
枯草菌変異株
【課題】タンパク質又はポリペプチドの生産性向上を可能とする枯草菌変異株、及び当該枯草菌を用いたタンパク質又はポリペプチドの製造方法の提供。
【解決手段】枯草菌変異株MGB874株に対して、rocRを導入せず且つrocD、rocE、及びrocFから選ばれる1以上の枯草菌遺伝子を導入するか、又は枯草菌遺伝子rocD及びrocRを導入した枯草菌変異株。
【解決手段】枯草菌変異株MGB874株に対して、rocRを導入せず且つrocD、rocE、及びrocFから選ばれる1以上の枯草菌遺伝子を導入するか、又は枯草菌遺伝子rocD及びrocRを導入した枯草菌変異株。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、有用なタンパク質又はポリペプチドの製造に用いる枯草菌変異株、及び当該枯草菌を用いたタンパク質又はポリペプチドの製造方法に関する。
【背景技術】
【0002】
微生物による有用物質の工業的生産は、アルコール飲料や味噌、醤油等の食品類をはじめとし、アミノ酸、有機酸、核酸関連物質、抗生物質、糖質、脂質、タンパク質等、その種類は多岐に渡っており、またその用途についても食品、医薬品、洗浄剤、化粧品等の日用品、或いは各種化成品原料に至るまで幅広い分野に広がっている。
【0003】
こうした微生物による有用物質の工業生産においては、その生産性の向上が重要な課題の一つであり、その手法として、突然変異等の遺伝学的手法による生産菌の育種が行われてきた。特に最近では、微生物遺伝学、バイオテクノロジーの発展により、遺伝子組換え技術等を用いたより効率的な生産菌の育種が行われるようになっている。さらに、近年のゲノム解析技術の急速な発展を受けて、対象とする微生物のゲノム情報を解読し、これらを積極的に産業に応用しようとする試みもなされている。
【0004】
近年、枯草菌については、枯草菌ゲノムに存在する約4100種類の遺伝子の破壊株が網羅的に研究され、271個の遺伝子が成育に必須であることが指摘されている(非特許文献1)。また、枯草菌等の胞子形成初期に関わる遺伝子やプロテアーゼ遺伝子、又は細胞壁或いは細胞膜中のテイコ酸へのD-アラニン付加に関わる遺伝子、更にはサーファクチンの生合成或いは分泌に関わる遺伝子を単独に欠失又は不活性化した菌株が構築されている(特許文献1〜献8参照)。
【0005】
しかるところ、本出願人は、枯草菌に由来する遺伝子破壊株を網羅的に解析し、枯草菌ゲノムの大量域を欠失させ、各種酵素の生産性に優れた変異株を見出すことに成功している(特許文献9)。
【0006】
しかしながら、限定された生産培地と安定な培養条件が用いられる工業的生産においては、タンパク質又はポリペプチドの生産にとって必要或いは不必要な遺伝子が存在すると考えられ、更なる研究が求められている。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】特開昭58-190390号公報
【特許文献2】特開昭61-1381号公報
【特許文献3】国際公開第89/04866号パンフレット
【特許文献4】特表平11-509096号公報
【特許文献5】特許第3210315号公報
【特許文献6】特表2001-527401号公報
【特許文献7】特表2002−520017号公報
【特許文献8】特表2001−503641号公報
【特許文献9】特開2007-130013号公報
【非特許文献】
【0008】
【非特許文献1】K. Kobayashi et al., Proc. Natl. Acad. Sci. USA., 100, 4678-4683, 2003
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明は、タンパク質又はポリペプチドの生産性向上を可能とする枯草菌変異株、及び当該枯草菌を用いたタンパク質又はポリペプチドの製造方法を提供することに関する。
【課題を解決するための手段】
【0010】
本発明者らは、枯草菌Bacillus subtilis Marburg No.168(枯草菌168株)を野生株とし、そのゲノムの合計874kbが削除された枯草菌変異株MGB874株を作製した。枯草菌変異株MGB874株は野生株である168株に比較し、目的タンパク質をコードする遺伝子が発現可能に導入された際に、その目的タンパク質の分泌生産性が有意に向上する変異株であった(前記特許文献9)。したがって削除した遺伝子は目的タンパク質の分泌生産においては不要あるいは有害な遺伝子であると推察された。しかし、種々検討したところ予想に反して、MGB874株において削除したpdp-rocR領域の96遺伝子のうちアルギニン分解系に関与する遺伝子rocRを導入せず且つrocF、rocE及びrocDから選ばれる遺伝子をMGB874株に導入した場合、又はrocD及びrocRをMGB874株に導入した場合に、目的のタンパク質又はポリペプチドの生産性が、導入前と比較して有意に向上することを見出した。
【0011】
すなわち本発明は、以下の1)〜7)に係るものである。
1)枯草菌変異株MGB874株に対して、rocRを導入せず且つrocD、rocE及びrocFから選ばれる1以上の枯草菌遺伝子を導入するか、又はrocD及びrocRを導入した枯草菌変異株。
2)上記1)の枯草菌変異株に、異種のタンパク質又はポリペプチドをコードする遺伝子を導入した組換え枯草菌。
3)異種のタンパク質又はポリペプチドをコードする遺伝子の上流に転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域から選ばれる1以上の領域を結合した上記1)又は2)の組換え枯草菌。
4)転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域を結合した上記3)の組換え枯草菌。
5)分泌シグナル領域がバチルス(Bacillus)属細菌のセルラーゼ遺伝子由来のものであり、転写開始制御領域及び翻訳開始制御領域が当該セルラーゼ遺伝子の開始コドンから始まる長さ0.6〜1kbの上流領域由来のものである上記3)又は4)の組換え枯草菌。
6)転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域が、配列番号1で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜659の塩基配列又は当該塩基配列のいずれかと70%以上の同一性を有する塩基配列からなるDNA断片、又は当該塩基配列の一部が欠失した塩基配列からなるDNA断片である上記4)の組換え枯草菌。
7)2)〜6)のいずれかの組換え枯草菌を用いるタンパク質又はポリペプチドの製造方法。
【発明の効果】
【0012】
本発明によれば、各種タンパク質又はポリペプチドの生産性に優れた枯草菌変異株が提供され、当該枯草菌変異株を用いることによって、生産性に優れた各種酵素の工業的生産が実現できる。また、当該枯草菌変異株は、各種酵素の産生メカニズム等の解明にも有用な生物材料となり得る。
【図面の簡単な説明】
【0013】
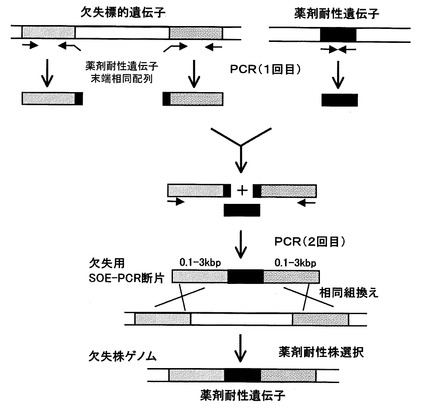
【図1】枯草菌のゲノム上から所定の領域を欠失させる方法の一例を説明するための模式図である。
【発明を実施するための形態】
【0014】
本発明の枯草菌変異株を構築するための親微生物には、枯草菌変異株MGB874株が用いられる。MGB874株は、枯草菌Bacillus subtilis Marburg No.168(枯草菌168株)を野生株とし、そのゲノムの大量域、すなわちprophage6 (yoaV-yobO)領域、prophage1 (ybbU-ybdE)領域、prophage4 (yjcM-yjdJ)領域、PBSX (ykdA-xlyA)領域、prophage5 (ynxB-dut)領域、prophage3 (ydiM-ydjC)領域、spb (yodU-ypqP)領域、pks (pksA-ymaC)領域、skin (spoIVCB-spoIIIC)領域、pps (ppsE-ppsA)領域、prophage2 (ydcL-ydeJ)領域、ydcL-ydeK-ydhU領域、yisB-yitD領域、yunA-yurT領域、cgeE-ypmQ領域、yeeK-yesX領域、pdp-rocR領域、ycxB-sipU領域、SKIN-Pro7 (spoIVCB-yraK)領域、sbo-ywhH領域、yybP-yyaJ領域及びyncM-fosB領域を欠失させたものである(前記特許文献9)。表1に示した当該欠失領域は、同表に示す一対のオリゴヌクレオチド・セットにより挟み込まれる領域として言い換えることができる。
【0015】
【表1】

【0016】
本発明において導入される遺伝子は、pdp-rocR領域の96遺伝子のうちアルギニン分解系の遺伝子である。アルギニンはアルギニントランスポーター[RocC、RocEおよびArtPQM(Genome Biol. 2: research: 0019.1)]により細胞内に取り込まれた後、RocFによりオルニチンとなり、RocD、RocAによってグルタミン酸まで変換された後、RocGにより2-オキソグルタル酸となることで分解される。RocBはカルバモイルリン酸とシトルリンからオルニチンを合成する酵素であると言われているが詳細な報告は論文として未発表である[J. Bacteriol. 186, 3399 (2004)]。転写因子RocRはアルギニン分解経路における遺伝子(rocABC, rocDEF, rocG)の転写を活性化し[Proc. Natl. Acad. Sci. USA 96, 10290 (1999), J. Bacteriol. 176, 1234 (1994)]、そのDNA結合活性はオルニチンで活性化すると推定されている[Mol. Microbiol. 24, 825 (1997), Nat. Rev. Microbiol. 5, 917 (2007)]。
rocD遺伝子は、ornithine aminotransferase、rocE 遺伝子は、arginine permease、rocF 遺伝子は、arginaseと推定されている[J. Mol. Biol. 249, 843(1995)]。
斯かる遺伝子は、枯草菌MGB874株の作製において削除されたpdp-rocR領域に属するものであり、従って、本発明の枯草菌変異株は、枯草菌MGB874株の作製において削除された遺伝子のうちrocRを復帰させることなく、少なくともrocD、rocE、及びrocFから選ばれる1以上の遺伝子を復帰させた株、又はrocD及びrocRを復帰させた株と言える。
