
新規PPARγプロモーター配列、及びその応用
【課題】PPARγの発現を直接調節することが可能な因子を含有する優れたPPARγ発現調節剤、並びに、前記因子を阻害することにより脂肪細胞の分化、脂質代謝の調節、破骨細胞機能の調節、及び炎症作用の調節が可能な、優れた脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤、及び前記PPARγ発現調節剤のスクリーニング方法の提供。
【解決手段】特定な配列からなる塩基配列を有する新規PPARγプロモーター配列、PPARγを直接調節可能な優れたPPARγ発現調節剤、DBP、TEF、HLF、及びE4BP4などの阻害を介した優れた脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤、前記PPARγ発現調節剤の効率的なスクリーニング方法である。
【解決手段】特定な配列からなる塩基配列を有する新規PPARγプロモーター配列、PPARγを直接調節可能な優れたPPARγ発現調節剤、DBP、TEF、HLF、及びE4BP4などの阻害を介した優れた脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤、前記PPARγ発現調節剤の効率的なスクリーニング方法である。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、新規PPARγプロモーター配列、並びに、PPARγ発現調節剤、脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤、及びスクリーニング方法に関する。
【背景技術】
【0002】
ライフスタイルの欧米化に伴い、糖尿病、高血圧、動脈硬化、脂質異常症などいわゆる生活習慣病が増加しており、その主要因としてメタボリックシンドロームがある。このメタボリックシンドロームは、内臓組織の脂肪細胞の増殖や脂肪の取り込みの増加などにより発症し、過剰な食事量による脂肪細胞への脂肪酸の取り込みや、それによる脂肪細胞の増殖や分化とも密接に関連している。したがって、これらを制御する因子を解明することが、生活習慣病の予防や治療に直接結びつくものと考えられる。
しかしながら、脂肪の摂取と、脂質代謝とを直接結びつける因子や、脂肪細胞の分化に関わる因子などは十分に解明されておらず、これらの分子機構の解明が強く求められている。
【0003】
ペルオキシソーム増殖剤応答性受容体(PPAR:Peroxisome Proliferator−Activated Receptor)は、ステロイドホルモン受容体ファミリーの1種であり、哺乳動物においてPPARα、PPARδ、PPARγの3種類のサブタイプが存在することが知られている。
PPARは、別の転写調節因子であるRXR(レチノイドX受容体)とヘテロダイマーを形成し、標的遺伝子の上流に存在する応答配列PPRE(PPAR response element)に結合し、標的遺伝子の転写を制御している。前記標的遺伝子としては、内分泌代謝、糖代謝、脂質代謝、マクロファージを介する炎症反応、骨代謝の制御、発ガン機構などの遺伝子が知られている。そのため、PPARは、これらの疾患の治療薬のターゲット分子として注目されている。
【0004】
これらの中でも、PPARγは、そのプロモーターに、数種のスプライスバリアントが存在することが知られており(非特許文献1参照)、数種のアイソフォームが存在する(非特許文献2〜3参照)。例えば、PPARγ1は、脂肪組織や免疫系臓器、副腎、小腸で発現し、PPARγ2は、脂肪細胞で特異的な発現が認められ、脂肪細胞の分化誘導や脂肪合成に重要な役割を担っていると考えられている(非特許文献4参照)。また、PPARγは、マウスの破骨細胞を制御するという報告(非特許文献5参照)や、炎症疾患の調節に有用であるという報告(非特許文献6参照)もある。
【0005】
従来、PPARγの作用機序に大きく関わる因子としては、PPARγのリガンドが報告されている(非特許文献7参照)。
しかしながら、PPARγの発現調節は多面的に制御されているため非常に複雑であり、PPARγの発現を直接調節することにより、脂肪細胞の増殖や分化を促進、停止、後退させる因子、破骨細胞機能を調節する因子、炎症を調節する因子などは未だ提供されておらず、PPARγを直接調節することが可能なPPARγ発現調節剤、PPARγの調節を介した、脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤の提供が強く望まれているのが現状である。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Fajas L, et al, J Biol Chem, 1997, 272, p.18779−18789
【非特許文献2】Fajas L, et al, FEBS Lett, 1998, 438, p.55−60
【非特許文献3】Sundvold H, Lien S, Biochem Biophys Res Commun, 2001, 28, p.383−90
【非特許文献4】Rosen ED, Walkey CJ, Puigserver P, Spiegelman BM, Genes Dev, 2000, 14, p.1293−307
【非特許文献5】Wan Y, Chong LW, Evans RM, Nature Medicine, 2007, 13, p.1496 − 1503
【非特許文献6】Ricote M, Li AC, Willson TM, Kelly CJ, Glass CK, Nature, 1998, 391, p.79−82
【非特許文献7】Rosen ED, Spiegelman BM, Nature, 2006, 444, p.847−853
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明は、従来における前記諸問題を解決し、以下の目的を達成することを課題とする。即ち、本発明は、PPARγの発現を直接調節することが可能な因子を含有する優れたPPARγ発現調節剤、並びに、前記因子を阻害することにより脂肪細胞の分化、脂質代謝の調節、破骨細胞機能の調節、及び炎症作用の調節が可能な、優れた脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤、及び前記PPARγ発現調節剤のスクリーニング方法を提供することを目的とする。
【課題を解決するための手段】
【0008】
前記課題を解決するため、本発明者らは、脂肪分化、脂質代謝、破骨細胞調節、炎症疾患などに関与するPPARγの発現調節に着目し、PPARγの発現調節に関与する因子について鋭意検討した結果、PPARγの発現制御に係る5’非翻訳領域(プロモーター領域)に新規スプライスバリアントが存在することを知見するとともに、その新規プロモーター配列を見出し、該プロモーター領域のプロモーターレポーターシステムを樹立した。また、PPARγの発現を直接調節することが可能なPPARγ発現調節剤のスクリーニング方法を確立し、該スクリーニング方法により、DBP、及びTEFがPPARγの遺伝子発現を促進すること、HLF、及びE4BP4がPPARγの発現を抑制すること、更に、DBP、TEF、HLF、及びE4BP4がPPARγのプロモーター領域に存在するD−ボックスに結合すること、DBP、及びTEFを阻害すると脂肪細胞への分化が阻害され、更にDBPの遺伝子発現は脂肪細胞の分化に伴い増加すること、E4BP4を阻害すると脂肪細胞への分化が促進し、更にE4PB4の遺伝子発現は脂肪細胞の分化に伴い減少することを知見し、本発明の完成に至った。なお、本発明者らは、前記5’非翻訳領域(プロモーター領域)に新規スプライスバリアントを有するPPARγを、「PPARγ6」と命名した。
【0009】
本発明は、本発明者らによる前記知見に基づくものであり、前記課題を解決するための手段としては、以下の通りである。即ち、
<1> 配列番号:1で表される塩基配列を有することを特徴とするPPARγプロモーター配列である。
<2> 配列番号:1で表される塩基配列のうち、塩基番号159〜166、塩基番号202〜209、及び塩基番号218〜225を少なくとも有する前記<1>に記載のPPARγプロモーター配列である。
<3> DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかを含有することを特徴とするPPARγ発現調節剤である。
<4> PPARγの発現促進作用を有する前記<3>に記載のPPARγ発現調節剤である。
<5> PPARγの発現促進作用を有する因子が、DBP、及びTEF、並びに、該DBP、及び該TEFをコードするcDNAの少なくともいずれかである前記<4>に記載のPPARγ発現調節剤である。
<6> PPARγの発現抑制作用を有する前記<3>に記載のPPARγ発現調節剤である。
<7> PPARγの発現抑制作用を有する因子が、HLF、及びE4BP4、並びに、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかである前記<6>に記載のPPARγ発現調節剤である。
<8> PPARγの発現調節作用が、PPARγプロモーター配列に結合することによる前記<3>から<7>のいずれかに記載のPPARγ発現調節剤である。
<9> PPARγプロモーター配列が、配列番号:1で表される塩基配列を有する前記<8>に記載のPPARγ発現調節剤である。
<10> PPARγの発現調節作用が、配列番号:1で表される塩基配列中のD−ボックスに結合することによる前記<3>から<9>のいずれかに記載のPPARγ発現調節剤である。
<11> DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかを阻害することを特徴とする脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤である。
<12> 阻害が、miRNAによる前記<11>に記載の脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤である。
<13> miRNAの配列が、配列番号:2〜33で表される塩基配列の少なくともいずれかを有する前記<12>に記載の脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤である。
<14> PPARγプロモーター配列を有するレポーター遺伝子を構築するプロモーターレポーター遺伝子構築工程と、該プロモーターレポーター遺伝子と、被験物質を含む被験試料とを細胞に導入する導入工程と、該プロモーターレポーター遺伝子のレポーター活性を指標にして、PPARγプロモーター活性を検出する検出工程と、該被験試料が、PPARγプロモーター活性の促進作用、及び抑制作用のいずれかの作用を有しているか否かを判断し、PPARγプロモーター活性の促進作用、及び抑制作用のいずれかの作用を有する該被験試料中の被験物質をPPARγ発現調節剤としてスクリーニングするスクリーニング工程と、を含むことを特徴とするPPARγ発現調節剤のスクリーニング方法である。
<15> PPARγプロモーター配列が、配列番号:1で表される塩基配列を有する前記<14>に記載のPPARγ発現調節剤のスクリーニング方法である。
<16> プロモーターレポーター遺伝子のPPARγプロモーター配列が、該PPARγプロモーター配列が有する3箇所のD−ボックスの少なくともいずれかを欠失した欠失変異体である前記<14>から<15>のいずれかに記載のPPARγ発現調節剤のスクリーニング方法である。
【発明の効果】
【0010】
本発明によれば、従来における前記諸問題を解決し、前記目的を達成することができ、
PPARγの発現を直接調節することが可能な因子を含有する優れたPPARγ発現調節剤、並びに、前記因子を阻害することにより脂肪細胞の分化、脂質代謝の調節、破骨細胞機能の調節、及び炎症作用の調節が可能な、優れた脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤、及び前記PPARγ発現調節剤のスクリーニング方法を提供することができる。
【図面の簡単な説明】
【0011】
【図1A】図1Aは、ヒトPPARγ遺伝子のエクソン・イントロン構造を模式的に表した図である。
【図1B】図1Bは、ヒトPPARγ遺伝子の6種のスプライスバリアントを模式的に表した図である。
【図2】図2は、PPARγ6のプロモーター構造、及びD−ボックスを模式的に表した図である。
【図3】図3は、DBP発現ベクターを一過性に発現させたCaco−2細胞における、DBP、及びPPARγ6のmRNA量を表した図である。
【図4A】図4Aは、DBPのPPARγ6プロモーターへの作用を、PPARγ6プロモーターレポーター遺伝子を用いたルシフェラーゼアッセイで解析した図である。
【図4B】図4Bは、TEFのPPARγ6プロモーターへの作用を、PPARγ6プロモーターレポーター遺伝子を用いたルシフェラーゼアッセイで解析した図である。
【図4C】図4Cは、HLFのPPARγ6プロモーターへの作用を、PPARγ6プロモーターレポーター遺伝子を用いたルシフェラーゼアッセイで解析した図である。
【図4D】図4Dは、E4BP4のPPARγ6プロモーターへの作用を、PPARγ6のプロモーターレポーター遺伝子を用いたルシフェラーゼアッセイで解析した図である。
【図5A】図5Aは、PPARγ6プロモーターの欠失変異体のコンストラクトを模式的に表した図である。
【図5B】図5Bは、DBPのPPARγ6のプロモーターへの作用を、PPARγ6プロモーターの欠失変異体のレポーター遺伝子を用いて、ルシフェラーゼアッセイで解析した図である。
【図5C】図5Cは、DBPのPPARγ6のプロモーターへの作用を、PPARγ6のプロモーター欠失変異体のレポーター遺伝子を用いて、ルシフェラーゼアッセイで解析した図である。
【図6A】図6Aは、PPARγ6プロモーターのD−ボックス欠失変異体のコンストラクトを模式的に表した図である。
【図6B】図6Bは、DBPのPPARγ6プロモーターへの作用を、PPARγ6プロモーターのD−ボックス欠失変異体のレポーター遺伝子を用いて、ルシフェラーゼアッセイで解析した図である。
【図6C】図6Cは、E4BP4のPPARγ6プロモーターへの作用を、PPARγ6プロモーターのD−ボックス欠失変異体のレポーター遺伝子を用いて、ルシフェラーゼアッセイで解析した図である。
【図7A】図7Aは、DBPに対するプローブ特異性をゲルシフトアッセイにより調べた図である。
【図7B】図7Bは、TEFに対するプローブ特異性をゲルシフトアッセイにより調べた図である。
【図7C】図7Cは、HLFに対するプローブ特異性をゲルシフトアッセイにより調べた図である。
【図7D】図7Dは、E4BP4に対するプローブ特異性をゲルシフトアッセイにより調べた図である。
【図7E】図7Eは、DBPの、PPARγ6プロモーターに存在するD−ボックスへの結合能をゲルシフトアッセイで解析した図である。
【図7F】図7Fは、TEFの、PPARγ6プロモーターに存在するD−ボックスへの結合能をゲルシフトアッセイで解析した図である。
【図7G】図7Gは、HLFの、PPARγ6プロモーターに存在するD−ボックスへの結合能をゲルシフトアッセイで解析した図である。
【図7H】図7Hは、E4BP4の、PPARγ6プロモーターに存在するD−ボックスへの結合能をゲルシフトアッセイで解析した図である。
【図8】図8は、DBP、TEF、及びE4BP4の脂肪細胞分化への影響を、RNAiで解析した図である。
【図9A】図9Aは、脂肪細胞分化におけるDBPのmRNAの発現量の変化を表した図である。
【図9B】図9Bは、脂肪細胞分化におけるE4BP4のmRNAの発現量の変化を表した図である。
【発明を実施するための形態】
【0012】
(新規PPARγプロモーター配列)
本発明の新規PPARγプロモーター配列は、配列番号:1で表される塩基配列を有する。
前記新規PPARγプロモーター配列は、5’非翻訳領域が、既知のPPARγの配列と異なる、本発明者らが見出したPPARγの新規スプライスバリアント(図1B参照)の発現を制御するプロモーター配列である。
前記新規PPARγプロモーター配列は、配列番号:1で表される塩基配列のみからなる塩基配列であることが好ましいが、そのプロモーター活性を有する限りは、特に制限はなく、目的に応じて適宜選択することができ、配列番号:1で表される塩基配列の一部からなる塩基配列であってもよく、配列番号:1で表される塩基配列に、更にその他の塩基配列が付加された塩基配列であってもよい。
前記新規PPARγプロモーター配列が、配列番号:1で表される塩基配列の一部からなる場合、その一部の塩基配列としては、PPARγのプロモーター活性を有する限りは、特に制限はなく、目的に応じて適宜選択することができるが、配列番号:1で表される塩基配列の、塩基番号159〜166、塩基番号202〜209、及び塩基番号218〜225を有することが好ましい。
前記その他の塩基配列としても、PPARγのプロモーター活性を有する限りは、特に制限はなく、目的に応じて適宜選択することができる。
また、前記新規PPARγプロモーター配列は、配列番号:1で表される塩基配列に、1若しくは数個の塩基が、欠失、置換、挿入された塩基配列からなるものであってもよい。
【0013】
<新規スプライスバリアント>
ヒトPPARγの遺伝子配列は、図1Aに示すように、10個のエクソン(A1、C1C2、A2、B、E1、E2、E3、E4、E5、及びE6)を有する。このうち、A1、C1C2、A2、及びBは、非翻訳領域であり、前記E1〜E6が翻訳されることによりPPARγのタンパク質を形成する。
前記ヒトPPARγの発現調節は、そのプロモーターにより制御されているが、前記ヒトPPARγの5’非翻訳領域に存在するプロモーターには、従来、5種のスプライスバリアント(PPARγ1〜5)が存在することが知られていた(図1B参照)。前記5’非翻訳領域には、4つのエクソン(A1、C1C2、A2、及びB)が存在し、これらの4つのエクソンが選択的スプライシングを受けることにより、スプライスバリアントが形成される(図1A、及び図1B参照)。
既知のスプライスバリアントとしては、図1Bに示すように、5’非翻訳領域がA1、及びA2からなるPPARγ1、5’非翻訳領域がBからなるPPARγ2、5’非翻訳領域がA2からなるPPARγ3、5’非翻訳領域にいずれのエクソンも有さないPPARγ4、及び5’非翻訳領域がC1C2からなるPPARγ5が存在する。
本発明者らは、これらの既知のスプライスバリアントとは全く異なる、5’非翻訳領域がC1C2、及びA2からなるヒトPPARγが存在することを見出し(図1B参照)、「PPARγ6」と命名した(以下、「PPARγ6」と称することがある)。
【0014】
前記PPARγ6が、新規スプライスバリアントを有するか否かを解析する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、エクソンC1C2からエクソンE1にかけてのPCR産物のサイズ、及び配列決定などにより解析する方法などが挙げられる。
【0015】
前記PPARγ6のプロモーター配列が、DNA結合配列を有するか否かを解析する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、MatInspector(Genomatix社製)により解析する方法などが挙げられる。
【0016】
前記MatInspector(Genomatix社製)による解析の結果、前記C1C2は、後述するDBP(Albumin gene D−site Binding Protein:D部位結合タンパク質)の結合配列であるD−ボックスが3箇所(D1、D2、及びD3)存在していた(図2参照)。
マウス及びヒトの、プロモーター領域及びエクソン1領域の配列についてDNA解析ソフトGeneWorksでアライメントを行い、前記領域の塩基配列に相同性が高いことを確認した。これらの解析結果より、前記D−ボックスは、種間で極めてよく保存されていることが明らかとなった。
【0017】
(PPARγ発現調節剤)
本発明のPPARγ発現調節剤は、DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかを含有し、必要に応じて、更にその他の成分を含有する。
【0018】
<DBP、TEF、HLF、及びE4BP4>
−DBP、TEF、及びHLF−
前記DBP、前記TEF(Thyrotroph Embryonic Factor:甲状腺刺激ホルモン分泌細胞胎児性因子)、及び前記HLF(Hepatic Leukemia Factor:肝白血病要因遺伝子)は、PAR−bZIP(proline and acidic amino acid−rich basic leucine zipper)転写因子ファミリーに属するタンパク質である(Wuarin J et al, J Cell Sci Suppl, 1992, 6, p.123−127)。前記PAR−bZIP転写因子ファミリーは、ホモあるいはヘテロダイマーを形成して、プロモーター領域にあるD−ボックス(RTTAYGTAAY)に結合し、転写活性化に働くことが知られている(Khatib ZA et al, Genomics, 1994, 123, p.344−51)。前記PAR−bZIP転写因子ファミリーは、哺乳類の概日リズムを制御する因子として知られている(Mitsui S et al, Genes Dev, 2001, 15, p.995−1006)が、脂肪細胞の分化などを含め、他の機能に関しては全く報告がない。
【0019】
−E4BP4−
前記E4BP4(アデノウイルスE4プロモーター結合蛋白4)は、bZIP転写因子ファミリーに属するタンパク質である。前記bZIPファミリーは、前記PAR−bZIP転写因子ファミリーと同様に、プロモーター領域にあるD−ボックスに結合し、転写抑制化に働くことが知られている(Mitsui S, Yamaguchi S, Matsuo T, Ishida Y, Okamura H, Genes Dev, 2001, 15, p.995−1006)。前記E4BP4も、前記PAR−bZIP転写因子ファミリーと同様に、哺乳類の概日リズムを制御する因子として知られている(Mitsui S, Yamaguchi S, Matsuo T, Ishida Y, Okamura H, Genes Dev, 2001, 15, p.995−1006)が、脂肪分化における作用などを含め、他の機能に関しては全く報告がない。
【0020】
−入手方法−
前記PPARγ発現調節剤中に含まれる、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの入手方法としては、特に制限はなく、目的に応じて適宜選択することができ、前記DBP、TEF、HLF、及びE4BP4がタンパク質の状態である場合、例えば、市販品を用いる方法、培養細胞より精製する方法、遺伝子組換え技術により組換体タンパク質を得る方法、無細胞系を用いた真核生物由来タンパク質合成の方法などが挙げられるが、これらの中でも無細胞系を用いた真核生物由来タンパク質合成の方法により得ることが、可溶性、及び機能タンパク質を迅速に合成できる点で好ましい。
前記DBP、TEF、HLF、及びE4BP4がcDNAの状態である場合、例えば、市販品を用いる方法、cDNAを合成する方法、cDNAライブラリーからクローニングする方法などが挙げられるが、これらの中でもcDNAを合成する方法により得ることが、迅速に取得できる点で好ましい。
【0021】
−含有量−
前記PPARγ発現調節剤中の、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかの含有量としては、特に制限はなく、目的に応じて適宜選択することができる。また、前記PPARγ発現調節剤は、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかそのものであってもよい。
前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかの含有比率としても特に制限はなく、目的に応じて適宜選択することができるが、PPARγの発現を促進する目的で用いられる場合と、PPARγの発現を抑制する目的で用いられる場合とで、前記含有比率を変えることが好ましい。
【0022】
−−PPARγの発現促進作用を有するPPARγ発現調節剤−−
前記PPARγ発現調節剤が、PPARγの発現を促進する目的で用いられる場合、前記PPARγ発現調節剤中の、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかの含有比率としては、前記DBP、及びTEF、並びに、該DBP、及び該TEFをコードするcDNAの少なくともいずれかが、前記HLF、及びE4BP4、並びに、該HLF、及び該E4BP4をコードするcDNAより多く含有されていることが好ましく、前記DBP、及びTEF、並びに、該DBP、及び該TEFをコードするcDNAの少なくともいずれかのみ含有されていることがより好ましい。
前記PPARγ発現調節剤が、前記DBP、及びTEF、並びに、該DBP、及び該TEFをコードするcDNAの少なくともいずれかであると、前記PPARγ発現調節剤によるPPARγの発現促進作用が強い点で有利である。
なお、PPARγの発現促進作用を有するPPARγ発現調節剤を、PPARγ発現促進剤として使用することができる。
【0023】
−−PPARγの発現抑制作用を有するPPARγ発現調節剤−−
前記PPARγ発現調節剤が、PPARγの発現を抑制する目的で用いられる場合、前記PPARγ発現調節剤中の、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかの含有比率としては、前記HLF、及びE4BP4、並びに、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかが、前記DBP、及びTEF、並びに、該DBP、及び該TEFをコードするcDNAより多く含有されていることが好ましく、前記HLF、及びE4BP4、並びに、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかのみ含有されていることがより好ましい。
前記PPARγ発現調節剤が、前記HLF、及びE4BP4、並びに、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかであると、前記PPARγ発現調節剤によるPPARγの発現抑制作用が強い点で有利である。
