
核酸配列の不均衡性の決定
生体試料中に核酸配列の不均衡が存在するか否かを判定するための、方法、システム、及び装置が提供される。不均衡を判定するための1つ以上のカットオフ値、例えば2つの配列(又は配列のセット)の比率が選択される。このカットオフ値は、母体血漿等の母体核酸配列のバックグラウンドを含む試料中の胎児DNAのパーセンテージに、少なくとも部分的に基づき決定され得る。また、このカットオフ値は、反応あたりの配列の平均濃度に基づいても決定され得る。一つの側面において、このカットオフ値は、特定の核酸配列を含むことが見込まれる情報的ウェルの割合から決定され、その割合は、上述のパーセンテージ及び/又は平均濃度に基づき決定される。このカットオフ値は、逐次確率比検定(SPRT)等の、多くの異なる種類の方法を使用して決定され得る。

【発明の詳細な説明】
【技術分野】
【0001】
優先権の主張
本出願は、「DETERMINING A NUCLEIC ACID SEQUENCE IMBALANCE」と題された、2007年7月23日に出願された米国仮出願第60/951438号(代理人整理番号016285−005200US)からの優先権を主張し、そして当該仮出願の正規の出願であり、当該仮出願の全ての内容は、全ての目的のために参照により本明細書に援用される。
【0002】
関連出願の相互参照
本出願はまた、「DIAGNOSING FETAL CHROMOSOMAL ANEUPLOIDY USING GENOMIC SEQUENCING,」と題された、同時に出願された正規の出願(代理人整理番号016285−005220US)に関し、そしてその全ての内容は全ての目的のために参照により本明細書に援用される。
【0003】
発明の技術分野
本発明は、一般的には、異なる核酸配列間における不均衡を決定することによる、遺伝子型及び疾患の診断的検査に関し、そしてより詳細には、母体血液試料の検査を通じての、胎児のダウン症候群、他の染色体の異数性、突然変異、及び遺伝子型に関する。また、本発明は、癌の検出、移植のモニタリング、及び感染性疾患のモニタリングに関する。
【背景技術】
【0004】
遺伝的疾患、癌及び他の病態は、2つの対応する染色体若しくは対立遺伝子、又は他の複数の核酸配列の不均衡の原因又は結果となる場合が多い。この場合、1つの遺伝子に対するもう1つの遺伝子の割合が、正常の場合より多く、又は少ない。通常、正常の場合の比率はちょうど50:50である。ダウン症候群(トリソミー21)は、第21番染色体が追加されて不均衡を有する。
【0005】
胎児染色体異数性、例えばトリソミー21の、出産前における従来の診断方法は、侵襲性の手順、例えば羊水穿刺又は絨毛膜検体採取による、胎児試料の採取を伴うものであり、そしてそれらは胎児が死亡する危険性がある。非侵襲性の手順、例えば超音波検査法及び生化学マーカーによるスクリーニングは、最終的な侵襲性診断手順に先立ってリスク層の妊婦に使用された。しかし、これらのスクリーニング法は通常、例えば核となる染色体の異常性の代わりに、トリソミー21と関わりのある付帯現象を測定するため、その診断精度は最適ではなく、また、妊娠年齢によって大きな影響を受ける等の短所を有する。
【0006】
1997年における、母体血漿中における循環性無細胞胎児DNAの発見は、非侵襲性出生前診断のための新しい可能性を提供した(Lo,YMD and Chiu,RWK 2007 Nat Rev Genet 8,71−77)。この方法は、伴性遺伝子疾患(Costa,JM et al.2002 N Engl J Med 346,1502)及び特定の単一遺伝子疾患(Lo,YMD et al.1998 N Engl J Med 339,1734−1738)の出生前診断に対して容易に応用されたが、胎児染色体異数性の出産前における検出へのその適用は、相当な困難を要した(Lo, YMD and Chiu, RWK 2007,上記参照)。第一に、胎児の核酸は通常、胎児の核酸の分析を阻害し得る、母体由来核酸の高いバックグラウンドを含む母体血漿中に存在する(Lo,YMD et al.1998 Am J Hum Genet 62,768−775)。第二に、胎児の核酸は母体血漿中において主に無細胞形態で循環しており、そしてそのことは、胎児ゲノム中の遺伝子量又は染色体量の情報を得ることを困難としている。
【0007】
これらの課題を解決する著しい発展が近年なされた(Benachi,A & Costa,JM 2007 Lancet 369,440−442)。一つの方法は、母体血漿中における胎児特異的な核酸を検出し、その結果、母体のバックグラウンド障害の問題を克服することである(Lo, YMD and Chiu,RWK 2007,上記参照)。第21番染色体の量は、胎盤由来のDNA/RNA分子中における多型対立遺伝子の比から推定された。しかしこの方法は、試料中の標的核酸の含有量が少ないほど精度が低くなる上に、標的とされた多型に関してヘテロ接合性である胎児にしか適用できない。一つの多型が使用されるのであれば、斯かる胎児は人口群の一部に過ぎない。
【0008】
Dhallan et al(Dhallan,R,et al.2007,上記参照、Dhallan,R,et al.2007 Lancet 369,474−481)は、母体血漿へホルムアルデヒドを加えることにより、循環性胎児DNAの割合を増やす代替的な方法について記載している。母体血漿中における胎児の第21番染色体配列の割合は、第21番染色体上の一塩基多型(SNPs)に関して、父系遺伝性の胎児特異的対立遺伝子と胎児特異的ではない対立遺伝子との比を評価することによって決定された。SNP比は、基準染色体に関して同様に計算された。胎児の第21番染色体の不均衡はその後、第21番染色体のSNP比と基準染色体のSNP比との間の統計的な有意差を検出することによって推測され、ここで有意性は、0.05以下の固定されたp値を使用して定義される。高い個体群範囲を確保するために、一つの染色体あたり500個超のSNPが標的とされた。しかし、胎児DNAを高い割合に増大させるためのホルムアルデヒドの有効性について議論があり(Chung,GTY,et al.2005 Clin Chem 51,655−658)、したがって当該方法の再現性はさらに評価される必要がある。同様に、胎児及び母体の各々が、各々の染色体について異なる数のSNPsに関する情報を提供するため、SNP比の比較に関する統計的検出力は、事例ごとに変化するだろう(Lo,YMD&Chiu,RWK.2007 Lancet 369,1997)。さらにこれらの方法は遺伝子多型の検出に依存するため、それらはこれらの多型に関してヘテロ接合性である胎児に限定される。
【0009】
トリソミー21及び正倍数体の胎児から得られる羊膜細胞培養中の、第21番染色体遺伝子座及び基準遺伝子座のポリメラーゼ連鎖反応(PCR)及びDNA定量化を使用することにより、Zimmermann他(2002 Clin Chem 48,362−363)は、前者の第21番染色体DNA配列における1.5倍の増加に基づいて、胎児の当該2つの群を区別することができた。DNA鋳型濃度における2倍の違いは、僅か一回の閾値サイクル(Ct)の違いに相当するため、1.5倍の違いの区別は、従来のリアルタイムPCRの限界であった。より精密な程度の定量的識別を達成するために、代替的な方法が必要とされる。
【0010】
デジタルPCRは、核酸試料中の対立遺伝子比率の不均衡を検出するために開発された(Chang,HW et al.2002 J Natl Cancer Inst 94,1697−1703)。臨床的に、それは腫瘍DNA試料中におけるヘテロ接合性の欠如(LOH)の検出において有用であることが示された(Zhou,W.et al.2002 Lancet 359,219−225)。実験結果が試料中におけるLOHの存在を示唆するものであるか否かを分類するため、先の試験においてデジタルPCRの結果分析のために逐次確率比検定(SPRT)が採用された(El Karoui at al.2006 Stat Med 25,3124−3133)。前記研究で使用された方法において、LOHを決定するカットオフ値には、2/3のDNAの2つの対立遺伝子の固定された参照比を使用した。羊水中の胎児性核酸の量、割合、及び濃度は様々であるため、これらの方法は、羊水中の母性核酸のバックグラウンド中の胎児性核酸を使用するトリソミー21の検出には適さない。
【0011】
循環性の(circulating)胎児核酸解析に基づく、特に遺伝的多型及び/又は胎児特異的マーカーの使用に依存しない、胎児のトリソミー21(及び他の不均衡)の非侵襲性検査が望ましい。また、正確を期するのに必要なデータのウェル数及び/又は母体血漿核酸分子の量を減少させるためには、カットオフ値の決定及び配列の計数を正確にするのが望ましく、それにより、効率及び費用対効果も増大する。また、非侵襲性検査は、診断ミスを最小にするべく高感度かつ特異的であることが望ましい。
【0012】
羊水中の胎児性DNAを検出する他の用途は、ベータサラセミア等の単独の遺伝子障害の出生前診断である。しかしながら、胎児性DNAは羊水中のDNAのごく一部に過ぎないため、このアプローチは、胎児が父親から受け継ぎ、母親には存在しない突然変異のみ検出することが出来ると考えられる。この突然変異の例として、ベータサラセミアを引き起こすベータグロビン遺伝子の41/42コドン中の4bpの欠失(Chiu RWK et al. 2002 Lancet, 360, 998-1000)、及び嚢胞性線維症を引き起こす嚢胞性線維症膜コンダクタンス制御因子遺伝子のQ890X突然変異(Gonzalez-Gonzalez et al 2002 Prenat Diagn, 22, 946-8)が挙げられる。しかしながら、ベータサラセミア及び嚢胞性線維症のいずれも、常染色体劣性条件であるため、それらの疾患が発症するには胎児がそれぞれの親から突然変異を受け継ぐ必要があり、父性遺伝突然変異の検出のみでは、胎児の罹患危険率を25%から50%まで上昇させ得るに過ぎない。これは、診断的に理想的なものではない。すなわち、既存のアプローチを診断に適用することは、羊水中に父親から受け継いだ突然変異が検出されず、故にその胎児がホモ接合性病態を有する可能性が排除されたときには、シナリオどおりに運用されたといえる。しかしながら、診断的に、このアプローチは、上記結果が、父性突然変異を検出しないことに基づくという短所を有する。故に、上記制限を有さず、羊水から、胎児の完全な遺伝子型(ホモ正常、ホモ突然変異、又はヘテロ接合)を決定することが出来るアプローチが極めて望ましい。
【発明の概要】
【課題を解決するための手段】
【0013】
簡潔な概要
本発明の態様は、生体試料中に核酸配列の不均衡(例えば、対立遺伝子の不均衡、突然変異の不均衡、又は染色体の不均衡)が存在するか否かを決定する方法、システム、及び装置を提供する。例えば2つの配列(又は二つのセットの配列)の量の比率等の、不均衡を決定する1つ以上のカットオフ値が選択される。
【0014】
一つの態様において、前記カットオフ値は、少なくとも部分的に、母体血漿又は血清又は尿等の、母性核酸配列のバックグラウンドを含有する生体試料中の胎児性(臨床関連核酸)配列のパーセンテージに基づき決定される。もう一つの態様において、前記カットオフ値は、複数の反応中の配列の平均濃度に基づき決定される。一つの側面において、前記カットオフ値は、特定の核酸配列を含有することが推定される情報的ウェル(informative well)の割合から決定される、ここで、該割合は、上記パーセンテージ及び/又は平均濃度に基づき決定される。
【0015】
前記カットオフ値は、SPRT、誤検出(false discovery)、信頼区間、受信者動作特性(ROC)等の、様々な種類の方法を使用して決定され得る。この方策は、更に、信頼分類(confident classification)が作成され得る前に必要な検査の量を最小化した。これは、通常鋳型の量が限定されている血漿核酸の解析において特に重要である。
【0016】
一つの例示的態様において、
生体試料内の核酸配列中の不均衡の存否を決定する方法であり、該方法は;
複数の反応からデータを取得し、ここで、該データが、
(1)臨床関連核酸配列の第一の量を示す第一のセットの定量データ;及び
(2)臨床関連核酸配列ではない基準核酸配列の第二の量を示す第二のセットの定量データ;
を含み;
該二つのセットのデータからパラメーターを決定し;
複数の反応それぞれにおける参照核酸配列の平均濃度から第一のカットオフ値を導き出し、ここで、該参照核酸配列は、臨床関連核酸配列又は基準核酸配列のいずれかであり;
該パラメーターと該第一のカットオフ値とを比較し;続いて、
該比較に基づき、核酸配列に不均衡が存在するか否かの分類を決定する;
ことを含む。
【0017】
もう一つの例示的態様において、
生体試料内の核酸配列中の不均衡の存否を決定する方法であり、該方法は;
複数の反応からデータを取得し、ここで、該データが、
(1)臨床関連核酸配列の第一の量を示す第一のセットの定量データ;及び
(2)臨床関連核酸配列ではない基準核酸配列の第二の量を示す第二のセットの定量データ;
を含み、ここで、該臨床関連核酸配列及び該基準核酸配列が、第一の種類の細胞及び1つ以上の第二の種類の細胞に由来し;
該二つのセットのデータからパラメーターを決定し;
生体試料中の第一の種類の細胞に由来する核酸配列の量の測定に由来する第一のパーセンテージから第一のカットオフ値を導き出し;
該パラメーターと該カットオフ値とを比較し;続いて、
該比較に基づき、核酸配列に不均衡が存在するか否かの分類を決定する;
ことを含む。
【0018】
本発明の他の実施形態は、本明細書に記載された方法と関連するシステム及びコンピュータで読み取り可能な媒体に関する。
【0019】
本発明の性質及び利点のさらなる理解は、以下の詳細な説明及び添付の図面を参照にして得られる。
【図面の簡単な説明】
【0020】
【図1】デジタルDNA実験を説明するフローチャートである。
【0021】
【図2A】本発明の一つの態様に係るデジタルRNA-SNP及びRCD法を説明する。
【0022】
【図2B】癌においてよく検出される染色体異常の一覧を示す。
【0023】
【図3】本発明の一つの態様に係る、ダウン症候群の決定に使用されるSPRT曲線を有するグラフを説明する。
【0024】
【図4】本発明の一つの態様に係る、胎児細胞のパーセンテージを使用する病態の決定方法を示す。
【0025】
【図5】本発明の一つの態様に係る、平均濃度を使用する病態の決定方法を示す。
【0026】
【図6】本発明の一つの態様に係る、予想されるデジタルRNA-SNP対立遺伝子比、並びにウェルあたりの平均参照鋳型濃度(mr)として表現した、広範な鋳型濃度におけるトリソミー21試料のPrの一覧を示す。
【0027】
【図7】本発明の一つの態様に係る、ウェルあたりの平均参照鋳型濃度(mr)として発現した広範な鋳型濃度におけるトリソミー21試料において、分画胎児性DNA濃度が10%、25%、50%、及び100%の場合の予想されるPrの一覧を示す
【0028】
【図8】本発明の一つの態様に係る、デジタルRNA-SNP解析における、mr値が0.1、0.5、及び1.0の場合の、SPRT曲線の相異の程度を説明するプロットを示す。
【0029】
【図9A】本発明の一つの態様に係る、96ウェルデジタルRNA-SNP解析における、正倍数体及びトリソミー21の症例を分類する新旧のSPRTアルゴリズムの有効性比較の表を示す。
【0030】
【図9B】本発明の一つの態様に係る、384ウェルデジタルRNA-SNP解析における、正倍数体及びトリソミー21の症例を分類する新旧のSPRTアルゴリズムの有効性比較の表を示す。
【0031】
【図10】本発明の一つの態様に係る、正しく、及び誤って分類された正倍数性又は異数性の、並びに所定の情報的カウント(informative count)で分類できない胎児のパーセンテージを示す表である。
【0032】
【図11】本発明の一つの態様に係る、純粋(100%)なDNA試料における、デジタルRCD解析のコンピューターシミュレーションを示す表1100である。
【0033】
【図12】本発明の一つの態様に係る、胎児性DNAの分画濃度が異なる、正倍数性又はトリソミー21胎児由来の試料を分類するための、mr=0.5でなされたデジタルRCD解析の正確性のコンピューターシミュレーションの結果を示す表1200である。
【0034】
【図13】図13Aは、本発明の一つの態様に係る、正倍数性及びトリソミー21妊婦由来の胎盤組織における田字たるRNA-SNP解析の表1300を示す。図13Bは、本発明の一つの態様に係る、正倍数性及びトリソミー21妊婦由来の母体血漿組織におけるデジタルRNA-SNP解析の表1350を示す。
【0035】
【図14】本発明の一つの態様に係る、RCD解析で得られたカットオフ曲線を説明するプロットを示す。
【0036】
【図15】図15Aは、本発明の一つの態様に係る、正倍数性及びトリソミー21妊婦由来の胎盤組織におけるデジタルRNA-SNP解析の表を示す。図15Bは、本発明の一つの態様に係る、1つの母体血漿試料に由来する12枚の反応パネルのデジタルRNA-SNPデータの表を示す。図15Cは、本発明の一つの態様に係る、正倍数性及びトリソミー21妊婦由来の母体血漿のデジタルRNA-SNP解析の表を示す。
【0037】
【図16】図16Aは、本発明の一つの態様に係る、正倍数性及びトリソミー18胎盤のデジタルRNA-SNP解析の表を示す。図16Bは、本発明の一つの態様に係る、正倍数性及びトリソミー18胎盤のデジタルRNA-SNPデータのSPRT解釈を示す。
【0038】
【図17】本発明の一つの態様に係る、正倍数性及びトリソミー21妊婦の50%胎盤/母体血球細胞DNA混合物の、デジタルRCD解析の表を示す。
【0039】
【図18】本発明の一つの態様に係る、分類の正誤の境界決定を描写する、SPRT曲線を示す。
【0040】
【図19】本発明の一つの態様に係る、正倍数性及びトリソミー21妊婦由来の羊水のデジタルRCD解析の表を示す。
【0041】
【図20】本発明の一つの態様に係る、正倍数性及びトリソミー18妊婦由来の胎盤DNA試料のデジタルRNC解析の表(E=正倍数性;T18=トリソミー18)を示す。
【0042】
【図21】本発明の一つの態様に係る、正倍数性及びトリソミー18妊婦由来の、50%胎盤/母体血球細胞DNA混合物の、多重デジタルRCD解析の表(E=正倍数性;T21=トリソミー21;U=分類不能)を示す。
【0043】
【図22A】本発明の一つの態様に係る、50%正倍数性又はトリソミー21胎盤ゲノムDNA/50%母体軟膜DNA混合物の、多重デジタルRCD解析の表を示す。
【0044】
【図22B】本発明の一つの態様に係る、50%正倍数性又はトリソミー21胎盤ゲノムDNA/50%母体軟膜DNA混合物の、多重デジタルRCD解析の表を示す。
【0045】
【図23】両親とも同一の突然変異を担持していた場合のシナリオを示す。
【0046】
【図24】図24Aは、本発明の一つの態様に係る、女性/男性、及び男性/男性のDNA混合物の、デジタルRMD解析の表を示す。図24Bは、本発明の一つの態様に係る、25%が女性由来、75%が男性由来のDNAの混合物の、デジタルRMD解析の表を示す。
【0047】
【図25】本発明の一つの態様に係る、HbE突然変異の母体血漿試料を模した15%〜50%DNA混合物の、デジタルRMD解析の表を示す。
【0048】
【図26】図26Aは、本発明の一つの態様に係る、CD41/42の突然変異の母体血漿試料を模した5%〜50%DNA混合物の、デジタルRMD解析の表を示す。図26Bは、本発明の一つの態様に係る、CD41/42の突然変異の母体血漿試料を模した20%DNA混合物の、デジタルRMD解析の表を示す。
【0049】
【図27】本発明の一つの態様に係るシステム及び方法に有用なコンピューター装置の一例のブロック図を示す。
【発明を実施するための形態】
【0050】
定義
本明細書において使用される用語「生体試料」は、対象(例えば、妊娠女性のようなヒト)から採取される任意の試料のことであり、そして1つ以上の目的の核酸分子を含む。
【0051】
用語「核酸」又は「ポリヌクレオチド」は、デオキシリボ核酸(DNA)又はリボ核酸(RNA)、及び一本又は二本鎖のいずれかの形態であるそれらのポリマーのことである。具体的に限定されない限り、当該用語は、基準核酸と同程度の結合特性を有する、既知の天然ヌクレオチドアナログを含む核酸を含み、そして天然に存在するヌクレオチドに同様の方法で代謝される。他に示されない限り、特定の核酸配列はまた、その保存的に改変された改変体(例えば、縮重コドン置換)、対立遺伝子、オルソログ、SNPs、及び相補的配列、ならびに明確に示された配列を暗黙的に含む。具体的には、縮重コドン置換は、一つ以上選択された(又は全ての)コドンの第三の位置が混合塩基及び/又はデオキシイノシン残基で置換されている配列を生じさせることによって達成され得る(Batzer et al.,Nucleic Acid Res.19:5081(1991);Ohtsuka et al,J.Biol.Chem.260:2605−2608(1985);及びRossolini et al, Mol.Cell.Probes 8:91−98(1994))。核酸という用語は、遺伝子、cDNA、mRNA、低分子ノンコーディングRNA、マイクロRNA(miRNA)、Piwi相互作用RNA、及び遺伝子又は遺伝子座によってコードされたショートヘアピンRNA(shRNA)にも、交換可能的に使用される。
【0052】
用語「遺伝子」は、ポリペプチド鎖を産生することに関与するDNA断片を意味する。それは、コード領域に先行する、及びコード領域に続く領域(リーダー及びトレイラー)、ならびに個々のコード断片(エキソン)間の介在配列を含み得る。
【0053】
本明細書において使用される用語「反応」は、特定の目的ポリヌクレオチド配列の存在又は不在を示す、化学的、酵素的、又は物理的作用に関与する任意の工程のことである。「反応」の例は、例えばポリメラーゼ連鎖反応(PCR)のような増幅反応である。「反応」の別の例は、合成又はライゲーションのいずれかによる配列決定の反応である。「情報的反応(informative reaction)」とは、一つ以上の特定の目的ポリヌクレオチド配列の存在を示す反応を意味し、そして場合によっては、唯一つの目的ポリヌクレオチドが存在することを示す反応を意味する。本明細書において使用される用語「ウェル」は、密閉された構造、例えばPCRアレイにおける、ウェル形状バイアル、セル、又はチャンバー内のあらかじめ決められた位置における反応のことである。
【0054】
本明細書において使用される用語「臨床関連核酸配列」は、その潜在的不均衡が試験される、より大きな遺伝子配列断片に相当するポリヌクレオチド配列のことか、又はそのより大きな遺伝子配列自体に相当するポリヌクレオチド配列のことであり得る。一つの実施例は、第21番染色体の配列である。他の例は、第18番、第13番、X及びY染色体を含む。さらに他の例は、胎児がその親の片方又は両方から受け継いているかもしれない変異遺伝子配列、又は遺伝子多型、又はコピー数変異を含む。さらに他の実施例は、悪性腫瘍中において変異した、欠失した、又は増幅された配列、例えばヘテロ接合性の欠損又は遺伝子重複が起こっている配列を含む。幾つかの態様において、複数の臨床関連核酸配列、又は臨床関連核酸配列の同等の複数のマーカーが、当該不均衡を検出するためのデータを提供するために使用され得る。例えば、第21番染色体上の5つの不連続配列からのデータは、可能性のある第21番染色体不均衡の決定のための付加的な方法において使用され得、効果的に試料の容量を1/5へと減少させる。
【0055】
本明細書において使用される用語「基準核酸配列」は、正常な場合、臨床関連核酸配列に対する比率が既知、例えば1:1の比率である核酸配列のことである。一つの例として、基準核酸配列及び臨床関連核酸配列は、ヘテロ接合性により明確に区別し得る、同一の染色体由来の2つの対立遺伝子である。別の例において、基準核酸配列は1つの対立遺伝子であり、これに対してヘテロ接合性である別の対立遺伝子が、臨床関連核酸配列である。さらに、各々の基準核酸配列及び臨床関連核酸配列の幾つかは、異なる個体由来であってもよい。
【0056】
本明細書において使用される用語「参照核酸配列」は、一反応あたりの平均濃度が既知又は同等と測定された核酸配列のことである。
【0057】
本明細書において使用される用語「過剰出現核酸配列」は、生体試料中における他の配列以上に豊富である、目的の2つの配列(例えば臨床関連配列及び基準配列)に混在する核酸配列のことである。
【0058】
本明細書において使用される用語「に基づく」は、「少なくとも一部に基づく」ことを意味し、そして、一つの方法の入力とその方法の出力との関係において生じる、一つの値の決定において使用される別の値(又は結果)のことをいう。本明細書において使用される用語「導く」はまた、一つの方法の入力とその方法の出力との関係のことであり、例えば誘導が公式計算であるときに起こる。
【0059】
本明細書において使用される用語「定量的データ」は、一つ以上の反応から得られ、そして一つ以上の数値を提供するデータを意味する。例えば、特定の配列のための蛍光マーカーを示すウェルの数が、定量的データであろう。
【0060】
本明細書において使用される用語「パラメーター」は、定量的データのセットを特徴づける数値、及び/又は定量的データセット間における数的関係を意味する。例えば、第一の核酸配列の第一の量と第二の核酸配列の第二の量との間における比率(又は比率の関数)はパラメーターである。
【0061】
本明細書において使用される用語「カットオフ値」は、生体試料の分類の2つ以上の状態(例えば罹患した状態と罹患していない状態)間を裁定するために使用される数値を意味する。例えば、パラメーターが当該カットオフ値よりも大きいならば、定量的データの第一の分類がなされ(例えば罹患した状態);又はパラメーターがカットオフ値よりも小さいならば、定量データの異なる分類がなされる(例えば罹患していない状態)。
【0062】
本明細書において使用される用語「不均衡」は、臨床関連核酸配列量における少なくとも一つのカットオフ値によって定義される、基準量からの任意の有意な偏差を意味する。例えば、参照量の比率が3/5のとき、測定比が1:1である場合、不均衡が生じているといえる。
【0063】
発明の詳細な説明
分発明は、生体試料中に存在する、臨床的関連核酸配列の量が、他の非臨床関連核酸配列との関連で、参照(例えば正常体)と比較して増大したか又は減少したか(例えば、染色体又は対立遺伝子の不均衡)を判定する方法、システム、及び装置を提供する。参照量と比較して、例えば2つの配列の量の比率等に関して変化が存在する(即ち不均衡)か否かを判定するために、1つ以上のカットオフ値が決定される。参照量において検出される変化は、臨床的に重要な核酸配列と、他の臨床的に重要ではない配列との関係における任意の偏差(上向きまたは下向き)であり得る。したがって、基準状態は、任意の比または他の量(例えば1−1対応以外)であり得、そして変化を示す測定された状態は、一つ以上のカットオフ値によって決定される基準量とは異なる任意の比または他の量であり得る。
【0064】
臨床的に重要な染色体領域(臨床的に重要な核酸配列とも呼ばれる領域)およびバックグラウンド核酸配列は、第一のタイプの細胞由来であり得、そして一つ以上の第二のタイプの細胞由来であり得る。例えば、胎児/胎盤細胞由来の胎児核酸配列は、母体細胞由来の母体核酸配列のバックグラウンドを含む生体試料、例えば母体血漿中において存在する。一つの実施形態において、カットオフ値は、生体試料中における第一のタイプの細胞のパーセンテージに少なくとも一部基づいて決定される。試料中における胎児配列のパーセンテージは、任意の胎児由来遺伝子座によって決定され得、そして臨床的に重要な核酸配列を測定するために限定されないことに留意すべきである。別の実施形態において、カットオフ値は、体内における非悪性細胞由来の核酸配列のバックグラウンドを含む生体試料、例えば血漿、血清、唾液、または尿における腫瘍配列のパーセンテージに少なくとも一部基づいて決定される。
【0065】
なおももう一つの態様において、前記カットオフ値は、複数の反応における配列の平均濃度に基づいて決定される。一つの側面において、該カットオフ値は、特定の核酸配列を含有することが推定される、情報的ウェルの割合から決定され、ここで、該割合は、上記パーセンテージ及び/又は平均濃度に基づき決定される。SPRT、誤検出、信頼区間、受信者操作特性(ROC)等の、様々な種類の方法を使用して、前記カットオフ値が決定され得る。この方策は、更に、信頼分類が作成され得る前に必要な検査の量を最小化する。これは、通常鋳型の量が限定されている血漿核酸の解析において特に重要である。デジタルPCRに関連して提示されているが、他の方法も使用され得る。
【0066】
デジタルPCRは、核酸を極限まで希釈して、殆どの陽性増幅が単一の鋳型分子に由来するシグナルを反映するように、複数のPCR解析がなされる。従って、デジタルPCRは、個々の鋳型分子の計数を可能とする。解析されたPCRの合計数中で、正の増殖が見られたものの割合から、元の試料あるいは希釈していない試料中の鋳型濃度を推定することができる。この技法は、様々な遺伝的現象の検出が可能なことが示されており(Vogelstein, B et al. 1999, supra)、既に、腫瘍試料中(Zhou, W. et al. 2002, supra)、及び癌患者血漿中(Chang, HW et al. 2002, supra)のヘテロ接合性の消失の検出に使用されている。デジタルPCRによる鋳型分子の定量は、レポーター色素及び核酸濃度との間で容量依存的応答関係にないので、その解析精度は、理論的には、リアルタイムPCRを超える。従って、デジタルPCRは、潜在的には、標的と参照遺伝子座との間のより微細な程度の量的差異の判別を可能とし得る。
【0067】
これを試験するために、発明者らはまず、デジタルPCRが、PLCA4 mRNA(Lo, YMD, et al. 2007 Nat Med 13, 218-223)の対立遺伝子の比率を判定可能であるかを検証した。該mRNAは第21番染色体由来の胎盤性転写産物であり、母体血漿中に存在するため、胎児のトリソミー21及び正倍数体を峻別することが出来る。このアプローチは、デジタルRNA-SNP法と称される。それから、発明者らは、デジタルPCRの精度増大により、遺伝的多型に依存しない胎児染色体の異数性の検出が可能となるか否かを検証した。発明者らは、これを、デジタル相対的染色体量(RCD)解析と称する。前者のアプローチは多型依存的であるものの定量的識別能の要求は低く、一方で後者のアプローチは、多型非依存的であるが、高精度の定量的識別能を要求する。
【0068】
I.デジタルRNA-SNP
A.概要
デジタルPCRは、DNA試料において2つの対立遺伝子における存在比の非対称性を検出することができる。例えば、腫瘍DNA試料におけるヘテロ接合性の消失(LOH)の検出の使用されている。DNA試料中に2つの対立遺伝子A及びGが存在しているとする。そして、対立遺伝子Aが、LOH細胞において消失し得るとする。LOHが腫瘍試料中の細胞の50%の発生するとき、該DNA試料における対立遺伝子比G:Aは、2:1となり得る。しかしながら、腫瘍試料でLOHが発生しない場合、対立遺伝子比G:Aは、1:1となり得る。
【0069】
図1は、デジタルPCR実験を説明したフローチャート100である。工程110において、DNA試料を希釈し、続いて個別のウェルに分注する。ここで留意すべきなのは、発明者らが、幾つかの種類の核酸は、元の試料中で既に多分に希釈されていることを特定したことである。このように、目的の鋳型が既に必要な濃度で存在しているときは、希釈を必要としない。以前の研究で、あるDNA試料は、特定の「鋳型DNA」の平均濃度が、ウェルあたり約0.5分子、即ちウェル2つに約1つの鋳型が入る程度にまで希釈されている(例えば、Zhou et al 2002を参照されたい)。ここで留意すべきなのは、鋳型DNAが、対立遺伝子A又はGのいずれかに相当し、この特異的濃度に与えられる論理的根拠は存在しないことである。
【0070】
工程120において、各ウェル中で、PCRプロセスが実行され、対立遺伝子A及び/又はBが同時に検出される。工程130において、各ウェル中のマーカー(蛍光等)が同定され、ウェル中にA若しくはGのいずれか一方が入っているか、又はそれらのいずれも入っているか、若しくはいずれも入っていないかが判定される。LOHが起こらない場合、DNA試料中の対立遺伝子A及びGの量は同一であろう(細胞1個に1コピー)。従って、対立遺伝子Aが陽性であるウェルの頻度及び対立遺伝子Gが陽性であるウェルの頻度は同一となるであろう。しかしながら、腫瘍細胞の50%以上でLOHが発生した場合、対立遺伝子G及びAの存在比率は、少なくとも2:1となるであろう。従来の方法によれば、この試料は、少なくとも50%は癌性であると単純に推定されていた。すなわち、対立遺伝子Gが陽性のウェルの出現頻度が、対立遺伝子Aが陽性のウェルの出現頻度より高い。結果として、対立遺伝子Gが陽性のウェルの個数は、対立遺伝子Aが陽性のウェルの個数より多くなる。
【0071】
工程140において、デジタルPCRの結果を分類するために、各対立遺伝子の一方が陽性で他方が陰性であるウェルの個数が使用され得る。上記例において、対立遺伝子Aが陽性であるが対立遺伝子Gが陰性であるウェルの個数、及び対立遺伝子Gが陽性であるが対立遺伝子Aが陰性であるウェルの個数が計数される。一つの態様において、陽性のウェルがより少ない対立遺伝子が、参照対立遺伝子とみなされる。
【0072】
工程150において、情報的ウェルの総数は、2つの対立遺伝子のいずれかが陽性のウェルの個数の合計として決定される。工程160において、より多くの陽性ウェルを有する対立遺伝子に関する情報的ウェルの割合(Pr)(パラメーターの一種)が計算される。Pr=(より多くの陽性のウェルを有する対立遺伝子のみが陽性のウェルの個数/1つの対立遺伝子(A又はG)のみが陽性のウェルの総数)となる。他の態様として、少なくとも1つの対立遺伝子を有する全てのウェルにより分別された対立遺伝子の1つを有するウェルを使用し得る。
【0073】
工程170において、Prの値が対立遺伝子の不均衡を示すか否かを決定する。正確性と効率性が求められるので、この課題は簡単なものではない。不均衡を決定する1つの方法として、Bayesian型尤度法、逐次確率比検定(SPRT)が使用される。SPRTは、2つの確率的仮説をデータの蓄積により比較することを可能とする方法である。言い換えると、これは、対立遺伝子の歪みの存否を示唆するものとして、デジタルPCRの結果を分類する統計学的方法でああある。これは、解析すべくウェルの個数を最少としつつ、所定の統計的鹸出力及び精度を達成するのに有利である。
【0074】
例示的SPRT解析において、実験結果を、帰無仮説及び対立仮説に対して検定され得る。対立仮説は、試料中で対立遺伝子比に歪みが存在するときに許容される。帰無仮説は、試料中で対立遺伝子比に歪みが存在しないときに許容される。前記Prは、帰無仮説又は対立仮説を許容する2つのカットオフ値と比較され得る。いずれの仮説も許容されなければ、そのサンプルは、分類不能としてマークされ、これは、観察されたデジタルPCRの結果が、所望の統計的信頼性を有する試料を分類するのに不十分であることを意味する。
【0075】
帰無仮説又は対立仮説を許容するカットオフ値は、典型的には、それらの仮説において主張される仮定の下の固定されたPr値に基づき計算されている。帰無仮説において、試料は対立遺伝子比の歪みを示していないと想定される。従って、対立遺伝子Aが陽性であるウェルの頻度及び対立遺伝子Gが陽性であるウェルの頻度は同一であり、従って、Pr値は1/2と予想される。対立仮説において、Pr値は、2/3、又は0.5と2/3の中間、例えば0.585と予想される。また、実験数が限定されるため、上限(.585+3/N)及び下限(.585-3/N)が選択され得る。
【0076】
B.ダウン症候群の検出
本発明の一つの態様において、妊婦血漿から胎児のダウン症候群を検出するために、デジタルSNPが使用される。胎児/胎盤細胞に特異的なマーカーを使用して、第21番染色体中の対立遺伝子の比率が測定され得る。例えば、観察されたPLAC4対立遺伝子の過剰出現の度合が統計的に有意か否かを決定するために、SPRTが使用される。
【0077】
一つの例示的な態様において、デジタルRNA-SNPは、第21番染色体から転写され、胎盤により発現されるPLAC4 mRNA上に位置する、A/G SNPの多型的対立遺伝子の比率の不均衡を決定する。ヘテロ接合性正倍数性胎児において、対立遺伝子A及びGは、胎児ゲノム中で等しく表示されるべきである(ゲノム比1:1)ところ、トリソミー21において、トリソミー第21染色体は、胎児ゲノム中でSNP対立遺伝子の一つを追加的に1コピー有し、これが2:1のゲノム比をもたらす。デジタルPCR解析の目的は、解析される試料中のPLAC4対立遺伝子の量が等しいか否かを決定することである。従って、PLRC4の対立遺伝子A及びGのいずれも標的の鋳型である。PLAC4 mRNAを増幅するためにリアルタイムPCRアッセイが設計され、TaqMan蛍光プローブにより、2つのSNP対立遺伝子が識別された。図2Aに、解析工程の模式図を示す。
【0078】
図2Aは、本発明の態様におけるデジタルRNA-SNP法200を説明する。工程210において、試料が調製される。工程220において、核酸配列、例えば抽出されたRNA試料中のPLAC4 mRNAが定量される。一つの態様において、この工程は、標的がデジタルPCR解析の「領域」に達するのにいかほどの希釈が必要であるかの知見を実施者に提供する。
【0079】
工程230において、試料が希釈される。工程240において、希釈された試料の濃度が測定される。希釈された試料の濃度が、〜1鋳型(即ち参照若しくは非参照配列、又はいずれかの対立遺伝子)/ウェルとなっていることが確認される。幾つかの態様は、この測定のために、第IV章に記載される技術を使用する。例えば、発明者らは、前記希釈された試料を、リアルタイムPCR用に96ウェルに分注し、所望の希釈が達成されていることを確認した。希釈濃度は未知のままであり得るので、この工程を省略している態様も後に記載する。
【0080】
工程250において、アレイの各ウェルにおいて、デジタルPCRが実行される。例えば、等しく希釈された試料が、リアルタイムPCR解析用384ウェルに分注された。PCRの結果から、各核酸配列用のマーカーの量及び情報的ウェルの個数が同定される。情報的ウェルは、対立遺伝子A又はBのいずれかが陽性であるが、両方が陽性ではないウェルを意味する。工程260において、予想されるRrの値の計算が実行される。これらの工程は、後で詳述する。前記計算は、工程250において決定された数値からパラメーターを決定することを含む。例えば、ウェルあたりの実際の平均鋳型濃度が計算され得る。
【0081】
工程270において、SPRT又は他の尤度比が実行され、不均衡の存否が決定される。正倍数体の場合、発明者らは、A陽性及びG陽性のウェルの個数が等しくなると予想する。しかしながら、トリソミー21胎児由来の鋳型分子を解析すると、一方の対立遺伝子を1つだけ含むウェルの個数は、他方の対立遺伝子を1つだけ含むウェルの個数よりも多くなるはずである。要するに、トリソミー21には、対立遺伝子の不均衡が予想される。
【0082】
上記のように、SPRTは、2つの確率的仮説をデータの蓄積として比較することを可能とするBayesian型尤度法である。トリソミー21検出用デジタルPCR解析において、対立遺伝子の武均衡が存在する(即ちトリソミー21が検出された)とき、対立仮説が許容され;対立遺伝子の不均衡が存在しない(即ちトリソミー21が検出されない)とき、帰無仮説が許容される。多数派の対立遺伝子は、潜在的過剰出現対立遺伝子と称され、情報的ウェル全ての中に占めるその割合(Pr)が計算され得る。SPRTは、Prがトリソミー21試料で予想される十分な程度の対立遺伝子の不均衡を同定するか否かを決定するのに応用される。
【0083】
操作上、SPRTは、いずれかの仮説を許容又は排除する確率的境界を規定するために構築されるSPRTカーブのペアを有するグラフの使用を通じて、応用及び解釈され得る。図3は、本発明の一つの態様における、ダウン症候群を決定するためのSPRT曲線を有するグラフを説明する。これらのSPRT曲線は、信頼区分が作成され得るとき、所定の情報的ウェルの総数(x軸)に対する、潜在的に過剰出現対立遺伝子が陽性である情報的ウェル数の割合Pr(y軸)を描画している。図3で描かれる如く、上方の曲線は、対立仮説を許容する確率的境界を設定し;下方の曲線は、帰無仮説を許容する確率的境界を設定する。
【0084】
実験的に導き出されたPr値は、予想されたPr値と比較され、いずれかの仮説を排除又は許容する。もし帰無仮説が許容されると、それらの試料が由来する妊婦は、正倍数体胎児を妊娠しているのとして分類される。もし対立仮説が許容されると、それらの試料が由来する妊婦は、トリソミー21胎児を妊娠しているものとして分類される。あるいは、もし、所定の情報的カウントのPrが、疾患の分類における統計的信頼に要求されるレベルに達していないとき、いずれの仮説も許容され得ない。これらのケースは、更にデータが入手出来なければ、分類不可能とみなされた。疾患分類が不可能である場合、データが蓄積されてSPRTにより分類可能となるまでは、追加的な384ウェルプレートが実行され得る。
【0085】
故に、SPRTは、他の統計的方法よりも、所定のレベルの信頼性を達成するのにより少数の検査で足りる点が有利である。実用語で言えば、SPRTは、所望の量のデータが蓄積され次第いずれかの仮説を許容又は排除することが出来るので、不要な追加的解析を最少限にすることが出来る。この特性は、入手可能な鋳型分子の数が限定される血漿中の、一般に低濃度で存在する核酸の解析において特に重要である。厳格な分類に加えて、分類は、パーセント精度のものが含まれる場合もある。例えば、カットオフ値との比較から得られた分類は、試料が特定のパーセンテージで核酸配列が不均衡である可能性を示し、又は同様に、決定された不均一性が、所定のパーセンテージ程度の正確性を有している。
【0086】
突然変異、あるいは遺伝的多形に関して胎児の遺伝子型を決定するために、母体血漿又は血清中の胎児核酸を使用して、同様のアプローチが適用され得る。ここで、胎児のゲノムの半分は母親から受け継いでいることを想起されたい。例として、2つの対立遺伝子A及びBを有する特定の遺伝子座を想定する。母親がヘテロ接合の遺伝子型ABであったとすると、その胎児の遺伝子型は、理論上、AA、BB、又はABとなる。胎児の遺伝型がAB、即ち母親の遺伝型と同一のとき、母体血漿中には、遺伝子型ABの核酸ばかり(母親及び胎児由来)が存在するだろう。よって、核酸又は対立遺伝子の均衡は、母体血漿において確認される。他方、胎児の遺伝型がAA又はBBであるとき、母体血清中で、対立遺伝子A又はBのいずれかが過剰出現し、対立遺伝子の不均衡が生じるだろう。この検討は、疾患を引き起こす突然変異(例えば、ベータサラセミア又は脊髄性筋萎縮症)にも適用可能であり、その場合、Aは、野生型対立遺伝子とみなされ得て、そしてBは、突然変異対立遺伝子とみなされ得る。
【0087】
II.デジタルRCD
デジタルRNA-SNPの短所は、解析されるSNPにおいてヘテロ接合性である場合に限り適用され得ることである。循環性胎児核酸解析に基づく、胎児トリソミー21又は他の胎児染色体異数性(例えば、トリソミー18、13、及び性染色体異数性等)用の非侵襲性検査が、遺伝的多形の使用と独立しているのが理想的であり得る点が改善点である。従って、一つの態様において、参照染色体(本文中では、第1番染色体と称する)上に位置する遺伝子座に対する非多型第21番染色体遺伝子座のデジタルPCR解析により、染色体量が決定される。第21番染色体と第1番染色体との比率は、正倍数体胎児のゲノムにおいては2:2であるが、トリソミー21の場合、これが変化する。トリソミー21検出用デジタルPCR解析において、比較されるべき2つの仮説は、染色体不均衡が存在しない(即ちトリソミー21が検出されない)帰無仮説、及び染色体不均衡が存在する(即ちトリソミー21が検出される)対立仮説であり得る。
【0088】
このアプローチは、他の染色体異数性、例えばトリソミー18における第18番染色体、トリソミー13における第13番染色体、ターナー症候群におけるX染色体等を含む、他の染色体に一般化され得る。加えて、第1番染色体と別に、異数性に関係の無い他の染色体も、参照染色体として使用され得る。通常癌において参照染色体と比較して部分的に起こる、染色体の欠失の比率の変化を解析することにより、同様のアプローチを、癌の検出に適用することも出来る。前者の例として、直腸結腸癌における染色体5q、肺癌における染色体3p、及び鼻咽腔癌における染色体9pを含む。図2Bは、配列の不均衡をもたらす、幾つかの通常の癌関連染色体異常を列挙したものである。
【0089】
また、図2Aは、本発明の態様におけるデジタルRCD法205を説明する。工程220〜230の一つの態様において、抽出されたDNAは、例えばナノドロップ技術により定量され、1つのウェル第21番染色体又は正規化(normalizing)染色体(第1番染色体等)のいずれかに由来する約1個の標的鋳型が含まれる濃度まで希釈される。工程240の一つの態様において、確認は、96ウェルの形式においてのみ、第1番染色体を使用したアッセイにより、希釈したDNA試料を解析することにより実行され得て、384ウェルプレートにおける両TaqManプローブを使用したRCD解析を進行する前に、陰性のウェルが37%未満であるか否かを確認する。37%の重要性は、第IV章で述べる。
【0090】
工程240の検査及び工程250の結果は、TaqManプローブのペアにより区別されるパラロガス配列変異により識別される両染色体上に存在するパラロガス配列を増幅するように設計されたリアルタイムPCRアッセイを用いてなされ得る。これに関連して、情報的ウェルは、第21番染色体又は第1番染色体遺伝子座のいずれかが陽性であるが、両方が陽性ではないものとして定義される。正倍数体の胎児において、いずれかの遺伝子座が陽性である情報的ウェルの個数は、概ね等しくあるべきである。トリソミー21胎児において、第21番染色体が陽性のウェルは、第1番染色体の場合よりも過剰出現すべきである。過剰出現の正確な割合は、後の章で述べる。
【0091】
III .胎児配列パーセンテージの組込み
上記方法200及び205の態様の短所は、胎児特異的なマーカーが必要なことである。従って、本発明の1つの態様において、非胎児特異的マーカーが使用される。そのような非胎児特異的マーカーを使用するために、本発明の態様は、母体血漿(即ち生体試料)中の胎児DNAの分別濃度(fractional concentration)を測定する。この情報を用いて、後述のように、Prのより有用な数値が計算される。
【0092】
母体血漿中で胎児DNAの分別パーセンテージが小さくとも、トリソミー21胎児は、胎児DNAのゲノム当量(GE)あたりの追加的な量の第21番染色体の、母体血漿中への放出に関与する。例えば、全DNAが50GE/mlで、その内胎児に由来するDNAが5GE/mlである(即ち胎児DNAの分別濃度は10%)、正倍数体の妊娠に由来する材料血漿試料は、材料血漿1mlあたり総数100コピーの第21番染色体(材料由来が90コピー+胎児由来が10コピー)配列を含有すべきである。トリソミー21妊娠において、各胎児性GEは、第21番染色体を3コピー有するため、母体血漿中の第21番染色体配列は、合計で105コピー(材料由来が90コピー+胎児由来が15コピー)となる。よって、胎児DNA濃度が10%において、三染色体性妊娠の母体血漿中の第21番染色体由来の配列の量は、正倍数体の場合の1.05倍になるべきである。故に、分析アプローチが小程度の量的層異を決定するように発達すれば、胎児トリソミー21の非侵襲性出生前診断用の、多型性独立的検査が達成され得る。
【0093】
従って、過剰出現の度合いは、解析されるDNA試料中の胎児DNA分別濃度に依存し得る。例えば、胎盤DNAが解析されたとき、胎児ゲノム中の理論上のRCD比率は3:2、即ち1.5倍異なるべきである。しかしながら、上記の如く、胎児性DNAを10%含有する母体血漿試料を解析した場合、理論上のRCD比率は1.05倍まで低下し得る。実験的に導き出されたPrは、情報的ウェルの総数で、第21番染色体遺伝子座のみ陽性であるウェルの個数を割ることにより計算される。この実験的に導き出されたPrは、計算されたPr及び前記理論上のRCD比率と共に、SPRT解析に供される。
