
核内ノンコーディングRNAの機能解析方法
【課題】核内に存在するncRNAを破壊することにより、該RNA分子の機能解析を行う方法を提供する。
【解決手段】標的ncRNAの二次構造における一本鎖領域に相補的な配列と実質的に同一の配列を含むアンチセンスオリゴ分子を細胞核内に導入して、該RNA分子を破壊することを含む、標的ncRNAの機能解析方法。
【解決手段】標的ncRNAの二次構造における一本鎖領域に相補的な配列と実質的に同一の配列を含むアンチセンスオリゴ分子を細胞核内に導入して、該RNA分子を破壊することを含む、標的ncRNAの機能解析方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明はノンコーディングRNAの機能を解析する方法に関する。より詳細には、本発明は、細胞核内に存在するノンコーディングRNAの機能を解析する方法に関する。
【背景技術】
【0002】
細胞の標的遺伝子の発現阻害方法のひとつとして2本鎖RNAを該細胞に導入することによりその配列に相同性を持つmRNAの分解を促進し、結果的にmRNAの鋳型となる遺伝子の発現を阻害する方法(以下、「RNAi法」とも称する)がある。細胞において発現している多くの遺伝子は未だその機能が未知であるものが多く、遺伝子発現阻害は遺伝子の機能解析において重要な方法である(非特許文献1)。
【0003】
またRNAi法以前から用いられている遺伝子発現抑制方法として1本鎖のRNAを用いるアンチセンスRNA法やRNAとRNase耐性を有するヌクレオチドとのハイブリッド分子を用いるキメラオリゴ法が知られているが、これらの手法はキメラオリゴ分子の導入が細胞毒性を誘導すること(非特許文献2)や遺伝子破壊がより効率的なRNAi法の出現により利用されなくなっている。
【0004】
従来、遺伝子は主としてタンパク質の設計図であるとの認識から、タンパク質に翻訳されるmRNAを標的として機能解析技術が開発されていた。しかし、ヒトを含む多くの生物のゲノム解析や発現遺伝子解析の進展によりタンパク質に翻訳されないRNA分子(以下、「ncRNA」: non-coding RNAと称する)が非常に多く存在すること、これらのncRNAが細胞の遺伝子発現調節に関わることにより、細胞の分化や状態の維持に重要な役割を果たしていることが理解されてきた。
【0005】
これらのncRNA中で、細胞質に分泌されるタンパク質翻訳型のmRNAと異なり、細胞核の中で機能する分子が多数存在することが知られており、これらのncRNAが遺伝子の転写制御やRNA分子の成熟過程に関与することを示す文献や特定の疾患に関わることを示す報告がされているが、まだそのほとんどの分子は機能未知である。これら核内で機能するRNA分子の機能解析は生物の生体反応を理解する上で重要な標的分子であるが、現在までにこれらの効率的な破壊方法は報告されていない。
【0006】
【非特許文献1】Fire et al., Nature. 391:806-811, 1998
【非特許文献2】Drygin et al., Nucleic Acids Res., 32: 6585-6594, 2004
【発明の開示】
【発明が解決しようとする課題】
【0007】
従来の遺伝子機能解析は、タンパク質に翻訳されるmRNA、すなわち細胞質に分泌されるRNAをターゲットとして開発されている。例えば、近年頻繁に利用されているRNAi法は、細胞質に存在するRNA分解機構を利用したRNA破壊を用いて遺伝子発現を抑制する。
【0008】
一方、核内に存在するRNA分子に対しては、細胞質に存在するRNA分解機構が利用できないため、従来のRNAi法等では核内RNA分子の破壊による機能解析を実施し得ない。
【0009】
そこで本発明は、核内に存在するncRNAを破壊することにより、該分子の機能を解析する方法を提供することを目的とする。
【課題を解決するための手段】
【0010】
本発明者らは、鋭意検討を重ねた結果、核内に存在する標的ncRNAに対するアンチセンスオリゴ分子を細胞核内に導入することにより、該RNA分子を破壊できることを見出し、本発明を完成させるに至った。
【0011】
すなわち、本発明は以下の特徴を包含する:
(1) 細胞核内に存在するノンコーディングRNA(non-coding RNA)の機能を解析する方法であって、該RNA分子に対するアンチセンスオリゴ分子を細胞核内に導入して、該RNA分子を破壊することを含む、前記方法。
【0012】
(2) 前記アンチセンスオリゴ分子は、前記RNA分子の二次構造における一本鎖領域に相補的な配列と少なくとも80%の同一性を有する塩基配列を含むことを特徴とする、上記(1)記載の方法。
【0013】
(3) 前記アンチセンスオリゴ分子は、前記RNA分子の二次構造における一本鎖領域に相補的な塩基配列を含むことを特徴とする上記(2)記載の方法。
【0014】
(4) 前記一本鎖領域は、前記RNA分子の二次構造解析により予め推定された領域であることを特徴とする上記(2)記載の方法。
【0015】
(5) 前記一本鎖領域は、前記RNA分子に特異的な配列領域であることを特徴とする上記(2)記載の方法。
【0016】
(6) 前記アンチセンスオリゴ分子は、核酸類似物質が導入されたキメラアンチセンスオリゴ分子であることを特徴とする上記(1)〜(5)のいずれか記載の方法。
【0017】
(7) 核酸類似物質は、2'-O-メチルリボヌクレオチドであることを特徴とする上記(6)記載の方法。
【0018】
(8) 前記アンチセンスオリゴ分子の細胞核内への導入をエレクトロポレーション法により実施することを特徴とする上記(1)〜(7)のいずれか記載の方法。
【発明の効果】
【0019】
本発明によれば、核内に存在する標的ncRNAを破壊することができ、該RNA分子を機能解析することができる。
【発明を実施するための最良の形態】
【0020】
以下、本発明について詳細に説明する。
本発明は、核内に存在するncRNAの機能を解析する方法に関する。より具体的に、本発明の方法は、細胞核内に存在する標的ncRNAを破壊することによって、該RNA分子を機能解析することを含む。ここで、ncRNAの機能解析としては、該RNA分子の機能を推定・同定すること等が挙げられる。
【0021】
本発明の方法において、細胞核内に存在する標的ncRNAの破壊は、該RNA分子に対するアンチセンスオリゴ分子を細胞核内に導入することにより行う。
【0022】
