
果実軟化に関連するα−マンノシダーゼのポリヌクレオチド配列および果実貯蔵寿命を延長するためのその使用
本発明は、α-マンノシダーゼをコードする単離されたポリヌクレオチド配列を提供する。本発明は、センスの向きまたはアンチセンスの向きでα-マンノシダーゼをコードするポリヌクレオチド配列を含むDNA構築物、RNAi構築物、該構築物を含む組換えベクター、および本発明において開示される組換えベクターを含む宿主細胞をさらに提供する。本発明は、前記ポリヌクレオチドを発現して、α-マンノシダーゼタンパク質の蓄積が低下し、果実貯蔵寿命が延長した、トランスジェニックの植物、植物細胞、トランスジェニック子孫および種子をさらに提供する。
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、トマト(ソラナム リコペルシカム(Solanum lycopersicum))およびトウガラシ(カプシカム アンニューム(Capsicum annuum))のα-マンノシダーゼをコードするポリヌクレオチドに関する。
【背景技術】
【0002】
発明の背景
今日の世界的流通の世界において、果実成熟の制御は戦略的に重要である(Causier B, Kieffer M and Davies B, 2002, Science 296: 275-276(非特許文献1))。果実の貯蔵寿命および貯蔵を制限する主因のうちの一つは、過度の軟化である。これに関して、多数の酵素を抑制する試みは、貯蔵寿命の延長または軟化の所望の低下において限られた成功しか収めていない。成熟は、消費される官能特性を達成するための、果実の発達および代謝における主要な変遷を含む複雑な過程である。これらのうち、テクスチャは、嗜好性、消費者受容性、貯蔵寿命、輸送能力、および収穫後疾患/病原体に対する抵抗性に関する主要な品質特性であり、これらは全て直接コストに影響を与える。これらのテクスチャの変化が主として細胞壁構造の変化に起因することは、自明であると見なされている。細胞壁は、炭水化物と、多重遺伝子ファミリーによってコードされたタンパク質とから主になる動的な成分である。これらの細胞壁多糖は、自然界に見出される最も豊富な有機化合物であり、その構造的および機能的な多様性は、合成(糖転移酵素)、修飾(炭水化物エステラーゼ)、および分解(糖質加水分解酵素および多糖リアーゼ)に関与する一連の多数の酵素によって反映される。生物の遺伝子のおよそ1〜2%がこれらの過程に関与しており、このことは、果実成熟におけるそれらの重要性を明らかにしている(Henrissat B, Coutinho PM and Davies GJ, 2001, Plant Mol Biol 47: 55-72(非特許文献2)およびJamet E, Canut H, Boudart G and Pont-Lezica RF, 2006, Trends Plant Sci 11: 33-39(非特許文献3))。一般に、果実の堅さの低下には、多糖加水分解酵素/糖類加水分解酵素、トランスグリコシラーゼ、リアーゼ、およびエクスパンシンのような、多数の細胞壁分解酵素の発現の増加が伴う(Brummell DA, 2006, Funct Plant Biol 33: 103-119(非特許文献4))。一般的な触媒活性は配列から推論され得るが、これらのタンパク質の大部分の正確な酵素機能および生物学的役割は未知である。炭水化物代謝に関与する一式の酵素のうち、加水分解酵素は主要な部分を占める。これらの加水分解酵素は、細胞壁またはその他の細胞器官に見出されるN-糖タンパク質を標的とし、N-複合糖質を分解して、遊離N-グリカン含有量を増加させる。さらに、これらの遊離N-グリカンは、トマトにおいて成熟を刺激する生物学的活性を有することが公知である(Priem B, Gitti R, Bush CA and Gross KC, 1993 Plant Physiol 102: 445-458(非特許文献5))。
【先行技術文献】
【非特許文献】
【0003】
【非特許文献1】Causier B, Kieffer M and Davies B, 2002, Science 296: 275-276
【非特許文献2】Henrissat B, Coutinho PM and Davies GJ, 2001, Plant Mol Biol 47: 55-72
【非特許文献3】Jamet E, Canut H, Boudart G and Pont-Lezica RF, 2006, Trends Plant Sci 11: 33-39
【非特許文献4】Brummell DA, 2006, Funct Plant Biol 33: 103-119
【非特許文献5】Priem B, Gitti R, Bush CA and Gross KC, 1993 Plant Physiol 102: 445-458
【発明の概要】
【0004】
本発明は、トマト(ソラナム リコペルシカム)およびトウガラシ(カプシカム アンニューム)の果実軟化に関連する酵素α-マンノシダーゼの精製および特徴決定に関する。本発明は、トマトおよびトウガラシのα-マンノシダーゼ遺伝子の同定、単離、およびクローニングにも関する。本発明は、α-マンノシダーゼタンパク質の蓄積が低下し、果実貯蔵寿命が延長した、トランスジェニック植物にさらに関する。
【0005】
本発明の一つの局面は、マンノシダーゼ活性を有するポリペプチドをコードする単離されたポリヌクレオチドを提供し、このポリヌクレオチドのヌクレオチド配列は、SEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードする。
【0006】
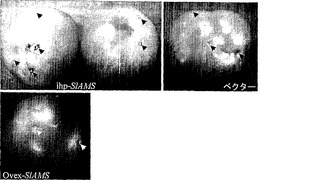
本発明の別の局面は、SEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列を有するポリペプチドを提供する。
【0007】
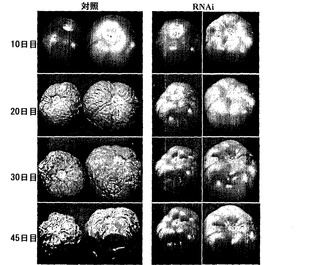
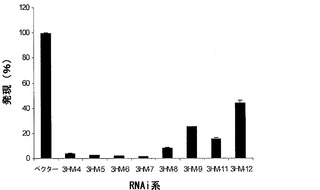
本発明のさらに別の局面は、SEQ ID NO:1またはSEQ ID NO:4に記載の配列に由来する少なくとも20個の連続ヌクレオチドを含むセンスポリヌクレオチド、および該センスポリヌクレオチド鎖とハイブリダイズするアンチセンスポリヌクレオチドを含み、このアンチセンスポリヌクレオチド鎖およびセンスポリヌクレオチド鎖が二重鎖を形成する、トランスジェニック植物においてマンノシダーゼの発現を抑制するためのRNAi構築物を提供する。
【0008】
本発明のさらに別の局面は、マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドを含み、このポリヌクレオチドのヌクレオチド配列がSEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードし、かつこのポリヌクレオチドがアンチセンスの向きである、組換えベクターによって、植物細胞、組織、またはそれらの一部を形質転換する工程を含む、植物における果実軟化を遅延させる方法を提供する。
【0009】
本発明のさらに別の局面は、マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドの少なくとも20個の連続ヌクレオチドの断片を少なくとも含むRNA干渉(RNAi)構築物の発現によって、トランスジェニック植物におけるマンノシダーゼのレベルを非トランスジェニック植物におけるマンノシダーゼのレベルと比較して減少させる工程であって、このポリヌクレオチドのヌクレオチド配列が、SEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードする、工程;および非トランスジェニック植物と比べて低下したマンノシダーゼのレベルについて、得られた植物をスクリーニングする工程を含む、植物における果実軟化を遅延させる方法を提供する。
【0010】
本発明のさらに別の局面は、SEQ ID NO:1またはSEQ ID NO:3に記載の配列に由来する少なくとも20個の連続ヌクレオチドを含むセンスポリヌクレオチド鎖、およびセンスポリヌクレオチド鎖にハイブリダイズするアンチセンスポリヌクレオチド鎖を含むRNA干渉(RNAi)構築物の発現によって、トランスジェニック植物におけるマンノシダーゼのレベルを非トランスジェニック植物におけるマンノシダーゼのレベルと比較して減少させる工程であって、このアンチセンスポリヌクレオチド鎖およびセンスポリヌクレオチド鎖が二重鎖を形成する、工程;ならびに非トランスジェニック植物と比べて低下したマンノシダーゼのレベルについて、得られた植物をスクリーニングする工程を含む、植物における果実軟化を遅延させる方法を提供する。
【0011】
本発明のさらに別の局面は、マンノシダーゼの発現が果実軟化を遅延させるために制御されている、マンノシダーゼをコードするポリヌクレオチドを含むトランスジェニック植物、その種子および子孫を提供する。
【0012】
本発明において開示されるトランスジェニック植物には、トマト、トウガラシ、パパイア、マンゴー、バナナ、モモ、セイヨウナシ、柑橘類、パイナップル、グアバ、アボカド、イチゴ、リンゴ、およびザクロが包含される。
【0013】
上記のおよびその他の主題の特色、局面、および利点は、以下の説明、添付の図面、および添付の特許請求の範囲に関してよりよく理解されるであろう。
【図面の簡単な説明】
【0014】
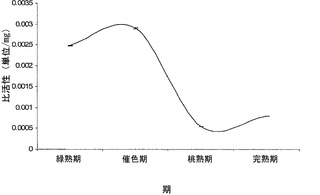
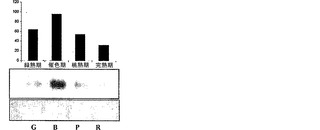
【図1a】異なる成熟期におけるトマト果実中のα-マンノシダーゼの比活性を示すグラフ。
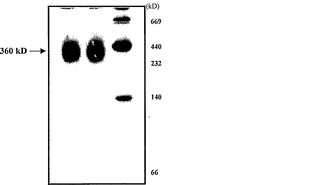
【図1b】6%非変性ゲル上で分離された、精製されたα-マンノシダーゼ。
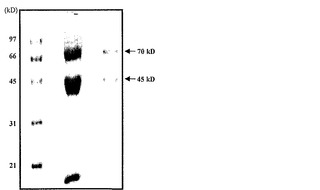
【図1c】12.5%SDS-PAGEで変性させられ分離された、精製されたタンパク質。
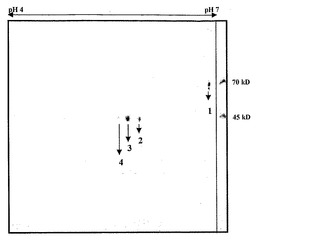
【図1d】一次元目に4-7 IPGストリップ上でフォーカシングされ、二次元目に12.5%SDS-PAGE上で分解されたα-マンノシダーゼタンパク質。
【図1e】未消化(PP)と比較してシフトを示す、α-マンノシダーゼ特異抗体によって検出された、精製されたタンパク質のEndoH消化(PP+EndoH)。
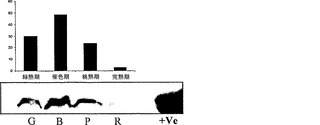
【図1f】異なる成熟期(G-緑熟期、B-催色期、P-桃熟期、およびR-完熟期)におけるα-マンノシダーゼのイムノブロット分析。
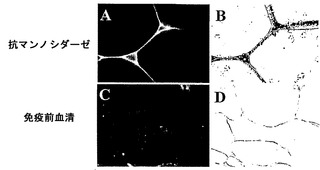
【図1g】トマト果実におけるα-マンノシダーゼの細胞内局在。
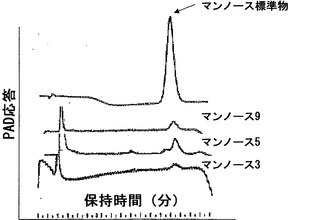
【図1h】α-マンノシダーゼと共にインキュベートした際の三つのN-結合型オリゴ糖のHPAEクロマトグラム。
【図2a】ノーザンブロット分析を使用した、トマトにおける異なる成熟期におけるα-マンノシダーゼ遺伝子の発現。
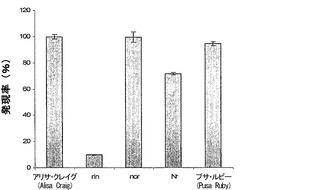
【図2b】野生型と比較した、成熟不全変異体rin、nor、およびNrにおけるα-マンノシダーゼの相対的な発現を示すRT-PCR分析。
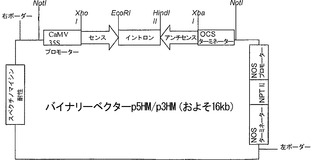
【図2c】組換えベクターp5HM/p3HMの模式図。
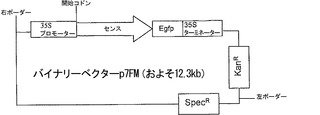
【図2d】組換えベクターp7FMの模式図。
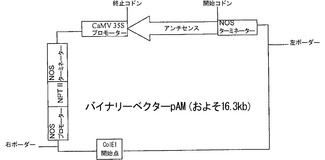
【図2e】組換えベクターpAMの模式図。
【図3a】サイレンシングベクター、ブランクバイナリーベクター、および過剰発現ベクターによるアグロインジェクションを受けたトマト果実。
【図3b】アグロインジェクションを受けた代表的なトマト果実を示す微速度撮影写真。
【図3c】マンノシダーゼ転写物レベルの低下を決定するためのRT-PCRによるRNAiトランスジェニック系の発現分析。データは平均値±SE(n=3)である。
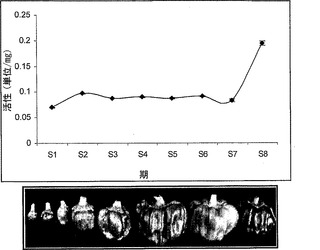
【図4a】トウガラシにおける期別のα-マンノシダーゼの比活性。
【図4b】6%非変性ゲル上で分離された、精製されたα-マンノシダーゼ。
【図4c】12.5%SDS-PAGEで変性させられ分離された、精製されたタンパク質。
【図4d】一次元目に4-7 IPGストリップ上でフォーカシングされ、二次元目に12.5%SDS-PAGE上で分解されたα-マンノシダーゼタンパク質。
【発明を実施するための形態】
【0015】
発明の詳細な説明
本発明は、トマトおよびトウガラシの果実軟化に関連するα-マンノシダーゼに関する。本発明は、特に、α-マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチド配列に関する。ここで、植物におけるRNAiおよびアンチセンスにより媒介されるα-マンノシダーゼのサイレンシングは果実軟化を遅延させ、過剰発現は果実軟化を増強する。
【0016】
本発明は、α-マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドを含むDNA構築物、組換えベクター、および組換え宿主細胞を提供する。本発明は、さらに、本発明において開示されるα-マンノシダーゼのポリヌクレオチドを使用して、植物における果実軟化を遅延させる方法を提供する。本発明は、さらに、本発明において開示されるポリヌクレオチドを使用した、植物における早期の果実軟化のための方法を提供する。さらに、本発明は、果実軟化に影響を与えるためにα-マンノシダーゼの発現が制御されているトランスジェニック植物、それらの子孫および種子を提供する。
【0017】
アンチセンス技術およびRNAi技術を使用してβ-D-N-アセチルマンノシダーゼ遺伝子の発現を抑制することにより得られたトランスジェニック植物は、非トランスジェニック果実より約1.5〜2.5倍堅い果実を生じた。さらに、RNAi技術によって作製されたトランスジェニック果実は、非トランスジェニック果実よりほぼ30日長く、収穫後45日目まで軟化の兆候を示さなかった。アンチセンス技術によって作製されたトランスジェニック果実は、非トランスジェニック果実よりほぼ20日長く、収穫後35日目まで軟化の兆候を示さなかった。これらの結果は、驚くべき予想外のものである。トランスジェニック植物における遺伝子のさらなる抑制は、栄養生長、開花および果実発達、成熟までの日数、および収量に対しては負の効果を及ぼさなかった。
【0018】
本発明は、植物における炭水化物代謝およびN-グリカンプロセシングを担うグリコシル加水分解酵素ファミリー遺伝子であるα-マンノシダーゼを、RNA干渉(RNAi)技術によって抑制することにより、トマトの貯蔵寿命を延長する方法を記載する。本発明は、さらに、トマト果皮由来のα-マンノシダーゼタンパク質複合体の精製および特徴決定を記載する。
【0019】
2DEフォーカシングされたタンパク質複合体のLC MS/MS分析は、相互作用パートナーの同定に役立ち、α-マンノシダーゼ遺伝子のクローニングにも役立った。α-マンノシダーゼ遺伝子の発現は、催色期に最も高く、成熟/軟化の開始と相関していた。トランスジェニック果実の分子的分析によって、α-マンノシダーゼ転写物が、99%特異的に分解され、30日延長された貯蔵寿命を有する、2.5倍堅い果実を与えることが示された。研究の過程で、本発明者らは、マンノシダーゼが、植物に存在するテルペノイドであるゲラニオールによって誘導され、rinによって調節されることを見出した。
【0020】
pNP-アルファ-D-マンノピリノシド(mannopyrinoside)を基質として使用して、本発明者らは、マンノシダーゼの活性が催色期に最大となり(図1a)、緑熟期、桃熟期、および完熟期がそれに続くことを見出した。呼吸またはエチレン生成、カロテノイドの蓄積、および軟化のような過程の開始を特徴付ける催色期におけるα-マンノシダーゼの蓄積は、それが成熟および/または軟化に関与しているという本発明者らの仮説を強化した。
【0021】
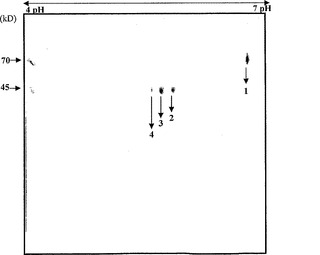
トマトα-マンノシダーゼを催色期トマトの果皮から精製し、さらに特徴決定した。トマト果皮を液体窒素中で破砕し、抽出緩衝液(0.25 NaCl、1mM PMSFを含む100mMトリス-Cl、pH7.8)に一夜懸濁させた。タンパク質を均質になるまで精製するため、硫酸アンモニウム沈殿(0〜70%)、イオン交換(DEAEセファロース)、およびゲルろ過クロマトグラフィ(Sephadex G-100)のような方法を利用した。精製された酵素は、6%非変性PAGE上で360kDの単一のタンパク質を構成し(図1b)、それはα-マンノシダーゼ活性-バンド相関によって確認された。しかしながら、これに反して、superdex 200分析用カラムを使用したゲルろ過によって決定されたタンパク質の分子量は、290kDであった。さらに、SDS-PAGE上で分離すると、タンパク質は70kDおよび45kDの二つのサブユニットに分解され、従って、それがヘテロ二量体であることが明らかとなった(図1c)。明瞭な画像を入手するため、タンパク質複合体を二次元ゲル電気泳動(2DE)上で分離したところ、70kDサブユニットは単一のタンパク質であり、45kDサブユニットは、異なるpIを有する三つのタンパク質へとさらに分解されることが明らかになった(図1d)。本発明者らはLC/MS-MS分析によってスポットを同定し、それらが各々異なるタンパク質であることを見出した。70kDスポットはα-マンノシダーゼ(スポット番号1、pI6.8)を表し、三つのタンパク質へ分けられる45kDのスポットは、推定単糖トランスポーター(スポット番号2、pI6.4)、遺伝子の一部であるゲラニオール応答因子(スポット番号3、pI6.3)、およびH+ ATPアーゼ(スポット番号4、pI6.2)を有していた。従って、本発明者らは、45kD複合体に見出された三つのタンパク質のうちの二つは相互作用タンパク質であるかもしれないと予測した。