rocD、rocE、及びrocFの各遺伝子は、1種又はそれ以上を適宜組み合わせて導入すればよく、3遺伝子を全て導入してもよく、rocD及びrocRを復帰させる場合は2遺伝子を導入する。さらに、rocRを復帰させることなく少なくともrocD、rocE、及びrocFから選ばれる1以上の遺伝子を復帰させる場合において、異種タンパク質又はポリペプチドの生産性を抑制しない他の遺伝子を導入してもよい。
【0017】
本発明の枯草菌株の構築方法は特に限定されないが、rocD、rocE、及びrocFから選ばれる1以上の遺伝子を含むDNA断片あるいはゲノムDNAを供与体DNAとして用いて、受容体DNAである枯草菌MGB874株ゲノムに対して、rocRを導入せず且つrocD、rocE、及びrocFから選ばれる1以上の遺伝子を含むDNAを導入する方法、又はrocD及びrocR遺伝子を導入する方法を実施することができる。
例えば、rocD、rocE、及びrocFから選ばれる1以上の遺伝子を含むゲノムDNAを供与体DNAとして利用する場合、アルギニン分解系の遺伝子rocF、rocE、rocD、rocRを含むpdp-rocR領域を有する枯草菌菌株例えばMGB625株のゲノムDNAを利用して、枯草菌MGB874株ゲノムにrocF、rocE、rocD、rocRを含むpdp-rocR領域全体を復帰させることもできる。MGB625株は、枯草菌168株を元に、表1に示された領域の一部であるprophage6 (yoaV-yobO)領域、prophage1 (ybbU-ybdE)領域、prophage4 (yjcM-yjdJ)領域、PBSX (ykdA-xlyA)領域、prophage5 (ynxB-dut)領域、prophage3 (ydiM-ydjC)領域、spb (yodU-ypqP)領域、pks (pksA-ymaC)領域、skin (spoIVCB-spoIIIC)領域、pps (ppsE-ppsA)領域、prophage2 (ydcL-ydeJ)領域、ydcL-ydeK-ydhU領域、yisB-yitD領域、yunA-yurT領域、cgeE-ypmQ領域、yeeK-yesX領域を削除して構築された株である(前記特許文献9参照)。
あるいは、MGB625株からpdp-rocR領域の一部を削除した菌株のゲノムDNAを供与体DNAとして、pdp-rocR領域のうちrocF、rocE、rocD、rocRの一部を含む領域のみを復帰させることもできる。
以下に、rocF、rocE、rocD、rocRの一部を含む領域のみを枯草菌MGB874株ゲノムに復帰させるために用いる供与体DNAを、MGB625株を利用して構築する例を説明する。
ただし、利用する枯草菌株は特にMGB625株に限定するものではなく、rocRを含まず且つrocF、rocE、rocD遺伝子から選ばれる1以上の遺伝子を含むか、又はrocD及びrocRを含む枯草菌であればよい。また、rocRのみを復帰させる実験は比較のために行う。
【0018】
まず、MGB625株から、以下に示す方法により薬剤耐性遺伝子をマーカー遺伝子として、pdp-rocR領域の一部にあたるpdp-phrG領域、pdp- rocF領域、pdp- rocE領域、pdp-rocD領域のいずれかを削除した菌株を構築する。その結果構築される菌株は、それぞれrocF-rocE-rocD-rocRがゲノム上に残存する菌株、rocE-rocD-rocRがゲノム上に残存する菌株、rocD-rocRがゲノム上に残存する菌株、rocRがゲノム上に残存する菌株となる。
【0019】
枯草菌ゲノム上から領域削除する方法としては、特に限定されないが、例えば図1に示す以下の方法を適用することができる。本方法で用いる欠失用DNA断片は、欠失対象領域の上流に隣接する約0.1〜3kb断片(上流断片と称する)と、薬剤耐性遺伝子マーカーを含む断片、同じく下流に隣接する約0.1〜3kb断片(下流断片と称する)が連結したDNA断片である。
例えば、薬剤耐性遺伝子マーカーとしてクロラムフェニコール耐性遺伝子を用いる場合、枯草菌168株から抽出したゲノムDNAを鋳型とし、後記表2に示す“pdpfw2”と“pdprv/Cm”のプライマーセットを用いて、ゲノム上の当該遺伝子の上流に隣接する断片(A)を調製する。また、“削除領域下流に隣接する遺伝子名fw/Cm”と“削除領域下流に隣接する遺伝子名rv2”のプライマーセットを用いて、下流に隣接する断片(B)をそれぞれ調製する。
【0020】
一方、プラスミドpC194[J. Bacteriol. 150 (2), 815 (1982)]のクロラムフェニコール耐性遺伝子をプラスミドpUC18のXbaI−BamHI切断点に挿入した組換えプラ
スミドpCBB31を鋳型とし、表2に示したCmFとCmRプライマーセットを用いて、クロラムフェニコール耐性遺伝子を含む1kb断片(C)を調製する。
次に、得られた(A)(B)(C)3断片を混合して鋳型とし、表2のプライマー“pdpfw1”と“削除領域下流に隣接する遺伝子名rv1”を用いたSOE−PCRを行うことによって、3断片を(A)(C)(B)の順になる様に結合させたDNA断片を得る(図1参照)。
すなわち、(B)がプライマー番号14と6のプラマーセット由来断片の場合はプライマー番号2、5を用いてSOE−PCRを行い、(B)がプライマー番号15と8のプライマーセット由来断片の場合はプライマー番号2、7を用いてSOE−PCRを行い、(B)がプライマー番号16と10のプライマーセット由来断片の場合はプライマー番号2、9を用いてSOE−PCRを行い、(B)がプライマー番号17と12のプライマーセット由来断片の場合はプライマー番号2、11を用いてSOE−PCRを行う。
これらのDNA断片のそれぞれを用いてコンピテントセル法[J.Bacteriol.,81,741(1960)]により枯草菌MGB625株の形質転換を行う。すなわち、枯草菌をSPI培地(0.20 % 硫酸アンモニウム、1.40 % リン酸水素二カリウム、0.60 % リン酸二水素カリウム、0.10 % クエン酸三ナトリウム二水和物、0.50 % グルコース、0.02 % カザミノ酸(Difco)、5 mM 硫酸マグネシウム、0.25μM 塩化マンガン、50μg/ml トリプトファン)において37℃で、生育度(OD 600)の値が1程度になるまで振盪培養する。振盪培養後、培養液の一部を9倍量のSPII培地(0.20 % 硫酸アンモニウム、1.40 % リン酸水素二カリウム、0.60 % リン酸二水素カリウム、0.10 % クエン酸三ナトリウム二水和物、0.50 % グルコース、0.01 % カザミノ酸(Difco)、5 mM 硫酸マグネシウム、0.4 0μM 塩化マンガン、5μg/ml トリプトファン)に接種し、更に生育度(OD 600)の値が0.4程度になるまで振盪培養することで、枯草菌のコンピテントセルを調製する。次いで調製したコンピテントセル懸濁液(SPII培地における培養液)100μLに上記DNA断片を含む溶液(上記SOE−PCRの反応液)5μLを添加し、37℃で1時間振盪培養後、クロラムフェニコール含むLB寒天培地(1 % トリプトン、0.5 % 酵母エキス、1 % NaCl、1.5 % 寒天) に全量を塗沫する。37℃における静置培養の後、生育したコロニーを形質転換体として分離する。
形質転換体のゲノムDNAを抽出し、これを鋳型DNAとしたPCRにより、目的の領域削除が行われていることを確認することができる。ゲノムDNAの抽出は、斉藤と三浦(Saito & Miura)の方法[Saito & Miura Biochem. Biophys. Acta. 72, 619-629 (1963)
]、あるいは、市販のゲノムDNA抽出キット、例えば、UltraClean Microbial DNA Isolation Kit(MO BIO Laboratories, Inc.)を用いて行うことができる。
【0021】
次に、抽出した上記ゲノムDNAのそれぞれを用いてコンピテントセル法によりMGB874株を形質転換する。得られた各形質転換体のゲノムを抽出し、MGB874株ゲノム上のpdp-rocR領域が削除された位置に、クロラムフェニコール耐性遺伝子とrocF-rocE-rocD-rocR、又は、クロラムフェニコール耐性遺伝子とrocE-rocD-rocR、又は、クロラムフェニコール耐性遺伝子とrocD-rocR、又は、rocRが挿入していること、および、MGB874株のpdp-rocR領域以外の削除領域に復帰が生じていないことをPCRにより確認する。斯くして、rocF-rocE-rocD-rocRが復帰した株(以降、MGB874+rocDEFR株)、rocE-rocD-rocRが復帰した株(以降、MGB874+rocEFR株)、rocD-rocRが復帰した株(以降、MGB874+rocDR株) 、rocRが復帰した株(以降、MGB874+rocR株)を取得できる。
更に、MGB874+rocDEFR株、MGB874+rocEFR株MGB874+rocDR株から以下の方法によりネオマイシン耐性遺伝子をマーカー遺伝子としてrocRを削除することで、MGB874株に対して、rocF-rocE-rocDが復帰した株、rocE-rocDが復帰した株、rocDが復帰した株を構築する。
【0022】
rocRの削除の方法としては、特に限定されないが、上述した領域削除の手順において用いたのとは異なる薬剤耐性遺伝子マーカーを用いて同様にして作製することができる(図1参照)。例えば、薬剤耐性遺伝子マーカーとしてネオマイシン耐性遺伝子を用いる場合、枯草菌168株から抽出したゲノムDNAを鋳型とし、表2に示したrocR-FWとrocR/neoR、及びrocR/neoFとrocR-RVの各プライマーセットを用いて、ゲノム上のrocR遺伝子の上流に隣接する断片(A)、及び下流に隣接する断片(B)をそれぞれ調製する。
一方、プラスミドpBEST501[Nucleic Acids Res. 17、4410(1989)]のネオマイシン耐性遺伝子を鋳型とし、表2に示したneofとneorプライマーセットを用いて、ネオマイシン耐性遺伝子を含む1.3kb断片(C)を調製する。次に、得られた(A)(B)(C)3断片を混合して鋳型とし、表のプライマーrocRFW2とrocR-RV2を用いたSOE−PCRを行うことによって、3断片を(A)(C)(B)の順になる様に結合させたDNA断片を得る(図1参照)。
このDNA断片を用いてコンピテントセル法によりMGB874+rocDEFR株、MGB874+rocEFR株、MGB874+rocDR株の形質転換を行い、得られた形質転換体のゲノムを抽出し、PCRによってrocR遺伝子が欠失され、ネオマイシン耐性遺伝子に置換していることを確認する。斯くして、rocF-rocE-rocDが復帰した株(以降、MGB874+rocDEF株)、rocE-rocDが復帰した株(以降、MGB874+rocDE株)、rocDが復帰した株(以降、MGB874+rocD株)を取得することができる。
