
架橋マトリックスの特性を調節するための酵素的架橋剤の改変
ポリマーを酵素架橋することにより形成される、改良されたマトリックスまたはヒドロゲルであって、
架橋化酵素分子が、マトリックスの架橋密度、力学的特性、もしくは他の特性を改善するため、かつ/または架橋速度および/もしくは架橋度の制御を改善するために改変されていること、ならびに、
架橋化酵素分子の改変が、基質ポリマーの架橋によってマトリックスが形成されるのに伴い、架橋マトリックス内における酵素分子の感知体積を変化させるものであること
を特徴とするマトリックスまたはヒドロゲル。
これらの製造方法および使用方法も提供される。

架橋化酵素分子が、マトリックスの架橋密度、力学的特性、もしくは他の特性を改善するため、かつ/または架橋速度および/もしくは架橋度の制御を改善するために改変されていること、ならびに、
架橋化酵素分子の改変が、基質ポリマーの架橋によってマトリックスが形成されるのに伴い、架橋マトリックス内における酵素分子の感知体積を変化させるものであること
を特徴とするマトリックスまたはヒドロゲル。
これらの製造方法および使用方法も提供される。
【発明の詳細な説明】
【背景技術】
【0001】
酵素架橋マトリックスの利用
酵素架橋マトリックスは、食品産業、化粧品産業、および医療産業において、様々な用途に利用されている。酵素架橋ヒドロゲルは、特に医療用途において、組織シーラント、組織接着剤、止血製剤、組織工学用マトリックス、薬物送達プラットフォームなどの様々な医療用途に広く利用されている。ゼラチンやポロキサマーなどのヒドロゲルのいくつかは、特定の条件下(たとえば温度変化など)においてポリマー鎖同士が物理的に相互作用することによって形成され得るが、一方、ポリマー溶液のヒドロゲルが形成されるには架橋を必要とすることが多い。埋め込み可能なヒドロゲルにおいては、固体ゲルが実際に形成されることに加えて、このゲルを使用する組織で一般的に見られる状態、たとえば、力学的ストレス、体温上昇、酵素的分解、化学的分解などに対する抵抗性が必要とされる。このように、多くの場合において、ヒドロゲルマトリックスの架橋が必要とされる。架橋は、ヒドロゲルをプレキャストまたは成型することによって体外で行ってもよい。この架橋方法は、主に、組織工学または薬物送達においてヒドロゲルを使用する場合に用いられる。あるいは、架橋を体内で行ってもよく(in situゲル化またはin situ架橋)、所望の部位に注入または塗布した溶液を架橋させてゲルを形成させる。
【0002】
ゲル形成は、様々な架橋アプローチによって開始することができる。化学的アプローチによるゲル形成としては、シアノアクリレートにおいて見られるような接触による重合開始、または光開始などの外部刺激による重合開始が挙げられる。あるいは、グルタルアルデヒドやカルボジイミドなどの低分子量架橋剤(Otani Y, Tabata Y, Ikada Y. Ann Thorac Surg 1999, 67, 922-6. Sung HW, Huang DM, Chang WH, Huang RN, Hsu JC. J Biomed Mater Res 1999, 46, 520-30. Otani, Y.; Tabata, Y.; Ikada, Y. Biomaterials 1998, 19, 2167-73. Lim, D. W.; Nettles, D. L.; Setton, L. A.; Chilkoti, A. Biomacromolecules 2008, 9, 222-30.)または活性化置換基(Iwata, H.; Matsuda, S.; Mitsuhashi, K.; Itoh,E.; Ikada, Y. Biomaterials 1998, 19, 1869-76)を用いて、あらかじめ形成されたポリマーを化学的に架橋することによってもゲルを形成することができる。
【0003】
しかしながら、化学的架橋剤は、毒性、発癌性、または刺激性を有することが多いため、食品、化粧品、または医薬品において化学的架橋を行うと問題が生じることがある。さらに、化学的架橋剤は、架橋マトリックス外へと容易に拡散し得る小型分子であり、局所的・全身的障害を引き起こし得る。
【0004】
化学的架橋に代わるもののひとつとして、酵素的架橋アプローチが挙げられる。ゲル形成を開始するための酵素的架橋アプローチは、種々の異なる架橋化酵素に基づいて研究がなされている。このような酵素的架橋アプローチとしては、たとえば、ムラサキイガイ接着剤などの接着剤による酵素的架橋(Strausberg RL, Link RP. Trends Biotechnol 1990, 8, 53-7)、またはフィブリンシーラントで利用されているような血液凝固による酵素的架橋(Jackson MR. Am J Surg 2001, 182, 1S-7S. Spotnitz WD. Am J Surg 2001, 182, 8S-14S Buchta C, Hedrich HC, Macher M, Hocker P, Redl H. Biomaterials 2005, 26, 6233-41. 27-30)が挙げられる。
【0005】
ムラサキイガイ接着剤による架橋は、接着性タンパク質のフェノール(すなわちドーパ)残基が反応性キノン残基へと酵素的に変換されることによって開始され、次いで、この反応性キノン残基においてタンパク質間架橋反応が起こる(Burzio LA, Waite JH. Biochemistry 2000, 39, 11147-53. McDowell LM, Burzio LA, Waite JH, Schaefer JJ. Biol Chem 1999, 274, 20293-5)。この種のシーラントには、チロシナーゼや、ラッカーゼおよびペルオキシダーゼが酵素として用いられている。チロシナーゼはチロシンから、ラッカーゼおよびペルオキシダーゼは他のフェノール化合物から、キノン類およびフリーラジカルを生成することによって作用する。次いで、キノン類およびフリーラジカルは、タンパク質の遊離アミン、またはタンパク質および多糖類に含まれる同様に修飾されたフェノール基を架橋することができる。
【0006】
技術モデルとなっている第2の架橋作用として、血液凝固において見られるトランスグルタミナーゼ触媒反応が挙げられる(Ehrbar M, Rizzi SC, Hlushchuk R, Djonov V, Zisch AH, Hubbell JA, Weber FE, Lutolf MP. Biomaterials 2007, 28, 3856-66)。in situゲル形成のためのバイオミメティックアプローチにおいては、第XIIIa因子や他の組織トランスグルタミナーゼの使用について研究がなされてきた(Sperinde J, Griffith L. Macromolecules 2000, 33, 5476-5480. Sanborn TJ, Messersmith PB, Barron AE. Biomaterials 2002, 23, 2703-10)。
【0007】
さらなるin situ架橋ゲル形成のうち特に興味深いものとして、カルシウム非依存性微生物トランスグルタミナーゼ(mTG)によるゼラチンの架橋が挙げられる。mTGは、第XIIIa因子と類似した架橋反応を触媒するが、活性化のためにトロンビンもカルシウムも必要としない。mTGに関する初期の研究は、食品産業において用いることを目的としていたが(Babin H, Dickinson E. Food Hydrocolloids 2001, 15, 271-276. Motoki M, Seguro K. Trends in Food Science & Technology 1998, 9, 204-210.)、後の研究では医療分野における利用可能性が検討された。過去のインビトロ研究において、mTGはゼラチンを架橋して数分以内にゲルを形成できること、ゼラチン−mTG接着剤は湿った組織にも濡れた組織にも接着することができること、およびその接着強度はフィブリンベースのシーラントと同等もしくはそれ以上であることが示されている(Chen TH, Payne GFら, Biomaterials 2003, 24, 2831-2841. McDermott MK, Payne GFら, Biomacromolecules 2004, 5, 1270-1279. Chen T, Payne GFら, J Biomed Mater Res B Appl Biomater 2006, 77, 416-22.)。ゼラチンとmTGの医療用接着剤としての使用は、国際公開第2008/076407号に記載されている。
【0008】
架橋マトリックスを形成するための架橋剤として酵素を使用した際の不都合のひとつとして、所望のゲル状態が形成された後でも酵素が架橋反応を継続し得ることが挙げられる。架橋反応の継続は望ましくない場合が多く、その理由のひとつとして過剰な架橋により硬くて脆く柔軟性に劣るゲルが生じ得ることが挙げられる。さらに、架橋マトリックスの力学的特性がゲルの寿命を通して変化し続け、一貫した特性を持たせることが難しい。所望の架橋密度を上回っても酵素的架橋が継続されることは、架橋マトリックスまたはヒドロゲルが一旦形成されても酵素が架橋反応を触媒し続けることができることに起因する。これは、溶液粘度が非常に増加しても、酵素がマトリックス全体に拡散し続けることができるかどうかに依存する。この見解は、ポリマー溶液の部分的架橋から生じるネットワーク形成の初期段階において、溶液粘度が急速に増加するとトランスグルタミナーゼの移動は急速に低下することを、ペプチドグラフト化合成ポリマー溶液についての研究に基づき示唆したHuら(Hu BH, Messersmith PB. J. Am. Chem. Soc., 2003, 125(47), pp 14298-14299)による報告と一致している。
【0009】
力学的特性の低下を生じる過剰な酵素的架橋ついては、過去にいくつかの文献において報告されている。
【0010】
Bauerらは、高レベルの微生物トランスグルタミナーゼ(mTG)がコムギグルテンタンパク質の過剰な架橋を引き起こし、これによって、グルテンネットワークの柔軟性の喪失および力学的損傷が引き起こされることを実証した(Bauer N, Koehler P, Wieser H, and Schieberle P. Studies on Effects of Microbial Transglutaminase on Gluten Proteins of Wheat II Rheological Properties. Cereal Chem. 80(6): 787-790)。
【0011】
Sakaiらは、フェノール間の共有結合性架橋が多いほど力学的安定性は効果的に増強されるが、フェノール間の架橋が多すぎると形成されるゲルが脆くなることを見出した(Sakai S, Kawakami K. Synthesis and characterization of both ionically and enzymatically crosslinkable alginate, Acta Biomater 3 (2007), pp. 495-501)。
【0012】
カルシウム依存性トランスグルタミナーゼなどの補因子依存性架橋化酵素を用いる場合、一定時間反応させた後、結合反応または他の方法によって補因子を除去すると、架橋度を制限することができる。しかしながら、補因子を除去することは、ヒドロゲルが補因子を捕捉し得るヒドロゲル形成においては技術的に不可能である場合が多い。微生物源から入手可能なトランスグルタミナーゼなどの補因子非依存性酵素を用いる場合、反応系で熱処理を行うことによって架橋度を制限することができる。しかしながら、熱処理はタンパク質機能に悪影響を及ぼすため行わない方が望ましい。さらに、あらゆる反応系に熱処理が適するとは限らない。
【0013】
架橋マトリックス内で過剰な架橋が生じることのほかに、所望の架橋状態が得られた後にマトリックス内において架橋化酵素が持続的に拡散することによって、酵素がゲル外へと速い速度で拡散し得る(これは酵素溶出としても知られている)。これによって、体内に放出された高レベルの架橋化酵素が体内組織と相互作用して、局所的または全身的障害を引き起こすという問題が生じ得る。
【発明の概要】
【発明が解決しようとする課題】
【0014】
種々の用途に用いることができる、改良された酵素により架橋された組成物が必要とされており、このような組成物は有用であると考えられる。
【0015】
したがって、固体マトリックスの形成が開始された後に、所望の力学的特性が得られた時点で架橋マトリックスの酵素的架橋を停止させ、かつ/または固体架橋マトリックスからの酵素の溶出度および溶出速度を低下させる機構が必要性とされており、このような機構は有用であると考えられる。
【0016】
少なくとも実施形態のいくつかにおいて、本発明は、上記の従来技術の欠点を克服するものであり、かつ、(多くの利点のうち、これに限定されるものではないが)上記の技術的問題を解決するための方法を提供するものである。これは、ポリマーを酵素架橋することにより形成されるマトリックスまたはヒドロゲルであって、架橋化酵素分子が、マトリックスの架橋密度、力学的特性、もしくは他の特性を改善するため、かつ/または架橋速度および/もしくは架橋度の制御を改善するために改変されていることを特徴とする、マトリックスまたはヒドロゲルを提供することによって達成される。
【図面の簡単な説明】
【0017】
【図1】PEG化生成物のサイズおよび分布に対する、反応pHおよび活性化PEG濃度の影響を示す画像である。
【図2】PEG化生成物のサイズおよび分布に対する、反応時間およびpHの影響を示す画像である。
【図3】種々の濃度のPEG−NHS(2kD)を用いたPEG化mTGのSDS分析を示す画像である。
【図4】同一の架橋ゼラチンゲルからのmTGおよびPEG化mTGの溶出を示す画像である。
【図5】種々の架橋ゼラチンゲルから溶出したmTG(左側)およびPEG化mTG(右側)を示す画像である。
【図6】非PEG化mTGまたは2種のPEG化mTGで作製したゼラチンシーラントにおける破壊圧力を示すグラフである。
【図7】mTGとデキストランとのコンジュゲーション生成物のSDS−PAGE分析を示す画像である。
【図8】西洋ワサビペルオキシダーゼ(HRP)のPEG化生成物のSDS−PAGE分析を示す画像である。
【図9】種々の反応条件を用いた、PEG化mTG生成物のSDS−PAGE分析を示す画像であり、種々のPEG化度がゲルに示されている。
【図10】反応性PEGが二官能基型PEG−NHS(10kD)である、PEG化mTG生成物のSDS−PAGE分析を示す画像である。
【図11】MALDI−TOF質量分析装置によって得られた、PEG化mTGの典型的なバッチの質量対電荷スペクトルである。
【図12】PEG試薬とアミンの比率を固定し、反応物の濃度を様々に変えたときの、PEG化mTG生成物のSDS−PAGE分析を示す画像である。
【発明を実施するための形態】
【0018】
酵素分子の改変は、形成される架橋マトリックスにおける酵素分子の感知体積を改変することによって行ってもよい。改変された酵素分子の感知体積は、ポリマーが架橋することによって形成されたマトリックスの架橋度によって決定することが好ましく、感知体積の増加は、改変していない酵素分子による架橋と比較して、架橋度が減少することによって示される。
【0019】
酵素分子の感知体積を増加させる方法のひとつとして、少なくとも1つの分子または部分を酵素分子に共有結合または非共有結合させることによって、分子のサイズおよび/または流体力学的体積を増加させる方法が挙げられる。本発明者らは、分子を酵素に共有結合させることによって、ヒドロゲルまたは架橋マトリックスの酵素的架橋度を制御することができ、この酵素分子の改変により最終的な架橋度を低下させることができることを実証している。このようにして、過剰な架橋を防ぐことができる。
【0020】
感知体積を増加させる別の方法として、酵素分子の正味の電荷がポリマー鎖またはコポリマー鎖の正味の電荷とは反対の符号に帯電するように、酵素分子の静電荷を改変する方法が挙げられる。これは、酵素の等電点(pI)を変化させることによって達成することができる。
【0021】
これに限定されるものではないが、仮説によれば、酵素分子の感知体積を増加させると、架橋マトリックスまたは架橋ヒドロゲルにおける酵素分子の移動または拡散が低下する。これによって、ヒドロゲルの所望の物質特性に有益な程度を越えて、酵素分子の架橋活性により架橋が継続されることを防ぐことができる。
【0022】
本明細書で定義されるように、「感知体積(perceived volume)」または「有効体積」は、架橋マトリックス内の架橋化酵素の有効な流体力学的体積を指す。感知体積は、架橋反応前または架橋反応中に、酵素が、別の分子、担体、ポリマー、タンパク質、多糖類などと共有結合または非共有結合することによって増加してもよい。
【0023】
本明細書で定義されるように、「拡散」または「移動」は、溶液、ヒドロゲル、またはマトリックスにおける架橋化酵素または他のタンパク質の、ブラウン運動に起因するランダムな分子運動を指す。
【0024】
本明細書で定義されるように、「拡散係数」とは、単一種の分子が特定の条件下で拡散する度合いを定量化したものを指す。酵素拡散の測定に代わるものとして、ヒドロゲルからの酵素の溶出を測定することが挙げられるが、これに限定されない。
【0025】
本明細書で定義されるように、「移動の低下」は、溶液中またはヒドロゲル内において、タンパク質または酵素の分子運動が遅いことまたはタンパク質または酵素の拡散係数が小さいことを指す。
【0026】
本明細書で定義されるように、「サイズ」は、分子の分子量、分子の流体力学的体積、または分子の感知体積を指す。
【0027】
本明細書で使用されているように、「分子量」(MWと略す)は、ダルトンまたはキロダルトンで表される、タンパク質またはポリマーの絶対重量を指す。たとえば、PEG化タンパク質(すなわち、1個以上のPEG(ポリエチレングリコール)分子が結合したタンパク質)の分子量は、そのPEG化タンパク質の構成成分の分子量をすべて合計したものである。
【0028】
本明細書で定義されるように、「流体力学的体積」は、一般的にサイズ排除クロマトグラフィーを用いて測定してもよいタンパク質または酵素の見かけの分子量を指す。構成成分の流体力学的体積は、構成成分が流体中で運動していると仮定したときの直径または体積を指す。
【0029】
本明細書で定義されるように、「マトリックス」は、架橋された材料の組成物を指す。一般的に、組成物中に含まれるマトリックス形成材料が架橋すると、この組成物は、液体状態からゲル状態へ遷移することによって、「ゲル」、「ヒドロゲル」、または「ゲル化組成物」を形成する。ゲルは、ある一定の粘弾特性およびレオロジー特性を有し得ることから、ある程度の耐久性および膨潤性を有する。このような材料はポリマーであることが多い。マトリックスは、コポリマーとも称される架橋していない材料を含んでもよい。
【0030】
本明細書で使用されているように、「ポリマー」は、反復単位を含む、天然分子、合成分子、または半合成分子を指す。
【0031】
本明細書で使用されているように、「コポリマー」は、架橋反応に関与しても関与しなくてもよいマトリックスの構成成分を指し、通常、マトリックスの主要な構成成分ではない。このようなコポリマーとしては、デキストランなどの多糖類および/またはカルボキシメチルセルロースなどのセルロースポリマーが挙げられるが、これらに限定されない。コポリマーは、酵素またはマトリックス材料(たとえば、マトリックスの主成分であるタンパク質など)に共有結合していないことが好ましい。
【0032】
本明細書で使用されているように、「担体」は、架橋反応前または架橋反応中に、架橋化酵素と共有結合または非共有結合する、ポリマー、タンパク質、多糖類、または他の構成成分を指す。
【0033】
本明細書で定義されるように、「架橋化酵素」は、ポリマーストランドの基質残基を(たとえば、トランスグルタミネーションによって)直接的または(たとえば、キノンまたはフリーラジカルの生成を通して)間接的に架橋して、緊密なマトリックス(coherent matrix)(たとえば、ヒドロゲルなど)を形成することのできる酵素または酵素の組み合わせを指す。
【0034】
本発明の少なくともいくつかの実施形態によれば、
架橋マトリックスであって、
改変酵素分子により架橋される基質ポリマーを含み、
改変酵素分子が、基質ポリマーの架橋によってマトリックスが形成されるのに伴い、架橋マトリックス内における酵素分子の感知体積が変化するように改変されたものであることを特徴とする、架橋マトリックスが提供される。
【0035】
改変酵素分子は、改変酵素分子の実際のサイズが増加するように改変されたものであってもよい。改変酵素分子は、改変酵素分子の流体力学的体積が増加するように改変されたものであってもよい。改変酵素分子は、改変酵素の等電点(pI)を、改変していない酵素の等電点と異なるように変化させることによって、改変酵素分子の静電荷が基質ポリマーの正味の電荷と反対の符号に帯電されるように改変されたものであってもよい。また、(無水コハク酸による)スクシニル化、(無水酢酸による)アセチル化、(シアネートによる)カルバミル化、(アルデヒドによる)還元的アルキル化、および無水マレイン酸処理からなる群から選択される工程によって、酵素のリシンのε−アミノ基が改変されていてもよい。さらに、負の荷電量が減少するように、酵素のカルボン酸含有側鎖の1つ以上が改変されていてもよい。
【0036】
上記改変は、少なくとも1つの分子または部分が改変酵素分子に共有結合または非共有結合されていることを含んでもよい。また、上記改変は、改変作用を有する分子が改変酵素分子に共有結合されていることを含んでもよい。改変酵素分子は、非改変酵素よりも低い拡散速度および架橋速度を有してもよく、非改変酵素と少なくとも類似した測定酵素活性を有してもよい。
【0037】
上記架橋速度は、非改変酵素の架橋速度よりも少なくとも10%遅くてもよい。
【0038】
上記改変作用を有する分子は、担体またはポリマーを含んでもよい。このポリマーは、合成ポリマー、セルロースポリマー、タンパク質、または多糖類を含んでもよい。このセルロースポリマーは、カルボキシメチルセルロース、ヒドロキシプロピルメチルセルロース、ヒドロキシエチルセルロース、またはメチルセルロースのうちの1種以上を含んでもよい。上記多糖類は、デキストラン、コンドロイチン硫酸、デルマタン硫酸、ケラタン硫酸、ヘパリン、ヘパラン硫酸、ヒアルロン酸、またはデンプン誘導体のうちの1種以上を含んでもよい。
【0039】
上記改変作用を有する分子は、PEG(ポリエチレングリコール)を含んでもよい。このPEGは、PEG誘導体を含んでもよい。このPEG誘導体は、活性化PEGを含んでもよい。この活性化PEGは、メトキシPEG(mPEG)、もしくはその誘導体、mPEG−NHS、mPEGのスクシンイミジル(NHS)エステル(mPEG−スクシナート−NHS)、mPEG−グルタレート−NHS、mPEG−バレラート−NHS、mPEG−カルボナート−NHS、mPEG−カルボキシメチル−NHS、mPEG−プロピオナート−NHS、mPEG−カルボキシペンチル−NHS)、mPEG−ニトロフェニルカルボナート、mPEG−プロピルアルデヒド、mPEG−トシラート、mPEG−カルボニルイミダゾール、mPEG−イソシアネート、mPEG−エポキシド、またはこれらの組み合わせのうちの1種以上を含んでもよい。上記活性化PEGは、上記酵素のアミン基またはチオール基と反応してもよい。上記活性化された酵素のリシン残基に対する上記活性化PEGのモル比は、0.5〜25の範囲であってもよい。上記活性化PEGは、一官能基型、ヘテロ二官能基型、ホモ二官能基型、または多官能基型であってもよい。上記活性化PEGは、分岐PEGまたはマルチアームPEGであってもよい。上記活性化PEGは、1000ダルトン〜40,000ダルトンの範囲のサイズであってもよい。
【0040】
上記マトリックスは、上記酵素または上記基質ポリマーに共有結合していないコポリマーをさらに含んでもよい。このコポリマーは、多糖類またはセルロースポリマーを含んでもよい。この多糖類は、デキストラン、コンドロイチン硫酸、デルマタン硫酸、ケラタン硫酸、ヘパリン、ヘパラン硫酸、ヒアルロン酸、またはデンプン誘導体を含んでもよい。上記セルロースポリマーは、カルボキシメチルセルロース、ヒドロキシプロピルメチルセルロース、ヒドロキシエチルセルロース、メチルセルロースを含んでもよい。
【0041】
上記改変酵素分子は、上記改変酵素分子と複数の酵素分子とが架橋することによって、複数の架橋化酵素分子が凝集体を形成するように改変されたものであってもよい。
【0042】
上記改変酵素分子の改変またはその改変の度合いは、マトリックスの少なくとも1種の特性に変化を与えてもよい。この少なくとも1種の特性は、引張り強さ、剛性、上記基質ポリマーの架橋度、粘度、弾性、可撓性、破断歪み、破断応力、ポアソン比、膨潤力、およびヤング率、およびこれらの組み合わせからなる群から選択されてもよい。
【0043】
上記改変酵素の改変の度合いによって、上記改変酵素のマトリックス内における移動または上記改変酵素のマトリックスからの拡散が決定されてもよい。上記改変酵素の改変によって、上記改変酵素と上記タンパク質との溶液またはマトリックスにおける前記改変酵素の拡散係数が、非改変酵素と上記タンパク質との溶液またはマトリックスにおける非改変酵素の拡散係数よりも低下してもよい。上記改変酵素の改変の度合いによって、マトリックスの1種以上の力学的特性が決定されてもよい。上記改変酵素分子によって溶液中で架橋ポリマーが形成される架橋速度が、非改変酵素分子と比較して速くてもよい。
【0044】
本発明の少なくともいくつかの実施形態によれば、
マトリックスの形成を調節する方法であって、
マトリックスが形成されるのに伴い、架橋マトリックス内における酵素分子の感知体積が変化するように酵素分子を改変すること;
改変した酵素分子と、改変した酵素分子の基質である少なくとも1種の基質ポリマーとを混合すること;および
改変した酵素分子で該少なくとも1種の基質ポリマーを架橋して、その形成の少なくとも一部が酵素分子の改変によって調節されたマトリックスを形成すること
を含む方法が提供される。上記改変によって、上記少なくとも1種の基質ポリマーの架橋度が増加するのに伴い、上記改変酵素分子による架橋の速度が低下してもよい。上記改変酵素分子と上記少なくとも1種の基質ポリマーとを溶液中で混合すると、上記の改変によって、溶液の粘度が増加するのに伴い、上記少なくとも1種の基質ポリマーの架橋度が調節されてもよい。上記改変は、pH7〜9において上記酵素をPEG化することを含んでもよい。このPEG化反応は、pH7.5〜8.5で行われてもよい。
【0045】
上述の方法および/またはマトリックスの少なくともいくつかの実施形態によれば、上記少なくとも1種の基質ポリマーは、元より架橋可能なポリマー、上記改変酵素により架橋可能な部分変性ポリマー、および上記改変酵素により架橋可能な官能基またはペプチドを含む改変ポリマーからなる群から選択される基質ポリマーを含む。上記少なくとも1種の基質ポリマーは、ゼラチン、コラーゲン、カゼイン、もしくはアルブミン、または改変ポリマーを含んでもよく、上記改変酵素分子は、改変トランスグルタミナーゼおよび/または改変酸化酵素を含んでもよい。上記少なくとも1種の基質ポリマーは、動物組織もしくは動物組織から得られるコラーゲンを部分的に加水分解することによって得られるゼラチン;細菌系、酵母系、動物系、昆虫系、植物系、もしくは任意の種類の細胞培養を用いて産生される組換えゼラチン;およびこれらの組み合わせからなる群から選択されるゼラチンを含んでもよく;該動物組織は、動物の皮、結合組織、枝角、角、骨、および魚鱗からなる群から選択されてもよい。上記ゼラチンは、哺乳動物由来または魚類由来であってよい。上記ゼラチンは、Aタイプ(酸処理)またはBタイプ(アルカリ処理)であってよい。上記ゼラチンは、250〜300ブルームであってよい。上記ゼラチンの平均分子量は、75〜150kdaであってよい。
【0046】
上記改変トランスグルタミナーゼは、改変微生物トランスグルタミナーゼを含んでもよい。上記改変ポリマーは、改変微生物トランスグルタミナーゼによる架橋が可能となるように改変されたものであってもよい。上記改変酸化酵素は、チロシナーゼ、ラッカーゼ、またはペルオキシダーゼのうちの1種以上を含んでもよい。上記マトリックスは、上記少なくとも1種の基質ポリマーとして、上記改変酸化酵素により架橋されるフェノール酸を含む炭水化物をさらに含んでもよい。上記炭水化物は、アラビノキシランまたはペクチンのうちの1種以上を含んでもよい。上記酵素分子をPEG化により改変し、このPEG化により、マトリックスが使用される宿主動物の免疫系から上記酵素分子がマスクされ、これによって、上記酵素分子の免疫原性がマスクされてもよい。この宿主動物はヒトであってもよい。
【0047】
少なくともいくつかの実施形態によれば、本明細書に記載のマトリックスを組織に塗布することを含む、体液が滲出した組織を密封するための方法が提供される。この体液は血液を含んでもよく、このマトリックスは止血剤であってもよい。
【0048】
少なくともいくつかの実施形態によれば、本明細書に記載のマトリックスを含む、止血剤または外科用シーラントが提供される。
【0049】
少なくともいくつかの実施形態によれば、本明細書に記載のマトリックスを含む、創傷を密封するための組成物が提供される。少なくともいくつかの実施形態によれば、組織における縫合線またはステープルラインを密封するための組成物の使用が提供される。
【0050】
少なくともいくつかの実施形態によれば、本明細書に記載のマトリックスを含む、局所的薬物送達のビヒクルのための組成物が提供される。
【0051】
少なくともいくつかの実施形態によれば、注入可能な足場として適した、本明細書に記載のマトリックスを含む、組織工学のための組成物が提供される。
【0052】
少なくともいくつかの実施形態によれば、
組成物を改変する方法であって、
架橋可能な官能基を有する改変酵素と、改変酵素によって架橋可能な少なくとも1種の部分を有するタンパク質とを提供すること;および
改変酵素とタンパク質を混合して、改変酵素によりタンパク質を架橋し、かつタンパク質により改変酵素自体を架橋可能な官能基において架橋すること
を含む方法が提供される。
【0053】
ポリマーストランドの基質残基を直接的に架橋する直接架橋化酵素としては、トランスグルタミナーゼおよび酸化酵素が挙げられるが、これらに限定されない。トランスグルタミナーゼとしては、微生物トランスグルタミナーゼ(mTG)、組織トランスグルタミナーゼ(tTG)、および第XIII因子が挙げられる。これらの酵素は、天然酵素由来または組換え酵素由来であってよい。ポリマーストランドに含まれるグルタミンのアミノ酸およびリシンのアミノ酸は、トランスグルタミナーゼによる架橋の基質である。
【0054】
上記酸化酵素としては、チロシナーゼ、ラッカーゼ、およびペルオキシダーゼが挙げられるが、これらに限定されない。これらの酵素は、キノン生成(チロシナーゼ)またはフリーラジカル生成(ラッカーゼ、ペルオキシダーゼ)によってポリマーを架橋する。次いで、キノンまたはフリーラジカルは、互いにまたは他のアミノ酸もしくはフェノール酸と相互作用してポリマーを架橋する。これらの酵素が架橋可能な基質は、チロシンまたは他の芳香族アミノ酸を含有する任意のタンパク質であってもよい。このような基質は、フェルラ酸などのフェノール酸を含む炭水化物であってもよい。このような炭水化物は、たとえば、アラビノキシランまたはペクチンであってよい。
【0055】
1種以上の適切な官能基を有する合成または部分合成のポリマーも、上記のいずれかの酵素の架橋可能な基質として機能することができる。
【0056】
本発明の別の実施形態において、酵素の組み合わせが用いられる。
【0057】
本明細書で定義されるように、「ポリマーストランド」または「ポリマー鎖」は、酵素架橋のための基質ポリマーを指す。本発明の少なくともいくつかの実施形態によれば、(例示に過ぎず、これらに限定されるものではない)以下の分類のうちの1つに属することが好ましい。
1)上記の酵素によって元より架橋可能な基質残基を有する任意のポリマー、および自体が上記の酵素によって元より架橋可能である任意のポリマー。たとえば、トランスグルタミナーゼを用いる場合、このようなポリマーとして、上記の酵素によって元より架橋可能な、ゼラチン、コラーゲン、およびカゼインなどのタンパク質またはポリペプチドが挙げられる。
