
昆虫細胞におけるAAVの生成に有用なAAV−REP78の翻訳の改変型開始コドンを有するベクター
本発明は、昆虫細胞における組換えパルボウイルス(例えばアデノ随伴ウイルス)ベクターの生成用の核酸構築体、このような構築体を含む昆虫細胞、及びこれらの細胞を使用して組換えパルボウイルスビリオンを生成する方法に関する。昆虫細胞はパルボウイルスrepタンパク質をコードする第1のヌクレオチド配列を含むことが好ましく、パルボウイルスRep78タンパク質の翻訳の開始コドンは、昆虫細胞における発現後に部分的なエキソンスキッピングを行う次善の開始コドンである。昆虫細胞は、少なくとも1つのパルボウイルス(AAV)逆方向末端反復(ITR)ヌクレオチド配列を含む第2のヌクレオチド配列、及びパルボウイルスカプシドタンパク質をコードする配列を含む第3のヌクレオチド配列をさらに含む。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、昆虫細胞におけるアデノ随伴ウイルスの生成、及び昆虫細胞におけるアデノ随伴ウイルスベクターの生産性を増大させるウイルスrepタンパク質の発現及び安定性が改善されたアデノ随伴ウイルスに関する。
【背景技術】
【0002】
アデノ随伴ウイルス(AAV)は、ヒト遺伝子治療用の最も有望なウイルスベクターの1つとして考えることができる。AAVは分裂及び非分裂ヒト細胞に効率よく感染する能力を有し、AAVウイルスのゲノムは宿主細胞のゲノム中の単一の染色体部位に組み込まれ、最も重要なことに、AAVは多くのヒトに存在するのに、それは如何なる疾患とも関連しない。これらの利点に鑑みて、組換えアデノ随伴ウイルス(rAAV)はB型血友病、悪性メラノーマ、嚢胞性線維症、及び他の疾患に関する遺伝子治療の臨床試験において評価されている。
【0003】
AAVの複製をin vitroで持続させる宿主細胞は、いずれも哺乳動物細胞型に由来する。したがって、遺伝子治療において使用するためのrAAVは、これまで主に例えば293細胞、COS細胞、HeLa細胞、KB細胞、及び他の哺乳動物細胞系などの哺乳動物細胞系で生成されている(例えば、米国特許第6,156,303号、米国特許第5,387,484号、米国特許第5,741,683号、米国特許第5,691,176号、米国特許第5,688,676号、US20020081721、WO00/47757、WO00/24916、及びWO96/17947を参照)。rAAVベクターは、AAVの複製起点(逆方向末端反復即ちITR)に隣接する治療遺伝子、AAV複製タンパク質Rep78、Rep68、Rep52、及びRep40に関する遺伝子、及びビリオン又は構造タンパク質VP1、VP2、及びVP3に関する遺伝子を含むDNAプラスミドを提供することによって、このような哺乳動物細胞培養系において典型的には生成される。さらに、アデノウイルス由来の初期遺伝子(E2A、E4ORF6、VARNA)を含むプラスミドを提供してAAV遺伝子の発現を増大させ、ベクター収率を改善する(例えば、Grimm et al.、1998、Hum.Gene Ther.9:2745〜2760を参照)。しかしながら、大部分のこれらの哺乳動物細胞培養系では、細胞当たりで生じるAAV粒子の数は約104個の粒子である(Clark、2002、Kidney Int.61(Suppl.1):9〜15中で総説されている)。臨床試験用には、1015個を超えるrAAVの粒子が必要とされる可能性がある。この数のrAAV粒子を生成するためには、175cm2フラスコの5,000個の細胞に相当する、約1011個の培養ヒト293細胞を用いたトランスフェクション及び培養が必要とされる可能性があり、これは最大1011個の293細胞のトランスフェクションを意味する。したがって、臨床試験用の物質を得るための哺乳動物細胞培養系を使用したrAAVの大規模な生成は既に問題があることが証明されており、商業規模での生成はさらに現実的でない可能性がある。さらに、哺乳動物細胞培養において生成される臨床用途のベクターは、哺乳動物宿主細胞中に存在する望ましくない、おそらくは病原性の物質で汚染されるリスクが常に存在する。
【0004】
哺乳動物生成系のこれらの問題を克服するために、近年、昆虫細胞を使用するAAV生成系が開発されてきている(Urabe et al.、2002、Hum.Gene Ther.13:1935〜1943;US20030148506及びUS20040197895)。昆虫細胞におけるAAVの生成には、3つのAAVカプシドタンパク質(VP1、VP2及びVP3)の正確な化学量論値を得るためのいくつかの改変が必要であった、これは2つのスプライス受容部位の代替的使用と昆虫細胞によって正確に再現されないVP2のACG開始コドンの次善的利用の組合せに頼るものである。昆虫細胞におけるカプシドタンパク質の正確な化学量論値を模倣するために、Urabe et al.(2002、上記)は、スプライシングを必要とせずに3つすべてのVPタンパク質を発現することができ、最上流開始コドンが次善の開始コドンACGによって置換されている、1つのポリシストロン性メッセンジャーに転写される構築体を使用する。同時係属出願(PCT/NL2005/050018)において、本発明者は、昆虫細胞におけるAAVカプシドタンパク質の化学量論値のさらなる最適化による生成に基づく、バキュロウイルス生成AAVベクターの感染性をさらに改良している。
【0005】
Urabe et al.(2002、上記)により最初に開発されたAAV昆虫細胞発現系におけるAAVRepタンパク質の発現用には、それぞれ別々の昆虫細胞プロモーター、それぞれΔIE1及びPo1Hプロモーターの制御下で、2つの独立したRep発現単位(1つはRep78用であり1つはRep52用)を有する、組換えバキュロウイルス構築体が使用される。この系では、ΔIE1プロモーター、Po1Hプロモーターよりはるかに弱いプロモーターがRep78発現を促進するために選択されたが、これは、哺乳動物細胞中では、Rep52と比較したRep78の少量の発現が高いベクター収率を助長することが知られているからである(Li et al.、1997、J Virol.71:5236〜43;Grimm et al.、1998、上記)。
【0006】
しかしながら、さらに近年、Kohlbrenner et al.(2005、Mol.Ther.12:1217〜25)は、Urabe et alによって使用された2つのRepタンパク質の発現用のバキュロウイルス構築体は、固有の不安定性という欠点を有することを報告した。Urabeの原型ベクターにおける2つのRep遺伝子のパリンドローム配向を分離し、Rep52及びRep78の発現用の2つの別個のバキュロウイルスベクターを設計することによって、Kohlbrenner et al.(2005、上記)はベクターの継代安定性を増大させた。しかしながら、少なくとも5継代の間の昆虫細胞における2つの独立したバキュロウイルス−Rep構築体からRep78及びRep52が一貫して発現されるにもかかわらず、rAAVベクターの収率は、Urabe et al.(2002、上記)により設計された原型バキュロウイルス−Rep構築体と比較して5〜10倍低い。
【発明の概要】
【発明が解決しようとする課題】
【0007】
したがって、昆虫細胞におけるAAVベクターの大規模な(商業的)生成の、前述の重大な限界を克服する必要性が依然として存在する。したがって本発明の目的は、昆虫細胞におけるAAVベクターの安定した高収率の(大規模な)生成をもたらす手段及び方法を提供することである。
【課題を解決するための手段】
【0008】
発明の説明
定義
本明細書で使用する用語「作動可能に連結した」は、機能的関係のポリヌクレオチド(又はポリペプチド)要素の連結を指す。それが他の核酸配列と機能的関係に置かれているとき、核酸は「作動可能に連結している」。例えば、それがコード配列の転写に影響を与える場合、転写制御配列はコード配列と作動可能に連結している。作動可能な連結は、連結したDNA配列は典型的には隣接しており、必要な場合2つのタンパク質コード領域が接合して、隣接しリーディングフレーム内に存在することを意味する。
【0009】
「発現制御配列」は、それが作動可能に連結しているヌクレオチド配列の発現を制御する核酸配列を指す。発現制御配列がヌクレオチド配列の転写及び/又は翻訳を制御及び調節するとき、発現制御配列はヌクレオチド配列と「作動可能に連結している」。したがって発現制御配列は、プロモーター、エンハンサー、内部リボソーム侵入部位(IRES)、転写ターミネーター、タンパク質コード遺伝子の前の開始コドン、イントロンのスプライシングシグナル、及び停止コドンを含み得る。用語「発現制御配列」は、その存在が発現に影響を与えるように設計されている配列を少なくとも含むものとし、他の有利な構成要素も含み得る。例えば、リーダー配列及び融合パートナー配列は発現制御配列である。この用語は、フレーム内外の望ましくない潜在的な開始コドンが配列から除去されているような核酸配列の設計も含み得る。この用語は、望ましくない潜在的なスプライス部位が除去されているような核酸配列の設計も含み得る。この用語は、ポリA尾部、即ちmRNAの3’端における一連のアデニン残基、ポリA配列と呼ばれる配列の付加を誘導する配列、即ちポリアデニル化配列(pA)を含む。それを設計してmRNAの安定性を増大させることも可能である。転写及び翻訳の安定性に影響を与える発現制御配列、例えばプロモーター、及び翻訳を行う配列、例えばコザック配列は、昆虫細胞において知られている。発現制御配列は、より低い発現レベル又はより高い発現レベルが得られるように、作動可能に連結しているヌクレオチド配列を調節するような性質の配列でよい。
【0010】
本明細書で使用する用語「プロモーター」又は「転写調節配列」は、1つ又は複数のコード配列の転写を制御するために機能し、コード配列の転写開始部位の転写の方向に対して上流に位置し、DNA依存性RNAポリメラーゼに関する結合部位、転写開始部位、及び転写因子結合部位、リプレッサー及びアクチベータータンパク質結合部位だけには限られないが、これらを含めた任意の他のDNA配列、及び直接的又は間接的に作用してプロモーターからの転写の量を調節することが当業者に知られている、任意の他のヌクレオチド配列の存在によって構造的に確認される核酸断片を指す。「構成的」プロモーターは、大部分の生理的条件及び発生条件下において大部分の組織中で活性があるプロモーターである。「誘導性」プロモーターは、例えば化学的誘導物質の適用によって、生理的又は発生的に調節されるプロモーターである。「組織特異的」プロモーターは、特定の型の組織又は細胞中のみで活性がある。
【0011】
用語「実質的に同一」、「実質的同一性」、又は「本質的に類似」又は「本質的類似性」は、2つのペプチド又は2つのヌクレオチド配列が、初期設定のパラメーターを使用してプログラムGAP又はBESTFITなどによって最適にアラインメントをとるとき、本明細書の他の箇所で定義する少なくともある程度の割合の配列同一性を共有することを意味する。GAPはNeedleman及びWunschの大域的アラインメントアルゴリズムを使用して、2つの配列のそれらの全長でアラインメントをとり、整合の数を最大にし、ギャップの数を最少にする。一般に、GAP初期設定のパラメーター、及びギャップ生成ペナルティー=50(ヌクレオチド)/8(タンパク質)及びギャップ伸長ペナルティー=3(ヌクレオチド)/2(タンパク質)が使用される。ヌクレオチドに関しては、使用する初期設定のスコア行列はnwsgapdnaであり、及びタンパク質に関しては、初期設定のスコア行列はBlosum62である(Henikoff & Henikoff、1992、PNAS89、915〜919)。RNA配列がDNA配列と本質的に類似しているか、或いはある程度の配列同一性を有すると言えるとき、DNA配列中のチミン(T)はRNA配列中のウラシル(U)と等しいと考えられることは明らかである。配列アラインメント及びパーセント配列同一性のスコアは、Accelrys Inc.、9685 Scranton Road、San Diego、CA 92121〜3752 USAから入手可能なGCG Wisconsin Package、バージョン10.3などのコンピュータプログラム、又はオープンソースソフトウェアEmboss for Windows(登録商標)(現バージョン2.7.1〜07)を使用して決定することができる。或いは、パーセント類似性又は同一性は、例えばFASTA、BLASTなどのデータベースを検索することによって決定することができる。
【0012】
本発明のパルボウイルスRepタンパク質をコードするヌクレオチド配列は、それぞれ適度、或いは好ましくは厳密なハイブリダイゼーション条件下において、配列番号10のヌクレオチド配列とハイブリダイズするそれらの能力によって定義することもできる。厳密なハイブリダイゼーション条件は、約1Mの塩、好ましくは6×SSCを含む溶液、又は同程度のイオン強度を有する任意の他の溶液中で約65℃の温度において、少なくとも約25、好ましくは約50ヌクレオチド、75又は100及び最も好ましくは約200以上のヌクレオチドの核酸配列をハイブリダイズさせ、約0.1Mの塩、或いはそれ未満、好ましくは0.2×SSCを含む溶液、又は同程度のイオン強度を有する任意の他の溶液中で65℃において洗浄することが可能である条件として本明細書では定義する。ハイブリダイゼーションは一晩、即ち少なくとも10時間実施することが好ましく、且つ洗浄は少なくとも1時間実施し、洗浄溶液は少なくとも2回交換することが好ましい。これらの条件は、約90%以上の配列同一性を有する配列の特異的なハイブリダイゼーションを通常可能にするはずである。
【0013】
適度な条件は、約1Mの塩、好ましくは6×SSCを含む溶液、又は同程度のイオン強度を有する任意の他の溶液中で約45℃の温度において、少なくとも50ヌクレオチド、好ましくは約200以上のヌクレオチドの核酸配列をハイブリダイズさせ、約1Mの塩、好ましくは6×SSCを含む溶液、又は同程度のイオン強度を有する任意の他の溶液中で室温において洗浄することが可能である条件として本明細書では定義する。ハイブリダイゼーションは一晩、即ち少なくとも10時間実施することが好ましく、且つ洗浄は少なくとも1時間実施し、洗浄溶液は少なくとも2回交換することが好ましい。これらの条件は、50%までの配列同一性を有する配列の特異的なハイブリダイゼーションを通常可能にするはずである。当業者はこれらのハイブリダイゼーション条件を改変して、50%と90%の間で同一性が異なる配列を特異的に同定することができるはずである。
【0014】
発明の詳細な説明
本発明は、哺乳動物細胞中の核酸の導入及び/又は発現用のベクターとして使用するための、動物パルボウイルス、具体的には感染性のヒト又はサルAAVなどのデペンドウイルス属、及びそれらの構成要素(例えば、動物パルボウイルスゲノム)の使用に関する。具体的には本発明は、昆虫細胞中での生成時のこのようなパルボウイルスベクターの生産性の改善に関する。
【0015】
パルボウイルス科のウイルスは小さなDNA動物ウイルスである。パルボウイルス科は、2つの亜科:脊椎動物に感染するパルボウイルス科、及び昆虫に感染するデンソウイルス科に分けることができる。パルボウイルス亜科のメンバーは本明細書ではパルボウイルスと呼び、デペンドウイルス属を含む。その属の名称から推測することができるように、デペンドウイルスのメンバーは、それらが細胞培養中の増殖感染にアデノウイルス又はヘルペスウイルスなどのヘルパーウイルスとの同時感染を通常必要とする点で独特である。デペンドウイルス属は、ヒト(例えば、血清型1、2、3A、3B、4、5、及び6)又は霊長類(例えば、血清型1及び4)に通常感染するAAV、及び他の温血動物(例えば、ウシ、イヌ、ウマ、及びヒツジアデノ随伴ウイルス)に感染する関連ウイルスを含む。パルボウイルス及びパルボウイルス科の他のメンバーに関する他の情報は、Kenneth I.Berns、「パルボウイルス科:ウイルス及びそれらの複製(Parvoviridae:The Viruses and Their Replication)」、Chapter 69 in Fields Virology(3dEd.1996)中に記載されている。便宜上本発明は、AAVを参照することによって本明細書でさらに例示し記載する。しかしながら、本発明はAAVに限られず、他のパルボウイルスに同様に適用することができることは理解される。
【0016】
すべての知られているAAV血清型のゲノムの構成は非常に類似している。AAVのゲノムは、約5,000ヌクレオチド(nt)長未満である線状の一本鎖DNA分子である。逆方向末端反復(ITR)は、非構造複製(Rep)タンパク質及び構造(VP)タンパク質に独特なコードヌクレオチド配列と隣接している。VPタンパク質(VP1、−2及び−3)はカプシドを形成する。末端の145ntは自己相補的であり、T形状ヘアピンを形成するエネルギー的に安定な分子内二本鎖を形成することができるように構成されている。これらのヘアピン構造はウイルスDNA複製の起点として機能し、細胞DNAポリメラーゼ複合体用のプライマーとして働く。哺乳動物細胞におけるwtAAV感染の後、Rep遺伝子(即ち、Rep78及びRep52)がそれぞれP5プロモーター及びP19プロモーターから発現され、両方のRepタンパク質がウイルスゲノムの複製において機能を有する。RepのORF中のスプライシング事象は、実際4つのRepタンパク質(即ち、Rep78、Rep68、Rep52及びRep40)の発現をもたらす。しかしながら、Rep78及びRep52タンパク質をコードする非スプライシングmRNAは、哺乳動物細胞中でのAAVベクター生成に十分であることが示されてきている。昆虫細胞中でも、Rep78及びRep52タンパク質はAAVベクター生成に十分である。
【0017】
「組換えパルボウイルス又はAAVベクター」(又は「rAAVベクター」)は、1つ又は複数の対象とするポリヌクレオチド配列、対象とする遺伝子、又はパルボウイルス又はAAV逆方向末端反復配列(ITRs)に隣接する「導入遺伝子」を含むベクターを本明細書では指す。AAVrep及びcap遺伝子産物(即ち、AAVRep及びCapタンパク質)を発現している昆虫宿主細胞中に存在するとき、このようなrAAVベクターを複製し感染性ウイルス粒子にパッケージ化することができる。rAAVベクターが大きな核酸構築体(例えば、染色体中、或いはクローニング又はトランスフェクションに使用されるプラスミド又はバキュロウイルスなどの他のベクター)に組み込まれているとき、したがってrAAVベクターは典型的には「プロベクター」と呼ばれ、これはAAVパッケージ化機能及び必要なヘルパー機能の存在下での複製及びカプセル化によって「レスキュー」することができる。
【0018】
第1の態様では、本発明は、動物のパルボウイルスRepタンパク質をコードするヌクレオチド配列を含むオープンリーディングフレームを含むヌクレオチド配列であって、パルボウイルスRep78タンパク質の翻訳の開始コドンが次善開始コドンであるヌクレオチド配列に関する。次善開始コドンは、部分的なエキソンスキッピングを行う開始コドンであることが好ましい。部分的なエキソンスキッピングは、少なくとも一部分のリボソームがさらに下流の開始コドンではなくRep78タンパク質の次善開始コドンで翻訳を開始せず、好ましくはさらに下流の開始コドンがRep52タンパク質の開始コドンであることを意味すると本明細書では理解される。次善開始コドンは、昆虫細胞におけるヌクレオチド配列の発現後に部分的なエキソンスキッピングを行うことが好ましい。次善開始コドンは昆虫細胞中で部分的なエキソンスキッピングを行い、バキュロウイルス発現を使用し、好ましくは感染後約20〜40時間、より好ましくは感染後約30〜40時間で1:10〜10:1、1:5〜5:1、又は1:3〜3:1の範囲のRep78とRep52のモル比を昆虫細胞においてもたらすことが好ましい。Rep78とRep52のモル比は、好ましくはRep78とRep52両方の共通のエピトープを認識するモノクローナル抗体を使用して、或いは実施例1.1.3中に記載した抗体を使用して、実施例1.1.3中に記載したのと同様にウェスタンブロッティングによって決定することができる。
【0019】
用語「次善開始コドン」は、本明細書では3ヌクレオチドの開始コドン自体のみでなく、その状況も指す。したがって、次善開始コドンは、次善の状況、例えばコザックでない状況での「最適」ATGコドンからなっていてよい。しかしながら、より好ましいのは、3ヌクレオチドの開始コドン自体が次善、即ちATGではない次善開始コドンである。次善は本明細書では、正常ATGコドンと比較して他の同一状況での翻訳の開始時に、コドンが有効性が低いことを意味すると理解される。次善コドンの有効性は、他の同一状況での正常ATGコドンの有効性の90、80、60、40又は20%未満であることが好ましい。翻訳開始の相対的有効性を比較するための方法は、本質的に当業者に知られている。好ましい次善開始コドンは、ACG、TTG、CTG、及びGTGから選択することができる。より好ましくはACGである。
【0020】
動物のパルボウイルスRepタンパク質をコードするヌクレオチド配列は、Rep78及びRep52タンパク質などの昆虫細胞におけるパルボウイルスベクター生成に必要とされ十分である非構造Repタンパク質をコードするヌクレオチド配列として本明細書では理解する。動物パルボウイルスのヌクレオチド配列はデペンドウイルスに由来することが好ましく、ヒト又はサルアデノ随伴ウイルス(AAV)に由来することがより好ましく、ヒト(例えば、血清型1、2、3A、3B、4、5、及び6)又は霊長類(例えば、血清型1及び4)に通常感染するAAVに由来することが最も好ましい。動物のパルボウイルスRepタンパク質をコードするヌクレオチド配列の一例は配列番号10で与え、これはRepタンパク質をコードするAAV血清型2の配列ゲノムの一部分を示す。Rep78コード配列はヌクレオチド11〜1876を含み、Rep52コード配列はヌクレオチド683〜1876を含む。Rep78及びRep52タンパク質の正確な分子量、及び翻訳開始コドンの正確な位置は、異なるパルボウイルス間で異なる可能性があることは理解される。しかしながら当業者は、AAV−2以外のパルボウイルス由来のヌクレオチド配列中の対応する位置の確認の仕方を知っているはずである。動物のパルボウイルスRepタンパク質をコードするヌクレオチド配列は、したがって:
