
核移行性ペプチド
【課題】 新規な核移行性ペプチドを提供する。
【解決手段】 複数個のアルギニン残基を含み、核移行性を有する第1のペプチド(例えば、アデノウイルス由来コアペプチドmu)と、複数個のリシン残基を含む第2のペプチド(例えば、シミアンウイルス40由来核移行性シグナル)とを融合させて得られる核移行性ペプチドを提供する。
【解決手段】 複数個のアルギニン残基を含み、核移行性を有する第1のペプチド(例えば、アデノウイルス由来コアペプチドmu)と、複数個のリシン残基を含む第2のペプチド(例えば、シミアンウイルス40由来核移行性シグナル)とを融合させて得られる核移行性ペプチドを提供する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、核移行性ペプチドに関する。
【背景技術】
【0002】
近年、核酸等の目的物質を標的部位に確実に送達するためのベクター又はキャリアーの開発が盛んに行われている。
ベクターの一例としては、リポソーム膜の外表面に、細胞膜の表面上に存在する受容体又は抗原と結合できる物質(例えば、トラスフェリン、インシュリン、葉酸、ヒアルロン酸、抗体又はその断片、糖鎖)が導入されたリポソーム(特許文献1及び2参照)が挙げられる。このリポソームによれば、リポソームのエンドサイトーシス効率を向上させることができる。
【0003】
ベクターの別の例としては、リポソーム膜の外表面にGALAが導入されたリポソーム(非特許文献1参照)が挙げられる。リポソームはエンドサイトーシスするとエンドソーム内に包含された状態となり、エンドソーム内のリポソームは、エンドソームがリソソームと融合することにより分解されるが、このリポソームによれば、リポソーム封入物質をエンドソームから脱出させ、細胞質中へ放出させることができる。
【0004】
キャリアーの一例としては、ポリアルギニン、アルギニンリッチペプチド等のポリカチオン性ペプチドキャリアーが開発されている(特許文献3参照)。また、アデノウイルス由来コアペプチドmu等のアルギニンリッチペプチドの核移行性について報告されている(非特許文献2参照)。
【0005】
ポリカチオン性ペプチドは静電的相互作用により核酸と複合体化する。ポリカチオン性ペプチドと複合体化した核酸は、外部環境から保護され、核酸分解酵素による分解から逃れることができる。細胞膜の表面上に存在する受容体又は抗原と結合できる物質をポリカチオン性ペプチドに共有結合させることにより、ポリカチオン性ペプチドと複合体形成した核酸を、エンドサイトーシスにより細胞内へ導入させることができる。ポリカチオン性ペプチドと複合体形成した核酸がエンドサイトーシスするとエンドソーム内に包含された状態となる。エンドソーム内のpHは、エンドソーム膜上のプロトンATPaseの作用により下がるが、ポリカチオン性ペプチドが生理pH以下でプロトン化する窒素原子を有していれば、エンドソーム内へ集積してきたプロトンをポリカチオン性物質が消費し、pHの低下を抑制する。そして、エンドソーム内へ塩化物イオンの流入を伴ったプロトンの集積が続くことにより、浸透圧が増大する。その結果、エンドソーム内外の浸透圧のバランスがくずれ、エンドソームが崩壊し、ポリカチオン性ペプチドと複合体形成した核酸は細胞質内へ放出されるものと考えられる。
【特許文献1】特開平4−346918号公報
【特許文献2】特開2004−10481号公報
【特許文献3】特開2001−199997号公報
【非特許文献1】T.Kakudo等,「バイオケミストリー(Biochemistry)」,2004年,第43巻,5618-5623頁
【非特許文献2】M. Keller等,「ケムバイオケム(ChemBiocChem)」,2003年,第4巻,296-298頁
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明は、新規な核移行性ペプチド、及び当該ペプチドと核酸とを含む複合体を細胞内に導入し、当該複合体に含まれる核酸を細胞内で発現させる方法を提供することを目的とする。
【課題を解決するための手段】
【0007】
本発明者は、アデノウイルス由来コアペプチドmu(配列番号1)等のアルギニンリッチペプチドが優れた核移行性を有する一方、当該ペプチドと複合体形成した核酸の転写を抑制することを見出すとともに、アデノウイルス由来コアペプチドmu等のアルギニンリッチペプチドにシミアンウイルス40由来核移行性シグナル(配列番号2)等のリシンリッチペプチドを付加することにより、アルギニンリッチペプチドの優れた核移行性を保持したまま、アルギニンリッチペプチドによる核酸の転写抑制を改善することができることを見出し、本発明を完成させるに至った。
【0008】
すなわち、本発明は、以下の発明を包含する。
(1)複数個のアルギニン残基を含み、核移行性を有する第1のペプチドと、複数個のリシン残基を含む第2のペプチドとを融合させて得られる核移行性ペプチド。
【0009】
(2)前記第1のペプチドにおけるアルギニン残基の含有率が35%以上である前記(1)記載の核移行性ペプチド。
【0010】
(3)前記第1のペプチドが、下記(a)及び(b)からなる群より選択される前記(2)記載の核移行性ペプチド。
(a)配列番号1記載のアミノ酸配列からなるペプチド
(b)配列番号1記載のアミノ酸配列において、1又は複数個のアミノ酸が欠失、置換、挿入又は付加されたアミノ酸配列からなり、核移行性を有するペプチド
【0011】
(4)前記第2のペプチドにおけるリシン残基の含有率が25%以上である前記(1)〜(3)のいずれかに記載の核移行性ペプチド。
【0012】
(5)前記第2のペプチドが核移行性を有する前記(4)記載の核移行性ペプチド。
【0013】
(6)前記第2のペプチドが下記(c)及び(d)からなる群より選択される前記(5)記載の核移行性ペプチド。
(c)配列番号2記載のアミノ酸配列からなるペプチド
(d)配列番号2記載のアミノ酸配列において、1又は複数個のアミノ酸が欠失、置換、挿入又は付加されたアミノ酸配列からなり、核移行性を有するペプチド
【0014】
(7)1又は複数個の前記第1のペプチドと1又は複数個の前記第2のペプチドとを融合させて得られる前記(1)〜(6)のいずれかに記載の核移行性ペプチド。
【0015】
(8)核酸と、前記核酸と静電的に結合した前記(1)〜(7)のいずれかに記載の核移行性ペプチドとを含む複合体。
【0016】
(9)前記(8)記載の複合体を細胞内に導入し、前記複合体に含まれる核酸を細胞内で発現させる方法。
【0017】
(10)次式:
【0018】
【数1】

【0019】
[式中、CPは複合体作製時に使用するペプチドの濃度(μg/μL)を表し、nKはペプチド1分子あたりのリシン残基数を表し、nRはペプチド1分子あたりのアルギニン残基数を表し、MWPはペプチドの分子量を表し、CDは複合体作製時に使用する核酸の濃度(μg/μL)を表し、MWDは核酸のヌクレオチド1残基あたりの平均分子量を表す。]
で定義される電荷比が1〜4である前記複合体を細胞内に導入する前記(9)記載の方法。
【発明の効果】
【0020】
本発明の核移行性ペプチドにおいては、第1のペプチドが有する核移行性を保持したまま、第1のペプチドによる核酸の転写抑制が改善されている。したがって、本発明の核移行性ペプチドと複合体形成した核酸は、核内へ効率よく移行することができるとともに、核内において効率よく発現することができる。
【発明を実施するための最良の形態】
【0021】
以下、本発明について詳細に説明する。
本発明において、「核移行性」とは、細胞質から核内へ移行する能力を意味する。
本発明のペプチドは、第1のペプチドと第2のペプチドとを融合して得られるペプチドと同一のアミノ酸配列からなる全てのペプチドを含む。したがって、本発明のペプチドの製法は、第1のペプチドと第2のペプチドとを融合する方法に限定されるものではない。
【0022】
本発明のペプチドは、複数個のアルギニン残基と複数個のリシン残基とを含むポリカチオン性ペプチドであるので、静電的結合を介して核酸と複合体を形成することができる。本発明のペプチドと核酸との複合体を細胞内に導入すると、当該複合体は核内に移行し、当該複合体に含まれる核酸は核内で発現する。本発明のペプチドと複合体形成した核酸の転写効率は、第1のペプチドと複合体形成した核酸の転写効率よりも高い。すなわち、本発明のペプチドにおいては、第2のペプチド部分の存在により、第1のペプチド部分による核酸の転写抑制が改善されている。
【0023】
第1のペプチドは、複数個のアルギニン残基を含み、核移行性を有するペプチドである。
第1のペプチドにおけるアルギニン残基の含有率は、第1のペプチドが核移行性を有する限り特に限定されるものではないが、通常35%以上、好ましくは40%以上、さらに好ましくは45%以上である。なお、第1のペプチドにおけるアルギニン残基の含有率の上限値は100%である。第1のペプチドと核酸との複合体を細胞内に導入すると、当該複合体は核内に移行し、当該複合体に含まれる核酸は核内で発現するが、第1のペプチドにおけるアルギニン残基の含有率が35%以上であると、核酸との静電的結合力が強固となるため、核酸への転写因子の結合阻害が顕著となり、核酸の転写抑制が顕著となる。
【0024】
第1のペプチドは、核移行性を有する限り、アルギニン残基のみから構成されていてもよいし、他のアミノ酸残基を含んでいてもよい。第1のペプチドにおけるリシン残基の含有率は、通常10%以下、好ましくは5%以下、さらに好ましくは0%である。第1のペプチドにおけるリシン残基の含有率が10%以下であると、第1のペプチドと核酸との複合体を細胞内に導入したときの核酸の転写抑制は顕著なままである。
【0025】
第1のペプチドを構成するアミノ酸残基の総数は、複数個のアルギニン残基を含み、核移行性を有する限り特に限定されるものではないが、通常5〜35個、好ましくは10〜30個、さらに好ましくは15〜20個である。
【0026】
第1のペプチドのアミノ酸配列は、複数個のアルギニン残基を含み、核移行性を有する限り特に限定されるものではない。
【0027】
第1のペプチドは、例えば、下記(a)及び(b)からなる群より選択することができる。
