
核酸配列の機能およびそれにより核酸配列にコードされる発現産物の決定方法
【課題】宿主細胞にRNAを導入することにより、核酸配列及び核酸発現産物の機能を調査及び決定する方法を提供する。
【解決手段】生命体における核酸および/または核酸発現産物の機能を調査する方法であって、(a)核酸に由来するRNAを、前記宿主細胞へ導入すること、(b)宿主細胞への当該RNAの導入の結果として生じる、宿主細胞に対する効果の調査、(c)(b)のステップで決定された宿主細胞における効果に基づいた核酸および/または発現産物の一つ以上の機能の同定、の各ステップを含む方法。
【解決手段】生命体における核酸および/または核酸発現産物の機能を調査する方法であって、(a)核酸に由来するRNAを、前記宿主細胞へ導入すること、(b)宿主細胞への当該RNAの導入の結果として生じる、宿主細胞に対する効果の調査、(c)(b)のステップで決定された宿主細胞における効果に基づいた核酸および/または発現産物の一つ以上の機能の同定、の各ステップを含む方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、一般的に核酸配列および核酸発現産物の機能を、宿主細胞にRNAを導入することにより、調査または決定する方法に関する。
【背景技術】
【0002】
ヒトゲノムの配列決定の最初のドラフトが完了し、次の課題は、記述的ゲノム研究から機能的ゲノム研究に移行することである(Fields,S.et al.(1999)Proc.Natl.Acad.Sci.U.S.A.96, 8825−8826)。
【0003】
アンチセンスRNAまたはRNAi(RNA干渉)の使用は、近年において、対象とする遺伝子発現のオフ切り替えのための広い用途が見出されている。
【0004】
アンチセンスRNAは配列特異的に、相補性の規則にしたがって、当然、自然に転写された標的配列のmRNAに結合する。この方法は原核生物において最初に記述された(Green,P.J.et al.(1986)Annu.Rev. Biochem.55,569−597)。今日では、それにより、非常に多くの型の真核細胞におけるいかなる所望される遺伝子の迅速で、束の間の、転写後サイレンシングを可能にし(Izant,J.G.and Weintraub,H.(1985)Science 229,345−352)、広く使用されている。
【0005】
RNAi(RNA干渉)において、標的配列と特異的に相互作用する短い二重鎖RNA(〜21bp)が細胞内に導入される。結果として、多タンパク質複合体が活性化されることで、転写されたmRNAが翻訳可能になる前に、mRNAの早期分解が引き起こされる (Hammond,S.M.et al.(2000)Nature, 404,293−296)。しかしながら、インターフェロン系の活性化を通じて、または、二重鎖RNAに依存するタンパク質キナーゼ応答(PKR)の誘導を通じて、(Elbashir S.M.et al. (2001) Nature,411,494−498;Caplen,N.J. et al.(2001)Proc.Natl.Acad.Sci.U.S.A. 98,9742−9747)、または、濃度依存的な刺激、または、非標的遺伝子の抑制を通じて(Persengiev,S.P.et al.(2004)RNA 10,12−18)siRNAによっても非特異的効果が引き起こされる。
【0006】
上記方法は、アンチセンスRNA、または、RNAiを用いて、翻訳を阻止し、したがって遺伝子の機能欠損を引き起こす。
【発明の概要】
【発明が解決しようとする課題】
【0007】
しかしながら、特定の遺伝子、特に異所的に活性化または過剰発現された遺伝子、たとえば腫瘍において選択的に発現した遺伝子を機能的特徴づけるために、機能獲得の効果をシミュレートすることはより有用である。
【0008】
数多くの遺伝子転換技術も、このために利用可能である。したがって、一過性の形質転換体および安定して形質転換された細胞クローンの両方が生産される。調査中の遺伝子は、DNAプラスミド構造体を用いて、または、レトロウイルスベクターあるいはアデノウイルスベクターの助けにより細胞内に挿入することができる。さらに、挿入遺伝子の発現を構成的にすることができ、または、制御可能なプロモーターにより制御することができる。この目的に利用される、全ての遺伝子転移形態は、固有のシステムの欠点を有している。プラスミドやウィルスによる一過性形質転換体は、ほとんどの細胞型に対して非常に非効率であり(特に、初代細胞)、結果として、多様な亜集団の形質転換をもたらし、これらは、発現レベルの十分正確な滴定をできなくしている。しかしながら、ひとつのおよびそれと同一の遺伝子産物が多様な機能を、異なった細胞、または、たとえ同じ細胞でも異なった濃度で、発揮しているため、これらの技術から得られた情報量は、非常に限られている。軽視できない他の問題は、プラスミドを用いた形質転換が、遅延後にしか転写がおきないことである。その転写まで時間は、形質転換された細胞の細胞周期のフェーズに依存し、かつ、細胞集団に応じて非常に多様である。結果、非同期な、転写および発現がおこる。かわりに、安定形質転換細胞を用いることが提案されている。しかしながら、安定形質転換細胞株の生産には、非常に時間がかかり、かつ、効率的に全ての細胞系に対して実行することはできない。したがって、ほとんどの哺乳類生命体の新鮮な単離細胞が安定して形質転換されない。他の短所は、形質転換後の、例えば抗体の使用や物理的措置(例えば細胞分類)による形質転換細胞の濃縮により、遺伝子産物が導入されたにもかかわらず、起源細胞型から多くの点で異なった亜集団の選別を引き起こすことがある。
【0009】
本発明は、遺伝子産物の機能の特徴づけのための方法に関し、インビトロで転写された長鎖RNAの転移に基づく。インビトロで転写されたRNAの細胞への転移は確立された方法であり、腫瘍抗原を抗原提示樹上細胞(DC)に導入するために、現在特に集中して、腫瘍免疫学において試験されている(Boczkowski,D.et al.(2000)Cancer Res.60,1028−1034;Heiser,A.et al.(2001)J.Immunol.166,2953−2960;Nair,S.K.et al.(1998)Nat.Biotechnol.16,364−369)。反対に、今までは遺伝子機能の系統的決定のために、インビトロで転写されたRNAの転移が利用されていなかった。
【課題を解決するための手段】
【0010】
本発明は、一般的に核酸および/または発現産物、特にペプチドやタンパク質の機能の特徴づけのための方法に関する。本発明による方法において、調査中の核酸および/または調査中の発現産物の機能は、完全に未知であってもよく、または、調査中の核酸および/または調査中の発現産物の一つ以上の機能は、既知であってもよく、この態様においては、本発明による方法は、さらなる機能の同定を目的としている。
【0011】
本発明の一つの特徴は、宿主細胞における核酸の発現の効果を調査する方法であって、(a)核酸に由来するRNAを、前記宿主細胞へ導入すること;および、(b)宿主細胞への当該RNAの導入の結果として生じる、宿主細胞に対する効果の調査、の段階を含む方法に関する。
【0012】
RNAの宿主細胞への導入の結果として生じる宿主細胞に対する効果は、核酸および/または発現産物の機能についての情報を提供することができる。
【0013】
したがって、本発明のもう一つの特徴は、生命体における核酸および/または核酸発現産物の機能を調査する方法であって(a)核酸および/または、発現産物の一部または全配列のコードに由来するRNAの宿主細胞への導入;(b)RNAの宿主細胞への導入の結果として生じる宿主細胞に対する効果の調査;(c)(b)のステップで決定された宿主細胞に対する効果に基づいた核酸および/または発現産物の一つ以上の機能の同定、の各ステップを含む方法に関する。
【0014】
本発明による方法は、特に殆ど時間消費しないこと、反応速度論的解析の可能なことおよび、殆ど全ての細胞系に適用可能なことに関して利点がある。特に、発現には、RNAが細胞の細胞質に入りさえすれば十分で、細胞核に入る必要はないため、調査中の核酸、たとえば遺伝子の導入は、DNA構造体の使用に比べ、RNAの形を取った方が利点がある。したがって、RNA転移は、形質転換される細胞の分裂活性に依存しない。さらに、RNAを用いて達成可能となる転換率は、比較的高く(Van Tendeloo,V.F.et al.(2001)Blood,98,49−56;Saeboe−Larssen,S.et al.(2002)J.Immunol.Methods,259,191−203)、得られるタンパク質量は、生理学的発現におけるその導入率に対応する。翻訳が完了したタンパク質は、数時間以内に検出可能であり、機能的に意味のあるものになるため、反応速度的な研究を非常に良好に行うことができる(Van Meirvenne,S.et al.(2002)Cancer Gene Ther.9,787−797)。
【0015】
細胞内へのRNA転移は、本発明によれば、実施例で実証された以下の特徴に基づいて、その後の機能的特徴づけに対して理想的である。
【0016】
・ RNA転移によって、腫瘍細胞や未転換一次細胞を含む、調査対象の全ての真核細胞型に、遺伝子産物を挿入できる(表1)。
・ 転換率は非常に高く、多くの細胞型に対して90%より大であり、したがって、選択する必要がない。
・ 発現レベルは、形質転換に用いるRNA量によって、非常に良好に滴定することができる。このようにして、細胞内の発現レベルの差異の効果をシミュレートすることが可能となる。
・ 形質転換後、同期翻訳が直ちに起こるため、年代順に生物学的余効を感度よく検出できる。
・ RNA形質転換は、トランスクリプトームに関して、形質転換細胞の方法依存変動を殆ど示さない。
【0017】
本発明による方法は、特に細胞および細胞株の転写調節因子、腫瘍遺伝子、潜在腫瘍遺伝子の異所性発現のシミュレーションに好適であり、したがって、それら遺伝子の機能的特徴付けに好適である。このような場合、宿主細胞に挿入されるRNAは、腫瘍遺伝子または、潜在腫瘍遺伝由来である。
【0018】
本発明による方法のひとつの実施の態様において、宿主細胞に挿入されるRNAは、それをエンコードする核酸をクローニングし、そのクローン核酸の転写によって、好ましくは、インビトロ転写によって得られる。転写調節プロモーターは、RNAポリメラーゼのためのプロモーターならどれでも良い。特にRNAポリメラーゼの例としては、T7、T3およびSP6のRNAポリメラーゼがある。好ましくは、本発明によるインビトロでの転写は、T7またはSP6プロモーターによって調節される。
【0019】
特定の実施形態において、RNAをコードする核酸は、ポリアデニルカセットと併せて存在する。これらの実施形態においては、核酸の発現により、好ましくはインビトロの転写により得られるRNAは、定義された長さのポリアデニル化部分を有することが好ましい。ポリAストレッチは、好ましくは10から500長、より好ましくは30から300長、さらに好ましくは65から200長、特には100から150長のアデノシン残基を有する。特に好ましい実施形態においては、ポリAストレッチは、およそ120長のアデノシン残基を有する。
【0020】
本発明のさらなる実施形態において、宿主細胞に挿入するRNAは、その5'末端にてCap構造または調節配列を有し、これが宿主細胞での翻訳をプロモートする。
【0021】
本発明に係る方法の特に好ましい実施形態において、RNAを宿主細胞に導入することにより、RNAによりエンコードされた発現産物の発現が引き起こされる。本発明の特定の実施形態においては、発現産物はペプチド、タンパク質、酵素、抗体および抗原からなる群より選択することができる。
【0022】
本発明によれば、機能調査について、宿主細胞に導入されるRNAの量を制御することが可能になり、したがって宿主細胞において発現した発現産物の量を制御することが可能になる。
【0023】
特定の実施形態において、本発明の宿主細胞に導入されるRNAは、RNAライブラリーの一部である。本発明のこの態様は、宿主細胞への導入における宿主細胞に対する特定の影響を起こすことを可能にする又は宿主細胞の特定の機能を制御あるいは特定の機能に影響を与えることを可能にするRNAの発生について、異なる核酸または核酸の変異体(定義:RNAライブラリー)の群をスクリーニングする方法にも関係する。
【0024】
したがって、本発明のこの態様は、特定の効果または機能を有する核酸および/または核酸発現産物を見つける方法であって、(a)宿主細胞へのRNAの導入;(b)RNAの宿主細胞への導入の結果として生じる宿主細胞に対する効果の調査;(c)宿主細胞に対する効果を引き起こすRNAの同定の各ステップを含む方法に関する。この方法は、宿主細胞に対する効果を引き起こすRNAのヌクレオチド配列の決定をも含む。前記ヌクレオチド配列の決定は、公知の方法、特にオリジナル核酸のシークエンシングにより行うことができる。
【0025】
本発明に係る方法は、RNAの宿主細胞への導入および宿主細胞中のRNAの可能性ある発現の結果として生じる宿主細胞に対する効果の調査および同定を含む。
【0026】
RNAの宿主細胞への導入が、当該宿主細胞に対する効果を示すには多くの様式があることは当業者にとって明らかだろう。したがって、本発明に係る方法においては、RNAの宿主細胞への導入の結果として生じる効果を調査することを可能にする様々な方法がある。このような方法は、特に限定されないが、以下の一つ以上のパラメータ:細胞増殖、細胞死、細胞移動、細胞接着、薬物に対して改変された耐性または改変された感受性、細胞老化、特定分子の生産または生産抑制、mRNAの量、タンパク質産物の量、タンパク質活性レベル、タンパク質のリン酸化に対する効果、タンパク質または核酸のプロセッシングに対する効果、RNA安定性に対する効果、シグナル伝達または二次メッセンジャーに対する効果、の決定を含む。
【0027】
RNAの宿主細胞への導入の結果として生じる宿主細胞に対する効果の調査は、好ましくは当該宿主細胞の表現型および/または生化学的特徴の調査を含む。
【0028】
特定の実施形態においては、RNAの宿主細胞への導入の結果として生じる宿主細胞に対する効果の調査が、宿主細胞のタンパク質発現プロファイルの調査(プロテオーム解析)および/またはRNA発現プロファイルの調査(トランスクリプトーム解析)を含む。
【0029】
さらなる実施形態において、RNAの宿主細胞への導入の結果として生じる宿主細胞に対する効果の調査が、宿主細胞の代謝の調査を含む。この代謝の調査は、公知の方法にしたがった酵素反応からの基質または産物の集積調査を含むことが好ましい。
【0030】
他の実施形態において、RNAの宿主細胞への導入の結果として生じる宿主細胞に対する効果の調査が、遺伝子調節の調査を含む。
【0031】
本発明に係る方法の特に好ましい実施形態において、宿主細胞に対する効果の調査は、RNAが導入されていない細胞(転移コントロール)との比較、または機能と関係のないRNAを導入した細胞(ネガティブコントロール)、あるいは機能を仲介するRNAを導入した細胞(ポジティブコントロール)との比較で行う。これら細胞は、コントロールまたは基準としての役割を果たす。また、これら細胞は用いられる宿主細胞と一致していることが好ましい。
