balse PM, 2000; Sawyer TK, in "Structure Based Drug Design", edited by Veerapandian P, Marcel Dekker Inc., pg. 557-663,1997)。
【0071】
本発明の別の目的は、IL−8様/TMEM9活性を有する本発明のポリペプチド、それらに対して生成する抗体又は結合タンパク質と結合するポリペプチド、相当の融合タンパク質、あるいは上文で開示したアンタゴニスト活性を有する突然変異体、をコードする単離された核酸にある。好ましくは、これらの核酸は、配列番号1又は配列番号3又は配列番号6から成る群から選択されるDNA配列、あるいは前記DNA配列のいずれかの相補体、を含んで成るべきである。
【0072】
あるいは、本発明の核酸は、配列番号1又は配列番号3又は配列番号6から成る核酸配列、あるいは前記DNA配列の相補体と、高ストリンジェントな条件下でハイブリダイズし、あるいは少なくとも一続きの約30個のヌクレオチドの範囲にわたり少なくとも約85%の同一性を示すはずである。
【0073】
用語「高ストリンジェントな条件」は、非常に類似の分子の会合を容易にするハイブリダイゼーション反応における条件であって、50%ホルムアルデヒド、5xSSC(150mMのNaCl、15mMのクエン酸三ナトリウム)、50mMのリン酸ナトリウム(pH7.6)、5xデンハルト溶液、10%硫酸デキストラン、及び20μg/mlの変性してせん断されたサケ精子DNAを含んで成る溶液中で60〜65℃一晩インキュベーションし、続いて0.1xSSC中フィルターを同一の温度で洗浄することに存するものを意味する。
【0074】
実質的に同一のヌクレオチド配列を含むこれらの核酸は、コードされたポリペプチドを維持し、修飾し、導入し、又は発現するために使用されうるプラスミド、ベクター及び任意な他のDNAコンストラクトに含まれることがある。特に、前記核酸分子が発現制御配列と作用可能に連結しているベクターは、原核又は真核宿主細胞における、コードされたポリペプチドの発現を可能にしうる。
【0075】
用語「実質的に同一のヌクレオチド配列」は、遺伝コードの縮重が原因で、所定のアミノ酸配列をもコードする全ての他の核酸配列を含む。この意味で、文献は、組換え体の発現にとって好ましい又は最適化されたコドンに対する示唆を提供している(Kane JF et al., 1995)。
【0076】
核酸及びベクターは、異なる目的で細胞内に導入されることがあり、その結果トランスジェニック細胞及び動物を生成せしめる。本発明のポリペプチドを発現することができる細胞の産生方法は、そのようなベクター及び核酸で細胞を遺伝子操作することを含んで成る。
【0077】
特に、宿主細胞(例えば、細菌細胞)は、本発明の核酸及びベクターによってコードされたポリペプチドの一過性又は安定性の発現を可能にする形質転換によって修飾されることがある。あるいは、前記分子は、正常な発現レベルと比較した場合に、本発明のポリペプチドの増強又は低下した発現レベルを有するトランスジェニック動物細胞又はヒト以外の動物を生成させるために(非相同組換え/相同組換え又はそれらの安定な組込み及び維持を可能にする任意な他の方法による)使用されることがある。そのような正確な修飾は、本発明の核酸及び、例えば遺伝子治療(Meth. Enzymol., vol. 346,2002)又は部位特異的なリコンビナーゼ(Kolb AF, 2002)に関連する技術、を利用することによって得ることができる。本願で開示するIL−8様/TMEM9ポリペプチドを基にしたそれらの機能の系統だった研究のためのモデル系も、ヒト細胞系へのジーンターゲティングによって生成されうる(Bunz F, 2002)。
【0078】
遺伝子サイレンシング法を実施して、本発明のポリペプチドをコードする遺伝子の内在性発現をダウンレギュレートすることもできる。RNA干渉(RNAi)(S.M. Elbashir ら. Nature 2001, 411, 494-498)は、使用可能な配列特異的転写後遺伝子サイレンシングのための1つの方法である。短いdsRNAオリゴヌクレオチドをin vitroで合成して細胞内に導入する。これらdsRNAオリゴヌクレオチドの配列特異的結合によって標的mRNAの分解が開始され、標的タンパク質の発現が減少又は阻害される。
【0079】
上述のとおり評価される遺伝子サイレンシング法の有効性は、ポリペプチド発現の測定を介して(例えばウェスタンブロットによる)、又はTaqManベースの方法を用いてRNAレベルで評価することができる。
【0080】
本発明のポリペプチドは、当業界で知られている任意な方法、例えば組換えDNA関連技術、及び化学合成技術、によって調製されることもある。特に、本発明のポリペプチドを生成する方法は、上述のように、核酸又はベクターが発現する条件下で宿主細胞又はトランスジェニック細胞を培養し、そして培養物から前記核酸又はベクターによってコードされるポリペプチドを回収することを含んで成ることもある。例えば、ベクターが、細胞外又はシグナルペプチド含有タンパク質との融合タンパク質としてポリペプチドを発現する場合、組換え産物は細胞外空間に分泌されることがあり、そして、培養細胞から、更なる処理の観点で更に容易に回収され、且つ精製されることがあり、あるいは、当該細胞は直接使用され、又は投与されることがある。
【0081】
本発明のタンパク質をコードするDNA配列は、任意の適当な手段(形質転換、トランスフェクション、接合、プロトプラスト融合、エレクトロポレーション、リン酸カルシウム沈殿、直接的なマイクロインジェクション等)によって適切な宿主細胞に導入されうる、適当なエピソームの又は非相同/相同組換えのベクター内に挿入され、そしてライゲーションされることがある。特定のプラスミド又はウイルスベクターを選択するのに重要な因子は、ベクターを含むレシピエント細胞が当該ベクターを含まないそれらのレシピエント細胞から容易に認識され、且つ選択されうることの容易さ;特定の宿主で望まれるベクターのコピー数;及び異なる主の宿主細胞間でベクターを「シャトル」し得ることが望ましいか否か、を含む。
【0082】
ベクターは、転写開始/終結制御配列の支配下での原核又は真核宿主細胞による本発明のポリペプチドを含む単離されたポリペプチド又は融合ポリペプチドの発現を可能にするはずであり、これらは、前記細胞において構成的に活性又は誘導性のものが選択される。前記細胞が実質的に豊富な細胞系は、続いて安定な細胞系を提供するよう単離されうる。
【0083】
異なる転写制御配列及び翻訳制御配列は、真核生物宿主(例えば、酵母、昆虫、植物、又は哺乳類の細胞)に対し、その宿主の性質に依存して利用することができる。それらは、ウイルス源、例えばアデノウイルス、ウシパピローマウイルス、シミアンウイルス等に由来してもよく、ここでは、制御シグナルが、高レベルで発現する特定の遺伝子と関連している。例としてはヘルペスウイルスのTKプロモーター、SV40初期プロモーター、酵母GAL4遺伝子プロモーター等がある。転写開始制御シグナルは、抑制及び活性を可能にして、その結果前記遺伝子発現が調節されうるようなものが選択されうる。導入されたDNAによって安定して形質転換された細胞は、前記発現ベクターを含む宿主細胞の選択を可能にする1又は複数のマーカーを導入することによって選択されうる。当該マーカーはまた、栄養要求宿主に対する光合成栄養、殺生物耐性、例えば抗生物質、又は重金属、例えば銅、等をも提供しうる。選択マーカー遺伝子は、発現されるDNA遺伝子配列に直接連結されるか、あるいは同時トランスフェクションによって同一の細胞内に導入されうる。
【0084】
宿主細胞は原核生物又は真核生物のいずれかのものでありうる。好ましいものは、真核生物宿主、例えば哺乳類細胞、例えばヒト、サル、マウス、及びチャイニーズハムスター卵巣(CHO)細胞であり、それは、それらがタンパク質に対する翻訳後修飾、例えば正確な折りたたみ及びグリコシル化を提供するためである。酵母もグリコシル化を含む翻訳後ペプチド修飾を実施しうる。多くの組換えDNAストラテジーが存在しており、これらは、酵母内で所望のタンパク質の産生に利用されうる強力なプロモーター配列及び高コピー数のプラスミドを利用する。酵母は、クローン化哺乳類遺伝子産物のリーダー配列を認識し、そしてリーダー配列を有するペプチド(すなわち、プレペプチド)を分泌する。
【0085】
本発明の上文で言及した態様は、新規IL−8様/TMEM9ポリペプチドの配列に対して本願が提示する開示を、一般的な分子生物学的技術の知識と組み合わせることによって達成されうる。
【0086】
多数の本及び概説が、ベクター及び原核生物又は真核生物宿主細胞を用いて組換えタンパク質をどのようにクローン化し、そして産生するかについての技術を提供しており、例えばオックスフォード大学出版局によって発行されている「A Practical Approach」のシリーズの中の幾つかのタイトルである ("DNA Cloning 2: Expression Systems", 1995;"DNA Cloning 4: Mammalian Systems", 1996;"Protein Expression", 1999;"Protein Purification Techniques", 2001).。
【0087】
更に、最新のより重点的な文献は、ハイスループットな様式でポリペプチドを発現する技術(Chambers SP, 2002; Coleman TA, et al., 1997)、治療的用途を有する組換えタンパク質の大規模産生のために産業上利用される細胞系及び方法(Andersen DC and Krummen L, 2002; Chu L and Robinson DK, 2001)、及び注目のポリペプチドを発現する別の真核生物発現系であって、所望のタンパク質の経済的な産生についてかなりの可能性を有すると考えられるもの、例えばトランスジェニック植物を基にしたもの(Giddings G, 2001)又は酵母ピキア・パストリス(Pichia pastors)を基にしたもの(Lin Cereghino GP et al., 2002)、の概論を提供する。組換えタンパク質産物は、発現したポリペプチドの量及び質を証明するため(Baker KN et al., 2002)、生物学的同等性及び免疫原性の問題があるか否かを調べるのと同様に(Schellekens H, 2002; Gendel SM, 2002)、種々の分析技術により迅速にモニタリングされうる。
【0088】
全体的に、合成IL−8様ポリペプチドは文献において開示されており、そして、短い長さで示した本発明のIL−8様/TMEM9ポリペプチドに有効に適用されうる多数の化学合成技術の例が、固相又は液相合成技術として文献にて利用可能となっている。例えば、合成されるポリペプチドのカルボキシ末端に相当するアミノ酸は、有機溶媒中で不溶の支持体に結合し、そして、適切な保護基で保護されたそれらのアミノ基及び側鎖の官能基を有するアミノ酸がカルボキシ末端からアミノ末端へと順番に1つずつ縮合するものと、樹脂又は前記ペプチドのアミノ基の保護基と結合したアミノ酸が放出されるものとの交互の反復反応によって、ペプチド鎖はその結果この様式で伸長される。固層合成法は、使用する保護基の型に依存して、tBoc法及びFmoc法に大きく分類される。典型的に使用される保護基には、tBoc(t−ブトキシカルボニル)、Cl−Z(2−クロロベンジルオキシカルボニル)、Br−Z(2−ブロモベンジルオキシカルボニル)、Bzl(ベンジル)、Fmoc(9−フルオレニルメトキシカルボニル)、Mbh(4,4’−ジメトキシジベンズヒドリル)、Mtr(4−メトキシ−2,3,6−トリメチルベンゼンスルホニル)、Trt(トリチル)、Tos(トシル)、Z(ベンジルオキシカルボニル)及びCl2−Bzl(2,6−ジクロロベンジル)がアミノ基の場合に含まれ;NO2(ニトロ)及びPmc(2,2,5,7,8−ペンタメチルクロマン−6−スルホニル)がグアミジノ基の場合に含まれ;そしてtBu(t−ブチル)がヒドロキシル基の場合に含まれる。所望のペプチドの合成後、それは脱保護反応にかけられ、そして固体の支持体から切り離される。そのようなペプチド切断反応は、Boc法の場合フッ化水素又はトリフルオロメタンスルホン酸を用いて、そしてFmoc法の場合TFAを用いて実施されうる。
【0089】
本発明のポリペプチドの精製は、この目的のために知られている方法、すなわち、抽出、沈殿、クロマトグラフィー、電気泳動等の任意な常用の方法、のうちのいずれか1つによって実施されうる。本発明のタンパク質を精製するために優先して使用されうる追加の精製手順は、標的タンパク質に結合し、且つ、産生され、そしてカラム内に含まれるゲルマトリックス上に固定される、モノクローナル抗体又は親和性の基を用いるアフィニティークロマトグラフィーである。タンパク質は、ヘパリン又は特異的抗体によりカラムと結合し、一方、不純物はカラムを通過する。洗浄後、タンパク質はpH又はイオン強度の変化によってゲルから溶出される。あるいは、HPLC(高性能液体クロマトグラフィー)を使用することができる。溶出は、タンパク質精製に一般的に利用される水−アセトニトリルベースの溶媒を用いて実施されうる。
【0090】
本発明の新規ポリペプチドの開示、及びそれらに関連して開示されている試薬(抗体、核酸、細胞)の開示はまた、細胞への又は動物内でのそれらの発現レベルを増強し、又は低下させる化合物をスクリーニングし、そして特徴付けることを可能にする。
【0091】
「オリゴヌクレオチド」は、化学合成されうる一本鎖ポリデオキシヌクレオチド又は二本の相補的なポリデオキシヌクレオチド鎖のいずれかを意味する。そのような合成オリゴヌクレオチドは、5’ホスフェートを有さず、そのためキナーゼの存在下ホスフェートをATPと一緒に添加しないことには別のオリゴヌクレオチドとライゲーションしないであろう。合成オリゴヌクレオチドは、脱リン酸されていないフラグメントにライゲーションするであろう。
【0092】
本発明は、本発明の化合物の精製された調製物(ポリペプチド、核酸、細胞等)を含む。精製された調製物とは、本明細書で使用する場合、乾燥重量当たり少なくとも1%、好ましくは少なくとも5%本発明の化合物を含む調製物を意味する。
【0093】
本願は、一連の新規IL−8様/TMEM9ポリペプチド及び複数の可能性のある用途を有する関連試薬を開示する。特に、本発明のポリペプチドのIL−8様/TMEM9活性の増大が疾患の治療又は予防において望まれる場合はいつでも、試薬、例えば開示されているIL−8様/TMEM9ポリペプチド、相当の融合タンパク質及びペプチドミメティクス、コード核酸、発現細胞、又はそれらの発現を増強する化合物が使用されうる。
【0094】
従って、本発明は、開示したIL−8様/TMEM9ポリペプチド、相当の融合タンパク質及びペプチドミメティクス、コード核酸、発現している細胞、又はそれらの発現を増強する化合物のうちの1つを活性成分として含む、本発明のポリペプチドのIL−8様/TMEM9活性の増加が必要である疾患の治療又は予防のための医薬組成物を開示している。これらの医薬組成物の調製方法は、開示したIL−8様/TMEM9ポリペプチド、相当の融合タンパク質及びペプチドミメティクス、コード核酸、発現細胞、又はそれらの発現を増強する化合物を、医薬として許容される担体と一緒に組み合わせることを含んで成る。本発明のポリペプチドのIL−8様/TMEM9活性の増加が必要である疾患を治療又は予防する方法は、治療として有効量の、開示したIL−8様/TMEM9ポリペプチド、相当の融合タンパク質及びペプチドミメティクス、コード核酸、発現細胞、又はそれらの発現を増強する化合物、の投与を含んで成る。
【0095】
本願で開示されている試薬の中でも、前記のリガンド、アンタゴニスト又は本発明のポリペプチドの発現又は活性を低下させる化合物は、複数の用途を有し、特に、これらは、本発明のポリペプチドの過剰なIL−8様/TMEM9活性に関連した疾患の治療又は診断において使用されうる。
【0096】
従って、本発明は、本発明のポリペプチドの過剰なIL−8様/TMEM9活性に関連する疾患の処置又は予防のための医薬組成物であって、そのようなポリペプチドの発現又は活性を低下させるリガンド、アンタゴニスト又は化合物のうちの1つを活性成分として含む医薬組成物を開示する。これらの医薬組成物の調製方法は、前記のリガンド、アンタゴニスト、又は化合物を、医薬として許容される担体と一緒に組み合わせることを含んで成る。本発明のポリペプチドの過剰なIL−8様/TMEM9活性に関連する疾患の処置又は予防のための方法は、治療的に有効量のアンタゴニスト、リガンド又は化合物の投与を含んで成る。
【0097】
本発明の医薬組成物は、IL−8様/TMEM9ポリペプチド又は関連試薬に加え、医薬として許容される適当な担体、生物学的に適合性のある媒体及び添加物を含んでもよく、これらは動物への投与に適したものであり(例えば、生理学的食塩水)、且つ、活性化合物を医薬として使用されうる調製物へと処理するのを容易にする助剤(賦形剤、安定化剤、佐剤、又は希釈剤のようなもの)を最終的に含んで成るものである。
【0098】
医薬組成物は、投与形態の要求に合致する任意の許容される方法で製剤化されうる。例えば、生体材料の中でも、糖−高分子接合体、ポリエチレングリコール及び他の天然又は合成ポリマーは、薬物送達の有効性に関して活性成分を改善するのに使用されうる。特定の投与形態を評価する技術及びモデルは文献に開示されている(Davis BG and Robinson MA, 2002; Gupta P et al., 2002; Luo B and Prestwich GD, 2001; Cleland JL et al., 2001; Pillai O and Panchagnula R, 2001)。
【0099】
これらの目的に適したポリマーは生体適合性があり、すなわち、それらは生物系に対して非毒性であり、そして多数のそのようなポリマーは既知である。そのようなポリマーは天然で疎水性又は親水性であっても、生分解性、非生分解性、又はそれらの組み合わせであってもよい。これらのポリマーには、天然ポリマー(例えばコラーゲン、ゼラチン、セルロース、ヒアルロン酸)、並びに合成ポリマー(例えばポリエステル、ポリオルトエステル、ポリ無水物)が含まれる。疎水性の非分解性ポリマーの例には、ポリジメチルシロキサン、ポリウレタン、ポリテトラフルオロエチレン、ポリエチレン、ポリ塩化ビニル、及びポリメチルメタエリラート(methaerylate)が含まれる。親水性の非分解性ポリマーの例には、ポリ(2−ヒドロキシエチルメタクリラート)、ポリビニルアルコール、ポリ(N−ビニルピロリドン)、ポリアルキレン、ポリアクリルアミド、及びそれらのコポリマーが含まれる。連続反復単位としてエチレンオキシドを含んで成る好ましいポリマーには、例えばポリエチレングリコール(PEG)がある。
【0100】
任意の許容される投与形態は、活性成分の所望の血液レベルを確立するために、当業者によって使用され、そして決定されうる。例えば、投与は、種々の非経口経路、例えば皮下、静脈内、皮内、筋肉内、腹腔内、経鼻、経皮、経口、又は頬側でなされることがある。本発明の医薬組成物はまた、維持又は制御放出剤形、例えば蓄積注射、浸透ポンプ等を含むものであって、既定の速度での前記ポリペプチドの長期投与のためのもので、好ましくは正確な投与量の単回投与に適した単位剤形で投与されてもよい。
【0101】
非経口投与は、ボーラス注入又は時間をかけての漸進的な潅流によってもよい。非経口投与のための調製物には、滅菌水溶液又は非水溶液、懸濁液、及び乳濁液が含まれ、これらは当業界で知られている佐剤又は賦形剤を含むことがあり、且つルーチンな方法に従い調製されうる。尚、適切な油性注射懸濁液としての活性化合物の懸濁液が投与されることがある。適当な親油性溶媒又は媒体には、脂肪油、例えばごま油、又は合成脂肪酸エステル、例えばごま油、例えば合成脂肪酸エステル、例えばオレイン酸エチル又はトリグリセリドが含まれる。懸濁液の粘性を増大させる物質を含みうる水性注射懸濁液は、例えばカルボキシメチルセルロースナトリウム、ソルビトール、及び/又はデキストランが含まれる。任意に、懸濁液はまた安定化剤を含んでもよい。医薬組成物は注射による投与に適した溶液を含み、且つ、賦形剤と一緒に約0.01〜99.99パーセント、好ましくは約20〜75パーセントの活性化合物を含む。
【0102】
用語「治療的に有効量」とは、疾患の経過及び重症度に影響を及ぼし、そのような病状の緩和又は寛解を導くのに十分な量の活性成分を意味する。有効量は、投与経路及び患者の症状に依存する。
【0103】
用語「医薬として許容される」は、活性成分の生物学的活性の有効性を妨げず、且つ投与される宿主にとって毒性でない任意の担体を包含することを意味する。例えば、非経口投与の場合、上記活性成分は、単位剤形で、生理食塩水、デキストロース溶液、血清アルブミン及びリンガー液のような媒体中で製剤化されうる。担体はまた、デンプン、セルロース、タルク、グルコース、ラクトース、スクロース、ゼラチン、麦芽、コメ、コムギ、チョーク、シリカゲル、ステアリン酸マグネシウム、ステアリン酸ナトリウム、モノステアリン酸グリセロール、塩化ナトリウム、乾燥スキムミルク、グリセロール、プロピレングリコール、水、エタノール、及び種々の油から選択することができ、これらは、石油、動物、植物又は合成起源のもの(ピーナッツ油、大豆油、鉱油、ごま油)を含む。
【0104】
投与される用量は、レシピエントの年齢、性別、健康、及び体重、併用の処置の種類、もしあるのであれば、処置の頻度、及び所望の効果の性質、に依存する。用量は、当業者によって理解され、且つ決定可能なように、個々の対象者に合わせられる。各処置に必要とされる総量は、複数回投与又は単回投与によって投与されうる。本発明の医薬組成物は、単独で、又はその症状に対する、又は当該症状のほかの症候に対する他の治療薬と一緒に投与されうる。通常、活性成分の一日量は、0.01〜100ミリグラム/キログラム体重/日含まれる。通常、分割量又は維持放出形態で与えると、1〜40ミリグラム/キログラム/日が所望の結果を得るのに有効である。第二回目又はその後の投与は、個体に対して投与された最初又は前回の用量と同一、それ未満、又はそれ以上の用量で実施されうる。
【0105】
治療目的又は製造目的を有する方法とは別に、複数のほかの方法が本願で開示されているIL−8様/TMEM9ポリペプチド及び関連試薬を利用することができる。
【0106】
最初の例において、本発明のIL−8様/TMEM9ポリペプチドに関連する疾患を処置するのに有効な候補化合物をスクリーニングする方法であって:
(a)そのようなポリペプチドを発現する宿主細胞、当該ポリペプチドの発現レベルが増大又は低下しているトランスジェニックなヒト以外の動物、又はトランスジェニック動物細胞を、候補化合物と接触させること;及び
(b)前記動物又は細胞に対する前記化合物の効果を決定すること、
を含んで成る方法が提供される。
【0107】
第二の例において、本発明のポリペプチドのアンタゴニスト/阻害物質又はアゴニスト/活性化物質としての候補化合物を同定する方法であって:
(a)前記ポリペプチドと、前記化合物、及び哺乳類細胞又は哺乳類細胞膜と
を接触させること;及び
(b)前記の分子が、前記ポリペプチドと前記哺乳類細胞又は哺乳類細胞膜との相互作用、あるいは当該相互作用から生じる応答、を阻止又は増強するか否かを決定すること、
を含んで成る方法が提供される。
【0108】
第三の例において、試料中の本発明のポリペプチドの活性及び/又は存在を決定する方法は、当該ポリペプチド又はそのコードRNA/DNAのいずれかを検出することができる。従って、そのような方法は:
(a)タンパク質含有試料を準備すること;
(b)前記試料と本発明のリガンドとを接触させること;及び
(c)前記ポリペプチドに結合した前記リガンドの存在を決定し、それにより前記試料中のポリペプチドの活性及び/又は存在を決定すること、
を含んで成る。
【0109】
あるいは、前記方法は:
(a)核酸含有試料を準備すること;
(b)前記試料と本発明の核酸とを接触させること;及び
(c)当該核酸と前記試料中の核酸とのハイブリダイゼーションを決定し、それにより前記試料中の核酸の存在を決定すること、
を含んで成る。
【0110】
この意味において、配列番号1又は配列番号3又は配列番号6に示すヌクレオチド配列に由来したプライマー配列を、更にポリメラーゼ連鎖反応増幅により、試料中の本発明のポリペプチドの転写産物又はこれをコードしている核酸の量を決定するためにも使用することができる。
【0111】
本発明の更なる目的は、試料中の本発明のIL−8様/TMEM9ポリペプチドの活性及び/又は存在を測定するためのキットであって、本願で開示されている1又は複数の試薬:本発明のIL−8様/TMEM9ポリペプチド、アンタゴニスト、リガンド又はペプチドミメティクス、単離された核酸又はベクター、医薬組成物、発現細胞、あるいは発現レベルを増大又は低下させる化合物、を含んで成るキットである。
【0112】
このようなキットは、in vitro診断法又はスクリーニング法において使用することができ、それらの実際の組成は、試料の具体的様式(例えば、患者由来の生物学的試料組織)、及び測定される分子種に適合されるべきである。例えば、IL−8様/TMEM9ポリペプチドの濃度を測定することが望ましい場合、キットは、ウェスタンブロットで得られるシグナルを比較するために、精製された形の抗体及び相当のタンパク質を含有してもよい。あるいは、IL−8様/TMEM9ポリペプチドの転写産物の濃度を測定することが望ましい場合は、このキットは、相当のORF配列でデザインされた特異的核酸プローブを含むか、又はそのようなプローブを含有する核酸アレイの形であってよい。これらのキットは、タンパク質−、ペプチドミメティクス−、又は細胞−ベースのマイクロアレイの形であっても良く(Templin MFら, 2002;Pellois JPら, 2002;Blagoev B及びPandey A, 2001)、本願で開示されているタンパク質、ペプチドミメティクス及び細胞を利用することにより、ハイ-スループットプロテオミクス試験が可能となる。
【0113】
治療的用途
本願は、複数の適用可能性を有するSCS0010ポリペプチドを開示する。特に、本発明のポリペプチドのIL−8様/TMEM9活性の増大が疾患の治療又は予防において所望とされる場合はいつでも、試薬、例えば開示したSCS0010ポリペプチド、相当の融合タンパク質及びペプチドミメティクス、コード核酸、発現細胞、又はそれらの発現を増強する化合物が使用されうる。
【0114】
従って、本発明は、開示したIL−8様/TMEM9ポリペプチド、相当の融合タンパク質及びペプチドミメティクス、コード核酸、発現している細胞、又はそれらの発現を増強する化合物のうちの1つを活性成分として含む、本発明のポリペプチドのIL−8様/TMEM9活性の増加が必要である疾患の治療又は予防のための医薬組成物を開示している。これらの医薬組成物の調製方法は、開示したIL−8様/TMEM9ポリペプチド、相当の融合タンパク質及びペプチドミメティクス、コード核酸、発現細胞、又はそれらの発現を増強する化合物を、医薬として許容される担体と一緒に組み合わせることを含んで成る。本発明のポリペプチドのIL−8様/TMEM9活性の増加が必要である疾患を処置又は予防する方法は、治療として有効量の、開示したIL−8様/TMEM9ポリペプチド、相当の融合タンパク質及びペプチドミメティクス、コード核酸、発現細胞、又はそれらの発現を増強する化合物、の投与を含んで成る。
【0115】
本願で開示されている試薬の中でも、前記のリガンド、アンタゴニスト又は本発明のポリペプチドの発現又は活性を低下させる化合物は、複数の用途を有し、特に、これらは、本発明のポリペプチドの過剰なIL−8様/TMEM9活性に関連した疾患の治療又は診断において使用されうる。
【0116】
従って、本発明は、本発明のポリペプチドの過剰なIL−8様/TMEM9活性に関連する疾患の処置又は予防のための医薬組成物であって、そのようなポリペプチドの発現又は活性を低下させるリガンド、アンタゴニスト又は化合物のうちの1つを活性成分として含む医薬組成物を開示する。これらの医薬組成物の調製方法は、前記のリガンド、アンタゴニスト、又は化合物を、医薬として許容される担体と一緒に組み合わせることを含んで成る。本発明のポリペプチドの過剰なIL−8様/TMEM9活性に関連する疾患の処置又は予防のための方法は、治療的に有効量のアンタゴニスト、リガンド又は化合物の投与を含んで成る。
【0117】
SCS0010ポリペプチドのアゴニスト及びアンタゴニストは、SCS0010ポリペプチドの活性を調節する分子、SCS0010ポリペプチドの成熟型の少なくとも1つの活性を増加又は減少するいずれかの分子を含む。アゴニスト又はアンタゴニストは、SCS0010ポリペプチドと相互作用するタンパク質、ペプチド、炭水化物、脂質、又は低分子量の分子のような、コファクターであってもよく、これによりそれらの活性を調節する。
【0118】
潜在的なポリペプチドアゴニスト又はアンタゴニストには、TMEM9ポリペプチドの可溶型(SCS0010)又は膜結合型(TMEM9)のいずれかであって、前記タンパク質の細胞外ドメインの一部又は全部を含んで成るものと反応する抗体が含まれる。SCS0010ポリペプチドの発現を制御する分子は、典型的には、発現のアンチセンスレギュレーターとして作用することができるSCS0010ポリペプチドをコードしている核酸を含む。
【0119】
従って、本願は、適切に調製された医薬組成物における活性成分として、IL−8様ポリペプチドが関与している疾患及び症状、例えば、免疫障害、転移、移植片拒絶、血管形成、アルツハイマー病、アテローム性動脈硬化症、亜致死性の内毒素血症、敗血症ショック、羊膜腔の微生物感染、回帰熱のヤーリッシュ・ヘルクスハイマー反応、中枢神経系の感染病、急性膵炎、潰瘍性大腸炎、膿胸、溶血性尿毒症症候群、髄膜炎菌性の疾患、胃の感染症、腹膜炎、乾癬、関節リウマチ、敗血症、喘息及び糸球体腎炎の予防及び処置において有用であり得る新規IL−8様/TMEM9ポリペプチド及び一連の関連試薬を開示する。
【0120】
これらの用途の幾つかは、Gene Expression Omnibus (GEO)データベース:http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=geoに由来する実験によって支持されている。
【0121】
GEOはハイスループットの実験データの広い範囲のための公共の貯蔵場所としての役割を果たしている。これらのデータは、mRNA、ゲノムDNA、及びタンパク質の存在量を測定するシングル及びデュアル・チャネルマイクロアレイベースの実験、並びにアレイ以外の技術、例えば連続的遺伝子発現解析(SAGE)、及び質量分析プロテオミクスデータ、を含む。
【0122】
血清に応答する初代線維芽細胞の時間的な転写プログラム上のGEOレコードGDS85において(デュアル・チャネルマイクロアレイ実験)、TMEM9が若干阻害を受け、ほぼ1時間でピークを迎えることが示された。線維芽細胞は、血管疾患、炎症、線維症、線維性障害、関節リウマチ、クローン病、硬皮症、呼吸器感染、喘息、アレルギー反応、気管支炎、間質性肺炎、糖尿病性のネフロパシー、心不整脈、心臓肥大、腫瘍、潰瘍、眼障害、脂肪異栄養症、全身性硬化症、骨関節症、神経病理学を含む多数の病気、並びに老化、組織修復及び形成外科に関係するもの、に関与していることが知られている。更に、GEOレコードGDS60における、遺伝子発現と気道への抗原チャレンジとの間の時間的関係の決定による肺の中のブタクサ花粉の免疫原性タンパク質に対するアレルギー反応の解析において(シングルチャネルマイクロアレイ実験)、アレルゲンチャレンジにより、TMEM9が3つのうち2つの試料中で一貫して低下した(TMEM9はごくわずかに発現している)ことが示された。
【0123】
GEOレコードGDS350における肺線維症モデル(A/J、ブレオマイシン耐性)において、気管内をブレオマイシン処理されたA/Jマウスの肺がコントロールと比較された(デュアルチャネルマイクロアレイ実験)。この研究は、TMEM9が肺線維症モデルの4つのうち3つの試料において、コントロールと比較して若干阻害されたことを示した。
【0124】
GEOレコードGDS61における、肺の高血圧の回復において、肺高血圧症後の血管リモデリングが、正常酸素圧及び低酸素圧で処理した動物の肺と比較して実施された(シングルチャネルマイクロアレイ実験)。低酸素は、血管収縮に続き、肺血管平滑筋の肥大/肥厚及び内皮細胞増殖を誘導する。10時間後、TMEM9の発現は、正常酸素圧のものと比較すると、低酸素圧処理した動物において一貫して低下したことが示された。
【0125】
IL−8についての概説において、Mukaida Nは、「種々のタイプの細胞が、広範な刺激、例えば炎症性サイトカイン、微生物及びそれらの産物、並びに環境的変化、例えば低酸素症、再潅流、及び酸素過剰症に応じて大量のIL−8/CXCL8を産生することができる」(Am J Physiol Lung Cell Mol Physiol. 2003 Apr; 284 (4): L566-77 "Pathophysiological roles ofinterleukin-8/CXCL8 in pulmonary diseases")ことを証明する証拠を公表している。更に、Mukaidaは「多数の観察が、好中球に対する強力な作用により好中球媒介型の急性炎症における鍵となる媒介因子としてIL−8/CXCL8を確立した」ことを言及している。最後にMukaidaは、「複数の一連の証拠が、IL−8/CXCL8が好中球以外の種々のタイプの細胞、例えばリンパ球、単球、内皮細胞、及び線維芽細胞に対する広範な作用を有する」ことに言及している。更に、IL−8の生物学的機能は、これが更に種々の病理学的症状、例えばアテローム性動脈硬化症、炎症、肺病理及びガンにおいても重要な役割を有することを示唆している。
【0126】
SCS0010はまた、以下の研究で示唆されているようなガン治療においても有用なことがある。
【0127】
Kurokawaらは、3つの異なるcDNAライブラリー由来の3072の遺伝子及び肝臓ガン形成に関与していることが疑われている298の追加の既知の遺伝子を含むPCRアレイ実験において、わずかに7つの遺伝子が、そのうちTMEM9が肝臓ガン形成における共通の重要な役割を果たしうることを示した(Kurokawa et al. J Exp Clin Cancer Res. 2004 Mar; 23(1) : 135-41. PCR-array gene expression profiling of hepatocellular carcinoma)。
【0128】
GEOレコードGDS88における、抗ガン剤についてのNational Cancer Instituteのスクリーニングに使用された細胞系の解析において(デュアルチャネルマイクロアレイ実験)、TMEM9が、参照プールと比較して、黒色腫、白血病及び卵巣ガン細胞系において若干阻害されることを示した。
【0129】
GEOレコードGDS143における、表皮ガン形成において、前ガン状態の皮膚病変(光線角化症)と正常な表皮と培養ケラチノサイトとの比較が実施された(SAGE実験)。TMEM9は、培養ケラチノサイト及び光線角化症の表皮において発現しないのと比較して、高度に発現することが示された。
【0130】
GEOレコードGDS103における、心筋細胞形成において、未分化の多能性状態における及び心筋細胞への分化の誘導後のP19胚性癌腫(EC)細胞のトランスクリプトームを特徴付けるために(SAGE実験)、TMEM9は、プレーティングから0.5日後のEC細胞又はTMEM9の発現が起こらなかった未分化のP19EC細胞と比較して、プレーティングから3.5日後のP19EC細胞において発現することが示された。
【0131】
GEOレコードGDS121において、エストロゲン又はタモキシフェン処理後の遺伝子発現プロファイルの網羅的解析が実施された(SAGE実験)。タモキシフェン処理したZR75−1ヒト乳ガン細胞系が未処理又はエストロゲン処理のZR75−1細胞と比較してTMEM9の発現を誘導したことが示された。
【0132】
GEOレコードGDS122における、細胞の恒常性及び腫瘍細胞の増殖の制御におけるJNK2のシグナル伝達の重要性を強調するc−Jun NH82末端キナーゼの標的の同定において(SAGE実験)、TMEM9の発現が、モック処理したPC3細胞と比較してJNK2アンチセンスオリゴヌクレオチドPC3処理細胞において高いことが示された。IL−8活性は、JNK2のドミナントネガティブ突然変異体に対して感受性があり、そしてNF−カッパBとJNKの協調的な活性が強力なIL−8転写に必要なことが示されている。
【0133】
複数のGEOレコードはまた、種々の組織におけるTMEM9の発現を提供している(GDS503, GDS8, GDS393, GDS217, GDS541, GDS546, GDS217, GDS381, GDS102, GDS541, GDS117, GDS541, GDS544, GDS580, GDS551, GDS545, GDS542及びGDS548)。TMEM9の発現は、腎臓(近位尿細管、皮質集合管、皮質の太い上行脚)、脳全体、側坐核、脳の上衣腫、脳の髄芽腫、リンパ節、乳腺上皮、胸部の筋上皮、乳ガン、腺管ガン(胸部)、白質、小児の皮質、周辺部網膜、中心網膜、腹膜、結腸(原発腫瘍)、胃ガン、卵巣の表面上皮、原発性悪性の皮膚メラノーマ、前立腺ガン(進行した腫瘍)において示されている。TMEM9のGenecard entryによると(http://bioinformatics.weizmann.ac.il/cards/)、TMEM9は骨髄、脾臓、胸腺、脳、脊髄、心臓、骨格筋、加増、膵臓、前立腺、腎臓及び肺において発現している。
【0134】
TMEM9の発現情報はまた、UniGene Cluster Hs. 181444(ホモサピエンス;http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=unigene)から入手可能である。
【0135】
UniGeneによると、TMEM9は、成人のホモサピエンスの神経腫瘍に由来するライブラリー5967NT0048において高度に発現している。尚、TMEM9のcDNA供給源は、深刻な乳頭ガン、高悪性度、2個体のプールされた腫瘍;心臓;中分化した子宮内膜腺ガン、3個体のプールされた腫瘍;グリア芽腫(プールされたもの);印環細胞の特徴を有する低分化腺ガン;低分化した子宮内膜腺ガン、2個体のプールされた腫瘍;未分化希突起グリオーマ;リンパ腫、濾胞性混合小細胞大細胞;高分化した子宮内膜腺ガン、7個体のプールされた腫瘍;扁平上皮細胞ガン、低分化(4個体のプールされた腫瘍。原発性及び転移性のものを含む):プールされたもの;精巣;NEUROBLASTOMA COT 50-NORMALIZED;胸腺、プールされたもの;子宮;骨;正常な子宮内膜、中間の分泌期、周期23日;下垂体;NEUROBLASTOMA COT 25-NORMALIZED; PLACENTA COT 25- NORMALIZED;初期の乳頭の深刻な癌腫;胎盤;腺癌腫瘍、5個体のプールされたもの(説明書を参照のこと);正常な鼻咽頭;クローン病を有する3人の患者由来の結腸粘膜;5個体のプールされた肉腫、例えば粘液性脂肪肉腫、単発性線維性腫瘍、悪性線維性組織球腫、消化管間質腫瘍及び中皮腫;脳全体;肝臓及び脾臓;精巣;肝臓;胎児肝臓;肺;カルチノイド;中分化した腺ガン;2個体のプールされたウイルムス腫瘍。1つは原発性、そして1つは脳に対して転移性;髄芽腫;CNCAP (3) T-225細胞系;胸部;結腸;2個体のプールされた扁平上皮細胞癌腫;精製された膵島;頭頸部;末梢神経系;胎盤;プールされた軟骨肉腫腫瘍細胞;ガン細胞系;腺ガン細胞系;膜芽細胞腫;腎細胞腺ガン;プールされた胚細胞性腫瘍;脳;脳;前立腺;扁平上皮細胞癌腫;子宮頸ガン細胞系;大細胞癌腫;小細胞癌腫;子宮内膜腺ガン細胞系;黒色性黒色腫;腎腫瘍;絨毛ガン;正常な胸部;大細胞癌腫、未分化;神経系腫瘍;膵臓;卵巣;副腎腫;胚性肝細胞、WA01、38回継代;臍帯血;EGFRの増幅を伴うグリア芽腫;1p/19qの損失を伴う未分化希突起グリオーマ;メラニン細胞;腺ガン細胞系;副腎皮質癌腫細胞系;筋肉(骨格);十二指腸腺ガン細胞系;網膜;上皮(細胞系);副腎腫細胞系;神経芽細胞腫;粘液性類表皮ガン;軟骨;海馬;多発性硬化症の病変;血液;白血球;未分化神経外胚葉;NEUROBLASTOMA COT 10-NORMALIZED;類上皮癌腫細胞系;移行上皮性乳頭腫細胞系;正常な肺;視床下部;プールされた結腸、腎臓、胃;プールされた膵臓及び脾臓;癌腫細胞系;骨髄;プールされた肺及び脾臓;膀胱腫瘍;プールされた脳、肺、前立腺;上皮小体腫瘍;視神経;卵巣(3個体のプール);ヒト網膜;神経芽細胞腫;胎児の眼、水晶体、前眼部、視神経、網膜、網膜の中心の且つ斑点のあるRPE及び脈絡;RPE及び脈絡胎児の眼;網膜の中心及び斑点;腹水;プールされたヒトメラニン細胞、胎児の心臓、及び妊娠子宮;前頭葉;胎児の脳;リンパ節に対して転移性の悪性黒色腫;全脳;2個体のプールされた腫瘍(明細胞型);腎臓;胃;眼;肺;B細胞、慢性リンパ球性白血病;原発性肺嚢胞性線維症上皮細胞;混合型;転移性軟骨肉腫;平滑筋肉腫;悪性度IIの軟骨肉腫;肺限局性線維症;軟骨肉腫;網膜;肝細胞癌腫細胞系;乳頭漿液性腺ガン;交感神経幹;恐らくTP53が突然変異しており、且つEGFR増幅していないグリア芽腫;後根神経節;肺ガン;混合型(40腫のRNAのプール);骨髄腫;細胞系;奇形癌腫細胞系;インスリノーマ;胚中心B細胞;水晶体;肺胞マクロファージ;RPE/脈絡膜;脂肪細胞;骨髄肉腫細胞系;内軟骨肉腫細胞系;軟骨下骨;子宮内膜;胚;副睾丸;卵巣腫瘍;類表皮癌腫細胞系;低級前立腺新生物;胚細胞腫瘍;副腎腺腫;正常前立腺;結腸腫瘍RER+;肝臓;松果体;転移性前立腺骨病変;類上皮癌腫、である。
【0136】
IL−8のレベルは再発寛解型多発性硬化症のクラドリビン処置の後脳脊髄液(CSF)中で有意に低下したことも指摘されうる(RR-MS; Bartosik-Psujek H et al. Acta Neurol Scand. 2004 Jun; 109 (6): 390-2. "Interleukin-8 and RANTES levels in patients with relapsing-remitting multiple sclerosis (RR-MS) treated with cladribine.")。
【0137】
尚、Kveineらは、TMEM9のmRNAが副腎、甲状腺、卵巣、精巣、及び前立腺に豊富に存在し、そしてそれには及ばないが、気管、脊髄、胃、結腸、小腸、胸腺、及び脾臓において発現していることを示した(Kveine et al. Biochem Biophys Res Commun. 2002 Oct 4; 297 (4): 912-7. "Characterization of the novel human transmembrane protein 9(TMEM9) that localizes to lysosomes and late endosomes.")。彼等はまた、骨髄、リンパ節、及び末梢血リンパ球(PBL)における発現が低いことも観察し、そしてTMEM9のmRNAがB細胞系起源、T細胞系起源、骨髄起源及び赤血球起源の造血細胞系において発現していることを証明している。
【0138】