【0022】
生化学的特徴決定は、酵素が、60℃まで、活性の考慮すべき低下なしに1時間安定していることを明らかにした。酵素にとって至適の温度およびpHは、それぞれ45℃およびpH6.0であった。p-ニトロフェノール-アルファ-D-マンノピラノシドに対して決定された酵素のKmは、4.6mMである。トマトα-マンノシダーゼの性質を同定するため、それをSDS-PAGE上で分解し、PAS染色に供したところ、タンパク質は、糖タンパク質染色陽性であった。上記の結果をさらに確認するため、タンパク質のEndoH消化を実施したところ、炭水化物/グリカン部分の切断による2〜3kDのシフトが示され、従って、上記の結果が立証された(図1e)。特定の成熟期における比活性をタンパク質蓄積と相関させるため、α-マンノシダーゼに相同なタンパク質の70kDサブユニットに対して産生されたポリクローナル抗体を使用して、全ての期のイムノブロット分析を行った。イムノブロット分析は、タンパク質蓄積が催色期に最大となり、緑熟期、桃熟期、および完熟期がそれに続くことを明らかにした(図1f)。さらに、細胞内の分布を決定するため、免疫局在性アッセイを行ったところ、細胞壁の最も内側の縁にシグナルが見られたことから、それが細胞壁タンパク質であることが明らかになった(図1g)。本発明者らの所見は、分泌のためのシグナルペプチドを有する細胞壁タンパク質に、α-マンノシダーゼホモログ(Q8LPJ3)を分類した、アラビドプシス(Arabidopsis)における報告によってさらに補強された。N-グリカンおよびそれらのプロセシングに対するα-マンノシダーゼの役割を判定するため、本発明者らは、果実果皮中に一般的に見出される、三つのN結合型オリゴ糖(オリゴマンノース-3、オリゴマンノース-5、およびオリゴマンノース-9)を標的とした。N-グリカンに対する作用を確証するため、α-マンノシダーゼを、37℃で一夜、N結合型オリゴ糖と共にインキュベートし、反応混合物をCarbopac PA-1カラム上で分解した。酵素は、全てのN結合型オリゴ糖から末端マンノース残基を切断することができ、従って、それが成熟/軟化に関与していることの間接的な証拠が得られた(図1h)
【0023】
ペプチドQHVADDYAKおよびSGAYVFRPに対応する縮重プライマーを使用して、cDNAを増幅した。このプライマーのオリゴヌクレオチド配列は、SEQ ID NO:5およびSEQ ID NO:6に記載されている。プライマーは、LC-MS/MSデータ、および多重アラインメントによって同定されたモチーフを使用して設計された。5'末および3'末の欠けている部分を、cDNA末端のランダム増幅(Random Amplification of cDNA Ends)(RACE)によって増幅し、全長にした。配列のインシリコ分析は、コーディング領域が3090bp長(SEQ ID NO:1)であることを示し、推定タンパク質は、114KDaの質量計算値および6.48のpI理論値を有する1029アミノ酸長(SEQ ID NO:2)であった。トマトα-マンノシダーゼタンパク質は、アラビドプシスおよびイネのα-マンノシダーゼタンパク質との、それぞれ66%および62%の同一性を示した。その進化的局面を洞察するため、系統学的分析を実施したところ、α-マンノシダーゼは、三つの公知のアラビドプシスα-マンノシダーゼタンパク質のうちの二つと最も近縁であることが示された。発現パターンを確証し、先の結果を実証するため、ノーザンブロット分析を実施したところ、マンノシダーゼ転写物が催色期に最も豊富であり、緑熟期、桃熟期、および完熟期がそれに続くことが明らかになった(図2a)。
左プライマー:

右プライマー:
ここで、KはGまたはTであり、RはAまたはGであり、MはAまたはCである。
【0024】
α-マンノシダーゼの発現を、トマト変異体rin(ripening inhibitor)、nor、およびNrにおいて分析した。その分析は、α-マンノシダーゼが、変異体rinにおいては90%、Nrにおいては30%阻害されており、nor変異体は野生型と比較可能であることを明らかにした(図2b)。果実成熟の発達調節のために必要とされるMADSボックス転写因子をコードする遺伝子に変異を保持している変異体のさらなる分析を実施した。本発明者らは、rin変異体および野生型において、マンノシダーゼを誘導するため、エチレンの前駆物質であるACCを使用した。ACCは、rinにおいても野生型においても対照と比較してα-マンノシダーゼをアップレギュレートすることができた。これらの観察は、α-マンノシダーゼがエチレンによってもrinによっても独立にまたは相乗的に調節されることを示した。
【0025】
多くの観察に支持されて、成熟および/またはそれに関連した軟化における役割を証明するため、サイレンシングによって、α-マンノシダーゼを機能的に特徴決定することが不可避となった。アグロインジェクションを使用したエンドジーン(Endogene)抑制は、遺伝子機能を分析するための便利なツールになっている(Orzaez DF, Mirabel S, Wieland WH and Granell A, 2006, Plant Physiol 140: 3-11)。このdsRNAにより媒介されるサイレンシング技術を使用して、本発明者らは、pHANNIBALベクターを使用してα-マンノシダーゼを一過性にサイレンシングした(Wesley SV, Helliwell CA, Smith NA, Wang MB, Rouse DT, Liu Q, Gooding PS, Singh SP, Abbott D, Stoutjesdijk PA, Robinson SP, Gleave AP, Green AG and Waterhouse PM, 2001, Plant J 27: 581-590)。その発現カセットは、発現時にdsRNAへと組み立てられるよう設計された、CaMV 35Sプロモーターの制御下にある、イントロンによって隔てられた、センスの向きおよびアンチセンスの向きにサブクローニングされたそれぞれのUTR領域を含む5'領域または3'領域のいずれかに由来する遺伝子の600bp断片からなっていた。RNAiカセットの構築において使用されたプライマーのオリゴヌクレオチド配列は、SEQ ID NO:7、SEQ ID NO:8、SEQ ID NO:9、SEQ ID NO:10、SEQ ID NO:11、SEQ ID NO:12、SEQ ID NO:13、およびSEQ ID NO:14に記載されている。このカセットを、さらに、バイナリーベクターpART 27へサブクローニングし、そのバイナリーベクターをp5HM/p3HMと名付けた(図2c)。各果実に、サイズに依って、花柱の先端の近くの表面上の2〜3箇所のスポットに注入した(図3a)。1週間後、RNAiによるアグロインジェクションを受けた果実は、対照(ブランクバイナリーベクター)のものと比較して、果実の周囲の最大面積をカバーする、顕著な緑色のセクターを注入スポットの周囲に発達させた(図3a)。分子レベルで抑制を確認し定量化するため、注入の2〜4日後に採取した果実を、リアルタイムRT-PCRによって、α-マンノシダーゼに対応するmRNAレベルの低下を決定するために使用した。オリゴヌクレオチドプライマーは、light cycler probe design 2プログラム(Roche)を使用して設計した。α-マンノシダーゼの増幅において使用されたプライマーのオリゴヌクレオチド配列は、SEQ ID NO:15およびSEQ ID NO:16に記載されている。内因性対照遺伝子アクチンの増幅のために使用されたプライマーは、SEQ ID NO:17およびSEQ ID NO:18に記載する。
【0026】
結果は、緑熟期の2日前に注入された場合には、mRNAレベルが対照と比べて70%低下し、緑熟期に注入された場合には、50%低下することを明らかにした。結果をバリデートするため、pK7FWG 2.0において調製され、p7FMと名付けられたα-マンノシダーゼの過剰発現構築物(図2d)の、果実へのアグロインジェクションを行ったところ、その果実は、初期の間は、対照に類似した挙動を示した。過剰発現の効果は後期に見られ、果実は対照(ブランクベクターおよび未注入)よりはるかに急速に劣化し、テクスチャを喪失した。効果を見るため、アンチセンス構築物もpBI121において調製し、pAMと名付けた(図2e)。アンチセンスを注入された果実は、過剰発現果実とサイレンシングを受けた果実との中間型であった。このmRNAレベルの低下のため、本発明者らは、21〜23残基の低分子量RNA(siRNA)の存在を調査することとした。この目的のため、本発明者らは、対照と共に、RNAiの、緑熟期および催色期の果実から可溶性の低分子RNAを抽出し、ノーザンブロット分析を実施した。本発明者らは、RNAiにより媒介されるサイレンシングの最大の特徴であるα-マンノシダーゼに特異的な低分子干渉(si)RNAを果実果皮において観察した。さらに、本発明者らの結果は、非ウイルスベクターが、ウイルスベクターと同一の効率および程度でサイレンシングを達成し得ることも証明している。
【0027】
アグロインジェクションの30日後、アグロインジェクションを受けた果実のテクスチャの堅さを定量化することを試みて、TA-XT2テクスチャアナライザーを使用した圧縮分析を実施した。成熟は均一の過程ではないため、果実の異なる面で5mmまで果実を圧縮し(2〜3箇所の圧縮/果実)、花柱の付根をテクスチャアナライザーのプレートに触れさせて単一の10mmの圧縮を行うことにより、果実の堅さを測定した。全ての例において、RNAiアグロインジェクションを受けた果実は、統計的に堅く、5mmおよび10mmの圧縮において、それぞれ2倍および1.6倍、カウンターパートより堅かった。さらに、アグロインジェクションを受けた果実の微速度撮影写真は、RNAi注入果実は、45日目までテクスチャおよび堅さを保持するのに対し、対照は15日後にテクスチャの縮みおよび緩みを開始することを明らかにした(図3b)。これらの有望な結果より、本発明者らは、同ベクターを使用して安定的なRNAi系を産生することにした。トマト子葉のアグロバクテリウムにより媒介される形質転換は、イントロンによって隔てられた遺伝子のセンス断片およびアンチセンス断片を発現するRNAi系をもたらした。子葉を、サイレンシングベクターによって形質転換されたアグロバクテリウム株EHA105と共培養し、カナマイシンプレート上で選択した。この研究において使用された株の他に、LBA 4404およびGV 3101のような株も同一の効果を得るために利用され得る。苗条が出現するまで、外植片を12〜15日毎に継代培養した。苗条が1〜2cm長になった時、それらを切断し、発根培地において生長させた。サイレンシングを受けたと推定される植物(T0)を、リアルタイムRT-PCRによって転写物レベルの低下について調査したところ、α-マンノシダーゼが催色期に99%抑制されることが明らかになった(図3c)。次いで、桃熟期に果実を採取し、テクスチャ分析のため室温で貯蔵した。アンチセンス技術によって作製されたトランスジェニック果実は、非トランスジェニック果実と比較して、マンノシダーゼの80%の抑制および1.5倍の堅さを示した。30日後、生長、発達、および種子生成に対する負の効果なしに、RNAi果実が対照より2.5倍堅いことが観察された。アンチセンス技術によって作製されたトランスジェニック果実は、20日の貯蔵寿命の延長を示した。
【0028】
トウガラシは非クライマクテリック型果実であり、果実の発達および成熟を8期に分割した。第8期から成熟果実を採取し、タンパク質抽出をトリス-Cl pH7において実施した。トウガラシの発達および成熟の全ての異なる期において活性アッセイを実施したところ、最大活性は第8期果実に見出された(図4a)。トウガラシマンノシダーゼを、イオン交換およびゲルろ過のような種々のクロマトグラフィ技術を利用することにより精製した。活性を含有している溶出画分を、濃縮し、ゲル上で分解した。精製されたタンパク質は、6%非変性ゲル上のおよそ500kDの単一のバンドであり(図4b)、SDS-PAGEでは、70kDおよび50kDの二つのサブユニットが分解された(図4c)。精製されたタンパク質の2DEエレクトロフォーカシングによって、それが複合体であることが明らかになり、相互作用タンパク質であることが暗示された。より低い50kDのバンドは、トマトマンノシダーゼに極めて類似した、異なるpIを有する三つのタンパク質に分離された(図4d)。免疫局在性から、それが細胞壁に局在していることが明らかになった。トウガラシα-マンノシダーゼはトマトα-マンノシダーゼと90%同一であった。トウガラシ由来のα-マンノシダーゼの分子クローニングは、コーディング領域が3093bp長(SEQ ID NO:3)であることを明らかにした。トウガラシ由来の推定タンパク質は1030アミノ酸長(SEQ ID NO:4)であった。トウガラシにおけるアグロインジェクションによるα-マンノシダーゼの一過性のサイレンシングは、それが成熟に関連した軟化に関与していることを明らかにした。
【0029】
ゲラニオールは、様々な癌細胞系に対するインビトロおよびインビボの抗腫瘍活性を有する、非環式の食事性モノテルペンであって、従って、癌化学防御のための薬剤の新たなクラスと見なされている。ゲラニオールは、カンジダ アルビカンス(Candida albicans)およびサッカロミセス セレビシエ(Saccharomyces cervisiae)の膜機能に干渉することも報告された。ゲラニオールは水に不溶性であるため、実生に供給するため、20%DMSOを使用した(Hendry GAF and Jones OTG, 1984, Induction of cytochrome p-450 in intact mung beans. New Phytol 96: 153-159)。本発明者らは、ゲラニオール処理の30分後、マンノシダーゼ遺伝子発現が、対照(DMSOのみ)と比較して3倍アップレギュレートされたことを観察した。このことから、本発明者らは、多面的なrin変異体において、α-マンノシダーゼ誘導に対するゲラニオールの役割をさらに分析することにした。rin実生に10%ゲラニオールを供給し、試料を収集した。相対遺伝子発現分析は、α-マンノシダーゼ転写物が、30分後、6倍アップレギュレートされることを明らかにし、先の結果を実証した。
【0030】
本発明の一つの態様は、α-マンノシダーゼ活性を有するポリペプチドをコードする、単離されたポリヌクレオチドを提供する。該ポリヌクレオチドのヌクレオチド配列は、
(a)SEQ ID NO:2に記載のアミノ酸配列との90%の類似性を有するポリペプチドをコードするヌクレオチド配列;
(b)SEQ ID NO:4に記載のアミノ酸配列との90%の同一性を有するポリペプチドをコードするヌクレオチド配列;
(c)(a)または(b)のヌクレオチド配列に相補的なヌクレオチド配列
(d)SEQ ID NO:1に記載のヌクレオチド配列;
(e)SEQ ID NO:3に記載のヌクレオチド配列;
(f)(d)または(e)のヌクレオチド配列に相補的なヌクレオチド配列
からなる群より選択される。
【0031】
本発明の別の態様は、ヌクレオチド配列がSEQ ID NO:1に記載の、トマトから単離される、本発明において開示されるα-マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドを提供する。
【0032】
本発明の別の態様は、ヌクレオチド配列がSEQ ID NO:3に記載の、トウガラシから単離される、本発明において開示されるα-マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドを提供する。
【0033】
本発明の一つの態様は、SEQ ID NO:1およびSEQ ID NO:3に記載のヌクレオチド配列を有する、単離されたポリヌクレオチドを提供する。
【0034】
本発明のさらなる態様は、SEQ ID NO:2およびSEQ ID NO:4に記載のアミノ酸配列を有する、α-マンノシダーゼ活性を有するポリペプチドを提供する。
【0035】
本発明のさらに別の態様は、本発明において開示されるα-マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドを含み、このポリヌクレオチド配列がプロモーター配列と機能的に連結されている、DNA構築物を提供する。
【0036】
本発明のさらなる態様は、イントロンによって隔てられたポリヌクレオチド配列の部分が、センスの向きおよびアンチセンスの向きである、本発明のDNA構築物を提供する。
【0037】
本発明のさらに別の態様は、α-マンノシダーゼ活性を有するポリペプチドをコードするSEQ ID NO:1に記載のヌクレオチド配列を有するポリヌクレオチドの部分を含み、イントロンによって隔てられたこのポリヌクレオチド配列が、センスの向きおよびアンチセンスの向きである、DNA構築物を提供する。
【0038】
本発明のさらに別の態様は、α-マンノシダーゼ活性を有するポリペプチドをコードするSEQ ID NO:3に記載のヌクレオチド配列を有するポリヌクレオチドの部分を含み、イントロンによって隔てられたこのポリヌクレオチド配列が、センスの向きおよびアンチセンスの向きである、DNA構築物を提供する。
【0039】
本発明は、本発明において開示されるDNA構築物を含む組換えベクターも提供する。
【0040】
本発明は、α-マンノシダーゼ活性を有するポリペプチドをコードするSEQ ID NO:1に記載のヌクレオチド配列を有するポリヌクレオチドの部分を含み、イントロンによって隔てられたこのポリヌクレオチド配列が、センスの向きおよびアンチセンスの向きである、組換えベクターをさらに提供する。
【0041】
本発明は、α-マンノシダーゼ活性を有するポリペプチドをコードするSEQ ID NO:3に記載のヌクレオチド配列を有するポリヌクレオチドの部分を含み、イントロンによって隔てられたこのポリヌクレオチド配列が、センスの向きおよびアンチセンスの向きである、組換えベクターをさらに提供する。
【0042】
一つの態様において、本発明は、本発明において開示されるベクターを含む組換え宿主細胞を提供する。
【0043】
