
核酸増幅法による単純ヘルペスウイルス1型および2型の検出

【課題】HSVの診断を助ける迅速且つ高感度なツールの開発の、臨床上の必要性が存在する。
【解決手段】本発明は、サンプル中の単純ヘルペスウイルス(HSV)の存在または非存在を、HSVの糖タンパク質G(US4)遺伝子の部分を増幅することによって検出し、および本明細書で開示するようなプライマーおよび検出プライマーを使用して増幅された核酸の存在を検出する方法に関する。本発明の方法は、さらに、サンプル中のHSVのタイプ、すなわちHSV−1またはHSV−2を同定する。本明細書に記載された増幅方法で使用されうるプライマーおよび検出プライマーを含むキットも本発明に包含される。
【解決手段】本発明は、サンプル中の単純ヘルペスウイルス(HSV)の存在または非存在を、HSVの糖タンパク質G(US4)遺伝子の部分を増幅することによって検出し、および本明細書で開示するようなプライマーおよび検出プライマーを使用して増幅された核酸の存在を検出する方法に関する。本発明の方法は、さらに、サンプル中のHSVのタイプ、すなわちHSV−1またはHSV−2を同定する。本明細書に記載された増幅方法で使用されうるプライマーおよび検出プライマーを含むキットも本発明に包含される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、核酸増幅法により単純ヘルペスウイルス(HSV)を同定するための診断方法と核酸配列に関する。
【背景技術】
【0002】
本出願は、特願2006−513315の分割出願である。
【0003】
単純ヘルペスは、ヒトにおける初感染および回帰感染の原因であり、伝染性単球増加症(エプスタイン−バーウイルス)、水痘および帯状疱疹(水痘帯状疱疹ウイルス)を起こすウイルスに関連する、エンベロープで包まれた二本鎖DNAウイルスである。単純ヘルペスウイルス感染の症状には、皮膚または粘膜上の小さな疱疹の出現が含まれる。疱疹の出現が収まった後、ウイルスは、感染した領域に神経繊維を供給する神経細胞群(神経節)の内側に休眠(潜伏)状態で生き残っている。周期的に、ウイルスは再活性化し、再度成長を開始し、神経繊維を通して皮膚に戻って行き、これによって先の感染と同じ皮膚領域に疱疹の出現を引き起こす。明確な疱疹がない場合であっても、ウイルスはしばしば、皮膚または粘膜に存在することがある。単純ヘルペスウイルス(HSV)は2種類のタイプ、HSV−1およびHSV−2に分類される。ヒトHSV−1およびHSV−2の完全なゲノムは配列決定されている(例えば、NCBI Accession Nos. X14112 and Z86099(非特許文献1)参照)。
【0004】
HSVは、失明および脳炎を含めた種々の疾患に寄与するかまたは疾患を引き起こすことが示されている。局所的な発生の他に、HSV−1およびHSV−2は脳炎に関連する。この脳炎の病体生理学は、ヒトでは理解が不十分である。動物モデルでは、このウイルスは末梢神経を通して中枢神経系に入り脳の炎症を引き起こすことが示唆されている。HSV−1は成人の脳炎の一般的な原因である。HSV−2は、母親の性器感染に関連する新生児脳炎の一般的な原因である。HSV−2は、社会における最も一般的な性感染症の1つである。HSVに関連する脳炎は、100万人あたり1から4人の年間発症率を有し、全てのタイプの脳炎のうち死亡率が最も高い。HSV脳炎は、全年齢の人を、一年のうちのいずれの時期においても冒す。成人では、HSVに関連する脳炎は潜伏したウイルスの再活性化によると考えられる。症状には、発熱、頭痛、痙攣(seizures)、意識レベルの変化および人格変化が含まれうる。他の病患に対するこれらの症状の類似点が臨床診断を困難にしている。治療しないままでいると、単純ヘルペス脳炎(HSE)の死亡率は、治療を受けたものの中での19パーセント程度の低さと比べ、70パーセント程度の高さになる。治療された患者の中では、約38パーセントが、最終的に正常機能に戻ることが報告されている。従って、早期にHSV感染を診断できることが非常に重要である。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】EP 0 497 272
【特許文献2】米国特許第5,010,183号明細書
【特許文献3】米国特許第5,860,937号明細書
【特許文献4】米国特許第5,728,822号明細書
【特許文献5】米国特許第5,990,301号明細書
【特許文献6】米国特許第6,261,785号明細書
【特許文献7】米国特許第5,455,166号明細書
【特許文献8】米国特許第5,270,184号明細書
【特許文献9】米国特許第5,547,861号明細書
【特許文献10】米国特許第5,928,869号明細書
【特許文献11】米国特許第5,593,867号明細書
【特許文献12】米国特許第5,550,025号明細書
【特許文献13】米国特許第5,935,791号明細書
【特許文献14】米国特許第5,888,739号明細書
【特許文献15】米国特許第5,846,726号明細書
【非特許文献】
【0006】
【非特許文献1】NCBI Accession Nos. X14112 and Z86099
【非特許文献2】Saiki et al., 1985, Science 230: 1350-1354
【非特許文献3】Wu et al., 1989, Genomics 4: 560-569
【非特許文献4】Barringer et al., 1990, Gene 89: 117-122
【非特許文献5】Barany, 1991, Proc. Natl. Acad. Sci. USA 88: 189-193
【非特許文献6】Kwoh et al., 1989, Proc. Natl. Acad. Sci. USA 86: 1173-1177
【非特許文献7】Guatelli et al., 1990, Proc. Natl. Acad. Sci. USA 87: 1874-1878
【非特許文献8】Lizardi et al., 1988, BioTechnology 6: 1197-1202
【非特許文献9】Walker et al., 1992, Proc. Natl. Acad. Sci. USA 89: 392-396
【非特許文献10】Walker et al., 1992, Nuc. Acids. Res. 20:1691-1696
【非特許文献11】T. Maniatis et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, 1982
【非特許文献12】J. Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, 1989
【非特許文献13】Current Protocols in Molecular Biology, Eds. F. M. Ausubel et al., Vol. 1, "Preparation and Analysis of DNA", John Wiley and Sons, Inc., 1994-1995, Suppls. 26, 29, 35 and 42; pp. 2.10.7-2.10.16
【非特許文献14】G. M. Wahl and S. L. Berger (1987; Methods Enzymol. 152: 399-407)
【非特許文献15】A. R. Kimmel, 1987; Methods of Enzymol. 152: 507-511
【非特許文献16】Nadeau, et al., 1999 Anal. Biochem. 276:177-187
【非特許文献17】J. Sambrook and D. W. Russell, Molecular Cloning: a Laboratory Manual, 3rd Edition, Cold Spring Harbor Laboratory Press, USA, (2001)
【発明の概要】
【発明が解決しようとする課題】
【0007】
HSV感染の診断は、一般に、適切な臨床サンプルについて細胞培養を用いて行われる。しかし、細胞培養物内容HSVを単離する能力は、古い病巣、宿主の免疫応答の存在、および再活性化の症状の出現で低下する。血清学的診断法、特に髄液(CSF)中のHSVの診断法は、感度または特異性が十分ではなく、脳炎の初期治療介入の選択を含む決定に使用するには時間がかかりすぎる。HSVは細胞培養を用いた脳脊髄液中では滅多に検出されず、僅かに4パーセントの場合が培養物陽性であった。血清学的方法はまた、初期感染後、抗体応答が現れるのに2から3週間の間遅れることにより、HSEの診断には適さない。脳生検を含む診断である「ゴールドスタンダード」法は、侵襲的であり、長期の罹病率の重大なリスクが問題となる。コンピューター断層撮影法および磁気共鳴映像法は、診断ツールとしては特異的でなく、感度が不足している。
【0008】
現時点では、HSVの検出のための免疫学的方法は、信頼できず且つ実施するのが困難である。分子法の検出は、感度を高め、結果に至る時間が従来の手段で可能であるよりも早まる可能性を提供する。迅速で、高感度で、且つ特異的なHSV疾患の診断が絶対必要であるという事実が存在する。従って、HSVの診断を助ける迅速且つ高感度なツールの開発の、臨床上の必要性が存在する。HSV感染の型を決めるためのツールの必要性も残っている。ウイルス感染に関連する特異的な原因物質の迅速な同定は、短い時間内で適切な治療法を決定するのに用いることができる情報を提供する。
【課題を解決するための手段】
【0009】
本発明は、哺乳動物内の単純ヘルペスウイルス(HSV)、特に単純ヘルペスウイルス1型(HSV−1)または単純ヘルペスウイルス2型(HSV−2)を検出する方法および組成物に関する。この方法は、プライマーを用いて単純ヘルペスウイルス標的配列を増幅し、検出することを含む。一実施形態は、鎖置換増幅(Strand Displacement Amplification)(SDA)の増幅技術を使用する。
【0010】
本発明の核酸プライマーは、HSV−1またはHSV−2中の標的配列を一義的に増幅し、これによってHSVの高感度な検出および型の同定を可能にする。本発明はまた、ユニバーサル蛍光エネルギー移動プローブを用いるリアルタイムな検出を含むHSVの検出のための鎖置換増幅(SDA)をベースとする方法を指向する。本発明のプローブおよびプライマーは、直接、迅速、且つ高感度なHSV核酸の検出を提供し、従って、免疫学的アッセイに、魅力的な代替法を提供する。
【発明の効果】
【0011】
本発明のプローブおよびプライマーは、培養された微生物の正体を確認するための手段として、サンプルを培養した後に使用されうる。別法として、これらは公知の増幅方法を用いてHSV核酸の検出および同定をするために、培養に先立って、または培養に代えて用いることができる。本発明のプローブ、プライマーおよび組成物、並びにこのプローブ、プライマーおよび組成物を用いたアッセイ法は、HSV−1とHSV−2の核酸標的配列を迅速に識別する手段を提供し、一般に依拠されている時間のかかる免疫学的および生化学的手順に頼ることなく、医者がHSVを同定し、診断し、治療することを可能にする。
【図面の簡単な説明】
【0012】
【図1】単純ヘルペスウイルス1型の糖タンパク質G(US4)遺伝子のコンセンサス配列(配列番号1)を示す図である。
【図2】HSV−1標的領域(配列番号2)のゲノム配列の位置、および、HSV−1DNAの特異的検出用に設計されたプライマー、バンパー、およびアダプタの配置を示すマップである。
【図3】「MOTA」発現の結果を示すグラフである。
【図4】BDプローブテック(商標)ETシステムを用いた「PAT」アルゴリズムを示すグラフである。
【図5】種々のHSV−1系統の希釈物についてのSDA法の分析感度を表す図である。
【図6】単純ヘルペスウイルス2型(HSV−2)の糖タンパク質G(US4)のフラグメントのコンセンサス配列(配列番号3)の図である。
【図7】HSV−2標的領域(配列番号4)のゲノム配列、および、HSV−2DNAの特異的検出用に設計されたプライマー、バンパー、およびアダプタの配置を示すマップである。
【発明を実施するための形態】
【0013】
本発明の種々の目的、利点、および新規な特徴は、添付の図面と組み合わせた以下の詳細な説明から容易に理解されるであろう。
【0014】
本発明は、核酸増幅反応における単純ヘルペスウイルス(HSV)の型の特異性を示す、単離され精製された核酸、ポリヌクレオチド、増幅プライマーおよびアッセイプローブを提供する。本発明のプローブおよびプライマーを用いてHSV核酸を検出および同定する方法も提供する。
【0015】
本発明の一実施形態は、標的核酸配列の存在を検出する増幅方法であって、標的結合配列を有する1以上の増幅プライマーを用い、増幅された標的配列を産生し、この標的配列を検出する増幅方法に関する。増幅方法の非制限的な例には、ポリメラーゼ連鎖反応(PCR;Saiki et al., 1985, Science 230: 1350-1354(非特許文献2)参照。参照文献として本明細書に取り込む)、リガーゼ連鎖反応(LCR;Wu et al., 1989, Genomics 4: 560-569; Barringer et al., 1990, Gene 89: 117-122; Barany, 1991, Proc. Natl. Acad. Sci. USA 88: 189-193(非特許文献3、4、5)参照。この全てを参照文献として本明細書に取り込む)、in situ ハイブリダイゼーション、転写介在増幅手法(TMA、Kwoh etal., 1989, Proc. Natl. Acad. Sci. USA 86 : 1173-1177(非特許文献6)参照。参照文献として本明細書に取り込む)、自己持続性配列複製法(Self-Sustaining Sequence Replication(3SR、Guatelli et al., 1990, Proc. Natl. Acad. Sci. USA 87: 1874-1878(非特許文献7)参照。参照文献として本明細書に取り込む)、ローリングサークル増幅法(Rolling Circle Amplification)(RCA)、核酸配列をベースとする増幅法(Nucleic Acid Sequence Based Amplification)(NASBA)、Qβレプリカーゼシステム(Lizardi et al., 1988, BioTechnology 6: 1197-1202(非特許文献8)。参照文献として本明細書に取り込む)、および、好熱性SDA(tSDA)を含めた鎖置換増幅法(SDA、Walker et al., 1992, Proc. Natl. Acad. Sci. USA 89: 392-396; Walker et al., 1992, Nuc. Acids. Res. 20:1691-1696(非特許文献9、10)およびEP 0 497 272(特許文献1)この全てを参照文献として本明細書に取り込む)が含まれる。
【0016】
本発明の他の実施形態は、HSV標的配列の指数関数的増幅によってサンプル中のHSV核酸配列の存在を検出する鎖置換増幅(SDA)法に関する。さらなる実施形態では、SDAは、選択された検出(detector)プライマーを用いて、米国特許第5,648,211号明細書(特許文献2)に記載されるように約52℃で実施され、米国特許第5,919,630号明細書;同5,928,869号明細書;同5,958,700号明細書および同6,261,785号明細書(特許文献3〜6)に記載されるように増幅の間に標的を検出する(これらの文献は全て、参照により本明細書に取り込む)。SDAで一般的なように、制限酵素およびポリメラーゼのような試薬、プライマー、酵素、および他の成分を反応マイクロウェル、容器、または、レセプタクルに添加する。SDAはサンプルからDNA配列を増幅するが、この方法では、一度全ての成分を互いに混合し、重要な成分が使い尽くされるまで反応を継続する。ポリメラーゼ連鎖反応(PCR)とは対照的に、SDAは等温反応プロセスであり、従って、一度反応が開始されると反応の進行中に外部から制御をしない。
【0017】
本発明のSDA法は、増幅を開始するのに少なくとも2種類のHSV増幅プライマーおよび2種のバンパープライマーを必要とする。増幅プライマーはHSV−1またはHSV−2に高度に特異的であるように設計されている。SDA法は、混合物中での同時増幅反応を含み、PCR増幅法で必要な温度サイクルのための別々のフェーズとサイクルを必要としない。本発明のSDAのさらなる利点は、指数関数的な増幅である。DNAポリメラーゼ伸張、ニッキング、置換およびニック部位の再生のステップが、どちらかの末端に部分的制限酵素部位(例えばBsoBI部位)を持つ、置き換えられた単鎖分子で起こり、この分子は続いて、培養され、増幅プライマーによって捕捉され、これによって、HSV標的配列が指数関数的に増幅される。SDA法はまた、特に高スループット法に対するワークフローを改善する。SDAは、マイクロアレイをベースとする適用例に組み込まれうる。この適用例では、小容積量(ナノリットル)のサンプルおよび試薬を使用して、単一のプラットホームで複数のSDAアッセイを実施することにより、HSV標的DNAを増幅し、マイクロチップアレイ上で増幅サンプルを検出する。等温増幅プロセスには、このプラットホームの設計および維持における技術的難題が極めて僅かしか存在しないので、サンプル中のHSVを検出するためのSDA法の主たる利点は、労働力の必要性を最小限にすることと、高スループットの可能性である。
【0018】
本明細書で使用する用語「標的」または「標的配列」とは、増幅および検出されるHSV核酸配列、HSV−1またはHSV−2をいう。これらには、増幅される本来のHSV核酸配列、増幅される本来のHSV核酸配列の相補性第二鎖、および増幅反応で産生される本来のHSV配列のコピーのどちらかの鎖が含まれる。これらのコピーは、増幅プライマーがアニールする配列のコピーを含有するので、増幅可能な標的として機能する。増幅反応中に生成される標的配列のコピーは、増幅産物、アンプリマーまたはアンプリコンと称される。HSV−1およびHSV−2標的配列は、HSV−1およびHSV−2のゲノム配列の遺伝子の糖タンパク質G(US4)に局在化している。HSV−1標的配列は、図1のコンセンサス配列の555から680塩基の間に位置する。HSV−2標的配列は、図6のコンセンサス配列の867から990塩基の間に位置する。糖タンパク質G(US24)遺伝子は、図2および図7のHSV−1ゲノム配列の136744から137460の間におよびHSV−2のゲノム配列の137878から139977の間の位置にそれぞれ位置する。
【0019】
本明細書で使用する「増幅プライマー」は、標的配列にアニールし、増幅によって伸張されうるプライマーである。標的配列に結合する増幅プライマーの領域は標的結合配列である。増幅技術には、好熱性SDA(tSDA)を含めた鎖置換増幅(SDA)、ポリメラーゼ連鎖反応(PCR)、リガーゼ連鎖反応(LCR)、in situ ハイブリダイゼーション、自己持続性配列複製法(Self-Sustaining Sequence Replication(3SR))、ローリングサークル増幅法(Rolling Circle Amplification)(RCA)、核酸配列をベースとする増幅法(Nucleic Acid Sequence Based Amplification)(NASBA)、および転写介在増幅手法(TMA)が含まれるが、これらに限定されない。
【0020】
一実施形態では、増幅プライマーは鎖置換増幅(SDA)法に使用される。増幅プライマーは、3’末端にHSV標的配列に結合する標的結合配列部分を含み、5'末端に標的配列に結合またはアニールしない部分を含む。標的配列に結合しないSDA増幅プライマーの部分はまた、米国特許第5,455,166号明細書(特許文献7)および米国特許第5,270,184号明細書(特許文献8)(これらは参照文献として本明細書に取り込む)に記載されているような、標的結合配列の上流の、テール、および制限エンドヌクレアーゼに対する認識部位を含む。この認識部位は、Walkerら(非特許文献9および10)に記載されているように、この認識部位が半修飾(hemimodified)されている場合に、DNA二本鎖の一方の鎖にニックを入れるエンドヌクレアーゼに特異的である。テールは、制限エンドヌクレアーゼ認識部位の配列の上流にあり、増幅プライマーの残りの部分がSDA中にニックを入れられ、置換されるときに、ポリメラーゼ再プライミング部位として機能する。テールの再プライミング機能は、SDA反応を維持し、単一の標的分子から多数のアンプリコンの合成を可能にする。テールの長さおよび配列は一般に重要ではなく、型通りに選択でき、且つ修飾することができる。
【0021】
本発明の一実施形態は、増幅プライマーに標的特異性を付与する標的結合配列に基づく。ここで、本発明で例示される標的結合配列は、HSVの検出のための他の種々の方法に使用できることも理解されるべきである。例えば、他の選択肢として、本明細書に開示されている標的結合配列は、事前の増幅を用いずに、または増幅後のアッセイにおいてHSVの直接の検出のためにハイブリダイゼーションプローブとして使用されうる。このようなハイブリダイゼーション法は当分野で周知であり、典型的には、ハイブリダイゼーションの検出を促進するために標的結合配列と会合された、またはこれに連結された検出可能な標識を使用する。さらに、表1および表2にプライマー配列(それぞれ配列番号:5〜25および36〜47)を列挙したが、これには大文字を使用しておよびアンダーラインを引いて示した標的結合配列が含まれている。これらの標的結合配列は、追加の特化された配列を必要としない増幅反応(例えばPCR)のプライマーとして使用できるか、または、NASBA、in situハイブリダイゼーション、TMA、3SR、プライマーの標的結合配列に連結されたRNAポリメラーゼプロモータを必要とする増幅プライマーをベースとする他の転写、または任意の他のプライマー伸長増副反応に追加されうる。プライマー中の特化された非標的結合配列を必要とするこれらの増幅法は、増幅反応を進行するのに必要であり、典型的には、標的配列に特化した配列を追加するように機能する。
例えば、制限酵素認識部位は、SDAで起こる指数関数的増幅に必要である(米国特許第5,455,166明細書および同5,270,184号明細書(特許文献7、8)参照)。対照的に、自己持続性配列複製法(3SR)および核酸配列をベースとする増幅法(NASBA)は、5’末端近傍にポリメラーゼプロモータを含む(3SRアッセイは、Guatelli et al., 1990, Proc. Natl. Acad. Sci. USA 87: 1874-1878(非特許文献7)に開示されている。)。このプロモータは、標的結合配列に追加され、テンプレートの複数のRNAコピーの転写を指示することによって増幅反応を推進する働きをする。選択された増幅反応に使用するために、このような特化した配列を標的結合配列に連結することは、通常行われることであり、当業者に周知である。
【0022】
対照的に、標的の末端に特化された配列を必要としないPCRのような増幅方法は、一般に、標的結合配列のみからなる増幅プライマーを使用する。これらの他の増幅方法における検出の目的に対しては、当業者によって理解されるように、プライマーが検出可能に標識されうる。
【0023】
核酸はアニールするためには完全な相補性を必要としないので、当業者は、本明細書に開示されたプローおよびプライマー配列がHSV−1およびHSV−2特異的プライマーおよびプローブとしての有用性を損なうことなくある程度まで修飾できることを理解するであろう。用語「相同性」は相補性の程度を言う。部分的相同性または完全相同性があり得、完全相同性は同一と同義である。同一の配列を標的核酸へハイブリダイゼーションするのを阻害する部分的相同性配列は、「実質的に相同性である」と称される。標的配列への完全に相補的な配列のハイブリダイゼーションの阻害は、低ストリンジェンシー条件下でハイブリダイゼーションアッセイ(例えばサザンまたはノーザンブロット、溶液ハイブリダイゼーションなど)を用いて試験することができる。実質的に相同性である配列またはプローブは、低ストリンジェンシー条件下で標的配列への、完全に相補的な配列またはプローブの結合(すなわちハイブリダイゼーション)に競合し且つこの結合を阻害する。それにもかかわらず、低ストリンジェンシー条件は、非特異的結合を許さず、低ストリンジェンシー条件は、2つの配列の他方への結合が特異的(すなわち選択的)相互作用であることを必要とする。
【0024】
