
本態性高血圧症におけるGタンパク質関連変異体
【課題】本態性高血圧症の素因を有する個体を同定するための診断試験、およびその試験の実施のための遺伝子学的、細胞学的、生化学的な手段を提供する。
【解決手段】D1受容体を有しGRK4を発現する腎細胞を得て、該D1受容体の翻訳後修飾の程度を測定する。また、GRK4活性を測定する。さらに、GRK4タンパク質を産生するトランスジーンを含有するニ倍性ゲノムを含むトランスジェニック動物と腎細胞が該GRK4タンパク質を発現しない正常血圧の動物と比較する。
【解決手段】D1受容体を有しGRK4を発現する腎細胞を得て、該D1受容体の翻訳後修飾の程度を測定する。また、GRK4活性を測定する。さらに、GRK4タンパク質を産生するトランスジーンを含有するニ倍性ゲノムを含むトランスジェニック動物と腎細胞が該GRK4タンパク質を発現しない正常血圧の動物と比較する。
【発明の詳細な説明】
【技術分野】
【0001】
本明細書に記載の本発明につながる研究は、アメリカ国立衛生研究所の補助金NIH:DK3930.HL.23081により一部資金を受けた。従って、米国政府が本発明になんらかの権利を有し得る。
【0002】
(技術分野)
本発明は、本態性高血圧、より詳細にはこの疾患を診断および治療する方法における遺伝マーカーの使用に関する。
【背景技術】
【0003】
(背景技術)
本態性高血圧すなわち原因不明の高血圧は、アメリカ合衆国における白人の25−30%が冒される疾患である。高血圧症を治療せずに放置すると、心臓疾患、卒中、心筋梗塞および末期腎臓病に至る。高血圧患者は、通常病気だと感じないので、末期の臓器不全が始まるまでしばしば診断を受けず、治療しないまま放置する。従って、高血圧はヒトにおける心臓血管の罹患率および死亡率の主な原因である。多くの高血圧患者は、高塩分の食事が血圧を上昇させ、またはすでに上昇した血圧を悪化させるという点で塩分に敏感である。高血圧を発症させる性質を測定する方法がわかれば、心臓血管疾患の減少に重要な影響を与えるだろう。
【0004】
ヒトにおいて遺伝的要因が血圧の変動性の30−40%を占めると考えられている(Ward R. (1990) In Hypertension: Pathophysiology, Diagnosis and Management, Laragh JH. And Brenner BM eds., (Raven Press, Ltd., New York, NY), pp81-100)。しかしながら、別の意見では、1つの主要遺伝子が40%を占めた状態で、高血圧の遺伝率は80%であると示唆された(Cavalli Sporza LL., Boomer WF. In The Genetics of Human Population (1971), (WH Freeman Co., South San Francisco, CA) pp534-536)。単一の主要遺伝子が、血圧に作用して、有意な程度に、血圧制御において副次的役割を果たす多くの他の遺伝子に優性であるのかも知れない。
【0005】
高血圧の発症と持続における腎臓の中心的役割は充分に確立された。正常な腎臓を高血圧ラットに移植すると、その血圧は正常化する。一方、高血圧のラットの腎臓を正常血圧のラットに移植すると、高血圧を発症する。従って、高血圧は腎臓に従うようである。また、大部分のヒトの高血圧遺伝子の型は、腎臓におけるナトリウムの高い再吸収に関連することがわかっている。腎臓のナトリウム排出および血圧を調節する多くのホルモン性システムがあるが、ドーパミンの腎臓パラクリン機能が長期の血圧調節における重要なメカニズムとして充分に確立されている。高血圧におけるナトリウムに対する近位尿細管の結合性の増加は、ドーパミンの欠陥的な腎臓のパラクリン作用により引き起こされる。ドーパミンはナトリウムの再吸収を減少させる。従って、ドーパミン作用の欠陥がナトリウム再吸収の増加および高血圧をまねく。
【0006】
ドーパミンは、Gタンパク質結合受容体のロドプシン様ファミリーに属する細胞表面受容体のある種類によってその機能を発揮する;これらの受容体は通常7つの膜貫通ドメインを有する。CNSおよびいくつかの内分泌器におけるドーパミン受容体は、2つの主な種類、D1様受容体およびD2様受容体に分類される。CNS以外の腎臓などの臓器においては、D1様受容体はDA1受容体と呼ばれ、D2様受容体はDA2受容体と呼ばれている。ドーパミン受容体がCNSの内または外で独占的に発現はしないので、これらの区別はおそらくもはや必要ではない。しかしながら、神経組織と腎臓組織とにおいてD1受容体の異なる調節がある。D1受容体遺伝子の2つのエキソンが神経組織では転写されるのに対し、第2エキソンのみが腎臓組織では転写される。短いおよび長いD1転写物の異なる発現は、活性化因子タンパク質の組織特異的発現がD1受容体遺伝子の5’非コード領域におけるプロモータからの転写を行うためである。D2様ドーパミン受容体のサブタイプは各々、いくつかのイソ形を有する。しかしながら、特定のイソ形が末梢神経で特異的に発現されることはない。参照、Jose, et al., Pharmac. Ther. 80:149-182(1998)。
【0007】
2つのD1様受容体:D1およびD5受容体は、哺乳動物において発現し、各々げっ歯類のD1AおよびD1Bとして知られている。2つの付加的D1様受容体、D1CおよびD1Dは、非哺乳類において発現する。D1様受容体はアデニリルシクラーゼの刺激に関係している。D1A受容体はまた、ホスホリパーゼC活性を刺激するが、アデニリルシクラーゼの刺激に対して補助的である。D1様受容体は、まだクローン化されてなく、百日咳毒素非感受性のGタンパク質、Gqを通してホスホリパーゼC(PLC)に結合し、D1およびD5受容体とは異なるようである(Jose et al, Pharmacol. Ther. 80:149-182(1998)。3つのD2様受容体は哺乳動物において発現する:D2、D3およびD4受容体である。D2様受容体はアデニリルシクラーゼおよびCa2+チャネルの阻害に関連する。D2およびD3受容体はNG108−15細胞における電位依存性カリウム電流を減少すると報告されているが、D2様受容体はまたK+チャネルを刺激する。シナプス前神経に存在するD2およびD3受容体の両方がまた、ドーパミンおよびノルエピネフリン両方の放出を減少するのに役立ち得る。
【0008】
すべての哺乳動物のドーパミン受容体は、最初に脳からクローン化され、腎臓および尿路において発現することがわかった。ドーパミン受容体サブタイプは、腎臓血管系、糸球体および尿細管に従って特徴的に発現をする。腎臓血管系、糸球体および尿細管において、このサブタイプは、腎臓血行動態、電解質および水の輸送、およびレニン分泌を調節する。外因性ドーパミンは低用量で、腎臓血管抵抗力を低下させ、腎臓の血流を増加するが、糸球体濾過値に変動的な効果を有する。付加的な腎臓効果には、血行動態および細管メカニズムにより生じる溶質および水の排出の増加がある。近位尿細管のドーパミンを産生する能力とこれらの細管における受容体の存在は、ドーパミンが自己分泌またはパラクリン方法で作用し得ることを示唆している。外因性腎臓ドーパミンはいくつかのネフロンセグメント(近位細管、ヘンレの髄質の厚い上行性の脚(mTAL)、皮質性集合管(CCD))における作用により溶質と水の排出を増加する。各ネフロンセグメントにおけるドーパミンの阻害効果の大きさは、穏やかであるが、ネフロンに沿った多数の部位の作用により、溶質および水の排出が著しく増加する。ドーパミンの腎臓効果は溶質(例えば、ナトリウム、リン酸塩)またはタンパク質負荷の条件下において最も明らかである。D1様受容体、おそらくD1サブタイプの受容体は、腎臓を血管拡張し、管腔膜におけるナトリウム/水素交換体活性の阻害および側底膜におけるナトリウム/カリウムATP分解酵素活性の阻害により近位尿細管におけるナトリウム輸送を阻害する。D1様受容体はまたmTALおよびCCDにおけるナトリウム輸送を減少させる。腎臓における主な機能的D1様受容体は、D1受容体である。シナプス前D2様受容体はまた血管拡張性である。シナプス後D2様受容体は単独で、腎臓近位ナトリウム輸送を刺激し、CCDにおけるバソプレシン作用を阻害する。しかしながら、D1様受容体とともに、シナプス後D2様受容体は相乗的に作用して、近位尿細管においてナトリウム輸送を阻害する。近位尿細管における主なD2様受容体は、D3受容体であり、CCDにおける主なD2様受容体は、D4受容体である。おそらくD3サブタイプのシナプス後D2受容体がレニン分泌を阻害する能力は、レニン分泌に対するD1様受容体の刺激効果を中和し、相乗的作用をもたらして、ナトリウム補充状態におけるナトリウム排出を増加する(Jose et al., 前出)。
【0009】
結果、長年の熱心な努力により、本態性高血圧の病因について多くのことがわかったにもかかわらず、血圧を制御する単一の主要遺伝子は発見されていない。従って、血圧の調節に関する主要遺伝子の発見が、本態性高血圧を引き起こすメカニズムを理解するのに重要であり、重要な新しい診断法および治療法を導くであろう。
【発明の概要】
【0010】
(本発明の要約)
キナーゼは、リン酸基のタンパク質への付加を触媒する酵素である。Gタンパク質結合受容体キナーゼ(GRK)は、セリンおよびスレオニン残基におけるGタンパク質結合受容体タンパク質をリン酸化するタンパク質キナーゼのファミリーである。GRKはアレスチンと呼ばれる他のタンパク質とともに、ホルモン性反応の相同的脱感作を媒介する。参照、Premont, et al., FASEB J. 9:175-182(1995)。6つのGRK、つまりGRK1−GRK6が同定された。参照、Premont, et al., 前出.;Palczewski, Protein Sci. 3:1355-1361(1994); および Inglese, et al, J. Biol. Chem. 268:23735-23738(1993)。GRK4は最もわかっていないGRKファミリーのメンバーである。Premont et al., J. Biol. Chem. 271:6403-6410(1996)は、精巣にその実質的存在を測定し、GRK1を除くいかなるGRKのなかで最も分布していない。Premontの文献は、どのような具体的な型の精巣細胞がGRK4を発現するのかわからないことを認めているが、GRK4が、LH/CG受容体、性腺刺激ホルモン放出ホルモン受容体、および卵胞刺激ホルモン受容体および種々の嗅覚受容体を含む多くの受容体のいずれにも結合し得ると推測する。後に、Gros, J. Clin. Invest. 99(9):2087-2093(1997)は、高血圧者のリンパ球におけるアデニリルシクラーゼ活性化の減少におけるGRK2の活性を示唆した。Grosはまた、GRK活性の増加がGRK2発現の増加のみに関連し、他のGRKの活性が変わらないことを観察した。
【0011】
出願人はいくつかの重要な発見をした。第1にGRK4イソ形発現は、腎臓、特に近位尿細管および皮質集合管細胞において有意な程度に生じる。第2に、いくつかの既知のGRK4の多形性型および3つの以前知られていなかった多形性高血圧者に多い。第3に、本態性高血圧者に関すると知られている近位尿細管細胞におけるD1受容体/アデニリルシクラーゼ共役欠損が、限定しないが、D1受容体の過剰リン酸化に関係することである。
【0012】
本発明の実際上の態様は、3つの基本的領域、つまり診断、薬剤の発見、治療に分類される。従って、本発明の第1の態様は、本態性高血圧にかかりやすい個体を同定する方法に関する。該方法は、個体から単離したD1受容体およびGRK4を発現する腎臓細胞のサンプルを用いて行い得る。該方法において、細胞をアッセイして、例えばリン酸化またはパルミトイル化などのD1受容体の翻訳後修飾の範囲を測定する。正常血圧者から単離した細胞に関する受容体の翻訳後修飾の変化は、本態性高血圧の素因を示唆している。別法として、GRK遺伝子またはその断片を分析して、本態性高血圧に関連のGRK4を検出するために、核酸サンプルを個体から単離する。本発明において本態性高血圧に関連するとして同定した具体的な変異体には、次のものが含まれる:R65L、A142V、A486V、2つ二重変異体R65LとA486VおよびR65LとA142Vおよび3重変異体R65LとA142VとA486V。本態性高血圧に関連の他の変異体GRK4をさらに同定することは、単に、本態性高血圧と診断された個体から単離したGRK4を分析し、GRK4遺伝子の配列を分析することにより行い得る。さらに本発明において、非腎臓細胞におけるこれらのGRK4の発現により、これらの非腎臓細胞がドーパミン作動性シグナルを正常というより“適切に”形質導入し得ないことが明らかになった。
【0013】
本発明の関連態様は、R65LとA142Vの二重変異、R65LとA486Vの二重変異、またはR65LとA142VとA486Vの3重変異を有するGRK4タンパク質をコードする単離および精製された核酸に関する。前出の変異を含有するGRK4遺伝子断片に特異的にハイブリダイズするオリゴヌクレオチドも開示されている。さらに本態性高血圧に関連の変異を含有するGRK4遺伝子の断片をハイブリダイズするオリゴヌクレオチドプライマー、またはプライマー対が開示されている。好ましいプライマーは、GRK4遺伝子のエキソン3、5、8、14または16に特異的にハイブリダイズし、GRK4遺伝子のヌクレオチド431−503(エキソン3)、594−697(エキソン5)、857−995(エキソン8)、1662−1798(エキソン14)または1937−1991(エキソン16)を含むDNA配列を増幅するのに有用である。
【0014】
本発明の別の態様は、抗高血圧活性のための物質を、GRK4の立体配座および/または活性において変化をもたらすそれらの能力により試験する様々なシステムに関する。これらのシステムは、例えば、本態性高血圧に関連の野生型またはイソ形または変異体などのGRK4タンパク質と、検出しようとする抗高血圧剤との相互作用があるとGRK4タンパク質の立体配座的変化を引き起こす薬剤との間の複合体から、GRK4およびGRK4基質を含有する再構築されたシステムまでの範囲に及ぶ。GRK4とGRK4基質間の相互作用を測定し得るいかなるシステムも、潜在的な抗高血圧剤のスクリーニングに使用し得る。従って、システムは、人工膜、例えば脂質ミセルなどの細胞様部分から全細胞までに及ぶ。好ましい全細胞には、D1受容体遺伝子(またはその機能的断片)で形質導入された細胞および野生型または変異体のGRK4遺伝子および不死化されたヒト近位細管細胞がある。これら種々のシステムに生じるGRK4活性の変化は、例えばいかなる2次メッセンジャー成分または(限定するわけではないが)アデニリルシクラーゼにより発生するcAMP、Gタンパク質活性、ナトリウム輸送体またはポンプ活性などの終末点およびリン酸化またはパルミトイル化などの翻訳後修飾などの細胞活性における卵管通気を測定することにより検出し得る。本態性高血圧に関連のGRK4タンパク質をコードするトランスジーンを含有するトランスジェニック動物などのインビボシステムも開示されている。該システムにおいて、トランスジーンは腎臓細胞において発現して、トランスジェニック動物に本態性高血圧の状態を起こす。
【0015】
本発明のさらに別の態様は、インビトロまたはインビボの近位尿細管細胞においてナトリウム輸送を減少(ナトリウム利尿を増加)する方法に関する。この治療を適用する基本的な目的は、GRK4活性を変えることである。一つの好ましい方法は、高血圧者の腎臓細胞においてGRK4の発現を減少または防止する1つまたは複数の薬剤を投与することを含む。GRK4のmRNAまたはDNAは、転写または翻訳を防止するアンチセンスRNAまたは優性阻害変異体などのオリゴヌクレオチドで攻撃される。GRK4のmRNAまたはプレmRNAを開裂するリボザイムも有用である。他の治療の適用には、GRK4活性(阻害または刺激のいずれか)を阻害するかまたは高めるなど、変更する薬剤が含まれる。
【0016】
いかなる特定の作用理論にも拘束されるわけではないが、出願人の考えでは、ヒト被験者において腎欠陥は高血圧のなんらかの原因であり、GRK4変異は特に、D1受容体の直接または間接のリガンド非依存性セリン過剰リン酸化を引き起こし、Gタンパク質/エフェクター複合体の結合をはずす。結果、ドーパミンのナトリウム利尿作用が低下され、腎臓がナトリウムと水分の適切な平衡を保てなくなり、ナトリウムの滞留と血圧の上昇をもたらす。特に、正常血圧の被験者からではなく、高血圧の被験者から得た近位尿細管は、ドーパミンD1受容体のアデニリルシクラーゼによる欠損結合を示す。欠損結合はD1受容体のリガンド非依存性リン酸化に関連する。出願人は、少なくとも6つの変異した遺伝子をキナーゼ型4(GRK4)関連Gタンパク質に発見し、該遺伝子は高血圧の被験者におけるD1受容体のリガンド非依存性リン酸化を調節する。
【図面の簡単な説明】
【0017】
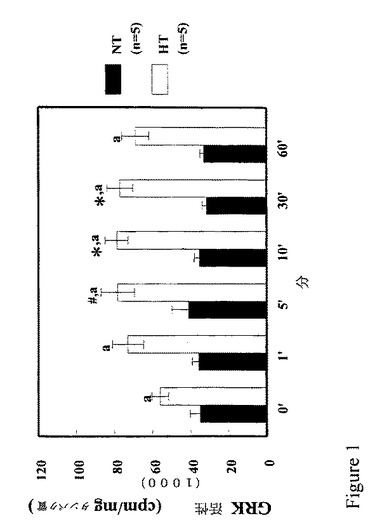
【図1】図1は、正常血圧者からではなく高血圧者からの近位尿細管細胞においてD1様アゴニストがGRK活性を刺激することを示すグラフである。
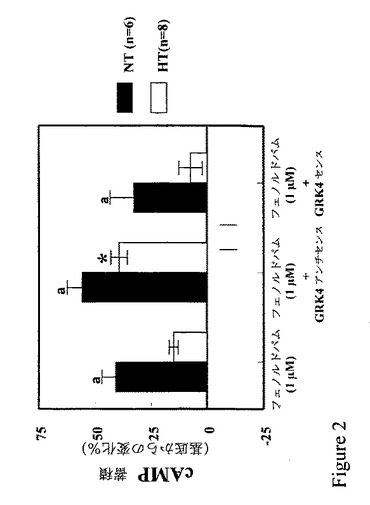
【図2】図2は、GRK4の発現を防止すると、高血圧者からの近位尿細管細胞の能力が正常値に回復し、D1様アゴニストによって生じるcAMP産生物を増加することを示すグラフである。
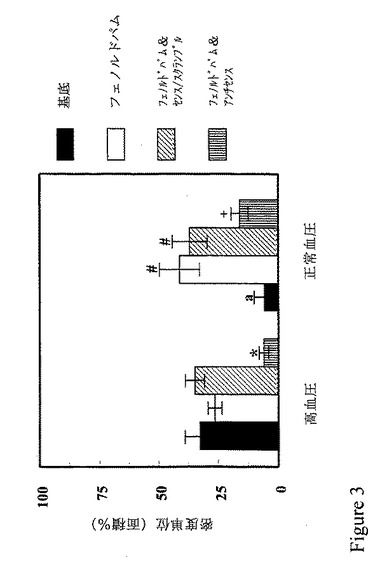
【図3】図3は、休眠近位尿細管細胞におけるD1受容体のリン酸化は、正常血圧の被験者より高血圧者において大きく、D1様アゴニスト刺激に反応しないことを示すグラフである。GRK4発現が防止される場合、D1受容体のリン酸化は停止する。
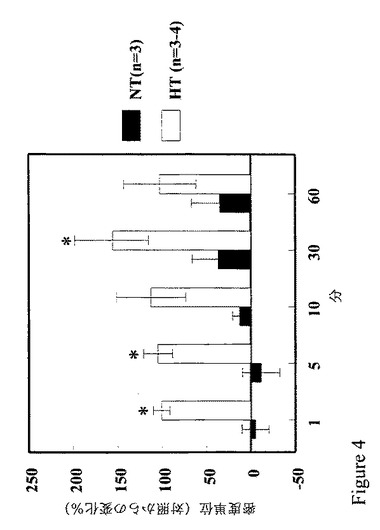
【図4】図4は、正常血圧者ではなく高血圧者でD1様アゴニスト刺激に対する反応において、近位尿細管におけるGRK4ガンマ/∂の発現が増加することを示すグラフである。
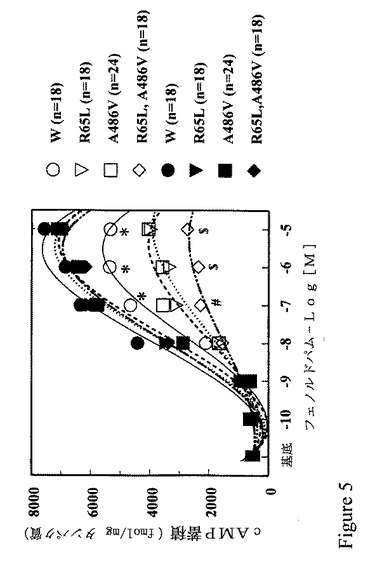
【図5】図5は、GRK4ガンマの変異により、GRK4ガンマおよびD1受容体を過発現するチャイニーズハムスターの卵巣細胞においてD1様アゴニスト刺激に反応するD1受容体の能力が減少することを示すグラフである。
【発明を実施するための形態】
【0018】
(本発明を実行する最適な方法)
ヒトGRK4遺伝子転写の構造は、多量の選択的スプライシングを受けて、GRK4タンパク質の4つの型をコードする4つの異なるGRK4のmRNAの型を産生する。選択的スプライシングはGRK4のアミノおよび/またはカルボキシル末端領域で生じ、4つのイソ形を産生する。
【0019】
GRK4は初め、Ambrose, et al., Hum. Mol. Genet. 1:697-703(1993)において報告され、次いでPremont et al., J. Biol. Chem. 271(11):6403-6410(1996)においてより広範囲に特徴付けられた。Premontにおいて、GRK4は精巣においてのみ非常に豊富で、GRK4のmRNAは脳細胞および骨格筋肉においてわずかに存在すると報告されている。GRK4遺伝子はプロモータ領域を除いて、約75キロベース(kDa)に及び、16エキソンから構成されている。GRK4の最長型は、無傷のアミノおよびカルボキシル末端の選択的エキソン配列を有し、GRK4アルファと呼ばれた。推測されるタンパク質配列は、予測分子量66.5kDaを有する578アミノ酸を含有する。次に短い型、GRK4ベータは、コドンから構成されるアミノ酸末端の選択的エキソンのみを欠失し、分子量62.5kDaを有する546アミノ酸を含有する。GRK4ガンマは、46コドンであるカルボキシル末端の選択的エキソンのみを欠失するイソ形である。従って、このイソ形は532アミノ酸を含有し、予測分子量61.2kDaを有する。GRK4ガンマは正式にはGRK4Aと呼ばれていた。参照、Sallese et al., Biochem. Biophys. Res. Commun. 199:848-854(1994)。GRK4デルタは予測分子量57.6kDaを有する500アミノ酸を含有し、最短のイソ形である。GRK4デルタは両方の選択的エキソンを欠失する。GRKデルタは初めIT11およびGRK4Bと呼ばれていた。参照、Sallese et al., 前出、およびAmbrose, et al., 前出。さらに最近、2つの付加的イソ形が発見された、つまり:エキソン13および15を欠失し、予測分子量53.6kDaを有する466アミノ酸を含有するGRK4イプシロンと、エキソン2、13および15を欠失し、予測分子量49.9kDaを有する434アミノ酸を含有するGRK4ゼータである。
【0020】
また、GRK4の5つの単一ヌクレオチド多形性、つまり:R65L(CGT−CTT);A142V(GCC−GTC);V247I(GTA−ATA);A486V(GCG−GTG)およびD562G(GAC−GGC)が知られている。参照、Premont, et al., 前出。出願人は、R65L、A142VおよびA486Vの多形性が本態性高血圧に関係することを発見した。出願人はまた、高血圧者に多い3つの付加的多形性を発見した、つまり:二重変異体、R65LとA142VおよびR65LとA486V;および3重変異体R65LとA142VとA486V。表1は、6つのGRK4イソ形のアミノ酸配列と対応するヌクレオチド配列を示す。本態性高血圧に関連の多形において変化するアミノ酸および対応するヌクレオチドは、太字で示されている。イプシロンおよびゼータのイソ形の5’非翻訳領域の配列は示されていない。
