
核磁気共鳴法を用いた、標的タンパク質のアゴニストおよびアンタゴニストの同定方法、およびこの方法に用いるプログラム。
本発明によって、任意の化合物が標的タンパク質と接触・結合した際に、その標的タンパク質にどのような構造変化が生じるのかを簡便かつ迅速に決定する方法、そのような方法を用いる化合物の選択・スクリーニング方法、およびそのような方法を実行するためのコンピュータプログラムが提供される。
本発明の1つの実施形態において、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量、ドメインの原子座標から、ドメインのSaupeオーダーマトリクスエレメントを得て、そのマトリクスを対角化して、ドメインの配向情報を得る。その配向情報の変化を構造変化の指標とすることによって、本発明の簡便かつ迅速な標的タンパク質の構造変化測定が行われる。
本発明の1つの実施形態において、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量、ドメインの原子座標から、ドメインのSaupeオーダーマトリクスエレメントを得て、そのマトリクスを対角化して、ドメインの配向情報を得る。その配向情報の変化を構造変化の指標とすることによって、本発明の簡便かつ迅速な標的タンパク質の構造変化測定が行われる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、核磁気共鳴を用いることによって、化合物がタンパク質と結合した場合のタンパク質の構造変化を測定する方法、その構造変化に基づいて化合物を選択する方法、およびその方法において使用されるプログラムに関する。
【背景技術】
【0002】
細胞が細胞外からの刺激に対して応答する場合、細胞内の種々のタンパク質が関与する。そのため、これらタンパク質を標的とするアゴニストおよび/またはアンタゴニストは、新規薬物のリード化合物として用いることができる。
【0003】
タンパク質の、アゴニストおよび/またはアンタゴニストの探索のための従来法としては、標的タンパク質に対する物理的結合を指標とする探索方法、および標的タンパク質が関与する生物学的反応を誘導する能力(機能誘導能)を指標とする探索方法が挙げられる。
【0004】
標的タンパク質に対する物理的結合を指標とする方法は、化合物ライブラリー(例えば、組み合わせライブラリー)由来の化合物が、標的タンパク質にどの程度の強さで結合するか、また何分子結合するかなどを調べ、ある評価基準以上の標的タンパク質に対する親和性を有する化合物を選び出すという方法である。
【0005】
一方、標的タンパク質の機能誘導能、すなわち、標的タンパク質が関与する生物学的反応を指標とする方法は、化合物ライブラリー(例えば、組み合わせライブラリー)由来の化合物を、標的タンパク質を発現する細胞と接触させ、その結果生じる生物学的反応の変化(例えば、生物学的反応の開始および終了のような質的変化、あるいは生物学的反応の程度の増加または減少のような量的変化)を指標とする方法である。機能誘導能を指標とする方法を行うには、標的タンパク質に応じたアッセイ系をそれぞれ立ち上げる必要がある。そのため、例えば、生きた細胞を対象として化合物の効果を解析する場合にはアッセイ系に依存して信頼性・実験精度が左右されるなどの制約がある。
【0006】
タンパク質のアゴニストおよび/またはアンタゴニストを同定し、次に同定された化合物を、新規薬物のリード化合物として用いることができる。新規薬物のスクリーニングにおいては、その候補化合物が、標的タンパク質と結合することによって、どのようにその標的タンパク質の立体構造が変化するか知ることが有用である。なぜなら、そのような立体構造変化の情報は、より適切な構造を有する候補薬物のデザインの指標となるからである。
【0007】
しかし、物理的結合・機能誘導能のいずれを指標とする場合にも、化合物が直接タンパク質に結合している状態を見ているわけではない。従って間接的に得た標的タンパク質への結合能あるいは機能誘導能から化合物の構造と機能を類推することになる。物理的結合の有無あるいは機能誘導能の有無だけでは、標的タンパク質と化合物の相互作用の実態を知ることは難しく、いつも明確な薬物デザインの指針を与えるとはかぎらない。
【0008】
このような従来のアッセイ法が持つ問題点を克服するために、核磁気共鳴法(NMR)により化合物が標的タンパク質の機能部位に結合していることを確認する核磁気共鳴法(NMR)を用いたアッセイ法が発表されている(Shuker S.B. et al. Science (1996) 274, 1531−1534頁)(特開2001−321192)。この先行技術では、標的タンパク質の機能とは無関係な場所に非特異的に化合物が結合するケースを除くことができるために、非特異的結合と特異的結合の区別が難しい従来の物理的結合アッセイよりも効率的にリード化合物を特定することが可能である。しかし、この技術は結合部位を特定することは可能であるが、タンパク質の構造変化自体を測定するのには適していない。そのため、この技術は、標的タンパク質の活性部位に結合する阻害剤の設計には有効であるが、逆にタンパク質の活性誘導を行う作動薬(例えば、アゴニスト)のデザインには効果が少ない。作動薬リード化合物の同定には、標的タンパク質の基質結合部位に正しく結合し、さらに標的タンパク質の活性構造を誘導する作用の有無を指標としたスクリーニングが必要であるため、従来技術は、作動薬リード化合物の同定には不十分であった。
【0009】
化合物が標的タンパク質に対して活性構造を誘導し得るかどうかはX線結晶構造解析を基にして決定することができる。しかし、X線結晶構造解析は、多大な労力と時間を必要とする方法であることから、多数の化合物に対して標的タンパク質との共結晶化を行い立体構造解析を繰り返すことは、効率が求められるスクリーニングには適当ではない。そこで、任意の化合物が標的タンパク質と接触・結合した際に、その標的タンパク質にどのような構造変化が生じるのかを簡便かつ迅速に決定する方法、およびそのような方法を実行するためのコンピュータプログラムが求められている。
【特許文献1】特開2001−321192
【非特許文献1】Shuker S.B. et al. Science (1996) 274, 1531−1534頁
【発明の開示】
【発明が解決しようとする課題】
【0010】
任意の化合物が標的タンパク質と接触・結合した際に、その標的タンパク質にどのような構造変化が生じるのかを簡便かつ迅速に決定する方法、そのような方法を用いる化合物の選択・スクリーニング方法、およびそのような方法を実行するためのコンピュータプログラムを提供することが、本発明の課題である。
【0011】
例えば、核磁気共鳴を用いることによって、任意の化合物がタンパク質と結合・接触した場合の、タンパク質の構造変化を測定する方法を提供することが、本発明の課題の1つである。さらに、任意の化合物がタンパク質と結合・接触した場合に、タンパク質の構造変化を生じる化合物を同定・スクリーニングする方法を提供することが、本発明の課題の1つである。また、例えば、リガンド、アゴニスト、などのように、タンパク質と結合した際にそのタンパク質の構造変化を生じることが公知の物質がそのタンパク質と結合した際の構造変化と、任意の化合物がそのタンパク質と結合した際の構造変化とを比較して、リガンド、アゴニストと類似の構造変化をもたらす物質を同定・スクリーニングする方法を提供することもまた、本発明の別の課題の1つである。
【0012】
また、これら方法において使用されるコンピュータプログラムを提供することも、本発明の課題の1つである。
【課題を解決するための手段】
【0013】
本発明者らは、標的タンパク質溶液に化合物を添加して誘導される標的タンパク質の立体構造変化をTROSY NMR相関スペクトル(例えば、1H/15N−NMRスペクトル)の変化から読み取る方法およびプログラムを開発し、本発明を完成した。本発明者らは、1)化合物の標的タンパク質上での結合部位、2)標的タンパク質が化合物によって誘導される構造変化の2つを同時に観測することができる新規な方法を開発し、本発明を完成した。
【0014】
従って、本発明は以下を提供する。
1.タンパク質と化合物とを接触させた場合の、該タンパク質の構造変化を測定する方法であって、以下:
(a)該タンパク質内のドメインを選択する工程;
(b)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を得る工程;
(c)該タンパク質と該化合物を接触させる条件下における、該ドメインの配向情報を以下
(i)該ドメインの既知の原子座標を得て、
(ii)液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(iii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(iv)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、
ことにより得る工程;および、
(d)工程(b)において得られた配向情報と、工程(c)において得られた配向情報との差異から、該タンパク質の構造変化を測定する工程、
を包含する、方法。
2.前記(b)工程が、
(b)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を以下
(v)該ドメインの既知の原子座標を得て、
(vi)液晶物質存在下において、該タンパク質を該化合物と接触させずに、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(vii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(viii)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、ことにより得る工程である、項目1に記載の測定方法。
3.前記(b)工程が、
(b)前記タンパク質が前記化合物と接触しない条件下において既に得られている原子座標から、前記ドメインの配向情報を得る工程;
である、項目1に記載の測定方法。
4.項目1に記載の測定方法であって、前記工程(c)において二次元TROSY NMRスペクトルを用いて得られる分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、方法。
5.項目4に記載の測定方法であって、ここで、前記(iii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、方法。
6.項目2に記載の測定方法であって、前記工程(b)において二次元TROSY NMRスペクトルを用いて得られる分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、方法。
7.項目6に記載の測定方法であって、ここで、前記(vii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させることなく、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、方法。
8.項目5または7に記載の方法であって、ここで、前記タンパク質と前記化合物とが接触した場合の、該タンパク質の構造変化が、以下:
(ix)該化合物と接触させる前の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【0015】
【化1】

を用いて示し、
(x)該化合物と接触させた後の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【0016】
【化2】
を用いて示し、
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、そして
(xii)以下の式によって分子配向変化度を求める
分子配向変化度=a2+b2+c2、
ことによって分子配向変化度として数値化される、方法。
9.項目2に記載の方法であって、さらに、前記化合物が前記タンパク質に結合した位置を特定する工程を包含する、方法。
10.項目9に記載の方法であって、前記化合物が前記タンパク質に結合した位置を特定する工程が、前記工程(b)において得られた二次元TROSY NMRスペクトルと、前記工程(c)において得られた二次元TROSY NMRスペクトルとを比較して、スペクトルの変化を検出し、そして、該スペクトルの変化を生じた、該タンパク質中のアミノ酸残基を同定すること、によって行われる、方法。
11.項目1に記載の方法であって、前記液晶物質が、以下からなる群から選択される混合物を含む、方法:
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とドデシル硫酸ナトリウム(SDS)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物、
1,2−ジ−O−ドデシル−sn−グリセロ−3−ホスフォコリン(DIODPC)と3−(コラミドプロピル)−ジメチルアンモニオ−2−ヒドロキシ−1−プロパンサルフェート(CHAPS)との混合物、
n−アルキル−ポリ(エチレングリコール)/n−アルキルアルコール混合物、
繊維状ファージ、
セチルピリジニウムクロライド(CPCl)−ヘキサノール−NaCl混合物、
セチルピリジニウムブロミド(CPBr)−ヘキサノール−NaCl混合物、
Halobacterium属の紫膜断片
微結晶セルロース、および
ポリアクリルアミドゲル。
12.前記液晶物質が、7.5%(w/v)のジミリストイルホスファチジルコリン(D
MPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物である、項目11に記載の方法。
13.接触の際に、タンパク質内のドメインの構造変化を生じる化合物を選択する方法であって、以下:
(a)該タンパク質内のドメインを選択する工程;
(b)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を得る工程;
(c)該タンパク質と該化合物を接触させる条件下における、該ドメインの配向情報を以下
(i)該ドメインの既知の原子座標を得て、
(ii)液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(iii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(iv)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、
ことにより得る工程;
(d)工程(b)において得られた配向情報と、工程(c)において得られた配向情報との比較から、該化合物が該タンパク質と接触した際に該ドメインの構造変化を生じる化合物であるか否かを決定する工程、
を包含する、方法。
14.前記(b)工程が、
(b)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を以下
(v)該ドメインの既知の原子座標を得て、
(vi)液晶物質存在下において、該タンパク質を該化合物と接触させずに、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(vii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(viii)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、ことにより得る工程である、項目13に記載の測定方法。
15.前記(b)工程が、
(b)前記タンパク質が前記化合物と接触しない条件下において既に得られている原子座標から、前記ドメインの配向情報を得る工程;
である、項目13に記載の測定方法。
16.項目13に記載の測定方法であって、前記工程(c)において二次元TROSY NMRスペクトルを用いて得られる分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、方法。
17.項目16に記載の測定方法であって、ここで、前記(iii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、方法。
18.項目14に記載の測定方法であって、前記工程(b)において二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、方法。
19.項目18に記載の測定方法であって、ここで、前記(vii)のSaupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させることなく、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、方法。
20.項目17または19に記載の方法であって、ここで、前記工程(c)における配向情報の比較が、以下:
(ix)該化合物と接触する前の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【0017】
【化3】
を用いて示し、
(x)該化合物と接触した後の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【0018】
【化4】
を用いて示し、
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、そして
(xii)以下の式によって分子配向変化度を求め、
分子配向変化度=a2+b2+c2、
該分子配向変化度を比較することによって行われる、方法。
21.項目14に記載の方法であって、さらに、前記化合物が前記タンパク質に結合した位置を特定する工程を包含する、方法。
22.項目21に記載の方法であって、前記化合物が前記タンパク質に結合した位置を特定する工程が、前記工程(b)において得られた二次元TROSY NMRスペクトルと、前記工程(c)において得られた二次元TROSY NMRスペクトルとを比較して、スペクトルの変化を検出し、そして、該スペクトルの変化を生じた、該タンパク質中のアミノ酸残基を同定すること、によって行われる、方法。
23.項目13に記載の方法であって、前記液晶物質が、以下からなる群から選択される混合物を含む、方法:
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とドデシル硫酸ナトリウム(SDS)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物、
1,2−ジ−O−ドデシル−sn−グリセロ−3−ホスフォコリン(DIODPC)と3−(コラミドプロピル)−ジメチルアンモニオ−2−ヒドロキシ−1−プロパンサルフェート(CHAPS)との混合物、
n−アルキル−ポリ(エチレングリコール)/n−アルキルアルコール混合物、
繊維状ファージ、
セチルピリジニウムクロライド(CPCl)−ヘキサノール−NaCl混合物、
セチルピリジニウムブロミド(CPBr)−ヘキサノール−NaCl混合物、
Halobacterium属の紫膜断片
微結晶セルロース、および
ポリアクリルアミドゲル。
24.前記液晶物質が、7.5%(w/v)のジミリストイルホスファチジルコリン(D
MPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物である、項目23に記載の方法。
25.タンパク質と接触することによって、第1の化合物と該タンパク質とが接触した場合の該タンパク質の構造変化と類似する該タンパク質の構造変化を生じる、第2の化合物を選択する方法であって、以下:
(a)該タンパク質内のドメインを選択する工程;
(b)該タンパク質と該第1の化合物を接触させる条件下における、該ドメインの配向情報を得る工程;
(c)該タンパク質と該第2の化合物を接触させる条件下における、該ドメインの配向情報を以下
(i)該ドメインの既知の原子座標を得て、
(ii)液晶物質存在下において該タンパク質と該第2の化合物を接触させ、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(iii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(iv)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、
ことにより得る工程;
(d)工程(b)において得られた配向情報と、工程(c)において得られた配向情報とを比較する工程;および
(e)工程(d)における比較の結果から、該第2の化合物が該タンパク質と接触することによる該タンパク質の構造変化が、該第1の化合物が該タンパク質と接触することによる該タンパク質の構造変化と類似するか否かを決定する工程、
を包含する、方法。
26.前記(b)工程が、
(b)該タンパク質が該第1の化合物と接触する条件下における、該ドメインの配向情報を以下
(v)該ドメインの既知の原子座標を得て、
(vi)液晶物質存在下において、該タンパク質を該第1の化合物と接触させた場合に二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(vii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(viii)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、ことにより得る工程である、項目25に記載の測定方法。
27.前記(b)工程が、
(b)前記タンパク質が前記第1の化合物を接触させる条件下において既に得られている原子座標から、前記ドメインの配向情報を得る工程;
である、項目25に記載の測定方法。
28.項目25に記載の測定方法であって、前記工程(c)において二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、方法。
29.項目28に記載の測定方法であって、ここで、前記(iii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、方法。
30.項目26に記載の測定方法であって、前記工程(b)において二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、方法。
31.項目30に記載の測定方法であって、ここで、前記(vii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該第2の化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、方法。
32.項目29または31に記載の方法であって、ここで、前記前記工程(d)における配向情報の比較が、以下:
(ix)該第1の化合物と接触した場合の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【0019】
【化5】
を用いて示し、
(x)該第2の化合物と接触させた場合の、該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【0020】
【化6】
を用いて示し、そして
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、該a、b、およびcを配向情報の比較のための指標とする、
ことによって行われる、方法。
33.項目29または31に記載の方法であって、ここで、前記工程(d)における配向情報の比較が、以下:
(ix)該第1の化合物と接触した場合の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【0021】
【化7】
を用いて示し、
(x)該第2の化合物と接触させた場合の、該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【0022】
【化8】
を用いて示し、そして
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、そして
(xii)以下の式によって活性構造一致度を求め
活性構造一致度=a2+b2+c2、
該活性構造一致度を配向情報の比較のための指標とする、
ことによって行われる、方法。
34.項目25に記載の方法であって、さらに、前記第1および第2の化合物の少なくとも1つが前記タンパク質に結合した位置を特定する工程を包含する、方法。
35.項目34に記載の方法であって、前記第1および第2の化合物の少なくとも1つが前記タンパク質に結合した位置を特定する工程が、該化合物の非存在下における該タンパク質の二次元TROSY NMRスペクトルと、該化合物の存在下における該タンパク質の二次元TROSY NMRスペクトルを比較して、スペクトルの変化を検出し、そして、該スペクトルの変化を生じた、該タンパク質中のアミノ酸残基を同定すること、
によって行われる、方法。
36.項目25に記載の方法であって、前記液晶物質が、以下からなる群から選択される混合物を含む、方法:
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とドデシル硫酸ナトリウム(SDS)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物、
1,2−ジ−O−ドデシル−sn−グリセロ−3−ホスフォコリン(DIODPC)と3−(コラミドプロピル)−ジメチルアンモニオ−2−ヒドロキシ−1−プロパンサルフェート(CHAPS)との混合物、
n−アルキル−ポリ(エチレングリコール)/n−アルキルアルコール混合物、
繊維状ファージ、
セチルピリジニウムクロライド(CPCl)−ヘキサノール−NaCl混合物、
セチルピリジニウムブロミド(CPBr)−ヘキサノール−NaCl混合物、
Halobacterium属の紫膜断片
微結晶セルロース、および
ポリアクリルアミドゲル。
37.前記液晶物質が、7.5%(w/v)のジミリストイルホスファチジルコリン(D
MPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物である、項目36に記載の方法。
38.タンパク質と化合物とを接触させた場合の、該タンパク質内の選択されたドメインの構造変化を数値化するプログラムであって、以下:
(a)該ドメインについて、該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を得る手段;
(b)該タンパク質と該化合物を接触させる条件下における、該ドメインの配向情報を以下
(i)該ドメインの既知の原子座標のデータを得て、
(ii)液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量のデータを得て、
(iii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(iv)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、
ことにより得る手段;および、
(c)手段(a)において得られた配向情報と、手段(b)において測定された配向情報との差異から、該タンパク質の構造変化を計算する手段、
とをコンピュータに実現させるための、プログラム。
39.前記(a)手段が、
(a)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を以下
(v)該ドメインの既知の原子座標を得て、
(vi)液晶物質存在下において、該タンパク質を該化合物と接触させずに、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(vii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(viii)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、ことにより得る手段である、項目38に記載のプログラム。
40.前記(a)手段が、
(a)前記タンパク質が前記化合物と接触しない条件下において既に得られている原子座標から、前記ドメインの配向情報を得る手段;
である、項目38に記載のプログラム。
41.項目38に記載のプログラムであって、前記工程(b)において二次元TROSY
NMRスペクトルを用いて得られる分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、プログラム。
42.項目41に記載のプログラムであって、ここで、前記(iii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、プログラム。
43.項目39に記載のプログラムであって、前記工程(a)において二次元TROSY
NMRスペクトルを用いて得られる分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、プログラム。
44.項目43に記載のプログラムであって、ここで、前記(vii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させることなく、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、プログラム。
45.項目42または44に記載のプログラムであって、ここで、前記タンパク質と前記化合物とが接触した場合の、該タンパク質の構造変化が、以下:
(ix)該化合物と接触させる前の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【0023】
【化9】
を用いて示し、
(x)該化合物と接触させた後の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【0024】
【化10】
を用いて示し、
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、そして
(xii)以下の式によって分子配向変化度を求める
分子配向変化度=a2+b2+c2、
ことによって分子配向変化度として数値化される、プログラム。
46.項目39に記載のプログラムであって、さらに、前記化合物が前記タンパク質に結合した位置を特定する手段を包含する、プログラム。
47.項目46に記載のプログラムであって、前記化合物が前記タンパク質に結合した位置を特定する手段が、前記工程(a)において得られた二次元TROSY NMRスペクトルと、前記工程(b)において得られた二次元TROSY NMRスペクトルとを比較して、スペクトルの変化を検出し、そして、該スペクトルの変化を生じた、該タンパク質中のアミノ酸残基を同定すること、によって行われる、プログラム。
48.接触の際に、タンパク質内のドメインの構造変化を生じる化合物を選択するプログラムであって、以下:
(a)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を得る手段;
(b)該タンパク質と該化合物を接触させる条件下における、該ドメインの配向情報を以下
(i)該ドメインの既知の原子座標を得て、
(ii)液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(iii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(iv)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、
ことにより得る手段;
(c)手段(a)において得られた配向情報と、手段(b)において得られた配向情報との比較から、該化合物が該タンパク質と接触した際に該ドメインの構造変化を生じる化合物であるか否かを決定する手段、
を包含する、プログラム。
49.前記(a)手段が、
(a)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を以下
(v)該ドメインの既知の原子座標を得て、
(vi)液晶物質存在下において、該タンパク質を該化合物と接触させずに、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(vii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(viii)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、ことにより得る手段である、項目48に記載のプログラム。
50.前記(a)手段が、
(a)前記タンパク質が前記化合物と接触しない条件下において既に得られている原子座標から、前記ドメインの配向情報を得る手段;
である、項目48に記載のプログラム。
51.項目48に記載のプログラムであって、前記工程(b)において二次元TROSY
NMRスペクトルを用いて得られる分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、プログラム。
52.項目51に記載のプログラムであって、ここで、前記(iii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、プログラム。
53.項目52に記載のプログラムであって、前記工程(a)において二次元TROSY
NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、プログラム。
54.項目53に記載のプログラムであって、ここで、前記(vii)のSaupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させることなく、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、プログラム。
55.項目52または54に記載のプログラムであって、ここで、前記工程(c)における配向情報の比較が、以下:
(ix)該化合物と接触する前の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【0025】
【化11】
を用いて示し、
(x)該化合物と接触した後の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【0026】
【化12】
を用いて示し、
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、そして
(xii)以下の式によって分子配向変化度を求め、
分子配向変化度=a2+b2+c2、
該分子配向変化度を比較することによって行われる、プログラム。
56.項目48に記載のプログラムであって、さらに、前記化合物が前記タンパク質に結合した位置を特定する手段を包含する、プログラム。
57.項目56に記載のプログラムであって、前記化合物が前記タンパク質に結合した位置を特定する手段が、前記工程(a)において得られた二次元TROSY NMRスペクトルと、前記工程(b)において得られた二次元TROSY NMRスペクトルとを比較して、スペクトルの変化を検出し、そして、該スペクトルの変化を生じた、該タンパク質中のアミノ酸残基を同定すること、によって行われる、プログラム。
58.タンパク質と接触することによって、第1の化合物と該タンパク質とが接触した場合の該タンパク質内のドメインの構造変化と類似する該タンパク質内のドメインの構造変化を生じる、第2の化合物を選択するプログラムであって、以下:
(a)該タンパク質と該第1の化合物を接触させる条件下における、該ドメインの配向情報を得る手段;
(b)該タンパク質と該第2の化合物を接触させる条件下における、該ドメインの配向情報を以下
(i)該ドメインの既知の原子座標を得て、
(ii)液晶物質存在下において該タンパク質と該第2の化合物を接触させ、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(iii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(iv)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、
ことにより得る手段;
(c)手段(a)において得られた配向情報と、手段(b)において得られた配向情報とを比較する手段;および
(d)手段(c)における比較の結果から、該第2の化合物が該タンパク質と接触することによる該タンパク質の構造変化が、該第1の化合物が該タンパク質と接触することによる該タンパク質の構造変化と類似するか否かを決定する手段、
を包含する、プログラム。
59.前記(a)手段が、
(a)該タンパク質が該第1の化合物と接触する条件下における、該ドメインの配向情報を以下
(v)該ドメインの既知の原子座標を得て、
(vi)液晶物質存在下において、該タンパク質を該第1の化合物と接触させた場合に二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(vii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(viii)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、ことにより得る手段である、項目25に記載のプログラム。
60.前記(a)手段が、
(a)前記タンパク質が前記第1の化合物を接触させる条件下において既に得られている原子座標から、前記ドメインの配向情報を得る手段;
である、項目58に記載のプログラム。
61.項目58に記載のプログラムであって、前記工程(b)において二次元TROSY
NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、プログラム。
62.項目61に記載のプログラムであって、ここで、前記(iii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、プログラム。
63.項目59に記載のプログラムであって、前記工程(a)において二次元TROSY
NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、プログラム。
64.項目63に記載のプログラムであって、ここで、前記(vii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該第2の化合物を接触させ、二次元TROSY
NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、プログラム。
65.項目62または64に記載のプログラムであって、ここで、前記工程(c)における配向情報の比較が、以下:
(ix)該第1の化合物と接触した場合の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【0027】
【化13】
を用いて示し、
(x)該第2の化合物と接触させた場合の、該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【0028】
【化14】
を用いて示し、そして
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、該a、b、およびcを配向情報の比較のための指標とする、
ことによって行われる、プログラム。
66.項目62または64に記載のプログラムであって、ここで、前記工程(c)における配向情報の比較が、以下:
(ix)該第1の化合物と接触した場合の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【0029】
【化15】
を用いて示し、
(x)該第2の化合物と接触させた場合の、該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【0030】
【化16】
を用いて示し、そして
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、そして
(xii)以下の式によって活性構造一致度を求め
活性構造一致度=a2+b2+c2、
該活性構造一致度を配向情報の比較のための指標とする、
ことによって行われる、プログラム。
67.項目58に記載のプログラムであって、さらに、前記第1および第2の化合物の少なくとも1つが前記タンパク質に結合した位置を特定する手段を包含する、プログラム。68.項目67に記載のプログラムであって、前記第1および第2の化合物の少なくとも1つが前記タンパク質に結合した位置を特定する手段が、該化合物の非存在下における該タンパク質の二次元TROSY NMRスペクトルと、該化合物の存在下における該タンパク質の二次元TROSY NMRスペクトルとを比較して、スペクトルの変化を検出し、そして該スペクトルの変化を生じた、該タンパク質中のアミノ酸残基を同定すること、によって行われる、プログラム。
【0031】
69.項目1、13、または25に記載の方法を実行し得る装置。
【0032】
70.項目38、48、または58に記載のプログラムを備える装置。
【0033】
71.項目38、48、または58に記載のプログラムを含む記憶媒体。
【発明の効果】
【0034】
本発明によって、標的タンパク質溶液に化合物を添加して誘導される標的タンパク質の立体構造変化をTROSY NMR相関スペクトル(例えば、1H/15N−NMRスペクトル)の変化から読み取る方法およびプログラムが提供される。本発明により、1)化合物の標的タンパク質上での結合部位、2)標的タンパク質が化合物によって誘導される構造変化の2つを同時に観測することができる新規な方法が提供される。
【図面の簡単な説明】
【0035】
【図1】図1は、ドメイン間の相対配向変化を基にして選択的に作動薬のリード化合物を同定する方法を模式的に示した図である。
【図2】図2は、タンパク質の分子配向角度の表現法を示す。
【図3】図3は、実施例3で用いた配向ゲルの写真である。右上の白棒は、10mmの長さを示す。
【図4】図4は、この配向ゲルを用いた、クロレラウイルスPBCV1のグアニリル転移酵素であるmRNA5’−キャッピング酵素のTROSYシグナルを示す。6%(w/w)の伸張させた50%アクリルアミド/50%アクリレートゲル(配向ゲル)を用いて、20℃(293K)で実験を行った。「I」は、等方的状態において収集されたデーを示す。「A」は、整列された状態において収集されたデータを示す。
【図5】図5は、オープン型結晶構造を基にした解析の結果を示す。配向テンソルは、N末端ドメイン(ラージドメイン)23残基、およびC末端ドメイン(スモールドメイン)28残基を用いて決定した。
【図6】図6は、オープン型結晶構造を基にした解析の結果を示す。配向テンソルは、N末端ドメイン(ラージドメイン)23残基、およびC末端ドメイン(スモールドメイン)28残基を用いて決定した。
【図7】図7は、クローズ型結晶構造を基にした解析の結果を示す。配向テンソルは、N末端ドメイン(ラージドメイン)21残基、およびC末端ドメイン(スモールドメイン)28残基を用いて決定した。
【図8】図8は、クローズ型結晶構造を基にした解析の結果を示す。配向テンソルは、N末端ドメイン(ラージドメイン)21残基、およびC末端ドメイン(スモールドメイン)28残基を用いて決定した。
【発明を実施するための最良の形態】
【0036】
以下、本発明を説明する。本明細書の全体にわたり、単数形の表現は、特に言及しない限り、その複数形の概念をも含むことが理解されるべきである。従って、単数形の冠詞(例えば、英語の場合は「a」、「an」、「the」など)は、特に言及しない限り、その複数形の概念をも含むことが理解されるべきである。また、本明細書において使用される用語は、特に言及しない限り、当該分野で通常用いられる意味で用いられることが理解されるべきである。したがって、他に定義されない限り、本明細書中で使用される全ての専門用語および科学技術用語は、本発明の属する分野の当業者によって一般的に理解されるのと同じ意味を有する。矛盾する場合、本明細書(定義を含めて)が優先する。