【0032】
サブトラクティブハイブリダイゼーション(subtractive hybridization)法は、RNAの宿主細胞への導入が宿主細胞のRNA発現プロファイルに対する作用を含むのか否か決定するために用いることができる。この実施形態においては、例えばコントロール細胞に存在するRNAを、着目しているRNAを挿入する宿主細胞により発現されたRNAから「差し引いた(subtracted)」としている。
【0033】
さらなる実施形態において、RNAの宿主細胞への導入の結果として生じる宿主細胞に対する効果の調査は、相補性解析を含む。この実施形態においては、宿主細胞に内在する遺伝子の機能を、RNAの宿主細胞への導入により置き換え、または増強させる。
【0034】
さらなる実施形態において、RNAの宿主細胞への導入の結果として生じる宿主細胞に対する効果の調査は、宿主細胞の表現型の特徴の調査、例えば形態学的、巨視的または微視的調査などの方法による調査を含む。
【0035】
また、宿主細胞は、RNAの宿主細胞への導入の結果として生じる宿主細胞に対する効果の調査の前に、さらなる処置または状態の変更をすることができる。これにより、ある実施形態においては、細胞の因子に対する効果を一時的に調査する。宿主細胞は、核酸および/または核酸発現産物の機能のさらなる決定のために、特定の生命体におけるインビボの状況下に組み込むこともできる。
【0036】
RNAの宿主細胞への導入の結果として生じる宿主細胞に対する効果の調査は、核酸および/または発現産物が有する、この核酸および/または発現産物が由来する生命体の中の一つ以上の機能を同定することを可能にする。
【0037】
本発明に係る方法は、着目する核酸および/または着目する発現産物に機能的に関連する一つ以上の細胞因子を同定することも可能にする。このような細胞因子の群には、着目するRNAの宿主細胞への導入に対する増加した発現を表示するものがあるだろう。もう一つのこのような細胞因子の群には、着目するRNAの宿主細胞への導入に対する減少した発現を表示するものがあるだろう。これにより、本発明は、細胞因子および着目する核酸および/または発現産物の間の機能的な関連性を同定する手段をも提供する。
【0038】
ある実施形態においては、宿主細胞が、調査する核酸および/または調査する発現産物と同様の生命体由来であることが好ましい、すなわち調査する核酸および/または調査する発現産物が宿主細胞と同種であることが好ましい。しかしながら、調査する核酸および/または調査する発現産物は、宿主細胞とは異種であることが可能である。
【0039】
着目する核酸および/または着目する発現産物は、少なくとも初めに、その核酸および/またはその発現産物を自然に発現することが知られている宿主細胞中で調査することが好ましい。
【0040】
しかしながら、ある実施形態においては、異種の核酸および/または発現産物を、宿主細胞、特に哺乳類細胞中で試験することが望まれることもある。これにより、例えば特定の配列、例えば真菌由来の配列が哺乳類の配列のための置換産物として機能する可能性があるか否かを決定することも可能である。もしそうである場合、この異種の配列は、例えば哺乳類の配列のための治療上の置換産物として用いることができる。
【0041】
さらに、本発明に係る方法において、核酸および/または発現産物の誘導体で一つ以上の機能について試験することが望まれることもある。これは、特定の核酸および/または特定の発現産物を改変すること、および改変された核酸および/または改変された発現産物がオリジナルの核酸および/またはオリジナルの発現産物の一つ以上の効果および/または機能を維持するか否かを試験することを可能にする。多くの場合において、オリジナルの核酸および/またはオリジナルの発現産物の一つ以上の効果および/または機能を維持する核酸および/または発現産物の誘導体を製造することが望まれる。一方で、それらはオリジナルの核酸および/またはオリジナルの発現産物の一つ以上の効果および/または機能を持たないものである。
【0042】
ある実施形態においては、高いスループットのシステムを、本発明に係る方法を行うために用いる。この態様の一つの実施態様において、システムは随意にコンピュータ化またはロボット化することができ、および多数のコンパートメントの使用を含むこともできる。特に好ましい実施形態においては、このコンパートメントをマルチウェルのプレートに配列させて、RNAおよび宿主細胞を多くのコンパートメントに配置させることが好ましい。
【0043】
RNAの宿主細胞への導入の結果として生じる宿主細胞に対する効果は、公知のマイクロアレイ技術および処理により調査することができる。
【0044】
(発明の詳細な説明)
本発明によれば、組み換え核酸の産物、細胞の培養、および核酸、特にRNAの細胞への導入、特にエレクトロポレーションおよびリポフェクションについて標準的な方法を使用することができる。酵素反応および精製技術は、製造者の指示または公知の手法により行う。
【0045】
「生命体」という用語は、本発明によれば、遺伝物質の増幅または伝達する能力を有し、かつ、植物および動物並びに微生物、例えばバクテリア、イースト、真菌およびウィルスを含む生物学上の単位に関する。
【0046】
「形質転換」という用語は、本発明によれば、一つ以上の核酸の生命体または細胞、または後者由来の細胞への導入に関する。
【0047】
「宿主細胞」という用語は、本発明によれば、外部の核酸、特にRNAを用いて形質の変換または転換が可能な細胞に関する。核酸は、宿主細胞に一つ以上のコピーにて存在することができ、およびある実施形態においては宿主細胞で発現される。
【0048】
本発明によれば、「宿主細胞」は、原核および真核細胞、特にヒトおよび動物細胞、植物細胞および微生物細胞を包含する。本発明で使用される特に好ましい宿主細胞は、真核細胞、特に哺乳類細胞、例えばヒト、マウス、ハムスター、ブタ、ヤギおよび霊長類由来の細胞である。これらの細胞は、多様な組織の型由来であり、および一次細胞および細胞株を包含することができる。
【0049】
宿主細胞は、ヒトの組織由来であることが好ましく、特に限定されないが、神経細胞、脳細胞、上皮細胞、結合組織細胞(例えば線維芽細胞、骨芽細胞)、血液細胞(例えば白血球、リンパ球、単球および好中球、特に樹状細胞、血小板)、感覚細胞、筋細胞、肺細胞、心臓細胞、肝細胞、皮膚細胞、膵臓細胞、乳腺細胞、腎臓細胞、腸細胞、胃細胞、結腸細胞、前立腺細胞、卵巣細胞、胚細胞および幹細胞が挙げられ、これらは多様な細胞型に分化する能力を有する。
【0050】
培養細胞株も使用することができる。本発明のある実施形態においては、例えば着目する核酸および/または着目する発現産物がこれら細胞でしか正常に発現しない場合に、部分的にまたは完全に分化した細胞を用いる。
【0051】
本発明によれば、「RNAの宿主細胞への導入の結果として生じる宿主細胞に対する効果」という用語は、RNAの宿主細胞への導入に起因する可能性のある、宿主細胞に対する知覚可能な効果のいずれも包含し、特に宿主細胞の成長、発達、増幅、遺伝的継承、生物学および/または表現型に対する効果を包含する。
【0052】
「表現型」または「表現型の特徴」という用語は、生命体または宿主細胞の外観に関する。表現型は、内的および外的な構造および機能の全てを含む。個々の発達の経過において、生命体およびそれに由来する細胞の表現型は変化することができる。生命体およびそれに由来する細胞の外的特徴は、その遺伝的情報(遺伝子型)により決定されるが、表現型はどの遺伝子が実際に発現されるのか(発現)に依存する。
【0053】
「生化学」または「生化学的特徴」という用語は、生命体またはそれに由来する細胞における基質の代謝および伝達に関する。
【0054】
本発明によれば、「生命体またはそれに由来する細胞における核酸および/または核酸発現産物の機能」という用語は、生命体またはそれに由来する細胞、好ましくはその核酸および/またはその核酸発現産物が由来する生命体またはそれに由来する細胞における、核酸および/または核酸発現産物の機能的役割に関する。ある実施形態においては、この用語は、発ガン現象における核酸および/または核酸発現産物の機能的役割に関する。核酸に関しては、この用語は、特に生命体またはそれに由来する細胞における核酸の発現または発現の欠如から生じる効果に関する。
【0055】
本発明によれば、「核酸および/または核酸発現産物の機能」という用語は、核酸および/または核酸発現産物が一つだけまたはいくつかの機能を有することを意味することがあり、後者の場合、一つ以上の機能が本発明に係る方法により調査される。
【0056】
「遺伝子」という用語は、本発明によれば、特定の核酸配列に関し、この配列は一つ以上の細胞の産物の産生、および/または一つ以上の細胞間または細胞内の機能の獲得に関与する。特にこの用語は、特定のタンパク質、または機能性または構造性RNA分子をコードするDNAセグメントに関する。
【0057】
本発明の核酸は、リボ核酸(RNA)またはRNAの産生に使用することができるデオキシリボ核酸(DNA)であることが好ましい。本発明によれば、核酸は、ゲノムDNA、cDNA、mRNA、組み換え技術により作り出された分子および化学的に合成された分子を包含する。また、本発明によれば、核酸は、一本鎖または二本鎖、および直鎖状または共有結合的に環状に閉じた分子の形態をとることができる。
【0058】
また、本発明の核酸、特にRNAは、核酸間でのホモロジー(相同性)がある程度あれば、すなわちこの核酸がヌクレオチド配列において直接的または相補的に十分な一致があれば、この他の核酸から誘導される。さらに、核酸は、一番目の核酸(当該核酸)が二番目の核酸(他の核酸)の誘導体である場合にも、この他の核酸から誘導される。また、「核酸から誘導されたRNA」という用語は、特にRNAが核酸またはそれから誘導される核酸の転写産物であること、および/または翻訳によって、そのRNAが由来する核酸の転写および翻訳に基づいて生じるタンパク質発現産物と、同様の配列を有するタンパク質発現産物を結果的にもたらす配列を有することを意味する。
【0059】
本発明によれば、核酸の「誘導体」は、その核酸において一つまたは複数のヌクレオチドの置換、削除および/または追加が存在することを意味する。さらに、「誘導体」という用語は、核酸の核酸塩基、糖またはリン酸に対する化学的誘導体も包含する。また、「誘導体」という用語は、非天然にて発生するヌクレオチドおよびヌクレオチド誘導体を含む核酸をも包含する。
【0060】
本発明によれば、核酸は単離することができる。本発明によれば、「単離された核酸」は、核酸が、(i)例えばポリメラーゼ連鎖反応(PCR)によりインビトロで増幅されること、(ii)クローニングにより組み換えにて作り出されること、(iii)例えば切断およびゲル電気泳動による分離により精製されること、または(iv)例えば化学合成により合成されることを意味する。単離された核酸は、組み換えDNA技術による操作が可能な核酸である。
【0061】
核酸は、二つの配列が互いにハイブリダイズ可能であり、安定した二重鎖を形成できる場合、他方の核酸と「相補的」となる。なお、このハイブリダイズは、好ましくはポリヌクレオチド間での特定のハイブリダイズを許容する条件(ストリンジェントな条件)下で生じるものである。ストリンジェントな条件は、例えばMolecular Cloning:A Laboratory Manual,J.Sambrook et al.,Publ.,2nd edition,Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989またはCurrent Protocols in Molecular Biology,F.M.Ausubel et al.,Publ.,John Wiley&Sons,Inc.,New Yorkに記載されており、および例えば65℃でハイブリダイゼーションバッファ(3.5×SSC、0.02%Ficoll、0.02%ポリビニルピロリドン、0.02%ウシ血清アルブミン、2.5mM NaH2PO4(pH7)、0.5%SDS,2mM EDTA)でのハイブリダイズに関する。SSCは、0.15Mの塩化ナトリウム/0.15Mのクエン酸ナトリウム、pH7である。ハイブリダイズ後、DNAが移動してきた膜を、例えば2×SSCで室温にて洗浄し、その後0.1−0.5×SSC/0.1×SDSで68℃までの温度で洗浄する。
【0062】
本発明によれば、相補的核酸は、少なくとも70%、少なくとも80%、少なくとも90%、および好ましくは少なくとも95%、少なくとも98%、または少なくとも99%のヌクレオチドの同一性を有する。
【0063】
本発明の方法により核酸と機能的に連結される発現制御配列または調節配列は、その核酸に対して同種または異種であってもよい。コード配列および調節配列は、それらが共有結合する場合、コード配列の転写または翻訳が調節配列の制御下または影響下にて行われるように、「機能的」に連結する。コード配列が機能的タンパク質に翻訳される場合、調節配列のこのコード配列との機能的連結した状態でのこの調節配列の提示が、コード配列での読み枠移動を引き起こしたり、または所望するタンパク質またはペプチドへのコード配列を翻訳不能としないように、コード配列の転写を導く。
【0064】
本発明によれば、「発現制御配列」または「調節配列」は、プロモーター、リボソーム結合配列および他の制御要素を包含し、この制御要素は遺伝子の転写または誘導されたRNAの翻訳を制御するものである。本発明のある実施形態においては、発現制御配列は制御され得る。調節配列の正確な構造は、種に依存して、または細胞型に依存して異なるが、転写または翻訳の開始に関与する、一般的に5'−未転写の配列、および5'および3'−未翻訳の配列、例えばTATAボックス、キャップ配列、CAAT配列などを包含する。特に、5'−未翻訳の調節配列は、機能的に結合した遺伝子の転写制御のためのプロモーター配列を有するプロモーター領域を包含する。調節配列は、エンハンサー配列または上流のアクチベータ配列も包含し得る。
【0065】
さらに、本発明の核酸は、この核酸によりエンコードされたタンパク質またはポリペプチドを特定の標的部位にガイドするポリペプチドをコードする他の核酸と連結することができる。特に、本発明の核酸は、エンコードされたタンパク質またはポリペプチドの転座または安定化を宿主細胞の細胞膜またはこの細胞の特定のオルガネラにおいてコンパートメント化された部分の細胞膜の上で引き起こすポリペプチドをコードする他の核酸と連結することができる。
【0066】
本発明の好ましい実施形態において、クローン核酸は、この核酸の発現を制御するプロモーターを随意に含むベクター内に存在する。「ベクター」という用語は、その最も一般的な意味において用いられ、例えば核酸を原核および/または真核細胞に挿入して、必要に応じてゲノムに組み込むことを可能にする、核酸のためのいかなる中間的な媒体(vehicle)をも包含する。このようなベクターは、細胞中で複製および/または発現されることが好ましい。中間的な媒体(vehicle)は、例えばエレクトロポレーション、微小粒子衝撃法(microprojectile bombardment)、リポソーム投与(liposomal administration)、アグロバクテリアを用いた転移、またはDNAまたはRNAウィルスを介した挿入における用途に適応させることができる。