なお、PPARγの発現抑制作用を有するPPARγ発現調節剤を、PPARγ発現抑制剤として使用することができる。
【0024】
<PPARγの発現調節作用>
前記PPARγ発現調節剤が、PPARγの発現を調節する方法としては、特に制限はなく、目的に応じて適宜選択することができるが、前記PPARγ発現調節剤が、PPARγのプロモーター配列に結合することによりPPARγの発現を調節することが好ましい。
前記PPARγ発現調節剤が結合するプロモーター配列としては、特に制限はなく、目的に応じて適宜選択することができるが、前記PPARγ6のプロモーター配列であることが好ましく、前記PPARγ6のプロモーター配列に存在する3箇所のD−ボックス(D1、D2、及びD3)の少なくともいずれかに結合することがより好ましく、前記D2に結合することが更に好ましく、前記3箇所のD−ボックス全てに結合することが特に好ましい。
前記3箇所のD−ボックス全てに結合すると、PPARγの発現を顕著に促進又は抑制できる点で好ましい。
前記結合を確認する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、ゲルシフトアッセイ、チップアッセイ(ChIP assay)などが挙げられる。
【0025】
<その他の成分>
前記その他の成分としては、特に制限はなく、薬理学的に許容される担体の中から目的に応じて適宜選択することができ、例えば、エタノール、水、デンプンなどが挙げられる。
前記PPARγ発現調節剤中の、その他の成分の含有量としては、特に制限はなく、目的に応じて適宜選択することができる。
【0026】
<使用>
前記PPARγ発現調節剤は、1種単独で使用されてもよいし、他の成分を有効成分とする医薬と併用してもよい。また、前記PPARγ発現調節剤は、他の成分を有効成分とする医薬中に、配合された状態で使用されてもよい。
【0027】
<剤型>
前記PPARγ発現調節剤の剤型としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、経口固形剤、経口液剤、注射剤、吸入散剤などが挙げられる。
【0028】
−経口固形剤−
前記経口固形剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、錠剤、被覆錠剤、顆粒剤、散剤、カプセル剤などが挙げられる。
前記経口固形剤の製造方法としては、特に制限はなく、常法を使用することができ、例えば、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかに、賦形剤、及び必要に応じて前記その他の成分、各種添加剤を加えることにより、製造することができる。ここで、前記賦形剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、乳糖、白糖、塩化ナトリウム、ブドウ糖、デンプン、炭酸カルシウム、カオリン、微結晶セルロース、珪酸などが挙げられる。また、前記添加剤としても、特に制限はなく、目的に応じて適宜選択することができ、例えば、結合剤、崩壊剤、滑沢剤、着色剤、矯味/矯臭剤などが挙げられる。
前記結合剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、水、エタノール、プロパノール、単シロップ、ブドウ糖液、デンプン液、ゼラチン液、カルボキシメチルセルロース、ヒドロキシプロピルセルロース、ヒドロキシプロピルスターチ、メチルセルロース、エチルセルロース、シェラック、リン酸カルシウム、ポリビニルピロリドンなどが挙げられる。
前記崩壊剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、乾燥デンプン、アルギン酸ナトリウム、カンテン末、炭酸水素ナトリウム、炭酸カルシウム、ラウリル硫酸ナトリウム、ステアリン酸モノグリセリド、乳糖などが挙げられる。
前記滑沢剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、精製タルク、ステアリン酸塩、ホウ砂、ポリエチレングリコールなどが挙げられる。
前記着色剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、酸化チタン、酸化鉄などが挙げられる。
前記矯味/矯臭剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、白糖、橙皮、クエン酸、酒石酸などが挙げられる。
【0029】
−経口液剤−
前記経口液剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、内服液剤、シロップ剤、エリキシル剤などが挙げられる。
前記経口液剤の製造方法としては、特に制限はなく、常法を使用することができ、例えば、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれか、及び必要に応じて前記その他の成分に、添加剤を加えることにより、製造することができる。ここで、前記添加剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、矯味/矯臭剤、緩衝剤、安定化剤などが挙げられる。
前記矯味/矯臭剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、白糖、橙皮、クエン酸、酒石酸などが挙げられる。
前記緩衝剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、クエン酸ナトリウムなどが挙げられる。
前記安定化剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、トラガント、アラビアゴム、ゼラチンなどが挙げられる。
【0030】
−注射剤−
前記注射剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、溶液、懸濁液、用事溶解用固形剤などが挙げられる。
前記注射剤の製造方法としては、特に制限はなく、常法を使用することができ、例えば、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれか、及び必要に応じて前記その他の成分に、pH調節剤、緩衝剤、安定化剤、等張化剤、局所麻酔剤などを添加することにより、製造することができる。ここで、前記pH調節剤及び前記緩衝剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、クエン酸ナトリウム、酢酸ナトリウム、リン酸ナトリウムなどが挙げられる。また、前記安定化剤としても、特に制限はなく、目的に応じて適宜選択することができ、例えば、ピロ亜硫酸ナトリウム、EDTA、チオグリコール酸、チオ乳酸などが挙げられる。前記等張化剤としても、特に制限はなく、目的に応じて適宜選択することができ、例えば、塩化ナトリウム、ブドウ糖などが挙げられる。前記局所麻酔剤としても、特に制限はなく、目的に応じて適宜選択することができ、例えば、塩酸プロカイン、塩酸リドカインなどが挙げられる。
【0031】
<投与>
前記PPARγ発現調節剤の投与方法、投与量、投与時期、及び投与対象としては、特に制限はなく、目的に応じて適宜選択することができる。
前記投与方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、経口投与法、注射による方法、吸入による方法などが挙げられる。
前記投与量としては、特に制限はなく、投与対象個体の年齢、体重、体質、症状、他の成分を有効成分とする医薬の投与の有無など、様々な要因を考慮して適宜選択することができる。
前記投与対象となる動物種としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、ヒト、サル、ブタ、ウシ、ヒツジ、ヤギ、イヌ、ネコ、マウス、ラット、トリなどが挙げられるが、これらの中でもヒトに好適に用いられる。
【0032】
<用途>
前記PPARγ発現調節剤は、PPARγの発現調節作用を有することから、脂肪細胞への分化の調節、脂質代謝の調節、破骨細胞機能の調節、炎症の調節などに好適に利用可能である。
【0033】
(脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤)
本発明の脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤は、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかを阻害する因子を含有し、必要に応じて、更にその他の成分を含有する。
前記脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤は、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかを阻害することにより、PPARγの発現を促進又は抑制し、脂肪分化調節作用、脂質代謝調節作用、破骨細胞機能調節作用、及び炎症調節作用を発揮することが好ましい。
【0034】
<阻害する因子>
前記阻害する因子としては、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかを阻害することができれば、特に制限はなく、目的に応じて適宜選択することができ、例えば、miRNA(micro RNA)、siRNA(smoll interfering RNA)、shRNA(short hairpin RNA)などが挙げられるが、これらの中でも、miRNAが、完全に相補的でない場合でも翻訳阻害を引き起こすことができる点、及びRNAi(RNA干渉)として利用できる配列の範囲が広い点で好ましい。
本発明において、前記miRNAは、本発明の効果を損なわない限り、特に制限はなく、目的に応じて適宜選択することができ、DNAに置換されているものであってもよく、RNAとDNAとのキメラであってもよく、前記miRNAを発現し得る分子であってもよい。
前記miRNAを発現し得る分子としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、前記miRNAをDNAに置換した塩基配列を有するベクターなどが挙げられる。
【0035】
−miRNA−
前記miRNAとしては、特に制限はなく、目的に応じて適宜選択することができるが、前記DBP、TEF、HLF、及びE4BP4の少なくともいずれかの遺伝子発現を抑制する塩基配列を有することが好ましい。
前記DBP、TEF、HLF、及びE4BP4の少なくともいずれかの遺伝子発現を抑制する塩基配列としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、下記配列番号:2〜33で表される塩基配列などが挙げられる。
前記阻害する因子中のmiRNAは、配列番号:2〜33で表される塩基配列全てを有していてもよいが、発現を阻害する遺伝子の種類によって、下記塩基配列を選択することが好ましい。
また、前記DBP、TEF、HLF、及びE4BP4の少なくともいずれかの遺伝子発現を抑制する塩基配列の由来する生物種としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、マウス、ヒトなどが挙げられるが、これらの中でも、ヒトの配列であることが、前記脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤をヒトに用いる場合、抗原性を有さない点で好ましい。
【0036】
前記DBPの遺伝子発現を抑制するマウス由来のmiRNA配列としては、例えば、下記配列番号:2〜5で表される塩基配列の少なくともいずれかを有することが好ましい。
AAGAGGAGCUGCAAGAGCCGGGUUUUGGCCACUGACUGACCCGGCUCUCAGCUCCUCUU (配列番号:2)
AAGAGGAGCUGAGAGCCGGGUCAGUCAGUGGCCAAAACCCGGCUCUUGCAGCUCCUCUU (配列番号:3)
AAGAGGAGCTGCAAGAGCCGGGTTTTGGCCACTGACTGACCCGGCTCTCAGCTCCTCTT (配列番号:4)
AAGAGGAGCTGAGAGCCGGGTCAGTCAGTGGCCAAAACCCGGCTCTTGCAGCTCCTCTT (配列番号:5)
【0037】
前記DBPの遺伝子発現を抑制するヒト由来のmiRNA配列としては、例えば、下記配列番号:6〜9で表される塩基配列の少なくともいずれかを有することが好ましい。
AAGCCGAGCCGCACGAACCCGGUUUUGGCCACUGACUGACCGGGUUCGCGGCUCGGCUU (配列番号:6)
AAGCCGAGCCGCGAACCCGGUCAGUCAGUGGCCAAAACCGGGUUCGUGCGGCUCGGCUU (配列番号:7)
AAGCCGAGCCGCACGAACCCGGTTTTGGCCACTGACTGACCGGGTTCGCGGCTCGGCTT (配列番号:8)
AAGCCGAGCCGCGAACCCGGTCAGTCAGTGGCCAAAACCGGGTTCGTGCGGCTCGGCTT (配列番号:9)
【0038】
前記TEFの遺伝子発現を抑制するマウス由来のmiRNA配列としては、例えば、下記配列番号:10〜13で表される塩基配列の少なくともいずれかを有することが好ましい。
UUUAGCUGCCACAUUGUUCUUGUUUUGGCCACUGACUGACAAGAACAAUGGCAGCUAAA (配列番号:10)
UUUAGCUGCCAUUGUUCUUGUCAGUCAGUGGCCAAAACAAGAACAAUGUGGCAGCUAAA (配列番号:11)
TTTAGCTGCCACATTGTTCTTGTTTTGGCCACTGACTGACAAGAACAATGGCAGCTAAA (配列番号:12)
TTTAGCTGCCATTGTTCTTGTCAGTCAGTGGCCAAAACAAGAACAATGTGGCAGCTAAA (配列番号:13)
【0039】
前記TEFの遺伝子発現を抑制するヒト由来のmiRNA配列としては、例えば、下記配列番号:14〜17で表される塩基配列の少なくともいずれかを有することが好ましい。
UUUAGCUGCCACGUUGUUCUUGUUUUGGCCACUGACUGACAAGAACAACGUCAGCUAAA (配列番号:14)
UUUAGCUGACGUUGUUCUUGUCAGUCAGUGGCCAAAACAAGAACAACGUGGCAGCUAAA (配列番号:15)
TTTAGCTGCCACGTTGTTCTTGTTTTGGCCACTGACTGACAAGAACAACGTCAGCTAAA (配列番号:16)
TTTAGCTGACGTTGTTCTTGTCAGTCAGTGGCCAAAACAAGAACAACGTGGCAGCTAAA (配列番号:17)
【0040】
前記HLFの遺伝子発現を抑制するマウス由来のmiRNA配列としては、例えば、下記配列番号:18〜21で表される塩基配列の少なくともいずれかを有することが好ましい。
CAUCCUGAAGACGCAUUUAGUGUUUUGGCCACUGACUGACACUAAAUGCGUUCAGGAUG (配列番号:18)
CAUCCUGAACGCAUUUAGUGUCAGUCAGUGGCCAAAACACUAAAUGCGUCUUCAGGAUG (配列番号:19)
CATCCTGAAGACGCATTTAGTGTTTTGGCCACTGACTGACACTAAATGCGTTCAGGATG (配列番号:20)
CATCCTGAACGCATTTAGTGTCAGTCAGTGGCCAAAACACTAAATGCGTCTTCAGGATG (配列番号:21)
【0041】
前記HLFの遺伝子発現を抑制するヒト由来のmiRNA配列としては、例えば、下記配列番号:22〜25で表される塩基配列の少なくともいずれかを有することが好ましい。
ACUAAAUGCGUCUUCGUGGUGGUUUUGGCCACUGACUGACCACCACGAACGCAUUUAGU (配列番号:22)
ACUAAAUGCGUUCGUGGUGGUCAGUCAGUGGCCAAAACCACCACGAAGACGCAUUUAGU (配列番号:23)
ACTAAATGCGTCTTCGTGGTGGTTTTGGCCACTGACTGACCACCACGAACGCATTTAGT (配列番号:24)
ACTAAATGCGTTCGTGGTGGTCAGTCAGTGGCCAAAACCACCACGAAGACGCATTTAGT (配列番号:25)
【0042】
前記E4BP4の遺伝子発現を抑制するマウス由来のmiRNA配列としては、例えば、下記配列番号:26〜29で表される塩基配列の少なくともいずれかを有することが好ましい。
AUCACAGGGAACUUGUUCUCAGUUUUGGCCACUGACUGACUGAGAACAUUCCCUGUGAU (配列番号:26)
AUCACAGGGAAUGUUCUCAGUCAGUCAGUGGCCAAAACUGAGAACAAGUUCCCUGUGAU (配列番号:27)
ATCACAGGGAACTTGTTCTCAGTTTTGGCCACTGACTGACTGAGAACATTCCCTGTGAT (配列番号:28)
ATCACAGGGAATGTTCTCAGTCAGTCAGTGGCCAAAACTGAGAACAAGTTCCCTGTGAT (配列番号:29)
【0043】
前記E4BP4の遺伝子発現を抑制するヒト由来のmiRNA配列としては、例えば、下記配列番号:30〜33で表される塩基配列の少なくともいずれかを有することが好ましい。
AUAAUCUGGAACUUGUUUUCAGUUUUGGCCACUGACUGACUGAAAACAAGUCAGAUUAU (配列番号:30)
AUAAUCUGACUUGUUUUCAGUCAGUCAGUGGCCAAAACUGAAAACAAGUUCCAGAUUAU (配列番号:31)
ATAATCTGGAACTTGTTTTCAGTTTTGGCCACTGACTGACTGAAAACAAGTCAGATTAT (配列番号:32)
ATAATCTGACTTGTTTTCAGTCAGTCAGTGGCCAAAACTGAAAACAAGTTCCAGATTAT (配列番号:33)
【0044】
前記配列番号:2〜33で表される塩基配列のいずれかを有する前記miRNAは、前記配列番号:2〜33で表される塩基配列の全体からなる塩基配列であることが、発現阻害効率が高い点で好ましいが、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかを阻害することができれば、特に制限はなく、目的に応じて適宜選択することができ、例えば、前記配列番号:2〜33で表される塩基配列の一部からなる配列であってもよい。
前記miRNAが、前記配列番号:2〜33で表される塩基配列の一部からなる配列である場合、その一部の塩基配列としては、前記miRNAが、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかを阻害する能力を有する限り、特に制限はなく、目的に応じて適宜選択することができる。
また、前記miRNAは、前記配列番号:2〜33で表される塩基配列のいずれかの1若しくは数個の塩基が、欠失、置換、挿入された塩基配列からなるものであってもよい。
【0045】
−−入手方法−−
前記配列番号:2〜33で表される塩基配列の少なくともいずれかを有する前記miRNAを入手する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、合成による方法、市販品を用いる方法などが挙げられる。
【0046】
−−含有量−−
前記脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤中の、前記miRNAの含有量としては、特に制限はなく、目的に応じて適宜選択することができる。
前記miRNAが、前記配列番号:2〜33で表される塩基配列の少なくともいずれかである場合、その含有比率としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、前記DBP、及び該DBPをコードするcDNAを抑制する目的で用いられる場合、前記TEF、及び該TEFをコードするcDNAを抑制する目的で用いられる場合、前記HLF、及び該HLFをコードするcDNAを抑制する目的で用いられる場合、前記E4BP4、及び該E4BP4をコードするcDNAを抑制する目的で用いられる場合で、前記含有比率を変えることが好ましい。
【0047】
−−−DBP、及び該DBPをコードするcDNAの抑制−−−
前記DBP、及び該DBPをコードするcDNAを抑制する目的で用いられる場合、前記miRNAの含有比率としては、配列番号:2〜9で表される塩基配列の少なくともいずれかを有する前記miRNAが、配列番号:10〜33で表される塩基配列を有する前記miRNAより多いことが好ましく、配列番号:2〜9で表される塩基配列の少なくともいずれかのみを有する前記miRNAがより好ましく、配列番号:6〜9で表される塩基配列の少なくともいずれかのみを有する前記miRNAが特に好ましい。
前記脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤が、配列番号:2〜9で表される塩基配列の少なくともいずれかのみを有する前記miRNAである場合、配列番号:2〜9で表される塩基配列それぞれの含有比率としては、特に制限はなく、目的に応じて適宜選択することができる。
【0048】
−−−TEF、及び該TEFをコードするcDNAの抑制−−−
前記TEF、及び該TEFをコードするcDNAを抑制する目的で用いられる場合、前記miRNAの含有比率としては、配列番号:10〜17で表される塩基配列の少なくともいずれかを有するmiRNAが、配列番号:2〜9で表される塩基配列を有するmiRNA、及び配列番号:18〜33で表される塩基配列を有するmiRNAより多いことが好ましく、配列番号:10〜17で表される塩基配列の少なくともいずれかのみを有するmiRNAがより好ましく、配列番号:14〜17で表される塩基配列の少なくともいずれかのみを有するmiRNAが特に好ましい。
前記脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤が、配列番号:10〜17で表される塩基配列の少なくともいずれかのみを有するmiRNAである場合、配列番号:10〜17で表される塩基配列それぞれの含有比率としては、特に制限はなく、目的に応じて適宜選択することができる。
【0049】
−−−HLF、及び該HLFをコードするcDNAの抑制−−−
前記HLF、及び該HLFをコードするcDNAを抑制する目的で用いられる場合、前記miRNAの含有比率としては、配列番号:18〜25で表される塩基配列の少なくともいずれかを有するmiRNAが、配列番号:2〜17で表される塩基配列を有するmiRNA、及び配列番号:26〜33で表される塩基配列を有するmiRNAより多いことが好ましく、配列番号:18〜25で表される塩基配列の少なくともいずれかのみを有するmiRNAがより好ましく、配列番号:22〜25で表される塩基配列の少なくともいずれかのみを有するmiRNAが特に好ましい。
前記脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤が、配列番号:18〜25で表される塩基配列の少なくともいずれかのみを有するmiRNAである場合、配列番号:18〜25で表される塩基配列それぞれの含有比率としては、特に制限はなく、目的に応じて適宜選択することができる。
【0050】
−−−E4BP4、及び該E4BP4をコードするcDNAの抑制−−−
前記E4BP4、及び該E4BP4をコードするcDNAを抑制する目的で用いられる場合、前記miRNAの含有比率としては、配列番号:26〜33で表される塩基配列の少なくともいずれかを有するmiRNAが、配列番号:2〜25で表される塩基配列を有するmiRNAより多いことが好ましく、配列番号:26〜33で表される塩基配列の少なくともいずれかを有するmiRNAそのものがより好ましく、配列番号:30〜33で表される塩基配列の少なくともいずれかのみを有するmiRNAが特に好ましい。
前記脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤が、配列番号:26〜33で表される塩基配列の少なくともいずれかのみを有するmiRNAである場合、配列番号:26〜33で表される塩基配列それぞれの含有比率としては、特に制限はなく、目的に応じて適宜選択することができる。
【0051】
<阻害活性>
前記脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤が、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかを阻害しているか否かを確認する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、前記配列番号:2〜33を用いたRNAiにより確認する方法などが挙げられる。
前記RNAiに用いる細胞としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、脂肪細胞前駆細胞株である3T3−L1細胞、マクロファージ由来の細胞株であるRAW264細胞、骨髄由来マスト細胞mBMMCsなどが挙げられる。
前記3T3−L1細胞を用いた場合、前記配列番号:2〜33で表される塩基配列のいずれかによりRNAiを行った前記3T3−L1細胞の脂肪滴の数と、前記配列番号:2〜33で表される塩基配列を導入していない前記3T3−L1細胞の脂肪滴の数とを比較することで、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかの阻害活性を確認することができる。
【0052】
<その他の成分>
前記その他の成分としては、特に制限はなく、薬理学的に許容される担体の中から目的に応じて適宜選択することができ、例えば、エタノール、水、デンプンなどが挙げられる。
前記脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤中の、その他の成分の含有量としては、特に制限はなく、目的に応じて適宜選択することができる。
【0053】
<使用>