ここで、本明細書で使用する「細胞核内に存在するncRNA」又は「標的ncRNA」とは、タンパク質に翻訳されないncRNAのうち、細胞核の中で機能するRNA分子を指し、該分子が本発明のアンチセンスオリゴ分子と二重鎖を形成する一本鎖領域を構成するものであれば特に制限されない。換言すると、本発明に係る機能解析方法で使用するアンチセンスオリゴ分子は、標的ncRNAの二次構造における一本鎖領域に相補的な配列と実質的に同一の塩基配列を含む。
【0023】
本明細書で使用する「実質的に同一」とは、標的ncRNAの二次構造における一本鎖領域に相補的な配列と、少なくとも80%、好ましくは少なくとも85%、より好ましくは少なくとも90%、より好ましくは少なくとも95%、最も好ましくは少なくとも97%、98%又は99%の同一性を有することを指す。なお、本明細書において「同一性」の値は、複数の塩基配列間の同一性を演算するソフトウェア(例えばBLASTP、BLASTN、FASTAなど)をデフォルトの設定で使用することにより算出した値を意味する。本発明のアンチセンスオリゴ分子が、前記一本鎖領域に相補的な配列を含むことが最も好ましい。
【0024】
本発明に用いるアンチセンスオリゴ分子において、標的ncRNAとの結合領域であるアンチセンス領域を構成する塩基数は特に制限されないが、該領域を構成する塩基数が少ないと標的ncRNAと二重鎖を形成することが困難になり、多すぎると非標的配列との非特異的結合が生じる点に留意すべきことは言うまでもない。最適なアンチセンス領域の長さは、当業者に公知であるか又は当業者が適宜設定することができ、これに限定されるものではないが、例えば10〜40塩基長、好ましくは15〜27塩基長、例えば20塩基長である。また上記アンチセンス領域は、アンチセンスオリゴ分子内において連続して存在する必要はない。すなわち上記アンチセンス領域は、標的ncRNAと二重鎖を形成することができる限り、1個又は数個の非相補的塩基を1又は数箇所に介在させてもよい。
【0025】
かかるアンチセンスオリゴ分子は、標的ncRNAに特異的な配列領域として予め同定された領域と相補的な配列と実質的に又は完全に同一の配列を含むように設計することができる。これにより、本発明のアンチセンスオリゴ分子による標的ncRNAの特異的な破壊の精度を向上させることができる。このような特異的配列領域は、例えば設計対象とする配列領域を遺伝子データベースに登録されている当該生物のゲノム情報と比較することにより決定することができる。
【0026】
また本発明のアンチセンスオリゴ分子による標的ncRNAの特異的な破壊の精度を向上させるその他の方法として、これに限定されるものではないが、例えば特表2003-500064号などに開示される手法を挙げることができ、本発明においてこれらの手法を使用することもできる。
【0027】
本発明のアンチセンスオリゴ分子は、当業者に周知の方法、例えば適当な配列のクローニング及び制限酵素による切断、ホスホトリエステル法(例えばNarangら,1979年,Meth.Enzymol.,第68巻,p90〜99参照)、ホスホジエステル法(例えばBrownら,1979年、Meth.Enzymol.,第68巻,p109〜151参照)、エチルホスホアミダイト法(例えばBeaucageら,1981年,Tetrahedron Lett.,第22巻,p1859〜1862参照)などの方法により、直接的に合成することができる。また市販の自動DNA合成装置を使用することによって合成してもよい。その際、機能解析の標的とするncRNAの塩基配列に基づいて、該ncRNAの二次構造における一本鎖領域を予め推定しておくことが好ましい。かかる二次構造解析は、これに限定されるものではないが、例えばRNAfold、RNAz、MFOLDなど、当業者に公知の二次構造予測プログラムを使用することによって行うことができる。
【0028】
本発明のアンチセンスオリゴ分子は、核酸類似物質の導入、ヘアピンループ構造を形成する塩基の導入等により、核内に存在する核酸分解酵素に対する耐性が付与されたものであることが好ましい。これにより、核内に存在する種々の核酸分解酵素による分解により本発明のアンチセンスオリゴ分子が標的ncRNAと結合する前に消失してしまうのを回避し、より効率的に標的ncRNAを破壊することができる。
【0029】
本発明のアンチセンスオリゴ分子は、核酸類似物質の導入により核酸分解酵素に対する耐性が付与されたキメラアンチセンスオリゴ分子であることが好ましい。具体的に、本発明のキメラアンチセンスオリゴ分子とは、アンチセンスオリゴ分子内の塩基の一部が核酸類似物質で置き換えられたものを指す。
【0030】
本発明に使用できる核酸類似物質として、これに限定されるものではないが、例えば2’-O-メチルリボヌクレオチド、モルフォリノオリゴ、2’-4’-BNA、Locked nucleic acid (LNA)などを挙げることができる。本発明では、核酸類似物質として2'-O-メチルリボヌクレオチドを使用することが好ましい。
【0031】
本発明のキメラアンチセンスオリゴ分子は当業者に公知の方法を用いて合成することができる。例えば、核酸類似物質として2’-O-メチルリボヌクレオチドを使用する場合には、5'-ジメトキシトリチル-2’-O-メチルリボヌクレオシド-3’-〔(2-シアノエチル)-(N,N-ジイソプロピル)〕-ホスホロアミダイトユニットを用いて、ホスホロアミダイト法によるDNA/RNA自動合成機を利用して目的のキメラアンチセンスオリゴ分子を合成することができる。
【0032】
核酸類似物質を導入するアンチセンスオリゴ分子内の塩基の最適な位置及び数は、アンチセンスオリゴ分子の全長及び配列に依存する二次構造やGC含量などによって変化するものであるが、標的ncRNAとの二重鎖形成前の分解を回避でき、かつ標的ncRNAとの二重鎖形成後に核酸分解酵素による分解が生じる限り特に限定されず、当業者は適宜導入塩基の位置及び数を決定することができる。本発明のキメラアンチセンスオリゴ分子の一例として、図1にその両末端の各5塩基に核酸類似物質を導入した20塩基長のアンチセンス領域からなる分子を示す。図中、Xは標的とする遺伝子に応じて設計される任意の塩基を表し、両末端各5塩基に核酸分解酵素に対する抵抗性付与を目的として2’-O-メチルリボヌクレオチドを用いている。かかる構造を有する例示の分子は、下記の実施例において、特定の標的ncRNAを特異的に破壊する分子として有効であることが示されている。
【0033】
上記のようにして合成した本発明のアンチセンスオリゴ分子を細胞核内に導入すると、核内に存在する標的ncRNAと該アンチセンスオリゴ分子とが二重鎖を形成し、核内に存在する核酸分解酵素(例えばRNAse)によって該二重鎖が分解されることで、標的ncRNAを破壊することができる。