当業者によって理解されるように、アニーリングのストリンジェンシーは同一または関連するポリヌクレオチド配列を同定または検出するために変更されうる。熟練した専門家によってさらに理解されるように、融解温度、Tmは、ヌクレオチドの数でのプライマーまたはプローブの長さ、またはアニーリングバッファの成分および条件のような多くのパラメータに依存して、当分野で公知のように式で近似することができる(例えば、T. Maniatis et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, 1982およびJ. Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, 1989; Current Protocols in Molecular Biology, Eds. F. M. Ausubel et al., Vol. 1, "Preparation and Analysis of DNA", John Wiley and Sons, Inc., 1994-1995, Suppls. 26, 29, 35 and 42; pp. 2.10.7-2.10.16; G. M. Wahl and S. L. Berger (1987; Methods Enzymol. 152: 399-407);およびA. R. Kimmel, 1987; Methods of Enzymol. 152: 507-511(非特許文献11〜15)参照)。一般的指針として、Tmは、配列相同性が1%減少するごとに約1℃から1.5℃低下する。温度範囲は約50℃から62℃で変化しうるが、増幅プライマーは52℃で最適化されるように設計されうる。しかし、50℃未満の温度はプライマーに特異性を欠如させ、一方62℃を超える温度はハイブリダイゼーションを引き起こさない。増幅プライマーを設計する場合にさらに考慮することは、グアニンとシトシンの含有量である。一般には、プライマーに対するGC含有量は、約60〜70%であるが、これはより少なくてもよく、当業者によって適切に調節することができる。標的結合配列のハイブリダイゼーション領域は、約42〜48℃のTmを有しうる。アニーリングの相同性および部分相同性核酸配列は、アニーリングの条件を変更してストリンジェンシーを増加または減少すること(すなわち、アニーリング温度またはバッファの塩の含有量を調節すること)によって得ることができる。このような開示された配列のマイナーな変更、およびHSV−1およびHSV−2の特異性を維持するための必要なアニーリング条件の調節には、決まりきった実験手順のみが必要であり、そのような変更および調節は当業者の範囲内にある。
【0025】
HSV−1およびHSV−2標的配列の検出用に設計された増幅プライマーは、それぞれ、配列番号7〜18および38〜43として表1および2に同定されている。これらの増幅プライマーは、標的配列が高相同性コンセンサス糖タンパク質G(US4)遺伝子領域にアニールするように設計されている(図1〜2および6〜7参照)。HSV標的DNA配列にアニールするか、またはこれらに対して相補的である増幅プライマー内のHSV標的結合配列領域は、アンダーラインを付し、大文字で示した(表1および2参照)。SDA検出プライマー配列の残っている5’部位は、SDA反応が進行するのに必要であるBsoBI制限エンドヌクレアーゼ認識部位(RERS)(小文字のイタリック体で示した)、並びに、ジェネリックな非標的特異的5’テール末端配列を含む。
【0026】
それぞれ配列番号7〜8および38のHSV−1およびHSV−2増幅プライマーは左側(「第一」)S1増幅プライマーであり、それぞれ配列番号9〜18および39〜43は右側(「第二」)S2増幅プライマーである。増幅の目的では、少なくとも1つの左側および右側プライマー対が反応中に存在するように、特異型の一対のHSV増幅プライマーを単独(すなわち、1つのHSV−1左側増幅プライマーおよび1つのHSV−1右側増幅プライマー)で使用するか、または組み合わせて(すなわち、1つのHSV−1SDA左側プライマーと2つのHSV−1SDA右側増幅プライマー)を使用する。多数の増幅プライマーを使用して、標的配列の幾つかの領域を増幅することができる。プライマーの濃度は、HSV−1の第1増幅プライマーが500nMの濃度で唯一の第一増幅プライマーとして使用され、2つのHSV−1右側増幅プライマーが併用して使用される場合には、各々250nMの濃度を有するように適切に調節されうる。
【0027】
用語「伸長生成物」は、一般に、ポリメラーゼのような酵素を用いてプライマーまたは標的配列を伸長することによって産生される配列を言う。一実施形態では、増幅プライマーのハイブリダイゼーションおよびテンプレートとしてHSV標的配列を用いるポリメラーゼによる増幅プライマーの伸長は、増幅プライマー伸長生成物を産生する。
【0028】
「バンパープライマー」または「外部プライマー(external primer)」は、バンパープライマーが下流の増幅プライマーおよびその伸張生成物を置換するように増幅プライマーの上流で標的配列にアニールするプライマーである。本明細書で使用する場合、用語「バンパープライマー」は、HSV標的結合配列を含むポリヌクレオチドをいう。有用なバンパープライマーは、表1および2に、それぞれ配列番号23〜25および46〜47として同定されている。左側または第一のHSV−1およびHSV−2バンパープライマーは、それぞれ配列番号23および46であり、一方右側または第二のHSV−1およびHSV−2バンパープライマーはそれぞれ配列番号24〜25および47である。バンパープライマーは、増幅プライマーの標的結合部位に十分近接し、バンパープライマーの伸張後に増幅プライマー伸張生成物の置換を可能にする増幅プライマーの上流の位置で増幅プライマーの側面に位置する配列の保存領域から誘導される。例えば、配列番号23(HSV1GGLB1.0)のHSV−1第一バンパープライマーの5’末端はHSV−1ゲノム配列の塩基137,256に位置する(図2)。配列番号25(HSV1GGRB1.1)のHSV−1第二バンパープライマーの5’末端は、HSV−1ゲノム配列の塩基137,382に位置する。SDAの初期のラウンドの間に、バンパープライマーはHSV標的配列にハイブリダイズし、ポリメラーゼ伸張、すなわち下流の増幅プライマー伸張生成物によって置換され、複製および/または指数関数的増幅のさらなるラウンドを受けうる単鎖DNAの生成を引き起こす。
【0029】
用語「アッセイプローブ」は、核酸の検出または同定を促進するために使用される任意の核酸を言う。例えば、本発明の一実施形態では、アッセイプローブはHSV核酸の検出または同定に使用される。以下に記載される検出プローブ、検出プライマー、キャプチャープローブおよびプライマーはアッセイプローブの例である。
【0030】
特に、特異的なHSV−型を検出および同定するのに有用な「検出プローブ」は標識化されるかまたはタグ付けされる。検出プローブの検出可能な標識は、直接的または間接的に検出され、標的核酸配列の存在を示しうる部分である。直接検出については、アッセイまたは検出プローブは放射性同位体でタグ付けされ、オートラジオグラフィーで検出されるか、または、蛍光部分でタグ付けされ、当分野で公知の通りの蛍光によって検出する。あるいは、アッセイプローブは、検出を可能にする追加の試薬で標識することによって間接的に検出することができる。間接的に検出可能な標識には、例えば、化学発光剤、視覚化または着色された反応生成物をもたらす酵素、および、リガンドで検出可能に標識されたリガンド結合パートナーであって、リガンド(例えばハプテン、抗体または抗原)を標識されたリガンド−特異的結合パートナーへ結合することによって検出できるものが含まれる。
【0031】
増幅生成物の検出に対しては、本明細書で開示された標的結合配列を含む増幅プライマーが当分野で公知の方法で標識されうるか、または、開示された標識結合配列を含む標識された検出プライマーを、増幅のリアルタイム均一検出に対して米国特許第5,547,861、米国特許第5,928,869、米国特許第5,593,867号明細書、米国特許第5,550,025号明細書、米国特許第5,935,791号明細書、米国特許第5,888,739号明細書、米国特許第5,846,726号明細書(特許文献9〜15に開示された増幅プライマーと共に使用することができる。このような検出プライマーは直接的にまたは間接的に検出可能な配列を含み、この検出可能な配列は、初期には標的にハイブリダイズしないが、一旦標的にハイブリダイズして伸長されると検出プライマーの検出を促進する。例えば、このような検出可能な配列は、制限部位を含む配列、または、ヘアピンおよびg−カルテット配列(これらに限定されない)のような、近傍にフルオロフォアおよびクエンチャー部分を結合する二次構造を形成する配列、または、当分野で公知である標識されたオリゴヌクレオチド(しばしばレポータープローブと言われる)へ線状配列の相補体をハイブリダイゼーションすることによって検出される線状配列でありうる。あるいは、増幅生成物は、リアルタイムに、または、挿入色素(intercalating dye)の使用を介した増幅後に、もしくは選択した一連の増幅プライマーの範囲に入る本明細書で開示された任意の標的結合配列から選択されるプローブのハイブリダイゼーションによる増幅後に検出されうる。
【0032】
末端および内部標識法は当分野で公知であり、検出プライマーのそれぞれの位置でドナー色素およびアクセプター色素を連結するのに使用される。5'-末端標識法の例には、a)5’−対−5’カップリングしたリボヌクレオチドの過ヨウ素酸塩酸化とこれに続くアミン含有標識との反応、b)エチレンジアミンと5’−ホスホリル化ポリヌクレオチドの縮合とこれに続くアミン反応性標識との反応、およびc)固相DNA合成でアミノヘキシルホスファイト試薬を用いた脂肪族アミン置換基の導入とこれに続くアミン反応性標識との反応が含まれる。標識はまた、特別な脂肪族アミン含有ヌクレオチドホスホラミダイト試薬を用いて特定の位置で、合成DNAオリゴヌクレオチドに連結される。検出プライマーへ選択された標識を連結し、連結した試薬を働かせるための適切な方法を選択することは、当分野ではごく普通の手順である。
【0033】
他の実施形態は、検出される標的配列に依存して多数の検出プライマーを必要とする、特異的標的配列にハイブリダイズする検出プライマーを使用する。しかし、特異的HSV型の検出および同定のための実施形態では、Nadeauら(1999)によって記載されているリアルタイムSDA検出法を修飾したユニバーサル検出システム(Universal detection system)を使用する。ユニバーサル検出システムは、多数のアッセイに対して同じ蛍光検出プライマーの対を使用することができ、経費、時間、および技術的な複雑さを低減するような幾つかの利点を提供する。
【0034】
「シグナル」または「アダプタ」プライマーは、HSV標的配列にハイブリダイズする標的結合部分およびジェネリックでHSV標的配列に結合しないテール部分を有する。アダプタプライマーはユニバーサル検出用の検出プライマーと共に使用される。検出プライマーは、相補性アダプタプライマーのテール部分(すなわち、非標的結合配列)にハイブリダイズする。シグナルまたはアダプタプライマーは、第一および第二増幅プライマーの間の介在領域に少なくとも部分的に位置する標的配列の領域にハイブリダイズし、シグナルまたはアダプタ配列が増幅反応中に置換されるように設計される。配列番号19〜22および44〜45を有するHSV−1およびHSV−2シグナルまたはアダプタプライマーは、それぞれ表1および2に示されている。
【0035】
検出プライマーは、「ユニバーサル検出プライマー」または「検出プライマー」であってもよく、検出可能に標識された5'テール末端部分と、相補性アダプタプライマーテール配列に結合する3'末端部分を有する。一般に、検出プライマーの3’末端は、HSVまたは内部増幅対照(Internal Amplification Control)(IAC)標的配列に対する任意の十分な相補性を有する配列を含まない。検出プライマーはまた、5'末端に制限酵素認識部位を有する。
【0036】
手短には、このユニバーサル検出システムは、増幅のためのSDA法と同時に、且つ同じ反応容器中で使用することができる。ユニバーサル検出システムは、未標識のアダプタプライマーの標的依存的伸長を含む。アダプタプライマーは、HSV−1またはHSV−2標的特異的3'配列および5’ジェネリックテールを含み、それぞれ配列番号19〜22および配列番号44〜45と、その相補体で例示される。アダプタプライマーは、S1増幅プライマーの下流で、増幅されたHSV標的配列にハイブリダイズする。DNAポリメラーゼは、アダプタ配列およびS1増幅プライマーの3’末端から伸長し、この場合、増幅プライマーの伸長はアダプタプライマー伸長生成物を置換する。S2増幅プライマーは、アダプタプライマー伸長生成物にアニールする。DNAポリメラーゼはS2増幅プライマーの3’末端を伸長し、アダプタプライマー伸長生成物およびその相補体を含み、ニッキング可能な制限酵素認識部位を有する二本鎖分子を産生する。ニッキングとは、DNA二重鎖の2つの鎖のうちの1つのみのホスホジエステル結合を切断することを言う。対応する制限酵素は、制限酵素認識部位で二本鎖分子をニッキングし、ニッキングされた短いテールを含む5’部分と、ニッキングされた長い相補性アダプタプライマー伸長生成物を含む3’部分を生じる。BsoBI酵素のような対応する制限酵素で制限酵素部位をニッキングし、ニッキングされた部位から鎖を伸長すると、アダプタプライマー相補体の単鎖コピーが置換される。DHAポリメラーゼはニッキングされたテールの3’末端を伸長し、これによってニッキングされた単鎖の相補性アダプタプライマー伸長生成物を置換する。S1増幅プライマー伸長生成物および伸長されたHSV標的配列は、さらにSDAによって指数関数的に増幅される。次に、置換された相補性アダプタプライマー伸長生成物は検出プライマーによって捕獲されるが、この捕獲では、検出プライマーの3'末端が相補性アダプタプライマー伸長生成物の5’部分にアニールする。検出プライマーは、検出可能な標識を含み、標的配列を検出する。検出プライマーの3'末端からのDNAポリメラーゼ伸長および相補性アダプタプライマー伸長生成物は、存在する場合にはヘアピンを開き、検出プライマー伸長生成物とその相補体を含む二本鎖検出分子を産生する。各鎖は、開裂可能な制限酵素認識部位を含み、開裂されたときに、ドナーおよびクエンチャー色素を切り離し、フルオロフォアおよびクエンチャー部分を切り離し、標的特異的蛍光を生じる。この切り離しによって、クエンチャーはもはやフルオロフォアによって発光された蛍光を抑制することができない。二本鎖化された検出プライマー制限酵素認識部位の完全な開裂は、フルオロフォアとクエンチャーを切り離すことによって蛍光シグナルを増加させる。
【0037】
本発明の一実施形態では、検出プライマーは、蛍光ドナー部分(またはフルオロフォア)およびクエンチャー部分であって、各部分が制限酵素認識部位に隣接しているものにより蛍光検出用にタグ付けされうる。表1および2は、配列番号30〜35を有する検出プライマー配列を示す。ユニバーサル検出では、標的配列を検出するための検出プライマーが、一般にアダプタプライマーと共に使用される。ドナー色素、すなわちローダミン(ROX)およびクエンチャー色素、すなわちP−(ジメチルアミノフェニルアゾ)安息香酸(DABCYL)で標識された、配列番号30〜33を有する検出プライマーは、本発明でHSV標的配列の検出に使用される。他のドナーおよびクエンチャー色素対は、SDAで使用するために、クエンチャー色素がドナー色素によって放出される蛍光を十分に吸収するように、当業者によって容易に選択されうる。例えば、ドナーおよびクエンチャー色素は異なる波長での吸収によって容易に検出され、識別される。ドナーおよびクエンチャー色素に依存して、クエンチャー色素はある場合にはクエンチャーとして、そして他の場合にはドナー色素として作用してもよい。
【0038】
この実施形態では、配列番号30〜35の検出プライマーは、検出プライマーの5'末端に位置する制限酵素認識部位によって切り離されるドナーおよびクエンチャー色素対を有する。さらに、配列番号30の検出プライマーは、ドナーおよびクエンチャー部分の間に位置するヘアピン構造配列であって、制限酵素認識部位がこの中に存在するヘアピン構造配列を含む配列を有する。ヘアピン構造は、ドナー色素によって放出される蛍光がクエンチャー色素によって抑制されるように2つの色素を近接した位置に持ってくる。しかし、配列番号31〜35の検出プライマーは、2つの色素の間に線状の配列を有し、この線状の配列は、クエンチャーがフルオロフォアによって放出される蛍光を吸収するのに十分短い長さである。
【0039】
当分野で知られている多くのドナー/クエンチャー色素対が本発明の実施形態で有用である。これらには、フルオレセイン(FAM(商標);Glen Research; Sterling, VA)/ローダミン(ROX(商標);Molecular Probes;Eugene,OR);ROX/P−(ジメチルアミノフェニルアゾ安息香酸(DABCYL(商標);Glen Research);FAM/DABCYL;フルオレセインイソチオシアネート(FITC)/テトラメチルローダミンイソチオシアネート(TRITC);FITC/テキサスレッド(Molecular Probes);FITC/N−ヒドロキシスクシンイミジル1−ピレンブチレート(PYB);FITC/エオシンイソチオシアネート(EITC);N−ヒドロキシスクシンイミジル1−ペンタンスルホネート(PYS)/FITC;FITC/ローダミン X;およびFITC/テトラメチルローダミン(TAMRA)が含まれるが、これらに限定されない。特定のドナー/クエンチャー対の選択は、重要ではない。
【0040】
しかし、エネルギー移動消光メカニズムについては、ドナーフルオロフォアの放出波長がクエンチャーの励起波長とオーバーラップしていること、すなわち2つの色素の間で、十分なスペクトルのオーバーラップが存在し、十分なエネルギー移動、電荷移動または蛍光消光が可能であることのみが必要である。ROXはEMmax=608nmを有し、FAMはEMmax=520nmを有する。当業者は、適切なドナーおよびクエンチャー色素対を選択することにおいて豊富な知識を有するであろう。P−(ジメチルアミノフェニルアゾ)安息香酸(DABCYL)は、非蛍光性クエンチャー色素であり、隣接するフルオロフォア、例えばFAMまたは5−(2’−アミノエチル)アミノナフタレン(EDANS)からの蛍光を効果的に消光する。特定のドナー/クエンチャー対をこの開示で例示するが、他のものも当業者には明かであり、本発明でも有用である。消光を起こすメカニズムにかかわらず、本発明の検出プライマーにおいて蛍光の消光をもたらす任意の色素対が本発明の方法で使用するのに適している。他のクエンチャーの非制限的な例には、ブラックホールクエンチャー(Black Hole Quencher)(商標)(Biosearch Technologies, Inc.;Novato,CA)およびアイオワブラック(Iowa Black)(商標)(Integrated DNA Technologies,Inc.;Corralville,IA)が含まれる。
【0041】
蛍光は、特定の増幅生成物の蓄積をモニタするために核酸増幅反応過程中に測定される。蛍光シグナルは、産生された特定のアンプリコンの量に比例する。HSV標的核酸配列の存在下では、蛍光は増強されるであろう。標的がない場合、蛍光は反応の間一貫して低いままであろう。蛍光の増加、または実質的に蛍光の変化がないことは、それぞれHSV標的配列の存在および非存在を示す。
【0042】
サンプルの蛍光は、典型的には、サンプルがHSV DNAを含むかどうかを決定するために所定時間にわたって測定される。一実施形態では、蛍光は1時間の過程で60パスモニターされる。手短には、ほぼ1分ごとに、サンプル容器中で測定された蛍光の量、補正値(必要であれば)、および各カラムに対するキャーリブレータに関するデータを集める。データを、グラフの曲線の下の領域に換算した結果を表す「MOTA」(メトリック・アザー・ザン・アクセラレーション(Metric Other Than Acceleration))法を用いて分析することができる。グラフは、パスの数(X−軸)対相対蛍光単位(Y−軸)(図3参照)を測定する。MOTA領域が大きければ大きいほど、生成される蛍光は大きく、且つ増幅された生成物の検出の効率がよい。
【0043】
さらに別の実施形態では、図4に示されるパス・アフター・スレッシュホールド(Passes After Threshold)(PAT)アルゴリズムを使用し、これはBDプローブテック(BD ProbeTec)(商標)ETシステムで使用するために特に開発されている。MOTAと同様に、より高いPATスコアーは、SDA反応がより効果的なことを示す。PATアルゴリズムを用いる場合、バックグラウンドで集められた蛍光強度のシグナルが予め決められた閾値を越える時点をT3(「タイム−トゥ−スレッシュホールド(Time-To-Threshold)」と表す。このグラフも、パスの数対相対蛍光単位を測定する。同じT3閾値を各サンプルに対して使用する。PATスコアーは60マイナスT3の値に等しい。陰性サンプルは、蛍光の最低閾値に達しておらず、従ってゼロのPAT値が割り当てられる。陽性サンプルは、アッセイおよび標的のレベルに依存して、0よりも大きなPAT値、好ましくは1から60の間、より好ましくは40〜55の間のPAT値を有する。より低いT3スコアーおよび対応するより高いPAT値が、より効果的なSDAと相関する。PATアルゴリズムは、最も再現性のある増幅曲線の領域にのみ利用する。結果として、PATアルゴリズム法は、ウェル間またはサンプル間の認識できる差を最小限にし、検出物を比較する他の方法よりも正確である。PATはBDプローブテック(商標)ETシステムによって自動で行うことができる。BDプローブテック(商標)ETの出力は、PATスコアーおよび報告できる結果を提供する。
【0044】
さらなる実施形態では、「内部増幅対照」(「IAC」)は、陰性の結果を確認するため、および潜在的な阻害検体を特定するため、またはサンプル中に入り込んだ微生物、例えばウイルス、細菌、および菌類(これらに限定されない)の定量化を促進するために、存在している方法に取り込むことができる。診断への応用では、2つの異なるDNA配列、すなわちHSV標的配列およびIAC標的配列の同時増幅と検出がIACの使用を可能にする。「IAC標的配列」または「IAC配列」は、配列番号26〜27および配列番号48〜49のIAC標的配列がHSV−1およびHSV−2標的配列と比べて約5〜10塩基ミスマッチしていることを除いて、HSV標的配列と同じである。これらの修飾塩基がIACアダプタプライマーの特異的なアニーリングを可能にするのに十分である。
【0045】
「IACアダプタプライマー」は、IACアダプタプライマーがIAC結合配列を介して「IAC標的配列」または「IAC配列」にハイブリダイズすることを除いて、シグナルまたはアダプタプライマーと同じように機能する。IACアダプタプライマーはまた、IAC標的配列とハイブリダイズしないジェネリックな配列を含む5'テール部分を有する。どちらかと言えば、検出プライマーはIACアダプタプライマー相補体のテール部分にハイブリダイズすることができる。HSV−1およびHSV−2のSDAアッセイで使用されるIACアダプタプライマーは、それぞれ配列番号28〜29および50〜51から選択でき、IAC標的配列の増幅に有用である。IACアダプタプライマーの3’末端に位置するIAC標的結合配列は、HSV−1またはHSV−2アダプタプライマーがハイブリダイズしないか、またはIAC標的配列の増幅を妨げるように十分にHSV標的配列と異なっている。表1および2において、IACアダプタプライマーの3’末端のIAC標的結合配列は小文字にアンダーラインを付すことによって示した。IACアダプタプライマーは、陰性の結果を確認し、反応を阻害する検体をモニタするのに有用である。定量SDAでは、IACと陰性の標的配列の間で律速試薬を競合させることも有用であるかもしれない(Nadeau, et al., 1999 Anal. Biochem. 276:177-187(非特許文献16)。
【0046】
本発明の検出プライマーは、HSV標的配列またはIAC標的配列のいずれかを検出するのに使用することができる。しかし、本発明の一実施形態では、HSV−1またはHSV−2標的配列を検出するために使用される検出プライマーは、配列番号30〜33のものである。IAC標的配列を検出するのに有用な検出プライマーは、ドナーおよびクエンチャー色素対がそれぞれフルオレセイン(FAM)およびDABCYLである配列番号34〜35から選択されるものであり、本明細書で「IAC検出プライマー」と称される。当業者は、IAC標的配列の同定がHSV−1またはHSV−2標的配列の検出から明確に区別できるように、標識を有する適切な検出プライマーを選択する知力を有するであろう。従って、HSV標的配列およびIAC標的配列の検出に使用される検出プライマーは交換することができ、その結果、配列番号30〜33の検出プライマーがIAC標的配列の検出に使用でき、配列番号34〜35がHSV標的配列の検出に使用できる。
【0047】