(a)配列番号1記載のアミノ酸配列からなるペプチド(以下「ペプチド(a)」という場合がある。)
(b)配列番号1記載のアミノ酸配列において、1又は複数個のアミノ酸が欠失、置換、挿入又は付加されたアミノ酸配列からなり、核移行性を有するペプチド(以下「ペプチド(b)」という場合がある。)
【0028】
ペプチド(a)は、アデノウイルス由来コアペプチドmuであり、核移行性を有する。ペプチド(a)を構成するアミノ酸残基の総数は19個であり、ペプチド(a)に含まれるアルギニン残基は9個(アルギニン残基の含有率は約47%)であり、リシン残基は0個(リシン残基の含有率は0%)である。
【0029】
ペプチド(b)には、ペプチド(a)に対して人為的に欠失、置換、挿入、付加等の変異を導入したペプチドの他、変異が導入された状態で天然に存在するペプチドや、それに対して人為的に変異を導入したペプチドも含まれる。変異が導入された状態で天然に存在するタンパク質としては、例えば、多型によって生じ得るペプチドが挙げられる。
【0030】
配列番号1記載のアミノ酸配列において欠失、置換、挿入又は付加されるアミノ酸の個数は、ペプチド(b)が核移行性を有する限り特に限定されるものではなく、その個数は1又は複数個である。欠失に関する具体的範囲は、通常1〜3個、好ましくは1〜2個、さらに好ましくは1個であり、置換に関する具体的範囲は、通常1〜3個、好ましくは1〜2個、さらに好ましくは1個であり、挿入に関する具体的範囲は、通常1〜15個、好ましくは1〜10個、さらに好ましくは1〜5個であり、付加に関する具体的範囲は、通常1〜15個、好ましくは1〜10個、さらに好ましくは1〜5個である。
【0031】
配列番号1記載のアミノ酸配列において欠失、置換、挿入又は付加されるアミノ酸の位置は、ペプチド(b)が核移行性を有する限り特に限定されるものではない。
【0032】
第2のペプチドは、複数個のリシン残基を含むペプチドである。
第2のペプチドにおけるリシン残基の含有率は特に限定されるものではないが、通常25%以上、好ましくは30%以上、さらに好ましくは35%以上である。なお、第2のペプチドにおけるリシン残基の含有率の上限値は100%である。第2のペプチドにおけるリシン残基の含有率が25%以上であると、第1のペプチドによる核酸の転写抑制を効果的に改善することができるので、本発明のペプチドと核酸との複合体を細胞内に導入すると、当該複合体は核内に移行し、当該複合体に含まれる核酸は核内で効率よく発現することができる。
【0033】
第2のペプチドは、リシン残基のみから構成されていてもよいし、他のアミノ酸残基を含んでいてもよい。第2のペプチドにおけるアルギニン残基の含有率は、通常20%以下、好ましくは15%以下、さらに好ましくは10%以下である。第2のペプチドにおけるアルギニン残基の含有率が20%以下であると、第1のペプチドによる核酸の転写抑制を効果的に改善することができるので、本発明のペプチドと核酸との複合体を細胞内に導入したときの、核酸の発現効率を向上させることができる。
【0034】
第2のペプチドを構成するアミノ酸残基の総数は、第1のペプチドの核移行性を抑制しない限り特に限定されるものではないが、通常5〜30個、好ましくは5〜20個、さらに好ましくは5〜15個である。
【0035】
第2のペプチドのアミノ酸配列は、複数個のリシン残基を含み、第1のペプチドの核移行性を抑制しない限り特に限定されるものではない。
【0036】
第2のペプチドは、核移行性を有するペプチドであることが好ましい。第2のペプチドが核移行性を有するペプチドであると、本発明のペプチドと核酸との複合体を細胞内に導入したときに、当該複合体が効率よく核内に移行することができる。
【0037】
第2のペプチドは、例えば、下記(c)及び(d)からなる群より選択することができる。
(c)配列番号2記載のアミノ酸配列からなるペプチド(以下「ペプチド(c)」という場合がある。)
(d)配列番号2記載のアミノ酸配列において、1又は複数個のアミノ酸が欠失、置換、挿入又は付加されたアミノ酸配列からなり、核移行性を有するペプチド(以下「ペプチド(d)」という場合がある。)
【0038】
ペプチド(c)は、シミアンウイルス40由来核移行性シグナル(SV40−NLS)であり、核移行性を有する。ペプチド(c)を構成するアミノ酸残基の総数は11個であり、ペプチド(c)に含まれるアルギニン残基の個数は1個(アルギニン残基の含有率は約9%)であり、リシン残基の個数は4個(リシン残基の含有率は約36%)である。
【0039】
ペプチド(d)には、ペプチド(c)に対して人為的に欠失、置換、挿入、付加等の変異を導入したペプチドの他、変異が導入された状態で天然に存在するペプチドや、それに対して人為的に変異を導入したペプチドも含まれる。変異が導入された状態で天然に存在するタンパク質としては、例えば、多型によって生じ得るペプチドが挙げられる。
【0040】
配列番号2記載のアミノ酸配列において欠失、置換、挿入又は付加されるアミノ酸の個数は、ペプチド(d)が核移行性を有する限り特に限定されるものではなく、その個数は1又は複数個である。欠失に関する具体的範囲は、通常1〜3個、好ましくは1〜2個、さらに好ましくは1個であり、置換に関する具体的範囲は、通常1〜3個、好ましくは1〜2個、さらに好ましくは1個であり、挿入に関する具体的範囲は、通常1〜15個、好ましくは1〜10個、さらに好ましくは1〜5個であり、付加に関する具体的範囲は、通常1〜15個、好ましくは1〜10個、さらに好ましくは1〜5個である。
【0041】
配列番号2記載のアミノ酸配列において欠失、置換、挿入又は付加されるアミノ酸の位置は、ペプチド(d)が核移行性を有する限り特に限定されるものではない。
【0042】
本発明のペプチドに含まれる第1のペプチド部分の個数は特に限定されるものではなく、1個であってもよいし複数個であってもよい。第1のペプチド部分の個数は、通常1〜10個、好ましくは1〜5個、さらに好ましくは1〜2個である。
【0043】
本発明のペプチドに含まれる第2のペプチド部分の個数は特に限定されるものではなく、1個であってもよいし複数個であってもよい。第2のペプチド部分の個数は、通常1〜10個、好ましくは1〜5個、さらに好ましくは1〜2個である。
【0044】
本発明のペプチドにおいて、第1のペプチド部分と第2のペプチド部分との配列順序は特に限定されるものではない。
【0045】
本発明のペプチドに第1のペプチド部分が複数個含まれる場合、第1のペプチド部分同士が連続していてもよいし、第1のペプチド部分間に第2のペプチド部分が介在していてもよい。
【0046】
本発明のペプチドに第2のペプチド部分が複数個含まれる場合、第2のペプチド部分同士が連続していてもよいし、第2のペプチド部分間に第1のペプチド部分が介在していてもよい。
【0047】
第1のペプチド部分と第2のペプチド部分とは、第1のペプチド部分のN末端と第2のペプチド部分のC末端とが結合していてもよいし、第1のペプチド部分のC末端と第2のペプチド部分のN末端とが結合していてもよい。
【0048】
第1のペプチド部分と第2のペプチド部分とは、直接結合していてもよいし、適当なリンカーを介して結合していてもよい。リンカーのアミノ酸配列は、本発明のペプチドの核内移行性を抑制しない限り特に限定されるものではない。リンカーを構成するアミノ酸残基の総数は、本発明のペプチドの核内移行性を抑制しない限り特に限定されるものではないが、通常1〜10個、好ましくは1〜7個、さらに好ましくは1〜5個である。
【0049】
本発明のペプチドは、例えば、本発明のペプチドを生産し得る形質転換体を培養することにより得ることができる。本発明のペプチドを生産し得る形質転換体は、第1のペプチドをコードするオープンリーディングフレームと第2のペプチドをコードするオープンリーディングフレームとが所望の順序で連結されたDNAを、発現ベクターのプロモーターの下流に挿入して組換えベクターを作製し、当該組換えベクターを宿主細胞に導入することにより得ることができる。
【0050】
ペプチド(a)をコードするDNAは、例えば、アデノウイルスを感染させた細胞から抽出したmRNAを用いてcDNAライブラリーを作製し、ペプチド(a)をコードするDNAの塩基配列に基づいて合成したプローブを用いて、cDNAライブラリーから目的のDNAを含むクローンをスクリーニングすることにより得ることができる。また、ペプチド(a)をコードするDNAは、その塩基配列に従って化学合成により得ることもできる。DNAの化学合成は、市販のDNA合成機、例えば、チオホスファイト法を利用したDNA合成機(島津製作所社製)、フォスフォアミダイト法を利用したDNA合成機(パーキン・エルマー社製)を用いて行うことができる。
【0051】
ペプチド(b)をコードするDNAは、ペプチド(a)をコードするDNAに、部位特異的変異誘発法等の公知の方法を用いて人為的に変異を導入することにより得ることができる。変異の導入は、例えば、変異導入用キット、例えば、Mutant-K(TAKARA社製)、Mutant-G(TAKARA社製)、TAKARA社のLA PCR in vitro Mutagenesis シリーズキットを用いて行うことができる。また、ペプチド(b)をコードするDNAは、その塩基配列に従って化学合成により得ることもできる。
【0052】
ペプチド(c)をコードするDNAは、例えば、シミアンウイルスを感染させた細胞から抽出したmRNAを用いてcDNAライブラリーを作製し、ペプチド(c)をコードするDNAの塩基配列に基づいて合成したプローブを用いて、cDNAライブラリーから目的のDNAを含むクローンをスクリーニングすることにより得ることができる。また、ペプチド(c)をコードするDNAは、その塩基配列に従って化学合成により得ることもできる。
【0053】
ペプチド(d)をコードするDNAは、ペプチド(c)をコードするDNAに、部位特異的変異誘発法等の公知の方法を用いて人為的に変異を導入することにより得ることができる。また、ペプチド(d)をコードするDNAは、その塩基配列に従って化学合成により得ることもできる。
【0054】