a)配列番号11のアミノ酸配列と少なくとも50、60、70、80、88、89、90、95、97、98、又は99%の配列同一性を有するアミノ酸配列を含むポリペプチドをコードするヌクレオチド配列、
b)配列番号10の11〜1876位のヌクレオチド配列と少なくとも50、60、70、80、81、82、85、90、95、97、98、又は99%の配列同一性を有するヌクレオチド配列、
c)その相補鎖が(a)又は(b)の核酸分子配列とハイブリダイズするヌクレオチド配列、
d)遺伝コードの縮重のために(c)の核酸分子の配列とその配列が異なるヌクレオチド配列
として定義することもできる。
【0021】
ヌクレオチド配列は、昆虫細胞におけるパルボウイルスベクター生成に必要とされ十分である動物のパルボウイルスRepタンパク質をコードすることが好ましい。
【0022】
本発明の他の好ましいヌクレオチド配列は、パルボウイルスRep78タンパク質をコードするヌクレオチド配列の開始コドンの上流に、配列番号7の9ヌクレオチドの配列、又は配列番号7と実質的に相同なヌクレオチド配列を含む発現制御配列を含む。配列番号7のヌクレオチド配列と実質的同一性を有し、パルボウイルスRep78タンパク質の発現の増大を助長するはずである配列は、例えば配列番号7の9ヌクレオチドの配列と少なくとも60%、70%、80%又は90%の同一性を有する配列である。
【0023】
他のパルボウイルスの、Rep78及びRep52翻訳開始部位以外の、Repタンパク質コード配列中の偽性と考えられる翻訳開始部位の除去は、昆虫細胞において認識され得る推定スプライス部位の除去と同様に、当業者によって十分理解されているはずである。昆虫細胞中での正確な発現のための野生型パルボウイルス配列の様々な改変は、例えばSambrook and Russell(2001)「分子クローニング:研究室用マニュアル(Molecular Cloning:A Laboratory Manual)」(第3版)、Cold Spring Harbor Laboratory、Cold Spring Harbor Laboratory Press、ニューヨーク中に記載された技術などの、よく知られている遺伝子工学技術の適用によって実施される。Repタンパク質コード領域の様々な他の改変は当業者に知られており、これらはRepタンパク質の収率を増大させ得る。これらの改変は本発明の範囲内にある。
【0024】
さらなる態様では、本発明は、上記で定義したパルボウイルスRepタンパク質をコードするヌクレオチド配列を含む核酸構築体に関する。構築体中では、パルボウイルスRepタンパク質をコードするヌクレオチド配列は、昆虫細胞中での発現用の発現制御配列と作動可能に連結していることが好ましい。これらの発現制御配列は、昆虫細胞中で活性があるプロモーターを少なくとも含むはずである。昆虫宿主細胞中で外来遺伝子を発現させるための当業者に知られている技術を使用して、本発明を実施することができる。昆虫細胞におけるポリペプチドの分子工学及び発現に関する方法は、例えばSummers and Smith.1986.「バキュロウイルスベクター及び昆虫培養手順に関する方法のマニュアル(A Manual of Methods for Baculovirus Vectors and Insect Culture Procedures)」、Texas Agricultural Experimental Station Bull.No.7555、College Station、Tex.;Luckow.1991.In Prokop et al.、「バキュロウイルスベクターを用いた昆虫細胞における異種遺伝子のクローニング及び発現(Cloning and Expression of Heterologous Genes in Insect Cells with Baculovirus Vectors)」「組換えDNA技術及び適用(Recombinant DNA Technology and Applications)」、97〜152;King、L.A.and R.D.Possee、1992、「バキュロウイルス発現系(The baculovirus expression system)」、Chapman and Hall、United Kingdom;O’Reilly、D.R.、L.K.Miller、V.A.Luckow、1992、「バキュロウイルス発現ベクター(Baculovirus Expression Vectors)」:「研究室用マニュアル(A Laboratory Manual)」、ニューヨーク;W.H.Freeman and Richardson、C.D.、1995、「バキュロウイルス発現のプロトコル、分子生物学における方法(Baculovirus Expression Protocols,Methods in Molecular Biology)」、volume39;米国特許第4,745,051号;US2003148506;及びWO03/074714中に記載されている。パルボウイルスRepタンパク質をコードする本発明のヌクレオチド配列の転写に特に適したプロモーターは、例えば多角体プロモーターである。しかしながら、昆虫細胞中で活性がある他のプロモーター、例えばpl0、p35、IE−1又はΔIE−1プロモーター、及び前述の参照文献中に記載された他のプロモーターが、当技術分野で知られている。
【0025】
昆虫細胞におけるパルボウイルスRepタンパク質の発現用の核酸構築体は、昆虫細胞適合性ベクターであることが好ましい。「昆虫細胞適合性ベクター」又は「ベクター」は、昆虫又は昆虫細胞の生産的な形質転換又はトランスフェクションが可能である核酸分子であると理解される。代表的な生物学的ベクターには、プラスミド、線状核酸分子、及び組換えウイルスがある。それが昆虫細胞適合性である限り、任意のベクターを利用することができる。ベクターを昆虫細胞ゲノムに組み込むことはできるが、昆虫細胞中のベクターの存在は不変的である必要はなく、一時的なエピソームベクターも含まれる。ベクターは知られている任意の手段によって、例えば細胞の化学処理、エレクトロポレーション、又は感染によって導入することができる。好ましい実施形態では、ベクターはバキュロウイルス、ウイルスベクター、又はプラスミドである。より好ましい実施形態では、ベクターはバキュロウイルスであり、即ち、構築体はバキュロウイルスベクターである。バキュロウイルスベクター及びそれらの使用法は、昆虫細胞の分子工学に関する前に引用した参照文献中に記載されている。
【0026】
他の態様では、本発明は、1つのパルボウイルスRepタンパク質をコードする単一のオープンリーディングフレームを含む唯一の型のヌクレオチド配列を含む昆虫細胞に関する。単一のオープンリーディングフレームは1つ又は複数のパルボウイルスRepタンパク質をコードすることが好ましく、オープンリーディングフレームはパルボウイルスRepタンパク質のすべてをコードすることがより好ましく、オープンリーディングフレームは、好ましくは昆虫細胞において少なくともRep52とRep78タンパク質の両方がそこから発現され得る完全長Rep78タンパク質をコードすることが最も好ましい。昆虫細胞は、例えば多コピーエピソームベクターでは、1つの型のヌクレオチド配列の2つ以上のコピーを含み得るが、これらは本質的に1つ及び同一の核酸分子、又は1つ及び同一のRepアミノ酸配列をコードする少なくともいくつかの核酸分子、例えば、遺伝コードの縮重のために相互間でわずかに異なる核酸分子の多コピーであることは、本明細書では理解される。パルボウイルスRepタンパク質をコードする唯一の型の核酸分子の存在は、昆虫細胞におけるパルボウイルス生成のレベル(安定性)に影響を与える欠陥Rep発現構築体を生成する可能性がある、Rep配列を含む異なる型のベクター中に存在し得る相同的配列間の組換えを回避する。好ましくは、昆虫細胞では、1つ又は複数のパルボウイルスRepタンパク質をコードする単一のオープンリーディングフレームを含むヌクレオチド配列が核酸構築体の一部分であり、ヌクレオチド配列が昆虫細胞中での発現用の発現制御配列と作動可能に連結していることが好ましい。さらに好ましい昆虫細胞は、パルボウイルスRepタンパク質をコードする、上記で定義したヌクレオチド配列、好ましくは上記で定義した次善開始コドンを有するコード配列、又は上記で定義した核酸構築体を、「第1の」ヌクレオチド配列として含み、或いは昆虫細胞は、このようなヌクレオチド配列を含む上記で定義した核酸構築体を、「第1の」核酸構築体として含む。
【0027】
組換えパルボウイルス(rAAV)ベクターの複製を可能にし、培養中に維持することができる任意の昆虫細胞を、本発明によって使用することができる。例えば、使用する細胞系は、スポドプテラフルギベルダ(Spodoptera frugiperda)、ショウジョウバエの細胞系、又は蚊細胞系、例えばアエデスアルボピクタス(Aedes albopictus)派生細胞系由来であってよい。好ましい昆虫細胞又は細胞系は、例えばSe301、SeIZD2109、SeUCR1、Sf9、Sf900+、Sf21、BTI−TN−5B1−4、MG−1、Tn368、HzAm1、Ha2302、Hz2E5、High Five(Invitrogen、CA、USA)及びexpresSF+(登録商標)(米国特許第6,103,526号;Protein Sciences Corp.、CT、USA)を含めた、バキュロウイルス感染に感受性である昆虫種由来の細胞である。
【0028】
本発明による好ましい昆虫細胞は、前に記載した「第1の」ヌクレオチド配列又は核酸構築体以外に、
a)少なくとも1つのパルボウイルス逆方向末端反復(ITR)ヌクレオチド配列を含む第2のヌクレオチド配列と、
b)昆虫細胞における発現用の発現制御配列と作動可能に連結しているパルボウイルスCapタンパク質コード配列を含む第3のヌクレオチド配列と
をさらに含む。
【0029】
本発明の文脈では、「少なくとも1つのパルボウイルスITRヌクレオチド配列」は、「A」、「B」、及び「C」領域とも呼ばれる、大部分が相補的であり、対称的に配列した配列を含むパリンドローム配列を意味すると理解される。ITRは複製起点、複製における「シス」の役割を有する部位、即ち、パリンドローム配列及びパリンドロームに固有の特異的配列を認識する、例えばRep78(又はRep68)などの複製タンパク質として作用するトランスの認識部位である部位として機能する。ITR配列の対称性の1つの例外はITRの「D」領域である。それは独特である(1つのITR内に相補配列を有していない)。一本鎖DNAの切断は、A領域とD領域の間の接合部で起こる。それは新たなDNA合成が始まる領域である。D領域は通常パリンドロームの片側に位置し、核酸複製ステップに対する方向性をもたらす。哺乳動物細胞において複製するパルボウイルスは、2つのITR配列を典型的には有する。しかしながら、結合部位がA領域の両鎖上に存在し、D領域は対称的に、1つがパリンドロームの各側に位置するように、ITRを工学処理することは可能である。二本鎖環状DNA鋳型(例えば、プラスミド)上では、Rep78又はRep68支援核酸複製が次いで両方向に進行し、1つのITRは環状ベクターのパルボウイルス複製に十分である。したがって、1つのITRヌクレオチド配列は本発明の文脈で使用することができる。しかしながら、2個又は他の偶数個の通常のITRを使用することが好ましい。2個のITR配列を使用することが最も好ましい。好ましいパルボウイルスのITRはAAVのITRである。安全上の理由で、細胞中への最初の導入後にさらに増殖することができない組換えパルボウイルス(rAAV)ベクターを構築することが望ましい可能性がある。レシピエント中の望ましくないベクター増殖を制限するためのこのような安全メカニズムは、US2003148506中に記載されたのと同様に、キメラITRを有するrAAVを使用することによって提供することができる。
【0030】
組換えパルボウイルス(rAAV)ベクターの生成用に昆虫細胞中で利用する核酸構築体の数は、本発明中では無制限である。例えば、1個、2個、3個、4個、5個、或いはそれより多くの別個の構築体を利用して、本発明の方法に従い昆虫細胞中でrAAVを生成することができる。5個の構築体を利用する場合、1個の構築体はAAVVP1をコードし、他の構築体はAAVVP2をコードし、さらに他の構築体はAAVVP3をコードし、さらに他の構築体は上記で定義したRepタンパク質をコードし、且つ最終構築体は少なくとも1個のAAVITRを含む。5個より少ない構築体を利用する場合、構築体は少なくとも1個のAAVITR、及びVP1、VP2、VP3、及びRepタンパク質コード配列の様々な組合せを含むことができる。2個の構築体又は3個の構築体を使用することが好ましく、前に記載したのと同様の2個の構築体がより好ましい。2個の構築体を使用する場合、昆虫細胞は(a)上記で定義したRepタンパク質の発現用の第1の核酸構築体であって、上記の(b)で定義した第3のヌクレオチド配列をさらに含む構築体(昆虫細胞中での発現のために少なくとも1個の発現制御配列と作動可能に連結しているパルボウイルスCapタンパク質コード配列を含む;以下も参照)、及び(c)上記の(a)で定義した第2のヌクレオチド配列を含む第2の核酸構築体(少なくとも1個のパルボウイルス/AAVITRヌクレオチド配列を含む)を含むことが好ましい。3個の構築体を使用する場合、カプシドタンパク質の発現とRepタンパク質の発現に別々の構築体を使用すること以外は、2個の構築体に使用するのと同じ形状を使用することが好ましい。それぞれの構築体における配列は、互いに対して任意の順序であってよい。例えば、1個の構築体がVPカプシドタンパク質をコードするヌクレオチド配列を含むITR及びORFを含む場合、ITR配列間のDNAの複製によって、VPのORFが複製されるか或いは複製されないように、構築体上にVPのORFが局在する可能性がある。他の例では、Repコード配列及び/又はVPカプシドタンパク質をコードするヌクレオチド配列を含むORFは、構築体上で任意の順序であってよい。さらに第2、第3及び他の核酸構築体は昆虫細胞適合性ベクターであることが好ましく、前に記載したバキュロウイルスベクターであることが好ましいことが理解される。或いは、本発明の昆虫細胞では、1つ又は複数の第1のヌクレオチド配列、第2のヌクレオチド配列、第3のヌクレオチド配列、及び第4のヌクレオチド配列及び任意選択の他のヌクレオチド配列を、昆虫細胞のゲノムに安定して組み込むことができる。当業者は、昆虫ゲノム中にヌクレオチド配列を安定して導入する方法、及びゲノム中にこのようなヌクレオチド配列を有する細胞を同定する方法を知っている。ゲノム中への組込みは、例えば昆虫ゲノムの領域と非常に相同性が高いヌクレオチド配列を含むベクターの使用によって助長することができる。トランスポゾンなどの特異的配列の使用は、ゲノム中にヌクレオチド配列を導入する他の方法である。
【0031】
本発明では、パルボウイルスカプシド(Cap)タンパク質コード配列を含む第3のヌクレオチド配列は、3つのパルボウイルスカプシドタンパク質、VP1、−2及び−3のそれぞれをコードする配列を含むと本明細書では理解される。カプシドタンパク質コード配列を含む第3のヌクレオチド配列は、様々な形で存在してよい。例えば、カプシドタンパク質、VP1、−2及び−3のそれぞれに関する別個のコード配列を使用することができ、それぞれのコード配列は昆虫細胞中での発現のために発現制御配列と作動可能に連結している。しかしながら、より好ましくは、第3のヌクレオチド配列は、全3個の動物パルボウイルス(AAV)VP1、VP2、及びVP3カプシドタンパク質をコードする1個のオープンリーディングフレームを含み、VP1カプシドタンパク質の翻訳の開始コドンは、例えばUrabe et al.(2002、上記)によって記載されたのと同様のATGではない次善開始コドンである。VP1カプシドタンパク質に関する次善開始コドンは、Rep78タンパク質に関して上記で定義したのと同様であってよい。VP1カプシドタンパク質に関するより好ましい次善開始コドンはACG、TTG、CTG及びGTGから選択することができ、この中ではCTG及びGTGが最も好ましい。カプシドタンパク質の発現に好ましい第3のヌクレオチド配列は、VP1カプシドタンパク質をコードするヌクレオチド配列の開始コドンの上流に、配列番号7の9ヌクレオチドの配列又は配列番号7と実質的に相同的なヌクレオチドの配列を含む発現制御配列をさらに含む。配列番号7のヌクレオチド配列と実質的同一性を有し、VP1の発現の増大を助長するはずである配列は、例えば配列番号7の9ヌクレオチドの配列と少なくとも60%、70%、80%又は90%の同一性を有する配列である。カプシドタンパク質の発現に好ましい他の第3のヌクレオチド配列は、12ヌクレオチド位におけるC、21ヌクレオチド位におけるA、及び24ヌクレオチド位におけるC(翻訳開始コドンの第1のヌクレオチドである位置1を基準とする;配列番号1を参照)から選択されるVP1カプシドタンパク質をコードするヌクレオチド配列の少なくとも1つの改変をさらに含むことが好ましい。他の血清型のVP1の翻訳の偽性と考えられる開始コドンの除去は、昆虫細胞において認識され得る推定スプライス部位の除去と同様に、当業者によって十分理解されているはずである。VPコード領域の様々な他の改変は当業者に知られており、それらはVP及びビリオンの収率を増大させるか、或いは指向性の変化などの他の望ましい影響を有するか、或いはビリオンの抗原性を低下させるいずれかの可能性がある。これらの改変は本発明の範囲内にある。好ましくは、パルボウイルスカプシドタンパク質をコードする本発明のヌクレオチド配列は、昆虫細胞中での発現のために発現制御配列と作動可能に連結しており、発現制御配列は昆虫細胞中で活性があるプロモーターを少なくとも含むはずである。昆虫宿主細胞中でパルボウイルスカプシドタンパク質を発現させるための、このような制御配列及び他の技術及び物質(例えばベクター)は、Repタンパク質について上記で既に記載している。
【0032】
本発明の好ましい実施形態では、本発明の昆虫細胞中に存在する第2のヌクレオチド配列、即ち少なくとも1つのパルボウイルス(AAV)ITRを含む配列は、対象とする遺伝子産物をコードする少なくとも1つのヌクレオチド配列をさらに含み、対象とする遺伝子産物をコードする少なくとも1つのヌクレオチド配列は、昆虫細胞において生成される組換えパルボウイルス(rAAV)ベクターのゲノムに組み込まれることが好ましい。対象とする遺伝子産物をコードする少なくとも1つのヌクレオチド配列は、哺乳動物細胞中での発現用の配列であることが好ましい。第2のヌクレオチド配列は2つのパルボウイルス(AAV)ITRヌクレオチド配列を含み、対象とする遺伝子産物をコードする少なくとも1つのヌクレオチド配列は、2つのパルボウイルス(AAV)ITRヌクレオチド配列間に位置することが好ましい。それが2つの通常のITRの間に位置するか、或いは2カ所のD領域で工学処理したITRの片側に位置する場合、(哺乳動物細胞中での発現用の)対象とする遺伝子産物をコードするヌクレオチド配列は、昆虫細胞において生成される組換えパルボウイルス(rAAV)ベクターに組み込まれることが好ましい。
【0033】
本明細書で上記で定義した第2のヌクレオチド配列は、昆虫細胞中で複製される組換えパルボウイルス(rAAV)ベクターに組み込まれるように位置する、昆虫細胞中での発現用の少なくとも1つの「対象とする遺伝子産物」をコードするヌクレオチド配列をしたがって含み得る。任意のヌクレオチド配列は、本発明の方法に従い生成した組換えパルボウイルス(rAAV)ベクターでトランスフェクトした哺乳動物細胞中での、後の発現用に組み込むことが可能である。ヌクレオチド配列は例えばタンパク質をコードすることができ、それがRNAi物質、即ちRNA干渉することができるRNA分子、例えばshRNA(低分子ヘアピンRNA)又はsiRNA(低分子干渉RNA)などを発現することができる。「siRNA」は、哺乳動物細胞中では無毒である短い長さの二本鎖RNAである小分子の干渉RNAを意味する(Elbashir et al.、2001、Nature411:494〜98;Caplen et al.、2001、Proc.Natl.Acad.Sci.USA98:9742〜47)。好ましい実施形態では、第2のヌクレオチド配列は2つのヌクレオチド配列を含むことができ、それぞれが哺乳動物細胞中での発現用の対象とする1つの遺伝子産物をコードする。対象とする産物をコードする2つのヌクレオチド配列のそれぞれは、昆虫細胞中で複製される組換えパルボウイルス(rAAV)ベクターに組み込まれるように位置する。
【0034】
哺乳動物細胞中での発現用の対象とする産物は、治療用遺伝子産物であってよい。治療用遺伝子産物は、標的細胞中で発現されると、例えば望ましくない活性の除去、例えば感染細胞の除去、又は例えば酵素活性の欠失を引き起こす遺伝的欠陥の補足などの望ましい治療効果をもたらす、ポリペプチド、又はRNA分子(siRNA)、又は他の遺伝子産物であってよい。治療用ポリペプチド遺伝子産物の例には、CFTR、因子IX、リポタンパク質リパーゼ(LPL、好ましくはLPL S447X;WO01/00220参照)、アポリポタンパク質Al、ウリジン二リン酸グルクロノシルトランスフェラーゼ(UGT)、網膜色素変性GTPaseレギュレーター相互作用タンパク質(RP−GRIP)、及びサイトカイン又は例えばIL−10のようなインターロイキンがある。
【0035】