【0037】
(用語の定義)
以下に本明細書において特に使用される用語の定義を列挙する。
本明細書において使用される用語「タンパク質」「ポリペプチド」、「オリゴペプチド」および「ペプチド」は、本明細書において同じ意味で使用され、任意の長さのアミノ酸のポリマーをいう。このポリマーは、直鎖であっても分岐していてもよく、環状であってもよい。アミノ酸は、天然のものであっても非天然のものであってもよく、改変されたアミノ酸であってもよい。この用語はまた、複数のポリペプチド鎖の複合体へとアセンブルされ得る。この用語はまた、天然または人工的に改変されたアミノ酸ポリマーも包含する。そのような改変としては、例えば、ジスルフィド結合形成、グリコシル化、脂質化、アセチル化、リン酸化または任意の他の操作もしくは改変(例えば、標識成分との結合体化)。この定義にはまた、例えば、アミノ酸の1または2以上のアナログを含むポリペプチド(例えば、非天然のアミノ酸などを含む)、ペプチド様化合物(例えば、ペプトイド)および当該分野において公知の他の改変が包含される。
【0038】
本明細書においてタンパク質の「ドメイン」とは、1残基のアミノ酸、または連続した少なくとも2つのアミノ酸残基を含む、タンパク質分子の特定の一部を意味する。本明細書においてドメインは、独立して特定の機能を有しても、有していなくてもよい。
【0039】
本明細書において、「アミノ酸」は、天然のものでも非天然のものでもよい。「誘導体アミノ酸」または「アミノ酸アナログ」とは、天然に存在するアミノ酸とは異なるがもとのアミノ酸と同様の機能を有するものをいう。そのような誘導体アミノ酸およびアミノ酸アナログは、当該分野において周知である。用語「天然のアミノ酸」とは、天然のアミノ酸のL−異性体を意味する。天然のアミノ酸は、グリシン、アラニン、バリン、ロイシン、イソロイシン、セリン、メチオニン、トレオニン、フェニルアラニン、チロシン、トリプトファン、システイン、プロリン、ヒスチジン、アスパラギン酸、アスパラギン、グルタミン酸、グルタミン、γ−カルボキシグルタミン酸、アルギニン、オルニチン、およびリジンである。特に示されない限り、本明細書でいう全てのアミノ酸はL体である。用語「非天然アミノ酸」とは、タンパク質中で通常は天然に見出されないアミノ酸を意味する。非天然アミノ酸の例として、ノルロイシン、パラ−ニトロフェニルアラニン、ホモフェニルアラニン、パラ−フルオロフェニルアラニン、3−アミノ−2−ベンジルプロピオン酸、ホモアルギニンのD体またはL体およびD−フェニルアラニンが挙げられる。「アミノ酸アナログ」とは、アミノ酸ではないが、アミノ酸の物性および/または機能に類似する分子をいう。アミノ酸アナログとしては、例えば、エチオニン、カナバニン、2−メチルグルタミンなどが挙げられる。アミノ酸模倣物とは、アミノ酸の一般的な化学構造とは異なる構造を有するが、天然に存在するアミノ酸と同様な様式で機能する化合物をいう。
【0040】
アミノ酸は、その一般に公知の3文字記号か、またはIUPAC−IUB Biochemical Nomenclature Commissionにより推奨される1文字記号のいずれかにより、本明細書中で言及され得る。ヌクレオチドも同様に、一般に受け入れられた1文字コードにより言及され得る。
【0041】
本明細書中において、「対応する」アミノ酸とは、あるタンパク質分子またはポリペプチド分子において、比較の基準となるタンパク質またはポリペプチドにおける所定のアミノ酸と同様の作用を有するか、または有することが予測されるアミノ酸をいい、特に酵素分子にあっては、活性部位中の同様の位置に存在し触媒活性に同様の寄与をするアミノ酸をいう。
【0042】
本明細書において使用される用語「受容体」とは、1個以上のリガンドと可逆的、かつ特異的に複合体化する1個以上の結合ドメインを備える生物学的な構造であって、ここで、この複合体化は生物学的な構造を有する。受容体は、完全に細胞の外部(細胞外の受容体)、細胞膜の中(しかし、受容体の部分を細胞外部の環境および細胞質ゾルに向けている)、または完全に細胞の中(細胞内の受容体)に存在し得る。これらはまた、細胞と独立的に機能し得る。細胞膜中の受容体は、細胞を、その境界の外部の空間と連絡(例えば、シグナル伝達)させ、そして細胞の内側および外側への分子およびイオンの輸送において機能させることを可能とする。本明細書において使用する場合、受容体は、受容体全長であっても、受容体のフラグメントであってもよい。
【0043】
受容体フラグメントを用いる場合には、例えば、受容体タンパク質のリガンド認識に関係する部位を用いることができる。受容体タンパク質のリガンド認識に関係する部位は、以下のように同定することができる。ホモロジーやドメイン検索により相同性や機能上の類似点の高いタンパク質の構造からリガンド認識領域を推定することができる。例えば、同一のリガンドに特異的に結合する、異なる受容体分子のアミノ酸配列をBLASTのデフォルトパラメータを用いて算出した場合、50%以上、好ましくは55%以上、より好ましくは60%以上、さらに好ましくは65%以上の相同性を示す領域が、リガンド認識領域として推定される。
さらに欠損変異やアミノ酸置換などを導入した変異受容体をコードする遺伝子を動物細胞などに一過性発現させ、その機能に必須の領域を決定することも、当業者は容易になし得る。
【0044】
本明細書において使用される用語「リガンド」とは、特異的な受容体または受容体のファミリーに対する結合パートナーである。リガンドは、受容体に対する内因性のリガンドであるか、またはその代わりに、薬剤、薬剤候補、もしくは薬理学的手段のような受容体に対する合成リガンドであり得る。
【0045】
本明細書において「アゴニスト」とは、ある生体作用物質(リガンド)の受容体に結合し、その物質のもつ作用と同じ(あるいは類似の)作用を現わす因子をいう。
【0046】
本明細書において「アンタゴニスト」とは、ある生体作用物質(リガンド)の受容体への結合に拮抗的に働き、それ自身はその受容体を介した生理作用を現わさない因子をいう。拮抗薬、遮断剤(ブロッカー)、阻害剤(インヒビター)などもこのアンタゴニストに包含される。
【0047】
本明細書において「NMR」とは、核磁気共鳴を意味する。磁気モーメントを持つ核の集団を静磁場B0の中に置くと、核ゼーマン効果によって、磁気モーメントの大きさと磁場の強さに従った不連続なエネルギー準位に分布する。この準位の間隔に相当する周波数をもつ電磁波を照射すると共鳴吸収が観測される現象を核磁気共鳴という。
【0048】
本明細書において「二次元NMR」とは、核磁気共鳴スペクトルを2つの周波数軸に展開する方法をいう。この測定には複数のパルスを用い、その時間間隔と観測パルス後の時間とを二つの時間軸としてフーリエ変換を行う。シグナル強度は、通常、等高線によって表される。二次元NMRとしては、代表的には、TROSY(transverse relaxation−optimized spectroscopy)、COSY(二次元シフト相関NMR法)、SECSY(二次元スピンエコー相関分光法)、FOCSY(二次元折り返し補正分光法)、HOHAHA(二次元同核ハートマン法)、NOESY(二次元NOE法)、2D−J法(二次元J分解分光法)、リレーCOSY(リレーコヒーレンス移動分光法)が挙げられるが、これらに限定されない。好ましいNMRはTROSYである。TROSYとは、Wuthrichらによって開発された、超高磁場NMRに適した方法である(Proc.Natl.Acad.Sci.USA vol.94、12366−12371頁(1997))。分子量の大きな分子のNMRにおいては、核磁気緩和を支配するのは、(1)磁気双極子相互作用と、(2)化学シフト異方性であるが、超高磁場では、化学シフト異方性が非常に大きくなるために、緩和時間が短くなり、線幅が広くなる。TROSYは、これら2つの相互作用を相殺することによって、緩和時間を長くし、線幅を狭くする方法である。TROSYによって高分子量のタンパク質(例えば、900kDa以上)について、二次元NMRスペクトルを得ることが可能になった。
【0049】
本明細書において二次元NMRを行う場合、蛋白質を標識するために使用される安定同位体の核種としては、15N, 13C, 2Hまたはこれらの組み合わせが挙げられるが、これらに限定されない。好ましい標識に用いられる安定同位体核の種類としては15N単標識、13C単標識、15N/2H二重標識、13C/2H二重標識、15N/13C/2H三重標識である。
【0050】
本明細書において、「液晶物質」とは、標的タンパク質の弱い配向を実現する物質である。液晶物質としては、 ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とドデシル硫酸ナトリウム(SDS)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物、
1,2−ジ−O−ドデシル−sn−グリセロ−3−ホスフォコリン(DIODPC)と3−(コラミドプロピル)−ジメチルアンモニオ−2−ヒドロキシ−1−プロパンサルフェート(CHAPS)との混合物、
n−アルキル−ポリ(エチレングリコール)/n−アルキルアルコール混合物、
繊維状ファージ、
セチルピリジニウムクロライド(CPCl)−ヘキサノール−NaCl混合物、
セチルピリジニウムブロミド(CPBr)−ヘキサノール−NaCl混合物、
Halobacterium属の紫膜断片
微結晶セルロース、および
ポリアクリルアミドゲル。
が挙げられるが、これらに限定されない。
【0051】
これら液晶物質が、脂質である場合、脂質からなるbicelleを標的タンパク質と混合し、混合物の温度を、相転移以上に設定することによって、標的タンパク質を弱く配向することができる。
【0052】
本明細書において「分子の配向状態」とは、Z軸を静磁場方向とした座標軸に対する分子の座標軸の傾きを示すオイラー角によって示される。
【0053】
本明細書において「配向情報」とは、タンパク質内のドメインの配向に関する情報であって、分子配向テンソルとして得られる情報をいう。本明細書において、配向情報は、例えば、お互いに直行する単位ベクトルによって表すことができる。
【0054】
本明細書において、「NMRシグナルの軸方向の変化量」とは、安定同位体元素について展開された二次元NMRの周波数軸について、軸方向での分子配向依存的な変化量をいう。例えば、1Hを用いる場合、1H軸方向での分子配向依存的な変化量をいい、また、13Cを用いる場合、13C軸方向での分子配向依存的な変化量をいう。
【0055】
本発明において試験される化合物としては、任意の化合物が利用可能であり、例えば、低分子有機化合物、低分子無機化合物、核酸、核酸アナログ、ペプチド、ペプチドアナログ、金属イオンなどが挙げられるが、これらに限定されない。
【0056】
本発明において、低分子有機化合物、または低分子無機化合物を合成する際には、コンビナトリアルケミストリーを用いることができる。コンビナトリアルケミストリーとは、ある基本骨格(scaffold)に、複数の置換基を組合せ(combinatorial)の手法で導入し、短期間で多数かつ多様のある分子の一群(library)を合成し、この分子群の中から目的とする化合物を見つけだすための有力な合成法である。コンビナトリアルケミストリーのための種々の手法が周知である。歴史的には、コンビナトリアルケミストリーは、Merrifield(米国)のビーズを用いた「ペプチド合成法」(1963年)を基盤とした合成技術の進展と、Furka(ハンガリー)らの「スプリット合成法」(1991年)に触発され、1992年以降盛んに研究開発が進められてきた技術である。特に固相合成法は、合成後の精製が不要なことがスピードアップ化のニーズにマッチして近年急速に発展してきている技術である。
【0057】
本明細書において使用される用語「ポリヌクレオチド」、「オリゴヌクレオチド」および「核酸」は、本明細書において同じ意味で使用され、任意の長さのヌクレオチドのポリマーをいう。この用語はまた、「誘導体オリゴヌクレオチド」または「誘導体ポリヌクレオチド」を含む。「誘導体オリゴヌクレオチド」または「誘導体ポリヌクレオチド」とは、ヌクレオチドの誘導体を含むか、またはヌクレオチド間の結合が通常とは異なるオリゴヌクレオチドまたはポリヌクレオチドをいい、互換的に使用される。そのようなオリゴヌクレオチドとして具体的には、例えば、2’−O−メチル−リボヌクレオチド、オリゴヌクレオチド中のリン酸ジエステル結合がホスホロチオエート結合に変換された誘導体オリゴヌクレオチド、オリゴヌクレオチド中のリン酸ジエステル結合がN3’−P5’ホスホロアミデート結合に変換された誘導体オリゴヌクレオチド、オリゴヌクレオチド中のリボースとリン酸ジエステル結合とがペプチド核酸結合に変換された誘導体オリゴヌクレオチド、オリゴヌクレオチド中のウラシルがC−5プロピニルウラシルで置換された誘導体オリゴヌクレオチド、オリゴヌクレオチド中のウラシルがC−5チアゾールウラシルで置換された誘導体オリゴヌクレオチド、オリゴヌクレオチド中のシトシンがC−5プロピニルシトシンで置換された誘導体オリゴヌクレオチド、オリゴヌクレオチド中のシトシンがフェノキサジン修飾シトシン(phenoxazine−modified cytosine)で置換された誘導体オリゴヌクレオチド、DNA中のリボースが2’−O−プロピルリボースで置換された誘導体オリゴヌクレオチドおよびオリゴヌクレオチド中のリボースが2’−メトキシエトキシリボースで置換された誘導体オリゴヌクレオチドなどが例示される。他にそうではないと示されなければ、特定の核酸配列はまた、明示的に示された配列と同様に、その保存的に改変された改変体(例えば、縮重コドン置換体)および相補配列を包含することが企図される。具体的には、縮重コドン置換体は、1またはそれ以上の選択された(または、すべての)コドンの3番目の位置が混合塩基および/またはデオキシイノシン残基で置換された配列を作成することにより達成され得る(Batzerら、Nucleic Acid Res.19:5081(1991);Ohtsukaら、J.Biol.Chem.260:2605−2608(1985);Rossoliniら、Mol.Cell.Probes 8:91−98(1994))。用語「核酸」はまた、本明細書において、遺伝子、cDNA、mRNA、オリゴヌクレオチド、およびポリヌクレオチドと互換可能に使用される。特定の核酸配列はまた、「スプライス改変体」を包含する。同様に、核酸によりコードされた特定のタンパク質は、その核酸のスプライス改変体によりコードされる任意のタンパク質を暗黙に包含する。その名が示唆するように「スプライス改変体」は、遺伝子のオルタナティブスプライシングの産物である。転写後、最初の核酸転写物は、異なる(別の)核酸スプライス産物が異なるポリペプチドをコードするようにスプライスされ得る。スプライス改変体の産生機構は変化するが、エキソンのオルタナティブスプライシングを含む。読み過し転写により同じ核酸に由来する別のポリペプチドもまた、この定義に包含される。スプライシング反応の任意の産物(組換え形態のスプライス産物を含む)がこの定義に含まれる。
【0058】
本明細書において「ヌクレオチド」は、天然のものでも非天然のものでもよい。「誘導体ヌクレオチド」または「ヌクレオチドアナログ」とは、天然に存在するヌクレオチドとは異なるがもとのヌクレオチドと同様の機能を有するものをいう。そのような誘導体ヌクレオチドおよびヌクレオチドアナログは、当該分野において周知である。そのような誘導体ヌクレオチドおよびヌクレオチドアナログの例としては、ホスホロチオエート、ホスホルアミデート、メチルホスホネート、キラルメチルホスホネート、2−O−メチルリボヌクレオチド、ペプチド−核酸(PNA)が含まれるが、これらに限定されない。
【0059】
本明細書において、「フラグメント」とは、全長のポリペプチドまたはポリヌクレオチド(長さがn)に対して、1〜n−1までの配列長さを有するポリペプチドまたはポリヌクレオチドをいう。フラグメントの長さは、その目的に応じて、適宜変更することができ、例えば、その長さの下限としては、ポリペプチドの場合、3、4、5、6、7、8、9、10、15,20、25、30、40、50およびそれ以上のアミノ酸が挙げられ、ここの具体的に列挙していない整数で表される長さ(例えば、11など)もまた、下限として適切であり得る。また、ポリヌクレオチドの場合、5、6、7、8、9、10、15,20、25、30、40、50、75、100およびそれ以上のヌクレオチドが挙げられ、ここの具体的に列挙していない整数で表される長さ(例えば、11など)もまた、下限として適切であり得る。本明細書において使用する場合、好ましくは、受容体「フラグメント」は、全長受容体が特異的に結合し得るリガンドに特異的に結合する。
【0060】
本発明において使用されるタンパク質は、天然の供給源から精製されたものであっても、遺伝子工学的に宿主細胞を形質転換して、組換え発現されたものであってもよい。遺伝子工学的にポリペプチドを製造する方法としては、例えば、そのポリペプチドを産生する原核生物である細菌を培養し、細菌中に組換え受容体タンパク質を封入体として蓄積させ、その宿主細菌を破壊することによって、そのポリペプチドを得る方法が挙げられる。
【0061】
「形質転換体」とは、宿主細胞を形質転換することによって作製された細胞などの生命体の全部または一部をいう。形質転換体としては、原核細胞が例示される。形質転換体は、その対象に依存して、形質転換細胞、形質転換組織、形質転換宿主などともいわれ、本明細書においてそれらの形態をすべて包含するが、特定の文脈において特定の形態を指し得る。
【0062】
形質転換体を得るための宿主細菌細胞は、生理活性を保持するポリペプチドを発現するものであれば、特に限定されず、従来から遺伝子操作において利用される各種の宿主細菌細胞を用いることができる。原核細胞としては、エシェリヒア属、セラチア属、バチルス属、ブレビバクテリウム属、コリネバクテリウム属、ミクロバクテリウム属、シュードモナス属等に属する原核細胞、例えば、Escherichia coli XL1−Blue、Escherichia coli XL2−Blue、Escherichia coli DH1、Escherichia coli MC1000、Escherichia coli KY3276、Escherichia coli W1485、Escherichia coli JM109、Escherichia coli
HB101、Escherichia coli No.49、Escherichia coli W3110、Escherichia coli NY49、Escherichia coli BL21(DE3)、Escherichia coli BL21(DE3)pLysS、Escherichia coli HMS174(DE3)、Escherichia coli HMS174(DE3)pLysS、Serratia ficaria、Serratia fonticola、Serratia liquefaciens、Serratia marcescens、Bacillus subtilis、Bacillus amyloliquefaciens、Brevibacterium ammmoniagenes、Brevibacterium immariophilum ATCC14068、Brevibacterium saccharolyticumATCC14066、Corynebacterium glutamicum ATCC13032、Corynebacterium glutamicum ATCC14067、Corynebacterium glutamicum ATCC13869、Corynebacterium acetoacidophilum ATCC13870、Microbacterium ammoniaphilum ATCC15354、Pseudomonas sp.D−0110などが例示される。
【0063】
本発明において得られた細胞に由来するポリペプチドは、天然型のポリペプチドと実質的に同一の作用を有する限り、アミノ酸配列中の1以上のアミノ酸が置換、付加および/または欠失していてもよく、糖鎖が置換、付加および/または欠失していてもよい。
【0064】
あるアミノ酸は、相互作用結合能力の明らかな低下または消失なしに、例えば、リガンド分子の結合部位のようなタンパク質構造において他のアミノ酸に置換され得る。あるタンパク質の生物学的機能を規定するのは、タンパク質の相互作用能力および性質である。従って、特定のアミノ酸の置換がアミノ酸配列において、またはそのDNAコード配列のレベルにおいて行われ得、置換後もなお、もとの性質を維持するタンパク質が生じ得る。従って、生物学的有用性の明らかな損失なしに、種々の改変が、本明細書において開示されたペプチドまたはこのペプチドをコードする対応するDNAにおいて行われ得る。
【0065】
上記のような改変を設計する際に、アミノ酸の疎水性指数が考慮され得る。タンパク質における相互作用的な生物学的機能を与える際の疎水性アミノ酸指数の重要性は、一般に当該分野で認められている(Kyte.JおよびDoolittle,R.F.J.Mol.Biol. 157(1):105−132,1982)。アミノ酸の疎水的性質は、生成したタンパク質の二次構造に寄与し、次いでそのタンパク質と他の分子(例えば、酵素、基質、受容体、DNA、抗体、抗原など)との相互作用を規定する。各アミノ酸は、それらの疎水性および電荷の性質に基づく疎水性指数を割り当てられる。それらは:イソロイシン(+4.5);バリン(+4.2);ロイシン(+3.8);フェニルアラニン(+2.8);システイン/シスチン(+2.5);メチオニン(+1.9);アラニン(+1.8);グリシン(−0.4);スレオニン(−0.7);セリン(−0.8);トリプトファン(−0.9);チロシン(−1.3);プロリン(−1.6);ヒスチジン(−3.2);グルタミン酸(−3.5);グルタミン(−3.5);アスパラギン酸(−3.5);アスパラギン(−3.5);リジン(−3.9);およびアルギニン(−4.5))である。
【0066】
あるアミノ酸を、同様の疎水性指数を有する他のアミノ酸により置換して、そして依然として同様の生物学的機能を有するタンパク質(例えば、リガンド結合能において等価なタンパク質)を生じさせ得ることが当該分野で周知である。このようなアミノ酸置換において、疎水性指数が±2以内であることが好ましく、±1以内であることがより好ましく、および±0.5以内であることがさらにより好ましい。疎水性に基づくこのようなアミノ酸の置換は効率的であることが当該分野において理解される。米国特許第4、554、101号に記載されるように、以下の親水性指数がアミノ酸残基に割り当てられている:アルギニン(+3.0);リジン(+3.0);アスパラギン酸(+3.0±1);グルタミン酸(+3.0±1);セリン(+0.3);アスパラギン(+0.2);グルタミン(+0.2);グリシン(0);スレオニン(−0.4);プロリン(−0.5±1);アラニン(−0.5);ヒスチジン(−0.5);システイン(−1.0);メチオニン(−1.3);バリン(−1.5);ロイシン(−1.8);イソロイシン(−1.8);チロシン(−2.3);フェニルアラニン(−2.5);およびトリプトファン(−3.4)。アミノ酸が同様の親水性指数を有しかつ依然として生物学的等価体を与え得る別のものに置換され得ることが理解される。このようなアミノ酸置換において、親水性指数が±2以内であることが好ましく、±1以内であることがより好ましく、および±0.5以内であることがさらにより好ましい。
【0067】
本発明において、「保存的置換」とは、アミノ酸置換において、元のアミノ酸と置換されるアミノ酸との親水性指数または/および疎水性指数が上記のように類似している置換をいう。保存的置換の例は、当業者に周知であり、例えば、次の各グループ内での置換:アルギニンおよびリジン;グルタミン酸およびアスパラギン酸;セリンおよびスレオニン;グルタミンおよびアスパラギン;ならびにバリン、ロイシン、およびイソロイシン、などが挙げられるがこれらに限定されない。
【0068】
本明細書において、「改変体」とは、もとのポリペプチドまたはポリヌクレオチドなどの物質に対して、一部が変更されているものをいう。そのような改変体としては、置換改変体、付加改変体、欠失改変体、短縮(truncated)改変体、対立遺伝子変異体などが挙げられる。対立遺伝子(allele)とは、同一遺伝子座に属し、互いに区別される遺伝的改変体のことをいう。従って、「対立遺伝子変異体」とは、ある遺伝子に対して、対立遺伝子の関係にある改変体をいう。「種相同体またはホモログ(homolog)」とは、ある種の中で、ある遺伝子とアミノ酸レベルまたはヌクレオチドレベルで、相同性(好ましくは、60%以上の相同性、より好ましくは、80%以上、85%以上、90%以上、95%以上の相同性)を有するものをいう。そのような種相同体を取得する方法は、本明細書の記載から明らかである。「オルソログ(ortholog)」とは、オルソロガス遺伝子(orthologous gene)ともいい、二つの遺伝子がある共通祖先からの種分化に由来する遺伝子をいう。例えば、多重遺伝子構造をもつヘモグロビン遺伝子ファミリーを例にとると、ヒトとマウスのαヘモグロビン遺伝子はオルソログであるが,ヒトのαヘモグロビン遺伝子とβヘモグロビン遺伝子はパラログ(遺伝子重複で生じた遺伝子)である。オルソログは、分子系統樹の推定に有用であることから、オルソログもまた、本発明において有用であり得る。
【0069】
「保存的(に改変された)改変体」は、アミノ酸配列および核酸配列の両方に適用される。特定の核酸配列に関して、保存的に改変された改変体とは、同一のまたは本質的に同一のアミノ酸配列をコードする核酸をいい、核酸がアミノ酸配列をコードしない場合には、本質的に同一な配列をいう。遺伝コードの縮重のため、多数の機能的に同一な核酸が任意の所定のタンパク質をコードする。例えば、コドンGCA、GCC、GCG、およびGCUはすべて、アミノ酸アラニンをコードする。したがって、アラニンがコドンにより特定される全ての位置で、そのコドンは、コードされたポリペプチドを変更することなく、記載された対応するコドンの任意のものに変更され得る。このような核酸の変動は、保存的に改変された変異の1つの種である「サイレント改変(変異)」である。ポリペプチドをコードする本明細書中のすべての核酸配列はまた、その核酸の可能なすべてのサイレント変異を記載する。当該分野において、核酸中の各コドン(通常メチオニンのための唯一のコドンであるAUG、および通常トリプトファンのための唯一のコドンであるTGGを除く)が、機能的に同一な分子を産生するために改変され得ることが理解される。したがって、ポリペプチドをコードする核酸の各サイレント変異は、記載された各配列において暗黙に含まれる。好ましくは、そのような改変は、ポリペプチドの高次構造に多大な影響を与えるアミノ酸であるシステインの置換を回避するようになされ得る。
【0070】
本明細書中において、機能的に等価なポリペプチドを作製するために、アミノ酸の置換のほかに、アミノ酸の付加、欠失、または修飾もまた行うことができる。アミノ酸の置換とは、もとのペプチドを1つ以上、例えば、1〜10個、好ましくは1〜5個、より好ましくは1〜3個のアミノ酸で置換することをいう。アミノ酸の付加とは、もとのペプチド鎖に1つ以上、例えば、1〜10個、好ましくは1〜5個、より好ましくは1〜3個のアミノ酸を付加することをいう。アミノ酸の欠失とは、もとのペプチドから1つ以上、例えば、1〜10個、好ましくは1〜5個、より好ましくは1〜3個のアミノ酸を欠失させることをいう。アミノ酸修飾は、アミド化、カルボキシル化、硫酸化、ハロゲン化、アルキル化、グリコシル化、リン酸化、水酸化、アシル化(例えば、アセチル化)などを含むが、これらに限定されない。置換、または付加されるアミノ酸は、天然のアミノ酸であってもよく、非天然のアミノ酸、またはアミノ酸アナログでもよい。天然のアミノ酸が好ましい。
【0071】
このような核酸は、周知のPCR法により得ることができ、化学的に合成することもできる。これらの方法に、例えば、部位特異的変位誘発法、ハイブリダイゼーション法などを組み合わせてもよい。
【0072】
本明細書において、ポリペプチドまたはポリヌクレオチドの「置換、付加または欠失」とは、もとのポリペプチドまたはポリヌクレオチドに対して、それぞれアミノ酸もしくはその代替物、またはヌクレオチドもしくはその代替物が、置き換わること、付け加わることまたは取り除かれることをいう。このような置換、付加または欠失の技術は、当該分野において周知であり、そのような技術の例としては、部位特異的変異誘発技術などが挙げられる。置換、付加または欠失は、1つ以上であれば任意の数でよく、そのような数は、その置換、付加または欠失を有する改変体において目的とする機能(例えば、癌マーカー、神経疾患マーカーなど)が保持される限り、多くすることができる。例えば、そのような数は、1または数個であり得、そして好ましくは、全体の長さの20%以内、10%以内、または100個以下、50個以下、25個以下などであり得る。
【0073】
高分子構造(例えば、ポリペプチド構造)は種々のレベルの構成に関して記述され得る。この構成の一般的な議論については、例えば、Albertsら、Molecular Biology of the Cell(第3版、1994)、ならびに、CantorおよびSchimmel、Biophysical Chemistry Part I:The Conformation of Biological Macromolecules(1980)を参照。「一次構造」とは、特定のペプチドのアミノ酸配列をいう。「二次構造」とは、ポリペプチド内の局所的に配置された三次元構造をいう。これらの構造はドメインとして一般に公知である。ドメインは、ポリペプチドの緻密単位を形成し、そして代表的には50〜350アミノ酸長であるそのポリペプチドの部分である。代表的なドメインは、βシート(βストランドなど)およびα−ヘリックスのストレッチ(stretch)のような、部分から作られる。「三次構造」とは、ポリペプチドモノマーの完全な三次元構造をいう。「四次構造」とは、独立した三次単位の非共有的会合により形成される三次元構造をいう。異方性に関する用語は、エネルギー分野において知られる用語と同様に使用される。
【0074】
本明細書において使用される用語「生体分子」とは、生体に関連する分子をいう。本明細書において「生体」とは、生物学的な有機体をいい、動物、植物、菌類、ウイルスなどを含むがそれらに限定されない。生体分子は、生体から抽出される分子を包含するが、それに限定されず、生体に影響を与え得る分子であれば生体分子の定義に入る。そのような生体分子には、タンパク質、ポリペプチド、オリゴペプチド、ペプチド、ポリヌクレオチド、オリゴヌクレオチド、ヌクレオチド、核酸(例えば、cDNA、ゲノムDNAのようなDNA、mRNAのようなRNAを含む)、ポリサッカリド、オリゴサッカリド、脂質、低分子(例えば、ホルモン、リガンド、情報伝達物質、有機低分子、コンビナトリアルライブラリ化合物など)、これらの複合分子などが包含されるがそれらに限定されない。本明細書において好ましい生体分子は、受容体および受容体フラグメント、ならびにそれらのリガンドである。
【0075】
本明細書においてタンパク質として受容体を使用する場合、受容体としては、レクチン様酸化LDL受容体(LOX−1)を含むスカベンジャー受容体、インスリン受容体ファミリーに属する受容体、EGF受容体ファミリーに属する受容体、PDGF受容体ファミリーに属する受容体、VEGF受容体ファミリーに属する受容体、FGF受容体ファミリーに属する受容体、NGF受容体ファミリーなどの増殖因子受容体、ならびに、TGF-βスーパーファミリー受容体、Toll-like受容体ファミリーの受容体、LDL受容体関連タンパク質ファミリーの受容体、およびGタンパク質共役型受容体ファミリーの受容体が挙げられるが、これらに限定されない。
【0076】
本発明は、標的タンパク質溶液に化合物を添加して誘導される標的タンパク質の立体構造変化をTROSY NMR相関スペクトル(例えば、1H/15N−NMRスペクトル)の変化から読み取る技術を提供する。この技術により、1)化合物の標的タンパク質上での結合部位、2)標的タンパク質が化合物によって誘導される構造変化の2つを同時に観測することができる。
【0077】
タンパク質の多くはリガンド、アゴニストなどの化合物によって活性な構造へ誘導されて機能発現を行う。そのため、標的タンパク質を活性構造に誘導する化合物は作動薬のリード化合物になり得る。その一方で、リガンド、アゴニストなどによるタンパク質の活性構造への誘導を阻害する物質は、新規なアンタゴニストをスクリーニングする際のリード化合物になり得る。従って、化合物が標的タンパク質と結合・接触した際に、そのタンパク質の構造がそのように変化するのかを検出することは、新規な薬物のスクリーニングにおいて重要である。
【0078】
しかし、既存のNMRによる化合物と標的タンパク質の相互作用解析技術は、タンパク質構造変化に関する定量的データを提供しない。そのため、化合物が標的タンパク質と結合・接触した際に、そのタンパク質の構造がそのように変化するのかを検出する方法が望まれている。
【0079】
本発明は、核磁気共鳴を用いることによって、化合物がタンパク質と結合した場合のタンパク質の構造変化を測定する方法、およびその方法およびその方法において使用されるプログラムを提供する。
【0080】
より具体的には、本発明は、核磁気共鳴を用いることによって、任意の化合物がタンパク質と結合・接触した場合の、タンパク質の構造変化を測定する方法に関する。さらに本発明は、任意の化合物がタンパク質と結合・接触した場合に、タンパク質の構造変化を生じる化合物を同定・スクリーニングする方法を提供する。また、本発明は、例えば、リガンド、アゴニスト、などのように、タンパク質と結合した際にそのタンパク質の構造変化を生じることが公知の物質がそのタンパク質と結合した際の構造変化と、任意の化合物がそのタンパク質と結合した際の構造変化とを比較して、リガンド、アゴニストと類似の構造変化をもたらす物質を同定・スクリーニングする方法を提供する。例えば、本発明によって、化合物により誘導されるタンパク質構造変化の定量的情報を提供するため作動薬リード化合物の同定を可能とするスクリーニング法が提供される。
【0081】
この方法は、タンパク質をNMR磁場中に弱く配向させることで観測される異方性スピン相互作用の利用を基にしている。異方性スピン相互作用のひとつである残余双極子効果によるタンパク質中の各結合軸の静磁場に対する配向情報の取得と、そのタンパク質構造解析への応用に関しては多くの研究例が報告されている。また、残余双極子効果を基にしたタンパク質を構成するドメイン間の相対配向を決定する技術も報告されている。ただし、残余双極子観測法は原理的に適用できる標的タンパク質の分子量に限界があり約60kDa(アミノ酸数では約500残基)以下のタンパク質に対してしか適用できない。薬物スクリーニングにおける標的タンパク質としては、通常60kDaよりもはるかに大きな分子量のタンパク質が用いられる。従って、創薬を目的とした化合物スクリーニング技術に適用するには、その汎用性に問題があった。
【0082】
本発明で鍵となる新しい技術は、従来法が持つ標的タンパク質分子量の適用限界を超えて、原理的には900kDa以上のタンパク質を対象としても化合物により誘導されるタンパク質中のドメイン間相対配向を定量的に観測し、その構造変化を測定することできるというものである。
【0083】
例えば、本発明は、新規薬物のリード化合物を探索するスクリーニング技術を提供する。具体的には、化合物との相互作用により誘導される標的タンパク質構造変化をTROSY 1H/15N相関NMRスペクトル、またはTROSY 2H/15N相関NMRスペクトルにより定量的に評価する技術を提供する。さらにこの技術を利用して、標的タンパク質の活性構造への変化の有無を指標とした動作薬(アゴニスト)リード化合物の同定を行うスクリーニング法も提供される。
【0084】
(NMR構造解析)
本明細書において用いられるNMR による高次構造解析に用いられるタンパク質試料は、通常、生合成により安定同位体標識することによって構造解析される。一般には、まず遺伝子操作した大腸菌等によって標的タンパク質の発現系を確立し、安定同位体標識した炭素源や窒素源(すべての炭素を13Cラベルしたグルコース、窒素を15Nラベルした塩化アンモニウムなど)を加えた培地で発現させることにより、すべての炭素および窒素を安定同位体標識したタンパク質を得ることができる。得られた標識タンパク質は、クロマトグラフィーなどで精製した後、限外濾過などで濃縮することによってNMR測定に供することができる。この際、以下のような条件に留意することが望ましい。
【0085】
使用する容器については、タンパク質の粘度が高いことを考慮してガラス壁面に付着、分散を抑えるために、NMR 試料管を含めて、使用する器具にはシリコンなどでコーティング処理を施すことが好ましい。また、試料の扱いには、パスツールピペットなどを用いることが望ましい。
【0086】
溶媒としては、通常、水溶液が用いられる。ただし、タンパク質は多くの交換性アミドプロトンを有しており、重水溶液にしてしまうと、スペクトルの解析で鍵となるアミドプロトンが重水素置換されることにより、多くの情報が失われることから、通常、軽水溶液で試料調製し、その後NMRロック用として10%程度の重水を添加することが行われている。必要に応じて、界面活性剤、還元剤などを加えていてもよい。
【0087】
試料濃度は、通常、1mM 程度(例えば、0.4mM〜2mM)にする。試料の量に余裕があり、溶解度が高いのであれば、5mM〜10mM でも測定可能であるが、濃度を高くすると溶液中で会合が起こる危険性があるので留意する。会合による予期しない解析結果を防ぐためにも、低濃度と高濃度の試料で線形を比較して会合の有無を確認することが好ましい。
【0088】
使用するpHとしては、通常、5〜7程度が好ましい。pH を下げることにより、アミド
プロトンなどの交換可能プロトンの交換速度を遅くなるからである。ただし、pH が低い
と高次構造に影響を与える懸念があるため、円二色性(CD)スペクトル等で確認しておくことが好ましい。
【0089】
緩衝剤としては、スペクトル解析の障害となることを防ぐため、プロトンを持たないものを使用することが望ましい。そのようなものとしては、リン酸緩衝液、重水素化酢酸緩衝液などが挙げられるがそれらに限定されない。
【0090】
使用する温度としては、 生体内の温度に近付けるため、30〜40 ℃程度にすることが望ましい。温度を高めに設定して溶液の粘性を下げることにより、タンパク質分子のT2が長くなることから、一般には温度が高い方が良好なスペクトルが得られる。
【0091】
試料溶液に溶存酸素が含まれていると、常磁性緩和のために緩和時間が短くなり、感度の損失を招き、TROSY相関の検出の障害となる場合もあることから、このことを防ぐために脱気をおこない、溶存酸素を取り除くことが好ましい。脱気操作は減圧による脱気、不活性ガスのバブリングなどがあり、好ましくは減圧脱気が使用される。また、気泡を立てないことが望ましい。
【0092】
溶液の保存のために、アジドを加えることが望ましいが、必ずしも加える必要はない。
【0093】
試料調製ののち、NMRのシム調整を行う。まず、NMRロックをかけて分解能調整を行う。タンパク質溶液NMR の測定は、原則として軽水溶液での測定であるため、巨大な水信号(標的タンパク質の数万倍〜数百万倍の1H 濃度)を消去することが好ましいからである。これは、分解能調整が目的であり、好ましくは、良質のスペクトルを得るため、測定試料ごとに充分な分解能調整を行うことが望ましい。タンパク質溶液NMRの測定で試料管は、通常回転させないことから、Z 軸だけでなく、X、Y軸および高次の項の調整も含めて分解能をしっかりと上げておくことが、良いスペクトルを得るために必要である。
【0094】
次に、パルス幅の測定を行う。水溶液で測定するタンパク質溶液NMRは、塩強度の違いにより試料によってパルス幅が変化する場合があることから、原則として測定試料ごとにパルス幅の測定をおこなうことが必要である。
【0095】
タンパク質溶液NMRで用いられるパルス・シーケンスは、数多くのRFパルスが配列されていることから、パルス幅のズレが全体ではかなり大きく影響してくるとされている。したがって、多次元NMR測定を実行する前に、標的タンパク質の信号でパルス幅を測定し、その値を用いて多次元NMR測定をおこなうことが望ましい。
【0096】
次に、NMRによるサンプルの確認を行う。試料調整してからしばらく保管した後など、タンパク質が劣化している可能性があるときなどにはNMRによる確認をおこなうことが望ましい。
【0097】
そこで、通常15N−1H HSQC測定を行い、2次元NMRスペクトルで確認する。信号のパターンがおかしいとき、位相が合わないようなときなどの不具合があるときは、標的タンパク質が壊れてしまっている可能性があることから、再度実験条件の調整が必要である。
【0098】
次に、スペクトル解析(信号の帰属)を行う。NOEスペクトルを例にとってスペクトルの解析法の概略を以下に説明する。NMRによって得られたNOE由来の距離情報をもとに1H信号の帰属を行い、高次構造を導き出すことができる。幾重にも重なり合った信号を、3次元・4次元といった測定を用いてことごとく分離し、既知のアミノ酸配列に割り当てていく。その原理には、スペクトルがアミノ酸ごとに特定のパターンを持っていること、化学シフトおよびスピン結合定数などに極めて特徴的な法則があることを利用する。このような特徴的な性質から、現在では帰属の自動化も行われている。以下に、タンパク質NMRスペクトルの特徴的な化学シフトを示す。
【0099】
タンパク質の1Hの化学シフトはその環境ごとにおおよそ限定されている。すなわち、アミドプロトン由来の信号(8ppm付近)、側鎖の芳香環由来の信号(7ppm付近)、α位のプロトン由来の信号(4ppm付近)、β位のプロトン由来の信号(2〜3ppm付近)、メチル基由来の信号(1ppm付近)が、それぞれ観測される。
【0100】
13Cの化学シフトにも同様の相関があり、13Cではそれぞれの領域を選択的に励起することで、α位やβ位の炭素、カルボニル炭素などをあたかも別核種のように取り扱うことにより、帰属のための様々な測定法のなかで活用される。
【0101】
アミノ酸配列の帰属に加えて、高次構造に由来する化学シフトの変化が見られる。たとえば、α位のプロトン由来の信号はαヘリックス構造の中では高磁場シフトし、βシート構造の中では低磁場シフトする。その他にも、側鎖の芳香環との位置関係により±1ppm程度シフトすることがある。また金属結合タンパク質などでは、金属と結合した部位でかなり大きなシフトが見られる。
【0102】
次に、同種核スピン結合も考慮すべきである。タンパク質を構成するアミノ酸は、ペプチド結合(アミド結合)で連結されていることから、カルボニル基によって1H−1H間のスピン結合が分断され、1H−1H間のスピン結合は同一アミノ酸残基内に限定される。したがって、1H−1H間のスピン結合のパターンは、アミノ酸の種類ごとに特徴的であり、この事実を利用して、アミノ酸の同定を行う。また、α位のプロトンとアミドプロトンの3JHαHNスピン結合定数から導かれる二面角φは主鎖の2次構造の決定に役立つ。
【0103】
また、異種核間スピン結合も考慮すべきである。主鎖に関連する1H−1H以外のスピン結合定数としては、例えば、H−N(90−100Hz)、N−Cα(11Hz)、Cα−H(140Hz)、Cα−Cβ(30−40Hz)、Cα−C(=O)(55Hz)、Cα−X−N(7Hz)、13C−15N(15Hz)などが挙げられる。