本発明の他の実施形態は、高スループットプロセスで、同時に複数のサンプルをアッセイすることに関する。サンプルには、脳脊髄液(CSF)、生殖器病変、口腔病変、粘膜病変、眼の病変、皮膚病変、直腸スワブ、膣スワブ、膣分泌物、尿、末梢血白血球および組織(例えば脳生検からのもの)から集めたものが含まれるが、これらに限定されない。サンプルは、プレート、スライド、ウェル、皿、ビーズ、粒子、カップ、ストランド、チップおよびストリップでアッセイすることができる。一実施形態では、この方法は、BDプローブテック(商標)ET GT/GCで増幅されたDNAのアッセイで使用するのに適合した形態の96マイクロウェルプレートで行われる。この方法は、乾燥されたマイクロウェルの形態であって、乾燥された組成物が多数のサンプルを同時にアッセイするのに使用するための、HSV−1またはHSV−2のSDA検出を行うのに必要な全てのプライマーおよびプローブを含む形態で実施される。
【0048】
本発明の方法によって選択された標的配列の存在を検出するアッセイは、溶液または固相で行うことができる。検出核酸がプライマーとして機能する、リアルタイムまたは終点均一アッセイ(endpoint homogeneous assay)は、典型的には、溶液で実施される。本発明の検出プライマーを用いるハイブリダイゼーションアッセイは、溶液で行うことができるだけでなく、標的のリアルタイムまたは終点検出のための固相アッセイに特に適切である。固相アッセイでは、検出オリゴヌクレオチドを、当分野で公知の方法を用いて内部または末端標識を介して固相(例えば、ビーズ、膜または反応容器)に固定する。例えば、ビオチン標識された検出オリゴヌクレオチドは、適切なハイブリダイゼーション条件下で標的に曝したときに蛍光の変化をもたらすアビジンで修飾された固相上に固定することができる。この方法では、標的の捕獲はサンプルから標的の分離を促進し、アッセイのシグナルまたは他の特徴の検出を妨げうるサンプル中の物質を除去することができる。
【0049】
HSV−1またはHSV−2標的配列を検出し同定するために使用されるプライマーおよびプローブを表1に列記した。特異的HSV標的結合配列にはアンダーラインを付し、大文字で示し、一方、制限酵素エンドヌクレアーゼ部位は小文字のイタリック体で示した。IACアダプタプライマーについては、IAC標的結合配列を小文字にアンダーラインを付すことにより示した。全てのプライマーは、5’→3’の方向に位置づけされている。
【0050】
【表1−1】

【0051】
【表1−2】
【0052】
HSV−2標的配列を検出し同定するために使用されるプライマーおよびプローブを表2に列記した。
【0053】
【表2−1】
【0054】
【表2−2】
【0055】
本発明の核酸プライマーは、種々の系統についてのHSV遺伝子の糖タンパク質G(Glycoprotein G)(US4)を分析することによって生じるコンセンサス配列に基づいて設計される(図1および6、表1および2参照)。SDAおよびユニバーサル検出法で使用するためのバンパープライマー、アダプタプライマー、および検出プライマーも示されている。設計されたHSV−1プライマーは、表4に例示されるように全ての系統に認められるHSV−1標的配列を特異的に増幅する。HSV−2プライマーは、表7に例示されるように全ての系統に認められるHSV−2標的配列を特異的に増幅する。HSV−1とHSV−2標的配列の間の相同性は約90%であるので、プライマーは、HSV−1およびHSV−2の間を特異的に明確に区別するために注意深く設計される。標的結合配列に実質的に相同である配列および表1および2に列挙されたこのような実質的に相同な標的結合配列を含むプライマーもまた本発明に包含される。
【0056】
本発明の一実施形態では、HSV−1標的領域は、まず、ヒトHSV−1の完全なHSV−1ゲノム配列、すなわち152,261塩基長を有する系統17(NCBIアクセス番号X14112)から選択される。糖タンパク質「US4」遺伝子は、136,744〜137,460塩基に位置する。HSV−1左側バンパープライマー(HSV1LB1.0)(5'末端)は、核酸137,256に位置する。HSV−1右側バンパープライマー(HSV1RB1.1)(5'末端)は、核酸137,382に位置する。全てのHSV−1SDAシステム用のプライマーは、これらのバンパープライマー座標(coordinates)の範囲内に位置する。
【0057】
本発明の他の実施形態はヒトHSV−2の完全なHSV−1ゲノム配列、すなわち154,746塩基長を有する系統HG52(NCBIアクセス番号Z86099)に関する。糖タンパク質G「US4」遺伝子は、137,878〜139,977塩基に位置する。HSV−2左側バンパープライマー(HSV2LB1.0)(5'末端)は、位置139,773に位置する。HSV−2右側バンパープライマー(HSV2RB1.0)(5'末端)は、位置139,896に位置する。全てのHSV−2SDAシステム用のプライマーは、このバンパープライマー座標(coordinates)の範囲内に位置する。
【0058】
PCR増幅プライマーは、HSV標的DNAをプラスミドベクターへクローニングするために設計される。配列番号5〜6および配列番号36〜37のHSV−1およびHSV−2PCR増幅プライマーは、それぞれ、HSVゲノムの高度に保存された標的配列領域に対して相補的である。PCR増幅プライマーは、HSVの糖タンパク質G(US4)遺伝子のDNAフラグメントを含むHSV標的配列領域を増幅する。HSV標的領域を含む単純ヘルペスウイルス(HSV)ゲノムの増幅されたフラグメントは、手頃な制限酵素部位を含むプラスミドベクターに指向性を持ってクローニングされる。HSVフラグメントは、当業者によって理解されるように任意のプラスミドベクターにクローニングすることができるが、一実施形態では、増幅されたHSV−1およびHSV−2フラグメントは、選択されたHSV標的領域に特異的なPCR増幅プライマーを用いて、Escherichia coliプラスミドベクター、pUC19(ジーンバンク/EMBLアクセス番号L09137)およびpUC18(ジーンバンク/EMBLアクセス番号L09136)にそれぞれクローニングされる。HSVフラグメントはHSV標的ストックと称される。標的HSV DNAは、ピコグリーン(PicoGreen)(登録商標)二本鎖DNA定量アッセイ(Molecular Probes,Inc.)を用いて定量されうる。表1および表2に列記されたプライマーの名前の中の「L」または「R」の存在は、増幅反応に使用されたときに、それぞれ「左側」または「右側」プライマーを示す。
【0059】
本発明の一実施形態では、PCR増幅プライマー、配列番号5〜6および36〜37は、最初に、それぞれHSV−1およびHSV−2遺伝子の糖タンパク質G遺伝子の152および254塩基対フラグメントを増幅する。それぞれ配列番号5および36のHSV−1およびHSV−2左側PCRプライマーは、各々EcoRI制限酵素部位を含めて設計される。それぞれ配列番号6および37のHSV−1およびHSV−2右側PCRプライマーは、各々BamHI制限酵素部位を有する。このフラグメントは、次に、適切な位置でpUCプラスミドベクターにクローニングされる。例示されたプラスミドベクターは、HSV−1およびHSV−2に対してそれぞれpUC19およびpUC18であり、これらは制限酵素部位EcoRIおよびBamHIを有する。制限酵素消化によって精製および線状化した後、続いて、HSV標的フラグメントをHSV増幅プライマーおよびバンパープライマーを用いて指数関数的に増幅する。
【0060】
本発明の標的結合配列およびプライマーは核酸増幅に有用である。一実施形態では、プライマーは鎖置換増幅(SDA)に特に有用である。これは、プライマーの伸長、半修飾制限エンドヌクレアーゼ認識/開裂部位のニッキング、単鎖伸長生成物の置換、伸長生成物(または本来の標的配列)へのプライマーのアニーリング、および引き続きのプライマーの伸長が反応混合物中で同時に起こる等温の核酸増幅法である。さらに、SDAは15分未満で1010倍を超える標的配列の複製を可能にする。一方、PCRでは、反応のステップは反応中の温度サイクルの結果として、別々のフェーズおよびサイクルで起こる。好熱性鎖置換増幅(tSDA)は、本質的に、熱に安定なポリメラーゼおよび熱に安定な制限エンドヌクレアーゼの置き換えを伴う、本明細書で、および、walkerら(1992, Proc. Natl. Acad. Sci USA. USA 89:392-396および1992, Nucl. Acids Res. 20:1691-1696(非特許文献9および10))によって記載された従来のSDA方法として実施することができる。温度は、置換された酵素に適したより高い温度に調整することができる。
【0061】
HSV増幅生成物または増幅された標的配列を検出する別の方法は、ポリアクリルアミドまたはアガロースゲル電気泳動であって、アガロースが臭化エチジウムで染色されるものによって特徴的なサイズを検出することによるものである。HSV−1またはHSV−2増幅プライマーを用いて生成された増幅生成物は、定量的ハイブリダイゼーション、または分子生物学の分野で熟練した者に公知の核酸検出のための等価な技術(Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring, NY (1989)(非特許文献12))によって検出することができる。
【0062】
表1および2に列記したプライマーは、サンプル中のHSV−1およびHSV−2の検出および同定に有用である。本明細書で使用される場合、S1およびS2増幅プライマーは、それぞれ第一および第二の増幅プライマーを表し、一方B1およびB2バンパープライマーは、それぞれ第一および第二のバンパープライマーを表す。手短には、SDA法では、S1増幅プライマーは単鎖HSV標的配列にハイブリダイズする。S1増幅プライマーのちょうど上流または5’で、第一のバンパープライマー、B1、が単鎖HSV標的配列にハイブリダイズする。DNAポリメラーゼは、B1バンパープライマーおよびS1増幅プライマーの3'末端を伸長するが、この場合、B1バンパープライマーの伸長は、最終的に、S1SDA伸長生成物を置換する。S1SDA伸長生成物は、S2増幅プライマー、およびS2増幅プライマーの上流でアニールするB2バンパープライマーによって捕獲される。DNAポリメラーゼはS2SDAおよびB2バンパープライマーの3’末端を伸長し、この場合、B2バンパープライマーの伸長は、下流のS2SDA増幅プライマー伸長生成物を置換する。S1増幅プライマーは置換されたS2増幅プライマー伸長生成物にアニールし、DNAポリメラーゼはハイブリダイズしたS1増幅プライマーの3’末端を伸長し、S2増幅プライマー伸長生成物およびその相補鎖を有する二本鎖分子を生じる。各鎖は、いずれかの末端でニック可能な制限酵素認識部位を有する。対応する制限酵素を添加して、チオレート化されたシトシンを含む修飾されたDNA鎖をニッキングし、ニック部位の3’に、短いニッキングされたテールおよび長い伸長産物を形成する。DNAポリメラーゼは、5’→3’方向に、ニッキングされた短いテールの3’からニッキングされた短いテールを伸長し、長い単鎖伸長生成物を置換する。手短には、S2増幅プライマー伸長生成物のニッキングされたテールとその相補体のニッキングされたテールは、それぞれ、ニッキングされた単鎖S2増幅プライマー伸長生成物と、ニッキングされた単鎖相補性S2増幅プライマー伸長生成物を置換する。一実施形態では、BsoBI酵素を使用して、配列番号52〜53および54〜55の配列をそれぞれ有する各鎖をニッキングし、且つ開裂または切断する。以下に示されるニック部位は、増幅プライマー配列に組み込まれ、半ホスホロチオエート化された認識配列(dCsTP、チオエート化されたシトシン)を必要とする。ニック部位であっても、配列番号53は、dCsTPの存在下でさえ、二本鎖開裂をする傾向があり、ニッキング可能な増幅プライマーを設計する場合には好ましい配列ではない。
【0063】
ニック部位:5’−CTCGGG−3’(配列番号52)および5’−CCCGGG−3’(配列番号53)
切断部位:5’−CTCGAG−3’(配列番号54)および5’−CCCGAG−3’(配列番号55)
【0064】
本発明のさらなる実施形態では、検出プライマーはHSV標的配列を検出するのに有用である。HSV標的配列に特異的なS1増幅プライマーおよび検出プライマーを使用することができ、この場合、検出プライマーはHSV標的結合配列を有する。DNAポリメラーゼは、S1プライマーおよび検出プライマーの3‘末端から伸長する。S1プライマーの伸長は、下流の検出プライマー伸長生成物を溶液中に置換し、この場合、これは捕獲されて相補性S2増幅プライマーにハイブリダイズする。DNAポリメラーゼは、S2増幅プライマーの3’末端を伸長し、検出プライマーの二次構造を開き、二本鎖制限酵素部位を形成し、2つの色素をクエンチャーの消光能力を無能力化して蛍光を生じるような距離まで引き離す。追加の蛍光は、制限酵素認識部位を開裂し、フルオロフォアとクエンチャーをさらに引き離すことによって生じる。
【0065】
SDA法に有用な酵素は、半ホスホロチオエート化認識配列に単鎖ニックを生じさせるものであり、この場合、ホスホロチオエート化された核酸の取り込みはニッキングおよび修復のさらなるラウンドを妨げない。これらの特徴を有する酵素の非制限的な例には、HincII、BsoBI、AvaI、NciI、およびFnu4HIが含まれる。有用なDNAポリメラーゼは、単鎖ニック部位でDNA合成を開始し、伸長した核酸鎖にホスホロチオエート化されたヌクレオチドを組み込み、そして5‘→3’エキソヌクレアーゼ活性を持たない鎖を置換するものである。開裂は、二本鎖または単鎖DNAのホスホジエステル結合の切断をいう。これらの特性を示すDNAポリメラーゼの非制限的な例には、エキソヌクレアーゼ欠損クレノウ、およびBstポリメラーゼおよびBcaポリメラーゼのエキソヌクレアーゼ欠損フラグメントが含まれる。他のDNAポリメラーゼおよび制限酵素はSDAに適している(Walker et al. Proc. Natl. Acad. Sci USA, Vol. 89, pp. 392-396, January 1992, Applied Biological Sciences(非特許文献9))が、エキソ−BstポリメラーゼおよびBsoBIがこれらの熱特性および互いの適合性によって選択された。本発明の一実施形態では、BsoBI制限エンドヌクレアーゼ認識部位が使用され、これはイタリック体で示されている(表1および2参照)。HSV標的結合配列を単独で使用して、専用の配列または構造を必要とすることなく反応中にHSV標的を増幅できること、および、SDA以外の増幅反応(たとえばRNAポリメラーゼプロモータ)で必要とされる他の専用の配列が、このシステムで、たとえば本明細書に記載されているRERS含有配列に置き換えられることは容易に認識できるであろう。
【0066】
次に、標的ストックは、増幅プライマーの存在下で、増幅プライマー単独またはバンパープライマー、ユニバーサル検出用のシグナル/アダプタプライマー、およびユニバーサル検出プライマーと組み合わせて増幅される。増幅反応については、1つの「左側」増幅プライマーを含む少なくとも1対が選択され、1つの「右側」増幅プライマーが選択されて、HSV標的ストック配列の各鎖を増幅する。左側および右側増幅プライマーに加えて、SDA反応では、1つの左側および右側バンパープライマー対を最初に使用する。さらに、検出用に、シグナル/アダプタプライマーおよび検出プライマーを、HSV標的ストック配列を検出し、同定するために使用する。
【0067】
HSV−1またはHSV−2DNAを特異的に増幅し、検出するいくつかのHSVシステムが本発明に包含される。例えば、HSV−1システムは以下のプライマーを含む:HSV1GGLP1.1、HSV1GGRP5.2、HSV1GGAD2.1、HSV1GGLB1.0、HSV1GGRB1.1、およびTBD16(D/R)、または、そのほかには、HSV1GGLP1.1、HSV1GGRP5.2、HSV1GGAD3.0またはHSV1GGAD3.1、HSV1GGLB1.0、HSV1GGRB1.1、MPC.DR、HSV1IAC AD8.1またはHSV1IACAD8.7、MPC2.FD。他の実施形態では、プライマーの種々の組み合わせを用いたHSV−2システムを表3に列記した。プライマーの他の組み合わせが意図されるが、当業者は、サンプル中のHSV−1またはHSV−2を検出するためにプライマーを組み合わせる知力を有するであろう。プライマーは、表1および2に列記されたものから選択され、サンプル中のHSV−1またはHSV−2を同定するために統計学的に設計された実験で試験される。あるいは、HSV−1またはHSV−2に特異的であり、表1および2に列記されたものと実質的に相同であるプライマーをHSV−1またはHSV−2標的配列の検出に使用することもできる。
【0068】
【表3】
【0069】
商業的な利便性のためには、核酸の特異的な検出および同定のための増幅プライマーはキットの形態でパッケージングすることができる。一般にこのようなキットは、少なくとも1対のHSV増幅プライマーを含有する。核酸増幅反応を実施するための試薬、例えば、バッファー、追加のプライマー、ヌクレオチドトリホスフェート、酵素などを標的特異的増幅プライマーに含むことができる。キットの成分は、本発明の方法の特定の実施形態を実施するための使用説明書を含む、一般的な容器内に一緒にパッケージングされる。他の任意成分、例えばアッセイプローブとして使用するのに適した標識をタグ付けしたプライマー、および/またはその標識を検出するための試薬または手段をキットに含むことができる。
【0070】
本発明の一実施形態では、第一の増幅プライマーすなわちS1SDA増幅プライマー、および第二の増幅プライマーすなわちS2SDA増幅プライマーを含むキットを提供する。このキットは、第一のB1バンパープライマーおよび第二のB2バンパプライマー、検出プライマー、並びに、任意選択的に、IACアダプタプライマーおよびIAC標的配列を含む内部増幅対照(IAC)標的配列を同時に検出するための試薬をさらに含むことができる。キットは、HSV−1またはHSV−2に特異的なプライマーから構成されていてもよく、または、キットは、HSV−1およびHSV−2の両方を指向するプライマーから構成されていてもよい。この場合、当業者は、HSV−1を検出し同定する増幅反応がHSV−1プライマーを利用し、HSV−2を検出し同定する増幅反応がHSV−2プライマーを利用することを理解するであろう。サンプルがHSV−1またはHSV−2DNAを含有するかどうかを同定するために、HSV−1およびHSV−2のためのプライマーは混合されるべきではない。
【0071】
さらに別の実施形態では、本発明のキットおよびプライマーは、臨床サンプルがHSV−1またはHSV−2を含有するかどうかを検出し診断するために使用することができる。臨床サンプルは、SDA増幅プライマーを用いて増幅および検出することができるか、またはバンパープライマー、アダプタプライマー、および検出プライマーをさらに含むSDA反応に使用することができる。本発明の一実施形態では、IACアダプタプライマーは、HSV−1またはHSV−2に対する陽性および陰性対照に添加して反応の内部増幅対照として使用することができる。当業者は、本明細書の記載を読むことによって、および、当分野で一般的な方法および技術から、サンプル中のHSV−1およびHSV−2の検出および同定用のプライマーをどのように作成し使用するかを理解するであろう。
【0072】
さらに、商業的な実施形態では、本発明のプライマーおよびSDA用試薬を含む組成物を乾燥形態または液体形態で提供することができる。組成物は乾燥形態の場合により安定で容易に取り扱うことができる。乾燥された組成物は、マイクロタイタープレート、マイクロアレイ、または他の適切な容器のような固相に添加されるか、またはそのような固相にあらかじめ処理することができる。この場合、サンプルおよびSDAバッファを加えることのみが必要である。この形態は、多数のサンプルを同時にアッセイすることを容易にし、高スループット法に有用である。本発明の一実施形態では、BDプローブテック(商標)ET装置を使用することができる。
【0073】
標的結合配列、および、任意選択的に、選択された増幅反応に必要な配列または選択された検出反応に必要な配列からなる本発明に従った核酸が、スペーサー、リンカー、酵素の標識または結合のための配列、または他のものとして機能する他の特定の配列を含んでもよいことは理解されるべきである。このような追加の配列は、選択された反応中の核酸の最適な機能を得るために必要であることは一般に知られている。
【0074】
本明細書で引用した、すべての特許、特許出願、刊行されたPCT出願および物品、本、参照文献、参照マニュアルおよび要約は、本発明が関係する分野の状態をより完全に説明するためにその全体を参照により本明細書に取り込む。
【0075】
本発明の範囲および精神から逸脱することなく、上記の主題に種々の変更を行うことができるので、上記説明に含まれるかまたは添付の特許請求の範囲で定義されるすべての主題は本発明の記述および例示として解釈されることが意図される。本発明の多くの修飾および変更が上記教示に照らして可能である。
【実施例】
【0076】
本明細書の実施例は、本発明を実施する種々の側面を例示することを意味し、本発明の範囲をいずれの方法においても制限することを意図しない。実施例には、ベクターの構築またはそのようなベクターへのcDNAの挿入のような、使用される従来法の詳細な説明は含まれない。このような方法は、当業者に周知であり、多くの刊行物、例えばJ. Sambrook and D. W. Russell, Molecular Cloning: a Laboratory Manual, 3rd Edition, Cold Spring Harbor Laboratory Press, USA, (2001)(非特許文献17)に説明されている。
【0077】
(実施例1)
HSV−1糖タンパク質−G鎖置換増幅標的領域のクローニング
表1に同定されている配列番号5および配列番号6のPCR増幅プライマーを用いてHSV−1(系統ATCC:VR−539)からのDNAについてPCRを行った。これらのプライマーを使用して、HSV−1の糖タンパク質G(US4)遺伝子内の152塩基対フラグメントを増幅した。PCR増幅されたDNAをpUC19プラスミドベクター(Gibco BRL; Grand Island,NY)に挿入した。組み換えクローンをHSVIGGプラスミド#1と命名した。プラスミド#1DNAをBamH1制限酵素で消化することによって精製および線状化した。次に、DNAをQIAクイック(Qiagen,Inc.;Valencia,CA)スピンカラムを用いて精製し、蛍光ピコグリーン(登録商標)試薬で分析することによって定量した。将来の実験のために標的HSV−1DNAの希釈物を10ng/μlのヒトDNAを含む水中で調製した。特異的HSV−1系統の希釈物および各希釈物中のHSV−1検出の結果を図5に示した。「プラス」の記号はサンプル中にHSV−1の存在を示し、「マイナス」の記号は、サンプル中にHSV−1がないことを示し、疑問符はサンプルの汚染の疑いを示す。すべての系統は、サンプルO−2526を除いて、ストックの1:10希釈で陽性であった。ストックの1:1,000希釈で23系統のうち20系統が陽性であった。ストックの1:100,000希釈では、23系統のうち15系統が陽性であった。
【0078】
(実施例2)
クローニングされたHSV−1DNAの増幅
HSV−1DNAを検出するためのアッセイの最初のステップとして、実施例1で説明した標的核酸法を用いてHSV−1DNAの増幅のためのSDAシステムを開発した。DNA増幅アッセイの分析感度はクローニングされたプラスミドの希釈物を用いて評価した。8つの複製SDA反応を各標的レベルで実施した。これら8つの反応は、反応あたり100、50、25、12.5、6.25、および0コピーの二本鎖DNAに等しい。増幅は、BDプローブテックET装置(BD Diagnostic System;Sparks,MD)を使用し、各50nMのHSV1GGLB1.0およびHSV1GGRB1.0バンパープライマー、100nMのHSV1GGLP1.0左側増幅プライマー、500nMのHSV1GGRP1.0右側増幅プライマー、250nMのHSV1GGAD1.0アダプタプライマー、500nMのTBD10.2 D/R検出プライマーを用いて実施した。これらのプライマーの配列は表1に列記されている。最終のバッファ条件は、以下の通りである:143mMビシン(Bicine)、82mMKOH、25MMKiPO4、12.5%DMSO、5mM酢酸マンガン、500mM2’−デオキシシトシン−5−o−(1−チオトリホスフェート)(dCsTP)、それぞれ100nMのdATP、dGTPおよびdTTP、100ng/μlBSA、約12単位のBstポリメラーゼ、および約30単位のBsoBI制限エンドヌクレアーゼ。
【0079】
