
核酸結合ポリペプチドライブラリー
【課題】各ポリペプチドが少なくとも部分的に無作為されている1つ以上のジンクフィンガーを含んでなるジンクフィンガーポリペプチドライブラリーの提供。
【解決手段】この発明は、各ポリペプチドが少なくとも部分的に無作為化されている1つ以上のジンクフィンガーを含んでなるジンクフィンガーポリペプチドライブラリー、およびオーバーラップするジンクフィンガーポリペプチドをコードするジンクフィンガーポリペプチドライブラリーセットに関し、このポリペプチドは選択の後、多重フィンガージンクフィンガーポリペプチドが形成されるように組み立てられることができる。
【解決手段】この発明は、各ポリペプチドが少なくとも部分的に無作為化されている1つ以上のジンクフィンガーを含んでなるジンクフィンガーポリペプチドライブラリー、およびオーバーラップするジンクフィンガーポリペプチドをコードするジンクフィンガーポリペプチドライブラリーセットに関し、このポリペプチドは選択の後、多重フィンガージンクフィンガーポリペプチドが形成されるように組み立てられることができる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ジンクフィンガー(zinc finger)ポリペプチドの選択のためのライブラリー系に関する。特に本発明は、ジンクフィンガーモチーフがオーバーラップする領域で無作為化(randomised)されたバイナリー系、および選択された位置に限定して生じさせた無作為化を組込むスマートなライブラリーに関する。
【背景技術】
【0002】
タンパク質‐核酸認識は、真核および原核細胞の機能を調節する大多数の生体分子的制御機構に中心的役割を果すありふれた現象である。例えば、タンパク質‐DNA相互作用は遺伝子発現の調節の基礎を成し、したがって、分子生物学者により最も広範に研究されている対象の1つである。
【0003】
豊富な生化学および構造的情報により多数の事例におけるタンパク質‐DNA認識の詳細が説明され、認識の一般原理が現われるに至っている。多くのDNA結合タンパク質は、DNA認識のための折りたたみドメインを独立して含有し、これらのドメインは、順に、ロイシンジッパー、「ヘリックス‐ターン‐ヘリックス」およびジンクフィンガーのような多数の構造ファミリーに属する。
【0004】
構造ドメインの大きな変化に拘らず、タンパク質およびDNAの間に現在まで観察された相互作用の特異性は、ほとんどタンパク質αヘリックス表面とDNAの主溝(main groove)の相補性に由来する(Klug, (1993) Gene 135:83-92)。αヘリックスと主溝の繰返し物理的相互作用に照らして、特定のアミノ酸とDNA塩基の間の接触は、規則の簡単なセット:要するに、結合部位配列選好に対するタンパク質一次構造に関係する立体化学的認識コードにより記述することができる可能性に気持が駆られる。
【0005】
しかし、全てのDNA結合タンパク質によっても、DNA認識を記述できるコードは見出されていないことは明らかである。異なる構造ファミリー由来のDNA結合タンパク質の認識αヘリックスはDNAの主溝と相互作用する方法に重要な相異を示すため、多数の複合体の構造は認識パターンに類似性を与えない。既知のDNA結合モチーフの大部分は特に応用自在なものでなく、現れうるどのコードも非常に少数の関係するDNA配列との結合を記述しがちである。
【0006】
その上、DNA結合タンパク質の各ファミリー内ですら、コードの解読はこれまでとらえどころがなかった。タンパク質DNA相互作用が複雑なため、タンパク質の一次構造と核酸の間にはアミノ酸と塩基の関係を直接に規定する簡単な「アルファベット」等価性がみられない。
【0007】
国際特許出願WO 96/06166はこの問題を研究して、ジンクフィンガー核酸結合タンパク質についてタンパク質‐DNA相互作用を説明する「シラブル(sillabic)」コードを提供する。シラブルコードは、特定塩基への結合を規定する結合タンパク質の1つ以上の特徴に基づくコードであり、その特徴は「シラブル」もしくはそれぞれの特異的な接触を定義する複合指示の形で組合わせ可能である。
【0008】
しかし、このコードは不完全であり、各3塩基連鎖(triplet)の5'位置のG以外のヌクレオチドの特異的選択を可能にする特定の指示を与えない。該方法は、他の特異性に対する核酸結合蛋白質を作製するために、無作為化とその後の選択に依存する。しかし、部分的無作為化と選択の助けをかりても、WO 96/06166に報じられた方法も先行技術の他のどの方法も、Zif268の第1フィンガーに基いて、5'塩基がGまたはT以外の3塩基連鎖と結合する能力のあるジンクフィンガーポリペプチドを単離することに成功してない。これはジンクフィンガータンパク質を設計する能力において重大な不足点である。
【0009】
さらに、この文書は、全ての先行技術と同じく、ジンクフィンガーが核酸3塩基連鎖(triplet)またはその多重体と結合するという概念に依存する。今回、本発明者らは、ジンクフィンガー結合部位がオーバーラップする4bpサブサイトにより決定されることおよびサブサイト間の境界の配列特異性は隣接フィンガー間の相乗作用から生じることを確認した。これは、新規のDNA結合特異性をもつジンクフィンガーの設計および選択に対して重要な示唆を与える。
【特許文献1】国際公報第WO 96/06166号パンフレット
【非特許文献1】Klug, (1993) Gene 135:83-92
【発明の概要】
【発明が解決しようとする課題】
【0010】
本発明は、各ポリペプチドが少なくとも部分的に無作為されている1つ以上のジンクフィンガーを含んでなるジンクフィンガーポリペプチドライブラリーを提供する。好ましくは、本発明は、オーバーラップするジンクフィンガーポリペプチドをコードし、各ポリペプチドが少なくとも部分的に無作為化されている1つ以上のジンクフィンガーを含んでなり、該ポリペプチドは選択の後に組み立てられて多重ジンクフィンガーポリペプチドを形成することができる、一群のジンクフィンガーポリペプチドライブラリーを提供する。
【課題を解決するための手段】
【0011】
本発明は、ジンクフィンガーポリペプチドの設計におけるオーバーラップする4bpサブサイト認識の重要性を認識する。各ライブラリー内でただ1つのジンクフィンガーを無作為化する古典的ジンクフィンガーライブラリー設計においてはジンクフィンガー間に起る相乗作用は見落されている。
【0012】
したがって、本発明は、各ポリペプチドが少なくとも部分的に無作為されている1つ以上のジンクフィンガーを含んでなるジンクフィンガーポリペプチドライブラリーを提供する。
【0013】
好ましくは、本発明は、オーバーラップするジンクフィンガーポリペプチドをコードし、各ポリペプチドが少なくとも部分的に無作為化されている1つ以上のジンクフィンガーを含んでなり、該ポリペプチドは選択の後に組み立てられて多重ジンクフィンガーポリペプチドを形成することができる、一群のジンクフィンガーポリペプチドライブラリーを提供する。
【0014】
他の様態では、本発明は、上に記載したとおり、無作為化が結合部位特異性における改変を指揮することが知られているアミノ酸を置換することに限定されるライブラリーに関する。本発明は、標的配列としての任意の核酸配列に対するライブラリーの選択、およびそれに結合する特異的核酸結合タンパク質の製造を可能にするアミノ酸位置の偏り(bias)を示すコードを提供する。さらに、本発明は、所与の核酸配列に特異的なジンクフィンガータンパク質を設計しかつ最適化する方法を提供する。したがって、本発明は古典的ジンクフィンガーと核酸との相互作用について明らかにされた認識の偏りに関する。この事例において、全ての核酸配列との結合をカバーするルールのパターンが提供される。
【発明の効果】
【0015】
本発明で開示されるコードは、隣接ジンクフィンガー間の相乗的相互作用を考慮し、それによって、任意の所望の結合部位の選択を可能にする。
【図面の簡単な説明】
【0016】
【図1】図1は、ジンクフィンガー‐DNA相互作用を図示したものである。A:Zif268のDNA塩基3塩基連鎖についての古典的な3塩基連鎖相互作用のモデル;B:同様な4塩基連鎖(quadruplet)相互作用を示すモデル;C:認識コード決定のためのライブラリー設計のモデル。
【図2】図2は、認識コードの決定においてファージ提示選択に使う3つのフィンガーのアミノ酸配列を示す。
【図3】図3には、ファージ選択から得られた配列特異的ジンクフィンガークローン、およびそれらの結合部位を示す記号を挙げる。
【図4】図4は、ファージ選択から単離したクローンの塩基/アミノ酸相関を示す。認識パターンを灰色に塗って強調してある。
【図5】図5は、αヘリックスの位置2に対して選択された4塩基連鎖の位置1と結合する配列特異的相互作用を図示する。
【図6】図6は、本発明によるライブラリーの構築の模式図である。
【発明を実施するための形態】
【0017】
以下、本発明を詳細に説明する。
【0018】
本発明はライブラリーに関係する。用語「ライブラリー」は当業界の通常の用法により使われ、ポリペプチド、または、好ましくはポリペプチドをコードする核酸の集合体を表す。本発明のポリペプチドは、無作為化(randomisation)領域を含み、このため各ライブラリーは、個々のポリペプチドがお互いに異なる配列をもつポリペプチドのレパートリーを含んでなるかまたはコードする。同じ原理は、ファージ提示によるような、選択のために開発される実質的に全てのライブラリーに存在する。
【0019】
本明細書で使う無作為化とは、異なるポリペプチドの任意の所与の位置に様々なアミノ酸が存在しうるような、ライブラリーを含んでなるポリペプチドの配列の改変を意味する。無作為化は、与えられた位置にどのアミノ酸も存在しうるような完全なものであっても、またある特定のアミノ酸のみが存在するような部分的なものであってもよい。好ましくは、無作為化は、核酸レベルでの変異誘発により、例えば変異タンパク質をコードする新規遺伝子を合成し、それらを発現させて各種の異なるタンパク質を得ることにより達成される。あるいは、所望の変異遺伝子を得る目的で、存在する遺伝子自身を部位指定または無作為変異誘発のような方法により変異させることができる。
【0020】
変異は当業者に周知の方法で実施することができる。しかし、目的のタンパク質をコードする核酸配列の部位特異的突然変異誘発が好ましい。多数の部位特異的突然変異誘発の方法が、M13のような1本鎖ファージを用いる方法からPCRに基く技術まで、当業界で公知である("PCR Protocols: A guide to methods and applications(PCRプロトコル:方法および応用のガイド)", M.A. Innis, D.H. Gelfand. J.J. Sninsky, T.J. White(編), Academic Press, New York, 1990を参照すること)。好ましくは、市販される変異誘発システム(Altered Site II Mutagenesis System (Promega))をメーカーにより与えられた指示に従って用いることができる。
【0021】
変異遺伝子により産生されるタンパク質のスクリーニングは、好ましくは、遺伝子を発現してタンパク質産物の結合能力を検定することにより実施される。これを達成しうる簡単でかつ有利で迅速な方法は、ファージ提示(phage display)による方法であり、この方法では、変異体ポリペプチドは、バクテリオファージm13のマイナーなコートタンパク質pIIまたはバクテリオファージFdの遺伝子IIIのような、繊維状バクテリオファージのコートタンパク質との融合タンパク質として発現され、変異遺伝子により形質転換されたバクテリオファージのカプシド上に提示される。標的核酸配列をプローブとして使ってファージ表面上のタンパク質に直接結合させ、アフィニティ精製により有利な変異体を持つファージを選択する。その後、該ファージを細菌宿主を通じての継代により増幅し、所望のファージの変異体プールを富化するためにさらなる回数の選択および増幅を行い、最終的に好ましいクローンを単離する。当業界で公知のファージ提示の詳細な手法は、例えば、米国特許第5,223,409号;ChooおよびKlug, (1995) Current Opinions in Biotechnology 6:431-436;Smith. (1985) Science 228:1315-1317;およびMcCafferyら, (1990) Nature 348:552-554;に開示されており、これらは全て、本明細書に参照により組み入れられる。ファージ提示のためのベクターシステムおよびキットは、例えばファルマシア(Pharmacia)社から市販されている。
【0022】
本発明によるライブラリーが含んでなるポリペプチドは、ジンクフィンガーポリペプチドである。言換えれば、それらは、Cys2-His2ジンクフィンガーモチーフを含んでなる。各ポリペプチドが1つ以上のジンクフィンガーを含んでなることが本発明の特徴であり、該ライブラリーはポリペプチド上の2つ以上のジンクフィンガー間の相互作用に基いて選択することができる。
【0023】
当業界で公知であるように、ジンクフィンガーは核酸と結合する分子である。各ジンクフィンガーは、標的核酸の4塩基連鎖(quadruplet)配列と、ジンクフィンガーのαヘリックスの特定アミノ酸残基と核酸との間の接触を通して結合する。本発明に規定される4塩基連鎖は、核酸の-(マイナス)鎖を3'から5'へ読んだとき、第1の4塩基連鎖の塩基4は第2の4塩基連鎖の塩基1であり、かつ以後もそのように、オーバーラップしている。したがって、本出願において、各4塩基連鎖の塩基は、1〜4の数字により参照され、1は3'塩基でありかつ4は5'塩基である。塩基4は古典的なジンクフィンガー3塩基連鎖の5'塩基と等価である。一般的に、塩基4はαヘリックスの位置+6における接触を通して、塩基3は位置+3における接触を通して、塩基2は位置-1における接触を通して、塩基1は二本鎖核酸の対立鎖への位置+2における接触を通して結合される。
【0024】
本明細書に言及するジンクフィンガーの全ての核酸結合残基位置は、フィンガーのαヘリックスにおける第1残基から数えられ、+1〜+9の範囲にある。「-1」は、Cys2-His2ジンクフィンガーポリペプチド中のαヘリックスのすぐ前のフレームワーク構造の残基を指す。
【0025】
「++2」と称する残基は、隣接する(C末端)フィンガーに存在する。これらは、(+鎖の)塩基1に対する位置+2と、先行(3')4塩基連鎖の塩基4(オーバーラップにより同じ塩基である)に対する先行(N末端)フィンガー位置+6との間の相乗的共同作用を反映する。C末端隣接フィンガーがない場合、「++」相互作用は機能しない。
【0026】
Cys2-His2ジンクフィンガー結合タンパク質は、当業界で周知のように、ジンクフィンガーとして知られるαヘリカルの亜鉛金属原子が配位した結合モチーフを経由して、標的核酸配列と結合する。ジンクフィンガー核酸結合タンパク質の各ジンクフィンガーは、核酸結合配列内の核酸4塩基連鎖との結合を決定することに関わる。好ましくは、各結合タンパク質には2つ以上のジンクフィンガー、例えば2,3,4,5または6つのジンクフィンガーがある。各ジンクフィンガー結合タンパク質に3つのジンクフィンガーがあるのが有利である。
【0027】
本発明により、本質的に人工の核酸結合タンパク質の産生が可能になる。これらのタンパク質については、所望の物性をもつまたは他の理由の蛋白質を与えるためにアミノ酸の人工的類似体を使うことができる。かくして、特定的に「任意のアミノ酸(any amino acid)」に言及する文脈において、用語「アミノ酸(amino acid)」は、当業界で公知の方法によりどの種類のタンパク質構築に用いうる天然または人工のアミノ酸もしくはアミノ酸類似体も意味する。さらに、本明細書で参照されるどの特定のアミノ酸も、その機能的類似体、特に人工の機能類似体により置換することができる。したがって、本明細書で使われる命名法は、定義されたアミノ酸の機能的類似体を特定的に本発明の範囲内に含む。
【0028】
ジンクフィンガー結合タンパク質のαヘリックスは核酸鎖に反平行に(antiparallel)に整列し、ジンクフィンガーのN末端からC末端への配列に対応するために、一次核酸配列は3'から5'へ配列する。核酸配列は通常、5'から3'に向けてかつアミノ酸配列はN末端からC末端に向けて書かれるので、その結果、核酸配列およびジンクフィンガータンパク質を慣例により整列すると、ジンクフィンガーは3'から5'に整列している核酸の-(マイナス)鎖と一次相互作用をもつことになる。これらの慣例は本明細書に使われる命名法で踏襲される。しかし、実際には、タンパク質GLIのフィンガー4のようなある特定のフィンガーは、核酸の+(プラス)鎖と結合することは注意すべきである(Suzukiら, (1994) NAR 22:3397-3405およびPavletichおよびPabo, (1993) Science 261:1701-1707)。このようなフィンガーの核酸結合分子への結合は、本発明により予想される。
【0029】
本発明のライブラリーにより、単一DNA標的に対する隣接フィンガーの同時選択を促進することにより隣接ジンクフィンガー間の相乗的共同作用に対する選択が可能になる。これは、同じジンクフィンガーポリペプチドにおいて、1つ以上のジンクフィンガーを無作為化することにより達成される。好ましい実施様態では、各ポリペプチド中のほぼ1個半のジンクフィンガーが無作為化されるが、これはライブラリー設計により変化する。
【0030】
本発明のライブラリーにコードされるジンクフィンガーポリペプチドは、1つ以上であれば、いくつの数のジンクフィンガーを含んでなってもよい。各ポリペプチドが3〜6つのジンクフィンガーをコードするのが有利である。各ライブラリーにおいて、無作為化の範囲は少なくとも1対のジンクフィンガーのオーバーラップをカバーする。好ましくは、1対のオーバーラップをカバーする。
【0031】
好ましくは、本発明のライブラリーはセットとして提供される。したがって、F1、F2およびF3を含んでなる3つのジンクフィンガーポリペプチドを、各ライブラリーが2つのジンクフィンガーポリペプチドを含んでなる2つのライブラリーの1セットで提供することができる。第1のライブラリーは本質的にF1およびF2からなるポリペプチドで構成され、第2のライブラリーは本質的にF2およびF3からなるポリペプチドで構成される。各ライブラリーの無作為化は、それぞれF1およびF2、ならびにF2およびF3の間のオーバーラップを含む。
【0032】
好ましくは、各ライブラリーは少なくとも第1のフィンガーの位置6および第2フィンガーの位置2での無作為化を含んでなるであろう。これらの残基は二本鎖核酸標的上の同じ塩基対と接触するので、それらを一緒に変えることが有利である。
【0033】
3つのジンクフィンガーポリペプチドの場合には、第1ライブラリーはフィンガーF1およびF2で無作為化されるであろうが、第2はF2およびF3で無作為化される。ポリペプチドは、F2配列において選択後に組替えられ、F1、F2およびF3を含有する単一ポリペプチドを作製する。このポリペプチドはF1およびF2、ならびにF2およびF3の間のオーバーラップを考慮して選択されているであろう。
【0034】
有利なように、各ジンクフィンガーでより多数の位置を変えることができる。好ましくは、第1ジンクフィンガーでは位置-1、1、2、3、5および6から選択された残基がそして、第2ジンクフィンガーでは位置-1、1、2および3が改変される。片方のライブラリーについては、第2フィンガーでは位置3、5および6が、そして第3フィンガーでは位置-1、1、2および3が改変されうる。最後のフィンガー(3つのフィンガータンパク質の場合にはこれは第3フィンガーであろう)では、残基5および6も改変されうる。
【0035】
ライブラリーを選択後に可能とするため、ポリペプチドは、好ましくは、2つのライブラリーにより共有されるジンクフィンガーをコードする核酸中に適切な制限酵素切断部位を含むように設計される。開裂部位の位置は、各ライブラリー中の共有されるジンクフィンガーに行われる改変の正確な部位を指令するであろう。したがって、3つのジンクフィンガータンパク質をコードする2つのライブラリーの1セット中に、もし開裂部位がαヘリックスの位置3と5の間にあれば、位置3と5は第1ライブラリーで、5と6とは第2ライブラリーで無作為化されうる。
【0036】
無作為化または改変のための残基は、位置-1、1、2、3、5および6から選択されるのが好ましいが、さらなる残基も無作為化することができる。例えば、位置8の無作為化は有利でありうる。さらに、与えられた全ての位置より少ない位置を無作為化することも予測される。
【0037】
好ましい実施様態では、3つのフィンガータンパク質の選択のための2つのライブラリー系は、第1ライブラリーにおいて、F1位置-1、1、2、3、5および6で改変され、F2位置-1、1、2および3で改変される。第2ライブラリーは、F2位置3および6で改変され、F3位置-1、1、2、3、5および6で改変される。この場合、開裂および組換え点は残基3および5の間、好ましくはF2のαヘリックスの残基4および5の間であろう。
【0038】
組換え操作の後、組替えられたポリペプチドをコードする核酸は、適切な発現系で発現されるか、またはさらなる選択のためにFdファージにクローニングされる。
【0039】
本発明の好ましい様態では、本発明のライブラリーは、全てのアミノ酸でなくある特定のアミノ酸がコードされるように、選択した位置で真に無作為化されるのでなく部分的に無作為化される。この方法は、2つの目的で使うことができる。
【0040】
第1の実施様態では、改変は、ジンクフィンガーのαヘリックスの所与の位置に組込まれたときに核酸標的配列の配列特異的結合を指令する能力のあることが知られているアミノ酸に制限される。標的配列にかかわりなく、ある特定のアミノ酸はある特定の位置での組込みに適切でないことが知られている。これらのアミノ酸は回避される。
【0041】
第2の実施様態では、改変は、ジンクフィンガーのαヘリックスの所与の位置に組込まれたときに核酸標的配列の配列特異的結合を指令する能力のあることが知られているアミノ酸に制限され、かつ改変は、所与の位置での特定の標的配列との結合を有利にすることが知られている残基を特定するように行われる。このようにして、本発明は、的の絞られたライブラリーの設計を可能にし、このライブラリーから特定の標的配列または一連の関係標的配列と結合する能力のあるポリペプチドを選択することができる。