【0021】
【表1】

【表2】
【表3】
【表4】
【表5】
【表6】
【表7】
【0022】
本発明の第1の態様は、本態性高血圧症になりやすいまたは素因がある危険性の個体、または誰がそうであるかをスクリーニングする方法に関する。本態性高血圧症は、未知の病因の高血圧症として定義される。完全に特徴付けされているいくつかの高血圧症と異なり、本態性高血圧症の既知の原因はない。GRK4遺伝子、その基本的機能およびD1受容体との相互作用と、本態性高血圧症との間の関連または関係の同定は、個体のスクリーニングを可能にし、彼等が測定された高血圧症に関する遺伝的基礎を有するかどうか、または彼等が正常血圧である場合その素因を有するかどうかを決定できる。患者が正常血圧であるが(偽の低血圧読取を導く種々の条件がある)、高血圧症の臨床的証拠もある場合(終末器官疾病のような)、高血圧症変異のスクリーニングが、高血圧症の存在の確認に使用できる。このように、本態性高血圧症の素因を有するとして同定された個体は、次いで血圧をより綿密にモニターし、食事療法などの方法で、疾病の初期の段階において処置される。
【0023】
このような診断法の一つは、個体からの、D1受容体を有しGRK4を発現する腎細胞の単離を必要とする。本方法の実施に有用な腎細胞は、腎近位尿細管細胞および皮質集合管細胞を含む。それらは、尿サンプルから簡便に得られる。次いで細胞内のD1受容体の翻訳後修飾の程度を測定する。正常血圧の個体から単離された細胞と比べたD1受容体の翻訳後修飾における変化は、GRK4活性の変化によりもたらされると考えられ、それは、本態性高血圧症の素因の指標である。パルミトイル化およびリン酸化のような数個の翻訳後事象が、このような細胞内で起こり得る。高血圧症個体から単離されたこのような細胞内のD1受容体は、過リン酸化として知られる事象を示す。この期間までに、結合したリン分子を有するD1受容体の量が増加することを意味する。翻訳後修飾は、D1受容体とD1受容体抗体による免疫沈降およびホスホセリン抗体に対する免疫ブロット、または放射活性パルミチン酸による細胞の標識およびD1受容体抗体との免疫沈降のような、標準法にしたがって検出および測定できる(Ng et al., Eur. J. Pharmacol. 267:7-19 (1994))。
【0024】
他のこのような方法は、核酸サンプル、例えば、DNAまたはRNAを個体から得、個体のGRK4遺伝子の核酸配列を変異に関して分析することを必要とする。それによる変異の存在が、個体の本態性高血圧症に対する素因の指標である。核酸サンプルは、GRK4のDNAが遍在するため、任意の細胞タイプから得ることができる。血液からのDNA抽出物は、特に適当な源である。GRK4α番号付けに関して、本方法で同定される好ましいGRK4変異体は、アミノ酸残基65でArg→Leu(R65L)、アミノ酸残基142でAla→Val(A142V)、アミノ酸残基486でAla→Val(A486V)、二重変異体R65L、A142VおよびR65L、A486V、および三重変異体R65L、A142V、A486Vを含む。GRK4対立遺伝子は、本態性高血圧症の変異に関して、直接またはクローニングに続いてスクリーニングし得る。クローニングは、例えば、ゲノムDNAの適当な断片サイズへの消化、および得られる断片のベクターへのライゲートによる、慣用法を使用して連続できる。一方、ポリメラーゼ連鎖反応(PCR)を、GRK4遺伝子の特異的エキソン、例えば、エキソン3、5、8、14および16で行い得る。このようなプライマーの例は、表2に示す。PCRは、野生型または変異のGRKの任意の配列で行うことができる。PCRはまた、GRK4のmRNAでも行うことができる。このように、当業者は、GRK4対立遺伝子の増幅のためのプライマーまたはプライマー対が、全てのイソ形および多形性(表1に示すような)において同一のヌクレオチド配列に基づいて設計し得るか、またはそれらは活性化変異をもたらす特異的ヌクレオチド置換を含む配列に基づき得る。本発明のこの態様の実施に有用な他のプライマーは、ヌクレオチド431−503(エキソン3)、ヌクレオチド594−697(エキソン5)、ヌクレオチド857−995(エキソン8)、ヌクレオチド1662−1798(エキソン14)、およびヌクレオチド1937−1991(エキソン16)を含むDNA配列を増幅する。
【0025】
【表8】
* GenBank Accession #U33153からU33168
GRK4対立遺伝子は、クローン化対立遺伝子または増幅断片のヌクレオチド配列を決定し、それを正常対立遺伝子のヌクレオチド配列と比較することにより、正常対立遺伝子と異なる核酸配列の存在に関して試験する。他の既知の方法は、活性化対立遺伝子の存在の確認について、より完全であるが、まだ幾分は間接的な試験を提供する。これらの方法は、一本鎖確認分析(SSCA)、変性勾配ゲル電気泳動(DGGE)、RNase保護アッセイ、対立遺伝子特異的オリゴヌクレオチド(ASO)、E. coli mutSタンパク質のようなヌクレオチドミスマッチを認識するタンパク質の使用、および対立遺伝子特異的PCRを含む。これらの方法は、Orita et al., Proc. Natl. Acad. Sci. USA 86:2766-2770 (1989); Sheffield et al., Proc. Nat. Acad. Sci. USA 86:232-236 (1989); Finkelstein et al., Genomics 7:167-172 (1990)およびKinszler et al., Science 251:1366-1370 (1991); Conner et al., Proc. Nat. Acad. Sci. USA 80:278-282 (1983); Modrich, Ann. Rev. Genet. 25:229-253 (1991); およびRano & Kidd, Nucl. Acids Res. 17:8392 (1989)に各々記載されている。対立遺伝子特異的PCRに関して、その3'末端で特定のGRK4変異にハイブリダイズするプライマーを使用する。GRK4変異が存在しない場合、増幅産物は検出されない。増幅産物の検出は、EPA0332435に記載のように、Amplification Refractory Mutation System (ARMS)により行い得る。
【0026】
最初の3つの方法(SSCA、DGGEおよびRNase保護アッセイ)において、新規電気泳動バンドが出現する。SSCAは、一本鎖分子内塩基対形成の変化によりもたらされる配列変化のために異なって移動するバンドを検出する。RNase保護は、2個またはそれ以上の小さい断片への変異ポリヌクレオチドの開裂を含む。DGGEは、変性勾配ゲルを使用して、野生型配列と比較した変異配列の移動速度の差異を検出する。対立遺伝子特異的オリゴヌクレオチドアッセイにおいて、特異的配列を検出するオリゴヌクレオチドを設計し、アッセイを、ハイブリダイゼーションシグナルの存在または非存在の検出により行う。mutSアッセイにおいて、タンパク質は、変異体と野生型配列の間のヘテロ二本鎖のヌクレオチドミスマッチを含む配列とのみ結合する。
【0027】
本発明のミスマッチは、二つの鎖が100%相補的でないハイブリダイズした核酸二本鎖である。完全な相同性の欠如は、欠失、挿入、反転または置換によるものであり得る。ミスマッチ検出は、遺伝子またはそのmRNA生産物における点突然変異の検出に使用できる。これらの方法は配列決定よりも感受性が低いが、多数のサンプルについて行うのに簡便である。ミスマッチ開裂法の例は、RNase保護法である。本発明の実施において、この方法は、ヒト野生型GRK4遺伝子コード配列に相補的な標識リボプローブの使用を含む。リボプローブおよび腫瘍組織から単離されたmRNAまたはDNAは、共にアニール(ハイブリダイズ)され、続いて二本鎖RNA構造内のあるミスマッチを検出できる酵素RNase Aで消化される。ミスマッチがRNase Aで検出された場合、それはミスマッチの部位を開裂する。このように、アニールRNA調製物が電気泳動ゲルマトリックスで分離するとき、ミスマッチがRNase Aにより検出および開裂されている場合、RNA生産物は、リボプローブの完全長二本鎖RNAおよびmRNAまたはDNAより短いように見える。リボプローブは、GRK4のmRNAまたは遺伝子の完全長である必要はなく、いずれかのセグメントであり得る。リボプローブがGRK4のmRNAまたは遺伝子のセグメントのみを含む場合、ミスマッチに関して全mRNAをスクリーンするために、これらのプローブの多くを使用することが望ましい。
【0028】
同様の形態で、DNAプローブは酵素的または化学的な開裂を介してミスマッチの検出に使用できる。例えば、Cotton et al., (1988), Proc. Natl. Acad. Sci. USA 85:4397; Shenk et al., (1975), Proc. Natl. Acad. Sci. USA 72:989; およびNovack et al., (1986), Proc. Natl. Acad. Sci. USA 83:586参照。
【0029】
あるいは、ミスマッチは、マッチした二本鎖と比べてミスマッチ二本鎖の電気泳動移動性のシフトにより検出できる。例えば、Cariello, (1988), Human Genetics 42:726参照。リボプローブまたはDNAプローブのいずれかで、変異を含んでいるはずの細胞性mRNAまたはDNAをPCRプローブハイブリダイゼーションを使用して増幅できる。GRK4遺伝子のDNAにおける変化は、特に、変化が欠失および挿入のような正味の再配置である場合、サザンハイブリダイゼーションを使用してまた検出できる。
【0030】
PCRの使用により増幅されているGRK4遺伝子のDNA配列は、また対立遺伝子特異的プローブを使用してスクリーニングし得る。これらのプローブは核酸オリゴマーであって、既知の変異を担持する各々GRK4遺伝子配列の領域を含む。例えば、一つのオリゴマーは約30ヌクレオチド長であり得、GRK4遺伝子配列の部分に対応する。このような対立特異的遺伝子プローブの1組を使用することにより、PCR増幅生産物をスクリーニングし、GRK4遺伝子における先に同定された変異の存在を同定する。対立遺伝子特異的プローブと増幅GRK4配列とのハイブリダイゼーションは、例えば、ナイロンフィルター上で行うことができる。ストリンジェントなハイブリダイゼーション条件下での特定のプローブへのハイブリダイゼーションは、DNAサンプル中の対立遺伝子特異的プローブと同じ変異の存在を示す。このような対立遺伝子特異的の例は、表3に示す。
【表9】
*GRK4、GenBank Accession # U33054に基づく
【0031】
GRK4のコード領域の外側の変異は、イントロンのような非コード領域およびGRK4遺伝子の近くまたはその中の調節配列の試験により検出できる。非コード領域の変異は重要であるという先の指摘は、ノーザンブロット実験によるものであって、対照個体と比較して、高血圧症患者における異常サイズまたは多数のメッセンジャーRNA分子が示される。
【0032】
GRK4のmRNA発現の変化は、当分野で既知の方法で検出できる。これらは、ノーザンブロット分析、PCR増幅およびRNase保護を含む。減少したmRNA発現は、野生型GRK4遺伝子の変化を示す。野生型GRK4遺伝子の変化は、また野生型アンギオテンシノーゲンの変化に関するスクリーニングにより検出できる。例えば、GRK4に免疫反応性のモノクローナル抗体は、組織のスクリーニングに使用できる。同源抗原の欠失は、GRK4遺伝子変異を示す。変異対立遺伝子に特異的な抗体は、また変異GRK4遺伝子生産物の検出にも使用する。このような免疫学的アッセイは、当分野で既知の簡便な形態で行うことができる。これらは、ウエスタンブロット、免疫組織学的アッセイおよびELISAアッセイを含む。変化したGRK4を検出する任意の手段を野生型GRK4遺伝子の変化の検出に使用できる。変異GRK4遺伝子産物の発見は、野生型GRK4遺伝子の変化を示す。
【0033】
出願人は、上記の6個のGRK4変異体以外のGRK4変異体が、本態性高血圧症に関連することを推測する。このような変異体のインビトロでの同定は、変異体挿入のD1受容体含有細胞がドーパミン作動性シグナルを伝達し得なくする変異体の能力を測定することによりなされる。このことは、ドーパミン受容体がGタンパク質ユニットの活性化をしないか、またはナトリウム輸送体を阻害するために必要な細胞質性二次メッセンジャーの産生をしないことを意味する。ドーパミン作動性の伝達の欠失は、特に、D1受容体/アデニリルサイクラーゼ(AC)またはGタンパク質結合欠陥、および上記のタイプの翻訳後修飾でとりわけ明白である。これらの減少の測定は、ドーパミンまたはそのアゴニストの刺激に対する能力:(a)アデニリルサイクラーゼ活性またはcANP産生または活性化タンパク質キナーゼA、(b)活性化ホスホリパーゼCまたは活性化タンパク質キナーゼC、(c)ホスホリパーゼA2活性および(d)G−タンパク質活性またはナトリウム/水素交換体またはナトリウム/カリウムATPaseのようなナトリウム輸送タンパク質の阻害の測定で行える。
【0034】
本態性高血圧症に関連する他のGRK4は、本態性高血圧症に罹患している個体から得られまたはクローン化されたGRK4遺伝子の配列決定により、簡単に同定できる。
【0035】
野生型GRK4または本態性高血圧症と関連するGRK4は、抗高血圧症活性について多くの異なるタイプの物質のスクリーニングのための種々のシステムに包含し得る。一般に、GRK4およびGRK4基質を含むシステム、およびGRK4の立体配座または活性(およびその変化)を測定できるシステムを、抗高血圧症活性の物質のスクリーニングのために使用し得る。このように、本発明のこの態様の最も広い意味においては、細胞全体を必要としない。システムは、本質として人工的であり得、例えば、脂質ミセル内に入れられている。分子相互作用を研究するための無細胞システムの説明としては、Hammond et al., Nature 327:730-732 (1987)を参照。しかし細胞全体が好ましく、GRK4基質として、D1受容体またはその機能的断片である。“機能的断片”なる用語は、リン酸化、パルミトイル化またはインビトロの他の手段で翻訳後修飾されている受容体の一部を意味する。本発明の好ましい方法は、GRK4核酸で形質転換した細胞の使用を含む。一般に、哺乳類、細菌および昆虫細胞を含む種々の細胞のタイプを使用できる。チャイニーズハムスター卵巣(CHO)細胞、ヒト胚腎(HEK)繊維芽(LTK)細胞、MDCKおよびLLCP細胞のような哺乳類細胞が好ましい。CHO細胞は、それらがインビボで近位尿細管と同じように行動すると予測されるため、より好ましい。GRK4およびD1受容体核酸での細胞の形質転換は、標準法にしたがって行い得る。例えば、Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY (1989)およびAusubel et al., Current Protocols in Molecular Biology, Wiley & Sons (1994)参照。
【0036】
本出願のこの態様のより好ましい具体的態様において、本方法は、正常血圧または高血圧症の動物、例えば、ヒトから単離した細管細胞を使用して調製した不死化腎近位尿細管を使用して行う。一般に、細管細胞は、皮質を小切片(例えば、1mm3)に切断して腎臓から単離し、容器の適当な生育表面(例えば、コラーゲン被覆T−フラスコ)に置く。接着が容器の転倒により認められたら(例えば、室温で約30分)、容器を起し、適当な培地を添加する。好ましい培地は、物質(wt/ml):インシュリン(5マイクログラム)、トランスフェリン(5マイクログラム)、セレニウム(5ナノグラム)、ヒドロコルチゾン(36ナノグラム)、トリヨードチロニン(4ピコグラム)および表皮成長因子(19ナノグラム)を添加したダルベッコ最小必須およびF−12培地である。組織をインキュベートし、約3日間、95%空気、5%CO2で分配しないままにした。Detrisac, et al., Kidney Int. 25:383-390 (1984)。あるいは、皮質の切片を、コラゲナーゼで消化させ、連続的に212および140マイクロメーターにふるって、培養前に40マイクロメーターふるいで濃縮し得る。Courjault-Gautier et al., J. Am. Soc. Nephrol. 5:1949-1963 (1994)参照。“不死化”なる用語は、細胞が培養においていつまでも生育することを意味する。単離の腎近位尿細管細胞を、SV40ウイルス、例えば、SV40tsA変異体ウイルスで感染させ、次いで、感染約7−8週間後の過成長を得ることにより不死化し得る。これらの細胞は、GRK4タンパク質が機能するインビボ環境をより密接に模倣している利点を提供する。高血圧症患者からの不死化細胞は、しばしば抗高血圧症活性のための薬剤のスクリーニングに使用できる細胞について殆ど無限の供給を提供する。
【0037】
抗高血圧症特性を有すると推定される物質または薬剤は、物質または薬剤のGRK4システムへの添加によるGRK4の立体配座または活性の変化の測定により同定し得る。GRK4活性は、アデニレートキナーゼ活性の測定のように間接的に、または培養に添加したリン酸化可能な物質のリン酸化の程度の測定のように直接的に測定し得る。GRK4活性化または不活性化変異体、例えば、各々GRK4活性の増加またはGRK4活性の減少を導くGRK4の変異体または多形性は興味深い。GRK4活性の変化は、D1受容体により例示されるGタンパク質結合受容体の機能の変化を導くことができる。GRK4は、レニン−アンギオテンシン系、カリクレイン−キニン、エンドセリン、房および脳のナトリウム排泄増加性ペプチド、セロトニン、バソプレッシン、カルシウム検出受容体および上皮ナトリウムチャンネルのような本態性高血圧症に関与する他のタンパク質の機能を調節し得る。
【0038】
他のタイプのスクリーニング薬剤は、GRK4タンパク質、例えば野生型またはイソ形または本態性高血圧症に関与する変異体と、検出する抗高血圧症剤との相互作用によりGRK4タンパク質の立体配座的変化をもたらす薬剤との間の複合体を含む。複合薬剤の選択は、立体配座的分析を行う方法に依存する。このような分析は、分光測光法、フルオレッセンス、核磁気共鳴、エバネッセント波法および原子間力顕微鏡により行い得る。
【0039】
さらに別のタイプのスクリーニング剤およびプロトコールは、本態性高血圧症のトランスジェニック動物モデルの使用を含み、動物は本発明の野生型GRK4または変異GRK4をコードする核酸を発現する。変異GRK4の発現は、高血圧症により特徴付けられる表現型を明らかにし、動物が急性または慢性のナトリウム負荷を排出する能力を減少させる。トランスジェニックモデルはまた、GRK4の発現および活性における、血圧を減少することが示されている高カルシウム、高カリウムおよび高マグネシウムのような食事操作の効果の試験にも使用できる。明らかに、排出システムを有する動物は、本態性高血圧症のモデルとして使用できる。マウスのような齧歯類が好ましい。
【0040】
トランスジェニック動物は、当分野で既知の方法により製造できる。トランスジェニック動物の製造に適用できる方法は、動物の核DNA内へのトランスジーンの挿入である。これらのトランスジェニック動物は、例えば、米国特許4,873,191;5,849,578;5,731,489;5,614,396;5,487,992;5,464,764;5,387,742;5,347,075;5,298,422;5,288,846;5,221,778;5,175,384;5,175,383;4,873,191および4,736,866ならびにBurke and Olson, Methods in Enzymology 194:251-270 (1991), Capecchi, Science 244:1288-1292 (1989), Davies et al., Nucleic Acids Research 20(11):2693-2698 (1992), Dickinson et al., Human Molecular Genetics 2(8):1299-1302 (1993), Huxley et al., Genomics 9:742-750 (1991), Jakobovits et al., Nature 362:255-261 (1993), Lamb et al., Nature Genetics 5:22-29 (1993), Pearson and Choi, Proc. Natl. Acad. Sci. 90:10578-10582 (1993), Rothstein, Methods in Enzymology 194:281-301 (1991), Schedl et al., Nature 362:258-261 (1993), およびStrauss et al., Science 259:1904-1907 (1993)に明示の当分野で既知の標準法を使用して構築される。更に、公開国際特許出願WO94/23049、WO93/14200、WO94/06908およびWO94/28123は、これらに関する更なる関連の教示を提供する。