前記脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤は、1種単独で使用されてもよいし、2種以上を併用してもよく、他の成分を有効成分とする医薬と併用されてもよい。また、前記脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤は、他の成分を有効成分とする医薬中に、配合された状態で使用されてもよい。
【0054】
<剤型>
前記脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤の剤型としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、経口固形剤、経口液剤、注射剤、吸入散剤などが挙げられる。
【0055】
−経口固形剤−
前記経口固形剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、錠剤、被覆錠剤、顆粒剤、散剤、カプセル剤などが挙げられる。
前記経口固形剤の製造方法としては、特に制限はなく、常法を使用することができ、例えば、前記miRNAに、賦形剤、及び必要に応じて前記その他の成分、各種添加剤を加えることにより、製造することができる。ここで、前記賦形剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、乳糖、白糖、塩化ナトリウム、ブドウ糖、デンプン、炭酸カルシウム、カオリン、微結晶セルロース、珪酸などが挙げられる。また、前記添加剤としても、特に制限はなく、目的に応じて適宜選択することができ、例えば、結合剤、崩壊剤、滑沢剤、着色剤、矯味/矯臭剤などが挙げられる。
前記結合剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、水、エタノール、プロパノール、単シロップ、ブドウ糖液、デンプン液、ゼラチン液、カルボキシメチルセルロース、ヒドロキシプロピルセルロース、ヒドロキシプロピルスターチ、メチルセルロース、エチルセルロース、シェラック、リン酸カルシウム、ポリビニルピロリドンなどが挙げられる。
前記崩壊剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、乾燥デンプン、アルギン酸ナトリウム、カンテン末、炭酸水素ナトリウム、炭酸カルシウム、ラウリル硫酸ナトリウム、ステアリン酸モノグリセリド、乳糖などが挙げられる。
前記滑沢剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、精製タルク、ステアリン酸塩、ホウ砂、ポリエチレングリコールなどが挙げられる。
前記着色剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、酸化チタン、酸化鉄などが挙げられる。
前記矯味/矯臭剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、白糖、橙皮、クエン酸、酒石酸などが挙げられる。
【0056】
−経口液剤−
前記経口液剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、内服液剤、シロップ剤、エリキシル剤などが挙げられる。
前記経口液剤の製造方法としては、特に制限はなく、常法を使用することができ、例えば、前記miRNA、及び必要に応じて前記その他の成分に、添加剤を加えることにより、製造することができる。ここで、前記添加剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、矯味/矯臭剤、緩衝剤、安定化剤などが挙げられる。
前記矯味/矯臭剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、白糖、橙皮、クエン酸、酒石酸などが挙げられる。
前記緩衝剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、クエン酸ナトリウムなどが挙げられる。
前記安定化剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、トラガント、アラビアゴム、ゼラチンなどが挙げられる。
【0057】
−注射剤−
前記注射剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、溶液、懸濁液、用事溶解用固形剤などが挙げられる。
前記注射剤の製造方法としては、特に制限はなく、常法を使用することができ、例えば、前記miRNA、及び必要に応じて前記その他の成分に、pH調節剤、緩衝剤、安定化剤、等張化剤、局所麻酔剤などを添加することにより、製造することができる。ここで、前記pH調節剤及び前記緩衝剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、クエン酸ナトリウム、酢酸ナトリウム、リン酸ナトリウムなどが挙げられる。また、前記安定化剤としても、特に制限はなく、目的に応じて適宜選択することができ、例えば、ピロ亜硫酸ナトリウム、EDTA、チオグリコール酸、チオ乳酸などが挙げられる。前記等張化剤としても、特に制限はなく、目的に応じて適宜選択することができ、例えば、塩化ナトリウム、ブドウ糖などが挙げられる。前記局所麻酔剤としても、特に制限はなく、目的に応じて適宜選択することができ、例えば、塩酸プロカイン、塩酸リドカインなどが挙げられる。
【0058】
<投与>
前記脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤の投与方法、投与量、投与時期、及び投与対象としては、特に制限はなく、目的に応じて適宜選択することができる。
前記投与方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、経口投与法、注射による方法、吸入による方法などが挙げられる。
前記投与量としては、特に制限はなく、投与対象個体の年齢、体重、体質、症状、他の成分を有効成分とする医薬の投与の有無など、様々な要因を考慮して適宜選択することができる。
前記投与対象となる動物種としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、ヒト、サル、ブタ、ウシ、ヒツジ、ヤギ、イヌ、ネコ、マウス、ラット、トリなどが挙げられるが、これらの中でもヒトに好適に用いられる。
【0059】
<用途>
前記脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤は、前記DBP、TEF、及びE4BP4、並びに、該DBP、該TEF、及び該E4BPをコードするcDNAの少なくともいずれかの阻害作用を有することから、医薬、健康食品、機能性食品などに好適に利用可能である。
【0060】
(スクリーニング方法)
本発明のスクリーニング方法は、PPARγプロモーター配列を有するレポーター遺伝子を構築するプロモーターレポーター遺伝子構築工程と、該プロモーターレポーター遺伝子と、被験物質を含む被験試料とを細胞に導入する導入工程と、該プロモーターレポーター遺伝子のレポーター活性を指標にして、PPARγプロモーター活性を検出する検出工程と、該被験試料が、PPARγプロモーター活性の促進作用、及び抑制作用のいずれかの作用を有しているか否かを判断し、PPARγプロモーター活性の促進作用、及び抑制作用のいずれかの作用を有する該被験試料中の被験物質をPPARγ発現調節剤としてスクリーニングするスクリーニング工程と、を含み、必要に応じて、更にその他の工程を含む。
【0061】
<レポーター遺伝子構築工程>
前記レポーター遺伝子構築工程は、PPARγプロモーター配列を有するプロモーターレポーター遺伝子を構築する工程である。
【0062】
−PPARγプロモーター配列−
前記PPARγプロモーター配列としては、特に制限はなく、目的に応じて適宜選択することができるが、前述した配列番号:1で表される塩基配列を有するPPARγ6プロモーター配列を用いることが好ましい。
なお、本発明のスクリーニング方法に用いる前記PPARγ6のプロモーター配列は、前述した配列番号:1で表される塩基配列の全体からなる塩基配列を用いてもよいし、配列番号:1で表される塩基配列の一部からなる塩基配列を用いてもよく、配列番号:1で表される塩基配列中に存在する3ヵ所のD−ボックスの少なくともいずれかを欠失させた欠失変異体を用いてもよい。
前記D−ボックスの欠失変異体を、前記スクリーニング方法に用いると、前記被験物質とD−ボックスとの関係が明らかにできる点で有利である。
【0063】
前記PPARγ6のプロモーター配列を入手する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、ヒトゲノミックDNAを鋳型に用い、下記配列番号:34〜35で表される塩基配列をプライマーとしてPCRにより増幅する方法などが挙げられる。
プライマー配列
GATGAGAGCTGGGGAGAAGG (配列番号:34)
CTAAGGAAAGGGAATTAAGCAGAA (配列番号:35)
【0064】
−レポーター遺伝子−
前記レポーター遺伝子としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、ルシフェラーゼなどが挙げられる。
前記ルシフェラーゼの遺伝子配列の入手方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、市販のベクター(例えば、pGL3−basic、Promega社製)から入手する方法、NCBIなどのデータベースから入手する方法などが挙げられる。
【0065】
−−構築方法−−
前記プロモーターレポーター遺伝子を構築する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、前記ルシフェラーゼ遺伝子を有する前記pGL3−basic(Promega社製)、及び前記PPARγ6のプロモーター配列のPCR産物を制限酵素処理し、前記PPARγ6のプロモーター配列のPCR産物をクローニングすることにより構築する方法などが挙げられる。
なお、前記ルシフェラーゼは安定性の高いタンパク質であるため、プロモーター活性の変動(増減)をモニターし難いことがある。そのため、前記pGL3−basicは、ルシフェラーゼ遺伝子のC末端にPEST配列(プロリン、グルタミン酸、セリン、及びスレオニンに富む配列)を導入させた配列であることが、細胞内におけるルシフェラーゼの分解を促進できる点で好ましい。
【0066】
<導入工程>
前記導入工程は、前記プロモーターレポーター遺伝子と、被験物質を含む被験試料とを細胞に導入する工程である。
【0067】
−被験試料−
前記被験試料とは、少なくとも被験物質を含有し、必要に応じて、更にその他の成分を含有する。
【0068】
−−被験物質−−
前記被験物質としては、特に制限はなく、目的に応じて適宜選択することができるが、発現ベクターにクローニングされた遺伝子であることが好ましい。
前記発現ベクターとしては、特に制限はなく、目的に応じて適宜選択することができ、例えば、pcDNA3.1(Invitrogen社製)などが挙げられる。
【0069】
−−その他の成分−−
前記その他の成分としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、水、PBS、TE(Tris−EDTA)などが挙げられる。
【0070】
−細胞−
前記細胞としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、NIH3T3細胞、HepG2細胞、Caco2細胞などが挙げられる。
なお、前記NIH3T3細胞は、マウス胎児皮膚由来細胞株であり、前記HepG2細胞は、ヒト肝ガン由来細胞株であり、前記Caco2細胞は、ヒト結腸由来細胞株である。
前記細胞の培地としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、DMEM、MEMなどが挙げられる。
前記細胞の培養条件としては、特に制限はなく、細胞の種類に応じて適宜選択することができるが、前記NIH3T3細胞、及び前記HepG2細胞の場合は、10質量%FBS(ウシ胎児結成)、DMEM(Dulbecco's modified Eagle medium)、37℃、5%CO2が好ましく、前記Caco2細胞の場合は、20質量%FBS、DMEM、37℃、5%CO2が好ましい。
【0071】
−導入方法−
前記プロモーターレポーター遺伝子と、前記被験物質を含む被験試料とを細胞に導入する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、リン酸カルシウム法、リポフェクション法、エレクトロポレーション法などが挙げられるが、これらの中でもリポフェクション法が、簡単で導入効率が高い点で好ましい。
前記プロモーターレポーター遺伝子の導入量としては、特に制限はなく、目的に応じて適宜選択することができるが、5ng〜20ngが好ましく、10ng〜15ngがより好ましい。
前記被験物質が、遺伝子である場合、その導入量としては、特に制限はなく、遺伝子の種類、発現ベクターの種類などに応じて適宜選択することができるが、62.5ng〜1,000ngが好ましく、100ng〜200ngがより好ましい。
前記細胞に、前記プロモーターレポーター遺伝子、及び被験物質を導入する時期としても、特に制限はなく、目的に応じて適宜選択することができる。
なお、前記遺伝子の導入量を一定にするため、pcDNA3.1(Invitrogen社製)を用いることが好ましい。
【0072】
<検出工程>
前記検出工程は、前記プロモーターレポーター遺伝子のレポーター活性を指標にして、PPARγプロモーター活性を検出する工程である。
【0073】
前記レポーター活性を検出する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、dual−luciferase reporter assay system(Promega社製)を用いて、前記ルシフェラーゼを反応させ、ルミノメーター(例えば、Model TD−20/20(Turner Designs社製))により検出する方法などが挙げられる。
前記ルシフェラーゼは、前記PPARγプロモーターの制御下で発現するため、前記ルシフェラーゼのレポーター活性を検出することで、前記PPARγのプロモーター活性を検出することができる。
【0074】
<スクリーニング工程>
前記スクリーニング工程は、前記被験物質を含む被験試料が、PPARγプロモーター活性の促進作用、及び抑制作用のいずれかの作用を有しているか否かを判断し、PPARγプロモーター活性の促進作用、及び抑制作用のいずれかの作用を有する該被験試料中の被験物質をPPARγ発現調節剤としてスクリーニングする工程である。
【0075】
前記PPARγプロモーター活性の促進作用、及び抑制作用のいずれかの作用を有しているか否かを判断する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、前記レポーター活性の強度を比較する方法などが挙げられる。
前記レポーター活性の強度を比較する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、前記被験試料を導入せず、プロモーターレポーター遺伝子のみを導入した細胞をコントロールとし、該コントロールと、被験試料、及びプロモーターレポーター遺伝子を導入した細胞とを比較して、前記被験試料、及びプロモーターレポーター遺伝子を導入した細胞のレポーター活性が、前記コントロールのレポーター活性より強い場合、前記被験試料中の被験物質をPPARγ発現調節剤としてスクリーニングする方法、前記コントロールと比較せず、前記被験物質を含む被験試料間のレポーター活性の強度の違いにより、レポーター活性が強い前記被験試料中の被験物質を、より強いPPARγ発現調節作用を有するPPARγ発現調節剤としてスクリーニングする方法などが挙げられる。
【実施例】
【0076】
以下に本発明の実施例を挙げて本発明を具体的に説明するが、本発明はこれらの実施例に何ら限定されるものではない。
【0077】
(実施例1:新規PPARγプロモーター配列の解析)
新規PPARγであるPPARγ6のプロモーター配列は、新規スプライスバリアントcDNAの5’配列を元に転写開始点を決定し、第1エクソン(C1C2)(図1B)を有する上流領域を、下記方法により解析した。
【0078】
<PPARγ6プロモーター配列の増幅>
ヒトゲノミックDNA(Novagen社製)を鋳型に用いて、下記配列番号:34〜35で表される塩基配列をプライマーとして用い、96℃・30秒間、62℃・30秒間、72℃・90秒間を30サイクルの条件でPCRを行い、PPARγ6プロモーター配列を増幅した。前記PPARγ6プロモーター領域の長さは、約1kbであった。
前記PPARγ6プロモーター配列のPCR産物は、pCR2.1ベクター(Invitrogen社製)にTA法でクローニングした(以下、「hPPARγ6−pCR2.1」と称することがある。)。
【0079】
プライマー配列
GATGAGAGCTGGGGAGAAGG (配列番号:34)
CTAAGGAAAGGGAATTAAGCAGAA (配列番号:35)
【0080】
<PPARγ6のプロモーター配列の解析>
前記hPPARγ6−pCR2.1を用い、ABI PRISM377 シーケンサー(アプライドバイオシステム社製)によりシーケンスをいった。シーケンスの結果から明らかになった配列について、NCBI BLAST cDNA、及びETSデータベースにて検索を行い、スプライシングパターンを決定した。その結果、PPARγ6プロモーター配列は、PPARγの新規スプライスバリアントである、配列番号:1で表される塩基配列を有することが明らかとなった。
【0081】
<PPARγ6プロモーター領域のDNA結合配列予測解析>
前記hPPARγ6−pCR2.1を用い、PPARγ6のプロモーター配列のDNA結合配列予測解析を、MatInspector(Genomatix社製)により行った。
その結果、PPARγ6のプロモーター配列は、C1C2領域に、3箇所のD−ボックス(D1、D2、及びD3)を有することが確認された(図2)。前記D−ボックスは、配列番号:1で表される塩基配列のうち、塩基番号159〜166(D1)、塩基番号202〜209(D2)、及び塩基番号218〜225(D3)に存在する。なお、図2中、TSSは、転写開始領域(Transcript Start Site)を示す。
【0082】
(実施例2:DBPによるPPARγ6の発現調節)
DBPがPPARγ6の発現を調節しているか否かを、以下の方法を用いて調べた。
【0083】
<方法>
−Caco−2細胞の培養−
Caco−2細胞(細胞株バンクATCC)を、24ウエルプレートに5×105細胞/プレート播種し、DMEM(Invitrogen社製)で、37℃、5%CO2の条件下で3日間培養した。
【0084】
−DBP発現ベクターの構築−
DBP遺伝子を、マウス肝臓由来cDNAを鋳型にしてPCRにより増幅した。前記PCR産物と、pcDNA3.1ベクター(Invitrogen社製)とをEcoRIで制限酵素処理し、前記pcDNA3.1ベクターに、前記DBP遺伝子をクローニングした(以下、「pcDNA3.1−DBP」と称することがある)。
【0085】
−トランスフェクション−
前記pcDNA3.1−DBP 125ng、及びコントロールとしてDBPをクローニングしていないpcDNA3.1ベクターのいずれかを、Lipofectamin2000(Invitrogen社製)を用いて前記Caco−2細胞へトランスフェクションし、前記DBPを、前記Caco−2細胞に一過性に過剰発現させた。前記Caco−2細胞は、後述するmRNAの定量までの間、DMEMで、37℃、5%CO2の条件下で1日間培養した。
【0086】
−mRNAの定量−
前記DBPを過剰発現させたCaco−2細胞、及びコントロールのCaco−2細胞から、ISOGEN((株)日本ジーン製)を用いてRNA抽出、及び精製を行い、DBPのmRNA量、及びPPARγ6のmRNA量をRT−PCR法により測定した。
【0087】
<結果>
図3に前記RT−PCRの結果を示す。
図3より、DBPのmRNAの発現量は、コントロールと比較して有意に高く、pcDNA3.1−DBPをトランスフェクションしたCaco−2細胞では、DBPが過剰発現されていることが確認された。
PPARγ6のmRNAの発現量は、pcDNA3.1−DBPをトランスフェクションし、DBPを過剰発現したCaco−2細胞では、コントロールのCaco−2細胞と比較して有意に増加していた。
これらの結果より、PPARγ6の発現は、DBPにより促進されたことが示唆される。
【0088】
(実施例3:DBP、TEF、HLF、及びE4BP4によるPPARγ6の転写制御)
DBP、TEF、HLF、及びE4BP4が、PPARγ6の転写を制御しているか否かについて、以下のレポーターシステムにより調べた。
【0089】
<方法>
−DBP発現ベクターの構築−
DBP遺伝子を、マウス肝臓由来cDNAを鋳型にしてPCRにより増幅した。前記PCR産物と、pcDNA3.1ベクター(Invitrogen社製)とをEcoRIで制限酵素処理し、前記pcDNA3.1ベクターに、前記DBP遺伝子をクローニングした(以下、「pcDNA3.1−DBP」と称することがある)。
【0090】
−TEF発現ベクターの構築−
TEF遺伝子を、ヒト肝臓由来cDNAを鋳型にしてPCRにより増幅した。前記PCR産物と、pcDNA3.1ベクター(Invitrogen社製)とをEcoRIで制限酵素処理し、前記pcDNA3.1ベクターに、前記TEF遺伝子をクローニングした(以下、「pcDNA3.1−TEF」と称することがある)。
【0091】
−HLF発現ベクターの構築−
HLF遺伝子を、ヒト肝臓由来cDNAを鋳型にしてPCRにより増幅した。前記PCR産物と、pCR3.1ベクター(Invitrogen社製)とをTAクローニング法によって、前記pCR3.1ベクターに、前記HLF遺伝子をクローニングした(以下、「pCR3.1−HLF」と称することがある)。
【0092】
−E4BP4発現ベクターの構築−
E4BP4遺伝子を、マウス肝臓由来cDNAを鋳型にしてPCRにより増幅した。前記PCR産物と、pcDNA3ベクター(Invitrogen社製)とをBamHI、及びKpnIで制限酵素処理し、前記pcDNA3ベクターに、前記E4BP4遺伝子をクローニングした(以下、「pcDNA3−E4BP4」と称することがある)。
【0093】
−プロモーターレポーター遺伝子の構築−
−−pGL3−dlucの構築−−
pGL3−basic(Promega社製)に、ルシフェラーゼをコードする遺伝子のC末端に相当する領域をEcoRV、及びXbaIで制限酵素処理することにより、pGL3−basic中のルシフェラーゼ遺伝子のC末端に、PEST配列を導入した(以下、「pGL3−dluc」と称することがある。)。
【0094】
−−pGL3−dlucへのPPARγ6プロモーター配列のサブクローニング−−
実施例1のhPPARγ6−pCR2.1と、前記pGL3−dlucとを、KpnI、及びBglII(共に(株)日本ジーン製)で制限酵素処理し、PPARγ6プロモーター配列をpGL3−dlucにサブクローニングすることにより、プロモーターレポーター遺伝子(以下、「hPPARγ6−prom−1.0k/pGL3−basic」と称することがある)を構築した。
【0095】
−細胞の培養−
NIH3T3細胞(理化学研究所提供)を、35mmプレートに1×105細胞/プレート播種し、10質量%FBSを含むDMEMで、37℃、5%CO2の条件下で2日間培養した。
【0096】
−導入−
前記hPPARγ6−prom−1.0k/pGL3−basicと、前記pcDNA3.1−DBP、前記pcDNA3.1−TEF、前記pCR3.1−HLF、及び前記pcDNA3−E4BP4のいずれかとを、前記NIH3T3細胞に、Tfx−50(Promega社製)を用いて、製造元の方法に従い、トランスフェクションした。
なお、前記hPPARγ6−prom−1.0k/pGL3−basicは、10ngトランスフェクションに用いた。前記pcDNA3.1−DBPは0ng〜1,000ngの濃度で、前記pcDNA3.1−TEFは0ng〜500ngの濃度で、前記pCR3.1−HLFは0ng〜500ngの濃度で、前記pcDNA3−E4BP4は0ng〜1,000ngの濃度でそれぞれトランスフェクションを行った。
トランスフェクション24時間後に、dual−luciferase reporter assay system(Promega社製)によりpRL−TK(Prommega社製)を導入した。導入遺伝子量を一定にする為、pcDNA3(Invitrogen社製)を用いた。
なお、トランスフェクション効率は、前記pRL−TKにより確認した。
導入後、10質量%FBSを含むDMEMで、37℃、5%CO2の条件下で2日間培養した。
【0097】
−検出−
前記dual−luciferase reporter assay system(Promega社製)を用いて、製造元の方法に従い、前記hPPARγ6−prom−1.0k/pGL3−basicより発現したルシフェラーゼを反応させた。
前記ルシフェラーゼ活性(レポーター活性)の測定は、ルミノメーターModel TD−20/20(Turner Designs社製)を用いて行った。
【0098】
<結果>
ルシフェラーゼ活性の測定結果を、図4A〜Dに示す。
図4Aより、DBPは、導入量依存的にルシフェラーゼ活性の増加が認められ、その活性は、DBPを500ng導入したときに最も高かった。また、図4Bより、TEFもDBPと同様に、導入量依存的にルシフェラーゼ活性の増加が認められた。これらの結果より、DBP、及びTEFは、PPARγ6プロモーター活性を促進することが認められた。
図4C〜Dより、HLF、及びE4BP4は、DBP、及びTEFとは反対に、導入量依存的にルシフェラーゼ活性の低下が認められた。これらの結果より、HLF、及びE4BP4は、PPARγ6プロモーター活性を抑制することが認められた。
DBP、TEF、及びHLFは、同じPAR−bZIPファミリーに属する転写因子であるが、PPARγ6に対する活性は異なるものであることが示唆された。
【0099】
(実施例4:DBPのPPARγ6プロモーターへの結合部位)
DBPが、PPARγ6のプロモーター領域のどの部位を介してPPARγ6の転写活性化を行っているか、PPARγ6プロモーター領域を欠失させた数種の欠失変異体を用いて、下記方法により検討を行った。