【0042】
一般的応用のためのライブラリー系を提供する第1の実施様態では、無作為化は好ましくは、上に示した位置の全てで行われる。好ましくは、それぞれの所与の位置で現れるように選択されるアミノ酸は表1に掲げたとおりである:
【表1】

【0043】
各フィンガーが、表1に与えられた各位置で無作為化されている必要はない。好ましい実施様態では、3フィンガータンパク質を選択するライブラリーは、表2に与えられた明細により構築される。
【表2】
【0044】
第2の実施様態では、特定の位置の各アミノ酸の同一性は、本明細書に提供されたジンクフィンガー認識ルールにより選択される。したがって、好ましい様態では、本発明は、標的核酸配列内の核酸4塩基連鎖への結合を可能にするCys2-His2ジンクフィンガークラスの核酸結合タンパク質を調製する方法を提供し、該方法において、タンパク質のαヘリカルジンクフィンガー核酸結合モチーフによる4塩基連鎖の各塩基との結合は次のとおり決定される:
a)もし4塩基連鎖の塩基4がGであれば、αヘリックスの位置+6はArgもしくはLysであり;
b)もし4塩基連鎖の塩基4がAであれば、αヘリックスの+6はGln、AsnもしくはValであり;
c)もし4塩基連鎖の塩基4がTであれば、αヘリックスの位置+6はSer、Thr、ValもしくはLysであり;
d)もし4塩基連鎖の塩基4がCであれば、αヘリックスの位置+6はSer、Thr、Val、Ala、GluもしくはAsnであり;
e)もし4塩基連鎖の塩基3がGであれば、αヘリックスの位置+3はHisであり;
f)もし4塩基連鎖の塩基3がAであれば、αヘリックスの位置+3はAsnであり;
g)もし4塩基連鎖の塩基3がTであれば、αヘリックスの位置+3はAla、SerもしくはValであり;もしそれがAlaであれば、-1または+6の残基の1つは小さい残基であり;
h)もし4塩基連鎖の塩基3がCであれば、αヘリックスの位置+3はSer、Asp、Glu、Leu、ThrもしくはValであり;
i)もし4塩基連鎖の塩基2がGであれば、αヘリックスの位置-1はArgであり;
j)もし4塩基連鎖の塩基2がAであれば、αヘリックスの位置-1はGlnであり;
k)もし4塩基連鎖の塩基2がTであれば、αヘリックスの位置-1はHisもしくはThrであり;
l)もし4塩基連鎖の塩基2がCであれば、αヘリックスの位置-1はAspもしくはHisであり;
m)もし4塩基連鎖の塩基1がGであれば、αヘリックスの位置+2はGluであり;
n)もし4塩基連鎖の塩基1がAであれば、αヘリックスの位置+2はAspもしくはGlnでなく;
o)もし4塩基連鎖の塩基1がCであれば、αヘリックスの位置+2はAsn、Gln、Arg、HisもしくはLysであり;
p)もし4塩基連鎖の塩基1がTであれば、αヘリックスの位置+2はSerもしくはThrである。
【0045】
以上は、所与の核酸配列に特異的なジンクフィンガー結合タンパク質の設計を可能にするルールのセットを表す。これに関係する新規の知見は、ヘリックスの位置+2は、4塩基連鎖の塩基1との結合を決定することに関与するということである。そうすることによって、塩基1および4が、隣接する4塩基連鎖にオーバーラップしているので、4塩基連鎖の塩基4の結合を決定する位置+6と相乗的に同時作動する。
【0046】
ジンクフィンガーポリペプチドは、オーバーラップする4塩基連鎖配列と結合すると考えられたが、本発明の方法は、ポリペプチドが多重のオーバーラップ4塩基連鎖でない標的配列と結合するように設計することが可能である。例えば、ジンクフィンガーポリペプチドをパリンドローム標的配列と結合するように設計することができる。このような配列は通常、例えば、制限酵素標的配列として見出される。
【0047】
好ましくは、3個より少ないヌクレオチドと結合するジンクフィンガーの作製が、ジンクフィンガー中に関係位置の核酸とのH結合を支持することができないアミノ酸を特定することにより達成される。
【0048】
有利には、これは、位置-1のGly(塩基2との接触をなくするため)、ならびに/または、位置+3および/もしくは+6のAla(それぞれ第3もしくは第4塩基との接触をなくするために)を置換することにより達成される。
【0049】
好ましくは、標的配列の最後の(3')塩基との接触は、もし必要があれば、核酸のリン酸主鎖と直接接触できる関係位置において残基を置換することにより強化されるべきである。
【0050】
これらおよび他の考慮は、本発明によるライブラリーセットに組込むことができる。
【0051】
ジンクフィンガー結合モチーフは当業者には周知の構造であり、例えば、ミラーらその他(Millerら, (1985) EMBO J. 4:1609-1614; Berg (1988) PNAS (USA) 85:99-102; Leeら, (1989) Science 245:635-637; USSN 08/422.107.に対応する国際特許出願WO96/06166およびWO96/32475を参照すること、これらは参照により本明細書に組入れられる)に定義されている。
【0052】
本明細書に使われる「核酸」は、天然の核酸塩基もしくは合成塩基、またはそれらの混合物から構築されたRNAおよびDNAの両方を意味する。しかし、好ましくは、本発明の結合タンパク質はDNA結合タンパク質である。
【0053】
一般的に、好ましいジンクフィンガーフレームワークは、構造:
(A) X0-2 C X1-5 C X9-14 H X3-6 (H/C)
(式中、Xは任意のアミノ酸であり、下付き数字はXで表す残基の可能な数を示す)を有する。
【0054】
本発明の好ましい様態では、ジンクフィンガー核酸結合モチーフは次の一次構造:
(B)
(式中、X(Xa、XbおよびXcを含む)は任意のアミノ酸であり、X2-4およびX2-3はそれぞれ2個もしくは4個、または2個もしくは3個の存在を意味する)を有するモチーフとして表現することができる。一緒に亜鉛金属原子に配位するCysおよびHis残基は太字で記されているが、αヘリックスの+4のLeu残基のように通常は不変である。
【0055】
この表現に対する修正は、ジンクフィンガー機能を無効にすることなく、アミノ酸の挿入、変異または欠失によって起りうるし、または行うことができる。例えば、第2His残基はCysで置換しうることが公知であり(Krizekら, (1991) J. Am. Chem. Soc. 113: 4518-4523)、+4のLeuはある場合にはArgにより置換しうる。Xcの前のPhe残基は、Trp以外のどの芳香族によって置換することもできる。さらに、実験は、ジンクフィンガーの好ましい構造および残基割当からの逸脱は、許容されかつある特定の核酸配列との結合に有利であることすら証しうることを示した。しかし、これを考慮しても、4つのCysまたはHis残基に接触する亜鉛原子が配位したαヘリックスに関わる一般構造は変らない。本明細書に使われる上の構造(A)および(B)は、Cys2-His2タイプの全てのジンクフィンガー構造を表す例示の構造とみなされる。
【0056】
好ましくは、Xaは(F/Y)-XまたはP-(F/Y)-Xである。この式中で、Xはアミノ酸である。好ましくは、この式中で、XはE、K、TまたはSである。好ましくは劣るが、Q、V、AおよびPも予測される。残りのアミノ酸は可能性を残す。
【0057】
好ましくは、X2-4は4個よりも2個のアミノ酸からなる。これらのアミノ酸の第1は、どのアミノ酸であってもよいが、S、E、K、T、PおよびRが好ましい。PまたはRであるのが有利である。これらのアミノ酸の第2は、好ましくはEであるが、どのアミノ酸でも使うことができる。
【0058】
好ましくは、XbはTまたはIである。
【0059】
好ましくは、XcはSまたはTである。
【0060】
好ましくは、X2-3はG-K-A、G-K-C、G-K-SまたはG-K-Gである。しかし、好ましい残基からの逸脱、例えばM-R-NまたはM-Rの形も可能である。
【0061】
好ましくは、リンカーはT-G-E-KまたはT-G-E-K-Pである。
【0062】
上に述べたように、主な結合相互作用は、アミノ酸-1、+2、+3および+6と生じる。アミノ酸+4および+7はほとんど不変である。残るアミノ酸は本質的にどのアミノ酸であってもよい。好ましくは、位置+9はArgもしくはLysにより占められる。位置+1、+5および+8は、疎水性アミノ酸ではない、すなわちPhe、TrpもしくはTyrでないことが有利である。
【0063】
したがって、ほとんどの好ましい様態で、以上を一緒に持ち込んで、本発明により、所与の核酸4塩基連鎖と特異的に結合するジンクフィンガー核酸結合モチーフ内のすべての残基の定義が可能になる。
【0064】
本発明により提供されるコードは完全に固定したものではない。ある特定の選択が提供される。例えば、位置+1、+5および+8はどのアミノ酸が配置されてもよく、また、他の位置はある特定の選択がされてもよい。例えば、本発明のルールは、中央T残基との結合に、Ala、SerもしくはValのどれか1つを+3に使いうることを提供する。したがって最も広範囲の意味では、本発明は、全ての定義された標的核酸4塩基連鎖と結合することができる非常に多数のタンパク質を提供する。
【0065】
しかし、好ましくは、可能性の数は大いに減らすことができる。例えば、決定的でない残基+1、+5および+8は、残基Lys、ThrおよびGlnによりそれぞれデフォールトオプション(default option)として占められてよい。他を選択する場合、例えば、最初に与えられるオプションをデフォールトとして用いてよい。したがって、本発明によるコードにより、標的4塩基連鎖と結合する単一の定義されたポリペプチド(「デフォールト」ポリペプチド)の設計が可能になる。
【0066】
本発明のさらなる様態では、標的核酸配列と結合することができるCys2-His2ジンクフィンガークラスの核酸結合タンパク質を調製する方法が提供され、その方法は、
a)天然のジンクフィンガーおよびコンセンサスジンクフィンガーからなる群から1つのモデルジンクフィンガードメインを選択すること;および
b)上に述べたルールにより必要とされるようにフィンガーの位置-1、+2、+3および+6の1つ以上を変異すること
の工程を含む。
【0067】
一般的に、天然のジンクフィンガーは、その核酸結合特異性が公知であるフィンガーから選択してよい。例えば、これらは、結晶構造が解明されているフィンガー:すなわちZif268(Elrod-Ericksonら, (1996) Structure 4:1171-1180)、GLI(PavletichおよびPabo, (1993) Science 261:1701-1707)、Tramtrack(Fairallら, (1993) Nature 366:483-487)およびYY1(Houbaviyら, (1996) PNAS (USA) 93:13577-13582)であってよい。
【0068】
天然のZif268中のジンクフィンガー2はジンクフィンガーを遺伝子操作できる優れた出発点となり、かつ、好ましい。
【0069】
コンセンサスジンクフィンガー構造物を、その結合ドメインが公知であるか否かに拘らず、公知のジンクフィンガーの配列と比較して調製してもよい。好ましくは、該コンセンサス構造は、コンセンサス構造PYKCPECGKSFSQKSDLVKHQRTHTGおよびコンセンサス構造PYKCSECGKAFSQKSNLTRHQRIHTGEKPからなる群から選ばれる。
【0070】
該コンセンサスは、クリゼクら(Krizekら, (1991) J. Am. Chem. Soc. 113:4518-4523)およびヤコフ(Jacobs, (1993) 博士論文, University of Cambridge, UK)により提供されたコンセンサスから誘導される。両ケースとも、上に記載した2つのジンクフィンガーモチーフを一緒に結合するためのリンカー配列すなわちTGEKまたはTGEKPを、コンセンサスの末端に形成することができる。したがって、Pは必要があれば除いてもよく、または、TGで終るコンセンサスの場合はEK(P)を加えることができる。
【0071】
選択したモデルフィンガーの核酸特異性がわかっているときは、標的核酸に対する結合特異性を修正する目的で、天然と所望標的とで相異のある塩基についてその塩基との結合に影響することがわかっている残基に、フィンガーの変異を指定することができる。そうでなければ、モデルフィンガーの変異は、前述のルールに与えられたように、残基-1、+2、+3および+6に集中すべきである。
【0072】
改良された結合を有する結合タンパク質を産生する目的で、さらに、残基選択を支援するために、本発明により提供されるルールを、タンパク質/核酸界面の物理的または仮想的モデルにより補うことができる。
【0073】
本発明により設計したジンクフィンガー結合モチーフは、多重のジンクフィンガーを有する核酸結合タンパク質に組み合わせることができる。好ましくは、タンパク質は少なくとも2つのジンクフィンガーを有する。もともとジンクフィンガー結合タンパク質は、Tramtrackのような2つのジンクフィンガータンパク質も知られているが、一般的には少なくも3つのジンクフィンガーを有する。少なくとも3つのジンクフィンガーの存在が好ましい。結合タンパク質は、所要フィンガーの末端と末端、N末端とC末端を接続することにより構築することができる。好ましくは、これは、ジンクフィンガーをコードする関連核酸コード配列を一緒に結合して、結合タンパク質全体をコードする複合コード配列を作製することによって達成される。したがって、本発明は、上に定義した核酸結合タンパク質を製造する方法を提供し、その方法では該核酸結合タンパク質は組換えDNA技術により構築され、その方法は、
a)上に記載したように、N末端からC末端へ配置された2個以上のジンクフィンガー結合モチーフをコードする核酸コード配列を製造し;
b)核酸配列を適切な発現ベクターに挿入し;および
c)宿主生物に核酸配列を発現させて核酸結合タンパク質を得る
工程を含んでなる。
【0074】
「リーダー(leader)」ペプチドをN末端フィンガーに付加することができる。好ましくは、該リーダーペプチドはMAEEKPである。
【0075】
本発明により核酸結合タンパク質をコードする核酸は、さらなる遺伝子操作のため、ベクターに組込むことができる。本明細書で使用する場合、ベクター(またはプラスミド)は、その発現または複製のために異種核酸を細胞に導入するために使われる分離したエレメントを意味する。このようなビヒクルの選択と使用は、当業界の通常技術者の技術範囲内である。多くのベクターを利用することができ、適切なベクターの選択は、ベクターの意図する用途、すなわち該ベクターをDNA増幅のためまたは核酸発現のためのいずれに使うか、該ベクターに挿入するDNAのサイズ、および該ベクターで形質転換する宿主細胞に依存するであろう。それぞれのベクターは、その機能(DNAの増幅またはDNAの発現)およびベクターが適合しうる宿主細胞に依存して様々な成分を含有する。ベクター成分は一般的に、限定されるものでないが、1つ以上の次のものを含有する:複製の起点、1つ以上のマーカー遺伝子、エンハンサーエレメント、プロモーター、転写終結配列およびシグナル配列。
【0076】
一般的に、発現およびクローニングベクターは両方とも、ベクターが1つ以上の選択された宿主細胞中で複製できるようにする核酸配列を含有する。典型的には、クローニングベクターにおいて、この配列は、ベクターに宿主染色体DNAと関係なく複製する能力をあたえ、複製起点または自律的複製配列を含む。このような配列は、各種の細菌、酵母およびウイルスにおいて周知である。プラスミドpBR322由来の複製起点はほとんどのグラム陰性細菌用に適切であり、2μプラスミド起点は酵母用に適切であり、そして様々なウイルス起点(例えば、SV40、ポリオーマ、アデノウイルス)は哺乳動物細胞のクローニングベクター用に有用である。一般的に、哺乳動物発現ベクター用の複製起点成分は、COS細胞のような高レベルDNA複製能力のある哺乳動物細胞に使うのでなければ不要である。
【0077】
ほとんどの発現ベクターはシャトルベクターであり、すなわち、少なくとも1クラスの生物の複製能力があるが、発現のために他のクラスの生物にトランスフェクトすることができる。例えば、あるベクターは大腸菌(E. coli)でクローニングされ、その後、その同じベクターは酵母または哺乳動物細胞にトランスフェクトされるが、宿主細胞染色体に依存せず複製する能力を持たない。DNAは宿主ゲノムへの挿入によって複製させることもできる。しかし、核酸結合タンパク質をコードするゲノムDNAの回収は、核酸結合タンパク質DNAを切除するために制限酵素消化を必要とするので、外因的に複製したベクターの回収よりもっと複雑である。DNAはPCRにより増幅することができ、またどのような複製成分もなしに、直接、宿主細胞にトランスフェクトすることができる。
【0078】
発現およびクローニングベクターは、選択マーカー(selectable marker)とも呼ばれる選択遺伝子を含有するのが有利である。この遺伝子は、選択培地で増殖する形質転換した宿主細胞の生存または増殖に必要なタンパク質をコードする。選択遺伝子を含有するベクターで形質転換されない宿主細胞は、この培地で生存しないであろう。典型的な選択遺伝子は、抗生物質および他の毒素、例えばアンピシリン、ネオマイシン、メトトレキセートもしくはテトラサイクリンへの耐性を与え、栄養要求欠乏を補い、または複合培地から利用できない重要な栄養を供給するタンパク質をコードする。
【0079】
酵母に適切な選択遺伝子マーカーについては、マーカー遺伝子の表現型発現により形質転換体の選択を容易にするどのマーカー遺伝子でも使うことができる。酵母に対する適切なマーカーは、例えば、抗生物質G418、ハイグロマイシンまたはブレオマイシン耐性を与えるものであるか、または、栄養要求性酵母突然変異体、例えばURA3、LEU2、LYS2、TRP1、またはHIS3遺伝子に原栄養(prototrophy)を与える。
【0080】
ベクターの複製は大腸菌で行うのが好都合であるので、大腸菌遺伝子マーカーおよび大腸菌複製起点を含むのが有利である。これらは、pBR322、Bluescript(登録商標)ベクターまたはpUCプラスミド、例えばpUC18もしくはpUC19のような大腸菌プラスミドから得ることができ、これらは大腸菌複製起点およびアンピシリンのような抗生物質への耐性を与える大腸菌遺伝子マーカーの両方を含有する。
【0081】
哺乳動物細胞に適切な選択マーカーは、核酸結合タンパク質核酸を取上げる能力のある(competent)細胞の確認を可能にするジヒドロ葉酸レダクターゼ(DHFR、メトトレキセート耐性)、チミジンキナーゼ、またはG418もしくはハイグロマイシン耐性を与える遺伝子のようなマーカーである。哺乳動物細胞形質転換体は、マーカーを取上げて発現する形質転換体だけがユニークに生存適合する選択圧力下に置かれる。DHFRまたはグルタミンシンターゼ(GS)マーカーの場合、選択圧力が次第に増加する条件下で形質転換体を培養することが可能であり、それにより選択遺伝子および核酸結合タンパク質をコードする連結されたDNAの両方を、(その染色体組込み部位で)増幅させる。増幅は、増殖に重要なタンパク質の産生のより大きな需要のある遺伝子ならびに所望のタンパク質をコードしうる密接に関連する遺伝子を一緒に、組換え細胞の染色体内で直列に反復させるプロセスである。所望タンパク質の増加量は、通常、このように増幅したDNAから合成される。
【0082】
発現およびクローニングベクターは、通常、宿主生物により認識され、核酸結合タンパク質をコードする核酸に機能しうる形で連結されたプロモーターを含有する。このようなプロモーターは、誘導性(inducible)であっても、構成的(constitutive)であっても良い。該プロモーターは、ソースDNAから制限酵素消化によりプロモーターを除去し、単離したプロモーター配列をベクターに挿入することにより、機能しうる形で核酸結合タンパク質をコードするDNAに連結される。元来の核酸結合タンパク質プロモーター配列および多くの異種プロモーターの両方を、核酸結合タンパク質をコードするDNAの直接増幅および/または発現に使うことができる。
【0083】
原核生物宿主での使用に適切なプロモーターは、例えば、β-ラクタマーゼおよびラクトースプロモーター系、アルカリホスファターゼ、トリプトファン(Trp)プロモーター系ならびにtacプロモーターのようなハイブリッドプロモーターを含む。これらのヌクレオチド配列は公表されており、熟練技術者は、必要な制限酵素切断部位を与えるためにリンカーまたはアダプターを使って、該プロモーターを核酸結合タンパク質をコードするDNAに機能しうる形でライゲートすることが可能である。細菌系に使用するためのプロモーターは、一般的に、核酸結合タンパク質をコードするDNAに機能しうる形で連結したシャイン・ダルガルノ配列も含むであろう。
【0084】
好ましい発現ベクターは、細菌内で機能する能力のあるファージx(phagex)またはT7のようなバクテリオファージのプロモーターを含んでなる細菌発現ベクターである。最も広く使われる発現系の1つでは、融合タンパク質をコードする核酸を、該ベクターからT7 RNAポリメラーゼにより転写することができる(Studierら, Methods in Enzymol. 185; 60-89, 1990)。pETベクターと共に使われる大腸菌BL21(DE3)宿主株では、T7 RNAポリメラーゼは宿主細菌のλ溶原菌DE3から産生され、その発現はIPTG誘導性lac UV5プロモーターの制御下にある。この系は成功裏に多くのタンパク質の過剰産生に用いられている。あるいは、ポリメラーゼ遺伝子は、市販されるCE6ファージ(Novagen, Madison, USA)のようなintファージでの感染によりλファージに導入することができる。他のベクターとして、PLEX(Invitrogen, NL)のようなλPLプロモーターを含有するベクター、pTrcHisXpressTm(Invitrogen)もしくはpTrc99(Pharmacia Biotech, SE)のようなtrcプロモーターを含有するベクター、またはpKK223-3(Pharmacia Biotech)もしくはPMAL(New England Biolabs, MA, USA)のようなtacプロモーターを含有するベクターが挙げられる。
【0085】