【0067】
組み換えRNAは、本発明によれば、インビトロでの適切なDNA鋳型の転写により産生することができる。さらに、組み換えRNAは、配列の安定化、キャッピングおよびポリアデニル化によって修飾することができる。
【0068】
本発明によれば、「発現」という用語は、その最も一般的な意味において用いられ、RNAの産生、またはRNAおよびタンパク質/ポリペプチドの産生を包含する。また、発現は、核酸の部分的な発現も包含する。さらに、発現は過渡的であっても、安定的であってもよい。RNAに関して、「発現」という用語は、特にタンパク質/ポリペプチドの産生に関する。
【0069】
発現産物、特にタンパク質あるいはポリペプチド、または本発明の意味におけるアミノ酸配列の「誘導体」は、アミノ酸挿入変異体、アミノ酸削除変異体、および/またはアミノ酸置換変異体を包含する。
【0070】
アミノ酸挿入変異体は、アミノ−および/またはカルボキシ−末端融合物、および特定のアミノ酸配列の中での個々のまたはいくつかのアミノ酸の挿入物を包含する。挿入を伴うアミノ酸配列変異体において、一つ以上のアミノ酸残基は、アミノ酸配列の所定の位置で挿入されるが、結果産物の好適なスクリーニングによるランダム挿入も可能である。アミノ酸削除変異体は、配列から一つ以上のアミノ酸を除くことにより特徴付けられる。アミノ酸置換変異体は、配列の少なくとも一つの残基を除き、かつ、その場所にもう一つの残基を挿入することにより特徴付けられる。この修飾は、同種のタンパク質間またはポリペプチド間で保存されていない、アミノ酸配列の位置でなされていることが好ましい。アミノ酸は、同様の特性、例えば疎水性、親水性、電気陰性度、側鎖の容積などを有する他のもので置換すること(同類置換)が好ましい。同類置換は、例えばあるアミノ酸をもう一つのアミノ酸で置換することに関する。以下にその置換対象となるアミノ酸と同じ群に挙げる。
【0071】
1.小さな脂肪族の、非極性あるいはわずかに極性の残基:Ala、Ser、Thr(Pro、Gly)
2.負のチャージを帯びた残基およびそれらのアミド:Asn、Asp、Glu、Gln
3.正のチャージを帯びた残基:His、Arg、Lys
4.大きな脂肪族の、非極性残基:Met、Leu、Ile、Val(Cys)
5.大きな芳香族残基:Phe、Tyr、Trp
【0072】
3残基が、タンパク質構造において特定の役割を果たすため、括弧内にて示されている。Glyは、側鎖を有さない唯一の残基であり、これにより鎖に柔軟性を付与する。Proは、独特の配置を有しており、これによりポリペプチド構造に大きな変化を導くことができる。Cysは、分子内、または他のCysとの分子間のジスルフィド架橋を形成することができる。
【0073】
上述したアミノ酸変異体は、組み換えDNA操作を用いて容易に作り出すことができる。全部公知または部分的に公知の配列を有するDNAの所定部位での置換変異を挿入するための技術は、公知であり、例えばM13突然変異生成が挙げられる。タンパク質産生のための置換、挿入または欠失といったDNA配列の操作は、例えばSambrook et al.(1989)に詳細に記載されている。
【0074】
本発明によれば、タンパク質またはポリペプチドの「誘導体」は、タンパク質またはポリペプチドに関連する糖質、脂質および/またはタンパク質またはポリペプチドに関連する単一のまたは複数の分子の置換、欠失および/または付加も包含する。さらに、「誘導体」は、タンパク質またはポリペプチドの機能的、化学的な等価物の全てに当てはまる。
【0075】
本発明によれば、発現産物の部分またはフラグメントは、好ましくは、それが由来する発現産物の機能的な特性を有する。この機能的な特性は、抗生物質との相互作用、他のポリペプチドまたはタンパク質との相互作用、核酸の選択的結合および酵素活性を包含する。
【0076】
本発明によれば、核酸の部分またはフラグメントは、好ましくは上記で定義したような発現産物の部分またはフラグメントをコードする核酸の部分に関する。
【0077】
本発明によれば、核酸の細胞への導入のために、多様な方法を用いることができる。このような方法は、核酸−CaPO4沈殿物の形質転換、DEAEに関連する核酸の形質転換、着目する核酸を担持したウィルスを用いた形質転換または感染、エレクトロポレーション、リポソーム媒介の形質転換などを包含する。核酸を特定細胞へ案内することは、ある実施形態においては好ましい。このような実施形態において、核酸を細胞に送達するために用いるキャリア(例えば、レトロウィルスまたはリポソーム)は、結合した案内分子(directing molecule)を備える。例えば、抗体のような分子は、標的細胞の表面膜タンパク質に特異的であったり、あるいは標的細胞上のレセプターのためのリガンドであったりするが、核酸キャリアに取り込んだり、または結合させることができる。核酸をリポソームにより導入することが望まれる場合には、エンドサイトーシスに関連する表面膜タンパク質に結合するタンパク質をリポソーム製剤に取り込んで、標的および/または吸収を可能とすることができる。このようなタンパク質は、カプシドタンパク質またはそのフラグメントを包含する。このタンパク質は、特定の細胞型、内在するタンパク質に対する抗体、細胞内部位にガイドするタンパク質などに特異的である。
【0078】
本発明によれば、「相補性解析」という用語は、特定の遺伝子を削除または変異させて、その遺伝子の役割にてもはや十分に機能しないようにした後に、核酸を生命体または細胞に導入したときにこの生命体または細胞にて得られる効果の調査に関する。相補的な遺伝子またはその発現産物が、欠損遺伝子または変異遺伝子を伴った生命体または細胞に導入されると、好ましくはそれらはその遺伝子の機能を再生する能力を有する。
【0079】
本発明によれば、「マイクロアレイ」という用語は、生物学上の材料、例えば核酸、ペプチドまたはタンパク質を高密度にて適用し、結合するために用いられるデバイスに関する。マイクロアレイは、多量のサンプルを制御された間隔にて適用することを許容し、それらがパラレルな実験に使用することができる固体キャリア支持体からなり、またはそのような固体キャリア支持体を有する。
【0080】
本発明によれば、「高密度」という用語は、様々なサンプル、例えば核酸サンプルを高密度で保持し、これにより多くの質問に対して回答を与えることを可能にするアレイに関連して用いられる。
【0081】
本発明は、以下に示す図面および実施例により詳細に説明されるが、これら図面および実施例は説明としてのみ用いられるのであり、限定として理解されるべきではない。当業者は、本記載および本実施例に基づいてさらなる実施形態に拡げることができるものであり、これらも本発明により保護される。
【図面の簡単な説明】
【0082】
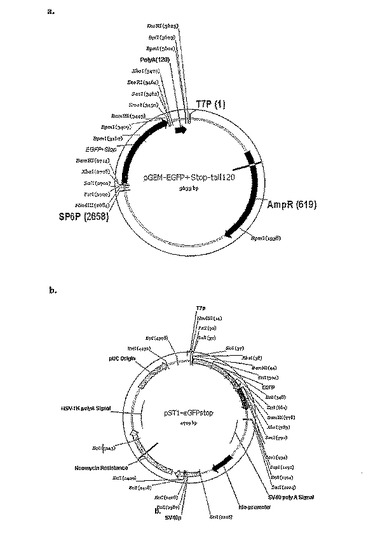
【図1】用いられたeGFPベクターの表示−a.pGEM3Z eGFPテール、b.pSTI eGFPテールである。これらのベクターは、続くクローニングのための、開始ベクターとして用いられた。
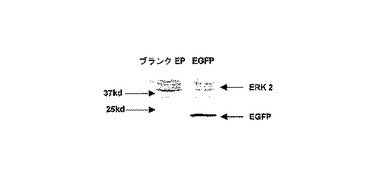
【図2】786−0細胞における、形質転換後24時間のeGFP IVT RNAの翻訳を検出するためのウエスタンブロット。eGFP−pGEM3ZテールIVT RNAの20μgが、200Vかつ250μFで形質転換された。一次抗体として、濃度1μg/mlのウサギ抗GFPを用いた。二次抗体として、濃度0.5μg/mlのヤギ抗ウサギHRPを用いた。マウス抗ERK2抗体を濃度0.2μg/mlのローディングコントロールとして用いた。ここで、二次抗体として、ヤギ抗マウスHRPを、1/5000希釈で用いられた。
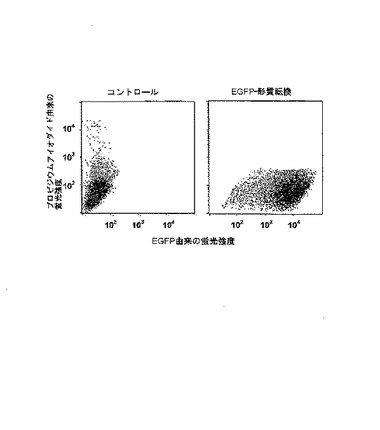
【図3】1×107個の786−0細胞に、eGFP IVT RNAの20μgが形質転換された後24時間の、又は、RNAなしでコントロール形質転換後の、FACSドットブロットの表示。
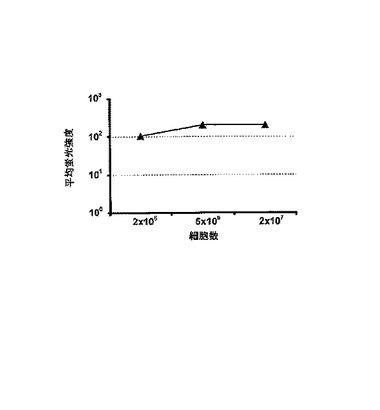
【図4】eGFPの平均蛍光強度を測定して、エレクトロポレーション効率に対する細胞カウントの影響の測定。eGFP−pGEM3ZテールIVT RNAの20μgが、上記数のK562細胞に形質転換された。細胞は形質転換後24時間培養された。平均蛍光強度は、FACS−Caliburを使用して測定した。
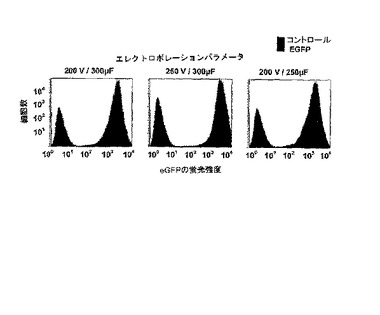
【図5】エレクトロポレーション効率や平均蛍光強度に対するエレクトロポレーション条件の影響の測定。MCF7細胞に、eGFP IVT RNAを20μg形質転換した。細胞は形質転換後24時間培養された。平均蛍光強度は、FACS−Caliburを使用して測定した。
【図6】eGFP IVT RNA又は、2dGFP IVT RNAの20μgを用いた、2×107個の786−0細胞の形質転換後、転写およびタンパク質量に対する時間の影響の測定。a.定量RT−PCR分析に基づく、相対転写量の測定。正規化は非形質転換細胞に対して行った。cDNA量の違いを考慮するために、18s特異的PCRを行い、その値で正規化に用いた。b.FACS−Caliburを使用したeGFPの平均蛍光強度の測定。
【図7】eGFP IVT RNAを用いた5×106個のMCF−7細胞の形質転換後24時間のeGFPタンパク質レベルに対するRNA量の影響の測定。a.非形質転換コントロールと比較したヒストグラムブロットでの蛍光強度の測定の表示。b.平均蛍光強度の表示。
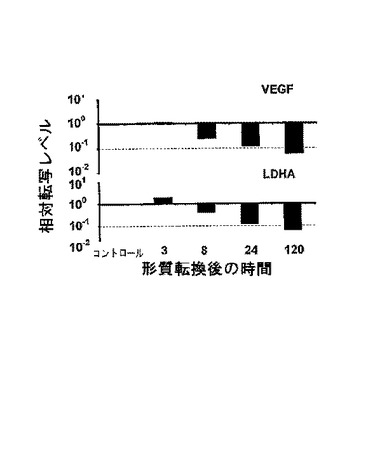
【図8】2×107個の786−0細胞形質転換後24時間のHIF1−α、LDHA、VEGF、および、Glut−1の相対転写量のリアルタイムRT−PCRによる定量測定。eGFP形質転換コントロールと比較した相対発現の表示。
【図9】VHL IVT RNAの20μgを用いた2×107個の786−0細胞の形質転換後、VEGFおよびLDHAの相対転写量の時間に対する変化。非形質転換細胞と比較して、相対転写量を測定した。正規化は18s rRNAによって行った。
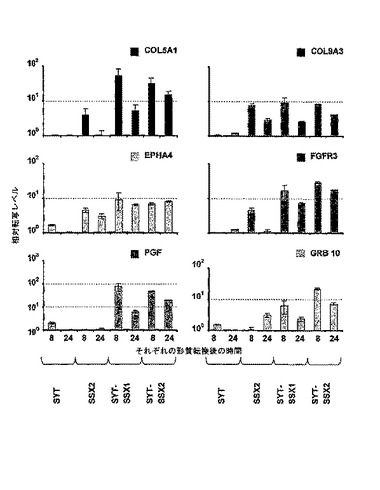
【図10】SYT、SSX2、SYT−SSX1、SYT−SSX2、または、コントロールとして、eGFP IVT RNAを用いた、1×107個のMCF−7細胞の形質転換後の相対転写量の表示。正規化はHPRTによって成立した。eGFP IVT RNAを用いて形質転換された細胞と比較して、相対転写量を測定した。
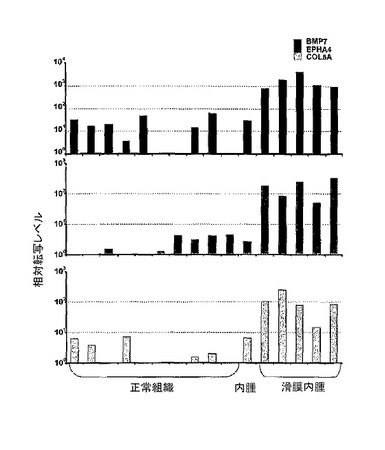
【図11】乳房、肺、腎臓、卵巣、睾丸、肝臓、リンパ節、胸腺、脾臓および副腎の正常組織と比較した、滑膜肉腫における、BMP7、EPHA4およびCOL5A1の相対転写量の表示。正規化はHPRTによって行った。BMP−7およびCOL5A1の発現の測定のため、試験された肉腫は、骨肉種であった。一方、EPHA4の調査のため、滑膜肉腫を試験し、SYT−SSX1またはSYT−SSX2に対して、ポジティブではなかった。転位の欠損によって、病理の診断は不確かなものである。
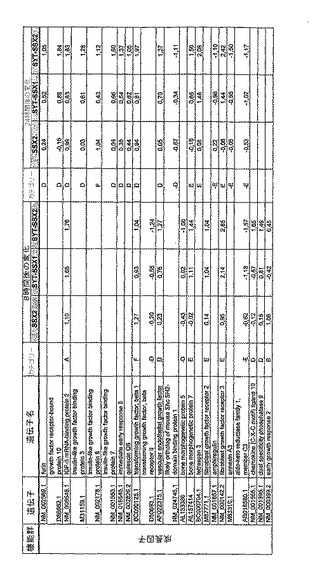
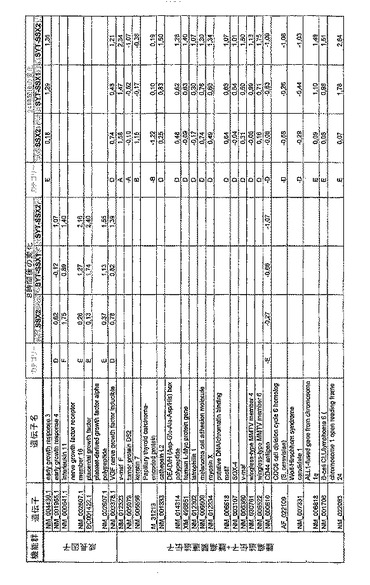
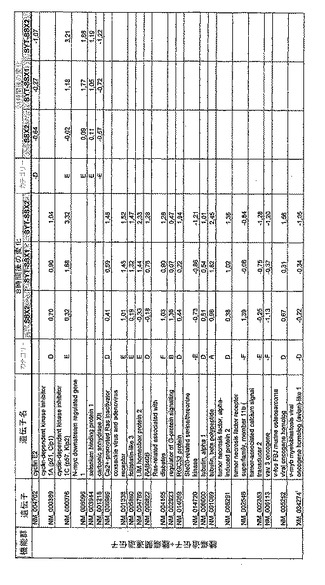
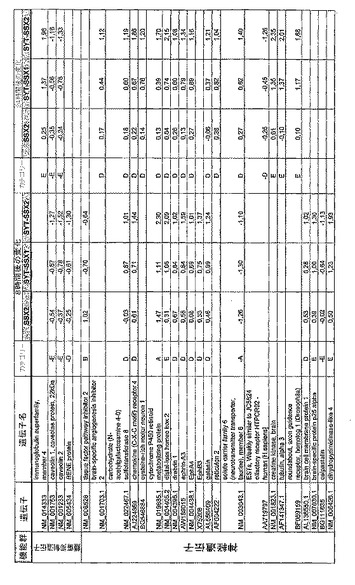
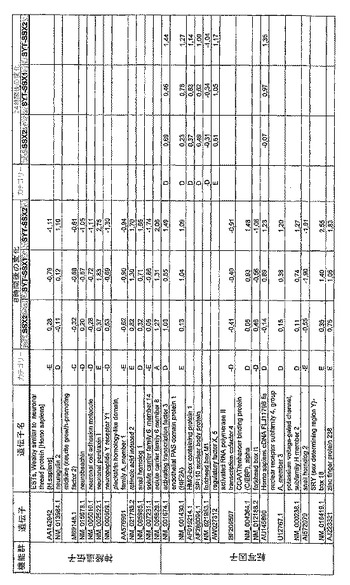
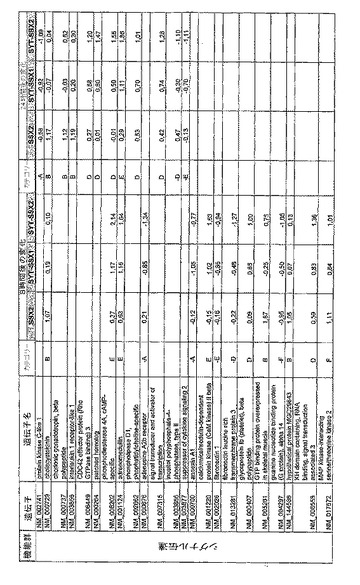
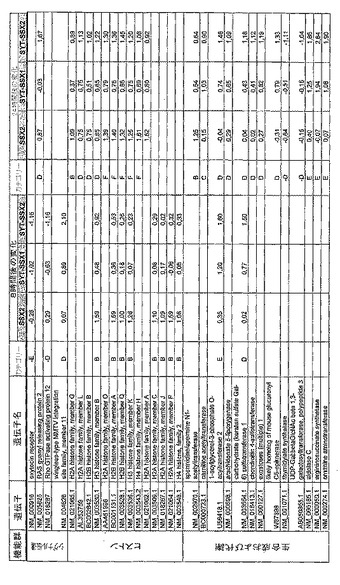
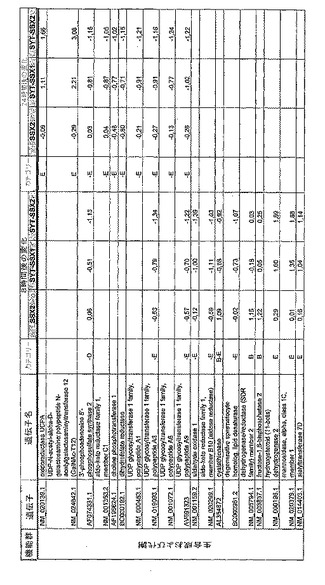
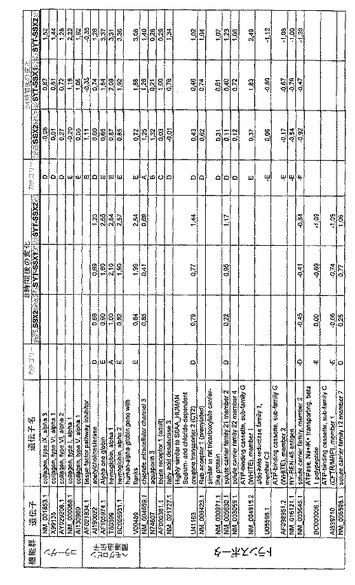
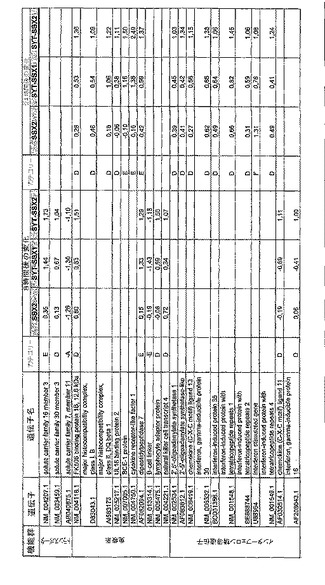
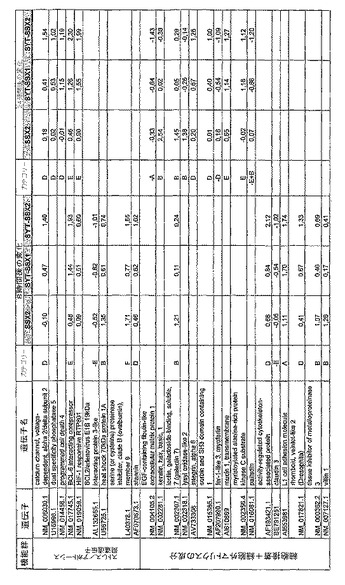
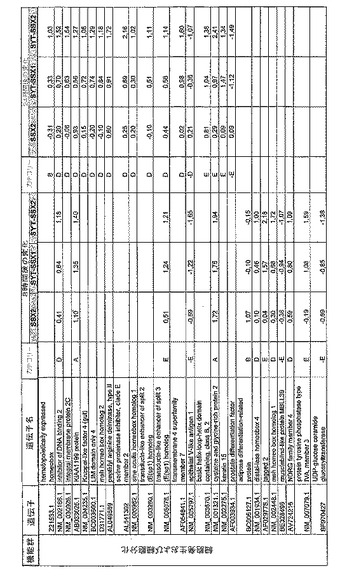
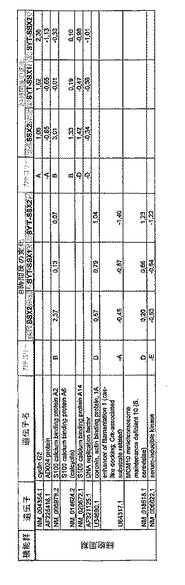
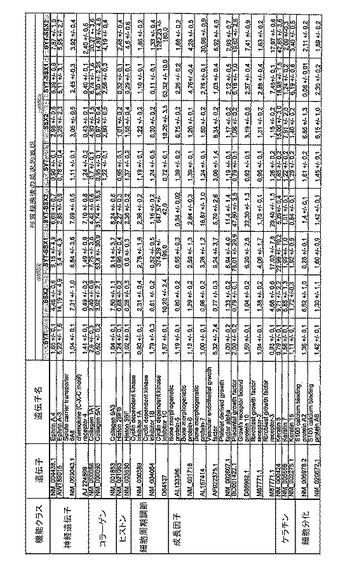
【図12】機能によってグループ化され、かつ、eGFPを用いた形質転換コントロールと比較して、SSX2、SYT−SSX1またはSYT−SSX2を用いたMCF−7細胞の形質転換体に対して、5%の有意性基準で、少なくとも21倍の差異で発現された、遺伝子一覧表。記載のない値は非有意調節領域に位置する、つまり<21の値である。表に示された値は、いずれの場合も、2が基底の指数である。つまり、結果数は差異発現のファクターを表している。
【図13】図12に同じ。
【図14】図12に同じ。
【図15】図12に同じ。
【図16】図12に同じ。
【図17】図12に同じ。
【図18】図12に同じ。
【図19】図12に同じ。
【図20】図12に同じ。
【図21】図12に同じ。
【図22】図12に同じ。
【図23】図12に同じ。
【図24】図12に同じ。
【図25】eGFP形質転換細胞と比較して、リアルタイムRT−PCRを用いて測定された、相対転写量の表示。正規化はHPRTによって行った。生物学的上のtriplicates(biological triplicates)間の標準偏差が記載されている。
【実施例】
【0083】
実施例1:ベクターの生産
【0084】
ベクターは、それぞれのコード配列を基礎ベクターであるpGEM3Zベクター(Invitrogen,San Diego,CA)または、pCMV Scriptベクター(Stratagene,Amsterdam,NL)にクローンして、生産した。
【0085】
pGEM3Zベクターは、開始コドンの前にSP6プロモーターを持ち、そこからインビトロで転写可能である。pCMV Scriptベクターは同じ部位にT7プロモーターを有する。
【0086】
これらの基礎ベクターから出発し、大幅な組み換えで導入する。これは、ポリアデニル化された、インビトロで転写された(IVT)RNAの生産のために必須である。
【0087】
120塩基のポリA伸長部分を、pGEM3ZベクターにXhoI切断部位を解して導入した。ポリA伸長直後、制限部位を導入し、これにより、ポリA伸長がリニア化する。後述するように、pGEM3Zテールベクターをこのベクターにしている(図1a)。
【0088】
120塩基対のポリA伸長部分をpCMV ScriptベクターにSacI切断部位の後ろで導入した。後述するように、pSTIテールベクターをこのベクターとしている(図1b)。
【0089】
eGFPまたは、各組織から増幅された遺伝子を、精製および各ベクターによる制限後、連結し、E.coli MRF1-に形質転換した。得られたプラスミドを配列決定し、その配列を分析した。