【0104】
また、NOE相関も考慮されるべきである。NOE相関は、高次構造解析に用いられるが、アミドプロトンとα位のプロトンとのカルボニル基をはさんだ1H同種核NOEは、その二つのプロトンが、アミド結合を介して隣り合ったアミノ酸残基に由来することを示しており、1H同種核実験によるアミノ酸配列の帰属で用いられる。この手法を、スピン結合や化学シフトをもとに決定したアミノ酸の種別情報とあわせて、配列特異的連鎖帰属法という。
【0105】
帰属作業が終わったら、必要に応じて、高次構造解析を行う。タンパク質の高次構造には、アミノ酸配列が立体的な規則性を持って形作るαヘリックス構造、βシート構造のような2次構造、いくつかの2次構造が空間的に配置された3次構造、3次構造で構築されたドメインが空間的に配置された4次構造があり、これらを明らかにすることがタンパク質溶液NMR測定において構造解析において重要であり、目的に応じて種々行われる。
【0106】
一次配列が既知のアミノ酸配列の場合、既知のポリペプチド鎖がどのように絡み合ってタンパク質を構築しているのかを、NMRから得られる情報を利用して解像する。この際利用されるのが、ディスタンス・ジオメトリー法、束縛条件付き分子動力学法と呼ばれる構造最適化計算アルゴリズムである。いずれの場合にも、NMRから得られる距離情報としてのNOE信号を集められる限り集めに集め、それをパラメータとして計算プログラムに与えることによって高次構造を導き出す手法である。
【0107】
本明細書において、「ディスタンス・ジオメトリー法」とは、距離情報と結合角(2面角)の集合とを空間的な位置情報としてみて、それをもとに構造を導き出す手法をいう。通常は、共有結合の結合長および結合角を一般的なタンパク質の標準値に固定し、2面角を変数として構造計算を進める。NOE由来の距離の束縛条件を満たす方向に収束した構造をいくつか取り出し、最も適切と思われる構造を最終構造とする。この手法は、変数が少ないために計算も速く、コンピュータに対する負荷も小さいが、原子の接触によって構造変化が制限される(共有結合同士のすり抜けができない)ため、真の構造に収束することが難しく、次に述べる束縛条件付き分子動力学法などの初期構造を求めるために利用される。
【0108】
本明細書において、「束縛条件付き分子動力学法」とは、分子運動をシミュレートするために一般的に用いられている手法として知られる分子動力学法(仮想的に分子を振動させ、構造を最適化する方法)の一つであり、NOE由来の距離の束縛条件をポテンシャルとして加え、構造を最適化していく手法をいう。この手法は位置情報としてデカルト(Cartesian)座標を使用しており、また変数も多いために膨大な計算量が必要とされ、コンピュータに対する負荷も大きいが、現在ではコンピュータの計算能力の飛躍的な進歩により計算時間が短縮され、頻繁に利用されている。
【0109】
本明細書において「シミュレーテッド・アニーリング法」とは、分子動力学計算の1手法であり、一般的な分子動力学計算と異なり、計算の初期段階で共有結合に寄与するパラメータを弱く設定し、系の温度を急激に上げることにより分子運動を激化させ、その後徐々に系の温度を下げて分子運動を遅くしつつ、それぞれのパラメータを強めて構造を収束させる手法をいう。これにより、原子の接触によって構造変化が妨げられることによる見かけの収束に構造が落ち着くことを防ぐことができる。
【0110】
上記で触れた3次元あるいは4次元展開されたNOEスペクトルの観測を行う場合、1H−15Nあるいは、1H−18Cという異種核スピン間での相関シグナルの検出が基本となる。高分子量タンパク質(30kDa以上)を解析対象とする場合、それぞれの核スピンのT2緩和時間が短くなるため生じる早いスピン緩和のために、この異種核相関シグナルを検出することが困難となる。高分子量タンパク質を対象とする場合に避けられない早いスピン緩和の問題を解決し、高分子量タンパク質でも十分な観測感度で異種核種の相関シグナルの観測を可能とする方法として、TROSYがWuthrichのグループによって開発された。この新しい異種核間相関シグナルの検出技術は、これまでタンパク質立体構造決定に用いられてきた既存の測定法に組み込むことが可能であり、上記のタンパク質構造解析過程はTROSYを利用したデータ測定法に切り替えることで、30kDaを超える分子量のタンパク質にも適用可能である。
【0111】
以上、本明細書において使用可能なNMR手法を説明したが、このような手法は、当該分野において公知であり、例えば、Kurt Wuthrich 著,京極好正・小林祐次訳,タンパク質と核酸のNMR:二次元NMR による構造解析,東京化学同人.(1991);M.Sattler,J.Schleucher,C.Griesinger,Progr.NMR Spectrosc.34,93−158.(1999);J.Cavanagh,W.J.Fairbrother,A.G.Palmer III,N.J.Skelton,Protein NMR Spectroscopy:Principles and Practice,Academic Press.(1995);T.L.James and N.J.Oppenheimer(eds.),Methods in Enzymology,Vol.239,Academic Press.(1994);荒田洋治,タンパク質のNMR:構造データの解釈と評価,共立出版.(1996); 荒田洋治,NMR の書,丸善.(2000); 根本暢明,吉田卓也,小林祐次,科学と工業,69 69(10),419−425.(1995);根本暢明,吉田卓也,小林祐次,科学と工業,70 70(2),48−55.(1996)などに記載されており、本明細書においてその内容を参考として援用する。
【0112】
(作動薬のリード化合物の同定)
作動薬のリード化合物の同定の1形態を以下に説明する。タンパク質は図1に楕円で示すようにドメインと呼ばれる部分構造体がつながって形作られる。多くのタンパク質は、機能発現の際にこのドメイン間の相対的配置が変化して、機能を発現するに最適な構造をとる。活性構造への誘導は、通常は低分子化合物のタンパク質への結合により行なわれる。阻害剤は、タンパク質へは結合するが活性構造へは誘導せず不活性な構造を維持するように作用する。一方、タンパク質を活性化する化合物の場合は、タンパク質に結合することにより機能発現に最適な立体構造変化を誘導する。化合物結合により誘導されるタンパク質構造が活性構造かどうかは、天然の基質が結合したことにより誘導される立体構造を雛形として判断することができる。本発明は、タンパク質のドメイン間の相対配向を定量的に決定するNMR解析技術を提供することで、TROSYスペクトル変化という簡便に観測可能な実験値からタンパク質が活性構造にあるかどうかを判定することを可能とする。この技術を利用することで、図1にイメージとして描いているようなドメイン間の相対配向変化を基にして選択的に作動薬のリード化合物を同定することができる。
【0113】
(タンパク質の分子配向の計算法)
NMRスペクトルを用いて、タンパク質の分子配向の決定に必要な計算を、例えば、以下の様に行うことが可能であるが、これに限定されない。
【0114】
TROSYスペクトル上で観測されるシグナルの15N軸方向での分子配向依存的な変化量Δδtrosy(k)は、1H−15N核スピン間の残余双極子効果Dnhと、15N核まわりの電子分布の空間的異方性に由来する分子配向依存的な15N軸方向でのシグナルの変化量Δδ15Nの2つの量により次の関係で結び付けられる。
【0115】
Δδtrosy=(1/2)Dnh+Δδ15N (1)
残余双極子効果Dnhはタンパク質中の主鎖にある各NH結合スペクトルのi番目(i=x、y、z)の分子座標軸に対する角度φiと分子の外部磁場に対する分子の配向を規定するSaupeオーダーマトリクス、S(Sijは、分子配向マトリクスの要素を表す。i,j=x,y,z)により次の関係により表現される。D0nhは1H−15N間の双極子間相互作用の強度を示す。
【0116】
Dknh=D0nhΣSijcosφkicosφkj (2)
i,j=x,y,z
一方15N核周りの電子分布の異方性のために生じる、分子の外部磁場に対する配向依存的な15N核の外部磁場に対する磁気遮蔽度の違いに由来する15N化学シフトの変化Δδ15Nは、各ペプチド結合に固有な化学シフト異方性テンソルを分子座標軸上で表現した要素σij i,j=x,y,zと分子配向を規定するSaupeオーダーマトリクスを用いて次の式で表現できる。
【0117】
Δδ15N=(2/3)ΣSijδij (3)
i,j=x,y,z
式(1)−(3)の関係から、分子の磁場中での配向角度に依存したTROSYシグナル15N軸方向での変化量Δδtrosyは、次の式で表現できる。
【0118】
Δδtrosy(k)=ΣSij{(1/2)D0nh cosφkicosφkj+(2/3)δij} (4)
i,j=x,y,z
スペクトル上で観測される複数のシグナルkについて得られる式(4)の関係を用い、特異値分解法(Singular Value Decomposition)により実験値Δδtrosyと、標的タンパク質のドメイン構造内の原子座標から計算される値{(1/2)D0nhcosφi
cosφj+(2/3)δij}から、分子配向を決定するために必要なSaupeオーダーマトリクスの全ての要素Sijを決定することができる。この場合、少なくとも5以上、好ましくは少なくとも6以上、より好ましくは少なくとも7以上、少なくとも8以上、少なくとも9以上、または少なくとも10以上のシグナルkについて得られる式(4)の関係を用いる。また、全てのシグナルkについて式(4)の関係を用いることも可能である。Saupeオーダーマトリクスを対角化することで、標的タンパク質または標的タンパク質内のドメイン構造の静磁場に対する配向情報を得ることができる。ここで、得られる物理量は、分子座標軸で表現される分子配向テンソル量として与えられる。この分子配向テンソル量は分子配向強度(配向テンソルという3次元直行座標系で表現される各軸方向の大きさであり、どの程度その軸に分子が配向しているか(存在確率に相当する)を表現する量である)と、分子の配向状態を記述する新たな座標軸(Z軸が静磁場方向となる)の分子座標に対するオイラー角(a、b、c)で表現する。なお、上記の式は、15Nを用いて例示的に示しているが、必ずしも15Nを用いる必要はなく、その他の安定同位体(例えば、13Cなど)を用いることもできる。
【0119】
(タンパク質の配向情報の決定)
タンパク質構造は、x、y、z軸上の原子の位置によって表現される。タンパク質が磁場に対して配向する場合には、配向状態にあるタンパク質を表現する座標軸(Dxx,Dyy,Dzz)がもとの座上軸からどの程度傾いているかで表現する。そのために、図に示すような3つの角度、α、β、γ、が定義される。これをオイラー角と呼ぶ。磁場の配向方向はDzz軸に一致すると仮定して表現される(図2)。
【0120】
本明細書において、ドメイン構造変化を表す指標としては、分子配向変化度(DOC)および活性構造一致度(DOS)が挙げられるがこれらに限定されない。
【0121】
本明細書において、2つの配向の間での「分子配向変化度」(DOC;degree of orientational change)は、例えば、
(i)化合物と結合・接触前のタンパク質ドメインの第1の配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【0122】
【化17】
を用いて示し、
(ii)化合物と結合・接触後のタンパク質の第2のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【0123】
【化18】
を用いて示し、
(iii)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、この角度をドメイン構造変化を表す指標とするか、または、以下の式
分子配向変化度=a2+b2+c2、
によって分子配向変化度を求め、この分子配向変化度を構造変化の比較のための指標とする。例えば、この角度a、b、およびcが小さい場合、またはDOCが小さい場合、化合物の結合・接触によって誘導されたタンパク質の構造変化が、化合物が存在しない場合と大きく違わないことを示す。また、この角度a、b、およびcが大きい場合、またはDOCが大きい場合、化合物の結合・接触によって誘導されたタンパク質の構造変化が、化合物が存在しない場合と大きく異なることを示す。
【0124】
また、アゴニスト化合物をスクリーニングする場合、リガンド結合によって誘導された構造変化が、どの程度活性構造に近いかを指標とすることができる。この場合、リガンドと結合・接触した後の活性構造のドメイン配向を規定する分子配向テンソルの各軸を表現する第3の単位ベクトルを
【0125】
【化19】
として、化合物と結合・接触した後の第2の単位ベクトルとを用いて、これら2つの単位ベクトルの各々の間の角度を、a、b、およびcとして表し、この角度をドメイン構造変化を表す指標とするか、または、活性構造一致度(degree of orientational similarity to the active conformation;DOS)を以下の式
活性構造一致度=a2+b2+c2、
によって求め、この分子配向変化度を構造変化の比較のための指標とする。例えば、この角度a、b、およびcが小さい場合、またはDOSが小さい場合、化合物の結合・接触によって誘導されたタンパク質の構造変化が、リガンドによって誘導されたタンパク質の構造変化に近い、すなわち構造変化が類似することを示す指標となる。
【0126】
また、タンパク質内のドメインの構造変化は、ドメインの比較する各々の状態での配向情報に基づくDOCおよび/またはDOSの差異として数値化される。
【0127】
従って、本明細書において、第1の化合物と該タンパク質とが接触した場合のタンパク質の構造変化と、第2の化合物と該タンパク質とが接触した場合のタンパク質の構造変化とを比較した場合において、タンパク質の構造変化が「類似する」とは、構造変化の指標としてのDOSの差が小さいことを意味する。例えば、タンパク質と接触することによって、第1の化合物と該タンパク質とが接触した場合のタンパク質の構造変化と類似するタンパク質の構造変化を生じる、第2の化合物の選択は、
(a)タンパク質内のドメインを選択する工程;
(b)タンパク質と該第1の化合物を接触させる条件下における、ドメインの配向情報を得る工程;
(c)タンパク質と第2の化合物を接触させる条件下における、ドメインの配向情報を以下
(i)ドメインの既知の原子座標を得て、
(ii)液晶物質存在下において該タンパク質と該第2の化合物を接触させ、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(iii)ドメインの原子座標、およびNMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(iv)決定されたマトリクスを対角化して、ドメインの配向情報とする、
ことにより得る工程;および
(d)工程(b)において得られた配向情報と、工程(c)において測定された配向情報について、DOSを比較する工程;
によって行なわれる。
【0128】
従って、第1の化合物によって生じる構造変化と第2の化合物によって生じる構造変化が同一の場合、a=b=c=0であり、従って、DOS=0である。第1の化合物によって生じる構造変化と第2の化合物によって生じる構造変化が「類似する」とは、例えば、DOSの値が、75(deg2)以下、48(deg2)以下、27(deg2)以下、12(deg2)以下、3(deg2)以下、0.75(deg2)以下、0.48(deg2)以下、0.27(deg2)以下、0.12(deg2)以下、または0.03(deg2)以下の場合をいう。
【0129】
あるいは、構造変化が類似するか否かは、例えば、複数の候補化合物の集団内の第2の化合物と第1の化合物との間でDOSを計算して、相対的に小さいDOSを生じるか、または最も小さいDOSを生じる第2の化合物が、第1の化合物によって生じる標的タンパク質の構造変化と類似する構造変化を生じるとして、判断することが可能である。
【0130】
(タンパク質の原子座標の入手)
タンパク質のドメイン間構造と機能について、米国Yale大学のM.Gresteinのグループがタンパク質の構造変化の様式をMolMovDBというデータベースにまとめている(MolMovDB、http://MolMovDB.org)。
【0131】
例えば、GroEL、アデニレートキナーゼ、カルモジュリン、HIV−1逆転写酵素、クエン酸シンターゼ、ジフテリアトキシン、HIV−1プロテアーゼ、EGF−レセプター、ヘキソキナーゼ、グルコキナーゼ、EPOレセプター、ホスホグリセレートキナーゼ、リコベリン(recoverin)、mRNAキャッピング酵素、DNAポリメラーゼβについての情報がデータベースから利用可能である。
【0132】
(タンパク質の原子座標から、タンパク質ドメインの配向情報を得る方法)
上記記載の方法によってタンパク質の原子座標を入手した場合、その原子座標から、タンパク質のドメインの配向情報を得ることが可能である。例えば、残余双極子効果Dnhはタンパク質中の主鎖にある各NH結合スペクトルのi番目(i=x、y、z)の分子座標軸に対する角度φiと分子の外部磁場に対する分子の配向を規定するSaupeオーダーマトリクス、S(Sijは、分子配向マトリクスの要素を表す。i,j=x,y,z)により次の関係により表現される。
【0133】
Dknh=D0nhΣSijcosφkicosφkj (5)
i,j=x,y,z
そのため、タンパク質のドメインの原子座標より計算されるcosφki cosφkjと、1H−15N核スピン間の残余双極子効果Dnhと、1H−15N間の双極子間相互作用の強度D0nhとを用いることによって、分子配向を決定するために必要なSaupeオーダーマトリクスの全ての要素Sijを決定することができる。
【0134】
(タンパク質に化合物が結合した際の、タンパク質上の結合位置を特定する方法)
化合物が、タンパク質に結合した場合に、そのタンパク質上の結合位置を、例えば、以下のようにして決定することが可能である。
・化合物の非存在下におけるタンパク質の二次元TROSY NMRスペクトルを得ること、
・化合物をタンパク質と接触させて、タンパク質の二次元TROSY NMRスペクトルを得ること、
・上記のにおいて得られた各々の二次元TROSY NMRスペクトルを比較して、スペクトルの変化を検出すること、
・スペクトルの変化を生じた原子がどのアミノ酸残基の原子であるのかを、二次元NMRの帰属から決定することにより、その化合物が結合したタンパク質の部位周辺のアミノ酸残基(好ましくは複数のアミノ酸残基)を特定すること、
・必要であれば、その化合物の立体構造と、上記において特定されたアミノ酸残基周辺のタンパク質の立体構造とを比較し、その化合物がタンパク質上の結合部位に結合した際の立体構造を推定すること、
によって行なわれる。
【0135】
(タンパク質内ドメインの構造変化の測定)
本発明の1つの局面に従って、例えば、以下の工程を行って、タンパク質内ドメインの構造変化を測定し、および/または構造変化(好ましくは、所望の構造変化)を生じる化合物を選択することが可能である。
【0136】
(a)該タンパク質内のドメインを選択する工程;
(b)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を得る工程;
(c)該タンパク質と該化合物を接触させる条件下における、該ドメインの配向情報を以下
(i)該ドメインの既知の原子座標を得て、
(ii)液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(iii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(iv)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、
ことにより得る工程;および、
(d)工程(b)において得られた配向情報と、工程(c)において得られた配向情報との差異から、該タンパク質の構造変化を測定する工程。
【0137】
本発明の方法の測定対象となるドメインとしては、測定対象となるタンパク質内の任意のドメインが利用可能である。しかし、測定対象としてのドメインとしては、例えば、化合物との結合の有無によって配向情報の変化が大きなドメイン、および/または異なる化合物との結合によって異なる配向情報の変化を生じるドメインが好ましい。
【0138】
上記測定方法における前記(b)工程は、必要に応じて
(b)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を以下
(v)該ドメインの既知の原子座標を得て、
(vi)液晶物質存在下において、該タンパク質を該化合物と接触させずに、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(vii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(viii)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、ことにより得る工程であっても、または
(b)前記タンパク質が前記化合物と接触しない条件下において既に得られている原子座標から、前記ドメインの配向情報を得る工程;
であってもよい。上記複数のSaupeオーダーマトリクスエレメントを決定するためのTROSYスペクトルは、同時にまたは別に測定され得る。
【0139】
また、前記工程(b)および/または(c)において二次元TROSY NMRスペクトルを用いて得られる分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量は、15Nを用いる場合は、15N軸方向の変化量であり得る。別の安定同位体の核種を用いる場合は、その核種の軸方向の変化量であってもよい。
【0140】
ここで、前記(iii)および/または(vii)Saupeオーダーマトリクスエレメントの決定は、15Nを用いる場合は、好ましくは、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定することによって行われる。15N以外の安定同位体の核種を用いる場合は、その核種の核スピン対について上記計算を行ってもよい。その場合、N−H結合の距離に対応する別の異種核間結合の距離および静的双極カップリングが公知であることから、その公知の値を上記計算式において用いることができる。
【0141】
タンパク質内ドメインの構造変化は、例えば、Saupeオーダーマトリクスエレメントを対角化して、ドメインの配向情報とすることによって、得られる。従って、ドメインの配向情報自体を相互に比較することによって、ドメインの構造変化の指標とすることが可能である。例えば、化合物の存在下と非存在下における所定のドメインの配向情報に変化がある場合、その化合物は、タンパク質と結合した場合に、その所定のドメインの構造変化をもたらしたことが判明する。化合物の存在下と非存在下の配向情報は、各々を実験的に求めることが可能であるが、例えば、化合物の非存在下での配向情報が、NMR実験またはX線解析などによって既に得られている場合、その情報を用いて、化合物存在下での実験によって得られた配向情報と比較することが可能である。
【0142】
さらに、構造変化の指標としては、DOCおよびDOSを用いることができる。そのため、任意の化合物がもたらす構造変化の大きさとしては、DOCを用いることができる。DOCの値が大きい場合、構造変化が大きいことを示す。また、任意の化合物が、所定のリガンドまたはアゴニストと類似の構造変化を生じるか否かについては、DOSを用いることができる。DOSの値がより小さい場合、構造変化がより類似することを示す。
【0143】
標的タンパク質の活性誘導を行う作動薬をスクリーニングは、例えば、以下の手順によって行うことが可能である:
・ドメインの構造変化をもたらすことが公知のリガンドまたはアゴニストが標的タンパク質に結合する条件下で、標的タンパク質内ドメインの配向情報(A)を得る;
・スクリーニングの候補化合物が標的タンパク質に結合する条件下で、標的タンパク質内ドメインの配向情報(B)を得る;
・これら配向情報(A)および(B)を比較して、これら配向情報が同一であるか、または類似するかを決定する;
・これら配向情報が同一または類似である場合、候補化合物は、標的タンパク質の活性誘導を行う作動薬と同様の構造変化を標的タンパク質にもたらす化合物であることが判明する。従って、このようにして選択された候補化合物は、有用な作動薬リード化合物である。
【0144】
本発明においては、必要に応じて、化合物がタンパク質に結合した位置を特定する工程を行うことも可能である。その位置の特定は、例えば、工程(b)において得られた二次元TROSY NMRスペクトルと、工程(c)において得られた二次元TROSY NMRスペクトルとを比較して、スペクトルの変化を検出し、そして、該スペクトルの変化を生じた、該タンパク質中のアミノ酸残基を同定すること、によって行われる。このアミノ酸残基の同定は、変化したスペクトルのアミノ酸配列への帰属に基づいて、行われる。
【0145】
本発明において使用される液晶物質としては、以下からなる群から選択される混合物が挙げられる:
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とドデシル硫酸ナトリウム(SDS)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物、
1,2−ジ−O−ドデシル−sn−グリセロ−3−ホスフォコリン(DIODPC)と3−(コラミドプロピル)−ジメチルアンモニオ−2−ヒドロキシ−1−プロパンサルフェート(CHAPS)との混合物、
n−アルキル−ポリ(エチレングリコール)/n−アルキルアルコール混合物、
繊維状ファージ、
セチルピリジニウムクロライド(CPCl)−ヘキサノール−NaCl混合物、
セチルピリジニウムブロミド(CPBr)−ヘキサノール−NaCl混合物、
Halobacterium属の紫膜断片
微結晶セルロース、および
ポリアクリルアミドゲル(例えば、アクリルアミド/アクリレートゲル)、
ならびに上記混合物の少なくとも2つを混合した混合物。
【0146】
例えば、液晶物質は、7.5%(w/v)のジミリストイルホスファチジルコリン
(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物である。
【0147】
本発明の別の局面において、本発明の方法を実行するプログラム、およびそのようなプログラムを含む記憶媒体も提供される。
【0148】
本発明のさらなる局面において、本発明の方法を実行するために使用し得る装置、および本発明のプログラムを含む装置が提供される。
【0149】
以下に実施例等により本発明を詳しく説明するが、本発明はこれらに限定されるものではない。
【実施例1】
【0150】
(液晶物質によるタンパク質の配向)
液晶物質によるタンパク質の配向は、例えば、以下の方法により行われる。
【0151】
(1.ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)との混合物を用いる、タンパク質の弱い配向化)
好ましくは6.8−8.0のpH、より好ましくは、6.5のpHで、DMPCとDHPCとを、好ましくは2.5−3.0の混合比、より好ましくは3.0の混合比で混合する。この混合物を、好ましくは5−20%(w/v)、より好ましくは7.5%(w/v)の濃度で使用し、好ましくは25−40℃、より好ましくは、30℃で分子配向を行う。(M.Ottinger and A.Bax、J.Biomol.NMR 12、361(1998))
(2.ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とドデシル硫酸ナトリウム(SDS)との混合物を用いる、タンパク質の弱い配向化)
上記1のDMPC/DHPC溶液に対して、例えば、SDSのような界面活性剤を添加して、リン脂質膜表面を負に帯電させ、負の表面電荷を持つタンパク質のリン脂質への不必要な吸着を防ぐ。SDSを添加した場合でも、pHを、好ましくは6.0−8.0、より好ましくは、pHを6.5として、好ましくは5−20%(w/v)の濃度で、より好ましくは、7.5%(w/v)で、タンパク質試料溶液と混合し、好ましくは20−35℃、より好ましくは30℃で分子配向を行う。この場合、DMPCとDHPCとを、好ましくは2.5−3.0の混合比、より好ましくは3.0の混合比で混合する。SDSは、DMPCに対して、好ましくは1−3%添加する。(J.A.Losonczi and
J.H.Prestigard、J.Biomol.NMR 12、447(1998))
(3.ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物を用いる、タンパク質の弱い配向化)
上記1のDMPC/DHPC溶液に対して、正の表面電荷を有するタンパク質のリン脂質への不要な吸着を避けるためには、例えば、CTABのような界面活性剤を添加する。CTABを添加した場合でも、pHを好ましくは6.0−8.0、より好ましくはpHを6.5として、好ましくは5−20%(w/v)の濃度で、より好ましくは7.5%(w/v)で、タンパク質試料溶液と混合し、好ましくは20−35℃、より好ましくは30℃で分子配向を行う。この場合、DMPCとDHPCとを、2.5−3.0の混合比、好ましくは3.0の混合比で混合する。CTABは、DMPCに対して、1−3%添加する。(J.A.Losonczi and J.H.Prestigard、J.Biomol.NMR 12、447(1998))
(4.1,2−ジ−O−ドデシル−sn−グリセロ−3−ホスフォコリン(DIODPC)と3−(コラミドプロピル)−ジメチルアンモニオ−2−ヒドロキシ−1−プロパンサルフェート(CHAPS)との混合物を用いる、タンパク質の弱い配向化)
DIODPC/CHAPSOの割合は、4.8が最適であるがこれに限定されない。試料中の最適なリン脂質膜濃度は、5%(w/v)であるがこれに限定されない。試料溶液のpHは、好ましくは1.0−6.5の範囲である。分子配向は、好ましくは10−60℃の範囲である(S.Cavagneroら、J.Biomol.NMR13、387(1999))。
【0152】
(5.n−アルキル−ポリ(エチレングリコール)/n−アルキルアルコール混合物を用いる、タンパク質の弱い配向化)
上記のアルコール混合物を、限定されることはないが、溶液に5%(w/v)添加することにより、好ましくは0−40℃の範囲で分子配向を誘導できる。アルコールの種類としては、n−ヘキサノールまたはn−オクタノールを用いる。n−アルキル−ポリ(エチレングリコール)/n−アルキルアルコールの割合は、好ましくは0.6−0.9の範囲である。pHは、任意のpHが使用可能である。(M.Ruckert and G.Otting、J.Am.Chem.Soc.122、7793(2000))
(6.繊維状ファージを用いる、タンパク質の弱い配向化)
pf1またはタバコモザイクウイルス(TMV)のような繊維状の形態を有するファージを試料溶液中に共存させることによって、磁場中での分子配向を実現できる。試料溶液中でのファージ濃度は、好ましくは10−50mg/mlの範囲で用いられる。より好ましくは25mg/mlの濃度である。pHは好ましくは6以下である。任意の温度が使用可能であるが、より好ましくは20−40℃の範囲で使用される。(Hansenら、Nat.Struct.Biol.5、1065(1998);Cloreら、J.Am.Chem.Soc.120、10571(1998))
(7.セチルピリジニウムクロライド(CPCl)−ヘキサノール−NaCl混合物、またはセチルピリジニウムブロミド(CPBr)−ヘキサノール−NaCl混合物を用いる、タンパク質の弱い配向化)
CPBr:ヘキサノール=1:1.33(w/w)〜1:1(w/w)を用いる。NaBrの濃度は、好ましくは25−40mMであり、NaClの濃度は、好ましくは200−500mMである。この液晶物質を使用する際の濃度は、好ましくは1−10%(w/v)であり、好ましくは15−50℃の範囲の温度で分子配向する。
【0153】
(8.Halobacterium属の紫膜断片を用いる、タンパク質の弱い配向化)
Halobacterium属の紫膜断片、好ましくは、Halobacterium salinariumの紫膜断片を、1−10mg/mlの濃度で用いる。任意の溶液温度、および任意のpHが利用可能である。溶液中のNaCl濃度は、好ましくは50mM以下である。(Prosserら、J.Am.Chem.Soc. 120、11010(1988))
(9.微結晶セルロースを用いる、タンパク質の弱い配向化)
好ましくは、80mg/mlの溶液を調製し、タンパク質を導入することによって、分子配向が可能である。任意のpH、温度が利用可能である。溶液のイオン濃度が上昇すると、分子配向を実現するために必要となる試料溶液に添加する。(Flemingら、J.Am.Chem.Soc.122、5224(2000))
(10.ポリアクリルアミドゲルを用いる、タンパク質の弱い配向化)
NMR試料管内に、縦方向、または横方向に圧縮したポリアクリルアミドゲル中に、タンパク質溶液を注入して、タンパク質を機械的に分子配向させる。任意のpH、イオン強度、測定温度が使用可能である。(Saasら、J.Biomol.NMR 18、303(2000);Tyckoら、J.Am.Chem.Soc.122、9340(2000))
(11.アクリルアミド/アクリレートゲルを用いる、タンパク質の配向化)
ポリアクリルアミドゲルの一例として、50%アクリルアミド/50%アクリレートゲルを2.5倍に伸長させることによって作製した配向ゲルを用いることができる。アクリルアミド/アクリレートを用いてゲルを生成する際に、標識したタンパク質を添加することによって、ゲル中でタンパク質を配向させることができる。
【実施例2】
【0154】
(TROSYシグナル変化から得た分子配向情報の従来法による解析結果との比較)
0.5mM ユビキチン(8.6kDa)を20mM リン酸ナトリウム緩衝液(pH6.4)に溶かして7.5%(w/v)のDMPC/DHPC/CTABを共存させた溶液を30℃で分子配向状態を作り、以下の手順で実験を行なった。
【0155】
(実験)
(NMR分光法)
本分析で使用された全てのスペクトルを、セチルトリメチルアンモニウムブロミド(CTAB)をドープされた、ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とのバイセル(bicelle)混合物が、2H NMRスペクトルにおいて、10Hzの残余四重極分裂を与えるように配列される条件下で、30℃で記録した。データ収集を、1H周波数が499.84MHzで作動するVarian INOVA500分光器上で実施した。マジック角度実験について、本発明者らは、15Nに対して調整されたXを備える1H検出コイルVarian gHX NanoProbeを使用した。サンプル回転速度を、2.6kHzに設定し、磁界に対してバイセルを配列したことに関するトルクを除去した(Courtieu,J.; Bayles, J. P.; Fung, B. M. Prog. NMR spectrosc. 1994,26, 141-169.、Kurita,J.; Shimahara, H.; Utsunomiya-Tate, N.; Tate, S. J. MagnReson. 2003,163,163-173.)。1H−15N双極子カップリングを、1H−15N IRAP−HSQCスペクトルにおいて測定した(Ottiger,M.;Delaglio, F.; Bax, A. J. Magn Reson. 1998, 131, 373-378.)。1H−15N TROSYスペクトル(Pervushin,K.;Riek, R.; Wider, G.; Wuthrich, K. Proc. Natl. Acad. Sci. U. S. A 1997,94,12366-12371.、Weigelt, J. J. Am. Chem. Soc. 1998, 120, 10778-10779.)を、7.5%(重量/体積)のDMPC:DHPC:CTABバイセルを含有する、50mMのリン酸ナトリウム(pH6.4)および2mM EDTA中に溶解された0.5mM 15N標識ユビキチンを含む同一の溶液をサンプル溶液を使用して、マジック角度サンプル回転法を用いずに、およびそれを用いて、それぞれ等方的状態および異方的状態において収集した。
【0156】
全てのデータを、NMRpipeプログラムソフトウェアパッケージ(Delaglio, F.; Grzesiek, S.; Vuister, G. W.;Zhu, G.; Pfeifer, J.;Bax, A. J. Biomol. NMR 1995, 6, 277-293.)を用いて処理した。ピーク位置を、プログラムPIPP(Garrett,D.S.; Powers, R.; Gronenborn, A. M.; Clore, G. M. J. Magn. Reson. 1991,95,214-220.)を用いてピーク最大値の60%と80%との間の各計算等高線に対して楕円をフィッティングすることによって決定した。各楕円の中心は、ピーク位置の基準を提供し、所定のピークの60〜80%の強度範囲における全ての等高線について得られた値を、ピーク位置を提供するために平均した。t1次元およびt2次元でのデータ収集のための取得時間は、それぞれ111msおよび128msであり、200(t1、15N)×1024(t2、1H)複素点のマトリクスサイズを有した。広範囲なゼロフィルをされた最終処理データは、1H次元および15N次元それぞれについて、2.0Hzおよび0.88Hzのデジタル分解能を示した。
【0157】
(配列テンソルの計算)
配列誘導性TROSYシフト変化およびその誤差評価に由来する整列テンソルの決定を、公開されたサブルーチンを組込んだ、組織内で書かれたC−プログラムによって実施した(Press,W. H.; Teukolsky, S. A.; Vetterling, W. T.; Flannery, B. P.Numerical recipes in C 2nd ed.; Cambridge university press: New York, 1992.)。配列テンソルを、実験データから計算し、そしてプログラムMOLMOL(Koradi,R.;Billeter, M.; Wuthrich, K. J. Mol. Graph. 1996, 14, 51-55.)によって付加された水素原子を有する1.8ÅのX線座標(PDB登録コード、1UBQ)(Vijay-Kumar,S.;Bugg, C. E.; Cook, W. J. J. Mol. Biol. 1987, 194, 531-544.)から生成された参照構造から計算した。TROSYシフトの実験不確定性の効果を、生成されたデータセットに対して、特異値分解(SVD)計算(Losonczi,J.A.; Andrec, M.; Fischer, M. W.; Prestegard, J. H. J. Magn. Reson. 1999,138,334-342.)を繰り返すことによって評価した。このデータセットは、ガウス雑音を実験データに付加することによって生成された。ガウス雑音を、相対的確率(−exp(x2/2δ2))で分布すると仮定した。ここで、xは、実験値であり、δは、TROSYシフトの連続的な実験の間で対rms偏位から見積もられたrmsノイズである。
【0158】
「構造ノイズ」を、文献(Zweckstetter, M.; Bax, A. J. Biomol. NMR, 2002, 23, 127-137.)に記載される方法と類似する方法で、ランダムな様式でNH結合ベクトルを再配向することによって、シミュレートした。オリジナルベクトルと再配向ベクトルとの間の偏位を、オリジナルの方向から5度の傾斜角を有する円錐で記述する。本構造ノイズのシミュレーションにおいて、オリジナルのNHベクトルと再配向NHベクトルとの間の傾斜角を、ガウス分布(−exp(α2/2θ2))にあると仮定した。ここでαは、傾斜角であり、θは、5度に固定された傾斜角に関する推定された標準偏差である。円錐中の再配向されたベクトルの横方向の回転角を、等しく分布されているとしてシミュレートした。シミュレートされたデータからの適切な値の選択は、実験データとシミュレートされたΔδtrosy値との間のrms偏位を、2.55Hz未満に維持することによって行った。この値は、シミュレートされた値の68%に対して受容される速度を示し、この値は、これらの値が、平均の1標準偏差内に存在することを意味する。
【0159】
30℃で7.5%(w/v)のDMPC/DHPC/CTABバイセルに溶解した15N標識ユビキチンについてのΔδtrosy値を以下の表に示す。
【0160】
【表1】
上記の結果を得た実験においては、それぞれ、異方性の状態、および整列された状態において収集されたデータを得るために、MASを適用せずに、またはMASを適用して、実験を行なった。MAS(マジック角試料回転)とは、magic angle sample spinningの略である。MASを適用してNMR測定を行うと、液晶物質共存下であっても、物理的に分子配向を消すことが可能である。そのため、液晶物質共存下であっても、液晶物質非共存下の場合と同じデータを得ることができる。
【0161】
一連の1H−15N TROSYデータから、Δδtrosy値を、
Δδtrosy値=δ15Ntrosy(MAS非適用)−δ15Ntrosy(MAS適用)
として求めた。
【0162】
Saupeオーダーマトリクスエレメントの決定を以下のとおりに行った。
【0163】
ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させることなく、二次元NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (6)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定した。
【0164】
このSijを対角化することによって、オイラー角α、β、γを得た。
【0165】
この分子配向により誘導されたTROSYシグナル変化から決定されたユビキチンの分子配向角度を従来の残余双極子効果観測から得た。
【0166】
残余双極子効果観測は、従来の残余双極子効果観測法:IPAP-HSQC法(Inphase-Antiphase mode Heteronuclear Single Quantum Coherence spectroscopy; Ottigerら、J.Magn.Reson. 131, 373 (1998))を用いて、液晶分子を利用して分子配向を誘導する前後において観測される各アミノ基に由来するシグナルの1Jnh(1H-15N核スピン間の単結合スピン結合定数)が示す強度変化を観測し、液晶共存下で分子配向状態で観測される1Jnh(aniso)と、液晶が共存せず蛋白質分子が配向状態にない時に観測される1Jnh(iso)を用いて、
Dnh = 1Jnh(aniso) - 1Jnh(iso)
として、各1H-15N核スピン間の残余双極子効果Dnhを求めることにより行なった。
【0167】
残余双極子効果観測から得られた分子配向角度と比較した結果は以下のとおりである。(分子座標軸に対する分子配向軸の傾きをオイラー角表示で示す)。
【0168】
【表2】
この結果から、TROSYシグナル変化に基づいて決定される分子配向角度情報は、従来の分子配向決定技術と整合性のある結果を与えることが確認された。すなわち高分子量タンパク質を対象として、化合物との結合により誘導される標的タンパク質の構造変化を定量的にTROSYシグナル変化だけから観測することが可能であると示された。