さらに、本発明による核酸結合タンパク質遺伝子は、好ましくは、細菌宿主からのポリペプチドの分泌を容易にするための分泌配列を含み、封入体中よりもむしろ可溶性の元来のペプチドとして産生させる。該ペプチドは細菌の周辺腔または培地から適当に回収することができる。
【0086】
酵母宿主での使用のための適切なプロモーター配列は、調節されるかまたは構成的であることができ、好ましくは、高度に発現した酵母遺伝子、特にSaccharomyces cerevisiae遺伝子由来のものである。したがって、TRP1遺伝子、ADHIもしくはADHII遺伝子、酸性ホスファターゼ遺伝子(PH05)のプロモーター、aもしくはα因子をコードする酵母接合フェロモン遺伝子のプロモーターまたはエノラーゼ、グリセルアルデヒド-3-リン酸デヒドロゲナーゼ(GAP)、3-ホスホグリセリン酸キナーゼ(PGK)、ヘキソキナーゼ、ピルビン酸デカルボキシラーゼ、ホスホフルクトキナーゼ、グルコース-6-リン酸イソメラーゼ、3-ホスホグリセリン酸ムターゼ、ピルビン酸キナーゼ、トリオースリン酸イソメラーゼ、ホスホグルコースイソメラーゼもしくはグルコキナーゼ遺伝子のプロモーターのような解糖系酵素をコードする遺伝子由来のプロモーター、またはTATA結合タンパク質(TBP)遺伝子由来のプロモーターを使うことができる。さらに、1つの酵母遺伝子の上流活性化配列(UAS)および他の酵母遺伝子の機能的TATAボックスを含む下流プロモーターエレメントを含んでなるハイブリッドプロモーター、例えば、酵母PH05遺伝子のUASおよび酵母GAP遺伝子の機能的TATAボックスを含む下流プロモーターエレメントを含むハイブリッドプロモーター(PH05-GAPハイブリッドプロモーター)を使うことができる。適切な構成的PH05プロモーターは、例えば、PH05遺伝子のヌクレオチド-173で開始しヌクレオチド-9で終結するPH05(-173)プロモーターエレメントのような、上流調節エレメント(UAS)を欠いた短縮酸性ホスファターゼPH05プロモーターである。
【0087】
哺乳動物宿主内でのベクターからの核酸結合タンパク質遺伝子転写は、プロモーターが宿主細胞系と適合しうることを条件として、ポリオーマウイルス、アデノウイルス、伝染性上皮腫ウイルス(fowlpox virus)、ウシ乳頭腫ウイルス、トリサルコーマウイルス、サイトメガロウイルス(CMV)、レトロウイルスおよびサルウイルス40(SV40)のようなウイルスのゲノム由来のプロモーター、アクチンプロモーターのような異種哺乳動物プロモーターまたは非常に強いプロモーター、例えばリボソームタンパク質プロモーター、および、核酸結合タンパク質配列に通常関連するプロモーターにより制御することができる。
【0088】
高等真核生物による核酸結合タンパク質をコードするDNAの転写は、ベクターにエンハンサー配列を挿入することにより増加することができる。エンハンサーは比較的配向および位置に依らない。哺乳動物遺伝子由来の多くのエンハンサー配列が公知である(例えば、エラスターゼおよびグロビン)。しかし、典型的には、真核細胞ウイルス由来のエンハンサーを用いるであろう。例として、複製起点の後期側(bp 100-270)のSV40エンハンサーおよびCMV初期プロモーターエンハンサーが挙げられる。該エンハンサーは、核酸結合タンパク質DNAの位置5'または3'でベクター中にスプライスされうるが、プロモーターから部位5'に位置することが好ましい。
【0089】
本発明による核酸結合タンパク質をコードする真核発現ベクターは、遺伝子座制御領域(LCR)を含んでなることが有利である。LCRは、宿主細胞クロマチンに組込まれた導入遺伝子の高レベルの組込み部位に依らない発現を指令することが可能であり、これは特に、ベクターの染色体組込みが起っている永久的トランスフェクトされた真核細胞系において、またはトランスジェニック動物において、核酸結合タンパク質遺伝子を発現させるときに重要である。
【0090】
真核ベクターは、転写の終結およびmRNAの安定化のために必要な配列も含有することができる。このような配列は、一般的に、真核生物またはウイルスのDNAまたはcDNAの5'および3'非翻訳領域から利用可能である。これらの領域は、核酸結合タンパク質をコードするmRNAの非翻訳部のポリアデニル化断片として転写されるヌクレオチドセグメントを含有する。
【0091】
発現ベクターは、かかるDNAを発現する能力のある、プロモーター領域のような調節配列と機能しうる形で連結された、核酸結合タンパク質核酸を発現する能力のあるベクターを含む。したがって、発現ベクターは、プラスミド、ファージ、組換えウイルスまたは他のベクターのような、適切な宿主細胞への導入によりクローン化DNAの発現をもたらす組換えDNAまたはRNA構築物を意味する。適切な発現ベクターは当業者には周知であり、真核および/または原核細胞で複製可能であるもの、およびエピソームのままのもの、または宿主細胞ゲノムに組込まれるものを含む。例えば、核酸結合タンパク質をコードするDNAを、哺乳動物細胞のcDNAの発現に適したベクター、例えばpEVRFのようなCMVエンハンサーに基くベクターに挿入することができる(Matthiasら, (1989) NAR 17, 6418)。
【0092】
本発明の実施に特に有用なのは、哺乳動物細胞の核酸結合タンパク質をコードするDNAの一過性発現を提供する発現ベクターである。一過性発現は、通常、宿主細胞内で効率的に複製が可能な発現ベクターの使用に関わり、宿主細胞が発現ベクターの多数のコピーを蓄積して、その結果、高レベルの核酸結合タンパク質を合成する。本発明の目的に対して、一過性発現系は、例えば核酸結合タンパク質変異体を確認するため、潜在ホスホリル化部位を確認するため、またはタンパク質の機能ドメインを特性決定するために有用である。
【0093】
本発明によるベクターの構築は、通常の連結反応技術を用いる。単離したプラスミドまたはDNA断片を開裂し、加工し、所望の形に再連結して所要のプラスミドを作製する。もし所望であれば、構築プラスミド内の正しい配列を確証する分析を公知の様式で実施する。発現ベクターを構築し、in vitro転写物を調製し、宿主細胞にDNAを導入し、そして核酸結合タンパク質発現および機能を評価する分析を実施する適切な方法は、当業者には公知である。サンプル中の遺伝子存在、増幅および/または発現は、例えば、本明細書に与えられた配列に基く適切に標識したプローブを使用して、通常のサザンブロット、mRNAの転写物を定量するためのノーザンブロット、ドットブロット(DNAもしくはRNA分析)、またはin situハイブリダイゼーションによって、直接に測定することができる。当業者は、もし所望であれば、これらの方法がいかに変更されうるかを容易に予測するであろう。
【0094】
本発明の他の実施態様によって、上記核酸を含有する細胞が提供される。原核生物、酵母および高等真核生物細胞のような宿主細胞を、DNAを複製しかつ核酸結合タンパク質を産生するために使うことができる。適切な原核生物として、グラム陰性またはグラム陽性菌、具体的には大腸菌(例えば大腸菌 K-12株、DH5aおよびHB101)、またはバチルスのような、真性細菌が挙げられる。核酸結合タンパク質をコードするベクターに適切なさらなる宿主は、繊維状真菌または酵母、例えばSaccharomyces cerevisiaeのような真核微生物を含む。高等真核細胞としては、昆虫および脊椎動物細胞、特に、ヒト細胞を含む哺乳動物細胞または他の多細胞生物由来の有核細胞が挙げられる。近年、培養(組織培養)中の脊椎動物細胞の増殖は日常的手法になっている。有用な哺乳動物宿主細胞系の例は、チャイニーズハムスター卵巣(CHO)細胞、NIH 3T3細胞、HeLa細胞もしくは293T細胞のような、上皮または繊維芽細胞系である。本発明の開示でいう宿主細胞は、in vitro培養の細胞ならびに宿主動物内の細胞を含んでなる。
【0095】
DNAは、細胞に安定に組み込まれていても、または当分野で公知の方法を使い一過的に発現されてもよい。安定にトランスフェクトされた哺乳動物細胞は、細胞を選択マーカー遺伝子を有する発現ベクターでトランスフェクトし、トランスフェクトされた細胞をマーカー遺伝子を発現する細胞を選択する条件下で増殖させることにより、調製することができる。一過性のトランスフェクタントを調製するため、哺乳動物細胞は、トランスフェクション効率をモニターするレポーター遺伝子でトランスフェクトされる。
【0096】
このような安定にまたは一過的にトランスフェクトされた細胞を産生するには、細胞を、核酸結合タンパク質を形成するに十分な量の核酸結合タンパク質をコードする核酸でトランスフェクトしなければならない。核酸結合タンパク質をコードするDNAの正確な量は、実験的に決定され、特定の細胞とアッセイに対して最適化される。
【0097】
宿主細胞は、本発明の上に述べた発現またはクローニングベクターでトランスフェクトされ、または好ましくは形質転換され、プロモーターを誘起し、形質転換体を選択し、または所望の配列をコードする遺伝子を増幅するために適切に修飾された通常の栄養培地で培養される。異種DNAの宿主細胞への導入は、リン酸カルシウム共沈技術によるまたはエレクトロポレーションによる異種DNAをコードするベクターでのトランスフェクションのような、当分野で公知の方法により実施することができる。多数のトランスフェクションの方法が当業者には公知である。トランスフェクションの成功は、一般的に、このベクターの作動の徴候が宿主細胞中で起るときに認められる。形質転換は、使用する特定の宿主細胞に適切な標準技術を使って達成される。
【0098】
クローン化DNAの適切な発現ベクターへの組込み、1つ以上の別個の遺伝子をそれぞれコードするプラスミドベクターもしくはプラスミドベクターの組合わせ、または線形DNAによる真核細胞のトランスフェクション、およびトランスフェクトされた細胞の選択は、当分野で周知である(例えば、Sambrookら, (1989) Molecular Cloning: A Laboratory Manual(分子クローニング:実験室マニュアル),第二版、Cold Spring Harbor Laboratory Pressを参照すること)。
【0099】
トランスフェクトされたまたは形質転換された細胞は、当業界で公知の培地および培養方法を使い、好ましくは、DNAによりコードされた核酸結合タンパク質が発現される条件下で培養する。適切な培地の組成は、当業者に公知であり、容易に調製することができる。適切な培地は市販もされている。
【0100】
本発明による核酸結合タンパク質は、診断および研究ツールを含む広範囲の応用に用いることができる。複雑な(complex)混合物中の核酸分子の存在を確認するための診断ツールとして有利に用いることができる。本発明による核酸結合分子は、標的核酸分子の単一塩基対変化を識別することができる。
【0101】
したがって、本発明は、標的核酸分子の存在を確認する方法を提供し、該方法は:
a)上に述べた方法により標的核酸分子に特異的である核酸結合タンパク質を調製すること;
b)結合を促進し、未結合で残る核酸結合タンパク質を除去する条件下で、標的核酸分子を含んでなる試験システムを核酸結合タンパク質にさらすこと;
c)試験システムの核酸結合タンパク質の存在を検出すること
の工程を含んでなる。
【0102】
好ましい実施態様では、本発明の核酸結合分子はELISAアッセイに組み込むことができる。例えば、本発明の分子を提示するファージを、標的核酸の存在を検出するために使うことができ、酵素結合抗ファージ抗体を使って可視化することができる。
【0103】
診断用のジンクフィンガーファージの使用のさらなる改良が、例えば、バクテリオファージのマイナーコートタンパク質(gVIII)と融合したマーカータンパク質を同時発現することにより行うことができる。抗ファージ抗体による検出はすたれている(obsolete)ので、それぞれの診断の時間と費用はさらに低減されよう。必要に応じて、提示のための適切なマーカーは、蛍光タンパク質(A. B. Cubittら, (1995) Trends Biochem Sci. 20, 448-455;T.T. Yangら, (1996) Gene 173, 19-23)、または、先にgIII上に提示されているアルカリホスファターゼ(J. McCafferty, R. H. Jackson, D. J. Chiswell, (1991) Protein Engineering 4, 955-961)のような酵素を含む。別個のマーカーによる様々なタイプの診断ファージの標識付けにより、単一核酸サンプルの多重スクリーニングが可能になる。このようなきめ細かさがなくとも、基本的なELISA技術は、信頼性があり、迅速で、簡単で、特に安価である。さらに、特別な装置も必要としないし、放射性同位体のような有害な試薬を用いないので、臨床での日常的使用に受入れられる。このプロトコルの主な利点は、ゲル電気泳動の必要性を除き、自動化核酸診断への道を開くことにある。
【0104】
本発明は、精巧な特異性で遺伝子操作することができる核酸結合タンパク質を提供する。したがって、本発明は、特異的な核酸結合を必要とする分子の設計に役立つ。例えば、本発明によるタンパク質は、本明細書に記載したジンクフィンガーを含んでなる核酸結合ドメインと融合した核酸開裂ドメインを有するキメラ制限酵素の製造に用いることができる。
【0105】
さらに、本発明は、本明細書に記載した核酸結合タンパク質の使用に関わる治療薬および治療法を提供する。特に、本発明は、ウイルスインテグラーゼのようなインテグラーゼ、および本発明によるin vivo核酸配列を標的とする核酸結合タンパク質を含んでなるポリペプチド融合体の使用を提供する(Bushman, (1994) PNAS (USA)91:9233-9237)。遺伝子治療用途において、本法は機能的遺伝子の欠陥遺伝子への送達、または望ましくない核酸を破壊する目的のナンセンス核酸の送達に応用することができる。あるいは、遺伝子をセントロメアのような既知の、核酸の繰返し範囲に、LCRのような活性化配列と一緒に送達することもできる。これは核酸のゲノムへの安全かつ予測可能な組込みへの経路を表すであろう。
【0106】
通常の治療用途では、本発明による核酸結合タンパク質は、変異した必須タンパク質(vital protein)を有する細胞を特異的にノックアウトするために使うことができる。例えば、もし変異rasをもつ細胞を標的とすれば、それらはrasが細胞生存に必須であるので、破壊されるであろう。あるいは、転写因子の作用を、細胞に転写因子に特異的な結合部位に結合する薬剤を投与することにより、調節、好ましくは抑制することができる。例えば、HIVtatの活性をHIV TARに特異的な結合タンパク質により抑制することができる。
【0107】
さらに、本発明による結合タンパク質は、不可逆的核酸破壊および細胞死を引き起す能力があるヌクレアーゼのような毒性分子と結合することができる。このような薬剤は、内因性核酸に変異を含む細胞を選択的に破壊する能力がある。
【0108】
以上に説明したこれらの核酸結合タンパク質および誘導体は感染などの治療にも、生物特異的抗生物質また抗ウイルス医薬の形態で応用することができる。このような応用では、結合タンパク質をヌクレアーゼまたは他の核毒素(nuclear toxin)と組合せて、微生物の核酸を特異的に標的とすることができる。
【0109】
本発明は、同様に、本発明による化合物または製薬上許容されるそれらの塩を有効成分として含有する医薬製剤およびそれらの製造方法に関する。
【0110】
本発明による化合物または製薬上許容されるそれらの塩を含有する本発明による医薬製剤は、温血動物への、経口等の経腸、さらに直腸、および非経口投与用の製剤であり、薬理学的有効成分が製剤自身にまたは製薬上許容される担体と一緒に存在する。有効成分の一日量は年齢および個体の症状に、また投与の方法に依存する。
【0111】
新規の医薬製剤は、例えば、約10%〜約80%、好ましくは約20%〜約60%の有効成分を含む。経腸または非経口投与用の本発明による医薬製剤は、例えば、糖衣錠、錠剤、カプセルまたは座薬、およびさらにアンプルのような単位用量形態の製剤である。これらは、それ自体公知の方法により、例えば通常の混合、造粒(granulating)、糖コーティング、溶解または凍結乾燥プロセスの方法により調製される。したがって、経口用途の医薬製剤は、有効成分を固体担体と組合せ、もし所望であれば、得た混合物を造粒し、そしてもし所望であるか必要であれば、錠剤もしくは糖衣錠コアを与える適切な賦形剤を添加した後に、混合物または造粒物を加工することによって、得ることができる。
【0112】
適切な担体は、特定的には、例えばラクトース、スクロース、マンニトールもしくはソルビトールのような糖類、セルロース調製物および/または例えばリン酸三カルシウムもしくはリン酸水素カルシウムのようなリン酸カルシウム等の充填剤、さらにはトウモロコシ、コムギ、コメまたはジャガイモデンプンを使うデンプンペースト、ゼラチン、トラガカント、メチルセルロースおよび/またはポリビニルピロリドンのような結合剤、もし所望であれば上述のデンプン、さらにカルボキシメチルデンプン、架橋ポリビニルピロリドン、寒天、アルギン酸またはアルギン酸ナトリウムのようなその塩等の崩壊剤であり;補助剤は、主に滑剤、流動調節剤および潤滑剤、例えば珪酸、タルク、ステアリン酸またはステアリン酸マグネシウムもしくはカルシウムのようなその塩、および/またはポリエチレングリコールである。糖衣錠コアは、もし所望であれば胃液耐性のある適切なコーティングにより提供され、なかんずくもし所望であれば、アラビアゴム、タルク、ポリビニルピロリドン、ポリエチレングリコールおよび/または二酸化チタンを含有する濃厚糖類溶液、適切な有機溶媒または溶媒混合物のコーティング溶液、または、胃液耐性コーティングの製剤にはアセチルセルロースフタレートもしくはヒドロキシプロピルメチルセルロースフタレートのような適切なセルロース調製物の溶液を使う。例えば有効成分の異なる用量を同定するかまたは示すために、着色剤または顔料を錠剤または糖衣錠コーティングに加えることができる。
【0113】
他の経口使用できる医薬製剤は、硬ゼラチンカプセル、ならびに、ゼラチンおよびグリセロールまたはソルビトールのような可塑剤で作られた軟閉鎖カプセルである。硬ゼラチンカプセルは、有効成分を顆粒の形態で、例えばラクトースのような充填剤、デンプンのような結合剤、および/またはタルクもしくはステアリン酸マグネシウムのような潤滑剤、および、所望であれば安定剤との混合物で含有することができる。軟カプセルでは、有効成分は、好ましくは脂肪油、パラフィン油または液体ポリエチレングリコールのような適切な液体に溶解または懸濁されており、安定剤を加えることもできる。
【0114】
適切な直腸経由で利用可能な医薬製剤は、例えば、有効成分と座薬基剤の組合わせからなる座薬である。適切な座薬基剤は、例えば、天然または合成トリグリセライド、パラフィン系炭化水素、ポリエチレングリコールまたは高級アルカノールである。さらに、有効成分と基剤との組合わせを含有するゼラチン直腸カプセルも使うことができる。適切な基剤は、例えば、液体トリグリセライド、ポリエチレングリコールまたはパラフィン系炭化水素である。
【0115】
非経口投与に適切な製剤は主に、有効成分の水溶性形態、例えば水溶性塩の水溶液、およびさらに、脂肪油、例えばゴマ油、または合成脂肪酸エステル、例えばオレイン酸エチルもしくはトリグリセライドのような適切な親油性溶媒またはビヒクルを使う、適切な油状注入懸濁液、または、増粘性物質、例えばカルボキシメチルセルロースナトリウム、ソルビトールおよび/またはデキストラン、かつ必要であれば安定剤も含有する水性注入懸濁液のような、有効成分の懸濁液である。
【0116】
有効成分用量は、温血動物種、年齢、および個体の症状ならびに投与方法による。通常の場合、ほぼ75kg体重の患者に対する経口投与の場合には、約10mg〜約250mgの一日量と見積られる。
【0117】
本発明は以下に、説明のみの目的で、次の実施例に記載される。
【0118】
実施例1
ジンクフィンガーの結合部位選好(preference)の決定
ジンクフィンガーファージ提示ライブラリーの設計
隣接DNAサブサイト間の界面におけるジンクフィンガー‐DNA認識を、ジンクフィンガーファージ提示ライブラリーを使い研究した。このライブラリーは、Zif268の3フィンガーDNA結合ドメインに基くが、フィンガー2(F2)およびフィンガー3(F3)は、それらのDNAサブサイトの界面を横切る接触ネットワークを形成しうる残基位置においてアミノ酸のランダム化が行われている。ライブラリーの詳細設計を、選択に使った総括的な(generic)DNA結合部位とともに、図1cに示す。簡単に説明すると、ライブラリーはF2残基位置6(以後、F2[+6]と記す)およびF3残基位置-1、+1、+2および+3(以後、F3[-1]、F3[+2]、などと記す)におけるランダム化を含有する。
【0119】
ライブラリー選択は、Zif268オペレーターに似るが、F2とF3のDNAサブサイト間の塩基ステップを形成するDNAダブレットの塩基の組織的組合わせを有するDNA結合部位を使って実施した。DNA結合部位は一般的形態が5'-GNX-XCG-GCG-3'であり、ここにX-XはDNAサブサイト間の界面における塩基の所与の組合わせを記し、Nは4種の塩基がDNA位置3において等しく発現されることを記す。したがって、この実験では、F3[+3]とヌクレオチド位置3Nの間の相互作用は完全な自由度が与えられている。このライブラリーの特徴により、ヌクレオチド位置4Xおよび5Xの塩基の所与の組合わせと結合する関連ジンクフィンガーの大きなファミリー(またはデータベース)の選択が可能になるが、これらは、F3の名目上のトリプレットサブサイトの中間塩基との異なる相互作用のために同一ではない。
【0120】
構築されるべき第1ライブラリー、LIB-Aは、F2残基位置6およびF3残基位置-1、+1、+2および+3におけるランダム化物を含有し(図2を参照すること)、DNA配列5'-GNX-XCG-GCG-3'を使ってソートされる。ここにX-XはDNA位置4Xおよび5Xにおける2つの塩基の既知の組合わせを記し、NはDNA位置3において等しい確率をもつ4種の塩基のいずれかを記す。第2ライブラリー、LIB-Bは、F2残基位置6およびF3残基位置-1および2におけるランダム化物を含有し、DNA配列5'-GCX-XCG-GCG-3'を使ってソートされる。ここにX-XはDNA位置4Xおよび5Xにおける2つの塩基の既知の組合わせを記す。
【0121】