代替的或いは追加的に、第2の遺伝子産物として、本明細書で上記で定義した第2のヌクレオチド配列は、細胞の形質転換及び発現を評価するためのマーカータンパク質として働くポリペプチドをコードするヌクレオチド配列を含むことができる。この目的に適したマーカータンパク質は、例えば蛍光タンパク質GFP、及び選択可能なマーカー遺伝子HSVチミジンキナーゼ(HAT培地における選択用)、細菌ハイグロマイシンBホスホトランスフェラーゼ(ハイグロマイシンBにおける選択用)、Tn5アミノグリコシドホスホトランスフェラーゼ(G418における選択用)、及びジヒドロ葉酸還元酵素(DHFR)(メトトレキセートにおける選択用)、CD20、低親和性の神経成長因子遺伝子である。これらのマーカー遺伝子を得るための供給源、及びそれらを使用するための方法は、Sambrook and Russell(2001)「分子クローニング:研究室用マニュアル(Molecular Cloning:A Laboratory Manual)」(第3版)、Cold Spring Harbor Laboratory、Cold Spring Harbor Laboratory Press、ニューヨーク中に示されている。さらに、本明細書で上記で定義した第2のヌクレオチド配列は、必要であると考えられる場合、本発明の組換えパルボウイルス(rAAV)ベクターで形質導入した細胞由来の対象を治癒することができるフェイルセーフ機構として働くことができる、ポリペプチドをコードするヌクレオチド配列を含むことができる。しばしば自殺遺伝子と呼ばれる、このようなヌクレオチド配列は、タンパク質が発現されるトランスジェニック細胞を死滅させることができる毒性物質にプロドラッグを変換する可能性があるタンパク質をコードする。このような自殺遺伝子の適切な例には、例えば大腸菌のシトシンデアミナーゼ遺伝子、又は単純ヘルペスウイルス、サイトメガロウイルス及び水痘帯状疱疹ウイルス由来のチミジンキナーゼ遺伝子の1つがあり、この場合ガンシクロビルをプロドラッグとして使用して、対象中のトランスジェニック細胞を死滅させることができる(例えば、Clair et al.、1987、Antimicrob.Agents Chemother.31:844〜849を参照)。
【0036】
他の実施形態では、対象とする遺伝子産物の1つはAAVタンパク質であり得る。特に、Rep78又はRep68などのRepタンパク質、又はその機能断片。Rep78及び/又はRep68をコードするヌクレオチド配列は、本発明の組換えパルボウイルス(rAAV)ベクターのゲノム上に存在し、ベクターで形質導入した哺乳動物細胞中で発現される場合、形質導入した哺乳動物細胞のゲノムへの組換えパルボウイルス(rAAV)ベクターの組込みを可能にする。rAAV形質導入又は感染哺乳動物細胞中でのRep78及び/又はRep68の発現は、ベクターによって細胞中に導入される対象とする任意の他の遺伝子産物の長期又は永続的な発現を可能にすることによって、組換えパルボウイルス(rAAV)ベクターのいくつかの用途に関する利点をもたらす可能性がある。
【0037】
本発明の組換えパルボウイルス(rAAV)ベクターでは、哺乳動物細胞中での発現用の対象とする遺伝子産物をコードする少なくとも1つのヌクレオチド配列は、少なくとも1つの哺乳動物細胞適合性の発現制御配列、例えばプロモーターと作動可能に連結していることが好ましい。多くのこのようなプロモーターは、当技術分野で知られている(Sambrook and Russel、2001、上記を参照)。CMVプロモーターなどの、多くの細胞型において広範囲で発現される構成的プロモーターを使用することができる。しかしながら、より好ましいのは、誘導性、組織特異的、細胞型特異的、又は細胞周期特異的であるプロモーターであるはずである。例えば、肝臓特異的な発現に関しては、α1−アンチトリプシンプロモーター、甲状腺ホルモン結合グロブリンプロモーター、アルブミンプロモーター、LPS(チロキシン結合グロブリン)プロモーター、HCR−ApoCIIハイブリッドプロモーター、HCR−hAATハイブリッドプロモーター及びアポリポタンパク質Eプロモーターからプロモーターを選択することができる。他の例には、腫瘍選択的、及び特に神経細胞腫瘍選択的発現用のE2Fプロモーター(Parr et al.、1997、Nat.Med.3:1145〜9)、又は単核血液細胞において使用するためのIL−2プロモーターがある(Hagenbaugh et al.、1997、J Exp Med;185:2101〜10)。
【0038】
AAVはいくつかの哺乳動物細胞に感染することができる。例えば、Tratschin et al.(1985、Mol.Cell Biol.5:3251〜3260)及びGrimm et al.(1999、Hum.Gene Ther.10:2445〜2450)を参照。しかしながら、ヒト滑膜繊維芽細胞のAAV形質導入は同様のネズミ細胞中より著しく有効であり、Jennings et al.、Arthritis Res、3:1(2001)、AAVの細胞指向性は血清型間で異なる。例えば、哺乳動物のCNS細胞指向性及び形質導入効率に関するAAV2、AAV4、及びAAV5の間の違いを論じている、Davidson et al.(2000、Proc.Natl.Acad.Sci.USA、97:3428〜3432)を参照されたい。
【0039】
昆虫細胞中で組換えAAVベクターを生成するための本発明の方法において使用することができるAAV配列は、任意のAAV血清型のゲノムに由来してよい。一般に、AAV血清型はアミノ酸及び核酸レベルで相当な相同性のゲノム配列を有し、一組の同一な遺伝的機能をもたらし、物理的及び機能的にほぼ等しいビリオンを生成し、実際同一であるメカニズムによって複製及び構築する。様々なAAV血清型のゲノム配列及びゲノム類似性の総説に関しては、例えば、GenBankアクセッション番号U89790;GenBankアクセッション番号J01901;GenBankアクセッション番号AF043303;GenBankアクセッション番号AF085716;Chlorini et al.(1997、J.Vir.71:6823〜33);Srivastava et al.(1983、J.Vir.45:555〜64);Chlorini et al.(1999、J.Vir.73:1309〜1319);Rutledge et al.(1998、J.Vir.72:309〜319);及びWu et al.(2000、J.Vir.74:8635〜47)を参照。AAV血清型1、2、3、4及び5は、本発明の文脈で使用するためのAAVヌクレオチド配列の好ましい供給源である。本発明の文脈で使用するためのAAVのITR配列は、AAV1、AAV2、及び/又はAAV4に由来することが好ましい。同様に、Rep(Rep78及びRep52)コード配列は、AAV1、AAV2、及び/又はAAV4に由来することが好ましい。本発明の文脈で使用するためのVP1、VP2、及びVP3カプシドタンパク質をコードする配列は、しかしながら、任意の知られている42の血清型から、より好ましくはAAV1、AAV2、AAV3、AAV4、AAV5、AAV6、AAV7、AAV8又はAAV9、或いは例えばカプシドシャッフリング技術及びAAVカプシドライブラリーによって入手される、新たに開発されたAAV様粒子から得ることができる。
【0040】
AAVのRep及びITR配列は、大部分の血清型の間で特に保存されている。様々なAAV血清型のRep78タンパク質は例えば89%を超えて同一であり、AAV2、AAV3A、AAV3B、及びAAV6の間のゲノムレベルでの全体のヌクレオチド配列の同一性は約82%である(Bantel−Schaal et al.、1999、J.Virol.、73(2):939〜947)。さらに、多くのAAV血清型のRep配列及びITRは、哺乳動物中でのAAV粒子の生成において、他の血清型由来の対応する配列と効率よく相互補完する(即ち、機能的に置換する)ことが知られている。US2003148506は、AAVのRep及びITR配列は、昆虫細胞中の他のAAVのRep及びITR配列とも効率よく相互補完することを報告している。
【0041】
AAVVPタンパク質は、AAVビリオンの細胞指向性を決定することが知られている。VPタンパク質コード配列は、異なるAAV血清型間のRepタンパク質及び遺伝子ほど有意に保存されていない。他の血清型の対応する配列と相互補完するRep及びITR配列の能力は、1つの血清型(例えば、AAV3)のカプシドタンパク質及び他のAAV血清型(例えば、AAV2)のRep及び/又はITR配列を含む偽型rAAV粒子の生成をもたらす。このような偽型rAAV粒子は、本発明の一部分である。
【0042】
改変「AAV」配列も、例えば昆虫細胞においてrAAVベクターを生成するために、本発明の文脈で使用することができる。このような改変配列は、例えばAAV1、AAV2、AAV3、AAV4、AAV5、AAV6、AAV7、AAV8又はAAV9のITR、Repと少なくとも約70%、少なくとも約75%、少なくとも約80%、少なくとも約85%、少なくとも約90%、少なくとも約95%、又はそれより高いヌクレオチド及び/又はアミノ酸配列同一性を有する配列(例えば、約75〜99%のヌクレオチド配列同一性を有する配列)を含み、或いはVPを野生型AAVのITR、Rep、又はVP配列の代わりに使用することができる。
【0043】
多くの点で他のAAV血清型と類似しているが、AAV5は他の知られているヒト及びサルの血清型より、他のヒト及びサルのAAV血清型と異なる。それを考慮すると、rAAV5の生成は昆虫細胞における他の血清型の生成と異なる可能性がある。本発明の方法を利用してrAAV5を生成する場合、1つ又は複数の構築体は、2つ以上の構築体の場合をひとまとめにして、AAV5ITRを含むヌクレオチド配列、AAV5Repコード配列を含むヌクレオチド配列(即ち、ヌクレオチド配列はAAV5Rep78を含む)を含むことが好ましい。このようなITR及びRep配列を望むように改変して、昆虫細胞におけるrAAV5又は偽型rAAV5ベクターの有効な生成を得ることができる。例えば、Rep配列の開始コドンを改変することができ、VPスプライス部位を改変又は除去することができ、且つ/或いはVP1開始コドン及び隣接ヌクレオチドを改変して、昆虫細胞におけるrAAV5ベクターの生成を改善することができる。
【0044】
したがって、他の態様では、本発明は昆虫細胞において(上記で定義した組換えパルボウイルス(rAAV)ベクターを含む)組換えパルボウイルス(rAAV)ビリオンを生成するための方法に関する。この方法は、(a)組換えパルボウイルス(rAAV)ベクターが生成するような条件下において本明細書で上記で定義した昆虫細胞を培養するステップと、(b)組換えパルボウイルス(rAAV)ベクターを回収するステップとを含むことが好ましい。この方法で生成される組換えパルボウイルス(rAAV)ベクターは、組換えパルボウイルス(rAAV)ベクター核酸を含む感染性パルボウイルス又はAAVビリオンであることが好ましいことは、本明細書において理解される。培養中の昆虫細胞に関する増殖条件、及び培養中の昆虫細胞における異種産物の生成は当技術分野でよく知られており、例えば昆虫細胞の分子工学に関する前に引用した参照文献中に記載されている。
【0045】
この方法は、抗AAV抗体、好ましくは固定化抗体を使用する、組換えパルボウイルス(rAAV)ベクター(を含むビリオン)の親和性精製のステップをさらに含むことが好ましい。抗AAV抗体は、モノクローナル抗体であることが好ましい。特に適切な抗体は、例えばラクダ又はラマなどから入手可能な単鎖ラクダ抗体又はその断片である(例えば、Muyldermans、2001、Biotechnol.74:277〜302を参照)。rAAVの親和性精製用の抗体は、AAVカプシドタンパク質上のエピトープと特異的に結合する抗体であることが好ましく、エピトープは2つ以上のAAV血清型のカプシドタンパク質上に存在するエピトープであることが好ましい。例えば抗体は、AAV2カプシドとの特異的結合に基づいて産生又は選択することができるが、さらに同時に、抗体はAAV1、AAV3及びAAV5カプシドとも特異的に結合することができる。
【0046】
他の態様では、本発明は、上記で定義した核酸構築体及び細胞を使用して、前に記載した本発明の方法において生成されるrAAVビリオンに関する。rAAVビリオンは、そのゲノム中に対象とする遺伝子産物をコードする少なくとも1つのヌクレオチド配列を含むことが好ましく、少なくとも1つのヌクレオチド配列は原型AAVヌクレオチド配列ではなく、AAVVP1、VP2、及びVP3カプシドタンパク質の化学量論では、VP1の量は:(a)VP2の量の少なくとも100、105、110、120、150、200又は400%であり、或いは(b)VP3の量の少なくとも8、10、10.5、11、12、15、20又は40%であり、或いは(c)少なくとも(a)と(b)の両方で定義した量である。VP1、VP2及びVP3の量は、VP1、VP2及びVP3のそれぞれに共通であるエピトープを認識する抗体を使用して測定することが好ましい。VP1、VP2及び/又はVP3の相対量の定量化を可能にするはずである、様々なイムノアッセイが当技術分野で利用可能である(例えば、「抗体の使用(Using Antibodies)」、E.Harlow and D.Lane、1999、Cold Spring Harbor Laboratory Press、ニューヨークを参照)。3つのカプシドタンパク質のそれぞれに共通であるエピトープを認識するのに適した抗体は、例えば(Progen、ドイツから市販されている)マウス抗CapB1抗体である。本発明による好ましいrAAVビリオンは、そのゲノム中に対象とする遺伝子産物をコードする少なくとも1つのヌクレオチド配列を含むビリオンであり、少なくとも1つのヌクレオチド配列は原型AAVヌクレオチド配列ではなく、AAVビリオンは1アミノ酸位にロイシン又はバリンを含むVP1カプシドタンパク質を含む。本発明によるさらに好ましいAAVビリオンは上記で定義したカプシドタンパク質の比を有しており、1アミノ酸位にロイシン又はバリンを含むVP1カプシドタンパク質を含む。
【0047】
本文書中及びその特許請求の範囲中において、動詞「含む」及びその活用は、その語句の後の項目が含まれるが、具体的に言及されていない項目が除外されるわけではないことを意味するように、非限定的な意味で使用する。さらに、不定冠詞「a」又は「an」による要素に対する言及は、1つ及びただ1つの要素が存在することを状況が明らかに必要としない限り、2つ以上の要素が存在する可能性を除外するわけではない。したがって不定冠詞「a」又は「an」は、「少なくとも1つ」を通常意味する。
【図面の簡単な説明】
【0048】
【図1A】Rep発現における野生型AAVゲノムの構成の図である。Rep78及びRep52遺伝子は、それぞれP5及びP19プロモーターから発現される。(それぞれRep78及びRep52のスプライス変異体である)Rep68及びRep40の発現は示さない。両方の発現単位がATG開始部位を含む。
【図1B】本発明の構築体が、1つのプロモーター(例えば、多角体(PolH)プロモーター)の制御下でRepのORFを有することを示す図である。このプロモーターはRep78とRep52両方の発現を誘導するが、これは、Rep78の開始コドンATGが他のACG開始コドンに変換され、リボソームによって一部分がスキップされるからである。
【図1C】Urabe et al(2002、上記)による原型構築体は、2つの異なるプロモーター(それぞれΔIE1及びpolH)から独立にRep78及びRep52を誘導することを示す図である。
【図2】昆虫細胞で5回継代した組換えバキュロウイルスから発現された、Repタンパク質のウェスタンブロット分析の図である。Urabe et al.、2002によって設計された原型バキュロウイルス(原型REP/Bac−to−Bac)は、5回の継代でRep78/52発現のゆっくりした低下をもたらす。バキュロウイルス骨格PSC(原型REP/PSC)に挿入した、Urabe et al.、2002によって設計されたRep78及び52の発現単位も、昆虫細胞での継代後にRep78/52発現の低下をもたらす。しかしながら、PSC骨格中にACG開始コドンを含むREP発現単位を有するバキュロウイルス(REP−ACG/PSC)は、少なくとも5回の継代でRep78/52の安定した発現をもたらす。ウェスタンブロット分析は、実施例1.1.3中に記載したように実施した。
【図3】グラフ中にプロットした表1の結果の図である。
【図4】昆虫細胞における様々なrAAV構築体の安定性の比較の図である。SF+細胞におけるrAAV生成は、実施例1中で前に記載したように実施した。すべての生成に関して、ITR含有バキュロウイルスとカプシド遺伝子含有バキュロウイルスは同一であり、継代数はRep遺伝子含有バキュロウイルスと同じであった。4つの異なるRep遺伝子含有バキュロウイルスを使用した:1)REP−ACG/PSC、2)SLR:Urabe et al.(2002、上記)による原型構築体、3)Rep52+Rep78(B2B):2つの別個のBac−to−Bacバキュロウイルス、1つはRep78遺伝子を含み、もう1つはRep52遺伝子を含む。4)Rep52+Rep78(PSC):2つの別個のプロテインサイエンスのバキュロウイルス、1つはRep78遺伝子を含み、もう1つはRep52遺伝子を含む。
【図5】第8代までのREP−ACG/PSCバキュロウイルス構築体の安定性の図である。SF+細胞におけるrAAV生成は、実施例1中で記載したように実施した。
【図6】Urabe et al.(2002、上記)からの原型構築体及び本発明によるREP−ACG/PSC構築体のrepタンパク質発現に対する、継代効果の影響の比較の図である。バキュロウイルスの継代及びウェスタンブロットは、実施例1中に記載したように実施した。repバキュロウイルスの正常な継代の間に、SF細胞にバキュロウイルスを加えた後40時間でサンプルを採取し、ウェスタンブロットを実施した。
【発明を実施するための形態】
【実施例】
【0049】
(実施例1)
Rep構築体
1.1.材料及び方法
1.1.1 バキュロウイルスプラスミドの構築
1つの2シストロン性メッセンジャーRNAからRep78及びRep52を発現させるために、発現ベクターpFastBacDualSLR(Urabe et al.、2002、上記)上に位置するRep78のATG開始コドンをACGに転換した。使用した上流プライマーは以下の通りであった:
BamHI
5'-cgcggatcctgttaagACGGCGGGGTTTTACGAGATTGTGATTAAGGTC-3'
(配列番号8)
プライマー配列、正方向
【0050】
PCR反応中で使用した3’プライマーはREP78遺伝子に隣接しており、XbaI部位(TCTAGA)を含む:
XbaI
5'-AGGCTCTAGATTCGAAAGCGGCCCG-3'
(配列番号9)
プライマー配列、逆方向
【0051】
前述のプライマーセット間の配列を、以下のプロトコル:95℃、5分間の1サイクル;95℃、15秒間の35サイクル;55℃、30秒間;72℃、2分間;72℃、10分間の1サイクル;4℃、恒久的を使用して、PCR(反応体積50μl;1×PfX Amp.バッファー、0.3mMのdNTP’s、1mMのMgSO4、150mMの正方向プライマー、150mMの逆方向プライマー、2×エンハンサー溶液、鋳型50ng(pFastBacDualSLR)、1UのPlatinum Pfx(Invitrogen、Carlsbad、CA、USA))によって増幅した。PCR産物は、Zero Blunt TOPO PCRクローニングキット(Invitrogen)を使用してPCR平滑末端II−TOPOでクローニングした。Rep78は制限部位SpeI及び×baIを使用してpFastBacDual(Invitrogen)にサブクローニングした。突然変異Rep発現カセットは、(制限酵素BstZ171及びAvrIIを使用して)、バキュロウイルス発現構築体(EcoRV及びXbaIで切断した)pPSC10(Protein Sciences Corporation、Meriden、CT、USA)に最終的にクローニングした。構築体の配列分析は、Baseclear、Leiden、オランダによって確認した。
【0052】
1.1.2 組換えバキュロウイルスの生成
オートグラファカリフォルニカ(Autographa californica)核多角体病ウイルス(AcNPV)由来の組換えバキュロウイルスを、GeneXpress BaculoKIT(Protein Sciences Corporation)を使用して生成した。トランスフェクションは以下のように実施した:丸底14m1チューブ内で200μlのGRACE培地を6μlのセルフェクチン(Invitrogen)と混合し、エッペンドルフチューブ内で200μlのGRACE培地を、50μlのウイルスDNA(プロテインサイエンス)及び2μgのトランスファープラスミド(REP)と混合した。エッペンドルフチューブからの内容物をチューブに加え、注意深く混合した。室温で30分間のインキュベーション後、1,300μlのGRACEをトランスフェクション混合物に加えた。T25フラスコ中の昆虫細胞はGRACE培地で洗浄し、トランスフェクション混合物は細胞層に一滴ずつ加えた。28℃で6時間のインキュベーション後、10%のFBSを補ったSF900II血清を注意深く加え、T25フラスコは5日間28℃のストーブ内に置き、その後組換えバキュロウイルスを採取した。
【0053】
1.1.3 ウェスタンブロット分析
バキュロウイルス−REPを用いて昆虫細胞(SF+)を感染させた。細胞感染後16、40、及び64時間でサンプルを採取し、0.1V10×TRIS溶解バッファー(1.5MのNaCl、0.5MのTRIS、0.01MのMgCl、1%のTRITON X−100、pH8.5、濾過滅菌済)を加えることによって細胞を溶かし、シェーカー(Innova 44、New Brunswick)内で28℃において30分間インキュベートした。30分間37℃でのベンゾナーゼとのインキュベーションによって、遊離DNA及びRNAを分解した。細胞溶解物は遠心分離にかけた(1,900×g;15分間;4℃)。NuPAGE LDSサンプルバッファー(4×、Invitrogen)を上清のサンプルに加え、4〜12%のBis−Trisゲル(120V)に載せた。タンパク質は、10V(半乾燥ブロッティング)で30分間PVDF膜(BioRad)にブロッティングした。ウェスタンブロットによる免疫化学測定法を、Superblock−PBSブロッキングバッファー(PIERCE)で膜をブロッキングし、マウス抗Rep(303.9、Progen、ドイツ;希釈1:50)及びウサギ抗マウスHRP(DAKO、希釈1:500)と後にインキュベートすることによって実施した。lumi−light plusウェスタンブロッティング基質(Roche)を用いた化学発光染色によって、Rep−タンパク質を視覚化した。