そのようなものとして、SCS0010核酸分子、それらのポリペプチド、アゴニスト又はアンタゴニスト(例えば、アゴニスト抗体又はアンタゴニスト抗体)は種々の疾患、線維症、線維性障害、関節リウマチ、クローン病、硬皮症、呼吸器感染、喘息、アレルギー反応、気管支炎、間質性肺炎、糖尿病性のネフロパシー、心不整脈、心臓肥大、腫瘍、潰瘍、眼障害、脂肪異栄養症、全身性硬化症、骨関節症、神経病理学、肺高血圧症、低酸素症、肺血管平滑筋の肥大/肥厚、黒色腫、白血病及び卵巣ガン、肺細胞癌腫の発生、前ガン状態の皮膚病変(光線角化症)、胚性癌腫、乳ガン、脳上衣腫、脳髄芽腫、乳腺癌、腺管癌(胸部)、結腸原発性腫瘍、胃ガン、原発性悪性の皮膚メラノーマ、前立腺ガン(進行した腫瘍)、神経腫瘍、漿液性乳頭ガン、子宮内膜腺ガン、グリア芽腫、印環細胞の特徴を有する低分化腺ガン、未分化希突起グリオーマ、リンパ腫、扁平上皮細胞癌腫、神経芽細胞腫、乳頭漿液性腺ガン、肉腫、例えば粘液性脂肪肉腫、単発性線維性腫瘍、悪性線維性組織球腫、消化管間質腫瘍及び中皮腫、中分化した腺ガン、2個体のプールされたウイルムス腫瘍(原発性、そして1つは脳に対して転移性)、髄芽腫、扁平上皮細胞癌腫、軟骨肉腫、癌腫、網膜芽腫、腎細胞腺ガン、胚細胞性腫瘍、扁平上皮細胞癌腫、子宮頸ガン細胞系、大細胞癌腫、小細胞癌腫、子宮内膜腺ガン細胞系、黒色性黒色腫、腎腫瘍、絨毛ガン、神経腫瘍、副腎腫、胚性肝細胞、グリア芽腫、急性骨髄性白血病、未分化希突起グリオーマ、副腎皮質癌腫、十二指腸腺ガン、副腎腫細胞系、粘液性類表皮ガン、多発性硬化症の病変、類上皮癌腫、移行上皮性乳頭腫、膀胱腫瘍、上皮小体腫瘍、悪性黒色腫、慢性リンパ球性白血病、原発性肺嚢胞性線維症、転移性軟骨肉腫、平滑筋肉腫、悪性度IIの軟骨肉腫、肺限局性線維症、軟骨肉腫、肝細胞癌腫、乳頭漿液性腺ガン細胞系、肺ガン、骨髄腫、奇形癌腫細胞系、インスリノーマ、内軟骨肉腫、副睾丸、卵巣腫瘍、類表皮癌腫細胞系、前立腺新生物、胚細胞腫瘍、副腎腺腫、結腸腫瘍、転移性前立腺骨病変、類上皮癌腫並びに老化、組織修復及び形成外科に関係するものを診断又は処置するのに有用でありうる。尚、SCS0010核酸分子、そのアゴニスト又はアンタゴニスト(例えば、アゴニスト抗体又はアンタゴニスト抗体)は、TMEM9が検出されている器官及び組織に由来する、又はそこに存在する疾患(例えば腫瘍)を診断又は処置するのに有用なことがある。SCS0010のアンタゴニスト又はアゴニスト(例えば抗体)は、実施例5及び/又は図11、12において決定されるようなSCS0010の部位及びドメインに対するものであってもよい。例えば、抗体は、SCS0010の3つの保存されたシステインリッチドメイン(内在性TMEM9にも存在する)のうちの少なくとも1つに対するものであってもよく、これは理論に拘束されることを望むものではないが、SCS0010とそれ自体との又は他のTMEM9ファミリーメンバーとの二量体化を阻止しうる。
【0139】
本発明のポリペプチド及び関連試薬の治療的用途は、創薬及び前臨床開発時の前駆脂肪細胞因子−1様ポリペプチド及び他の生物学的産物の評価に関して、動物細胞、組織を使用するin vivo/in vitroアッセイ手段によるか、又は公知のin silico/コンピュータ支援の方法(Johnson DE及びWolfgang GH, 2000)により、評価(安全性、薬物動態及び有効性に関して)される。
【0140】
本発明は、以降、以下の実施例により特定の態様を参照して説明されるが、これらは本発明を何ら限定するものとして解されるべきではない。当該説明の内容は、上文の教示に照らして、そしてそれ故に、特許請求の範囲の意味及び目的を超えて広げることなく、当業者によって実施されうる全ての改変及び置換を含んで成る。
【0141】
【表1】

【表2】
【実施例1】
【0142】
ASTRALデータベース(Brenner SEら, "The ASTRAL compendium for protein structure and sequence analysis", Nucleic Acids Res., 2000 Jan 1; 28(1): 254-6)から得たIL−8様タンパク質ドメイン配列を用い、ヒトゲノム配列(Celeraバージョン)から予測された遺伝子の相同タンパク質配列を検索した。
【0143】
これらのドメイン配列を使用して、ホモログを含む配列プロファイルを作成した。更に、ジスルフィド結合に関与する保存されたシステインに相当する各プロファイルの位置は、これらの位置での厳密なAA保存を要求するように修飾された。IL−8様ドメインの配列プロファイルは、PIMAII(Profile Induced Multiple Alignment;ボストン大学ソフトウェア、バージョンII, Das S及びSmith TF, 2000)を用いて作成した。PIMAIIアルゴリズムは繰り返し相同配列を並置し、そして配列プロファイルを作成する。
【0144】
クエリータンパク質配列は、下記の3種のプログラムのひとつにより作成された、遺伝子予測及びそれらの翻訳(ヒトゲノムR27のCELERAバージョン)から得た:Genescan (Burge C, Karlin S., "Prediction of complete gene structures in human genomic DNA”, J Mol Biol., 1997 Apr 25; 268(1): 78-94)、Grail(Xu Y, Uberbacher EC., “Automated gene identification in large-scale genomic sequences", J Comput Biol., 1997 Fall; 4(3): 325-38)及びFgenesh(Celera社独自のソフトウェア)。
【0145】
相同性は、プロファイルとクエリー配列の間のグローバル-ローカルアラインメントを作成するPIMAIIを用いて検出した。この場合、このアルゴリズムは、クエリーとしてのEGF機能ドメインのプロファイルと共に使用した。PIMAIIは、クエリープロファイルを、タンパク質配列に翻訳された遺伝子予測のデータベースと比較し、その結果そのドメインを含むDNA配列との合致を同定することができる。
【0146】
更にこの配列の既知のIL−8様含有タンパク質とのBLAST(Basic Local Alignment Search Tool;NCBI ver2)による比較は、一番近いホモログを同定した(Gish W, States DJ., "Identification of protein coding regions by database similarity search", Nat Genet., 1993 Mar; 3(3): 266-72;Pearson WR, Miller W., "Dynamic programming algorithms for biological sequence comparison", Methods Enzymol., 1992; 210: 575-601;Altschul SFら,"Basic local alignment search tool", J Mol Biol., 1990 Oct 5; 215(3): 403-10)。
【0147】
検出に使用したPIMAIIパラメータは、PIMA先行(prior)アミノ酸確率行列及びZ-カットオフスコア10であった。使用したBLASTパラメータは以下のとおりである:比較行列=BLOSUM62;ワードレングス=3;カットオフE値=10;ギャップオープニング及びイクステンション=デフォルト;フィルターなし。
【0148】
一旦機能ドメインがこの配列において同定されたならば、これらの遺伝子は、一番近いホモログの配列を用いgenewiseアルゴリズムで再度予測した(Birney Eら,"PairWise and Search Wise: finding the optimal alignment in a simultaneous comparison of a protein profile against all DNA translation frames", Nucleic Acids Res., 1996 Jul 15; 24(14): 2730-9)。
【0149】
IL−8様の相同性ドメインのプロファイルは、PERL(Practical Extraction and Report Language)及びPIMAIIに記載されたPSI-BLAST(Altshulら 1997)スクリプトを用い自動的に作成した。
【0150】
合計51の(IL−8様)の18種の予測された遺伝子は、新規である可能性があると判断されたので選択した。
【0151】
前記タンパク質配列の新規性は、最終的には、BLASTを用いるタンパク質データベース(SwissProt/Trembl, Human IPI and Derwent GENESEQ)の検索により評価した。機能的ドメインに対する整合性及び新規性は、選択した新規候補についての「人の手による」検査で調べた。
【実施例2】
【0152】
実施例1に記載の方法により単離されたひとつの配列は、本明細書においてSCS0010ポリペプチド配列と称されるものである。最も類似の、既知のポリペプチド配列は、アクセッション番号ABP61498のヒトNF−κB活性化タンパク質であるが、第三のエキソンが異なる。図2は、SCS0010をコードする遺伝子のイントロン/エキソン構造を示す(64000111830404)。
【実施例3】
【0153】
SCS0010のスプライス変異体の同定及びクローニング
1.SCS0010のクローニング
1.1 cDNAライブラリー
ヒト胎盤cDNAライブラリー(バクテリオファージラムダ(λ)GT10)は、Clontech(カタログ番号HL1075b)から購入した。バクテリオファージλDNAは、Wizard Lambda Preps DNA精製システムを用いて、メーカーの使用説明書に従い(Promega, Corporation, Madison WI)、感染したE.コリ宿主株の小規模培養物から調製した。生じたファージDNAは、50ng/μlの濃度で再懸濁し、そして後のPCR反応に使用した。
【0154】
1.2 PCR用の遺伝子特異的クローニングプライマー
18〜25塩基の長さを有する一対のPCRプライマーを、Primer Designer Software(Scientific & Educational Software, PO Box 72045, Durham, NC 27722-2045, USA)を用いて仮のcDNAのコード配列を増幅するために設計した。PCRプライマーは最適化された結果、55±10℃付近のTM及び40〜60%のGC含量を有する。標的配列について高い選択性を有していたプライマーが選択された(ほとんど又は全く非特異的プライミング無し)。
【0155】
1.3 ヒト胎盤ファージcDNAライブラリー由来のSCS0010cDNAのPCR
遺伝子特異的PCR増幅プライマー(SCS0010−CP1及びSCS0010−CP2、図3及び表4)は、SCS0010のほぼ予測されたコード配列を含むことが予想される299bpの産物を増幅するよう設計された。PCRは1xAmplitaq(登録商標)バッファー、200μMのdNTP、50ピコモルの各クローニングプライマー、2.5ユニットのAmplitaq(登録商標)(Perkin Elmer)及び100ngの各ファージライブラリープールDNAを含む50μlの終量で、以下の通りプログラムしたMJ Research DNA Engineを用いて実施した:94℃、1分;94℃、1分、χ℃、及び1分、72度を40サイクル;続いて72℃、7分を1サイクル、そして4℃の保持サイクル。
【0156】
増幅産物を0.8%アガロースゲル/1xTAEバッファー(Invitrogen)上で可視化し、そしてWizard Lambda Preps DNA精製システム(Promega)を用いてゲルから精製した。50Lの滅菌水で溶出したPCR産物は、直接サブクローニングするか、又は−20℃で保存した。
【0157】
1.4 PCR産物のサブクローニング
PCR産物は、Invitrogen Corporationから購入したTOPOクローニングキットをメーカー指定の条件を使用して用いることでトポイソメラーゼI修飾クローニングベクター(pCR4−TOPO)内にサブクローニングした。要約すると、4μlのゲル精製PCR産物を1μlのTOPTベクター及び1μlの塩溶液と一緒に15分間室温でインキュベートした。続いて、反応混合物でE.コリ株TOP10(Invitrogen)を以下の通り形質転換した:50μlのOne Shot TOP10細胞のアリコートを氷上で融解し、そして2μlのTOPO反応液を添加した。混合物を15分間氷上でインキュベートし、そして次に42℃で正確に30秒間インキュベートすることで熱ショックを与えた。試料を氷に戻し、そして250μlの温めたSOC培地(室温)を添加した。試料を振とうしながら(220rpm)1時間37℃でインキュベートした。続いて、アンピシリン(100μg/ml)を含むLブロス(LB)プレート上に形質転換混合物をプレーティングし、そして一晩37℃でインキュベートした。インサートを含むアンピシリン耐性コロニーをコロニーPCRで同定した。
【0158】
1.4 コロニーPCR
コロニーを、滅菌した爪楊枝を用い、50μl滅菌水中に接種した。播種材料のうち10μlのアリコートが、使用したプライマーがT7及びT3であることを除き上述のように20μlの合計の反応液量中でPCRにかけられた。サイクリング条件は以下の通りである:94℃で2分間;94℃で30秒間、48℃で30秒間及び72℃で1分間を30サイクル。試料は、次の解析まで4℃で維持した(保持サイクル)
【0159】
PCR反応産物を、1X TAE緩衝液中1%アガロースゲル上で分析した。予想されたPCR産物サイズ(約299bp+マルチクローニングサイト又はMCSに起因する105bp)を生じたコロニーを、振盪しながら(220rpm)で、アンピシリン(100μg/ml)を含有する5ml L-ブロス(LB)中で一晩37℃で増殖した。
【0160】
1.5 プラスミドDNAの調製及び配列決定
ミニプレッププラスミドDNAは、Qiaprep Turbo 9600 ロボット型システム(Qiagen)又はWizard Plus SMミニプレップキット(Promegaカタログ番号1460)をメーカーの使用説明書に従い用いることで5mlの培養液から調製した。プラスミドDNAは、100μlの滅菌水で溶出した。DNA濃度は、Eppendorf BO光度計を用いて測定した。プラスミドDNA(200〜500ng)を、T7プライマー及びT3プライマーにより、Big DyeTerminatorシステム(Applied Biosystemsカタログ番号4390246)をメーカーの使用説明書に従い使用し、DNA配列決定にかけた。プライマー配列は表3に示すとおりである。配列決定反応液を、Dye-Exカラム(Qiagen)又はMontage SEQ 96クリーナッププレート(Millipore カタログ番号LSKS09624)を使用し精製し、その後Applied Biosystems 3700シークエンサーで解析した。
【0161】
配列解析は、予測されたSCS0010配列との100%のマッチを含むクローンを同定した(図3)。クローニングしたPCR産物(pCR4−TOPO−SCS0010)(プラスミドID.14613)のプラスミドマップを図4に示す。
【0162】
2.SCS0010の哺乳類細胞発現ベクターの構築
続いて、DNA配列決定で同定したSCS0010のコード配列を含むpCR4−TOPOクローン(pCR4−TOPO−SCS0010、プラスミドID.14613)(図4)を使用して、インサートをGateway(登録商標)クローニング法(Invitrogen)で哺乳類細胞発現ベクターpEAK12d(図6)及びpDEST12.2(図7)内にサブクローニングした。
【0163】
2.1 インフレームで6HISタグ配列と融合したGateway適合SCS0010ORFの生成
Gatewayクローニング法の第一段階は、attB1組換え部位及びコザック配列が5’末端に隣接し、並びにインフレームで6ヒスチジン(6HIS)タグ、停止コドン及びattB2組換え部位をコードしている配列(Gateway適合cDNA)が3’末端で隣接した、SCS0010のORFを生成する2段階のPCR反応を伴う。第一のPCR反応液(終量50μl)は、1μl(40ng)のpCR4−TOPO−SCS0010(プラスミドID.14613)、1.5μlのdNTP(10mM)、10μlの10X Pfxポリメラーゼ緩衝液、1μlのMgSO4(50mM)、各0.5μlの遺伝子特異的プライマー(100μM)(SCS0010−EX1及びSCS0010−EX2)、及び0.5μlのPlatinum Pfx DNAポリメラーゼ(Invitrogen)を含む。このPCR反応は、95℃で2分間の最初の変性工程、それに続く94℃で15秒;55℃で30秒及び68℃で2分間の12サイクル;並びに、4℃の保持サイクルを用い行った。この増幅産物を、1X TAE緩衝液(Invitrogen)中の0.8%アガロースゲル上で可視化し、予測された分子量に移動した産物を、メーカーの使用説明書に従い、Wizard PCRプレップDNA精製システム(Promega)を用いゲルから精製し、50μl滅菌水中に回収した。
【0164】
第二のPCR反応液(終量50μl)は、10μlの精製されたPCR1産物、1.5μlのdNTP(10mM)、5μlの10X Pfxポリメラーゼ緩衝液、1μlのMgSO4(50mM)、0.5μlの各Gateway変換プライマー(100μM)(GCPフォワード及びGCPリバース)、及び0.5μlのPlatinum Pfx DNA ポリメラーゼを含んだ。第2のPCR反応条件は:95℃で1分間;94℃で15秒、50℃で30秒及び68℃で2分間を4サイクル;94℃で15秒、55℃で30秒及び68℃で2分間を25サイクル;引き続き4℃の保持サイクルとした。PCR産物は、メーカーの使用説明書に従い、Wizard PCRプレップDNA精製システム(Promega)を用いゲル精製した。
【0165】
2.2 Gateway適合SCS0010ORFの、GatewayエントリーベクターpDONR221並びに発現ベクターpEAK12d及びpDEST12.2へのサブクローニング
Gatewayクローニング法の第二段階は、下記のような、Gateway修飾PCR産物のGatewayエントリーベクターpDONR221(Invitrogen、図3)へのサブクローニングを伴う:5μlのPCR2由来の精製産物を、1.5μlのpDONR221ベクター(0.1μg/μl)、2μlのBPバッファー及び1.5μlのBPクロナーゼ酵素混合物(Invitrogen)と共に、終量10μlで室温で1時間インキュベーションした。この反応をプロテイナーゼK 1μl(2μg/μl)の添加により停止させ、37℃で更に10分間インキュベーションした。この反応液のアリコート(1μl)を用い、E.コリDH10B細胞を、以下の通りエレクトロポレーションにより形質転換した:DH10Bエレクトロコンピテント細胞(Invitrogen)25μlアリコートを氷上で解凍し、1μlのBP反応混合液を添加した。混合物を、冷却した0.1cmエレクトロポレーション用キュベットに移し、細胞をBioRad Gene-Pulser(登録商標)をメーカーの推奨プロトコールに従い使用しエレクトロポレーションした。予め室温に温めたSOC培地(0.5ml)を、エレクトロポレーション直後に添加した。この混合物を、15mlのスナップ-キャップチューブに移し、振盪しながら(220rpm)37℃で1時間インキュベーションした。その後形質転換混合物のアリコート(10μl及び50μl)を、カナマイシン(40μg/ml)を含有するL-ブロス(LB)プレート上にプレーティングし、37℃で一晩インキュベーションした。
【0166】
プラスミドミニ-プレップDNAを、Qiaprep Turbo 9600 ロボット型システム(Qiagen)を用い、生じたコロニーのうちの6つに由来する5ml培養物から調製した。プラスミドDNA(150-200ng)を、BigDyeTerminatorシステム(Applied Biosystems カタログ番号4390246)をメーカーの使用説明書に従い用い、21M13及びM13RevプライマーによるDNA配列決定にかけた。これらのプライマー配列を表3に示す。配列決定反応は、Dye-Exカラム(Qiagen)又はMontage SEQ 96クリーンアッププレート(Millipore カタログ番号LSKS09624)を用いて精製し、その後Applied Biosystems 3700シークエンサーで解析した。
【0167】
正確な配列を含むこれらのクローンのひとつ(pENTR−SCS0010−6HIS、プラスミドID14691、図8)のプラスミド溶出液(2μl又は約150ng)を、終量10μl中に1.5μlのpEAK12dベクター又はpDEST12.2ベクター(図4及び5)のいずれか(0.1μg/μl)、2μlのLRバッファー及び1.5μlのLRクロナーゼ(Invitrogen)を含有する組換え反応液において使用した。この混合物を、室温で1時間インキュベーションし、プロテイナーゼK(2μg)の添加により停止し、37℃で更に10分間インキュベーションした。この反応液のアリコート(1μl)を用い、E.コリDH10B細胞を、以下の通りエレクトロポレーションにより形質転換した:DH10Bエレクトロコンピテント細胞(Invitrogen)の25μlアリコートを氷上で解凍し、1μlのBP反応混合液を添加した。混合物を、冷却した0.1cmエレクトロポレーション用キュベットに移し、細胞をBioRad Gene-Pulser(登録商標)をメーカーの推奨プロトコールに従い使用しエレクトロポレーションした。予め室温に温めたSOC培地(0.5ml)を、エレクトロポレーション直後に添加した。この混合物を、15mlのスナップ-キャップチューブに移し、振盪しながら(220rpm)37℃で1時間インキュベーションした。その後形質転換混合物のアリコート(10μl及び50μl)を、アンピシリン(100μg/ml)を含有するL-ブロス(LB)プレート上にプレーティングし、37℃で一晩インキュベーションした。
【0168】
プラスミドミニ-プレップDNAを、Qiaprep Turbo 9600 ロボット型システム(Qiagen)を用い、各ベクターでサブクローニングして生じたコロニーのうちの6つに由来する5ml培養物から調製した。pEAK12dベクター中のプラスミドDNA(200-500ng)を、先に説明したように、pEAK12F及びpEAK12Rプライマーにより、DNA配列決定にかけた。pDEST12.2ベクター内のプラスミドDNA(200-500ng)を、先に説明したように、21M13及びM13Revプライマーにより、DNA配列決定にかけた。プライマー配列は表3に示す。
【0169】
Sambrook J.らの説明した方法(1989 (Molecular Cloning, a Laboratory Manual, 第2版, Cold Spring Harbor Laboratory Press))により、CsCl勾配で精製されたマキシ-プレップDNAを、配列を確認した各クローン(pEAK12d−SCS0010−6HIS、プラスミドID14699、図9、及びpDEST12.2−SCS0010−6HIS、プラスミドID14700、図10)のひとつの500ml培養物から調製した。プラスミドDNAは、濃度1μg/μlで滅菌水(又は10mM Tris-HCl、pH8.5)中に再懸濁し、-20℃で保存した。
【0170】
【表3】
下線の配列=コザック配列
太字=停止コドン
イタリックの配列=Hisタグ
【実施例4】
【0171】
SCS0010の発現及び精製
1.哺乳類細胞における発現及びクローニングしたHisタグ付きプラスミドの精製を通じての機能ゲノミクス
エプスタインバーウイルス核抗原を発現しているヒト胎児腎臓293細胞(HEK293−EBNA、Invitrogen)をEx-cell VPRO無血清培地(シードストック、維持培地、JRH)の懸濁液中で維持した。トランスフェクションの日に細胞を計数し、遠心し(低速)、そしてペレットを、1%FCSを補足した所望の量のFEME(下文を参照のこと)培地内で再懸濁して1XE6生存細胞/mlの細胞濃度を生成させた。cDNAをFEME(1Lの体積当たり200ml)中1Lの体積当たり2mgに希釈した(2%のeGFPを同時トランスフェクションする)。ポリエチレンイミントランスフェクション試薬(1Lの体積当たり4mg)を続いてcDNA溶液に添加し、ボルテックスにかけ、そして室温で10分間インキュベートした(トランスフェクション混合物の生成)。
【0172】
続いて、このトランスフェクション混合物をスピナーに添加し、そして90分間CO2インキュベーター(5%CO2及び37℃)内でインキュベートした。新鮮なFEME培地(1%FCS)を、例えば、最初のスピナーの体積を二倍にするために90分後に添加した。続いて、スピナーを6日間インキュベートした。6日目(収集日)に、スピナーの上清(500ml)を遠心(4℃、400g)、そしてプラスミド番号及び発酵番号と一緒に独特の識別子を有するポット内に据えた。
【0173】
精製方法
C末端6Hisタグを有する組換えタンパク質を含む500mlの培養液試料を1倍量の冷たいバッファーA(50mMのNaH2PO4;600mMのNaCl;8.7%(w/v)のグリセロール、pH7.5)で1000mlの終量に希釈した。試料を0.22μmの滅菌フィルター(Millipore, 500mlのフィルターユニット)を通じて濾過し、そして1Lの滅菌した四角い培地ボトル(Nalgene)中4℃で維持した。
【0174】
精製は、自動サンプルローダー(Labomatic)と接続されたVISIONワークステーション(Applied Biosystems)上で4℃で実施された。精製手順は、ニッケルイオンがチャージされたPoros 20MC(Applied Biosystems)上での金属アフィニティークロマトグラフィー(10x50mm、3.93ml)、続いて、Sephadex G-25 medium(Amersham Pharmacia)ゲル濾過カラム(1.0x15cm)上でのバッファー交換、の2連続工程から構成された。
【0175】
第一のクロマトグラフィー工程のために、金属アフィニティーカラムを30倍のカラム体積のEDTA溶液(100mMのEDTA;1MのNaCl;pH8.0)で再生し、15倍のカラム体積の100mMNiSO4溶液で洗浄することでニッケルイオンを再チャージし、そして10倍のカラム体積のバッファーA、続いて7倍のカラム体積のバッファーB(50mMのNaH2PO4;600mMのNaCl;8.7%(w/v)のグリセロール、400mM;イミダゾール、pH7.5)で洗浄し、そして最後に、15mMのイミダゾールを含む15倍のカラム体積のバッファーAで平衡化した。試料をLabomaticサンプルローダーで200mlのサンプルループ内に移し、そして次にニッケル金属アフィニティーカラム上に20ml/分の流速でチャージした。チャージ手順を5回繰り返すことで全試料(1000ml)をニッケルカラム上に移した。次に、カラムは、12倍のカラム体積のバッファーA、続いて20mMのイミダゾールを含む28倍のカラム体積のバッファーAで洗浄した。20mMのイミダゾール洗浄の間に、緩やかに付着した混入タンパク質をカラムから溶出した。組換えHisタグ付きタンパク質を最後に10倍のカラム体積のバッファーBを用い2ml/分の流速で溶出し、そして溶出したタンパク質を2.7mlの画分で回収した。
【0176】
第二のクロマトグラフィー工程のために、Sephadex G-25ゲル濾過カラムを2mlのバッファーD(1.137MのNaCl;2.7mMのKCl;1.5mMのKH2PO4;8mMのNa2HPO4;pH7.4)で再生し、そして次に4倍のカラム体積のバッファーC(137mMのNaCl;2.7mMのKCl;1.5mMのKH2PO4;8mMのNa2HPO4;20%(w/v)のグリセロール;pH7.2)で平衡化した。ニッケルカラムから溶出したピーク画分は、VISION上に組み込まれたサンプルローダーを通じて、Sephadex G-25カラム上に自動的に載せられ、そしてタンパク質はバッファーCで2ml/分の流速で溶出した。脱塩した試料を2.7mlの画分で回収した。当該画分を0.22μmの滅菌遠心フィルター(Millipore)を通じて濾過し、等分し、凍結し、そして−80℃で保存した。試料のアリコートをSDS−PAGE(4〜12%NuPAGEゲル;Novex)上でクーマシーブルー染色及び抗His抗体を用いるウェスタンブロットにより解析した。
【0177】
クーマシーブルー染色。NuPageゲルを0.1%クーマシーブルーR250染色溶液(30%メタノール、10%酢酸)中で室温で1時間染色し、そして次に20%メタノール、7.5%酢酸中でバックグラウンドがクリアーになり、そしてタンパク質のバンドがはっきりと見えるようになるまで脱染した。
【0178】
ウェスタンブロット
電気泳動の後、タンパク質を290mAで1時間、4℃でゲルからニトロセルロース膜にエレクトロトランスファーした。膜を5%粉ミルク/バッファーE(137mMのNaCl;2.7mMKCl;1.5mMのKH2PO4;8mMのNa2HPO4;0.1%Tween20、pH7.4)を用いて1時間室温でブロッキングし、そして次に2つのウサギポリクローナル抗His抗体混合物(G−18及びH−15。各0.2μg/ml;Santa Cruz)/2.5%粉ミルク/バッファーEと一緒にインキュベートした。更に室温で1時間インキュベートした後、膜をバッファーEで洗浄し(3x10分)、そして次に、2.5%粉ミルクを含むバッファーE中で1/3000希釈した二次HRP複合抗ウサギ抗体(DAKO, HRP 0399)と一緒に2時間室温でインキュベートした。バッファーEで洗浄した後(3x10分)、膜をECLキット(Amersham Pharmacia)で1分間展開した。次に膜をHyperfilm(Amersham Pharmacia)に曝露し、フィルムを現像し、そしてウェスタンブロットのイメージを視覚解析した。
【0179】
タンパク質アッセイ
タンパク質濃度はBCAタンパク質アッセイキット(Pierce)とスタンダードとしてウシ血清アルブミンを用いて決定した。回収量は420μgであった。
【実施例5】
【0180】
SCS0010の特徴づけ
Blastを用いて、SCS0010は、皮膚乳頭由来タンパク質4(DERP4)、HSPC186又はUNQ631/PRO1248(SwissProtエントリーQ9P0T7、TME9_HUMAN)としても知られている膜貫通タンパク質9のスプライス変異体であることが決定された。TME9_HUMANとSCS0010との間のアラインメントを図11に示し、これはSCS0010が3’末端で異なることを示している。TMEM9の既知の特徴は:
・183アミノ酸長(20574Da)
・後期エンドソーム及びリソソームに局在しているI型膜タンパク質
・3つのグリコシル化型
・二量体化の可能性
・TMEM9ファミリーに属すること、及び
・細胞内輸送において提案される役割、
である。
【0181】
TMEM9は、Kveineらによって特徴付けられている(Kveine et al. Biochem Biophys Res Commun. 2002 Oct 4 ; 297 (4): 912-7. "Characterization of the novel human transmembrane protein 9 (TMEM9) that localizes to lysosomes and late endosomes.")。TMEM9ファミリーは、Interpro (http://www.ebi.ac.uk/interpro)において以下のように説明されている:「このファミリーは、ホモサピエンス膜貫通タンパク質9SwissProtQ9P0T7と相同な複数の真核生物膜貫通タンパク質を含む。TMEM9遺伝子は、N末端シグナルペプチド、1つの膜貫通領域、3つの潜在的なNグリコシル化部位及び3つの保存されたN末端のシステインリッチドメインを含む183アミノ酸のタンパク質をコードする。当該タンパク質は、線虫(C.エレガンス)からホモサピエンスまでの種で高度に保存されており、そして膜貫通タンパク質の新規ファミリーに属する。TMEM9の正確な機能は知られてないが、広範に発現しており、そして後期エンドソーム及びリソソームに局在することがわかっている。MEDLINE:。このファミリーのメンバーはCXCXCリピートINTERPRO:IPR004153をそれらのN末端領域に含む」。インターロイキン−8(IL−8)は、CXCケモカインサブファミリーのメンバーであり、そしてSCS0010はCXCXリピートを含む、IL−8に対する共通のパターンを示している。
【0182】
バイオインフォマティクスツール、例えばSMART(http://smart.embl-heidelberg.de/)、を用い、TMEM9及びスプライシング変異体SCS0010の推定のドメインを同定した。SMARTの結果を図12に示す。この解析は、SCS0010がTMEM9の可溶性変異体であることを示し、その結果SCS0010が膜結合型のTMEM9と比較して独特の機能を示すことを示唆している。そのようなものとして、分泌型のSCS0010タンパク質は内在性TMEM9のアンタゴニストとしてin vitroで働くようである。Kveineらは、TMEM9がN末端領域に見られる3つの保存されたシステインリッチドメインを有することを示唆している。彼等は更に、これらの領域がタンパク質の折りたたみ、タンパク質の相互作用、及び多量体化に関与しうることに言及している。これらの領域はSC0010において完全に保存されている(図12において矢印で示す)。そのようなものとして、そして理論に拘束されることを望むものではないが、可溶性SCS0010は、二量体化によるTMEM9の制御に関与しているようである。従って、SCS0010は、治療的用途の項目に記載のような疾患の診断及び処置において特に有用であることを示しているようである。本明細書で同定したSCS0010のモチーフ及び部位(例えば、グリコシル化部位、プロテオグリカン部位、SH2ドメイン、保存されたシステインリッチドメイン)は、処置及び/又は診断のために(アゴニスト又はアンタゴニスト、例えば抗体によって)修飾及び/又は標的化されうる。
【0183】
理論に拘束されることを望むものではないが、プロテオグリカンのためのTMEM9部位における突然変異(後述のELMの結果を参照のこと)は、ガンになる遺伝的素因に関与しているようである。Kveineらによって同定されたTMEM9のN末端部分の3つのNグリコシル化部位は、このタンパク質の構造及び機能にとって潜在的に重要なものであることが示唆された。彼等は異なる機能及び/又は親和性の特性を有すると思われる3つのグリコシル化形態を同定した。TMEM9とガンの関連も、Kurokawaらの刊行物(Kurokawa et al. J Exp Clin Cancer Res. 2004 Mar; 23(1) : 135-41. PCR-array gene expression profiling of hepatocellular carcinoma)、並びにマイクロアレイベースの実験及び他の発現実験(治療的用途の項目を参照のこと)によって支持されている。従って、可溶性SC0010は、TMEM9との結合によって、多数の疾患、例えばガンを軽減及び/又は予防することができるようである。
【0184】
尚、Prosite(http://us.expasy.org/prosite/)及びELM(http://elm.eu.org/basicELM/)も、これらの配列について行った。
【0185】
Prositeの結果
> PDOC00001 PS00001 ASN_GLYCOSYLATION Nグリコシル化部位[パターン][警告:発生の可能性が高いパターン]
25-28 NKSS
42-45 NISG
51-54 NVSQ
【0186】
> PDOC00005 PS00005 PKC_PHOSPHO_SITE プロテインキナーゼCリン酸化部位[パターン][警告:発生の可能性が高いパターン]
53-55 SqK
91-93 TiK
【0187】
> PDOC00006 PS00006 CK2_PHOSPHO_SITE カゼインキナーゼIIリン酸化部位[パターン][警告:発生の可能性が高いパターン]
27-30 SseD
53-56 SqkD
【0188】
【表4】
【0189】
ELMの結果の説明(ELMによるもの)
MOD_GlcNHglycan:プロテオグリカンは細胞表面及び細胞外マトリックス内で見られる。それらは細胞コミュニケーションにとって重要であり、例えば形態形成及び発生における役割を担っている。幾つかのプロテオグリカンにおける突然変異は、ガンになる遺伝的素因と関連している。コアタンパク質は、露出しているセリン残基上のグリコサミノグリカン鎖の付着によって修飾されている。ヘパラン硫酸の場合、この過程は、ゴルジ層板においてプロテインキシロシルトランスフェラーゼ(EC2.4.2.26)がUDP−キシロース由来のキシロースをセリンのヒドロキシル基に転移させることによって開始する。この系は後生動物において進化してきたようである。
【0190】
MOD N-GLC 1:Nグリコシル化は、真核細胞における分泌型及び膜結合型タンパク質の最も一般的な修飾である。Nグリコシル化の全工程は、100超の酵素及び輸送タンパク質を含んで成る。全てのN結合型オリゴ糖の生合成は、ERにおいて大きな前駆体オリゴ糖から開始する。このオリゴ糖[(Glc)3(Man)9(GIcNAc)2]の構造は植物、動物、単細胞真核生物において同一である。この前駆体は、前記オリゴ糖の担体として働く長鎖ポリイソプレノイド脂質であるドリコールと結合している。続いて、前記オリゴ糖は、ER酵素によって、ドリコール担体から新生タンパク質上のアスパラギン酸残基に転移する。オリゴ糖鎖は、続いて、前記糖タンパク質がゴルジ装置を通り抜ける場合にプロセシングされる。場合によっては、この修飾は、より多くのマンノース基の付着を伴い、他の場合にはより複雑な構造の型が付着する。
【0191】
LIG SH2 STAT5:Srcホモロジー2(SH2)ドメインは、異なるシグナル伝達経路に関与する大多数のタンパク質において見られる小さいモジュラードメインである。それらはリン酸化チロシン残基を含む特異的なモチーフと結合することができ、シグナルを下流に伝播してタンパク質間相互作用を促進し、そして/あるいは酵素活性を修飾する。SH2ドメインの異なるファミリーは、異なる結合特異性を有することがあり、これは通常、pYに対してC末端側の数残基(位置+1、+2及び+3)によって決定される。リン酸化されてないペプチドはSH2ドメインと結合しない。少なくとも3つの異なる結合モチーフが知られている。pYEEI(SrcファミリーSH2ドメイン)、pY[IV].[VILP](SH−PTP2、ホスホリパーゼC−ガンマ)、pY.[EN](GRB2)。しかしながら、SH2ドメインとそれらの基質との相互作用は、他の表面領域の協調的な接触とも独立している。
【実施例6】
【0192】
ケモカイン様ポリペプチドの確証及び特徴づけのための細胞−及び動物−ベースのアッセイ
構造−活性の相関関係に対する研究は、ケモカインがアミノ末端領域を利用することで受容体と結合し、そして受容体を活性化していることを示唆している。この領域のアミノ酸に対するタンパク質分解的な消化、突然変異、又は化学的修飾は、アンタゴニスト的な活性を有する化合物を生成しうる(Loetscher P and Clark-Lewis I, J Leukoc Biol, 69: 881-884,2001 Lambeir A, et al. J Biol Chem, 276: 29839-29845, 2001, Proost P, et al. Blood, 98 (13): 3554-3561,2001)。従って、相当のケモカインのアミノ末端領域又は他の領域の1又は複数の残基の特異的な修飾(欠失、非保存的な置換)から生じるアンタゴニスト的な分子は、炎症及び自己免疫疾患についての治療的な可能性を有していると考えられる(WO 02/28419; WO 00/27880; WO 99/33989; Schwarz MK and Wells TN, Curr Opin Chem Biol, 3: 407-17,1999)。それ故に、本願の更なる目的は、本発明のポリペプチドの修飾によって生じたそのような種類のアンタゴニストによって表される。
【0193】
本発明のポリペプチド及び関連試薬の治療上の応用は、創薬及び前臨床段階の開発の間のケモカイン及び他の生物学的産物の確証のために知られている、動物細胞、組織及びモデルを利用するin vivo/in vitroアッセイによって(Coleman RA et al., Drug Discov Today, 6: 1116-1126,2001 ; Li AP, Drug Discov Today, 6: 357-366,2001 ; Methods Mol. Biol vol. 138, "Chemokines Protocols", edited by Proudfoot AI et al. , Humana Press Inc. , 2000; Methods Enzymol, vol. 287 and 288, Academic Press, 1997)、又はin silico/コンピューターを利用したアプローチによって、(安全性、薬物動態及び有効性の観点から)評価されうる。
【0194】
本願は、疾患、例えば細胞増殖障害、自己免疫/炎症性障害、心臓血管障害、神経障害、発達障害、代謝障害、感染及び他の病状の処置又は予防において、適切に製剤化された医薬組成物における活性成分として有用でありうる新規ケモカイン様ポリペプチド及び一連の関連試薬を開示する。特に、既知のケモカインの特性を考慮すると、開示するポリペプチド及び試薬は、異常な又は欠陥のある細胞遊走に関与する症状に対処するはずである。そのような症状の非限定的な例は以下の通りである:関節炎、関節リウマチ(RA)、乾癬性関節炎、骨関節炎、全身性エリテマトーデス(SLE)、全身性硬化症、強皮症、多発性筋炎、糸球体腎炎、線維症、肺線維症及び肺炎、アレルギー性疾患及び過敏性疾患、皮膚炎、喘息、慢性閉塞性肺疾患(COPD)、炎症性腸疾患(IBD)、クローン病、潰瘍性大腸炎、多発性硬化症、敗血症ショック、HIV感染、移植片拒絶、創傷治癒、転移、子宮内膜症、肝炎、肝硬変、ガン、無痛覚症、並びにアテローム性動脈硬化症に関連する血管炎症。
【0195】
細胞培養物又は動物モデルを用いてケモカインの特異性、能力、及び有効性を試験するための複数のアッセイが開発されており、例えば、in vitroでの走化性アッセイ (Proudfoot A, et al. J Biol Chem 276: 10620-10626,2001 ;Lusti-Narasimhan M et al. , J Biol Chem, 270: 2716-21,1995)、又はマウス耳介膨脹(Garrigue JL et al., Contact Dermatitis, 30: 231-7,1994)である。有用なツール及び生成物(抗体、トランスジェニック動物、放射性標識タンパク質等)をもたらすための多数の他のアッセイ及び技術は、ケモカインに対して貢献する概説及び本において記載されており(Methods Mol. Biol vol. 138, "Chemokines Protocols", edited by Proudfoot Al et al., Humana Press Inc., 2000; Methods Enzymol, vol. 287 and 288, Academic Press, 1997)、そして、より正確には、本発明のケモカイン様ポリペプチド及び関連試薬の生物学的活性を、可能性のある治療又は診断方法及び使用と関連して証明するために使用されうる。
【0196】
サイトカイン発現調節アッセイ
1 序論
以下のin vitroの細胞ベースの三回一組のアッセイは、IL−2、IFN−γ、TNF−α、IL−5、IL−4及びIL−10についてのサイトカインビーズアレイ(CBA)アッセイによって測定した場合の異なるヒト末梢血単核細胞(hPBMC)に対して作用するコンカナバリンA(ConA)によって誘導されるサイトカイン分泌に対する本発明のタンパク質の作用を測定する。
【0197】
最適な条件は、96ウェルプレート1ウェル当たり100,000個の細胞であり、そして2%のグリセロール中最終的に100μlである。
【0198】
分裂促進因子(ConA)の最適濃度は5ng/mlである。
【0199】
当該アッセイに最適な時間は48時間である。
【0200】
読み取りの選択はCBAである。
【0201】
2 装置及びソフトウェア
・96ウェルマイクロタイタープレートフォトメーターEX(Labsystem).