【0100】
−欠失変異体の作製−
前記欠失変異体は、実施例2で作製したhPPARγ6−prom−1.0k/pGL3−basicに、QuikChange XL site−directed mutagenesis kit(Stratagene社製)を用いて、製造元の方法に従い、下記表1に示すコンストラクト1〜10を作製した(図5A)。
下記表1は、PPARγ6プロモーターのC1C2の5’末端を、転写開始点、即ち「0」とし、C1C2の転写開始点より上流の塩基数を「−」で示し、C1C2の転写開始点より下流の塩基数を「+」で示した。なお、C1C2は、全長297bである。
コンストラクト1は、PPARγ6プロモーター配列全長を有する野生型である。コンストラクト2〜5は、PPARγ6プロモーター配列の上流、即ち5’末端を欠失させた欠失変異体である。コンストラクト6は、PPARγ6プロモーター配列に存在する3箇所のD−ボックスを欠失させた欠失変異体である。コンストラクト7〜9は、PPARγ6プロモーター配列に存在する3箇所のD−ボックス、及び5’末端を欠失させた欠失変異体である。コンストラクト10(ネガティブコントロール)は、PPARγ6プロモーター配列を導入していない、pGL3−basicのみのコンストラクトである。
【0101】
【表1】

【0102】
−NIH3T3細胞の培養−
NIH3T3細胞(理化学研究所提供)を、35mmディッシュに、1×106細胞/ディッシュ播種し、DMEMを用い、37℃、5%CO2の条件下で2日間培養した。
【0103】
−導入−
前記コンストラクト1〜10のいずれかをそれぞれ10ngと、pcDNA3.1−DBP 125ngとを、PolyFect(Qiagen社製)を用いて、前記NIH3T3細胞にトランスフェクションし、24時間後に、100nMデキサメタゾン(Sigma社製)を添加したDMEMに交換し、更に2時間後に、0.1mMルシフェリン(Promega社製)を含むDMEMに交換し、Kronos(Atto社製)を用いて、ルシフェラーゼ活性を10分間ごとに測定した。
なお、コントロールとしては、pcDNA3.1−DBPを導入せず、コンストラクト1〜10のいずれかのみを導入したサンプルを用いた。前記コントロールのルシフェラーゼ活性が、NIH3T3細胞におけるPPARγ6プロモーターの基礎転写活性を示すものとする。
【0104】
−結果−
結果を、図5B〜Cに示す。
図5B〜Cより、コンストラクト1のルシフェラーゼ活性は、pcDNA3.1−DBPにより基礎転写活性の約2倍になっており、PPARγ6のプロモーター活性がDBPにより顕著に活性化されたことが認められた。このDBPによるPPARγ6の転写活性化は、C1C2より上流を欠失したコンストラクト2〜5においても認められた(図5B)。C1C2が一部欠失したコンストラクト4〜5では、コンストラクト1〜3と比較して、わずかにPPARγ6のプロモーター活性が低かったが、いずれも各コントロール(DBP(−))と比較して、PPARγ6の転写は活性化されていることが認められた。
一方、C1C2より上流は欠失せず、C1C2に存在するD−ボックス部位のみを欠失したコンストラクト6では、PPARγ6プロモーターの基礎転写活性が完全に消失し、そのレベルは、コンストラクト10(ネガティブコントロール)と同程度であった。更に、DBPによるPPARγ6の転写活性化も認められなかった(図5C)。また、コンストラクト7〜9において、C1C2のD−ボックス部位を欠失させ、更にC1C2より上流を欠失させた場合も、PPARγ6プロモーターの基礎転写活性は完全に消失し、DBPによるPPARγ6の転写活性化も認められなかった(図5C)。
【0105】
これらの結果より、DBPによるPPARγ6プロモーターの活性化には、PPARγ6プロモーター配列のC1C2に存在するD−ボックスが重要であり、DBPは、D−ボックスを介してPPARγ6プロモーターの活性化、即ちPPARγ6の発現を促進していることが示唆された。
【0106】
(実施例5:DBP、及びE4BP4による、D−ボックスを介したPPARγ6の転写制御)
実施例4の結果、DBPはPPARγ6プロモーター配列に存在するD−ボックスに作用していることが認められた。実施例1のPPARγ6プロモーター配列解析の結果、PPARγ6プロモーターには3箇所D−ボックスが存在することが明らかとなっていることから、これらの3箇所のD−ボックスと、DBP、及びE4BP4との関係について、下記方法により検討を行った。
【0107】
<方法>
実施例4のコンストラクト5(5’末端:+126、3’末端:+297)を用い、実施例4と同様の方法により、QuikChange XL site−directed mutagenesis kit(Stratagene社製)を用いて、PPARγ6プロモーターの3箇所のD−ボックスの全部又は一部を欠失させた下記表2、及び図6Aに示すコンストラクト11〜15を作製した。
下記表2の「○」は、D−ボックスを有する箇所を示し、「×」は、D−ボックスを欠失した箇所を示す。
前記コンストラクト11〜15のいずれかをそれぞれ10ngと、pcDNA3.1−DBP 250ng、及びpcDNA3−E4BP4 400ngのいずれかとを、実施例4と同様の方法で、前記NIH3T3細胞に導入し、ルシフェラーゼ活性を測定した。
なお、コントロールとしては、pcDNA3.1−DBP、及びpcDNA3−E4BP4を導入せず、コンストラクト11〜15のいずれかのみを導入したサンプルを用いた。前記コントロールのルシフェラーゼ活性が、NIH3T3細胞におけるPPARγ6プロモーターの基礎転写活性を示すものとする。
【0108】
【表2】
【0109】
<結果>
図6BにDBPの結果を、図6CにE4BP4の結果を示す。なお、図6B、及び図6C中、「**」は、t−検定によるp値が、p<0.01を示し、「***」は、p<0.001を示す。
【0110】
−DBP−
PPARγ6プロモーター配列に存在する3箇所のD−ボックスを有するコンストラクト11は、DBPによりPPARγ6プロモーターを活性化し、その活性は、コントロール(基礎転写活性)の約2倍であった(図6B)。
D−ボックスのD2、及びD3の両方を欠失させたコンストラクト12では、コンストラクト11と比較して、基礎転写活性に変化はなかったが、DBP存在下では、コンストラクト11と比較して、PPARγ6プロモーターの活性化の強度が弱かった。また、コントロール(基礎転写活性)と比較して、DBPによるPPARγ6プロモーターの活性化は認められたものの、その活性化強度は弱かった(図6B)。
D−ボックスのD1、及びD3の両方を欠失させたコンストラクト13では、コンストラクト11と比較して、基礎転写活性が低下し、DBP存在下のコンストラクト11と比較しても、DBPによるPPARγ6プロモーターの活性化の強度は弱かった。しかし、コンストラクト13のコントロールと比較して、ルシフェラーゼの発現量は約2倍以上であり、DBPによるPPARγ6プロモーターの強い活性化が認められた(図6B)。
D−ボックスのD1、及びD2の両方を欠失させたコンストラクト14も、コンストラクト13の結果と同様であり、コンストラクト11と比較して、基礎転写活性が低下し、DBP存在下のコンストラクト11と比較しても、DBPによるPPARγ6の活性化の強度は弱かった。しかし、コンストラクト14のコントロールと比較して、ルシフェラーゼの発現量は約2倍以上であり、DBPによるPPARγ6のプロモーターの活性化が認められた(図6B)。
D−ボックスのD1〜D3の全てを欠失させたコンストラクト15では、コンストラクト11と比較して、基礎転写活性が低下し、DBP存在下でもルシフェラーゼの発現レベルは、コンストラクト15のコントロールと同レベルであったことから、DBPによるPPARγプロモーターの活性化が完全に消失していることが認められた(図6B)。
【0111】
−E4BP4−
PPARγ6プロモーター配列に存在する3箇所のD−ボックスを有するコンストラクト11は、E4BP4によりPPARγ6プロモーター活性を抑制し、その活性は、コントロール(基礎転写活性)の約1/2であった(図6C)。
D−ボックスのD2、及びD3の両方を欠失させたコンストラクト12では、コンストラクト11と比較して、基礎転写活性に変化はなく、E4BP4存在下でも、コンストラクト11と比較して変化は認められなかった。即ち、E4BP4によるPPARγ6プロモーター活性の抑制が認められ、その活性は、コンストラクト12のコントロールの約1/2であった。(図6C)。
D−ボックスのD1、及びD3の両方を欠失させたコンストラクト13では、コンストラクト11と比較して、基礎転写活性が低下し、コンストラクト13のコントロールと比較して、E4BP4によるPPARγ6プロモーター活性の抑制が弱かった(図6C)。
D−ボックスのD1、及びD2の両方を欠失させたコンストラクト14も、コンストラクト13の結果と同様であり、コンストラクト11と比較して、基礎転写活性が低下し、コンストラクト14のコントロールと比較して、E4BP4によるPPARγ6プロモーター活性の抑制が弱かった(図6C)。
D−ボックスのD1〜D3の全てを欠失させたコンストラクト15では、コンストラクト11と比較して、基礎転写活性が低下し、E4BP4存在下でもルシフェラーゼの発現レベルは、コンストラクト15のコントロールと同レベルであったことから、E4BP4によるPPARγプロモーター活性の抑制が完全に消失していることが認められた(図6C)。
【0112】
実施例5の結果から、PPARγ6の転写調節は、主に、基礎転写活性に係る転写開始点上流配列約700bと、PAR−bZIP転写因子ファミリーによって転写調節を受ける転写開始点より下流の3箇所のD−ボックスを有する配列とによって行われていることが明らかになった。
更に、DBPによって正に、E4BP4によって負に制御を受ける、D−ボックスを介した転写調節システムを有することが示された。
また、3箇所のD−ボックスによって、DBP、及びE4BP4による転写調節が行われていることが明らかになったことから、PPARγ6プロモーター配列に存在するD−ボックスの有無によって、PPARγ6の転写調節作用がDBP、及びE4BP4を介するものであるかどうかを判定出来る可能性が示唆された。
【0113】
(実施例6:DBP、TEF、HLF、及びE4BP4のPPARγ6のD−ボックスへの結合能)
実施例2〜3より、DBP、TEF、HLF、及びE4BP4がPPARγ6の転写活性に関与することが示唆され、更に実施例4〜5より、DBP、及びE4BP4によるPPARγ6の転写制御は、PPARγ6のプロモーター領域に存在するD−ボックスが重要であることが示唆された。
そこで、DBP、TEF、HLF、及びE4BP4が、PPARγ6のプロモーター領域に存在するD−ボックスに直接結合するか否かについて、ゲルシフトアッセイにより検討を行った。
【0114】
<方法>
−プローブの作製−
ゲルシフトアッセイは、PPARγ6プロモーターに存在する3箇所のD−ボックス(D1、D2、及びD3)の配列をもとに、D1に対するプローブとしては、下記配列番号:36で表される2本鎖プローブを、D2に対するプローブとしては、下記配列番号:37で表される2本鎖プローブを、D3に対するプローブとしては、下記配列番号:38で表される2本鎖プローブを作製した。配列番号:36〜38で表されるプローブの下線は、D−ボックスを示す。前記プローブに、T4ポリヌクレオチドキナーゼを用いてγ−32P ATPを5’末端に導入して標識した。
AGGGAGGAGCCTAAGGTAAGGAGTCAGAAA (配列番号:36)
AGCTGCGGCTTTTATATAAGGTCAGTGGTA (配列番号:37)
TAAGGTCAGTGGTAGGTAAGGAAGGGGCCT (配列番号:38)
【0115】
−タンパク質の合成−
ゲルシフトアッセイに用いるタンパク質は、実施例3で構築したpcDNA3.1−DBP、pcDNA3.1−TEF、pCR3.1−HLF、及びpcDNA3−E4BP4を鋳型として、TNT T7 Quick Coupled Transcription/Translation System (Promega社製)を用い、製造元の方法に従って合成した。
前記DBP、TEF、HLF、及びE4BP4の合成タンパク質は、Gel Shift Binding Buffer (Takara Bio社製)に懸濁した。
【0116】
−ゲルシフトアッセイ−
−−プローブ特異性の確認−−
前記DBP合成タンパク質、TEF合成タンパク質、HLF合成タンパク質、及びE4BP4合成タンパク質をそれぞれ4μg、前記標識した各プローブ(配列番号:36〜38)0.0175pmol、特異的コンペティターとして、前記各プローブ(配列番号:36〜38)の非標識のプローブ1.75pmol、及び、非特異的コンペティター(合成委託)として、1.75pmolを用い、下記表3〜6に従い、サンプル1〜32を調製し、4質量%ポリアクリルアミドゲルにアプライし、150V、2時間の条件で電気泳動を行った。ゲルはゲルドライヤー(バイオラド社製)で乾燥した後、ゲルイメージを、BAS5000ゲルイメージ解析装置(Fuji Film社製)を用いて取得した。
なお、表1中、「+」は添加、「−」は非添加を示す。
【0117】
【表3】
【0118】
【表4】
【0119】
【表5】
【0120】
【表6】
【0121】
図7AにDBP合成タンパク質に対するプローブ特異性の確認の結果を、図7BにTEF合成タンパク質に対するプローブ特異性の確認の結果を、図7CにHLF合成タンパク質に対するプローブ特異性の確認の結果を、図7DにE4BP4合成タンパク質に対するプローブ特異性の確認の結果を示す。
図7A〜D中、「WT」は特異的コンペティターを添加したサンプル、「MT」は非特異的コンペティターを添加したサンプルを示す。ポジティブコントロールとしては、それぞれの合成タンパク質のプローブのみを添加したもの(サンプルNo.1、5、9、13、17、21、25、及び29)を用い、ネガティブコントロールとしては、プローブのみを添加したもの(サンプルNo.4、8、12、16、20、24、28、及び32)を用いた。
これらの結果より、ポジティブコントロールとしてコンペティターを添加しなかったサンプルは、DBP、TEF、HLF、及びE4BP4のどのタンパク質においても、それぞれバンドのシフトが認められた(図7A〜D:サンプル1、5、9、13、17、21、25、及び29)。また、ネガティブコントロールとしてプローブのみを添加したサンプルは、DBP、TEF、HLF、及びE4BP4のどのタンパク質においても、バンドのシフトが認められなかった(図7A〜D:サンプル4、8、12、16、20、24、28、及び32)。
特異的コンペティターを添加した場合、シフトしたバンドが消失したことから、前記バンドは、配列番号:36〜38に特異的な配列を認識していることが確認された(図7A〜D:サンプル2、6、10、14、18、22、26、及び30)。非特異的コンペティターを用いた場合、コンペティターを添加しなかった場合と同程度のバンドが検出されることを確認した(図7A〜D:サンプル3、7、11、15、19、23、27、及び31)。
【0122】
−−DBP、TEF、HLF、及びE4BP4のD−ボックスへの結合の確認−−
前記DBP合成タンパク質、TEF合成タンパク質、HLF合成タンパク質、及びE4BP4合成タンパク質をそれぞれ4μg、及び前記標識した各プローブ(配列番号:36〜38)0.0175pmolを使用して、下記表7に従い、サンプル33〜48を調製し、4質量%ポリアクリルアミドゲルにアプライし、150V、2時間の条件で電気泳動を行った。ゲルはゲルドライヤー(バイオラド社製)で乾燥した後、ゲルイメージを、BAS5000(Fuji Film社製)を用いて取得した。
なお、ポジティブコントロールとしては、下記配列番号:39で表される塩基配列を有する、PAR(proline and acidic amino acid−rich)、及びE4BP4のコンセンサス配列を用いた(Mitsui S, et al., Genes Dev, 2001, 15(8), p.995−1006)。
GCCCGCTACATATTACGTAACAAGCGTTCGC (配列番号:39)
【0123】
【表7】
【0124】
<結果>
図7EにDBPのゲルシフトアッセイの結果を、図7GにTEFのゲルシフトアッセイの結果を図7FにHLFのゲルシフトアッセイの結果を、図7HにE4BP4のゲルシフトアッセイの結果を示す。
図7E〜Hより、PPARγ6プロモーター領域に存在する3箇所のD−ボックスへの、DBP、及びE4BP4の結合は、D1、D2、及びD3の全てにおいて結合が認められた(図7E:サンプルNo.33〜36、及び図7H:サンプルNo.45〜48)。特に、D2に関して強い特異的バンドが検出された(図7E:サンプルNo.34、及び図7H:サンプルNo.47)。また、TEF、及びHLFは、D2への結合が認められた(図7F:サンプルNo.39、図7G:サンプルNo.43)。
これらの結果より、DBP、TEF、HLF、及びE4BP4によるPPARγ6プロモーターの転写調節は、DBP、TEF、HLF、及びE4BP4が、直接PPARγ6プロモーターに存在するD−ボックスに結合して行われている可能性が示唆された。
【0125】
(実施例7:脂肪細胞分化過程におけるDBP、TEF、及びE4BP4の役割)
脂肪細胞分化過程におけるDBP、TEF、及びE4BP4の役割について、下記方法により検討を行った。
【0126】
<oil red O染色による検討>
−方法−
−−miRNA配列の合成−−
Invitrogen社より提供されたRNAi配列情報を元に、下記配列番号:4、5、16、17、28、及び29で表される2本鎖miRNA(マイクロRNA)配列を合成した。
DBPに対するmiRNAは、下記配列番号:4で表される塩基配列、及び下記配列番号:5で表される塩基配列である。
TEFに対するmiRNAは、下記配列番号:16で表される塩基配列、及び下記配列番号:17で表される塩基配列である。
E4BP4に対するmiRNAは、下記配列番号:28で表される塩基配列、及び下記配列番号:29で表される塩基配列である。
前記miRNA(配列番号:4、5、16、17、28、及び29)をアニーリングした後、Block−itベクター(Invitrogen社製)に導入した。
AAGAGGAGCTGCAAGAGCCGGGTTTTGGCCACTGACTGACCCGGCTCTCAGCTCCTCTT (配列番号:4)
AAGAGGAGCTGAGAGCCGGGTCAGTCAGTGGCCAAAACCCGGCTCTTGCAGCTCCTCTT (配列番号:5)
TTTAGCTGCCACGTTGTTCTTGTTTTGGCCACTGACTGACAAGAACAACGTCAGCTAAA (配列番号:16)
TTTAGCTGACGTTGTTCTTGTCAGTCAGTGGCCAAAACAAGAACAACGTGGCAGCTAAA (配列番号:17)
ATCACAGGGAACTTGTTCTCAGTTTTGGCCACTGACTGACTGAGAACATTCCCTGTGAT (配列番号:28)
ATCACAGGGAATGTTCTCAGTCAGTCAGTGGCCAAAACTGAGAACAAGTTCCCTGTGAT (配列番号:29)
【0127】
−−細胞の培養−−
3T3−L1細胞(細胞株バンクATCC)を、24ウエルプレートに5×104細胞/プレート播種し、脂肪前駆細胞培養用培地(DMEM+10%CS)で、37℃、5%CO2の条件下で70%コンフルエントになるまで培養した。
【0128】
−−トランスフェクション−−
前記DBP、TEF、及びE4BP4のmiRNAの配列を有するベクター(pcDNA 6.2−GW/EmGFP−miR、Invitrogen社製)0.8μgを、リポフェクタミン2000(Invitrogen社製)を用いてトランスフェクションした。前記pcDNA 6.2−GW/EmGFP−miRは、ブラストシジン遺伝子配列を有することから、ブラストシジン(Invitrogen社製)5μg/mLによって導入細胞のセレクションを行なった。この細胞が、85%コンフルエントになった段階で、インスリン(ナカライテスク(株)製)10μg/mL、デキサメタゾン(ナカライテスク(株)製)0.25μM、及びIBMX(ナカライテスク(株)製)0.5mMにより分化誘導をかけ、分化誘導後7日目にoil red Oを用いた染色を行った。
【0129】
−結果−
結果を図8に示す。図8中、スケールバーは50μmである。
脂肪細胞への分化の評価は、脂肪滴数を指標に行った。DBP、及びTEFのmiRNAを、単独あるいは両者を導入した細胞では脂肪滴数の減少が認められ、脂肪細胞への分化が阻害された。一方、E4BP4 miRNAを導入した細胞では脂肪滴が増加する傾向が認められた。
【0130】
<定量PCRによる検討>
−方法−
−−細胞−−
3T3−L1細胞(ATCC社製)を、24ウエルプレートに5×104細胞/プレート播種し、脂肪前駆細胞培養用培地で、37℃、5%CO2の条件下で70%コンフルエントになるまで培養した。
【0131】
−−トランスフェクション−−
前記DBP、及びE4BP4 発現ベクター(Invitrogen社製)を、リポフェクタミン2000(Invitrogen社製)を用いてトランスフェクションし、ブラストシジン(Invitrogen社製)5μg/mLによってセレクションを行なった。この細胞が、85%コンフルエントになった段階で、インスリン(ナカライテスク(株)製)10μg/mL、デキサメタゾン(ナカライテスク(株)製)0.25μM、及びIBMX(ナカライテスク(株)製)0.5mMにより分化誘導をかけ、分化誘導後7日目にRNA抽出を行った。
【0132】
−−mRNAの抽出、及び精製−−
前記脂肪細胞の分化の検討を行った3T3−L1細胞より、トライゾール(Invitrogen社製)を用いてRNA抽出、及び精製を行い、DBPのmRNA量、及びE4BP4のmRNA量をqPCR(50℃2分間を1サイクル、95℃10分間を1サイクル、及び95℃15秒間、及び60℃1分間を40サイクル)により測定した。
【0133】
結果を図9A〜Bに示す。
DBPを発現させ、脂肪細胞へ分化した3T3−L1細胞は、分化に伴い、DBPの発現量が増加した(図9A)。
一方、E4BP4を発現させ、脂肪細胞へ分化した3T3−L1細胞は、分化に伴い、DBPの発現量が減少した(図9B)。
【0134】
実施例7の結果、DBP、TEF、及びE4BP4の発現は、脂肪分化と関連のあることが示唆された。その作用としては、DBP、及びTEFが、脂肪分化を促進し、E4BP4は、脂肪分化を抑制するものと考えられる。
【産業上の利用可能性】
【0135】
本発明のPPARγ発現調節剤は、PPARγの発現調節作用を有することから、脂肪細胞への分化の調節、脂質代謝の調節、破骨細胞機能の調節、炎症の調節などに好適に利用可能である。
また、本発明の脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤は、DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BPをコードするcDNAの少なくともいずれかの阻害作用を有することから、医薬、健康食品、機能性食品などに好適に利用可能である。
更に、本発明のスクリーニング方法は、PPARγの発現調節作用を有する因子を効率よくスクリーニングできることから、PPARγ発現調節剤のスクリーニングに好適に利用可能である。
【技術分野】
【0001】
本発明は、新規PPARγプロモーター配列、並びに、PPARγ発現調節剤、脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤、及びスクリーニング方法に関する。
【背景技術】
【0002】
ライフスタイルの欧米化に伴い、糖尿病、高血圧、動脈硬化、脂質異常症などいわゆる生活習慣病が増加しており、その主要因としてメタボリックシンドロームがある。このメタボリックシンドロームは、内臓組織の脂肪細胞の増殖や脂肪の取り込みの増加などにより発症し、過剰な食事量による脂肪細胞への脂肪酸の取り込みや、それによる脂肪細胞の増殖や分化とも密接に関連している。したがって、これらを制御する因子を解明することが、生活習慣病の予防や治療に直接結びつくものと考えられる。
しかしながら、脂肪の摂取と、脂質代謝とを直接結びつける因子や、脂肪細胞の分化に関わる因子などは十分に解明されておらず、これらの分子機構の解明が強く求められている。
【0003】
ペルオキシソーム増殖剤応答性受容体(PPAR:Peroxisome Proliferator−Activated Receptor)は、ステロイドホルモン受容体ファミリーの1種であり、哺乳動物においてPPARα、PPARδ、PPARγの3種類のサブタイプが存在することが知られている。
PPARは、別の転写調節因子であるRXR(レチノイドX受容体)とヘテロダイマーを形成し、標的遺伝子の上流に存在する応答配列PPRE(PPAR response element)に結合し、標的遺伝子の転写を制御している。前記標的遺伝子としては、内分泌代謝、糖代謝、脂質代謝、マクロファージを介する炎症反応、骨代謝の制御、発ガン機構などの遺伝子が知られている。そのため、PPARは、これらの疾患の治療薬のターゲット分子として注目されている。
【0004】
これらの中でも、PPARγは、そのプロモーターに、数種のスプライスバリアントが存在することが知られており(非特許文献1参照)、数種のアイソフォームが存在する(非特許文献2〜3参照)。例えば、PPARγ1は、脂肪組織や免疫系臓器、副腎、小腸で発現し、PPARγ2は、脂肪細胞で特異的な発現が認められ、脂肪細胞の分化誘導や脂肪合成に重要な役割を担っていると考えられている(非特許文献4参照)。また、PPARγは、マウスの破骨細胞を制御するという報告(非特許文献5参照)や、炎症疾患の調節に有用であるという報告(非特許文献6参照)もある。
【0005】
従来、PPARγの作用機序に大きく関わる因子としては、PPARγのリガンドが報告されている(非特許文献7参照)。
しかしながら、PPARγの発現調節は多面的に制御されているため非常に複雑であり、PPARγの発現を直接調節することにより、脂肪細胞の増殖や分化を促進、停止、後退させる因子、破骨細胞機能を調節する因子、炎症を調節する因子などは未だ提供されておらず、PPARγを直接調節することが可能なPPARγ発現調節剤、PPARγの調節を介した、脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤の提供が強く望まれているのが現状である。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Fajas L, et al, J Biol Chem, 1997, 272, p.18779−18789