【0023】
次に、斯くして得られた枯草変異株に、目的とするタンパク質又はポリペプチドをコードする遺伝子を導入することによって、本発明の組換え枯草菌を得ることができる。
目的タンパク質又はポリペプチド遺伝子は特に限定されず、洗浄剤用、食品用、繊維処理用、飼料処理用、化粧品用、医薬品用、診断薬用など各種産業用酵素や、生理活性ペプチドなどが含まれる。また、産業用酵素の機能別には、酸化還元酵素 (Oxidoreductase) 、転移酵素 (Transferase) 、加水分解酵素 (Hydrolase) 、脱離酵素 (Lyase)、異性化酵素 (Isomerase) 、合成酵素 (Ligase/Synthetase) 等が含まれるが、好適にはセルラーゼ、α-アミラーゼ、プロテアーゼ等の加水分解酵素の遺伝子が挙げられる。
【0024】
セルラーゼとしては、例えば、多糖加水分解酵素の分類(Biochem.J.,280,309,1991)
中でファミリー5に属するセルラーゼが挙げられ、中でも微生物由来、特にBacillus属細菌由来のセルラーゼが挙げられる。より具体的な例として、配列番号2で示されるアミノ酸配列からなるBacillus属細菌KSM-S237株(FERM BP-7875)由来のアルカリセルラーゼ、又は、配列番号4で示されるアミノ酸配列からなるBacillus属細菌KSM-64株(FERM BP-2886)由来のアルカリセルラーゼ、或いは、当該アミノ酸配列と70%、好ましくは80%、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有するアミノ酸配列からなるセルラーゼ、又は配列番号2又は配列番号4で示されるアミノ酸配列において1若しくは数個のアミノ酸が置換、欠失若しくは付加されたアミノ酸配列からなるアルカリセルラーゼが挙げられる。
【0025】
α−アミラーゼの具体例としては、微生物由来のα−アミラーゼが挙げられ、特にBacillus属細菌由来の液化型アミラーゼが好ましい。より具体的な例として、配列番号5で示されるアミノ酸配列からなるBacillus属細菌KSM-K38株(FERM BP-6946)由来のアルカリアミラーゼ、或いは当該アミノ酸配列と70%、好ましくは80%、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有するアミノ酸配列からなるアミラーゼ、又は配列番号5で示されるアミノ酸配列において1若しくは数個のアミノ酸が置換、欠失若しくは付加されたアミノ酸配列からなるアルカリアミラーゼが挙げられる。
【0026】
プロテアーゼの具体例としては、微生物由来、特にBacillus属細菌由来のセリンプロテアーゼや金属プロテアーゼ等が挙げられる。より具体的な例として、配列番号6で示されるアミノ酸配列からなるバチルス クラウジ(Bacillus clausii)KSM-K16株(FERM BP-3376)由来のアルカリプロテアーゼ、或いは当該アミノ酸配列と70%、好ましくは80%、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有するアミノ酸配列からなるプロテアーゼ、又は配列番号6で示されるアミノ酸配列において1若しくは数個のアミノ酸が置換、欠失若しくは付加されたアミノ酸配列からなるアルカリプロテアーゼが挙げられる。
【0027】
尚、本発明においてアミノ酸配列および塩基配列の同一性はLipman-Pearson法 (Science,227,1435,1985)によって計算される。具体的には、遺伝情報処理ソフトウェアGenetyx-Win(ソフトウェア開発)のホモロジー解析(Search homology)プログラムを用いて、Unit size to compare(ktup)を2として解析を行うことにより算出される。
また、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列としては、1乃至10個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列が好ましく、また、付加には、両末端への1〜数個のアミノ酸の付加が含まれる。
【0028】
本発明の微生物に導入される目的タンパク質又はポリペプチドの遺伝子は、その上流に当該遺伝子の転写、翻訳、分泌に関わる制御領域、即ち、プロモーター及び転写開始点を含む転写開始制御領域、リボソーム結合部位及び開始コドンを含む翻訳開始領域並びに分泌シグナルペプチド領域から選ばれる1以上の領域が適正な形で結合されていることが望ましい。特に、転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域が結合されていることが好ましく、更に分泌シグナルペプチド領域がバチルス(Bacillus)属細菌のセルラーゼ遺伝子由来のものであり、転写開始制御領域及び翻訳開始制御領域が当該セルラーゼ遺伝子の開始コドンから始まる長さ0.6〜1kbの上流領域であるものが、目的のタンパク質又はポリペプチド遺伝子と適正な形で結合されていることが望ましい。例えば、特開2000-210081号公報や特開平4-190793号公報等に記載されているバチルス(Bacillus)属細菌、すなわちKSM-S237株(FERM BP-7875)、KSM-64株(FERM BP-2886)由来のセルラーゼ遺伝子の転写開始制御領域、翻訳開始領域及び分泌シグナルペプチド領域が目的のタンパク質又はポリペプチドの構造遺伝子と適正に結合されていることが望ましい。より具体的には配列番号1で示される塩基配列の塩基番号1〜659の塩基配列、配列番号3で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜696の塩基配列、また当該塩基配列に対して70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有する塩基配列からなるDNA断片、又は上記いずれかの塩基配列からなるDNAとストリンジェントの条件でハイブリダイズし且つ遺伝子の転写、翻訳、分泌に関わる機能を有するDNA、或いは上記いずれかの塩基配列の一部が欠失した塩基配列からなるDNA断片が、目的のタンパク質又はポリペプチドの構造遺伝子と適正に結合されていることが望ましい。尚、ここで、上記塩基配列の一部が欠失した塩基配列からなるDNA断片とは、上記塩基配列の一部を欠失しているが、遺伝子の転写、翻訳、分泌に関わる機能を保持しているDNA断片を意味する。また此処で言うストリンジェントな条件とは、例えば[Molecular cloning-a Laboratory manual 2nd edition(Sambrookら、1989)]に記載の条件等が挙げられる。例えば、6XSSC(1XSSCの組成:0.15M塩化ナトリウム、0.015Mクエン酸ナトリウム、pH7.0)、0.5%SDS、5Xデンハート及び100mg/mLニシン精子DNAを含む溶液にプローブとともに65℃で8〜16時間恒温し、ハイブリダイズさせる条件が挙げられる。
【0029】
上記の目的タンパク質又はポリペプチド遺伝子を含むDNA断片と適当なプラスミドベクターを結合させた組換えプラスミドを、一般的な形質転換法によって上記枯草菌変異株に取り込ませることによって、本発明の組換え枯草菌を得ることができる。また、当該DNA断片に枯草菌変異株のゲノムとの適当な相同領域を結合したDNA断片を用い、枯草菌変異株ゲノムに直接組み込むことによっても本発明の組換え枯草菌を得ることができる。
【0030】
本発明の組換え枯草菌を用いた目的のタンパク質又はポリペプチドの生産は、当該菌株を同化性の炭素源、窒素源、その他の必須成分を含む培地に接種し、通常の微生物培養法にて培養し、培養終了後、タンパク質又はポリペプチドを採取・精製することにより行えばよい。そして、後記実施例に示すように、目的のタンパク質又はポリペプチドの生産性は、本発明の遺伝子を導入していない枯草菌変異株MGB874株を用いた場合と比較して、その向上が達成されている。
【0031】
以下に、当該発明の組換え微生物の構築方法と及び当該組換え微生物を用いたセルラーゼ生産方法について具体的に説明する。
【実施例】
【0032】
実施例1 本発明に係る微生物株の構築
まず、MGB874株にrocF、rocE、rocD、rocRを復帰させるために利用する菌株を、MGB625株のゲノム構造を改変して構築した。すなわち、MGB625株から、以下に示す方法によりクロラムフェニコール耐性遺伝子をマーカー遺伝子としてpdp-rocR領域の一部にあたるpdp-phrG領域を削除して、rocF-rocE-rocD-rocRがゲノム上に残存する菌株を構築した。同様にpdp- rocF領域を削除してrocE-rocD-rocRがゲノム上に残存する菌株、pdp-rocE領域を削除してrocD-rocRがゲノム上に残存する菌株、pdp-rocF領域を削除してrocRがゲノム上に残存する菌株を構築した。
【0033】
領域削除において行うPCR(Polymerase Chain Reaction)にはGeneAmp PCR System(アプライドバイオシステムズ)を使用し、Pyrobest DNA Polymerase(タカラバイオ)と付属の試薬類を用いて行った。PCRの反応液組成は、適宜希釈した鋳型DNAを1μLとし、センス及びアンチセンスプライマーを各々20pmolとし、Pyrobest DNA Polymeraseを2.5U添加して、反応液総量を50μLとした。PCRの反応条件は、98℃で10秒間、55℃で30秒間及び72℃で1〜5分間(目的増幅産物に応じて調整。目安は1kbあたり1分間)の3段階の温度変化を30回繰り返した後、72℃で5分間反応させることにより行った。
領域削除の方法としては、枯草菌168株から抽出したゲノムDNAを鋳型とし、表2に示した“pdpfw2”と“pdprv/Cm” のプライマーセットを用いて、ゲノム上の当該遺伝子の上流に隣接する1.0kb断片(A)をPCRにより調製した。
【0034】
“削除領域下流に隣接する遺伝子名fw/Cm”と“削除領域下流に隣接する遺伝子名rv2”のプライマーセットを用いて、下流に隣接する1.0kb断片(B)をそれぞれ調製した。一方、プラスミドpC194[J. Bacteriol. 150 (2), 815 (1982)]のクロラムフェニコール耐性遺伝子をプラスミドpUC18のXbaI−BamHI切断点に挿入した組換えプ