実施例2:IVT RNAの生産
【0090】
IVT RNAの生産の第一段階は、各プラスミドのリニア化を含む。用いられる制限酵素は、pGEM3Zテールベクターについては、BpiIであり、pSTIテールベクターについてはSapIだった。両方とも、ベクターの中を一度だけ、つまり挿入テールの後ろが切断された。リニア化後、酵素は、フェノール−クロロホルム沈殿によって不活性化し、除去した。この為に、フェノールおよびクロロホルムの混合物の等容量(isovolume)が制限チャージに追加され、完全に混合された。10000×gの手短な遠心分離により、下部の有機層およびDNAを含む上部の水層への分離が生じた。後者を新しい反応槽に移した。その後、水層を等容量の純クロロホルムで混合し、全てのフェノール残留物を除去した。遠心分離後、水層を除去し、2倍等容量のエタノールとpH4.5の10%v/v3M酢酸ナトリウムと−20℃にて加えて2時間沈殿させた。
【0091】
DNAは、10000×gで、4℃の遠心分離で45分間沈降し、塩を除去するために70%エタノールで洗浄し、適量のRNAse−free水中で取っておいた。リニア化が成功かつ完了であることを確かめるために、ゲル電気泳動を行なった。DNA濃度は、260nmの光度測定法で、測定した。DNA純度の測定のため、さらに280nmの吸光度を測定してOD260/280比を得た。
【0092】
10μgのリニア化DNAをインビトロ転写に用いた。これについては、4/5のdGTPにさらにキャップ構造を付与した40μlのdNTPと、l0μlの10×バッファと、20μlのdTTと、10μlのT7またはSP6ポリメラーゼを、37℃で2時間でインキュベートした。ポリメラーゼは、転写されるORFから5'方向に位置する、T7またはSP6認識配列に結合し、相補的RNA鎖を合成する。
【0093】
IVT RNAは、MegaClear Kitを用いて精製した。これについては、RNAをシリカ膜に最適に結合させるために必要な塩を含んだ結合緩衝濃縮物に、そのRNAを取っておいた。エタノールの添加により、RNA水和殻から水が除去した。その混合物を、シリカカラムにロードし、10000×gで2分遠心分離させた。RNAをそのカラムに結合させ、一方不純物、たとえば、酵素残留物を洗い落とした。何度かの洗浄後、精製されたRNAを溶出した。溶出緩衝液を、95℃まで予熱して効率的に溶出した。
【0094】
質管理および定量化は、ゲル電気泳動および光度測定によって、行った。
実施例3:細胞へのエレクトロポレーション
【0095】
エレクトロポレーション法の原理は、短い電流パルスによって、細胞の膜電位を乱すことに基づいている。外部刺激による膜電位の変化は、次の式で表される。
【数1】

Vmは膜電位であり、fは細胞外の電界分布に対する細胞の影響を表す形状因子である。fEexは適用された電界を表し、rは細胞半径を表し、φは外部から適用された電界に対する角度を表す。fは、通常1.5として与えられるが、それは他の多くの因子に依存している。適用された電界が、細胞膜の容量を越えるとき、すなわち、ΔVmが1Vとして与えられる閾値ΔVsより大きいとき、細胞へのエレクトロポレーションは成功する(Kinosita,K.,Jr.and Tsong,T.Y.(1977)Nature,268,438−441)。二分子膜としての細胞膜の構造は、真核細胞に共通の特徴であるため、この値は異なった細胞株に対して、ほとんど変動を示さない。
【0096】
絶縁破壊の膜電位になると、一時的に親水性穿孔が形成され、そこを通じて水が細胞内に浸透して、たとえば核酸等の分子の細胞内への移動が生じる(Weaver,J.C.(1995)Methods Mol.Biol.55,3−28;Neumann,E.et al.(1999)Bioelectrochem.Bioenerg.48,3−16)。
【0097】
エレクトロポレーションの前に、用いる接着細胞をセミコンフルエントの状態まで培養し、PBSで洗浄後、トリプシンを用いて、細胞培養フラスコから取り外した。細胞を10%FCS(ウシ胎仔血清)添加培地に移し、500×gで8分遠心分離した。ペレットを血清フリー培地X−Vivoのなかで再懸濁し、再び、500×gで8分遠心分離した。この洗浄操作を続くエレクトロポレーションに影響するFCSの残留物を除去するためにさらに2回行なった。
【0098】
洗浄後、細胞を250μlにて所望の細胞密度になるように調整し、エレクトロポレーションキュベットに移し、氷上で静置した。適量のインビトロで転写されたRNAを添加し、十分攪拌した後、エレクトロポレーションを、200Vかつ250μFで行った。その後、細胞を速やかに適当な培養液に移し、37℃、5%CO2下でインキュベートした。
実施例4:実験条件の確立と実験系の特徴付け
【0099】
eGFP−pGEM3Zテール発現ベクターを、実験条件の確立のために用いた。5'に位置するRNAポリメラーゼプロモーターSP6によって、インビトロでのRNAの発現が可能になる。
【0100】
インビトロで転写されたeGFP RNAは、以下のように、多様な細胞型の形質転換効率の試験のために用いた。形質転換効率は、eGFPの蛍光強度の測定によりFACS−Caliburを用いて、測定した。細胞内のRNA量は、定量RT−PCRによって測定した。腫瘍細胞株に90%より高い効率で形質転換可能であるだけでなく、初代樹状細胞にも70から80%の効率で形質転換可能であることが示された(図2および3並びに表1)。
【0101】
【表1】
表1:20μgのeGFP IVT RNAを用いた5×106個の細胞の形質転換後に測定した多様な細胞の形質転換効率。形質転換後24時間にFACS−Caliburを用いての蛍光強度の測定。
形質転換条件:200Vかつ250μFのエレクトロポレーション。
【0102】
さらに、形質転換効率による細胞の影響は2×106から2×107細胞の範囲において除外することができた(図4)。また、エレクトロポレーション条件は、試験範囲内での形質転換効率に対して決定的な影響を及ぼさない(図5)。
【0103】
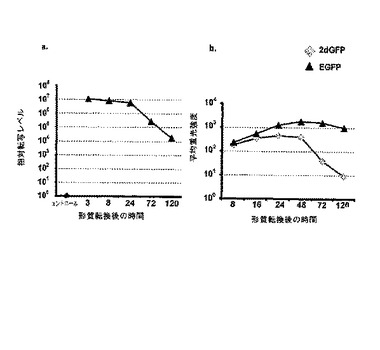
RNAの安定性は24時間にわたって立証された(図6a)。一方、その半減期に依存するが48から72時間にわたって、細胞内でタンパク質を検出することができる(図6b)。
【0104】
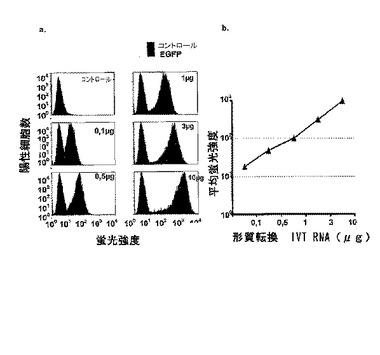
転写されたRNA量および得られるタンパク質量の間には直接的な依存性があることが示すことができた(図7)。
【0105】
この方法に因る分子変化は、cDNAマイクロアレイを用いて測定された。プロテアソーム関連遺伝子、ヒートショック遺伝子およびアポトーシス関連遺伝子の分子変化は、8時間後および24時間後に見られただけであった(表2)。
【0106】
【表2】
表2:非形質転換コントロール細胞と比較して、20μgのeGFP IVT RNAで形質転換した2×107個の786−0細胞における十分に調節された遺伝子の数。変化は、>2または<0.5の調節で有意であるだけである。
【0107】
siRNAの使用の後に記載されてきたような(Elbashir,S.M.et al.(2001)Nature,411,494−498;Caplen,N.J.et al.(2001)Proc.Natl.Acad.Sci.U.S.A.98,9742−9747)インターフェロン系の活性はない。
実施例5:モデル遺伝子のIVT RNA形質転換
【0108】
さらなる、IVT RNAを用いた細胞の形質転換の可能性および限界の詳細な検討のため、多種のモデル遺伝子を異種発現したIVT RNAとして細胞に導入した。よく特徴づけされた遺伝子HIF1−α(低酸素症誘導因子)およびVHL(von−Hippel Lindau)をモデル遺伝子として用いた。これらの遺伝子のコード領域を睾丸組織由来のcDNAを用いて増幅し、eGFPのために記載されたベクター系へクローンした。
【0109】
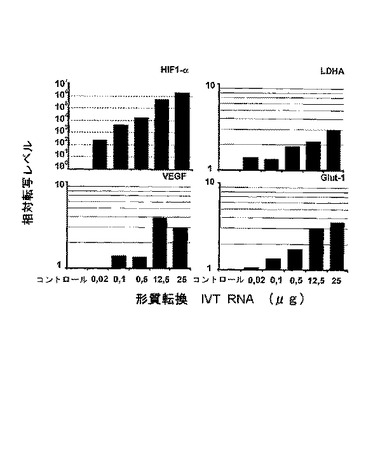
HIF1−αやVHLについては、多種な標的遺伝子が記載されており、それらの発現は、不完全細胞株の形質転換後に測定した。HIF1−α転写因子の形質転換後、標的遺伝子であるLDHA(乳酸脱水素酵素A)、VEGF(血管内皮増殖因子1α)や、Glut−1(グルコーストランスポーター1)の発現増加を検出した(図8)。それらの誘導の程度は、転移HIF1−α IVT RNA量に基づいて滴定することができる。HIF1−α転写因子の転移によって、その標的遺伝子の強固な調節が可能である。用いられたIVT RNA量の滴定を通じて、HIF1−αにより誘導された遺伝子の転写量を滴定することもできる。
【0110】
転写因子の分子効果は、ほとんど直接かつ即座に検出することができる。したがって、HIF1はその標的遺伝子のプロモーター領域の共通配列に直接結合し(Semenza,G.L.(2000)Genes Dev.14,1983−1991;Semenza,G.L.(2000)J.Appl.Physiol 88,1474−1480)、速やかにそれらの転写を開始させる。間接的かつ副次的効果もIVT RNA転移により、明らかにできるか否かを調査するために、HIF1−αよりも上位であるpVHL(von−Hippel−Lidau)タンパク質を第2モデル遺伝子として選択した。pVHLは、ユビキチンリガーゼE3複合体に属し、HIF1のαサブユニットの分解を引き起こす。したがって、VHLの発現増加は、細胞内のHIF1濃度の減少を引き起こし、HIF1標的遺伝子の発現の減少に関係している。VEGFおよびLDHA発現における時間に依存した減少は、VHLでの細胞の形質転換後、明確に実証された(図9)。
実施例6:例としてSYT−SSX1およびSYT−SSX2を用いた推定発癌転写物の従来未知の機能の同定。
【0111】
RNA転移技術は、腫瘍生物学に対する遺伝子の関連性の迅速な解読を可能にする。証明のために普遍的に発現する遺伝子である、SYT遺伝子およびCG遺伝子であり、睾丸や腫瘍でのみ発現するSSX1またはSSX2遺伝子の融合産物を選択した。これらの遺伝子を選択した理由としては、一方においては、滑膜肉腫において90%を超える(Sreekantaiah,C.et al.(1994)Am.J.Pathol.144,1121−1134)転座t(X;18)(p11.2;q11.2)(Clark,J.et al.(1994)Nat.Genet.7,502−508;Crew,A.J.(1995)EMBO J.14,2333−2340)の頻度であり、この転座頻度は、腫瘍生物学の関連性を示唆し、したがって、発癌能を示唆するからである。また、他方では、この融合産物が転写活性化QPGYドメインを含有していることから、転座の結果として、直接分子変化が期待されるためである。腫瘍の発達及び体系における、転座の分子効果及びそれらの重要性は未だ大部分が知られていない。SYT−SSX1及びSYT−SSX2 IVT RNAの形質転換体により誘導された分子変化をAffymetrix社のオリゴヌクレオチドマイクロアレイを用いて、解析した。SYT−SSX1及びSYT−SSX2を用いた細胞に形質転換後、多数の遺伝子発現の増加が検出できた。形質転換後、検出できた調節遺伝子としては、第1成長因子、神経遺伝子、腫瘍遺伝子または腫瘍関連遺伝子が挙げられ、また、これらは、細胞周期の調節またはシグナル伝達に関与する(図12〜24)。これら遺伝子のいくつかの差次的発現は、定量RT−PCRにより、実証された(図25;図10)。滑膜肉腫における、これら遺伝子の機能的関連性を検出するために、さらにそれらの発現を、正常組織と比較して滑膜肉腫において測定した(図11)。
【0112】
実施例としてSYT−SSX1およびSYT−SSX2の形質転換体を用いて、RNA転移およびそれに続くAffymetrix社のオリゴヌクレオチドマイクロアレイを用いた分子変化の測定により、発癌現象における遺伝子の有意性を決定することが可能であることを明らかに示すことができた。形質転換細胞の直接的および間接的な標的遺伝子は、決定され、遺伝子機能の我々の理解に寄与する。
【0113】
IVT RNAの薬物動態的特徴と併せて、IVT RNAはシグナル伝達カスケードを解読するための良好な基礎を構成する。様々な時点での遺伝子発現プロファイルの構成および適切なソフトウェアを用いたそれらの解析は、定義された遺伝子群の単位を記述することにより、結果として起こる活性化された調節遺伝子のカスケードを発見することを可能にする。このように、RNA転位技術を用いて、発癌現象において、今まで殆ど特徴付けられていない遺伝子の機能的有意性を解明することが可能となる。さらに、新規に同定された標的遺伝子の転移により、形質転換遺伝子の直接的および間接的な効果を発見することが可能となる。
【技術分野】
【0001】
本発明は、一般的に核酸配列および核酸発現産物の機能を、宿主細胞にRNAを導入することにより、調査または決定する方法に関する。
【背景技術】
【0002】
ヒトゲノムの配列決定の最初のドラフトが完了し、次の課題は、記述的ゲノム研究から機能的ゲノム研究に移行することである(Fields,S.et al.(1999)Proc.Natl.Acad.Sci.U.S.A.96, 8825−8826)。
【0003】
アンチセンスRNAまたはRNAi(RNA干渉)の使用は、近年において、対象とする遺伝子発現のオフ切り替えのための広い用途が見出されている。
【0004】
アンチセンスRNAは配列特異的に、相補性の規則にしたがって、当然、自然に転写された標的配列のmRNAに結合する。この方法は原核生物において最初に記述された(Green,P.J.et al.(1986)Annu.Rev. Biochem.55,569−597)。今日では、それにより、非常に多くの型の真核細胞におけるいかなる所望される遺伝子の迅速で、束の間の、転写後サイレンシングを可能にし(Izant,J.G.and Weintraub,H.(1985)Science 229,345−352)、広く使用されている。
【0005】
RNAi(RNA干渉)において、標的配列と特異的に相互作用する短い二重鎖RNA(〜21bp)が細胞内に導入される。結果として、多タンパク質複合体が活性化されることで、転写されたmRNAが翻訳可能になる前に、mRNAの早期分解が引き起こされる (Hammond,S.M.et al.(2000)Nature, 404,293−296)。しかしながら、インターフェロン系の活性化を通じて、または、二重鎖RNAに依存するタンパク質キナーゼ応答(PKR)の誘導を通じて、(Elbashir S.M.et al. (2001) Nature,411,494−498;Caplen,N.J. et al.(2001)Proc.Natl.Acad.Sci.U.S.A. 98,9742−9747)、または、濃度依存的な刺激、または、非標的遺伝子の抑制を通じて(Persengiev,S.P.et al.(2004)RNA 10,12−18)siRNAによっても非特異的効果が引き起こされる。
【0006】
上記方法は、アンチセンスRNA、または、RNAiを用いて、翻訳を阻止し、したがって遺伝子の機能欠損を引き起こす。