・Graph Pad Prismソフトウェア
・エクセルソフトウェア
・フローサイトメーター Becton-Dickinson
・CBA解析ソフトウェア
・細胞培養のためのフード
・細胞培養のためのインキュベーター
・遠心機
・ピペット
【0202】
3 材料と試薬
・軟膜
・DMEM GIBCO
・ヒト血清AB型 SIGMA
・L−グルタミン GIBCO
・ペニシリン−ストレプトマイシン GIBCO
・フィコール PHARMACIA
・細胞培養のための96ウェルマイクロタイタープレート COSTAR
・コンカナバリンA SIGMA
・ヒトTh1/Th2サイトカインCBAキット Becton-Dickinson
・PBS GIBCO
・FALCON 50ml滅菌済み Becton-Dickinson
・BSA SIGMA
・グリセロール MERCK
・DMSO SIGMA
・96ウェルマイクロタイタープレートコニカルボトム NUNC
【0203】
4 方法
4.1 軟膜由来のヒトPBMCの精製
軟膜1〜2をDMEMで希釈する。25mlの希釈した血液をその後ゆっくりと50mlのファルコンチューブ中の15mlのフィコール層に添加し、そしてチューブを遠心する(2000rpm、20分、室温、中断無し)。中間層(リング)を続いて回収し、そして細胞を25mlのDMEMで洗浄し、その後遠心段階を行う(1200rpm、5分)。この手順を3回繰り返す。軟膜は合計約600x106個の細胞をもたらす。
【0204】
4.2 スクリーニング
80μlの1.25x106個の細胞/mlをDMEM+2.5%ヒト血清+1%L−グルタミン+1%ペニシリン−ストレプトマイシン中で希釈し、そしてその後96ウェルマイクロタイタープレートに添加する。
【0205】
ウェル当たり10μl添加する(ウェル当たり1つの条件):タンパク質をPBS+20%グリセロール中で希釈する(最終的なタンパク質の希釈は1/10である)。
【0206】
ウェル当たり10μlのConA刺激物を続いて添加する(ウェル当たり1つの条件):ConA 50μg/ml(ConAの終濃度は5μg/mlである)。
【0207】
48時間後、細胞上清を回収し、そしてヒトサイトカインをヒトTh1/Th2サイトカインCBAキット(Becton-Dickinson)で測定する。
【0208】
4.3 CBA解析
(より具体的には、CBAキットの小冊子を参照のこと)
i)混合ヒトTh1/Th2捕獲ビーズ
当該実験に必要なアッセイチューブの数を決定する。
【0209】
各捕獲ビーズの懸濁液を混合する前に激しく数秒間ボルテックスにかける。
【0210】
解析を行う各アッセイにつき、各捕獲ビーズの10μlのアリコートを「混合捕獲ビーズ」と標識を付した一本のチューブに添加する。ビーズ混合物を徹底的にボルテックスにかける。
【0211】
ii)試験試料の調製
上清は、アッセイ希釈物を用いて希釈した(1:4)(20μlの上清+60μlのアッセイ希釈物)。試料の希釈物は、96ウェルマイクロタイタープレートコニカルボトム(NUNC)中に試料を移す前に混合する。
【0212】
iii)ヒトTh1/Th2サイトカインCBAアッセイ手順
50μlの希釈した上清を96ウェルマイクロタイタープレートコニカルボトム(NUNC)中に添加する。50μlの混合した捕獲ビーズを添加し、続いて50μlのヒトTh1/Th2PE検出試薬を添加する。プレートを続いて3時間室温でインキュベートし、そして直接的な光への曝露から保護し、続いて1500rpmで5分間遠心する。上清を続いて慎重に捨てる。次の段階において、200μlの洗浄バッファーを2回各ウェルに添加し、1500rpmで5分間延伸し、そして上清を慎重に捨てる。130μlの洗浄バッファーをその後各ウェルに添加し、ビーズペレットを再懸濁する。試料は最終的にフローサイトメーター上で解析する。データはCBAアプリケーションソフト、Activity Base及びマイクロソフトエクセルソフトウェアを用いて解析する。
【0213】
アッセイの読み取りから、in vitroで、本発明のタンパク質が試験した全てのサイトカイン(IFN−γ、TNF−α、IL−2、IL−4、IL−5、IL−10)に対し一貫した阻害効果を有するか否かが評価されうる。
【0214】
更にEC50値に基づき、どれが最もよく阻害されたサイトカインであるか、そしてそのようなサイトカインに特に関連していることが知られている特定の自己免疫疾患/炎症性疾患に達しているかが容易に評価されうる。
【実施例7】
【0215】
自己免疫/炎症アッセイ
Tリンパ球応答を標的とするアッセイ
・Fas−リガンド誘導型T細胞死
このアッセイは、受容体媒介型の細胞死の新規モジュレーターを明らかにする。
【0216】
このアッセイにおいて、T細胞のアポトーシスはジャーカット細胞(ヒトT細胞系)を。組換え6ヒスチジンタグ付きFasリガンドとモノクローナル抗6−His抗体とを組み合わせて刺激することで誘導される。死亡は、細胞が死んでいる場合に培養液中に放出される細胞質酵素LDHの放出で定量される。読み取りは、490nmで読まれる比色アッセイで行う。T細胞は、多数の自己免疫疾患において病原性であることが証明されており、抗原特異的T細胞死を制御することができるということは治療上の戦略となる(例えば、クローン病患者における抗TNFα治療)。
【0217】
・ヒト−MLR:増殖及びサイトカイン分泌
この細胞ベースのアッセイは、リンパ球の増殖及びサイトカインの分泌又は別のドナー由来のPBMCによる刺激時の阻害(アロ反応性)に対する新規タンパク質の効果を測定する。これらのアッセイは、あらゆる自己免疫疾患における必須の細胞性応答である抗原特異的T細胞及び抗原提示細胞の機能について対処する。分泌型サイトカイン(IL−2,4,5,10,TNF−α及びIFN−γ)はCBAで定量される。注記:増殖とサイトカイン分泌は独立の反応である。
【0218】
・マウス−MLR:増殖
この細胞ベースのアッセイは、別のドナー(マウス株)由来の脾臓細胞による刺激後のマウス脾臓細胞のリンパ球増殖又は阻害に対する新規タンパク質の効果を測定する。この細胞ベースのアッセイは、Tリンパ球及び抗原提示細胞の応答に対する新規タンパク質の効果を測定し、そしてh−MLRアッセイにおけるポジティブの活性とヒットの同一性を確認するために使用されるであろう。このアッセイは、ヒト疾患のマウスモデルにおいて試験されるタンパク質を選択するために使用されるであろう。
【0219】
・スーパー抗原TSSTで刺激したヒトPBMC
スーパー抗原は、T細胞に影響を及ぼす免疫系の強力なモジュレーターである。スーパー抗原は、免疫介在疾患、例えばIBD、炎症性皮膚疾患、例えばアトピー性皮膚炎及び乾癬に影響を及ぼす。この細胞性アッセイにおいて、我々は、TCRを介するが、古典的な抗原に対するT細胞応答、特に共刺激分子に関するものと異なる要件を有するTリンパ球活性化を特異的にターゲティングする。
【0220】
・ConA又はPHAのいずれかで刺激したヒトPBMC
これらの細胞ベースのアッセイは、サイトカインビーズアレイ(CBA)アッセイ(IL−2、IFN−γ、TNF−α、IL−5、IL−4及びIL−10)によって測定した場合の異なる細胞に対する2つの異なる刺激作用によって誘導されるサイトカイン分泌に対する新規タンパク質の効果を測定する。
【0221】
サイトカインの多くが、損傷、環境及び細胞内標的に依存して、前炎症性又は抗炎症性という二重の作用を有することがある。サイトカイン分泌を調節する能力を有するあらゆるタンパク質が、治療上の潜在能力を有するようである(例えば、IFN−γ及びTNF−αの低下はTh1介在性自己免疫疾患において有益なようであり、一方、IL−4、IL−5の低下はTh2介在性の疾患において有益なようであり、IL−10の誘導はMS及びSLEにおいて興味深い)。
【0222】
単球/マクロファージと顆粒球の反応をターゲティングするアッセイ
・LPSで刺激したヒトPBMC
この細胞ベースのアッセイは、単球/マクロファージ及び顆粒球に対して作用するLPSによって誘導されるサイトカイン分泌(IFN−γ、TNF−α)に対する新規タンパク質の効果を測定する。
【0223】
IFN−γ及びTNF−α分泌を調節する能力を有するあらゆるタンパク質がTh1介在性の自己免疫疾患において有益なようである。
【0224】
好中球反応をターゲティングするアッセイ
好中球は、炎症性疾患及び自己免疫疾患、例えば関節リウマチにおいて重要である。白血球走化性因子、例えばIL−8は、細胞と微小血管内皮細胞との間の一連の接着性の相互作用を開始させて好中球の活性化、接着、そして最終的に遊走をもたらす。好中球の組織浸潤は、これらの細胞の細胞形態の特定の変化を伴う細胞骨格エレメントの再構成に依存する。
【0225】
この細胞ベースのアッセイは、ヒト好中球の細胞骨格再構成に対する新規タンパク質の効果を測定する。
【0226】
Bリンパ球反応をターゲティングするアッセイ
自己抗体並びに浸潤B細胞は、種々の自己免疫疾患、例えば全身性エリテマトーデス(SLE)、関節リウマチ(RA)、シェーグレン症候群及び重症筋無力症の病原において重要であると考えられている。B細胞の恒常性の異常調節が免疫寛容に影響を及ぼして、病原性抗体を産生し、且つ持続性の炎症を引き起こす自己反応性B細胞の不適切な生存をもたらしうるという有力な証拠がある。B細胞受容体のトリガリング後のB細胞の増殖、生存及び分化の制御において必須の役割を果たす新規因子の同定は、新規治療の開発に高度に関連するものである。
【0227】
・B細胞の増殖
この細胞ベースのアッセイは、B細胞の生存に対する新規タンパク質の効果を測定する。
【0228】
・B細胞の同時刺激
この細胞ベースのアッセイは、B細胞の同時刺激に対する新規タンパク質の効果を測定する。
【0229】
単球及びミクログリアの反応をターゲティングするアッセイ
・THP−1カルシウム流入
THP−1細胞アッセイにおけるCa+流入は、小胞体からの細胞内カルシウム放出(一般的な二次メッセンジャーの事象)を引き起こす能力に対する新規タンパク質の効果を測定する。
【0230】
ミクログリア細胞増殖(次のIACに提示されうるもの)
ミクログリア前駆細胞の増殖の間、多数のコロニー刺激因子、例えば幾つかのサイトカインが、重要な役割を果たすことが知られている。それらの中でも、M−CSFはマクロファージ/ミクログリアの成熟の最終段階に必須であり、そして他のどの因子によっても代替不可能である。この生物学的反応の評価は、ミクログリア活性に影響を及ぼす方法を代表することがあり、そしてそれ故にMSから治療上の潜在性を有する分子を同定する機会を代表することがある。
【0231】
M−CSFに対するミクログリア細胞系の増殖反応を測定する細胞ベースのアッセイが開発された。その実現可能性及び頑健性のフェーズは最適な結果を示した。このアッセイは96ウェルプレートで行われ;非放射性の基質が必要とされ、容易に自動化される。
【実施例8】
【0232】
タンパク質機能の生物学的関連性の調査に適した神経学的アッセイ
多数の神経学的アッセイが本出願人によって開発されており、そしてそれらはタンパク質の生物学的関連性の研究において使用するものである。本出願人によって開発された神経学的アッセイの例には、4つのタイプのアッセイがある。これらを以下論じる。
【0233】
i.乏突起膠細胞ベースのアッセイ
乏突起膠細胞はCNSにおけるミエリン形成にとって重要である。多発性硬化症において、それらは最初に攻撃される細胞であり、それらの損失は主要な行動障害を引き起こす。炎症の抑制に加え、MSで生じる病変の不完全な再ミエリン化の増強は、MSの治療上のストラテジーとして提唱されている。ニューロンのように、成熟した乏突起膠細胞は分割しないが、新規乏突起膠細胞は前駆細胞から生じることがある。成体の脳にはこれらの前駆細胞はほとんど存在せず、そして胚においてでさえも、前駆細胞の数はHTSにとって不適切である。
【0234】
Oli−neuは、t−neu発ガン遺伝子による乏突起膠細胞の不死化によって得られるマウスの細胞系である。それらは十分に研究されており、若い乏突起膠細胞の細胞学を研究するための代表的な細胞系として受け入れられている。
【0235】
これらの細胞は2つのタイプのアッセイにおいて使用されうる。
【0236】
一方は、乏突起膠細胞の増殖を刺激する因子を同定するためのものであり、そして他方は、それらの分化を促進する因子を発見するためのものである。それらの事象は、再生を助け、且つ脱髄疾患を治療する観点から重要である。
【0237】
別の可能性のある細胞系はヒト細胞系MO3−13である。MO3−13は、横紋筋肉腫細胞と成人ヒト乏突起膠細胞との融合から生じる。しかしながら、これらの細胞は乏突起膠細胞に分化する能力が低下しており、そしてそれらの増殖速度は増殖アッセイを可能にするのに十分ではない。それにも関わらず、それらは乏突起膠細胞の幾つかの特徴を有しており、それらの形態は、核転座の研究によく適合している。従って、この細胞系は、3つの転写因子、それぞれNF−κB、Stat−1及びStat−2の核転座に基づいたアッセイにおいて使用されることがある。Jak/Stat転写経路は多数の因子、例えばIFN−α、β、γ、サイトカイン(例えば、IL−2、IL−6;IL−5)又はホルモン(例えば、GH、TPO、EPO)によって活性化される複雑な経路である。反応の特異性は活性化されたStatの組み合わせに依存する。例えば、IFN−βはStat1、2及び3の核転座を活性化し、一方、IFN−γはStat1だけ活性化することは注目に値する。同様に、多数のサイトカイン及び増殖因子がNF−κBの転座を誘導した。これらのアッセイの目的は、所定のタンパク質についての活性化経路の写真を撮ることである。
【0238】
ii.星状細胞ベースのアッセイ
星状細胞の生物学は非常に複雑であるが、2つの一般的な状態が認識されている。静止状態と称される1つの状態において、星状細胞は、グルタミン酸をポンピングし、そしてエネルギー性の下層をニューロン及び乏突起膠細胞に提供することでニューロンの代謝性及び興奮性のレベルを制御する。活性化された状態において、星状細胞はケモカイン及びサイトカイン並びに一酸化窒素を産生する。最初の状態は正常な健康状態と見なされうるが、第二の状態は炎症、発作又は神経変性疾患の間に生じる。
【0239】
多数の因子及び多数の経路が星状細胞の活性化を調節することが知られている。星状細胞の活性化を調節する新規因子を同定するために、アストログリオーマ起源のヒト細胞系であるU373細胞が使用されうる。NF−κB、c−Jun並びにStatは、星状細胞活性化において中心的な役割を果たすことが知られているシグナル伝達分子である。
【0240】
NF−κB、c−Jun並びにStat1、2及び3の核転座に基づく一連のスクリーニングが実施されうる。これらの経路のプロトタイプの活性化因子はIL−1b、IFN−ベータ又はIFN−ガンマである。この目的は、CNS疾患の処置における治療物質として使用されうるタンパク質を同定することである。
【0241】
C.ニューロンベースのアッセイ
ニューロンは非常に複雑且つ多様な細胞であるが、それらは全て共通の2つの事を有している。第一にそれらは分裂終了細胞であり、第二にそれらは他の細胞を神経支配している。それらの生存は、神経支配された標的細胞がしばしば産生する栄養性因子の存在と関連している。多数の神経変性疾患において、標的の神経支配が失われると細胞体の萎縮及びアポトーシス性の細胞死に至る。従って、標的の欠損を補足するする栄養性因子の同定は神経変性疾患の処置において非常に重要である。
【0242】
この観点から、ラットPC12細胞のサブクローンであるNS1細胞を用いる生存アッセイが実施されうる。これらの細胞は何年も使用されており、そして多くの神経生物学の知識が、一次ニューロンに対して確認する前に、これらの細胞に対して最初に獲得されており、例えばニューロンの生存及び分化に関与する経路である(MEK、PI3K、CREB)。対照的に、マウス神経芽細胞腫であるマウスN2A細胞は、古典的な神経栄養因子には反応しないが、Junキナーゼ阻害剤は血清枯渇によって誘導されるアポトーシスを防ぐ。従って、これらの細胞系に対するアッセイは、異なるタイプの「生存促進」タンパク質を発見する助けとなるであろう。
【0243】
これまでのアッセイにおいて、本発明者が増殖と分化両方を促進する因子を同定しうることに注目することは重要である。ニューロンの分化を特異的に促進する因子を同定するために、神経突起伸長に基づいたNS1分化アッセイが使用されうる。神経変性疾患における軸策及び樹状突起の発芽を促進させることは、2つの理由から有利なことがある。最初に、それは変性ニューロンが再成長し、そして標的細胞との接触を再確立するのを助ける。第二に、それは、神経変性の末期、例えばパーキンソン又はADを遅らせる代償的な現象である、健康な線維からのいわゆる側枝の発芽を増強する。
【0244】
1.内皮細胞ベースのアッセイ
脳と血管の間の血液脳関門(BBB)は、皮質の髄液と血清の間の組成の差異に重要である。BBBは、内皮細胞と星状細胞との密接な接着から生じる。それは、脳における白血球の透過を防ぐことで免疫寛容状態を維持し、そして同一の細胞内シグナル伝達経路を用いる2つの平行した内分泌系の発達を可能にする。しかしながら、多くの疾患又は外傷において、BBBの完全性は変化し、そして白血球並びに血清タンパク質は脳に進入して神経炎症を誘導する。BBBのin vitroモデルは簡単ではないが、一次内皮細胞、例えばヒト臍帯静脈内皮細胞(HUVEC)は、BBB生物学の幾つかの側面を模倣することができる。例えば、BBBの漏出は、細胞なカルシウム放出を刺激するタンパク質によって誘導することができる。BBBの完全性を調節するタンパク質を同定する観点から、トロンビンを用いての又は用いないカルシウム動員アッセイがHUVECで実施されうる。
【実施例9】
【0245】
前記タンパク質機能の生物学的関連性の調査に適した線維芽細胞アッセイ
多数の線維芽細胞アッセイが本出願人によって開発されており、これはタンパク質機能の生物学的関連性の研究において使用するためのものである。本出願人によって開発された線維芽細胞アッセイの例には8つのタイプのアッセイが含まれる。これらを以下論じる。
【0246】
線維芽細胞の活性化及び病理学的増殖は、線維症として知られている現象を引き起こす重要な段階である。線維症は、細胞外マトリックス、例えばコラーゲンの過剰な沈着を特徴とする。間質細胞、例えば線維芽細胞は、特異的な線維化誘発性(pro-fibrotic)タンパク質及び抗線維化タンパク質を発現している。ケラチノサイト増殖因子(KGF)は抗線維化分子によってよく特徴付けられている。更に、酸化的損傷及び炎症誘発性の刺激は、主要な事象の中でも、筋線維芽細胞の表現型、そして最終的には線維症を引き起こすことが提唱されている。NF−κBは酸化的ストレス及び炎症反応のメディエーターである。線維芽細胞の生物学に基づき、本発明者は4つの細胞ベースのアッセイ、すなわち線維芽細胞増殖、コラーゲン産生、NF−κB活性化及びKFG産生アッセイを開発した。
【0247】
A.ヒト線維芽細胞増殖アッセイ
線維芽細胞の活性化及び病理学的な増殖は、線維症として知られている表現型を引き起こす重要な段階である。当該アッセイは、細胞性の核酸に結合したCyQuant GR Dyeを介する蛍光増強に基づくものであり、新規タンパク質及び小分子に対するヒト皮膚由来線維芽細胞の増殖反応を測定するものである。
【0248】
B.ヒト線維芽細胞によるI型コラーゲン産生
線維症は、細胞外マトリックス、特にコラーゲンの過剰な蓄積を特徴としている。I型コラーゲンの過剰産生は全身性硬化症の主要な徴候である。TGFβは、in vitro及びin vivoでコラーゲンの産生をアップレギュレートすることが知られている。本発明者は、新規な線維化誘発性(pro-fibrotic)分子及び抗線維化分子がヒト皮膚由来線維芽細胞によるI型コラーゲンの産生の基礎レベル又はTGFβ1刺激レベルを調節する能力を試験するための細胞ベースのアッセイを開発した。
【0249】
C.ヒト線維芽細胞によるケラチノサイト増殖因子(KGF)の産生
KGFは、正常及び病的な成長発達の間の、多くの器官(例えば、肺、膵臓、腎臓、前立腺、乳房、腺、子宮)における間質と上皮の相互作用の重要なメディエーターである。KGFは特に間質細胞により産生され、そしてその受容体は特に上皮細胞で発現している。KGFは線維症の病理学的反応の間に重要な役割を果たしているようであり、その結果これらの反応のマーカーとして使用することができると提唱されている。KGFのELISAアッセイが開発されており、ヒトの肺由来線維芽細胞を用いた結果、KGFの産生が有意にIL−1β及びTNFαによってアップレギュレートされることがあり、そしてTGFβによってダウンレギュレートされることがあることが証明された。これらのサイトカインは、KGF産生を誘導することができる新規タンパク質をスクリーニングする際の参照分子として使用されるであろう。
【0250】
D.線維芽細胞におけるNF−κB転写活性
酸化的損傷及び炎症誘発性の刺激は、主要な事象の中でも、筋線維芽細胞の表現型、そして最終的には線維症を引き起こすことが提唱されている。NF−κBは酸化的ストレス及び炎症反応のメディエーターである。Swiss3T3線維芽細胞は、安定的に組み込まれたNF−κB−SEAP(分泌型アルカリホスファターゼ)コンストラクトを用いて作出された。NF−κB−SEAPは、κエンハンサーに対する転写因子の結合を測定してNF−κB経路の活性化の直接的な測定を可能にするよう設計された。SEAP酵素は培養液中に分泌され、その結果試料は、細胞を回収することなく、転写活性についてのアッセイに対しての種々の時点で回収することができる。Swiss3T3−NF−κB−SEAP線維芽細胞系を細胞ベースのアッセイとして使用して新規な機能ゲノミクスタンパク質を試験することができ、そしてこれは小分子、特に炎症誘発性/抗炎症性の活性を有するものを試験するのに非常に見込みがある。
【0251】
E.線維芽細胞における結合組織成長因子(CTGF)プロモーターの活性化/抑制
38kDのシステインリッチなタンパク質であるCTGFは、線維芽細胞による細胞外マトリックス成分の産生を刺激する。CTGFの過剰発現は、伝えられるところによると、多くの線維性ヒト組織、例えば肺、皮膚、肝臓、腎臓及び血管において見出されている。in vitroでは、TGFβは、ヒト肺線維芽細胞におけるCTGF遺伝子の転写を活性化する。CTGFプロモーター−レポーターは、レポーターとして分泌型アルカリホスファターゼ(SEAP)を用いて構築され、そして3T3線維芽細胞は安定的に組み込まれたCTGF−SEAPコンストラクトを用いて作出された。これらの線維芽細胞を用いて、CTGFプロモーターがSARP−1、OPG及びFSGによってダウンレギュレートされ、そしてTGFβによってアップレギュレートされることが証明された。
【0252】
F.KL−6産生
最初に肺腺癌関連タンパク質として発見され、そしてその後MUC−1と称されることとなったKL−6は、高分子量糖タンパク質であり、現在ではCD9抗原として分類されている。KL−6は特発性肺線維症(IPF)及び他の肺間質性疾患を有する患者の血清及びBALFの両方で増大する。IPFに罹患している患者由来の肺組織において、KL−6抗体で標識された細胞の多くが再生II型肺細胞である。KL−6に対するポリクローナル抗体を産生する2つのペプチドを設計した。KL−6ELISAを使用して、ヒト肺由来II型肺細胞によるKL−6の産生を測定することができる。
【0253】
G.可溶性組換えTRAIL(TNF−関連アポトーシス誘導リガンド)で処理したL−929線維芽細胞のアポトーシスの中和
TRAILはオステオプロテジェリン(OPG)の細胞性リガンドのうちの1つであることが証明されている。このアッセイを使用してOPGの生物学的活性を測定することができる。
【0254】
H.ヒト線維芽細胞によるRANKL(NF−κBリガンドの受容体活性化因子)の産生
RANKLはOPGの別のリガンドである。このアッセイもOPGの生物学的活性を測定するために使用することができる。
【実施例10】
【0255】
タンパク質機能の生物学的関連性の調査に適したリプロダクティブ・ヘルスアッセイ
多くのリプロダクティブ・ヘルス関連アッセイが本出願人により開発されており、これらはSCS0010タンパク質機能の生物学的関連性の調査に使用するものである。男性不妊におけるSCS0010の予想される意味を考慮すると(治療用途を参照のこと)、本アッセイは特に関連性があるように見える。本出願人によって開発されたリプロダクティブ・ヘルス関連アッセイの例は、リプロダクティブ・ヘルスについての18の細胞ベースアッセイを含む。これらについて以下論じる。
【0256】
1.一次ヒト子宮平滑筋増殖アッセイ:
子宮平滑筋細胞の増殖は、女性における子宮筋腫疾患における腫瘍の発達のための前駆体である。このアッセイにおける目的は、一次ヒト子宮平滑筋細胞の増殖を阻害するタンパク質を同定することである。
【0257】
2.JEG−3着床アッセイ
JEG−3細胞は、着床の間の胚盤胞のモデルとして使用される栄養絨毛膜ヒトガン細胞系である。Ishikawa細胞は、脱落膜のモデルとして使用される比較的未分化のヒト子宮内膜ガン細胞系である。JEG−3細胞は、ヒト脱落膜組織に「着床」する。このアッセイにおいては2チャンバーシステムを使用し、ここで、蛍光標識したJEG−3細胞は、Ishikawa細胞又はIshikawa条件培地がロアチャンバー内に据えられた場合にマトリゲルコーティングした多孔性膜を通じてアッパーチャンバーからロアチャンバーへ進入する。遊走する細胞は、プレートリーダー内で定量される。in vivoでの着床を幇助するのに使用するためのJEG−3細胞の侵入を増大させるタンパク質を同定することがこの目的である。
【0258】
C.オステオポンチンビーズアッセイ(Ishikawa細胞):
Ishikawaヒト子宮内膜ガン細胞を着床のモデルとして使用する。ヒトにおける着床時に、種々のインテグリンが、胚盤胞によって発現するタンパク質と結合すると考えられている子宮内膜により発現する。Ishikawa細胞は、文献にてavb3を発現することが証明されており、avb3は「着床有効期間(window of implantation)」の間子宮内膜で発現するインテグリンである。このインテグリンは、栄養膜で発現するオステオポンチンと結合すると考えられている。このアッセイにおいて、オステオポンチンでコーティングした蛍光ビーズは線維芽細胞を表し、そしてIshikawa細胞は、エストラジオールでそれらを処理することによりそれらが結合することを受け入れるようプライミングされる。この目的は、着床の時期に子宮内膜の受容性を増大させるための補助として、Ishikawa細胞がオステオポンチンと結合する能力を増大させるタンパク質を同定することである。
【0259】
D.HuF6アッセイ
HuF6細胞は一次ヒト子宮線維芽細胞である。これらの細胞をIL−1βで処理することにより脱落膜が起こるように誘導することができる。脱落膜のマーカーは、PGE2の産生であり、これはELISAによって測定される。この目的は、妊娠初期の間に脱落膜を増強する手段としてHuF6細胞によるPGE2の産生を増大するタンパク質を同定することである。
【0260】
2.子宮内膜症アッセイ:
腹膜のTNFαは、子宮から脱落した子宮内膜細胞が腹膜の中皮細胞に接着して増殖するように誘導することで子宮内膜症における役割を果たしている。このアッセイにおいて、BEND細胞は、子宮内膜症の間の接着についてのアッセイとして、それらがフィブロネクチンでコーティングした蛍光ビーズに結合する能力を増大させるTNFαで処理される。この目的は、TNFαが前記細胞のビーズ結合能を刺激する能力を低下させ又は刺激するタンパク質を同定することである。
【0261】
F.hLHRで安定トランスフェクションしたJC−410ブタ顆粒膜細胞を用いるサイクリックAMPアッセイ
多嚢胞性卵巣症候群において、下垂体由来のLHは比較的多く、そして卵巣の莢膜細胞からのアンドロゲンの産生を誘導する。このアッセイは、PCOSの間の卵巣でのLHの作用を低下させるのに使用することができる、LHのシグナル伝達の阻害剤を探索するために使用される。JC−410ブタ顆粒膜細胞系は、ヒトLH受容体を安定トランスフェクションされる。LHによる処理は、cAMPの産生をもたらす。
【0262】
G.hFSHRで安定トランスフェクションしたJC−410ブタ顆粒膜細胞を用いるサイクリックAMPアッセイ
JC−410ブタ顆粒膜細胞をヒトFSHRで安定トランスフェクションした。FSHによる処理はcAMP産生を刺激し、これをこのアッセイで測定する。この目的は、顆粒膜細胞におけるFSH作用を増強するタンパク質を同定することである。
【0263】
H.LβT2(マウス)下垂体細胞アッセイ:
LβT2は、不死化マウス性腺刺激ホルモン分泌細胞系である。アクチビン単独又はGnRH+アクチビンによる刺激は、FSHの分泌をもたらす(GnRH単独による刺激はLHの分泌をもたらす)。前記細胞は、GnRHと協調してFSH産生を刺激するよう働くタンパク質を発見するためにGnRH+バイオスクリーニングタンパク質で処理してもよく、あるいはそれらは、アクチビン単独のようにFSH分泌を刺激することのできるタンパク質を発見するためにバイオスクリーニングタンパク質で処理してもよい。
【0264】
I.卵丘膨化アッセイ:
マウス卵丘−卵母細胞複合体(2/ウェル)を用いた卵丘膨化アッセイは、卵母細胞の成熟に影響を及ぼすタンパク質についてアッセイする96ウェルフォーマットにおいて確認されている(卵丘膨化によって測定)。アッセイ当たり2つの96ウェルプレートを処理することができ、そして一週当たり2回のアッセイを実施することができる。バイオスクリーニングタンパク質を一種類の濃度だけでアッセイする場合、全てのバイオクスクリーニングIタンパク質は一ヶ月でアッセイすることができる。読み取りは膨化についてのイエス/ノーアンサーで行ってもよく、あるいは画像解析プログラムを使用して定量的に膨化を測定することもできる。
【0265】
J.RWPE増殖アッセイ:
良性前立腺肥大症は前立腺の上皮及び間質の増殖を特徴とし、これはアポトーシスでバランスを失わず、その結果器官の拡大がもたらされる。RWPEはHPV−18で不死化された正常なヒト前立腺上細胞系であり、これは一次ヒト前立腺上皮細胞の代わりに使用されることがある。
【0266】
K.HT−1080線維肉腫浸潤アッセイ:
このアッセイは、JEG−3着床アッセイ(上記)のポジティブな細胞のコントロールとして開発された。これはガンの転移のためのモデルとしてよく確立されたアッセイである。蛍光標識HT−1080ヒト線維肉腫細胞は、2−チャンバーシステムのアッパーチャンバーにおいて培養され、そして、多孔性のマトリゲルコーティングした膜を通じてボトムチャンバー内に浸潤するよう刺激することができ、ここで、それらが定量される。この目的は、浸潤を阻害するタンパク質を同定することである。前記細胞は、血清をボトムチャンバーに添加することにより浸潤するよう刺激され、そしてドキシサイクリンで阻害される。
【0267】
L.一次ヒト子宮平滑筋細胞アッセイ
子宮筋腫疾患の特徴の1つは、平滑筋腫となっている子宮平滑筋細胞によるコラーゲン沈着である。一次ヒト子宮平滑筋細胞はTGFβ処理でコラーゲンを産生するよう刺激され、これはRebifで阻止される。この目的は、この線維化の表現型を阻害するタンパク質を発見することである。
【0268】
M.ヒト平滑筋腫細胞増殖アッセイ:
ヒト平滑筋腫細胞系は、増殖アッセイにおける子宮筋腫のモデルとして使用されうる。当該細胞は非常にゆっくりと生育し、そして本発明者はエストラジオール及び増殖因子でそれらを処理することによりそれらを刺激してより速い速度で生育させる。この目的は、平滑筋腫細胞のエストラジオール依存性の増殖を阻害するタンパク質を同定することである。
【0269】
N.937遊走アッセイ
子宮内膜症の病変は、免疫細胞を腹腔に補充するサイトカインを分泌する。これらの免疫細胞(特に活性化マクロファージ及びTリンパ球)は、子宮内膜症に共通の炎症の症候を媒介する。RANTESは子宮内膜症の間質細胞によって産生されることが証明されており、そして腹水中に存在している。このアッセイにおいて、活性化マクロファージのモデルとして使用される単球細胞系であるU937は、2−チャンバーシステムのより下の位置で処理してアッパーチャンバーから遊走させることで誘導されうる。当該細胞を蛍光色素で前処理する場合、それらをロアチャンバー内で定量することができる。この目的は、U937細胞の遊走を阻害するタンパク質を同定することである。
【0270】
O.JEG3ヒト栄養膜アッセイ:
胚盤胞の栄養膜はクラスIのHLA分子であるHLA−Gを産生し、これは、母体による胚の免疫拒絶を防ぐのに重要であると考えられている。子癇前症の間、HLA−Gレベルは低いか全くなく、その結果恐らく特徴的な症候が生じ、例えば、母体の免疫学的な干渉による子宮内膜及びらせん動脈への栄養膜の浸潤が乏しいことが挙げられる。JEG−3ヒト栄養膜細胞系はHLA−Gを産生し、これはIL−10又はLIFによる処理で増大することができる。ELISAを使用してJEG−3によるHLA−G産生を測定することができ、これは、HLA−G産生を増大することができる他のタンパク質の発見を目的とするものである。
【0271】
P.一次ラット卵巣分散体(dispersate)アッセイ:
JC−410FSHR/LHR細胞系由来の大量のステロイドを測定することが難しいため、未成熟ラットから採取した全卵巣由来の一次細胞を用いるアッセイを開発した。最初に、これらの培養物からのエストラジオールの産生をFSH及び/又はLH処理後に測定する。この目的は、ゴナドトロピン刺激型のステロイド産生を増強するタンパク質、又はこれらの培養物によりステロイド産生を増大させるよう単独で働くタンパク質を同定することである。
【0272】
Q.マウスIVFアッセイ:
このアッセイにおいて、卵母細胞を受精する能力により測定された精子の機能は、精子の受精能力を刺激するタンパク質を発見する目的でアッセイされる。
【0273】
R.一次ヒト前立腺間質細胞増殖アッセイ:
BPHの上皮成分についてのアッセイは既に上文で説明している(上記RWPEアッセイを参照のこと)。このアッセイは、BPHの間これらの細胞の増殖についてのモデルとして一次ヒト前立腺間質細胞を使用する。この目的は、これらの細胞の増殖を阻害するタンパク質を同定することである。
【0274】
【表5】
【表6】
【図面の簡単な説明】
【0275】
【図1】SCS0010とIL−8様ファミリーのメンバーとのアラインメント。
【図2】SCS0010のイントロン/エキソン構造。
【図3】SCS0010のヌクレオチド配列と翻訳。
【図4】pCR4−TOPO−SCS0010のマップ
【図5】pDONR221のマップ。
【図6】発現ベクターpEAK12dのマップ。
【図7】発現ベクターpDEST12.2のマップ。
【図8】pENTR−SCS0010−6HISのマップ。
【図9】pEAK12d−SCS0010−6HISのマップ。
【図10】pDEST12.2−SCS0010−6HISのマップ。
【図11】TMEM9とSCS0010とのアラインメント
【図12】TMEM9とSCS0010のSMARTドメイン。
【技術分野】
【0001】
本発明は、新規ポリペプチド、更に具体的にはIL−8様/TMEM9ポリペプチドをコードするものとしてヒトゲノムにおいて同定された核酸配列に関する。
【0002】
本明細書で引用する全ての刊行物、特許及び特許出願は、引用により全体が組み入れられる。
【背景技術】
【0003】
既に多くの新規ポリペプチドが、同じファミリーの公知のポリペプチドに対する厳密な相同性の基準を適用することにより同定されている。しかしIL−8様ポリペプチド(及び任意な他のタンパク質ファミリー)についてのヒトゲノム中のポリペプチド-コード配列中の実際の内容は依然不明であるので、ヒトゲノムに存在するオープンリーディングフレーム(ORF、すなわち、ヌクレオチドの連続的にコードしているトリプレットを含み、終止コドンによって中断されておらず、且つ潜在的にポリペプチドに翻訳可能なDNA配列)全体に、別の、そしてあまり厳密でない相同性/構造の基準を適用することにより、IL−8様ポリペプチド活性を有するポリペプチドをコードするDNA配列を同定することの可能性は尚も存在している。
【0004】
哺乳類の免疫応答は、一連の複雑なネットワーク様の相互作用に基づいており、これらには、細胞活動(移動、増殖、分化等)を調節することができる細胞成分(例えばリンパ球又は顆粒球)及び可溶性タンパク質が関与している。従って、ヒトの障害、特に免疫系に関連するものの診断、予防、及び治療における重大な進歩を提供する目的での、細胞調節因子の単離及び特徴づけに大きな関心が存在している。
【0005】
ケモカインは、それらが細胞の方向付けられた遊走及び活性化に関与しているため、これらの可溶性タンパク質の中に含まれる。この小さい(70〜130アミノ酸)分泌型のヘパリン結合炎症誘発性タンパク質のスーパーファミリーは、これらの細胞の補充を必要とする、血液から組織への白血球の血管外遊走における役割が特に知られている(Fernandez EJ and Lolis E, Annu. Rev. Pharmacol. Toxicol.. 42:469-499. 2002)。
【0006】