【0054】
1.2 結果
本発明の新たに設計したRep−構築体(REP−ACG/PSC)の成績を、1)PSCバキュロウイルス骨格と2)Bac−to−Bacバキュロウイルス骨格(Urabe et al.、2002)の両方の原型Rep構築体と比較した。全3個の構築体を第5代まで連続して継代した。AAV1−LPL生成の実験は、それぞれ第2、3、4及び5代のAAV−LPL及びAAV−Cap組換えバキュロウイルスと組合せた、第2、3、4及び5代のRep−構築体を使用して実施した(ここで使用したAAV−LPL及びAAV−Cap組換えバキュロウイルスは、実施例2中で以下に記載する)。AAV1−LPLの生成収率はqPCRによって測定し、表1中に示す。Urabe et al.、2002によって設計された原型バキュロウイルス(原型REP/Bac−to−Bac)は、5回の継代でAAV生成の急速な低下をもたらす。バキュロウイルス骨格PSC(原型REP/PSC)に挿入した、Urabe et al.、2002によって設計されたRepの発現単位も、昆虫細胞での継代後にAAV生成の低下をもたらす。しかしながら、PSC骨格中にACG開始コドンを含むRep発現単位を有するバキュロウイルス(REP−ACG/PSC)は、少なくとも5回の継代で安定したAAV生成をもたらす。したがって、数回の継代(例えば、第2代〜第5代)におけるAAV−LPLの再現性のある生成収率は、REP−ACG構築体を含むバキュロウイルスを使用してのみ得られた。
【0055】
【表1】
表1:数回継代したバキュロウイルス構築体を使用したrAAVビリオンの生成:Sf9細胞を、第2、3、4又は5代のLPL−ベクター単位、第2、3、4又は5代のRep−発現単位、及び第2、3、4又は5代のCap−発現単位をコードする3つの組換えバキュロウイルスで感染させた。3日後に細胞を採取し、AAV収率(1ml当たりのベクターゲノム;vg/ml)をqPCRによって測定した。
【0056】
【表2】
表2:昆虫細胞の継代(第2代〜第5代)後に様々なBac−Rep構築体で実施したQ−PCR。
【0057】
表2は、組換えバキュロウイルスにおけるRep−発現単位用、及び隣接するバキュロウイルスORF用に設計した、定量PCR(Q−PCR)アッセイの結果を示す(1ml当たりの遺伝子コピー数;gc/ml)。Q−PCRの値の間の比は、Rep−バキュロウイルス中の欠失の存在を決定する。1という比は、バッチ中のすべてのバキュロウイルスが組換えRep78又は52配列を含むことを理論上意味する。Urabe et al.、2002によって設計された原型バキュロウイルス(原型REP/Bac−to−Bac)は、第5代での相当量の組換えバキュロウイルスがRep配列中の欠失を有することを示す。バキュロウイルス骨格PSC(原型REP/PSC)に挿入した、Urabe et al.、2002によって設計されたRep78及び52用の発現単位は、組換えバキュロウイルスの非常に初期の劇的な消失を示す。しかしながら、PSC骨格中にACG開始コドンを含むRep発現単位を有するバキュロウイルス(REP−ACG/PSC)(クローンC4及びA3)は、少なくとも5回の継代で安定した組換えバキュロウイルスを示す。
【0058】
(実施例2)
Cap構築体
2.1.1 バキュロウイルスプラスミドの構築
1つのポリシストロン性メッセンジャーRNAからVP1、2、3を発現させるために、(Urabe et al.、2002、上記)によって記載されたのと同様にバキュロウイルス−AAV−Cap構築体を設計した。簡単に言うと、VP1のATG開始コドンをACGに突然変異させた。11位の考えられるATG開始コドンはACGに変わった。VP1開始コドンの下流のスプライス受容体部位を破壊した(21及び24位における突然変異)。突然変異Cap発現カセットは、BamH1/StuI制限部位を有するバキュロウイルス発現構築体;pfastBacDual(pFBDAAV1VPm11)にクローニングした。このプラスミド(pFBDAAV1VPm11)は、VP1用の他の開始コドンを導入するための出発物質であった。前述の突然変異を導入するためにUrabe et al.(2002、上記)によって使用された正方向プライマーは、以下の通りであった:
BamHI 1 11 21 24
5'-cgcggatcctgttaagACGGCTGCCGACGGTTATCTACCCGATTGGCTC-3'
(配列番号1)
【0059】
出発物質としてpFBDAAV1VPm11(Urabe et al.、2002、上記)を使用して発現構築体を作製するために、以下の正方向プライマーを使用した:
5'-cgcggatcctgttaagTTGGCTGCCGACGGTTATCTACCCGATTGGCTC-3'
(配列番号2)
5'-cgcggatcctgttaagATTGCTGCCGACGGTTATCTACCCGATTGGCTC-3'
(配列番号3)
5'-cgcggatcctgttaagGTGGCTGCCGACGGTTATCTACCCGATTGGCTC-3'
(配列番号4)
5'-cgcggatcctgttaagCTGGCTGCCGACGGTTATCTACCCGATTGGCTC-3'
(配列番号5)
【0060】
前述の正方向プライマーと共にPCR反応中で使用した逆方向プライマーは、VP1開始コドンの下流230bpまでの位置を対象とし、特有のStuI部位(AGGCCT)を含む。
5'-GTCGTAGGCCTTGTCGTGCTCGAGGGCCGC-3'
(配列番号6)
【0061】
PCRにより正方向と逆方向プライマー対の前述のセットを用いて断片を増幅した。BamHI及びStuIを用いたPCR産物の消化後、PCR産物はpFBDAAV1vpm11のBamHI/StuI部位にサブクローニングし、様々な試験用バキュロウイルス−AAV−Cap構築体を生成した。DNA構築体はBaseclear、Leiden、オランダでの配列分析によって確認した。
【0062】
2.1.2 組換えバキュロウイルスの生成
オートグラファカリフォルニカ(Autographa californica)核多角体病ウイルス(AcNPV)由来の組換えバキュロウイルスを、Bac−to−Bacバキュロウイルス発現系(Invitrogen)を使用して生成した。0.1のmoiで1ml当たり2×106個のSf9細胞を感染させることによってrBac−Capを増幅した。感染後3日で細胞を遠心し、ウイルスを含む上清を回収した。
【0063】
2.1.3 組換えAAVの生成
Urabe et al.、2002による3つの組換えバキュロウイルスを使用してrAAVバッチを生成した。しかしながら、この試験に関しては、1つのバキュロウイルスはLPLS447X−導入遺伝子の発現構築体を有していた。この導入遺伝子から発現される治療活性物質は、ヒトリポタンパク質リパーゼの天然に存在する変異体、448アミノ酸の単鎖ポリペプチドである。LPLS447Xの変異体は、タンパク質のC末端に2アミノ酸の欠失を有する。第2のバキュロウイルスはAAV複製遺伝子、Rep78及びRep52の発現構築体を有していた。第3のバキュロウイルスはAAV1カプシド配列及びVP1のACG又はTTG、CTG、GTG開始コドンのいずれかを有していた。
【0064】
プラスミド−トランスフェクション系を用いて生成した哺乳動物rAAVのバッチは、Grimm et al.、1998(「組換えアデノ随伴ウイルスベクターを生成及び精製するための新規のツール(Novel tools for production and purification of recombinant adeno−associated virus vectors)」Hum Gene Ther.1998 Dec10;9(18):2745〜60)に従い生成した。
【0065】
2.1.3 ウェスタンブロット分析
バキュロウイルス−Capを用いて昆虫細胞を感染させた。感染後3日で、細胞を遠心分離にかけた(3,000g;15分間)。上清は0.22umのMillexフィルターを介して濾過した。NuPAGE LDSサンプルバッファー(Invitrogen)を上清のサンプルに加え、4〜12%のBis−Trisゲル(120V)に載せた。ゲルは100Vで施した。タンパク質は1時間、100V、350mAでニトロセルロース膜(BioRad)にブロッティングした。ウェスタンブロットによる免疫化学測定法を、1%マーヴェル、乾燥スキムミルクで膜をブロッキングし、マウス抗Cap(Progen、ドイツからのB1;希釈1:50)及びウサギ抗マウス−HRP(DAKO、希釈1:100)と後にインキュベートすることによって実施した。発光及びウェスタンブロッティング基質(Roche)を用いた化学発光染色によって、VP1、2及び3を視覚化した。
【0066】
2.1.4 生化学的測定
放射性トリオレオイルグリセロールエマルジョン基質を使用して以前に記載されたのと同様に(Nilsson−Ehle and Scholtz、1976)、ヒトLPLS447Xの活性をアッセイした。ヒトLPLS447Xの免疫反応体を、ニワトリIgY及びマウス5D2抗hLPL抗体(Liu et at.、2000)を用いたサンドウィッチELISAを使用してアッセイした。血漿トリグリセリドレベルは、製造者のプロトコル(Boehringer Mannheim、#450032)に従い市販のキットを使用することによって測定した。
【0067】
2.2 結果
2.2.1 組換えバキュロウイルスの構築
Urabe et al.(2002、上記)により設計されたバキュロウイルスプラスミド中でのVP1の発現用の異なる他の開始コドンを導入するために、BamHI制限部位及びVP1のACG開始コドンの代わりにTTG、ATT、GTG又はCTGコドンのいずれかを含む一連の上流プライマーを設計した。StuI部位を含む下流プライマーと組合せてこれらのプライマーを使用したPCRによって、いくつかの増幅断片を生成し、これらはpFBDVPm11(Bac−Cap)のBamHI/StuI部位にサブクローニングした。生成したバキュロウイルスプラスミドは、Bac−to−Bacバキュロウイルス発現系を使用する組換えバキュロウイルスの調製に使用した。調製した組換えバキュロウイルスは昆虫細胞に感染させて、AAVカプシドを生成した。感染後3日で、異なるバキュロウイルスバッチのウイルスタンパク質の発現を、ウェスタンブロットで測定した。ウェスタンブロットから、VP1のTTG開始コドンを含むバキュロウイルス構築体は、以前に使用したACG開始コドンと比較して高いレベルで、このタンパク質を発現したことが明らかになった。TTGコドンを使用したVP1とVP2の間の比は、野生型AAVに関して報告される比と同様に1:1であったことが分かった(示さず)。
【0068】
2.2.2 培養中の細胞におけるrAAVバッチの感染
TTG開始コドンを有する組換えバキュロウイルス由来のAAVカプシドの感染性を調べるために、rAAVを作製した。さらにrAAVバッチを、哺乳動物HEK293細胞におけるプラスミドトランスフェクションによって作製した。両rAAVバッチのベクターゲノム力価をqPCRによって測定した。この力価を使用して、高いmoiでマイクロタイタープレート中のHEK293細胞を感染させた。感染後2日で、導入遺伝子産物(LPLS447X)に関する定量アッセイ(LPLS447X−質量アッセイ)を、感染細胞の培地で実施した。このアッセイは、バキュロウイルス生成rAAVによって生成したLPLS447Xの量は、プラスミド生成rAAVによって生成したLPLと同等であったことを示した(示さず)。
【0069】
2.2.3 マウスにおけるrAAVバッチの注射
バキュロウイルス生成系、及び従来の哺乳動物プラスミド生成系によって生成したrAAVバッチをマウスの筋肉内に注射して、in vivoでのLPLS447X−タンパク質活性及びトリグリセリド非摂取を追跡した。注射後第3日、第7日及び第2週で、血液サンプルを採取し評価した。ウイルス投与後第3日と第7日の間に、哺乳動物rAAVを注射した両マウス及びバキュロrAAVを注射した1マウスからサンプル採取した血液−血漿は、乳濁状から完全に透明な状態に変わった。1匹のバキュロrAAV注射マウス由来の血液血漿は依然として比較的乳濁状であったが、しかしながら脂肪レベルは明らかに低下した。トリグリセリドレベルは、すべての治療マウスのそれぞれで低下した(示さず)。第14日に、両方の哺乳動物rAAV及びバキュロウイルス−(TTG)−AAV治療マウスにおけるTGレベルは、TGレベルが96%低下した。ウイルス投与後2週間で採取した血漿サンプルは、バキュロウイルス−AAVと哺乳動物−AAVで治療したマウスのLPLS447X−活性は、同等であったことを示した(示さず)。
【0070】
(実施例3)
昆虫細胞における改変型Rep78開始コドンを有するrAAV構築体の安定性
3.1 昆虫細胞における様々なrAAV構築体の安定性の比較
SF+細胞におけるrAAV生成は、実施例1中に前に記載したのと同様に実施した。すべての生成に関して、ITR含有バキュロウイルスとカプシド遺伝子含有バキュロウイルスは同一であり、継代数はRep遺伝子含有バキュロウイルスと同じであった。4つの異なるRep遺伝子含有バキュロウイルス:1)REP−ACG/PSC、2)SLR:Urabe et al.(2002、上記)による原型構築体、3)Rep52+Rep78(B2B):1つがRep78遺伝子を含みもう1つがRep52遺伝子を含む、2つの別個のBac−to−Bacバキュロウイルス、4)Rep52+Rep78(PSC):1つがRep78遺伝子を含みもう1つがRep52遺伝子を含む、2つの別個のプロテインサイエンスのバキュロウイルスを使用した。
【0071】
結果は図4中に示す。5回目のバキュロウイルス継代したrAAV生成は、原型Rep構築体と比較して、及び分岐Rep構築体と比較して、本発明によるREP−ACG/PSCを使用して10倍を超えて既に改善されている。
【0072】
3.2 第8代までのバキュロウイルス構築体の安定性
SF+細胞におけるrAAV生成は、実施例1中に記載したのと同様に実施した。すべての生成に関して、ITR含有バキュロウイルスとカプシド遺伝子含有バキュロウイルスは同一であり、継代数はREP−ACG/PSCバキュロウイルスと同じであった。結果は図5中に示す。REP−ACG/PSCバキュロウイルスは少なくとも第8代まで安定性がある。REP−ACG/PSCのrAAV生成力価は、少なくとも第8代のバキュロウイルスまで安定している。
【0073】
3.3 repタンパク質発現に対する継代の影響
Urabe et al.(2002、上記)からの原型構築体Repタンパク質の発現に対する継代数の影響を、本発明によるREP−ACG/PSC構築体と比較した。バキュロウイルスの継代及びウェスタンブロットは、実施例1中に記載したように実施した。repバキュロウイルスの正常な継代の間に、SF細胞にバキュロウイルスを加えた後40時間でサンプルを採取し、ウェスタンブロットを実施した。図6は、原型Urabe構築体(SLR)の初期継代と比較した多数回継代の低下したRep発現を明らかに示すが、一方REP−ACG/PSC構築体におけるRep発現は、少数回の継代と比較して多数回の継代で同じ状態である。
【技術分野】
【0001】
本発明は、昆虫細胞におけるアデノ随伴ウイルスの生成、及び昆虫細胞におけるアデノ随伴ウイルスベクターの生産性を増大させるウイルスrepタンパク質の発現及び安定性が改善されたアデノ随伴ウイルスに関する。
【背景技術】
【0002】
アデノ随伴ウイルス(AAV)は、ヒト遺伝子治療用の最も有望なウイルスベクターの1つとして考えることができる。AAVは分裂及び非分裂ヒト細胞に効率よく感染する能力を有し、AAVウイルスのゲノムは宿主細胞のゲノム中の単一の染色体部位に組み込まれ、最も重要なことに、AAVは多くのヒトに存在するのに、それは如何なる疾患とも関連しない。これらの利点に鑑みて、組換えアデノ随伴ウイルス(rAAV)はB型血友病、悪性メラノーマ、嚢胞性線維症、及び他の疾患に関する遺伝子治療の臨床試験において評価されている。
【0003】
AAVの複製をin vitroで持続させる宿主細胞は、いずれも哺乳動物細胞型に由来する。したがって、遺伝子治療において使用するためのrAAVは、これまで主に例えば293細胞、COS細胞、HeLa細胞、KB細胞、及び他の哺乳動物細胞系などの哺乳動物細胞系で生成されている(例えば、米国特許第6,156,303号、米国特許第5,387,484号、米国特許第5,741,683号、米国特許第5,691,176号、米国特許第5,688,676号、US20020081721、WO00/47757、WO00/24916、及びWO96/17947を参照)。rAAVベクターは、AAVの複製起点(逆方向末端反復即ちITR)に隣接する治療遺伝子、AAV複製タンパク質Rep78、Rep68、Rep52、及びRep40に関する遺伝子、及びビリオン又は構造タンパク質VP1、VP2、及びVP3に関する遺伝子を含むDNAプラスミドを提供することによって、このような哺乳動物細胞培養系において典型的には生成される。さらに、アデノウイルス由来の初期遺伝子(E2A、E4ORF6、VARNA)を含むプラスミドを提供してAAV遺伝子の発現を増大させ、ベクター収率を改善する(例えば、Grimm et al.、1998、Hum.Gene Ther.9:2745〜2760を参照)。しかしながら、大部分のこれらの哺乳動物細胞培養系では、細胞当たりで生じるAAV粒子の数は約104個の粒子である(Clark、2002、Kidney Int.61(Suppl.1):9〜15中で総説されている)。臨床試験用には、1015個を超えるrAAVの粒子が必要とされる可能性がある。この数のrAAV粒子を生成するためには、175cm2フラスコの5,000個の細胞に相当する、約1011個の培養ヒト293細胞を用いたトランスフェクション及び培養が必要とされる可能性があり、これは最大1011個の293細胞のトランスフェクションを意味する。したがって、臨床試験用の物質を得るための哺乳動物細胞培養系を使用したrAAVの大規模な生成は既に問題があることが証明されており、商業規模での生成はさらに現実的でない可能性がある。さらに、哺乳動物細胞培養において生成される臨床用途のベクターは、哺乳動物宿主細胞中に存在する望ましくない、おそらくは病原性の物質で汚染されるリスクが常に存在する。
【0004】
哺乳動物生成系のこれらの問題を克服するために、近年、昆虫細胞を使用するAAV生成系が開発されてきている(Urabe et al.、2002、Hum.Gene Ther.13:1935〜1943;US20030148506及びUS20040197895)。昆虫細胞におけるAAVの生成には、3つのAAVカプシドタンパク質(VP1、VP2及びVP3)の正確な化学量論値を得るためのいくつかの改変が必要であった、これは2つのスプライス受容部位の代替的使用と昆虫細胞によって正確に再現されないVP2のACG開始コドンの次善的利用の組合せに頼るものである。昆虫細胞におけるカプシドタンパク質の正確な化学量論値を模倣するために、Urabe et al.(2002、上記)は、スプライシングを必要とせずに3つすべてのVPタンパク質を発現することができ、最上流開始コドンが次善の開始コドンACGによって置換されている、1つのポリシストロン性メッセンジャーに転写される構築体を使用する。同時係属出願(PCT/NL2005/050018)において、本発明者は、昆虫細胞におけるAAVカプシドタンパク質の化学量論値のさらなる最適化による生成に基づく、バキュロウイルス生成AAVベクターの感染性をさらに改良している。
【0005】
Urabe et al.(2002、上記)により最初に開発されたAAV昆虫細胞発現系におけるAAVRepタンパク質の発現用には、それぞれ別々の昆虫細胞プロモーター、それぞれΔIE1及びPo1Hプロモーターの制御下で、2つの独立したRep発現単位(1つはRep78用であり1つはRep52用)を有する、組換えバキュロウイルス構築体が使用される。この系では、ΔIE1プロモーター、Po1Hプロモーターよりはるかに弱いプロモーターがRep78発現を促進するために選択されたが、これは、哺乳動物細胞中では、Rep52と比較したRep78の少量の発現が高いベクター収率を助長することが知られているからである(Li et al.、1997、J Virol.71:5236〜43;Grimm et al.、1998、上記)。
【0006】
しかしながら、さらに近年、Kohlbrenner et al.(2005、Mol.Ther.12:1217〜25)は、Urabe et alによって使用された2つのRepタンパク質の発現用のバキュロウイルス構築体は、固有の不安定性という欠点を有することを報告した。Urabeの原型ベクターにおける2つのRep遺伝子のパリンドローム配向を分離し、Rep52及びRep78の発現用の2つの別個のバキュロウイルスベクターを設計することによって、Kohlbrenner et al.(2005、上記)はベクターの継代安定性を増大させた。しかしながら、少なくとも5継代の間の昆虫細胞における2つの独立したバキュロウイルス−Rep構築体からRep78及びRep52が一貫して発現されるにもかかわらず、rAAVベクターの収率は、Urabe et al.(2002、上記)により設計された原型バキュロウイルス−Rep構築体と比較して5〜10倍低い。
【発明の概要】
【発明が解決しようとする課題】
【0007】
したがって、昆虫細胞におけるAAVベクターの大規模な(商業的)生成の、前述の重大な限界を克服する必要性が依然として存在する。したがって本発明の目的は、昆虫細胞におけるAAVベクターの安定した高収率の(大規模な)生成をもたらす手段及び方法を提供することである。