2)上記の酵素によって架橋可能な基質残基を含むが、自体の構造によってそのままでは上記の酵素により架橋できないポリマー。この場合、酵素で架橋する前にポリマー構造を改変しなければならない。たとえば、トランスグルタミナーゼを用いる場合、このようなポリマーとして、酵素の結合を妨げる球状構造を有するために上記の酵素の天然基質ではない、アルブミン、ラクトグロブリンなどのタンパク質が挙げられる。これらのタンパク質は、還元剤、変性剤、または熱を用いて部分的に変性することによって、基質へと変換することができる。
3)酵素架橋のための基質ポリマーではないが、上記の酵素の基質であるペプチドまたは官能基で改変することによって、上記の酵素で架橋可能となる天然または合成のポリマー。
【0058】
このようなポリマーとして、適切な種類のタンパク質が挙げられるがこれには限定されず、このようなタンパク質は、たとえば、上述したようにゼラチンを含んでもよい。ゼラチンは、当技術分野において公知のタンパク質を含む任意の種類のゼラチンを含んでもよい。このようなゼラチンとして、動物組織および/もしくは動物組織から得られるコラーゲンを部分的に加水分解することによって得られるゼラチン(動物組織として、動物の皮;靱帯、軟骨など(これらに限定されない)の結合組織;枝角、角など;および/または骨;および/または魚鱗および/または魚骨;または他の成分が挙げられるが、これらに限定されない);ならびに/または細菌系、酵母系、動物系、昆虫系、植物系、もしくは任意の種類の細胞培養を用いて産生される組換えゼラチンが好ましいが、これらに限定されない。
【0059】
本発明の好ましい実施形態によれば、動物源由来のゼラチンは、哺乳動物源由来のゼラチンを含むことが好ましく、豚皮、豚骨、牛骨、牛皮革片(ハイド)、および他のブタ由来物もしくはウシ由来物のうちの1種以上を含むことがより好ましい。また、ブタのゼラチンはアナフィラキシーを起こしにくいことから、動物源由来のゼラチンはブタのゼラチンを含むことがより好ましい。動物源由来のゼラチンは、Aタイプ(酸処理)でもBタイプ(アルカリ処理)でもよいが、Aタイプであることが好ましい。
【0060】
動物源由来のゼラチンは、一般的に低温度(50〜60℃であるが、この温度範囲に必ずしも限定されない)で行われる1回目の抽出で得られるゼラチンを含むことが好ましい。この温度の抽出により得られるゼラチンは、250〜300ブルームの範囲であり、少なくとも約95〜100kDaの高い分子量を有する。275〜300ブルームのゼラチンを用いることが好ましい。
【0061】
このようなゼラチンの製造会社として、PB Gelatins(Tessenderlo Group(ベルギー))が挙げられるが、これに限定されない。
【0062】
本発明の実施形態のいくつかによれば、動物源由来のゼラチンは魚類由来のゼラチンを含んでもよい。いずれの魚類由来であってもよく、コイ、タラ、カワカマス、マグロなどの冷水魚が好ましい。このようなゼラチンのpH(10%溶液で測定)は、4〜6の範囲であることが好ましい。
【0063】
冷水魚ゼラチンは10℃の水中で溶液となることから、冷水魚ゼラチンはすべて0ブルームであると考えられる。本発明において、高分子量の冷水魚ゼラチンを用いてもよく、またこれを用いることが好ましく、少なくとも約95〜115kDaの平均分子量を有する冷水魚ゼラチンがより好ましいものとして挙げられる。この平均分子量は、250〜300ブルームの動物ゼラチンの分子量に等しい。冷水魚ゼラチンは、プロリンおよびヒドロキシプロリンの含量が少ないことから、動物ゼラチンよりもかなり低い温度で熱可逆性のゲル化を起こす。アミノ酸残基1000個あたり、動物ゼラチンは135〜145個のプロリン残基および90〜100個のヒドロキシプロリン残基を有するのに対し、冷水魚ゼラチンは100〜130個のプロリン残基および50〜75個のヒドロキシプロリン残基を有する(Haug IJ, Draget KI, Smidsrod O.(2004).Food Hydrocolloids. 18:203-213)。
【0064】
このようなゼラチンの製造会社として、Norland Products(クランベリー(ニュージャージー))が挙げられるが、これに限定されない。
【0065】
本発明の実施形態のいくつかにおいては、低エンドトキシンゼラチンを用いて、ゼラチン−mTG組成物のゼラチン溶液成分を形成する。このようなゼラチンは、Gelita(登録商標)(エーバーバッハ(ドイツ))などの供給業者から市販されている。低エンドトキシンゼラチンは、1グラムあたりのエンドトキシン単位(EU)が1000を下回るゼラチンとして定義される。エンドトキシンが500EU/グラムを下回るゼラチンを用いることがより好ましい。
【0066】
脊椎または脳と接触する材料とともに使用する場合などの非常に慎重さが要求される用途においては、エンドトキシンが100EU/gを下回るゼラチンが好ましく、50EU/gを下回るゼラチンがより好ましい。エンドトキシンが10EU/gを下回るゼラチンは非常に高価であるが、慎重さが要求される用途において、本発明の少なくともいくつかの実施形態の一部として用いることもできる。
【0067】
本発明の実施形態のいくつかによれば、架橋されるタンパク質成分としてのゼラチンの代わりに、I型、II型、もしくは他の型の加水分解コラーゲンまたは非加水分解コラーゲンを使用する。種々の型のコラーゲンが熱的に安定なmTG架橋ゲルを形成する能力を有することが実証されている。
【0068】
本発明の実施形態のいくつかによれば、組換えヒトゼラチンが用いられる。このようなゼラチンは、Fibrogen(登録商標)(サンフランシスコ(カリフォルニア))などの供給業者から市販されている。組換えゼラチンの純度は、少なくとも約90%が好ましく、少なくとも約95%であることがより好ましい。いくつかの組換えゼラチンは、10℃においてゲル化しないことから、0ブルームであると考えられる。本発明の実施形態のいくつかにおいて、高分子量の組換えゼラチンを用いることが好ましく、少なくとも約95〜100kDaの分子量の組換えゼラチンがより好ましいものとして挙げられる。
【0069】
上述したように、架橋可能なタンパク質はゼラチンを含むことが好ましいが、追加的にまたはそれに代わるものとして、別の種類のタンパク質を含んでもよい。本発明の実施形態のいくつかによれば、このようなタンパク質はトランスグルタミナーゼの基質でもあり、適切なトランスグルタミナーゼ特異的ポリペプチド配列およびポリマー配列を有することが好ましい。このタンパク質は、単独で生体接着剤を形成する特性を有する合成ポリマー配列、または、トランスグルタミナーゼによる材料の架橋を増強するトランスグルタミナーゼ特異的基質で修飾されていることがより好ましいポリマー配列を含んでいてもよいが、これらに限定されない。いずれの種類の材料も下記に例示されるが、これらの例示に限定されない。
【0070】
トランスグルタミナーゼの適切な架橋標的を有し、かつ好ましくは約20℃〜約40℃の転移温度を有する合成ポリペプチド配列およびポリマー配列が開発されている。好ましい物理的特性として、組織結合能および線維形成能が挙げられるが、これらに限定されない。(上記の)ゲル化ゼラチンと同様に、このようなポリペプチドは、転移温度を低下させる1種以上の物質を有する組成物に用いてもよい。
【0071】
上記のペプチドは、ZymoGenetics Incにより出願された米国特許第5,428,014号および第5,939,385号に例示されているがこれらには限定されず、これらの特許文献はいずれも、あたかもその全体が本明細書に記載されているかのように参照により本明細書に組み込まれる。いずれの特許文献においても、トランスグルタミナーゼにより架橋可能な、生体適合性かつ生体接着性のポリペプチドが記載されており、トランスグルタミナーゼは、タンパク質結合グルタミニル残基のγ−カルボキサミド基とLys残基のε−アミノ基との間でのアシル転移反応を触媒し、その結果、ε−(γ−グルタミル)リシンイソペプチド結合を形成することが知られている。
【0072】
実施形態のいくつかによれば、得られた組成物は局所的薬物送達のためのビヒクルとして使用される。
【0073】
実施形態のいくつかによれば、得られた組成物は組織工学のための注入可能な足場である。
【0074】
実施形態のいくつかによれば、本発明の組成物は止血組成物である。実施形態のいくつかによれば、本発明の組成物は体液封止組成物である。
【0075】
本発明の組成物は、出血を速やかに止めることによって、負傷後または手術後の血液の喪失を最小限に留めるものであることが好ましい。
【0076】
本明細書で使用されているように、「創傷」は、循環系からの血液の喪失、または生理学的導管(たとえば、いかなる種類の脈管など)からの他の体液の喪失を引き起こす、患者のいかなる組織における損傷を指す。組織は、臓器や血管などの内部組織、または皮膚などの外部組織であり得る。血液喪失または体液の喪失は、破裂した臓器などからの体内における喪失、または裂傷などからの体の外面からの喪失であり得る。創傷は、臓器などの柔組織における創傷、または骨などの硬組織における創傷であり得る。損傷は、いかなる作因または原因によって引き起こされたものであってもよく、たとえば、外傷、感染、または外科的介入が挙げられる。損傷は、致命的なもの、または致命的ではないものであり得る。
【0077】
外科的創傷閉鎖は、現在、組織を引き合わせることによって治癒を促進する縫合およびステープルによって行われている。しかしながら、これらの方法は、体液の滲出を防ぐのに十分な密閉性を得ることができない場合が非常に多い。したがって、ステープルラインおよび縫合線に沿って生じることが多い体液滲出に挙げられるような、手術後の体液滲出を防ぐための装置および方法が医療分野において一層必要とされており、いまだにその要求は満たされていない。このような装置および方法は、末梢血管再構築、硬膜再構築、胸部手術、心臓血管手術、肺手術、神経手術、および消化管手術において、縫合またはステープルを補助して、出血または他の体液の滲出を止めるためのものとして必要とされている。現在市販されている高圧止血装置のほとんどは、たとえ接着性を有したとしても名ばかりのものである。したがって、少なくともいくつかの実施形態による本発明の組成物は、これらの欠点を克服するものであり、止血に用いてもよい。
【0078】
本明細書で使用されているように、「約」は、示された値のプラスマイナス約10パーセントの値を意味する。
【0079】
本発明の種々の実施形態における他の特徴および利点は、以下の詳細な説明および特許請求の範囲から明らかになるであろう。
【0080】
本明細書において、添付された図面を参照して本発明を例示としてのみ説明する。ここで、特に詳細図に関していえば、示された具体例は、本発明の好ましい実施形態を説明することのみを目的とした例示に過ぎず、本発明の原理および概念的見地を最も有用にかつ容易に理解されるように説明することを目的として示されるものであることが強調される。この点において、本発明の構造の詳細は、本発明を基本的に理解するのに必要とされる以上には詳しく説明されておらず、本発明を実際に使用する際の態様のいくつかが当業者にとって明らかとなるように図面を用いて説明がなされている。
【0081】
以下の各項目の見出しは、説明を容易にすることのみを目的として記載されている。これらの見出しは、いかなる方法によっても限定されることをなんら意図するものではないことが理解される。また、特に定義されない限り、本明細書で使用される技術用語および科学用語はすべて、本発明が属する分野の当業者によって一般に理解されるのと同じ意味を有する。本明細書に記載されたものと類似または同一の方法および原料を、本発明の実施または実験に用いることができるが、適切な方法および原料は以下に記載されているものである。矛盾点がある場合、定義を包含する本発明の明細書の記載が優先する。さらに、原料、方法、および実施例は、例示にとどまりなんら限定するものではないことが意図される。
【0082】
ヒドロゲルにおける酵素的架橋剤の感知体積の増加
架橋マトリックス内の架橋化酵素による触媒反応の速度の調節は、酵素含有ポリマー溶液の粘度および部分的に架橋した溶液の架橋密度(反応基の利用可能度)のほかに、架橋化酵素分子の感知体積を調節することにより達成できることが見出された。
【0083】
本発明の少なくともいくつかの実施形態によれば、架橋反応の開始前または架橋反応中に酵素分子の感知体積を増加させることによって、マトリックスの力学的状態が所望のものに近づくのに伴い触媒による架橋の速度が低下するように上記の調節を行ってもよく、かつ、このように調節することが好ましい。このようにすると、所望の架橋密度において凝固マトリックスがサイズの増した酵素を捕捉し、さらなる架橋が阻止される。検出酵素体積は、他の要因のうちでもとりわけ、酵素分子量および流体力学的体積の関数である。
【0084】
架橋マトリックスの最終的な架橋度は、マトリックスが形成されるのに伴い架橋マトリックス内の酵素分子の感知体積が増加するように、酵素分子、マトリックス材料、架橋環境、またはこれらの要因のいくつかの組み合わせを調整することによって、制限することができる。単一の仮説によって限定されることを望むものではないが、検出酵素の体積を増加させることにより、架橋マトリックス内の酵素の移動を低下させることができる。酵素の移動を低下させることによって最終的な架橋密度を調節することは、溶液粘度が低い状態の架橋反応の初期段階においては酵素分子が移動度を保持しているが、架橋が進行して溶液粘度が増加するのに伴い移動度が低下し、初期の固体マトリックスまたはヒドロゲルが形成されるとより顕著に移動度が失われる場合において最も効果的である。当然のことながら、特定の用途に望ましい架橋プロファイルおよび架橋度が得られるように、マトリックス内における酵素の移動は正確に制御されるべきである。
【0085】
単一の仮説によって限定されることを望むものではないが、サイズまたは流体力学的体積の増した酵素は、改変していない酵素と比較して、架橋マトリックス内における拡散係数または移動度が低いため、架橋可能な基質へと到達しにくくなる。
【0086】
サイズおよび/または流体力学的体積の増した酵素分子
架橋マトリックスにおける酵素分子の移動を低下させる方法として、酵素分子の有効サイズを増加させることが好ましい。これは、酵素分子の分子量(MW)、流体力学的体積、またはMWと流体力学的体積のいずれもを増加させることによって達成することができる。この方法は、架橋マトリックスの構成成分に影響を与えないと考えられる点で好ましい。
【0087】
本発明の明細書に記載された実施形態を効果的なものとするために、酵素活性を取り除かないように、または所望のポリマー基質を架橋して固体マトリックスもしくはヒドロゲルを形成する能力を取り除かないように、酵素分子のサイズを増加させることが好ましい。酵素は、適切な時間内にマトリックスを形成するのに十分な活性を保持していることが好ましい。さらに、サイズの増した酵素分子は、マトリックス内で移動を停止する前に所望の架橋度までの架橋を触媒するのに十分な移動度を架橋マトリックスにおいて保持することが好ましい。
【0088】
架橋マトリックスまたは架橋ヒドロゲルにおいて酵素分子サイズを増加させるための方法がいくつか同定されている。
1.酵素同士を架橋(分子間架橋)させて可溶性マルチユニットコンジュゲートを形成させる方法。この例のひとつが、下記の実施例18に記載されている。
2.酵素を担体に共有結合(固定化)させる方法。
I.可溶性タンパク質(たとえば、アルブミン)への固定化;(Allen TMら, 1985, JPET 234: 250-254, alpha-Glucosidase-albumin conjugates: effect of chronic administration in mice)。
II.可溶性ポリマーへの固定化。ポリマー担体が酵素よりも大きく、1つ以上の酵素分子が、それぞれのポリマー分子に固定化されることが好ましい。1つの酵素分子を、2つ以上の付着部位を介して2つ以上のポリマー分子に結合させることも可能である。担体は天然、合成、または半合成であってもよい。このような方法の多くは、酵素のインビボ安定性の増加または免疫原性の低下のために開発された。このようなポリマーのファミリーのうちのひとつとしてセルロースエーテルが挙げられ、これには、たとえば、カルボキシメチルセルロース、ヒドロキシプロピルメチルセルロース、ヒドロキシエチルセルロース、メチルセルロースなどが挙げられるが、これらに限定されない。このような固定化は、トリプシン(Villaongaら, 2000, Journal of Molecular Catalysis B: 10, 483-490 Enzymatic Preparation and functional properties of trypsin modified by carboxymethylcellulose)およびリゾチーム(Chen SHら, 2003, Enzyme and Microbial Technology 33, 643-649, Reversible immobilization of lysozyme via coupling to reversibly soluble polymer)などの酵素を用いて過去に行われているが、酵素架橋ヒドロゲルまたはマトリックスの力学的特性に変化を与えるためにこのような酵素固定化が用いられたことは過去にはない。
III.グリコサミノグリカン(GAG)への結合。グリコサミノグリカンとしては、コンドロイチン硫酸、デルマタン硫酸、ケラタン硫酸、ヘパリン、ヘパラン硫酸、およびヒアルロン酸が挙げられるが、これらに限定されない。上記のように、このような結合は、過去に行われているが(Luchter-Wasylewska Eら, 1991, Biotechnology and applied biochemistry 13: 36-47, Stabilization of human prostatic acid phosphatase by coupling with chondroitin sulfate)、酵素架橋ヒドロゲルまたはマトリックスの力学的特性に変化を与えるのに用いられたことはない。
IV.酵素は多糖類に結合させることもできる。多糖類としては、たとえば、デキストラン、およびヒドロキシエチルデンプンなどのデンプン誘導体などが挙げられる。この例のひとつは、酸化デキストランに酵素を結合させた実施例13に記載されている。
3.共有結合性改変によって1つの酵素分子に1つ以上の部分を付加する方法。必ずしもそうではないが、これらの部分は酵素よりも小さいことが多い。このような改変の例のひとつとして、架橋化酵素のPEG化が挙げられ、下記の複数の実施例に広く記載されている。
4.他の種類の共有結合。たとえば、酵素の表面にビオチン分子をグラフト化(ビオチン化)し、このビオチン化した酵素を、アビジンもしくはストレプトアビジンを含有する分子またはポリマー上に固定化して行う。担体は、架橋反応前または架橋反応中に架橋化酵素を捕捉する機能を有する架橋不能な可溶性ポリマーであってもよい。あるいは、架橋可能なポリマー自体に捕捉群(たとえば、アビジンまたはストレプトアビジン)をグラフト化し、その結果、架橋可能なポリマーの架橋反応中に架橋化酵素が徐々に固定化されるようにしてもよい。
5.担体またはポリマーに酵素を非共有結合させる方法。たとえば、酵素の正味の電荷が担体の正味の電荷と反対の符号に帯電している場合、酵素と担体またはポリマーとの間の静電気的相互作用によって安定な非共有結合性結合が形成される。
【0089】
酵素分子のサイズの増加に関する技術
酵素のサイズの増加については過去に数回にわたり開示されているが、酵素架橋マトリックスまたはヒドロゲルの形成および/またはその形成の調節についてはいままでに考慮されたことはない。サイズの増した酵素による架橋マトリックスにおける架橋反応性が過去に一度も明らかされたことがないように、サイズの増した酵素を架橋マトリックスに使用することは全く新規である。さらに、驚くべきことに、本発明の発明者らは、比色法による酵素活性アッセイを用いて酵素活性を単独で検討することにより、サイズの増した酵素の酵素活性と、ゲル化速度によって示されるこの酵素のヒドロゲル形成における架橋活性とが明らかに異なるものであることを実証した。たとえば、実施例5において、酵素触媒によるゲル化の速度と、比色アッセイを用いて測定した酵素活性値との比較が記載されている。この実施例において記載されているPEG化は、酵素のサイズを増加させるための方法として例示され、なんらこれに限定するものではない。
【0090】
PEG化は、酵素分子にポリエチレングリコール(PEG)分子を共有結合により付着させることであり、酵素分子のサイズを増加させる方法として好ましい。1つ以上のPEG分子を付加する操作は、「PEG化」として知られている。
【0091】
PEGは、生体に対して不活性であり、かつ、埋め込み分子または注入分子をPEG化することにより、これらに対する免疫原性応答を制限できる能力を有することから、酵素サイズを増加させるための材料として望ましい。このような免疫原性のマスキング(免疫原性応答の制限)が、本明細書に記載されているように酵素をPEG化することによって達成できるかは不明だが、単一の仮説によって限定されることを望まずとも、酵素をPEG化することにより、酵素に対する免疫原性応答を実際に制限することが可能であると考えられ、さらに、拡大解釈すれば、架橋マトリックスに対する免疫原性応答も制限することが可能であると考えられる。
【0092】
酵素をPEG化する方法のひとつとして、酵素のアミン基と反応する活性化メトキシルPEG(mPEG)を酵素と反応させること(アミンPEG化)が挙げられる。活性化mPEGとしては、mPEGのスクシンイミジル(NHS)エステル(mPEG−スクシナート−NHS、mPEG−グルタレート−NHS、mPEG−バレラート−NHS、mPEG−カルボナート−NHS、mPEG−カルボキシメチル−NHS、mPEG−プロピオナート−NHS、mPEG−カルボキシペンチル−NHS)、mPEG−ニトロフェニルカルボナート、mPEG−プロピルアルデヒド、mPEG−トシラート、mPEG−カルボニルイミダゾール、mPEG−イソシアネート、およびmPEG−エポキシドが挙げられるが、これらに限定されない。
【0093】
活性化mPEGは、酵素のチオール基と反応するものであってもよい(チオールPEG化)。
【0094】
活性化PEGは、一官能基型、ヘテロ二官能基型、またはホモ二官能基型であってもよい。
【0095】
活性化PEGは、分岐PEGまたはマルチアームPEGであってもよい。
【0096】
活性化PEGのサイズは、1000ダルトン〜40,000ダルトンの範囲であってもよい。
【0097】
活性化PEGと酵素のリシン基とのモル比は、0.1:1〜100:1、好ましくは、0.5:1〜10:1である。
【0098】
PEG化反応物のpHは、7〜9であることが好ましい。反応物のpHが、7.5〜8.5であることがより好ましい。
【0099】
好ましい実施形態によれば、PEG化されなかった酵素を取り除くため、または、PEG化生成物のサイズを縮小するために、PEG化酵素をさらに精製してもよい。精製は、サイズ排除クロマトグラフィーを用いて行ってもよい。あるいは、これに代わるものとしてまたは追加的に、SPセファロース、Qセファロース、SMセファロース、DEAEセファロースなどのイオンクロマトグラフィーを用いて精製を行ってもよい。あるいは、これに代わるものとしてまたは追加的に、PEG化されなかった酵素を取り除く精製は、透析、限外濾過、または硫酸アンモニウム分画を用いて行ってもよい。
【0100】
下記に示される種々の実施例は、架橋ヒドロゲル形成を調節するためのトランスグルタミナーゼのPEG化について記載されている。実施例1においては、PEG−NHS(5kD)を用いたmTGのPEG化反応が記載されている。PEG化生成物のサイズおよび分布は、PEGとmTGとの比率、および反応物のpHに依存する。
【0101】
実施例2においては、PEG−NHS(5kD)を用いたmTGのPEG化反応が記載されている。PEG化生成物のサイズおよび分布は、反応の継続時間およびpHに依存する。
【0102】
実施例3においては、PEG−NHS(2kD)を用いたmTGのPEG化反応が記載されている。PEG化生成物のサイズおよび分布は、PEGとmTGとの比率に依存する。
【0103】
実施例4においては、種々のPEG化mTG(5kD PEG)調製物のPEG化度を決定するためのTNBSアッセイが記載されている。実験結果より、PEG化度が、反応における活性化PEG:mTGの比率に依存することが示唆されている。
【0104】
実施例5においては、PEG化mTGの活性を測定するためのアッセイが記載されている。実験結果より、PEG化mTGは、ヒドロキシルアミンおよびCBZ−Gln−Glyなどの小さい基質に対して活性の大部分を有するが、ゼラチンなどの大きい基質に対しては活性の大部分が喪失されることが示唆されている。
【0105】
実施例6および7においては、架橋ゼラチンゲルからのmTGおよびPEG化mTGの溶出プロファイルのSDS−PAGE分析が記載されている。実験結果より、PEG化mTGは、非PEG化mTGよりも、ゆっくりとかつ小さい移動度でゲルから溶出したことが示唆され、これはおそらくPEG化mTGのサイズまたは流体力学的体積がより大きいことに起因すると考えられる。
【0106】
実施例8においては、架橋ゼラチンゲルから溶出したmTGの活性の測定が記載されている。実験結果より、架橋ゼラチンゲルから溶出した非PEG化mTGは、その活性の大部分(算出した最大活性:86%)を保持していることが示唆される。
【0107】
実施例9においては、PEG化mTGまたは非PEG化mTGで架橋したゼラチンゲルの力学的試験が記載されている。実験結果より、PEG化mTGで架橋したゼラチンゲルは、非PEG化mTGで架橋したゲルと比較して、より強くかつより柔軟なことが示されている。
【0108】
実施例10においては、種々のゼラチンシーラント調製物の破壊圧力試験が記載されている。実験結果より、PEG化mTGで作製したゼラチンシーラントは、破壊圧力試験において、非PEG化mTGで作製したシーラントに匹敵することが示唆されている。
【0109】
実施例11においては、インビボブタモデルのステープルラインを補強するためのシーラントの使用が記載されている。
【0110】
実施例12においては、架橋化酵素を不溶性担体に非共有結合させた効果が記載されている。
【0111】
実施例13においては、酸化デキストランで酵素を改変した効果が記載されている。
【0112】
実施例14においては、PEG化によって西洋ワサビペルオキシダーゼ(別の架橋化酵素)を改変することにより、ペルオキシダーゼ架橋により形成されるマトリックスを改変できることが実証されている。
【0113】
実施例15においては、架橋化酵素の部分的PEG化の効果が実証されている。
【0114】
実施例16においては、遊離PEG(架橋化酵素と共に溶液中に存在するが、酵素には共有結合されていないPEG分子)はゲル化になんら効果を与えないことが実証されている。
【0115】
実施例17においては、非改変酵素と改変酵素との種々の混合物のゲル化に対する影響が示されている。
【0116】
実施例18においては、二官能基型PEG−酵素ブリッジのゲル化に対する影響が実証されている。
【0117】
実施例19は、PEG化mTG(微生物トランスグルタミナーゼ)の質量分析に関する。
【0118】
実施例20においては、PEGとアミンの比率を固定したときの、種々の濃度の反応物によるmTGのPEG化が記載されており、反応物の総濃度がPEG化度に大きな影響を与えることが実証されている。
【0119】
驚くべきことに、PEG化は、微生物トランスグルタミナーゼ(mTG)がゼラチンを架橋する速度を低下させるが、トランスグルタミナーゼの最も標準的な活性アッセイであるヒドロキサマートアッセイにおける活性を低下させないことがこれらの実施例において見出された。この結果は、サイズの増した酵素はヒドロゲルを形成する効果が著しく低いと考えられることからヒドロゲル形成に用いるのは望ましくないであろうとする従来技術の教示と相反するものである。
【0120】
従来技術において、TGアーゼ(トランスグルタミナーゼ)がPEG化と関連してときおり言及されていることには注意しておかなければならない。しかしながら、このことに言及した文献は、(PEG化の基質としてTGアーゼを使用するのではなく)タンパク質の部位特異的PEG化を可能にするためのまたはこれを増強するためのツールとしてTGアーゼを使用することを教示するものであり、この使用は、タンパク質のグルタミル残基とPEG分子に付着させた一級アミン基とのトランスグルタミネーション反応を触媒することによって行われる。しかしながら、このような従来技術は、TGアーゼ自体をPEG化することによって、TGアーゼの架橋活性を変化もしくは調節すること、またはTGアーゼによって架橋されたヒドロゲルマトリックスの力学的特性を変化もしくは調節することを教示または示唆するものではない。
【0121】
架橋マトリックスへの結合による架橋化酵素の移動の低下
架橋マトリックスにおいて酵素の移動を低下させる別の実施形態において、酵素は、架橋反応を触媒するのと同時に架橋マトリックス自体への結合反応を起こす。酵素がポリマー溶液中を移動してポリマーを架橋しマトリックスを形成するのに伴い、酵素は徐々にポリマー自体に結合することによりマトリックス内に固定化される。たとえば、ビオチン化酵素は、アビジンまたはストレプトアビジンでコーティングされたポリマーを含む架橋可能なポリマー成分と混合することができる。米国特許第6046024号(Method of producing a fibrin monomer using a biotinylated enzyme and immobilized avidin)には、アビジン修飾アガロースを添加することによって、フィブリノゲン溶液からビオチン化トロンビンを捕捉する方法が記載されている。この方法では、アガロースは可溶性ではないが、米国特許第5026785号(Avidin and streptavidin modified water-soluble polymers such as polyacrylamide, and the use thereof in the construction of soluble multivalent macromolecular conjugates)によって記載されているように、アビジンまたはストレプトアビジンを水溶性ポリマーに結合させることも可能である。トランスグルタミナーゼをビオチン化し、次いで、アビジン処理した表面に吸着させることは、実施可能であることが示されている(Huang XLら, J. Agric. Food Chem., 1995, 43(4), pp 895-901)。あるいは、架橋化酵素をアビジンまたはストレプトアビジンに共有結合させ、得られたコンジュゲートを、ビオチン化ポリマーを含む架橋反応に添加してもよい。ビオチン化されるものは、架橋可能なポリマー自体(たとえば、ゼラチンなど)、または架橋不能なコポリマー(たとえば、デキストランなど)であってもよい。分子量が500,000ダルトン以下のデキストラン−ビオチンコンジュゲートは、供給業者から入手できる。
【0122】
架橋マトリックスにおける静電気的相互作用による架橋化酵素の移動の低下
本発明の別の実施形態において、酵素の移動は、酵素の正味の電荷がポリマー担体の正味の電荷と反対の符号に帯電した、酵素とポリマー担体との間の静電気的相互作用に基づいた可逆性結合によって低減される。酵素は、担体とプレインキュベートしてから架橋反応に添加してもよいし、架橋反応中に担体に結合させてもよい。たとえば、架橋化酵素が中性pHにおいて正に荷電している場合、この架橋化酵素を、負に荷電した担体(たとえば、カルボキシメチルセルロース(CMC))に静電気的に結合させてもよい。酵素をCMCとインキュベートして結合させ、得られた複合体を架橋反応に添加してもよいし、あるいは、酵素とCMCとを別々に添加してもよい。後者の場合、架橋反応中に酵素がCMCに徐々に結合するであろう。架橋可能なポリマーが架橋化酵素と反対の符号の電荷を有する場合、架橋反応中に酵素を架橋可能なポリマーストランド自体に結合させることも可能である。あるいは、架橋可能なポリマーまたは担体の電荷と反対の符号の電荷を酵素が有するように、架橋化酵素の等電点(pI)を変更することもできる。
【0123】