蛍光を、1時間の過程で60パスモニターした。結果を曲線の下の領域の換算、すなわち「MOTA」スコアーで表した。陽性のMOTAスコアーは、決まりきった実験によって容易に決定することができる。本発明の目的では、3500以上のMOTAスコアーを「陽性」とみなした。アッセイが100%の陽性の結果を与える最も低いレベルのHSV1GG標的DNAは反応あたり100コピーである。8つの複製のうちの7つ(87.5%)が反応あたり50コピーの標的DNAでも陽性であった。
【0080】
(実施例3)
SDAによる単純ヘルペス1ウイルス粒子の検出
HSV−1を検出するための本発明のプライマーおよびプローブの能力を確認するために、アメリカンタイプカルチャーコレクション(ATCC;Manassas,VA)から得たHSV−1の4系統、Quest Diagnostic(Baltimore,MD)から得た10系統、およびオハイオ州立大学(OSU)からの14の分類されていないHSVサンプルについてSDAを実施した。
【0081】
HSVの分類されていない系統は、PCRによってDNAポリメラーゼ遺伝子の領域を増幅することにより特徴づけた。1セットの増幅プライマーを設計して、HSV−1およびHSV−2の両方の同じ領域を増幅した。この2つのタイプのウイルスは、HSV−1の系統から生成されるPCR産物中には存在しないHSV−2PCRフラグメント中のApal制限エンドヌクレアーゼ認識部位の存在によって明確に区別できた。Apal制限酵素でインキュベートした場合、HSV−2由来の増幅生成物を2つの短いフラグメントに開裂し、一方HSV−1から得られるものは無傷のままである。制限されたフラグメントは、適切な対照とともにアガロースゲル電気泳動で解析した。
【0082】
SDAシステムでHSVの存在および非存在を評価するために使用されたウイルスストックの濃度は未知であった。ウイルスストックをリン酸塩緩衝食塩水で1:10に希釈し、この懸濁液の10μlをSDAで試験した。結果を表4に示す。HSV−1のすべての系統がHSV−1増幅プライマーを用いて検出され、多様な起源からのHSV−1の系統を検出するための、開示されたプライマーおよびプローブの能力が実証された。Apal制限消化物によって分類されたHSVの、以前には分類されていない14の系統のうち、9つがHSV−1であると決定され、4つがHSV−2であると決定され、1つがPCRによって増幅されなかった(表4および5参照)。
【0083】
【表4】
【0084】
(実施例4)
SDA法の分析感度
本発明で開示されたプライマーおよびプローブを使用したHSV−1アッセイの検出限界を決定するために、クローニングした標的核酸の希釈物およびウイルス粒子の一連の希釈物についてSDA反応を実施した。ウイルス粒子のストックを電子顕微鏡(Electron Microscopy Bioservices)で数えた。16の複製を各標的レベルで試験した。
【0085】
アッセイの感度および特異性を確認するために、種々の地理的な位置から得た23系統のHSV−1を、微生物ストックの1:10、1:1,000および1:100,000希釈で試験した。以前に分類されていない系統から、およびQuest Diagnositicsからのサンプルの力価は未知であった。ATCCからのHSV−1の2つのストックの力価はほぼ以下の通りであった:VR260、1.5×104TCID/μlおよびVR−539、2.0×105TCID/μl。結果を図5に示す。系統#0−2526を除いて、すべての系統がストック懸濁液の1:10希釈で陽性であった。23系統のうち、試験した20系統がストックの1:1000希釈で陽性であり、試験した15の系統が1:100,000の希釈で陽性であった。
【0086】
(実施例5)
HSV−1DNAに対するSDAの特異性
HSV−2の16の系統をHSV1GGSDAシステムで試験した。HSV−2希釈物の10ミリリットルの各懸濁液を反応あたり添加した。試験した17のストックのうちの1つがHSV1GGシステムで陽性であった。結果を表5に示す。加えて、23の他の微生物を、開示された本発明の方法のプライマーおよびプローブを用いて試験した。これらの微生物には、細菌、酵母および臨床標本中で遭遇するであろう他のウイルスが含まれる。試験された微生物のいずれもHSV−1に対して陽性ではなかった。結果を表6に示す。
【0087】
【表5】
【0088】
【表6】
【0089】
(実施例6)
HSV−2糖タンパク質−GSDA標的領域のクローニング
プライマーHSV2PCRRおよびHDV2PCRLを用いてHSV−2系統のATCC VR−540からのDNAについて、69℃のアニーリング温度でPCRを実施した。これらのプライマーは、HSV−2の糖タンパク質G(US4)遺伝子の範囲内の254塩基対フラグメントを増幅する。pUC18プラスミドベクター(invitrogen)に増幅したDNAを挿入した。組み換えクローンにはpHSV2−NT#9−1という名を付けた。プラスミドDNAを、BamHI制限酵素を用いて消化することにより精製し線状化した。DNAをQIAGEN QIAクイックスピンカラムを用いて精製し、蛍光ピコグリーン(登録商標)試薬による分析で定量した。将来の実験のための標的DNAの希釈物を7ng/μlのサーモン精子DNAを含有する水中で調製した。
【0090】
(実施例7)
クローニングされたHSV−2DNAの増幅
DNA増幅アッセイの分析感度はクローニングされたプラスミドの希釈物を用いて評価した。8つの複製SDA反応をステム2.0および5.2を用いて各標的レベルで実施した。これら8つの反応は、反応あたり500、250、100、50、25、10、および0コピーの二本鎖DNAに等しい。増幅は、BDプローブテックET装置を使用し、各50nMのHSV2GGLB1.0およびHSV2GGRB1.0、100nMのHSV2GGLP1.0、500nMのHSV2GGRP2.0もしくは5.2、250nMのHSV2GGAD1.0、500nMのMPC.D/Rを用いて実施した。これらのプライマーの配列は表2に列記されている。最終のバッファ条件は、以下の通りである:71mMビシン(Bicine)、56.6mMKOH、23.9mMKPO4、15.4%DMSO、5mM酢酸マンガン、500mM2’−デオキシシトシン−5−o−(1−チオトリホスフェート)(dCsTP)、それぞれ100nMのdATP、dGTPおよびdTTP、100μg/μlBSA、約3.515単位のBstポリメラーゼ、および約30単位のBsoBI制限エンドヌクレアーゼ。
【0091】
蛍光を、1時間の過程で60パスモニターした。結果を曲線の下の領域の換算、すなわち「MOTA」スコアーおよびPATスコアー(パス・アフター・スレッシュホールド)で表した。本発明の目的では、3500以上のMOTAスコアーおよび0を超えるPATスコアーを「陽性」とみなした。アッセイが100%の陽性の結果を与える最も低いレベルのHSV2GG標的DNAは、システム2.0および5.2の両方に対して反応あたり50コピーである。8つの複製のうちの7つが、システム2.0に対して25コピーで陽性であり、8つの複製のうちの6つが、システム5.2に対して25コピーで陽性であった。
【0092】
(実施例8)
SDAによるHSV−2ウイルス粒子の検出
HSV−2を検出するための本発明のプライマーおよびプローブの能力を確認するために、アメリカンタイプカルチャーコレクション(ATCC)から得たHSV−2の2系統、Quest Diagnostic(Baltimore,MD)から得た9系統、およびApaI分析によってHSV−2として分類されたオハイオ州立大学(OSU)からの5系統についてSDAを実施した。
【0093】
OSUから得た系統は、PCRによってDNAポリメラーゼ遺伝子の領域を増幅することにより特徴づけた。1セットのPCRプライマーを設計して、HSV−1およびHSV−2の両方の同じ領域を増幅した。この2つのタイプのウイルスは、HSV−1の系統から生成されるPCR産物中には存在しないHSV−2PCRフラグメント中のApaI制限エンドヌクレアーゼ認識部位の存在によって明確に区別できた。ApaI制限酵素でインキュベートした場合、HSV−2由来のPCR生成物を2つの短いフラグメントに開裂し、一方HSV−1から得られるものは無傷のままである。制限されたフラグメントは、適切な対照とともにアガロースゲル電気泳動で解析した。
【0094】
SDAシステムを評価するために使用されたウイルスストックの濃度は未知であった。ウイルスストックをリン酸塩緩衝食塩水で1:10に希釈し、この懸濁液の10μlをSDAで試験した。結果を表7に示す。HSV−2のすべての系統がシステム1.0、2.0および5.2からの増幅プライマーを用いて検出され、多様な起源からのHSV−1の系統を検出するための、開示されたプライマーおよびプローブの能力が実証された。
【0095】
【表7−1】
【0096】
【表7−2】
【0097】
(実施例9)
HSV−2DNAに対するSDAの特異性
HSV−1の25の系統をHSV2GGSDAシステム1.0、2.0および5.2で試験した。HSV−1希釈物の10ミリリットルの各懸濁液を反応あたり添加した。試験した25の系統のいずれもHSV−2システムで検出されなかった。結果を表8に示す。加えて、他の微生物のパネルを、開示された本発明の方法のプライマーおよびプローブを用いて試験した。これらの微生物には、細菌、酵母および臨床標本中で遭遇するであろう他のウイルスが含まれる。試験された微生物のいずれもHSV−1に対して陽性ではなかった。結果を表9に示す。
【0098】
【表8−1】
【0099】
【表8−2】
【0100】
【表9】
【技術分野】
【0001】
本発明は、核酸増幅法により単純ヘルペスウイルス(HSV)を同定するための診断方法と核酸配列に関する。
【背景技術】
【0002】
本出願は、特願2006−513315の分割出願である。
【0003】
単純ヘルペスは、ヒトにおける初感染および回帰感染の原因であり、伝染性単球増加症(エプスタイン−バーウイルス)、水痘および帯状疱疹(水痘帯状疱疹ウイルス)を起こすウイルスに関連する、エンベロープで包まれた二本鎖DNAウイルスである。単純ヘルペスウイルス感染の症状には、皮膚または粘膜上の小さな疱疹の出現が含まれる。疱疹の出現が収まった後、ウイルスは、感染した領域に神経繊維を供給する神経細胞群(神経節)の内側に休眠(潜伏)状態で生き残っている。周期的に、ウイルスは再活性化し、再度成長を開始し、神経繊維を通して皮膚に戻って行き、これによって先の感染と同じ皮膚領域に疱疹の出現を引き起こす。明確な疱疹がない場合であっても、ウイルスはしばしば、皮膚または粘膜に存在することがある。単純ヘルペスウイルス(HSV)は2種類のタイプ、HSV−1およびHSV−2に分類される。ヒトHSV−1およびHSV−2の完全なゲノムは配列決定されている(例えば、NCBI Accession Nos. X14112 and Z86099(非特許文献1)参照)。
【0004】
HSVは、失明および脳炎を含めた種々の疾患に寄与するかまたは疾患を引き起こすことが示されている。局所的な発生の他に、HSV−1およびHSV−2は脳炎に関連する。この脳炎の病体生理学は、ヒトでは理解が不十分である。動物モデルでは、このウイルスは末梢神経を通して中枢神経系に入り脳の炎症を引き起こすことが示唆されている。HSV−1は成人の脳炎の一般的な原因である。HSV−2は、母親の性器感染に関連する新生児脳炎の一般的な原因である。HSV−2は、社会における最も一般的な性感染症の1つである。HSVに関連する脳炎は、100万人あたり1から4人の年間発症率を有し、全てのタイプの脳炎のうち死亡率が最も高い。HSV脳炎は、全年齢の人を、一年のうちのいずれの時期においても冒す。成人では、HSVに関連する脳炎は潜伏したウイルスの再活性化によると考えられる。症状には、発熱、頭痛、痙攣(seizures)、意識レベルの変化および人格変化が含まれうる。他の病患に対するこれらの症状の類似点が臨床診断を困難にしている。治療しないままでいると、単純ヘルペス脳炎(HSE)の死亡率は、治療を受けたものの中での19パーセント程度の低さと比べ、70パーセント程度の高さになる。治療された患者の中では、約38パーセントが、最終的に正常機能に戻ることが報告されている。従って、早期にHSV感染を診断できることが非常に重要である。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】EP 0 497 272
【特許文献2】米国特許第5,010,183号明細書
【特許文献3】米国特許第5,860,937号明細書
【特許文献4】米国特許第5,728,822号明細書
【特許文献5】米国特許第5,990,301号明細書
【特許文献6】米国特許第6,261,785号明細書
【特許文献7】米国特許第5,455,166号明細書
【特許文献8】米国特許第5,270,184号明細書
【特許文献9】米国特許第5,547,861号明細書
【特許文献10】米国特許第5,928,869号明細書
【特許文献11】米国特許第5,593,867号明細書
【特許文献12】米国特許第5,550,025号明細書
【特許文献13】米国特許第5,935,791号明細書
【特許文献14】米国特許第5,888,739号明細書
【特許文献15】米国特許第5,846,726号明細書
【非特許文献】
【0006】
【非特許文献1】NCBI Accession Nos. X14112 and Z86099
【非特許文献2】Saiki et al., 1985, Science 230: 1350-1354
【非特許文献3】Wu et al., 1989, Genomics 4: 560-569
【非特許文献4】Barringer et al., 1990, Gene 89: 117-122
【非特許文献5】Barany, 1991, Proc. Natl. Acad. Sci. USA 88: 189-193
【非特許文献6】Kwoh et al., 1989, Proc. Natl. Acad. Sci. USA 86: 1173-1177
【非特許文献7】Guatelli et al., 1990, Proc. Natl. Acad. Sci. USA 87: 1874-1878
【非特許文献8】Lizardi et al., 1988, BioTechnology 6: 1197-1202
【非特許文献9】Walker et al., 1992, Proc. Natl. Acad. Sci. USA 89: 392-396
【非特許文献10】Walker et al., 1992, Nuc. Acids. Res. 20:1691-1696
【非特許文献11】T. Maniatis et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, 1982
【非特許文献12】J. Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, 1989
【非特許文献13】Current Protocols in Molecular Biology, Eds. F. M. Ausubel et al., Vol. 1, "Preparation and Analysis of DNA", John Wiley and Sons, Inc., 1994-1995, Suppls. 26, 29, 35 and 42; pp. 2.10.7-2.10.16
【非特許文献14】G. M. Wahl and S. L. Berger (1987; Methods Enzymol. 152: 399-407)
【非特許文献15】A. R. Kimmel, 1987; Methods of Enzymol. 152: 507-511
【非特許文献16】Nadeau, et al., 1999 Anal. Biochem. 276:177-187
【非特許文献17】J. Sambrook and D. W. Russell, Molecular Cloning: a Laboratory Manual, 3rd Edition, Cold Spring Harbor Laboratory Press, USA, (2001)
【発明の概要】
【発明が解決しようとする課題】
【0007】
HSV感染の診断は、一般に、適切な臨床サンプルについて細胞培養を用いて行われる。しかし、細胞培養物内容HSVを単離する能力は、古い病巣、宿主の免疫応答の存在、および再活性化の症状の出現で低下する。血清学的診断法、特に髄液(CSF)中のHSVの診断法は、感度または特異性が十分ではなく、脳炎の初期治療介入の選択を含む決定に使用するには時間がかかりすぎる。HSVは細胞培養を用いた脳脊髄液中では滅多に検出されず、僅かに4パーセントの場合が培養物陽性であった。血清学的方法はまた、初期感染後、抗体応答が現れるのに2から3週間の間遅れることにより、HSEの診断には適さない。脳生検を含む診断である「ゴールドスタンダード」法は、侵襲的であり、長期の罹病率の重大なリスクが問題となる。コンピューター断層撮影法および磁気共鳴映像法は、診断ツールとしては特異的でなく、感度が不足している。
【0008】
現時点では、HSVの検出のための免疫学的方法は、信頼できず且つ実施するのが困難である。分子法の検出は、感度を高め、結果に至る時間が従来の手段で可能であるよりも早まる可能性を提供する。迅速で、高感度で、且つ特異的なHSV疾患の診断が絶対必要であるという事実が存在する。従って、HSVの診断を助ける迅速且つ高感度なツールの開発の、臨床上の必要性が存在する。HSV感染の型を決めるためのツールの必要性も残っている。ウイルス感染に関連する特異的な原因物質の迅速な同定は、短い時間内で適切な治療法を決定するのに用いることができる情報を提供する。
【課題を解決するための手段】
【0009】
本発明は、哺乳動物内の単純ヘルペスウイルス(HSV)、特に単純ヘルペスウイルス1型(HSV−1)または単純ヘルペスウイルス2型(HSV−2)を検出する方法および組成物に関する。この方法は、プライマーを用いて単純ヘルペスウイルス標的配列を増幅し、検出することを含む。一実施形態は、鎖置換増幅(Strand Displacement Amplification)(SDA)の増幅技術を使用する。
【0010】
本発明の核酸プライマーは、HSV−1またはHSV−2中の標的配列を一義的に増幅し、これによってHSVの高感度な検出および型の同定を可能にする。本発明はまた、ユニバーサル蛍光エネルギー移動プローブを用いるリアルタイムな検出を含むHSVの検出のための鎖置換増幅(SDA)をベースとする方法を指向する。本発明のプローブおよびプライマーは、直接、迅速、且つ高感度なHSV核酸の検出を提供し、従って、免疫学的アッセイに、魅力的な代替法を提供する。
【発明の効果】
【0011】
本発明のプローブおよびプライマーは、培養された微生物の正体を確認するための手段として、サンプルを培養した後に使用されうる。別法として、これらは公知の増幅方法を用いてHSV核酸の検出および同定をするために、培養に先立って、または培養に代えて用いることができる。本発明のプローブ、プライマーおよび組成物、並びにこのプローブ、プライマーおよび組成物を用いたアッセイ法は、HSV−1とHSV−2の核酸標的配列を迅速に識別する手段を提供し、一般に依拠されている時間のかかる免疫学的および生化学的手順に頼ることなく、医者がHSVを同定し、診断し、治療することを可能にする。
【図面の簡単な説明】
【0012】
【図1】単純ヘルペスウイルス1型の糖タンパク質G(US4)遺伝子のコンセンサス配列(配列番号1)を示す図である。
【図2】HSV−1標的領域(配列番号2)のゲノム配列の位置、および、HSV−1DNAの特異的検出用に設計されたプライマー、バンパー、およびアダプタの配置を示すマップである。
【図3】「MOTA」発現の結果を示すグラフである。
【図4】BDプローブテック(商標)ETシステムを用いた「PAT」アルゴリズムを示すグラフである。
【図5】種々のHSV−1系統の希釈物についてのSDA法の分析感度を表す図である。
【図6】単純ヘルペスウイルス2型(HSV−2)の糖タンパク質G(US4)のフラグメントのコンセンサス配列(配列番号3)の図である。
【図7】HSV−2標的領域(配列番号4)のゲノム配列、および、HSV−2DNAの特異的検出用に設計されたプライマー、バンパー、およびアダプタの配置を示すマップである。
【発明を実施するための形態】
【0013】
本発明の種々の目的、利点、および新規な特徴は、添付の図面と組み合わせた以下の詳細な説明から容易に理解されるであろう。
【0014】
本発明は、核酸増幅反応における単純ヘルペスウイルス(HSV)の型の特異性を示す、単離され精製された核酸、ポリヌクレオチド、増幅プライマーおよびアッセイプローブを提供する。本発明のプローブおよびプライマーを用いてHSV核酸を検出および同定する方法も提供する。
【0015】
本発明の一実施形態は、標的核酸配列の存在を検出する増幅方法であって、標的結合配列を有する1以上の増幅プライマーを用い、増幅された標的配列を産生し、この標的配列を検出する増幅方法に関する。増幅方法の非制限的な例には、ポリメラーゼ連鎖反応(PCR;Saiki et al., 1985, Science 230: 1350-1354(非特許文献2)参照。参照文献として本明細書に取り込む)、リガーゼ連鎖反応(LCR;Wu et al., 1989, Genomics 4: 560-569; Barringer et al., 1990, Gene 89: 117-122; Barany, 1991, Proc. Natl. Acad. Sci. USA 88: 189-193(非特許文献3、4、5)参照。この全てを参照文献として本明細書に取り込む)、in situ ハイブリダイゼーション、転写介在増幅手法(TMA、Kwoh etal., 1989, Proc. Natl. Acad. Sci. USA 86 : 1173-1177(非特許文献6)参照。参照文献として本明細書に取り込む)、自己持続性配列複製法(Self-Sustaining Sequence Replication(3SR、Guatelli et al., 1990, Proc. Natl. Acad. Sci. USA 87: 1874-1878(非特許文献7)参照。参照文献として本明細書に取り込む)、ローリングサークル増幅法(Rolling Circle Amplification)(RCA)、核酸配列をベースとする増幅法(Nucleic Acid Sequence Based Amplification)(NASBA)、Qβレプリカーゼシステム(Lizardi et al., 1988, BioTechnology 6: 1197-1202(非特許文献8)。参照文献として本明細書に取り込む)、および、好熱性SDA(tSDA)を含めた鎖置換増幅法(SDA、Walker et al., 1992, Proc. Natl. Acad. Sci. USA 89: 392-396; Walker et al., 1992, Nuc. Acids. Res. 20:1691-1696(非特許文献9、10)およびEP 0 497 272(特許文献1)この全てを参照文献として本明細書に取り込む)が含まれる。
【0016】
本発明の他の実施形態は、HSV標的配列の指数関数的増幅によってサンプル中のHSV核酸配列の存在を検出する鎖置換増幅(SDA)法に関する。さらなる実施形態では、SDAは、選択された検出(detector)プライマーを用いて、米国特許第5,648,211号明細書(特許文献2)に記載されるように約52℃で実施され、米国特許第5,919,630号明細書;同5,928,869号明細書;同5,958,700号明細書および同6,261,785号明細書(特許文献3〜6)に記載されるように増幅の間に標的を検出する(これらの文献は全て、参照により本明細書に取り込む)。SDAで一般的なように、制限酵素およびポリメラーゼのような試薬、プライマー、酵素、および他の成分を反応マイクロウェル、容器、または、レセプタクルに添加する。SDAはサンプルからDNA配列を増幅するが、この方法では、一度全ての成分を互いに混合し、重要な成分が使い尽くされるまで反応を継続する。ポリメラーゼ連鎖反応(PCR)とは対照的に、SDAは等温反応プロセスであり、従って、一度反応が開始されると反応の進行中に外部から制御をしない。
【0017】