ラスミドpCBB31を鋳型とし、表2に示したCmFとCmRプライマーセットを用いて、クロラムフェニコール耐性遺伝子を含む1kb断片(C)をPCRにより調製した。次に、得られた(A)(B)(C)3断片を混合して鋳型とし、表2のプライマー“pdpfw1”と“削除領域下流に隣接する遺伝子名rv1”を用いたSOE−PCRを行うことによって、3断片を(A)(C)(B)の順になる様に結合させ、3.0kbのDNA断片を得た(図1参照)。すなわち、(B)がプライマー番号14と6のプラマーセット由来断片の場合はプライマー番号2、5を用いてSOE−PCRを行い、(B)がプライマー番号15と8のプライマーセット由来断片の場合はプライマー番号2、7を用いてSOE−PCRを行い、(B)がプライマー番号16と10のプライマーセット由来断片の場合はプライマー番号2、9を用いてSOE−PCRを行い、(B)がプライマー番号17と12のプライマーセット由来断片の場合はプライマー番号2、11を用いてSOE−PCRを行った。
【0035】
これらのDNA断片のそれぞれを用いてコンピテントセル法[J.Bacteriol.,81,741(1960)]により枯草菌MGB625株の形質転換を行った。すなわち、枯草菌をSPI培地(0.20 % 硫酸アンモニウム、1.40 % リン酸水素二カリウム、0.60 % リン酸二水素カリウム、0 . 10 % クエン酸三ナトリウム二水和物、0.50 % グルコース、0.02 % カザミノ酸(Difco )、5 mM 硫酸マグネシウム、0.25μM 塩化マンガン、50μg/ml トリプトファン)において37℃で、生育度(OD 600)の値が1程度になるまで振盪培養する。振盪培養後、培養液の一部を9倍量のSPII培地(0.20 % 硫酸アンモニウム、1.40 %リン酸水素二カリウム、0.60 % リン酸二水素カリウム、0.10 % クエン酸三ナトリウム二水和物、0.50 % グルコース、0.01 % カザミノ酸 (Difco)、5 mM 硫酸マグネシウム、0.40μM 塩化マンガン、5μg/ml トリプトファン)に接種し、更に生育度(OD 600)の値が0.4程度になるまで振盪培養することで、枯草菌のコンピテントセルを調製した。次いで調製したコンピテントセル懸濁液(SPII培地における培養液)100μLに上記DNA断片を含む溶液(上記SOE−PCRの反応液)5 μLを添加し、37℃で1時間振盪培養後、クロラムフェニコール含むLB寒天培地(1 % トリプトン、0.5 % 酵母エキス、1 % NaCl、1.5 % 寒天) に全量を塗沫する。37℃における静置培養の後、生育したコロニーを形質転換体として分離した。
市販のゲノムDNA抽出キット、UltraClean Microbial DNA Isolation Kit(MO BIO Laboratories,Inc.)を用いて形質転換体のゲノムDNAを抽出し、これを鋳型DNAとしたPCRにより、目的の領域がクロラムフェニコール耐性遺伝子に置換していることを確認した。
【0036】
【表2】
【0037】
次に、MGB625株から、クロラムフェニコール耐性遺伝子をマーカー遺伝子としてpdp-rocR領域の一部にあたるpdp-phrG領域を削除してrocF-rocE-rocD-rocRがゲノム上に残存する菌株、同様にpdp- rocF領域を削除してrocE-rocD-rocRがゲノム上に残存する菌株、pdp-rocE領域を削除してrocD-rocRがゲノム上に残存する菌株、pdp-rocF領域を削除してrocRがゲノム上に残存する菌株のゲノムDNAをそれぞれ調製した。
各ゲノムDNAを用いてコンピテントセル法によりMGB874株を形質転換した。クロラムフェニコールを含むLB寒天培地上に生育したコロニーを形質転換体として分離した。得られた各形質転換体のゲノムを抽出し、ゲノム上のpdp-rocR領域が削除された位置に、クロラムフェニコール耐性遺伝子とrocF-rocE-rocD-rocRが挿入していること、又は、クロラムフェニコール耐性遺伝子とrocE-rocD-rocRが挿入していること、又は、クロラムフェニコール耐性遺伝子とrocD-rocRが挿入していること、、又は、クロラムフェニコール耐性遺伝子とrocRが挿入していることをPCRにより確認した。
【0038】
次に、MGB625株からMGB874株を構築するまでに削除されている6個の領域の内、pdp-rocR領域以外の5個の削除領域であるycxB-sipU領域、SKIN-Pro7 (spoIVCB-yraK)領域、sbo-ywhH領域、yybP-yyaJ領域及びyncM-fosB領域に復帰が生じていないことをPCRにより確認した。このようにして、rocF-rocE-rocD-rocRが復帰した株(以降、MGB874+rocDEFR株)、rocE-rocD-rocRが復帰した株(以降、MGB874+rocEFR株)、rocD-rocRが復帰した株(以降、MGB874+rocDR株) 、rocRが復帰した株(以降、MGB874+rocR株)を取得できた。
更に、MGB874+rocDEFR株、MGB874+rocEFR株MGB874+rocDR株から以下の方法によりネオマイシン耐性遺伝子をマーカー遺伝子としてrocRを削除することで、MGB874株に対して、rocF-rocE-rocDが復帰した株、rocE-rocDが復帰した株、rocDが復帰した株を構築した。
【0039】
rocRの削除の方法としては、上述した領域削除の手順においてクロラムフェニコール耐性遺伝子の代わりにネオマイシン耐性遺伝子を用いた以外は同様にして作製した。すなわち、枯草菌168株から抽出したゲノムDNAを鋳型とし、表2に示したrocR-FWとrocR/neoR、及びrocR/neoFとrocR-RVの各プライマーセットを用いて、ゲノム上のrocR遺伝子の上流に隣接する1.0kb断片(A)、及び下流に隣接する1.0kb断片(B)をそれぞれ調製した。一方、プラスミドpBEST501[Nucleic Acids Res., 17,4410(1989)]のネオマイシン耐性遺伝子を鋳型とし、表2に示したneofとneorプライマーセットを用いて、ネオマイシン耐性遺伝子を含む1.3kb断片(C)を調製した。次に、得られた(A)(B)(C)3断片を混合して鋳型とし、表のプライマーrocRFW2とrocR-RV2を用いたSOE−PCRを行うことによって、3断片を(A)(C)(B)の順になる様に結合させ、3.0kbのDNA断片を得た(図1参照)。このDNA断片を用いてコンピテントセル法によりMGB874+rocDEFR株、MGB874+rocEFR株、MGB874+rocDR株の形質転換を行い、ネオマイシンを含むLB寒天培地上に生育したコロニーを形質転換体として分離した。得られた形質転換体のゲノムを抽出し、PCRによってrocR遺伝子が欠失され、ネオマイシン耐性遺伝子に置換していることを確認した。これらの結果、rocF-rocE-rocDが復帰した株(以降、MGB874+rocDEF株)、rocE-rocDが復帰した株(以降、MGB874+rocDE株)、rocDが復帰した株(以降、MGB874+rocD株)を取得した。
【0040】
実施例2 アルカリセルラーゼ分泌生産評価
アルカリセルラーゼ分泌生産性評価は以下の様に行った。即ち、バチルス エスピー(Bacillus sp.)KSM-S237株(FERM BP-7875)由来のアルカリセルラーゼ遺伝子(特開2000-210081号公報)断片(3.1 kb)がシャトルベクターpHY300PLKのBamHI制限酵素切断点に挿入された組換えプラスミドpHY-S237を、プロトプラスト形質転換法によって各菌株に導入した。これによって得られた組換え菌株を10 mLのLB培地で一夜37℃で振盪培養を行い、更にこの培養液0.05 mLを50 mLの2×L−マルトース培地(2% トリプトン、1% 酵母エキス、1% NaCl、7.5% マルトース、7.5 ppm硫酸マンガン4-5水和物、15 ppmテトラサイクリン)に接種し、30℃にて3日間振盪培養を行った。遠心分離によって菌体を除いた培養液上清のアルカリセルラーゼ活性を測定し、培養によって菌体外に分泌生産されたアルカリセルラーゼの量を求めた。
【0041】
セルラーゼ活性測定については、1/7.5M リン酸緩衝液(pH7.4 和光純薬)で適宜希釈したサンプル溶液50μLに0.4mM p-nitrophenyl-β-D-cellotrioside(生化学工業)を50μL加えて混和し、30℃にて反応を行った際に遊離するp-ニトロフェノール量を420nmにおける吸光度(OD420nm)変化により定量した。1分間に1μmolのp-ニトロフェノールを遊離させる酵素量を1Uとした。
【0042】
【表3】
【0043】
表3に示したように、MGB874+rocDR株、MGB874+rocDEF株、MGB874+rocDE株、MGB874+rocD株において、MGB874よりアルカリセルラーゼの分泌生産性が向上していることが明らかとなった。
【技術分野】
【0001】
本発明は、有用なタンパク質又はポリペプチドの製造に用いる枯草菌変異株、及び当該枯草菌を用いたタンパク質又はポリペプチドの製造方法に関する。
【背景技術】
【0002】
微生物による有用物質の工業的生産は、アルコール飲料や味噌、醤油等の食品類をはじめとし、アミノ酸、有機酸、核酸関連物質、抗生物質、糖質、脂質、タンパク質等、その種類は多岐に渡っており、またその用途についても食品、医薬品、洗浄剤、化粧品等の日用品、或いは各種化成品原料に至るまで幅広い分野に広がっている。
【0003】
こうした微生物による有用物質の工業生産においては、その生産性の向上が重要な課題の一つであり、その手法として、突然変異等の遺伝学的手法による生産菌の育種が行われてきた。特に最近では、微生物遺伝学、バイオテクノロジーの発展により、遺伝子組換え技術等を用いたより効率的な生産菌の育種が行われるようになっている。さらに、近年のゲノム解析技術の急速な発展を受けて、対象とする微生物のゲノム情報を解読し、これらを積極的に産業に応用しようとする試みもなされている。
【0004】
近年、枯草菌については、枯草菌ゲノムに存在する約4100種類の遺伝子の破壊株が網羅的に研究され、271個の遺伝子が成育に必須であることが指摘されている(非特許文献1)。また、枯草菌等の胞子形成初期に関わる遺伝子やプロテアーゼ遺伝子、又は細胞壁或いは細胞膜中のテイコ酸へのD-アラニン付加に関わる遺伝子、更にはサーファクチンの生合成或いは分泌に関わる遺伝子を単独に欠失又は不活性化した菌株が構築されている(特許文献1〜献8参照)。
【0005】