【0034】
本発明のアンチセンスオリゴ分子の導入の対象となる細胞としては、これに限定されるものではないが、例えば動物(例えばヒト)又は植物由来の生細胞、或いはそれらの培養細胞などを挙げることができ、該細胞が由来する器官、組織等も制限されない。
【0035】
本発明のアンチセンスオリゴ分子の細胞核内への導入は、核酸分子を細胞内に導入することができる当業者に公知の方法を用いて実施することができる。かかる方法として、これに限定されるものではないが、例えばエレクトロポレーション法、リポフェクション法、リン酸カルシウム法、マイクロインジェクション法、ウイルスベクター法等を挙げることができる。
【0036】
アンチセンスオリゴ分子の細胞核内への導入を、エレクトロポレーション法を用いて実施する(本明細書中、「ヌクレオフェクション」ともいう)ことが特に好ましい。エレクトロポレーション法は、高電圧パルスで一時的に脂質二重層の細胞膜構造を不安定化して孔を生じさせ、そこから核酸分子を取り込ませる方法であり、一般的に、導入効率に優れ、導入が困難な細胞にも導入が可能であるという利点を有する。本発明においても、エレクトロポレーション法を用いた細胞へのアンチセンスオリゴ分子の導入は、細胞種を問わず核内標的ncRNAの高い破壊効率を達成していることから(例えば図5、図7参照)、アンチセンスオリゴ分子の核内への効率的な導入にエレクトロポレーションが有効であることが示されている。またエレクトロポレーションによるアンチセンスオリゴ分子の導入は、導入後の高い細胞生存率をも実現しており(下記実施例参照)、細胞毒性の観点からも好ましい手法であることが示されている。またエレクトロポレーションに採用されるべき電圧、パルスの長さは、標的細胞の種類、培養条件、エレクトロポレーションに用いる溶液の組成などに応じて当業者が適宜設定することができ、これに限定されるものではないが、それぞれ500〜800V/cm、10〜25ミリ秒、例えば550V/cm、25ミリ秒で行うことができる。本発明において、エレクトロポレーションに使用する培養条件、エレクトロポレーション用溶液の組成は、細胞内への核酸の導入の際に当業者に一般的に使用されているものを用いればよく、特別な条件及び組成を用いる必要はない。
【0037】
本発明において、エレクトロポレーションに当業者に公知の核酸導入装置を使用することができる。かかる核酸導入装置として、これに限定されるものではないが、例えばヌクレオフェクター(アマクサ社製)、ジーンパルサー(バイオラッド社製)などを使用することができる。
【0038】
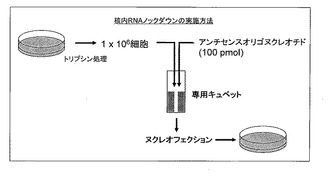
図2に、エレクトロポレーションの原理を利用する核酸導入装置を用いた、培養細胞に対する核酸分子導入方法の概略図を示す。培養細胞が接着性を有する場合はトリプシン処理等によって細胞を浮遊化したのち、核酸導入装置を用いて本発明のアンチセンスオリゴ分子を細胞に導入し、その後、導入処理を行った細胞を細胞種に適した培地上で培養して、標的ncRNAの機能解析を行う。
【0039】
標的ncRNAの機能は、本発明のアンチセンスオリゴ分子の細胞核内への導入後、例えば導入していない細胞に比較して発現量が増加した遺伝子を同定すること、該細胞に現れる表現形の変化を解析すること、顕微鏡等を用いて細胞の構造等を観察すること等によって同定・推定することができる。
【0040】
以下、実施例により本発明を詳細に説明するが、本発明の範囲はこの実施例に限定されるものではない。
【実施例】
【0041】
(1)アンチセンスオリゴ分子の設計と合成
核内で機能するncRNAであるU7 RNAを破壊の標的分子とした。U7 RNAの塩基配列情報を用いて該RNA分子の二次構造解析をMFOLDを用いて行い、機能部位である1本鎖領域を算出した。U7 RNAの1本鎖領域に対して相補的な20塩基のアンチセンスオリゴ配列を設計した。このアンチセンスオリゴ配列に基づいて、細胞内における非特異的な分解に抵抗性を持たせる目的で両末端の各5塩基に2’-O-メチルリボヌクレオチドを持つキメラアンチセンスオリゴ核酸を化学合成した。合成したオリゴ配列は以下の通りである:
U7用キメラオリゴ:5'-mUmUmCmUmAAAAGAGCTGTmAmAmCmAmC-3'
次にU7 RNA同様に核内で機能するncRNAとして知られるU84 RNA、及び対照実験用としてヒトには存在しないGFP RNAに対しても同様にキメラアンチセンスオリゴ核酸の合成を行った。合成したキメラアンチセンスオリゴ核酸の配列を以下に示す:
U84用キメラオリゴ:5'-mCmAmAmGmGGTGATAGATGmAmGmGmGmU-3'
GFP用キメラオリゴ:5'-mUmCmAmCmCTTCACCCTCTmCmCmAmCmU-3'
【0042】
(2)U7 RNAの破壊実験
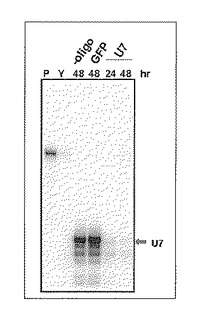
培養して得たヒト由来の培養細胞であるHeLa細胞にトリプシン処理を行って細胞を浮遊化し、約百万個の細胞群を得た。これらの細胞群に対して合成したU7 RNAに相補的な塩基配列を有するキメラアンチセンスオリゴ核酸を、上記非特許文献2に記載の遺伝子破壊に必要なオリゴヌクレオチドの50%有効量(2.2μM)より低濃度の1μMの濃度で添加し、エレクトロポレーション法を用いた核酸導入装置であるヌクレオフェクター(アマクサ社製)を説明書にしたがって使用し核酸導入を行った。核酸導入を行った細胞を引き続いて培地上で培養を行い、これらの細胞中に存在するU7 RNA分子の量をRNaseプロテクションアッセイ法(例えばHirose & Steitz, PNAS 98: 12914-12919, 2001参照)によって観察した。対照実験として行ったキメラオリゴを加えなかった実験区(-oligo)及びGFP用キメラオリゴを加えた実験区では核酸導入処理後48時間経過した時点においてU7 RNAの存在が確認された。しかし、U7 RNA用キメラオリゴを加えた実験区では培養時間24時間及び48時間のいずれの時間においてもU7 RNAの顕著な減少が確認された(図3)。本結果から本発明者らの合成したU7用キメラオリゴをヌクレオフェクターによってHeLa細胞に導入することにより、低濃度のキメラオリゴ分子でも効率的にU7 RNAの破壊が誘導されることが明らかになった。また本RNaseプロテクションアッセイ法でU7 RNAの顕著な減少が観察された事実から、本発明者らの開発したncRNAの破壊方法によって、細胞内に存在する標的ncRNAが破壊されること、また標的ncRNA破壊処理を行った細胞群の全体にその効果が見られることが確認された。