本発明のSDA法は、増幅を開始するのに少なくとも2種類のHSV増幅プライマーおよび2種のバンパープライマーを必要とする。増幅プライマーはHSV−1またはHSV−2に高度に特異的であるように設計されている。SDA法は、混合物中での同時増幅反応を含み、PCR増幅法で必要な温度サイクルのための別々のフェーズとサイクルを必要としない。本発明のSDAのさらなる利点は、指数関数的な増幅である。DNAポリメラーゼ伸張、ニッキング、置換およびニック部位の再生のステップが、どちらかの末端に部分的制限酵素部位(例えばBsoBI部位)を持つ、置き換えられた単鎖分子で起こり、この分子は続いて、培養され、増幅プライマーによって捕捉され、これによって、HSV標的配列が指数関数的に増幅される。SDA法はまた、特に高スループット法に対するワークフローを改善する。SDAは、マイクロアレイをベースとする適用例に組み込まれうる。この適用例では、小容積量(ナノリットル)のサンプルおよび試薬を使用して、単一のプラットホームで複数のSDAアッセイを実施することにより、HSV標的DNAを増幅し、マイクロチップアレイ上で増幅サンプルを検出する。等温増幅プロセスには、このプラットホームの設計および維持における技術的難題が極めて僅かしか存在しないので、サンプル中のHSVを検出するためのSDA法の主たる利点は、労働力の必要性を最小限にすることと、高スループットの可能性である。
【0018】
本明細書で使用する用語「標的」または「標的配列」とは、増幅および検出されるHSV核酸配列、HSV−1またはHSV−2をいう。これらには、増幅される本来のHSV核酸配列、増幅される本来のHSV核酸配列の相補性第二鎖、および増幅反応で産生される本来のHSV配列のコピーのどちらかの鎖が含まれる。これらのコピーは、増幅プライマーがアニールする配列のコピーを含有するので、増幅可能な標的として機能する。増幅反応中に生成される標的配列のコピーは、増幅産物、アンプリマーまたはアンプリコンと称される。HSV−1およびHSV−2標的配列は、HSV−1およびHSV−2のゲノム配列の遺伝子の糖タンパク質G(US4)に局在化している。HSV−1標的配列は、図1のコンセンサス配列の555から680塩基の間に位置する。HSV−2標的配列は、図6のコンセンサス配列の867から990塩基の間に位置する。糖タンパク質G(US24)遺伝子は、図2および図7のHSV−1ゲノム配列の136744から137460の間におよびHSV−2のゲノム配列の137878から139977の間の位置にそれぞれ位置する。
【0019】
本明細書で使用する「増幅プライマー」は、標的配列にアニールし、増幅によって伸張されうるプライマーである。標的配列に結合する増幅プライマーの領域は標的結合配列である。増幅技術には、好熱性SDA(tSDA)を含めた鎖置換増幅(SDA)、ポリメラーゼ連鎖反応(PCR)、リガーゼ連鎖反応(LCR)、in situ ハイブリダイゼーション、自己持続性配列複製法(Self-Sustaining Sequence Replication(3SR))、ローリングサークル増幅法(Rolling Circle Amplification)(RCA)、核酸配列をベースとする増幅法(Nucleic Acid Sequence Based Amplification)(NASBA)、および転写介在増幅手法(TMA)が含まれるが、これらに限定されない。
【0020】
一実施形態では、増幅プライマーは鎖置換増幅(SDA)法に使用される。増幅プライマーは、3’末端にHSV標的配列に結合する標的結合配列部分を含み、5'末端に標的配列に結合またはアニールしない部分を含む。標的配列に結合しないSDA増幅プライマーの部分はまた、米国特許第5,455,166号明細書(特許文献7)および米国特許第5,270,184号明細書(特許文献8)(これらは参照文献として本明細書に取り込む)に記載されているような、標的結合配列の上流の、テール、および制限エンドヌクレアーゼに対する認識部位を含む。この認識部位は、Walkerら(非特許文献9および10)に記載されているように、この認識部位が半修飾(hemimodified)されている場合に、DNA二本鎖の一方の鎖にニックを入れるエンドヌクレアーゼに特異的である。テールは、制限エンドヌクレアーゼ認識部位の配列の上流にあり、増幅プライマーの残りの部分がSDA中にニックを入れられ、置換されるときに、ポリメラーゼ再プライミング部位として機能する。テールの再プライミング機能は、SDA反応を維持し、単一の標的分子から多数のアンプリコンの合成を可能にする。テールの長さおよび配列は一般に重要ではなく、型通りに選択でき、且つ修飾することができる。
【0021】
本発明の一実施形態は、増幅プライマーに標的特異性を付与する標的結合配列に基づく。ここで、本発明で例示される標的結合配列は、HSVの検出のための他の種々の方法に使用できることも理解されるべきである。例えば、他の選択肢として、本明細書に開示されている標的結合配列は、事前の増幅を用いずに、または増幅後のアッセイにおいてHSVの直接の検出のためにハイブリダイゼーションプローブとして使用されうる。このようなハイブリダイゼーション法は当分野で周知であり、典型的には、ハイブリダイゼーションの検出を促進するために標的結合配列と会合された、またはこれに連結された検出可能な標識を使用する。さらに、表1および表2にプライマー配列(それぞれ配列番号:5〜25および36〜47)を列挙したが、これには大文字を使用しておよびアンダーラインを引いて示した標的結合配列が含まれている。これらの標的結合配列は、追加の特化された配列を必要としない増幅反応(例えばPCR)のプライマーとして使用できるか、または、NASBA、in situハイブリダイゼーション、TMA、3SR、プライマーの標的結合配列に連結されたRNAポリメラーゼプロモータを必要とする増幅プライマーをベースとする他の転写、または任意の他のプライマー伸長増副反応に追加されうる。プライマー中の特化された非標的結合配列を必要とするこれらの増幅法は、増幅反応を進行するのに必要であり、典型的には、標的配列に特化した配列を追加するように機能する。
例えば、制限酵素認識部位は、SDAで起こる指数関数的増幅に必要である(米国特許第5,455,166明細書および同5,270,184号明細書(特許文献7、8)参照)。対照的に、自己持続性配列複製法(3SR)および核酸配列をベースとする増幅法(NASBA)は、5’末端近傍にポリメラーゼプロモータを含む(3SRアッセイは、Guatelli et al., 1990, Proc. Natl. Acad. Sci. USA 87: 1874-1878(非特許文献7)に開示されている。)。このプロモータは、標的結合配列に追加され、テンプレートの複数のRNAコピーの転写を指示することによって増幅反応を推進する働きをする。選択された増幅反応に使用するために、このような特化した配列を標的結合配列に連結することは、通常行われることであり、当業者に周知である。
【0022】
対照的に、標的の末端に特化された配列を必要としないPCRのような増幅方法は、一般に、標的結合配列のみからなる増幅プライマーを使用する。これらの他の増幅方法における検出の目的に対しては、当業者によって理解されるように、プライマーが検出可能に標識されうる。
【0023】
核酸はアニールするためには完全な相補性を必要としないので、当業者は、本明細書に開示されたプローおよびプライマー配列がHSV−1およびHSV−2特異的プライマーおよびプローブとしての有用性を損なうことなくある程度まで修飾できることを理解するであろう。用語「相同性」は相補性の程度を言う。部分的相同性または完全相同性があり得、完全相同性は同一と同義である。同一の配列を標的核酸へハイブリダイゼーションするのを阻害する部分的相同性配列は、「実質的に相同性である」と称される。標的配列への完全に相補的な配列のハイブリダイゼーションの阻害は、低ストリンジェンシー条件下でハイブリダイゼーションアッセイ(例えばサザンまたはノーザンブロット、溶液ハイブリダイゼーションなど)を用いて試験することができる。実質的に相同性である配列またはプローブは、低ストリンジェンシー条件下で標的配列への、完全に相補的な配列またはプローブの結合(すなわちハイブリダイゼーション)に競合し且つこの結合を阻害する。それにもかかわらず、低ストリンジェンシー条件は、非特異的結合を許さず、低ストリンジェンシー条件は、2つの配列の他方への結合が特異的(すなわち選択的)相互作用であることを必要とする。
【0024】
当業者によって理解されるように、アニーリングのストリンジェンシーは同一または関連するポリヌクレオチド配列を同定または検出するために変更されうる。熟練した専門家によってさらに理解されるように、融解温度、Tmは、ヌクレオチドの数でのプライマーまたはプローブの長さ、またはアニーリングバッファの成分および条件のような多くのパラメータに依存して、当分野で公知のように式で近似することができる(例えば、T. Maniatis et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, 1982およびJ. Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, 1989; Current Protocols in Molecular Biology, Eds. F. M. Ausubel et al., Vol. 1, "Preparation and Analysis of DNA", John Wiley and Sons, Inc., 1994-1995, Suppls. 26, 29, 35 and 42; pp. 2.10.7-2.10.16; G. M. Wahl and S. L. Berger (1987; Methods Enzymol. 152: 399-407);およびA. R. Kimmel, 1987; Methods of Enzymol. 152: 507-511(非特許文献11〜15)参照)。一般的指針として、Tmは、配列相同性が1%減少するごとに約1℃から1.5℃低下する。温度範囲は約50℃から62℃で変化しうるが、増幅プライマーは52℃で最適化されるように設計されうる。しかし、50℃未満の温度はプライマーに特異性を欠如させ、一方62℃を超える温度はハイブリダイゼーションを引き起こさない。増幅プライマーを設計する場合にさらに考慮することは、グアニンとシトシンの含有量である。一般には、プライマーに対するGC含有量は、約60〜70%であるが、これはより少なくてもよく、当業者によって適切に調節することができる。標的結合配列のハイブリダイゼーション領域は、約42〜48℃のTmを有しうる。アニーリングの相同性および部分相同性核酸配列は、アニーリングの条件を変更してストリンジェンシーを増加または減少すること(すなわち、アニーリング温度またはバッファの塩の含有量を調節すること)によって得ることができる。このような開示された配列のマイナーな変更、およびHSV−1およびHSV−2の特異性を維持するための必要なアニーリング条件の調節には、決まりきった実験手順のみが必要であり、そのような変更および調節は当業者の範囲内にある。
【0025】
HSV−1およびHSV−2標的配列の検出用に設計された増幅プライマーは、それぞれ、配列番号7〜18および38〜43として表1および2に同定されている。これらの増幅プライマーは、標的配列が高相同性コンセンサス糖タンパク質G(US4)遺伝子領域にアニールするように設計されている(図1〜2および6〜7参照)。HSV標的DNA配列にアニールするか、またはこれらに対して相補的である増幅プライマー内のHSV標的結合配列領域は、アンダーラインを付し、大文字で示した(表1および2参照)。SDA検出プライマー配列の残っている5’部位は、SDA反応が進行するのに必要であるBsoBI制限エンドヌクレアーゼ認識部位(RERS)(小文字のイタリック体で示した)、並びに、ジェネリックな非標的特異的5’テール末端配列を含む。
【0026】
それぞれ配列番号7〜8および38のHSV−1およびHSV−2増幅プライマーは左側(「第一」)S1増幅プライマーであり、それぞれ配列番号9〜18および39〜43は右側(「第二」)S2増幅プライマーである。増幅の目的では、少なくとも1つの左側および右側プライマー対が反応中に存在するように、特異型の一対のHSV増幅プライマーを単独(すなわち、1つのHSV−1左側増幅プライマーおよび1つのHSV−1右側増幅プライマー)で使用するか、または組み合わせて(すなわち、1つのHSV−1SDA左側プライマーと2つのHSV−1SDA右側増幅プライマー)を使用する。多数の増幅プライマーを使用して、標的配列の幾つかの領域を増幅することができる。プライマーの濃度は、HSV−1の第1増幅プライマーが500nMの濃度で唯一の第一増幅プライマーとして使用され、2つのHSV−1右側増幅プライマーが併用して使用される場合には、各々250nMの濃度を有するように適切に調節されうる。
【0027】
用語「伸長生成物」は、一般に、ポリメラーゼのような酵素を用いてプライマーまたは標的配列を伸長することによって産生される配列を言う。一実施形態では、増幅プライマーのハイブリダイゼーションおよびテンプレートとしてHSV標的配列を用いるポリメラーゼによる増幅プライマーの伸長は、増幅プライマー伸長生成物を産生する。
【0028】
「バンパープライマー」または「外部プライマー(external primer)」は、バンパープライマーが下流の増幅プライマーおよびその伸張生成物を置換するように増幅プライマーの上流で標的配列にアニールするプライマーである。本明細書で使用する場合、用語「バンパープライマー」は、HSV標的結合配列を含むポリヌクレオチドをいう。有用なバンパープライマーは、表1および2に、それぞれ配列番号23〜25および46〜47として同定されている。左側または第一のHSV−1およびHSV−2バンパープライマーは、それぞれ配列番号23および46であり、一方右側または第二のHSV−1およびHSV−2バンパープライマーはそれぞれ配列番号24〜25および47である。バンパープライマーは、増幅プライマーの標的結合部位に十分近接し、バンパープライマーの伸張後に増幅プライマー伸張生成物の置換を可能にする増幅プライマーの上流の位置で増幅プライマーの側面に位置する配列の保存領域から誘導される。例えば、配列番号23(HSV1GGLB1.0)のHSV−1第一バンパープライマーの5’末端はHSV−1ゲノム配列の塩基137,256に位置する(図2)。配列番号25(HSV1GGRB1.1)のHSV−1第二バンパープライマーの5’末端は、HSV−1ゲノム配列の塩基137,382に位置する。SDAの初期のラウンドの間に、バンパープライマーはHSV標的配列にハイブリダイズし、ポリメラーゼ伸張、すなわち下流の増幅プライマー伸張生成物によって置換され、複製および/または指数関数的増幅のさらなるラウンドを受けうる単鎖DNAの生成を引き起こす。
【0029】
用語「アッセイプローブ」は、核酸の検出または同定を促進するために使用される任意の核酸を言う。例えば、本発明の一実施形態では、アッセイプローブはHSV核酸の検出または同定に使用される。以下に記載される検出プローブ、検出プライマー、キャプチャープローブおよびプライマーはアッセイプローブの例である。
【0030】
特に、特異的なHSV−型を検出および同定するのに有用な「検出プローブ」は標識化されるかまたはタグ付けされる。検出プローブの検出可能な標識は、直接的または間接的に検出され、標的核酸配列の存在を示しうる部分である。直接検出については、アッセイまたは検出プローブは放射性同位体でタグ付けされ、オートラジオグラフィーで検出されるか、または、蛍光部分でタグ付けされ、当分野で公知の通りの蛍光によって検出する。あるいは、アッセイプローブは、検出を可能にする追加の試薬で標識することによって間接的に検出することができる。間接的に検出可能な標識には、例えば、化学発光剤、視覚化または着色された反応生成物をもたらす酵素、および、リガンドで検出可能に標識されたリガンド結合パートナーであって、リガンド(例えばハプテン、抗体または抗原)を標識されたリガンド−特異的結合パートナーへ結合することによって検出できるものが含まれる。
【0031】
増幅生成物の検出に対しては、本明細書で開示された標的結合配列を含む増幅プライマーが当分野で公知の方法で標識されうるか、または、開示された標識結合配列を含む標識された検出プライマーを、増幅のリアルタイム均一検出に対して米国特許第5,547,861、米国特許第5,928,869、米国特許第5,593,867号明細書、米国特許第5,550,025号明細書、米国特許第5,935,791号明細書、米国特許第5,888,739号明細書、米国特許第5,846,726号明細書(特許文献9〜15に開示された増幅プライマーと共に使用することができる。このような検出プライマーは直接的にまたは間接的に検出可能な配列を含み、この検出可能な配列は、初期には標的にハイブリダイズしないが、一旦標的にハイブリダイズして伸長されると検出プライマーの検出を促進する。例えば、このような検出可能な配列は、制限部位を含む配列、または、ヘアピンおよびg−カルテット配列(これらに限定されない)のような、近傍にフルオロフォアおよびクエンチャー部分を結合する二次構造を形成する配列、または、当分野で公知である標識されたオリゴヌクレオチド(しばしばレポータープローブと言われる)へ線状配列の相補体をハイブリダイゼーションすることによって検出される線状配列でありうる。あるいは、増幅生成物は、リアルタイムに、または、挿入色素(intercalating dye)の使用を介した増幅後に、もしくは選択した一連の増幅プライマーの範囲に入る本明細書で開示された任意の標的結合配列から選択されるプローブのハイブリダイゼーションによる増幅後に検出されうる。
【0032】
末端および内部標識法は当分野で公知であり、検出プライマーのそれぞれの位置でドナー色素およびアクセプター色素を連結するのに使用される。5'-末端標識法の例には、a)5’−対−5’カップリングしたリボヌクレオチドの過ヨウ素酸塩酸化とこれに続くアミン含有標識との反応、b)エチレンジアミンと5’−ホスホリル化ポリヌクレオチドの縮合とこれに続くアミン反応性標識との反応、およびc)固相DNA合成でアミノヘキシルホスファイト試薬を用いた脂肪族アミン置換基の導入とこれに続くアミン反応性標識との反応が含まれる。標識はまた、特別な脂肪族アミン含有ヌクレオチドホスホラミダイト試薬を用いて特定の位置で、合成DNAオリゴヌクレオチドに連結される。検出プライマーへ選択された標識を連結し、連結した試薬を働かせるための適切な方法を選択することは、当分野ではごく普通の手順である。
【0033】
他の実施形態は、検出される標的配列に依存して多数の検出プライマーを必要とする、特異的標的配列にハイブリダイズする検出プライマーを使用する。しかし、特異的HSV型の検出および同定のための実施形態では、Nadeauら(1999)によって記載されているリアルタイムSDA検出法を修飾したユニバーサル検出システム(Universal detection system)を使用する。ユニバーサル検出システムは、多数のアッセイに対して同じ蛍光検出プライマーの対を使用することができ、経費、時間、および技術的な複雑さを低減するような幾つかの利点を提供する。
【0034】
「シグナル」または「アダプタ」プライマーは、HSV標的配列にハイブリダイズする標的結合部分およびジェネリックでHSV標的配列に結合しないテール部分を有する。アダプタプライマーはユニバーサル検出用の検出プライマーと共に使用される。検出プライマーは、相補性アダプタプライマーのテール部分(すなわち、非標的結合配列)にハイブリダイズする。シグナルまたはアダプタプライマーは、第一および第二増幅プライマーの間の介在領域に少なくとも部分的に位置する標的配列の領域にハイブリダイズし、シグナルまたはアダプタ配列が増幅反応中に置換されるように設計される。配列番号19〜22および44〜45を有するHSV−1およびHSV−2シグナルまたはアダプタプライマーは、それぞれ表1および2に示されている。
【0035】
検出プライマーは、「ユニバーサル検出プライマー」または「検出プライマー」であってもよく、検出可能に標識された5'テール末端部分と、相補性アダプタプライマーテール配列に結合する3'末端部分を有する。一般に、検出プライマーの3’末端は、HSVまたは内部増幅対照(Internal Amplification Control)(IAC)標的配列に対する任意の十分な相補性を有する配列を含まない。検出プライマーはまた、5'末端に制限酵素認識部位を有する。
【0036】
手短には、このユニバーサル検出システムは、増幅のためのSDA法と同時に、且つ同じ反応容器中で使用することができる。ユニバーサル検出システムは、未標識のアダプタプライマーの標的依存的伸長を含む。アダプタプライマーは、HSV−1またはHSV−2標的特異的3'配列および5’ジェネリックテールを含み、それぞれ配列番号19〜22および配列番号44〜45と、その相補体で例示される。アダプタプライマーは、S1増幅プライマーの下流で、増幅されたHSV標的配列にハイブリダイズする。DNAポリメラーゼは、アダプタ配列およびS1増幅プライマーの3’末端から伸長し、この場合、増幅プライマーの伸長はアダプタプライマー伸長生成物を置換する。S2増幅プライマーは、アダプタプライマー伸長生成物にアニールする。DNAポリメラーゼはS2増幅プライマーの3’末端を伸長し、アダプタプライマー伸長生成物およびその相補体を含み、ニッキング可能な制限酵素認識部位を有する二本鎖分子を産生する。ニッキングとは、DNA二重鎖の2つの鎖のうちの1つのみのホスホジエステル結合を切断することを言う。対応する制限酵素は、制限酵素認識部位で二本鎖分子をニッキングし、ニッキングされた短いテールを含む5’部分と、ニッキングされた長い相補性アダプタプライマー伸長生成物を含む3’部分を生じる。BsoBI酵素のような対応する制限酵素で制限酵素部位をニッキングし、ニッキングされた部位から鎖を伸長すると、アダプタプライマー相補体の単鎖コピーが置換される。DHAポリメラーゼはニッキングされたテールの3’末端を伸長し、これによってニッキングされた単鎖の相補性アダプタプライマー伸長生成物を置換する。S1増幅プライマー伸長生成物および伸長されたHSV標的配列は、さらにSDAによって指数関数的に増幅される。次に、置換された相補性アダプタプライマー伸長生成物は検出プライマーによって捕獲されるが、この捕獲では、検出プライマーの3'末端が相補性アダプタプライマー伸長生成物の5’部分にアニールする。検出プライマーは、検出可能な標識を含み、標的配列を検出する。検出プライマーの3'末端からのDNAポリメラーゼ伸長および相補性アダプタプライマー伸長生成物は、存在する場合にはヘアピンを開き、検出プライマー伸長生成物とその相補体を含む二本鎖検出分子を産生する。各鎖は、開裂可能な制限酵素認識部位を含み、開裂されたときに、ドナーおよびクエンチャー色素を切り離し、フルオロフォアおよびクエンチャー部分を切り離し、標的特異的蛍光を生じる。この切り離しによって、クエンチャーはもはやフルオロフォアによって発光された蛍光を抑制することができない。二本鎖化された検出プライマー制限酵素認識部位の完全な開裂は、フルオロフォアとクエンチャーを切り離すことによって蛍光シグナルを増加させる。
【0037】
本発明の一実施形態では、検出プライマーは、蛍光ドナー部分(またはフルオロフォア)およびクエンチャー部分であって、各部分が制限酵素認識部位に隣接しているものにより蛍光検出用にタグ付けされうる。表1および2は、配列番号30〜35を有する検出プライマー配列を示す。ユニバーサル検出では、標的配列を検出するための検出プライマーが、一般にアダプタプライマーと共に使用される。ドナー色素、すなわちローダミン(ROX)およびクエンチャー色素、すなわちP−(ジメチルアミノフェニルアゾ)安息香酸(DABCYL)で標識された、配列番号30〜33を有する検出プライマーは、本発明でHSV標的配列の検出に使用される。他のドナーおよびクエンチャー色素対は、SDAで使用するために、クエンチャー色素がドナー色素によって放出される蛍光を十分に吸収するように、当業者によって容易に選択されうる。例えば、ドナーおよびクエンチャー色素は異なる波長での吸収によって容易に検出され、識別される。ドナーおよびクエンチャー色素に依存して、クエンチャー色素はある場合にはクエンチャーとして、そして他の場合にはドナー色素として作用してもよい。
【0038】
この実施形態では、配列番号30〜35の検出プライマーは、検出プライマーの5'末端に位置する制限酵素認識部位によって切り離されるドナーおよびクエンチャー色素対を有する。さらに、配列番号30の検出プライマーは、ドナーおよびクエンチャー部分の間に位置するヘアピン構造配列であって、制限酵素認識部位がこの中に存在するヘアピン構造配列を含む配列を有する。ヘアピン構造は、ドナー色素によって放出される蛍光がクエンチャー色素によって抑制されるように2つの色素を近接した位置に持ってくる。しかし、配列番号31〜35の検出プライマーは、2つの色素の間に線状の配列を有し、この線状の配列は、クエンチャーがフルオロフォアによって放出される蛍光を吸収するのに十分短い長さである。
【0039】