一つの態様において、本発明は、α-マンノシダーゼ活性を有するポリペプチドをコードするSEQ ID NO:1に記載のヌクレオチド配列を有するポリヌクレオチドの部分を含み、イントロンによって隔てられたこのポリヌクレオチド配列が、センスの向きおよびアンチセンスの向きである、組換え宿主細胞を提供する。
【0044】
一つの態様において、本発明は、マンノシダーゼ活性を有するポリペプチドをコードするSEQ ID NO:3に記載のヌクレオチド配列を有するポリヌクレオチドの部分を含み、イントロンによって隔てられたこのポリヌクレオチド配列が、センスの向きおよびアンチセンスの向きである、組換え宿主細胞を提供する。
【0045】
別の態様において、本発明は、アグロバクテリウム、大腸菌、および酵母からなる群より選択される宿主細胞に関する。
【0046】
別の態様において、本発明は、植物細胞、組織、またはそれらの一部を本発明の組換えベクターによって形質転換する工程を含み、該ベクターが、イントロンによって隔てられた、センスの向きおよびアンチセンスの向きであるα-マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドの部分を含む、植物において果実軟化を遅延させる方法を提供する。
【0047】
別の態様において、本発明は、植物細胞、組織、またはそれらの一部を本発明の組換えベクターによって形質転換する工程を含み、該ベクターが、アンチセンスの向きであるα-マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドを含む、植物において果実軟化を遅延させる方法を提供する。
【0048】
別の態様において、本発明は、植物細胞、組織、またはそれらの一部を本発明の組換えベクターによって形質転換する工程を含み、該ベクターが、センスの向きであるα-マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドを含む、植物における早期果実軟化のための方法を提供する。
【0049】
別の態様において、本発明は、植物細胞、組織、またはそれらの一部を本発明の組換えベクターによって形質転換する工程を含み、該ベクターが、アンチセンスの向きであるα-マンノシダーゼ活性を有するポリペプチドをコードするSEQ ID NO:1に記載のヌクレオチド配列を有するポリヌクレオチドを含む、植物において果実軟化を遅延させる方法を提供する。
【0050】
別の態様において、本発明は、植物細胞、組織、またはそれらの一部を本発明の組換えベクターによって形質転換する工程を含み、該ベクターが、センスの向きであるα-マンノシダーゼ活性を有するポリペプチドをコードするSEQ ID NO:3に記載のヌクレオチド配列を有するポリヌクレオチドを含む、植物における早期果実軟化のための方法を提供する。
【0051】
本発明において提供される植物における果実軟化の遅延または早期果実軟化のための方法において、形質転換は、アグロバクテリウムにより媒介される形質転換、遺伝子銃、電気穿孔、およびインプランタ(in planta)形質転換のような、当技術分野において公知の方法によって実施され得る。
【0052】
一つの態様において、本発明は、α-マンノシダーゼの発現が果実軟化を引き起こし最終的には果実成熟をもたらすよう制御されている、本発明において開示される方法によって作製されたトランスジェニック植物を提供し、該植物は、トマト、トウガラシ、マンゴー、バナナ、パパイア、柑橘類、グアバ、アボカド、ブドウ、パイナップル、イチゴ、リンゴ、ザクロ、およびその他の果実収穫植物からなる群より選択される。
【0053】
本発明において開示される方法によって作製されるトランスジェニック植物は、トマトである。
【0054】
本発明は、本発明において開示されるトランスジェニック植物の種子または子孫も提供する。
【0055】
本発明の一つの態様において、マンノシダーゼ活性を有するポリペプチドをコードする単離されたポリヌクレオチドが提供され、該ポリヌクレオチドのヌクレオチド配列は、SEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約88%の同一性を有するポリペプチドをコードする。
【0056】
本発明の別の態様において、マンノシダーゼ活性を有するポリペプチドをコードする単離されたポリヌクレオチドが提供され、該ポリヌクレオチドのヌクレオチド配列は、SEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約90%の同一性を有するポリペプチドをコードする。
【0057】
本発明の別の態様は、マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドに由来する少なくとも20個の連続ヌクレオチドを含み、該ポリヌクレオチドのヌクレオチド配列は、SEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードし、これらのヌクレオチドがヘアピン構造のみを形成し全域において二次的なループを形成しないよう選択されている、RNAi構築物を提供する。
【0058】
本発明の別の態様は、マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドに由来する少なくとも20個の連続ヌクレオチドを含み、該ポリヌクレオチドのヌクレオチド配列は、SEQ ID NO:1またはSEQ ID NO:3に記載のものであり、これらのヌクレオチドがヘアピン構造のみを形成し全域において二次的なループを形成しないよう選択されている、RNAi構築物を提供する。
【0059】
本発明において開示されるRNAi構築物は、植物において転写後にヘアピン構造を形成する。
【0060】
本発明において開示されるRNAi構築物は、ヘアピン核酸である。
【0061】
本発明において開示されるRNAi構築物は、植物において転写後にヘアピン構造を形成し、二次的なループ構造を形成しない。
【0062】
別の態様は、植物において転写後にヘアピン構造を形成する、本発明において開示されるポリヌクレオチド配列を含むRNAi構築物を提供する。
【0063】
本発明のさらに別の局面は、マンノシダーゼ活性を有するポリペプチドをコードする単離されたポリヌクレオチドを提供し、このポリヌクレオチドのヌクレオチド配列は、SEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードする。
【0064】
本発明のさらに別の局面は、マンノシダーゼ活性を有するポリペプチドをコードする単離されたポリヌクレオチドを提供し、このポリヌクレオチドのヌクレオチド配列は、SEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列を有するポリペプチドをコードする。
【0065】
本発明のさらに別の局面は、マンノシダーゼ活性を有するポリペプチドをコードする単離されたポリヌクレオチドを提供し、このポリヌクレオチドのヌクレオチド配列は、SEQ ID NO:1またはSEQ ID NO:3に記載されている。
【0066】
本発明のさらに別の態様は、SEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列を有するポリペプチドを提供する。
【0067】
本発明のさらに別の局面は、マンノシダーゼ活性を有するポリペプチドをコードする単離されたポリヌクレオチドを含み、このポリヌクレオチドのヌクレオチド配列がSEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードし、かつこのポリヌクレオチドがプロモーター配列に機能的に連結されている、DNA構築物を提供する。
【0068】
本発明のさらに別の局面は、マンノシダーゼ活性を有するポリペプチドをコードする単離されたポリヌクレオチドを含み、このポリヌクレオチドのヌクレオチド配列がSEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列を有するポリペプチドをコードし、かつこのポリヌクレオチドがプロモーター配列に機能的に連結されている、DNA構築物を提供する。
【0069】
本発明のさらに別の局面は、マンノシダーゼ活性を有するポリペプチドをコードする単離されたポリヌクレオチドを含み、このポリヌクレオチドのヌクレオチド配列がSEQ ID NO:1またはSEQ ID NO:3に記載されており、かつこのポリヌクレオチドがプロモーター配列に機能的に連結されている、DNA構築物を提供する。
【0070】
本発明のさらに別の局面は、マンノシダーゼ活性を有するポリペプチドをコードする単離されたポリヌクレオチドを含み、このポリヌクレオチドのヌクレオチド配列がSEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードし、このポリヌクレオチドがプロモーター配列に機能的に連結されており、かつこのポリヌクレオチド配列がセンスの向きである、DNA構築物を提供する。
【0071】
本発明のさらに別の局面は、マンノシダーゼ活性を有するポリペプチドをコードする単離されたポリヌクレオチドを含み、このポリヌクレオチドのヌクレオチド配列がSEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードし、このポリヌクレオチドがプロモーター配列に機能的に連結されており、かつこのポリヌクレオチド配列がアンチセンスの向きである、DNA構築物を提供する。
【0072】
本発明の別の態様は、SEQ ID NO:1またはSEQ ID NO:4に記載の配列に由来する少なくとも20個の連続ヌクレオチドを含むセンスポリヌクレオチド鎖、および該センスポリヌクレオチド鎖にハイブリダイズするアンチセンスポリヌクレオチド鎖を含み、このアンチセンスポリヌクレオチド鎖およびセンスポリヌクレオチド鎖が二重鎖を形成する、トランスジェニック植物においてマンノシダーゼの発現を抑制するためのRNAi構築物を提供する。
【0073】
本発明において開示されるRNAi構築物において、RNAi構築物はヘアピン核酸である。
【0074】
本発明において開示されるRNAi構築物において、センス鎖は100〜600個のヌクレオチドを含む。
【0075】
本発明の別の態様は、本発明において開示されるDNA構築物を含む組換えベクターを提供する。
【0076】
本発明の別の態様は、アグロバクテリウム(Agrobacterium)、大腸菌(E.coli)、および酵母からなる群より選択される組換え宿主細胞を提供する。
【0077】
本発明のさらに別の態様は、マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドを含む組換えベクターによって、植物細胞、組織、またはそれらの一部を形質転換する工程を含み、該ポリヌクレオチドのヌクレオチド配列がSEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードし、かつこのポリヌクレオチドがアンチセンスの向きである、植物において果実軟化を遅延させる方法を提供する。
【0078】
本発明のさらに別の態様は、マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドを含む組換えベクターによって、植物細胞、組織、またはそれらの一部を形質転換する工程を含み、該ポリヌクレオチドのヌクレオチド配列がSEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードし、このポリヌクレオチドがアンチセンスの向きであり、かつこのポリヌクレオチドのヌクレオチド配列がSEQ ID NO:1またはSEQ ID NO:3に記載されている、植物において果実軟化を遅延させる方法を提供する。
【0079】
本発明のさらに別の態様は、マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドの少なくとも20個の連続ヌクレオチドの断片を少なくとも含み、このポリヌクレオチドのヌクレオチド配列がSEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードする、RNA干渉(RNAi)構築物の発現によって、トランスジェニック植物におけるマンノシダーゼのレベルを非トランスジェニック植物におけるマンノシダーゼのレベルと比較して減少させる工程;および非トランスジェニック植物と比べて低下したマンノシダーゼのレベルについて、得られた植物をスクリーニングする工程を含む、植物における果実軟化を遅延させる方法を提供する。
【0080】
本発明のさらに別の態様は、マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドの少なくとも20個の連続ヌクレオチドの断片を少なくとも含み、このポリヌクレオチドのヌクレオチド配列がSEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードし、かつポリヌクレオチドのヌクレオチド配列がSEQ ID NO:1またはSEQ ID NO:3に記載されている、RNA干渉(RNAi)構築物の発現によって、トランスジェニック植物におけるマンノシダーゼのレベルを非トランスジェニック植物におけるマンノシダーゼのレベルと比較して減少させる工程;および非トランスジェニック植物と比べて低下したマンノシダーゼのレベルについて、得られた植物をスクリーニングする工程を含む、植物における果実軟化を遅延させる方法を提供する。
【0081】
本発明のさらに別の態様は、SEQ ID NO:1またはSEQ ID NO:3に記載の配列に由来する少なくとも20個の連続ヌクレオチドを含むセンスポリヌクレオチド鎖、およびセンスポリヌクレオチド鎖にハイブリダイズするアンチセンスポリヌクレオチド鎖を含み、このアンチセンスポリヌクレオチド鎖およびセンスポリヌクレオチド鎖が二重鎖を形成する、RNA干渉(RNAi)構築物の発現によって、トランスジェニック植物におけるマンノシダーゼのレベルを非トランスジェニック植物におけるマンノシダーゼのレベルと比較して減少させる工程;ならびに非トランスジェニック植物と比べて低下したマンノシダーゼのレベルについて、得られた植物をスクリーニングする工程を含む、植物における果実軟化を遅延させる方法を提供する。
【0082】
本発明のさらに別の態様は、SEQ ID NO:1またはSEQ ID NO:3に記載の配列に由来する少なくとも20個の連続ヌクレオチドを含むセンスポリヌクレオチド鎖、およびセンスポリヌクレオチド鎖にハイブリダイズするアンチセンスポリヌクレオチド鎖を含み、このアンチセンスポリヌクレオチド鎖およびセンスポリヌクレオチド鎖が二重鎖を形成する、ヘアピン核酸であるRNA干渉(RNAi)構築物の発現によって、トランスジェニック植物におけるマンノシダーゼのレベルを非トランスジェニック植物におけるマンノシダーゼのレベルと比較して減少させる工程;ならびに非トランスジェニック植物と比べて低下したマンノシダーゼのレベルについて、得られた植物をスクリーニングする工程を含む、植物における果実軟化を遅延させる方法を提供する。
【0083】
本発明のさらに別の態様は、マンノシダーゼの発現が果実軟化を遅延させるために制御されている、マンノシダーゼをコードするポリヌクレオチドを含むトランスジェニック植物、その種子または子孫を提供する。
【0084】
本発明において開示されるトランスジェニック植物には、トマト、トウガラシ、パパイア、マンゴー、バナナ、モモ、セイヨウナシ、柑橘類、パイナップル、グアバ、アボカド、イチゴ、リンゴ、およびザクロが包含される。
【0085】
本発明において開示されるトランスジェニック植物はトマトである。
【0086】
本発明において開示されるトランスジェニック植物はトウガラシである。
【実施例】
【0087】
本明細書に記載された以下の実施例は、例示を目的とするものに過ぎず、それを考慮すれば、様々な修飾または変化が当業者に示唆されるであろうこと、そしてそれらが本願の本旨および範囲ならびに添付の特許請求の範囲の範囲内に含まれることが理解されるべきである。
【0088】
実施例1
トマト由来のα-マンノシダーゼの精製および酵素アッセイ
トマト種子(cv.プサ ルビー(Pusa Ruby)および変異体)を、予め滅菌した土壌で発芽させ、その後、実生を、およそ25℃の温度、70%の湿度、および14/10hの明/暗管理の温室に移植した。分析のため、開花期に花にタグを付した後、緑熟期、催色期、桃熟期、および完熟期に果実を採取した。研究において使用された変異体は、Tomato Genetics Resource Center(TGRC)より取得された。
【0089】
果皮を、液体窒素中で粉砕し、1/4容量の抽出緩衝液(0.25M NaClおよび1mM PMSFを含む100mMトリス-Cl pH7.8)に懸濁させた。遠心分離後の上清を0〜70%硫酸アンモニウム沈殿に供し、10分間10,000gで遠心分離した。このようにして入手されたペレットを、25mMトリス-Cl pH7.8に対して一夜透析した。透析物をDEAEセファロースpH7.8上でのクロマトグラフィに供し、増加する塩勾配により溶出させた(補足図2A〜B)。100mMの塩濃度で溶出した画分をプールし、0〜80%硫酸アンモニウム沈殿に供した。ペレットを、少量の抽出緩衝液に溶解させ、sepadex G-100ゲルろ過カラム上に直接負荷した。
【0090】
酵素を欠くブランクと共に、940μlのMQ、20μlの20mM基質(pNP-α-D-マンノピリノシド)、および一定分量20μlの酵素からなる反応混合物1mlにおいて、37℃で15分間、マンノシダーゼをアッセイした。40μlの0.5M Na2CO3を添加することにより、反応を中止させた。pNP放出の結果として現れた色を、405nmで記録し、標準曲線を使用して定量化した。
【0091】
実施例2
二次元ゲル電気泳動、SDS-PAGE、およびイムノブロット分析