【0043】
(3)U7 RNAの機能解析実験
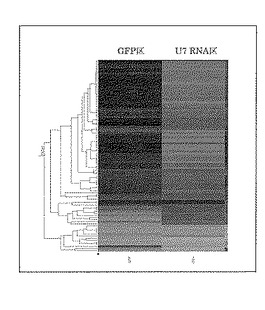
機能解析手法として標的ncRNAの破壊が細胞の遺伝子発現に与える影響を解析する目的でマイクロアレイを用いた遺伝子発現解析を行った。U7用キメラオリゴの導入によってU7 RNAの破壊を行ったHeLa細胞より全RNAを抽出精製してAgilent社製のマイクロアレイ(44k Whole Human Genome Oligo Microarray)を用いて遺伝子発現解析を行い、対象実験としてGFP用キメラオリゴの導入を行ったHeLa細胞の発現遺伝子と比較し、U7 RNAの破壊の影響を観察した。解析を行ったヒト遺伝子22,833遺伝子中、U7 RNAの遺伝子破壊によって細胞内に遺伝子量に2倍以上の変化が観察された遺伝子は93遺伝子であった(図4)。この93遺伝子の内50遺伝子は細胞内においてヒストン遺伝子の合成に関連がある遺伝子群であったことから、U7 RNAは核内においてヒストン遺伝子合成に関連する機能を有することが推察された。
【0044】
(4)核酸導入方法の比較
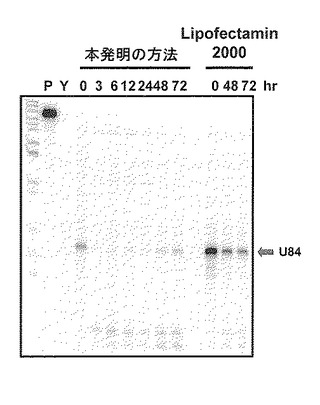
核酸導入方法の比較として、既知の遺伝子導入方法であるエレクトロポレーション法とリポフェクタミン法とを比較した。標的ncRNA分子としてU84分子を選択し、先に合成したU84用キメラオリゴを用いて実験を行った。エレクトロポレーション法を用いた方法は標的ncRNA分子及び使用したキメラオリゴを除き、上記(2)と同様に行った。リポフェクタミンを用いた方法は、Invitogen社製Lipofectamine2000試薬をマニュアルに従いキメラオリゴと混合したものを細胞培養液に加えることにより行った。エレクトロポレーション法を用いた方法では核酸導入処理後3時間から72時間に至るまでU84分子が破壊されていることがRNaseプロテクションアッセイ法で確認されたが、リポフェクタミン法を用いた方法では十分なU84分子破壊が観察されなかった(図5)。また、エレクトロポレーション法を用いた方法では核酸導入処理後の培養において細胞はほぼ100%の生存率が観察されたが、リポフェクタミン法を用いた方法では一部の細胞が死滅する減少が観察された。この比較により、アンチセンスオリゴ分子の導入におけるエレクトロポレーションの使用は、細胞毒性が低く、効率的な標的分子の破壊を実現する有効な方法であることがわかった。
【0045】
(5)導入する核酸構造の比較
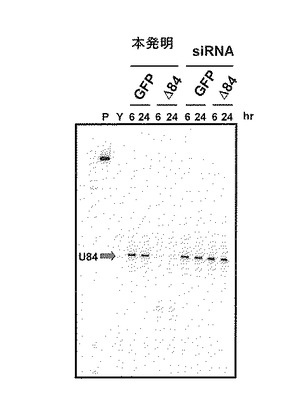
遺伝子破壊を目的とする実験において多用される2重鎖RNA分子(siRNA)を比較対象として、ncRNA破壊効率の比較を行った。なお、本実験は、核内への導入に使用した核酸分子を除き、上記(4)と同様に行った。siRNAは細胞質に存在する遺伝子を標的としたRNAの破壊に効果的であるが、本実験の標的としたU84 RNAでは破壊の誘導が観察されなかった(図6)。本結果からキメラアンチセンスオリゴの利用が効果的であることが確認された。
【0046】
(6)種々の培養細胞を用いたncRNA破壊実験
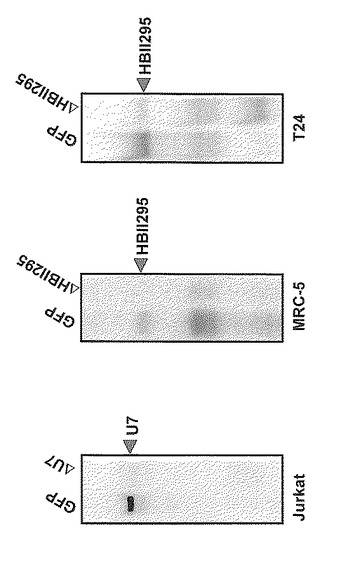
本発明のncRNA破壊方法をT細胞白血病由来のJurkat細胞、肺がん由来のMCR5、膀胱ガン由来のT24細胞に対して実施し、破壊の効果を検証した。Jurkat細胞ではU7 RNA、その他の細胞種ではHBII295 RNAを破壊の標的分子とし、上記(2)と同様の方法で行った。この検証実験により、本発明の方法は細胞種と標的分子とを限定することなく用いることが可能であることが確認された(図7)。
【図面の簡単な説明】
【0047】
【図1】図1は、本発明に用いるキメラアンチセンスオリゴ分子の構造の一例を示す。
【図2】図2は、エレクトロポレーションの原理を利用する核酸導入装置を用いた、核酸分子導入方法の概略図を示す。
【図3】図3は、U7 RNA分子破壊実験の結果を示す。
【図4】図4は、U7 RNA分子の破壊が及ぼす影響の解析結果を示す。
【図5】図5は、核酸導入方法による効果の比較を示す。
【図6】図6は、導入する核酸構造による効果の比較を示す。
【図7】図7は、種々の細胞種に対するncRNA破壊の検証結果を示す。
【技術分野】
【0001】
本発明はノンコーディングRNAの機能を解析する方法に関する。より詳細には、本発明は、細胞核内に存在するノンコーディングRNAの機能を解析する方法に関する。
【背景技術】
【0002】
細胞の標的遺伝子の発現阻害方法のひとつとして2本鎖RNAを該細胞に導入することによりその配列に相同性を持つmRNAの分解を促進し、結果的にmRNAの鋳型となる遺伝子の発現を阻害する方法(以下、「RNAi法」とも称する)がある。細胞において発現している多くの遺伝子は未だその機能が未知であるものが多く、遺伝子発現阻害は遺伝子の機能解析において重要な方法である(非特許文献1)。
【0003】
またRNAi法以前から用いられている遺伝子発現抑制方法として1本鎖のRNAを用いるアンチセンスRNA法やRNAとRNase耐性を有するヌクレオチドとのハイブリッド分子を用いるキメラオリゴ法が知られているが、これらの手法はキメラオリゴ分子の導入が細胞毒性を誘導すること(非特許文献2)や遺伝子破壊がより効率的なRNAi法の出現により利用されなくなっている。
【0004】
従来、遺伝子は主としてタンパク質の設計図であるとの認識から、タンパク質に翻訳されるmRNAを標的として機能解析技術が開発されていた。しかし、ヒトを含む多くの生物のゲノム解析や発現遺伝子解析の進展によりタンパク質に翻訳されないRNA分子(以下、「ncRNA」: non-coding RNAと称する)が非常に多く存在すること、これらのncRNAが細胞の遺伝子発現調節に関わることにより、細胞の分化や状態の維持に重要な役割を果たしていることが理解されてきた。
【0005】
これらのncRNA中で、細胞質に分泌されるタンパク質翻訳型のmRNAと異なり、細胞核の中で機能する分子が多数存在することが知られており、これらのncRNAが遺伝子の転写制御やRNA分子の成熟過程に関与することを示す文献や特定の疾患に関わることを示す報告がされているが、まだそのほとんどの分子は機能未知である。これら核内で機能するRNA分子の機能解析は生物の生体反応を理解する上で重要な標的分子であるが、現在までにこれらの効率的な破壊方法は報告されていない。