【0094】
図4は、本発明の一つの態様における、胎児核酸のパーセンテージを使用した病態の判定方法400を示す。工程410において、胎児由来分子の分別パーセンテージが測定される。一つの態様において、分別パーセンテージは、非胎児特異的マーカー(即ち母親及び胎児のいずれにも存在する遺伝子配列)との関係における、胎児特異的マーカー(例えば、Y染色体、遺伝的多型(SNP等)、胎盤エピジェネティックサイン(placental epigenetic signature)等)の量を測定することにより判定される。リアルタイムPCR、デジタルPCR、シークエンシング反応(超並列ゲノムシークエンシングを含む)又は他の任意の定量法を使用して、正確な測定がなされ得る。一つの側面において、潜在的に測定自体により対立遺伝子の不均衡を生じさせ得る遺伝子ターゲティングは、使用しないのが好ましい。
【0095】
工程420において、デジタルPCR又は他の測定方法が実行され、この工程は、試料の希釈、希釈された試料のウェルへの分注、及び各ウェル内の反応の測定を含む。工程430において、相異する参照核酸配列(染色体又は対立遺伝子)のマーカーを同定するのに、PCRの結果が使用される。工程440において、過剰出現配列の正確な割合(Pr)が計算される。工程450において、病態を判定するためのカットオフ値が、試料中の胎児由来分子のパーセンテージを使用して計算される。工程460において、前記正確なPr及びカットオフ値から、不均衡の存否が判定される。
【0096】
一つの態様において、参照核酸配列の分別パーセンテージは、デジタルRNA-SNP法に組み込まれる。よって、癌細胞によるLOHを調査するとき、これは、腫瘍試料中癌細胞が50%未満の状態で行われ得る。より正確なPrを取得するのに、50%以上が癌細胞である腫瘍試料も使用され得て、これは、誤診断をもたらす擬陽性の数を減少させる。もう一つの態様において、胎児核酸のパーセンテージは、胎児が親の遺伝子突然変異(例えば、嚢胞性線維症、ベータサラセミア、又は脊髄性筋萎縮症を引き起こすもの等)又は多型を受け継いでいるか否かを、母体血漿核酸の解析から判定するために、デジタルPCR法に組み込まれる。
【0097】
IV.ウェルあたりの平均濃度の組込み
従来の方法(例えば、上記Zhou, W. et al. 2002を参照されたい)のもう一つの短所は、鋳型の濃度を、1ウェルに1個となるようにする必要があることである。正確な濃度を求めるのが困難である場合、これは誤差を生じる場合がある。更に、ウェルあたり鋳型1個となるように正確な濃度に調整しても、従来の方法、即ち古いアルゴリズムでは、対立仮説を許容するために予想されたPr値は、対立遺伝子の比率であり、ウェルあたりの平均の鋳型濃度から独立している。
【0098】
しかしながら、希釈された試料中の鋳型の自然の統計的変動のために、実際にはウェルあたり1個の鋳型が存在するとは限らない。本発明の態様は、少なくとも1つの配列の経筋濃度を測定し、そしてその測定値がカットオフ値、即ち予想されるPrを計算するのに使用される。一つの態様において、この計算は、異なる複数の核酸配列を含むウェルの確率を決定するための統計的分布を含み、そしてこれは、予想されるPrを決定するのに使用される。
【0099】
一つの態様に置いて、平均濃度は、1つの参照核酸配列から得られ、この配列は、例えば、DNA試料中でより低い濃度で存在する核酸配列である。不均衡が存在しない場合、試料中の2つの配列の濃度は同一のはずであり、いずれか1つが、参照対立遺伝子とみなされる。試料が、例えばLOHを有している場合、癌細胞中で欠失した対立遺伝子が、参照対立遺伝子とみなされ得る。参照対立遺伝子の平均濃度は、mrと表記される。もう一つの態様において、より高濃度で存在する配列を、参照配列として扱う。
【0100】
A.デジタルSNP.SPRT及びデジタルPCRの使用例
図5は、本発明の一つの態様における、平均鋳型濃度を使用した、病態判定方法500を示す。工程510において、異なる複数の配列の量が測定される。これは、例えば、上記の如く、デジタルPCR実験においてマーカーを計数すること等によりなされ得る。しかしながら、本方法は、増幅工程を含まない、又は蛍光マーカーを使用せず、質量等の物理的特性、特定の光学的特性、又は塩基対特性等の他の特性を使用することが出来る他の方法によりなされ得る。
【0101】
工程520において、過剰出現配列の実際の割合が決定される。これは、上記のように、その配列のみを示すウェルの個数をとり、情報的ウェルの個数により区別されることによりなされ得る。工程530において、少なくとも1つの配列(参照配列)の平均濃度が測定される。一つの態様において、この参照配列は、過剰出現配列である。もう一つの態様において、この参照配列は、過小表示した(underrepresented)配列である。測定は、デジタルPCR実験において参照配列が陰性であるウェルの個数を計数することによりなされ得る。陰性のウェルの割合と平均鋳型濃度との間の関係は、次の小区分に記載のポアソン分布により記述されている。
【0102】
工程540において、異なる複数の配列が陽性であるウェルの予想量が計算され、例えばポアソン分布が使用される。この予想量は、ウェルあたりの配列の確率(probability)、ウェルあたりの平均配列、その配列を含むウェルの個数、又は他の任意の適切な量としてあり得る。工程550において、予想されるPrが、前記予想量から計算される。工程560において、カットオフ値が銭予想されるPrから計算され、例えば、SPRTを使用して計算される。工程570において、核酸配列の不均衡の分類が判定される。方法500の特別の側面が、ここで記載される。
【0103】
配列の予想量の決定
工程530からウェルあたりの平均濃度が知られれば、工程540において、その配列を示すウェルの予想される個数が計算される。この量は、%、少数値、又は整数値で表現され得る。説明のため、参照鋳型配列の平均濃度(mr)が0.5であり、PLAC4 SNP、rs8130833におけるトリソミー21の胎児の遺伝子型はAGGとする。故に、参照鋳型は対立遺伝子Aとし、過剰出現鋳型は対立遺伝子Gとする。
【0104】
一つの態様において、ポアソン分布は、デジタルPCR等の測定手順のウェルの反応混合物中の、対立遺伝子Aの分布であるとする。他の態様において、二項分布等の他の分布関数が使用される。
【0105】
ポアソン方程式は:
【化1】
で表され、ここで、nは、ウェルあたりの鋳型分子の個数;P(n)は、特定のウェル中の鋳型分子の確率;そしてmは、特定のデジタルPCR実験における1つのウェル中の鋳型分子の平均数である。
【0106】
従って、対立遺伝子Aを平均濃度0.5である場合、対立遺伝子Aの分子がウェルに含まれない確率は:
【化2】
となる。
【0107】
故に、任意のウェルが対立遺伝子Aを少なくとも1分子含有する確率は、1−0.6065=0.3935となるであろう。従って、約39%のウェルは、対立遺伝子Aを少なくとも1分子含有すると予想される。
【0108】
非参照核酸配列に関しては、トリソミー21胎児の各細胞において、AとGとのゲノム比率は1:2である。抽出されたRNA又はDNA試料においてこのAとGとの比率が変化していないと仮定すると、ウェルあたりの対立遺伝子Gの平均濃度は、対立遺伝子Aの2倍、即ち2x0.5=1である。
【0109】
従って、対立遺伝子Gの平均濃度が1の場合、対立遺伝子Gの分子がウェルに含まれない確率は:
【化3】
となる。
【0110】
故に、任意のウェルが対立遺伝子Gを少なくとも1分子含有する確率は、1−0.3679=0.6321となるであろう。従って、約63%のウェルは、対立遺伝子Gを少なくとも1分子含有すると予想される。
【0111】
2.過剰出現配列の割合の決定
予想量が計算された後、過剰出現核酸配列の割合が決定され得る。ウェルに充填された対立遺伝子A及びBは独立であると仮定すると、両方の対立遺伝子を含有するウェルの確率は、0.3935x0.6321=0.2487となる、よって、約25%のウェルが、両方の対立遺伝子を含有すると予想される。
【0112】
対立遺伝子Aを含有するが対立遺伝子Gを含有しないウェルの割合は、少なくとも一つの対立遺伝子Aを含有する確率から、対立遺伝子A及びBのいずれも含有する確率を引く:0.3935−0.2487=0.1488となる。同様に、対立遺伝子Gを含有するが対立遺伝子Aを含有しないウェルの割合は:0.6321−0.2487=0.3834となる。情報的ウェルは、対立遺伝子A又はBのいずれかが陽性であるが、両方陽性ではないウェルと定義される。
【0113】
故に、デジタルRNA-SNPにおいて、対立遺伝子A又は対立遺伝子Gのいずれかを含むウェルの期待される割合は、0.1488/0.3834である。換言すると、対立遺伝子Gのみ陽性の場合、Aのみ陽性の場合と比較して、ウェルの個数は2.65倍となる。これは、過剰出現対立遺伝子が他の対立遺伝子の2倍となる胎児ゲノムの比率と対照的である。
【0114】
SPRT解析において、過剰出現対立遺伝子が陽性である情報的ウェルの割合(Pr)は、SPRT曲線を使用して計算及び解釈される。ここまでの実施例において、情報的ウェルの割合は:0.1448+0.3834=0.5852となる。故に、mrが0.5の場合に、トリソミー21の予想されるPrは:0.3834/0.5282=0.73となる。
【0115】
平均の鋳型濃度(m)がポアソン方程式の肝要なパラメーターであるから、Prは、mに伴って変化する。図6は、本発明の一つの態様に従い、ウェルあたりの平均の参照鋳型濃度(mr)として表された、予想されるデジタルRNA-SNP対立遺伝子比率、及び様々な範囲の鋳型濃度のトリソミー21試料のPrを列記する表600を示す。表600は、一連のウェルあたりの平均の参照鋳型濃度(mr)における、過剰出現アリルが陽性である情報的ウェルの比率及び割合(Pr)、及び予想される対立遺伝子比を示す。
【0116】
予想されるPr値は、ウェルあたりの参照対立遺伝子の平均濃度(mr)に従い非線形的に変化する。表600に示すように、対立仮説を許容するためのPrの予想される値は、mrに従い増大する。帰無仮説を許容するための予想されるPrの値は、0.5で固定され、対立遺伝子の不均衡を有する又は有しない試料は、mrが増大すると、Prの数値においてより大きく隔てられる。他の態様において、帰無仮説を許容するための数値は、0.5以外になり得ることに留意されたい。これは、正常の比率が1:1ではなく、例えば5:3であり、そのため5:3の比率が変化するときに不均衡が生じたことになる場合にあり得る。そのため、2つの異なる核酸配列の量の相異は、ケースバイケースで判定され得る。
【0117】
しかしながら、従来の方法(例えば、上記Zhou, W. et al. 2002を参照されたい)は、LOH試料に固定した期待されるPr値を使用していたため、それらはLOHを有する(対立仮説を許容する)試料のPr値よりも低く見積もられていた。見積もりの低さは、mrに従い増大し得る。換言すると、DNA試料中の参照対立遺伝子の平均濃度が高くなるにつれ、旧来の方法は不正確になっていく。この、対立仮説を許容するためのPrの低い見積もりは、帰無仮説及び対立仮説を許容するためのカットオフ値の計算を両方とも不正確にし得る。
【0118】
3.予想されるPrに基づくカットオフ値の計算
SPRTを使用する態様において、El Karoui at al. (2006)のSPRT曲線の境界の上端及び下端を計算するための方程式を使用し得る。更に、帰無仮説又は対立仮説を許容するために好ましい統計的信頼性のレベルは、それらの方程式の閾値尤度比(threshold likelihood ratio)を調整することにより変化し得る。本文中、閾値尤度比は8とする。なぜなら、この値は、癌検出において、対立遺伝子の不均衡の有無で試料を峻別するのに良好な成績を提供することが示されているからである。故に、一つの態様において、SPRT曲線の境界の上端及び下端を計算するための方程式は:
境界上端=[(ln8)/n-lnδ]/lnγ
境界下端=[(ln1/8)/n-lnδ]/lnγ
で表され、式中、
δ=-(1-θ1)/(1-θ0)
γ=-(θ1(1-θ0)/θ0(1-θ1)
θ0=帰無仮説が真であるときの非参照対立遺伝子を含む情報的ウェルの割合=0.5(下記参照)
θ1=対立仮説が真であるときの、非参照(即ち過剰出現)対立遺伝子を含む情報的ウェルの割合
N=情報的ウェルの個数=いずれか1つの対立遺伝子のみが陽性であるウェルの個数
で表される。ここで、lnは、自然対数、即ちlogeを表す数学記号である。
【0119】
帰無仮説を許容するためのθ0を決定するために、試料が正倍数体胎児を妊娠した妊婦から取得されたと仮定する。この仮定の下、いずれかの鋳型が陽性であるウェルの予想される個数は、1:1となるべきであり、そのため非参照対立遺伝子を含む情報的ウェルの予想される割合は、0.5となるべきである。
【0120】
対立仮説を許容するためのθ1を決定するために、試料がトリソミー21胎児を妊娠した妊婦から取得されたと仮定する。デジタルRNA-SNP解析のためにされる、トリソミー21の場合の予想されるPr の計算は、表600に詳述される。故に、デジタルRNA-SNP解析のためのθ1は、表600の最後のカラムに示すデータを参照する。
【0121】
4.平均濃度の測定
mrの測定は、当業者に知られる、又は今後知られ得る様々なメカニズムを通じて実行され得る。一つの態様において、mrの値は、デジタルPCR解析の実験プロセスの間に決定される。mrの値と参照対立遺伝子が陽性であるウェルの総数との間の関係は分布(例えばポアソン分布)により支配されるので、mrは、参照対立遺伝子が陽性であるウェルの個数から計算され得て、ここで、mr=-ln(1-参照対立遺伝子が陽性であるウェルの割合)の計算式が使用される。ここで、lnは、自然対数、即ちlogeを表す数学記号である。このアプローチは、デジタルPCR実験に使用されるDNA試料におけるmrの直接的かつ正確な見積もりを提供する。
【0122】
この方法は、所望の濃度を実現するのに使用され得る。例えば、方法200の工程240と同じく、抽出された試料の核酸は特定の濃度、例えば反応ウェルあたり鋳型分子1個の濃度まで希釈され得る。ポアソン分布を使用する一つの態様において、鋳型が入らないウェルの予想される割合は、e-mと計算され得て、ここでmは、ウェルあたりの平均鋳型分子濃度である。例えば、ウェルあたり鋳型分子1個の平均濃度において、鋳型分子が入らないウェルの予想される割合は、e-1、即ち0.37(37%)である。残りの63%のウェルは、1個以上の鋳型分子が含まれ得る。典型的には、デジタルPCRを実行して、陽性かつ情報的ウェルの個数が計数される。情報的ウェルの定義及びデジタルPCRデータを解釈する方法は、装置に依存する。
【0123】
他の態様において、ウェルあたりの平均濃度のmrは、他の定量方法、例えば定量リアルタイムPCR、半定量競合PCR、質量分析法を用いる実競合PCR等により測定される。
【0124】
B.デジタルRCD
平均濃度を使用するデジタルRCDは、上記デジタルSNP法と同様の方式で実行され得る。参照染色体(第21番染色体ではない)マーカー、第21番染色体マーカー、及びそれら両マーカーが陽性であるウェルの個数が、デジタルPCRにより決定され得る。ウェルあたりの参照マーカーの平均濃度(mr)は、デジタルSNP解析におけるmrの計算の場合と同じく、ポアソン分布の関数に従い、第21番染色体マーカーの陽性と無関係に、参照マーカーが陰性であるウェルの総数から計算される。
【0125】
それから、SPRT解析は、正倍数体又はトリソミー21の胎児を妊娠した妊婦から取得した血漿試料を分類するのに使用され得る。このシナリオにおいて、参照マーカー及び第21番染色体マーカーが陽性であるウェルの予想される比率は1:1であり、そのため、第21番染色体の陽性シグナルを示す情報的ウェルの予想される割合は0.5となる。胎児が第21番染色体においてトリソミーであるとき、対立仮説が許容され得る。このシナリオにおいて、試料DNAが胎児由来のもののみであれば、各ウェル中の第21番染色体の平均濃度は、参照マーカーの平均濃度(mr)の3/2倍であり得る。
【0126】
デジタルRCDは、胎児特異的マーカー、例えば胎盤エピジェネティックサイン(Chim, SSC. et al. 2005 Proc Natl Acad Sci USA 102, 14753-14758)等の検出を通じて、染色体量を決定するのに使用され得るが、このデジタルRCD解析の一つの態様は、非胎児特異的マーカーを使用する。そのため、非胎児特異的マーカーが使用されるとき、胎児由来分子のパーセンテージを測定する追加的な工程が加えられる。従って、ウェルあたりの第21番染色体の平均濃度は、試料中の胎児DNAの割合に依存し、そして、mr[(200%+胎児DNAパーセンテージ)/200%]を使用して計算され得る。
【0127】
説明のために、もう一度特定の例を使用する。ウェルあたりの参照染色体、第1番染色体の平均濃度()を0.5と仮定し、試料中50%のDNAが胎児由来で、DNAの50%が母親由来と仮定する。
【0128】
従って、ポアソン分布を使用すると、ウェルあたりの平均濃度が0.5のときの第1番染色体の遺伝子座のいずれの分子も含有しないウェルの割合は:
【化4】
となる。
【0129】
故に、第1番染色体の遺伝子座の少なくとも1つの分子をウェルが含む確率は:1-0.6065=0.3935である。従って、約39%のウェルは、少なくとも1個の遺伝子座の分子を含有すると予想され得る。
【0130】
このトリソミー21胎児の各細胞において、第21番染色体と第1番染色体とのゲノム比率は、3:2となる。DNA試料中の第21番染色体と第1番染色体との間の比率は、分別胎児DNA濃度(胎児DNA%)に依存し:3x胎児DNA%+2(1-胎児DNA%):2x胎児DNA%+2x(1-胎児DNA%)となる。よって、分別胎児DNA濃度が50%であるこの場合、比率は:(3x50%+2x50%)/(2x50%+2x50%)=1.25となる。シグナルSNP法が胎児特異的マーカーを使用していなければ、このような計算を、非参照配列の平均濃度を計算するのにも使用することが出来ていた。
【0131】
故に、ウェルあたりの第1番染色体の遺伝子座の平均濃度が0.5であるとき、ウェルあたりの第21番染色体の平均濃度は:1.25x0.5=0.625となる。よって、ウェルあたりの平均濃度が0.625であるときの、第21番染色体遺伝子座の分子がウェルに入らない確率は:
【化5】
となる。
【0132】
故に、任意のウェルが少なくとも1個の第21番染色体の遺伝子座を含む確率は:1-0.5353=0.4647となる。よって、約46%のウェルは、少なくとも1個の前記遺伝子座の分子を含むと期待される。ウェルにいずれの遺伝子座が入るかが独立していると仮定すると、ウェルに両方の遺伝子座が入る確率は:0.3935x0.4647=0.1829となる。よって、約18%のウェルは、両方の遺伝子座を含むと期待される。
【0133】
第1番染色体の遺伝子座を含むが第21番染色体の遺伝子座を含まないと期待されるウェルの割合は、少なくとも1個の第1番染色体の遺伝子座を含むウェルの個数から、両方の遺伝子座を含むウェルの個数を引いた値であり:0.3935-0.1829=0.2126となる。同様に、第21番染色体の遺伝子座を含むが両方の遺伝子座を含まないと期待されるウェルの割合は:0.4647-0.1829=0.2818となる。情報的ウェルは、第1番染色体の遺伝子座又は第21番染色体の遺伝子座のいずれかが陽性であるが、それらの両方が両性ではないウェルとして定義される。
【0134】
故に、デジタルRCDにおいて予想される第21番染色体と第1番染色体との比率は、0.2818/0.2106=1.34となる。換言すると、第21番染色体遺伝子座のみ陽性であるウェルの割合は、第1番染色体遺伝子座のみ陽性であるウェルの割合の1.34倍となる。これは、DNA試料における1.25の比率と対照的である。
【0135】
SPRT解析において、第21番染色体遺伝子座が陽性である情報的ウェルの割合(Pr)は、SPRT曲線を使用して計算及び解釈される必要がある。今回の事例では、情報的ウェルの割合は:0.2106+0.2818=0.4924となる。故に、mrが0.5で、胎児DNAが50%の場合のトリソミー21の予想されるPrは:0.2818/0.4924=0.57となる。
【0136】
平均鋳型濃度(m)がポアソン方程式の肝要なパラメーターなので、Prはmに応じて変化する。図7は、本発明の一つの態様における、ウェルあたりの平均参照鋳型濃度(mr)として表現される範囲の鋳型濃度で、分別胎児DNA濃度が10%、25%、50%及び100%のトリソミー21試料における、予想されるPrを列記する、表700を示す。デジタルRCD解析のためのトリソミー21の検体の予想されるPrの計算は、表700に詳述している。故に、胎児DNA分別濃度が様々な試料のデジタルRCD解析用のθ1は、表700に記載の対応する予想されるPr値を示すカラムから取得され得る。
【0137】
C.結果
1.異なるmrの比較
対立遺伝子又は染色体の不均衡を理論的(胎児ゲノム中の如く)に導き出した場合と実験的に予想した場合との間の程度の違いの基準、及びその後者の場合のmr値を決定する計算を、表600及び表700に示す。トリソミー21試料のデジタルRNA-SNP解析において、mr=0.5のとき、参照対立遺伝子のみを含むウェルに対する、過剰出現対立遺伝子のみを含むウェルの比率、即ちデジタルRNA-SNP比率は、2.65となる(表600)。100%胎児検体からなる検体のデジタルRCD解析において、mr=0.5のとき、第1番染色体遺伝子座のみが陽性であるウェルに対する、第1番染色体遺伝子座のみが陽性であるウェルの比率、即ちデジタルRCD比率は、0.63/(1-0.63)=1.7).となる。分別胎児DNA濃度が低下すると、mrが同一となるまでデジタルRCD比率も低下する(表700)
【0138】
表600及び700に示されるように、対立遺伝子又は染色体の過剰出現の程度は、mrに応じて増大する。しかしながら、情報的ウェルのパーセンテージは、mr=0.5付近で頭打ちとなり、続いてmrが更に増大するにつれて、徐々に低下していく。実際に、情報的ウェルの割合の低下は、もし検体の鋳型分子の量が無制限であるならば、ウェルの総数を増大することで補償され得る。しかし、ウェルを追加すると、試薬のコストが増大する。故に、最適のデジタルPCR効率は、鋳型濃度と、試料あたり検査されるウェルの総数との間のトレードオフにより決定される。
【0139】
2.SPRT曲線を使用する実例
上に考察したように、デジタルPCR実験のための対立遺伝子又は染色体の不均衡の予想される程度は、反応混合物(例えばウェル)あたりの実際の鋳型濃度に依存する。発明者らは、参照対立遺伝子に基づく鋳型濃度、即ちウェルあたりの平均参照鋳型濃度(mr)を記載する。上記公式に示されるように、予想されるPrは、上方及び下方のSPTR曲線を描画するのに使用され得る。同じく、予想されるPrは、mrの値に依存し、SPRT曲線の描画は、本質的に、mrの値に依存し得る。よって、実際には、特定の実験から得られたPrを解釈するのに、デジタルPCRデータセットの実際のmrに関連するSPRT曲線のセットを使用する必要があろう。
【0140】
図8は、本発明の一つの態様における、デジタルRNA-SNP解析のための、mrの値が0.1、0.5、及び1.0の場合の、SPRT曲線の相異の程度を説明するプロット800を示す。デジタルPCRデータの各セットは、特定の実験の正確なmrの値に関する特定の曲線を使用して解釈されるべきである。デジタルRNA-SNP及びRCDアプローチのための、対立遺伝子又は染色体の不均衡の予想される程度は異なる(前者は2:1、後者は3:2である)ため、2つのデジタルPCRシステムのために、異なるセットのSPRT曲線が必要となる。実験的に導き出されたPrは、デジタルPCR反応の対応するmrにより選択される適切なSPRT曲線を用いて解釈される。これは、従来報告されていた、固定されたセットの曲線を使用する、デジタルPCRによるLOHの分子検出のためのSPRTの使用と対照的である。
【0141】
SPRTを使用してデジタルPCRデータを解釈する実際の方法は、仮想のデジタルRNA-SNP反応を使用して、下で説明される。それぞれの場合のデジタルRNA-SNP解析の後、対立遺伝子Aのみが陽性であるウェルの個数、対立遺伝子Gのみが陽性であるウェルの個数、又は両方の対立遺伝子が陽性であるウェルの個数が計数される。参照対立遺伝子は、陽性ウェルの個数がより少数の対立遺伝子として定義される。mrの値は、ポアソン確率密度関数に従い、他方の対立遺伝子が陽性か否かに関係なく、参照対立遺伝子が陰性のウェルの総数を使用して計算される。発明者らの仮説例のデータを以下に示す。
【0142】
96ウェルの反応において、20ウェルが対立遺伝子Aのみ陽性であり、24ウェルが対立遺伝子Gのみ陽性であり、そして33ウェルが両方とも陽性である。G陽性ウェルよりもA陽性ウェルの方が少ないので、対立遺伝子Aが参照対立遺伝子とみなされる。参照対立遺伝子が陰性のウェルの個数は、96-20-33=43である。よって、ポアソン方程式を使用してmrが計算され:-ln(43/96)=0.80となる。このケースの実験的に決定されたPrは:24/(20+24)=0.55となる。
【0143】
表600によると、mr=0.8で、トリソミー21試料の予想されるPrは、0.76となる。よって、このケースでは、θ1は0.76となる。θ1=0.76に基づくSPRT曲線が、実験的に導き出されたPr(このケースでは0.55)を解釈するのに使用され得る。Pr=0.55が、関連するSPRT曲線上に収まったとき、そのデータ点は、下方の曲線に分類される。故に、本ケースは、正倍数体として分類される。図3を参照されたい。
【0144】
3.従来の手法との比較
図9Aは、96ウェルデジタルRNA-SNP解析における正倍数体及びトリソミー21の検体を分類するための、新旧のSPRTアルゴリズムの有効性を比較する、表900を示す。図9Bは、384ウェルデジタルRNA-SNP解析における正倍数体及びトリソミー21の検体を分類するための、新旧のSPRTアルゴリズムの有効性を比較する、表950を示す。新しいアルゴリズムは、デジタルPCRデータから導き出されたmrに対して特異的なSPRT曲線を選択することを意味する。古いアルゴリズムは、全てのデジタルPCR反応に対して固定されたセットのSPRT曲線を使用することを意味する。分級精度におけるカットオフ値の計算が不正確であることの効果は、表900に示すシミュレーション解析により明らかにされる。
【0145】
表900及び950に示されるように、従来の研究における固定されたセットのSPRT曲線を使用する場合と比較して、分類不能データの割合は、発明者らのアプローチの方が相当に低い。例えば、mr =0.5で発明者らのアプローチを使用した場合、96ウェル及び384ウェルデジタルRNA-SNP解析において、それぞれ14%及び0%のトリソミー21試料が分類不能であったが、固定した曲線を使用した場合、それぞれ62%及び10%が分類不能であった(表900)。故に、発明者らのアプローチは、より少数の情報的ウェルの個数で、疾患分類を可能とする。
【0146】
表900に示されるように、前記新しいアルゴリズムは、mrが0.1〜2.0の全ての値である場合において、対立遺伝子比率の歪みの有無をもって、より正確に試料を分類することが出来る。例えば、mr=1.0で、96ウェルデジタルRNA-SNP反応を実行する場合、前記新しいアルゴリズムは、対立遺伝子比率の歪みを有する又は有しない試料を、それぞれ88%及び92%の正確性で分類した。一方、前記古いアルゴリズムを使用した場合、対立遺伝子比率の歪みを有する又は有しない試料を正確に分類したパーセンテージは、それぞれ僅か19%及び36%であった。
【0147】
この新しいアルゴリズムを使用することにより、対立遺伝子比率の歪みを有する試料及び有しない試料の分離の精度は、mrに応じて増大し得る。結果として、分類精度は、mrに応じて増大し得る。mrが2.0を超えて増大すると、試料を2つの群に分離する場合の分類精度の向上の作用は低下する。なぜなら、情報的ウェルのパーセンテージが減少するからである。一方、古いアルゴリズムを使用した場合、分類精度はmrが増大すると顕著に低下する。なぜなら、予想されるP値の真の値からの逸脱が増大するからである。
【0148】
発明者らの実験データ及びシミュレーションデータは、デジタルRNA-SNPは、トリソミー21検出の有効かつ正確な方法であることを示す。母体血漿中のPLAC4 mRNAは純粋に胎児に由来するため、13個の母体血漿サンプル中12個は、正確な分類のために1回の384ウェルデジタルPCR実験で十分であった(図13Bの表1350)。このホモ接合性の、リアルタイムデジタルPCRに基づくアプローチは、RNA-SNP解析のための、質量分析に基づくアプローチ(上記Lo, YMD, et al. 2007 Nat Med,)の代替手法を提案する。胎盤特異的mRNA転写産物は別として、発明者らは、母体血清中の他の種類の胎児特異的核酸種が、デジタルPCRに基づく胎児染色体の異数性検出に使用可能であることをも想定する。一例として、エピジェネティック対立遺伝子比率(EAR)アプローチ(Tong, YK et al. (2006) Clin Chem 52, 2194-2202)を使用する、トリソミー18の非侵襲的な出生前検出に最近使用されている、胎児エピジェネティックマーカー(Chim, SSC et al. (2005) Proc Natl Acad Sci USA 102, 14753-14758; Chan, KCA et al. (2006) Clin Chem 52, 2211 -2218)が挙げられる。これにより、発明者らは、デジタルEARが、可能な解析技術であると予測する。
【0149】
V.%の増大、複数のマーカー、及びPCR代替手法
上記のように、本発明の態様を母体血漿から抽出したDNAに応用することは、妊娠11週〜17週の、母体血漿中の胎児DNAの分別濃度が約3%という僅かな割合でしか存在しないとき、複雑なものとなり得る。しかしながら、本明細書中に記載されるように、デジタルRCDは、異数性DNAが少数集団として存在するときであっても、異数性検出を可能とする。妊娠初期の如く胎児DNAの分別濃度が低いものほど、デジタルRCDにより多数の情報的カウントが必要となる。本研究の重要性は、図12の表1200に要約したように、発明者らが、診断的実験の基礎となる、例えば分別胎児DNA及び全鋳型分子等のベンチマークパラメーターのセットを提供したことである。発明者らの見解では、分別胎児DNA濃度が25%のとき、総数で7680の反応が、ベンチマークパラメーターの特に魅力的なセットである。これらのパラメーターによれば、表1200に示すように、分別胎児DNA濃度が25%のとき、正倍数体及びトリソミー21試料を、97%の正確性で分類することが可能となる。
【0150】
単位体積の母体血漿あたりに存在する血漿DNA分子の個数は限定される(Lo, YMD. et al. 1998 Am J Hum Genet 62, 768-7758)。例えば、妊娠早期において、常染色体遺伝子座(β-グロビン)の中間母体血漿濃度は、986コピー/mlであり、胎児と母親の両方の遺伝子座が算入されている(Lo, YMD. et al. 1998 Am J Hum Genet 62, 768-7758)。7680分子を捕らえるのに、約8mlの母体血漿から抽出したDNAが必要とされ得る。この血漿の体積は、約15mlの母体血液から取得され得て、これは通常の診療における限界である。しかしながら、発明者らは、複数のセットのchr21及び参照染色体標的を、デジタルRCD解析のために組み合わせ得ることを想定する。chr21及び参照染色体標的の5個のペアにおいて、分析に必要な数の鋳型分子を得るのに、ちょうど1.6mLの材料血漿が必要となり得る。多重単一分子PCRが実行され得る。この多重単一分子解析のロバスト性は、単一分子ハプロタイピングにおいて既に実証されている(Ding, C. and Cantor, CR. 2003 Proc Natl Acad Sci USA 100, 7449-7453)。
【0151】
あるいは、25%の分別胎児DNA濃度を達成するために、母体血漿中の胎児DNAを選択的に増幅する方法(Li, Y. et al. 2004 Clin Chem 50, 1002-1011)、若しくは母体DNAバックグラウンドを抑制する方法(Dhallan, R et al. 2004 JAMA 291, 1114- 1119)、又はそれらの両方が採用され得る。そうした胎児DNAの増幅や母体DNAの抑制等の物理的手法とは別に、特定のDNAメチル化パターンを示す胎児DNA分子を標的とする(Chim, SSC et al, 2005 Proc Natl Acad Sci USA 102,14753-14758, Chan, KCA et al. 2006 Clin Chem 52, 2211-2218; Chiu, RWK et al 2007 Am J Pathol 170, 941-950.)等による、分子増幅法を使用することも可能であり得る。
【0152】
加えて、現在は、デジタルPCRを実施する研究において使用される、デジタルリアルタイムPCR解析を手動で調整する多くの代替的なアプローチが存在する。これらの代替的アプローチは、マイクロ流体工学デジタルPCRチップ(Warren, L et al. 2006 Proc Natl Acad Sci USA 103, 17807-17812; Ottesen, EA et al. 2006 Science 314, 1464-1467)、エマルジョンPCR(Dressman, D et al. 2003 Proc Natl Acad Sci USA 100, 8817-8822)、並びにRoche 454プラットフォーム、Illumina Solexaプラットフォーム、及びApplied BiosystemsのSOLiD(商標)システム等を使用する超並列ゲノムシークエンシング(Margulies, M. et al. 2005 Nature 437, 376-380)等を含む。後者に関しては、発明者らの手法は、単一DNA分子に対する超並列シークエンシング法にも応用可能であり、これは、Helicos True Single Molecule DNAシークエンシング技術、Pacific Biosciencesの単一分子リアルタイム(SMRT(商標))技術、及びナノポアシークエンシング(Soni GV及びMeller A. 2007 Clin Chem 53, 1996-2001)等の増幅工程を要しない。これらの方法を使用することにより、デジタルRNA-SNP及びデジタルRCDを、大量の試料に対して迅速に実行することが出来て、それにより、非侵襲性出生前診断に提案される方法の臨床的柔軟性が向上する。
【実施例】
【0153】
以下の実施例は、例示のために提供されるものであり、主張する発明を限定することを目的としない。
I.コンピューターシミュレーション
SPRTアプローチを使用してのトリソミー21診断の精度を見積もるために、コンピューターシミュレーションを実行した。このコンピューターシミュレーションは、Microsoft Excel 2003ソフトウェア(Microsoft Corp., USA)及びSAS 9.1 for Windows(登録商標)ソフトウェア(SAS Institute Inc., NC, USA)を用いて行われた。デジタルPCRの性能は、参照鋳型濃度(mr)、情報的カウントの個数と、対立遺伝子又は染色体の不均衡の推定される程度(Pr)との間の相互作用である。これらの変数を各々、様々に変化させてシミュレーションを実行した。デジタルRNA-SNPとデジタルRCDとでSPRT曲線の決定境界が異なるため、それら2つのシミュレーション解析は別個に実行された。
【0154】
想定した各デジタルPCR条件(即ち胎児DNA分別濃度、ウェル総数)について、2ラウンドのシミュレーションを実行した。第一のラウンドでは、正倍数体胎児を妊娠した妊婦から検査試料を取得したというシナリオを想定した。第二のラウンドでは、トリソミー21胎児を妊娠した妊婦から検査試料を取得したというシナリオを想定した。各ラウンドにおいて、胎児5000人分の検査を想定した。
【0155】
A.RNA-SNP
デジタルRNA-SNPにおいて、mr=0.1〜mr=2.0である384ウェル実験のシミュレーションを実行した。各mr値で、発明者らは、正倍数体胎児5000人及びトリソミー21胎児5000人を試験したというシナリオを想定した。10000人の胎児を分類するために、所定のmrに適したSPRT曲線を使用した。図10は、本発明の態様における所定の情報的カウントにより、正倍数体又は異数体として正しく及び誤って分類された、並びに分類不能であった胎児のパーセンテージを示す、表1000である。mrが0.5〜2.0の場合、性倍数体と異数体の診断の正確性は、いずれも100%である。mr=0.5のとき、384ウェルの解析後に正確に分類できた性倍数体及びトリソミー21胎児は、それぞれ僅か57%及び88%である。
【0156】
これらのシミュレーションデータは、以下に記載される工程により生成された。
【0157】
工程1において、各ウェルについて、SASプログラム(www.sas.com/technologies/analytics/statistics/index.html)のランダム(ポアソン)関数を使用して、それぞれ対立遺伝子A及びGを表す2つのランダムな数字を生成した。ランダム(ポアソン)関数は、0から始まる正の整数(即ち0、1、2、3・・・)を生成し、各整数が生成される確率は、ウェルあたりの対立遺伝子の平均濃度を表す所定の平均値についてのポアソン確率密度関数に従い、各整数の確率に応じて決定した。対立遺伝子Aを表すランダム数字がゼロより大きとき、そのウェルは対立遺伝子Aが陽性、即ち対立遺伝子Aの分子を1つ以上含むとみなされた。同様に、対立遺伝子Gを表すランダム数字がゼロより大きいとき、そのウェルは対立遺伝子Gが陽性とみなされた。
【0158】
正倍数体胎児を妊娠した妊婦のシナリオを想定するにあたり、対立遺伝子A及び対立遺伝子Gにランダム数字を生産するために、同一の平均値が使用された。例えば、mr=0.5でのデジタルRNA-SNP解析を想定する解析において、対立遺伝子A及びGのいずれの平均値も0.5に設定した。これは、いずれの対立遺伝子も平均濃度がウェルあたり0.5分子であることを意味する、ポアソン方程式を使用して、平均濃度が0.5で、対立遺伝子A又はBが陽性であるウェルの割合は同一であり得て、0.3935であった。表600を参照されたい。
【0159】
トリソミー21を妊娠した妊婦のmr=0.5でのデジタルRNA-SNP解析を想定するとき、過剰出現対立遺伝子のウェルあたりの平均濃度は、参照対立遺伝子の値の2倍、即ち1と期待され得る。この条件下で、過剰出現対立遺伝子が陽性であるウェルの確率は、0.6321であった。表600を参照されたい。
【0160】
デジタルPCRウェルのランダム数字を生成した後、ウェルは、以下の状態のいずれかに分類され得る:
a.対立遺伝子A及びGのいずれも陰性
b.対立遺伝子A及びGのいずれも陽性
c.対立遺伝子Aは陽性であるがGは陰性
d.対立遺伝子Gは陽性であるがAは陰性
【0161】
工程2において、所望のウェル数(本シミュレーションでは384ウェル)に達するまで工程1が反復された。対立遺伝子Aのみ、又は対立遺伝子Gのみが陽性であるウェルの子数が計数された。陽性ウェルが少ない方の対立遺伝子を参照対立遺伝子とみなし、陽性ウェルが多い方の対立遺伝子を潜在的に過剰出現対立遺伝子とみなした。情報的ウェルの個数は、いずれかの対立遺伝子が陽性であるがそれらの両方が陽性ではないウェルの総数とした。そして、潜在的に過剰出現対立遺伝子を含む情報的ウェルの割合(Pr)を計算した。本発明の一つの態様に従い、帰無仮説又は対立仮説を許容するためのSPRT曲線における境界の上端及び下端を計算した。
【0162】
工程3において、正倍数体又はトリソミー21胎児を妊娠した妊婦の2つの各シナリオについて、5000回シミュレーションを実行した。各シミュレーションを、妊婦から取得した独立の生体試料とみなし得る。表1000において、正倍数体の検体の正しい分類は、帰無仮説が許容された正倍数体の検体を意味し、そして、正倍数体の検体の誤った分類は、対立仮説が許容された正倍数体の検体を意味する。同様に、対立仮説が許容されたトリソミー21の検体は正しく分類されたものとみなされ、そして、帰無仮説が許容されたトリソミー21の検体は誤って分類されたものとみなされる。いずれの群においても、事前に指定されたウェルの総数がシミュレーションされた後に、帰無仮説及び対立仮説のいずれも許容されない検体は、分類不能とみなした。
【0163】
工程4において、mrを0.1〜2.0の範囲で0.1ずつ増大させて設定し、工程1〜3を実行した。
【0164】
B.RCD
図11は、本発明の一つの態様における、mrが0.1〜2.0の範囲で、純粋(100%)な胎児DNA試料についてなされるデジタルRCDのコンピューターシミュレーションを示す、表1100である。分別胎児DNA濃度が低下するに従い、第21番染色体の過剰出現の度合は低下し、それにより疾患分類を確定するのにより多数の情報的ウェルが必要となる。故に、シミュレーションは、mr=0.5で、384〜7680ウェルのウェル総数の範囲において、50%、25%、及び10%の胎児DNA濃度において、更に実行された。
【0165】
図12は、本発明の一つの態様における、胎児DNAの分別濃度が異なる、正倍数体又はトリソミー21胎児由来のサンプルを分類するための、mr=0.5における、デジタルRCD解析の精度のコンピューターシミュレーションの結果を示す、表1200である。デジタルRCDの効率は、検体のDNA分別濃度が高くなるに従い良好になる。胎児DNA濃度が25%で、PCR解析の総数が7680回で、正倍数体及び異数体の検体のいずれも、分類を誤らず97%が分類可能であった。検体の残りの3%については、分類を達成するのに更なる解析を必要とする。
【0166】
デジタルRCD解析をシミュレーションする手順は、デジタルRNA-SNP解析において記載した物と同様であった。このシミュレーションの工程を、以下に記載する。
【0167】
工程1において、参照遺伝子座、第1番染色体、及び第21番染色体遺伝子座を表現するために、ポアソン確率密度関数の下で2つのランダムな数字が生成された。正倍数体の胎児を妊娠した対象において、第1番染色体及び第21番染色体の平均濃度は、いずれも等しかった。このシミュレーション解析において、各遺伝子座のウェルあたりの平均鋳型濃度には、0.5を採用した。トリソミー21胎児を妊娠した対象において、このシミュレーションにおけるmrは0.5であったが、ウェルあたりの第21番染色体の平均濃度は、表700に示されるように、検査される使用中の分別DNA濃度に依存していた。参照遺伝子座及び/又は第21番染色体遺伝子座の1個のウェルへの分配は、ウェルあたり遺伝子座の適切な平均濃度で、ポアソン確率密度関数に従い生成された各遺伝子座を表すランダムな数字により決定された。
【0168】
工程2において、所望のウェル数、例えば384ウェルプレートの実験であれば384ウェルに達するまで、工程1が反復された。第1番染色体のみ、及び第21番染色体のみが陽性であるウェルの子数が計数された。情報的ウェルの個数は、いずれかの染色体が陽性であるがそれらの両方が陽性ではないウェルの総数とした。そして、第21番染色体が陽性である情報的ウェルの割合(Pr)を計算した。前記SPRT解析の章で記載したのと同様に、本発明の一つの態様に従い、帰無仮説又は対立仮説を許容するためのSPRT曲線における境界の上端及び下端を計算した。
【0169】
工程3において、正倍数体又はトリソミー21胎児を妊娠した妊婦の2つの各シナリオについて、5000回シミュレーションを実行した。各シミュレーションを、妊婦から取得した独立の生体試料とみなし得る。表1100において、正倍数体の検体の正しい分類は、帰無仮説が許容された正倍数体の検体を意味し、そして、正倍数体の検体の誤った分類は、対立仮説が許容された正倍数体の検体を意味する。同様に、対立仮説が許容されたトリソミー21の検体は正しく分類されたものとみなされ、そして、帰無仮説が許容されたトリソミー21の検体は誤って分類されたものとみなされる。いずれの群においても、事前に指定されたウェルの総数がシミュレーションされた後に、帰無仮説及び対立仮説のいずれも許容されない検体は、分類不能とみなした。
【0170】
工程4において、384〜7680ウェルのウェル総数の範囲において、10%、25%、50%及び100%の胎児DNA濃度のサンプルにおいて、工程1〜3を反復した。
【0171】
II.トリソミー21検出の変法
A.PLAC4用RNA-SNP
第21番染色体上のPLAC4のrs8130833 SNP(Lo, YMD et al. 2007 Nat Med 13, 218-223)を使用して、デジタルRNA-SNPの実際の実行可能性を実証した。2つの正倍数体及び2つのトリソミー21ヘテロ接合体の胎盤に由来する胎盤DNA及びRNAを解析した。これらの胎盤DNA試料を、デジタルRNA-SNPプロトコールを用いて分析したが、逆転写工程を省略したので、本質的には、その手順を、デジタルDNA-SNP解析に転換したものであった。検体を正しく分類する可能性と情報的ウェルの割合との間のバランスを取るために、発明者らは、ウェルあたり任意の種類の対立遺伝子が1個入るように試料を希釈し、そしてそれを96ウェルデジタルPCR解析により検証した。これは、384ウェルデジタルRNA-SNP実験により追認された。Pr及びmrが計算され、このmrに対するSPRT曲線が、データ解釈に使用された。
【0172】
図13Aは、本発明の一つの態様における、正倍数体及びトリソミー21胎児を妊娠した妊婦の胎盤組織をデジタルRNA-SNP解析の結果である、表1300を示す。質量分析アッセイにより、遺伝子型を決定した。「正倍数体」は、実験的に取得したPrが分類不能領域(unclassifiable region)を下回ったときに割り当てられ;トリソミー21を意味する「T21」は、実験的に取得したPrが分類不能領域を上回ったときに割り当てられる。これらの各検体は、1回の384ウェル実験で、DNA及びRNA試料のいずれを使用しても、正しく分類された。
【0173】
発明者らは、更に、正倍数体胎児を妊娠した妊婦9人及びトリソミー21胎児を妊娠した妊婦4人から採取した血漿RNAを検査した。図13Bは、本発明の一つの態様における、正倍数体及びトリソミー21を妊娠した妊婦から採取した母体血漿のデジタルRNA-SNP解析の結果である、表1350を示す。検体は全て、正しく分類された。1つのトリソミー21検体(M2272P)の最初の結果は、SPRT曲線の分類不能領域内に収まった。そこで、追加的な384ウェル実験が実行された。計768ウェルから収集したデータから、新たなmr及びPrが計算され、このmr値に基づいて選択された新たなセットのSPRT曲線を使用して、分類が実行された。その結果、上記検体は、異数体として正しく評点された。
【0174】
発明者らの実験的データ及びシミュレーションデータは、デジタルRNA-SNPが、トリソミー21を検出する有効かつ正確な方法であることを示す。母体血漿中のPLAC4 mRNAは純粋に胎児に由来するので、13個中12個の母体血漿試料を正しく分類するのに、僅か1回の384ウェルデジタルPCR実験で十分である。故に、この均質な、リアルタイムデジタルPCRに基づくアプローチは、RNA-SNP解析のための、質量分析に基づくアプローチという代替手法を提案する。胎盤特異的mRNA転写産物とは別に、発明者らは、母体血清中の他の種類の胎児特異的核酸種が、胎児染色体の異数性のデジタルPCRに基づく検出に使用され得ることを想定する。一例として、近頃エピジェネティック対立遺伝子比率(EAR)アプローチTong YK et al. 2006 Clin Chem, 52, 2194-2202)を使用したトリソミー18の非侵襲性出生前検出に使用されている、胎児エピジェネティックマーカーが挙げられる。そのため、発明者らは、デジタルEARが、可能な解析技術になり得ると予想している。