【発明の概要】
【発明が解決しようとする課題】
【0007】
しかしながら、特定の遺伝子、特に異所的に活性化または過剰発現された遺伝子、たとえば腫瘍において選択的に発現した遺伝子を機能的特徴づけるために、機能獲得の効果をシミュレートすることはより有用である。
【0008】
数多くの遺伝子転換技術も、このために利用可能である。したがって、一過性の形質転換体および安定して形質転換された細胞クローンの両方が生産される。調査中の遺伝子は、DNAプラスミド構造体を用いて、または、レトロウイルスベクターあるいはアデノウイルスベクターの助けにより細胞内に挿入することができる。さらに、挿入遺伝子の発現を構成的にすることができ、または、制御可能なプロモーターにより制御することができる。この目的に利用される、全ての遺伝子転移形態は、固有のシステムの欠点を有している。プラスミドやウィルスによる一過性形質転換体は、ほとんどの細胞型に対して非常に非効率であり(特に、初代細胞)、結果として、多様な亜集団の形質転換をもたらし、これらは、発現レベルの十分正確な滴定をできなくしている。しかしながら、ひとつのおよびそれと同一の遺伝子産物が多様な機能を、異なった細胞、または、たとえ同じ細胞でも異なった濃度で、発揮しているため、これらの技術から得られた情報量は、非常に限られている。軽視できない他の問題は、プラスミドを用いた形質転換が、遅延後にしか転写がおきないことである。その転写まで時間は、形質転換された細胞の細胞周期のフェーズに依存し、かつ、細胞集団に応じて非常に多様である。結果、非同期な、転写および発現がおこる。かわりに、安定形質転換細胞を用いることが提案されている。しかしながら、安定形質転換細胞株の生産には、非常に時間がかかり、かつ、効率的に全ての細胞系に対して実行することはできない。したがって、ほとんどの哺乳類生命体の新鮮な単離細胞が安定して形質転換されない。他の短所は、形質転換後の、例えば抗体の使用や物理的措置(例えば細胞分類)による形質転換細胞の濃縮により、遺伝子産物が導入されたにもかかわらず、起源細胞型から多くの点で異なった亜集団の選別を引き起こすことがある。
【0009】
本発明は、遺伝子産物の機能の特徴づけのための方法に関し、インビトロで転写された長鎖RNAの転移に基づく。インビトロで転写されたRNAの細胞への転移は確立された方法であり、腫瘍抗原を抗原提示樹上細胞(DC)に導入するために、現在特に集中して、腫瘍免疫学において試験されている(Boczkowski,D.et al.(2000)Cancer Res.60,1028−1034;Heiser,A.et al.(2001)J.Immunol.166,2953−2960;Nair,S.K.et al.(1998)Nat.Biotechnol.16,364−369)。反対に、今までは遺伝子機能の系統的決定のために、インビトロで転写されたRNAの転移が利用されていなかった。
【課題を解決するための手段】
【0010】
本発明は、一般的に核酸および/または発現産物、特にペプチドやタンパク質の機能の特徴づけのための方法に関する。本発明による方法において、調査中の核酸および/または調査中の発現産物の機能は、完全に未知であってもよく、または、調査中の核酸および/または調査中の発現産物の一つ以上の機能は、既知であってもよく、この態様においては、本発明による方法は、さらなる機能の同定を目的としている。
【0011】
本発明の一つの特徴は、宿主細胞における核酸の発現の効果を調査する方法であって、(a)核酸に由来するRNAを、前記宿主細胞へ導入すること;および、(b)宿主細胞への当該RNAの導入の結果として生じる、宿主細胞に対する効果の調査、の段階を含む方法に関する。
【0012】
RNAの宿主細胞への導入の結果として生じる宿主細胞に対する効果は、核酸および/または発現産物の機能についての情報を提供することができる。
【0013】
したがって、本発明のもう一つの特徴は、生命体における核酸および/または核酸発現産物の機能を調査する方法であって(a)核酸および/または、発現産物の一部または全配列のコードに由来するRNAの宿主細胞への導入;(b)RNAの宿主細胞への導入の結果として生じる宿主細胞に対する効果の調査;(c)(b)のステップで決定された宿主細胞に対する効果に基づいた核酸および/または発現産物の一つ以上の機能の同定、の各ステップを含む方法に関する。
【0014】
本発明による方法は、特に殆ど時間消費しないこと、反応速度論的解析の可能なことおよび、殆ど全ての細胞系に適用可能なことに関して利点がある。特に、発現には、RNAが細胞の細胞質に入りさえすれば十分で、細胞核に入る必要はないため、調査中の核酸、たとえば遺伝子の導入は、DNA構造体の使用に比べ、RNAの形を取った方が利点がある。したがって、RNA転移は、形質転換される細胞の分裂活性に依存しない。さらに、RNAを用いて達成可能となる転換率は、比較的高く(Van Tendeloo,V.F.et al.(2001)Blood,98,49−56;Saeboe−Larssen,S.et al.(2002)J.Immunol.Methods,259,191−203)、得られるタンパク質量は、生理学的発現におけるその導入率に対応する。翻訳が完了したタンパク質は、数時間以内に検出可能であり、機能的に意味のあるものになるため、反応速度的な研究を非常に良好に行うことができる(Van Meirvenne,S.et al.(2002)Cancer Gene Ther.9,787−797)。
【0015】
細胞内へのRNA転移は、本発明によれば、実施例で実証された以下の特徴に基づいて、その後の機能的特徴づけに対して理想的である。
【0016】
・ RNA転移によって、腫瘍細胞や未転換一次細胞を含む、調査対象の全ての真核細胞型に、遺伝子産物を挿入できる(表1)。
・ 転換率は非常に高く、多くの細胞型に対して90%より大であり、したがって、選択する必要がない。
・ 発現レベルは、形質転換に用いるRNA量によって、非常に良好に滴定することができる。このようにして、細胞内の発現レベルの差異の効果をシミュレートすることが可能となる。
・ 形質転換後、同期翻訳が直ちに起こるため、年代順に生物学的余効を感度よく検出できる。
・ RNA形質転換は、トランスクリプトームに関して、形質転換細胞の方法依存変動を殆ど示さない。
【0017】
本発明による方法は、特に細胞および細胞株の転写調節因子、腫瘍遺伝子、潜在腫瘍遺伝子の異所性発現のシミュレーションに好適であり、したがって、それら遺伝子の機能的特徴付けに好適である。このような場合、宿主細胞に挿入されるRNAは、腫瘍遺伝子または、潜在腫瘍遺伝由来である。
【0018】
本発明による方法のひとつの実施の態様において、宿主細胞に挿入されるRNAは、それをエンコードする核酸をクローニングし、そのクローン核酸の転写によって、好ましくは、インビトロ転写によって得られる。転写調節プロモーターは、RNAポリメラーゼのためのプロモーターならどれでも良い。特にRNAポリメラーゼの例としては、T7、T3およびSP6のRNAポリメラーゼがある。好ましくは、本発明によるインビトロでの転写は、T7またはSP6プロモーターによって調節される。
【0019】
特定の実施形態において、RNAをコードする核酸は、ポリアデニルカセットと併せて存在する。これらの実施形態においては、核酸の発現により、好ましくはインビトロの転写により得られるRNAは、定義された長さのポリアデニル化部分を有することが好ましい。ポリAストレッチは、好ましくは10から500長、より好ましくは30から300長、さらに好ましくは65から200長、特には100から150長のアデノシン残基を有する。特に好ましい実施形態においては、ポリAストレッチは、およそ120長のアデノシン残基を有する。
【0020】
本発明のさらなる実施形態において、宿主細胞に挿入するRNAは、その5'末端にてCap構造または調節配列を有し、これが宿主細胞での翻訳をプロモートする。
【0021】
本発明に係る方法の特に好ましい実施形態において、RNAを宿主細胞に導入することにより、RNAによりエンコードされた発現産物の発現が引き起こされる。本発明の特定の実施形態においては、発現産物はペプチド、タンパク質、酵素、抗体および抗原からなる群より選択することができる。
【0022】
本発明によれば、機能調査について、宿主細胞に導入されるRNAの量を制御することが可能になり、したがって宿主細胞において発現した発現産物の量を制御することが可能になる。
【0023】
特定の実施形態において、本発明の宿主細胞に導入されるRNAは、RNAライブラリーの一部である。本発明のこの態様は、宿主細胞への導入における宿主細胞に対する特定の影響を起こすことを可能にする又は宿主細胞の特定の機能を制御あるいは特定の機能に影響を与えることを可能にするRNAの発生について、異なる核酸または核酸の変異体(定義:RNAライブラリー)の群をスクリーニングする方法にも関係する。
【0024】
したがって、本発明のこの態様は、特定の効果または機能を有する核酸および/または核酸発現産物を見つける方法であって、(a)宿主細胞へのRNAの導入;(b)RNAの宿主細胞への導入の結果として生じる宿主細胞に対する効果の調査;(c)宿主細胞に対する効果を引き起こすRNAの同定の各ステップを含む方法に関する。この方法は、宿主細胞に対する効果を引き起こすRNAのヌクレオチド配列の決定をも含む。前記ヌクレオチド配列の決定は、公知の方法、特にオリジナル核酸のシークエンシングにより行うことができる。
【0025】
本発明に係る方法は、RNAの宿主細胞への導入および宿主細胞中のRNAの可能性ある発現の結果として生じる宿主細胞に対する効果の調査および同定を含む。
【0026】
RNAの宿主細胞への導入が、当該宿主細胞に対する効果を示すには多くの様式があることは当業者にとって明らかだろう。したがって、本発明に係る方法においては、RNAの宿主細胞への導入の結果として生じる効果を調査することを可能にする様々な方法がある。このような方法は、特に限定されないが、以下の一つ以上のパラメータ:細胞増殖、細胞死、細胞移動、細胞接着、薬物に対して改変された耐性または改変された感受性、細胞老化、特定分子の生産または生産抑制、mRNAの量、タンパク質産物の量、タンパク質活性レベル、タンパク質のリン酸化に対する効果、タンパク質または核酸のプロセッシングに対する効果、RNA安定性に対する効果、シグナル伝達または二次メッセンジャーに対する効果、の決定を含む。
【0027】
RNAの宿主細胞への導入の結果として生じる宿主細胞に対する効果の調査は、好ましくは当該宿主細胞の表現型および/または生化学的特徴の調査を含む。
【0028】
特定の実施形態においては、RNAの宿主細胞への導入の結果として生じる宿主細胞に対する効果の調査が、宿主細胞のタンパク質発現プロファイルの調査(プロテオーム解析)および/またはRNA発現プロファイルの調査(トランスクリプトーム解析)を含む。
【0029】
さらなる実施形態において、RNAの宿主細胞への導入の結果として生じる宿主細胞に対する効果の調査が、宿主細胞の代謝の調査を含む。この代謝の調査は、公知の方法にしたがった酵素反応からの基質または産物の集積調査を含むことが好ましい。
【0030】
他の実施形態において、RNAの宿主細胞への導入の結果として生じる宿主細胞に対する効果の調査が、遺伝子調節の調査を含む。
【0031】
本発明に係る方法の特に好ましい実施形態において、宿主細胞に対する効果の調査は、RNAが導入されていない細胞(転移コントロール)との比較、または機能と関係のないRNAを導入した細胞(ネガティブコントロール)、あるいは機能を仲介するRNAを導入した細胞(ポジティブコントロール)との比較で行う。これら細胞は、コントロールまたは基準としての役割を果たす。また、これら細胞は用いられる宿主細胞と一致していることが好ましい。
【0032】
サブトラクティブハイブリダイゼーション(subtractive hybridization)法は、RNAの宿主細胞への導入が宿主細胞のRNA発現プロファイルに対する作用を含むのか否か決定するために用いることができる。この実施形態においては、例えばコントロール細胞に存在するRNAを、着目しているRNAを挿入する宿主細胞により発現されたRNAから「差し引いた(subtracted)」としている。
【0033】
さらなる実施形態において、RNAの宿主細胞への導入の結果として生じる宿主細胞に対する効果の調査は、相補性解析を含む。この実施形態においては、宿主細胞に内在する遺伝子の機能を、RNAの宿主細胞への導入により置き換え、または増強させる。
【0034】
さらなる実施形態において、RNAの宿主細胞への導入の結果として生じる宿主細胞に対する効果の調査は、宿主細胞の表現型の特徴の調査、例えば形態学的、巨視的または微視的調査などの方法による調査を含む。
【0035】
また、宿主細胞は、RNAの宿主細胞への導入の結果として生じる宿主細胞に対する効果の調査の前に、さらなる処置または状態の変更をすることができる。これにより、ある実施形態においては、細胞の因子に対する効果を一時的に調査する。宿主細胞は、核酸および/または核酸発現産物の機能のさらなる決定のために、特定の生命体におけるインビボの状況下に組み込むこともできる。
【0036】
RNAの宿主細胞への導入の結果として生じる宿主細胞に対する効果の調査は、核酸および/または発現産物が有する、この核酸および/または発現産物が由来する生命体の中の一つ以上の機能を同定することを可能にする。
【0037】
本発明に係る方法は、着目する核酸および/または着目する発現産物に機能的に関連する一つ以上の細胞因子を同定することも可能にする。このような細胞因子の群には、着目するRNAの宿主細胞への導入に対する増加した発現を表示するものがあるだろう。もう一つのこのような細胞因子の群には、着目するRNAの宿主細胞への導入に対する減少した発現を表示するものがあるだろう。これにより、本発明は、細胞因子および着目する核酸および/または発現産物の間の機能的な関連性を同定する手段をも提供する。
【0038】
ある実施形態においては、宿主細胞が、調査する核酸および/または調査する発現産物と同様の生命体由来であることが好ましい、すなわち調査する核酸および/または調査する発現産物が宿主細胞と同種であることが好ましい。しかしながら、調査する核酸および/または調査する発現産物は、宿主細胞とは異種であることが可能である。
【0039】
着目する核酸および/または着目する発現産物は、少なくとも初めに、その核酸および/またはその発現産物を自然に発現することが知られている宿主細胞中で調査することが好ましい。
【0040】
しかしながら、ある実施形態においては、異種の核酸および/または発現産物を、宿主細胞、特に哺乳類細胞中で試験することが望まれることもある。これにより、例えば特定の配列、例えば真菌由来の配列が哺乳類の配列のための置換産物として機能する可能性があるか否かを決定することも可能である。もしそうである場合、この異種の配列は、例えば哺乳類の配列のための治療上の置換産物として用いることができる。
【0041】