しかるところ、本出願人は、枯草菌に由来する遺伝子破壊株を網羅的に解析し、枯草菌ゲノムの大量域を欠失させ、各種酵素の生産性に優れた変異株を見出すことに成功している(特許文献9)。
【0006】
しかしながら、限定された生産培地と安定な培養条件が用いられる工業的生産においては、タンパク質又はポリペプチドの生産にとって必要或いは不必要な遺伝子が存在すると考えられ、更なる研究が求められている。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】特開昭58-190390号公報
【特許文献2】特開昭61-1381号公報
【特許文献3】国際公開第89/04866号パンフレット
【特許文献4】特表平11-509096号公報
【特許文献5】特許第3210315号公報
【特許文献6】特表2001-527401号公報
【特許文献7】特表2002−520017号公報
【特許文献8】特表2001−503641号公報
【特許文献9】特開2007-130013号公報
【非特許文献】
【0008】
【非特許文献1】K. Kobayashi et al., Proc. Natl. Acad. Sci. USA., 100, 4678-4683, 2003
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明は、タンパク質又はポリペプチドの生産性向上を可能とする枯草菌変異株、及び当該枯草菌を用いたタンパク質又はポリペプチドの製造方法を提供することに関する。
【課題を解決するための手段】
【0010】
本発明者らは、枯草菌Bacillus subtilis Marburg No.168(枯草菌168株)を野生株とし、そのゲノムの合計874kbが削除された枯草菌変異株MGB874株を作製した。枯草菌変異株MGB874株は野生株である168株に比較し、目的タンパク質をコードする遺伝子が発現可能に導入された際に、その目的タンパク質の分泌生産性が有意に向上する変異株であった(前記特許文献9)。したがって削除した遺伝子は目的タンパク質の分泌生産においては不要あるいは有害な遺伝子であると推察された。しかし、種々検討したところ予想に反して、MGB874株において削除したpdp-rocR領域の96遺伝子のうちアルギニン分解系に関与する遺伝子rocRを導入せず且つrocF、rocE及びrocDから選ばれる遺伝子をMGB874株に導入した場合、又はrocD及びrocRをMGB874株に導入した場合に、目的のタンパク質又はポリペプチドの生産性が、導入前と比較して有意に向上することを見出した。
【0011】
すなわち本発明は、以下の1)〜7)に係るものである。
1)枯草菌変異株MGB874株に対して、rocRを導入せず且つrocD、rocE及びrocFから選ばれる1以上の枯草菌遺伝子を導入するか、又はrocD及びrocRを導入した枯草菌変異株。
2)上記1)の枯草菌変異株に、異種のタンパク質又はポリペプチドをコードする遺伝子を導入した組換え枯草菌。
3)異種のタンパク質又はポリペプチドをコードする遺伝子の上流に転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域から選ばれる1以上の領域を結合した上記1)又は2)の組換え枯草菌。
4)転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域を結合した上記3)の組換え枯草菌。
5)分泌シグナル領域がバチルス(Bacillus)属細菌のセルラーゼ遺伝子由来のものであり、転写開始制御領域及び翻訳開始制御領域が当該セルラーゼ遺伝子の開始コドンから始まる長さ0.6〜1kbの上流領域由来のものである上記3)又は4)の組換え枯草菌。
6)転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域が、配列番号1で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜659の塩基配列又は当該塩基配列のいずれかと70%以上の同一性を有する塩基配列からなるDNA断片、又は当該塩基配列の一部が欠失した塩基配列からなるDNA断片である上記4)の組換え枯草菌。
7)2)〜6)のいずれかの組換え枯草菌を用いるタンパク質又はポリペプチドの製造方法。
【発明の効果】
【0012】
本発明によれば、各種タンパク質又はポリペプチドの生産性に優れた枯草菌変異株が提供され、当該枯草菌変異株を用いることによって、生産性に優れた各種酵素の工業的生産が実現できる。また、当該枯草菌変異株は、各種酵素の産生メカニズム等の解明にも有用な生物材料となり得る。
【図面の簡単な説明】
【0013】
【図1】枯草菌のゲノム上から所定の領域を欠失させる方法の一例を説明するための模式図である。
【発明を実施するための形態】
【0014】
本発明の枯草菌変異株を構築するための親微生物には、枯草菌変異株MGB874株が用いられる。MGB874株は、枯草菌Bacillus subtilis Marburg No.168(枯草菌168株)を野生株とし、そのゲノムの大量域、すなわちprophage6 (yoaV-yobO)領域、prophage1 (ybbU-ybdE)領域、prophage4 (yjcM-yjdJ)領域、PBSX (ykdA-xlyA)領域、prophage5 (ynxB-dut)領域、prophage3 (ydiM-ydjC)領域、spb (yodU-ypqP)領域、pks (pksA-ymaC)領域、skin (spoIVCB-spoIIIC)領域、pps (ppsE-ppsA)領域、prophage2 (ydcL-ydeJ)領域、ydcL-ydeK-ydhU領域、yisB-yitD領域、yunA-yurT領域、cgeE-ypmQ領域、yeeK-yesX領域、pdp-rocR領域、ycxB-sipU領域、SKIN-Pro7 (spoIVCB-yraK)領域、sbo-ywhH領域、yybP-yyaJ領域及びyncM-fosB領域を欠失させたものである(前記特許文献9)。表1に示した当該欠失領域は、同表に示す一対のオリゴヌクレオチド・セットにより挟み込まれる領域として言い換えることができる。
【0015】
【表1】
【0016】
本発明において導入される遺伝子は、pdp-rocR領域の96遺伝子のうちアルギニン分解系の遺伝子である。アルギニンはアルギニントランスポーター[RocC、RocEおよびArtPQM(Genome Biol. 2: research: 0019.1)]により細胞内に取り込まれた後、RocFによりオルニチンとなり、RocD、RocAによってグルタミン酸まで変換された後、RocGにより2-オキソグルタル酸となることで分解される。RocBはカルバモイルリン酸とシトルリンからオルニチンを合成する酵素であると言われているが詳細な報告は論文として未発表である[J. Bacteriol. 186, 3399 (2004)]。転写因子RocRはアルギニン分解経路における遺伝子(rocABC, rocDEF, rocG)の転写を活性化し[Proc. Natl. Acad. Sci. USA 96, 10290 (1999), J. Bacteriol. 176, 1234 (1994)]、そのDNA結合活性はオルニチンで活性化すると推定されている[Mol. Microbiol. 24, 825 (1997), Nat. Rev. Microbiol. 5, 917 (2007)]。
rocD遺伝子は、ornithine aminotransferase、rocE 遺伝子は、arginine permease、rocF 遺伝子は、arginaseと推定されている[J. Mol. Biol. 249, 843(1995)]。
斯かる遺伝子は、枯草菌MGB874株の作製において削除されたpdp-rocR領域に属するものであり、従って、本発明の枯草菌変異株は、枯草菌MGB874株の作製において削除された遺伝子のうちrocRを復帰させることなく、少なくともrocD、rocE、及びrocFから選ばれる1以上の遺伝子を復帰させた株、又はrocD及びrocRを復帰させた株と言える。
rocD、rocE、及びrocFの各遺伝子は、1種又はそれ以上を適宜組み合わせて導入すればよく、3遺伝子を全て導入してもよく、rocD及びrocRを復帰させる場合は2遺伝子を導入する。さらに、rocRを復帰させることなく少なくともrocD、rocE、及びrocFから選ばれる1以上の遺伝子を復帰させる場合において、異種タンパク質又はポリペプチドの生産性を抑制しない他の遺伝子を導入してもよい。
【0017】
本発明の枯草菌株の構築方法は特に限定されないが、rocD、rocE、及びrocFから選ばれる1以上の遺伝子を含むDNA断片あるいはゲノムDNAを供与体DNAとして用いて、受容体DNAである枯草菌MGB874株ゲノムに対して、rocRを導入せず且つrocD、rocE、及びrocFから選ばれる1以上の遺伝子を含むDNAを導入する方法、又はrocD及びrocR遺伝子を導入する方法を実施することができる。
例えば、rocD、rocE、及びrocFから選ばれる1以上の遺伝子を含むゲノムDNAを供与体DNAとして利用する場合、アルギニン分解系の遺伝子rocF、rocE、rocD、rocRを含むpdp-rocR領域を有する枯草菌菌株例えばMGB625株のゲノムDNAを利用して、枯草菌MGB874株ゲノムにrocF、rocE、rocD、rocRを含むpdp-rocR領域全体を復帰させることもできる。MGB625株は、枯草菌168株を元に、表1に示された領域の一部であるprophage6 (yoaV-yobO)領域、prophage1 (ybbU-ybdE)領域、prophage4 (yjcM-yjdJ)領域、PBSX (ykdA-xlyA)領域、prophage5 (ynxB-dut)領域、prophage3 (ydiM-ydjC)領域、spb (yodU-ypqP)領域、pks (pksA-ymaC)領域、skin (spoIVCB-spoIIIC)領域、pps (ppsE-ppsA)領域、prophage2 (ydcL-ydeJ)領域、ydcL-ydeK-ydhU領域、yisB-yitD領域、yunA-yurT領域、cgeE-ypmQ領域、yeeK-yesX領域を削除して構築された株である(前記特許文献9参照)。
あるいは、MGB625株からpdp-rocR領域の一部を削除した菌株のゲノムDNAを供与体DNAとして、pdp-rocR領域のうちrocF、rocE、rocD、rocRの一部を含む領域のみを復帰させることもできる。
以下に、rocF、rocE、rocD、rocRの一部を含む領域のみを枯草菌MGB874株ゲノムに復帰させるために用いる供与体DNAを、MGB625株を利用して構築する例を説明する。