当分野で知られている多くのドナー/クエンチャー色素対が本発明の実施形態で有用である。これらには、フルオレセイン(FAM(商標);Glen Research; Sterling, VA)/ローダミン(ROX(商標);Molecular Probes;Eugene,OR);ROX/P−(ジメチルアミノフェニルアゾ安息香酸(DABCYL(商標);Glen Research);FAM/DABCYL;フルオレセインイソチオシアネート(FITC)/テトラメチルローダミンイソチオシアネート(TRITC);FITC/テキサスレッド(Molecular Probes);FITC/N−ヒドロキシスクシンイミジル1−ピレンブチレート(PYB);FITC/エオシンイソチオシアネート(EITC);N−ヒドロキシスクシンイミジル1−ペンタンスルホネート(PYS)/FITC;FITC/ローダミン X;およびFITC/テトラメチルローダミン(TAMRA)が含まれるが、これらに限定されない。特定のドナー/クエンチャー対の選択は、重要ではない。
【0040】
しかし、エネルギー移動消光メカニズムについては、ドナーフルオロフォアの放出波長がクエンチャーの励起波長とオーバーラップしていること、すなわち2つの色素の間で、十分なスペクトルのオーバーラップが存在し、十分なエネルギー移動、電荷移動または蛍光消光が可能であることのみが必要である。ROXはEMmax=608nmを有し、FAMはEMmax=520nmを有する。当業者は、適切なドナーおよびクエンチャー色素対を選択することにおいて豊富な知識を有するであろう。P−(ジメチルアミノフェニルアゾ)安息香酸(DABCYL)は、非蛍光性クエンチャー色素であり、隣接するフルオロフォア、例えばFAMまたは5−(2’−アミノエチル)アミノナフタレン(EDANS)からの蛍光を効果的に消光する。特定のドナー/クエンチャー対をこの開示で例示するが、他のものも当業者には明かであり、本発明でも有用である。消光を起こすメカニズムにかかわらず、本発明の検出プライマーにおいて蛍光の消光をもたらす任意の色素対が本発明の方法で使用するのに適している。他のクエンチャーの非制限的な例には、ブラックホールクエンチャー(Black Hole Quencher)(商標)(Biosearch Technologies, Inc.;Novato,CA)およびアイオワブラック(Iowa Black)(商標)(Integrated DNA Technologies,Inc.;Corralville,IA)が含まれる。
【0041】
蛍光は、特定の増幅生成物の蓄積をモニタするために核酸増幅反応過程中に測定される。蛍光シグナルは、産生された特定のアンプリコンの量に比例する。HSV標的核酸配列の存在下では、蛍光は増強されるであろう。標的がない場合、蛍光は反応の間一貫して低いままであろう。蛍光の増加、または実質的に蛍光の変化がないことは、それぞれHSV標的配列の存在および非存在を示す。
【0042】
サンプルの蛍光は、典型的には、サンプルがHSV DNAを含むかどうかを決定するために所定時間にわたって測定される。一実施形態では、蛍光は1時間の過程で60パスモニターされる。手短には、ほぼ1分ごとに、サンプル容器中で測定された蛍光の量、補正値(必要であれば)、および各カラムに対するキャーリブレータに関するデータを集める。データを、グラフの曲線の下の領域に換算した結果を表す「MOTA」(メトリック・アザー・ザン・アクセラレーション(Metric Other Than Acceleration))法を用いて分析することができる。グラフは、パスの数(X−軸)対相対蛍光単位(Y−軸)(図3参照)を測定する。MOTA領域が大きければ大きいほど、生成される蛍光は大きく、且つ増幅された生成物の検出の効率がよい。
【0043】
さらに別の実施形態では、図4に示されるパス・アフター・スレッシュホールド(Passes After Threshold)(PAT)アルゴリズムを使用し、これはBDプローブテック(BD ProbeTec)(商標)ETシステムで使用するために特に開発されている。MOTAと同様に、より高いPATスコアーは、SDA反応がより効果的なことを示す。PATアルゴリズムを用いる場合、バックグラウンドで集められた蛍光強度のシグナルが予め決められた閾値を越える時点をT3(「タイム−トゥ−スレッシュホールド(Time-To-Threshold)」と表す。このグラフも、パスの数対相対蛍光単位を測定する。同じT3閾値を各サンプルに対して使用する。PATスコアーは60マイナスT3の値に等しい。陰性サンプルは、蛍光の最低閾値に達しておらず、従ってゼロのPAT値が割り当てられる。陽性サンプルは、アッセイおよび標的のレベルに依存して、0よりも大きなPAT値、好ましくは1から60の間、より好ましくは40〜55の間のPAT値を有する。より低いT3スコアーおよび対応するより高いPAT値が、より効果的なSDAと相関する。PATアルゴリズムは、最も再現性のある増幅曲線の領域にのみ利用する。結果として、PATアルゴリズム法は、ウェル間またはサンプル間の認識できる差を最小限にし、検出物を比較する他の方法よりも正確である。PATはBDプローブテック(商標)ETシステムによって自動で行うことができる。BDプローブテック(商標)ETの出力は、PATスコアーおよび報告できる結果を提供する。
【0044】
さらなる実施形態では、「内部増幅対照」(「IAC」)は、陰性の結果を確認するため、および潜在的な阻害検体を特定するため、またはサンプル中に入り込んだ微生物、例えばウイルス、細菌、および菌類(これらに限定されない)の定量化を促進するために、存在している方法に取り込むことができる。診断への応用では、2つの異なるDNA配列、すなわちHSV標的配列およびIAC標的配列の同時増幅と検出がIACの使用を可能にする。「IAC標的配列」または「IAC配列」は、配列番号26〜27および配列番号48〜49のIAC標的配列がHSV−1およびHSV−2標的配列と比べて約5〜10塩基ミスマッチしていることを除いて、HSV標的配列と同じである。これらの修飾塩基がIACアダプタプライマーの特異的なアニーリングを可能にするのに十分である。
【0045】
「IACアダプタプライマー」は、IACアダプタプライマーがIAC結合配列を介して「IAC標的配列」または「IAC配列」にハイブリダイズすることを除いて、シグナルまたはアダプタプライマーと同じように機能する。IACアダプタプライマーはまた、IAC標的配列とハイブリダイズしないジェネリックな配列を含む5'テール部分を有する。どちらかと言えば、検出プライマーはIACアダプタプライマー相補体のテール部分にハイブリダイズすることができる。HSV−1およびHSV−2のSDAアッセイで使用されるIACアダプタプライマーは、それぞれ配列番号28〜29および50〜51から選択でき、IAC標的配列の増幅に有用である。IACアダプタプライマーの3’末端に位置するIAC標的結合配列は、HSV−1またはHSV−2アダプタプライマーがハイブリダイズしないか、またはIAC標的配列の増幅を妨げるように十分にHSV標的配列と異なっている。表1および2において、IACアダプタプライマーの3’末端のIAC標的結合配列は小文字にアンダーラインを付すことによって示した。IACアダプタプライマーは、陰性の結果を確認し、反応を阻害する検体をモニタするのに有用である。定量SDAでは、IACと陰性の標的配列の間で律速試薬を競合させることも有用であるかもしれない(Nadeau, et al., 1999 Anal. Biochem. 276:177-187(非特許文献16)。
【0046】
本発明の検出プライマーは、HSV標的配列またはIAC標的配列のいずれかを検出するのに使用することができる。しかし、本発明の一実施形態では、HSV−1またはHSV−2標的配列を検出するために使用される検出プライマーは、配列番号30〜33のものである。IAC標的配列を検出するのに有用な検出プライマーは、ドナーおよびクエンチャー色素対がそれぞれフルオレセイン(FAM)およびDABCYLである配列番号34〜35から選択されるものであり、本明細書で「IAC検出プライマー」と称される。当業者は、IAC標的配列の同定がHSV−1またはHSV−2標的配列の検出から明確に区別できるように、標識を有する適切な検出プライマーを選択する知力を有するであろう。従って、HSV標的配列およびIAC標的配列の検出に使用される検出プライマーは交換することができ、その結果、配列番号30〜33の検出プライマーがIAC標的配列の検出に使用でき、配列番号34〜35がHSV標的配列の検出に使用できる。
【0047】
本発明の他の実施形態は、高スループットプロセスで、同時に複数のサンプルをアッセイすることに関する。サンプルには、脳脊髄液(CSF)、生殖器病変、口腔病変、粘膜病変、眼の病変、皮膚病変、直腸スワブ、膣スワブ、膣分泌物、尿、末梢血白血球および組織(例えば脳生検からのもの)から集めたものが含まれるが、これらに限定されない。サンプルは、プレート、スライド、ウェル、皿、ビーズ、粒子、カップ、ストランド、チップおよびストリップでアッセイすることができる。一実施形態では、この方法は、BDプローブテック(商標)ET GT/GCで増幅されたDNAのアッセイで使用するのに適合した形態の96マイクロウェルプレートで行われる。この方法は、乾燥されたマイクロウェルの形態であって、乾燥された組成物が多数のサンプルを同時にアッセイするのに使用するための、HSV−1またはHSV−2のSDA検出を行うのに必要な全てのプライマーおよびプローブを含む形態で実施される。
【0048】
本発明の方法によって選択された標的配列の存在を検出するアッセイは、溶液または固相で行うことができる。検出核酸がプライマーとして機能する、リアルタイムまたは終点均一アッセイ(endpoint homogeneous assay)は、典型的には、溶液で実施される。本発明の検出プライマーを用いるハイブリダイゼーションアッセイは、溶液で行うことができるだけでなく、標的のリアルタイムまたは終点検出のための固相アッセイに特に適切である。固相アッセイでは、検出オリゴヌクレオチドを、当分野で公知の方法を用いて内部または末端標識を介して固相(例えば、ビーズ、膜または反応容器)に固定する。例えば、ビオチン標識された検出オリゴヌクレオチドは、適切なハイブリダイゼーション条件下で標的に曝したときに蛍光の変化をもたらすアビジンで修飾された固相上に固定することができる。この方法では、標的の捕獲はサンプルから標的の分離を促進し、アッセイのシグナルまたは他の特徴の検出を妨げうるサンプル中の物質を除去することができる。
【0049】
HSV−1またはHSV−2標的配列を検出し同定するために使用されるプライマーおよびプローブを表1に列記した。特異的HSV標的結合配列にはアンダーラインを付し、大文字で示し、一方、制限酵素エンドヌクレアーゼ部位は小文字のイタリック体で示した。IACアダプタプライマーについては、IAC標的結合配列を小文字にアンダーラインを付すことにより示した。全てのプライマーは、5’→3’の方向に位置づけされている。
【0050】
【表1−1】
【0051】
【表1−2】
【0052】
HSV−2標的配列を検出し同定するために使用されるプライマーおよびプローブを表2に列記した。
【0053】
【表2−1】
【0054】
【表2−2】
【0055】
本発明の核酸プライマーは、種々の系統についてのHSV遺伝子の糖タンパク質G(Glycoprotein G)(US4)を分析することによって生じるコンセンサス配列に基づいて設計される(図1および6、表1および2参照)。SDAおよびユニバーサル検出法で使用するためのバンパープライマー、アダプタプライマー、および検出プライマーも示されている。設計されたHSV−1プライマーは、表4に例示されるように全ての系統に認められるHSV−1標的配列を特異的に増幅する。HSV−2プライマーは、表7に例示されるように全ての系統に認められるHSV−2標的配列を特異的に増幅する。HSV−1とHSV−2標的配列の間の相同性は約90%であるので、プライマーは、HSV−1およびHSV−2の間を特異的に明確に区別するために注意深く設計される。標的結合配列に実質的に相同である配列および表1および2に列挙されたこのような実質的に相同な標的結合配列を含むプライマーもまた本発明に包含される。
【0056】
本発明の一実施形態では、HSV−1標的領域は、まず、ヒトHSV−1の完全なHSV−1ゲノム配列、すなわち152,261塩基長を有する系統17(NCBIアクセス番号X14112)から選択される。糖タンパク質「US4」遺伝子は、136,744〜137,460塩基に位置する。HSV−1左側バンパープライマー(HSV1LB1.0)(5'末端)は、核酸137,256に位置する。HSV−1右側バンパープライマー(HSV1RB1.1)(5'末端)は、核酸137,382に位置する。全てのHSV−1SDAシステム用のプライマーは、これらのバンパープライマー座標(coordinates)の範囲内に位置する。
【0057】
本発明の他の実施形態はヒトHSV−2の完全なHSV−1ゲノム配列、すなわち154,746塩基長を有する系統HG52(NCBIアクセス番号Z86099)に関する。糖タンパク質G「US4」遺伝子は、137,878〜139,977塩基に位置する。HSV−2左側バンパープライマー(HSV2LB1.0)(5'末端)は、位置139,773に位置する。HSV−2右側バンパープライマー(HSV2RB1.0)(5'末端)は、位置139,896に位置する。全てのHSV−2SDAシステム用のプライマーは、このバンパープライマー座標(coordinates)の範囲内に位置する。
【0058】
PCR増幅プライマーは、HSV標的DNAをプラスミドベクターへクローニングするために設計される。配列番号5〜6および配列番号36〜37のHSV−1およびHSV−2PCR増幅プライマーは、それぞれ、HSVゲノムの高度に保存された標的配列領域に対して相補的である。PCR増幅プライマーは、HSVの糖タンパク質G(US4)遺伝子のDNAフラグメントを含むHSV標的配列領域を増幅する。HSV標的領域を含む単純ヘルペスウイルス(HSV)ゲノムの増幅されたフラグメントは、手頃な制限酵素部位を含むプラスミドベクターに指向性を持ってクローニングされる。HSVフラグメントは、当業者によって理解されるように任意のプラスミドベクターにクローニングすることができるが、一実施形態では、増幅されたHSV−1およびHSV−2フラグメントは、選択されたHSV標的領域に特異的なPCR増幅プライマーを用いて、Escherichia coliプラスミドベクター、pUC19(ジーンバンク/EMBLアクセス番号L09137)およびpUC18(ジーンバンク/EMBLアクセス番号L09136)にそれぞれクローニングされる。HSVフラグメントはHSV標的ストックと称される。標的HSV DNAは、ピコグリーン(PicoGreen)(登録商標)二本鎖DNA定量アッセイ(Molecular Probes,Inc.)を用いて定量されうる。表1および表2に列記されたプライマーの名前の中の「L」または「R」の存在は、増幅反応に使用されたときに、それぞれ「左側」または「右側」プライマーを示す。
【0059】
本発明の一実施形態では、PCR増幅プライマー、配列番号5〜6および36〜37は、最初に、それぞれHSV−1およびHSV−2遺伝子の糖タンパク質G遺伝子の152および254塩基対フラグメントを増幅する。それぞれ配列番号5および36のHSV−1およびHSV−2左側PCRプライマーは、各々EcoRI制限酵素部位を含めて設計される。それぞれ配列番号6および37のHSV−1およびHSV−2右側PCRプライマーは、各々BamHI制限酵素部位を有する。このフラグメントは、次に、適切な位置でpUCプラスミドベクターにクローニングされる。例示されたプラスミドベクターは、HSV−1およびHSV−2に対してそれぞれpUC19およびpUC18であり、これらは制限酵素部位EcoRIおよびBamHIを有する。制限酵素消化によって精製および線状化した後、続いて、HSV標的フラグメントをHSV増幅プライマーおよびバンパープライマーを用いて指数関数的に増幅する。
【0060】
本発明の標的結合配列およびプライマーは核酸増幅に有用である。一実施形態では、プライマーは鎖置換増幅(SDA)に特に有用である。これは、プライマーの伸長、半修飾制限エンドヌクレアーゼ認識/開裂部位のニッキング、単鎖伸長生成物の置換、伸長生成物(または本来の標的配列)へのプライマーのアニーリング、および引き続きのプライマーの伸長が反応混合物中で同時に起こる等温の核酸増幅法である。さらに、SDAは15分未満で1010倍を超える標的配列の複製を可能にする。一方、PCRでは、反応のステップは反応中の温度サイクルの結果として、別々のフェーズおよびサイクルで起こる。好熱性鎖置換増幅(tSDA)は、本質的に、熱に安定なポリメラーゼおよび熱に安定な制限エンドヌクレアーゼの置き換えを伴う、本明細書で、および、walkerら(1992, Proc. Natl. Acad. Sci USA. USA 89:392-396および1992, Nucl. Acids Res. 20:1691-1696(非特許文献9および10))によって記載された従来のSDA方法として実施することができる。温度は、置換された酵素に適したより高い温度に調整することができる。
【0061】
HSV増幅生成物または増幅された標的配列を検出する別の方法は、ポリアクリルアミドまたはアガロースゲル電気泳動であって、アガロースが臭化エチジウムで染色されるものによって特徴的なサイズを検出することによるものである。HSV−1またはHSV−2増幅プライマーを用いて生成された増幅生成物は、定量的ハイブリダイゼーション、または分子生物学の分野で熟練した者に公知の核酸検出のための等価な技術(Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring, NY (1989)(非特許文献12))によって検出することができる。
【0062】
表1および2に列記したプライマーは、サンプル中のHSV−1およびHSV−2の検出および同定に有用である。本明細書で使用される場合、S1およびS2増幅プライマーは、それぞれ第一および第二の増幅プライマーを表し、一方B1およびB2バンパープライマーは、それぞれ第一および第二のバンパープライマーを表す。手短には、SDA法では、S1増幅プライマーは単鎖HSV標的配列にハイブリダイズする。S1増幅プライマーのちょうど上流または5’で、第一のバンパープライマー、B1、が単鎖HSV標的配列にハイブリダイズする。DNAポリメラーゼは、B1バンパープライマーおよびS1増幅プライマーの3'末端を伸長するが、この場合、B1バンパープライマーの伸長は、最終的に、S1SDA伸長生成物を置換する。S1SDA伸長生成物は、S2増幅プライマー、およびS2増幅プライマーの上流でアニールするB2バンパープライマーによって捕獲される。DNAポリメラーゼはS2SDAおよびB2バンパープライマーの3’末端を伸長し、この場合、B2バンパープライマーの伸長は、下流のS2SDA増幅プライマー伸長生成物を置換する。S1増幅プライマーは置換されたS2増幅プライマー伸長生成物にアニールし、DNAポリメラーゼはハイブリダイズしたS1増幅プライマーの3’末端を伸長し、S2増幅プライマー伸長生成物およびその相補鎖を有する二本鎖分子を生じる。各鎖は、いずれかの末端でニック可能な制限酵素認識部位を有する。対応する制限酵素を添加して、チオレート化されたシトシンを含む修飾されたDNA鎖をニッキングし、ニック部位の3’に、短いニッキングされたテールおよび長い伸長産物を形成する。DNAポリメラーゼは、5’→3’方向に、ニッキングされた短いテールの3’からニッキングされた短いテールを伸長し、長い単鎖伸長生成物を置換する。手短には、S2増幅プライマー伸長生成物のニッキングされたテールとその相補体のニッキングされたテールは、それぞれ、ニッキングされた単鎖S2増幅プライマー伸長生成物と、ニッキングされた単鎖相補性S2増幅プライマー伸長生成物を置換する。一実施形態では、BsoBI酵素を使用して、配列番号52〜53および54〜55の配列をそれぞれ有する各鎖をニッキングし、且つ開裂または切断する。以下に示されるニック部位は、増幅プライマー配列に組み込まれ、半ホスホロチオエート化された認識配列(dCsTP、チオエート化されたシトシン)を必要とする。ニック部位であっても、配列番号53は、dCsTPの存在下でさえ、二本鎖開裂をする傾向があり、ニッキング可能な増幅プライマーを設計する場合には好ましい配列ではない。
【0063】
ニック部位:5’−CTCGGG−3’(配列番号52)および5’−CCCGGG−3’(配列番号53)
切断部位:5’−CTCGAG−3’(配列番号54)および5’−CCCGAG−3’(配列番号55)
【0064】
本発明のさらなる実施形態では、検出プライマーはHSV標的配列を検出するのに有用である。HSV標的配列に特異的なS1増幅プライマーおよび検出プライマーを使用することができ、この場合、検出プライマーはHSV標的結合配列を有する。DNAポリメラーゼは、S1プライマーおよび検出プライマーの3‘末端から伸長する。S1プライマーの伸長は、下流の検出プライマー伸長生成物を溶液中に置換し、この場合、これは捕獲されて相補性S2増幅プライマーにハイブリダイズする。DNAポリメラーゼは、S2増幅プライマーの3’末端を伸長し、検出プライマーの二次構造を開き、二本鎖制限酵素部位を形成し、2つの色素をクエンチャーの消光能力を無能力化して蛍光を生じるような距離まで引き離す。追加の蛍光は、制限酵素認識部位を開裂し、フルオロフォアとクエンチャーをさらに引き離すことによって生じる。
【0065】
SDA法に有用な酵素は、半ホスホロチオエート化認識配列に単鎖ニックを生じさせるものであり、この場合、ホスホロチオエート化された核酸の取り込みはニッキングおよび修復のさらなるラウンドを妨げない。これらの特徴を有する酵素の非制限的な例には、HincII、BsoBI、AvaI、NciI、およびFnu4HIが含まれる。有用なDNAポリメラーゼは、単鎖ニック部位でDNA合成を開始し、伸長した核酸鎖にホスホロチオエート化されたヌクレオチドを組み込み、そして5‘→3’エキソヌクレアーゼ活性を持たない鎖を置換するものである。開裂は、二本鎖または単鎖DNAのホスホジエステル結合の切断をいう。これらの特性を示すDNAポリメラーゼの非制限的な例には、エキソヌクレアーゼ欠損クレノウ、およびBstポリメラーゼおよびBcaポリメラーゼのエキソヌクレアーゼ欠損フラグメントが含まれる。他のDNAポリメラーゼおよび制限酵素はSDAに適している(Walker et al. Proc. Natl. Acad. Sci USA, Vol. 89, pp. 392-396, January 1992, Applied Biological Sciences(非特許文献9))が、エキソ−BstポリメラーゼおよびBsoBIがこれらの熱特性および互いの適合性によって選択された。本発明の一実施形態では、BsoBI制限エンドヌクレアーゼ認識部位が使用され、これはイタリック体で示されている(表1および2参照)。HSV標的結合配列を単独で使用して、専用の配列または構造を必要とすることなく反応中にHSV標的を増幅できること、および、SDA以外の増幅反応(たとえばRNAポリメラーゼプロモータ)で必要とされる他の専用の配列が、このシステムで、たとえば本明細書に記載されているRERS含有配列に置き換えられることは容易に認識できるであろう。
【0066】
次に、標的ストックは、増幅プライマーの存在下で、増幅プライマー単独またはバンパープライマー、ユニバーサル検出用のシグナル/アダプタプライマー、およびユニバーサル検出プライマーと組み合わせて増幅される。増幅反応については、1つの「左側」増幅プライマーを含む少なくとも1対が選択され、1つの「右側」増幅プライマーが選択されて、HSV標的ストック配列の各鎖を増幅する。左側および右側増幅プライマーに加えて、SDA反応では、1つの左側および右側バンパープライマー対を最初に使用する。さらに、検出用に、シグナル/アダプタプライマーおよび検出プライマーを、HSV標的ストック配列を検出し、同定するために使用する。
【0067】
HSV−1またはHSV−2DNAを特異的に増幅し、検出するいくつかのHSVシステムが本発明に包含される。例えば、HSV−1システムは以下のプライマーを含む:HSV1GGLP1.1、HSV1GGRP5.2、HSV1GGAD2.1、HSV1GGLB1.0、HSV1GGRB1.1、およびTBD16(D/R)、または、そのほかには、HSV1GGLP1.1、HSV1GGRP5.2、HSV1GGAD3.0またはHSV1GGAD3.1、HSV1GGLB1.0、HSV1GGRB1.1、MPC.DR、HSV1IAC AD8.1またはHSV1IACAD8.7、MPC2.FD。他の実施形態では、プライマーの種々の組み合わせを用いたHSV−2システムを表3に列記した。プライマーの他の組み合わせが意図されるが、当業者は、サンプル中のHSV−1またはHSV−2を検出するためにプライマーを組み合わせる知力を有するであろう。プライマーは、表1および2に列記されたものから選択され、サンプル中のHSV−1またはHSV−2を同定するために統計学的に設計された実験で試験される。あるいは、HSV−1またはHSV−2に特異的であり、表1および2に列記されたものと実質的に相同であるプライマーをHSV−1またはHSV−2標的配列の検出に使用することもできる。