【非特許文献2】Fajas L, et al, FEBS Lett, 1998, 438, p.55−60
【非特許文献3】Sundvold H, Lien S, Biochem Biophys Res Commun, 2001, 28, p.383−90
【非特許文献4】Rosen ED, Walkey CJ, Puigserver P, Spiegelman BM, Genes Dev, 2000, 14, p.1293−307
【非特許文献5】Wan Y, Chong LW, Evans RM, Nature Medicine, 2007, 13, p.1496 − 1503
【非特許文献6】Ricote M, Li AC, Willson TM, Kelly CJ, Glass CK, Nature, 1998, 391, p.79−82
【非特許文献7】Rosen ED, Spiegelman BM, Nature, 2006, 444, p.847−853
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明は、従来における前記諸問題を解決し、以下の目的を達成することを課題とする。即ち、本発明は、PPARγの発現を直接調節することが可能な因子を含有する優れたPPARγ発現調節剤、並びに、前記因子を阻害することにより脂肪細胞の分化、脂質代謝の調節、破骨細胞機能の調節、及び炎症作用の調節が可能な、優れた脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤、及び前記PPARγ発現調節剤のスクリーニング方法を提供することを目的とする。
【課題を解決するための手段】
【0008】
前記課題を解決するため、本発明者らは、脂肪分化、脂質代謝、破骨細胞調節、炎症疾患などに関与するPPARγの発現調節に着目し、PPARγの発現調節に関与する因子について鋭意検討した結果、PPARγの発現制御に係る5’非翻訳領域(プロモーター領域)に新規スプライスバリアントが存在することを知見するとともに、その新規プロモーター配列を見出し、該プロモーター領域のプロモーターレポーターシステムを樹立した。また、PPARγの発現を直接調節することが可能なPPARγ発現調節剤のスクリーニング方法を確立し、該スクリーニング方法により、DBP、及びTEFがPPARγの遺伝子発現を促進すること、HLF、及びE4BP4がPPARγの発現を抑制すること、更に、DBP、TEF、HLF、及びE4BP4がPPARγのプロモーター領域に存在するD−ボックスに結合すること、DBP、及びTEFを阻害すると脂肪細胞への分化が阻害され、更にDBPの遺伝子発現は脂肪細胞の分化に伴い増加すること、E4BP4を阻害すると脂肪細胞への分化が促進し、更にE4PB4の遺伝子発現は脂肪細胞の分化に伴い減少することを知見し、本発明の完成に至った。なお、本発明者らは、前記5’非翻訳領域(プロモーター領域)に新規スプライスバリアントを有するPPARγを、「PPARγ6」と命名した。
【0009】
本発明は、本発明者らによる前記知見に基づくものであり、前記課題を解決するための手段としては、以下の通りである。即ち、
<1> 配列番号:1で表される塩基配列を有することを特徴とするPPARγプロモーター配列である。
<2> 配列番号:1で表される塩基配列のうち、塩基番号159〜166、塩基番号202〜209、及び塩基番号218〜225を少なくとも有する前記<1>に記載のPPARγプロモーター配列である。
<3> DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかを含有することを特徴とするPPARγ発現調節剤である。
<4> PPARγの発現促進作用を有する前記<3>に記載のPPARγ発現調節剤である。
<5> PPARγの発現促進作用を有する因子が、DBP、及びTEF、並びに、該DBP、及び該TEFをコードするcDNAの少なくともいずれかである前記<4>に記載のPPARγ発現調節剤である。
<6> PPARγの発現抑制作用を有する前記<3>に記載のPPARγ発現調節剤である。
<7> PPARγの発現抑制作用を有する因子が、HLF、及びE4BP4、並びに、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかである前記<6>に記載のPPARγ発現調節剤である。
<8> PPARγの発現調節作用が、PPARγプロモーター配列に結合することによる前記<3>から<7>のいずれかに記載のPPARγ発現調節剤である。
<9> PPARγプロモーター配列が、配列番号:1で表される塩基配列を有する前記<8>に記載のPPARγ発現調節剤である。
<10> PPARγの発現調節作用が、配列番号:1で表される塩基配列中のD−ボックスに結合することによる前記<3>から<9>のいずれかに記載のPPARγ発現調節剤である。
<11> DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかを阻害することを特徴とする脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤である。
<12> 阻害が、miRNAによる前記<11>に記載の脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤である。
<13> miRNAの配列が、配列番号:2〜33で表される塩基配列の少なくともいずれかを有する前記<12>に記載の脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤である。
<14> PPARγプロモーター配列を有するレポーター遺伝子を構築するプロモーターレポーター遺伝子構築工程と、該プロモーターレポーター遺伝子と、被験物質を含む被験試料とを細胞に導入する導入工程と、該プロモーターレポーター遺伝子のレポーター活性を指標にして、PPARγプロモーター活性を検出する検出工程と、該被験試料が、PPARγプロモーター活性の促進作用、及び抑制作用のいずれかの作用を有しているか否かを判断し、PPARγプロモーター活性の促進作用、及び抑制作用のいずれかの作用を有する該被験試料中の被験物質をPPARγ発現調節剤としてスクリーニングするスクリーニング工程と、を含むことを特徴とするPPARγ発現調節剤のスクリーニング方法である。
<15> PPARγプロモーター配列が、配列番号:1で表される塩基配列を有する前記<14>に記載のPPARγ発現調節剤のスクリーニング方法である。
<16> プロモーターレポーター遺伝子のPPARγプロモーター配列が、該PPARγプロモーター配列が有する3箇所のD−ボックスの少なくともいずれかを欠失した欠失変異体である前記<14>から<15>のいずれかに記載のPPARγ発現調節剤のスクリーニング方法である。
【発明の効果】
【0010】
本発明によれば、従来における前記諸問題を解決し、前記目的を達成することができ、
PPARγの発現を直接調節することが可能な因子を含有する優れたPPARγ発現調節剤、並びに、前記因子を阻害することにより脂肪細胞の分化、脂質代謝の調節、破骨細胞機能の調節、及び炎症作用の調節が可能な、優れた脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤、及び前記PPARγ発現調節剤のスクリーニング方法を提供することができる。
【図面の簡単な説明】
【0011】
【図1A】図1Aは、ヒトPPARγ遺伝子のエクソン・イントロン構造を模式的に表した図である。
【図1B】図1Bは、ヒトPPARγ遺伝子の6種のスプライスバリアントを模式的に表した図である。
【図2】図2は、PPARγ6のプロモーター構造、及びD−ボックスを模式的に表した図である。
【図3】図3は、DBP発現ベクターを一過性に発現させたCaco−2細胞における、DBP、及びPPARγ6のmRNA量を表した図である。
【図4A】図4Aは、DBPのPPARγ6プロモーターへの作用を、PPARγ6プロモーターレポーター遺伝子を用いたルシフェラーゼアッセイで解析した図である。
【図4B】図4Bは、TEFのPPARγ6プロモーターへの作用を、PPARγ6プロモーターレポーター遺伝子を用いたルシフェラーゼアッセイで解析した図である。
【図4C】図4Cは、HLFのPPARγ6プロモーターへの作用を、PPARγ6プロモーターレポーター遺伝子を用いたルシフェラーゼアッセイで解析した図である。
【図4D】図4Dは、E4BP4のPPARγ6プロモーターへの作用を、PPARγ6のプロモーターレポーター遺伝子を用いたルシフェラーゼアッセイで解析した図である。
【図5A】図5Aは、PPARγ6プロモーターの欠失変異体のコンストラクトを模式的に表した図である。
【図5B】図5Bは、DBPのPPARγ6のプロモーターへの作用を、PPARγ6プロモーターの欠失変異体のレポーター遺伝子を用いて、ルシフェラーゼアッセイで解析した図である。
【図5C】図5Cは、DBPのPPARγ6のプロモーターへの作用を、PPARγ6のプロモーター欠失変異体のレポーター遺伝子を用いて、ルシフェラーゼアッセイで解析した図である。
【図6A】図6Aは、PPARγ6プロモーターのD−ボックス欠失変異体のコンストラクトを模式的に表した図である。
【図6B】図6Bは、DBPのPPARγ6プロモーターへの作用を、PPARγ6プロモーターのD−ボックス欠失変異体のレポーター遺伝子を用いて、ルシフェラーゼアッセイで解析した図である。
【図6C】図6Cは、E4BP4のPPARγ6プロモーターへの作用を、PPARγ6プロモーターのD−ボックス欠失変異体のレポーター遺伝子を用いて、ルシフェラーゼアッセイで解析した図である。
【図7A】図7Aは、DBPに対するプローブ特異性をゲルシフトアッセイにより調べた図である。
【図7B】図7Bは、TEFに対するプローブ特異性をゲルシフトアッセイにより調べた図である。
【図7C】図7Cは、HLFに対するプローブ特異性をゲルシフトアッセイにより調べた図である。
【図7D】図7Dは、E4BP4に対するプローブ特異性をゲルシフトアッセイにより調べた図である。
【図7E】図7Eは、DBPの、PPARγ6プロモーターに存在するD−ボックスへの結合能をゲルシフトアッセイで解析した図である。
【図7F】図7Fは、TEFの、PPARγ6プロモーターに存在するD−ボックスへの結合能をゲルシフトアッセイで解析した図である。
【図7G】図7Gは、HLFの、PPARγ6プロモーターに存在するD−ボックスへの結合能をゲルシフトアッセイで解析した図である。
【図7H】図7Hは、E4BP4の、PPARγ6プロモーターに存在するD−ボックスへの結合能をゲルシフトアッセイで解析した図である。
【図8】図8は、DBP、TEF、及びE4BP4の脂肪細胞分化への影響を、RNAiで解析した図である。
【図9A】図9Aは、脂肪細胞分化におけるDBPのmRNAの発現量の変化を表した図である。
【図9B】図9Bは、脂肪細胞分化におけるE4BP4のmRNAの発現量の変化を表した図である。
【発明を実施するための形態】
【0012】
(新規PPARγプロモーター配列)
本発明の新規PPARγプロモーター配列は、配列番号:1で表される塩基配列を有する。
前記新規PPARγプロモーター配列は、5’非翻訳領域が、既知のPPARγの配列と異なる、本発明者らが見出したPPARγの新規スプライスバリアント(図1B参照)の発現を制御するプロモーター配列である。
前記新規PPARγプロモーター配列は、配列番号:1で表される塩基配列のみからなる塩基配列であることが好ましいが、そのプロモーター活性を有する限りは、特に制限はなく、目的に応じて適宜選択することができ、配列番号:1で表される塩基配列の一部からなる塩基配列であってもよく、配列番号:1で表される塩基配列に、更にその他の塩基配列が付加された塩基配列であってもよい。
前記新規PPARγプロモーター配列が、配列番号:1で表される塩基配列の一部からなる場合、その一部の塩基配列としては、PPARγのプロモーター活性を有する限りは、特に制限はなく、目的に応じて適宜選択することができるが、配列番号:1で表される塩基配列の、塩基番号159〜166、塩基番号202〜209、及び塩基番号218〜225を有することが好ましい。
前記その他の塩基配列としても、PPARγのプロモーター活性を有する限りは、特に制限はなく、目的に応じて適宜選択することができる。
また、前記新規PPARγプロモーター配列は、配列番号:1で表される塩基配列に、1若しくは数個の塩基が、欠失、置換、挿入された塩基配列からなるものであってもよい。
【0013】
<新規スプライスバリアント>
ヒトPPARγの遺伝子配列は、図1Aに示すように、10個のエクソン(A1、C1C2、A2、B、E1、E2、E3、E4、E5、及びE6)を有する。このうち、A1、C1C2、A2、及びBは、非翻訳領域であり、前記E1〜E6が翻訳されることによりPPARγのタンパク質を形成する。
前記ヒトPPARγの発現調節は、そのプロモーターにより制御されているが、前記ヒトPPARγの5’非翻訳領域に存在するプロモーターには、従来、5種のスプライスバリアント(PPARγ1〜5)が存在することが知られていた(図1B参照)。前記5’非翻訳領域には、4つのエクソン(A1、C1C2、A2、及びB)が存在し、これらの4つのエクソンが選択的スプライシングを受けることにより、スプライスバリアントが形成される(図1A、及び図1B参照)。
既知のスプライスバリアントとしては、図1Bに示すように、5’非翻訳領域がA1、及びA2からなるPPARγ1、5’非翻訳領域がBからなるPPARγ2、5’非翻訳領域がA2からなるPPARγ3、5’非翻訳領域にいずれのエクソンも有さないPPARγ4、及び5’非翻訳領域がC1C2からなるPPARγ5が存在する。
本発明者らは、これらの既知のスプライスバリアントとは全く異なる、5’非翻訳領域がC1C2、及びA2からなるヒトPPARγが存在することを見出し(図1B参照)、「PPARγ6」と命名した(以下、「PPARγ6」と称することがある)。
【0014】
前記PPARγ6が、新規スプライスバリアントを有するか否かを解析する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、エクソンC1C2からエクソンE1にかけてのPCR産物のサイズ、及び配列決定などにより解析する方法などが挙げられる。
【0015】
前記PPARγ6のプロモーター配列が、DNA結合配列を有するか否かを解析する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、MatInspector(Genomatix社製)により解析する方法などが挙げられる。
【0016】
前記MatInspector(Genomatix社製)による解析の結果、前記C1C2は、後述するDBP(Albumin gene D−site Binding Protein:D部位結合タンパク質)の結合配列であるD−ボックスが3箇所(D1、D2、及びD3)存在していた(図2参照)。
マウス及びヒトの、プロモーター領域及びエクソン1領域の配列についてDNA解析ソフトGeneWorksでアライメントを行い、前記領域の塩基配列に相同性が高いことを確認した。これらの解析結果より、前記D−ボックスは、種間で極めてよく保存されていることが明らかとなった。
【0017】
(PPARγ発現調節剤)
本発明のPPARγ発現調節剤は、DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかを含有し、必要に応じて、更にその他の成分を含有する。
【0018】
<DBP、TEF、HLF、及びE4BP4>
−DBP、TEF、及びHLF−
前記DBP、前記TEF(Thyrotroph Embryonic Factor:甲状腺刺激ホルモン分泌細胞胎児性因子)、及び前記HLF(Hepatic Leukemia Factor:肝白血病要因遺伝子)は、PAR−bZIP(proline and acidic amino acid−rich basic leucine zipper)転写因子ファミリーに属するタンパク質である(Wuarin J et al, J Cell Sci Suppl, 1992, 6, p.123−127)。前記PAR−bZIP転写因子ファミリーは、ホモあるいはヘテロダイマーを形成して、プロモーター領域にあるD−ボックス(RTTAYGTAAY)に結合し、転写活性化に働くことが知られている(Khatib ZA et al, Genomics, 1994, 123, p.344−51)。前記PAR−bZIP転写因子ファミリーは、哺乳類の概日リズムを制御する因子として知られている(Mitsui S et al, Genes Dev, 2001, 15, p.995−1006)が、脂肪細胞の分化などを含め、他の機能に関しては全く報告がない。
【0019】
−E4BP4−
前記E4BP4(アデノウイルスE4プロモーター結合蛋白4)は、bZIP転写因子ファミリーに属するタンパク質である。前記bZIPファミリーは、前記PAR−bZIP転写因子ファミリーと同様に、プロモーター領域にあるD−ボックスに結合し、転写抑制化に働くことが知られている(Mitsui S, Yamaguchi S, Matsuo T, Ishida Y, Okamura H, Genes Dev, 2001, 15, p.995−1006)。前記E4BP4も、前記PAR−bZIP転写因子ファミリーと同様に、哺乳類の概日リズムを制御する因子として知られている(Mitsui S, Yamaguchi S, Matsuo T, Ishida Y, Okamura H, Genes Dev, 2001, 15, p.995−1006)が、脂肪分化における作用などを含め、他の機能に関しては全く報告がない。
【0020】
−入手方法−
前記PPARγ発現調節剤中に含まれる、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの入手方法としては、特に制限はなく、目的に応じて適宜選択することができ、前記DBP、TEF、HLF、及びE4BP4がタンパク質の状態である場合、例えば、市販品を用いる方法、培養細胞より精製する方法、遺伝子組換え技術により組換体タンパク質を得る方法、無細胞系を用いた真核生物由来タンパク質合成の方法などが挙げられるが、これらの中でも無細胞系を用いた真核生物由来タンパク質合成の方法により得ることが、可溶性、及び機能タンパク質を迅速に合成できる点で好ましい。
前記DBP、TEF、HLF、及びE4BP4がcDNAの状態である場合、例えば、市販品を用いる方法、cDNAを合成する方法、cDNAライブラリーからクローニングする方法などが挙げられるが、これらの中でもcDNAを合成する方法により得ることが、迅速に取得できる点で好ましい。
【0021】
−含有量−
前記PPARγ発現調節剤中の、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかの含有量としては、特に制限はなく、目的に応じて適宜選択することができる。また、前記PPARγ発現調節剤は、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかそのものであってもよい。
前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかの含有比率としても特に制限はなく、目的に応じて適宜選択することができるが、PPARγの発現を促進する目的で用いられる場合と、PPARγの発現を抑制する目的で用いられる場合とで、前記含有比率を変えることが好ましい。
【0022】
−−PPARγの発現促進作用を有するPPARγ発現調節剤−−
前記PPARγ発現調節剤が、PPARγの発現を促進する目的で用いられる場合、前記PPARγ発現調節剤中の、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかの含有比率としては、前記DBP、及びTEF、並びに、該DBP、及び該TEFをコードするcDNAの少なくともいずれかが、前記HLF、及びE4BP4、並びに、該HLF、及び該E4BP4をコードするcDNAより多く含有されていることが好ましく、前記DBP、及びTEF、並びに、該DBP、及び該TEFをコードするcDNAの少なくともいずれかのみ含有されていることがより好ましい。
前記PPARγ発現調節剤が、前記DBP、及びTEF、並びに、該DBP、及び該TEFをコードするcDNAの少なくともいずれかであると、前記PPARγ発現調節剤によるPPARγの発現促進作用が強い点で有利である。
なお、PPARγの発現促進作用を有するPPARγ発現調節剤を、PPARγ発現促進剤として使用することができる。
【0023】
−−PPARγの発現抑制作用を有するPPARγ発現調節剤−−
前記PPARγ発現調節剤が、PPARγの発現を抑制する目的で用いられる場合、前記PPARγ発現調節剤中の、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかの含有比率としては、前記HLF、及びE4BP4、並びに、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかが、前記DBP、及びTEF、並びに、該DBP、及び該TEFをコードするcDNAより多く含有されていることが好ましく、前記HLF、及びE4BP4、並びに、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかのみ含有されていることがより好ましい。
前記PPARγ発現調節剤が、前記HLF、及びE4BP4、並びに、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかであると、前記PPARγ発現調節剤によるPPARγの発現抑制作用が強い点で有利である。
なお、PPARγの発現抑制作用を有するPPARγ発現調節剤を、PPARγ発現抑制剤として使用することができる。
【0024】
<PPARγの発現調節作用>
前記PPARγ発現調節剤が、PPARγの発現を調節する方法としては、特に制限はなく、目的に応じて適宜選択することができるが、前記PPARγ発現調節剤が、PPARγのプロモーター配列に結合することによりPPARγの発現を調節することが好ましい。
前記PPARγ発現調節剤が結合するプロモーター配列としては、特に制限はなく、目的に応じて適宜選択することができるが、前記PPARγ6のプロモーター配列であることが好ましく、前記PPARγ6のプロモーター配列に存在する3箇所のD−ボックス(D1、D2、及びD3)の少なくともいずれかに結合することがより好ましく、前記D2に結合することが更に好ましく、前記3箇所のD−ボックス全てに結合することが特に好ましい。
前記3箇所のD−ボックス全てに結合すると、PPARγの発現を顕著に促進又は抑制できる点で好ましい。
前記結合を確認する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、ゲルシフトアッセイ、チップアッセイ(ChIP assay)などが挙げられる。
【0025】
<その他の成分>
前記その他の成分としては、特に制限はなく、薬理学的に許容される担体の中から目的に応じて適宜選択することができ、例えば、エタノール、水、デンプンなどが挙げられる。
前記PPARγ発現調節剤中の、その他の成分の含有量としては、特に制限はなく、目的に応じて適宜選択することができる。
【0026】
<使用>
前記PPARγ発現調節剤は、1種単独で使用されてもよいし、他の成分を有効成分とする医薬と併用してもよい。また、前記PPARγ発現調節剤は、他の成分を有効成分とする医薬中に、配合された状態で使用されてもよい。
【0027】
<剤型>
前記PPARγ発現調節剤の剤型としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、経口固形剤、経口液剤、注射剤、吸入散剤などが挙げられる。
【0028】
−経口固形剤−
前記経口固形剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、錠剤、被覆錠剤、顆粒剤、散剤、カプセル剤などが挙げられる。
前記経口固形剤の製造方法としては、特に制限はなく、常法を使用することができ、例えば、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかに、賦形剤、及び必要に応じて前記その他の成分、各種添加剤を加えることにより、製造することができる。ここで、前記賦形剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、乳糖、白糖、塩化ナトリウム、ブドウ糖、デンプン、炭酸カルシウム、カオリン、微結晶セルロース、珪酸などが挙げられる。また、前記添加剤としても、特に制限はなく、目的に応じて適宜選択することができ、例えば、結合剤、崩壊剤、滑沢剤、着色剤、矯味/矯臭剤などが挙げられる。
前記結合剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、水、エタノール、プロパノール、単シロップ、ブドウ糖液、デンプン液、ゼラチン液、カルボキシメチルセルロース、ヒドロキシプロピルセルロース、ヒドロキシプロピルスターチ、メチルセルロース、エチルセルロース、シェラック、リン酸カルシウム、ポリビニルピロリドンなどが挙げられる。
前記崩壊剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、乾燥デンプン、アルギン酸ナトリウム、カンテン末、炭酸水素ナトリウム、炭酸カルシウム、ラウリル硫酸ナトリウム、ステアリン酸モノグリセリド、乳糖などが挙げられる。
前記滑沢剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、精製タルク、ステアリン酸塩、ホウ砂、ポリエチレングリコールなどが挙げられる。
前記着色剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、酸化チタン、酸化鉄などが挙げられる。