発現ベクターとしては、宿主細胞において自立複製が可能なものであれば特に限定されず、例えば、プラスミドベクター、ファージベクター、ウイルスベクター等を使用することができる。
【0055】
宿主細胞としては、目的とする遺伝子を発現し得る限り、原核細胞、酵母、動物細胞、昆虫細胞、植物細胞等のいずれを使用してもよい。また、動物個体、植物個体、カイコ虫体等を使用してもよい。
【0056】
宿主細胞への組換えベクターの導入方法は、エレクトロポレーション法、リン酸カルシウム法、リポフェクション法等を使用することができる。
【0057】
形質転換体の培養は、宿主細胞の培養に用いられる通常の方法に従って行うことができる。
形質転換体の培養物より本発明のペプチドを採取することにより、本発明のペプチドを得ることができる。培養物には、培養上清、培養細胞、培養菌体、細胞又は菌体の破砕物のいずれもが含まれる。
【0058】
本発明のペプチドが形質転換体の細胞内に蓄積される場合には、培養物を遠心分離することにより、培養物中の細胞を集め、該細胞を洗浄した後に細胞を破砕して、本発明のペプチドを抽出する。本発明のペプチドが形質転換体の細胞外に分泌される場合には、培養上清をそのまま使用するか、遠心分離等により培養上清から細胞又は菌体を除去する。
【0059】
得られたペプチドは、溶媒抽出法、硫安等による塩析法脱塩法、有機溶媒による沈殿法、ジエチルアミノエチル(DEAE)−セファロース、イオン交換クロマトグラフィー法、疎水性クロマトグラフィー法、ゲルろ過法、アフィニティークロマトグラフィー法等により精製することができる。
【0060】
本発明のペプチドは、そのアミノ酸配列に基づいて、Fmoc法(フルオレニルメチルオキシカルボニル法)、tBoc法(t−ブチルオキシカルボニル法)等の化学合成法によって製造することができる。この際、市販のペプチド合成機を使用することができる。
【0061】
本発明のペプチドは、複数個のアルギニン残基と複数個のリシン残基とを含むポリカチオン性ペプチドであるので、静電的結合を介して核酸と複合体を形成することができる。本発明のペプチドと核酸との複合体は、本発明のペプチドと核酸とを例えば4〜37℃で0.5〜3時間インキュベーションすることにより作製することができる。本発明のペプチドと複合体形成させる核酸としては、例えば、目的のタンパク質をコードするDNA、当該DNAが組み込まれたプラスミド等を使用することができる。
【0062】
本発明のペプチドと核酸との複合体は、脂質を含んでいてもよい。脂質としては、例えば、リン脂質、糖脂質、ステロール、飽和又は不飽和の脂肪酸等が挙げられる。リン脂質としては、例えば、ホスファチジルコリン、ホスファチジルグリセロール、ホスファチジルエタノールアミン、ホスファチジルセリン、ホスファチジルイノシトール、ホスファチジン酸、カルジオリピン、スフィンゴミエリン、卵黄レシチン、大豆レシチン、これらの水素添加物等が挙げられる。糖脂質としては、例えば、グリセロ糖脂質、スフィンゴ糖脂質等が挙げられる。ステロールとしては、例えば、動物由来のステロール(例えば、コレステロール、コレステロールコハク酸、ラノステロール、ジヒドロラノステロール、デスモステロール、ジヒドロコレステロール)、植物由来のステロール(フィトステロール)(例えば、スチグマステロール、シトステロール、カンペステロール、ブラシカステロール)、微生物由来のステロール(例えば、チモステロール、エルゴステロール)等が挙げられる。飽和又は不飽和の脂肪酸としては、例えば、パルミチン酸、オレイン酸、ステアリン酸、アラキドン酸、ミリスチン酸等の炭素数12〜20の飽和又は不飽和の脂肪酸が挙げられる。
【0063】
本発明のペプチドと核酸との複合体は、リポソーム内に封入してもよい。本発明のペプチドと核酸との複合体の存在下、脂質膜を水和し、次いで攪拌又は超音波処理することにより、本発明のペプチドと核酸との複合体が封入されたリポソームを製造することができる。
【0064】
本発明のペプチドと核酸との複合体の粒子径は特に限定されるものではないが、通常30〜105nm、好ましくは30〜100nm、さらに好ましくは30〜95nmである。本発明のペプチドと核酸との複合体の粒子径が上記範囲にあると、当該複合体が効率よく核内に移行することができる。
【0065】
本発明のペプチドと核酸との複合体を細胞内に導入する方法は特に限定されるものではなく、常法に従って行うことができる。
本発明のペプチドと核酸との複合体を細胞内に導入する方法としては、例えば、エンドサイトーシスを利用した方法、細胞質内へのインジェクション、核内へのインジェクション、エレクトロポレーション、ハイドロダイナミックインジェクション等が挙げられる。
【0066】
エンドサイトーシスを利用する場合、本発明のペプチドと核酸との複合体又は当該複合体を封入したリポソームを、細胞膜の表面上に存在する受容体又は抗原と結合できる物質で修飾することにより、当該複合体を細胞内へ効率よく導入することができる。
【0067】
エンドサイトーシスによる細胞内導入は、本発明のペプチドと核酸との複合体又は当該複合体を封入したリポソームを、細胞とともに例えば4〜37℃で0.5〜48時間インキュベーションすることにより行うことができる。
【0068】
細胞膜の表面上に存在する受容体又は抗原と結合できる物質の種類は特に限定されるものではなく、細胞膜結合性物質としては、例えば、トラスフェリン、インシュリン、葉酸、ヒアルロン酸、抗体又はその断片、糖鎖、成長因子、アポリポタンパク質等が挙げられる。成長因子としては、例えば、上皮成長因子(EGF)、インシュリン様成長因子(IGF)、繊維芽細胞増殖因子(FGF)等が挙げられる。アポリポタンパク質としては、例えば、アポA−1、アポB−48、アポB−100、アポE等が挙げられる。抗体の断片としては、例えば、Fab断片、F(ab)'2断片、単鎖抗体(scFv)等が挙げられる。
【0069】
本発明のペプチドと核酸との複合体において、次式:
【0070】
【数2】
【0071】
[式中、CPは複合体作製時に使用するペプチドの濃度(μg/μL)を表し、nKはペプチド1分子あたりのリジン残基数を表し、nRはペプチド1分子あたりのアルギニン残基数を表し、MWPはペプチドの分子量を表し、CDは複合体作製時に使用する核酸の濃度(μg/μL)を表し、MWDは核酸のヌクレオチド1残基あたりの平均分子量を表す。]
で定義される電荷比は1〜4であることが好ましく、2〜3であることがさらに好ましい。本発明のペプチドと核酸との複合体の電荷比が1〜4であると、当該複合体に含まれる核酸を効率よく核内で発現させることができる。
【0072】
本発明のペプチドと核酸との複合体を導入する細胞が由来する生物種は特に限定されるものではなく、動物、植物、微生物等のいずれであってもよいが、動物であることが好ましく、哺乳動物であることがさらに好ましい。哺乳動物としては、例えば、ヒト、サル、ウシ、ヒツジ、ヤギ、ウマ、ブタ、ウサギ、イヌ、ネコ、ラット、マウス、モルモット等が挙げられる。また、目的物質を送達すべき細胞の種類は特に限定されるものではなく、例えば、体細胞、生殖細胞、幹細胞又はこれらの培養細胞等が挙げられる。
【実施例】
【0073】
1.ポリカチオン性ペプチドと核酸との複合体の作製
ポリカチオン性ペプチドとして、アデノウイルス由来コアペプチドmu(以下「mu」という。)、シミアンウイルス40由来核移行性シグナル(SV40−NLS)とmuとの融合ペプチド(以下「NLS−mu」という。)、及びポリL−リシン(以下「PLL」という。)を使用した。muのアミノ酸配列を配列番号1に示し、NLS−muのアミノ酸配列を配列番号2に示し、融合ペプチドのアミノ酸配列を配列番号3に示す。muの分子量は2440.89であり、NLS−muの分子量は3810.48であり、PLLの分子量は25700である。なお、ポリカチオン性ペプチドの合成は、ペプチド合成業者に委託した。
【0074】
核酸としては、EGFP(enhanced Green Fluorescent Protein)をコードするDNA(クロンテック社製)を組み込んだプラスミドpcDNA3.1(+)(インビトロジェン社製)(以下「pcDNA3.1(+)−EGFP」という。)を使用した。
【0075】
電荷比(charge ratio (+/-))を1,2,5,10の4種類に設定し、mu又はNLS−muと核酸との複合体を作製した。
【0076】
mu又はNLS−muと核酸との複合体に関する電荷比は、ペプチド中の1個のリジン残基、1個のアルギニン残基をそれぞれ+1の電荷、DNA中の1個のリン酸基を−1の電荷として次式により算出した。
【0077】
【数3】
【0078】
[式中、CPは複合体作製時に使用するペプチドの濃度(μg/μL)を表し、nKはペプチド1分子あたりのリジン残基数を表し、nRはペプチド1分子あたりのアルギニン残基数を表し、MWPはペプチドの分子量を表し、CDは複合体作製時に使用する核酸の濃度(μg/μL)を表し、MWDは核酸のヌクレオチド1残基あたりの平均分子量を表す。]
【0079】
なお、muに関し、nKは0であり、nRは9であり、MWPは2440.89であり、NLS−muに関し、nKは4であり、nRは10であり、MWPは3810.48である。また、pcDNA3.1(+)−EGFPに関し、MWDは308であり、CDは0.1に設定した。
【0080】
電荷比が1のとき、muに関するCPは0.0875に設定し、NLS−muに関するCPは0.0885に設定した。電荷比が2のとき、muに関するCPは0.175に設定し、NLS−muに関するCPは0.177に設定した。電荷比が5のとき、muに関するCPは0.4375に設定し、NLS−muに関するCPは0.4425に設定した。電荷比が10のとき、muに関するCPは0.875に設定し、NLS−muに関するCPは0.885に設定した。
【0081】
電荷比(charge ratio (+/-))を2に設定し、PLLと核酸との複合体を作製した。
PLLと核酸との複合体に関する電荷比は次式により計算した。
【0082】
【数4】
【0083】
[式中、CPは複合体作製時に使用するペプチドの濃度(μg/μL)を表し、MWKはリジン1残基あたりの分子量を表し、CDは複合体作製時に使用する核酸の濃度(μg/μL)を表し、MWDは核酸のヌクレオチド1残基あたりの平均分子量を表す。]