【課題を解決するための手段】
【0008】
発明の説明
定義
本明細書で使用する用語「作動可能に連結した」は、機能的関係のポリヌクレオチド(又はポリペプチド)要素の連結を指す。それが他の核酸配列と機能的関係に置かれているとき、核酸は「作動可能に連結している」。例えば、それがコード配列の転写に影響を与える場合、転写制御配列はコード配列と作動可能に連結している。作動可能な連結は、連結したDNA配列は典型的には隣接しており、必要な場合2つのタンパク質コード領域が接合して、隣接しリーディングフレーム内に存在することを意味する。
【0009】
「発現制御配列」は、それが作動可能に連結しているヌクレオチド配列の発現を制御する核酸配列を指す。発現制御配列がヌクレオチド配列の転写及び/又は翻訳を制御及び調節するとき、発現制御配列はヌクレオチド配列と「作動可能に連結している」。したがって発現制御配列は、プロモーター、エンハンサー、内部リボソーム侵入部位(IRES)、転写ターミネーター、タンパク質コード遺伝子の前の開始コドン、イントロンのスプライシングシグナル、及び停止コドンを含み得る。用語「発現制御配列」は、その存在が発現に影響を与えるように設計されている配列を少なくとも含むものとし、他の有利な構成要素も含み得る。例えば、リーダー配列及び融合パートナー配列は発現制御配列である。この用語は、フレーム内外の望ましくない潜在的な開始コドンが配列から除去されているような核酸配列の設計も含み得る。この用語は、望ましくない潜在的なスプライス部位が除去されているような核酸配列の設計も含み得る。この用語は、ポリA尾部、即ちmRNAの3’端における一連のアデニン残基、ポリA配列と呼ばれる配列の付加を誘導する配列、即ちポリアデニル化配列(pA)を含む。それを設計してmRNAの安定性を増大させることも可能である。転写及び翻訳の安定性に影響を与える発現制御配列、例えばプロモーター、及び翻訳を行う配列、例えばコザック配列は、昆虫細胞において知られている。発現制御配列は、より低い発現レベル又はより高い発現レベルが得られるように、作動可能に連結しているヌクレオチド配列を調節するような性質の配列でよい。
【0010】
本明細書で使用する用語「プロモーター」又は「転写調節配列」は、1つ又は複数のコード配列の転写を制御するために機能し、コード配列の転写開始部位の転写の方向に対して上流に位置し、DNA依存性RNAポリメラーゼに関する結合部位、転写開始部位、及び転写因子結合部位、リプレッサー及びアクチベータータンパク質結合部位だけには限られないが、これらを含めた任意の他のDNA配列、及び直接的又は間接的に作用してプロモーターからの転写の量を調節することが当業者に知られている、任意の他のヌクレオチド配列の存在によって構造的に確認される核酸断片を指す。「構成的」プロモーターは、大部分の生理的条件及び発生条件下において大部分の組織中で活性があるプロモーターである。「誘導性」プロモーターは、例えば化学的誘導物質の適用によって、生理的又は発生的に調節されるプロモーターである。「組織特異的」プロモーターは、特定の型の組織又は細胞中のみで活性がある。
【0011】
用語「実質的に同一」、「実質的同一性」、又は「本質的に類似」又は「本質的類似性」は、2つのペプチド又は2つのヌクレオチド配列が、初期設定のパラメーターを使用してプログラムGAP又はBESTFITなどによって最適にアラインメントをとるとき、本明細書の他の箇所で定義する少なくともある程度の割合の配列同一性を共有することを意味する。GAPはNeedleman及びWunschの大域的アラインメントアルゴリズムを使用して、2つの配列のそれらの全長でアラインメントをとり、整合の数を最大にし、ギャップの数を最少にする。一般に、GAP初期設定のパラメーター、及びギャップ生成ペナルティー=50(ヌクレオチド)/8(タンパク質)及びギャップ伸長ペナルティー=3(ヌクレオチド)/2(タンパク質)が使用される。ヌクレオチドに関しては、使用する初期設定のスコア行列はnwsgapdnaであり、及びタンパク質に関しては、初期設定のスコア行列はBlosum62である(Henikoff & Henikoff、1992、PNAS89、915〜919)。RNA配列がDNA配列と本質的に類似しているか、或いはある程度の配列同一性を有すると言えるとき、DNA配列中のチミン(T)はRNA配列中のウラシル(U)と等しいと考えられることは明らかである。配列アラインメント及びパーセント配列同一性のスコアは、Accelrys Inc.、9685 Scranton Road、San Diego、CA 92121〜3752 USAから入手可能なGCG Wisconsin Package、バージョン10.3などのコンピュータプログラム、又はオープンソースソフトウェアEmboss for Windows(登録商標)(現バージョン2.7.1〜07)を使用して決定することができる。或いは、パーセント類似性又は同一性は、例えばFASTA、BLASTなどのデータベースを検索することによって決定することができる。
【0012】
本発明のパルボウイルスRepタンパク質をコードするヌクレオチド配列は、それぞれ適度、或いは好ましくは厳密なハイブリダイゼーション条件下において、配列番号10のヌクレオチド配列とハイブリダイズするそれらの能力によって定義することもできる。厳密なハイブリダイゼーション条件は、約1Mの塩、好ましくは6×SSCを含む溶液、又は同程度のイオン強度を有する任意の他の溶液中で約65℃の温度において、少なくとも約25、好ましくは約50ヌクレオチド、75又は100及び最も好ましくは約200以上のヌクレオチドの核酸配列をハイブリダイズさせ、約0.1Mの塩、或いはそれ未満、好ましくは0.2×SSCを含む溶液、又は同程度のイオン強度を有する任意の他の溶液中で65℃において洗浄することが可能である条件として本明細書では定義する。ハイブリダイゼーションは一晩、即ち少なくとも10時間実施することが好ましく、且つ洗浄は少なくとも1時間実施し、洗浄溶液は少なくとも2回交換することが好ましい。これらの条件は、約90%以上の配列同一性を有する配列の特異的なハイブリダイゼーションを通常可能にするはずである。
【0013】
適度な条件は、約1Mの塩、好ましくは6×SSCを含む溶液、又は同程度のイオン強度を有する任意の他の溶液中で約45℃の温度において、少なくとも50ヌクレオチド、好ましくは約200以上のヌクレオチドの核酸配列をハイブリダイズさせ、約1Mの塩、好ましくは6×SSCを含む溶液、又は同程度のイオン強度を有する任意の他の溶液中で室温において洗浄することが可能である条件として本明細書では定義する。ハイブリダイゼーションは一晩、即ち少なくとも10時間実施することが好ましく、且つ洗浄は少なくとも1時間実施し、洗浄溶液は少なくとも2回交換することが好ましい。これらの条件は、50%までの配列同一性を有する配列の特異的なハイブリダイゼーションを通常可能にするはずである。当業者はこれらのハイブリダイゼーション条件を改変して、50%と90%の間で同一性が異なる配列を特異的に同定することができるはずである。
【0014】
発明の詳細な説明
本発明は、哺乳動物細胞中の核酸の導入及び/又は発現用のベクターとして使用するための、動物パルボウイルス、具体的には感染性のヒト又はサルAAVなどのデペンドウイルス属、及びそれらの構成要素(例えば、動物パルボウイルスゲノム)の使用に関する。具体的には本発明は、昆虫細胞中での生成時のこのようなパルボウイルスベクターの生産性の改善に関する。
【0015】
パルボウイルス科のウイルスは小さなDNA動物ウイルスである。パルボウイルス科は、2つの亜科:脊椎動物に感染するパルボウイルス科、及び昆虫に感染するデンソウイルス科に分けることができる。パルボウイルス亜科のメンバーは本明細書ではパルボウイルスと呼び、デペンドウイルス属を含む。その属の名称から推測することができるように、デペンドウイルスのメンバーは、それらが細胞培養中の増殖感染にアデノウイルス又はヘルペスウイルスなどのヘルパーウイルスとの同時感染を通常必要とする点で独特である。デペンドウイルス属は、ヒト(例えば、血清型1、2、3A、3B、4、5、及び6)又は霊長類(例えば、血清型1及び4)に通常感染するAAV、及び他の温血動物(例えば、ウシ、イヌ、ウマ、及びヒツジアデノ随伴ウイルス)に感染する関連ウイルスを含む。パルボウイルス及びパルボウイルス科の他のメンバーに関する他の情報は、Kenneth I.Berns、「パルボウイルス科:ウイルス及びそれらの複製(Parvoviridae:The Viruses and Their Replication)」、Chapter 69 in Fields Virology(3dEd.1996)中に記載されている。便宜上本発明は、AAVを参照することによって本明細書でさらに例示し記載する。しかしながら、本発明はAAVに限られず、他のパルボウイルスに同様に適用することができることは理解される。
【0016】
すべての知られているAAV血清型のゲノムの構成は非常に類似している。AAVのゲノムは、約5,000ヌクレオチド(nt)長未満である線状の一本鎖DNA分子である。逆方向末端反復(ITR)は、非構造複製(Rep)タンパク質及び構造(VP)タンパク質に独特なコードヌクレオチド配列と隣接している。VPタンパク質(VP1、−2及び−3)はカプシドを形成する。末端の145ntは自己相補的であり、T形状ヘアピンを形成するエネルギー的に安定な分子内二本鎖を形成することができるように構成されている。これらのヘアピン構造はウイルスDNA複製の起点として機能し、細胞DNAポリメラーゼ複合体用のプライマーとして働く。哺乳動物細胞におけるwtAAV感染の後、Rep遺伝子(即ち、Rep78及びRep52)がそれぞれP5プロモーター及びP19プロモーターから発現され、両方のRepタンパク質がウイルスゲノムの複製において機能を有する。RepのORF中のスプライシング事象は、実際4つのRepタンパク質(即ち、Rep78、Rep68、Rep52及びRep40)の発現をもたらす。しかしながら、Rep78及びRep52タンパク質をコードする非スプライシングmRNAは、哺乳動物細胞中でのAAVベクター生成に十分であることが示されてきている。昆虫細胞中でも、Rep78及びRep52タンパク質はAAVベクター生成に十分である。
【0017】
「組換えパルボウイルス又はAAVベクター」(又は「rAAVベクター」)は、1つ又は複数の対象とするポリヌクレオチド配列、対象とする遺伝子、又はパルボウイルス又はAAV逆方向末端反復配列(ITRs)に隣接する「導入遺伝子」を含むベクターを本明細書では指す。AAVrep及びcap遺伝子産物(即ち、AAVRep及びCapタンパク質)を発現している昆虫宿主細胞中に存在するとき、このようなrAAVベクターを複製し感染性ウイルス粒子にパッケージ化することができる。rAAVベクターが大きな核酸構築体(例えば、染色体中、或いはクローニング又はトランスフェクションに使用されるプラスミド又はバキュロウイルスなどの他のベクター)に組み込まれているとき、したがってrAAVベクターは典型的には「プロベクター」と呼ばれ、これはAAVパッケージ化機能及び必要なヘルパー機能の存在下での複製及びカプセル化によって「レスキュー」することができる。
【0018】
第1の態様では、本発明は、動物のパルボウイルスRepタンパク質をコードするヌクレオチド配列を含むオープンリーディングフレームを含むヌクレオチド配列であって、パルボウイルスRep78タンパク質の翻訳の開始コドンが次善開始コドンであるヌクレオチド配列に関する。次善開始コドンは、部分的なエキソンスキッピングを行う開始コドンであることが好ましい。部分的なエキソンスキッピングは、少なくとも一部分のリボソームがさらに下流の開始コドンではなくRep78タンパク質の次善開始コドンで翻訳を開始せず、好ましくはさらに下流の開始コドンがRep52タンパク質の開始コドンであることを意味すると本明細書では理解される。次善開始コドンは、昆虫細胞におけるヌクレオチド配列の発現後に部分的なエキソンスキッピングを行うことが好ましい。次善開始コドンは昆虫細胞中で部分的なエキソンスキッピングを行い、バキュロウイルス発現を使用し、好ましくは感染後約20〜40時間、より好ましくは感染後約30〜40時間で1:10〜10:1、1:5〜5:1、又は1:3〜3:1の範囲のRep78とRep52のモル比を昆虫細胞においてもたらすことが好ましい。Rep78とRep52のモル比は、好ましくはRep78とRep52両方の共通のエピトープを認識するモノクローナル抗体を使用して、或いは実施例1.1.3中に記載した抗体を使用して、実施例1.1.3中に記載したのと同様にウェスタンブロッティングによって決定することができる。
【0019】
用語「次善開始コドン」は、本明細書では3ヌクレオチドの開始コドン自体のみでなく、その状況も指す。したがって、次善開始コドンは、次善の状況、例えばコザックでない状況での「最適」ATGコドンからなっていてよい。しかしながら、より好ましいのは、3ヌクレオチドの開始コドン自体が次善、即ちATGではない次善開始コドンである。次善は本明細書では、正常ATGコドンと比較して他の同一状況での翻訳の開始時に、コドンが有効性が低いことを意味すると理解される。次善コドンの有効性は、他の同一状況での正常ATGコドンの有効性の90、80、60、40又は20%未満であることが好ましい。翻訳開始の相対的有効性を比較するための方法は、本質的に当業者に知られている。好ましい次善開始コドンは、ACG、TTG、CTG、及びGTGから選択することができる。より好ましくはACGである。
【0020】
動物のパルボウイルスRepタンパク質をコードするヌクレオチド配列は、Rep78及びRep52タンパク質などの昆虫細胞におけるパルボウイルスベクター生成に必要とされ十分である非構造Repタンパク質をコードするヌクレオチド配列として本明細書では理解する。動物パルボウイルスのヌクレオチド配列はデペンドウイルスに由来することが好ましく、ヒト又はサルアデノ随伴ウイルス(AAV)に由来することがより好ましく、ヒト(例えば、血清型1、2、3A、3B、4、5、及び6)又は霊長類(例えば、血清型1及び4)に通常感染するAAVに由来することが最も好ましい。動物のパルボウイルスRepタンパク質をコードするヌクレオチド配列の一例は配列番号10で与え、これはRepタンパク質をコードするAAV血清型2の配列ゲノムの一部分を示す。Rep78コード配列はヌクレオチド11〜1876を含み、Rep52コード配列はヌクレオチド683〜1876を含む。Rep78及びRep52タンパク質の正確な分子量、及び翻訳開始コドンの正確な位置は、異なるパルボウイルス間で異なる可能性があることは理解される。しかしながら当業者は、AAV−2以外のパルボウイルス由来のヌクレオチド配列中の対応する位置の確認の仕方を知っているはずである。動物のパルボウイルスRepタンパク質をコードするヌクレオチド配列は、したがって:
a)配列番号11のアミノ酸配列と少なくとも50、60、70、80、88、89、90、95、97、98、又は99%の配列同一性を有するアミノ酸配列を含むポリペプチドをコードするヌクレオチド配列、
b)配列番号10の11〜1876位のヌクレオチド配列と少なくとも50、60、70、80、81、82、85、90、95、97、98、又は99%の配列同一性を有するヌクレオチド配列、
c)その相補鎖が(a)又は(b)の核酸分子配列とハイブリダイズするヌクレオチド配列、
d)遺伝コードの縮重のために(c)の核酸分子の配列とその配列が異なるヌクレオチド配列
として定義することもできる。
【0021】
ヌクレオチド配列は、昆虫細胞におけるパルボウイルスベクター生成に必要とされ十分である動物のパルボウイルスRepタンパク質をコードすることが好ましい。
【0022】
本発明の他の好ましいヌクレオチド配列は、パルボウイルスRep78タンパク質をコードするヌクレオチド配列の開始コドンの上流に、配列番号7の9ヌクレオチドの配列、又は配列番号7と実質的に相同なヌクレオチド配列を含む発現制御配列を含む。配列番号7のヌクレオチド配列と実質的同一性を有し、パルボウイルスRep78タンパク質の発現の増大を助長するはずである配列は、例えば配列番号7の9ヌクレオチドの配列と少なくとも60%、70%、80%又は90%の同一性を有する配列である。
【0023】
他のパルボウイルスの、Rep78及びRep52翻訳開始部位以外の、Repタンパク質コード配列中の偽性と考えられる翻訳開始部位の除去は、昆虫細胞において認識され得る推定スプライス部位の除去と同様に、当業者によって十分理解されているはずである。昆虫細胞中での正確な発現のための野生型パルボウイルス配列の様々な改変は、例えばSambrook and Russell(2001)「分子クローニング:研究室用マニュアル(Molecular Cloning:A Laboratory Manual)」(第3版)、Cold Spring Harbor Laboratory、Cold Spring Harbor Laboratory Press、ニューヨーク中に記載された技術などの、よく知られている遺伝子工学技術の適用によって実施される。Repタンパク質コード領域の様々な他の改変は当業者に知られており、これらはRepタンパク質の収率を増大させ得る。これらの改変は本発明の範囲内にある。
【0024】
さらなる態様では、本発明は、上記で定義したパルボウイルスRepタンパク質をコードするヌクレオチド配列を含む核酸構築体に関する。構築体中では、パルボウイルスRepタンパク質をコードするヌクレオチド配列は、昆虫細胞中での発現用の発現制御配列と作動可能に連結していることが好ましい。これらの発現制御配列は、昆虫細胞中で活性があるプロモーターを少なくとも含むはずである。昆虫宿主細胞中で外来遺伝子を発現させるための当業者に知られている技術を使用して、本発明を実施することができる。昆虫細胞におけるポリペプチドの分子工学及び発現に関する方法は、例えばSummers and Smith.1986.「バキュロウイルスベクター及び昆虫培養手順に関する方法のマニュアル(A Manual of Methods for Baculovirus Vectors and Insect Culture Procedures)」、Texas Agricultural Experimental Station Bull.No.7555、College Station、Tex.;Luckow.1991.In Prokop et al.、「バキュロウイルスベクターを用いた昆虫細胞における異種遺伝子のクローニング及び発現(Cloning and Expression of Heterologous Genes in Insect Cells with Baculovirus Vectors)」「組換えDNA技術及び適用(Recombinant DNA Technology and Applications)」、97〜152;King、L.A.and R.D.Possee、1992、「バキュロウイルス発現系(The baculovirus expression system)」、Chapman and Hall、United Kingdom;O’Reilly、D.R.、L.K.Miller、V.A.Luckow、1992、「バキュロウイルス発現ベクター(Baculovirus Expression Vectors)」:「研究室用マニュアル(A Laboratory Manual)」、ニューヨーク;W.H.Freeman and Richardson、C.D.、1995、「バキュロウイルス発現のプロトコル、分子生物学における方法(Baculovirus Expression Protocols,Methods in Molecular Biology)」、volume39;米国特許第4,745,051号;US2003148506;及びWO03/074714中に記載されている。パルボウイルスRepタンパク質をコードする本発明のヌクレオチド配列の転写に特に適したプロモーターは、例えば多角体プロモーターである。しかしながら、昆虫細胞中で活性がある他のプロモーター、例えばpl0、p35、IE−1又はΔIE−1プロモーター、及び前述の参照文献中に記載された他のプロモーターが、当技術分野で知られている。
【0025】
昆虫細胞におけるパルボウイルスRepタンパク質の発現用の核酸構築体は、昆虫細胞適合性ベクターであることが好ましい。「昆虫細胞適合性ベクター」又は「ベクター」は、昆虫又は昆虫細胞の生産的な形質転換又はトランスフェクションが可能である核酸分子であると理解される。代表的な生物学的ベクターには、プラスミド、線状核酸分子、及び組換えウイルスがある。それが昆虫細胞適合性である限り、任意のベクターを利用することができる。ベクターを昆虫細胞ゲノムに組み込むことはできるが、昆虫細胞中のベクターの存在は不変的である必要はなく、一時的なエピソームベクターも含まれる。ベクターは知られている任意の手段によって、例えば細胞の化学処理、エレクトロポレーション、又は感染によって導入することができる。好ましい実施形態では、ベクターはバキュロウイルス、ウイルスベクター、又はプラスミドである。より好ましい実施形態では、ベクターはバキュロウイルスであり、即ち、構築体はバキュロウイルスベクターである。バキュロウイルスベクター及びそれらの使用法は、昆虫細胞の分子工学に関する前に引用した参照文献中に記載されている。
【0026】