13cmのゲルストリップで、250μLの2-D再水和緩衝液で、精製されたタンパク質試料20μgを用いて、等電点フォーカシングを実施した。まず、タンパク質を、13cm長のイモビリン(immobiline)ドライストリップ(pH4-7)上で、受動的な再水和後に、電荷に従って分離した。エレクトロフォーカシングは、20℃で、20,000Vhr、IPGphorシステム(Amersham Biosciences)を使用して実施された。フォーカシング後のストリップを、10mlの平衡緩衝液において、1%(w/v)DTTにより還元し、続いて2.5%(w/v)ヨードアセトアミドによりアルキル化した。次いで、SDS-PAGE上での二次元目分離のため、ストリップを12.5%ポリアクリルアミドゲル上に負荷した。電気泳動されたタンパク質を、silver stain plusキット(Bio-Rad, CA, USA)により染色した。スポットをゲルから切り出し、エレクトロスプレーイオントラップ飛行時間型質量分析(LC-MS/MS)(Q-Star Pulsar i, Applied Biosystems)によって分析した。スペクトルを、緑色植物亜界(Viridiplantae)(緑色植物)データベースに対して、Mascot配列マッチングソフトウェア(www.matrixscience.com)によって分析した。
【0092】
イムノブロット分析のため、異なる成熟期からの全タンパク質50μgを定量化し、12.5%SDS-PAGEゲル上へ負荷した。電気泳動後、ゲルを150mAの定電流でHybond-C(Amersham)メンブレンに転写した。メンブレン上の非特異的な部位を、1時間、トリス緩衝生理食塩水(TBS)によりブロッキングし、4℃で一夜、一次抗体と共にインキュベートした。西洋ワサビペルオキシダーゼ結合抗ウサギ抗血清を二次抗体として用いて、免疫検出を1時間実施し、化学発光基質(Pierce Biotechnology)にブロットを曝した。
【0093】
SDS PSGEで、トマトマンノシダーゼは、70kDおよび40kDの二つのサブユニットへ分解された。しかしながら、非変性PAGE上でネイティブタンパク質のサイズは360kDであることが見出された。
【0094】
トマトの異なる成熟期におけるイムノブロット分析は、トマトのマンノシダーゼタンパク質蓄積が催色期に最大となることを確認した。
【0095】
実施例3
糖タンパク質染色およびEndoH消化
糖タンパク質染色を、製造業者の指示(GelCode(登録商標)Glycoprotein Staining Kit, PIERCE biotechnology)に従って実施した。EndoH消化を、製造業者の指示(New England Biolabs)に従って実施した。反応後、それを12.5%SDS PAGEで分解し、ニトロセルロースメンブレン上へブロットし、マンノシダーゼ抗体により検出した。
【0096】
染色実験は、マンノシダーゼが2〜3kDのグリカン部分を有する糖タンパク質であることを確認した。
【0097】
実施例3
クローニング、特徴決定、およびノーザンハイブリダイゼーション
多重配列アラインメントによって同定されたペプチドタグおよびモチーフを使用して、縮重プライマーを設計した。増幅断片を、pGEM T-Easyベクターにクローニングし配列決定した。次いで、残りの5'領域および3'領域を、RACE(Invitrogen)を使用して増幅した。
【0098】
Dynabeads(登録商標)mRNA purificationキット(DYNAL biotech)を使用して、全ての成熟期からの75マイクログラムの全RNAからメッセンジャーRNAを単離した。mRNAを一定の50ボルトで1.2%ホルムアルデヒドゲル/1×MOPSで分解し、Hybond-Nメンブレン(Amersham)にブロットし、リン酸緩衝液(0.5M NaPi pH7.2、7%SDS)中でハイブリダイズさせた。ハイブリダイゼーションを12時間実施し、メンブレンを-80℃で72時間hyperfilm(Amersham)に曝した。
【0099】
トマトマンノシダーゼおよびトウガラシマンノシダーゼをコードするポリペプチドを、pGEM-T Easyベクターにクローニングし、それぞれ、pGMTおよびpGMCと命名した。
【0100】
実施例4
免疫局在性
緑熟期および催色期の果実を採取し、果皮の5mm×5mm切片を4%パラホルムアルデヒドで一夜固定した。翌日、切片を1×PBSにより洗浄し、クリオスタット(Lieca CM1510S)上に乗せた。凍結組織を切片化し(10〜15μm)、1時間室温でスライド上で乾燥させた。スライドを1×PBSにより洗浄し、続いて、非特異的な部位をブロッキングするため室温で1時間3%BSA中でインキュベートし、1×PBSにより3回洗浄した。果実果皮切片を有するスライドを、マンノシダーゼのポリクローナル抗体と共に4℃で一夜インキュベートした。翌日、切片を、1×PBSにより3回洗浄し、FITCにより標識された二次抗体と共に3時間インキュベートした。次いで、10分間隔で8回、1×PBSにより切片を洗浄した。次いで、試料を適切なフィルターを有する蛍光顕微鏡において観察した。これにより、マンノシダーゼが細胞壁タンパク質であることが確認された。
【0101】
実施例5
高性能陰イオン交換クロマトグラフィ
タンパク質1μgを、各々100ngのN結合型オリゴ糖(Dextra, U.K)と共に37℃で一夜インキュベートし、酵素を除去するため、試料をPVDFメンブレンでろ過した。ろ過された試料10μlを分析のために使用した。勾配ポンプ(GP 40)、陰イオン交換カラム(Carbopac PA-I、4×250mm)、アルゴンによって溶出液を加圧するためのeluant degas module(EDM-2)が装備されたHPAE-PADシステム(Dionex DX 500 BioLC)を、単糖の分析のために使用した。分離された単糖を、金電極およびAg/AgCl基準電極が装備されたED 40検出器によって検出した。得られたクロマトグラフィデータを、PCに基づくoracle 2データ取得システム(Indtech Analytical, Bombay)を使用して、積分し、プロットした。結果を確認するため、各試料の分析の前後に、標準物(マンノース)を100 nmolの濃度で注入した。これにより、マンノシダーゼのN-グリカンプロセシング能力が確認された。
【0102】
実施例6
RNAi/アンチセンス/過剰発現/MYMIVプラスミドの構築およびアグロバクテリウムに基づく一過性形質転換
pHANNIBAL(細菌におけるアンピシリン耐性を有する)ベクターを使用して、マンノシダーゼをサイレンシングした。アンチセンス構築物は、GUS遺伝子を交換して、マンノシダーゼの全長を逆向きにクローニングすることにより、pBI 121で調製された。VIGSベクターについては、GFP配列を交換することにより600bpをMYMIVベクターにクローニングした。過剰発現構築物は、ゲートウェイ(gateway)クローニング法によってPK7FWG2ベクターにクローニングされた全長マンノシダーゼ遺伝子からなっていた。アグロインジェクションのため、アグロバクテリウム予備培養物(3ml)を、抗生物質を含むYEP(酵母抽出物1%、ペプトン1%、およびNaCl 0.5%)培地において28℃で個々のコロニーから24時間増殖させた。この培養物の1/10を、抗生物質を含む50mlの誘導培地(0.5%牛肉抽出物、0.1%酵母抽出物、0.5%ペプトン、0.5%ショ糖、2mM MgSO4、20mMアセトシリンゴン、10mM MES、pH5.6)に移し、一夜増殖させた。翌日、培養物を遠心分離によって回収し、浸透培地(10mM MgCl2、10mM MES、200mMアセトシリンゴン、pH5.6)に再懸濁させ、4時間、温和に撹拌しながら室温でインキュベートした。次いで、0.5mmの針を有する1mlの注射器を使用して、花柱の先端の近くで1〜2mmの深さにまで針を導入し、穏和に注入することにより、果実への培養物のアグロインジェクションを行った。
【0103】
実施例7
RNA単離および定量的リアルタイムRT-PCR
RNAを塩化リチウム法に従って単離し、nanodrop(ND-100)を使用して定量化した。5μgの全RNAをsuperscript II(Invitrogen)を使用してcDNAへと逆転写した。サイバーグリーン(cyber green)を用い、light Cycler Version 4.05(Roche diagnostics)を使用して、定量的RT-PCRを実施した。サイクルは、95℃で10分の初期Taq活性化、それに続く、94℃10s、55℃10s、および72℃10sからなる45サイクルからなっていた。次いで、増幅の正確さを確証するため融解曲線分析を実施した。2-ΔΔCT法を使用して、データは、内因性参照遺伝子に対して規準化された、対照と比べた遺伝子発現の変化倍率として提示される。アクチンをコードする遺伝子を、内因性対照として使用した。
【0104】
実施例8
低分子RNAの単離およびノーザンハイブリダイゼーション
全RNAを、以前に記載されたようにして単離した。LiClによりRNAを沈殿させた後、それを70%エタノールにより洗浄し、ペレットを空気乾燥させた。次いで、ペレットをDEPC水1mlに溶解させ、5分間65℃に加熱し、2分間氷上で冷却した。高分子量RNAを沈殿させるため、ポリエチレングリコール(分子量8000)およびNaClを、それぞれ5%および0.5Mの最終濃度で添加した。氷上で30分のインキュベーションの後、RNAを30分間13,000rpmで遠心分離した。上清を分離し、3倍容量のエタノールおよび1/10容量の3M酢酸ナトリウム(pH5.2)により沈殿させた。チューブを-20℃に一夜置いた。翌日、低分子量RNAを、10分間の13,000rpmでの遠心分離によって沈殿させた。ペレットを乾燥させ、DEPC水50μlに溶解させ、70Vで4〜5時間15%尿素PAGE上で分解した。次いで、ゲルを一定の100Vで1時間0.5×TBEを使用してナイロンメンブレンに転写した。メンブレンをUV架橋し、40℃で、50%ホルムアミド、7%SDS、および50mM Na2HPO4/NaH2PO4 pH7.2においてプレハイブリダイズさせた。4時間後、変性プローブを添加し、30℃で14〜16時間ハイブリダイズさせた。ブロットを、RTで2分間、2×SSCおよび1%SDSを使用して洗浄し、次いで、0.5×SSCおよび0.1%SDSにより洗浄し、フィルムに曝した。21〜23ヌクレオチドの遺伝子特異的なsiRNAが、RNAi果実において検出され、RNAiによって媒介されたマンノシダーゼのサイレンシングが確認された。
【0105】
実施例9
トマト形質転換
トランスジェニックトマト植物を生成するため、2週齢の実生からの子葉を、記載されるようにして使用した。トマト種子を4%市販漂白剤を使用して滅菌し、ムラシゲ スクーグ(Murashige and Skoog)(MS)培地上で発芽させた。発芽の2週間後、子葉を切り取り、異なる構築物を含有しているA.ツメファシエンス(tumefaciens)株EHA105と、30分間、共培養した。次いで、子葉を、選択のため、50mg/lカナマイシンを含有しているMSプレート上に収集した。小植物が再生した時、それらを発根培地に移した。発根後、植物を温室に移動させ、成熟するまで生長させた。
【0106】
実施例10
テクスチャ分析
果実の堅さを、TA-XT Plus(Stable Microsystems UK)を使用して決定した。各果実を、幅75mmのP75圧縮プレートにより分析し、1mm s-1の試験スピードで垂直の方向に5mmおよび10mmまで圧縮した。堅さは、5gの適用された力に対する応答力として定義された。値を、統計的有意性を決定するため、t検定に供した。これらの結果は、RNAi果実が非トランスジェニック果実より2.5倍大きい堅さを有することを示した。
【0107】
実施例11
実生に対するゲラニオール/ACC処理
プサ ルビー種子を滅菌し、MS培地上で発芽させた。15日後、実生を、20%DMSO中の10mMゲラニオールまたは水中の1mM ACC(1-アミノシクロプロパン-1-カルボン酸)を含有している液体培地に移した。採取された試料からRNAを単離し、リアルタイムRT-PCR分析のためcDNAへと逆転写した。発現を、対照実生(0時間)に対して相対的に計算した。結果は、マンノシダーゼがゲラニオールおよびエチレンによってアップレギュレートされることを示した。
【0108】
実施例12
染色および顕微鏡検
切片をクリオスタット付きミクロトーム(Leica 1050)で切断し、スライド上で乾燥させた。スライドを、0.1Mリン酸緩衝液(pH6.8)中の0.05%トルイジンブルー(sigma)の水性溶液に2分間浸漬し、水で2分間洗浄した。染色された切片を、カバーガラス下に水で乗せ、Nikon 80i落射蛍光/位相差/明視野顕微鏡を使用して、100倍の拡大率で撮影した。壁構造を調査するため、切片を、多糖に結合する蛍光増白剤である0.05%カルコフロールに浸し、蒸留水で洗浄した。切片を適切なフィルターにより調査した。細胞壁の成分に結合したカルコフロールによって、強い水色蛍光が生じた。
【0109】
SEQ ID NO:1 5'および3'のUTR領域を含むトマトα-マンノシダーゼのcDNA
SEQ ID NO:2 トマトα-マンノシダーゼのタンパク質配列
SEQ ID NO:3 5'および3'のUTR領域を含むトウガラシα-マンノシダーゼのcDNA配列
SEQ ID NO:4 トウガラシα-マンノシダーゼのタンパク質配列
【技術分野】
【0001】
発明の分野
本発明は、トマト(ソラナム リコペルシカム(Solanum lycopersicum))およびトウガラシ(カプシカム アンニューム(Capsicum annuum))のα-マンノシダーゼをコードするポリヌクレオチドに関する。
【背景技術】
【0002】
発明の背景
今日の世界的流通の世界において、果実成熟の制御は戦略的に重要である(Causier B, Kieffer M and Davies B, 2002, Science 296: 275-276(非特許文献1))。果実の貯蔵寿命および貯蔵を制限する主因のうちの一つは、過度の軟化である。これに関して、多数の酵素を抑制する試みは、貯蔵寿命の延長または軟化の所望の低下において限られた成功しか収めていない。成熟は、消費される官能特性を達成するための、果実の発達および代謝における主要な変遷を含む複雑な過程である。これらのうち、テクスチャは、嗜好性、消費者受容性、貯蔵寿命、輸送能力、および収穫後疾患/病原体に対する抵抗性に関する主要な品質特性であり、これらは全て直接コストに影響を与える。これらのテクスチャの変化が主として細胞壁構造の変化に起因することは、自明であると見なされている。細胞壁は、炭水化物と、多重遺伝子ファミリーによってコードされたタンパク質とから主になる動的な成分である。これらの細胞壁多糖は、自然界に見出される最も豊富な有機化合物であり、その構造的および機能的な多様性は、合成(糖転移酵素)、修飾(炭水化物エステラーゼ)、および分解(糖質加水分解酵素および多糖リアーゼ)に関与する一連の多数の酵素によって反映される。生物の遺伝子のおよそ1〜2%がこれらの過程に関与しており、このことは、果実成熟におけるそれらの重要性を明らかにしている(Henrissat B, Coutinho PM and Davies GJ, 2001, Plant Mol Biol 47: 55-72(非特許文献2)およびJamet E, Canut H, Boudart G and Pont-Lezica RF, 2006, Trends Plant Sci 11: 33-39(非特許文献3))。一般に、果実の堅さの低下には、多糖加水分解酵素/糖類加水分解酵素、トランスグリコシラーゼ、リアーゼ、およびエクスパンシンのような、多数の細胞壁分解酵素の発現の増加が伴う(Brummell DA, 2006, Funct Plant Biol 33: 103-119(非特許文献4))。一般的な触媒活性は配列から推論され得るが、これらのタンパク質の大部分の正確な酵素機能および生物学的役割は未知である。炭水化物代謝に関与する一式の酵素のうち、加水分解酵素は主要な部分を占める。これらの加水分解酵素は、細胞壁またはその他の細胞器官に見出されるN-糖タンパク質を標的とし、N-複合糖質を分解して、遊離N-グリカン含有量を増加させる。さらに、これらの遊離N-グリカンは、トマトにおいて成熟を刺激する生物学的活性を有することが公知である(Priem B, Gitti R, Bush CA and Gross KC, 1993 Plant Physiol 102: 445-458(非特許文献5))。
【先行技術文献】
【非特許文献】
【0003】
【非特許文献1】Causier B, Kieffer M and Davies B, 2002, Science 296: 275-276
【非特許文献2】Henrissat B, Coutinho PM and Davies GJ, 2001, Plant Mol Biol 47: 55-72
【非特許文献3】Jamet E, Canut H, Boudart G and Pont-Lezica RF, 2006, Trends Plant Sci 11: 33-39
【非特許文献4】Brummell DA, 2006, Funct Plant Biol 33: 103-119
【非特許文献5】Priem B, Gitti R, Bush CA and Gross KC, 1993 Plant Physiol 102: 445-458
【発明の概要】
【0004】
本発明は、トマト(ソラナム リコペルシカム)およびトウガラシ(カプシカム アンニューム)の果実軟化に関連する酵素α-マンノシダーゼの精製および特徴決定に関する。本発明は、トマトおよびトウガラシのα-マンノシダーゼ遺伝子の同定、単離、およびクローニングにも関する。本発明は、α-マンノシダーゼタンパク質の蓄積が低下し、果実貯蔵寿命が延長した、トランスジェニック植物にさらに関する。
【0005】
本発明の一つの局面は、マンノシダーゼ活性を有するポリペプチドをコードする単離されたポリヌクレオチドを提供し、このポリヌクレオチドのヌクレオチド配列は、SEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードする。
【0006】
本発明の別の局面は、SEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列を有するポリペプチドを提供する。
【0007】