【0084】
なお、MWKは128であり、MWDは308であり、CPは0.083に設定し、CDは0.1に設定した。
【0085】
ポリカチオン性ペプチド溶液(100μL)を攪拌し、0.1μg/μLのpcDNA3.1(+)−EGFP/H2Oを、20μLずつを5回滴下して加えた。滴下後、30秒間攪拌を継続し、室温で30分間静置した。静置後、ELS−8000を用いて、ポリカチオン性ペプチドと核酸との複合体の粒子径測定を行った。なお、H2Oとしては、DISMIC−13CP(cellulose acetate, pore size = 0.20μm)を用いて濾過したH2Oを用いた。
各複合体の粒子径(nm)は表1に示すとおりである。
【0086】
【表1】
【0087】
2.各複合体に関する核膜透過及び核内転写効率の評価
各複合体について、核膜透過及び核内転写効率の評価を行った。なお、PLLと核酸との複合体(電荷比=2)はコントロールとして使用した。
【0088】
(1)マイクロインジェクション
インジェクションを行う前日に、glass base dish(IWAKI)上に5×104細胞/mLの濃度でHeLa細胞を培養した。各複合体を、濾過済み0.5% ローダミン(Rh)/H2Oを用いて、pcDNA換算で33.2fmol/μL(= 1000 copies/cell)、0.33fmol/μL(= 10 copies/cell)となるように希釈した。33.2fmol/μLのものを細胞質インジェクションに、0.33fmol/μLのものを核インジェクションに使用した。
【0089】
細胞質インジェクションにおいては、HeLa細胞の細胞質に1000 copies/cellの濃度のサンプルをインジェクションした(インジェクション条件:Pi = 50-70 hPa, Pc = 30 hPa, Ti = 0.2 s)。インジェクションは400個の細胞に行った。インジェクション後、新しいD’MEMに培地を交換し、37℃、5%CO2下でインキュベートした。インジェクション24時間後、蛍光顕微鏡にて細胞を観察し、Rh発色細胞数及びEGFP発現細胞数をカウントし、遺伝子発現率をEGFP発現細胞数/Rh発色細胞数より算出した。
【0090】
核内インジェクションにおいては、HeLa細胞の核内に10 copies/cellの濃度のサンプルをインジェクションした(インジェクション条件:Pi = 50 hPa, Pc = 30 hPa, Ti = 0.2 s)。インジェクションは400〜450個の細胞に行った。インジェクション後、新しいD’MEMに培地を交換し、37℃、5%CO2下でインキュベートした。インジェクション24時間後、蛍光顕微鏡にて細胞を観察し、Rh発色細胞数及びEGFP発現細胞数をカウントし、遺伝子発現率をEGFP発現細胞数/Rh発色細胞数より算出した。
【0091】
核内インジェクションにおける遺伝子発現が、核内における転写効率を反映するパラメーターと考えられる。また、核インジェクション時の発現率を細胞質インジェクション時の発現率で除することにより、核移行性の指標となる核移行スコアー(nuclear transfer score : NT score)を算出した。
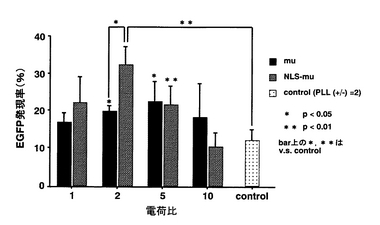
各複合体の細胞質インジェクション時におけるEGFP発現率を図1に示す。
図中、各値は平均値±標準偏差(n=3)を表す。
【0092】
図1に示すように、電荷比が1のとき、各複合体のEGFP発現率はいずれも20%前後であった。電荷比が2のとき、NLS−muと核酸との複合体が最も高いEGFP発現率を示し(32.6%)、コントロールであるPLLと核酸との複合体のEGFP発現率(12.3%)の2.7倍、muと核酸との複合体のEGFP発現率(20.1%)の1.6倍であり、各複合体のEGFP発現率に顕著な違いが認められた。NLS−muと核酸との複合体のEGFP発現率は、電荷比が2よりも大きくなるに従って減少した。
【0093】
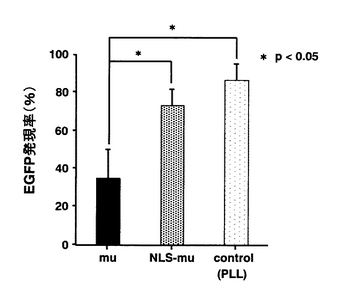
核インジェクション時における各複合体のEGFP発現率の比較は、細胞質インジェクション時における各複合体のEGFP発現率に顕著な違いが認められた電荷比が2の場合にて行った。各複合体の核インジェクション時におけるEGFP発現率を図2に示す。
【0094】
図2に示すように、PLLと核酸との複合体のEGFP発現率が最も高く(86.7%)、NLS−muと核酸との複合体のEGFP発現率(73.6%)も同程度であったが、muと核酸との複合体のEGFP発現率は低かった(35.0%)。
以上の結果から、電荷比が2のときの各複合体の核移行スコアーを算出し、PLLと核酸との複合体の核移行スコアーを1としたときの相対値を表2に示す。
【0095】
【表2】
【0096】
表中、各値は平均値±標準偏差を表す。
電荷比が2のとき、muと核酸との複合体の核移行スコアー及びNLS−muと核酸との複合体の核移行スコアーは、PLLと核酸との複合体の核移行スコアーの3〜4倍であった。
【0097】
以上の結果から、SV40−NLSをmuに付加することにより、muの核移行性を保持しつつ、muによる核酸の転写抑制を緩和することができ、これにより、NLS−muと核酸との複合体を細胞内に導入したときの、核酸の発現を亢進することができることが示された。
【0098】
アルギニンに富むペプチドは、リシンに富むペプチドと比較して、核酸と強固な複合体を形成することが示唆されている(Plank. C. et al., Hum Gene Ther 10 : 319-322 (1999))。muはアルギニンに富むペプチド(アルギニン残基の含有率は約47%、リシン残基の含有率は0%)であるので、muと核酸との強い相互作用により、転写因子の核酸への結合が阻害され、muと複合体形成した核酸の転写が抑制されたものと考えられる。一方、SV40−NLSは、リシンに富むペプチド(リシン残基の含有率は約36%、アルギニン残基の含有率は約9%)であるので、SV40−NLSをmuに付加することにより(融合ペプチドにおけるアルギニン残基の含有率は約33%、リシン残基の含有率は約13%)、muと核酸との強い相互作用が緩和され、muによる核酸の転写抑制を緩和されたものと考えられる。
【0099】
通常、ポリカチオン性ペプチドと核酸との複合体は、核膜孔を透過して核内へ移行するものと考えられており、核膜孔を透過し得る粒子サイズの上限値は39nmであると考えられている(N. Psnte et al., Mol Biol Cell 13: 425-434 (2002))。NLS−muと核酸との複合体の粒子サイズは、表1に示すように39nmを超えており、NLS−muと核酸との複合体の場合は、柔軟性に富んだ構造であるため核膜孔を透過することができるものと考えられる。但し、電荷比が10のとき、NLS−muと核酸との複合体の核移行スコアーは1.56(細胞質インジェクション時のEGFP発現率は10.5%、核内インジェクション時のEGFP発現率は47.3%)であり、電荷比の増大とともに核移行スコアーの低下が認められた。電荷比の増大とともに粒子サイズも増大するので、核膜孔を透過し難くなり、核移行スコアーが低下したものと考えられる。
【図面の簡単な説明】
【0100】
【図1】各複合体の細胞質インジェクション時におけるEGFP発現率を示す図である。
【図2】各複合体の核インジェクション時におけるEGFP発現率を示す図である。
【技術分野】
【0001】
本発明は、核移行性ペプチドに関する。
【背景技術】
【0002】
近年、核酸等の目的物質を標的部位に確実に送達するためのベクター又はキャリアーの開発が盛んに行われている。
ベクターの一例としては、リポソーム膜の外表面に、細胞膜の表面上に存在する受容体又は抗原と結合できる物質(例えば、トラスフェリン、インシュリン、葉酸、ヒアルロン酸、抗体又はその断片、糖鎖)が導入されたリポソーム(特許文献1及び2参照)が挙げられる。このリポソームによれば、リポソームのエンドサイトーシス効率を向上させることができる。
【0003】
ベクターの別の例としては、リポソーム膜の外表面にGALAが導入されたリポソーム(非特許文献1参照)が挙げられる。リポソームはエンドサイトーシスするとエンドソーム内に包含された状態となり、エンドソーム内のリポソームは、エンドソームがリソソームと融合することにより分解されるが、このリポソームによれば、リポソーム封入物質をエンドソームから脱出させ、細胞質中へ放出させることができる。
【0004】
キャリアーの一例としては、ポリアルギニン、アルギニンリッチペプチド等のポリカチオン性ペプチドキャリアーが開発されている(特許文献3参照)。また、アデノウイルス由来コアペプチドmu等のアルギニンリッチペプチドの核移行性について報告されている(非特許文献2参照)。
【0005】
ポリカチオン性ペプチドは静電的相互作用により核酸と複合体化する。ポリカチオン性ペプチドと複合体化した核酸は、外部環境から保護され、核酸分解酵素による分解から逃れることができる。細胞膜の表面上に存在する受容体又は抗原と結合できる物質をポリカチオン性ペプチドに共有結合させることにより、ポリカチオン性ペプチドと複合体形成した核酸を、エンドサイトーシスにより細胞内へ導入させることができる。ポリカチオン性ペプチドと複合体形成した核酸がエンドサイトーシスするとエンドソーム内に包含された状態となる。エンドソーム内のpHは、エンドソーム膜上のプロトンATPaseの作用により下がるが、ポリカチオン性ペプチドが生理pH以下でプロトン化する窒素原子を有していれば、エンドソーム内へ集積してきたプロトンをポリカチオン性物質が消費し、pHの低下を抑制する。そして、エンドソーム内へ塩化物イオンの流入を伴ったプロトンの集積が続くことにより、浸透圧が増大する。その結果、エンドソーム内外の浸透圧のバランスがくずれ、エンドソームが崩壊し、ポリカチオン性ペプチドと複合体形成した核酸は細胞質内へ放出されるものと考えられる。
【特許文献1】特開平4−346918号公報