他の態様では、本発明は、1つのパルボウイルスRepタンパク質をコードする単一のオープンリーディングフレームを含む唯一の型のヌクレオチド配列を含む昆虫細胞に関する。単一のオープンリーディングフレームは1つ又は複数のパルボウイルスRepタンパク質をコードすることが好ましく、オープンリーディングフレームはパルボウイルスRepタンパク質のすべてをコードすることがより好ましく、オープンリーディングフレームは、好ましくは昆虫細胞において少なくともRep52とRep78タンパク質の両方がそこから発現され得る完全長Rep78タンパク質をコードすることが最も好ましい。昆虫細胞は、例えば多コピーエピソームベクターでは、1つの型のヌクレオチド配列の2つ以上のコピーを含み得るが、これらは本質的に1つ及び同一の核酸分子、又は1つ及び同一のRepアミノ酸配列をコードする少なくともいくつかの核酸分子、例えば、遺伝コードの縮重のために相互間でわずかに異なる核酸分子の多コピーであることは、本明細書では理解される。パルボウイルスRepタンパク質をコードする唯一の型の核酸分子の存在は、昆虫細胞におけるパルボウイルス生成のレベル(安定性)に影響を与える欠陥Rep発現構築体を生成する可能性がある、Rep配列を含む異なる型のベクター中に存在し得る相同的配列間の組換えを回避する。好ましくは、昆虫細胞では、1つ又は複数のパルボウイルスRepタンパク質をコードする単一のオープンリーディングフレームを含むヌクレオチド配列が核酸構築体の一部分であり、ヌクレオチド配列が昆虫細胞中での発現用の発現制御配列と作動可能に連結していることが好ましい。さらに好ましい昆虫細胞は、パルボウイルスRepタンパク質をコードする、上記で定義したヌクレオチド配列、好ましくは上記で定義した次善開始コドンを有するコード配列、又は上記で定義した核酸構築体を、「第1の」ヌクレオチド配列として含み、或いは昆虫細胞は、このようなヌクレオチド配列を含む上記で定義した核酸構築体を、「第1の」核酸構築体として含む。
【0027】
組換えパルボウイルス(rAAV)ベクターの複製を可能にし、培養中に維持することができる任意の昆虫細胞を、本発明によって使用することができる。例えば、使用する細胞系は、スポドプテラフルギベルダ(Spodoptera frugiperda)、ショウジョウバエの細胞系、又は蚊細胞系、例えばアエデスアルボピクタス(Aedes albopictus)派生細胞系由来であってよい。好ましい昆虫細胞又は細胞系は、例えばSe301、SeIZD2109、SeUCR1、Sf9、Sf900+、Sf21、BTI−TN−5B1−4、MG−1、Tn368、HzAm1、Ha2302、Hz2E5、High Five(Invitrogen、CA、USA)及びexpresSF+(登録商標)(米国特許第6,103,526号;Protein Sciences Corp.、CT、USA)を含めた、バキュロウイルス感染に感受性である昆虫種由来の細胞である。
【0028】
本発明による好ましい昆虫細胞は、前に記載した「第1の」ヌクレオチド配列又は核酸構築体以外に、
a)少なくとも1つのパルボウイルス逆方向末端反復(ITR)ヌクレオチド配列を含む第2のヌクレオチド配列と、
b)昆虫細胞における発現用の発現制御配列と作動可能に連結しているパルボウイルスCapタンパク質コード配列を含む第3のヌクレオチド配列と
をさらに含む。
【0029】
本発明の文脈では、「少なくとも1つのパルボウイルスITRヌクレオチド配列」は、「A」、「B」、及び「C」領域とも呼ばれる、大部分が相補的であり、対称的に配列した配列を含むパリンドローム配列を意味すると理解される。ITRは複製起点、複製における「シス」の役割を有する部位、即ち、パリンドローム配列及びパリンドロームに固有の特異的配列を認識する、例えばRep78(又はRep68)などの複製タンパク質として作用するトランスの認識部位である部位として機能する。ITR配列の対称性の1つの例外はITRの「D」領域である。それは独特である(1つのITR内に相補配列を有していない)。一本鎖DNAの切断は、A領域とD領域の間の接合部で起こる。それは新たなDNA合成が始まる領域である。D領域は通常パリンドロームの片側に位置し、核酸複製ステップに対する方向性をもたらす。哺乳動物細胞において複製するパルボウイルスは、2つのITR配列を典型的には有する。しかしながら、結合部位がA領域の両鎖上に存在し、D領域は対称的に、1つがパリンドロームの各側に位置するように、ITRを工学処理することは可能である。二本鎖環状DNA鋳型(例えば、プラスミド)上では、Rep78又はRep68支援核酸複製が次いで両方向に進行し、1つのITRは環状ベクターのパルボウイルス複製に十分である。したがって、1つのITRヌクレオチド配列は本発明の文脈で使用することができる。しかしながら、2個又は他の偶数個の通常のITRを使用することが好ましい。2個のITR配列を使用することが最も好ましい。好ましいパルボウイルスのITRはAAVのITRである。安全上の理由で、細胞中への最初の導入後にさらに増殖することができない組換えパルボウイルス(rAAV)ベクターを構築することが望ましい可能性がある。レシピエント中の望ましくないベクター増殖を制限するためのこのような安全メカニズムは、US2003148506中に記載されたのと同様に、キメラITRを有するrAAVを使用することによって提供することができる。
【0030】
組換えパルボウイルス(rAAV)ベクターの生成用に昆虫細胞中で利用する核酸構築体の数は、本発明中では無制限である。例えば、1個、2個、3個、4個、5個、或いはそれより多くの別個の構築体を利用して、本発明の方法に従い昆虫細胞中でrAAVを生成することができる。5個の構築体を利用する場合、1個の構築体はAAVVP1をコードし、他の構築体はAAVVP2をコードし、さらに他の構築体はAAVVP3をコードし、さらに他の構築体は上記で定義したRepタンパク質をコードし、且つ最終構築体は少なくとも1個のAAVITRを含む。5個より少ない構築体を利用する場合、構築体は少なくとも1個のAAVITR、及びVP1、VP2、VP3、及びRepタンパク質コード配列の様々な組合せを含むことができる。2個の構築体又は3個の構築体を使用することが好ましく、前に記載したのと同様の2個の構築体がより好ましい。2個の構築体を使用する場合、昆虫細胞は(a)上記で定義したRepタンパク質の発現用の第1の核酸構築体であって、上記の(b)で定義した第3のヌクレオチド配列をさらに含む構築体(昆虫細胞中での発現のために少なくとも1個の発現制御配列と作動可能に連結しているパルボウイルスCapタンパク質コード配列を含む;以下も参照)、及び(c)上記の(a)で定義した第2のヌクレオチド配列を含む第2の核酸構築体(少なくとも1個のパルボウイルス/AAVITRヌクレオチド配列を含む)を含むことが好ましい。3個の構築体を使用する場合、カプシドタンパク質の発現とRepタンパク質の発現に別々の構築体を使用すること以外は、2個の構築体に使用するのと同じ形状を使用することが好ましい。それぞれの構築体における配列は、互いに対して任意の順序であってよい。例えば、1個の構築体がVPカプシドタンパク質をコードするヌクレオチド配列を含むITR及びORFを含む場合、ITR配列間のDNAの複製によって、VPのORFが複製されるか或いは複製されないように、構築体上にVPのORFが局在する可能性がある。他の例では、Repコード配列及び/又はVPカプシドタンパク質をコードするヌクレオチド配列を含むORFは、構築体上で任意の順序であってよい。さらに第2、第3及び他の核酸構築体は昆虫細胞適合性ベクターであることが好ましく、前に記載したバキュロウイルスベクターであることが好ましいことが理解される。或いは、本発明の昆虫細胞では、1つ又は複数の第1のヌクレオチド配列、第2のヌクレオチド配列、第3のヌクレオチド配列、及び第4のヌクレオチド配列及び任意選択の他のヌクレオチド配列を、昆虫細胞のゲノムに安定して組み込むことができる。当業者は、昆虫ゲノム中にヌクレオチド配列を安定して導入する方法、及びゲノム中にこのようなヌクレオチド配列を有する細胞を同定する方法を知っている。ゲノム中への組込みは、例えば昆虫ゲノムの領域と非常に相同性が高いヌクレオチド配列を含むベクターの使用によって助長することができる。トランスポゾンなどの特異的配列の使用は、ゲノム中にヌクレオチド配列を導入する他の方法である。
【0031】
本発明では、パルボウイルスカプシド(Cap)タンパク質コード配列を含む第3のヌクレオチド配列は、3つのパルボウイルスカプシドタンパク質、VP1、−2及び−3のそれぞれをコードする配列を含むと本明細書では理解される。カプシドタンパク質コード配列を含む第3のヌクレオチド配列は、様々な形で存在してよい。例えば、カプシドタンパク質、VP1、−2及び−3のそれぞれに関する別個のコード配列を使用することができ、それぞれのコード配列は昆虫細胞中での発現のために発現制御配列と作動可能に連結している。しかしながら、より好ましくは、第3のヌクレオチド配列は、全3個の動物パルボウイルス(AAV)VP1、VP2、及びVP3カプシドタンパク質をコードする1個のオープンリーディングフレームを含み、VP1カプシドタンパク質の翻訳の開始コドンは、例えばUrabe et al.(2002、上記)によって記載されたのと同様のATGではない次善開始コドンである。VP1カプシドタンパク質に関する次善開始コドンは、Rep78タンパク質に関して上記で定義したのと同様であってよい。VP1カプシドタンパク質に関するより好ましい次善開始コドンはACG、TTG、CTG及びGTGから選択することができ、この中ではCTG及びGTGが最も好ましい。カプシドタンパク質の発現に好ましい第3のヌクレオチド配列は、VP1カプシドタンパク質をコードするヌクレオチド配列の開始コドンの上流に、配列番号7の9ヌクレオチドの配列又は配列番号7と実質的に相同的なヌクレオチドの配列を含む発現制御配列をさらに含む。配列番号7のヌクレオチド配列と実質的同一性を有し、VP1の発現の増大を助長するはずである配列は、例えば配列番号7の9ヌクレオチドの配列と少なくとも60%、70%、80%又は90%の同一性を有する配列である。カプシドタンパク質の発現に好ましい他の第3のヌクレオチド配列は、12ヌクレオチド位におけるC、21ヌクレオチド位におけるA、及び24ヌクレオチド位におけるC(翻訳開始コドンの第1のヌクレオチドである位置1を基準とする;配列番号1を参照)から選択されるVP1カプシドタンパク質をコードするヌクレオチド配列の少なくとも1つの改変をさらに含むことが好ましい。他の血清型のVP1の翻訳の偽性と考えられる開始コドンの除去は、昆虫細胞において認識され得る推定スプライス部位の除去と同様に、当業者によって十分理解されているはずである。VPコード領域の様々な他の改変は当業者に知られており、それらはVP及びビリオンの収率を増大させるか、或いは指向性の変化などの他の望ましい影響を有するか、或いはビリオンの抗原性を低下させるいずれかの可能性がある。これらの改変は本発明の範囲内にある。好ましくは、パルボウイルスカプシドタンパク質をコードする本発明のヌクレオチド配列は、昆虫細胞中での発現のために発現制御配列と作動可能に連結しており、発現制御配列は昆虫細胞中で活性があるプロモーターを少なくとも含むはずである。昆虫宿主細胞中でパルボウイルスカプシドタンパク質を発現させるための、このような制御配列及び他の技術及び物質(例えばベクター)は、Repタンパク質について上記で既に記載している。
【0032】
本発明の好ましい実施形態では、本発明の昆虫細胞中に存在する第2のヌクレオチド配列、即ち少なくとも1つのパルボウイルス(AAV)ITRを含む配列は、対象とする遺伝子産物をコードする少なくとも1つのヌクレオチド配列をさらに含み、対象とする遺伝子産物をコードする少なくとも1つのヌクレオチド配列は、昆虫細胞において生成される組換えパルボウイルス(rAAV)ベクターのゲノムに組み込まれることが好ましい。対象とする遺伝子産物をコードする少なくとも1つのヌクレオチド配列は、哺乳動物細胞中での発現用の配列であることが好ましい。第2のヌクレオチド配列は2つのパルボウイルス(AAV)ITRヌクレオチド配列を含み、対象とする遺伝子産物をコードする少なくとも1つのヌクレオチド配列は、2つのパルボウイルス(AAV)ITRヌクレオチド配列間に位置することが好ましい。それが2つの通常のITRの間に位置するか、或いは2カ所のD領域で工学処理したITRの片側に位置する場合、(哺乳動物細胞中での発現用の)対象とする遺伝子産物をコードするヌクレオチド配列は、昆虫細胞において生成される組換えパルボウイルス(rAAV)ベクターに組み込まれることが好ましい。
【0033】
本明細書で上記で定義した第2のヌクレオチド配列は、昆虫細胞中で複製される組換えパルボウイルス(rAAV)ベクターに組み込まれるように位置する、昆虫細胞中での発現用の少なくとも1つの「対象とする遺伝子産物」をコードするヌクレオチド配列をしたがって含み得る。任意のヌクレオチド配列は、本発明の方法に従い生成した組換えパルボウイルス(rAAV)ベクターでトランスフェクトした哺乳動物細胞中での、後の発現用に組み込むことが可能である。ヌクレオチド配列は例えばタンパク質をコードすることができ、それがRNAi物質、即ちRNA干渉することができるRNA分子、例えばshRNA(低分子ヘアピンRNA)又はsiRNA(低分子干渉RNA)などを発現することができる。「siRNA」は、哺乳動物細胞中では無毒である短い長さの二本鎖RNAである小分子の干渉RNAを意味する(Elbashir et al.、2001、Nature411:494〜98;Caplen et al.、2001、Proc.Natl.Acad.Sci.USA98:9742〜47)。好ましい実施形態では、第2のヌクレオチド配列は2つのヌクレオチド配列を含むことができ、それぞれが哺乳動物細胞中での発現用の対象とする1つの遺伝子産物をコードする。対象とする産物をコードする2つのヌクレオチド配列のそれぞれは、昆虫細胞中で複製される組換えパルボウイルス(rAAV)ベクターに組み込まれるように位置する。
【0034】
哺乳動物細胞中での発現用の対象とする産物は、治療用遺伝子産物であってよい。治療用遺伝子産物は、標的細胞中で発現されると、例えば望ましくない活性の除去、例えば感染細胞の除去、又は例えば酵素活性の欠失を引き起こす遺伝的欠陥の補足などの望ましい治療効果をもたらす、ポリペプチド、又はRNA分子(siRNA)、又は他の遺伝子産物であってよい。治療用ポリペプチド遺伝子産物の例には、CFTR、因子IX、リポタンパク質リパーゼ(LPL、好ましくはLPL S447X;WO01/00220参照)、アポリポタンパク質Al、ウリジン二リン酸グルクロノシルトランスフェラーゼ(UGT)、網膜色素変性GTPaseレギュレーター相互作用タンパク質(RP−GRIP)、及びサイトカイン又は例えばIL−10のようなインターロイキンがある。
【0035】
代替的或いは追加的に、第2の遺伝子産物として、本明細書で上記で定義した第2のヌクレオチド配列は、細胞の形質転換及び発現を評価するためのマーカータンパク質として働くポリペプチドをコードするヌクレオチド配列を含むことができる。この目的に適したマーカータンパク質は、例えば蛍光タンパク質GFP、及び選択可能なマーカー遺伝子HSVチミジンキナーゼ(HAT培地における選択用)、細菌ハイグロマイシンBホスホトランスフェラーゼ(ハイグロマイシンBにおける選択用)、Tn5アミノグリコシドホスホトランスフェラーゼ(G418における選択用)、及びジヒドロ葉酸還元酵素(DHFR)(メトトレキセートにおける選択用)、CD20、低親和性の神経成長因子遺伝子である。これらのマーカー遺伝子を得るための供給源、及びそれらを使用するための方法は、Sambrook and Russell(2001)「分子クローニング:研究室用マニュアル(Molecular Cloning:A Laboratory Manual)」(第3版)、Cold Spring Harbor Laboratory、Cold Spring Harbor Laboratory Press、ニューヨーク中に示されている。さらに、本明細書で上記で定義した第2のヌクレオチド配列は、必要であると考えられる場合、本発明の組換えパルボウイルス(rAAV)ベクターで形質導入した細胞由来の対象を治癒することができるフェイルセーフ機構として働くことができる、ポリペプチドをコードするヌクレオチド配列を含むことができる。しばしば自殺遺伝子と呼ばれる、このようなヌクレオチド配列は、タンパク質が発現されるトランスジェニック細胞を死滅させることができる毒性物質にプロドラッグを変換する可能性があるタンパク質をコードする。このような自殺遺伝子の適切な例には、例えば大腸菌のシトシンデアミナーゼ遺伝子、又は単純ヘルペスウイルス、サイトメガロウイルス及び水痘帯状疱疹ウイルス由来のチミジンキナーゼ遺伝子の1つがあり、この場合ガンシクロビルをプロドラッグとして使用して、対象中のトランスジェニック細胞を死滅させることができる(例えば、Clair et al.、1987、Antimicrob.Agents Chemother.31:844〜849を参照)。
【0036】
他の実施形態では、対象とする遺伝子産物の1つはAAVタンパク質であり得る。特に、Rep78又はRep68などのRepタンパク質、又はその機能断片。Rep78及び/又はRep68をコードするヌクレオチド配列は、本発明の組換えパルボウイルス(rAAV)ベクターのゲノム上に存在し、ベクターで形質導入した哺乳動物細胞中で発現される場合、形質導入した哺乳動物細胞のゲノムへの組換えパルボウイルス(rAAV)ベクターの組込みを可能にする。rAAV形質導入又は感染哺乳動物細胞中でのRep78及び/又はRep68の発現は、ベクターによって細胞中に導入される対象とする任意の他の遺伝子産物の長期又は永続的な発現を可能にすることによって、組換えパルボウイルス(rAAV)ベクターのいくつかの用途に関する利点をもたらす可能性がある。
【0037】
本発明の組換えパルボウイルス(rAAV)ベクターでは、哺乳動物細胞中での発現用の対象とする遺伝子産物をコードする少なくとも1つのヌクレオチド配列は、少なくとも1つの哺乳動物細胞適合性の発現制御配列、例えばプロモーターと作動可能に連結していることが好ましい。多くのこのようなプロモーターは、当技術分野で知られている(Sambrook and Russel、2001、上記を参照)。CMVプロモーターなどの、多くの細胞型において広範囲で発現される構成的プロモーターを使用することができる。しかしながら、より好ましいのは、誘導性、組織特異的、細胞型特異的、又は細胞周期特異的であるプロモーターであるはずである。例えば、肝臓特異的な発現に関しては、α1−アンチトリプシンプロモーター、甲状腺ホルモン結合グロブリンプロモーター、アルブミンプロモーター、LPS(チロキシン結合グロブリン)プロモーター、HCR−ApoCIIハイブリッドプロモーター、HCR−hAATハイブリッドプロモーター及びアポリポタンパク質Eプロモーターからプロモーターを選択することができる。他の例には、腫瘍選択的、及び特に神経細胞腫瘍選択的発現用のE2Fプロモーター(Parr et al.、1997、Nat.Med.3:1145〜9)、又は単核血液細胞において使用するためのIL−2プロモーターがある(Hagenbaugh et al.、1997、J Exp Med;185:2101〜10)。
【0038】
AAVはいくつかの哺乳動物細胞に感染することができる。例えば、Tratschin et al.(1985、Mol.Cell Biol.5:3251〜3260)及びGrimm et al.(1999、Hum.Gene Ther.10:2445〜2450)を参照。しかしながら、ヒト滑膜繊維芽細胞のAAV形質導入は同様のネズミ細胞中より著しく有効であり、Jennings et al.、Arthritis Res、3:1(2001)、AAVの細胞指向性は血清型間で異なる。例えば、哺乳動物のCNS細胞指向性及び形質導入効率に関するAAV2、AAV4、及びAAV5の間の違いを論じている、Davidson et al.(2000、Proc.Natl.Acad.Sci.USA、97:3428〜3432)を参照されたい。
【0039】
昆虫細胞中で組換えAAVベクターを生成するための本発明の方法において使用することができるAAV配列は、任意のAAV血清型のゲノムに由来してよい。一般に、AAV血清型はアミノ酸及び核酸レベルで相当な相同性のゲノム配列を有し、一組の同一な遺伝的機能をもたらし、物理的及び機能的にほぼ等しいビリオンを生成し、実際同一であるメカニズムによって複製及び構築する。様々なAAV血清型のゲノム配列及びゲノム類似性の総説に関しては、例えば、GenBankアクセッション番号U89790;GenBankアクセッション番号J01901;GenBankアクセッション番号AF043303;GenBankアクセッション番号AF085716;Chlorini et al.(1997、J.Vir.71:6823〜33);Srivastava et al.(1983、J.Vir.45:555〜64);Chlorini et al.(1999、J.Vir.73:1309〜1319);Rutledge et al.(1998、J.Vir.72:309〜319);及びWu et al.(2000、J.Vir.74:8635〜47)を参照。AAV血清型1、2、3、4及び5は、本発明の文脈で使用するためのAAVヌクレオチド配列の好ましい供給源である。本発明の文脈で使用するためのAAVのITR配列は、AAV1、AAV2、及び/又はAAV4に由来することが好ましい。同様に、Rep(Rep78及びRep52)コード配列は、AAV1、AAV2、及び/又はAAV4に由来することが好ましい。本発明の文脈で使用するためのVP1、VP2、及びVP3カプシドタンパク質をコードする配列は、しかしながら、任意の知られている42の血清型から、より好ましくはAAV1、AAV2、AAV3、AAV4、AAV5、AAV6、AAV7、AAV8又はAAV9、或いは例えばカプシドシャッフリング技術及びAAVカプシドライブラリーによって入手される、新たに開発されたAAV様粒子から得ることができる。
【0040】
AAVのRep及びITR配列は、大部分の血清型の間で特に保存されている。様々なAAV血清型のRep78タンパク質は例えば89%を超えて同一であり、AAV2、AAV3A、AAV3B、及びAAV6の間のゲノムレベルでの全体のヌクレオチド配列の同一性は約82%である(Bantel−Schaal et al.、1999、J.Virol.、73(2):939〜947)。さらに、多くのAAV血清型のRep配列及びITRは、哺乳動物中でのAAV粒子の生成において、他の血清型由来の対応する配列と効率よく相互補完する(即ち、機能的に置換する)ことが知られている。US2003148506は、AAVのRep及びITR配列は、昆虫細胞中の他のAAVのRep及びITR配列とも効率よく相互補完することを報告している。