本発明のさらに別の局面は、SEQ ID NO:1またはSEQ ID NO:4に記載の配列に由来する少なくとも20個の連続ヌクレオチドを含むセンスポリヌクレオチド、および該センスポリヌクレオチド鎖とハイブリダイズするアンチセンスポリヌクレオチドを含み、このアンチセンスポリヌクレオチド鎖およびセンスポリヌクレオチド鎖が二重鎖を形成する、トランスジェニック植物においてマンノシダーゼの発現を抑制するためのRNAi構築物を提供する。
【0008】
本発明のさらに別の局面は、マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドを含み、このポリヌクレオチドのヌクレオチド配列がSEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードし、かつこのポリヌクレオチドがアンチセンスの向きである、組換えベクターによって、植物細胞、組織、またはそれらの一部を形質転換する工程を含む、植物における果実軟化を遅延させる方法を提供する。
【0009】
本発明のさらに別の局面は、マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドの少なくとも20個の連続ヌクレオチドの断片を少なくとも含むRNA干渉(RNAi)構築物の発現によって、トランスジェニック植物におけるマンノシダーゼのレベルを非トランスジェニック植物におけるマンノシダーゼのレベルと比較して減少させる工程であって、このポリヌクレオチドのヌクレオチド配列が、SEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードする、工程;および非トランスジェニック植物と比べて低下したマンノシダーゼのレベルについて、得られた植物をスクリーニングする工程を含む、植物における果実軟化を遅延させる方法を提供する。
【0010】
本発明のさらに別の局面は、SEQ ID NO:1またはSEQ ID NO:3に記載の配列に由来する少なくとも20個の連続ヌクレオチドを含むセンスポリヌクレオチド鎖、およびセンスポリヌクレオチド鎖にハイブリダイズするアンチセンスポリヌクレオチド鎖を含むRNA干渉(RNAi)構築物の発現によって、トランスジェニック植物におけるマンノシダーゼのレベルを非トランスジェニック植物におけるマンノシダーゼのレベルと比較して減少させる工程であって、このアンチセンスポリヌクレオチド鎖およびセンスポリヌクレオチド鎖が二重鎖を形成する、工程;ならびに非トランスジェニック植物と比べて低下したマンノシダーゼのレベルについて、得られた植物をスクリーニングする工程を含む、植物における果実軟化を遅延させる方法を提供する。
【0011】
本発明のさらに別の局面は、マンノシダーゼの発現が果実軟化を遅延させるために制御されている、マンノシダーゼをコードするポリヌクレオチドを含むトランスジェニック植物、その種子および子孫を提供する。
【0012】
本発明において開示されるトランスジェニック植物には、トマト、トウガラシ、パパイア、マンゴー、バナナ、モモ、セイヨウナシ、柑橘類、パイナップル、グアバ、アボカド、イチゴ、リンゴ、およびザクロが包含される。
【0013】
上記のおよびその他の主題の特色、局面、および利点は、以下の説明、添付の図面、および添付の特許請求の範囲に関してよりよく理解されるであろう。
【図面の簡単な説明】
【0014】
【図1a】異なる成熟期におけるトマト果実中のα-マンノシダーゼの比活性を示すグラフ。
【図1b】6%非変性ゲル上で分離された、精製されたα-マンノシダーゼ。
【図1c】12.5%SDS-PAGEで変性させられ分離された、精製されたタンパク質。
【図1d】一次元目に4-7 IPGストリップ上でフォーカシングされ、二次元目に12.5%SDS-PAGE上で分解されたα-マンノシダーゼタンパク質。
【図1e】未消化(PP)と比較してシフトを示す、α-マンノシダーゼ特異抗体によって検出された、精製されたタンパク質のEndoH消化(PP+EndoH)。
【図1f】異なる成熟期(G-緑熟期、B-催色期、P-桃熟期、およびR-完熟期)におけるα-マンノシダーゼのイムノブロット分析。
【図1g】トマト果実におけるα-マンノシダーゼの細胞内局在。
【図1h】α-マンノシダーゼと共にインキュベートした際の三つのN-結合型オリゴ糖のHPAEクロマトグラム。
【図2a】ノーザンブロット分析を使用した、トマトにおける異なる成熟期におけるα-マンノシダーゼ遺伝子の発現。
【図2b】野生型と比較した、成熟不全変異体rin、nor、およびNrにおけるα-マンノシダーゼの相対的な発現を示すRT-PCR分析。
【図2c】組換えベクターp5HM/p3HMの模式図。
【図2d】組換えベクターp7FMの模式図。
【図2e】組換えベクターpAMの模式図。
【図3a】サイレンシングベクター、ブランクバイナリーベクター、および過剰発現ベクターによるアグロインジェクションを受けたトマト果実。
【図3b】アグロインジェクションを受けた代表的なトマト果実を示す微速度撮影写真。
【図3c】マンノシダーゼ転写物レベルの低下を決定するためのRT-PCRによるRNAiトランスジェニック系の発現分析。データは平均値±SE(n=3)である。
【図4a】トウガラシにおける期別のα-マンノシダーゼの比活性。
【図4b】6%非変性ゲル上で分離された、精製されたα-マンノシダーゼ。
【図4c】12.5%SDS-PAGEで変性させられ分離された、精製されたタンパク質。
【図4d】一次元目に4-7 IPGストリップ上でフォーカシングされ、二次元目に12.5%SDS-PAGE上で分解されたα-マンノシダーゼタンパク質。
【発明を実施するための形態】
【0015】
発明の詳細な説明
本発明は、トマトおよびトウガラシの果実軟化に関連するα-マンノシダーゼに関する。本発明は、特に、α-マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチド配列に関する。ここで、植物におけるRNAiおよびアンチセンスにより媒介されるα-マンノシダーゼのサイレンシングは果実軟化を遅延させ、過剰発現は果実軟化を増強する。
【0016】
本発明は、α-マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドを含むDNA構築物、組換えベクター、および組換え宿主細胞を提供する。本発明は、さらに、本発明において開示されるα-マンノシダーゼのポリヌクレオチドを使用して、植物における果実軟化を遅延させる方法を提供する。本発明は、さらに、本発明において開示されるポリヌクレオチドを使用した、植物における早期の果実軟化のための方法を提供する。さらに、本発明は、果実軟化に影響を与えるためにα-マンノシダーゼの発現が制御されているトランスジェニック植物、それらの子孫および種子を提供する。
【0017】
アンチセンス技術およびRNAi技術を使用してβ-D-N-アセチルマンノシダーゼ遺伝子の発現を抑制することにより得られたトランスジェニック植物は、非トランスジェニック果実より約1.5〜2.5倍堅い果実を生じた。さらに、RNAi技術によって作製されたトランスジェニック果実は、非トランスジェニック果実よりほぼ30日長く、収穫後45日目まで軟化の兆候を示さなかった。アンチセンス技術によって作製されたトランスジェニック果実は、非トランスジェニック果実よりほぼ20日長く、収穫後35日目まで軟化の兆候を示さなかった。これらの結果は、驚くべき予想外のものである。トランスジェニック植物における遺伝子のさらなる抑制は、栄養生長、開花および果実発達、成熟までの日数、および収量に対しては負の効果を及ぼさなかった。
【0018】
本発明は、植物における炭水化物代謝およびN-グリカンプロセシングを担うグリコシル加水分解酵素ファミリー遺伝子であるα-マンノシダーゼを、RNA干渉(RNAi)技術によって抑制することにより、トマトの貯蔵寿命を延長する方法を記載する。本発明は、さらに、トマト果皮由来のα-マンノシダーゼタンパク質複合体の精製および特徴決定を記載する。
【0019】
2DEフォーカシングされたタンパク質複合体のLC MS/MS分析は、相互作用パートナーの同定に役立ち、α-マンノシダーゼ遺伝子のクローニングにも役立った。α-マンノシダーゼ遺伝子の発現は、催色期に最も高く、成熟/軟化の開始と相関していた。トランスジェニック果実の分子的分析によって、α-マンノシダーゼ転写物が、99%特異的に分解され、30日延長された貯蔵寿命を有する、2.5倍堅い果実を与えることが示された。研究の過程で、本発明者らは、マンノシダーゼが、植物に存在するテルペノイドであるゲラニオールによって誘導され、rinによって調節されることを見出した。
【0020】
pNP-アルファ-D-マンノピリノシド(mannopyrinoside)を基質として使用して、本発明者らは、マンノシダーゼの活性が催色期に最大となり(図1a)、緑熟期、桃熟期、および完熟期がそれに続くことを見出した。呼吸またはエチレン生成、カロテノイドの蓄積、および軟化のような過程の開始を特徴付ける催色期におけるα-マンノシダーゼの蓄積は、それが成熟および/または軟化に関与しているという本発明者らの仮説を強化した。
【0021】
トマトα-マンノシダーゼを催色期トマトの果皮から精製し、さらに特徴決定した。トマト果皮を液体窒素中で破砕し、抽出緩衝液(0.25 NaCl、1mM PMSFを含む100mMトリス-Cl、pH7.8)に一夜懸濁させた。タンパク質を均質になるまで精製するため、硫酸アンモニウム沈殿(0〜70%)、イオン交換(DEAEセファロース)、およびゲルろ過クロマトグラフィ(Sephadex G-100)のような方法を利用した。精製された酵素は、6%非変性PAGE上で360kDの単一のタンパク質を構成し(図1b)、それはα-マンノシダーゼ活性-バンド相関によって確認された。しかしながら、これに反して、superdex 200分析用カラムを使用したゲルろ過によって決定されたタンパク質の分子量は、290kDであった。さらに、SDS-PAGE上で分離すると、タンパク質は70kDおよび45kDの二つのサブユニットに分解され、従って、それがヘテロ二量体であることが明らかとなった(図1c)。明瞭な画像を入手するため、タンパク質複合体を二次元ゲル電気泳動(2DE)上で分離したところ、70kDサブユニットは単一のタンパク質であり、45kDサブユニットは、異なるpIを有する三つのタンパク質へとさらに分解されることが明らかになった(図1d)。本発明者らはLC/MS-MS分析によってスポットを同定し、それらが各々異なるタンパク質であることを見出した。70kDスポットはα-マンノシダーゼ(スポット番号1、pI6.8)を表し、三つのタンパク質へ分けられる45kDのスポットは、推定単糖トランスポーター(スポット番号2、pI6.4)、遺伝子の一部であるゲラニオール応答因子(スポット番号3、pI6.3)、およびH+ ATPアーゼ(スポット番号4、pI6.2)を有していた。従って、本発明者らは、45kD複合体に見出された三つのタンパク質のうちの二つは相互作用タンパク質であるかもしれないと予測した。
【0022】
生化学的特徴決定は、酵素が、60℃まで、活性の考慮すべき低下なしに1時間安定していることを明らかにした。酵素にとって至適の温度およびpHは、それぞれ45℃およびpH6.0であった。p-ニトロフェノール-アルファ-D-マンノピラノシドに対して決定された酵素のKmは、4.6mMである。トマトα-マンノシダーゼの性質を同定するため、それをSDS-PAGE上で分解し、PAS染色に供したところ、タンパク質は、糖タンパク質染色陽性であった。上記の結果をさらに確認するため、タンパク質のEndoH消化を実施したところ、炭水化物/グリカン部分の切断による2〜3kDのシフトが示され、従って、上記の結果が立証された(図1e)。特定の成熟期における比活性をタンパク質蓄積と相関させるため、α-マンノシダーゼに相同なタンパク質の70kDサブユニットに対して産生されたポリクローナル抗体を使用して、全ての期のイムノブロット分析を行った。イムノブロット分析は、タンパク質蓄積が催色期に最大となり、緑熟期、桃熟期、および完熟期がそれに続くことを明らかにした(図1f)。さらに、細胞内の分布を決定するため、免疫局在性アッセイを行ったところ、細胞壁の最も内側の縁にシグナルが見られたことから、それが細胞壁タンパク質であることが明らかになった(図1g)。本発明者らの所見は、分泌のためのシグナルペプチドを有する細胞壁タンパク質に、α-マンノシダーゼホモログ(Q8LPJ3)を分類した、アラビドプシス(Arabidopsis)における報告によってさらに補強された。N-グリカンおよびそれらのプロセシングに対するα-マンノシダーゼの役割を判定するため、本発明者らは、果実果皮中に一般的に見出される、三つのN結合型オリゴ糖(オリゴマンノース-3、オリゴマンノース-5、およびオリゴマンノース-9)を標的とした。N-グリカンに対する作用を確証するため、α-マンノシダーゼを、37℃で一夜、N結合型オリゴ糖と共にインキュベートし、反応混合物をCarbopac PA-1カラム上で分解した。酵素は、全てのN結合型オリゴ糖から末端マンノース残基を切断することができ、従って、それが成熟/軟化に関与していることの間接的な証拠が得られた(図1h)
【0023】
ペプチドQHVADDYAKおよびSGAYVFRPに対応する縮重プライマーを使用して、cDNAを増幅した。このプライマーのオリゴヌクレオチド配列は、SEQ ID NO:5およびSEQ ID NO:6に記載されている。プライマーは、LC-MS/MSデータ、および多重アラインメントによって同定されたモチーフを使用して設計された。5'末および3'末の欠けている部分を、cDNA末端のランダム増幅(Random Amplification of cDNA Ends)(RACE)によって増幅し、全長にした。配列のインシリコ分析は、コーディング領域が3090bp長(SEQ ID NO:1)であることを示し、推定タンパク質は、114KDaの質量計算値および6.48のpI理論値を有する1029アミノ酸長(SEQ ID NO:2)であった。トマトα-マンノシダーゼタンパク質は、アラビドプシスおよびイネのα-マンノシダーゼタンパク質との、それぞれ66%および62%の同一性を示した。その進化的局面を洞察するため、系統学的分析を実施したところ、α-マンノシダーゼは、三つの公知のアラビドプシスα-マンノシダーゼタンパク質のうちの二つと最も近縁であることが示された。発現パターンを確証し、先の結果を実証するため、ノーザンブロット分析を実施したところ、マンノシダーゼ転写物が催色期に最も豊富であり、緑熟期、桃熟期、および完熟期がそれに続くことが明らかになった(図2a)。
左プライマー:
右プライマー:
ここで、KはGまたはTであり、RはAまたはGであり、MはAまたはCである。
【0024】
α-マンノシダーゼの発現を、トマト変異体rin(ripening inhibitor)、nor、およびNrにおいて分析した。その分析は、α-マンノシダーゼが、変異体rinにおいては90%、Nrにおいては30%阻害されており、nor変異体は野生型と比較可能であることを明らかにした(図2b)。果実成熟の発達調節のために必要とされるMADSボックス転写因子をコードする遺伝子に変異を保持している変異体のさらなる分析を実施した。本発明者らは、rin変異体および野生型において、マンノシダーゼを誘導するため、エチレンの前駆物質であるACCを使用した。ACCは、rinにおいても野生型においても対照と比較してα-マンノシダーゼをアップレギュレートすることができた。これらの観察は、α-マンノシダーゼがエチレンによってもrinによっても独立にまたは相乗的に調節されることを示した。
【0025】
多くの観察に支持されて、成熟および/またはそれに関連した軟化における役割を証明するため、サイレンシングによって、α-マンノシダーゼを機能的に特徴決定することが不可避となった。アグロインジェクションを使用したエンドジーン(Endogene)抑制は、遺伝子機能を分析するための便利なツールになっている(Orzaez DF, Mirabel S, Wieland WH and Granell A, 2006, Plant Physiol 140: 3-11)。このdsRNAにより媒介されるサイレンシング技術を使用して、本発明者らは、pHANNIBALベクターを使用してα-マンノシダーゼを一過性にサイレンシングした(Wesley SV, Helliwell CA, Smith NA, Wang MB, Rouse DT, Liu Q, Gooding PS, Singh SP, Abbott D, Stoutjesdijk PA, Robinson SP, Gleave AP, Green AG and Waterhouse PM, 2001, Plant J 27: 581-590)。その発現カセットは、発現時にdsRNAへと組み立てられるよう設計された、CaMV 35Sプロモーターの制御下にある、イントロンによって隔てられた、センスの向きおよびアンチセンスの向きにサブクローニングされたそれぞれのUTR領域を含む5'領域または3'領域のいずれかに由来する遺伝子の600bp断片からなっていた。RNAiカセットの構築において使用されたプライマーのオリゴヌクレオチド配列は、SEQ ID NO:7、SEQ ID NO:8、SEQ ID NO:9、SEQ ID NO:10、SEQ ID NO:11、SEQ ID NO:12、SEQ ID NO:13、およびSEQ ID NO:14に記載されている。このカセットを、さらに、バイナリーベクターpART 27へサブクローニングし、そのバイナリーベクターをp5HM/p3HMと名付けた(図2c)。各果実に、サイズに依って、花柱の先端の近くの表面上の2〜3箇所のスポットに注入した(図3a)。1週間後、RNAiによるアグロインジェクションを受けた果実は、対照(ブランクバイナリーベクター)のものと比較して、果実の周囲の最大面積をカバーする、顕著な緑色のセクターを注入スポットの周囲に発達させた(図3a)。分子レベルで抑制を確認し定量化するため、注入の2〜4日後に採取した果実を、リアルタイムRT-PCRによって、α-マンノシダーゼに対応するmRNAレベルの低下を決定するために使用した。オリゴヌクレオチドプライマーは、light cycler probe design 2プログラム(Roche)を使用して設計した。α-マンノシダーゼの増幅において使用されたプライマーのオリゴヌクレオチド配列は、SEQ ID NO:15およびSEQ ID NO:16に記載されている。内因性対照遺伝子アクチンの増幅のために使用されたプライマーは、SEQ ID NO:17およびSEQ ID NO:18に記載する。