別の実施形態において、架橋化酵素は改変されて、架橋化酵素の等電点(pI)が変化し、任意のpHにおいて異なる正味の電荷に帯電される。酵素のpIを低下させる方法として、(無水コハク酸による)スクシニル化、(無水酢酸による)アセチル化、(シアネートによる)カルバミル化、(アルデヒドによる)還元的アルキル化、無水マレイン酸処理などの工程(これらに限定されない)によってリシンのε−アミノ基を改変することが挙げられる。これにより、1個の改変アミノ酸あたり1価以下の電荷が減少する(スクシニル化では、2価以下の正の正味の電荷が減少する)ことによりタンパク質の正の正味の電荷が減少し、pIも減少する。これとは逆に、グルタミン酸およびアスパラギン酸などのカルボン酸を含有する側鎖を改変し、タンパク質の負の荷電量を減少させ、pIを増加させてもよい。たとえば、酵素をEDC 1−エチル−3−[3−ジメチルアミノプロピル]カルボジイミド)およびエチレンジアミン(EDA)で処理することができる。EDCはカルボン酸基を活性化し、これらのカルボン酸基とEDAとの間にアミド結合を形成する。その結果、タンパク質の正の正味の電荷が増加し、pIも増加する。
【0124】
ヒドロゲルからのタンパク質の遊離は、ヒドロゲルポリマー鎖と捕捉されたタンパク質との間の静電気的引力および静電気的斥力に関連づけられている。組換えゼラチンマトリックスからのタンパク質遊離実験において、静電気的斥力は、捕捉されたタンパク質の拡散係数を増加させ、逆に、静電気的引力は拡散係数を減少させることが示唆されている(Marc Sutter, Juergen Siepmann, Wim E. Hennink and Wim Jiskoot, Recombinant gelatin hydrogels for the sustained release of proteins, Journal of Controlled Release Volume 119, Issue 3, 22 June 2007, Pages 301-312)。
【0125】
ヒドロゲルポリマー鎖自体のpIを変化させることは、ヒドロゲルからのタンパク質の遊離を調節する方法として示唆されている。しかしながら、ヒドロゲル内に捕捉されたタンパク質とヒドロゲル鎖との間の静電気的相互作用を調整することを含む従来技術は、ヒドロゲルからの治療用タンパク質の遊離速度を調節する方法に関係しており、このタンパク質自体はヒドロゲルの形成に関与していない。本発明の少なくともいくつかの実施形態において、静電気的相互作用は、ヒドロゲルの力学的特性を向上させるために改変され、この力学的特性は、酵素による架橋過程のヒドロゲルマトリックスにおける酵素の移動および拡散係数に関連し得る。
【0126】
したがって、架橋可能なマトリックスにおける架橋化酵素の拡散または移動が、ポリマーヒドロゲルではなく捕捉された酵素のpIを改変することによって強力に制限されるため、捕捉された架橋化酵素のpIを変化させることは架橋を阻止する新規な手法である。
【実施例】
【0127】
実施例1:PEG化生成物のサイズおよび分布に対する、反応pHおよびPEG:mTG比の影響
材料:
活性化PEG:mPEG−グルタレート−NHS 5kDa(SunBright ME−050GS、日油株式会社(日本))。
mTG:
SPセファロースイオン交換クロマトグラフィーを用いてさらに10%精製した味の素アクティバ。活性:0.2Mクエン酸ナトリウム(pH6)において604ユニット/ml。
クエン酸ナトリウム、Hepes、SDS、およびβ−メルカプトエタノールはシグマアルドリッチから入手した。
30%アクリルアミド/Bis 29:1およびBio−Safe Coomassie G−250ステインはバイオ・ラッドから入手した。
分子量マーカーは、Precision Plus Dual Color(バイオ・ラッド)であった。
【0128】
1ユニットのmTG活性は、pH6.0、37℃において、N−CBZ−Gln−Glyおよびヒドロキシルアミンから、1分間あたり1.0μmolのヒドロキサマートの形成を触媒する。一連の反応は、それぞれの反応物が0.2mlの容量になるように設定した。それぞれの反応物は、15U/ml mTG、適切な反応緩衝液(90mM クエン酸ナトリウム(pH6)または100mM Hepes(pH7))、および種々の量の活性化PEGを含有した。PEG−NHSは、タンパク質の一級アミン、リシン残基の側鎖上のε−アミン、およびタンパク質のアミノ末端と反応する。反応混合物におけるPEGのリシン残基に対する比率は以下に詳細に記載する。
【0129】
反応物を、37℃で1:36時間インキュベートし、最終濃度が110mMになるまでグリシンを添加して、酵素と反応しなかった余分な活性化PEG分子を中和した。
【0130】
各反応物から得られた試料を、SDSおよびβ−メルカプトエタノールの存在下、90℃で加熱して変性させ、SDS−PAGE(8%分離ゲル、4%濃縮ゲル、ミニプロティアン電気泳動システム、バイオ・ラッド)を用いて分析した。タンパク質を可視化するために、Bio−Safe Coomassie G−250ステインでゲルを染色し、水で脱色した。CanoScan 8800Fスキャナでゲルをスキャンし、画像(図1に示す)を得た。この画像は、PEG化生成物のサイズおよび分布に対する、反応pHおよび活性化PEG濃度の影響を示している。レーンは以下のように割り当てた。
レーン1:mTG(対照)
レーン2:分子量マーカー(上から下へ順に:250kD、150kD、100kD、75kD、50kD、37kD、25kD)
レーン3:53.3mg/ml活性化PEG;90mMクエン酸Na、pH6;PEGのリシンに対する比率9.15
レーン4:26.6mg/ml活性化PEG;90mMクエン酸Na、pH6;PEGのリシンに対する比率4.59
レーン5:13.3mg/ml活性化PEG;90mMクエン酸Na、pH6;PEGのリシンに対する比率2.30
レーン6:53.3mg/ml活性化PEG;100mM Hepes、pH7;PEGのリシンに対する比率9.15
レーン7:26.6mg/ml活性化PEG;100mM Hepes、pH7;PEGのリシンに対する比率4.59
レーン8:13.3mg/ml活性化PEG;100mM Hepes、pH7;PEGのリシンに対する比率2.30
【0131】
図1から明らかなように、PEG量を増加しpHを高くすると、ゲル上に見かけの分子量が増した酵素が見られた。
【0132】
実施例2:PEG化生成物のサイズおよび分布に対する、反応pHおよび継続時間の影響
それぞれの反応物は15U/ml mTGを含有した。
材料:
活性化PEG:mPEG−グルタレート−NHS 5kDa(SunBright ME−050GS、日油株式会社(日本))。
mTG:SPセファロースイオン交換クロマトグラフィーを用いてさらに10%精製した味の素アクティバ。活性:0.2Mクエン酸ナトリウム(pH6)において604ユニット/ml。
クエン酸ナトリウム、Hepes、SDS、およびβ−メルカプトエタノールはシグマアルドリッチから入手した。
30%アクリルアミド/Bis 29:1およびBio−Safe Coomassie G−250ステインはバイオ・ラッドから入手した。
分子量マーカーは、Precision Plus Dual Color(バイオ・ラッド)であった。
【0133】
1ユニットのmTG活性は、pH6.0、37℃において、N−CBZ−Gln−Glyおよびヒドロキシルアミンから、1分間あたり1.0μmolのヒドロキサマートの形成を触媒する。
【0134】
一連の反応は、それぞれの反応物が0.2mlの容量になるように設定した。それぞれの反応物は、15U/ml mTG、適切な反応緩衝液(100mM Hepes(pH7)または100mM Hepes(pH8))、および25mg/ml PEG−NHSを含有した。反応混合物におけるPEGのリシン残基に対する比率は4.59であった。
【0135】
反応物を室温で2時間インキュベートした。下記の種々の時点で試料を採取し、最終濃度が110mMになるまでグリシンを添加し、酵素と反応しなかった余分な活性化PEG分子を中和した。
【0136】
各反応物から得られた試料を、SDSおよびβ−メルカプトエタノールの存在下、90℃で加熱して変性させ、SDS−PAGE(8%分離ゲル、4%濃縮ゲル、ミニプロティアン電気泳動システム、バイオ・ラッド)を用いて分析した。タンパク質を可視化するために、Bio−Safe Coomassie G−250ステインでゲルを染色し、水で脱色した。CanoScan 8800Fスキャナでゲルをスキャンし、画像(図2に示す)を得た。この画像は、PEG化生成物のサイズおよび分布に対する、反応時間およびpHの影響を示している。レーンは以下のように割り当てた。
レーン1:25mg/ml活性化PEG;100mM Hepes、pH8;反応時間15分
レーン2:25mg/ml活性化PEG;100mM Hepes、pH8;反応時間30分
レーン3:25mg/ml活性化PEG;100mM Hepes、pH8;反応時間60分
レーン4:25mg/ml活性化PEG;100mM Hepes、pH8;反応時間120分
レーン5:分子量マーカー(上から下へ順に:250kD、150kD、100kD、75kD、50kD、37kD、25kD)
レーン6:25mg/ml活性化PEG;100mM Hepes、pH7;反応時間15分
レーン7:25mg/ml活性化PEG;100mM Hepes、pH7;反応時間30分
レーン8:25mg/ml活性化PEG;100mM Hepes、pH7;反応時間60分
レーン9:25mg/ml活性化PEG;100mM Hepes、pH7;反応時間120分
【0137】
図2に示されているように、反応時間を延長しpHを高くすると、ゲル上に見かけの分子量が増した酵素が見られた。
【0138】
実施例3:PEG−NHS(2kD)を用いたmTGのPEG化:
PEG化生成物のサイズおよび分布に対する、PEGとmTGの比率の影響
材料:
活性化PEG:mPEG−グルタレート−NHS 2kDa(SunBright ME−020CS、日油株式会社(日本))。
mTG:SPセファロースイオン交換クロマトグラフィーを用いてさらに10%精製した味の素アクティバ。活性:0.2Mクエン酸ナトリウム(pH6)において604ユニット/ml。
クエン酸ナトリウム、Hepes、SDS、およびβ−メルカプトエタノールはシグマアルドリッチから入手した。
30%アクリルアミド/Bis 29:1およびBio−Safe Coomassie G−250ステインはバイオ・ラッドから入手した。
分子量マーカーは、Precision Plus Dual Color(バイオ・ラッド)であった。
【0139】
1ユニットのmTG活性は、pH6.0、37℃において、N−CBZ−Gln−Glyおよびヒドロキシルアミンから、1分間あたり1.0μmolのヒドロキサマートの形成を触媒する。反応物(200μl)は、15U/ml mTG、100mM Hepes(pH8)、および種々の濃度のPEG NHS(2kD)を含有した。反応物を、37℃で2時間インキュベートし、1.5Mグリシン(最終濃度71mM)10μlを添加して、酵素と反応しなかったPEG−NHS分子を中和した。各反応物から得た試料を、SDSおよびβ−メルカプトエタノールの存在下、90℃で加熱することにより変性させ、SDS−PAGE(8%分離ゲル、4%濃縮ゲル、ミニプロティアン電気泳動システム、バイオ・ラッド)を用いて分析した。タンパク質を可視化するために、Bio−Safe Coomassie G−250ステインでゲルを染色し、水で脱色した。CanoScan 8800Fスキャナでゲルをスキャンし、画像(図3に示す)を得た。この画像は、種々の濃度のPEG−NHS(2kD)を用いたPEG化mTGのSDS分析を示している。レーンは以下のように割り当てた。
レーン1:分子量マーカー
レーン2:1.75mg/ml PEG−NHS 2kD;PEGのリシンに対する比率0.74
レーン3:3.5mg/ml PEG−NHS 2kD;PEGのリシンに対する比率1.48
レーン4:7mg/ml PEG−NHS 2kD;PEGのリシンに対する比率2.97
レーン5:14mg/ml PEG−NHS 2kD;PEGのリシンに対する比率5.93
レーン6:28mg/ml PEG−NHS 2kD;PEGのリシンに対する比率11.86
レーン7:56mg/ml PEG−NHS 2kD;PEGのリシンに対する比率23.72
【0140】
図3に示されているように、PEG−NHS量を増加させると、ゲル上に見かけの分子量が増した酵素が見られた。
【0141】
実施例4:PEG化度を測定するためのTNBSアッセイ
材料:
グリシンおよび5%TNBS溶液(ピクリルスルホン酸)はシグマアルドリッチから入手した。
重炭酸ナトリウムはFrutarom(イスラエル)から入手した。
希釈TNBS溶液は、5%TNBSと重炭酸緩衝液(pH8.5)とを1:500の比率で混合することにより調製した。
分光光度計はAnthelie Advanced(Secomam)であった。
【0142】
検量線を作成するため、以下の濃度のグリシン溶液を重炭酸緩衝液(pH8.5)で調製した:1μg/ml、2μg/ml、4μg/ml、8μg/ml。
【0143】
希釈TNBS溶液0.5mlを、標準グリシン溶液または試料1mlと混合した。混合物を37℃で2時間インキュベートした。次に、10%SDS溶液0.5mlと1M HCl 0.25mlとを添加して反応を停止した。溶液をキュベットに移し、分光光度計を用いて335nmでのO.D.を読み取った。
【0144】
グリシンについて設定した検量線に基づいて、各PEG化mTGの遊離NH2基のパーセンテージを決定した。
%PEG化=100−遊離NH2のパーセンテージ
算出したPEG化mTGの平均MW:38,000+(%PEG化:100×18×5000)
【0145】
結果を以下の表1に示す。
【0146】
【表1】
【0147】
上記の表により、PEG化を増加させることによって、mTGの見かけの(算出された)分子量が増加したことが示された。さらに、PEG化度は遊離リシン基のパーセンテージの減少と相関し、これによって、予想されたようにリシン基がPEG化されていたことが示された。
【0148】
実施例5:PEG化mTGの活性を測定するためのアッセイ
材料:
尿素、クエン酸Na、酢酸Na、および塩化カルシウムはシグマアルドリッチから入手した。
ゼラチン(豚皮、Aタイプ、275ブルーム)はGelitaより入手した。
【0149】
PEG化反応物(8ml)は、15U/ml mTG、100mM HEPES(pH7)、および14mg/ml PEG−NHS(5kD)を含有した。反応条件は図1のレーン8と同様であった。
【0150】
反応物を37℃で1:50時間インキュベートし、2.34Mグリシン0.4mlを添加して(最終濃度100mM)反応していない活性化PEGを中和した。室温で15分後、Amicon Ultra−4 Centrifugal Filter Unit MWCO 30,000(ミリポア)を用いて、反応混合物を2mlに濃縮し、反応緩衝液を0.2M クエン酸ナトリウムに交換した。濃縮したPEG化mTGを4×と称し、これをクエン酸緩衝液で2倍希釈および4倍希釈したものをそれぞれ2×および1×と称す。
【0151】
ゼラチンを基質として用いる活性アッセイ
mTG 0.5mlをゼラチン調製物(25%ゼラチン、3.8M尿素、0.15M CaCl2、0.1M酢酸Na、pH6)1mlと混合し、37℃でインキュベートし、ゲル化時間を記録した。ゲル化時間は、反応チューブを逆さにしても液体が流れ落ちない時間と定義される。
【0152】
ヒドロキサマートアッセイを用いる活性アッセイ
反応物A 15μl 1×非PEG化mTG(15U/ml)+135μlクエン酸緩衝液
反応物B 15μl 1×PEG化mTG+135μlクエン酸緩衝液
反応物C 15μl 2×PEG化mTG+135μlクエン酸緩衝液
反応物D 15μl 4×PEG化mTG+135μlクエン酸緩衝液
反応物A〜Dのそれぞれに反応カクテル1mLを添加し、37℃で10分間または20分間インキュベートした。各時点において、反応物0.23mlをTCA0.5mLと共に試験管に加えた。
【0153】
ヒドロキサマート反応基質カクテル(20ml、pH6)
240mg CBZ−Glu−Gly(シグマアルドリッチ)
139mg 塩酸ヒドロキシルアミン(シグマアルドリッチ)
61.5mg 還元型グルタチオン(シグマアルドリッチ)
4ml 0.2Mクエン酸Na緩衝液、pH6
20mlになるまで加えた水
結果を下記の表2に示す。
【0154】
【表2】
【0155】
上記の表2によると、トランスグルタミナーゼをPEG化することにより、1×PEG化においてゲル化時間が延長したが、ヒドロキサマートアッセイ(ヒドロゲルは形成されず、遊離溶液中で行われるアッセイ)においては酵素活性にほとんど影響を与えなかったことが示された。実際、PEG化量が増加したことによりゲル化時間が延長したが、これはおそらく、形成過程の架橋ポリマーネットワーク内の基質分子と接触するのに必要な酵素の移動度が低下したことによると考えられる。あるいは、PEG化が酵素の活性部位に構造的変化を与え、その結果、基質に対して非PEG化酵素と同程度に効率的な結合ができなくなったか、または酵素の活性部位の近傍に挿入された1つ以上のPEG分子が、結合しようとする基質分子に対する立体障害となったとも説明することができる。ヒドロキサマートアッセイにおいてPEG化酵素活性が低下しなかったのは、このアッセイにおける基質のサイズが小さいこと、または架橋ポリマーネットワークを形成させなかったことが理由として考えられ得る。これらの説明はすべて、単一の仮説によって限定されることを望むことなく示されている。
【0156】
この実験結果より、PEG化がヒドロゲルの形成にもたらす効果は、溶液中の酵素活性に対する効果とはまったく異なることが裏付けられた。単一の仮説によって限定されることを望むものではないが、これらの異なる効果は、酵素の見かけのサイズが増したことおよび/または(サイズが増したこと以外による)感知体積が増したことにより引き起こされ、その結果、ヒドロゲルの形成を調節するのに有益な効果がもたられたと考えられる。いかなる場合においても、本発明の少なくともいくつかの実施形態によれば、PEG化することによりもたらされる差別的な効果により、形成過程のヒドロゲルにおける架橋を調節することが可能となる。
【0157】
実施例6:ゼラチンゲルからのPEG化mTGおよび非PEG化mTGの溶出プロファイル
材料:
ゼラチン(豚皮、Aタイプ、275ブルーム)はGelitaより入手した。
30%アクリルアミド/Bis 29:1およびBio−Safe Coomassie G−250ステインはバイオ・ラッドから入手した。
分子量マーカーは、Precision Plus Dual Color(バイオ・ラッド)であった。
【0158】
PEG化mTG(反応条件は図1のレーン6におけるものと同様であった)と20U/ml mTGとを1:1で含む酵素混合物0.67mlと、ゼラチン溶液(25%ゼラチン、3.8M尿素、0.15M CaCl2、0.1M酢酸Na、pH6)1.33mlと混合することによって、架橋ゼラチンゲルを作製した。得られたゲルをサランラップで包み、37℃で2時間インキュベートした。次に、生理食塩水を10ml含む試験管にゲルを入れ、恒温振とう培養機を用いて37℃で4時間インキュベートした。1時間ごとに試料を採取した。Amicon Ultra−4 Centrifugal Filter Unit MWCO 30,000(ミリポア)を用いて試料を濃縮し、SDSおよびβ−メルカプトエタノールの存在下90℃で加熱することにより変性させ、SDS−PAGE(8%分離ゲル、4%濃縮ゲル、ミニプロティアン電気泳動システム、バイオ・ラッド)を用いて分析した。タンパク質を可視化するため、Bio−Safe Coomassie G−250ステインでゲルを染色し、水で脱色した。SDS−PAGEのバンド強度を定量化するため、CanoScan 8800Fスキャナでゲルをスキャンし、得られた画像(図4に示す)をQuantity Oneソフトウェア(バイオ・ラッド)を用いて解析した。レーンは以下のように割り当てた。
レーン1:t=1時間に採取された10μlの試料
レーン2:t=1時間に採取された20μlの試料
レーン3:t=4時間に採取された10μlの試料
レーン4:t=4時間に採取された20μlの試料
レーン5:分子量マーカー
レーン6:7μl mTG+PEG化mTG混合物
レーン7:3μl mTG+PEG化mTG混合物
レーン8:1.5μl mTG+PEG化mTG混合物
【0159】
図4は、同一の架橋ゼラチンゲルからのmTGおよびPEG化mTGの溶出を示す。表3は、ゲルから溶出したトランスグルタミナーゼの相対量を示す。
【0160】
【表3】
【0161】
実施例7:架橋ゼラチンゲルからの種々のPEG化mTGの溶出
異なる濃度(2kDまたは5kD)のPEG−NHSを用いたmTGの大規模なPEG化
活性化PEG:
試薬:
mPEG−グルタリル−NHS、MW 5000(SunBright ME−050GS、日油株式会社(日本))。
mPEG−スクシニル−NHS、MW 2000(SunBright ME−020CS、日油株式会社(日本))。
mTG:SPセファロースイオン交換クロマトグラフィーを用いてさらに10%精製した味の素アクティバ。
30%アクリルアミド/Bis 29:1およびBio−Safe Coomassie G−250ステインはバイオ・ラッドから入手した。
SDSおよびβ−メルカプトエタノールはシグマアルドリッチから入手した。
【0162】
PEG化反応物(32ml)は、15U/ml mTG、100mM HEPES(pH8)、および種々の濃度のPEG−NHS(2kDまたは5kD)を含有した。反応物を室温で2.5時間インキュベートし、1.5Mグリシン(最終濃度97mM)2.2mlを添加して、反応しなかった活性化PEGを中和した。室温でさらに15分間インキュベーション後、ビバスピン20(ザルトリウス)を用いて反応混合物を8mlに濃縮し、それと同時に、反応緩衝液を0.2Mクエン酸Na(pH6)に交換した。
【0163】
1倍量の種々の濃度のPEG化mTG(上記)をそれぞれ、2倍量のゼラチン調製物(25%ゼラチン、3.8M尿素、0.15M CaCl2、0.1M酢酸Na、pH6)と混合した。混合物を、テフロン加工のイヌ用の骨の形をした型へ流し込んだ。ゲルが形成された後、ゲルを型から取り出し、重量を計り、乾燥を防ぐために密閉型試験管へ入れ、37℃で3時間インキュベートした。次に、ゲル1グラムあたり正確に5mlのクエン酸Na(pH6)を加え、エアシェーカーを用いて試験管を37℃、100rpmでインキュベートした。1時間後、2時間後、3時間後、および30℃、18時間でのさらなるインキュベーション後に、各時間点において試料をそれぞれ0.5mlずつ採取した。
【0164】
各時間点で得られた試料を、SDSおよびβ−メルカプトエタノールの存在下、90℃で加熱して変性させ、SDS−PAGE(8%分離ゲル、4%濃縮ゲル、ミニプロティアン電気泳動システム、バイオ・ラッド)を用いて分析した。タンパク質を可視化するために、Bio−Safe Coomassie G−250ステインでゲルを染色し、水で脱色した。SDS−PAGEにおけるバンドの強度を定量化するために、CanoScan 8800Fスキャナでゲルをスキャンし、得られた画像(図5に示す)をQuantity Oneソフトウェア(バイオ・ラッド)を用いて解析した。
【0165】
遊離すると考えられる酵素の最大理論量も同様にSDS−PAGEに供し、これを100%の遊離と見なした。実際に溶出した試料を横一列に並べて流し、100%遊離に対するバンド強度を算出した。SDS−PAGEにおけるバンドの強度を定量化するために、CanoScan 8800Fスキャナでゲルをスキャンし、得られた画像を、Quantity Oneソフトウェア(バイオ・ラッド)を用いて解析した。
【0166】
図5は、種々の架橋ゼラチンゲルから溶出したmTG(左側)およびPEG化mTG(右側)を示す。レーンは以下のように割り当てた。
レーン1:mTG、100%遊離対照
レーン2:架橋ゼラチンゲルから遊離したmTG、1時間時点
レーン3:架橋ゼラチンゲルから遊離したmTG、2時間時点
レーン4:架橋ゼラチンゲルから遊離したmTG、3時間時点
レーン5:架橋ゼラチンゲルから遊離したmTG、18時間時点
レーン6:PEG化mTG(7mg/ml PEG−NHS、5kD)、100%遊離対照
レーン7:ゼラチンゲルから遊離したPEG化mTG(7mg/ml PEG−NHS、5kD)、1時間時点
レーン8:ゼラチンゲルから遊離したPEG化mTG(7mg/ml PEG−NHS、5kD)、2時間時点
レーン9:ゼラチンゲルから遊離したPEG化mTG(7mg/ml PEG−NHS、5kD)、3時間時点
レーン10:ゼラチンゲルから遊離したPEG化mTG(7mg/ml PEG−NHS、5kD)、18時間時点
【0167】
総遊離量を以下の表4に示す。
【0168】
【表4】
【0169】
実施例8:架橋ゲルから溶出したmTGの活性
18時間後にゼラチンゲルから溶出した非PEG化mTG(実施例7を参照)9mlを、ビバスピン20(MWCO 30,000(ザルトリウス))を用いて0.47mlに濃縮した。濃縮した酵素の活性を、実施例5に記載されているのと同様にヒドロキサマートアッセイを用いて測定した。
【0170】
測定した活性は、3.65U/mlであった。
【0171】
算出した活性(ゲルにおける最初の活性5U/ml、図5におけるSDS−PAGEによる18時間後の遊離%、および表4におけるその定量化値26.7%に基づく)は、4.24U/mlであった。
【0172】
実施例9:PEG化mTGまたは非PEG化mTGで架橋したゼラチンゲルの力学的試験
尿素、クエン酸Na、酢酸Na、および塩化カルシウムはシグマアルドリッチから入手した。
ゼラチン(豚皮、Aタイプ、275ブルーム)はGelitaより入手した。
SPセファロースイオン交換クロマトグラフィーを用いてさらに10%精製した味の素アクティバをmTGとして用いた。活性:0.2Mクエン酸ナトリウム(pH6)において604ユニット/ml。
種々のPEG化度のPEG化mTG(2kDまたは5kDのPEG−NHSを使用)を、実施例7の記載と同様に調製した。
【0173】
PEG化mTG溶液1部と、ゼラチン溶液(25%ゼラチン、3.8M尿素、0.15M CaCl2、0.1M酢酸Na、pH6)2部とを混合した。混合物を、テフロン加工のイヌ用の骨の形をした型へ流し込んだ。ゲルが形成された後、ゲルを型から取り出し、生理食塩水中に浸して37℃で4時間インキュベートした。次いで、デジタルカリパスを用いて、イヌ用の骨の形をしたゲルの寸法を測定した。15U/ml非PEG化mTG1部とゼラチン溶液2部とを用いて対照試料を作製した。いずれの試料に対しても以下の試験プロトコルを行った。
【0174】
約12mm(幅)×1.9mm(厚さ)×20mm(長さ)の試料を、引張試験システム(3343モデル;インストロン)のクランプに固定した。引張試験の直前に各試料の正確な寸法を測定し、これらの測定した数値を用いて各試料の物質特性を算出した。測定および固定後、予荷重が0.025Nになるまで、各試料に0.25mm/秒の速度で引張力を加えた。これを0%の歪み点とした。予荷重後、試料が破断するまで試料に引張歪を0.5mm/秒の速度で連続的に加えた。
【0175】
破断した時点で最大の歪みおよび応力が生じ、生じた最大引張歪および最大引張応力を、破断時点における破断歪み(%)および破断応力(kPa)として記録した。各試料について、弾性率を10%歪みと30%歪みの間の直線領域から算出した。
【0176】
各種の架橋ゼラチンゲルを5回繰り返して試験し、得られた平均および標準偏差を表5に要約した。
【0177】
【表5】
【0178】
表5は、種々のPEG化mTGの力学的試験の結果を示す。最も左の列は、PEG化の条件を「A−B」で示している。すなわち、Aが7の場合7mg/ml PEGを指し、Aが14の場合14mg/ml PEGを指し、Aが28の場合28mg/ml PEGを指す。Bが2の場合2kD PEGを指し、Bが5の場合5kD PEGを指す。表に示されているように、PEG量を増加させると、ゲル化時間は延長し、ヤング率は低下した。これに対し、PEGサイズを増加させると、得られたゲルの引張り強さおよび可撓性が増加した。
【0179】
実施例10:破壊圧力試験により評価した生体組織におけるシーラントの性能
ブタの小腸組織から残留物を取り除き、10cm四方に切断した。各組織片を14ゲージ針で穿刺した。次いで、各組織を生理食塩水中に浸漬し、37℃でインキュベートした。実施例7に記載の方法で調製したシーラント材を塗布する前に、組織を平らにし、ガーゼパッドを用いて各組織の塗布部位の水分を拭き取った。1mLシリンジを用いて、試験シーラント約0.1〜0.2mLを各塗布部位に塗布した。塗布から5分以内に組織を生理食塩水で洗浄し、37℃で4時間インキュベートした。各試験は3回以上繰り返した。
【0180】
破壊圧力試験を行うため、片側が(クランプを用いて)密閉され、反対側が圧力メーターおよび手動ポンプに(プラスチック固定具を用いて)接続されたパースペックス箱に組織を入れた。パースペックス箱を生理食塩水で満たし、組織試料を完全に浸漬した。手動ポンプを用いて一定速度(20mL/分)で空気を注入した。泡の発生によって破壊圧力を決定した。
【0181】
結果を図6に示す。非PEG化mTGまたは2種のPEG化mTGを用いて作製したゼラチンシーラントについての破壊圧力が示されている。図に示されているように、いずれのPEG化mTGについても破壊圧力強度の中央値は増加したが、PEG化度がより低い酵素においてやや高い効果が示されている。
【0182】
実施例11:インビボブタモデルのステープルラインを補強するためのシーラントの使用
コヴィディエンEEA輪状吻合用外科用ステープラーを用いて、ブタの直腸の輪状吻合を行った。
【0183】
外科用シーラントはゼラチン溶液とPEG化TGとで構成した。反応容量72ml中、28mg/ml 5kDa PEG−NHSを用いて、実施例7と同様にPEG化TGを調製した。PEG化酵素の活性が40U/mlの非PEG化酵素と等しくなるように、ビバスピン20(MWCO 30,000(ザルトリウス))を用いて反応混合物を3mlに濃縮した。(ゼラチン溶液2.66mlおよびPEG化酵素溶液1.33で構成した)シーラント4mLを、直腸のステープルラインの周囲に均一に塗布し、4分間かけて硬化させた。次いで、ブタの傷口を縫合した。
【0184】
手術から14日後にブタを屠殺した。密封した吻合領域を肉眼での病理観察により評価し、シーラントを触診してその力学的特性を定性的に評価した。
【0185】
結果:
シーラントは、14日間の埋め込み期間にわたって顕著な分解を起こさなかった。シーラントは、ステープルラインに強く接着しており、ステープルラインの全長にわたって100%完全な状態で維持されていた。シーラント材は、柔軟性および可撓性を有し、輪状ステープルラインの形状および動きに適合した。
【0186】
シーラントが塗布された領域またはステープルライン領域において、炎症または腹部癒着は見られなかった。吻合は完全に治癒し、滲出の兆候はなかった。直腸において狭窄は観察されなかった。
【0187】
実施例12:架橋化酵素の不溶性担体への非共有結合
SPセファロースを、mTG(微生物トランスグルタミナーゼ)に結合させ、ゲルを作製した。この固定化トランスグルタミナーゼを用いると16〜23分でゲルが形成されたが、可溶性酵素は6分未満でゲルを形成した。したがって、固定化により、ゲル化に必要な時間が延長した。
【0188】
洗浄したSPセファロースビーズ(GEヘルスケア)500μlを、13.5mg/ml精製mTG2.7mlと、50mM酢酸Na(pH5.5)11.55mlと共に混合した(合計15ml)。
【0189】
この混合物を、シェーカーを用いて、室温で20分間インキュベートした。次いで、50mM酢酸Na(pH5.5)11.5mlを用いて、ビーズを3回洗浄(1回の洗浄あたり3分間)した。洗浄工程後、タンパク質の70%がビーズに結合していた。50mM酢酸Na(pH5.5)9.5mlにビーズを再懸濁し、最終容量を10mlとした。最終容量が600μlの以下の種々の組成を有するように、mTG担持ビーズと50mM酢酸Na(pH5.5)とを混合した(括弧内には、結合したmTGの量、および結合していないmTG1mgを用いて測定された活性(33ヒドロキサマートユニット)に基づいて算出されたmTG活性の理論値を示す)。
A:292μlビーズ+308酢酸Na(1.244mg/ml=41U/ml)
B:400μlビーズ+200μl酢酸Na(1.704mg/ml=56.2U/ml)
C:500μlビーズ+100μl酢酸Na(2.13mg/ml=70.3U/ml)
D:550μlビーズ+50μl酢酸Na(2.34mg/ml=77.2U/ml)
【0190】
各反応物A〜D500μlと、4.5M尿素を含む酢酸ナトリウム緩衝液中の25%ゼラチン溶液1mlとを、シリンジからシリンジへと往復させて混合した。ゼラチンが流動しなくなった時間を目視で確認してゲル化時間を決定した。
ゲル化時間:
A:約23分
B:約21分
C:約20分