【0175】
B.RCD
また、トリソミー21を検出するためのデジタルRCDの実際の実行可能性は、第21番染色体及び第1番染色体上のパラロガス配列を標的にするPCRアッセイを使用して調査された。パラロガス遺伝子座は、実施例の方法により、ここで使用された。第21番染色体上の非パラロガス配列、及び他の任意の参照配列も、RCDに使用され得る。2つの正倍数体及び2つのトリソミー21胎盤に由来する胎盤DNA試料を、いずれかの染色体由来の標的鋳型がウェルあたりに約1個入る濃度に希釈し、それを、96ウェルデジタルPCR解析により確認した。確認された各試料を、384ウェルデジタルRCD実験で解析し、そしてPr及びmrの値を計算した。デジタルRCDにおいて、第1番染色体のパラログが、参照鋳型であった。このmr値を、データ解釈のための対応するセットのSPRT曲線の選択に使用した。図14Aに示されるように、全ての胎盤試料が正しく分類された。
【0176】
母体血漿中の胎児DNAの如く、過剰の正倍数性DNAと混在しているトリソミー21DNAを検出するのに、デジタルRCDアプローチが使用され得ることを明示するために、正倍数性母体血球細胞DNAのバックグラウンド中に、50%及び25%のトリソミー21胎盤DNAを含む混合物を解析した。トリソミー21検体10個、及び正倍数体の検体10個に由来する胎盤DNAのそれぞれが、同量の正倍数性血球細胞DNAと混合され、これで20個の50%DNA混合物が出来た。図14Bは、本発明の一つの態様における、それら50%胎児DNA混合物のRCD解析のためのSPRT解釈を説明する、プロット1440を示す。同様に、トリソミー21検体5個、及び正倍数体の検体5個に由来する胎盤DNAのそれぞれが、3倍量の正倍数性血球細胞DNAと混合され、これで10個の25%DNA混合物が出来た。図14Cは、それら25%胎児DNA混合物のRCD解析のためのSPRT解釈を説明する、プロット1470を示す。図14B及び図14Cに示すように、全ての正倍数性DNA混合物及び異数性DNA混合物は、正しく分類された。
【0177】
各試料は、図14B及び14Cにマークされるように、多くの384ウェルデジタルPCR解析の後に、解析可能な点に達した。50%DNA混合物において、必要とされる384ウェルプレートの数は、1〜5枚の範囲であった。25%DNA混合物において、必要とされる384ウェルプレートの数は、1〜7枚の範囲であった。全384デジタルPCR解析を追加して正しく分類された検体の累積的割合は、表1200に表されるコンピューターシミュレーションにより予測されたものと同等であった。
【0178】
III.デジタルPCRを用いる方法
A.デジタルRNA-SNP
全てのRNA試料は、まず、ThermoScript逆転写逆転写酵素(Invitrogen)を使用して、遺伝子特異的逆転写プライマーを用いて逆転写された。逆転写プライマーの配列は、5'-AGTATATAGAACCATGTTTAGGCCAGA-3'(Integrated DNA Technologies, Coralville, IA)であった。以後、デジタルRNA-SNPのための逆転写されたRNA(即ちcDNA)試料とDNA試料(例えば胎盤DNA)とは、本質的に同じ処理がされる。デジタルDNA解析に先立ち、DNA及びcDNA試料は、まず、PLAC4に付いてのリアルタイムPCRアッセイを使用して定量された。ここで、プライマーとして、5'-CCGCTAGGGTGTCTTTTAAGC-3'、5'-GTGTTGCAATACAAAATGAGTTTCT-3'、及び蛍光プローブとして、5'-(FAM)ATTGGAGCAAATTC(MGBNFQ)-3'が用いられた(Applied Biosystems, Foster City, CA)。なお、FAMは、6-カルボキシフルオレセインであり、MGBNFQは、無蛍光クエンチャーが結合するマイナーグルーブである。
【0179】
単位複製配列をコードする、HPLC精製単鎖合成DNAオリゴヌクレオチドの連続希釈により、検量線が容易された。前記配列は、5'-CGCCGCTAGGGTGTCTTTTAAGCTATTGGAGCAAATTCAAATTTGGCTTAAAGAA AAAGAAACTCATTTTGTATTGCAACACCAGGAGTATCCCAAGGGACTCG-3'であった。反応体積を25μlとして、2X TaqMan Universal PCR Master Mix (Applied Biosystems)を使用して、反応を準備した。各反応において、各プライマーを400nL、及びプローブを80nM使用した。50℃5分で反応を開始し、続いて95℃10分、そして95℃15秒及び65℃1分を45サイクル、以上をABI PRISM 7900HT Sequence Detection System (Applied Biosystems)で行った。そして、その後のデジタルPCR増幅が、ウェルあたり鋳型分子が約1個で実行され得るように、DNA又はcDNA試料の連続希釈が行われた。そのような濃度では、約37%の反応ウェルが、増幅において陰性であることが予想され、まずは96ウェルデジタルリアルタイムPCR解析を行うことによりそのことが確認された。それから、イントロンをまたがないプライマー(順プライマーが5'-TTTGTATTGCAACACCATTTGG-3'であり、遺伝子特異的な逆プライマーは前に記載した)のセットを使用して、384ウェルプレートで行われるデジタルRNA-SNP解析が行われた。
【0180】
PLAC4上のrs8130833 SNPの2つの各対立遺伝子を標的とする、2つの対立遺伝子特異的なTaqManプローブが設計された。それらの配列は、対立遺伝子A及びGについて、それぞれ、5'-(FAM)TCGTCGTCTAACTTG(MGBNFQ)-3'及び5'-(VIC)ATTCGTCATCTAACTTG(MGBNFQ)とした。反応体積5μlの、2X TaqMan Universal PCR Master Mixを使用して、反応を準備した。各反応は、1xTaqMan Universal PCR Master Mix、各プライマーを572nM、対立遺伝子G特異的プローブを107nM、及び対立遺伝子A特異的プローブを357nM含んでいた。ABI PRISM 7900HT Sequence Detection System中で、反応を遂行した。反応は、50℃2分で始まり、そして95℃10分、更に45サイクルの95℃15秒及び57℃1分が続いた。反応の間、SDS 2.2.2ソフトウェアの「Absolute Quantification」アプリケーション(Applied Biosystems)により、蛍光データが収集された。このソフトウェアが、基準値及び閾値を自動的に計算した。対立遺伝子A又はGのいずれかが陽性であるウェルの個数が記録され、そしてその結果がSPRT解析の用に供された。
【0181】
B.デジタルRCD解析
本研究で使用された全ての胎盤及び母体軟膜のDNA試料は、まずNanoDrop分光光度計(NanoDrop Technology, Wilmington, DE)により定量された。DNA濃度は、6.6pg/細胞の転換を使用して、コピー/mlに転換される。ウェルあたり約1個の鋳型が入る濃度に相当するDNA濃度は、DNA試料の連続希釈により決定され、そして96ウェルフォーマットのリアルタイムPCRで、約37%のウェルが増幅において陽性であることにより確認された。確認プレート用のPCRは、参照染色体のプローブのみが添加される点を除いて、下記と同様に準備された。デジタルRCD解析において、第21番及び第1番染色体上のパラロガス遺伝子座(Deutsch, S. et al. 2004 J Med Genet 41, 908-915)を、最初に、順プライマー5'-GTTGTTCTGCAAAAAACCTTCGA-3'及び逆プライマー5'-CTTGGCCAGAAATACTTCATTACCATAT-3'により、同時に増幅させる。2つの染色体特異的TaqManプローブは、第21番及び第1番染色体パラログを標的にするように設計されており、それらの配列は、それぞれ、5'-(FAM)TACCTCCATAATGAGTAA A(MGBNFQ)-3'及び5'-(VIC)CGTACCTCTGTAATGTGTAA(MGBNFQ)-3'である。各反応は、1xTaqMan Universal PCR Master Mix(Applied Biosystems)、各プライマー450nM、及び各プローブ125nMを含んでいた。反応体積の合計は、5μL/ウェルであった。この反応は、50℃2分で始まり、そして95℃10分、更に50サイクルの95℃15秒及び60℃1分が続く。リアルタイムPCR実験は全て、ABI PRISM 7900HT Sequence Detection System(Applied Biosystems)で遂行され、蛍光データは、SDS 2.2.2ソフトウェア(Applied Biosystems)の「Absolute Quantification」アプリケーションで収集した。既定の基準値及び手入力の閾値が使用された。第21番又は第1番のいずれかの染色体が陽性であるウェルの個数が記録され、そしてSPRT解析の用に供された。SPRTにより疾患分類が可能となるまでに、1枚以上の384ウェルプレートが解析されるべきである。
【0182】
IV.マイクロ流体工学に基づくデジタルPCRの使用
A.デジタルRNA-SNP
本実施例は、マイクロ流体工学に基づくデジタルPCRを使用した、デジタルPCR解析の性能を明示する。本章には、ここに示される一つの方法はFluidigm BioMark(商標)システムを使用するが、これは例示を目的としており、限定を目的とするものではない。このシステムは、1回の反応で9000個以上のデジタルPCRを実行することが可能である。
【0183】
正倍数性胎児又はトリソミー21胎児を妊娠した妊婦から、胎盤組織及び母体末梢血試料が得られた。胎盤DNA試料において、プライマー伸長に続いて質量分析を行うことにより、PLAC4遺伝子上のrs8130833 SNPのジェノタイピングが遂行された。RNAは、胎盤及び母体血漿試料から抽出された。
【0184】
全てのRNA試料は、遺伝子特異的逆転写プライマー(5'-AGTATATAGAACCATGTTTAGGCCAGA-3')で、ThermoScript逆転写酵素(Invitrogen)を使用して、逆転写された。胎盤cDNA試料において、ウェルあたり約1個の鋳型分子が入る濃度でその後のデジタルPCR増幅が遂行されるように、連続希釈が遂行された。
【0185】
デジタルPCRは、12.765デジタルアレイ(Fluidigm)を有するBioMark System(商標)(Fluidigm)で行われる。各デジタルアレイは、12種類の試料アッセイ混合物に対応するための、12枚のパネルから構成される。各パネルは、更に、7-nL反応/ウェルを実行するために、765ウェルに区切られている。前記PLAC4遺伝子上のrs8130833 SNP領域は、順プライマー(5'-TTTGTATTGCAACACCATTTGG-3')及び上記遺伝子特異的逆転写プライマーにより増幅された。rs8130833 SNPの2つの各対立遺伝子を標的とする、2つの対立遺伝子特異的TaqManプローブが設計された。これらの配列は、対立遺伝子Gに対するものを5'-(FAM)TCGTCGTCTAACTTG(MGBNFQ)-3'、そしてAに対するものを5'-(VIC)ATTCGTCATCTAACTTG(MGBNFQ)とした。1枚のアレイパネルにおける反応は、反応体積10μL中のTaqMan Universal PCR Master Mixを使用して準備された。各反応は、1x TaqMan Universal PCR Master Mix、572nMの各プライマー、53.5nMの対立遺伝子G特異的プローブ、178.5 nMの対立遺伝子A特異的プローブ及び3.5 μLのcDNA鋳型を含む。各胎盤cDNA試料に1枚の反応パネルが使用されたが、各母体血漿試料には、12枚のパネルが使用された。試料アッセイ混合物は、NanoFlex(商標)IFCコントローラー (Fluidigm)によりデジタルアレイに注入された。反応はBioMark(商標)System中で実行された。その反応は、50℃2分で始まり、そして95℃10分、更に40サイクルの95℃15秒及び57℃1分が続いた。
【0186】
1つの正倍数体及び2つのT21ヘテロ接合体の胎盤に由来する胎盤RNA試料が、765ウェルの反応パネル中で解析された。各資料において、対立遺伝子A又はGが陽性(だが両方陽性ではない)である情報的ウェルが計数された。全ての情報的ウェル中の過剰出現対立遺伝子の割合(Pr)が決定された。これらの反応の、ウェルあたりの平均参照鋳型濃度(mr)に適したSPRT曲線は、実験的に取得されたPrが、倍数体又はT21試料のいずれを示唆するかを決定するのに利用された。
【0187】
発明者らは、更に、正倍数体の胎児を妊娠した女性4人及びトリソミー21胎児を妊娠した女性1人に由来する血漿RNA試料を検査した。各資料は、血漿RNA試料あたり12枚の765ウェル反応パネル中で、即ち9180回の反応で解析された。図15Bは、この血漿RNA試料における、12枚の各パネルにおける、情報的ウェルの個数を示す。この表に示すように、血漿資料中の鋳型濃度は、いずれかの反応パネル中の情報的ウェルが、SPRT分類に十分でない程度に希釈された。この試料を正倍数体試料と分類するのに、3つの反応パネルからの情報的ウェルを組み合わせなければならなかった(図15C)。図15Cは、2〜12枚のパネルからのデータを集積して使用することにより、全ての血漿検体が正しく分類され得ることを示す。
【0188】
デジタルPCRを手作業で実行する方法と比較して、このマイクロ流体工学に基づく方法は、格段に迅速で、労働集約的(labor-intensive)である。全てのプロセスが、2時間半以内に完了し得る。
【0189】
トリソミー18の出生前検出のためのデジタルRNA-SNP解析
本実施例において、第18番染色体中胎盤で発現する転写産物である、セルピンプロテアーゼ阻害剤クレードB(オボアルブミン)メンバー2(SERPINB2)mRNAに関する、デジタルPCRに基づく対立遺伝子区別アッセイを使用して、トリソミー18胎児の多型対立遺伝子の比率の不均衡を検出した。胎盤組織試料からのDNA及びRNAの抽出は、QIAamp DNA Mini Kit(Qiagen, Hilden, Germany)を使用して、取扱説明書に従い遂行された。抽出されたRNA試料は、ゲノムDNAのコンタミネーションを排除するために、DNA分解酵素I(Invitrogen)処理に付された。SERPINB2遺伝子上のrs6098 SNPのジェノタイピングは、上記のように、MassARRAY Compact(Sequenom, San Diego)を使用した均質MassEXTEND (hME)アッセイで、胎盤組織DNA試料において遂行された。
【0190】
SERPINB2転写産物の逆転写は、ThermoScript逆転写酵素(Invitrogen)を使用して、遺伝子特異的プライマー5'-CGCAGACTTCTCACCAAACA-3'(Integrated DNA Technologies, Coralville, IA)を用いて、胎盤組織RNA試料について実行された。全てのcDNA試料は、その後のデジタルPCR増幅が、反応ウェルあたり鋳型分子が1個入る平均濃度で実行され得るような濃度まで希釈された。デジタルPCRは、TaqMan Universal PCR Master Mix(Applied Biosystems, Foster City, CA)及びBiomarkTM PCR Reagents(Fluidigm, San Francisco)を使用して準備された。順プライマー5'-CTCAGCTCTGCAATCAATGC-3'(Integrated DNA Technologies)及び逆プライマー(前記逆転写用遺伝子特異的プライマーと同一)が、600nMの濃度で使用された。SERPINB2上のrs6098SNPの対立遺伝子A又はG を標的とする2つのTaqManプローブは、5'-(FAM)CCACAGGGAATTATTT(MGBNFQ)-3'及び5'-(FAM)CCACAGGGGATTATTT(MGBNFQ)-3'(Applied Biosystems)であった。FAMは、6-カルボキシフルオレセインであり、MGBNFQは、マイナーグルーブ結合無蛍光クエンチャーであり、それらはそれぞれ300nM及び500nMの濃度で使用された。各試料-試薬混合物は、NanoflexTM IFC Controller (Fluidigm)を使用するBiomarkTM 12.765 Digital Array上の756反応ウェルに分注された。その後、そのアレイは、加熱増幅及び蛍光検出のために、BiomarkTM Real-time PCR System (Fluidigm)中に置かれた。その反応は、50℃2分で始まり、そして95℃5分、更に45サイクルの95℃15秒及び59℃1分が続いた。増幅の後、情報的ウェル(対立遺伝子A又はGのいずれか一方のみが陽性であるウェル)の個数及び両方の対立遺伝子が陽性であるウェルの個数が計数され、そして、逐時確率比検定(SPRT)解析に付された。
【0191】
ヘテロ接合性倍数体胎児において、対立遺伝子A及びGは、胎児ゲノム中に等しく存在する(1:1)はずであり、一方トリソミー18において、一つの対立遺伝子に追加的なコピーが存在するため、胎児ゲノム中の比率が2:1となる。異なる試料を解釈するために、一連のDPRT曲線が作成された。これらの曲線は、分類に必要となる、所定の情報的ウェルの総数(x軸)に対する、過剰出現対立遺伝子が陽性である情報的ウェル数の割合Pr(y軸)を描画している。各試料において、実験的に導き出されたPrは、予想されたPr値と比較された。上方の曲線を上回った試料はトリソミー18として分類され、他方下方の曲線を下回った試料は、正倍数体として分類された。2つの曲線の間の領域は、分類不能領域である。
【0192】
胎児トリソミー18を検出するためのデジタルRNA-SNP解析の実現可能性は、SERPINB2遺伝子中のrs6098 SNPを使用することで実証された。正倍数体及びトリソミー18胎児を妊娠した対象に由来する胎盤組織DNA試料を、まず、ヘテロ接合の検体を同定するために、質量分析によりジェノタイピングした。その結果、前記胎盤試料の内9個が正倍数体、そして3つがトリソミー18ヘテロ接合体であることがわかり、それらはデジタルRNA-SNP解析に付された。各試料において、Pr及びmrが計算され、そして、このmr値に対応するSPRT曲線が、疾患分類に使用された。図16Aに示されるように、全ての試料が正しく分類された。トリソミー18胎盤のPr値は分類不能領域を上回り、他方正倍数体の胎盤の場合は、この領域を下回った。
【0193】
mr=0.1、0.2及び0.3に基づくSPRT曲線を用いた試料は、図16Bに説明される。これらのデータは、このデジタルRNA-SNP法が、トリソミー18の妊娠を診断するための有用な診断方法であることを示唆する。データ点が上方の曲線を上回る試料は異数体として分類され、一方データポイントが下方の曲線を下回る試料は、正倍数体として分類された。
【0194】
C.デジタルRCD解析
本実施例は、マイクロ流体工学に基づくデジタルPCRを使用するデジタルRCD解析の効率を明示する。Fluidigm BioMark(商標)Systemを使用するこのアプローチの一種がここに記載されるが、これは例示を目的とするものであり、限定を目的とするものではない。このシステムは、1回の反応でデジタルPCRを9000回以上実行することが出来る。
【0195】
正倍数体又はトリソミー21(T21)胎児を妊娠した妊婦から、胎盤組織、母体血球細胞、及び羊水試料が採取された。10個のT21及び10個の正倍数体由来の胎盤DNAを、それぞれ等量の正倍数体母体血球細胞DNAと混合することにより、20個の50%DNA混合物を作り出した。それらの混合試料中の胎児の割合の正確さを保証するため、まずは抽出されたDNAが、260nmで光学密度(OD)を測定することにより定量された。それからそれらは、12.765 Digital Arrays (Fluidigm)を使用して、BioMark(商標)System (Fluidigm)により、デジタル的に(digitally)定量された。それらの試料を定量するアッセイは、参照染色体に対するプローブが使用された点を除き、下記と同様に行われた。
【0196】
前記50%DNA混合物及び羊水試料中の染色体量は、第1番染色体と第21番染色体との非多型遺伝子座を比較する、デジタルPCR解析により決定された。第21番及び第1番の染色体上の一対のパラロガス遺伝子座の101bpのアンプリコンを、まず、順プライマー5'-GTTGTTCTGCAAAAAACCTTCGA-3'及び逆プライマー5'-CTTGGCCAGAAATACTTCATTACCATAT-3'により同時に増幅した。2つの染色体特異的TaqManプローブが、第21番と第1番染色体のパラログを区別するように設計され、それらの配列は、それぞれ、5'-(FAM)TACCTCCATA ATGAGTAAA(MGBNFQ)-3'、及び5'-(VIC)CGTACCTCTGTAATGTGTAA(MGBNFQ)-3'であった。パラロガス遺伝子座は、ここでは、例示のみを目的として使用される。言い換えると、このような解析に非パラロガス遺伝子座も使用され得る。
【0197】
トリソミー18(T18)を検出するためにデジタルRCDアプローチの使用を明示するため、第21番及び第18番染色体上のパラロガス配列を標的とするもう一つのアッセイが設計された。第21番及び第18番の染色体上のパラロガス遺伝子座の128bpのアンプリコンを、まず、順プライマー5'-GTACAGAAACCACAAACTGATCGG-3'及び逆プライマー5'-GTCCAGGCTGTGGGCCT-3'により同時に増幅した。2つの染色体特異的TaqManプローブが、第21番と第18番染色体のパラログを区別するように設計され、それらの配列は、それぞれ、5'-(FAM)AAGAGGCGAGGCAA(MGBNFQ)-3'及び5'-(VIC)AAGAGGACAGGCAAC(MGBNFQ)-3'であった。パラロガス遺伝子座の使用は、ここでは、例示のみを目的とする。言い換えると、このような解析に非パラロガス遺伝子座も使用され得る。
【0198】
全ての実験は、BioMark(商標)System (Fluidigm)上で、12.765 Digital Arrays (Fluidigm)を使用して遂行された。1枚のパネル上での反応は、反応体積10μlで、2x TaqMan Universal PCR Master Mix (Applied Biosystems)を使用して準備された。各反応は、1x TaqMan Universal PCR Master Mix、各プライマーを900nL、及びプローブを125nM、及び50%胎盤/母体血球細胞DNA試料を3.5μl含んで行われた。試料/アッセイ混合物は、NanoFlex(商標)IFCコントローラー(Fluidigm)により、デジタルアレイに注入された。反応は、検出用BioMark(商標)System上で実行された。この反応は、50℃2分で始まり、そして95℃10分、更に40サイクルの95℃15秒及び57℃1分が続いた。
【0199】
正倍数体及びT21の50%胎盤/母体血球細胞DNA試料は、chr21/chr1アッセイを用いて、デジタルアレイ上で解析された。各試料において、chr21又はchr1マーカーのいずれか1方のみ(両方ではない)が陽性であるものを含む情報的ウェルの個数が計数された。全ての情報的ウェル中の、過剰出現マーカーの割合(Pr)が決定された。ウェルあたりの正確な平均参照鋳型濃度(mr)に対応するSPRT曲線が、実験的に取得されたPrが正倍数体又はT21のいずれの試料を示唆するものかを判定するのに使用された。分類不能の試料が残っているときは、分類が可能となるまで、追加的なパネルからのデータが蓄積された。図17に示すように、全ての50%胎盤/母体血球細胞DNA試料は、このアプローチを使用して正しく分類され、それらの分類には、1枚〜4枚の範囲のパネルが必要であった。図18に示すように、正しい分類のための決定境界を示すために、SPRT曲線の描画もなされた。
【0200】
発明者らは、更に、RCD解析を、正倍数体胎児を妊娠した23人の妊婦及びT21胎児を妊娠した6人の妊婦から得られた羊水試料に適用した。各試料は、chr21/chr1アッセイと共に、単一の765ウェル中で解析された。図19は、このSPRT分類を要約して示した。図19のように、29個の試料は、全て正しく分類された。よって、デジタルRCD法は、羊水及び絨毛膜絨毛生検等の、出生前診断に使用される様々な試料において、マイクロサテライト(Levett LJ, et al. A large-scale evaluation of amnio-PCR for the rapid prenatal diagnosis of fetal trisomy. Ultrasound Obstet Gynecol 2001; 17: 115-8)若しくは単一ヌクレオチド多型(SNP)マーカー(Tsui NB, et al. Detection of trisomy 21 by quantitative mass spectrometric analysis of single-nucleotide polymorphisms. Clin Chem 2005; 51 : 2358-62)、又はリアルタイム非デジタルPCRを使用してトリソミーを検出すする、代替的なアプローチである。
【0201】
T18の検体を検出するという試みにおいて、発明者らは、3個の正倍数体及び5個のT18の胎盤DNA試料において、chr21/chr18アッセイを利用した。全ての情報的ウェル中の過剰出現マーカーの割合(Pr)が計算された。1個のT18検体が正倍数体と誤分類したのを除き、他の全ての試料は、正しく分類された。それらの結果は、図20に概説された。
【0202】
V.質量分析プラットフォーム上の多重デジタルRCDアッセイの使用
単位体積の母体血漿あたりに存在する血漿DNA分子の数は限られている(Lo YMD. et al. 1998 Am J Hum Genet 62, 768-7758)。例えば、妊娠早期において、常染色体遺伝子座、β-グロビン遺伝子の中間母体血漿濃度は、母子両方由来のものを合わせても、986コピー/mlであることが示されている(Lo YMD. et al. 1998 Am J Hum Genet 62, 768-7758)。7680分子を確保するためには、約8mlの母体血漿からDNAを抽出することが必要な場合がある。この血漿の体積は、約15mlの母体血液から取得される量であり、この量は日常的な実施の限界に近い。しかしながら、発明者らは、福数のセットのchr21及び参照染色体標的が、デジタルRCD解析に組み合わせられ得ることを想定する。5ペアのchr21及び参照染色体標的において、解析に必要な鋳型分子の個数を提供するのに必要となり得る母体血漿は、僅か1.6mlである。多重単一分子PCRが遂行され得る。この多重単一分子解析のロバストネスは、以前単一分子ハプロタイピングにおいて明示されている(Ding, C. and Cantor, CR. 2003 Proc Natl Acad Sci USA lW, 7449-7453)。
【0203】
一例において、胎盤組織及び母体血球細胞試料は、正倍数体又はトリソミー21(T21)胎児を妊娠した妊婦から取得された。5個の正倍数体及び5個のT21胎盤DNA試料を、それぞれ等量の母体血球細胞DNAと混合し、50%胎児DNAを含む血漿試料を模した、10個のDNA混合物を作り出した。それらの混合試料中の胎児の割合の正確さを保証するため、まずは抽出されたDNAが、260nmで光学密度(OD)を測定することにより定量された。それからそれらは、384ウェルの形式で、リアルタイムPCRにより、デジタル的に定量された。それらの試料を定量するアッセイは、前記デジタルRCD解析の例と同様に行われた。
【0204】
前記50%混合物中の染色体量は、第1番染色体と第21番染色体との非多型遺伝子座を比較する、デジタルPCR解析により決定された。この方法は、デジタル相対的染色体量(RCD)解析と呼ばれる。
第21番及び第1番の染色体上の一対のパラロガス遺伝子座の121bp(各プライマー上の10bpを含む)のアンプリコンを、まず、順プライマー5'-ACGTTGGATGGTTGTTCTGCAAAAAACCTTCGA-3'及び逆プライマー5'-ACGTTGGATGCTTGGCCAGAAATACTTCATTACCATAT-3'により同時に増幅した。第21番染色体と第1番染色体との間の塩基の相異を標的にする延長プライマーが設計され、その配列は、
5'-CTCATCCTCACTTCGTACCTC-3'であった。
【0205】
T21検体を検出するための多重デジタルPCRアッセイの利用を示すために、第21番及び第18番染色体のパラロガス配列を標的とするもう一つのデジタルRCDアッセイが設計された。第21番及び第18番の染色体上の一対のパラロガス遺伝子座の148bp(各プライマー上の10bpを含む)のアンプリコンを、まず、順プライマー5'-ACGTTGGATGGTACAGAAACCACAAACTGATCGG-S'及び逆プライマー5'-ACGTTGGATGGTCC AGGCTGTGGGCCT-3'により同時に増幅した。第21番染色体と第18番染色体との間の塩基の相異を標的にする延長プライマーが設計され、その配列は、
5'-ACAAAAGGGGGAAGAGG-3'であった。
【0206】
多重デジタルRCD解析は、プライマー伸長プロトコールを使用して遂行された。PCR反応は、5μlの反応体積で、GeneAmp PCR Core Reagent Kit (Applied Biosystems)を使用して準備された。各反応は、1xBufferII、2 mMのMgCl2、200μMのdNTP混合物、0.2UのAmpliTaq Gold、各200nMの4種類のプライマー、及び50% DNA混合物を含むものとした。このアッセイ/試料混合物を384ウェルPCRプレートに充填し、そして反応は、50℃2分で始まり、そして95℃10分、更に40サイクルの95℃15秒及び57℃1分が続いた。
【0207】
PCR生産物は、シュリンプアルカリホスファターゼ(SAP)で処理され、遊離dNTPが除去された。この混合物を37℃で40分、続いて85℃で5分インキュベーションした。プライマー伸長反応は、それから実行された。手短に説明すると、伸長カクテル中のchr21/chr1アッセイ用の伸長プライマー771nM、chr21/chr18アッセイ用の伸長プライマー1.54nM、0.67UのThermosequenase (Sequenom)、並びにそれぞれ64μMのddCTP、 ddGTP、dATP、及びdTTPが、SAP処理されたPCR産物に添加された。反応条件は、94℃2分、続いて94℃5秒、50℃5秒、及び72℃5秒を80サイクルとした。最終洗浄として、16μlの水及び3mgのClean Resin (Sequenom)が前記伸長生産物に添加された。この混合物をローテーター中で20〜30分間混合し、続いて351gで5分間遠心分離した。MassARRAY Nanodispenser S (Sequenom)により、15〜25nLの最終生産物が、SpectroCHIP (Sequenom)上に分注された。SpectroCHIPからのデータの取得は、MassARRAY Analyzer Compact Mass Spectrometer (Sequenom)中で行われた。質量データは、解析のため、MassARRAY Typer (Sequenom)ソフトウェア中に移された。
【0208】
5個の正倍数体及び5個のT21の50%胎盤/母体DNA試料は、二重RCDアッセイを用いて解析された。各試料において、個別のアッセイからの情報的ウェル、即ちchr21又はchr1又はchr18マーカーのいずれか1つのみ陽性であるウェルの個数が計数された。全ての情報的ウェル中のchr21マーカーが陽性のウェルの割合(Pr)は、各RCDアッセイについて別個に計算された。それから、Prが正倍数体又はT21試料のいずれを示唆するものであるかを決定するため、逐時確率比検定(SPRT)が利用された。こうすることにより、各ウェルが2回計数されたことになるので、必要なウェルの個数が減少した。
【0209】
Chr21/chr1アッセイは、通常最初に行われる。試料に分類不可能なものが残っていた場合は、chr21/chr18アッセイから得た数値を更なる計算のために追加する場合がある。追加のプレートは、分類不可能なものが残っている試料に、分類が可能となるまで使用され得る。図21に示されるように、全ての正倍数体50%混合試料は、単一の384ウェルプレートの使用で、正しく分類された。幾つかのT21検体は、正しい分類のために、2枚以上のプレートを必要とした。もし1つのアッセイしか使用されないとしたら、分類が達成されるのに必要な数の情報的ウェルを確保するのに、より多くのプレートが必要であった。例えば、検体N0230は、いずれかのRCDアッセイのみを使用しても分類できなかった。しかしながら、2つのアッセイを組み合わせてデータを得たとき、正しい分類が達成された。もしこの二重RCDアッセイが使用されないとすると、プレートを追加しての解析が必要となっていた。発明者らは、より高いレベルのアッセイの多重化を用いることで、更にウェル数を減少させることを意図する。
【0210】
もう一つの実施例において、発明者らは、第21番染色体上の4つの異なるアンプリコンと、第21番染色体以外の常染色体上に位置するそれらの対応するパラロガスパートナーを標的とする、4重アッセイを開発した。この4重アッセイは、正倍数体及びトリソミー21妊娠対象由来の試料のSPRT分類の前の、デジタルRCD解析において使用された。胎盤試料からのDNAの抽出は、QIAamp組織キット(Qiagen, Hilden, Germany)を使用して遂行された。
【0211】
本研究で使用される全ての胎盤及び母体軟膜DNA使用は、まずNanoDrop 分光光度計(NanoDrop Technology, Wilmington, DE)で定量された。DNA濃度を、6.6 pg/細胞の転換を使用して、ゲノム当量(GE)/μLに転換した。ウェルあたり約1個の鋳型が入る濃度に相当するDNA濃度は、DNA試料の連続希釈により決定された。その条件下で、発明者らは、約37%のウェルが増幅において陰性であると予想し得る。多重デジタルRCD解析において、4セットのパラロガス配列標的が選択され:
第21番及び第1番染色体上のパラロガス遺伝子座は、順プライマー5'-ACGTTGGATGTTGATGAAGTCTCATCTCTACTTCG 3'及び逆プライマー5'-ACGTTGGATGCAATAAGCTTGGCCAGAAATACT-3'により同時に増幅され、81bpのアンプリコンが生じる。
第21番及び第7番染色体上のパラロガス遺伝子座は、順プライマー5'-ACGTTGGATGGAATTTAAGCTAAATCAGCCTGAACTG-3'及び逆プライマー5'-ACGTTGGATGGTTTCTCATAGTTCATCGTAGGCTTAT-3'により同時に増幅され、82bpのアンプリコンが生じる。
第21番及び第2番染色体上のパラロガス遺伝子座は、順プライマー5'-ACGTTGGATGTCAGGCAGGGTTCTATGCAG-3'及び逆プライマー5'-ACGTTGGATGAGGCGGCTTCCTGGCTCTT-3'により同時に増幅され、101bpのアンプリコンが生じる。
第21番及び第6番染色体上のパラロガス遺伝子座は、順プライマー5'-ACGTTGGATGGCTCGTCTCAGGCTCGTAGTT-3'及び逆プライマー5'-ACGTTGGATGTTTCTTCGAGCCCTTCTTGG-3'により同時に増幅され、102bpのアンプリコンが生じる。
各反応は、10x緩衝剤II(Applied Biosystems)、MgCl2、及び100nMの各プライマーを含んでいた。反応体積の合計は、5μL/ウェルであった。この反応は、95℃5分で始まり、そして45サイクルの95℃30秒、62℃30秒及び72℃30秒が続き、最後に72℃7分が追加された。公知のPCR増幅は全て、GeneAmp PCR Core Reagent Kit (Applied Biosystems)で遂行された。遊離ヌクレオチドは、シュリンプアルカリホスファターゼ(SAP)により不活性化された。各反応は、10xSAP緩衝剤(Sequenom)及びSAP酵素(Sequenom)を含むものであった。各PCRに、2μlのSAP混合物が添加された。このSAP反応物は、37℃で40分、及び85℃で5分インキュベートされた。SAP処理の後、iPLEX Gold kit (Sequenom)を使用して、PCR生産物のプライマー伸長反応が行われた。第21番及び第1番染色体上のパラロガス配列ミスマッチ(PSM)が、伸長プライマー5'-GTCTCATCTCTACTTCGTACCTC-3'により探索(interrogate)された。第21番及び第7番染色体上のPSMは、伸長プライマー5'-TTTTACGCTGTCCCCATTT-3'により探索された。第21番及び第2番染色体上のPSMは、伸長プライマー5'-GGTCTATGCAGGAGCCGAC-3'により探索された。第21番及び第6番染色体上のPSMは、伸長プライマー5'-TGGGCGCGGGAGCGGACTTCGCTGG-3'により探索された。各反応は、10xPLEX緩衝剤(Sequenom)、iPLEX停止混合物(Sequenom)、iPLEX酵素(Sequenom)及び343nMの各伸長プライマーを含むものであった。但し、第21番及び第6番染色体上のPSMに対する伸長プライマーは、1.3μMで使用した。2μlのiPLEX混合物が5μlのPCR線産物に添加された。このiPLEX反応は、200ショートサイクルプログラムに従いサイクル反応に付された(cycled)。要するに、前記試料は、まず94℃で35秒変性し、続いて52℃で5秒アニーリングし、そして80℃で5秒伸長した。アニーリング及び伸長サイクルは、5サイクルが4回以上反復され、それから94℃5秒の変性工程に戻り、その後、再び前記5サイクルのアニーリング及び伸長ループに戻された。前記5回のアニーリング及び伸長サイクル及び1回の変性工程が、合計40と成るように39回繰り返された。最後に、72℃で3分、伸長が行われた。iPLEX反応生産物は、16μlの水で希釈され、各PCRのために、6mgの樹脂で脱塩した。384ウェルを1600gで3分間遠心して、SpectroCHIP (Sequenom)に分注して、マトリックス支援レーザー脱離/イオン化-飛行時間(MALDI-TOF)質量分析MS Analysis (Sequenom)に付した。
【0212】
各4つのアッセイにおいて、第21番又は参照染色体のいずれか1つのみ陽性であるウェルの数が、独立して記録された。各アッセイにおいて、ポアソン補正した第21番染色体及び参照染色体の数が計算された。4つのアッセイ全ての第21番染色体及び参照染色体のポアソン補正した数の合計が計算され、4重のアッセイの情報的カウントとみなされた。Pr値は、4重のアッセイの第21番染色体のカウントを、4重のアッセイの第21番染色体及び参照染色体のカウントの合計で割った値とした。実験的に導き出したPr値は、SPRT解析に供された。1枚以上の384ウェルプレートが、SPRTによる疾患分類が可能となるまで解析された。2つの50%胎盤ゲノムDNA/50%母体軟膜DNA混合物と、2つの50%トリソミー21胎盤ゲノムDNA/50%母体軟膜DNA混合物との合計が解析された。
【0213】
前記実験的に導き出されたPr値を、予想されたPr値と比較して、帰無仮説又は対立仮説を検査する。あるいは、所定の情報的カウントのPrが疾患分類のための統計的信頼度の要求されるレベルにまだ達していなければ、帰無仮説及び対立仮説のいずれも許容され得ない。これらの検体は、更にデータが取得されない限り、分類不可能とみなされた。
【0214】
それらの結果及び各試料のSPRT分類は、図22A及び22Bに示す。2つの正倍数体試料は、SPRT分類が可能となるまでに、それぞれ2回及び5回の384ウェルデジタルRCD解析を必要とした。4重アッセイの各構成要素の独立したデータは、それらのいずれもSPRTによる疾患分類が不可能であった。両トリソミー21試料は、それぞれ僅か1重の384ウェルデジタルRCD解析を用いて正しく分類された。同様に、4重アッセイの各構成要素の独立したデータは、それらのいずれもSPRTによる疾患分類が不可能であった。しかしながら、4重アッセイから得た合成カウントは、正しいSPRT分類を可能とした。これらのデータは、多重デジタルRCDを使用することにより、所定の回数実行されるデジタルPCR解析において、1重デジタルRCDアッセイの使用と比較して、効果的な数の情報的カウントが実質的に増大することを示すものであった。
【0215】
VI.デジタルエピジェネティック相対的染色体量の使用
ここで、発明者らは、デジタルエピジェネティック相対的染色体量(デジタルERCD)と称されるアプローチを紹介する。ERCDにおいて、染色体の異数性に関与する染色体(例えばトリソミー21における第21番染色体)及び参照染色体上に、胎児特異的DNAメチル化パターン又は他のエピジェネティックな変化を呈するエピジェネティックマーカーが、デジタルPCR解析に供される。正常な胎児を妊娠した妊婦から抽出した血漿DNA中の、第21番染色体エピジェネティックマーカーが陽性であるウェルの数と、参照染色体エピジェネティックマーカーが陽性であるウェルの数との比率は、参照範囲(reference range)を提供し得る。この比率は、胎児がトリソミー21を有すると増大するものと予想される。この解析において、1つ以上の第21番染色体マーカー及び1つ以上の参照染色体マーカーが使用され得ることは、当業者に明らかである。
【0216】
胎児(胎盤)特異的メチル化パターンを呈する第21番染色体上の遺伝子の一例として、ホロカルボキシラーゼ合成酵素(HLCS)遺伝子が挙げられる。HLCSは、胎盤中で高メチル化されるが、母体血球細胞では低メチル化されている。これは米国特許出願第11/784499号の範囲内であり、本明細書中に参照により援用される。胎児(胎盤)特異的メチル化パターンを呈する参照染色体上の遺伝子の一例として、第3番染色体上のRASSF1Aが挙げられる[10]。RASSF1Aは、胎盤中で高メチル化されるが、母体血球細胞では低メチル化されている。本明細書に参照により援用される米国特許出願第11/784501号を参照されたい。
【0217】
母体血漿細胞を使用して行う胎児のトリソミー21を検出するデジタルPCRに、高メチル化HLCS及び高メチル化RASSF1Aを適用するとき、母体末梢血細胞がまず採取される。それから、その血液は、遠心分離にかけられ、血漿が回収される。更にその血漿からDNAが、QIAamp Blood kit (Qiagen)等の当業者に周知の技術を使用して抽出される。この血漿DNAは、更に、HpaII及びBstUI等の1つ以上のメチル化感受性制限酵素を使用して消化される。これらのメチル化感受性制限酵素は、メチル化されていない母体由来の遺伝子を切断するが、胎児由来の高メチル化された遺伝子を切断しない。消化された血漿DNA試料は、それから、制限酵素処理されても無傷のHLCS又はRASSF1A配列のいずれかが反応ウェル中に、平均で約0.2〜1分子検出され得る程度に希釈される。この希釈されたDNAを増幅するのに、2つのリアルタイムPCRシステムが使用され得る。一つは、HLCS遺伝子がメチル化していなければ前記制限酵素により切断されるべき領域を網羅する、2つのプライマー、及びHLCS遺伝子に特異的な1つのTaqManプローブが用いられる。もう一つは、同様に2つのプライマー及び1つのプローブであってRASSF1Aに対するものが用いられる。校舎のRASSF1Aのプライマー/プローブセットに関して、Chan et al 2006, Clin Chem 52, 2211-2218に、一つの実施例が記載されている。HLCS及びRASSFlAを標的とするTaqManプローブは、それぞれFAM及びVIC等、異なる蛍光レポーターを有する場合がある。よって、このデジタルPCR実験を実行するのに、1枚の384ウェルプレートで足りる場合がある。HLCS又はRASSF1Aのいずれか1つのみが陽性と評点されるウェルの個数が計数され、これらのカウントの比率が取得され得る。このHLCS:RASSF1A比率は、正常な正倍数体の胎児を妊娠した妊婦から採取した母体血漿と比較して、トリソミー21の胎児を妊娠した妊婦から採取した母体血漿のほうが高くなると予想される。その過剰出現の程度は、デジタルPCR反応におけるウェルあたりの平均参照鋳型濃度に依存し得る。
【0218】
これらの結果を評点するのに、例えば、RASSF1Aの陽性に無関係にHLCSが陽性であるウェルの数を計数し;及び逆にHLCSの陽性に無関係にRASSF1Aが陽性であるウェルの数を計数する等の、他の方法を採用することも可能である。更に、胎児のトリソミー21状態を示すために、前記比率を計算する代わりに、HLCS及びRASSF1Aカウントの合計又は差異のいずれかを使用することも出来る。
【0219】
プレート中のデジタルPCR実行とは別に、当業者には、他のデジタルPCRの変法、例えばマイクロ流体工学チップ、ナノリットルPCRマイクロプレートシステム、エマルジョンPCR、ポロニー(polony)PCR及びローリングサークル増幅、プライマー伸長、及び質量分析等が使用され得ることも明らかである。これらのデジタルPCRの変法は例示を目的として列挙されもので、限定を目的とするものではない。
【0220】
リアルタイムPCRとは別に、当業者には、デジタルPCRの結果を評点するために、質量分析等の方法が使用され得ることも明らかである。
【0221】
HLCS及びRASSF1Aの胎児型と母親型とを区別するためのメチル化感受性制限酵素の使用とは別に、当業者には、例えば亜硫酸水素修飾、メチル化特異的PCR、抗メチル化シトシン抗体を使用する免疫沈降法、質量分析等の、メチル化状態を確認する他の方法も適用可能であることは明らかである。
【0222】
また、当業者には、本実施例及び本発明の他の実施例に説明されたアプローチが、胎児DNAが見出され得る、母体尿、羊水、経頸管流出物(transcervical washing)、絨毛膜、母体唾液等を含む他の生体液においても使用され得ることは明らかである。
【0223】
VII.エマルジョンPCR及びその他の方策を使用する、超並列ゲノムシークエンシング
ここで、発明者らは、核酸分子のデジタル読出しが、母体血漿中のトリソミー21等の胎児染色体異数性の検出に使用され得るというもう一つの例を記載する。胎児染色体の異数性は、染色体又は染色体領域の量が異常であることにより引き起こされる。誤診断を最少化するために、非侵襲的検査は、高い感受性及び特異性を有することが望ましい。しかしながら、胎児DNAは母体血漿及び血清中で絶対的に低濃度で存在し、全てのDNA中ごく一部を占めるに過ぎない。よって、特定の遺伝子座を標的とするデジタルPCRサンプリングの数は、同一の標本中で無限に増大させることは出来ない。故に、実行されるデジタルPCRサンプリングの数を増大させずに、1つの標本から取得され得るデータの量を増大させるのに、特異的な標的遺伝子座の複数のセットの解析が用いられ得る。
【0224】
従って、態様によっては、母体バックグラウンド核酸を含有する生体試料中に僅かに存在する限定的な量の胎児核酸から推察され得る遺伝的情報の量を最大化することによる、胎児染色体の異数性の非侵襲的検出が可能となる。一つの側面において、取得される遺伝的情報の量は、正しい診断をするのに十分であるが、必要とされるコスト及び生体試料のインプット量はそれほど多くはない。
【0225】