【特許文献2】特開2004−10481号公報
【特許文献3】特開2001−199997号公報
【非特許文献1】T.Kakudo等,「バイオケミストリー(Biochemistry)」,2004年,第43巻,5618-5623頁
【非特許文献2】M. Keller等,「ケムバイオケム(ChemBiocChem)」,2003年,第4巻,296-298頁
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明は、新規な核移行性ペプチド、及び当該ペプチドと核酸とを含む複合体を細胞内に導入し、当該複合体に含まれる核酸を細胞内で発現させる方法を提供することを目的とする。
【課題を解決するための手段】
【0007】
本発明者は、アデノウイルス由来コアペプチドmu(配列番号1)等のアルギニンリッチペプチドが優れた核移行性を有する一方、当該ペプチドと複合体形成した核酸の転写を抑制することを見出すとともに、アデノウイルス由来コアペプチドmu等のアルギニンリッチペプチドにシミアンウイルス40由来核移行性シグナル(配列番号2)等のリシンリッチペプチドを付加することにより、アルギニンリッチペプチドの優れた核移行性を保持したまま、アルギニンリッチペプチドによる核酸の転写抑制を改善することができることを見出し、本発明を完成させるに至った。
【0008】
すなわち、本発明は、以下の発明を包含する。
(1)複数個のアルギニン残基を含み、核移行性を有する第1のペプチドと、複数個のリシン残基を含む第2のペプチドとを融合させて得られる核移行性ペプチド。
【0009】
(2)前記第1のペプチドにおけるアルギニン残基の含有率が35%以上である前記(1)記載の核移行性ペプチド。
【0010】
(3)前記第1のペプチドが、下記(a)及び(b)からなる群より選択される前記(2)記載の核移行性ペプチド。
(a)配列番号1記載のアミノ酸配列からなるペプチド
(b)配列番号1記載のアミノ酸配列において、1又は複数個のアミノ酸が欠失、置換、挿入又は付加されたアミノ酸配列からなり、核移行性を有するペプチド
【0011】
(4)前記第2のペプチドにおけるリシン残基の含有率が25%以上である前記(1)〜(3)のいずれかに記載の核移行性ペプチド。
【0012】
(5)前記第2のペプチドが核移行性を有する前記(4)記載の核移行性ペプチド。
【0013】
(6)前記第2のペプチドが下記(c)及び(d)からなる群より選択される前記(5)記載の核移行性ペプチド。
(c)配列番号2記載のアミノ酸配列からなるペプチド
(d)配列番号2記載のアミノ酸配列において、1又は複数個のアミノ酸が欠失、置換、挿入又は付加されたアミノ酸配列からなり、核移行性を有するペプチド
【0014】
(7)1又は複数個の前記第1のペプチドと1又は複数個の前記第2のペプチドとを融合させて得られる前記(1)〜(6)のいずれかに記載の核移行性ペプチド。
【0015】
(8)核酸と、前記核酸と静電的に結合した前記(1)〜(7)のいずれかに記載の核移行性ペプチドとを含む複合体。
【0016】
(9)前記(8)記載の複合体を細胞内に導入し、前記複合体に含まれる核酸を細胞内で発現させる方法。
【0017】
(10)次式:
【0018】
【数1】
【0019】
[式中、CPは複合体作製時に使用するペプチドの濃度(μg/μL)を表し、nKはペプチド1分子あたりのリシン残基数を表し、nRはペプチド1分子あたりのアルギニン残基数を表し、MWPはペプチドの分子量を表し、CDは複合体作製時に使用する核酸の濃度(μg/μL)を表し、MWDは核酸のヌクレオチド1残基あたりの平均分子量を表す。]
で定義される電荷比が1〜4である前記複合体を細胞内に導入する前記(9)記載の方法。
【発明の効果】
【0020】
本発明の核移行性ペプチドにおいては、第1のペプチドが有する核移行性を保持したまま、第1のペプチドによる核酸の転写抑制が改善されている。したがって、本発明の核移行性ペプチドと複合体形成した核酸は、核内へ効率よく移行することができるとともに、核内において効率よく発現することができる。
【発明を実施するための最良の形態】
【0021】
以下、本発明について詳細に説明する。
本発明において、「核移行性」とは、細胞質から核内へ移行する能力を意味する。
本発明のペプチドは、第1のペプチドと第2のペプチドとを融合して得られるペプチドと同一のアミノ酸配列からなる全てのペプチドを含む。したがって、本発明のペプチドの製法は、第1のペプチドと第2のペプチドとを融合する方法に限定されるものではない。
【0022】
本発明のペプチドは、複数個のアルギニン残基と複数個のリシン残基とを含むポリカチオン性ペプチドであるので、静電的結合を介して核酸と複合体を形成することができる。本発明のペプチドと核酸との複合体を細胞内に導入すると、当該複合体は核内に移行し、当該複合体に含まれる核酸は核内で発現する。本発明のペプチドと複合体形成した核酸の転写効率は、第1のペプチドと複合体形成した核酸の転写効率よりも高い。すなわち、本発明のペプチドにおいては、第2のペプチド部分の存在により、第1のペプチド部分による核酸の転写抑制が改善されている。
【0023】
第1のペプチドは、複数個のアルギニン残基を含み、核移行性を有するペプチドである。
第1のペプチドにおけるアルギニン残基の含有率は、第1のペプチドが核移行性を有する限り特に限定されるものではないが、通常35%以上、好ましくは40%以上、さらに好ましくは45%以上である。なお、第1のペプチドにおけるアルギニン残基の含有率の上限値は100%である。第1のペプチドと核酸との複合体を細胞内に導入すると、当該複合体は核内に移行し、当該複合体に含まれる核酸は核内で発現するが、第1のペプチドにおけるアルギニン残基の含有率が35%以上であると、核酸との静電的結合力が強固となるため、核酸への転写因子の結合阻害が顕著となり、核酸の転写抑制が顕著となる。
【0024】
第1のペプチドは、核移行性を有する限り、アルギニン残基のみから構成されていてもよいし、他のアミノ酸残基を含んでいてもよい。第1のペプチドにおけるリシン残基の含有率は、通常10%以下、好ましくは5%以下、さらに好ましくは0%である。第1のペプチドにおけるリシン残基の含有率が10%以下であると、第1のペプチドと核酸との複合体を細胞内に導入したときの核酸の転写抑制は顕著なままである。
【0025】
第1のペプチドを構成するアミノ酸残基の総数は、複数個のアルギニン残基を含み、核移行性を有する限り特に限定されるものではないが、通常5〜35個、好ましくは10〜30個、さらに好ましくは15〜20個である。
【0026】
第1のペプチドのアミノ酸配列は、複数個のアルギニン残基を含み、核移行性を有する限り特に限定されるものではない。
【0027】
第1のペプチドは、例えば、下記(a)及び(b)からなる群より選択することができる。
(a)配列番号1記載のアミノ酸配列からなるペプチド(以下「ペプチド(a)」という場合がある。)
(b)配列番号1記載のアミノ酸配列において、1又は複数個のアミノ酸が欠失、置換、挿入又は付加されたアミノ酸配列からなり、核移行性を有するペプチド(以下「ペプチド(b)」という場合がある。)
【0028】
ペプチド(a)は、アデノウイルス由来コアペプチドmuであり、核移行性を有する。ペプチド(a)を構成するアミノ酸残基の総数は19個であり、ペプチド(a)に含まれるアルギニン残基は9個(アルギニン残基の含有率は約47%)であり、リシン残基は0個(リシン残基の含有率は0%)である。
【0029】
ペプチド(b)には、ペプチド(a)に対して人為的に欠失、置換、挿入、付加等の変異を導入したペプチドの他、変異が導入された状態で天然に存在するペプチドや、それに対して人為的に変異を導入したペプチドも含まれる。変異が導入された状態で天然に存在するタンパク質としては、例えば、多型によって生じ得るペプチドが挙げられる。
【0030】
配列番号1記載のアミノ酸配列において欠失、置換、挿入又は付加されるアミノ酸の個数は、ペプチド(b)が核移行性を有する限り特に限定されるものではなく、その個数は1又は複数個である。欠失に関する具体的範囲は、通常1〜3個、好ましくは1〜2個、さらに好ましくは1個であり、置換に関する具体的範囲は、通常1〜3個、好ましくは1〜2個、さらに好ましくは1個であり、挿入に関する具体的範囲は、通常1〜15個、好ましくは1〜10個、さらに好ましくは1〜5個であり、付加に関する具体的範囲は、通常1〜15個、好ましくは1〜10個、さらに好ましくは1〜5個である。
【0031】
配列番号1記載のアミノ酸配列において欠失、置換、挿入又は付加されるアミノ酸の位置は、ペプチド(b)が核移行性を有する限り特に限定されるものではない。
【0032】
第2のペプチドは、複数個のリシン残基を含むペプチドである。
第2のペプチドにおけるリシン残基の含有率は特に限定されるものではないが、通常25%以上、好ましくは30%以上、さらに好ましくは35%以上である。なお、第2のペプチドにおけるリシン残基の含有率の上限値は100%である。第2のペプチドにおけるリシン残基の含有率が25%以上であると、第1のペプチドによる核酸の転写抑制を効果的に改善することができるので、本発明のペプチドと核酸との複合体を細胞内に導入すると、当該複合体は核内に移行し、当該複合体に含まれる核酸は核内で効率よく発現することができる。
【0033】
第2のペプチドは、リシン残基のみから構成されていてもよいし、他のアミノ酸残基を含んでいてもよい。第2のペプチドにおけるアルギニン残基の含有率は、通常20%以下、好ましくは15%以下、さらに好ましくは10%以下である。第2のペプチドにおけるアルギニン残基の含有率が20%以下であると、第1のペプチドによる核酸の転写抑制を効果的に改善することができるので、本発明のペプチドと核酸との複合体を細胞内に導入したときの、核酸の発現効率を向上させることができる。
【0034】
第2のペプチドを構成するアミノ酸残基の総数は、第1のペプチドの核移行性を抑制しない限り特に限定されるものではないが、通常5〜30個、好ましくは5〜20個、さらに好ましくは5〜15個である。
【0035】