【0041】
当分野で既知の方法を、標的遺伝子トランスジーンを動物に挿入し、トランスジェニック動物の創始系を製造する。このような方法は、前核マイクロインジェクション(Hoppe, P. C. and Wagner, T. E., 1989, 米国特許4,873,191);生殖細胞系へのレトロウイルス仲介遺伝子移入(Van der Putten et al., Proc. Natl. Acad. Sci., USA 82:6148-6152 (1985));胚幹細胞への遺伝子標的化(Thompson et al., Cell 56:313-321 (1989));胚のエレクトロポレーション(Lo. Mol. Cell. Biol. 3:1803-1814 (1983))および精子仲介遺伝子移入(Lavitrano et al., 1989, Cell 57:717-723 (1989))を含むが、これらに限定されない。これらの方法に関する一般的総説は、Gordon, Transgenic Animals, Intl. Rev. Cytol. 115:171-229 (1989)参照。
【0042】
本発明は、全細胞にGRK4トランスジーンを担持するトランスジェニック動物、および全てではないが、いくつかの細胞にトランスジーンを担持する動物、即ち、モザイク動物を提供する。トランスジーンは、単一トランスジーンまたは、例えば、頭と頭のタンデム、頭と尾のタンデムのコンカタマーとして統合され得る。トランスジーンはまた、例えば、Lasko et al., Proc. Natl. Acad. Sci. USA 89:6232-6236 (1992)の教示にしたがって、特定の細胞タイプに選択的に挿入し、その中で活性化され得る。当業者は、このような細胞タイプ特異的活性化に必要な調節配列は、目的の特定の細胞タイプに依存することを認める。標的遺伝子トランスジーンが内因性標的遺伝子の染色体部位に統合されるのが望ましいとき、遺伝子標的化が好ましい。簡単に述べると、このような方法を用いるとき、目的の内因性標的遺伝子と相同的なあるヌクレオチド配列を含むベクターを、染色体配列との相同的組換えを介して、内因性標的遺伝子のヌクレオチド配列に統合し、機能を中断する目的で設計する。トランスジーンはまた、例えば、Gu et al., Science 265:103-106 (1994)の教示にしたがって、特定の細胞タイプに選択的に挿入し、そしてその細胞タイプのみで目的の内因性遺伝子を不活化し得る。このような細胞タイプ特異的不活性化に必要な調節配列は、目的の特定の細胞タイプに依存し、当業者には明らかである。
【0043】
トランスジェニック動物を産生したら、組換え標的遺伝子およびタンパク質の発現を、標準法を使用してアッセイし得る。最初のスクリーニングは、動物組織の分析のためにサザンブロット分析またはPCR法を使用して達成し得、トランスジーンの統合が起きているかをアッセイする。トランスジェニック動物の組織内のトランスジーンのmRNA発現のレベルは、また動物から得た組織のノーザンブロット分析、インシチュハイブリダイゼージョン、およびRT−PCRを含むが、これらに限定されない方法を使用して評価し得る。標的遺伝子発現組織のサンプルは、また、目的の標的遺伝子トランスジーン遺伝子生産物に特異的な抗体を使用して、標的細胞化学的に評価し得る。
【0044】
標的遺伝子mRNAまたは標的遺伝子トランスジーンペプチド(標的遺伝子産物エピトープを指向する抗体を使用して、免疫細胞化学的に検出)を容易に検出可能なレベルで発現する標的遺伝子トランスジェニック動物を、次いで更に評価し、本態性高血圧症の特徴的症状を示す動物を同定する。
【0045】
好ましい具体的態様において、GRK4トランスジーンを適当なベクターに挿入し、テトラサイクリン感受性プロモーターに操作不可能的に結合し、次いで、胚性幹(ES)細胞に導入する。ES細胞は、次いで遺伝的に変えたES細胞を宿主胚盤胞にマイクロインジェクションすることにより、または桑実胚共培養により再導入する。創始系動物を得、次いで、GRK4トランスジーンに同型接合の動物を使用する。Thompson, et al., Am. J. Physiol. 269:E793-E803 (1995)参照。
【0046】
治療様相は、標的GRK4活性をナトリウム尿排泄亢進まで増加させることを伴うか、またはそうでなければナトリウムと水の適当なバランスに関する正常性に近づく。例えば、GRK4発現は、RNAレベルまたはDNAレベルで、腎細胞のGRK4の発現を変える薬を投与することにより予防できる。このような薬剤は、好ましくは、アンチセンスオリゴヌクレオチド、優性阻害変異体DNA分子、およびGRK4 mRNA、プレmRNA、またはGRK4 DNAの結合によりGRK4発現を減少するまたは防止するリボザイムのようなオリゴヌクレオチド分子である。アンチセンスオリゴヌクレオチドの高血圧者への投与は、米国特許5,856,099;5,856,103;5,783,683;5,840,708;および5,591,600;5,849,903;5,135,917;5,098,890;および5,087,617に記載の製剤および媒体にしたがって行うことができる。現在当分野で既知のアンチセンス法は、また、Uhlmann et al., Chem. Rev. 90:543-584 (1994); Oligodeoxynucleotides: Antisense Inhibitors of Gene Expression (Cohen, ed. 1989); Delivery Strategies for Antisense Oligonucleotide Therapeutics, CRC press (Saghir Akhtar, et. 1995); およびStein, C. A., and Cohen, Jack S., "Oligodeoxynucleotides as Inhibitors of Gene Expression: A Review," Cancer Research, 48:2659-2668 (1988)にも記載されている。
【0047】
合成アンチセンスオリゴヌクレオチドは、標的ヌクレオチド配列とハイブリダイズし、所望の効果、例えば、mRNA分子の翻訳の遮断を発揮するのに十分な長さでなければならない。しかし、相対的に小さいオリゴヌクレオチドを使用するのが有利である。なぜなら、それらは、細胞にインビボでより効率的に取りこまれ、より多い数のアンチセンスオリゴヌクレオチドが標的mRNAの位置に送達されるようである。好ましくは、十分な特異性を達するために、アンチセンスオリゴヌクレオチドは少なくとも15ヌクレオチド長、好ましくは20ヌクレオチド長でなければならない。好ましいアンチセンスは、5'CAC GAT GTT CTC GAG CTC CAT3'(塩基255−275に相補的)および5'CTC CAT GTC CTG GCG CCG3')塩基243−260に相補的)である。
【0048】
上記のような小さいオリゴヌクレオチドは、雑多なヌクレアーゼの分解に非常に感受性である。更に、このような分子は、不十分な膜貫通のために、細胞内に入ることができないことがある。この理由のために、当業者は、一般に、安定性および膜貫通を増加する種々の方法で修飾したオリゴヌクレオチドを合成する。修飾アンチセンスオリゴヌクレオチドの使用は、本発明で好ましい。“アンチセンスオリゴヌクレオチドアナログ”なる用語は、下記のような、このような修飾オリゴヌクレオチドを意味する。
【0049】
本発明のオリゴヌクレオチドは、簡便には既知の方法の固相法を使用して合成し、本明細書に記載の配列をコードする標的GRKのDNAまたはRNAの予め選択した配列と相補的および/または特異的にハイブリダイズできるように設計する。核酸合成装置は商品として入手可能であり、その使用は、妥当な長さの所望のオリゴヌクレオチドの製造に有効であるとして当業者に理解されている。
【0050】
GRK4 mRNAまたはプレmRNAの分解を触媒する、例えば、ハンマーヘッドまたはハリピンタイプのリボザイムを設計でき、標準法にしたがって製造できる。治療的使用のリボザイムの設計、製造および製剤に関する詳細な教示は、例えば、米国特許5,856,463(およびその中に引用されている刊行物)参照。
【0051】
GRK4活性はまた、タンパク質に直接作用することにより完全にまたは部分的に発現されたGRK4タンパク質の触媒活性、例えば、リン酸化または非リン酸化作用を変化(例えば、阻害または促進)する薬理的アンタゴニストのような薬剤の投与により標的化できる。他の治療的作用は、GRK4タンパク質とペプチド様薬剤の直接の結合を伴う。これらの方法および薬剤の全ては、GRK4を発現する腎細胞におけるD1受容体/AC結合の正常化をもたらし、その結果、腎近位尿細管におけるナトリウム輸送を減少させる。
【0052】
本発明を、以下の詳細な実施例を参照して更に記載する。これらの実施例は説明の目的のみで提供され、特記しない限り、本明細書に記載の本発明の限定を意図するものではない。
【実施例1】
【0053】
実施例
組織培養
ヒト腎を、腎癌腫のため片側の腎を摘出した患者から新鮮な外科的標本として得た。患者記録を調べて、正常血圧(n=9)と本態性高血圧症(n=14)とに分類した。収縮期血圧が140mmHg未満で弛緩期血圧が90mmHg未満の者を正常血圧者とした。収縮期血圧が140mmHg以上で弛緩期血圧が90mmHg以上の者および/または抗高血圧剤服用者を高血圧者とした。
【0054】
組織学的に確認された正常な腎切片からの腎近位尿細管細胞の培養物(0.075%タイプIコラーゲンで被覆した24ウエルプラスチック管における5x105細胞/ウエル)を、37℃で95%O2/5%CO2中でインキュベートし、デルベッコ修飾イーグル培地とハムF12培地との1:1の混合物からなる血清を含まない培地で生育した。この培地には、セレニウム(5ng/ml)、インスリン(5μg/ml)、トランスフェリン(5μg/ml)、ヒドロコルチゾン(36ng/ml)、トリヨードチロシン(4pg/ml)、表皮成長因子(10ng/ml)(5)が補充されている。半コンフルエント(90−95%)のときは、細胞を継代培養して(6−8継代)、トリプシンEDTA(0.05%、0.02%)を使用する実験プロトコールに用いた。この培養条件は、腎近位尿細管細胞の特性を保持するヒト腎近位尿細管の成長に役立つ。Sanada, H. et al., J. Invest. Med. 45: 277A (1997)。
【0055】
光学顕微鏡による免疫組織化学
HISTOCHOICE中に不死した培養物における腎の組織および細胞について免疫組織化学的検査を、Sanada, H. et al.(前出)に記載のように行った。アフィニティー・カラム精製ポリクローナル・ヒトD1受容体抗体が合成ペプチド配列GSGETQPFC(アミノ酸299−307)に対して生じる。参照、Sanada, H. et al.(前出)。2種の市販(Santa Cruz Biotechnology, Inc, Santa Cruz, CA)のGRK4イソ形抗体を使用した。1つのGRK4抗体がαイソ形とβイソ形の両方を認識し、他の1つが両αβイソ形を認識した。これらの抗体の特異性はすでに報告されている。Sanada, H. et al.(前出)および Guyton A. C. Circulatory Physiology III, Arterial Pressure and Hypertension, W. B. Saunders Co., Philadelphia, PA (1980)。
【0056】
免疫組織化学的検査によると、GRK4α/βおよびGRK4α/σのイソ形発現が腎の近位および遠位の尿曲細管のみでみられた(ヘレン係蹄、皮質および髄質の集合管、糸球体、腎動脈ではみられなかった)。GRK4α/σは管腔膜および基底膜の両方でみられたが、GRK4α/βは管腔膜のみでみられた。これら2種のGRK4の腎発現について、高血圧者と正常者とで差異がなかった(表示せず)。GRK4α/βおよびGRK4α/σの発現は、培養物の腎近位尿細管細胞において持続した(写真は示さず)。
【0057】
GRK活性の測定
GRK活性を Benovic, Methods Enzymol. 200: 351-362 (1991)に従って測定した。腎近位尿細管抽出物の調製は、氷冷の融解緩衝液中での均質化によった。この緩衝液は、25トリスHCl、pH7.5、5EDTA、5EGTA、ロイペプシン(10μg/ml)、アプロチニン(20μg/ml)および1PMSFを含む。粗均質物を30,000gで30分間遠心分離した。ペレットの抽出を200mMのNaClで30分間行い、30,000gで30分間遠心分離した。上澄み液をGRKアッセイおよびイムノブロッティグに用いた。20μgのタンパク質抽出液を、ロドプシン強化桿外側セグメントとともに、10mMのMgCl2および0.1mMのATP(γ32P−ATP含有)中でインキュベートした。白色光で15分間室温でのインキュベーション後、反応を氷冷の融解緩衝液で停止し、30,000gで15分間遠心分離した。ペレットをラエムリ緩衝液に再懸濁し、12%SDS−PAGEに掛けた。ゲルにオートラジオグラフィを行い、リン酸化ロドプシンの定量を、密度計測と適当なサイズでの切除バンドの放射活性計測で行った。また、GRK活性の測定をGRKイソ形抗体の存在および不存在で行った。
【0058】
図1は、D1様アゴニストであるフェノルドパムがGRK活性に対する作用を有さないことを示す。これは、正常血圧者からの腎近位尿細管細胞におけるロドプシンのリン酸化により調べられた。このデータによると、基質としてロドプシンを使用し得るGRK(すなわち、GRK2、GRK3、GRK4α、GRK5、GRK6)は、血圧が正常であるとき、腎近位尿細管細胞におけるD1受容体の脱感作に関与しない。また、培養物中の腎近位尿細管細胞におけるD1受容体およびGRK4の発現は、高血圧者と正常血圧者とで類似していた(データは示さず)。しかし、高血圧者からの腎近位尿細管細胞において、フェノルドパムがGRK活性を増加した。さらに、腎近位尿細管細胞における基本的GRK活性は、高血圧者において正常血圧者におけるよりも大きかった。これらの研究から高血圧における腎近位尿細管細胞でのGRKの異常な機能がわかる。
【0059】
フェノルドパムによりつくられたGRK活性の増加(高血圧における)は、GRK2、GRK3およびGRK4α/σに対する抗体によって遮断され、これらのGRKの1つまたはすべての活性がGRK4活性のフェノルドパム仲介増加に関与し得ることを表す。Tiberi et al., J. Biol. Chem. 271: 3771-3778 (1996)。しかし、GRK2およびGRK3の偏在的発現は、げっ歯類およびヒトの本態性高血圧の病因において認識されている腎の特徴と適合しない。Guyton, W. B. Saunders Co. Phil., PA (1980); Guidi et al., J. Am. Soc. Nephrol 7: 1131-1138 (1996)。GRK2のコード領域の配列には、高血圧者と正常血圧者との間に差異がみられなかった(データは表示せず)。このことから、高血圧患者のリンパ球のおけるGRK2活性の増加が高血圧にとって二次的であり(Gros et al., J. Clin. Invest. 99: 2087-2093 (1997))、遺伝的および誘発された高血圧のげっ歯類おけるGRK5の活性および発現の増加について示唆されていたことと同様である。Ishizaka et al., J. Biol. Chem. 272: 32482-32488 (1997)。
【0060】
cAMP蓄積の測定
細胞をダルベッコ・リン酸緩衝液(D−PBS)で2回洗い、次いで各ウエルに1mMの3−イソブチル−1−メチルキサンチンを加えた。細胞を37℃で30分間、薬剤とともに、または薬剤なしでインキュベートした。薬剤は、ドパミンおよびD1様受容体アゴニスト、フェノルドパム、D1様受容体アンタゴニスト、SCH23390(Research Biochemicals International, Natick, MA)、およびフォルスコリン(Sigma Chemical Co., St. Louis, MO)である。次いで、細胞をD−PBSで2回洗い、−80℃に冷凍して、さらに0.1NのHClで分解した。cAMPの濃度を放射免疫検定法で測定した。Sanada, H. et al.,(前出)、Kinoshita, S. J. Chlin. Invest. 84: 1849-1856 (1989)。タンパク質の濃度をBCAタンパク質アッセイ・キットで測定した。
【0061】
GRK4活性の増加が高血圧での腎近位尿細管細胞におけるD1受容体の非共役に関与しているかどうかを確認するために、アンチセンス・オリゴヌクレオチドによるGRK4翻訳の阻害後のcAMPに対するD1様アゴニスト刺激作用を調べた。図2からわかるように、D1様アゴニストのフェノルドパムによるcAMPの蓄積は、高血圧者よりも正常血圧者の腎近位尿細管細胞において大きい。センス/スクランブルまたはアンチセンスのいずれのGRK4オリゴヌクレオチドも基底性またはフォルスコリン刺激cAMP産生に作用しなかった。フェノルドパムのみに比べると、センスまたはスクランブルのいずれのGRK4オリゴヌクレオチドもいずれのグループでのcAMP蓄積に作用しなかった。しかし、アンチセンスGRK4オリゴヌクレオチドはフェノルドパムの能力を高め、高血圧者由来の細胞でのcAMPの蓄積を促進し(正常血圧者からでは、しない)、その値は、フェノルドパムで処置された正常血圧者の細胞で見られる値とほぼ同じであった。
【0062】
免疫沈降
近位管細胞を、運搬物、フェノルドパム、センス、スクランブルまたはアンチセンスのプロピン/ホスホロチオエート・オリゴヌクレオチド(5nM)とともに、上記のようにインキュベートした。膜を氷冷融解緩衝液(PBSと、1%NP40、0.5%デオキシコール酸ナトリウム、0.1%SDS,1mMのEDTA、1mMのEGTA、1mMバナジン酸ナトリウム、1mMのPMSF、10μg/mlアプロチニン、10μg/mlロイペプシン)で溶解した。溶解物を、IgG精製アンチD1受容体抗体とともに氷上1時間、そしてタンパク質Aアガロースとともに12時間ゆすりながら4℃でインキュベートした。SDSポリアクリルアミド・ゲル電気泳動により分離したタンパク質を、ニトロセルロース膜上に電気泳動で移した。移行ブロット・シートを、10mMトリスHCl、pH7.5、150mMのNaCl、0.1%ツィン20中で5−10%の脂肪なし乾燥乳で阻害し、希釈の親和性精製ポリクローナル・アンチホスホセリン抗体(Zymed Lab, San Francisco, CA)とともにインキュベートした。Sanada, H. et al.(前出)。オートラジオグラムおよび免疫ブロットをECLシステム(Amersham, Arlington Heights, Il)で可視化し、エンシトメトリーで定量した。Sanada, H. et al.(前出)。
【0063】
次の検討は、アンチセンスGRK4オリゴヌクレオチドの示差作用がD1受容体のリン酸化まで及ぶかどうかである。図3からわかるように、腎近位尿細管細胞でのセリン−リン酸化D1受容体の基底レベルは、正常血圧者よりも高血圧者で高く、高血圧者でのGRK活性の基底レベルの増加に相関する(図1に示す)。フェノルドパムは、セリン−リン酸化D1受容体の量を正常血圧者で増加せしめたが、高血圧者では増加せしめない。このことは、以前の報告 Sanada, H. et al.(前出)に一致する。センスまたはスクランブルのいずれのGRK4オリゴヌクレオチドは、両グループの対象者のフェノルドパム処置細胞におけるD1受容体のリン酸化に作用しなかった。一方、GRK4アンチセンス処置は、高血圧者からのフェノルドパム処置腎近位尿細管細胞におけるD1受容体のリン酸化を、基底値より低いレベルまでほとんど消失せしめた。GRK4アンチセンス処置は、正常血圧者からのフェノルドパム処置腎近位尿細管細胞におけるD1受容体のリン酸化も下げたが、基底値よりは高かった。高血圧者の腎近位尿細管細胞においてGRK4に対するアンチセンス・オリゴヌクレオチドによりD1受容体のリン酸化がほとんど完全に抑制されることから、高血圧のおけるD1受容体のリン酸化および脱感作に関与する主要なGKRがGRK4であって、このネフロン切片で発現される他のGRKでないことがわかる。
【0064】
遺伝子型決定
ホモ接合性GRK4遺伝子変異体の発生率が高血圧者で約60%および一般人で16%であるという当初の観察に基づくと、能力分析(能力:0.8、α:0.05、効果:45%)がグループにつき14−21のサンプルサイズで、グループ間の有意な差違が検出できる。従って、さらに高血圧者18名および正常血圧者11名の末梢血によるDNAを入手した。少なくとも2人の研究者で全志願者を検査し、医療記録を検討した。高血圧症の病歴および潜在的高血圧症の臨床徴候をもたず、降圧剤薬物治療を行なっておらず、血管拡張療法または血圧に影響し得る他の薬剤を受けず、および最近の3回の診療訪問で収縮期血圧が140mmHgより低く、弛緩期血圧が90mmHより低い状態であった者を正常圧として、被検者を分類した。高血圧症の患者らは、顕著であり持続的な血圧上昇(収縮期血圧160mmHg以上で弛緩期血圧が95mmHg以上)を少なくとも3回の機会で有していた。高血圧者 (腎臓由来のDNA:n=14、末梢血由来のDNA:n=18)は全員20歳以上であった。各個体の本態性高血圧の後期発症についての先天性な問題を除去するため、正常血圧者(腎臓由来のDNA:n=9、末梢血由来のDNA:n=11)は全員45歳以上であった。