454プラットフォーム(Roche)(Margulies,M.et al.2005 Nature 437,376−380)、Illumina Genome Analyzer (またはSolexaプラットフォーム)、またはSOLiDシステム(Applied Biosystems)、またはHelicos True Single Molecule DNAシークエンシング技術(Harris TD et al.2008 Science,320,106−109)、Pacific Biosciencesのsingle molecule,real−time(SMRT(商標))技術、およびナノポアシークエンシング(Soni GV and Meller A.2007 Clin Chem 53:1996−2001)等により達成され得る超並列シークエンシングは、並行方式において高次に多重化している標本から単離された多くの核酸分子のシークエンシングを可能とする(Dear Brief Funct Genomic Proteomic 2003;1:397−416)。これらのプラットフォームの各々は、クローン的に増大された、または増幅さえされていない核酸断片の単一分子をシークエンシングする。
【0226】
各反応において、多数のシークエンスデータが、10万〜100万または場合により数百万、または10億個のオーダーで各々の試料から生じるので、得られる配列決定されたデータは、元の標本における核酸種の混合物の代表的な特性を形成する。例えば、シークエンシングされたデータのハプロタイプ、トランスクリプトーム、およびメチル化プロファイルは、元の標本のものと類似する(Brenner et al Nat Biotech 2000;18:630−634;Taylor et al Cancer Res 2007;67:8511−8518)。各標本から大量の配列をサンプリングするために、同一配列の数、例えば数倍の範囲または高い冗長性での核酸プールのシークエンシングから生じたものはまた、元の試料中の特定の核酸種または遺伝子座のカウントの優れた定量的表示である。
【0227】
一つの態様において、ランダムシークエンシングは、妊婦の血漿中に存在するDNA断片に対して行われ、これにより母親又は胎児のいずれかを起源とするゲノム配列が得られる。ランダムシークエンシングは、生体試料中に存在する核酸分子のランダムな部分をサンプリング(シークエンシング)することを含む。配列がランダムであるから、核酸分子(即ちそのゲノム)の異なる小集団(分画)は、各解析においてシークエンシングされ得る。態様によっては、同一の試料を使用しているにも拘らず試料毎に、及び解析毎にこの小集団が変化するときであっても作用し得る。分画の例は、ゲノムの約0.1%、0.5%、又は1%である。他の態様において、この分画は、少なくともこれらの数値のいずれか1つである。
【0228】
バイオインフォマティクスの手法がその後、これらのDNA配列の各々をヒトゲノムへと配置するために使用され得る。かかる配列は、ヒトゲノムの繰り返し領域、または個体間変動、例えばコピー数変動を受ける領域において存在するため、かかる配列比率は、その後の解析から排除される可能性がある。目的の染色体の量、および一つ以上の他の染色体の量がこのようにして決定され得る。
【0229】
一つの態様において、染色体異数性に潜在的に関与する染色体、例えば第21番染色体、第18番染色体、または第13番染色体のパラメーター(例えば分数表示(fractional representation))はしたがって、バイオインフォマティクスの手順の結果から計算され得る。当該分数表示は、全ての染色体配列(例えば臨床的に重要な染色体を含む全ての染色体の幾つかの測定)、または特定の染色体小集団(例えば試験されるもの以外のちょうど一つのもの)の量に基づいて得られる。
【0230】
一つの態様において、パラメーター(例えば、臨床的に重要な染色体の分数表示)はその後、正常の(すなわち正倍数体の)胎児に関与する妊娠において確立された基準範囲と比較される。当該手順における幾つかの変法において、基準範囲(すなわちカットオフ値)は、特定の母体血漿試料中における胎児DNAの分別濃度(f)に従って調節される可能性がある。fの値は、シークエンシングのデータセットから、例えば胎児が男である場合に、Y染色体にマッピング可能な配列を使用して決定され得る。fの値はまた、分離した分析、例えば胎児のエピジェネティックマーカーを使用して(Chan KCA et al 2006 Clin Chem 52,2211−8)、または一塩基多型の分析から決定されてもよい。
【0231】
一つの態様において、標本中における核酸のプールが、100%未満の遺伝子範囲でシークエンシングされるときでさえ、捕獲された核酸分子の比率中において、大部分の各々の核酸種が一回のみシークエンシングされる。同様に、特定の遺伝子座または染色体の量の不均衡は、定量的に決定され得る。すなわち、遺伝子座または染色体の量の不均衡は、当該標本のマッピング可能な、他のシークエンシングされたタグ中における前記遺伝子座のパーセンテージ表示から推測される。
【0232】
超並列ゲノムシークエンスアプローチの一つの側面において、全ての染色体から代表的データが、同時に生産され得る。特定の断片の起源は、予め選択されない。シークエンシングはランダムに行われ、それからデータベース検索が行われ、特定の断片がどこに由来するかを確認する。これを、第21番染色体由来の特定の断片と、第1番染色体由来のもう一つが増幅された状況と対比する。
【0233】
一つの実施例において、かかる配列の割合は、異数性に関与する染色体、例えば本実例において第21番染色体から得られる。かかるシークエンシングの実行により得られたさらに別の配列は、他の染色体に由来し得る。他の染色体と比較した、第21番染色体の相対的大きさを考慮することによって、基準範囲内における、かかるシークエンシングから第21番染色体特異的配列を引き当てる正規化頻度を得ることができる。胎児がトリソミー21を有するならば、かかるシークエンシングから、第21番染色体由来配列を引き当てる正規化頻度は増加し、したがってトリソミー21の検出を可能とするだろう。正規化頻度における変化の程度は、分析試料中における胎児核酸の分別濃度に依存するだろう。
【0234】
一つの態様において、ヒトゲノムおよびヒト血漿DNA試料の単一端シークエンシングのために、我々はIlluminaゲノムアナライザーを使用した。Illuminaゲノムアナライザーは、フローセルと呼ばれる固体表面上に捕獲された、クローン的に増大された単一のDNA分子をシークエンシングする。各々のフローセルは、8個の個々の標本または標本のプールをシークエンシングするための8つのレーンを有する。各々のレーンは、ヒトゲノム中における30億塩基対の配列の一部のみである、約200Mbの配列を生じさせることが可能である。各々のゲノムDNAまたは血漿DNA試料は、一つのフローセルのレーンを使用してシークエンシングされた。生じた短い配列タグは、ヒト基準ゲノム配列と整列され、そして染色体の複製起点が示された。各々の染色体に整列された、個々のシークエンシングされたタグの総数は表にされ、そして基準ヒトゲノムまたは非疾患性の代表的標本から期待される各々の染色体の相対的なサイズと比較された。染色体の獲得または喪失はその後同定された。
【0235】
記載された手法は、本明細書において記載された遺伝子/染色体用量の方法の一つの例示にすぎない。代替的に、ペアエンドシークエンシング(paired end sequencing)が行われ得る。シークエンシングされた断片の長さとCampbell他(Nat Genet 2008;40:722−729)によって記載された基準ゲノムにおいて期待されたものとを比較する代わりに、整列され、シークエンシングされたタグ数が数えられ、そして染色体の位置に従ってソートされる。染色体領域、または染色体全体の獲得または喪失は、タグの数と基準ゲノムにおいて期待される染色体の大きさ、または非疾患性の代表的な標本のものとを比較することによって決定された。
【0236】
別の態様において、一回の反応においてシークエンシングされる核酸プール画分は、シークエンシング前にさらに補助的に選択される。例えば、ハイブリダイゼーション型の技術、例えばオリゴヌクレオチドアレイは、特定の染色体、例えば潜在的に異数性の染色体および試験された異数性に関与しない他の染色体(単数または複数)から、核酸配列を初めに補助的に選択するために使用され得る。別の例は、試料プールからの核酸配列の特定の下位群が、シークエンシング前に補助的に選択されるか、または増大される。例えば上で議論されたように、母体血漿中における胎児DNA分子は、母体のバックグラウンドDNA分子よりも短い断片を含むことが報告された(Chan et al Clin Chem 2004;50:88−92)。したがって、例えばゲル電気泳動、もしくはサイズ排除カラムによって、またはマイクロ流体工学的手法によって、分子サイズに従って試料中における核酸配列を分画するために、当業者に既知の一つ以上の方法を使用し得る。さらに、代替的には、母体血漿中の無細胞胎児DNAを分析する例において、胎児核酸部分は、母体バックグラウンドを抑制する方法によって、例えばホルムアルデヒドの添加によって増幅され得る(Dhallan et al JAMA 2004;291:1114−9)。一つの態様において、補助的に選択された核酸のプールの一部分または小集団は、ランダムにシークエンシングされる。
【0237】
他の単一分子シークエンシングの方法、例えばRoche454プラットフォーム、Applied Biosystems SOLiDプラットフォーム、Helicos True Single Molecule DNAシークエンシング技術、Pacific Biosciencesの単一分子リアルタイム(SMRT(登録商標))技術、およびナノポアシークエンシングによる方法が、本出願において同様に使用され得る。
【0238】
結果及び更なる考察の実施例(例えばシークエンシング及び計算パラメーター)は、同時に出願した、"DIAGNOSING FETAL CHROMOSOMAL ANEUPLOIDY USING GENOMIC SEQUENCING," (Attorney Docket No. 016285-005220US)に見出される。この出願は、参照により援用される。カットオフ値を決定するための本明細書中に記載の方法は、本章に記載のように、その反応がシークエンシングであったときに運用され得る。
【0239】
母体血漿中における胎児DNAの分別濃度の決定はまた、シークエンシングの実施とは別になされ得る。例えば、Y染色体DNA濃度は、リアルタイムPCR、マイクロ流体工学PCR、または質量分析を使用してあらかじめ決定され得る。実際、胎児DNA濃度は、Y染色体以外の女性胎児において適用可能な遺伝子座を使用して決定され得る。例えば、Chanらは、胎児由来のメチル化されたRASSF1A配列が、母体由来のメチル化されていないRASSF1A配列のバックグラウンドにおける妊娠女性の血漿中において検出されることを示した(Chan et al,Clin Chem 2006;52:2211−8)。したがって、分別胎児DNA濃度は、メチル化されたRASSF1A配列の量を全RASSF1A(メチル化されたもの、およびメチル化されていないもの)配列で割ることによって決定され得る。
【0240】
血液凝固の間に、DNAが母体の血液細胞から放出されるため、本発明を実施するにあたり、母体血清よりも母体血漿が好ましいことが予想される。したがって、血清が使用されるならば、胎児DNAの分別濃度は、母体血漿中において母体血清中よりも低いだろうと予想される。すなわち、母体血清が使用されるならば、同時に、同一の妊娠女性から得られる血漿試料と比較して、胎児染色体異数性を診断するためにより多くの配列が必要となると予想される。
【0241】
胎児DNAの分別濃度を決定するためのさらに別の代替的な方法は、妊娠女性と胎児との間の多型の違いの定量を通じてなされ得る(Dhallan R, et al.2007 Lancet,369,474−481)。この方法の例は、妊娠女性がホモ接合性であり、そして胎児がヘテロ接合性である多型部位を標的とし得る。胎児特異的対立遺伝子の量は、胎児DNAの分別濃度を決定するために、共通の対立遺伝子の量と比較し得る。
【0242】
一つ以上の特異的配列を検出及び定量する、比較ゲノムハイブリダイゼーション、マイクロアレイ比較ゲノムハイブリダイゼーション、定量的リアルタイムポリメラーゼ連鎖反応を含む、染色体異常を検出するための既存の技術とは対照的に、超並列シークエンシングは、あらかじめ決定もしくは定義されたDNA配列のセットの検出または分析に依存しない。標本プールからの、DNA分子のランダムな代表的断片がシークエンシングされる。様々な染色体領域に整列された異なる配列タグの数を、腫瘍性DNAを含む標本または含まない標本間で比較した。染色体異常は、標本中の所定の染色体領域に整列された配列の数(またはパーセンテージ)の相異によって明らかとされ得る。
【0243】
別の例において、血漿の無細胞DNAにおけるシークエンシング技術を、特定の癌の検出のために、血漿DNA中の染色体異常を検出するために使用してもよい。異なる癌は、典型的な染色体異常のセットを有する。複数の染色体領域における変化(増幅および欠失)が使用され得る。したがって、増幅された領域に整列された配列の割合が増大し、現象した整列された配列の割合が減少し得る。染色体あたりのパーセンテージ表示は、全ゲノムと関連する、与えられた任意の染色体のゲノム表示のパーセンテージとして表現される参照ゲノム中における各々の対応する染色体に関するサイズと比較され得る。直接的な比較、または参照染色体との比較が同様に使用され得る。
【0244】
VIII.突然変異検出
母体血漿中の胎児DNAは、母体血漿DNAの平均3〜6%を占める、胎児由来の少数の集団として存在する。そのため、当該技術分野の従来技術は、胎児が父親から受け継いで、母体血漿における大勢の母体DNAバックグラウンドから区別できるDNA標的の検出に焦点を当てていた。そのような従来検出されていた標的の例として、Y染色体上のSRF遺伝子(Lo YMD et al. 1998 Am J Hum Genet, 62, 768-775)、及び母親がRhD陰性であればRHD遺伝子(Lo YMD et al. 1998 N Engl J Med, 339, 1734- 1738)が挙げられる。
【0245】
胎児突然変異を検出するための従来の母体血漿を使用する方策は、父親が保因者である常染色体優性の条件に限定されており、父親と母親が異なる突然変異を保有していた場合の、直接突然変異を検出することによる、又は連鎖解析による、常染色体劣性疾患を除外していた(Ding C. et al 2004 Proc Natl Acad Sci USA l0l, 10762- 10767)。これらの従来の方策は、顕著に限定されていた。例えば、父親と母親とがいずれも同一の突然変異を保有していた場合、母体血漿中の直接の突然変異検出による有意な出生前診断を遂行することは不可能となる。
【0246】
そのようなシナリオが、図23に説明されている。このシナリオにおいて、NN、NM、及びMMという、3つの可能な胎児遺伝子型が存在し得て、ここで、Nは正常対立遺伝子を表し、Mは突然変異対立遺伝子を表す。突然変異対立遺伝子の例としては、嚢胞性線維症、ベータサラセミア、アルファサラセミア、鎌状赤血球貧血症、脊髄性筋萎縮症、先天性副腎過形成の原因遺伝子等が挙げられる。そのような障害の他の例は、Online Mendelian Inheritance in Man (OMIM) www.ncbi.nlm.nih.gov/sites/entrez?db=OMIM&itool=toolbarで閲覧できる。母体血漿中で、殆どのDNAは母体由来であり、NMであり得る。3つの胎児遺伝子型において、母体血漿中で特異的に検出され得る特異的な胎児対立遺伝子は存在し得ない。故に、従来の方策は、この場合は利用し得ない。
【0247】
本明細書に記載の態様は、そのようなシナリオに対処することが出来る。母親及び胎児がいずれもNMとなるシナリオにおいて、対立遺伝子N及びMは、両対立遺伝子は均衡を保っている。しかしながら、母親がNMで胎児がNNである場合、母体血漿中で、対立遺伝子Nが過剰出現して、対立遺伝子の不均衡が生じる。他方、母親がNMで胎児がMMである場合、母体血漿中で、対立遺伝子Mが過剰出現して、対立遺伝子の不均衡が生じる。故に、胎児突然変異の検出において、帰無仮説は、胎児の遺伝子型がNMであるときに、対立遺伝子の不均衡が存在しないことを意味する。対立仮説は、対立遺伝子の不均衡が存在し、対立遺伝子N又はMのいずれが過剰出現しているかに依存して、胎児の遺伝子型がNN又はMMであることを意味する。
【0248】
対立遺伝子の不均衡の存否は、本明細書中に記載のデジタルPCRを使用することにより決定され得る。第一のシナリオにおいて、特定の体積の母体血漿が100個の細胞から放出されたDNAを含み、それらの細胞の内50個が母体由来で、50個が胎児由来であるとする。そうすると、この体積の血漿中の胎児DNAの分別濃度は50%となる。母親の遺伝子型がNMであるとき、母親由来の対立遺伝子Nが50個、及び対立遺伝子Mが50個存在し得る。胎児の遺伝子型がNMであるとき、胎児母親由来の対立遺伝子Nが50個、及び対立遺伝子Mが50個存在し得る。よって、対立遺伝子NとMと間に対立遺伝子の不均衡は存在せず、それらは各100コピーずつ存在する。一方、胎児の遺伝子型がNNであるとき、この体積の血漿中に、胎児由来の対立遺伝子Nが100個存在し得る。そのため、全体で対立遺伝子Nが150個に対し、Mが50個存在し得る。言い換えると、NとMとの間で対立遺伝子の不均衡が存在し、NはMに対し、3:1の比率で過剰出現している。
【0249】
逆に、胎児の遺伝子型がMMであるとき、この体積の血漿中に、胎児由来の対立遺伝子Mが100個存在し得る。そのため、全体で対立遺伝子Mが150個に対し、Nが50個存在し得る。言い換えると、NとMとの間で対立遺伝子の不均衡が存在し、MはNに対し、3:1の比率で過剰出現している。このような対立遺伝子の不均衡は、デジタルPCRにより測定され得る。この対立遺伝子は、参照鋳型とみなされる。デジタルRNA-SNP及びデジタルRCD解析と同様に、田字たるPCR実験における対立遺伝子の実際の区別は、ポアソン確率密度関数に支配される。よって、本シナリオにおける理論上の対立遺伝子の不均衡の程度が3:1であっても、予想される対立遺伝子の不均衡の程度は、デジタルPCR解析の間の、ウェルあたりの平均鋳型濃度に依存し得る。よって、ウェルあたりの平均参照鋳型濃度(mr)にふさわしいSPRT解析等のカットオフの解釈が、検体の分類に使用されるべきである。
【0250】
更に、測定されるべき対立遺伝子の不均衡の程度は、分別DNA濃度に依存する。上記実施例と対照的に、特定の体積の母体血漿が100個の細胞から放出されたDNAを含有し、それらの細胞の90個が母親由来で、10個が胎児由来であることを想定されたい。そうすると、その体積の結晶中の胎児DNAの分別濃度は、10%である。母親の遺伝型がNMであるとすると、母親由来の対立遺伝子Nが90個、及び対立遺伝子Mが90個存在し得る。胎児の遺伝型がNMであるとすると、胎児由来の対立遺伝子Nが10個、及び対立遺伝子Mが10個存在し得る。よって、この場合、対立遺伝子N及びMの間に対立遺伝子の不均衡は存在せず、それぞれが合計100コピーを有している。他方、胎児の遺伝型がNNであるとすると、この体積の血漿中に、胎児由来の対立遺伝子Nが20個存在し得る。そうすると、対立遺伝子Nが合計で110個、対立遺伝子Mが合計で90個存在することとなる。
【0251】
言い換えると、NとMとの間に対立遺伝子の不均等が存在しており、Nが過剰出現している。逆に、状況を逆にして、胎児の遺伝子型がMMであるとすると、この体積の血漿中に、胎児由来の対立遺伝子Mが20個存在することとなる。そうすると、対立遺伝子Mが110個、対立遺伝子Nが90個存在することとなる。言い換えると、NとMとの間に対立遺伝子の不均衡が存在しており、Mが過剰出現している。胎児DNA分別濃度が10%の場合の理論上の対立遺伝子の不均衡の程度は、110:90であり、これは、上述の例で示した、胎児DNAが50%存在するときの比率3:1とは異なっている。よって、胎児DNA分別濃度に相応のSPRT解析等のカットオフの解釈が、検体の分類に使用されるべきである。
【0252】
故に、血漿DNAは抽出されて、例えば従来技術のリアルタイムPCRアッセイ(Lo, et al. 1998 Am J Hum Genet 62, 768-775)により、又は当業者に周知の他の種類の定量要素(quantifier)、例えばSNPマーカー(Dhallan R et al. 2007 Lancet, 369, 474- 481)、及び胎児エピジェネティックマーカー(Chan KCA et al. 2006 Clin Chem, 52, 2211-2218)等により、血漿試料中の母体及び胎児のDNAの量が定量され得る。胎児DNAパーセンテージが計算され得る。それから、デジタルPCR解析の間、各反応ウェルが平均1分子の鋳型(対立遺伝子N又はMのいずれかであり得る)を含み得るように、定量された血漿DNA試料が調製(例えば希釈又は濃縮)される。デジタルPCR解析は、一対のプライマー及び1つが対立遺伝子Nに特異的で、もう1つが対立遺伝子Nに特異的な2つのTaqManプローブを使用して遂行され得る。Mのみ陽性であるウェルの個数及びNのみ陽性であるウェルの個数が計数され得る。対立遺伝子の不均衡の証拠の存否を判定するために、これらのウェルの比率が使用され得る。対立遺伝子の不均衡の統計的な証拠は、当業者に周知の方法、例えばSPRTの使用等により検査され得る。本解析の一つの変法において、Mのみが陽性である、又はM及びNが陽性であるウェルの数を計数することが可能であり;並びにNのみが陽性である、又はM及びNが陽性であるウェルの数を計数することが可能であり;そしてそれらの計数の比率を導き出すことが出来る。繰り返すが、対立遺伝子の不均衡の統計的な証拠は、当業者に周知の方法、例えばSPRTの使用等により検査され得る。
【0253】
デジタル相対突然変異量(relative mutant dosage)(RMD)と称される、胎児遺伝子の突然変異の量の決定は、男性/女性(XX/XY)DNA混合物を使用して実証された。図24Aに記載されるように、男性又は女性由来の血球細胞DNAを男性DNAと混合し、XYのバックグラウンド中に、XX又はXY遺伝子型が、25%及び50%の分別濃度で存在する試料を調製した。
【0254】
加えて、血液細胞試料は、12人の男性対象及び12人の女性対象からも取得された。女性血球細胞DNA(遺伝子型XX)を、それぞれ3倍過剰量の男性血球細胞DNA(遺伝子型XY)と混合し、こうして、図24Bに記載されるように、遺伝子型XYのバックグラウンド75%中に、XX遺伝子型を有するDNAが25%混合された12個のDNA試料を調製した。
【0255】
SPRTの目的は、バックグラウンドDNA中に存在する少数の遺伝子型を判定することであった。75%XY遺伝子型DNAをバックグラウンドとし、25%XX遺伝子型DNAを含むDNA混合物において、少数の対立遺伝子は、75%のDNA由来のYであり得る。試料中のDNAの25%は遺伝子型XXに由来するため、この試料中にDNAが200分子存在しているとすると、150分子が、XY個体を起源としている。故に、対立遺伝子Yの個数は、75個と期待される。また、男性DNAに(遺伝子型XY)由来する対立遺伝子Xの個数も75個である。女性DNAに(遺伝子型XX)由来する対立遺伝子Xの個数は、50個(25の2倍)である。よって、X:Yの比率は、125/75=(1+25%)/(1−25%)=5/3である。
【0256】
この研究の第二の部分において、ベータグロビン遺伝子、即ちヘモグロビンベータ(HBB)遺伝子上に、HbE(G->A)及びCD41/42(CTTT/-)突然変異を担持する、男性及び女性の対象から、血液細胞試料を取得した。全ての可能な遺伝子型(MM、MN、又はNN)を有する男性胎児を妊娠した、ヘテロ接合体(MN、ここでM=突然変異、N=野生型とする)の母親から得られた母体血漿試料を模倣するために、野生型対立遺伝子のホモ接合体(NN)又は前記2つの突然変異の1つに関してのヘテロ接合体(MN)のいずれかである男性に由来する血球細胞DNAのそれぞれを、前記のものと同じ突然変異に関するヘテロ接合体(MN)の女性から採取した血球細胞DNA試料と混合した。こうして、様々な男性/女性分別DNA濃度のDNA混合物を調製した。CD41/42に関するホモ接合体(MM)女性由来の血球細胞DNA使用も、上記DNA混合物の調製に使用された。SPRT分類に使用される実際の男性DNAの比率を確認するため、各DNA混合物の分別男性DNA濃度を、ZFY/Xアッセイを使用して決定した。
【0257】
デジタルZFY/Xアッセイは、SPRTを実証するために、及びDNA混合物中の分別男性DNA濃度を決定するために使用された。X染色体及びY染色体上のジンクフィンガータンパク質配列(ZFX及びZFY)の量を、デジタルPCR解析により決定した。最初に、順プライマー5'-CAAGTGCTGGACTCAGATGTAACTG-3'及び逆プライマー5'-TGAAGTAATGTCAGAAGCTAAAACATCA-3'により、ZFX及びZFY遺伝子座の87bpのアンプリコンを同時に増幅した。X染色体とY染色体との間のパラログを区別するために2つの染色体特異的TaqManプローブが設計され、それらの配列は、それぞれ5'-(VIC)TCTTTAGCACATTGCA(MGBNFQ)-3'及び5'-(FAM)TCTTTACCACACTGCAC(MGBNFQ)-3'とした。
【0258】
突然変異対立遺伝子に対する正常の対立遺伝子のデジタルPCR解析により、DNA混合物中の突然変異の量を決定した。HbE突然変異において、まず、順プライマー5'-GGGCAAGGTGAACGTGGAT -3'及び逆プライマー5'-CTATTGGTCTCCTTAAACCTGTCTTGTAA-3'により、正常及び突然変異対立遺伝子が、同時に増幅された。正常(G)及び突然変異(A)対立遺伝子を区別するために2つの対立遺伝子特異的TaqManプローブが設計され、それらの配列は、それぞれ、5'-(VIC)TTGGTGGTGAGGCC(MGBNFQ)-3'及び5'-(FAM)TTGGTGGTAAGGCC(MGBNFQ)-3'とした。
【0259】
CD41/42欠失突然変異において、まず、順プライマー5'-TTTTCCCACCCTTAGGCTGC-3'及び逆プライマー5'-ACAGCATCAGGAGTGGACAGATC-3'により、正常及び突然変異対立遺伝子の87bp及び83bpのアンプリコンが、同時に増幅された。正常及び突然変異対立遺伝子(欠失無し及び欠失有り)の間を区別する2つの対立遺伝子特異的TaqManプローブが設計され、それらの配列は、それぞれ5'-(VIC)CAGAGGTTCTTTGAGTCCT(MGBNFQ)-3'及び5'-(FAM)AGAGGTTGAGTCCTT(MGBNFQ)-3'とされた。HbE突然変異の結果は、図26A及び26Bに記載される。
【0260】
これらの実験は、12.765 Digital Arrays (Fluidigm)を使用した、BioMark(商標)System (Fluidigm)により遂行された。1つのパネルの反応が、反応体積10μlで、2x TaqMan Universal PCR Master Mix (Applied Biosystems)を使用して準備された。CD41/42及びZFY/Xアッセイにおいて、各反応は、1xTaqMan Universal PCR Master Mix、各プライマー900 nM、各プローブ125nM、及び1 ng/μLのDNA混合物3.5 μLを含むものであった。HbEアッセイにおいて、正常(G)及び突然変異体(A)対立遺伝子を標的とするプローブを、それぞれ250nM及び125nM添加した。この試料/アッセイ混合物を、NanoFlex(商標)IFCコントローラー(Fluidigm)により、デジタルアレイに注入した。反応は、シグナル検出用のBioMark(商標)System上で遂行された。この反応は、50℃2分で始まり、続いて95℃10分、更に、95℃15秒及び57℃(ZFX/Y及びCD41/42用)又は56℃(HbE用)1分を50サイクルで行われた。各検体について少なくとも1枚の反応パネルが使用され、分類不可能なままの試料がある場合、判定を行えるようになるまで追加的なパネルからデータを蓄積した。
【0261】
また、例えばマイクロ流体工学チップ、ナノリットルPCRマイクロプレートシステム、エマルジョンPCR、ポロニーPCR、ローリングサークル増幅、プライマー伸長及び質量分析等の、当業者に周知の方法を使用して、前記デジタルPCRを実行することが出来ることは、当業者にとって明白であり得る。
【0262】
IX.癌を用いた例
一つの態様において、本発明は、癌性腫瘍において生じ得る、対立遺伝子比率の歪みの有無で試料を分類するために実行され得る。一つの態様において、各検体における、対立遺伝子Aのみ、対立遺伝子Gのみ、及びそれらの対立遺伝子両方の陽性シグナルを有するウェルの数が、デジタルPCRにより決定された。参照配列は、陽性ウェルの数がより少ない対立遺伝子として定義された(万が一両対立遺伝子の陽性ウェル数が同一であった場合は、両方とも参照対立遺伝子として使用される)。ウェルあたりの参照対立遺伝子の推定平均濃度(mr)は、ポアソン確率密度関数に従い、他方の対立遺伝子が陽性か否かにかかわらず、参照対立遺伝子が陰性であるウェルの合計を使用して計算された。発明者らは、この計算を説明するため、一つの仮想的な例を使用する。
【0263】
96ウェル反応において、20ウェルは対立遺伝子Aが陽性であり、24ウェルは対立遺伝子Gが陽性であり、そして28ウェルはそれらの対立遺伝子のいずれも陽性である。前記対立遺伝子Aは、陽性のウェルが他方より少ないので、参照対立遺伝子とみなされる。参照対立遺伝子が陰性であるウェルの数は、96−20−28=48となる。よって、mrがポアソン分布を使用して計算され、-ln(48/96)=0.693となる。
【0264】
LOH検出との関連で、帰無仮説は、1つの対立遺伝子の欠失が起こったことにより引き起こされる、対立遺伝子比率の歪みの喪失が推定される試料を意味する。この推定の下、2つの対立遺伝子の陽性ウェルの個数の予想される比率は1:1となり、そのため、潜在的に過剰出現対立遺伝子を含む情報的ウェル(1つの対立遺伝子のみ陽性であるウェル)の割合は、0.5となる。
【0265】
LOH検出との関連で、対立仮説は、試料の細胞の50%において1つの対立遺伝子の欠失が起こったことにより引き起こされる、対立遺伝子比率の歪みが推定される試料を意味する。過剰出現対立遺伝子と参照対立遺伝子との間の比率は2:1であるから、ウェルあたりの過剰出現対立遺伝子の平均濃度は、参照対立遺伝子の濃度の2倍となり得る。しかしながら、過剰出現対立遺伝子が陽性であるウェルの数は、単純に参照対立遺伝子の陽性ウェル数の2倍とはならず、ポアソン分布に従い得る。
【0266】
情報的ウェルは、対立遺伝子A又はGのいずれかが陽性であるが、両対立遺伝子とも陽性ではないウェルとして定義される。対立遺伝子比の歪みを有する試料の過剰出現対立遺伝子を含むウェルの数の予想される比率の計算は、表600で示したものと同様である。上記例において、LOHが腫瘍細胞の50%に出現するとしたら、ウェルあたりの対立遺伝子Gの平均の独活は、0.693の2倍=1.386となり得る。LOHが腫瘍細胞の50%を越えて出現するとしたら、ウェルあたりの対立遺伝子Gの平均濃度は、公式:1/[1−(LOHの割合)]x mrに従い得る。
【0267】
対立遺伝子Gが陽性であるウェルの予想される割合は、1−C-1.386=0.75(即ち75%又は72ウェル)対立遺伝子A又はGにおけるウェルの陽性判定が独立していると仮定すると、それらのウェルの0.5x0.75=0.375が、対立遺伝子A及びGの両方とも陽性であり得る。故に、0.5-0.375=0.125のウェルは、対立遺伝子Aのみ陽性で、そして0.75-0.375=0.375のウェルは、対立遺伝子Gのみ陽性であり得る。よって、情報的ウェルの割合は、0.125+0.375=0.5となり得る。対立遺伝子Gを担持する情報的ウェルの予想される比率は、0.375/0.5=0.75となり得る。このPrの予想される値は、試料中の対立遺伝子比率の歪み(ここでは即ちLOH)の存非を変低するための適切なSPRT曲線の構築に使用される。
【0268】
更に、デジタルPCR解析により実験的に決定された、非参照対立遺伝子を担持する情報的ウェルの実際の比率が、帰無仮説若しくは対立仮説のいずれが許容されるか、又はより多くのウェルを用いた更なる解析が必要となり得るか否かを判定するために使用された。帰無仮説又は対立仮説を許容するPrの決定境界は、閾値尤度比8に基づいて計算された。この閾値尤度比の値は、癌検出との関係で、対立遺伝子不均衡を有する試料と有しない試料とを峻別するのに、良好な成績を提供することが示されている(Zhou, W, et al. (2001) Nat Biotechnol 19, 78-81; Zhou et al 2002,上記)からである。上記例において、情報的ウェルの数は、20+24=44となり、実験的に得られたPrは、24/44=0.5455となり得る。決定境界は、<0.5879で虚無仮説が許容され、>0.6379で対立仮説が許容される。よって、この例における試料は、対立遺伝子比の歪みを有しないものと分類され得る。
【0269】
結果として、発明者らは、試料中の配列の不均衡を検出するためのアプローチを概説した。一つの態様において、この発明は、母体血漿中の胎児核酸の分析により、トリソミー21等の胎児染色体の異数性を、非侵襲的に検出するのに使用され得る。このアプローチは、羊水、絨毛膜試料、母体尿、頸管内試料、母体唾液等を含む、胎児核酸を含有する他の生体材料にも適用することが出来る。第一に、発明者らは、トリソミー21胎児を妊娠した妊婦の母体血漿中の、第21番染色体によって胎盤特異的に発現するPLAC4 mRNAのSNPの対立遺伝子の不均衡の判定のための、この発明の使用を示した。第二に、発明者らは、トリソミー21の非侵襲的出生前検出のための、相対的染色体量(RCD)解析による、多型に基づかない方法として、本発明が使用され得ることを示した。そのようなデジタルRCDに基づくアプローチは、胎児DNAを含有する試料中の第21番染色体の合計のコピー数が、参照染色体との関係で過剰出現しているか否かを直接評価することを含む。複雑な装置を要せずに、デジタルRCDは、25%の胎児DNAを賀乳する試料中のトリソミー21の検出を可能とする。発明者らは、前記デジタルPCRデータを解釈するために、逐時確率比検定(SPRT)を適用した。コンピューターシミュレーション解析により、本疾患分類アルゴリズムの高い精度が確認された。
【0270】
発明者らは、更に、本アプローチが、母体血漿中の胎児の突然変異又は多型検出、及び血漿中の腫瘍由来核酸の解析による悪性細胞のゲノム中の局所的な重複及び欠失の検出等の、染色体異常以外の他の形態の核酸配列不均衡の判定に応用され得ることを更に概説した。
【0271】
この適用において記載された任意のソフトウェア・コンポーネントまたはソフトウェア関数は、好適なコンピュータ言語、例えばJava(登録商標)、C++またはPerlを使用して、例えば従来の、またはオブジェクト指向の技術を使用して実行され得る。ソフトウェアコードは、記憶および/または伝達のためのコンピュータ読み取り可能媒体、ランダムアクセスメモリー(RAM)、読み取り専用メモリー(ROM)、磁気媒体、例えばハードドライブもしくはフロッピー(登録商標)ディスク、または光学媒体、例えばコンパクトディスク(CD)もしくはDVD(デジタル多用途ディスク)、フラッシュメモリーなどを含む好適な媒体における、一連の指示またはコマンドとして蓄積され得る。コンピュータ読み取り可能媒体は、かかる記憶または伝達装置の任意の組み合わせであり得る。
【0272】
かかるプログラムはまた、コード化され、そしてインターネットを含む種々のプロトコールに適合する有線、光、および/または無線ネットワークを経由した伝達のために適用されるキャリア信号を使用して伝達され得る。したがって、本発明の態様のコンピュータ読み取り可能媒体は、かかるプログラムでコード化されたデータ信号を使用して作られ得る。かかるプログラムでコード化されたコンピュータ読み取り可能媒体は、互換性のある装置と共にパッケージされるか、または他の装置からは分離して(例えばインターネットダウンロードを通じて)提供され得る。任意のかかるコンピュータ読み取り可能媒体は、単一のプログラム製品(例えばハードドライブもしくは全コンピュータシステム)上にまたはその中に装備されてもよく、そしてシステムもしくはネットワーク内における異なるコンピュータプログラム製品上もしくはその中に存在してもよい。コンピュータシステムは、ユーザーに本明細書において言及された任意の結果を提供するためのモニター、プリンター、または他の好適なディスプレイを含んでもよい。
【0273】
コンピュータシステムの例を図27に示す。図27に示されたサブシステムは、システムバス2775を通じて相互接続される。さらなるサブシステム、例えばプリンター2774、キーボード2778、固定ディスク2779、ディスプレイアダプター2782と連結されたモニター2776、およびその他のものが示される。I/Oコントローラー2771と連結する、周辺機器および入力/出力(I/O)装置は、当技術分野において既知である任意の数の手段、例えばシリアルポート2777によってコンピュータシステムと接続され得る。例えば、シリアルポート2777または外部インターフェース981は、コンピュータ装置をインターネットのような広域ネットワーク、マウス入力装置、またはスキャナーと接続するために使用され得る。システムバス経由の相互接続により、セントラルプロセッサ2773を個々のサブシステムと通信させ、そしてシステムメモリー2772または固定ディスク2779からの指示の実行、およびサブシステム間の情報の交換をさせることが可能である。システムメモリー2772および/または固定ディスク2779は、コンピュータ読み取り可能媒体を具現化し得る。
【0274】
本発明の例示的態様である上の記載は、例証および説明を目的として存在する。記載された明確な形態は、本発明を網羅しているものではなく、または限定することは意図されておらず、そして多くの変更および変形は、上の教示の観点において可能である。当該態様は、本発明の原理、及びその実用的な利用を最もよく説明するために選択かつ記載され、それにより他の当業者は、意図される特定の使用に適するように、様々な態様で、及び様々に変形させて、本発明を、最適な形で利用することが出来る。
【0275】
本明細書において引用された全ての出願、特許および特許出願は、全ての目的で、それら全てが参照により本明細書に援用される。
【技術分野】
【0001】
優先権の主張
本出願は、「DETERMINING A NUCLEIC ACID SEQUENCE IMBALANCE」と題された、2007年7月23日に出願された米国仮出願第60/951438号(代理人整理番号016285−005200US)からの優先権を主張し、そして当該仮出願の正規の出願であり、当該仮出願の全ての内容は、全ての目的のために参照により本明細書に援用される。
【0002】
関連出願の相互参照
本出願はまた、「DIAGNOSING FETAL CHROMOSOMAL ANEUPLOIDY USING GENOMIC SEQUENCING,」と題された、同時に出願された正規の出願(代理人整理番号016285−005220US)に関し、そしてその全ての内容は全ての目的のために参照により本明細書に援用される。
【0003】
発明の技術分野
本発明は、一般的には、異なる核酸配列間における不均衡を決定することによる、遺伝子型及び疾患の診断的検査に関し、そしてより詳細には、母体血液試料の検査を通じての、胎児のダウン症候群、他の染色体の異数性、突然変異、及び遺伝子型に関する。また、本発明は、癌の検出、移植のモニタリング、及び感染性疾患のモニタリングに関する。
【背景技術】
【0004】
遺伝的疾患、癌及び他の病態は、2つの対応する染色体若しくは対立遺伝子、又は他の複数の核酸配列の不均衡の原因又は結果となる場合が多い。この場合、1つの遺伝子に対するもう1つの遺伝子の割合が、正常の場合より多く、又は少ない。通常、正常の場合の比率はちょうど50:50である。ダウン症候群(トリソミー21)は、第21番染色体が追加されて不均衡を有する。
【0005】
胎児染色体異数性、例えばトリソミー21の、出産前における従来の診断方法は、侵襲性の手順、例えば羊水穿刺又は絨毛膜検体採取による、胎児試料の採取を伴うものであり、そしてそれらは胎児が死亡する危険性がある。非侵襲性の手順、例えば超音波検査法及び生化学マーカーによるスクリーニングは、最終的な侵襲性診断手順に先立ってリスク層の妊婦に使用された。しかし、これらのスクリーニング法は通常、例えば核となる染色体の異常性の代わりに、トリソミー21と関わりのある付帯現象を測定するため、その診断精度は最適ではなく、また、妊娠年齢によって大きな影響を受ける等の短所を有する。
【0006】
1997年における、母体血漿中における循環性無細胞胎児DNAの発見は、非侵襲性出生前診断のための新しい可能性を提供した(Lo,YMD and Chiu,RWK 2007 Nat Rev Genet 8,71−77)。この方法は、伴性遺伝子疾患(Costa,JM et al.2002 N Engl J Med 346,1502)及び特定の単一遺伝子疾患(Lo,YMD et al.1998 N Engl J Med 339,1734−1738)の出生前診断に対して容易に応用されたが、胎児染色体異数性の出産前における検出へのその適用は、相当な困難を要した(Lo, YMD and Chiu, RWK 2007,上記参照)。第一に、胎児の核酸は通常、胎児の核酸の分析を阻害し得る、母体由来核酸の高いバックグラウンドを含む母体血漿中に存在する(Lo,YMD et al.1998 Am J Hum Genet 62,768−775)。第二に、胎児の核酸は母体血漿中において主に無細胞形態で循環しており、そしてそのことは、胎児ゲノム中の遺伝子量又は染色体量の情報を得ることを困難としている。
【0007】
これらの課題を解決する著しい発展が近年なされた(Benachi,A & Costa,JM 2007 Lancet 369,440−442)。一つの方法は、母体血漿中における胎児特異的な核酸を検出し、その結果、母体のバックグラウンド障害の問題を克服することである(Lo, YMD and Chiu,RWK 2007,上記参照)。第21番染色体の量は、胎盤由来のDNA/RNA分子中における多型対立遺伝子の比から推定された。しかしこの方法は、試料中の標的核酸の含有量が少ないほど精度が低くなる上に、標的とされた多型に関してヘテロ接合性である胎児にしか適用できない。一つの多型が使用されるのであれば、斯かる胎児は人口群の一部に過ぎない。
【0008】
Dhallan et al(Dhallan,R,et al.2007,上記参照、Dhallan,R,et al.2007 Lancet 369,474−481)は、母体血漿へホルムアルデヒドを加えることにより、循環性胎児DNAの割合を増やす代替的な方法について記載している。母体血漿中における胎児の第21番染色体配列の割合は、第21番染色体上の一塩基多型(SNPs)に関して、父系遺伝性の胎児特異的対立遺伝子と胎児特異的ではない対立遺伝子との比を評価することによって決定された。SNP比は、基準染色体に関して同様に計算された。胎児の第21番染色体の不均衡はその後、第21番染色体のSNP比と基準染色体のSNP比との間の統計的な有意差を検出することによって推測され、ここで有意性は、0.05以下の固定されたp値を使用して定義される。高い個体群範囲を確保するために、一つの染色体あたり500個超のSNPが標的とされた。しかし、胎児DNAを高い割合に増大させるためのホルムアルデヒドの有効性について議論があり(Chung,GTY,et al.2005 Clin Chem 51,655−658)、したがって当該方法の再現性はさらに評価される必要がある。同様に、胎児及び母体の各々が、各々の染色体について異なる数のSNPsに関する情報を提供するため、SNP比の比較に関する統計的検出力は、事例ごとに変化するだろう(Lo,YMD&Chiu,RWK.2007 Lancet 369,1997)。さらにこれらの方法は遺伝子多型の検出に依存するため、それらはこれらの多型に関してヘテロ接合性である胎児に限定される。
【0009】
トリソミー21及び正倍数体の胎児から得られる羊膜細胞培養中の、第21番染色体遺伝子座及び基準遺伝子座のポリメラーゼ連鎖反応(PCR)及びDNA定量化を使用することにより、Zimmermann他(2002 Clin Chem 48,362−363)は、前者の第21番染色体DNA配列における1.5倍の増加に基づいて、胎児の当該2つの群を区別することができた。DNA鋳型濃度における2倍の違いは、僅か一回の閾値サイクル(Ct)の違いに相当するため、1.5倍の違いの区別は、従来のリアルタイムPCRの限界であった。より精密な程度の定量的識別を達成するために、代替的な方法が必要とされる。
【0010】
デジタルPCRは、核酸試料中の対立遺伝子比率の不均衡を検出するために開発された(Chang,HW et al.2002 J Natl Cancer Inst 94,1697−1703)。臨床的に、それは腫瘍DNA試料中におけるヘテロ接合性の欠如(LOH)の検出において有用であることが示された(Zhou,W.et al.2002 Lancet 359,219−225)。実験結果が試料中におけるLOHの存在を示唆するものであるか否かを分類するため、先の試験においてデジタルPCRの結果分析のために逐次確率比検定(SPRT)が採用された(El Karoui at al.2006 Stat Med 25,3124−3133)。前記研究で使用された方法において、LOHを決定するカットオフ値には、2/3のDNAの2つの対立遺伝子の固定された参照比を使用した。羊水中の胎児性核酸の量、割合、及び濃度は様々であるため、これらの方法は、羊水中の母性核酸のバックグラウンド中の胎児性核酸を使用するトリソミー21の検出には適さない。
【0011】
循環性の(circulating)胎児核酸解析に基づく、特に遺伝的多型及び/又は胎児特異的マーカーの使用に依存しない、胎児のトリソミー21(及び他の不均衡)の非侵襲性検査が望ましい。また、正確を期するのに必要なデータのウェル数及び/又は母体血漿核酸分子の量を減少させるためには、カットオフ値の決定及び配列の計数を正確にするのが望ましく、それにより、効率及び費用対効果も増大する。また、非侵襲性検査は、診断ミスを最小にするべく高感度かつ特異的であることが望ましい。
【0012】
羊水中の胎児性DNAを検出する他の用途は、ベータサラセミア等の単独の遺伝子障害の出生前診断である。しかしながら、胎児性DNAは羊水中のDNAのごく一部に過ぎないため、このアプローチは、胎児が父親から受け継ぎ、母親には存在しない突然変異のみ検出することが出来ると考えられる。この突然変異の例として、ベータサラセミアを引き起こすベータグロビン遺伝子の41/42コドン中の4bpの欠失(Chiu RWK et al. 2002 Lancet, 360, 998-1000)、及び嚢胞性線維症を引き起こす嚢胞性線維症膜コンダクタンス制御因子遺伝子のQ890X突然変異(Gonzalez-Gonzalez et al 2002 Prenat Diagn, 22, 946-8)が挙げられる。しかしながら、ベータサラセミア及び嚢胞性線維症のいずれも、常染色体劣性条件であるため、それらの疾患が発症するには胎児がそれぞれの親から突然変異を受け継ぐ必要があり、父性遺伝突然変異の検出のみでは、胎児の罹患危険率を25%から50%まで上昇させ得るに過ぎない。これは、診断的に理想的なものではない。すなわち、既存のアプローチを診断に適用することは、羊水中に父親から受け継いだ突然変異が検出されず、故にその胎児がホモ接合性病態を有する可能性が排除されたときには、シナリオどおりに運用されたといえる。しかしながら、診断的に、このアプローチは、上記結果が、父性突然変異を検出しないことに基づくという短所を有する。故に、上記制限を有さず、羊水から、胎児の完全な遺伝子型(ホモ正常、ホモ突然変異、又はヘテロ接合)を決定することが出来るアプローチが極めて望ましい。