2つの異なるジンクフィンガーファージ提示ライブラリー用の遺伝子は、4種の合成DNAオリゴヌクレオチドから、方向性末端対末端(directional end to end)連結反応により3種の短い相補的DNAリンカーを使って構築する。該オリゴヌクレオチドは、フィンガー2および3の適切なアミノ酸位置に選択的にランダム化された(配列NNSの;N=A/C/G/G、S=G/C)コドンを有する。該構築物をNot IおよびSfi I制限酵素切断部位を含有するプライマーを使ってPCRにより増幅し、上記のエンドヌクレアーゼで消化してクローニングオーバーハング(overhang)を作り、ファージベクターFd-Tet-SN中にライゲートした。電気コンピテント(electrocompetent)大腸菌(E. coli)TG 1細胞を組換えベクターにより形質転換し、15μg/mlテトラサイクリンを含有するTYE培地(1.5%寒天、1%バクトトリプトン、0.5%バクト酵母抽出物、0.8%NaCl)上にプレーティングした。
【0122】
未研究のいくつかのタンパク質‐DNA相互作用にこの自由度を認めることは、いずれか1つの選択実験から得られるクローンの多様性を増加する有用な方法である。しかし同時に、どの時点でも前後の(contextual)自由度が認められる接触数を限定することが重要であり、そうしなければ、特に強い分子間相互作用のサブセットが選択を支配する危険がある。この可能性を予測して、位置F2[+6]およびF3[-1および+2]にのみランダム化残基を含有し、したがって選択において前後の自由度を認めない、より小さいサブライブラリーも作製した。このライブラリーから選択されたクローンを本明細書で記すときは、星印でマークした。
【0123】
実験方法
2つのジンクフィンガーライブラリーからのファージ選択は、各ライブラリーのメンバーにより特異的に結合されうるDNA配列の多様性を決定する目的で別々に実施する。各ライブラリーについて、ヌクレオチド位置4Xおよび5Xにおける塩基の全16通りの可能な組合わせに対応する異なるDNA結合部位を使用して、16回の選択を実施する。特異的結合ファージを選択するために使われるDNA結合部位は固相表面に固定し、一方、他の15種のDNA部位はそれぞれ10倍過剰で特異性競合剤として溶液中に存在させる。
【0124】
ファージ選択
テトラサイクリン耐性コロニーをプレートから、50μM ZnCl2および15μg/mlテトラサイクリンを含有する2xTY培地(16g/リットルバクトトリプトン、10g/リットルバクト酵母抽出物、5g/リットルNaCl)に移し、一夜、30℃で振とう培養器で培養した。300gで5分間遠心分離して、ファージ粒子を含有する清澄な培養上清を得た。
【0125】
ビオチン化DNA標的部位(1pmol)をストレプトアビジン被覆チューブ(Boehringer Mannheim)に結合させた。ファージ上清溶液をPBS選択バッファー(50μM ZnCl2、2% Marvel、1% Tween、20μg/ml音波処理サケ精子DNA、10pmol/mlの他の15種の可能な非ビオチン化DNA部位のそれぞれを含有するPBS)中に1:10に希釈し、1mlを各チューブにアプライし、1時間、20℃でおいた。この後、チューブを空にし、50μM ZnCl2、2% Marvel、1% Tweenを含有するPBSにより20回洗浄した。保持されたファージを0.1mlの0.1Mトリエチルアミン中に溶出させ、等容積の1M Tris(pH7.4)で中和した。対数期の大腸菌TG 1(0.5ml)を溶出ファージ(50μl)で感染させ、続くラウンドの選択のためのファージ上清を調製するために使った。3ラウンドの選択の後、選択ファージで感染させた大腸菌をプレーティングし、個々のコロニーを拾上げて結合部位特徴(signature)アッセイおよびDNA配列決定用のファージを増殖するために使った。
【0126】
特定のDNA結合部位に対する3ラウンドのファージ選択の後、個々のジンクフィンガークローンを回収し、そして各クローンのDNA結合特異性を結合部位サイン法により決定した。これは、各ライブラリーが位置4Xおよび5Xのいずれかで1つの固定塩基および1つのランダム化塩基を含有する(すなわち、ライブラリーGN、AN、TN、CN、およびNG、NA、NT、NC)ように設計された8つの異なるDNA結合部位ライブラリーに対する、各ジンクフィンガーファージの結合スクリーニングに関わる。したがって、選択実験に使われた16のDNA結合部位のそれぞれは、ユニークな2つのライブラリーの組合わせにより規定される。例えば、4G5Gを含有するDNA結合部位は、8つのライブラリー中、関連するダブレットが1つのランダム化ヌクレオチドおよび他のグアニンとして固定されたヌクレオチドを有する2つ、すなわちライブラリー4G5Nおよび4N5Gにのみ存在する。結合部位特徴に使われた8つのDNAライブラリーを、マイクロタイタープレートに横に(across)配列し、ジンクフィンガーファージ結合をファージELISAにより検出した。8つのDNAライブラリーとの結合パターンは、各ファージクローンのDNA配列特異性(または選好性)を表し、そして比較的特異性のあるとみられるクローンのみを続いて配列決定して、ランダム化ジンクフィンガー残基位置に存在するアミノ酸の同一性を明らかにした。
【0127】
方法は、先に記載されたとおりである(Choo, Y.およびKlug, A. (1994) Proc. Natl. Acad. Sci. U.S.A. 91, 11163-11167;Choo, Y.およびKlug, A. (1994) Proc. Natl. Acad. Sci. U.S.A. 91, 11168-11172)。簡単に説明すると、Zif268オペレーター変異型を含有する、5'ビオチン化した位置的にランダム化したオリゴヌクレオチドライブラリーを、記載したとおり、プライマー伸長により合成する。DNAライブラリー(LIB-Aに対して0.4pmol/ウエルおよびLIB-Bに対して1.2pmol/ウエル)を、50μM ZnCl2を含有するPBS(PBS/Zn)の入ったストレプトアビジン被覆ELISAウエル(Boehringer-Mannheim)に加える。ファージ溶液(一晩細菌培養した上清を2% Marvel、1% Tweenおよび20μg/ml音波処理サケ精子DNAを含有するPBS/Zn中に1:10で希釈したもの)を各ウエル(50μl/ウエル)にアプライする。結合は、20°で1時間進める。未結合ファージは、1%Tweenを含有するPBS/Znで6回洗浄し、その後PBS/Znで3回洗浄する。結合したファージをELISAにより、西洋わさびペルオキシダーゼとコンジュゲートした抗M13 IgG(Pharmacia Biotech)で検出し、比色シグナルをSOFFMAX 2.32(Molecular Devices)を使い定量する。
【0128】
個々のジンクフィンガークローンのコード配列は、ファージ配列に相補的な外部プライマーを使ってPCRにより増幅する。これらのPCR産物をThermo Sequenase cycle sequencing(Amersham Life Science)を使って手動で配列決定する。
【0129】
ファージ選択ジンクフィンガーの分析
図3は、フィンガー2および3のDNAサブサイト間の塩基ステップを形成する16の異なるDNAダブレットを使い、両方のライブラリーから選択した比較的配列特異性のあるジンクフィンガーファージの結合部位特徴を示す。これらの結果は、5'位置(モデルシステムのヌクレオチド5X)がグアニン以外の塩基で固定された3塩基連鎖を含む、ほとんど全てのサブサイトに特異的に結合するジンクフィンガークローンが選択されたことを示す。結局、該選択物は、4種の塩基のいずれも名目的トリプレットの5'および3'位置の両方に特異的に結合することが可能であることを示す。結果を図4に総括する。
【0130】
大きなライブラリーのメンバーは16のダブレットから15のダブレットを認識するフィンガーを生じるが、より小さいサブライブラリーからの選択は、16のダブレットの8つとだけ特異的に結合しうるフィンガーを生じる。この効率の相異が大きなライブラリー中のよりランダム化された位置の包含に由来するか、大きなライブラリーで設計された前後の自由度により与えられるコンフォメーションのフレキシビリティーに由来するのか、またはその両方であるのかはわかっていない。特異的なジンクフィンガーを生じないただ1つの塩基ステップは、4G5Aである。このジヌクレオチドは、選択に使うDNA結合部位のコンテキスト(context)に好ましくないDNA変形を誘起するのかも知れない。
【0131】
実施例2
位置1に対する+2特異性の決定
ジンクフィンガーのαヘリックス位置2に存在するアミノ酸は、2つのオーバーラップDNA4塩基連鎖(quadruplet)サブサイト(図1Bを参照すること;上に説明した4塩基連鎖の位置1または4に対応する、位置5/5')の界面における塩基対に対する特異性の決定を助けることができる。野生型Zif268のF3[+2]に存在するAsp残基は、DNA認識にある役割を果すことが示されており、さらなる例が本発明のファージ提示実験により作製されている(詳細は実施例1および図5Aを参照すること)。
【0132】
次の実験プロトコルは実施例1のそれである。図5AはDNA結合特異性に与えるαヘリックス位置2の影響を示す関連するジンクフィンガークローンの1例を示す。この場合、フィンガー2の位置6は不変(Asn)で、この場合の変化として、この塩基との接触を選択するためのジンクフィンガーの特異性は、フィンガー3の位置+2により指揮される。
【0133】
ジンクフィンガーのこのファミリーは、4T5Aまたは4T5Cサブサイト界面を含有するDNA結合部位を使う選択物から誘導される。5X-5'X塩基対に対する塩基選好性はF3[+2]に存在するアミノ酸により、恐らくは交差鎖接触の形成により、決定される。
【0134】
図5Bは、F3[+2]で選択されたある特定のアミノ酸と位置5'Xに存在する塩基の同一性の関係の例を示す。選択は5つのアミノ酸(Asn、Gln、Arg、LysおよびHis)からのDNA接触の可能性を表し、これらのアミノ酸は全てヌクレオチド位置5'Xでグアニン(O6)またはチミン(O4)のいずれかの環外酸素原子に水素結合を与えることができる。この図には、F3[+2]でこれらのアミノ酸により単離されたクローンを、位置5'Xで塩基選好性を示す結合部位特徴と一緒に掲げた。全体的に、Serは、全ての既知のジンクフィンガーの半分を越える位置2での存在により、38%の出現率(occurrence)で選択を支配する。トレオニン、AlaおよびGlyはしばしば選択物に存在する(それぞれ、15%、15%および9%)が、識別の区別可能なパターンを示さなかった。ある種のアミノ酸(Cys、Asp、Phe、Ile、Leu、Met、Pro、ValおよびTrp)は位置2に決して選択されない。しかし、これらのある特定の状況下で結合する能力は排除されるべきでない。
【0135】
F3[+2]で選択されるアミノ酸の小さいサブセットは、位置5'Xにおける塩基対の同一性と有意な関係を示し(図5B)、これらの間の交差鎖相互作用はDNA認識の一般的機構でありうることを示唆する。これらの関係のほとんどは、Zif268モデルの枠組に従い、F3[+2]の水素結合供与体とDNA位置5'Xのグアニンまたはチミンの間の対形成として理論付けすることができる。位置2に決して選択されないアミノ酸、または選択されるが有意な相関を示さないアミノ酸とは対照的に、一貫してこの位置からDNA認識にある役割を果すように思われるアミノ酸は、多重水素結合基を側鎖に有する。これらの残基はネットワーク支持への参加により一層大きな特異性を達成するので、塩基認識にある役割を果しうる可能性がある。
【0136】
実施例3
一般目的ライブラリーの構築
この実施例で構築される2成分ライブラリーシステムは、それぞれZif268の3つのフィンガーをコードするがいくつかのアミノ酸位置では選択的にランダム化された、ライブラリーLIB1/2およびLIB2/3を含む。新しいライブラリーは、モジュールジンクフィンガーのモデルに固執する代りに、隣接ジンクフィンガーのある特定のアミノ酸位置において協奏的(concerted)変異を有する。したがって、LIB1/2はF1位置-1、2、3、5および6ならびにF2位置-1、1、2および3に、同時に変異を有する。LIB2/3は、F2位置3および6ならびにF3位置-1、1、2、3および5、6に同時に変異を含有する。各ライブラリーの残りのアミノ酸は、野生型(WT)Zif268配列のとおりである。この2つのライブラリーをFdファージにGIII融合物として標準技術によりクローニングした。
【0137】
それぞれの変異位置に許容されるアミノ酸は、次のとおりである。
【0138】
選択および組換え
選択は、DNA配列GCG-GMN-OPQをLIB1/2に対して、およびDNA配列IJK-LMG-GCGをLIB2/3に対して使って実施した(式中、下線を引いた塩基は野生型 Zif268残基により結合され、他の文字は所与のヌクレオチドを表す)。Zif268結合部位の保存されたヌクレオチドは、Zif268 DNA結合ドメインの保存部分との結合により相互作用の記録(register)を固定する役割を果す。2成分ファージ提示ライブラリーを混合してこれら2つの共通(generic)部位を使う選択を単一のチューブで実施することができるし、または、該選択を別々に実施することができる。数ラウンドの選択の後に、2つのライブラリーを組替えて配列IJK-LMN-OPQを認識するキメラDNA結合ドメインを作製する。
【0139】
組換え反応は、選択した3つのフィンガードメインをPCRにより増幅し、PCR産物を制限酵素Ddelを使い切断することにより実施する。これにより、両方のジンクフィンガーライブラリーの遺伝子を、F2αヘリックス位置4および5をコードするDNA配列で切断する。消化した産物をランダムに再度ライゲートして、キメラDNA結合ドメイン(および再構成した野生型 Zif268を含む他の産物)をコードする組換え遺伝子を作製する。キメラDNA結合ドメインは、PCRによる混合産物から、野生型遺伝子よりむしろ組換えF1およびF3遺伝子を認識する選択的プライマーを使い選択的に増幅し、そして(さらなる選択のために)Fdファージにまたは(例えば大腸菌での発現のために)他のベクターにクローニングされる。
【0140】
2成分系ライブラリーからの最初の選択を完成まで押し進めることができ、かくして組換えによる単一クローンの構築が可能になる。あるいは、もし最初の選択のストリンジェンシーを低くすれば、多くの候補を組換え後の様々なキメラドメインの構築に利用できるであろう。後者の場合、最良の組換えタンパク質は、ファージ上の選択のさらなる繰返しにより、選択することができる。
【技術分野】
【0001】
本発明は、ジンクフィンガー(zinc finger)ポリペプチドの選択のためのライブラリー系に関する。特に本発明は、ジンクフィンガーモチーフがオーバーラップする領域で無作為化(randomised)されたバイナリー系、および選択された位置に限定して生じさせた無作為化を組込むスマートなライブラリーに関する。
【背景技術】
【0002】
タンパク質‐核酸認識は、真核および原核細胞の機能を調節する大多数の生体分子的制御機構に中心的役割を果すありふれた現象である。例えば、タンパク質‐DNA相互作用は遺伝子発現の調節の基礎を成し、したがって、分子生物学者により最も広範に研究されている対象の1つである。
【0003】
豊富な生化学および構造的情報により多数の事例におけるタンパク質‐DNA認識の詳細が説明され、認識の一般原理が現われるに至っている。多くのDNA結合タンパク質は、DNA認識のための折りたたみドメインを独立して含有し、これらのドメインは、順に、ロイシンジッパー、「ヘリックス‐ターン‐ヘリックス」およびジンクフィンガーのような多数の構造ファミリーに属する。
【0004】
構造ドメインの大きな変化に拘らず、タンパク質およびDNAの間に現在まで観察された相互作用の特異性は、ほとんどタンパク質αヘリックス表面とDNAの主溝(main groove)の相補性に由来する(Klug, (1993) Gene 135:83-92)。αヘリックスと主溝の繰返し物理的相互作用に照らして、特定のアミノ酸とDNA塩基の間の接触は、規則の簡単なセット:要するに、結合部位配列選好に対するタンパク質一次構造に関係する立体化学的認識コードにより記述することができる可能性に気持が駆られる。
【0005】
しかし、全てのDNA結合タンパク質によっても、DNA認識を記述できるコードは見出されていないことは明らかである。異なる構造ファミリー由来のDNA結合タンパク質の認識αヘリックスはDNAの主溝と相互作用する方法に重要な相異を示すため、多数の複合体の構造は認識パターンに類似性を与えない。既知のDNA結合モチーフの大部分は特に応用自在なものでなく、現れうるどのコードも非常に少数の関係するDNA配列との結合を記述しがちである。
【0006】
その上、DNA結合タンパク質の各ファミリー内ですら、コードの解読はこれまでとらえどころがなかった。タンパク質DNA相互作用が複雑なため、タンパク質の一次構造と核酸の間にはアミノ酸と塩基の関係を直接に規定する簡単な「アルファベット」等価性がみられない。
【0007】
国際特許出願WO 96/06166はこの問題を研究して、ジンクフィンガー核酸結合タンパク質についてタンパク質‐DNA相互作用を説明する「シラブル(sillabic)」コードを提供する。シラブルコードは、特定塩基への結合を規定する結合タンパク質の1つ以上の特徴に基づくコードであり、その特徴は「シラブル」もしくはそれぞれの特異的な接触を定義する複合指示の形で組合わせ可能である。
【0008】
しかし、このコードは不完全であり、各3塩基連鎖(triplet)の5'位置のG以外のヌクレオチドの特異的選択を可能にする特定の指示を与えない。該方法は、他の特異性に対する核酸結合蛋白質を作製するために、無作為化とその後の選択に依存する。しかし、部分的無作為化と選択の助けをかりても、WO 96/06166に報じられた方法も先行技術の他のどの方法も、Zif268の第1フィンガーに基いて、5'塩基がGまたはT以外の3塩基連鎖と結合する能力のあるジンクフィンガーポリペプチドを単離することに成功してない。これはジンクフィンガータンパク質を設計する能力において重大な不足点である。
【0009】
さらに、この文書は、全ての先行技術と同じく、ジンクフィンガーが核酸3塩基連鎖(triplet)またはその多重体と結合するという概念に依存する。今回、本発明者らは、ジンクフィンガー結合部位がオーバーラップする4bpサブサイトにより決定されることおよびサブサイト間の境界の配列特異性は隣接フィンガー間の相乗作用から生じることを確認した。これは、新規のDNA結合特異性をもつジンクフィンガーの設計および選択に対して重要な示唆を与える。
【特許文献1】国際公報第WO 96/06166号パンフレット
【非特許文献1】Klug, (1993) Gene 135:83-92
【発明の概要】
【発明が解決しようとする課題】
【0010】
本発明は、各ポリペプチドが少なくとも部分的に無作為されている1つ以上のジンクフィンガーを含んでなるジンクフィンガーポリペプチドライブラリーを提供する。好ましくは、本発明は、オーバーラップするジンクフィンガーポリペプチドをコードし、各ポリペプチドが少なくとも部分的に無作為化されている1つ以上のジンクフィンガーを含んでなり、該ポリペプチドは選択の後に組み立てられて多重ジンクフィンガーポリペプチドを形成することができる、一群のジンクフィンガーポリペプチドライブラリーを提供する。
【課題を解決するための手段】
【0011】
本発明は、ジンクフィンガーポリペプチドの設計におけるオーバーラップする4bpサブサイト認識の重要性を認識する。各ライブラリー内でただ1つのジンクフィンガーを無作為化する古典的ジンクフィンガーライブラリー設計においてはジンクフィンガー間に起る相乗作用は見落されている。
【0012】
したがって、本発明は、各ポリペプチドが少なくとも部分的に無作為されている1つ以上のジンクフィンガーを含んでなるジンクフィンガーポリペプチドライブラリーを提供する。
【0013】
好ましくは、本発明は、オーバーラップするジンクフィンガーポリペプチドをコードし、各ポリペプチドが少なくとも部分的に無作為化されている1つ以上のジンクフィンガーを含んでなり、該ポリペプチドは選択の後に組み立てられて多重ジンクフィンガーポリペプチドを形成することができる、一群のジンクフィンガーポリペプチドライブラリーを提供する。
【0014】
他の様態では、本発明は、上に記載したとおり、無作為化が結合部位特異性における改変を指揮することが知られているアミノ酸を置換することに限定されるライブラリーに関する。本発明は、標的配列としての任意の核酸配列に対するライブラリーの選択、およびそれに結合する特異的核酸結合タンパク質の製造を可能にするアミノ酸位置の偏り(bias)を示すコードを提供する。さらに、本発明は、所与の核酸配列に特異的なジンクフィンガータンパク質を設計しかつ最適化する方法を提供する。したがって、本発明は古典的ジンクフィンガーと核酸との相互作用について明らかにされた認識の偏りに関する。この事例において、全ての核酸配列との結合をカバーするルールのパターンが提供される。
【発明の効果】
【0015】
本発明で開示されるコードは、隣接ジンクフィンガー間の相乗的相互作用を考慮し、それによって、任意の所望の結合部位の選択を可能にする。
【図面の簡単な説明】
【0016】
【図1】図1は、ジンクフィンガー‐DNA相互作用を図示したものである。A:Zif268のDNA塩基3塩基連鎖についての古典的な3塩基連鎖相互作用のモデル;B:同様な4塩基連鎖(quadruplet)相互作用を示すモデル;C:認識コード決定のためのライブラリー設計のモデル。
【図2】図2は、認識コードの決定においてファージ提示選択に使う3つのフィンガーのアミノ酸配列を示す。
【図3】図3には、ファージ選択から得られた配列特異的ジンクフィンガークローン、およびそれらの結合部位を示す記号を挙げる。
【図4】図4は、ファージ選択から単離したクローンの塩基/アミノ酸相関を示す。認識パターンを灰色に塗って強調してある。
【図5】図5は、αヘリックスの位置2に対して選択された4塩基連鎖の位置1と結合する配列特異的相互作用を図示する。
【図6】図6は、本発明によるライブラリーの構築の模式図である。
【発明を実施するための形態】
【0017】
以下、本発明を詳細に説明する。
【0018】