【0006】
【非特許文献1】Fire et al., Nature. 391:806-811, 1998
【非特許文献2】Drygin et al., Nucleic Acids Res., 32: 6585-6594, 2004
【発明の開示】
【発明が解決しようとする課題】
【0007】
従来の遺伝子機能解析は、タンパク質に翻訳されるmRNA、すなわち細胞質に分泌されるRNAをターゲットとして開発されている。例えば、近年頻繁に利用されているRNAi法は、細胞質に存在するRNA分解機構を利用したRNA破壊を用いて遺伝子発現を抑制する。
【0008】
一方、核内に存在するRNA分子に対しては、細胞質に存在するRNA分解機構が利用できないため、従来のRNAi法等では核内RNA分子の破壊による機能解析を実施し得ない。
【0009】
そこで本発明は、核内に存在するncRNAを破壊することにより、該分子の機能を解析する方法を提供することを目的とする。
【課題を解決するための手段】
【0010】
本発明者らは、鋭意検討を重ねた結果、核内に存在する標的ncRNAに対するアンチセンスオリゴ分子を細胞核内に導入することにより、該RNA分子を破壊できることを見出し、本発明を完成させるに至った。
【0011】
すなわち、本発明は以下の特徴を包含する:
(1) 細胞核内に存在するノンコーディングRNA(non-coding RNA)の機能を解析する方法であって、該RNA分子に対するアンチセンスオリゴ分子を細胞核内に導入して、該RNA分子を破壊することを含む、前記方法。
【0012】
(2) 前記アンチセンスオリゴ分子は、前記RNA分子の二次構造における一本鎖領域に相補的な配列と少なくとも80%の同一性を有する塩基配列を含むことを特徴とする、上記(1)記載の方法。
【0013】
(3) 前記アンチセンスオリゴ分子は、前記RNA分子の二次構造における一本鎖領域に相補的な塩基配列を含むことを特徴とする上記(2)記載の方法。
【0014】
(4) 前記一本鎖領域は、前記RNA分子の二次構造解析により予め推定された領域であることを特徴とする上記(2)記載の方法。
【0015】
(5) 前記一本鎖領域は、前記RNA分子に特異的な配列領域であることを特徴とする上記(2)記載の方法。
【0016】
(6) 前記アンチセンスオリゴ分子は、核酸類似物質が導入されたキメラアンチセンスオリゴ分子であることを特徴とする上記(1)〜(5)のいずれか記載の方法。
【0017】
(7) 核酸類似物質は、2'-O-メチルリボヌクレオチドであることを特徴とする上記(6)記載の方法。
【0018】
(8) 前記アンチセンスオリゴ分子の細胞核内への導入をエレクトロポレーション法により実施することを特徴とする上記(1)〜(7)のいずれか記載の方法。
【発明の効果】
【0019】
本発明によれば、核内に存在する標的ncRNAを破壊することができ、該RNA分子を機能解析することができる。
【発明を実施するための最良の形態】
【0020】
以下、本発明について詳細に説明する。
本発明は、核内に存在するncRNAの機能を解析する方法に関する。より具体的に、本発明の方法は、細胞核内に存在する標的ncRNAを破壊することによって、該RNA分子を機能解析することを含む。ここで、ncRNAの機能解析としては、該RNA分子の機能を推定・同定すること等が挙げられる。
【0021】
本発明の方法において、細胞核内に存在する標的ncRNAの破壊は、該RNA分子に対するアンチセンスオリゴ分子を細胞核内に導入することにより行う。
【0022】
ここで、本明細書で使用する「細胞核内に存在するncRNA」又は「標的ncRNA」とは、タンパク質に翻訳されないncRNAのうち、細胞核の中で機能するRNA分子を指し、該分子が本発明のアンチセンスオリゴ分子と二重鎖を形成する一本鎖領域を構成するものであれば特に制限されない。換言すると、本発明に係る機能解析方法で使用するアンチセンスオリゴ分子は、標的ncRNAの二次構造における一本鎖領域に相補的な配列と実質的に同一の塩基配列を含む。
【0023】
本明細書で使用する「実質的に同一」とは、標的ncRNAの二次構造における一本鎖領域に相補的な配列と、少なくとも80%、好ましくは少なくとも85%、より好ましくは少なくとも90%、より好ましくは少なくとも95%、最も好ましくは少なくとも97%、98%又は99%の同一性を有することを指す。なお、本明細書において「同一性」の値は、複数の塩基配列間の同一性を演算するソフトウェア(例えばBLASTP、BLASTN、FASTAなど)をデフォルトの設定で使用することにより算出した値を意味する。本発明のアンチセンスオリゴ分子が、前記一本鎖領域に相補的な配列を含むことが最も好ましい。
【0024】
本発明に用いるアンチセンスオリゴ分子において、標的ncRNAとの結合領域であるアンチセンス領域を構成する塩基数は特に制限されないが、該領域を構成する塩基数が少ないと標的ncRNAと二重鎖を形成することが困難になり、多すぎると非標的配列との非特異的結合が生じる点に留意すべきことは言うまでもない。最適なアンチセンス領域の長さは、当業者に公知であるか又は当業者が適宜設定することができ、これに限定されるものではないが、例えば10〜40塩基長、好ましくは15〜27塩基長、例えば20塩基長である。また上記アンチセンス領域は、アンチセンスオリゴ分子内において連続して存在する必要はない。すなわち上記アンチセンス領域は、標的ncRNAと二重鎖を形成することができる限り、1個又は数個の非相補的塩基を1又は数箇所に介在させてもよい。
【0025】
かかるアンチセンスオリゴ分子は、標的ncRNAに特異的な配列領域として予め同定された領域と相補的な配列と実質的に又は完全に同一の配列を含むように設計することができる。これにより、本発明のアンチセンスオリゴ分子による標的ncRNAの特異的な破壊の精度を向上させることができる。このような特異的配列領域は、例えば設計対象とする配列領域を遺伝子データベースに登録されている当該生物のゲノム情報と比較することにより決定することができる。
【0026】
また本発明のアンチセンスオリゴ分子による標的ncRNAの特異的な破壊の精度を向上させるその他の方法として、これに限定されるものではないが、例えば特表2003-500064号などに開示される手法を挙げることができ、本発明においてこれらの手法を使用することもできる。
【0027】