ただし、利用する枯草菌株は特にMGB625株に限定するものではなく、rocRを含まず且つrocF、rocE、rocD遺伝子から選ばれる1以上の遺伝子を含むか、又はrocD及びrocRを含む枯草菌であればよい。また、rocRのみを復帰させる実験は比較のために行う。
【0018】
まず、MGB625株から、以下に示す方法により薬剤耐性遺伝子をマーカー遺伝子として、pdp-rocR領域の一部にあたるpdp-phrG領域、pdp- rocF領域、pdp- rocE領域、pdp-rocD領域のいずれかを削除した菌株を構築する。その結果構築される菌株は、それぞれrocF-rocE-rocD-rocRがゲノム上に残存する菌株、rocE-rocD-rocRがゲノム上に残存する菌株、rocD-rocRがゲノム上に残存する菌株、rocRがゲノム上に残存する菌株となる。
【0019】
枯草菌ゲノム上から領域削除する方法としては、特に限定されないが、例えば図1に示す以下の方法を適用することができる。本方法で用いる欠失用DNA断片は、欠失対象領域の上流に隣接する約0.1〜3kb断片(上流断片と称する)と、薬剤耐性遺伝子マーカーを含む断片、同じく下流に隣接する約0.1〜3kb断片(下流断片と称する)が連結したDNA断片である。
例えば、薬剤耐性遺伝子マーカーとしてクロラムフェニコール耐性遺伝子を用いる場合、枯草菌168株から抽出したゲノムDNAを鋳型とし、後記表2に示す“pdpfw2”と“pdprv/Cm”のプライマーセットを用いて、ゲノム上の当該遺伝子の上流に隣接する断片(A)を調製する。また、“削除領域下流に隣接する遺伝子名fw/Cm”と“削除領域下流に隣接する遺伝子名rv2”のプライマーセットを用いて、下流に隣接する断片(B)をそれぞれ調製する。
【0020】
一方、プラスミドpC194[J. Bacteriol. 150 (2), 815 (1982)]のクロラムフェニコール耐性遺伝子をプラスミドpUC18のXbaI−BamHI切断点に挿入した組換えプラ
スミドpCBB31を鋳型とし、表2に示したCmFとCmRプライマーセットを用いて、クロラムフェニコール耐性遺伝子を含む1kb断片(C)を調製する。
次に、得られた(A)(B)(C)3断片を混合して鋳型とし、表2のプライマー“pdpfw1”と“削除領域下流に隣接する遺伝子名rv1”を用いたSOE−PCRを行うことによって、3断片を(A)(C)(B)の順になる様に結合させたDNA断片を得る(図1参照)。
すなわち、(B)がプライマー番号14と6のプラマーセット由来断片の場合はプライマー番号2、5を用いてSOE−PCRを行い、(B)がプライマー番号15と8のプライマーセット由来断片の場合はプライマー番号2、7を用いてSOE−PCRを行い、(B)がプライマー番号16と10のプライマーセット由来断片の場合はプライマー番号2、9を用いてSOE−PCRを行い、(B)がプライマー番号17と12のプライマーセット由来断片の場合はプライマー番号2、11を用いてSOE−PCRを行う。
これらのDNA断片のそれぞれを用いてコンピテントセル法[J.Bacteriol.,81,741(1960)]により枯草菌MGB625株の形質転換を行う。すなわち、枯草菌をSPI培地(0.20 % 硫酸アンモニウム、1.40 % リン酸水素二カリウム、0.60 % リン酸二水素カリウム、0.10 % クエン酸三ナトリウム二水和物、0.50 % グルコース、0.02 % カザミノ酸(Difco)、5 mM 硫酸マグネシウム、0.25μM 塩化マンガン、50μg/ml トリプトファン)において37℃で、生育度(OD 600)の値が1程度になるまで振盪培養する。振盪培養後、培養液の一部を9倍量のSPII培地(0.20 % 硫酸アンモニウム、1.40 % リン酸水素二カリウム、0.60 % リン酸二水素カリウム、0.10 % クエン酸三ナトリウム二水和物、0.50 % グルコース、0.01 % カザミノ酸(Difco)、5 mM 硫酸マグネシウム、0.4 0μM 塩化マンガン、5μg/ml トリプトファン)に接種し、更に生育度(OD 600)の値が0.4程度になるまで振盪培養することで、枯草菌のコンピテントセルを調製する。次いで調製したコンピテントセル懸濁液(SPII培地における培養液)100μLに上記DNA断片を含む溶液(上記SOE−PCRの反応液)5μLを添加し、37℃で1時間振盪培養後、クロラムフェニコール含むLB寒天培地(1 % トリプトン、0.5 % 酵母エキス、1 % NaCl、1.5 % 寒天) に全量を塗沫する。37℃における静置培養の後、生育したコロニーを形質転換体として分離する。
形質転換体のゲノムDNAを抽出し、これを鋳型DNAとしたPCRにより、目的の領域削除が行われていることを確認することができる。ゲノムDNAの抽出は、斉藤と三浦(Saito & Miura)の方法[Saito & Miura Biochem. Biophys. Acta. 72, 619-629 (1963)
]、あるいは、市販のゲノムDNA抽出キット、例えば、UltraClean Microbial DNA Isolation Kit(MO BIO Laboratories, Inc.)を用いて行うことができる。
【0021】
次に、抽出した上記ゲノムDNAのそれぞれを用いてコンピテントセル法によりMGB874株を形質転換する。得られた各形質転換体のゲノムを抽出し、MGB874株ゲノム上のpdp-rocR領域が削除された位置に、クロラムフェニコール耐性遺伝子とrocF-rocE-rocD-rocR、又は、クロラムフェニコール耐性遺伝子とrocE-rocD-rocR、又は、クロラムフェニコール耐性遺伝子とrocD-rocR、又は、rocRが挿入していること、および、MGB874株のpdp-rocR領域以外の削除領域に復帰が生じていないことをPCRにより確認する。斯くして、rocF-rocE-rocD-rocRが復帰した株(以降、MGB874+rocDEFR株)、rocE-rocD-rocRが復帰した株(以降、MGB874+rocEFR株)、rocD-rocRが復帰した株(以降、MGB874+rocDR株) 、rocRが復帰した株(以降、MGB874+rocR株)を取得できる。
更に、MGB874+rocDEFR株、MGB874+rocEFR株MGB874+rocDR株から以下の方法によりネオマイシン耐性遺伝子をマーカー遺伝子としてrocRを削除することで、MGB874株に対して、rocF-rocE-rocDが復帰した株、rocE-rocDが復帰した株、rocDが復帰した株を構築する。
【0022】
rocRの削除の方法としては、特に限定されないが、上述した領域削除の手順において用いたのとは異なる薬剤耐性遺伝子マーカーを用いて同様にして作製することができる(図1参照)。例えば、薬剤耐性遺伝子マーカーとしてネオマイシン耐性遺伝子を用いる場合、枯草菌168株から抽出したゲノムDNAを鋳型とし、表2に示したrocR-FWとrocR/neoR、及びrocR/neoFとrocR-RVの各プライマーセットを用いて、ゲノム上のrocR遺伝子の上流に隣接する断片(A)、及び下流に隣接する断片(B)をそれぞれ調製する。
一方、プラスミドpBEST501[Nucleic Acids Res. 17、4410(1989)]のネオマイシン耐性遺伝子を鋳型とし、表2に示したneofとneorプライマーセットを用いて、ネオマイシン耐性遺伝子を含む1.3kb断片(C)を調製する。次に、得られた(A)(B)(C)3断片を混合して鋳型とし、表のプライマーrocRFW2とrocR-RV2を用いたSOE−PCRを行うことによって、3断片を(A)(C)(B)の順になる様に結合させたDNA断片を得る(図1参照)。
このDNA断片を用いてコンピテントセル法によりMGB874+rocDEFR株、MGB874+rocEFR株、MGB874+rocDR株の形質転換を行い、得られた形質転換体のゲノムを抽出し、PCRによってrocR遺伝子が欠失され、ネオマイシン耐性遺伝子に置換していることを確認する。斯くして、rocF-rocE-rocDが復帰した株(以降、MGB874+rocDEF株)、rocE-rocDが復帰した株(以降、MGB874+rocDE株)、rocDが復帰した株(以降、MGB874+rocD株)を取得することができる。
【0023】
次に、斯くして得られた枯草変異株に、目的とするタンパク質又はポリペプチドをコードする遺伝子を導入することによって、本発明の組換え枯草菌を得ることができる。
目的タンパク質又はポリペプチド遺伝子は特に限定されず、洗浄剤用、食品用、繊維処理用、飼料処理用、化粧品用、医薬品用、診断薬用など各種産業用酵素や、生理活性ペプチドなどが含まれる。また、産業用酵素の機能別には、酸化還元酵素 (Oxidoreductase) 、転移酵素 (Transferase) 、加水分解酵素 (Hydrolase) 、脱離酵素 (Lyase)、異性化酵素 (Isomerase) 、合成酵素 (Ligase/Synthetase) 等が含まれるが、好適にはセルラーゼ、α-アミラーゼ、プロテアーゼ等の加水分解酵素の遺伝子が挙げられる。
【0024】
セルラーゼとしては、例えば、多糖加水分解酵素の分類(Biochem.J.,280,309,1991)
中でファミリー5に属するセルラーゼが挙げられ、中でも微生物由来、特にBacillus属細菌由来のセルラーゼが挙げられる。より具体的な例として、配列番号2で示されるアミノ酸配列からなるBacillus属細菌KSM-S237株(FERM BP-7875)由来のアルカリセルラーゼ、又は、配列番号4で示されるアミノ酸配列からなるBacillus属細菌KSM-64株(FERM BP-2886)由来のアルカリセルラーゼ、或いは、当該アミノ酸配列と70%、好ましくは80%、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有するアミノ酸配列からなるセルラーゼ、又は配列番号2又は配列番号4で示されるアミノ酸配列において1若しくは数個のアミノ酸が置換、欠失若しくは付加されたアミノ酸配列からなるアルカリセルラーゼが挙げられる。
【0025】
α−アミラーゼの具体例としては、微生物由来のα−アミラーゼが挙げられ、特にBacillus属細菌由来の液化型アミラーゼが好ましい。より具体的な例として、配列番号5で示されるアミノ酸配列からなるBacillus属細菌KSM-K38株(FERM BP-6946)由来のアルカリアミラーゼ、或いは当該アミノ酸配列と70%、好ましくは80%、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有するアミノ酸配列からなるアミラーゼ、又は配列番号5で示されるアミノ酸配列において1若しくは数個のアミノ酸が置換、欠失若しくは付加されたアミノ酸配列からなるアルカリアミラーゼが挙げられる。