第2のペプチドのアミノ酸配列は、複数個のリシン残基を含み、第1のペプチドの核移行性を抑制しない限り特に限定されるものではない。
【0036】
第2のペプチドは、核移行性を有するペプチドであることが好ましい。第2のペプチドが核移行性を有するペプチドであると、本発明のペプチドと核酸との複合体を細胞内に導入したときに、当該複合体が効率よく核内に移行することができる。
【0037】
第2のペプチドは、例えば、下記(c)及び(d)からなる群より選択することができる。
(c)配列番号2記載のアミノ酸配列からなるペプチド(以下「ペプチド(c)」という場合がある。)
(d)配列番号2記載のアミノ酸配列において、1又は複数個のアミノ酸が欠失、置換、挿入又は付加されたアミノ酸配列からなり、核移行性を有するペプチド(以下「ペプチド(d)」という場合がある。)
【0038】
ペプチド(c)は、シミアンウイルス40由来核移行性シグナル(SV40−NLS)であり、核移行性を有する。ペプチド(c)を構成するアミノ酸残基の総数は11個であり、ペプチド(c)に含まれるアルギニン残基の個数は1個(アルギニン残基の含有率は約9%)であり、リシン残基の個数は4個(リシン残基の含有率は約36%)である。
【0039】
ペプチド(d)には、ペプチド(c)に対して人為的に欠失、置換、挿入、付加等の変異を導入したペプチドの他、変異が導入された状態で天然に存在するペプチドや、それに対して人為的に変異を導入したペプチドも含まれる。変異が導入された状態で天然に存在するタンパク質としては、例えば、多型によって生じ得るペプチドが挙げられる。
【0040】
配列番号2記載のアミノ酸配列において欠失、置換、挿入又は付加されるアミノ酸の個数は、ペプチド(d)が核移行性を有する限り特に限定されるものではなく、その個数は1又は複数個である。欠失に関する具体的範囲は、通常1〜3個、好ましくは1〜2個、さらに好ましくは1個であり、置換に関する具体的範囲は、通常1〜3個、好ましくは1〜2個、さらに好ましくは1個であり、挿入に関する具体的範囲は、通常1〜15個、好ましくは1〜10個、さらに好ましくは1〜5個であり、付加に関する具体的範囲は、通常1〜15個、好ましくは1〜10個、さらに好ましくは1〜5個である。
【0041】
配列番号2記載のアミノ酸配列において欠失、置換、挿入又は付加されるアミノ酸の位置は、ペプチド(d)が核移行性を有する限り特に限定されるものではない。
【0042】
本発明のペプチドに含まれる第1のペプチド部分の個数は特に限定されるものではなく、1個であってもよいし複数個であってもよい。第1のペプチド部分の個数は、通常1〜10個、好ましくは1〜5個、さらに好ましくは1〜2個である。
【0043】
本発明のペプチドに含まれる第2のペプチド部分の個数は特に限定されるものではなく、1個であってもよいし複数個であってもよい。第2のペプチド部分の個数は、通常1〜10個、好ましくは1〜5個、さらに好ましくは1〜2個である。
【0044】
本発明のペプチドにおいて、第1のペプチド部分と第2のペプチド部分との配列順序は特に限定されるものではない。
【0045】
本発明のペプチドに第1のペプチド部分が複数個含まれる場合、第1のペプチド部分同士が連続していてもよいし、第1のペプチド部分間に第2のペプチド部分が介在していてもよい。
【0046】
本発明のペプチドに第2のペプチド部分が複数個含まれる場合、第2のペプチド部分同士が連続していてもよいし、第2のペプチド部分間に第1のペプチド部分が介在していてもよい。
【0047】
第1のペプチド部分と第2のペプチド部分とは、第1のペプチド部分のN末端と第2のペプチド部分のC末端とが結合していてもよいし、第1のペプチド部分のC末端と第2のペプチド部分のN末端とが結合していてもよい。
【0048】
第1のペプチド部分と第2のペプチド部分とは、直接結合していてもよいし、適当なリンカーを介して結合していてもよい。リンカーのアミノ酸配列は、本発明のペプチドの核内移行性を抑制しない限り特に限定されるものではない。リンカーを構成するアミノ酸残基の総数は、本発明のペプチドの核内移行性を抑制しない限り特に限定されるものではないが、通常1〜10個、好ましくは1〜7個、さらに好ましくは1〜5個である。
【0049】
本発明のペプチドは、例えば、本発明のペプチドを生産し得る形質転換体を培養することにより得ることができる。本発明のペプチドを生産し得る形質転換体は、第1のペプチドをコードするオープンリーディングフレームと第2のペプチドをコードするオープンリーディングフレームとが所望の順序で連結されたDNAを、発現ベクターのプロモーターの下流に挿入して組換えベクターを作製し、当該組換えベクターを宿主細胞に導入することにより得ることができる。
【0050】
ペプチド(a)をコードするDNAは、例えば、アデノウイルスを感染させた細胞から抽出したmRNAを用いてcDNAライブラリーを作製し、ペプチド(a)をコードするDNAの塩基配列に基づいて合成したプローブを用いて、cDNAライブラリーから目的のDNAを含むクローンをスクリーニングすることにより得ることができる。また、ペプチド(a)をコードするDNAは、その塩基配列に従って化学合成により得ることもできる。DNAの化学合成は、市販のDNA合成機、例えば、チオホスファイト法を利用したDNA合成機(島津製作所社製)、フォスフォアミダイト法を利用したDNA合成機(パーキン・エルマー社製)を用いて行うことができる。
【0051】
ペプチド(b)をコードするDNAは、ペプチド(a)をコードするDNAに、部位特異的変異誘発法等の公知の方法を用いて人為的に変異を導入することにより得ることができる。変異の導入は、例えば、変異導入用キット、例えば、Mutant-K(TAKARA社製)、Mutant-G(TAKARA社製)、TAKARA社のLA PCR in vitro Mutagenesis シリーズキットを用いて行うことができる。また、ペプチド(b)をコードするDNAは、その塩基配列に従って化学合成により得ることもできる。
【0052】
ペプチド(c)をコードするDNAは、例えば、シミアンウイルスを感染させた細胞から抽出したmRNAを用いてcDNAライブラリーを作製し、ペプチド(c)をコードするDNAの塩基配列に基づいて合成したプローブを用いて、cDNAライブラリーから目的のDNAを含むクローンをスクリーニングすることにより得ることができる。また、ペプチド(c)をコードするDNAは、その塩基配列に従って化学合成により得ることもできる。
【0053】
ペプチド(d)をコードするDNAは、ペプチド(c)をコードするDNAに、部位特異的変異誘発法等の公知の方法を用いて人為的に変異を導入することにより得ることができる。また、ペプチド(d)をコードするDNAは、その塩基配列に従って化学合成により得ることもできる。
【0054】
発現ベクターとしては、宿主細胞において自立複製が可能なものであれば特に限定されず、例えば、プラスミドベクター、ファージベクター、ウイルスベクター等を使用することができる。
【0055】
宿主細胞としては、目的とする遺伝子を発現し得る限り、原核細胞、酵母、動物細胞、昆虫細胞、植物細胞等のいずれを使用してもよい。また、動物個体、植物個体、カイコ虫体等を使用してもよい。
【0056】
宿主細胞への組換えベクターの導入方法は、エレクトロポレーション法、リン酸カルシウム法、リポフェクション法等を使用することができる。
【0057】
形質転換体の培養は、宿主細胞の培養に用いられる通常の方法に従って行うことができる。
形質転換体の培養物より本発明のペプチドを採取することにより、本発明のペプチドを得ることができる。培養物には、培養上清、培養細胞、培養菌体、細胞又は菌体の破砕物のいずれもが含まれる。
【0058】
本発明のペプチドが形質転換体の細胞内に蓄積される場合には、培養物を遠心分離することにより、培養物中の細胞を集め、該細胞を洗浄した後に細胞を破砕して、本発明のペプチドを抽出する。本発明のペプチドが形質転換体の細胞外に分泌される場合には、培養上清をそのまま使用するか、遠心分離等により培養上清から細胞又は菌体を除去する。
【0059】
得られたペプチドは、溶媒抽出法、硫安等による塩析法脱塩法、有機溶媒による沈殿法、ジエチルアミノエチル(DEAE)−セファロース、イオン交換クロマトグラフィー法、疎水性クロマトグラフィー法、ゲルろ過法、アフィニティークロマトグラフィー法等により精製することができる。
【0060】
本発明のペプチドは、そのアミノ酸配列に基づいて、Fmoc法(フルオレニルメチルオキシカルボニル法)、tBoc法(t−ブチルオキシカルボニル法)等の化学合成法によって製造することができる。この際、市販のペプチド合成機を使用することができる。
【0061】
本発明のペプチドは、複数個のアルギニン残基と複数個のリシン残基とを含むポリカチオン性ペプチドであるので、静電的結合を介して核酸と複合体を形成することができる。本発明のペプチドと核酸との複合体は、本発明のペプチドと核酸とを例えば4〜37℃で0.5〜3時間インキュベーションすることにより作製することができる。本発明のペプチドと複合体形成させる核酸としては、例えば、目的のタンパク質をコードするDNA、当該DNAが組み込まれたプラスミド等を使用することができる。
【0062】
本発明のペプチドと核酸との複合体は、脂質を含んでいてもよい。脂質としては、例えば、リン脂質、糖脂質、ステロール、飽和又は不飽和の脂肪酸等が挙げられる。リン脂質としては、例えば、ホスファチジルコリン、ホスファチジルグリセロール、ホスファチジルエタノールアミン、ホスファチジルセリン、ホスファチジルイノシトール、ホスファチジン酸、カルジオリピン、スフィンゴミエリン、卵黄レシチン、大豆レシチン、これらの水素添加物等が挙げられる。糖脂質としては、例えば、グリセロ糖脂質、スフィンゴ糖脂質等が挙げられる。ステロールとしては、例えば、動物由来のステロール(例えば、コレステロール、コレステロールコハク酸、ラノステロール、ジヒドロラノステロール、デスモステロール、ジヒドロコレステロール)、植物由来のステロール(フィトステロール)(例えば、スチグマステロール、シトステロール、カンペステロール、ブラシカステロール)、微生物由来のステロール(例えば、チモステロール、エルゴステロール)等が挙げられる。飽和又は不飽和の脂肪酸としては、例えば、パルミチン酸、オレイン酸、ステアリン酸、アラキドン酸、ミリスチン酸等の炭素数12〜20の飽和又は不飽和の脂肪酸が挙げられる。