本発明のアンチセンスオリゴ分子は、当業者に周知の方法、例えば適当な配列のクローニング及び制限酵素による切断、ホスホトリエステル法(例えばNarangら,1979年,Meth.Enzymol.,第68巻,p90〜99参照)、ホスホジエステル法(例えばBrownら,1979年、Meth.Enzymol.,第68巻,p109〜151参照)、エチルホスホアミダイト法(例えばBeaucageら,1981年,Tetrahedron Lett.,第22巻,p1859〜1862参照)などの方法により、直接的に合成することができる。また市販の自動DNA合成装置を使用することによって合成してもよい。その際、機能解析の標的とするncRNAの塩基配列に基づいて、該ncRNAの二次構造における一本鎖領域を予め推定しておくことが好ましい。かかる二次構造解析は、これに限定されるものではないが、例えばRNAfold、RNAz、MFOLDなど、当業者に公知の二次構造予測プログラムを使用することによって行うことができる。
【0028】
本発明のアンチセンスオリゴ分子は、核酸類似物質の導入、ヘアピンループ構造を形成する塩基の導入等により、核内に存在する核酸分解酵素に対する耐性が付与されたものであることが好ましい。これにより、核内に存在する種々の核酸分解酵素による分解により本発明のアンチセンスオリゴ分子が標的ncRNAと結合する前に消失してしまうのを回避し、より効率的に標的ncRNAを破壊することができる。
【0029】
本発明のアンチセンスオリゴ分子は、核酸類似物質の導入により核酸分解酵素に対する耐性が付与されたキメラアンチセンスオリゴ分子であることが好ましい。具体的に、本発明のキメラアンチセンスオリゴ分子とは、アンチセンスオリゴ分子内の塩基の一部が核酸類似物質で置き換えられたものを指す。
【0030】
本発明に使用できる核酸類似物質として、これに限定されるものではないが、例えば2’-O-メチルリボヌクレオチド、モルフォリノオリゴ、2’-4’-BNA、Locked nucleic acid (LNA)などを挙げることができる。本発明では、核酸類似物質として2'-O-メチルリボヌクレオチドを使用することが好ましい。
【0031】
本発明のキメラアンチセンスオリゴ分子は当業者に公知の方法を用いて合成することができる。例えば、核酸類似物質として2’-O-メチルリボヌクレオチドを使用する場合には、5'-ジメトキシトリチル-2’-O-メチルリボヌクレオシド-3’-〔(2-シアノエチル)-(N,N-ジイソプロピル)〕-ホスホロアミダイトユニットを用いて、ホスホロアミダイト法によるDNA/RNA自動合成機を利用して目的のキメラアンチセンスオリゴ分子を合成することができる。
【0032】
核酸類似物質を導入するアンチセンスオリゴ分子内の塩基の最適な位置及び数は、アンチセンスオリゴ分子の全長及び配列に依存する二次構造やGC含量などによって変化するものであるが、標的ncRNAとの二重鎖形成前の分解を回避でき、かつ標的ncRNAとの二重鎖形成後に核酸分解酵素による分解が生じる限り特に限定されず、当業者は適宜導入塩基の位置及び数を決定することができる。本発明のキメラアンチセンスオリゴ分子の一例として、図1にその両末端の各5塩基に核酸類似物質を導入した20塩基長のアンチセンス領域からなる分子を示す。図中、Xは標的とする遺伝子に応じて設計される任意の塩基を表し、両末端各5塩基に核酸分解酵素に対する抵抗性付与を目的として2’-O-メチルリボヌクレオチドを用いている。かかる構造を有する例示の分子は、下記の実施例において、特定の標的ncRNAを特異的に破壊する分子として有効であることが示されている。
【0033】
上記のようにして合成した本発明のアンチセンスオリゴ分子を細胞核内に導入すると、核内に存在する標的ncRNAと該アンチセンスオリゴ分子とが二重鎖を形成し、核内に存在する核酸分解酵素(例えばRNAse)によって該二重鎖が分解されることで、標的ncRNAを破壊することができる。
【0034】
本発明のアンチセンスオリゴ分子の導入の対象となる細胞としては、これに限定されるものではないが、例えば動物(例えばヒト)又は植物由来の生細胞、或いはそれらの培養細胞などを挙げることができ、該細胞が由来する器官、組織等も制限されない。
【0035】
本発明のアンチセンスオリゴ分子の細胞核内への導入は、核酸分子を細胞内に導入することができる当業者に公知の方法を用いて実施することができる。かかる方法として、これに限定されるものではないが、例えばエレクトロポレーション法、リポフェクション法、リン酸カルシウム法、マイクロインジェクション法、ウイルスベクター法等を挙げることができる。
【0036】
アンチセンスオリゴ分子の細胞核内への導入を、エレクトロポレーション法を用いて実施する(本明細書中、「ヌクレオフェクション」ともいう)ことが特に好ましい。エレクトロポレーション法は、高電圧パルスで一時的に脂質二重層の細胞膜構造を不安定化して孔を生じさせ、そこから核酸分子を取り込ませる方法であり、一般的に、導入効率に優れ、導入が困難な細胞にも導入が可能であるという利点を有する。本発明においても、エレクトロポレーション法を用いた細胞へのアンチセンスオリゴ分子の導入は、細胞種を問わず核内標的ncRNAの高い破壊効率を達成していることから(例えば図5、図7参照)、アンチセンスオリゴ分子の核内への効率的な導入にエレクトロポレーションが有効であることが示されている。またエレクトロポレーションによるアンチセンスオリゴ分子の導入は、導入後の高い細胞生存率をも実現しており(下記実施例参照)、細胞毒性の観点からも好ましい手法であることが示されている。またエレクトロポレーションに採用されるべき電圧、パルスの長さは、標的細胞の種類、培養条件、エレクトロポレーションに用いる溶液の組成などに応じて当業者が適宜設定することができ、これに限定されるものではないが、それぞれ500〜800V/cm、10〜25ミリ秒、例えば550V/cm、25ミリ秒で行うことができる。本発明において、エレクトロポレーションに使用する培養条件、エレクトロポレーション用溶液の組成は、細胞内への核酸の導入の際に当業者に一般的に使用されているものを用いればよく、特別な条件及び組成を用いる必要はない。
【0037】
本発明において、エレクトロポレーションに当業者に公知の核酸導入装置を使用することができる。かかる核酸導入装置として、これに限定されるものではないが、例えばヌクレオフェクター(アマクサ社製)、ジーンパルサー(バイオラッド社製)などを使用することができる。
【0038】