【0065】
培養中の腎臓近位尿細管細胞および腎組織または任意の末梢白血球から、ゲノムDNAを抽出した(塩析法)。多形性ヌクレオチドを含むGRK4のエキソンを、表2で示すプライマーで増幅した。反応混合物 20μlは、それぞれ、1XPCR緩衝液、0.2mMの各dNTP、1.25mM MgCl2、0.2μMの各プライマー、TaqDNAポリメラーゼ 0.5単位およびゲノムDNA 50ngを含んでいた。反応混合物を94℃で5分間変性し、次いで、94℃の30秒間変性、55℃のリアニール 30秒間および72℃の30秒伸長を30サイクル行なった。72℃での5分間の最終伸長でPCRが完了した。PCR産物 2μlをバイオダインB+膜上にスポットした。下記の野生型および変異体の対立遺伝子特異的オリゴヌクレオチドプローブそれぞれについてドットブロットを行った(表4)。プローブ標識化、膜調製、ハイブリダイゼーション、および洗浄条件は公知の手順である。Song et al., Clin. Chem. 43:1857-1861 (1997)参照。任意の被検者250人の1801位ヌクレオチドは変異体(G)であった。993位の多形性ヌクレオチドの頻度は高血圧者と正常血圧者で違いがないことも判明した。従って、調査の結果、448、679、1711位の3つのみ(表4)の多形性部位が存在する。サンガー・ジデオキシ連鎖停止法でcDNA配列を決定した。
【0066】
表4.正常血圧者および高血圧者のGRK4変異体
【表10】
【0067】
対立遺伝子特異的オリゴヌクレオチドを使用したドットブロット分析で遺伝子型を決定した。高血圧者の4名は2つの部位(65位および142位アミノ酸)で相同性があった。高血圧者のR65L、A142Vおよび/またはA486Vのホモ接合性変異体の頻度(53%、32中の17)は、正常血圧者で示されたもの(5%、20中の1)と有意に異なっていた(X2=10.56、P=0.0012)。ホモ接合性変異体A142Vの頻度もまた、高血圧者(34%、32中の11)および正常血圧者(0%、20中の0)で有意に異なっていた(X2=6.78、P=0.0092)。
【0068】
ヒト腎臓由来GRK4のcDNAの配列決定、さらに、腎臓および末梢白血球細胞由来のDNA中の5つの多形性部位の遺伝子型決定から、3つの変異体が明らかになった:ヌクレオチド448、CGTからCTT(アミノ酸R65L)、ヌクレオチド679、GCCからGTC(アミノ酸A142V)、およびヌクレオチド1711、GCGからGTG(アミノ酸A486V)(オートラジオグラフは示していない)の変異体が、正常血圧者より高血圧者で頻繁に現われた(表4)。高血圧者におけるR65L、A142V、および/またはA486Vのホモ接合性変異の頻度(53%,32中の17)は、正常血圧者で示されたもの(5%,20中の1)と有意な違いがあり(X2=10.56,P=0.0012)(表4)、無作為成人被検者50人でみられたものと異なっていた(X2=10.99,P=0.0009)。この血圧不明の無作為集団の中で、16%がR65Lおよび/またはA486Vでホモ接合性であり、および50%がR65LまたはA486Vの何れかでヘテロ接合性であった;ホモ接合性対立遺伝子の頻度16%は、本態性高血圧症の発生率に近い(Lifton R P. Science 272:676-680 (1996))。GRK4 A142V位のホモ接合性変異も、それ自体は、高血圧症 (34%、32中の11) の方が正常血圧者(0%、20中の0)より頻繁であった(X2=6.78,P=0.0092)。
【0069】
GRK4αは、リン酸化ロドプシンにレポートされる唯一のGRK4イソ形である(Sallese et al., J. Biol. Chem. 272:10188-10195 1997))が、我々の研究では、フェノルドパムによるD1アゴニスト刺激は、正常血圧者の腎臓近位尿細管細胞中のGRK活性を増強しなかった(図1)。従って、GRK4αはD1受容体の脱感作に関与していないと結論した。その確信によると、ロドプシンを普通はリン酸化しないGRK4イソ形(例えばGRK4γ)(Premont et al., J. Biol. Chem. 271:6403-6410 (1996); Sallese et al., supra.; and Virlon et al., Endocrinol. 139:2784-2795 (1998))は、高血圧症で活性となり得る。たしかにGRK活性中のD1様アゴニスト介在増加は、高血圧者の腎臓近位尿細管細胞中のGRK4αδの膜発現の増加と関連があることが分かったが、正常血圧者(図4)では関連しない。
【0070】
トランスフェクションおよび細胞培養
ラットD1(rD1)またはヒトD1(hD1)受容体cDNAを、発現ベクターpPUR(Clontech, Palo Alto, CA)またはpcDNA3.1/Zeo(Invitrogen, Carlsbad, CA)中で、それぞれ、EcoR1およびXbaI部位でサブクローン化した。得られた作成物を使用し、pTeT−オフ レギュレータープラスミドを発現するCHO細胞を、安定してトランスフェクトした(Clontech, Palo Alto, CA)。この操作にリン酸化カルシウムを使用した。Yamaguchi et al., Mol. Pharmacol. 49:373-378 (1996)参照。ヒト腎臓皮質由来mRNAのRT/PCRから得られた、GRK4γおよびGRK4δのcDNAを、pTet-オフ 応答プラスミド中へ入れサブクローン化した(pTRE−rD1またはpTRE−hD1とpTK−Hygをそれぞれ20:1の割合で混合した)(Clontech, Palo Alto, CA)。
【0071】
GRK4α遺伝子の変異が他の機能的結果を有するかどうか測定するために、D1様アゴニストの効果を、D1受容体および野生型または変異体GRK4α cDNAの両方をトランスフェクトする、チャイニーズハムスター卵巣(CHO)細胞中のcAMP産物について調べた。 GRK4δを対照として使用した。GRK4α欠損下におけるCHO細胞中の投与応答カーブは、HEK-293細胞(低い内因性GRK活性を有する細胞)で示されるものと類似していた。Premont et al, 前出。野生型GRK4αの発現は、D1アゴニストのcAMP産物を刺激する能力を減少させた(図5)。しかし、D1アゴニスト活性の阻害は、GRK4α変異体R65Lおよび/またはA486Vよりも大きくなった。野生型または変異体のGRK4αの効果は、D1受容体またはGRK4αの何れかの発現量の違いからではない(データは示していない)。野生型GRK4αまたはその変異体は、フォルスコリンの能力に作用せず、GRK4αとD1受容体の相互作用の特異性を表わすcAMP蓄積を刺激する。フェノルドパムの作用はD1受容体に対して選択的であり、フェノルドパム効果は、D1様アンタゴニストSCH23390によって遮蔽されたことによるからである(データは示していない)。他の研究では、野生型GRK4δは、野生型GRK4αによって誘発されたD1受容体の脱感作と比較し、D1様アゴニスト介在のcAMP蓄積に対して効果がない(データは示していない)。腎臓近位尿細管細胞の機能的研究およびCHO細胞の発現研究では、GRK4αの活性増加が、D1受容体リガンドの能力の減少の原因であり、高血圧症でエフェクター酵素およびイオン輸送タンパク質と結合することを提唱する。言いかえると、高血圧者における腎臓近位尿細管中のD1受容体の脱感作は、腎臓の能力増強につながり、塩化ナトリウムの負荷を除去する。腎臓の塩化ナトリウム排出の不足は、高血圧症の進行に極めて重要であると考えられる。 Guyton, A.C., Circulatory Physiology III. Arterial Pressure and Hypertension, W.B. Saunders Co.,, Philadelphia, PA (1980); Guidi et al., J. Am. Soc. Nephrol. 7:1131-1138 (1996)。確かに、腎臓ナトリウム輸送を調節する遺伝子は、血圧の調製に重要であると記されている。Lipton, R.P. Science 272:676-680 (1996)およびKaret & Lifton, Recent Prog. Horm. Res. 52:263-276 (1997)。
【0072】
物質または薬剤を生物体中へ注入し、引き起こされたGRK4活性の減少が抗高血圧療法として役立ち得るかどうかを調べるために、自発性高血圧症ラット(SHR)でさらなる実験を行なった。6匹の雌ラット(4週齢、体重100g)を左一側性腎摘出し、2週間そのままにしておいて、手術から回復させた。回復後、単独のアウトレットカテーテルを装備する30日浸透圧ミニポンプを、ホスホロチオネート/プロピン介在アンチセンスGRK4オリゴヌクレオチド(5nM、1マイクロリットル/時間)またはスクランブルGRK4オリゴヌクレオチドで充填し、残された左腎臓の腎皮質中へ移植する。カテーテルのアウトレットを残された腎臓の腎皮質中のほぼ1mm深さに挿入し、スーパーグルーで固定した。ラットを手術から回復させ、血圧と尿出量を毎日測定した(容量と電解液)。30日後、ラットを屠殺し、残された腎臓をGRK4のウエスタンブロット分析に使用した。我々の研究によると、血圧は、スクランブルGRK4オリゴヌクレオチド(n=3)を処置したラットのときと比較して、GRK4(n=3)にアンチセンスオリゴヌクレオチドを処置したラットで減少した。さらに、ウエスタンブロット分析で、アンチセンスオリゴヌクレオチドは腎臓GRK4の発現を減少させることを証明した。
【0073】
結果として、この例では、本態性高血圧者由来の腎臓近位尿細管細胞におけるD1受容体/アデニリルシクラーゼ共役で欠損が示された。ヒト本態性高血圧症の腎臓近位尿細管細胞のGRK活性の増加は、GRK4の活性化ミスセンス変異によるものであり、この効果はトランスフェクトされた細胞モデルで再現された。その上、GRK4の翻訳を防止し、高血圧症におけるD1受容体のアデニリルシクラーゼに対する結合を正常化した。さらに何れの特定の理論に拘束されることを意図することなく、出願人は、ホモ接合性アミノ酸変異は、D1受容体のリガンドに依存しないセリン-リン酸化を引き起こし、Gタンパク質/エフェクター複合体からその結合解離をもたらすと考える。腎臓近位尿細管中のD1受容体の脱感作は、ドーパミンのナトリウム排泄作用を低下し、最終的にナトリウム保持および高血圧症となる。これらの結論は、GRK4に対するアンチセンスオリゴヌクレオチドを自発性高血圧症ラットの腎臓内に注入すると、腎臓内でGRK4濃度が減少し、平均動脈血圧が低下するという上記実験の結果に対応している。従って、GRK4の濃度または活性を変える物質または薬剤は、降圧性薬物治療の新規段階を表わす。
【0074】
ドーパミンDIA受容体とGプロテイン/エフェクター酵素複合体とのネフロンセグメント特異的欠損結合は、自発性高血圧症ラット(SHR)の腎臓ナトリウム保持の原因となり得る。腎臓近位尿細管中でナトリウム輸送を阻害する外因性および腎臓内因性のドーパミンの能力減少は、SHRと正常圧対照(ウィスター-キョート(WKY)ラット)とのF2交配において高血圧症を同様におこす。同様の欠損がダール塩過敏性ラットで判明した。さらに重要なことに、本態性高血圧症のヒトでも判明した。従って、高血圧症ヒト由来の腎臓近位尿細管の初期の培養は、腎臓D1様受容体とアデニリルシクラーゼ(AC)の欠損結合を有しており、同様な結合欠損が高血圧症齧歯動物でもみられた。これらのインビトロデータは、欠損D1様受容体を証明するインビボ研究(Gタンパク質/エフェクター酵素複合体は、相同性または異種性脱感作、受容体下方制御、Gタンパク質またはエフェクター酵素“欠損”またはD1様受容体の1次配列における変異によらない)に一致する。むしろ、D1様受容体の脱共役は、D-l受容体(腎臓中の主要なD1様受容体)のリガンドに依存しない過リン酸化の結果であり、限られた器官およびネフロンの発現を伴うGRK4イソ形のホモ接合性変異体の結果である。
【産業上の利用可能性】
【0075】
産業上の利用可能性
本発明の診断検査は、個体をスクリーニングし、その本態性高血圧症の素因を同定できるであろう。これらの検査の実施に関する、遺伝、細胞および生化学的手段も提供する。本発明は、薬剤を発見するための手段および方法も提供し、および抗高血圧性の活性または特性を有する物質を同定する。組成物および方法は、それぞれ本態性高血圧症を有する個体の腎細胞中のナトリウム輸送を正常化するものであり、この疾患の処置手段を提供する。
【0076】
本明細書で引用したすべての特許および非特許の刊行物は、本発明の属する当業者の技術のレベルを示している。すべての出版物または特許出願は、出典明示により本明細書の一部とすることを具体的に個々に示したのと同様に、出典表示より本明細書の一部とする。
【技術分野】
【0001】
本明細書に記載の本発明につながる研究は、アメリカ国立衛生研究所の補助金NIH:DK3930.HL.23081により一部資金を受けた。従って、米国政府が本発明になんらかの権利を有し得る。
【0002】
(技術分野)
本発明は、本態性高血圧、より詳細にはこの疾患を診断および治療する方法における遺伝マーカーの使用に関する。
【背景技術】
【0003】
(背景技術)
本態性高血圧すなわち原因不明の高血圧は、アメリカ合衆国における白人の25−30%が冒される疾患である。高血圧症を治療せずに放置すると、心臓疾患、卒中、心筋梗塞および末期腎臓病に至る。高血圧患者は、通常病気だと感じないので、末期の臓器不全が始まるまでしばしば診断を受けず、治療しないまま放置する。従って、高血圧はヒトにおける心臓血管の罹患率および死亡率の主な原因である。多くの高血圧患者は、高塩分の食事が血圧を上昇させ、またはすでに上昇した血圧を悪化させるという点で塩分に敏感である。高血圧を発症させる性質を測定する方法がわかれば、心臓血管疾患の減少に重要な影響を与えるだろう。
【0004】
ヒトにおいて遺伝的要因が血圧の変動性の30−40%を占めると考えられている(Ward R. (1990) In Hypertension: Pathophysiology, Diagnosis and Management, Laragh JH. And Brenner BM eds., (Raven Press, Ltd., New York, NY), pp81-100)。しかしながら、別の意見では、1つの主要遺伝子が40%を占めた状態で、高血圧の遺伝率は80%であると示唆された(Cavalli Sporza LL., Boomer WF. In The Genetics of Human Population (1971), (WH Freeman Co., South San Francisco, CA) pp534-536)。単一の主要遺伝子が、血圧に作用して、有意な程度に、血圧制御において副次的役割を果たす多くの他の遺伝子に優性であるのかも知れない。
【0005】
高血圧の発症と持続における腎臓の中心的役割は充分に確立された。正常な腎臓を高血圧ラットに移植すると、その血圧は正常化する。一方、高血圧のラットの腎臓を正常血圧のラットに移植すると、高血圧を発症する。従って、高血圧は腎臓に従うようである。また、大部分のヒトの高血圧遺伝子の型は、腎臓におけるナトリウムの高い再吸収に関連することがわかっている。腎臓のナトリウム排出および血圧を調節する多くのホルモン性システムがあるが、ドーパミンの腎臓パラクリン機能が長期の血圧調節における重要なメカニズムとして充分に確立されている。高血圧におけるナトリウムに対する近位尿細管の結合性の増加は、ドーパミンの欠陥的な腎臓のパラクリン作用により引き起こされる。ドーパミンはナトリウムの再吸収を減少させる。従って、ドーパミン作用の欠陥がナトリウム再吸収の増加および高血圧をまねく。
【0006】
ドーパミンは、Gタンパク質結合受容体のロドプシン様ファミリーに属する細胞表面受容体のある種類によってその機能を発揮する;これらの受容体は通常7つの膜貫通ドメインを有する。CNSおよびいくつかの内分泌器におけるドーパミン受容体は、2つの主な種類、D1様受容体およびD2様受容体に分類される。CNS以外の腎臓などの臓器においては、D1様受容体はDA1受容体と呼ばれ、D2様受容体はDA2受容体と呼ばれている。ドーパミン受容体がCNSの内または外で独占的に発現はしないので、これらの区別はおそらくもはや必要ではない。しかしながら、神経組織と腎臓組織とにおいてD1受容体の異なる調節がある。D1受容体遺伝子の2つのエキソンが神経組織では転写されるのに対し、第2エキソンのみが腎臓組織では転写される。短いおよび長いD1転写物の異なる発現は、活性化因子タンパク質の組織特異的発現がD1受容体遺伝子の5’非コード領域におけるプロモータからの転写を行うためである。D2様ドーパミン受容体のサブタイプは各々、いくつかのイソ形を有する。しかしながら、特定のイソ形が末梢神経で特異的に発現されることはない。参照、Jose, et al., Pharmac. Ther. 80:149-182(1998)。
【0007】
2つのD1様受容体:D1およびD5受容体は、哺乳動物において発現し、各々げっ歯類のD1AおよびD1Bとして知られている。2つの付加的D1様受容体、D1CおよびD1Dは、非哺乳類において発現する。D1様受容体はアデニリルシクラーゼの刺激に関係している。D1A受容体はまた、ホスホリパーゼC活性を刺激するが、アデニリルシクラーゼの刺激に対して補助的である。D1様受容体は、まだクローン化されてなく、百日咳毒素非感受性のGタンパク質、Gqを通してホスホリパーゼC(PLC)に結合し、D1およびD5受容体とは異なるようである(Jose et al, Pharmacol. Ther. 80:149-182(1998)。3つのD2様受容体は哺乳動物において発現する:D2、D3およびD4受容体である。D2様受容体はアデニリルシクラーゼおよびCa2+チャネルの阻害に関連する。D2およびD3受容体はNG108−15細胞における電位依存性カリウム電流を減少すると報告されているが、D2様受容体はまたK+チャネルを刺激する。シナプス前神経に存在するD2およびD3受容体の両方がまた、ドーパミンおよびノルエピネフリン両方の放出を減少するのに役立ち得る。
【0008】
すべての哺乳動物のドーパミン受容体は、最初に脳からクローン化され、腎臓および尿路において発現することがわかった。ドーパミン受容体サブタイプは、腎臓血管系、糸球体および尿細管に従って特徴的に発現をする。腎臓血管系、糸球体および尿細管において、このサブタイプは、腎臓血行動態、電解質および水の輸送、およびレニン分泌を調節する。外因性ドーパミンは低用量で、腎臓血管抵抗力を低下させ、腎臓の血流を増加するが、糸球体濾過値に変動的な効果を有する。付加的な腎臓効果には、血行動態および細管メカニズムにより生じる溶質および水の排出の増加がある。近位尿細管のドーパミンを産生する能力とこれらの細管における受容体の存在は、ドーパミンが自己分泌またはパラクリン方法で作用し得ることを示唆している。外因性腎臓ドーパミンはいくつかのネフロンセグメント(近位細管、ヘンレの髄質の厚い上行性の脚(mTAL)、皮質性集合管(CCD))における作用により溶質と水の排出を増加する。各ネフロンセグメントにおけるドーパミンの阻害効果の大きさは、穏やかであるが、ネフロンに沿った多数の部位の作用により、溶質および水の排出が著しく増加する。ドーパミンの腎臓効果は溶質(例えば、ナトリウム、リン酸塩)またはタンパク質負荷の条件下において最も明らかである。D1様受容体、おそらくD1サブタイプの受容体は、腎臓を血管拡張し、管腔膜におけるナトリウム/水素交換体活性の阻害および側底膜におけるナトリウム/カリウムATP分解酵素活性の阻害により近位尿細管におけるナトリウム輸送を阻害する。D1様受容体はまたmTALおよびCCDにおけるナトリウム輸送を減少させる。腎臓における主な機能的D1様受容体は、D1受容体である。シナプス前D2様受容体はまた血管拡張性である。シナプス後D2様受容体は単独で、腎臓近位ナトリウム輸送を刺激し、CCDにおけるバソプレシン作用を阻害する。しかしながら、D1様受容体とともに、シナプス後D2様受容体は相乗的に作用して、近位尿細管においてナトリウム輸送を阻害する。近位尿細管における主なD2様受容体は、D3受容体であり、CCDにおける主なD2様受容体は、D4受容体である。おそらくD3サブタイプのシナプス後D2受容体がレニン分泌を阻害する能力は、レニン分泌に対するD1様受容体の刺激効果を中和し、相乗的作用をもたらして、ナトリウム補充状態におけるナトリウム排出を増加する(Jose et al., 前出)。
【0009】
結果、長年の熱心な努力により、本態性高血圧の病因について多くのことがわかったにもかかわらず、血圧を制御する単一の主要遺伝子は発見されていない。従って、血圧の調節に関する主要遺伝子の発見が、本態性高血圧を引き起こすメカニズムを理解するのに重要であり、重要な新しい診断法および治療法を導くであろう。
【発明の概要】
【0010】
(本発明の要約)