【0026】
プロテアーゼの具体例としては、微生物由来、特にBacillus属細菌由来のセリンプロテアーゼや金属プロテアーゼ等が挙げられる。より具体的な例として、配列番号6で示されるアミノ酸配列からなるバチルス クラウジ(Bacillus clausii)KSM-K16株(FERM BP-3376)由来のアルカリプロテアーゼ、或いは当該アミノ酸配列と70%、好ましくは80%、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有するアミノ酸配列からなるプロテアーゼ、又は配列番号6で示されるアミノ酸配列において1若しくは数個のアミノ酸が置換、欠失若しくは付加されたアミノ酸配列からなるアルカリプロテアーゼが挙げられる。
【0027】
尚、本発明においてアミノ酸配列および塩基配列の同一性はLipman-Pearson法 (Science,227,1435,1985)によって計算される。具体的には、遺伝情報処理ソフトウェアGenetyx-Win(ソフトウェア開発)のホモロジー解析(Search homology)プログラムを用いて、Unit size to compare(ktup)を2として解析を行うことにより算出される。
また、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列としては、1乃至10個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列が好ましく、また、付加には、両末端への1〜数個のアミノ酸の付加が含まれる。
【0028】
本発明の微生物に導入される目的タンパク質又はポリペプチドの遺伝子は、その上流に当該遺伝子の転写、翻訳、分泌に関わる制御領域、即ち、プロモーター及び転写開始点を含む転写開始制御領域、リボソーム結合部位及び開始コドンを含む翻訳開始領域並びに分泌シグナルペプチド領域から選ばれる1以上の領域が適正な形で結合されていることが望ましい。特に、転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域が結合されていることが好ましく、更に分泌シグナルペプチド領域がバチルス(Bacillus)属細菌のセルラーゼ遺伝子由来のものであり、転写開始制御領域及び翻訳開始制御領域が当該セルラーゼ遺伝子の開始コドンから始まる長さ0.6〜1kbの上流領域であるものが、目的のタンパク質又はポリペプチド遺伝子と適正な形で結合されていることが望ましい。例えば、特開2000-210081号公報や特開平4-190793号公報等に記載されているバチルス(Bacillus)属細菌、すなわちKSM-S237株(FERM BP-7875)、KSM-64株(FERM BP-2886)由来のセルラーゼ遺伝子の転写開始制御領域、翻訳開始領域及び分泌シグナルペプチド領域が目的のタンパク質又はポリペプチドの構造遺伝子と適正に結合されていることが望ましい。より具体的には配列番号1で示される塩基配列の塩基番号1〜659の塩基配列、配列番号3で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜696の塩基配列、また当該塩基配列に対して70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有する塩基配列からなるDNA断片、又は上記いずれかの塩基配列からなるDNAとストリンジェントの条件でハイブリダイズし且つ遺伝子の転写、翻訳、分泌に関わる機能を有するDNA、或いは上記いずれかの塩基配列の一部が欠失した塩基配列からなるDNA断片が、目的のタンパク質又はポリペプチドの構造遺伝子と適正に結合されていることが望ましい。尚、ここで、上記塩基配列の一部が欠失した塩基配列からなるDNA断片とは、上記塩基配列の一部を欠失しているが、遺伝子の転写、翻訳、分泌に関わる機能を保持しているDNA断片を意味する。また此処で言うストリンジェントな条件とは、例えば[Molecular cloning-a Laboratory manual 2nd edition(Sambrookら、1989)]に記載の条件等が挙げられる。例えば、6XSSC(1XSSCの組成:0.15M塩化ナトリウム、0.015Mクエン酸ナトリウム、pH7.0)、0.5%SDS、5Xデンハート及び100mg/mLニシン精子DNAを含む溶液にプローブとともに65℃で8〜16時間恒温し、ハイブリダイズさせる条件が挙げられる。
【0029】
上記の目的タンパク質又はポリペプチド遺伝子を含むDNA断片と適当なプラスミドベクターを結合させた組換えプラスミドを、一般的な形質転換法によって上記枯草菌変異株に取り込ませることによって、本発明の組換え枯草菌を得ることができる。また、当該DNA断片に枯草菌変異株のゲノムとの適当な相同領域を結合したDNA断片を用い、枯草菌変異株ゲノムに直接組み込むことによっても本発明の組換え枯草菌を得ることができる。
【0030】
本発明の組換え枯草菌を用いた目的のタンパク質又はポリペプチドの生産は、当該菌株を同化性の炭素源、窒素源、その他の必須成分を含む培地に接種し、通常の微生物培養法にて培養し、培養終了後、タンパク質又はポリペプチドを採取・精製することにより行えばよい。そして、後記実施例に示すように、目的のタンパク質又はポリペプチドの生産性は、本発明の遺伝子を導入していない枯草菌変異株MGB874株を用いた場合と比較して、その向上が達成されている。
【0031】
以下に、当該発明の組換え微生物の構築方法と及び当該組換え微生物を用いたセルラーゼ生産方法について具体的に説明する。
【実施例】
【0032】
実施例1 本発明に係る微生物株の構築
まず、MGB874株にrocF、rocE、rocD、rocRを復帰させるために利用する菌株を、MGB625株のゲノム構造を改変して構築した。すなわち、MGB625株から、以下に示す方法によりクロラムフェニコール耐性遺伝子をマーカー遺伝子としてpdp-rocR領域の一部にあたるpdp-phrG領域を削除して、rocF-rocE-rocD-rocRがゲノム上に残存する菌株を構築した。同様にpdp- rocF領域を削除してrocE-rocD-rocRがゲノム上に残存する菌株、pdp-rocE領域を削除してrocD-rocRがゲノム上に残存する菌株、pdp-rocF領域を削除してrocRがゲノム上に残存する菌株を構築した。
【0033】
領域削除において行うPCR(Polymerase Chain Reaction)にはGeneAmp PCR System(アプライドバイオシステムズ)を使用し、Pyrobest DNA Polymerase(タカラバイオ)と付属の試薬類を用いて行った。PCRの反応液組成は、適宜希釈した鋳型DNAを1μLとし、センス及びアンチセンスプライマーを各々20pmolとし、Pyrobest DNA Polymeraseを2.5U添加して、反応液総量を50μLとした。PCRの反応条件は、98℃で10秒間、55℃で30秒間及び72℃で1〜5分間(目的増幅産物に応じて調整。目安は1kbあたり1分間)の3段階の温度変化を30回繰り返した後、72℃で5分間反応させることにより行った。
領域削除の方法としては、枯草菌168株から抽出したゲノムDNAを鋳型とし、表2に示した“pdpfw2”と“pdprv/Cm” のプライマーセットを用いて、ゲノム上の当該遺伝子の上流に隣接する1.0kb断片(A)をPCRにより調製した。
【0034】
“削除領域下流に隣接する遺伝子名fw/Cm”と“削除領域下流に隣接する遺伝子名rv2”のプライマーセットを用いて、下流に隣接する1.0kb断片(B)をそれぞれ調製した。一方、プラスミドpC194[J. Bacteriol. 150 (2), 815 (1982)]のクロラムフェニコール耐性遺伝子をプラスミドpUC18のXbaI−BamHI切断点に挿入した組換えプ
ラスミドpCBB31を鋳型とし、表2に示したCmFとCmRプライマーセットを用いて、クロラムフェニコール耐性遺伝子を含む1kb断片(C)をPCRにより調製した。次に、得られた(A)(B)(C)3断片を混合して鋳型とし、表2のプライマー“pdpfw1”と“削除領域下流に隣接する遺伝子名rv1”を用いたSOE−PCRを行うことによって、3断片を(A)(C)(B)の順になる様に結合させ、3.0kbのDNA断片を得た(図1参照)。すなわち、(B)がプライマー番号14と6のプラマーセット由来断片の場合はプライマー番号2、5を用いてSOE−PCRを行い、(B)がプライマー番号15と8のプライマーセット由来断片の場合はプライマー番号2、7を用いてSOE−PCRを行い、(B)がプライマー番号16と10のプライマーセット由来断片の場合はプライマー番号2、9を用いてSOE−PCRを行い、(B)がプライマー番号17と12のプライマーセット由来断片の場合はプライマー番号2、11を用いてSOE−PCRを行った。
【0035】
これらのDNA断片のそれぞれを用いてコンピテントセル法[J.Bacteriol.,81,741(1960)]により枯草菌MGB625株の形質転換を行った。すなわち、枯草菌をSPI培地(0.20 % 硫酸アンモニウム、1.40 % リン酸水素二カリウム、0.60 % リン酸二水素カリウム、0 . 10 % クエン酸三ナトリウム二水和物、0.50 % グルコース、0.02 % カザミノ酸(Difco )、5 mM 硫酸マグネシウム、0.25μM 塩化マンガン、50μg/ml トリプトファン)において37℃で、生育度(OD 600)の値が1程度になるまで振盪培養する。振盪培養後、培養液の一部を9倍量のSPII培地(0.20 % 硫酸アンモニウム、1.40 %リン酸水素二カリウム、0.60 % リン酸二水素カリウム、0.10 % クエン酸三ナトリウム二水和物、0.50 % グルコース、0.01 % カザミノ酸 (Difco)、5 mM 硫酸マグネシウム、0.40μM 塩化マンガン、5μg/ml トリプトファン)に接種し、更に生育度(OD 600)の値が0.4程度になるまで振盪培養することで、枯草菌のコンピテントセルを調製した。次いで調製したコンピテントセル懸濁液(SPII培地における培養液)100μLに上記DNA断片を含む溶液(上記SOE−PCRの反応液)5 μLを添加し、37℃で1時間振盪培養後、クロラムフェニコール含むLB寒天培地(1 % トリプトン、0.5 % 酵母エキス、1 % NaCl、1.5 % 寒天) に全量を塗沫する。37℃における静置培養の後、生育したコロニーを形質転換体として分離した。