D:約16分
対照(結合していないmTG 10U/ml):5.5分
【0191】
結合したmTGのゲル化時間は、遊離酵素と比較して顕著に延長された。これは、酵素を大きな足場または不溶性担体に結合させると、ヒドロゲルマトリックス内における酵素の移動が低下し、その結果、結合した酵素の架橋によるゲル化(力学的剛性が増したことを示す)は、遊離(結合していない)酵素によるものと比較して、時間が延長されることを示唆する。したがって、酵素を結合させると、ヒドロゲルマトリックスの力学的特性が改変される。
【0192】
実施例13:酸化デキストランでの酵素改変
この実験は、可溶性大型分子と結合させることによって酵素を改変することにより、その力学的特性が改変されることを実証している。
【0193】
方法
デキストラン1グラムを精製水20mlに溶解した。過ヨウ素酸ナトリウム1.3グラムを添加し、反応物をアルミホイルによって光から保護しながら、室温で80分間(9:50〜11:10)撹拌した。
【0194】
グリセロール2グラムを添加して、反応しなかった過ヨウ素酸塩をクエンチした。
【0195】
反応物を1Lの精製水で2:00時間かけて3回透析し、1回ごとに精製水を交換した。
【0196】
【表6】
【0197】
反応物を室温で一晩インキュベートし、次いで、ビバスピン20(ザルトリウス)を用いたダイアフィルトレーションによって精製した。
【0198】
結果
図7は、コンジュゲーション反応物A〜DのSDS−PAGE分析を示す。4〜15%ミニプロティアン TGXゲル(バイオ・ラッド)上に、4.35μg(反応物A)、4.38μg(反応物B)、1.98μg(反応物C)、および3μg(反応物D)のデキストランコンジュゲート化mTGを載せた。それぞれの試料は、0.1%SDSを含有していたが、還元剤は含有しておらず、ゲル上に載せる前に85℃で10分間加熱した。ゲルに一定電圧(200V)をかけて試料を流し、Bio−Safe Coomassie G−250溶液(バイオ・ラッド)でタンパク質バンドを染色することによって可視化した。分子量マーカーはPrecision Plus(バイオ・ラッド)であった。この実施例では、架橋化酵素(この場合、mTG(微生物トランスグルタミナーゼ))を可溶性ポリマー上に固定化できることが示されている。さらに、デキストラン:mTG比においてデキストランの比率が高いと、より多くの遊離mTG分子が、デキストランとの結合により高分子量コンジュゲートに変換された。
【0199】
実施例14:西洋ワサビペルオキシダーゼのPEG化
この実験では、西洋ワサビペルオキシダーゼ(別の架橋化酵素)をPEG化して改変することにより、ペルオキシダーゼ架橋により形成されるマトリックスを改変できることが実証されている。本発明の別の実施形態において、架橋化酵素は西洋ワサビペルオキシダーゼ(HRP)であり、PEG分子をHRP分子に付着させることによってHRPを改変し、これによって、HRP架橋により形成されるゼラチンヒドロゲルの力学的特性を改変することができる。
【0200】
方法
フェノール修飾ゼラチン(ゼラチン−Ph)の調製:
高分子量ゼラチン(Aタイプ)2グラムを、50mM MES(2−(N−モルホリノ)エタンスルホン酸(シグマアルドリッチ))緩衝液(pH6)100mlに溶解した。この2%w/w溶液に、チラミン(シグマアルドリッチ)0.984グラム、NHS(N−ヒドロキシスクシンイミド(シグマアルドリッチ))0.218グラム、およびEDC(1−エチル−3−[3−ジメチルアミノプロピル]カルボジイミド(シグマアルドリッチ))0.72グラムを添加した。反応物を室温で16時間撹拌し、次いで、蒸留水で十分に透析した。透析物を凍結乾燥し、得られた乾燥物を0.1Mリン酸緩衝液(pH6.0)に溶解して、最終容積が16mlの12.5%w/wゼラチンを得た。
【0201】
HRPのPEG化:
100mM Hepes(pH8.0)中において、2mg/ml HRP I型(シグマ、セントルイス(ミズーリ))と、60mg/ml PEG−NHS 5kDとを2時間反応させ、次いで、110mMグリシンを添加して反応しなかったPEG−NHSをクエンチし、さらに30分間インキュベーションした。PEG化HRPを、25mMリン酸緩衝液(pH6.0)で十分に透析することによって精製した。
【0202】
ゼラチン−PhのHRP依存性およびPEG化HRP依存性ゲル化:
ゼラチン成分:ゼラチン−Ph 5ml+20mM H2O2 0.5mlをガラスバイアル中で混合し、このうち4.4mlをシリンジAに移した。
HRP/PEG化HRP成分:0.035mg/ml HRPまたはPEG化HRP 1mlをシリンジBに入れた。
ゼラチン成分と酵素成分とをシリンジからシリンジへと往復させて混合し、次いで、上下反転させながら37℃でインキュベートしゲル化時間を決定した。
【0203】
20分後、ゲルの重量を計り、生理食塩水10mlに浸漬させて37℃で16時間インキュベートし、次いで、ゲルの重量を再び計って膨潤率を決定した。
【0204】
結果
混合後、ゼラチンおよび酵素の混合物は3分以内にゲルを形成した。HRPタンパク質およびPEG化HRPタンパク質のSDS−PAGE分析を図8に示す。HRPおよびPEG化HRP(各20μg)を4〜15%ミニプロティアンTGXゲル(バイオ・ラッド)上に載せた。それぞれの試料は0.1%SDSを含有していたが、還元剤は含有しておらず、ゲル上に載せる前に85℃で10分間加熱した。ゲルに一定電圧(200V)をかけて試料を流し、Bio−Safe Coomassie G−250溶液(バイオ・ラッド)でタンパク質バンドを染色することによって可視化した。分子量マーカーはPrecision Plus(バイオ・ラッド)であった。
【0205】
測定した膨潤率を以下の表7に詳細に示す。
【0206】
【表7】
【0207】
上記の表7に示されているように、PEG化HRPで作製したゲルは、非PEG化HRPで作製したゲルよりも大きく膨潤した。これは、PEG化(改変型)HRPにより形成されたゼラチンヒドロゲルの力学的特性が、遊離(非改変型)HRPにより形成されたヒドロゲルの力学的特性とは有意に異なることを実証している。いずれのゲルも熱に強く、80℃で1時間加熱しても溶解しなかった。
【0208】
実施例15:部分的PEG化の効果
異なるPEG化度を有する架橋化酵素により、異なる力学的特性がもたらされた。この実施例においては、流体力学的体積(この場合、PEG化度)を調節することによって、酵素的に架橋されたヒドロゲルの力学的特性がどのように特異的に調節され得るかが実証されており、流体力学的体積が大きい(すなわち、PEG化度が高い)場合、弾性の高いマトリックスが形成され、流体力学的体積が小さい(すなわち、PEG化度が低い)場合、弾性の低いマトリックスが形成される。当然のことながら、流体力学的体積が改変されていない(すなわち、PEG化なし)場合は、最も弾性の低いマトリックスが形成される。インストロン社製試験器によるデータおよびSDS−Pageゲルによるデータを、これらの効果に関して以下に記載する。
【0209】
方法
3種のPEG化反応を並列に実施した。反応は、PEG−NHS 5Kを用い、100mM HEPES(pH8.0)中において、室温で2.5時間行った。反応後、反応しなかった余分なPEGを110mMグリシンで中和し、さらに30分間インキュベーションを続けた。
【0210】
反応Aおよび反応Bは、いずれも同じPEG:アミン比を有したが、AのmTGおよびPEGはいずれも、Bよりも3倍高い濃度であった。反応Cは、反応Aと類似しているが、PEG:アミン比はAの半分であった。結果を表8に示す。
【0211】
【表8】
【0212】
これらの反応の終了後、得られたPEG化mTG溶液それぞれを、mTG溶液とゼラチン溶液の比が1:2になるように、(4.5M尿素を含む酢酸ナトリウム緩衝液中の)25%ゼラチン溶液と反応させて、ゼラチンヒドロゲルを形成させた。ゼラチンとの反応時間を各溶液において統一することによって、各PEG化mTG溶液のmTG活性レベルを標準化した。各ヒドロゲルが形成された後、37℃で2時間インキュベートし、引張試験システムを用いて力学的試験を行った。
【0213】
結果を、PEG化mTGをSDS−PAGE分析した画像である図9に示す。反応A、B、およびCから得られたPEG化mTG(それぞれ5μg)を、6%ポリアクリルアミドゲル上に載せ、SDS−PAGEに供した。それぞれの試料は、0.1%SDSを含有していたが、還元剤は含有しておらず、ゲル上に載せる前に85℃で10分間加熱した。ゲルに一定電圧(200V)をかけて試料を流し、Bio−Safe Coomassie G−250溶液(バイオ・ラッド)でタンパク質バンドを染色することによって可視化した。分子量マーカーはPrecision Plus(バイオ・ラッド)であった。
【0214】
SDS−PAGEプロファイルにより、反応A、B、およびCにおいて、PEG分子とmTG分子とがどのように異なる度合いで結合したかが示されており、Aにおいては多数のPEG分子がmTGに結合し、Bにおいてはより少ないPEG分子がmTGに結合し、Cにおいてはさらに少ないPEG分子がmTGに結合した。
【0215】
以下の結果に示されるように、PEG化度の差異は、ゼラチンマトリックスの力学的特性に顕著な影響を与える。すなわち、PEG化度が最も高かったmTGであるAは、弾性の最も高い(破断時点における引張歪が最も高い)ヒドロゲルを形成し、非PEG化mTGは弾性の最も低いヒドロゲルを形成し、部分的にPEG化されたmTGであるBおよびCは、それぞれのPEG化度に相関して、PEG化度が最も高かったmTGと非PEG化mTGとの間の力学的特性を有した。
【0216】
【表9】
【0217】
実施例16:遊離PEGはゲル化に効果を与えない
対照として、インストロン社製試験器によるゲルの力学的特性に関する結果を、遊離PEGを用いてまたは用いずに評価した。「遊離」とは、PEG分子が、架橋化酵素と共に溶液中に存在するが、酵素には共有結合されていないことを意味する。実験結果より、遊離PEGを用いても、PEGが架橋化酵素に共有結合すること(すなわち、PEGによる酵素自体の改変)によってもたらされる力学的特性の改変は見られなかったことが示された。
【0218】
方法
20%PEG6000を含むまたは含まないゼラチン溶液(酢酸ナトリウム緩衝液中の25%ゼラチンおよび4.5M尿素)のそれぞれのアリコート4mlと、15U/ml mTGのアリコート2mlとを混合した。それぞれから得られた溶液2mlを、実施例9の記載と同様に、イヌ用の骨の形をした型へ流し込んだ。実施例9の記載と同様に、得られたゲルを型から取り出し、生理食塩水中で37℃、2時間インキュベートし、次いで、引張試験を行った。
【0219】
結果
【表10】
【0220】
表10の結果より、遊離PEG(可塑剤)の添加により、酵素架橋ヒドロゲルマトリックスの力学的特性にわずかな効果が見られたか、またはまったく効果が見られず、さらに、このような力学的特性の改変は、PEGを酵素に付着させて酵素分子の流体力学的体積を増加させることによる改変と比較して、わずかなものであることが実証されている。特に、遊離PEGを添加することによって、マトリックスの弾性(破断歪み)は全く向上しなかった。これに対し、別の実施例のいくつかに示されているように、酵素分子をPEG化すると、マトリックス弾性が有意に増加する。
【0221】
実施例17:混合した改変/非改変架橋化酵素
非改変酵素と改変酵素との種々の混合物を試験した。特定の割合で混合することにより、種々の異なる力学的特性を有するように改変することができる。
【0222】
本発明の別の実施形態において、酵素架橋マトリックスの力学的特性を改変するために、改変酵素は非改変(遊離)酵素と共に用いられる。
【0223】
方法
20%PEG6000を含むまたは含まないゼラチン溶液(25%ゼラチン、4.5M尿素、酢酸ナトリウム緩衝液)のそれぞれのアリコート4mlと、非PEG化mTGを加えたまたは加えていない55U/ml PEG化mTG 2mlと混合した。それぞれから得られた溶液2mlを、実施例9の記載と同様に、イヌ用の骨の形をした型へ流し込んだ。実施例9の記載と同様に、得られたゲルを型から取り出し、生理食塩水中で37℃、24時間インキュベートし、次いで、引張試験を行った。
【0224】
結果
【表11】
【0225】
表11の結果より、酵素架橋ヒドロゲルの力学的特性は、改変酵素のみを用いることによっても、また、低い程度にとどまるが、改変酵素と遊離酵素の混合物を用いることによっても、改変できることが示されている。
【0226】
実施例18:二官能基型PEG−酵素ブリッジ
この実験では、二官能基型PEGブリッジによって酵素同士を架橋できることが実証された。この実施例では、2個以上の酵素分子を互いに結合させて、酵素凝集体全体の流体力学的体積を増加させることができる。これを達成する方法のひとつとして、酵素分子間にブリッジを形成することのできる二官能基型分子を用いることが挙げられる。
【0227】
方法
100mM Hepes(pH8)中において、mTG(15U/ml、0.5mg/ml)と、種々の濃度の二官能基型PEG−NHS(10kD)とを、室温で2時間インキュベートした。次いで、110mMグリシンを30分を上回る時間をかけて添加し、反応しなかった余分なPEGを中和した。具体的な条件を以下の表12に示す。
【0228】
【表12】
【0229】
反応終了後、各反応組成の酵素5μgを7.5%ポリアクリルアミドゲル上に載せ、SDS−PAGE分析に供した。それぞれの試料は、0.1%SDSを含有していたが、還元剤は含有しておらず、ゲル上に載せる前に85℃で10分間加熱した。ゲルに一定電圧(200V)をかけて試料を流し、Bio−Safe Coomassie G−250溶液(バイオ・ラッド)でタンパク質バンドを染色することによって可視化した。分子量マーカーはPrecision Plus(バイオ・ラッド)であった。
【0230】
力学的特性試験を行うために、ゼラチン溶液(25%ゼラチン、4.5M尿素、酢酸ナトリウム緩衝液)のアリコート4mlと、15U/mlの遊離mTGまたはPEG化mTG(反応C)のアリコート2mlと混合した。得られた溶液2mlを、実施例9の記載と同様に、イヌ用の骨の形をした型へ流し込んだ。実施例9の記載と同様に、得られたゲルを型から取り出し、生理食塩水中で37℃、24時間インキュベートし、次いで、引張試験を行った。
【0231】
結果
図10のSDS−PAGE結果により、相対的に低いPEG:mTG比において、mTGのいくらかは、類似の濃度の一官能基型PEG(5kD)を含む反応において得られるPEG化生成物よりも大きな、非常に高い分子量の生成物に変換されたことが示されている。これにより、この高分子量生成物が、二官能基型PEGブリッジによって互いに架橋された酵素分子のマルチマーからなること、および、二官能基型PEGの使用は、架橋化酵素分子同士を結合させることによって架橋化酵素分子を改変する効果を有することが実証されている。
【0232】
遊離酵素分子による架橋により形成されたゼラチンヒドロゲルと比較した以下の力学的試験の結果により、架橋化酵素分子同士を結合させると、この連結した酵素分子による架橋により形成されるゼラチンヒドロゲルを顕著に改変できることが示されている。結果を表13に示す。
【0233】
【表13】
【0234】
実施例19:PEG化mTGの質量分析
PEG化mTG(PEG−NHS−5kDで改変した微生物トランスグルタミナーゼ酵素)の3種のバッチを、MALDI−TOF質量分析法によって分析した。図11は、これらのうちの1バッチのm/zスペクトルを示す。
【0235】
質量分析
無傷の分子の質量測定を、遅延イオン抽出、反射器、および337nmの窒素レーザーを用いたBruker Reflex IIIマトリックス支援レーザー脱離/イオン化(MALDI)飛行時間型(TOF)質量分析装置(Bruker、ブレーメン(ドイツ))で実施した。各質量スペクトルは、レーザーを200回照射した累積データから作成した。タンパク質に対する外部較正は、BSAおよびミオグロビンタンパク質(シグマ、セントルイス(ミズーリ))を用いて行った。
【0236】
MALDI−TOF MSのための試料調製−dried droplet法
2,5−ジヒドロキシ安息香酸(DHB)マトリックス剤と、0.1%トリフルオロ酢酸(TFA)−アセトニトリル(ACN)とを2:1の割合で混合したもの0.5Lを、ギ酸/イソプロパノール/H20(1:3:2)に溶解した試料溶液0.5Lとターゲット上で混合し、空気乾燥させた。溶媒が蒸発した後、試料を0.1%TFAで1〜3回再洗浄した。
【0237】
【表14】
【0238】
表14の結果は、5kDaのPEG−NHS試薬を用いてmTG架橋化酵素をPEG化したことにより、1個の酵素分子に複数個のPEG分子が結合されたことを示している。
【0239】
実施例20:PEGとアミンの比率を固定したときの、種々の濃度の反応物によるmTGのPEG化
この実施例により、反応物の総濃度がPEG化度に大きな影響を与えることが実証されている。PEG:アミン比を固定した場合、反応物(PEGおよびmTG)の濃度とPEG化度とに相関性があることが実証された。
【0240】
方法
100mM Hepes(pH8.0)中において、PEG−NHS−5kDを用いたmTGのPEG化を室温で2.5時間かけて行い、次いで、110mMグリシンを添加して反応しなかったPEG−NHSを中和した。反応後、各反応組成の酵素5μgを、6.0%ポリアクリルアミドゲル上に載せ、SDS−PAGE分析に供した。それぞれの試料は、0.1%SDSを含有していたが、還元剤は含有しておらず、ゲル上に載せる前に85℃で10分間加熱した。ゲルに一定電圧(200V)をかけて試料を流し、Bio−Safe Coomassie G−250溶液(バイオ・ラッド)でタンパク質バンドを染色することによって可視化した。分子量マーカーはPrecision Plus(バイオ・ラッド)であった。
【0241】
反応は以下の条件(表15)に従って実施した。
【0242】
【表15】
【0243】
結果
図12に示したSDS−PAGEレーンより、PEG化mTG分子が得られたことが確認できる。画像に示されているように、PEG:アミン比を固定した場合でも、反応物の濃度が高いほど、PEG化度の高い生成物が得られた。
【0244】
本発明の選択された実施形態は個別に記載かつ説明されているが、記載された実施形態の適切な態様は組み合わされてもよく、さらに、複数の実施形態を組み合わせてもよいことが理解される。
【0245】
加えて、本発明の選択された実施形態が記載かつ説明されているが、本発明は、記載された実施形態に限定されないことが理解される。むしろ、本発明の原理および精神、ならびに特許請求の範囲およびその等価物によって定義される範囲から逸脱することなく、これらの実施形態に変更を加えてもよいことが理解される。
【背景技術】
【0001】
酵素架橋マトリックスの利用
酵素架橋マトリックスは、食品産業、化粧品産業、および医療産業において、様々な用途に利用されている。酵素架橋ヒドロゲルは、特に医療用途において、組織シーラント、組織接着剤、止血製剤、組織工学用マトリックス、薬物送達プラットフォームなどの様々な医療用途に広く利用されている。ゼラチンやポロキサマーなどのヒドロゲルのいくつかは、特定の条件下(たとえば温度変化など)においてポリマー鎖同士が物理的に相互作用することによって形成され得るが、一方、ポリマー溶液のヒドロゲルが形成されるには架橋を必要とすることが多い。埋め込み可能なヒドロゲルにおいては、固体ゲルが実際に形成されることに加えて、このゲルを使用する組織で一般的に見られる状態、たとえば、力学的ストレス、体温上昇、酵素的分解、化学的分解などに対する抵抗性が必要とされる。このように、多くの場合において、ヒドロゲルマトリックスの架橋が必要とされる。架橋は、ヒドロゲルをプレキャストまたは成型することによって体外で行ってもよい。この架橋方法は、主に、組織工学または薬物送達においてヒドロゲルを使用する場合に用いられる。あるいは、架橋を体内で行ってもよく(in situゲル化またはin situ架橋)、所望の部位に注入または塗布した溶液を架橋させてゲルを形成させる。
【0002】
ゲル形成は、様々な架橋アプローチによって開始することができる。化学的アプローチによるゲル形成としては、シアノアクリレートにおいて見られるような接触による重合開始、または光開始などの外部刺激による重合開始が挙げられる。あるいは、グルタルアルデヒドやカルボジイミドなどの低分子量架橋剤(Otani Y, Tabata Y, Ikada Y. Ann Thorac Surg 1999, 67, 922-6. Sung HW, Huang DM, Chang WH, Huang RN, Hsu JC. J Biomed Mater Res 1999, 46, 520-30. Otani, Y.; Tabata, Y.; Ikada, Y. Biomaterials 1998, 19, 2167-73. Lim, D. W.; Nettles, D. L.; Setton, L. A.; Chilkoti, A. Biomacromolecules 2008, 9, 222-30.)または活性化置換基(Iwata, H.; Matsuda, S.; Mitsuhashi, K.; Itoh,E.; Ikada, Y. Biomaterials 1998, 19, 1869-76)を用いて、あらかじめ形成されたポリマーを化学的に架橋することによってもゲルを形成することができる。
【0003】
しかしながら、化学的架橋剤は、毒性、発癌性、または刺激性を有することが多いため、食品、化粧品、または医薬品において化学的架橋を行うと問題が生じることがある。さらに、化学的架橋剤は、架橋マトリックス外へと容易に拡散し得る小型分子であり、局所的・全身的障害を引き起こし得る。
【0004】
化学的架橋に代わるもののひとつとして、酵素的架橋アプローチが挙げられる。ゲル形成を開始するための酵素的架橋アプローチは、種々の異なる架橋化酵素に基づいて研究がなされている。このような酵素的架橋アプローチとしては、たとえば、ムラサキイガイ接着剤などの接着剤による酵素的架橋(Strausberg RL, Link RP. Trends Biotechnol 1990, 8, 53-7)、またはフィブリンシーラントで利用されているような血液凝固による酵素的架橋(Jackson MR. Am J Surg 2001, 182, 1S-7S. Spotnitz WD. Am J Surg 2001, 182, 8S-14S Buchta C, Hedrich HC, Macher M, Hocker P, Redl H. Biomaterials 2005, 26, 6233-41. 27-30)が挙げられる。
【0005】
ムラサキイガイ接着剤による架橋は、接着性タンパク質のフェノール(すなわちドーパ)残基が反応性キノン残基へと酵素的に変換されることによって開始され、次いで、この反応性キノン残基においてタンパク質間架橋反応が起こる(Burzio LA, Waite JH. Biochemistry 2000, 39, 11147-53. McDowell LM, Burzio LA, Waite JH, Schaefer JJ. Biol Chem 1999, 274, 20293-5)。この種のシーラントには、チロシナーゼや、ラッカーゼおよびペルオキシダーゼが酵素として用いられている。チロシナーゼはチロシンから、ラッカーゼおよびペルオキシダーゼは他のフェノール化合物から、キノン類およびフリーラジカルを生成することによって作用する。次いで、キノン類およびフリーラジカルは、タンパク質の遊離アミン、またはタンパク質および多糖類に含まれる同様に修飾されたフェノール基を架橋することができる。
【0006】
技術モデルとなっている第2の架橋作用として、血液凝固において見られるトランスグルタミナーゼ触媒反応が挙げられる(Ehrbar M, Rizzi SC, Hlushchuk R, Djonov V, Zisch AH, Hubbell JA, Weber FE, Lutolf MP. Biomaterials 2007, 28, 3856-66)。in situゲル形成のためのバイオミメティックアプローチにおいては、第XIIIa因子や他の組織トランスグルタミナーゼの使用について研究がなされてきた(Sperinde J, Griffith L. Macromolecules 2000, 33, 5476-5480. Sanborn TJ, Messersmith PB, Barron AE. Biomaterials 2002, 23, 2703-10)。
【0007】
さらなるin situ架橋ゲル形成のうち特に興味深いものとして、カルシウム非依存性微生物トランスグルタミナーゼ(mTG)によるゼラチンの架橋が挙げられる。mTGは、第XIIIa因子と類似した架橋反応を触媒するが、活性化のためにトロンビンもカルシウムも必要としない。mTGに関する初期の研究は、食品産業において用いることを目的としていたが(Babin H, Dickinson E. Food Hydrocolloids 2001, 15, 271-276. Motoki M, Seguro K. Trends in Food Science & Technology 1998, 9, 204-210.)、後の研究では医療分野における利用可能性が検討された。過去のインビトロ研究において、mTGはゼラチンを架橋して数分以内にゲルを形成できること、ゼラチン−mTG接着剤は湿った組織にも濡れた組織にも接着することができること、およびその接着強度はフィブリンベースのシーラントと同等もしくはそれ以上であることが示されている(Chen TH, Payne GFら, Biomaterials 2003, 24, 2831-2841. McDermott MK, Payne GFら, Biomacromolecules 2004, 5, 1270-1279. Chen T, Payne GFら, J Biomed Mater Res B Appl Biomater 2006, 77, 416-22.)。ゼラチンとmTGの医療用接着剤としての使用は、国際公開第2008/076407号に記載されている。
【0008】
架橋マトリックスを形成するための架橋剤として酵素を使用した際の不都合のひとつとして、所望のゲル状態が形成された後でも酵素が架橋反応を継続し得ることが挙げられる。架橋反応の継続は望ましくない場合が多く、その理由のひとつとして過剰な架橋により硬くて脆く柔軟性に劣るゲルが生じ得ることが挙げられる。さらに、架橋マトリックスの力学的特性がゲルの寿命を通して変化し続け、一貫した特性を持たせることが難しい。所望の架橋密度を上回っても酵素的架橋が継続されることは、架橋マトリックスまたはヒドロゲルが一旦形成されても酵素が架橋反応を触媒し続けることができることに起因する。これは、溶液粘度が非常に増加しても、酵素がマトリックス全体に拡散し続けることができるかどうかに依存する。この見解は、ポリマー溶液の部分的架橋から生じるネットワーク形成の初期段階において、溶液粘度が急速に増加するとトランスグルタミナーゼの移動は急速に低下することを、ペプチドグラフト化合成ポリマー溶液についての研究に基づき示唆したHuら(Hu BH, Messersmith PB. J. Am. Chem. Soc., 2003, 125(47), pp 14298-14299)による報告と一致している。
【0009】
力学的特性の低下を生じる過剰な酵素的架橋ついては、過去にいくつかの文献において報告されている。
【0010】
Bauerらは、高レベルの微生物トランスグルタミナーゼ(mTG)がコムギグルテンタンパク質の過剰な架橋を引き起こし、これによって、グルテンネットワークの柔軟性の喪失および力学的損傷が引き起こされることを実証した(Bauer N, Koehler P, Wieser H, and Schieberle P. Studies on Effects of Microbial Transglutaminase on Gluten Proteins of Wheat II Rheological Properties. Cereal Chem. 80(6): 787-790)。
【0011】
Sakaiらは、フェノール間の共有結合性架橋が多いほど力学的安定性は効果的に増強されるが、フェノール間の架橋が多すぎると形成されるゲルが脆くなることを見出した(Sakai S, Kawakami K. Synthesis and characterization of both ionically and enzymatically crosslinkable alginate, Acta Biomater 3 (2007), pp. 495-501)。
【0012】
カルシウム依存性トランスグルタミナーゼなどの補因子依存性架橋化酵素を用いる場合、一定時間反応させた後、結合反応または他の方法によって補因子を除去すると、架橋度を制限することができる。しかしながら、補因子を除去することは、ヒドロゲルが補因子を捕捉し得るヒドロゲル形成においては技術的に不可能である場合が多い。微生物源から入手可能なトランスグルタミナーゼなどの補因子非依存性酵素を用いる場合、反応系で熱処理を行うことによって架橋度を制限することができる。しかしながら、熱処理はタンパク質機能に悪影響を及ぼすため行わない方が望ましい。さらに、あらゆる反応系に熱処理が適するとは限らない。
【0013】
架橋マトリックス内で過剰な架橋が生じることのほかに、所望の架橋状態が得られた後にマトリックス内において架橋化酵素が持続的に拡散することによって、酵素がゲル外へと速い速度で拡散し得る(これは酵素溶出としても知られている)。これによって、体内に放出された高レベルの架橋化酵素が体内組織と相互作用して、局所的または全身的障害を引き起こすという問題が生じ得る。
【発明の概要】
【発明が解決しようとする課題】
【0014】
種々の用途に用いることができる、改良された酵素により架橋された組成物が必要とされており、このような組成物は有用であると考えられる。
【0015】
したがって、固体マトリックスの形成が開始された後に、所望の力学的特性が得られた時点で架橋マトリックスの酵素的架橋を停止させ、かつ/または固体架橋マトリックスからの酵素の溶出度および溶出速度を低下させる機構が必要性とされており、このような機構は有用であると考えられる。
【0016】
少なくとも実施形態のいくつかにおいて、本発明は、上記の従来技術の欠点を克服するものであり、かつ、(多くの利点のうち、これに限定されるものではないが)上記の技術的問題を解決するための方法を提供するものである。これは、ポリマーを酵素架橋することにより形成されるマトリックスまたはヒドロゲルであって、架橋化酵素分子が、マトリックスの架橋密度、力学的特性、もしくは他の特性を改善するため、かつ/または架橋速度および/もしくは架橋度の制御を改善するために改変されていることを特徴とする、マトリックスまたはヒドロゲルを提供することによって達成される。
【図面の簡単な説明】
【0017】
【図1】PEG化生成物のサイズおよび分布に対する、反応pHおよび活性化PEG濃度の影響を示す画像である。
【図2】PEG化生成物のサイズおよび分布に対する、反応時間およびpHの影響を示す画像である。
【図3】種々の濃度のPEG−NHS(2kD)を用いたPEG化mTGのSDS分析を示す画像である。
【図4】同一の架橋ゼラチンゲルからのmTGおよびPEG化mTGの溶出を示す画像である。
【図5】種々の架橋ゼラチンゲルから溶出したmTG(左側)およびPEG化mTG(右側)を示す画像である。
【図6】非PEG化mTGまたは2種のPEG化mTGで作製したゼラチンシーラントにおける破壊圧力を示すグラフである。
【図7】mTGとデキストランとのコンジュゲーション生成物のSDS−PAGE分析を示す画像である。
【図8】西洋ワサビペルオキシダーゼ(HRP)のPEG化生成物のSDS−PAGE分析を示す画像である。
【図9】種々の反応条件を用いた、PEG化mTG生成物のSDS−PAGE分析を示す画像であり、種々のPEG化度がゲルに示されている。
【図10】反応性PEGが二官能基型PEG−NHS(10kD)である、PEG化mTG生成物のSDS−PAGE分析を示す画像である。
【図11】MALDI−TOF質量分析装置によって得られた、PEG化mTGの典型的なバッチの質量対電荷スペクトルである。
【図12】PEG試薬とアミンの比率を固定し、反応物の濃度を様々に変えたときの、PEG化mTG生成物のSDS−PAGE分析を示す画像である。
【発明を実施するための形態】
【0018】