【0026】
結果は、緑熟期の2日前に注入された場合には、mRNAレベルが対照と比べて70%低下し、緑熟期に注入された場合には、50%低下することを明らかにした。結果をバリデートするため、pK7FWG 2.0において調製され、p7FMと名付けられたα-マンノシダーゼの過剰発現構築物(図2d)の、果実へのアグロインジェクションを行ったところ、その果実は、初期の間は、対照に類似した挙動を示した。過剰発現の効果は後期に見られ、果実は対照(ブランクベクターおよび未注入)よりはるかに急速に劣化し、テクスチャを喪失した。効果を見るため、アンチセンス構築物もpBI121において調製し、pAMと名付けた(図2e)。アンチセンスを注入された果実は、過剰発現果実とサイレンシングを受けた果実との中間型であった。このmRNAレベルの低下のため、本発明者らは、21〜23残基の低分子量RNA(siRNA)の存在を調査することとした。この目的のため、本発明者らは、対照と共に、RNAiの、緑熟期および催色期の果実から可溶性の低分子RNAを抽出し、ノーザンブロット分析を実施した。本発明者らは、RNAiにより媒介されるサイレンシングの最大の特徴であるα-マンノシダーゼに特異的な低分子干渉(si)RNAを果実果皮において観察した。さらに、本発明者らの結果は、非ウイルスベクターが、ウイルスベクターと同一の効率および程度でサイレンシングを達成し得ることも証明している。
【0027】
アグロインジェクションの30日後、アグロインジェクションを受けた果実のテクスチャの堅さを定量化することを試みて、TA-XT2テクスチャアナライザーを使用した圧縮分析を実施した。成熟は均一の過程ではないため、果実の異なる面で5mmまで果実を圧縮し(2〜3箇所の圧縮/果実)、花柱の付根をテクスチャアナライザーのプレートに触れさせて単一の10mmの圧縮を行うことにより、果実の堅さを測定した。全ての例において、RNAiアグロインジェクションを受けた果実は、統計的に堅く、5mmおよび10mmの圧縮において、それぞれ2倍および1.6倍、カウンターパートより堅かった。さらに、アグロインジェクションを受けた果実の微速度撮影写真は、RNAi注入果実は、45日目までテクスチャおよび堅さを保持するのに対し、対照は15日後にテクスチャの縮みおよび緩みを開始することを明らかにした(図3b)。これらの有望な結果より、本発明者らは、同ベクターを使用して安定的なRNAi系を産生することにした。トマト子葉のアグロバクテリウムにより媒介される形質転換は、イントロンによって隔てられた遺伝子のセンス断片およびアンチセンス断片を発現するRNAi系をもたらした。子葉を、サイレンシングベクターによって形質転換されたアグロバクテリウム株EHA105と共培養し、カナマイシンプレート上で選択した。この研究において使用された株の他に、LBA 4404およびGV 3101のような株も同一の効果を得るために利用され得る。苗条が出現するまで、外植片を12〜15日毎に継代培養した。苗条が1〜2cm長になった時、それらを切断し、発根培地において生長させた。サイレンシングを受けたと推定される植物(T0)を、リアルタイムRT-PCRによって転写物レベルの低下について調査したところ、α-マンノシダーゼが催色期に99%抑制されることが明らかになった(図3c)。次いで、桃熟期に果実を採取し、テクスチャ分析のため室温で貯蔵した。アンチセンス技術によって作製されたトランスジェニック果実は、非トランスジェニック果実と比較して、マンノシダーゼの80%の抑制および1.5倍の堅さを示した。30日後、生長、発達、および種子生成に対する負の効果なしに、RNAi果実が対照より2.5倍堅いことが観察された。アンチセンス技術によって作製されたトランスジェニック果実は、20日の貯蔵寿命の延長を示した。
【0028】
トウガラシは非クライマクテリック型果実であり、果実の発達および成熟を8期に分割した。第8期から成熟果実を採取し、タンパク質抽出をトリス-Cl pH7において実施した。トウガラシの発達および成熟の全ての異なる期において活性アッセイを実施したところ、最大活性は第8期果実に見出された(図4a)。トウガラシマンノシダーゼを、イオン交換およびゲルろ過のような種々のクロマトグラフィ技術を利用することにより精製した。活性を含有している溶出画分を、濃縮し、ゲル上で分解した。精製されたタンパク質は、6%非変性ゲル上のおよそ500kDの単一のバンドであり(図4b)、SDS-PAGEでは、70kDおよび50kDの二つのサブユニットが分解された(図4c)。精製されたタンパク質の2DEエレクトロフォーカシングによって、それが複合体であることが明らかになり、相互作用タンパク質であることが暗示された。より低い50kDのバンドは、トマトマンノシダーゼに極めて類似した、異なるpIを有する三つのタンパク質に分離された(図4d)。免疫局在性から、それが細胞壁に局在していることが明らかになった。トウガラシα-マンノシダーゼはトマトα-マンノシダーゼと90%同一であった。トウガラシ由来のα-マンノシダーゼの分子クローニングは、コーディング領域が3093bp長(SEQ ID NO:3)であることを明らかにした。トウガラシ由来の推定タンパク質は1030アミノ酸長(SEQ ID NO:4)であった。トウガラシにおけるアグロインジェクションによるα-マンノシダーゼの一過性のサイレンシングは、それが成熟に関連した軟化に関与していることを明らかにした。
【0029】
ゲラニオールは、様々な癌細胞系に対するインビトロおよびインビボの抗腫瘍活性を有する、非環式の食事性モノテルペンであって、従って、癌化学防御のための薬剤の新たなクラスと見なされている。ゲラニオールは、カンジダ アルビカンス(Candida albicans)およびサッカロミセス セレビシエ(Saccharomyces cervisiae)の膜機能に干渉することも報告された。ゲラニオールは水に不溶性であるため、実生に供給するため、20%DMSOを使用した(Hendry GAF and Jones OTG, 1984, Induction of cytochrome p-450 in intact mung beans. New Phytol 96: 153-159)。本発明者らは、ゲラニオール処理の30分後、マンノシダーゼ遺伝子発現が、対照(DMSOのみ)と比較して3倍アップレギュレートされたことを観察した。このことから、本発明者らは、多面的なrin変異体において、α-マンノシダーゼ誘導に対するゲラニオールの役割をさらに分析することにした。rin実生に10%ゲラニオールを供給し、試料を収集した。相対遺伝子発現分析は、α-マンノシダーゼ転写物が、30分後、6倍アップレギュレートされることを明らかにし、先の結果を実証した。
【0030】
本発明の一つの態様は、α-マンノシダーゼ活性を有するポリペプチドをコードする、単離されたポリヌクレオチドを提供する。該ポリヌクレオチドのヌクレオチド配列は、
(a)SEQ ID NO:2に記載のアミノ酸配列との90%の類似性を有するポリペプチドをコードするヌクレオチド配列;
(b)SEQ ID NO:4に記載のアミノ酸配列との90%の同一性を有するポリペプチドをコードするヌクレオチド配列;
(c)(a)または(b)のヌクレオチド配列に相補的なヌクレオチド配列
(d)SEQ ID NO:1に記載のヌクレオチド配列;
(e)SEQ ID NO:3に記載のヌクレオチド配列;
(f)(d)または(e)のヌクレオチド配列に相補的なヌクレオチド配列
からなる群より選択される。
【0031】
本発明の別の態様は、ヌクレオチド配列がSEQ ID NO:1に記載の、トマトから単離される、本発明において開示されるα-マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドを提供する。
【0032】
本発明の別の態様は、ヌクレオチド配列がSEQ ID NO:3に記載の、トウガラシから単離される、本発明において開示されるα-マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドを提供する。
【0033】
本発明の一つの態様は、SEQ ID NO:1およびSEQ ID NO:3に記載のヌクレオチド配列を有する、単離されたポリヌクレオチドを提供する。
【0034】
本発明のさらなる態様は、SEQ ID NO:2およびSEQ ID NO:4に記載のアミノ酸配列を有する、α-マンノシダーゼ活性を有するポリペプチドを提供する。
【0035】
本発明のさらに別の態様は、本発明において開示されるα-マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドを含み、このポリヌクレオチド配列がプロモーター配列と機能的に連結されている、DNA構築物を提供する。
【0036】
本発明のさらなる態様は、イントロンによって隔てられたポリヌクレオチド配列の部分が、センスの向きおよびアンチセンスの向きである、本発明のDNA構築物を提供する。
【0037】
本発明のさらに別の態様は、α-マンノシダーゼ活性を有するポリペプチドをコードするSEQ ID NO:1に記載のヌクレオチド配列を有するポリヌクレオチドの部分を含み、イントロンによって隔てられたこのポリヌクレオチド配列が、センスの向きおよびアンチセンスの向きである、DNA構築物を提供する。
【0038】
本発明のさらに別の態様は、α-マンノシダーゼ活性を有するポリペプチドをコードするSEQ ID NO:3に記載のヌクレオチド配列を有するポリヌクレオチドの部分を含み、イントロンによって隔てられたこのポリヌクレオチド配列が、センスの向きおよびアンチセンスの向きである、DNA構築物を提供する。
【0039】
本発明は、本発明において開示されるDNA構築物を含む組換えベクターも提供する。
【0040】
本発明は、α-マンノシダーゼ活性を有するポリペプチドをコードするSEQ ID NO:1に記載のヌクレオチド配列を有するポリヌクレオチドの部分を含み、イントロンによって隔てられたこのポリヌクレオチド配列が、センスの向きおよびアンチセンスの向きである、組換えベクターをさらに提供する。
【0041】
本発明は、α-マンノシダーゼ活性を有するポリペプチドをコードするSEQ ID NO:3に記載のヌクレオチド配列を有するポリヌクレオチドの部分を含み、イントロンによって隔てられたこのポリヌクレオチド配列が、センスの向きおよびアンチセンスの向きである、組換えベクターをさらに提供する。
【0042】
一つの態様において、本発明は、本発明において開示されるベクターを含む組換え宿主細胞を提供する。
【0043】
一つの態様において、本発明は、α-マンノシダーゼ活性を有するポリペプチドをコードするSEQ ID NO:1に記載のヌクレオチド配列を有するポリヌクレオチドの部分を含み、イントロンによって隔てられたこのポリヌクレオチド配列が、センスの向きおよびアンチセンスの向きである、組換え宿主細胞を提供する。
【0044】
一つの態様において、本発明は、マンノシダーゼ活性を有するポリペプチドをコードするSEQ ID NO:3に記載のヌクレオチド配列を有するポリヌクレオチドの部分を含み、イントロンによって隔てられたこのポリヌクレオチド配列が、センスの向きおよびアンチセンスの向きである、組換え宿主細胞を提供する。
【0045】
別の態様において、本発明は、アグロバクテリウム、大腸菌、および酵母からなる群より選択される宿主細胞に関する。
【0046】
別の態様において、本発明は、植物細胞、組織、またはそれらの一部を本発明の組換えベクターによって形質転換する工程を含み、該ベクターが、イントロンによって隔てられた、センスの向きおよびアンチセンスの向きであるα-マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドの部分を含む、植物において果実軟化を遅延させる方法を提供する。
【0047】
別の態様において、本発明は、植物細胞、組織、またはそれらの一部を本発明の組換えベクターによって形質転換する工程を含み、該ベクターが、アンチセンスの向きであるα-マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドを含む、植物において果実軟化を遅延させる方法を提供する。
【0048】
別の態様において、本発明は、植物細胞、組織、またはそれらの一部を本発明の組換えベクターによって形質転換する工程を含み、該ベクターが、センスの向きであるα-マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドを含む、植物における早期果実軟化のための方法を提供する。
【0049】
別の態様において、本発明は、植物細胞、組織、またはそれらの一部を本発明の組換えベクターによって形質転換する工程を含み、該ベクターが、アンチセンスの向きであるα-マンノシダーゼ活性を有するポリペプチドをコードするSEQ ID NO:1に記載のヌクレオチド配列を有するポリヌクレオチドを含む、植物において果実軟化を遅延させる方法を提供する。
【0050】
別の態様において、本発明は、植物細胞、組織、またはそれらの一部を本発明の組換えベクターによって形質転換する工程を含み、該ベクターが、センスの向きであるα-マンノシダーゼ活性を有するポリペプチドをコードするSEQ ID NO:3に記載のヌクレオチド配列を有するポリヌクレオチドを含む、植物における早期果実軟化のための方法を提供する。
【0051】
本発明において提供される植物における果実軟化の遅延または早期果実軟化のための方法において、形質転換は、アグロバクテリウムにより媒介される形質転換、遺伝子銃、電気穿孔、およびインプランタ(in planta)形質転換のような、当技術分野において公知の方法によって実施され得る。
【0052】
一つの態様において、本発明は、α-マンノシダーゼの発現が果実軟化を引き起こし最終的には果実成熟をもたらすよう制御されている、本発明において開示される方法によって作製されたトランスジェニック植物を提供し、該植物は、トマト、トウガラシ、マンゴー、バナナ、パパイア、柑橘類、グアバ、アボカド、ブドウ、パイナップル、イチゴ、リンゴ、ザクロ、およびその他の果実収穫植物からなる群より選択される。
【0053】
本発明において開示される方法によって作製されるトランスジェニック植物は、トマトである。
【0054】
本発明は、本発明において開示されるトランスジェニック植物の種子または子孫も提供する。
【0055】
本発明の一つの態様において、マンノシダーゼ活性を有するポリペプチドをコードする単離されたポリヌクレオチドが提供され、該ポリヌクレオチドのヌクレオチド配列は、SEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約88%の同一性を有するポリペプチドをコードする。
【0056】
本発明の別の態様において、マンノシダーゼ活性を有するポリペプチドをコードする単離されたポリヌクレオチドが提供され、該ポリヌクレオチドのヌクレオチド配列は、SEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約90%の同一性を有するポリペプチドをコードする。
【0057】
本発明の別の態様は、マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドに由来する少なくとも20個の連続ヌクレオチドを含み、該ポリヌクレオチドのヌクレオチド配列は、SEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードし、これらのヌクレオチドがヘアピン構造のみを形成し全域において二次的なループを形成しないよう選択されている、RNAi構築物を提供する。
【0058】
本発明の別の態様は、マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドに由来する少なくとも20個の連続ヌクレオチドを含み、該ポリヌクレオチドのヌクレオチド配列は、SEQ ID NO:1またはSEQ ID NO:3に記載のものであり、これらのヌクレオチドがヘアピン構造のみを形成し全域において二次的なループを形成しないよう選択されている、RNAi構築物を提供する。
【0059】
本発明において開示されるRNAi構築物は、植物において転写後にヘアピン構造を形成する。
【0060】
本発明において開示されるRNAi構築物は、ヘアピン核酸である。
【0061】
本発明において開示されるRNAi構築物は、植物において転写後にヘアピン構造を形成し、二次的なループ構造を形成しない。
【0062】
別の態様は、植物において転写後にヘアピン構造を形成する、本発明において開示されるポリヌクレオチド配列を含むRNAi構築物を提供する。
【0063】
本発明のさらに別の局面は、マンノシダーゼ活性を有するポリペプチドをコードする単離されたポリヌクレオチドを提供し、このポリヌクレオチドのヌクレオチド配列は、SEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードする。
【0064】
本発明のさらに別の局面は、マンノシダーゼ活性を有するポリペプチドをコードする単離されたポリヌクレオチドを提供し、このポリヌクレオチドのヌクレオチド配列は、SEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列を有するポリペプチドをコードする。
【0065】
本発明のさらに別の局面は、マンノシダーゼ活性を有するポリペプチドをコードする単離されたポリヌクレオチドを提供し、このポリヌクレオチドのヌクレオチド配列は、SEQ ID NO:1またはSEQ ID NO:3に記載されている。
【0066】
本発明のさらに別の態様は、SEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列を有するポリペプチドを提供する。
【0067】
本発明のさらに別の局面は、マンノシダーゼ活性を有するポリペプチドをコードする単離されたポリヌクレオチドを含み、このポリヌクレオチドのヌクレオチド配列がSEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードし、かつこのポリヌクレオチドがプロモーター配列に機能的に連結されている、DNA構築物を提供する。