ケモカインは、それらが全て、保存されたシステインが分子内結合を形成している中心領域を含んでいるため、機能的に関連しているだけでなく、構造的に関連している。特に、成熟ポリペプチドのN末端配列における保存されたシステイン残基の数及び位置は、一般的に認識されているケモカイン分類の基礎的な基準であり、これらは孤立した又は隣接するシステイン残基、あるいは1〜3個のアミノ酸で分離されたシステイン残基を含むケモカイン間で本質的に分類される。
【0007】
一連の膜受容体は、全て7回膜貫通Gタンパク質共役受容体であって、ケモカインに対して、それらの生物学的活性を標的細胞に発揮させるようにする結合パートナーである。ケモカインの生理学的作用は、同時発生の相互作用の複雑で且つ統合された系に起因する。異なる細胞は、それらの状態及び/又は型に従い、受容体の特定の組み合わせを提示することができる。更に、ケモカイン受容体は、しばしば重複しているリガンド特異性を有し、その結果、単一の受容体は、単一のケモカインが異なる受容体に結合することができるのと同様に、異なるケモカインにより高い親和性で結合することができる。
【0008】
通常ケモカインは、損傷、炎症、又は他の組織変化の部位で産生され、そしてパラ分泌又は自己分泌の様式でそれらの活性を発揮する。しかしながら、炎症及び免疫反応における細胞型特異的な遊走及び活性化はケモカイン単独での活性ではない。他の生理学的活性、例えば造血又は血管形成、及び病理学的症状、例えば転移、移植片拒絶、アルツハイマー病又はアテローム性動脈硬化症は、少なくとも、これらのタンパク質のうちの幾つかによって制御されているようである。これは、ケモカイン及び/又はそれらの受容体は、複数の動物モデル又は臨床試料においてかなりアップレギュレートされ、そして/あるいは活性化されていることが明らかとなっているためである(Haskell CA et al., Curr. Opin. Invest. Drugs, 3: 399-455,2002 ; Lucas AD and Greaves DR, Exp. Rev. Mol. Med. 2001 ; Frederick MJ and Clayman GL, Exp. Rev. Mol. Med. 2001; Godessart N and Kunkel SL, Curr. Opin. Immunol., 13: 670-675, 2001 ; Reape TJ and Groot PH, Atherosclerosis,147 : 213- 25,1999)。
【0009】
治療物質としてケモカインを用いることには潜在的な欠点(特に、凝集する傾向及び無差別な結合)があるが、ケモカインに対してアンタゴニスト的な特性を有する分子は、過剰なケモカイン活性に関連する障害における治療的介入の価値のある機会を提供すると広く考えられる。特異的なケモカイン及びそれらの受容体の阻害は、不所望な又は制御されない細胞過程、例えば補充又は活性化を防ぐ解決策とみなされている(Baggiolini M, J. Intern. Med., 250: 91-104,2001 ; Proudfoot A, et al. Immunol. Rev., 177: 246-256,2000 ; Rossi D and Zlotnik A, Annu. Rev. Immunol., 18: 217-42,2000)。
【0010】
現在利用可能なヒトゲノム及び生理学に対する技術及び情報も新規で且つ有用な治療分子及び治療標的を提供するかもしれない新規なケモカイン及び受容体を発見するために使用された。最初に、ケモカイン遺伝子は、ヒトゲノムのジーンリッチ領域の、第4及び第17染色体上に規則正しくマッピングされたが(Nomiyama H et al., Genes Immun, 2: 110-113,2001)、この文献は、それぞれ特徴的なパターンでリンパ様組織及び他の組織において発現しており、且つ伝統的な遺伝子クラスターと異なる染色体の座位にマッピングされている転写物のバイオインフォマティクス解析を利用することにより新規なケモカインを特徴付けるための異なるアプローチの多数の例を提供している。ケモカインは、通常リンパ様組織及び他の組織において発現しているが、新規ケモカインは特異的な発現パターンを有することがあり、そして伝統的な遺伝子クラスターと異なる染色体の座位にマッピングされうる(WO 02/70706; Wells TN. and Peitsch MC. Methods Mol. Biol., 138: 65-73,2000 ; Chantry DF et al., J Leukoc Biol, 64: 49-54, 1998; Rossi D et al., J. Immunol, 158: 1033-1036,1997)。
【0011】
インターロイキン−8(IL−8)は、好中球活性化ペプチド−1又はSCYB8とも称される、炎症性刺激に応じて複数の細胞型によって分泌される組織由来のペプチドである。
【0012】
インターロイキン−8は、13のヒトCXCケモカインから成るファミリーのうちの1つである。これらの小さい塩基性のヘパリン結合タンパク質は炎症誘発性であり、そして主に好中球の活性化及び末梢血から組織への好中球の遊走を媒介する。IL−8は、呼吸器合胞体ウイルス(RSV)によって引き起こされるウイルス性の下気道感染気管支炎の発病に関与している。この疾患は毎年主要な伝染病に関与しており、何千人もの乳児が入院治療を必要としている。高レベルのIL−8がRSV気管支炎を有する乳児の鼻汁及び気管の吸引物で見られ、そしてIL−8のレベルは疾患の重症度と関連しているようである(Smyth, R. L. et al., Arch. Dis. Child. 82 (Suppl. 1) : A4-A5, 2000)。
【0013】
IL−8は造血前駆細胞(HPC)の迅速な動員を誘導する。動員は、マウスにおいて、ベータ−2−インテグリンLfa1(CD11A)に対する中和抗体による前処理によって完全に防止されうる。尚、マウスHPCはLfa1を発現せず、これは動員が補助細胞集団を必要とすることを示唆している。
【0014】
Pruijt, J. F. Mらは(Proc. Nat. Acad. Sci. 99: 6228-6233,2002)、多形核細胞(PMN)がIL−8誘導型のHPC動員における主要な制御因子としての役割を果たすことを示した。PMNの役割は、単回投与の抗好中球抗体の投与により好中球減少性となったマウスにおいて研究された。完全な好中球減少症が最大3〜5日で観察され、但し、7日目にリバウンドして好中球が増大する。IL−8誘導型の動員能力は、好中球減少期の間に有意に低下し、PMNの循環と共に再び現れ、そして好中球期(neutrophilic phase)の間に比例して増大した。このデータは、HPCのIL−8誘導型の動員が循環しているPMNのin vivoでの活性化を必要とすることを示した。
【0015】
多数の新規ケモカインが既知のケモカインに対し厳しいホモロジー基準を適用することによって同定されている。しかしながら、ケモカインについてのヒトゲノムにおけるポリペプチドコード配列の実際の内容(及びあらゆるほかのタンパク質ファミリーに関してのもの)は未だに知られておらず、オープンリーディングフレーム(ORF、すなわち、ヌクレオチドの連続的にコードしているトリプレットを含み、終止コドンによって中断されておらず、且つ潜在的にポリペプチドに翻訳可能なDNA配列)の解析において別の基準を適用することにより、アミノ酸をコードするDNA配列を同定することの可能性は尚も存在している。
【0016】
IL−8用タンパク質が疾患において有する役割は現在研究中である。しかしながら、新規IL−8様タンパク質の同定は、これらのタンパク質が関与しているとある病状に至る基礎的な経路の理解を深め、且つ、これらの障害を処置するのにより有効な遺伝子又は薬物治療を開発するのにかなり重要であるのは明らかである。
【発明の開示】
【発明が解決しようとする課題】
【0017】
本発明は、新規IL−8様/TMEM9ポリペプチドをコードするヒトゲノムのオープンリーディングフレーム(ORF)の同定に基づいている。このポリペプチドは、本明細書ではSCS0010ポリペプチドと称する。
【課題を解決するための手段】
【0018】
従って、本発明は、配列番号2で示すアミノ酸配列を有する、単離されたSCS0010ポリペプチド、並びに、IL−8様/TMEM9ポリペプチドの活性を有するポリペプチドとしての、それらの成熟型、それらのヒスチジンタグ付き型、変異体、及びフラグメント、を提供する。本発明はまた、それらをコードする核酸、当該核酸を含むベクター、及びこれらのベクター又は核酸を含む細胞、並びに他の関連する試薬、例えば融合タンパク質、リガンド、及びアンタゴニスト、を含む。
【0019】
本発明は、これらの分子を同定し、且つ生成するための方法、それらを含む医薬組成物を調製するための方法、並びに、疾患の診断、予防及び処置においてそれらを使用するための方法、を提供する。
【0020】
本発明の詳細な説明
本発明の第一の側面に従い、IL−8様/TMEM9活性を有する単離されたポリペプチドであって:
a)配列番号2に記載のアミノ酸配列;
b)配列番号2に記載の配列のポリペプチドの成熟型(配列番号4又は配列番号7);
c)配列番号4に記載の配列のポリペプチドのヒスチジンタグ付き型(配列番号5)
d)配列番号2に記載のアミノ酸配列の変異体であって、選択した配列において特定されている任意のアミノ酸が非保存的に置換されており、但し、当該配列中の15%未満のアミノ酸残基がそのように変化している、変異体;
e) a)〜c)に示すアミノ酸配列の活性フラグメント、前駆体、塩、又は誘導体、
から成る群から選択されるポリペプチド、が提供される。
【0021】
本明細書に記載の新規ポリペプチドは、クエリー配列としてIL−8様タンパク質を用いて同定され、そして最終的なアノテーションは、アミノ酸配列のホモロジーに基づいて行った。新規ポリペプチドはまた、内在性膜貫通タンパク質9のスプライス変異体としても同定された(TMEM9;実施例を参照のこと)。
【0022】
ヒトゲノムにおける既知のORF配列を翻訳することによって得られるアミノ酸配列の全体が、この共通配列を用いて挑戦され、そしてポジティブなヒットは、予測される構造的且つ機能的な「サイン」(N末端のシグナル配列及びC末端のアルファヘリックス)の存在について更にスクリーニングされ、そして最終的に、配列の特徴を既知のIL−8様ポリペプチドと比較することによって選択される。従って、本発明の新規ポリペプチドはIL−8様活性及びTMEM9活性を有することが予想されうる。
【0023】
用語「活性な」及び「活性」は、本願において配列番号2に提示するアミノ酸配列のIL−8様/TMEM9ポリペプチドについて予測されるIL−8様/TMEM9の特性を意味する。IL−8及び関連タンパク質は、炎症細胞、線維芽細胞及びケラチノサイトを創傷部位に引き付ける走化性活性を有する;それらはマイトジェンとして働いて細胞増殖を刺激する;サイトカインは血管形成、すなわち新規血管が創傷内部に増殖することを刺激することができる;それらはECMの産生及び分解に対しての著しい作用を有する;それらはまた、隣接細胞による他のサイトカイン及び増殖因子の合成に影響を及ぼす。IL−8様ケモカインとして機能する能力は、70pg/ml程度の低さのIL−8濃度を検出することができるヒトIL−8ELISAのようなアッセイキット(IBL, Hamburg)を用いて測定されうる。TMEM9は実施例で説明する。
【0024】
第二の側面において、本発明は、本発明の第一の側面のポリペプチドをコードしている精製された核酸分子を提供する。好ましくは、精製された核酸分子は、配列番号1に記載の核酸配列(配列番号2に記載のアミノ酸配列のIL−8様/TMEM9ポリペプチドをコードしている)又は配列番号3に記載の核酸配列(前記ポリペプチドの成熟型であって、アミノ酸配列が配列番号4に記載のものをコードしている)又は配列番号6に記載の核酸配列(前記ポリペプチドの成熟型であって、アミノ酸配列が配列番号7に記載のものをコードしている)を有する。好ましい成熟ポリヌクレオチド配列は、配列番号4に記載の好ましい成熟ポリペプチド型をコードしている配列番号3である。
【0025】
第三の側面において、本発明は、高ストリンジェントな条件下で、本発明の第二の側面の核酸分子とハイブリダイズする、精製された核酸分子を提供する。
【0026】
第四の側面において、本発明は、本発明の第二又は第三の側面の核酸分子を含む、発現ベクターなどの、ベクターを提供する。
【0027】
第五の側面において、本発明は、本発明の第四の側面のベクターにより形質転換された宿主細胞を提供する。
【0028】
第六の側面において、本発明は、本発明の第一の側面のポリペプチドと特異的に結合し、且つ好ましくは本発明の第一の側面のポリペプチドのIL−8様/TMEM9活性を阻害するリガンドを提供する。本発明のポリペプチドに対するリガンドは、天然の又は修飾された基質、酵素、受容体、最大2000Da、好ましくは800Da又はそれ未満の小型の天然又は合成の有機分子のような小型有機分子、ペプチドミメティクス、無機分子、ペプチド、ポリペプチド、抗体、前述のものの構造的又は機能的ミメティクスを含む、様々な形であってよい。
【0029】
第七の側面において、本発明は、本発明の第一の側面のポリペプチドをコードしている天然の遺伝子の発現を変更するか、又は本発明の第一の側面のポリペプチドの活性を制御するのに有効である化合物を提供する。
【0030】
本発明の第七の側面の化合物は、遺伝子の発現のレベル又はポリペプチドの活性を増加する(アゴニスト作用)又は減少する(アンタゴニスト作用)のいずれかであってもよい。重要なことは、本発明のIL−8様/TMEM9ポリペプチドの機能の同定は、疾患の治療及び/又は診断に有効である化合物を同定することが可能であるスクリーニング法のデザインを可能にする。
【0031】
第八の側面において、本発明は、治療又は診断において使用するための、本発明の第一の側面のポリペプチド、又は本発明の第二もしくは第三の側面の核酸分子、又は本発明の第四の側面のベクター、又は本発明の第五の側面の宿主細胞、又は本発明の第六の側面のリガンド、又は本発明の第七の側面の化合物を提供する。これらの分子は、IL−8様ポリペプチドが関与している疾患及び症状、例えば、免疫障害、転移、移植片拒絶、血管形成、アルツハイマー病、アテローム性動脈硬化症、亜致死性の内毒素血症、敗血症ショック、羊膜腔の微生物感染、回帰熱のヤーリッシュ・ヘルクスハイマー反応、中枢神経系の感染病、急性膵炎、潰瘍性大腸炎、膿胸、溶血性尿毒症症候群、髄膜炎菌性の疾患、胃の感染症、腹膜炎、乾癬、関節リウマチ、敗血症、喘息及び糸球体腎炎、の予防及び処置のための薬物の製造においても使用されうる。
【0032】
第九の側面において、本発明は、患者の疾患を診断する方法であって、当該患者の組織において、本発明の第一の側面のポリペプチドをコードしている天然の遺伝子の発現レベル又は本発明の第一の側面のポリペプチドの活性を評価すること、及び当該発現のレベル又は活性を対照レベルと比較することを含む方法を提供し、ここで当該対照レベルとは異なるレベルは、疾患の指標である。このような方法は、in vitroで実行されることが好ましいであろう。同様の方法を用い、患者における疾患の治療的処置をモニタリングすることができ、ここで経時的に対照レベルへと向かうポリペプチド又は核酸分子の発現のレベル又は活性の変化は、疾患の回復の指標である。
【0033】
本発明の第一の側面のポリペプチドを検出する好ましい方法は:(a)本発明の第六の側面のリガンド、例えば抗体を、生物学的試料と、リガンド−ポリペプチド複合体を形成するのに適した条件下で接触する工程;及び(b)当該複合体を検出する工程、を含んで成る。
【0034】
本発明の第九の観点に従う多数の異なるそのような方法が存在するのは当業者にとって自明であり、例えば、短いプローブによる核酸ハイブリダイゼーション法、点変異解析、ポリメラーゼ連鎖反応(PCR)増幅、及び、抗体を用いて異常なタンパク質レベルを検出する方法である。同様の方法を短期又は長期的に用いて、疾患の治療的処置が患者においてモニタリングされることが可能となりうる。本発明はまた、これらの疾患診断方法に有用なキットも提供する。
【0035】
第十の側面において、IL−8様/TMEM9タンパク質として本発明の第一の側面のポリペプチドの使用を提供する。適当な使用には、細胞増殖、代謝又は分化の制御因子としての使用、受容体/リガンド対の一部としての使用及び上記リストから選択される生理学的又は病理学的症状の診断用マーカーとしての使用が含まれる。
【0036】
第十一の側面において、本発明は、本発明の第一の側面のポリペプチド、又は本発明の第二もしくは第三の側面の核酸分子、又は本発明の第四の側面のベクター、又は本発明の第五の側面の宿主細胞、又は本発明の第六の側面のリガンド、あるいは本発明の第七の側面の化合物を、医薬として許容される担体と組合わせて含んで成る医薬組成物を提供する。
【0037】
第十二の側面において、本発明は、IL−8様ポリペプチドが関与している疾患及び症状、例えば、免疫障害、転移、移植片拒絶、血管形成、アルツハイマー病、アテローム性動脈硬化症、亜致死性の内毒素血症、敗血症ショック、羊膜腔の微生物感染、回帰熱のヤーリッシュ・ヘルクスハイマー反応、中枢神経系の感染病、急性膵炎、潰瘍性大腸炎、膿胸、溶血性尿毒症症候群、髄膜炎菌性の疾患、胃の感染症、腹膜炎、乾癬、関節リウマチ、敗血症、喘息及び糸球体腎炎、の診断又は処置のための薬物の製造における使用のための、本発明の第一の側面のポリペプチド、又は本発明の第二もしくは第三の側面の核酸分子、又は本発明の第四の側面のベクター、又は本発明の第五の側面の宿主細胞、又は本発明の第六の側面のリガンド、あるいは本発明の第七の側面の化合物を提供する。
【0038】
第十三の側面において、本発明は、患者の疾患を処置する方法であって、当該患者へ、本発明の第一の側面のポリペプチド、又は本発明の第二もしくは第三の側面の核酸分子、又は本発明の第四の側面のベクター、又は本発明の第五の側面の宿主細胞、又は本発明の第六の側面のリガンド、あるいは本発明の第七の側面の化合物を投与することを含んで成る方法を提供する。
【0039】
本発明の第一の側面のポリペプチドをコードする天然の遺伝子の発現、又は本発明の第一の側面のポリペプチドの活性が、健常な対象者の発現のレベル又は活性と比較したとき罹患患者で低下する疾患については、この患者に投与される前記ポリペプチド、核酸分子、リガンド又は化合物はアゴニストであるはずである。逆に、前記天然の遺伝子の発現又はポリペプチドの活性が、健常な対象者の発現のレベル又は活性と比較したとき罹患患者で上昇する疾患については、この患者に投与される前記ポリペプチド、核酸分子、リガンド又は化合物はアンタゴニストであるはずである。このようなアンタゴニストの例にはアンチセンス核酸分子、リボザイム及びリガンド、例えば抗体が含まれる。
【0040】
第十四の側面において、本発明は、本発明の第一の側面のポリペプチドを高レベル、低レベルで発現、又は発現しないように形質転換したトランスジェニック又はノックアウト非ヒト動物を提供する。このようなトランスジェニック動物は、疾患の研究用モデルとして非常に有用であり、さらに前記疾患の治療又は診断に有効な化合物の同定を目的とするスクリーニング方法で用いることもできる。
【0041】
本発明を利用するために用いることができる標準的な技術及び手法の要約は後述する。本発明は、記載される特定の方法論、プロトコール、細胞系、ベクター及び試薬に限定されないことは理解されよう。本明細書で用いられる専門用語は単に特定の態様を説明するためのものであり、この用語によって本発明の範囲を限定しようとするものではないこともまた理解されよう。本発明の範囲は特許請求の範囲の用語によってのみ限定される。
【0042】
本明細書では、ヌクレオチド及びアミノ酸についての標準的な略語が用いられる。
【0043】
本発明の実施では別に指示がなければ、分子生物学、微生物学、組換えDNA技術及び免疫学の通常の技術が用いられ、これらの技術は当業者の技術範囲内である。
【0044】
このような技術は、文献で十分に説明されている。特に適切な解説書の例には以下が含まれる:Sambrook Molecular Cloning; A Laboratory Manual, Second Edition (1989);DNA Cloning, Vol.I及びII (D.N. Glover編集、1985);Oligonucleotide Synthesis (M.J. Gait編集、1984);Nucleic Acid Hybridization (B.D. Hames及びS.J. Higgins編集、1984);Transcription and Translation (B.D. Hames及びS.J. Higgins編集、1984);Animal Cell Culture (R.I. Freshney編集、1986);Immobilized Cells and Enzymes (IRL Press, 1986);B. Perbal, A Practical Guide to Molecular Cloning (1984);the Methods in Enzymology series (Academic Press, Inc.)特にVol. 154及び155;Gene Transfer Vectors for Mammalian Cells (J.H. Miller and M.P. Calos編集、1987, Cold Spring Harbor Laboratory);Immunochemical Methods in Cell and Molecular Biology (Mayer及びWalker編集、1987, Academic Press, London);Scopes, (1987) Protein Purification: Principles and Practice, Second Edition (Springer Verlag, NY);及びHandbook of Experimental Immunology, Vols. I-IV (D.M. Weir and C.C. Blackwell編集、1986)。
【0045】
本発明の第一の側面は、配列番号2又は配列番号4又は配列番号5又は配列番号6に記載のアミノ酸配列の変異体を含み、ここで、選択された配列で特定された任意のアミノ酸は、非保存的に置換され、但し当該配列中の15%未満のアミノ酸残基がそのように変化している。指定された数の非保存的置換を有するタンパク質配列は、通常利用可能なバイオインフォマティクスツールを用いて同定することができる(Mulder NJ及びApweiler R, 2002; Rehm BH, 2001)。
【0046】
このような配列に加え、一連のポリペプチドは、本発明の開示の一部を形成する。IL−8様ポリペプチドはN末端配列のタンパク質分解性の除去(シグナルペプチダーゼ及び他のタンパク質分解酵素による)を含む成熟過程を進行することが分かっているので、本願は、配列番号2に記載の配列のポリペプチドの成熟型も特許請求する。これらのポリペプチド配列は配列番号4又は配列番号7に記載されている。好ましい成熟型は配列番号4に記載されている。成熟型は、IL−8様/TMEM9活性を示し、且つin vivo(発現している細胞又は動物によるもの)又はin vitro(精製されたポリペプチドの特異的酵素による修飾によるもの)での翻訳後成熟過程から生じるあらゆるポリペプチドを含むことが意図されている。他の選択的(alternative)成熟型も、糖又はリン酸のような化学基の付加から生じ得る。
【0047】
本願はまた、配列番号4に記載の配列のポリペプチドのヒスチジンタグ付き型も特許請求する。このポリペプチド配列は配列番号5に記載されている。
【0048】
特許請求する他のポリペプチドは、配列番号2、配列番号4又は配列番号5又は配列番号7で示すアミノ酸配列の活性変異体であり、ここで、選択した配列において特定されている任意のアミノ酸が非保存的に置換されており、但し、当該配列中の15%未満、好ましくは10%、5%、3%、又は1%未満のアミノ酸残基がそのように変化している。指定のパーセンテージは、開示した新規アミノ酸配列全体について測定されされなければならない。
【0049】
本発明によると、あらゆる置換が、分子の構造及び生物学的機能を保存するために、好ましくは「保存的」置換又は「安全」置換であるべきであり、これは、十分に類似の化学的特性を有するアミノ酸を導入する、一般的に定義されている置換のことである(例えば、塩基性正電荷アミノ酸は別の塩基性正電荷アミノ酸によって置換されるべきである)。
【0050】
文献は、保存的アミノ酸置換の選択が、タンパク質の配列及び/又は構造に対する統計的及び/又は物理化学的研究に基づいて実施されうる多数のモデルを提供している(Rogov Si and Nekrasov AN, 2001)。タンパク質デザイン実験は、アミノ酸の特異的なサブセットの使用が、折りたたみ可能で且つ活性なタンパク質を生成することができ、その結果、タンパク質構造内に容易に適合され且つ、機能的且つ構造的ホモログ及びパラログを検出するために使用されうるアミノ酸の「同義」置換の分類に役立つ(Murphy LR et al., 2000)。同義アミノ酸群及び更に好ましい同義アミノ酸群を表1に示す。
【0051】
相当のIL−8様/TMEM9ポリペプチドに対し、同等の、又は改善された活性を有する活性変異体は、コードDNAの常用の突然変異誘発技術、コードDNA配列のレベルでのコンビナトリアル技術(例えば、DNAシャッフリング、ファージディスプレー/セレクション)から、あるいはコンピューター支援設計研究、続く従来技術において説明されているような所望の活性についてのバリデーション、から生じることがある。
【0052】
特異的な非保存的突然変異も、異なる目的で本発明のポリペプチド内に導入されることがある。IL−8様/TMEM9ポリペプチドの親和性を低下する突然変異は、その再使用能及び再循環能を高め、これはその治療効能を高める可能性がある(Robinson CR, 2002)。最終的には本発明のポリペプチド中に存在する免疫原性エピトープが、ワクチン開発のために活用されるか(Stevanovic S, 2002)、もしくはタンパク質安定性を増大する変異を選択し、それらを補正するための公知の方法に従いそれらの配列を修飾することにより排除され得る(van den Burg B及びEijsink V, 2002;国際公開公報第02/05146号、第00/34317号、第98/52976号)。
【0053】
本発明の更に別のポリペプチドは、上述の配列のアミノ酸配列の活性なフラグメント、前駆体、塩、又は機能的に等価の誘導体である。
【0054】
フラグメントは、それらの機能を変化することなく末端又は内部のアミノ酸の欠失を提示すべきであり、そして、前記タンパク質の機能的な高次構造に必須のアミノ酸を除去又は置換することなく、一般的に数個のアミノ酸、例えば10個以下、好ましくは3個以下のものを包含すべきである。小さいフラグメントは、抗原決定基を形成することがある。
【0055】
「前駆体」は、細胞又は身体への投与前又は後に代謝的で且つ酵素的なプロセシングにより本発明の化合物へと変換されうる化合物である。
【0056】
用語「塩」は、本明細書では、本発明のポリペプチドのカルボキシル基の塩及びアミノ基の酸付加塩の両方を意味する。カルボキシル基の塩は、当業界で知られている手段によって形成することができ、そして無機塩、例えばナトリウム、カルシウム、アンモニウム、第二鉄又は亜鉛の塩など、及び、例えばアミンで形成されるような有機性塩、例えばトリエタノールアミン、アルギニン又はリジン、ピペリジン、プロカインなどを含む。酸付加塩には、例えば、鉱酸、例えば塩酸又は硫酸との塩、及び有機酸、例えば酢酸又はシュウ酸との塩、が含まれる。そのような塩のいずれも、本発明のペプチド及びポリペプチド又はそれらの類似体と実質的に同様の活性を有するはずである。
【0057】
用語「誘導体」は、本明細書で使用する場合、アミノ酸部分の側鎖又はアミノ末端若しくはカルボキシ末端の基の上に存在する官能基から、既知の方法に従い調製されうる誘導体を意味する。そのような分子は、通常一次配列を変化させない他の修飾、例えば、ポリペプチドのin vivo又はin vitroでの化学的誘導体化(アセチル化又はカルボキシル化)であって、ペプチドの合成及びプロセシングの間に又は更なるプロセシング段階において、当該ペプチドのリン酸化(ホスホチロシン、ホスホセリン、又はホスホスレオニン残基の導入)又はグリコシル化(哺乳類のグリコシル化酵素にポリペプチドを曝露することによるもの)のパターンを修飾することによって生成されるもの、からも生じることがある。あるいは、誘導体は、前記カルボキシル基のエステル又は脂肪族アミド及び遊離アミノ基のN−アシル誘導体又は遊離ヒドロキシル基のO−アシル誘導体を含み、且つ、例えばアルカノイル基又はアリール基として、アシル基により形成される。
【0058】
前記誘導体の生成は、内部又は末端の位置において、適切な残基の部位指定の修飾を包含してもよい。付着に使用する残基は、ポリマーの付着が可能な側鎖(すなわち、官能基を有する側鎖、例えばリジン、アスパラギン酸、グルタミン酸、システイン、ヒスチジン等)を有するはずである。あるいは、ポリマーの付着が可能な側鎖を有する残基は、前記ポリペプチドのアミノ酸を置換することができ、あるいは前記ポリペプチドの内部又は末端の位置で付加することができる。また、遺伝学的にコードされているアミノ酸の側鎖は、ポリマー付着のために化学的に修飾されることがあり、あるいは適切な側鎖の官能基を有する非天然アミノ酸も利用されることがある。好ましい付着方法は、ペプチド合成と化学的ライゲーションの組み合わせを利用する。有利には、水溶性ポリマーの付着が、生分解性リンカーを介して、特にタンパク質のアミノ末端領域でなされる。そのような修飾は、前記リンカーの分解時にポリマーの修飾無しに前記タンパク質を放出する前駆体(又は「プロドラッグ」)型のタンパク質を提供するように働く。
【0059】
ポリマー付着は、アンタゴニストの特定の位置で自然に発生するアミノ酸の側鎖又はアンタゴニストの特定の位置で自然に発生するアミノ酸を置換する天然若しくは非天然の側鎖に対してだけでなく、標的の位置でアミノ酸の側鎖に付着する炭水化物又は他の部分に対してもなされうる。希少な又は非天然のアミノ酸も、特異的に操作された菌種においてタンパク質を発現することによって導入されうる(Bock A, 2001)。
【0060】
上文で示した変異体は全て、自然に発生するものでも、ヒト以外の生物において同定されるものでも、あるいは、化学合成、部位指定突然変異誘発技術、又は任意な他の適当な既知の技術により調製される人工的なもの、であってもよく、これは当業界で提示されている技術を用いて当業者によってルーチンに得ることができ、そして試験することができる、実質的に相当の突然変異型又は短縮型のペプチドの有限集合を提供する。
【0061】
本願で開示されている新規アミノ酸配列は、異なる種類の試薬及び分子を提供するために使用されうる。これらの化合物の例は、それらの完全配列又は特異的フラグメント、例えば抗原決定基を用いて同定されうる結合タンパク質又は抗体である。ペプチドライブラリーは、特許請求の範囲のアミノ酸配列に結合する抗体又は他のタンパク質をスクリーニングし、そして特徴付けるための、そして同様の結合特性を有する前記ポリペプチドの別の型を同定するための既知の方法(Tribbick G, 2002)において使用されうる。
【0062】
本願は、上述のポリペプチドのいずれかを含む融合タンパク質も開示している。これらのポリペプチドは、ポリペプチドのIL−8様/TMEM9活性を著しく損なうことも、追加の特性を提供する可能性もなく、本願で開示されたタンパク質配列とは異種のタンパク質配列を含むべきである。このような特性の例は、より容易な精製手法、より長い体液中の半減期の維持、追加の結合部分、末端タンパク質分解性の消化による成熟、又は細胞外局在化がある。この後者の特徴は、先の定義に含まれる融合タンパク質又はキメラタンパク質の特定群を定義するために特に重要であり、その理由は、特許請求する分子が、これらのポリペプチドの単離及び精製が促進される空間のみではなく、一般にIL−8様/TMEM9ポリペプチド及びそれらの受容体が相互作用する空間にも、局在化することを可能にするからである。
【0063】
前記部分、リガンド及びリンカーの設計は、融合タンパク質の構築、精製、検出及び使用のための方法及び戦略同様、文献において開示されている(Nilsson J et al., 1997; Methods Enzymol, Vol. 326-328, Academic Press, 2000)。本発明の融合タンパク質に含まれうる好ましいタンパク質配列は、これらのタンパク質配列:膜結合タンパク質、イムノグロブリンの定常領域、多量体化ドメイン、細胞外タンパク質、シグナルペプチド含有タンパク質、エクスポートシグナル含有タンパク質、に属する。これらの配列の特徴及びそれらの特異的な使用は詳細に開示されており、例えばアルブミン融合タンパク質(WO01/77137)、多量体化ドメインを含む融合タンパク質(WO01/02240、WO00/24782)、免疫複合体(Garnett MC, 2001)、又はアフィニティークロマトグラフィーによる組み換え産物の精製を可能にする配列を含む融合タンパク質(Constans A, 2002; Burgess RR and Thompson NE, 2002; Lowe CR et al., 2001; J. Bioch. Biophy. Meth., vol. 49 (1-3),2001; Sheibani N, 1999)である。
【0064】
本発明のポリペプチドは、それらに特異的に結合するリガンドを生成し、そして特徴付けるために使用されうる。これらの分子は、天然又は人工の、化学的観点(結合タンパク質、抗体、分子インプリンティングポリマー)から非常に異なるものであってもよく、そして当業界の技術(WO 02/74938 ; Kuroiwa Y et al., 2002; Haupt K, 2002; van Dijk MA and van de Winkel JG, 2001; Gavilondo JV and Larrick JW, 2000)を適用することによって生成されてもよい。そのようなリガンドは、それらを生成せしめたポリペプチドのIL−8様活性を拮抗化し、又は阻害することができる。特に、共通で且つ効率的なリガンドは、膜結合タンパク質又は抗体の細胞外ドメインで表され、これは、モノクローナル抗体、ポリクローナル抗体、ヒト化抗体、又は抗原結合フラグメントの形態であってもよい。
【0065】
上述のポリペプチド及びポリペプチドベースの誘導体化された試薬は、所望の使用方法及び/又は製造方法に従い、別の型、例えば、放射性標識、蛍光標識、ビオチン、又は細胞毒性物質の中から選択される分子との活性のある接合体又は複合体であってもよい。
【0066】
特定の分子、例えばペプチドミメティクス(peptide mimetic)も、本発明のポリペプチドの配列及び/又は構造を基に設計されることがある。ペプチドミメティクス(peptidomimeticとも称される)は、アミノ酸の側鎖の、アミノ酸のキラリティーの、及び/又はペプチド主鎖のレベルで化学的に修飾されたペプチドである。これらの変更は、改善された調製、性能及び/又は薬物動態の特徴を有する本発明のポリペプチドのアゴニスト又はアンタゴニストを提供することが意図される。
【0067】
例えば、前記ペプチドが、問題のある対象へのインジェクションの後にペプチダーゼによる開裂を受けやすい場合、非開裂性のペプチドミメティクスによる特に感受性のあるペプチド結合の置換は、より安定で且つその結果治療薬として更に有用なペプチドを提供することができる。同様に、L−アミノ酸残基の置換は、タンパク質分解に対してほとんど感受性がないペプチドであって、最終的にペプチド以外の有機化合物により類似のペプチドを賦与する標準的な方法である。アミノ末端のブロッキング基、例えばt−ブチルオキシカルボニル、アセチル、テイル(theyl)、スクシニル、メトキシスクシニル、スベリル(suberyl)、アジピル、アゼライル(azelayl)、ダンシル、ベンジルオキシカルボニル、フルオレニルメトキシカルボニル、メトキシアゼライル、メトキシアジピル、メトキシスベリル、及び2,4−ジニトロフェニルも有用である。増大した性能、延長された活性、精製の容易さ、及び/又は増大した半減期を提供する多くの他の修飾が従来技術で開示されている(WO 02/10195; Villain M et al., 2001)。
【0068】
ペプチドミメティクスに含まれるアミノ酸誘導体にとって好ましい別の同義群は、表IIに規定されているものである。アミノ酸誘導体の非限定的なリストには、アミノイソ酪酸(Aib)、ヒドロキシプロリン(Hyp)、1,2,3,4−テトラヒドロ−イソキノリン−3−COOH、インドリン−2カルボン酸、4−ジフルオロ−プロリン、L−チアゾリジン−4−カルボン酸、L−ホモプロリン、3,4−デヒドロプロリン、3,4−ジヒドロキシ−フェニルアラニン、シクロヘキシル−グリシン、及びフェニルグリシンも含まれる。
【0069】