酵素分子の改変は、形成される架橋マトリックスにおける酵素分子の感知体積を改変することによって行ってもよい。改変された酵素分子の感知体積は、ポリマーが架橋することによって形成されたマトリックスの架橋度によって決定することが好ましく、感知体積の増加は、改変していない酵素分子による架橋と比較して、架橋度が減少することによって示される。
【0019】
酵素分子の感知体積を増加させる方法のひとつとして、少なくとも1つの分子または部分を酵素分子に共有結合または非共有結合させることによって、分子のサイズおよび/または流体力学的体積を増加させる方法が挙げられる。本発明者らは、分子を酵素に共有結合させることによって、ヒドロゲルまたは架橋マトリックスの酵素的架橋度を制御することができ、この酵素分子の改変により最終的な架橋度を低下させることができることを実証している。このようにして、過剰な架橋を防ぐことができる。
【0020】
感知体積を増加させる別の方法として、酵素分子の正味の電荷がポリマー鎖またはコポリマー鎖の正味の電荷とは反対の符号に帯電するように、酵素分子の静電荷を改変する方法が挙げられる。これは、酵素の等電点(pI)を変化させることによって達成することができる。
【0021】
これに限定されるものではないが、仮説によれば、酵素分子の感知体積を増加させると、架橋マトリックスまたは架橋ヒドロゲルにおける酵素分子の移動または拡散が低下する。これによって、ヒドロゲルの所望の物質特性に有益な程度を越えて、酵素分子の架橋活性により架橋が継続されることを防ぐことができる。
【0022】
本明細書で定義されるように、「感知体積(perceived volume)」または「有効体積」は、架橋マトリックス内の架橋化酵素の有効な流体力学的体積を指す。感知体積は、架橋反応前または架橋反応中に、酵素が、別の分子、担体、ポリマー、タンパク質、多糖類などと共有結合または非共有結合することによって増加してもよい。
【0023】
本明細書で定義されるように、「拡散」または「移動」は、溶液、ヒドロゲル、またはマトリックスにおける架橋化酵素または他のタンパク質の、ブラウン運動に起因するランダムな分子運動を指す。
【0024】
本明細書で定義されるように、「拡散係数」とは、単一種の分子が特定の条件下で拡散する度合いを定量化したものを指す。酵素拡散の測定に代わるものとして、ヒドロゲルからの酵素の溶出を測定することが挙げられるが、これに限定されない。
【0025】
本明細書で定義されるように、「移動の低下」は、溶液中またはヒドロゲル内において、タンパク質または酵素の分子運動が遅いことまたはタンパク質または酵素の拡散係数が小さいことを指す。
【0026】
本明細書で定義されるように、「サイズ」は、分子の分子量、分子の流体力学的体積、または分子の感知体積を指す。
【0027】
本明細書で使用されているように、「分子量」(MWと略す)は、ダルトンまたはキロダルトンで表される、タンパク質またはポリマーの絶対重量を指す。たとえば、PEG化タンパク質(すなわち、1個以上のPEG(ポリエチレングリコール)分子が結合したタンパク質)の分子量は、そのPEG化タンパク質の構成成分の分子量をすべて合計したものである。
【0028】
本明細書で定義されるように、「流体力学的体積」は、一般的にサイズ排除クロマトグラフィーを用いて測定してもよいタンパク質または酵素の見かけの分子量を指す。構成成分の流体力学的体積は、構成成分が流体中で運動していると仮定したときの直径または体積を指す。
【0029】
本明細書で定義されるように、「マトリックス」は、架橋された材料の組成物を指す。一般的に、組成物中に含まれるマトリックス形成材料が架橋すると、この組成物は、液体状態からゲル状態へ遷移することによって、「ゲル」、「ヒドロゲル」、または「ゲル化組成物」を形成する。ゲルは、ある一定の粘弾特性およびレオロジー特性を有し得ることから、ある程度の耐久性および膨潤性を有する。このような材料はポリマーであることが多い。マトリックスは、コポリマーとも称される架橋していない材料を含んでもよい。
【0030】
本明細書で使用されているように、「ポリマー」は、反復単位を含む、天然分子、合成分子、または半合成分子を指す。
【0031】
本明細書で使用されているように、「コポリマー」は、架橋反応に関与しても関与しなくてもよいマトリックスの構成成分を指し、通常、マトリックスの主要な構成成分ではない。このようなコポリマーとしては、デキストランなどの多糖類および/またはカルボキシメチルセルロースなどのセルロースポリマーが挙げられるが、これらに限定されない。コポリマーは、酵素またはマトリックス材料(たとえば、マトリックスの主成分であるタンパク質など)に共有結合していないことが好ましい。
【0032】
本明細書で使用されているように、「担体」は、架橋反応前または架橋反応中に、架橋化酵素と共有結合または非共有結合する、ポリマー、タンパク質、多糖類、または他の構成成分を指す。
【0033】
本明細書で定義されるように、「架橋化酵素」は、ポリマーストランドの基質残基を(たとえば、トランスグルタミネーションによって)直接的または(たとえば、キノンまたはフリーラジカルの生成を通して)間接的に架橋して、緊密なマトリックス(coherent matrix)(たとえば、ヒドロゲルなど)を形成することのできる酵素または酵素の組み合わせを指す。
【0034】
本発明の少なくともいくつかの実施形態によれば、
架橋マトリックスであって、
改変酵素分子により架橋される基質ポリマーを含み、
改変酵素分子が、基質ポリマーの架橋によってマトリックスが形成されるのに伴い、架橋マトリックス内における酵素分子の感知体積が変化するように改変されたものであることを特徴とする、架橋マトリックスが提供される。
【0035】
改変酵素分子は、改変酵素分子の実際のサイズが増加するように改変されたものであってもよい。改変酵素分子は、改変酵素分子の流体力学的体積が増加するように改変されたものであってもよい。改変酵素分子は、改変酵素の等電点(pI)を、改変していない酵素の等電点と異なるように変化させることによって、改変酵素分子の静電荷が基質ポリマーの正味の電荷と反対の符号に帯電されるように改変されたものであってもよい。また、(無水コハク酸による)スクシニル化、(無水酢酸による)アセチル化、(シアネートによる)カルバミル化、(アルデヒドによる)還元的アルキル化、および無水マレイン酸処理からなる群から選択される工程によって、酵素のリシンのε−アミノ基が改変されていてもよい。さらに、負の荷電量が減少するように、酵素のカルボン酸含有側鎖の1つ以上が改変されていてもよい。
【0036】
上記改変は、少なくとも1つの分子または部分が改変酵素分子に共有結合または非共有結合されていることを含んでもよい。また、上記改変は、改変作用を有する分子が改変酵素分子に共有結合されていることを含んでもよい。改変酵素分子は、非改変酵素よりも低い拡散速度および架橋速度を有してもよく、非改変酵素と少なくとも類似した測定酵素活性を有してもよい。
【0037】
上記架橋速度は、非改変酵素の架橋速度よりも少なくとも10%遅くてもよい。
【0038】
上記改変作用を有する分子は、担体またはポリマーを含んでもよい。このポリマーは、合成ポリマー、セルロースポリマー、タンパク質、または多糖類を含んでもよい。このセルロースポリマーは、カルボキシメチルセルロース、ヒドロキシプロピルメチルセルロース、ヒドロキシエチルセルロース、またはメチルセルロースのうちの1種以上を含んでもよい。上記多糖類は、デキストラン、コンドロイチン硫酸、デルマタン硫酸、ケラタン硫酸、ヘパリン、ヘパラン硫酸、ヒアルロン酸、またはデンプン誘導体のうちの1種以上を含んでもよい。
【0039】
上記改変作用を有する分子は、PEG(ポリエチレングリコール)を含んでもよい。このPEGは、PEG誘導体を含んでもよい。このPEG誘導体は、活性化PEGを含んでもよい。この活性化PEGは、メトキシPEG(mPEG)、もしくはその誘導体、mPEG−NHS、mPEGのスクシンイミジル(NHS)エステル(mPEG−スクシナート−NHS)、mPEG−グルタレート−NHS、mPEG−バレラート−NHS、mPEG−カルボナート−NHS、mPEG−カルボキシメチル−NHS、mPEG−プロピオナート−NHS、mPEG−カルボキシペンチル−NHS)、mPEG−ニトロフェニルカルボナート、mPEG−プロピルアルデヒド、mPEG−トシラート、mPEG−カルボニルイミダゾール、mPEG−イソシアネート、mPEG−エポキシド、またはこれらの組み合わせのうちの1種以上を含んでもよい。上記活性化PEGは、上記酵素のアミン基またはチオール基と反応してもよい。上記活性化された酵素のリシン残基に対する上記活性化PEGのモル比は、0.5〜25の範囲であってもよい。上記活性化PEGは、一官能基型、ヘテロ二官能基型、ホモ二官能基型、または多官能基型であってもよい。上記活性化PEGは、分岐PEGまたはマルチアームPEGであってもよい。上記活性化PEGは、1000ダルトン〜40,000ダルトンの範囲のサイズであってもよい。
【0040】
上記マトリックスは、上記酵素または上記基質ポリマーに共有結合していないコポリマーをさらに含んでもよい。このコポリマーは、多糖類またはセルロースポリマーを含んでもよい。この多糖類は、デキストラン、コンドロイチン硫酸、デルマタン硫酸、ケラタン硫酸、ヘパリン、ヘパラン硫酸、ヒアルロン酸、またはデンプン誘導体を含んでもよい。上記セルロースポリマーは、カルボキシメチルセルロース、ヒドロキシプロピルメチルセルロース、ヒドロキシエチルセルロース、メチルセルロースを含んでもよい。
【0041】
上記改変酵素分子は、上記改変酵素分子と複数の酵素分子とが架橋することによって、複数の架橋化酵素分子が凝集体を形成するように改変されたものであってもよい。
【0042】
上記改変酵素分子の改変またはその改変の度合いは、マトリックスの少なくとも1種の特性に変化を与えてもよい。この少なくとも1種の特性は、引張り強さ、剛性、上記基質ポリマーの架橋度、粘度、弾性、可撓性、破断歪み、破断応力、ポアソン比、膨潤力、およびヤング率、およびこれらの組み合わせからなる群から選択されてもよい。
【0043】
上記改変酵素の改変の度合いによって、上記改変酵素のマトリックス内における移動または上記改変酵素のマトリックスからの拡散が決定されてもよい。上記改変酵素の改変によって、上記改変酵素と上記タンパク質との溶液またはマトリックスにおける前記改変酵素の拡散係数が、非改変酵素と上記タンパク質との溶液またはマトリックスにおける非改変酵素の拡散係数よりも低下してもよい。上記改変酵素の改変の度合いによって、マトリックスの1種以上の力学的特性が決定されてもよい。上記改変酵素分子によって溶液中で架橋ポリマーが形成される架橋速度が、非改変酵素分子と比較して速くてもよい。
【0044】
本発明の少なくともいくつかの実施形態によれば、
マトリックスの形成を調節する方法であって、
マトリックスが形成されるのに伴い、架橋マトリックス内における酵素分子の感知体積が変化するように酵素分子を改変すること;
改変した酵素分子と、改変した酵素分子の基質である少なくとも1種の基質ポリマーとを混合すること;および
改変した酵素分子で該少なくとも1種の基質ポリマーを架橋して、その形成の少なくとも一部が酵素分子の改変によって調節されたマトリックスを形成すること
を含む方法が提供される。上記改変によって、上記少なくとも1種の基質ポリマーの架橋度が増加するのに伴い、上記改変酵素分子による架橋の速度が低下してもよい。上記改変酵素分子と上記少なくとも1種の基質ポリマーとを溶液中で混合すると、上記の改変によって、溶液の粘度が増加するのに伴い、上記少なくとも1種の基質ポリマーの架橋度が調節されてもよい。上記改変は、pH7〜9において上記酵素をPEG化することを含んでもよい。このPEG化反応は、pH7.5〜8.5で行われてもよい。
【0045】
上述の方法および/またはマトリックスの少なくともいくつかの実施形態によれば、上記少なくとも1種の基質ポリマーは、元より架橋可能なポリマー、上記改変酵素により架橋可能な部分変性ポリマー、および上記改変酵素により架橋可能な官能基またはペプチドを含む改変ポリマーからなる群から選択される基質ポリマーを含む。上記少なくとも1種の基質ポリマーは、ゼラチン、コラーゲン、カゼイン、もしくはアルブミン、または改変ポリマーを含んでもよく、上記改変酵素分子は、改変トランスグルタミナーゼおよび/または改変酸化酵素を含んでもよい。上記少なくとも1種の基質ポリマーは、動物組織もしくは動物組織から得られるコラーゲンを部分的に加水分解することによって得られるゼラチン;細菌系、酵母系、動物系、昆虫系、植物系、もしくは任意の種類の細胞培養を用いて産生される組換えゼラチン;およびこれらの組み合わせからなる群から選択されるゼラチンを含んでもよく;該動物組織は、動物の皮、結合組織、枝角、角、骨、および魚鱗からなる群から選択されてもよい。上記ゼラチンは、哺乳動物由来または魚類由来であってよい。上記ゼラチンは、Aタイプ(酸処理)またはBタイプ(アルカリ処理)であってよい。上記ゼラチンは、250〜300ブルームであってよい。上記ゼラチンの平均分子量は、75〜150kdaであってよい。
【0046】
上記改変トランスグルタミナーゼは、改変微生物トランスグルタミナーゼを含んでもよい。上記改変ポリマーは、改変微生物トランスグルタミナーゼによる架橋が可能となるように改変されたものであってもよい。上記改変酸化酵素は、チロシナーゼ、ラッカーゼ、またはペルオキシダーゼのうちの1種以上を含んでもよい。上記マトリックスは、上記少なくとも1種の基質ポリマーとして、上記改変酸化酵素により架橋されるフェノール酸を含む炭水化物をさらに含んでもよい。上記炭水化物は、アラビノキシランまたはペクチンのうちの1種以上を含んでもよい。上記酵素分子をPEG化により改変し、このPEG化により、マトリックスが使用される宿主動物の免疫系から上記酵素分子がマスクされ、これによって、上記酵素分子の免疫原性がマスクされてもよい。この宿主動物はヒトであってもよい。
【0047】
少なくともいくつかの実施形態によれば、本明細書に記載のマトリックスを組織に塗布することを含む、体液が滲出した組織を密封するための方法が提供される。この体液は血液を含んでもよく、このマトリックスは止血剤であってもよい。
【0048】
少なくともいくつかの実施形態によれば、本明細書に記載のマトリックスを含む、止血剤または外科用シーラントが提供される。
【0049】
少なくともいくつかの実施形態によれば、本明細書に記載のマトリックスを含む、創傷を密封するための組成物が提供される。少なくともいくつかの実施形態によれば、組織における縫合線またはステープルラインを密封するための組成物の使用が提供される。
【0050】
少なくともいくつかの実施形態によれば、本明細書に記載のマトリックスを含む、局所的薬物送達のビヒクルのための組成物が提供される。
【0051】
少なくともいくつかの実施形態によれば、注入可能な足場として適した、本明細書に記載のマトリックスを含む、組織工学のための組成物が提供される。
【0052】
少なくともいくつかの実施形態によれば、
組成物を改変する方法であって、
架橋可能な官能基を有する改変酵素と、改変酵素によって架橋可能な少なくとも1種の部分を有するタンパク質とを提供すること;および
改変酵素とタンパク質を混合して、改変酵素によりタンパク質を架橋し、かつタンパク質により改変酵素自体を架橋可能な官能基において架橋すること
を含む方法が提供される。
【0053】
ポリマーストランドの基質残基を直接的に架橋する直接架橋化酵素としては、トランスグルタミナーゼおよび酸化酵素が挙げられるが、これらに限定されない。トランスグルタミナーゼとしては、微生物トランスグルタミナーゼ(mTG)、組織トランスグルタミナーゼ(tTG)、および第XIII因子が挙げられる。これらの酵素は、天然酵素由来または組換え酵素由来であってよい。ポリマーストランドに含まれるグルタミンのアミノ酸およびリシンのアミノ酸は、トランスグルタミナーゼによる架橋の基質である。
【0054】
上記酸化酵素としては、チロシナーゼ、ラッカーゼ、およびペルオキシダーゼが挙げられるが、これらに限定されない。これらの酵素は、キノン生成(チロシナーゼ)またはフリーラジカル生成(ラッカーゼ、ペルオキシダーゼ)によってポリマーを架橋する。次いで、キノンまたはフリーラジカルは、互いにまたは他のアミノ酸もしくはフェノール酸と相互作用してポリマーを架橋する。これらの酵素が架橋可能な基質は、チロシンまたは他の芳香族アミノ酸を含有する任意のタンパク質であってもよい。このような基質は、フェルラ酸などのフェノール酸を含む炭水化物であってもよい。このような炭水化物は、たとえば、アラビノキシランまたはペクチンであってよい。
【0055】
1種以上の適切な官能基を有する合成または部分合成のポリマーも、上記のいずれかの酵素の架橋可能な基質として機能することができる。
【0056】
本発明の別の実施形態において、酵素の組み合わせが用いられる。
【0057】
本明細書で定義されるように、「ポリマーストランド」または「ポリマー鎖」は、酵素架橋のための基質ポリマーを指す。本発明の少なくともいくつかの実施形態によれば、(例示に過ぎず、これらに限定されるものではない)以下の分類のうちの1つに属することが好ましい。
1)上記の酵素によって元より架橋可能な基質残基を有する任意のポリマー、および自体が上記の酵素によって元より架橋可能である任意のポリマー。たとえば、トランスグルタミナーゼを用いる場合、このようなポリマーとして、上記の酵素によって元より架橋可能な、ゼラチン、コラーゲン、およびカゼインなどのタンパク質またはポリペプチドが挙げられる。
2)上記の酵素によって架橋可能な基質残基を含むが、自体の構造によってそのままでは上記の酵素により架橋できないポリマー。この場合、酵素で架橋する前にポリマー構造を改変しなければならない。たとえば、トランスグルタミナーゼを用いる場合、このようなポリマーとして、酵素の結合を妨げる球状構造を有するために上記の酵素の天然基質ではない、アルブミン、ラクトグロブリンなどのタンパク質が挙げられる。これらのタンパク質は、還元剤、変性剤、または熱を用いて部分的に変性することによって、基質へと変換することができる。
3)酵素架橋のための基質ポリマーではないが、上記の酵素の基質であるペプチドまたは官能基で改変することによって、上記の酵素で架橋可能となる天然または合成のポリマー。
【0058】
このようなポリマーとして、適切な種類のタンパク質が挙げられるがこれには限定されず、このようなタンパク質は、たとえば、上述したようにゼラチンを含んでもよい。ゼラチンは、当技術分野において公知のタンパク質を含む任意の種類のゼラチンを含んでもよい。このようなゼラチンとして、動物組織および/もしくは動物組織から得られるコラーゲンを部分的に加水分解することによって得られるゼラチン(動物組織として、動物の皮;靱帯、軟骨など(これらに限定されない)の結合組織;枝角、角など;および/または骨;および/または魚鱗および/または魚骨;または他の成分が挙げられるが、これらに限定されない);ならびに/または細菌系、酵母系、動物系、昆虫系、植物系、もしくは任意の種類の細胞培養を用いて産生される組換えゼラチンが好ましいが、これらに限定されない。
【0059】
本発明の好ましい実施形態によれば、動物源由来のゼラチンは、哺乳動物源由来のゼラチンを含むことが好ましく、豚皮、豚骨、牛骨、牛皮革片(ハイド)、および他のブタ由来物もしくはウシ由来物のうちの1種以上を含むことがより好ましい。また、ブタのゼラチンはアナフィラキシーを起こしにくいことから、動物源由来のゼラチンはブタのゼラチンを含むことがより好ましい。動物源由来のゼラチンは、Aタイプ(酸処理)でもBタイプ(アルカリ処理)でもよいが、Aタイプであることが好ましい。
【0060】
動物源由来のゼラチンは、一般的に低温度(50〜60℃であるが、この温度範囲に必ずしも限定されない)で行われる1回目の抽出で得られるゼラチンを含むことが好ましい。この温度の抽出により得られるゼラチンは、250〜300ブルームの範囲であり、少なくとも約95〜100kDaの高い分子量を有する。275〜300ブルームのゼラチンを用いることが好ましい。
【0061】
このようなゼラチンの製造会社として、PB Gelatins(Tessenderlo Group(ベルギー))が挙げられるが、これに限定されない。
【0062】
本発明の実施形態のいくつかによれば、動物源由来のゼラチンは魚類由来のゼラチンを含んでもよい。いずれの魚類由来であってもよく、コイ、タラ、カワカマス、マグロなどの冷水魚が好ましい。このようなゼラチンのpH(10%溶液で測定)は、4〜6の範囲であることが好ましい。
【0063】
冷水魚ゼラチンは10℃の水中で溶液となることから、冷水魚ゼラチンはすべて0ブルームであると考えられる。本発明において、高分子量の冷水魚ゼラチンを用いてもよく、またこれを用いることが好ましく、少なくとも約95〜115kDaの平均分子量を有する冷水魚ゼラチンがより好ましいものとして挙げられる。この平均分子量は、250〜300ブルームの動物ゼラチンの分子量に等しい。冷水魚ゼラチンは、プロリンおよびヒドロキシプロリンの含量が少ないことから、動物ゼラチンよりもかなり低い温度で熱可逆性のゲル化を起こす。アミノ酸残基1000個あたり、動物ゼラチンは135〜145個のプロリン残基および90〜100個のヒドロキシプロリン残基を有するのに対し、冷水魚ゼラチンは100〜130個のプロリン残基および50〜75個のヒドロキシプロリン残基を有する(Haug IJ, Draget KI, Smidsrod O.(2004).Food Hydrocolloids. 18:203-213)。
【0064】
このようなゼラチンの製造会社として、Norland Products(クランベリー(ニュージャージー))が挙げられるが、これに限定されない。
【0065】
本発明の実施形態のいくつかにおいては、低エンドトキシンゼラチンを用いて、ゼラチン−mTG組成物のゼラチン溶液成分を形成する。このようなゼラチンは、Gelita(登録商標)(エーバーバッハ(ドイツ))などの供給業者から市販されている。低エンドトキシンゼラチンは、1グラムあたりのエンドトキシン単位(EU)が1000を下回るゼラチンとして定義される。エンドトキシンが500EU/グラムを下回るゼラチンを用いることがより好ましい。
【0066】
脊椎または脳と接触する材料とともに使用する場合などの非常に慎重さが要求される用途においては、エンドトキシンが100EU/gを下回るゼラチンが好ましく、50EU/gを下回るゼラチンがより好ましい。エンドトキシンが10EU/gを下回るゼラチンは非常に高価であるが、慎重さが要求される用途において、本発明の少なくともいくつかの実施形態の一部として用いることもできる。
【0067】
本発明の実施形態のいくつかによれば、架橋されるタンパク質成分としてのゼラチンの代わりに、I型、II型、もしくは他の型の加水分解コラーゲンまたは非加水分解コラーゲンを使用する。種々の型のコラーゲンが熱的に安定なmTG架橋ゲルを形成する能力を有することが実証されている。
【0068】
本発明の実施形態のいくつかによれば、組換えヒトゼラチンが用いられる。このようなゼラチンは、Fibrogen(登録商標)(サンフランシスコ(カリフォルニア))などの供給業者から市販されている。組換えゼラチンの純度は、少なくとも約90%が好ましく、少なくとも約95%であることがより好ましい。いくつかの組換えゼラチンは、10℃においてゲル化しないことから、0ブルームであると考えられる。本発明の実施形態のいくつかにおいて、高分子量の組換えゼラチンを用いることが好ましく、少なくとも約95〜100kDaの分子量の組換えゼラチンがより好ましいものとして挙げられる。
【0069】
上述したように、架橋可能なタンパク質はゼラチンを含むことが好ましいが、追加的にまたはそれに代わるものとして、別の種類のタンパク質を含んでもよい。本発明の実施形態のいくつかによれば、このようなタンパク質はトランスグルタミナーゼの基質でもあり、適切なトランスグルタミナーゼ特異的ポリペプチド配列およびポリマー配列を有することが好ましい。このタンパク質は、単独で生体接着剤を形成する特性を有する合成ポリマー配列、または、トランスグルタミナーゼによる材料の架橋を増強するトランスグルタミナーゼ特異的基質で修飾されていることがより好ましいポリマー配列を含んでいてもよいが、これらに限定されない。いずれの種類の材料も下記に例示されるが、これらの例示に限定されない。
【0070】
トランスグルタミナーゼの適切な架橋標的を有し、かつ好ましくは約20℃〜約40℃の転移温度を有する合成ポリペプチド配列およびポリマー配列が開発されている。好ましい物理的特性として、組織結合能および線維形成能が挙げられるが、これらに限定されない。(上記の)ゲル化ゼラチンと同様に、このようなポリペプチドは、転移温度を低下させる1種以上の物質を有する組成物に用いてもよい。
【0071】
上記のペプチドは、ZymoGenetics Incにより出願された米国特許第5,428,014号および第5,939,385号に例示されているがこれらには限定されず、これらの特許文献はいずれも、あたかもその全体が本明細書に記載されているかのように参照により本明細書に組み込まれる。いずれの特許文献においても、トランスグルタミナーゼにより架橋可能な、生体適合性かつ生体接着性のポリペプチドが記載されており、トランスグルタミナーゼは、タンパク質結合グルタミニル残基のγ−カルボキサミド基とLys残基のε−アミノ基との間でのアシル転移反応を触媒し、その結果、ε−(γ−グルタミル)リシンイソペプチド結合を形成することが知られている。
【0072】
実施形態のいくつかによれば、得られた組成物は局所的薬物送達のためのビヒクルとして使用される。
【0073】
実施形態のいくつかによれば、得られた組成物は組織工学のための注入可能な足場である。
【0074】
実施形態のいくつかによれば、本発明の組成物は止血組成物である。実施形態のいくつかによれば、本発明の組成物は体液封止組成物である。
【0075】
本発明の組成物は、出血を速やかに止めることによって、負傷後または手術後の血液の喪失を最小限に留めるものであることが好ましい。
【0076】
本明細書で使用されているように、「創傷」は、循環系からの血液の喪失、または生理学的導管(たとえば、いかなる種類の脈管など)からの他の体液の喪失を引き起こす、患者のいかなる組織における損傷を指す。組織は、臓器や血管などの内部組織、または皮膚などの外部組織であり得る。血液喪失または体液の喪失は、破裂した臓器などからの体内における喪失、または裂傷などからの体の外面からの喪失であり得る。創傷は、臓器などの柔組織における創傷、または骨などの硬組織における創傷であり得る。損傷は、いかなる作因または原因によって引き起こされたものであってもよく、たとえば、外傷、感染、または外科的介入が挙げられる。損傷は、致命的なもの、または致命的ではないものであり得る。
【0077】
外科的創傷閉鎖は、現在、組織を引き合わせることによって治癒を促進する縫合およびステープルによって行われている。しかしながら、これらの方法は、体液の滲出を防ぐのに十分な密閉性を得ることができない場合が非常に多い。したがって、ステープルラインおよび縫合線に沿って生じることが多い体液滲出に挙げられるような、手術後の体液滲出を防ぐための装置および方法が医療分野において一層必要とされており、いまだにその要求は満たされていない。このような装置および方法は、末梢血管再構築、硬膜再構築、胸部手術、心臓血管手術、肺手術、神経手術、および消化管手術において、縫合またはステープルを補助して、出血または他の体液の滲出を止めるためのものとして必要とされている。現在市販されている高圧止血装置のほとんどは、たとえ接着性を有したとしても名ばかりのものである。したがって、少なくともいくつかの実施形態による本発明の組成物は、これらの欠点を克服するものであり、止血に用いてもよい。
【0078】
本明細書で使用されているように、「約」は、示された値のプラスマイナス約10パーセントの値を意味する。
【0079】
本発明の種々の実施形態における他の特徴および利点は、以下の詳細な説明および特許請求の範囲から明らかになるであろう。
【0080】
本明細書において、添付された図面を参照して本発明を例示としてのみ説明する。ここで、特に詳細図に関していえば、示された具体例は、本発明の好ましい実施形態を説明することのみを目的とした例示に過ぎず、本発明の原理および概念的見地を最も有用にかつ容易に理解されるように説明することを目的として示されるものであることが強調される。この点において、本発明の構造の詳細は、本発明を基本的に理解するのに必要とされる以上には詳しく説明されておらず、本発明を実際に使用する際の態様のいくつかが当業者にとって明らかとなるように図面を用いて説明がなされている。
【0081】
以下の各項目の見出しは、説明を容易にすることのみを目的として記載されている。これらの見出しは、いかなる方法によっても限定されることをなんら意図するものではないことが理解される。また、特に定義されない限り、本明細書で使用される技術用語および科学用語はすべて、本発明が属する分野の当業者によって一般に理解されるのと同じ意味を有する。本明細書に記載されたものと類似または同一の方法および原料を、本発明の実施または実験に用いることができるが、適切な方法および原料は以下に記載されているものである。矛盾点がある場合、定義を包含する本発明の明細書の記載が優先する。さらに、原料、方法、および実施例は、例示にとどまりなんら限定するものではないことが意図される。
【0082】
ヒドロゲルにおける酵素的架橋剤の感知体積の増加
架橋マトリックス内の架橋化酵素による触媒反応の速度の調節は、酵素含有ポリマー溶液の粘度および部分的に架橋した溶液の架橋密度(反応基の利用可能度)のほかに、架橋化酵素分子の感知体積を調節することにより達成できることが見出された。
【0083】
本発明の少なくともいくつかの実施形態によれば、架橋反応の開始前または架橋反応中に酵素分子の感知体積を増加させることによって、マトリックスの力学的状態が所望のものに近づくのに伴い触媒による架橋の速度が低下するように上記の調節を行ってもよく、かつ、このように調節することが好ましい。このようにすると、所望の架橋密度において凝固マトリックスがサイズの増した酵素を捕捉し、さらなる架橋が阻止される。検出酵素体積は、他の要因のうちでもとりわけ、酵素分子量および流体力学的体積の関数である。
【0084】
架橋マトリックスの最終的な架橋度は、マトリックスが形成されるのに伴い架橋マトリックス内の酵素分子の感知体積が増加するように、酵素分子、マトリックス材料、架橋環境、またはこれらの要因のいくつかの組み合わせを調整することによって、制限することができる。単一の仮説によって限定されることを望むものではないが、検出酵素の体積を増加させることにより、架橋マトリックス内の酵素の移動を低下させることができる。酵素の移動を低下させることによって最終的な架橋密度を調節することは、溶液粘度が低い状態の架橋反応の初期段階においては酵素分子が移動度を保持しているが、架橋が進行して溶液粘度が増加するのに伴い移動度が低下し、初期の固体マトリックスまたはヒドロゲルが形成されるとより顕著に移動度が失われる場合において最も効果的である。当然のことながら、特定の用途に望ましい架橋プロファイルおよび架橋度が得られるように、マトリックス内における酵素の移動は正確に制御されるべきである。
【0085】
単一の仮説によって限定されることを望むものではないが、サイズまたは流体力学的体積の増した酵素は、改変していない酵素と比較して、架橋マトリックス内における拡散係数または移動度が低いため、架橋可能な基質へと到達しにくくなる。
【0086】
サイズおよび/または流体力学的体積の増した酵素分子
架橋マトリックスにおける酵素分子の移動を低下させる方法として、酵素分子の有効サイズを増加させることが好ましい。これは、酵素分子の分子量(MW)、流体力学的体積、またはMWと流体力学的体積のいずれもを増加させることによって達成することができる。この方法は、架橋マトリックスの構成成分に影響を与えないと考えられる点で好ましい。
【0087】
本発明の明細書に記載された実施形態を効果的なものとするために、酵素活性を取り除かないように、または所望のポリマー基質を架橋して固体マトリックスもしくはヒドロゲルを形成する能力を取り除かないように、酵素分子のサイズを増加させることが好ましい。酵素は、適切な時間内にマトリックスを形成するのに十分な活性を保持していることが好ましい。さらに、サイズの増した酵素分子は、マトリックス内で移動を停止する前に所望の架橋度までの架橋を触媒するのに十分な移動度を架橋マトリックスにおいて保持することが好ましい。
【0088】
架橋マトリックスまたは架橋ヒドロゲルにおいて酵素分子サイズを増加させるための方法がいくつか同定されている。
1.酵素同士を架橋(分子間架橋)させて可溶性マルチユニットコンジュゲートを形成させる方法。この例のひとつが、下記の実施例18に記載されている。