【実施例3】
【0169】
(高分子量タンパク質について、本発明を用いる構造変化の測定)
従来のNMRでは測定することができなかった30kDaを超える分子量を有するタンパク質の配向情報を、本発明を用いて以下のように得て、構造変化を測定した。
【0170】
対象のタンパク質として、35kDaの分子量を有する、クロレラウイルスPBCV1のグアニリル転移酵素であるmRNA5’−キャッピング酵素(PBCV1-CE)を用いた。35kDaという分子量は、従来の測定法による分子配向解析では困難になる分子量域である。このタンパク質はリガンドにより誘導される立体構造の変化に依存して活性が制御されることが公知である。結晶構造解析により、リガンド結合していないPBCV1-CEは結晶中で2つの構造(オープン型およびクローズ型)を取ることが明らかにされている。本実施例では活性型構造を誘導するGTPの結合阻害剤であるピロリン酸共存下でのPBCV1-CEの溶液中での構造を解析し、リガンドを持たない構造からの変化を解析した。
【0171】
(主鎖の帰属)
TROSY測定のデータ解析には、主鎖の帰属データが必要であるので、2H/13C/15Nで三重標識したPBCV1−CEを用いて約50%の主鎖の帰属を行った。
(TROSY測定条件)
2H/15N標識した0.5mMのPBCV1-CE(200mM リン酸カリウム,pH7.6中)を、6%(w/w)の約2.5倍に伸張させた50%アクリルアミド/50%アクリレートゲル(配向ゲル)(図3)に入れることで、PBCV1-CEを磁場に対して配向させた。サンプル溶液には、活性阻害剤であるピロリン酸を5mMを入れた。TROSY測定を20℃で行った(図4)。分子配向依存的なTROSYシフト変化を、配向ゲル中のPBCV1-CEのTROSYシグナルの、通常溶液の状態のTROSYシグナルからの15N軸側のずれとして観測した。十分なシグナルの分離を示す51残基分(N末端ドメイン、23残基、C末端ドメイン、28残基)のTROSYシフト変化に基づいて、各ドメインの配向テンソルを決定した。
得られたドメイン相対配向の結果の精度を確認するために、結晶構造中で見つかっているPBCV1-CEの2つの異なるドメイン配向を持つ構造(オープン型およびクローズ型)をもとにして解析を行った。なお、クローズ型の結晶構造を基にした解析では、部分的なドメイン内構造変化の影響を除くためにN末端ドメインの2残基分のデータを解析からはずした。
(分子配向解析)
本発明の方法に従って、オープン型構造のデータを用いて、ドメイン間分子配向解析を行った。Δδtrosy値を得て、実施例2と同様にSaupeオーダーマトリクスエレメントの決定を行い、ヌクレオチド非存在下でのオイラー角α、β、およびγを決定した。
PBCV1-CEは立体構造上明確なドメイン構造を持つ。アミノ酸残基番号で15-222からなるドメイン(ラージドメイン)と同じく239−311からなるドメイン(スモールドメイン)のそれぞれに由来するTROSYシグナルの配向依存的な変化から、各ドメインの配向テンソルを決定した。その結果、表3に示すように、ラージドメイン(N末端)の分子配向軸の分子座標軸からの傾きを表現するオイラー角は(α=95.9°、β=123.1°、γ=96.2°、Q−因子=0.38)であるのに対し、スモールドメイン(C末端)の方は(α=98.4°、β=-114.7°、γ=165.4°、Q−因子=0.45)となった。
【表3】
それぞれのドメインの分子配向軸は図5および図6に示すような関係になった。相対的なドメインの配向は、2つの配向軸を一致させるようにスモールドメインを回転させることで決定されるが、各ドメインの配向テンソルからスモールドメインは結晶構造に比べて約65°回転していることが明らかとなった。
オープン型の構造を用いて得られたドメイン相対配向の結果の精度を確認するために、結晶構造中で見つかっているPBCV1-CEの別のドメイン配向(クローズ型)を持つ構造を用いた。その結果を、以下の表4、ならびに図7および図8に示す。
【表4】
本発明の解析方法の結果決定されたドメイン間の相対配向は、オープン型およびクローズ型で同様となり、決定されたドメイン間相対配向の精度が確認された。
【0172】
同様の方法を用いて、GTP存在下およびATP存在下の各々についてのオイラー角α、β、およびγを決定する。さらに、このオイラー角から、GTP添加時、およびATP添加時のDOCを求める。このDOCは、GTP添加時、およびATP添加時の構造変化の指標となる。
【実施例4】
【0173】
(タンパク質の作動薬リード化合物のスクリーニング)
タンパク質の新規のアゴニストおよび/またはアンタゴニストの探索は、新規薬物の製造のために重要である。特に新規のアゴニストは、作動薬リード化合物として利用され得るが、従来技術は、作動薬リード化合物の同定には不十分であった。
【0174】
そこで、本発明を用いて、作動薬リード化合物のスクリーニングを行う。モデルとして、実施例3において用いられるmRNA5’−キャッピング酵素を用いる。
【0175】
作動薬リード化合物は、mRNA5’−キャッピング酵素を活性化するGTPと同様の構造変化をmRNA5’−キャッピング酵素に生じると考えられるため、以下のようにスクリーニングを行う。
【0176】
GTPの代わりに候補化合物を添加した実験を行う以外は、実施例3と同様の条件下で、mRNA5’−キャッピング酵素の配向情報を得る。候補化合物としては、ヌクレオチドアナログ、特にGTPアナログ、およびATPアナログを用いることができる。GTP存在下での配向情報と、候補化合物存在下での配向情報とから、DOSを計算する。
【0177】
複数の候補化合物の中から、DOS値が小さい化合物を、作動薬リード化合物として選択する。
【実施例5】
【0178】
(候補化合物から選択された作動薬リード化合物の結合部位の同定)
作動薬リード化合物を選択した場合、その作動薬リード化合物の結合部位を同定することが可能である。本実施例においては、実施例4において最も小さいDOS値を生じた化合物を用いて、以下のように実験を行う。
・実施例4において用いる候補化合物非存在下での二次元TROSY NMRスペクトルのアミノ酸残基への帰属を常法に従って行う。
・化合物存在下でのタンパク質内ドメインの二次元TROSY NMRスペクトルと、化合物非存在下での二次元TROSY NMRスペクトルとを比較する。その比較において、スペクトルの変化を検出する。変化したスペクトルに対応するアミノ酸残基を、上記帰属の結果から、作動薬リード化合物の結合部位として同定する。
【実施例6】
【0179】
(標的タンパク質に対する化合物の親和性の決定)
本発明において、例えば、以下の方法を用いて標的タンパク質に対する化合物の親和性を決定することが可能である。
・実施例4において用いる候補化合物非存在下での二次元TROSY NMRスペクトルのアミノ酸残基への帰属を常法に従って行う。
・化合物存在下でのタンパク質内ドメインの二次元TROSY NMRスペクトルと、化合物非存在下での二次元TROSY NMRスペクトルとを比較する。その比較から、スペクトルの変化を検出する。変化したスペクトルに対応するアミノ酸残基を、上記帰属の結果から、同定する。
・同定されたアミノ酸に対応するスペクトルは、化合物非存在下の場合と、化合物存在下の場合とで、異なる位置に出現する。しかし、用いる化合物の種類によっては、化合物存在下であっても、化合物非存在下において検出される同定されたアミノ酸に対応するスペクトルが残存する場合がある。このように単一のアミノ酸残基に対応するスペクトルが二重に存在するのは、二次元TROSY NMR測定の溶液中に、化合物と結合したタンパク質と、未結合のタンパク質が混在するためである。この二重に存在する単一のアミノ酸残基に対応するスペクトルの強度の比は、化合物結合タンパク質と化合物未結合タンパク質との存在比と相関する。従って、この二重のスペクトルの強度の比は、試験した化合物とタンパク質との親和性の指標となる。この原理を用いて、以下のように化合物の相対的親和性を測定する。
・実施例5に従って、化合物の結合するアミノ酸残基を同定する。
・化合物A存在下においてNMR測定を行い、化合物の結合するアミノ酸に対応するスペクトルについて、
(化合物A存在下においてのみ検出される位置に出現するスペクトル)/(化合物非存在下において検出される位置に残存するスペクトル)
の比を求める。
・化合物B存在下においてNMR測定を行い、化合物の結合するアミノ酸に対応するスペクトルについて、
(化合物B存在下においてのみ検出される位置に出現するスペクトル)/(化合物非存在下において検出される位置に残存するスペクトル)
の比を求める。
・化合物AおよびBについて、これらの比を比較し、この比の大きい化合物が、標的タンパク質に対して、より親和性が大きい化合物であると判断される。
【産業上の利用可能性】
【0180】
化合物の結合により生じるタンパク質の構造変化を検出することによって、タンパク質のリガンド、アゴニスト、アンタゴニストのスクリーニングが迅速かつ容易になる。さらに、構造変化の検出は、標的タンパク質の活性構造を誘導するか否かを指標として、新規薬物の候補化合物をスクリーニングすることを可能にする。
【技術分野】
【0001】
本発明は、核磁気共鳴を用いることによって、化合物がタンパク質と結合した場合のタンパク質の構造変化を測定する方法、その構造変化に基づいて化合物を選択する方法、およびその方法において使用されるプログラムに関する。
【背景技術】
【0002】
細胞が細胞外からの刺激に対して応答する場合、細胞内の種々のタンパク質が関与する。そのため、これらタンパク質を標的とするアゴニストおよび/またはアンタゴニストは、新規薬物のリード化合物として用いることができる。
【0003】
タンパク質の、アゴニストおよび/またはアンタゴニストの探索のための従来法としては、標的タンパク質に対する物理的結合を指標とする探索方法、および標的タンパク質が関与する生物学的反応を誘導する能力(機能誘導能)を指標とする探索方法が挙げられる。
【0004】
標的タンパク質に対する物理的結合を指標とする方法は、化合物ライブラリー(例えば、組み合わせライブラリー)由来の化合物が、標的タンパク質にどの程度の強さで結合するか、また何分子結合するかなどを調べ、ある評価基準以上の標的タンパク質に対する親和性を有する化合物を選び出すという方法である。
【0005】
一方、標的タンパク質の機能誘導能、すなわち、標的タンパク質が関与する生物学的反応を指標とする方法は、化合物ライブラリー(例えば、組み合わせライブラリー)由来の化合物を、標的タンパク質を発現する細胞と接触させ、その結果生じる生物学的反応の変化(例えば、生物学的反応の開始および終了のような質的変化、あるいは生物学的反応の程度の増加または減少のような量的変化)を指標とする方法である。機能誘導能を指標とする方法を行うには、標的タンパク質に応じたアッセイ系をそれぞれ立ち上げる必要がある。そのため、例えば、生きた細胞を対象として化合物の効果を解析する場合にはアッセイ系に依存して信頼性・実験精度が左右されるなどの制約がある。
【0006】
タンパク質のアゴニストおよび/またはアンタゴニストを同定し、次に同定された化合物を、新規薬物のリード化合物として用いることができる。新規薬物のスクリーニングにおいては、その候補化合物が、標的タンパク質と結合することによって、どのようにその標的タンパク質の立体構造が変化するか知ることが有用である。なぜなら、そのような立体構造変化の情報は、より適切な構造を有する候補薬物のデザインの指標となるからである。
【0007】
しかし、物理的結合・機能誘導能のいずれを指標とする場合にも、化合物が直接タンパク質に結合している状態を見ているわけではない。従って間接的に得た標的タンパク質への結合能あるいは機能誘導能から化合物の構造と機能を類推することになる。物理的結合の有無あるいは機能誘導能の有無だけでは、標的タンパク質と化合物の相互作用の実態を知ることは難しく、いつも明確な薬物デザインの指針を与えるとはかぎらない。
【0008】
このような従来のアッセイ法が持つ問題点を克服するために、核磁気共鳴法(NMR)により化合物が標的タンパク質の機能部位に結合していることを確認する核磁気共鳴法(NMR)を用いたアッセイ法が発表されている(Shuker S.B. et al. Science (1996) 274, 1531−1534頁)(特開2001−321192)。この先行技術では、標的タンパク質の機能とは無関係な場所に非特異的に化合物が結合するケースを除くことができるために、非特異的結合と特異的結合の区別が難しい従来の物理的結合アッセイよりも効率的にリード化合物を特定することが可能である。しかし、この技術は結合部位を特定することは可能であるが、タンパク質の構造変化自体を測定するのには適していない。そのため、この技術は、標的タンパク質の活性部位に結合する阻害剤の設計には有効であるが、逆にタンパク質の活性誘導を行う作動薬(例えば、アゴニスト)のデザインには効果が少ない。作動薬リード化合物の同定には、標的タンパク質の基質結合部位に正しく結合し、さらに標的タンパク質の活性構造を誘導する作用の有無を指標としたスクリーニングが必要であるため、従来技術は、作動薬リード化合物の同定には不十分であった。
【0009】
化合物が標的タンパク質に対して活性構造を誘導し得るかどうかはX線結晶構造解析を基にして決定することができる。しかし、X線結晶構造解析は、多大な労力と時間を必要とする方法であることから、多数の化合物に対して標的タンパク質との共結晶化を行い立体構造解析を繰り返すことは、効率が求められるスクリーニングには適当ではない。そこで、任意の化合物が標的タンパク質と接触・結合した際に、その標的タンパク質にどのような構造変化が生じるのかを簡便かつ迅速に決定する方法、およびそのような方法を実行するためのコンピュータプログラムが求められている。
【特許文献1】特開2001−321192
【非特許文献1】Shuker S.B. et al. Science (1996) 274, 1531−1534頁
【発明の開示】
【発明が解決しようとする課題】
【0010】
任意の化合物が標的タンパク質と接触・結合した際に、その標的タンパク質にどのような構造変化が生じるのかを簡便かつ迅速に決定する方法、そのような方法を用いる化合物の選択・スクリーニング方法、およびそのような方法を実行するためのコンピュータプログラムを提供することが、本発明の課題である。
【0011】
例えば、核磁気共鳴を用いることによって、任意の化合物がタンパク質と結合・接触した場合の、タンパク質の構造変化を測定する方法を提供することが、本発明の課題の1つである。さらに、任意の化合物がタンパク質と結合・接触した場合に、タンパク質の構造変化を生じる化合物を同定・スクリーニングする方法を提供することが、本発明の課題の1つである。また、例えば、リガンド、アゴニスト、などのように、タンパク質と結合した際にそのタンパク質の構造変化を生じることが公知の物質がそのタンパク質と結合した際の構造変化と、任意の化合物がそのタンパク質と結合した際の構造変化とを比較して、リガンド、アゴニストと類似の構造変化をもたらす物質を同定・スクリーニングする方法を提供することもまた、本発明の別の課題の1つである。
【0012】
また、これら方法において使用されるコンピュータプログラムを提供することも、本発明の課題の1つである。
【課題を解決するための手段】
【0013】
本発明者らは、標的タンパク質溶液に化合物を添加して誘導される標的タンパク質の立体構造変化をTROSY NMR相関スペクトル(例えば、1H/15N−NMRスペクトル)の変化から読み取る方法およびプログラムを開発し、本発明を完成した。本発明者らは、1)化合物の標的タンパク質上での結合部位、2)標的タンパク質が化合物によって誘導される構造変化の2つを同時に観測することができる新規な方法を開発し、本発明を完成した。
【0014】
従って、本発明は以下を提供する。
1.タンパク質と化合物とを接触させた場合の、該タンパク質の構造変化を測定する方法であって、以下:
(a)該タンパク質内のドメインを選択する工程;
(b)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を得る工程;
(c)該タンパク質と該化合物を接触させる条件下における、該ドメインの配向情報を以下
(i)該ドメインの既知の原子座標を得て、
(ii)液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(iii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(iv)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、
ことにより得る工程;および、
(d)工程(b)において得られた配向情報と、工程(c)において得られた配向情報との差異から、該タンパク質の構造変化を測定する工程、
を包含する、方法。
2.前記(b)工程が、
(b)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を以下
(v)該ドメインの既知の原子座標を得て、
(vi)液晶物質存在下において、該タンパク質を該化合物と接触させずに、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(vii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(viii)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、ことにより得る工程である、項目1に記載の測定方法。
3.前記(b)工程が、
(b)前記タンパク質が前記化合物と接触しない条件下において既に得られている原子座標から、前記ドメインの配向情報を得る工程;
である、項目1に記載の測定方法。
4.項目1に記載の測定方法であって、前記工程(c)において二次元TROSY NMRスペクトルを用いて得られる分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、方法。
5.項目4に記載の測定方法であって、ここで、前記(iii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、方法。
6.項目2に記載の測定方法であって、前記工程(b)において二次元TROSY NMRスペクトルを用いて得られる分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、方法。
7.項目6に記載の測定方法であって、ここで、前記(vii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させることなく、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、方法。
8.項目5または7に記載の方法であって、ここで、前記タンパク質と前記化合物とが接触した場合の、該タンパク質の構造変化が、以下:
(ix)該化合物と接触させる前の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【0015】
【化1】
を用いて示し、
(x)該化合物と接触させた後の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【0016】
【化2】
を用いて示し、
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、そして
(xii)以下の式によって分子配向変化度を求める
分子配向変化度=a2+b2+c2、
ことによって分子配向変化度として数値化される、方法。
9.項目2に記載の方法であって、さらに、前記化合物が前記タンパク質に結合した位置を特定する工程を包含する、方法。
10.項目9に記載の方法であって、前記化合物が前記タンパク質に結合した位置を特定する工程が、前記工程(b)において得られた二次元TROSY NMRスペクトルと、前記工程(c)において得られた二次元TROSY NMRスペクトルとを比較して、スペクトルの変化を検出し、そして、該スペクトルの変化を生じた、該タンパク質中のアミノ酸残基を同定すること、によって行われる、方法。
11.項目1に記載の方法であって、前記液晶物質が、以下からなる群から選択される混合物を含む、方法:
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とドデシル硫酸ナトリウム(SDS)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物、
1,2−ジ−O−ドデシル−sn−グリセロ−3−ホスフォコリン(DIODPC)と3−(コラミドプロピル)−ジメチルアンモニオ−2−ヒドロキシ−1−プロパンサルフェート(CHAPS)との混合物、
n−アルキル−ポリ(エチレングリコール)/n−アルキルアルコール混合物、
繊維状ファージ、
セチルピリジニウムクロライド(CPCl)−ヘキサノール−NaCl混合物、
セチルピリジニウムブロミド(CPBr)−ヘキサノール−NaCl混合物、
Halobacterium属の紫膜断片
微結晶セルロース、および
ポリアクリルアミドゲル。
12.前記液晶物質が、7.5%(w/v)のジミリストイルホスファチジルコリン(D
MPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物である、項目11に記載の方法。
13.接触の際に、タンパク質内のドメインの構造変化を生じる化合物を選択する方法であって、以下:
(a)該タンパク質内のドメインを選択する工程;
(b)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を得る工程;
(c)該タンパク質と該化合物を接触させる条件下における、該ドメインの配向情報を以下
(i)該ドメインの既知の原子座標を得て、
(ii)液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(iii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(iv)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、
ことにより得る工程;
(d)工程(b)において得られた配向情報と、工程(c)において得られた配向情報との比較から、該化合物が該タンパク質と接触した際に該ドメインの構造変化を生じる化合物であるか否かを決定する工程、
を包含する、方法。
14.前記(b)工程が、
(b)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を以下
(v)該ドメインの既知の原子座標を得て、
(vi)液晶物質存在下において、該タンパク質を該化合物と接触させずに、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(vii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(viii)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、ことにより得る工程である、項目13に記載の測定方法。
15.前記(b)工程が、
(b)前記タンパク質が前記化合物と接触しない条件下において既に得られている原子座標から、前記ドメインの配向情報を得る工程;
である、項目13に記載の測定方法。
16.項目13に記載の測定方法であって、前記工程(c)において二次元TROSY NMRスペクトルを用いて得られる分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、方法。
17.項目16に記載の測定方法であって、ここで、前記(iii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、方法。
18.項目14に記載の測定方法であって、前記工程(b)において二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、方法。
19.項目18に記載の測定方法であって、ここで、前記(vii)のSaupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させることなく、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、方法。
20.項目17または19に記載の方法であって、ここで、前記工程(c)における配向情報の比較が、以下:
(ix)該化合物と接触する前の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【0017】
【化3】
を用いて示し、
(x)該化合物と接触した後の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【0018】
【化4】
を用いて示し、
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、そして
(xii)以下の式によって分子配向変化度を求め、
分子配向変化度=a2+b2+c2、
該分子配向変化度を比較することによって行われる、方法。
21.項目14に記載の方法であって、さらに、前記化合物が前記タンパク質に結合した位置を特定する工程を包含する、方法。
22.項目21に記載の方法であって、前記化合物が前記タンパク質に結合した位置を特定する工程が、前記工程(b)において得られた二次元TROSY NMRスペクトルと、前記工程(c)において得られた二次元TROSY NMRスペクトルとを比較して、スペクトルの変化を検出し、そして、該スペクトルの変化を生じた、該タンパク質中のアミノ酸残基を同定すること、によって行われる、方法。
23.項目13に記載の方法であって、前記液晶物質が、以下からなる群から選択される混合物を含む、方法:
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とドデシル硫酸ナトリウム(SDS)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物、
1,2−ジ−O−ドデシル−sn−グリセロ−3−ホスフォコリン(DIODPC)と3−(コラミドプロピル)−ジメチルアンモニオ−2−ヒドロキシ−1−プロパンサルフェート(CHAPS)との混合物、
n−アルキル−ポリ(エチレングリコール)/n−アルキルアルコール混合物、
繊維状ファージ、
セチルピリジニウムクロライド(CPCl)−ヘキサノール−NaCl混合物、
セチルピリジニウムブロミド(CPBr)−ヘキサノール−NaCl混合物、
Halobacterium属の紫膜断片
微結晶セルロース、および
ポリアクリルアミドゲル。
24.前記液晶物質が、7.5%(w/v)のジミリストイルホスファチジルコリン(D
MPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物である、項目23に記載の方法。
25.タンパク質と接触することによって、第1の化合物と該タンパク質とが接触した場合の該タンパク質の構造変化と類似する該タンパク質の構造変化を生じる、第2の化合物を選択する方法であって、以下:
(a)該タンパク質内のドメインを選択する工程;
(b)該タンパク質と該第1の化合物を接触させる条件下における、該ドメインの配向情報を得る工程;
(c)該タンパク質と該第2の化合物を接触させる条件下における、該ドメインの配向情報を以下
(i)該ドメインの既知の原子座標を得て、
(ii)液晶物質存在下において該タンパク質と該第2の化合物を接触させ、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(iii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(iv)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、
ことにより得る工程;
(d)工程(b)において得られた配向情報と、工程(c)において得られた配向情報とを比較する工程;および
(e)工程(d)における比較の結果から、該第2の化合物が該タンパク質と接触することによる該タンパク質の構造変化が、該第1の化合物が該タンパク質と接触することによる該タンパク質の構造変化と類似するか否かを決定する工程、
を包含する、方法。
26.前記(b)工程が、
(b)該タンパク質が該第1の化合物と接触する条件下における、該ドメインの配向情報を以下
(v)該ドメインの既知の原子座標を得て、
(vi)液晶物質存在下において、該タンパク質を該第1の化合物と接触させた場合に二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(vii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(viii)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、ことにより得る工程である、項目25に記載の測定方法。
27.前記(b)工程が、
(b)前記タンパク質が前記第1の化合物を接触させる条件下において既に得られている原子座標から、前記ドメインの配向情報を得る工程;
である、項目25に記載の測定方法。
28.項目25に記載の測定方法であって、前記工程(c)において二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、方法。
29.項目28に記載の測定方法であって、ここで、前記(iii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、方法。
30.項目26に記載の測定方法であって、前記工程(b)において二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、方法。
31.項目30に記載の測定方法であって、ここで、前記(vii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該第2の化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、方法。
32.項目29または31に記載の方法であって、ここで、前記前記工程(d)における配向情報の比較が、以下:
(ix)該第1の化合物と接触した場合の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【0019】
【化5】
を用いて示し、
(x)該第2の化合物と接触させた場合の、該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【0020】
【化6】
を用いて示し、そして
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、該a、b、およびcを配向情報の比較のための指標とする、
ことによって行われる、方法。
33.項目29または31に記載の方法であって、ここで、前記工程(d)における配向情報の比較が、以下:
(ix)該第1の化合物と接触した場合の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【0021】
【化7】
を用いて示し、
(x)該第2の化合物と接触させた場合の、該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【0022】
【化8】
を用いて示し、そして
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、そして
(xii)以下の式によって活性構造一致度を求め
活性構造一致度=a2+b2+c2、
該活性構造一致度を配向情報の比較のための指標とする、
ことによって行われる、方法。
34.項目25に記載の方法であって、さらに、前記第1および第2の化合物の少なくとも1つが前記タンパク質に結合した位置を特定する工程を包含する、方法。
35.項目34に記載の方法であって、前記第1および第2の化合物の少なくとも1つが前記タンパク質に結合した位置を特定する工程が、該化合物の非存在下における該タンパク質の二次元TROSY NMRスペクトルと、該化合物の存在下における該タンパク質の二次元TROSY NMRスペクトルを比較して、スペクトルの変化を検出し、そして、該スペクトルの変化を生じた、該タンパク質中のアミノ酸残基を同定すること、
によって行われる、方法。
36.項目25に記載の方法であって、前記液晶物質が、以下からなる群から選択される混合物を含む、方法:
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とドデシル硫酸ナトリウム(SDS)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物、
1,2−ジ−O−ドデシル−sn−グリセロ−3−ホスフォコリン(DIODPC)と3−(コラミドプロピル)−ジメチルアンモニオ−2−ヒドロキシ−1−プロパンサルフェート(CHAPS)との混合物、
n−アルキル−ポリ(エチレングリコール)/n−アルキルアルコール混合物、
繊維状ファージ、
セチルピリジニウムクロライド(CPCl)−ヘキサノール−NaCl混合物、
セチルピリジニウムブロミド(CPBr)−ヘキサノール−NaCl混合物、
Halobacterium属の紫膜断片
微結晶セルロース、および
ポリアクリルアミドゲル。
37.前記液晶物質が、7.5%(w/v)のジミリストイルホスファチジルコリン(D
MPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物である、項目36に記載の方法。
38.タンパク質と化合物とを接触させた場合の、該タンパク質内の選択されたドメインの構造変化を数値化するプログラムであって、以下:
(a)該ドメインについて、該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を得る手段;
(b)該タンパク質と該化合物を接触させる条件下における、該ドメインの配向情報を以下
(i)該ドメインの既知の原子座標のデータを得て、
(ii)液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量のデータを得て、
(iii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(iv)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、
ことにより得る手段;および、
(c)手段(a)において得られた配向情報と、手段(b)において測定された配向情報との差異から、該タンパク質の構造変化を計算する手段、
とをコンピュータに実現させるための、プログラム。
39.前記(a)手段が、
(a)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を以下
(v)該ドメインの既知の原子座標を得て、
(vi)液晶物質存在下において、該タンパク質を該化合物と接触させずに、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(vii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(viii)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、ことにより得る手段である、項目38に記載のプログラム。
40.前記(a)手段が、
(a)前記タンパク質が前記化合物と接触しない条件下において既に得られている原子座標から、前記ドメインの配向情報を得る手段;
である、項目38に記載のプログラム。
41.項目38に記載のプログラムであって、前記工程(b)において二次元TROSY
NMRスペクトルを用いて得られる分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、プログラム。
42.項目41に記載のプログラムであって、ここで、前記(iii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、プログラム。
43.項目39に記載のプログラムであって、前記工程(a)において二次元TROSY
NMRスペクトルを用いて得られる分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、プログラム。
44.項目43に記載のプログラムであって、ここで、前記(vii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させることなく、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、プログラム。
45.項目42または44に記載のプログラムであって、ここで、前記タンパク質と前記化合物とが接触した場合の、該タンパク質の構造変化が、以下:
(ix)該化合物と接触させる前の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【0023】
【化9】
を用いて示し、
(x)該化合物と接触させた後の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【0024】
【化10】
を用いて示し、
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、そして
(xii)以下の式によって分子配向変化度を求める
分子配向変化度=a2+b2+c2、
ことによって分子配向変化度として数値化される、プログラム。
46.項目39に記載のプログラムであって、さらに、前記化合物が前記タンパク質に結合した位置を特定する手段を包含する、プログラム。
47.項目46に記載のプログラムであって、前記化合物が前記タンパク質に結合した位置を特定する手段が、前記工程(a)において得られた二次元TROSY NMRスペクトルと、前記工程(b)において得られた二次元TROSY NMRスペクトルとを比較して、スペクトルの変化を検出し、そして、該スペクトルの変化を生じた、該タンパク質中のアミノ酸残基を同定すること、によって行われる、プログラム。
48.接触の際に、タンパク質内のドメインの構造変化を生じる化合物を選択するプログラムであって、以下:
(a)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を得る手段;
(b)該タンパク質と該化合物を接触させる条件下における、該ドメインの配向情報を以下
(i)該ドメインの既知の原子座標を得て、
(ii)液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(iii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(iv)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、
ことにより得る手段;
(c)手段(a)において得られた配向情報と、手段(b)において得られた配向情報との比較から、該化合物が該タンパク質と接触した際に該ドメインの構造変化を生じる化合物であるか否かを決定する手段、
を包含する、プログラム。
49.前記(a)手段が、
(a)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を以下
(v)該ドメインの既知の原子座標を得て、
(vi)液晶物質存在下において、該タンパク質を該化合物と接触させずに、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(vii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(viii)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、ことにより得る手段である、項目48に記載のプログラム。
50.前記(a)手段が、
(a)前記タンパク質が前記化合物と接触しない条件下において既に得られている原子座標から、前記ドメインの配向情報を得る手段;
である、項目48に記載のプログラム。
51.項目48に記載のプログラムであって、前記工程(b)において二次元TROSY
NMRスペクトルを用いて得られる分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、プログラム。
52.項目51に記載のプログラムであって、ここで、前記(iii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、プログラム。
53.項目52に記載のプログラムであって、前記工程(a)において二次元TROSY
NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、プログラム。
54.項目53に記載のプログラムであって、ここで、前記(vii)のSaupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させることなく、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、プログラム。
55.項目52または54に記載のプログラムであって、ここで、前記工程(c)における配向情報の比較が、以下:
(ix)該化合物と接触する前の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【0025】
【化11】
を用いて示し、
(x)該化合物と接触した後の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【0026】
【化12】
を用いて示し、
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、そして
(xii)以下の式によって分子配向変化度を求め、
分子配向変化度=a2+b2+c2、
該分子配向変化度を比較することによって行われる、プログラム。
56.項目48に記載のプログラムであって、さらに、前記化合物が前記タンパク質に結合した位置を特定する手段を包含する、プログラム。
57.項目56に記載のプログラムであって、前記化合物が前記タンパク質に結合した位置を特定する手段が、前記工程(a)において得られた二次元TROSY NMRスペクトルと、前記工程(b)において得られた二次元TROSY NMRスペクトルとを比較して、スペクトルの変化を検出し、そして、該スペクトルの変化を生じた、該タンパク質中のアミノ酸残基を同定すること、によって行われる、プログラム。
58.タンパク質と接触することによって、第1の化合物と該タンパク質とが接触した場合の該タンパク質内のドメインの構造変化と類似する該タンパク質内のドメインの構造変化を生じる、第2の化合物を選択するプログラムであって、以下:
(a)該タンパク質と該第1の化合物を接触させる条件下における、該ドメインの配向情報を得る手段;
(b)該タンパク質と該第2の化合物を接触させる条件下における、該ドメインの配向情報を以下
(i)該ドメインの既知の原子座標を得て、