前記矯味/矯臭剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、白糖、橙皮、クエン酸、酒石酸などが挙げられる。
【0029】
−経口液剤−
前記経口液剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、内服液剤、シロップ剤、エリキシル剤などが挙げられる。
前記経口液剤の製造方法としては、特に制限はなく、常法を使用することができ、例えば、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれか、及び必要に応じて前記その他の成分に、添加剤を加えることにより、製造することができる。ここで、前記添加剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、矯味/矯臭剤、緩衝剤、安定化剤などが挙げられる。
前記矯味/矯臭剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、白糖、橙皮、クエン酸、酒石酸などが挙げられる。
前記緩衝剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、クエン酸ナトリウムなどが挙げられる。
前記安定化剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、トラガント、アラビアゴム、ゼラチンなどが挙げられる。
【0030】
−注射剤−
前記注射剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、溶液、懸濁液、用事溶解用固形剤などが挙げられる。
前記注射剤の製造方法としては、特に制限はなく、常法を使用することができ、例えば、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれか、及び必要に応じて前記その他の成分に、pH調節剤、緩衝剤、安定化剤、等張化剤、局所麻酔剤などを添加することにより、製造することができる。ここで、前記pH調節剤及び前記緩衝剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、クエン酸ナトリウム、酢酸ナトリウム、リン酸ナトリウムなどが挙げられる。また、前記安定化剤としても、特に制限はなく、目的に応じて適宜選択することができ、例えば、ピロ亜硫酸ナトリウム、EDTA、チオグリコール酸、チオ乳酸などが挙げられる。前記等張化剤としても、特に制限はなく、目的に応じて適宜選択することができ、例えば、塩化ナトリウム、ブドウ糖などが挙げられる。前記局所麻酔剤としても、特に制限はなく、目的に応じて適宜選択することができ、例えば、塩酸プロカイン、塩酸リドカインなどが挙げられる。
【0031】
<投与>
前記PPARγ発現調節剤の投与方法、投与量、投与時期、及び投与対象としては、特に制限はなく、目的に応じて適宜選択することができる。
前記投与方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、経口投与法、注射による方法、吸入による方法などが挙げられる。
前記投与量としては、特に制限はなく、投与対象個体の年齢、体重、体質、症状、他の成分を有効成分とする医薬の投与の有無など、様々な要因を考慮して適宜選択することができる。
前記投与対象となる動物種としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、ヒト、サル、ブタ、ウシ、ヒツジ、ヤギ、イヌ、ネコ、マウス、ラット、トリなどが挙げられるが、これらの中でもヒトに好適に用いられる。
【0032】
<用途>
前記PPARγ発現調節剤は、PPARγの発現調節作用を有することから、脂肪細胞への分化の調節、脂質代謝の調節、破骨細胞機能の調節、炎症の調節などに好適に利用可能である。
【0033】
(脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤)
本発明の脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤は、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかを阻害する因子を含有し、必要に応じて、更にその他の成分を含有する。
前記脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤は、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかを阻害することにより、PPARγの発現を促進又は抑制し、脂肪分化調節作用、脂質代謝調節作用、破骨細胞機能調節作用、及び炎症調節作用を発揮することが好ましい。
【0034】
<阻害する因子>
前記阻害する因子としては、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかを阻害することができれば、特に制限はなく、目的に応じて適宜選択することができ、例えば、miRNA(micro RNA)、siRNA(smoll interfering RNA)、shRNA(short hairpin RNA)などが挙げられるが、これらの中でも、miRNAが、完全に相補的でない場合でも翻訳阻害を引き起こすことができる点、及びRNAi(RNA干渉)として利用できる配列の範囲が広い点で好ましい。
本発明において、前記miRNAは、本発明の効果を損なわない限り、特に制限はなく、目的に応じて適宜選択することができ、DNAに置換されているものであってもよく、RNAとDNAとのキメラであってもよく、前記miRNAを発現し得る分子であってもよい。
前記miRNAを発現し得る分子としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、前記miRNAをDNAに置換した塩基配列を有するベクターなどが挙げられる。
【0035】
−miRNA−
前記miRNAとしては、特に制限はなく、目的に応じて適宜選択することができるが、前記DBP、TEF、HLF、及びE4BP4の少なくともいずれかの遺伝子発現を抑制する塩基配列を有することが好ましい。
前記DBP、TEF、HLF、及びE4BP4の少なくともいずれかの遺伝子発現を抑制する塩基配列としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、下記配列番号:2〜33で表される塩基配列などが挙げられる。
前記阻害する因子中のmiRNAは、配列番号:2〜33で表される塩基配列全てを有していてもよいが、発現を阻害する遺伝子の種類によって、下記塩基配列を選択することが好ましい。
また、前記DBP、TEF、HLF、及びE4BP4の少なくともいずれかの遺伝子発現を抑制する塩基配列の由来する生物種としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、マウス、ヒトなどが挙げられるが、これらの中でも、ヒトの配列であることが、前記脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤をヒトに用いる場合、抗原性を有さない点で好ましい。
【0036】
前記DBPの遺伝子発現を抑制するマウス由来のmiRNA配列としては、例えば、下記配列番号:2〜5で表される塩基配列の少なくともいずれかを有することが好ましい。
AAGAGGAGCUGCAAGAGCCGGGUUUUGGCCACUGACUGACCCGGCUCUCAGCUCCUCUU (配列番号:2)
AAGAGGAGCUGAGAGCCGGGUCAGUCAGUGGCCAAAACCCGGCUCUUGCAGCUCCUCUU (配列番号:3)
AAGAGGAGCTGCAAGAGCCGGGTTTTGGCCACTGACTGACCCGGCTCTCAGCTCCTCTT (配列番号:4)
AAGAGGAGCTGAGAGCCGGGTCAGTCAGTGGCCAAAACCCGGCTCTTGCAGCTCCTCTT (配列番号:5)
【0037】
前記DBPの遺伝子発現を抑制するヒト由来のmiRNA配列としては、例えば、下記配列番号:6〜9で表される塩基配列の少なくともいずれかを有することが好ましい。
AAGCCGAGCCGCACGAACCCGGUUUUGGCCACUGACUGACCGGGUUCGCGGCUCGGCUU (配列番号:6)
AAGCCGAGCCGCGAACCCGGUCAGUCAGUGGCCAAAACCGGGUUCGUGCGGCUCGGCUU (配列番号:7)
AAGCCGAGCCGCACGAACCCGGTTTTGGCCACTGACTGACCGGGTTCGCGGCTCGGCTT (配列番号:8)
AAGCCGAGCCGCGAACCCGGTCAGTCAGTGGCCAAAACCGGGTTCGTGCGGCTCGGCTT (配列番号:9)
【0038】
前記TEFの遺伝子発現を抑制するマウス由来のmiRNA配列としては、例えば、下記配列番号:10〜13で表される塩基配列の少なくともいずれかを有することが好ましい。
UUUAGCUGCCACAUUGUUCUUGUUUUGGCCACUGACUGACAAGAACAAUGGCAGCUAAA (配列番号:10)
UUUAGCUGCCAUUGUUCUUGUCAGUCAGUGGCCAAAACAAGAACAAUGUGGCAGCUAAA (配列番号:11)
TTTAGCTGCCACATTGTTCTTGTTTTGGCCACTGACTGACAAGAACAATGGCAGCTAAA (配列番号:12)
TTTAGCTGCCATTGTTCTTGTCAGTCAGTGGCCAAAACAAGAACAATGTGGCAGCTAAA (配列番号:13)
【0039】
前記TEFの遺伝子発現を抑制するヒト由来のmiRNA配列としては、例えば、下記配列番号:14〜17で表される塩基配列の少なくともいずれかを有することが好ましい。
UUUAGCUGCCACGUUGUUCUUGUUUUGGCCACUGACUGACAAGAACAACGUCAGCUAAA (配列番号:14)
UUUAGCUGACGUUGUUCUUGUCAGUCAGUGGCCAAAACAAGAACAACGUGGCAGCUAAA (配列番号:15)
TTTAGCTGCCACGTTGTTCTTGTTTTGGCCACTGACTGACAAGAACAACGTCAGCTAAA (配列番号:16)
TTTAGCTGACGTTGTTCTTGTCAGTCAGTGGCCAAAACAAGAACAACGTGGCAGCTAAA (配列番号:17)
【0040】
前記HLFの遺伝子発現を抑制するマウス由来のmiRNA配列としては、例えば、下記配列番号:18〜21で表される塩基配列の少なくともいずれかを有することが好ましい。
CAUCCUGAAGACGCAUUUAGUGUUUUGGCCACUGACUGACACUAAAUGCGUUCAGGAUG (配列番号:18)
CAUCCUGAACGCAUUUAGUGUCAGUCAGUGGCCAAAACACUAAAUGCGUCUUCAGGAUG (配列番号:19)
CATCCTGAAGACGCATTTAGTGTTTTGGCCACTGACTGACACTAAATGCGTTCAGGATG (配列番号:20)
CATCCTGAACGCATTTAGTGTCAGTCAGTGGCCAAAACACTAAATGCGTCTTCAGGATG (配列番号:21)
【0041】
前記HLFの遺伝子発現を抑制するヒト由来のmiRNA配列としては、例えば、下記配列番号:22〜25で表される塩基配列の少なくともいずれかを有することが好ましい。
ACUAAAUGCGUCUUCGUGGUGGUUUUGGCCACUGACUGACCACCACGAACGCAUUUAGU (配列番号:22)
ACUAAAUGCGUUCGUGGUGGUCAGUCAGUGGCCAAAACCACCACGAAGACGCAUUUAGU (配列番号:23)
ACTAAATGCGTCTTCGTGGTGGTTTTGGCCACTGACTGACCACCACGAACGCATTTAGT (配列番号:24)
ACTAAATGCGTTCGTGGTGGTCAGTCAGTGGCCAAAACCACCACGAAGACGCATTTAGT (配列番号:25)
【0042】
前記E4BP4の遺伝子発現を抑制するマウス由来のmiRNA配列としては、例えば、下記配列番号:26〜29で表される塩基配列の少なくともいずれかを有することが好ましい。
AUCACAGGGAACUUGUUCUCAGUUUUGGCCACUGACUGACUGAGAACAUUCCCUGUGAU (配列番号:26)
AUCACAGGGAAUGUUCUCAGUCAGUCAGUGGCCAAAACUGAGAACAAGUUCCCUGUGAU (配列番号:27)
ATCACAGGGAACTTGTTCTCAGTTTTGGCCACTGACTGACTGAGAACATTCCCTGTGAT (配列番号:28)
ATCACAGGGAATGTTCTCAGTCAGTCAGTGGCCAAAACTGAGAACAAGTTCCCTGTGAT (配列番号:29)
【0043】
前記E4BP4の遺伝子発現を抑制するヒト由来のmiRNA配列としては、例えば、下記配列番号:30〜33で表される塩基配列の少なくともいずれかを有することが好ましい。
AUAAUCUGGAACUUGUUUUCAGUUUUGGCCACUGACUGACUGAAAACAAGUCAGAUUAU (配列番号:30)
AUAAUCUGACUUGUUUUCAGUCAGUCAGUGGCCAAAACUGAAAACAAGUUCCAGAUUAU (配列番号:31)
ATAATCTGGAACTTGTTTTCAGTTTTGGCCACTGACTGACTGAAAACAAGTCAGATTAT (配列番号:32)
ATAATCTGACTTGTTTTCAGTCAGTCAGTGGCCAAAACTGAAAACAAGTTCCAGATTAT (配列番号:33)
【0044】
前記配列番号:2〜33で表される塩基配列のいずれかを有する前記miRNAは、前記配列番号:2〜33で表される塩基配列の全体からなる塩基配列であることが、発現阻害効率が高い点で好ましいが、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかを阻害することができれば、特に制限はなく、目的に応じて適宜選択することができ、例えば、前記配列番号:2〜33で表される塩基配列の一部からなる配列であってもよい。
前記miRNAが、前記配列番号:2〜33で表される塩基配列の一部からなる配列である場合、その一部の塩基配列としては、前記miRNAが、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかを阻害する能力を有する限り、特に制限はなく、目的に応じて適宜選択することができる。
また、前記miRNAは、前記配列番号:2〜33で表される塩基配列のいずれかの1若しくは数個の塩基が、欠失、置換、挿入された塩基配列からなるものであってもよい。
【0045】
−−入手方法−−
前記配列番号:2〜33で表される塩基配列の少なくともいずれかを有する前記miRNAを入手する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、合成による方法、市販品を用いる方法などが挙げられる。
【0046】
−−含有量−−
前記脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤中の、前記miRNAの含有量としては、特に制限はなく、目的に応じて適宜選択することができる。
前記miRNAが、前記配列番号:2〜33で表される塩基配列の少なくともいずれかである場合、その含有比率としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、前記DBP、及び該DBPをコードするcDNAを抑制する目的で用いられる場合、前記TEF、及び該TEFをコードするcDNAを抑制する目的で用いられる場合、前記HLF、及び該HLFをコードするcDNAを抑制する目的で用いられる場合、前記E4BP4、及び該E4BP4をコードするcDNAを抑制する目的で用いられる場合で、前記含有比率を変えることが好ましい。
【0047】
−−−DBP、及び該DBPをコードするcDNAの抑制−−−
前記DBP、及び該DBPをコードするcDNAを抑制する目的で用いられる場合、前記miRNAの含有比率としては、配列番号:2〜9で表される塩基配列の少なくともいずれかを有する前記miRNAが、配列番号:10〜33で表される塩基配列を有する前記miRNAより多いことが好ましく、配列番号:2〜9で表される塩基配列の少なくともいずれかのみを有する前記miRNAがより好ましく、配列番号:6〜9で表される塩基配列の少なくともいずれかのみを有する前記miRNAが特に好ましい。
前記脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤が、配列番号:2〜9で表される塩基配列の少なくともいずれかのみを有する前記miRNAである場合、配列番号:2〜9で表される塩基配列それぞれの含有比率としては、特に制限はなく、目的に応じて適宜選択することができる。
【0048】
−−−TEF、及び該TEFをコードするcDNAの抑制−−−
前記TEF、及び該TEFをコードするcDNAを抑制する目的で用いられる場合、前記miRNAの含有比率としては、配列番号:10〜17で表される塩基配列の少なくともいずれかを有するmiRNAが、配列番号:2〜9で表される塩基配列を有するmiRNA、及び配列番号:18〜33で表される塩基配列を有するmiRNAより多いことが好ましく、配列番号:10〜17で表される塩基配列の少なくともいずれかのみを有するmiRNAがより好ましく、配列番号:14〜17で表される塩基配列の少なくともいずれかのみを有するmiRNAが特に好ましい。
前記脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤が、配列番号:10〜17で表される塩基配列の少なくともいずれかのみを有するmiRNAである場合、配列番号:10〜17で表される塩基配列それぞれの含有比率としては、特に制限はなく、目的に応じて適宜選択することができる。
【0049】
−−−HLF、及び該HLFをコードするcDNAの抑制−−−
前記HLF、及び該HLFをコードするcDNAを抑制する目的で用いられる場合、前記miRNAの含有比率としては、配列番号:18〜25で表される塩基配列の少なくともいずれかを有するmiRNAが、配列番号:2〜17で表される塩基配列を有するmiRNA、及び配列番号:26〜33で表される塩基配列を有するmiRNAより多いことが好ましく、配列番号:18〜25で表される塩基配列の少なくともいずれかのみを有するmiRNAがより好ましく、配列番号:22〜25で表される塩基配列の少なくともいずれかのみを有するmiRNAが特に好ましい。
前記脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤が、配列番号:18〜25で表される塩基配列の少なくともいずれかのみを有するmiRNAである場合、配列番号:18〜25で表される塩基配列それぞれの含有比率としては、特に制限はなく、目的に応じて適宜選択することができる。
【0050】
−−−E4BP4、及び該E4BP4をコードするcDNAの抑制−−−
前記E4BP4、及び該E4BP4をコードするcDNAを抑制する目的で用いられる場合、前記miRNAの含有比率としては、配列番号:26〜33で表される塩基配列の少なくともいずれかを有するmiRNAが、配列番号:2〜25で表される塩基配列を有するmiRNAより多いことが好ましく、配列番号:26〜33で表される塩基配列の少なくともいずれかを有するmiRNAそのものがより好ましく、配列番号:30〜33で表される塩基配列の少なくともいずれかのみを有するmiRNAが特に好ましい。
前記脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤が、配列番号:26〜33で表される塩基配列の少なくともいずれかのみを有するmiRNAである場合、配列番号:26〜33で表される塩基配列それぞれの含有比率としては、特に制限はなく、目的に応じて適宜選択することができる。
【0051】
<阻害活性>
前記脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤が、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかを阻害しているか否かを確認する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、前記配列番号:2〜33を用いたRNAiにより確認する方法などが挙げられる。
前記RNAiに用いる細胞としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、脂肪細胞前駆細胞株である3T3−L1細胞、マクロファージ由来の細胞株であるRAW264細胞、骨髄由来マスト細胞mBMMCsなどが挙げられる。
前記3T3−L1細胞を用いた場合、前記配列番号:2〜33で表される塩基配列のいずれかによりRNAiを行った前記3T3−L1細胞の脂肪滴の数と、前記配列番号:2〜33で表される塩基配列を導入していない前記3T3−L1細胞の脂肪滴の数とを比較することで、前記DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかの阻害活性を確認することができる。
【0052】
<その他の成分>
前記その他の成分としては、特に制限はなく、薬理学的に許容される担体の中から目的に応じて適宜選択することができ、例えば、エタノール、水、デンプンなどが挙げられる。
前記脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤中の、その他の成分の含有量としては、特に制限はなく、目的に応じて適宜選択することができる。
【0053】
<使用>
前記脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤は、1種単独で使用されてもよいし、2種以上を併用してもよく、他の成分を有効成分とする医薬と併用されてもよい。また、前記脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤は、他の成分を有効成分とする医薬中に、配合された状態で使用されてもよい。
【0054】
<剤型>
前記脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤の剤型としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、経口固形剤、経口液剤、注射剤、吸入散剤などが挙げられる。
【0055】
−経口固形剤−
前記経口固形剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、錠剤、被覆錠剤、顆粒剤、散剤、カプセル剤などが挙げられる。
前記経口固形剤の製造方法としては、特に制限はなく、常法を使用することができ、例えば、前記miRNAに、賦形剤、及び必要に応じて前記その他の成分、各種添加剤を加えることにより、製造することができる。ここで、前記賦形剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、乳糖、白糖、塩化ナトリウム、ブドウ糖、デンプン、炭酸カルシウム、カオリン、微結晶セルロース、珪酸などが挙げられる。また、前記添加剤としても、特に制限はなく、目的に応じて適宜選択することができ、例えば、結合剤、崩壊剤、滑沢剤、着色剤、矯味/矯臭剤などが挙げられる。
前記結合剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、水、エタノール、プロパノール、単シロップ、ブドウ糖液、デンプン液、ゼラチン液、カルボキシメチルセルロース、ヒドロキシプロピルセルロース、ヒドロキシプロピルスターチ、メチルセルロース、エチルセルロース、シェラック、リン酸カルシウム、ポリビニルピロリドンなどが挙げられる。
前記崩壊剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、乾燥デンプン、アルギン酸ナトリウム、カンテン末、炭酸水素ナトリウム、炭酸カルシウム、ラウリル硫酸ナトリウム、ステアリン酸モノグリセリド、乳糖などが挙げられる。
前記滑沢剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、精製タルク、ステアリン酸塩、ホウ砂、ポリエチレングリコールなどが挙げられる。
前記着色剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、酸化チタン、酸化鉄などが挙げられる。
前記矯味/矯臭剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、白糖、橙皮、クエン酸、酒石酸などが挙げられる。
【0056】
−経口液剤−
前記経口液剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、内服液剤、シロップ剤、エリキシル剤などが挙げられる。
前記経口液剤の製造方法としては、特に制限はなく、常法を使用することができ、例えば、前記miRNA、及び必要に応じて前記その他の成分に、添加剤を加えることにより、製造することができる。ここで、前記添加剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、矯味/矯臭剤、緩衝剤、安定化剤などが挙げられる。
前記矯味/矯臭剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、白糖、橙皮、クエン酸、酒石酸などが挙げられる。
前記緩衝剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、クエン酸ナトリウムなどが挙げられる。
前記安定化剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、トラガント、アラビアゴム、ゼラチンなどが挙げられる。
【0057】
−注射剤−
前記注射剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、溶液、懸濁液、用事溶解用固形剤などが挙げられる。