2.酵素を担体に共有結合(固定化)させる方法。
I.可溶性タンパク質(たとえば、アルブミン)への固定化;(Allen TMら, 1985, JPET 234: 250-254, alpha-Glucosidase-albumin conjugates: effect of chronic administration in mice)。
II.可溶性ポリマーへの固定化。ポリマー担体が酵素よりも大きく、1つ以上の酵素分子が、それぞれのポリマー分子に固定化されることが好ましい。1つの酵素分子を、2つ以上の付着部位を介して2つ以上のポリマー分子に結合させることも可能である。担体は天然、合成、または半合成であってもよい。このような方法の多くは、酵素のインビボ安定性の増加または免疫原性の低下のために開発された。このようなポリマーのファミリーのうちのひとつとしてセルロースエーテルが挙げられ、これには、たとえば、カルボキシメチルセルロース、ヒドロキシプロピルメチルセルロース、ヒドロキシエチルセルロース、メチルセルロースなどが挙げられるが、これらに限定されない。このような固定化は、トリプシン(Villaongaら, 2000, Journal of Molecular Catalysis B: 10, 483-490 Enzymatic Preparation and functional properties of trypsin modified by carboxymethylcellulose)およびリゾチーム(Chen SHら, 2003, Enzyme and Microbial Technology 33, 643-649, Reversible immobilization of lysozyme via coupling to reversibly soluble polymer)などの酵素を用いて過去に行われているが、酵素架橋ヒドロゲルまたはマトリックスの力学的特性に変化を与えるためにこのような酵素固定化が用いられたことは過去にはない。
III.グリコサミノグリカン(GAG)への結合。グリコサミノグリカンとしては、コンドロイチン硫酸、デルマタン硫酸、ケラタン硫酸、ヘパリン、ヘパラン硫酸、およびヒアルロン酸が挙げられるが、これらに限定されない。上記のように、このような結合は、過去に行われているが(Luchter-Wasylewska Eら, 1991, Biotechnology and applied biochemistry 13: 36-47, Stabilization of human prostatic acid phosphatase by coupling with chondroitin sulfate)、酵素架橋ヒドロゲルまたはマトリックスの力学的特性に変化を与えるのに用いられたことはない。
IV.酵素は多糖類に結合させることもできる。多糖類としては、たとえば、デキストラン、およびヒドロキシエチルデンプンなどのデンプン誘導体などが挙げられる。この例のひとつは、酸化デキストランに酵素を結合させた実施例13に記載されている。
3.共有結合性改変によって1つの酵素分子に1つ以上の部分を付加する方法。必ずしもそうではないが、これらの部分は酵素よりも小さいことが多い。このような改変の例のひとつとして、架橋化酵素のPEG化が挙げられ、下記の複数の実施例に広く記載されている。
4.他の種類の共有結合。たとえば、酵素の表面にビオチン分子をグラフト化(ビオチン化)し、このビオチン化した酵素を、アビジンもしくはストレプトアビジンを含有する分子またはポリマー上に固定化して行う。担体は、架橋反応前または架橋反応中に架橋化酵素を捕捉する機能を有する架橋不能な可溶性ポリマーであってもよい。あるいは、架橋可能なポリマー自体に捕捉群(たとえば、アビジンまたはストレプトアビジン)をグラフト化し、その結果、架橋可能なポリマーの架橋反応中に架橋化酵素が徐々に固定化されるようにしてもよい。
5.担体またはポリマーに酵素を非共有結合させる方法。たとえば、酵素の正味の電荷が担体の正味の電荷と反対の符号に帯電している場合、酵素と担体またはポリマーとの間の静電気的相互作用によって安定な非共有結合性結合が形成される。
【0089】
酵素分子のサイズの増加に関する技術
酵素のサイズの増加については過去に数回にわたり開示されているが、酵素架橋マトリックスまたはヒドロゲルの形成および/またはその形成の調節についてはいままでに考慮されたことはない。サイズの増した酵素による架橋マトリックスにおける架橋反応性が過去に一度も明らかされたことがないように、サイズの増した酵素を架橋マトリックスに使用することは全く新規である。さらに、驚くべきことに、本発明の発明者らは、比色法による酵素活性アッセイを用いて酵素活性を単独で検討することにより、サイズの増した酵素の酵素活性と、ゲル化速度によって示されるこの酵素のヒドロゲル形成における架橋活性とが明らかに異なるものであることを実証した。たとえば、実施例5において、酵素触媒によるゲル化の速度と、比色アッセイを用いて測定した酵素活性値との比較が記載されている。この実施例において記載されているPEG化は、酵素のサイズを増加させるための方法として例示され、なんらこれに限定するものではない。
【0090】
PEG化は、酵素分子にポリエチレングリコール(PEG)分子を共有結合により付着させることであり、酵素分子のサイズを増加させる方法として好ましい。1つ以上のPEG分子を付加する操作は、「PEG化」として知られている。
【0091】
PEGは、生体に対して不活性であり、かつ、埋め込み分子または注入分子をPEG化することにより、これらに対する免疫原性応答を制限できる能力を有することから、酵素サイズを増加させるための材料として望ましい。このような免疫原性のマスキング(免疫原性応答の制限)が、本明細書に記載されているように酵素をPEG化することによって達成できるかは不明だが、単一の仮説によって限定されることを望まずとも、酵素をPEG化することにより、酵素に対する免疫原性応答を実際に制限することが可能であると考えられ、さらに、拡大解釈すれば、架橋マトリックスに対する免疫原性応答も制限することが可能であると考えられる。
【0092】
酵素をPEG化する方法のひとつとして、酵素のアミン基と反応する活性化メトキシルPEG(mPEG)を酵素と反応させること(アミンPEG化)が挙げられる。活性化mPEGとしては、mPEGのスクシンイミジル(NHS)エステル(mPEG−スクシナート−NHS、mPEG−グルタレート−NHS、mPEG−バレラート−NHS、mPEG−カルボナート−NHS、mPEG−カルボキシメチル−NHS、mPEG−プロピオナート−NHS、mPEG−カルボキシペンチル−NHS)、mPEG−ニトロフェニルカルボナート、mPEG−プロピルアルデヒド、mPEG−トシラート、mPEG−カルボニルイミダゾール、mPEG−イソシアネート、およびmPEG−エポキシドが挙げられるが、これらに限定されない。
【0093】
活性化mPEGは、酵素のチオール基と反応するものであってもよい(チオールPEG化)。
【0094】
活性化PEGは、一官能基型、ヘテロ二官能基型、またはホモ二官能基型であってもよい。
【0095】
活性化PEGは、分岐PEGまたはマルチアームPEGであってもよい。
【0096】
活性化PEGのサイズは、1000ダルトン〜40,000ダルトンの範囲であってもよい。
【0097】
活性化PEGと酵素のリシン基とのモル比は、0.1:1〜100:1、好ましくは、0.5:1〜10:1である。
【0098】
PEG化反応物のpHは、7〜9であることが好ましい。反応物のpHが、7.5〜8.5であることがより好ましい。
【0099】
好ましい実施形態によれば、PEG化されなかった酵素を取り除くため、または、PEG化生成物のサイズを縮小するために、PEG化酵素をさらに精製してもよい。精製は、サイズ排除クロマトグラフィーを用いて行ってもよい。あるいは、これに代わるものとしてまたは追加的に、SPセファロース、Qセファロース、SMセファロース、DEAEセファロースなどのイオンクロマトグラフィーを用いて精製を行ってもよい。あるいは、これに代わるものとしてまたは追加的に、PEG化されなかった酵素を取り除く精製は、透析、限外濾過、または硫酸アンモニウム分画を用いて行ってもよい。
【0100】
下記に示される種々の実施例は、架橋ヒドロゲル形成を調節するためのトランスグルタミナーゼのPEG化について記載されている。実施例1においては、PEG−NHS(5kD)を用いたmTGのPEG化反応が記載されている。PEG化生成物のサイズおよび分布は、PEGとmTGとの比率、および反応物のpHに依存する。
【0101】
実施例2においては、PEG−NHS(5kD)を用いたmTGのPEG化反応が記載されている。PEG化生成物のサイズおよび分布は、反応の継続時間およびpHに依存する。
【0102】
実施例3においては、PEG−NHS(2kD)を用いたmTGのPEG化反応が記載されている。PEG化生成物のサイズおよび分布は、PEGとmTGとの比率に依存する。
【0103】
実施例4においては、種々のPEG化mTG(5kD PEG)調製物のPEG化度を決定するためのTNBSアッセイが記載されている。実験結果より、PEG化度が、反応における活性化PEG:mTGの比率に依存することが示唆されている。
【0104】
実施例5においては、PEG化mTGの活性を測定するためのアッセイが記載されている。実験結果より、PEG化mTGは、ヒドロキシルアミンおよびCBZ−Gln−Glyなどの小さい基質に対して活性の大部分を有するが、ゼラチンなどの大きい基質に対しては活性の大部分が喪失されることが示唆されている。
【0105】
実施例6および7においては、架橋ゼラチンゲルからのmTGおよびPEG化mTGの溶出プロファイルのSDS−PAGE分析が記載されている。実験結果より、PEG化mTGは、非PEG化mTGよりも、ゆっくりとかつ小さい移動度でゲルから溶出したことが示唆され、これはおそらくPEG化mTGのサイズまたは流体力学的体積がより大きいことに起因すると考えられる。
【0106】
実施例8においては、架橋ゼラチンゲルから溶出したmTGの活性の測定が記載されている。実験結果より、架橋ゼラチンゲルから溶出した非PEG化mTGは、その活性の大部分(算出した最大活性:86%)を保持していることが示唆される。
【0107】
実施例9においては、PEG化mTGまたは非PEG化mTGで架橋したゼラチンゲルの力学的試験が記載されている。実験結果より、PEG化mTGで架橋したゼラチンゲルは、非PEG化mTGで架橋したゲルと比較して、より強くかつより柔軟なことが示されている。
【0108】
実施例10においては、種々のゼラチンシーラント調製物の破壊圧力試験が記載されている。実験結果より、PEG化mTGで作製したゼラチンシーラントは、破壊圧力試験において、非PEG化mTGで作製したシーラントに匹敵することが示唆されている。
【0109】
実施例11においては、インビボブタモデルのステープルラインを補強するためのシーラントの使用が記載されている。
【0110】
実施例12においては、架橋化酵素を不溶性担体に非共有結合させた効果が記載されている。
【0111】
実施例13においては、酸化デキストランで酵素を改変した効果が記載されている。
【0112】
実施例14においては、PEG化によって西洋ワサビペルオキシダーゼ(別の架橋化酵素)を改変することにより、ペルオキシダーゼ架橋により形成されるマトリックスを改変できることが実証されている。
【0113】
実施例15においては、架橋化酵素の部分的PEG化の効果が実証されている。
【0114】
実施例16においては、遊離PEG(架橋化酵素と共に溶液中に存在するが、酵素には共有結合されていないPEG分子)はゲル化になんら効果を与えないことが実証されている。
【0115】
実施例17においては、非改変酵素と改変酵素との種々の混合物のゲル化に対する影響が示されている。
【0116】
実施例18においては、二官能基型PEG−酵素ブリッジのゲル化に対する影響が実証されている。
【0117】
実施例19は、PEG化mTG(微生物トランスグルタミナーゼ)の質量分析に関する。
【0118】
実施例20においては、PEGとアミンの比率を固定したときの、種々の濃度の反応物によるmTGのPEG化が記載されており、反応物の総濃度がPEG化度に大きな影響を与えることが実証されている。
【0119】
驚くべきことに、PEG化は、微生物トランスグルタミナーゼ(mTG)がゼラチンを架橋する速度を低下させるが、トランスグルタミナーゼの最も標準的な活性アッセイであるヒドロキサマートアッセイにおける活性を低下させないことがこれらの実施例において見出された。この結果は、サイズの増した酵素はヒドロゲルを形成する効果が著しく低いと考えられることからヒドロゲル形成に用いるのは望ましくないであろうとする従来技術の教示と相反するものである。
【0120】
従来技術において、TGアーゼ(トランスグルタミナーゼ)がPEG化と関連してときおり言及されていることには注意しておかなければならない。しかしながら、このことに言及した文献は、(PEG化の基質としてTGアーゼを使用するのではなく)タンパク質の部位特異的PEG化を可能にするためのまたはこれを増強するためのツールとしてTGアーゼを使用することを教示するものであり、この使用は、タンパク質のグルタミル残基とPEG分子に付着させた一級アミン基とのトランスグルタミネーション反応を触媒することによって行われる。しかしながら、このような従来技術は、TGアーゼ自体をPEG化することによって、TGアーゼの架橋活性を変化もしくは調節すること、またはTGアーゼによって架橋されたヒドロゲルマトリックスの力学的特性を変化もしくは調節することを教示または示唆するものではない。
【0121】
架橋マトリックスへの結合による架橋化酵素の移動の低下
架橋マトリックスにおいて酵素の移動を低下させる別の実施形態において、酵素は、架橋反応を触媒するのと同時に架橋マトリックス自体への結合反応を起こす。酵素がポリマー溶液中を移動してポリマーを架橋しマトリックスを形成するのに伴い、酵素は徐々にポリマー自体に結合することによりマトリックス内に固定化される。たとえば、ビオチン化酵素は、アビジンまたはストレプトアビジンでコーティングされたポリマーを含む架橋可能なポリマー成分と混合することができる。米国特許第6046024号(Method of producing a fibrin monomer using a biotinylated enzyme and immobilized avidin)には、アビジン修飾アガロースを添加することによって、フィブリノゲン溶液からビオチン化トロンビンを捕捉する方法が記載されている。この方法では、アガロースは可溶性ではないが、米国特許第5026785号(Avidin and streptavidin modified water-soluble polymers such as polyacrylamide, and the use thereof in the construction of soluble multivalent macromolecular conjugates)によって記載されているように、アビジンまたはストレプトアビジンを水溶性ポリマーに結合させることも可能である。トランスグルタミナーゼをビオチン化し、次いで、アビジン処理した表面に吸着させることは、実施可能であることが示されている(Huang XLら, J. Agric. Food Chem., 1995, 43(4), pp 895-901)。あるいは、架橋化酵素をアビジンまたはストレプトアビジンに共有結合させ、得られたコンジュゲートを、ビオチン化ポリマーを含む架橋反応に添加してもよい。ビオチン化されるものは、架橋可能なポリマー自体(たとえば、ゼラチンなど)、または架橋不能なコポリマー(たとえば、デキストランなど)であってもよい。分子量が500,000ダルトン以下のデキストラン−ビオチンコンジュゲートは、供給業者から入手できる。
【0122】
架橋マトリックスにおける静電気的相互作用による架橋化酵素の移動の低下
本発明の別の実施形態において、酵素の移動は、酵素の正味の電荷がポリマー担体の正味の電荷と反対の符号に帯電した、酵素とポリマー担体との間の静電気的相互作用に基づいた可逆性結合によって低減される。酵素は、担体とプレインキュベートしてから架橋反応に添加してもよいし、架橋反応中に担体に結合させてもよい。たとえば、架橋化酵素が中性pHにおいて正に荷電している場合、この架橋化酵素を、負に荷電した担体(たとえば、カルボキシメチルセルロース(CMC))に静電気的に結合させてもよい。酵素をCMCとインキュベートして結合させ、得られた複合体を架橋反応に添加してもよいし、あるいは、酵素とCMCとを別々に添加してもよい。後者の場合、架橋反応中に酵素がCMCに徐々に結合するであろう。架橋可能なポリマーが架橋化酵素と反対の符号の電荷を有する場合、架橋反応中に酵素を架橋可能なポリマーストランド自体に結合させることも可能である。あるいは、架橋可能なポリマーまたは担体の電荷と反対の符号の電荷を酵素が有するように、架橋化酵素の等電点(pI)を変更することもできる。
【0123】
別の実施形態において、架橋化酵素は改変されて、架橋化酵素の等電点(pI)が変化し、任意のpHにおいて異なる正味の電荷に帯電される。酵素のpIを低下させる方法として、(無水コハク酸による)スクシニル化、(無水酢酸による)アセチル化、(シアネートによる)カルバミル化、(アルデヒドによる)還元的アルキル化、無水マレイン酸処理などの工程(これらに限定されない)によってリシンのε−アミノ基を改変することが挙げられる。これにより、1個の改変アミノ酸あたり1価以下の電荷が減少する(スクシニル化では、2価以下の正の正味の電荷が減少する)ことによりタンパク質の正の正味の電荷が減少し、pIも減少する。これとは逆に、グルタミン酸およびアスパラギン酸などのカルボン酸を含有する側鎖を改変し、タンパク質の負の荷電量を減少させ、pIを増加させてもよい。たとえば、酵素をEDC 1−エチル−3−[3−ジメチルアミノプロピル]カルボジイミド)およびエチレンジアミン(EDA)で処理することができる。EDCはカルボン酸基を活性化し、これらのカルボン酸基とEDAとの間にアミド結合を形成する。その結果、タンパク質の正の正味の電荷が増加し、pIも増加する。
【0124】
ヒドロゲルからのタンパク質の遊離は、ヒドロゲルポリマー鎖と捕捉されたタンパク質との間の静電気的引力および静電気的斥力に関連づけられている。組換えゼラチンマトリックスからのタンパク質遊離実験において、静電気的斥力は、捕捉されたタンパク質の拡散係数を増加させ、逆に、静電気的引力は拡散係数を減少させることが示唆されている(Marc Sutter, Juergen Siepmann, Wim E. Hennink and Wim Jiskoot, Recombinant gelatin hydrogels for the sustained release of proteins, Journal of Controlled Release Volume 119, Issue 3, 22 June 2007, Pages 301-312)。
【0125】
ヒドロゲルポリマー鎖自体のpIを変化させることは、ヒドロゲルからのタンパク質の遊離を調節する方法として示唆されている。しかしながら、ヒドロゲル内に捕捉されたタンパク質とヒドロゲル鎖との間の静電気的相互作用を調整することを含む従来技術は、ヒドロゲルからの治療用タンパク質の遊離速度を調節する方法に関係しており、このタンパク質自体はヒドロゲルの形成に関与していない。本発明の少なくともいくつかの実施形態において、静電気的相互作用は、ヒドロゲルの力学的特性を向上させるために改変され、この力学的特性は、酵素による架橋過程のヒドロゲルマトリックスにおける酵素の移動および拡散係数に関連し得る。
【0126】
したがって、架橋可能なマトリックスにおける架橋化酵素の拡散または移動が、ポリマーヒドロゲルではなく捕捉された酵素のpIを改変することによって強力に制限されるため、捕捉された架橋化酵素のpIを変化させることは架橋を阻止する新規な手法である。
【実施例】
【0127】
実施例1:PEG化生成物のサイズおよび分布に対する、反応pHおよびPEG:mTG比の影響
材料:
活性化PEG:mPEG−グルタレート−NHS 5kDa(SunBright ME−050GS、日油株式会社(日本))。
mTG:
SPセファロースイオン交換クロマトグラフィーを用いてさらに10%精製した味の素アクティバ。活性:0.2Mクエン酸ナトリウム(pH6)において604ユニット/ml。
クエン酸ナトリウム、Hepes、SDS、およびβ−メルカプトエタノールはシグマアルドリッチから入手した。
30%アクリルアミド/Bis 29:1およびBio−Safe Coomassie G−250ステインはバイオ・ラッドから入手した。
分子量マーカーは、Precision Plus Dual Color(バイオ・ラッド)であった。
【0128】
1ユニットのmTG活性は、pH6.0、37℃において、N−CBZ−Gln−Glyおよびヒドロキシルアミンから、1分間あたり1.0μmolのヒドロキサマートの形成を触媒する。一連の反応は、それぞれの反応物が0.2mlの容量になるように設定した。それぞれの反応物は、15U/ml mTG、適切な反応緩衝液(90mM クエン酸ナトリウム(pH6)または100mM Hepes(pH7))、および種々の量の活性化PEGを含有した。PEG−NHSは、タンパク質の一級アミン、リシン残基の側鎖上のε−アミン、およびタンパク質のアミノ末端と反応する。反応混合物におけるPEGのリシン残基に対する比率は以下に詳細に記載する。
【0129】
反応物を、37℃で1:36時間インキュベートし、最終濃度が110mMになるまでグリシンを添加して、酵素と反応しなかった余分な活性化PEG分子を中和した。
【0130】
各反応物から得られた試料を、SDSおよびβ−メルカプトエタノールの存在下、90℃で加熱して変性させ、SDS−PAGE(8%分離ゲル、4%濃縮ゲル、ミニプロティアン電気泳動システム、バイオ・ラッド)を用いて分析した。タンパク質を可視化するために、Bio−Safe Coomassie G−250ステインでゲルを染色し、水で脱色した。CanoScan 8800Fスキャナでゲルをスキャンし、画像(図1に示す)を得た。この画像は、PEG化生成物のサイズおよび分布に対する、反応pHおよび活性化PEG濃度の影響を示している。レーンは以下のように割り当てた。
レーン1:mTG(対照)
レーン2:分子量マーカー(上から下へ順に:250kD、150kD、100kD、75kD、50kD、37kD、25kD)
レーン3:53.3mg/ml活性化PEG;90mMクエン酸Na、pH6;PEGのリシンに対する比率9.15
レーン4:26.6mg/ml活性化PEG;90mMクエン酸Na、pH6;PEGのリシンに対する比率4.59
レーン5:13.3mg/ml活性化PEG;90mMクエン酸Na、pH6;PEGのリシンに対する比率2.30
レーン6:53.3mg/ml活性化PEG;100mM Hepes、pH7;PEGのリシンに対する比率9.15
レーン7:26.6mg/ml活性化PEG;100mM Hepes、pH7;PEGのリシンに対する比率4.59
レーン8:13.3mg/ml活性化PEG;100mM Hepes、pH7;PEGのリシンに対する比率2.30
【0131】
図1から明らかなように、PEG量を増加しpHを高くすると、ゲル上に見かけの分子量が増した酵素が見られた。
【0132】
実施例2:PEG化生成物のサイズおよび分布に対する、反応pHおよび継続時間の影響
それぞれの反応物は15U/ml mTGを含有した。
材料:
活性化PEG:mPEG−グルタレート−NHS 5kDa(SunBright ME−050GS、日油株式会社(日本))。
mTG:SPセファロースイオン交換クロマトグラフィーを用いてさらに10%精製した味の素アクティバ。活性:0.2Mクエン酸ナトリウム(pH6)において604ユニット/ml。
クエン酸ナトリウム、Hepes、SDS、およびβ−メルカプトエタノールはシグマアルドリッチから入手した。
30%アクリルアミド/Bis 29:1およびBio−Safe Coomassie G−250ステインはバイオ・ラッドから入手した。
分子量マーカーは、Precision Plus Dual Color(バイオ・ラッド)であった。
【0133】
1ユニットのmTG活性は、pH6.0、37℃において、N−CBZ−Gln−Glyおよびヒドロキシルアミンから、1分間あたり1.0μmolのヒドロキサマートの形成を触媒する。
【0134】
一連の反応は、それぞれの反応物が0.2mlの容量になるように設定した。それぞれの反応物は、15U/ml mTG、適切な反応緩衝液(100mM Hepes(pH7)または100mM Hepes(pH8))、および25mg/ml PEG−NHSを含有した。反応混合物におけるPEGのリシン残基に対する比率は4.59であった。
【0135】
反応物を室温で2時間インキュベートした。下記の種々の時点で試料を採取し、最終濃度が110mMになるまでグリシンを添加し、酵素と反応しなかった余分な活性化PEG分子を中和した。
【0136】
各反応物から得られた試料を、SDSおよびβ−メルカプトエタノールの存在下、90℃で加熱して変性させ、SDS−PAGE(8%分離ゲル、4%濃縮ゲル、ミニプロティアン電気泳動システム、バイオ・ラッド)を用いて分析した。タンパク質を可視化するために、Bio−Safe Coomassie G−250ステインでゲルを染色し、水で脱色した。CanoScan 8800Fスキャナでゲルをスキャンし、画像(図2に示す)を得た。この画像は、PEG化生成物のサイズおよび分布に対する、反応時間およびpHの影響を示している。レーンは以下のように割り当てた。
レーン1:25mg/ml活性化PEG;100mM Hepes、pH8;反応時間15分
レーン2:25mg/ml活性化PEG;100mM Hepes、pH8;反応時間30分
レーン3:25mg/ml活性化PEG;100mM Hepes、pH8;反応時間60分
レーン4:25mg/ml活性化PEG;100mM Hepes、pH8;反応時間120分
レーン5:分子量マーカー(上から下へ順に:250kD、150kD、100kD、75kD、50kD、37kD、25kD)
レーン6:25mg/ml活性化PEG;100mM Hepes、pH7;反応時間15分
レーン7:25mg/ml活性化PEG;100mM Hepes、pH7;反応時間30分
レーン8:25mg/ml活性化PEG;100mM Hepes、pH7;反応時間60分
レーン9:25mg/ml活性化PEG;100mM Hepes、pH7;反応時間120分
【0137】
図2に示されているように、反応時間を延長しpHを高くすると、ゲル上に見かけの分子量が増した酵素が見られた。
【0138】
実施例3:PEG−NHS(2kD)を用いたmTGのPEG化:
PEG化生成物のサイズおよび分布に対する、PEGとmTGの比率の影響
材料:
活性化PEG:mPEG−グルタレート−NHS 2kDa(SunBright ME−020CS、日油株式会社(日本))。
mTG:SPセファロースイオン交換クロマトグラフィーを用いてさらに10%精製した味の素アクティバ。活性:0.2Mクエン酸ナトリウム(pH6)において604ユニット/ml。
クエン酸ナトリウム、Hepes、SDS、およびβ−メルカプトエタノールはシグマアルドリッチから入手した。
30%アクリルアミド/Bis 29:1およびBio−Safe Coomassie G−250ステインはバイオ・ラッドから入手した。
分子量マーカーは、Precision Plus Dual Color(バイオ・ラッド)であった。
【0139】
1ユニットのmTG活性は、pH6.0、37℃において、N−CBZ−Gln−Glyおよびヒドロキシルアミンから、1分間あたり1.0μmolのヒドロキサマートの形成を触媒する。反応物(200μl)は、15U/ml mTG、100mM Hepes(pH8)、および種々の濃度のPEG NHS(2kD)を含有した。反応物を、37℃で2時間インキュベートし、1.5Mグリシン(最終濃度71mM)10μlを添加して、酵素と反応しなかったPEG−NHS分子を中和した。各反応物から得た試料を、SDSおよびβ−メルカプトエタノールの存在下、90℃で加熱することにより変性させ、SDS−PAGE(8%分離ゲル、4%濃縮ゲル、ミニプロティアン電気泳動システム、バイオ・ラッド)を用いて分析した。タンパク質を可視化するために、Bio−Safe Coomassie G−250ステインでゲルを染色し、水で脱色した。CanoScan 8800Fスキャナでゲルをスキャンし、画像(図3に示す)を得た。この画像は、種々の濃度のPEG−NHS(2kD)を用いたPEG化mTGのSDS分析を示している。レーンは以下のように割り当てた。
レーン1:分子量マーカー
レーン2:1.75mg/ml PEG−NHS 2kD;PEGのリシンに対する比率0.74
レーン3:3.5mg/ml PEG−NHS 2kD;PEGのリシンに対する比率1.48
レーン4:7mg/ml PEG−NHS 2kD;PEGのリシンに対する比率2.97
レーン5:14mg/ml PEG−NHS 2kD;PEGのリシンに対する比率5.93
レーン6:28mg/ml PEG−NHS 2kD;PEGのリシンに対する比率11.86
レーン7:56mg/ml PEG−NHS 2kD;PEGのリシンに対する比率23.72
【0140】
図3に示されているように、PEG−NHS量を増加させると、ゲル上に見かけの分子量が増した酵素が見られた。
【0141】
実施例4:PEG化度を測定するためのTNBSアッセイ
材料:
グリシンおよび5%TNBS溶液(ピクリルスルホン酸)はシグマアルドリッチから入手した。
重炭酸ナトリウムはFrutarom(イスラエル)から入手した。
希釈TNBS溶液は、5%TNBSと重炭酸緩衝液(pH8.5)とを1:500の比率で混合することにより調製した。
分光光度計はAnthelie Advanced(Secomam)であった。
【0142】
検量線を作成するため、以下の濃度のグリシン溶液を重炭酸緩衝液(pH8.5)で調製した:1μg/ml、2μg/ml、4μg/ml、8μg/ml。
【0143】
希釈TNBS溶液0.5mlを、標準グリシン溶液または試料1mlと混合した。混合物を37℃で2時間インキュベートした。次に、10%SDS溶液0.5mlと1M HCl 0.25mlとを添加して反応を停止した。溶液をキュベットに移し、分光光度計を用いて335nmでのO.D.を読み取った。
【0144】
グリシンについて設定した検量線に基づいて、各PEG化mTGの遊離NH2基のパーセンテージを決定した。
%PEG化=100−遊離NH2のパーセンテージ
算出したPEG化mTGの平均MW:38,000+(%PEG化:100×18×5000)
【0145】
結果を以下の表1に示す。
【0146】
【表1】
【0147】
上記の表により、PEG化を増加させることによって、mTGの見かけの(算出された)分子量が増加したことが示された。さらに、PEG化度は遊離リシン基のパーセンテージの減少と相関し、これによって、予想されたようにリシン基がPEG化されていたことが示された。
【0148】
実施例5:PEG化mTGの活性を測定するためのアッセイ
材料:
尿素、クエン酸Na、酢酸Na、および塩化カルシウムはシグマアルドリッチから入手した。
ゼラチン(豚皮、Aタイプ、275ブルーム)はGelitaより入手した。
【0149】
PEG化反応物(8ml)は、15U/ml mTG、100mM HEPES(pH7)、および14mg/ml PEG−NHS(5kD)を含有した。反応条件は図1のレーン8と同様であった。
【0150】
反応物を37℃で1:50時間インキュベートし、2.34Mグリシン0.4mlを添加して(最終濃度100mM)反応していない活性化PEGを中和した。室温で15分後、Amicon Ultra−4 Centrifugal Filter Unit MWCO 30,000(ミリポア)を用いて、反応混合物を2mlに濃縮し、反応緩衝液を0.2M クエン酸ナトリウムに交換した。濃縮したPEG化mTGを4×と称し、これをクエン酸緩衝液で2倍希釈および4倍希釈したものをそれぞれ2×および1×と称す。
【0151】
ゼラチンを基質として用いる活性アッセイ
mTG 0.5mlをゼラチン調製物(25%ゼラチン、3.8M尿素、0.15M CaCl2、0.1M酢酸Na、pH6)1mlと混合し、37℃でインキュベートし、ゲル化時間を記録した。ゲル化時間は、反応チューブを逆さにしても液体が流れ落ちない時間と定義される。
【0152】
ヒドロキサマートアッセイを用いる活性アッセイ