【0063】
本発明のペプチドと核酸との複合体は、リポソーム内に封入してもよい。本発明のペプチドと核酸との複合体の存在下、脂質膜を水和し、次いで攪拌又は超音波処理することにより、本発明のペプチドと核酸との複合体が封入されたリポソームを製造することができる。
【0064】
本発明のペプチドと核酸との複合体の粒子径は特に限定されるものではないが、通常30〜105nm、好ましくは30〜100nm、さらに好ましくは30〜95nmである。本発明のペプチドと核酸との複合体の粒子径が上記範囲にあると、当該複合体が効率よく核内に移行することができる。
【0065】
本発明のペプチドと核酸との複合体を細胞内に導入する方法は特に限定されるものではなく、常法に従って行うことができる。
本発明のペプチドと核酸との複合体を細胞内に導入する方法としては、例えば、エンドサイトーシスを利用した方法、細胞質内へのインジェクション、核内へのインジェクション、エレクトロポレーション、ハイドロダイナミックインジェクション等が挙げられる。
【0066】
エンドサイトーシスを利用する場合、本発明のペプチドと核酸との複合体又は当該複合体を封入したリポソームを、細胞膜の表面上に存在する受容体又は抗原と結合できる物質で修飾することにより、当該複合体を細胞内へ効率よく導入することができる。
【0067】
エンドサイトーシスによる細胞内導入は、本発明のペプチドと核酸との複合体又は当該複合体を封入したリポソームを、細胞とともに例えば4〜37℃で0.5〜48時間インキュベーションすることにより行うことができる。
【0068】
細胞膜の表面上に存在する受容体又は抗原と結合できる物質の種類は特に限定されるものではなく、細胞膜結合性物質としては、例えば、トラスフェリン、インシュリン、葉酸、ヒアルロン酸、抗体又はその断片、糖鎖、成長因子、アポリポタンパク質等が挙げられる。成長因子としては、例えば、上皮成長因子(EGF)、インシュリン様成長因子(IGF)、繊維芽細胞増殖因子(FGF)等が挙げられる。アポリポタンパク質としては、例えば、アポA−1、アポB−48、アポB−100、アポE等が挙げられる。抗体の断片としては、例えば、Fab断片、F(ab)'2断片、単鎖抗体(scFv)等が挙げられる。
【0069】
本発明のペプチドと核酸との複合体において、次式:
【0070】
【数2】
【0071】
[式中、CPは複合体作製時に使用するペプチドの濃度(μg/μL)を表し、nKはペプチド1分子あたりのリジン残基数を表し、nRはペプチド1分子あたりのアルギニン残基数を表し、MWPはペプチドの分子量を表し、CDは複合体作製時に使用する核酸の濃度(μg/μL)を表し、MWDは核酸のヌクレオチド1残基あたりの平均分子量を表す。]
で定義される電荷比は1〜4であることが好ましく、2〜3であることがさらに好ましい。本発明のペプチドと核酸との複合体の電荷比が1〜4であると、当該複合体に含まれる核酸を効率よく核内で発現させることができる。
【0072】
本発明のペプチドと核酸との複合体を導入する細胞が由来する生物種は特に限定されるものではなく、動物、植物、微生物等のいずれであってもよいが、動物であることが好ましく、哺乳動物であることがさらに好ましい。哺乳動物としては、例えば、ヒト、サル、ウシ、ヒツジ、ヤギ、ウマ、ブタ、ウサギ、イヌ、ネコ、ラット、マウス、モルモット等が挙げられる。また、目的物質を送達すべき細胞の種類は特に限定されるものではなく、例えば、体細胞、生殖細胞、幹細胞又はこれらの培養細胞等が挙げられる。
【実施例】
【0073】
1.ポリカチオン性ペプチドと核酸との複合体の作製
ポリカチオン性ペプチドとして、アデノウイルス由来コアペプチドmu(以下「mu」という。)、シミアンウイルス40由来核移行性シグナル(SV40−NLS)とmuとの融合ペプチド(以下「NLS−mu」という。)、及びポリL−リシン(以下「PLL」という。)を使用した。muのアミノ酸配列を配列番号1に示し、NLS−muのアミノ酸配列を配列番号2に示し、融合ペプチドのアミノ酸配列を配列番号3に示す。muの分子量は2440.89であり、NLS−muの分子量は3810.48であり、PLLの分子量は25700である。なお、ポリカチオン性ペプチドの合成は、ペプチド合成業者に委託した。
【0074】
核酸としては、EGFP(enhanced Green Fluorescent Protein)をコードするDNA(クロンテック社製)を組み込んだプラスミドpcDNA3.1(+)(インビトロジェン社製)(以下「pcDNA3.1(+)−EGFP」という。)を使用した。
【0075】
電荷比(charge ratio (+/-))を1,2,5,10の4種類に設定し、mu又はNLS−muと核酸との複合体を作製した。
【0076】
mu又はNLS−muと核酸との複合体に関する電荷比は、ペプチド中の1個のリジン残基、1個のアルギニン残基をそれぞれ+1の電荷、DNA中の1個のリン酸基を−1の電荷として次式により算出した。
【0077】
【数3】
【0078】
[式中、CPは複合体作製時に使用するペプチドの濃度(μg/μL)を表し、nKはペプチド1分子あたりのリジン残基数を表し、nRはペプチド1分子あたりのアルギニン残基数を表し、MWPはペプチドの分子量を表し、CDは複合体作製時に使用する核酸の濃度(μg/μL)を表し、MWDは核酸のヌクレオチド1残基あたりの平均分子量を表す。]
【0079】
なお、muに関し、nKは0であり、nRは9であり、MWPは2440.89であり、NLS−muに関し、nKは4であり、nRは10であり、MWPは3810.48である。また、pcDNA3.1(+)−EGFPに関し、MWDは308であり、CDは0.1に設定した。
【0080】
電荷比が1のとき、muに関するCPは0.0875に設定し、NLS−muに関するCPは0.0885に設定した。電荷比が2のとき、muに関するCPは0.175に設定し、NLS−muに関するCPは0.177に設定した。電荷比が5のとき、muに関するCPは0.4375に設定し、NLS−muに関するCPは0.4425に設定した。電荷比が10のとき、muに関するCPは0.875に設定し、NLS−muに関するCPは0.885に設定した。
【0081】
電荷比(charge ratio (+/-))を2に設定し、PLLと核酸との複合体を作製した。
PLLと核酸との複合体に関する電荷比は次式により計算した。
【0082】
【数4】
【0083】
[式中、CPは複合体作製時に使用するペプチドの濃度(μg/μL)を表し、MWKはリジン1残基あたりの分子量を表し、CDは複合体作製時に使用する核酸の濃度(μg/μL)を表し、MWDは核酸のヌクレオチド1残基あたりの平均分子量を表す。]
【0084】
なお、MWKは128であり、MWDは308であり、CPは0.083に設定し、CDは0.1に設定した。
【0085】
ポリカチオン性ペプチド溶液(100μL)を攪拌し、0.1μg/μLのpcDNA3.1(+)−EGFP/H2Oを、20μLずつを5回滴下して加えた。滴下後、30秒間攪拌を継続し、室温で30分間静置した。静置後、ELS−8000を用いて、ポリカチオン性ペプチドと核酸との複合体の粒子径測定を行った。なお、H2Oとしては、DISMIC−13CP(cellulose acetate, pore size = 0.20μm)を用いて濾過したH2Oを用いた。
各複合体の粒子径(nm)は表1に示すとおりである。
【0086】
【表1】
【0087】
2.各複合体に関する核膜透過及び核内転写効率の評価
各複合体について、核膜透過及び核内転写効率の評価を行った。なお、PLLと核酸との複合体(電荷比=2)はコントロールとして使用した。
【0088】
(1)マイクロインジェクション
インジェクションを行う前日に、glass base dish(IWAKI)上に5×104細胞/mLの濃度でHeLa細胞を培養した。各複合体を、濾過済み0.5% ローダミン(Rh)/H2Oを用いて、pcDNA換算で33.2fmol/μL(= 1000 copies/cell)、0.33fmol/μL(= 10 copies/cell)となるように希釈した。33.2fmol/μLのものを細胞質インジェクションに、0.33fmol/μLのものを核インジェクションに使用した。
【0089】
細胞質インジェクションにおいては、HeLa細胞の細胞質に1000 copies/cellの濃度のサンプルをインジェクションした(インジェクション条件:Pi = 50-70 hPa, Pc = 30 hPa, Ti = 0.2 s)。インジェクションは400個の細胞に行った。インジェクション後、新しいD’MEMに培地を交換し、37℃、5%CO2下でインキュベートした。インジェクション24時間後、蛍光顕微鏡にて細胞を観察し、Rh発色細胞数及びEGFP発現細胞数をカウントし、遺伝子発現率をEGFP発現細胞数/Rh発色細胞数より算出した。
【0090】
核内インジェクションにおいては、HeLa細胞の核内に10 copies/cellの濃度のサンプルをインジェクションした(インジェクション条件:Pi = 50 hPa, Pc = 30 hPa, Ti = 0.2 s)。インジェクションは400〜450個の細胞に行った。インジェクション後、新しいD’MEMに培地を交換し、37℃、5%CO2下でインキュベートした。インジェクション24時間後、蛍光顕微鏡にて細胞を観察し、Rh発色細胞数及びEGFP発現細胞数をカウントし、遺伝子発現率をEGFP発現細胞数/Rh発色細胞数より算出した。
【0091】
核内インジェクションにおける遺伝子発現が、核内における転写効率を反映するパラメーターと考えられる。また、核インジェクション時の発現率を細胞質インジェクション時の発現率で除することにより、核移行性の指標となる核移行スコアー(nuclear transfer score : NT score)を算出した。
各複合体の細胞質インジェクション時におけるEGFP発現率を図1に示す。