(ii)液晶物質存在下において該タンパク質と該第2の化合物を接触させ、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(iii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(iv)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、
ことにより得る手段;
(c)手段(a)において得られた配向情報と、手段(b)において得られた配向情報とを比較する手段;および
(d)手段(c)における比較の結果から、該第2の化合物が該タンパク質と接触することによる該タンパク質の構造変化が、該第1の化合物が該タンパク質と接触することによる該タンパク質の構造変化と類似するか否かを決定する手段、
を包含する、プログラム。
59.前記(a)手段が、
(a)該タンパク質が該第1の化合物と接触する条件下における、該ドメインの配向情報を以下
(v)該ドメインの既知の原子座標を得て、
(vi)液晶物質存在下において、該タンパク質を該第1の化合物と接触させた場合に二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(vii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(viii)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、ことにより得る手段である、項目25に記載のプログラム。
60.前記(a)手段が、
(a)前記タンパク質が前記第1の化合物を接触させる条件下において既に得られている原子座標から、前記ドメインの配向情報を得る手段;
である、項目58に記載のプログラム。
61.項目58に記載のプログラムであって、前記工程(b)において二次元TROSY
NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、プログラム。
62.項目61に記載のプログラムであって、ここで、前記(iii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、プログラム。
63.項目59に記載のプログラムであって、前記工程(a)において二次元TROSY
NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、プログラム。
64.項目63に記載のプログラムであって、ここで、前記(vii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該第2の化合物を接触させ、二次元TROSY
NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、プログラム。
65.項目62または64に記載のプログラムであって、ここで、前記工程(c)における配向情報の比較が、以下:
(ix)該第1の化合物と接触した場合の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【0027】
【化13】
を用いて示し、
(x)該第2の化合物と接触させた場合の、該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【0028】
【化14】
を用いて示し、そして
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、該a、b、およびcを配向情報の比較のための指標とする、
ことによって行われる、プログラム。
66.項目62または64に記載のプログラムであって、ここで、前記工程(c)における配向情報の比較が、以下:
(ix)該第1の化合物と接触した場合の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【0029】
【化15】
を用いて示し、
(x)該第2の化合物と接触させた場合の、該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【0030】
【化16】
を用いて示し、そして
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、そして
(xii)以下の式によって活性構造一致度を求め
活性構造一致度=a2+b2+c2、
該活性構造一致度を配向情報の比較のための指標とする、
ことによって行われる、プログラム。
67.項目58に記載のプログラムであって、さらに、前記第1および第2の化合物の少なくとも1つが前記タンパク質に結合した位置を特定する手段を包含する、プログラム。68.項目67に記載のプログラムであって、前記第1および第2の化合物の少なくとも1つが前記タンパク質に結合した位置を特定する手段が、該化合物の非存在下における該タンパク質の二次元TROSY NMRスペクトルと、該化合物の存在下における該タンパク質の二次元TROSY NMRスペクトルとを比較して、スペクトルの変化を検出し、そして該スペクトルの変化を生じた、該タンパク質中のアミノ酸残基を同定すること、によって行われる、プログラム。
【0031】
69.項目1、13、または25に記載の方法を実行し得る装置。
【0032】
70.項目38、48、または58に記載のプログラムを備える装置。
【0033】
71.項目38、48、または58に記載のプログラムを含む記憶媒体。
【発明の効果】
【0034】
本発明によって、標的タンパク質溶液に化合物を添加して誘導される標的タンパク質の立体構造変化をTROSY NMR相関スペクトル(例えば、1H/15N−NMRスペクトル)の変化から読み取る方法およびプログラムが提供される。本発明により、1)化合物の標的タンパク質上での結合部位、2)標的タンパク質が化合物によって誘導される構造変化の2つを同時に観測することができる新規な方法が提供される。
【図面の簡単な説明】
【0035】
【図1】図1は、ドメイン間の相対配向変化を基にして選択的に作動薬のリード化合物を同定する方法を模式的に示した図である。
【図2】図2は、タンパク質の分子配向角度の表現法を示す。
【図3】図3は、実施例3で用いた配向ゲルの写真である。右上の白棒は、10mmの長さを示す。
【図4】図4は、この配向ゲルを用いた、クロレラウイルスPBCV1のグアニリル転移酵素であるmRNA5’−キャッピング酵素のTROSYシグナルを示す。6%(w/w)の伸張させた50%アクリルアミド/50%アクリレートゲル(配向ゲル)を用いて、20℃(293K)で実験を行った。「I」は、等方的状態において収集されたデーを示す。「A」は、整列された状態において収集されたデータを示す。
【図5】図5は、オープン型結晶構造を基にした解析の結果を示す。配向テンソルは、N末端ドメイン(ラージドメイン)23残基、およびC末端ドメイン(スモールドメイン)28残基を用いて決定した。
【図6】図6は、オープン型結晶構造を基にした解析の結果を示す。配向テンソルは、N末端ドメイン(ラージドメイン)23残基、およびC末端ドメイン(スモールドメイン)28残基を用いて決定した。
【図7】図7は、クローズ型結晶構造を基にした解析の結果を示す。配向テンソルは、N末端ドメイン(ラージドメイン)21残基、およびC末端ドメイン(スモールドメイン)28残基を用いて決定した。
【図8】図8は、クローズ型結晶構造を基にした解析の結果を示す。配向テンソルは、N末端ドメイン(ラージドメイン)21残基、およびC末端ドメイン(スモールドメイン)28残基を用いて決定した。
【発明を実施するための最良の形態】
【0036】
以下、本発明を説明する。本明細書の全体にわたり、単数形の表現は、特に言及しない限り、その複数形の概念をも含むことが理解されるべきである。従って、単数形の冠詞(例えば、英語の場合は「a」、「an」、「the」など)は、特に言及しない限り、その複数形の概念をも含むことが理解されるべきである。また、本明細書において使用される用語は、特に言及しない限り、当該分野で通常用いられる意味で用いられることが理解されるべきである。したがって、他に定義されない限り、本明細書中で使用される全ての専門用語および科学技術用語は、本発明の属する分野の当業者によって一般的に理解されるのと同じ意味を有する。矛盾する場合、本明細書(定義を含めて)が優先する。
【0037】
(用語の定義)
以下に本明細書において特に使用される用語の定義を列挙する。
本明細書において使用される用語「タンパク質」「ポリペプチド」、「オリゴペプチド」および「ペプチド」は、本明細書において同じ意味で使用され、任意の長さのアミノ酸のポリマーをいう。このポリマーは、直鎖であっても分岐していてもよく、環状であってもよい。アミノ酸は、天然のものであっても非天然のものであってもよく、改変されたアミノ酸であってもよい。この用語はまた、複数のポリペプチド鎖の複合体へとアセンブルされ得る。この用語はまた、天然または人工的に改変されたアミノ酸ポリマーも包含する。そのような改変としては、例えば、ジスルフィド結合形成、グリコシル化、脂質化、アセチル化、リン酸化または任意の他の操作もしくは改変(例えば、標識成分との結合体化)。この定義にはまた、例えば、アミノ酸の1または2以上のアナログを含むポリペプチド(例えば、非天然のアミノ酸などを含む)、ペプチド様化合物(例えば、ペプトイド)および当該分野において公知の他の改変が包含される。
【0038】
本明細書においてタンパク質の「ドメイン」とは、1残基のアミノ酸、または連続した少なくとも2つのアミノ酸残基を含む、タンパク質分子の特定の一部を意味する。本明細書においてドメインは、独立して特定の機能を有しても、有していなくてもよい。
【0039】
本明細書において、「アミノ酸」は、天然のものでも非天然のものでもよい。「誘導体アミノ酸」または「アミノ酸アナログ」とは、天然に存在するアミノ酸とは異なるがもとのアミノ酸と同様の機能を有するものをいう。そのような誘導体アミノ酸およびアミノ酸アナログは、当該分野において周知である。用語「天然のアミノ酸」とは、天然のアミノ酸のL−異性体を意味する。天然のアミノ酸は、グリシン、アラニン、バリン、ロイシン、イソロイシン、セリン、メチオニン、トレオニン、フェニルアラニン、チロシン、トリプトファン、システイン、プロリン、ヒスチジン、アスパラギン酸、アスパラギン、グルタミン酸、グルタミン、γ−カルボキシグルタミン酸、アルギニン、オルニチン、およびリジンである。特に示されない限り、本明細書でいう全てのアミノ酸はL体である。用語「非天然アミノ酸」とは、タンパク質中で通常は天然に見出されないアミノ酸を意味する。非天然アミノ酸の例として、ノルロイシン、パラ−ニトロフェニルアラニン、ホモフェニルアラニン、パラ−フルオロフェニルアラニン、3−アミノ−2−ベンジルプロピオン酸、ホモアルギニンのD体またはL体およびD−フェニルアラニンが挙げられる。「アミノ酸アナログ」とは、アミノ酸ではないが、アミノ酸の物性および/または機能に類似する分子をいう。アミノ酸アナログとしては、例えば、エチオニン、カナバニン、2−メチルグルタミンなどが挙げられる。アミノ酸模倣物とは、アミノ酸の一般的な化学構造とは異なる構造を有するが、天然に存在するアミノ酸と同様な様式で機能する化合物をいう。
【0040】
アミノ酸は、その一般に公知の3文字記号か、またはIUPAC−IUB Biochemical Nomenclature Commissionにより推奨される1文字記号のいずれかにより、本明細書中で言及され得る。ヌクレオチドも同様に、一般に受け入れられた1文字コードにより言及され得る。
【0041】
本明細書中において、「対応する」アミノ酸とは、あるタンパク質分子またはポリペプチド分子において、比較の基準となるタンパク質またはポリペプチドにおける所定のアミノ酸と同様の作用を有するか、または有することが予測されるアミノ酸をいい、特に酵素分子にあっては、活性部位中の同様の位置に存在し触媒活性に同様の寄与をするアミノ酸をいう。
【0042】
本明細書において使用される用語「受容体」とは、1個以上のリガンドと可逆的、かつ特異的に複合体化する1個以上の結合ドメインを備える生物学的な構造であって、ここで、この複合体化は生物学的な構造を有する。受容体は、完全に細胞の外部(細胞外の受容体)、細胞膜の中(しかし、受容体の部分を細胞外部の環境および細胞質ゾルに向けている)、または完全に細胞の中(細胞内の受容体)に存在し得る。これらはまた、細胞と独立的に機能し得る。細胞膜中の受容体は、細胞を、その境界の外部の空間と連絡(例えば、シグナル伝達)させ、そして細胞の内側および外側への分子およびイオンの輸送において機能させることを可能とする。本明細書において使用する場合、受容体は、受容体全長であっても、受容体のフラグメントであってもよい。
【0043】
受容体フラグメントを用いる場合には、例えば、受容体タンパク質のリガンド認識に関係する部位を用いることができる。受容体タンパク質のリガンド認識に関係する部位は、以下のように同定することができる。ホモロジーやドメイン検索により相同性や機能上の類似点の高いタンパク質の構造からリガンド認識領域を推定することができる。例えば、同一のリガンドに特異的に結合する、異なる受容体分子のアミノ酸配列をBLASTのデフォルトパラメータを用いて算出した場合、50%以上、好ましくは55%以上、より好ましくは60%以上、さらに好ましくは65%以上の相同性を示す領域が、リガンド認識領域として推定される。
さらに欠損変異やアミノ酸置換などを導入した変異受容体をコードする遺伝子を動物細胞などに一過性発現させ、その機能に必須の領域を決定することも、当業者は容易になし得る。
【0044】
本明細書において使用される用語「リガンド」とは、特異的な受容体または受容体のファミリーに対する結合パートナーである。リガンドは、受容体に対する内因性のリガンドであるか、またはその代わりに、薬剤、薬剤候補、もしくは薬理学的手段のような受容体に対する合成リガンドであり得る。
【0045】
本明細書において「アゴニスト」とは、ある生体作用物質(リガンド)の受容体に結合し、その物質のもつ作用と同じ(あるいは類似の)作用を現わす因子をいう。
【0046】
本明細書において「アンタゴニスト」とは、ある生体作用物質(リガンド)の受容体への結合に拮抗的に働き、それ自身はその受容体を介した生理作用を現わさない因子をいう。拮抗薬、遮断剤(ブロッカー)、阻害剤(インヒビター)などもこのアンタゴニストに包含される。
【0047】
本明細書において「NMR」とは、核磁気共鳴を意味する。磁気モーメントを持つ核の集団を静磁場B0の中に置くと、核ゼーマン効果によって、磁気モーメントの大きさと磁場の強さに従った不連続なエネルギー準位に分布する。この準位の間隔に相当する周波数をもつ電磁波を照射すると共鳴吸収が観測される現象を核磁気共鳴という。
【0048】
本明細書において「二次元NMR」とは、核磁気共鳴スペクトルを2つの周波数軸に展開する方法をいう。この測定には複数のパルスを用い、その時間間隔と観測パルス後の時間とを二つの時間軸としてフーリエ変換を行う。シグナル強度は、通常、等高線によって表される。二次元NMRとしては、代表的には、TROSY(transverse relaxation−optimized spectroscopy)、COSY(二次元シフト相関NMR法)、SECSY(二次元スピンエコー相関分光法)、FOCSY(二次元折り返し補正分光法)、HOHAHA(二次元同核ハートマン法)、NOESY(二次元NOE法)、2D−J法(二次元J分解分光法)、リレーCOSY(リレーコヒーレンス移動分光法)が挙げられるが、これらに限定されない。好ましいNMRはTROSYである。TROSYとは、Wuthrichらによって開発された、超高磁場NMRに適した方法である(Proc.Natl.Acad.Sci.USA vol.94、12366−12371頁(1997))。分子量の大きな分子のNMRにおいては、核磁気緩和を支配するのは、(1)磁気双極子相互作用と、(2)化学シフト異方性であるが、超高磁場では、化学シフト異方性が非常に大きくなるために、緩和時間が短くなり、線幅が広くなる。TROSYは、これら2つの相互作用を相殺することによって、緩和時間を長くし、線幅を狭くする方法である。TROSYによって高分子量のタンパク質(例えば、900kDa以上)について、二次元NMRスペクトルを得ることが可能になった。
【0049】
本明細書において二次元NMRを行う場合、蛋白質を標識するために使用される安定同位体の核種としては、15N, 13C, 2Hまたはこれらの組み合わせが挙げられるが、これらに限定されない。好ましい標識に用いられる安定同位体核の種類としては15N単標識、13C単標識、15N/2H二重標識、13C/2H二重標識、15N/13C/2H三重標識である。
【0050】
本明細書において、「液晶物質」とは、標的タンパク質の弱い配向を実現する物質である。液晶物質としては、 ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とドデシル硫酸ナトリウム(SDS)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物、
1,2−ジ−O−ドデシル−sn−グリセロ−3−ホスフォコリン(DIODPC)と3−(コラミドプロピル)−ジメチルアンモニオ−2−ヒドロキシ−1−プロパンサルフェート(CHAPS)との混合物、
n−アルキル−ポリ(エチレングリコール)/n−アルキルアルコール混合物、
繊維状ファージ、
セチルピリジニウムクロライド(CPCl)−ヘキサノール−NaCl混合物、
セチルピリジニウムブロミド(CPBr)−ヘキサノール−NaCl混合物、
Halobacterium属の紫膜断片
微結晶セルロース、および
ポリアクリルアミドゲル。
が挙げられるが、これらに限定されない。
【0051】
これら液晶物質が、脂質である場合、脂質からなるbicelleを標的タンパク質と混合し、混合物の温度を、相転移以上に設定することによって、標的タンパク質を弱く配向することができる。
【0052】
本明細書において「分子の配向状態」とは、Z軸を静磁場方向とした座標軸に対する分子の座標軸の傾きを示すオイラー角によって示される。
【0053】
本明細書において「配向情報」とは、タンパク質内のドメインの配向に関する情報であって、分子配向テンソルとして得られる情報をいう。本明細書において、配向情報は、例えば、お互いに直行する単位ベクトルによって表すことができる。
【0054】
本明細書において、「NMRシグナルの軸方向の変化量」とは、安定同位体元素について展開された二次元NMRの周波数軸について、軸方向での分子配向依存的な変化量をいう。例えば、1Hを用いる場合、1H軸方向での分子配向依存的な変化量をいい、また、13Cを用いる場合、13C軸方向での分子配向依存的な変化量をいう。
【0055】
本発明において試験される化合物としては、任意の化合物が利用可能であり、例えば、低分子有機化合物、低分子無機化合物、核酸、核酸アナログ、ペプチド、ペプチドアナログ、金属イオンなどが挙げられるが、これらに限定されない。
【0056】
本発明において、低分子有機化合物、または低分子無機化合物を合成する際には、コンビナトリアルケミストリーを用いることができる。コンビナトリアルケミストリーとは、ある基本骨格(scaffold)に、複数の置換基を組合せ(combinatorial)の手法で導入し、短期間で多数かつ多様のある分子の一群(library)を合成し、この分子群の中から目的とする化合物を見つけだすための有力な合成法である。コンビナトリアルケミストリーのための種々の手法が周知である。歴史的には、コンビナトリアルケミストリーは、Merrifield(米国)のビーズを用いた「ペプチド合成法」(1963年)を基盤とした合成技術の進展と、Furka(ハンガリー)らの「スプリット合成法」(1991年)に触発され、1992年以降盛んに研究開発が進められてきた技術である。特に固相合成法は、合成後の精製が不要なことがスピードアップ化のニーズにマッチして近年急速に発展してきている技術である。
【0057】
本明細書において使用される用語「ポリヌクレオチド」、「オリゴヌクレオチド」および「核酸」は、本明細書において同じ意味で使用され、任意の長さのヌクレオチドのポリマーをいう。この用語はまた、「誘導体オリゴヌクレオチド」または「誘導体ポリヌクレオチド」を含む。「誘導体オリゴヌクレオチド」または「誘導体ポリヌクレオチド」とは、ヌクレオチドの誘導体を含むか、またはヌクレオチド間の結合が通常とは異なるオリゴヌクレオチドまたはポリヌクレオチドをいい、互換的に使用される。そのようなオリゴヌクレオチドとして具体的には、例えば、2’−O−メチル−リボヌクレオチド、オリゴヌクレオチド中のリン酸ジエステル結合がホスホロチオエート結合に変換された誘導体オリゴヌクレオチド、オリゴヌクレオチド中のリン酸ジエステル結合がN3’−P5’ホスホロアミデート結合に変換された誘導体オリゴヌクレオチド、オリゴヌクレオチド中のリボースとリン酸ジエステル結合とがペプチド核酸結合に変換された誘導体オリゴヌクレオチド、オリゴヌクレオチド中のウラシルがC−5プロピニルウラシルで置換された誘導体オリゴヌクレオチド、オリゴヌクレオチド中のウラシルがC−5チアゾールウラシルで置換された誘導体オリゴヌクレオチド、オリゴヌクレオチド中のシトシンがC−5プロピニルシトシンで置換された誘導体オリゴヌクレオチド、オリゴヌクレオチド中のシトシンがフェノキサジン修飾シトシン(phenoxazine−modified cytosine)で置換された誘導体オリゴヌクレオチド、DNA中のリボースが2’−O−プロピルリボースで置換された誘導体オリゴヌクレオチドおよびオリゴヌクレオチド中のリボースが2’−メトキシエトキシリボースで置換された誘導体オリゴヌクレオチドなどが例示される。他にそうではないと示されなければ、特定の核酸配列はまた、明示的に示された配列と同様に、その保存的に改変された改変体(例えば、縮重コドン置換体)および相補配列を包含することが企図される。具体的には、縮重コドン置換体は、1またはそれ以上の選択された(または、すべての)コドンの3番目の位置が混合塩基および/またはデオキシイノシン残基で置換された配列を作成することにより達成され得る(Batzerら、Nucleic Acid Res.19:5081(1991);Ohtsukaら、J.Biol.Chem.260:2605−2608(1985);Rossoliniら、Mol.Cell.Probes 8:91−98(1994))。用語「核酸」はまた、本明細書において、遺伝子、cDNA、mRNA、オリゴヌクレオチド、およびポリヌクレオチドと互換可能に使用される。特定の核酸配列はまた、「スプライス改変体」を包含する。同様に、核酸によりコードされた特定のタンパク質は、その核酸のスプライス改変体によりコードされる任意のタンパク質を暗黙に包含する。その名が示唆するように「スプライス改変体」は、遺伝子のオルタナティブスプライシングの産物である。転写後、最初の核酸転写物は、異なる(別の)核酸スプライス産物が異なるポリペプチドをコードするようにスプライスされ得る。スプライス改変体の産生機構は変化するが、エキソンのオルタナティブスプライシングを含む。読み過し転写により同じ核酸に由来する別のポリペプチドもまた、この定義に包含される。スプライシング反応の任意の産物(組換え形態のスプライス産物を含む)がこの定義に含まれる。
【0058】
本明細書において「ヌクレオチド」は、天然のものでも非天然のものでもよい。「誘導体ヌクレオチド」または「ヌクレオチドアナログ」とは、天然に存在するヌクレオチドとは異なるがもとのヌクレオチドと同様の機能を有するものをいう。そのような誘導体ヌクレオチドおよびヌクレオチドアナログは、当該分野において周知である。そのような誘導体ヌクレオチドおよびヌクレオチドアナログの例としては、ホスホロチオエート、ホスホルアミデート、メチルホスホネート、キラルメチルホスホネート、2−O−メチルリボヌクレオチド、ペプチド−核酸(PNA)が含まれるが、これらに限定されない。
【0059】
本明細書において、「フラグメント」とは、全長のポリペプチドまたはポリヌクレオチド(長さがn)に対して、1〜n−1までの配列長さを有するポリペプチドまたはポリヌクレオチドをいう。フラグメントの長さは、その目的に応じて、適宜変更することができ、例えば、その長さの下限としては、ポリペプチドの場合、3、4、5、6、7、8、9、10、15,20、25、30、40、50およびそれ以上のアミノ酸が挙げられ、ここの具体的に列挙していない整数で表される長さ(例えば、11など)もまた、下限として適切であり得る。また、ポリヌクレオチドの場合、5、6、7、8、9、10、15,20、25、30、40、50、75、100およびそれ以上のヌクレオチドが挙げられ、ここの具体的に列挙していない整数で表される長さ(例えば、11など)もまた、下限として適切であり得る。本明細書において使用する場合、好ましくは、受容体「フラグメント」は、全長受容体が特異的に結合し得るリガンドに特異的に結合する。
【0060】
本発明において使用されるタンパク質は、天然の供給源から精製されたものであっても、遺伝子工学的に宿主細胞を形質転換して、組換え発現されたものであってもよい。遺伝子工学的にポリペプチドを製造する方法としては、例えば、そのポリペプチドを産生する原核生物である細菌を培養し、細菌中に組換え受容体タンパク質を封入体として蓄積させ、その宿主細菌を破壊することによって、そのポリペプチドを得る方法が挙げられる。
【0061】
「形質転換体」とは、宿主細胞を形質転換することによって作製された細胞などの生命体の全部または一部をいう。形質転換体としては、原核細胞が例示される。形質転換体は、その対象に依存して、形質転換細胞、形質転換組織、形質転換宿主などともいわれ、本明細書においてそれらの形態をすべて包含するが、特定の文脈において特定の形態を指し得る。
【0062】
形質転換体を得るための宿主細菌細胞は、生理活性を保持するポリペプチドを発現するものであれば、特に限定されず、従来から遺伝子操作において利用される各種の宿主細菌細胞を用いることができる。原核細胞としては、エシェリヒア属、セラチア属、バチルス属、ブレビバクテリウム属、コリネバクテリウム属、ミクロバクテリウム属、シュードモナス属等に属する原核細胞、例えば、Escherichia coli XL1−Blue、Escherichia coli XL2−Blue、Escherichia coli DH1、Escherichia coli MC1000、Escherichia coli KY3276、Escherichia coli W1485、Escherichia coli JM109、Escherichia coli
HB101、Escherichia coli No.49、Escherichia coli W3110、Escherichia coli NY49、Escherichia coli BL21(DE3)、Escherichia coli BL21(DE3)pLysS、Escherichia coli HMS174(DE3)、Escherichia coli HMS174(DE3)pLysS、Serratia ficaria、Serratia fonticola、Serratia liquefaciens、Serratia marcescens、Bacillus subtilis、Bacillus amyloliquefaciens、Brevibacterium ammmoniagenes、Brevibacterium immariophilum ATCC14068、Brevibacterium saccharolyticumATCC14066、Corynebacterium glutamicum ATCC13032、Corynebacterium glutamicum ATCC14067、Corynebacterium glutamicum ATCC13869、Corynebacterium acetoacidophilum ATCC13870、Microbacterium ammoniaphilum ATCC15354、Pseudomonas sp.D−0110などが例示される。
【0063】
本発明において得られた細胞に由来するポリペプチドは、天然型のポリペプチドと実質的に同一の作用を有する限り、アミノ酸配列中の1以上のアミノ酸が置換、付加および/または欠失していてもよく、糖鎖が置換、付加および/または欠失していてもよい。
【0064】
あるアミノ酸は、相互作用結合能力の明らかな低下または消失なしに、例えば、リガンド分子の結合部位のようなタンパク質構造において他のアミノ酸に置換され得る。あるタンパク質の生物学的機能を規定するのは、タンパク質の相互作用能力および性質である。従って、特定のアミノ酸の置換がアミノ酸配列において、またはそのDNAコード配列のレベルにおいて行われ得、置換後もなお、もとの性質を維持するタンパク質が生じ得る。従って、生物学的有用性の明らかな損失なしに、種々の改変が、本明細書において開示されたペプチドまたはこのペプチドをコードする対応するDNAにおいて行われ得る。
【0065】
上記のような改変を設計する際に、アミノ酸の疎水性指数が考慮され得る。タンパク質における相互作用的な生物学的機能を与える際の疎水性アミノ酸指数の重要性は、一般に当該分野で認められている(Kyte.JおよびDoolittle,R.F.J.Mol.Biol. 157(1):105−132,1982)。アミノ酸の疎水的性質は、生成したタンパク質の二次構造に寄与し、次いでそのタンパク質と他の分子(例えば、酵素、基質、受容体、DNA、抗体、抗原など)との相互作用を規定する。各アミノ酸は、それらの疎水性および電荷の性質に基づく疎水性指数を割り当てられる。それらは:イソロイシン(+4.5);バリン(+4.2);ロイシン(+3.8);フェニルアラニン(+2.8);システイン/シスチン(+2.5);メチオニン(+1.9);アラニン(+1.8);グリシン(−0.4);スレオニン(−0.7);セリン(−0.8);トリプトファン(−0.9);チロシン(−1.3);プロリン(−1.6);ヒスチジン(−3.2);グルタミン酸(−3.5);グルタミン(−3.5);アスパラギン酸(−3.5);アスパラギン(−3.5);リジン(−3.9);およびアルギニン(−4.5))である。
【0066】
あるアミノ酸を、同様の疎水性指数を有する他のアミノ酸により置換して、そして依然として同様の生物学的機能を有するタンパク質(例えば、リガンド結合能において等価なタンパク質)を生じさせ得ることが当該分野で周知である。このようなアミノ酸置換において、疎水性指数が±2以内であることが好ましく、±1以内であることがより好ましく、および±0.5以内であることがさらにより好ましい。疎水性に基づくこのようなアミノ酸の置換は効率的であることが当該分野において理解される。米国特許第4、554、101号に記載されるように、以下の親水性指数がアミノ酸残基に割り当てられている:アルギニン(+3.0);リジン(+3.0);アスパラギン酸(+3.0±1);グルタミン酸(+3.0±1);セリン(+0.3);アスパラギン(+0.2);グルタミン(+0.2);グリシン(0);スレオニン(−0.4);プロリン(−0.5±1);アラニン(−0.5);ヒスチジン(−0.5);システイン(−1.0);メチオニン(−1.3);バリン(−1.5);ロイシン(−1.8);イソロイシン(−1.8);チロシン(−2.3);フェニルアラニン(−2.5);およびトリプトファン(−3.4)。アミノ酸が同様の親水性指数を有しかつ依然として生物学的等価体を与え得る別のものに置換され得ることが理解される。このようなアミノ酸置換において、親水性指数が±2以内であることが好ましく、±1以内であることがより好ましく、および±0.5以内であることがさらにより好ましい。
【0067】
本発明において、「保存的置換」とは、アミノ酸置換において、元のアミノ酸と置換されるアミノ酸との親水性指数または/および疎水性指数が上記のように類似している置換をいう。保存的置換の例は、当業者に周知であり、例えば、次の各グループ内での置換:アルギニンおよびリジン;グルタミン酸およびアスパラギン酸;セリンおよびスレオニン;グルタミンおよびアスパラギン;ならびにバリン、ロイシン、およびイソロイシン、などが挙げられるがこれらに限定されない。
【0068】
本明細書において、「改変体」とは、もとのポリペプチドまたはポリヌクレオチドなどの物質に対して、一部が変更されているものをいう。そのような改変体としては、置換改変体、付加改変体、欠失改変体、短縮(truncated)改変体、対立遺伝子変異体などが挙げられる。対立遺伝子(allele)とは、同一遺伝子座に属し、互いに区別される遺伝的改変体のことをいう。従って、「対立遺伝子変異体」とは、ある遺伝子に対して、対立遺伝子の関係にある改変体をいう。「種相同体またはホモログ(homolog)」とは、ある種の中で、ある遺伝子とアミノ酸レベルまたはヌクレオチドレベルで、相同性(好ましくは、60%以上の相同性、より好ましくは、80%以上、85%以上、90%以上、95%以上の相同性)を有するものをいう。そのような種相同体を取得する方法は、本明細書の記載から明らかである。「オルソログ(ortholog)」とは、オルソロガス遺伝子(orthologous gene)ともいい、二つの遺伝子がある共通祖先からの種分化に由来する遺伝子をいう。例えば、多重遺伝子構造をもつヘモグロビン遺伝子ファミリーを例にとると、ヒトとマウスのαヘモグロビン遺伝子はオルソログであるが,ヒトのαヘモグロビン遺伝子とβヘモグロビン遺伝子はパラログ(遺伝子重複で生じた遺伝子)である。オルソログは、分子系統樹の推定に有用であることから、オルソログもまた、本発明において有用であり得る。
【0069】
「保存的(に改変された)改変体」は、アミノ酸配列および核酸配列の両方に適用される。特定の核酸配列に関して、保存的に改変された改変体とは、同一のまたは本質的に同一のアミノ酸配列をコードする核酸をいい、核酸がアミノ酸配列をコードしない場合には、本質的に同一な配列をいう。遺伝コードの縮重のため、多数の機能的に同一な核酸が任意の所定のタンパク質をコードする。例えば、コドンGCA、GCC、GCG、およびGCUはすべて、アミノ酸アラニンをコードする。したがって、アラニンがコドンにより特定される全ての位置で、そのコドンは、コードされたポリペプチドを変更することなく、記載された対応するコドンの任意のものに変更され得る。このような核酸の変動は、保存的に改変された変異の1つの種である「サイレント改変(変異)」である。ポリペプチドをコードする本明細書中のすべての核酸配列はまた、その核酸の可能なすべてのサイレント変異を記載する。当該分野において、核酸中の各コドン(通常メチオニンのための唯一のコドンであるAUG、および通常トリプトファンのための唯一のコドンであるTGGを除く)が、機能的に同一な分子を産生するために改変され得ることが理解される。したがって、ポリペプチドをコードする核酸の各サイレント変異は、記載された各配列において暗黙に含まれる。好ましくは、そのような改変は、ポリペプチドの高次構造に多大な影響を与えるアミノ酸であるシステインの置換を回避するようになされ得る。
【0070】
本明細書中において、機能的に等価なポリペプチドを作製するために、アミノ酸の置換のほかに、アミノ酸の付加、欠失、または修飾もまた行うことができる。アミノ酸の置換とは、もとのペプチドを1つ以上、例えば、1〜10個、好ましくは1〜5個、より好ましくは1〜3個のアミノ酸で置換することをいう。アミノ酸の付加とは、もとのペプチド鎖に1つ以上、例えば、1〜10個、好ましくは1〜5個、より好ましくは1〜3個のアミノ酸を付加することをいう。アミノ酸の欠失とは、もとのペプチドから1つ以上、例えば、1〜10個、好ましくは1〜5個、より好ましくは1〜3個のアミノ酸を欠失させることをいう。アミノ酸修飾は、アミド化、カルボキシル化、硫酸化、ハロゲン化、アルキル化、グリコシル化、リン酸化、水酸化、アシル化(例えば、アセチル化)などを含むが、これらに限定されない。置換、または付加されるアミノ酸は、天然のアミノ酸であってもよく、非天然のアミノ酸、またはアミノ酸アナログでもよい。天然のアミノ酸が好ましい。
【0071】
このような核酸は、周知のPCR法により得ることができ、化学的に合成することもできる。これらの方法に、例えば、部位特異的変位誘発法、ハイブリダイゼーション法などを組み合わせてもよい。
【0072】
本明細書において、ポリペプチドまたはポリヌクレオチドの「置換、付加または欠失」とは、もとのポリペプチドまたはポリヌクレオチドに対して、それぞれアミノ酸もしくはその代替物、またはヌクレオチドもしくはその代替物が、置き換わること、付け加わることまたは取り除かれることをいう。このような置換、付加または欠失の技術は、当該分野において周知であり、そのような技術の例としては、部位特異的変異誘発技術などが挙げられる。置換、付加または欠失は、1つ以上であれば任意の数でよく、そのような数は、その置換、付加または欠失を有する改変体において目的とする機能(例えば、癌マーカー、神経疾患マーカーなど)が保持される限り、多くすることができる。例えば、そのような数は、1または数個であり得、そして好ましくは、全体の長さの20%以内、10%以内、または100個以下、50個以下、25個以下などであり得る。
【0073】
高分子構造(例えば、ポリペプチド構造)は種々のレベルの構成に関して記述され得る。この構成の一般的な議論については、例えば、Albertsら、Molecular Biology of the Cell(第3版、1994)、ならびに、CantorおよびSchimmel、Biophysical Chemistry Part I:The Conformation of Biological Macromolecules(1980)を参照。「一次構造」とは、特定のペプチドのアミノ酸配列をいう。「二次構造」とは、ポリペプチド内の局所的に配置された三次元構造をいう。これらの構造はドメインとして一般に公知である。ドメインは、ポリペプチドの緻密単位を形成し、そして代表的には50〜350アミノ酸長であるそのポリペプチドの部分である。代表的なドメインは、βシート(βストランドなど)およびα−ヘリックスのストレッチ(stretch)のような、部分から作られる。「三次構造」とは、ポリペプチドモノマーの完全な三次元構造をいう。「四次構造」とは、独立した三次単位の非共有的会合により形成される三次元構造をいう。異方性に関する用語は、エネルギー分野において知られる用語と同様に使用される。
【0074】
本明細書において使用される用語「生体分子」とは、生体に関連する分子をいう。本明細書において「生体」とは、生物学的な有機体をいい、動物、植物、菌類、ウイルスなどを含むがそれらに限定されない。生体分子は、生体から抽出される分子を包含するが、それに限定されず、生体に影響を与え得る分子であれば生体分子の定義に入る。そのような生体分子には、タンパク質、ポリペプチド、オリゴペプチド、ペプチド、ポリヌクレオチド、オリゴヌクレオチド、ヌクレオチド、核酸(例えば、cDNA、ゲノムDNAのようなDNA、mRNAのようなRNAを含む)、ポリサッカリド、オリゴサッカリド、脂質、低分子(例えば、ホルモン、リガンド、情報伝達物質、有機低分子、コンビナトリアルライブラリ化合物など)、これらの複合分子などが包含されるがそれらに限定されない。本明細書において好ましい生体分子は、受容体および受容体フラグメント、ならびにそれらのリガンドである。
【0075】
本明細書においてタンパク質として受容体を使用する場合、受容体としては、レクチン様酸化LDL受容体(LOX−1)を含むスカベンジャー受容体、インスリン受容体ファミリーに属する受容体、EGF受容体ファミリーに属する受容体、PDGF受容体ファミリーに属する受容体、VEGF受容体ファミリーに属する受容体、FGF受容体ファミリーに属する受容体、NGF受容体ファミリーなどの増殖因子受容体、ならびに、TGF-βスーパーファミリー受容体、Toll-like受容体ファミリーの受容体、LDL受容体関連タンパク質ファミリーの受容体、およびGタンパク質共役型受容体ファミリーの受容体が挙げられるが、これらに限定されない。
【0076】
本発明は、標的タンパク質溶液に化合物を添加して誘導される標的タンパク質の立体構造変化をTROSY NMR相関スペクトル(例えば、1H/15N−NMRスペクトル)の変化から読み取る技術を提供する。この技術により、1)化合物の標的タンパク質上での結合部位、2)標的タンパク質が化合物によって誘導される構造変化の2つを同時に観測することができる。
【0077】
タンパク質の多くはリガンド、アゴニストなどの化合物によって活性な構造へ誘導されて機能発現を行う。そのため、標的タンパク質を活性構造に誘導する化合物は作動薬のリード化合物になり得る。その一方で、リガンド、アゴニストなどによるタンパク質の活性構造への誘導を阻害する物質は、新規なアンタゴニストをスクリーニングする際のリード化合物になり得る。従って、化合物が標的タンパク質と結合・接触した際に、そのタンパク質の構造がそのように変化するのかを検出することは、新規な薬物のスクリーニングにおいて重要である。
【0078】
しかし、既存のNMRによる化合物と標的タンパク質の相互作用解析技術は、タンパク質構造変化に関する定量的データを提供しない。そのため、化合物が標的タンパク質と結合・接触した際に、そのタンパク質の構造がそのように変化するのかを検出する方法が望まれている。
【0079】
本発明は、核磁気共鳴を用いることによって、化合物がタンパク質と結合した場合のタンパク質の構造変化を測定する方法、およびその方法およびその方法において使用されるプログラムを提供する。
【0080】
より具体的には、本発明は、核磁気共鳴を用いることによって、任意の化合物がタンパク質と結合・接触した場合の、タンパク質の構造変化を測定する方法に関する。さらに本発明は、任意の化合物がタンパク質と結合・接触した場合に、タンパク質の構造変化を生じる化合物を同定・スクリーニングする方法を提供する。また、本発明は、例えば、リガンド、アゴニスト、などのように、タンパク質と結合した際にそのタンパク質の構造変化を生じることが公知の物質がそのタンパク質と結合した際の構造変化と、任意の化合物がそのタンパク質と結合した際の構造変化とを比較して、リガンド、アゴニストと類似の構造変化をもたらす物質を同定・スクリーニングする方法を提供する。例えば、本発明によって、化合物により誘導されるタンパク質構造変化の定量的情報を提供するため作動薬リード化合物の同定を可能とするスクリーニング法が提供される。
【0081】
この方法は、タンパク質をNMR磁場中に弱く配向させることで観測される異方性スピン相互作用の利用を基にしている。異方性スピン相互作用のひとつである残余双極子効果によるタンパク質中の各結合軸の静磁場に対する配向情報の取得と、そのタンパク質構造解析への応用に関しては多くの研究例が報告されている。また、残余双極子効果を基にしたタンパク質を構成するドメイン間の相対配向を決定する技術も報告されている。ただし、残余双極子観測法は原理的に適用できる標的タンパク質の分子量に限界があり約60kDa(アミノ酸数では約500残基)以下のタンパク質に対してしか適用できない。薬物スクリーニングにおける標的タンパク質としては、通常60kDaよりもはるかに大きな分子量のタンパク質が用いられる。従って、創薬を目的とした化合物スクリーニング技術に適用するには、その汎用性に問題があった。
【0082】
本発明で鍵となる新しい技術は、従来法が持つ標的タンパク質分子量の適用限界を超えて、原理的には900kDa以上のタンパク質を対象としても化合物により誘導されるタンパク質中のドメイン間相対配向を定量的に観測し、その構造変化を測定することできるというものである。
【0083】
例えば、本発明は、新規薬物のリード化合物を探索するスクリーニング技術を提供する。具体的には、化合物との相互作用により誘導される標的タンパク質構造変化をTROSY 1H/15N相関NMRスペクトル、またはTROSY 2H/15N相関NMRスペクトルにより定量的に評価する技術を提供する。さらにこの技術を利用して、標的タンパク質の活性構造への変化の有無を指標とした動作薬(アゴニスト)リード化合物の同定を行うスクリーニング法も提供される。