「アミノ酸誘導体」とは、20個の遺伝子でコードされる天然のアミノ酸とは異なるアミノ酸又はアミノ酸様の化学的存在物を意図する。特に、アミノ酸誘導体は、置換型又は非置換型の直鎖若しくは分枝、又は環状のアルキル部分を含むことがあり、且つ1又は複数のヘテロ原子を含むことがある。アミノ酸誘導体は、新規に生成されてもよく、又は商業的供給源(Calbiochem-Novabiochem AG, Switzerland; Bachem, USA)から入手してもよい。
【0070】
タンパク質の構造及び機能をプローブし、そして/あるいは改善させるために、in vitro及びin vivo両方の翻訳系を用いて非天然アミノ酸誘導体をタンパク質に組み込む種々の方法論が文献において開示されている(Dougherty DA, 2000)。非ペプチドミメティクスと同様、ペプチドミメティクスの合成及び開発のための技術も当業界で周知である(Golebiowski A et al., 2001; Hruby VJ and Balse PM, 2000; Sawyer TK, in "Structure Based Drug Design", edited by Veerapandian P, Marcel Dekker Inc., pg. 557-663,1997)。
【0071】
本発明の別の目的は、IL−8様/TMEM9活性を有する本発明のポリペプチド、それらに対して生成する抗体又は結合タンパク質と結合するポリペプチド、相当の融合タンパク質、あるいは上文で開示したアンタゴニスト活性を有する突然変異体、をコードする単離された核酸にある。好ましくは、これらの核酸は、配列番号1又は配列番号3又は配列番号6から成る群から選択されるDNA配列、あるいは前記DNA配列のいずれかの相補体、を含んで成るべきである。
【0072】
あるいは、本発明の核酸は、配列番号1又は配列番号3又は配列番号6から成る核酸配列、あるいは前記DNA配列の相補体と、高ストリンジェントな条件下でハイブリダイズし、あるいは少なくとも一続きの約30個のヌクレオチドの範囲にわたり少なくとも約85%の同一性を示すはずである。
【0073】
用語「高ストリンジェントな条件」は、非常に類似の分子の会合を容易にするハイブリダイゼーション反応における条件であって、50%ホルムアルデヒド、5xSSC(150mMのNaCl、15mMのクエン酸三ナトリウム)、50mMのリン酸ナトリウム(pH7.6)、5xデンハルト溶液、10%硫酸デキストラン、及び20μg/mlの変性してせん断されたサケ精子DNAを含んで成る溶液中で60〜65℃一晩インキュベーションし、続いて0.1xSSC中フィルターを同一の温度で洗浄することに存するものを意味する。
【0074】
実質的に同一のヌクレオチド配列を含むこれらの核酸は、コードされたポリペプチドを維持し、修飾し、導入し、又は発現するために使用されうるプラスミド、ベクター及び任意な他のDNAコンストラクトに含まれることがある。特に、前記核酸分子が発現制御配列と作用可能に連結しているベクターは、原核又は真核宿主細胞における、コードされたポリペプチドの発現を可能にしうる。
【0075】
用語「実質的に同一のヌクレオチド配列」は、遺伝コードの縮重が原因で、所定のアミノ酸配列をもコードする全ての他の核酸配列を含む。この意味で、文献は、組換え体の発現にとって好ましい又は最適化されたコドンに対する示唆を提供している(Kane JF et al., 1995)。
【0076】
核酸及びベクターは、異なる目的で細胞内に導入されることがあり、その結果トランスジェニック細胞及び動物を生成せしめる。本発明のポリペプチドを発現することができる細胞の産生方法は、そのようなベクター及び核酸で細胞を遺伝子操作することを含んで成る。
【0077】
特に、宿主細胞(例えば、細菌細胞)は、本発明の核酸及びベクターによってコードされたポリペプチドの一過性又は安定性の発現を可能にする形質転換によって修飾されることがある。あるいは、前記分子は、正常な発現レベルと比較した場合に、本発明のポリペプチドの増強又は低下した発現レベルを有するトランスジェニック動物細胞又はヒト以外の動物を生成させるために(非相同組換え/相同組換え又はそれらの安定な組込み及び維持を可能にする任意な他の方法による)使用されることがある。そのような正確な修飾は、本発明の核酸及び、例えば遺伝子治療(Meth. Enzymol., vol. 346,2002)又は部位特異的なリコンビナーゼ(Kolb AF, 2002)に関連する技術、を利用することによって得ることができる。本願で開示するIL−8様/TMEM9ポリペプチドを基にしたそれらの機能の系統だった研究のためのモデル系も、ヒト細胞系へのジーンターゲティングによって生成されうる(Bunz F, 2002)。
【0078】
遺伝子サイレンシング法を実施して、本発明のポリペプチドをコードする遺伝子の内在性発現をダウンレギュレートすることもできる。RNA干渉(RNAi)(S.M. Elbashir ら. Nature 2001, 411, 494-498)は、使用可能な配列特異的転写後遺伝子サイレンシングのための1つの方法である。短いdsRNAオリゴヌクレオチドをin vitroで合成して細胞内に導入する。これらdsRNAオリゴヌクレオチドの配列特異的結合によって標的mRNAの分解が開始され、標的タンパク質の発現が減少又は阻害される。
【0079】
上述のとおり評価される遺伝子サイレンシング法の有効性は、ポリペプチド発現の測定を介して(例えばウェスタンブロットによる)、又はTaqManベースの方法を用いてRNAレベルで評価することができる。
【0080】
本発明のポリペプチドは、当業界で知られている任意な方法、例えば組換えDNA関連技術、及び化学合成技術、によって調製されることもある。特に、本発明のポリペプチドを生成する方法は、上述のように、核酸又はベクターが発現する条件下で宿主細胞又はトランスジェニック細胞を培養し、そして培養物から前記核酸又はベクターによってコードされるポリペプチドを回収することを含んで成ることもある。例えば、ベクターが、細胞外又はシグナルペプチド含有タンパク質との融合タンパク質としてポリペプチドを発現する場合、組換え産物は細胞外空間に分泌されることがあり、そして、培養細胞から、更なる処理の観点で更に容易に回収され、且つ精製されることがあり、あるいは、当該細胞は直接使用され、又は投与されることがある。
【0081】
本発明のタンパク質をコードするDNA配列は、任意の適当な手段(形質転換、トランスフェクション、接合、プロトプラスト融合、エレクトロポレーション、リン酸カルシウム沈殿、直接的なマイクロインジェクション等)によって適切な宿主細胞に導入されうる、適当なエピソームの又は非相同/相同組換えのベクター内に挿入され、そしてライゲーションされることがある。特定のプラスミド又はウイルスベクターを選択するのに重要な因子は、ベクターを含むレシピエント細胞が当該ベクターを含まないそれらのレシピエント細胞から容易に認識され、且つ選択されうることの容易さ;特定の宿主で望まれるベクターのコピー数;及び異なる主の宿主細胞間でベクターを「シャトル」し得ることが望ましいか否か、を含む。
【0082】
ベクターは、転写開始/終結制御配列の支配下での原核又は真核宿主細胞による本発明のポリペプチドを含む単離されたポリペプチド又は融合ポリペプチドの発現を可能にするはずであり、これらは、前記細胞において構成的に活性又は誘導性のものが選択される。前記細胞が実質的に豊富な細胞系は、続いて安定な細胞系を提供するよう単離されうる。
【0083】
異なる転写制御配列及び翻訳制御配列は、真核生物宿主(例えば、酵母、昆虫、植物、又は哺乳類の細胞)に対し、その宿主の性質に依存して利用することができる。それらは、ウイルス源、例えばアデノウイルス、ウシパピローマウイルス、シミアンウイルス等に由来してもよく、ここでは、制御シグナルが、高レベルで発現する特定の遺伝子と関連している。例としてはヘルペスウイルスのTKプロモーター、SV40初期プロモーター、酵母GAL4遺伝子プロモーター等がある。転写開始制御シグナルは、抑制及び活性を可能にして、その結果前記遺伝子発現が調節されうるようなものが選択されうる。導入されたDNAによって安定して形質転換された細胞は、前記発現ベクターを含む宿主細胞の選択を可能にする1又は複数のマーカーを導入することによって選択されうる。当該マーカーはまた、栄養要求宿主に対する光合成栄養、殺生物耐性、例えば抗生物質、又は重金属、例えば銅、等をも提供しうる。選択マーカー遺伝子は、発現されるDNA遺伝子配列に直接連結されるか、あるいは同時トランスフェクションによって同一の細胞内に導入されうる。
【0084】
宿主細胞は原核生物又は真核生物のいずれかのものでありうる。好ましいものは、真核生物宿主、例えば哺乳類細胞、例えばヒト、サル、マウス、及びチャイニーズハムスター卵巣(CHO)細胞であり、それは、それらがタンパク質に対する翻訳後修飾、例えば正確な折りたたみ及びグリコシル化を提供するためである。酵母もグリコシル化を含む翻訳後ペプチド修飾を実施しうる。多くの組換えDNAストラテジーが存在しており、これらは、酵母内で所望のタンパク質の産生に利用されうる強力なプロモーター配列及び高コピー数のプラスミドを利用する。酵母は、クローン化哺乳類遺伝子産物のリーダー配列を認識し、そしてリーダー配列を有するペプチド(すなわち、プレペプチド)を分泌する。
【0085】
本発明の上文で言及した態様は、新規IL−8様/TMEM9ポリペプチドの配列に対して本願が提示する開示を、一般的な分子生物学的技術の知識と組み合わせることによって達成されうる。
【0086】
多数の本及び概説が、ベクター及び原核生物又は真核生物宿主細胞を用いて組換えタンパク質をどのようにクローン化し、そして産生するかについての技術を提供しており、例えばオックスフォード大学出版局によって発行されている「A Practical Approach」のシリーズの中の幾つかのタイトルである ("DNA Cloning 2: Expression Systems", 1995;"DNA Cloning 4: Mammalian Systems", 1996;"Protein Expression", 1999;"Protein Purification Techniques", 2001).。
【0087】
更に、最新のより重点的な文献は、ハイスループットな様式でポリペプチドを発現する技術(Chambers SP, 2002; Coleman TA, et al., 1997)、治療的用途を有する組換えタンパク質の大規模産生のために産業上利用される細胞系及び方法(Andersen DC and Krummen L, 2002; Chu L and Robinson DK, 2001)、及び注目のポリペプチドを発現する別の真核生物発現系であって、所望のタンパク質の経済的な産生についてかなりの可能性を有すると考えられるもの、例えばトランスジェニック植物を基にしたもの(Giddings G, 2001)又は酵母ピキア・パストリス(Pichia pastors)を基にしたもの(Lin Cereghino GP et al., 2002)、の概論を提供する。組換えタンパク質産物は、発現したポリペプチドの量及び質を証明するため(Baker KN et al., 2002)、生物学的同等性及び免疫原性の問題があるか否かを調べるのと同様に(Schellekens H, 2002; Gendel SM, 2002)、種々の分析技術により迅速にモニタリングされうる。
【0088】
全体的に、合成IL−8様ポリペプチドは文献において開示されており、そして、短い長さで示した本発明のIL−8様/TMEM9ポリペプチドに有効に適用されうる多数の化学合成技術の例が、固相又は液相合成技術として文献にて利用可能となっている。例えば、合成されるポリペプチドのカルボキシ末端に相当するアミノ酸は、有機溶媒中で不溶の支持体に結合し、そして、適切な保護基で保護されたそれらのアミノ基及び側鎖の官能基を有するアミノ酸がカルボキシ末端からアミノ末端へと順番に1つずつ縮合するものと、樹脂又は前記ペプチドのアミノ基の保護基と結合したアミノ酸が放出されるものとの交互の反復反応によって、ペプチド鎖はその結果この様式で伸長される。固層合成法は、使用する保護基の型に依存して、tBoc法及びFmoc法に大きく分類される。典型的に使用される保護基には、tBoc(t−ブトキシカルボニル)、Cl−Z(2−クロロベンジルオキシカルボニル)、Br−Z(2−ブロモベンジルオキシカルボニル)、Bzl(ベンジル)、Fmoc(9−フルオレニルメトキシカルボニル)、Mbh(4,4’−ジメトキシジベンズヒドリル)、Mtr(4−メトキシ−2,3,6−トリメチルベンゼンスルホニル)、Trt(トリチル)、Tos(トシル)、Z(ベンジルオキシカルボニル)及びCl2−Bzl(2,6−ジクロロベンジル)がアミノ基の場合に含まれ;NO2(ニトロ)及びPmc(2,2,5,7,8−ペンタメチルクロマン−6−スルホニル)がグアミジノ基の場合に含まれ;そしてtBu(t−ブチル)がヒドロキシル基の場合に含まれる。所望のペプチドの合成後、それは脱保護反応にかけられ、そして固体の支持体から切り離される。そのようなペプチド切断反応は、Boc法の場合フッ化水素又はトリフルオロメタンスルホン酸を用いて、そしてFmoc法の場合TFAを用いて実施されうる。
【0089】
本発明のポリペプチドの精製は、この目的のために知られている方法、すなわち、抽出、沈殿、クロマトグラフィー、電気泳動等の任意な常用の方法、のうちのいずれか1つによって実施されうる。本発明のタンパク質を精製するために優先して使用されうる追加の精製手順は、標的タンパク質に結合し、且つ、産生され、そしてカラム内に含まれるゲルマトリックス上に固定される、モノクローナル抗体又は親和性の基を用いるアフィニティークロマトグラフィーである。タンパク質は、ヘパリン又は特異的抗体によりカラムと結合し、一方、不純物はカラムを通過する。洗浄後、タンパク質はpH又はイオン強度の変化によってゲルから溶出される。あるいは、HPLC(高性能液体クロマトグラフィー)を使用することができる。溶出は、タンパク質精製に一般的に利用される水−アセトニトリルベースの溶媒を用いて実施されうる。
【0090】
本発明の新規ポリペプチドの開示、及びそれらに関連して開示されている試薬(抗体、核酸、細胞)の開示はまた、細胞への又は動物内でのそれらの発現レベルを増強し、又は低下させる化合物をスクリーニングし、そして特徴付けることを可能にする。
【0091】
「オリゴヌクレオチド」は、化学合成されうる一本鎖ポリデオキシヌクレオチド又は二本の相補的なポリデオキシヌクレオチド鎖のいずれかを意味する。そのような合成オリゴヌクレオチドは、5’ホスフェートを有さず、そのためキナーゼの存在下ホスフェートをATPと一緒に添加しないことには別のオリゴヌクレオチドとライゲーションしないであろう。合成オリゴヌクレオチドは、脱リン酸されていないフラグメントにライゲーションするであろう。
【0092】
本発明は、本発明の化合物の精製された調製物(ポリペプチド、核酸、細胞等)を含む。精製された調製物とは、本明細書で使用する場合、乾燥重量当たり少なくとも1%、好ましくは少なくとも5%本発明の化合物を含む調製物を意味する。
【0093】
本願は、一連の新規IL−8様/TMEM9ポリペプチド及び複数の可能性のある用途を有する関連試薬を開示する。特に、本発明のポリペプチドのIL−8様/TMEM9活性の増大が疾患の治療又は予防において望まれる場合はいつでも、試薬、例えば開示されているIL−8様/TMEM9ポリペプチド、相当の融合タンパク質及びペプチドミメティクス、コード核酸、発現細胞、又はそれらの発現を増強する化合物が使用されうる。
【0094】
従って、本発明は、開示したIL−8様/TMEM9ポリペプチド、相当の融合タンパク質及びペプチドミメティクス、コード核酸、発現している細胞、又はそれらの発現を増強する化合物のうちの1つを活性成分として含む、本発明のポリペプチドのIL−8様/TMEM9活性の増加が必要である疾患の治療又は予防のための医薬組成物を開示している。これらの医薬組成物の調製方法は、開示したIL−8様/TMEM9ポリペプチド、相当の融合タンパク質及びペプチドミメティクス、コード核酸、発現細胞、又はそれらの発現を増強する化合物を、医薬として許容される担体と一緒に組み合わせることを含んで成る。本発明のポリペプチドのIL−8様/TMEM9活性の増加が必要である疾患を治療又は予防する方法は、治療として有効量の、開示したIL−8様/TMEM9ポリペプチド、相当の融合タンパク質及びペプチドミメティクス、コード核酸、発現細胞、又はそれらの発現を増強する化合物、の投与を含んで成る。
【0095】
本願で開示されている試薬の中でも、前記のリガンド、アンタゴニスト又は本発明のポリペプチドの発現又は活性を低下させる化合物は、複数の用途を有し、特に、これらは、本発明のポリペプチドの過剰なIL−8様/TMEM9活性に関連した疾患の治療又は診断において使用されうる。
【0096】
従って、本発明は、本発明のポリペプチドの過剰なIL−8様/TMEM9活性に関連する疾患の処置又は予防のための医薬組成物であって、そのようなポリペプチドの発現又は活性を低下させるリガンド、アンタゴニスト又は化合物のうちの1つを活性成分として含む医薬組成物を開示する。これらの医薬組成物の調製方法は、前記のリガンド、アンタゴニスト、又は化合物を、医薬として許容される担体と一緒に組み合わせることを含んで成る。本発明のポリペプチドの過剰なIL−8様/TMEM9活性に関連する疾患の処置又は予防のための方法は、治療的に有効量のアンタゴニスト、リガンド又は化合物の投与を含んで成る。
【0097】
本発明の医薬組成物は、IL−8様/TMEM9ポリペプチド又は関連試薬に加え、医薬として許容される適当な担体、生物学的に適合性のある媒体及び添加物を含んでもよく、これらは動物への投与に適したものであり(例えば、生理学的食塩水)、且つ、活性化合物を医薬として使用されうる調製物へと処理するのを容易にする助剤(賦形剤、安定化剤、佐剤、又は希釈剤のようなもの)を最終的に含んで成るものである。
【0098】
医薬組成物は、投与形態の要求に合致する任意の許容される方法で製剤化されうる。例えば、生体材料の中でも、糖−高分子接合体、ポリエチレングリコール及び他の天然又は合成ポリマーは、薬物送達の有効性に関して活性成分を改善するのに使用されうる。特定の投与形態を評価する技術及びモデルは文献に開示されている(Davis BG and Robinson MA, 2002; Gupta P et al., 2002; Luo B and Prestwich GD, 2001; Cleland JL et al., 2001; Pillai O and Panchagnula R, 2001)。
【0099】
これらの目的に適したポリマーは生体適合性があり、すなわち、それらは生物系に対して非毒性であり、そして多数のそのようなポリマーは既知である。そのようなポリマーは天然で疎水性又は親水性であっても、生分解性、非生分解性、又はそれらの組み合わせであってもよい。これらのポリマーには、天然ポリマー(例えばコラーゲン、ゼラチン、セルロース、ヒアルロン酸)、並びに合成ポリマー(例えばポリエステル、ポリオルトエステル、ポリ無水物)が含まれる。疎水性の非分解性ポリマーの例には、ポリジメチルシロキサン、ポリウレタン、ポリテトラフルオロエチレン、ポリエチレン、ポリ塩化ビニル、及びポリメチルメタエリラート(methaerylate)が含まれる。親水性の非分解性ポリマーの例には、ポリ(2−ヒドロキシエチルメタクリラート)、ポリビニルアルコール、ポリ(N−ビニルピロリドン)、ポリアルキレン、ポリアクリルアミド、及びそれらのコポリマーが含まれる。連続反復単位としてエチレンオキシドを含んで成る好ましいポリマーには、例えばポリエチレングリコール(PEG)がある。
【0100】
任意の許容される投与形態は、活性成分の所望の血液レベルを確立するために、当業者によって使用され、そして決定されうる。例えば、投与は、種々の非経口経路、例えば皮下、静脈内、皮内、筋肉内、腹腔内、経鼻、経皮、経口、又は頬側でなされることがある。本発明の医薬組成物はまた、維持又は制御放出剤形、例えば蓄積注射、浸透ポンプ等を含むものであって、既定の速度での前記ポリペプチドの長期投与のためのもので、好ましくは正確な投与量の単回投与に適した単位剤形で投与されてもよい。
【0101】
非経口投与は、ボーラス注入又は時間をかけての漸進的な潅流によってもよい。非経口投与のための調製物には、滅菌水溶液又は非水溶液、懸濁液、及び乳濁液が含まれ、これらは当業界で知られている佐剤又は賦形剤を含むことがあり、且つルーチンな方法に従い調製されうる。尚、適切な油性注射懸濁液としての活性化合物の懸濁液が投与されることがある。適当な親油性溶媒又は媒体には、脂肪油、例えばごま油、又は合成脂肪酸エステル、例えばごま油、例えば合成脂肪酸エステル、例えばオレイン酸エチル又はトリグリセリドが含まれる。懸濁液の粘性を増大させる物質を含みうる水性注射懸濁液は、例えばカルボキシメチルセルロースナトリウム、ソルビトール、及び/又はデキストランが含まれる。任意に、懸濁液はまた安定化剤を含んでもよい。医薬組成物は注射による投与に適した溶液を含み、且つ、賦形剤と一緒に約0.01〜99.99パーセント、好ましくは約20〜75パーセントの活性化合物を含む。
【0102】
用語「治療的に有効量」とは、疾患の経過及び重症度に影響を及ぼし、そのような病状の緩和又は寛解を導くのに十分な量の活性成分を意味する。有効量は、投与経路及び患者の症状に依存する。
【0103】
用語「医薬として許容される」は、活性成分の生物学的活性の有効性を妨げず、且つ投与される宿主にとって毒性でない任意の担体を包含することを意味する。例えば、非経口投与の場合、上記活性成分は、単位剤形で、生理食塩水、デキストロース溶液、血清アルブミン及びリンガー液のような媒体中で製剤化されうる。担体はまた、デンプン、セルロース、タルク、グルコース、ラクトース、スクロース、ゼラチン、麦芽、コメ、コムギ、チョーク、シリカゲル、ステアリン酸マグネシウム、ステアリン酸ナトリウム、モノステアリン酸グリセロール、塩化ナトリウム、乾燥スキムミルク、グリセロール、プロピレングリコール、水、エタノール、及び種々の油から選択することができ、これらは、石油、動物、植物又は合成起源のもの(ピーナッツ油、大豆油、鉱油、ごま油)を含む。
【0104】
投与される用量は、レシピエントの年齢、性別、健康、及び体重、併用の処置の種類、もしあるのであれば、処置の頻度、及び所望の効果の性質、に依存する。用量は、当業者によって理解され、且つ決定可能なように、個々の対象者に合わせられる。各処置に必要とされる総量は、複数回投与又は単回投与によって投与されうる。本発明の医薬組成物は、単独で、又はその症状に対する、又は当該症状のほかの症候に対する他の治療薬と一緒に投与されうる。通常、活性成分の一日量は、0.01〜100ミリグラム/キログラム体重/日含まれる。通常、分割量又は維持放出形態で与えると、1〜40ミリグラム/キログラム/日が所望の結果を得るのに有効である。第二回目又はその後の投与は、個体に対して投与された最初又は前回の用量と同一、それ未満、又はそれ以上の用量で実施されうる。
【0105】
治療目的又は製造目的を有する方法とは別に、複数のほかの方法が本願で開示されているIL−8様/TMEM9ポリペプチド及び関連試薬を利用することができる。
【0106】
最初の例において、本発明のIL−8様/TMEM9ポリペプチドに関連する疾患を処置するのに有効な候補化合物をスクリーニングする方法であって:
(a)そのようなポリペプチドを発現する宿主細胞、当該ポリペプチドの発現レベルが増大又は低下しているトランスジェニックなヒト以外の動物、又はトランスジェニック動物細胞を、候補化合物と接触させること;及び
(b)前記動物又は細胞に対する前記化合物の効果を決定すること、
を含んで成る方法が提供される。
【0107】
第二の例において、本発明のポリペプチドのアンタゴニスト/阻害物質又はアゴニスト/活性化物質としての候補化合物を同定する方法であって:
(a)前記ポリペプチドと、前記化合物、及び哺乳類細胞又は哺乳類細胞膜と
を接触させること;及び
(b)前記の分子が、前記ポリペプチドと前記哺乳類細胞又は哺乳類細胞膜との相互作用、あるいは当該相互作用から生じる応答、を阻止又は増強するか否かを決定すること、
を含んで成る方法が提供される。
【0108】
第三の例において、試料中の本発明のポリペプチドの活性及び/又は存在を決定する方法は、当該ポリペプチド又はそのコードRNA/DNAのいずれかを検出することができる。従って、そのような方法は:
(a)タンパク質含有試料を準備すること;
(b)前記試料と本発明のリガンドとを接触させること;及び
(c)前記ポリペプチドに結合した前記リガンドの存在を決定し、それにより前記試料中のポリペプチドの活性及び/又は存在を決定すること、
を含んで成る。
【0109】
あるいは、前記方法は:
(a)核酸含有試料を準備すること;
(b)前記試料と本発明の核酸とを接触させること;及び
(c)当該核酸と前記試料中の核酸とのハイブリダイゼーションを決定し、それにより前記試料中の核酸の存在を決定すること、
を含んで成る。
【0110】
この意味において、配列番号1又は配列番号3又は配列番号6に示すヌクレオチド配列に由来したプライマー配列を、更にポリメラーゼ連鎖反応増幅により、試料中の本発明のポリペプチドの転写産物又はこれをコードしている核酸の量を決定するためにも使用することができる。
【0111】
本発明の更なる目的は、試料中の本発明のIL−8様/TMEM9ポリペプチドの活性及び/又は存在を測定するためのキットであって、本願で開示されている1又は複数の試薬:本発明のIL−8様/TMEM9ポリペプチド、アンタゴニスト、リガンド又はペプチドミメティクス、単離された核酸又はベクター、医薬組成物、発現細胞、あるいは発現レベルを増大又は低下させる化合物、を含んで成るキットである。
【0112】
このようなキットは、in vitro診断法又はスクリーニング法において使用することができ、それらの実際の組成は、試料の具体的様式(例えば、患者由来の生物学的試料組織)、及び測定される分子種に適合されるべきである。例えば、IL−8様/TMEM9ポリペプチドの濃度を測定することが望ましい場合、キットは、ウェスタンブロットで得られるシグナルを比較するために、精製された形の抗体及び相当のタンパク質を含有してもよい。あるいは、IL−8様/TMEM9ポリペプチドの転写産物の濃度を測定することが望ましい場合は、このキットは、相当のORF配列でデザインされた特異的核酸プローブを含むか、又はそのようなプローブを含有する核酸アレイの形であってよい。これらのキットは、タンパク質−、ペプチドミメティクス−、又は細胞−ベースのマイクロアレイの形であっても良く(Templin MFら, 2002;Pellois JPら, 2002;Blagoev B及びPandey A, 2001)、本願で開示されているタンパク質、ペプチドミメティクス及び細胞を利用することにより、ハイ-スループットプロテオミクス試験が可能となる。
【0113】
治療的用途
本願は、複数の適用可能性を有するSCS0010ポリペプチドを開示する。特に、本発明のポリペプチドのIL−8様/TMEM9活性の増大が疾患の治療又は予防において所望とされる場合はいつでも、試薬、例えば開示したSCS0010ポリペプチド、相当の融合タンパク質及びペプチドミメティクス、コード核酸、発現細胞、又はそれらの発現を増強する化合物が使用されうる。
【0114】
従って、本発明は、開示したIL−8様/TMEM9ポリペプチド、相当の融合タンパク質及びペプチドミメティクス、コード核酸、発現している細胞、又はそれらの発現を増強する化合物のうちの1つを活性成分として含む、本発明のポリペプチドのIL−8様/TMEM9活性の増加が必要である疾患の治療又は予防のための医薬組成物を開示している。これらの医薬組成物の調製方法は、開示したIL−8様/TMEM9ポリペプチド、相当の融合タンパク質及びペプチドミメティクス、コード核酸、発現細胞、又はそれらの発現を増強する化合物を、医薬として許容される担体と一緒に組み合わせることを含んで成る。本発明のポリペプチドのIL−8様/TMEM9活性の増加が必要である疾患を処置又は予防する方法は、治療として有効量の、開示したIL−8様/TMEM9ポリペプチド、相当の融合タンパク質及びペプチドミメティクス、コード核酸、発現細胞、又はそれらの発現を増強する化合物、の投与を含んで成る。
【0115】
本願で開示されている試薬の中でも、前記のリガンド、アンタゴニスト又は本発明のポリペプチドの発現又は活性を低下させる化合物は、複数の用途を有し、特に、これらは、本発明のポリペプチドの過剰なIL−8様/TMEM9活性に関連した疾患の治療又は診断において使用されうる。
【0116】
従って、本発明は、本発明のポリペプチドの過剰なIL−8様/TMEM9活性に関連する疾患の処置又は予防のための医薬組成物であって、そのようなポリペプチドの発現又は活性を低下させるリガンド、アンタゴニスト又は化合物のうちの1つを活性成分として含む医薬組成物を開示する。これらの医薬組成物の調製方法は、前記のリガンド、アンタゴニスト、又は化合物を、医薬として許容される担体と一緒に組み合わせることを含んで成る。本発明のポリペプチドの過剰なIL−8様/TMEM9活性に関連する疾患の処置又は予防のための方法は、治療的に有効量のアンタゴニスト、リガンド又は化合物の投与を含んで成る。
【0117】
SCS0010ポリペプチドのアゴニスト及びアンタゴニストは、SCS0010ポリペプチドの活性を調節する分子、SCS0010ポリペプチドの成熟型の少なくとも1つの活性を増加又は減少するいずれかの分子を含む。アゴニスト又はアンタゴニストは、SCS0010ポリペプチドと相互作用するタンパク質、ペプチド、炭水化物、脂質、又は低分子量の分子のような、コファクターであってもよく、これによりそれらの活性を調節する。
【0118】
潜在的なポリペプチドアゴニスト又はアンタゴニストには、TMEM9ポリペプチドの可溶型(SCS0010)又は膜結合型(TMEM9)のいずれかであって、前記タンパク質の細胞外ドメインの一部又は全部を含んで成るものと反応する抗体が含まれる。SCS0010ポリペプチドの発現を制御する分子は、典型的には、発現のアンチセンスレギュレーターとして作用することができるSCS0010ポリペプチドをコードしている核酸を含む。
【0119】
従って、本願は、適切に調製された医薬組成物における活性成分として、IL−8様ポリペプチドが関与している疾患及び症状、例えば、免疫障害、転移、移植片拒絶、血管形成、アルツハイマー病、アテローム性動脈硬化症、亜致死性の内毒素血症、敗血症ショック、羊膜腔の微生物感染、回帰熱のヤーリッシュ・ヘルクスハイマー反応、中枢神経系の感染病、急性膵炎、潰瘍性大腸炎、膿胸、溶血性尿毒症症候群、髄膜炎菌性の疾患、胃の感染症、腹膜炎、乾癬、関節リウマチ、敗血症、喘息及び糸球体腎炎の予防及び処置において有用であり得る新規IL−8様/TMEM9ポリペプチド及び一連の関連試薬を開示する。
【0120】
これらの用途の幾つかは、Gene Expression Omnibus (GEO)データベース:http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=geoに由来する実験によって支持されている。
【0121】
GEOはハイスループットの実験データの広い範囲のための公共の貯蔵場所としての役割を果たしている。これらのデータは、mRNA、ゲノムDNA、及びタンパク質の存在量を測定するシングル及びデュアル・チャネルマイクロアレイベースの実験、並びにアレイ以外の技術、例えば連続的遺伝子発現解析(SAGE)、及び質量分析プロテオミクスデータ、を含む。
【0122】
血清に応答する初代線維芽細胞の時間的な転写プログラム上のGEOレコードGDS85において(デュアル・チャネルマイクロアレイ実験)、TMEM9が若干阻害を受け、ほぼ1時間でピークを迎えることが示された。線維芽細胞は、血管疾患、炎症、線維症、線維性障害、関節リウマチ、クローン病、硬皮症、呼吸器感染、喘息、アレルギー反応、気管支炎、間質性肺炎、糖尿病性のネフロパシー、心不整脈、心臓肥大、腫瘍、潰瘍、眼障害、脂肪異栄養症、全身性硬化症、骨関節症、神経病理学を含む多数の病気、並びに老化、組織修復及び形成外科に関係するもの、に関与していることが知られている。更に、GEOレコードGDS60における、遺伝子発現と気道への抗原チャレンジとの間の時間的関係の決定による肺の中のブタクサ花粉の免疫原性タンパク質に対するアレルギー反応の解析において(シングルチャネルマイクロアレイ実験)、アレルゲンチャレンジにより、TMEM9が3つのうち2つの試料中で一貫して低下した(TMEM9はごくわずかに発現している)ことが示された。
【0123】
GEOレコードGDS350における肺線維症モデル(A/J、ブレオマイシン耐性)において、気管内をブレオマイシン処理されたA/Jマウスの肺がコントロールと比較された(デュアルチャネルマイクロアレイ実験)。この研究は、TMEM9が肺線維症モデルの4つのうち3つの試料において、コントロールと比較して若干阻害されたことを示した。
【0124】
GEOレコードGDS61における、肺の高血圧の回復において、肺高血圧症後の血管リモデリングが、正常酸素圧及び低酸素圧で処理した動物の肺と比較して実施された(シングルチャネルマイクロアレイ実験)。低酸素は、血管収縮に続き、肺血管平滑筋の肥大/肥厚及び内皮細胞増殖を誘導する。10時間後、TMEM9の発現は、正常酸素圧のものと比較すると、低酸素圧処理した動物において一貫して低下したことが示された。
【0125】
IL−8についての概説において、Mukaida Nは、「種々のタイプの細胞が、広範な刺激、例えば炎症性サイトカイン、微生物及びそれらの産物、並びに環境的変化、例えば低酸素症、再潅流、及び酸素過剰症に応じて大量のIL−8/CXCL8を産生することができる」(Am J Physiol Lung Cell Mol Physiol. 2003 Apr; 284 (4): L566-77 "Pathophysiological roles ofinterleukin-8/CXCL8 in pulmonary diseases")ことを証明する証拠を公表している。更に、Mukaidaは「多数の観察が、好中球に対する強力な作用により好中球媒介型の急性炎症における鍵となる媒介因子としてIL−8/CXCL8を確立した」ことを言及している。最後にMukaidaは、「複数の一連の証拠が、IL−8/CXCL8が好中球以外の種々のタイプの細胞、例えばリンパ球、単球、内皮細胞、及び線維芽細胞に対する広範な作用を有する」ことに言及している。更に、IL−8の生物学的機能は、これが更に種々の病理学的症状、例えばアテローム性動脈硬化症、炎症、肺病理及びガンにおいても重要な役割を有することを示唆している。
【0126】
SCS0010はまた、以下の研究で示唆されているようなガン治療においても有用なことがある。
【0127】
Kurokawaらは、3つの異なるcDNAライブラリー由来の3072の遺伝子及び肝臓ガン形成に関与していることが疑われている298の追加の既知の遺伝子を含むPCRアレイ実験において、わずかに7つの遺伝子が、そのうちTMEM9が肝臓ガン形成における共通の重要な役割を果たしうることを示した(Kurokawa et al. J Exp Clin Cancer Res. 2004 Mar; 23(1) : 135-41. PCR-array gene expression profiling of hepatocellular carcinoma)。
【0128】
GEOレコードGDS88における、抗ガン剤についてのNational Cancer Instituteのスクリーニングに使用された細胞系の解析において(デュアルチャネルマイクロアレイ実験)、TMEM9が、参照プールと比較して、黒色腫、白血病及び卵巣ガン細胞系において若干阻害されることを示した。
【0129】
GEOレコードGDS143における、表皮ガン形成において、前ガン状態の皮膚病変(光線角化症)と正常な表皮と培養ケラチノサイトとの比較が実施された(SAGE実験)。TMEM9は、培養ケラチノサイト及び光線角化症の表皮において発現しないのと比較して、高度に発現することが示された。
【0130】
GEOレコードGDS103における、心筋細胞形成において、未分化の多能性状態における及び心筋細胞への分化の誘導後のP19胚性癌腫(EC)細胞のトランスクリプトームを特徴付けるために(SAGE実験)、TMEM9は、プレーティングから0.5日後のEC細胞又はTMEM9の発現が起こらなかった未分化のP19EC細胞と比較して、プレーティングから3.5日後のP19EC細胞において発現することが示された。
【0131】
GEOレコードGDS121において、エストロゲン又はタモキシフェン処理後の遺伝子発現プロファイルの網羅的解析が実施された(SAGE実験)。タモキシフェン処理したZR75−1ヒト乳ガン細胞系が未処理又はエストロゲン処理のZR75−1細胞と比較してTMEM9の発現を誘導したことが示された。
【0132】