さらに、本発明に係る方法において、核酸および/または発現産物の誘導体で一つ以上の機能について試験することが望まれることもある。これは、特定の核酸および/または特定の発現産物を改変すること、および改変された核酸および/または改変された発現産物がオリジナルの核酸および/またはオリジナルの発現産物の一つ以上の効果および/または機能を維持するか否かを試験することを可能にする。多くの場合において、オリジナルの核酸および/またはオリジナルの発現産物の一つ以上の効果および/または機能を維持する核酸および/または発現産物の誘導体を製造することが望まれる。一方で、それらはオリジナルの核酸および/またはオリジナルの発現産物の一つ以上の効果および/または機能を持たないものである。
【0042】
ある実施形態においては、高いスループットのシステムを、本発明に係る方法を行うために用いる。この態様の一つの実施態様において、システムは随意にコンピュータ化またはロボット化することができ、および多数のコンパートメントの使用を含むこともできる。特に好ましい実施形態においては、このコンパートメントをマルチウェルのプレートに配列させて、RNAおよび宿主細胞を多くのコンパートメントに配置させることが好ましい。
【0043】
RNAの宿主細胞への導入の結果として生じる宿主細胞に対する効果は、公知のマイクロアレイ技術および処理により調査することができる。
【0044】
(発明の詳細な説明)
本発明によれば、組み換え核酸の産物、細胞の培養、および核酸、特にRNAの細胞への導入、特にエレクトロポレーションおよびリポフェクションについて標準的な方法を使用することができる。酵素反応および精製技術は、製造者の指示または公知の手法により行う。
【0045】
「生命体」という用語は、本発明によれば、遺伝物質の増幅または伝達する能力を有し、かつ、植物および動物並びに微生物、例えばバクテリア、イースト、真菌およびウィルスを含む生物学上の単位に関する。
【0046】
「形質転換」という用語は、本発明によれば、一つ以上の核酸の生命体または細胞、または後者由来の細胞への導入に関する。
【0047】
「宿主細胞」という用語は、本発明によれば、外部の核酸、特にRNAを用いて形質の変換または転換が可能な細胞に関する。核酸は、宿主細胞に一つ以上のコピーにて存在することができ、およびある実施形態においては宿主細胞で発現される。
【0048】
本発明によれば、「宿主細胞」は、原核および真核細胞、特にヒトおよび動物細胞、植物細胞および微生物細胞を包含する。本発明で使用される特に好ましい宿主細胞は、真核細胞、特に哺乳類細胞、例えばヒト、マウス、ハムスター、ブタ、ヤギおよび霊長類由来の細胞である。これらの細胞は、多様な組織の型由来であり、および一次細胞および細胞株を包含することができる。
【0049】
宿主細胞は、ヒトの組織由来であることが好ましく、特に限定されないが、神経細胞、脳細胞、上皮細胞、結合組織細胞(例えば線維芽細胞、骨芽細胞)、血液細胞(例えば白血球、リンパ球、単球および好中球、特に樹状細胞、血小板)、感覚細胞、筋細胞、肺細胞、心臓細胞、肝細胞、皮膚細胞、膵臓細胞、乳腺細胞、腎臓細胞、腸細胞、胃細胞、結腸細胞、前立腺細胞、卵巣細胞、胚細胞および幹細胞が挙げられ、これらは多様な細胞型に分化する能力を有する。
【0050】
培養細胞株も使用することができる。本発明のある実施形態においては、例えば着目する核酸および/または着目する発現産物がこれら細胞でしか正常に発現しない場合に、部分的にまたは完全に分化した細胞を用いる。
【0051】
本発明によれば、「RNAの宿主細胞への導入の結果として生じる宿主細胞に対する効果」という用語は、RNAの宿主細胞への導入に起因する可能性のある、宿主細胞に対する知覚可能な効果のいずれも包含し、特に宿主細胞の成長、発達、増幅、遺伝的継承、生物学および/または表現型に対する効果を包含する。
【0052】
「表現型」または「表現型の特徴」という用語は、生命体または宿主細胞の外観に関する。表現型は、内的および外的な構造および機能の全てを含む。個々の発達の経過において、生命体およびそれに由来する細胞の表現型は変化することができる。生命体およびそれに由来する細胞の外的特徴は、その遺伝的情報(遺伝子型)により決定されるが、表現型はどの遺伝子が実際に発現されるのか(発現)に依存する。
【0053】
「生化学」または「生化学的特徴」という用語は、生命体またはそれに由来する細胞における基質の代謝および伝達に関する。
【0054】
本発明によれば、「生命体またはそれに由来する細胞における核酸および/または核酸発現産物の機能」という用語は、生命体またはそれに由来する細胞、好ましくはその核酸および/またはその核酸発現産物が由来する生命体またはそれに由来する細胞における、核酸および/または核酸発現産物の機能的役割に関する。ある実施形態においては、この用語は、発ガン現象における核酸および/または核酸発現産物の機能的役割に関する。核酸に関しては、この用語は、特に生命体またはそれに由来する細胞における核酸の発現または発現の欠如から生じる効果に関する。
【0055】
本発明によれば、「核酸および/または核酸発現産物の機能」という用語は、核酸および/または核酸発現産物が一つだけまたはいくつかの機能を有することを意味することがあり、後者の場合、一つ以上の機能が本発明に係る方法により調査される。
【0056】
「遺伝子」という用語は、本発明によれば、特定の核酸配列に関し、この配列は一つ以上の細胞の産物の産生、および/または一つ以上の細胞間または細胞内の機能の獲得に関与する。特にこの用語は、特定のタンパク質、または機能性または構造性RNA分子をコードするDNAセグメントに関する。
【0057】
本発明の核酸は、リボ核酸(RNA)またはRNAの産生に使用することができるデオキシリボ核酸(DNA)であることが好ましい。本発明によれば、核酸は、ゲノムDNA、cDNA、mRNA、組み換え技術により作り出された分子および化学的に合成された分子を包含する。また、本発明によれば、核酸は、一本鎖または二本鎖、および直鎖状または共有結合的に環状に閉じた分子の形態をとることができる。
【0058】
また、本発明の核酸、特にRNAは、核酸間でのホモロジー(相同性)がある程度あれば、すなわちこの核酸がヌクレオチド配列において直接的または相補的に十分な一致があれば、この他の核酸から誘導される。さらに、核酸は、一番目の核酸(当該核酸)が二番目の核酸(他の核酸)の誘導体である場合にも、この他の核酸から誘導される。また、「核酸から誘導されたRNA」という用語は、特にRNAが核酸またはそれから誘導される核酸の転写産物であること、および/または翻訳によって、そのRNAが由来する核酸の転写および翻訳に基づいて生じるタンパク質発現産物と、同様の配列を有するタンパク質発現産物を結果的にもたらす配列を有することを意味する。
【0059】
本発明によれば、核酸の「誘導体」は、その核酸において一つまたは複数のヌクレオチドの置換、削除および/または追加が存在することを意味する。さらに、「誘導体」という用語は、核酸の核酸塩基、糖またはリン酸に対する化学的誘導体も包含する。また、「誘導体」という用語は、非天然にて発生するヌクレオチドおよびヌクレオチド誘導体を含む核酸をも包含する。
【0060】
本発明によれば、核酸は単離することができる。本発明によれば、「単離された核酸」は、核酸が、(i)例えばポリメラーゼ連鎖反応(PCR)によりインビトロで増幅されること、(ii)クローニングにより組み換えにて作り出されること、(iii)例えば切断およびゲル電気泳動による分離により精製されること、または(iv)例えば化学合成により合成されることを意味する。単離された核酸は、組み換えDNA技術による操作が可能な核酸である。
【0061】
核酸は、二つの配列が互いにハイブリダイズ可能であり、安定した二重鎖を形成できる場合、他方の核酸と「相補的」となる。なお、このハイブリダイズは、好ましくはポリヌクレオチド間での特定のハイブリダイズを許容する条件(ストリンジェントな条件)下で生じるものである。ストリンジェントな条件は、例えばMolecular Cloning:A Laboratory Manual,J.Sambrook et al.,Publ.,2nd edition,Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989またはCurrent Protocols in Molecular Biology,F.M.Ausubel et al.,Publ.,John Wiley&Sons,Inc.,New Yorkに記載されており、および例えば65℃でハイブリダイゼーションバッファ(3.5×SSC、0.02%Ficoll、0.02%ポリビニルピロリドン、0.02%ウシ血清アルブミン、2.5mM NaH2PO4(pH7)、0.5%SDS,2mM EDTA)でのハイブリダイズに関する。SSCは、0.15Mの塩化ナトリウム/0.15Mのクエン酸ナトリウム、pH7である。ハイブリダイズ後、DNAが移動してきた膜を、例えば2×SSCで室温にて洗浄し、その後0.1−0.5×SSC/0.1×SDSで68℃までの温度で洗浄する。
【0062】
本発明によれば、相補的核酸は、少なくとも70%、少なくとも80%、少なくとも90%、および好ましくは少なくとも95%、少なくとも98%、または少なくとも99%のヌクレオチドの同一性を有する。
【0063】
本発明の方法により核酸と機能的に連結される発現制御配列または調節配列は、その核酸に対して同種または異種であってもよい。コード配列および調節配列は、それらが共有結合する場合、コード配列の転写または翻訳が調節配列の制御下または影響下にて行われるように、「機能的」に連結する。コード配列が機能的タンパク質に翻訳される場合、調節配列のこのコード配列との機能的連結した状態でのこの調節配列の提示が、コード配列での読み枠移動を引き起こしたり、または所望するタンパク質またはペプチドへのコード配列を翻訳不能としないように、コード配列の転写を導く。
【0064】
本発明によれば、「発現制御配列」または「調節配列」は、プロモーター、リボソーム結合配列および他の制御要素を包含し、この制御要素は遺伝子の転写または誘導されたRNAの翻訳を制御するものである。本発明のある実施形態においては、発現制御配列は制御され得る。調節配列の正確な構造は、種に依存して、または細胞型に依存して異なるが、転写または翻訳の開始に関与する、一般的に5'−未転写の配列、および5'および3'−未翻訳の配列、例えばTATAボックス、キャップ配列、CAAT配列などを包含する。特に、5'−未翻訳の調節配列は、機能的に結合した遺伝子の転写制御のためのプロモーター配列を有するプロモーター領域を包含する。調節配列は、エンハンサー配列または上流のアクチベータ配列も包含し得る。
【0065】
さらに、本発明の核酸は、この核酸によりエンコードされたタンパク質またはポリペプチドを特定の標的部位にガイドするポリペプチドをコードする他の核酸と連結することができる。特に、本発明の核酸は、エンコードされたタンパク質またはポリペプチドの転座または安定化を宿主細胞の細胞膜またはこの細胞の特定のオルガネラにおいてコンパートメント化された部分の細胞膜の上で引き起こすポリペプチドをコードする他の核酸と連結することができる。
【0066】
本発明の好ましい実施形態において、クローン核酸は、この核酸の発現を制御するプロモーターを随意に含むベクター内に存在する。「ベクター」という用語は、その最も一般的な意味において用いられ、例えば核酸を原核および/または真核細胞に挿入して、必要に応じてゲノムに組み込むことを可能にする、核酸のためのいかなる中間的な媒体(vehicle)をも包含する。このようなベクターは、細胞中で複製および/または発現されることが好ましい。中間的な媒体(vehicle)は、例えばエレクトロポレーション、微小粒子衝撃法(microprojectile bombardment)、リポソーム投与(liposomal administration)、アグロバクテリアを用いた転移、またはDNAまたはRNAウィルスを介した挿入における用途に適応させることができる。
【0067】
組み換えRNAは、本発明によれば、インビトロでの適切なDNA鋳型の転写により産生することができる。さらに、組み換えRNAは、配列の安定化、キャッピングおよびポリアデニル化によって修飾することができる。
【0068】
本発明によれば、「発現」という用語は、その最も一般的な意味において用いられ、RNAの産生、またはRNAおよびタンパク質/ポリペプチドの産生を包含する。また、発現は、核酸の部分的な発現も包含する。さらに、発現は過渡的であっても、安定的であってもよい。RNAに関して、「発現」という用語は、特にタンパク質/ポリペプチドの産生に関する。
【0069】
発現産物、特にタンパク質あるいはポリペプチド、または本発明の意味におけるアミノ酸配列の「誘導体」は、アミノ酸挿入変異体、アミノ酸削除変異体、および/またはアミノ酸置換変異体を包含する。
【0070】
アミノ酸挿入変異体は、アミノ−および/またはカルボキシ−末端融合物、および特定のアミノ酸配列の中での個々のまたはいくつかのアミノ酸の挿入物を包含する。挿入を伴うアミノ酸配列変異体において、一つ以上のアミノ酸残基は、アミノ酸配列の所定の位置で挿入されるが、結果産物の好適なスクリーニングによるランダム挿入も可能である。アミノ酸削除変異体は、配列から一つ以上のアミノ酸を除くことにより特徴付けられる。アミノ酸置換変異体は、配列の少なくとも一つの残基を除き、かつ、その場所にもう一つの残基を挿入することにより特徴付けられる。この修飾は、同種のタンパク質間またはポリペプチド間で保存されていない、アミノ酸配列の位置でなされていることが好ましい。アミノ酸は、同様の特性、例えば疎水性、親水性、電気陰性度、側鎖の容積などを有する他のもので置換すること(同類置換)が好ましい。同類置換は、例えばあるアミノ酸をもう一つのアミノ酸で置換することに関する。以下にその置換対象となるアミノ酸と同じ群に挙げる。
【0071】
1.小さな脂肪族の、非極性あるいはわずかに極性の残基:Ala、Ser、Thr(Pro、Gly)
2.負のチャージを帯びた残基およびそれらのアミド:Asn、Asp、Glu、Gln
3.正のチャージを帯びた残基:His、Arg、Lys
4.大きな脂肪族の、非極性残基:Met、Leu、Ile、Val(Cys)
5.大きな芳香族残基:Phe、Tyr、Trp
【0072】
3残基が、タンパク質構造において特定の役割を果たすため、括弧内にて示されている。Glyは、側鎖を有さない唯一の残基であり、これにより鎖に柔軟性を付与する。Proは、独特の配置を有しており、これによりポリペプチド構造に大きな変化を導くことができる。Cysは、分子内、または他のCysとの分子間のジスルフィド架橋を形成することができる。
【0073】
上述したアミノ酸変異体は、組み換えDNA操作を用いて容易に作り出すことができる。全部公知または部分的に公知の配列を有するDNAの所定部位での置換変異を挿入するための技術は、公知であり、例えばM13突然変異生成が挙げられる。タンパク質産生のための置換、挿入または欠失といったDNA配列の操作は、例えばSambrook et al.(1989)に詳細に記載されている。
【0074】