【0041】
AAVVPタンパク質は、AAVビリオンの細胞指向性を決定することが知られている。VPタンパク質コード配列は、異なるAAV血清型間のRepタンパク質及び遺伝子ほど有意に保存されていない。他の血清型の対応する配列と相互補完するRep及びITR配列の能力は、1つの血清型(例えば、AAV3)のカプシドタンパク質及び他のAAV血清型(例えば、AAV2)のRep及び/又はITR配列を含む偽型rAAV粒子の生成をもたらす。このような偽型rAAV粒子は、本発明の一部分である。
【0042】
改変「AAV」配列も、例えば昆虫細胞においてrAAVベクターを生成するために、本発明の文脈で使用することができる。このような改変配列は、例えばAAV1、AAV2、AAV3、AAV4、AAV5、AAV6、AAV7、AAV8又はAAV9のITR、Repと少なくとも約70%、少なくとも約75%、少なくとも約80%、少なくとも約85%、少なくとも約90%、少なくとも約95%、又はそれより高いヌクレオチド及び/又はアミノ酸配列同一性を有する配列(例えば、約75〜99%のヌクレオチド配列同一性を有する配列)を含み、或いはVPを野生型AAVのITR、Rep、又はVP配列の代わりに使用することができる。
【0043】
多くの点で他のAAV血清型と類似しているが、AAV5は他の知られているヒト及びサルの血清型より、他のヒト及びサルのAAV血清型と異なる。それを考慮すると、rAAV5の生成は昆虫細胞における他の血清型の生成と異なる可能性がある。本発明の方法を利用してrAAV5を生成する場合、1つ又は複数の構築体は、2つ以上の構築体の場合をひとまとめにして、AAV5ITRを含むヌクレオチド配列、AAV5Repコード配列を含むヌクレオチド配列(即ち、ヌクレオチド配列はAAV5Rep78を含む)を含むことが好ましい。このようなITR及びRep配列を望むように改変して、昆虫細胞におけるrAAV5又は偽型rAAV5ベクターの有効な生成を得ることができる。例えば、Rep配列の開始コドンを改変することができ、VPスプライス部位を改変又は除去することができ、且つ/或いはVP1開始コドン及び隣接ヌクレオチドを改変して、昆虫細胞におけるrAAV5ベクターの生成を改善することができる。
【0044】
したがって、他の態様では、本発明は昆虫細胞において(上記で定義した組換えパルボウイルス(rAAV)ベクターを含む)組換えパルボウイルス(rAAV)ビリオンを生成するための方法に関する。この方法は、(a)組換えパルボウイルス(rAAV)ベクターが生成するような条件下において本明細書で上記で定義した昆虫細胞を培養するステップと、(b)組換えパルボウイルス(rAAV)ベクターを回収するステップとを含むことが好ましい。この方法で生成される組換えパルボウイルス(rAAV)ベクターは、組換えパルボウイルス(rAAV)ベクター核酸を含む感染性パルボウイルス又はAAVビリオンであることが好ましいことは、本明細書において理解される。培養中の昆虫細胞に関する増殖条件、及び培養中の昆虫細胞における異種産物の生成は当技術分野でよく知られており、例えば昆虫細胞の分子工学に関する前に引用した参照文献中に記載されている。
【0045】
この方法は、抗AAV抗体、好ましくは固定化抗体を使用する、組換えパルボウイルス(rAAV)ベクター(を含むビリオン)の親和性精製のステップをさらに含むことが好ましい。抗AAV抗体は、モノクローナル抗体であることが好ましい。特に適切な抗体は、例えばラクダ又はラマなどから入手可能な単鎖ラクダ抗体又はその断片である(例えば、Muyldermans、2001、Biotechnol.74:277〜302を参照)。rAAVの親和性精製用の抗体は、AAVカプシドタンパク質上のエピトープと特異的に結合する抗体であることが好ましく、エピトープは2つ以上のAAV血清型のカプシドタンパク質上に存在するエピトープであることが好ましい。例えば抗体は、AAV2カプシドとの特異的結合に基づいて産生又は選択することができるが、さらに同時に、抗体はAAV1、AAV3及びAAV5カプシドとも特異的に結合することができる。
【0046】
他の態様では、本発明は、上記で定義した核酸構築体及び細胞を使用して、前に記載した本発明の方法において生成されるrAAVビリオンに関する。rAAVビリオンは、そのゲノム中に対象とする遺伝子産物をコードする少なくとも1つのヌクレオチド配列を含むことが好ましく、少なくとも1つのヌクレオチド配列は原型AAVヌクレオチド配列ではなく、AAVVP1、VP2、及びVP3カプシドタンパク質の化学量論では、VP1の量は:(a)VP2の量の少なくとも100、105、110、120、150、200又は400%であり、或いは(b)VP3の量の少なくとも8、10、10.5、11、12、15、20又は40%であり、或いは(c)少なくとも(a)と(b)の両方で定義した量である。VP1、VP2及びVP3の量は、VP1、VP2及びVP3のそれぞれに共通であるエピトープを認識する抗体を使用して測定することが好ましい。VP1、VP2及び/又はVP3の相対量の定量化を可能にするはずである、様々なイムノアッセイが当技術分野で利用可能である(例えば、「抗体の使用(Using Antibodies)」、E.Harlow and D.Lane、1999、Cold Spring Harbor Laboratory Press、ニューヨークを参照)。3つのカプシドタンパク質のそれぞれに共通であるエピトープを認識するのに適した抗体は、例えば(Progen、ドイツから市販されている)マウス抗CapB1抗体である。本発明による好ましいrAAVビリオンは、そのゲノム中に対象とする遺伝子産物をコードする少なくとも1つのヌクレオチド配列を含むビリオンであり、少なくとも1つのヌクレオチド配列は原型AAVヌクレオチド配列ではなく、AAVビリオンは1アミノ酸位にロイシン又はバリンを含むVP1カプシドタンパク質を含む。本発明によるさらに好ましいAAVビリオンは上記で定義したカプシドタンパク質の比を有しており、1アミノ酸位にロイシン又はバリンを含むVP1カプシドタンパク質を含む。
【0047】
本文書中及びその特許請求の範囲中において、動詞「含む」及びその活用は、その語句の後の項目が含まれるが、具体的に言及されていない項目が除外されるわけではないことを意味するように、非限定的な意味で使用する。さらに、不定冠詞「a」又は「an」による要素に対する言及は、1つ及びただ1つの要素が存在することを状況が明らかに必要としない限り、2つ以上の要素が存在する可能性を除外するわけではない。したがって不定冠詞「a」又は「an」は、「少なくとも1つ」を通常意味する。
【図面の簡単な説明】
【0048】
【図1A】Rep発現における野生型AAVゲノムの構成の図である。Rep78及びRep52遺伝子は、それぞれP5及びP19プロモーターから発現される。(それぞれRep78及びRep52のスプライス変異体である)Rep68及びRep40の発現は示さない。両方の発現単位がATG開始部位を含む。
【図1B】本発明の構築体が、1つのプロモーター(例えば、多角体(PolH)プロモーター)の制御下でRepのORFを有することを示す図である。このプロモーターはRep78とRep52両方の発現を誘導するが、これは、Rep78の開始コドンATGが他のACG開始コドンに変換され、リボソームによって一部分がスキップされるからである。
【図1C】Urabe et al(2002、上記)による原型構築体は、2つの異なるプロモーター(それぞれΔIE1及びpolH)から独立にRep78及びRep52を誘導することを示す図である。
【図2】昆虫細胞で5回継代した組換えバキュロウイルスから発現された、Repタンパク質のウェスタンブロット分析の図である。Urabe et al.、2002によって設計された原型バキュロウイルス(原型REP/Bac−to−Bac)は、5回の継代でRep78/52発現のゆっくりした低下をもたらす。バキュロウイルス骨格PSC(原型REP/PSC)に挿入した、Urabe et al.、2002によって設計されたRep78及び52の発現単位も、昆虫細胞での継代後にRep78/52発現の低下をもたらす。しかしながら、PSC骨格中にACG開始コドンを含むREP発現単位を有するバキュロウイルス(REP−ACG/PSC)は、少なくとも5回の継代でRep78/52の安定した発現をもたらす。ウェスタンブロット分析は、実施例1.1.3中に記載したように実施した。
【図3】グラフ中にプロットした表1の結果の図である。
【図4】昆虫細胞における様々なrAAV構築体の安定性の比較の図である。SF+細胞におけるrAAV生成は、実施例1中で前に記載したように実施した。すべての生成に関して、ITR含有バキュロウイルスとカプシド遺伝子含有バキュロウイルスは同一であり、継代数はRep遺伝子含有バキュロウイルスと同じであった。4つの異なるRep遺伝子含有バキュロウイルスを使用した:1)REP−ACG/PSC、2)SLR:Urabe et al.(2002、上記)による原型構築体、3)Rep52+Rep78(B2B):2つの別個のBac−to−Bacバキュロウイルス、1つはRep78遺伝子を含み、もう1つはRep52遺伝子を含む。4)Rep52+Rep78(PSC):2つの別個のプロテインサイエンスのバキュロウイルス、1つはRep78遺伝子を含み、もう1つはRep52遺伝子を含む。
【図5】第8代までのREP−ACG/PSCバキュロウイルス構築体の安定性の図である。SF+細胞におけるrAAV生成は、実施例1中で記載したように実施した。
【図6】Urabe et al.(2002、上記)からの原型構築体及び本発明によるREP−ACG/PSC構築体のrepタンパク質発現に対する、継代効果の影響の比較の図である。バキュロウイルスの継代及びウェスタンブロットは、実施例1中に記載したように実施した。repバキュロウイルスの正常な継代の間に、SF細胞にバキュロウイルスを加えた後40時間でサンプルを採取し、ウェスタンブロットを実施した。
【発明を実施するための形態】
【実施例】
【0049】
(実施例1)
Rep構築体
1.1.材料及び方法
1.1.1 バキュロウイルスプラスミドの構築
1つの2シストロン性メッセンジャーRNAからRep78及びRep52を発現させるために、発現ベクターpFastBacDualSLR(Urabe et al.、2002、上記)上に位置するRep78のATG開始コドンをACGに転換した。使用した上流プライマーは以下の通りであった:
BamHI
5'-cgcggatcctgttaagACGGCGGGGTTTTACGAGATTGTGATTAAGGTC-3'
(配列番号8)
プライマー配列、正方向
【0050】
PCR反応中で使用した3’プライマーはREP78遺伝子に隣接しており、XbaI部位(TCTAGA)を含む:
XbaI
5'-AGGCTCTAGATTCGAAAGCGGCCCG-3'
(配列番号9)
プライマー配列、逆方向
【0051】
前述のプライマーセット間の配列を、以下のプロトコル:95℃、5分間の1サイクル;95℃、15秒間の35サイクル;55℃、30秒間;72℃、2分間;72℃、10分間の1サイクル;4℃、恒久的を使用して、PCR(反応体積50μl;1×PfX Amp.バッファー、0.3mMのdNTP’s、1mMのMgSO4、150mMの正方向プライマー、150mMの逆方向プライマー、2×エンハンサー溶液、鋳型50ng(pFastBacDualSLR)、1UのPlatinum Pfx(Invitrogen、Carlsbad、CA、USA))によって増幅した。PCR産物は、Zero Blunt TOPO PCRクローニングキット(Invitrogen)を使用してPCR平滑末端II−TOPOでクローニングした。Rep78は制限部位SpeI及び×baIを使用してpFastBacDual(Invitrogen)にサブクローニングした。突然変異Rep発現カセットは、(制限酵素BstZ171及びAvrIIを使用して)、バキュロウイルス発現構築体(EcoRV及びXbaIで切断した)pPSC10(Protein Sciences Corporation、Meriden、CT、USA)に最終的にクローニングした。構築体の配列分析は、Baseclear、Leiden、オランダによって確認した。
【0052】
1.1.2 組換えバキュロウイルスの生成
オートグラファカリフォルニカ(Autographa californica)核多角体病ウイルス(AcNPV)由来の組換えバキュロウイルスを、GeneXpress BaculoKIT(Protein Sciences Corporation)を使用して生成した。トランスフェクションは以下のように実施した:丸底14m1チューブ内で200μlのGRACE培地を6μlのセルフェクチン(Invitrogen)と混合し、エッペンドルフチューブ内で200μlのGRACE培地を、50μlのウイルスDNA(プロテインサイエンス)及び2μgのトランスファープラスミド(REP)と混合した。エッペンドルフチューブからの内容物をチューブに加え、注意深く混合した。室温で30分間のインキュベーション後、1,300μlのGRACEをトランスフェクション混合物に加えた。T25フラスコ中の昆虫細胞はGRACE培地で洗浄し、トランスフェクション混合物は細胞層に一滴ずつ加えた。28℃で6時間のインキュベーション後、10%のFBSを補ったSF900II血清を注意深く加え、T25フラスコは5日間28℃のストーブ内に置き、その後組換えバキュロウイルスを採取した。
【0053】
1.1.3 ウェスタンブロット分析
バキュロウイルス−REPを用いて昆虫細胞(SF+)を感染させた。細胞感染後16、40、及び64時間でサンプルを採取し、0.1V10×TRIS溶解バッファー(1.5MのNaCl、0.5MのTRIS、0.01MのMgCl、1%のTRITON X−100、pH8.5、濾過滅菌済)を加えることによって細胞を溶かし、シェーカー(Innova 44、New Brunswick)内で28℃において30分間インキュベートした。30分間37℃でのベンゾナーゼとのインキュベーションによって、遊離DNA及びRNAを分解した。細胞溶解物は遠心分離にかけた(1,900×g;15分間;4℃)。NuPAGE LDSサンプルバッファー(4×、Invitrogen)を上清のサンプルに加え、4〜12%のBis−Trisゲル(120V)に載せた。タンパク質は、10V(半乾燥ブロッティング)で30分間PVDF膜(BioRad)にブロッティングした。ウェスタンブロットによる免疫化学測定法を、Superblock−PBSブロッキングバッファー(PIERCE)で膜をブロッキングし、マウス抗Rep(303.9、Progen、ドイツ;希釈1:50)及びウサギ抗マウスHRP(DAKO、希釈1:500)と後にインキュベートすることによって実施した。lumi−light plusウェスタンブロッティング基質(Roche)を用いた化学発光染色によって、Rep−タンパク質を視覚化した。
【0054】
1.2 結果
本発明の新たに設計したRep−構築体(REP−ACG/PSC)の成績を、1)PSCバキュロウイルス骨格と2)Bac−to−Bacバキュロウイルス骨格(Urabe et al.、2002)の両方の原型Rep構築体と比較した。全3個の構築体を第5代まで連続して継代した。AAV1−LPL生成の実験は、それぞれ第2、3、4及び5代のAAV−LPL及びAAV−Cap組換えバキュロウイルスと組合せた、第2、3、4及び5代のRep−構築体を使用して実施した(ここで使用したAAV−LPL及びAAV−Cap組換えバキュロウイルスは、実施例2中で以下に記載する)。AAV1−LPLの生成収率はqPCRによって測定し、表1中に示す。Urabe et al.、2002によって設計された原型バキュロウイルス(原型REP/Bac−to−Bac)は、5回の継代でAAV生成の急速な低下をもたらす。バキュロウイルス骨格PSC(原型REP/PSC)に挿入した、Urabe et al.、2002によって設計されたRepの発現単位も、昆虫細胞での継代後にAAV生成の低下をもたらす。しかしながら、PSC骨格中にACG開始コドンを含むRep発現単位を有するバキュロウイルス(REP−ACG/PSC)は、少なくとも5回の継代で安定したAAV生成をもたらす。したがって、数回の継代(例えば、第2代〜第5代)におけるAAV−LPLの再現性のある生成収率は、REP−ACG構築体を含むバキュロウイルスを使用してのみ得られた。
【0055】
【表1】
表1:数回継代したバキュロウイルス構築体を使用したrAAVビリオンの生成:Sf9細胞を、第2、3、4又は5代のLPL−ベクター単位、第2、3、4又は5代のRep−発現単位、及び第2、3、4又は5代のCap−発現単位をコードする3つの組換えバキュロウイルスで感染させた。3日後に細胞を採取し、AAV収率(1ml当たりのベクターゲノム;vg/ml)をqPCRによって測定した。
【0056】
【表2】
表2:昆虫細胞の継代(第2代〜第5代)後に様々なBac−Rep構築体で実施したQ−PCR。
【0057】
表2は、組換えバキュロウイルスにおけるRep−発現単位用、及び隣接するバキュロウイルスORF用に設計した、定量PCR(Q−PCR)アッセイの結果を示す(1ml当たりの遺伝子コピー数;gc/ml)。Q−PCRの値の間の比は、Rep−バキュロウイルス中の欠失の存在を決定する。1という比は、バッチ中のすべてのバキュロウイルスが組換えRep78又は52配列を含むことを理論上意味する。Urabe et al.、2002によって設計された原型バキュロウイルス(原型REP/Bac−to−Bac)は、第5代での相当量の組換えバキュロウイルスがRep配列中の欠失を有することを示す。バキュロウイルス骨格PSC(原型REP/PSC)に挿入した、Urabe et al.、2002によって設計されたRep78及び52用の発現単位は、組換えバキュロウイルスの非常に初期の劇的な消失を示す。しかしながら、PSC骨格中にACG開始コドンを含むRep発現単位を有するバキュロウイルス(REP−ACG/PSC)(クローンC4及びA3)は、少なくとも5回の継代で安定した組換えバキュロウイルスを示す。
【0058】
(実施例2)
Cap構築体
2.1.1 バキュロウイルスプラスミドの構築
1つのポリシストロン性メッセンジャーRNAからVP1、2、3を発現させるために、(Urabe et al.、2002、上記)によって記載されたのと同様にバキュロウイルス−AAV−Cap構築体を設計した。簡単に言うと、VP1のATG開始コドンをACGに突然変異させた。11位の考えられるATG開始コドンはACGに変わった。VP1開始コドンの下流のスプライス受容体部位を破壊した(21及び24位における突然変異)。突然変異Cap発現カセットは、BamH1/StuI制限部位を有するバキュロウイルス発現構築体;pfastBacDual(pFBDAAV1VPm11)にクローニングした。このプラスミド(pFBDAAV1VPm11)は、VP1用の他の開始コドンを導入するための出発物質であった。前述の突然変異を導入するためにUrabe et al.(2002、上記)によって使用された正方向プライマーは、以下の通りであった:
BamHI 1 11 21 24
5'-cgcggatcctgttaagACGGCTGCCGACGGTTATCTACCCGATTGGCTC-3'
(配列番号1)
【0059】
出発物質としてpFBDAAV1VPm11(Urabe et al.、2002、上記)を使用して発現構築体を作製するために、以下の正方向プライマーを使用した:
5'-cgcggatcctgttaagTTGGCTGCCGACGGTTATCTACCCGATTGGCTC-3'
(配列番号2)
5'-cgcggatcctgttaagATTGCTGCCGACGGTTATCTACCCGATTGGCTC-3'
(配列番号3)
5'-cgcggatcctgttaagGTGGCTGCCGACGGTTATCTACCCGATTGGCTC-3'
(配列番号4)
5'-cgcggatcctgttaagCTGGCTGCCGACGGTTATCTACCCGATTGGCTC-3'
(配列番号5)
【0060】
前述の正方向プライマーと共にPCR反応中で使用した逆方向プライマーは、VP1開始コドンの下流230bpまでの位置を対象とし、特有のStuI部位(AGGCCT)を含む。
5'-GTCGTAGGCCTTGTCGTGCTCGAGGGCCGC-3'
(配列番号6)
【0061】
PCRにより正方向と逆方向プライマー対の前述のセットを用いて断片を増幅した。BamHI及びStuIを用いたPCR産物の消化後、PCR産物はpFBDAAV1vpm11のBamHI/StuI部位にサブクローニングし、様々な試験用バキュロウイルス−AAV−Cap構築体を生成した。DNA構築体はBaseclear、Leiden、オランダでの配列分析によって確認した。
【0062】
2.1.2 組換えバキュロウイルスの生成
オートグラファカリフォルニカ(Autographa californica)核多角体病ウイルス(AcNPV)由来の組換えバキュロウイルスを、Bac−to−Bacバキュロウイルス発現系(Invitrogen)を使用して生成した。0.1のmoiで1ml当たり2×106個のSf9細胞を感染させることによってrBac−Capを増幅した。感染後3日で細胞を遠心し、ウイルスを含む上清を回収した。
【0063】
2.1.3 組換えAAVの生成
Urabe et al.、2002による3つの組換えバキュロウイルスを使用してrAAVバッチを生成した。しかしながら、この試験に関しては、1つのバキュロウイルスはLPLS447X−導入遺伝子の発現構築体を有していた。この導入遺伝子から発現される治療活性物質は、ヒトリポタンパク質リパーゼの天然に存在する変異体、448アミノ酸の単鎖ポリペプチドである。LPLS447Xの変異体は、タンパク質のC末端に2アミノ酸の欠失を有する。第2のバキュロウイルスはAAV複製遺伝子、Rep78及びRep52の発現構築体を有していた。第3のバキュロウイルスはAAV1カプシド配列及びVP1のACG又はTTG、CTG、GTG開始コドンのいずれかを有していた。