本発明はライブラリーに関係する。用語「ライブラリー」は当業界の通常の用法により使われ、ポリペプチド、または、好ましくはポリペプチドをコードする核酸の集合体を表す。本発明のポリペプチドは、無作為化(randomisation)領域を含み、このため各ライブラリーは、個々のポリペプチドがお互いに異なる配列をもつポリペプチドのレパートリーを含んでなるかまたはコードする。同じ原理は、ファージ提示によるような、選択のために開発される実質的に全てのライブラリーに存在する。
【0019】
本明細書で使う無作為化とは、異なるポリペプチドの任意の所与の位置に様々なアミノ酸が存在しうるような、ライブラリーを含んでなるポリペプチドの配列の改変を意味する。無作為化は、与えられた位置にどのアミノ酸も存在しうるような完全なものであっても、またある特定のアミノ酸のみが存在するような部分的なものであってもよい。好ましくは、無作為化は、核酸レベルでの変異誘発により、例えば変異タンパク質をコードする新規遺伝子を合成し、それらを発現させて各種の異なるタンパク質を得ることにより達成される。あるいは、所望の変異遺伝子を得る目的で、存在する遺伝子自身を部位指定または無作為変異誘発のような方法により変異させることができる。
【0020】
変異は当業者に周知の方法で実施することができる。しかし、目的のタンパク質をコードする核酸配列の部位特異的突然変異誘発が好ましい。多数の部位特異的突然変異誘発の方法が、M13のような1本鎖ファージを用いる方法からPCRに基く技術まで、当業界で公知である("PCR Protocols: A guide to methods and applications(PCRプロトコル:方法および応用のガイド)", M.A. Innis, D.H. Gelfand. J.J. Sninsky, T.J. White(編), Academic Press, New York, 1990を参照すること)。好ましくは、市販される変異誘発システム(Altered Site II Mutagenesis System (Promega))をメーカーにより与えられた指示に従って用いることができる。
【0021】
変異遺伝子により産生されるタンパク質のスクリーニングは、好ましくは、遺伝子を発現してタンパク質産物の結合能力を検定することにより実施される。これを達成しうる簡単でかつ有利で迅速な方法は、ファージ提示(phage display)による方法であり、この方法では、変異体ポリペプチドは、バクテリオファージm13のマイナーなコートタンパク質pIIまたはバクテリオファージFdの遺伝子IIIのような、繊維状バクテリオファージのコートタンパク質との融合タンパク質として発現され、変異遺伝子により形質転換されたバクテリオファージのカプシド上に提示される。標的核酸配列をプローブとして使ってファージ表面上のタンパク質に直接結合させ、アフィニティ精製により有利な変異体を持つファージを選択する。その後、該ファージを細菌宿主を通じての継代により増幅し、所望のファージの変異体プールを富化するためにさらなる回数の選択および増幅を行い、最終的に好ましいクローンを単離する。当業界で公知のファージ提示の詳細な手法は、例えば、米国特許第5,223,409号;ChooおよびKlug, (1995) Current Opinions in Biotechnology 6:431-436;Smith. (1985) Science 228:1315-1317;およびMcCafferyら, (1990) Nature 348:552-554;に開示されており、これらは全て、本明細書に参照により組み入れられる。ファージ提示のためのベクターシステムおよびキットは、例えばファルマシア(Pharmacia)社から市販されている。
【0022】
本発明によるライブラリーが含んでなるポリペプチドは、ジンクフィンガーポリペプチドである。言換えれば、それらは、Cys2-His2ジンクフィンガーモチーフを含んでなる。各ポリペプチドが1つ以上のジンクフィンガーを含んでなることが本発明の特徴であり、該ライブラリーはポリペプチド上の2つ以上のジンクフィンガー間の相互作用に基いて選択することができる。
【0023】
当業界で公知であるように、ジンクフィンガーは核酸と結合する分子である。各ジンクフィンガーは、標的核酸の4塩基連鎖(quadruplet)配列と、ジンクフィンガーのαヘリックスの特定アミノ酸残基と核酸との間の接触を通して結合する。本発明に規定される4塩基連鎖は、核酸の-(マイナス)鎖を3'から5'へ読んだとき、第1の4塩基連鎖の塩基4は第2の4塩基連鎖の塩基1であり、かつ以後もそのように、オーバーラップしている。したがって、本出願において、各4塩基連鎖の塩基は、1〜4の数字により参照され、1は3'塩基でありかつ4は5'塩基である。塩基4は古典的なジンクフィンガー3塩基連鎖の5'塩基と等価である。一般的に、塩基4はαヘリックスの位置+6における接触を通して、塩基3は位置+3における接触を通して、塩基2は位置-1における接触を通して、塩基1は二本鎖核酸の対立鎖への位置+2における接触を通して結合される。
【0024】
本明細書に言及するジンクフィンガーの全ての核酸結合残基位置は、フィンガーのαヘリックスにおける第1残基から数えられ、+1〜+9の範囲にある。「-1」は、Cys2-His2ジンクフィンガーポリペプチド中のαヘリックスのすぐ前のフレームワーク構造の残基を指す。
【0025】
「++2」と称する残基は、隣接する(C末端)フィンガーに存在する。これらは、(+鎖の)塩基1に対する位置+2と、先行(3')4塩基連鎖の塩基4(オーバーラップにより同じ塩基である)に対する先行(N末端)フィンガー位置+6との間の相乗的共同作用を反映する。C末端隣接フィンガーがない場合、「++」相互作用は機能しない。
【0026】
Cys2-His2ジンクフィンガー結合タンパク質は、当業界で周知のように、ジンクフィンガーとして知られるαヘリカルの亜鉛金属原子が配位した結合モチーフを経由して、標的核酸配列と結合する。ジンクフィンガー核酸結合タンパク質の各ジンクフィンガーは、核酸結合配列内の核酸4塩基連鎖との結合を決定することに関わる。好ましくは、各結合タンパク質には2つ以上のジンクフィンガー、例えば2,3,4,5または6つのジンクフィンガーがある。各ジンクフィンガー結合タンパク質に3つのジンクフィンガーがあるのが有利である。
【0027】
本発明により、本質的に人工の核酸結合タンパク質の産生が可能になる。これらのタンパク質については、所望の物性をもつまたは他の理由の蛋白質を与えるためにアミノ酸の人工的類似体を使うことができる。かくして、特定的に「任意のアミノ酸(any amino acid)」に言及する文脈において、用語「アミノ酸(amino acid)」は、当業界で公知の方法によりどの種類のタンパク質構築に用いうる天然または人工のアミノ酸もしくはアミノ酸類似体も意味する。さらに、本明細書で参照されるどの特定のアミノ酸も、その機能的類似体、特に人工の機能類似体により置換することができる。したがって、本明細書で使われる命名法は、定義されたアミノ酸の機能的類似体を特定的に本発明の範囲内に含む。
【0028】
ジンクフィンガー結合タンパク質のαヘリックスは核酸鎖に反平行に(antiparallel)に整列し、ジンクフィンガーのN末端からC末端への配列に対応するために、一次核酸配列は3'から5'へ配列する。核酸配列は通常、5'から3'に向けてかつアミノ酸配列はN末端からC末端に向けて書かれるので、その結果、核酸配列およびジンクフィンガータンパク質を慣例により整列すると、ジンクフィンガーは3'から5'に整列している核酸の-(マイナス)鎖と一次相互作用をもつことになる。これらの慣例は本明細書に使われる命名法で踏襲される。しかし、実際には、タンパク質GLIのフィンガー4のようなある特定のフィンガーは、核酸の+(プラス)鎖と結合することは注意すべきである(Suzukiら, (1994) NAR 22:3397-3405およびPavletichおよびPabo, (1993) Science 261:1701-1707)。このようなフィンガーの核酸結合分子への結合は、本発明により予想される。
【0029】
本発明のライブラリーにより、単一DNA標的に対する隣接フィンガーの同時選択を促進することにより隣接ジンクフィンガー間の相乗的共同作用に対する選択が可能になる。これは、同じジンクフィンガーポリペプチドにおいて、1つ以上のジンクフィンガーを無作為化することにより達成される。好ましい実施様態では、各ポリペプチド中のほぼ1個半のジンクフィンガーが無作為化されるが、これはライブラリー設計により変化する。
【0030】
本発明のライブラリーにコードされるジンクフィンガーポリペプチドは、1つ以上であれば、いくつの数のジンクフィンガーを含んでなってもよい。各ポリペプチドが3〜6つのジンクフィンガーをコードするのが有利である。各ライブラリーにおいて、無作為化の範囲は少なくとも1対のジンクフィンガーのオーバーラップをカバーする。好ましくは、1対のオーバーラップをカバーする。
【0031】
好ましくは、本発明のライブラリーはセットとして提供される。したがって、F1、F2およびF3を含んでなる3つのジンクフィンガーポリペプチドを、各ライブラリーが2つのジンクフィンガーポリペプチドを含んでなる2つのライブラリーの1セットで提供することができる。第1のライブラリーは本質的にF1およびF2からなるポリペプチドで構成され、第2のライブラリーは本質的にF2およびF3からなるポリペプチドで構成される。各ライブラリーの無作為化は、それぞれF1およびF2、ならびにF2およびF3の間のオーバーラップを含む。
【0032】
好ましくは、各ライブラリーは少なくとも第1のフィンガーの位置6および第2フィンガーの位置2での無作為化を含んでなるであろう。これらの残基は二本鎖核酸標的上の同じ塩基対と接触するので、それらを一緒に変えることが有利である。
【0033】
3つのジンクフィンガーポリペプチドの場合には、第1ライブラリーはフィンガーF1およびF2で無作為化されるであろうが、第2はF2およびF3で無作為化される。ポリペプチドは、F2配列において選択後に組替えられ、F1、F2およびF3を含有する単一ポリペプチドを作製する。このポリペプチドはF1およびF2、ならびにF2およびF3の間のオーバーラップを考慮して選択されているであろう。
【0034】
有利なように、各ジンクフィンガーでより多数の位置を変えることができる。好ましくは、第1ジンクフィンガーでは位置-1、1、2、3、5および6から選択された残基がそして、第2ジンクフィンガーでは位置-1、1、2および3が改変される。片方のライブラリーについては、第2フィンガーでは位置3、5および6が、そして第3フィンガーでは位置-1、1、2および3が改変されうる。最後のフィンガー(3つのフィンガータンパク質の場合にはこれは第3フィンガーであろう)では、残基5および6も改変されうる。
【0035】
ライブラリーを選択後に可能とするため、ポリペプチドは、好ましくは、2つのライブラリーにより共有されるジンクフィンガーをコードする核酸中に適切な制限酵素切断部位を含むように設計される。開裂部位の位置は、各ライブラリー中の共有されるジンクフィンガーに行われる改変の正確な部位を指令するであろう。したがって、3つのジンクフィンガータンパク質をコードする2つのライブラリーの1セット中に、もし開裂部位がαヘリックスの位置3と5の間にあれば、位置3と5は第1ライブラリーで、5と6とは第2ライブラリーで無作為化されうる。
【0036】
無作為化または改変のための残基は、位置-1、1、2、3、5および6から選択されるのが好ましいが、さらなる残基も無作為化することができる。例えば、位置8の無作為化は有利でありうる。さらに、与えられた全ての位置より少ない位置を無作為化することも予測される。
【0037】
好ましい実施様態では、3つのフィンガータンパク質の選択のための2つのライブラリー系は、第1ライブラリーにおいて、F1位置-1、1、2、3、5および6で改変され、F2位置-1、1、2および3で改変される。第2ライブラリーは、F2位置3および6で改変され、F3位置-1、1、2、3、5および6で改変される。この場合、開裂および組換え点は残基3および5の間、好ましくはF2のαヘリックスの残基4および5の間であろう。
【0038】
組換え操作の後、組替えられたポリペプチドをコードする核酸は、適切な発現系で発現されるか、またはさらなる選択のためにFdファージにクローニングされる。
【0039】
本発明の好ましい様態では、本発明のライブラリーは、全てのアミノ酸でなくある特定のアミノ酸がコードされるように、選択した位置で真に無作為化されるのでなく部分的に無作為化される。この方法は、2つの目的で使うことができる。
【0040】
第1の実施様態では、改変は、ジンクフィンガーのαヘリックスの所与の位置に組込まれたときに核酸標的配列の配列特異的結合を指令する能力のあることが知られているアミノ酸に制限される。標的配列にかかわりなく、ある特定のアミノ酸はある特定の位置での組込みに適切でないことが知られている。これらのアミノ酸は回避される。
【0041】
第2の実施様態では、改変は、ジンクフィンガーのαヘリックスの所与の位置に組込まれたときに核酸標的配列の配列特異的結合を指令する能力のあることが知られているアミノ酸に制限され、かつ改変は、所与の位置での特定の標的配列との結合を有利にすることが知られている残基を特定するように行われる。このようにして、本発明は、的の絞られたライブラリーの設計を可能にし、このライブラリーから特定の標的配列または一連の関係標的配列と結合する能力のあるポリペプチドを選択することができる。
【0042】
一般的応用のためのライブラリー系を提供する第1の実施様態では、無作為化は好ましくは、上に示した位置の全てで行われる。好ましくは、それぞれの所与の位置で現れるように選択されるアミノ酸は表1に掲げたとおりである:
【表1】
【0043】
各フィンガーが、表1に与えられた各位置で無作為化されている必要はない。好ましい実施様態では、3フィンガータンパク質を選択するライブラリーは、表2に与えられた明細により構築される。
【表2】
【0044】
第2の実施様態では、特定の位置の各アミノ酸の同一性は、本明細書に提供されたジンクフィンガー認識ルールにより選択される。したがって、好ましい様態では、本発明は、標的核酸配列内の核酸4塩基連鎖への結合を可能にするCys2-His2ジンクフィンガークラスの核酸結合タンパク質を調製する方法を提供し、該方法において、タンパク質のαヘリカルジンクフィンガー核酸結合モチーフによる4塩基連鎖の各塩基との結合は次のとおり決定される:
a)もし4塩基連鎖の塩基4がGであれば、αヘリックスの位置+6はArgもしくはLysであり;
b)もし4塩基連鎖の塩基4がAであれば、αヘリックスの+6はGln、AsnもしくはValであり;
c)もし4塩基連鎖の塩基4がTであれば、αヘリックスの位置+6はSer、Thr、ValもしくはLysであり;
d)もし4塩基連鎖の塩基4がCであれば、αヘリックスの位置+6はSer、Thr、Val、Ala、GluもしくはAsnであり;
e)もし4塩基連鎖の塩基3がGであれば、αヘリックスの位置+3はHisであり;
f)もし4塩基連鎖の塩基3がAであれば、αヘリックスの位置+3はAsnであり;
g)もし4塩基連鎖の塩基3がTであれば、αヘリックスの位置+3はAla、SerもしくはValであり;もしそれがAlaであれば、-1または+6の残基の1つは小さい残基であり;
h)もし4塩基連鎖の塩基3がCであれば、αヘリックスの位置+3はSer、Asp、Glu、Leu、ThrもしくはValであり;
i)もし4塩基連鎖の塩基2がGであれば、αヘリックスの位置-1はArgであり;
j)もし4塩基連鎖の塩基2がAであれば、αヘリックスの位置-1はGlnであり;
k)もし4塩基連鎖の塩基2がTであれば、αヘリックスの位置-1はHisもしくはThrであり;
l)もし4塩基連鎖の塩基2がCであれば、αヘリックスの位置-1はAspもしくはHisであり;
m)もし4塩基連鎖の塩基1がGであれば、αヘリックスの位置+2はGluであり;
n)もし4塩基連鎖の塩基1がAであれば、αヘリックスの位置+2はAspもしくはGlnでなく;
o)もし4塩基連鎖の塩基1がCであれば、αヘリックスの位置+2はAsn、Gln、Arg、HisもしくはLysであり;
p)もし4塩基連鎖の塩基1がTであれば、αヘリックスの位置+2はSerもしくはThrである。
【0045】
以上は、所与の核酸配列に特異的なジンクフィンガー結合タンパク質の設計を可能にするルールのセットを表す。これに関係する新規の知見は、ヘリックスの位置+2は、4塩基連鎖の塩基1との結合を決定することに関与するということである。そうすることによって、塩基1および4が、隣接する4塩基連鎖にオーバーラップしているので、4塩基連鎖の塩基4の結合を決定する位置+6と相乗的に同時作動する。
【0046】
ジンクフィンガーポリペプチドは、オーバーラップする4塩基連鎖配列と結合すると考えられたが、本発明の方法は、ポリペプチドが多重のオーバーラップ4塩基連鎖でない標的配列と結合するように設計することが可能である。例えば、ジンクフィンガーポリペプチドをパリンドローム標的配列と結合するように設計することができる。このような配列は通常、例えば、制限酵素標的配列として見出される。
【0047】
好ましくは、3個より少ないヌクレオチドと結合するジンクフィンガーの作製が、ジンクフィンガー中に関係位置の核酸とのH結合を支持することができないアミノ酸を特定することにより達成される。
【0048】
有利には、これは、位置-1のGly(塩基2との接触をなくするため)、ならびに/または、位置+3および/もしくは+6のAla(それぞれ第3もしくは第4塩基との接触をなくするために)を置換することにより達成される。
【0049】
好ましくは、標的配列の最後の(3')塩基との接触は、もし必要があれば、核酸のリン酸主鎖と直接接触できる関係位置において残基を置換することにより強化されるべきである。
【0050】
これらおよび他の考慮は、本発明によるライブラリーセットに組込むことができる。
【0051】
ジンクフィンガー結合モチーフは当業者には周知の構造であり、例えば、ミラーらその他(Millerら, (1985) EMBO J. 4:1609-1614; Berg (1988) PNAS (USA) 85:99-102; Leeら, (1989) Science 245:635-637; USSN 08/422.107.に対応する国際特許出願WO96/06166およびWO96/32475を参照すること、これらは参照により本明細書に組入れられる)に定義されている。
【0052】
本明細書に使われる「核酸」は、天然の核酸塩基もしくは合成塩基、またはそれらの混合物から構築されたRNAおよびDNAの両方を意味する。しかし、好ましくは、本発明の結合タンパク質はDNA結合タンパク質である。
【0053】
一般的に、好ましいジンクフィンガーフレームワークは、構造:
(A) X0-2 C X1-5 C X9-14 H X3-6 (H/C)
(式中、Xは任意のアミノ酸であり、下付き数字はXで表す残基の可能な数を示す)を有する。
【0054】
本発明の好ましい様態では、ジンクフィンガー核酸結合モチーフは次の一次構造:
(B)
(式中、X(Xa、XbおよびXcを含む)は任意のアミノ酸であり、X2-4およびX2-3はそれぞれ2個もしくは4個、または2個もしくは3個の存在を意味する)を有するモチーフとして表現することができる。一緒に亜鉛金属原子に配位するCysおよびHis残基は太字で記されているが、αヘリックスの+4のLeu残基のように通常は不変である。
【0055】
この表現に対する修正は、ジンクフィンガー機能を無効にすることなく、アミノ酸の挿入、変異または欠失によって起りうるし、または行うことができる。例えば、第2His残基はCysで置換しうることが公知であり(Krizekら, (1991) J. Am. Chem. Soc. 113: 4518-4523)、+4のLeuはある場合にはArgにより置換しうる。Xcの前のPhe残基は、Trp以外のどの芳香族によって置換することもできる。さらに、実験は、ジンクフィンガーの好ましい構造および残基割当からの逸脱は、許容されかつある特定の核酸配列との結合に有利であることすら証しうることを示した。しかし、これを考慮しても、4つのCysまたはHis残基に接触する亜鉛原子が配位したαヘリックスに関わる一般構造は変らない。本明細書に使われる上の構造(A)および(B)は、Cys2-His2タイプの全てのジンクフィンガー構造を表す例示の構造とみなされる。
【0056】
好ましくは、Xaは(F/Y)-XまたはP-(F/Y)-Xである。この式中で、Xはアミノ酸である。好ましくは、この式中で、XはE、K、TまたはSである。好ましくは劣るが、Q、V、AおよびPも予測される。残りのアミノ酸は可能性を残す。
【0057】
好ましくは、X2-4は4個よりも2個のアミノ酸からなる。これらのアミノ酸の第1は、どのアミノ酸であってもよいが、S、E、K、T、PおよびRが好ましい。PまたはRであるのが有利である。これらのアミノ酸の第2は、好ましくはEであるが、どのアミノ酸でも使うことができる。
【0058】
好ましくは、XbはTまたはIである。
【0059】
好ましくは、XcはSまたはTである。
【0060】
好ましくは、X2-3はG-K-A、G-K-C、G-K-SまたはG-K-Gである。しかし、好ましい残基からの逸脱、例えばM-R-NまたはM-Rの形も可能である。
【0061】
好ましくは、リンカーはT-G-E-KまたはT-G-E-K-Pである。
【0062】
上に述べたように、主な結合相互作用は、アミノ酸-1、+2、+3および+6と生じる。アミノ酸+4および+7はほとんど不変である。残るアミノ酸は本質的にどのアミノ酸であってもよい。好ましくは、位置+9はArgもしくはLysにより占められる。位置+1、+5および+8は、疎水性アミノ酸ではない、すなわちPhe、TrpもしくはTyrでないことが有利である。
【0063】
したがって、ほとんどの好ましい様態で、以上を一緒に持ち込んで、本発明により、所与の核酸4塩基連鎖と特異的に結合するジンクフィンガー核酸結合モチーフ内のすべての残基の定義が可能になる。
【0064】
本発明により提供されるコードは完全に固定したものではない。ある特定の選択が提供される。例えば、位置+1、+5および+8はどのアミノ酸が配置されてもよく、また、他の位置はある特定の選択がされてもよい。例えば、本発明のルールは、中央T残基との結合に、Ala、SerもしくはValのどれか1つを+3に使いうることを提供する。したがって最も広範囲の意味では、本発明は、全ての定義された標的核酸4塩基連鎖と結合することができる非常に多数のタンパク質を提供する。