【0084】
(NMR構造解析)
本明細書において用いられるNMR による高次構造解析に用いられるタンパク質試料は、通常、生合成により安定同位体標識することによって構造解析される。一般には、まず遺伝子操作した大腸菌等によって標的タンパク質の発現系を確立し、安定同位体標識した炭素源や窒素源(すべての炭素を13Cラベルしたグルコース、窒素を15Nラベルした塩化アンモニウムなど)を加えた培地で発現させることにより、すべての炭素および窒素を安定同位体標識したタンパク質を得ることができる。得られた標識タンパク質は、クロマトグラフィーなどで精製した後、限外濾過などで濃縮することによってNMR測定に供することができる。この際、以下のような条件に留意することが望ましい。
【0085】
使用する容器については、タンパク質の粘度が高いことを考慮してガラス壁面に付着、分散を抑えるために、NMR 試料管を含めて、使用する器具にはシリコンなどでコーティング処理を施すことが好ましい。また、試料の扱いには、パスツールピペットなどを用いることが望ましい。
【0086】
溶媒としては、通常、水溶液が用いられる。ただし、タンパク質は多くの交換性アミドプロトンを有しており、重水溶液にしてしまうと、スペクトルの解析で鍵となるアミドプロトンが重水素置換されることにより、多くの情報が失われることから、通常、軽水溶液で試料調製し、その後NMRロック用として10%程度の重水を添加することが行われている。必要に応じて、界面活性剤、還元剤などを加えていてもよい。
【0087】
試料濃度は、通常、1mM 程度(例えば、0.4mM〜2mM)にする。試料の量に余裕があり、溶解度が高いのであれば、5mM〜10mM でも測定可能であるが、濃度を高くすると溶液中で会合が起こる危険性があるので留意する。会合による予期しない解析結果を防ぐためにも、低濃度と高濃度の試料で線形を比較して会合の有無を確認することが好ましい。
【0088】
使用するpHとしては、通常、5〜7程度が好ましい。pH を下げることにより、アミド
プロトンなどの交換可能プロトンの交換速度を遅くなるからである。ただし、pH が低い
と高次構造に影響を与える懸念があるため、円二色性(CD)スペクトル等で確認しておくことが好ましい。
【0089】
緩衝剤としては、スペクトル解析の障害となることを防ぐため、プロトンを持たないものを使用することが望ましい。そのようなものとしては、リン酸緩衝液、重水素化酢酸緩衝液などが挙げられるがそれらに限定されない。
【0090】
使用する温度としては、 生体内の温度に近付けるため、30〜40 ℃程度にすることが望ましい。温度を高めに設定して溶液の粘性を下げることにより、タンパク質分子のT2が長くなることから、一般には温度が高い方が良好なスペクトルが得られる。
【0091】
試料溶液に溶存酸素が含まれていると、常磁性緩和のために緩和時間が短くなり、感度の損失を招き、TROSY相関の検出の障害となる場合もあることから、このことを防ぐために脱気をおこない、溶存酸素を取り除くことが好ましい。脱気操作は減圧による脱気、不活性ガスのバブリングなどがあり、好ましくは減圧脱気が使用される。また、気泡を立てないことが望ましい。
【0092】
溶液の保存のために、アジドを加えることが望ましいが、必ずしも加える必要はない。
【0093】
試料調製ののち、NMRのシム調整を行う。まず、NMRロックをかけて分解能調整を行う。タンパク質溶液NMR の測定は、原則として軽水溶液での測定であるため、巨大な水信号(標的タンパク質の数万倍〜数百万倍の1H 濃度)を消去することが好ましいからである。これは、分解能調整が目的であり、好ましくは、良質のスペクトルを得るため、測定試料ごとに充分な分解能調整を行うことが望ましい。タンパク質溶液NMRの測定で試料管は、通常回転させないことから、Z 軸だけでなく、X、Y軸および高次の項の調整も含めて分解能をしっかりと上げておくことが、良いスペクトルを得るために必要である。
【0094】
次に、パルス幅の測定を行う。水溶液で測定するタンパク質溶液NMRは、塩強度の違いにより試料によってパルス幅が変化する場合があることから、原則として測定試料ごとにパルス幅の測定をおこなうことが必要である。
【0095】
タンパク質溶液NMRで用いられるパルス・シーケンスは、数多くのRFパルスが配列されていることから、パルス幅のズレが全体ではかなり大きく影響してくるとされている。したがって、多次元NMR測定を実行する前に、標的タンパク質の信号でパルス幅を測定し、その値を用いて多次元NMR測定をおこなうことが望ましい。
【0096】
次に、NMRによるサンプルの確認を行う。試料調整してからしばらく保管した後など、タンパク質が劣化している可能性があるときなどにはNMRによる確認をおこなうことが望ましい。
【0097】
そこで、通常15N−1H HSQC測定を行い、2次元NMRスペクトルで確認する。信号のパターンがおかしいとき、位相が合わないようなときなどの不具合があるときは、標的タンパク質が壊れてしまっている可能性があることから、再度実験条件の調整が必要である。
【0098】
次に、スペクトル解析(信号の帰属)を行う。NOEスペクトルを例にとってスペクトルの解析法の概略を以下に説明する。NMRによって得られたNOE由来の距離情報をもとに1H信号の帰属を行い、高次構造を導き出すことができる。幾重にも重なり合った信号を、3次元・4次元といった測定を用いてことごとく分離し、既知のアミノ酸配列に割り当てていく。その原理には、スペクトルがアミノ酸ごとに特定のパターンを持っていること、化学シフトおよびスピン結合定数などに極めて特徴的な法則があることを利用する。このような特徴的な性質から、現在では帰属の自動化も行われている。以下に、タンパク質NMRスペクトルの特徴的な化学シフトを示す。
【0099】
タンパク質の1Hの化学シフトはその環境ごとにおおよそ限定されている。すなわち、アミドプロトン由来の信号(8ppm付近)、側鎖の芳香環由来の信号(7ppm付近)、α位のプロトン由来の信号(4ppm付近)、β位のプロトン由来の信号(2〜3ppm付近)、メチル基由来の信号(1ppm付近)が、それぞれ観測される。
【0100】
13Cの化学シフトにも同様の相関があり、13Cではそれぞれの領域を選択的に励起することで、α位やβ位の炭素、カルボニル炭素などをあたかも別核種のように取り扱うことにより、帰属のための様々な測定法のなかで活用される。
【0101】
アミノ酸配列の帰属に加えて、高次構造に由来する化学シフトの変化が見られる。たとえば、α位のプロトン由来の信号はαヘリックス構造の中では高磁場シフトし、βシート構造の中では低磁場シフトする。その他にも、側鎖の芳香環との位置関係により±1ppm程度シフトすることがある。また金属結合タンパク質などでは、金属と結合した部位でかなり大きなシフトが見られる。
【0102】
次に、同種核スピン結合も考慮すべきである。タンパク質を構成するアミノ酸は、ペプチド結合(アミド結合)で連結されていることから、カルボニル基によって1H−1H間のスピン結合が分断され、1H−1H間のスピン結合は同一アミノ酸残基内に限定される。したがって、1H−1H間のスピン結合のパターンは、アミノ酸の種類ごとに特徴的であり、この事実を利用して、アミノ酸の同定を行う。また、α位のプロトンとアミドプロトンの3JHαHNスピン結合定数から導かれる二面角φは主鎖の2次構造の決定に役立つ。
【0103】
また、異種核間スピン結合も考慮すべきである。主鎖に関連する1H−1H以外のスピン結合定数としては、例えば、H−N(90−100Hz)、N−Cα(11Hz)、Cα−H(140Hz)、Cα−Cβ(30−40Hz)、Cα−C(=O)(55Hz)、Cα−X−N(7Hz)、13C−15N(15Hz)などが挙げられる。
【0104】
また、NOE相関も考慮されるべきである。NOE相関は、高次構造解析に用いられるが、アミドプロトンとα位のプロトンとのカルボニル基をはさんだ1H同種核NOEは、その二つのプロトンが、アミド結合を介して隣り合ったアミノ酸残基に由来することを示しており、1H同種核実験によるアミノ酸配列の帰属で用いられる。この手法を、スピン結合や化学シフトをもとに決定したアミノ酸の種別情報とあわせて、配列特異的連鎖帰属法という。
【0105】
帰属作業が終わったら、必要に応じて、高次構造解析を行う。タンパク質の高次構造には、アミノ酸配列が立体的な規則性を持って形作るαヘリックス構造、βシート構造のような2次構造、いくつかの2次構造が空間的に配置された3次構造、3次構造で構築されたドメインが空間的に配置された4次構造があり、これらを明らかにすることがタンパク質溶液NMR測定において構造解析において重要であり、目的に応じて種々行われる。
【0106】
一次配列が既知のアミノ酸配列の場合、既知のポリペプチド鎖がどのように絡み合ってタンパク質を構築しているのかを、NMRから得られる情報を利用して解像する。この際利用されるのが、ディスタンス・ジオメトリー法、束縛条件付き分子動力学法と呼ばれる構造最適化計算アルゴリズムである。いずれの場合にも、NMRから得られる距離情報としてのNOE信号を集められる限り集めに集め、それをパラメータとして計算プログラムに与えることによって高次構造を導き出す手法である。
【0107】
本明細書において、「ディスタンス・ジオメトリー法」とは、距離情報と結合角(2面角)の集合とを空間的な位置情報としてみて、それをもとに構造を導き出す手法をいう。通常は、共有結合の結合長および結合角を一般的なタンパク質の標準値に固定し、2面角を変数として構造計算を進める。NOE由来の距離の束縛条件を満たす方向に収束した構造をいくつか取り出し、最も適切と思われる構造を最終構造とする。この手法は、変数が少ないために計算も速く、コンピュータに対する負荷も小さいが、原子の接触によって構造変化が制限される(共有結合同士のすり抜けができない)ため、真の構造に収束することが難しく、次に述べる束縛条件付き分子動力学法などの初期構造を求めるために利用される。
【0108】
本明細書において、「束縛条件付き分子動力学法」とは、分子運動をシミュレートするために一般的に用いられている手法として知られる分子動力学法(仮想的に分子を振動させ、構造を最適化する方法)の一つであり、NOE由来の距離の束縛条件をポテンシャルとして加え、構造を最適化していく手法をいう。この手法は位置情報としてデカルト(Cartesian)座標を使用しており、また変数も多いために膨大な計算量が必要とされ、コンピュータに対する負荷も大きいが、現在ではコンピュータの計算能力の飛躍的な進歩により計算時間が短縮され、頻繁に利用されている。
【0109】
本明細書において「シミュレーテッド・アニーリング法」とは、分子動力学計算の1手法であり、一般的な分子動力学計算と異なり、計算の初期段階で共有結合に寄与するパラメータを弱く設定し、系の温度を急激に上げることにより分子運動を激化させ、その後徐々に系の温度を下げて分子運動を遅くしつつ、それぞれのパラメータを強めて構造を収束させる手法をいう。これにより、原子の接触によって構造変化が妨げられることによる見かけの収束に構造が落ち着くことを防ぐことができる。
【0110】
上記で触れた3次元あるいは4次元展開されたNOEスペクトルの観測を行う場合、1H−15Nあるいは、1H−18Cという異種核スピン間での相関シグナルの検出が基本となる。高分子量タンパク質(30kDa以上)を解析対象とする場合、それぞれの核スピンのT2緩和時間が短くなるため生じる早いスピン緩和のために、この異種核相関シグナルを検出することが困難となる。高分子量タンパク質を対象とする場合に避けられない早いスピン緩和の問題を解決し、高分子量タンパク質でも十分な観測感度で異種核種の相関シグナルの観測を可能とする方法として、TROSYがWuthrichのグループによって開発された。この新しい異種核間相関シグナルの検出技術は、これまでタンパク質立体構造決定に用いられてきた既存の測定法に組み込むことが可能であり、上記のタンパク質構造解析過程はTROSYを利用したデータ測定法に切り替えることで、30kDaを超える分子量のタンパク質にも適用可能である。
【0111】
以上、本明細書において使用可能なNMR手法を説明したが、このような手法は、当該分野において公知であり、例えば、Kurt Wuthrich 著,京極好正・小林祐次訳,タンパク質と核酸のNMR:二次元NMR による構造解析,東京化学同人.(1991);M.Sattler,J.Schleucher,C.Griesinger,Progr.NMR Spectrosc.34,93−158.(1999);J.Cavanagh,W.J.Fairbrother,A.G.Palmer III,N.J.Skelton,Protein NMR Spectroscopy:Principles and Practice,Academic Press.(1995);T.L.James and N.J.Oppenheimer(eds.),Methods in Enzymology,Vol.239,Academic Press.(1994);荒田洋治,タンパク質のNMR:構造データの解釈と評価,共立出版.(1996); 荒田洋治,NMR の書,丸善.(2000); 根本暢明,吉田卓也,小林祐次,科学と工業,69 69(10),419−425.(1995);根本暢明,吉田卓也,小林祐次,科学と工業,70 70(2),48−55.(1996)などに記載されており、本明細書においてその内容を参考として援用する。
【0112】
(作動薬のリード化合物の同定)
作動薬のリード化合物の同定の1形態を以下に説明する。タンパク質は図1に楕円で示すようにドメインと呼ばれる部分構造体がつながって形作られる。多くのタンパク質は、機能発現の際にこのドメイン間の相対的配置が変化して、機能を発現するに最適な構造をとる。活性構造への誘導は、通常は低分子化合物のタンパク質への結合により行なわれる。阻害剤は、タンパク質へは結合するが活性構造へは誘導せず不活性な構造を維持するように作用する。一方、タンパク質を活性化する化合物の場合は、タンパク質に結合することにより機能発現に最適な立体構造変化を誘導する。化合物結合により誘導されるタンパク質構造が活性構造かどうかは、天然の基質が結合したことにより誘導される立体構造を雛形として判断することができる。本発明は、タンパク質のドメイン間の相対配向を定量的に決定するNMR解析技術を提供することで、TROSYスペクトル変化という簡便に観測可能な実験値からタンパク質が活性構造にあるかどうかを判定することを可能とする。この技術を利用することで、図1にイメージとして描いているようなドメイン間の相対配向変化を基にして選択的に作動薬のリード化合物を同定することができる。
【0113】
(タンパク質の分子配向の計算法)
NMRスペクトルを用いて、タンパク質の分子配向の決定に必要な計算を、例えば、以下の様に行うことが可能であるが、これに限定されない。
【0114】
TROSYスペクトル上で観測されるシグナルの15N軸方向での分子配向依存的な変化量Δδtrosy(k)は、1H−15N核スピン間の残余双極子効果Dnhと、15N核まわりの電子分布の空間的異方性に由来する分子配向依存的な15N軸方向でのシグナルの変化量Δδ15Nの2つの量により次の関係で結び付けられる。
【0115】
Δδtrosy=(1/2)Dnh+Δδ15N (1)
残余双極子効果Dnhはタンパク質中の主鎖にある各NH結合スペクトルのi番目(i=x、y、z)の分子座標軸に対する角度φiと分子の外部磁場に対する分子の配向を規定するSaupeオーダーマトリクス、S(Sijは、分子配向マトリクスの要素を表す。i,j=x,y,z)により次の関係により表現される。D0nhは1H−15N間の双極子間相互作用の強度を示す。
【0116】
Dknh=D0nhΣSijcosφkicosφkj (2)
i,j=x,y,z
一方15N核周りの電子分布の異方性のために生じる、分子の外部磁場に対する配向依存的な15N核の外部磁場に対する磁気遮蔽度の違いに由来する15N化学シフトの変化Δδ15Nは、各ペプチド結合に固有な化学シフト異方性テンソルを分子座標軸上で表現した要素σij i,j=x,y,zと分子配向を規定するSaupeオーダーマトリクスを用いて次の式で表現できる。
【0117】
Δδ15N=(2/3)ΣSijδij (3)
i,j=x,y,z
式(1)−(3)の関係から、分子の磁場中での配向角度に依存したTROSYシグナル15N軸方向での変化量Δδtrosyは、次の式で表現できる。
【0118】
Δδtrosy(k)=ΣSij{(1/2)D0nh cosφkicosφkj+(2/3)δij} (4)
i,j=x,y,z
スペクトル上で観測される複数のシグナルkについて得られる式(4)の関係を用い、特異値分解法(Singular Value Decomposition)により実験値Δδtrosyと、標的タンパク質のドメイン構造内の原子座標から計算される値{(1/2)D0nhcosφi
cosφj+(2/3)δij}から、分子配向を決定するために必要なSaupeオーダーマトリクスの全ての要素Sijを決定することができる。この場合、少なくとも5以上、好ましくは少なくとも6以上、より好ましくは少なくとも7以上、少なくとも8以上、少なくとも9以上、または少なくとも10以上のシグナルkについて得られる式(4)の関係を用いる。また、全てのシグナルkについて式(4)の関係を用いることも可能である。Saupeオーダーマトリクスを対角化することで、標的タンパク質または標的タンパク質内のドメイン構造の静磁場に対する配向情報を得ることができる。ここで、得られる物理量は、分子座標軸で表現される分子配向テンソル量として与えられる。この分子配向テンソル量は分子配向強度(配向テンソルという3次元直行座標系で表現される各軸方向の大きさであり、どの程度その軸に分子が配向しているか(存在確率に相当する)を表現する量である)と、分子の配向状態を記述する新たな座標軸(Z軸が静磁場方向となる)の分子座標に対するオイラー角(a、b、c)で表現する。なお、上記の式は、15Nを用いて例示的に示しているが、必ずしも15Nを用いる必要はなく、その他の安定同位体(例えば、13Cなど)を用いることもできる。
【0119】
(タンパク質の配向情報の決定)
タンパク質構造は、x、y、z軸上の原子の位置によって表現される。タンパク質が磁場に対して配向する場合には、配向状態にあるタンパク質を表現する座標軸(Dxx,Dyy,Dzz)がもとの座上軸からどの程度傾いているかで表現する。そのために、図に示すような3つの角度、α、β、γ、が定義される。これをオイラー角と呼ぶ。磁場の配向方向はDzz軸に一致すると仮定して表現される(図2)。
【0120】
本明細書において、ドメイン構造変化を表す指標としては、分子配向変化度(DOC)および活性構造一致度(DOS)が挙げられるがこれらに限定されない。
【0121】
本明細書において、2つの配向の間での「分子配向変化度」(DOC;degree of orientational change)は、例えば、
(i)化合物と結合・接触前のタンパク質ドメインの第1の配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【0122】
【化17】
を用いて示し、
(ii)化合物と結合・接触後のタンパク質の第2のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【0123】
【化18】
を用いて示し、
(iii)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、この角度をドメイン構造変化を表す指標とするか、または、以下の式
分子配向変化度=a2+b2+c2、
によって分子配向変化度を求め、この分子配向変化度を構造変化の比較のための指標とする。例えば、この角度a、b、およびcが小さい場合、またはDOCが小さい場合、化合物の結合・接触によって誘導されたタンパク質の構造変化が、化合物が存在しない場合と大きく違わないことを示す。また、この角度a、b、およびcが大きい場合、またはDOCが大きい場合、化合物の結合・接触によって誘導されたタンパク質の構造変化が、化合物が存在しない場合と大きく異なることを示す。
【0124】
また、アゴニスト化合物をスクリーニングする場合、リガンド結合によって誘導された構造変化が、どの程度活性構造に近いかを指標とすることができる。この場合、リガンドと結合・接触した後の活性構造のドメイン配向を規定する分子配向テンソルの各軸を表現する第3の単位ベクトルを
【0125】
【化19】
として、化合物と結合・接触した後の第2の単位ベクトルとを用いて、これら2つの単位ベクトルの各々の間の角度を、a、b、およびcとして表し、この角度をドメイン構造変化を表す指標とするか、または、活性構造一致度(degree of orientational similarity to the active conformation;DOS)を以下の式
活性構造一致度=a2+b2+c2、
によって求め、この分子配向変化度を構造変化の比較のための指標とする。例えば、この角度a、b、およびcが小さい場合、またはDOSが小さい場合、化合物の結合・接触によって誘導されたタンパク質の構造変化が、リガンドによって誘導されたタンパク質の構造変化に近い、すなわち構造変化が類似することを示す指標となる。
【0126】
また、タンパク質内のドメインの構造変化は、ドメインの比較する各々の状態での配向情報に基づくDOCおよび/またはDOSの差異として数値化される。
【0127】
従って、本明細書において、第1の化合物と該タンパク質とが接触した場合のタンパク質の構造変化と、第2の化合物と該タンパク質とが接触した場合のタンパク質の構造変化とを比較した場合において、タンパク質の構造変化が「類似する」とは、構造変化の指標としてのDOSの差が小さいことを意味する。例えば、タンパク質と接触することによって、第1の化合物と該タンパク質とが接触した場合のタンパク質の構造変化と類似するタンパク質の構造変化を生じる、第2の化合物の選択は、
(a)タンパク質内のドメインを選択する工程;
(b)タンパク質と該第1の化合物を接触させる条件下における、ドメインの配向情報を得る工程;
(c)タンパク質と第2の化合物を接触させる条件下における、ドメインの配向情報を以下
(i)ドメインの既知の原子座標を得て、
(ii)液晶物質存在下において該タンパク質と該第2の化合物を接触させ、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(iii)ドメインの原子座標、およびNMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(iv)決定されたマトリクスを対角化して、ドメインの配向情報とする、
ことにより得る工程;および
(d)工程(b)において得られた配向情報と、工程(c)において測定された配向情報について、DOSを比較する工程;
によって行なわれる。
【0128】
従って、第1の化合物によって生じる構造変化と第2の化合物によって生じる構造変化が同一の場合、a=b=c=0であり、従って、DOS=0である。第1の化合物によって生じる構造変化と第2の化合物によって生じる構造変化が「類似する」とは、例えば、DOSの値が、75(deg2)以下、48(deg2)以下、27(deg2)以下、12(deg2)以下、3(deg2)以下、0.75(deg2)以下、0.48(deg2)以下、0.27(deg2)以下、0.12(deg2)以下、または0.03(deg2)以下の場合をいう。
【0129】
あるいは、構造変化が類似するか否かは、例えば、複数の候補化合物の集団内の第2の化合物と第1の化合物との間でDOSを計算して、相対的に小さいDOSを生じるか、または最も小さいDOSを生じる第2の化合物が、第1の化合物によって生じる標的タンパク質の構造変化と類似する構造変化を生じるとして、判断することが可能である。
【0130】
(タンパク質の原子座標の入手)
タンパク質のドメイン間構造と機能について、米国Yale大学のM.Gresteinのグループがタンパク質の構造変化の様式をMolMovDBというデータベースにまとめている(MolMovDB、http://MolMovDB.org)。
【0131】
例えば、GroEL、アデニレートキナーゼ、カルモジュリン、HIV−1逆転写酵素、クエン酸シンターゼ、ジフテリアトキシン、HIV−1プロテアーゼ、EGF−レセプター、ヘキソキナーゼ、グルコキナーゼ、EPOレセプター、ホスホグリセレートキナーゼ、リコベリン(recoverin)、mRNAキャッピング酵素、DNAポリメラーゼβについての情報がデータベースから利用可能である。
【0132】
(タンパク質の原子座標から、タンパク質ドメインの配向情報を得る方法)
上記記載の方法によってタンパク質の原子座標を入手した場合、その原子座標から、タンパク質のドメインの配向情報を得ることが可能である。例えば、残余双極子効果Dnhはタンパク質中の主鎖にある各NH結合スペクトルのi番目(i=x、y、z)の分子座標軸に対する角度φiと分子の外部磁場に対する分子の配向を規定するSaupeオーダーマトリクス、S(Sijは、分子配向マトリクスの要素を表す。i,j=x,y,z)により次の関係により表現される。
【0133】
Dknh=D0nhΣSijcosφkicosφkj (5)
i,j=x,y,z
そのため、タンパク質のドメインの原子座標より計算されるcosφki cosφkjと、1H−15N核スピン間の残余双極子効果Dnhと、1H−15N間の双極子間相互作用の強度D0nhとを用いることによって、分子配向を決定するために必要なSaupeオーダーマトリクスの全ての要素Sijを決定することができる。
【0134】
(タンパク質に化合物が結合した際の、タンパク質上の結合位置を特定する方法)
化合物が、タンパク質に結合した場合に、そのタンパク質上の結合位置を、例えば、以下のようにして決定することが可能である。
・化合物の非存在下におけるタンパク質の二次元TROSY NMRスペクトルを得ること、
・化合物をタンパク質と接触させて、タンパク質の二次元TROSY NMRスペクトルを得ること、
・上記のにおいて得られた各々の二次元TROSY NMRスペクトルを比較して、スペクトルの変化を検出すること、
・スペクトルの変化を生じた原子がどのアミノ酸残基の原子であるのかを、二次元NMRの帰属から決定することにより、その化合物が結合したタンパク質の部位周辺のアミノ酸残基(好ましくは複数のアミノ酸残基)を特定すること、
・必要であれば、その化合物の立体構造と、上記において特定されたアミノ酸残基周辺のタンパク質の立体構造とを比較し、その化合物がタンパク質上の結合部位に結合した際の立体構造を推定すること、
によって行なわれる。
【0135】
(タンパク質内ドメインの構造変化の測定)
本発明の1つの局面に従って、例えば、以下の工程を行って、タンパク質内ドメインの構造変化を測定し、および/または構造変化(好ましくは、所望の構造変化)を生じる化合物を選択することが可能である。
【0136】
(a)該タンパク質内のドメインを選択する工程;
(b)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を得る工程;
(c)該タンパク質と該化合物を接触させる条件下における、該ドメインの配向情報を以下
(i)該ドメインの既知の原子座標を得て、
(ii)液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(iii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(iv)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、
ことにより得る工程;および、
(d)工程(b)において得られた配向情報と、工程(c)において得られた配向情報との差異から、該タンパク質の構造変化を測定する工程。
【0137】
本発明の方法の測定対象となるドメインとしては、測定対象となるタンパク質内の任意のドメインが利用可能である。しかし、測定対象としてのドメインとしては、例えば、化合物との結合の有無によって配向情報の変化が大きなドメイン、および/または異なる化合物との結合によって異なる配向情報の変化を生じるドメインが好ましい。
【0138】
上記測定方法における前記(b)工程は、必要に応じて
(b)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を以下
(v)該ドメインの既知の原子座標を得て、
(vi)液晶物質存在下において、該タンパク質を該化合物と接触させずに、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(vii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(viii)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、ことにより得る工程であっても、または
(b)前記タンパク質が前記化合物と接触しない条件下において既に得られている原子座標から、前記ドメインの配向情報を得る工程;
であってもよい。上記複数のSaupeオーダーマトリクスエレメントを決定するためのTROSYスペクトルは、同時にまたは別に測定され得る。
【0139】
また、前記工程(b)および/または(c)において二次元TROSY NMRスペクトルを用いて得られる分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量は、15Nを用いる場合は、15N軸方向の変化量であり得る。別の安定同位体の核種を用いる場合は、その核種の軸方向の変化量であってもよい。
【0140】
ここで、前記(iii)および/または(vii)Saupeオーダーマトリクスエレメントの決定は、15Nを用いる場合は、好ましくは、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定することによって行われる。15N以外の安定同位体の核種を用いる場合は、その核種の核スピン対について上記計算を行ってもよい。その場合、N−H結合の距離に対応する別の異種核間結合の距離および静的双極カップリングが公知であることから、その公知の値を上記計算式において用いることができる。
【0141】
タンパク質内ドメインの構造変化は、例えば、Saupeオーダーマトリクスエレメントを対角化して、ドメインの配向情報とすることによって、得られる。従って、ドメインの配向情報自体を相互に比較することによって、ドメインの構造変化の指標とすることが可能である。例えば、化合物の存在下と非存在下における所定のドメインの配向情報に変化がある場合、その化合物は、タンパク質と結合した場合に、その所定のドメインの構造変化をもたらしたことが判明する。化合物の存在下と非存在下の配向情報は、各々を実験的に求めることが可能であるが、例えば、化合物の非存在下での配向情報が、NMR実験またはX線解析などによって既に得られている場合、その情報を用いて、化合物存在下での実験によって得られた配向情報と比較することが可能である。
【0142】
さらに、構造変化の指標としては、DOCおよびDOSを用いることができる。そのため、任意の化合物がもたらす構造変化の大きさとしては、DOCを用いることができる。DOCの値が大きい場合、構造変化が大きいことを示す。また、任意の化合物が、所定のリガンドまたはアゴニストと類似の構造変化を生じるか否かについては、DOSを用いることができる。DOSの値がより小さい場合、構造変化がより類似することを示す。
【0143】
標的タンパク質の活性誘導を行う作動薬をスクリーニングは、例えば、以下の手順によって行うことが可能である:
・ドメインの構造変化をもたらすことが公知のリガンドまたはアゴニストが標的タンパク質に結合する条件下で、標的タンパク質内ドメインの配向情報(A)を得る;
・スクリーニングの候補化合物が標的タンパク質に結合する条件下で、標的タンパク質内ドメインの配向情報(B)を得る;
・これら配向情報(A)および(B)を比較して、これら配向情報が同一であるか、または類似するかを決定する;
・これら配向情報が同一または類似である場合、候補化合物は、標的タンパク質の活性誘導を行う作動薬と同様の構造変化を標的タンパク質にもたらす化合物であることが判明する。従って、このようにして選択された候補化合物は、有用な作動薬リード化合物である。
【0144】
本発明においては、必要に応じて、化合物がタンパク質に結合した位置を特定する工程を行うことも可能である。その位置の特定は、例えば、工程(b)において得られた二次元TROSY NMRスペクトルと、工程(c)において得られた二次元TROSY NMRスペクトルとを比較して、スペクトルの変化を検出し、そして、該スペクトルの変化を生じた、該タンパク質中のアミノ酸残基を同定すること、によって行われる。このアミノ酸残基の同定は、変化したスペクトルのアミノ酸配列への帰属に基づいて、行われる。
【0145】
本発明において使用される液晶物質としては、以下からなる群から選択される混合物が挙げられる:
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とドデシル硫酸ナトリウム(SDS)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物、
1,2−ジ−O−ドデシル−sn−グリセロ−3−ホスフォコリン(DIODPC)と3−(コラミドプロピル)−ジメチルアンモニオ−2−ヒドロキシ−1−プロパンサルフェート(CHAPS)との混合物、
n−アルキル−ポリ(エチレングリコール)/n−アルキルアルコール混合物、
繊維状ファージ、
セチルピリジニウムクロライド(CPCl)−ヘキサノール−NaCl混合物、
セチルピリジニウムブロミド(CPBr)−ヘキサノール−NaCl混合物、
Halobacterium属の紫膜断片
微結晶セルロース、および
ポリアクリルアミドゲル(例えば、アクリルアミド/アクリレートゲル)、
ならびに上記混合物の少なくとも2つを混合した混合物。
【0146】
例えば、液晶物質は、7.5%(w/v)のジミリストイルホスファチジルコリン
(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物である。
【0147】
本発明の別の局面において、本発明の方法を実行するプログラム、およびそのようなプログラムを含む記憶媒体も提供される。
【0148】
本発明のさらなる局面において、本発明の方法を実行するために使用し得る装置、および本発明のプログラムを含む装置が提供される。
【0149】
以下に実施例等により本発明を詳しく説明するが、本発明はこれらに限定されるものではない。
【実施例1】
【0150】
(液晶物質によるタンパク質の配向)
液晶物質によるタンパク質の配向は、例えば、以下の方法により行われる。
【0151】
(1.ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)との混合物を用いる、タンパク質の弱い配向化)
好ましくは6.8−8.0のpH、より好ましくは、6.5のpHで、DMPCとDHPCとを、好ましくは2.5−3.0の混合比、より好ましくは3.0の混合比で混合する。この混合物を、好ましくは5−20%(w/v)、より好ましくは7.5%(w/v)の濃度で使用し、好ましくは25−40℃、より好ましくは、30℃で分子配向を行う。(M.Ottinger and A.Bax、J.Biomol.NMR 12、361(1998))
(2.ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とドデシル硫酸ナトリウム(SDS)との混合物を用いる、タンパク質の弱い配向化)
上記1のDMPC/DHPC溶液に対して、例えば、SDSのような界面活性剤を添加して、リン脂質膜表面を負に帯電させ、負の表面電荷を持つタンパク質のリン脂質への不必要な吸着を防ぐ。SDSを添加した場合でも、pHを、好ましくは6.0−8.0、より好ましくは、pHを6.5として、好ましくは5−20%(w/v)の濃度で、より好ましくは、7.5%(w/v)で、タンパク質試料溶液と混合し、好ましくは20−35℃、より好ましくは30℃で分子配向を行う。この場合、DMPCとDHPCとを、好ましくは2.5−3.0の混合比、より好ましくは3.0の混合比で混合する。SDSは、DMPCに対して、好ましくは1−3%添加する。(J.A.Losonczi and
J.H.Prestigard、J.Biomol.NMR 12、447(1998))
(3.ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物を用いる、タンパク質の弱い配向化)
上記1のDMPC/DHPC溶液に対して、正の表面電荷を有するタンパク質のリン脂質への不要な吸着を避けるためには、例えば、CTABのような界面活性剤を添加する。CTABを添加した場合でも、pHを好ましくは6.0−8.0、より好ましくはpHを6.5として、好ましくは5−20%(w/v)の濃度で、より好ましくは7.5%(w/v)で、タンパク質試料溶液と混合し、好ましくは20−35℃、より好ましくは30℃で分子配向を行う。この場合、DMPCとDHPCとを、2.5−3.0の混合比、好ましくは3.0の混合比で混合する。CTABは、DMPCに対して、1−3%添加する。(J.A.Losonczi and J.H.Prestigard、J.Biomol.NMR 12、447(1998))
(4.1,2−ジ−O−ドデシル−sn−グリセロ−3−ホスフォコリン(DIODPC)と3−(コラミドプロピル)−ジメチルアンモニオ−2−ヒドロキシ−1−プロパンサルフェート(CHAPS)との混合物を用いる、タンパク質の弱い配向化)
DIODPC/CHAPSOの割合は、4.8が最適であるがこれに限定されない。試料中の最適なリン脂質膜濃度は、5%(w/v)であるがこれに限定されない。試料溶液のpHは、好ましくは1.0−6.5の範囲である。分子配向は、好ましくは10−60℃の範囲である(S.Cavagneroら、J.Biomol.NMR13、387(1999))。
【0152】
(5.n−アルキル−ポリ(エチレングリコール)/n−アルキルアルコール混合物を用いる、タンパク質の弱い配向化)
上記のアルコール混合物を、限定されることはないが、溶液に5%(w/v)添加することにより、好ましくは0−40℃の範囲で分子配向を誘導できる。アルコールの種類としては、n−ヘキサノールまたはn−オクタノールを用いる。n−アルキル−ポリ(エチレングリコール)/n−アルキルアルコールの割合は、好ましくは0.6−0.9の範囲である。pHは、任意のpHが使用可能である。(M.Ruckert and G.Otting、J.Am.Chem.Soc.122、7793(2000))
(6.繊維状ファージを用いる、タンパク質の弱い配向化)
pf1またはタバコモザイクウイルス(TMV)のような繊維状の形態を有するファージを試料溶液中に共存させることによって、磁場中での分子配向を実現できる。試料溶液中でのファージ濃度は、好ましくは10−50mg/mlの範囲で用いられる。より好ましくは25mg/mlの濃度である。pHは好ましくは6以下である。任意の温度が使用可能であるが、より好ましくは20−40℃の範囲で使用される。(Hansenら、Nat.Struct.Biol.5、1065(1998);Cloreら、J.Am.Chem.Soc.120、10571(1998))
(7.セチルピリジニウムクロライド(CPCl)−ヘキサノール−NaCl混合物、またはセチルピリジニウムブロミド(CPBr)−ヘキサノール−NaCl混合物を用いる、タンパク質の弱い配向化)
CPBr:ヘキサノール=1:1.33(w/w)〜1:1(w/w)を用いる。NaBrの濃度は、好ましくは25−40mMであり、NaClの濃度は、好ましくは200−500mMである。この液晶物質を使用する際の濃度は、好ましくは1−10%(w/v)であり、好ましくは15−50℃の範囲の温度で分子配向する。
【0153】
(8.Halobacterium属の紫膜断片を用いる、タンパク質の弱い配向化)
Halobacterium属の紫膜断片、好ましくは、Halobacterium salinariumの紫膜断片を、1−10mg/mlの濃度で用いる。任意の溶液温度、および任意のpHが利用可能である。溶液中のNaCl濃度は、好ましくは50mM以下である。(Prosserら、J.Am.Chem.Soc. 120、11010(1988))
(9.微結晶セルロースを用いる、タンパク質の弱い配向化)
好ましくは、80mg/mlの溶液を調製し、タンパク質を導入することによって、分子配向が可能である。任意のpH、温度が利用可能である。溶液のイオン濃度が上昇すると、分子配向を実現するために必要となる試料溶液に添加する。(Flemingら、J.Am.Chem.Soc.122、5224(2000))
(10.ポリアクリルアミドゲルを用いる、タンパク質の弱い配向化)
NMR試料管内に、縦方向、または横方向に圧縮したポリアクリルアミドゲル中に、タンパク質溶液を注入して、タンパク質を機械的に分子配向させる。任意のpH、イオン強度、測定温度が使用可能である。(Saasら、J.Biomol.NMR 18、303(2000);Tyckoら、J.Am.Chem.Soc.122、9340(2000))
(11.アクリルアミド/アクリレートゲルを用いる、タンパク質の配向化)
ポリアクリルアミドゲルの一例として、50%アクリルアミド/50%アクリレートゲルを2.5倍に伸長させることによって作製した配向ゲルを用いることができる。アクリルアミド/アクリレートを用いてゲルを生成する際に、標識したタンパク質を添加することによって、ゲル中でタンパク質を配向させることができる。
【実施例2】
【0154】
(TROSYシグナル変化から得た分子配向情報の従来法による解析結果との比較)
0.5mM ユビキチン(8.6kDa)を20mM リン酸ナトリウム緩衝液(pH6.4)に溶かして7.5%(w/v)のDMPC/DHPC/CTABを共存させた溶液を30℃で分子配向状態を作り、以下の手順で実験を行なった。
【0155】
(実験)
(NMR分光法)
本分析で使用された全てのスペクトルを、セチルトリメチルアンモニウムブロミド(CTAB)をドープされた、ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とのバイセル(bicelle)混合物が、2H NMRスペクトルにおいて、10Hzの残余四重極分裂を与えるように配列される条件下で、30℃で記録した。データ収集を、1H周波数が499.84MHzで作動するVarian INOVA500分光器上で実施した。マジック角度実験について、本発明者らは、15Nに対して調整されたXを備える1H検出コイルVarian gHX NanoProbeを使用した。サンプル回転速度を、2.6kHzに設定し、磁界に対してバイセルを配列したことに関するトルクを除去した(Courtieu,J.; Bayles, J. P.; Fung, B. M. Prog. NMR spectrosc. 1994,26, 141-169.、Kurita,J.; Shimahara, H.; Utsunomiya-Tate, N.; Tate, S. J. MagnReson. 2003,163,163-173.)。1H−15N双極子カップリングを、1H−15N IRAP−HSQCスペクトルにおいて測定した(Ottiger,M.;Delaglio, F.; Bax, A. J. Magn Reson. 1998, 131, 373-378.)。1H−15N TROSYスペクトル(Pervushin,K.;Riek, R.; Wider, G.; Wuthrich, K. Proc. Natl. Acad. Sci. U. S. A 1997,94,12366-12371.、Weigelt, J. J. Am. Chem. Soc. 1998, 120, 10778-10779.)を、7.5%(重量/体積)のDMPC:DHPC:CTABバイセルを含有する、50mMのリン酸ナトリウム(pH6.4)および2mM EDTA中に溶解された0.5mM 15N標識ユビキチンを含む同一の溶液をサンプル溶液を使用して、マジック角度サンプル回転法を用いずに、およびそれを用いて、それぞれ等方的状態および異方的状態において収集した。