【発明の概要】
【課題を解決するための手段】
【0013】
簡潔な概要
本発明の態様は、生体試料中に核酸配列の不均衡(例えば、対立遺伝子の不均衡、突然変異の不均衡、又は染色体の不均衡)が存在するか否かを決定する方法、システム、及び装置を提供する。例えば2つの配列(又は二つのセットの配列)の量の比率等の、不均衡を決定する1つ以上のカットオフ値が選択される。
【0014】
一つの態様において、前記カットオフ値は、少なくとも部分的に、母体血漿又は血清又は尿等の、母性核酸配列のバックグラウンドを含有する生体試料中の胎児性(臨床関連核酸)配列のパーセンテージに基づき決定される。もう一つの態様において、前記カットオフ値は、複数の反応中の配列の平均濃度に基づき決定される。一つの側面において、前記カットオフ値は、特定の核酸配列を含有することが推定される情報的ウェル(informative well)の割合から決定される、ここで、該割合は、上記パーセンテージ及び/又は平均濃度に基づき決定される。
【0015】
前記カットオフ値は、SPRT、誤検出(false discovery)、信頼区間、受信者動作特性(ROC)等の、様々な種類の方法を使用して決定され得る。この方策は、更に、信頼分類(confident classification)が作成され得る前に必要な検査の量を最小化した。これは、通常鋳型の量が限定されている血漿核酸の解析において特に重要である。
【0016】
一つの例示的態様において、
生体試料内の核酸配列中の不均衡の存否を決定する方法であり、該方法は;
複数の反応からデータを取得し、ここで、該データが、
(1)臨床関連核酸配列の第一の量を示す第一のセットの定量データ;及び
(2)臨床関連核酸配列ではない基準核酸配列の第二の量を示す第二のセットの定量データ;
を含み;
該二つのセットのデータからパラメーターを決定し;
複数の反応それぞれにおける参照核酸配列の平均濃度から第一のカットオフ値を導き出し、ここで、該参照核酸配列は、臨床関連核酸配列又は基準核酸配列のいずれかであり;
該パラメーターと該第一のカットオフ値とを比較し;続いて、
該比較に基づき、核酸配列に不均衡が存在するか否かの分類を決定する;
ことを含む。
【0017】
もう一つの例示的態様において、
生体試料内の核酸配列中の不均衡の存否を決定する方法であり、該方法は;
複数の反応からデータを取得し、ここで、該データが、
(1)臨床関連核酸配列の第一の量を示す第一のセットの定量データ;及び
(2)臨床関連核酸配列ではない基準核酸配列の第二の量を示す第二のセットの定量データ;
を含み、ここで、該臨床関連核酸配列及び該基準核酸配列が、第一の種類の細胞及び1つ以上の第二の種類の細胞に由来し;
該二つのセットのデータからパラメーターを決定し;
生体試料中の第一の種類の細胞に由来する核酸配列の量の測定に由来する第一のパーセンテージから第一のカットオフ値を導き出し;
該パラメーターと該カットオフ値とを比較し;続いて、
該比較に基づき、核酸配列に不均衡が存在するか否かの分類を決定する;
ことを含む。
【0018】
本発明の他の実施形態は、本明細書に記載された方法と関連するシステム及びコンピュータで読み取り可能な媒体に関する。
【0019】
本発明の性質及び利点のさらなる理解は、以下の詳細な説明及び添付の図面を参照にして得られる。
【図面の簡単な説明】
【0020】
【図1】デジタルDNA実験を説明するフローチャートである。
【0021】
【図2A】本発明の一つの態様に係るデジタルRNA-SNP及びRCD法を説明する。
【0022】
【図2B】癌においてよく検出される染色体異常の一覧を示す。
【0023】
【図3】本発明の一つの態様に係る、ダウン症候群の決定に使用されるSPRT曲線を有するグラフを説明する。
【0024】
【図4】本発明の一つの態様に係る、胎児細胞のパーセンテージを使用する病態の決定方法を示す。
【0025】
【図5】本発明の一つの態様に係る、平均濃度を使用する病態の決定方法を示す。
【0026】
【図6】本発明の一つの態様に係る、予想されるデジタルRNA-SNP対立遺伝子比、並びにウェルあたりの平均参照鋳型濃度(mr)として表現した、広範な鋳型濃度におけるトリソミー21試料のPrの一覧を示す。
【0027】
【図7】本発明の一つの態様に係る、ウェルあたりの平均参照鋳型濃度(mr)として発現した広範な鋳型濃度におけるトリソミー21試料において、分画胎児性DNA濃度が10%、25%、50%、及び100%の場合の予想されるPrの一覧を示す
【0028】
【図8】本発明の一つの態様に係る、デジタルRNA-SNP解析における、mr値が0.1、0.5、及び1.0の場合の、SPRT曲線の相異の程度を説明するプロットを示す。
【0029】
【図9A】本発明の一つの態様に係る、96ウェルデジタルRNA-SNP解析における、正倍数体及びトリソミー21の症例を分類する新旧のSPRTアルゴリズムの有効性比較の表を示す。
【0030】
【図9B】本発明の一つの態様に係る、384ウェルデジタルRNA-SNP解析における、正倍数体及びトリソミー21の症例を分類する新旧のSPRTアルゴリズムの有効性比較の表を示す。
【0031】
【図10】本発明の一つの態様に係る、正しく、及び誤って分類された正倍数性又は異数性の、並びに所定の情報的カウント(informative count)で分類できない胎児のパーセンテージを示す表である。
【0032】
【図11】本発明の一つの態様に係る、純粋(100%)なDNA試料における、デジタルRCD解析のコンピューターシミュレーションを示す表1100である。
【0033】
【図12】本発明の一つの態様に係る、胎児性DNAの分画濃度が異なる、正倍数性又はトリソミー21胎児由来の試料を分類するための、mr=0.5でなされたデジタルRCD解析の正確性のコンピューターシミュレーションの結果を示す表1200である。
【0034】
【図13】図13Aは、本発明の一つの態様に係る、正倍数性及びトリソミー21妊婦由来の胎盤組織における田字たるRNA-SNP解析の表1300を示す。図13Bは、本発明の一つの態様に係る、正倍数性及びトリソミー21妊婦由来の母体血漿組織におけるデジタルRNA-SNP解析の表1350を示す。
【0035】
【図14】本発明の一つの態様に係る、RCD解析で得られたカットオフ曲線を説明するプロットを示す。
【0036】
【図15】図15Aは、本発明の一つの態様に係る、正倍数性及びトリソミー21妊婦由来の胎盤組織におけるデジタルRNA-SNP解析の表を示す。図15Bは、本発明の一つの態様に係る、1つの母体血漿試料に由来する12枚の反応パネルのデジタルRNA-SNPデータの表を示す。図15Cは、本発明の一つの態様に係る、正倍数性及びトリソミー21妊婦由来の母体血漿のデジタルRNA-SNP解析の表を示す。
【0037】
【図16】図16Aは、本発明の一つの態様に係る、正倍数性及びトリソミー18胎盤のデジタルRNA-SNP解析の表を示す。図16Bは、本発明の一つの態様に係る、正倍数性及びトリソミー18胎盤のデジタルRNA-SNPデータのSPRT解釈を示す。
【0038】
【図17】本発明の一つの態様に係る、正倍数性及びトリソミー21妊婦の50%胎盤/母体血球細胞DNA混合物の、デジタルRCD解析の表を示す。
【0039】
【図18】本発明の一つの態様に係る、分類の正誤の境界決定を描写する、SPRT曲線を示す。
【0040】
【図19】本発明の一つの態様に係る、正倍数性及びトリソミー21妊婦由来の羊水のデジタルRCD解析の表を示す。
【0041】
【図20】本発明の一つの態様に係る、正倍数性及びトリソミー18妊婦由来の胎盤DNA試料のデジタルRNC解析の表(E=正倍数性;T18=トリソミー18)を示す。
【0042】
【図21】本発明の一つの態様に係る、正倍数性及びトリソミー18妊婦由来の、50%胎盤/母体血球細胞DNA混合物の、多重デジタルRCD解析の表(E=正倍数性;T21=トリソミー21;U=分類不能)を示す。
【0043】
【図22A】本発明の一つの態様に係る、50%正倍数性又はトリソミー21胎盤ゲノムDNA/50%母体軟膜DNA混合物の、多重デジタルRCD解析の表を示す。
【0044】
【図22B】本発明の一つの態様に係る、50%正倍数性又はトリソミー21胎盤ゲノムDNA/50%母体軟膜DNA混合物の、多重デジタルRCD解析の表を示す。
【0045】
【図23】両親とも同一の突然変異を担持していた場合のシナリオを示す。
【0046】
【図24】図24Aは、本発明の一つの態様に係る、女性/男性、及び男性/男性のDNA混合物の、デジタルRMD解析の表を示す。図24Bは、本発明の一つの態様に係る、25%が女性由来、75%が男性由来のDNAの混合物の、デジタルRMD解析の表を示す。
【0047】
【図25】本発明の一つの態様に係る、HbE突然変異の母体血漿試料を模した15%〜50%DNA混合物の、デジタルRMD解析の表を示す。
【0048】
【図26】図26Aは、本発明の一つの態様に係る、CD41/42の突然変異の母体血漿試料を模した5%〜50%DNA混合物の、デジタルRMD解析の表を示す。図26Bは、本発明の一つの態様に係る、CD41/42の突然変異の母体血漿試料を模した20%DNA混合物の、デジタルRMD解析の表を示す。
【0049】
【図27】本発明の一つの態様に係るシステム及び方法に有用なコンピューター装置の一例のブロック図を示す。
【発明を実施するための形態】
【0050】
定義
本明細書において使用される用語「生体試料」は、対象(例えば、妊娠女性のようなヒト)から採取される任意の試料のことであり、そして1つ以上の目的の核酸分子を含む。
【0051】
用語「核酸」又は「ポリヌクレオチド」は、デオキシリボ核酸(DNA)又はリボ核酸(RNA)、及び一本又は二本鎖のいずれかの形態であるそれらのポリマーのことである。具体的に限定されない限り、当該用語は、基準核酸と同程度の結合特性を有する、既知の天然ヌクレオチドアナログを含む核酸を含み、そして天然に存在するヌクレオチドに同様の方法で代謝される。他に示されない限り、特定の核酸配列はまた、その保存的に改変された改変体(例えば、縮重コドン置換)、対立遺伝子、オルソログ、SNPs、及び相補的配列、ならびに明確に示された配列を暗黙的に含む。具体的には、縮重コドン置換は、一つ以上選択された(又は全ての)コドンの第三の位置が混合塩基及び/又はデオキシイノシン残基で置換されている配列を生じさせることによって達成され得る(Batzer et al.,Nucleic Acid Res.19:5081(1991);Ohtsuka et al,J.Biol.Chem.260:2605−2608(1985);及びRossolini et al, Mol.Cell.Probes 8:91−98(1994))。核酸という用語は、遺伝子、cDNA、mRNA、低分子ノンコーディングRNA、マイクロRNA(miRNA)、Piwi相互作用RNA、及び遺伝子又は遺伝子座によってコードされたショートヘアピンRNA(shRNA)にも、交換可能的に使用される。
【0052】
用語「遺伝子」は、ポリペプチド鎖を産生することに関与するDNA断片を意味する。それは、コード領域に先行する、及びコード領域に続く領域(リーダー及びトレイラー)、ならびに個々のコード断片(エキソン)間の介在配列を含み得る。
【0053】
本明細書において使用される用語「反応」は、特定の目的ポリヌクレオチド配列の存在又は不在を示す、化学的、酵素的、又は物理的作用に関与する任意の工程のことである。「反応」の例は、例えばポリメラーゼ連鎖反応(PCR)のような増幅反応である。「反応」の別の例は、合成又はライゲーションのいずれかによる配列決定の反応である。「情報的反応(informative reaction)」とは、一つ以上の特定の目的ポリヌクレオチド配列の存在を示す反応を意味し、そして場合によっては、唯一つの目的ポリヌクレオチドが存在することを示す反応を意味する。本明細書において使用される用語「ウェル」は、密閉された構造、例えばPCRアレイにおける、ウェル形状バイアル、セル、又はチャンバー内のあらかじめ決められた位置における反応のことである。
【0054】
本明細書において使用される用語「臨床関連核酸配列」は、その潜在的不均衡が試験される、より大きな遺伝子配列断片に相当するポリヌクレオチド配列のことか、又はそのより大きな遺伝子配列自体に相当するポリヌクレオチド配列のことであり得る。一つの実施例は、第21番染色体の配列である。他の例は、第18番、第13番、X及びY染色体を含む。さらに他の例は、胎児がその親の片方又は両方から受け継いているかもしれない変異遺伝子配列、又は遺伝子多型、又はコピー数変異を含む。さらに他の実施例は、悪性腫瘍中において変異した、欠失した、又は増幅された配列、例えばヘテロ接合性の欠損又は遺伝子重複が起こっている配列を含む。幾つかの態様において、複数の臨床関連核酸配列、又は臨床関連核酸配列の同等の複数のマーカーが、当該不均衡を検出するためのデータを提供するために使用され得る。例えば、第21番染色体上の5つの不連続配列からのデータは、可能性のある第21番染色体不均衡の決定のための付加的な方法において使用され得、効果的に試料の容量を1/5へと減少させる。
【0055】
本明細書において使用される用語「基準核酸配列」は、正常な場合、臨床関連核酸配列に対する比率が既知、例えば1:1の比率である核酸配列のことである。一つの例として、基準核酸配列及び臨床関連核酸配列は、ヘテロ接合性により明確に区別し得る、同一の染色体由来の2つの対立遺伝子である。別の例において、基準核酸配列は1つの対立遺伝子であり、これに対してヘテロ接合性である別の対立遺伝子が、臨床関連核酸配列である。さらに、各々の基準核酸配列及び臨床関連核酸配列の幾つかは、異なる個体由来であってもよい。
【0056】
本明細書において使用される用語「参照核酸配列」は、一反応あたりの平均濃度が既知又は同等と測定された核酸配列のことである。
【0057】
本明細書において使用される用語「過剰出現核酸配列」は、生体試料中における他の配列以上に豊富である、目的の2つの配列(例えば臨床関連配列及び基準配列)に混在する核酸配列のことである。
【0058】
本明細書において使用される用語「に基づく」は、「少なくとも一部に基づく」ことを意味し、そして、一つの方法の入力とその方法の出力との関係において生じる、一つの値の決定において使用される別の値(又は結果)のことをいう。本明細書において使用される用語「導く」はまた、一つの方法の入力とその方法の出力との関係のことであり、例えば誘導が公式計算であるときに起こる。
【0059】
本明細書において使用される用語「定量的データ」は、一つ以上の反応から得られ、そして一つ以上の数値を提供するデータを意味する。例えば、特定の配列のための蛍光マーカーを示すウェルの数が、定量的データであろう。
【0060】
本明細書において使用される用語「パラメーター」は、定量的データのセットを特徴づける数値、及び/又は定量的データセット間における数的関係を意味する。例えば、第一の核酸配列の第一の量と第二の核酸配列の第二の量との間における比率(又は比率の関数)はパラメーターである。
【0061】
本明細書において使用される用語「カットオフ値」は、生体試料の分類の2つ以上の状態(例えば罹患した状態と罹患していない状態)間を裁定するために使用される数値を意味する。例えば、パラメーターが当該カットオフ値よりも大きいならば、定量的データの第一の分類がなされ(例えば罹患した状態);又はパラメーターがカットオフ値よりも小さいならば、定量データの異なる分類がなされる(例えば罹患していない状態)。
【0062】
本明細書において使用される用語「不均衡」は、臨床関連核酸配列量における少なくとも一つのカットオフ値によって定義される、基準量からの任意の有意な偏差を意味する。例えば、参照量の比率が3/5のとき、測定比が1:1である場合、不均衡が生じているといえる。
【0063】
発明の詳細な説明
分発明は、生体試料中に存在する、臨床的関連核酸配列の量が、他の非臨床関連核酸配列との関連で、参照(例えば正常体)と比較して増大したか又は減少したか(例えば、染色体又は対立遺伝子の不均衡)を判定する方法、システム、及び装置を提供する。参照量と比較して、例えば2つの配列の量の比率等に関して変化が存在する(即ち不均衡)か否かを判定するために、1つ以上のカットオフ値が決定される。参照量において検出される変化は、臨床的に重要な核酸配列と、他の臨床的に重要ではない配列との関係における任意の偏差(上向きまたは下向き)であり得る。したがって、基準状態は、任意の比または他の量(例えば1−1対応以外)であり得、そして変化を示す測定された状態は、一つ以上のカットオフ値によって決定される基準量とは異なる任意の比または他の量であり得る。
【0064】
臨床的に重要な染色体領域(臨床的に重要な核酸配列とも呼ばれる領域)およびバックグラウンド核酸配列は、第一のタイプの細胞由来であり得、そして一つ以上の第二のタイプの細胞由来であり得る。例えば、胎児/胎盤細胞由来の胎児核酸配列は、母体細胞由来の母体核酸配列のバックグラウンドを含む生体試料、例えば母体血漿中において存在する。一つの実施形態において、カットオフ値は、生体試料中における第一のタイプの細胞のパーセンテージに少なくとも一部基づいて決定される。試料中における胎児配列のパーセンテージは、任意の胎児由来遺伝子座によって決定され得、そして臨床的に重要な核酸配列を測定するために限定されないことに留意すべきである。別の実施形態において、カットオフ値は、体内における非悪性細胞由来の核酸配列のバックグラウンドを含む生体試料、例えば血漿、血清、唾液、または尿における腫瘍配列のパーセンテージに少なくとも一部基づいて決定される。
【0065】
なおももう一つの態様において、前記カットオフ値は、複数の反応における配列の平均濃度に基づいて決定される。一つの側面において、該カットオフ値は、特定の核酸配列を含有することが推定される、情報的ウェルの割合から決定され、ここで、該割合は、上記パーセンテージ及び/又は平均濃度に基づき決定される。SPRT、誤検出、信頼区間、受信者操作特性(ROC)等の、様々な種類の方法を使用して、前記カットオフ値が決定され得る。この方策は、更に、信頼分類が作成され得る前に必要な検査の量を最小化する。これは、通常鋳型の量が限定されている血漿核酸の解析において特に重要である。デジタルPCRに関連して提示されているが、他の方法も使用され得る。
【0066】
デジタルPCRは、核酸を極限まで希釈して、殆どの陽性増幅が単一の鋳型分子に由来するシグナルを反映するように、複数のPCR解析がなされる。従って、デジタルPCRは、個々の鋳型分子の計数を可能とする。解析されたPCRの合計数中で、正の増殖が見られたものの割合から、元の試料あるいは希釈していない試料中の鋳型濃度を推定することができる。この技法は、様々な遺伝的現象の検出が可能なことが示されており(Vogelstein, B et al. 1999, supra)、既に、腫瘍試料中(Zhou, W. et al. 2002, supra)、及び癌患者血漿中(Chang, HW et al. 2002, supra)のヘテロ接合性の消失の検出に使用されている。デジタルPCRによる鋳型分子の定量は、レポーター色素及び核酸濃度との間で容量依存的応答関係にないので、その解析精度は、理論的には、リアルタイムPCRを超える。従って、デジタルPCRは、潜在的には、標的と参照遺伝子座との間のより微細な程度の量的差異の判別を可能とし得る。
【0067】
これを試験するために、発明者らはまず、デジタルPCRが、PLCA4 mRNA(Lo, YMD, et al. 2007 Nat Med 13, 218-223)の対立遺伝子の比率を判定可能であるかを検証した。該mRNAは第21番染色体由来の胎盤性転写産物であり、母体血漿中に存在するため、胎児のトリソミー21及び正倍数体を峻別することが出来る。このアプローチは、デジタルRNA-SNP法と称される。それから、発明者らは、デジタルPCRの精度増大により、遺伝的多型に依存しない胎児染色体の異数性の検出が可能となるか否かを検証した。発明者らは、これを、デジタル相対的染色体量(RCD)解析と称する。前者のアプローチは多型依存的であるものの定量的識別能の要求は低く、一方で後者のアプローチは、多型非依存的であるが、高精度の定量的識別能を要求する。
【0068】
I.デジタルRNA-SNP
A.概要
デジタルPCRは、DNA試料において2つの対立遺伝子における存在比の非対称性を検出することができる。例えば、腫瘍DNA試料におけるヘテロ接合性の消失(LOH)の検出の使用されている。DNA試料中に2つの対立遺伝子A及びGが存在しているとする。そして、対立遺伝子Aが、LOH細胞において消失し得るとする。LOHが腫瘍試料中の細胞の50%の発生するとき、該DNA試料における対立遺伝子比G:Aは、2:1となり得る。しかしながら、腫瘍試料でLOHが発生しない場合、対立遺伝子比G:Aは、1:1となり得る。
【0069】
図1は、デジタルPCR実験を説明したフローチャート100である。工程110において、DNA試料を希釈し、続いて個別のウェルに分注する。ここで留意すべきなのは、発明者らが、幾つかの種類の核酸は、元の試料中で既に多分に希釈されていることを特定したことである。このように、目的の鋳型が既に必要な濃度で存在しているときは、希釈を必要としない。以前の研究で、あるDNA試料は、特定の「鋳型DNA」の平均濃度が、ウェルあたり約0.5分子、即ちウェル2つに約1つの鋳型が入る程度にまで希釈されている(例えば、Zhou et al 2002を参照されたい)。ここで留意すべきなのは、鋳型DNAが、対立遺伝子A又はGのいずれかに相当し、この特異的濃度に与えられる論理的根拠は存在しないことである。
【0070】
工程120において、各ウェル中で、PCRプロセスが実行され、対立遺伝子A及び/又はBが同時に検出される。工程130において、各ウェル中のマーカー(蛍光等)が同定され、ウェル中にA若しくはGのいずれか一方が入っているか、又はそれらのいずれも入っているか、若しくはいずれも入っていないかが判定される。LOHが起こらない場合、DNA試料中の対立遺伝子A及びGの量は同一であろう(細胞1個に1コピー)。従って、対立遺伝子Aが陽性であるウェルの頻度及び対立遺伝子Gが陽性であるウェルの頻度は同一となるであろう。しかしながら、腫瘍細胞の50%以上でLOHが発生した場合、対立遺伝子G及びAの存在比率は、少なくとも2:1となるであろう。従来の方法によれば、この試料は、少なくとも50%は癌性であると単純に推定されていた。すなわち、対立遺伝子Gが陽性のウェルの出現頻度が、対立遺伝子Aが陽性のウェルの出現頻度より高い。結果として、対立遺伝子Gが陽性のウェルの個数は、対立遺伝子Aが陽性のウェルの個数より多くなる。
【0071】
工程140において、デジタルPCRの結果を分類するために、各対立遺伝子の一方が陽性で他方が陰性であるウェルの個数が使用され得る。上記例において、対立遺伝子Aが陽性であるが対立遺伝子Gが陰性であるウェルの個数、及び対立遺伝子Gが陽性であるが対立遺伝子Aが陰性であるウェルの個数が計数される。一つの態様において、陽性のウェルがより少ない対立遺伝子が、参照対立遺伝子とみなされる。
【0072】
工程150において、情報的ウェルの総数は、2つの対立遺伝子のいずれかが陽性のウェルの個数の合計として決定される。工程160において、より多くの陽性ウェルを有する対立遺伝子に関する情報的ウェルの割合(Pr)(パラメーターの一種)が計算される。Pr=(より多くの陽性のウェルを有する対立遺伝子のみが陽性のウェルの個数/1つの対立遺伝子(A又はG)のみが陽性のウェルの総数)となる。他の態様として、少なくとも1つの対立遺伝子を有する全てのウェルにより分別された対立遺伝子の1つを有するウェルを使用し得る。
【0073】
工程170において、Prの値が対立遺伝子の不均衡を示すか否かを決定する。正確性と効率性が求められるので、この課題は簡単なものではない。不均衡を決定する1つの方法として、Bayesian型尤度法、逐次確率比検定(SPRT)が使用される。SPRTは、2つの確率的仮説をデータの蓄積により比較することを可能とする方法である。言い換えると、これは、対立遺伝子の歪みの存否を示唆するものとして、デジタルPCRの結果を分類する統計学的方法でああある。これは、解析すべくウェルの個数を最少としつつ、所定の統計的鹸出力及び精度を達成するのに有利である。
【0074】
例示的SPRT解析において、実験結果を、帰無仮説及び対立仮説に対して検定され得る。対立仮説は、試料中で対立遺伝子比に歪みが存在するときに許容される。帰無仮説は、試料中で対立遺伝子比に歪みが存在しないときに許容される。前記Prは、帰無仮説又は対立仮説を許容する2つのカットオフ値と比較され得る。いずれの仮説も許容されなければ、そのサンプルは、分類不能としてマークされ、これは、観察されたデジタルPCRの結果が、所望の統計的信頼性を有する試料を分類するのに不十分であることを意味する。
【0075】
帰無仮説又は対立仮説を許容するカットオフ値は、典型的には、それらの仮説において主張される仮定の下の固定されたPr値に基づき計算されている。帰無仮説において、試料は対立遺伝子比の歪みを示していないと想定される。従って、対立遺伝子Aが陽性であるウェルの頻度及び対立遺伝子Gが陽性であるウェルの頻度は同一であり、従って、Pr値は1/2と予想される。対立仮説において、Pr値は、2/3、又は0.5と2/3の中間、例えば0.585と予想される。また、実験数が限定されるため、上限(.585+3/N)及び下限(.585-3/N)が選択され得る。
【0076】
B.ダウン症候群の検出
本発明の一つの態様において、妊婦血漿から胎児のダウン症候群を検出するために、デジタルSNPが使用される。胎児/胎盤細胞に特異的なマーカーを使用して、第21番染色体中の対立遺伝子の比率が測定され得る。例えば、観察されたPLAC4対立遺伝子の過剰出現の度合が統計的に有意か否かを決定するために、SPRTが使用される。
【0077】
一つの例示的な態様において、デジタルRNA-SNPは、第21番染色体から転写され、胎盤により発現されるPLAC4 mRNA上に位置する、A/G SNPの多型的対立遺伝子の比率の不均衡を決定する。ヘテロ接合性正倍数性胎児において、対立遺伝子A及びGは、胎児ゲノム中で等しく表示されるべきである(ゲノム比1:1)ところ、トリソミー21において、トリソミー第21染色体は、胎児ゲノム中でSNP対立遺伝子の一つを追加的に1コピー有し、これが2:1のゲノム比をもたらす。デジタルPCR解析の目的は、解析される試料中のPLAC4対立遺伝子の量が等しいか否かを決定することである。従って、PLRC4の対立遺伝子A及びGのいずれも標的の鋳型である。PLAC4 mRNAを増幅するためにリアルタイムPCRアッセイが設計され、TaqMan蛍光プローブにより、2つのSNP対立遺伝子が識別された。図2Aに、解析工程の模式図を示す。
【0078】
図2Aは、本発明の態様におけるデジタルRNA-SNP法200を説明する。工程210において、試料が調製される。工程220において、核酸配列、例えば抽出されたRNA試料中のPLAC4 mRNAが定量される。一つの態様において、この工程は、標的がデジタルPCR解析の「領域」に達するのにいかほどの希釈が必要であるかの知見を実施者に提供する。
【0079】
工程230において、試料が希釈される。工程240において、希釈された試料の濃度が測定される。希釈された試料の濃度が、〜1鋳型(即ち参照若しくは非参照配列、又はいずれかの対立遺伝子)/ウェルとなっていることが確認される。幾つかの態様は、この測定のために、第IV章に記載される技術を使用する。例えば、発明者らは、前記希釈された試料を、リアルタイムPCR用に96ウェルに分注し、所望の希釈が達成されていることを確認した。希釈濃度は未知のままであり得るので、この工程を省略している態様も後に記載する。
【0080】
工程250において、アレイの各ウェルにおいて、デジタルPCRが実行される。例えば、等しく希釈された試料が、リアルタイムPCR解析用384ウェルに分注された。PCRの結果から、各核酸配列用のマーカーの量及び情報的ウェルの個数が同定される。情報的ウェルは、対立遺伝子A又はBのいずれかが陽性であるが、両方が陽性ではないウェルを意味する。工程260において、予想されるRrの値の計算が実行される。これらの工程は、後で詳述する。前記計算は、工程250において決定された数値からパラメーターを決定することを含む。例えば、ウェルあたりの実際の平均鋳型濃度が計算され得る。
【0081】
工程270において、SPRT又は他の尤度比が実行され、不均衡の存否が決定される。正倍数体の場合、発明者らは、A陽性及びG陽性のウェルの個数が等しくなると予想する。しかしながら、トリソミー21胎児由来の鋳型分子を解析すると、一方の対立遺伝子を1つだけ含むウェルの個数は、他方の対立遺伝子を1つだけ含むウェルの個数よりも多くなるはずである。要するに、トリソミー21には、対立遺伝子の不均衡が予想される。
【0082】
上記のように、SPRTは、2つの確率的仮説をデータの蓄積として比較することを可能とするBayesian型尤度法である。トリソミー21検出用デジタルPCR解析において、対立遺伝子の武均衡が存在する(即ちトリソミー21が検出された)とき、対立仮説が許容され;対立遺伝子の不均衡が存在しない(即ちトリソミー21が検出されない)とき、帰無仮説が許容される。多数派の対立遺伝子は、潜在的過剰出現対立遺伝子と称され、情報的ウェル全ての中に占めるその割合(Pr)が計算され得る。SPRTは、Prがトリソミー21試料で予想される十分な程度の対立遺伝子の不均衡を同定するか否かを決定するのに応用される。
【0083】
操作上、SPRTは、いずれかの仮説を許容又は排除する確率的境界を規定するために構築されるSPRTカーブのペアを有するグラフの使用を通じて、応用及び解釈され得る。図3は、本発明の一つの態様における、ダウン症候群を決定するためのSPRT曲線を有するグラフを説明する。これらのSPRT曲線は、信頼区分が作成され得るとき、所定の情報的ウェルの総数(x軸)に対する、潜在的に過剰出現対立遺伝子が陽性である情報的ウェル数の割合Pr(y軸)を描画している。図3で描かれる如く、上方の曲線は、対立仮説を許容する確率的境界を設定し;下方の曲線は、帰無仮説を許容する確率的境界を設定する。
【0084】
実験的に導き出されたPr値は、予想されたPr値と比較され、いずれかの仮説を排除又は許容する。もし帰無仮説が許容されると、それらの試料が由来する妊婦は、正倍数体胎児を妊娠しているのとして分類される。もし対立仮説が許容されると、それらの試料が由来する妊婦は、トリソミー21胎児を妊娠しているものとして分類される。あるいは、もし、所定の情報的カウントのPrが、疾患の分類における統計的信頼に要求されるレベルに達していないとき、いずれの仮説も許容され得ない。これらのケースは、更にデータが入手出来なければ、分類不可能とみなされた。疾患分類が不可能である場合、データが蓄積されてSPRTにより分類可能となるまでは、追加的な384ウェルプレートが実行され得る。
【0085】
故に、SPRTは、他の統計的方法よりも、所定のレベルの信頼性を達成するのにより少数の検査で足りる点が有利である。実用語で言えば、SPRTは、所望の量のデータが蓄積され次第いずれかの仮説を許容又は排除することが出来るので、不要な追加的解析を最少限にすることが出来る。この特性は、入手可能な鋳型分子の数が限定される血漿中の、一般に低濃度で存在する核酸の解析において特に重要である。厳格な分類に加えて、分類は、パーセント精度のものが含まれる場合もある。例えば、カットオフ値との比較から得られた分類は、試料が特定のパーセンテージで核酸配列が不均衡である可能性を示し、又は同様に、決定された不均一性が、所定のパーセンテージ程度の正確性を有している。
【0086】
突然変異、あるいは遺伝的多形に関して胎児の遺伝子型を決定するために、母体血漿又は血清中の胎児核酸を使用して、同様のアプローチが適用され得る。ここで、胎児のゲノムの半分は母親から受け継いでいることを想起されたい。例として、2つの対立遺伝子A及びBを有する特定の遺伝子座を想定する。母親がヘテロ接合の遺伝子型ABであったとすると、その胎児の遺伝子型は、理論上、AA、BB、又はABとなる。胎児の遺伝型がAB、即ち母親の遺伝型と同一のとき、母体血漿中には、遺伝子型ABの核酸ばかり(母親及び胎児由来)が存在するだろう。よって、核酸又は対立遺伝子の均衡は、母体血漿において確認される。他方、胎児の遺伝型がAA又はBBであるとき、母体血清中で、対立遺伝子A又はBのいずれかが過剰出現し、対立遺伝子の不均衡が生じるだろう。この検討は、疾患を引き起こす突然変異(例えば、ベータサラセミア又は脊髄性筋萎縮症)にも適用可能であり、その場合、Aは、野生型対立遺伝子とみなされ得て、そしてBは、突然変異対立遺伝子とみなされ得る。
【0087】
II.デジタルRCD
デジタルRNA-SNPの短所は、解析されるSNPにおいてヘテロ接合性である場合に限り適用され得ることである。循環性胎児核酸解析に基づく、胎児トリソミー21又は他の胎児染色体異数性(例えば、トリソミー18、13、及び性染色体異数性等)用の非侵襲性検査が、遺伝的多形の使用と独立しているのが理想的であり得る点が改善点である。従って、一つの態様において、参照染色体(本文中では、第1番染色体と称する)上に位置する遺伝子座に対する非多型第21番染色体遺伝子座のデジタルPCR解析により、染色体量が決定される。第21番染色体と第1番染色体との比率は、正倍数体胎児のゲノムにおいては2:2であるが、トリソミー21の場合、これが変化する。トリソミー21検出用デジタルPCR解析において、比較されるべき2つの仮説は、染色体不均衡が存在しない(即ちトリソミー21が検出されない)帰無仮説、及び染色体不均衡が存在する(即ちトリソミー21が検出される)対立仮説であり得る。
【0088】
このアプローチは、他の染色体異数性、例えばトリソミー18における第18番染色体、トリソミー13における第13番染色体、ターナー症候群におけるX染色体等を含む、他の染色体に一般化され得る。加えて、第1番染色体と別に、異数性に関係の無い他の染色体も、参照染色体として使用され得る。通常癌において参照染色体と比較して部分的に起こる、染色体の欠失の比率の変化を解析することにより、同様のアプローチを、癌の検出に適用することも出来る。前者の例として、直腸結腸癌における染色体5q、肺癌における染色体3p、及び鼻咽腔癌における染色体9pを含む。図2Bは、配列の不均衡をもたらす、幾つかの通常の癌関連染色体異常を列挙したものである。
【0089】
また、図2Aは、本発明の態様におけるデジタルRCD法205を説明する。工程220〜230の一つの態様において、抽出されたDNAは、例えばナノドロップ技術により定量され、1つのウェル第21番染色体又は正規化(normalizing)染色体(第1番染色体等)のいずれかに由来する約1個の標的鋳型が含まれる濃度まで希釈される。工程240の一つの態様において、確認は、96ウェルの形式においてのみ、第1番染色体を使用したアッセイにより、希釈したDNA試料を解析することにより実行され得て、384ウェルプレートにおける両TaqManプローブを使用したRCD解析を進行する前に、陰性のウェルが37%未満であるか否かを確認する。37%の重要性は、第IV章で述べる。
【0090】
工程240の検査及び工程250の結果は、TaqManプローブのペアにより区別されるパラロガス配列変異により識別される両染色体上に存在するパラロガス配列を増幅するように設計されたリアルタイムPCRアッセイを用いてなされ得る。これに関連して、情報的ウェルは、第21番染色体又は第1番染色体遺伝子座のいずれかが陽性であるが、両方が陽性ではないものとして定義される。正倍数体の胎児において、いずれかの遺伝子座が陽性である情報的ウェルの個数は、概ね等しくあるべきである。トリソミー21胎児において、第21番染色体が陽性のウェルは、第1番染色体の場合よりも過剰出現すべきである。過剰出現の正確な割合は、後の章で述べる。
【0091】
III .胎児配列パーセンテージの組込み
上記方法200及び205の態様の短所は、胎児特異的なマーカーが必要なことである。従って、本発明の1つの態様において、非胎児特異的マーカーが使用される。そのような非胎児特異的マーカーを使用するために、本発明の態様は、母体血漿(即ち生体試料)中の胎児DNAの分別濃度(fractional concentration)を測定する。この情報を用いて、後述のように、Prのより有用な数値が計算される。
【0092】
母体血漿中で胎児DNAの分別パーセンテージが小さくとも、トリソミー21胎児は、胎児DNAのゲノム当量(GE)あたりの追加的な量の第21番染色体の、母体血漿中への放出に関与する。例えば、全DNAが50GE/mlで、その内胎児に由来するDNAが5GE/mlである(即ち胎児DNAの分別濃度は10%)、正倍数体の妊娠に由来する材料血漿試料は、材料血漿1mlあたり総数100コピーの第21番染色体(材料由来が90コピー+胎児由来が10コピー)配列を含有すべきである。トリソミー21妊娠において、各胎児性GEは、第21番染色体を3コピー有するため、母体血漿中の第21番染色体配列は、合計で105コピー(材料由来が90コピー+胎児由来が15コピー)となる。よって、胎児DNA濃度が10%において、三染色体性妊娠の母体血漿中の第21番染色体由来の配列の量は、正倍数体の場合の1.05倍になるべきである。故に、分析アプローチが小程度の量的層異を決定するように発達すれば、胎児トリソミー21の非侵襲性出生前診断用の、多型性独立的検査が達成され得る。
【0093】
従って、過剰出現の度合いは、解析されるDNA試料中の胎児DNA分別濃度に依存し得る。例えば、胎盤DNAが解析されたとき、胎児ゲノム中の理論上のRCD比率は3:2、即ち1.5倍異なるべきである。しかしながら、上記の如く、胎児性DNAを10%含有する母体血漿試料を解析した場合、理論上のRCD比率は1.05倍まで低下し得る。実験的に導き出されたPrは、情報的ウェルの総数で、第21番染色体遺伝子座のみ陽性であるウェルの個数を割ることにより計算される。この実験的に導き出されたPrは、計算されたPr及び前記理論上のRCD比率と共に、SPRT解析に供される。
【0094】
図4は、本発明の一つの態様における、胎児核酸のパーセンテージを使用した病態の判定方法400を示す。工程410において、胎児由来分子の分別パーセンテージが測定される。一つの態様において、分別パーセンテージは、非胎児特異的マーカー(即ち母親及び胎児のいずれにも存在する遺伝子配列)との関係における、胎児特異的マーカー(例えば、Y染色体、遺伝的多型(SNP等)、胎盤エピジェネティックサイン(placental epigenetic signature)等)の量を測定することにより判定される。リアルタイムPCR、デジタルPCR、シークエンシング反応(超並列ゲノムシークエンシングを含む)又は他の任意の定量法を使用して、正確な測定がなされ得る。一つの側面において、潜在的に測定自体により対立遺伝子の不均衡を生じさせ得る遺伝子ターゲティングは、使用しないのが好ましい。
【0095】
工程420において、デジタルPCR又は他の測定方法が実行され、この工程は、試料の希釈、希釈された試料のウェルへの分注、及び各ウェル内の反応の測定を含む。工程430において、相異する参照核酸配列(染色体又は対立遺伝子)のマーカーを同定するのに、PCRの結果が使用される。工程440において、過剰出現配列の正確な割合(Pr)が計算される。工程450において、病態を判定するためのカットオフ値が、試料中の胎児由来分子のパーセンテージを使用して計算される。工程460において、前記正確なPr及びカットオフ値から、不均衡の存否が判定される。
【0096】
一つの態様において、参照核酸配列の分別パーセンテージは、デジタルRNA-SNP法に組み込まれる。よって、癌細胞によるLOHを調査するとき、これは、腫瘍試料中癌細胞が50%未満の状態で行われ得る。より正確なPrを取得するのに、50%以上が癌細胞である腫瘍試料も使用され得て、これは、誤診断をもたらす擬陽性の数を減少させる。もう一つの態様において、胎児核酸のパーセンテージは、胎児が親の遺伝子突然変異(例えば、嚢胞性線維症、ベータサラセミア、又は脊髄性筋萎縮症を引き起こすもの等)又は多型を受け継いでいるか否かを、母体血漿核酸の解析から判定するために、デジタルPCR法に組み込まれる。
【0097】
IV.ウェルあたりの平均濃度の組込み
従来の方法(例えば、上記Zhou, W. et al. 2002を参照されたい)のもう一つの短所は、鋳型の濃度を、1ウェルに1個となるようにする必要があることである。正確な濃度を求めるのが困難である場合、これは誤差を生じる場合がある。更に、ウェルあたり鋳型1個となるように正確な濃度に調整しても、従来の方法、即ち古いアルゴリズムでは、対立仮説を許容するために予想されたPr値は、対立遺伝子の比率であり、ウェルあたりの平均の鋳型濃度から独立している。
【0098】
しかしながら、希釈された試料中の鋳型の自然の統計的変動のために、実際にはウェルあたり1個の鋳型が存在するとは限らない。本発明の態様は、少なくとも1つの配列の経筋濃度を測定し、そしてその測定値がカットオフ値、即ち予想されるPrを計算するのに使用される。一つの態様において、この計算は、異なる複数の核酸配列を含むウェルの確率を決定するための統計的分布を含み、そしてこれは、予想されるPrを決定するのに使用される。
【0099】
一つの態様に置いて、平均濃度は、1つの参照核酸配列から得られ、この配列は、例えば、DNA試料中でより低い濃度で存在する核酸配列である。不均衡が存在しない場合、試料中の2つの配列の濃度は同一のはずであり、いずれか1つが、参照対立遺伝子とみなされる。試料が、例えばLOHを有している場合、癌細胞中で欠失した対立遺伝子が、参照対立遺伝子とみなされ得る。参照対立遺伝子の平均濃度は、mrと表記される。もう一つの態様において、より高濃度で存在する配列を、参照配列として扱う。
【0100】
A.デジタルSNP.SPRT及びデジタルPCRの使用例
図5は、本発明の一つの態様における、平均鋳型濃度を使用した、病態判定方法500を示す。工程510において、異なる複数の配列の量が測定される。これは、例えば、上記の如く、デジタルPCR実験においてマーカーを計数すること等によりなされ得る。しかしながら、本方法は、増幅工程を含まない、又は蛍光マーカーを使用せず、質量等の物理的特性、特定の光学的特性、又は塩基対特性等の他の特性を使用することが出来る他の方法によりなされ得る。
【0101】
工程520において、過剰出現配列の実際の割合が決定される。これは、上記のように、その配列のみを示すウェルの個数をとり、情報的ウェルの個数により区別されることによりなされ得る。工程530において、少なくとも1つの配列(参照配列)の平均濃度が測定される。一つの態様において、この参照配列は、過剰出現配列である。もう一つの態様において、この参照配列は、過小表示した(underrepresented)配列である。測定は、デジタルPCR実験において参照配列が陰性であるウェルの個数を計数することによりなされ得る。陰性のウェルの割合と平均鋳型濃度との間の関係は、次の小区分に記載のポアソン分布により記述されている。
【0102】
工程540において、異なる複数の配列が陽性であるウェルの予想量が計算され、例えばポアソン分布が使用される。この予想量は、ウェルあたりの配列の確率(probability)、ウェルあたりの平均配列、その配列を含むウェルの個数、又は他の任意の適切な量としてあり得る。工程550において、予想されるPrが、前記予想量から計算される。工程560において、カットオフ値が銭予想されるPrから計算され、例えば、SPRTを使用して計算される。工程570において、核酸配列の不均衡の分類が判定される。方法500の特別の側面が、ここで記載される。
【0103】
配列の予想量の決定
工程530からウェルあたりの平均濃度が知られれば、工程540において、その配列を示すウェルの予想される個数が計算される。この量は、%、少数値、又は整数値で表現され得る。説明のため、参照鋳型配列の平均濃度(mr)が0.5であり、PLAC4 SNP、rs8130833におけるトリソミー21の胎児の遺伝子型はAGGとする。故に、参照鋳型は対立遺伝子Aとし、過剰出現鋳型は対立遺伝子Gとする。
【0104】
一つの態様において、ポアソン分布は、デジタルPCR等の測定手順のウェルの反応混合物中の、対立遺伝子Aの分布であるとする。他の態様において、二項分布等の他の分布関数が使用される。
【0105】
ポアソン方程式は:
【化1】
で表され、ここで、nは、ウェルあたりの鋳型分子の個数;P(n)は、特定のウェル中の鋳型分子の確率;そしてmは、特定のデジタルPCR実験における1つのウェル中の鋳型分子の平均数である。
【0106】
従って、対立遺伝子Aを平均濃度0.5である場合、対立遺伝子Aの分子がウェルに含まれない確率は:
【化2】
となる。
【0107】
故に、任意のウェルが対立遺伝子Aを少なくとも1分子含有する確率は、1−0.6065=0.3935となるであろう。従って、約39%のウェルは、対立遺伝子Aを少なくとも1分子含有すると予想される。
【0108】
非参照核酸配列に関しては、トリソミー21胎児の各細胞において、AとGとのゲノム比率は1:2である。抽出されたRNA又はDNA試料においてこのAとGとの比率が変化していないと仮定すると、ウェルあたりの対立遺伝子Gの平均濃度は、対立遺伝子Aの2倍、即ち2x0.5=1である。
【0109】
従って、対立遺伝子Gの平均濃度が1の場合、対立遺伝子Gの分子がウェルに含まれない確率は:
【化3】
となる。
【0110】
故に、任意のウェルが対立遺伝子Gを少なくとも1分子含有する確率は、1−0.3679=0.6321となるであろう。従って、約63%のウェルは、対立遺伝子Gを少なくとも1分子含有すると予想される。
【0111】
2.過剰出現配列の割合の決定
予想量が計算された後、過剰出現核酸配列の割合が決定され得る。ウェルに充填された対立遺伝子A及びBは独立であると仮定すると、両方の対立遺伝子を含有するウェルの確率は、0.3935x0.6321=0.2487となる、よって、約25%のウェルが、両方の対立遺伝子を含有すると予想される。
【0112】
対立遺伝子Aを含有するが対立遺伝子Gを含有しないウェルの割合は、少なくとも一つの対立遺伝子Aを含有する確率から、対立遺伝子A及びBのいずれも含有する確率を引く:0.3935−0.2487=0.1488となる。同様に、対立遺伝子Gを含有するが対立遺伝子Aを含有しないウェルの割合は:0.6321−0.2487=0.3834となる。情報的ウェルは、対立遺伝子A又はBのいずれかが陽性であるが、両方陽性ではないウェルと定義される。
【0113】
故に、デジタルRNA-SNPにおいて、対立遺伝子A又は対立遺伝子Gのいずれかを含むウェルの期待される割合は、0.1488/0.3834である。換言すると、対立遺伝子Gのみ陽性の場合、Aのみ陽性の場合と比較して、ウェルの個数は2.65倍となる。これは、過剰出現対立遺伝子が他の対立遺伝子の2倍となる胎児ゲノムの比率と対照的である。
【0114】
SPRT解析において、過剰出現対立遺伝子が陽性である情報的ウェルの割合(Pr)は、SPRT曲線を使用して計算及び解釈される。ここまでの実施例において、情報的ウェルの割合は:0.1448+0.3834=0.5852となる。故に、mrが0.5の場合に、トリソミー21の予想されるPrは:0.3834/0.5282=0.73となる。
【0115】
平均の鋳型濃度(m)がポアソン方程式の肝要なパラメーターであるから、Prは、mに伴って変化する。図6は、本発明の一つの態様に従い、ウェルあたりの平均の参照鋳型濃度(mr)として表された、予想されるデジタルRNA-SNP対立遺伝子比率、及び様々な範囲の鋳型濃度のトリソミー21試料のPrを列記する表600を示す。表600は、一連のウェルあたりの平均の参照鋳型濃度(mr)における、過剰出現アリルが陽性である情報的ウェルの比率及び割合(Pr)、及び予想される対立遺伝子比を示す。
【0116】
予想されるPr値は、ウェルあたりの参照対立遺伝子の平均濃度(mr)に従い非線形的に変化する。表600に示すように、対立仮説を許容するためのPrの予想される値は、mrに従い増大する。帰無仮説を許容するための予想されるPrの値は、0.5で固定され、対立遺伝子の不均衡を有する又は有しない試料は、mrが増大すると、Prの数値においてより大きく隔てられる。他の態様において、帰無仮説を許容するための数値は、0.5以外になり得ることに留意されたい。これは、正常の比率が1:1ではなく、例えば5:3であり、そのため5:3の比率が変化するときに不均衡が生じたことになる場合にあり得る。そのため、2つの異なる核酸配列の量の相異は、ケースバイケースで判定され得る。