前記注射剤の製造方法としては、特に制限はなく、常法を使用することができ、例えば、前記miRNA、及び必要に応じて前記その他の成分に、pH調節剤、緩衝剤、安定化剤、等張化剤、局所麻酔剤などを添加することにより、製造することができる。ここで、前記pH調節剤及び前記緩衝剤としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、クエン酸ナトリウム、酢酸ナトリウム、リン酸ナトリウムなどが挙げられる。また、前記安定化剤としても、特に制限はなく、目的に応じて適宜選択することができ、例えば、ピロ亜硫酸ナトリウム、EDTA、チオグリコール酸、チオ乳酸などが挙げられる。前記等張化剤としても、特に制限はなく、目的に応じて適宜選択することができ、例えば、塩化ナトリウム、ブドウ糖などが挙げられる。前記局所麻酔剤としても、特に制限はなく、目的に応じて適宜選択することができ、例えば、塩酸プロカイン、塩酸リドカインなどが挙げられる。
【0058】
<投与>
前記脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤の投与方法、投与量、投与時期、及び投与対象としては、特に制限はなく、目的に応じて適宜選択することができる。
前記投与方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、経口投与法、注射による方法、吸入による方法などが挙げられる。
前記投与量としては、特に制限はなく、投与対象個体の年齢、体重、体質、症状、他の成分を有効成分とする医薬の投与の有無など、様々な要因を考慮して適宜選択することができる。
前記投与対象となる動物種としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、ヒト、サル、ブタ、ウシ、ヒツジ、ヤギ、イヌ、ネコ、マウス、ラット、トリなどが挙げられるが、これらの中でもヒトに好適に用いられる。
【0059】
<用途>
前記脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤は、前記DBP、TEF、及びE4BP4、並びに、該DBP、該TEF、及び該E4BPをコードするcDNAの少なくともいずれかの阻害作用を有することから、医薬、健康食品、機能性食品などに好適に利用可能である。
【0060】
(スクリーニング方法)
本発明のスクリーニング方法は、PPARγプロモーター配列を有するレポーター遺伝子を構築するプロモーターレポーター遺伝子構築工程と、該プロモーターレポーター遺伝子と、被験物質を含む被験試料とを細胞に導入する導入工程と、該プロモーターレポーター遺伝子のレポーター活性を指標にして、PPARγプロモーター活性を検出する検出工程と、該被験試料が、PPARγプロモーター活性の促進作用、及び抑制作用のいずれかの作用を有しているか否かを判断し、PPARγプロモーター活性の促進作用、及び抑制作用のいずれかの作用を有する該被験試料中の被験物質をPPARγ発現調節剤としてスクリーニングするスクリーニング工程と、を含み、必要に応じて、更にその他の工程を含む。
【0061】
<レポーター遺伝子構築工程>
前記レポーター遺伝子構築工程は、PPARγプロモーター配列を有するプロモーターレポーター遺伝子を構築する工程である。
【0062】
−PPARγプロモーター配列−
前記PPARγプロモーター配列としては、特に制限はなく、目的に応じて適宜選択することができるが、前述した配列番号:1で表される塩基配列を有するPPARγ6プロモーター配列を用いることが好ましい。
なお、本発明のスクリーニング方法に用いる前記PPARγ6のプロモーター配列は、前述した配列番号:1で表される塩基配列の全体からなる塩基配列を用いてもよいし、配列番号:1で表される塩基配列の一部からなる塩基配列を用いてもよく、配列番号:1で表される塩基配列中に存在する3ヵ所のD−ボックスの少なくともいずれかを欠失させた欠失変異体を用いてもよい。
前記D−ボックスの欠失変異体を、前記スクリーニング方法に用いると、前記被験物質とD−ボックスとの関係が明らかにできる点で有利である。
【0063】
前記PPARγ6のプロモーター配列を入手する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、ヒトゲノミックDNAを鋳型に用い、下記配列番号:34〜35で表される塩基配列をプライマーとしてPCRにより増幅する方法などが挙げられる。
プライマー配列
GATGAGAGCTGGGGAGAAGG (配列番号:34)
CTAAGGAAAGGGAATTAAGCAGAA (配列番号:35)
【0064】
−レポーター遺伝子−
前記レポーター遺伝子としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、ルシフェラーゼなどが挙げられる。
前記ルシフェラーゼの遺伝子配列の入手方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、市販のベクター(例えば、pGL3−basic、Promega社製)から入手する方法、NCBIなどのデータベースから入手する方法などが挙げられる。
【0065】
−−構築方法−−
前記プロモーターレポーター遺伝子を構築する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、前記ルシフェラーゼ遺伝子を有する前記pGL3−basic(Promega社製)、及び前記PPARγ6のプロモーター配列のPCR産物を制限酵素処理し、前記PPARγ6のプロモーター配列のPCR産物をクローニングすることにより構築する方法などが挙げられる。
なお、前記ルシフェラーゼは安定性の高いタンパク質であるため、プロモーター活性の変動(増減)をモニターし難いことがある。そのため、前記pGL3−basicは、ルシフェラーゼ遺伝子のC末端にPEST配列(プロリン、グルタミン酸、セリン、及びスレオニンに富む配列)を導入させた配列であることが、細胞内におけるルシフェラーゼの分解を促進できる点で好ましい。
【0066】
<導入工程>
前記導入工程は、前記プロモーターレポーター遺伝子と、被験物質を含む被験試料とを細胞に導入する工程である。
【0067】
−被験試料−
前記被験試料とは、少なくとも被験物質を含有し、必要に応じて、更にその他の成分を含有する。
【0068】
−−被験物質−−
前記被験物質としては、特に制限はなく、目的に応じて適宜選択することができるが、発現ベクターにクローニングされた遺伝子であることが好ましい。
前記発現ベクターとしては、特に制限はなく、目的に応じて適宜選択することができ、例えば、pcDNA3.1(Invitrogen社製)などが挙げられる。
【0069】
−−その他の成分−−
前記その他の成分としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、水、PBS、TE(Tris−EDTA)などが挙げられる。
【0070】
−細胞−
前記細胞としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、NIH3T3細胞、HepG2細胞、Caco2細胞などが挙げられる。
なお、前記NIH3T3細胞は、マウス胎児皮膚由来細胞株であり、前記HepG2細胞は、ヒト肝ガン由来細胞株であり、前記Caco2細胞は、ヒト結腸由来細胞株である。
前記細胞の培地としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、DMEM、MEMなどが挙げられる。
前記細胞の培養条件としては、特に制限はなく、細胞の種類に応じて適宜選択することができるが、前記NIH3T3細胞、及び前記HepG2細胞の場合は、10質量%FBS(ウシ胎児結成)、DMEM(Dulbecco's modified Eagle medium)、37℃、5%CO2が好ましく、前記Caco2細胞の場合は、20質量%FBS、DMEM、37℃、5%CO2が好ましい。
【0071】
−導入方法−
前記プロモーターレポーター遺伝子と、前記被験物質を含む被験試料とを細胞に導入する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、リン酸カルシウム法、リポフェクション法、エレクトロポレーション法などが挙げられるが、これらの中でもリポフェクション法が、簡単で導入効率が高い点で好ましい。
前記プロモーターレポーター遺伝子の導入量としては、特に制限はなく、目的に応じて適宜選択することができるが、5ng〜20ngが好ましく、10ng〜15ngがより好ましい。
前記被験物質が、遺伝子である場合、その導入量としては、特に制限はなく、遺伝子の種類、発現ベクターの種類などに応じて適宜選択することができるが、62.5ng〜1,000ngが好ましく、100ng〜200ngがより好ましい。
前記細胞に、前記プロモーターレポーター遺伝子、及び被験物質を導入する時期としても、特に制限はなく、目的に応じて適宜選択することができる。
なお、前記遺伝子の導入量を一定にするため、pcDNA3.1(Invitrogen社製)を用いることが好ましい。
【0072】
<検出工程>
前記検出工程は、前記プロモーターレポーター遺伝子のレポーター活性を指標にして、PPARγプロモーター活性を検出する工程である。
【0073】
前記レポーター活性を検出する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、dual−luciferase reporter assay system(Promega社製)を用いて、前記ルシフェラーゼを反応させ、ルミノメーター(例えば、Model TD−20/20(Turner Designs社製))により検出する方法などが挙げられる。
前記ルシフェラーゼは、前記PPARγプロモーターの制御下で発現するため、前記ルシフェラーゼのレポーター活性を検出することで、前記PPARγのプロモーター活性を検出することができる。
【0074】
<スクリーニング工程>
前記スクリーニング工程は、前記被験物質を含む被験試料が、PPARγプロモーター活性の促進作用、及び抑制作用のいずれかの作用を有しているか否かを判断し、PPARγプロモーター活性の促進作用、及び抑制作用のいずれかの作用を有する該被験試料中の被験物質をPPARγ発現調節剤としてスクリーニングする工程である。
【0075】
前記PPARγプロモーター活性の促進作用、及び抑制作用のいずれかの作用を有しているか否かを判断する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、前記レポーター活性の強度を比較する方法などが挙げられる。
前記レポーター活性の強度を比較する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、前記被験試料を導入せず、プロモーターレポーター遺伝子のみを導入した細胞をコントロールとし、該コントロールと、被験試料、及びプロモーターレポーター遺伝子を導入した細胞とを比較して、前記被験試料、及びプロモーターレポーター遺伝子を導入した細胞のレポーター活性が、前記コントロールのレポーター活性より強い場合、前記被験試料中の被験物質をPPARγ発現調節剤としてスクリーニングする方法、前記コントロールと比較せず、前記被験物質を含む被験試料間のレポーター活性の強度の違いにより、レポーター活性が強い前記被験試料中の被験物質を、より強いPPARγ発現調節作用を有するPPARγ発現調節剤としてスクリーニングする方法などが挙げられる。
【実施例】
【0076】
以下に本発明の実施例を挙げて本発明を具体的に説明するが、本発明はこれらの実施例に何ら限定されるものではない。
【0077】
(実施例1:新規PPARγプロモーター配列の解析)
新規PPARγであるPPARγ6のプロモーター配列は、新規スプライスバリアントcDNAの5’配列を元に転写開始点を決定し、第1エクソン(C1C2)(図1B)を有する上流領域を、下記方法により解析した。
【0078】
<PPARγ6プロモーター配列の増幅>
ヒトゲノミックDNA(Novagen社製)を鋳型に用いて、下記配列番号:34〜35で表される塩基配列をプライマーとして用い、96℃・30秒間、62℃・30秒間、72℃・90秒間を30サイクルの条件でPCRを行い、PPARγ6プロモーター配列を増幅した。前記PPARγ6プロモーター領域の長さは、約1kbであった。
前記PPARγ6プロモーター配列のPCR産物は、pCR2.1ベクター(Invitrogen社製)にTA法でクローニングした(以下、「hPPARγ6−pCR2.1」と称することがある。)。
【0079】
プライマー配列
GATGAGAGCTGGGGAGAAGG (配列番号:34)
CTAAGGAAAGGGAATTAAGCAGAA (配列番号:35)
【0080】
<PPARγ6のプロモーター配列の解析>
前記hPPARγ6−pCR2.1を用い、ABI PRISM377 シーケンサー(アプライドバイオシステム社製)によりシーケンスをいった。シーケンスの結果から明らかになった配列について、NCBI BLAST cDNA、及びETSデータベースにて検索を行い、スプライシングパターンを決定した。その結果、PPARγ6プロモーター配列は、PPARγの新規スプライスバリアントである、配列番号:1で表される塩基配列を有することが明らかとなった。
【0081】
<PPARγ6プロモーター領域のDNA結合配列予測解析>
前記hPPARγ6−pCR2.1を用い、PPARγ6のプロモーター配列のDNA結合配列予測解析を、MatInspector(Genomatix社製)により行った。
その結果、PPARγ6のプロモーター配列は、C1C2領域に、3箇所のD−ボックス(D1、D2、及びD3)を有することが確認された(図2)。前記D−ボックスは、配列番号:1で表される塩基配列のうち、塩基番号159〜166(D1)、塩基番号202〜209(D2)、及び塩基番号218〜225(D3)に存在する。なお、図2中、TSSは、転写開始領域(Transcript Start Site)を示す。
【0082】
(実施例2:DBPによるPPARγ6の発現調節)
DBPがPPARγ6の発現を調節しているか否かを、以下の方法を用いて調べた。
【0083】
<方法>
−Caco−2細胞の培養−
Caco−2細胞(細胞株バンクATCC)を、24ウエルプレートに5×105細胞/プレート播種し、DMEM(Invitrogen社製)で、37℃、5%CO2の条件下で3日間培養した。
【0084】
−DBP発現ベクターの構築−
DBP遺伝子を、マウス肝臓由来cDNAを鋳型にしてPCRにより増幅した。前記PCR産物と、pcDNA3.1ベクター(Invitrogen社製)とをEcoRIで制限酵素処理し、前記pcDNA3.1ベクターに、前記DBP遺伝子をクローニングした(以下、「pcDNA3.1−DBP」と称することがある)。
【0085】
−トランスフェクション−
前記pcDNA3.1−DBP 125ng、及びコントロールとしてDBPをクローニングしていないpcDNA3.1ベクターのいずれかを、Lipofectamin2000(Invitrogen社製)を用いて前記Caco−2細胞へトランスフェクションし、前記DBPを、前記Caco−2細胞に一過性に過剰発現させた。前記Caco−2細胞は、後述するmRNAの定量までの間、DMEMで、37℃、5%CO2の条件下で1日間培養した。
【0086】
−mRNAの定量−
前記DBPを過剰発現させたCaco−2細胞、及びコントロールのCaco−2細胞から、ISOGEN((株)日本ジーン製)を用いてRNA抽出、及び精製を行い、DBPのmRNA量、及びPPARγ6のmRNA量をRT−PCR法により測定した。
【0087】
<結果>
図3に前記RT−PCRの結果を示す。
図3より、DBPのmRNAの発現量は、コントロールと比較して有意に高く、pcDNA3.1−DBPをトランスフェクションしたCaco−2細胞では、DBPが過剰発現されていることが確認された。
PPARγ6のmRNAの発現量は、pcDNA3.1−DBPをトランスフェクションし、DBPを過剰発現したCaco−2細胞では、コントロールのCaco−2細胞と比較して有意に増加していた。
これらの結果より、PPARγ6の発現は、DBPにより促進されたことが示唆される。
【0088】
(実施例3:DBP、TEF、HLF、及びE4BP4によるPPARγ6の転写制御)
DBP、TEF、HLF、及びE4BP4が、PPARγ6の転写を制御しているか否かについて、以下のレポーターシステムにより調べた。
【0089】
<方法>
−DBP発現ベクターの構築−
DBP遺伝子を、マウス肝臓由来cDNAを鋳型にしてPCRにより増幅した。前記PCR産物と、pcDNA3.1ベクター(Invitrogen社製)とをEcoRIで制限酵素処理し、前記pcDNA3.1ベクターに、前記DBP遺伝子をクローニングした(以下、「pcDNA3.1−DBP」と称することがある)。
【0090】
−TEF発現ベクターの構築−
TEF遺伝子を、ヒト肝臓由来cDNAを鋳型にしてPCRにより増幅した。前記PCR産物と、pcDNA3.1ベクター(Invitrogen社製)とをEcoRIで制限酵素処理し、前記pcDNA3.1ベクターに、前記TEF遺伝子をクローニングした(以下、「pcDNA3.1−TEF」と称することがある)。
【0091】
−HLF発現ベクターの構築−
HLF遺伝子を、ヒト肝臓由来cDNAを鋳型にしてPCRにより増幅した。前記PCR産物と、pCR3.1ベクター(Invitrogen社製)とをTAクローニング法によって、前記pCR3.1ベクターに、前記HLF遺伝子をクローニングした(以下、「pCR3.1−HLF」と称することがある)。
【0092】
−E4BP4発現ベクターの構築−
E4BP4遺伝子を、マウス肝臓由来cDNAを鋳型にしてPCRにより増幅した。前記PCR産物と、pcDNA3ベクター(Invitrogen社製)とをBamHI、及びKpnIで制限酵素処理し、前記pcDNA3ベクターに、前記E4BP4遺伝子をクローニングした(以下、「pcDNA3−E4BP4」と称することがある)。
【0093】
−プロモーターレポーター遺伝子の構築−
−−pGL3−dlucの構築−−
pGL3−basic(Promega社製)に、ルシフェラーゼをコードする遺伝子のC末端に相当する領域をEcoRV、及びXbaIで制限酵素処理することにより、pGL3−basic中のルシフェラーゼ遺伝子のC末端に、PEST配列を導入した(以下、「pGL3−dluc」と称することがある。)。
【0094】
−−pGL3−dlucへのPPARγ6プロモーター配列のサブクローニング−−
実施例1のhPPARγ6−pCR2.1と、前記pGL3−dlucとを、KpnI、及びBglII(共に(株)日本ジーン製)で制限酵素処理し、PPARγ6プロモーター配列をpGL3−dlucにサブクローニングすることにより、プロモーターレポーター遺伝子(以下、「hPPARγ6−prom−1.0k/pGL3−basic」と称することがある)を構築した。
【0095】
−細胞の培養−
NIH3T3細胞(理化学研究所提供)を、35mmプレートに1×105細胞/プレート播種し、10質量%FBSを含むDMEMで、37℃、5%CO2の条件下で2日間培養した。
【0096】
−導入−
前記hPPARγ6−prom−1.0k/pGL3−basicと、前記pcDNA3.1−DBP、前記pcDNA3.1−TEF、前記pCR3.1−HLF、及び前記pcDNA3−E4BP4のいずれかとを、前記NIH3T3細胞に、Tfx−50(Promega社製)を用いて、製造元の方法に従い、トランスフェクションした。
なお、前記hPPARγ6−prom−1.0k/pGL3−basicは、10ngトランスフェクションに用いた。前記pcDNA3.1−DBPは0ng〜1,000ngの濃度で、前記pcDNA3.1−TEFは0ng〜500ngの濃度で、前記pCR3.1−HLFは0ng〜500ngの濃度で、前記pcDNA3−E4BP4は0ng〜1,000ngの濃度でそれぞれトランスフェクションを行った。
トランスフェクション24時間後に、dual−luciferase reporter assay system(Promega社製)によりpRL−TK(Prommega社製)を導入した。導入遺伝子量を一定にする為、pcDNA3(Invitrogen社製)を用いた。
なお、トランスフェクション効率は、前記pRL−TKにより確認した。
導入後、10質量%FBSを含むDMEMで、37℃、5%CO2の条件下で2日間培養した。
【0097】
−検出−
前記dual−luciferase reporter assay system(Promega社製)を用いて、製造元の方法に従い、前記hPPARγ6−prom−1.0k/pGL3−basicより発現したルシフェラーゼを反応させた。
前記ルシフェラーゼ活性(レポーター活性)の測定は、ルミノメーターModel TD−20/20(Turner Designs社製)を用いて行った。
【0098】
<結果>
ルシフェラーゼ活性の測定結果を、図4A〜Dに示す。
図4Aより、DBPは、導入量依存的にルシフェラーゼ活性の増加が認められ、その活性は、DBPを500ng導入したときに最も高かった。また、図4Bより、TEFもDBPと同様に、導入量依存的にルシフェラーゼ活性の増加が認められた。これらの結果より、DBP、及びTEFは、PPARγ6プロモーター活性を促進することが認められた。
図4C〜Dより、HLF、及びE4BP4は、DBP、及びTEFとは反対に、導入量依存的にルシフェラーゼ活性の低下が認められた。これらの結果より、HLF、及びE4BP4は、PPARγ6プロモーター活性を抑制することが認められた。
DBP、TEF、及びHLFは、同じPAR−bZIPファミリーに属する転写因子であるが、PPARγ6に対する活性は異なるものであることが示唆された。
【0099】
(実施例4:DBPのPPARγ6プロモーターへの結合部位)
DBPが、PPARγ6のプロモーター領域のどの部位を介してPPARγ6の転写活性化を行っているか、PPARγ6プロモーター領域を欠失させた数種の欠失変異体を用いて、下記方法により検討を行った。
【0100】
−欠失変異体の作製−
前記欠失変異体は、実施例2で作製したhPPARγ6−prom−1.0k/pGL3−basicに、QuikChange XL site−directed mutagenesis kit(Stratagene社製)を用いて、製造元の方法に従い、下記表1に示すコンストラクト1〜10を作製した(図5A)。
下記表1は、PPARγ6プロモーターのC1C2の5’末端を、転写開始点、即ち「0」とし、C1C2の転写開始点より上流の塩基数を「−」で示し、C1C2の転写開始点より下流の塩基数を「+」で示した。なお、C1C2は、全長297bである。
コンストラクト1は、PPARγ6プロモーター配列全長を有する野生型である。コンストラクト2〜5は、PPARγ6プロモーター配列の上流、即ち5’末端を欠失させた欠失変異体である。コンストラクト6は、PPARγ6プロモーター配列に存在する3箇所のD−ボックスを欠失させた欠失変異体である。コンストラクト7〜9は、PPARγ6プロモーター配列に存在する3箇所のD−ボックス、及び5’末端を欠失させた欠失変異体である。コンストラクト10(ネガティブコントロール)は、PPARγ6プロモーター配列を導入していない、pGL3−basicのみのコンストラクトである。
【0101】
【表1】
【0102】
−NIH3T3細胞の培養−
NIH3T3細胞(理化学研究所提供)を、35mmディッシュに、1×106細胞/ディッシュ播種し、DMEMを用い、37℃、5%CO2の条件下で2日間培養した。
【0103】
−導入−
前記コンストラクト1〜10のいずれかをそれぞれ10ngと、pcDNA3.1−DBP 125ngとを、PolyFect(Qiagen社製)を用いて、前記NIH3T3細胞にトランスフェクションし、24時間後に、100nMデキサメタゾン(Sigma社製)を添加したDMEMに交換し、更に2時間後に、0.1mMルシフェリン(Promega社製)を含むDMEMに交換し、Kronos(Atto社製)を用いて、ルシフェラーゼ活性を10分間ごとに測定した。
なお、コントロールとしては、pcDNA3.1−DBPを導入せず、コンストラクト1〜10のいずれかのみを導入したサンプルを用いた。前記コントロールのルシフェラーゼ活性が、NIH3T3細胞におけるPPARγ6プロモーターの基礎転写活性を示すものとする。
【0104】
−結果−
結果を、図5B〜Cに示す。
図5B〜Cより、コンストラクト1のルシフェラーゼ活性は、pcDNA3.1−DBPにより基礎転写活性の約2倍になっており、PPARγ6のプロモーター活性がDBPにより顕著に活性化されたことが認められた。このDBPによるPPARγ6の転写活性化は、C1C2より上流を欠失したコンストラクト2〜5においても認められた(図5B)。C1C2が一部欠失したコンストラクト4〜5では、コンストラクト1〜3と比較して、わずかにPPARγ6のプロモーター活性が低かったが、いずれも各コントロール(DBP(−))と比較して、PPARγ6の転写は活性化されていることが認められた。
一方、C1C2より上流は欠失せず、C1C2に存在するD−ボックス部位のみを欠失したコンストラクト6では、PPARγ6プロモーターの基礎転写活性が完全に消失し、そのレベルは、コンストラクト10(ネガティブコントロール)と同程度であった。更に、DBPによるPPARγ6の転写活性化も認められなかった(図5C)。また、コンストラクト7〜9において、C1C2のD−ボックス部位を欠失させ、更にC1C2より上流を欠失させた場合も、PPARγ6プロモーターの基礎転写活性は完全に消失し、DBPによるPPARγ6の転写活性化も認められなかった(図5C)。
【0105】
これらの結果より、DBPによるPPARγ6プロモーターの活性化には、PPARγ6プロモーター配列のC1C2に存在するD−ボックスが重要であり、DBPは、D−ボックスを介してPPARγ6プロモーターの活性化、即ちPPARγ6の発現を促進していることが示唆された。
【0106】
(実施例5:DBP、及びE4BP4による、D−ボックスを介したPPARγ6の転写制御)
実施例4の結果、DBPはPPARγ6プロモーター配列に存在するD−ボックスに作用していることが認められた。実施例1のPPARγ6プロモーター配列解析の結果、PPARγ6プロモーターには3箇所D−ボックスが存在することが明らかとなっていることから、これらの3箇所のD−ボックスと、DBP、及びE4BP4との関係について、下記方法により検討を行った。