市販のゲノムDNA抽出キット、UltraClean Microbial DNA Isolation Kit(MO BIO Laboratories,Inc.)を用いて形質転換体のゲノムDNAを抽出し、これを鋳型DNAとしたPCRにより、目的の領域がクロラムフェニコール耐性遺伝子に置換していることを確認した。
【0036】
【表2】
【0037】
次に、MGB625株から、クロラムフェニコール耐性遺伝子をマーカー遺伝子としてpdp-rocR領域の一部にあたるpdp-phrG領域を削除してrocF-rocE-rocD-rocRがゲノム上に残存する菌株、同様にpdp- rocF領域を削除してrocE-rocD-rocRがゲノム上に残存する菌株、pdp-rocE領域を削除してrocD-rocRがゲノム上に残存する菌株、pdp-rocF領域を削除してrocRがゲノム上に残存する菌株のゲノムDNAをそれぞれ調製した。
各ゲノムDNAを用いてコンピテントセル法によりMGB874株を形質転換した。クロラムフェニコールを含むLB寒天培地上に生育したコロニーを形質転換体として分離した。得られた各形質転換体のゲノムを抽出し、ゲノム上のpdp-rocR領域が削除された位置に、クロラムフェニコール耐性遺伝子とrocF-rocE-rocD-rocRが挿入していること、又は、クロラムフェニコール耐性遺伝子とrocE-rocD-rocRが挿入していること、又は、クロラムフェニコール耐性遺伝子とrocD-rocRが挿入していること、、又は、クロラムフェニコール耐性遺伝子とrocRが挿入していることをPCRにより確認した。
【0038】
次に、MGB625株からMGB874株を構築するまでに削除されている6個の領域の内、pdp-rocR領域以外の5個の削除領域であるycxB-sipU領域、SKIN-Pro7 (spoIVCB-yraK)領域、sbo-ywhH領域、yybP-yyaJ領域及びyncM-fosB領域に復帰が生じていないことをPCRにより確認した。このようにして、rocF-rocE-rocD-rocRが復帰した株(以降、MGB874+rocDEFR株)、rocE-rocD-rocRが復帰した株(以降、MGB874+rocEFR株)、rocD-rocRが復帰した株(以降、MGB874+rocDR株) 、rocRが復帰した株(以降、MGB874+rocR株)を取得できた。
更に、MGB874+rocDEFR株、MGB874+rocEFR株MGB874+rocDR株から以下の方法によりネオマイシン耐性遺伝子をマーカー遺伝子としてrocRを削除することで、MGB874株に対して、rocF-rocE-rocDが復帰した株、rocE-rocDが復帰した株、rocDが復帰した株を構築した。
【0039】
rocRの削除の方法としては、上述した領域削除の手順においてクロラムフェニコール耐性遺伝子の代わりにネオマイシン耐性遺伝子を用いた以外は同様にして作製した。すなわち、枯草菌168株から抽出したゲノムDNAを鋳型とし、表2に示したrocR-FWとrocR/neoR、及びrocR/neoFとrocR-RVの各プライマーセットを用いて、ゲノム上のrocR遺伝子の上流に隣接する1.0kb断片(A)、及び下流に隣接する1.0kb断片(B)をそれぞれ調製した。一方、プラスミドpBEST501[Nucleic Acids Res., 17,4410(1989)]のネオマイシン耐性遺伝子を鋳型とし、表2に示したneofとneorプライマーセットを用いて、ネオマイシン耐性遺伝子を含む1.3kb断片(C)を調製した。次に、得られた(A)(B)(C)3断片を混合して鋳型とし、表のプライマーrocRFW2とrocR-RV2を用いたSOE−PCRを行うことによって、3断片を(A)(C)(B)の順になる様に結合させ、3.0kbのDNA断片を得た(図1参照)。このDNA断片を用いてコンピテントセル法によりMGB874+rocDEFR株、MGB874+rocEFR株、MGB874+rocDR株の形質転換を行い、ネオマイシンを含むLB寒天培地上に生育したコロニーを形質転換体として分離した。得られた形質転換体のゲノムを抽出し、PCRによってrocR遺伝子が欠失され、ネオマイシン耐性遺伝子に置換していることを確認した。これらの結果、rocF-rocE-rocDが復帰した株(以降、MGB874+rocDEF株)、rocE-rocDが復帰した株(以降、MGB874+rocDE株)、rocDが復帰した株(以降、MGB874+rocD株)を取得した。
【0040】
実施例2 アルカリセルラーゼ分泌生産評価
アルカリセルラーゼ分泌生産性評価は以下の様に行った。即ち、バチルス エスピー(Bacillus sp.)KSM-S237株(FERM BP-7875)由来のアルカリセルラーゼ遺伝子(特開2000-210081号公報)断片(3.1 kb)がシャトルベクターpHY300PLKのBamHI制限酵素切断点に挿入された組換えプラスミドpHY-S237を、プロトプラスト形質転換法によって各菌株に導入した。これによって得られた組換え菌株を10 mLのLB培地で一夜37℃で振盪培養を行い、更にこの培養液0.05 mLを50 mLの2×L−マルトース培地(2% トリプトン、1% 酵母エキス、1% NaCl、7.5% マルトース、7.5 ppm硫酸マンガン4-5水和物、15 ppmテトラサイクリン)に接種し、30℃にて3日間振盪培養を行った。遠心分離によって菌体を除いた培養液上清のアルカリセルラーゼ活性を測定し、培養によって菌体外に分泌生産されたアルカリセルラーゼの量を求めた。
【0041】
セルラーゼ活性測定については、1/7.5M リン酸緩衝液(pH7.4 和光純薬)で適宜希釈したサンプル溶液50μLに0.4mM p-nitrophenyl-β-D-cellotrioside(生化学工業)を50μL加えて混和し、30℃にて反応を行った際に遊離するp-ニトロフェノール量を420nmにおける吸光度(OD420nm)変化により定量した。1分間に1μmolのp-ニトロフェノールを遊離させる酵素量を1Uとした。
【0042】
【表3】
【0043】
表3に示したように、MGB874+rocDR株、MGB874+rocDEF株、MGB874+rocDE株、MGB874+rocD株において、MGB874よりアルカリセルラーゼの分泌生産性が向上していることが明らかとなった。
【特許請求の範囲】
【請求項1】
枯草菌変異株MGB874株に対して、rocRを導入せず且つrocD、rocE及びrocFから選ばれる1以上の枯草菌遺伝子を導入するか、又は枯草菌遺伝子rocD及びrocRを導入した枯草菌変異株。
【請求項2】
請求項1記載の枯草菌変異株に、異種のタンパク質又はポリペプチドをコードする遺伝子を導入した組換え枯草菌。
【請求項3】
異種のタンパク質又はポリペプチドをコードする遺伝子の上流に転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域から選ばれる1以上の領域を結合した請求項2記載の組換え枯草菌。
【請求項4】
転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域を結合した請求項3記載の組換え枯草菌。
【請求項5】
分泌シグナル領域がバチルス(Bacillus)属細菌のセルラーゼ遺伝子由来のものであり、転写開始制御領域及び翻訳開始制御領域が当該セルラーゼ遺伝子の開始コドンから始まる長さ0.6〜1kbの上流領域由来のものである請求項3又は4記載の組換え枯草菌。
【請求項6】
転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域が、配列番号1で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜659の塩基配列又は当該塩基配列のいずれかと70%以上の同一性を有する塩基配列からなるDNA断片、又は当該塩基配列の一部が欠失した塩基配列からなるDNA断片である請求項4記載の組換え枯草菌。
【請求項7】
請求項2〜6のいずれか1項記載の組換え枯草菌を用いるタンパク質又はポリペプチドの製造方法。
【請求項1】
枯草菌変異株MGB874株に対して、rocRを導入せず且つrocD、rocE及びrocFから選ばれる1以上の枯草菌遺伝子を導入するか、又は枯草菌遺伝子rocD及びrocRを導入した枯草菌変異株。
【請求項2】
請求項1記載の枯草菌変異株に、異種のタンパク質又はポリペプチドをコードする遺伝子を導入した組換え枯草菌。
【請求項3】
異種のタンパク質又はポリペプチドをコードする遺伝子の上流に転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域から選ばれる1以上の領域を結合した請求項2記載の組換え枯草菌。
【請求項4】
転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域を結合した請求項3記載の組換え枯草菌。
【請求項5】
分泌シグナル領域がバチルス(Bacillus)属細菌のセルラーゼ遺伝子由来のものであり、転写開始制御領域及び翻訳開始制御領域が当該セルラーゼ遺伝子の開始コドンから始まる長さ0.6〜1kbの上流領域由来のものである請求項3又は4記載の組換え枯草菌。
【請求項6】
転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域が、配列番号1で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜659の塩基配列又は当該塩基配列のいずれかと70%以上の同一性を有する塩基配列からなるDNA断片、又は当該塩基配列の一部が欠失した塩基配列からなるDNA断片である請求項4記載の組換え枯草菌。
【請求項7】
請求項2〜6のいずれか1項記載の組換え枯草菌を用いるタンパク質又はポリペプチドの製造方法。
【図1】

【公開番号】特開2010−178705(P2010−178705A)
【公開日】平成22年8月19日(2010.8.19)
【国際特許分類】
【出願番号】特願2009−26676(P2009−26676)
【出願日】平成21年2月6日(2009.2.6)
【出願人】(000000918)花王株式会社 (8,290)
【Fターム(参考)】
【公開日】平成22年8月19日(2010.8.19)
【国際特許分類】
【出願日】平成21年2月6日(2009.2.6)
【出願人】(000000918)花王株式会社 (8,290)
【Fターム(参考)】
[ Back to top ]