【0156】
全てのデータを、NMRpipeプログラムソフトウェアパッケージ(Delaglio, F.; Grzesiek, S.; Vuister, G. W.;Zhu, G.; Pfeifer, J.;Bax, A. J. Biomol. NMR 1995, 6, 277-293.)を用いて処理した。ピーク位置を、プログラムPIPP(Garrett,D.S.; Powers, R.; Gronenborn, A. M.; Clore, G. M. J. Magn. Reson. 1991,95,214-220.)を用いてピーク最大値の60%と80%との間の各計算等高線に対して楕円をフィッティングすることによって決定した。各楕円の中心は、ピーク位置の基準を提供し、所定のピークの60〜80%の強度範囲における全ての等高線について得られた値を、ピーク位置を提供するために平均した。t1次元およびt2次元でのデータ収集のための取得時間は、それぞれ111msおよび128msであり、200(t1、15N)×1024(t2、1H)複素点のマトリクスサイズを有した。広範囲なゼロフィルをされた最終処理データは、1H次元および15N次元それぞれについて、2.0Hzおよび0.88Hzのデジタル分解能を示した。
【0157】
(配列テンソルの計算)
配列誘導性TROSYシフト変化およびその誤差評価に由来する整列テンソルの決定を、公開されたサブルーチンを組込んだ、組織内で書かれたC−プログラムによって実施した(Press,W. H.; Teukolsky, S. A.; Vetterling, W. T.; Flannery, B. P.Numerical recipes in C 2nd ed.; Cambridge university press: New York, 1992.)。配列テンソルを、実験データから計算し、そしてプログラムMOLMOL(Koradi,R.;Billeter, M.; Wuthrich, K. J. Mol. Graph. 1996, 14, 51-55.)によって付加された水素原子を有する1.8ÅのX線座標(PDB登録コード、1UBQ)(Vijay-Kumar,S.;Bugg, C. E.; Cook, W. J. J. Mol. Biol. 1987, 194, 531-544.)から生成された参照構造から計算した。TROSYシフトの実験不確定性の効果を、生成されたデータセットに対して、特異値分解(SVD)計算(Losonczi,J.A.; Andrec, M.; Fischer, M. W.; Prestegard, J. H. J. Magn. Reson. 1999,138,334-342.)を繰り返すことによって評価した。このデータセットは、ガウス雑音を実験データに付加することによって生成された。ガウス雑音を、相対的確率(−exp(x2/2δ2))で分布すると仮定した。ここで、xは、実験値であり、δは、TROSYシフトの連続的な実験の間で対rms偏位から見積もられたrmsノイズである。
【0158】
「構造ノイズ」を、文献(Zweckstetter, M.; Bax, A. J. Biomol. NMR, 2002, 23, 127-137.)に記載される方法と類似する方法で、ランダムな様式でNH結合ベクトルを再配向することによって、シミュレートした。オリジナルベクトルと再配向ベクトルとの間の偏位を、オリジナルの方向から5度の傾斜角を有する円錐で記述する。本構造ノイズのシミュレーションにおいて、オリジナルのNHベクトルと再配向NHベクトルとの間の傾斜角を、ガウス分布(−exp(α2/2θ2))にあると仮定した。ここでαは、傾斜角であり、θは、5度に固定された傾斜角に関する推定された標準偏差である。円錐中の再配向されたベクトルの横方向の回転角を、等しく分布されているとしてシミュレートした。シミュレートされたデータからの適切な値の選択は、実験データとシミュレートされたΔδtrosy値との間のrms偏位を、2.55Hz未満に維持することによって行った。この値は、シミュレートされた値の68%に対して受容される速度を示し、この値は、これらの値が、平均の1標準偏差内に存在することを意味する。
【0159】
30℃で7.5%(w/v)のDMPC/DHPC/CTABバイセルに溶解した15N標識ユビキチンについてのΔδtrosy値を以下の表に示す。
【0160】
【表1】
上記の結果を得た実験においては、それぞれ、異方性の状態、および整列された状態において収集されたデータを得るために、MASを適用せずに、またはMASを適用して、実験を行なった。MAS(マジック角試料回転)とは、magic angle sample spinningの略である。MASを適用してNMR測定を行うと、液晶物質共存下であっても、物理的に分子配向を消すことが可能である。そのため、液晶物質共存下であっても、液晶物質非共存下の場合と同じデータを得ることができる。
【0161】
一連の1H−15N TROSYデータから、Δδtrosy値を、
Δδtrosy値=δ15Ntrosy(MAS非適用)−δ15Ntrosy(MAS適用)
として求めた。
【0162】
Saupeオーダーマトリクスエレメントの決定を以下のとおりに行った。
【0163】
ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させることなく、二次元NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (6)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定した。
【0164】
このSijを対角化することによって、オイラー角α、β、γを得た。
【0165】
この分子配向により誘導されたTROSYシグナル変化から決定されたユビキチンの分子配向角度を従来の残余双極子効果観測から得た。
【0166】
残余双極子効果観測は、従来の残余双極子効果観測法:IPAP-HSQC法(Inphase-Antiphase mode Heteronuclear Single Quantum Coherence spectroscopy; Ottigerら、J.Magn.Reson. 131, 373 (1998))を用いて、液晶分子を利用して分子配向を誘導する前後において観測される各アミノ基に由来するシグナルの1Jnh(1H-15N核スピン間の単結合スピン結合定数)が示す強度変化を観測し、液晶共存下で分子配向状態で観測される1Jnh(aniso)と、液晶が共存せず蛋白質分子が配向状態にない時に観測される1Jnh(iso)を用いて、
Dnh = 1Jnh(aniso) - 1Jnh(iso)
として、各1H-15N核スピン間の残余双極子効果Dnhを求めることにより行なった。
【0167】
残余双極子効果観測から得られた分子配向角度と比較した結果は以下のとおりである。(分子座標軸に対する分子配向軸の傾きをオイラー角表示で示す)。
【0168】
【表2】
この結果から、TROSYシグナル変化に基づいて決定される分子配向角度情報は、従来の分子配向決定技術と整合性のある結果を与えることが確認された。すなわち高分子量タンパク質を対象として、化合物との結合により誘導される標的タンパク質の構造変化を定量的にTROSYシグナル変化だけから観測することが可能であると示された。
【実施例3】
【0169】
(高分子量タンパク質について、本発明を用いる構造変化の測定)
従来のNMRでは測定することができなかった30kDaを超える分子量を有するタンパク質の配向情報を、本発明を用いて以下のように得て、構造変化を測定した。
【0170】
対象のタンパク質として、35kDaの分子量を有する、クロレラウイルスPBCV1のグアニリル転移酵素であるmRNA5’−キャッピング酵素(PBCV1-CE)を用いた。35kDaという分子量は、従来の測定法による分子配向解析では困難になる分子量域である。このタンパク質はリガンドにより誘導される立体構造の変化に依存して活性が制御されることが公知である。結晶構造解析により、リガンド結合していないPBCV1-CEは結晶中で2つの構造(オープン型およびクローズ型)を取ることが明らかにされている。本実施例では活性型構造を誘導するGTPの結合阻害剤であるピロリン酸共存下でのPBCV1-CEの溶液中での構造を解析し、リガンドを持たない構造からの変化を解析した。
【0171】
(主鎖の帰属)
TROSY測定のデータ解析には、主鎖の帰属データが必要であるので、2H/13C/15Nで三重標識したPBCV1−CEを用いて約50%の主鎖の帰属を行った。
(TROSY測定条件)
2H/15N標識した0.5mMのPBCV1-CE(200mM リン酸カリウム,pH7.6中)を、6%(w/w)の約2.5倍に伸張させた50%アクリルアミド/50%アクリレートゲル(配向ゲル)(図3)に入れることで、PBCV1-CEを磁場に対して配向させた。サンプル溶液には、活性阻害剤であるピロリン酸を5mMを入れた。TROSY測定を20℃で行った(図4)。分子配向依存的なTROSYシフト変化を、配向ゲル中のPBCV1-CEのTROSYシグナルの、通常溶液の状態のTROSYシグナルからの15N軸側のずれとして観測した。十分なシグナルの分離を示す51残基分(N末端ドメイン、23残基、C末端ドメイン、28残基)のTROSYシフト変化に基づいて、各ドメインの配向テンソルを決定した。
得られたドメイン相対配向の結果の精度を確認するために、結晶構造中で見つかっているPBCV1-CEの2つの異なるドメイン配向を持つ構造(オープン型およびクローズ型)をもとにして解析を行った。なお、クローズ型の結晶構造を基にした解析では、部分的なドメイン内構造変化の影響を除くためにN末端ドメインの2残基分のデータを解析からはずした。
(分子配向解析)
本発明の方法に従って、オープン型構造のデータを用いて、ドメイン間分子配向解析を行った。Δδtrosy値を得て、実施例2と同様にSaupeオーダーマトリクスエレメントの決定を行い、ヌクレオチド非存在下でのオイラー角α、β、およびγを決定した。
PBCV1-CEは立体構造上明確なドメイン構造を持つ。アミノ酸残基番号で15-222からなるドメイン(ラージドメイン)と同じく239−311からなるドメイン(スモールドメイン)のそれぞれに由来するTROSYシグナルの配向依存的な変化から、各ドメインの配向テンソルを決定した。その結果、表3に示すように、ラージドメイン(N末端)の分子配向軸の分子座標軸からの傾きを表現するオイラー角は(α=95.9°、β=123.1°、γ=96.2°、Q−因子=0.38)であるのに対し、スモールドメイン(C末端)の方は(α=98.4°、β=-114.7°、γ=165.4°、Q−因子=0.45)となった。
【表3】
それぞれのドメインの分子配向軸は図5および図6に示すような関係になった。相対的なドメインの配向は、2つの配向軸を一致させるようにスモールドメインを回転させることで決定されるが、各ドメインの配向テンソルからスモールドメインは結晶構造に比べて約65°回転していることが明らかとなった。
オープン型の構造を用いて得られたドメイン相対配向の結果の精度を確認するために、結晶構造中で見つかっているPBCV1-CEの別のドメイン配向(クローズ型)を持つ構造を用いた。その結果を、以下の表4、ならびに図7および図8に示す。
【表4】
本発明の解析方法の結果決定されたドメイン間の相対配向は、オープン型およびクローズ型で同様となり、決定されたドメイン間相対配向の精度が確認された。
【0172】
同様の方法を用いて、GTP存在下およびATP存在下の各々についてのオイラー角α、β、およびγを決定する。さらに、このオイラー角から、GTP添加時、およびATP添加時のDOCを求める。このDOCは、GTP添加時、およびATP添加時の構造変化の指標となる。
【実施例4】
【0173】
(タンパク質の作動薬リード化合物のスクリーニング)
タンパク質の新規のアゴニストおよび/またはアンタゴニストの探索は、新規薬物の製造のために重要である。特に新規のアゴニストは、作動薬リード化合物として利用され得るが、従来技術は、作動薬リード化合物の同定には不十分であった。
【0174】
そこで、本発明を用いて、作動薬リード化合物のスクリーニングを行う。モデルとして、実施例3において用いられるmRNA5’−キャッピング酵素を用いる。
【0175】
作動薬リード化合物は、mRNA5’−キャッピング酵素を活性化するGTPと同様の構造変化をmRNA5’−キャッピング酵素に生じると考えられるため、以下のようにスクリーニングを行う。
【0176】
GTPの代わりに候補化合物を添加した実験を行う以外は、実施例3と同様の条件下で、mRNA5’−キャッピング酵素の配向情報を得る。候補化合物としては、ヌクレオチドアナログ、特にGTPアナログ、およびATPアナログを用いることができる。GTP存在下での配向情報と、候補化合物存在下での配向情報とから、DOSを計算する。
【0177】
複数の候補化合物の中から、DOS値が小さい化合物を、作動薬リード化合物として選択する。
【実施例5】
【0178】
(候補化合物から選択された作動薬リード化合物の結合部位の同定)
作動薬リード化合物を選択した場合、その作動薬リード化合物の結合部位を同定することが可能である。本実施例においては、実施例4において最も小さいDOS値を生じた化合物を用いて、以下のように実験を行う。
・実施例4において用いる候補化合物非存在下での二次元TROSY NMRスペクトルのアミノ酸残基への帰属を常法に従って行う。
・化合物存在下でのタンパク質内ドメインの二次元TROSY NMRスペクトルと、化合物非存在下での二次元TROSY NMRスペクトルとを比較する。その比較において、スペクトルの変化を検出する。変化したスペクトルに対応するアミノ酸残基を、上記帰属の結果から、作動薬リード化合物の結合部位として同定する。
【実施例6】
【0179】
(標的タンパク質に対する化合物の親和性の決定)
本発明において、例えば、以下の方法を用いて標的タンパク質に対する化合物の親和性を決定することが可能である。
・実施例4において用いる候補化合物非存在下での二次元TROSY NMRスペクトルのアミノ酸残基への帰属を常法に従って行う。
・化合物存在下でのタンパク質内ドメインの二次元TROSY NMRスペクトルと、化合物非存在下での二次元TROSY NMRスペクトルとを比較する。その比較から、スペクトルの変化を検出する。変化したスペクトルに対応するアミノ酸残基を、上記帰属の結果から、同定する。
・同定されたアミノ酸に対応するスペクトルは、化合物非存在下の場合と、化合物存在下の場合とで、異なる位置に出現する。しかし、用いる化合物の種類によっては、化合物存在下であっても、化合物非存在下において検出される同定されたアミノ酸に対応するスペクトルが残存する場合がある。このように単一のアミノ酸残基に対応するスペクトルが二重に存在するのは、二次元TROSY NMR測定の溶液中に、化合物と結合したタンパク質と、未結合のタンパク質が混在するためである。この二重に存在する単一のアミノ酸残基に対応するスペクトルの強度の比は、化合物結合タンパク質と化合物未結合タンパク質との存在比と相関する。従って、この二重のスペクトルの強度の比は、試験した化合物とタンパク質との親和性の指標となる。この原理を用いて、以下のように化合物の相対的親和性を測定する。
・実施例5に従って、化合物の結合するアミノ酸残基を同定する。
・化合物A存在下においてNMR測定を行い、化合物の結合するアミノ酸に対応するスペクトルについて、
(化合物A存在下においてのみ検出される位置に出現するスペクトル)/(化合物非存在下において検出される位置に残存するスペクトル)
の比を求める。
・化合物B存在下においてNMR測定を行い、化合物の結合するアミノ酸に対応するスペクトルについて、
(化合物B存在下においてのみ検出される位置に出現するスペクトル)/(化合物非存在下において検出される位置に残存するスペクトル)
の比を求める。
・化合物AおよびBについて、これらの比を比較し、この比の大きい化合物が、標的タンパク質に対して、より親和性が大きい化合物であると判断される。
【産業上の利用可能性】
【0180】
化合物の結合により生じるタンパク質の構造変化を検出することによって、タンパク質のリガンド、アゴニスト、アンタゴニストのスクリーニングが迅速かつ容易になる。さらに、構造変化の検出は、標的タンパク質の活性構造を誘導するか否かを指標として、新規薬物の候補化合物をスクリーニングすることを可能にする。
【特許請求の範囲】
【請求項1】
タンパク質と化合物とを接触させた場合の、該タンパク質の構造変化を測定する方法であって、以下:
(a)該タンパク質内のドメインを選択する工程;
(b)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を得る工程;
(c)該タンパク質と該化合物を接触させる条件下における、該ドメインの配向情報を以下
(i)該ドメインの既知の原子座標を得て、
(ii)液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(iii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(iv)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、
ことにより得る工程;および、
(d)工程(b)において得られた配向情報と、工程(c)において得られた配向情報との差異から、該タンパク質の構造変化を測定する工程、
を包含する、方法。
【請求項2】
前記(b)工程が、
(b)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を以下
(v)該ドメインの既知の原子座標を得て、
(vi)液晶物質存在下において、該タンパク質を該化合物と接触させずに、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(vii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(viii)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、ことにより得る工程である、請求項1に記載の測定方法。
【請求項3】
前記(b)工程が、
(b)前記タンパク質が前記化合物と接触しない条件下において既に得られている原子座標から、前記ドメインの配向情報を得る工程;
である、請求項1に記載の測定方法。
【請求項4】
請求項1に記載の測定方法であって、前記工程(c)において二次元TROSY NMRスペクトルを用いて得られる分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、方法。
【請求項5】
請求項4に記載の測定方法であって、ここで、前記(iii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、方法。
【請求項6】
請求項2に記載の測定方法であって、前記工程(b)において二次元TROSY NMRスペクトルを用いて得られる分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、方法。
【請求項7】
請求項6に記載の測定方法であって、ここで、前記(vii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させることなく、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、方法。
【請求項8】
請求項5または7に記載の方法であって、ここで、前記タンパク質と前記化合物とが接触した場合の、該タンパク質の構造変化が、以下:
(ix)該化合物と接触させる前の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【化20】
を用いて示し、
(x)該化合物と接触させた後の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【化21】
を用いて示し、
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、そして
(xii)以下の式によって分子配向変化度を求める
分子配向変化度=a2+b2+c2、
ことによって分子配向変化度として数値化される、方法。
【請求項9】
請求項2に記載の方法であって、さらに、前記化合物が前記タンパク質に結合した位置を特定する工程を包含する、方法。
【請求項10】
請求項9に記載の方法であって、前記化合物が前記タンパク質に結合した位置を特定する工程が、前記工程(b)において得られた二次元TROSY NMRスペクトルと、前記工程(c)において得られた二次元TROSY NMRスペクトルとを比較して、スペクトルの変化を検出し、そして、該スペクトルの変化を生じた、該タンパク質中のアミノ酸残基を同定すること、によって行われる、方法。
【請求項11】
請求項1に記載の方法であって、前記液晶物質が、以下からなる群から選択される混合物を含む、方法:
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とドデシル硫酸ナトリウム(SDS)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物、
1,2−ジ−O−ドデシル−sn−グリセロ−3−ホスフォコリン(DIODPC)と3−(コラミドプロピル)−ジメチルアンモニオ−2−ヒドロキシ−1−プロパンサルフェート(CHAPS)との混合物、
n−アルキル−ポリ(エチレングリコール)/n−アルキルアルコール混合物、
繊維状ファージ、
セチルピリジニウムクロライド(CPCl)−ヘキサノール−NaCl混合物、
セチルピリジニウムブロミド(CPBr)−ヘキサノール−NaCl混合物、
Halobacterium属の紫膜断片
微結晶セルロース、および
ポリアクリルアミドゲル。
【請求項12】
前記液晶物質が、7.5%(w/v)のジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物である、請求項11に記載の方法。
【請求項13】
接触の際に、タンパク質内のドメインの構造変化を生じる化合物を選択する方法であって、以下:
(a)該タンパク質内のドメインを選択する工程;
(b)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を得る工程;
(c)該タンパク質と該化合物を接触させる条件下における、該ドメインの配向情報を以下
(i)該ドメインの既知の原子座標を得て、
(ii)液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(iii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(iv)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、
ことにより得る工程;
(d)工程(b)において得られた配向情報と、工程(c)において得られた配向情報との比較から、該化合物が該タンパク質と接触した際に該ドメインの構造変化を生じる化合物であるか否かを決定する工程、
を包含する、方法。
【請求項14】
前記(b)工程が、
(b)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を以下
(v)該ドメインの既知の原子座標を得て、
(vi)液晶物質存在下において、該タンパク質を該化合物と接触させずに、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(vii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(viii)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、ことにより得る工程である、請求項13に記載の測定方法。
【請求項15】
前記(b)工程が、
(b)前記タンパク質が前記化合物と接触しない条件下において既に得られている原子座標から、前記ドメインの配向情報を得る工程;
である、請求項13に記載の測定方法。
【請求項16】
請求項13に記載の測定方法であって、前記工程(c)において二次元TROSY NMRスペクトルを用いて得られる分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、方法。
【請求項17】
請求項16に記載の測定方法であって、ここで、前記(iii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、方法。
【請求項18】
請求項14に記載の測定方法であって、前記工程(b)において二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、方法。
【請求項19】
請求項18に記載の測定方法であって、ここで、前記(vii)のSaupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させることなく、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、方法。
【請求項20】
請求項17または19に記載の方法であって、ここで、前記工程(c)における配向情報の比較が、以下:
(ix)該化合物と接触する前の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【化22】
を用いて示し、
(x)該化合物と接触した後の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【化23】
を用いて示し、
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、そして
(xii)以下の式によって分子配向変化度を求め、
分子配向変化度=a2+b2+c2、
該分子配向変化度を比較することによって行われる、方法。
【請求項21】
請求項14に記載の方法であって、さらに、前記化合物が前記タンパク質に結合した位置を特定する工程を包含する、方法。
【請求項22】
請求項21に記載の方法であって、前記化合物が前記タンパク質に結合した位置を特定する工程が、前記工程(b)において得られた二次元TROSY NMRスペクトルと、前記工程(c)において得られた二次元TROSY NMRスペクトルとを比較して、スペクトルの変化を検出し、そして、該スペクトルの変化を生じた、該タンパク質中のアミノ酸残基を同定すること、によって行われる、方法。
【請求項23】
請求項13に記載の方法であって、前記液晶物質が、以下からなる群から選択される混合物を含む、方法:
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とドデシル硫酸ナトリウム(SDS)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物、
1,2−ジ−O−ドデシル−sn−グリセロ−3−ホスフォコリン(DIODPC)と3−(コラミドプロピル)−ジメチルアンモニオ−2−ヒドロキシ−1−プロパンサルフェート(CHAPS)との混合物、
n−アルキル−ポリ(エチレングリコール)/n−アルキルアルコール混合物、
繊維状ファージ、
セチルピリジニウムクロライド(CPCl)−ヘキサノール−NaCl混合物、
セチルピリジニウムブロミド(CPBr)−ヘキサノール−NaCl混合物、
Halobacterium属の紫膜断片
微結晶セルロース、および
ポリアクリルアミドゲル。
【請求項24】
前記液晶物質が、7.5%(w/v)のジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物である、請求項23に記載の方法。
【請求項25】
タンパク質と接触することによって、第1の化合物と該タンパク質とが接触した場合の該タンパク質の構造変化と類似する該タンパク質の構造変化を生じる、第2の化合物を選択する方法であって、以下:
(a)該タンパク質内のドメインを選択する工程;
(b)該タンパク質と該第1の化合物を接触させる条件下における、該ドメインの配向情報を得る工程;
(c)該タンパク質と該第2の化合物を接触させる条件下における、該ドメインの配向情報を以下
(i)該ドメインの既知の原子座標を得て、
(ii)液晶物質存在下において該タンパク質と該第2の化合物を接触させ、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(iii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(iv)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、
ことにより得る工程;
(d)工程(b)において得られた配向情報と、工程(c)において得られた配向情報とを比較する工程;および
(e)工程(d)における比較の結果から、該第2の化合物が該タンパク質と接触することによる該タンパク質の構造変化が、該第1の化合物が該タンパク質と接触することによる該タンパク質の構造変化と類似するか否かを決定する工程、
を包含する、方法。
【請求項26】
前記(b)工程が、
(b)該タンパク質が該第1の化合物と接触する条件下における、該ドメインの配向情報を以下
(v)該ドメインの既知の原子座標を得て、
(vi)液晶物質存在下において、該タンパク質を該第1の化合物と接触させた場合に二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(vii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(viii)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、ことにより得る工程である、請求項25に記載の測定方法。
【請求項27】
前記(b)工程が、
(b)前記タンパク質が前記第1の化合物を接触させる条件下において既に得られている原子座標から、前記ドメインの配向情報を得る工程;
である、請求項25に記載の測定方法。
【請求項28】
請求項25に記載の測定方法であって、前記工程(c)において二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、方法。
【請求項29】
請求項28に記載の測定方法であって、ここで、前記(iii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、方法。
【請求項30】
請求項26に記載の測定方法であって、前記工程(b)において二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、方法。
【請求項31】
請求項30に記載の測定方法であって、ここで、前記(vii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該第2の化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、方法。
【請求項32】
請求項29または31に記載の方法であって、ここで、前記前記工程(d)における配向情報の比較が、以下:
(ix)該第1の化合物と接触した場合の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【化24】
を用いて示し、
(x)該第2の化合物と接触させた場合の、該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【化25】
を用いて示し、そして
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、該a、b、およびcを配向情報の比較のための指標とする、
ことによって行われる、方法。
【請求項33】
請求項29または31に記載の方法であって、ここで、前記工程(d)における配向情報の比較が、以下:
(ix)該第1の化合物と接触した場合の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【化26】
を用いて示し、
(x)該第2の化合物と接触させた場合の、該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【化27】
を用いて示し、そして
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、そして
(xii)以下の式によって活性構造一致度を求め
活性構造一致度=a2+b2+c2、
該活性構造一致度を配向情報の比較のための指標とする、
ことによって行われる、方法。
【請求項34】
請求項25に記載の方法であって、さらに、前記第1および第2の化合物の少なくとも1つが前記タンパク質に結合した位置を特定する工程を包含する、方法。
【請求項35】
請求項34に記載の方法であって、前記第1および第2の化合物の少なくとも1つが前記タンパク質に結合した位置を特定する工程が、該化合物の非存在下における該タンパク質の二次元TROSY NMRスペクトルと、該化合物の存在下における該タンパク質の二次元TROSY NMRスペクトルを比較して、スペクトルの変化を検出し、そして、該スペクトルの変化を生じた、該タンパク質中のアミノ酸残基を同定すること、
によって行われる、方法。
【請求項36】
請求項25に記載の方法であって、前記液晶物質が、以下からなる群から選択される混合物を含む、方法:
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とドデシル硫酸ナトリウム(SDS)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物、
1,2−ジ−O−ドデシル−sn−グリセロ−3−ホスフォコリン(DIODPC)と3−(コラミドプロピル)−ジメチルアンモニオ−2−ヒドロキシ−1−プロパンサルフェート(CHAPS)との混合物、
n−アルキル−ポリ(エチレングリコール)/n−アルキルアルコール混合物、
繊維状ファージ、
セチルピリジニウムクロライド(CPCl)−ヘキサノール−NaCl混合物、
セチルピリジニウムブロミド(CPBr)−ヘキサノール−NaCl混合物、
Halobacterium属の紫膜断片
微結晶セルロース、および
ポリアクリルアミドゲル。
【請求項37】
前記液晶物質が、7.5%(w/v)のジミリストイルホスファチジルコリン(DMPC
)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物である、請求項36に記載の方法。
【請求項38】
タンパク質と化合物とを接触させた場合の、該タンパク質内の選択されたドメインの構造変化を数値化するプログラムであって、以下:
(a)該ドメインについて、該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を得る手段;
(b)該タンパク質と該化合物を接触させる条件下における、該ドメインの配向情報を以下
(i)該ドメインの既知の原子座標のデータを得て、
(ii)液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量のデータを得て、
(iii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(iv)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、
ことにより得る手段;および、
(c)手段(a)において得られた配向情報と、手段(b)において測定された配向情報との差異から、該タンパク質の構造変化を計算する手段、
とをコンピュータに実現させるための、プログラム。
【請求項39】
前記(a)手段が、
(a)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を以下
(v)該ドメインの既知の原子座標を得て、
(vi)液晶物質存在下において、該タンパク質を該化合物と接触させずに、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(vii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(viii)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、ことにより得る手段である、請求項38に記載のプログラム。
【請求項40】
前記(a)手段が、
(a)前記タンパク質が前記化合物と接触しない条件下において既に得られている原子座標から、前記ドメインの配向情報を得る手段;
である、請求項38に記載のプログラム。
【請求項41】
請求項38に記載のプログラムであって、前記工程(b)において二次元TROSY NMRスペクトルを用いて得られる分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、プログラム。
【請求項42】
請求項41に記載のプログラムであって、ここで、前記(iii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、プログラム。
【請求項43】
請求項39に記載のプログラムであって、前記工程(a)において二次元TROSY NMRスペクトルを用いて得られる分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、プログラム。
【請求項44】
請求項43に記載のプログラムであって、ここで、前記(vii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させることなく、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、プログラム。
【請求項45】
請求項42または44に記載のプログラムであって、ここで、前記タンパク質と前記化合物とが接触した場合の、該タンパク質の構造変化が、以下:
(ix)該化合物と接触させる前の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【化28】
を用いて示し、
(x)該化合物と接触させた後の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【化29】
を用いて示し、
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、そして
(xii)以下の式によって分子配向変化度を求める
分子配向変化度=a2+b2+c2、
ことによって分子配向変化度として数値化される、プログラム。
【請求項46】
請求項39に記載のプログラムであって、さらに、前記化合物が前記タンパク質に結合した位置を特定する手段を包含する、プログラム。
【請求項47】
請求項46に記載のプログラムであって、前記化合物が前記タンパク質に結合した位置を特定する手段が、前記工程(a)において得られた二次元TROSY NMRスペクトルと、前記工程(b)において得られた二次元TROSY NMRスペクトルとを比較して、スペクトルの変化を検出し、そして、該スペクトルの変化を生じた、該タンパク質中のアミノ酸残基を同定すること、によって行われる、プログラム。
【請求項48】
接触の際に、タンパク質内のドメインの構造変化を生じる化合物を選択するプログラムであって、以下:
(a)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を得る手段;
(b)該タンパク質と該化合物を接触させる条件下における、該ドメインの配向情報を以下
(i)該ドメインの既知の原子座標を得て、
(ii)液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(iii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(iv)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、
ことにより得る手段;
(c)手段(a)において得られた配向情報と、手段(b)において得られた配向情報との比較から、該化合物が該タンパク質と接触した際に該ドメインの構造変化を生じる化合物であるか否かを決定する手段、
を包含する、プログラム。
【請求項49】
前記(a)手段が、
(a)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を以下
(v)該ドメインの既知の原子座標を得て、
(vi)液晶物質存在下において、該タンパク質を該化合物と接触させずに、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(vii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(viii)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、ことにより得る手段である、請求項48に記載のプログラム。
【請求項50】
前記(a)手段が、
(a)前記タンパク質が前記化合物と接触しない条件下において既に得られている原子座標から、前記ドメインの配向情報を得る手段;
である、請求項48に記載のプログラム。
【請求項51】
請求項48に記載のプログラムであって、前記工程(b)において二次元TROSY NMRスペクトルを用いて得られる分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、プログラム。
【請求項52】
請求項51に記載のプログラムであって、ここで、前記(iii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、プログラム。
【請求項53】
請求項52に記載のプログラムであって、前記工程(a)において二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、プログラム。
【請求項54】
請求項53に記載のプログラムであって、ここで、前記(vii)のSaupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させることなく、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5D0nh cosφki cosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、プログラム。
【請求項55】
請求項52または54に記載のプログラムであって、ここで、前記工程(c)における配向情報の比較が、以下:
(ix)該化合物と接触する前の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【化30】
を用いて示し、