GEOレコードGDS122における、細胞の恒常性及び腫瘍細胞の増殖の制御におけるJNK2のシグナル伝達の重要性を強調するc−Jun NH82末端キナーゼの標的の同定において(SAGE実験)、TMEM9の発現が、モック処理したPC3細胞と比較してJNK2アンチセンスオリゴヌクレオチドPC3処理細胞において高いことが示された。IL−8活性は、JNK2のドミナントネガティブ突然変異体に対して感受性があり、そしてNF−カッパBとJNKの協調的な活性が強力なIL−8転写に必要なことが示されている。
【0133】
複数のGEOレコードはまた、種々の組織におけるTMEM9の発現を提供している(GDS503, GDS8, GDS393, GDS217, GDS541, GDS546, GDS217, GDS381, GDS102, GDS541, GDS117, GDS541, GDS544, GDS580, GDS551, GDS545, GDS542及びGDS548)。TMEM9の発現は、腎臓(近位尿細管、皮質集合管、皮質の太い上行脚)、脳全体、側坐核、脳の上衣腫、脳の髄芽腫、リンパ節、乳腺上皮、胸部の筋上皮、乳ガン、腺管ガン(胸部)、白質、小児の皮質、周辺部網膜、中心網膜、腹膜、結腸(原発腫瘍)、胃ガン、卵巣の表面上皮、原発性悪性の皮膚メラノーマ、前立腺ガン(進行した腫瘍)において示されている。TMEM9のGenecard entryによると(http://bioinformatics.weizmann.ac.il/cards/)、TMEM9は骨髄、脾臓、胸腺、脳、脊髄、心臓、骨格筋、加増、膵臓、前立腺、腎臓及び肺において発現している。
【0134】
TMEM9の発現情報はまた、UniGene Cluster Hs. 181444(ホモサピエンス;http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=unigene)から入手可能である。
【0135】
UniGeneによると、TMEM9は、成人のホモサピエンスの神経腫瘍に由来するライブラリー5967NT0048において高度に発現している。尚、TMEM9のcDNA供給源は、深刻な乳頭ガン、高悪性度、2個体のプールされた腫瘍;心臓;中分化した子宮内膜腺ガン、3個体のプールされた腫瘍;グリア芽腫(プールされたもの);印環細胞の特徴を有する低分化腺ガン;低分化した子宮内膜腺ガン、2個体のプールされた腫瘍;未分化希突起グリオーマ;リンパ腫、濾胞性混合小細胞大細胞;高分化した子宮内膜腺ガン、7個体のプールされた腫瘍;扁平上皮細胞ガン、低分化(4個体のプールされた腫瘍。原発性及び転移性のものを含む):プールされたもの;精巣;NEUROBLASTOMA COT 50-NORMALIZED;胸腺、プールされたもの;子宮;骨;正常な子宮内膜、中間の分泌期、周期23日;下垂体;NEUROBLASTOMA COT 25-NORMALIZED; PLACENTA COT 25- NORMALIZED;初期の乳頭の深刻な癌腫;胎盤;腺癌腫瘍、5個体のプールされたもの(説明書を参照のこと);正常な鼻咽頭;クローン病を有する3人の患者由来の結腸粘膜;5個体のプールされた肉腫、例えば粘液性脂肪肉腫、単発性線維性腫瘍、悪性線維性組織球腫、消化管間質腫瘍及び中皮腫;脳全体;肝臓及び脾臓;精巣;肝臓;胎児肝臓;肺;カルチノイド;中分化した腺ガン;2個体のプールされたウイルムス腫瘍。1つは原発性、そして1つは脳に対して転移性;髄芽腫;CNCAP (3) T-225細胞系;胸部;結腸;2個体のプールされた扁平上皮細胞癌腫;精製された膵島;頭頸部;末梢神経系;胎盤;プールされた軟骨肉腫腫瘍細胞;ガン細胞系;腺ガン細胞系;膜芽細胞腫;腎細胞腺ガン;プールされた胚細胞性腫瘍;脳;脳;前立腺;扁平上皮細胞癌腫;子宮頸ガン細胞系;大細胞癌腫;小細胞癌腫;子宮内膜腺ガン細胞系;黒色性黒色腫;腎腫瘍;絨毛ガン;正常な胸部;大細胞癌腫、未分化;神経系腫瘍;膵臓;卵巣;副腎腫;胚性肝細胞、WA01、38回継代;臍帯血;EGFRの増幅を伴うグリア芽腫;1p/19qの損失を伴う未分化希突起グリオーマ;メラニン細胞;腺ガン細胞系;副腎皮質癌腫細胞系;筋肉(骨格);十二指腸腺ガン細胞系;網膜;上皮(細胞系);副腎腫細胞系;神経芽細胞腫;粘液性類表皮ガン;軟骨;海馬;多発性硬化症の病変;血液;白血球;未分化神経外胚葉;NEUROBLASTOMA COT 10-NORMALIZED;類上皮癌腫細胞系;移行上皮性乳頭腫細胞系;正常な肺;視床下部;プールされた結腸、腎臓、胃;プールされた膵臓及び脾臓;癌腫細胞系;骨髄;プールされた肺及び脾臓;膀胱腫瘍;プールされた脳、肺、前立腺;上皮小体腫瘍;視神経;卵巣(3個体のプール);ヒト網膜;神経芽細胞腫;胎児の眼、水晶体、前眼部、視神経、網膜、網膜の中心の且つ斑点のあるRPE及び脈絡;RPE及び脈絡胎児の眼;網膜の中心及び斑点;腹水;プールされたヒトメラニン細胞、胎児の心臓、及び妊娠子宮;前頭葉;胎児の脳;リンパ節に対して転移性の悪性黒色腫;全脳;2個体のプールされた腫瘍(明細胞型);腎臓;胃;眼;肺;B細胞、慢性リンパ球性白血病;原発性肺嚢胞性線維症上皮細胞;混合型;転移性軟骨肉腫;平滑筋肉腫;悪性度IIの軟骨肉腫;肺限局性線維症;軟骨肉腫;網膜;肝細胞癌腫細胞系;乳頭漿液性腺ガン;交感神経幹;恐らくTP53が突然変異しており、且つEGFR増幅していないグリア芽腫;後根神経節;肺ガン;混合型(40腫のRNAのプール);骨髄腫;細胞系;奇形癌腫細胞系;インスリノーマ;胚中心B細胞;水晶体;肺胞マクロファージ;RPE/脈絡膜;脂肪細胞;骨髄肉腫細胞系;内軟骨肉腫細胞系;軟骨下骨;子宮内膜;胚;副睾丸;卵巣腫瘍;類表皮癌腫細胞系;低級前立腺新生物;胚細胞腫瘍;副腎腺腫;正常前立腺;結腸腫瘍RER+;肝臓;松果体;転移性前立腺骨病変;類上皮癌腫、である。
【0136】
IL−8のレベルは再発寛解型多発性硬化症のクラドリビン処置の後脳脊髄液(CSF)中で有意に低下したことも指摘されうる(RR-MS; Bartosik-Psujek H et al. Acta Neurol Scand. 2004 Jun; 109 (6): 390-2. "Interleukin-8 and RANTES levels in patients with relapsing-remitting multiple sclerosis (RR-MS) treated with cladribine.")。
【0137】
尚、Kveineらは、TMEM9のmRNAが副腎、甲状腺、卵巣、精巣、及び前立腺に豊富に存在し、そしてそれには及ばないが、気管、脊髄、胃、結腸、小腸、胸腺、及び脾臓において発現していることを示した(Kveine et al. Biochem Biophys Res Commun. 2002 Oct 4; 297 (4): 912-7. "Characterization of the novel human transmembrane protein 9(TMEM9) that localizes to lysosomes and late endosomes.")。彼等はまた、骨髄、リンパ節、及び末梢血リンパ球(PBL)における発現が低いことも観察し、そしてTMEM9のmRNAがB細胞系起源、T細胞系起源、骨髄起源及び赤血球起源の造血細胞系において発現していることを証明している。
【0138】
そのようなものとして、SCS0010核酸分子、それらのポリペプチド、アゴニスト又はアンタゴニスト(例えば、アゴニスト抗体又はアンタゴニスト抗体)は種々の疾患、線維症、線維性障害、関節リウマチ、クローン病、硬皮症、呼吸器感染、喘息、アレルギー反応、気管支炎、間質性肺炎、糖尿病性のネフロパシー、心不整脈、心臓肥大、腫瘍、潰瘍、眼障害、脂肪異栄養症、全身性硬化症、骨関節症、神経病理学、肺高血圧症、低酸素症、肺血管平滑筋の肥大/肥厚、黒色腫、白血病及び卵巣ガン、肺細胞癌腫の発生、前ガン状態の皮膚病変(光線角化症)、胚性癌腫、乳ガン、脳上衣腫、脳髄芽腫、乳腺癌、腺管癌(胸部)、結腸原発性腫瘍、胃ガン、原発性悪性の皮膚メラノーマ、前立腺ガン(進行した腫瘍)、神経腫瘍、漿液性乳頭ガン、子宮内膜腺ガン、グリア芽腫、印環細胞の特徴を有する低分化腺ガン、未分化希突起グリオーマ、リンパ腫、扁平上皮細胞癌腫、神経芽細胞腫、乳頭漿液性腺ガン、肉腫、例えば粘液性脂肪肉腫、単発性線維性腫瘍、悪性線維性組織球腫、消化管間質腫瘍及び中皮腫、中分化した腺ガン、2個体のプールされたウイルムス腫瘍(原発性、そして1つは脳に対して転移性)、髄芽腫、扁平上皮細胞癌腫、軟骨肉腫、癌腫、網膜芽腫、腎細胞腺ガン、胚細胞性腫瘍、扁平上皮細胞癌腫、子宮頸ガン細胞系、大細胞癌腫、小細胞癌腫、子宮内膜腺ガン細胞系、黒色性黒色腫、腎腫瘍、絨毛ガン、神経腫瘍、副腎腫、胚性肝細胞、グリア芽腫、急性骨髄性白血病、未分化希突起グリオーマ、副腎皮質癌腫、十二指腸腺ガン、副腎腫細胞系、粘液性類表皮ガン、多発性硬化症の病変、類上皮癌腫、移行上皮性乳頭腫、膀胱腫瘍、上皮小体腫瘍、悪性黒色腫、慢性リンパ球性白血病、原発性肺嚢胞性線維症、転移性軟骨肉腫、平滑筋肉腫、悪性度IIの軟骨肉腫、肺限局性線維症、軟骨肉腫、肝細胞癌腫、乳頭漿液性腺ガン細胞系、肺ガン、骨髄腫、奇形癌腫細胞系、インスリノーマ、内軟骨肉腫、副睾丸、卵巣腫瘍、類表皮癌腫細胞系、前立腺新生物、胚細胞腫瘍、副腎腺腫、結腸腫瘍、転移性前立腺骨病変、類上皮癌腫並びに老化、組織修復及び形成外科に関係するものを診断又は処置するのに有用でありうる。尚、SCS0010核酸分子、そのアゴニスト又はアンタゴニスト(例えば、アゴニスト抗体又はアンタゴニスト抗体)は、TMEM9が検出されている器官及び組織に由来する、又はそこに存在する疾患(例えば腫瘍)を診断又は処置するのに有用なことがある。SCS0010のアンタゴニスト又はアゴニスト(例えば抗体)は、実施例5及び/又は図11、12において決定されるようなSCS0010の部位及びドメインに対するものであってもよい。例えば、抗体は、SCS0010の3つの保存されたシステインリッチドメイン(内在性TMEM9にも存在する)のうちの少なくとも1つに対するものであってもよく、これは理論に拘束されることを望むものではないが、SCS0010とそれ自体との又は他のTMEM9ファミリーメンバーとの二量体化を阻止しうる。
【0139】
本発明のポリペプチド及び関連試薬の治療的用途は、創薬及び前臨床開発時の前駆脂肪細胞因子−1様ポリペプチド及び他の生物学的産物の評価に関して、動物細胞、組織を使用するin vivo/in vitroアッセイ手段によるか、又は公知のin silico/コンピュータ支援の方法(Johnson DE及びWolfgang GH, 2000)により、評価(安全性、薬物動態及び有効性に関して)される。
【0140】
本発明は、以降、以下の実施例により特定の態様を参照して説明されるが、これらは本発明を何ら限定するものとして解されるべきではない。当該説明の内容は、上文の教示に照らして、そしてそれ故に、特許請求の範囲の意味及び目的を超えて広げることなく、当業者によって実施されうる全ての改変及び置換を含んで成る。
【0141】
【表1】
【表2】
【実施例1】
【0142】
ASTRALデータベース(Brenner SEら, "The ASTRAL compendium for protein structure and sequence analysis", Nucleic Acids Res., 2000 Jan 1; 28(1): 254-6)から得たIL−8様タンパク質ドメイン配列を用い、ヒトゲノム配列(Celeraバージョン)から予測された遺伝子の相同タンパク質配列を検索した。
【0143】
これらのドメイン配列を使用して、ホモログを含む配列プロファイルを作成した。更に、ジスルフィド結合に関与する保存されたシステインに相当する各プロファイルの位置は、これらの位置での厳密なAA保存を要求するように修飾された。IL−8様ドメインの配列プロファイルは、PIMAII(Profile Induced Multiple Alignment;ボストン大学ソフトウェア、バージョンII, Das S及びSmith TF, 2000)を用いて作成した。PIMAIIアルゴリズムは繰り返し相同配列を並置し、そして配列プロファイルを作成する。
【0144】
クエリータンパク質配列は、下記の3種のプログラムのひとつにより作成された、遺伝子予測及びそれらの翻訳(ヒトゲノムR27のCELERAバージョン)から得た:Genescan (Burge C, Karlin S., "Prediction of complete gene structures in human genomic DNA”, J Mol Biol., 1997 Apr 25; 268(1): 78-94)、Grail(Xu Y, Uberbacher EC., “Automated gene identification in large-scale genomic sequences", J Comput Biol., 1997 Fall; 4(3): 325-38)及びFgenesh(Celera社独自のソフトウェア)。
【0145】
相同性は、プロファイルとクエリー配列の間のグローバル-ローカルアラインメントを作成するPIMAIIを用いて検出した。この場合、このアルゴリズムは、クエリーとしてのEGF機能ドメインのプロファイルと共に使用した。PIMAIIは、クエリープロファイルを、タンパク質配列に翻訳された遺伝子予測のデータベースと比較し、その結果そのドメインを含むDNA配列との合致を同定することができる。
【0146】
更にこの配列の既知のIL−8様含有タンパク質とのBLAST(Basic Local Alignment Search Tool;NCBI ver2)による比較は、一番近いホモログを同定した(Gish W, States DJ., "Identification of protein coding regions by database similarity search", Nat Genet., 1993 Mar; 3(3): 266-72;Pearson WR, Miller W., "Dynamic programming algorithms for biological sequence comparison", Methods Enzymol., 1992; 210: 575-601;Altschul SFら,"Basic local alignment search tool", J Mol Biol., 1990 Oct 5; 215(3): 403-10)。
【0147】
検出に使用したPIMAIIパラメータは、PIMA先行(prior)アミノ酸確率行列及びZ-カットオフスコア10であった。使用したBLASTパラメータは以下のとおりである:比較行列=BLOSUM62;ワードレングス=3;カットオフE値=10;ギャップオープニング及びイクステンション=デフォルト;フィルターなし。
【0148】
一旦機能ドメインがこの配列において同定されたならば、これらの遺伝子は、一番近いホモログの配列を用いgenewiseアルゴリズムで再度予測した(Birney Eら,"PairWise and Search Wise: finding the optimal alignment in a simultaneous comparison of a protein profile against all DNA translation frames", Nucleic Acids Res., 1996 Jul 15; 24(14): 2730-9)。
【0149】
IL−8様の相同性ドメインのプロファイルは、PERL(Practical Extraction and Report Language)及びPIMAIIに記載されたPSI-BLAST(Altshulら 1997)スクリプトを用い自動的に作成した。
【0150】
合計51の(IL−8様)の18種の予測された遺伝子は、新規である可能性があると判断されたので選択した。
【0151】
前記タンパク質配列の新規性は、最終的には、BLASTを用いるタンパク質データベース(SwissProt/Trembl, Human IPI and Derwent GENESEQ)の検索により評価した。機能的ドメインに対する整合性及び新規性は、選択した新規候補についての「人の手による」検査で調べた。
【実施例2】
【0152】
実施例1に記載の方法により単離されたひとつの配列は、本明細書においてSCS0010ポリペプチド配列と称されるものである。最も類似の、既知のポリペプチド配列は、アクセッション番号ABP61498のヒトNF−κB活性化タンパク質であるが、第三のエキソンが異なる。図2は、SCS0010をコードする遺伝子のイントロン/エキソン構造を示す(64000111830404)。
【実施例3】
【0153】
SCS0010のスプライス変異体の同定及びクローニング
1.SCS0010のクローニング
1.1 cDNAライブラリー
ヒト胎盤cDNAライブラリー(バクテリオファージラムダ(λ)GT10)は、Clontech(カタログ番号HL1075b)から購入した。バクテリオファージλDNAは、Wizard Lambda Preps DNA精製システムを用いて、メーカーの使用説明書に従い(Promega, Corporation, Madison WI)、感染したE.コリ宿主株の小規模培養物から調製した。生じたファージDNAは、50ng/μlの濃度で再懸濁し、そして後のPCR反応に使用した。
【0154】
1.2 PCR用の遺伝子特異的クローニングプライマー
18〜25塩基の長さを有する一対のPCRプライマーを、Primer Designer Software(Scientific & Educational Software, PO Box 72045, Durham, NC 27722-2045, USA)を用いて仮のcDNAのコード配列を増幅するために設計した。PCRプライマーは最適化された結果、55±10℃付近のTM及び40〜60%のGC含量を有する。標的配列について高い選択性を有していたプライマーが選択された(ほとんど又は全く非特異的プライミング無し)。
【0155】
1.3 ヒト胎盤ファージcDNAライブラリー由来のSCS0010cDNAのPCR
遺伝子特異的PCR増幅プライマー(SCS0010−CP1及びSCS0010−CP2、図3及び表4)は、SCS0010のほぼ予測されたコード配列を含むことが予想される299bpの産物を増幅するよう設計された。PCRは1xAmplitaq(登録商標)バッファー、200μMのdNTP、50ピコモルの各クローニングプライマー、2.5ユニットのAmplitaq(登録商標)(Perkin Elmer)及び100ngの各ファージライブラリープールDNAを含む50μlの終量で、以下の通りプログラムしたMJ Research DNA Engineを用いて実施した:94℃、1分;94℃、1分、χ℃、及び1分、72度を40サイクル;続いて72℃、7分を1サイクル、そして4℃の保持サイクル。
【0156】
増幅産物を0.8%アガロースゲル/1xTAEバッファー(Invitrogen)上で可視化し、そしてWizard Lambda Preps DNA精製システム(Promega)を用いてゲルから精製した。50Lの滅菌水で溶出したPCR産物は、直接サブクローニングするか、又は−20℃で保存した。
【0157】
1.4 PCR産物のサブクローニング
PCR産物は、Invitrogen Corporationから購入したTOPOクローニングキットをメーカー指定の条件を使用して用いることでトポイソメラーゼI修飾クローニングベクター(pCR4−TOPO)内にサブクローニングした。要約すると、4μlのゲル精製PCR産物を1μlのTOPTベクター及び1μlの塩溶液と一緒に15分間室温でインキュベートした。続いて、反応混合物でE.コリ株TOP10(Invitrogen)を以下の通り形質転換した:50μlのOne Shot TOP10細胞のアリコートを氷上で融解し、そして2μlのTOPO反応液を添加した。混合物を15分間氷上でインキュベートし、そして次に42℃で正確に30秒間インキュベートすることで熱ショックを与えた。試料を氷に戻し、そして250μlの温めたSOC培地(室温)を添加した。試料を振とうしながら(220rpm)1時間37℃でインキュベートした。続いて、アンピシリン(100μg/ml)を含むLブロス(LB)プレート上に形質転換混合物をプレーティングし、そして一晩37℃でインキュベートした。インサートを含むアンピシリン耐性コロニーをコロニーPCRで同定した。
【0158】
1.4 コロニーPCR
コロニーを、滅菌した爪楊枝を用い、50μl滅菌水中に接種した。播種材料のうち10μlのアリコートが、使用したプライマーがT7及びT3であることを除き上述のように20μlの合計の反応液量中でPCRにかけられた。サイクリング条件は以下の通りである:94℃で2分間;94℃で30秒間、48℃で30秒間及び72℃で1分間を30サイクル。試料は、次の解析まで4℃で維持した(保持サイクル)
【0159】
PCR反応産物を、1X TAE緩衝液中1%アガロースゲル上で分析した。予想されたPCR産物サイズ(約299bp+マルチクローニングサイト又はMCSに起因する105bp)を生じたコロニーを、振盪しながら(220rpm)で、アンピシリン(100μg/ml)を含有する5ml L-ブロス(LB)中で一晩37℃で増殖した。
【0160】
1.5 プラスミドDNAの調製及び配列決定
ミニプレッププラスミドDNAは、Qiaprep Turbo 9600 ロボット型システム(Qiagen)又はWizard Plus SMミニプレップキット(Promegaカタログ番号1460)をメーカーの使用説明書に従い用いることで5mlの培養液から調製した。プラスミドDNAは、100μlの滅菌水で溶出した。DNA濃度は、Eppendorf BO光度計を用いて測定した。プラスミドDNA(200〜500ng)を、T7プライマー及びT3プライマーにより、Big DyeTerminatorシステム(Applied Biosystemsカタログ番号4390246)をメーカーの使用説明書に従い使用し、DNA配列決定にかけた。プライマー配列は表3に示すとおりである。配列決定反応液を、Dye-Exカラム(Qiagen)又はMontage SEQ 96クリーナッププレート(Millipore カタログ番号LSKS09624)を使用し精製し、その後Applied Biosystems 3700シークエンサーで解析した。
【0161】
配列解析は、予測されたSCS0010配列との100%のマッチを含むクローンを同定した(図3)。クローニングしたPCR産物(pCR4−TOPO−SCS0010)(プラスミドID.14613)のプラスミドマップを図4に示す。
【0162】
2.SCS0010の哺乳類細胞発現ベクターの構築
続いて、DNA配列決定で同定したSCS0010のコード配列を含むpCR4−TOPOクローン(pCR4−TOPO−SCS0010、プラスミドID.14613)(図4)を使用して、インサートをGateway(登録商標)クローニング法(Invitrogen)で哺乳類細胞発現ベクターpEAK12d(図6)及びpDEST12.2(図7)内にサブクローニングした。
【0163】
2.1 インフレームで6HISタグ配列と融合したGateway適合SCS0010ORFの生成
Gatewayクローニング法の第一段階は、attB1組換え部位及びコザック配列が5’末端に隣接し、並びにインフレームで6ヒスチジン(6HIS)タグ、停止コドン及びattB2組換え部位をコードしている配列(Gateway適合cDNA)が3’末端で隣接した、SCS0010のORFを生成する2段階のPCR反応を伴う。第一のPCR反応液(終量50μl)は、1μl(40ng)のpCR4−TOPO−SCS0010(プラスミドID.14613)、1.5μlのdNTP(10mM)、10μlの10X Pfxポリメラーゼ緩衝液、1μlのMgSO4(50mM)、各0.5μlの遺伝子特異的プライマー(100μM)(SCS0010−EX1及びSCS0010−EX2)、及び0.5μlのPlatinum Pfx DNAポリメラーゼ(Invitrogen)を含む。このPCR反応は、95℃で2分間の最初の変性工程、それに続く94℃で15秒;55℃で30秒及び68℃で2分間の12サイクル;並びに、4℃の保持サイクルを用い行った。この増幅産物を、1X TAE緩衝液(Invitrogen)中の0.8%アガロースゲル上で可視化し、予測された分子量に移動した産物を、メーカーの使用説明書に従い、Wizard PCRプレップDNA精製システム(Promega)を用いゲルから精製し、50μl滅菌水中に回収した。
【0164】
第二のPCR反応液(終量50μl)は、10μlの精製されたPCR1産物、1.5μlのdNTP(10mM)、5μlの10X Pfxポリメラーゼ緩衝液、1μlのMgSO4(50mM)、0.5μlの各Gateway変換プライマー(100μM)(GCPフォワード及びGCPリバース)、及び0.5μlのPlatinum Pfx DNA ポリメラーゼを含んだ。第2のPCR反応条件は:95℃で1分間;94℃で15秒、50℃で30秒及び68℃で2分間を4サイクル;94℃で15秒、55℃で30秒及び68℃で2分間を25サイクル;引き続き4℃の保持サイクルとした。PCR産物は、メーカーの使用説明書に従い、Wizard PCRプレップDNA精製システム(Promega)を用いゲル精製した。
【0165】
2.2 Gateway適合SCS0010ORFの、GatewayエントリーベクターpDONR221並びに発現ベクターpEAK12d及びpDEST12.2へのサブクローニング
Gatewayクローニング法の第二段階は、下記のような、Gateway修飾PCR産物のGatewayエントリーベクターpDONR221(Invitrogen、図3)へのサブクローニングを伴う:5μlのPCR2由来の精製産物を、1.5μlのpDONR221ベクター(0.1μg/μl)、2μlのBPバッファー及び1.5μlのBPクロナーゼ酵素混合物(Invitrogen)と共に、終量10μlで室温で1時間インキュベーションした。この反応をプロテイナーゼK 1μl(2μg/μl)の添加により停止させ、37℃で更に10分間インキュベーションした。この反応液のアリコート(1μl)を用い、E.コリDH10B細胞を、以下の通りエレクトロポレーションにより形質転換した:DH10Bエレクトロコンピテント細胞(Invitrogen)25μlアリコートを氷上で解凍し、1μlのBP反応混合液を添加した。混合物を、冷却した0.1cmエレクトロポレーション用キュベットに移し、細胞をBioRad Gene-Pulser(登録商標)をメーカーの推奨プロトコールに従い使用しエレクトロポレーションした。予め室温に温めたSOC培地(0.5ml)を、エレクトロポレーション直後に添加した。この混合物を、15mlのスナップ-キャップチューブに移し、振盪しながら(220rpm)37℃で1時間インキュベーションした。その後形質転換混合物のアリコート(10μl及び50μl)を、カナマイシン(40μg/ml)を含有するL-ブロス(LB)プレート上にプレーティングし、37℃で一晩インキュベーションした。
【0166】
プラスミドミニ-プレップDNAを、Qiaprep Turbo 9600 ロボット型システム(Qiagen)を用い、生じたコロニーのうちの6つに由来する5ml培養物から調製した。プラスミドDNA(150-200ng)を、BigDyeTerminatorシステム(Applied Biosystems カタログ番号4390246)をメーカーの使用説明書に従い用い、21M13及びM13RevプライマーによるDNA配列決定にかけた。これらのプライマー配列を表3に示す。配列決定反応は、Dye-Exカラム(Qiagen)又はMontage SEQ 96クリーンアッププレート(Millipore カタログ番号LSKS09624)を用いて精製し、その後Applied Biosystems 3700シークエンサーで解析した。
【0167】
正確な配列を含むこれらのクローンのひとつ(pENTR−SCS0010−6HIS、プラスミドID14691、図8)のプラスミド溶出液(2μl又は約150ng)を、終量10μl中に1.5μlのpEAK12dベクター又はpDEST12.2ベクター(図4及び5)のいずれか(0.1μg/μl)、2μlのLRバッファー及び1.5μlのLRクロナーゼ(Invitrogen)を含有する組換え反応液において使用した。この混合物を、室温で1時間インキュベーションし、プロテイナーゼK(2μg)の添加により停止し、37℃で更に10分間インキュベーションした。この反応液のアリコート(1μl)を用い、E.コリDH10B細胞を、以下の通りエレクトロポレーションにより形質転換した:DH10Bエレクトロコンピテント細胞(Invitrogen)の25μlアリコートを氷上で解凍し、1μlのBP反応混合液を添加した。混合物を、冷却した0.1cmエレクトロポレーション用キュベットに移し、細胞をBioRad Gene-Pulser(登録商標)をメーカーの推奨プロトコールに従い使用しエレクトロポレーションした。予め室温に温めたSOC培地(0.5ml)を、エレクトロポレーション直後に添加した。この混合物を、15mlのスナップ-キャップチューブに移し、振盪しながら(220rpm)37℃で1時間インキュベーションした。その後形質転換混合物のアリコート(10μl及び50μl)を、アンピシリン(100μg/ml)を含有するL-ブロス(LB)プレート上にプレーティングし、37℃で一晩インキュベーションした。
【0168】
プラスミドミニ-プレップDNAを、Qiaprep Turbo 9600 ロボット型システム(Qiagen)を用い、各ベクターでサブクローニングして生じたコロニーのうちの6つに由来する5ml培養物から調製した。pEAK12dベクター中のプラスミドDNA(200-500ng)を、先に説明したように、pEAK12F及びpEAK12Rプライマーにより、DNA配列決定にかけた。pDEST12.2ベクター内のプラスミドDNA(200-500ng)を、先に説明したように、21M13及びM13Revプライマーにより、DNA配列決定にかけた。プライマー配列は表3に示す。
【0169】
Sambrook J.らの説明した方法(1989 (Molecular Cloning, a Laboratory Manual, 第2版, Cold Spring Harbor Laboratory Press))により、CsCl勾配で精製されたマキシ-プレップDNAを、配列を確認した各クローン(pEAK12d−SCS0010−6HIS、プラスミドID14699、図9、及びpDEST12.2−SCS0010−6HIS、プラスミドID14700、図10)のひとつの500ml培養物から調製した。プラスミドDNAは、濃度1μg/μlで滅菌水(又は10mM Tris-HCl、pH8.5)中に再懸濁し、-20℃で保存した。
【0170】
【表3】
下線の配列=コザック配列
太字=停止コドン
イタリックの配列=Hisタグ
【実施例4】
【0171】
SCS0010の発現及び精製
1.哺乳類細胞における発現及びクローニングしたHisタグ付きプラスミドの精製を通じての機能ゲノミクス
エプスタインバーウイルス核抗原を発現しているヒト胎児腎臓293細胞(HEK293−EBNA、Invitrogen)をEx-cell VPRO無血清培地(シードストック、維持培地、JRH)の懸濁液中で維持した。トランスフェクションの日に細胞を計数し、遠心し(低速)、そしてペレットを、1%FCSを補足した所望の量のFEME(下文を参照のこと)培地内で再懸濁して1XE6生存細胞/mlの細胞濃度を生成させた。cDNAをFEME(1Lの体積当たり200ml)中1Lの体積当たり2mgに希釈した(2%のeGFPを同時トランスフェクションする)。ポリエチレンイミントランスフェクション試薬(1Lの体積当たり4mg)を続いてcDNA溶液に添加し、ボルテックスにかけ、そして室温で10分間インキュベートした(トランスフェクション混合物の生成)。
【0172】
続いて、このトランスフェクション混合物をスピナーに添加し、そして90分間CO2インキュベーター(5%CO2及び37℃)内でインキュベートした。新鮮なFEME培地(1%FCS)を、例えば、最初のスピナーの体積を二倍にするために90分後に添加した。続いて、スピナーを6日間インキュベートした。6日目(収集日)に、スピナーの上清(500ml)を遠心(4℃、400g)、そしてプラスミド番号及び発酵番号と一緒に独特の識別子を有するポット内に据えた。
【0173】
精製方法
C末端6Hisタグを有する組換えタンパク質を含む500mlの培養液試料を1倍量の冷たいバッファーA(50mMのNaH2PO4;600mMのNaCl;8.7%(w/v)のグリセロール、pH7.5)で1000mlの終量に希釈した。試料を0.22μmの滅菌フィルター(Millipore, 500mlのフィルターユニット)を通じて濾過し、そして1Lの滅菌した四角い培地ボトル(Nalgene)中4℃で維持した。
【0174】
精製は、自動サンプルローダー(Labomatic)と接続されたVISIONワークステーション(Applied Biosystems)上で4℃で実施された。精製手順は、ニッケルイオンがチャージされたPoros 20MC(Applied Biosystems)上での金属アフィニティークロマトグラフィー(10x50mm、3.93ml)、続いて、Sephadex G-25 medium(Amersham Pharmacia)ゲル濾過カラム(1.0x15cm)上でのバッファー交換、の2連続工程から構成された。
【0175】
第一のクロマトグラフィー工程のために、金属アフィニティーカラムを30倍のカラム体積のEDTA溶液(100mMのEDTA;1MのNaCl;pH8.0)で再生し、15倍のカラム体積の100mMNiSO4溶液で洗浄することでニッケルイオンを再チャージし、そして10倍のカラム体積のバッファーA、続いて7倍のカラム体積のバッファーB(50mMのNaH2PO4;600mMのNaCl;8.7%(w/v)のグリセロール、400mM;イミダゾール、pH7.5)で洗浄し、そして最後に、15mMのイミダゾールを含む15倍のカラム体積のバッファーAで平衡化した。試料をLabomaticサンプルローダーで200mlのサンプルループ内に移し、そして次にニッケル金属アフィニティーカラム上に20ml/分の流速でチャージした。チャージ手順を5回繰り返すことで全試料(1000ml)をニッケルカラム上に移した。次に、カラムは、12倍のカラム体積のバッファーA、続いて20mMのイミダゾールを含む28倍のカラム体積のバッファーAで洗浄した。20mMのイミダゾール洗浄の間に、緩やかに付着した混入タンパク質をカラムから溶出した。組換えHisタグ付きタンパク質を最後に10倍のカラム体積のバッファーBを用い2ml/分の流速で溶出し、そして溶出したタンパク質を2.7mlの画分で回収した。
【0176】
第二のクロマトグラフィー工程のために、Sephadex G-25ゲル濾過カラムを2mlのバッファーD(1.137MのNaCl;2.7mMのKCl;1.5mMのKH2PO4;8mMのNa2HPO4;pH7.4)で再生し、そして次に4倍のカラム体積のバッファーC(137mMのNaCl;2.7mMのKCl;1.5mMのKH2PO4;8mMのNa2HPO4;20%(w/v)のグリセロール;pH7.2)で平衡化した。ニッケルカラムから溶出したピーク画分は、VISION上に組み込まれたサンプルローダーを通じて、Sephadex G-25カラム上に自動的に載せられ、そしてタンパク質はバッファーCで2ml/分の流速で溶出した。脱塩した試料を2.7mlの画分で回収した。当該画分を0.22μmの滅菌遠心フィルター(Millipore)を通じて濾過し、等分し、凍結し、そして−80℃で保存した。試料のアリコートをSDS−PAGE(4〜12%NuPAGEゲル;Novex)上でクーマシーブルー染色及び抗His抗体を用いるウェスタンブロットにより解析した。
【0177】
クーマシーブルー染色。NuPageゲルを0.1%クーマシーブルーR250染色溶液(30%メタノール、10%酢酸)中で室温で1時間染色し、そして次に20%メタノール、7.5%酢酸中でバックグラウンドがクリアーになり、そしてタンパク質のバンドがはっきりと見えるようになるまで脱染した。
【0178】
ウェスタンブロット
電気泳動の後、タンパク質を290mAで1時間、4℃でゲルからニトロセルロース膜にエレクトロトランスファーした。膜を5%粉ミルク/バッファーE(137mMのNaCl;2.7mMKCl;1.5mMのKH2PO4;8mMのNa2HPO4;0.1%Tween20、pH7.4)を用いて1時間室温でブロッキングし、そして次に2つのウサギポリクローナル抗His抗体混合物(G−18及びH−15。各0.2μg/ml;Santa Cruz)/2.5%粉ミルク/バッファーEと一緒にインキュベートした。更に室温で1時間インキュベートした後、膜をバッファーEで洗浄し(3x10分)、そして次に、2.5%粉ミルクを含むバッファーE中で1/3000希釈した二次HRP複合抗ウサギ抗体(DAKO, HRP 0399)と一緒に2時間室温でインキュベートした。バッファーEで洗浄した後(3x10分)、膜をECLキット(Amersham Pharmacia)で1分間展開した。次に膜をHyperfilm(Amersham Pharmacia)に曝露し、フィルムを現像し、そしてウェスタンブロットのイメージを視覚解析した。