【0065】
しかし、好ましくは、可能性の数は大いに減らすことができる。例えば、決定的でない残基+1、+5および+8は、残基Lys、ThrおよびGlnによりそれぞれデフォールトオプション(default option)として占められてよい。他を選択する場合、例えば、最初に与えられるオプションをデフォールトとして用いてよい。したがって、本発明によるコードにより、標的4塩基連鎖と結合する単一の定義されたポリペプチド(「デフォールト」ポリペプチド)の設計が可能になる。
【0066】
本発明のさらなる様態では、標的核酸配列と結合することができるCys2-His2ジンクフィンガークラスの核酸結合タンパク質を調製する方法が提供され、その方法は、
a)天然のジンクフィンガーおよびコンセンサスジンクフィンガーからなる群から1つのモデルジンクフィンガードメインを選択すること;および
b)上に述べたルールにより必要とされるようにフィンガーの位置-1、+2、+3および+6の1つ以上を変異すること
の工程を含む。
【0067】
一般的に、天然のジンクフィンガーは、その核酸結合特異性が公知であるフィンガーから選択してよい。例えば、これらは、結晶構造が解明されているフィンガー:すなわちZif268(Elrod-Ericksonら, (1996) Structure 4:1171-1180)、GLI(PavletichおよびPabo, (1993) Science 261:1701-1707)、Tramtrack(Fairallら, (1993) Nature 366:483-487)およびYY1(Houbaviyら, (1996) PNAS (USA) 93:13577-13582)であってよい。
【0068】
天然のZif268中のジンクフィンガー2はジンクフィンガーを遺伝子操作できる優れた出発点となり、かつ、好ましい。
【0069】
コンセンサスジンクフィンガー構造物を、その結合ドメインが公知であるか否かに拘らず、公知のジンクフィンガーの配列と比較して調製してもよい。好ましくは、該コンセンサス構造は、コンセンサス構造PYKCPECGKSFSQKSDLVKHQRTHTGおよびコンセンサス構造PYKCSECGKAFSQKSNLTRHQRIHTGEKPからなる群から選ばれる。
【0070】
該コンセンサスは、クリゼクら(Krizekら, (1991) J. Am. Chem. Soc. 113:4518-4523)およびヤコフ(Jacobs, (1993) 博士論文, University of Cambridge, UK)により提供されたコンセンサスから誘導される。両ケースとも、上に記載した2つのジンクフィンガーモチーフを一緒に結合するためのリンカー配列すなわちTGEKまたはTGEKPを、コンセンサスの末端に形成することができる。したがって、Pは必要があれば除いてもよく、または、TGで終るコンセンサスの場合はEK(P)を加えることができる。
【0071】
選択したモデルフィンガーの核酸特異性がわかっているときは、標的核酸に対する結合特異性を修正する目的で、天然と所望標的とで相異のある塩基についてその塩基との結合に影響することがわかっている残基に、フィンガーの変異を指定することができる。そうでなければ、モデルフィンガーの変異は、前述のルールに与えられたように、残基-1、+2、+3および+6に集中すべきである。
【0072】
改良された結合を有する結合タンパク質を産生する目的で、さらに、残基選択を支援するために、本発明により提供されるルールを、タンパク質/核酸界面の物理的または仮想的モデルにより補うことができる。
【0073】
本発明により設計したジンクフィンガー結合モチーフは、多重のジンクフィンガーを有する核酸結合タンパク質に組み合わせることができる。好ましくは、タンパク質は少なくとも2つのジンクフィンガーを有する。もともとジンクフィンガー結合タンパク質は、Tramtrackのような2つのジンクフィンガータンパク質も知られているが、一般的には少なくも3つのジンクフィンガーを有する。少なくとも3つのジンクフィンガーの存在が好ましい。結合タンパク質は、所要フィンガーの末端と末端、N末端とC末端を接続することにより構築することができる。好ましくは、これは、ジンクフィンガーをコードする関連核酸コード配列を一緒に結合して、結合タンパク質全体をコードする複合コード配列を作製することによって達成される。したがって、本発明は、上に定義した核酸結合タンパク質を製造する方法を提供し、その方法では該核酸結合タンパク質は組換えDNA技術により構築され、その方法は、
a)上に記載したように、N末端からC末端へ配置された2個以上のジンクフィンガー結合モチーフをコードする核酸コード配列を製造し;
b)核酸配列を適切な発現ベクターに挿入し;および
c)宿主生物に核酸配列を発現させて核酸結合タンパク質を得る
工程を含んでなる。
【0074】
「リーダー(leader)」ペプチドをN末端フィンガーに付加することができる。好ましくは、該リーダーペプチドはMAEEKPである。
【0075】
本発明により核酸結合タンパク質をコードする核酸は、さらなる遺伝子操作のため、ベクターに組込むことができる。本明細書で使用する場合、ベクター(またはプラスミド)は、その発現または複製のために異種核酸を細胞に導入するために使われる分離したエレメントを意味する。このようなビヒクルの選択と使用は、当業界の通常技術者の技術範囲内である。多くのベクターを利用することができ、適切なベクターの選択は、ベクターの意図する用途、すなわち該ベクターをDNA増幅のためまたは核酸発現のためのいずれに使うか、該ベクターに挿入するDNAのサイズ、および該ベクターで形質転換する宿主細胞に依存するであろう。それぞれのベクターは、その機能(DNAの増幅またはDNAの発現)およびベクターが適合しうる宿主細胞に依存して様々な成分を含有する。ベクター成分は一般的に、限定されるものでないが、1つ以上の次のものを含有する:複製の起点、1つ以上のマーカー遺伝子、エンハンサーエレメント、プロモーター、転写終結配列およびシグナル配列。
【0076】
一般的に、発現およびクローニングベクターは両方とも、ベクターが1つ以上の選択された宿主細胞中で複製できるようにする核酸配列を含有する。典型的には、クローニングベクターにおいて、この配列は、ベクターに宿主染色体DNAと関係なく複製する能力をあたえ、複製起点または自律的複製配列を含む。このような配列は、各種の細菌、酵母およびウイルスにおいて周知である。プラスミドpBR322由来の複製起点はほとんどのグラム陰性細菌用に適切であり、2μプラスミド起点は酵母用に適切であり、そして様々なウイルス起点(例えば、SV40、ポリオーマ、アデノウイルス)は哺乳動物細胞のクローニングベクター用に有用である。一般的に、哺乳動物発現ベクター用の複製起点成分は、COS細胞のような高レベルDNA複製能力のある哺乳動物細胞に使うのでなければ不要である。
【0077】
ほとんどの発現ベクターはシャトルベクターであり、すなわち、少なくとも1クラスの生物の複製能力があるが、発現のために他のクラスの生物にトランスフェクトすることができる。例えば、あるベクターは大腸菌(E. coli)でクローニングされ、その後、その同じベクターは酵母または哺乳動物細胞にトランスフェクトされるが、宿主細胞染色体に依存せず複製する能力を持たない。DNAは宿主ゲノムへの挿入によって複製させることもできる。しかし、核酸結合タンパク質をコードするゲノムDNAの回収は、核酸結合タンパク質DNAを切除するために制限酵素消化を必要とするので、外因的に複製したベクターの回収よりもっと複雑である。DNAはPCRにより増幅することができ、またどのような複製成分もなしに、直接、宿主細胞にトランスフェクトすることができる。
【0078】
発現およびクローニングベクターは、選択マーカー(selectable marker)とも呼ばれる選択遺伝子を含有するのが有利である。この遺伝子は、選択培地で増殖する形質転換した宿主細胞の生存または増殖に必要なタンパク質をコードする。選択遺伝子を含有するベクターで形質転換されない宿主細胞は、この培地で生存しないであろう。典型的な選択遺伝子は、抗生物質および他の毒素、例えばアンピシリン、ネオマイシン、メトトレキセートもしくはテトラサイクリンへの耐性を与え、栄養要求欠乏を補い、または複合培地から利用できない重要な栄養を供給するタンパク質をコードする。
【0079】
酵母に適切な選択遺伝子マーカーについては、マーカー遺伝子の表現型発現により形質転換体の選択を容易にするどのマーカー遺伝子でも使うことができる。酵母に対する適切なマーカーは、例えば、抗生物質G418、ハイグロマイシンまたはブレオマイシン耐性を与えるものであるか、または、栄養要求性酵母突然変異体、例えばURA3、LEU2、LYS2、TRP1、またはHIS3遺伝子に原栄養(prototrophy)を与える。
【0080】
ベクターの複製は大腸菌で行うのが好都合であるので、大腸菌遺伝子マーカーおよび大腸菌複製起点を含むのが有利である。これらは、pBR322、Bluescript(登録商標)ベクターまたはpUCプラスミド、例えばpUC18もしくはpUC19のような大腸菌プラスミドから得ることができ、これらは大腸菌複製起点およびアンピシリンのような抗生物質への耐性を与える大腸菌遺伝子マーカーの両方を含有する。
【0081】
哺乳動物細胞に適切な選択マーカーは、核酸結合タンパク質核酸を取上げる能力のある(competent)細胞の確認を可能にするジヒドロ葉酸レダクターゼ(DHFR、メトトレキセート耐性)、チミジンキナーゼ、またはG418もしくはハイグロマイシン耐性を与える遺伝子のようなマーカーである。哺乳動物細胞形質転換体は、マーカーを取上げて発現する形質転換体だけがユニークに生存適合する選択圧力下に置かれる。DHFRまたはグルタミンシンターゼ(GS)マーカーの場合、選択圧力が次第に増加する条件下で形質転換体を培養することが可能であり、それにより選択遺伝子および核酸結合タンパク質をコードする連結されたDNAの両方を、(その染色体組込み部位で)増幅させる。増幅は、増殖に重要なタンパク質の産生のより大きな需要のある遺伝子ならびに所望のタンパク質をコードしうる密接に関連する遺伝子を一緒に、組換え細胞の染色体内で直列に反復させるプロセスである。所望タンパク質の増加量は、通常、このように増幅したDNAから合成される。
【0082】
発現およびクローニングベクターは、通常、宿主生物により認識され、核酸結合タンパク質をコードする核酸に機能しうる形で連結されたプロモーターを含有する。このようなプロモーターは、誘導性(inducible)であっても、構成的(constitutive)であっても良い。該プロモーターは、ソースDNAから制限酵素消化によりプロモーターを除去し、単離したプロモーター配列をベクターに挿入することにより、機能しうる形で核酸結合タンパク質をコードするDNAに連結される。元来の核酸結合タンパク質プロモーター配列および多くの異種プロモーターの両方を、核酸結合タンパク質をコードするDNAの直接増幅および/または発現に使うことができる。
【0083】
原核生物宿主での使用に適切なプロモーターは、例えば、β-ラクタマーゼおよびラクトースプロモーター系、アルカリホスファターゼ、トリプトファン(Trp)プロモーター系ならびにtacプロモーターのようなハイブリッドプロモーターを含む。これらのヌクレオチド配列は公表されており、熟練技術者は、必要な制限酵素切断部位を与えるためにリンカーまたはアダプターを使って、該プロモーターを核酸結合タンパク質をコードするDNAに機能しうる形でライゲートすることが可能である。細菌系に使用するためのプロモーターは、一般的に、核酸結合タンパク質をコードするDNAに機能しうる形で連結したシャイン・ダルガルノ配列も含むであろう。
【0084】
好ましい発現ベクターは、細菌内で機能する能力のあるファージx(phagex)またはT7のようなバクテリオファージのプロモーターを含んでなる細菌発現ベクターである。最も広く使われる発現系の1つでは、融合タンパク質をコードする核酸を、該ベクターからT7 RNAポリメラーゼにより転写することができる(Studierら, Methods in Enzymol. 185; 60-89, 1990)。pETベクターと共に使われる大腸菌BL21(DE3)宿主株では、T7 RNAポリメラーゼは宿主細菌のλ溶原菌DE3から産生され、その発現はIPTG誘導性lac UV5プロモーターの制御下にある。この系は成功裏に多くのタンパク質の過剰産生に用いられている。あるいは、ポリメラーゼ遺伝子は、市販されるCE6ファージ(Novagen, Madison, USA)のようなintファージでの感染によりλファージに導入することができる。他のベクターとして、PLEX(Invitrogen, NL)のようなλPLプロモーターを含有するベクター、pTrcHisXpressTm(Invitrogen)もしくはpTrc99(Pharmacia Biotech, SE)のようなtrcプロモーターを含有するベクター、またはpKK223-3(Pharmacia Biotech)もしくはPMAL(New England Biolabs, MA, USA)のようなtacプロモーターを含有するベクターが挙げられる。
【0085】
さらに、本発明による核酸結合タンパク質遺伝子は、好ましくは、細菌宿主からのポリペプチドの分泌を容易にするための分泌配列を含み、封入体中よりもむしろ可溶性の元来のペプチドとして産生させる。該ペプチドは細菌の周辺腔または培地から適当に回収することができる。
【0086】
酵母宿主での使用のための適切なプロモーター配列は、調節されるかまたは構成的であることができ、好ましくは、高度に発現した酵母遺伝子、特にSaccharomyces cerevisiae遺伝子由来のものである。したがって、TRP1遺伝子、ADHIもしくはADHII遺伝子、酸性ホスファターゼ遺伝子(PH05)のプロモーター、aもしくはα因子をコードする酵母接合フェロモン遺伝子のプロモーターまたはエノラーゼ、グリセルアルデヒド-3-リン酸デヒドロゲナーゼ(GAP)、3-ホスホグリセリン酸キナーゼ(PGK)、ヘキソキナーゼ、ピルビン酸デカルボキシラーゼ、ホスホフルクトキナーゼ、グルコース-6-リン酸イソメラーゼ、3-ホスホグリセリン酸ムターゼ、ピルビン酸キナーゼ、トリオースリン酸イソメラーゼ、ホスホグルコースイソメラーゼもしくはグルコキナーゼ遺伝子のプロモーターのような解糖系酵素をコードする遺伝子由来のプロモーター、またはTATA結合タンパク質(TBP)遺伝子由来のプロモーターを使うことができる。さらに、1つの酵母遺伝子の上流活性化配列(UAS)および他の酵母遺伝子の機能的TATAボックスを含む下流プロモーターエレメントを含んでなるハイブリッドプロモーター、例えば、酵母PH05遺伝子のUASおよび酵母GAP遺伝子の機能的TATAボックスを含む下流プロモーターエレメントを含むハイブリッドプロモーター(PH05-GAPハイブリッドプロモーター)を使うことができる。適切な構成的PH05プロモーターは、例えば、PH05遺伝子のヌクレオチド-173で開始しヌクレオチド-9で終結するPH05(-173)プロモーターエレメントのような、上流調節エレメント(UAS)を欠いた短縮酸性ホスファターゼPH05プロモーターである。
【0087】
哺乳動物宿主内でのベクターからの核酸結合タンパク質遺伝子転写は、プロモーターが宿主細胞系と適合しうることを条件として、ポリオーマウイルス、アデノウイルス、伝染性上皮腫ウイルス(fowlpox virus)、ウシ乳頭腫ウイルス、トリサルコーマウイルス、サイトメガロウイルス(CMV)、レトロウイルスおよびサルウイルス40(SV40)のようなウイルスのゲノム由来のプロモーター、アクチンプロモーターのような異種哺乳動物プロモーターまたは非常に強いプロモーター、例えばリボソームタンパク質プロモーター、および、核酸結合タンパク質配列に通常関連するプロモーターにより制御することができる。
【0088】
高等真核生物による核酸結合タンパク質をコードするDNAの転写は、ベクターにエンハンサー配列を挿入することにより増加することができる。エンハンサーは比較的配向および位置に依らない。哺乳動物遺伝子由来の多くのエンハンサー配列が公知である(例えば、エラスターゼおよびグロビン)。しかし、典型的には、真核細胞ウイルス由来のエンハンサーを用いるであろう。例として、複製起点の後期側(bp 100-270)のSV40エンハンサーおよびCMV初期プロモーターエンハンサーが挙げられる。該エンハンサーは、核酸結合タンパク質DNAの位置5'または3'でベクター中にスプライスされうるが、プロモーターから部位5'に位置することが好ましい。
【0089】
本発明による核酸結合タンパク質をコードする真核発現ベクターは、遺伝子座制御領域(LCR)を含んでなることが有利である。LCRは、宿主細胞クロマチンに組込まれた導入遺伝子の高レベルの組込み部位に依らない発現を指令することが可能であり、これは特に、ベクターの染色体組込みが起っている永久的トランスフェクトされた真核細胞系において、またはトランスジェニック動物において、核酸結合タンパク質遺伝子を発現させるときに重要である。
【0090】
真核ベクターは、転写の終結およびmRNAの安定化のために必要な配列も含有することができる。このような配列は、一般的に、真核生物またはウイルスのDNAまたはcDNAの5'および3'非翻訳領域から利用可能である。これらの領域は、核酸結合タンパク質をコードするmRNAの非翻訳部のポリアデニル化断片として転写されるヌクレオチドセグメントを含有する。
【0091】
発現ベクターは、かかるDNAを発現する能力のある、プロモーター領域のような調節配列と機能しうる形で連結された、核酸結合タンパク質核酸を発現する能力のあるベクターを含む。したがって、発現ベクターは、プラスミド、ファージ、組換えウイルスまたは他のベクターのような、適切な宿主細胞への導入によりクローン化DNAの発現をもたらす組換えDNAまたはRNA構築物を意味する。適切な発現ベクターは当業者には周知であり、真核および/または原核細胞で複製可能であるもの、およびエピソームのままのもの、または宿主細胞ゲノムに組込まれるものを含む。例えば、核酸結合タンパク質をコードするDNAを、哺乳動物細胞のcDNAの発現に適したベクター、例えばpEVRFのようなCMVエンハンサーに基くベクターに挿入することができる(Matthiasら, (1989) NAR 17, 6418)。
【0092】
本発明の実施に特に有用なのは、哺乳動物細胞の核酸結合タンパク質をコードするDNAの一過性発現を提供する発現ベクターである。一過性発現は、通常、宿主細胞内で効率的に複製が可能な発現ベクターの使用に関わり、宿主細胞が発現ベクターの多数のコピーを蓄積して、その結果、高レベルの核酸結合タンパク質を合成する。本発明の目的に対して、一過性発現系は、例えば核酸結合タンパク質変異体を確認するため、潜在ホスホリル化部位を確認するため、またはタンパク質の機能ドメインを特性決定するために有用である。
【0093】
本発明によるベクターの構築は、通常の連結反応技術を用いる。単離したプラスミドまたはDNA断片を開裂し、加工し、所望の形に再連結して所要のプラスミドを作製する。もし所望であれば、構築プラスミド内の正しい配列を確証する分析を公知の様式で実施する。発現ベクターを構築し、in vitro転写物を調製し、宿主細胞にDNAを導入し、そして核酸結合タンパク質発現および機能を評価する分析を実施する適切な方法は、当業者には公知である。サンプル中の遺伝子存在、増幅および/または発現は、例えば、本明細書に与えられた配列に基く適切に標識したプローブを使用して、通常のサザンブロット、mRNAの転写物を定量するためのノーザンブロット、ドットブロット(DNAもしくはRNA分析)、またはin situハイブリダイゼーションによって、直接に測定することができる。当業者は、もし所望であれば、これらの方法がいかに変更されうるかを容易に予測するであろう。
【0094】
本発明の他の実施態様によって、上記核酸を含有する細胞が提供される。原核生物、酵母および高等真核生物細胞のような宿主細胞を、DNAを複製しかつ核酸結合タンパク質を産生するために使うことができる。適切な原核生物として、グラム陰性またはグラム陽性菌、具体的には大腸菌(例えば大腸菌 K-12株、DH5aおよびHB101)、またはバチルスのような、真性細菌が挙げられる。核酸結合タンパク質をコードするベクターに適切なさらなる宿主は、繊維状真菌または酵母、例えばSaccharomyces cerevisiaeのような真核微生物を含む。高等真核細胞としては、昆虫および脊椎動物細胞、特に、ヒト細胞を含む哺乳動物細胞または他の多細胞生物由来の有核細胞が挙げられる。近年、培養(組織培養)中の脊椎動物細胞の増殖は日常的手法になっている。有用な哺乳動物宿主細胞系の例は、チャイニーズハムスター卵巣(CHO)細胞、NIH 3T3細胞、HeLa細胞もしくは293T細胞のような、上皮または繊維芽細胞系である。本発明の開示でいう宿主細胞は、in vitro培養の細胞ならびに宿主動物内の細胞を含んでなる。
【0095】
DNAは、細胞に安定に組み込まれていても、または当分野で公知の方法を使い一過的に発現されてもよい。安定にトランスフェクトされた哺乳動物細胞は、細胞を選択マーカー遺伝子を有する発現ベクターでトランスフェクトし、トランスフェクトされた細胞をマーカー遺伝子を発現する細胞を選択する条件下で増殖させることにより、調製することができる。一過性のトランスフェクタントを調製するため、哺乳動物細胞は、トランスフェクション効率をモニターするレポーター遺伝子でトランスフェクトされる。
【0096】
このような安定にまたは一過的にトランスフェクトされた細胞を産生するには、細胞を、核酸結合タンパク質を形成するに十分な量の核酸結合タンパク質をコードする核酸でトランスフェクトしなければならない。核酸結合タンパク質をコードするDNAの正確な量は、実験的に決定され、特定の細胞とアッセイに対して最適化される。
【0097】