【0068】
【表3】
【0069】
商業的な利便性のためには、核酸の特異的な検出および同定のための増幅プライマーはキットの形態でパッケージングすることができる。一般にこのようなキットは、少なくとも1対のHSV増幅プライマーを含有する。核酸増幅反応を実施するための試薬、例えば、バッファー、追加のプライマー、ヌクレオチドトリホスフェート、酵素などを標的特異的増幅プライマーに含むことができる。キットの成分は、本発明の方法の特定の実施形態を実施するための使用説明書を含む、一般的な容器内に一緒にパッケージングされる。他の任意成分、例えばアッセイプローブとして使用するのに適した標識をタグ付けしたプライマー、および/またはその標識を検出するための試薬または手段をキットに含むことができる。
【0070】
本発明の一実施形態では、第一の増幅プライマーすなわちS1SDA増幅プライマー、および第二の増幅プライマーすなわちS2SDA増幅プライマーを含むキットを提供する。このキットは、第一のB1バンパープライマーおよび第二のB2バンパプライマー、検出プライマー、並びに、任意選択的に、IACアダプタプライマーおよびIAC標的配列を含む内部増幅対照(IAC)標的配列を同時に検出するための試薬をさらに含むことができる。キットは、HSV−1またはHSV−2に特異的なプライマーから構成されていてもよく、または、キットは、HSV−1およびHSV−2の両方を指向するプライマーから構成されていてもよい。この場合、当業者は、HSV−1を検出し同定する増幅反応がHSV−1プライマーを利用し、HSV−2を検出し同定する増幅反応がHSV−2プライマーを利用することを理解するであろう。サンプルがHSV−1またはHSV−2DNAを含有するかどうかを同定するために、HSV−1およびHSV−2のためのプライマーは混合されるべきではない。
【0071】
さらに別の実施形態では、本発明のキットおよびプライマーは、臨床サンプルがHSV−1またはHSV−2を含有するかどうかを検出し診断するために使用することができる。臨床サンプルは、SDA増幅プライマーを用いて増幅および検出することができるか、またはバンパープライマー、アダプタプライマー、および検出プライマーをさらに含むSDA反応に使用することができる。本発明の一実施形態では、IACアダプタプライマーは、HSV−1またはHSV−2に対する陽性および陰性対照に添加して反応の内部増幅対照として使用することができる。当業者は、本明細書の記載を読むことによって、および、当分野で一般的な方法および技術から、サンプル中のHSV−1およびHSV−2の検出および同定用のプライマーをどのように作成し使用するかを理解するであろう。
【0072】
さらに、商業的な実施形態では、本発明のプライマーおよびSDA用試薬を含む組成物を乾燥形態または液体形態で提供することができる。組成物は乾燥形態の場合により安定で容易に取り扱うことができる。乾燥された組成物は、マイクロタイタープレート、マイクロアレイ、または他の適切な容器のような固相に添加されるか、またはそのような固相にあらかじめ処理することができる。この場合、サンプルおよびSDAバッファを加えることのみが必要である。この形態は、多数のサンプルを同時にアッセイすることを容易にし、高スループット法に有用である。本発明の一実施形態では、BDプローブテック(商標)ET装置を使用することができる。
【0073】
標的結合配列、および、任意選択的に、選択された増幅反応に必要な配列または選択された検出反応に必要な配列からなる本発明に従った核酸が、スペーサー、リンカー、酵素の標識または結合のための配列、または他のものとして機能する他の特定の配列を含んでもよいことは理解されるべきである。このような追加の配列は、選択された反応中の核酸の最適な機能を得るために必要であることは一般に知られている。
【0074】
本明細書で引用した、すべての特許、特許出願、刊行されたPCT出願および物品、本、参照文献、参照マニュアルおよび要約は、本発明が関係する分野の状態をより完全に説明するためにその全体を参照により本明細書に取り込む。
【0075】
本発明の範囲および精神から逸脱することなく、上記の主題に種々の変更を行うことができるので、上記説明に含まれるかまたは添付の特許請求の範囲で定義されるすべての主題は本発明の記述および例示として解釈されることが意図される。本発明の多くの修飾および変更が上記教示に照らして可能である。
【実施例】
【0076】
本明細書の実施例は、本発明を実施する種々の側面を例示することを意味し、本発明の範囲をいずれの方法においても制限することを意図しない。実施例には、ベクターの構築またはそのようなベクターへのcDNAの挿入のような、使用される従来法の詳細な説明は含まれない。このような方法は、当業者に周知であり、多くの刊行物、例えばJ. Sambrook and D. W. Russell, Molecular Cloning: a Laboratory Manual, 3rd Edition, Cold Spring Harbor Laboratory Press, USA, (2001)(非特許文献17)に説明されている。
【0077】
(実施例1)
HSV−1糖タンパク質−G鎖置換増幅標的領域のクローニング
表1に同定されている配列番号5および配列番号6のPCR増幅プライマーを用いてHSV−1(系統ATCC:VR−539)からのDNAについてPCRを行った。これらのプライマーを使用して、HSV−1の糖タンパク質G(US4)遺伝子内の152塩基対フラグメントを増幅した。PCR増幅されたDNAをpUC19プラスミドベクター(Gibco BRL; Grand Island,NY)に挿入した。組み換えクローンをHSVIGGプラスミド#1と命名した。プラスミド#1DNAをBamH1制限酵素で消化することによって精製および線状化した。次に、DNAをQIAクイック(Qiagen,Inc.;Valencia,CA)スピンカラムを用いて精製し、蛍光ピコグリーン(登録商標)試薬で分析することによって定量した。将来の実験のために標的HSV−1DNAの希釈物を10ng/μlのヒトDNAを含む水中で調製した。特異的HSV−1系統の希釈物および各希釈物中のHSV−1検出の結果を図5に示した。「プラス」の記号はサンプル中にHSV−1の存在を示し、「マイナス」の記号は、サンプル中にHSV−1がないことを示し、疑問符はサンプルの汚染の疑いを示す。すべての系統は、サンプルO−2526を除いて、ストックの1:10希釈で陽性であった。ストックの1:1,000希釈で23系統のうち20系統が陽性であった。ストックの1:100,000希釈では、23系統のうち15系統が陽性であった。
【0078】
(実施例2)
クローニングされたHSV−1DNAの増幅
HSV−1DNAを検出するためのアッセイの最初のステップとして、実施例1で説明した標的核酸法を用いてHSV−1DNAの増幅のためのSDAシステムを開発した。DNA増幅アッセイの分析感度はクローニングされたプラスミドの希釈物を用いて評価した。8つの複製SDA反応を各標的レベルで実施した。これら8つの反応は、反応あたり100、50、25、12.5、6.25、および0コピーの二本鎖DNAに等しい。増幅は、BDプローブテックET装置(BD Diagnostic System;Sparks,MD)を使用し、各50nMのHSV1GGLB1.0およびHSV1GGRB1.0バンパープライマー、100nMのHSV1GGLP1.0左側増幅プライマー、500nMのHSV1GGRP1.0右側増幅プライマー、250nMのHSV1GGAD1.0アダプタプライマー、500nMのTBD10.2 D/R検出プライマーを用いて実施した。これらのプライマーの配列は表1に列記されている。最終のバッファ条件は、以下の通りである:143mMビシン(Bicine)、82mMKOH、25MMKiPO4、12.5%DMSO、5mM酢酸マンガン、500mM2’−デオキシシトシン−5−o−(1−チオトリホスフェート)(dCsTP)、それぞれ100nMのdATP、dGTPおよびdTTP、100ng/μlBSA、約12単位のBstポリメラーゼ、および約30単位のBsoBI制限エンドヌクレアーゼ。
【0079】
蛍光を、1時間の過程で60パスモニターした。結果を曲線の下の領域の換算、すなわち「MOTA」スコアーで表した。陽性のMOTAスコアーは、決まりきった実験によって容易に決定することができる。本発明の目的では、3500以上のMOTAスコアーを「陽性」とみなした。アッセイが100%の陽性の結果を与える最も低いレベルのHSV1GG標的DNAは反応あたり100コピーである。8つの複製のうちの7つ(87.5%)が反応あたり50コピーの標的DNAでも陽性であった。
【0080】
(実施例3)
SDAによる単純ヘルペス1ウイルス粒子の検出
HSV−1を検出するための本発明のプライマーおよびプローブの能力を確認するために、アメリカンタイプカルチャーコレクション(ATCC;Manassas,VA)から得たHSV−1の4系統、Quest Diagnostic(Baltimore,MD)から得た10系統、およびオハイオ州立大学(OSU)からの14の分類されていないHSVサンプルについてSDAを実施した。
【0081】
HSVの分類されていない系統は、PCRによってDNAポリメラーゼ遺伝子の領域を増幅することにより特徴づけた。1セットの増幅プライマーを設計して、HSV−1およびHSV−2の両方の同じ領域を増幅した。この2つのタイプのウイルスは、HSV−1の系統から生成されるPCR産物中には存在しないHSV−2PCRフラグメント中のApal制限エンドヌクレアーゼ認識部位の存在によって明確に区別できた。Apal制限酵素でインキュベートした場合、HSV−2由来の増幅生成物を2つの短いフラグメントに開裂し、一方HSV−1から得られるものは無傷のままである。制限されたフラグメントは、適切な対照とともにアガロースゲル電気泳動で解析した。
【0082】
SDAシステムでHSVの存在および非存在を評価するために使用されたウイルスストックの濃度は未知であった。ウイルスストックをリン酸塩緩衝食塩水で1:10に希釈し、この懸濁液の10μlをSDAで試験した。結果を表4に示す。HSV−1のすべての系統がHSV−1増幅プライマーを用いて検出され、多様な起源からのHSV−1の系統を検出するための、開示されたプライマーおよびプローブの能力が実証された。Apal制限消化物によって分類されたHSVの、以前には分類されていない14の系統のうち、9つがHSV−1であると決定され、4つがHSV−2であると決定され、1つがPCRによって増幅されなかった(表4および5参照)。
【0083】
【表4】
【0084】
(実施例4)
SDA法の分析感度
本発明で開示されたプライマーおよびプローブを使用したHSV−1アッセイの検出限界を決定するために、クローニングした標的核酸の希釈物およびウイルス粒子の一連の希釈物についてSDA反応を実施した。ウイルス粒子のストックを電子顕微鏡(Electron Microscopy Bioservices)で数えた。16の複製を各標的レベルで試験した。
【0085】
アッセイの感度および特異性を確認するために、種々の地理的な位置から得た23系統のHSV−1を、微生物ストックの1:10、1:1,000および1:100,000希釈で試験した。以前に分類されていない系統から、およびQuest Diagnositicsからのサンプルの力価は未知であった。ATCCからのHSV−1の2つのストックの力価はほぼ以下の通りであった:VR260、1.5×104TCID/μlおよびVR−539、2.0×105TCID/μl。結果を図5に示す。系統#0−2526を除いて、すべての系統がストック懸濁液の1:10希釈で陽性であった。23系統のうち、試験した20系統がストックの1:1000希釈で陽性であり、試験した15の系統が1:100,000の希釈で陽性であった。
【0086】
(実施例5)
HSV−1DNAに対するSDAの特異性
HSV−2の16の系統をHSV1GGSDAシステムで試験した。HSV−2希釈物の10ミリリットルの各懸濁液を反応あたり添加した。試験した17のストックのうちの1つがHSV1GGシステムで陽性であった。結果を表5に示す。加えて、23の他の微生物を、開示された本発明の方法のプライマーおよびプローブを用いて試験した。これらの微生物には、細菌、酵母および臨床標本中で遭遇するであろう他のウイルスが含まれる。試験された微生物のいずれもHSV−1に対して陽性ではなかった。結果を表6に示す。
【0087】
【表5】
【0088】
【表6】
【0089】
(実施例6)
HSV−2糖タンパク質−GSDA標的領域のクローニング
プライマーHSV2PCRRおよびHDV2PCRLを用いてHSV−2系統のATCC VR−540からのDNAについて、69℃のアニーリング温度でPCRを実施した。これらのプライマーは、HSV−2の糖タンパク質G(US4)遺伝子の範囲内の254塩基対フラグメントを増幅する。pUC18プラスミドベクター(invitrogen)に増幅したDNAを挿入した。組み換えクローンにはpHSV2−NT#9−1という名を付けた。プラスミドDNAを、BamHI制限酵素を用いて消化することにより精製し線状化した。DNAをQIAGEN QIAクイックスピンカラムを用いて精製し、蛍光ピコグリーン(登録商標)試薬による分析で定量した。将来の実験のための標的DNAの希釈物を7ng/μlのサーモン精子DNAを含有する水中で調製した。
【0090】
(実施例7)
クローニングされたHSV−2DNAの増幅
DNA増幅アッセイの分析感度はクローニングされたプラスミドの希釈物を用いて評価した。8つの複製SDA反応をステム2.0および5.2を用いて各標的レベルで実施した。これら8つの反応は、反応あたり500、250、100、50、25、10、および0コピーの二本鎖DNAに等しい。増幅は、BDプローブテックET装置を使用し、各50nMのHSV2GGLB1.0およびHSV2GGRB1.0、100nMのHSV2GGLP1.0、500nMのHSV2GGRP2.0もしくは5.2、250nMのHSV2GGAD1.0、500nMのMPC.D/Rを用いて実施した。これらのプライマーの配列は表2に列記されている。最終のバッファ条件は、以下の通りである:71mMビシン(Bicine)、56.6mMKOH、23.9mMKPO4、15.4%DMSO、5mM酢酸マンガン、500mM2’−デオキシシトシン−5−o−(1−チオトリホスフェート)(dCsTP)、それぞれ100nMのdATP、dGTPおよびdTTP、100μg/μlBSA、約3.515単位のBstポリメラーゼ、および約30単位のBsoBI制限エンドヌクレアーゼ。
【0091】
蛍光を、1時間の過程で60パスモニターした。結果を曲線の下の領域の換算、すなわち「MOTA」スコアーおよびPATスコアー(パス・アフター・スレッシュホールド)で表した。本発明の目的では、3500以上のMOTAスコアーおよび0を超えるPATスコアーを「陽性」とみなした。アッセイが100%の陽性の結果を与える最も低いレベルのHSV2GG標的DNAは、システム2.0および5.2の両方に対して反応あたり50コピーである。8つの複製のうちの7つが、システム2.0に対して25コピーで陽性であり、8つの複製のうちの6つが、システム5.2に対して25コピーで陽性であった。
【0092】
(実施例8)
SDAによるHSV−2ウイルス粒子の検出
HSV−2を検出するための本発明のプライマーおよびプローブの能力を確認するために、アメリカンタイプカルチャーコレクション(ATCC)から得たHSV−2の2系統、Quest Diagnostic(Baltimore,MD)から得た9系統、およびApaI分析によってHSV−2として分類されたオハイオ州立大学(OSU)からの5系統についてSDAを実施した。
【0093】
OSUから得た系統は、PCRによってDNAポリメラーゼ遺伝子の領域を増幅することにより特徴づけた。1セットのPCRプライマーを設計して、HSV−1およびHSV−2の両方の同じ領域を増幅した。この2つのタイプのウイルスは、HSV−1の系統から生成されるPCR産物中には存在しないHSV−2PCRフラグメント中のApaI制限エンドヌクレアーゼ認識部位の存在によって明確に区別できた。ApaI制限酵素でインキュベートした場合、HSV−2由来のPCR生成物を2つの短いフラグメントに開裂し、一方HSV−1から得られるものは無傷のままである。制限されたフラグメントは、適切な対照とともにアガロースゲル電気泳動で解析した。
【0094】
SDAシステムを評価するために使用されたウイルスストックの濃度は未知であった。ウイルスストックをリン酸塩緩衝食塩水で1:10に希釈し、この懸濁液の10μlをSDAで試験した。結果を表7に示す。HSV−2のすべての系統がシステム1.0、2.0および5.2からの増幅プライマーを用いて検出され、多様な起源からのHSV−1の系統を検出するための、開示されたプライマーおよびプローブの能力が実証された。
【0095】
【表7−1】
【0096】
【表7−2】
【0097】
(実施例9)
HSV−2DNAに対するSDAの特異性
HSV−1の25の系統をHSV2GGSDAシステム1.0、2.0および5.2で試験した。HSV−1希釈物の10ミリリットルの各懸濁液を反応あたり添加した。試験した25の系統のいずれもHSV−2システムで検出されなかった。結果を表8に示す。加えて、他の微生物のパネルを、開示された本発明の方法のプライマーおよびプローブを用いて試験した。これらの微生物には、細菌、酵母および臨床標本中で遭遇するであろう他のウイルスが含まれる。試験された微生物のいずれもHSV−1に対して陽性ではなかった。結果を表9に示す。
【0098】
【表8−1】
【0099】
【表8−2】
【0100】
【表9】
【特許請求の範囲】
【請求項1】
配列番号7〜8のいずれか1つの標的結合配列からなる配列、5’BsoBI制限エンドヌクレアーゼ認識部位、および5’非標的特異的5’テール末端配列を含むことを特徴とするポリヌクレオチド。
【請求項2】
配列番号7または8の配列からなることを特徴とする請求項1に記載のポリヌクレオチド。
【請求項3】
単純ヘルペスウイルス1型(HSV−1)標的配列を検出する方法であって、
(a)請求項1または2に記載のポリヌクレオチドをハイブリダイズする工程、および(b)HSV−1標的配列を検出する工程を含むことを特徴とする方法。
【請求項4】
(a)配列番号8又は7のいずれか1つの標的結合配列からなるHSV−1標的結合部位を有する配列、5’BsoBI制限エンドヌクレアーゼ認識部位、および5’非標的特異的5’テール末端配列を含む第一の増幅プライマーを用いて、HSV−1標的配列を増幅する工程、
(b)配列番号18を含むか、または配列番号9〜17のいずれか1つの標的結合配列からなるHSV−1標的結合部位を有する配列を含む第二の増幅プライマー配列を用いて、HSV−1標的配列を増幅する工程、および
(c)増幅されたHSV−1標的配列を検出する工程
をさらに含むことを特徴とする請求項3に記載の方法。
【請求項5】
前記第一のプライマーが配列番号7または配列番号8の配列を含むことを特徴とする請求項4に記載の方法。
【請求項6】
請求項4または5に記載の方法であって、
(i) 増幅反応が鎖置換増幅(SDA)反応であるか、または
(ii)検出方法が、直接検出、ポリメラーゼ連鎖反応(PCR)、リガーゼ連鎖反応(LCR)、in situ ハイブリダイゼーション、転写介在増幅手法(TMA)、自己持続性配列複製法(Self-Sustaining Sequence Replication)(3SR)、ローリングサークル増幅法(Rolling Circle Amplification)(RCA)、Qβレプリカーゼシステム、および核酸配列をベースとする増幅法(Nucleic Acid Sequence Based Amplification)(NASBA)からなる群から選択されるか、または
(iii)第一の増幅プライマーが、ヘアピン、g−カルテット、およびアッセイプローブに結合する配列から選択される配列をさらに含むか、または
(iv)第一の増幅プライマーが、好ましくは蛍光部分である検出可能な標識をさらに含むか、または
(v)第一の増幅プライマーが、制限酵素認識部位およびRNAポリメラーゼプロモータから選択される配列を含むか、または
(vi)前記方法が、内部増幅対照(IAC)を増幅する工程をさらに含む
ことを特徴とする方法。
【請求項7】
(i)IACが配列番号26〜27から選択されるか、または
(ii)前記方法が、増幅されたIACを検出する工程、好ましくは、増幅されたIACが増幅された標的配列とは異なる手段で検出される工程をさらに含む
ことを特徴とする請求項6に記載の方法。
【請求項8】
請求項7に記載の方法であって、
(a)配列番号18を含むか、または配列番号9〜17のいずれか1つの標的結合配列からなるHSV−1標的結合部位を有する配列を含む1以上のポリヌクレオチドをハイブリダイズする工程、および
(b)HSV−1標的配列を検出する工程であって、好ましくは、前記1以上のポリヌクレオチドが、好ましくは蛍光部分である検出可能な標識をさらに含む工程
をさらに含むことを特徴とする方法。
【請求項9】
請求項4または5において規定された1以上の第一のプライマーを含むことを特徴とする配列を検出するためのキット。
【請求項10】
請求項9に記載のキットであって、
(I)(i)増幅プライマー、好ましくは配列番号9〜18から選択されるもの、または
(ii)バンパープライマー、好ましくは配列番号23〜25から選択されるもの、または
(iii)アダプタプライマー、好ましくは配列番号19〜22から選択されるもの、および
(iv)検出プライマー、好ましくは配列番号30〜35から選択されるもの
をさらに含むか、或いは
(II)前記キットはHSV−1標的配列を検出するためのものであり、配列番号7〜8からの1つ、および配列番号9〜18からの少なくとも1つから選択される増幅プライマーの1つ以上;および配列番号23〜25から選択されるバンパープライマーを含み、好ましくは、前記キットは、(i)配列番号19〜22から選択される1以上のアダプタプライマー;および(ii)配列番号30〜35からなる群から選択される1以上の検出プライマー;および/または(iii)配列番号26〜27から選択される1以上のIAC標的配列;および(iv)配列番号28〜29からなる群から選択される1以上のIACアダプタプライマーをさらに含む
ことを特徴とするキット。
【請求項11】
請求項4または5において定義された1以上の第一のプライマーを含むことを特徴とするHSV−1を検出するための組成物。
【請求項12】
(I)配列番号9〜18および配列番号23〜25のいずれか1つの標的結合配列からなる1以上のプライマーをさらに含むこと、および/または
(II)(a)アダプタプライマーの3’末端に位置する標的結合配列を介して標的配列にハイブリダイズすることができるアダプタプライマー配列であって、このアダプタプライマーが5’ジェネリックテールを含み、かつこのアダプタプライマーが配列番号19〜22からなる群から選択されるもの;および
(b)検出プライマーの3’部分を介してアダプタプライマーの5’テールにハイブリダイズすることができる検出プライマー配列であって、この検出プライマー配列が5’制限酵素認識部位、ならびに、蛍光部分、放射性同位体、化学発光剤、目で見える反応生成物を生じることができる酵素基質、およびリガンドで検出可能に標識されたリガンド結合パートナーからなる群から選択される検出可能な標識を含むもの
をさらに含み、好ましくは、
(i)前記検出プライマー配列がヘアピンおよびカルテットからなる群から選択される構造部分を含むか、または
(ii)前記検出可能な標識が蛍光部分であり、好ましくはこの蛍光部分が、フルオロセイン(FAM)/ローダミン(ROX);FAM/P−(ジメチルアミノフェニルアゾ)安息香酸(DABCYL);ROX/DABCYL;フルオロセインイソチオシアネート(FITC)/テトラメチルローダミンイソチオシアネート(TRITC);FITC/テキサスレッド(商標);FITC/N−ヒドロキシスクシンイミジル1−ピレンブチレート(PYB);FITC/エオシンイソチオシアネート(EITC);N−ヒドロキシスクシンイミジル1−ペンタンスルホネート(PYS)/FITC;FITC/ローダミン X;およびFITC/テトラメチルローダミン(TAMRA)からなる群から選択されるドナーおよびクエンチャー色素対を含むか、または
(iii)前記検出プライマーが配列番号30〜35からなる群から選択されるものであること;および/または
(III)増幅反応によってサンプル中のHSV−1標的配列の検出のためのプライマーを含み、このプライマーは、
(a)HSV−1標的配列にハイブリダイズできる第一の増幅プライマー配列であって、第一の増幅プライマーが配列番号7〜8からなる群から選択されるもの、
(b)HSV−1標的配列の相補体にハイブリダイズできる第二の増幅プライマー配列であって、第二の増幅プライマーが配列番号9〜18からなる群から選択されるもの、