【0068】
本発明のさらに別の局面は、マンノシダーゼ活性を有するポリペプチドをコードする単離されたポリヌクレオチドを含み、このポリヌクレオチドのヌクレオチド配列がSEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列を有するポリペプチドをコードし、かつこのポリヌクレオチドがプロモーター配列に機能的に連結されている、DNA構築物を提供する。
【0069】
本発明のさらに別の局面は、マンノシダーゼ活性を有するポリペプチドをコードする単離されたポリヌクレオチドを含み、このポリヌクレオチドのヌクレオチド配列がSEQ ID NO:1またはSEQ ID NO:3に記載されており、かつこのポリヌクレオチドがプロモーター配列に機能的に連結されている、DNA構築物を提供する。
【0070】
本発明のさらに別の局面は、マンノシダーゼ活性を有するポリペプチドをコードする単離されたポリヌクレオチドを含み、このポリヌクレオチドのヌクレオチド配列がSEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードし、このポリヌクレオチドがプロモーター配列に機能的に連結されており、かつこのポリヌクレオチド配列がセンスの向きである、DNA構築物を提供する。
【0071】
本発明のさらに別の局面は、マンノシダーゼ活性を有するポリペプチドをコードする単離されたポリヌクレオチドを含み、このポリヌクレオチドのヌクレオチド配列がSEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードし、このポリヌクレオチドがプロモーター配列に機能的に連結されており、かつこのポリヌクレオチド配列がアンチセンスの向きである、DNA構築物を提供する。
【0072】
本発明の別の態様は、SEQ ID NO:1またはSEQ ID NO:4に記載の配列に由来する少なくとも20個の連続ヌクレオチドを含むセンスポリヌクレオチド鎖、および該センスポリヌクレオチド鎖にハイブリダイズするアンチセンスポリヌクレオチド鎖を含み、このアンチセンスポリヌクレオチド鎖およびセンスポリヌクレオチド鎖が二重鎖を形成する、トランスジェニック植物においてマンノシダーゼの発現を抑制するためのRNAi構築物を提供する。
【0073】
本発明において開示されるRNAi構築物において、RNAi構築物はヘアピン核酸である。
【0074】
本発明において開示されるRNAi構築物において、センス鎖は100〜600個のヌクレオチドを含む。
【0075】
本発明の別の態様は、本発明において開示されるDNA構築物を含む組換えベクターを提供する。
【0076】
本発明の別の態様は、アグロバクテリウム(Agrobacterium)、大腸菌(E.coli)、および酵母からなる群より選択される組換え宿主細胞を提供する。
【0077】
本発明のさらに別の態様は、マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドを含む組換えベクターによって、植物細胞、組織、またはそれらの一部を形質転換する工程を含み、該ポリヌクレオチドのヌクレオチド配列がSEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードし、かつこのポリヌクレオチドがアンチセンスの向きである、植物において果実軟化を遅延させる方法を提供する。
【0078】
本発明のさらに別の態様は、マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドを含む組換えベクターによって、植物細胞、組織、またはそれらの一部を形質転換する工程を含み、該ポリヌクレオチドのヌクレオチド配列がSEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードし、このポリヌクレオチドがアンチセンスの向きであり、かつこのポリヌクレオチドのヌクレオチド配列がSEQ ID NO:1またはSEQ ID NO:3に記載されている、植物において果実軟化を遅延させる方法を提供する。
【0079】
本発明のさらに別の態様は、マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドの少なくとも20個の連続ヌクレオチドの断片を少なくとも含み、このポリヌクレオチドのヌクレオチド配列がSEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードする、RNA干渉(RNAi)構築物の発現によって、トランスジェニック植物におけるマンノシダーゼのレベルを非トランスジェニック植物におけるマンノシダーゼのレベルと比較して減少させる工程;および非トランスジェニック植物と比べて低下したマンノシダーゼのレベルについて、得られた植物をスクリーニングする工程を含む、植物における果実軟化を遅延させる方法を提供する。
【0080】
本発明のさらに別の態様は、マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドの少なくとも20個の連続ヌクレオチドの断片を少なくとも含み、このポリヌクレオチドのヌクレオチド配列がSEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードし、かつポリヌクレオチドのヌクレオチド配列がSEQ ID NO:1またはSEQ ID NO:3に記載されている、RNA干渉(RNAi)構築物の発現によって、トランスジェニック植物におけるマンノシダーゼのレベルを非トランスジェニック植物におけるマンノシダーゼのレベルと比較して減少させる工程;および非トランスジェニック植物と比べて低下したマンノシダーゼのレベルについて、得られた植物をスクリーニングする工程を含む、植物における果実軟化を遅延させる方法を提供する。
【0081】
本発明のさらに別の態様は、SEQ ID NO:1またはSEQ ID NO:3に記載の配列に由来する少なくとも20個の連続ヌクレオチドを含むセンスポリヌクレオチド鎖、およびセンスポリヌクレオチド鎖にハイブリダイズするアンチセンスポリヌクレオチド鎖を含み、このアンチセンスポリヌクレオチド鎖およびセンスポリヌクレオチド鎖が二重鎖を形成する、RNA干渉(RNAi)構築物の発現によって、トランスジェニック植物におけるマンノシダーゼのレベルを非トランスジェニック植物におけるマンノシダーゼのレベルと比較して減少させる工程;ならびに非トランスジェニック植物と比べて低下したマンノシダーゼのレベルについて、得られた植物をスクリーニングする工程を含む、植物における果実軟化を遅延させる方法を提供する。
【0082】
本発明のさらに別の態様は、SEQ ID NO:1またはSEQ ID NO:3に記載の配列に由来する少なくとも20個の連続ヌクレオチドを含むセンスポリヌクレオチド鎖、およびセンスポリヌクレオチド鎖にハイブリダイズするアンチセンスポリヌクレオチド鎖を含み、このアンチセンスポリヌクレオチド鎖およびセンスポリヌクレオチド鎖が二重鎖を形成する、ヘアピン核酸であるRNA干渉(RNAi)構築物の発現によって、トランスジェニック植物におけるマンノシダーゼのレベルを非トランスジェニック植物におけるマンノシダーゼのレベルと比較して減少させる工程;ならびに非トランスジェニック植物と比べて低下したマンノシダーゼのレベルについて、得られた植物をスクリーニングする工程を含む、植物における果実軟化を遅延させる方法を提供する。
【0083】
本発明のさらに別の態様は、マンノシダーゼの発現が果実軟化を遅延させるために制御されている、マンノシダーゼをコードするポリヌクレオチドを含むトランスジェニック植物、その種子または子孫を提供する。
【0084】
本発明において開示されるトランスジェニック植物には、トマト、トウガラシ、パパイア、マンゴー、バナナ、モモ、セイヨウナシ、柑橘類、パイナップル、グアバ、アボカド、イチゴ、リンゴ、およびザクロが包含される。
【0085】
本発明において開示されるトランスジェニック植物はトマトである。
【0086】
本発明において開示されるトランスジェニック植物はトウガラシである。
【実施例】
【0087】
本明細書に記載された以下の実施例は、例示を目的とするものに過ぎず、それを考慮すれば、様々な修飾または変化が当業者に示唆されるであろうこと、そしてそれらが本願の本旨および範囲ならびに添付の特許請求の範囲の範囲内に含まれることが理解されるべきである。
【0088】
実施例1
トマト由来のα-マンノシダーゼの精製および酵素アッセイ
トマト種子(cv.プサ ルビー(Pusa Ruby)および変異体)を、予め滅菌した土壌で発芽させ、その後、実生を、およそ25℃の温度、70%の湿度、および14/10hの明/暗管理の温室に移植した。分析のため、開花期に花にタグを付した後、緑熟期、催色期、桃熟期、および完熟期に果実を採取した。研究において使用された変異体は、Tomato Genetics Resource Center(TGRC)より取得された。
【0089】
果皮を、液体窒素中で粉砕し、1/4容量の抽出緩衝液(0.25M NaClおよび1mM PMSFを含む100mMトリス-Cl pH7.8)に懸濁させた。遠心分離後の上清を0〜70%硫酸アンモニウム沈殿に供し、10分間10,000gで遠心分離した。このようにして入手されたペレットを、25mMトリス-Cl pH7.8に対して一夜透析した。透析物をDEAEセファロースpH7.8上でのクロマトグラフィに供し、増加する塩勾配により溶出させた(補足図2A〜B)。100mMの塩濃度で溶出した画分をプールし、0〜80%硫酸アンモニウム沈殿に供した。ペレットを、少量の抽出緩衝液に溶解させ、sepadex G-100ゲルろ過カラム上に直接負荷した。
【0090】
酵素を欠くブランクと共に、940μlのMQ、20μlの20mM基質(pNP-α-D-マンノピリノシド)、および一定分量20μlの酵素からなる反応混合物1mlにおいて、37℃で15分間、マンノシダーゼをアッセイした。40μlの0.5M Na2CO3を添加することにより、反応を中止させた。pNP放出の結果として現れた色を、405nmで記録し、標準曲線を使用して定量化した。
【0091】
実施例2
二次元ゲル電気泳動、SDS-PAGE、およびイムノブロット分析
13cmのゲルストリップで、250μLの2-D再水和緩衝液で、精製されたタンパク質試料20μgを用いて、等電点フォーカシングを実施した。まず、タンパク質を、13cm長のイモビリン(immobiline)ドライストリップ(pH4-7)上で、受動的な再水和後に、電荷に従って分離した。エレクトロフォーカシングは、20℃で、20,000Vhr、IPGphorシステム(Amersham Biosciences)を使用して実施された。フォーカシング後のストリップを、10mlの平衡緩衝液において、1%(w/v)DTTにより還元し、続いて2.5%(w/v)ヨードアセトアミドによりアルキル化した。次いで、SDS-PAGE上での二次元目分離のため、ストリップを12.5%ポリアクリルアミドゲル上に負荷した。電気泳動されたタンパク質を、silver stain plusキット(Bio-Rad, CA, USA)により染色した。スポットをゲルから切り出し、エレクトロスプレーイオントラップ飛行時間型質量分析(LC-MS/MS)(Q-Star Pulsar i, Applied Biosystems)によって分析した。スペクトルを、緑色植物亜界(Viridiplantae)(緑色植物)データベースに対して、Mascot配列マッチングソフトウェア(www.matrixscience.com)によって分析した。
【0092】
イムノブロット分析のため、異なる成熟期からの全タンパク質50μgを定量化し、12.5%SDS-PAGEゲル上へ負荷した。電気泳動後、ゲルを150mAの定電流でHybond-C(Amersham)メンブレンに転写した。メンブレン上の非特異的な部位を、1時間、トリス緩衝生理食塩水(TBS)によりブロッキングし、4℃で一夜、一次抗体と共にインキュベートした。西洋ワサビペルオキシダーゼ結合抗ウサギ抗血清を二次抗体として用いて、免疫検出を1時間実施し、化学発光基質(Pierce Biotechnology)にブロットを曝した。
【0093】
SDS PSGEで、トマトマンノシダーゼは、70kDおよび40kDの二つのサブユニットへ分解された。しかしながら、非変性PAGE上でネイティブタンパク質のサイズは360kDであることが見出された。
【0094】
トマトの異なる成熟期におけるイムノブロット分析は、トマトのマンノシダーゼタンパク質蓄積が催色期に最大となることを確認した。
【0095】
実施例3
糖タンパク質染色およびEndoH消化
糖タンパク質染色を、製造業者の指示(GelCode(登録商標)Glycoprotein Staining Kit, PIERCE biotechnology)に従って実施した。EndoH消化を、製造業者の指示(New England Biolabs)に従って実施した。反応後、それを12.5%SDS PAGEで分解し、ニトロセルロースメンブレン上へブロットし、マンノシダーゼ抗体により検出した。
【0096】
染色実験は、マンノシダーゼが2〜3kDのグリカン部分を有する糖タンパク質であることを確認した。
【0097】
実施例3
クローニング、特徴決定、およびノーザンハイブリダイゼーション
多重配列アラインメントによって同定されたペプチドタグおよびモチーフを使用して、縮重プライマーを設計した。増幅断片を、pGEM T-Easyベクターにクローニングし配列決定した。次いで、残りの5'領域および3'領域を、RACE(Invitrogen)を使用して増幅した。
【0098】
Dynabeads(登録商標)mRNA purificationキット(DYNAL biotech)を使用して、全ての成熟期からの75マイクログラムの全RNAからメッセンジャーRNAを単離した。mRNAを一定の50ボルトで1.2%ホルムアルデヒドゲル/1×MOPSで分解し、Hybond-Nメンブレン(Amersham)にブロットし、リン酸緩衝液(0.5M NaPi pH7.2、7%SDS)中でハイブリダイズさせた。ハイブリダイゼーションを12時間実施し、メンブレンを-80℃で72時間hyperfilm(Amersham)に曝した。
【0099】
トマトマンノシダーゼおよびトウガラシマンノシダーゼをコードするポリペプチドを、pGEM-T Easyベクターにクローニングし、それぞれ、pGMTおよびpGMCと命名した。
【0100】
実施例4
免疫局在性
緑熟期および催色期の果実を採取し、果皮の5mm×5mm切片を4%パラホルムアルデヒドで一夜固定した。翌日、切片を1×PBSにより洗浄し、クリオスタット(Lieca CM1510S)上に乗せた。凍結組織を切片化し(10〜15μm)、1時間室温でスライド上で乾燥させた。スライドを1×PBSにより洗浄し、続いて、非特異的な部位をブロッキングするため室温で1時間3%BSA中でインキュベートし、1×PBSにより3回洗浄した。果実果皮切片を有するスライドを、マンノシダーゼのポリクローナル抗体と共に4℃で一夜インキュベートした。翌日、切片を、1×PBSにより3回洗浄し、FITCにより標識された二次抗体と共に3時間インキュベートした。次いで、10分間隔で8回、1×PBSにより切片を洗浄した。次いで、試料を適切なフィルターを有する蛍光顕微鏡において観察した。これにより、マンノシダーゼが細胞壁タンパク質であることが確認された。
【0101】
実施例5
高性能陰イオン交換クロマトグラフィ
タンパク質1μgを、各々100ngのN結合型オリゴ糖(Dextra, U.K)と共に37℃で一夜インキュベートし、酵素を除去するため、試料をPVDFメンブレンでろ過した。ろ過された試料10μlを分析のために使用した。勾配ポンプ(GP 40)、陰イオン交換カラム(Carbopac PA-I、4×250mm)、アルゴンによって溶出液を加圧するためのeluant degas module(EDM-2)が装備されたHPAE-PADシステム(Dionex DX 500 BioLC)を、単糖の分析のために使用した。分離された単糖を、金電極およびAg/AgCl基準電極が装備されたED 40検出器によって検出した。得られたクロマトグラフィデータを、PCに基づくoracle 2データ取得システム(Indtech Analytical, Bombay)を使用して、積分し、プロットした。結果を確認するため、各試料の分析の前後に、標準物(マンノース)を100 nmolの濃度で注入した。これにより、マンノシダーゼのN-グリカンプロセシング能力が確認された。
【0102】
実施例6
RNAi/アンチセンス/過剰発現/MYMIVプラスミドの構築およびアグロバクテリウムに基づく一過性形質転換
pHANNIBAL(細菌におけるアンピシリン耐性を有する)ベクターを使用して、マンノシダーゼをサイレンシングした。アンチセンス構築物は、GUS遺伝子を交換して、マンノシダーゼの全長を逆向きにクローニングすることにより、pBI 121で調製された。VIGSベクターについては、GFP配列を交換することにより600bpをMYMIVベクターにクローニングした。過剰発現構築物は、ゲートウェイ(gateway)クローニング法によってPK7FWG2ベクターにクローニングされた全長マンノシダーゼ遺伝子からなっていた。アグロインジェクションのため、アグロバクテリウム予備培養物(3ml)を、抗生物質を含むYEP(酵母抽出物1%、ペプトン1%、およびNaCl 0.5%)培地において28℃で個々のコロニーから24時間増殖させた。この培養物の1/10を、抗生物質を含む50mlの誘導培地(0.5%牛肉抽出物、0.1%酵母抽出物、0.5%ペプトン、0.5%ショ糖、2mM MgSO4、20mMアセトシリンゴン、10mM MES、pH5.6)に移し、一夜増殖させた。翌日、培養物を遠心分離によって回収し、浸透培地(10mM MgCl2、10mM MES、200mMアセトシリンゴン、pH5.6)に再懸濁させ、4時間、温和に撹拌しながら室温でインキュベートした。次いで、0.5mmの針を有する1mlの注射器を使用して、花柱の先端の近くで1〜2mmの深さにまで針を導入し、穏和に注入することにより、果実への培養物のアグロインジェクションを行った。