キナーゼは、リン酸基のタンパク質への付加を触媒する酵素である。Gタンパク質結合受容体キナーゼ(GRK)は、セリンおよびスレオニン残基におけるGタンパク質結合受容体タンパク質をリン酸化するタンパク質キナーゼのファミリーである。GRKはアレスチンと呼ばれる他のタンパク質とともに、ホルモン性反応の相同的脱感作を媒介する。参照、Premont, et al., FASEB J. 9:175-182(1995)。6つのGRK、つまりGRK1−GRK6が同定された。参照、Premont, et al., 前出.;Palczewski, Protein Sci. 3:1355-1361(1994); および Inglese, et al, J. Biol. Chem. 268:23735-23738(1993)。GRK4は最もわかっていないGRKファミリーのメンバーである。Premont et al., J. Biol. Chem. 271:6403-6410(1996)は、精巣にその実質的存在を測定し、GRK1を除くいかなるGRKのなかで最も分布していない。Premontの文献は、どのような具体的な型の精巣細胞がGRK4を発現するのかわからないことを認めているが、GRK4が、LH/CG受容体、性腺刺激ホルモン放出ホルモン受容体、および卵胞刺激ホルモン受容体および種々の嗅覚受容体を含む多くの受容体のいずれにも結合し得ると推測する。後に、Gros, J. Clin. Invest. 99(9):2087-2093(1997)は、高血圧者のリンパ球におけるアデニリルシクラーゼ活性化の減少におけるGRK2の活性を示唆した。Grosはまた、GRK活性の増加がGRK2発現の増加のみに関連し、他のGRKの活性が変わらないことを観察した。
【0011】
出願人はいくつかの重要な発見をした。第1にGRK4イソ形発現は、腎臓、特に近位尿細管および皮質集合管細胞において有意な程度に生じる。第2に、いくつかの既知のGRK4の多形性型および3つの以前知られていなかった多形性高血圧者に多い。第3に、本態性高血圧者に関すると知られている近位尿細管細胞におけるD1受容体/アデニリルシクラーゼ共役欠損が、限定しないが、D1受容体の過剰リン酸化に関係することである。
【0012】
本発明の実際上の態様は、3つの基本的領域、つまり診断、薬剤の発見、治療に分類される。従って、本発明の第1の態様は、本態性高血圧にかかりやすい個体を同定する方法に関する。該方法は、個体から単離したD1受容体およびGRK4を発現する腎臓細胞のサンプルを用いて行い得る。該方法において、細胞をアッセイして、例えばリン酸化またはパルミトイル化などのD1受容体の翻訳後修飾の範囲を測定する。正常血圧者から単離した細胞に関する受容体の翻訳後修飾の変化は、本態性高血圧の素因を示唆している。別法として、GRK遺伝子またはその断片を分析して、本態性高血圧に関連のGRK4を検出するために、核酸サンプルを個体から単離する。本発明において本態性高血圧に関連するとして同定した具体的な変異体には、次のものが含まれる:R65L、A142V、A486V、2つ二重変異体R65LとA486VおよびR65LとA142Vおよび3重変異体R65LとA142VとA486V。本態性高血圧に関連の他の変異体GRK4をさらに同定することは、単に、本態性高血圧と診断された個体から単離したGRK4を分析し、GRK4遺伝子の配列を分析することにより行い得る。さらに本発明において、非腎臓細胞におけるこれらのGRK4の発現により、これらの非腎臓細胞がドーパミン作動性シグナルを正常というより“適切に”形質導入し得ないことが明らかになった。
【0013】
本発明の関連態様は、R65LとA142Vの二重変異、R65LとA486Vの二重変異、またはR65LとA142VとA486Vの3重変異を有するGRK4タンパク質をコードする単離および精製された核酸に関する。前出の変異を含有するGRK4遺伝子断片に特異的にハイブリダイズするオリゴヌクレオチドも開示されている。さらに本態性高血圧に関連の変異を含有するGRK4遺伝子の断片をハイブリダイズするオリゴヌクレオチドプライマー、またはプライマー対が開示されている。好ましいプライマーは、GRK4遺伝子のエキソン3、5、8、14または16に特異的にハイブリダイズし、GRK4遺伝子のヌクレオチド431−503(エキソン3)、594−697(エキソン5)、857−995(エキソン8)、1662−1798(エキソン14)または1937−1991(エキソン16)を含むDNA配列を増幅するのに有用である。
【0014】
本発明の別の態様は、抗高血圧活性のための物質を、GRK4の立体配座および/または活性において変化をもたらすそれらの能力により試験する様々なシステムに関する。これらのシステムは、例えば、本態性高血圧に関連の野生型またはイソ形または変異体などのGRK4タンパク質と、検出しようとする抗高血圧剤との相互作用があるとGRK4タンパク質の立体配座的変化を引き起こす薬剤との間の複合体から、GRK4およびGRK4基質を含有する再構築されたシステムまでの範囲に及ぶ。GRK4とGRK4基質間の相互作用を測定し得るいかなるシステムも、潜在的な抗高血圧剤のスクリーニングに使用し得る。従って、システムは、人工膜、例えば脂質ミセルなどの細胞様部分から全細胞までに及ぶ。好ましい全細胞には、D1受容体遺伝子(またはその機能的断片)で形質導入された細胞および野生型または変異体のGRK4遺伝子および不死化されたヒト近位細管細胞がある。これら種々のシステムに生じるGRK4活性の変化は、例えばいかなる2次メッセンジャー成分または(限定するわけではないが)アデニリルシクラーゼにより発生するcAMP、Gタンパク質活性、ナトリウム輸送体またはポンプ活性などの終末点およびリン酸化またはパルミトイル化などの翻訳後修飾などの細胞活性における卵管通気を測定することにより検出し得る。本態性高血圧に関連のGRK4タンパク質をコードするトランスジーンを含有するトランスジェニック動物などのインビボシステムも開示されている。該システムにおいて、トランスジーンは腎臓細胞において発現して、トランスジェニック動物に本態性高血圧の状態を起こす。
【0015】
本発明のさらに別の態様は、インビトロまたはインビボの近位尿細管細胞においてナトリウム輸送を減少(ナトリウム利尿を増加)する方法に関する。この治療を適用する基本的な目的は、GRK4活性を変えることである。一つの好ましい方法は、高血圧者の腎臓細胞においてGRK4の発現を減少または防止する1つまたは複数の薬剤を投与することを含む。GRK4のmRNAまたはDNAは、転写または翻訳を防止するアンチセンスRNAまたは優性阻害変異体などのオリゴヌクレオチドで攻撃される。GRK4のmRNAまたはプレmRNAを開裂するリボザイムも有用である。他の治療の適用には、GRK4活性(阻害または刺激のいずれか)を阻害するかまたは高めるなど、変更する薬剤が含まれる。
【0016】
いかなる特定の作用理論にも拘束されるわけではないが、出願人の考えでは、ヒト被験者において腎欠陥は高血圧のなんらかの原因であり、GRK4変異は特に、D1受容体の直接または間接のリガンド非依存性セリン過剰リン酸化を引き起こし、Gタンパク質/エフェクター複合体の結合をはずす。結果、ドーパミンのナトリウム利尿作用が低下され、腎臓がナトリウムと水分の適切な平衡を保てなくなり、ナトリウムの滞留と血圧の上昇をもたらす。特に、正常血圧の被験者からではなく、高血圧の被験者から得た近位尿細管は、ドーパミンD1受容体のアデニリルシクラーゼによる欠損結合を示す。欠損結合はD1受容体のリガンド非依存性リン酸化に関連する。出願人は、少なくとも6つの変異した遺伝子をキナーゼ型4(GRK4)関連Gタンパク質に発見し、該遺伝子は高血圧の被験者におけるD1受容体のリガンド非依存性リン酸化を調節する。
【図面の簡単な説明】
【0017】
【図1】図1は、正常血圧者からではなく高血圧者からの近位尿細管細胞においてD1様アゴニストがGRK活性を刺激することを示すグラフである。
【図2】図2は、GRK4の発現を防止すると、高血圧者からの近位尿細管細胞の能力が正常値に回復し、D1様アゴニストによって生じるcAMP産生物を増加することを示すグラフである。
【図3】図3は、休眠近位尿細管細胞におけるD1受容体のリン酸化は、正常血圧の被験者より高血圧者において大きく、D1様アゴニスト刺激に反応しないことを示すグラフである。GRK4発現が防止される場合、D1受容体のリン酸化は停止する。
【図4】図4は、正常血圧者ではなく高血圧者でD1様アゴニスト刺激に対する反応において、近位尿細管におけるGRK4ガンマ/∂の発現が増加することを示すグラフである。
【図5】図5は、GRK4ガンマの変異により、GRK4ガンマおよびD1受容体を過発現するチャイニーズハムスターの卵巣細胞においてD1様アゴニスト刺激に反応するD1受容体の能力が減少することを示すグラフである。
【発明を実施するための形態】
【0018】
(本発明を実行する最適な方法)
ヒトGRK4遺伝子転写の構造は、多量の選択的スプライシングを受けて、GRK4タンパク質の4つの型をコードする4つの異なるGRK4のmRNAの型を産生する。選択的スプライシングはGRK4のアミノおよび/またはカルボキシル末端領域で生じ、4つのイソ形を産生する。
【0019】
GRK4は初め、Ambrose, et al., Hum. Mol. Genet. 1:697-703(1993)において報告され、次いでPremont et al., J. Biol. Chem. 271(11):6403-6410(1996)においてより広範囲に特徴付けられた。Premontにおいて、GRK4は精巣においてのみ非常に豊富で、GRK4のmRNAは脳細胞および骨格筋肉においてわずかに存在すると報告されている。GRK4遺伝子はプロモータ領域を除いて、約75キロベース(kDa)に及び、16エキソンから構成されている。GRK4の最長型は、無傷のアミノおよびカルボキシル末端の選択的エキソン配列を有し、GRK4アルファと呼ばれた。推測されるタンパク質配列は、予測分子量66.5kDaを有する578アミノ酸を含有する。次に短い型、GRK4ベータは、コドンから構成されるアミノ酸末端の選択的エキソンのみを欠失し、分子量62.5kDaを有する546アミノ酸を含有する。GRK4ガンマは、46コドンであるカルボキシル末端の選択的エキソンのみを欠失するイソ形である。従って、このイソ形は532アミノ酸を含有し、予測分子量61.2kDaを有する。GRK4ガンマは正式にはGRK4Aと呼ばれていた。参照、Sallese et al., Biochem. Biophys. Res. Commun. 199:848-854(1994)。GRK4デルタは予測分子量57.6kDaを有する500アミノ酸を含有し、最短のイソ形である。GRK4デルタは両方の選択的エキソンを欠失する。GRKデルタは初めIT11およびGRK4Bと呼ばれていた。参照、Sallese et al., 前出、およびAmbrose, et al., 前出。さらに最近、2つの付加的イソ形が発見された、つまり:エキソン13および15を欠失し、予測分子量53.6kDaを有する466アミノ酸を含有するGRK4イプシロンと、エキソン2、13および15を欠失し、予測分子量49.9kDaを有する434アミノ酸を含有するGRK4ゼータである。
【0020】
また、GRK4の5つの単一ヌクレオチド多形性、つまり:R65L(CGT−CTT);A142V(GCC−GTC);V247I(GTA−ATA);A486V(GCG−GTG)およびD562G(GAC−GGC)が知られている。参照、Premont, et al., 前出。出願人は、R65L、A142VおよびA486Vの多形性が本態性高血圧に関係することを発見した。出願人はまた、高血圧者に多い3つの付加的多形性を発見した、つまり:二重変異体、R65LとA142VおよびR65LとA486V;および3重変異体R65LとA142VとA486V。表1は、6つのGRK4イソ形のアミノ酸配列と対応するヌクレオチド配列を示す。本態性高血圧に関連の多形において変化するアミノ酸および対応するヌクレオチドは、太字で示されている。イプシロンおよびゼータのイソ形の5’非翻訳領域の配列は示されていない。
【0021】
【表1】
【表2】
【表3】
【表4】
【表5】
【表6】
【表7】
【0022】
本発明の第1の態様は、本態性高血圧症になりやすいまたは素因がある危険性の個体、または誰がそうであるかをスクリーニングする方法に関する。本態性高血圧症は、未知の病因の高血圧症として定義される。完全に特徴付けされているいくつかの高血圧症と異なり、本態性高血圧症の既知の原因はない。GRK4遺伝子、その基本的機能およびD1受容体との相互作用と、本態性高血圧症との間の関連または関係の同定は、個体のスクリーニングを可能にし、彼等が測定された高血圧症に関する遺伝的基礎を有するかどうか、または彼等が正常血圧である場合その素因を有するかどうかを決定できる。患者が正常血圧であるが(偽の低血圧読取を導く種々の条件がある)、高血圧症の臨床的証拠もある場合(終末器官疾病のような)、高血圧症変異のスクリーニングが、高血圧症の存在の確認に使用できる。このように、本態性高血圧症の素因を有するとして同定された個体は、次いで血圧をより綿密にモニターし、食事療法などの方法で、疾病の初期の段階において処置される。
【0023】
このような診断法の一つは、個体からの、D1受容体を有しGRK4を発現する腎細胞の単離を必要とする。本方法の実施に有用な腎細胞は、腎近位尿細管細胞および皮質集合管細胞を含む。それらは、尿サンプルから簡便に得られる。次いで細胞内のD1受容体の翻訳後修飾の程度を測定する。正常血圧の個体から単離された細胞と比べたD1受容体の翻訳後修飾における変化は、GRK4活性の変化によりもたらされると考えられ、それは、本態性高血圧症の素因の指標である。パルミトイル化およびリン酸化のような数個の翻訳後事象が、このような細胞内で起こり得る。高血圧症個体から単離されたこのような細胞内のD1受容体は、過リン酸化として知られる事象を示す。この期間までに、結合したリン分子を有するD1受容体の量が増加することを意味する。翻訳後修飾は、D1受容体とD1受容体抗体による免疫沈降およびホスホセリン抗体に対する免疫ブロット、または放射活性パルミチン酸による細胞の標識およびD1受容体抗体との免疫沈降のような、標準法にしたがって検出および測定できる(Ng et al., Eur. J. Pharmacol. 267:7-19 (1994))。
【0024】
他のこのような方法は、核酸サンプル、例えば、DNAまたはRNAを個体から得、個体のGRK4遺伝子の核酸配列を変異に関して分析することを必要とする。それによる変異の存在が、個体の本態性高血圧症に対する素因の指標である。核酸サンプルは、GRK4のDNAが遍在するため、任意の細胞タイプから得ることができる。血液からのDNA抽出物は、特に適当な源である。GRK4α番号付けに関して、本方法で同定される好ましいGRK4変異体は、アミノ酸残基65でArg→Leu(R65L)、アミノ酸残基142でAla→Val(A142V)、アミノ酸残基486でAla→Val(A486V)、二重変異体R65L、A142VおよびR65L、A486V、および三重変異体R65L、A142V、A486Vを含む。GRK4対立遺伝子は、本態性高血圧症の変異に関して、直接またはクローニングに続いてスクリーニングし得る。クローニングは、例えば、ゲノムDNAの適当な断片サイズへの消化、および得られる断片のベクターへのライゲートによる、慣用法を使用して連続できる。一方、ポリメラーゼ連鎖反応(PCR)を、GRK4遺伝子の特異的エキソン、例えば、エキソン3、5、8、14および16で行い得る。このようなプライマーの例は、表2に示す。PCRは、野生型または変異のGRKの任意の配列で行うことができる。PCRはまた、GRK4のmRNAでも行うことができる。このように、当業者は、GRK4対立遺伝子の増幅のためのプライマーまたはプライマー対が、全てのイソ形および多形性(表1に示すような)において同一のヌクレオチド配列に基づいて設計し得るか、またはそれらは活性化変異をもたらす特異的ヌクレオチド置換を含む配列に基づき得る。本発明のこの態様の実施に有用な他のプライマーは、ヌクレオチド431−503(エキソン3)、ヌクレオチド594−697(エキソン5)、ヌクレオチド857−995(エキソン8)、ヌクレオチド1662−1798(エキソン14)、およびヌクレオチド1937−1991(エキソン16)を含むDNA配列を増幅する。
【0025】
【表8】
* GenBank Accession #U33153からU33168
GRK4対立遺伝子は、クローン化対立遺伝子または増幅断片のヌクレオチド配列を決定し、それを正常対立遺伝子のヌクレオチド配列と比較することにより、正常対立遺伝子と異なる核酸配列の存在に関して試験する。他の既知の方法は、活性化対立遺伝子の存在の確認について、より完全であるが、まだ幾分は間接的な試験を提供する。これらの方法は、一本鎖確認分析(SSCA)、変性勾配ゲル電気泳動(DGGE)、RNase保護アッセイ、対立遺伝子特異的オリゴヌクレオチド(ASO)、E. coli mutSタンパク質のようなヌクレオチドミスマッチを認識するタンパク質の使用、および対立遺伝子特異的PCRを含む。これらの方法は、Orita et al., Proc. Natl. Acad. Sci. USA 86:2766-2770 (1989); Sheffield et al., Proc. Nat. Acad. Sci. USA 86:232-236 (1989); Finkelstein et al., Genomics 7:167-172 (1990)およびKinszler et al., Science 251:1366-1370 (1991); Conner et al., Proc. Nat. Acad. Sci. USA 80:278-282 (1983); Modrich, Ann. Rev. Genet. 25:229-253 (1991); およびRano & Kidd, Nucl. Acids Res. 17:8392 (1989)に各々記載されている。対立遺伝子特異的PCRに関して、その3'末端で特定のGRK4変異にハイブリダイズするプライマーを使用する。GRK4変異が存在しない場合、増幅産物は検出されない。増幅産物の検出は、EPA0332435に記載のように、Amplification Refractory Mutation System (ARMS)により行い得る。
【0026】
最初の3つの方法(SSCA、DGGEおよびRNase保護アッセイ)において、新規電気泳動バンドが出現する。SSCAは、一本鎖分子内塩基対形成の変化によりもたらされる配列変化のために異なって移動するバンドを検出する。RNase保護は、2個またはそれ以上の小さい断片への変異ポリヌクレオチドの開裂を含む。DGGEは、変性勾配ゲルを使用して、野生型配列と比較した変異配列の移動速度の差異を検出する。対立遺伝子特異的オリゴヌクレオチドアッセイにおいて、特異的配列を検出するオリゴヌクレオチドを設計し、アッセイを、ハイブリダイゼーションシグナルの存在または非存在の検出により行う。mutSアッセイにおいて、タンパク質は、変異体と野生型配列の間のヘテロ二本鎖のヌクレオチドミスマッチを含む配列とのみ結合する。
【0027】
本発明のミスマッチは、二つの鎖が100%相補的でないハイブリダイズした核酸二本鎖である。完全な相同性の欠如は、欠失、挿入、反転または置換によるものであり得る。ミスマッチ検出は、遺伝子またはそのmRNA生産物における点突然変異の検出に使用できる。これらの方法は配列決定よりも感受性が低いが、多数のサンプルについて行うのに簡便である。ミスマッチ開裂法の例は、RNase保護法である。本発明の実施において、この方法は、ヒト野生型GRK4遺伝子コード配列に相補的な標識リボプローブの使用を含む。リボプローブおよび腫瘍組織から単離されたmRNAまたはDNAは、共にアニール(ハイブリダイズ)され、続いて二本鎖RNA構造内のあるミスマッチを検出できる酵素RNase Aで消化される。ミスマッチがRNase Aで検出された場合、それはミスマッチの部位を開裂する。このように、アニールRNA調製物が電気泳動ゲルマトリックスで分離するとき、ミスマッチがRNase Aにより検出および開裂されている場合、RNA生産物は、リボプローブの完全長二本鎖RNAおよびmRNAまたはDNAより短いように見える。リボプローブは、GRK4のmRNAまたは遺伝子の完全長である必要はなく、いずれかのセグメントであり得る。リボプローブがGRK4のmRNAまたは遺伝子のセグメントのみを含む場合、ミスマッチに関して全mRNAをスクリーンするために、これらのプローブの多くを使用することが望ましい。
【0028】
同様の形態で、DNAプローブは酵素的または化学的な開裂を介してミスマッチの検出に使用できる。例えば、Cotton et al., (1988), Proc. Natl. Acad. Sci. USA 85:4397; Shenk et al., (1975), Proc. Natl. Acad. Sci. USA 72:989; およびNovack et al., (1986), Proc. Natl. Acad. Sci. USA 83:586参照。
【0029】
あるいは、ミスマッチは、マッチした二本鎖と比べてミスマッチ二本鎖の電気泳動移動性のシフトにより検出できる。例えば、Cariello, (1988), Human Genetics 42:726参照。リボプローブまたはDNAプローブのいずれかで、変異を含んでいるはずの細胞性mRNAまたはDNAをPCRプローブハイブリダイゼーションを使用して増幅できる。GRK4遺伝子のDNAにおける変化は、特に、変化が欠失および挿入のような正味の再配置である場合、サザンハイブリダイゼーションを使用してまた検出できる。