本発明によれば、タンパク質またはポリペプチドの「誘導体」は、タンパク質またはポリペプチドに関連する糖質、脂質および/またはタンパク質またはポリペプチドに関連する単一のまたは複数の分子の置換、欠失および/または付加も包含する。さらに、「誘導体」は、タンパク質またはポリペプチドの機能的、化学的な等価物の全てに当てはまる。
【0075】
本発明によれば、発現産物の部分またはフラグメントは、好ましくは、それが由来する発現産物の機能的な特性を有する。この機能的な特性は、抗生物質との相互作用、他のポリペプチドまたはタンパク質との相互作用、核酸の選択的結合および酵素活性を包含する。
【0076】
本発明によれば、核酸の部分またはフラグメントは、好ましくは上記で定義したような発現産物の部分またはフラグメントをコードする核酸の部分に関する。
【0077】
本発明によれば、核酸の細胞への導入のために、多様な方法を用いることができる。このような方法は、核酸−CaPO4沈殿物の形質転換、DEAEに関連する核酸の形質転換、着目する核酸を担持したウィルスを用いた形質転換または感染、エレクトロポレーション、リポソーム媒介の形質転換などを包含する。核酸を特定細胞へ案内することは、ある実施形態においては好ましい。このような実施形態において、核酸を細胞に送達するために用いるキャリア(例えば、レトロウィルスまたはリポソーム)は、結合した案内分子(directing molecule)を備える。例えば、抗体のような分子は、標的細胞の表面膜タンパク質に特異的であったり、あるいは標的細胞上のレセプターのためのリガンドであったりするが、核酸キャリアに取り込んだり、または結合させることができる。核酸をリポソームにより導入することが望まれる場合には、エンドサイトーシスに関連する表面膜タンパク質に結合するタンパク質をリポソーム製剤に取り込んで、標的および/または吸収を可能とすることができる。このようなタンパク質は、カプシドタンパク質またはそのフラグメントを包含する。このタンパク質は、特定の細胞型、内在するタンパク質に対する抗体、細胞内部位にガイドするタンパク質などに特異的である。
【0078】
本発明によれば、「相補性解析」という用語は、特定の遺伝子を削除または変異させて、その遺伝子の役割にてもはや十分に機能しないようにした後に、核酸を生命体または細胞に導入したときにこの生命体または細胞にて得られる効果の調査に関する。相補的な遺伝子またはその発現産物が、欠損遺伝子または変異遺伝子を伴った生命体または細胞に導入されると、好ましくはそれらはその遺伝子の機能を再生する能力を有する。
【0079】
本発明によれば、「マイクロアレイ」という用語は、生物学上の材料、例えば核酸、ペプチドまたはタンパク質を高密度にて適用し、結合するために用いられるデバイスに関する。マイクロアレイは、多量のサンプルを制御された間隔にて適用することを許容し、それらがパラレルな実験に使用することができる固体キャリア支持体からなり、またはそのような固体キャリア支持体を有する。
【0080】
本発明によれば、「高密度」という用語は、様々なサンプル、例えば核酸サンプルを高密度で保持し、これにより多くの質問に対して回答を与えることを可能にするアレイに関連して用いられる。
【0081】
本発明は、以下に示す図面および実施例により詳細に説明されるが、これら図面および実施例は説明としてのみ用いられるのであり、限定として理解されるべきではない。当業者は、本記載および本実施例に基づいてさらなる実施形態に拡げることができるものであり、これらも本発明により保護される。
【図面の簡単な説明】
【0082】
【図1】用いられたeGFPベクターの表示−a.pGEM3Z eGFPテール、b.pSTI eGFPテールである。これらのベクターは、続くクローニングのための、開始ベクターとして用いられた。
【図2】786−0細胞における、形質転換後24時間のeGFP IVT RNAの翻訳を検出するためのウエスタンブロット。eGFP−pGEM3ZテールIVT RNAの20μgが、200Vかつ250μFで形質転換された。一次抗体として、濃度1μg/mlのウサギ抗GFPを用いた。二次抗体として、濃度0.5μg/mlのヤギ抗ウサギHRPを用いた。マウス抗ERK2抗体を濃度0.2μg/mlのローディングコントロールとして用いた。ここで、二次抗体として、ヤギ抗マウスHRPを、1/5000希釈で用いられた。
【図3】1×107個の786−0細胞に、eGFP IVT RNAの20μgが形質転換された後24時間の、又は、RNAなしでコントロール形質転換後の、FACSドットブロットの表示。
【図4】eGFPの平均蛍光強度を測定して、エレクトロポレーション効率に対する細胞カウントの影響の測定。eGFP−pGEM3ZテールIVT RNAの20μgが、上記数のK562細胞に形質転換された。細胞は形質転換後24時間培養された。平均蛍光強度は、FACS−Caliburを使用して測定した。
【図5】エレクトロポレーション効率や平均蛍光強度に対するエレクトロポレーション条件の影響の測定。MCF7細胞に、eGFP IVT RNAを20μg形質転換した。細胞は形質転換後24時間培養された。平均蛍光強度は、FACS−Caliburを使用して測定した。
【図6】eGFP IVT RNA又は、2dGFP IVT RNAの20μgを用いた、2×107個の786−0細胞の形質転換後、転写およびタンパク質量に対する時間の影響の測定。a.定量RT−PCR分析に基づく、相対転写量の測定。正規化は非形質転換細胞に対して行った。cDNA量の違いを考慮するために、18s特異的PCRを行い、その値で正規化に用いた。b.FACS−Caliburを使用したeGFPの平均蛍光強度の測定。
【図7】eGFP IVT RNAを用いた5×106個のMCF−7細胞の形質転換後24時間のeGFPタンパク質レベルに対するRNA量の影響の測定。a.非形質転換コントロールと比較したヒストグラムブロットでの蛍光強度の測定の表示。b.平均蛍光強度の表示。
【図8】2×107個の786−0細胞形質転換後24時間のHIF1−α、LDHA、VEGF、および、Glut−1の相対転写量のリアルタイムRT−PCRによる定量測定。eGFP形質転換コントロールと比較した相対発現の表示。
【図9】VHL IVT RNAの20μgを用いた2×107個の786−0細胞の形質転換後、VEGFおよびLDHAの相対転写量の時間に対する変化。非形質転換細胞と比較して、相対転写量を測定した。正規化は18s rRNAによって行った。
【図10】SYT、SSX2、SYT−SSX1、SYT−SSX2、または、コントロールとして、eGFP IVT RNAを用いた、1×107個のMCF−7細胞の形質転換後の相対転写量の表示。正規化はHPRTによって成立した。eGFP IVT RNAを用いて形質転換された細胞と比較して、相対転写量を測定した。
【図11】乳房、肺、腎臓、卵巣、睾丸、肝臓、リンパ節、胸腺、脾臓および副腎の正常組織と比較した、滑膜肉腫における、BMP7、EPHA4およびCOL5A1の相対転写量の表示。正規化はHPRTによって行った。BMP−7およびCOL5A1の発現の測定のため、試験された肉腫は、骨肉種であった。一方、EPHA4の調査のため、滑膜肉腫を試験し、SYT−SSX1またはSYT−SSX2に対して、ポジティブではなかった。転位の欠損によって、病理の診断は不確かなものである。
【図12】機能によってグループ化され、かつ、eGFPを用いた形質転換コントロールと比較して、SSX2、SYT−SSX1またはSYT−SSX2を用いたMCF−7細胞の形質転換体に対して、5%の有意性基準で、少なくとも21倍の差異で発現された、遺伝子一覧表。記載のない値は非有意調節領域に位置する、つまり<21の値である。表に示された値は、いずれの場合も、2が基底の指数である。つまり、結果数は差異発現のファクターを表している。
【図13】図12に同じ。
【図14】図12に同じ。
【図15】図12に同じ。
【図16】図12に同じ。
【図17】図12に同じ。
【図18】図12に同じ。
【図19】図12に同じ。
【図20】図12に同じ。
【図21】図12に同じ。
【図22】図12に同じ。
【図23】図12に同じ。
【図24】図12に同じ。
【図25】eGFP形質転換細胞と比較して、リアルタイムRT−PCRを用いて測定された、相対転写量の表示。正規化はHPRTによって行った。生物学的上のtriplicates(biological triplicates)間の標準偏差が記載されている。
【実施例】
【0083】
実施例1:ベクターの生産
【0084】
ベクターは、それぞれのコード配列を基礎ベクターであるpGEM3Zベクター(Invitrogen,San Diego,CA)または、pCMV Scriptベクター(Stratagene,Amsterdam,NL)にクローンして、生産した。
【0085】
pGEM3Zベクターは、開始コドンの前にSP6プロモーターを持ち、そこからインビトロで転写可能である。pCMV Scriptベクターは同じ部位にT7プロモーターを有する。
【0086】
これらの基礎ベクターから出発し、大幅な組み換えで導入する。これは、ポリアデニル化された、インビトロで転写された(IVT)RNAの生産のために必須である。
【0087】
120塩基のポリA伸長部分を、pGEM3ZベクターにXhoI切断部位を解して導入した。ポリA伸長直後、制限部位を導入し、これにより、ポリA伸長がリニア化する。後述するように、pGEM3Zテールベクターをこのベクターにしている(図1a)。
【0088】
120塩基対のポリA伸長部分をpCMV ScriptベクターにSacI切断部位の後ろで導入した。後述するように、pSTIテールベクターをこのベクターとしている(図1b)。
【0089】
eGFPまたは、各組織から増幅された遺伝子を、精製および各ベクターによる制限後、連結し、E.coli MRF1-に形質転換した。得られたプラスミドを配列決定し、その配列を分析した。
実施例2:IVT RNAの生産
【0090】
IVT RNAの生産の第一段階は、各プラスミドのリニア化を含む。用いられる制限酵素は、pGEM3Zテールベクターについては、BpiIであり、pSTIテールベクターについてはSapIだった。両方とも、ベクターの中を一度だけ、つまり挿入テールの後ろが切断された。リニア化後、酵素は、フェノール−クロロホルム沈殿によって不活性化し、除去した。この為に、フェノールおよびクロロホルムの混合物の等容量(isovolume)が制限チャージに追加され、完全に混合された。10000×gの手短な遠心分離により、下部の有機層およびDNAを含む上部の水層への分離が生じた。後者を新しい反応槽に移した。その後、水層を等容量の純クロロホルムで混合し、全てのフェノール残留物を除去した。遠心分離後、水層を除去し、2倍等容量のエタノールとpH4.5の10%v/v3M酢酸ナトリウムと−20℃にて加えて2時間沈殿させた。
【0091】
DNAは、10000×gで、4℃の遠心分離で45分間沈降し、塩を除去するために70%エタノールで洗浄し、適量のRNAse−free水中で取っておいた。リニア化が成功かつ完了であることを確かめるために、ゲル電気泳動を行なった。DNA濃度は、260nmの光度測定法で、測定した。DNA純度の測定のため、さらに280nmの吸光度を測定してOD260/280比を得た。
【0092】
10μgのリニア化DNAをインビトロ転写に用いた。これについては、4/5のdGTPにさらにキャップ構造を付与した40μlのdNTPと、l0μlの10×バッファと、20μlのdTTと、10μlのT7またはSP6ポリメラーゼを、37℃で2時間でインキュベートした。ポリメラーゼは、転写されるORFから5'方向に位置する、T7またはSP6認識配列に結合し、相補的RNA鎖を合成する。
【0093】
IVT RNAは、MegaClear Kitを用いて精製した。これについては、RNAをシリカ膜に最適に結合させるために必要な塩を含んだ結合緩衝濃縮物に、そのRNAを取っておいた。エタノールの添加により、RNA水和殻から水が除去した。その混合物を、シリカカラムにロードし、10000×gで2分遠心分離させた。RNAをそのカラムに結合させ、一方不純物、たとえば、酵素残留物を洗い落とした。何度かの洗浄後、精製されたRNAを溶出した。溶出緩衝液を、95℃まで予熱して効率的に溶出した。
【0094】
質管理および定量化は、ゲル電気泳動および光度測定によって、行った。
実施例3:細胞へのエレクトロポレーション
【0095】
エレクトロポレーション法の原理は、短い電流パルスによって、細胞の膜電位を乱すことに基づいている。外部刺激による膜電位の変化は、次の式で表される。
【数1】
Vmは膜電位であり、fは細胞外の電界分布に対する細胞の影響を表す形状因子である。fEexは適用された電界を表し、rは細胞半径を表し、φは外部から適用された電界に対する角度を表す。fは、通常1.5として与えられるが、それは他の多くの因子に依存している。適用された電界が、細胞膜の容量を越えるとき、すなわち、ΔVmが1Vとして与えられる閾値ΔVsより大きいとき、細胞へのエレクトロポレーションは成功する(Kinosita,K.,Jr.and Tsong,T.Y.(1977)Nature,268,438−441)。二分子膜としての細胞膜の構造は、真核細胞に共通の特徴であるため、この値は異なった細胞株に対して、ほとんど変動を示さない。
【0096】
絶縁破壊の膜電位になると、一時的に親水性穿孔が形成され、そこを通じて水が細胞内に浸透して、たとえば核酸等の分子の細胞内への移動が生じる(Weaver,J.C.(1995)Methods Mol.Biol.55,3−28;Neumann,E.et al.(1999)Bioelectrochem.Bioenerg.48,3−16)。
【0097】
エレクトロポレーションの前に、用いる接着細胞をセミコンフルエントの状態まで培養し、PBSで洗浄後、トリプシンを用いて、細胞培養フラスコから取り外した。細胞を10%FCS(ウシ胎仔血清)添加培地に移し、500×gで8分遠心分離した。ペレットを血清フリー培地X−Vivoのなかで再懸濁し、再び、500×gで8分遠心分離した。この洗浄操作を続くエレクトロポレーションに影響するFCSの残留物を除去するためにさらに2回行なった。
【0098】
洗浄後、細胞を250μlにて所望の細胞密度になるように調整し、エレクトロポレーションキュベットに移し、氷上で静置した。適量のインビトロで転写されたRNAを添加し、十分攪拌した後、エレクトロポレーションを、200Vかつ250μFで行った。その後、細胞を速やかに適当な培養液に移し、37℃、5%CO2下でインキュベートした。
実施例4:実験条件の確立と実験系の特徴付け
【0099】
eGFP−pGEM3Zテール発現ベクターを、実験条件の確立のために用いた。5'に位置するRNAポリメラーゼプロモーターSP6によって、インビトロでのRNAの発現が可能になる。
【0100】
インビトロで転写されたeGFP RNAは、以下のように、多様な細胞型の形質転換効率の試験のために用いた。形質転換効率は、eGFPの蛍光強度の測定によりFACS−Caliburを用いて、測定した。細胞内のRNA量は、定量RT−PCRによって測定した。腫瘍細胞株に90%より高い効率で形質転換可能であるだけでなく、初代樹状細胞にも70から80%の効率で形質転換可能であることが示された(図2および3並びに表1)。