図2に、エレクトロポレーションの原理を利用する核酸導入装置を用いた、培養細胞に対する核酸分子導入方法の概略図を示す。培養細胞が接着性を有する場合はトリプシン処理等によって細胞を浮遊化したのち、核酸導入装置を用いて本発明のアンチセンスオリゴ分子を細胞に導入し、その後、導入処理を行った細胞を細胞種に適した培地上で培養して、標的ncRNAの機能解析を行う。
【0039】
標的ncRNAの機能は、本発明のアンチセンスオリゴ分子の細胞核内への導入後、例えば導入していない細胞に比較して発現量が増加した遺伝子を同定すること、該細胞に現れる表現形の変化を解析すること、顕微鏡等を用いて細胞の構造等を観察すること等によって同定・推定することができる。
【0040】
以下、実施例により本発明を詳細に説明するが、本発明の範囲はこの実施例に限定されるものではない。
【実施例】
【0041】
(1)アンチセンスオリゴ分子の設計と合成
核内で機能するncRNAであるU7 RNAを破壊の標的分子とした。U7 RNAの塩基配列情報を用いて該RNA分子の二次構造解析をMFOLDを用いて行い、機能部位である1本鎖領域を算出した。U7 RNAの1本鎖領域に対して相補的な20塩基のアンチセンスオリゴ配列を設計した。このアンチセンスオリゴ配列に基づいて、細胞内における非特異的な分解に抵抗性を持たせる目的で両末端の各5塩基に2’-O-メチルリボヌクレオチドを持つキメラアンチセンスオリゴ核酸を化学合成した。合成したオリゴ配列は以下の通りである:
U7用キメラオリゴ:5'-mUmUmCmUmAAAAGAGCTGTmAmAmCmAmC-3'
次にU7 RNA同様に核内で機能するncRNAとして知られるU84 RNA、及び対照実験用としてヒトには存在しないGFP RNAに対しても同様にキメラアンチセンスオリゴ核酸の合成を行った。合成したキメラアンチセンスオリゴ核酸の配列を以下に示す:
U84用キメラオリゴ:5'-mCmAmAmGmGGTGATAGATGmAmGmGmGmU-3'
GFP用キメラオリゴ:5'-mUmCmAmCmCTTCACCCTCTmCmCmAmCmU-3'
【0042】
(2)U7 RNAの破壊実験
培養して得たヒト由来の培養細胞であるHeLa細胞にトリプシン処理を行って細胞を浮遊化し、約百万個の細胞群を得た。これらの細胞群に対して合成したU7 RNAに相補的な塩基配列を有するキメラアンチセンスオリゴ核酸を、上記非特許文献2に記載の遺伝子破壊に必要なオリゴヌクレオチドの50%有効量(2.2μM)より低濃度の1μMの濃度で添加し、エレクトロポレーション法を用いた核酸導入装置であるヌクレオフェクター(アマクサ社製)を説明書にしたがって使用し核酸導入を行った。核酸導入を行った細胞を引き続いて培地上で培養を行い、これらの細胞中に存在するU7 RNA分子の量をRNaseプロテクションアッセイ法(例えばHirose & Steitz, PNAS 98: 12914-12919, 2001参照)によって観察した。対照実験として行ったキメラオリゴを加えなかった実験区(-oligo)及びGFP用キメラオリゴを加えた実験区では核酸導入処理後48時間経過した時点においてU7 RNAの存在が確認された。しかし、U7 RNA用キメラオリゴを加えた実験区では培養時間24時間及び48時間のいずれの時間においてもU7 RNAの顕著な減少が確認された(図3)。本結果から本発明者らの合成したU7用キメラオリゴをヌクレオフェクターによってHeLa細胞に導入することにより、低濃度のキメラオリゴ分子でも効率的にU7 RNAの破壊が誘導されることが明らかになった。また本RNaseプロテクションアッセイ法でU7 RNAの顕著な減少が観察された事実から、本発明者らの開発したncRNAの破壊方法によって、細胞内に存在する標的ncRNAが破壊されること、また標的ncRNA破壊処理を行った細胞群の全体にその効果が見られることが確認された。
【0043】
(3)U7 RNAの機能解析実験
機能解析手法として標的ncRNAの破壊が細胞の遺伝子発現に与える影響を解析する目的でマイクロアレイを用いた遺伝子発現解析を行った。U7用キメラオリゴの導入によってU7 RNAの破壊を行ったHeLa細胞より全RNAを抽出精製してAgilent社製のマイクロアレイ(44k Whole Human Genome Oligo Microarray)を用いて遺伝子発現解析を行い、対象実験としてGFP用キメラオリゴの導入を行ったHeLa細胞の発現遺伝子と比較し、U7 RNAの破壊の影響を観察した。解析を行ったヒト遺伝子22,833遺伝子中、U7 RNAの遺伝子破壊によって細胞内に遺伝子量に2倍以上の変化が観察された遺伝子は93遺伝子であった(図4)。この93遺伝子の内50遺伝子は細胞内においてヒストン遺伝子の合成に関連がある遺伝子群であったことから、U7 RNAは核内においてヒストン遺伝子合成に関連する機能を有することが推察された。
【0044】
(4)核酸導入方法の比較
核酸導入方法の比較として、既知の遺伝子導入方法であるエレクトロポレーション法とリポフェクタミン法とを比較した。標的ncRNA分子としてU84分子を選択し、先に合成したU84用キメラオリゴを用いて実験を行った。エレクトロポレーション法を用いた方法は標的ncRNA分子及び使用したキメラオリゴを除き、上記(2)と同様に行った。リポフェクタミンを用いた方法は、Invitogen社製Lipofectamine2000試薬をマニュアルに従いキメラオリゴと混合したものを細胞培養液に加えることにより行った。エレクトロポレーション法を用いた方法では核酸導入処理後3時間から72時間に至るまでU84分子が破壊されていることがRNaseプロテクションアッセイ法で確認されたが、リポフェクタミン法を用いた方法では十分なU84分子破壊が観察されなかった(図5)。また、エレクトロポレーション法を用いた方法では核酸導入処理後の培養において細胞はほぼ100%の生存率が観察されたが、リポフェクタミン法を用いた方法では一部の細胞が死滅する減少が観察された。この比較により、アンチセンスオリゴ分子の導入におけるエレクトロポレーションの使用は、細胞毒性が低く、効率的な標的分子の破壊を実現する有効な方法であることがわかった。
【0045】
(5)導入する核酸構造の比較
遺伝子破壊を目的とする実験において多用される2重鎖RNA分子(siRNA)を比較対象として、ncRNA破壊効率の比較を行った。なお、本実験は、核内への導入に使用した核酸分子を除き、上記(4)と同様に行った。siRNAは細胞質に存在する遺伝子を標的としたRNAの破壊に効果的であるが、本実験の標的としたU84 RNAでは破壊の誘導が観察されなかった(図6)。本結果からキメラアンチセンスオリゴの利用が効果的であることが確認された。