【0030】
PCRの使用により増幅されているGRK4遺伝子のDNA配列は、また対立遺伝子特異的プローブを使用してスクリーニングし得る。これらのプローブは核酸オリゴマーであって、既知の変異を担持する各々GRK4遺伝子配列の領域を含む。例えば、一つのオリゴマーは約30ヌクレオチド長であり得、GRK4遺伝子配列の部分に対応する。このような対立特異的遺伝子プローブの1組を使用することにより、PCR増幅生産物をスクリーニングし、GRK4遺伝子における先に同定された変異の存在を同定する。対立遺伝子特異的プローブと増幅GRK4配列とのハイブリダイゼーションは、例えば、ナイロンフィルター上で行うことができる。ストリンジェントなハイブリダイゼーション条件下での特定のプローブへのハイブリダイゼーションは、DNAサンプル中の対立遺伝子特異的プローブと同じ変異の存在を示す。このような対立遺伝子特異的の例は、表3に示す。
【表9】
*GRK4、GenBank Accession # U33054に基づく
【0031】
GRK4のコード領域の外側の変異は、イントロンのような非コード領域およびGRK4遺伝子の近くまたはその中の調節配列の試験により検出できる。非コード領域の変異は重要であるという先の指摘は、ノーザンブロット実験によるものであって、対照個体と比較して、高血圧症患者における異常サイズまたは多数のメッセンジャーRNA分子が示される。
【0032】
GRK4のmRNA発現の変化は、当分野で既知の方法で検出できる。これらは、ノーザンブロット分析、PCR増幅およびRNase保護を含む。減少したmRNA発現は、野生型GRK4遺伝子の変化を示す。野生型GRK4遺伝子の変化は、また野生型アンギオテンシノーゲンの変化に関するスクリーニングにより検出できる。例えば、GRK4に免疫反応性のモノクローナル抗体は、組織のスクリーニングに使用できる。同源抗原の欠失は、GRK4遺伝子変異を示す。変異対立遺伝子に特異的な抗体は、また変異GRK4遺伝子生産物の検出にも使用する。このような免疫学的アッセイは、当分野で既知の簡便な形態で行うことができる。これらは、ウエスタンブロット、免疫組織学的アッセイおよびELISAアッセイを含む。変化したGRK4を検出する任意の手段を野生型GRK4遺伝子の変化の検出に使用できる。変異GRK4遺伝子産物の発見は、野生型GRK4遺伝子の変化を示す。
【0033】
出願人は、上記の6個のGRK4変異体以外のGRK4変異体が、本態性高血圧症に関連することを推測する。このような変異体のインビトロでの同定は、変異体挿入のD1受容体含有細胞がドーパミン作動性シグナルを伝達し得なくする変異体の能力を測定することによりなされる。このことは、ドーパミン受容体がGタンパク質ユニットの活性化をしないか、またはナトリウム輸送体を阻害するために必要な細胞質性二次メッセンジャーの産生をしないことを意味する。ドーパミン作動性の伝達の欠失は、特に、D1受容体/アデニリルサイクラーゼ(AC)またはGタンパク質結合欠陥、および上記のタイプの翻訳後修飾でとりわけ明白である。これらの減少の測定は、ドーパミンまたはそのアゴニストの刺激に対する能力:(a)アデニリルサイクラーゼ活性またはcANP産生または活性化タンパク質キナーゼA、(b)活性化ホスホリパーゼCまたは活性化タンパク質キナーゼC、(c)ホスホリパーゼA2活性および(d)G−タンパク質活性またはナトリウム/水素交換体またはナトリウム/カリウムATPaseのようなナトリウム輸送タンパク質の阻害の測定で行える。
【0034】
本態性高血圧症に関連する他のGRK4は、本態性高血圧症に罹患している個体から得られまたはクローン化されたGRK4遺伝子の配列決定により、簡単に同定できる。
【0035】
野生型GRK4または本態性高血圧症と関連するGRK4は、抗高血圧症活性について多くの異なるタイプの物質のスクリーニングのための種々のシステムに包含し得る。一般に、GRK4およびGRK4基質を含むシステム、およびGRK4の立体配座または活性(およびその変化)を測定できるシステムを、抗高血圧症活性の物質のスクリーニングのために使用し得る。このように、本発明のこの態様の最も広い意味においては、細胞全体を必要としない。システムは、本質として人工的であり得、例えば、脂質ミセル内に入れられている。分子相互作用を研究するための無細胞システムの説明としては、Hammond et al., Nature 327:730-732 (1987)を参照。しかし細胞全体が好ましく、GRK4基質として、D1受容体またはその機能的断片である。“機能的断片”なる用語は、リン酸化、パルミトイル化またはインビトロの他の手段で翻訳後修飾されている受容体の一部を意味する。本発明の好ましい方法は、GRK4核酸で形質転換した細胞の使用を含む。一般に、哺乳類、細菌および昆虫細胞を含む種々の細胞のタイプを使用できる。チャイニーズハムスター卵巣(CHO)細胞、ヒト胚腎(HEK)繊維芽(LTK)細胞、MDCKおよびLLCP細胞のような哺乳類細胞が好ましい。CHO細胞は、それらがインビボで近位尿細管と同じように行動すると予測されるため、より好ましい。GRK4およびD1受容体核酸での細胞の形質転換は、標準法にしたがって行い得る。例えば、Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY (1989)およびAusubel et al., Current Protocols in Molecular Biology, Wiley & Sons (1994)参照。
【0036】
本出願のこの態様のより好ましい具体的態様において、本方法は、正常血圧または高血圧症の動物、例えば、ヒトから単離した細管細胞を使用して調製した不死化腎近位尿細管を使用して行う。一般に、細管細胞は、皮質を小切片(例えば、1mm3)に切断して腎臓から単離し、容器の適当な生育表面(例えば、コラーゲン被覆T−フラスコ)に置く。接着が容器の転倒により認められたら(例えば、室温で約30分)、容器を起し、適当な培地を添加する。好ましい培地は、物質(wt/ml):インシュリン(5マイクログラム)、トランスフェリン(5マイクログラム)、セレニウム(5ナノグラム)、ヒドロコルチゾン(36ナノグラム)、トリヨードチロニン(4ピコグラム)および表皮成長因子(19ナノグラム)を添加したダルベッコ最小必須およびF−12培地である。組織をインキュベートし、約3日間、95%空気、5%CO2で分配しないままにした。Detrisac, et al., Kidney Int. 25:383-390 (1984)。あるいは、皮質の切片を、コラゲナーゼで消化させ、連続的に212および140マイクロメーターにふるって、培養前に40マイクロメーターふるいで濃縮し得る。Courjault-Gautier et al., J. Am. Soc. Nephrol. 5:1949-1963 (1994)参照。“不死化”なる用語は、細胞が培養においていつまでも生育することを意味する。単離の腎近位尿細管細胞を、SV40ウイルス、例えば、SV40tsA変異体ウイルスで感染させ、次いで、感染約7−8週間後の過成長を得ることにより不死化し得る。これらの細胞は、GRK4タンパク質が機能するインビボ環境をより密接に模倣している利点を提供する。高血圧症患者からの不死化細胞は、しばしば抗高血圧症活性のための薬剤のスクリーニングに使用できる細胞について殆ど無限の供給を提供する。
【0037】
抗高血圧症特性を有すると推定される物質または薬剤は、物質または薬剤のGRK4システムへの添加によるGRK4の立体配座または活性の変化の測定により同定し得る。GRK4活性は、アデニレートキナーゼ活性の測定のように間接的に、または培養に添加したリン酸化可能な物質のリン酸化の程度の測定のように直接的に測定し得る。GRK4活性化または不活性化変異体、例えば、各々GRK4活性の増加またはGRK4活性の減少を導くGRK4の変異体または多形性は興味深い。GRK4活性の変化は、D1受容体により例示されるGタンパク質結合受容体の機能の変化を導くことができる。GRK4は、レニン−アンギオテンシン系、カリクレイン−キニン、エンドセリン、房および脳のナトリウム排泄増加性ペプチド、セロトニン、バソプレッシン、カルシウム検出受容体および上皮ナトリウムチャンネルのような本態性高血圧症に関与する他のタンパク質の機能を調節し得る。
【0038】
他のタイプのスクリーニング薬剤は、GRK4タンパク質、例えば野生型またはイソ形または本態性高血圧症に関与する変異体と、検出する抗高血圧症剤との相互作用によりGRK4タンパク質の立体配座的変化をもたらす薬剤との間の複合体を含む。複合薬剤の選択は、立体配座的分析を行う方法に依存する。このような分析は、分光測光法、フルオレッセンス、核磁気共鳴、エバネッセント波法および原子間力顕微鏡により行い得る。
【0039】
さらに別のタイプのスクリーニング剤およびプロトコールは、本態性高血圧症のトランスジェニック動物モデルの使用を含み、動物は本発明の野生型GRK4または変異GRK4をコードする核酸を発現する。変異GRK4の発現は、高血圧症により特徴付けられる表現型を明らかにし、動物が急性または慢性のナトリウム負荷を排出する能力を減少させる。トランスジェニックモデルはまた、GRK4の発現および活性における、血圧を減少することが示されている高カルシウム、高カリウムおよび高マグネシウムのような食事操作の効果の試験にも使用できる。明らかに、排出システムを有する動物は、本態性高血圧症のモデルとして使用できる。マウスのような齧歯類が好ましい。
【0040】
トランスジェニック動物は、当分野で既知の方法により製造できる。トランスジェニック動物の製造に適用できる方法は、動物の核DNA内へのトランスジーンの挿入である。これらのトランスジェニック動物は、例えば、米国特許4,873,191;5,849,578;5,731,489;5,614,396;5,487,992;5,464,764;5,387,742;5,347,075;5,298,422;5,288,846;5,221,778;5,175,384;5,175,383;4,873,191および4,736,866ならびにBurke and Olson, Methods in Enzymology 194:251-270 (1991), Capecchi, Science 244:1288-1292 (1989), Davies et al., Nucleic Acids Research 20(11):2693-2698 (1992), Dickinson et al., Human Molecular Genetics 2(8):1299-1302 (1993), Huxley et al., Genomics 9:742-750 (1991), Jakobovits et al., Nature 362:255-261 (1993), Lamb et al., Nature Genetics 5:22-29 (1993), Pearson and Choi, Proc. Natl. Acad. Sci. 90:10578-10582 (1993), Rothstein, Methods in Enzymology 194:281-301 (1991), Schedl et al., Nature 362:258-261 (1993), およびStrauss et al., Science 259:1904-1907 (1993)に明示の当分野で既知の標準法を使用して構築される。更に、公開国際特許出願WO94/23049、WO93/14200、WO94/06908およびWO94/28123は、これらに関する更なる関連の教示を提供する。
【0041】
当分野で既知の方法を、標的遺伝子トランスジーンを動物に挿入し、トランスジェニック動物の創始系を製造する。このような方法は、前核マイクロインジェクション(Hoppe, P. C. and Wagner, T. E., 1989, 米国特許4,873,191);生殖細胞系へのレトロウイルス仲介遺伝子移入(Van der Putten et al., Proc. Natl. Acad. Sci., USA 82:6148-6152 (1985));胚幹細胞への遺伝子標的化(Thompson et al., Cell 56:313-321 (1989));胚のエレクトロポレーション(Lo. Mol. Cell. Biol. 3:1803-1814 (1983))および精子仲介遺伝子移入(Lavitrano et al., 1989, Cell 57:717-723 (1989))を含むが、これらに限定されない。これらの方法に関する一般的総説は、Gordon, Transgenic Animals, Intl. Rev. Cytol. 115:171-229 (1989)参照。
【0042】
本発明は、全細胞にGRK4トランスジーンを担持するトランスジェニック動物、および全てではないが、いくつかの細胞にトランスジーンを担持する動物、即ち、モザイク動物を提供する。トランスジーンは、単一トランスジーンまたは、例えば、頭と頭のタンデム、頭と尾のタンデムのコンカタマーとして統合され得る。トランスジーンはまた、例えば、Lasko et al., Proc. Natl. Acad. Sci. USA 89:6232-6236 (1992)の教示にしたがって、特定の細胞タイプに選択的に挿入し、その中で活性化され得る。当業者は、このような細胞タイプ特異的活性化に必要な調節配列は、目的の特定の細胞タイプに依存することを認める。標的遺伝子トランスジーンが内因性標的遺伝子の染色体部位に統合されるのが望ましいとき、遺伝子標的化が好ましい。簡単に述べると、このような方法を用いるとき、目的の内因性標的遺伝子と相同的なあるヌクレオチド配列を含むベクターを、染色体配列との相同的組換えを介して、内因性標的遺伝子のヌクレオチド配列に統合し、機能を中断する目的で設計する。トランスジーンはまた、例えば、Gu et al., Science 265:103-106 (1994)の教示にしたがって、特定の細胞タイプに選択的に挿入し、そしてその細胞タイプのみで目的の内因性遺伝子を不活化し得る。このような細胞タイプ特異的不活性化に必要な調節配列は、目的の特定の細胞タイプに依存し、当業者には明らかである。
【0043】
トランスジェニック動物を産生したら、組換え標的遺伝子およびタンパク質の発現を、標準法を使用してアッセイし得る。最初のスクリーニングは、動物組織の分析のためにサザンブロット分析またはPCR法を使用して達成し得、トランスジーンの統合が起きているかをアッセイする。トランスジェニック動物の組織内のトランスジーンのmRNA発現のレベルは、また動物から得た組織のノーザンブロット分析、インシチュハイブリダイゼージョン、およびRT−PCRを含むが、これらに限定されない方法を使用して評価し得る。標的遺伝子発現組織のサンプルは、また、目的の標的遺伝子トランスジーン遺伝子生産物に特異的な抗体を使用して、標的細胞化学的に評価し得る。
【0044】
標的遺伝子mRNAまたは標的遺伝子トランスジーンペプチド(標的遺伝子産物エピトープを指向する抗体を使用して、免疫細胞化学的に検出)を容易に検出可能なレベルで発現する標的遺伝子トランスジェニック動物を、次いで更に評価し、本態性高血圧症の特徴的症状を示す動物を同定する。
【0045】
好ましい具体的態様において、GRK4トランスジーンを適当なベクターに挿入し、テトラサイクリン感受性プロモーターに操作不可能的に結合し、次いで、胚性幹(ES)細胞に導入する。ES細胞は、次いで遺伝的に変えたES細胞を宿主胚盤胞にマイクロインジェクションすることにより、または桑実胚共培養により再導入する。創始系動物を得、次いで、GRK4トランスジーンに同型接合の動物を使用する。Thompson, et al., Am. J. Physiol. 269:E793-E803 (1995)参照。
【0046】
治療様相は、標的GRK4活性をナトリウム尿排泄亢進まで増加させることを伴うか、またはそうでなければナトリウムと水の適当なバランスに関する正常性に近づく。例えば、GRK4発現は、RNAレベルまたはDNAレベルで、腎細胞のGRK4の発現を変える薬を投与することにより予防できる。このような薬剤は、好ましくは、アンチセンスオリゴヌクレオチド、優性阻害変異体DNA分子、およびGRK4 mRNA、プレmRNA、またはGRK4 DNAの結合によりGRK4発現を減少するまたは防止するリボザイムのようなオリゴヌクレオチド分子である。アンチセンスオリゴヌクレオチドの高血圧者への投与は、米国特許5,856,099;5,856,103;5,783,683;5,840,708;および5,591,600;5,849,903;5,135,917;5,098,890;および5,087,617に記載の製剤および媒体にしたがって行うことができる。現在当分野で既知のアンチセンス法は、また、Uhlmann et al., Chem. Rev. 90:543-584 (1994); Oligodeoxynucleotides: Antisense Inhibitors of Gene Expression (Cohen, ed. 1989); Delivery Strategies for Antisense Oligonucleotide Therapeutics, CRC press (Saghir Akhtar, et. 1995); およびStein, C. A., and Cohen, Jack S., "Oligodeoxynucleotides as Inhibitors of Gene Expression: A Review," Cancer Research, 48:2659-2668 (1988)にも記載されている。
【0047】
合成アンチセンスオリゴヌクレオチドは、標的ヌクレオチド配列とハイブリダイズし、所望の効果、例えば、mRNA分子の翻訳の遮断を発揮するのに十分な長さでなければならない。しかし、相対的に小さいオリゴヌクレオチドを使用するのが有利である。なぜなら、それらは、細胞にインビボでより効率的に取りこまれ、より多い数のアンチセンスオリゴヌクレオチドが標的mRNAの位置に送達されるようである。好ましくは、十分な特異性を達するために、アンチセンスオリゴヌクレオチドは少なくとも15ヌクレオチド長、好ましくは20ヌクレオチド長でなければならない。好ましいアンチセンスは、5'CAC GAT GTT CTC GAG CTC CAT3'(塩基255−275に相補的)および5'CTC CAT GTC CTG GCG CCG3')塩基243−260に相補的)である。
【0048】
上記のような小さいオリゴヌクレオチドは、雑多なヌクレアーゼの分解に非常に感受性である。更に、このような分子は、不十分な膜貫通のために、細胞内に入ることができないことがある。この理由のために、当業者は、一般に、安定性および膜貫通を増加する種々の方法で修飾したオリゴヌクレオチドを合成する。修飾アンチセンスオリゴヌクレオチドの使用は、本発明で好ましい。“アンチセンスオリゴヌクレオチドアナログ”なる用語は、下記のような、このような修飾オリゴヌクレオチドを意味する。
【0049】
本発明のオリゴヌクレオチドは、簡便には既知の方法の固相法を使用して合成し、本明細書に記載の配列をコードする標的GRKのDNAまたはRNAの予め選択した配列と相補的および/または特異的にハイブリダイズできるように設計する。核酸合成装置は商品として入手可能であり、その使用は、妥当な長さの所望のオリゴヌクレオチドの製造に有効であるとして当業者に理解されている。
【0050】
GRK4 mRNAまたはプレmRNAの分解を触媒する、例えば、ハンマーヘッドまたはハリピンタイプのリボザイムを設計でき、標準法にしたがって製造できる。治療的使用のリボザイムの設計、製造および製剤に関する詳細な教示は、例えば、米国特許5,856,463(およびその中に引用されている刊行物)参照。
【0051】
GRK4活性はまた、タンパク質に直接作用することにより完全にまたは部分的に発現されたGRK4タンパク質の触媒活性、例えば、リン酸化または非リン酸化作用を変化(例えば、阻害または促進)する薬理的アンタゴニストのような薬剤の投与により標的化できる。他の治療的作用は、GRK4タンパク質とペプチド様薬剤の直接の結合を伴う。これらの方法および薬剤の全ては、GRK4を発現する腎細胞におけるD1受容体/AC結合の正常化をもたらし、その結果、腎近位尿細管におけるナトリウム輸送を減少させる。
【0052】
本発明を、以下の詳細な実施例を参照して更に記載する。これらの実施例は説明の目的のみで提供され、特記しない限り、本明細書に記載の本発明の限定を意図するものではない。
【実施例1】
【0053】
実施例
組織培養
ヒト腎を、腎癌腫のため片側の腎を摘出した患者から新鮮な外科的標本として得た。患者記録を調べて、正常血圧(n=9)と本態性高血圧症(n=14)とに分類した。収縮期血圧が140mmHg未満で弛緩期血圧が90mmHg未満の者を正常血圧者とした。収縮期血圧が140mmHg以上で弛緩期血圧が90mmHg以上の者および/または抗高血圧剤服用者を高血圧者とした。
【0054】