【0117】
しかしながら、従来の方法(例えば、上記Zhou, W. et al. 2002を参照されたい)は、LOH試料に固定した期待されるPr値を使用していたため、それらはLOHを有する(対立仮説を許容する)試料のPr値よりも低く見積もられていた。見積もりの低さは、mrに従い増大し得る。換言すると、DNA試料中の参照対立遺伝子の平均濃度が高くなるにつれ、旧来の方法は不正確になっていく。この、対立仮説を許容するためのPrの低い見積もりは、帰無仮説及び対立仮説を許容するためのカットオフ値の計算を両方とも不正確にし得る。
【0118】
3.予想されるPrに基づくカットオフ値の計算
SPRTを使用する態様において、El Karoui at al. (2006)のSPRT曲線の境界の上端及び下端を計算するための方程式を使用し得る。更に、帰無仮説又は対立仮説を許容するために好ましい統計的信頼性のレベルは、それらの方程式の閾値尤度比(threshold likelihood ratio)を調整することにより変化し得る。本文中、閾値尤度比は8とする。なぜなら、この値は、癌検出において、対立遺伝子の不均衡の有無で試料を峻別するのに良好な成績を提供することが示されているからである。故に、一つの態様において、SPRT曲線の境界の上端及び下端を計算するための方程式は:
境界上端=[(ln8)/n-lnδ]/lnγ
境界下端=[(ln1/8)/n-lnδ]/lnγ
で表され、式中、
δ=-(1-θ1)/(1-θ0)
γ=-(θ1(1-θ0)/θ0(1-θ1)
θ0=帰無仮説が真であるときの非参照対立遺伝子を含む情報的ウェルの割合=0.5(下記参照)
θ1=対立仮説が真であるときの、非参照(即ち過剰出現)対立遺伝子を含む情報的ウェルの割合
N=情報的ウェルの個数=いずれか1つの対立遺伝子のみが陽性であるウェルの個数
で表される。ここで、lnは、自然対数、即ちlogeを表す数学記号である。
【0119】
帰無仮説を許容するためのθ0を決定するために、試料が正倍数体胎児を妊娠した妊婦から取得されたと仮定する。この仮定の下、いずれかの鋳型が陽性であるウェルの予想される個数は、1:1となるべきであり、そのため非参照対立遺伝子を含む情報的ウェルの予想される割合は、0.5となるべきである。
【0120】
対立仮説を許容するためのθ1を決定するために、試料がトリソミー21胎児を妊娠した妊婦から取得されたと仮定する。デジタルRNA-SNP解析のためにされる、トリソミー21の場合の予想されるPr の計算は、表600に詳述される。故に、デジタルRNA-SNP解析のためのθ1は、表600の最後のカラムに示すデータを参照する。
【0121】
4.平均濃度の測定
mrの測定は、当業者に知られる、又は今後知られ得る様々なメカニズムを通じて実行され得る。一つの態様において、mrの値は、デジタルPCR解析の実験プロセスの間に決定される。mrの値と参照対立遺伝子が陽性であるウェルの総数との間の関係は分布(例えばポアソン分布)により支配されるので、mrは、参照対立遺伝子が陽性であるウェルの個数から計算され得て、ここで、mr=-ln(1-参照対立遺伝子が陽性であるウェルの割合)の計算式が使用される。ここで、lnは、自然対数、即ちlogeを表す数学記号である。このアプローチは、デジタルPCR実験に使用されるDNA試料におけるmrの直接的かつ正確な見積もりを提供する。
【0122】
この方法は、所望の濃度を実現するのに使用され得る。例えば、方法200の工程240と同じく、抽出された試料の核酸は特定の濃度、例えば反応ウェルあたり鋳型分子1個の濃度まで希釈され得る。ポアソン分布を使用する一つの態様において、鋳型が入らないウェルの予想される割合は、e-mと計算され得て、ここでmは、ウェルあたりの平均鋳型分子濃度である。例えば、ウェルあたり鋳型分子1個の平均濃度において、鋳型分子が入らないウェルの予想される割合は、e-1、即ち0.37(37%)である。残りの63%のウェルは、1個以上の鋳型分子が含まれ得る。典型的には、デジタルPCRを実行して、陽性かつ情報的ウェルの個数が計数される。情報的ウェルの定義及びデジタルPCRデータを解釈する方法は、装置に依存する。
【0123】
他の態様において、ウェルあたりの平均濃度のmrは、他の定量方法、例えば定量リアルタイムPCR、半定量競合PCR、質量分析法を用いる実競合PCR等により測定される。
【0124】
B.デジタルRCD
平均濃度を使用するデジタルRCDは、上記デジタルSNP法と同様の方式で実行され得る。参照染色体(第21番染色体ではない)マーカー、第21番染色体マーカー、及びそれら両マーカーが陽性であるウェルの個数が、デジタルPCRにより決定され得る。ウェルあたりの参照マーカーの平均濃度(mr)は、デジタルSNP解析におけるmrの計算の場合と同じく、ポアソン分布の関数に従い、第21番染色体マーカーの陽性と無関係に、参照マーカーが陰性であるウェルの総数から計算される。
【0125】
それから、SPRT解析は、正倍数体又はトリソミー21の胎児を妊娠した妊婦から取得した血漿試料を分類するのに使用され得る。このシナリオにおいて、参照マーカー及び第21番染色体マーカーが陽性であるウェルの予想される比率は1:1であり、そのため、第21番染色体の陽性シグナルを示す情報的ウェルの予想される割合は0.5となる。胎児が第21番染色体においてトリソミーであるとき、対立仮説が許容され得る。このシナリオにおいて、試料DNAが胎児由来のもののみであれば、各ウェル中の第21番染色体の平均濃度は、参照マーカーの平均濃度(mr)の3/2倍であり得る。
【0126】
デジタルRCDは、胎児特異的マーカー、例えば胎盤エピジェネティックサイン(Chim, SSC. et al. 2005 Proc Natl Acad Sci USA 102, 14753-14758)等の検出を通じて、染色体量を決定するのに使用され得るが、このデジタルRCD解析の一つの態様は、非胎児特異的マーカーを使用する。そのため、非胎児特異的マーカーが使用されるとき、胎児由来分子のパーセンテージを測定する追加的な工程が加えられる。従って、ウェルあたりの第21番染色体の平均濃度は、試料中の胎児DNAの割合に依存し、そして、mr[(200%+胎児DNAパーセンテージ)/200%]を使用して計算され得る。
【0127】
説明のために、もう一度特定の例を使用する。ウェルあたりの参照染色体、第1番染色体の平均濃度()を0.5と仮定し、試料中50%のDNAが胎児由来で、DNAの50%が母親由来と仮定する。
【0128】
従って、ポアソン分布を使用すると、ウェルあたりの平均濃度が0.5のときの第1番染色体の遺伝子座のいずれの分子も含有しないウェルの割合は:
【化4】
となる。
【0129】
故に、第1番染色体の遺伝子座の少なくとも1つの分子をウェルが含む確率は:1-0.6065=0.3935である。従って、約39%のウェルは、少なくとも1個の遺伝子座の分子を含有すると予想され得る。
【0130】
このトリソミー21胎児の各細胞において、第21番染色体と第1番染色体とのゲノム比率は、3:2となる。DNA試料中の第21番染色体と第1番染色体との間の比率は、分別胎児DNA濃度(胎児DNA%)に依存し:3x胎児DNA%+2(1-胎児DNA%):2x胎児DNA%+2x(1-胎児DNA%)となる。よって、分別胎児DNA濃度が50%であるこの場合、比率は:(3x50%+2x50%)/(2x50%+2x50%)=1.25となる。シグナルSNP法が胎児特異的マーカーを使用していなければ、このような計算を、非参照配列の平均濃度を計算するのにも使用することが出来ていた。
【0131】
故に、ウェルあたりの第1番染色体の遺伝子座の平均濃度が0.5であるとき、ウェルあたりの第21番染色体の平均濃度は:1.25x0.5=0.625となる。よって、ウェルあたりの平均濃度が0.625であるときの、第21番染色体遺伝子座の分子がウェルに入らない確率は:
【化5】
となる。
【0132】
故に、任意のウェルが少なくとも1個の第21番染色体の遺伝子座を含む確率は:1-0.5353=0.4647となる。よって、約46%のウェルは、少なくとも1個の前記遺伝子座の分子を含むと期待される。ウェルにいずれの遺伝子座が入るかが独立していると仮定すると、ウェルに両方の遺伝子座が入る確率は:0.3935x0.4647=0.1829となる。よって、約18%のウェルは、両方の遺伝子座を含むと期待される。
【0133】
第1番染色体の遺伝子座を含むが第21番染色体の遺伝子座を含まないと期待されるウェルの割合は、少なくとも1個の第1番染色体の遺伝子座を含むウェルの個数から、両方の遺伝子座を含むウェルの個数を引いた値であり:0.3935-0.1829=0.2126となる。同様に、第21番染色体の遺伝子座を含むが両方の遺伝子座を含まないと期待されるウェルの割合は:0.4647-0.1829=0.2818となる。情報的ウェルは、第1番染色体の遺伝子座又は第21番染色体の遺伝子座のいずれかが陽性であるが、それらの両方が両性ではないウェルとして定義される。
【0134】
故に、デジタルRCDにおいて予想される第21番染色体と第1番染色体との比率は、0.2818/0.2106=1.34となる。換言すると、第21番染色体遺伝子座のみ陽性であるウェルの割合は、第1番染色体遺伝子座のみ陽性であるウェルの割合の1.34倍となる。これは、DNA試料における1.25の比率と対照的である。
【0135】
SPRT解析において、第21番染色体遺伝子座が陽性である情報的ウェルの割合(Pr)は、SPRT曲線を使用して計算及び解釈される必要がある。今回の事例では、情報的ウェルの割合は:0.2106+0.2818=0.4924となる。故に、mrが0.5で、胎児DNAが50%の場合のトリソミー21の予想されるPrは:0.2818/0.4924=0.57となる。
【0136】
平均鋳型濃度(m)がポアソン方程式の肝要なパラメーターなので、Prはmに応じて変化する。図7は、本発明の一つの態様における、ウェルあたりの平均参照鋳型濃度(mr)として表現される範囲の鋳型濃度で、分別胎児DNA濃度が10%、25%、50%及び100%のトリソミー21試料における、予想されるPrを列記する、表700を示す。デジタルRCD解析のためのトリソミー21の検体の予想されるPrの計算は、表700に詳述している。故に、胎児DNA分別濃度が様々な試料のデジタルRCD解析用のθ1は、表700に記載の対応する予想されるPr値を示すカラムから取得され得る。
【0137】
C.結果
1.異なるmrの比較
対立遺伝子又は染色体の不均衡を理論的(胎児ゲノム中の如く)に導き出した場合と実験的に予想した場合との間の程度の違いの基準、及びその後者の場合のmr値を決定する計算を、表600及び表700に示す。トリソミー21試料のデジタルRNA-SNP解析において、mr=0.5のとき、参照対立遺伝子のみを含むウェルに対する、過剰出現対立遺伝子のみを含むウェルの比率、即ちデジタルRNA-SNP比率は、2.65となる(表600)。100%胎児検体からなる検体のデジタルRCD解析において、mr=0.5のとき、第1番染色体遺伝子座のみが陽性であるウェルに対する、第1番染色体遺伝子座のみが陽性であるウェルの比率、即ちデジタルRCD比率は、0.63/(1-0.63)=1.7).となる。分別胎児DNA濃度が低下すると、mrが同一となるまでデジタルRCD比率も低下する(表700)
【0138】
表600及び700に示されるように、対立遺伝子又は染色体の過剰出現の程度は、mrに応じて増大する。しかしながら、情報的ウェルのパーセンテージは、mr=0.5付近で頭打ちとなり、続いてmrが更に増大するにつれて、徐々に低下していく。実際に、情報的ウェルの割合の低下は、もし検体の鋳型分子の量が無制限であるならば、ウェルの総数を増大することで補償され得る。しかし、ウェルを追加すると、試薬のコストが増大する。故に、最適のデジタルPCR効率は、鋳型濃度と、試料あたり検査されるウェルの総数との間のトレードオフにより決定される。
【0139】
2.SPRT曲線を使用する実例
上に考察したように、デジタルPCR実験のための対立遺伝子又は染色体の不均衡の予想される程度は、反応混合物(例えばウェル)あたりの実際の鋳型濃度に依存する。発明者らは、参照対立遺伝子に基づく鋳型濃度、即ちウェルあたりの平均参照鋳型濃度(mr)を記載する。上記公式に示されるように、予想されるPrは、上方及び下方のSPTR曲線を描画するのに使用され得る。同じく、予想されるPrは、mrの値に依存し、SPRT曲線の描画は、本質的に、mrの値に依存し得る。よって、実際には、特定の実験から得られたPrを解釈するのに、デジタルPCRデータセットの実際のmrに関連するSPRT曲線のセットを使用する必要があろう。
【0140】
図8は、本発明の一つの態様における、デジタルRNA-SNP解析のための、mrの値が0.1、0.5、及び1.0の場合の、SPRT曲線の相異の程度を説明するプロット800を示す。デジタルPCRデータの各セットは、特定の実験の正確なmrの値に関する特定の曲線を使用して解釈されるべきである。デジタルRNA-SNP及びRCDアプローチのための、対立遺伝子又は染色体の不均衡の予想される程度は異なる(前者は2:1、後者は3:2である)ため、2つのデジタルPCRシステムのために、異なるセットのSPRT曲線が必要となる。実験的に導き出されたPrは、デジタルPCR反応の対応するmrにより選択される適切なSPRT曲線を用いて解釈される。これは、従来報告されていた、固定されたセットの曲線を使用する、デジタルPCRによるLOHの分子検出のためのSPRTの使用と対照的である。
【0141】
SPRTを使用してデジタルPCRデータを解釈する実際の方法は、仮想のデジタルRNA-SNP反応を使用して、下で説明される。それぞれの場合のデジタルRNA-SNP解析の後、対立遺伝子Aのみが陽性であるウェルの個数、対立遺伝子Gのみが陽性であるウェルの個数、又は両方の対立遺伝子が陽性であるウェルの個数が計数される。参照対立遺伝子は、陽性ウェルの個数がより少数の対立遺伝子として定義される。mrの値は、ポアソン確率密度関数に従い、他方の対立遺伝子が陽性か否かに関係なく、参照対立遺伝子が陰性のウェルの総数を使用して計算される。発明者らの仮説例のデータを以下に示す。
【0142】
96ウェルの反応において、20ウェルが対立遺伝子Aのみ陽性であり、24ウェルが対立遺伝子Gのみ陽性であり、そして33ウェルが両方とも陽性である。G陽性ウェルよりもA陽性ウェルの方が少ないので、対立遺伝子Aが参照対立遺伝子とみなされる。参照対立遺伝子が陰性のウェルの個数は、96-20-33=43である。よって、ポアソン方程式を使用してmrが計算され:-ln(43/96)=0.80となる。このケースの実験的に決定されたPrは:24/(20+24)=0.55となる。
【0143】
表600によると、mr=0.8で、トリソミー21試料の予想されるPrは、0.76となる。よって、このケースでは、θ1は0.76となる。θ1=0.76に基づくSPRT曲線が、実験的に導き出されたPr(このケースでは0.55)を解釈するのに使用され得る。Pr=0.55が、関連するSPRT曲線上に収まったとき、そのデータ点は、下方の曲線に分類される。故に、本ケースは、正倍数体として分類される。図3を参照されたい。
【0144】
3.従来の手法との比較
図9Aは、96ウェルデジタルRNA-SNP解析における正倍数体及びトリソミー21の検体を分類するための、新旧のSPRTアルゴリズムの有効性を比較する、表900を示す。図9Bは、384ウェルデジタルRNA-SNP解析における正倍数体及びトリソミー21の検体を分類するための、新旧のSPRTアルゴリズムの有効性を比較する、表950を示す。新しいアルゴリズムは、デジタルPCRデータから導き出されたmrに対して特異的なSPRT曲線を選択することを意味する。古いアルゴリズムは、全てのデジタルPCR反応に対して固定されたセットのSPRT曲線を使用することを意味する。分級精度におけるカットオフ値の計算が不正確であることの効果は、表900に示すシミュレーション解析により明らかにされる。
【0145】
表900及び950に示されるように、従来の研究における固定されたセットのSPRT曲線を使用する場合と比較して、分類不能データの割合は、発明者らのアプローチの方が相当に低い。例えば、mr =0.5で発明者らのアプローチを使用した場合、96ウェル及び384ウェルデジタルRNA-SNP解析において、それぞれ14%及び0%のトリソミー21試料が分類不能であったが、固定した曲線を使用した場合、それぞれ62%及び10%が分類不能であった(表900)。故に、発明者らのアプローチは、より少数の情報的ウェルの個数で、疾患分類を可能とする。
【0146】
表900に示されるように、前記新しいアルゴリズムは、mrが0.1〜2.0の全ての値である場合において、対立遺伝子比率の歪みの有無をもって、より正確に試料を分類することが出来る。例えば、mr=1.0で、96ウェルデジタルRNA-SNP反応を実行する場合、前記新しいアルゴリズムは、対立遺伝子比率の歪みを有する又は有しない試料を、それぞれ88%及び92%の正確性で分類した。一方、前記古いアルゴリズムを使用した場合、対立遺伝子比率の歪みを有する又は有しない試料を正確に分類したパーセンテージは、それぞれ僅か19%及び36%であった。
【0147】
この新しいアルゴリズムを使用することにより、対立遺伝子比率の歪みを有する試料及び有しない試料の分離の精度は、mrに応じて増大し得る。結果として、分類精度は、mrに応じて増大し得る。mrが2.0を超えて増大すると、試料を2つの群に分離する場合の分類精度の向上の作用は低下する。なぜなら、情報的ウェルのパーセンテージが減少するからである。一方、古いアルゴリズムを使用した場合、分類精度はmrが増大すると顕著に低下する。なぜなら、予想されるP値の真の値からの逸脱が増大するからである。
【0148】
発明者らの実験データ及びシミュレーションデータは、デジタルRNA-SNPは、トリソミー21検出の有効かつ正確な方法であることを示す。母体血漿中のPLAC4 mRNAは純粋に胎児に由来するため、13個の母体血漿サンプル中12個は、正確な分類のために1回の384ウェルデジタルPCR実験で十分であった(図13Bの表1350)。このホモ接合性の、リアルタイムデジタルPCRに基づくアプローチは、RNA-SNP解析のための、質量分析に基づくアプローチ(上記Lo, YMD, et al. 2007 Nat Med,)の代替手法を提案する。胎盤特異的mRNA転写産物は別として、発明者らは、母体血清中の他の種類の胎児特異的核酸種が、デジタルPCRに基づく胎児染色体の異数性検出に使用可能であることをも想定する。一例として、エピジェネティック対立遺伝子比率(EAR)アプローチ(Tong, YK et al. (2006) Clin Chem 52, 2194-2202)を使用する、トリソミー18の非侵襲的な出生前検出に最近使用されている、胎児エピジェネティックマーカー(Chim, SSC et al. (2005) Proc Natl Acad Sci USA 102, 14753-14758; Chan, KCA et al. (2006) Clin Chem 52, 2211 -2218)が挙げられる。これにより、発明者らは、デジタルEARが、可能な解析技術であると予測する。
【0149】
V.%の増大、複数のマーカー、及びPCR代替手法
上記のように、本発明の態様を母体血漿から抽出したDNAに応用することは、妊娠11週〜17週の、母体血漿中の胎児DNAの分別濃度が約3%という僅かな割合でしか存在しないとき、複雑なものとなり得る。しかしながら、本明細書中に記載されるように、デジタルRCDは、異数性DNAが少数集団として存在するときであっても、異数性検出を可能とする。妊娠初期の如く胎児DNAの分別濃度が低いものほど、デジタルRCDにより多数の情報的カウントが必要となる。本研究の重要性は、図12の表1200に要約したように、発明者らが、診断的実験の基礎となる、例えば分別胎児DNA及び全鋳型分子等のベンチマークパラメーターのセットを提供したことである。発明者らの見解では、分別胎児DNA濃度が25%のとき、総数で7680の反応が、ベンチマークパラメーターの特に魅力的なセットである。これらのパラメーターによれば、表1200に示すように、分別胎児DNA濃度が25%のとき、正倍数体及びトリソミー21試料を、97%の正確性で分類することが可能となる。
【0150】
単位体積の母体血漿あたりに存在する血漿DNA分子の個数は限定される(Lo, YMD. et al. 1998 Am J Hum Genet 62, 768-7758)。例えば、妊娠早期において、常染色体遺伝子座(β-グロビン)の中間母体血漿濃度は、986コピー/mlであり、胎児と母親の両方の遺伝子座が算入されている(Lo, YMD. et al. 1998 Am J Hum Genet 62, 768-7758)。7680分子を捕らえるのに、約8mlの母体血漿から抽出したDNAが必要とされ得る。この血漿の体積は、約15mlの母体血液から取得され得て、これは通常の診療における限界である。しかしながら、発明者らは、複数のセットのchr21及び参照染色体標的を、デジタルRCD解析のために組み合わせ得ることを想定する。chr21及び参照染色体標的の5個のペアにおいて、分析に必要な数の鋳型分子を得るのに、ちょうど1.6mLの材料血漿が必要となり得る。多重単一分子PCRが実行され得る。この多重単一分子解析のロバスト性は、単一分子ハプロタイピングにおいて既に実証されている(Ding, C. and Cantor, CR. 2003 Proc Natl Acad Sci USA 100, 7449-7453)。
【0151】
あるいは、25%の分別胎児DNA濃度を達成するために、母体血漿中の胎児DNAを選択的に増幅する方法(Li, Y. et al. 2004 Clin Chem 50, 1002-1011)、若しくは母体DNAバックグラウンドを抑制する方法(Dhallan, R et al. 2004 JAMA 291, 1114- 1119)、又はそれらの両方が採用され得る。そうした胎児DNAの増幅や母体DNAの抑制等の物理的手法とは別に、特定のDNAメチル化パターンを示す胎児DNA分子を標的とする(Chim, SSC et al, 2005 Proc Natl Acad Sci USA 102,14753-14758, Chan, KCA et al. 2006 Clin Chem 52, 2211-2218; Chiu, RWK et al 2007 Am J Pathol 170, 941-950.)等による、分子増幅法を使用することも可能であり得る。
【0152】
加えて、現在は、デジタルPCRを実施する研究において使用される、デジタルリアルタイムPCR解析を手動で調整する多くの代替的なアプローチが存在する。これらの代替的アプローチは、マイクロ流体工学デジタルPCRチップ(Warren, L et al. 2006 Proc Natl Acad Sci USA 103, 17807-17812; Ottesen, EA et al. 2006 Science 314, 1464-1467)、エマルジョンPCR(Dressman, D et al. 2003 Proc Natl Acad Sci USA 100, 8817-8822)、並びにRoche 454プラットフォーム、Illumina Solexaプラットフォーム、及びApplied BiosystemsのSOLiD(商標)システム等を使用する超並列ゲノムシークエンシング(Margulies, M. et al. 2005 Nature 437, 376-380)等を含む。後者に関しては、発明者らの手法は、単一DNA分子に対する超並列シークエンシング法にも応用可能であり、これは、Helicos True Single Molecule DNAシークエンシング技術、Pacific Biosciencesの単一分子リアルタイム(SMRT(商標))技術、及びナノポアシークエンシング(Soni GV及びMeller A. 2007 Clin Chem 53, 1996-2001)等の増幅工程を要しない。これらの方法を使用することにより、デジタルRNA-SNP及びデジタルRCDを、大量の試料に対して迅速に実行することが出来て、それにより、非侵襲性出生前診断に提案される方法の臨床的柔軟性が向上する。
【実施例】
【0153】
以下の実施例は、例示のために提供されるものであり、主張する発明を限定することを目的としない。
I.コンピューターシミュレーション
SPRTアプローチを使用してのトリソミー21診断の精度を見積もるために、コンピューターシミュレーションを実行した。このコンピューターシミュレーションは、Microsoft Excel 2003ソフトウェア(Microsoft Corp., USA)及びSAS 9.1 for Windows(登録商標)ソフトウェア(SAS Institute Inc., NC, USA)を用いて行われた。デジタルPCRの性能は、参照鋳型濃度(mr)、情報的カウントの個数と、対立遺伝子又は染色体の不均衡の推定される程度(Pr)との間の相互作用である。これらの変数を各々、様々に変化させてシミュレーションを実行した。デジタルRNA-SNPとデジタルRCDとでSPRT曲線の決定境界が異なるため、それら2つのシミュレーション解析は別個に実行された。
【0154】
想定した各デジタルPCR条件(即ち胎児DNA分別濃度、ウェル総数)について、2ラウンドのシミュレーションを実行した。第一のラウンドでは、正倍数体胎児を妊娠した妊婦から検査試料を取得したというシナリオを想定した。第二のラウンドでは、トリソミー21胎児を妊娠した妊婦から検査試料を取得したというシナリオを想定した。各ラウンドにおいて、胎児5000人分の検査を想定した。
【0155】
A.RNA-SNP
デジタルRNA-SNPにおいて、mr=0.1〜mr=2.0である384ウェル実験のシミュレーションを実行した。各mr値で、発明者らは、正倍数体胎児5000人及びトリソミー21胎児5000人を試験したというシナリオを想定した。10000人の胎児を分類するために、所定のmrに適したSPRT曲線を使用した。図10は、本発明の態様における所定の情報的カウントにより、正倍数体又は異数体として正しく及び誤って分類された、並びに分類不能であった胎児のパーセンテージを示す、表1000である。mrが0.5〜2.0の場合、性倍数体と異数体の診断の正確性は、いずれも100%である。mr=0.5のとき、384ウェルの解析後に正確に分類できた性倍数体及びトリソミー21胎児は、それぞれ僅か57%及び88%である。
【0156】
これらのシミュレーションデータは、以下に記載される工程により生成された。
【0157】
工程1において、各ウェルについて、SASプログラム(www.sas.com/technologies/analytics/statistics/index.html)のランダム(ポアソン)関数を使用して、それぞれ対立遺伝子A及びGを表す2つのランダムな数字を生成した。ランダム(ポアソン)関数は、0から始まる正の整数(即ち0、1、2、3・・・)を生成し、各整数が生成される確率は、ウェルあたりの対立遺伝子の平均濃度を表す所定の平均値についてのポアソン確率密度関数に従い、各整数の確率に応じて決定した。対立遺伝子Aを表すランダム数字がゼロより大きとき、そのウェルは対立遺伝子Aが陽性、即ち対立遺伝子Aの分子を1つ以上含むとみなされた。同様に、対立遺伝子Gを表すランダム数字がゼロより大きいとき、そのウェルは対立遺伝子Gが陽性とみなされた。
【0158】
正倍数体胎児を妊娠した妊婦のシナリオを想定するにあたり、対立遺伝子A及び対立遺伝子Gにランダム数字を生産するために、同一の平均値が使用された。例えば、mr=0.5でのデジタルRNA-SNP解析を想定する解析において、対立遺伝子A及びGのいずれの平均値も0.5に設定した。これは、いずれの対立遺伝子も平均濃度がウェルあたり0.5分子であることを意味する、ポアソン方程式を使用して、平均濃度が0.5で、対立遺伝子A又はBが陽性であるウェルの割合は同一であり得て、0.3935であった。表600を参照されたい。
【0159】
トリソミー21を妊娠した妊婦のmr=0.5でのデジタルRNA-SNP解析を想定するとき、過剰出現対立遺伝子のウェルあたりの平均濃度は、参照対立遺伝子の値の2倍、即ち1と期待され得る。この条件下で、過剰出現対立遺伝子が陽性であるウェルの確率は、0.6321であった。表600を参照されたい。
【0160】
デジタルPCRウェルのランダム数字を生成した後、ウェルは、以下の状態のいずれかに分類され得る:
a.対立遺伝子A及びGのいずれも陰性
b.対立遺伝子A及びGのいずれも陽性
c.対立遺伝子Aは陽性であるがGは陰性
d.対立遺伝子Gは陽性であるがAは陰性
【0161】
工程2において、所望のウェル数(本シミュレーションでは384ウェル)に達するまで工程1が反復された。対立遺伝子Aのみ、又は対立遺伝子Gのみが陽性であるウェルの子数が計数された。陽性ウェルが少ない方の対立遺伝子を参照対立遺伝子とみなし、陽性ウェルが多い方の対立遺伝子を潜在的に過剰出現対立遺伝子とみなした。情報的ウェルの個数は、いずれかの対立遺伝子が陽性であるがそれらの両方が陽性ではないウェルの総数とした。そして、潜在的に過剰出現対立遺伝子を含む情報的ウェルの割合(Pr)を計算した。本発明の一つの態様に従い、帰無仮説又は対立仮説を許容するためのSPRT曲線における境界の上端及び下端を計算した。
【0162】
工程3において、正倍数体又はトリソミー21胎児を妊娠した妊婦の2つの各シナリオについて、5000回シミュレーションを実行した。各シミュレーションを、妊婦から取得した独立の生体試料とみなし得る。表1000において、正倍数体の検体の正しい分類は、帰無仮説が許容された正倍数体の検体を意味し、そして、正倍数体の検体の誤った分類は、対立仮説が許容された正倍数体の検体を意味する。同様に、対立仮説が許容されたトリソミー21の検体は正しく分類されたものとみなされ、そして、帰無仮説が許容されたトリソミー21の検体は誤って分類されたものとみなされる。いずれの群においても、事前に指定されたウェルの総数がシミュレーションされた後に、帰無仮説及び対立仮説のいずれも許容されない検体は、分類不能とみなした。
【0163】
工程4において、mrを0.1〜2.0の範囲で0.1ずつ増大させて設定し、工程1〜3を実行した。
【0164】
B.RCD
図11は、本発明の一つの態様における、mrが0.1〜2.0の範囲で、純粋(100%)な胎児DNA試料についてなされるデジタルRCDのコンピューターシミュレーションを示す、表1100である。分別胎児DNA濃度が低下するに従い、第21番染色体の過剰出現の度合は低下し、それにより疾患分類を確定するのにより多数の情報的ウェルが必要となる。故に、シミュレーションは、mr=0.5で、384〜7680ウェルのウェル総数の範囲において、50%、25%、及び10%の胎児DNA濃度において、更に実行された。
【0165】
図12は、本発明の一つの態様における、胎児DNAの分別濃度が異なる、正倍数体又はトリソミー21胎児由来のサンプルを分類するための、mr=0.5における、デジタルRCD解析の精度のコンピューターシミュレーションの結果を示す、表1200である。デジタルRCDの効率は、検体のDNA分別濃度が高くなるに従い良好になる。胎児DNA濃度が25%で、PCR解析の総数が7680回で、正倍数体及び異数体の検体のいずれも、分類を誤らず97%が分類可能であった。検体の残りの3%については、分類を達成するのに更なる解析を必要とする。
【0166】
デジタルRCD解析をシミュレーションする手順は、デジタルRNA-SNP解析において記載した物と同様であった。このシミュレーションの工程を、以下に記載する。
【0167】
工程1において、参照遺伝子座、第1番染色体、及び第21番染色体遺伝子座を表現するために、ポアソン確率密度関数の下で2つのランダムな数字が生成された。正倍数体の胎児を妊娠した対象において、第1番染色体及び第21番染色体の平均濃度は、いずれも等しかった。このシミュレーション解析において、各遺伝子座のウェルあたりの平均鋳型濃度には、0.5を採用した。トリソミー21胎児を妊娠した対象において、このシミュレーションにおけるmrは0.5であったが、ウェルあたりの第21番染色体の平均濃度は、表700に示されるように、検査される使用中の分別DNA濃度に依存していた。参照遺伝子座及び/又は第21番染色体遺伝子座の1個のウェルへの分配は、ウェルあたり遺伝子座の適切な平均濃度で、ポアソン確率密度関数に従い生成された各遺伝子座を表すランダムな数字により決定された。
【0168】
工程2において、所望のウェル数、例えば384ウェルプレートの実験であれば384ウェルに達するまで、工程1が反復された。第1番染色体のみ、及び第21番染色体のみが陽性であるウェルの子数が計数された。情報的ウェルの個数は、いずれかの染色体が陽性であるがそれらの両方が陽性ではないウェルの総数とした。そして、第21番染色体が陽性である情報的ウェルの割合(Pr)を計算した。前記SPRT解析の章で記載したのと同様に、本発明の一つの態様に従い、帰無仮説又は対立仮説を許容するためのSPRT曲線における境界の上端及び下端を計算した。
【0169】
工程3において、正倍数体又はトリソミー21胎児を妊娠した妊婦の2つの各シナリオについて、5000回シミュレーションを実行した。各シミュレーションを、妊婦から取得した独立の生体試料とみなし得る。表1100において、正倍数体の検体の正しい分類は、帰無仮説が許容された正倍数体の検体を意味し、そして、正倍数体の検体の誤った分類は、対立仮説が許容された正倍数体の検体を意味する。同様に、対立仮説が許容されたトリソミー21の検体は正しく分類されたものとみなされ、そして、帰無仮説が許容されたトリソミー21の検体は誤って分類されたものとみなされる。いずれの群においても、事前に指定されたウェルの総数がシミュレーションされた後に、帰無仮説及び対立仮説のいずれも許容されない検体は、分類不能とみなした。
【0170】
工程4において、384〜7680ウェルのウェル総数の範囲において、10%、25%、50%及び100%の胎児DNA濃度のサンプルにおいて、工程1〜3を反復した。
【0171】
II.トリソミー21検出の変法
A.PLAC4用RNA-SNP
第21番染色体上のPLAC4のrs8130833 SNP(Lo, YMD et al. 2007 Nat Med 13, 218-223)を使用して、デジタルRNA-SNPの実際の実行可能性を実証した。2つの正倍数体及び2つのトリソミー21ヘテロ接合体の胎盤に由来する胎盤DNA及びRNAを解析した。これらの胎盤DNA試料を、デジタルRNA-SNPプロトコールを用いて分析したが、逆転写工程を省略したので、本質的には、その手順を、デジタルDNA-SNP解析に転換したものであった。検体を正しく分類する可能性と情報的ウェルの割合との間のバランスを取るために、発明者らは、ウェルあたり任意の種類の対立遺伝子が1個入るように試料を希釈し、そしてそれを96ウェルデジタルPCR解析により検証した。これは、384ウェルデジタルRNA-SNP実験により追認された。Pr及びmrが計算され、このmrに対するSPRT曲線が、データ解釈に使用された。
【0172】
図13Aは、本発明の一つの態様における、正倍数体及びトリソミー21胎児を妊娠した妊婦の胎盤組織をデジタルRNA-SNP解析の結果である、表1300を示す。質量分析アッセイにより、遺伝子型を決定した。「正倍数体」は、実験的に取得したPrが分類不能領域(unclassifiable region)を下回ったときに割り当てられ;トリソミー21を意味する「T21」は、実験的に取得したPrが分類不能領域を上回ったときに割り当てられる。これらの各検体は、1回の384ウェル実験で、DNA及びRNA試料のいずれを使用しても、正しく分類された。
【0173】
発明者らは、更に、正倍数体胎児を妊娠した妊婦9人及びトリソミー21胎児を妊娠した妊婦4人から採取した血漿RNAを検査した。図13Bは、本発明の一つの態様における、正倍数体及びトリソミー21を妊娠した妊婦から採取した母体血漿のデジタルRNA-SNP解析の結果である、表1350を示す。検体は全て、正しく分類された。1つのトリソミー21検体(M2272P)の最初の結果は、SPRT曲線の分類不能領域内に収まった。そこで、追加的な384ウェル実験が実行された。計768ウェルから収集したデータから、新たなmr及びPrが計算され、このmr値に基づいて選択された新たなセットのSPRT曲線を使用して、分類が実行された。その結果、上記検体は、異数体として正しく評点された。
【0174】
発明者らの実験的データ及びシミュレーションデータは、デジタルRNA-SNPが、トリソミー21を検出する有効かつ正確な方法であることを示す。母体血漿中のPLAC4 mRNAは純粋に胎児に由来するので、13個中12個の母体血漿試料を正しく分類するのに、僅か1回の384ウェルデジタルPCR実験で十分である。故に、この均質な、リアルタイムデジタルPCRに基づくアプローチは、RNA-SNP解析のための、質量分析に基づくアプローチという代替手法を提案する。胎盤特異的mRNA転写産物とは別に、発明者らは、母体血清中の他の種類の胎児特異的核酸種が、胎児染色体の異数性のデジタルPCRに基づく検出に使用され得ることを想定する。一例として、近頃エピジェネティック対立遺伝子比率(EAR)アプローチTong YK et al. 2006 Clin Chem, 52, 2194-2202)を使用したトリソミー18の非侵襲性出生前検出に使用されている、胎児エピジェネティックマーカーが挙げられる。そのため、発明者らは、デジタルEARが、可能な解析技術になり得ると予想している。
【0175】
B.RCD
また、トリソミー21を検出するためのデジタルRCDの実際の実行可能性は、第21番染色体及び第1番染色体上のパラロガス配列を標的にするPCRアッセイを使用して調査された。パラロガス遺伝子座は、実施例の方法により、ここで使用された。第21番染色体上の非パラロガス配列、及び他の任意の参照配列も、RCDに使用され得る。2つの正倍数体及び2つのトリソミー21胎盤に由来する胎盤DNA試料を、いずれかの染色体由来の標的鋳型がウェルあたりに約1個入る濃度に希釈し、それを、96ウェルデジタルPCR解析により確認した。確認された各試料を、384ウェルデジタルRCD実験で解析し、そしてPr及びmrの値を計算した。デジタルRCDにおいて、第1番染色体のパラログが、参照鋳型であった。このmr値を、データ解釈のための対応するセットのSPRT曲線の選択に使用した。図14Aに示されるように、全ての胎盤試料が正しく分類された。
【0176】
母体血漿中の胎児DNAの如く、過剰の正倍数性DNAと混在しているトリソミー21DNAを検出するのに、デジタルRCDアプローチが使用され得ることを明示するために、正倍数性母体血球細胞DNAのバックグラウンド中に、50%及び25%のトリソミー21胎盤DNAを含む混合物を解析した。トリソミー21検体10個、及び正倍数体の検体10個に由来する胎盤DNAのそれぞれが、同量の正倍数性血球細胞DNAと混合され、これで20個の50%DNA混合物が出来た。図14Bは、本発明の一つの態様における、それら50%胎児DNA混合物のRCD解析のためのSPRT解釈を説明する、プロット1440を示す。同様に、トリソミー21検体5個、及び正倍数体の検体5個に由来する胎盤DNAのそれぞれが、3倍量の正倍数性血球細胞DNAと混合され、これで10個の25%DNA混合物が出来た。図14Cは、それら25%胎児DNA混合物のRCD解析のためのSPRT解釈を説明する、プロット1470を示す。図14B及び図14Cに示すように、全ての正倍数性DNA混合物及び異数性DNA混合物は、正しく分類された。
【0177】
各試料は、図14B及び14Cにマークされるように、多くの384ウェルデジタルPCR解析の後に、解析可能な点に達した。50%DNA混合物において、必要とされる384ウェルプレートの数は、1〜5枚の範囲であった。25%DNA混合物において、必要とされる384ウェルプレートの数は、1〜7枚の範囲であった。全384デジタルPCR解析を追加して正しく分類された検体の累積的割合は、表1200に表されるコンピューターシミュレーションにより予測されたものと同等であった。
【0178】
III.デジタルPCRを用いる方法
A.デジタルRNA-SNP
全てのRNA試料は、まず、ThermoScript逆転写逆転写酵素(Invitrogen)を使用して、遺伝子特異的逆転写プライマーを用いて逆転写された。逆転写プライマーの配列は、5'-AGTATATAGAACCATGTTTAGGCCAGA-3'(Integrated DNA Technologies, Coralville, IA)であった。以後、デジタルRNA-SNPのための逆転写されたRNA(即ちcDNA)試料とDNA試料(例えば胎盤DNA)とは、本質的に同じ処理がされる。デジタルDNA解析に先立ち、DNA及びcDNA試料は、まず、PLAC4に付いてのリアルタイムPCRアッセイを使用して定量された。ここで、プライマーとして、5'-CCGCTAGGGTGTCTTTTAAGC-3'、5'-GTGTTGCAATACAAAATGAGTTTCT-3'、及び蛍光プローブとして、5'-(FAM)ATTGGAGCAAATTC(MGBNFQ)-3'が用いられた(Applied Biosystems, Foster City, CA)。なお、FAMは、6-カルボキシフルオレセインであり、MGBNFQは、無蛍光クエンチャーが結合するマイナーグルーブである。
【0179】
単位複製配列をコードする、HPLC精製単鎖合成DNAオリゴヌクレオチドの連続希釈により、検量線が容易された。前記配列は、5'-CGCCGCTAGGGTGTCTTTTAAGCTATTGGAGCAAATTCAAATTTGGCTTAAAGAA AAAGAAACTCATTTTGTATTGCAACACCAGGAGTATCCCAAGGGACTCG-3'であった。反応体積を25μlとして、2X TaqMan Universal PCR Master Mix (Applied Biosystems)を使用して、反応を準備した。各反応において、各プライマーを400nL、及びプローブを80nM使用した。50℃5分で反応を開始し、続いて95℃10分、そして95℃15秒及び65℃1分を45サイクル、以上をABI PRISM 7900HT Sequence Detection System (Applied Biosystems)で行った。そして、その後のデジタルPCR増幅が、ウェルあたり鋳型分子が約1個で実行され得るように、DNA又はcDNA試料の連続希釈が行われた。そのような濃度では、約37%の反応ウェルが、増幅において陰性であることが予想され、まずは96ウェルデジタルリアルタイムPCR解析を行うことによりそのことが確認された。それから、イントロンをまたがないプライマー(順プライマーが5'-TTTGTATTGCAACACCATTTGG-3'であり、遺伝子特異的な逆プライマーは前に記載した)のセットを使用して、384ウェルプレートで行われるデジタルRNA-SNP解析が行われた。
【0180】
PLAC4上のrs8130833 SNPの2つの各対立遺伝子を標的とする、2つの対立遺伝子特異的なTaqManプローブが設計された。それらの配列は、対立遺伝子A及びGについて、それぞれ、5'-(FAM)TCGTCGTCTAACTTG(MGBNFQ)-3'及び5'-(VIC)ATTCGTCATCTAACTTG(MGBNFQ)とした。反応体積5μlの、2X TaqMan Universal PCR Master Mixを使用して、反応を準備した。各反応は、1xTaqMan Universal PCR Master Mix、各プライマーを572nM、対立遺伝子G特異的プローブを107nM、及び対立遺伝子A特異的プローブを357nM含んでいた。ABI PRISM 7900HT Sequence Detection System中で、反応を遂行した。反応は、50℃2分で始まり、そして95℃10分、更に45サイクルの95℃15秒及び57℃1分が続いた。反応の間、SDS 2.2.2ソフトウェアの「Absolute Quantification」アプリケーション(Applied Biosystems)により、蛍光データが収集された。このソフトウェアが、基準値及び閾値を自動的に計算した。対立遺伝子A又はGのいずれかが陽性であるウェルの個数が記録され、そしてその結果がSPRT解析の用に供された。
【0181】
B.デジタルRCD解析
本研究で使用された全ての胎盤及び母体軟膜のDNA試料は、まずNanoDrop分光光度計(NanoDrop Technology, Wilmington, DE)により定量された。DNA濃度は、6.6pg/細胞の転換を使用して、コピー/mlに転換される。ウェルあたり約1個の鋳型が入る濃度に相当するDNA濃度は、DNA試料の連続希釈により決定され、そして96ウェルフォーマットのリアルタイムPCRで、約37%のウェルが増幅において陽性であることにより確認された。確認プレート用のPCRは、参照染色体のプローブのみが添加される点を除いて、下記と同様に準備された。デジタルRCD解析において、第21番及び第1番染色体上のパラロガス遺伝子座(Deutsch, S. et al. 2004 J Med Genet 41, 908-915)を、最初に、順プライマー5'-GTTGTTCTGCAAAAAACCTTCGA-3'及び逆プライマー5'-CTTGGCCAGAAATACTTCATTACCATAT-3'により、同時に増幅させる。2つの染色体特異的TaqManプローブは、第21番及び第1番染色体パラログを標的にするように設計されており、それらの配列は、それぞれ、5'-(FAM)TACCTCCATAATGAGTAA A(MGBNFQ)-3'及び5'-(VIC)CGTACCTCTGTAATGTGTAA(MGBNFQ)-3'である。各反応は、1xTaqMan Universal PCR Master Mix(Applied Biosystems)、各プライマー450nM、及び各プローブ125nMを含んでいた。反応体積の合計は、5μL/ウェルであった。この反応は、50℃2分で始まり、そして95℃10分、更に50サイクルの95℃15秒及び60℃1分が続く。リアルタイムPCR実験は全て、ABI PRISM 7900HT Sequence Detection System(Applied Biosystems)で遂行され、蛍光データは、SDS 2.2.2ソフトウェア(Applied Biosystems)の「Absolute Quantification」アプリケーションで収集した。既定の基準値及び手入力の閾値が使用された。第21番又は第1番のいずれかの染色体が陽性であるウェルの個数が記録され、そしてSPRT解析の用に供された。SPRTにより疾患分類が可能となるまでに、1枚以上の384ウェルプレートが解析されるべきである。
【0182】
IV.マイクロ流体工学に基づくデジタルPCRの使用
A.デジタルRNA-SNP
本実施例は、マイクロ流体工学に基づくデジタルPCRを使用した、デジタルPCR解析の性能を明示する。本章には、ここに示される一つの方法はFluidigm BioMark(商標)システムを使用するが、これは例示を目的としており、限定を目的とするものではない。このシステムは、1回の反応で9000個以上のデジタルPCRを実行することが可能である。
【0183】
正倍数性胎児又はトリソミー21胎児を妊娠した妊婦から、胎盤組織及び母体末梢血試料が得られた。胎盤DNA試料において、プライマー伸長に続いて質量分析を行うことにより、PLAC4遺伝子上のrs8130833 SNPのジェノタイピングが遂行された。RNAは、胎盤及び母体血漿試料から抽出された。
【0184】
全てのRNA試料は、遺伝子特異的逆転写プライマー(5'-AGTATATAGAACCATGTTTAGGCCAGA-3')で、ThermoScript逆転写酵素(Invitrogen)を使用して、逆転写された。胎盤cDNA試料において、ウェルあたり約1個の鋳型分子が入る濃度でその後のデジタルPCR増幅が遂行されるように、連続希釈が遂行された。