【0179】
タンパク質アッセイ
タンパク質濃度はBCAタンパク質アッセイキット(Pierce)とスタンダードとしてウシ血清アルブミンを用いて決定した。回収量は420μgであった。
【実施例5】
【0180】
SCS0010の特徴づけ
Blastを用いて、SCS0010は、皮膚乳頭由来タンパク質4(DERP4)、HSPC186又はUNQ631/PRO1248(SwissProtエントリーQ9P0T7、TME9_HUMAN)としても知られている膜貫通タンパク質9のスプライス変異体であることが決定された。TME9_HUMANとSCS0010との間のアラインメントを図11に示し、これはSCS0010が3’末端で異なることを示している。TMEM9の既知の特徴は:
・183アミノ酸長(20574Da)
・後期エンドソーム及びリソソームに局在しているI型膜タンパク質
・3つのグリコシル化型
・二量体化の可能性
・TMEM9ファミリーに属すること、及び
・細胞内輸送において提案される役割、
である。
【0181】
TMEM9は、Kveineらによって特徴付けられている(Kveine et al. Biochem Biophys Res Commun. 2002 Oct 4 ; 297 (4): 912-7. "Characterization of the novel human transmembrane protein 9 (TMEM9) that localizes to lysosomes and late endosomes.")。TMEM9ファミリーは、Interpro (http://www.ebi.ac.uk/interpro)において以下のように説明されている:「このファミリーは、ホモサピエンス膜貫通タンパク質9SwissProtQ9P0T7と相同な複数の真核生物膜貫通タンパク質を含む。TMEM9遺伝子は、N末端シグナルペプチド、1つの膜貫通領域、3つの潜在的なNグリコシル化部位及び3つの保存されたN末端のシステインリッチドメインを含む183アミノ酸のタンパク質をコードする。当該タンパク質は、線虫(C.エレガンス)からホモサピエンスまでの種で高度に保存されており、そして膜貫通タンパク質の新規ファミリーに属する。TMEM9の正確な機能は知られてないが、広範に発現しており、そして後期エンドソーム及びリソソームに局在することがわかっている。MEDLINE:。このファミリーのメンバーはCXCXCリピートINTERPRO:IPR004153をそれらのN末端領域に含む」。インターロイキン−8(IL−8)は、CXCケモカインサブファミリーのメンバーであり、そしてSCS0010はCXCXリピートを含む、IL−8に対する共通のパターンを示している。
【0182】
バイオインフォマティクスツール、例えばSMART(http://smart.embl-heidelberg.de/)、を用い、TMEM9及びスプライシング変異体SCS0010の推定のドメインを同定した。SMARTの結果を図12に示す。この解析は、SCS0010がTMEM9の可溶性変異体であることを示し、その結果SCS0010が膜結合型のTMEM9と比較して独特の機能を示すことを示唆している。そのようなものとして、分泌型のSCS0010タンパク質は内在性TMEM9のアンタゴニストとしてin vitroで働くようである。Kveineらは、TMEM9がN末端領域に見られる3つの保存されたシステインリッチドメインを有することを示唆している。彼等は更に、これらの領域がタンパク質の折りたたみ、タンパク質の相互作用、及び多量体化に関与しうることに言及している。これらの領域はSC0010において完全に保存されている(図12において矢印で示す)。そのようなものとして、そして理論に拘束されることを望むものではないが、可溶性SCS0010は、二量体化によるTMEM9の制御に関与しているようである。従って、SCS0010は、治療的用途の項目に記載のような疾患の診断及び処置において特に有用であることを示しているようである。本明細書で同定したSCS0010のモチーフ及び部位(例えば、グリコシル化部位、プロテオグリカン部位、SH2ドメイン、保存されたシステインリッチドメイン)は、処置及び/又は診断のために(アゴニスト又はアンタゴニスト、例えば抗体によって)修飾及び/又は標的化されうる。
【0183】
理論に拘束されることを望むものではないが、プロテオグリカンのためのTMEM9部位における突然変異(後述のELMの結果を参照のこと)は、ガンになる遺伝的素因に関与しているようである。Kveineらによって同定されたTMEM9のN末端部分の3つのNグリコシル化部位は、このタンパク質の構造及び機能にとって潜在的に重要なものであることが示唆された。彼等は異なる機能及び/又は親和性の特性を有すると思われる3つのグリコシル化形態を同定した。TMEM9とガンの関連も、Kurokawaらの刊行物(Kurokawa et al. J Exp Clin Cancer Res. 2004 Mar; 23(1) : 135-41. PCR-array gene expression profiling of hepatocellular carcinoma)、並びにマイクロアレイベースの実験及び他の発現実験(治療的用途の項目を参照のこと)によって支持されている。従って、可溶性SC0010は、TMEM9との結合によって、多数の疾患、例えばガンを軽減及び/又は予防することができるようである。
【0184】
尚、Prosite(http://us.expasy.org/prosite/)及びELM(http://elm.eu.org/basicELM/)も、これらの配列について行った。
【0185】
Prositeの結果
> PDOC00001 PS00001 ASN_GLYCOSYLATION Nグリコシル化部位[パターン][警告:発生の可能性が高いパターン]
25-28 NKSS
42-45 NISG
51-54 NVSQ
【0186】
> PDOC00005 PS00005 PKC_PHOSPHO_SITE プロテインキナーゼCリン酸化部位[パターン][警告:発生の可能性が高いパターン]
53-55 SqK
91-93 TiK
【0187】
> PDOC00006 PS00006 CK2_PHOSPHO_SITE カゼインキナーゼIIリン酸化部位[パターン][警告:発生の可能性が高いパターン]
27-30 SseD
53-56 SqkD
【0188】
【表4】
【0189】
ELMの結果の説明(ELMによるもの)
MOD_GlcNHglycan:プロテオグリカンは細胞表面及び細胞外マトリックス内で見られる。それらは細胞コミュニケーションにとって重要であり、例えば形態形成及び発生における役割を担っている。幾つかのプロテオグリカンにおける突然変異は、ガンになる遺伝的素因と関連している。コアタンパク質は、露出しているセリン残基上のグリコサミノグリカン鎖の付着によって修飾されている。ヘパラン硫酸の場合、この過程は、ゴルジ層板においてプロテインキシロシルトランスフェラーゼ(EC2.4.2.26)がUDP−キシロース由来のキシロースをセリンのヒドロキシル基に転移させることによって開始する。この系は後生動物において進化してきたようである。
【0190】
MOD N-GLC 1:Nグリコシル化は、真核細胞における分泌型及び膜結合型タンパク質の最も一般的な修飾である。Nグリコシル化の全工程は、100超の酵素及び輸送タンパク質を含んで成る。全てのN結合型オリゴ糖の生合成は、ERにおいて大きな前駆体オリゴ糖から開始する。このオリゴ糖[(Glc)3(Man)9(GIcNAc)2]の構造は植物、動物、単細胞真核生物において同一である。この前駆体は、前記オリゴ糖の担体として働く長鎖ポリイソプレノイド脂質であるドリコールと結合している。続いて、前記オリゴ糖は、ER酵素によって、ドリコール担体から新生タンパク質上のアスパラギン酸残基に転移する。オリゴ糖鎖は、続いて、前記糖タンパク質がゴルジ装置を通り抜ける場合にプロセシングされる。場合によっては、この修飾は、より多くのマンノース基の付着を伴い、他の場合にはより複雑な構造の型が付着する。
【0191】
LIG SH2 STAT5:Srcホモロジー2(SH2)ドメインは、異なるシグナル伝達経路に関与する大多数のタンパク質において見られる小さいモジュラードメインである。それらはリン酸化チロシン残基を含む特異的なモチーフと結合することができ、シグナルを下流に伝播してタンパク質間相互作用を促進し、そして/あるいは酵素活性を修飾する。SH2ドメインの異なるファミリーは、異なる結合特異性を有することがあり、これは通常、pYに対してC末端側の数残基(位置+1、+2及び+3)によって決定される。リン酸化されてないペプチドはSH2ドメインと結合しない。少なくとも3つの異なる結合モチーフが知られている。pYEEI(SrcファミリーSH2ドメイン)、pY[IV].[VILP](SH−PTP2、ホスホリパーゼC−ガンマ)、pY.[EN](GRB2)。しかしながら、SH2ドメインとそれらの基質との相互作用は、他の表面領域の協調的な接触とも独立している。
【実施例6】
【0192】
ケモカイン様ポリペプチドの確証及び特徴づけのための細胞−及び動物−ベースのアッセイ
構造−活性の相関関係に対する研究は、ケモカインがアミノ末端領域を利用することで受容体と結合し、そして受容体を活性化していることを示唆している。この領域のアミノ酸に対するタンパク質分解的な消化、突然変異、又は化学的修飾は、アンタゴニスト的な活性を有する化合物を生成しうる(Loetscher P and Clark-Lewis I, J Leukoc Biol, 69: 881-884,2001 Lambeir A, et al. J Biol Chem, 276: 29839-29845, 2001, Proost P, et al. Blood, 98 (13): 3554-3561,2001)。従って、相当のケモカインのアミノ末端領域又は他の領域の1又は複数の残基の特異的な修飾(欠失、非保存的な置換)から生じるアンタゴニスト的な分子は、炎症及び自己免疫疾患についての治療的な可能性を有していると考えられる(WO 02/28419; WO 00/27880; WO 99/33989; Schwarz MK and Wells TN, Curr Opin Chem Biol, 3: 407-17,1999)。それ故に、本願の更なる目的は、本発明のポリペプチドの修飾によって生じたそのような種類のアンタゴニストによって表される。
【0193】
本発明のポリペプチド及び関連試薬の治療上の応用は、創薬及び前臨床段階の開発の間のケモカイン及び他の生物学的産物の確証のために知られている、動物細胞、組織及びモデルを利用するin vivo/in vitroアッセイによって(Coleman RA et al., Drug Discov Today, 6: 1116-1126,2001 ; Li AP, Drug Discov Today, 6: 357-366,2001 ; Methods Mol. Biol vol. 138, "Chemokines Protocols", edited by Proudfoot AI et al. , Humana Press Inc. , 2000; Methods Enzymol, vol. 287 and 288, Academic Press, 1997)、又はin silico/コンピューターを利用したアプローチによって、(安全性、薬物動態及び有効性の観点から)評価されうる。
【0194】
本願は、疾患、例えば細胞増殖障害、自己免疫/炎症性障害、心臓血管障害、神経障害、発達障害、代謝障害、感染及び他の病状の処置又は予防において、適切に製剤化された医薬組成物における活性成分として有用でありうる新規ケモカイン様ポリペプチド及び一連の関連試薬を開示する。特に、既知のケモカインの特性を考慮すると、開示するポリペプチド及び試薬は、異常な又は欠陥のある細胞遊走に関与する症状に対処するはずである。そのような症状の非限定的な例は以下の通りである:関節炎、関節リウマチ(RA)、乾癬性関節炎、骨関節炎、全身性エリテマトーデス(SLE)、全身性硬化症、強皮症、多発性筋炎、糸球体腎炎、線維症、肺線維症及び肺炎、アレルギー性疾患及び過敏性疾患、皮膚炎、喘息、慢性閉塞性肺疾患(COPD)、炎症性腸疾患(IBD)、クローン病、潰瘍性大腸炎、多発性硬化症、敗血症ショック、HIV感染、移植片拒絶、創傷治癒、転移、子宮内膜症、肝炎、肝硬変、ガン、無痛覚症、並びにアテローム性動脈硬化症に関連する血管炎症。
【0195】
細胞培養物又は動物モデルを用いてケモカインの特異性、能力、及び有効性を試験するための複数のアッセイが開発されており、例えば、in vitroでの走化性アッセイ (Proudfoot A, et al. J Biol Chem 276: 10620-10626,2001 ;Lusti-Narasimhan M et al. , J Biol Chem, 270: 2716-21,1995)、又はマウス耳介膨脹(Garrigue JL et al., Contact Dermatitis, 30: 231-7,1994)である。有用なツール及び生成物(抗体、トランスジェニック動物、放射性標識タンパク質等)をもたらすための多数の他のアッセイ及び技術は、ケモカインに対して貢献する概説及び本において記載されており(Methods Mol. Biol vol. 138, "Chemokines Protocols", edited by Proudfoot Al et al., Humana Press Inc., 2000; Methods Enzymol, vol. 287 and 288, Academic Press, 1997)、そして、より正確には、本発明のケモカイン様ポリペプチド及び関連試薬の生物学的活性を、可能性のある治療又は診断方法及び使用と関連して証明するために使用されうる。
【0196】
サイトカイン発現調節アッセイ
1 序論
以下のin vitroの細胞ベースの三回一組のアッセイは、IL−2、IFN−γ、TNF−α、IL−5、IL−4及びIL−10についてのサイトカインビーズアレイ(CBA)アッセイによって測定した場合の異なるヒト末梢血単核細胞(hPBMC)に対して作用するコンカナバリンA(ConA)によって誘導されるサイトカイン分泌に対する本発明のタンパク質の作用を測定する。
【0197】
最適な条件は、96ウェルプレート1ウェル当たり100,000個の細胞であり、そして2%のグリセロール中最終的に100μlである。
【0198】
分裂促進因子(ConA)の最適濃度は5ng/mlである。
【0199】
当該アッセイに最適な時間は48時間である。
【0200】
読み取りの選択はCBAである。
【0201】
2 装置及びソフトウェア
・96ウェルマイクロタイタープレートフォトメーターEX(Labsystem).
・Graph Pad Prismソフトウェア
・エクセルソフトウェア
・フローサイトメーター Becton-Dickinson
・CBA解析ソフトウェア
・細胞培養のためのフード
・細胞培養のためのインキュベーター
・遠心機
・ピペット
【0202】
3 材料と試薬
・軟膜
・DMEM GIBCO
・ヒト血清AB型 SIGMA
・L−グルタミン GIBCO
・ペニシリン−ストレプトマイシン GIBCO
・フィコール PHARMACIA
・細胞培養のための96ウェルマイクロタイタープレート COSTAR
・コンカナバリンA SIGMA
・ヒトTh1/Th2サイトカインCBAキット Becton-Dickinson
・PBS GIBCO
・FALCON 50ml滅菌済み Becton-Dickinson
・BSA SIGMA
・グリセロール MERCK
・DMSO SIGMA
・96ウェルマイクロタイタープレートコニカルボトム NUNC
【0203】
4 方法
4.1 軟膜由来のヒトPBMCの精製
軟膜1〜2をDMEMで希釈する。25mlの希釈した血液をその後ゆっくりと50mlのファルコンチューブ中の15mlのフィコール層に添加し、そしてチューブを遠心する(2000rpm、20分、室温、中断無し)。中間層(リング)を続いて回収し、そして細胞を25mlのDMEMで洗浄し、その後遠心段階を行う(1200rpm、5分)。この手順を3回繰り返す。軟膜は合計約600x106個の細胞をもたらす。
【0204】
4.2 スクリーニング
80μlの1.25x106個の細胞/mlをDMEM+2.5%ヒト血清+1%L−グルタミン+1%ペニシリン−ストレプトマイシン中で希釈し、そしてその後96ウェルマイクロタイタープレートに添加する。
【0205】
ウェル当たり10μl添加する(ウェル当たり1つの条件):タンパク質をPBS+20%グリセロール中で希釈する(最終的なタンパク質の希釈は1/10である)。
【0206】
ウェル当たり10μlのConA刺激物を続いて添加する(ウェル当たり1つの条件):ConA 50μg/ml(ConAの終濃度は5μg/mlである)。
【0207】
48時間後、細胞上清を回収し、そしてヒトサイトカインをヒトTh1/Th2サイトカインCBAキット(Becton-Dickinson)で測定する。
【0208】
4.3 CBA解析
(より具体的には、CBAキットの小冊子を参照のこと)
i)混合ヒトTh1/Th2捕獲ビーズ
当該実験に必要なアッセイチューブの数を決定する。
【0209】
各捕獲ビーズの懸濁液を混合する前に激しく数秒間ボルテックスにかける。
【0210】
解析を行う各アッセイにつき、各捕獲ビーズの10μlのアリコートを「混合捕獲ビーズ」と標識を付した一本のチューブに添加する。ビーズ混合物を徹底的にボルテックスにかける。
【0211】
ii)試験試料の調製
上清は、アッセイ希釈物を用いて希釈した(1:4)(20μlの上清+60μlのアッセイ希釈物)。試料の希釈物は、96ウェルマイクロタイタープレートコニカルボトム(NUNC)中に試料を移す前に混合する。
【0212】
iii)ヒトTh1/Th2サイトカインCBAアッセイ手順
50μlの希釈した上清を96ウェルマイクロタイタープレートコニカルボトム(NUNC)中に添加する。50μlの混合した捕獲ビーズを添加し、続いて50μlのヒトTh1/Th2PE検出試薬を添加する。プレートを続いて3時間室温でインキュベートし、そして直接的な光への曝露から保護し、続いて1500rpmで5分間遠心する。上清を続いて慎重に捨てる。次の段階において、200μlの洗浄バッファーを2回各ウェルに添加し、1500rpmで5分間延伸し、そして上清を慎重に捨てる。130μlの洗浄バッファーをその後各ウェルに添加し、ビーズペレットを再懸濁する。試料は最終的にフローサイトメーター上で解析する。データはCBAアプリケーションソフト、Activity Base及びマイクロソフトエクセルソフトウェアを用いて解析する。
【0213】
アッセイの読み取りから、in vitroで、本発明のタンパク質が試験した全てのサイトカイン(IFN−γ、TNF−α、IL−2、IL−4、IL−5、IL−10)に対し一貫した阻害効果を有するか否かが評価されうる。
【0214】
更にEC50値に基づき、どれが最もよく阻害されたサイトカインであるか、そしてそのようなサイトカインに特に関連していることが知られている特定の自己免疫疾患/炎症性疾患に達しているかが容易に評価されうる。
【実施例7】
【0215】
自己免疫/炎症アッセイ
Tリンパ球応答を標的とするアッセイ
・Fas−リガンド誘導型T細胞死
このアッセイは、受容体媒介型の細胞死の新規モジュレーターを明らかにする。
【0216】
このアッセイにおいて、T細胞のアポトーシスはジャーカット細胞(ヒトT細胞系)を。組換え6ヒスチジンタグ付きFasリガンドとモノクローナル抗6−His抗体とを組み合わせて刺激することで誘導される。死亡は、細胞が死んでいる場合に培養液中に放出される細胞質酵素LDHの放出で定量される。読み取りは、490nmで読まれる比色アッセイで行う。T細胞は、多数の自己免疫疾患において病原性であることが証明されており、抗原特異的T細胞死を制御することができるということは治療上の戦略となる(例えば、クローン病患者における抗TNFα治療)。
【0217】
・ヒト−MLR:増殖及びサイトカイン分泌
この細胞ベースのアッセイは、リンパ球の増殖及びサイトカインの分泌又は別のドナー由来のPBMCによる刺激時の阻害(アロ反応性)に対する新規タンパク質の効果を測定する。これらのアッセイは、あらゆる自己免疫疾患における必須の細胞性応答である抗原特異的T細胞及び抗原提示細胞の機能について対処する。分泌型サイトカイン(IL−2,4,5,10,TNF−α及びIFN−γ)はCBAで定量される。注記:増殖とサイトカイン分泌は独立の反応である。
【0218】
・マウス−MLR:増殖
この細胞ベースのアッセイは、別のドナー(マウス株)由来の脾臓細胞による刺激後のマウス脾臓細胞のリンパ球増殖又は阻害に対する新規タンパク質の効果を測定する。この細胞ベースのアッセイは、Tリンパ球及び抗原提示細胞の応答に対する新規タンパク質の効果を測定し、そしてh−MLRアッセイにおけるポジティブの活性とヒットの同一性を確認するために使用されるであろう。このアッセイは、ヒト疾患のマウスモデルにおいて試験されるタンパク質を選択するために使用されるであろう。
【0219】
・スーパー抗原TSSTで刺激したヒトPBMC
スーパー抗原は、T細胞に影響を及ぼす免疫系の強力なモジュレーターである。スーパー抗原は、免疫介在疾患、例えばIBD、炎症性皮膚疾患、例えばアトピー性皮膚炎及び乾癬に影響を及ぼす。この細胞性アッセイにおいて、我々は、TCRを介するが、古典的な抗原に対するT細胞応答、特に共刺激分子に関するものと異なる要件を有するTリンパ球活性化を特異的にターゲティングする。
【0220】
・ConA又はPHAのいずれかで刺激したヒトPBMC
これらの細胞ベースのアッセイは、サイトカインビーズアレイ(CBA)アッセイ(IL−2、IFN−γ、TNF−α、IL−5、IL−4及びIL−10)によって測定した場合の異なる細胞に対する2つの異なる刺激作用によって誘導されるサイトカイン分泌に対する新規タンパク質の効果を測定する。
【0221】
サイトカインの多くが、損傷、環境及び細胞内標的に依存して、前炎症性又は抗炎症性という二重の作用を有することがある。サイトカイン分泌を調節する能力を有するあらゆるタンパク質が、治療上の潜在能力を有するようである(例えば、IFN−γ及びTNF−αの低下はTh1介在性自己免疫疾患において有益なようであり、一方、IL−4、IL−5の低下はTh2介在性の疾患において有益なようであり、IL−10の誘導はMS及びSLEにおいて興味深い)。
【0222】
単球/マクロファージと顆粒球の反応をターゲティングするアッセイ
・LPSで刺激したヒトPBMC
この細胞ベースのアッセイは、単球/マクロファージ及び顆粒球に対して作用するLPSによって誘導されるサイトカイン分泌(IFN−γ、TNF−α)に対する新規タンパク質の効果を測定する。
【0223】
IFN−γ及びTNF−α分泌を調節する能力を有するあらゆるタンパク質がTh1介在性の自己免疫疾患において有益なようである。
【0224】
好中球反応をターゲティングするアッセイ
好中球は、炎症性疾患及び自己免疫疾患、例えば関節リウマチにおいて重要である。白血球走化性因子、例えばIL−8は、細胞と微小血管内皮細胞との間の一連の接着性の相互作用を開始させて好中球の活性化、接着、そして最終的に遊走をもたらす。好中球の組織浸潤は、これらの細胞の細胞形態の特定の変化を伴う細胞骨格エレメントの再構成に依存する。
【0225】
この細胞ベースのアッセイは、ヒト好中球の細胞骨格再構成に対する新規タンパク質の効果を測定する。
【0226】
Bリンパ球反応をターゲティングするアッセイ
自己抗体並びに浸潤B細胞は、種々の自己免疫疾患、例えば全身性エリテマトーデス(SLE)、関節リウマチ(RA)、シェーグレン症候群及び重症筋無力症の病原において重要であると考えられている。B細胞の恒常性の異常調節が免疫寛容に影響を及ぼして、病原性抗体を産生し、且つ持続性の炎症を引き起こす自己反応性B細胞の不適切な生存をもたらしうるという有力な証拠がある。B細胞受容体のトリガリング後のB細胞の増殖、生存及び分化の制御において必須の役割を果たす新規因子の同定は、新規治療の開発に高度に関連するものである。
【0227】
・B細胞の増殖
この細胞ベースのアッセイは、B細胞の生存に対する新規タンパク質の効果を測定する。
【0228】
・B細胞の同時刺激
この細胞ベースのアッセイは、B細胞の同時刺激に対する新規タンパク質の効果を測定する。
【0229】
単球及びミクログリアの反応をターゲティングするアッセイ
・THP−1カルシウム流入
THP−1細胞アッセイにおけるCa+流入は、小胞体からの細胞内カルシウム放出(一般的な二次メッセンジャーの事象)を引き起こす能力に対する新規タンパク質の効果を測定する。
【0230】
ミクログリア細胞増殖(次のIACに提示されうるもの)
ミクログリア前駆細胞の増殖の間、多数のコロニー刺激因子、例えば幾つかのサイトカインが、重要な役割を果たすことが知られている。それらの中でも、M−CSFはマクロファージ/ミクログリアの成熟の最終段階に必須であり、そして他のどの因子によっても代替不可能である。この生物学的反応の評価は、ミクログリア活性に影響を及ぼす方法を代表することがあり、そしてそれ故にMSから治療上の潜在性を有する分子を同定する機会を代表することがある。
【0231】
M−CSFに対するミクログリア細胞系の増殖反応を測定する細胞ベースのアッセイが開発された。その実現可能性及び頑健性のフェーズは最適な結果を示した。このアッセイは96ウェルプレートで行われ;非放射性の基質が必要とされ、容易に自動化される。
【実施例8】
【0232】
タンパク質機能の生物学的関連性の調査に適した神経学的アッセイ
多数の神経学的アッセイが本出願人によって開発されており、そしてそれらはタンパク質の生物学的関連性の研究において使用するものである。本出願人によって開発された神経学的アッセイの例には、4つのタイプのアッセイがある。これらを以下論じる。
【0233】
i.乏突起膠細胞ベースのアッセイ
乏突起膠細胞はCNSにおけるミエリン形成にとって重要である。多発性硬化症において、それらは最初に攻撃される細胞であり、それらの損失は主要な行動障害を引き起こす。炎症の抑制に加え、MSで生じる病変の不完全な再ミエリン化の増強は、MSの治療上のストラテジーとして提唱されている。ニューロンのように、成熟した乏突起膠細胞は分割しないが、新規乏突起膠細胞は前駆細胞から生じることがある。成体の脳にはこれらの前駆細胞はほとんど存在せず、そして胚においてでさえも、前駆細胞の数はHTSにとって不適切である。
【0234】
Oli−neuは、t−neu発ガン遺伝子による乏突起膠細胞の不死化によって得られるマウスの細胞系である。それらは十分に研究されており、若い乏突起膠細胞の細胞学を研究するための代表的な細胞系として受け入れられている。
【0235】
これらの細胞は2つのタイプのアッセイにおいて使用されうる。
【0236】
一方は、乏突起膠細胞の増殖を刺激する因子を同定するためのものであり、そして他方は、それらの分化を促進する因子を発見するためのものである。それらの事象は、再生を助け、且つ脱髄疾患を治療する観点から重要である。
【0237】
別の可能性のある細胞系はヒト細胞系MO3−13である。MO3−13は、横紋筋肉腫細胞と成人ヒト乏突起膠細胞との融合から生じる。しかしながら、これらの細胞は乏突起膠細胞に分化する能力が低下しており、そしてそれらの増殖速度は増殖アッセイを可能にするのに十分ではない。それにも関わらず、それらは乏突起膠細胞の幾つかの特徴を有しており、それらの形態は、核転座の研究によく適合している。従って、この細胞系は、3つの転写因子、それぞれNF−κB、Stat−1及びStat−2の核転座に基づいたアッセイにおいて使用されることがある。Jak/Stat転写経路は多数の因子、例えばIFN−α、β、γ、サイトカイン(例えば、IL−2、IL−6;IL−5)又はホルモン(例えば、GH、TPO、EPO)によって活性化される複雑な経路である。反応の特異性は活性化されたStatの組み合わせに依存する。例えば、IFN−βはStat1、2及び3の核転座を活性化し、一方、IFN−γはStat1だけ活性化することは注目に値する。同様に、多数のサイトカイン及び増殖因子がNF−κBの転座を誘導した。これらのアッセイの目的は、所定のタンパク質についての活性化経路の写真を撮ることである。
【0238】
ii.星状細胞ベースのアッセイ
星状細胞の生物学は非常に複雑であるが、2つの一般的な状態が認識されている。静止状態と称される1つの状態において、星状細胞は、グルタミン酸をポンピングし、そしてエネルギー性の下層をニューロン及び乏突起膠細胞に提供することでニューロンの代謝性及び興奮性のレベルを制御する。活性化された状態において、星状細胞はケモカイン及びサイトカイン並びに一酸化窒素を産生する。最初の状態は正常な健康状態と見なされうるが、第二の状態は炎症、発作又は神経変性疾患の間に生じる。
【0239】
多数の因子及び多数の経路が星状細胞の活性化を調節することが知られている。星状細胞の活性化を調節する新規因子を同定するために、アストログリオーマ起源のヒト細胞系であるU373細胞が使用されうる。NF−κB、c−Jun並びにStatは、星状細胞活性化において中心的な役割を果たすことが知られているシグナル伝達分子である。
【0240】
NF−κB、c−Jun並びにStat1、2及び3の核転座に基づく一連のスクリーニングが実施されうる。これらの経路のプロトタイプの活性化因子はIL−1b、IFN−ベータ又はIFN−ガンマである。この目的は、CNS疾患の処置における治療物質として使用されうるタンパク質を同定することである。
【0241】
C.ニューロンベースのアッセイ
ニューロンは非常に複雑且つ多様な細胞であるが、それらは全て共通の2つの事を有している。第一にそれらは分裂終了細胞であり、第二にそれらは他の細胞を神経支配している。それらの生存は、神経支配された標的細胞がしばしば産生する栄養性因子の存在と関連している。多数の神経変性疾患において、標的の神経支配が失われると細胞体の萎縮及びアポトーシス性の細胞死に至る。従って、標的の欠損を補足するする栄養性因子の同定は神経変性疾患の処置において非常に重要である。
【0242】
この観点から、ラットPC12細胞のサブクローンであるNS1細胞を用いる生存アッセイが実施されうる。これらの細胞は何年も使用されており、そして多くの神経生物学の知識が、一次ニューロンに対して確認する前に、これらの細胞に対して最初に獲得されており、例えばニューロンの生存及び分化に関与する経路である(MEK、PI3K、CREB)。対照的に、マウス神経芽細胞腫であるマウスN2A細胞は、古典的な神経栄養因子には反応しないが、Junキナーゼ阻害剤は血清枯渇によって誘導されるアポトーシスを防ぐ。従って、これらの細胞系に対するアッセイは、異なるタイプの「生存促進」タンパク質を発見する助けとなるであろう。
【0243】
これまでのアッセイにおいて、本発明者が増殖と分化両方を促進する因子を同定しうることに注目することは重要である。ニューロンの分化を特異的に促進する因子を同定するために、神経突起伸長に基づいたNS1分化アッセイが使用されうる。神経変性疾患における軸策及び樹状突起の発芽を促進させることは、2つの理由から有利なことがある。最初に、それは変性ニューロンが再成長し、そして標的細胞との接触を再確立するのを助ける。第二に、それは、神経変性の末期、例えばパーキンソン又はADを遅らせる代償的な現象である、健康な線維からのいわゆる側枝の発芽を増強する。
【0244】
1.内皮細胞ベースのアッセイ
脳と血管の間の血液脳関門(BBB)は、皮質の髄液と血清の間の組成の差異に重要である。BBBは、内皮細胞と星状細胞との密接な接着から生じる。それは、脳における白血球の透過を防ぐことで免疫寛容状態を維持し、そして同一の細胞内シグナル伝達経路を用いる2つの平行した内分泌系の発達を可能にする。しかしながら、多くの疾患又は外傷において、BBBの完全性は変化し、そして白血球並びに血清タンパク質は脳に進入して神経炎症を誘導する。BBBのin vitroモデルは簡単ではないが、一次内皮細胞、例えばヒト臍帯静脈内皮細胞(HUVEC)は、BBB生物学の幾つかの側面を模倣することができる。例えば、BBBの漏出は、細胞なカルシウム放出を刺激するタンパク質によって誘導することができる。BBBの完全性を調節するタンパク質を同定する観点から、トロンビンを用いての又は用いないカルシウム動員アッセイがHUVECで実施されうる。
【実施例9】
【0245】
前記タンパク質機能の生物学的関連性の調査に適した線維芽細胞アッセイ
多数の線維芽細胞アッセイが本出願人によって開発されており、これはタンパク質機能の生物学的関連性の研究において使用するためのものである。本出願人によって開発された線維芽細胞アッセイの例には8つのタイプのアッセイが含まれる。これらを以下論じる。
【0246】
線維芽細胞の活性化及び病理学的増殖は、線維症として知られている現象を引き起こす重要な段階である。線維症は、細胞外マトリックス、例えばコラーゲンの過剰な沈着を特徴とする。間質細胞、例えば線維芽細胞は、特異的な線維化誘発性(pro-fibrotic)タンパク質及び抗線維化タンパク質を発現している。ケラチノサイト増殖因子(KGF)は抗線維化分子によってよく特徴付けられている。更に、酸化的損傷及び炎症誘発性の刺激は、主要な事象の中でも、筋線維芽細胞の表現型、そして最終的には線維症を引き起こすことが提唱されている。NF−κBは酸化的ストレス及び炎症反応のメディエーターである。線維芽細胞の生物学に基づき、本発明者は4つの細胞ベースのアッセイ、すなわち線維芽細胞増殖、コラーゲン産生、NF−κB活性化及びKFG産生アッセイを開発した。
【0247】
A.ヒト線維芽細胞増殖アッセイ
線維芽細胞の活性化及び病理学的な増殖は、線維症として知られている表現型を引き起こす重要な段階である。当該アッセイは、細胞性の核酸に結合したCyQuant GR Dyeを介する蛍光増強に基づくものであり、新規タンパク質及び小分子に対するヒト皮膚由来線維芽細胞の増殖反応を測定するものである。
【0248】
B.ヒト線維芽細胞によるI型コラーゲン産生
線維症は、細胞外マトリックス、特にコラーゲンの過剰な蓄積を特徴としている。I型コラーゲンの過剰産生は全身性硬化症の主要な徴候である。TGFβは、in vitro及びin vivoでコラーゲンの産生をアップレギュレートすることが知られている。本発明者は、新規な線維化誘発性(pro-fibrotic)分子及び抗線維化分子がヒト皮膚由来線維芽細胞によるI型コラーゲンの産生の基礎レベル又はTGFβ1刺激レベルを調節する能力を試験するための細胞ベースのアッセイを開発した。
【0249】
C.ヒト線維芽細胞によるケラチノサイト増殖因子(KGF)の産生
KGFは、正常及び病的な成長発達の間の、多くの器官(例えば、肺、膵臓、腎臓、前立腺、乳房、腺、子宮)における間質と上皮の相互作用の重要なメディエーターである。KGFは特に間質細胞により産生され、そしてその受容体は特に上皮細胞で発現している。KGFは線維症の病理学的反応の間に重要な役割を果たしているようであり、その結果これらの反応のマーカーとして使用することができると提唱されている。KGFのELISAアッセイが開発されており、ヒトの肺由来線維芽細胞を用いた結果、KGFの産生が有意にIL−1β及びTNFαによってアップレギュレートされることがあり、そしてTGFβによってダウンレギュレートされることがあることが証明された。これらのサイトカインは、KGF産生を誘導することができる新規タンパク質をスクリーニングする際の参照分子として使用されるであろう。
【0250】
D.線維芽細胞におけるNF−κB転写活性
酸化的損傷及び炎症誘発性の刺激は、主要な事象の中でも、筋線維芽細胞の表現型、そして最終的には線維症を引き起こすことが提唱されている。NF−κBは酸化的ストレス及び炎症反応のメディエーターである。Swiss3T3線維芽細胞は、安定的に組み込まれたNF−κB−SEAP(分泌型アルカリホスファターゼ)コンストラクトを用いて作出された。NF−κB−SEAPは、κエンハンサーに対する転写因子の結合を測定してNF−κB経路の活性化の直接的な測定を可能にするよう設計された。SEAP酵素は培養液中に分泌され、その結果試料は、細胞を回収することなく、転写活性についてのアッセイに対しての種々の時点で回収することができる。Swiss3T3−NF−κB−SEAP線維芽細胞系を細胞ベースのアッセイとして使用して新規な機能ゲノミクスタンパク質を試験することができ、そしてこれは小分子、特に炎症誘発性/抗炎症性の活性を有するものを試験するのに非常に見込みがある。
【0251】
E.線維芽細胞における結合組織成長因子(CTGF)プロモーターの活性化/抑制