組織学的に確認された正常な腎切片からの腎近位尿細管細胞の培養物(0.075%タイプIコラーゲンで被覆した24ウエルプラスチック管における5x105細胞/ウエル)を、37℃で95%O2/5%CO2中でインキュベートし、デルベッコ修飾イーグル培地とハムF12培地との1:1の混合物からなる血清を含まない培地で生育した。この培地には、セレニウム(5ng/ml)、インスリン(5μg/ml)、トランスフェリン(5μg/ml)、ヒドロコルチゾン(36ng/ml)、トリヨードチロシン(4pg/ml)、表皮成長因子(10ng/ml)(5)が補充されている。半コンフルエント(90−95%)のときは、細胞を継代培養して(6−8継代)、トリプシンEDTA(0.05%、0.02%)を使用する実験プロトコールに用いた。この培養条件は、腎近位尿細管細胞の特性を保持するヒト腎近位尿細管の成長に役立つ。Sanada, H. et al., J. Invest. Med. 45: 277A (1997)。
【0055】
光学顕微鏡による免疫組織化学
HISTOCHOICE中に不死した培養物における腎の組織および細胞について免疫組織化学的検査を、Sanada, H. et al.(前出)に記載のように行った。アフィニティー・カラム精製ポリクローナル・ヒトD1受容体抗体が合成ペプチド配列GSGETQPFC(アミノ酸299−307)に対して生じる。参照、Sanada, H. et al.(前出)。2種の市販(Santa Cruz Biotechnology, Inc, Santa Cruz, CA)のGRK4イソ形抗体を使用した。1つのGRK4抗体がαイソ形とβイソ形の両方を認識し、他の1つが両αβイソ形を認識した。これらの抗体の特異性はすでに報告されている。Sanada, H. et al.(前出)および Guyton A. C. Circulatory Physiology III, Arterial Pressure and Hypertension, W. B. Saunders Co., Philadelphia, PA (1980)。
【0056】
免疫組織化学的検査によると、GRK4α/βおよびGRK4α/σのイソ形発現が腎の近位および遠位の尿曲細管のみでみられた(ヘレン係蹄、皮質および髄質の集合管、糸球体、腎動脈ではみられなかった)。GRK4α/σは管腔膜および基底膜の両方でみられたが、GRK4α/βは管腔膜のみでみられた。これら2種のGRK4の腎発現について、高血圧者と正常者とで差異がなかった(表示せず)。GRK4α/βおよびGRK4α/σの発現は、培養物の腎近位尿細管細胞において持続した(写真は示さず)。
【0057】
GRK活性の測定
GRK活性を Benovic, Methods Enzymol. 200: 351-362 (1991)に従って測定した。腎近位尿細管抽出物の調製は、氷冷の融解緩衝液中での均質化によった。この緩衝液は、25トリスHCl、pH7.5、5EDTA、5EGTA、ロイペプシン(10μg/ml)、アプロチニン(20μg/ml)および1PMSFを含む。粗均質物を30,000gで30分間遠心分離した。ペレットの抽出を200mMのNaClで30分間行い、30,000gで30分間遠心分離した。上澄み液をGRKアッセイおよびイムノブロッティグに用いた。20μgのタンパク質抽出液を、ロドプシン強化桿外側セグメントとともに、10mMのMgCl2および0.1mMのATP(γ32P−ATP含有)中でインキュベートした。白色光で15分間室温でのインキュベーション後、反応を氷冷の融解緩衝液で停止し、30,000gで15分間遠心分離した。ペレットをラエムリ緩衝液に再懸濁し、12%SDS−PAGEに掛けた。ゲルにオートラジオグラフィを行い、リン酸化ロドプシンの定量を、密度計測と適当なサイズでの切除バンドの放射活性計測で行った。また、GRK活性の測定をGRKイソ形抗体の存在および不存在で行った。
【0058】
図1は、D1様アゴニストであるフェノルドパムがGRK活性に対する作用を有さないことを示す。これは、正常血圧者からの腎近位尿細管細胞におけるロドプシンのリン酸化により調べられた。このデータによると、基質としてロドプシンを使用し得るGRK(すなわち、GRK2、GRK3、GRK4α、GRK5、GRK6)は、血圧が正常であるとき、腎近位尿細管細胞におけるD1受容体の脱感作に関与しない。また、培養物中の腎近位尿細管細胞におけるD1受容体およびGRK4の発現は、高血圧者と正常血圧者とで類似していた(データは示さず)。しかし、高血圧者からの腎近位尿細管細胞において、フェノルドパムがGRK活性を増加した。さらに、腎近位尿細管細胞における基本的GRK活性は、高血圧者において正常血圧者におけるよりも大きかった。これらの研究から高血圧における腎近位尿細管細胞でのGRKの異常な機能がわかる。
【0059】
フェノルドパムによりつくられたGRK活性の増加(高血圧における)は、GRK2、GRK3およびGRK4α/σに対する抗体によって遮断され、これらのGRKの1つまたはすべての活性がGRK4活性のフェノルドパム仲介増加に関与し得ることを表す。Tiberi et al., J. Biol. Chem. 271: 3771-3778 (1996)。しかし、GRK2およびGRK3の偏在的発現は、げっ歯類およびヒトの本態性高血圧の病因において認識されている腎の特徴と適合しない。Guyton, W. B. Saunders Co. Phil., PA (1980); Guidi et al., J. Am. Soc. Nephrol 7: 1131-1138 (1996)。GRK2のコード領域の配列には、高血圧者と正常血圧者との間に差異がみられなかった(データは表示せず)。このことから、高血圧患者のリンパ球のおけるGRK2活性の増加が高血圧にとって二次的であり(Gros et al., J. Clin. Invest. 99: 2087-2093 (1997))、遺伝的および誘発された高血圧のげっ歯類おけるGRK5の活性および発現の増加について示唆されていたことと同様である。Ishizaka et al., J. Biol. Chem. 272: 32482-32488 (1997)。
【0060】
cAMP蓄積の測定
細胞をダルベッコ・リン酸緩衝液(D−PBS)で2回洗い、次いで各ウエルに1mMの3−イソブチル−1−メチルキサンチンを加えた。細胞を37℃で30分間、薬剤とともに、または薬剤なしでインキュベートした。薬剤は、ドパミンおよびD1様受容体アゴニスト、フェノルドパム、D1様受容体アンタゴニスト、SCH23390(Research Biochemicals International, Natick, MA)、およびフォルスコリン(Sigma Chemical Co., St. Louis, MO)である。次いで、細胞をD−PBSで2回洗い、−80℃に冷凍して、さらに0.1NのHClで分解した。cAMPの濃度を放射免疫検定法で測定した。Sanada, H. et al.,(前出)、Kinoshita, S. J. Chlin. Invest. 84: 1849-1856 (1989)。タンパク質の濃度をBCAタンパク質アッセイ・キットで測定した。
【0061】
GRK4活性の増加が高血圧での腎近位尿細管細胞におけるD1受容体の非共役に関与しているかどうかを確認するために、アンチセンス・オリゴヌクレオチドによるGRK4翻訳の阻害後のcAMPに対するD1様アゴニスト刺激作用を調べた。図2からわかるように、D1様アゴニストのフェノルドパムによるcAMPの蓄積は、高血圧者よりも正常血圧者の腎近位尿細管細胞において大きい。センス/スクランブルまたはアンチセンスのいずれのGRK4オリゴヌクレオチドも基底性またはフォルスコリン刺激cAMP産生に作用しなかった。フェノルドパムのみに比べると、センスまたはスクランブルのいずれのGRK4オリゴヌクレオチドもいずれのグループでのcAMP蓄積に作用しなかった。しかし、アンチセンスGRK4オリゴヌクレオチドはフェノルドパムの能力を高め、高血圧者由来の細胞でのcAMPの蓄積を促進し(正常血圧者からでは、しない)、その値は、フェノルドパムで処置された正常血圧者の細胞で見られる値とほぼ同じであった。
【0062】
免疫沈降
近位管細胞を、運搬物、フェノルドパム、センス、スクランブルまたはアンチセンスのプロピン/ホスホロチオエート・オリゴヌクレオチド(5nM)とともに、上記のようにインキュベートした。膜を氷冷融解緩衝液(PBSと、1%NP40、0.5%デオキシコール酸ナトリウム、0.1%SDS,1mMのEDTA、1mMのEGTA、1mMバナジン酸ナトリウム、1mMのPMSF、10μg/mlアプロチニン、10μg/mlロイペプシン)で溶解した。溶解物を、IgG精製アンチD1受容体抗体とともに氷上1時間、そしてタンパク質Aアガロースとともに12時間ゆすりながら4℃でインキュベートした。SDSポリアクリルアミド・ゲル電気泳動により分離したタンパク質を、ニトロセルロース膜上に電気泳動で移した。移行ブロット・シートを、10mMトリスHCl、pH7.5、150mMのNaCl、0.1%ツィン20中で5−10%の脂肪なし乾燥乳で阻害し、希釈の親和性精製ポリクローナル・アンチホスホセリン抗体(Zymed Lab, San Francisco, CA)とともにインキュベートした。Sanada, H. et al.(前出)。オートラジオグラムおよび免疫ブロットをECLシステム(Amersham, Arlington Heights, Il)で可視化し、エンシトメトリーで定量した。Sanada, H. et al.(前出)。
【0063】
次の検討は、アンチセンスGRK4オリゴヌクレオチドの示差作用がD1受容体のリン酸化まで及ぶかどうかである。図3からわかるように、腎近位尿細管細胞でのセリン−リン酸化D1受容体の基底レベルは、正常血圧者よりも高血圧者で高く、高血圧者でのGRK活性の基底レベルの増加に相関する(図1に示す)。フェノルドパムは、セリン−リン酸化D1受容体の量を正常血圧者で増加せしめたが、高血圧者では増加せしめない。このことは、以前の報告 Sanada, H. et al.(前出)に一致する。センスまたはスクランブルのいずれのGRK4オリゴヌクレオチドは、両グループの対象者のフェノルドパム処置細胞におけるD1受容体のリン酸化に作用しなかった。一方、GRK4アンチセンス処置は、高血圧者からのフェノルドパム処置腎近位尿細管細胞におけるD1受容体のリン酸化を、基底値より低いレベルまでほとんど消失せしめた。GRK4アンチセンス処置は、正常血圧者からのフェノルドパム処置腎近位尿細管細胞におけるD1受容体のリン酸化も下げたが、基底値よりは高かった。高血圧者の腎近位尿細管細胞においてGRK4に対するアンチセンス・オリゴヌクレオチドによりD1受容体のリン酸化がほとんど完全に抑制されることから、高血圧のおけるD1受容体のリン酸化および脱感作に関与する主要なGKRがGRK4であって、このネフロン切片で発現される他のGRKでないことがわかる。
【0064】
遺伝子型決定
ホモ接合性GRK4遺伝子変異体の発生率が高血圧者で約60%および一般人で16%であるという当初の観察に基づくと、能力分析(能力:0.8、α:0.05、効果:45%)がグループにつき14−21のサンプルサイズで、グループ間の有意な差違が検出できる。従って、さらに高血圧者18名および正常血圧者11名の末梢血によるDNAを入手した。少なくとも2人の研究者で全志願者を検査し、医療記録を検討した。高血圧症の病歴および潜在的高血圧症の臨床徴候をもたず、降圧剤薬物治療を行なっておらず、血管拡張療法または血圧に影響し得る他の薬剤を受けず、および最近の3回の診療訪問で収縮期血圧が140mmHgより低く、弛緩期血圧が90mmHより低い状態であった者を正常圧として、被検者を分類した。高血圧症の患者らは、顕著であり持続的な血圧上昇(収縮期血圧160mmHg以上で弛緩期血圧が95mmHg以上)を少なくとも3回の機会で有していた。高血圧者 (腎臓由来のDNA:n=14、末梢血由来のDNA:n=18)は全員20歳以上であった。各個体の本態性高血圧の後期発症についての先天性な問題を除去するため、正常血圧者(腎臓由来のDNA:n=9、末梢血由来のDNA:n=11)は全員45歳以上であった。
【0065】
培養中の腎臓近位尿細管細胞および腎組織または任意の末梢白血球から、ゲノムDNAを抽出した(塩析法)。多形性ヌクレオチドを含むGRK4のエキソンを、表2で示すプライマーで増幅した。反応混合物 20μlは、それぞれ、1XPCR緩衝液、0.2mMの各dNTP、1.25mM MgCl2、0.2μMの各プライマー、TaqDNAポリメラーゼ 0.5単位およびゲノムDNA 50ngを含んでいた。反応混合物を94℃で5分間変性し、次いで、94℃の30秒間変性、55℃のリアニール 30秒間および72℃の30秒伸長を30サイクル行なった。72℃での5分間の最終伸長でPCRが完了した。PCR産物 2μlをバイオダインB+膜上にスポットした。下記の野生型および変異体の対立遺伝子特異的オリゴヌクレオチドプローブそれぞれについてドットブロットを行った(表4)。プローブ標識化、膜調製、ハイブリダイゼーション、および洗浄条件は公知の手順である。Song et al., Clin. Chem. 43:1857-1861 (1997)参照。任意の被検者250人の1801位ヌクレオチドは変異体(G)であった。993位の多形性ヌクレオチドの頻度は高血圧者と正常血圧者で違いがないことも判明した。従って、調査の結果、448、679、1711位の3つのみ(表4)の多形性部位が存在する。サンガー・ジデオキシ連鎖停止法でcDNA配列を決定した。
【0066】
表4.正常血圧者および高血圧者のGRK4変異体
【表10】
【0067】
対立遺伝子特異的オリゴヌクレオチドを使用したドットブロット分析で遺伝子型を決定した。高血圧者の4名は2つの部位(65位および142位アミノ酸)で相同性があった。高血圧者のR65L、A142Vおよび/またはA486Vのホモ接合性変異体の頻度(53%、32中の17)は、正常血圧者で示されたもの(5%、20中の1)と有意に異なっていた(X2=10.56、P=0.0012)。ホモ接合性変異体A142Vの頻度もまた、高血圧者(34%、32中の11)および正常血圧者(0%、20中の0)で有意に異なっていた(X2=6.78、P=0.0092)。
【0068】
ヒト腎臓由来GRK4のcDNAの配列決定、さらに、腎臓および末梢白血球細胞由来のDNA中の5つの多形性部位の遺伝子型決定から、3つの変異体が明らかになった:ヌクレオチド448、CGTからCTT(アミノ酸R65L)、ヌクレオチド679、GCCからGTC(アミノ酸A142V)、およびヌクレオチド1711、GCGからGTG(アミノ酸A486V)(オートラジオグラフは示していない)の変異体が、正常血圧者より高血圧者で頻繁に現われた(表4)。高血圧者におけるR65L、A142V、および/またはA486Vのホモ接合性変異の頻度(53%,32中の17)は、正常血圧者で示されたもの(5%,20中の1)と有意な違いがあり(X2=10.56,P=0.0012)(表4)、無作為成人被検者50人でみられたものと異なっていた(X2=10.99,P=0.0009)。この血圧不明の無作為集団の中で、16%がR65Lおよび/またはA486Vでホモ接合性であり、および50%がR65LまたはA486Vの何れかでヘテロ接合性であった;ホモ接合性対立遺伝子の頻度16%は、本態性高血圧症の発生率に近い(Lifton R P. Science 272:676-680 (1996))。GRK4 A142V位のホモ接合性変異も、それ自体は、高血圧症 (34%、32中の11) の方が正常血圧者(0%、20中の0)より頻繁であった(X2=6.78,P=0.0092)。
【0069】
GRK4αは、リン酸化ロドプシンにレポートされる唯一のGRK4イソ形である(Sallese et al., J. Biol. Chem. 272:10188-10195 1997))が、我々の研究では、フェノルドパムによるD1アゴニスト刺激は、正常血圧者の腎臓近位尿細管細胞中のGRK活性を増強しなかった(図1)。従って、GRK4αはD1受容体の脱感作に関与していないと結論した。その確信によると、ロドプシンを普通はリン酸化しないGRK4イソ形(例えばGRK4γ)(Premont et al., J. Biol. Chem. 271:6403-6410 (1996); Sallese et al., supra.; and Virlon et al., Endocrinol. 139:2784-2795 (1998))は、高血圧症で活性となり得る。たしかにGRK活性中のD1様アゴニスト介在増加は、高血圧者の腎臓近位尿細管細胞中のGRK4αδの膜発現の増加と関連があることが分かったが、正常血圧者(図4)では関連しない。
【0070】
トランスフェクションおよび細胞培養
ラットD1(rD1)またはヒトD1(hD1)受容体cDNAを、発現ベクターpPUR(Clontech, Palo Alto, CA)またはpcDNA3.1/Zeo(Invitrogen, Carlsbad, CA)中で、それぞれ、EcoR1およびXbaI部位でサブクローン化した。得られた作成物を使用し、pTeT−オフ レギュレータープラスミドを発現するCHO細胞を、安定してトランスフェクトした(Clontech, Palo Alto, CA)。この操作にリン酸化カルシウムを使用した。Yamaguchi et al., Mol. Pharmacol. 49:373-378 (1996)参照。ヒト腎臓皮質由来mRNAのRT/PCRから得られた、GRK4γおよびGRK4δのcDNAを、pTet-オフ 応答プラスミド中へ入れサブクローン化した(pTRE−rD1またはpTRE−hD1とpTK−Hygをそれぞれ20:1の割合で混合した)(Clontech, Palo Alto, CA)。
【0071】
GRK4α遺伝子の変異が他の機能的結果を有するかどうか測定するために、D1様アゴニストの効果を、D1受容体および野生型または変異体GRK4α cDNAの両方をトランスフェクトする、チャイニーズハムスター卵巣(CHO)細胞中のcAMP産物について調べた。 GRK4δを対照として使用した。GRK4α欠損下におけるCHO細胞中の投与応答カーブは、HEK-293細胞(低い内因性GRK活性を有する細胞)で示されるものと類似していた。Premont et al, 前出。野生型GRK4αの発現は、D1アゴニストのcAMP産物を刺激する能力を減少させた(図5)。しかし、D1アゴニスト活性の阻害は、GRK4α変異体R65Lおよび/またはA486Vよりも大きくなった。野生型または変異体のGRK4αの効果は、D1受容体またはGRK4αの何れかの発現量の違いからではない(データは示していない)。野生型GRK4αまたはその変異体は、フォルスコリンの能力に作用せず、GRK4αとD1受容体の相互作用の特異性を表わすcAMP蓄積を刺激する。フェノルドパムの作用はD1受容体に対して選択的であり、フェノルドパム効果は、D1様アンタゴニストSCH23390によって遮蔽されたことによるからである(データは示していない)。他の研究では、野生型GRK4δは、野生型GRK4αによって誘発されたD1受容体の脱感作と比較し、D1様アゴニスト介在のcAMP蓄積に対して効果がない(データは示していない)。腎臓近位尿細管細胞の機能的研究およびCHO細胞の発現研究では、GRK4αの活性増加が、D1受容体リガンドの能力の減少の原因であり、高血圧症でエフェクター酵素およびイオン輸送タンパク質と結合することを提唱する。言いかえると、高血圧者における腎臓近位尿細管中のD1受容体の脱感作は、腎臓の能力増強につながり、塩化ナトリウムの負荷を除去する。腎臓の塩化ナトリウム排出の不足は、高血圧症の進行に極めて重要であると考えられる。 Guyton, A.C., Circulatory Physiology III. Arterial Pressure and Hypertension, W.B. Saunders Co.,, Philadelphia, PA (1980); Guidi et al., J. Am. Soc. Nephrol. 7:1131-1138 (1996)。確かに、腎臓ナトリウム輸送を調節する遺伝子は、血圧の調製に重要であると記されている。Lipton, R.P. Science 272:676-680 (1996)およびKaret & Lifton, Recent Prog. Horm. Res. 52:263-276 (1997)。
【0072】
物質または薬剤を生物体中へ注入し、引き起こされたGRK4活性の減少が抗高血圧療法として役立ち得るかどうかを調べるために、自発性高血圧症ラット(SHR)でさらなる実験を行なった。6匹の雌ラット(4週齢、体重100g)を左一側性腎摘出し、2週間そのままにしておいて、手術から回復させた。回復後、単独のアウトレットカテーテルを装備する30日浸透圧ミニポンプを、ホスホロチオネート/プロピン介在アンチセンスGRK4オリゴヌクレオチド(5nM、1マイクロリットル/時間)またはスクランブルGRK4オリゴヌクレオチドで充填し、残された左腎臓の腎皮質中へ移植する。カテーテルのアウトレットを残された腎臓の腎皮質中のほぼ1mm深さに挿入し、スーパーグルーで固定した。ラットを手術から回復させ、血圧と尿出量を毎日測定した(容量と電解液)。30日後、ラットを屠殺し、残された腎臓をGRK4のウエスタンブロット分析に使用した。我々の研究によると、血圧は、スクランブルGRK4オリゴヌクレオチド(n=3)を処置したラットのときと比較して、GRK4(n=3)にアンチセンスオリゴヌクレオチドを処置したラットで減少した。さらに、ウエスタンブロット分析で、アンチセンスオリゴヌクレオチドは腎臓GRK4の発現を減少させることを証明した。