【0185】
デジタルPCRは、12.765デジタルアレイ(Fluidigm)を有するBioMark System(商標)(Fluidigm)で行われる。各デジタルアレイは、12種類の試料アッセイ混合物に対応するための、12枚のパネルから構成される。各パネルは、更に、7-nL反応/ウェルを実行するために、765ウェルに区切られている。前記PLAC4遺伝子上のrs8130833 SNP領域は、順プライマー(5'-TTTGTATTGCAACACCATTTGG-3')及び上記遺伝子特異的逆転写プライマーにより増幅された。rs8130833 SNPの2つの各対立遺伝子を標的とする、2つの対立遺伝子特異的TaqManプローブが設計された。これらの配列は、対立遺伝子Gに対するものを5'-(FAM)TCGTCGTCTAACTTG(MGBNFQ)-3'、そしてAに対するものを5'-(VIC)ATTCGTCATCTAACTTG(MGBNFQ)とした。1枚のアレイパネルにおける反応は、反応体積10μL中のTaqMan Universal PCR Master Mixを使用して準備された。各反応は、1x TaqMan Universal PCR Master Mix、572nMの各プライマー、53.5nMの対立遺伝子G特異的プローブ、178.5 nMの対立遺伝子A特異的プローブ及び3.5 μLのcDNA鋳型を含む。各胎盤cDNA試料に1枚の反応パネルが使用されたが、各母体血漿試料には、12枚のパネルが使用された。試料アッセイ混合物は、NanoFlex(商標)IFCコントローラー (Fluidigm)によりデジタルアレイに注入された。反応はBioMark(商標)System中で実行された。その反応は、50℃2分で始まり、そして95℃10分、更に40サイクルの95℃15秒及び57℃1分が続いた。
【0186】
1つの正倍数体及び2つのT21ヘテロ接合体の胎盤に由来する胎盤RNA試料が、765ウェルの反応パネル中で解析された。各資料において、対立遺伝子A又はGが陽性(だが両方陽性ではない)である情報的ウェルが計数された。全ての情報的ウェル中の過剰出現対立遺伝子の割合(Pr)が決定された。これらの反応の、ウェルあたりの平均参照鋳型濃度(mr)に適したSPRT曲線は、実験的に取得されたPrが、倍数体又はT21試料のいずれを示唆するかを決定するのに利用された。
【0187】
発明者らは、更に、正倍数体の胎児を妊娠した女性4人及びトリソミー21胎児を妊娠した女性1人に由来する血漿RNA試料を検査した。各資料は、血漿RNA試料あたり12枚の765ウェル反応パネル中で、即ち9180回の反応で解析された。図15Bは、この血漿RNA試料における、12枚の各パネルにおける、情報的ウェルの個数を示す。この表に示すように、血漿資料中の鋳型濃度は、いずれかの反応パネル中の情報的ウェルが、SPRT分類に十分でない程度に希釈された。この試料を正倍数体試料と分類するのに、3つの反応パネルからの情報的ウェルを組み合わせなければならなかった(図15C)。図15Cは、2〜12枚のパネルからのデータを集積して使用することにより、全ての血漿検体が正しく分類され得ることを示す。
【0188】
デジタルPCRを手作業で実行する方法と比較して、このマイクロ流体工学に基づく方法は、格段に迅速で、労働集約的(labor-intensive)である。全てのプロセスが、2時間半以内に完了し得る。
【0189】
トリソミー18の出生前検出のためのデジタルRNA-SNP解析
本実施例において、第18番染色体中胎盤で発現する転写産物である、セルピンプロテアーゼ阻害剤クレードB(オボアルブミン)メンバー2(SERPINB2)mRNAに関する、デジタルPCRに基づく対立遺伝子区別アッセイを使用して、トリソミー18胎児の多型対立遺伝子の比率の不均衡を検出した。胎盤組織試料からのDNA及びRNAの抽出は、QIAamp DNA Mini Kit(Qiagen, Hilden, Germany)を使用して、取扱説明書に従い遂行された。抽出されたRNA試料は、ゲノムDNAのコンタミネーションを排除するために、DNA分解酵素I(Invitrogen)処理に付された。SERPINB2遺伝子上のrs6098 SNPのジェノタイピングは、上記のように、MassARRAY Compact(Sequenom, San Diego)を使用した均質MassEXTEND (hME)アッセイで、胎盤組織DNA試料において遂行された。
【0190】
SERPINB2転写産物の逆転写は、ThermoScript逆転写酵素(Invitrogen)を使用して、遺伝子特異的プライマー5'-CGCAGACTTCTCACCAAACA-3'(Integrated DNA Technologies, Coralville, IA)を用いて、胎盤組織RNA試料について実行された。全てのcDNA試料は、その後のデジタルPCR増幅が、反応ウェルあたり鋳型分子が1個入る平均濃度で実行され得るような濃度まで希釈された。デジタルPCRは、TaqMan Universal PCR Master Mix(Applied Biosystems, Foster City, CA)及びBiomarkTM PCR Reagents(Fluidigm, San Francisco)を使用して準備された。順プライマー5'-CTCAGCTCTGCAATCAATGC-3'(Integrated DNA Technologies)及び逆プライマー(前記逆転写用遺伝子特異的プライマーと同一)が、600nMの濃度で使用された。SERPINB2上のrs6098SNPの対立遺伝子A又はG を標的とする2つのTaqManプローブは、5'-(FAM)CCACAGGGAATTATTT(MGBNFQ)-3'及び5'-(FAM)CCACAGGGGATTATTT(MGBNFQ)-3'(Applied Biosystems)であった。FAMは、6-カルボキシフルオレセインであり、MGBNFQは、マイナーグルーブ結合無蛍光クエンチャーであり、それらはそれぞれ300nM及び500nMの濃度で使用された。各試料-試薬混合物は、NanoflexTM IFC Controller (Fluidigm)を使用するBiomarkTM 12.765 Digital Array上の756反応ウェルに分注された。その後、そのアレイは、加熱増幅及び蛍光検出のために、BiomarkTM Real-time PCR System (Fluidigm)中に置かれた。その反応は、50℃2分で始まり、そして95℃5分、更に45サイクルの95℃15秒及び59℃1分が続いた。増幅の後、情報的ウェル(対立遺伝子A又はGのいずれか一方のみが陽性であるウェル)の個数及び両方の対立遺伝子が陽性であるウェルの個数が計数され、そして、逐時確率比検定(SPRT)解析に付された。
【0191】
ヘテロ接合性倍数体胎児において、対立遺伝子A及びGは、胎児ゲノム中に等しく存在する(1:1)はずであり、一方トリソミー18において、一つの対立遺伝子に追加的なコピーが存在するため、胎児ゲノム中の比率が2:1となる。異なる試料を解釈するために、一連のDPRT曲線が作成された。これらの曲線は、分類に必要となる、所定の情報的ウェルの総数(x軸)に対する、過剰出現対立遺伝子が陽性である情報的ウェル数の割合Pr(y軸)を描画している。各試料において、実験的に導き出されたPrは、予想されたPr値と比較された。上方の曲線を上回った試料はトリソミー18として分類され、他方下方の曲線を下回った試料は、正倍数体として分類された。2つの曲線の間の領域は、分類不能領域である。
【0192】
胎児トリソミー18を検出するためのデジタルRNA-SNP解析の実現可能性は、SERPINB2遺伝子中のrs6098 SNPを使用することで実証された。正倍数体及びトリソミー18胎児を妊娠した対象に由来する胎盤組織DNA試料を、まず、ヘテロ接合の検体を同定するために、質量分析によりジェノタイピングした。その結果、前記胎盤試料の内9個が正倍数体、そして3つがトリソミー18ヘテロ接合体であることがわかり、それらはデジタルRNA-SNP解析に付された。各試料において、Pr及びmrが計算され、そして、このmr値に対応するSPRT曲線が、疾患分類に使用された。図16Aに示されるように、全ての試料が正しく分類された。トリソミー18胎盤のPr値は分類不能領域を上回り、他方正倍数体の胎盤の場合は、この領域を下回った。
【0193】
mr=0.1、0.2及び0.3に基づくSPRT曲線を用いた試料は、図16Bに説明される。これらのデータは、このデジタルRNA-SNP法が、トリソミー18の妊娠を診断するための有用な診断方法であることを示唆する。データ点が上方の曲線を上回る試料は異数体として分類され、一方データポイントが下方の曲線を下回る試料は、正倍数体として分類された。
【0194】
C.デジタルRCD解析
本実施例は、マイクロ流体工学に基づくデジタルPCRを使用するデジタルRCD解析の効率を明示する。Fluidigm BioMark(商標)Systemを使用するこのアプローチの一種がここに記載されるが、これは例示を目的とするものであり、限定を目的とするものではない。このシステムは、1回の反応でデジタルPCRを9000回以上実行することが出来る。
【0195】
正倍数体又はトリソミー21(T21)胎児を妊娠した妊婦から、胎盤組織、母体血球細胞、及び羊水試料が採取された。10個のT21及び10個の正倍数体由来の胎盤DNAを、それぞれ等量の正倍数体母体血球細胞DNAと混合することにより、20個の50%DNA混合物を作り出した。それらの混合試料中の胎児の割合の正確さを保証するため、まずは抽出されたDNAが、260nmで光学密度(OD)を測定することにより定量された。それからそれらは、12.765 Digital Arrays (Fluidigm)を使用して、BioMark(商標)System (Fluidigm)により、デジタル的に(digitally)定量された。それらの試料を定量するアッセイは、参照染色体に対するプローブが使用された点を除き、下記と同様に行われた。
【0196】
前記50%DNA混合物及び羊水試料中の染色体量は、第1番染色体と第21番染色体との非多型遺伝子座を比較する、デジタルPCR解析により決定された。第21番及び第1番の染色体上の一対のパラロガス遺伝子座の101bpのアンプリコンを、まず、順プライマー5'-GTTGTTCTGCAAAAAACCTTCGA-3'及び逆プライマー5'-CTTGGCCAGAAATACTTCATTACCATAT-3'により同時に増幅した。2つの染色体特異的TaqManプローブが、第21番と第1番染色体のパラログを区別するように設計され、それらの配列は、それぞれ、5'-(FAM)TACCTCCATA ATGAGTAAA(MGBNFQ)-3'、及び5'-(VIC)CGTACCTCTGTAATGTGTAA(MGBNFQ)-3'であった。パラロガス遺伝子座は、ここでは、例示のみを目的として使用される。言い換えると、このような解析に非パラロガス遺伝子座も使用され得る。
【0197】
トリソミー18(T18)を検出するためにデジタルRCDアプローチの使用を明示するため、第21番及び第18番染色体上のパラロガス配列を標的とするもう一つのアッセイが設計された。第21番及び第18番の染色体上のパラロガス遺伝子座の128bpのアンプリコンを、まず、順プライマー5'-GTACAGAAACCACAAACTGATCGG-3'及び逆プライマー5'-GTCCAGGCTGTGGGCCT-3'により同時に増幅した。2つの染色体特異的TaqManプローブが、第21番と第18番染色体のパラログを区別するように設計され、それらの配列は、それぞれ、5'-(FAM)AAGAGGCGAGGCAA(MGBNFQ)-3'及び5'-(VIC)AAGAGGACAGGCAAC(MGBNFQ)-3'であった。パラロガス遺伝子座の使用は、ここでは、例示のみを目的とする。言い換えると、このような解析に非パラロガス遺伝子座も使用され得る。
【0198】
全ての実験は、BioMark(商標)System (Fluidigm)上で、12.765 Digital Arrays (Fluidigm)を使用して遂行された。1枚のパネル上での反応は、反応体積10μlで、2x TaqMan Universal PCR Master Mix (Applied Biosystems)を使用して準備された。各反応は、1x TaqMan Universal PCR Master Mix、各プライマーを900nL、及びプローブを125nM、及び50%胎盤/母体血球細胞DNA試料を3.5μl含んで行われた。試料/アッセイ混合物は、NanoFlex(商標)IFCコントローラー(Fluidigm)により、デジタルアレイに注入された。反応は、検出用BioMark(商標)System上で実行された。この反応は、50℃2分で始まり、そして95℃10分、更に40サイクルの95℃15秒及び57℃1分が続いた。
【0199】
正倍数体及びT21の50%胎盤/母体血球細胞DNA試料は、chr21/chr1アッセイを用いて、デジタルアレイ上で解析された。各試料において、chr21又はchr1マーカーのいずれか1方のみ(両方ではない)が陽性であるものを含む情報的ウェルの個数が計数された。全ての情報的ウェル中の、過剰出現マーカーの割合(Pr)が決定された。ウェルあたりの正確な平均参照鋳型濃度(mr)に対応するSPRT曲線が、実験的に取得されたPrが正倍数体又はT21のいずれの試料を示唆するものかを判定するのに使用された。分類不能の試料が残っているときは、分類が可能となるまで、追加的なパネルからのデータが蓄積された。図17に示すように、全ての50%胎盤/母体血球細胞DNA試料は、このアプローチを使用して正しく分類され、それらの分類には、1枚〜4枚の範囲のパネルが必要であった。図18に示すように、正しい分類のための決定境界を示すために、SPRT曲線の描画もなされた。
【0200】
発明者らは、更に、RCD解析を、正倍数体胎児を妊娠した23人の妊婦及びT21胎児を妊娠した6人の妊婦から得られた羊水試料に適用した。各試料は、chr21/chr1アッセイと共に、単一の765ウェル中で解析された。図19は、このSPRT分類を要約して示した。図19のように、29個の試料は、全て正しく分類された。よって、デジタルRCD法は、羊水及び絨毛膜絨毛生検等の、出生前診断に使用される様々な試料において、マイクロサテライト(Levett LJ, et al. A large-scale evaluation of amnio-PCR for the rapid prenatal diagnosis of fetal trisomy. Ultrasound Obstet Gynecol 2001; 17: 115-8)若しくは単一ヌクレオチド多型(SNP)マーカー(Tsui NB, et al. Detection of trisomy 21 by quantitative mass spectrometric analysis of single-nucleotide polymorphisms. Clin Chem 2005; 51 : 2358-62)、又はリアルタイム非デジタルPCRを使用してトリソミーを検出すする、代替的なアプローチである。
【0201】
T18の検体を検出するという試みにおいて、発明者らは、3個の正倍数体及び5個のT18の胎盤DNA試料において、chr21/chr18アッセイを利用した。全ての情報的ウェル中の過剰出現マーカーの割合(Pr)が計算された。1個のT18検体が正倍数体と誤分類したのを除き、他の全ての試料は、正しく分類された。それらの結果は、図20に概説された。
【0202】
V.質量分析プラットフォーム上の多重デジタルRCDアッセイの使用
単位体積の母体血漿あたりに存在する血漿DNA分子の数は限られている(Lo YMD. et al. 1998 Am J Hum Genet 62, 768-7758)。例えば、妊娠早期において、常染色体遺伝子座、β-グロビン遺伝子の中間母体血漿濃度は、母子両方由来のものを合わせても、986コピー/mlであることが示されている(Lo YMD. et al. 1998 Am J Hum Genet 62, 768-7758)。7680分子を確保するためには、約8mlの母体血漿からDNAを抽出することが必要な場合がある。この血漿の体積は、約15mlの母体血液から取得される量であり、この量は日常的な実施の限界に近い。しかしながら、発明者らは、福数のセットのchr21及び参照染色体標的が、デジタルRCD解析に組み合わせられ得ることを想定する。5ペアのchr21及び参照染色体標的において、解析に必要な鋳型分子の個数を提供するのに必要となり得る母体血漿は、僅か1.6mlである。多重単一分子PCRが遂行され得る。この多重単一分子解析のロバストネスは、以前単一分子ハプロタイピングにおいて明示されている(Ding, C. and Cantor, CR. 2003 Proc Natl Acad Sci USA lW, 7449-7453)。
【0203】
一例において、胎盤組織及び母体血球細胞試料は、正倍数体又はトリソミー21(T21)胎児を妊娠した妊婦から取得された。5個の正倍数体及び5個のT21胎盤DNA試料を、それぞれ等量の母体血球細胞DNAと混合し、50%胎児DNAを含む血漿試料を模した、10個のDNA混合物を作り出した。それらの混合試料中の胎児の割合の正確さを保証するため、まずは抽出されたDNAが、260nmで光学密度(OD)を測定することにより定量された。それからそれらは、384ウェルの形式で、リアルタイムPCRにより、デジタル的に定量された。それらの試料を定量するアッセイは、前記デジタルRCD解析の例と同様に行われた。
【0204】
前記50%混合物中の染色体量は、第1番染色体と第21番染色体との非多型遺伝子座を比較する、デジタルPCR解析により決定された。この方法は、デジタル相対的染色体量(RCD)解析と呼ばれる。
第21番及び第1番の染色体上の一対のパラロガス遺伝子座の121bp(各プライマー上の10bpを含む)のアンプリコンを、まず、順プライマー5'-ACGTTGGATGGTTGTTCTGCAAAAAACCTTCGA-3'及び逆プライマー5'-ACGTTGGATGCTTGGCCAGAAATACTTCATTACCATAT-3'により同時に増幅した。第21番染色体と第1番染色体との間の塩基の相異を標的にする延長プライマーが設計され、その配列は、
5'-CTCATCCTCACTTCGTACCTC-3'であった。
【0205】
T21検体を検出するための多重デジタルPCRアッセイの利用を示すために、第21番及び第18番染色体のパラロガス配列を標的とするもう一つのデジタルRCDアッセイが設計された。第21番及び第18番の染色体上の一対のパラロガス遺伝子座の148bp(各プライマー上の10bpを含む)のアンプリコンを、まず、順プライマー5'-ACGTTGGATGGTACAGAAACCACAAACTGATCGG-S'及び逆プライマー5'-ACGTTGGATGGTCC AGGCTGTGGGCCT-3'により同時に増幅した。第21番染色体と第18番染色体との間の塩基の相異を標的にする延長プライマーが設計され、その配列は、
5'-ACAAAAGGGGGAAGAGG-3'であった。
【0206】
多重デジタルRCD解析は、プライマー伸長プロトコールを使用して遂行された。PCR反応は、5μlの反応体積で、GeneAmp PCR Core Reagent Kit (Applied Biosystems)を使用して準備された。各反応は、1xBufferII、2 mMのMgCl2、200μMのdNTP混合物、0.2UのAmpliTaq Gold、各200nMの4種類のプライマー、及び50% DNA混合物を含むものとした。このアッセイ/試料混合物を384ウェルPCRプレートに充填し、そして反応は、50℃2分で始まり、そして95℃10分、更に40サイクルの95℃15秒及び57℃1分が続いた。
【0207】
PCR生産物は、シュリンプアルカリホスファターゼ(SAP)で処理され、遊離dNTPが除去された。この混合物を37℃で40分、続いて85℃で5分インキュベーションした。プライマー伸長反応は、それから実行された。手短に説明すると、伸長カクテル中のchr21/chr1アッセイ用の伸長プライマー771nM、chr21/chr18アッセイ用の伸長プライマー1.54nM、0.67UのThermosequenase (Sequenom)、並びにそれぞれ64μMのddCTP、 ddGTP、dATP、及びdTTPが、SAP処理されたPCR産物に添加された。反応条件は、94℃2分、続いて94℃5秒、50℃5秒、及び72℃5秒を80サイクルとした。最終洗浄として、16μlの水及び3mgのClean Resin (Sequenom)が前記伸長生産物に添加された。この混合物をローテーター中で20〜30分間混合し、続いて351gで5分間遠心分離した。MassARRAY Nanodispenser S (Sequenom)により、15〜25nLの最終生産物が、SpectroCHIP (Sequenom)上に分注された。SpectroCHIPからのデータの取得は、MassARRAY Analyzer Compact Mass Spectrometer (Sequenom)中で行われた。質量データは、解析のため、MassARRAY Typer (Sequenom)ソフトウェア中に移された。
【0208】
5個の正倍数体及び5個のT21の50%胎盤/母体DNA試料は、二重RCDアッセイを用いて解析された。各試料において、個別のアッセイからの情報的ウェル、即ちchr21又はchr1又はchr18マーカーのいずれか1つのみ陽性であるウェルの個数が計数された。全ての情報的ウェル中のchr21マーカーが陽性のウェルの割合(Pr)は、各RCDアッセイについて別個に計算された。それから、Prが正倍数体又はT21試料のいずれを示唆するものであるかを決定するため、逐時確率比検定(SPRT)が利用された。こうすることにより、各ウェルが2回計数されたことになるので、必要なウェルの個数が減少した。
【0209】
Chr21/chr1アッセイは、通常最初に行われる。試料に分類不可能なものが残っていた場合は、chr21/chr18アッセイから得た数値を更なる計算のために追加する場合がある。追加のプレートは、分類不可能なものが残っている試料に、分類が可能となるまで使用され得る。図21に示されるように、全ての正倍数体50%混合試料は、単一の384ウェルプレートの使用で、正しく分類された。幾つかのT21検体は、正しい分類のために、2枚以上のプレートを必要とした。もし1つのアッセイしか使用されないとしたら、分類が達成されるのに必要な数の情報的ウェルを確保するのに、より多くのプレートが必要であった。例えば、検体N0230は、いずれかのRCDアッセイのみを使用しても分類できなかった。しかしながら、2つのアッセイを組み合わせてデータを得たとき、正しい分類が達成された。もしこの二重RCDアッセイが使用されないとすると、プレートを追加しての解析が必要となっていた。発明者らは、より高いレベルのアッセイの多重化を用いることで、更にウェル数を減少させることを意図する。
【0210】
もう一つの実施例において、発明者らは、第21番染色体上の4つの異なるアンプリコンと、第21番染色体以外の常染色体上に位置するそれらの対応するパラロガスパートナーを標的とする、4重アッセイを開発した。この4重アッセイは、正倍数体及びトリソミー21妊娠対象由来の試料のSPRT分類の前の、デジタルRCD解析において使用された。胎盤試料からのDNAの抽出は、QIAamp組織キット(Qiagen, Hilden, Germany)を使用して遂行された。
【0211】
本研究で使用される全ての胎盤及び母体軟膜DNA使用は、まずNanoDrop 分光光度計(NanoDrop Technology, Wilmington, DE)で定量された。DNA濃度を、6.6 pg/細胞の転換を使用して、ゲノム当量(GE)/μLに転換した。ウェルあたり約1個の鋳型が入る濃度に相当するDNA濃度は、DNA試料の連続希釈により決定された。その条件下で、発明者らは、約37%のウェルが増幅において陰性であると予想し得る。多重デジタルRCD解析において、4セットのパラロガス配列標的が選択され:
第21番及び第1番染色体上のパラロガス遺伝子座は、順プライマー5'-ACGTTGGATGTTGATGAAGTCTCATCTCTACTTCG 3'及び逆プライマー5'-ACGTTGGATGCAATAAGCTTGGCCAGAAATACT-3'により同時に増幅され、81bpのアンプリコンが生じる。
第21番及び第7番染色体上のパラロガス遺伝子座は、順プライマー5'-ACGTTGGATGGAATTTAAGCTAAATCAGCCTGAACTG-3'及び逆プライマー5'-ACGTTGGATGGTTTCTCATAGTTCATCGTAGGCTTAT-3'により同時に増幅され、82bpのアンプリコンが生じる。
第21番及び第2番染色体上のパラロガス遺伝子座は、順プライマー5'-ACGTTGGATGTCAGGCAGGGTTCTATGCAG-3'及び逆プライマー5'-ACGTTGGATGAGGCGGCTTCCTGGCTCTT-3'により同時に増幅され、101bpのアンプリコンが生じる。
第21番及び第6番染色体上のパラロガス遺伝子座は、順プライマー5'-ACGTTGGATGGCTCGTCTCAGGCTCGTAGTT-3'及び逆プライマー5'-ACGTTGGATGTTTCTTCGAGCCCTTCTTGG-3'により同時に増幅され、102bpのアンプリコンが生じる。
各反応は、10x緩衝剤II(Applied Biosystems)、MgCl2、及び100nMの各プライマーを含んでいた。反応体積の合計は、5μL/ウェルであった。この反応は、95℃5分で始まり、そして45サイクルの95℃30秒、62℃30秒及び72℃30秒が続き、最後に72℃7分が追加された。公知のPCR増幅は全て、GeneAmp PCR Core Reagent Kit (Applied Biosystems)で遂行された。遊離ヌクレオチドは、シュリンプアルカリホスファターゼ(SAP)により不活性化された。各反応は、10xSAP緩衝剤(Sequenom)及びSAP酵素(Sequenom)を含むものであった。各PCRに、2μlのSAP混合物が添加された。このSAP反応物は、37℃で40分、及び85℃で5分インキュベートされた。SAP処理の後、iPLEX Gold kit (Sequenom)を使用して、PCR生産物のプライマー伸長反応が行われた。第21番及び第1番染色体上のパラロガス配列ミスマッチ(PSM)が、伸長プライマー5'-GTCTCATCTCTACTTCGTACCTC-3'により探索(interrogate)された。第21番及び第7番染色体上のPSMは、伸長プライマー5'-TTTTACGCTGTCCCCATTT-3'により探索された。第21番及び第2番染色体上のPSMは、伸長プライマー5'-GGTCTATGCAGGAGCCGAC-3'により探索された。第21番及び第6番染色体上のPSMは、伸長プライマー5'-TGGGCGCGGGAGCGGACTTCGCTGG-3'により探索された。各反応は、10xPLEX緩衝剤(Sequenom)、iPLEX停止混合物(Sequenom)、iPLEX酵素(Sequenom)及び343nMの各伸長プライマーを含むものであった。但し、第21番及び第6番染色体上のPSMに対する伸長プライマーは、1.3μMで使用した。2μlのiPLEX混合物が5μlのPCR線産物に添加された。このiPLEX反応は、200ショートサイクルプログラムに従いサイクル反応に付された(cycled)。要するに、前記試料は、まず94℃で35秒変性し、続いて52℃で5秒アニーリングし、そして80℃で5秒伸長した。アニーリング及び伸長サイクルは、5サイクルが4回以上反復され、それから94℃5秒の変性工程に戻り、その後、再び前記5サイクルのアニーリング及び伸長ループに戻された。前記5回のアニーリング及び伸長サイクル及び1回の変性工程が、合計40と成るように39回繰り返された。最後に、72℃で3分、伸長が行われた。iPLEX反応生産物は、16μlの水で希釈され、各PCRのために、6mgの樹脂で脱塩した。384ウェルを1600gで3分間遠心して、SpectroCHIP (Sequenom)に分注して、マトリックス支援レーザー脱離/イオン化-飛行時間(MALDI-TOF)質量分析MS Analysis (Sequenom)に付した。
【0212】
各4つのアッセイにおいて、第21番又は参照染色体のいずれか1つのみ陽性であるウェルの数が、独立して記録された。各アッセイにおいて、ポアソン補正した第21番染色体及び参照染色体の数が計算された。4つのアッセイ全ての第21番染色体及び参照染色体のポアソン補正した数の合計が計算され、4重のアッセイの情報的カウントとみなされた。Pr値は、4重のアッセイの第21番染色体のカウントを、4重のアッセイの第21番染色体及び参照染色体のカウントの合計で割った値とした。実験的に導き出したPr値は、SPRT解析に供された。1枚以上の384ウェルプレートが、SPRTによる疾患分類が可能となるまで解析された。2つの50%胎盤ゲノムDNA/50%母体軟膜DNA混合物と、2つの50%トリソミー21胎盤ゲノムDNA/50%母体軟膜DNA混合物との合計が解析された。
【0213】
前記実験的に導き出されたPr値を、予想されたPr値と比較して、帰無仮説又は対立仮説を検査する。あるいは、所定の情報的カウントのPrが疾患分類のための統計的信頼度の要求されるレベルにまだ達していなければ、帰無仮説及び対立仮説のいずれも許容され得ない。これらの検体は、更にデータが取得されない限り、分類不可能とみなされた。
【0214】
それらの結果及び各試料のSPRT分類は、図22A及び22Bに示す。2つの正倍数体試料は、SPRT分類が可能となるまでに、それぞれ2回及び5回の384ウェルデジタルRCD解析を必要とした。4重アッセイの各構成要素の独立したデータは、それらのいずれもSPRTによる疾患分類が不可能であった。両トリソミー21試料は、それぞれ僅か1重の384ウェルデジタルRCD解析を用いて正しく分類された。同様に、4重アッセイの各構成要素の独立したデータは、それらのいずれもSPRTによる疾患分類が不可能であった。しかしながら、4重アッセイから得た合成カウントは、正しいSPRT分類を可能とした。これらのデータは、多重デジタルRCDを使用することにより、所定の回数実行されるデジタルPCR解析において、1重デジタルRCDアッセイの使用と比較して、効果的な数の情報的カウントが実質的に増大することを示すものであった。
【0215】
VI.デジタルエピジェネティック相対的染色体量の使用
ここで、発明者らは、デジタルエピジェネティック相対的染色体量(デジタルERCD)と称されるアプローチを紹介する。ERCDにおいて、染色体の異数性に関与する染色体(例えばトリソミー21における第21番染色体)及び参照染色体上に、胎児特異的DNAメチル化パターン又は他のエピジェネティックな変化を呈するエピジェネティックマーカーが、デジタルPCR解析に供される。正常な胎児を妊娠した妊婦から抽出した血漿DNA中の、第21番染色体エピジェネティックマーカーが陽性であるウェルの数と、参照染色体エピジェネティックマーカーが陽性であるウェルの数との比率は、参照範囲(reference range)を提供し得る。この比率は、胎児がトリソミー21を有すると増大するものと予想される。この解析において、1つ以上の第21番染色体マーカー及び1つ以上の参照染色体マーカーが使用され得ることは、当業者に明らかである。
【0216】
胎児(胎盤)特異的メチル化パターンを呈する第21番染色体上の遺伝子の一例として、ホロカルボキシラーゼ合成酵素(HLCS)遺伝子が挙げられる。HLCSは、胎盤中で高メチル化されるが、母体血球細胞では低メチル化されている。これは米国特許出願第11/784499号の範囲内であり、本明細書中に参照により援用される。胎児(胎盤)特異的メチル化パターンを呈する参照染色体上の遺伝子の一例として、第3番染色体上のRASSF1Aが挙げられる[10]。RASSF1Aは、胎盤中で高メチル化されるが、母体血球細胞では低メチル化されている。本明細書に参照により援用される米国特許出願第11/784501号を参照されたい。
【0217】
母体血漿細胞を使用して行う胎児のトリソミー21を検出するデジタルPCRに、高メチル化HLCS及び高メチル化RASSF1Aを適用するとき、母体末梢血細胞がまず採取される。それから、その血液は、遠心分離にかけられ、血漿が回収される。更にその血漿からDNAが、QIAamp Blood kit (Qiagen)等の当業者に周知の技術を使用して抽出される。この血漿DNAは、更に、HpaII及びBstUI等の1つ以上のメチル化感受性制限酵素を使用して消化される。これらのメチル化感受性制限酵素は、メチル化されていない母体由来の遺伝子を切断するが、胎児由来の高メチル化された遺伝子を切断しない。消化された血漿DNA試料は、それから、制限酵素処理されても無傷のHLCS又はRASSF1A配列のいずれかが反応ウェル中に、平均で約0.2〜1分子検出され得る程度に希釈される。この希釈されたDNAを増幅するのに、2つのリアルタイムPCRシステムが使用され得る。一つは、HLCS遺伝子がメチル化していなければ前記制限酵素により切断されるべき領域を網羅する、2つのプライマー、及びHLCS遺伝子に特異的な1つのTaqManプローブが用いられる。もう一つは、同様に2つのプライマー及び1つのプローブであってRASSF1Aに対するものが用いられる。校舎のRASSF1Aのプライマー/プローブセットに関して、Chan et al 2006, Clin Chem 52, 2211-2218に、一つの実施例が記載されている。HLCS及びRASSFlAを標的とするTaqManプローブは、それぞれFAM及びVIC等、異なる蛍光レポーターを有する場合がある。よって、このデジタルPCR実験を実行するのに、1枚の384ウェルプレートで足りる場合がある。HLCS又はRASSF1Aのいずれか1つのみが陽性と評点されるウェルの個数が計数され、これらのカウントの比率が取得され得る。このHLCS:RASSF1A比率は、正常な正倍数体の胎児を妊娠した妊婦から採取した母体血漿と比較して、トリソミー21の胎児を妊娠した妊婦から採取した母体血漿のほうが高くなると予想される。その過剰出現の程度は、デジタルPCR反応におけるウェルあたりの平均参照鋳型濃度に依存し得る。
【0218】
これらの結果を評点するのに、例えば、RASSF1Aの陽性に無関係にHLCSが陽性であるウェルの数を計数し;及び逆にHLCSの陽性に無関係にRASSF1Aが陽性であるウェルの数を計数する等の、他の方法を採用することも可能である。更に、胎児のトリソミー21状態を示すために、前記比率を計算する代わりに、HLCS及びRASSF1Aカウントの合計又は差異のいずれかを使用することも出来る。
【0219】
プレート中のデジタルPCR実行とは別に、当業者には、他のデジタルPCRの変法、例えばマイクロ流体工学チップ、ナノリットルPCRマイクロプレートシステム、エマルジョンPCR、ポロニー(polony)PCR及びローリングサークル増幅、プライマー伸長、及び質量分析等が使用され得ることも明らかである。これらのデジタルPCRの変法は例示を目的として列挙されもので、限定を目的とするものではない。
【0220】
リアルタイムPCRとは別に、当業者には、デジタルPCRの結果を評点するために、質量分析等の方法が使用され得ることも明らかである。
【0221】
HLCS及びRASSF1Aの胎児型と母親型とを区別するためのメチル化感受性制限酵素の使用とは別に、当業者には、例えば亜硫酸水素修飾、メチル化特異的PCR、抗メチル化シトシン抗体を使用する免疫沈降法、質量分析等の、メチル化状態を確認する他の方法も適用可能であることは明らかである。
【0222】
また、当業者には、本実施例及び本発明の他の実施例に説明されたアプローチが、胎児DNAが見出され得る、母体尿、羊水、経頸管流出物(transcervical washing)、絨毛膜、母体唾液等を含む他の生体液においても使用され得ることは明らかである。
【0223】
VII.エマルジョンPCR及びその他の方策を使用する、超並列ゲノムシークエンシング
ここで、発明者らは、核酸分子のデジタル読出しが、母体血漿中のトリソミー21等の胎児染色体異数性の検出に使用され得るというもう一つの例を記載する。胎児染色体の異数性は、染色体又は染色体領域の量が異常であることにより引き起こされる。誤診断を最少化するために、非侵襲的検査は、高い感受性及び特異性を有することが望ましい。しかしながら、胎児DNAは母体血漿及び血清中で絶対的に低濃度で存在し、全てのDNA中ごく一部を占めるに過ぎない。よって、特定の遺伝子座を標的とするデジタルPCRサンプリングの数は、同一の標本中で無限に増大させることは出来ない。故に、実行されるデジタルPCRサンプリングの数を増大させずに、1つの標本から取得され得るデータの量を増大させるのに、特異的な標的遺伝子座の複数のセットの解析が用いられ得る。
【0224】
従って、態様によっては、母体バックグラウンド核酸を含有する生体試料中に僅かに存在する限定的な量の胎児核酸から推察され得る遺伝的情報の量を最大化することによる、胎児染色体の異数性の非侵襲的検出が可能となる。一つの側面において、取得される遺伝的情報の量は、正しい診断をするのに十分であるが、必要とされるコスト及び生体試料のインプット量はそれほど多くはない。
【0225】
454プラットフォーム(Roche)(Margulies,M.et al.2005 Nature 437,376−380)、Illumina Genome Analyzer (またはSolexaプラットフォーム)、またはSOLiDシステム(Applied Biosystems)、またはHelicos True Single Molecule DNAシークエンシング技術(Harris TD et al.2008 Science,320,106−109)、Pacific Biosciencesのsingle molecule,real−time(SMRT(商標))技術、およびナノポアシークエンシング(Soni GV and Meller A.2007 Clin Chem 53:1996−2001)等により達成され得る超並列シークエンシングは、並行方式において高次に多重化している標本から単離された多くの核酸分子のシークエンシングを可能とする(Dear Brief Funct Genomic Proteomic 2003;1:397−416)。これらのプラットフォームの各々は、クローン的に増大された、または増幅さえされていない核酸断片の単一分子をシークエンシングする。
【0226】
各反応において、多数のシークエンスデータが、10万〜100万または場合により数百万、または10億個のオーダーで各々の試料から生じるので、得られる配列決定されたデータは、元の標本における核酸種の混合物の代表的な特性を形成する。例えば、シークエンシングされたデータのハプロタイプ、トランスクリプトーム、およびメチル化プロファイルは、元の標本のものと類似する(Brenner et al Nat Biotech 2000;18:630−634;Taylor et al Cancer Res 2007;67:8511−8518)。各標本から大量の配列をサンプリングするために、同一配列の数、例えば数倍の範囲または高い冗長性での核酸プールのシークエンシングから生じたものはまた、元の試料中の特定の核酸種または遺伝子座のカウントの優れた定量的表示である。
【0227】
一つの態様において、ランダムシークエンシングは、妊婦の血漿中に存在するDNA断片に対して行われ、これにより母親又は胎児のいずれかを起源とするゲノム配列が得られる。ランダムシークエンシングは、生体試料中に存在する核酸分子のランダムな部分をサンプリング(シークエンシング)することを含む。配列がランダムであるから、核酸分子(即ちそのゲノム)の異なる小集団(分画)は、各解析においてシークエンシングされ得る。態様によっては、同一の試料を使用しているにも拘らず試料毎に、及び解析毎にこの小集団が変化するときであっても作用し得る。分画の例は、ゲノムの約0.1%、0.5%、又は1%である。他の態様において、この分画は、少なくともこれらの数値のいずれか1つである。
【0228】
バイオインフォマティクスの手法がその後、これらのDNA配列の各々をヒトゲノムへと配置するために使用され得る。かかる配列は、ヒトゲノムの繰り返し領域、または個体間変動、例えばコピー数変動を受ける領域において存在するため、かかる配列比率は、その後の解析から排除される可能性がある。目的の染色体の量、および一つ以上の他の染色体の量がこのようにして決定され得る。
【0229】
一つの態様において、染色体異数性に潜在的に関与する染色体、例えば第21番染色体、第18番染色体、または第13番染色体のパラメーター(例えば分数表示(fractional representation))はしたがって、バイオインフォマティクスの手順の結果から計算され得る。当該分数表示は、全ての染色体配列(例えば臨床的に重要な染色体を含む全ての染色体の幾つかの測定)、または特定の染色体小集団(例えば試験されるもの以外のちょうど一つのもの)の量に基づいて得られる。
【0230】
一つの態様において、パラメーター(例えば、臨床的に重要な染色体の分数表示)はその後、正常の(すなわち正倍数体の)胎児に関与する妊娠において確立された基準範囲と比較される。当該手順における幾つかの変法において、基準範囲(すなわちカットオフ値)は、特定の母体血漿試料中における胎児DNAの分別濃度(f)に従って調節される可能性がある。fの値は、シークエンシングのデータセットから、例えば胎児が男である場合に、Y染色体にマッピング可能な配列を使用して決定され得る。fの値はまた、分離した分析、例えば胎児のエピジェネティックマーカーを使用して(Chan KCA et al 2006 Clin Chem 52,2211−8)、または一塩基多型の分析から決定されてもよい。
【0231】
一つの態様において、標本中における核酸のプールが、100%未満の遺伝子範囲でシークエンシングされるときでさえ、捕獲された核酸分子の比率中において、大部分の各々の核酸種が一回のみシークエンシングされる。同様に、特定の遺伝子座または染色体の量の不均衡は、定量的に決定され得る。すなわち、遺伝子座または染色体の量の不均衡は、当該標本のマッピング可能な、他のシークエンシングされたタグ中における前記遺伝子座のパーセンテージ表示から推測される。
【0232】
超並列ゲノムシークエンスアプローチの一つの側面において、全ての染色体から代表的データが、同時に生産され得る。特定の断片の起源は、予め選択されない。シークエンシングはランダムに行われ、それからデータベース検索が行われ、特定の断片がどこに由来するかを確認する。これを、第21番染色体由来の特定の断片と、第1番染色体由来のもう一つが増幅された状況と対比する。
【0233】
一つの実施例において、かかる配列の割合は、異数性に関与する染色体、例えば本実例において第21番染色体から得られる。かかるシークエンシングの実行により得られたさらに別の配列は、他の染色体に由来し得る。他の染色体と比較した、第21番染色体の相対的大きさを考慮することによって、基準範囲内における、かかるシークエンシングから第21番染色体特異的配列を引き当てる正規化頻度を得ることができる。胎児がトリソミー21を有するならば、かかるシークエンシングから、第21番染色体由来配列を引き当てる正規化頻度は増加し、したがってトリソミー21の検出を可能とするだろう。正規化頻度における変化の程度は、分析試料中における胎児核酸の分別濃度に依存するだろう。
【0234】
一つの態様において、ヒトゲノムおよびヒト血漿DNA試料の単一端シークエンシングのために、我々はIlluminaゲノムアナライザーを使用した。Illuminaゲノムアナライザーは、フローセルと呼ばれる固体表面上に捕獲された、クローン的に増大された単一のDNA分子をシークエンシングする。各々のフローセルは、8個の個々の標本または標本のプールをシークエンシングするための8つのレーンを有する。各々のレーンは、ヒトゲノム中における30億塩基対の配列の一部のみである、約200Mbの配列を生じさせることが可能である。各々のゲノムDNAまたは血漿DNA試料は、一つのフローセルのレーンを使用してシークエンシングされた。生じた短い配列タグは、ヒト基準ゲノム配列と整列され、そして染色体の複製起点が示された。各々の染色体に整列された、個々のシークエンシングされたタグの総数は表にされ、そして基準ヒトゲノムまたは非疾患性の代表的標本から期待される各々の染色体の相対的なサイズと比較された。染色体の獲得または喪失はその後同定された。
【0235】
記載された手法は、本明細書において記載された遺伝子/染色体用量の方法の一つの例示にすぎない。代替的に、ペアエンドシークエンシング(paired end sequencing)が行われ得る。シークエンシングされた断片の長さとCampbell他(Nat Genet 2008;40:722−729)によって記載された基準ゲノムにおいて期待されたものとを比較する代わりに、整列され、シークエンシングされたタグ数が数えられ、そして染色体の位置に従ってソートされる。染色体領域、または染色体全体の獲得または喪失は、タグの数と基準ゲノムにおいて期待される染色体の大きさ、または非疾患性の代表的な標本のものとを比較することによって決定された。
【0236】
別の態様において、一回の反応においてシークエンシングされる核酸プール画分は、シークエンシング前にさらに補助的に選択される。例えば、ハイブリダイゼーション型の技術、例えばオリゴヌクレオチドアレイは、特定の染色体、例えば潜在的に異数性の染色体および試験された異数性に関与しない他の染色体(単数または複数)から、核酸配列を初めに補助的に選択するために使用され得る。別の例は、試料プールからの核酸配列の特定の下位群が、シークエンシング前に補助的に選択されるか、または増大される。例えば上で議論されたように、母体血漿中における胎児DNA分子は、母体のバックグラウンドDNA分子よりも短い断片を含むことが報告された(Chan et al Clin Chem 2004;50:88−92)。したがって、例えばゲル電気泳動、もしくはサイズ排除カラムによって、またはマイクロ流体工学的手法によって、分子サイズに従って試料中における核酸配列を分画するために、当業者に既知の一つ以上の方法を使用し得る。さらに、代替的には、母体血漿中の無細胞胎児DNAを分析する例において、胎児核酸部分は、母体バックグラウンドを抑制する方法によって、例えばホルムアルデヒドの添加によって増幅され得る(Dhallan et al JAMA 2004;291:1114−9)。一つの態様において、補助的に選択された核酸のプールの一部分または小集団は、ランダムにシークエンシングされる。
【0237】
他の単一分子シークエンシングの方法、例えばRoche454プラットフォーム、Applied Biosystems SOLiDプラットフォーム、Helicos True Single Molecule DNAシークエンシング技術、Pacific Biosciencesの単一分子リアルタイム(SMRT(登録商標))技術、およびナノポアシークエンシングによる方法が、本出願において同様に使用され得る。
【0238】
結果及び更なる考察の実施例(例えばシークエンシング及び計算パラメーター)は、同時に出願した、"DIAGNOSING FETAL CHROMOSOMAL ANEUPLOIDY USING GENOMIC SEQUENCING," (Attorney Docket No. 016285-005220US)に見出される。この出願は、参照により援用される。カットオフ値を決定するための本明細書中に記載の方法は、本章に記載のように、その反応がシークエンシングであったときに運用され得る。
【0239】