【0064】
プラスミド−トランスフェクション系を用いて生成した哺乳動物rAAVのバッチは、Grimm et al.、1998(「組換えアデノ随伴ウイルスベクターを生成及び精製するための新規のツール(Novel tools for production and purification of recombinant adeno−associated virus vectors)」Hum Gene Ther.1998 Dec10;9(18):2745〜60)に従い生成した。
【0065】
2.1.3 ウェスタンブロット分析
バキュロウイルス−Capを用いて昆虫細胞を感染させた。感染後3日で、細胞を遠心分離にかけた(3,000g;15分間)。上清は0.22umのMillexフィルターを介して濾過した。NuPAGE LDSサンプルバッファー(Invitrogen)を上清のサンプルに加え、4〜12%のBis−Trisゲル(120V)に載せた。ゲルは100Vで施した。タンパク質は1時間、100V、350mAでニトロセルロース膜(BioRad)にブロッティングした。ウェスタンブロットによる免疫化学測定法を、1%マーヴェル、乾燥スキムミルクで膜をブロッキングし、マウス抗Cap(Progen、ドイツからのB1;希釈1:50)及びウサギ抗マウス−HRP(DAKO、希釈1:100)と後にインキュベートすることによって実施した。発光及びウェスタンブロッティング基質(Roche)を用いた化学発光染色によって、VP1、2及び3を視覚化した。
【0066】
2.1.4 生化学的測定
放射性トリオレオイルグリセロールエマルジョン基質を使用して以前に記載されたのと同様に(Nilsson−Ehle and Scholtz、1976)、ヒトLPLS447Xの活性をアッセイした。ヒトLPLS447Xの免疫反応体を、ニワトリIgY及びマウス5D2抗hLPL抗体(Liu et at.、2000)を用いたサンドウィッチELISAを使用してアッセイした。血漿トリグリセリドレベルは、製造者のプロトコル(Boehringer Mannheim、#450032)に従い市販のキットを使用することによって測定した。
【0067】
2.2 結果
2.2.1 組換えバキュロウイルスの構築
Urabe et al.(2002、上記)により設計されたバキュロウイルスプラスミド中でのVP1の発現用の異なる他の開始コドンを導入するために、BamHI制限部位及びVP1のACG開始コドンの代わりにTTG、ATT、GTG又はCTGコドンのいずれかを含む一連の上流プライマーを設計した。StuI部位を含む下流プライマーと組合せてこれらのプライマーを使用したPCRによって、いくつかの増幅断片を生成し、これらはpFBDVPm11(Bac−Cap)のBamHI/StuI部位にサブクローニングした。生成したバキュロウイルスプラスミドは、Bac−to−Bacバキュロウイルス発現系を使用する組換えバキュロウイルスの調製に使用した。調製した組換えバキュロウイルスは昆虫細胞に感染させて、AAVカプシドを生成した。感染後3日で、異なるバキュロウイルスバッチのウイルスタンパク質の発現を、ウェスタンブロットで測定した。ウェスタンブロットから、VP1のTTG開始コドンを含むバキュロウイルス構築体は、以前に使用したACG開始コドンと比較して高いレベルで、このタンパク質を発現したことが明らかになった。TTGコドンを使用したVP1とVP2の間の比は、野生型AAVに関して報告される比と同様に1:1であったことが分かった(示さず)。
【0068】
2.2.2 培養中の細胞におけるrAAVバッチの感染
TTG開始コドンを有する組換えバキュロウイルス由来のAAVカプシドの感染性を調べるために、rAAVを作製した。さらにrAAVバッチを、哺乳動物HEK293細胞におけるプラスミドトランスフェクションによって作製した。両rAAVバッチのベクターゲノム力価をqPCRによって測定した。この力価を使用して、高いmoiでマイクロタイタープレート中のHEK293細胞を感染させた。感染後2日で、導入遺伝子産物(LPLS447X)に関する定量アッセイ(LPLS447X−質量アッセイ)を、感染細胞の培地で実施した。このアッセイは、バキュロウイルス生成rAAVによって生成したLPLS447Xの量は、プラスミド生成rAAVによって生成したLPLと同等であったことを示した(示さず)。
【0069】
2.2.3 マウスにおけるrAAVバッチの注射
バキュロウイルス生成系、及び従来の哺乳動物プラスミド生成系によって生成したrAAVバッチをマウスの筋肉内に注射して、in vivoでのLPLS447X−タンパク質活性及びトリグリセリド非摂取を追跡した。注射後第3日、第7日及び第2週で、血液サンプルを採取し評価した。ウイルス投与後第3日と第7日の間に、哺乳動物rAAVを注射した両マウス及びバキュロrAAVを注射した1マウスからサンプル採取した血液−血漿は、乳濁状から完全に透明な状態に変わった。1匹のバキュロrAAV注射マウス由来の血液血漿は依然として比較的乳濁状であったが、しかしながら脂肪レベルは明らかに低下した。トリグリセリドレベルは、すべての治療マウスのそれぞれで低下した(示さず)。第14日に、両方の哺乳動物rAAV及びバキュロウイルス−(TTG)−AAV治療マウスにおけるTGレベルは、TGレベルが96%低下した。ウイルス投与後2週間で採取した血漿サンプルは、バキュロウイルス−AAVと哺乳動物−AAVで治療したマウスのLPLS447X−活性は、同等であったことを示した(示さず)。
【0070】
(実施例3)
昆虫細胞における改変型Rep78開始コドンを有するrAAV構築体の安定性
3.1 昆虫細胞における様々なrAAV構築体の安定性の比較
SF+細胞におけるrAAV生成は、実施例1中に前に記載したのと同様に実施した。すべての生成に関して、ITR含有バキュロウイルスとカプシド遺伝子含有バキュロウイルスは同一であり、継代数はRep遺伝子含有バキュロウイルスと同じであった。4つの異なるRep遺伝子含有バキュロウイルス:1)REP−ACG/PSC、2)SLR:Urabe et al.(2002、上記)による原型構築体、3)Rep52+Rep78(B2B):1つがRep78遺伝子を含みもう1つがRep52遺伝子を含む、2つの別個のBac−to−Bacバキュロウイルス、4)Rep52+Rep78(PSC):1つがRep78遺伝子を含みもう1つがRep52遺伝子を含む、2つの別個のプロテインサイエンスのバキュロウイルスを使用した。
【0071】
結果は図4中に示す。5回目のバキュロウイルス継代したrAAV生成は、原型Rep構築体と比較して、及び分岐Rep構築体と比較して、本発明によるREP−ACG/PSCを使用して10倍を超えて既に改善されている。
【0072】
3.2 第8代までのバキュロウイルス構築体の安定性
SF+細胞におけるrAAV生成は、実施例1中に記載したのと同様に実施した。すべての生成に関して、ITR含有バキュロウイルスとカプシド遺伝子含有バキュロウイルスは同一であり、継代数はREP−ACG/PSCバキュロウイルスと同じであった。結果は図5中に示す。REP−ACG/PSCバキュロウイルスは少なくとも第8代まで安定性がある。REP−ACG/PSCのrAAV生成力価は、少なくとも第8代のバキュロウイルスまで安定している。
【0073】
3.3 repタンパク質発現に対する継代の影響
Urabe et al.(2002、上記)からの原型構築体Repタンパク質の発現に対する継代数の影響を、本発明によるREP−ACG/PSC構築体と比較した。バキュロウイルスの継代及びウェスタンブロットは、実施例1中に記載したように実施した。repバキュロウイルスの正常な継代の間に、SF細胞にバキュロウイルスを加えた後40時間でサンプルを採取し、ウェスタンブロットを実施した。図6は、原型Urabe構築体(SLR)の初期継代と比較した多数回継代の低下したRep発現を明らかに示すが、一方REP−ACG/PSC構築体におけるRep発現は、少数回の継代と比較して多数回の継代で同じ状態である。
【特許請求の範囲】
【請求項1】
パルボウイルスRepタンパク質をコードするヌクレオチド配列を含むオープンリーディングフレームを含むヌクレオチド配列であって、パルボウイルスRep78タンパク質の翻訳の開始コドンが、昆虫細胞における発現後に部分的なエキソンスキッピングを行う開始コドンであるヌクレオチド配列。
【請求項2】
開始コドンがACG、TTG、CTG、及びGTGから選択される、請求項1に記載のヌクレオチド配列。
【請求項3】
AAVRep78タンパク質をコードするヌクレオチド配列の開始コドンの上流に、配列番号7の9ヌクレオチドの配列、又は配列番号7と実質的に相同なヌクレオチド配列を含む発現制御配列を含む、請求項1又は2に記載のヌクレオチド配列。
【請求項4】
パルボウイルスRepタンパク質がアデノ随伴ウイルス(AAV)Repタンパク質である、請求項1から3までのいずれか一項に記載のヌクレオチド配列。
【請求項5】
ヌクレオチド配列が昆虫細胞における発現用の発現制御配列と作動可能に連結している、請求項1から4までのいずれか一項に記載のヌクレオチド配列を含む核酸構築体。
【請求項6】
ヌクレオチド配列が多角体プロモーターと作動可能に連結している、請求項5に記載の核酸構築体。
【請求項7】
昆虫細胞適合性ベクター、好ましくはバキュロウイルスベクターである、請求項5又は6に記載の核酸構築体。
【請求項8】
1つ又は複数のパルボウイルスRepタンパク質をコードする単一のオープンリーディングフレームを含む唯一の型のヌクレオチド配列を含む昆虫細胞。
【請求項9】
単一のオープンリーディングフレームが完全長Rep78タンパク質をコードする、請求項8に記載の昆虫細胞。
【請求項10】
1つ又は複数のパルボウイルスRepタンパク質をコードする単一のオープンリーディングフレームを含むヌクレオチド配列が核酸構築体の一部分であり、ヌクレオチド配列が昆虫細胞における発現用の発現制御配列と作動可能に連結している、請求項8又は9に記載の昆虫細胞。
【請求項11】
請求項1から4までのいずれか一項に記載の1つ又は複数のパルボウイルスRepタンパク質をコードする単一のオープンリーディングフレームを含む第1のヌクレオチド配列、又は請求項5から7までのいずれか一項に記載の、ヌクレオチド配列が昆虫細胞における発現用の発現制御配列と作動可能に連結している第1の核酸構築体を含む、請求項8から10までのいずれか一項に記載の昆虫細胞。
【請求項12】
a)少なくとも1つのパルボウイルス逆方向末端反復(ITR)ヌクレオチド配列を含む第2のヌクレオチド配列と、
b)昆虫細胞における発現用の発現制御配列と作動可能に連結している配列をコードするパルボウイルスカプシドタンパク質を含む第3のヌクレオチド配列と
をさらに含む、請求項11に記載の昆虫細胞。
【請求項13】
a)請求項9の(b)に記載の第3のヌクレオチド配列をさらに含む、請求項5から7までのいずれか一項に記載の第1の核酸構築体と、
b)請求項9の(a)に記載の第2のヌクレオチド配列を含む第2の核酸構築体と
を含む、請求項12に記載の昆虫細胞。
【請求項14】
第2の核酸構築体が昆虫細胞適合性ベクター、好ましくはバキュロウイルスベクターである、請求項13に記載の昆虫細胞。
【請求項15】
第2のヌクレオチド配列が対象とする遺伝子産物をコードする少なくとも1つのヌクレオチド配列をさらに含み、前記対象とする遺伝子産物をコードする少なくとも1つのヌクレオチド配列が、昆虫細胞において生成されるパルボウイルスベクターのゲノムに組み込まれる、請求項12から14までのいずれか一項に記載の昆虫細胞。
【請求項16】
第2のヌクレオチド配列が2つのパルボウイルスITRヌクレオチド配列を含み、対象とする遺伝子産物をコードする少なくとも1つのヌクレオチド配列が、前記2つのパルボウイルスITRヌクレオチド配列間に位置する、請求項15に記載の昆虫細胞。
【請求項17】
第3のヌクレオチド配列がパルボウイルスVP1、VP2、及びVP3カプシドタンパク質をコードするヌクレオチド配列を含むオープンリーディングフレームを含み、パルボウイルスVP1カプシドタンパク質の翻訳の開始コドンがACG、TTG、CTG、及びGTGから選択される、請求項12から16までのいずれか一項に記載の昆虫細胞。
【請求項18】
第3のヌクレオチド配列が、パルボウイルスVP1カプシドタンパク質をコードするヌクレオチド配列の開始コドンの上流に、配列番号7の9ヌクレオチドの配列、又は配列番号7と実質的に相同なヌクレオチド配列を含む発現制御配列を含む、請求項17に記載の昆虫細胞。
【請求項19】
第3のヌクレオチド配列が、12ヌクレオチド位におけるC、21ヌクレオチド位におけるA、及び24ヌクレオチド位におけるCから選択されるパルボウイルスVP1カプシドタンパク質をコードするヌクレオチド配列の少なくとも1つの改変をさらに含む、請求項17又は18に記載の昆虫細胞。
【請求項20】
少なくとも1つの第1のヌクレオチド配列、第2のヌクレオチド配列、及び第3のヌクレオチド配列が昆虫細胞のゲノムに安定して組み込まれている、請求項12から19までのいずれか一項に記載の昆虫細胞。
【請求項21】
パルボウイルスがAAVである、請求項8から20までのいずれか一項に記載の昆虫細胞。
【請求項22】
請求項12、15及び16のいずれか一項に記載の第2のヌクレオチド配列を含む組換えパルボウイルスビリオンを昆虫細胞において生成するための方法であって、
a)組換えパルボウイルスビリオンが生成するような条件下で請求項12から21までのいずれか一項に記載の昆虫細胞を培養するステップと、
b)組換えパルボウイルスビリオンを回収するステップと
を含む方法。
【請求項23】
抗パルボウイルス抗体、好ましくは固定化抗体を使用する、ビリオンの親和性精製のステップをさらに含む、請求項22に記載の方法。
【請求項24】
抗パルボウイルス抗体が単鎖ラクダ抗体又はその断片である、請求項23に記載の方法。
【請求項25】
組換えパルボウイルスビリオンが組換えAAVビリオンである、請求項22から24までのいずれか一項に記載の方法。
【請求項1】
パルボウイルスRepタンパク質をコードするヌクレオチド配列を含むオープンリーディングフレームを含むヌクレオチド配列であって、パルボウイルスRep78タンパク質の翻訳の開始コドンが、昆虫細胞における発現後に部分的なエキソンスキッピングを行う開始コドンであるヌクレオチド配列。
【請求項2】
開始コドンがACG、TTG、CTG、及びGTGから選択される、請求項1に記載のヌクレオチド配列。
【請求項3】
AAVRep78タンパク質をコードするヌクレオチド配列の開始コドンの上流に、配列番号7の9ヌクレオチドの配列、又は配列番号7と実質的に相同なヌクレオチド配列を含む発現制御配列を含む、請求項1又は2に記載のヌクレオチド配列。
【請求項4】
パルボウイルスRepタンパク質がアデノ随伴ウイルス(AAV)Repタンパク質である、請求項1から3までのいずれか一項に記載のヌクレオチド配列。
【請求項5】
ヌクレオチド配列が昆虫細胞における発現用の発現制御配列と作動可能に連結している、請求項1から4までのいずれか一項に記載のヌクレオチド配列を含む核酸構築体。
【請求項6】
ヌクレオチド配列が多角体プロモーターと作動可能に連結している、請求項5に記載の核酸構築体。
【請求項7】
昆虫細胞適合性ベクター、好ましくはバキュロウイルスベクターである、請求項5又は6に記載の核酸構築体。
【請求項8】
1つ又は複数のパルボウイルスRepタンパク質をコードする単一のオープンリーディングフレームを含む唯一の型のヌクレオチド配列を含む昆虫細胞。
【請求項9】
単一のオープンリーディングフレームが完全長Rep78タンパク質をコードする、請求項8に記載の昆虫細胞。
【請求項10】
1つ又は複数のパルボウイルスRepタンパク質をコードする単一のオープンリーディングフレームを含むヌクレオチド配列が核酸構築体の一部分であり、ヌクレオチド配列が昆虫細胞における発現用の発現制御配列と作動可能に連結している、請求項8又は9に記載の昆虫細胞。
【請求項11】
請求項1から4までのいずれか一項に記載の1つ又は複数のパルボウイルスRepタンパク質をコードする単一のオープンリーディングフレームを含む第1のヌクレオチド配列、又は請求項5から7までのいずれか一項に記載の、ヌクレオチド配列が昆虫細胞における発現用の発現制御配列と作動可能に連結している第1の核酸構築体を含む、請求項8から10までのいずれか一項に記載の昆虫細胞。
【請求項12】
a)少なくとも1つのパルボウイルス逆方向末端反復(ITR)ヌクレオチド配列を含む第2のヌクレオチド配列と、
b)昆虫細胞における発現用の発現制御配列と作動可能に連結している配列をコードするパルボウイルスカプシドタンパク質を含む第3のヌクレオチド配列と
をさらに含む、請求項11に記載の昆虫細胞。
【請求項13】
a)請求項9の(b)に記載の第3のヌクレオチド配列をさらに含む、請求項5から7までのいずれか一項に記載の第1の核酸構築体と、
b)請求項9の(a)に記載の第2のヌクレオチド配列を含む第2の核酸構築体と
を含む、請求項12に記載の昆虫細胞。
【請求項14】
第2の核酸構築体が昆虫細胞適合性ベクター、好ましくはバキュロウイルスベクターである、請求項13に記載の昆虫細胞。
【請求項15】
第2のヌクレオチド配列が対象とする遺伝子産物をコードする少なくとも1つのヌクレオチド配列をさらに含み、前記対象とする遺伝子産物をコードする少なくとも1つのヌクレオチド配列が、昆虫細胞において生成されるパルボウイルスベクターのゲノムに組み込まれる、請求項12から14までのいずれか一項に記載の昆虫細胞。
【請求項16】
第2のヌクレオチド配列が2つのパルボウイルスITRヌクレオチド配列を含み、対象とする遺伝子産物をコードする少なくとも1つのヌクレオチド配列が、前記2つのパルボウイルスITRヌクレオチド配列間に位置する、請求項15に記載の昆虫細胞。
【請求項17】
第3のヌクレオチド配列がパルボウイルスVP1、VP2、及びVP3カプシドタンパク質をコードするヌクレオチド配列を含むオープンリーディングフレームを含み、パルボウイルスVP1カプシドタンパク質の翻訳の開始コドンがACG、TTG、CTG、及びGTGから選択される、請求項12から16までのいずれか一項に記載の昆虫細胞。
【請求項18】
第3のヌクレオチド配列が、パルボウイルスVP1カプシドタンパク質をコードするヌクレオチド配列の開始コドンの上流に、配列番号7の9ヌクレオチドの配列、又は配列番号7と実質的に相同なヌクレオチド配列を含む発現制御配列を含む、請求項17に記載の昆虫細胞。
【請求項19】
第3のヌクレオチド配列が、12ヌクレオチド位におけるC、21ヌクレオチド位におけるA、及び24ヌクレオチド位におけるCから選択されるパルボウイルスVP1カプシドタンパク質をコードするヌクレオチド配列の少なくとも1つの改変をさらに含む、請求項17又は18に記載の昆虫細胞。
【請求項20】
少なくとも1つの第1のヌクレオチド配列、第2のヌクレオチド配列、及び第3のヌクレオチド配列が昆虫細胞のゲノムに安定して組み込まれている、請求項12から19までのいずれか一項に記載の昆虫細胞。
【請求項21】
パルボウイルスがAAVである、請求項8から20までのいずれか一項に記載の昆虫細胞。
【請求項22】
請求項12、15及び16のいずれか一項に記載の第2のヌクレオチド配列を含む組換えパルボウイルスビリオンを昆虫細胞において生成するための方法であって、
a)組換えパルボウイルスビリオンが生成するような条件下で請求項12から21までのいずれか一項に記載の昆虫細胞を培養するステップと、
b)組換えパルボウイルスビリオンを回収するステップと
を含む方法。
【請求項23】
抗パルボウイルス抗体、好ましくは固定化抗体を使用する、ビリオンの親和性精製のステップをさらに含む、請求項22に記載の方法。
【請求項24】
抗パルボウイルス抗体が単鎖ラクダ抗体又はその断片である、請求項23に記載の方法。
【請求項25】
組換えパルボウイルスビリオンが組換えAAVビリオンである、請求項22から24までのいずれか一項に記載の方法。
【図1A】

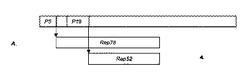
【図1B】
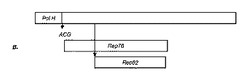
【図1C】
【図3】
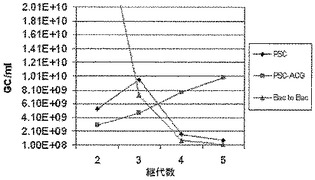
【図4】
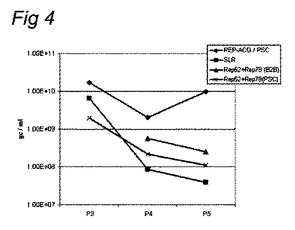
【図5】
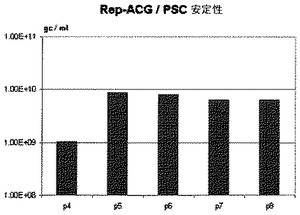
【図6】
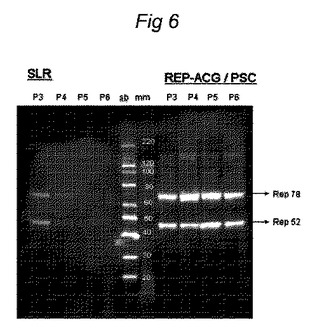
【図2】
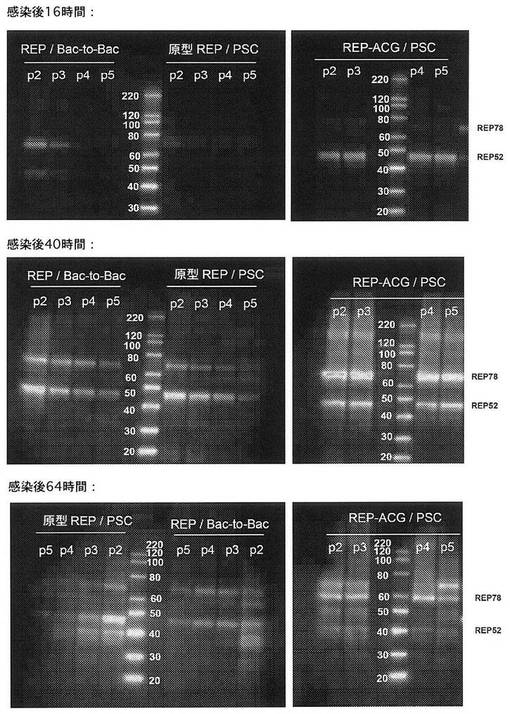
【図1B】
【図1C】
【図3】
【図4】
【図5】
【図6】
【図2】
【公表番号】特表2009−540823(P2009−540823A)
【公表日】平成21年11月26日(2009.11.26)
【国際特許分類】
【出願番号】特願2009−516420(P2009−516420)
【出願日】平成19年6月20日(2007.6.20)
【国際出願番号】PCT/NL2007/050298
【国際公開番号】WO2007/148971
【国際公開日】平成19年12月27日(2007.12.27)
【出願人】(506421611)アムステルダム モレキュラー セラピューティクス ビー.ブイ. (5)
【Fターム(参考)】
【公表日】平成21年11月26日(2009.11.26)
【国際特許分類】
【出願日】平成19年6月20日(2007.6.20)
【国際出願番号】PCT/NL2007/050298
【国際公開番号】WO2007/148971
【国際公開日】平成19年12月27日(2007.12.27)
【出願人】(506421611)アムステルダム モレキュラー セラピューティクス ビー.ブイ. (5)
【Fターム(参考)】
[ Back to top ]