【0073】
結果として、この例では、本態性高血圧者由来の腎臓近位尿細管細胞におけるD1受容体/アデニリルシクラーゼ共役で欠損が示された。ヒト本態性高血圧症の腎臓近位尿細管細胞のGRK活性の増加は、GRK4の活性化ミスセンス変異によるものであり、この効果はトランスフェクトされた細胞モデルで再現された。その上、GRK4の翻訳を防止し、高血圧症におけるD1受容体のアデニリルシクラーゼに対する結合を正常化した。さらに何れの特定の理論に拘束されることを意図することなく、出願人は、ホモ接合性アミノ酸変異は、D1受容体のリガンドに依存しないセリン-リン酸化を引き起こし、Gタンパク質/エフェクター複合体からその結合解離をもたらすと考える。腎臓近位尿細管中のD1受容体の脱感作は、ドーパミンのナトリウム排泄作用を低下し、最終的にナトリウム保持および高血圧症となる。これらの結論は、GRK4に対するアンチセンスオリゴヌクレオチドを自発性高血圧症ラットの腎臓内に注入すると、腎臓内でGRK4濃度が減少し、平均動脈血圧が低下するという上記実験の結果に対応している。従って、GRK4の濃度または活性を変える物質または薬剤は、降圧性薬物治療の新規段階を表わす。
【0074】
ドーパミンDIA受容体とGプロテイン/エフェクター酵素複合体とのネフロンセグメント特異的欠損結合は、自発性高血圧症ラット(SHR)の腎臓ナトリウム保持の原因となり得る。腎臓近位尿細管中でナトリウム輸送を阻害する外因性および腎臓内因性のドーパミンの能力減少は、SHRと正常圧対照(ウィスター-キョート(WKY)ラット)とのF2交配において高血圧症を同様におこす。同様の欠損がダール塩過敏性ラットで判明した。さらに重要なことに、本態性高血圧症のヒトでも判明した。従って、高血圧症ヒト由来の腎臓近位尿細管の初期の培養は、腎臓D1様受容体とアデニリルシクラーゼ(AC)の欠損結合を有しており、同様な結合欠損が高血圧症齧歯動物でもみられた。これらのインビトロデータは、欠損D1様受容体を証明するインビボ研究(Gタンパク質/エフェクター酵素複合体は、相同性または異種性脱感作、受容体下方制御、Gタンパク質またはエフェクター酵素“欠損”またはD1様受容体の1次配列における変異によらない)に一致する。むしろ、D1様受容体の脱共役は、D-l受容体(腎臓中の主要なD1様受容体)のリガンドに依存しない過リン酸化の結果であり、限られた器官およびネフロンの発現を伴うGRK4イソ形のホモ接合性変異体の結果である。
【産業上の利用可能性】
【0075】
産業上の利用可能性
本発明の診断検査は、個体をスクリーニングし、その本態性高血圧症の素因を同定できるであろう。これらの検査の実施に関する、遺伝、細胞および生化学的手段も提供する。本発明は、薬剤を発見するための手段および方法も提供し、および抗高血圧性の活性または特性を有する物質を同定する。組成物および方法は、それぞれ本態性高血圧症を有する個体の腎細胞中のナトリウム輸送を正常化するものであり、この疾患の処置手段を提供する。
【0076】
本明細書で引用したすべての特許および非特許の刊行物は、本発明の属する当業者の技術のレベルを示している。すべての出版物または特許出願は、出典明示により本明細書の一部とすることを具体的に個々に示したのと同様に、出典表示より本明細書の一部とする。
【特許請求の範囲】
【請求項1】
R65L変異、A142V変異、R65LとA486Vの二重変異またはR65LとA142とA486Vの三重変異を有するGRK4タンパク質をコードする単離された精製核酸。
【請求項2】
R65L変異、A142V変異、A486V変異、R65LとA142Vの二重変異、R65LとA486の二重変異またはR65LとA142とA486Vの三重変異をコードする配列を有するGRK4遺伝子に特異的にハイブリダイズするオリゴヌクレオチド。
【請求項3】
GRK4遺伝子のエキソン3、5、8、14または16にハイブリダイズし、そして、該遺伝子のヌクレオチド431−503(エキソン3)、594−697(エキソン5)、857−995(エキソン8)、1662−1798(エキソン14)、1937−1991(エキソン16)を含むDNA配列を増幅するのに有用であるオリゴヌクレオチド。
【請求項4】
本態性高血圧症の素因を有する個体を同定する方法であって、
該個体からD1受容体を有しGRK4を発現する腎細胞を得て、
該細胞をアッセイして、概D1受容体の翻訳後修飾の程度を測定する(なお、該D1受容体の翻訳後修飾における変化が、正常血圧の個体から単離され、D1受容体を有しGRK4を発現する腎細胞におけるD1受容体の翻訳後修飾の程度と比較して、本態性高血圧症の素因の指標となる)、
ことを含む方法。
【請求項5】
該細胞を該D1受容体のパルミトイル化の程度についてアッセイする、請求項4の方法。
【請求項6】
該細胞を該D1受容体のリン酸化の程度についてアッセイする、請求項4の方法。
【請求項7】
該細胞を該D1受容体の過リン酸化についてアッセイする、請求項4の方法。
【請求項8】
該腎細胞が腎近位尿細管細胞または皮質集合管細胞である、請求項4の方法。
【請求項9】
本態性高血圧症の素因を有する個体を同定する方法であって、
個体から核酸のサンプルを得て、
該サンプルからの、GRK4をコードする核酸またはその断片を、GRK4発現細胞にドーパミン作動性シグナルを翻訳せしめないようなGRK4変異について、分析する(なお、GRK4の該変異は本態性高血圧症の素因についての指標である)、
ことを含む方法。
【請求項10】
GRK4の該変異が、GRK4の該変異を発現する該細胞におけるD1受容体/アデニリルシクラーゼ共役欠損を起す、請求項9の方法。
【請求項11】
GRK4の該変異が、GRK4の該変異を発現する該細胞におけるD1受容体/Gタンパク質共役欠損を起す、請求項9の方法。
【請求項12】
該核酸のサンプルがDNAのサンプルである、請求項9の方法。
【請求項13】
該核酸のサンプルがRNAのサンプルである、請求項9の方法。
【請求項14】
該核酸のサンプルがゲノムDNAのサンプルである、請求項9の方法。
【請求項15】
該核酸のサンプルがcDNAのサンプルである、請求項9の方法。
【請求項16】
GRK4をコードする核酸の断片を分析する、請求項9の方法。
【請求項17】
GRK4核酸を変異R65Lについて分析する、請求項9の方法。
【請求項18】
GRK4核酸を変異A142Vについて分析する、請求項9の方法。
【請求項19】
GRK4核酸を変異A486Vについて分析する、請求項9の方法。
【請求項20】
GRK4核酸を変異R65L、A486Vについて分析する、請求項9の方法。
【請求項21】
GRK4核酸を変異R65L、A142Vについて分析する、請求項9の方法。
【請求項22】
GRK4核酸を変異R65L、A142、A486Vについて分析する、請求項9の方法。
【請求項23】
該検出工程がPCRでなされる、請求項9の方法。
【請求項24】
本態性高血圧症に関連するGRK4の変異を検出する方法であって、
高血圧の個体からの核酸サンプルを得て、
該サンプル由来のGRK4をコードする遺伝子の配列を決定する、
ことを含む方法。
【請求項25】
GRK4およびGRK4基質を含み、GRK活性を測定する再構築システム。
【請求項26】
該GRK4基質がD1受容体またはその機能的断片である、請求項25の再構築システム。
【請求項27】
該GRK4および該GRK4基質を発現する全細胞である、請求項26の再構築システム。
【請求項28】
該全細胞が、D1受容体をコードする第1異種性遺伝子および高血圧に関連のGRK4タンパク質をコードする第2異種性遺伝子でトランスフェクトされたチャイニーズハムスター卵巣細胞である、請求項27の再構築システム。
【請求項29】
該GRK4タンパク質が本態性高血圧症と関連している、請求項25の再構築システム。
【請求項30】
高血圧に関連するGRK4タンパク質と、抗高血圧活性について分析する物質との相互作用において概GRK4タンパク質の検出可能な立体配座的変化を提供する薬剤との複合体。
【請求項31】
不死化ヒト腎近位尿細管細胞。
【請求項32】
高血圧のヒトから得られる単離された精製腎近位尿細管細胞。
【請求項33】
不死化された、請求項32の高血圧のヒトから得られる単離された精製腎近位尿細管細胞。
【請求項34】
腎細胞中で発現されるGRK4タンパク質をコードし、該GRK4タンパク質を産生するトランスジーンを含有するニ倍性ゲノムを含むトランスジェニック動物(なお、該トランスジーンの発現によって、概トランスジェニック動物において発揮される本態性高血圧症の状態が、腎細胞が該GRK4タンパク質を発現しない正常血圧の動物と比較される)。
【請求項35】
該腎細胞が、該GRK4タンパク質を発現しない腎細胞の正常血圧の動物に比して、低いナトリウム拒否能力を持つ、請求項34のトランスジェニック動物。
【請求項36】
げっ歯類である、請求項34のトランスジェニック動物。
【請求項37】
マウスである、請求項34のトランスジェニック動物。
【請求項38】
推定の抗高血圧剤を同定する方法であって、
少なくとも1つの候補薬剤を請求項25の再構築システムに加え、
GRK4活性を検出する(該活性は推定の抗高血圧物質の指標である)、
ことを含む方法。
【請求項39】
GRK4活性を検出する該工程が、アデニレート・シクラーゼ活性を測定することを含む、請求項38の方法。
【請求項40】
GRK4活性を検出する該工程が、リン酸塩を付加し得る基質およびリン酸塩源を該培地に加えて該基質のリン酸化を測定することを含む、請求項38の方法。
【請求項41】
推定の抗高血圧剤を同定する方法であって、
少なくとも1つの候補薬剤を請求項30の複合体に接触せしめ、
該GRK4の立体配座的変化が起きたかどうかを検出する(該活性は推定の降圧物質の指標である)、
ことを含む方法。
【請求項42】
該検出が、分光測定、蛍光、核磁気共鳴、エバネセント波法または原子間力顕微鏡で行う、請求項41の方法。
【請求項43】
推定の抗高血圧剤を同定する方法であって、
少なくとも1つの候補薬剤を、高血圧動物から単離の、D1受容体およびGRK4を発現する不死化腎細胞に加え、
該細胞のドーパミン作動性シグナルの形質導入の変化を検出する(ドーパミン作動性シグナルの形質導入の変化は推定の抗高血活性の指標である)、
ことを含む方法。
【請求項44】
推定の抗高血圧剤を同定する方法であって、
該薬剤を投与した請求項34の第1トランスジェニック動物と、該薬剤を投与していない請求項34の第2トランスジェニック動物との電解質出力を比較する(推定の抗高血圧剤の同定は、該第1トランスジェニック動物の電解質出力が該第2トランスジェニック動物に比して増加することでなされる)、
ことを含む方法。
【請求項45】
ナトリウム利尿を増加する方法であって、本態性高血圧症の個体に、GRK4と相互作用する薬剤を投与して、該個体におけるナトリウム利尿を増加することを含む方法。
【請求項46】
該薬剤が、該高血圧個体の腎細胞におけるGRK4の発現を変化する、請求項45の方法。
【請求項47】
該薬剤が、GRK4のmRNAまたはDNAに結合するアンチセンスRNAを含む、請求項45の方法。
【請求項48】
該薬剤が、GRK4のmRNAまたはプレmRNAを開裂するリボチームを含む、請求項45の方法。
【請求項49】
該薬剤が、優性阻害変異DNA分子を含む、請求項45の方法。
【請求項50】
該薬剤がGRK4タンパク質に結合する、請求項45の方法。
【請求項51】
インビトロおよびインビボでGRK4のmRNAに特異的にハイブリダイズするオリゴヌクレオチド。
【請求項52】
アンチセンスRNA分子である、請求項51のオリゴヌクレオチド。
【請求項53】
優性阻害変異DNA分子である、請求項51のオリゴヌクレオチド。
【請求項54】
GRK4のmRNAまたはプレmRNAを開裂するリボチーム。
【請求項1】
R65L変異、A142V変異、R65LとA486Vの二重変異またはR65LとA142とA486Vの三重変異を有するGRK4タンパク質をコードする単離された精製核酸。
【請求項2】
R65L変異、A142V変異、A486V変異、R65LとA142Vの二重変異、R65LとA486の二重変異またはR65LとA142とA486Vの三重変異をコードする配列を有するGRK4遺伝子に特異的にハイブリダイズするオリゴヌクレオチド。
【請求項3】
GRK4遺伝子のエキソン3、5、8、14または16にハイブリダイズし、そして、該遺伝子のヌクレオチド431−503(エキソン3)、594−697(エキソン5)、857−995(エキソン8)、1662−1798(エキソン14)、1937−1991(エキソン16)を含むDNA配列を増幅するのに有用であるオリゴヌクレオチド。
【請求項4】
本態性高血圧症の素因を有する個体を同定する方法であって、
該個体からD1受容体を有しGRK4を発現する腎細胞を得て、
該細胞をアッセイして、概D1受容体の翻訳後修飾の程度を測定する(なお、該D1受容体の翻訳後修飾における変化が、正常血圧の個体から単離され、D1受容体を有しGRK4を発現する腎細胞におけるD1受容体の翻訳後修飾の程度と比較して、本態性高血圧症の素因の指標となる)、
ことを含む方法。
【請求項5】
該細胞を該D1受容体のパルミトイル化の程度についてアッセイする、請求項4の方法。
【請求項6】
該細胞を該D1受容体のリン酸化の程度についてアッセイする、請求項4の方法。
【請求項7】
該細胞を該D1受容体の過リン酸化についてアッセイする、請求項4の方法。
【請求項8】
該腎細胞が腎近位尿細管細胞または皮質集合管細胞である、請求項4の方法。
【請求項9】
本態性高血圧症の素因を有する個体を同定する方法であって、
個体から核酸のサンプルを得て、
該サンプルからの、GRK4をコードする核酸またはその断片を、GRK4発現細胞にドーパミン作動性シグナルを翻訳せしめないようなGRK4変異について、分析する(なお、GRK4の該変異は本態性高血圧症の素因についての指標である)、
ことを含む方法。
【請求項10】
GRK4の該変異が、GRK4の該変異を発現する該細胞におけるD1受容体/アデニリルシクラーゼ共役欠損を起す、請求項9の方法。
【請求項11】
GRK4の該変異が、GRK4の該変異を発現する該細胞におけるD1受容体/Gタンパク質共役欠損を起す、請求項9の方法。
【請求項12】
該核酸のサンプルがDNAのサンプルである、請求項9の方法。
【請求項13】
該核酸のサンプルがRNAのサンプルである、請求項9の方法。
【請求項14】
該核酸のサンプルがゲノムDNAのサンプルである、請求項9の方法。
【請求項15】
該核酸のサンプルがcDNAのサンプルである、請求項9の方法。
【請求項16】
GRK4をコードする核酸の断片を分析する、請求項9の方法。
【請求項17】
GRK4核酸を変異R65Lについて分析する、請求項9の方法。
【請求項18】
GRK4核酸を変異A142Vについて分析する、請求項9の方法。
【請求項19】
GRK4核酸を変異A486Vについて分析する、請求項9の方法。
【請求項20】
GRK4核酸を変異R65L、A486Vについて分析する、請求項9の方法。
【請求項21】
GRK4核酸を変異R65L、A142Vについて分析する、請求項9の方法。
【請求項22】
GRK4核酸を変異R65L、A142、A486Vについて分析する、請求項9の方法。
【請求項23】
該検出工程がPCRでなされる、請求項9の方法。
【請求項24】
本態性高血圧症に関連するGRK4の変異を検出する方法であって、
高血圧の個体からの核酸サンプルを得て、
該サンプル由来のGRK4をコードする遺伝子の配列を決定する、
ことを含む方法。
【請求項25】
GRK4およびGRK4基質を含み、GRK活性を測定する再構築システム。
【請求項26】
該GRK4基質がD1受容体またはその機能的断片である、請求項25の再構築システム。
【請求項27】
該GRK4および該GRK4基質を発現する全細胞である、請求項26の再構築システム。
【請求項28】
該全細胞が、D1受容体をコードする第1異種性遺伝子および高血圧に関連のGRK4タンパク質をコードする第2異種性遺伝子でトランスフェクトされたチャイニーズハムスター卵巣細胞である、請求項27の再構築システム。
【請求項29】
該GRK4タンパク質が本態性高血圧症と関連している、請求項25の再構築システム。
【請求項30】
高血圧に関連するGRK4タンパク質と、抗高血圧活性について分析する物質との相互作用において概GRK4タンパク質の検出可能な立体配座的変化を提供する薬剤との複合体。
【請求項31】
不死化ヒト腎近位尿細管細胞。
【請求項32】
高血圧のヒトから得られる単離された精製腎近位尿細管細胞。
【請求項33】
不死化された、請求項32の高血圧のヒトから得られる単離された精製腎近位尿細管細胞。
【請求項34】
腎細胞中で発現されるGRK4タンパク質をコードし、該GRK4タンパク質を産生するトランスジーンを含有するニ倍性ゲノムを含むトランスジェニック動物(なお、該トランスジーンの発現によって、概トランスジェニック動物において発揮される本態性高血圧症の状態が、腎細胞が該GRK4タンパク質を発現しない正常血圧の動物と比較される)。
【請求項35】
該腎細胞が、該GRK4タンパク質を発現しない腎細胞の正常血圧の動物に比して、低いナトリウム拒否能力を持つ、請求項34のトランスジェニック動物。
【請求項36】
げっ歯類である、請求項34のトランスジェニック動物。
【請求項37】
マウスである、請求項34のトランスジェニック動物。
【請求項38】
推定の抗高血圧剤を同定する方法であって、
少なくとも1つの候補薬剤を請求項25の再構築システムに加え、
GRK4活性を検出する(該活性は推定の抗高血圧物質の指標である)、
ことを含む方法。
【請求項39】
GRK4活性を検出する該工程が、アデニレート・シクラーゼ活性を測定することを含む、請求項38の方法。
【請求項40】
GRK4活性を検出する該工程が、リン酸塩を付加し得る基質およびリン酸塩源を該培地に加えて該基質のリン酸化を測定することを含む、請求項38の方法。
【請求項41】
推定の抗高血圧剤を同定する方法であって、
少なくとも1つの候補薬剤を請求項30の複合体に接触せしめ、
該GRK4の立体配座的変化が起きたかどうかを検出する(該活性は推定の降圧物質の指標である)、
ことを含む方法。
【請求項42】
該検出が、分光測定、蛍光、核磁気共鳴、エバネセント波法または原子間力顕微鏡で行う、請求項41の方法。
【請求項43】
推定の抗高血圧剤を同定する方法であって、
少なくとも1つの候補薬剤を、高血圧動物から単離の、D1受容体およびGRK4を発現する不死化腎細胞に加え、
該細胞のドーパミン作動性シグナルの形質導入の変化を検出する(ドーパミン作動性シグナルの形質導入の変化は推定の抗高血活性の指標である)、
ことを含む方法。
【請求項44】
推定の抗高血圧剤を同定する方法であって、
該薬剤を投与した請求項34の第1トランスジェニック動物と、該薬剤を投与していない請求項34の第2トランスジェニック動物との電解質出力を比較する(推定の抗高血圧剤の同定は、該第1トランスジェニック動物の電解質出力が該第2トランスジェニック動物に比して増加することでなされる)、
ことを含む方法。
【請求項45】
ナトリウム利尿を増加する方法であって、本態性高血圧症の個体に、GRK4と相互作用する薬剤を投与して、該個体におけるナトリウム利尿を増加することを含む方法。
【請求項46】
該薬剤が、該高血圧個体の腎細胞におけるGRK4の発現を変化する、請求項45の方法。
【請求項47】
該薬剤が、GRK4のmRNAまたはDNAに結合するアンチセンスRNAを含む、請求項45の方法。
【請求項48】
該薬剤が、GRK4のmRNAまたはプレmRNAを開裂するリボチームを含む、請求項45の方法。
【請求項49】
該薬剤が、優性阻害変異DNA分子を含む、請求項45の方法。
【請求項50】
該薬剤がGRK4タンパク質に結合する、請求項45の方法。
【請求項51】
インビトロおよびインビボでGRK4のmRNAに特異的にハイブリダイズするオリゴヌクレオチド。
【請求項52】
アンチセンスRNA分子である、請求項51のオリゴヌクレオチド。
【請求項53】
優性阻害変異DNA分子である、請求項51のオリゴヌクレオチド。
【請求項54】
GRK4のmRNAまたはプレmRNAを開裂するリボチーム。
【図1】

【図2】
【図3】
【図4】
【図5】
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2011−36253(P2011−36253A)
【公開日】平成23年2月24日(2011.2.24)
【国際特許分類】
【外国語出願】
【出願番号】特願2010−199161(P2010−199161)
【出願日】平成22年9月6日(2010.9.6)
【分割の表示】特願2000−527662(P2000−527662)の分割
【原出願日】平成11年1月12日(1999.1.12)
【出願人】(500330636)
【出願人】(500330647)
【Fターム(参考)】
【公開日】平成23年2月24日(2011.2.24)
【国際特許分類】
【出願番号】特願2010−199161(P2010−199161)
【出願日】平成22年9月6日(2010.9.6)
【分割の表示】特願2000−527662(P2000−527662)の分割
【原出願日】平成11年1月12日(1999.1.12)
【出願人】(500330636)
【出願人】(500330647)
【Fターム(参考)】
[ Back to top ]