(c)第一の増幅プライマーの上流でHSV−1標的配列にハイブリダイズできる第一のバンパープライマー配列であって、第一のバンパープライマーが配列番号23であるもの、および
(d)第二の増幅プライマーの上流でHSV−1標的配列の相補体にハイブリダイズできる第二のバンパープライマー配列であって、第二の増幅プライマーが配列番号24〜25からなる群から選択されるもの
を含むこと
を特徴とする請求項11に記載の組成物。
【請求項13】
サンプル中のHSV−1標的配列の存在を検出するための請求項3に記載の方法であって、前記方法が、
(a)配列番号7〜8からなる群から選択される第一の増幅プライマー、および配列番号9〜18からなる群から選択される第二の増幅プライマーにサンプルを添加し、増幅されたHSV−1核酸生成物を産生する工程、および
(b)増幅されたHSV−1核酸生成物を検出する工程であって、増幅生成物の検出がサンプル中のHSV−1の存在を示す工程
を含むことを特徴とする方法。
【請求項14】
請求項13に記載の方法であって、
(i)前記増幅されたHSV−1核酸生成物が、ユニバーサル検出、ゲル電気泳動、および定量的ハイブリダイゼーションからなる群から選択される方法によって検出されること、または
この方法が、
(c)配列番号23の第一のバンパープライマーおよび配列番号25または配列番号26の第二のバンパーをサンプルに添加する工程をさらに含むこと
を特徴とする方法。
【請求項15】
検出工程が、
(d)第一の増幅プライマーおよび配列番号19〜22からなる群から選択されるアダプタプライマーを増幅されたHSV−1標的配列にハイブリダイズする工程であって、アダプタプライマーが第一の増幅プライマーの下流でハイブリダイズする工程、
(e)第一の増幅プライマーおよびアダプタプライマーの3’末端を伸長し、アダプタプライマー伸長生成物を産生する工程であって、第一の増幅プライマーの伸長がアダプタプライマー伸長生成物を置換する工程、
(f)第二の増幅プライマーをアダプタプライマー伸長生成物へハイブリダイズする工程、
(g)第二の増幅プライマーの3’末端を伸長し、アダプタプライマー伸長生成物およびその相補体を含む二本鎖分子を産生する工程であって、各鎖がニック可能な制限酵素部位を含む工程、
(h)二本鎖分子をニッキングし、5’部分および3’部分を造り出す工程であって、5’部分がニッキングされたテールを含み、3’部分がニッキングされた相補性アダプタプライマー伸長生成物を含む工程、
(i)ニッキングされたテールの3’末端を伸長し、ニッキングされた相補性アダプタプライマー伸長生成物を置換する工程であって、置換され、ニッキングされた相補性アダプタプライマー伸長生成物が一本鎖である工程、
(j)一本鎖相補性アダプタプライマー伸長生成物へ検出プライマーをハイブリダイズする工程であって、検出プライマーが検出可能な標識を含み、HSV−1標的配列を検出する工程、
(k)検出プライマーおよび相補性アダプタプライマー伸長生成物の3’末端を伸長し、検出プライマー伸長生成物およびその相補体を含む二本鎖検出分子を産生する工程であって、各鎖が開裂可能な制限酵素認識部位を含む工程、
(l)二本鎖検出分子を開裂する工程、および
(m)検出可能な標識を検出する工程
をさらに含むことを特徴とする請求項14に記載の方法。
【請求項16】
(n)配列番号26〜27からなる群から選択される内部増幅対照(IAC)標的配列を増幅する工程、
(o)第一の増幅プライマーおよび配列番号28〜29からなる群から選択されるIACアダプタプライマーをIACにハイブリダイズする工程であって、アダプタプライマーが第一の増幅プライマーの下流でハイブリダイズする工程、
(p)第一の増幅プライマーおよびIACアダプタプライマーの3’末端を伸長し、IACアダプタプライマー伸長生成物を産生する工程であって、第一の増幅プライマーの伸長がIACアダプタプライマー伸長生成物を置換する工程、
(q)IACアダプタプライマー伸長生成物へ第二の増幅プライマーをハイブリダイズする工程、
(r)第二の増幅プライマーの3'末端を伸長し、IACアダプタプライマー伸長生成物およびその相補体を含む二本鎖分子を産生する工程であって、各鎖がニック可能な制限酵素部位を含む工程、
(s)二本鎖分子をニッキングする工程、
(t)相補性IACアダプタプライマー伸長生成物のニッキングされたテール末端を伸長し、一本鎖でニッキングされた相補性アダプタプライマー伸長生成物を置換する工程、
(u)一本鎖相補性IACアダプタプライマー伸長生成物にIAC検出プライマーをハイブリダイズする工程であって、IAC検出プライマーが検出可能な標識を含み、IACを検出する工程、
(v)IAC検出プライマーおよび一本鎖相補性IACアダプタプライマー伸長生成物の3’末端を伸長し、検出プライマー伸長生成物およびその相補体を含む二本鎖検出分子を産生する工程であって、各鎖が開裂可能な制限酵素認識部位を含む工程、
(w)二本鎖検出分子を開裂する工程、および
(x)検出可能な標識を検出する工程
をさらに含むことを特徴とする請求項15に記載の方法。
【請求項17】
HSV−1標的配列をサンプル中で検出するための請求項3に記載の方法であって、前記方法が、
(a)配列番号7〜8からなる群から選択される配列を有する第一の増幅プライマーをHSV−1標的配列へハイブリダイズする工程、
(b)配列番号23の第一のバンパープライマーをHSV−1標的配列へハイブリダイズする工程であって、第一のバンパープライマーが第一の増幅プライマーの上流でHSV−1標的配列にハイブリダイズし、第二のバンパープライマーが第二の増幅プライマーの上流で相補性HSV−1標的配列にハイブリダイズする工程、
(c)第一のバンパープライマーおよび第一の増幅プライマーの3’末端を伸長し、第一の増幅プライマー伸長生成物を産生する工程であって、第一のバンパープライマーの伸長が第一の増幅プライマー伸長生成物を置換する工程、
(d)配列番号9〜18からなる群から選択される配列を有する第二の増幅プライマーを第一の増幅プライマー伸長生成物へハイブリダイズする工程、
(e)配列番号24〜25からなる群から選択される配列を有する第二のバンパープライマーを第一の増幅プライマー伸長生成物へハイブリダイズする工程、
(f)第二のバンパープライマーおよび第二の増幅プライマーの3’末端を伸長し、第二の増幅プライマー伸長生成物を産生する工程であって、第二のバンパープライマーの伸長が第二の増幅プライマー伸長生成物を置換する工程、
(g)第一の増幅プライマーを第二の増幅プライマー伸長生成物へハイブリダイズする工程、
(h)工程(g)の第一の増幅プライマーの3’末端を伸長し、第二の増幅プライマー伸長生成物およびその相補体を含む二本鎖分子を産生する工程であって、各鎖がニック可能な制限酵素部位を含む工程、
(i)二本鎖分子をニッキングし、5’部分と3’部分を造り出す工程であって、5’部分はニッキングされたテールを含み、3’部分はニッキングされた第二の増幅プライマー伸長生成物およびその相補体を含む工程、
(j)ニッキングされたテールの3’末端を伸長し、ニッキングされた第二増幅プライマー伸長生成物を置換する工程であって、ニッキングされた相補性第二増幅プライマー伸長生成物が一本鎖である工程、
(k)ニッキングされたテールの3’末端を伸長し、ニッキングされた相補性第二増幅プライマー伸長生成物を置換する工程であって、ニッキングされた相補性第二増幅プライマー伸長生成物が一本鎖である工程、
(l)第一の増幅プライマーおよび第二の増幅プライマーを、各一本鎖分子に別々にハイブリダイズし、工程(h)〜(k)を繰り返し、これによってHSV−1標的配列を増幅することによって2種類の一本鎖分子を指数関数的に増幅する工程、
(m)増幅されたHSV−1標的配列を検出する工程
を含むことを特徴とする方法。
【請求項1】
配列番号7〜8のいずれか1つの標的結合配列からなる配列、5’BsoBI制限エンドヌクレアーゼ認識部位、および5’非標的特異的5’テール末端配列を含むことを特徴とするポリヌクレオチド。
【請求項2】
配列番号7または8の配列からなることを特徴とする請求項1に記載のポリヌクレオチド。
【請求項3】
単純ヘルペスウイルス1型(HSV−1)標的配列を検出する方法であって、
(a)請求項1または2に記載のポリヌクレオチドをハイブリダイズする工程、および(b)HSV−1標的配列を検出する工程を含むことを特徴とする方法。
【請求項4】
(a)配列番号8又は7のいずれか1つの標的結合配列からなるHSV−1標的結合部位を有する配列、5’BsoBI制限エンドヌクレアーゼ認識部位、および5’非標的特異的5’テール末端配列を含む第一の増幅プライマーを用いて、HSV−1標的配列を増幅する工程、
(b)配列番号18を含むか、または配列番号9〜17のいずれか1つの標的結合配列からなるHSV−1標的結合部位を有する配列を含む第二の増幅プライマー配列を用いて、HSV−1標的配列を増幅する工程、および
(c)増幅されたHSV−1標的配列を検出する工程
をさらに含むことを特徴とする請求項3に記載の方法。
【請求項5】
前記第一のプライマーが配列番号7または配列番号8の配列を含むことを特徴とする請求項4に記載の方法。
【請求項6】
請求項4または5に記載の方法であって、
(i) 増幅反応が鎖置換増幅(SDA)反応であるか、または
(ii)検出方法が、直接検出、ポリメラーゼ連鎖反応(PCR)、リガーゼ連鎖反応(LCR)、in situ ハイブリダイゼーション、転写介在増幅手法(TMA)、自己持続性配列複製法(Self-Sustaining Sequence Replication)(3SR)、ローリングサークル増幅法(Rolling Circle Amplification)(RCA)、Qβレプリカーゼシステム、および核酸配列をベースとする増幅法(Nucleic Acid Sequence Based Amplification)(NASBA)からなる群から選択されるか、または
(iii)第一の増幅プライマーが、ヘアピン、g−カルテット、およびアッセイプローブに結合する配列から選択される配列をさらに含むか、または
(iv)第一の増幅プライマーが、好ましくは蛍光部分である検出可能な標識をさらに含むか、または
(v)第一の増幅プライマーが、制限酵素認識部位およびRNAポリメラーゼプロモータから選択される配列を含むか、または
(vi)前記方法が、内部増幅対照(IAC)を増幅する工程をさらに含む
ことを特徴とする方法。
【請求項7】
(i)IACが配列番号26〜27から選択されるか、または
(ii)前記方法が、増幅されたIACを検出する工程、好ましくは、増幅されたIACが増幅された標的配列とは異なる手段で検出される工程をさらに含む
ことを特徴とする請求項6に記載の方法。
【請求項8】
請求項7に記載の方法であって、
(a)配列番号18を含むか、または配列番号9〜17のいずれか1つの標的結合配列からなるHSV−1標的結合部位を有する配列を含む1以上のポリヌクレオチドをハイブリダイズする工程、および
(b)HSV−1標的配列を検出する工程であって、好ましくは、前記1以上のポリヌクレオチドが、好ましくは蛍光部分である検出可能な標識をさらに含む工程
をさらに含むことを特徴とする方法。
【請求項9】
請求項4または5において規定された1以上の第一のプライマーを含むことを特徴とする配列を検出するためのキット。
【請求項10】
請求項9に記載のキットであって、
(I)(i)増幅プライマー、好ましくは配列番号9〜18から選択されるもの、または
(ii)バンパープライマー、好ましくは配列番号23〜25から選択されるもの、または
(iii)アダプタプライマー、好ましくは配列番号19〜22から選択されるもの、および
(iv)検出プライマー、好ましくは配列番号30〜35から選択されるもの
をさらに含むか、或いは
(II)前記キットはHSV−1標的配列を検出するためのものであり、配列番号7〜8からの1つ、および配列番号9〜18からの少なくとも1つから選択される増幅プライマーの1つ以上;および配列番号23〜25から選択されるバンパープライマーを含み、好ましくは、前記キットは、(i)配列番号19〜22から選択される1以上のアダプタプライマー;および(ii)配列番号30〜35からなる群から選択される1以上の検出プライマー;および/または(iii)配列番号26〜27から選択される1以上のIAC標的配列;および(iv)配列番号28〜29からなる群から選択される1以上のIACアダプタプライマーをさらに含む
ことを特徴とするキット。
【請求項11】
請求項4または5において定義された1以上の第一のプライマーを含むことを特徴とするHSV−1を検出するための組成物。
【請求項12】
(I)配列番号9〜18および配列番号23〜25のいずれか1つの標的結合配列からなる1以上のプライマーをさらに含むこと、および/または
(II)(a)アダプタプライマーの3’末端に位置する標的結合配列を介して標的配列にハイブリダイズすることができるアダプタプライマー配列であって、このアダプタプライマーが5’ジェネリックテールを含み、かつこのアダプタプライマーが配列番号19〜22からなる群から選択されるもの;および
(b)検出プライマーの3’部分を介してアダプタプライマーの5’テールにハイブリダイズすることができる検出プライマー配列であって、この検出プライマー配列が5’制限酵素認識部位、ならびに、蛍光部分、放射性同位体、化学発光剤、目で見える反応生成物を生じることができる酵素基質、およびリガンドで検出可能に標識されたリガンド結合パートナーからなる群から選択される検出可能な標識を含むもの
をさらに含み、好ましくは、
(i)前記検出プライマー配列がヘアピンおよびカルテットからなる群から選択される構造部分を含むか、または
(ii)前記検出可能な標識が蛍光部分であり、好ましくはこの蛍光部分が、フルオロセイン(FAM)/ローダミン(ROX);FAM/P−(ジメチルアミノフェニルアゾ)安息香酸(DABCYL);ROX/DABCYL;フルオロセインイソチオシアネート(FITC)/テトラメチルローダミンイソチオシアネート(TRITC);FITC/テキサスレッド(商標);FITC/N−ヒドロキシスクシンイミジル1−ピレンブチレート(PYB);FITC/エオシンイソチオシアネート(EITC);N−ヒドロキシスクシンイミジル1−ペンタンスルホネート(PYS)/FITC;FITC/ローダミン X;およびFITC/テトラメチルローダミン(TAMRA)からなる群から選択されるドナーおよびクエンチャー色素対を含むか、または
(iii)前記検出プライマーが配列番号30〜35からなる群から選択されるものであること;および/または
(III)増幅反応によってサンプル中のHSV−1標的配列の検出のためのプライマーを含み、このプライマーは、
(a)HSV−1標的配列にハイブリダイズできる第一の増幅プライマー配列であって、第一の増幅プライマーが配列番号7〜8からなる群から選択されるもの、
(b)HSV−1標的配列の相補体にハイブリダイズできる第二の増幅プライマー配列であって、第二の増幅プライマーが配列番号9〜18からなる群から選択されるもの、
(c)第一の増幅プライマーの上流でHSV−1標的配列にハイブリダイズできる第一のバンパープライマー配列であって、第一のバンパープライマーが配列番号23であるもの、および
(d)第二の増幅プライマーの上流でHSV−1標的配列の相補体にハイブリダイズできる第二のバンパープライマー配列であって、第二の増幅プライマーが配列番号24〜25からなる群から選択されるもの
を含むこと
を特徴とする請求項11に記載の組成物。
【請求項13】
サンプル中のHSV−1標的配列の存在を検出するための請求項3に記載の方法であって、前記方法が、
(a)配列番号7〜8からなる群から選択される第一の増幅プライマー、および配列番号9〜18からなる群から選択される第二の増幅プライマーにサンプルを添加し、増幅されたHSV−1核酸生成物を産生する工程、および
(b)増幅されたHSV−1核酸生成物を検出する工程であって、増幅生成物の検出がサンプル中のHSV−1の存在を示す工程
を含むことを特徴とする方法。
【請求項14】
請求項13に記載の方法であって、
(i)前記増幅されたHSV−1核酸生成物が、ユニバーサル検出、ゲル電気泳動、および定量的ハイブリダイゼーションからなる群から選択される方法によって検出されること、または
この方法が、
(c)配列番号23の第一のバンパープライマーおよび配列番号25または配列番号26の第二のバンパーをサンプルに添加する工程をさらに含むこと
を特徴とする方法。
【請求項15】
検出工程が、
(d)第一の増幅プライマーおよび配列番号19〜22からなる群から選択されるアダプタプライマーを増幅されたHSV−1標的配列にハイブリダイズする工程であって、アダプタプライマーが第一の増幅プライマーの下流でハイブリダイズする工程、
(e)第一の増幅プライマーおよびアダプタプライマーの3’末端を伸長し、アダプタプライマー伸長生成物を産生する工程であって、第一の増幅プライマーの伸長がアダプタプライマー伸長生成物を置換する工程、
(f)第二の増幅プライマーをアダプタプライマー伸長生成物へハイブリダイズする工程、
(g)第二の増幅プライマーの3’末端を伸長し、アダプタプライマー伸長生成物およびその相補体を含む二本鎖分子を産生する工程であって、各鎖がニック可能な制限酵素部位を含む工程、
(h)二本鎖分子をニッキングし、5’部分および3’部分を造り出す工程であって、5’部分がニッキングされたテールを含み、3’部分がニッキングされた相補性アダプタプライマー伸長生成物を含む工程、
(i)ニッキングされたテールの3’末端を伸長し、ニッキングされた相補性アダプタプライマー伸長生成物を置換する工程であって、置換され、ニッキングされた相補性アダプタプライマー伸長生成物が一本鎖である工程、
(j)一本鎖相補性アダプタプライマー伸長生成物へ検出プライマーをハイブリダイズする工程であって、検出プライマーが検出可能な標識を含み、HSV−1標的配列を検出する工程、
(k)検出プライマーおよび相補性アダプタプライマー伸長生成物の3’末端を伸長し、検出プライマー伸長生成物およびその相補体を含む二本鎖検出分子を産生する工程であって、各鎖が開裂可能な制限酵素認識部位を含む工程、
(l)二本鎖検出分子を開裂する工程、および
(m)検出可能な標識を検出する工程
をさらに含むことを特徴とする請求項14に記載の方法。
【請求項16】
(n)配列番号26〜27からなる群から選択される内部増幅対照(IAC)標的配列を増幅する工程、
(o)第一の増幅プライマーおよび配列番号28〜29からなる群から選択されるIACアダプタプライマーをIACにハイブリダイズする工程であって、アダプタプライマーが第一の増幅プライマーの下流でハイブリダイズする工程、
(p)第一の増幅プライマーおよびIACアダプタプライマーの3’末端を伸長し、IACアダプタプライマー伸長生成物を産生する工程であって、第一の増幅プライマーの伸長がIACアダプタプライマー伸長生成物を置換する工程、
(q)IACアダプタプライマー伸長生成物へ第二の増幅プライマーをハイブリダイズする工程、
(r)第二の増幅プライマーの3'末端を伸長し、IACアダプタプライマー伸長生成物およびその相補体を含む二本鎖分子を産生する工程であって、各鎖がニック可能な制限酵素部位を含む工程、
(s)二本鎖分子をニッキングする工程、
(t)相補性IACアダプタプライマー伸長生成物のニッキングされたテール末端を伸長し、一本鎖でニッキングされた相補性アダプタプライマー伸長生成物を置換する工程、
(u)一本鎖相補性IACアダプタプライマー伸長生成物にIAC検出プライマーをハイブリダイズする工程であって、IAC検出プライマーが検出可能な標識を含み、IACを検出する工程、
(v)IAC検出プライマーおよび一本鎖相補性IACアダプタプライマー伸長生成物の3’末端を伸長し、検出プライマー伸長生成物およびその相補体を含む二本鎖検出分子を産生する工程であって、各鎖が開裂可能な制限酵素認識部位を含む工程、
(w)二本鎖検出分子を開裂する工程、および
(x)検出可能な標識を検出する工程
をさらに含むことを特徴とする請求項15に記載の方法。
【請求項17】
HSV−1標的配列をサンプル中で検出するための請求項3に記載の方法であって、前記方法が、
(a)配列番号7〜8からなる群から選択される配列を有する第一の増幅プライマーをHSV−1標的配列へハイブリダイズする工程、
(b)配列番号23の第一のバンパープライマーをHSV−1標的配列へハイブリダイズする工程であって、第一のバンパープライマーが第一の増幅プライマーの上流でHSV−1標的配列にハイブリダイズし、第二のバンパープライマーが第二の増幅プライマーの上流で相補性HSV−1標的配列にハイブリダイズする工程、
(c)第一のバンパープライマーおよび第一の増幅プライマーの3’末端を伸長し、第一の増幅プライマー伸長生成物を産生する工程であって、第一のバンパープライマーの伸長が第一の増幅プライマー伸長生成物を置換する工程、
(d)配列番号9〜18からなる群から選択される配列を有する第二の増幅プライマーを第一の増幅プライマー伸長生成物へハイブリダイズする工程、
(e)配列番号24〜25からなる群から選択される配列を有する第二のバンパープライマーを第一の増幅プライマー伸長生成物へハイブリダイズする工程、
(f)第二のバンパープライマーおよび第二の増幅プライマーの3’末端を伸長し、第二の増幅プライマー伸長生成物を産生する工程であって、第二のバンパープライマーの伸長が第二の増幅プライマー伸長生成物を置換する工程、
(g)第一の増幅プライマーを第二の増幅プライマー伸長生成物へハイブリダイズする工程、
(h)工程(g)の第一の増幅プライマーの3’末端を伸長し、第二の増幅プライマー伸長生成物およびその相補体を含む二本鎖分子を産生する工程であって、各鎖がニック可能な制限酵素部位を含む工程、
(i)二本鎖分子をニッキングし、5’部分と3’部分を造り出す工程であって、5’部分はニッキングされたテールを含み、3’部分はニッキングされた第二の増幅プライマー伸長生成物およびその相補体を含む工程、
(j)ニッキングされたテールの3’末端を伸長し、ニッキングされた第二増幅プライマー伸長生成物を置換する工程であって、ニッキングされた相補性第二増幅プライマー伸長生成物が一本鎖である工程、
(k)ニッキングされたテールの3’末端を伸長し、ニッキングされた相補性第二増幅プライマー伸長生成物を置換する工程であって、ニッキングされた相補性第二増幅プライマー伸長生成物が一本鎖である工程、
(l)第一の増幅プライマーおよび第二の増幅プライマーを、各一本鎖分子に別々にハイブリダイズし、工程(h)〜(k)を繰り返し、これによってHSV−1標的配列を増幅することによって2種類の一本鎖分子を指数関数的に増幅する工程、
(m)増幅されたHSV−1標的配列を検出する工程
を含むことを特徴とする方法。
【図1】


【図2】
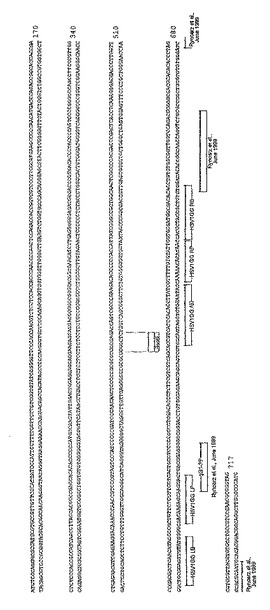
【図3】
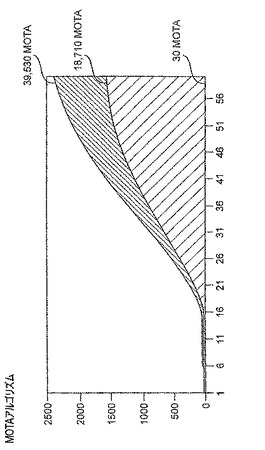
【図4】
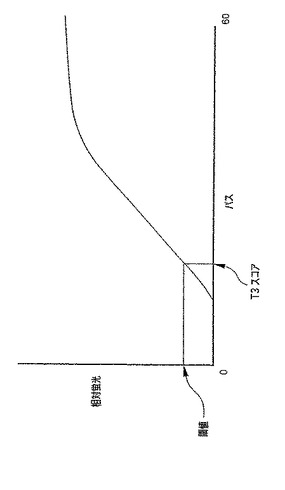
【図5】
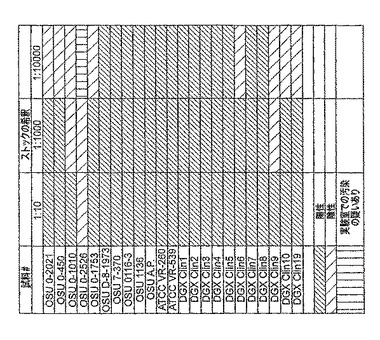
【図6】

【図7】
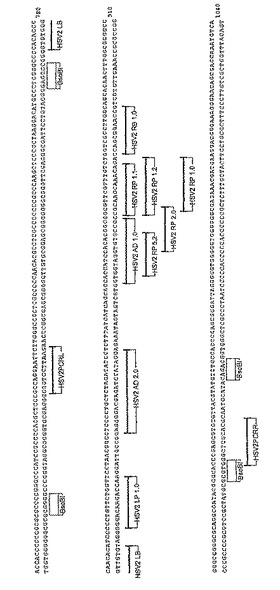
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【公開番号】特開2012−24109(P2012−24109A)
【公開日】平成24年2月9日(2012.2.9)
【国際特許分類】
【出願番号】特願2011−246483(P2011−246483)
【出願日】平成23年11月10日(2011.11.10)
【分割の表示】特願2006−513315(P2006−513315)の分割
【原出願日】平成16年4月26日(2004.4.26)
【出願人】(595117091)ベクトン・ディキンソン・アンド・カンパニー (539)
【氏名又は名称原語表記】BECTON, DICKINSON AND COMPANY
【住所又は居所原語表記】1 BECTON DRIVE, FRANKLIN LAKES, NEW JERSEY 07417−1880, UNITED STATES OF AMERICA
【Fターム(参考)】
【公開日】平成24年2月9日(2012.2.9)
【国際特許分類】
【出願日】平成23年11月10日(2011.11.10)
【分割の表示】特願2006−513315(P2006−513315)の分割
【原出願日】平成16年4月26日(2004.4.26)
【出願人】(595117091)ベクトン・ディキンソン・アンド・カンパニー (539)
【氏名又は名称原語表記】BECTON, DICKINSON AND COMPANY
【住所又は居所原語表記】1 BECTON DRIVE, FRANKLIN LAKES, NEW JERSEY 07417−1880, UNITED STATES OF AMERICA
【Fターム(参考)】
[ Back to top ]