【0101】
【表1】
表1:20μgのeGFP IVT RNAを用いた5×106個の細胞の形質転換後に測定した多様な細胞の形質転換効率。形質転換後24時間にFACS−Caliburを用いての蛍光強度の測定。
形質転換条件:200Vかつ250μFのエレクトロポレーション。
【0102】
さらに、形質転換効率による細胞の影響は2×106から2×107細胞の範囲において除外することができた(図4)。また、エレクトロポレーション条件は、試験範囲内での形質転換効率に対して決定的な影響を及ぼさない(図5)。
【0103】
RNAの安定性は24時間にわたって立証された(図6a)。一方、その半減期に依存するが48から72時間にわたって、細胞内でタンパク質を検出することができる(図6b)。
【0104】
転写されたRNA量および得られるタンパク質量の間には直接的な依存性があることが示すことができた(図7)。
【0105】
この方法に因る分子変化は、cDNAマイクロアレイを用いて測定された。プロテアソーム関連遺伝子、ヒートショック遺伝子およびアポトーシス関連遺伝子の分子変化は、8時間後および24時間後に見られただけであった(表2)。
【0106】
【表2】
表2:非形質転換コントロール細胞と比較して、20μgのeGFP IVT RNAで形質転換した2×107個の786−0細胞における十分に調節された遺伝子の数。変化は、>2または<0.5の調節で有意であるだけである。
【0107】
siRNAの使用の後に記載されてきたような(Elbashir,S.M.et al.(2001)Nature,411,494−498;Caplen,N.J.et al.(2001)Proc.Natl.Acad.Sci.U.S.A.98,9742−9747)インターフェロン系の活性はない。
実施例5:モデル遺伝子のIVT RNA形質転換
【0108】
さらなる、IVT RNAを用いた細胞の形質転換の可能性および限界の詳細な検討のため、多種のモデル遺伝子を異種発現したIVT RNAとして細胞に導入した。よく特徴づけされた遺伝子HIF1−α(低酸素症誘導因子)およびVHL(von−Hippel Lindau)をモデル遺伝子として用いた。これらの遺伝子のコード領域を睾丸組織由来のcDNAを用いて増幅し、eGFPのために記載されたベクター系へクローンした。
【0109】
HIF1−αやVHLについては、多種な標的遺伝子が記載されており、それらの発現は、不完全細胞株の形質転換後に測定した。HIF1−α転写因子の形質転換後、標的遺伝子であるLDHA(乳酸脱水素酵素A)、VEGF(血管内皮増殖因子1α)や、Glut−1(グルコーストランスポーター1)の発現増加を検出した(図8)。それらの誘導の程度は、転移HIF1−α IVT RNA量に基づいて滴定することができる。HIF1−α転写因子の転移によって、その標的遺伝子の強固な調節が可能である。用いられたIVT RNA量の滴定を通じて、HIF1−αにより誘導された遺伝子の転写量を滴定することもできる。
【0110】
転写因子の分子効果は、ほとんど直接かつ即座に検出することができる。したがって、HIF1はその標的遺伝子のプロモーター領域の共通配列に直接結合し(Semenza,G.L.(2000)Genes Dev.14,1983−1991;Semenza,G.L.(2000)J.Appl.Physiol 88,1474−1480)、速やかにそれらの転写を開始させる。間接的かつ副次的効果もIVT RNA転移により、明らかにできるか否かを調査するために、HIF1−αよりも上位であるpVHL(von−Hippel−Lidau)タンパク質を第2モデル遺伝子として選択した。pVHLは、ユビキチンリガーゼE3複合体に属し、HIF1のαサブユニットの分解を引き起こす。したがって、VHLの発現増加は、細胞内のHIF1濃度の減少を引き起こし、HIF1標的遺伝子の発現の減少に関係している。VEGFおよびLDHA発現における時間に依存した減少は、VHLでの細胞の形質転換後、明確に実証された(図9)。
実施例6:例としてSYT−SSX1およびSYT−SSX2を用いた推定発癌転写物の従来未知の機能の同定。
【0111】
RNA転移技術は、腫瘍生物学に対する遺伝子の関連性の迅速な解読を可能にする。証明のために普遍的に発現する遺伝子である、SYT遺伝子およびCG遺伝子であり、睾丸や腫瘍でのみ発現するSSX1またはSSX2遺伝子の融合産物を選択した。これらの遺伝子を選択した理由としては、一方においては、滑膜肉腫において90%を超える(Sreekantaiah,C.et al.(1994)Am.J.Pathol.144,1121−1134)転座t(X;18)(p11.2;q11.2)(Clark,J.et al.(1994)Nat.Genet.7,502−508;Crew,A.J.(1995)EMBO J.14,2333−2340)の頻度であり、この転座頻度は、腫瘍生物学の関連性を示唆し、したがって、発癌能を示唆するからである。また、他方では、この融合産物が転写活性化QPGYドメインを含有していることから、転座の結果として、直接分子変化が期待されるためである。腫瘍の発達及び体系における、転座の分子効果及びそれらの重要性は未だ大部分が知られていない。SYT−SSX1及びSYT−SSX2 IVT RNAの形質転換体により誘導された分子変化をAffymetrix社のオリゴヌクレオチドマイクロアレイを用いて、解析した。SYT−SSX1及びSYT−SSX2を用いた細胞に形質転換後、多数の遺伝子発現の増加が検出できた。形質転換後、検出できた調節遺伝子としては、第1成長因子、神経遺伝子、腫瘍遺伝子または腫瘍関連遺伝子が挙げられ、また、これらは、細胞周期の調節またはシグナル伝達に関与する(図12〜24)。これら遺伝子のいくつかの差次的発現は、定量RT−PCRにより、実証された(図25;図10)。滑膜肉腫における、これら遺伝子の機能的関連性を検出するために、さらにそれらの発現を、正常組織と比較して滑膜肉腫において測定した(図11)。
【0112】
実施例としてSYT−SSX1およびSYT−SSX2の形質転換体を用いて、RNA転移およびそれに続くAffymetrix社のオリゴヌクレオチドマイクロアレイを用いた分子変化の測定により、発癌現象における遺伝子の有意性を決定することが可能であることを明らかに示すことができた。形質転換細胞の直接的および間接的な標的遺伝子は、決定され、遺伝子機能の我々の理解に寄与する。
【0113】
IVT RNAの薬物動態的特徴と併せて、IVT RNAはシグナル伝達カスケードを解読するための良好な基礎を構成する。様々な時点での遺伝子発現プロファイルの構成および適切なソフトウェアを用いたそれらの解析は、定義された遺伝子群の単位を記述することにより、結果として起こる活性化された調節遺伝子のカスケードを発見することを可能にする。このように、RNA転位技術を用いて、発癌現象において、今まで殆ど特徴付けられていない遺伝子の機能的有意性を解明することが可能となる。さらに、新規に同定された標的遺伝子の転移により、形質転換遺伝子の直接的および間接的な効果を発見することが可能となる。
【特許請求の範囲】
【請求項1】
生命体における核酸および/または核酸発現産物の機能を調査する方法であって、
(a)発現産物の一部または全配列の核酸および/またはコードに由来するRNAの宿主細胞への導入、
(b)前記RNAの宿主細胞への導入の結果として生じる宿主細胞に対する効果の調査、および
(c)(b)のステップで決定された宿主細胞における効果に基づいた核酸および/または発現産物の一つ以上の機能の同定、
の各ステップを含む方法。
【請求項2】
前記RNAがそれをエンコードする核酸をクローニングし、インビトロでの転写によって得られることを特徴とする請求項1に記載の方法。
【請求項3】
前記インビトロでの転写は、T7、T3またはSP6プロモーターによって調節されることを特徴とする請求項2に記載の方法。
【請求項4】
前記核酸はポリアデニルカセットと組み合わせて存在することを特徴とする請求項2または3に記載の方法。
【請求項5】
前記RNAが定義された長さのポリアデニル化部分を有すること特徴とする請求項1から4のいずれかに記載の方法。
【請求項6】
前記RNAがCap構造を有することを特徴とする請求項1から5のいずれかに記載の方法。
【請求項7】
前記RNAの宿主細胞への導入は、前記RNAによりエンコードされた発現産物の発現を引き起こすことを特徴とする請求項1から6のいずれかに記載の方法。
【請求項8】
前記発現産物が、ペプチド、タンパク質、酵素、ホルモン、抗体からなる群から選択されることを特徴とする請求項7に記載の方法。
【請求項9】
前記宿主細胞に対する効果の調査が、前記宿主細胞の表現型および/または生化学的特徴の調査を含むことを特徴とする請求項1から8のいずれかに記載の方法。
【請求項10】
前記宿主細胞に対する効果の調査が、前記宿主細胞のタンパク質発現プロファイルの調査(プロテオーム解析)を含むことを特徴とする請求項1から9のいずれかに記載の方法。
【請求項11】
前記宿主細胞に対する効果の調査が、前記宿主細胞の前記RNA発現プロファイルの調査(トランスクリプトーム解析)を含むことを特徴とする、請求項1から10のいずれかに記載の方法。
【請求項12】
前記宿主細胞に対する効果の調査が、前記宿主細胞の代謝の調査を含むことを特徴とする請求項1から11のいずれかに記載の方法。
【請求項13】
前記代謝の調査が、酵素反応の基質または産物の集積の調査を含むことを特徴とする、請求項12に記載の方法。
【請求項14】
前記宿主細胞に対する効果の調査が、遺伝子調節の調査を含むことを特徴とする請求項1から13のいずれかに記載の方法。
【請求項15】
前記宿主細胞に対する効果が、RNAが導入されていない細胞と比較して調査されることを特徴とする請求項1から14のいずれかに記載の方法。
【請求項16】
前記宿主細胞に対する効果の調査が、相補性解析を含むことを特徴とする、請求項1から15のいずれかに記載の方法。
【請求項17】
前記宿主細胞が、原生または真核細胞であることを特徴とする、請求項1から16のいずれかに記載の方法。
【請求項18】
前記宿主細胞へのRNA導入が、エレクトロポレーション法またはリポソームを用いて、行なわれることを特徴とする、請求項1から17のいずれかに記載の方法。
【請求項19】
前記宿主細胞に対する効果の調査が、高密度マイクロアレイ処理を用いて行なわれることを特徴とする請求項1から18のいずれかに記載の方法。
【請求項1】
生命体における核酸および/または核酸発現産物の機能を調査する方法であって、
(a)発現産物の一部または全配列の核酸および/またはコードに由来するRNAの宿主細胞への導入、
(b)前記RNAの宿主細胞への導入の結果として生じる宿主細胞に対する効果の調査、および
(c)(b)のステップで決定された宿主細胞における効果に基づいた核酸および/または発現産物の一つ以上の機能の同定、
の各ステップを含む方法。
【請求項2】
前記RNAがそれをエンコードする核酸をクローニングし、インビトロでの転写によって得られることを特徴とする請求項1に記載の方法。
【請求項3】
前記インビトロでの転写は、T7、T3またはSP6プロモーターによって調節されることを特徴とする請求項2に記載の方法。
【請求項4】
前記核酸はポリアデニルカセットと組み合わせて存在することを特徴とする請求項2または3に記載の方法。
【請求項5】
前記RNAが定義された長さのポリアデニル化部分を有すること特徴とする請求項1から4のいずれかに記載の方法。
【請求項6】
前記RNAがCap構造を有することを特徴とする請求項1から5のいずれかに記載の方法。
【請求項7】
前記RNAの宿主細胞への導入は、前記RNAによりエンコードされた発現産物の発現を引き起こすことを特徴とする請求項1から6のいずれかに記載の方法。
【請求項8】
前記発現産物が、ペプチド、タンパク質、酵素、ホルモン、抗体からなる群から選択されることを特徴とする請求項7に記載の方法。
【請求項9】
前記宿主細胞に対する効果の調査が、前記宿主細胞の表現型および/または生化学的特徴の調査を含むことを特徴とする請求項1から8のいずれかに記載の方法。
【請求項10】
前記宿主細胞に対する効果の調査が、前記宿主細胞のタンパク質発現プロファイルの調査(プロテオーム解析)を含むことを特徴とする請求項1から9のいずれかに記載の方法。
【請求項11】
前記宿主細胞に対する効果の調査が、前記宿主細胞の前記RNA発現プロファイルの調査(トランスクリプトーム解析)を含むことを特徴とする、請求項1から10のいずれかに記載の方法。
【請求項12】
前記宿主細胞に対する効果の調査が、前記宿主細胞の代謝の調査を含むことを特徴とする請求項1から11のいずれかに記載の方法。
【請求項13】
前記代謝の調査が、酵素反応の基質または産物の集積の調査を含むことを特徴とする、請求項12に記載の方法。
【請求項14】
前記宿主細胞に対する効果の調査が、遺伝子調節の調査を含むことを特徴とする請求項1から13のいずれかに記載の方法。
【請求項15】
前記宿主細胞に対する効果が、RNAが導入されていない細胞と比較して調査されることを特徴とする請求項1から14のいずれかに記載の方法。
【請求項16】
前記宿主細胞に対する効果の調査が、相補性解析を含むことを特徴とする、請求項1から15のいずれかに記載の方法。
【請求項17】
前記宿主細胞が、原生または真核細胞であることを特徴とする、請求項1から16のいずれかに記載の方法。
【請求項18】
前記宿主細胞へのRNA導入が、エレクトロポレーション法またはリポソームを用いて、行なわれることを特徴とする、請求項1から17のいずれかに記載の方法。
【請求項19】
前記宿主細胞に対する効果の調査が、高密度マイクロアレイ処理を用いて行なわれることを特徴とする請求項1から18のいずれかに記載の方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【公開番号】特開2012−179049(P2012−179049A)
【公開日】平成24年9月20日(2012.9.20)
【国際特許分類】
【出願番号】特願2012−62748(P2012−62748)
【出願日】平成24年3月19日(2012.3.19)
【分割の表示】特願2007−548736(P2007−548736)の分割
【原出願日】平成17年12月22日(2005.12.22)
【出願人】(509146023)バイオエヌテック アーゲー (3)
【氏名又は名称原語表記】BioNTech AG
【住所又は居所原語表記】Hoelderlinstrasse 8 55131 Mainz Germany
【Fターム(参考)】
【公開日】平成24年9月20日(2012.9.20)
【国際特許分類】
【出願日】平成24年3月19日(2012.3.19)
【分割の表示】特願2007−548736(P2007−548736)の分割
【原出願日】平成17年12月22日(2005.12.22)
【出願人】(509146023)バイオエヌテック アーゲー (3)
【氏名又は名称原語表記】BioNTech AG
【住所又は居所原語表記】Hoelderlinstrasse 8 55131 Mainz Germany
【Fターム(参考)】
[ Back to top ]