【0107】
<方法>
実施例4のコンストラクト5(5’末端:+126、3’末端:+297)を用い、実施例4と同様の方法により、QuikChange XL site−directed mutagenesis kit(Stratagene社製)を用いて、PPARγ6プロモーターの3箇所のD−ボックスの全部又は一部を欠失させた下記表2、及び図6Aに示すコンストラクト11〜15を作製した。
下記表2の「○」は、D−ボックスを有する箇所を示し、「×」は、D−ボックスを欠失した箇所を示す。
前記コンストラクト11〜15のいずれかをそれぞれ10ngと、pcDNA3.1−DBP 250ng、及びpcDNA3−E4BP4 400ngのいずれかとを、実施例4と同様の方法で、前記NIH3T3細胞に導入し、ルシフェラーゼ活性を測定した。
なお、コントロールとしては、pcDNA3.1−DBP、及びpcDNA3−E4BP4を導入せず、コンストラクト11〜15のいずれかのみを導入したサンプルを用いた。前記コントロールのルシフェラーゼ活性が、NIH3T3細胞におけるPPARγ6プロモーターの基礎転写活性を示すものとする。
【0108】
【表2】
【0109】
<結果>
図6BにDBPの結果を、図6CにE4BP4の結果を示す。なお、図6B、及び図6C中、「**」は、t−検定によるp値が、p<0.01を示し、「***」は、p<0.001を示す。
【0110】
−DBP−
PPARγ6プロモーター配列に存在する3箇所のD−ボックスを有するコンストラクト11は、DBPによりPPARγ6プロモーターを活性化し、その活性は、コントロール(基礎転写活性)の約2倍であった(図6B)。
D−ボックスのD2、及びD3の両方を欠失させたコンストラクト12では、コンストラクト11と比較して、基礎転写活性に変化はなかったが、DBP存在下では、コンストラクト11と比較して、PPARγ6プロモーターの活性化の強度が弱かった。また、コントロール(基礎転写活性)と比較して、DBPによるPPARγ6プロモーターの活性化は認められたものの、その活性化強度は弱かった(図6B)。
D−ボックスのD1、及びD3の両方を欠失させたコンストラクト13では、コンストラクト11と比較して、基礎転写活性が低下し、DBP存在下のコンストラクト11と比較しても、DBPによるPPARγ6プロモーターの活性化の強度は弱かった。しかし、コンストラクト13のコントロールと比較して、ルシフェラーゼの発現量は約2倍以上であり、DBPによるPPARγ6プロモーターの強い活性化が認められた(図6B)。
D−ボックスのD1、及びD2の両方を欠失させたコンストラクト14も、コンストラクト13の結果と同様であり、コンストラクト11と比較して、基礎転写活性が低下し、DBP存在下のコンストラクト11と比較しても、DBPによるPPARγ6の活性化の強度は弱かった。しかし、コンストラクト14のコントロールと比較して、ルシフェラーゼの発現量は約2倍以上であり、DBPによるPPARγ6のプロモーターの活性化が認められた(図6B)。
D−ボックスのD1〜D3の全てを欠失させたコンストラクト15では、コンストラクト11と比較して、基礎転写活性が低下し、DBP存在下でもルシフェラーゼの発現レベルは、コンストラクト15のコントロールと同レベルであったことから、DBPによるPPARγプロモーターの活性化が完全に消失していることが認められた(図6B)。
【0111】
−E4BP4−
PPARγ6プロモーター配列に存在する3箇所のD−ボックスを有するコンストラクト11は、E4BP4によりPPARγ6プロモーター活性を抑制し、その活性は、コントロール(基礎転写活性)の約1/2であった(図6C)。
D−ボックスのD2、及びD3の両方を欠失させたコンストラクト12では、コンストラクト11と比較して、基礎転写活性に変化はなく、E4BP4存在下でも、コンストラクト11と比較して変化は認められなかった。即ち、E4BP4によるPPARγ6プロモーター活性の抑制が認められ、その活性は、コンストラクト12のコントロールの約1/2であった。(図6C)。
D−ボックスのD1、及びD3の両方を欠失させたコンストラクト13では、コンストラクト11と比較して、基礎転写活性が低下し、コンストラクト13のコントロールと比較して、E4BP4によるPPARγ6プロモーター活性の抑制が弱かった(図6C)。
D−ボックスのD1、及びD2の両方を欠失させたコンストラクト14も、コンストラクト13の結果と同様であり、コンストラクト11と比較して、基礎転写活性が低下し、コンストラクト14のコントロールと比較して、E4BP4によるPPARγ6プロモーター活性の抑制が弱かった(図6C)。
D−ボックスのD1〜D3の全てを欠失させたコンストラクト15では、コンストラクト11と比較して、基礎転写活性が低下し、E4BP4存在下でもルシフェラーゼの発現レベルは、コンストラクト15のコントロールと同レベルであったことから、E4BP4によるPPARγプロモーター活性の抑制が完全に消失していることが認められた(図6C)。
【0112】
実施例5の結果から、PPARγ6の転写調節は、主に、基礎転写活性に係る転写開始点上流配列約700bと、PAR−bZIP転写因子ファミリーによって転写調節を受ける転写開始点より下流の3箇所のD−ボックスを有する配列とによって行われていることが明らかになった。
更に、DBPによって正に、E4BP4によって負に制御を受ける、D−ボックスを介した転写調節システムを有することが示された。
また、3箇所のD−ボックスによって、DBP、及びE4BP4による転写調節が行われていることが明らかになったことから、PPARγ6プロモーター配列に存在するD−ボックスの有無によって、PPARγ6の転写調節作用がDBP、及びE4BP4を介するものであるかどうかを判定出来る可能性が示唆された。
【0113】
(実施例6:DBP、TEF、HLF、及びE4BP4のPPARγ6のD−ボックスへの結合能)
実施例2〜3より、DBP、TEF、HLF、及びE4BP4がPPARγ6の転写活性に関与することが示唆され、更に実施例4〜5より、DBP、及びE4BP4によるPPARγ6の転写制御は、PPARγ6のプロモーター領域に存在するD−ボックスが重要であることが示唆された。
そこで、DBP、TEF、HLF、及びE4BP4が、PPARγ6のプロモーター領域に存在するD−ボックスに直接結合するか否かについて、ゲルシフトアッセイにより検討を行った。
【0114】
<方法>
−プローブの作製−
ゲルシフトアッセイは、PPARγ6プロモーターに存在する3箇所のD−ボックス(D1、D2、及びD3)の配列をもとに、D1に対するプローブとしては、下記配列番号:36で表される2本鎖プローブを、D2に対するプローブとしては、下記配列番号:37で表される2本鎖プローブを、D3に対するプローブとしては、下記配列番号:38で表される2本鎖プローブを作製した。配列番号:36〜38で表されるプローブの下線は、D−ボックスを示す。前記プローブに、T4ポリヌクレオチドキナーゼを用いてγ−32P ATPを5’末端に導入して標識した。
AGGGAGGAGCCTAAGGTAAGGAGTCAGAAA (配列番号:36)
AGCTGCGGCTTTTATATAAGGTCAGTGGTA (配列番号:37)
TAAGGTCAGTGGTAGGTAAGGAAGGGGCCT (配列番号:38)
【0115】
−タンパク質の合成−
ゲルシフトアッセイに用いるタンパク質は、実施例3で構築したpcDNA3.1−DBP、pcDNA3.1−TEF、pCR3.1−HLF、及びpcDNA3−E4BP4を鋳型として、TNT T7 Quick Coupled Transcription/Translation System (Promega社製)を用い、製造元の方法に従って合成した。
前記DBP、TEF、HLF、及びE4BP4の合成タンパク質は、Gel Shift Binding Buffer (Takara Bio社製)に懸濁した。
【0116】
−ゲルシフトアッセイ−
−−プローブ特異性の確認−−
前記DBP合成タンパク質、TEF合成タンパク質、HLF合成タンパク質、及びE4BP4合成タンパク質をそれぞれ4μg、前記標識した各プローブ(配列番号:36〜38)0.0175pmol、特異的コンペティターとして、前記各プローブ(配列番号:36〜38)の非標識のプローブ1.75pmol、及び、非特異的コンペティター(合成委託)として、1.75pmolを用い、下記表3〜6に従い、サンプル1〜32を調製し、4質量%ポリアクリルアミドゲルにアプライし、150V、2時間の条件で電気泳動を行った。ゲルはゲルドライヤー(バイオラド社製)で乾燥した後、ゲルイメージを、BAS5000ゲルイメージ解析装置(Fuji Film社製)を用いて取得した。
なお、表1中、「+」は添加、「−」は非添加を示す。
【0117】
【表3】
【0118】
【表4】
【0119】
【表5】
【0120】
【表6】
【0121】
図7AにDBP合成タンパク質に対するプローブ特異性の確認の結果を、図7BにTEF合成タンパク質に対するプローブ特異性の確認の結果を、図7CにHLF合成タンパク質に対するプローブ特異性の確認の結果を、図7DにE4BP4合成タンパク質に対するプローブ特異性の確認の結果を示す。
図7A〜D中、「WT」は特異的コンペティターを添加したサンプル、「MT」は非特異的コンペティターを添加したサンプルを示す。ポジティブコントロールとしては、それぞれの合成タンパク質のプローブのみを添加したもの(サンプルNo.1、5、9、13、17、21、25、及び29)を用い、ネガティブコントロールとしては、プローブのみを添加したもの(サンプルNo.4、8、12、16、20、24、28、及び32)を用いた。
これらの結果より、ポジティブコントロールとしてコンペティターを添加しなかったサンプルは、DBP、TEF、HLF、及びE4BP4のどのタンパク質においても、それぞれバンドのシフトが認められた(図7A〜D:サンプル1、5、9、13、17、21、25、及び29)。また、ネガティブコントロールとしてプローブのみを添加したサンプルは、DBP、TEF、HLF、及びE4BP4のどのタンパク質においても、バンドのシフトが認められなかった(図7A〜D:サンプル4、8、12、16、20、24、28、及び32)。
特異的コンペティターを添加した場合、シフトしたバンドが消失したことから、前記バンドは、配列番号:36〜38に特異的な配列を認識していることが確認された(図7A〜D:サンプル2、6、10、14、18、22、26、及び30)。非特異的コンペティターを用いた場合、コンペティターを添加しなかった場合と同程度のバンドが検出されることを確認した(図7A〜D:サンプル3、7、11、15、19、23、27、及び31)。
【0122】
−−DBP、TEF、HLF、及びE4BP4のD−ボックスへの結合の確認−−
前記DBP合成タンパク質、TEF合成タンパク質、HLF合成タンパク質、及びE4BP4合成タンパク質をそれぞれ4μg、及び前記標識した各プローブ(配列番号:36〜38)0.0175pmolを使用して、下記表7に従い、サンプル33〜48を調製し、4質量%ポリアクリルアミドゲルにアプライし、150V、2時間の条件で電気泳動を行った。ゲルはゲルドライヤー(バイオラド社製)で乾燥した後、ゲルイメージを、BAS5000(Fuji Film社製)を用いて取得した。
なお、ポジティブコントロールとしては、下記配列番号:39で表される塩基配列を有する、PAR(proline and acidic amino acid−rich)、及びE4BP4のコンセンサス配列を用いた(Mitsui S, et al., Genes Dev, 2001, 15(8), p.995−1006)。
GCCCGCTACATATTACGTAACAAGCGTTCGC (配列番号:39)
【0123】
【表7】
【0124】
<結果>
図7EにDBPのゲルシフトアッセイの結果を、図7GにTEFのゲルシフトアッセイの結果を図7FにHLFのゲルシフトアッセイの結果を、図7HにE4BP4のゲルシフトアッセイの結果を示す。
図7E〜Hより、PPARγ6プロモーター領域に存在する3箇所のD−ボックスへの、DBP、及びE4BP4の結合は、D1、D2、及びD3の全てにおいて結合が認められた(図7E:サンプルNo.33〜36、及び図7H:サンプルNo.45〜48)。特に、D2に関して強い特異的バンドが検出された(図7E:サンプルNo.34、及び図7H:サンプルNo.47)。また、TEF、及びHLFは、D2への結合が認められた(図7F:サンプルNo.39、図7G:サンプルNo.43)。
これらの結果より、DBP、TEF、HLF、及びE4BP4によるPPARγ6プロモーターの転写調節は、DBP、TEF、HLF、及びE4BP4が、直接PPARγ6プロモーターに存在するD−ボックスに結合して行われている可能性が示唆された。
【0125】
(実施例7:脂肪細胞分化過程におけるDBP、TEF、及びE4BP4の役割)
脂肪細胞分化過程におけるDBP、TEF、及びE4BP4の役割について、下記方法により検討を行った。
【0126】
<oil red O染色による検討>
−方法−
−−miRNA配列の合成−−
Invitrogen社より提供されたRNAi配列情報を元に、下記配列番号:4、5、16、17、28、及び29で表される2本鎖miRNA(マイクロRNA)配列を合成した。
DBPに対するmiRNAは、下記配列番号:4で表される塩基配列、及び下記配列番号:5で表される塩基配列である。
TEFに対するmiRNAは、下記配列番号:16で表される塩基配列、及び下記配列番号:17で表される塩基配列である。
E4BP4に対するmiRNAは、下記配列番号:28で表される塩基配列、及び下記配列番号:29で表される塩基配列である。
前記miRNA(配列番号:4、5、16、17、28、及び29)をアニーリングした後、Block−itベクター(Invitrogen社製)に導入した。
AAGAGGAGCTGCAAGAGCCGGGTTTTGGCCACTGACTGACCCGGCTCTCAGCTCCTCTT (配列番号:4)
AAGAGGAGCTGAGAGCCGGGTCAGTCAGTGGCCAAAACCCGGCTCTTGCAGCTCCTCTT (配列番号:5)
TTTAGCTGCCACGTTGTTCTTGTTTTGGCCACTGACTGACAAGAACAACGTCAGCTAAA (配列番号:16)
TTTAGCTGACGTTGTTCTTGTCAGTCAGTGGCCAAAACAAGAACAACGTGGCAGCTAAA (配列番号:17)
ATCACAGGGAACTTGTTCTCAGTTTTGGCCACTGACTGACTGAGAACATTCCCTGTGAT (配列番号:28)
ATCACAGGGAATGTTCTCAGTCAGTCAGTGGCCAAAACTGAGAACAAGTTCCCTGTGAT (配列番号:29)
【0127】
−−細胞の培養−−
3T3−L1細胞(細胞株バンクATCC)を、24ウエルプレートに5×104細胞/プレート播種し、脂肪前駆細胞培養用培地(DMEM+10%CS)で、37℃、5%CO2の条件下で70%コンフルエントになるまで培養した。
【0128】
−−トランスフェクション−−
前記DBP、TEF、及びE4BP4のmiRNAの配列を有するベクター(pcDNA 6.2−GW/EmGFP−miR、Invitrogen社製)0.8μgを、リポフェクタミン2000(Invitrogen社製)を用いてトランスフェクションした。前記pcDNA 6.2−GW/EmGFP−miRは、ブラストシジン遺伝子配列を有することから、ブラストシジン(Invitrogen社製)5μg/mLによって導入細胞のセレクションを行なった。この細胞が、85%コンフルエントになった段階で、インスリン(ナカライテスク(株)製)10μg/mL、デキサメタゾン(ナカライテスク(株)製)0.25μM、及びIBMX(ナカライテスク(株)製)0.5mMにより分化誘導をかけ、分化誘導後7日目にoil red Oを用いた染色を行った。
【0129】
−結果−
結果を図8に示す。図8中、スケールバーは50μmである。
脂肪細胞への分化の評価は、脂肪滴数を指標に行った。DBP、及びTEFのmiRNAを、単独あるいは両者を導入した細胞では脂肪滴数の減少が認められ、脂肪細胞への分化が阻害された。一方、E4BP4 miRNAを導入した細胞では脂肪滴が増加する傾向が認められた。
【0130】
<定量PCRによる検討>
−方法−
−−細胞−−
3T3−L1細胞(ATCC社製)を、24ウエルプレートに5×104細胞/プレート播種し、脂肪前駆細胞培養用培地で、37℃、5%CO2の条件下で70%コンフルエントになるまで培養した。
【0131】
−−トランスフェクション−−
前記DBP、及びE4BP4 発現ベクター(Invitrogen社製)を、リポフェクタミン2000(Invitrogen社製)を用いてトランスフェクションし、ブラストシジン(Invitrogen社製)5μg/mLによってセレクションを行なった。この細胞が、85%コンフルエントになった段階で、インスリン(ナカライテスク(株)製)10μg/mL、デキサメタゾン(ナカライテスク(株)製)0.25μM、及びIBMX(ナカライテスク(株)製)0.5mMにより分化誘導をかけ、分化誘導後7日目にRNA抽出を行った。
【0132】
−−mRNAの抽出、及び精製−−
前記脂肪細胞の分化の検討を行った3T3−L1細胞より、トライゾール(Invitrogen社製)を用いてRNA抽出、及び精製を行い、DBPのmRNA量、及びE4BP4のmRNA量をqPCR(50℃2分間を1サイクル、95℃10分間を1サイクル、及び95℃15秒間、及び60℃1分間を40サイクル)により測定した。
【0133】
結果を図9A〜Bに示す。
DBPを発現させ、脂肪細胞へ分化した3T3−L1細胞は、分化に伴い、DBPの発現量が増加した(図9A)。
一方、E4BP4を発現させ、脂肪細胞へ分化した3T3−L1細胞は、分化に伴い、DBPの発現量が減少した(図9B)。
【0134】
実施例7の結果、DBP、TEF、及びE4BP4の発現は、脂肪分化と関連のあることが示唆された。その作用としては、DBP、及びTEFが、脂肪分化を促進し、E4BP4は、脂肪分化を抑制するものと考えられる。
【産業上の利用可能性】
【0135】
本発明のPPARγ発現調節剤は、PPARγの発現調節作用を有することから、脂肪細胞への分化の調節、脂質代謝の調節、破骨細胞機能の調節、炎症の調節などに好適に利用可能である。
また、本発明の脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤は、DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BPをコードするcDNAの少なくともいずれかの阻害作用を有することから、医薬、健康食品、機能性食品などに好適に利用可能である。
更に、本発明のスクリーニング方法は、PPARγの発現調節作用を有する因子を効率よくスクリーニングできることから、PPARγ発現調節剤のスクリーニングに好適に利用可能である。
【特許請求の範囲】
【請求項1】
配列番号:1で表される塩基配列を有することを特徴とするPPARγプロモーター配列。
【請求項2】
DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BPをコードするcDNAの少なくともいずれかを含有することを特徴とするPPARγ発現調節剤。
【請求項3】
PPARγの発現調節作用が、PPARγプロモーター配列に結合することによる請求項2に記載のPPARγ発現調節剤。
【請求項4】
DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかを阻害することを特徴とする脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤。
【請求項5】
阻害が、miRNAによる請求項4に記載の脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤。
【請求項6】
miRNAの配列が、配列番号:2〜33で表される塩基配列の少なくともいずれかを有する請求項5に記載の脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤。
【請求項7】
PPARγプロモーター配列を有するレポーター遺伝子を構築するプロモーターレポーター遺伝子構築工程と、
該プロモーターレポーター遺伝子と、被験物質を含む被験試料とを細胞に導入する導入工程と、
該プロモーターレポーター遺伝子のレポーター活性を指標にして、PPARγプロモーター活性を検出する検出工程と、
該被験試料が、PPARγプロモーター活性の促進作用、及び抑制作用のいずれかの作用を有しているか否かを判断し、PPARγプロモーター活性の促進作用、及び抑制作用のいずれかの作用を有する該被験試料中の被験物質をPPARγ発現調節剤としてスクリーニングするスクリーニング工程と、
を含むことを特徴とするPPARγ発現調節剤のスクリーニング方法。
【請求項8】
プロモーターレポーター遺伝子のPPARγプロモーター配列が、該PPARγプロモーター配列が有する3箇所のD−ボックスの少なくともいずれかを欠失した欠失変異体である請求項7に記載のPPARγ発現調節剤のスクリーニング方法。
【請求項1】
配列番号:1で表される塩基配列を有することを特徴とするPPARγプロモーター配列。
【請求項2】
DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BPをコードするcDNAの少なくともいずれかを含有することを特徴とするPPARγ発現調節剤。
【請求項3】
PPARγの発現調節作用が、PPARγプロモーター配列に結合することによる請求項2に記載のPPARγ発現調節剤。
【請求項4】
DBP、TEF、HLF、及びE4BP4、並びに、該DBP、該TEF、該HLF、及び該E4BP4をコードするcDNAの少なくともいずれかを阻害することを特徴とする脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤。
【請求項5】
阻害が、miRNAによる請求項4に記載の脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤。
【請求項6】
miRNAの配列が、配列番号:2〜33で表される塩基配列の少なくともいずれかを有する請求項5に記載の脂肪分化調節剤、脂質代謝調節剤、破骨細胞機能調節剤、及び炎症調節剤。
【請求項7】
PPARγプロモーター配列を有するレポーター遺伝子を構築するプロモーターレポーター遺伝子構築工程と、
該プロモーターレポーター遺伝子と、被験物質を含む被験試料とを細胞に導入する導入工程と、
該プロモーターレポーター遺伝子のレポーター活性を指標にして、PPARγプロモーター活性を検出する検出工程と、
該被験試料が、PPARγプロモーター活性の促進作用、及び抑制作用のいずれかの作用を有しているか否かを判断し、PPARγプロモーター活性の促進作用、及び抑制作用のいずれかの作用を有する該被験試料中の被験物質をPPARγ発現調節剤としてスクリーニングするスクリーニング工程と、
を含むことを特徴とするPPARγ発現調節剤のスクリーニング方法。
【請求項8】
プロモーターレポーター遺伝子のPPARγプロモーター配列が、該PPARγプロモーター配列が有する3箇所のD−ボックスの少なくともいずれかを欠失した欠失変異体である請求項7に記載のPPARγ発現調節剤のスクリーニング方法。
【図1A】


【図1B】
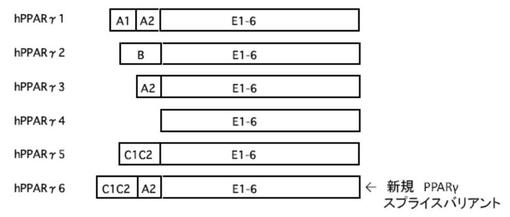
【図2】
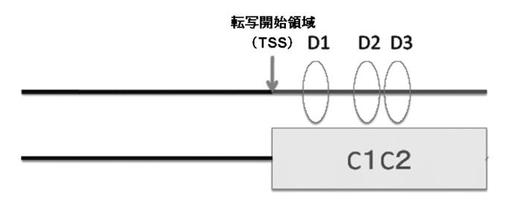
【図3】
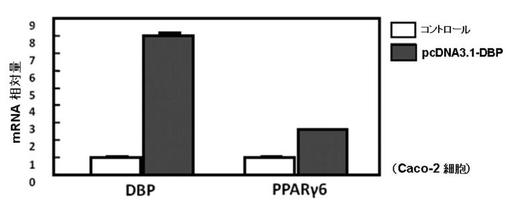
【図4A】

【図4B】
【図4C】

【図4D】
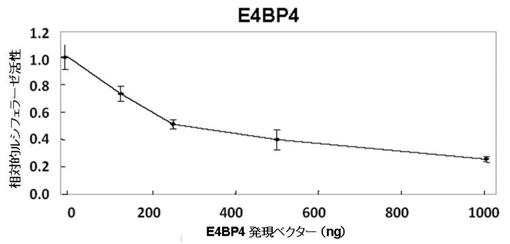
【図5A】
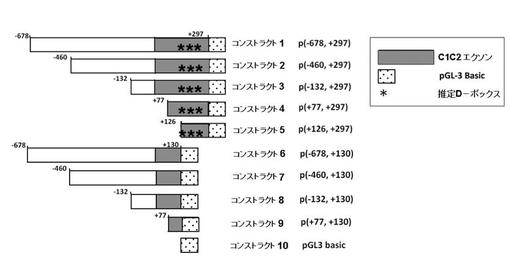
【図5B】
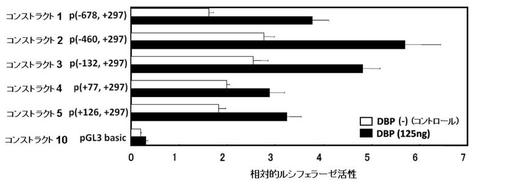
【図5C】

【図6A】

【図6B】
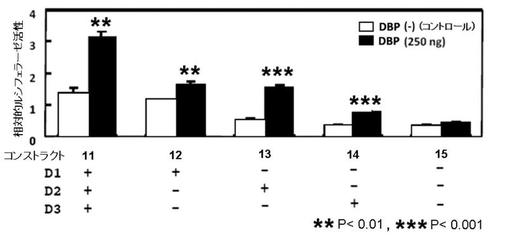
【図6C】

【図7A】

【図7B】

【図7C】
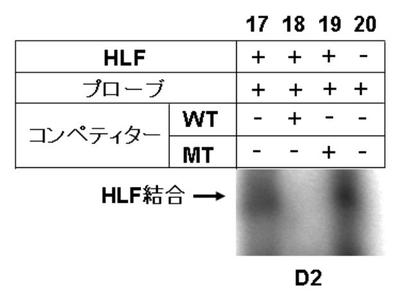
【図7D】
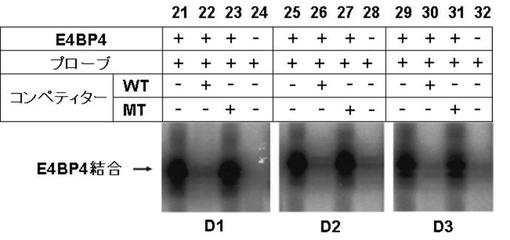
【図7E】
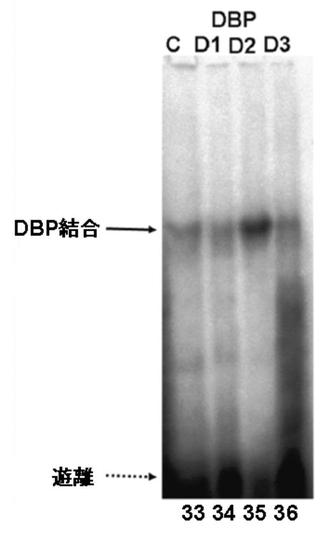
【図7F】
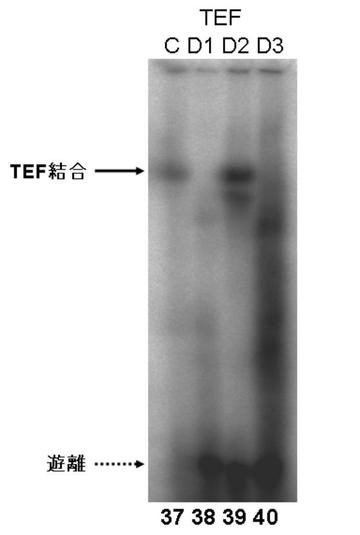
【図7G】
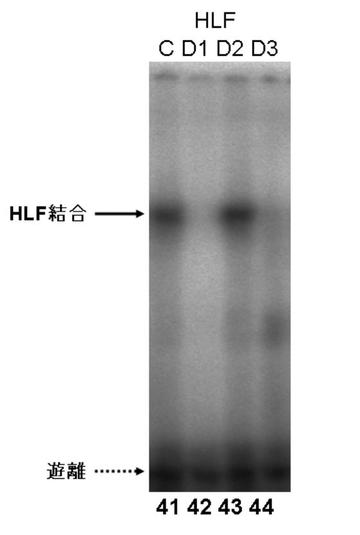
【図7H】
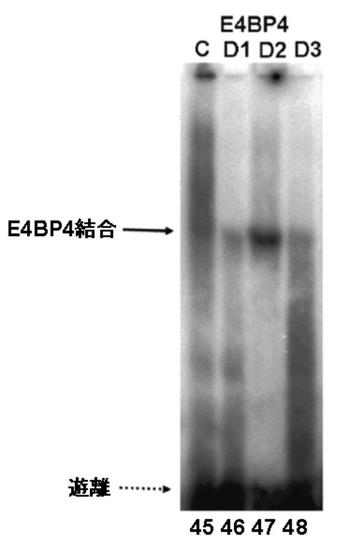
【図8】

【図9A】
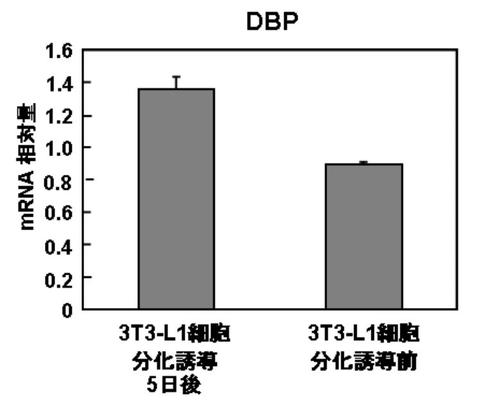
【図9B】
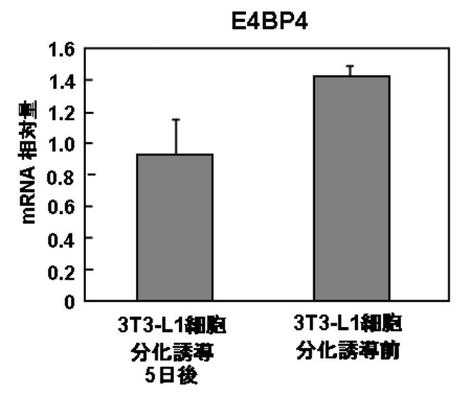
【図1B】
【図2】
【図3】
【図4A】
【図4B】
【図4C】
【図4D】
【図5A】
【図5B】
【図5C】
【図6A】
【図6B】
【図6C】
【図7A】
【図7B】
【図7C】
【図7D】
【図7E】
【図7F】
【図7G】
【図7H】
【図8】
【図9A】
【図9B】
【公開番号】特開2010−263818(P2010−263818A)
【公開日】平成22年11月25日(2010.11.25)
【国際特許分類】
【出願番号】特願2009−116845(P2009−116845)
【出願日】平成21年5月13日(2009.5.13)
【出願人】(504013775)学校法人 埼玉医科大学 (39)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
【公開日】平成22年11月25日(2010.11.25)
【国際特許分類】
【出願日】平成21年5月13日(2009.5.13)
【出願人】(504013775)学校法人 埼玉医科大学 (39)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
[ Back to top ]