(x)該化合物と接触した後の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【化31】
を用いて示し、
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、そして
(xii)以下の式によって分子配向変化度を求め、
分子配向変化度=a2+b2+c2、
該分子配向変化度を比較することによって行われる、プログラム。
【請求項56】
請求項48に記載のプログラムであって、さらに、前記化合物が前記タンパク質に結合した位置を特定する手段を包含する、プログラム。
【請求項57】
請求項56に記載のプログラムであって、前記化合物が前記タンパク質に結合した位置を特定する手段が、前記工程(a)において得られた二次元TROSY NMRスペクトルと、前記工程(b)において得られた二次元TROSY NMRスペクトルとを比較して、スペクトルの変化を検出し、そして、該スペクトルの変化を生じた、該タンパク質中のアミノ酸残基を同定すること、によって行われる、プログラム。
【請求項58】
タンパク質と接触することによって、第1の化合物と該タンパク質とが接触した場合の該タンパク質内のドメインの構造変化と類似する該タンパク質内のドメインの構造変化を生じる、第2の化合物を選択するプログラムであって、以下:
(a)該タンパク質と該第1の化合物を接触させる条件下における、該ドメインの配向情報を得る手段;
(b)該タンパク質と該第2の化合物を接触させる条件下における、該ドメインの配向情報を以下
(i)該ドメインの既知の原子座標を得て、
(ii)液晶物質存在下において該タンパク質と該第2の化合物を接触させ、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(iii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(iv)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、
ことにより得る手段;
(c)手段(a)において得られた配向情報と、手段(b)において得られた配向情報とを比較する手段;および
(d)手段(c)における比較の結果から、該第2の化合物が該タンパク質と接触することによる該タンパク質の構造変化が、該第1の化合物が該タンパク質と接触することによる該タンパク質の構造変化と類似するか否かを決定する手段、
を包含する、プログラム。
【請求項59】
前記(a)手段が、
(a)該タンパク質が該第1の化合物と接触する条件下における、該ドメインの配向情報を以下
(v)該ドメインの既知の原子座標を得て、
(vi)液晶物質存在下において、該タンパク質を該第1の化合物と接触させた場合に二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(vii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(viii)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、ことにより得る手段である、請求項25に記載のプログラム。
【請求項60】
前記(a)手段が、
(a)前記タンパク質が前記第1の化合物を接触させる条件下において既に得られている原子座標から、前記ドメインの配向情報を得る手段;
である、請求項58に記載のプログラム。
【請求項61】
請求項58に記載のプログラムであって、前記工程(b)において二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、プログラム。
【請求項62】
請求項61に記載のプログラムであって、ここで、前記(iii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定することによって行われる、プログラム。
【請求項63】
請求項59に記載のプログラムであって、前記工程(a)において二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、プログラム。
【請求項64】
請求項63に記載のプログラムであって、ここで、前記(vii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該第2の化合物を接触させ、二次元TROSY
NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、プログラム。
【請求項65】
請求項62または64に記載のプログラムであって、ここで、前記工程(c)における配向情報の比較が、以下:
(ix)該第1の化合物と接触した場合の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【化32】
を用いて示し、
(x)該第2の化合物と接触させた場合の、該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【化33】
を用いて示し、そして
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、該a、b、およびcを配向情報の比較のための指標とする、
ことによって行われる、プログラム。
【請求項66】
請求項62または64に記載のプログラムであって、ここで、前記工程(c)における配向情報の比較が、以下:
(ix)該第1の化合物と接触した場合の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【化34】
を用いて示し、
(x)該第2の化合物と接触させた場合の、該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【化35】
を用いて示し、そして
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、そして
(xii)以下の式によって活性構造一致度を求め
活性構造一致度=a2+b2+c2、
該活性構造一致度を配向情報の比較のための指標とする、
ことによって行われる、プログラム。
【請求項67】
請求項58に記載のプログラムであって、さらに、前記第1および第2の化合物の少なくとも1つが前記タンパク質に結合した位置を特定する手段を包含する、プログラム。
【請求項68】
請求項67に記載のプログラムであって、前記第1および第2の化合物の少なくとも1つが前記タンパク質に結合した位置を特定する手段が、該化合物の非存在下における該タンパク質の二次元TROSY NMRスペクトルと、該化合物の存在下における該タンパク質の二次元TROSY NMRスペクトルとを比較して、スペクトルの変化を検出し、そして該スペクトルの変化を生じた、該タンパク質中のアミノ酸残基を同定すること、によって行われる、プログラム。
【請求項69】
請求項1、13、または25に記載の方法を実行し得る装置。
【請求項70】
請求項38、48、または58に記載のプログラムを備える装置。
【請求項71】
請求項38、48、または58に記載のプログラムを含む記憶媒体。
【請求項1】
タンパク質と化合物とを接触させた場合の、該タンパク質の構造変化を測定する方法であって、以下:
(a)該タンパク質内のドメインを選択する工程;
(b)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を得る工程;
(c)該タンパク質と該化合物を接触させる条件下における、該ドメインの配向情報を以下
(i)該ドメインの既知の原子座標を得て、
(ii)液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(iii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(iv)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、
ことにより得る工程;および、
(d)工程(b)において得られた配向情報と、工程(c)において得られた配向情報との差異から、該タンパク質の構造変化を測定する工程、
を包含する、方法。
【請求項2】
前記(b)工程が、
(b)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を以下
(v)該ドメインの既知の原子座標を得て、
(vi)液晶物質存在下において、該タンパク質を該化合物と接触させずに、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(vii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(viii)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、ことにより得る工程である、請求項1に記載の測定方法。
【請求項3】
前記(b)工程が、
(b)前記タンパク質が前記化合物と接触しない条件下において既に得られている原子座標から、前記ドメインの配向情報を得る工程;
である、請求項1に記載の測定方法。
【請求項4】
請求項1に記載の測定方法であって、前記工程(c)において二次元TROSY NMRスペクトルを用いて得られる分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、方法。
【請求項5】
請求項4に記載の測定方法であって、ここで、前記(iii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、方法。
【請求項6】
請求項2に記載の測定方法であって、前記工程(b)において二次元TROSY NMRスペクトルを用いて得られる分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、方法。
【請求項7】
請求項6に記載の測定方法であって、ここで、前記(vii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させることなく、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、方法。
【請求項8】
請求項5または7に記載の方法であって、ここで、前記タンパク質と前記化合物とが接触した場合の、該タンパク質の構造変化が、以下:
(ix)該化合物と接触させる前の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【化20】
を用いて示し、
(x)該化合物と接触させた後の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【化21】
を用いて示し、
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、そして
(xii)以下の式によって分子配向変化度を求める
分子配向変化度=a2+b2+c2、
ことによって分子配向変化度として数値化される、方法。
【請求項9】
請求項2に記載の方法であって、さらに、前記化合物が前記タンパク質に結合した位置を特定する工程を包含する、方法。
【請求項10】
請求項9に記載の方法であって、前記化合物が前記タンパク質に結合した位置を特定する工程が、前記工程(b)において得られた二次元TROSY NMRスペクトルと、前記工程(c)において得られた二次元TROSY NMRスペクトルとを比較して、スペクトルの変化を検出し、そして、該スペクトルの変化を生じた、該タンパク質中のアミノ酸残基を同定すること、によって行われる、方法。
【請求項11】
請求項1に記載の方法であって、前記液晶物質が、以下からなる群から選択される混合物を含む、方法:
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とドデシル硫酸ナトリウム(SDS)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物、
1,2−ジ−O−ドデシル−sn−グリセロ−3−ホスフォコリン(DIODPC)と3−(コラミドプロピル)−ジメチルアンモニオ−2−ヒドロキシ−1−プロパンサルフェート(CHAPS)との混合物、
n−アルキル−ポリ(エチレングリコール)/n−アルキルアルコール混合物、
繊維状ファージ、
セチルピリジニウムクロライド(CPCl)−ヘキサノール−NaCl混合物、
セチルピリジニウムブロミド(CPBr)−ヘキサノール−NaCl混合物、
Halobacterium属の紫膜断片
微結晶セルロース、および
ポリアクリルアミドゲル。
【請求項12】
前記液晶物質が、7.5%(w/v)のジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物である、請求項11に記載の方法。
【請求項13】
接触の際に、タンパク質内のドメインの構造変化を生じる化合物を選択する方法であって、以下:
(a)該タンパク質内のドメインを選択する工程;
(b)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を得る工程;
(c)該タンパク質と該化合物を接触させる条件下における、該ドメインの配向情報を以下
(i)該ドメインの既知の原子座標を得て、
(ii)液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(iii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(iv)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、
ことにより得る工程;
(d)工程(b)において得られた配向情報と、工程(c)において得られた配向情報との比較から、該化合物が該タンパク質と接触した際に該ドメインの構造変化を生じる化合物であるか否かを決定する工程、
を包含する、方法。
【請求項14】
前記(b)工程が、
(b)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を以下
(v)該ドメインの既知の原子座標を得て、
(vi)液晶物質存在下において、該タンパク質を該化合物と接触させずに、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(vii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(viii)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、ことにより得る工程である、請求項13に記載の測定方法。
【請求項15】
前記(b)工程が、
(b)前記タンパク質が前記化合物と接触しない条件下において既に得られている原子座標から、前記ドメインの配向情報を得る工程;
である、請求項13に記載の測定方法。
【請求項16】
請求項13に記載の測定方法であって、前記工程(c)において二次元TROSY NMRスペクトルを用いて得られる分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、方法。
【請求項17】
請求項16に記載の測定方法であって、ここで、前記(iii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、方法。
【請求項18】
請求項14に記載の測定方法であって、前記工程(b)において二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、方法。
【請求項19】
請求項18に記載の測定方法であって、ここで、前記(vii)のSaupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させることなく、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、方法。
【請求項20】
請求項17または19に記載の方法であって、ここで、前記工程(c)における配向情報の比較が、以下:
(ix)該化合物と接触する前の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【化22】
を用いて示し、
(x)該化合物と接触した後の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【化23】
を用いて示し、
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、そして
(xii)以下の式によって分子配向変化度を求め、
分子配向変化度=a2+b2+c2、
該分子配向変化度を比較することによって行われる、方法。
【請求項21】
請求項14に記載の方法であって、さらに、前記化合物が前記タンパク質に結合した位置を特定する工程を包含する、方法。
【請求項22】
請求項21に記載の方法であって、前記化合物が前記タンパク質に結合した位置を特定する工程が、前記工程(b)において得られた二次元TROSY NMRスペクトルと、前記工程(c)において得られた二次元TROSY NMRスペクトルとを比較して、スペクトルの変化を検出し、そして、該スペクトルの変化を生じた、該タンパク質中のアミノ酸残基を同定すること、によって行われる、方法。
【請求項23】
請求項13に記載の方法であって、前記液晶物質が、以下からなる群から選択される混合物を含む、方法:
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とドデシル硫酸ナトリウム(SDS)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物、
1,2−ジ−O−ドデシル−sn−グリセロ−3−ホスフォコリン(DIODPC)と3−(コラミドプロピル)−ジメチルアンモニオ−2−ヒドロキシ−1−プロパンサルフェート(CHAPS)との混合物、
n−アルキル−ポリ(エチレングリコール)/n−アルキルアルコール混合物、
繊維状ファージ、
セチルピリジニウムクロライド(CPCl)−ヘキサノール−NaCl混合物、
セチルピリジニウムブロミド(CPBr)−ヘキサノール−NaCl混合物、
Halobacterium属の紫膜断片
微結晶セルロース、および
ポリアクリルアミドゲル。
【請求項24】
前記液晶物質が、7.5%(w/v)のジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物である、請求項23に記載の方法。
【請求項25】
タンパク質と接触することによって、第1の化合物と該タンパク質とが接触した場合の該タンパク質の構造変化と類似する該タンパク質の構造変化を生じる、第2の化合物を選択する方法であって、以下:
(a)該タンパク質内のドメインを選択する工程;
(b)該タンパク質と該第1の化合物を接触させる条件下における、該ドメインの配向情報を得る工程;
(c)該タンパク質と該第2の化合物を接触させる条件下における、該ドメインの配向情報を以下
(i)該ドメインの既知の原子座標を得て、
(ii)液晶物質存在下において該タンパク質と該第2の化合物を接触させ、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(iii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(iv)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、
ことにより得る工程;
(d)工程(b)において得られた配向情報と、工程(c)において得られた配向情報とを比較する工程;および
(e)工程(d)における比較の結果から、該第2の化合物が該タンパク質と接触することによる該タンパク質の構造変化が、該第1の化合物が該タンパク質と接触することによる該タンパク質の構造変化と類似するか否かを決定する工程、
を包含する、方法。
【請求項26】
前記(b)工程が、
(b)該タンパク質が該第1の化合物と接触する条件下における、該ドメインの配向情報を以下
(v)該ドメインの既知の原子座標を得て、
(vi)液晶物質存在下において、該タンパク質を該第1の化合物と接触させた場合に二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(vii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(viii)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、ことにより得る工程である、請求項25に記載の測定方法。
【請求項27】
前記(b)工程が、
(b)前記タンパク質が前記第1の化合物を接触させる条件下において既に得られている原子座標から、前記ドメインの配向情報を得る工程;
である、請求項25に記載の測定方法。
【請求項28】
請求項25に記載の測定方法であって、前記工程(c)において二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、方法。
【請求項29】
請求項28に記載の測定方法であって、ここで、前記(iii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、方法。
【請求項30】
請求項26に記載の測定方法であって、前記工程(b)において二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、方法。
【請求項31】
請求項30に記載の測定方法であって、ここで、前記(vii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該第2の化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、方法。
【請求項32】
請求項29または31に記載の方法であって、ここで、前記前記工程(d)における配向情報の比較が、以下:
(ix)該第1の化合物と接触した場合の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【化24】
を用いて示し、
(x)該第2の化合物と接触させた場合の、該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【化25】
を用いて示し、そして
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、該a、b、およびcを配向情報の比較のための指標とする、
ことによって行われる、方法。
【請求項33】
請求項29または31に記載の方法であって、ここで、前記工程(d)における配向情報の比較が、以下:
(ix)該第1の化合物と接触した場合の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【化26】
を用いて示し、
(x)該第2の化合物と接触させた場合の、該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【化27】
を用いて示し、そして
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、そして
(xii)以下の式によって活性構造一致度を求め
活性構造一致度=a2+b2+c2、
該活性構造一致度を配向情報の比較のための指標とする、
ことによって行われる、方法。
【請求項34】
請求項25に記載の方法であって、さらに、前記第1および第2の化合物の少なくとも1つが前記タンパク質に結合した位置を特定する工程を包含する、方法。
【請求項35】
請求項34に記載の方法であって、前記第1および第2の化合物の少なくとも1つが前記タンパク質に結合した位置を特定する工程が、該化合物の非存在下における該タンパク質の二次元TROSY NMRスペクトルと、該化合物の存在下における該タンパク質の二次元TROSY NMRスペクトルを比較して、スペクトルの変化を検出し、そして、該スペクトルの変化を生じた、該タンパク質中のアミノ酸残基を同定すること、
によって行われる、方法。
【請求項36】
請求項25に記載の方法であって、前記液晶物質が、以下からなる群から選択される混合物を含む、方法:
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とドデシル硫酸ナトリウム(SDS)との混合物、
ジミリストイルホスファチジルコリン(DMPC)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物、
1,2−ジ−O−ドデシル−sn−グリセロ−3−ホスフォコリン(DIODPC)と3−(コラミドプロピル)−ジメチルアンモニオ−2−ヒドロキシ−1−プロパンサルフェート(CHAPS)との混合物、
n−アルキル−ポリ(エチレングリコール)/n−アルキルアルコール混合物、
繊維状ファージ、
セチルピリジニウムクロライド(CPCl)−ヘキサノール−NaCl混合物、
セチルピリジニウムブロミド(CPBr)−ヘキサノール−NaCl混合物、
Halobacterium属の紫膜断片
微結晶セルロース、および
ポリアクリルアミドゲル。
【請求項37】
前記液晶物質が、7.5%(w/v)のジミリストイルホスファチジルコリン(DMPC
)とジヘキサノイルホスファチジルコリン(DHPC)とセチルトリメチルアンモニウムブロミド(CTAB)との混合物である、請求項36に記載の方法。
【請求項38】
タンパク質と化合物とを接触させた場合の、該タンパク質内の選択されたドメインの構造変化を数値化するプログラムであって、以下:
(a)該ドメインについて、該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を得る手段;
(b)該タンパク質と該化合物を接触させる条件下における、該ドメインの配向情報を以下
(i)該ドメインの既知の原子座標のデータを得て、
(ii)液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量のデータを得て、
(iii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(iv)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、
ことにより得る手段;および、
(c)手段(a)において得られた配向情報と、手段(b)において測定された配向情報との差異から、該タンパク質の構造変化を計算する手段、
とをコンピュータに実現させるための、プログラム。
【請求項39】
前記(a)手段が、
(a)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を以下
(v)該ドメインの既知の原子座標を得て、
(vi)液晶物質存在下において、該タンパク質を該化合物と接触させずに、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(vii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(viii)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、ことにより得る手段である、請求項38に記載のプログラム。
【請求項40】
前記(a)手段が、
(a)前記タンパク質が前記化合物と接触しない条件下において既に得られている原子座標から、前記ドメインの配向情報を得る手段;
である、請求項38に記載のプログラム。
【請求項41】
請求項38に記載のプログラムであって、前記工程(b)において二次元TROSY NMRスペクトルを用いて得られる分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、プログラム。
【請求項42】
請求項41に記載のプログラムであって、ここで、前記(iii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、プログラム。
【請求項43】
請求項39に記載のプログラムであって、前記工程(a)において二次元TROSY NMRスペクトルを用いて得られる分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、プログラム。
【請求項44】
請求項43に記載のプログラムであって、ここで、前記(vii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させることなく、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、プログラム。
【請求項45】
請求項42または44に記載のプログラムであって、ここで、前記タンパク質と前記化合物とが接触した場合の、該タンパク質の構造変化が、以下:
(ix)該化合物と接触させる前の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【化28】
を用いて示し、
(x)該化合物と接触させた後の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【化29】
を用いて示し、
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、そして
(xii)以下の式によって分子配向変化度を求める
分子配向変化度=a2+b2+c2、
ことによって分子配向変化度として数値化される、プログラム。
【請求項46】
請求項39に記載のプログラムであって、さらに、前記化合物が前記タンパク質に結合した位置を特定する手段を包含する、プログラム。
【請求項47】
請求項46に記載のプログラムであって、前記化合物が前記タンパク質に結合した位置を特定する手段が、前記工程(a)において得られた二次元TROSY NMRスペクトルと、前記工程(b)において得られた二次元TROSY NMRスペクトルとを比較して、スペクトルの変化を検出し、そして、該スペクトルの変化を生じた、該タンパク質中のアミノ酸残基を同定すること、によって行われる、プログラム。
【請求項48】
接触の際に、タンパク質内のドメインの構造変化を生じる化合物を選択するプログラムであって、以下:
(a)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を得る手段;
(b)該タンパク質と該化合物を接触させる条件下における、該ドメインの配向情報を以下
(i)該ドメインの既知の原子座標を得て、
(ii)液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(iii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(iv)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、
ことにより得る手段;
(c)手段(a)において得られた配向情報と、手段(b)において得られた配向情報との比較から、該化合物が該タンパク質と接触した際に該ドメインの構造変化を生じる化合物であるか否かを決定する手段、
を包含する、プログラム。
【請求項49】
前記(a)手段が、
(a)該タンパク質が該化合物と接触しない条件下における、該ドメインの配向情報を以下
(v)該ドメインの既知の原子座標を得て、
(vi)液晶物質存在下において、該タンパク質を該化合物と接触させずに、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(vii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(viii)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、ことにより得る手段である、請求項48に記載のプログラム。
【請求項50】
前記(a)手段が、
(a)前記タンパク質が前記化合物と接触しない条件下において既に得られている原子座標から、前記ドメインの配向情報を得る手段;
である、請求項48に記載のプログラム。
【請求項51】
請求項48に記載のプログラムであって、前記工程(b)において二次元TROSY NMRスペクトルを用いて得られる分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、プログラム。
【請求項52】
請求項51に記載のプログラムであって、ここで、前記(iii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、プログラム。
【請求項53】
請求項52に記載のプログラムであって、前記工程(a)において二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、プログラム。
【請求項54】
請求項53に記載のプログラムであって、ここで、前記(vii)のSaupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させることなく、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5D0nh cosφki cosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、プログラム。
【請求項55】
請求項52または54に記載のプログラムであって、ここで、前記工程(c)における配向情報の比較が、以下:
(ix)該化合物と接触する前の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【化30】
を用いて示し、
(x)該化合物と接触した後の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【化31】
を用いて示し、
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、そして
(xii)以下の式によって分子配向変化度を求め、
分子配向変化度=a2+b2+c2、
該分子配向変化度を比較することによって行われる、プログラム。
【請求項56】
請求項48に記載のプログラムであって、さらに、前記化合物が前記タンパク質に結合した位置を特定する手段を包含する、プログラム。
【請求項57】
請求項56に記載のプログラムであって、前記化合物が前記タンパク質に結合した位置を特定する手段が、前記工程(a)において得られた二次元TROSY NMRスペクトルと、前記工程(b)において得られた二次元TROSY NMRスペクトルとを比較して、スペクトルの変化を検出し、そして、該スペクトルの変化を生じた、該タンパク質中のアミノ酸残基を同定すること、によって行われる、プログラム。
【請求項58】
タンパク質と接触することによって、第1の化合物と該タンパク質とが接触した場合の該タンパク質内のドメインの構造変化と類似する該タンパク質内のドメインの構造変化を生じる、第2の化合物を選択するプログラムであって、以下:
(a)該タンパク質と該第1の化合物を接触させる条件下における、該ドメインの配向情報を得る手段;
(b)該タンパク質と該第2の化合物を接触させる条件下における、該ドメインの配向情報を以下
(i)該ドメインの既知の原子座標を得て、
(ii)液晶物質存在下において該タンパク質と該第2の化合物を接触させ、二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(iii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(iv)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、
ことにより得る手段;
(c)手段(a)において得られた配向情報と、手段(b)において得られた配向情報とを比較する手段;および
(d)手段(c)における比較の結果から、該第2の化合物が該タンパク質と接触することによる該タンパク質の構造変化が、該第1の化合物が該タンパク質と接触することによる該タンパク質の構造変化と類似するか否かを決定する手段、
を包含する、プログラム。
【請求項59】
前記(a)手段が、
(a)該タンパク質が該第1の化合物と接触する条件下における、該ドメインの配向情報を以下
(v)該ドメインの既知の原子座標を得て、
(vi)液晶物質存在下において、該タンパク質を該第1の化合物と接触させた場合に二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量を得て、
(vii)該ドメインの原子座標、および該NMRシグナルの軸方向の変化量から、該ドメインのSaupeオーダーマトリクスエレメントを決定し、そして
(viii)該決定されたマトリクスを対角化して、該ドメインの配向情報とする、ことにより得る手段である、請求項25に記載のプログラム。
【請求項60】
前記(a)手段が、
(a)前記タンパク質が前記第1の化合物を接触させる条件下において既に得られている原子座標から、前記ドメインの配向情報を得る手段;
である、請求項58に記載のプログラム。
【請求項61】
請求項58に記載のプログラムであって、前記工程(b)において二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、プログラム。
【請求項62】
請求項61に記載のプログラムであって、ここで、前記(iii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該化合物を接触させ、二次元TROSY NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定することによって行われる、プログラム。
【請求項63】
請求項59に記載のプログラムであって、前記工程(a)において二次元TROSY NMRスペクトルを用いて得られる、分子の磁場中での配向角度に依存したNMRシグナルの軸方向の変化量が、15N軸方向の変化量である、プログラム。
【請求項64】
請求項63に記載のプログラムであって、ここで、前記(vii)Saupeオーダーマトリクスエレメントの決定が、以下:
前記ドメイン内のk番目の15N核スピン対について、
i番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkiを得て、
j番目の分子軸に対する、該ドメイン内のk番目のスピン対のN−H結合のベクトル角としてφkjを得て、
N−H結合の距離が1.02Åの場合、静的双極カップリングD0nhを23.0kHzとするか、またはN−H結合の距離が1.04Åの場合、静的双極カップリングD0nhを21.7kHzとして、
該タンパク質内の任意の残基の15N核の化学シフト異方性のテンソル値をδijとして決定し、そして
液晶物質存在下において該タンパク質と該第2の化合物を接触させ、二次元TROSY
NMRスペクトルを用いて、k番目の15N核スピン対について得られたΔδtrosy(k)とともに、以下の式(1):
Δδtrosy(k)=ΣSij{0.5 D0nh cosφkicosφkj+(2/3)δij} (1)
i,j=x,y,z
を用いて、磁場に対する分子配向を規定するSaupeオーダーマトリクスエレメントであるSijを決定すること
によって行われる、プログラム。
【請求項65】
請求項62または64に記載のプログラムであって、ここで、前記工程(c)における配向情報の比較が、以下:
(ix)該第1の化合物と接触した場合の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【化32】
を用いて示し、
(x)該第2の化合物と接触させた場合の、該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【化33】
を用いて示し、そして
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、該a、b、およびcを配向情報の比較のための指標とする、
ことによって行われる、プログラム。
【請求項66】
請求項62または64に記載のプログラムであって、ここで、前記工程(c)における配向情報の比較が、以下:
(ix)該第1の化合物と接触した場合の該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第1の単位ベクトル
【化34】
を用いて示し、
(x)該第2の化合物と接触させた場合の、該タンパク質のドメインの配向情報から、配向テンソル軸の方向を3つのお互いに直交する第2の単位ベクトル
【化35】
を用いて示し、そして
(xi)第1の単位ベクトルの各々と第2の単位ベクトルの各々との間の角度を、a、b、およびcとして表し、そして
(xii)以下の式によって活性構造一致度を求め
活性構造一致度=a2+b2+c2、
該活性構造一致度を配向情報の比較のための指標とする、
ことによって行われる、プログラム。
【請求項67】
請求項58に記載のプログラムであって、さらに、前記第1および第2の化合物の少なくとも1つが前記タンパク質に結合した位置を特定する手段を包含する、プログラム。
【請求項68】
請求項67に記載のプログラムであって、前記第1および第2の化合物の少なくとも1つが前記タンパク質に結合した位置を特定する手段が、該化合物の非存在下における該タンパク質の二次元TROSY NMRスペクトルと、該化合物の存在下における該タンパク質の二次元TROSY NMRスペクトルとを比較して、スペクトルの変化を検出し、そして該スペクトルの変化を生じた、該タンパク質中のアミノ酸残基を同定すること、によって行われる、プログラム。
【請求項69】
請求項1、13、または25に記載の方法を実行し得る装置。
【請求項70】
請求項38、48、または58に記載のプログラムを備える装置。
【請求項71】
請求項38、48、または58に記載のプログラムを含む記憶媒体。
【図1】

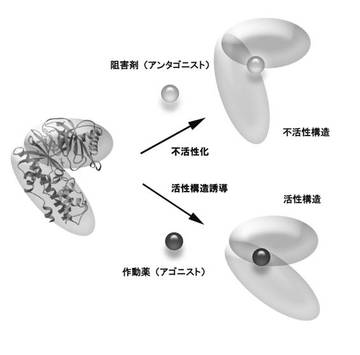
【図2】
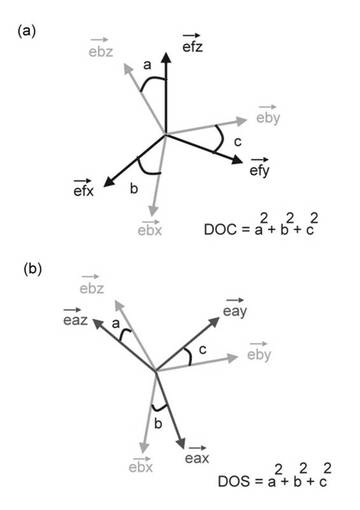
【図3】

【図4】
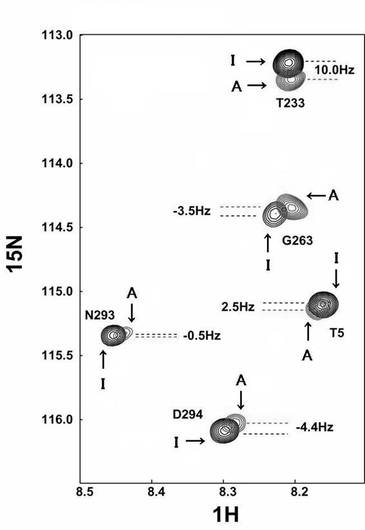
【図5】
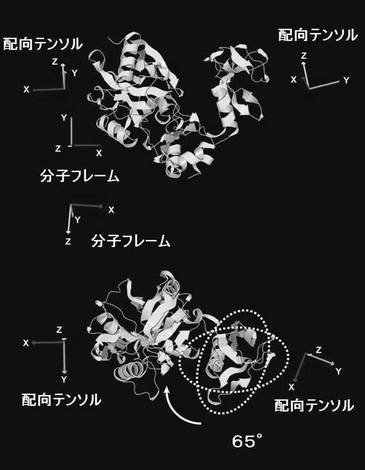
【図6】
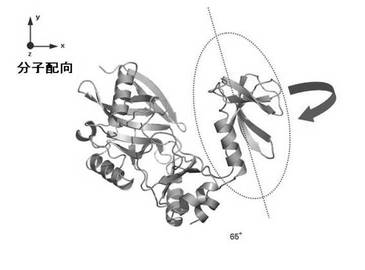
【図7】
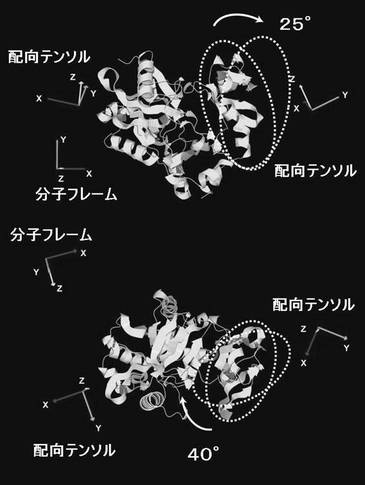
【図8】
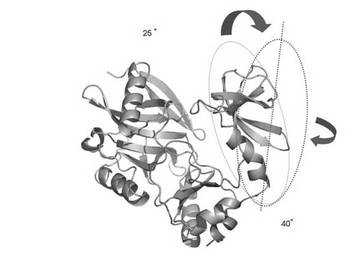
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【国際公開番号】WO2005/052620
【国際公開日】平成17年6月9日(2005.6.9)
【発行日】平成19年6月21日(2007.6.21)
【国際特許分類】
【出願番号】特願2005−515827(P2005−515827)
【国際出願番号】PCT/JP2004/017626
【国際出願日】平成16年11月26日(2004.11.26)
【出願人】(000005108)株式会社日立製作所 (27,607)
【Fターム(参考)】
【国際公開日】平成17年6月9日(2005.6.9)
【発行日】平成19年6月21日(2007.6.21)
【国際特許分類】
【国際出願番号】PCT/JP2004/017626
【国際出願日】平成16年11月26日(2004.11.26)
【出願人】(000005108)株式会社日立製作所 (27,607)
【Fターム(参考)】
[ Back to top ]