図中、各値は平均値±標準偏差(n=3)を表す。
【0092】
図1に示すように、電荷比が1のとき、各複合体のEGFP発現率はいずれも20%前後であった。電荷比が2のとき、NLS−muと核酸との複合体が最も高いEGFP発現率を示し(32.6%)、コントロールであるPLLと核酸との複合体のEGFP発現率(12.3%)の2.7倍、muと核酸との複合体のEGFP発現率(20.1%)の1.6倍であり、各複合体のEGFP発現率に顕著な違いが認められた。NLS−muと核酸との複合体のEGFP発現率は、電荷比が2よりも大きくなるに従って減少した。
【0093】
核インジェクション時における各複合体のEGFP発現率の比較は、細胞質インジェクション時における各複合体のEGFP発現率に顕著な違いが認められた電荷比が2の場合にて行った。各複合体の核インジェクション時におけるEGFP発現率を図2に示す。
【0094】
図2に示すように、PLLと核酸との複合体のEGFP発現率が最も高く(86.7%)、NLS−muと核酸との複合体のEGFP発現率(73.6%)も同程度であったが、muと核酸との複合体のEGFP発現率は低かった(35.0%)。
以上の結果から、電荷比が2のときの各複合体の核移行スコアーを算出し、PLLと核酸との複合体の核移行スコアーを1としたときの相対値を表2に示す。
【0095】
【表2】
【0096】
表中、各値は平均値±標準偏差を表す。
電荷比が2のとき、muと核酸との複合体の核移行スコアー及びNLS−muと核酸との複合体の核移行スコアーは、PLLと核酸との複合体の核移行スコアーの3〜4倍であった。
【0097】
以上の結果から、SV40−NLSをmuに付加することにより、muの核移行性を保持しつつ、muによる核酸の転写抑制を緩和することができ、これにより、NLS−muと核酸との複合体を細胞内に導入したときの、核酸の発現を亢進することができることが示された。
【0098】
アルギニンに富むペプチドは、リシンに富むペプチドと比較して、核酸と強固な複合体を形成することが示唆されている(Plank. C. et al., Hum Gene Ther 10 : 319-322 (1999))。muはアルギニンに富むペプチド(アルギニン残基の含有率は約47%、リシン残基の含有率は0%)であるので、muと核酸との強い相互作用により、転写因子の核酸への結合が阻害され、muと複合体形成した核酸の転写が抑制されたものと考えられる。一方、SV40−NLSは、リシンに富むペプチド(リシン残基の含有率は約36%、アルギニン残基の含有率は約9%)であるので、SV40−NLSをmuに付加することにより(融合ペプチドにおけるアルギニン残基の含有率は約33%、リシン残基の含有率は約13%)、muと核酸との強い相互作用が緩和され、muによる核酸の転写抑制を緩和されたものと考えられる。
【0099】
通常、ポリカチオン性ペプチドと核酸との複合体は、核膜孔を透過して核内へ移行するものと考えられており、核膜孔を透過し得る粒子サイズの上限値は39nmであると考えられている(N. Psnte et al., Mol Biol Cell 13: 425-434 (2002))。NLS−muと核酸との複合体の粒子サイズは、表1に示すように39nmを超えており、NLS−muと核酸との複合体の場合は、柔軟性に富んだ構造であるため核膜孔を透過することができるものと考えられる。但し、電荷比が10のとき、NLS−muと核酸との複合体の核移行スコアーは1.56(細胞質インジェクション時のEGFP発現率は10.5%、核内インジェクション時のEGFP発現率は47.3%)であり、電荷比の増大とともに核移行スコアーの低下が認められた。電荷比の増大とともに粒子サイズも増大するので、核膜孔を透過し難くなり、核移行スコアーが低下したものと考えられる。
【図面の簡単な説明】
【0100】
【図1】各複合体の細胞質インジェクション時におけるEGFP発現率を示す図である。
【図2】各複合体の核インジェクション時におけるEGFP発現率を示す図である。
【特許請求の範囲】
【請求項1】
複数個のアルギニン残基を含み、核移行性を有する第1のペプチドと、複数個のリシン残基を含む第2のペプチドとを融合させて得られる核移行性ペプチド。
【請求項2】
前記第1のペプチドにおけるアルギニン残基の含有率が35%以上である請求項1記載の核移行性ペプチド。
【請求項3】
前記第1のペプチドが、下記(a)及び(b)からなる群より選択される請求項2記載の核移行性ペプチド。
(a)配列番号1記載のアミノ酸配列からなるペプチド
(b)配列番号1記載のアミノ酸配列において、1又は複数個のアミノ酸が欠失、置換、挿入又は付加されたアミノ酸配列からなり、核移行性を有するペプチド
【請求項4】
前記第2のペプチドにおけるリシン残基の含有率が25%以上である請求項1〜3のいずれかに記載の核移行性ペプチド。
【請求項5】
前記第2のペプチドが核移行性を有する請求項4記載の核移行性ペプチド。
【請求項6】
前記第2のペプチドが下記(c)及び(d)からなる群より選択される請求項5記載の核移行性ペプチド。
(c)配列番号2記載のアミノ酸配列からなるペプチド
(d)配列番号2記載のアミノ酸配列において、1又は複数個のアミノ酸が欠失、置換、挿入又は付加されたアミノ酸配列からなり、核移行性を有するペプチド
【請求項7】
1又は複数個の前記第1のペプチドと1又は複数個の前記第2のペプチドとを融合させて得られる請求項1〜6のいずれかに記載の核移行性ペプチド。
【請求項8】
核酸と、前記核酸と静電的に結合した請求項1〜7のいずれかに記載の核移行性ペプチドとを含む複合体。
【請求項9】
請求項8記載の複合体を細胞内に導入し、前記複合体に含まれる核酸を細胞内で発現させる方法。
【請求項10】
次式:
【数1】
[式中、CPは複合体作製時に使用するペプチドの濃度(μg/μL)を表し、nKはペプチド1分子あたりのリシン残基数を表し、nRはペプチド1分子あたりのアルギニン残基数を表し、MWPはペプチドの分子量を表し、CDは複合体作製時に使用する核酸の濃度(μg/μL)を表し、MWDは核酸のヌクレオチド1残基あたりの平均分子量を表す。]
で定義される電荷比が1〜4である前記複合体を細胞内に導入する請求項9記載の方法。
【請求項1】
複数個のアルギニン残基を含み、核移行性を有する第1のペプチドと、複数個のリシン残基を含む第2のペプチドとを融合させて得られる核移行性ペプチド。
【請求項2】
前記第1のペプチドにおけるアルギニン残基の含有率が35%以上である請求項1記載の核移行性ペプチド。
【請求項3】
前記第1のペプチドが、下記(a)及び(b)からなる群より選択される請求項2記載の核移行性ペプチド。
(a)配列番号1記載のアミノ酸配列からなるペプチド
(b)配列番号1記載のアミノ酸配列において、1又は複数個のアミノ酸が欠失、置換、挿入又は付加されたアミノ酸配列からなり、核移行性を有するペプチド
【請求項4】
前記第2のペプチドにおけるリシン残基の含有率が25%以上である請求項1〜3のいずれかに記載の核移行性ペプチド。
【請求項5】
前記第2のペプチドが核移行性を有する請求項4記載の核移行性ペプチド。
【請求項6】
前記第2のペプチドが下記(c)及び(d)からなる群より選択される請求項5記載の核移行性ペプチド。
(c)配列番号2記載のアミノ酸配列からなるペプチド
(d)配列番号2記載のアミノ酸配列において、1又は複数個のアミノ酸が欠失、置換、挿入又は付加されたアミノ酸配列からなり、核移行性を有するペプチド
【請求項7】
1又は複数個の前記第1のペプチドと1又は複数個の前記第2のペプチドとを融合させて得られる請求項1〜6のいずれかに記載の核移行性ペプチド。
【請求項8】
核酸と、前記核酸と静電的に結合した請求項1〜7のいずれかに記載の核移行性ペプチドとを含む複合体。
【請求項9】
請求項8記載の複合体を細胞内に導入し、前記複合体に含まれる核酸を細胞内で発現させる方法。
【請求項10】
次式:
【数1】
[式中、CPは複合体作製時に使用するペプチドの濃度(μg/μL)を表し、nKはペプチド1分子あたりのリシン残基数を表し、nRはペプチド1分子あたりのアルギニン残基数を表し、MWPはペプチドの分子量を表し、CDは複合体作製時に使用する核酸の濃度(μg/μL)を表し、MWDは核酸のヌクレオチド1残基あたりの平均分子量を表す。]
で定義される電荷比が1〜4である前記複合体を細胞内に導入する請求項9記載の方法。
【図1】

【図2】
【図2】
【公開番号】特開2006−67891(P2006−67891A)
【公開日】平成18年3月16日(2006.3.16)
【国際特許分類】
【出願番号】特願2004−254911(P2004−254911)
【出願日】平成16年9月1日(2004.9.1)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 平成16年3月3日発行の刊行物「平成15年度 修士論文要旨集 北海道大学大学院薬学研究科」に発表。 平成16年3月16日開催の修士論文発表会(北海道大学主催)にて文書をもって発表。 平成16年6月30日発行の刊行物「第20回日本DDS学会 プログラム予稿集」に発表。 平成16年7月16日開催の第20回日本DDS学会にて文書をもって発表。
【出願人】(504173471)国立大学法人 北海道大学 (971)
【Fターム(参考)】
【公開日】平成18年3月16日(2006.3.16)
【国際特許分類】
【出願日】平成16年9月1日(2004.9.1)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 平成16年3月3日発行の刊行物「平成15年度 修士論文要旨集 北海道大学大学院薬学研究科」に発表。 平成16年3月16日開催の修士論文発表会(北海道大学主催)にて文書をもって発表。 平成16年6月30日発行の刊行物「第20回日本DDS学会 プログラム予稿集」に発表。 平成16年7月16日開催の第20回日本DDS学会にて文書をもって発表。
【出願人】(504173471)国立大学法人 北海道大学 (971)
【Fターム(参考)】
[ Back to top ]