反応物A 15μl 1×非PEG化mTG(15U/ml)+135μlクエン酸緩衝液
反応物B 15μl 1×PEG化mTG+135μlクエン酸緩衝液
反応物C 15μl 2×PEG化mTG+135μlクエン酸緩衝液
反応物D 15μl 4×PEG化mTG+135μlクエン酸緩衝液
反応物A〜Dのそれぞれに反応カクテル1mLを添加し、37℃で10分間または20分間インキュベートした。各時点において、反応物0.23mlをTCA0.5mLと共に試験管に加えた。
【0153】
ヒドロキサマート反応基質カクテル(20ml、pH6)
240mg CBZ−Glu−Gly(シグマアルドリッチ)
139mg 塩酸ヒドロキシルアミン(シグマアルドリッチ)
61.5mg 還元型グルタチオン(シグマアルドリッチ)
4ml 0.2Mクエン酸Na緩衝液、pH6
20mlになるまで加えた水
結果を下記の表2に示す。
【0154】
【表2】
【0155】
上記の表2によると、トランスグルタミナーゼをPEG化することにより、1×PEG化においてゲル化時間が延長したが、ヒドロキサマートアッセイ(ヒドロゲルは形成されず、遊離溶液中で行われるアッセイ)においては酵素活性にほとんど影響を与えなかったことが示された。実際、PEG化量が増加したことによりゲル化時間が延長したが、これはおそらく、形成過程の架橋ポリマーネットワーク内の基質分子と接触するのに必要な酵素の移動度が低下したことによると考えられる。あるいは、PEG化が酵素の活性部位に構造的変化を与え、その結果、基質に対して非PEG化酵素と同程度に効率的な結合ができなくなったか、または酵素の活性部位の近傍に挿入された1つ以上のPEG分子が、結合しようとする基質分子に対する立体障害となったとも説明することができる。ヒドロキサマートアッセイにおいてPEG化酵素活性が低下しなかったのは、このアッセイにおける基質のサイズが小さいこと、または架橋ポリマーネットワークを形成させなかったことが理由として考えられ得る。これらの説明はすべて、単一の仮説によって限定されることを望むことなく示されている。
【0156】
この実験結果より、PEG化がヒドロゲルの形成にもたらす効果は、溶液中の酵素活性に対する効果とはまったく異なることが裏付けられた。単一の仮説によって限定されることを望むものではないが、これらの異なる効果は、酵素の見かけのサイズが増したことおよび/または(サイズが増したこと以外による)感知体積が増したことにより引き起こされ、その結果、ヒドロゲルの形成を調節するのに有益な効果がもたられたと考えられる。いかなる場合においても、本発明の少なくともいくつかの実施形態によれば、PEG化することによりもたらされる差別的な効果により、形成過程のヒドロゲルにおける架橋を調節することが可能となる。
【0157】
実施例6:ゼラチンゲルからのPEG化mTGおよび非PEG化mTGの溶出プロファイル
材料:
ゼラチン(豚皮、Aタイプ、275ブルーム)はGelitaより入手した。
30%アクリルアミド/Bis 29:1およびBio−Safe Coomassie G−250ステインはバイオ・ラッドから入手した。
分子量マーカーは、Precision Plus Dual Color(バイオ・ラッド)であった。
【0158】
PEG化mTG(反応条件は図1のレーン6におけるものと同様であった)と20U/ml mTGとを1:1で含む酵素混合物0.67mlと、ゼラチン溶液(25%ゼラチン、3.8M尿素、0.15M CaCl2、0.1M酢酸Na、pH6)1.33mlと混合することによって、架橋ゼラチンゲルを作製した。得られたゲルをサランラップで包み、37℃で2時間インキュベートした。次に、生理食塩水を10ml含む試験管にゲルを入れ、恒温振とう培養機を用いて37℃で4時間インキュベートした。1時間ごとに試料を採取した。Amicon Ultra−4 Centrifugal Filter Unit MWCO 30,000(ミリポア)を用いて試料を濃縮し、SDSおよびβ−メルカプトエタノールの存在下90℃で加熱することにより変性させ、SDS−PAGE(8%分離ゲル、4%濃縮ゲル、ミニプロティアン電気泳動システム、バイオ・ラッド)を用いて分析した。タンパク質を可視化するため、Bio−Safe Coomassie G−250ステインでゲルを染色し、水で脱色した。SDS−PAGEのバンド強度を定量化するため、CanoScan 8800Fスキャナでゲルをスキャンし、得られた画像(図4に示す)をQuantity Oneソフトウェア(バイオ・ラッド)を用いて解析した。レーンは以下のように割り当てた。
レーン1:t=1時間に採取された10μlの試料
レーン2:t=1時間に採取された20μlの試料
レーン3:t=4時間に採取された10μlの試料
レーン4:t=4時間に採取された20μlの試料
レーン5:分子量マーカー
レーン6:7μl mTG+PEG化mTG混合物
レーン7:3μl mTG+PEG化mTG混合物
レーン8:1.5μl mTG+PEG化mTG混合物
【0159】
図4は、同一の架橋ゼラチンゲルからのmTGおよびPEG化mTGの溶出を示す。表3は、ゲルから溶出したトランスグルタミナーゼの相対量を示す。
【0160】
【表3】
【0161】
実施例7:架橋ゼラチンゲルからの種々のPEG化mTGの溶出
異なる濃度(2kDまたは5kD)のPEG−NHSを用いたmTGの大規模なPEG化
活性化PEG:
試薬:
mPEG−グルタリル−NHS、MW 5000(SunBright ME−050GS、日油株式会社(日本))。
mPEG−スクシニル−NHS、MW 2000(SunBright ME−020CS、日油株式会社(日本))。
mTG:SPセファロースイオン交換クロマトグラフィーを用いてさらに10%精製した味の素アクティバ。
30%アクリルアミド/Bis 29:1およびBio−Safe Coomassie G−250ステインはバイオ・ラッドから入手した。
SDSおよびβ−メルカプトエタノールはシグマアルドリッチから入手した。
【0162】
PEG化反応物(32ml)は、15U/ml mTG、100mM HEPES(pH8)、および種々の濃度のPEG−NHS(2kDまたは5kD)を含有した。反応物を室温で2.5時間インキュベートし、1.5Mグリシン(最終濃度97mM)2.2mlを添加して、反応しなかった活性化PEGを中和した。室温でさらに15分間インキュベーション後、ビバスピン20(ザルトリウス)を用いて反応混合物を8mlに濃縮し、それと同時に、反応緩衝液を0.2Mクエン酸Na(pH6)に交換した。
【0163】
1倍量の種々の濃度のPEG化mTG(上記)をそれぞれ、2倍量のゼラチン調製物(25%ゼラチン、3.8M尿素、0.15M CaCl2、0.1M酢酸Na、pH6)と混合した。混合物を、テフロン加工のイヌ用の骨の形をした型へ流し込んだ。ゲルが形成された後、ゲルを型から取り出し、重量を計り、乾燥を防ぐために密閉型試験管へ入れ、37℃で3時間インキュベートした。次に、ゲル1グラムあたり正確に5mlのクエン酸Na(pH6)を加え、エアシェーカーを用いて試験管を37℃、100rpmでインキュベートした。1時間後、2時間後、3時間後、および30℃、18時間でのさらなるインキュベーション後に、各時間点において試料をそれぞれ0.5mlずつ採取した。
【0164】
各時間点で得られた試料を、SDSおよびβ−メルカプトエタノールの存在下、90℃で加熱して変性させ、SDS−PAGE(8%分離ゲル、4%濃縮ゲル、ミニプロティアン電気泳動システム、バイオ・ラッド)を用いて分析した。タンパク質を可視化するために、Bio−Safe Coomassie G−250ステインでゲルを染色し、水で脱色した。SDS−PAGEにおけるバンドの強度を定量化するために、CanoScan 8800Fスキャナでゲルをスキャンし、得られた画像(図5に示す)をQuantity Oneソフトウェア(バイオ・ラッド)を用いて解析した。
【0165】
遊離すると考えられる酵素の最大理論量も同様にSDS−PAGEに供し、これを100%の遊離と見なした。実際に溶出した試料を横一列に並べて流し、100%遊離に対するバンド強度を算出した。SDS−PAGEにおけるバンドの強度を定量化するために、CanoScan 8800Fスキャナでゲルをスキャンし、得られた画像を、Quantity Oneソフトウェア(バイオ・ラッド)を用いて解析した。
【0166】
図5は、種々の架橋ゼラチンゲルから溶出したmTG(左側)およびPEG化mTG(右側)を示す。レーンは以下のように割り当てた。
レーン1:mTG、100%遊離対照
レーン2:架橋ゼラチンゲルから遊離したmTG、1時間時点
レーン3:架橋ゼラチンゲルから遊離したmTG、2時間時点
レーン4:架橋ゼラチンゲルから遊離したmTG、3時間時点
レーン5:架橋ゼラチンゲルから遊離したmTG、18時間時点
レーン6:PEG化mTG(7mg/ml PEG−NHS、5kD)、100%遊離対照
レーン7:ゼラチンゲルから遊離したPEG化mTG(7mg/ml PEG−NHS、5kD)、1時間時点
レーン8:ゼラチンゲルから遊離したPEG化mTG(7mg/ml PEG−NHS、5kD)、2時間時点
レーン9:ゼラチンゲルから遊離したPEG化mTG(7mg/ml PEG−NHS、5kD)、3時間時点
レーン10:ゼラチンゲルから遊離したPEG化mTG(7mg/ml PEG−NHS、5kD)、18時間時点
【0167】
総遊離量を以下の表4に示す。
【0168】
【表4】
【0169】
実施例8:架橋ゲルから溶出したmTGの活性
18時間後にゼラチンゲルから溶出した非PEG化mTG(実施例7を参照)9mlを、ビバスピン20(MWCO 30,000(ザルトリウス))を用いて0.47mlに濃縮した。濃縮した酵素の活性を、実施例5に記載されているのと同様にヒドロキサマートアッセイを用いて測定した。
【0170】
測定した活性は、3.65U/mlであった。
【0171】
算出した活性(ゲルにおける最初の活性5U/ml、図5におけるSDS−PAGEによる18時間後の遊離%、および表4におけるその定量化値26.7%に基づく)は、4.24U/mlであった。
【0172】
実施例9:PEG化mTGまたは非PEG化mTGで架橋したゼラチンゲルの力学的試験
尿素、クエン酸Na、酢酸Na、および塩化カルシウムはシグマアルドリッチから入手した。
ゼラチン(豚皮、Aタイプ、275ブルーム)はGelitaより入手した。
SPセファロースイオン交換クロマトグラフィーを用いてさらに10%精製した味の素アクティバをmTGとして用いた。活性:0.2Mクエン酸ナトリウム(pH6)において604ユニット/ml。
種々のPEG化度のPEG化mTG(2kDまたは5kDのPEG−NHSを使用)を、実施例7の記載と同様に調製した。
【0173】
PEG化mTG溶液1部と、ゼラチン溶液(25%ゼラチン、3.8M尿素、0.15M CaCl2、0.1M酢酸Na、pH6)2部とを混合した。混合物を、テフロン加工のイヌ用の骨の形をした型へ流し込んだ。ゲルが形成された後、ゲルを型から取り出し、生理食塩水中に浸して37℃で4時間インキュベートした。次いで、デジタルカリパスを用いて、イヌ用の骨の形をしたゲルの寸法を測定した。15U/ml非PEG化mTG1部とゼラチン溶液2部とを用いて対照試料を作製した。いずれの試料に対しても以下の試験プロトコルを行った。
【0174】
約12mm(幅)×1.9mm(厚さ)×20mm(長さ)の試料を、引張試験システム(3343モデル;インストロン)のクランプに固定した。引張試験の直前に各試料の正確な寸法を測定し、これらの測定した数値を用いて各試料の物質特性を算出した。測定および固定後、予荷重が0.025Nになるまで、各試料に0.25mm/秒の速度で引張力を加えた。これを0%の歪み点とした。予荷重後、試料が破断するまで試料に引張歪を0.5mm/秒の速度で連続的に加えた。
【0175】
破断した時点で最大の歪みおよび応力が生じ、生じた最大引張歪および最大引張応力を、破断時点における破断歪み(%)および破断応力(kPa)として記録した。各試料について、弾性率を10%歪みと30%歪みの間の直線領域から算出した。
【0176】
各種の架橋ゼラチンゲルを5回繰り返して試験し、得られた平均および標準偏差を表5に要約した。
【0177】
【表5】
【0178】
表5は、種々のPEG化mTGの力学的試験の結果を示す。最も左の列は、PEG化の条件を「A−B」で示している。すなわち、Aが7の場合7mg/ml PEGを指し、Aが14の場合14mg/ml PEGを指し、Aが28の場合28mg/ml PEGを指す。Bが2の場合2kD PEGを指し、Bが5の場合5kD PEGを指す。表に示されているように、PEG量を増加させると、ゲル化時間は延長し、ヤング率は低下した。これに対し、PEGサイズを増加させると、得られたゲルの引張り強さおよび可撓性が増加した。
【0179】
実施例10:破壊圧力試験により評価した生体組織におけるシーラントの性能
ブタの小腸組織から残留物を取り除き、10cm四方に切断した。各組織片を14ゲージ針で穿刺した。次いで、各組織を生理食塩水中に浸漬し、37℃でインキュベートした。実施例7に記載の方法で調製したシーラント材を塗布する前に、組織を平らにし、ガーゼパッドを用いて各組織の塗布部位の水分を拭き取った。1mLシリンジを用いて、試験シーラント約0.1〜0.2mLを各塗布部位に塗布した。塗布から5分以内に組織を生理食塩水で洗浄し、37℃で4時間インキュベートした。各試験は3回以上繰り返した。
【0180】
破壊圧力試験を行うため、片側が(クランプを用いて)密閉され、反対側が圧力メーターおよび手動ポンプに(プラスチック固定具を用いて)接続されたパースペックス箱に組織を入れた。パースペックス箱を生理食塩水で満たし、組織試料を完全に浸漬した。手動ポンプを用いて一定速度(20mL/分)で空気を注入した。泡の発生によって破壊圧力を決定した。
【0181】
結果を図6に示す。非PEG化mTGまたは2種のPEG化mTGを用いて作製したゼラチンシーラントについての破壊圧力が示されている。図に示されているように、いずれのPEG化mTGについても破壊圧力強度の中央値は増加したが、PEG化度がより低い酵素においてやや高い効果が示されている。
【0182】
実施例11:インビボブタモデルのステープルラインを補強するためのシーラントの使用
コヴィディエンEEA輪状吻合用外科用ステープラーを用いて、ブタの直腸の輪状吻合を行った。
【0183】
外科用シーラントはゼラチン溶液とPEG化TGとで構成した。反応容量72ml中、28mg/ml 5kDa PEG−NHSを用いて、実施例7と同様にPEG化TGを調製した。PEG化酵素の活性が40U/mlの非PEG化酵素と等しくなるように、ビバスピン20(MWCO 30,000(ザルトリウス))を用いて反応混合物を3mlに濃縮した。(ゼラチン溶液2.66mlおよびPEG化酵素溶液1.33で構成した)シーラント4mLを、直腸のステープルラインの周囲に均一に塗布し、4分間かけて硬化させた。次いで、ブタの傷口を縫合した。
【0184】
手術から14日後にブタを屠殺した。密封した吻合領域を肉眼での病理観察により評価し、シーラントを触診してその力学的特性を定性的に評価した。
【0185】
結果:
シーラントは、14日間の埋め込み期間にわたって顕著な分解を起こさなかった。シーラントは、ステープルラインに強く接着しており、ステープルラインの全長にわたって100%完全な状態で維持されていた。シーラント材は、柔軟性および可撓性を有し、輪状ステープルラインの形状および動きに適合した。
【0186】
シーラントが塗布された領域またはステープルライン領域において、炎症または腹部癒着は見られなかった。吻合は完全に治癒し、滲出の兆候はなかった。直腸において狭窄は観察されなかった。
【0187】
実施例12:架橋化酵素の不溶性担体への非共有結合
SPセファロースを、mTG(微生物トランスグルタミナーゼ)に結合させ、ゲルを作製した。この固定化トランスグルタミナーゼを用いると16〜23分でゲルが形成されたが、可溶性酵素は6分未満でゲルを形成した。したがって、固定化により、ゲル化に必要な時間が延長した。
【0188】
洗浄したSPセファロースビーズ(GEヘルスケア)500μlを、13.5mg/ml精製mTG2.7mlと、50mM酢酸Na(pH5.5)11.55mlと共に混合した(合計15ml)。
【0189】
この混合物を、シェーカーを用いて、室温で20分間インキュベートした。次いで、50mM酢酸Na(pH5.5)11.5mlを用いて、ビーズを3回洗浄(1回の洗浄あたり3分間)した。洗浄工程後、タンパク質の70%がビーズに結合していた。50mM酢酸Na(pH5.5)9.5mlにビーズを再懸濁し、最終容量を10mlとした。最終容量が600μlの以下の種々の組成を有するように、mTG担持ビーズと50mM酢酸Na(pH5.5)とを混合した(括弧内には、結合したmTGの量、および結合していないmTG1mgを用いて測定された活性(33ヒドロキサマートユニット)に基づいて算出されたmTG活性の理論値を示す)。
A:292μlビーズ+308酢酸Na(1.244mg/ml=41U/ml)
B:400μlビーズ+200μl酢酸Na(1.704mg/ml=56.2U/ml)
C:500μlビーズ+100μl酢酸Na(2.13mg/ml=70.3U/ml)
D:550μlビーズ+50μl酢酸Na(2.34mg/ml=77.2U/ml)
【0190】
各反応物A〜D500μlと、4.5M尿素を含む酢酸ナトリウム緩衝液中の25%ゼラチン溶液1mlとを、シリンジからシリンジへと往復させて混合した。ゼラチンが流動しなくなった時間を目視で確認してゲル化時間を決定した。
ゲル化時間:
A:約23分
B:約21分
C:約20分
D:約16分
対照(結合していないmTG 10U/ml):5.5分
【0191】
結合したmTGのゲル化時間は、遊離酵素と比較して顕著に延長された。これは、酵素を大きな足場または不溶性担体に結合させると、ヒドロゲルマトリックス内における酵素の移動が低下し、その結果、結合した酵素の架橋によるゲル化(力学的剛性が増したことを示す)は、遊離(結合していない)酵素によるものと比較して、時間が延長されることを示唆する。したがって、酵素を結合させると、ヒドロゲルマトリックスの力学的特性が改変される。
【0192】
実施例13:酸化デキストランでの酵素改変
この実験は、可溶性大型分子と結合させることによって酵素を改変することにより、その力学的特性が改変されることを実証している。
【0193】
方法
デキストラン1グラムを精製水20mlに溶解した。過ヨウ素酸ナトリウム1.3グラムを添加し、反応物をアルミホイルによって光から保護しながら、室温で80分間(9:50〜11:10)撹拌した。
【0194】
グリセロール2グラムを添加して、反応しなかった過ヨウ素酸塩をクエンチした。
【0195】
反応物を1Lの精製水で2:00時間かけて3回透析し、1回ごとに精製水を交換した。
【0196】
【表6】
【0197】
反応物を室温で一晩インキュベートし、次いで、ビバスピン20(ザルトリウス)を用いたダイアフィルトレーションによって精製した。
【0198】
結果
図7は、コンジュゲーション反応物A〜DのSDS−PAGE分析を示す。4〜15%ミニプロティアン TGXゲル(バイオ・ラッド)上に、4.35μg(反応物A)、4.38μg(反応物B)、1.98μg(反応物C)、および3μg(反応物D)のデキストランコンジュゲート化mTGを載せた。それぞれの試料は、0.1%SDSを含有していたが、還元剤は含有しておらず、ゲル上に載せる前に85℃で10分間加熱した。ゲルに一定電圧(200V)をかけて試料を流し、Bio−Safe Coomassie G−250溶液(バイオ・ラッド)でタンパク質バンドを染色することによって可視化した。分子量マーカーはPrecision Plus(バイオ・ラッド)であった。この実施例では、架橋化酵素(この場合、mTG(微生物トランスグルタミナーゼ))を可溶性ポリマー上に固定化できることが示されている。さらに、デキストラン:mTG比においてデキストランの比率が高いと、より多くの遊離mTG分子が、デキストランとの結合により高分子量コンジュゲートに変換された。
【0199】
実施例14:西洋ワサビペルオキシダーゼのPEG化
この実験では、西洋ワサビペルオキシダーゼ(別の架橋化酵素)をPEG化して改変することにより、ペルオキシダーゼ架橋により形成されるマトリックスを改変できることが実証されている。本発明の別の実施形態において、架橋化酵素は西洋ワサビペルオキシダーゼ(HRP)であり、PEG分子をHRP分子に付着させることによってHRPを改変し、これによって、HRP架橋により形成されるゼラチンヒドロゲルの力学的特性を改変することができる。
【0200】
方法
フェノール修飾ゼラチン(ゼラチン−Ph)の調製:
高分子量ゼラチン(Aタイプ)2グラムを、50mM MES(2−(N−モルホリノ)エタンスルホン酸(シグマアルドリッチ))緩衝液(pH6)100mlに溶解した。この2%w/w溶液に、チラミン(シグマアルドリッチ)0.984グラム、NHS(N−ヒドロキシスクシンイミド(シグマアルドリッチ))0.218グラム、およびEDC(1−エチル−3−[3−ジメチルアミノプロピル]カルボジイミド(シグマアルドリッチ))0.72グラムを添加した。反応物を室温で16時間撹拌し、次いで、蒸留水で十分に透析した。透析物を凍結乾燥し、得られた乾燥物を0.1Mリン酸緩衝液(pH6.0)に溶解して、最終容積が16mlの12.5%w/wゼラチンを得た。
【0201】
HRPのPEG化:
100mM Hepes(pH8.0)中において、2mg/ml HRP I型(シグマ、セントルイス(ミズーリ))と、60mg/ml PEG−NHS 5kDとを2時間反応させ、次いで、110mMグリシンを添加して反応しなかったPEG−NHSをクエンチし、さらに30分間インキュベーションした。PEG化HRPを、25mMリン酸緩衝液(pH6.0)で十分に透析することによって精製した。
【0202】
ゼラチン−PhのHRP依存性およびPEG化HRP依存性ゲル化:
ゼラチン成分:ゼラチン−Ph 5ml+20mM H2O2 0.5mlをガラスバイアル中で混合し、このうち4.4mlをシリンジAに移した。
HRP/PEG化HRP成分:0.035mg/ml HRPまたはPEG化HRP 1mlをシリンジBに入れた。
ゼラチン成分と酵素成分とをシリンジからシリンジへと往復させて混合し、次いで、上下反転させながら37℃でインキュベートしゲル化時間を決定した。
【0203】
20分後、ゲルの重量を計り、生理食塩水10mlに浸漬させて37℃で16時間インキュベートし、次いで、ゲルの重量を再び計って膨潤率を決定した。
【0204】
結果
混合後、ゼラチンおよび酵素の混合物は3分以内にゲルを形成した。HRPタンパク質およびPEG化HRPタンパク質のSDS−PAGE分析を図8に示す。HRPおよびPEG化HRP(各20μg)を4〜15%ミニプロティアンTGXゲル(バイオ・ラッド)上に載せた。それぞれの試料は0.1%SDSを含有していたが、還元剤は含有しておらず、ゲル上に載せる前に85℃で10分間加熱した。ゲルに一定電圧(200V)をかけて試料を流し、Bio−Safe Coomassie G−250溶液(バイオ・ラッド)でタンパク質バンドを染色することによって可視化した。分子量マーカーはPrecision Plus(バイオ・ラッド)であった。
【0205】
測定した膨潤率を以下の表7に詳細に示す。
【0206】
【表7】
【0207】
上記の表7に示されているように、PEG化HRPで作製したゲルは、非PEG化HRPで作製したゲルよりも大きく膨潤した。これは、PEG化(改変型)HRPにより形成されたゼラチンヒドロゲルの力学的特性が、遊離(非改変型)HRPにより形成されたヒドロゲルの力学的特性とは有意に異なることを実証している。いずれのゲルも熱に強く、80℃で1時間加熱しても溶解しなかった。
【0208】
実施例15:部分的PEG化の効果
異なるPEG化度を有する架橋化酵素により、異なる力学的特性がもたらされた。この実施例においては、流体力学的体積(この場合、PEG化度)を調節することによって、酵素的に架橋されたヒドロゲルの力学的特性がどのように特異的に調節され得るかが実証されており、流体力学的体積が大きい(すなわち、PEG化度が高い)場合、弾性の高いマトリックスが形成され、流体力学的体積が小さい(すなわち、PEG化度が低い)場合、弾性の低いマトリックスが形成される。当然のことながら、流体力学的体積が改変されていない(すなわち、PEG化なし)場合は、最も弾性の低いマトリックスが形成される。インストロン社製試験器によるデータおよびSDS−Pageゲルによるデータを、これらの効果に関して以下に記載する。
【0209】
方法
3種のPEG化反応を並列に実施した。反応は、PEG−NHS 5Kを用い、100mM HEPES(pH8.0)中において、室温で2.5時間行った。反応後、反応しなかった余分なPEGを110mMグリシンで中和し、さらに30分間インキュベーションを続けた。
【0210】
反応Aおよび反応Bは、いずれも同じPEG:アミン比を有したが、AのmTGおよびPEGはいずれも、Bよりも3倍高い濃度であった。反応Cは、反応Aと類似しているが、PEG:アミン比はAの半分であった。結果を表8に示す。
【0211】
【表8】
【0212】
これらの反応の終了後、得られたPEG化mTG溶液それぞれを、mTG溶液とゼラチン溶液の比が1:2になるように、(4.5M尿素を含む酢酸ナトリウム緩衝液中の)25%ゼラチン溶液と反応させて、ゼラチンヒドロゲルを形成させた。ゼラチンとの反応時間を各溶液において統一することによって、各PEG化mTG溶液のmTG活性レベルを標準化した。各ヒドロゲルが形成された後、37℃で2時間インキュベートし、引張試験システムを用いて力学的試験を行った。
【0213】
結果を、PEG化mTGをSDS−PAGE分析した画像である図9に示す。反応A、B、およびCから得られたPEG化mTG(それぞれ5μg)を、6%ポリアクリルアミドゲル上に載せ、SDS−PAGEに供した。それぞれの試料は、0.1%SDSを含有していたが、還元剤は含有しておらず、ゲル上に載せる前に85℃で10分間加熱した。ゲルに一定電圧(200V)をかけて試料を流し、Bio−Safe Coomassie G−250溶液(バイオ・ラッド)でタンパク質バンドを染色することによって可視化した。分子量マーカーはPrecision Plus(バイオ・ラッド)であった。
【0214】
SDS−PAGEプロファイルにより、反応A、B、およびCにおいて、PEG分子とmTG分子とがどのように異なる度合いで結合したかが示されており、Aにおいては多数のPEG分子がmTGに結合し、Bにおいてはより少ないPEG分子がmTGに結合し、Cにおいてはさらに少ないPEG分子がmTGに結合した。
【0215】
以下の結果に示されるように、PEG化度の差異は、ゼラチンマトリックスの力学的特性に顕著な影響を与える。すなわち、PEG化度が最も高かったmTGであるAは、弾性の最も高い(破断時点における引張歪が最も高い)ヒドロゲルを形成し、非PEG化mTGは弾性の最も低いヒドロゲルを形成し、部分的にPEG化されたmTGであるBおよびCは、それぞれのPEG化度に相関して、PEG化度が最も高かったmTGと非PEG化mTGとの間の力学的特性を有した。
【0216】
【表9】
【0217】
実施例16:遊離PEGはゲル化に効果を与えない
対照として、インストロン社製試験器によるゲルの力学的特性に関する結果を、遊離PEGを用いてまたは用いずに評価した。「遊離」とは、PEG分子が、架橋化酵素と共に溶液中に存在するが、酵素には共有結合されていないことを意味する。実験結果より、遊離PEGを用いても、PEGが架橋化酵素に共有結合すること(すなわち、PEGによる酵素自体の改変)によってもたらされる力学的特性の改変は見られなかったことが示された。
【0218】
方法
20%PEG6000を含むまたは含まないゼラチン溶液(酢酸ナトリウム緩衝液中の25%ゼラチンおよび4.5M尿素)のそれぞれのアリコート4mlと、15U/ml mTGのアリコート2mlとを混合した。それぞれから得られた溶液2mlを、実施例9の記載と同様に、イヌ用の骨の形をした型へ流し込んだ。実施例9の記載と同様に、得られたゲルを型から取り出し、生理食塩水中で37℃、2時間インキュベートし、次いで、引張試験を行った。
【0219】
結果
【表10】
【0220】
表10の結果より、遊離PEG(可塑剤)の添加により、酵素架橋ヒドロゲルマトリックスの力学的特性にわずかな効果が見られたか、またはまったく効果が見られず、さらに、このような力学的特性の改変は、PEGを酵素に付着させて酵素分子の流体力学的体積を増加させることによる改変と比較して、わずかなものであることが実証されている。特に、遊離PEGを添加することによって、マトリックスの弾性(破断歪み)は全く向上しなかった。これに対し、別の実施例のいくつかに示されているように、酵素分子をPEG化すると、マトリックス弾性が有意に増加する。
【0221】
実施例17:混合した改変/非改変架橋化酵素
非改変酵素と改変酵素との種々の混合物を試験した。特定の割合で混合することにより、種々の異なる力学的特性を有するように改変することができる。
【0222】
本発明の別の実施形態において、酵素架橋マトリックスの力学的特性を改変するために、改変酵素は非改変(遊離)酵素と共に用いられる。
【0223】
方法
20%PEG6000を含むまたは含まないゼラチン溶液(25%ゼラチン、4.5M尿素、酢酸ナトリウム緩衝液)のそれぞれのアリコート4mlと、非PEG化mTGを加えたまたは加えていない55U/ml PEG化mTG 2mlと混合した。それぞれから得られた溶液2mlを、実施例9の記載と同様に、イヌ用の骨の形をした型へ流し込んだ。実施例9の記載と同様に、得られたゲルを型から取り出し、生理食塩水中で37℃、24時間インキュベートし、次いで、引張試験を行った。
【0224】
結果
【表11】
【0225】
表11の結果より、酵素架橋ヒドロゲルの力学的特性は、改変酵素のみを用いることによっても、また、低い程度にとどまるが、改変酵素と遊離酵素の混合物を用いることによっても、改変できることが示されている。
【0226】
実施例18:二官能基型PEG−酵素ブリッジ
この実験では、二官能基型PEGブリッジによって酵素同士を架橋できることが実証された。この実施例では、2個以上の酵素分子を互いに結合させて、酵素凝集体全体の流体力学的体積を増加させることができる。これを達成する方法のひとつとして、酵素分子間にブリッジを形成することのできる二官能基型分子を用いることが挙げられる。
【0227】
方法
100mM Hepes(pH8)中において、mTG(15U/ml、0.5mg/ml)と、種々の濃度の二官能基型PEG−NHS(10kD)とを、室温で2時間インキュベートした。次いで、110mMグリシンを30分を上回る時間をかけて添加し、反応しなかった余分なPEGを中和した。具体的な条件を以下の表12に示す。
【0228】
【表12】
【0229】
反応終了後、各反応組成の酵素5μgを7.5%ポリアクリルアミドゲル上に載せ、SDS−PAGE分析に供した。それぞれの試料は、0.1%SDSを含有していたが、還元剤は含有しておらず、ゲル上に載せる前に85℃で10分間加熱した。ゲルに一定電圧(200V)をかけて試料を流し、Bio−Safe Coomassie G−250溶液(バイオ・ラッド)でタンパク質バンドを染色することによって可視化した。分子量マーカーはPrecision Plus(バイオ・ラッド)であった。
【0230】
力学的特性試験を行うために、ゼラチン溶液(25%ゼラチン、4.5M尿素、酢酸ナトリウム緩衝液)のアリコート4mlと、15U/mlの遊離mTGまたはPEG化mTG(反応C)のアリコート2mlと混合した。得られた溶液2mlを、実施例9の記載と同様に、イヌ用の骨の形をした型へ流し込んだ。実施例9の記載と同様に、得られたゲルを型から取り出し、生理食塩水中で37℃、24時間インキュベートし、次いで、引張試験を行った。
【0231】
結果
図10のSDS−PAGE結果により、相対的に低いPEG:mTG比において、mTGのいくらかは、類似の濃度の一官能基型PEG(5kD)を含む反応において得られるPEG化生成物よりも大きな、非常に高い分子量の生成物に変換されたことが示されている。これにより、この高分子量生成物が、二官能基型PEGブリッジによって互いに架橋された酵素分子のマルチマーからなること、および、二官能基型PEGの使用は、架橋化酵素分子同士を結合させることによって架橋化酵素分子を改変する効果を有することが実証されている。
【0232】
遊離酵素分子による架橋により形成されたゼラチンヒドロゲルと比較した以下の力学的試験の結果により、架橋化酵素分子同士を結合させると、この連結した酵素分子による架橋により形成されるゼラチンヒドロゲルを顕著に改変できることが示されている。結果を表13に示す。