【0046】
(6)種々の培養細胞を用いたncRNA破壊実験
本発明のncRNA破壊方法をT細胞白血病由来のJurkat細胞、肺がん由来のMCR5、膀胱ガン由来のT24細胞に対して実施し、破壊の効果を検証した。Jurkat細胞ではU7 RNA、その他の細胞種ではHBII295 RNAを破壊の標的分子とし、上記(2)と同様の方法で行った。この検証実験により、本発明の方法は細胞種と標的分子とを限定することなく用いることが可能であることが確認された(図7)。
【図面の簡単な説明】
【0047】
【図1】図1は、本発明に用いるキメラアンチセンスオリゴ分子の構造の一例を示す。
【図2】図2は、エレクトロポレーションの原理を利用する核酸導入装置を用いた、核酸分子導入方法の概略図を示す。
【図3】図3は、U7 RNA分子破壊実験の結果を示す。
【図4】図4は、U7 RNA分子の破壊が及ぼす影響の解析結果を示す。
【図5】図5は、核酸導入方法による効果の比較を示す。
【図6】図6は、導入する核酸構造による効果の比較を示す。
【図7】図7は、種々の細胞種に対するncRNA破壊の検証結果を示す。
【特許請求の範囲】
【請求項1】
細胞核内に存在するノンコーディングRNA(non-coding RNA)の機能を解析する方法であって、該RNA分子に対するアンチセンスオリゴ分子を細胞核内に導入して、該RNA分子を破壊することを含む、前記方法。
【請求項2】
前記アンチセンスオリゴ分子は、前記RNA分子の二次構造における一本鎖領域に相補的な配列と少なくとも80%の同一性を有する塩基配列を含むことを特徴とする、請求項1記載の方法。
【請求項3】
前記アンチセンスオリゴ分子は、前記RNA分子の二次構造における一本鎖領域に相補的な塩基配列を含むことを特徴とする請求項2記載の方法。
【請求項4】
前記一本鎖領域は、前記RNA分子の二次構造解析により予め推定された領域であることを特徴とする請求項2記載の方法。
【請求項5】
前記一本鎖領域は、前記RNA分子に特異的な配列領域であることを特徴とする請求項2記載の方法。
【請求項6】
前記アンチセンスオリゴ分子は、核酸類似物質が導入されたキメラアンチセンスオリゴ分子であることを特徴とする請求項1〜5のいずれか1項記載の方法。
【請求項7】
核酸類似物質は、2'-O-メチルリボヌクレオチドであることを特徴とする請求項6記載の方法。
【請求項8】
前記アンチセンスオリゴ分子の細胞核内への導入をエレクトロポレーション法により実施することを特徴とする請求項1〜7のいずれか1項記載の方法。
【請求項1】
細胞核内に存在するノンコーディングRNA(non-coding RNA)の機能を解析する方法であって、該RNA分子に対するアンチセンスオリゴ分子を細胞核内に導入して、該RNA分子を破壊することを含む、前記方法。
【請求項2】
前記アンチセンスオリゴ分子は、前記RNA分子の二次構造における一本鎖領域に相補的な配列と少なくとも80%の同一性を有する塩基配列を含むことを特徴とする、請求項1記載の方法。
【請求項3】
前記アンチセンスオリゴ分子は、前記RNA分子の二次構造における一本鎖領域に相補的な塩基配列を含むことを特徴とする請求項2記載の方法。
【請求項4】
前記一本鎖領域は、前記RNA分子の二次構造解析により予め推定された領域であることを特徴とする請求項2記載の方法。
【請求項5】
前記一本鎖領域は、前記RNA分子に特異的な配列領域であることを特徴とする請求項2記載の方法。
【請求項6】
前記アンチセンスオリゴ分子は、核酸類似物質が導入されたキメラアンチセンスオリゴ分子であることを特徴とする請求項1〜5のいずれか1項記載の方法。
【請求項7】
核酸類似物質は、2'-O-メチルリボヌクレオチドであることを特徴とする請求項6記載の方法。
【請求項8】
前記アンチセンスオリゴ分子の細胞核内への導入をエレクトロポレーション法により実施することを特徴とする請求項1〜7のいずれか1項記載の方法。
【図1】



【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【公開番号】特開2009−171895(P2009−171895A)
【公開日】平成21年8月6日(2009.8.6)
【国際特許分類】
【出願番号】特願2008−14036(P2008−14036)
【出願日】平成20年1月24日(2008.1.24)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 発行者:第9回日本RNA学会年会事務局 刊行物名:第9回日本RNA学会年会(第9回RNAミーティング)要旨集 掲載頁:50頁(講演番号:O−18) 刊行物発行年月日:平成19年7月28日
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成19年度、独立行政法人新エネルギー・産業技術総合開発機構、機能性RNAプロジェクト委託研究、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(000233055)日立ソフトウエアエンジニアリング株式会社 (1,610)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【出願人】(500535301)社団法人バイオ産業情報化コンソーシアム (22)
【Fターム(参考)】
【公開日】平成21年8月6日(2009.8.6)
【国際特許分類】
【出願日】平成20年1月24日(2008.1.24)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 発行者:第9回日本RNA学会年会事務局 刊行物名:第9回日本RNA学会年会(第9回RNAミーティング)要旨集 掲載頁:50頁(講演番号:O−18) 刊行物発行年月日:平成19年7月28日
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成19年度、独立行政法人新エネルギー・産業技術総合開発機構、機能性RNAプロジェクト委託研究、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(000233055)日立ソフトウエアエンジニアリング株式会社 (1,610)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【出願人】(500535301)社団法人バイオ産業情報化コンソーシアム (22)
【Fターム(参考)】
[ Back to top ]