宿主細胞は、本発明の上に述べた発現またはクローニングベクターでトランスフェクトされ、または好ましくは形質転換され、プロモーターを誘起し、形質転換体を選択し、または所望の配列をコードする遺伝子を増幅するために適切に修飾された通常の栄養培地で培養される。異種DNAの宿主細胞への導入は、リン酸カルシウム共沈技術によるまたはエレクトロポレーションによる異種DNAをコードするベクターでのトランスフェクションのような、当分野で公知の方法により実施することができる。多数のトランスフェクションの方法が当業者には公知である。トランスフェクションの成功は、一般的に、このベクターの作動の徴候が宿主細胞中で起るときに認められる。形質転換は、使用する特定の宿主細胞に適切な標準技術を使って達成される。
【0098】
クローン化DNAの適切な発現ベクターへの組込み、1つ以上の別個の遺伝子をそれぞれコードするプラスミドベクターもしくはプラスミドベクターの組合わせ、または線形DNAによる真核細胞のトランスフェクション、およびトランスフェクトされた細胞の選択は、当分野で周知である(例えば、Sambrookら, (1989) Molecular Cloning: A Laboratory Manual(分子クローニング:実験室マニュアル),第二版、Cold Spring Harbor Laboratory Pressを参照すること)。
【0099】
トランスフェクトされたまたは形質転換された細胞は、当業界で公知の培地および培養方法を使い、好ましくは、DNAによりコードされた核酸結合タンパク質が発現される条件下で培養する。適切な培地の組成は、当業者に公知であり、容易に調製することができる。適切な培地は市販もされている。
【0100】
本発明による核酸結合タンパク質は、診断および研究ツールを含む広範囲の応用に用いることができる。複雑な(complex)混合物中の核酸分子の存在を確認するための診断ツールとして有利に用いることができる。本発明による核酸結合分子は、標的核酸分子の単一塩基対変化を識別することができる。
【0101】
したがって、本発明は、標的核酸分子の存在を確認する方法を提供し、該方法は:
a)上に述べた方法により標的核酸分子に特異的である核酸結合タンパク質を調製すること;
b)結合を促進し、未結合で残る核酸結合タンパク質を除去する条件下で、標的核酸分子を含んでなる試験システムを核酸結合タンパク質にさらすこと;
c)試験システムの核酸結合タンパク質の存在を検出すること
の工程を含んでなる。
【0102】
好ましい実施態様では、本発明の核酸結合分子はELISAアッセイに組み込むことができる。例えば、本発明の分子を提示するファージを、標的核酸の存在を検出するために使うことができ、酵素結合抗ファージ抗体を使って可視化することができる。
【0103】
診断用のジンクフィンガーファージの使用のさらなる改良が、例えば、バクテリオファージのマイナーコートタンパク質(gVIII)と融合したマーカータンパク質を同時発現することにより行うことができる。抗ファージ抗体による検出はすたれている(obsolete)ので、それぞれの診断の時間と費用はさらに低減されよう。必要に応じて、提示のための適切なマーカーは、蛍光タンパク質(A. B. Cubittら, (1995) Trends Biochem Sci. 20, 448-455;T.T. Yangら, (1996) Gene 173, 19-23)、または、先にgIII上に提示されているアルカリホスファターゼ(J. McCafferty, R. H. Jackson, D. J. Chiswell, (1991) Protein Engineering 4, 955-961)のような酵素を含む。別個のマーカーによる様々なタイプの診断ファージの標識付けにより、単一核酸サンプルの多重スクリーニングが可能になる。このようなきめ細かさがなくとも、基本的なELISA技術は、信頼性があり、迅速で、簡単で、特に安価である。さらに、特別な装置も必要としないし、放射性同位体のような有害な試薬を用いないので、臨床での日常的使用に受入れられる。このプロトコルの主な利点は、ゲル電気泳動の必要性を除き、自動化核酸診断への道を開くことにある。
【0104】
本発明は、精巧な特異性で遺伝子操作することができる核酸結合タンパク質を提供する。したがって、本発明は、特異的な核酸結合を必要とする分子の設計に役立つ。例えば、本発明によるタンパク質は、本明細書に記載したジンクフィンガーを含んでなる核酸結合ドメインと融合した核酸開裂ドメインを有するキメラ制限酵素の製造に用いることができる。
【0105】
さらに、本発明は、本明細書に記載した核酸結合タンパク質の使用に関わる治療薬および治療法を提供する。特に、本発明は、ウイルスインテグラーゼのようなインテグラーゼ、および本発明によるin vivo核酸配列を標的とする核酸結合タンパク質を含んでなるポリペプチド融合体の使用を提供する(Bushman, (1994) PNAS (USA)91:9233-9237)。遺伝子治療用途において、本法は機能的遺伝子の欠陥遺伝子への送達、または望ましくない核酸を破壊する目的のナンセンス核酸の送達に応用することができる。あるいは、遺伝子をセントロメアのような既知の、核酸の繰返し範囲に、LCRのような活性化配列と一緒に送達することもできる。これは核酸のゲノムへの安全かつ予測可能な組込みへの経路を表すであろう。
【0106】
通常の治療用途では、本発明による核酸結合タンパク質は、変異した必須タンパク質(vital protein)を有する細胞を特異的にノックアウトするために使うことができる。例えば、もし変異rasをもつ細胞を標的とすれば、それらはrasが細胞生存に必須であるので、破壊されるであろう。あるいは、転写因子の作用を、細胞に転写因子に特異的な結合部位に結合する薬剤を投与することにより、調節、好ましくは抑制することができる。例えば、HIVtatの活性をHIV TARに特異的な結合タンパク質により抑制することができる。
【0107】
さらに、本発明による結合タンパク質は、不可逆的核酸破壊および細胞死を引き起す能力があるヌクレアーゼのような毒性分子と結合することができる。このような薬剤は、内因性核酸に変異を含む細胞を選択的に破壊する能力がある。
【0108】
以上に説明したこれらの核酸結合タンパク質および誘導体は感染などの治療にも、生物特異的抗生物質また抗ウイルス医薬の形態で応用することができる。このような応用では、結合タンパク質をヌクレアーゼまたは他の核毒素(nuclear toxin)と組合せて、微生物の核酸を特異的に標的とすることができる。
【0109】
本発明は、同様に、本発明による化合物または製薬上許容されるそれらの塩を有効成分として含有する医薬製剤およびそれらの製造方法に関する。
【0110】
本発明による化合物または製薬上許容されるそれらの塩を含有する本発明による医薬製剤は、温血動物への、経口等の経腸、さらに直腸、および非経口投与用の製剤であり、薬理学的有効成分が製剤自身にまたは製薬上許容される担体と一緒に存在する。有効成分の一日量は年齢および個体の症状に、また投与の方法に依存する。
【0111】
新規の医薬製剤は、例えば、約10%〜約80%、好ましくは約20%〜約60%の有効成分を含む。経腸または非経口投与用の本発明による医薬製剤は、例えば、糖衣錠、錠剤、カプセルまたは座薬、およびさらにアンプルのような単位用量形態の製剤である。これらは、それ自体公知の方法により、例えば通常の混合、造粒(granulating)、糖コーティング、溶解または凍結乾燥プロセスの方法により調製される。したがって、経口用途の医薬製剤は、有効成分を固体担体と組合せ、もし所望であれば、得た混合物を造粒し、そしてもし所望であるか必要であれば、錠剤もしくは糖衣錠コアを与える適切な賦形剤を添加した後に、混合物または造粒物を加工することによって、得ることができる。
【0112】
適切な担体は、特定的には、例えばラクトース、スクロース、マンニトールもしくはソルビトールのような糖類、セルロース調製物および/または例えばリン酸三カルシウムもしくはリン酸水素カルシウムのようなリン酸カルシウム等の充填剤、さらにはトウモロコシ、コムギ、コメまたはジャガイモデンプンを使うデンプンペースト、ゼラチン、トラガカント、メチルセルロースおよび/またはポリビニルピロリドンのような結合剤、もし所望であれば上述のデンプン、さらにカルボキシメチルデンプン、架橋ポリビニルピロリドン、寒天、アルギン酸またはアルギン酸ナトリウムのようなその塩等の崩壊剤であり;補助剤は、主に滑剤、流動調節剤および潤滑剤、例えば珪酸、タルク、ステアリン酸またはステアリン酸マグネシウムもしくはカルシウムのようなその塩、および/またはポリエチレングリコールである。糖衣錠コアは、もし所望であれば胃液耐性のある適切なコーティングにより提供され、なかんずくもし所望であれば、アラビアゴム、タルク、ポリビニルピロリドン、ポリエチレングリコールおよび/または二酸化チタンを含有する濃厚糖類溶液、適切な有機溶媒または溶媒混合物のコーティング溶液、または、胃液耐性コーティングの製剤にはアセチルセルロースフタレートもしくはヒドロキシプロピルメチルセルロースフタレートのような適切なセルロース調製物の溶液を使う。例えば有効成分の異なる用量を同定するかまたは示すために、着色剤または顔料を錠剤または糖衣錠コーティングに加えることができる。
【0113】
他の経口使用できる医薬製剤は、硬ゼラチンカプセル、ならびに、ゼラチンおよびグリセロールまたはソルビトールのような可塑剤で作られた軟閉鎖カプセルである。硬ゼラチンカプセルは、有効成分を顆粒の形態で、例えばラクトースのような充填剤、デンプンのような結合剤、および/またはタルクもしくはステアリン酸マグネシウムのような潤滑剤、および、所望であれば安定剤との混合物で含有することができる。軟カプセルでは、有効成分は、好ましくは脂肪油、パラフィン油または液体ポリエチレングリコールのような適切な液体に溶解または懸濁されており、安定剤を加えることもできる。
【0114】
適切な直腸経由で利用可能な医薬製剤は、例えば、有効成分と座薬基剤の組合わせからなる座薬である。適切な座薬基剤は、例えば、天然または合成トリグリセライド、パラフィン系炭化水素、ポリエチレングリコールまたは高級アルカノールである。さらに、有効成分と基剤との組合わせを含有するゼラチン直腸カプセルも使うことができる。適切な基剤は、例えば、液体トリグリセライド、ポリエチレングリコールまたはパラフィン系炭化水素である。
【0115】
非経口投与に適切な製剤は主に、有効成分の水溶性形態、例えば水溶性塩の水溶液、およびさらに、脂肪油、例えばゴマ油、または合成脂肪酸エステル、例えばオレイン酸エチルもしくはトリグリセライドのような適切な親油性溶媒またはビヒクルを使う、適切な油状注入懸濁液、または、増粘性物質、例えばカルボキシメチルセルロースナトリウム、ソルビトールおよび/またはデキストラン、かつ必要であれば安定剤も含有する水性注入懸濁液のような、有効成分の懸濁液である。
【0116】
有効成分用量は、温血動物種、年齢、および個体の症状ならびに投与方法による。通常の場合、ほぼ75kg体重の患者に対する経口投与の場合には、約10mg〜約250mgの一日量と見積られる。
【0117】
本発明は以下に、説明のみの目的で、次の実施例に記載される。
【0118】
実施例1
ジンクフィンガーの結合部位選好(preference)の決定
ジンクフィンガーファージ提示ライブラリーの設計
隣接DNAサブサイト間の界面におけるジンクフィンガー‐DNA認識を、ジンクフィンガーファージ提示ライブラリーを使い研究した。このライブラリーは、Zif268の3フィンガーDNA結合ドメインに基くが、フィンガー2(F2)およびフィンガー3(F3)は、それらのDNAサブサイトの界面を横切る接触ネットワークを形成しうる残基位置においてアミノ酸のランダム化が行われている。ライブラリーの詳細設計を、選択に使った総括的な(generic)DNA結合部位とともに、図1cに示す。簡単に説明すると、ライブラリーはF2残基位置6(以後、F2[+6]と記す)およびF3残基位置-1、+1、+2および+3(以後、F3[-1]、F3[+2]、などと記す)におけるランダム化を含有する。
【0119】
ライブラリー選択は、Zif268オペレーターに似るが、F2とF3のDNAサブサイト間の塩基ステップを形成するDNAダブレットの塩基の組織的組合わせを有するDNA結合部位を使って実施した。DNA結合部位は一般的形態が5'-GNX-XCG-GCG-3'であり、ここにX-XはDNAサブサイト間の界面における塩基の所与の組合わせを記し、Nは4種の塩基がDNA位置3において等しく発現されることを記す。したがって、この実験では、F3[+3]とヌクレオチド位置3Nの間の相互作用は完全な自由度が与えられている。このライブラリーの特徴により、ヌクレオチド位置4Xおよび5Xの塩基の所与の組合わせと結合する関連ジンクフィンガーの大きなファミリー(またはデータベース)の選択が可能になるが、これらは、F3の名目上のトリプレットサブサイトの中間塩基との異なる相互作用のために同一ではない。
【0120】
構築されるべき第1ライブラリー、LIB-Aは、F2残基位置6およびF3残基位置-1、+1、+2および+3におけるランダム化物を含有し(図2を参照すること)、DNA配列5'-GNX-XCG-GCG-3'を使ってソートされる。ここにX-XはDNA位置4Xおよび5Xにおける2つの塩基の既知の組合わせを記し、NはDNA位置3において等しい確率をもつ4種の塩基のいずれかを記す。第2ライブラリー、LIB-Bは、F2残基位置6およびF3残基位置-1および2におけるランダム化物を含有し、DNA配列5'-GCX-XCG-GCG-3'を使ってソートされる。ここにX-XはDNA位置4Xおよび5Xにおける2つの塩基の既知の組合わせを記す。
【0121】
2つの異なるジンクフィンガーファージ提示ライブラリー用の遺伝子は、4種の合成DNAオリゴヌクレオチドから、方向性末端対末端(directional end to end)連結反応により3種の短い相補的DNAリンカーを使って構築する。該オリゴヌクレオチドは、フィンガー2および3の適切なアミノ酸位置に選択的にランダム化された(配列NNSの;N=A/C/G/G、S=G/C)コドンを有する。該構築物をNot IおよびSfi I制限酵素切断部位を含有するプライマーを使ってPCRにより増幅し、上記のエンドヌクレアーゼで消化してクローニングオーバーハング(overhang)を作り、ファージベクターFd-Tet-SN中にライゲートした。電気コンピテント(electrocompetent)大腸菌(E. coli)TG 1細胞を組換えベクターにより形質転換し、15μg/mlテトラサイクリンを含有するTYE培地(1.5%寒天、1%バクトトリプトン、0.5%バクト酵母抽出物、0.8%NaCl)上にプレーティングした。
【0122】
未研究のいくつかのタンパク質‐DNA相互作用にこの自由度を認めることは、いずれか1つの選択実験から得られるクローンの多様性を増加する有用な方法である。しかし同時に、どの時点でも前後の(contextual)自由度が認められる接触数を限定することが重要であり、そうしなければ、特に強い分子間相互作用のサブセットが選択を支配する危険がある。この可能性を予測して、位置F2[+6]およびF3[-1および+2]にのみランダム化残基を含有し、したがって選択において前後の自由度を認めない、より小さいサブライブラリーも作製した。このライブラリーから選択されたクローンを本明細書で記すときは、星印でマークした。
【0123】
実験方法
2つのジンクフィンガーライブラリーからのファージ選択は、各ライブラリーのメンバーにより特異的に結合されうるDNA配列の多様性を決定する目的で別々に実施する。各ライブラリーについて、ヌクレオチド位置4Xおよび5Xにおける塩基の全16通りの可能な組合わせに対応する異なるDNA結合部位を使用して、16回の選択を実施する。特異的結合ファージを選択するために使われるDNA結合部位は固相表面に固定し、一方、他の15種のDNA部位はそれぞれ10倍過剰で特異性競合剤として溶液中に存在させる。
【0124】
ファージ選択
テトラサイクリン耐性コロニーをプレートから、50μM ZnCl2および15μg/mlテトラサイクリンを含有する2xTY培地(16g/リットルバクトトリプトン、10g/リットルバクト酵母抽出物、5g/リットルNaCl)に移し、一夜、30℃で振とう培養器で培養した。300gで5分間遠心分離して、ファージ粒子を含有する清澄な培養上清を得た。
【0125】
ビオチン化DNA標的部位(1pmol)をストレプトアビジン被覆チューブ(Boehringer Mannheim)に結合させた。ファージ上清溶液をPBS選択バッファー(50μM ZnCl2、2% Marvel、1% Tween、20μg/ml音波処理サケ精子DNA、10pmol/mlの他の15種の可能な非ビオチン化DNA部位のそれぞれを含有するPBS)中に1:10に希釈し、1mlを各チューブにアプライし、1時間、20℃でおいた。この後、チューブを空にし、50μM ZnCl2、2% Marvel、1% Tweenを含有するPBSにより20回洗浄した。保持されたファージを0.1mlの0.1Mトリエチルアミン中に溶出させ、等容積の1M Tris(pH7.4)で中和した。対数期の大腸菌TG 1(0.5ml)を溶出ファージ(50μl)で感染させ、続くラウンドの選択のためのファージ上清を調製するために使った。3ラウンドの選択の後、選択ファージで感染させた大腸菌をプレーティングし、個々のコロニーを拾上げて結合部位特徴(signature)アッセイおよびDNA配列決定用のファージを増殖するために使った。
【0126】
特定のDNA結合部位に対する3ラウンドのファージ選択の後、個々のジンクフィンガークローンを回収し、そして各クローンのDNA結合特異性を結合部位サイン法により決定した。これは、各ライブラリーが位置4Xおよび5Xのいずれかで1つの固定塩基および1つのランダム化塩基を含有する(すなわち、ライブラリーGN、AN、TN、CN、およびNG、NA、NT、NC)ように設計された8つの異なるDNA結合部位ライブラリーに対する、各ジンクフィンガーファージの結合スクリーニングに関わる。したがって、選択実験に使われた16のDNA結合部位のそれぞれは、ユニークな2つのライブラリーの組合わせにより規定される。例えば、4G5Gを含有するDNA結合部位は、8つのライブラリー中、関連するダブレットが1つのランダム化ヌクレオチドおよび他のグアニンとして固定されたヌクレオチドを有する2つ、すなわちライブラリー4G5Nおよび4N5Gにのみ存在する。結合部位特徴に使われた8つのDNAライブラリーを、マイクロタイタープレートに横に(across)配列し、ジンクフィンガーファージ結合をファージELISAにより検出した。8つのDNAライブラリーとの結合パターンは、各ファージクローンのDNA配列特異性(または選好性)を表し、そして比較的特異性のあるとみられるクローンのみを続いて配列決定して、ランダム化ジンクフィンガー残基位置に存在するアミノ酸の同一性を明らかにした。
【0127】
方法は、先に記載されたとおりである(Choo, Y.およびKlug, A. (1994) Proc. Natl. Acad. Sci. U.S.A. 91, 11163-11167;Choo, Y.およびKlug, A. (1994) Proc. Natl. Acad. Sci. U.S.A. 91, 11168-11172)。簡単に説明すると、Zif268オペレーター変異型を含有する、5'ビオチン化した位置的にランダム化したオリゴヌクレオチドライブラリーを、記載したとおり、プライマー伸長により合成する。DNAライブラリー(LIB-Aに対して0.4pmol/ウエルおよびLIB-Bに対して1.2pmol/ウエル)を、50μM ZnCl2を含有するPBS(PBS/Zn)の入ったストレプトアビジン被覆ELISAウエル(Boehringer-Mannheim)に加える。ファージ溶液(一晩細菌培養した上清を2% Marvel、1% Tweenおよび20μg/ml音波処理サケ精子DNAを含有するPBS/Zn中に1:10で希釈したもの)を各ウエル(50μl/ウエル)にアプライする。結合は、20°で1時間進める。未結合ファージは、1%Tweenを含有するPBS/Znで6回洗浄し、その後PBS/Znで3回洗浄する。結合したファージをELISAにより、西洋わさびペルオキシダーゼとコンジュゲートした抗M13 IgG(Pharmacia Biotech)で検出し、比色シグナルをSOFFMAX 2.32(Molecular Devices)を使い定量する。
【0128】
個々のジンクフィンガークローンのコード配列は、ファージ配列に相補的な外部プライマーを使ってPCRにより増幅する。これらのPCR産物をThermo Sequenase cycle sequencing(Amersham Life Science)を使って手動で配列決定する。
【0129】
ファージ選択ジンクフィンガーの分析
図3は、フィンガー2および3のDNAサブサイト間の塩基ステップを形成する16の異なるDNAダブレットを使い、両方のライブラリーから選択した比較的配列特異性のあるジンクフィンガーファージの結合部位特徴を示す。これらの結果は、5'位置(モデルシステムのヌクレオチド5X)がグアニン以外の塩基で固定された3塩基連鎖を含む、ほとんど全てのサブサイトに特異的に結合するジンクフィンガークローンが選択されたことを示す。結局、該選択物は、4種の塩基のいずれも名目的トリプレットの5'および3'位置の両方に特異的に結合することが可能であることを示す。結果を図4に総括する。
【0130】
大きなライブラリーのメンバーは16のダブレットから15のダブレットを認識するフィンガーを生じるが、より小さいサブライブラリーからの選択は、16のダブレットの8つとだけ特異的に結合しうるフィンガーを生じる。この効率の相異が大きなライブラリー中のよりランダム化された位置の包含に由来するか、大きなライブラリーで設計された前後の自由度により与えられるコンフォメーションのフレキシビリティーに由来するのか、またはその両方であるのかはわかっていない。特異的なジンクフィンガーを生じないただ1つの塩基ステップは、4G5Aである。このジヌクレオチドは、選択に使うDNA結合部位のコンテキスト(context)に好ましくないDNA変形を誘起するのかも知れない。
【0131】
実施例2
位置1に対する+2特異性の決定
ジンクフィンガーのαヘリックス位置2に存在するアミノ酸は、2つのオーバーラップDNA4塩基連鎖(quadruplet)サブサイト(図1Bを参照すること;上に説明した4塩基連鎖の位置1または4に対応する、位置5/5')の界面における塩基対に対する特異性の決定を助けることができる。野生型Zif268のF3[+2]に存在するAsp残基は、DNA認識にある役割を果すことが示されており、さらなる例が本発明のファージ提示実験により作製されている(詳細は実施例1および図5Aを参照すること)。