【0103】
実施例7
RNA単離および定量的リアルタイムRT-PCR
RNAを塩化リチウム法に従って単離し、nanodrop(ND-100)を使用して定量化した。5μgの全RNAをsuperscript II(Invitrogen)を使用してcDNAへと逆転写した。サイバーグリーン(cyber green)を用い、light Cycler Version 4.05(Roche diagnostics)を使用して、定量的RT-PCRを実施した。サイクルは、95℃で10分の初期Taq活性化、それに続く、94℃10s、55℃10s、および72℃10sからなる45サイクルからなっていた。次いで、増幅の正確さを確証するため融解曲線分析を実施した。2-ΔΔCT法を使用して、データは、内因性参照遺伝子に対して規準化された、対照と比べた遺伝子発現の変化倍率として提示される。アクチンをコードする遺伝子を、内因性対照として使用した。
【0104】
実施例8
低分子RNAの単離およびノーザンハイブリダイゼーション
全RNAを、以前に記載されたようにして単離した。LiClによりRNAを沈殿させた後、それを70%エタノールにより洗浄し、ペレットを空気乾燥させた。次いで、ペレットをDEPC水1mlに溶解させ、5分間65℃に加熱し、2分間氷上で冷却した。高分子量RNAを沈殿させるため、ポリエチレングリコール(分子量8000)およびNaClを、それぞれ5%および0.5Mの最終濃度で添加した。氷上で30分のインキュベーションの後、RNAを30分間13,000rpmで遠心分離した。上清を分離し、3倍容量のエタノールおよび1/10容量の3M酢酸ナトリウム(pH5.2)により沈殿させた。チューブを-20℃に一夜置いた。翌日、低分子量RNAを、10分間の13,000rpmでの遠心分離によって沈殿させた。ペレットを乾燥させ、DEPC水50μlに溶解させ、70Vで4〜5時間15%尿素PAGE上で分解した。次いで、ゲルを一定の100Vで1時間0.5×TBEを使用してナイロンメンブレンに転写した。メンブレンをUV架橋し、40℃で、50%ホルムアミド、7%SDS、および50mM Na2HPO4/NaH2PO4 pH7.2においてプレハイブリダイズさせた。4時間後、変性プローブを添加し、30℃で14〜16時間ハイブリダイズさせた。ブロットを、RTで2分間、2×SSCおよび1%SDSを使用して洗浄し、次いで、0.5×SSCおよび0.1%SDSにより洗浄し、フィルムに曝した。21〜23ヌクレオチドの遺伝子特異的なsiRNAが、RNAi果実において検出され、RNAiによって媒介されたマンノシダーゼのサイレンシングが確認された。
【0105】
実施例9
トマト形質転換
トランスジェニックトマト植物を生成するため、2週齢の実生からの子葉を、記載されるようにして使用した。トマト種子を4%市販漂白剤を使用して滅菌し、ムラシゲ スクーグ(Murashige and Skoog)(MS)培地上で発芽させた。発芽の2週間後、子葉を切り取り、異なる構築物を含有しているA.ツメファシエンス(tumefaciens)株EHA105と、30分間、共培養した。次いで、子葉を、選択のため、50mg/lカナマイシンを含有しているMSプレート上に収集した。小植物が再生した時、それらを発根培地に移した。発根後、植物を温室に移動させ、成熟するまで生長させた。
【0106】
実施例10
テクスチャ分析
果実の堅さを、TA-XT Plus(Stable Microsystems UK)を使用して決定した。各果実を、幅75mmのP75圧縮プレートにより分析し、1mm s-1の試験スピードで垂直の方向に5mmおよび10mmまで圧縮した。堅さは、5gの適用された力に対する応答力として定義された。値を、統計的有意性を決定するため、t検定に供した。これらの結果は、RNAi果実が非トランスジェニック果実より2.5倍大きい堅さを有することを示した。
【0107】
実施例11
実生に対するゲラニオール/ACC処理
プサ ルビー種子を滅菌し、MS培地上で発芽させた。15日後、実生を、20%DMSO中の10mMゲラニオールまたは水中の1mM ACC(1-アミノシクロプロパン-1-カルボン酸)を含有している液体培地に移した。採取された試料からRNAを単離し、リアルタイムRT-PCR分析のためcDNAへと逆転写した。発現を、対照実生(0時間)に対して相対的に計算した。結果は、マンノシダーゼがゲラニオールおよびエチレンによってアップレギュレートされることを示した。
【0108】
実施例12
染色および顕微鏡検
切片をクリオスタット付きミクロトーム(Leica 1050)で切断し、スライド上で乾燥させた。スライドを、0.1Mリン酸緩衝液(pH6.8)中の0.05%トルイジンブルー(sigma)の水性溶液に2分間浸漬し、水で2分間洗浄した。染色された切片を、カバーガラス下に水で乗せ、Nikon 80i落射蛍光/位相差/明視野顕微鏡を使用して、100倍の拡大率で撮影した。壁構造を調査するため、切片を、多糖に結合する蛍光増白剤である0.05%カルコフロールに浸し、蒸留水で洗浄した。切片を適切なフィルターにより調査した。細胞壁の成分に結合したカルコフロールによって、強い水色蛍光が生じた。
【0109】
SEQ ID NO:1 5'および3'のUTR領域を含むトマトα-マンノシダーゼのcDNA
SEQ ID NO:2 トマトα-マンノシダーゼのタンパク質配列
SEQ ID NO:3 5'および3'のUTR領域を含むトウガラシα-マンノシダーゼのcDNA配列
SEQ ID NO:4 トウガラシα-マンノシダーゼのタンパク質配列
【特許請求の範囲】
【請求項1】
マンノシダーゼ活性を有するポリペプチドをコードする単離されたポリヌクレオチドであって、該ポリヌクレオチドのヌクレオチド配列が、SEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードする、ポリヌクレオチド。
【請求項2】
SEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列を有するポリペプチドをコードする、請求項1記載のポリヌクレオチド。
【請求項3】
前記ポリヌクレオチドのヌクレオチド配列が、SEQ ID NO:1またはSEQ ID NO:3に記載される、請求項1記載のポリヌクレオチド。
【請求項4】
SEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列を有するポリペプチド。
【請求項5】
プロモーター配列に機能的に連結されている請求項1〜3のいずれか一項記載のポリヌクレオチドを含むDNA構築物。
【請求項6】
ポリヌクレオチド配列がセンスの向きである、請求項5記載のDNA構築物。
【請求項7】
ポリヌクレオチド配列がアンチセンスの向きである、請求項5記載のDNA構築物。
【請求項8】
トランスジェニック植物においてマンノシダーゼの発現を抑制するためのRNAi構築物であって、
SEQ ID NO:1またはSEQ ID NO:4に記載の配列に由来する少なくとも20個の連続ヌクレオチドを含むセンスポリヌクレオチド鎖、および
該センスポリヌクレオチド鎖とハイブリダイズするアンチセンスポリヌクレオチド鎖であって、該アンチセンスポリヌクレオチド鎖およびセンスポリヌクレオチド鎖が二重鎖を形成する、アンチセンスポリヌクレオチド鎖
を含むRNAi構築物。
【請求項9】
ヘアピン核酸である、請求項8記載のRNAi構築物。
【請求項10】
センス鎖が100〜600個のヌクレオチドを含む、請求項8記載のRNAi構築物。
【請求項11】
請求項5〜9のいずれか一項記載のDNA構築物を含む組換えベクター。
【請求項12】
請求項11記載の組換えベクターを含む組換え宿主細胞。
【請求項13】
アグロバクテリウム(Agrobacterium)、大腸菌(E.coli)、および酵母からなる群より選択される、請求項12記載の組換え宿主細胞。
【請求項14】
植物細胞、組織、またはそれらの一部を、マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドを含む組換えベクターによって形質転換する工程を含む、植物における果実軟化を遅延させる方法であって、該ポリヌクレオチドのヌクレオチド配列が、SEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードし、該ポリヌクレオチドがアンチセンスの向きである、方法。
【請求項15】
前記ポリヌクレオチドのヌクレオチド配列が、SEQ ID NO:1およびSEQ ID NO:3に記載される、請求項14記載の方法。
【請求項16】
植物における果実軟化を遅延させる方法であって、
マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドの少なくとも20個の連続ヌクレオチドの断片を少なくとも含むRNA干渉(RNAi)構築物の発現によって、トランスジェニック植物におけるマンノシダーゼのレベルを非トランスジェニック植物におけるマンノシダーゼのレベルと比較して減少させ、該ポリヌクレオチドのヌクレオチド配列が、SEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードする、工程;および
該非トランスジェニック植物と比べて低下したマンノシダーゼのレベルについて、得られた植物をスクリーニングする工程
を含む、方法。
【請求項17】
前記ポリヌクレオチドのヌクレオチド配列がSEQ ID NO:1およびSEQ ID NO:3に記載される、請求項16記載の方法。
【請求項18】
植物における果実軟化を遅延させる方法であって、
SEQ ID NO:1またはSEQ ID NO:3に記載の配列に由来する少なくとも20個の連続ヌクレオチドを含むセンスポリヌクレオチド鎖、および該センスポリヌクレオチド鎖にハイブリダイズするアンチセンスポリヌクレオチド鎖を含み、該アンチセンスポリヌクレオチド鎖およびセンスポリヌクレオチド鎖が二重鎖を形成する、RNA干渉(RNAi)構築物の発現によって、トランスジェニック植物におけるマンノシダーゼのレベルを非トランスジェニック植物におけるマンノシダーゼのレベルと比較して減少させる工程;ならびに
該非トランスジェニック植物と比べて低下したマンノシダーゼのレベルについて、得られた植物をスクリーニングする工程
を含む、方法。
【請求項19】
RNAi構築物がヘアピン核酸である、請求項18記載の方法。
【請求項20】
マンノシダーゼの発現が果実軟化を遅延させるために制御されている、請求項14〜19のいずれか一項記載の方法によって作製されたトランスジェニック植物。
【請求項21】
トマト、トウガラシ、パパイア、マンゴー、バナナ、モモ、セイヨウナシ、柑橘類、パイナップル、グアバ、アボカド、イチゴ、リンゴ、およびザクロからなる群より選択される、請求項20記載のトランスジェニック植物。
【請求項22】
トマトまたはトウガラシである、請求項20記載のトランスジェニック植物。
【請求項23】
請求項18〜22のいずれか一項記載のトランスジェニック植物の、トランスジェニック種子または子孫。
【請求項1】
マンノシダーゼ活性を有するポリペプチドをコードする単離されたポリヌクレオチドであって、該ポリヌクレオチドのヌクレオチド配列が、SEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードする、ポリヌクレオチド。
【請求項2】
SEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列を有するポリペプチドをコードする、請求項1記載のポリヌクレオチド。
【請求項3】
前記ポリヌクレオチドのヌクレオチド配列が、SEQ ID NO:1またはSEQ ID NO:3に記載される、請求項1記載のポリヌクレオチド。
【請求項4】
SEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列を有するポリペプチド。
【請求項5】
プロモーター配列に機能的に連結されている請求項1〜3のいずれか一項記載のポリヌクレオチドを含むDNA構築物。
【請求項6】
ポリヌクレオチド配列がセンスの向きである、請求項5記載のDNA構築物。
【請求項7】
ポリヌクレオチド配列がアンチセンスの向きである、請求項5記載のDNA構築物。
【請求項8】
トランスジェニック植物においてマンノシダーゼの発現を抑制するためのRNAi構築物であって、
SEQ ID NO:1またはSEQ ID NO:4に記載の配列に由来する少なくとも20個の連続ヌクレオチドを含むセンスポリヌクレオチド鎖、および
該センスポリヌクレオチド鎖とハイブリダイズするアンチセンスポリヌクレオチド鎖であって、該アンチセンスポリヌクレオチド鎖およびセンスポリヌクレオチド鎖が二重鎖を形成する、アンチセンスポリヌクレオチド鎖
を含むRNAi構築物。
【請求項9】
ヘアピン核酸である、請求項8記載のRNAi構築物。
【請求項10】
センス鎖が100〜600個のヌクレオチドを含む、請求項8記載のRNAi構築物。
【請求項11】
請求項5〜9のいずれか一項記載のDNA構築物を含む組換えベクター。
【請求項12】
請求項11記載の組換えベクターを含む組換え宿主細胞。
【請求項13】
アグロバクテリウム(Agrobacterium)、大腸菌(E.coli)、および酵母からなる群より選択される、請求項12記載の組換え宿主細胞。
【請求項14】
植物細胞、組織、またはそれらの一部を、マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドを含む組換えベクターによって形質転換する工程を含む、植物における果実軟化を遅延させる方法であって、該ポリヌクレオチドのヌクレオチド配列が、SEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードし、該ポリヌクレオチドがアンチセンスの向きである、方法。
【請求項15】
前記ポリヌクレオチドのヌクレオチド配列が、SEQ ID NO:1およびSEQ ID NO:3に記載される、請求項14記載の方法。
【請求項16】
植物における果実軟化を遅延させる方法であって、
マンノシダーゼ活性を有するポリペプチドをコードするポリヌクレオチドの少なくとも20個の連続ヌクレオチドの断片を少なくとも含むRNA干渉(RNAi)構築物の発現によって、トランスジェニック植物におけるマンノシダーゼのレベルを非トランスジェニック植物におけるマンノシダーゼのレベルと比較して減少させ、該ポリヌクレオチドのヌクレオチド配列が、SEQ ID NO:2またはSEQ ID NO:4に記載のアミノ酸配列と少なくとも約85%の同一性を有するポリペプチドをコードする、工程;および
該非トランスジェニック植物と比べて低下したマンノシダーゼのレベルについて、得られた植物をスクリーニングする工程
を含む、方法。
【請求項17】
前記ポリヌクレオチドのヌクレオチド配列がSEQ ID NO:1およびSEQ ID NO:3に記載される、請求項16記載の方法。
【請求項18】
植物における果実軟化を遅延させる方法であって、
SEQ ID NO:1またはSEQ ID NO:3に記載の配列に由来する少なくとも20個の連続ヌクレオチドを含むセンスポリヌクレオチド鎖、および該センスポリヌクレオチド鎖にハイブリダイズするアンチセンスポリヌクレオチド鎖を含み、該アンチセンスポリヌクレオチド鎖およびセンスポリヌクレオチド鎖が二重鎖を形成する、RNA干渉(RNAi)構築物の発現によって、トランスジェニック植物におけるマンノシダーゼのレベルを非トランスジェニック植物におけるマンノシダーゼのレベルと比較して減少させる工程;ならびに
該非トランスジェニック植物と比べて低下したマンノシダーゼのレベルについて、得られた植物をスクリーニングする工程
を含む、方法。
【請求項19】
RNAi構築物がヘアピン核酸である、請求項18記載の方法。
【請求項20】
マンノシダーゼの発現が果実軟化を遅延させるために制御されている、請求項14〜19のいずれか一項記載の方法によって作製されたトランスジェニック植物。
【請求項21】
トマト、トウガラシ、パパイア、マンゴー、バナナ、モモ、セイヨウナシ、柑橘類、パイナップル、グアバ、アボカド、イチゴ、リンゴ、およびザクロからなる群より選択される、請求項20記載のトランスジェニック植物。
【請求項22】
トマトまたはトウガラシである、請求項20記載のトランスジェニック植物。
【請求項23】
請求項18〜22のいずれか一項記載のトランスジェニック植物の、トランスジェニック種子または子孫。
【図1a】

【図1b】
【図1c】
【図1d】
【図1e】

【図1f】
【図1g】
【図1h】
【図2a】
【図2b】
【図2c】
【図2d】
【図2e】
【図3a】
【図3b】
【図3c】
【図4a】
【図4b】

【図4c】

【図4d】
【図1b】
【図1c】
【図1d】
【図1e】
【図1f】
【図1g】
【図1h】
【図2a】
【図2b】
【図2c】
【図2d】
【図2e】
【図3a】
【図3b】
【図3c】
【図4a】
【図4b】
【図4c】
【図4d】
【公表番号】特表2011−527563(P2011−527563A)
【公表日】平成23年11月4日(2011.11.4)
【国際特許分類】
【出願番号】特願2011−517315(P2011−517315)
【出願日】平成21年7月9日(2009.7.9)
【国際出願番号】PCT/IN2009/000387
【国際公開番号】WO2010/004582
【国際公開日】平成22年1月14日(2010.1.14)
【出願人】(510311274)
【出願人】(510311285)
【出願人】(510311296)
【出願人】(510311300)
【出願人】(510311311)
【Fターム(参考)】
【公表日】平成23年11月4日(2011.11.4)
【国際特許分類】
【出願日】平成21年7月9日(2009.7.9)
【国際出願番号】PCT/IN2009/000387
【国際公開番号】WO2010/004582
【国際公開日】平成22年1月14日(2010.1.14)
【出願人】(510311274)
【出願人】(510311285)
【出願人】(510311296)
【出願人】(510311300)
【出願人】(510311311)
【Fターム(参考)】
[ Back to top ]