【0233】
【表13】
【0234】
実施例19:PEG化mTGの質量分析
PEG化mTG(PEG−NHS−5kDで改変した微生物トランスグルタミナーゼ酵素)の3種のバッチを、MALDI−TOF質量分析法によって分析した。図11は、これらのうちの1バッチのm/zスペクトルを示す。
【0235】
質量分析
無傷の分子の質量測定を、遅延イオン抽出、反射器、および337nmの窒素レーザーを用いたBruker Reflex IIIマトリックス支援レーザー脱離/イオン化(MALDI)飛行時間型(TOF)質量分析装置(Bruker、ブレーメン(ドイツ))で実施した。各質量スペクトルは、レーザーを200回照射した累積データから作成した。タンパク質に対する外部較正は、BSAおよびミオグロビンタンパク質(シグマ、セントルイス(ミズーリ))を用いて行った。
【0236】
MALDI−TOF MSのための試料調製−dried droplet法
2,5−ジヒドロキシ安息香酸(DHB)マトリックス剤と、0.1%トリフルオロ酢酸(TFA)−アセトニトリル(ACN)とを2:1の割合で混合したもの0.5Lを、ギ酸/イソプロパノール/H20(1:3:2)に溶解した試料溶液0.5Lとターゲット上で混合し、空気乾燥させた。溶媒が蒸発した後、試料を0.1%TFAで1〜3回再洗浄した。
【0237】
【表14】
【0238】
表14の結果は、5kDaのPEG−NHS試薬を用いてmTG架橋化酵素をPEG化したことにより、1個の酵素分子に複数個のPEG分子が結合されたことを示している。
【0239】
実施例20:PEGとアミンの比率を固定したときの、種々の濃度の反応物によるmTGのPEG化
この実施例により、反応物の総濃度がPEG化度に大きな影響を与えることが実証されている。PEG:アミン比を固定した場合、反応物(PEGおよびmTG)の濃度とPEG化度とに相関性があることが実証された。
【0240】
方法
100mM Hepes(pH8.0)中において、PEG−NHS−5kDを用いたmTGのPEG化を室温で2.5時間かけて行い、次いで、110mMグリシンを添加して反応しなかったPEG−NHSを中和した。反応後、各反応組成の酵素5μgを、6.0%ポリアクリルアミドゲル上に載せ、SDS−PAGE分析に供した。それぞれの試料は、0.1%SDSを含有していたが、還元剤は含有しておらず、ゲル上に載せる前に85℃で10分間加熱した。ゲルに一定電圧(200V)をかけて試料を流し、Bio−Safe Coomassie G−250溶液(バイオ・ラッド)でタンパク質バンドを染色することによって可視化した。分子量マーカーはPrecision Plus(バイオ・ラッド)であった。
【0241】
反応は以下の条件(表15)に従って実施した。
【0242】
【表15】
【0243】
結果
図12に示したSDS−PAGEレーンより、PEG化mTG分子が得られたことが確認できる。画像に示されているように、PEG:アミン比を固定した場合でも、反応物の濃度が高いほど、PEG化度の高い生成物が得られた。
【0244】
本発明の選択された実施形態は個別に記載かつ説明されているが、記載された実施形態の適切な態様は組み合わされてもよく、さらに、複数の実施形態を組み合わせてもよいことが理解される。
【0245】
加えて、本発明の選択された実施形態が記載かつ説明されているが、本発明は、記載された実施形態に限定されないことが理解される。むしろ、本発明の原理および精神、ならびに特許請求の範囲およびその等価物によって定義される範囲から逸脱することなく、これらの実施形態に変更を加えてもよいことが理解される。
【特許請求の範囲】
【請求項1】
架橋マトリックスであって、
改変酵素分子により架橋される基質ポリマーを含み、
改変酵素分子が、基質ポリマーの架橋によってマトリックスが形成されるのに伴い、架橋マトリックス内における酵素分子の感知体積が変化するように改変されたものであることを特徴とする、架橋マトリックス。
【請求項2】
前記改変酵素分子が、前記改変酵素分子の実際のサイズが増加するように改変されたものであることを特徴とする、請求項1に記載のマトリックス。
【請求項3】
前記改変酵素分子が、前記改変酵素分子の流体力学的体積が増加するように改変されたものであることを特徴とする、請求項1に記載のマトリックス。
【請求項4】
前記改変酵素分子が、前記改変酵素の等電点(pI)を、改変していない酵素の等電点と異なるように変化させることによって、前記改変酵素分子の静電荷が前記基質ポリマーの正味の電荷と反対の符号に帯電されるように改変されたものであることを特徴とする、請求項1に記載のマトリックス。
【請求項5】
(無水コハク酸による)スクシニル化、(無水酢酸による)アセチル化、(シアネートによる)カルバミル化、(アルデヒドによる)還元的アルキル化、および無水マレイン酸処理からなる群から選択される工程によって、前記酵素のリシンのε−アミノ基が改変されていることを特徴とする、請求項4に記載のマトリックス。
【請求項6】
負の荷電量が減少するように、前記酵素のカルボン酸含有側鎖の1つ以上が改変されていることを特徴とする、請求項4に記載のマトリックス。
【請求項7】
前記改変が、少なくとも1つの分子または部分が前記改変酵素分子に共有結合または非共有結合されていることを含むことを特徴とする、請求項2〜6のいずれか一項に記載のマトリックス。
【請求項8】
前記改変が、改変作用を有する分子が前記改変酵素分子に共有結合されていることを含むことを特徴とする、請求項7に記載のマトリックス。
【請求項9】
前記改変酵素分子が、非改変酵素よりも低い拡散速度および架橋速度を有するが、非改変酵素と少なくとも類似した測定酵素活性を有することを特徴とする、請求項8に記載のマトリックス。
【請求項10】
前記架橋速度が、非改変酵素の架橋速度よりも少なくとも10%遅いことを特徴とする、請求項9に記載のマトリックス。
【請求項11】
前記改変作用を有する分子が、担体またはポリマーを含むことを特徴とする、請求項9に記載のマトリックス。
【請求項12】
前記ポリマーが、合成ポリマー、セルロースポリマー、タンパク質、または多糖類を含むことを特徴とする、請求項11に記載のマトリックス。
【請求項13】
前記セルロースポリマーが、カルボキシメチルセルロース、ヒドロキシプロピルメチルセルロース、ヒドロキシエチルセルロース、またはメチルセルロースのうちの1種以上を含むことを特徴とする、請求項12に記載のマトリックス。
【請求項14】
前記多糖類が、デキストラン、コンドロイチン硫酸、デルマタン硫酸、ケラタン硫酸、ヘパリン、ヘパラン硫酸、ヒアルロン酸、またはデンプン誘導体のうちの1種以上を含むことを特徴とする、請求項12に記載のマトリックス。
【請求項15】
前記改変作用を有する分子が、PEG(ポリエチレングリコール)を含むことを特徴とする、請求項12に記載のマトリックス。
【請求項16】
前記PEGがPEG誘導体を含むことを特徴とする、請求項15に記載のマトリックス。
【請求項17】
前記PEG誘導体が活性化PEGを含むことを特徴とする、請求項16に記載のマトリックス。
【請求項18】
前記活性化PEGが、メトキシPEG(mPEG)、もしくはその誘導体、mPEG−NHS、mPEGのスクシンイミジル(NHS)エステル(mPEG−スクシナート−NHS)、mPEG−グルタレート−NHS、mPEG−バレラート−NHS、mPEG−カルボナート−NHS、mPEG−カルボキシメチル−NHS、mPEG−プロピオナート−NHS、mPEG−カルボキシペンチル−NHS)、mPEG−ニトロフェニルカルボナート、mPEG−プロピルアルデヒド、mPEG−トシラート、mPEG−カルボニルイミダゾール、mPEG−イソシアネート、mPEG−エポキシド、またはこれらの組み合わせのうちの1種以上を含むことを特徴とする、請求項17に記載のマトリックス。
【請求項19】
前記活性化PEGが、前記酵素のアミン基またはチオール基と反応することを特徴とする、請求項17または18に記載のマトリックス。
【請求項20】
前記活性化された酵素のリシン残基に対する前記活性化PEGのモル比が、0.5〜25の範囲であることを特徴とする、請求項17〜19のいずれか一項に記載のマトリックス。
【請求項21】
前記活性化PEGが、一官能基型、ヘテロ二官能基型、ホモ二官能基型、または多官能基型であることを特徴とする、請求項17〜20のいずれか一項に記載のマトリックス。
【請求項22】
前記活性化PEGが、分岐PEGまたはマルチアームPEGであることを特徴とする、請求項17〜21のいずれか一項に記載のマトリックス。
【請求項23】
前記活性化PEGが、1000ダルトン〜40,000ダルトンの範囲のサイズであることを特徴とする、請求項17〜22のいずれか一項に記載のマトリックス。
【請求項24】
前記酵素または前記基質ポリマーのいずれにも共有結合していないコポリマーをさらに含むことを特徴とする、前記請求項のいずれか一項に記載のマトリックス。
【請求項25】
前記コポリマーが、多糖類またはセルロースポリマーを含むことを特徴とする、請求項24に記載のマトリックス。
【請求項26】
前記多糖類が、デキストラン、コンドロイチン硫酸、デルマタン硫酸、ケラタン硫酸、ヘパリン、ヘパラン硫酸、ヒアルロン酸、またはデンプン誘導体を含むことを特徴とする、請求項25に記載のマトリックス。
【請求項27】
前記セルロースポリマーが、カルボキシメチルセルロース、ヒドロキシプロピルメチルセルロース、ヒドロキシエチルセルロース、メチルセルロースを含むことを特徴とする、請求項25に記載のマトリックス。
【請求項28】
前記改変酵素分子が、前記改変酵素分子と複数の酵素分子とが架橋することによって、複数の架橋化酵素分子が凝集体を形成するように改変されたものであることを特徴とする、前記請求項のいずれか一項に記載のマトリックス。
【請求項29】
前記改変酵素分子の改変またはその改変の度合いが、マトリックスの少なくとも1種の特性に変化を与えることを特徴とする、前記請求項のいずれか一項に記載のマトリックス。
【請求項30】
前記少なくとも1種の特性が、引張り強さ、剛性、前記基質ポリマーの架橋度、粘度、弾性、可撓性、破断歪み、破断応力、ポアソン比、膨潤力、およびヤング率、およびこれらの組み合わせからなる群から選択されることを特徴とする、請求項29に記載のマトリックス。
【請求項31】
前記改変酵素の改変の度合いによって、前記改変酵素のマトリックス内における移動または前記改変酵素のマトリックスからの拡散が決定されることを特徴とする、前記請求項のいずれか一項に記載のマトリックス。
【請求項32】
前記改変酵素の前記改変によって、前記改変酵素と前記タンパク質との溶液またはマトリックスにおける前記改変酵素の拡散係数が、非改変酵素と前記タンパク質との溶液またはマトリックスにおける非改変酵素の拡散係数よりも低下することを特徴とする、請求項31に記載のマトリックス。
【請求項33】
前記改変酵素の改変の度合いによって、マトリックスの1種以上の力学的特性が決定されることを特徴とする、前記請求項のいずれか一項に記載のマトリックス。
【請求項34】
前記改変酵素分子によって溶液中で架橋ポリマーが形成される架橋速度が、非改変酵素分子と比較して速いことを特徴とする、前記請求項のいずれか一項に記載のマトリックス。
【請求項35】
マトリックスの形成を調節する方法であって、
マトリックスが形成されるのに伴い、架橋マトリックス内における酵素分子の感知体積が変化するように酵素分子を改変すること;
改変した酵素分子と、改変した酵素分子の基質である少なくとも1種の基質ポリマーとを混合すること;および
改変した酵素分子で該少なくとも1種の基質ポリマーを架橋して、その形成の少なくとも一部が酵素分子の改変によって調節されたマトリックスを形成すること
を含む方法。
【請求項36】
前記改変によって、前記少なくとも1種の基質ポリマーの架橋度が増加するのに伴い、前記改変酵素分子による架橋の速度が低下することを特徴とする、請求項35に記載の方法。
【請求項37】
前記改変酵素分子と前記少なくとも1種の基質ポリマーとを溶液中で混合すると、前記改変によって、溶液の粘度が増加するのに伴い、前記少なくとも1種の基質ポリマーの架橋度が調節されることを特徴とする、請求項36に記載の方法。
【請求項38】
前記改変が、pH7〜9において前記酵素をPEG化することを含むことを特徴とする、請求項37に記載の方法。
【請求項39】
前記PEG化反応が、pH7.5〜8.5で行われることを特徴とする、請求項38に記載の方法。
【請求項40】
前記少なくとも1種の基質ポリマーが、元より架橋可能なポリマー、前記改変酵素により架橋可能な部分変性ポリマー、および前記改変酵素により架橋可能な官能基またはペプチドを含む改変ポリマーからなる群から選択される基質ポリマーを含むことを特徴とする、前記請求項のいずれか一項に記載の方法またはマトリックス。
【請求項41】
前記少なくとも1種の基質ポリマーが、ゼラチン、コラーゲン、カゼイン、もしくはアルブミン、または改変ポリマーを含み、前記改変酵素分子が、改変トランスグルタミナーゼおよび/または改変酸化酵素を含むことを特徴とする、請求項40に記載の方法またはマトリックス。
【請求項42】
前記の少なくとも1種の基質ポリマーが、動物組織もしくは動物組織から得られるコラーゲンを部分的に加水分解することによって得られるゼラチン;細菌系、酵母系、動物系、昆虫系、植物系、もしくは任意の種類の細胞培養を用いて産生される組換えゼラチン;およびこれらの組み合わせからなる群から選択されるゼラチンを含み、
該動物組織が、動物の皮、結合組織、枝角、角、骨、および魚鱗からなる群から選択されることを特徴とする、請求項41に記載の方法またはマトリックス。
【請求項43】
前記ゼラチンが、哺乳動物由来または魚類由来であることを特徴とする、請求項42に記載の方法またはマトリックス。
【請求項44】
前記ゼラチンが、Aタイプ(酸処理)またはBタイプ(アルカリ処理)であることを特徴とする、請求項43に記載の方法またはマトリックス。
【請求項45】
前記ゼラチンが、250〜300ブルームであることを特徴とする、請求項44に記載の方法またはマトリックス。
【請求項46】
前記ゼラチンの平均分子量が、75〜150kdaであることを特徴とする、請求項43に記載の方法またはマトリックス。
【請求項47】
前記改変トランスグルタミナーゼが、改変された微生物トランスグルタミナーゼを含むことを特徴とする、請求項41〜46のいずれか一項に記載の方法またはマトリックス。
【請求項48】
前記改変ポリマーが、前記改変された微生物トランスグルタミナーゼによる架橋が可能となるように改変されたものであることを特徴とする、請求項47に記載の方法またはマトリックス。
【請求項49】
前記改変酸化酵素が、チロシナーゼ、ラッカーゼ、またはペルオキシダーゼのうちの1種以上を含むことを特徴とする、請求項41〜46に記載の方法またはマトリックス。
【請求項50】
前記マトリックスが、前記少なくとも1種の基質ポリマーとして、前記改変酸化酵素により架橋されるフェノール酸を含む炭水化物をさらに含むことを特徴とする、請求項49に記載の方法またはマトリックス。
【請求項51】
前記炭水化物が、アラビノキシランまたはペクチンのうちの1種以上を含むことを特徴とする、請求項50に記載の方法またはマトリックス。
【請求項52】
前記酵素分子をPEG化により改変し、このPEG化により、マトリックスが使用される宿主動物の免疫系から前記酵素分子がマスクされ、これによって、前記酵素分子の免疫原性がマスクされることを特徴とする、前記請求項のいずれか一項に記載の方法またはマトリックス。
【請求項53】
前記宿主動物がヒトであることを特徴とする、請求項52に記載の方法またはマトリックス。
【請求項54】
前記請求項のいずれか一項に記載のマトリックスを組織に塗布することを含む、体液が滲出した組織を密封するための方法。
【請求項55】
前記体液が血液を含み、前記マトリックスが止血剤であることを特徴とする、請求項54に記載の方法。
【請求項56】
前記請求項のいずれか一項に記載のマトリックスを含む、止血剤または外科用シーラント。
【請求項57】
前記請求項のいずれか一項に記載のマトリックスを含む、創傷を密封するための組成物。
【請求項58】
組織における縫合線またはステープルラインを密封するための、請求項57に記載の組成物の使用。
【請求項59】
前記請求項のいずれか一項に記載のマトリックスを含む、局所的薬物送達のビヒクルのための組成物。
【請求項60】
注入可能な足場として適した、前記請求項のいずれか一項に記載のマトリックスを含む、組織工学のための組成物。
【請求項61】
組成物を改変する方法であって、
架橋可能な官能基を有する改変酵素と、改変酵素によって架橋可能な少なくとも1種の部分を有するタンパク質とを提供すること;および
改変酵素とタンパク質を混合して、改変酵素によりタンパク質を架橋し、かつタンパク質により改変酵素自体を架橋可能な官能基において架橋すること
を含む方法。
【請求項1】
架橋マトリックスであって、
改変酵素分子により架橋される基質ポリマーを含み、
改変酵素分子が、基質ポリマーの架橋によってマトリックスが形成されるのに伴い、架橋マトリックス内における酵素分子の感知体積が変化するように改変されたものであることを特徴とする、架橋マトリックス。
【請求項2】
前記改変酵素分子が、前記改変酵素分子の実際のサイズが増加するように改変されたものであることを特徴とする、請求項1に記載のマトリックス。
【請求項3】
前記改変酵素分子が、前記改変酵素分子の流体力学的体積が増加するように改変されたものであることを特徴とする、請求項1に記載のマトリックス。
【請求項4】
前記改変酵素分子が、前記改変酵素の等電点(pI)を、改変していない酵素の等電点と異なるように変化させることによって、前記改変酵素分子の静電荷が前記基質ポリマーの正味の電荷と反対の符号に帯電されるように改変されたものであることを特徴とする、請求項1に記載のマトリックス。
【請求項5】
(無水コハク酸による)スクシニル化、(無水酢酸による)アセチル化、(シアネートによる)カルバミル化、(アルデヒドによる)還元的アルキル化、および無水マレイン酸処理からなる群から選択される工程によって、前記酵素のリシンのε−アミノ基が改変されていることを特徴とする、請求項4に記載のマトリックス。
【請求項6】
負の荷電量が減少するように、前記酵素のカルボン酸含有側鎖の1つ以上が改変されていることを特徴とする、請求項4に記載のマトリックス。
【請求項7】
前記改変が、少なくとも1つの分子または部分が前記改変酵素分子に共有結合または非共有結合されていることを含むことを特徴とする、請求項2〜6のいずれか一項に記載のマトリックス。
【請求項8】
前記改変が、改変作用を有する分子が前記改変酵素分子に共有結合されていることを含むことを特徴とする、請求項7に記載のマトリックス。
【請求項9】
前記改変酵素分子が、非改変酵素よりも低い拡散速度および架橋速度を有するが、非改変酵素と少なくとも類似した測定酵素活性を有することを特徴とする、請求項8に記載のマトリックス。
【請求項10】
前記架橋速度が、非改変酵素の架橋速度よりも少なくとも10%遅いことを特徴とする、請求項9に記載のマトリックス。
【請求項11】
前記改変作用を有する分子が、担体またはポリマーを含むことを特徴とする、請求項9に記載のマトリックス。
【請求項12】
前記ポリマーが、合成ポリマー、セルロースポリマー、タンパク質、または多糖類を含むことを特徴とする、請求項11に記載のマトリックス。
【請求項13】
前記セルロースポリマーが、カルボキシメチルセルロース、ヒドロキシプロピルメチルセルロース、ヒドロキシエチルセルロース、またはメチルセルロースのうちの1種以上を含むことを特徴とする、請求項12に記載のマトリックス。
【請求項14】
前記多糖類が、デキストラン、コンドロイチン硫酸、デルマタン硫酸、ケラタン硫酸、ヘパリン、ヘパラン硫酸、ヒアルロン酸、またはデンプン誘導体のうちの1種以上を含むことを特徴とする、請求項12に記載のマトリックス。
【請求項15】
前記改変作用を有する分子が、PEG(ポリエチレングリコール)を含むことを特徴とする、請求項12に記載のマトリックス。
【請求項16】
前記PEGがPEG誘導体を含むことを特徴とする、請求項15に記載のマトリックス。
【請求項17】
前記PEG誘導体が活性化PEGを含むことを特徴とする、請求項16に記載のマトリックス。
【請求項18】
前記活性化PEGが、メトキシPEG(mPEG)、もしくはその誘導体、mPEG−NHS、mPEGのスクシンイミジル(NHS)エステル(mPEG−スクシナート−NHS)、mPEG−グルタレート−NHS、mPEG−バレラート−NHS、mPEG−カルボナート−NHS、mPEG−カルボキシメチル−NHS、mPEG−プロピオナート−NHS、mPEG−カルボキシペンチル−NHS)、mPEG−ニトロフェニルカルボナート、mPEG−プロピルアルデヒド、mPEG−トシラート、mPEG−カルボニルイミダゾール、mPEG−イソシアネート、mPEG−エポキシド、またはこれらの組み合わせのうちの1種以上を含むことを特徴とする、請求項17に記載のマトリックス。
【請求項19】
前記活性化PEGが、前記酵素のアミン基またはチオール基と反応することを特徴とする、請求項17または18に記載のマトリックス。
【請求項20】
前記活性化された酵素のリシン残基に対する前記活性化PEGのモル比が、0.5〜25の範囲であることを特徴とする、請求項17〜19のいずれか一項に記載のマトリックス。
【請求項21】
前記活性化PEGが、一官能基型、ヘテロ二官能基型、ホモ二官能基型、または多官能基型であることを特徴とする、請求項17〜20のいずれか一項に記載のマトリックス。
【請求項22】
前記活性化PEGが、分岐PEGまたはマルチアームPEGであることを特徴とする、請求項17〜21のいずれか一項に記載のマトリックス。
【請求項23】
前記活性化PEGが、1000ダルトン〜40,000ダルトンの範囲のサイズであることを特徴とする、請求項17〜22のいずれか一項に記載のマトリックス。
【請求項24】
前記酵素または前記基質ポリマーのいずれにも共有結合していないコポリマーをさらに含むことを特徴とする、前記請求項のいずれか一項に記載のマトリックス。
【請求項25】
前記コポリマーが、多糖類またはセルロースポリマーを含むことを特徴とする、請求項24に記載のマトリックス。
【請求項26】
前記多糖類が、デキストラン、コンドロイチン硫酸、デルマタン硫酸、ケラタン硫酸、ヘパリン、ヘパラン硫酸、ヒアルロン酸、またはデンプン誘導体を含むことを特徴とする、請求項25に記載のマトリックス。
【請求項27】
前記セルロースポリマーが、カルボキシメチルセルロース、ヒドロキシプロピルメチルセルロース、ヒドロキシエチルセルロース、メチルセルロースを含むことを特徴とする、請求項25に記載のマトリックス。
【請求項28】
前記改変酵素分子が、前記改変酵素分子と複数の酵素分子とが架橋することによって、複数の架橋化酵素分子が凝集体を形成するように改変されたものであることを特徴とする、前記請求項のいずれか一項に記載のマトリックス。
【請求項29】
前記改変酵素分子の改変またはその改変の度合いが、マトリックスの少なくとも1種の特性に変化を与えることを特徴とする、前記請求項のいずれか一項に記載のマトリックス。
【請求項30】
前記少なくとも1種の特性が、引張り強さ、剛性、前記基質ポリマーの架橋度、粘度、弾性、可撓性、破断歪み、破断応力、ポアソン比、膨潤力、およびヤング率、およびこれらの組み合わせからなる群から選択されることを特徴とする、請求項29に記載のマトリックス。
【請求項31】
前記改変酵素の改変の度合いによって、前記改変酵素のマトリックス内における移動または前記改変酵素のマトリックスからの拡散が決定されることを特徴とする、前記請求項のいずれか一項に記載のマトリックス。
【請求項32】
前記改変酵素の前記改変によって、前記改変酵素と前記タンパク質との溶液またはマトリックスにおける前記改変酵素の拡散係数が、非改変酵素と前記タンパク質との溶液またはマトリックスにおける非改変酵素の拡散係数よりも低下することを特徴とする、請求項31に記載のマトリックス。
【請求項33】
前記改変酵素の改変の度合いによって、マトリックスの1種以上の力学的特性が決定されることを特徴とする、前記請求項のいずれか一項に記載のマトリックス。
【請求項34】
前記改変酵素分子によって溶液中で架橋ポリマーが形成される架橋速度が、非改変酵素分子と比較して速いことを特徴とする、前記請求項のいずれか一項に記載のマトリックス。
【請求項35】
マトリックスの形成を調節する方法であって、
マトリックスが形成されるのに伴い、架橋マトリックス内における酵素分子の感知体積が変化するように酵素分子を改変すること;
改変した酵素分子と、改変した酵素分子の基質である少なくとも1種の基質ポリマーとを混合すること;および
改変した酵素分子で該少なくとも1種の基質ポリマーを架橋して、その形成の少なくとも一部が酵素分子の改変によって調節されたマトリックスを形成すること
を含む方法。
【請求項36】
前記改変によって、前記少なくとも1種の基質ポリマーの架橋度が増加するのに伴い、前記改変酵素分子による架橋の速度が低下することを特徴とする、請求項35に記載の方法。
【請求項37】
前記改変酵素分子と前記少なくとも1種の基質ポリマーとを溶液中で混合すると、前記改変によって、溶液の粘度が増加するのに伴い、前記少なくとも1種の基質ポリマーの架橋度が調節されることを特徴とする、請求項36に記載の方法。
【請求項38】
前記改変が、pH7〜9において前記酵素をPEG化することを含むことを特徴とする、請求項37に記載の方法。
【請求項39】
前記PEG化反応が、pH7.5〜8.5で行われることを特徴とする、請求項38に記載の方法。
【請求項40】
前記少なくとも1種の基質ポリマーが、元より架橋可能なポリマー、前記改変酵素により架橋可能な部分変性ポリマー、および前記改変酵素により架橋可能な官能基またはペプチドを含む改変ポリマーからなる群から選択される基質ポリマーを含むことを特徴とする、前記請求項のいずれか一項に記載の方法またはマトリックス。
【請求項41】
前記少なくとも1種の基質ポリマーが、ゼラチン、コラーゲン、カゼイン、もしくはアルブミン、または改変ポリマーを含み、前記改変酵素分子が、改変トランスグルタミナーゼおよび/または改変酸化酵素を含むことを特徴とする、請求項40に記載の方法またはマトリックス。
【請求項42】
前記の少なくとも1種の基質ポリマーが、動物組織もしくは動物組織から得られるコラーゲンを部分的に加水分解することによって得られるゼラチン;細菌系、酵母系、動物系、昆虫系、植物系、もしくは任意の種類の細胞培養を用いて産生される組換えゼラチン;およびこれらの組み合わせからなる群から選択されるゼラチンを含み、
該動物組織が、動物の皮、結合組織、枝角、角、骨、および魚鱗からなる群から選択されることを特徴とする、請求項41に記載の方法またはマトリックス。
【請求項43】
前記ゼラチンが、哺乳動物由来または魚類由来であることを特徴とする、請求項42に記載の方法またはマトリックス。
【請求項44】
前記ゼラチンが、Aタイプ(酸処理)またはBタイプ(アルカリ処理)であることを特徴とする、請求項43に記載の方法またはマトリックス。
【請求項45】
前記ゼラチンが、250〜300ブルームであることを特徴とする、請求項44に記載の方法またはマトリックス。
【請求項46】
前記ゼラチンの平均分子量が、75〜150kdaであることを特徴とする、請求項43に記載の方法またはマトリックス。
【請求項47】
前記改変トランスグルタミナーゼが、改変された微生物トランスグルタミナーゼを含むことを特徴とする、請求項41〜46のいずれか一項に記載の方法またはマトリックス。
【請求項48】
前記改変ポリマーが、前記改変された微生物トランスグルタミナーゼによる架橋が可能となるように改変されたものであることを特徴とする、請求項47に記載の方法またはマトリックス。
【請求項49】
前記改変酸化酵素が、チロシナーゼ、ラッカーゼ、またはペルオキシダーゼのうちの1種以上を含むことを特徴とする、請求項41〜46に記載の方法またはマトリックス。
【請求項50】
前記マトリックスが、前記少なくとも1種の基質ポリマーとして、前記改変酸化酵素により架橋されるフェノール酸を含む炭水化物をさらに含むことを特徴とする、請求項49に記載の方法またはマトリックス。
【請求項51】
前記炭水化物が、アラビノキシランまたはペクチンのうちの1種以上を含むことを特徴とする、請求項50に記載の方法またはマトリックス。
【請求項52】
前記酵素分子をPEG化により改変し、このPEG化により、マトリックスが使用される宿主動物の免疫系から前記酵素分子がマスクされ、これによって、前記酵素分子の免疫原性がマスクされることを特徴とする、前記請求項のいずれか一項に記載の方法またはマトリックス。
【請求項53】
前記宿主動物がヒトであることを特徴とする、請求項52に記載の方法またはマトリックス。
【請求項54】
前記請求項のいずれか一項に記載のマトリックスを組織に塗布することを含む、体液が滲出した組織を密封するための方法。
【請求項55】
前記体液が血液を含み、前記マトリックスが止血剤であることを特徴とする、請求項54に記載の方法。
【請求項56】
前記請求項のいずれか一項に記載のマトリックスを含む、止血剤または外科用シーラント。
【請求項57】
前記請求項のいずれか一項に記載のマトリックスを含む、創傷を密封するための組成物。
【請求項58】
組織における縫合線またはステープルラインを密封するための、請求項57に記載の組成物の使用。
【請求項59】
前記請求項のいずれか一項に記載のマトリックスを含む、局所的薬物送達のビヒクルのための組成物。
【請求項60】
注入可能な足場として適した、前記請求項のいずれか一項に記載のマトリックスを含む、組織工学のための組成物。
【請求項61】
組成物を改変する方法であって、
架橋可能な官能基を有する改変酵素と、改変酵素によって架橋可能な少なくとも1種の部分を有するタンパク質とを提供すること;および
改変酵素とタンパク質を混合して、改変酵素によりタンパク質を架橋し、かつタンパク質により改変酵素自体を架橋可能な官能基において架橋すること
を含む方法。
【図1】

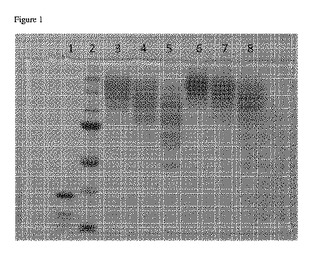
【図2】
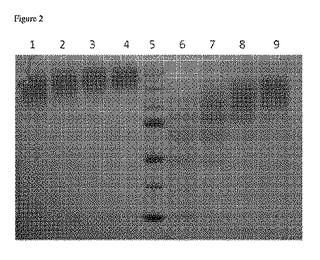
【図3】

【図11】
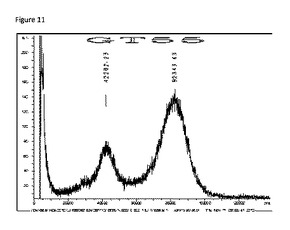
【図4】
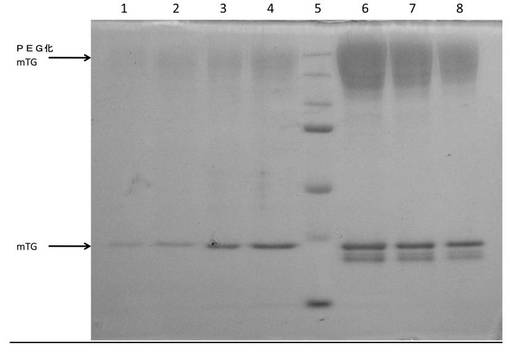
【図5】
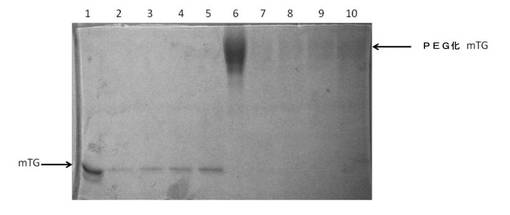
【図6】
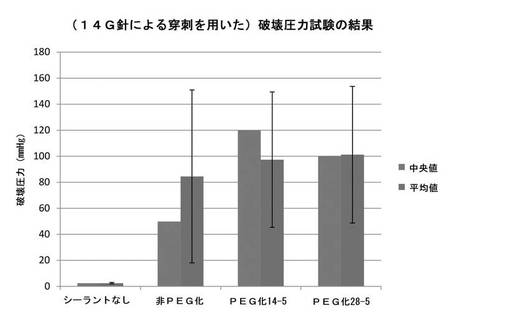
【図7】
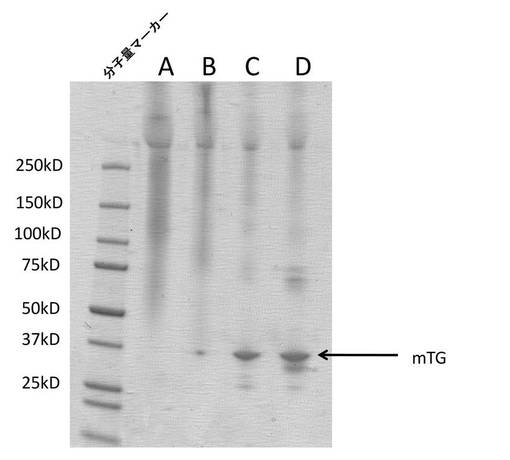
【図8】

【図9】
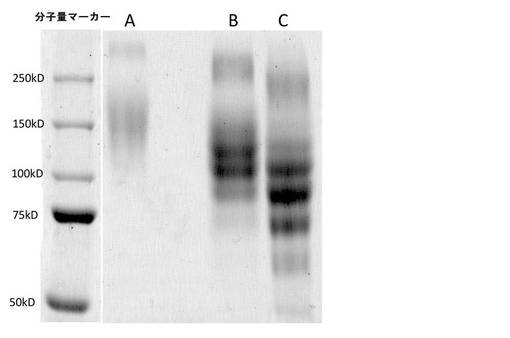
【図10】
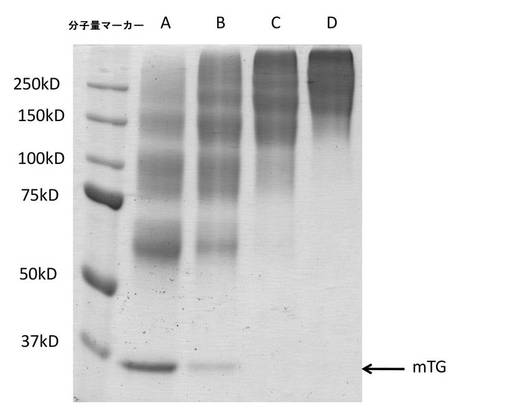
【図12】

【図2】
【図3】
【図11】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図12】
【公表番号】特表2013−515128(P2013−515128A)
【公表日】平成25年5月2日(2013.5.2)
【国際特許分類】
【出願番号】特願2012−545517(P2012−545517)
【出願日】平成22年12月22日(2010.12.22)
【国際出願番号】PCT/IB2010/056008
【国際公開番号】WO2011/077388
【国際公開日】平成23年6月30日(2011.6.30)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.サランラップ
2.テフロン
【出願人】(510332981)ライフボンド リミテッド (2)
【氏名又は名称原語表記】LIFEBOND LTD
【Fターム(参考)】
【公表日】平成25年5月2日(2013.5.2)
【国際特許分類】
【出願日】平成22年12月22日(2010.12.22)
【国際出願番号】PCT/IB2010/056008
【国際公開番号】WO2011/077388
【国際公開日】平成23年6月30日(2011.6.30)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.サランラップ
2.テフロン
【出願人】(510332981)ライフボンド リミテッド (2)
【氏名又は名称原語表記】LIFEBOND LTD
【Fターム(参考)】
[ Back to top ]