38kDのシステインリッチなタンパク質であるCTGFは、線維芽細胞による細胞外マトリックス成分の産生を刺激する。CTGFの過剰発現は、伝えられるところによると、多くの線維性ヒト組織、例えば肺、皮膚、肝臓、腎臓及び血管において見出されている。in vitroでは、TGFβは、ヒト肺線維芽細胞におけるCTGF遺伝子の転写を活性化する。CTGFプロモーター−レポーターは、レポーターとして分泌型アルカリホスファターゼ(SEAP)を用いて構築され、そして3T3線維芽細胞は安定的に組み込まれたCTGF−SEAPコンストラクトを用いて作出された。これらの線維芽細胞を用いて、CTGFプロモーターがSARP−1、OPG及びFSGによってダウンレギュレートされ、そしてTGFβによってアップレギュレートされることが証明された。
【0252】
F.KL−6産生
最初に肺腺癌関連タンパク質として発見され、そしてその後MUC−1と称されることとなったKL−6は、高分子量糖タンパク質であり、現在ではCD9抗原として分類されている。KL−6は特発性肺線維症(IPF)及び他の肺間質性疾患を有する患者の血清及びBALFの両方で増大する。IPFに罹患している患者由来の肺組織において、KL−6抗体で標識された細胞の多くが再生II型肺細胞である。KL−6に対するポリクローナル抗体を産生する2つのペプチドを設計した。KL−6ELISAを使用して、ヒト肺由来II型肺細胞によるKL−6の産生を測定することができる。
【0253】
G.可溶性組換えTRAIL(TNF−関連アポトーシス誘導リガンド)で処理したL−929線維芽細胞のアポトーシスの中和
TRAILはオステオプロテジェリン(OPG)の細胞性リガンドのうちの1つであることが証明されている。このアッセイを使用してOPGの生物学的活性を測定することができる。
【0254】
H.ヒト線維芽細胞によるRANKL(NF−κBリガンドの受容体活性化因子)の産生
RANKLはOPGの別のリガンドである。このアッセイもOPGの生物学的活性を測定するために使用することができる。
【実施例10】
【0255】
タンパク質機能の生物学的関連性の調査に適したリプロダクティブ・ヘルスアッセイ
多くのリプロダクティブ・ヘルス関連アッセイが本出願人により開発されており、これらはSCS0010タンパク質機能の生物学的関連性の調査に使用するものである。男性不妊におけるSCS0010の予想される意味を考慮すると(治療用途を参照のこと)、本アッセイは特に関連性があるように見える。本出願人によって開発されたリプロダクティブ・ヘルス関連アッセイの例は、リプロダクティブ・ヘルスについての18の細胞ベースアッセイを含む。これらについて以下論じる。
【0256】
1.一次ヒト子宮平滑筋増殖アッセイ:
子宮平滑筋細胞の増殖は、女性における子宮筋腫疾患における腫瘍の発達のための前駆体である。このアッセイにおける目的は、一次ヒト子宮平滑筋細胞の増殖を阻害するタンパク質を同定することである。
【0257】
2.JEG−3着床アッセイ
JEG−3細胞は、着床の間の胚盤胞のモデルとして使用される栄養絨毛膜ヒトガン細胞系である。Ishikawa細胞は、脱落膜のモデルとして使用される比較的未分化のヒト子宮内膜ガン細胞系である。JEG−3細胞は、ヒト脱落膜組織に「着床」する。このアッセイにおいては2チャンバーシステムを使用し、ここで、蛍光標識したJEG−3細胞は、Ishikawa細胞又はIshikawa条件培地がロアチャンバー内に据えられた場合にマトリゲルコーティングした多孔性膜を通じてアッパーチャンバーからロアチャンバーへ進入する。遊走する細胞は、プレートリーダー内で定量される。in vivoでの着床を幇助するのに使用するためのJEG−3細胞の侵入を増大させるタンパク質を同定することがこの目的である。
【0258】
C.オステオポンチンビーズアッセイ(Ishikawa細胞):
Ishikawaヒト子宮内膜ガン細胞を着床のモデルとして使用する。ヒトにおける着床時に、種々のインテグリンが、胚盤胞によって発現するタンパク質と結合すると考えられている子宮内膜により発現する。Ishikawa細胞は、文献にてavb3を発現することが証明されており、avb3は「着床有効期間(window of implantation)」の間子宮内膜で発現するインテグリンである。このインテグリンは、栄養膜で発現するオステオポンチンと結合すると考えられている。このアッセイにおいて、オステオポンチンでコーティングした蛍光ビーズは線維芽細胞を表し、そしてIshikawa細胞は、エストラジオールでそれらを処理することによりそれらが結合することを受け入れるようプライミングされる。この目的は、着床の時期に子宮内膜の受容性を増大させるための補助として、Ishikawa細胞がオステオポンチンと結合する能力を増大させるタンパク質を同定することである。
【0259】
D.HuF6アッセイ
HuF6細胞は一次ヒト子宮線維芽細胞である。これらの細胞をIL−1βで処理することにより脱落膜が起こるように誘導することができる。脱落膜のマーカーは、PGE2の産生であり、これはELISAによって測定される。この目的は、妊娠初期の間に脱落膜を増強する手段としてHuF6細胞によるPGE2の産生を増大するタンパク質を同定することである。
【0260】
2.子宮内膜症アッセイ:
腹膜のTNFαは、子宮から脱落した子宮内膜細胞が腹膜の中皮細胞に接着して増殖するように誘導することで子宮内膜症における役割を果たしている。このアッセイにおいて、BEND細胞は、子宮内膜症の間の接着についてのアッセイとして、それらがフィブロネクチンでコーティングした蛍光ビーズに結合する能力を増大させるTNFαで処理される。この目的は、TNFαが前記細胞のビーズ結合能を刺激する能力を低下させ又は刺激するタンパク質を同定することである。
【0261】
F.hLHRで安定トランスフェクションしたJC−410ブタ顆粒膜細胞を用いるサイクリックAMPアッセイ
多嚢胞性卵巣症候群において、下垂体由来のLHは比較的多く、そして卵巣の莢膜細胞からのアンドロゲンの産生を誘導する。このアッセイは、PCOSの間の卵巣でのLHの作用を低下させるのに使用することができる、LHのシグナル伝達の阻害剤を探索するために使用される。JC−410ブタ顆粒膜細胞系は、ヒトLH受容体を安定トランスフェクションされる。LHによる処理は、cAMPの産生をもたらす。
【0262】
G.hFSHRで安定トランスフェクションしたJC−410ブタ顆粒膜細胞を用いるサイクリックAMPアッセイ
JC−410ブタ顆粒膜細胞をヒトFSHRで安定トランスフェクションした。FSHによる処理はcAMP産生を刺激し、これをこのアッセイで測定する。この目的は、顆粒膜細胞におけるFSH作用を増強するタンパク質を同定することである。
【0263】
H.LβT2(マウス)下垂体細胞アッセイ:
LβT2は、不死化マウス性腺刺激ホルモン分泌細胞系である。アクチビン単独又はGnRH+アクチビンによる刺激は、FSHの分泌をもたらす(GnRH単独による刺激はLHの分泌をもたらす)。前記細胞は、GnRHと協調してFSH産生を刺激するよう働くタンパク質を発見するためにGnRH+バイオスクリーニングタンパク質で処理してもよく、あるいはそれらは、アクチビン単独のようにFSH分泌を刺激することのできるタンパク質を発見するためにバイオスクリーニングタンパク質で処理してもよい。
【0264】
I.卵丘膨化アッセイ:
マウス卵丘−卵母細胞複合体(2/ウェル)を用いた卵丘膨化アッセイは、卵母細胞の成熟に影響を及ぼすタンパク質についてアッセイする96ウェルフォーマットにおいて確認されている(卵丘膨化によって測定)。アッセイ当たり2つの96ウェルプレートを処理することができ、そして一週当たり2回のアッセイを実施することができる。バイオスクリーニングタンパク質を一種類の濃度だけでアッセイする場合、全てのバイオクスクリーニングIタンパク質は一ヶ月でアッセイすることができる。読み取りは膨化についてのイエス/ノーアンサーで行ってもよく、あるいは画像解析プログラムを使用して定量的に膨化を測定することもできる。
【0265】
J.RWPE増殖アッセイ:
良性前立腺肥大症は前立腺の上皮及び間質の増殖を特徴とし、これはアポトーシスでバランスを失わず、その結果器官の拡大がもたらされる。RWPEはHPV−18で不死化された正常なヒト前立腺上細胞系であり、これは一次ヒト前立腺上皮細胞の代わりに使用されることがある。
【0266】
K.HT−1080線維肉腫浸潤アッセイ:
このアッセイは、JEG−3着床アッセイ(上記)のポジティブな細胞のコントロールとして開発された。これはガンの転移のためのモデルとしてよく確立されたアッセイである。蛍光標識HT−1080ヒト線維肉腫細胞は、2−チャンバーシステムのアッパーチャンバーにおいて培養され、そして、多孔性のマトリゲルコーティングした膜を通じてボトムチャンバー内に浸潤するよう刺激することができ、ここで、それらが定量される。この目的は、浸潤を阻害するタンパク質を同定することである。前記細胞は、血清をボトムチャンバーに添加することにより浸潤するよう刺激され、そしてドキシサイクリンで阻害される。
【0267】
L.一次ヒト子宮平滑筋細胞アッセイ
子宮筋腫疾患の特徴の1つは、平滑筋腫となっている子宮平滑筋細胞によるコラーゲン沈着である。一次ヒト子宮平滑筋細胞はTGFβ処理でコラーゲンを産生するよう刺激され、これはRebifで阻止される。この目的は、この線維化の表現型を阻害するタンパク質を発見することである。
【0268】
M.ヒト平滑筋腫細胞増殖アッセイ:
ヒト平滑筋腫細胞系は、増殖アッセイにおける子宮筋腫のモデルとして使用されうる。当該細胞は非常にゆっくりと生育し、そして本発明者はエストラジオール及び増殖因子でそれらを処理することによりそれらを刺激してより速い速度で生育させる。この目的は、平滑筋腫細胞のエストラジオール依存性の増殖を阻害するタンパク質を同定することである。
【0269】
N.937遊走アッセイ
子宮内膜症の病変は、免疫細胞を腹腔に補充するサイトカインを分泌する。これらの免疫細胞(特に活性化マクロファージ及びTリンパ球)は、子宮内膜症に共通の炎症の症候を媒介する。RANTESは子宮内膜症の間質細胞によって産生されることが証明されており、そして腹水中に存在している。このアッセイにおいて、活性化マクロファージのモデルとして使用される単球細胞系であるU937は、2−チャンバーシステムのより下の位置で処理してアッパーチャンバーから遊走させることで誘導されうる。当該細胞を蛍光色素で前処理する場合、それらをロアチャンバー内で定量することができる。この目的は、U937細胞の遊走を阻害するタンパク質を同定することである。
【0270】
O.JEG3ヒト栄養膜アッセイ:
胚盤胞の栄養膜はクラスIのHLA分子であるHLA−Gを産生し、これは、母体による胚の免疫拒絶を防ぐのに重要であると考えられている。子癇前症の間、HLA−Gレベルは低いか全くなく、その結果恐らく特徴的な症候が生じ、例えば、母体の免疫学的な干渉による子宮内膜及びらせん動脈への栄養膜の浸潤が乏しいことが挙げられる。JEG−3ヒト栄養膜細胞系はHLA−Gを産生し、これはIL−10又はLIFによる処理で増大することができる。ELISAを使用してJEG−3によるHLA−G産生を測定することができ、これは、HLA−G産生を増大することができる他のタンパク質の発見を目的とするものである。
【0271】
P.一次ラット卵巣分散体(dispersate)アッセイ:
JC−410FSHR/LHR細胞系由来の大量のステロイドを測定することが難しいため、未成熟ラットから採取した全卵巣由来の一次細胞を用いるアッセイを開発した。最初に、これらの培養物からのエストラジオールの産生をFSH及び/又はLH処理後に測定する。この目的は、ゴナドトロピン刺激型のステロイド産生を増強するタンパク質、又はこれらの培養物によりステロイド産生を増大させるよう単独で働くタンパク質を同定することである。
【0272】
Q.マウスIVFアッセイ:
このアッセイにおいて、卵母細胞を受精する能力により測定された精子の機能は、精子の受精能力を刺激するタンパク質を発見する目的でアッセイされる。
【0273】
R.一次ヒト前立腺間質細胞増殖アッセイ:
BPHの上皮成分についてのアッセイは既に上文で説明している(上記RWPEアッセイを参照のこと)。このアッセイは、BPHの間これらの細胞の増殖についてのモデルとして一次ヒト前立腺間質細胞を使用する。この目的は、これらの細胞の増殖を阻害するタンパク質を同定することである。
【0274】
【表5】
【表6】
【図面の簡単な説明】
【0275】
【図1】SCS0010とIL−8様ファミリーのメンバーとのアラインメント。
【図2】SCS0010のイントロン/エキソン構造。
【図3】SCS0010のヌクレオチド配列と翻訳。
【図4】pCR4−TOPO−SCS0010のマップ
【図5】pDONR221のマップ。
【図6】発現ベクターpEAK12dのマップ。
【図7】発現ベクターpDEST12.2のマップ。
【図8】pENTR−SCS0010−6HISのマップ。
【図9】pEAK12d−SCS0010−6HISのマップ。
【図10】pDEST12.2−SCS0010−6HISのマップ。
【図11】TMEM9とSCS0010とのアラインメント
【図12】TMEM9とSCS0010のSMARTドメイン。
【特許請求の範囲】
【請求項1】
IL−8様/TMEM9活性を有する単離されたポリペプチドであって:
a.配列番号2に記載のアミノ酸配列;
b.配列番号2に記載の配列のポリペプチドの成熟型(配列番号4又は配列番号7);
c.配列番号4に記載の配列のポリペプチドのヒスチジンタグ型(配列番号5);
d.配列番号2で示すアミノ酸配列の活性変異体であって、選択した配列において特定されている任意のアミノ酸が非保存的に置換されており、但し、当該配列中の15%未満のアミノ酸残基がそのように変化している、変異体;
e.a)〜c)に示すアミノ酸配列の活性フラグメント、前駆体、塩、又は誘導体、
から成る群から選択されるポリペプチド。
【請求項2】
配列番号2又は配列番号4又は配列番号5又は配列番号7に示す配列の天然の対立遺伝子変異体である、請求項1に記載のポリペプチド。
【請求項3】
変異体が一塩基多型の翻訳である、請求項2に記載のポリペプチド。
【請求項4】
配列番号2又は配列番号4又は配列番号5又は配列番号7あるいはそれらのフラグメントに対して生成する抗体又は結合タンパク質と特異的に結合する、請求項1〜3のいずれか1項に記載のポリペプチド。
【請求項5】
請求項1〜4のいずれか1項に記載のポリペプチドを含んで成る融合タンパク質。
【請求項6】
以下のタンパク質配列:膜結合タンパク質、イムノグロブリン定常領域、多量体化ドメイン、細胞外タンパク質、シグナルペプチド含有タンパク質、エクスポートシグナル含有タンパク質、に属する1又は複数のアミノ酸配列、を更に含んで成る、請求項6に記載の融合タンパク質。
【請求項7】
相当のポリペプチドの1又は複数の残基の非保存的置換及び/又は欠失から生じるアミノ酸配列を含んで成る、請求項1〜4のいずれか1項に記載のポリペプチドのアンタゴニスト。
【請求項8】
請求項1〜4のいずれか1項に記載のポリペプチドに特異的に結合するリガンド。
【請求項9】
請求項1〜4のいずれか1項に記載のポリペプチドのIL−8様/TMEM9活性を拮抗又は阻害する、請求項8に記載のリガンド。
【請求項10】
モノクローナル抗体、ポリクローナル抗体、ヒト化抗体、抗原結合フラグメント、又は膜結合タンパク質の細胞外ドメイン、である、請求項9に記載のリガンド。
【請求項11】
放射性標識、蛍光標識、ビオチン、又は細胞毒性物質の中から選択される分子との活性な接合体(conjugate)又は複合体(complex)の形態の、請求項1〜6のいずれか1項に記載のポリペプチド。
【請求項12】
請求項1〜4のいずれか1項に記載のポリペプチドの配列及び/又は構造に基づいて設計されるペプチドミメティクス。
【請求項13】
b)請求項1〜4のいずれか1項に記載のIL−8様/TMEM9活性を有するポリペプチド;
c)請求項5又は6に記載の融合タンパク質;あるいは
d)請求項7に記載のアンタゴニスト、
から成る群から選択される単離されたポリペプチドをコードする単離された核酸。
【請求項14】
配列番号1又は配列番号3又は配列番号6から成るDNA配列、あるいは当該DNA配列の相補体、を含んで成る、請求項13に記載の核酸。
【請求項15】
配列番号1又は配列番号3又は配列番号6から成る群から選択される核酸、あるいは前記DNA配列の相補体と、
b)高ストリンジェントな条件下でハイブリダイズし;又は
c)少なくとも約30個のヌクレオチドの長さにわたり少なくとも約85%の同一性を示す、
精製された核酸。
【請求項16】
請求項13〜15のいずれか1項に記載の核酸を含んで成るベクター。
【請求項17】
前記核酸分子が、原核又は真核宿主細胞におけるコードポリペプチドの発現を可能にする発現制御配列と作用可能に連結している、請求項16に記載のベクター。
【請求項18】
請求項13〜15に記載の精製された核酸によりコードされるポリペプチド。
【請求項19】
請求項1〜7又は請求項18のいずれか1項に記載のポリペプチドを発現することができる細胞を産生する方法であって、請求項13〜17のいずれか1項に記載のベクター又は核酸で細胞を遺伝子操作することを含んで成る方法。
【請求項20】
請求項13〜17のいずれか1項に記載のベクター又は核酸で形質転換した宿主細胞。
【請求項21】
請求項13〜17のいずれか1項に記載のベクター又は核酸で形質転換されたトランスジェニック動物細胞であって、請求項1〜4のいずれか1項に記載のポリペプチドの増強又は低下した発現レベルを有する細胞。
【請求項22】
請求項1〜4のいずれか1項に記載のポリペプチドの増強又は低下した発現レベルを有するよう形質転換された、トランスジェニックなヒト以外の動物。
【請求項23】
請求項1〜7のいずれか1項に記載のポリペプチドを生成するための方法であって、請求項20又は21に記載の細胞を前記核酸又はベクターが発現する条件下で培養し、そして前記核酸又はベクターによってコードされるポリペプチドを培養物から回収することを含んで成る方法。
【請求項24】
細胞又は動物における請求項1〜4のいずれか1項に記載のポリペプチドの発現レベルを増強させる化合物。
【請求項25】
細胞又は動物における請求項1〜4のいずれか1項に記載のポリペプチドの発現レベルを低下させる化合物。
【請求項26】
アンチセンスオリゴヌクレオチド又は低分子干渉RNAである、請求項24に記載の化合物。
【請求項27】
請求項1〜6又は18のいずれか1項に記載のポリペプチド、請求項7に記載のアンタゴニスト、請求項8〜10のいずれか1項に記載のリガンド、請求項12に記載のペプチドミメティクス、請求項13〜17のいずれか1項に記載の核酸、請求項20又は21に記載の細胞、あるいは請求項24〜26のいずれか1項に記載の化合物を含む、精製された調製物。
【請求項28】
請求項1〜4のいずれか1項に記載のポリペプチドのIL−8様/TMEM9活性の増大が必要とされる場合の疾患の治療又は予防における、請求項1〜6又は18のいずれか1項に記載のポリペプチド、請求項12に記載のペプチドミメティクス、請求項13〜17のいずれか1項に記載の核酸、請求項20又は21に記載の細胞、あるいは請求項24に記載の化合物、の使用。
【請求項29】
IL−8様/TMEM9活性の増大を必要とする疾患の処置又は予防のための医薬組成物であって、請求項1〜6又は18のいずれか1項に記載のポリペプチド、請求項12に記載のペプチドミメティクス、請求項13〜17のいずれか1項に記載の核酸、請求項20又は21に記載の細胞、あるいは請求項24に記載の化合物を活性成分として含む医薬組成物。
【請求項30】
医薬組成物の調製方法であって、請求項1〜6又は18のいずれか1項に記載のポリペプチド、請求項12に記載のペプチドミメティクス、請求項13〜17のいずれか1項に記載の核酸、請求項20又は21に記載の細胞、あるいは請求項24に記載の化合物を、医薬として許容される担体と組み合わせることを含んで成る調製方法。
【請求項31】
請求項1〜4のいずれか1項に記載のポリペプチドのIL−8様/TMEM9活性の増大を必要とする疾患の処置又は予防のための方法であって、治療的に有効量の、請求項1〜6又は18のいずれか1項に記載のポリペプチド、請求項12に記載のペプチドミメティクス、請求項13〜17のいずれか1項に記載の核酸、請求項20又は21に記載の細胞、あるいは請求項24に記載の化合物の投与を含んで成る方法。
【請求項32】
請求項1〜4のいずれか1項に記載のポリペプチドの過剰なIL−8様/TMEM9活性に関連する疾患の治療又は予防における、請求項7に記載のアンタゴニスト、請求項8〜10のいずれか1項に記載のリガンド、あるいは請求項25又は26に記載の化合物の使用。
【請求項33】
請求項1〜4のいずれか1項に記載のポリペプチドの過剰なIL−8様/TMEM9活性に関連する疾患の治療又は予防のための医薬組成物であって、請求項7に記載のアンタゴニスト、請求項8〜10のいずれか1項に記載のリガンド、あるいは請求項25又は26に記載の化合物を活性成分として含む医薬組成物。
【請求項34】
請求項1〜4のいずれか1項に記載のポリペプチドの過剰なIL−8様/TMEM9活性に関連する疾患の処置又は予防のための医薬組成物の調製のための方法であって、請求項7に記載のアンタゴニスト、請求項8〜10のいずれか1項に記載のリガンド、あるいは請求項25又は26に記載の化合物を、医薬として許容される担体と組み合わせることを含んで成る方法。
【請求項35】
請求項1〜4のいずれか1項に記載のポリペプチドに関連する疾患の処置又は予防のための方法であって、治療的に有効量の、請求項7に記載のアンタゴニスト、請求項8〜10のいずれか1項に記載のリガンド、あるいは請求項25又は26に記載の化合物の投与を含んで成る方法。
【請求項36】
請求項1〜4のいずれか1項に記載のIL−8様/TMEM9ポリペプチドに関連する疾患を処置するのに有効な候補化合物をスクリーニングするための方法であって:
b)前記ポリペプチドの増強又は低下した発現レベルを有する、請求項20に記載の細胞、請求項21に記載のトランスジェニック動物細胞、又は請求項22に記載のトランスジェニックなヒト以外の動物を、候補化合物と接触させること、及び
c)前記動物又は細胞に対する前記化合物の効果を決定すること、
を含んで成る方法。
【請求項37】
請求項1〜4のいずれか1項に記載のポリペプチドのアンタゴニスト/阻害物質又はアゴニスト/活性化物質としての候補化合物を同定するための方法であって:
a)前記ポリペプチドと、前記化合物、及び前記ポリペプチドに結合することができる哺乳類細胞又は哺乳類細胞膜とを接触させること;及び
b)前記の分子が、前記ポリペプチドと前記哺乳類細胞又は哺乳類細胞膜との相互作用、又は当該相互作用から生じる応答、を阻止又は増強するか否かを測定すること、
を含んで成る方法。
【請求項38】
試料中の請求項1〜4のいずれか1項に記載のポリペプチドの活性及び/又は存在を決定するための方法であって:
a)タンパク質含有試料を準備すること;
b)前記試料と請求項8〜10のいずれか1項に記載のリガンドとを接触させること;及び
c)前記ポリペプチドに結合した前記リガンドの存在を決定すること、
を含んで成る方法。
【請求項39】
試料中の請求項1〜4のいずれか1項に記載のポリペプチドをコードする転写物又は核酸の存在又は量を決定するための方法であって:
a)核酸含有試料を準備すること;
b)前記試料と請求項13〜17のいずれか1項に記載の核酸とを接触させること;及び
c)前記核酸と前記試料中の核酸とのハイブリダイゼーションを決定すること、
を含んで成る方法。
【請求項40】
ポリメラーゼ連鎖反応により試料中の請求項1〜4のいずれか1項に記載のポリペプチドをコードする転写物又は核酸の存在又は量を決定するための配列番号1又は配列番号3又は配列番号6に記載のヌクレオチド配列由来のプライマーの使用。
【請求項41】
試料中の請求項1〜4のいずれか1項に記載のIL−8様/TMEM9ポリペプチドの活性及び/又は存在を測定するためのキットであって、1又は複数の以下の試薬:請求項1〜6又は18のいずれか1項に記載のポリペプチド、請求項7に記載のアンタゴニスト、請求項8〜10のいずれか1項に記載のリガンド、請求項11に記載のポリペプチド、請求項12に記載のペプチドミメティクス、請求項13〜17のいずれか1項に記載の核酸、請求項20又は21に記載の細胞、請求項24〜26のいずれか1項に記載の化合物、請求項29又は33に記載の医薬組成物、を含んで成るキット。
【請求項1】
IL−8様/TMEM9活性を有する単離されたポリペプチドであって:
a.配列番号2に記載のアミノ酸配列;
b.配列番号2に記載の配列のポリペプチドの成熟型(配列番号4又は配列番号7);
c.配列番号4に記載の配列のポリペプチドのヒスチジンタグ型(配列番号5);
d.配列番号2で示すアミノ酸配列の活性変異体であって、選択した配列において特定されている任意のアミノ酸が非保存的に置換されており、但し、当該配列中の15%未満のアミノ酸残基がそのように変化している、変異体;
e.a)〜c)に示すアミノ酸配列の活性フラグメント、前駆体、塩、又は誘導体、
から成る群から選択されるポリペプチド。
【請求項2】
配列番号2又は配列番号4又は配列番号5又は配列番号7に示す配列の天然の対立遺伝子変異体である、請求項1に記載のポリペプチド。
【請求項3】
変異体が一塩基多型の翻訳である、請求項2に記載のポリペプチド。
【請求項4】
配列番号2又は配列番号4又は配列番号5又は配列番号7あるいはそれらのフラグメントに対して生成する抗体又は結合タンパク質と特異的に結合する、請求項1〜3のいずれか1項に記載のポリペプチド。
【請求項5】
請求項1〜4のいずれか1項に記載のポリペプチドを含んで成る融合タンパク質。
【請求項6】
以下のタンパク質配列:膜結合タンパク質、イムノグロブリン定常領域、多量体化ドメイン、細胞外タンパク質、シグナルペプチド含有タンパク質、エクスポートシグナル含有タンパク質、に属する1又は複数のアミノ酸配列、を更に含んで成る、請求項6に記載の融合タンパク質。
【請求項7】
相当のポリペプチドの1又は複数の残基の非保存的置換及び/又は欠失から生じるアミノ酸配列を含んで成る、請求項1〜4のいずれか1項に記載のポリペプチドのアンタゴニスト。
【請求項8】
請求項1〜4のいずれか1項に記載のポリペプチドに特異的に結合するリガンド。
【請求項9】
請求項1〜4のいずれか1項に記載のポリペプチドのIL−8様/TMEM9活性を拮抗又は阻害する、請求項8に記載のリガンド。
【請求項10】
モノクローナル抗体、ポリクローナル抗体、ヒト化抗体、抗原結合フラグメント、又は膜結合タンパク質の細胞外ドメイン、である、請求項9に記載のリガンド。
【請求項11】
放射性標識、蛍光標識、ビオチン、又は細胞毒性物質の中から選択される分子との活性な接合体(conjugate)又は複合体(complex)の形態の、請求項1〜6のいずれか1項に記載のポリペプチド。
【請求項12】
請求項1〜4のいずれか1項に記載のポリペプチドの配列及び/又は構造に基づいて設計されるペプチドミメティクス。
【請求項13】
b)請求項1〜4のいずれか1項に記載のIL−8様/TMEM9活性を有するポリペプチド;
c)請求項5又は6に記載の融合タンパク質;あるいは
d)請求項7に記載のアンタゴニスト、
から成る群から選択される単離されたポリペプチドをコードする単離された核酸。
【請求項14】
配列番号1又は配列番号3又は配列番号6から成るDNA配列、あるいは当該DNA配列の相補体、を含んで成る、請求項13に記載の核酸。
【請求項15】
配列番号1又は配列番号3又は配列番号6から成る群から選択される核酸、あるいは前記DNA配列の相補体と、
b)高ストリンジェントな条件下でハイブリダイズし;又は
c)少なくとも約30個のヌクレオチドの長さにわたり少なくとも約85%の同一性を示す、
精製された核酸。
【請求項16】
請求項13〜15のいずれか1項に記載の核酸を含んで成るベクター。
【請求項17】
前記核酸分子が、原核又は真核宿主細胞におけるコードポリペプチドの発現を可能にする発現制御配列と作用可能に連結している、請求項16に記載のベクター。
【請求項18】
請求項13〜15に記載の精製された核酸によりコードされるポリペプチド。
【請求項19】
請求項1〜7又は請求項18のいずれか1項に記載のポリペプチドを発現することができる細胞を産生する方法であって、請求項13〜17のいずれか1項に記載のベクター又は核酸で細胞を遺伝子操作することを含んで成る方法。
【請求項20】
請求項13〜17のいずれか1項に記載のベクター又は核酸で形質転換した宿主細胞。
【請求項21】
請求項13〜17のいずれか1項に記載のベクター又は核酸で形質転換されたトランスジェニック動物細胞であって、請求項1〜4のいずれか1項に記載のポリペプチドの増強又は低下した発現レベルを有する細胞。
【請求項22】
請求項1〜4のいずれか1項に記載のポリペプチドの増強又は低下した発現レベルを有するよう形質転換された、トランスジェニックなヒト以外の動物。
【請求項23】
請求項1〜7のいずれか1項に記載のポリペプチドを生成するための方法であって、請求項20又は21に記載の細胞を前記核酸又はベクターが発現する条件下で培養し、そして前記核酸又はベクターによってコードされるポリペプチドを培養物から回収することを含んで成る方法。
【請求項24】
細胞又は動物における請求項1〜4のいずれか1項に記載のポリペプチドの発現レベルを増強させる化合物。
【請求項25】
細胞又は動物における請求項1〜4のいずれか1項に記載のポリペプチドの発現レベルを低下させる化合物。
【請求項26】
アンチセンスオリゴヌクレオチド又は低分子干渉RNAである、請求項24に記載の化合物。
【請求項27】
請求項1〜6又は18のいずれか1項に記載のポリペプチド、請求項7に記載のアンタゴニスト、請求項8〜10のいずれか1項に記載のリガンド、請求項12に記載のペプチドミメティクス、請求項13〜17のいずれか1項に記載の核酸、請求項20又は21に記載の細胞、あるいは請求項24〜26のいずれか1項に記載の化合物を含む、精製された調製物。
【請求項28】
請求項1〜4のいずれか1項に記載のポリペプチドのIL−8様/TMEM9活性の増大が必要とされる場合の疾患の治療又は予防における、請求項1〜6又は18のいずれか1項に記載のポリペプチド、請求項12に記載のペプチドミメティクス、請求項13〜17のいずれか1項に記載の核酸、請求項20又は21に記載の細胞、あるいは請求項24に記載の化合物、の使用。
【請求項29】
IL−8様/TMEM9活性の増大を必要とする疾患の処置又は予防のための医薬組成物であって、請求項1〜6又は18のいずれか1項に記載のポリペプチド、請求項12に記載のペプチドミメティクス、請求項13〜17のいずれか1項に記載の核酸、請求項20又は21に記載の細胞、あるいは請求項24に記載の化合物を活性成分として含む医薬組成物。
【請求項30】
医薬組成物の調製方法であって、請求項1〜6又は18のいずれか1項に記載のポリペプチド、請求項12に記載のペプチドミメティクス、請求項13〜17のいずれか1項に記載の核酸、請求項20又は21に記載の細胞、あるいは請求項24に記載の化合物を、医薬として許容される担体と組み合わせることを含んで成る調製方法。
【請求項31】
請求項1〜4のいずれか1項に記載のポリペプチドのIL−8様/TMEM9活性の増大を必要とする疾患の処置又は予防のための方法であって、治療的に有効量の、請求項1〜6又は18のいずれか1項に記載のポリペプチド、請求項12に記載のペプチドミメティクス、請求項13〜17のいずれか1項に記載の核酸、請求項20又は21に記載の細胞、あるいは請求項24に記載の化合物の投与を含んで成る方法。
【請求項32】
請求項1〜4のいずれか1項に記載のポリペプチドの過剰なIL−8様/TMEM9活性に関連する疾患の治療又は予防における、請求項7に記載のアンタゴニスト、請求項8〜10のいずれか1項に記載のリガンド、あるいは請求項25又は26に記載の化合物の使用。
【請求項33】
請求項1〜4のいずれか1項に記載のポリペプチドの過剰なIL−8様/TMEM9活性に関連する疾患の治療又は予防のための医薬組成物であって、請求項7に記載のアンタゴニスト、請求項8〜10のいずれか1項に記載のリガンド、あるいは請求項25又は26に記載の化合物を活性成分として含む医薬組成物。
【請求項34】
請求項1〜4のいずれか1項に記載のポリペプチドの過剰なIL−8様/TMEM9活性に関連する疾患の処置又は予防のための医薬組成物の調製のための方法であって、請求項7に記載のアンタゴニスト、請求項8〜10のいずれか1項に記載のリガンド、あるいは請求項25又は26に記載の化合物を、医薬として許容される担体と組み合わせることを含んで成る方法。
【請求項35】
請求項1〜4のいずれか1項に記載のポリペプチドに関連する疾患の処置又は予防のための方法であって、治療的に有効量の、請求項7に記載のアンタゴニスト、請求項8〜10のいずれか1項に記載のリガンド、あるいは請求項25又は26に記載の化合物の投与を含んで成る方法。
【請求項36】
請求項1〜4のいずれか1項に記載のIL−8様/TMEM9ポリペプチドに関連する疾患を処置するのに有効な候補化合物をスクリーニングするための方法であって:
b)前記ポリペプチドの増強又は低下した発現レベルを有する、請求項20に記載の細胞、請求項21に記載のトランスジェニック動物細胞、又は請求項22に記載のトランスジェニックなヒト以外の動物を、候補化合物と接触させること、及び
c)前記動物又は細胞に対する前記化合物の効果を決定すること、
を含んで成る方法。
【請求項37】
請求項1〜4のいずれか1項に記載のポリペプチドのアンタゴニスト/阻害物質又はアゴニスト/活性化物質としての候補化合物を同定するための方法であって:
a)前記ポリペプチドと、前記化合物、及び前記ポリペプチドに結合することができる哺乳類細胞又は哺乳類細胞膜とを接触させること;及び
b)前記の分子が、前記ポリペプチドと前記哺乳類細胞又は哺乳類細胞膜との相互作用、又は当該相互作用から生じる応答、を阻止又は増強するか否かを測定すること、
を含んで成る方法。
【請求項38】
試料中の請求項1〜4のいずれか1項に記載のポリペプチドの活性及び/又は存在を決定するための方法であって:
a)タンパク質含有試料を準備すること;
b)前記試料と請求項8〜10のいずれか1項に記載のリガンドとを接触させること;及び
c)前記ポリペプチドに結合した前記リガンドの存在を決定すること、
を含んで成る方法。
【請求項39】
試料中の請求項1〜4のいずれか1項に記載のポリペプチドをコードする転写物又は核酸の存在又は量を決定するための方法であって:
a)核酸含有試料を準備すること;
b)前記試料と請求項13〜17のいずれか1項に記載の核酸とを接触させること;及び
c)前記核酸と前記試料中の核酸とのハイブリダイゼーションを決定すること、
を含んで成る方法。
【請求項40】
ポリメラーゼ連鎖反応により試料中の請求項1〜4のいずれか1項に記載のポリペプチドをコードする転写物又は核酸の存在又は量を決定するための配列番号1又は配列番号3又は配列番号6に記載のヌクレオチド配列由来のプライマーの使用。
【請求項41】
試料中の請求項1〜4のいずれか1項に記載のIL−8様/TMEM9ポリペプチドの活性及び/又は存在を測定するためのキットであって、1又は複数の以下の試薬:請求項1〜6又は18のいずれか1項に記載のポリペプチド、請求項7に記載のアンタゴニスト、請求項8〜10のいずれか1項に記載のリガンド、請求項11に記載のポリペプチド、請求項12に記載のペプチドミメティクス、請求項13〜17のいずれか1項に記載の核酸、請求項20又は21に記載の細胞、請求項24〜26のいずれか1項に記載の化合物、請求項29又は33に記載の医薬組成物、を含んで成るキット。
【図1】

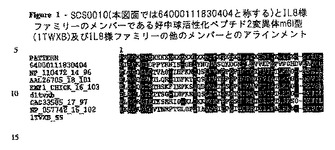
【図2】

【図3】

【図4】
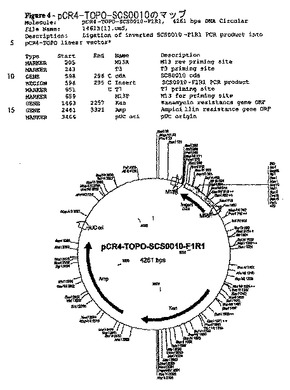
【図5】
【図6】

【図7】
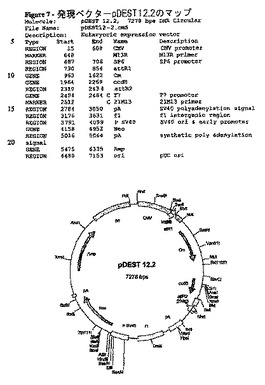
【図8】
【図9】
【図10】
【図11】

【図12】
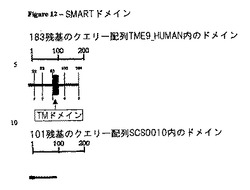
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【公表番号】特表2007−512803(P2007−512803A)
【公表日】平成19年5月24日(2007.5.24)
【国際特許分類】
【出願番号】特願2006−529939(P2006−529939)
【出願日】平成16年5月27日(2004.5.27)
【国際出願番号】PCT/EP2004/005739
【国際公開番号】WO2004/106503
【国際公開日】平成16年12月9日(2004.12.9)
【出願人】(599177396)アプライド リサーチ システムズ エーアールエス ホールディング ナームロゼ フェンノートシャップ (70)
【Fターム(参考)】
【公表日】平成19年5月24日(2007.5.24)
【国際特許分類】
【出願日】平成16年5月27日(2004.5.27)
【国際出願番号】PCT/EP2004/005739
【国際公開番号】WO2004/106503
【国際公開日】平成16年12月9日(2004.12.9)
【出願人】(599177396)アプライド リサーチ システムズ エーアールエス ホールディング ナームロゼ フェンノートシャップ (70)
【Fターム(参考)】
[ Back to top ]