母体血漿中における胎児DNAの分別濃度の決定はまた、シークエンシングの実施とは別になされ得る。例えば、Y染色体DNA濃度は、リアルタイムPCR、マイクロ流体工学PCR、または質量分析を使用してあらかじめ決定され得る。実際、胎児DNA濃度は、Y染色体以外の女性胎児において適用可能な遺伝子座を使用して決定され得る。例えば、Chanらは、胎児由来のメチル化されたRASSF1A配列が、母体由来のメチル化されていないRASSF1A配列のバックグラウンドにおける妊娠女性の血漿中において検出されることを示した(Chan et al,Clin Chem 2006;52:2211−8)。したがって、分別胎児DNA濃度は、メチル化されたRASSF1A配列の量を全RASSF1A(メチル化されたもの、およびメチル化されていないもの)配列で割ることによって決定され得る。
【0240】
血液凝固の間に、DNAが母体の血液細胞から放出されるため、本発明を実施するにあたり、母体血清よりも母体血漿が好ましいことが予想される。したがって、血清が使用されるならば、胎児DNAの分別濃度は、母体血漿中において母体血清中よりも低いだろうと予想される。すなわち、母体血清が使用されるならば、同時に、同一の妊娠女性から得られる血漿試料と比較して、胎児染色体異数性を診断するためにより多くの配列が必要となると予想される。
【0241】
胎児DNAの分別濃度を決定するためのさらに別の代替的な方法は、妊娠女性と胎児との間の多型の違いの定量を通じてなされ得る(Dhallan R, et al.2007 Lancet,369,474−481)。この方法の例は、妊娠女性がホモ接合性であり、そして胎児がヘテロ接合性である多型部位を標的とし得る。胎児特異的対立遺伝子の量は、胎児DNAの分別濃度を決定するために、共通の対立遺伝子の量と比較し得る。
【0242】
一つ以上の特異的配列を検出及び定量する、比較ゲノムハイブリダイゼーション、マイクロアレイ比較ゲノムハイブリダイゼーション、定量的リアルタイムポリメラーゼ連鎖反応を含む、染色体異常を検出するための既存の技術とは対照的に、超並列シークエンシングは、あらかじめ決定もしくは定義されたDNA配列のセットの検出または分析に依存しない。標本プールからの、DNA分子のランダムな代表的断片がシークエンシングされる。様々な染色体領域に整列された異なる配列タグの数を、腫瘍性DNAを含む標本または含まない標本間で比較した。染色体異常は、標本中の所定の染色体領域に整列された配列の数(またはパーセンテージ)の相異によって明らかとされ得る。
【0243】
別の例において、血漿の無細胞DNAにおけるシークエンシング技術を、特定の癌の検出のために、血漿DNA中の染色体異常を検出するために使用してもよい。異なる癌は、典型的な染色体異常のセットを有する。複数の染色体領域における変化(増幅および欠失)が使用され得る。したがって、増幅された領域に整列された配列の割合が増大し、現象した整列された配列の割合が減少し得る。染色体あたりのパーセンテージ表示は、全ゲノムと関連する、与えられた任意の染色体のゲノム表示のパーセンテージとして表現される参照ゲノム中における各々の対応する染色体に関するサイズと比較され得る。直接的な比較、または参照染色体との比較が同様に使用され得る。
【0244】
VIII.突然変異検出
母体血漿中の胎児DNAは、母体血漿DNAの平均3〜6%を占める、胎児由来の少数の集団として存在する。そのため、当該技術分野の従来技術は、胎児が父親から受け継いで、母体血漿における大勢の母体DNAバックグラウンドから区別できるDNA標的の検出に焦点を当てていた。そのような従来検出されていた標的の例として、Y染色体上のSRF遺伝子(Lo YMD et al. 1998 Am J Hum Genet, 62, 768-775)、及び母親がRhD陰性であればRHD遺伝子(Lo YMD et al. 1998 N Engl J Med, 339, 1734- 1738)が挙げられる。
【0245】
胎児突然変異を検出するための従来の母体血漿を使用する方策は、父親が保因者である常染色体優性の条件に限定されており、父親と母親が異なる突然変異を保有していた場合の、直接突然変異を検出することによる、又は連鎖解析による、常染色体劣性疾患を除外していた(Ding C. et al 2004 Proc Natl Acad Sci USA l0l, 10762- 10767)。これらの従来の方策は、顕著に限定されていた。例えば、父親と母親とがいずれも同一の突然変異を保有していた場合、母体血漿中の直接の突然変異検出による有意な出生前診断を遂行することは不可能となる。
【0246】
そのようなシナリオが、図23に説明されている。このシナリオにおいて、NN、NM、及びMMという、3つの可能な胎児遺伝子型が存在し得て、ここで、Nは正常対立遺伝子を表し、Mは突然変異対立遺伝子を表す。突然変異対立遺伝子の例としては、嚢胞性線維症、ベータサラセミア、アルファサラセミア、鎌状赤血球貧血症、脊髄性筋萎縮症、先天性副腎過形成の原因遺伝子等が挙げられる。そのような障害の他の例は、Online Mendelian Inheritance in Man (OMIM) www.ncbi.nlm.nih.gov/sites/entrez?db=OMIM&itool=toolbarで閲覧できる。母体血漿中で、殆どのDNAは母体由来であり、NMであり得る。3つの胎児遺伝子型において、母体血漿中で特異的に検出され得る特異的な胎児対立遺伝子は存在し得ない。故に、従来の方策は、この場合は利用し得ない。
【0247】
本明細書に記載の態様は、そのようなシナリオに対処することが出来る。母親及び胎児がいずれもNMとなるシナリオにおいて、対立遺伝子N及びMは、両対立遺伝子は均衡を保っている。しかしながら、母親がNMで胎児がNNである場合、母体血漿中で、対立遺伝子Nが過剰出現して、対立遺伝子の不均衡が生じる。他方、母親がNMで胎児がMMである場合、母体血漿中で、対立遺伝子Mが過剰出現して、対立遺伝子の不均衡が生じる。故に、胎児突然変異の検出において、帰無仮説は、胎児の遺伝子型がNMであるときに、対立遺伝子の不均衡が存在しないことを意味する。対立仮説は、対立遺伝子の不均衡が存在し、対立遺伝子N又はMのいずれが過剰出現しているかに依存して、胎児の遺伝子型がNN又はMMであることを意味する。
【0248】
対立遺伝子の不均衡の存否は、本明細書中に記載のデジタルPCRを使用することにより決定され得る。第一のシナリオにおいて、特定の体積の母体血漿が100個の細胞から放出されたDNAを含み、それらの細胞の内50個が母体由来で、50個が胎児由来であるとする。そうすると、この体積の血漿中の胎児DNAの分別濃度は50%となる。母親の遺伝子型がNMであるとき、母親由来の対立遺伝子Nが50個、及び対立遺伝子Mが50個存在し得る。胎児の遺伝子型がNMであるとき、胎児母親由来の対立遺伝子Nが50個、及び対立遺伝子Mが50個存在し得る。よって、対立遺伝子NとMと間に対立遺伝子の不均衡は存在せず、それらは各100コピーずつ存在する。一方、胎児の遺伝子型がNNであるとき、この体積の血漿中に、胎児由来の対立遺伝子Nが100個存在し得る。そのため、全体で対立遺伝子Nが150個に対し、Mが50個存在し得る。言い換えると、NとMとの間で対立遺伝子の不均衡が存在し、NはMに対し、3:1の比率で過剰出現している。
【0249】
逆に、胎児の遺伝子型がMMであるとき、この体積の血漿中に、胎児由来の対立遺伝子Mが100個存在し得る。そのため、全体で対立遺伝子Mが150個に対し、Nが50個存在し得る。言い換えると、NとMとの間で対立遺伝子の不均衡が存在し、MはNに対し、3:1の比率で過剰出現している。このような対立遺伝子の不均衡は、デジタルPCRにより測定され得る。この対立遺伝子は、参照鋳型とみなされる。デジタルRNA-SNP及びデジタルRCD解析と同様に、田字たるPCR実験における対立遺伝子の実際の区別は、ポアソン確率密度関数に支配される。よって、本シナリオにおける理論上の対立遺伝子の不均衡の程度が3:1であっても、予想される対立遺伝子の不均衡の程度は、デジタルPCR解析の間の、ウェルあたりの平均鋳型濃度に依存し得る。よって、ウェルあたりの平均参照鋳型濃度(mr)にふさわしいSPRT解析等のカットオフの解釈が、検体の分類に使用されるべきである。
【0250】
更に、測定されるべき対立遺伝子の不均衡の程度は、分別DNA濃度に依存する。上記実施例と対照的に、特定の体積の母体血漿が100個の細胞から放出されたDNAを含有し、それらの細胞の90個が母親由来で、10個が胎児由来であることを想定されたい。そうすると、その体積の結晶中の胎児DNAの分別濃度は、10%である。母親の遺伝型がNMであるとすると、母親由来の対立遺伝子Nが90個、及び対立遺伝子Mが90個存在し得る。胎児の遺伝型がNMであるとすると、胎児由来の対立遺伝子Nが10個、及び対立遺伝子Mが10個存在し得る。よって、この場合、対立遺伝子N及びMの間に対立遺伝子の不均衡は存在せず、それぞれが合計100コピーを有している。他方、胎児の遺伝型がNNであるとすると、この体積の血漿中に、胎児由来の対立遺伝子Nが20個存在し得る。そうすると、対立遺伝子Nが合計で110個、対立遺伝子Mが合計で90個存在することとなる。
【0251】
言い換えると、NとMとの間に対立遺伝子の不均等が存在しており、Nが過剰出現している。逆に、状況を逆にして、胎児の遺伝子型がMMであるとすると、この体積の血漿中に、胎児由来の対立遺伝子Mが20個存在することとなる。そうすると、対立遺伝子Mが110個、対立遺伝子Nが90個存在することとなる。言い換えると、NとMとの間に対立遺伝子の不均衡が存在しており、Mが過剰出現している。胎児DNA分別濃度が10%の場合の理論上の対立遺伝子の不均衡の程度は、110:90であり、これは、上述の例で示した、胎児DNAが50%存在するときの比率3:1とは異なっている。よって、胎児DNA分別濃度に相応のSPRT解析等のカットオフの解釈が、検体の分類に使用されるべきである。
【0252】
故に、血漿DNAは抽出されて、例えば従来技術のリアルタイムPCRアッセイ(Lo, et al. 1998 Am J Hum Genet 62, 768-775)により、又は当業者に周知の他の種類の定量要素(quantifier)、例えばSNPマーカー(Dhallan R et al. 2007 Lancet, 369, 474- 481)、及び胎児エピジェネティックマーカー(Chan KCA et al. 2006 Clin Chem, 52, 2211-2218)等により、血漿試料中の母体及び胎児のDNAの量が定量され得る。胎児DNAパーセンテージが計算され得る。それから、デジタルPCR解析の間、各反応ウェルが平均1分子の鋳型(対立遺伝子N又はMのいずれかであり得る)を含み得るように、定量された血漿DNA試料が調製(例えば希釈又は濃縮)される。デジタルPCR解析は、一対のプライマー及び1つが対立遺伝子Nに特異的で、もう1つが対立遺伝子Nに特異的な2つのTaqManプローブを使用して遂行され得る。Mのみ陽性であるウェルの個数及びNのみ陽性であるウェルの個数が計数され得る。対立遺伝子の不均衡の証拠の存否を判定するために、これらのウェルの比率が使用され得る。対立遺伝子の不均衡の統計的な証拠は、当業者に周知の方法、例えばSPRTの使用等により検査され得る。本解析の一つの変法において、Mのみが陽性である、又はM及びNが陽性であるウェルの数を計数することが可能であり;並びにNのみが陽性である、又はM及びNが陽性であるウェルの数を計数することが可能であり;そしてそれらの計数の比率を導き出すことが出来る。繰り返すが、対立遺伝子の不均衡の統計的な証拠は、当業者に周知の方法、例えばSPRTの使用等により検査され得る。
【0253】
デジタル相対突然変異量(relative mutant dosage)(RMD)と称される、胎児遺伝子の突然変異の量の決定は、男性/女性(XX/XY)DNA混合物を使用して実証された。図24Aに記載されるように、男性又は女性由来の血球細胞DNAを男性DNAと混合し、XYのバックグラウンド中に、XX又はXY遺伝子型が、25%及び50%の分別濃度で存在する試料を調製した。
【0254】
加えて、血液細胞試料は、12人の男性対象及び12人の女性対象からも取得された。女性血球細胞DNA(遺伝子型XX)を、それぞれ3倍過剰量の男性血球細胞DNA(遺伝子型XY)と混合し、こうして、図24Bに記載されるように、遺伝子型XYのバックグラウンド75%中に、XX遺伝子型を有するDNAが25%混合された12個のDNA試料を調製した。
【0255】
SPRTの目的は、バックグラウンドDNA中に存在する少数の遺伝子型を判定することであった。75%XY遺伝子型DNAをバックグラウンドとし、25%XX遺伝子型DNAを含むDNA混合物において、少数の対立遺伝子は、75%のDNA由来のYであり得る。試料中のDNAの25%は遺伝子型XXに由来するため、この試料中にDNAが200分子存在しているとすると、150分子が、XY個体を起源としている。故に、対立遺伝子Yの個数は、75個と期待される。また、男性DNAに(遺伝子型XY)由来する対立遺伝子Xの個数も75個である。女性DNAに(遺伝子型XX)由来する対立遺伝子Xの個数は、50個(25の2倍)である。よって、X:Yの比率は、125/75=(1+25%)/(1−25%)=5/3である。
【0256】
この研究の第二の部分において、ベータグロビン遺伝子、即ちヘモグロビンベータ(HBB)遺伝子上に、HbE(G->A)及びCD41/42(CTTT/-)突然変異を担持する、男性及び女性の対象から、血液細胞試料を取得した。全ての可能な遺伝子型(MM、MN、又はNN)を有する男性胎児を妊娠した、ヘテロ接合体(MN、ここでM=突然変異、N=野生型とする)の母親から得られた母体血漿試料を模倣するために、野生型対立遺伝子のホモ接合体(NN)又は前記2つの突然変異の1つに関してのヘテロ接合体(MN)のいずれかである男性に由来する血球細胞DNAのそれぞれを、前記のものと同じ突然変異に関するヘテロ接合体(MN)の女性から採取した血球細胞DNA試料と混合した。こうして、様々な男性/女性分別DNA濃度のDNA混合物を調製した。CD41/42に関するホモ接合体(MM)女性由来の血球細胞DNA使用も、上記DNA混合物の調製に使用された。SPRT分類に使用される実際の男性DNAの比率を確認するため、各DNA混合物の分別男性DNA濃度を、ZFY/Xアッセイを使用して決定した。
【0257】
デジタルZFY/Xアッセイは、SPRTを実証するために、及びDNA混合物中の分別男性DNA濃度を決定するために使用された。X染色体及びY染色体上のジンクフィンガータンパク質配列(ZFX及びZFY)の量を、デジタルPCR解析により決定した。最初に、順プライマー5'-CAAGTGCTGGACTCAGATGTAACTG-3'及び逆プライマー5'-TGAAGTAATGTCAGAAGCTAAAACATCA-3'により、ZFX及びZFY遺伝子座の87bpのアンプリコンを同時に増幅した。X染色体とY染色体との間のパラログを区別するために2つの染色体特異的TaqManプローブが設計され、それらの配列は、それぞれ5'-(VIC)TCTTTAGCACATTGCA(MGBNFQ)-3'及び5'-(FAM)TCTTTACCACACTGCAC(MGBNFQ)-3'とした。
【0258】
突然変異対立遺伝子に対する正常の対立遺伝子のデジタルPCR解析により、DNA混合物中の突然変異の量を決定した。HbE突然変異において、まず、順プライマー5'-GGGCAAGGTGAACGTGGAT -3'及び逆プライマー5'-CTATTGGTCTCCTTAAACCTGTCTTGTAA-3'により、正常及び突然変異対立遺伝子が、同時に増幅された。正常(G)及び突然変異(A)対立遺伝子を区別するために2つの対立遺伝子特異的TaqManプローブが設計され、それらの配列は、それぞれ、5'-(VIC)TTGGTGGTGAGGCC(MGBNFQ)-3'及び5'-(FAM)TTGGTGGTAAGGCC(MGBNFQ)-3'とした。
【0259】
CD41/42欠失突然変異において、まず、順プライマー5'-TTTTCCCACCCTTAGGCTGC-3'及び逆プライマー5'-ACAGCATCAGGAGTGGACAGATC-3'により、正常及び突然変異対立遺伝子の87bp及び83bpのアンプリコンが、同時に増幅された。正常及び突然変異対立遺伝子(欠失無し及び欠失有り)の間を区別する2つの対立遺伝子特異的TaqManプローブが設計され、それらの配列は、それぞれ5'-(VIC)CAGAGGTTCTTTGAGTCCT(MGBNFQ)-3'及び5'-(FAM)AGAGGTTGAGTCCTT(MGBNFQ)-3'とされた。HbE突然変異の結果は、図26A及び26Bに記載される。
【0260】
これらの実験は、12.765 Digital Arrays (Fluidigm)を使用した、BioMark(商標)System (Fluidigm)により遂行された。1つのパネルの反応が、反応体積10μlで、2x TaqMan Universal PCR Master Mix (Applied Biosystems)を使用して準備された。CD41/42及びZFY/Xアッセイにおいて、各反応は、1xTaqMan Universal PCR Master Mix、各プライマー900 nM、各プローブ125nM、及び1 ng/μLのDNA混合物3.5 μLを含むものであった。HbEアッセイにおいて、正常(G)及び突然変異体(A)対立遺伝子を標的とするプローブを、それぞれ250nM及び125nM添加した。この試料/アッセイ混合物を、NanoFlex(商標)IFCコントローラー(Fluidigm)により、デジタルアレイに注入した。反応は、シグナル検出用のBioMark(商標)System上で遂行された。この反応は、50℃2分で始まり、続いて95℃10分、更に、95℃15秒及び57℃(ZFX/Y及びCD41/42用)又は56℃(HbE用)1分を50サイクルで行われた。各検体について少なくとも1枚の反応パネルが使用され、分類不可能なままの試料がある場合、判定を行えるようになるまで追加的なパネルからデータを蓄積した。
【0261】
また、例えばマイクロ流体工学チップ、ナノリットルPCRマイクロプレートシステム、エマルジョンPCR、ポロニーPCR、ローリングサークル増幅、プライマー伸長及び質量分析等の、当業者に周知の方法を使用して、前記デジタルPCRを実行することが出来ることは、当業者にとって明白であり得る。
【0262】
IX.癌を用いた例
一つの態様において、本発明は、癌性腫瘍において生じ得る、対立遺伝子比率の歪みの有無で試料を分類するために実行され得る。一つの態様において、各検体における、対立遺伝子Aのみ、対立遺伝子Gのみ、及びそれらの対立遺伝子両方の陽性シグナルを有するウェルの数が、デジタルPCRにより決定された。参照配列は、陽性ウェルの数がより少ない対立遺伝子として定義された(万が一両対立遺伝子の陽性ウェル数が同一であった場合は、両方とも参照対立遺伝子として使用される)。ウェルあたりの参照対立遺伝子の推定平均濃度(mr)は、ポアソン確率密度関数に従い、他方の対立遺伝子が陽性か否かにかかわらず、参照対立遺伝子が陰性であるウェルの合計を使用して計算された。発明者らは、この計算を説明するため、一つの仮想的な例を使用する。
【0263】
96ウェル反応において、20ウェルは対立遺伝子Aが陽性であり、24ウェルは対立遺伝子Gが陽性であり、そして28ウェルはそれらの対立遺伝子のいずれも陽性である。前記対立遺伝子Aは、陽性のウェルが他方より少ないので、参照対立遺伝子とみなされる。参照対立遺伝子が陰性であるウェルの数は、96−20−28=48となる。よって、mrがポアソン分布を使用して計算され、-ln(48/96)=0.693となる。
【0264】
LOH検出との関連で、帰無仮説は、1つの対立遺伝子の欠失が起こったことにより引き起こされる、対立遺伝子比率の歪みの喪失が推定される試料を意味する。この推定の下、2つの対立遺伝子の陽性ウェルの個数の予想される比率は1:1となり、そのため、潜在的に過剰出現対立遺伝子を含む情報的ウェル(1つの対立遺伝子のみ陽性であるウェル)の割合は、0.5となる。
【0265】
LOH検出との関連で、対立仮説は、試料の細胞の50%において1つの対立遺伝子の欠失が起こったことにより引き起こされる、対立遺伝子比率の歪みが推定される試料を意味する。過剰出現対立遺伝子と参照対立遺伝子との間の比率は2:1であるから、ウェルあたりの過剰出現対立遺伝子の平均濃度は、参照対立遺伝子の濃度の2倍となり得る。しかしながら、過剰出現対立遺伝子が陽性であるウェルの数は、単純に参照対立遺伝子の陽性ウェル数の2倍とはならず、ポアソン分布に従い得る。
【0266】
情報的ウェルは、対立遺伝子A又はGのいずれかが陽性であるが、両対立遺伝子とも陽性ではないウェルとして定義される。対立遺伝子比の歪みを有する試料の過剰出現対立遺伝子を含むウェルの数の予想される比率の計算は、表600で示したものと同様である。上記例において、LOHが腫瘍細胞の50%に出現するとしたら、ウェルあたりの対立遺伝子Gの平均の独活は、0.693の2倍=1.386となり得る。LOHが腫瘍細胞の50%を越えて出現するとしたら、ウェルあたりの対立遺伝子Gの平均濃度は、公式:1/[1−(LOHの割合)]x mrに従い得る。
【0267】
対立遺伝子Gが陽性であるウェルの予想される割合は、1−C-1.386=0.75(即ち75%又は72ウェル)対立遺伝子A又はGにおけるウェルの陽性判定が独立していると仮定すると、それらのウェルの0.5x0.75=0.375が、対立遺伝子A及びGの両方とも陽性であり得る。故に、0.5-0.375=0.125のウェルは、対立遺伝子Aのみ陽性で、そして0.75-0.375=0.375のウェルは、対立遺伝子Gのみ陽性であり得る。よって、情報的ウェルの割合は、0.125+0.375=0.5となり得る。対立遺伝子Gを担持する情報的ウェルの予想される比率は、0.375/0.5=0.75となり得る。このPrの予想される値は、試料中の対立遺伝子比率の歪み(ここでは即ちLOH)の存非を変低するための適切なSPRT曲線の構築に使用される。
【0268】
更に、デジタルPCR解析により実験的に決定された、非参照対立遺伝子を担持する情報的ウェルの実際の比率が、帰無仮説若しくは対立仮説のいずれが許容されるか、又はより多くのウェルを用いた更なる解析が必要となり得るか否かを判定するために使用された。帰無仮説又は対立仮説を許容するPrの決定境界は、閾値尤度比8に基づいて計算された。この閾値尤度比の値は、癌検出との関係で、対立遺伝子不均衡を有する試料と有しない試料とを峻別するのに、良好な成績を提供することが示されている(Zhou, W, et al. (2001) Nat Biotechnol 19, 78-81; Zhou et al 2002,上記)からである。上記例において、情報的ウェルの数は、20+24=44となり、実験的に得られたPrは、24/44=0.5455となり得る。決定境界は、<0.5879で虚無仮説が許容され、>0.6379で対立仮説が許容される。よって、この例における試料は、対立遺伝子比の歪みを有しないものと分類され得る。
【0269】
結果として、発明者らは、試料中の配列の不均衡を検出するためのアプローチを概説した。一つの態様において、この発明は、母体血漿中の胎児核酸の分析により、トリソミー21等の胎児染色体の異数性を、非侵襲的に検出するのに使用され得る。このアプローチは、羊水、絨毛膜試料、母体尿、頸管内試料、母体唾液等を含む、胎児核酸を含有する他の生体材料にも適用することが出来る。第一に、発明者らは、トリソミー21胎児を妊娠した妊婦の母体血漿中の、第21番染色体によって胎盤特異的に発現するPLAC4 mRNAのSNPの対立遺伝子の不均衡の判定のための、この発明の使用を示した。第二に、発明者らは、トリソミー21の非侵襲的出生前検出のための、相対的染色体量(RCD)解析による、多型に基づかない方法として、本発明が使用され得ることを示した。そのようなデジタルRCDに基づくアプローチは、胎児DNAを含有する試料中の第21番染色体の合計のコピー数が、参照染色体との関係で過剰出現しているか否かを直接評価することを含む。複雑な装置を要せずに、デジタルRCDは、25%の胎児DNAを賀乳する試料中のトリソミー21の検出を可能とする。発明者らは、前記デジタルPCRデータを解釈するために、逐時確率比検定(SPRT)を適用した。コンピューターシミュレーション解析により、本疾患分類アルゴリズムの高い精度が確認された。
【0270】
発明者らは、更に、本アプローチが、母体血漿中の胎児の突然変異又は多型検出、及び血漿中の腫瘍由来核酸の解析による悪性細胞のゲノム中の局所的な重複及び欠失の検出等の、染色体異常以外の他の形態の核酸配列不均衡の判定に応用され得ることを更に概説した。
【0271】
この適用において記載された任意のソフトウェア・コンポーネントまたはソフトウェア関数は、好適なコンピュータ言語、例えばJava(登録商標)、C++またはPerlを使用して、例えば従来の、またはオブジェクト指向の技術を使用して実行され得る。ソフトウェアコードは、記憶および/または伝達のためのコンピュータ読み取り可能媒体、ランダムアクセスメモリー(RAM)、読み取り専用メモリー(ROM)、磁気媒体、例えばハードドライブもしくはフロッピー(登録商標)ディスク、または光学媒体、例えばコンパクトディスク(CD)もしくはDVD(デジタル多用途ディスク)、フラッシュメモリーなどを含む好適な媒体における、一連の指示またはコマンドとして蓄積され得る。コンピュータ読み取り可能媒体は、かかる記憶または伝達装置の任意の組み合わせであり得る。
【0272】
かかるプログラムはまた、コード化され、そしてインターネットを含む種々のプロトコールに適合する有線、光、および/または無線ネットワークを経由した伝達のために適用されるキャリア信号を使用して伝達され得る。したがって、本発明の態様のコンピュータ読み取り可能媒体は、かかるプログラムでコード化されたデータ信号を使用して作られ得る。かかるプログラムでコード化されたコンピュータ読み取り可能媒体は、互換性のある装置と共にパッケージされるか、または他の装置からは分離して(例えばインターネットダウンロードを通じて)提供され得る。任意のかかるコンピュータ読み取り可能媒体は、単一のプログラム製品(例えばハードドライブもしくは全コンピュータシステム)上にまたはその中に装備されてもよく、そしてシステムもしくはネットワーク内における異なるコンピュータプログラム製品上もしくはその中に存在してもよい。コンピュータシステムは、ユーザーに本明細書において言及された任意の結果を提供するためのモニター、プリンター、または他の好適なディスプレイを含んでもよい。
【0273】
コンピュータシステムの例を図27に示す。図27に示されたサブシステムは、システムバス2775を通じて相互接続される。さらなるサブシステム、例えばプリンター2774、キーボード2778、固定ディスク2779、ディスプレイアダプター2782と連結されたモニター2776、およびその他のものが示される。I/Oコントローラー2771と連結する、周辺機器および入力/出力(I/O)装置は、当技術分野において既知である任意の数の手段、例えばシリアルポート2777によってコンピュータシステムと接続され得る。例えば、シリアルポート2777または外部インターフェース981は、コンピュータ装置をインターネットのような広域ネットワーク、マウス入力装置、またはスキャナーと接続するために使用され得る。システムバス経由の相互接続により、セントラルプロセッサ2773を個々のサブシステムと通信させ、そしてシステムメモリー2772または固定ディスク2779からの指示の実行、およびサブシステム間の情報の交換をさせることが可能である。システムメモリー2772および/または固定ディスク2779は、コンピュータ読み取り可能媒体を具現化し得る。
【0274】
本発明の例示的態様である上の記載は、例証および説明を目的として存在する。記載された明確な形態は、本発明を網羅しているものではなく、または限定することは意図されておらず、そして多くの変更および変形は、上の教示の観点において可能である。当該態様は、本発明の原理、及びその実用的な利用を最もよく説明するために選択かつ記載され、それにより他の当業者は、意図される特定の使用に適するように、様々な態様で、及び様々に変形させて、本発明を、最適な形で利用することが出来る。
【0275】
本明細書において引用された全ての出願、特許および特許出願は、全ての目的で、それら全てが参照により本明細書に援用される。
【特許請求の範囲】
【請求項1】
生体試料内の核酸配列中の不均衡の存否を決定する方法であり、該方法が;
複数の反応からデータを取得し、ここで、該データが、
(1)臨床関連核酸配列の第一の量を示す第一のセットの定量データ;及び
(2)臨床関連核酸配列ではない基準核酸配列の第二の量を示す第二のセットの定量データ;
を含み;
該二つのセットのデータからパラメーターを決定し;
複数の反応それぞれにおける参照核酸配列の平均濃度から第一のカットオフ値を導き出し、ここで、該参照核酸配列は、臨床関連核酸配列又は基準核酸配列のいずれかであり;
該パラメーターと該第一のカットオフ値とを比較し;続いて、
該比較に基づき、核酸配列に不均衡が存在するか否かの分類を決定する;
ことを含む前記方法。
【請求項2】
前記第一のセットのデータが、それぞれが反応中の臨床関連核酸配列の一部の存在を検出する1つ以上の第一のマーカーから取得され、更に、前記第二のセットのデータが、それぞれが反応中の基準核酸配列の一部の存在を検出する1つ以上の第二のマーカーから取得される、請求項1に記載の方法。
【請求項3】
更に:
参照核酸配列のデータから得られた数値の入力を有する確率分布の逆数を使用した、複数の反応それぞれにおける参照核酸配列の平均濃度の決定
を含む、請求項1に記載の方法。
【請求項4】
前記臨床関連核酸配列が21番染色体由来であり、そして前記基準核酸配列が21番染色体以外の染色体由来である、請求項1に記載の方法。
【請求項5】
前記臨床関連核酸配列が18番又は13番染色体由来であり、そして前記基準核酸配列が18番又は13番染色体以外の染色体由来である、請求項1に記載の方法。
【請求項6】
前記臨床関連核酸配列が遺伝的多形の一つの対立遺伝子であり、そして前記基準核酸配列が該遺伝的多形のもう一つの対立遺伝子である、請求項1に記載の方法。
【請求項7】
前記臨床関連核酸配列が、嚢胞性線維症膜コンダクタンス制御因子(CFTR)遺伝子、ベータグロビン遺伝子、又はアルファグロビン遺伝子の突然変異コピーであり、前記基準核酸配列が、対応する遺伝子の野生型コピーである、請求項1に記載の方法。
【請求項8】
前記生体試料が、母体血漿又は血清である、請求項1に記載の方法。
【請求項9】
前記反応が増幅反応である、請求項1に記載の方法。
【請求項10】
前記反応がデジタルPCRプロセスの一部である、請求項1に記載の方法。
【請求項11】
前記反応がシークエンシング反応である、請求項1に記載の方法。
【請求項12】
臨床関連核酸配列及び基準核酸配列の第一の部分が第一の個体に由来し、そして臨床関連核酸配列及び基準核酸配列の第二の部分が第二の個体に由来する、請求項1に記載の方法。
【請求項13】
前記カットオフ値が、前記第一の部分の1つの測定値、又は前記第二の部分の1つの測定値に基づく、請求項12に記載の方法。
【請求項14】
更に、前記パラメーターを第二のカットオフ値と比較することを含む、請求項1に記載の方法。
【請求項15】
前記分類が、病態(disease state)、非病態(non-disease state)、及び分類不能(non-classifiable)を含む、請求項14に記載の方法。
【請求項16】
前記分類が、ホモ接合、ヘテロ接合、及び分類不能を含む、請求項14に記載の方法。
【請求項17】
前記第二のカットオフ値が、非病態における第二の基準核酸配列の量に対する、第一の臨床関連核酸配列の量の比率に基づく、請求項14に記載の方法。
【請求項18】
前記パラメーターが、第二の基準核酸配列の量に対する、第一の臨床関連核酸配列の量の比率に基づく、請求項1に記載の方法。
【請求項19】
前記第一のカットオフ値の計算が:逐次確率比の検定、誤検出比(false discovery rate)、信頼区間、及び受信者動作特性曲線(receiver operating characteristic curve)の少なくとも1つの使用を含む、請求項1に記載の方法。
【請求項20】
第一のカットオフ値の導出が:
過剰出現(overrepresented)核酸配列を含む情報的反応(informative reaction)の比率P1を決定し、ここで該核酸配列が前記参照又は非参照核酸配列のいずれかであり;そして
第一の比率P1から第一のカットオフ値を計算する
ことを含む、請求項1に記載の方法。
【請求項21】
前記比率P1の決定が:
少なくとも1つの過剰出現核酸配列を含む反応の第一の確率を決定し;
反応が情報的である第二の確率を計算し;そして
該第一及び第二の確率を使用して前記比率P1を計算する
ことを含む、請求項20に記載の方法。
【請求項22】
前記第一の確率が、参照核酸配列の平均濃度に、非参照核酸配列に対する予測割合を乗算することにより決定される、請求項21に記載の方法。
【請求項23】
前記第一の確率が、複数の反応のそれぞれにおける過剰出現核酸配列の平均濃度を有するポアソン分布をインプットとして使用して決定される、請求項21に記載の方法。
【請求項24】
更に:
少なくとも1つの過剰出現核酸配列を含む反応の第三の確率を決定する
工程を含み、ここで、該反応が情報的である第二の確率の計算が、第一の確率と第二の確率とが独立であると仮定することを含む、請求項21に記載の方法。
【請求項25】
生体試料内の核酸配列中の不均衡の存否を決定する方法であり、該方法が;
複数の反応からデータを取得し、ここで、該データが、
(1)臨床関連核酸配列の第一の量を示す第一のセットの定量データ;及び
(2)臨床関連核酸配列ではない基準核酸配列の第二の量を示す第二のセットの定量データ;
を含み、ここで、該臨床関連核酸配列及び該基準核酸配列が、第一の種類の細胞及び1種又は2種以上の第二の種類の細胞から取得されたものであり;
二つのセットのデータからパラメーターを決定し;
該生体試料中の該第一の種類の細胞から取得された核酸配列の量の測定により得られた第一のパーセンテージから第一のカットオフ値を導き出し;
該パラメーターと該第一のカットオフ値とを比較し;続いて、
該比較に基づき、核酸配列に不均衡が存在するか否かの分類を決定する;
ことを含む前記方法。
【請求項26】
前記第一の種類の細胞が第一の生体に由来し、そして前記第二の細胞が第二の生体に由来する、請求項25に記載の方法。
【請求項27】
第一のカットオフ値の導出が:
反応あたりの参照核酸配列の第一の平均濃度を決定することを含み、ここで、該参照核酸配列が、過小出現臨床関連核酸配列又は基準核酸配列であり;そして
前記第一のパーセンテージにより導き出された要素により、第一の平均濃度を増大させ、前記参照核酸配列ではない核酸配列の第二の平均濃度を取得する
ことを含む、前記方法。
【請求項28】
更に:
参照核酸配列のデータから導きだした数値のインプットを有する確率分布の逆数を使用して、複数の反応それぞれにおける参照核酸配列の平均濃度を決定する
ことを含む、請求項27に記載の方法。
【請求項29】
前記確率分布がポアソン分布である、請求項28に記載の方法。
【請求項30】
定量的リアルタイムPCR、デジタルPCR、半定量競合PCR、真競合PCR、又は質量分析を使用して、胎児特異的マーカーの量を決定することにより前記パーセンテージが測定される、請求項25に記載の方法。
【請求項31】
コンピュータープログラム製品であり、生体試料中に核酸配列の不均衡が存在するか否かを決定する処理を実行する電子計算システムをコントロールする複数の指令がコードされた読取り可能な媒体を含み、該処理が:
複数の反応からデータを受信し、ここで該データが:
(1)臨床関連核酸配列の第一の量を示す第一のセットの定量データ;及び
(2)臨床関連核酸配列ではない基準核酸配列の第二の量を示す第二のセットの定量データ;
を含み;
該二つのセットのデータからパラメーターを決定し;
複数の反応それぞれにおける参照核酸配列の平均濃度から第一のカットオフ値を導き出し、ここで、該参照核酸配列は、臨床関連核酸配列又は基準核酸配列のいずれかであり;
該パラメーターと該第一のカットオフ値とを比較し;続いて、
該比較に基づき、核酸配列に不均衡が存在するか否かの分類を決定する;
ことを含む前記コンピュータープログラム製品。
【請求項1】
生体試料内の核酸配列中の不均衡の存否を決定する方法であり、該方法が;
複数の反応からデータを取得し、ここで、該データが、
(1)臨床関連核酸配列の第一の量を示す第一のセットの定量データ;及び
(2)臨床関連核酸配列ではない基準核酸配列の第二の量を示す第二のセットの定量データ;
を含み;
該二つのセットのデータからパラメーターを決定し;
複数の反応それぞれにおける参照核酸配列の平均濃度から第一のカットオフ値を導き出し、ここで、該参照核酸配列は、臨床関連核酸配列又は基準核酸配列のいずれかであり;
該パラメーターと該第一のカットオフ値とを比較し;続いて、
該比較に基づき、核酸配列に不均衡が存在するか否かの分類を決定する;
ことを含む前記方法。
【請求項2】
前記第一のセットのデータが、それぞれが反応中の臨床関連核酸配列の一部の存在を検出する1つ以上の第一のマーカーから取得され、更に、前記第二のセットのデータが、それぞれが反応中の基準核酸配列の一部の存在を検出する1つ以上の第二のマーカーから取得される、請求項1に記載の方法。
【請求項3】
更に:
参照核酸配列のデータから得られた数値の入力を有する確率分布の逆数を使用した、複数の反応それぞれにおける参照核酸配列の平均濃度の決定
を含む、請求項1に記載の方法。
【請求項4】
前記臨床関連核酸配列が21番染色体由来であり、そして前記基準核酸配列が21番染色体以外の染色体由来である、請求項1に記載の方法。
【請求項5】
前記臨床関連核酸配列が18番又は13番染色体由来であり、そして前記基準核酸配列が18番又は13番染色体以外の染色体由来である、請求項1に記載の方法。
【請求項6】
前記臨床関連核酸配列が遺伝的多形の一つの対立遺伝子であり、そして前記基準核酸配列が該遺伝的多形のもう一つの対立遺伝子である、請求項1に記載の方法。
【請求項7】
前記臨床関連核酸配列が、嚢胞性線維症膜コンダクタンス制御因子(CFTR)遺伝子、ベータグロビン遺伝子、又はアルファグロビン遺伝子の突然変異コピーであり、前記基準核酸配列が、対応する遺伝子の野生型コピーである、請求項1に記載の方法。
【請求項8】
前記生体試料が、母体血漿又は血清である、請求項1に記載の方法。
【請求項9】
前記反応が増幅反応である、請求項1に記載の方法。
【請求項10】
前記反応がデジタルPCRプロセスの一部である、請求項1に記載の方法。
【請求項11】
前記反応がシークエンシング反応である、請求項1に記載の方法。
【請求項12】
臨床関連核酸配列及び基準核酸配列の第一の部分が第一の個体に由来し、そして臨床関連核酸配列及び基準核酸配列の第二の部分が第二の個体に由来する、請求項1に記載の方法。
【請求項13】
前記カットオフ値が、前記第一の部分の1つの測定値、又は前記第二の部分の1つの測定値に基づく、請求項12に記載の方法。
【請求項14】
更に、前記パラメーターを第二のカットオフ値と比較することを含む、請求項1に記載の方法。
【請求項15】
前記分類が、病態(disease state)、非病態(non-disease state)、及び分類不能(non-classifiable)を含む、請求項14に記載の方法。
【請求項16】
前記分類が、ホモ接合、ヘテロ接合、及び分類不能を含む、請求項14に記載の方法。
【請求項17】
前記第二のカットオフ値が、非病態における第二の基準核酸配列の量に対する、第一の臨床関連核酸配列の量の比率に基づく、請求項14に記載の方法。
【請求項18】
前記パラメーターが、第二の基準核酸配列の量に対する、第一の臨床関連核酸配列の量の比率に基づく、請求項1に記載の方法。
【請求項19】
前記第一のカットオフ値の計算が:逐次確率比の検定、誤検出比(false discovery rate)、信頼区間、及び受信者動作特性曲線(receiver operating characteristic curve)の少なくとも1つの使用を含む、請求項1に記載の方法。
【請求項20】
第一のカットオフ値の導出が:
過剰出現(overrepresented)核酸配列を含む情報的反応(informative reaction)の比率P1を決定し、ここで該核酸配列が前記参照又は非参照核酸配列のいずれかであり;そして
第一の比率P1から第一のカットオフ値を計算する
ことを含む、請求項1に記載の方法。
【請求項21】
前記比率P1の決定が:
少なくとも1つの過剰出現核酸配列を含む反応の第一の確率を決定し;
反応が情報的である第二の確率を計算し;そして
該第一及び第二の確率を使用して前記比率P1を計算する
ことを含む、請求項20に記載の方法。
【請求項22】
前記第一の確率が、参照核酸配列の平均濃度に、非参照核酸配列に対する予測割合を乗算することにより決定される、請求項21に記載の方法。
【請求項23】
前記第一の確率が、複数の反応のそれぞれにおける過剰出現核酸配列の平均濃度を有するポアソン分布をインプットとして使用して決定される、請求項21に記載の方法。
【請求項24】
更に:
少なくとも1つの過剰出現核酸配列を含む反応の第三の確率を決定する
工程を含み、ここで、該反応が情報的である第二の確率の計算が、第一の確率と第二の確率とが独立であると仮定することを含む、請求項21に記載の方法。
【請求項25】
生体試料内の核酸配列中の不均衡の存否を決定する方法であり、該方法が;
複数の反応からデータを取得し、ここで、該データが、
(1)臨床関連核酸配列の第一の量を示す第一のセットの定量データ;及び
(2)臨床関連核酸配列ではない基準核酸配列の第二の量を示す第二のセットの定量データ;
を含み、ここで、該臨床関連核酸配列及び該基準核酸配列が、第一の種類の細胞及び1種又は2種以上の第二の種類の細胞から取得されたものであり;
二つのセットのデータからパラメーターを決定し;
該生体試料中の該第一の種類の細胞から取得された核酸配列の量の測定により得られた第一のパーセンテージから第一のカットオフ値を導き出し;
該パラメーターと該第一のカットオフ値とを比較し;続いて、
該比較に基づき、核酸配列に不均衡が存在するか否かの分類を決定する;
ことを含む前記方法。
【請求項26】
前記第一の種類の細胞が第一の生体に由来し、そして前記第二の細胞が第二の生体に由来する、請求項25に記載の方法。
【請求項27】
第一のカットオフ値の導出が:
反応あたりの参照核酸配列の第一の平均濃度を決定することを含み、ここで、該参照核酸配列が、過小出現臨床関連核酸配列又は基準核酸配列であり;そして
前記第一のパーセンテージにより導き出された要素により、第一の平均濃度を増大させ、前記参照核酸配列ではない核酸配列の第二の平均濃度を取得する
ことを含む、前記方法。
【請求項28】
更に:
参照核酸配列のデータから導きだした数値のインプットを有する確率分布の逆数を使用して、複数の反応それぞれにおける参照核酸配列の平均濃度を決定する
ことを含む、請求項27に記載の方法。
【請求項29】
前記確率分布がポアソン分布である、請求項28に記載の方法。
【請求項30】
定量的リアルタイムPCR、デジタルPCR、半定量競合PCR、真競合PCR、又は質量分析を使用して、胎児特異的マーカーの量を決定することにより前記パーセンテージが測定される、請求項25に記載の方法。
【請求項31】
コンピュータープログラム製品であり、生体試料中に核酸配列の不均衡が存在するか否かを決定する処理を実行する電子計算システムをコントロールする複数の指令がコードされた読取り可能な媒体を含み、該処理が:
複数の反応からデータを受信し、ここで該データが:
(1)臨床関連核酸配列の第一の量を示す第一のセットの定量データ;及び
(2)臨床関連核酸配列ではない基準核酸配列の第二の量を示す第二のセットの定量データ;
を含み;
該二つのセットのデータからパラメーターを決定し;
複数の反応それぞれにおける参照核酸配列の平均濃度から第一のカットオフ値を導き出し、ここで、該参照核酸配列は、臨床関連核酸配列又は基準核酸配列のいずれかであり;
該パラメーターと該第一のカットオフ値とを比較し;続いて、
該比較に基づき、核酸配列に不均衡が存在するか否かの分類を決定する;
ことを含む前記コンピュータープログラム製品。
【図1】

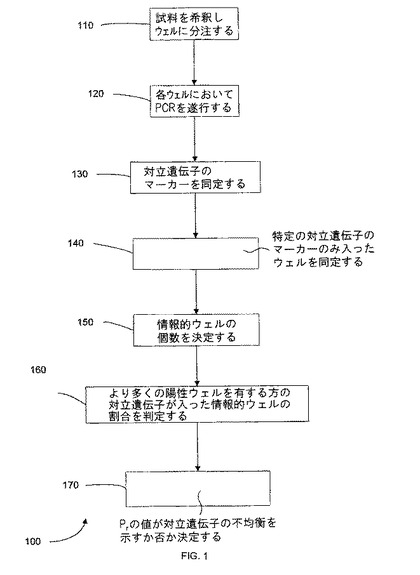
【図2A】

【図2B】

【図3】
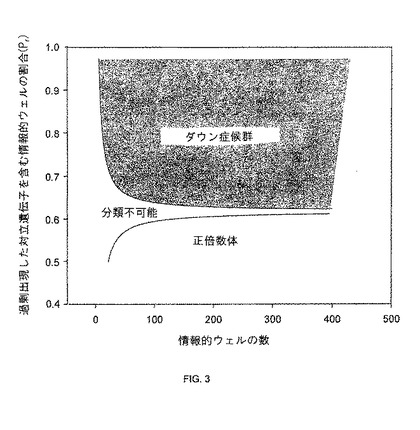
【図4】
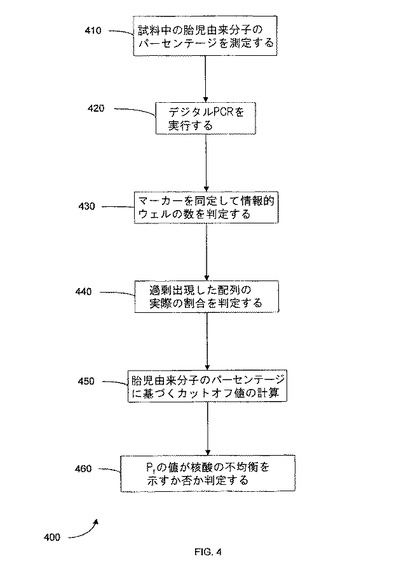
【図5】
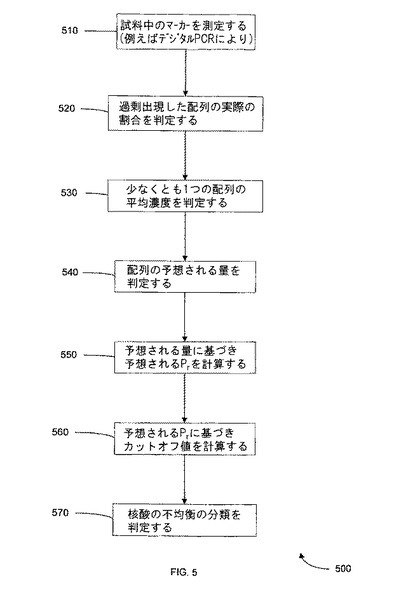
【図6】
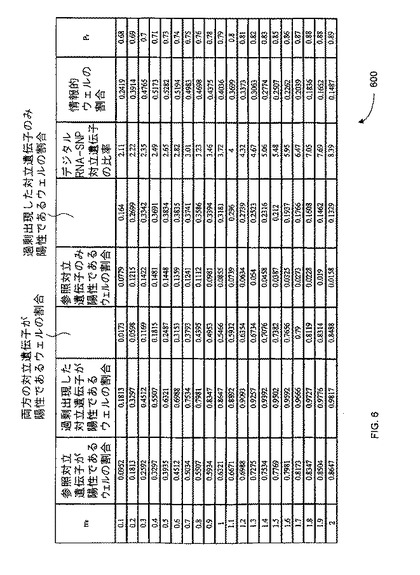
【図7】
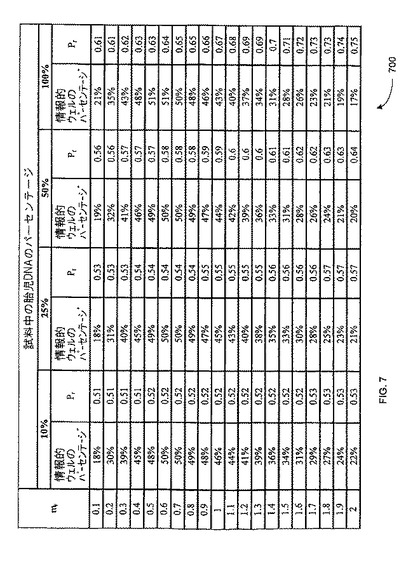
【図8】
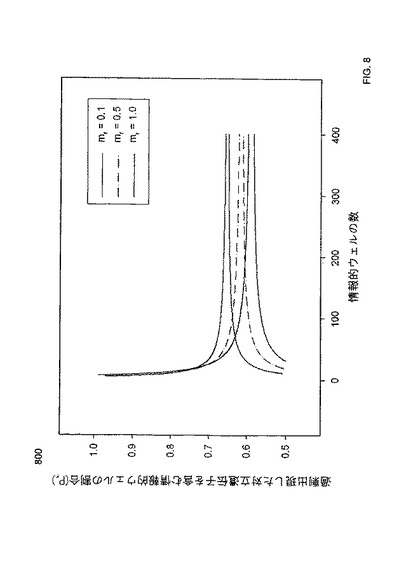
【図9A】
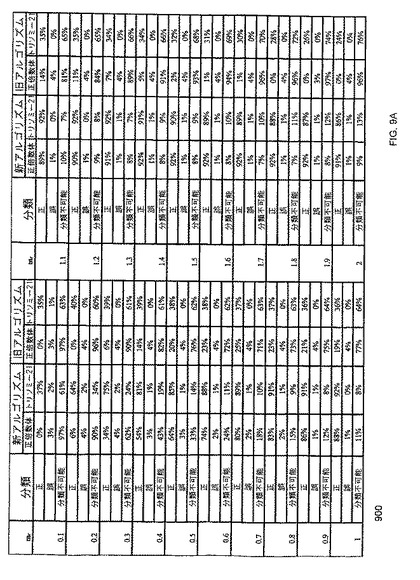
【図9B】
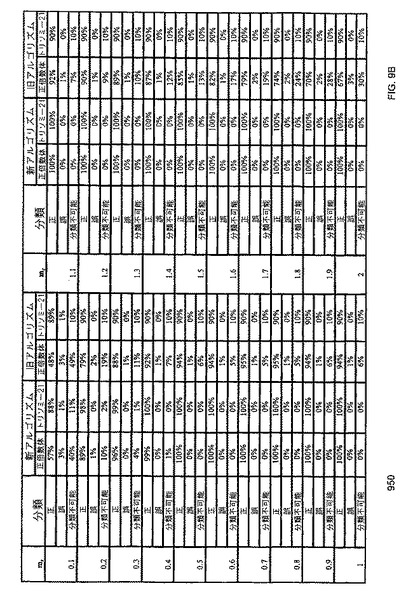
【図10】
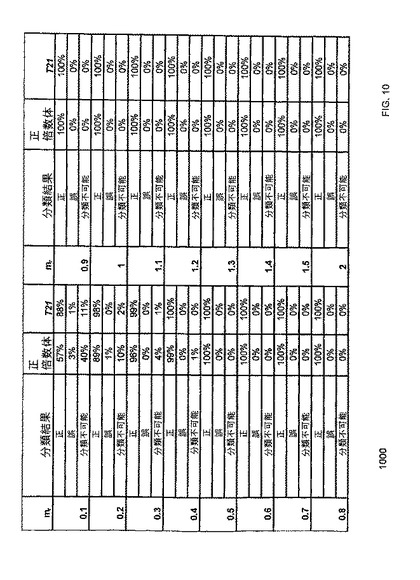
【図11】
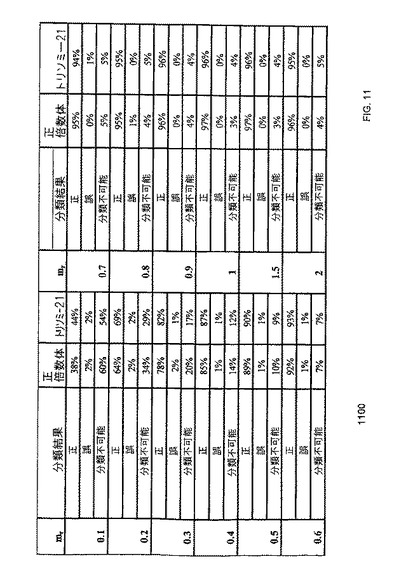
【図12】
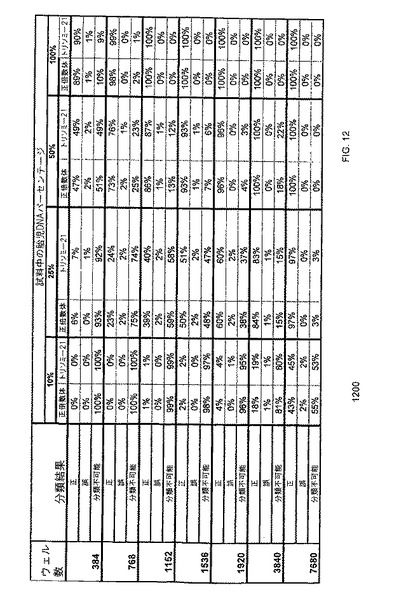
【図13】
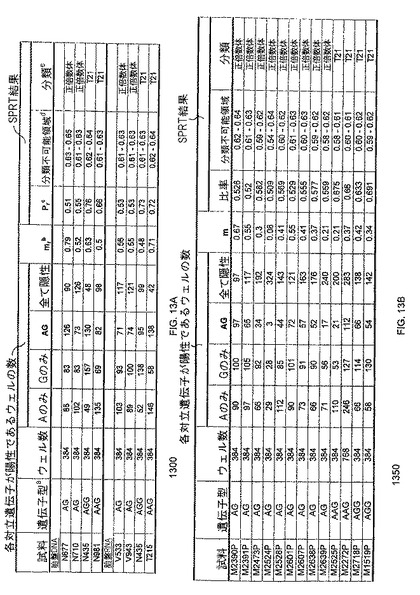
【図14】
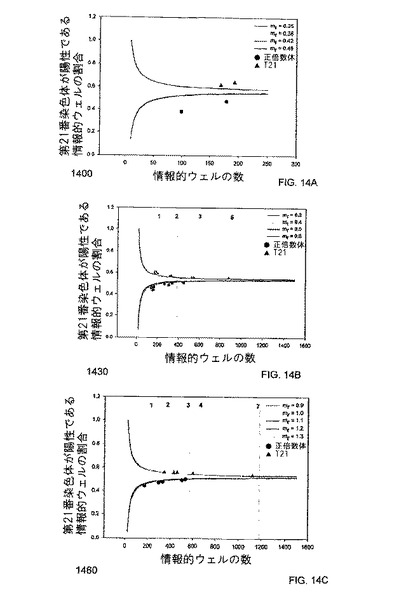
【図15】
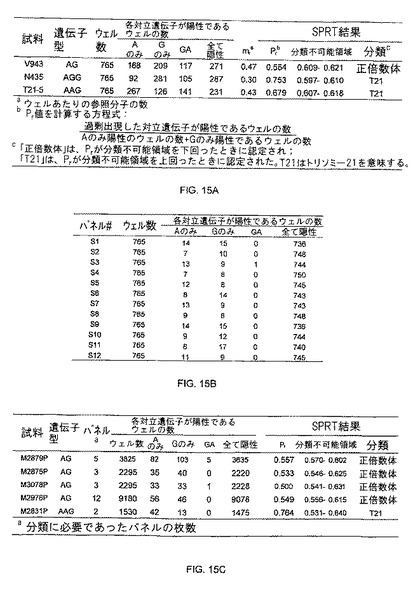
【図16】
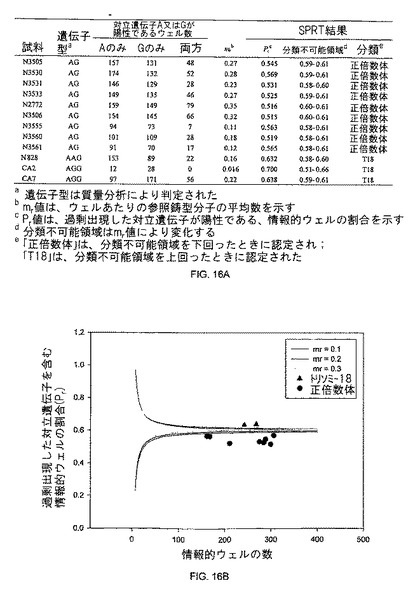
【図17】
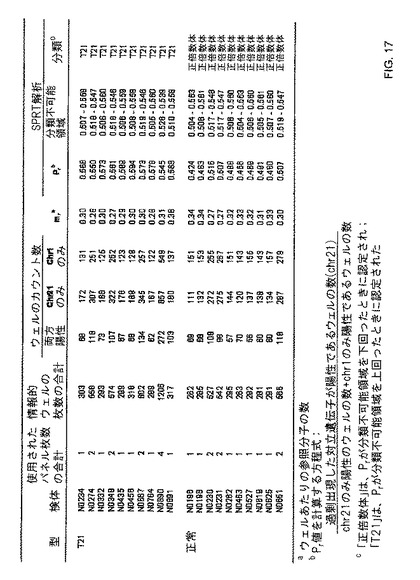
【図18】
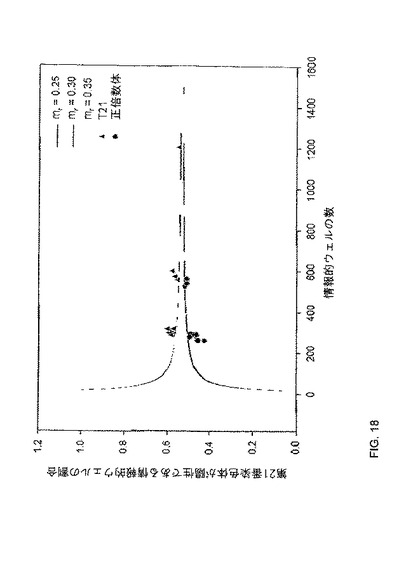
【図19】
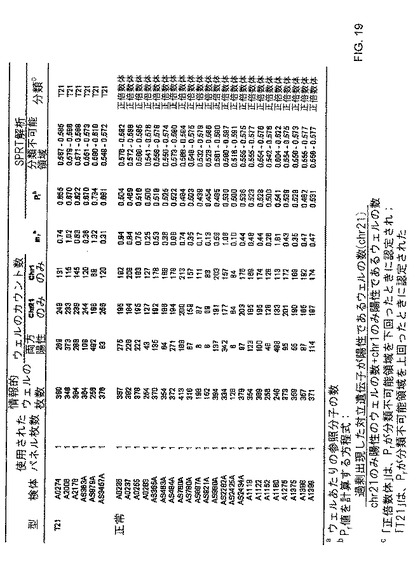
【図20】
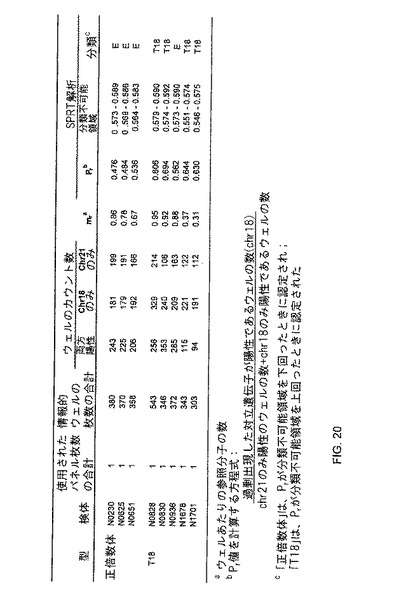
【図21】
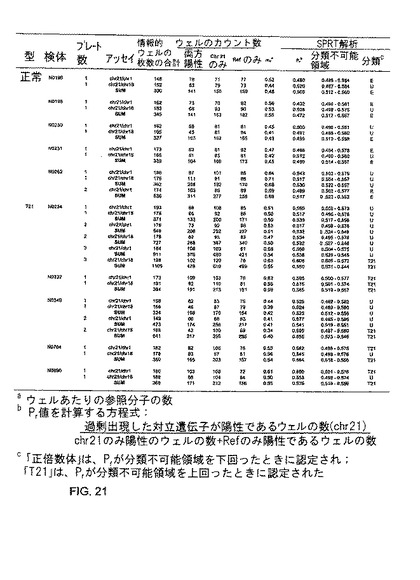
【図22A】

【図22B】

【図23】

【図24】
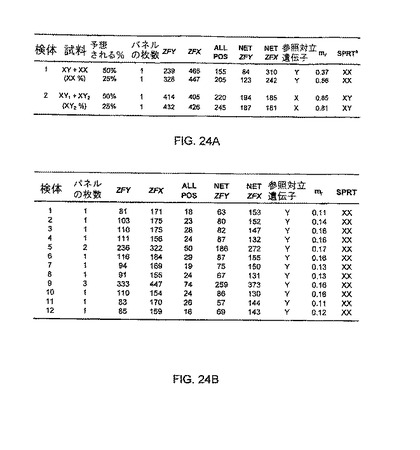
【図25】
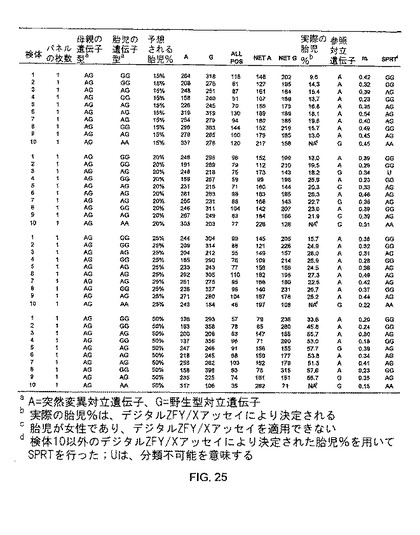
【図26】
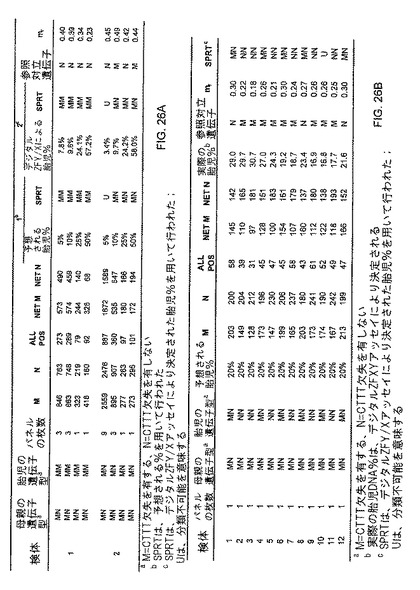
【図27】

【図2A】
【図2B】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9A】
【図9B】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22A】
【図22B】
【図23】
【図24】
【図25】
【図26】
【図27】
【公表番号】特表2010−534068(P2010−534068A)
【公表日】平成22年11月4日(2010.11.4)
【国際特許分類】
【出願番号】特願2010−517480(P2010−517480)
【出願日】平成20年7月23日(2008.7.23)
【国際出願番号】PCT/GB2008/002524
【国際公開番号】WO2009/013492
【国際公開日】平成21年1月29日(2009.1.29)
【出願人】(509229212)ザ チャイニーズ ユニバーシティー オブ ホンコン (4)
【Fターム(参考)】
【公表日】平成22年11月4日(2010.11.4)
【国際特許分類】
【出願日】平成20年7月23日(2008.7.23)
【国際出願番号】PCT/GB2008/002524
【国際公開番号】WO2009/013492
【国際公開日】平成21年1月29日(2009.1.29)
【出願人】(509229212)ザ チャイニーズ ユニバーシティー オブ ホンコン (4)
【Fターム(参考)】
[ Back to top ]