【0132】
次の実験プロトコルは実施例1のそれである。図5AはDNA結合特異性に与えるαヘリックス位置2の影響を示す関連するジンクフィンガークローンの1例を示す。この場合、フィンガー2の位置6は不変(Asn)で、この場合の変化として、この塩基との接触を選択するためのジンクフィンガーの特異性は、フィンガー3の位置+2により指揮される。
【0133】
ジンクフィンガーのこのファミリーは、4T5Aまたは4T5Cサブサイト界面を含有するDNA結合部位を使う選択物から誘導される。5X-5'X塩基対に対する塩基選好性はF3[+2]に存在するアミノ酸により、恐らくは交差鎖接触の形成により、決定される。
【0134】
図5Bは、F3[+2]で選択されたある特定のアミノ酸と位置5'Xに存在する塩基の同一性の関係の例を示す。選択は5つのアミノ酸(Asn、Gln、Arg、LysおよびHis)からのDNA接触の可能性を表し、これらのアミノ酸は全てヌクレオチド位置5'Xでグアニン(O6)またはチミン(O4)のいずれかの環外酸素原子に水素結合を与えることができる。この図には、F3[+2]でこれらのアミノ酸により単離されたクローンを、位置5'Xで塩基選好性を示す結合部位特徴と一緒に掲げた。全体的に、Serは、全ての既知のジンクフィンガーの半分を越える位置2での存在により、38%の出現率(occurrence)で選択を支配する。トレオニン、AlaおよびGlyはしばしば選択物に存在する(それぞれ、15%、15%および9%)が、識別の区別可能なパターンを示さなかった。ある種のアミノ酸(Cys、Asp、Phe、Ile、Leu、Met、Pro、ValおよびTrp)は位置2に決して選択されない。しかし、これらのある特定の状況下で結合する能力は排除されるべきでない。
【0135】
F3[+2]で選択されるアミノ酸の小さいサブセットは、位置5'Xにおける塩基対の同一性と有意な関係を示し(図5B)、これらの間の交差鎖相互作用はDNA認識の一般的機構でありうることを示唆する。これらの関係のほとんどは、Zif268モデルの枠組に従い、F3[+2]の水素結合供与体とDNA位置5'Xのグアニンまたはチミンの間の対形成として理論付けすることができる。位置2に決して選択されないアミノ酸、または選択されるが有意な相関を示さないアミノ酸とは対照的に、一貫してこの位置からDNA認識にある役割を果すように思われるアミノ酸は、多重水素結合基を側鎖に有する。これらの残基はネットワーク支持への参加により一層大きな特異性を達成するので、塩基認識にある役割を果しうる可能性がある。
【0136】
実施例3
一般目的ライブラリーの構築
この実施例で構築される2成分ライブラリーシステムは、それぞれZif268の3つのフィンガーをコードするがいくつかのアミノ酸位置では選択的にランダム化された、ライブラリーLIB1/2およびLIB2/3を含む。新しいライブラリーは、モジュールジンクフィンガーのモデルに固執する代りに、隣接ジンクフィンガーのある特定のアミノ酸位置において協奏的(concerted)変異を有する。したがって、LIB1/2はF1位置-1、2、3、5および6ならびにF2位置-1、1、2および3に、同時に変異を有する。LIB2/3は、F2位置3および6ならびにF3位置-1、1、2、3および5、6に同時に変異を含有する。各ライブラリーの残りのアミノ酸は、野生型(WT)Zif268配列のとおりである。この2つのライブラリーをFdファージにGIII融合物として標準技術によりクローニングした。
【0137】
それぞれの変異位置に許容されるアミノ酸は、次のとおりである。
【0138】
選択および組換え
選択は、DNA配列GCG-GMN-OPQをLIB1/2に対して、およびDNA配列IJK-LMG-GCGをLIB2/3に対して使って実施した(式中、下線を引いた塩基は野生型 Zif268残基により結合され、他の文字は所与のヌクレオチドを表す)。Zif268結合部位の保存されたヌクレオチドは、Zif268 DNA結合ドメインの保存部分との結合により相互作用の記録(register)を固定する役割を果す。2成分ファージ提示ライブラリーを混合してこれら2つの共通(generic)部位を使う選択を単一のチューブで実施することができるし、または、該選択を別々に実施することができる。数ラウンドの選択の後に、2つのライブラリーを組替えて配列IJK-LMN-OPQを認識するキメラDNA結合ドメインを作製する。
【0139】
組換え反応は、選択した3つのフィンガードメインをPCRにより増幅し、PCR産物を制限酵素Ddelを使い切断することにより実施する。これにより、両方のジンクフィンガーライブラリーの遺伝子を、F2αヘリックス位置4および5をコードするDNA配列で切断する。消化した産物をランダムに再度ライゲートして、キメラDNA結合ドメイン(および再構成した野生型 Zif268を含む他の産物)をコードする組換え遺伝子を作製する。キメラDNA結合ドメインは、PCRによる混合産物から、野生型遺伝子よりむしろ組換えF1およびF3遺伝子を認識する選択的プライマーを使い選択的に増幅し、そして(さらなる選択のために)Fdファージにまたは(例えば大腸菌での発現のために)他のベクターにクローニングされる。
【0140】
2成分系ライブラリーからの最初の選択を完成まで押し進めることができ、かくして組換えによる単一クローンの構築が可能になる。あるいは、もし最初の選択のストリンジェンシーを低くすれば、多くの候補を組換え後の様々なキメラドメインの構築に利用できるであろう。後者の場合、最良の組換えタンパク質は、ファージ上の選択のさらなる繰返しにより、選択することができる。
【特許請求の範囲】
【請求項1】
ジンクフィンガーポリペプチドをコードするライブラリーセットであって、該ライブラリーはオーバーラップするジンクフィンガーポリペプチドをコードし、各ポリペプチドは1つ以上のジンクフィンガーを含んでなり、各ポリペプチドは少なくとも部分的に無作為化され、該無作為化の範囲は1対の第1及び第2のジンクフィンガーのオーバーラップをカバーし、前記無作為化は少なくとも第1のジンクフィンガーの位置6及び第2のジンクフィンガーの位置2での無作為化を含み、前記ポリペプチドは選択の後に組み立てられて多重フィンガーのジンクフィンガーポリペプチドを形成することができ、ジンクフィンガーは核酸結合配列内の核酸4塩基連鎖との結合を決定することに関わり、オーバーラップするジンクフィンガーはオーバーラップする4塩基連鎖に結合するジンクフィンガーであり、核酸の-(マイナス鎖)を3'から5'へと読んだとき、第1の4塩基連鎖の塩基4は第2の4塩基連鎖の塩基1である、ライブラリーセット。
【請求項2】
各ポリペプチドが3から6のジンクフィンガーを含む、請求項1記載のライブラリーセット。
【請求項3】
各ポリペプチドにおいて、1個半のジンクフィンガーが無作為化される、請求項1又は2に記載のライブラリーセット。
【請求項4】
無作為化位置が位置-1、1、2、3、5および6から選択される、請求項1〜3のいずれかに記載のライブラリーセット。
【請求項5】
3ジンクフィンガーのポリペプチドをコードするライブラリーの1対を含んでなる、請求項1記載のライブラリーセット。
【請求項6】
アミノ酸残基の無作為化は次のアミノ酸が所与の位置に現れうるように制限される請求項1〜5のいずれかに記載のライブラリーセット:
位置 可能なアミノ酸
-1 R、Q、H、N、D、A、T;
1 S、R、K、N;
2 D、A、R、Q、H、K、S、N;
3 H、N、S、T、V、A、D;
5 I、T、K;
6 R、Q、V、A、E、K、N、T。
【請求項7】
次のアミノ酸が所与の位置に現れることができる3フィンガーのジンクフィンガータンパク質を選択するための請求項6に記載の2つのライブラリーセット:
【請求項8】
位置-1、2、3および6のアミノ酸が次のとおり選択される請求項1に記載のライブラリーセット:
a) もし4塩基連鎖の塩基4がGであれば、αヘリックスの位置+6はArgもしくはLysであり;
b)もし4塩基連鎖の塩基4がAであれば、αヘリックスの+6はGlu、AsnもしくはValであり;
c)もし4塩基連鎖の塩基4がTであれば、αヘリックスの位置+6はSer、Thr、ValもしくはLysであり;
d)もし4塩基連鎖の塩基4がCであれば、αヘリックスの+6はSer、Thr、Val、Ala、GluもしくはAsnであり;
e)もし4塩基連鎖の塩基3がGであれば、αヘリックスの位置+3はHisであり;
f)もし4塩基連鎖の塩基3がAであれば、αヘリックスの位置+3はAsnであり;
g)もし4塩基連鎖の塩基3がTであれば、αヘリックスの位置+3はAla、SerもしくはValであり;但し、もしそれがAlaであれば、-1または+6の残基の1つは小さい残基であり;
h)もし4塩基連鎖の塩基3がCであれば、αヘリックスの位置+3はSer、Asp、Glu、Leu、ThrもしくはValであり;
i)もし4塩基連鎖の塩基2がGであれば、αヘリックスの位置-1はArgであり;
j)もし4塩基連鎖の塩基2がAであれば、αヘリックスの位置-1はGlnであり;
k)もし4塩基連鎖の塩基2がTであれば、αヘリックスの位置-1はHisもしくはThrであり;
l)もし4塩基連鎖の塩基2がCであれば、αヘリックスの位置-1はAspもしくはHisであり;
m)もし4塩基連鎖の塩基1がGであれば、位置+2はGluであり;
n)もし4塩基連鎖の塩基1がAであれば、位置+2はArgもしくはGlnであり;
o)もし4塩基連鎖の塩基1がCであれば、位置+2はAsn、Gln、Arg、HisもしくはLysであり;
p)もし4塩基連鎖の塩基1がTであれば、位置+2はSerもしくはThrである。
【請求項9】
各ジンクフィンガーが一般的一次構造:
(式中、X(Xa、XbおよびXcを含む)はアミノ酸である)を有する、請求項1〜8のいずれかに記載のライブラリーセット。
【請求項10】
Xaが(F/Y)-XまたはP-(F/Y)-Xである請求項9に記載のライブラリーセット。
【請求項11】
X2-4がS-X、E-X、K-X、T-X、P-XおよびR-Xのいずれか1つから選ばれる請求項9または請求項10に記載のライブラリーセット。
【請求項12】
XbがTまたはIである請求項9〜11のいずれかに記載のライブラリーセット。
【請求項13】
X2-3がG-K-A、G-K-C、G-K-S、G-K-G、M-R-NまたはM-Rである請求項9〜12のいずれかに記載のライブラリーセット。
【請求項14】
リンカーがT-G-E-KまたはT-G-E-K-Pである請求項9〜13のいずれかに記載のライブラリーセット。
【請求項15】
位置+9がRまたはKである請求項9〜14のいずれかに記載のライブラリーセット。
【請求項16】
位置+1、+5および+8が、疎水性アミノ酸F、WまたはYのいずれか1つにより占められない請求項9〜15のいずれかに記載のライブラリーセット。
【請求項17】
位置+1、+5および+8が、それぞれ残基K、TおよびQにより占められる請求項16に記載のライブラリーセット。
【請求項18】
標的核酸配列と結合することができるCys2-His2ジンクフィンガークラスの核酸結合タンパク質のライブラリーを製造する方法であって、
a)天然のジンクフィンガーポリペプチドおよびコンセンサスジンクフィンガーポリペプチドからなる群からモデルジンクフィンガーポリペプチドを選択し;および
b)請求項1〜8のいずれかに記載のその中の1つ以上のフィンガーを無作為化する工程を含んでなる方法。
【請求項19】
モデルジンクフィンガーがコンセンサスジンクフィンガーであり、その構造はコンセンサス構造PYKCPECGKSFSQKSDLVKHQRTHTGおよびコンセンサス構造PYKCSECGKAFSQKSNLTRHQRIHTGEKPからなる群から選ばれる請求項18に記載の方法。
【請求項20】
モデルジンクフィンガーが天然のジンクフィンガーであり、その構造はZif268(Elrod-Ericksonら, (1996) Structure 4:1171-1180)、GLI(PavletichおよびPabo, (1993) Science 261:1701-1707)、Tramtrack(Fairallら, (1993) Nature 366:483-487)およびYY1(Houbaviyら, (1996) PNAS (USA) 93:13577-13582)からなる群から選ばれるタンパク質の1つのフィンガーから選ばれる請求項18に記載の方法。
【請求項21】
モデルジンクフィンガーがZif268のフィンガー2である請求項20に記載の方法。
【請求項22】
a)請求項18〜21のいずれかに記載の方法により標的核酸分子に特異的である核酸結合タンパク質を製造し;
b)結合を促進し未結合で残るいずれの核酸結合タンパク質をも除去する条件下で、標的核酸分子を含んでなる試験系を核酸結合タンパク質にさらし;
c)試験系の核酸結合タンパク質の存在を検出するステップを含んでなる標的核酸分子の有無を決定する方法。
【請求項23】
試験系中の核酸結合タンパク質の有無を抗体により検出する請求項22に記載の方法。
【請求項24】
使用する核酸結合タンパク質を繊維状バクテリオファージの表面上に提示して核酸結合タンパク質の有無をバクテリオファージまたはその成分を検出することにより検出する請求項22または請求項23に記載の方法。
【請求項1】
ジンクフィンガーポリペプチドをコードするライブラリーセットであって、該ライブラリーはオーバーラップするジンクフィンガーポリペプチドをコードし、各ポリペプチドは1つ以上のジンクフィンガーを含んでなり、各ポリペプチドは少なくとも部分的に無作為化され、該無作為化の範囲は1対の第1及び第2のジンクフィンガーのオーバーラップをカバーし、前記無作為化は少なくとも第1のジンクフィンガーの位置6及び第2のジンクフィンガーの位置2での無作為化を含み、前記ポリペプチドは選択の後に組み立てられて多重フィンガーのジンクフィンガーポリペプチドを形成することができ、ジンクフィンガーは核酸結合配列内の核酸4塩基連鎖との結合を決定することに関わり、オーバーラップするジンクフィンガーはオーバーラップする4塩基連鎖に結合するジンクフィンガーであり、核酸の-(マイナス鎖)を3'から5'へと読んだとき、第1の4塩基連鎖の塩基4は第2の4塩基連鎖の塩基1である、ライブラリーセット。
【請求項2】
各ポリペプチドが3から6のジンクフィンガーを含む、請求項1記載のライブラリーセット。
【請求項3】
各ポリペプチドにおいて、1個半のジンクフィンガーが無作為化される、請求項1又は2に記載のライブラリーセット。
【請求項4】
無作為化位置が位置-1、1、2、3、5および6から選択される、請求項1〜3のいずれかに記載のライブラリーセット。
【請求項5】
3ジンクフィンガーのポリペプチドをコードするライブラリーの1対を含んでなる、請求項1記載のライブラリーセット。
【請求項6】
アミノ酸残基の無作為化は次のアミノ酸が所与の位置に現れうるように制限される請求項1〜5のいずれかに記載のライブラリーセット:
位置 可能なアミノ酸
-1 R、Q、H、N、D、A、T;
1 S、R、K、N;
2 D、A、R、Q、H、K、S、N;
3 H、N、S、T、V、A、D;
5 I、T、K;
6 R、Q、V、A、E、K、N、T。
【請求項7】
次のアミノ酸が所与の位置に現れることができる3フィンガーのジンクフィンガータンパク質を選択するための請求項6に記載の2つのライブラリーセット:
【請求項8】
位置-1、2、3および6のアミノ酸が次のとおり選択される請求項1に記載のライブラリーセット:
a) もし4塩基連鎖の塩基4がGであれば、αヘリックスの位置+6はArgもしくはLysであり;
b)もし4塩基連鎖の塩基4がAであれば、αヘリックスの+6はGlu、AsnもしくはValであり;
c)もし4塩基連鎖の塩基4がTであれば、αヘリックスの位置+6はSer、Thr、ValもしくはLysであり;
d)もし4塩基連鎖の塩基4がCであれば、αヘリックスの+6はSer、Thr、Val、Ala、GluもしくはAsnであり;
e)もし4塩基連鎖の塩基3がGであれば、αヘリックスの位置+3はHisであり;
f)もし4塩基連鎖の塩基3がAであれば、αヘリックスの位置+3はAsnであり;
g)もし4塩基連鎖の塩基3がTであれば、αヘリックスの位置+3はAla、SerもしくはValであり;但し、もしそれがAlaであれば、-1または+6の残基の1つは小さい残基であり;
h)もし4塩基連鎖の塩基3がCであれば、αヘリックスの位置+3はSer、Asp、Glu、Leu、ThrもしくはValであり;
i)もし4塩基連鎖の塩基2がGであれば、αヘリックスの位置-1はArgであり;
j)もし4塩基連鎖の塩基2がAであれば、αヘリックスの位置-1はGlnであり;
k)もし4塩基連鎖の塩基2がTであれば、αヘリックスの位置-1はHisもしくはThrであり;
l)もし4塩基連鎖の塩基2がCであれば、αヘリックスの位置-1はAspもしくはHisであり;
m)もし4塩基連鎖の塩基1がGであれば、位置+2はGluであり;
n)もし4塩基連鎖の塩基1がAであれば、位置+2はArgもしくはGlnであり;
o)もし4塩基連鎖の塩基1がCであれば、位置+2はAsn、Gln、Arg、HisもしくはLysであり;
p)もし4塩基連鎖の塩基1がTであれば、位置+2はSerもしくはThrである。
【請求項9】
各ジンクフィンガーが一般的一次構造:
(式中、X(Xa、XbおよびXcを含む)はアミノ酸である)を有する、請求項1〜8のいずれかに記載のライブラリーセット。
【請求項10】
Xaが(F/Y)-XまたはP-(F/Y)-Xである請求項9に記載のライブラリーセット。
【請求項11】
X2-4がS-X、E-X、K-X、T-X、P-XおよびR-Xのいずれか1つから選ばれる請求項9または請求項10に記載のライブラリーセット。
【請求項12】
XbがTまたはIである請求項9〜11のいずれかに記載のライブラリーセット。
【請求項13】
X2-3がG-K-A、G-K-C、G-K-S、G-K-G、M-R-NまたはM-Rである請求項9〜12のいずれかに記載のライブラリーセット。
【請求項14】
リンカーがT-G-E-KまたはT-G-E-K-Pである請求項9〜13のいずれかに記載のライブラリーセット。
【請求項15】
位置+9がRまたはKである請求項9〜14のいずれかに記載のライブラリーセット。
【請求項16】
位置+1、+5および+8が、疎水性アミノ酸F、WまたはYのいずれか1つにより占められない請求項9〜15のいずれかに記載のライブラリーセット。
【請求項17】
位置+1、+5および+8が、それぞれ残基K、TおよびQにより占められる請求項16に記載のライブラリーセット。
【請求項18】
標的核酸配列と結合することができるCys2-His2ジンクフィンガークラスの核酸結合タンパク質のライブラリーを製造する方法であって、
a)天然のジンクフィンガーポリペプチドおよびコンセンサスジンクフィンガーポリペプチドからなる群からモデルジンクフィンガーポリペプチドを選択し;および
b)請求項1〜8のいずれかに記載のその中の1つ以上のフィンガーを無作為化する工程を含んでなる方法。
【請求項19】
モデルジンクフィンガーがコンセンサスジンクフィンガーであり、その構造はコンセンサス構造PYKCPECGKSFSQKSDLVKHQRTHTGおよびコンセンサス構造PYKCSECGKAFSQKSNLTRHQRIHTGEKPからなる群から選ばれる請求項18に記載の方法。
【請求項20】
モデルジンクフィンガーが天然のジンクフィンガーであり、その構造はZif268(Elrod-Ericksonら, (1996) Structure 4:1171-1180)、GLI(PavletichおよびPabo, (1993) Science 261:1701-1707)、Tramtrack(Fairallら, (1993) Nature 366:483-487)およびYY1(Houbaviyら, (1996) PNAS (USA) 93:13577-13582)からなる群から選ばれるタンパク質の1つのフィンガーから選ばれる請求項18に記載の方法。
【請求項21】
モデルジンクフィンガーがZif268のフィンガー2である請求項20に記載の方法。
【請求項22】
a)請求項18〜21のいずれかに記載の方法により標的核酸分子に特異的である核酸結合タンパク質を製造し;
b)結合を促進し未結合で残るいずれの核酸結合タンパク質をも除去する条件下で、標的核酸分子を含んでなる試験系を核酸結合タンパク質にさらし;
c)試験系の核酸結合タンパク質の存在を検出するステップを含んでなる標的核酸分子の有無を決定する方法。
【請求項23】
試験系中の核酸結合タンパク質の有無を抗体により検出する請求項22に記載の方法。
【請求項24】
使用する核酸結合タンパク質を繊維状バクテリオファージの表面上に提示して核酸結合タンパク質の有無をバクテリオファージまたはその成分を検出することにより検出する請求項22または請求項23に記載の方法。
【図1】

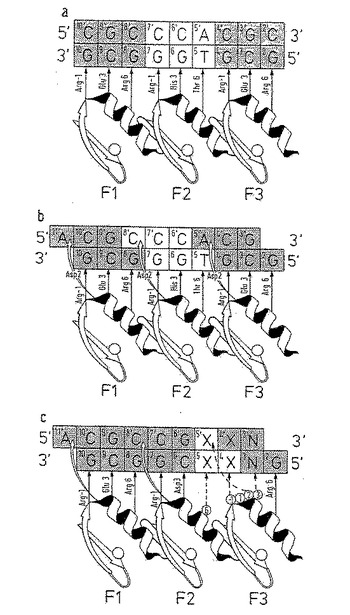
【図2】
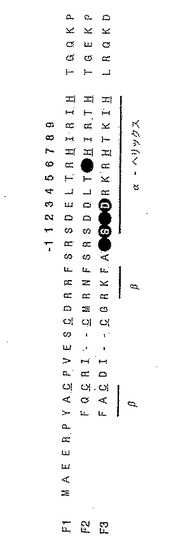
【図3】

【図4】
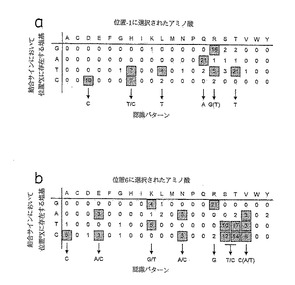
【図5】
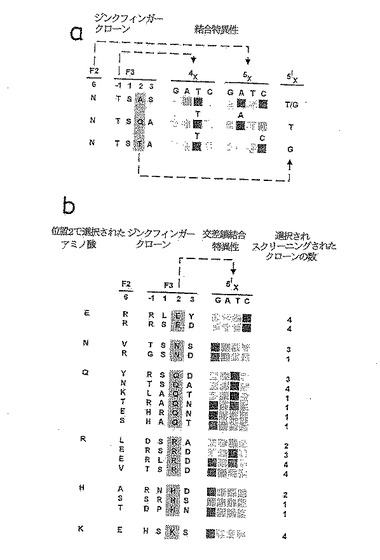
【図6】
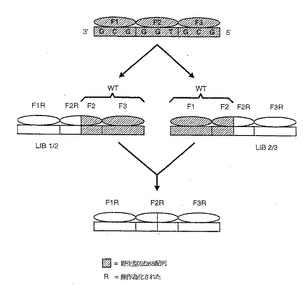
【図2】
【図3】
【図4】
【図5】
【図6】
【公開番号】特開2012−175982(P2012−175982A)
【公開日】平成24年9月13日(2012.9.13)
【国際特許分類】
【出願番号】特願2012−129566(P2012−129566)
【出願日】平成24年6月7日(2012.6.7)
【分割の表示】特願2008−278701(P2008−278701)の分割
【原出願日】平成10年5月26日(1998.5.26)
【出願人】(503415482)ジェンダック・リミテッド (2)
【Fターム(参考)】
【公開日】平成24年9月13日(2012.9.13)
【国際特許分類】
【出願日】平成24年6月7日(2012.6.7)
【分割の表示】特願2008−278701(P2008−278701)の分割
【原出願日】平成10年5月26日(1998.5.26)
【出願人】(503415482)ジェンダック・リミテッド (2)
【Fターム(参考)】
[ Back to top ]